Embed Size (px)
Citation preview

UNIVERSIDAD DE COSTA RICA
SISTEMA DE ESTUDIOS DE POSGRADO
DIAGNÓSTICO DE INFECCIONES CAUSADAS
POR BRUCELLA CETI EN CETÁCEOS
Tesis sometida a la consideración de la Comisión del Programa de Estudios de
Posgrado en Microbiología, Parasitología y Química Clínica para optar al
grado de Magister Scientiae en Microbiología
MARÍA GABRIELA HERNÁNDEZ MORA
Ciudad Universitaria Rodrigo Facio, Costa Rica
2011

ii
DEDICATORIA
Este trabajo está dedicado a mi madre Olga Hernández Mora, a mi abuelita Cecilia Mora
Borloz, mi hermano Alexander Ramos Hernández, a Edgardo Ramos Roque y a toda mi
familia y mis amigos en especial a Rocío González, Adriana Villegas y Charles Manire
quienes con sus consejos y palabras de aliento apoyaron mi carrera y mi sueño de trabajar
con delfines.

iii
AGRADECIMIENTOS
Al Dr. Edgardo Moreno Robles por brindarme su apoyo incondicional, guía personal y
académica además de su interés y visión desde el inicio de las investigaciones con delfines.
A mis asesores Dra. Caterina Guzmán Verri y Dr. Esteban Chaves Olarte por brindarme su
apoyo y guía desinteresada durante estos años de investigación.
A todo el personal del Programa de Investigación en Enfermedades Tropicales, de la
Escuela Medicina Veterinaria (EMV) de la Universidad Nacional (UNA), especialmente a
la señoras Daphne Garita Vargas y Mayra Bolaños Quijano; así como a los señores
Fernando Retana y Adrián Zamora Arce y del Laboratorio de Bacteriología al Dr. Elías
Barquero Calvo, Lic. Adriana Blanco Rojas y Laura Orozco Sánchez por su apoyo
incondicional, aun en horario fuera de trabajo, para el éxito en las investigaciones que
dieron origen a este trabajo. Además a todos los estudiantes y amigos de grado y posgrado
que me ayudaron durante este proceso.
Al personal del Departamento de Patología de la EMV, UNA en especial al Dr. Juan
Alberto Morales Acuña, Dra. Rocío González Barrientos, Bernal Valerio Alfaro y Laura
Alvarado Guzmán por todo su apoyo durante los años de investigación con delfines en esa
unidad académica.
Al Dr. Charles Manire y Ruth Manire, Lynn Byrd, Petra Cunningam- Smith, David Smith,
y al Dr. Jim Gelsleichter y Stephanie Leggett del Hospital de Delfines y Ballenas y del
laboratorio de Inmunología Marina del Mote Marine Laboratory, Sarasota Florida por
permitirme ser parte de su equipo de trabajo, así como tener acceso a su banco de sueros y
utilizar las instalaciones para el desarrollo de las investigaciones que hicieron posible este
trabajo.
Al Dr. José-María Blasco, Gobierno de Aragón, España, por sus consejos durante la
estandarización de iELISA.

iv
Al Dr. Juan José Romero de la EMV, UNA, por su colaboración y recomendación del
paquete estadístico.
A la Dra. Frances Gulland del Marine Mammal Center, Sausalito, California, Estados
Unidos, por suministrar las muestras de foca.
También agradecer al Lic. José David Palacios, MSc. Andrea Montero y MSc. Damián
Martínez miembros de la Fundación KETO que gracias a su trabajo en las zonas costeras
del Pacífico costarricense han hecho posible la concientización de los habitantes de estas
zonas en la importancia del apoyo y atención profesional durante un evento de
encallamiento de cetáceos.
Agradecer a FEES CONARE que permitió financiar parcialmente este trabajo a través de
los proyectos “Evaluación del impacto de la neurobrucelosis en el fenómeno de
encallamiento de delfines” de la UNA y Universidad de Costa Rica (UCR) y "Elaboración
de un protocolo para la identificación de bacterias del género Brucella que representan un
riesgo para la salud pública, veterinaria y la vigilancia epidemiológica en Costa Rica" del
Tecnológico de Costa Rica (TEC), UNA y UCR. También fue financiado con fondos del
Consejo Nacional para Investigaciones Científicas y Tecnológicas (CONICIT) en el
proyecto “ Desarrollo y validación de ensayos diagnósticos con alta sensibilidad y
especificidad para el estudio epidemiológico de la brucelosis en cetáceos que habitan los
litorales marinos de Costa Rica.”
Por último agradecer al Ministerio de Ciencia y Tecnología (MICIT) y al CONICIT por el
financiamiento otorgado para cursar la Maestría del Sistema de Posgrado de la Facultad de
Microbiología de la UCR que concluye con la presentación de esta tesis.

v
Esta tesis fue aceptada por la Comisión del Programa de Estudios de Posgrado en
Microbiología, Parasitología y Química Clínica de la Universidad de Costa Rica, como
requisito parcial para optar al grado de Magister Scientiae en Microbiología
___________________________________________
Eugenia Corrales Aguilar
Representante de la Decana
Sistema de Estudios de Posgrado
___________________________________________
Edgardo Moreno Robles, PhD
Director de tesis
___________________________________________
Esteban Chaves Olarte, PhD
Asesor
___________________________________________
Caterina Guzmán Verri, PhD
Asesora
___________________________________________’
Fernando García Santamaría PhD
Director
Programa de Posgrado en Microbiología, Parasitología y Química Clínica
___________________________________________
Lic. María Gabriela Hernández Mora
Candidata

vi
INDICE
Página
DEDICATORIA ii
AGRADECIMIENTOS iii
HOJA DE APROBACIÓN v
INDICE vi
RESUMEN viii
ABSTRACT ix
LISTA DE CUADROS x
LISTA DE FIGURAS xi
LISTA DE ABREVIATURAS xiii
INTRODUCCIÓN 1
OBJETIVOS 9
MATERIALES Y MÉTODOS 10
Aislamientos de B.ceti 10
Identificación de patologías 10
Sueros de Odontocetos 11
Antígenos de Brucella y generación de anticuerpos y conjugados 12
Ensayos serológicos e inmunoquímicos 12
Análisis estadísticos 15
RESULTADOS 17
DISCUSIÓN 24
CONCLUSIONES 32
REFERENCIAS 34
APÉNDICE
Publicaciones generadas de esta tesis
70
1. Hernández- Mora G., R. González-Barrientos, J. A.
Morales, E. Chaves-Olarte, C. Guzmán-Verri, E. Baquero-
Calvo, M.J. De-Miguel, C.M. Marín, J.M. Blasco, E.
Moreno. 2008 Neurobrucellosis in stranded dolphins, Costa

vii
Rica. Emerg. Infect. Dis. 14 (9): 1429- 1433.
2. Hernández- Mora G., C.A. Manire , R. González-
Barrientos , E. Baquero-Calvo, C. Guzmán-Verri E. Chaves-
Olarte, E. Moreno. 2009. Serological Diagnosis of Brucella
Infections in Odontocetes. Clin. Vaccine Immunol. 16(6):
906–915
3. González-Barrientos R., J.A. Morales, G. Hernández-
Mora, C. Guzmán Verri, E. Chaves Olarte, E.
Moreno.2010. Pathology of Striped Dolphins (Stenella
coeruleoalba) Infected with Brucella ceti. J Comp
Pathol;142(4):347-52.

viii
RESUMEN
La brucelosis en cetáceos es causada por Brucella ceti y debido a la biología de estos
animales, el acceso a nivel mundial a individuos positivos y en los cuales se haya logrado
aislar esta bacteria es limitado. A nivel nacional, de 10 delfines rayados (Stenella
coeruleoalba) encallados vivos o muertos en las costas del Pacífico, se logró aislar B.ceti a
partir de tejidos del sistema nervioso central, líquido cefalorraquídeo, tejido del sistema
retículoendotelial y reproductivo por lo que se pudo estudiar los hallazgos patológicos y
por primera vez describir la sintomatología clínica presentada por estos animales
confirmados como positivos. Así se determinó que esta bacteria además de producir
problemas reproductivos, tal y como se ha descrito para otras especies del género Brucella,
es capaz de causar la muerte en estos animales por meningoencefalomielitis y endocarditis,
lo que no es usual en otros hospederos terrestres, excepto en humanos. Además, se logró
diseñar y desarrollar un inmunoensayo ligado a enzimas indirecto (iELISA) en el que se
utilizaron los sueros control obtenidos de estos animales. Tomando en cuenta la diversidad
de especies existentes de cetáceos a nivel mundial, se desarrolló un conjugado con
peroxidasa que reconociera las inmunoglobulinas G (G+L) de 17 especies diferentes como
un único grupo. Para la estandarización se utilizaron los lipopolisacáridos de Brucella
melitensis y Brucella abortus, muestras de suero de 7 odontocetos en cautiverio sin historia
de enfermedad infecciosa, negativos en la prueba de Rosa de Bengala (RBT) y muestras de
7 delfines infectados con B. ceti. Debido a la ausencia de una prueba de oro estándar, se
comparó el rendimiento del iELISA desarrollado con inmunoensayos que usan proteína G
(gELISA), ELISA competitivo (cELISA), Inmunofluorescencia (IF) y prueba Dot Blot
(DB), usando como referencia el RBT en 179 muestras de sueros de odontocetos. La
correlación y concordancia entre el iELISA y el gELISA fue buena, mientras la correlación
y concordancia de estos dos ELISAs con el cELISA fue baja. A pesar de usar el mismo
anticuerpo anti-IgG de odontoceto, el iELISA fue más específico que las pruebas de IF y
DB. Además con los animales estudiados en Costa Rica de la especie Stenella
coeruleoalba y un caso de Tursiops truncatus de Estados Unidos se logró describir que
existe una asociación entre los animales con sintomatología nerviosa, aislamiento de B.ceti,
presencia de meningoencefalomielitis y altos títulos de anticuerpos en el iELISA
desarrollado.

ix
ABSTRACT
Brucellosis in cetaceans is caused by Brucella ceti and due to the biology of these animals,
the access to positive individuals where the bacteria have been successfully isolated is
limited worldwide. B. ceti was isolated from the central nervous system (CNS),
cerebrospinal fluid, reticuloendothelial and reproductive tissue from ten striped dolphins
(Stenella coeruleoalba) stranded alive or dead in the Pacific coast of Costa Rica. From
these animals, the pathological findings were described as well as the clinical signs which
have never been characterized before from confirmed positive animals. As other Brucella
species within the genus, this bacterium induces reproductive problems. It is also able to
cause death in these animals due to meningoencephalomielitis developed by the invasion of
the CNS, a feature that is not common in other hosts, with the exception of humans. Also
an indirect enzyme-linked immunosorbent assay (iELISA) were designed and developed.
It uses as positive controls the sera obtained from the animals described before and
knowing the diversity of existing species of cetaceans worldwide a conjugated of
peroxidase that recognized immunoglobulins G (H+L) from 17 odontocete species as a
single group. For standardization purposes, it used Brucella melitensis and Brucella
abortus lipopolysaccharides, serum samples from seven resident odontocetes with no
history of infectious disease displaying negative rose bengal test (RBT) reactions, and
serum samples from seven dolphins infected with B. ceti. Without a “gold” standard, the
performance of the iELISA developed were compared with those of the protein G ELISA
(gELISA), the competitive ELISA (cELISA), and the immunofluorescence (IF) and dot
blot (DB) tests, using 179 odontocete serum samples and RBT as reference. The diagnostic
potential based on sensitivity and specificity of the iELISA was superior to that of gELISA
and cELISA. The correlation and agreement between the iELISA and the gELISA were
good, while the correlation and agreement of the ELISAs with cELISA were low. In spite
of using the same anti-odontocete IgG antibody, the iELISA was more specific than were
the IF and DB tests. An association was observed between high antibody titers in the
iELISA developed and the presence of neurological symptoms as well as
meningoencephalomylities and the isolation of B.ceti in the striped dolphins and a bottle
nose dolphin (Tursiop truncatus) from Costa Rica and United States respectably.

x
LISTA DE CUADROS
Página
Cuadro 1. Determinación de las infecciones por Brucella en cetáceos
a nivel mundial según el ensayo serológico utilizado.
47
Cuadro 2. Valores Kappa de las pruebas diagnósticas para la detección
de anticuerpos anti-Brucella en odontocetos
50
Cuadro 3. Distribución de las muestras de suero positivas de las
especies de Odontocetos en los diferentes ensayos diagnósticos
51
Cuadro 4. Distribución de las muestras de suero positivas de acuerdo
al sexo y la edad de los diferentes odontocetos
52
Cuadro 5. Correlación del resultado serológico en el iELISA, Rosa de
Bengala, aislamiento de B.ceti, presencia de meningoencefalomielitis
de las muestras de suero de cetáceos estudiadas
53

xi
LISTA DE FIGURAS
Página
Figura 1. Distribución oceánica del diagnóstico serológico de las
infecciones de Brucella en cetáceos
54
Figura 2. Hallazgos clínicos y macroscópicos a nivel de sistema
nervioso central en los delfines rayados (S. coeruleoalba) encallados en
las costas del océano Pacífico nacional.
55
Figura 3. Hallazgos microscópicos principales de neurobrucelosis en
Stenella coeruleoalba.
56
Figura 4. Hallazgos macroscópicos y microscópico de la placentitis
brucelar presentada por una hembra de delfín rayado (S. coeruleoalba).
57
Figura 5. Hallazgos macroscópicos y microscópico de la endocarditis
de origen brucelar presentada por una hembra de delfín rayado (S.
coeruleoalba).
58
Figura 6. Electroforesis del PCR con los imprimidores para L7/L12
ribosomal específica del género Brucella spp, Master Mix 2X y ADN
de B.ceti cepa 1
59
Figura 7. Determinación de la reactividad del conjugado y anticuerpos
contra los sueros de las diferentes especies de odontocetos.
60
Figura 8. Rendimiento de la proteína G-HRP contra los sueros de
odontocetos y la calibración de los controles positivos y negativos.
61
Figura 9. Proporcionalidad de las lecturas de los ELISAs con muestras
de sueros de un T. truncatus y 4 S. coeruleoalba con cultivo de B. ceti.
62
Figura 10. Distribución de las muestras de suero positivas y negativas
en relación con los valores de ELISA y la correlación dentro de los tres
diferentes ELISAs.
63
Figura 11. Relación entre los valores de ELISA y los títulos de RBT
64

xii
Figura 12. Determinación de las sensibilidades y las especificidades
usando RBT como ensayo de referencia
65
Figura 13. Rendimiento comparativo del iELISA, gELISA y cELISA
con muestras de suero de Kogia spp. y S. bredanensis, usando RBT
como referencia.
66
Figura 14. Relación filogenética entre los mayores linajes de cetáceos
67
Figura 15. Contacto entre cetáceos encallados, animales domésticos y
humanos en Playa Matapalo, Puntarenas, Costa Rica.
68
Figura 16. Distribución mundial del delfín rayado (Stenella
coeruleoalba).
69

xiii
LISTA DE ABREVIATURAS
ABTS: 2.2’: Azino-bis (3- ethylbenz-thiazoline-6- sulfonic acid)
ADN: ácido desoxirribonucléico
D.O.: Densidad óptica
DB: Dot blot
ELISA: Prueba inmunoabsorbente ligado a enzimas
EMV: Escuela de Medicina Veterinaria
FITC: Isiotiocianato de fluoresceína
HRPO: Peroxidasa
HS: Suero de caballo
IF: Inmunofluorescencia
ITCR: Instituto Tecnológico de Costa Rica
LPS: Lipopolisacárido
PBS: Solución amortiguadora 0.1M de fosfatos con salina pH 7.2
PBS-T: PBS con 0.05% de Tween 20
PBS-T-BSA: PBS-T con 1% albúmina sérica bovina
PCR: Reacción en cadena de polimerasa
RBT: Prueba de aglutinación Rosa de Bengala
SNC: Sistema nervioso central
UCR: Universidad de Costa Rica
UNA: Universidad Nacional de Costa Rica
WB: Western blot

1
INTRODUCCIÓN
La brucelosis es una enfermedad causada por bacterias Gram negativas
intracelulares del género Brucella que afectan a animales terrestres, marinos y al ser
humano (Moreno et al., 2002). Se han descrito hasta el momento ocho especies que
infectan a mamíferos terrestres: Brucella abortus (bovinos), B. suis (suinos), B. melitensis
(caprinos), B. canis (perros), B. ovis (ovinos), B. neotomae (ratas del desierto) (Moreno et
al.,2002), B. microti (topillos comunes, zorro rojo y suelo) (Scholz et al., 2008a, Scholz et
al., 2008b, Scholz et al., 2009) y B. innopinata. Esta última se describió recientemente y
fue aislada de un implante de seno en una mujer, representando la especie más distante de
las especies de Brucella tanto a nivel molecular como fenotípica. A la fecha, su reservorio
se desconoce (Scholz et al., 2010).
Los primeros reportes de brucelas aisladas de mamíferos marinos se remontan a la
década de 1990 (Ross et al., 1994, Ewalt et al., 1994). Inicialmente se describió a dichos
aislamientos como integrantes de una única especie llamada “Brucella maris”, nombre que
se reemplazó más adelante por Brucella cetaceae y Brucella pinnipediae. En el año 2007
se reconoce que ambas especies difieren tanto genotípica como fenotípicamente de las que
parasitan mamíferos terrestres y se les asigna el nombre de Brucella pinnipedialis a
aquellas bacterias que infectan pinnípedos (focas principalmente) y la segunda, B. ceti, que
afecta principalmente cetáceos (delfines, ballenas y marsopas) (Foster et al., 2007).
Actualmente estas dos especies marinas pueden ser divididas en varias subgrupos indicando
su heterogeneidad según el genotipeo molecular utilizado (Bricker et al.,2000, Bricker et
al.,2003, Bourg et al.,2007,Clavareau et al., 1998, Cloeckaert et al.,2001, Cloeckaert et
al.,2003, Dawson et al,2008a, Groussaud et al.,2007, Jahans et al., 1997, Jensen et
al.,1999, Maquart et al.,2009a, Vizcaíno et al.,2004, Whatmore et al., 2007, Whatmore, et
al.,2008, Whatmore et al., 2009). El subgrupo que infecta a delfines es distinto al que
afecta ballenas y marsopas, por lo cual está actualmente en discusión la posibilidad de que
este grupo represente una nueva especie, B. delphini (Bourg et al.,2007, Groussaud et
al.,2007, Whatmore et al., 2007, Whatmore, 2009, Maquart et al., 2009a).
La experiencia clínica y el conocimiento de la patobiología de este microorganismo
marino en cetáceos es limitado debido a su biología. Sin embargo, al igual que en
mamíferos terrestres se han documentado desórdenes reproductivos y también patologías a

2
nivel de sistema nervioso central (SNC), que podrían causar eventualmente la muerte del
animal. Esto tiene implicaciones importantes en especies en peligro de extinción como el
delfín de Hector (Cephalorhynchus hectori hectori) y delfín de Maui (C. hectori maui)
(Duignan et al. 2005) o poblaciones no expuestas (Harwood y Hall, 1990). Por ejemplo, se
ha descrito placentitis brucelar, causando aborto en dos hembras nariz de botella (Tursiops
truncatus) en cautiverio (Ewalt et al., 1994, Miller et al., 1999). Los resultados de
necropsias en otros casos han mostrado absedación del “sub-blubber” (capa de grasa que
recubre su cuerpo), necrosis esplénica y hepática, linfoadenitis, infiltración de macrófagos
en el hígado y bazo, epididimitis, orquitis, mastitis, discoespondilitis espinal y meningitis
(Ross et al., 1994,Rhyan et al.,1997, Foster et al., 2002; González et al., 2002; Muñoz et
al., 2006, Dagleish et al., 2008, Hernández-Mora et al., 2009; González-Barrientos et al.,
2010).
El mecanismo de transmisión de Brucella en ambientes marinos es incierto hasta el
momento, sin embargo, existe evidencia que al igual que en especies terrestres, las bacterias
marinas podrían utilizar las rutas de transmisión por ingestión u exposición a productos
abortados, placenta, líquido amniótico y alantoideo, así como secreciones genitales o
transmisión venérea y leche. Lo anterior se basa en reportes donde Brucella ha sido aislada
de órganos reproductivos tanto de machos como de hembras, así como la transmisión
vertical (Miller et al., 1999, Dagleish et al., 2008, Hernández-Mora et al., 2008; González-
Barrientos et al., 2010). También se ha propuesto una vía alternativa de trasmisión en la
cual pulmonares de las especies Parafilaroides spp y Pseudalius inflexus podrían actuar
como posibles vectores (Garner et al., 1997, Perret et al., 2004, Dawson et al., 2008b).
La prevalencia de brucelosis en humanos proveniente de cetáceos puede estar
significativamente subestimada en países en desarrollo de África, Suramérica y el Sureste
de Asia, así como en regiones aborígenes del Ártico donde los cetáceos recién muertos son
frecuentemente manipulados para ser utilizados en el consumo humano o como carnada por
pescadores artesanales sin precauciones higiénicas (Van Waerebeek et al., 1997, Ofori-
Danson et al., 2003, Godfroid et al., 2005, Clapham & Van Waerebbek 2007, Alfaro-
Shigueto et al., 2008, Van Bressem et al., 2009).
En 1999 se reportó el primer caso de un ser humano afectado por exposición
ocupacional en donde se presentó enfermedad, seroconversión y aislamiento de una cepa

3
marina de Brucella (Brew et al., 1999). Posteriormente, Sohn et al., (2003), describieron la
infección con brucellas marinas en dos pacientes provenientes de Perú, que presentaban
neurobrucelosis y granulomas intracerebrales. En el 2006 se reportó el cuarto caso en una
persona infectada con brucellas marinas, que provenía de Nueva Zelanda y presentó
osteomielitis espinal (McDonald et al., 2006). En los casos de Perú y Nueva Zelanda no se
ha podido determinar la fuente de infección. Las brucellas aisladas correspondían a B.
pinnipedialis según la caracterización genotípica de ambos estudios (Sohn et al., 2003,
(McDonald et al., 2006), sin embargo, Zygmunt et al., 2010 y Whatmore et al., 2007,
afirman que el aislamiento de Nueva Zelanda posee un genotipo único que corresponde a
una cepa de origen marino pero diferente a la de los grupos de pinnípedos y cetáceos.
Según Van Bressem et al., (2009), en su artículo “Enfermedades infecciosas
emergentes en cetáceos a nivel mundial y el posible rol de factores de estrés ambiental”,
mencionan que uno de los autores de este artículo sufrió una enfermedad debilitante crónica
sin llegar a un diagnóstico en los años de 1990 a 1992, período donde el investigador
realizó la necropsia de cientos de cetáceos muertos frescos en los mercados de pescado en
Perú. Las manifestaciones clínicas fueron severas y sistémicas consistentes con brucelosis
tales como convulsiones asociadas a pérdida de la conciencia, severas mialgias y dolores de
espalda, fiebre ondulante, sudoraciones profusas en la noche, dolores de cabeza, fatiga
crónica, anorexia y pérdida de peso. También se reportó en este artículo, que para el
mismo período de tiempo, una mujer de edad media, quien distribuía carne de cetáceos para
consumo humano tuvo igual sintomatología en el puerto de Pucusana en el centro de Perú.
En ninguno de estos casos, a pesar de numerosas pruebas hematológicas, parasitológicas y
enfermedades comunes en Perú, se logró llegar al diagnóstico bacteriológico definitivo, por
lo que se mantuvo solamente un diagnóstico presuntivo.
Maquart et al., (2009b) analizó el comportamiento intracelular de Brucellas de
origen marino en macrófagos humanos. Los resultados demostraron tres patrones de
infección, la Brucella aislada del ser humano en Nueva Zelanda y otros dos grupos de cepas
de B. ceti y B. pinnipedialis fueron capaces de infectar a los macrófagos humanos THP-1
con la misma virulencia de B. melitensis 16M o B. suis 1330. Otros grupos de cepas de B.
ceti y B. pinnipedialis fueron capaces de invadir macrófagos de manera similar a las cepas
clásicas virulentas, pero fueron eliminadas después de 48 horas. El último patrón de

