Embed Size (px)
Citation preview

Dissertação de Mestrado
CARACTERIZAÇÃO ECOLÓGICA DOS ANFÍBIOS ANUROS DE ÁREAS
REFLORESTADAS DO ENTORNO DA REPRESA DE VOLTA GRANDE, NO RIO
GRANDE, MG/SP.
JOICE PAIVA VIDIGAL MARTINS
Ouro Preto, Dezembro de 2014
UNIVERSIDADE FEDERAL DE OURO PRETO
INSTITUTO DE CIÊNCIAS EXATAS E BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM
ECOLOGIA DE BIOMAS TROPICAIS

UNIVERSIDADE FEDERAL DE OURO PRETO
INSTITUTO DE CIÊNCIAS EXATAS E BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM
ECOLOGIA DE BIOMAS TROPICAIS
CARACTERIZAÇÃO ECOLÓGICA DOS ANFÍBIOS ANUROS DE ÁREAS
REFLORESTADAS DO ENTORNO DA REPRESA DE VOLTA GRANDE, NO RIO
GRANDE, MG/SP.
JOICE PAIVA VIDIGAL MARTINS
Dissertação apresentada ao Programa de Pós-
graduação em Ecologia de Biomas Tropicais da
Universidade Federal de Ouro Preto como parte dos
requisitos necessários para a obtenção do título de
Mestre em Ecologia de Biomas Tropicais.
ORIENTADORA: Dra. Maria Rita Silvério Pires



“Sejam alegres. A felicidade está em fazer alguém alegre, alguém feliz.”
Águila Bernardes Martins

AGRADECIMENTOS
Agradeço a minha mãe pela confiança que me passou. Sempre pareceu tranquila de
que eu fazia a minha parte e sabia que mesmo que fosse aos últimos minutos do
segundo tempo eu iria dar conta. A confiança que ela tem em mim sempre foi um
conforto e me pôs pra frente.
Agradeço ao meu pai pelas orações e pela força em todos os momentos do mestrado.
Também agradeço à Cibele, por me deixar tranquila de que meu pai está em boa
companhia.
Sou muito grata a vocês, meus pais, Diva e Daniel, pela educação que me deram, pelo
respeito à vida que me ensinaram ter. Prepararam-me bem para essa nova fase.
Agradeço ao meu irmão Cauê, por sempre me lembrar do índio jhow, me faz bem
saber que posso contar com você para o que precisar.
Agradeço também à vó Águila, uma guerreira, sempre divertida e espirituosa. Ela faz
parte de cada um que viveu muitas histórias com ela.
À vó Maria do Carmo, amorosa, aventureira e com muitos planos e sonhos ainda para
realizar. Muito do que sou hoje conquistei da minha infância brincando no sítio dela. E
muitas das tarefas que eu tive que realizar nos meus campos de biologia, aprendi
brincando no sítio dela. Afinal, quando se tem como brinquedos: facão, marreta,
cavadeira, enxada, dá pra se fazer muita coisa!
Agradeço aos amigos Natys e Rafa, que juntos viramos “o trio que o Marcão uniu”,
agora não tem como separar! Obrigada por tudo!
Aos meus amigos de campo. Foi dureza! Trabalhamos muito, cavamos, martelamos,
capinamos, abrimos valas, carregamos peso, abrimos trilha, revisamos armadilhas,
fizemos coletas diurnas e noturnas, seguimos fezes de guariba, procuramos o bendito
jacaré, e também ficamos impregnados com o cheiro de rola bosta. Foi uma das fazes
mais felizes de Ouro Preto. Fui muito feliz! Isto tudo só foi possível por causa da equipe
herpeto/masto. O elenco desta equipe são eles: Fifi, o garoto TPM; Adriele Prisca, a
garota que AMA o mundo e Little tets, o patrãozinho que nas horas vagas é velho do
rio. Agradeço muito a alegria que vivi ao lado de vocês! Agradeço também ao Gui

