Embed Size (px)
Citation preview

DECREASED EXPRESSION OF SMOOTH MUSCLE �-ACTIN RESULTS INDECREASED CONTRACTILE FUNCTION OF THE MOUSE BLADDER
R. A. ZIMMERMAN, J. J. TOMASEK, J. McRAE, C. J. HAAKSMA, R. J. SCHWARTZ, H. K. LIN,R. L. COWAN, A. N. JONES AND B. P. KROPP*
From the Departments of Urology (RAZ, HKL, RLC, ANJ, BPK) and Cell Biology (JJT, JM, CJH), University of Oklahoma HealthSciences Center, Oklahoma City, Oklahoma, and Department of Molecular and Cellular Biology (RJS), Baylor College of Medicine,
Houston, Texas
ABSTRACT
Purpose: Smooth muscle �-actin (SM�A) is an important actin isoform for functional contrac-tility in the mouse bladder. Alterations in the expression of SM�A have been associated with avariety of bladder pathological conditions. Recently, a SM�A-null mouse was generated anddifferences in vascular tone and contractility were observed between wild-type and SM�A-nullmice suggesting alterations in function of vascular smooth muscle. We used SM�A-null mice toexplore the hypothesis that SM�A is necessary for normal bladder function.
Materials and Methods: Reverse transcriptase polymerase chain reaction, Western blottingand immunohistochemical staining were used to confirm the absence of SM�A transcript andprotein in the bladder of SM�A-null mice. In vitro bladder contractility compared betweenbladder rings harvested from wild-type and SM�A-null mice was determined by force meas-urement following electrical field stimulation (EFS), and exposure to chemical agonists andantagonists including KCl, carbachol, atropine and tetrodotoxin. Resulting force generationprofiles for each tissue and agent were analyzed.
Results: There was no detectable SM�A transcript and protein expression in the bladder ofSM�A-null mice. Nine wild-type and 9 SM�A-null mice were used in the contractility study.Bladders from SM�A-null mice generated significantly less force than wild-type mice in responseto EFS after KCl. Similarly, bladders from SM�A-null mice generated less force than wild-typemice in response to pretreatment EFS, and EFS after carbachol and atropine, although thedifference was not significant. Surprisingly, the bladders in SM�A-null mice appeared to functionnormally and showed no gross or histological abnormalities.
Conclusions: SM�A appears to be necessary for the bladder to be able to generate normal levelsof contractile force. No functional deficits were observed in the bladders of these animals but nostress was placed on these bladders. To our knowledge this study represents the first report todemonstrate the importance of expression of SM�A in force generation in the bladder.
KEY WORDS: muscle, smooth; actins, bladder
Actins are highly conserved proteins that serve as compo-nents of the contractile apparatus and cytoskeleton of smoothmuscle (SM) cells. Functionally, they contribute to the con-tractility of these cells.1 In higher mammals the actin familyis composed of at least 6 different isoforms, including skele-tal, cardiac and SM �-actin (SM�A), SM �-actin, and cyto-plasmic � and �-actins.2 There is a high degree of sequenceconservation among the different isoforms suggesting thatthis multigene family arose from a common ancestral gene.3The various actin isoforms are conserved across species andconsidered tissue specific.2 Conservation of the various actinisoforms implies distinct functions in different tissues or celltypes.4
SM cells express SM�A and SM �-actin in addition tocytoplasmic � and �-actins.5 The dominant isoform in the SMof mammalian visceral organs is SM �-actin,6–8 while SM�Ais the major isoform in vascular smooth muscle.2, 9 In variousmammalian bladders the �-actin isoforms contribute nearly25% to 45%, while �-actin isoforms contribute 37% to 63% ofthe total actin (see table).10–13
The SM actins serve as molecular markers for the differ-
entiation, maturation and function of SM cells.8, 14 In earlybladder development SM�A marks the transition from un-differentiated mesenchyme to synthetic SM cells. Similarly,SM �-actin marks the transition from synthetic to immatureSM cells. Coordinate expression of both actins continues asthe SM cell matures.8, 15 It is reported that there is minimal(7%) increase in SM�A expression from 3-day-old to adult ratbladder.11
The bladder acts in storage and expulsion of urine. Duringfilling stages the bladder increases to large volumes whilemaintaining low intravesical pressures due to the visco-elastic properties. During emptying, coordinated SM contrac-tion occurs. Physiological and pharmacological changes havebeen documented in normal and physiologically altered blad-ders, mainly obstructed and obstruction-induced hypertro-phied bladders. Bladder outlet obstruction results in de-creased bladder emptying, decreased contractile response ofSM tissue, and decreased bladder compliance and capacity.16Actin isoforms, in particular SM�A, have been shown toserve as markers for normal and pathological SM and fibro-blastic tissues.6 Several studies have demonstrated changesin actin isoforms in obstructed and hypertrophied bladders(see table).7, 17 The altered expression of actin isoforms inresponse to pathology suggests functional significance.