4
infección lo presentó un aislamiento proveniente de una foca capuchina (Cystophora
cristata), el cual fue incapaz de entrar e infectar a los macrófagos humanos. Estos
resultados sugieren que existe diversidad en la patogenicidad de las cepas marinas.
La mayoría de los casos confirmados de brucelosis en cetáceos han sido
odontocetos encallados en las costas (Hernández-Mora et al., 2008). Estos animales
enfermos atraen la atención de las personas que entran en contacto con ellos para ayudarlos,
sin embargo, estos animales pueden servir como fuente de infección para humanos y otros
animales. Además, dependiendo de la región geográfica del encallamiento y condición del
animal, algunos de estos cetáceos encallados, luego de recibir tratamiento y recuperación de
diversas enfermedades son mantenidos en acuarios para exhibición, investigación o
establecimientos de terapia (Alaniz & Rojas., 2007, Antonioli & Reveley., 2005). Sin
embargo, una posible infección por Brucella y su diseminación en estos animales en
cautiverio no se investigan regularmente, ya sea por que la enfermedad como causa del
encallamiento es ignorada o bien debido al limitado número de ensayos diagnósticos
validados para la identificación presuntiva de esta enfermedad en cetáceos (Hernández-
Mora et al., 2009, Meegan et al.,2010).
La prevalencia de la infección en cetáceos ha sido evaluada serológicamente, debido
al limitado número de aislamientos de B. ceti existentes y se ha determinado que esta
enfermedad está ampliamente distribuida a nivel mundial, variando según la prueba
diagnóstica utilizada y la especie, la localización geográfica y población analizada
(animales encallados o de vida libre). De la familia Balaenide solamente la ballena de
cabeza arqueada Balaena mysticetus, especie localizada en el Ártico, ha sido analizada
serológicamente para esta enfermedad. En todos los casos, el resultado fue negativo
(Nielsen et al., 2001). De las 4 especies de balaenoptéridos que se han analizado (ballena
Minke, ballena Sei, ballena Bryde y ballena Fin) han resultado positivos individuos de
todas las especies. El mayor porcentaje de seropositividad se obtuvo en la ballena Minke
en el Pacífico Noreste, en donde 15 animales de 40 analizados resultaron positivos y
aunque no se lograron aislamientos, 10 animales resultaron positivos con la técnica de
PCR (Oshini et al., 2003). De las 3 especies de fisitéridos existentes, cachalote, cachalote
pigmeo y cachalote enano, se obtuvieron individuos seropositivos solamente en la especie
de cachalote pigmeo, tanto en animales del Golfo de México como en el océano Pacífico

5
Noroeste en Japón (Hernández Mora et al., 2009 y Ohishi et al., 2007). Las dos especies
pertenecientes a la familia Monodontidae (Narvales y Belugas) han sido estudiadas por
Nielsen et al., (2001), en el océano Ártico de Canadá. Para ambas especies se encontraron
aproximadamente un 6% de animales seropositivos. Hasta el momento de la familia
Ziphiidae, solamente 3 individuos han sido analizados: uno perteneciente a la especie de
Hyperoodon ampullatus o ballena nariz de botella del norte del océano Ártico Nielsen et
al., (2001), otro zifio de Blainville o Mesoplodon densirostris reportado de Gales,
Inglaterra por Jepson et al.,1997 y otro de la especie de zifio de Gervais o Mesoplodon
eurapeus del Golfo de México (Hernández Mora et al., 2009). Todos los casos fueron
seronegativos, adicionalmente a nivel nacional se registró en el año 2009 el encallamiento y
posterior muerte de un individuo de la especie de zifio de Cuvier o Ziphius cavisrostris, una
hembra preñada, serológicamente positiva sin embargo, no se logró aislar Brucella spp. De
las 36 especies de delfines existentes a nivel mundial, se han podido analizar 21, resultando
individuos seropositivos en las especies de delfín común de rostro largo (Delphinus
capensis) en Pacífico Sur en Perú (Van Bressem et al., 2001) delfines comunes (Delphinus
delphis) en Gales Inglaterra (Jepson et al., 1997), orca pigmea (Feresa attenuata) en el
Golfo de México (Hernández Mora et al., 2009), calderón común (Globicephala melas) en
Gales, Inglaterra (Jepson et al., 1997), delfín de Risso (Grampus griseus) en el Golfo de
México (Hernández Mora et al., 2009), delfín de Fraser (Lagenodelphis hosei) en el Golfo
de México (Hernández Mora et al., 2009), Delfín obscuro (Lagenorhynchus obscurus) en
Pacífico Sur en Perú (Van Bressem et al.,2001), Orca (Orcinus orca) en Gales Inglaterra,
(Jepson et al., 1997), delfín rayado (Stenella coeruleoalba) en el Mar Mediterráneo, en
Gales Inglaterra, en el Golfo de México y en el Pacífico de Costa Rica, (Jepson et al., 1997,
Van Bressem et al.,2001, Hernández Mora et al., 2009), delfín girador (Stenella
longirostris) en el Golfo de México, (Hernández Mora et al., 2009), delfín dientes rugosos
(Steno bredanensis) en el Golfo de México (Hernández Mora et al., 2009), delfín mular del
océano Indico (Tursiops aduncus) en las Islas Solomon (Tachibana et al.,2006) y delfín
nariz de botella (Tursiops truncatus) en Inglaterra, Golfo de México, Mar Mediterráneo,
Perú, Mar Negro y en el Pacífico de Costa Rica (Dawson et al., 2006, Hernández Mora et
al., 2009, Jepson et al., 1997, Miller et al.,1999, Van Bressem et al.,2001, Alekseev et
al.,2007). Las especies de delfines que han sido estudiadas pero han resultado negativos

6
los individuos analizados incluyen el calderón de aletas cortas (Globicephala
macrorhynchus) (Maratea et al., 2003), delfín de flancos blancos (Lagenorhynchus acutus)
(Jepson et al., 1997, Maratea et al., 2003, Nielsen et al., 2001), delfín de hocico blanco
(Lagenorhynchus albirostris) (Jepson et al., 1997), calderón pequeño (Peponocephala
electra) (Hernández Mora et al., 2009), delfín manchado (Stenella attenuata) (Hernández
Mora et al., 2009), delfín clymene (Stenella clymene) (Hernández Mora et al., 2009) y
delfín pintado del Atlántico (Stenella frontalis) (Hernández Mora et al., 2009). De la
familia Phocoenidae o focénidos solamente 2 especies han sido analizada para conocer la
seroprevalencia de esta enfermedad. La primera especie, la marsopa espinosa o Phocoena
spinipinnis resultó negativa. La segunda especie, la marsopa común o Phocoena phocoena
resultó positiva en 5 de los 7 estudios publicados hasta el momento realizados es Escocia,
Inglaterra, Connecticut, USA, Atlántico Noroeste y Ártico en Canadá, Mar del Norte y
Pacífico Sur en Perú (Foster et al., 2002, Jepson et. al., 1997, Maratea et al.,2003,
Neimanis et al., 2008, Nielsen et al., 2001) (Cuadro 1 y Figura 1).
Los estudios anteriormente citados fueron realizados utilizando ensayos
inmunoabsorbentes competitivos ligados a enzimas (cELISA) e indirectos con proteína G
(gELISA) y otros análisis serológicos desarrollados y validados satisfactoriamente para
brucelosis bovina y caprina (Jepson et al., 1997, Nielsen et al., 2001, Tachibana et al.,
2006, Tryland et al., 1999, Van Bressem et al., 2001). Sin embargo, estas pruebas
diagnósticas no han sido validadas satisfactoriamente para el diagnóstico serológico de
brucelosis en cetáceos. Esto se debe a la dificultad para obtener un número significativo de
muestras de suero de animales con infección probada, así como controles negativos de
animales libres de Brucella. Lo anterior es difícil de lograr ya que son especies de vida
silvestre, que migran en forma solitaria o en poblaciones en mar abierto, desconociéndose
en algunos casos incluso su biología. Además de las dificultades antes descritas para
validar los ensayos diagnósticos para esta enfermedad, se debe considerar que existen dos
diferentes órdenes de cetáceos con muchas familias y sus especies correspondientes. Por
ejemplo, el orden Odontoceti que incluye a los delfines, ballenas dentadas y marsopas,
contiene 9 familias, no todas monofiléticas y aproximadamente 73 especies diferentes
(May-Collado et al., 2006).

7
En Costa Rica se han reportado hasta el momento 29 especies de cetáceos (Sáenz et
al., 2004, May-Collado et al., 2005) y el estudio de esta enfermedad comenzó en el 2004,
con el análisis serológico de cetáceos encallados en su mayoría delfines rayados (Stenella
coeruleoalba) en la costa del océano Pacífico. A nivel histológico, en todos los casos de
esta especie se describía una meningitis no supurativa (Anexo 1 y 3) y por estas
características se decidió entonces remitir algunos cortes histológicos a España y Alemania
para el diagnóstico con inmunohistoquímica de Morbillivirus, enfermedad responsable de
altas mortalidades en el Mar Mediterráneo en la década de 1990 y que histológicamente
era similar a lo observado a nivel nacional. Sin embargo, en todos los casos el resultado fue
negativo. Este estudio patológico descartando Morbillivirus y confirmando la infección
con Brucella ceti se realizó de manera más detallada en 17 delfines rayados (Stenella
coeruleoalba) por González-Barrientos, R. (2008) y González-Barrientos,et al.,2010.
Dentro de los análisis microbiológicos realizados en estos animales se logró aislar B. ceti,
estos aislamientos permitieron utilizar los sueros de delfines confirmados positivos como
base para iniciar con el desarrollo de un ELISA indirecto (ELISAi) con alta sensibilidad y
especificidad tanto química como serológica.
Este trabajo de posgrado es la continuación y extensión del trabajo de Licenciatura
“Desarrollo y validación química y serológica de un ensayo inmunoenzimático ELISA para
el diagnóstico de la brucelosis en odontocetos” (Hernández-Mora G., 2006), lográndose en
esta segunda etapa la validación del iELISA con respecto a pruebas serológicas
convencionales de diagnóstico en animales terrestres como Rosa de Bengala y ELISA
competitivo y gELISA. Además gracias al desarrollo de los conjugados anti IgG de delfín,
se utilizaron para esta validación, pruebas serológicas como fluorescencia indirecta y Dot
Blot, pruebas no reportadas hasta el momento como utilizadas en el diagnóstico de esta
enfermedad en cetáceos. También se logró analizar un total de 179 muestras de 17
diferentes especies de odontocetos, aumentándose el número de muestras utilizadas
inicialmente, ya que además de muestras de animales encallados en las costas
costarricenses y de Estados Unidos, se obtuvieron muestras de animales residentes de
acuarios en este país y de un animal confirmado con aislamiento de B.ceti de España.
También se evalúo la reactividad cruzada del conjugado anti IgG de delfín peroxidasa
creado para las especies incluidas. Finalmente se realizaron los análisis estadísticos para

8
evaluar el rendimiento de todas las pruebas que permitieron establecer la sensibilidad y
especificidad y los valores predictivos del iELISA desarrollado. Por último se colaboró en
los estudios de diagnóstico patológico con el grupo de patología de la Escuela de Medicina
Veterinaria, en especial con la Dra. Rocío González Barrientos y el Dr. Juan Alberto
Morales, y con el aislamiento y caracterización de las cepas de B. ceti con investigadores
del PIET y con el grupo del Dr. José María Blasco, en Zaragoza España.

9
OBJETIVOS
GENERAL
Establecer el diagnóstico directo e indirecto de brucelosis en delfines encallados en las
costas del Pacífico de Costa Rica.
ESPECÍFICOS
1. Describir desde el punto de vista clínico y patológico los animales seropositivos y
con aislamiento confirmado de B. ceti.
2. Diseñar y estandarizar pruebas de ELISA indirectos, inmunofluorescencia y dot blot
usando la prueba de aglutinación de Rosa de Bengala como estándar, y sueros de
delfines con aislamiento de B. ceti positivos y compararlos con la prueba de ELISA
competitivo y G-ELISA usadas para el diagnóstico de la brucelosis en ganado

10
MATERIALES Y MÉTODOS
Aislamientos de B. ceti
Los cetáceos encallados y reportados por los habitantes de la costa Pacífico nacional
fueron atendidos desde enero del 2004 a julio del 2010 para este estudio. En total se
analizaron 25 cetáceos, 17 de ellos pertenecen a la especie de Stenella coeruleoalba o
delfín rayado, los 8 individuos restantes pertenecen a las especies de cachalote enano
(Kogia sima), delfín girador (Stenella longirostris), ballena jorobada (Megaptera
novaengliae), delfín nariz de botella (Tursiops truncatus) y 2 individuos de delfín
manchado (Stenella attenuata), zifio de cuvier (Ziphius cavirostris). A todos los animales
se les realizó necropsia. En total 17 animales fueron muestreados para el cultivo de B. ceti
y las muestras incluyeron tejido cerebral, líquido cefalorraquídeo, linfonodos, bazo, hígado,
riñones, y en las 2 hembras preñadas de las especies de Stenella coeruleoalba y Ziphius
cavirostris se incluyó las muestras de placenta, leche, líquidos y tejidos fetales. Todas las
muestras se cultivaron en el Departamento de Bacteriología de la Escuela Medicina
Veterinaria. Los restantes 8 animales no fueron muestreados para el cultivo de Brucella ya
que 3 individuos (la ballena Jorobada y 2 delfines rayados) se encontraban en avanzado
estado de descomposición por lo que las muestras no eran las adecuadas para estudios
microbiológicos y 5 animales muestreados durante el año 2004 y 2005 no se remitieron
muestras para cultivo específico de Brucella spp.
De 10 animales fue posible el aislamiento de Brucella y la caracterización
bioquímica convencional de los aislamientos fue respaldada por la identificación molecular
utilizando PCR con los imprimadores 5′GGC TGA TCT CGC AAA GAT-3′ y 5′-CCA
GGT CCT TGG CTT CCT TGAG-3′ (Invitrogen Corporation, Carlsbad, CA, USA) para el
gen L7 y L12 ribosomal específico para el género.
Identificación de las patologías
Desde el año 2001 al 2009, 17 delfines rayados (Stenella coeruleoalba) y 7
cetáceos más de especies como Stenella attenuata, S longirostris, Megaptera novaengliae,
Kogia sima, Tursiops truncatus y Ziphius cavirostris fueron estudiados en el Departamento
de Patología de la EMV. La realización de la necropsia y posterior recolección de los

11
tejidos fue realizada en el mismo Departamento y en el caso de la ballena jorobada y los
zifios debido a su tamaño, se les realizó la necropsia y recolección de tejidos en campo. Al
igual que para los estudios microbiológicos los tejidos de la ballena jorobada y los 2
delfines rayados no pudieron ser analizados debido a la autolisis. Todos los tejidos
recolectados se fijaron en formalina amortiguada al 10% y luego se incluyeron en cera de
parafina para ser seccionados y teñidos con hematoxilina eosina según Kiernan, (2003),
González-Barrientos R., (2008), González-Barrientos R. et al., (2010).
Sueros de Odontocetos
De un total de 179 muestras de suero de odontocetos de 17 especies diferentes, 145
pertenecen a animales atendidos en el Hospital de Delfines y Ballenas del Mote Marine
Laboratory, Sarasota, Florida, Estados Unidos. Estos sueros fueron facilitados por el
Dr. Charles A. Manire para este trabajo de investigación, los mismos se recolectaron desde
el año 1993 al 2007 por el personal del mismo Hospital y fueron mantenidos a -80°C. En
total, 14 sueros de animales residentes fueron facilitados por el personal veterinario del Six
Flags Marine World, Vallejo, California, Estados Unidos, 7 se recolectaron de animales
residentes del Gulf World, Panama City Beach, Florida, Estados Unidos, 12 animales
encallados en la costa del Pacífico de Costa Rica, fueron muestreados específicamente para
este trabajo y uno se obtuvo de un animal encallado de la costa del Atlántico de España.
Para los controles negativos se utilizaron sueros seleccionaron de 7 odontocetos residentes
(un miembro de Kogia breviceps, 2 Steno bredanensis, 3 Tursiops truncatus y un Stenella
coeruleoalba) sin síntomas clínicos de enfermedades infecciosas, todos con reacciones
negativas en la prueba de aglutinación de rosa de bengala (RBT) (Alton, G. et al., 1988).
Los controles positivos se obtuvieron de 7 delfines (6 S. coeruleoalba y un T. truncatus), de
los cuales se aisló B. ceti. Las muestras de suero se mantuvieron congeladas a -80°C o en
50% glicerol a -20°C. De estas muestras, un suero de S. coeruleoalba con un título de
aglutinación de Rosa de Bengala de 1/16 y lecturas altas (con una densidad óptica de 460
nm >1.6) en ELISA se utilizó como control positivo en los ensayos. Las muestras de suero
de los bovinos infectados en Costa Rica se obtuvieron de fincas con historial de múltiples
abortos y con aislamientos positivos de B. abortus biotipo 1. Los sueros de los bovinos

12
sanos se obtuvieron de fincas de ganado de carne principalmente, machos no vacunados
que se usaban para exportación (Gall et al., 1998).
Antígenos de Brucella y generación de anticuerpos y conjugados
Los lipopolisacáridos (LPS) de B. abortus 2308 (epitopo tipo AC) y B. melitensis 16
M (epitopo tipo MC) se obtuvieron del Laboratorio de Inmunología de la Escuela Medicina
Veterinaria, Universidad Nacional, previamente extraídos, purificados y mantenidos en
alícuotas en una concentración de 1 mg/ml y almacenados a -20°C según Aragón et al.,
1996 y Moreno et al., 1979. Antes de usarse, las soluciones de LPS se sonicaron y se
diluyeron. El aislamiento de inmunoglobulina G (IgG) de S. bredanensis se realizó según
los protocolos establecidos utilizando la combinación de precipitación en ácido caprílico,
sulfato de amonio y cromatografía (McKinney et al., 1987, Page & Thorpe, 1998, Reik.,
1987) y la fracción purificada se evaluó por geles con tinción de plata (Harlow & Lane,
1988). La preparación purificada de inmunoglobulina de delfín demostró solo 2 bandas
correspondientes a la cadena liviana y pesada de IgG. Los anticuerpos contra IgG de S.
bredanensis se desarrollaron en conejos como se describió previamente (Harlow &
Lane,1988, Lane & Harlow 1999, Lomonte, 2000). Las IgG de conejo se purificaron y
conjugaron con fluoresceína Isiotiocianato (FITC; Sigma Aldrich) o peroxidasa (HRPO;
Sigma Aldrich), siguiendo los protocolos descritos (Hudson & Hay, 1976, y Page &
Thorpe, 1988). La proteína G-HRPO recombinante se compró de Pierce Biotechnology,
Inc. Los conjugados se mantuvieron en la oscuridad y almacenados en alícuotas a -20°C en
presencia de 1% de albúmina bovina libre de lípidos (BSA) y 50 % glicerol.
Ensayos serológicos e inmunoquímicos
El RBT se realizó y evaluó según Alton, G. et al., 1988 utilizando 30 μl de suero y
30 μl del reactivo, con un período de incubación de 4 minutos, se observó la presencia o
ausencia de aglutinación. El cELISA para el diagnóstico de brucelosis (Svanova Biotech,
Uppsala, Sweden) se realizó de acuerdo con las instrucciones del fabricante, usando las
condiciones establecidas para el diagnóstico de los sueros bovinos. Brevemente, utilizando
las placas cubiertas con LPS de B.abortus, los anticuerpos presentes en las muestras
compiten con el anticuerpo monoclonal (mAb) por el epitopo C de la cadena O (Nielsen et

13
al.,2001). Luego el mAb se detecta con un conjugado con peroxidasa anti IgG de ratón. La
reacción se midió por la reducción de la densidad óptica con respecto a los controles. En
todos los ensayos, los sueros de delfín positivos y negativos se incluyeron en el análisis en
paralelo con los controles comerciales incluidos en el paquete comercial. El gELISA se
efectuó como se reporta para los sueros de bovinos (Saegerman et al.,2004), con las
siguientes modificaciones. Cien microlitros de las muestras control y los sueros se
diluyeron 1/250 en amortiguador salino de fosfatos o PBS-Tween BSA y se agregaron a los
hoyos de las microplacas de poliestireno Immunolon II 96 (Nalge Nunc International),
previamente cubiertos con LPS de Brucella. Luego de la incubación, las placas se lavaron
y 100 μl de volumen de la solución de 250 ng/ml de las proteína G- HRP diluido en PBS-
Tween conteniendo 0.1% BSA o en 0.1 M amortiguador de acetato (pH5) se añadió a cada
placa; luego, las placas se incubaron por una hora a 37°C en un agitador orbital. Las placas
se lavaron con PBS-Tween conteniendo 0.1% BSA; luego 100 μl del sustrato cromógeno
ABTS (Sigma- Aldrich) se añadió en cada hoyo. Después de incubar a 37°C, las placas se
leyeron con un lector de ELISA a 405 nm.
El ELISA indirecto (iELISA) se realizó como se describió anteriormente por Marín
et al., (1999), con modificaciones. Brevemente, las placas de poliestireno Immunolon II 96
hoyos, se cubrieron con 100 μl/ hoyo de la solución de LPS (25 a 200 μg/ml) por 18 horas a
temperatura ambiente, se sellaron con cobertores de plástico adherentes y se mantuvieron a
-20°C hasta ser usados. Luego de 5 lavados con 200 μl de PBS-Tween 20 por hoyo, los
controles y las muestras se diluyeron en PBS-Tween conteniendo 1% BSA, y 100 μl de la
solución se añadió a cada hoyo. Las placas se incubaron por una hora a 37°C en un
agitador orbital y luego se lavaron otra vez como se describió anteriormente. Cien
microlitros de anti IgG de delfín - HRP se diluyó en PBS-Tween conteniendo 1% BSA y se
añadió a cada hoyo. Las placas se incubaron otra vez por una hora a 37°C en un agitador
orbital. Después de la incubación, las placas se lavaron con PBS, y se agregó 100 μl del
sustrato cromógeno ABTS (Sigma-Aldrich) y tras 1 hora de incubación a 37°C, las placas
se leyeron en un lector de ELISA a 405 nm.
El ensayo inmunoenzimático dot blot (DB) se realizó utilizando las instrucciones
del fabricante de un sistema de microfiltración Bio-Dot (Bio-Rad Laboratories).
Brevemente, 100 μl de la solución del LPS de B. melitensis se dispensó en cada celda del

14
sistema de microfiltración ubicada sobre una membrana de nitrocelulosa (Bio-Rad). Esta
solución de LPS se filtró con la ayuda de una bomba de vacío. Después de la filtración, la
membrana se bloqueó con una solución de PBS con 1% de leche descremada (Sigma-
Aldrich), y la membrana se lavó 3 veces (cada lavado de 10 minutos) con PBS-Tween 20.
Se añadieron a cada hueco 50 μl de cada una de las muestras de suero y los controles
diluidos 1/5,000 en PBS-Tween - 0.1% BSA, y se incubaron por una hora a temperatura
ambiente en un agitador orbital. La membrana se lavó como se describió anteriormente y
se agregó a cada hueco 50 μl de anti IgG de delfín- HRP diluido en PBS-Tween
conteniendo 1% BSA (10 ng/ml). La membrana con el conjugado se incubó por 2 horas a
temperatura ambiente y se lavó 3 veces (cada lavado de 10 minutos) con PBS-Tween 20.
Ésta se sacó del bloque de microfiltración Bio-Dot y los puntos se revelaron en un film de
autoradiografía (Eastman Kodak Co.) con sustrato de quimioluminiscencia SuperSignal
(Thermo Fisher Scientific, Inc.), siguiendo las instrucciones del fabricante.
Para la inmunofluorescencia indirecta (IF), inicialmente se utilizaron células
completas de B. ceti como antígeno, sin embargo no se observó ninguna diferencia en la
sensibilidad o especificidad de la IF comparadas con las células completas de B. abortus
S19. Por razones de seguridad, se utilizó como antígeno en todos los experimentos de IF B.
abortus S19, que es una cepa atenuada y de menor riesgo para humanos. Las bacterias
vivas se secaron a 37°C en cubreobjetos y después se fijaron con paraformaldehído al 3.5%
a temperatura ambiente por 15 minutos. Posteriormente, se agregó 50 mM NH4Cl a los
cubreobjetos y se incubó por 10 minutos a temperatura ambiente. Los cubreobjetos se
lavaron con PBS y se les agregó 50 μl de suero diluido 1/50 en PBS conteniendo 10% de
suero de caballo (HS). Después de la incubación por 30 minutos a temperatura ambiente,
los cubreobjetos se lavaron 3 veces con PBS y una última vez con PBS conteniendo 0.1%
Triton X-100, a cada uno de ellos se les agregó 50 μl del anti IgG-FITC de delfín diluido en
PBS-HS y se incubó por 30 minutos a temperatura ambiente. Por último, los cubreobjetos
se lavaron como se describió anteriormente y a cada uno de estos se les se añadió 10 μl de
Mowiol (Merck) y se colocaron sobre un portaobjeto. Los portaobjetos se dejaron durante
la noche para su polimerización, y las muestras se evaluaron el próximo día por
microscopía de fluorescencia (Olympus BH-2).