(insônia) que, mesmo não sendo propriamente dito da equipe, considero como se
fosse. A gente falava mal de você principalmente quando estávamos entediados nas
viagens de campo. Logo, você também foi responsável pela nossa diversão.
Agradeço à Maria Rita, pois orientadora igual não há! Mãezona, divertida e soube
puxar a orelha da orientada quando preciso. Muito obrigada pelos ensinamentos, não
apenas os da escrita, mas por assumir uma postura de uma profissional que enxerga
as pessoas à sua volta como iguais e nunca com um olhar de superioridade.
Ao pessoal do LZV, pelas conversas sobre os campos e por aturarem a bagunça que a
equipe herpeto/masto fazia no laboratório, mas que fique claro que a bagunça foi
principalmente da masto.
Queria agradecer a comunidade horto dos contos, por ter me acolhido quando eu
precisei. É ótimo e essencial se sentir em casa. E foi assim que me senti morando com
vocês. Ric, Ed, Chris, Rafa, Tulinha, Nina, Galo, Galinha, Pintinho, Tartarugas e
Aranha, foi um prazer morar com vocês!
Um agradecimento especial ao brow brow, que me passou todos os betas pra entrar no
mestrado em Ouro Preto, por ser amigo e me ajudar SEMPRE que eu precisei.
Agradeço ao meu namorado, Giordano, por fazer Ouro Preto ainda melhor. E me
desculpa por deixar de te acompanhar na escalada e outros programas, mas agora
está ai o resultado. Valeu a pena me ausentar um pouco. Te bom dio!
Ao pessoal da escalada, que se tornaram amigos queridos.
Agradeço também à Grazi por me salvar com as estatísticas.
Agradeço a Dyana pela parceria na hora de trabalhar com os bichos. E valeu pela
paciência, tentarei ser uma co-orientadora melhor!
Aos amigos Lib e Josi, pelas boas vibrações enviadas à distância...rs...pela força e
pelos artigos. É sempre bom ter a quem recorrer quando as dúvidas aparecem e
quando bate o desespero do final de mestrado.
Agradeço às meninas que moraram comigo Samy, Cris, Fabíola, Lu, Flávia e Alice.
Valeu por estarem ao meu lado nos bons e maus momentos.

À Companhia Energética de Minas Gerais S.A. (CEMIG), Fundação Gorceix e
Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG), pelo apoio
logístico e concessão de bolsa.
À Yasmine, quem conquistou a participação da UFOP neste projeto, e chefiou a
batalha para o sucesso de todos os grupos que participaram.
À turma do projeto Cemig, por quebrarem o galho quando preciso, pelas conversas nos
intervalos dos campos e por compartilharem o pesar dos carrapatos.
Ao programa de pós-graduação em Ecologia de Biomas Tropicais pela oportunidade e
à UFOP, pela estrutura e apoio.
A estadia em Ouro Preto me surpreendeu, percebi que a vida é como a mata ciliar no
entorno de um rio. À medida que o rio corre a mata vai se diferenciando e se
diversificando. Eu ainda estou no começo desse rio, mas ele já não é o mesmo do que
era na nascente, são vários fragmentos de histórias se juntando num mosaico.

SUMÁRIO
RESUMO...........................................................................................................8
INTRODUÇÃO..................................................................................................9
MATERIAL E MÉTODOS................................................................................12
Área de estudo......................................................................................12
Coleta dos dados..................................................................................21
Análise dos dados................................................................................23
RESULTADOS................................................................................................24
DISCUSSÃO...................................................................................................31
CONCLUSÃO.................................................................................................35
REFERÊNCIA BIBLIOGRÁFICA....................................................................36
ANEXO 1........................................................................................................44
ANEXO 2........................................................................................................46