Functional differences have been demonstrated using a
* Correspondence and requests for reprints: Department of Urol-ogy, University of Oklahoma Health Sciences Center, 920 Stanton L.Young Blvd., Oklahoma City, Oklahoma 73104 (e-mail: [email protected]).
0022-5347/04/1724-1667/0 Vol. 172, 1667–1672, October 2004THE JOURNAL OF UROLOGY® Printed in U.S.A.Copyright © 2004 by AMERICAN UROLOGICAL ASSOCIATION DOI: 10.1097/01.ju.0000139874.48574.1b
1667

SM�A-null mouse. These animals have reduced vascular con-tractility, tone and blood flow, although there is no apparentfeeding or reproductive compromise.18 Surprisingly, skeletal�-actin is expressed in the vascular SM cells of these SM�A-null mice. These results suggest that SM�A is important inSM cell contractility and that skeletal �-actin may be able topartially compensate for the loss of SM�A in the vascular SMcells of these mice.
As SM�A represents a significant proportion of the totalactins expressed in most mammalian bladders, we hypothe-sized that bladder contractility and function may be compro-mised in SM�A-null mice. We investigated what histological,contractile and functional changes occur in the SM�A-nullmouse to further understand the role of these contractileproteins in the normal and pathological bladder.
MATERIALS AND METHODS
SM �-actin null mice. Adult SM�A-null and wild-type micewere obtained from a breeding colony at University of Okla-homa Health Sciences Center. The SM�A-null mice weregenerated by Dr. Robert J. Schwartz (Department of Molec-ular and Cellular Biology, Baylor College of Medicine, Hous-ton, Texas).18 Duplicate samples of genomic DNA were ob-tained for genotyping from tail snips, quantified byspectrophotometry and analyzed by polymerase chain reac-tion (PCR). To perform PCR amplification, an aliquot (2 �l) ofgenomic DNA (�100 ng) was added to 0.5 �M each of eitherwild-type or SM�A-null specific primers, 0.2 mM deoxynucle-otide triphosphate and 0.74 units Taq DNA polymerase in 1XPromega buffer B (Promega, Madison, Wisconsin) and 3.0mM MgCl2 in a 25 �l reaction. Temperature was cycledthrough 94C for 2.5 minutes initially, and then at 94C for 45seconds, 54C for 45 seconds and 72C for 1 minute for a totalof 30 cycles with a 7-minutes 72C terminal extension. PCRproducts were electrophoresed on 1.5% agarose gels andstained with ethidium bromide.Reverse transcriptase polymerase chain reaction (RT-PCR)
analysis. PCR analysis was performed to determine the lev-els of SM�A and glyceraldehyde-3-phosphate dehydrogenase(GAPDH) in SM�A-null and wild-type mouse bladders. Du-plicate frozen bladder specimens (approximately 200 mg)were frozen in liquid nitrogen, crushed and RNA extracted byguanidinium hydrochloride.19 Isolated total RNA (10 �g) wasreverse transcribed to a first-strand cDNA library using theSuperscript II RT kit (Invitrogen, Carlsbad, California). PCRamplification was performed using 33 ng cDNA in 1� Pro-mega buffer, 15 mM MgCl2, 0.2 mM deoxynucleotide triphos-phate, 0.5 �M oligonucleotide primers and 0.7 units TaqDNA polymerase in a 20 �l reaction. SM�A specific primersand GAPDH specific primers were used. Temperature was
cycled through 94C for 1.5 minutes initially, and then at 94Cfor 45 seconds, 52C for 45 seconds and 72C for 1 minute forthe appropriate number of cycles. PCR products were elec-trophoresed on 1.5% agarose gels and stained with ethidiumbromide.Western immunoblot analysis. Western immunoblotting