15
Los ensayos de inmunodifusión para la estimación de la reacción cruzada del
antisuero y los sueros de odontocetos, se realizó utilizando geles de 1% de agarosa (Hudson
& Hay,1976). Alternativamente, se realizó el Western Blot (WB) de las IgG purificadas y
sueros en membranas de difluoruro de polivinil difluoride (Pall Biosciences, Dreieich,
Germany) como se describió por Harlow & Lane (1988), y los resultados se revelaron con
IgG-HRP anti delfín seguido por la utilización del sustrato de quimioluminiscencia Super
Signal en un film de autoradiografía. El nivel de reacción del conjugado anti IgG de delfín-
HRP de las 17 especies diferentes de odontocetos se estimó con fotografías por
densitometría de pixeles de la cadena pesada de la IgG, usando el programa de dominio
público ImageJ (http://rsb.info.nih.gov/ij/). La IgG de la especie homóloga S. bredanensis
se utilizó como estándar interno (100% del valor) y en cada experimento el valor se expresó
como un porcentaje del estándar. El experimento de WB se realizó al menos 4 veces, los
valores relativos de cada cadena pesada de IgG se promediaron, y se calculó el error
estándar.
Análisis estadístico
El análisis estadístico comparativo se realizó utilizando el programa Win Episcope
2.0 (Thrusfield et al., 2001), de acceso público
(http://www.epidemiologia.vet.ulpgc.es/software.html). La interpretación del área bajo la
curva ROC (receiver operating characteristics) se realizó según Hanley & McNeil, (1983).
Los parámetros utilizados para establecer los valores del punto de corte y la interpretación
de los resultados diagnósticos de brucelosis estuvieron basados en trabajos previos (Marín
et al., 1999). Brevemente, los valores de punto de corte de los cELISA, gELISA e iELISA
se ajustaron con los límites del menor porcentaje de positividad obtenido por un suero
positivo y el mayor porcentaje de positividad obtenido por un suero negativo. Con estos
límites, el valor de punto de corte para cada ELISA se estableció empíricamente como los
valores de intersección que excluían el mínimo número de falsos positivos y falsos
negativos, tomando el RBT como referencia. Este ensayo de aglutinación se usó como
referencia debido a su rendimiento independiente de anticuerpos secundarios y por su
competencia en discriminar anticuerpos en el suero de animales infectados y no infectados
con Brucella (Alton, G. et al., 1988). Bajo estas condiciones, los valores del punto de corte

16
para el cELISA, gELISA e iELISA se establecieron empíricamente en 30.6%, 6.0% y 7.5%
de positividad respectivamente.

17
RESULTADOS
A nivel mundial existen 86 especies de cetáceos y tanto misticetos como
odontocetos han sido analizados serológicamente para el diagnóstico de brucelosis (Cuadro
1). Dentro de las 29 especies de cetáceos reportadas para el océano Pacífico de Costa Rica,
(Wehrtmann y Cortés., 2009) la especie de delfín rayado (Stenella coeruleoalba), es la que
con mayor frecuencia ha encallado de noviembre de 1999 a julio de 2010, presentándose en
21 de 30 casos estudiados en el Departamento de Patología de la EMV-UNA. Los 9
individuos restantes pertenecen a las especies de cachalote enano (Kogia sima), delfín
girador (Stenella longirostris), ballena jorobada (Megaptera novaengliae), delfín nariz de
botella (Tursiops truncatus), 2 individuos de zifio de cuvier (Ziphius cavirostris) y 3
individuos de delfín manchado (Stenella attenuata). De los 21 delfines rayados encallados,
17 fueron sometidos a necropsia completa y se determinó que las escoriaciones y lesiones
macroscópicas encontradas en estos animales se debían al mismo encallamiento en playas
rocosas o con fuerte oleaje y a la sintomatología clínica presentada (Figura 2A y 2B). En
general se concluyó que muchos de los hallazgos patológicos generales no se relacionaron
con brucelosis, pero los cambios detectados en el sistema nervioso central tales como
congestión a nivel de vasos sanguíneos con opacidad en las meninges y aumento en la
cantidad de líquido cefalorraquídeo (Figura 2 C), sí eran debidos a la infección con
Brucella. También un macho juvenil presentó hidrocéfalo interno secundario al proceso
inflamatorio por Brucella, involucrando los ventrículos laterales (Figura 2D). En la
mayoría de los casos hubo infiltración mononuclear perivascular de la materia gris y blanca
del cerebro, cerebelo y tallo encefálico. Los infiltrados fueron mayormente de células
plasmáticas, linfocitos y macrófagos (Figura 3). Un macho juvenil que tuvo
meningoencefalomielitis también tuvo osteoartritis fibrinopurulenta con una infiltración
severa de macrófagos y neutrófilos invadiendo el fluido sinovial afectando la articulación
escapulo humeral como se describió anteriormente por otros autores (Dagleish et al., 2008,
Galatius et al., 2009). Nueve delfines con meningoencefalomielitis (6 con serología
positiva y 4 con cultivo positivo de B.ceti) también tuvieron neumonía intersticial no
supurativa y otros 5 mostraron hepatitis linfocítica periportal (González- Barrientos et al.,
2010).

18
En los 17 animales se realizaron estudios de inmunohistoquímica para la detección
de antígeno de Brucella y Morvillivirus en cerebro, cerebelo, médula espinal resultando en
todos los casos negativos para ambos agentes etiológicos a pesar del aislamiento de
Brucella ceti de LCR de estos animales. Por lo tanto se determinó que la inmunohistoquímica
no fue una técnica suficientemente sensible para el diagnóstico de la neurobrucelosis en los
cetáceos analizados y se le atribuyó a las características del patrón inflamatorio presente, donde
la cantidad de macrófagos observada fue baja, disminuyendo de esta manera las probabilidades
de observar una inmuno reacción positiva para este agente (González-Barrientos, 2008) La
única hembra preñada de delfín rayado presentó una severa placentitis con múltiples focos
necróticos y se estimó por el tamaño del feto una gestación de aproximadamente 7 meses
(Figura 4A) (Hernández-Mora et al.,2008, González Barrientos et al., 2010). A nivel
microscópico se confirmó la presencia de necrosis diseminada y severa en la placenta con
infiltración abundante de células mononucleares y polimorfonucleares en los trofloblastos
(Figura 4B, 4C, 4D). Se detectó Brucella por IHC en el infiltrado inflamatorio así como
en algunas células coriónicas en las regiones necróticas, y la intensidad del marcaje fue
consistente con la presencia de grandes cantidades de bacteria (Figura 4D). A pesar de las
lesiones a nivel de placenta, no se encontraron cambios patológicos significativos en el feto
(no se muestra). Una hembra adulta tuvo una endocarditis severa con engrosamiento y
presencia de un nódulo vegetativo prominente en la válvula mitral (Figura 5A). La
endocarditis se caracterizó por la presencia de fibrina adyacente a la superficie de la válvula
mitral, con una infiltración predominantemente no supurativa de linfocitos, macrófagos,
células plasmáticas y células gigantes multinucleadas (Figura 5B). En la misma área se
observó una degeneración focal de las fibras del miocardio que se rodeaban con un
infiltrado linfocítico moderado y edema perivascular. También se observó fibrosis
pericárdica con infiltración de linfocitos y células plasmáticas. Por medio de IHC se
evidenció la presencia de Brucella asociado con el infiltrado células inflamatorias y
colonias de bacterias (Figura 5C y 5D). (González – Barrientos et al., 2009).
A excepción de un neonato de delfín rayado, todos los delfines rayados estudiados
fueron clasificados como positivos fuertes en el iELISA desarrollado y tuvieron una clara
aglutinación en la prueba de Rosa de Bengala. La sintomatología mostrada en estos
animales, que en algunos casos encallaron aún vivos, fue consistente con problemas

19
nerviosos, mostrando problemas de nado y flotabilidad con tremores corporales,
opistotonos y convulsiones (Figura 2A, Anexo 1). Estos síntomas clínicos fueron
compatibles con los hallazgos de histopatología a nivel de sistema nervioso central y los 10
aislamientos de B. ceti logrados del LCR de estos animales. Estos aislamientos fueron
analizados bajo la técnica de PCR para los genes L7 y L12 ribosomal que es específico para
el género Brucella y en todos los casos dieron como resultado el fragmento correspondiente
a 290 pb (Figura 6). Además de los aislamientos de LCR, la hembra preñada mostró
concomitantemente meningoencefalomielitis y placentitis y se logró aislar la bacteria
además de placenta, leche, líquidos y tejidos fetales (Figura 4).
De las otras especies de cetáceos que encallaron, a excepción de la ballena jorobada
y los 2 delfines rayados que debido a su nivel de descomposición no fue posible obtener ni
suero ni tejido para ser analizado, las especies de cachalote enano, delfín girador, delfín
nariz de botella, zifio de Cuvier y delfín manchado no se reportaron hallazgos similares a
los encontrados en la especie de delfín rayado a nivel de sistema nervioso central, ni
cardíaco; tampoco en la placenta de la otra hembra preñada de la especie de zifio de
Cuvier, a pesar que tuvo un resultado serológico positivo en el iELISA y una reacción de
aglutinación en Rosa de Bengala. De ningún animal de estas especies, se logró aislar B.
ceti aunque en algunos casos se clasificaron como positivos o positivos débiles
serológicamente tanto en el iELISA como en la prueba Rosa de Bengala (Cuadro 5).
Un problema de los inmunoensayos para el diagnóstico de la brucelosis en cetáceos
que dependen de un anticuerpo anti-delfín, es que la Brucella infecta a diferentes especies
de odontocetos. Tentativamente, esto restringiría el uso del conjugado anti-IgG de S.
bredanensis- HRP y –FITC desarrollados para este trabajo de investigación. Por lo tanto,
se decidió probar la reactividad del anticuerpo y del conjugado contra el suero de las 17
especies de odontocetos que se logró obtener suero tanto a nivel nacional como de Estados
Unidos y España. Como se muestra en la Figura 7 A, se generaron líneas de precipitación
contra las inmunoglobulinas de las 17 especies de odontocetos probadas por
inmunodifusión. Sin embargo, contra los sueros de Kogia sima, Kogia breviceps, y
Mesoplodon europaeus, se observó identidad parcial, lo que sugirió que el anticuerpo anti-
IgG de S. bredanensis tenía una menor especificidad y potencia para las inmunoglobulinas
de estas 3 especies. En los WBs, el conjugado de IgG-HRP reaccionó igualmente de forma

20
cruzada contra las cadenas pesadas y livianas de las IgG de las 17 especies, confirmándose
también en estos ensayos que las especies del género Kogiidae poseen menor reactividad
(Figura 7B). Se presentaron dos excepciones en la reactividad del conjugado de IgG-HRP.
La primera de ellas contra la cadena pesada de IgG de M. europaeus, la cual mostró una
buena reacción cruzada en WB y la segunda contra la cadena pesada de IgG de Stenella
longirostris, que reaccionó más débil que las otras especies del mismo género (Figura 7B).
No se observó reactividad cruzada del anticuerpo purificado o su respectivo conjugado anti-
IgG de S. bredanensis- HRP contra inmunoglobulinas de bovino (Figuras 7A y 7B), foca
común, cabra, humano o perro (no se muestra). El conjugado comercial de Bethyl
Laboratories, Inc. anti-IgG (H+L)- HRP de T. truncatus, también demostró amplia reacción
cruzada contra los sueros de odontocetos en el iELISA (no se muestra).
Debido a que el LPS de las Brucellas marinas tiene extensa reacción cruzada con el
LPS de otras Brucellas lisas al compartir diversos epitopos (Baucheron et al., 2002), se
evaluaron diferentes concentraciones de LPS de Brucella tipo AC o MC para la
estandarización del iELISA. Se determinó que las condiciones óptimas son las
concentraciones de 10 μg/ml de LPS tipo MC purificado, 10 ng/ml para el conjugado de
conejo IgG-HRP y una dilución de 1/1000 para el suero de delfín.
Según los fabricantes de gELISA (Manual protein G; Pierce Biotechnology), la
buena interacción de la proteína G y los anticuerpos de las diferentes especies depende
estrictamente del pH para su buen funcionamiento. Basado en lo anterior y que este ensayo
ha sido usado extensamente en el diagnóstico de las infecciones de Brucella en mamíferos
marinos (Cuadro 1) (Jepson et al., 1997, Tachibana et al., 2006, Tryland et al., 1999., Van
Bressem et al., 2001), se comparó la unión de la proteína G-HRP con las inmunoglobulinas
de odontocetos en dos soluciones tampón recomendadas. Este ensayo se realizó también
utilizando inmunoglobulinas de bovinos en las mismas condiciones. Mientras que los
anticuerpos contra Brucella del suero positivo de bovino se unieron eficientemente a la
proteína G-HRP en el gELISA en diluciones desde 1/250 a 1/ 4,000 y usando PBS-Tween
(pH 7.2) o solución tampón de acetato 0.1 M (pH 5), las inmunoglobulinas de los
odontocetos infectados con Brucella (dilución de suero en 1/250) no se unieron a la
proteína G-HRP usando la solución tampón PBS-Tween. Además, se observó que en el
gELISA, la cinética de unión de la proteína G a los anticuerpos de odontocetos fue más

21
lenta que con los anticuerpos bovinos (Figura 8A). Los anticuerpos bovinos reaccionaron
más fuertemente al LPS en comparación a los sueros de delfín, a pesar de que los animales
mostraron el mismo título de aglutinación en la prueba de RB (1/16). Igualmente, en el
iELISA ya fuera utilizando los conjugados anti IgG-HRP de bovino o anti IgG- HRP de
delfín, las densidades ópticas obtenidas fueron similares (~1.6). Por lo tanto, en todos los
ensayos de gELISA con sueros de odontocetos se usó solución tampón de acetato al 0.1 M
(pH 5).
Luego se procedió a ajustar las muestras de sueros positivos y negativos de
odontocetos en los diferentes ELISAs y se determinó que los valores de densidad óptica
obtenidos no se apartaron significativamente de la media y con un nivel de confianza de un
99%, los límites superiores e inferiores de variación de los ELISAs fueron cercanos al 10%,
por lo tanto, se llegó a la conclusión que los límites se mantuvieron sin sobreponer la
variación durante este tiempo de prueba. De esta forma, las muestras de suero de los
odontocetos con lecturas de densidad óptica altas, medias o bajas se seleccionaron como los
controles positivos para ser usados a través de los experimentos en los diferentes ensayos
serológicos (Figura 8B). Tal y como se esperaba, la intensidad de los puntos de los sueros
controles obtenidos en el análisis de DB fueron los mismos que en los ELISAs (Figura 8B).
Posteriormente, se determinó la proporcionalidad de los tres ELISAs, encontrándose que
las muestras de suero negativas variaban menos de 0.02 en unidades de densidad óptica en
correspondencia con las diluciones seriadas (no se muestra). Los valores de 5 muestras de
sueros positivos también se midieron de forma continua decreciente en el iELISA y el
gELISA (Figuras 9 A y 9 B) y de forma creciente en el cELISA (Figura 9 C). Sin embargo,
en el iELSA y el gELISA no todos los sueros siguieron la misma cinética, por ejemplo, dos
muestras positivas (ambas de S. coeruleoalba) mostraron densidades ópticas mayores, en
diluciones menores y tuvieron perfiles muy diferentes en las subsecuentes diluciones. Esto
sugiere que pueden ocurrir variaciones significativas en la avidez en sueros de animales
infectados. Además, mientras algunas muestras permanecieron en las mismas posiciones
relativas en los tres ELISAs, la posición de otras muestras varió (Figura 9).
Una vez que el rendimiento de los ensayos se ajustó ópticamente, se procedió a
comparar sus diagnósticos. La correlación y concordancia entre el iELISA, gELISA y
cELISA con respecto al RBT, se evaluó en las 179 muestras de sueros de los odontocetos

22
de las 17 especies diferentes (Figura 10). Mientras la correlación entre el iELISA y
gELISA fue relativamente alta (Rg/i2 = 0. 65), la correlación entre estos dos ELISAs y el
cELISA fue significativamente menor (Rc/i2 = 0.46 y Rc/g
2 = 0.37). Como se esperaba, los
altos títulos de RBT estuvieron asociados a porcentajes altos de positividad en los tres
ELISAs (Figura 11). Sin embargo, una muestra de suero de un delfín (S. coeruleoalba) con
cultivo de B. ceti y un título de 1/2 en RBT, cayó en el rango negativo del gELISA (Figura
11B). Esta misma muestra cayó en el rango positivo en el iELISA y cELISA, junto con las
otras 6 muestras con cultivo positivo de B. ceti. Esto indudablemente indica que las
infecciones de Brucella no necesariamente se correlacionan con altas lecturas en el ELISA
y sugiere una baja sensibilidad para el gELISA.
La sensibilidad y especificidad para los ELISAs se estimó utilizando el RBT como
referencia en el análisis de curvas ROC (Figura 12). El punto de corte teórico de 30.7%
para el cELISA y 5.9% para el gELISA, que correspondían a las intersecciones de las
sensibilidades y especificidades más altas de las curvas ROC, correspondieron con los
valores establecidos empíricamente de 30.6% y 6.0% respectivamente. Sin embargo, este
no fue el caso del iELISA, en el cual el valor del punto de corte (7.5%) establecido
empíricamente, favoreció la especificidad sobre la sensibilidad, afectando el valor kappa
levemente. De hecho, en todas las comparaciones realizadas, el rendimiento del iELISA
fue superior al rendimiento del gELISA y el cELISA, de acuerdo con la estimación de los
valores kappa (Cuadro 2). Además, en el iELISA se observó una transición de la curva
ROC más suave y una distribución más estrecha de las reacciones positivas comparado con
las reacciones en el gELISA y cELISA, indicando un mayor rendimiento para el iELISA.
Como se esperaba, se obtuvieron mayores valores kappa cuando los tres ensayos se usaron
como referencia, sin embargo, el número de muestras de suero que se probaron de manera
acumulada (consistentemente positivos o negativos de forma combinada en los diferentes
ensayos) se redujo de 179 a 130, indicando que el rendimiento de IF o DB es bajo si se
compara a los resultados del iELISA. El uso del mismo anticuerpo anti-delfín en el
iELISA, IF y DB favoreció el resultado del iELISA, cuando los últimos dos ensayos se
incluyeron como los ensayos de referencia.
El cELISA y gELISA no dependen de los conjugados anti-delfín. El cELISA utiliza
el conjugado de anticuerpo monoclonal HRP reaccionando contra el LPS de Brucella,

23
mismo epitopo que utiliza para competir y el gELISA usa la proteína G-HRP, que es una
proteína de unión “universal” a la cadena pesada de las IgG (Gall et al., 1998, Saegerman et
al., 2004). El conjugado anti IgG–HRP creado contra S. bredanensis mostró menor
reactividad contra las inmunoglobulinas Kogia spp. (Figura 7), por lo tanto, fue necesario
comparar el rendimiento de los ELISAs contra ambos tipos de sueros. Como se muestra en
la Figura 13, el iELISA mostró valores kappa predictivos más altos que el gELISA y
cELISA, siendo las muestras de suero de las Kogia spp. o su homólogo S. bredanensis.
Disminuyendo las diluciones de los sueros de Kogia spp. en una vez, aumentaron las
densidades ópticas de los sueros asignados como positivos sin incrementar el ruido de
fondo o la densidad óptica de los sueros negativos (no se muestra). Finalmente, el
conjugado HRP en el iELISA también dio como resultado altas lecturas (con porcentajes de
positividad >82%), en las muestras de suero heterólogas de los 6 S. coeruleoalba y un T.
truncatus con cultivos positivos (Figura 10), confirmando el rango amplio de reacciones
cruzadas de este anticuerpo anti IgG contra inmunoglobulinas G de odontocetos.
La frecuencia de reacciones positivas en los diferentes ensayos serológicos se
determinó para las 179 muestras de sueros en las 17 especies (Cuadro 3). Las reacciones
serológicas positivas se encontraron más frecuentemente en miembros de los géneros
Stenella, Steno, y Tursiops, pero no se evidenció una correlación fuerte entre el sexo y la
edad (Cuadro 4). Las reacciones positivas se observaron en mayor proporción en la prueba
de DB, seguida de la IF.

24
DISCUSIÓN
Las pruebas de aglutinación, así como los cELISA y gELISA han sido utilizados en
la detección de anticuerpos de cetáceos para el diagnóstico presuntivo de infecciones de
Brucella spp. La utilización a priori de estos ensayos se fundamenta en el hecho de que
estos no dependen de un conjugado específico dirigido directamente contra las
inmunoglobulinas de las especies analizadas. Aunque estos trabajos tienen importancia en
muchos aspectos para describir la enfermedad en ambientes marinos, ninguno de ellos ha
evaluado críticamente el rendimiento de estos ensayos serológicos en cetáceos y como se
mencionó anteriormente, uno de los mayores problemas que se debe enfrentar para
determinar la especificidad y sensibilidad de los inmunoensayos, es la ausencia de una
colección importante de muestras de sueros positivos y negativos de cetáceos a nivel
mundial.
Recientemente Meegan y colaboradores (2010) desarrollaron un cELISA para la
detección de anticuerpos contra la bacteria completa de B. pinnipedialis en mamíferos
marinos, con una sensibilidad y especificidad para cetáceos del 100% y 73% y en
pinnípedos del 77% y 67%. Este ensayo es una herramienta útil para el diagnóstico de la
brucelosis en diversas especies de pinnípedos y cetáceos, sin embargo, no toma en
consideración las relaciones filogenéticas existentes entre las especies de Brucella
involucradas y sus hospedadores. En este sentido se encuentra bien establecido que la clase
Artiodáctila (ungulados como vacas, cabras e hipopótamos) compartió el mismo ancestro
evolutivo que la clase Cetácea hace unos 50-65 millones de años y esto se refleja en la
relación evolutiva entre B.ceti y B.abortus o B.melitensis. Por otra parte la evidencia tanto
molecular como morfológica sostiene que los pinnípedos se encuentran mayormente
emparentados con Arctoides carnívoros como los úrsidos y mustélidos y no con mamíferos
marinos como los cetáceos (Berta A. et al., 2006, Arnason et al., 2006). Lo anterior puede
comprometer la sensibilidad y especificidad del ensayo utilizado tanto para cetáceos como
para pinnípedos. Los autores también justifican el uso de la bacteria completa de
B.pinnipedialis versus un componente específico de membrana ya que predicen una mejor
sensibilidad por que permite la unión de los anticuerpos a los diferentes epitopos
antigénicos, sin embargo, es este mismo factor el que compromete la especificidad de este
ensayo que de hecho tiene una mejor especificidad para cetáceos que para pinnípedos,

25
especie que se esperaría tuviera un mejor rendimiento en general al utilizar B.pinnipedialis.
Esto se puede explicar ya que dentro del proceso de adhesión del antígeno a la placa y
durante el ensayo, se pueden exponer diversos componentes de membrana e incluso
componentes citoplasmáticos de la bacteria que no son exclusivos del género Brucella y
por lo tanto podrían ser reconocidos de forma cruzada por los anticuerpos presentes en el
suero.
Dentro de las ventajas del RBT se incluyen su simplicidad, consistencia e
interpretación en los resultados como ensayo de referencia en las poblaciones infectadas
naturalmente (Alton, G. et al., 1988). Las reacciones falsas positivas en el RBT se dan
comúnmente debido al uso de plasma en vez de suero, en sueros muy hemolizados o por
reacciones cruzadas y los falsos negativos usualmente se dan por una baja avidez o títulos
reducidos de anticuerpos aglutinantes. A pesar de lo anterior, el RBT permanece robusto
en ausencia de otros ensayos independientes y su utilización como ensayo de referencia se
justifica (Alton, G. et al., 1988, Mainar-Jaime et al., 2005). Además, el hecho de que 7
muestras de suero de animales con cultivo de B. ceti presenten títulos de aglutinación ≥1/2
provee apoyo adicional para el uso de RBT como ensayo de referencia y punto inicial para
entender las reacciones serológicas en cetáceos infectados con Brucella.
Las reacciones “falsas negativas” observadas en iELISA y gELISA pueden ser
debidas a tres condiciones principales. La primera de ellas es la presencia de pequeñas
cantidades de anticuerpos aglutinantes IgM o IgA que pueden escapar a la detección de los
conjugados anti-IgG-HRP y proteína G-HRP. La segunda condición consiste en la
existencia de anticuerpos aglutinantes desarrollados contra epitopos diferentes al LPS por
ejemplo, proteínas que son excluidas en los ELISAs y la última posibilidad es que estén
presentes ambas condiciones. Sin embargo, en contraste a la proteína G, la cual se une
principalmente a la cadena pesada de las IgG, el antidelfín IgG-HRP usado en el iELISA
también reacciona contra la cadena liviana, incrementando la probabilidad de detectar otros
isotipos de inmunoglobulinas y por lo tanto, aumentar la sensibilidad serológica. Mainar-
Jaime y colaboradores (2005) han documentado que el cELISA posee menor sensibilidad
en relación con el iELISA y lo atribuyen al desplazamiento de los anticuerpos de baja
avidez por los anticuerpos monoclonales competitivos contra el epitopo C del LPS usado en
este ensayo.