9
RESUMO
As espécies colonizadoras precisam ter características que permitam que
cheguem ao local e se mantenham com os recursos e condições ali existentes. Dentre
os anfíbios, ocorrem espécies com diferentes capacidades para colonizar ambientes
alterados por ação antrópica. Esses animais podem ser utilizados para avaliar a
efetividade de áreas a partir de suas características ecológicas. O presente estudo foi
realizado em matas ciliares reflorestadas no Cerrado, localizadas no entorno do
reservatório da usina hidrelétrica de Volta Grande, entre os estados de São Paulo e
Minas Gerais. A mata ciliar nativa foi inundada para a formação da represa. Entre 10 e
30 anos, o entorno das áreas inundadas foi reflorestado e, atualmente, estas áreas de
mata ciliar reflorestadas estão isoladas de remanescentes florestais por matrizes
agrícolas. O objetivo do presente estudo foi analisar as espécies de anuros que
conseguiram colonizar as matas ciliares reflorestadas, utilizando como parâmetros
aspectos da paisagem local, bem como da ecologia e da história natural das espécies.
Foram amostrados cinco fragmentos ciliares reflorestados entre março de 2013 e
janeiro de 2014. As coletas se deram por armadilhas de pitfall, instaladas no interior
das matas ciliares, e por procura ativa no período noturno associada à zoofonia, nas
áreas do entorno desses fragmentos. Como resultados, as matas ciliares reflorestadas
da represa Volta Grande contribuem para a conservação de 21 espécies. Não foram
encontradas espécies endêmicas ou dependentes de matas ciliares. As espécies
capazes de colonizar estas áreas são predominantemente resistentes à alteração
antrópica, com ampla distribuição geográfica; habitat generalistas e apresentam modo
reprodutivo característico de áreas abertas e ambientes lênticos. Sendo assim, os
aspectos predominantes da ecologia e história natural das espécies colonizadoras
podem ser utilizados para comparações em estudos futuros em matas ciliares
reflorestadas. O tipo de matriz exerceu influência somente sobre a abundância da
anurofauna na área com matriz seringal, enquanto a composição diferiu entre as cinco
áreas. Nas menores áreas reflorestadas, foram encontrados os menores valores de
equitabilidade, provavelmente em função do efeito de borda. Dessa forma, é reforçada
a importância da largura da mata ciliar e conectividade com matas ciliares nativas para
que apresentem melhor distribuição da abundância entre as espécies e tenham um
ambiente que seja fonte de espécies típicas de mata ciliar.
Palavras chaves: Anfíbios; áreas reflorestadas; matriz; reservatório; usina
hidrelétrica; Cerrado.

10
INTRODUÇÃO
O Cerrado ocupa no território brasileiro 2.039.386 km² e vem sofrendo intensa e
acelerada ocupação antrópica, principalmente nos últimos 50 anos (Brasil 2011).
Devido ao elevado endemismo e à intensa redução de suas áreas originais, esse
bioma é considerado hotspot mundial para conservação da biodiversidade (Myers et al.
2000). O cerrado abriga aproximadamente 10.000 espécies de plantas, sendo 4.400
endêmicas, e 1268 espécies de vertebrados, sendo 117 endêmicas (Myers et al. 2000).
Até o ano de 2010, 49,16% de sua cobertura original já havia sido desmatada (Brasil
2011).
A conversão das áreas naturais do bioma cerrado em campos agrícolas, assim
como a poluição e o represamento das águas, tem reduzido a área de ocorrência de
muitas espécies (Dias 1994; Colli et al. 2002). Estas alterações de habitat geram
eventos de extinção de vários grupos faunísticos (Cosson et al. 1999), deixando os
ambientes mais suscetíveis à invasão por espécies exóticas (Laurance & Vasconcelos
2009). Em 2005, no Cerrado, as áreas de pastagem e monoculturas totalizavam
600.000km², enquanto a parcela de Unidade de conservação era de cerca de
33.000km² (aproximadamente 3%) (Klink & Machado 2005).
Em janeiro de 2002 haviam 433 centrais hidrelétricas em operação no Brasil, das
quais 304 eram empreendimentos de micro e pequenas centrais hidrelétricas (Aneel
2002). Estudos sobre os impactos causados por empreendimentos hidrelétricos alertam
para questões como a perda e fragmentação de habitat (Zhong & Power 1996),
alteração da biodiversidade pela redução de espécies nativas e introdução de espécies
exóticas (Power et al. 1996), isolamento populacional (Dynesius & Nilsson 1994) e
erosão genética (Rosenberg et al. 1997).
As áreas mais afetadas pela implantação de hidrelétricas são os habitats fluviais e
as matas ciliares, os quais são inundados pela construção da represa (Pavan 2007).
De acordo com Ribeiro & Walter (2001), matas ciliares são a vegetação florestal que
acompanha as margens dos rios de médio e grande porte e dificilmente ultrapassam
100m de largura. Estes autores diferenciam estas florestas das matas de galeria pelo
porte dos corpos d’água que acompanham. Sendo assim, as matas de galeria
envolvem riachos de pequeno porte. Apesar destas matas ocuparem pequena