was performed in duplicate to determine the levels of totalactin and SM�A in SM�A-null and wild-type mouse blad-ders. Bladder was harvested and total cellular proteins weresolubilized by homogenizing the tissue in 2� sample buffer �2% Dulbecco’s modified Eagle’s medium followed by boilingfor 3 minutes. The concentration of total cellular protein wasdetermined and 10 �g of protein were electrophoresed on a12.5% sodium dodecyl sulfate gel. Separated proteins weretransferred to polyvinylidene fluoride membranes (micronseparations). Membranes were blocked for 1 hour in blockingbuffer (1� TTBS buffer [0.3 M NaCl, 20 mM Tris/Cl, pH 7.8,0.1% v/v Tween-20 and 0.01% NaN3] � 2% casein) followedby incubation for 1 hour at room temperature (RT) with amouse anti-SM�A monoclonal antibody or a rabbit polyclonalanti-actin antibody followed by a goat anti-mouse or a goatanti-rabbit alkaline phosphatase (Sigma, St. Louis, Missouri)for 1 hour at RT and subsequently developed with Immun-Star chemiluminescence substrate (Bio-Rad) for 5 minutes atRT. X-ray film was exposed to chemiluminescence reactionfor 5 to 120 seconds at 25C and developed.Immunohistochemical staining. Immunohistochemical
staining for SM�A and general histology was performed onSM�A-null, heterozygous and wild-type mouse bladders.Bladders were fixed with 4% paraformaldehyde in 0.1 Msodium phosphate buffer (pH 7.4), dehydrated and embeddedin paraffin. Immunohistochemical staining for SM�A wasperformed using an avidin-biotin complex method. Briefly, 5�m sections were incubated in 10% goat serum blockingnonspecific binding followed by incubation with mouse anti-SM�A antibody conjugated to fluorescein (FITC). Sectionswere incubated in rabbit anti-FITC antibody (MolecularProbes, Eugene, Oregon) followed by biotinylated goat-anti-rabbit IgG (Vector Labs, Burlingame, California) and treatedwith avidin-biotin-peroxidase complex (Vector Labs). Colorwas developed in the presence of Vector Black Alkaline Phos-phatase Substrate (Vector Labs) and examined using a pho-tomicroscope.In vitro contractility. Adult wild-type and SM�A-null mice
were killed using CO2 asphyxiation and the bladders wereremoved promptly. Investigators were blinded to genotype ofthe individual animals. Bladder rings (bladders with domeand trigone removed) were suspended in 22 ml tissue baths(Radnoti) and bathed in oxygenated modified Krebs physio-logical salt solution (mmol/l NaCl 134, KCl 3.4, KH2PO4 1.3,
Previously reported actin isoform contributions of various species, ages and pathological states of mammalian bladders
References Species (tissue)Mean � SD
% �-Actin % �-Actin % �-Actin*
Matthew et al12 Mouse (bladder, wild-type) 44 � 3.5 10 � 1.1 46 � 2.3Skalli et al6 Rat (bladder) 33 � 2 16 � 1 51 � 3Malmqvist et al13 Rat (bladder 10 days) 40.9 � 2.6 18.8 � 1.1 40.3 � 2.9
Rat (bladder 10 days obstructed) 24.3 � 2.3 19.6 � 2.8 56.3 � 2.7Rat (bladder 7 wks) 43.1 � 1.7 19.6 � 0.9 37.4 � 1.0Rat (bladder 7 wks obstructed) 30.7 � 1.0 16.2 � 1.0 53.8 � 1.1Rat (bladder 7 wks) 42.0 � 0.7 20.5 � 1.0 37.9 � 1.0Rat (bladder 7 wks unobstructed) 35.5 � 1.0 20.3 � 0.9 44.2 � 1.1
Eddinger and Murphy11 Rat (3 days) 30 44Rat (21 days) 31 46Rat (adult) 37 40
Kim et al7 Rabbit (bladder normal) 23.7 � 2.2 21.3 � 4.0 55.0 � 4.3Rabbit (bladder obstructed) 22.8 � 3.9 13.4 � 3.6 63.8 � 5.3
Skalli et al6 Human (bladder normal) 34 � 1 11 � 1 55 � 1Malmqvist et al13 Human (bladder normal) 32.8 � 1.7 24.8 � 1.3 42.4 � 1.0
Human (bladder obstructed) 38.0 � 1.6 19.3 � 2.2 42.7 � 1.5* Represents �-smooth muscle actin plus �-cytoplasmic actin.
