Embed Size (px)
Citation preview

De
Ca
b
a
ARRAA
KADCMV
1
fraaplNpa
tfdmoims
MT
h0
Acta Tropica 149 (2015) 8–14
Contents lists available at ScienceDirect
Acta Tropica
jo u r n al homep age: www.elsev ier .com/ locate /ac ta t ropica
ifferent serotypes of dengue viruses differently regulate thexpression of the host cell antigen processing machinery
hye Sheng Gana,b,∗, Rohana Yusofa,b, Shatrah Othmana,b
Department of Molecular Medicine, Faculty of Medicine, University of Malaya, 50603 Kuala Lumpur, MalaysiaDrug Design and Development Research Group, University of Malaya, 50603 Kuala Lumpur, Malaysia
r t i c l e i n f o
rticle history:eceived 19 September 2014eceived in revised form 14 April 2015ccepted 6 May 2015vailable online 14 May 2015
a b s t r a c t
Dengue virus (DV) infection demonstrates an intriguing virus-induced intracellular membrane alterationthat results in the augmentation of major histocompatibility complex (MHC) class I-restricted antigenpresentation. As oppose to its biological function in attracting CD8+ T-cells, this phenomenon appears tofacilitate the immune evasion. However, the molecular events that attribute to the dysregulation of theantigen presenting mechanism (APM) by DV remain obscure. In this study, we aimed to characterize the
eywords:ntigen presentation mechanismenguelass Iajor histocompatibility complex
host cell APM upon infection with all serotypes of whole DV. Cellular RNA were isolated from infectedcells and the gene expressions of LMP2, LMP7, TAP1, TAP2, TAPBP, CALR, CANX, PDIA3, HLA-A and HLA-B were analyzed via quantitative PCR. The profiles of the gene expression were further validated. Weshowed that all four DV serotypes modulate host APM at the proteasomal level with DV2 showing themost prominent expression profile.
irus
. Introduction
Dengue virus (DV) belongs to the Flaviviridae family, comprisingour different serotypes that share similar genome organization andeplication cycle. DV consists of positive single-stranded RNA with
size of approximately 11 kb, flanked by two untranslated regionst its respective ends. The viral genome encodes three structuralroteins: capsid protein, precursor membrane protein and enve-
ope protein; and seven non-structural (NS) proteins: NS1, NS2A,S2B, NS3, NS4A, NS4B and NS5. The virus is transmitted to humansredominantly by the mosquito bites of Aedes aegypti and Aedeslbopictus (Clyde et al., 2006).
Despite the antigenically distinct serotypes, clinical manifesta-ions of DV infection remain the same, ranging from asymptomaticever to severe conditions such as dengue hemorrhagic fever andengue shock syndrome (Guzman and Kouri, 2002). Although theeticulous classification of dengue diseases has been well devel-
ped, the limited understanding of the molecular mechanisms
nvolved in the interactions between DV and the antigen processingachinery (APM) of infected cells impinges on the therapeutictrategies. Previous studies of DV infection have demonstrated the
∗ Corresponding author at: Department of Molecular Medicine, Faculty ofedicine, University of Malaya, 50603 Kuala Lumpur, Malaysia.
el.: +60 379674716.E-mail address: [email protected] (C.S. Gan).
ttp://dx.doi.org/10.1016/j.actatropica.2015.05.005001-706X/© 2015 Elsevier B.V. All rights reserved.
© 2015 Elsevier B.V. All rights reserved.
induction and enhancement of major histocompatibility complex(MHC) class I antigen presentation protein, human leukocyte anti-gen (HLA) (Hershkovitz et al., 2008; Othman et al., 2010). Theenhancement of HLA was subsequently demonstrated to possessprotective effect in immune response thus, unraveled anotheralternative mechanism of immune escape (Hershkovitz et al.,2008). Nevertheless, the complete molecular event of this immuneescape is currently not well characterized.
The MHC class I antigen presenting pathway plays a pivotal rolein signaling the patrolling immune cells in times of viral infec-tion. MHC class I molecules are found on every nucleated cell ofthe body and these complexes present peptide fragments found inthe cytoplasm. In the case of viral infection, where both the viralproteins and viral genome are abundant, peptides that originatefrom the virus may also be presented. Specific cytotoxic T lym-phocytes (CTL) continuously monitor for cell surface MHC class Imolecules that present viral peptides and this eventually leads toeradication of infected cells (Hewitt, 2003). As it has been reviewed,the MHC class I molecules are assembled in the endoplasmic reti-culum (ER) with the peptides derived from a myriad of availablecytosolic proteins such as the multicatalytic proteasome complex.These peptides were then percolated into the ER by the ATP-dependent peptide transporter associated with antigen processing
(TAP). Present in two subunits, TAP1 and TAP2, the ambulant pep-tides undergo selective pressure of specific motifs and lengths. Inthe ER, cofactors orchestrate the loading of these peptides onto theHLA class I molecules. Upon proper loading, the complex is directed
Tropic
t(ulwferi
iioPei
2
2
tbih1alg1p
2
mMpc1psaubtroa
ssvtu5gNtrg45
Results are expressed as mean ± standard error. Compari-
C.S. Gan et al. / Acta
o the trans-Golgi apparatus and traverses to the cell membraneSeliger et al., 2000). Alas, viruses were shown to have developednique mechanisms to hijack the APM, impeding the pathway that
eads to immune response. As an example, cells that were infectedith HIV demonstrated interference in MHC presentation. This is
ound to be brought about by the Nef protein. Nef is a proteinxpressed by the HIV virus and its expression resulted in the down-egulation of HLA molecules. This phenomenon has caused thenefficiency in CTL function in eradicating the virus (Petersen et al.,2003).