11
extensão na paisagem do cerrado (Ribeiro et al. 2001), elas exercem importantes
funções. Elas garantem a manutenção dos ecossistemas aquáticos e terrestres e o
equilíbrio geobiohidrológico do ambiente, além disso, funcionam como corredores para
dispersão de animais permitindo o fluxo gênico entre as populações (Fonseca et al.
2001).
Frente aos grandes empreendimentos, como as hidrelétricas, o Decreto Nº 97.632,
de 10 de abril de 1989 (Brasil 1989), prevê a recuperação das áreas degradadas,
visando a obtenção de estabilidade do meio ambiente. Esta recuperação deve ser feita
a partir de um plano de recuperação de áreas degradadas (PRAD). De acordo com a
Lei do Sistema Nacional de Unidades de Conservação Nº 9.985, de 18 de julho de
2000 (Brasil 2000), Art. 2, entende-se por recuperação a restituição de um ecossistema
ou de uma população silvestre degradada à uma condição não degradada, que pode
ser diferente de sua condição original. Com a construção de hidrelétricas, as represas
artificiais tornam-se um novo elemento na paisagem, enquanto que as áreas
adjacentes à área alagada tornam-se áreas de preservação permanente (Brasil 2002).
Portanto, as áreas no entorno dos reservatórios, devem ser recuperadas por
reflorestamentos.
O reflorestamento é uma medida para auxiliar a recuperação e, assim, iniciar os
estádios de sucessão nas áreas (Cullen 1998). À medida que este processo evolui, vão
se estabelecendo novas condições e recursos que irão propiciar a sucessão faunística
(Mexia 2008). Portanto, quando os fatores do habitat atingem as necessidades
particulares de cada espécie, a área vai sendo colonizada (Fox 1982). Com o tempo,
as condições e os recursos da área passam a não atender estas espécies da mesma
maneira, sendo assim, por competição ou outra interação, elas saem dos seus limiares
ótimos, deixam a área ou tornam-se menos abundantes (Mexia 2008).
A sucessão faunística também depende das áreas adjacentes (Majer 1989). O
padrão espacial da paisagem é composto basicamente por um mosaico de manchas e
corredores inseridos em uma matriz (Forman & Godron 1986). Estes elementos são
responsáveis pela distribuição dos organismos, matéria e energia (Forman & Godron
1986), pois podem representar diferentes funções para cada organismo, como barreira,
hábitat ou passagem. Nesse sentido, as matrizes apresentam diferentes níveis de
percolação, o que define a facilidade com que indivíduos das diversas espécies se

12
movimentam entre as manchas de florestas (Gardner & O’neil 1991). Portanto, algumas
matrizes são barreiras à movimentação dos organismos (Gascon et al. 1999; Ricketts
2001).
As espécies que irão colonizar determinada área precisam ter características que
permitam que elas cheguem ao local e se mantenham com os recursos e condições ali
existentes. As espécies generalistas, por exemplo, tem grande capacidade de
colonização, ocorrendo em mais de um habitat (Brandão & Araújo 2001). A partir das
informações sobre a ecologia e distribuição das espécies, é possível fazer uma
avaliação da área (Crema 2008; Magalhães 2012). Nesse sentido, no presente estudo,
os anfíbios foram utilizados para indicar a efetividade das áreas reflorestadas em
abrigar fauna típica de matas ciliares.
Segundo Azevedo-Ramos et al. (2005), um grupo bioindicador deve apresentar
alguns requisitos, tais como diversidade de formas, reunindo espécies mais exigentes e
outras mais adaptáveis, ocupação da maioria dos ambientes e nichos ecológicos e
mobilidade para responder às agressões ambientais. Os anfíbios anuros atendem bem
a estes requisitos. Dentre os anfíbios ocorrem espécies com diferentes capacidades
para colonizar em ambientes alterados por ação antrópica, variados níveis de
tolerância fisiológica, quanto às condições ambientais e quanto à dependência de
microambientes específicos para a reprodução (Moraes et al. 2007), sendo bons
bioindicadores (Feio et al. 1998).
Valdujo et al. (2011) registraram no Cerrado 209 espécies de anfíbios, sendo 150
consideradas típicas e 51,7%, endêmicas. Desta forma a fauna de anfíbios do Cerrado
é composta por diferentes grupos de espécies: 1) espécies endêmicas do Cerrado; 2)
espécies compartilhadas com domínios vegetacionais vizinhos e 3) espécies
generalistas, com distribuição em todo Brasil (Valdujo et al. 2011)
As espécies típicas de biomas florestais estão, em sua maioria, presentes nas matas
de galeria do Cerrado (Colli et al. 2002). São nestas matas que se encontram a maioria
dos anuros habitat-especialistas do Cerrado (Brandão & Araújo 2001). Contudo, esta
fisionomia não constitui barreira para a herpetofauna adaptada aos ambientes abertos
(Colli et al. 2002). Sendo assim, as matas ciliares são passíveis de apresentarem
anurofauna tanto de áreas abertas, quanto habitat-especialistas.