SMOOTH MUSCLE �-ACTIN IN BLADDER CONTRACTILITY1668
MgSO4 0.6, CaCl2 2.8, NaHCO3 16, glucose, 7.7) at pH of 7.4and 37C. Bladder rings were attached with 4-zero silk ties toforce displacement transducers (Kent Scientific Corp., Tor-rington, Connecticut) and changes in isometric tension wererecorded real time on a computer based data collection sys-tem (Strawberry Tree v4.01, Sunnyvale, California). An ini-tial passive force (tissue tension) of 1 gm20 was applied 3times at 15-minute intervals. This tension stretched thestrips to approximately 2.5 times the original length as de-fined by Zderic et al to provide an adequate length-tensionrelationship.21 Tissues were allowed to equilibrate for 1 hour,during which the Krebs solution was changed and replacedwith fresh buffer every 15 minutes. Tissues were subjected toelectrical field stimulation (EFS) with parameters of 100volts, 1.0 millisecond in duration, and 5 second trains at 5,10, 20, 30 and 40 Hz with 5 minutes between each stimula-tion. Bladder rings were exposed sequentially to potassiumchloride (2.7 mM), carbachol (1 �M), atropine (1 �M) andtetrodotoxin (3 �M) (Sigma). Tissue was rinsed and allowedto equilibrate with fresh Krebs solution for at least 30 min-utes after each treatment. Following each pharmacologicaltreatment, EFS stimulation was repeated. Frequency-dependent response curves normalized to tissue weight weregenerated, and data were analyzed for each treatment withrespect to total grams of tension generated and as a percent-age of response to carbachol.Statistical analysis. Data are represented as mean � SEM.
Statistical analysis and significance were determined usingthe Student t test with p �0.05 considered statistically sig-nificant and a rejection of the null hypothesis.
RESULTS
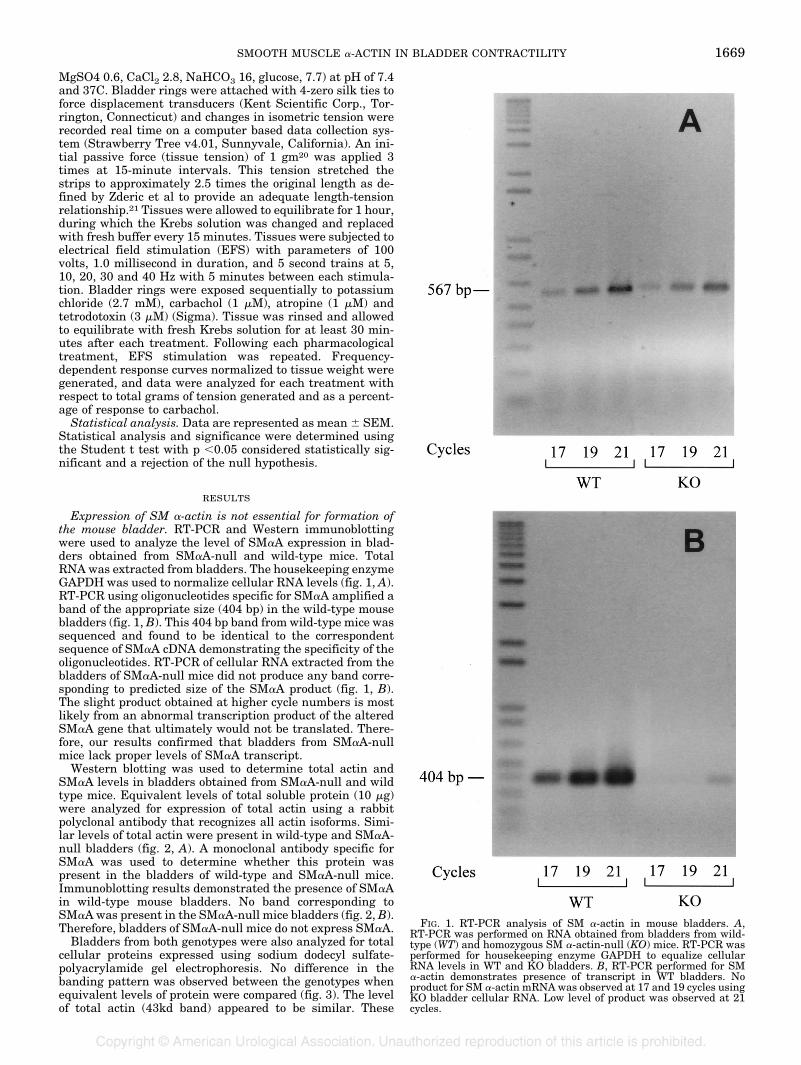
Expression of SM �-actin is not essential for formation ofthe mouse bladder. RT-PCR and Western immunoblottingwere used to analyze the level of SM�A expression in blad-ders obtained from SM�A-null and wild-type mice. TotalRNA was extracted from bladders. The housekeeping enzymeGAPDH was used to normalize cellular RNA levels (fig. 1, A).RT-PCR using oligonucleotides specific for SM�A amplified aband of the appropriate size (404 bp) in the wild-type mousebladders (fig. 1, B). This 404 bp band from wild-type mice wassequenced and found to be identical to the correspondentsequence of SM�A cDNA demonstrating the specificity of theoligonucleotides. RT-PCR of cellular RNA extracted from thebladders of SM�A-null mice did not produce any band corre-sponding to predicted size of the SM�A product (fig. 1, B).The slight product obtained at higher cycle numbers is mostlikely from an abnormal transcription product of the alteredSM�A gene that ultimately would not be translated. There-fore, our results confirmed that bladders from SM�A-nullmice lack proper levels of SM�A transcript.