In this study we characterized the APM of the host cell upon DVnfection. Human liver carcinoma cell line, HepG2, were employedn order to interrogate post-infection changes in the expressionf genes and proteins involved in APM by quantitative real-timeCR and Western blot, respectively. We therefore present here thexpression profiles of genes involved in the host cell APM upon DVnfection.
. Materials and methods
.1. Cell culture
HepG2 and C6/36 cells were obtained from American Type Cul-ure Collection (ATCC, USA). The HepG2 cell line was maintainedy regular sub-cultivating in 1× growth media of Dulbecco’s mod-
fied Eagle medium, DMEM (Gibco, USA), supplemented with 10%eat-inactivated fetal bovine serum (FBS), 100 U/mL of penicillin,00 U/mL of streptomycin and 20 mM of HEPES buffer at 37 ◦C in
humidified incubator in the presence of 5% CO2. The C6/36 celline was maintained by regular sub-cultivation in a 1× Leibovitz’srowth medium L-15 (Gibco, USA), supplemented with 10% FBS,00 U/mL of penicillin, 100 U/mL of streptomycin and 10% tryptosehosphate broth (Sigma, USA) at room temperature.
.2. Virus stocks
All four dengue virus strains were kind gifts from the Depart-ent of Medical Microbiology, Faculty of Medicine, University ofalaya. The virus stocks were prepared by promoting infection on
reformed monolayer of C6/36 cells, in a standard 75 cm2 tissueulture flask, in 3 mL of Leibovitz’s L-15 medium containing 2% FBS,00 U/mL of penicillin, 100 U/mL of streptomycin and 10% tryptosehosphate broth. The flask were left for 2 h for viral adsorption. Sub-equently, to encourage viral propagation, cell culture medium wasdded to 10 mL and incubated at room temperature for 6–7 days orntil the cytophatic effect (CPE) was observed. Virus was harvestedy one cycle of freeze-thaw, centrifuged at 1500 × g for 5 min andhe supernatant was finally filtered through polyvinyldene fluo-ide, PVDF, membrane (Sartorius AG, Germany) with the pore sizef 0.1 �m. The virus, present in the filtrate, was collected in smallliquots and stored at −80 ◦C until further use.
Serotyping of the virus strain was performed via reverse tran-cription PCR (RT-PCT) technique of the viral RNA using strainpecific primers. Total viral RNA was extracted using QIAampiral RNA mini kit (Qiagen, Germany) adhering to manufac-urer’s instruction. Serotyping of the virus stocks were confirmedsing the following primers: dengue virus serotype-1 NS3 region:′-atctataggtaccggcggtggaggttcaggagtgctatcc-3′ and 5′-attgatcaa-ctttaagtttcttttccta-3′, 700 base pairs; dengue virus serotype-2S2B-NS3 region: 5′-ccgggatccgccgatttggaactg-3′ and 5′-cccaagct-
caattttctctttcg-3′, 820 base pairs; dengue virus serotype-3 NS3
egion: 5′-atctataggtaccggcggtggaggttccggcgttttatgg-3′ and 5′-att-atcaagcttcaggtttcgcttttt-3′, 200 base pairs; dengue virus serotype-NS2 region: 5′-atctataggtaccggcggtggaggttcaggagccctgtgg-3′ and′-attgatcaagctttaatcttttctttcg-3′, 600 base pairs.
a 149 (2015) 8–14 9
Titers of the virus stocks were determined as previouslydescribed (Payne et al., 2006).
2.3. Analysis of mRNA expression
Preformed monolayer of HepG2 cell were either mock or denguevirus infected at a multiplicity of infection (m.o.i.) of one. Similarly,infections were performed on the cells at a ratio of 1 mL/25 cm2
of surface area. After 2 h of incubation at humidified incubatorfor effective viral adsorption, four times volume of 1× DMEMsupplemented with 2% FBS, 100 U/mL of penicillin, 100 U/mL ofstreptomycin and 20 mM of HEPES buffer were added into theflask and further incubated for an additional 48 h. The sameprocedure were repeated for each distinct serotypes of denguevirus.
Cellular RNA were isolated using the RNeasy mini kit (Qia-gen, Germany) adhering to the manufacturer’s instructions. Thepurified RNA was then subjected to quantification by spectropho-tometer and accessed for integrity by 2% denaturing agarosegel electrophoresis. Compliance to the recommended protocol,600 ng of RNA were reverse transcribed using high capacityRNA-to-cDNA kit (ABI, USA). TaqMan gene expression assays(ABI, USA) were used to assess the gene expression of the follow-ing genes: LMP2: HS00160610 m1, LMP7: HS00544760 g1, TAP1:HS00388675 m1, TAP2: HS00241060 m1, TAPBP: HS00917451 g1,PDIA3: HS00607126 m1, CALR: HS00189032 m1, CANX:HS01558409 m1, HLA-A: HS01058806 g1 and custom madeprobe set for HLA-B (Forward: 5′-AGCCCCGCTTCATCTCAGT-3′;Reverse: 5′-CGCTGTCGAACCTCACGAA-3′ and Probe: 5′-NEDTA-CGTGGACGACACCCAMGBNFQ-3′). In all assays, 40 ng of cDNAwere used and GAPDH were referred as the housekeeping gene.