13
O presente estudo foi realizado em matas ciliares reflorestadas localizadas no
entorno do reservatório da usina hidrelétrica de Volta Grande, entre os estados de São
Paulo e Minas Gerais. Toda vegetação da área foi removida na ocasião da construção
da hidrelétrica, que entrou em operação em 1974. Entre 10 e 30 anos, foi realizado um
reflorestamento linear no entorno das áreas inundadas, de modo que estas se
caracterizam atualmente como fragmentos não contínuos de mata ciliar. Estas novas
matas são circundadas por matriz aquática e por matrizes agrícolas como canavial,
pasto, seringal e cerrado, incluindo formações campestres e cerradão.
O objetivo do presente estudo foi registrar as espécies de anuros presentes nas
matas ciliares reflorestadas, utilizando como parâmetros aspectos do uso e ocupação
do entorno das áreas e dados de ecologia e história natural das espécies encontradas,
discutindo a capacidade de colonização das mesmas. Nesse sentido, buscamos
responder às seguintes questões:
Qual a composição da anurofauna das áreas de matas ciliares reflorestadas? Seria
essa fauna composta por espécies típicas das matas ciliares do cerrado?
Quais são as características das espécies dos anuros que colonizaram os ambientes
de matas ciliares reflorestadas e cercadas por matrizes agrícolas?
Quais dos seguintes fatores exercem maior influência para a efetividade na
manutenção da diversidade de anuros em áreas reflorestadas: tamanho do fragmento?
Tempo de reflorestamento? Ou o tipo de matriz circundante?
Material e Métodos
1. Área de estudo
O estudo foi conduzido em cinco matas ciliares reflorestadas, denominadas
Nativa, Noboro, Santa Bárbara, Figueira e Delta (figura 1, tabela 1). As áreas Delta e
Figueira são as mais próximas entre si, sendo, contudo, separadas pela rodovia
Anhanguera (SP330). Estas áreas estão localizadas às margens do reservatório da
usina hidrelétrica (UHE) de Volta Grande (22k 0800295; utm 7768185; elevação 462m),
ao longo do rio Grande, na divisa dos Estados de Minas Gerais (margem direita) e São
Paulo (margem esquerda). A área inundada do reservatório é de 221,7km² e o volume