26
Las reacciones “falsas positivas” detectadas en el iELISA y en el gELISA puede
deberse a anticuerpos dirigidos contra determinantes profundos del LPS de Brucella que no
están expuestos fácilmente en el RBT, pero que son accesibles en los LPS purificados
utilizados para cubrir las placas. De hecho, en bovinos y otros animales ha sido
demostrado que son capaces de desarrollar anticuerpos contra determinantes de LPS como
porciones del core y lípido A (Rojas et al., 2001), que son compartidos por otras alfa-
Proteobacterias relacionadas con Brucella (Delpino et al., 2004 y Velasco et al., 1997), por
ejemplo Bartonella, que al igual que los animales terrestres también infectan odontocetos
(Harms et al., 2008), por lo que las reacciones cruzadas contra estos epitopos profundos de
LPS pueden crearse. La mayor especificidad relativa del cELISA parece estar ligada a los
anticuerpos monoclonales competentes que específicamente reconocen el epitopo C de
LPS. Sin embargo, si el punto de corte es ajustado, el iELISA muestra una especificidad
comparable o mejor que el cELISA sin alcanzar los niveles de baja sensibilidad del ensayo.
Con respecto a la relación entre el iELISA y el gELISA relativamente buena (Rg/i2 = 0.65),
el primer ensayo muestra una sensibilidad significativamente mayor (SeiELISA = 86% versus
SegELISA = 58%). Cabe enfatizar, que una muestra de suero de delfín con cultivo positivo
de B. ceti y un número significativo de muestras positivas en el RBT fueron dadas por
negativas igualmente por el gELISA, esto sugiere que la proteína G-HRP falla en detectar
bajos títulos de anticuerpos o anticuerpos con baja avidez. Además a la fecha, los estudios
reportados por otros, no han tomado en consideración el cuidado con el amortiguador usado
para los ensayos con la proteína G en el diagnóstico de brucelosis en odontocetos, ya que
en condiciones regulares neutrales no trabaja. Esto puede deberse a que al usarse
amortiguadores con pH acídico, los anticuerpos de baja afinidad pueden ser removidos y
esto contribuye a reducir la sensibilidad del gELISA. En pequeños rumiantes, el gELISA
ha mostrado ser menos sensible que los otros ELISAs que usan conjugados anti IgG u otras
pruebas serológicas para el diagnóstico de infecciones de Brucella (Díaz-Aparicio et al.,
1994, y Ficapal et al., 1995). Por lo tanto, en odontocetos el uso sin restricción del gELISA
en pruebas de brucelosis debe hacerse con precaución, debido a que su sensibilidad parece
ser menor en estos animales que en bovinos.
En odontocetos la correlación y concordancia entre el iELISA, gELISA y cELISA
fue mucho menor de lo esperado, ya que en general, los tres ensayos muestran

27
relativamente buena correlación en bovinos (Díaz-Aparicio et al., 1993, Gall et al., 1998,
Marín et al., 1999, Saegerman et al., 2004) y es común que el ensayo de RBT muestre
buena correlación con los tres ELISAs en la detección de los anticuerpos anti-Brucella
(valores kappa variando entre 0.70 y 0.75). Basados en nuestra experiencia con el iELISA
desarrollado se puede afirmar, que éste fue altamente reproducible y su rendimiento fue
muy similar a lo que se ha observado en ganado usando el RBT. Sin embargo, a diferencia
de los resultados obtenidos con el iELISA, los resultados utilizando el cELISA mostraron
un comportamiento mucho más variable entre ensayo y ensayo, siendo el cELISA muy
sensible a las condiciones del suero analizado (incluyendo el almacenamiento), conjugado y
los controles. Aunque las últimas dos variables pueden ser controladas, la calidad del suero
depende de la recolección, que no siempre es controlado en el momento del sangrado de los
odontocetos encallados (Anexo 1). También la variabilidad del cELISA en comparación
con los resultados del iELISA, se ilustra en la forma irregular bajo las curvas ROC y en la
distribución más amplia de las muestras de suero en los odontocetos positivos y negativos.
Esta variabilidad entre los ELISAs podría explicarse con diferencias en la avidez o
en la forma de trabajar de los anticuerpos contra el LPS de Brucella según el ELISA
utilizado o la combinación de ambas condiciones. Para explicar un poco más este
fenómeno, se graficó las diferentes posiciones relativas de los sueros positivos en las curvas
de titulación (Figura 9).
Finalmente, la interpretación de las muestras de suero que poseían reacciones
cercanas al punto de corte fueron menos obvias en las pruebas de IF y DB que en los
ELISAs o en RBT. Este hecho puede explicarse con la mayor proporción de falsos
positivos observados en los ensayos de DB y IF con respecto al iELISA, a pesar de usar el
mismo conjugado anti-delfín IgG.
En brucelosis bovina, caprina y ovina el tipo de LPS de Brucella liso para fines
diagnósticos parece tener poca relevancia (Moreno & Moriyón, 2006). En el caso de sueros
de odontocetos, parece que el rendimiento del LPS tipo MC es ligeramente mejor que el
LPS tipo AC en el iELISA, con respecto al tiempo de lectura, densidades ópticas, y
diferencias entre sueros positivos y negativos. Sin embargo, los resultados finales de
“positivo” y “negativos” usando el LPS tipo AC en vez del tipo MC no divergió
significativamente, excepto en las lecturas cercanas al punto de corte (no se muestra). El

28
hecho que la densidad y el contenido de epitopos comunes del LPS de B.ceti varía con
respecto a los LPSs clásicos de las brucelas lisas terrestres (Baucheron et al., 2002), puede
influenciar la generación de anticuerpos en los odontocetos infectados con Brucella, los
cuales puede reconocer más fácilmente los epitopos comunes en los LPSs tipo MC que en
los de tipo AC. Sin embargo, dudamos que esta variable cuente significativamente en las
diferencias observadas entre los varios ELISAs. En todo caso, los resultados obtenidos con
este trabajo ilustran la relevancia en probar los LPSs como antígeno para la estandarización
de ensayos inmunoenzimáticos en cetáceos.
En diferentes estudios se ha demostrado que los anticuerpos hechos contra Bos
taurus, reaccionan de forma extensa contra las inmunoglobulinas de otras especies de
bóvidos de esta subfamilia y por lo tanto, son útiles en pruebas serológicas en
enfermedades incluyendo brucelosis (Henning et al., 1992). En odontocetos la reacción
cruzada de inmunoglobulinas es extensa, como quedó demostrado en este estudio y por
Nollens y colaboradores (2008), por lo tanto permite el uso de una sola especie de
anticuerpos anti IgG para discriminar la presencia de anticuerpos anti LPS de Brucella. El
hecho que los valores kappa obtenidos en el iELISA fueran significativamente mayores, sin
importar que la muestra de suero pertenecía a la especie homóloga S. bredanensis o a la
especie heteróloga Kogia spp. que los de los gELISA y cELISA refuerza este concepto.
A pesar que el cELISA no depende del conjugado anti IgG de odontoceto, la
congruencia con el RBT fue más bajo que con las muestras de suero de Kogia spp. y S.
bredanensis que las observadas con el gELISA y iELISA. Además, el conjugado anti IgG
de odontoceto, mostró buena reactividad contra Physeter macrocephalus (Cachalote), un
miembro de la familia Physeteridae, el cual es filogenéticamente más cercano a Kogiidae
que Delphinidae. Por lo tanto, la predicción es que la reacción cruzada de las
inmunoglobulinas puede ser utilizada para detectar anticuerpos de otras especies no
incluidas en este estudio, por ejemplo, Monodontidae (narvales) y los delfines de agua
dulce de las familias Iniidae, Pontoporiidae, Lipotidae, y Platanistidae. De hecho, todas
estas familias están filogenéticamente más cercanas a la familia Delphinidae, de la cual es
miembro S. bredanensis (Nikaido et al., 2001). Nollens y colaboradores (2008) reportan
una baja reactividad cruzada entre inmunoglobulinas de odontocetos y los anticuerpos de la
ballena gris (Eschrichtius robustus) y ballena jorobada (Megaptera novaeangliae), por lo

29
tanto, pareciera que el mismo conjugado anti-delfín IgG no trabajará para los misticetos o
ballenas, un orden que pertenece a una rama filogenética distante a los odontocetos
(Nikaido et al., 2001). Finalmente, predecimos que la extensa reacción cruzada con sueros
de odontocetos será observada igualmente con los anticuerpos comerciales anti delfines
IgG (H+L), por lo tanto, serán útiles para establecer otros iELISAs y otros ensayos
diagnósticos.
Una observación importante de este estudio fue la correlación entre los altos títulos
de anticuerpos contra los antígenos de Brucella y la presencia de síntomas neurológicos en
delfines. Del total de 179 muestras de suero obtenidas de Estados Unidos, España y Costa
Rica y analizadas para este estudio, 19 delfines que encallaron con el diagnóstico de
patologías neurológicas tales como meningitis no supurativa 17 de Costa Rica, uno de USA
y otro de España (Anexo 1 y 2), 18 mostraron reacciones positivas en RBT y iELISA por
encima del 82% de positividad. De éstos, 16 fueron S. coeruleoalba, 3 fueron T. truncatus
y 12 tuvieron cultivo positivo de B. ceti. En un T. truncatus, el aislamiento fue realizado
de forma retrospectiva de tejido congelado a -80 °C luego del diagnóstico serológico,
manteniendo el valor predictivo potencial cuando se utiliza en conjunto los ensayos iELISA
y el RBT.
Con respecto a la descripción de los hallazgos histopatológicos en los delfines
rayados analizados, éstos indican que el encallamiento de estos animales está asociado con
la meningoencefalomielitis causada por la infección con B. ceti (Anexo 1). Aunque estos
cambios patológicos se han descrito en el ser humano y otras especies de delfines
infectados con Brucella spp. (Foster et al., 2002; González et al., 2002, Pappas et al., 2005,
Davison et al., 2009, González-Barrientos et al., 2010), éstos son raramente vistos en
hospederos terrestres como ganado, cabras, ovejas o cerdos. En estos hospederos los
síntomas principales están relacionados con infecciones en el tracto reproductivo tales
como retención de placenta, mastitis intersticial, abortos, epididimitis y en algunos casos
higromas (Hagemoser et al., 1988; Meador et al., 1989; Musa et al., 1990, Campero et
al.,1994). Es importante notar que los hallazgos histopatológicos encontrados a nivel
cardíaco (endocarditis nodular) o a nivel neurológico (meningoencefalomielitis) en los
delfines S. coeruleoalba encallados y asociados a la infección comprobada de Brucella ceti
no han sido documentados en animales domésticos que sufren brucelosis y han sido solo

30
reportados en humanos (Shakir et al., 1987; Al-Harthi., 1989, Vinod et al., 2007, Je &
Song, 2008, Cay et al., 2009). Además las lesiones presentes en corazón, hígado,
pulmones, articulaciones y placenta de los animales sugieren que B. ceti tiene la habilidad
de causar infecciones crónicas en múltiples órganos antes de cruzar la barrera
hematoencefálica. La placentitis observada en la única hembra preñada de delfín a la que
se tuvo acceso fue similar a las reportadas en dos casos anteriores (Miller et al., 1999); sin
embargo, a pesar de las lesiones a nivel de placenta, no se detectaron cambios patológicos a
nivel del feto, a diferencia de como sucede en brucelosis bovina en donde una severa
placentitis se asocia con aborto y con patologías severas a nivel de sistema nervioso central
e inflamación importante a nivel pulmonar fetal (Hong et al., 1991).
Las descripciones de los hallazgos patológicos en humanos como hospedero
secundario accidental de brucelosis adquirida naturalmente son escasas en la literatura
(Hunt & Bothwell, 1967; Pappas et al., 2005, Özbek et al., 2007) por lo tanto la
descripción de los hallazgos encontrados en estos animales ayuda al conocimiento del curso
natural de esta enfermedad.
Rhyan y colaboradores (2001), realizaron infecciones experimentales con Brucella
spp aislada de marsopa común (Phocoena phocoena) y reportan que causó seroconversión
y aborto en ganado, por lo tanto a nivel nacional se desconoce el impacto real de la
Brucella de origen marino en las fincas costeras donde esta enfermedad no es monitoreada
ni por los propietarios ni por entes estatales y los cetáceos encallados entran en contacto
con animales domésticos por ejemplo perros y vacas al igual que seres humanos (Figura
15).
Según Jefferson et al., (2008) la distribución de los delfines rayados a nivel mundial
es amplia ya que se encuentran en los océanos Atlántico, Pacífico e Indico así como áreas
adyacentes (Figura 16) y aunque es considerada una especie de aguas templadas, el rango
en el cual es posible encontrarlos se extiende a regiones más lejanas y por lo tanto menos
templadas. Aunque los límites naturales reportados para esta especie va de 50°N a 40°S
rutinariamente esta especie logra alcanzar las aguas del Norte Europeo y existen reportes en
la Península de Kamchatka, parte sur de Groenlandia, Islas del Príncipe Eduardo en Canadá
e Islas Feroe en Dinamarca en donde son masacrados masivamente como parte de su
cultura. Además los delfines rayados son considerados animales pelágicos o de aguas

31
oceánicas y son vistos cerca de la costa solamente cuando aguas profundas se acercan a
éstas y en algunas áreas como el Pacífico Tropical Oriental (PTO) que incluye el Pacífico
nacional, se asocian mayormente a zonas de convergencia y de afloramientos marinos.
A pesar de la amplia distribución a nivel mundial y los encallamientos que ocurren
de esta y otras especies, la brucelosis no se asocia como causa de muerte de estos animales,
según la literatura disponible hasta el momento. La presencia de meningoencefalomielitis
asociada al aislamiento de Brucella ha sido lograda en los delfines rayados a nivel nacional
y en un único caso en España, suero que sirvió de control positivo también para este
trabajo. Por lo tanto es importante la educación y el adecuado manejo de los animales
encallados con sintomatología nerviosa en la región centroamericana en donde no se está
atendiendo ni estudiando las causas de los encallamientos de cetáceos. También es
necesario mayor investigación en la población silvestre del PTO en donde se estima que la
población de delfines rayados es de un millón de individuos y la infección por Brucella no
necesariamente compromete la vida del animal por lo tanto puede estarse presentando otras
patologías incluyendo problemas reproductivos que podría ser un factor crítico en la
sostenibilidad a largo plazo de la especie en conjunto con otros factores antropogénicos
tales como niveles inmunosupresores de contaminantes en los océanos.

32
CONCLUSIONES
Los ensayos iELISA y el RBT tienen alto valor potencial predictivo en el
diagnóstico de brucelosis en odontocetos cuando se usan en conjunto
La interpretación de resultados del gELISA en el diagnóstico de brucelosis en
odontocetos debe hacerse con precaución.
Basados en los resultados de este trabajo se recomienda que para la
estandarización de ensayos inmunoenzimáticos en el diagnóstico de brucelosis
en cetáceos, se deben probar el rendimiento tanto los LPS de B. abortus y de B.
melitensis en los ensayos, pues se pueden obtener mejores resultados utilizando
uno u otro.
La amplia reacción cruzada del conjugado anti IgG de S. bredanensis para
detectar inmunoglobulinas de otras especies, lo convierte en una herramienta
útil para el diagnóstico de brucelosis y otras enfermedades en odontocetos.
Los hallazgos patológicos que se observaron en los delfines rayados encallados
a nivel nacional puede ayudar a la comprensión del curso natural de la
enfermedad de la brucelosis tanto en humanos como en animales.
La identificación rápida y correcta de los animales infectados resulta en una
práctica relevante para los servicios de fauna acuática, acuarios, redes de rescate
y pescaderías.
La brucelosis en cetáceos ha mostrado ser una causa importante de
encallamientos debido a la meningoencefalomielitis desarrollada.
Se debe descartar la brucelosis antes de decidir una rehabilitación en cetáceos
encallados presentando problemas de nado y flotabilidad así como
sintomatología nerviosa.

33
La severidad de la brucelosis en delfines rayados o S. coeruleoalba indica que
esta especie de cetáceo es altamente susceptible a la infección y es
potencialmente reservorio de B. ceti en el océano Pacífico Tropical Oriental.
Se desconoce hasta el momento el impacto de esta enfermedad en la salud
pública mundial, ya que el diagnóstico de brucelosis en cetáceos no se realiza de
rutina en acuarios, ni establecimientos donde se ponen en contacto humanos y
delfines procedentes de vida libre. Por lo tanto se recomienda no entrar en
contacto con sangre ni con secreciones de estos animales.

34
REFERENCIAS
Alekseev, Y., E. I. Rozanova, E. N. Ustinova1, Y. Tumanov, I. N. Kuvshinova1 & A. M.
Shestopalov. 2007. The prevalence of antibodies to morbilliviruses, Brucella, and
Toxoplasma in the Black Sea bottlenose dolphin Tursiops truncatus ponticus
maintained in captivity. Russian Journal of Marine Biology.33:425-428.
Alaniz P. Y., & L. Rojas. 2007. Delfinarios. 1rst .ed. AGT Ed. DF, México.
Alfaro-Shigueto J.A, J.C. Mangel, K.Van Waerebeek . 2008. Small cetacean captures &
CPUE estimates in artisanal fisheries operating from a port in northern Peru, 2005–
2007. Paper SC/60/SM19 presented to IWC/60 Annual Meeting, Santiago, Chile,
June 2008. (Available from IWC Secretariat, Cambridge).
Alton, G. G., L. M. Jones, R. D. Angus, & J. M. Verger. 1988. Techniques for the
brucellosis laboratory. Institut National de la Recherche Agronomique, Paris,
France.
Al-Harthi, S.S. 1989. The morbidity and mortality pattern of Brucella endocarditis. Int J
Cardiol. 25:321-324.
Antonioli, C., & M. A. Reveley. 2005. Randomised controlled trial of animal facilitated
therapy with dolphins in the treatment of depression. BMJ. 331:1231.
Aragón, V., R. Díaz, E. Moreno, & I. Moriyón. 1996. Characterization of Brucella abortus
and Brucella melitensis native haptens as outer membrane O-type polysaccharides
independent from smooth lipopolysaccharide. J. Bacteriol. 178:1070-1079.
Arnason, U., A. Gullberg. A. Janke, M. Kullberg, N. Lehman, E. A. Petrov & R. Väinölä.
2006. Pinniped phylogeny and a new hypothesis for their origin and dispersal.
Molecular Phylogenetics and Evolution 41:345-354.
Baucheron, S., M. Grayon, S. Zygmunt, & A. Cloeckaert. 2002. Lipopolysaccharide
heterogeneity in Brucella strains isolated from marine mammals. Res. Microbiol.
153:277-280.
Berta A., J.L.Sumich & K.M. Kovacs. 2006. Marine Mammals: Evolutionary Biology.2nd
Ed. Elsevier.USA
Brew, S. D., L. L. Perrett, J. A. Stack, & A. P. MacMillan. 1999. Human exposure to
Brucella recovered from a sea mammal. Vet. Rec. 144: 483.

35
Bricker, B. J., D. R. Ewalt, A. P. MacMillan, G. Foster, & S. Brew. 2000. Molecular
characterization of Brucella strains isolated from marine mammals. J. Clin.
Microbiol. 38: 1258-1262.
Bricker, B., D. R Ewalt, & S. Halling. 2003. Brucella 'HOOF-Prints': strain typing by
multi-locus analysis of variable number tandem repeats (VNTRs).BMC Microbiol.
3: 15.
Bourg, G., D. O’Callaghan, & M.L. Boschiroli. 2007. The genomic structure of Brucella
strains isolated from marine mammals gives clues to evolutionary history within the
genus. Vet. Microbiol. 125:375-380.
Campero, C.M., P.W. Ladds, D. Hoffmann, B. Duffield, D. Watson & G. Fordyce. 1994.
Immunopathology of experimental Brucella abortus strain 19 infection of the
genitalia of bulls. Vet. Immunol. Immunopathol. 24:235-246.
Cay, S., G. Cagirci, O. Maden, Y. Balbay & S. Aydogdu. 2009. Brucella endocarditis: a
registry study. Kardiol Pol. 67:274-280.
Clapham P, K. & Van Waerebeek.2007. Bushmeat, the sum of the parts. Mol Ecol
16:2607–2609.
Clavareau, C., V. K.Wellemans, M. Walravens, J.M. Tryland, M. Verger, O. Grayon, A.
Cloeckaert, J. J. Letesson, & J. Godfroid. 1998. Phenotypic and molecular
characterization of a Brucella strain isolated from a minke whale (Balaenoptera
acutorostrata). Microbiol. 144: 3267-3273.
Cloeckaert, A., J.M. Verger, M. Grayon, J.Y. Paquet, B. Garin-Bastuji, G. Foster, & J.
Godfroid. 2001. Classification of Brucella spp. isolated from marine mammals by
DNA polymorphism at the omp2 locus. Microbes Infect. 3:729-738.
Cloeckaert, A., M. Grayon, O. Grépinet, & K. Sidi Boumedine. 2003. Classification of
Brucella strains isolated from marine mammals by infrequent restriction site PCR
and development of specific PCR identification tests. Microbes Infect. 5:593-602.
Dagleish M.P, J. Barley , J. Finlayso, R.J Reid, & G. Foster. 2008. Brucella ceti associated
pathology in the testicle of a harbour porpoise (Phocoena phocoena). J Comp
Pathol.139(1):54-59
Davison, N.J., M. P. Cranwell, L. L. Perrett, C. E. Dawson, R. Deaville, E. J. Stubberfield,
D. S. Jarvis & P. D. Jepson. 2009. Meningoencephalitis associated with Brucella

36
species in a live-stranded striped dolphin (Stenella coeruleoalba) in south-west
England. Vet Rec. 165:86-89.
Dawson, C. E., L. L. Perrett, E. J.Young, N. J. Davison & R. J. Monies. 2006. Isolation of
Brucella species from a bottlenosed dolphin (Tursiops truncatus). Vet Rec.158:831-
832.
Dawson C.E, E.J Stubberfield, L.L Perret, A.C. King, A.M. Whatmore, J.B. Bashiruddin,
J.A Stack, & A.P Macmillan. 2008a. Phenotypic and molecular characterisation of
Brucella isolates from marine mammals. BMC Microbiol. 8:224.
Dawson C.E., L.L Perret, E.J Stubberfield, & J.A Stack. 2008b. Isolation and
characterization of Brucella from lungworms of a harbor porpoise (Phocoena
phocoena) J. Wildl Dis 44: 237-246.
Delpino, M. V., C. A. Fossati, & P. C. Baldi. 2004. Occurrence and potential diagnostic
applications of serological cross-reactivities between Brucella and other alpha-
proteobacteria. Clin. Diagn. Lab. Immunol. 11868-873.
Díaz-Aparicio, E., C. Marín, B. Alonso-Urmeneta, V. Aragón, S. Pérez-Ortiz, M. Pardo, J.
M. Blasco, R. Díaz, & I. Moriyón. 1994. Evaluation of serological tests for
diagnosis of Brucella melitensis infection of goats. J. Clin. Microbiol. 32:1159-
1165.
Díaz-Aparicio, E., V. Aragón, C. Marín, B. Alonso, M. Font, E. Moreno, S. Pérez-Ortiz, J.
M. Blasco, R. Díaz, & I. Moriyón. 1993. Comparative analysis of Brucella serotype
A and M Yersinia enterocolitica O:9 polysaccharides for serological diagnosis of
brucellosis in cattle, sheep and goats. J. Clin. Microbiol. 31:3136-3141.
Duignan P.J., W. McDonald, G. Mackereth, Jamaluddin. 2005. Recent discovery of
Brucella infection in New Zeland Hector’s dolphins (Cephalorhychus hectori
hectori) a potential cause of low fecundity and high perinatal mortality. Wildlife
Disease Association International Conference, Cairns.
Ewalt, D. R., J. B. Payeur, B. M. Martin, D. R. Cummins, & W. G. Miller. 1994.
Characterization of a Brucella species from a bottlenose dolphin (Tursiops
truncatus). J.Vet. Diagn. Invest. 6: 448-452.