2.4. Analysis of proteins expression by Western blot
Respective proteins were analyzed by Western blot using thefollowing antibodies: anti-tapasin antibody ab13518 (Abcam, USA),anti-MHC class I + HLA B [EP2624] ab110645 (Abcam, USA), anti-proteasome 20S LMP2 antibody ab3328 (Abcam, USA), anti-TAP1antibody ab13516 (Abcam, USA), GAPDH (A-3): sc-137179 (SantaCruz Biotechnology, Inc., USA), goat anti-mouse IgG1-HRP: sc-2060(Santa Cruz Biotechnology, Inc., USA) and anti-rabbit IgG-HRP (CST,Germany). Total cellular proteins were isolated and quantified withBradford reagent (Bio Rad, USA). Eighty micrograms of protein wereloaded on a 10% acrylamide gel. Subsequently, the well-separatedproteins were transferred to a nitrocellulose blotting membraneby wet transfer apparatus (Bio Rad, USA) at 100 V for 75 min. Priorincubation with each primary antibody, the membrane was blockedwith 5% skimmed milk (Bio Rad, USA) in TBST for 1 h at 4 ◦C. Theprimary antibodies were incubated in concentration recommendedin the presence of 5% skimmed milk and TBST overnight at 4 ◦C.This is followed by the incubation with secondary antibody thatis conjugated with horseradish peroxidase for another 1 h at 4 ◦C.The membrane was extensively washed with TBST at every step.TMB stabilized substrate (Promega, USA) was used to develop themembrane. Band intensities were analyzed with Image-J software(http://rsbweb.nih.gov/ij/).
2.5. Statistical analysis
son between groups was made using non-parametric test, theKruskal–Wallis test. A p value <0.05 was considered statistically sig-nificant. All calculations were performed using statistical computerpackage SPSS version 17.0 software for Windows (IBM, USA).
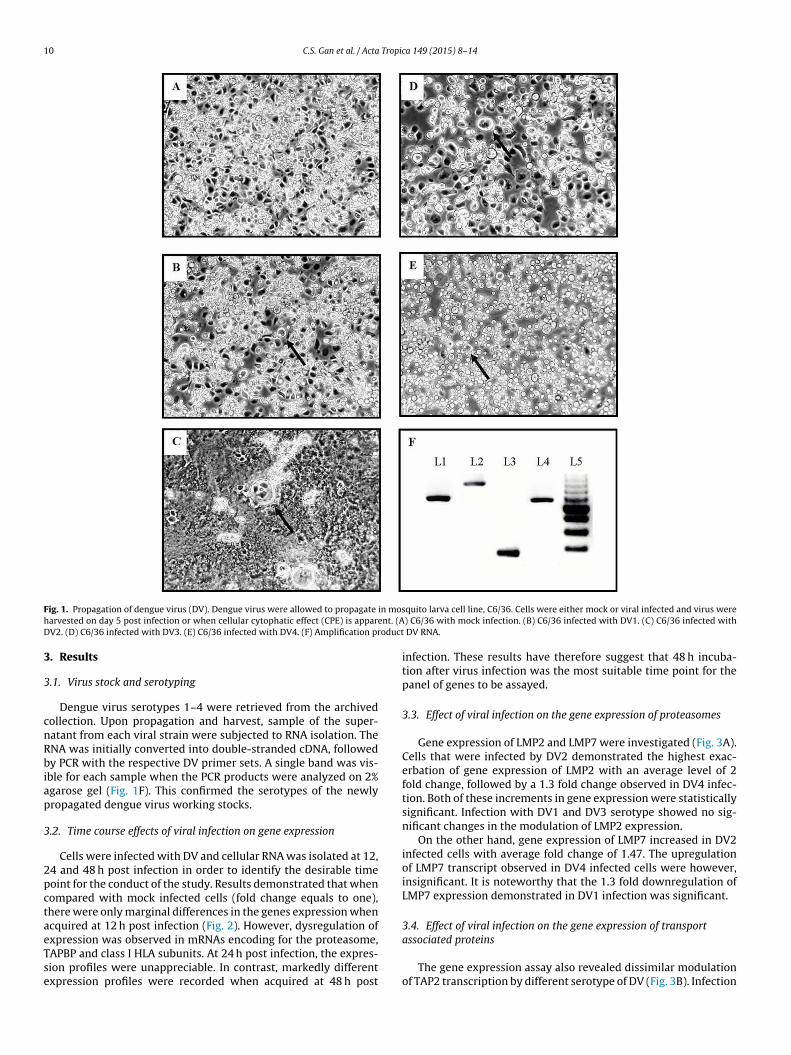
10 C.S. Gan et al. / Acta Tropica 149 (2015) 8–14
F in mosh ent. (AD oduct
3
3
cnRbiap
3
2pctaeTse
ig. 1. Propagation of dengue virus (DV). Dengue virus were allowed to propagate
arvested on day 5 post infection or when cellular cytophatic effect (CPE) is apparV2. (D) C6/36 infected with DV3. (E) C6/36 infected with DV4. (F) Amplification pr
. Results
.1. Virus stock and serotyping
Dengue virus serotypes 1–4 were retrieved from the archivedollection. Upon propagation and harvest, sample of the super-atant from each viral strain were subjected to RNA isolation. TheNA was initially converted into double-stranded cDNA, followedy PCR with the respective DV primer sets. A single band was vis-
ble for each sample when the PCR products were analyzed on 2%garose gel (Fig. 1F). This confirmed the serotypes of the newlyropagated dengue virus working stocks.