Western blotting was used to determine total actin andSM�A levels in bladders obtained from SM�A-null and wildtype mice. Equivalent levels of total soluble protein (10 �g)were analyzed for expression of total actin using a rabbitpolyclonal antibody that recognizes all actin isoforms. Simi-lar levels of total actin were present in wild-type and SM�A-null bladders (fig. 2, A). A monoclonal antibody specific forSM�A was used to determine whether this protein waspresent in the bladders of wild-type and SM�A-null mice.Immunoblotting results demonstrated the presence of SM�Ain wild-type mouse bladders. No band corresponding toSM�Awas present in the SM�A-null mice bladders (fig. 2, B).Therefore, bladders of SM�A-null mice do not express SM�A.
Bladders from both genotypes were also analyzed for totalcellular proteins expressed using sodium dodecyl sulfate-polyacrylamide gel electrophoresis. No difference in thebanding pattern was observed between the genotypes whenequivalent levels of protein were compared (fig. 3). The levelof total actin (43kd band) appeared to be similar. These
FIG. 1. RT-PCR analysis of SM �-actin in mouse bladders. A,RT-PCR was performed on RNA obtained from bladders from wild-type (WT) and homozygous SM �-actin-null (KO) mice. RT-PCR wasperformed for housekeeping enzyme GAPDH to equalize cellularRNA levels in WT and KO bladders. B, RT-PCR performed for SM�-actin demonstrates presence of transcript in WT bladders. Noproduct for SM �-actin mRNAwas observed at 17 and 19 cycles usingKO bladder cellular RNA. Low level of product was observed at 21cycles.
SMOOTH MUSCLE �-ACTIN IN BLADDER CONTRACTILITY 1669

results suggest that there are no major differences in pro-teins expressed, except for lack of SM�A, in SM�A-null andwild-type mouse bladders.
Immunohistochemical staining was performed to examinethe distribution of SM�A in the bladders. Bladder and vas-cular smooth muscle cells from wild-type mice stained withthe anti-SM�A antibody (fig. 4, B). An adjacent sectionshowed no staining in the absence of primary FITC conju-gated anti-SM�A antibody (fig. 4, A). No staining with theanti-SM�A antibody was observed in the bladders fromSM�A-null mice (fig. 4, C). Bladders from SM�A-null andwild-type mice had a similar histological appearance (fig. 4).Also, no urogenital disorders, such as hydronephrosis or hy-droureter, were observed in the SM�A-null mice. These re-sults suggest that lack of SM�A expression does not result inany obvious structural defects in the mouse bladder.SM �-actin is essential for contractile function of the mouse
bladder. Bladders from 9 adult (older than 72 days) wild-typeand 9 SM�A-null mice were subjected to contractility exper-
iments. Bladders were weighed after contractility assays. Nosignificant weight differences were observed between wild-type and SM�A-null mouse bladders (1.6 � 0.01 and 1.4 �0.01 mg, respectively). Frequency-dependent responses wereobserved upon subjecting bladder rings to EFS after phar-macological treatments (fig. 5).
Upon exposure to EFS before any pharmacological treat-ment wild-type bladders displayed a trend to generate moreforce than SM�A-null mouse bladders, although these differ-ences were just beyond the limit of significance (fig. 5, A).This finding may be reflective of the power of the study andmay be significant with an increased number of animals.Both bladder genotypes responded in a frequency-dependentmanner.
A significant difference in force generation between the
FIG. 2. Western immunoblot analysis of actin expression in mousebladders. A, western immunoblot analysis performed for total actindemonstrates similar levels in wild-type (WT) and SM �-actin-null(KO) bladders. B, western immunoblot analysis performed for SM�-actin reveals expression in WT but not KO bladders.
FIG. 3. Sodium dodecyl sulfate-polyacrylamide gel electrophoresiswas performed on total protein from wild-type (WT) and SM �-actin-null (KO) bladders. No difference was observed between WT and KObladders. Molecular weight standards are illustrated in left lane.
FIG. 4. Immunocytochemical staining of mouse bladders for SM�-actin. A, wild-type bladder smooth muscle and vascular smoothmuscle cells stain for SM �-actin. B, SM �-actin null bladder andvascular smooth muscle cells do not stain for SM �-actin. C, wild-type bladder smooth muscle and vascular smooth muscle cells do notstain if primary anti-SM �-actin antibody is not present duringstaining procedure. Staining present in urothelium in this controlsimilar to wild-type and in SM �-actin-null bladders demonstratesnonspecific background staining. Reduced from �100.