37
Ficapal, A., B. Alonso-Urmeneta, J. Velasco, I. Moriyón, & J. M. Blasco. 1995. Diagnosis
of Brucella ovis infection of rams with an ELISA using protein G as conjugate. Vet.
Rec. 137:145-147.
Foster, G., A. P. MacMillan, J. Godfroid, F. Howie, H. M. Ross, A. Cloeckaert, R. J. Reid,
S. Brew, & I. A. P Patterson. 2002. A review of Brucella spp. infection of sea
mammals with particular emphasis on insolates from Scotland. Vet. Microbiol. 90:
563-580.
Foster G., B.S.Osterman, J. Godfroid, I. Jacques, A. Cloeckaert. 2007. Brucella ceti sp. nov.
and Brucella pinnipedialis sp. nov. for Brucella strains with cetaceans and seals as
their preferred hosts. Int J Syst Evol Microbiol. 57:2688-93
Galatius, A., C. Sonne, C. C. Kinze, R. Dietz, & J. E. Jensen. 2009. Occurrence of vertebral
osteophytosis in a museum sample of white-beaked dolphins (Lagenorhynchus
albirostris) from Danish waters. J. Wildl. Dis. 45:19-28.
Gall, D., A. Colling, O. Mariño, E. Moreno, K. Nielsen, B. Pérez, & L. Samartino. 1998.
Enzyme immunoassays for the serological diagnosis of bovine brucellosis. Trial in
Latin America. Clin. Diag. Lab. Immunol. 5: 654-661.
Garner, M. M., D. M. Lambourn, S. J. Jeffries, P. B. Hall, J. C. Rhyan, D. R. Ewalt, L. M.
Polzin, & N. F. Cheville. 1997. Evidence of Brucella infection in Parafilaroides
lungworms in a Pacific harbor seal (Phoca vitulina richardsi). J. Vet. Diagn. Invest.
9: 298-303.
Godfroid, J., A. Cloeckaert, J. P. Liautard, S. Kohler, D. Fretin, K. Walravens, B. Garin-
Bastujl, & J. J. Letesson. 2005. From the discovery of the Malta fever’s agent to the
discovery of a marine mammal reservoir, brucellosis has continuously been a re-
emerging zoonosis. Vet. Res. 36: 313–326.
González, L., I. A. Patterson, R. J. Reid, G. Foster, M. Barberán, J. M. Blasco, S. Kennedy,
F. E. Howie, J. Godfroid, A. P. MacMillan, A. Schock & D. Buxton. 2002. Chronic
meningoencephalitis associated with Brucella sp. infection in live-stranded striped
dolphins (Stenella coeruleoalba). J. Comp. Pathol. 126: 147-152.
González-Barrientos R. 2008 Estudios anátomo-histopatológicos e inmunohistoquímicos
sobre las patologías de los cetáceos encallados en Costa Rica durante el período

38
2001-2007. Con énfasis especial en el diagnóstico de Brucella y Morbillivirus. Tesis
de Licenciatura, Universidad Nacional, Costa Rica
González-Barrientos R., J.A. Morales, G. Hernández-Mora, C. Guzmán Verri, E. Chaves
Olarte,& E. Moreno. 2009. Pathology of Striped Dolphins (Stenella coeruleoalba)
Infected with Brucella ceti. J Comp Pathol. 2010 May;142(4):347-52.
Groussaud, P., S.J. Shankster, M.S. Koylass, & A.M. Whatmore. 2007. Molecular 17
typing divides marine mammal strains of Brucella into at least three groups with
distinct host preferences. J. Med. Microbiol. 56:1512-1518.
Hagemoser W.A, B.L. Deyoe, & V.P. Meador. 1988. Histopathologic findings in Brucella
abortus infected, pregnant goats. American Journal of Veterinary Research, 49,274-
280.
Hanley, J. A., & B. J. McNeil. 1983. A method of comparing the 1 areas under receiver
operating characteristic curves derived from the same cases. Radiology. 148:839-
843.
Harlow, E., & D. Lane. 1988. Antibodies: a laboratory manual, 1st ed. Cold Spring Harbor
Laboratory, Cold Spring Harbor, NY
Harwood, J., & A. Hall. 1990. Mass mortality in marine mammals: Its implications for
population dynamics and genetics. Trends. Ecol. Evol. 5: 254-257.
Harms, C. A., R. G. Maggi, E. B. Breitschwerdt, C. L. Clemons-Chevis, M. Solangi, D. S.
Rotstein, P. A. Fair, L. J. Hansen, A. A. Hohn, G. N. Lovewell, W. A. McLellan, D.
A. Pabst, T. K. Rowles, L. H. Schwacke, F. I. Townsend, & R. S. Wells. 2008.
Bartonella species detection in captive, stranded and free-ranging cetaceans. Vet.
Res. 39:59.
Henning, D., & K. Nielsen. 1992. Cross-reactivity of monoclonal antibodies to bovine
immunoglobulins with immunoglobulins of other species. Vet. Immunol.
Immunopathol. 34235-243.
Hernández-Mora G. 2006 Desarrollo y validación química y serológica de un ensayo
inmunoenzimático ELISA para el diagnóstico de la brucelosis en odontocetos. Tesis
de Licenciatura, Universidad Nacional, Costa Rica.

39
Hernández- Mora G., R. González-Barrientos, J. A. Morales, E. Chaves-Olarte, C.
Guzmán-Verri, E. Baquero-Calvo, M.J. De-Miguel, C.M. Marín, J.M. Blasco, E.
Moreno.2008.Neurobrucellosis in stranded dolphins, Costa Rica. Emerg. Infect.
Diseas. 14 (9): 1429- 1433.
Hernández-Mora G, C. A. Manire, R. González-Barrientos, E. Barquero-Calvo, C.
Guzmán-Verri, L. Staggs, R. Thompson, E. Chaves-Olarte & E. Moreno. 2009.
Serological diagnosis of Brucella infections in odontocetes. Clin. Vaccine Immunol.
16:906-915.
Hong C.B., J.M. Jr Donahue, C.R. Giles, K.B. Poonacha, & P.A Tuttle.1991. Brucella
abortus-associated meningitis in aborted bovine fetuses. Veterinary Pathology, 28,
492-496.
Hudson, L., & F. C. Hay. 1976. Practical immunology. Blackwell Scientific, Oxford,
England.
Hunt A.C., & P.W. Bothwell.1967. Histological findings in human brucellosis. Journal of
Clinical Pathology, 20,267-272.
Jahans, K. L., G. Foster, & E. S. Broughton. 1997. The characterization of Brucella strains
isolated from marine mammals. Vet. Microbiol. 57: 373-382.
Je H.G., & H. Song.2008. Brucella endocarditis in a non-endemic country: first reported
case in East Asia. Circulation Journal, 72, 500-501.
Jefferson T.A, M.A. Webber, R.L. Pitman. 2008. Marine Mammals of the World : A
comprehensive guide to their identification. 1rst ed. Elseiver. Canadá.
Jensen, A. E., N. F. Cheville, C. O. Thoen, A. P. MacMillan, & G. W. Miller. 1999.
Genomic fingerprinting and development of a dendrogram for Brucella spp.
insolated from seals, porpoises and dolphins. J. Vet. Diafn. Invest. 11: 152-157.
Jepson, P. D., S. Brew, A. P. MacMillan, J. R. Baker, J. Barnett, J. K. Kirkwood, T.
Kuiken, I. R. Robinson, & V. R. Simpson. 1997. Antibodies to Brucella in marine
mammals around the coast of England and Wales. Vet. Rec. 141: 513-515.
Kiernan JA .2003. Histological and Histochemical Methods: Theory and Practice. Oxford
University Press, NewYork. pp. 1-477.
Lane, D. & E. Harlow. 1999. Using antibodies: a laboratory manual. 1st. ed. Cold spring
harbor, Laboratory, N. Y.

40
Lomonte, B. 2000. Manual de laboratorio de inmunología general. 2a. ed. Universidad de
Costa Rica, C.R.
Mainar-Jaime, R. C., P. M. Muñoz, M. J. de Miguel, M. J. Grilló, C. M. Marín, I. Moriyón,
& J. M. Blasco. 2005. Specificity dependence between serological tests for
diagnosing bovine brucellosis in Brucella-free farms showing false positive
serological reactions due to Yersinia enterocolitica O:9. Can. Vet. J. 46:913-916.
Marín, C. M., E. Moreno, I. Moriyón, R. Díaz, & J. M. Blasco. 1999. Performance of
competitive and indirect enzyme-linked immunosorbent assay, gel
immunoprecipitation with native hapten polysaccharide and standard serological
tests in the diagnosis of sheep brucellosis. Clin. Diag. Lab. Immunol. 6:269-272.
Maquart, M., P. Le Flèche, G. Foster, M. Tryland, F. Ramisse, 1 B. Djonne, S. Al Dahouk,
I. Jacques, H. Neubauer, K. Walravens, J. Godfroid, A. Cloeckaert, & G. Vergnaud.
2009a. MLVA-16 typing of 295 marine mammal Brucella isolates from different
animal and geographic origins identifies 7 major groups within Brucella ceti and
Brucella pinnipedialis. BMC Microbiol. 9:145.
Maquart, M., M. S. Zygmunt, & A. Cloeckaert. 2009b. Marine mammal Brucella isolates
with different genomic characteristics display a differential response when infecting
human macrophages in culture. Microbes Infect. 11:361-366.
Maratea, J., D. R. Ewalt, S. Frasca Jr., J. L. Dunn, S. De Guise, L. Szkudlarek, S.J. St.
Aubin & R. A. French. 2003. Evidence of Brucella sp. infection in marine mammals
stranded along the coast of southern. New England Journal of Zoo and Wildlife
Medicine 34:256-261.
May-Collado L, Gerrodette T, Calambokidis J, Rasmussen, & K, I. Sereg. 2005. Patterns of
cetacean sighting distribution in the Pacific exclusive economic zone of Costa Rica
based on data collected from 1979-2001. Rev Biol Trop.53(1-2):249-63.
May-Collado, L. J., & I. Agnarsson. 2006. Cytochrome b and Bayesian inference of whale
phylogeny. Mol. Phylogenet. Evol. 38:344-354
McDonald W.L., R. Jamaludin, G. Macereth, M. Hansen, S. Humphrey, P. Short, T.
Taylor, J. Swingler, C.E. Dawson, A.M.Whatmore, E. Stubberfield, L.L. Perrett, &
G. Simmons. 2006. Characterization of a Brucella sp. strain as a marine-

41
mammal type despite isolation from a patient with spinal osteomyelitis in
New Zealand. J Clin Microbiol, 44: 4363-4370.
McKinney, M. M., & A. Parkinson. 1987. A simple non-chromatographic procedure to
purify immunoglobulins from serum and ascites fluid. J. Immunol. Methods 96271-
278.
Meador V.P, B.L. Deyoe, & N.F. Cheville.1989. Pathogenesis of Brucella abortus
infection of the mammary gland and supramammary lymph node of the goat.
Veterinary Pathology, 26, 357-368
Meegan J, Field C, Sidor I, Romano T, Casinghino S, Smith CR, Kashinsky L, Fair PA,
Bossart G, Wells R, Dunn JL Development, validation, and utilization of a
competitive enzyme-linked immunosorbent assay for the detection of antibodies
against Brucella species in marine mammals. J Vet Diagn Invest. 2010
Nov;22(6):856-62.
Miller, W. G., L. G. Adams, T. A. Ficht, N. F. Cheville, J. P. Payeur, D. R. Harley, C.
House, & S. H. Ridgway. 1999. Brucella-induced abortions and infection in
bottlenose dolphins (Tursiops truncatus). J. Zoo. Wildl. Med. 30: 100-110.
Moreno, E., M. W. Pitt, L. M. Jones, G. G. Schuring, & D. T. Berman. 1979. Purification
and characterization of smooth and rough lipopolysaccharides from Brucella
abortus. J. Bacteriol. 138361-369
Moreno, E., A. Cloeckaert, & I. Moriyón. 2002. Brucella evolution and taxonomy. Vet.
Microbiol. 90: 209-227.
Moreno, E., & I. Moriyón. 2006. The genus Brucella, p. 315-456. In M. Dworkin, S.
Falkow, E. Rosenberg, K.-H. Schleifer, and E. Stackebrandt (ed.), The prokaryotes,
vol. 5. Springer-Verlag, New York, NY
Muñoz, P. M., G. García-Castrillo, P. López-García, J. C. González-Cueli, M. J. De
Miguel, C. M. Marín, M. Barberán, & J. M. Blasco. 2006. Isolation of Brucella
species from a live-stranded striped dolphin (Stenella coeruleoalba) in Spain. Vet.
Rec. 158: 450-451.
Musa M.T., K.L Jahans, & M.E Fadalla. 1990 Clinical manifestations of brucellosis in
cattle of the southern Darfur Province, western Sudan. Journal of Comparative
Pathology,103, 95-99.

42
Neimanis, A. S., H. N. Koopman, A. J. Westgate, K. Nielsen & F. A. Leighton. 2008.
Evidence of Exposure to Brucella sp. in Harbor Porpoises (Phocoena phocoena) from
the Bay of Fundy, Canada Journal of Wildlife Diseases, 44:480-485.
Nielsen O., K. Nielsen, & R. Stewart.1996. Serological evidence of Brucella spp. Exposure
in Atlantic walruses (Odobenus rosmarus rosmarus) and ringed seals (Phoca
hispida) of Arctic Canada. Arctic. 49: 383-386.
Nielsen O., R.E.A Stewart, K. Nielsen, L. Measures & P. Duignan. 2001. Serological
survey of Brucella spp. antibodies in some marine mammals of North America.
J Wildlife Dis. 37: 89-100.
Nikaido, M., F. Matsuno, H. Hamilton, R. L. Brownell, Jr., Y. Cao, W. Ding, Z. Zuoyan, A.
M. Shedlock, R. E. Fordyce, M. Hasegawa, & N. Okada. 2001. Retroposon analysis
of major cetacean lineages: the monophyly of toothed whales and the paraphyly of
river dolphins. Proc. Natl. Acad. Sci. USA 987384-7389.
Nollens, H. H., C. Ruiz, M. T. Walsh, F. M. Gulland, G. Bossart, E. D. Jensen, J. F.
McBain, & J. F. Wellehan. 2008. Cross-reactivity between immunoglobulin G
antibodies of whales and dolphins correlates with evolutionary distance. Clin.
Vaccine Immunol. 151547-1554.
Ofori-Danson P.K, K. Van Waerebeek, & S. Debrah.2003. A survey for the conservation of
dolphins in Ghanaian coastal waters. J Ghana Sci Assoc 5:45–54
Ohishi, K., R. Zenitani, T. Bando, Y. Goto, K. Uchida, T. Maruyama, S. Yamamoto, N.
Miyazaki & Y. Fujise. 2003. Pathological and serological evidence of Brucella-
infection in baleen whales (Mysticeti) in the western North Pacific. Comp. Immunol.
Microbiol. Infect. Dis. 26: 125-136.
Ohishi, K., K. Takishita, M. Kawato, R. Zenitani, T. Bando, Y. Fujise, Y. Goto, S.
Yamamoto, & T. Maruyama. 2004 Molecular evidence of new variant Brucella in
North Pacific common minke whales. Microbes Infect. 6:199-204.
Ohishi, K., E. Katsumata, K. Uchida & T. Maruyama. 2007. Two stranded pygmy with
anti- Brucella antibodies in Japan. Vet Rec. 160:628-629.
Ohishi, K., Y. Fujise, & T. Maruyama. 2008. Brucella spp. in the western north pacific and
antarctic cetaceans: A review. Journal of Cetacean Research and Management. 10:67-
72.

43
Özbek C., U. Yetkın, I. Yürekli, M. Bademcı & A. Gürbüz 2007. Development of
commissural perforation of an aortic valve in Brucella endocarditis with giant
vegetation . The Internet Journal of Thoracic and Cardiovascular Surgery. 10:1.
Page, M., & R. Thorpe. 1998. IgG purification. Methods Mol. Biol. 80:95-111.
Pappas G., N. Akritidis, M. Bosilkovski, & E. Tsianos. 2005 Brucellosis. New England
Journal of Medicine, 352, 2325-2336.
Perrett, L. L., C. E. Dawson, N. Davison, & S. Quinney. 2004. Brucella infection of
lungworms from a harbour porpoise. Vet. Rec. 154:800.
Reik, L. M. 1987. A simple non-chromatographic purification procedure for monoclonal
antibodies. J. Immunol. Meth. 100: 123-130.
Rhyan, J. C, S. D. Holland, T. Gidlewski, D. A. Saari, A. E. Jense, D. R. Ewalt, S. G.
Hennager, S. C. Olsen, & N. F. Cheville. 1997. Seminal vesiculitis and orchitis
caused by Brucella abortus biovar 1 in young bison bulls from South Dakota. J Vet
Diagn Invest. 9:368-374
Rhyan J. C., T. Gidlewski, D. R. Ewalt, S. G. Hennager, D. M. Lambourne & S. C. Olsen.
2001. Seroconversion and abortion in cattle experimentally infected with Brucella
sp. isolated from a Pacific harbor seal (Phoca vitulina richardsi). J. Vet. Diagn.
Invest. 13: 379–382.
Rojas, N., O. Zamora, J. Cascante, D. Garita, & E. Moreno. 2001. Comparison of the
antibody response in adult cattle against different epitopes of Brucella abortus
lipopolysaccharide. J. Vet. Med. B Infect. Dis. Vet. Public Health 48623-629.
Ross, H. M., G. Foster, R. J. Reid, K. L. Jahans, & A. P. MacMillan. 1994. Brucella species
infection in sea- mammals. Vet. Rec. 134: 359.
Ross, H. M., K. L. Jahans, A. P. MacMillan, R. J. Reid, P. M. Thompson, & G. Foster.
1996. Brucella species infection in North Sea seal and cetacean populations. Vet.
Rec. 138: 647-648.
Saegerman, C., L. De Waele, D. Gilson, J. Godfroid, P. Thiange, P. Michel, B. Limbourg,
T. K. Vo, J. Limet, J. J. Letesson, & D. Berkvens. 2004. Evaluation of three serum
i-ELISAs using monoclonal antibodies and protein G as peroxidase conjugate for
the diagnosis of bovine brucellosis. Vet. Microbiol. 10091-105.

44
Sáenz, J., G. Wong, & E. Carrillo. 2004. Ballenas y delfines de América Central:
descripción de los subórdenes de los cetáceos. 1a. ed. INBio, C.R.
Shakir R.A, A.S.N. Al-din, G.F. Araj, A.R. Lulu, & A. Mousa A. 1987. Clinical categories
of neurobrucellosis a report on 19 cases. Brain, 110, 213-223.
Sohn, A. H., W. S. Probert, C. A. Glaser, N. Gupta, A. W. Bollen, J. D. Wong, E. M.
Grace, & W. C. Mc Donald. 2003. Human neurobrucellosis with intracerebral
granuloma caused by a marine mammal Brucella spp. Emerg. Infect. Dis. 9: 485–
488.
Scholz, H.C., Z. Hubalek, J. Nesvadbova, H. Tomaso, G. Vergnaud, P. Le Flèche, A.M.
Whatmore, S. Al Dahouk, M. Krüger, C. Lodri, & M. Pfeffer. 2008a. Isolation of
Brucella microti from soil. Emerg. Infect. Dis. 14:1316-1317.
Scholz, H.C., Z. Hubalek, I. Sedlacek, G. Vergnaud, H. Tomaso, S. Al Dahouk, F. Melzer,
P. Kampfer, H. Neubauer, A. Cloeckaert, M. Maquart, M.S. Zygmunt, A.
Whatmore, E. Falsen, P. Bahn, C. Gollner, M. Pfeffer, B. Huber, H.J. Busse, & K.
Nockler. 2008b. Brucella microti sp. nov., isolated from the common 1 vole
Microtus arvalis.2 Int. J. Syst. Evol. Microbiol. 58:375-382.
Scholz, H.C., E. Hofer, G. Vergnaud, P. Le Fleche, A.M. Whatmore, S. Al Dahouk, M.
Pfeffer , M. Krüger, A. Cloeckaert, & H. Tomaso. 2009. Isolation of Brucella
microti from mandibular lymph nodes of red foxes, Vulpes vulpes, in Lower
Austria. Vector Borne Zoonotic Dis. 9:153-156.
Scholz, HC; K, Nöckler, C. Göllner, P. Bahn, G. Vergnaud, H. Tomaso, S. Al Dahouk, P.
Kämpfer, A. Cloeckaert, M. Maquart, M.S. Zygmunt, A.M. Whatmore, M. Pfeffer,
B. Huber, H.J. Busse,& B.K. De. 2010. Brucella inopinata sp. nov., isolated from a
breast implant infection. Int J Syst Evol Microbiol (60), Pt 4 801-808.
Tachibana M., K. Watanabe, S. Kim, Y. Omata, K. Murata, T. Hammond, & M. Watarai.
2006. Antibodies to Brucella spp. in Pacific Bottlenonse Dolphins from the
Solomon Islands. J Wildlife Dis. 42: 412-414.
Thrusfield M., C. Ortega, I. de Blas, J.P. Noordhuizen & K. Frankena. 2001. Win Episcope
2.0: improved epidemiological software for veterinary medicine. Vet. Rec. 148:567-
572.

45
Tryland, M., L. Kleivane, A. Alfredsson, M. Kjeld, A. Arnason, S. Stuen & J. Godfroid.
1999. Evidence of Brucella infection in marine mammals in the North Atlantic
Ocean. Vet. Rec. 144: 588-592.
Van Bressem, M. F., K. Van Waerebeek, J. A. Raga, J. Godfroid, S. D. Brew & A. P. Mac
Millan. 2001. Serological evidence of Brucella species infection in odontocetes
from the South Pacific and the Mediterranean. Vet. Rec. 148: 657-661.
Van Bressem, M.F., J.A. Raga, G. Di Guardo, P.D. Jepson, P.J. Duignan, U.Siebert,
T.Barrett, M.C.O.Santos, I.B.Moreno, S. Siciliano, A.Aguilar, & K.V.Waerebeek.
2009. Emerging infectious diseases in cetaceans worldwide and the possible role of
environmental stressors. Vol. 86: 143-157.
Van Waerebeek K., M.F. Van Bressem, F. Félix, J. Alfaro- Shigeto, A. García-Godos, L.
Chávez- Lisambart, K. Ontón, D. Montes, & R. Bell.1997. Mortality of dolphins
and porpoises in coastal fisheries off Peru and southern Ecuador in 1994. Biol
Conserv 81: 43- 49.
Velasco, J., R. Díaz, M. J. Grilló, M. Barberán, C. Marín, J. M. Blasco, & I. Moriyón.
1997. Antibody and delayed-type hypersensitivity responses to Ochrobactrum
anthropi cytosolic and outer membrane antigens in infections by smooth and rough
Brucella spp. Clin. Diagn. Lab. Immunol. 4279-284.
Vinod P., M.K. Singh, M.K. Garg & A. Agarwal.2007.Extensive meningoencephalitis,
retrobulbar neuritis and pulmonary involvement in a patient with
neurobrucellosis.Neurology India, 55, 157-159
Vizcaino, N., P. Caro-Hernandez, A. Cloeckaert, & L. Fernandez-Lago. 2004. DNA
polymorphism in the omp25/omp31 family of Brucella spp.: identification of a 1.7-
kb inversion in Brucella cetaceae and of a 15.1-kb genomic island, absent from
Brucella ovis,related to the synthesis of smooth lipopolysaccharide. Microbes
Infect. 6:821-834.
Wehrtmann I.S, J. Cortés.2009. Marine Biodiversity of Costa Rica, Central America. 1rst
Ed. Springer Science.USA
Whatmore A.M, L.P Lorraine, & A. P. MacMillan. 2007. Characterisation of the genetic
diversity of Brucella by multilocus Sequencing. BMC Microbiology, 7:34.