.2. Time course effects of viral infection on gene expression
Cells were infected with DV and cellular RNA was isolated at 12,4 and 48 h post infection in order to identify the desirable timeoint for the conduct of the study. Results demonstrated that whenompared with mock infected cells (fold change equals to one),here were only marginal differences in the genes expression whencquired at 12 h post infection (Fig. 2). However, dysregulation of
xpression was observed in mRNAs encoding for the proteasome,APBP and class I HLA subunits. At 24 h post infection, the expres-ion profiles were unappreciable. In contrast, markedly differentxpression profiles were recorded when acquired at 48 h postquito larva cell line, C6/36. Cells were either mock or viral infected and virus were) C6/36 with mock infection. (B) C6/36 infected with DV1. (C) C6/36 infected with
DV RNA.
infection. These results have therefore suggest that 48 h incuba-tion after virus infection was the most suitable time point for thepanel of genes to be assayed.
3.3. Effect of viral infection on the gene expression of proteasomes
Gene expression of LMP2 and LMP7 were investigated (Fig. 3A).Cells that were infected by DV2 demonstrated the highest exac-erbation of gene expression of LMP2 with an average level of 2fold change, followed by a 1.3 fold change observed in DV4 infec-tion. Both of these increments in gene expression were statisticallysignificant. Infection with DV1 and DV3 serotype showed no sig-nificant changes in the modulation of LMP2 expression.
On the other hand, gene expression of LMP7 increased in DV2infected cells with average fold change of 1.47. The upregulationof LMP7 transcript observed in DV4 infected cells were however,insignificant. It is noteworthy that the 1.3 fold downregulation ofLMP7 expression demonstrated in DV1 infection was significant.
3.4. Effect of viral infection on the gene expression of transport
associated proteinsThe gene expression assay also revealed dissimilar modulationof TAP2 transcription by different serotype of DV (Fig. 3B). Infection
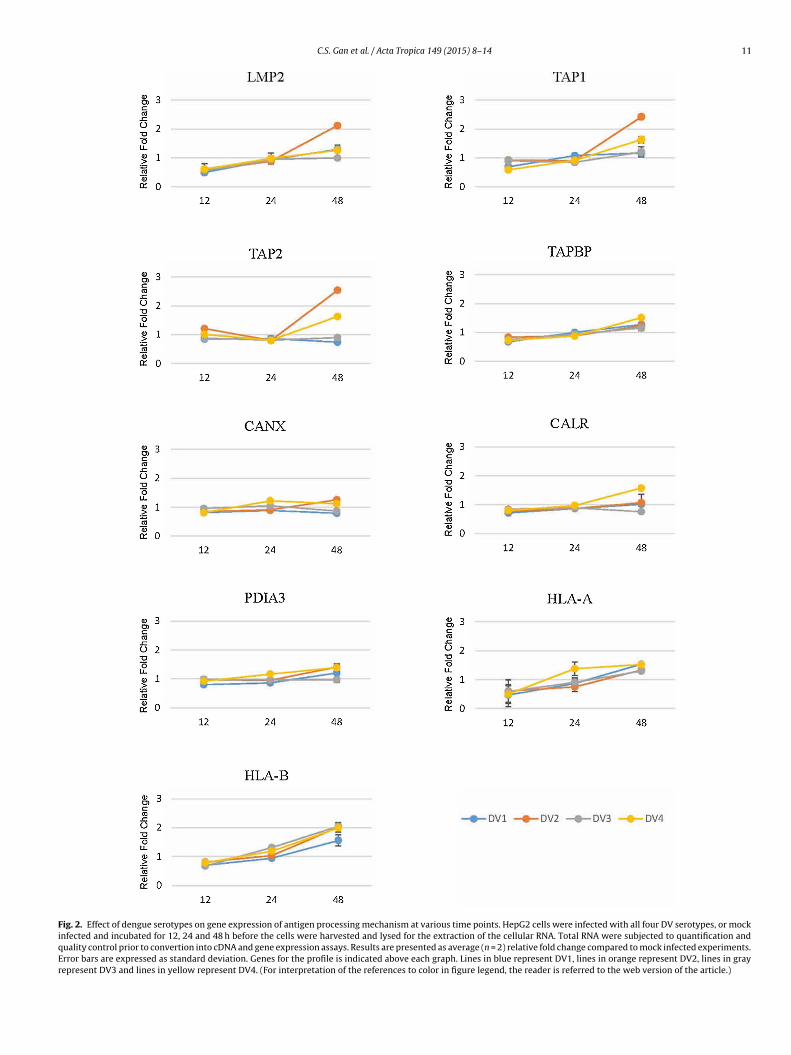
C.S. Gan et al. / Acta Tropica 149 (2015) 8–14 11
Fig. 2. Effect of dengue serotypes on gene expression of antigen processing mechanism at various time points. HepG2 cells were infected with all four DV serotypes, or mockinfected and incubated for 12, 24 and 48 h before the cells were harvested and lysed for the extraction of the cellular RNA. Total RNA were subjected to quantification andquality control prior to convertion into cDNA and gene expression assays. Results are presented as average (n = 2) relative fold change compared to mock infected experiments.Error bars are expressed as standard deviation. Genes for the profile is indicated above each graph. Lines in blue represent DV1, lines in orange represent DV2, lines in grayrepresent DV3 and lines in yellow represent DV4. (For interpretation of the references to color in figure legend, the reader is referred to the web version of the article.)