SMOOTH MUSCLE �-ACTIN IN BLADDER CONTRACTILITY1670
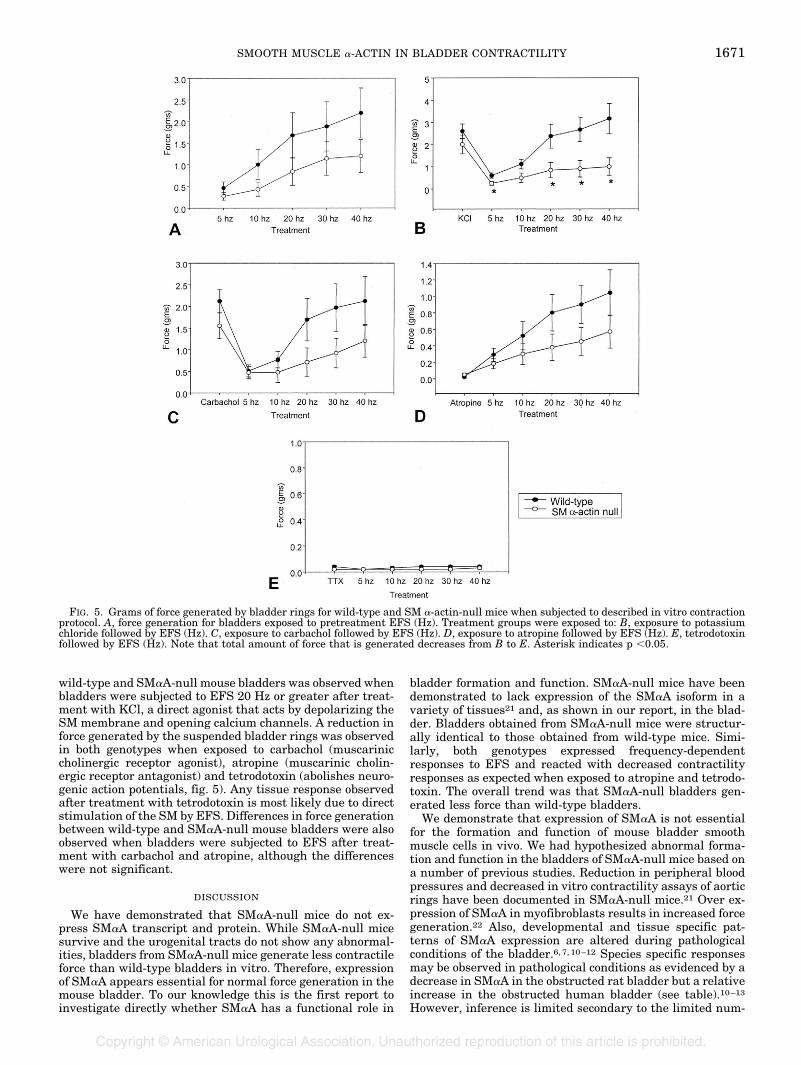
wild-type and SM�A-null mouse bladders was observed whenbladders were subjected to EFS 20 Hz or greater after treat-ment with KCl, a direct agonist that acts by depolarizing theSM membrane and opening calcium channels. A reduction inforce generated by the suspended bladder rings was observedin both genotypes when exposed to carbachol (muscariniccholinergic receptor agonist), atropine (muscarinic cholin-ergic receptor antagonist) and tetrodotoxin (abolishes neuro-genic action potentials, fig. 5). Any tissue response observedafter treatment with tetrodotoxin is most likely due to directstimulation of the SM by EFS. Differences in force generationbetween wild-type and SM�A-null mouse bladders were alsoobserved when bladders were subjected to EFS after treat-ment with carbachol and atropine, although the differenceswere not significant.
DISCUSSION
We have demonstrated that SM�A-null mice do not ex-press SM�A transcript and protein. While SM�A-null micesurvive and the urogenital tracts do not show any abnormal-ities, bladders from SM�A-null mice generate less contractileforce than wild-type bladders in vitro. Therefore, expressionof SM�A appears essential for normal force generation in themouse bladder. To our knowledge this is the first report toinvestigate directly whether SM�A has a functional role in
bladder formation and function. SM�A-null mice have beendemonstrated to lack expression of the SM�A isoform in avariety of tissues21 and, as shown in our report, in the blad-der. Bladders obtained from SM�A-null mice were structur-ally identical to those obtained from wild-type mice. Simi-larly, both genotypes expressed frequency-dependentresponses to EFS and reacted with decreased contractilityresponses as expected when exposed to atropine and tetrodo-toxin. The overall trend was that SM�A-null bladders gen-erated less force than wild-type bladders.