46
Whatmore, A. M., C. E. Dawson, P. Groussaud, M. S. Koylass, A. C. King, S. J.
Shankster, A. H. Sohn, W. S. Probert, & W. L. McDonald. 2008. Marine mammal
Brucella genotype associated with zoonotic infection. Emerg. Infect. Dis. 14:517-518.
Whatmore, A.M. 2009.Current understanding of the genetic diversity of Brucella, an
expanding genus of zoonotic pathogens. Infect. Genet. Evol. 9:1168-1184.
Zygmun M. S, M. Maquart, N. Bernardet, B. Doublet, & A. Cloeckaert.2010. Novel IS711-
Specific Chromosomal Locations in Marine Mammal Brucella Strains Useful for
their Identification and Classification. J. Clin. Microbiol. doi:10.1128/JCM.01069-
10.
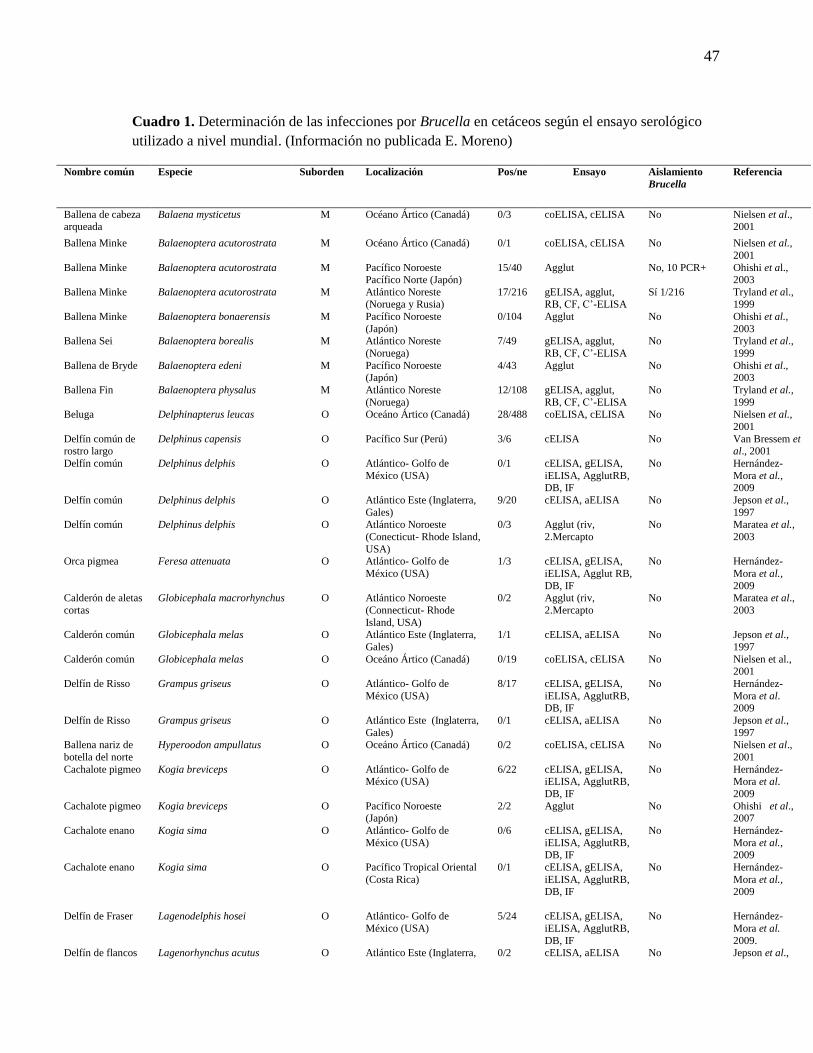
47
Cuadro 1. Determinación de las infecciones por Brucella en cetáceos según el ensayo serológico
utilizado a nivel mundial. (Información no publicada E. Moreno)
Nombre común Especie
Suborden Localización Pos/ne Ensayo Aislamiento
Brucella
Referencia
Ballena de cabeza arqueada
Balaena mysticetus M Océano Ártico (Canadá) 0/3 coELISA, cELISA No Nielsen et al., 2001
Ballena Minke Balaenoptera acutorostrata M Océano Ártico (Canadá) 0/1 coELISA, cELISA No Nielsen et al.,
2001
Ballena Minke Balaenoptera acutorostrata M Pacífico Noroeste Pacífico Norte (Japón)
15/40 Agglut No, 10 PCR+ Ohishi et al., 2003
Ballena Minke Balaenoptera acutorostrata M Atlántico Noreste
(Noruega y Rusia)
17/216 gELISA, agglut,
RB, CF, C’-ELISA
Sí 1/216 Tryland et al.,
1999
Ballena Minke Balaenoptera bonaerensis M Pacífico Noroeste
(Japón)
0/104 Agglut No Ohishi et al.,
2003 Ballena Sei Balaenoptera borealis M Atlántico Noreste
(Noruega)
7/49 gELISA, agglut,
RB, CF, C’-ELISA
No Tryland et al.,
1999
Ballena de Bryde Balaenoptera edeni M Pacífico Noroeste (Japón)
4/43 Agglut No Ohishi et al., 2003
Ballena Fin Balaenoptera physalus M Atlántico Noreste
(Noruega)
12/108 gELISA, agglut,
RB, CF, C’-ELISA
No Tryland et al.,
1999 Beluga Delphinapterus leucas O Oceáno Ártico (Canadá) 28/488 coELISA, cELISA No Nielsen et al.,
2001
Delfín común de rostro largo
Delphinus capensis O Pacífico Sur (Perú) 3/6 cELISA No Van Bressem et al., 2001
Delfín común Delphinus delphis O Atlántico- Golfo de
México (USA)
0/1 cELISA, gELISA,
iELISA, AgglutRB, DB, IF
No Hernández-
Mora et al., 2009
Delfín común Delphinus delphis O Atlántico Este (Inglaterra,
Gales)
9/20 cELISA, aELISA No Jepson et al.,
1997 Delfín común Delphinus delphis O Atlántico Noroeste
(Conecticut- Rhode Island,
USA)
0/3 Agglut (riv,
2.Mercapto
No Maratea et al.,
2003
Orca pigmea Feresa attenuata O Atlántico- Golfo de
México (USA)
1/3 cELISA, gELISA,
iELISA, Agglut RB,
DB, IF
No Hernández-
Mora et al.,
2009 Calderón de aletas
cortas
Globicephala macrorhynchus O Atlántico Noroeste
(Connecticut- Rhode
Island, USA)
0/2 Agglut (riv,
2.Mercapto
No Maratea et al.,
2003
Calderón común Globicephala melas O Atlántico Este (Inglaterra,
Gales)
1/1 cELISA, aELISA No Jepson et al.,
1997
Calderón común Globicephala melas O Oceáno Ártico (Canadá) 0/19 coELISA, cELISA No Nielsen et al., 2001
Delfín de Risso Grampus griseus O Atlántico- Golfo de
México (USA)
8/17 cELISA, gELISA,
iELISA, AgglutRB, DB, IF
No Hernández-
Mora et al. 2009
Delfín de Risso Grampus griseus O Atlántico Este (Inglaterra,
Gales)
0/1 cELISA, aELISA No Jepson et al.,
1997 Ballena nariz de
botella del norte
Hyperoodon ampullatus O Oceáno Ártico (Canadá) 0/2 coELISA, cELISA No Nielsen et al.,
2001
Cachalote pigmeo Kogia breviceps O Atlántico- Golfo de México (USA)
6/22 cELISA, gELISA, iELISA, AgglutRB,
DB, IF
No Hernández-Mora et al.
2009
Cachalote pigmeo Kogia breviceps O Pacífico Noroeste (Japón)
2/2 Agglut No Ohishi et al., 2007
Cachalote enano Kogia sima O Atlántico- Golfo de
México (USA)
0/6 cELISA, gELISA,
iELISA, AgglutRB, DB, IF
No Hernández-
Mora et al., 2009
Cachalote enano Kogia sima O Pacífico Tropical Oriental
(Costa Rica)
0/1 cELISA, gELISA,
iELISA, AgglutRB,
DB, IF
No Hernández-
Mora et al., 2009
Delfín de Fraser
Lagenodelphis hosei
O
Atlántico- Golfo de
México (USA)
5/24
cELISA, gELISA,
iELISA, AgglutRB,
DB, IF
No
Hernández-
Mora et al.
2009. Delfín de flancos Lagenorhynchus acutus O Atlántico Este (Inglaterra, 0/2 cELISA, aELISA No Jepson et al.,
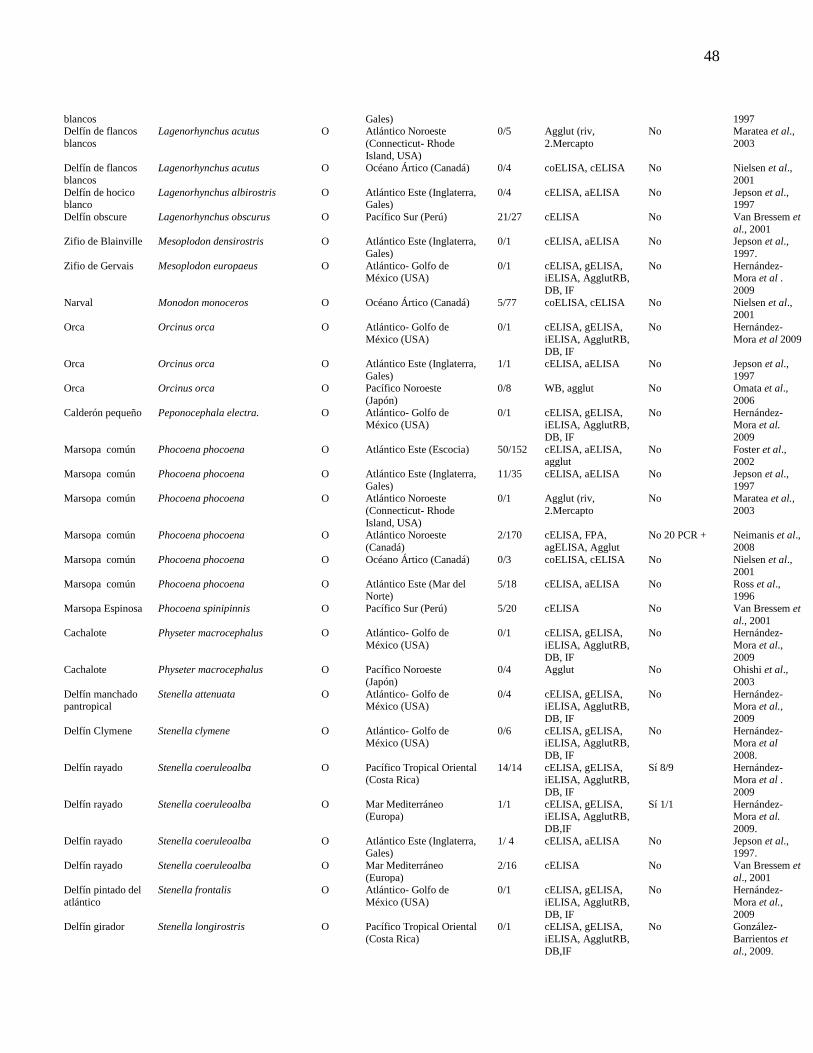
48
blancos Gales) 1997 Delfín de flancos
blancos
Lagenorhynchus acutus O Atlántico Noroeste
(Connecticut- Rhode
Island, USA)
0/5 Agglut (riv,
2.Mercapto
No Maratea et al.,
2003
Delfín de flancos
blancos
Lagenorhynchus acutus O Océano Ártico (Canadá) 0/4 coELISA, cELISA No Nielsen et al.,
2001
Delfín de hocico blanco
Lagenorhynchus albirostris O Atlántico Este (Inglaterra, Gales)
0/4 cELISA, aELISA No Jepson et al., 1997
Delfín obscure Lagenorhynchus obscurus O Pacífico Sur (Perú) 21/27 cELISA No Van Bressem et
al., 2001 Zifio de Blainville Mesoplodon densirostris O Atlántico Este (Inglaterra,
Gales)
0/1 cELISA, aELISA No Jepson et al.,
1997.
Zifio de Gervais Mesoplodon europaeus O Atlántico- Golfo de México (USA)
0/1 cELISA, gELISA, iELISA, AgglutRB,
DB, IF
No Hernández-Mora et al .
2009
Narval Monodon monoceros O Océano Ártico (Canadá) 5/77 coELISA, cELISA No Nielsen et al., 2001
Orca Orcinus orca O Atlántico- Golfo de
México (USA)
0/1 cELISA, gELISA,
iELISA, AgglutRB, DB, IF
No Hernández-
Mora et al 2009
Orca Orcinus orca O Atlántico Este (Inglaterra,
Gales)
1/1 cELISA, aELISA No Jepson et al.,
1997 Orca Orcinus orca O Pacífico Noroeste
(Japón)
0/8 WB, agglut No Omata et al.,
2006
Calderón pequeño Peponocephala electra. O Atlántico- Golfo de México (USA)
0/1 cELISA, gELISA, iELISA, AgglutRB,
DB, IF
No Hernández-Mora et al.
2009
Marsopa común Phocoena phocoena O Atlántico Este (Escocia) 50/152 cELISA, aELISA, agglut
No Foster et al., 2002
Marsopa común Phocoena phocoena O Atlántico Este (Inglaterra,
Gales)
11/35 cELISA, aELISA No Jepson et al.,
1997
Marsopa común Phocoena phocoena O Atlántico Noroeste
(Connecticut- Rhode
Island, USA)
0/1 Agglut (riv,
2.Mercapto
No Maratea et al.,
2003
Marsopa común Phocoena phocoena O Atlántico Noroeste
(Canadá)
2/170 cELISA, FPA,
agELISA, Agglut
No 20 PCR + Neimanis et al.,
2008
Marsopa común Phocoena phocoena O Océano Ártico (Canadá) 0/3 coELISA, cELISA No Nielsen et al., 2001
Marsopa común Phocoena phocoena O Atlántico Este (Mar del Norte)
5/18 cELISA, aELISA No Ross et al., 1996
Marsopa Espinosa Phocoena spinipinnis O Pacífico Sur (Perú) 5/20 cELISA No Van Bressem et
al., 2001 Cachalote Physeter macrocephalus O Atlántico- Golfo de
México (USA)
0/1 cELISA, gELISA,
iELISA, AgglutRB,
DB, IF
No Hernández-
Mora et al.,
2009 Cachalote Physeter macrocephalus O Pacífico Noroeste
(Japón)
0/4 Agglut No Ohishi et al.,
2003
Delfín manchado pantropical
Stenella attenuata O Atlántico- Golfo de México (USA)
0/4 cELISA, gELISA, iELISA, AgglutRB,
DB, IF
No Hernández-Mora et al.,
2009
Delfín Clymene Stenella clymene O Atlántico- Golfo de México (USA)
0/6 cELISA, gELISA, iELISA, AgglutRB,
DB, IF
No Hernández-Mora et al
2008.
Delfín rayado Stenella coeruleoalba O Pacífico Tropical Oriental (Costa Rica)
14/14 cELISA, gELISA, iELISA, AgglutRB,
DB, IF
Sí 8/9 Hernández-Mora et al .
2009
Delfín rayado Stenella coeruleoalba O Mar Mediterráneo (Europa)
1/1 cELISA, gELISA, iELISA, AgglutRB,
DB,IF
Sí 1/1 Hernández-Mora et al.
2009.
Delfín rayado Stenella coeruleoalba O Atlántico Este (Inglaterra, Gales)
1/ 4 cELISA, aELISA No Jepson et al., 1997.
Delfín rayado Stenella coeruleoalba O Mar Mediterráneo
(Europa)
2/16 cELISA No Van Bressem et
al., 2001
Delfín pintado del
atlántico
Stenella frontalis O Atlántico- Golfo de
México (USA)
0/1 cELISA, gELISA,
iELISA, AgglutRB,
DB, IF
No Hernández-
Mora et al.,
2009 Delfín girador Stenella longirostris O Pacífico Tropical Oriental
(Costa Rica)
0/1 cELISA, gELISA,
iELISA, AgglutRB,
DB,IF
No González-
Barrientos et
al., 2009.
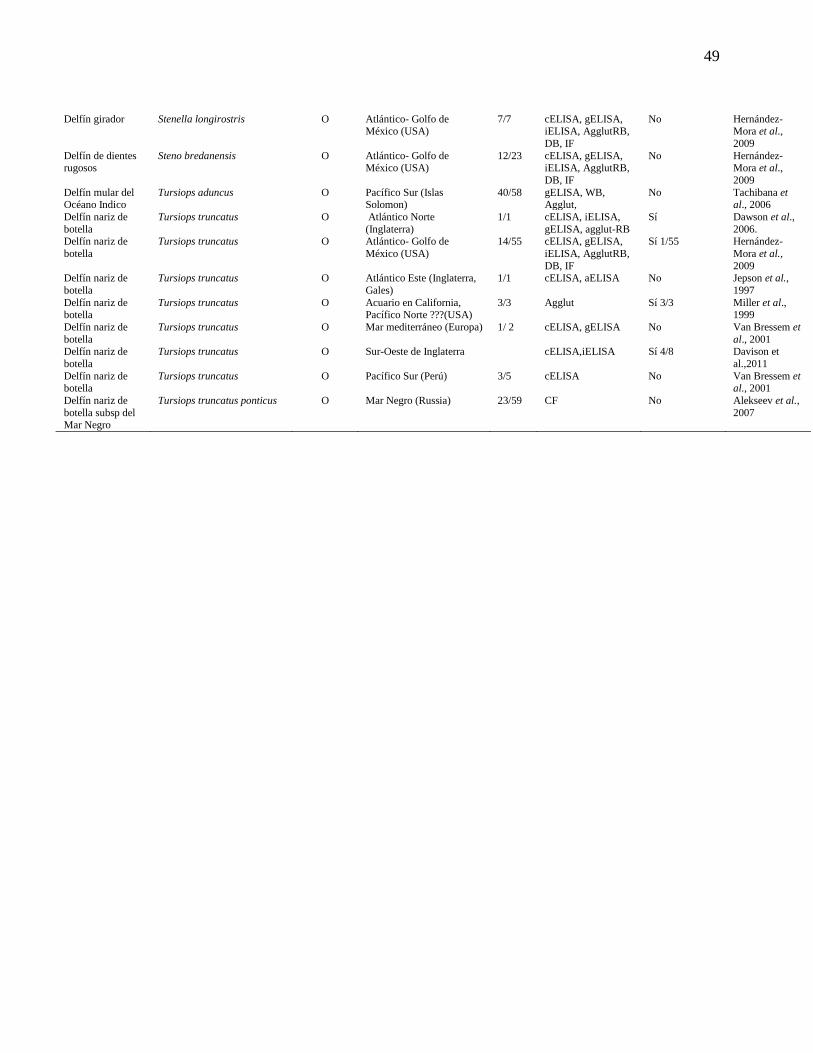
49
Delfín girador Stenella longirostris O Atlántico- Golfo de México (USA)
7/7 cELISA, gELISA, iELISA, AgglutRB,
DB, IF
No Hernández-Mora et al.,
2009
Delfín de dientes rugosos
Steno bredanensis O Atlántico- Golfo de México (USA)
12/23 cELISA, gELISA, iELISA, AgglutRB,
DB, IF
No Hernández-Mora et al.,
2009
Delfín mular del Océano Indico
Tursiops aduncus O Pacífico Sur (Islas Solomon)
40/58 gELISA, WB, Agglut,
No Tachibana et al., 2006
Delfín nariz de
botella
Tursiops truncatus O Atlántico Norte
(Inglaterra)
1/1 cELISA, iELISA,
gELISA, agglut-RB
Sí Dawson et al.,
2006. Delfín nariz de
botella
Tursiops truncatus O Atlántico- Golfo de
México (USA)
14/55 cELISA, gELISA,
iELISA, AgglutRB,
DB, IF
Sí 1/55 Hernández-
Mora et al.,
2009 Delfín nariz de
botella
Tursiops truncatus O Atlántico Este (Inglaterra,
Gales)
1/1 cELISA, aELISA No Jepson et al.,
1997
Delfín nariz de botella
Tursiops truncatus O Acuario en California, Pacífico Norte ???(USA)
3/3 Agglut Sí 3/3 Miller et al., 1999
Delfín nariz de
botella
Tursiops truncatus O Mar mediterráneo (Europa) 1/ 2 cELISA, gELISA No Van Bressem et
al., 2001 Delfín nariz de
botella
Tursiops truncatus O Sur-Oeste de Inglaterra cELISA,iELISA Sí 4/8 Davison et
al.,2011
Delfín nariz de botella
Tursiops truncatus O Pacífico Sur (Perú) 3/5 cELISA No Van Bressem et al., 2001
Delfín nariz de
botella subsp del Mar Negro
Tursiops truncatus ponticus O Mar Negro (Russia) 23/59 CF No Alekseev et al.,
2007
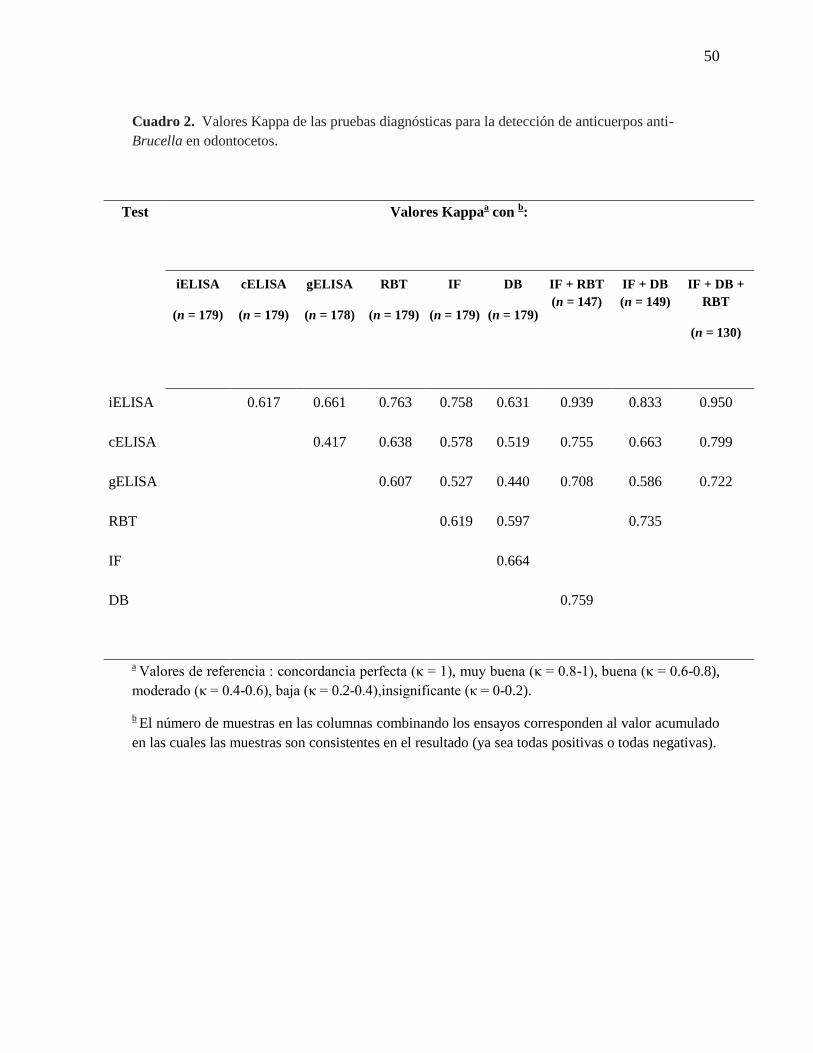
50
Cuadro 2. Valores Kappa de las pruebas diagnósticas para la detección de anticuerpos anti-
Brucella en odontocetos.
Test Valores Kappaa con
b:
iELISA
(n = 179)
cELISA
(n = 179)
gELISA
(n = 178)
RBT
(n = 179)
IF
(n = 179)
DB
(n = 179)
IF + RBT
(n = 147)
IF + DB
(n = 149)
IF + DB +
RBT
(n = 130)
iELISA 0.617 0.661 0.763 0.758 0.631 0.939 0.833 0.950
cELISA 0.417 0.638 0.578 0.519 0.755 0.663 0.799
gELISA 0.607 0.527 0.440 0.708 0.586 0.722
RBT 0.619 0.597 0.735
IF 0.664
DB 0.759
a Valores de referencia : concordancia perfecta (κ = 1), muy buena (κ = 0.8-1), buena (κ = 0.6-0.8),
moderado (κ = 0.4-0.6), baja (κ = 0.2-0.4),insignificante (κ = 0-0.2).
b El número de muestras en las columnas combinando los ensayos corresponden al valor acumulado
en las cuales las muestras son consistentes en el resultado (ya sea todas positivas o todas negativas).
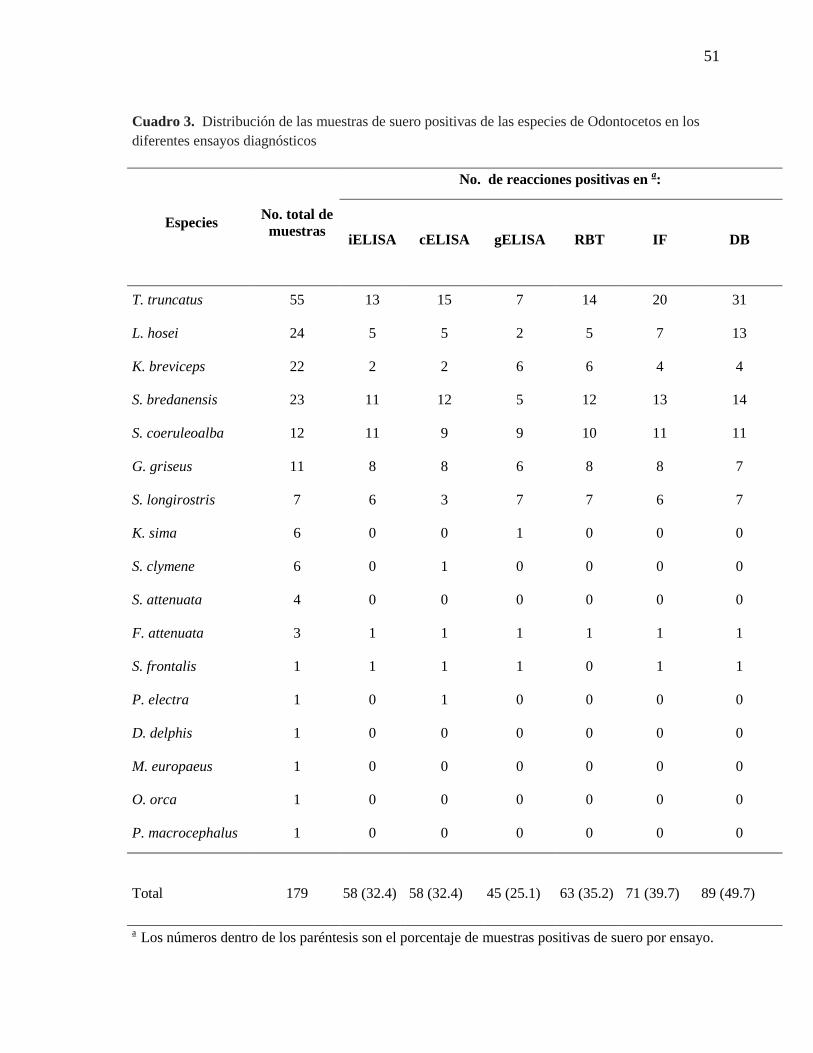
51
Cuadro 3. Distribución de las muestras de suero positivas de las especies de Odontocetos en los
diferentes ensayos diagnósticos
Especies No. total de
muestras
No. de reacciones positivas en a:
iELISA
cELISA
gELISA
RBT
IF
DB
T. truncatus 55 13 15 7 14 20 31
L. hosei 24 5 5 2 5 7 13
K. breviceps 22 2 2 6 6 4 4
S. bredanensis 23 11 12 5 12 13 14
S. coeruleoalba 12 11 9 9 10 11 11
G. griseus 11 8 8 6 8 8 7
S. longirostris 7 6 3 7 7 6 7
K. sima 6 0 0 1 0 0 0
S. clymene 6 0 1 0 0 0 0
S. attenuata 4 0 0 0 0 0 0
F. attenuata 3 1 1 1 1 1 1
S. frontalis 1 1 1 1 0 1 1
P. electra 1 0 1 0 0 0 0
D. delphis 1 0 0 0 0 0 0
M. europaeus 1 0 0 0 0 0 0
O. orca 1 0 0 0 0 0 0
P. macrocephalus 1 0 0 0 0 0 0
Total
179
58 (32.4)
58 (32.4)
45 (25.1)
63 (35.2)
71 (39.7)
89 (49.7)
a Los números dentro de los paréntesis son el porcentaje de muestras positivas de suero por ensayo.
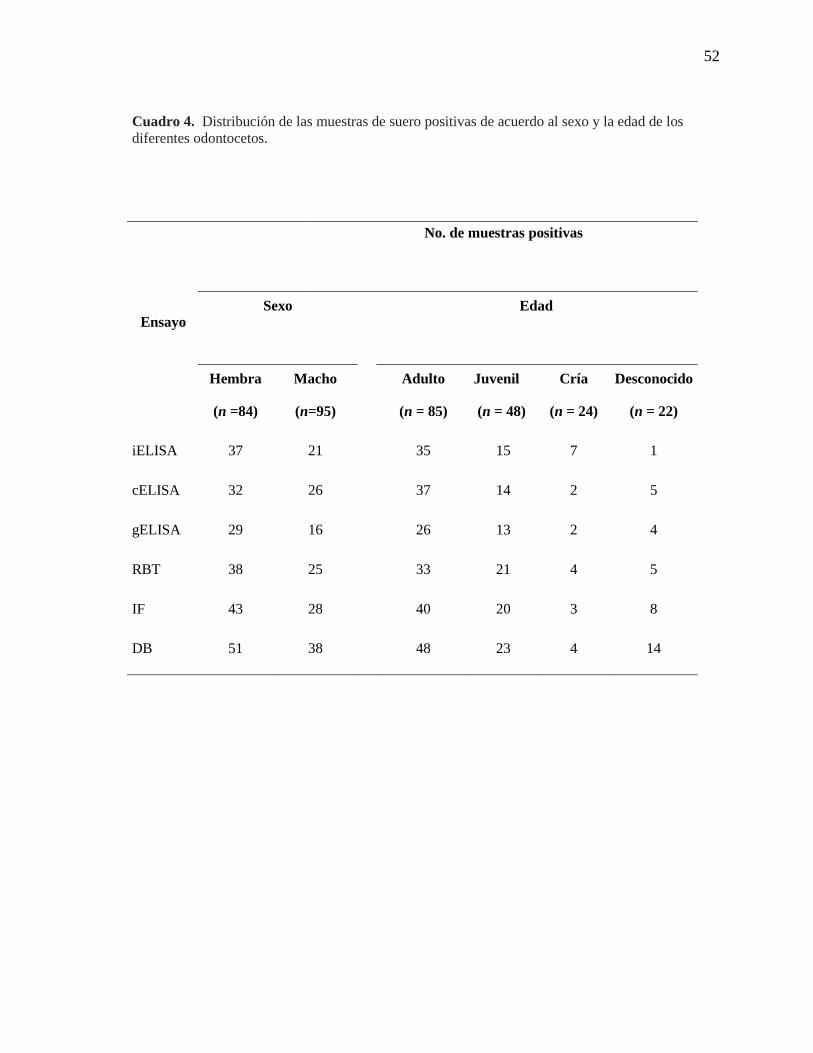
52
Cuadro 4. Distribución de las muestras de suero positivas de acuerdo al sexo y la edad de los
diferentes odontocetos.
Ensayo
No. de muestras positivas
Sexo
Edad
Hembra
(n =84)
Macho
(n=95)
Adulto
(n = 85)
Juvenil
(n = 48)
Cría
(n = 24)
Desconocido
(n = 22)
iELISA 37 21 35 15 7 1
cELISA 32 26 37 14 2 5
gELISA 29 16 26 13 2 4
RBT 38 25 33 21 4 5
IF 43 28 40 20 3 8
DB 51 38 48 23 4 14
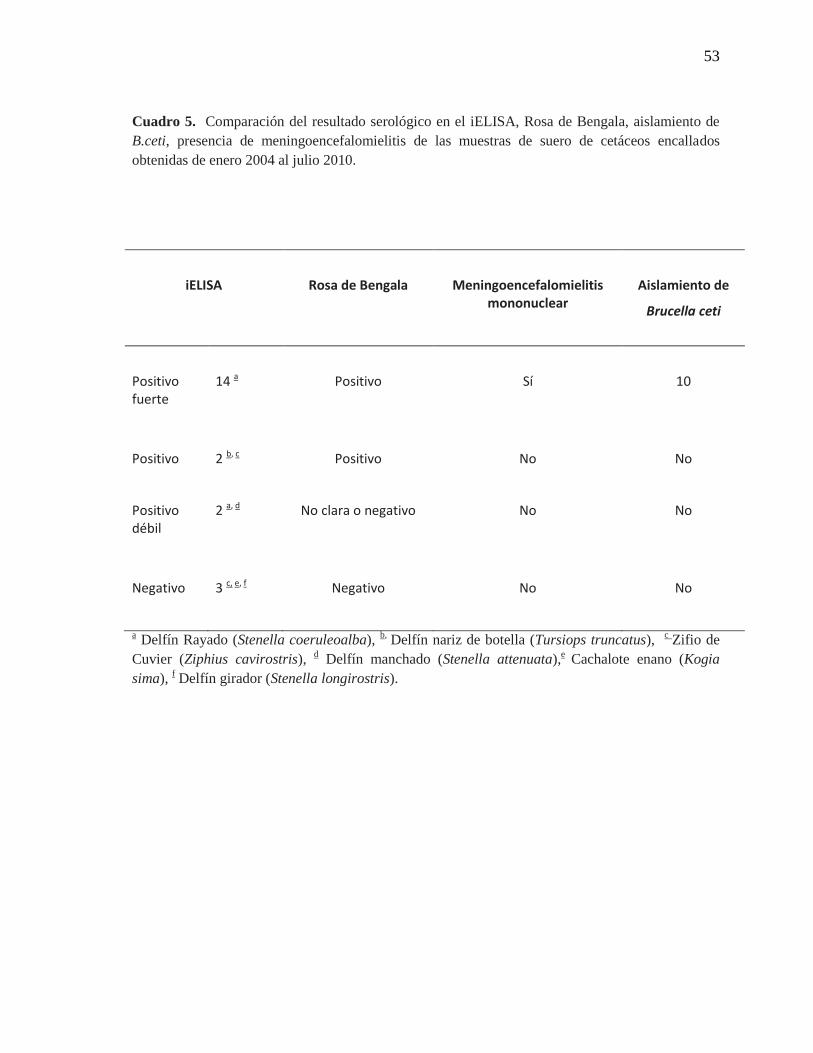
53
Cuadro 5. Comparación del resultado serológico en el iELISA, Rosa de Bengala, aislamiento de
B.ceti, presencia de meningoencefalomielitis de las muestras de suero de cetáceos encallados
obtenidas de enero 2004 al julio 2010.
iELISA
Rosa de Bengala
Meningoencefalomielitis mononuclear
Aislamiento de
Brucella ceti
Positivo fuerte
14 a
Positivo
Sí
10
Positivo 2 b, c Positivo No No
Positivo débil
2 a, d No clara o negativo No No
Negativo 3 c, e, f Negativo No No
a Delfín Rayado (Stenella coeruleoalba),
b, Delfín nariz de botella (Tursiops truncatus),
c Zifio de
Cuvier (Ziphius cavirostris), d Delfín manchado (Stenella attenuata),
e Cachalote enano (Kogia
sima), f Delfín girador (Stenella longirostris).