12 C.S. Gan et al. / Acta Tropica 149 (2015) 8–14
Fig. 3. Effect of dengue serotypes on genes expression of antigen procesing mechanism. HepG2 cells were infected with all four DV serotypes, or mock infected and incubatedfor 48 h before the cells were harvested and lysed for the extraction of the cellular RNA. Total RNA were subjected to quantification and quality control prior to convertioni elativs Exprea ssion
wgtda
sif
3i
pwoscwib
3
scic1De
nto cDNA and gene expression assays. Results are presented as the average (n = 3) rtandard deviation. Statistical significance, p < 0.05, * is indicated above the bar. (A)ssociated proteins. (C) Expression profiles of class I cheaperons proteins. (D) Expre
ith DV2 and DV4 has resulted in the upregulation of the TAP2ene, marking the average fold change of 2.24 and 1.61, respec-ively. In contrast, infection with DV1 and DV3 resulted in theownregulation of the gene with an average fold change of 1.46nd 1.31, respectively.
Similar profiles were detected in the expression of TAP1 tran-cript of those cells infected with DV2 and DV4. Nevertheless, onlynfection with DV2 marked a significant upregulation with averageold change of 2.65.
.5. Effect of viral infection on the gene expression of proteinsnvolved in peptide assembly onto the MHC class-I molecules
Amongst the four chaperone proteins involved in the loading ofeptide on the HLA complex, the differential expression of TAPBPas the most notable when cells were infected with all serotype
f DV (Fig. 3C). Despite the small increments in TAPBP expression,ignificant changes were observed in DV1-, DV2- and DV3-infectedells with 1.18 fold, 1.3 fold and 1.28 fold, respectively. Infectionsith DV4 also resulted in a 1.29 fold increment but was statically
nsignificant. CANX and CALR genes were significantly upregulatedy DV2 but not by other serotypes.
.6. Effect of viral infection on the gene expression of HLAs
Gene expression of HLA-A and HLA-B were investigated in thistudy (Fig. 3D). Cells infected by either DV1 or DV2 demonstratedonsistent upregulation of both HLA-A and HLA-B transcripts. Cellsnfected by DV1 has resulted in upregulation with average fold
hange of 1.43 in HLA-A expression and average fold change of.54 in HLA-B expression. On the other hand, cells infected byV2 marked an increment of average fold change of 1.52 in HLA-Axpression and average fold change of 2.12 in HLA-B expression.e fold change compared to mock infected expreiments. Error bars are expressed asssion profiles of immunoproteosome subunits. (B) Expression profiles of transport
profiles of human leukocyte antigen proteins.
Unlike DV3 that showed insignificant increase in the HLA-Aexpression, cells infected with DV4 marked a significant upregu-lation of the gene by 1.41 fold. However, the expression of HLA-Btranscript were unaffected by the infection of both DV3 and DV4.
3.7. Validation of gene expression profile by Western blot
Expression of the selected translational products was assessedto validate the genes expression profiles. One protein from eachfunctional stage of APM, which are peptide generation, peptidetransportation into ER, peptide loading and MHC class-I itself, wasrandomly selected for the validation process. The protein expres-sion profiles were congruent with that of the gene expressionprofiles (Fig. 4). Amongst the four dengue serotypes, only DV2demonstrated significantly altered expression in LMP2 and TAP1protein with respective average fold change of 5 and 2.9. The pro-tein expression profile for TAPBP was similar to that of the geneexpression profile where DV1, DV2 and DV3 demonstrated sig-nificant augmentation. All four dengue serotypes demonstratedsignificant upregulation of the MHC class I protein expression.