We demonstrate that expression of SM�A is not essentialfor the formation and function of mouse bladder smoothmuscle cells in vivo. We had hypothesized abnormal forma-tion and function in the bladders of SM�A-null mice based ona number of previous studies. Reduction in peripheral bloodpressures and decreased in vitro contractility assays of aorticrings have been documented in SM�A-null mice.21 Over ex-pression of SM�A in myofibroblasts results in increased forcegeneration.22 Also, developmental and tissue specific pat-terns of SM�A expression are altered during pathologicalconditions of the bladder.6, 7, 10–12 Species specific responsesmay be observed in pathological conditions as evidenced by adecrease in SM�A in the obstructed rat bladder but a relativeincrease in the obstructed human bladder (see table).10–13However, inference is limited secondary to the limited num-
FIG. 5. Grams of force generated by bladder rings for wild-type and SM �-actin-null mice when subjected to described in vitro contractionprotocol. A, force generation for bladders exposed to pretreatment EFS (Hz). Treatment groups were exposed to: B, exposure to potassiumchloride followed by EFS (Hz). C, exposure to carbachol followed by EFS (Hz). D, exposure to atropine followed by EFS (Hz). E, tetrodotoxinfollowed by EFS (Hz). Note that total amount of force that is generated decreases from B to E. Asterisk indicates p �0.05.
SMOOTH MUSCLE �-ACTIN IN BLADDER CONTRACTILITY 1671

ber of studies reporting on such trends. Therefore, it wasinteresting that loss of SM�A had no apparent effect on invivo bladder formation and function.
The decreased force generation that was observed in the invitro contractility studies most likely is a direct result of thelack of smooth muscle �-actin expression. Although the trendwas nonsignificant in many cases, increased power to thestudy may yield significance. It is plausible that the lack ofSM�A and the resulting compensatory mechanism(s) of thebladder may decrease the contractility enough to be demon-strated through in vitro studies but not enough to be offunctional consequence resulting in bladder distention, hy-dronephrosis or hydroureter.
Several possibilities as to why lack of expression of SM�Adid not result in abnormal in vivo bladder formation andfunction and a more dramatic alteration in force generationexist. One possibility is that other �-actin isoforms may beover expressed in the bladder to compensate for the loss ofSM�A. In the vascular smooth muscle cells of the SM�A-nullmouse skeletal �-actin expression is observed and suggestedto compensate for the lack of SM�A.21 However, it is unclearwhether this occurs in the bladder. Skeletal � and cardiac�-actins have not been described as being expressed in thebladder under normal or pathological conditions. Furtheranalysis of the expression patterns of these �-actin isoformsin the bladder needs to be carefully performed.
It is also possible that expression of SM �-actin is sufficientfor development and function of bladder smooth muscle cells.The ratio of �-actins-to-�-actins is different in vascular andbladder smooth muscle. SM�A is the predominant actin iso-form in vascular smooth muscle2, 9 and, in contrast, SM�-actin is highly expressed in the bladder (see table).10–13Interestingly, cardiac �-actin deficient mice die shortly afterbirth despite increased expression of SM�A in the heart.Ectopic expression of SM �-actin in these hearts using thecardiac �-myosin heavy chain promoter rescues these ani-mals although the hearts remain hypodynamic.23 These re-sults suggest functional differences between SM�A and�-actin isoforms and that only SM �-actin may be sufficientfor in vivo bladder formation and function.
CONCLUSIONS
SM�A expression correlates with bladder smooth musclecell differentiation and expression changes with variousbladder pathological conditions suggesting an important rolein bladder physiology. Expression of this actin isoform is notessential and cells must have a compensation mechanism forthe lack of SM�A expression. We have demonstrated theimportance of SM�A for normal contractile force generationin the mouse bladder. Future studies on the role of SM�A andcompensation mechanisms for lack of expression should helpin understanding the normal and pathological developmentand function of the bladder.