54
Figura 1. Distribución oceánica del diagnóstico serológico de las infecciones de Brucella
en cetáceos. Las reacciones positivas (numerador) en relación con el número total de
cetáceos analizados (denominador) está indicado luego del nombre de la especie. Datos
obtenidos de Alekseev et al., 2007; Dawson et al., 2006; Foster et al., 2002; González-
Barrientos et al., 2009; Hernández-Mora et al., 2008; Jepsone et al., 1997; Maratea et al.,
2003; Miller et al., 1999; Neimanis et al., 2008; Nielsen et al., 2001; Ohishi et al., 2007,
2003; Omata et al., 2006; Ross et al., 1996; Tachibana et al., 2006; Tryland et al., 1999;
Van Bressem et al., 2001. Los círculos corresponden al diagnóstico realizado para esta
investigación (Información no publicada E.Moreno)
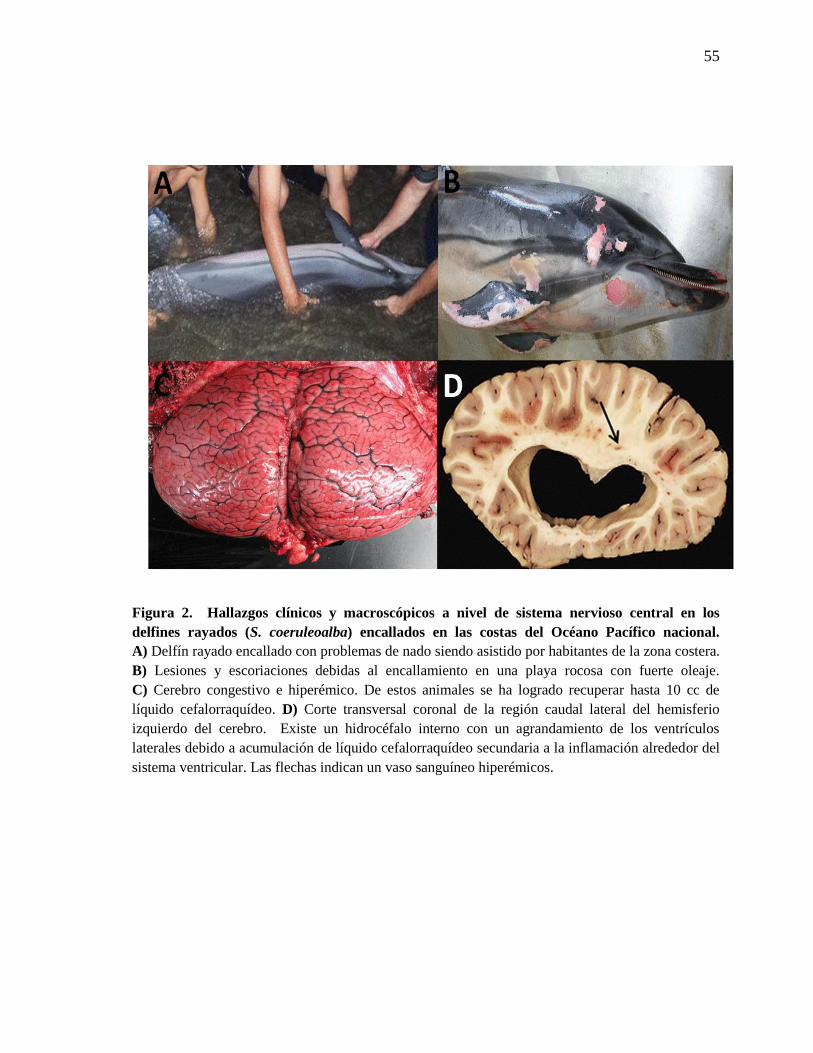
55
Figura 2. Hallazgos clínicos y macroscópicos a nivel de sistema nervioso central en los
delfines rayados (S. coeruleoalba) encallados en las costas del Océano Pacífico nacional.
A) Delfín rayado encallado con problemas de nado siendo asistido por habitantes de la zona costera.
B) Lesiones y escoriaciones debidas al encallamiento en una playa rocosa con fuerte oleaje.
C) Cerebro congestivo e hiperémico. De estos animales se ha logrado recuperar hasta 10 cc de
líquido cefalorraquídeo. D) Corte transversal coronal de la región caudal lateral del hemisferio
izquierdo del cerebro. Existe un hidrocéfalo interno con un agrandamiento de los ventrículos
laterales debido a acumulación de líquido cefalorraquídeo secundaria a la inflamación alrededor del
sistema ventricular. Las flechas indican un vaso sanguíneo hiperémicos.
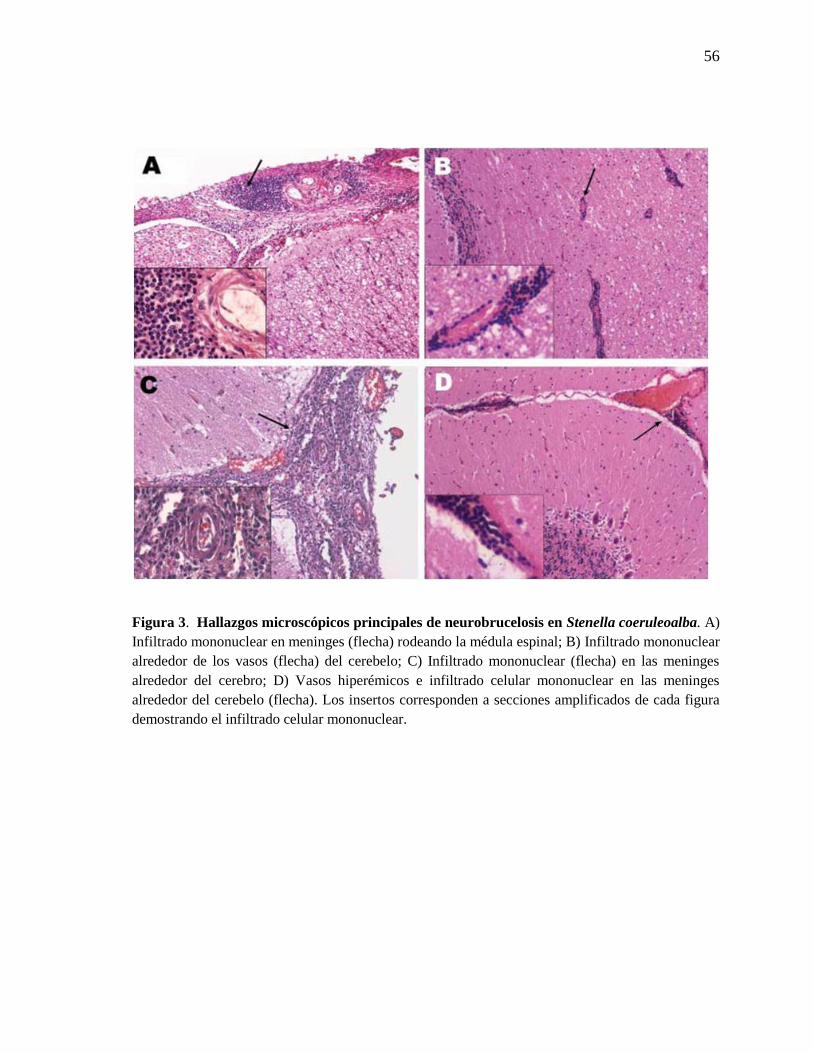
56
Figura 3. Hallazgos microscópicos principales de neurobrucelosis en Stenella coeruleoalba. A)
Infiltrado mononuclear en meninges (flecha) rodeando la médula espinal; B) Infiltrado mononuclear
alrededor de los vasos (flecha) del cerebelo; C) Infiltrado mononuclear (flecha) en las meninges
alrededor del cerebro; D) Vasos hiperémicos e infiltrado celular mononuclear en las meninges
alrededor del cerebelo (flecha). Los insertos corresponden a secciones amplificados de cada figura
demostrando el infiltrado celular mononuclear.
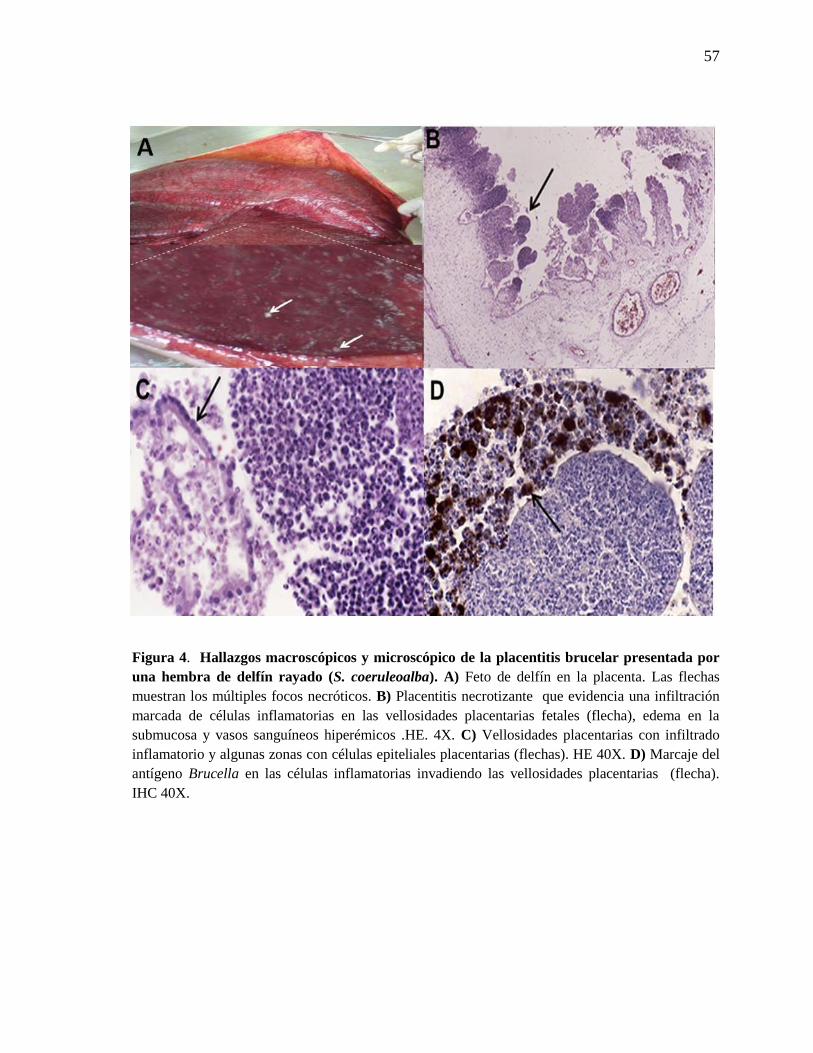
57
Figura 4. Hallazgos macroscópicos y microscópico de la placentitis brucelar presentada por
una hembra de delfín rayado (S. coeruleoalba). A) Feto de delfín en la placenta. Las flechas
muestran los múltiples focos necróticos. B) Placentitis necrotizante que evidencia una infiltración
marcada de células inflamatorias en las vellosidades placentarias fetales (flecha), edema en la
submucosa y vasos sanguíneos hiperémicos .HE. 4X. C) Vellosidades placentarias con infiltrado
inflamatorio y algunas zonas con células epiteliales placentarias (flechas). HE 40X. D) Marcaje del
antígeno Brucella en las células inflamatorias invadiendo las vellosidades placentarias (flecha).
IHC 40X.
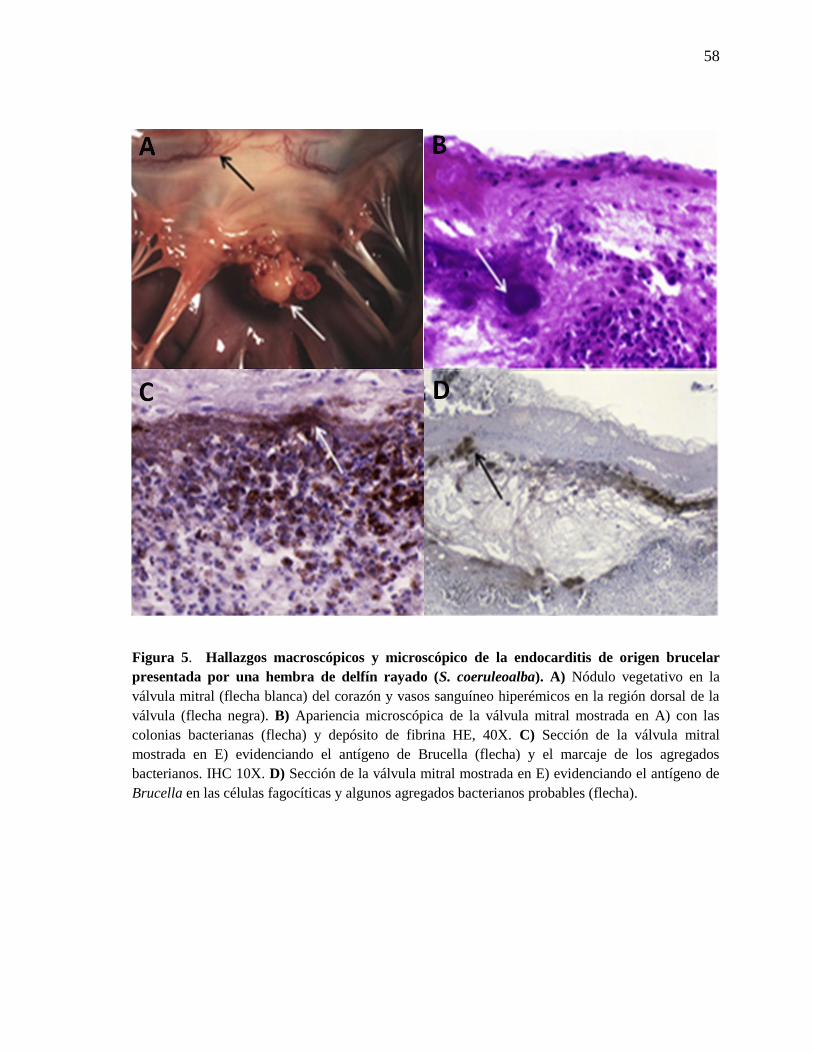
58
Figura 5. Hallazgos macroscópicos y microscópico de la endocarditis de origen brucelar
presentada por una hembra de delfín rayado (S. coeruleoalba). A) Nódulo vegetativo en la
válvula mitral (flecha blanca) del corazón y vasos sanguíneo hiperémicos en la región dorsal de la
válvula (flecha negra). B) Apariencia microscópica de la válvula mitral mostrada en A) con las
colonias bacterianas (flecha) y depósito de fibrina HE, 40X. C) Sección de la válvula mitral
mostrada en E) evidenciando el antígeno de Brucella (flecha) y el marcaje de los agregados
bacterianos. IHC 10X. D) Sección de la válvula mitral mostrada en E) evidenciando el antígeno de
Brucella en las células fagocíticas y algunos agregados bacterianos probables (flecha).
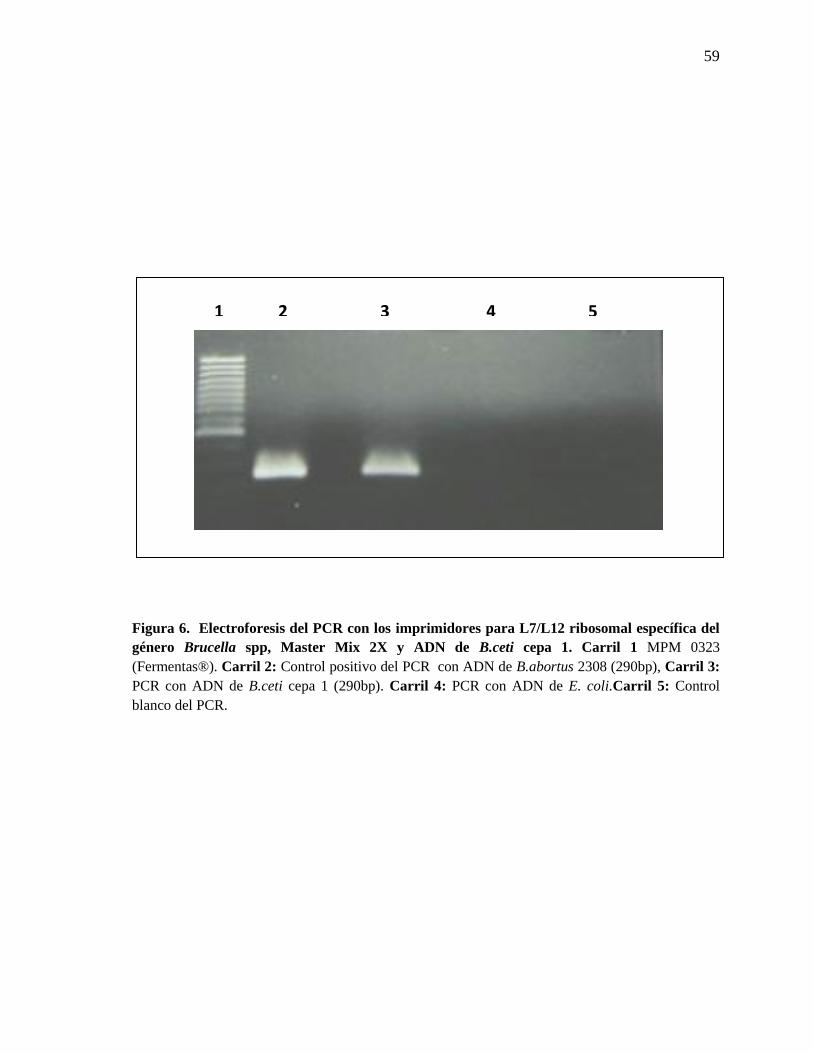
59
Figura 6. Electroforesis del PCR con los imprimidores para L7/L12 ribosomal específica del
género Brucella spp, Master Mix 2X y ADN de B.ceti cepa 1. Carril 1 MPM 0323
(Fermentas®). Carril 2: Control positivo del PCR con ADN de B.abortus 2308 (290bp), Carril 3:
PCR con ADN de B.ceti cepa 1 (290bp). Carril 4: PCR con ADN de E. coli.Carril 5: Control
blanco del PCR.
1 2 3 4 5
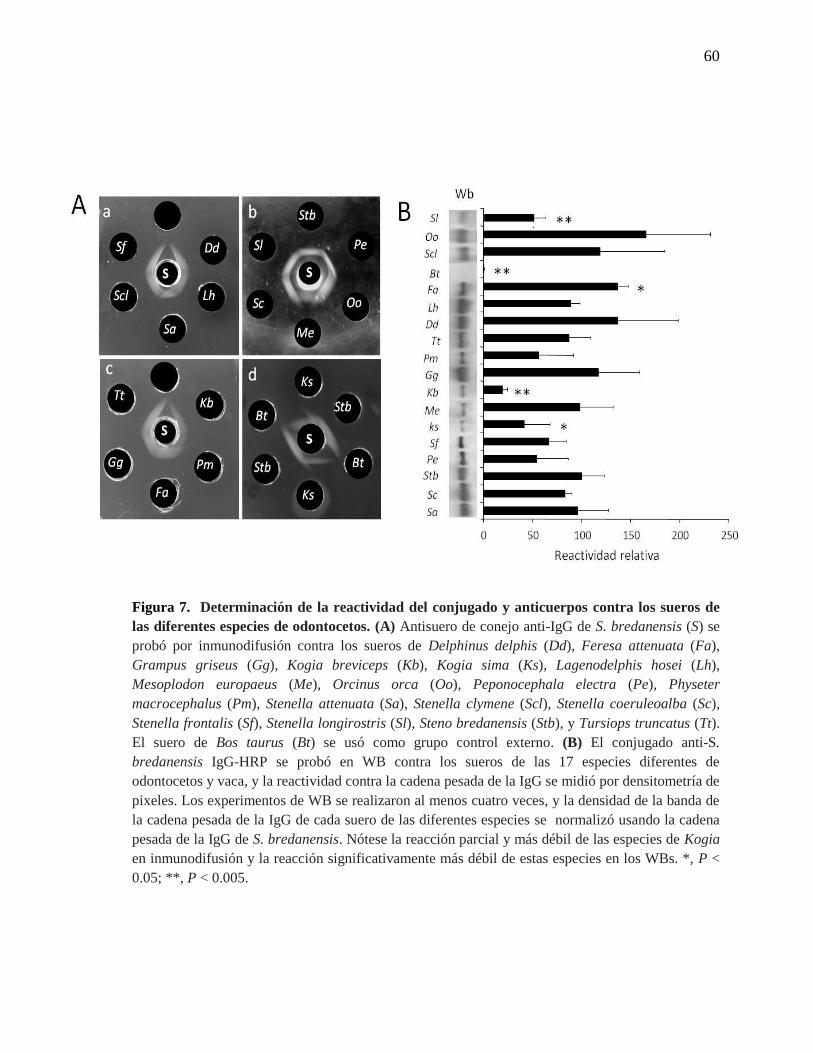
60
Figura 7. Determinación de la reactividad del conjugado y anticuerpos contra los sueros de
las diferentes especies de odontocetos. (A) Antisuero de conejo anti-IgG de S. bredanensis (S) se
probó por inmunodifusión contra los sueros de Delphinus delphis (Dd), Feresa attenuata (Fa),
Grampus griseus (Gg), Kogia breviceps (Kb), Kogia sima (Ks), Lagenodelphis hosei (Lh),
Mesoplodon europaeus (Me), Orcinus orca (Oo), Peponocephala electra (Pe), Physeter
macrocephalus (Pm), Stenella attenuata (Sa), Stenella clymene (Scl), Stenella coeruleoalba (Sc),
Stenella frontalis (Sf), Stenella longirostris (Sl), Steno bredanensis (Stb), y Tursiops truncatus (Tt).
El suero de Bos taurus (Bt) se usó como grupo control externo. (B) El conjugado anti-S.
bredanensis IgG-HRP se probó en WB contra los sueros de las 17 especies diferentes de
odontocetos y vaca, y la reactividad contra la cadena pesada de la IgG se midió por densitometría de
pixeles. Los experimentos de WB se realizaron al menos cuatro veces, y la densidad de la banda de
la cadena pesada de la IgG de cada suero de las diferentes especies se normalizó usando la cadena
pesada de la IgG de S. bredanensis. Nótese la reacción parcial y más débil de las especies de Kogia
en inmunodifusión y la reacción significativamente más débil de estas especies en los WBs. *, P <
0.05; **, P < 0.005.
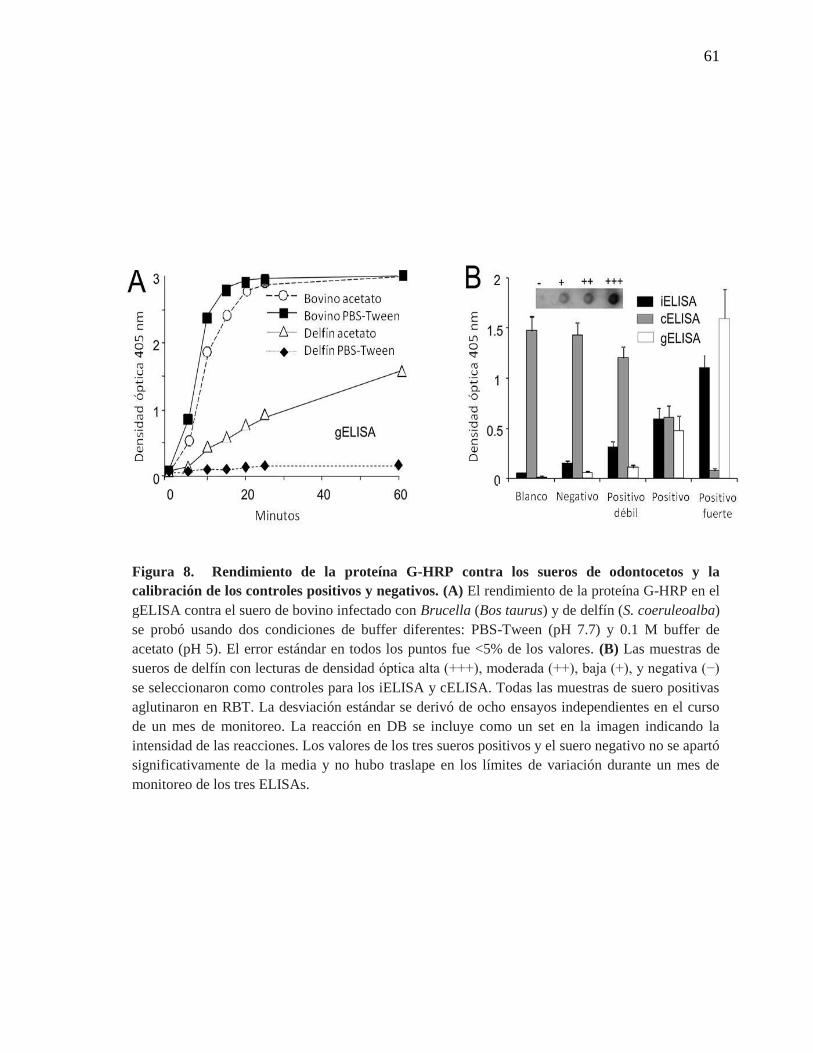
61
Figura 8. Rendimiento de la proteína G-HRP contra los sueros de odontocetos y la
calibración de los controles positivos y negativos. (A) El rendimiento de la proteína G-HRP en el
gELISA contra el suero de bovino infectado con Brucella (Bos taurus) y de delfín (S. coeruleoalba)
se probó usando dos condiciones de buffer diferentes: PBS-Tween (pH 7.7) y 0.1 M buffer de
acetato (pH 5). El error estándar en todos los puntos fue <5% de los valores. (B) Las muestras de
sueros de delfín con lecturas de densidad óptica alta (+++), moderada (++), baja (+), y negativa (−)
se seleccionaron como controles para los iELISA y cELISA. Todas las muestras de suero positivas
aglutinaron en RBT. La desviación estándar se derivó de ocho ensayos independientes en el curso
de un mes de monitoreo. La reacción en DB se incluye como un set en la imagen indicando la
intensidad de las reacciones. Los valores de los tres sueros positivos y el suero negativo no se apartó
significativamente de la media y no hubo traslape en los límites de variación durante un mes de
monitoreo de los tres ELISAs.
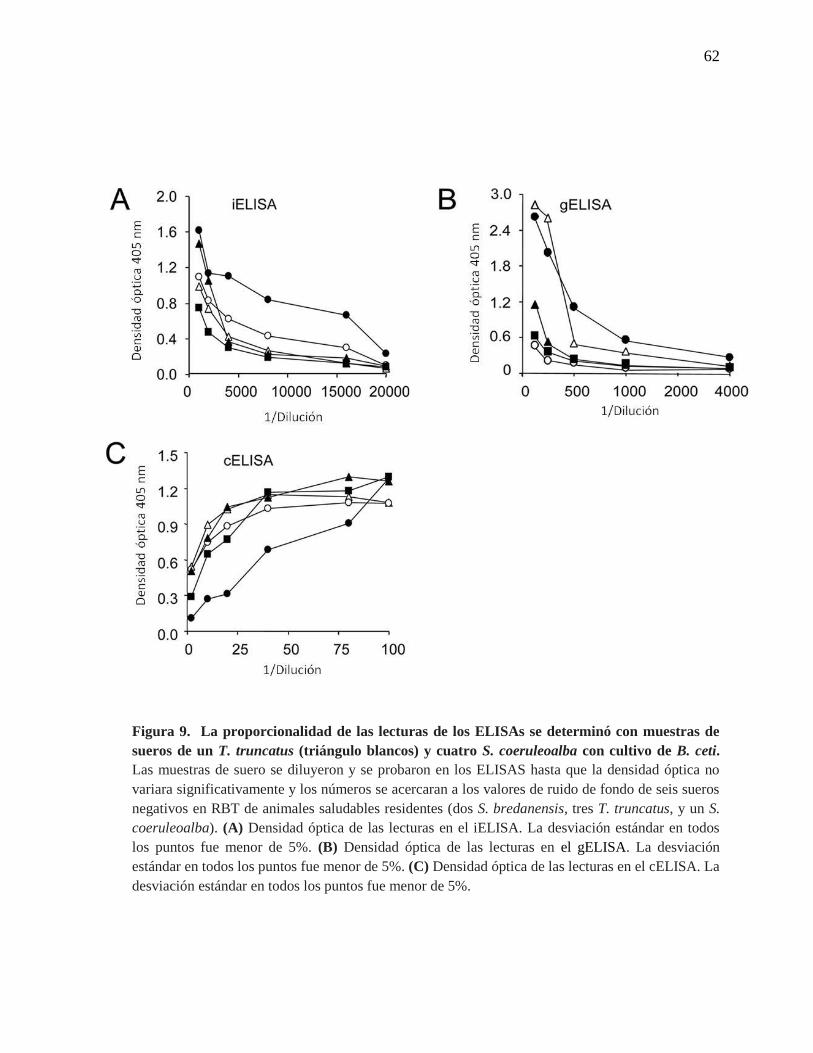
62
Figura 9. La proporcionalidad de las lecturas de los ELISAs se determinó con muestras de
sueros de un T. truncatus (triángulo blancos) y cuatro S. coeruleoalba con cultivo de B. ceti.
Las muestras de suero se diluyeron y se probaron en los ELISAS hasta que la densidad óptica no
variara significativamente y los números se acercaran a los valores de ruido de fondo de seis sueros
negativos en RBT de animales saludables residentes (dos S. bredanensis, tres T. truncatus, y un S.
coeruleoalba). (A) Densidad óptica de las lecturas en el iELISA. La desviación estándar en todos
los puntos fue menor de 5%. (B) Densidad óptica de las lecturas en el gELISA. La desviación
estándar en todos los puntos fue menor de 5%. (C) Densidad óptica de las lecturas en el cELISA. La
desviación estándar en todos los puntos fue menor de 5%.
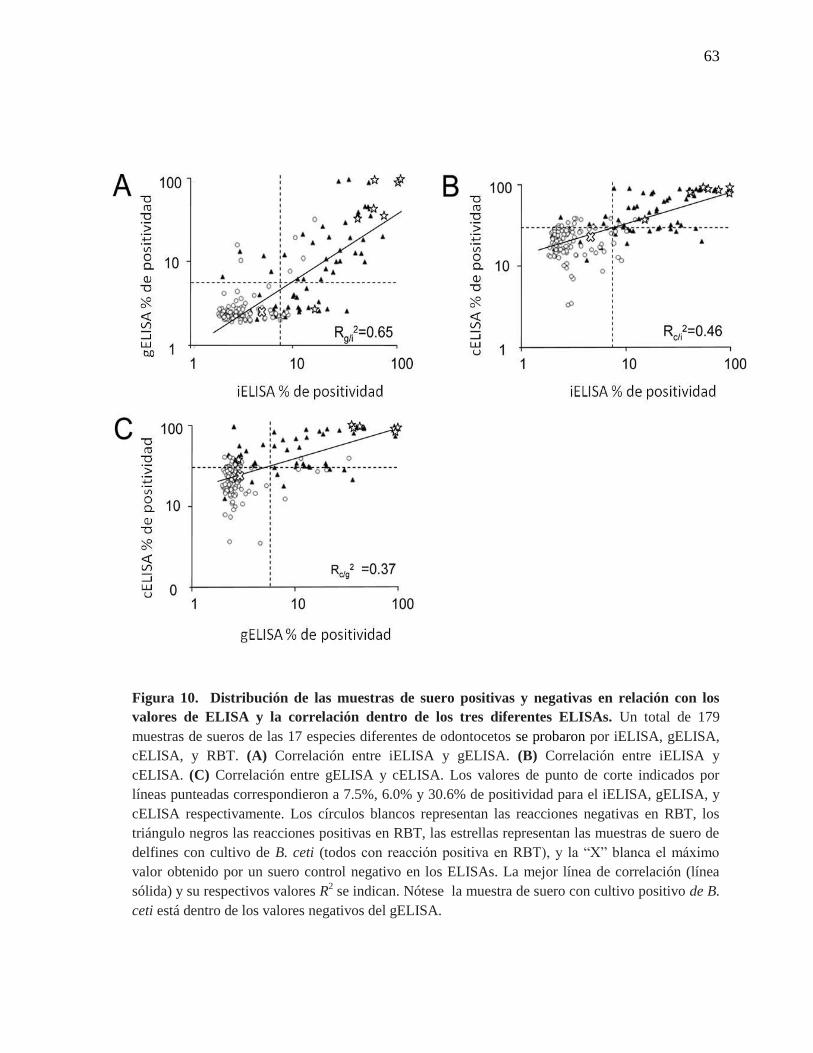
63
Figura 10. Distribución de las muestras de suero positivas y negativas en relación con los
valores de ELISA y la correlación dentro de los tres diferentes ELISAs. Un total de 179
muestras de sueros de las 17 especies diferentes de odontocetos se probaron por iELISA, gELISA,
cELISA, y RBT. (A) Correlación entre iELISA y gELISA. (B) Correlación entre iELISA y
cELISA. (C) Correlación entre gELISA y cELISA. Los valores de punto de corte indicados por
líneas punteadas correspondieron a 7.5%, 6.0% y 30.6% de positividad para el iELISA, gELISA, y
cELISA respectivamente. Los círculos blancos representan las reacciones negativas en RBT, los
triángulo negros las reacciones positivas en RBT, las estrellas representan las muestras de suero de
delfines con cultivo de B. ceti (todos con reacción positiva en RBT), y la “X” blanca el máximo
valor obtenido por un suero control negativo en los ELISAs. La mejor línea de correlación (línea
sólida) y su respectivos valores R2 se indican. Nótese la muestra de suero con cultivo positivo de B.
ceti está dentro de los valores negativos del gELISA.
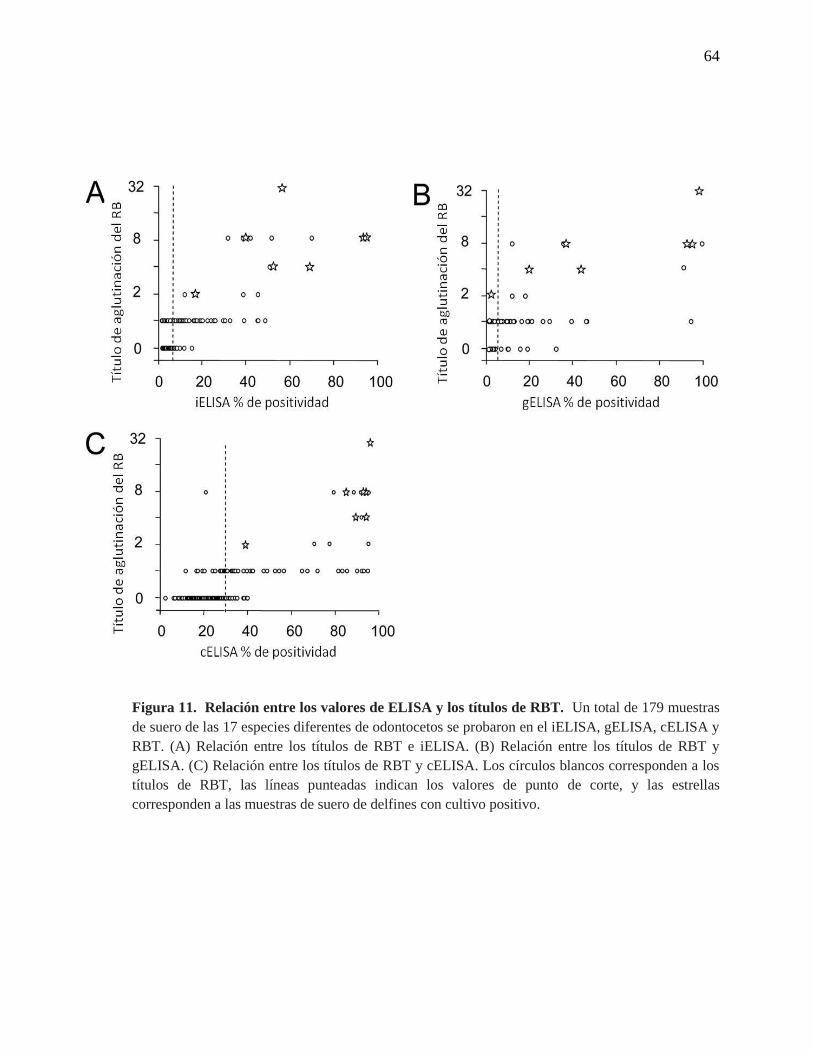
64
Figura 11. Relación entre los valores de ELISA y los títulos de RBT. Un total de 179 muestras
de suero de las 17 especies diferentes de odontocetos se probaron en el iELISA, gELISA, cELISA y
RBT. (A) Relación entre los títulos de RBT e iELISA. (B) Relación entre los títulos de RBT y
gELISA. (C) Relación entre los títulos de RBT y cELISA. Los círculos blancos corresponden a los
títulos de RBT, las líneas punteadas indican los valores de punto de corte, y las estrellas
corresponden a las muestras de suero de delfines con cultivo positivo.