4. Discussion
Conserved within its genus flavivirus, DV demonstrates simi-lar characteristics of the MHC class I manipulation of the host cell.In light of the augmentation of MHC class I in flavivirus-infectedcells, the mechanism for the increased abundance of MHC class Imolecules and its cognate of cofactor proteins was postulated asthe results of virus-induced increase of biosynthesis of the class
I molecules and/or ancillary molecules of the pathway, or a pro-longed half-life of the MHC class I molecules of those infectedcells (Lobigs et al., 2003). In this study, whole virus were usedand all DV serotypes, DV1 to DV4, demonstrated upregulation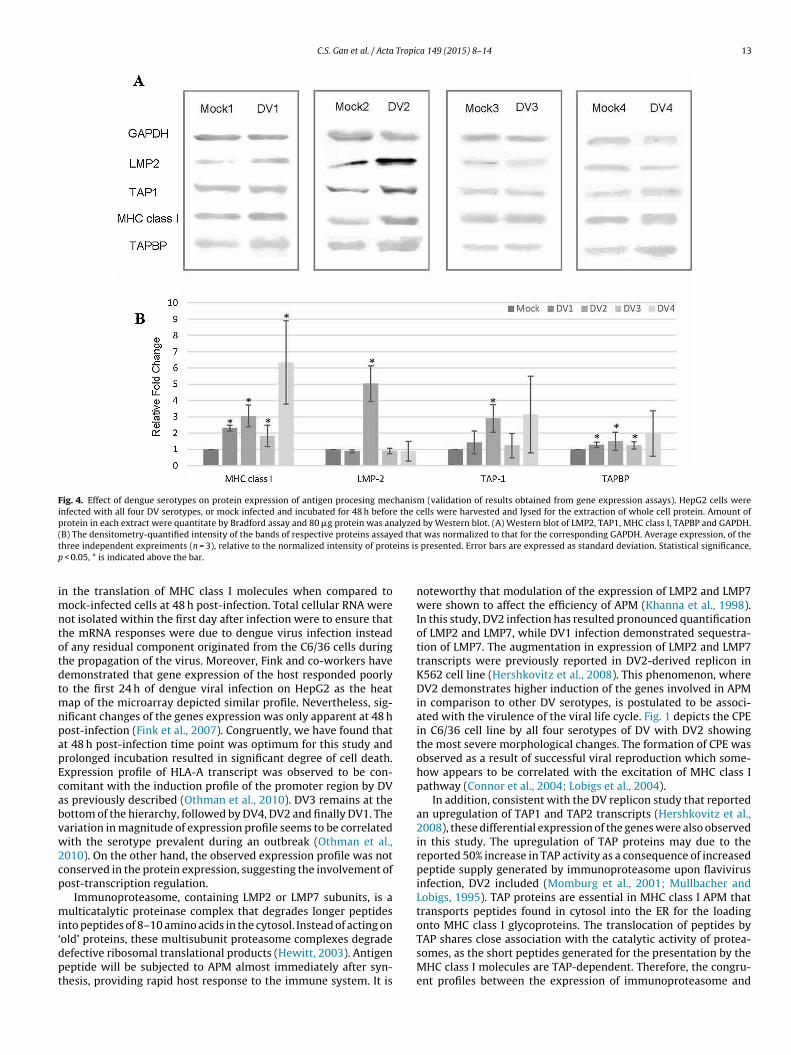
C.S. Gan et al. / Acta Tropica 149 (2015) 8–14 13
Fig. 4. Effect of dengue serotypes on protein expression of antigen procesing mechanism (validation of results obtained from gene expression assays). HepG2 cells wereinfected with all four DV serotypes, or mock infected and incubated for 48 h before the cells were harvested and lysed for the extraction of whole cell protein. Amount ofprotein in each extract were quantitate by Bradford assay and 80 �g protein was analyzed by Western blot. (A) Western blot of LMP2, TAP1, MHC class I, TAPBP and GAPDH.( ed that eins isp
imntotdtmnpapEcabvw2cp
mi‘dpt
B) The densitometry-quantified intensity of the bands of respective proteins assayhree independent expreiments (n = 3), relative to the normalized intensity of prot
< 0.05, * is indicated above the bar.
n the translation of MHC class I molecules when compared toock-infected cells at 48 h post-infection. Total cellular RNA were
ot isolated within the first day after infection were to ensure thathe mRNA responses were due to dengue virus infection insteadf any residual component originated from the C6/36 cells duringhe propagation of the virus. Moreover, Fink and co-workers haveemonstrated that gene expression of the host responded poorlyo the first 24 h of dengue viral infection on HepG2 as the heat
ap of the microarray depicted similar profile. Nevertheless, sig-ificant changes of the genes expression was only apparent at 48 host-infection (Fink et al., 2007). Congruently, we have found thatt 48 h post-infection time point was optimum for this study androlonged incubation resulted in significant degree of cell death.xpression profile of HLA-A transcript was observed to be con-omitant with the induction profile of the promoter region by DVs previously described (Othman et al., 2010). DV3 remains at theottom of the hierarchy, followed by DV4, DV2 and finally DV1. Theariation in magnitude of expression profile seems to be correlatedith the serotype prevalent during an outbreak (Othman et al.,
010). On the other hand, the observed expression profile was notonserved in the protein expression, suggesting the involvement ofost-transcription regulation.
Immunoproteasome, containing LMP2 or LMP7 subunits, is aulticatalytic proteinase complex that degrades longer peptides
nto peptides of 8–10 amino acids in the cytosol. Instead of acting on
old’ proteins, these multisubunit proteasome complexes degradeefective ribosomal translational products (Hewitt, 2003). Antigeneptide will be subjected to APM almost immediately after syn-hesis, providing rapid host response to the immune system. It ist was normalized to that for the corresponding GAPDH. Average expression, of the presented. Error bars are expressed as standard deviation. Statistical significance,
noteworthy that modulation of the expression of LMP2 and LMP7were shown to affect the efficiency of APM (Khanna et al., 1998).In this study, DV2 infection has resulted pronounced quantificationof LMP2 and LMP7, while DV1 infection demonstrated sequestra-tion of LMP7. The augmentation in expression of LMP2 and LMP7transcripts were previously reported in DV2-derived replicon inK562 cell line (Hershkovitz et al., 2008). This phenomenon, whereDV2 demonstrates higher induction of the genes involved in APMin comparison to other DV serotypes, is postulated to be associ-ated with the virulence of the viral life cycle. Fig. 1 depicts the CPEin C6/36 cell line by all four serotypes of DV with DV2 showingthe most severe morphological changes. The formation of CPE wasobserved as a result of successful viral reproduction which some-how appears to be correlated with the excitation of MHC class Ipathway (Connor et al., 2004; Lobigs et al., 2004).