REFERENCES
1. Pollard, T. D. and Cooper, J. A.: Actin and actin-binding pro-teins; a critical evaluation of mechanisms and functions. AnnRev Biochem, 55: 987, 1986
2. Vandekerckhove, J. and Weber, K.: At least six different actinsare expressed in a higher mammal: an analysis based on theamino acid sequence of the amino-terminal tryptic peptide. JMol Biol, 126: 783, 1978
3. Vandekerckhove, J. and Weber, K.: Mammalian cytoplasmic ac-tins are the products of at least two genes and differ in primarystructure in at least 25 identified positions from skeletal mus-
cle actins. Proc Natl Acad Sci USA, 75: 1106, 19784. Rubenstein, P. A.: The functional importance of multiple actin
isoforms. BioEssays, 12: 309, 19905. Fatigati, V. and Murphy, R. A.: Actin and tropomyosin variants
in smooth muscles; dependence on tissue type. J Biol Chem,259: 14383, 1984
6. Skalli, O., Vandekerckhove, J. and Gabbiani, G.: Actin-isoformpattern as a marker of normal or pathological smooth muscleand fibroblastic tissues. Differentiation, 33: 232, 1987
7. Kim, Y. S., Wang, Z., Levin, R. M. and Chacko, S.: Alterations inthe expression of the �-cytoplasmic and the �-smooth muscleactins in the hypertrophied urinary bladder smooth muscle.Mol Cell Biochem, 131: 115, 1994
8. Rivas, D. A., Chancellor, M. B., Shupp-Byrne, D., Shenot, P. J.,McHugh, K. and McCue, P.: A molecular marker for the de-velopment of interstitial cystitis in a rat model: isoactin geneexpression. J Urol, 157: 1937, 1997
9. Vandekerckhove, J. and Weber, K.: The complete amino acidsequence of actins from bovine aorta, bovine heart, bovine fastskeletal muscle and rabbit slow skeletal muscle. Differentia-tion, 14: 123, 1979
10. Malmqvist, U., Arner, A. and Uvelius, B.: Contractile and cy-toskeletal proteins in smooth muscle during hypertrophy andits reversal. Am J Physiol, 260: C1085, 1991
11. Eddinger, T. J. and Murphy, R. A.: Developmental changes inactin and myosin heavy chain isoform expression in smoothmuscle. Arch Biochem Biophys, 284: 232, 1991
12. Matthew, J. D., Khromov, A. S., McDuffie, M. J., Somlyo, A. V.,Somlyo, A. P., Taniguchi, S. et al: Contractile properties andproteins of smooth muscles of a calponin knockout mouse.J Physiol, 529: 811, 2000
13. Malmqvist, U., Arner, A. and Uvelius, B.: Cytoskeletal and con-tractile proteins in detrusor smooth muscle from bladders withoutlet obstruction—a comparative study in rat and man.Scand J Urol Nephrol, 25: 261, 1991
14. McHugh, K., Crawford, K. and Lessard, J.: A comprehensive anal-ysis of the developmental and tissue specific expression of theisoactin multigene family in the rat. Dev Biol, 148: 442, 1991
15. Saga, H., Kimura, K., Hayashi, K., Gotow, T., Uchiyama, Y.,Momiyama, T. et al: Phenotype-dependent expression of�-smooth muscle actin in visceral smooth muscle cells. ExpCell Res, 181: 256, 1999
16. Levin, R. M., Longhurst, P. A., Monson, F. C., Kato, K. andWein,A. J.: Effect of bladder outlet obstruction on the morphology,physiology, and pharmacology of the bladder. Prostate, 3: 9,1990
17. Chiavegato, A., Scatena, M., Roelofs, M., Ferrarese, P., Pauletto,P., Passerini-Glazel, G. et al: Cytoskeletal and cytocontractileprotein composition of smooth muscle cells in developing andobstructed rabbit bladder. Exp Cell Res, 207: 310, 1993
18. Schildmeyer, L. A., Braun, R., Taffet, G., Debiasi, M., Burns,A. E., Bradley, A. et al: Impaired vascular contractility andblood pressure homeostasis in the smooth muscle �-actin nullmouse. FASEB J, 14: 2213, 2000
19. Chomczynski, P. and Sacchi, N.: Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction. Anal Biochem, 162: 156, 1987
20. Busser, B. W., Hammond, T. G., Bjorling, D. E. and Saban, R.:Lipopolysaccharide up-regulates bradykinin 1 receptors in theisolated mouse bladder. J Urol, 160: 2267, 1998
21. Zderic, S. A., Liu, G. H., Haab, J. P., Monson, F. C., Gong, C. andLevin, R. M.: What is the most accurate way to study the activeproperties of bladder smooth muscle? Pharmacology, 48: 380,1994
22. Hinz, B., Celetta, G., Tomasek, J. J., Gabbiani, G. andChaponnier, C.: Alpha-smooth muscle actin expression up-regulates fibroblast contractile activity. Mol Biol Cell, 12:2730, 2001
23. Kumar, A., Crawford, K., Close, L., Madison, M., Lorenz, J.,Doetschman, T. et al: Rescue of cardiac alpha-actin-deficientmice by enteric smooth muscle gamma-actin. Proc Natl AcadSci USA, 94: 4406, 1997
SMOOTH MUSCLE �-ACTIN IN BLADDER CONTRACTILITY1672