65
Figura 12. Determinación de las sensibilidades y las especificidades usando RBT como ensayo
de referencia. Los valores de punto de corte se expresan como porcentaje de positividad y se
determinaron teóricamente bajo las curvas ROC de acuerdo a los valores aproximados de mayor
sensibilidad y especificidad con referencia a los porcentajes predictivos. Se estimaron los valores
Kappa (κ) para cada experimento. Las barras negras son las muestras de suero negativas, y las
barras blancas indican las muestras de suero positivas. Se, sensibilidad; Sp, especificidad; cut off,
punto de corte.
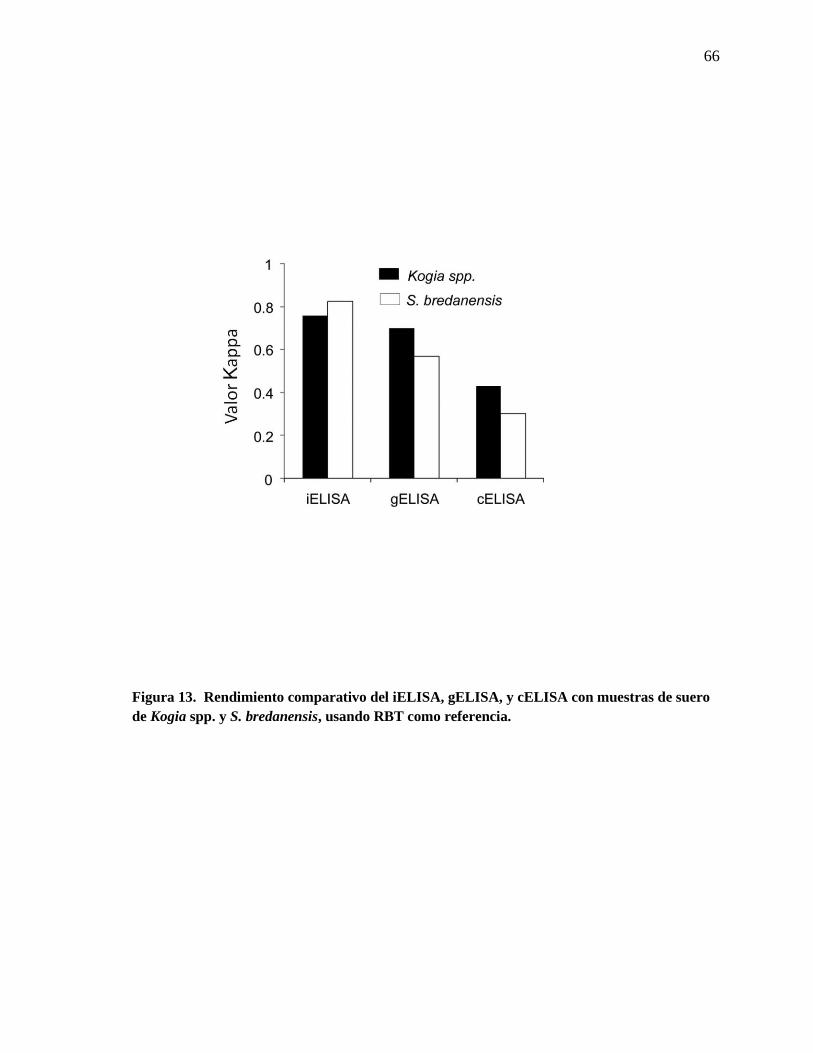
66
Figura 13. Rendimiento comparativo del iELISA, gELISA, y cELISA con muestras de suero
de Kogia spp. y S. bredanensis, usando RBT como referencia.
67
Figura 14. Relación filogenética entre los mayores linajes de cetáceos según Nikaido M. et al.,
(2001).

68
Figura 15. Contacto entre cetáceos encallados, animales domésticos y humanos en playa
Matapalo, Puntarenas, Costa Rica. El traslado del ganado por la playa de una finca a otra es una
práctica diaria, siendo ésta la playa de donde B. ceti ha sido aislada más frecuentemente de los
cetáceos encallados desde el año 2006.
69
Figura 16. Distribución mundial del delfín rayado (Stenella coeruleoalba). Aunque el rango
natural reportado para esta especie es 50° N a 40°S existen avistamientos en zonas menos templadas
según Jefferson T.A et al., 2008.

70
APÉNDICE





