In addition, consistent with the DV replicon study that reportedan upregulation of TAP1 and TAP2 transcripts (Hershkovitz et al.,2008), these differential expression of the genes were also observedin this study. The upregulation of TAP proteins may due to thereported 50% increase in TAP activity as a consequence of increasedpeptide supply generated by immunoproteasome upon flavivirusinfection, DV2 included (Momburg et al., 2001; Mullbacher andLobigs, 1995). TAP proteins are essential in MHC class I APM thattransports peptides found in cytosol into the ER for the loadingonto MHC class I glycoproteins. The translocation of peptides by
TAP shares close association with the catalytic activity of protea-somes, as the short peptides generated for the presentation by theMHC class I molecules are TAP-dependent. Therefore, the congru-ent profiles between the expression of immunoproteasome and
1 Tropi
twmrbmLievesowDbIatbu
bpoaifbpuakau2iiettedc(
thwHmlpDbccsSc
10.1016/S0167-5699(00)01692-3
4 C.S. Gan et al. / Acta
ransporter evident in this study suggest that the interaction of DVith the host cells APM occurs upstream of the pathway. Further-ore, previous studies have revealed the bidirectional promoter
egion of the LMP2 and TAP1 genes and the capability of inducingoth of the genes with the same stimuli (Wright et al., 1995). Thisay therefore explain the congruent expression profile observed in
MP2 and TAP1 gene. Nevertheless, the putative virus protein thats responsible and how is it interacting with host APM remains to belucidated. Manipulation of TAP activity was also shown by otheriruses in the flavivirus genus such as West Nile virus (Momburgt al., 2001). In this study, we demonstrated the differential expres-ion of LMP2, LMP7, TAP1 and TAP2 profiles by different serotypesf DV. The profile patterns can be clustered into three groups;here DV2 showing the pronounced induction of gene expression,V4 showing the intermediate induction of gene expression andoth DV1 and DV3 demonstrating suppression of gene expression.
nterestingly, this clustering is in fact coherent with the denguentigenic cartography; where DV2 and DV4 were antigenically dis-inct from DV1 and DV3 that share some similarities but distinctetween serotypes (http://www.whocc.infectiousdisease.cam.ac.k/antigenic-cartography/dengue).
Chaperon proteins are equally important for MHC class I assem-ly and maturation (Paulsson and Wang, 2003). Generally, theseroteins orchestrate the peptide optimization and quality controlsf the mature complex prior to its migration from the ER. Calnexinnd calreticulin are lectin-binding proteins that play pivotal rolesn facilitating the correct folding configuration of the short peptidesor proper loading (Parodi, 2000). Therefore, the congruent profilesetween the expressions level of calnexin, calreticulin and the TAProteins observed in this study may represent the homeostatic reg-lation of genes involved in APM, especially during the assemblynd maturation stages of the class I complex, with TAP as the gate-eeper. Interestingly, the expression level of tapasin was shownssociated with the expression level of HLA. Increase of tapasinpon DV infection was also obtained by others (Hershkovitz et al.,008). The upregulation of tapasin may due to its role in maintain-
ng the stability of the preformed (upregulated) class I complexes ints both peptide-free form and peptide-bound form. It was shownssential for the class I complexes to be bound by tapasin untilhey are cued to traverse off to the Golgi apparatus, otherwisehe complexes would be subjected to degradation (Schneeweisst al., 2009). Moreover, the absence of tapasin in a knock-out modelemonstrated reduced cell surface expression of MHC class I andonsequently impaired the elicitation of T cell-mediated responsesPaulsson and Wang, 2003).
Taken together, these results provide insights in understandinghe relationship between infection of all serotypes of DV and theost cell APM. Using the HepG2 cell line as a host, all serotypes ofhole DV (DV1–DV4) were shown to augment the expression ofLA class I molecules. In addition, DV2 infection demonstrated aore pronounced regulation of host’s APM; this may be the under-
ying reason for the acquisition of DV2 as the preferred denguerototype in laboratory research. We have provided evidence thatV affects the regulation of AMP at the proteasome level. This isecause, the expression profiles of TAP proteins and the dedicatedhaperones for assuring proper peptide loading, i.e. the calnexin,
alreticulin and PDIA3 proteins were congruent with the expres-ion profiles of the immunoproteosome subunits, LMP2 and LMP7.ubstantial further experimentation will be required and is beingarried out to clarify this hypothesis.ca 149 (2015) 8–14
Acknowledgements
This work was supported by the grants from Malaysian Ministerof Education-FRGS; FP019-2013A, University of Malaya ResearchGrant (UMRG); RP002-2012C and University of Malaya Postgradu-ate Grant; PG034-2014B.
References
Clyde, K., Kyle, J.L., Harris, E., 2006. Recent advances in deciphering viral and hostdeterminants of dengue virus replication and pathogenesis. J. Virol. 80 (23),11418–11431, http://dx.doi.org/10.1128/Jvi.01257-06
Connor, J.H., Naczki, C., Koumenis, C., Lyles, D.S., 2004. Replication and cytopathiceffect of oncolytic vesicular stomatitis virus in hypoxic tumor cells in vitro andin vivo. J. Virol. 78 (17), 8960–8970, http://dx.doi.org/10.1128/Jvi.78.17.8960-8970.2004
Fink, J., Gu, F., Ling, L., Tolfvenstam, T., Olfat, F., Chin, K.C., Aw, P., George, J., Kuznetsov,V.A., Schreiber, M., Vasudevan, S.G., Hibberd, M.L., 2007. Host gene expressionprofiling of dengue virus infection in cell lines and patients. PLoS Negl. Trop. Dis.1 (2), http://dx.doi.org/10.1371/journal.pntd.0000086, ARTN e86.
Guzman, M.G., Kouri, G., 2002. Dengue: an update. Lancet Infect. Dis. 2 (1), 33–42,http://dx.doi.org/10.1016/S1473-3099(01)00171-2
Hershkovitz, O., Zilka, A., Bar-Ilan, A., Abutbul, S., Davidson, A., Mazzon, M., Küm-merer, B.M., Monsoengo, A., Jacobs, M., Porgador, A., 2008. Dengue virus repliconexpressing the nonstructural proteins suffices to enhance membrane expressionof HLA class I and inhibit lysis by human NK cells. J. Virol. 82 (15), 7666–7676,http://dx.doi.org/10.1128/Jvi.01257-06
Hewitt, E.W., 2003. The MHC class I antigen presentation pathway: strategiesfor viral immune evasion. Immunology 110 (2), 163–169, http://dx.doi.org/10.1046/j.1365-2567.2003.01738.x
Khanna, R., Busson, P., Burrows, S.R., Raffoux, C., Moss, D.J., Nicholls, J.M.,Cooper, L., 1998. Molecular characterization of antigen-processing functionin nasopharyngeal carcinoma (NPC): evidence for efficient presentation ofEpstein-Barr virus cytotoxic T-cell epitopes by NPC cells. Cancer Res. 58 (2),310–314.
Lobigs, M., Mullbacher, A., Lee, E., 2004. Evidence that a mechanism for efficientflavivirus budding upregulates MHC class I. Immunol. Cell Biol. 82 (2), 184–188,http://dx.doi.org/10.1046/j.0818-9641.2004.01218.x
Lobigs, M., Mullbacher, A., Regner, M., 2003. MHC class I up-regulation by fla-viviruses: immune interaction with unknown advantage to host or pathogen.Immunol. Cell Biol. 81 (3), 217–223, http://dx.doi.org/10.1046/j.1440-1711.2003.01161.x
Momburg, F., Mullbacher, A., Lobigs, M., 2001. Modulation of transporter associatedwith antigen processing (TAP)-mediated peptide import into the endoplasmicreticulum by flavivirus infection. J. Virol. 75 (12), 5663–5671, http://dx.doi.org/10.1128/Jvi.75.12.5663-5671.2001
Mullbacher, A., Lobigs, M., 1995. Up-regulation of MHC class-I by flavivirus-inducedpeptide translocation into the endoplasmic-reticulum. Immunity 3 (2), 207–214,http://dx.doi.org/10.1016/1074-7613(95)90090-X
Othman, S., Rahman, N.A., Yusof, R., 2010. All serotypes of dengue virus induce HLA-A2 major histocompatibility complex class I promoter activity in human livercells. Trans. R. Soc. Trop. Med. Hyg. 104 (12), 806–808, http://dx.doi.org/10.1016/j.trstmh.2010.07.004
Parodi, A.J., 2000. Role of N-oligosaccharide endoplasmic reticulum processing reac-tions in glycoprotein folding and degradation. Biochem. J. 348, 1–13, http://dx.doi.org/10.1042/0264-6021:3480001
Paulsson, K., Wang, P., 2003. Chaperones and folding of MHC class I molecules in theendoplasmic reticulum. Biochim. Biophys. Acta – Mol. Cell Res. 1641 (1), 1–12,http://dx.doi.org/10.1016/S0167-4889(03)00048-X
Payne, A.F., Binduga-Gajewska, I., Kauffman, E.B., Kramer, L.D., 2006. Quantitationof flaviviruses by fluorescent focus assay. J. Virol. Methods 134 (1–2), 183–189,http://dx.doi.org/10.1016/j.jviromet.2006.01.003
Petersen, J.L., Morris, C.R., Solheim, J.C., 2003. Virus evasion of MHC class I moleculepresentation. J. Immunol. 171 (9), 4473–4478.
Schneeweiss, C., Garstka, M., Smith, J., Hutt, M.T., Springer, S., 2009. The mechanismof action of tapasin in the peptide exchange on MHC class I molecules deter-mined from kinetics simulation studies. Mol. Immunol. 46 (10), 2054–2063,http://dx.doi.org/10.1016/j.molimm.2009.02.032
Seliger, B., Maeurer, M.J., Ferrone, S., 2000. Antigen-processing machinery break-down and tumor growth. Immunol. Today 21 (9), 455–464, http://dx.doi.org/
Wright, K.L., White, L.C., Kelly, A., Beck, S., Trowsdale, J., Ting, J.P.Y., 1995. Coordi-nate regulation of the human TAP1 and LMP2 genes from a shared bidirectionalpromoter. J. Exp. Med. 181 (4), 1459–1471, http://dx.doi.org/10.1084/jem.181.4.1459.





