14
de água é de 2.268km³ (Braga 2001). A região tem uma estação seca bem definida
entre maio e outubro e a estação chuvosa ocorre entre novembro abril, com
precipitação média anual atingindo 1.506 milímetros (Martins 2014). A temperatura
média anual varia entre 22ºC e 24ºC (Filardi et al. 2007).
A região da UHE de Volta Grande é conhecida como Triangulo Mineiro e Norte do
estado de São Paulo e está inserida no bioma Cerrado. Com o enchimento do
reservatório em 1973, a mata ciliar original foi inundada, restando no entorno da
represa um cenário de solo exposto e compactado. Neste período, estas áreas de solo
exposto eram margeadas por cultivos agrícolas, com predomínio da cana-de-açúcar
(Ferreira 2009). No cumprimento da legislação, a Companhia Energética de Minas
Gerais S.A. (Cemig), que tem a concessão do aproveitamento da UHE de Volta Grande
desde 1967, reflorestou as margens da represa. No entanto, não foi usado um critério
padrão para esta recuperação, de modo que cada área apresenta larguras, tipos
vegetacionais e idades de reflorestamento diferentes.
Atualmente, a paisagem do entorno do Reservatório Volta Grande (RVG) é
representada por áreas de matas ciliares reflorestas, intercaladas por fisionomias do
cerrado em regeneração secundária, lagos, extensas plantações de cana, sorgo, pasto
ou seringal, além de fazendas, usinas e outras áreas urbanas (figuras 2 a 6). As cinco
áreas selecionadas para o presente estudo representam as diferentes fisionomias das
matrizes em que estão inseridas, bem como diferentes larguras e idades de
reflorestamento (tabela 1). Santa Bárbara e Delta foram reflorestadas mais
recentemente, há 10 anos, enquanto que Noboro e Figueira foram reflorestadas há 20
anos. Na área denominada Nativa ocorreu sucessão natural há cerca de 30 anos e é a
maior faixa de vegetação.
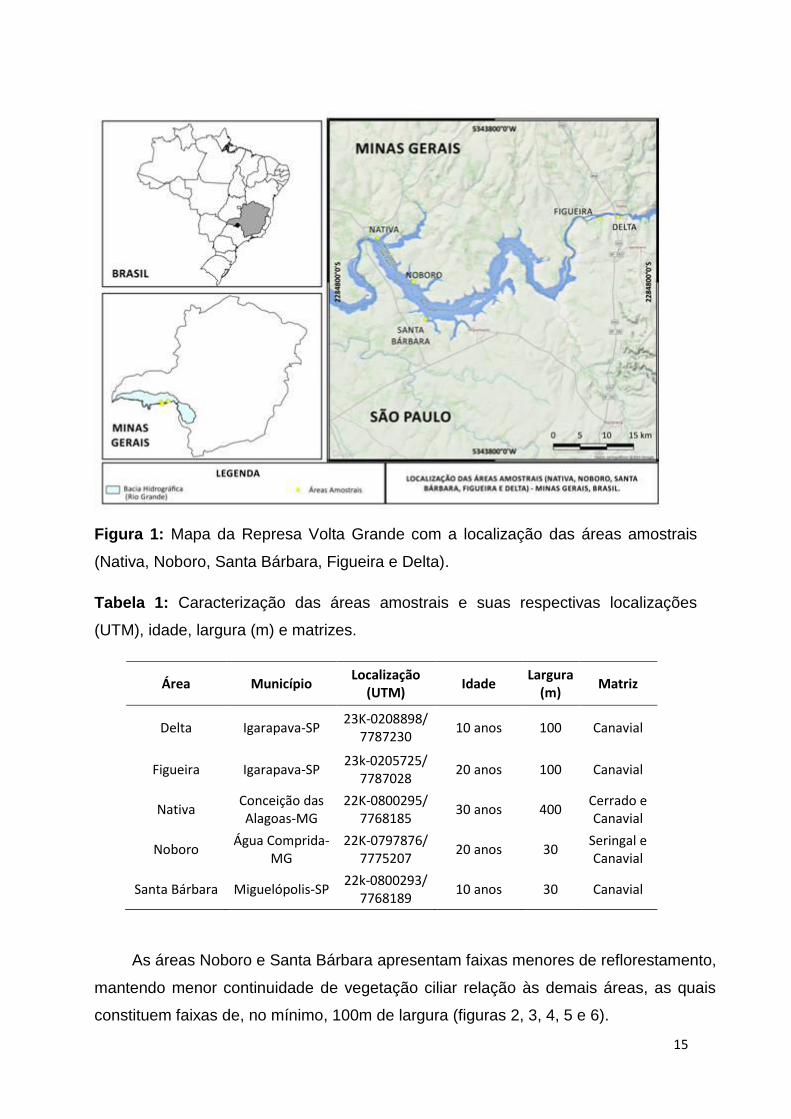
15
Figura 1: Mapa da Represa Volta Grande com a localização das áreas amostrais
(Nativa, Noboro, Santa Bárbara, Figueira e Delta).
Tabela 1: Caracterização das áreas amostrais e suas respectivas localizações
(UTM), idade, largura (m) e matrizes.
Área Município Localização
(UTM) Idade
Largura (m)
Matriz
Delta Igarapava-SP 23K-0208898/
7787230 10 anos 100 Canavial
Figueira Igarapava-SP 23k-0205725/
7787028 20 anos 100 Canavial
Nativa Conceição das Alagoas-MG
22K-0800295/ 7768185
30 anos 400 Cerrado e Canavial
Noboro Água Comprida-
MG 22K-0797876/
7775207 20 anos 30
Seringal e Canavial
Santa Bárbara Miguelópolis-SP 22k-0800293/
7768189 10 anos 30 Canavial
As áreas Noboro e Santa Bárbara apresentam faixas menores de reflorestamento,
mantendo menor continuidade de vegetação ciliar relação às demais áreas, as quais
constituem faixas de, no mínimo, 100m de largura (figuras 2, 3, 4, 5 e 6).

16
As áreas foram caracterizadas quanto ao uso e ocupação utilizando o programa
ArcGis. Para isso foram gerados buffers de três quilômetros para cada área, tendo
como centro as matas ciliares reflorestadas. Nas cinco áreas a matriz predominante foi
a monocultura de cana-de-açúcar, cobrindo no mínimo 65% destas. Sendo a área de
Santa Bárbara com o maior percentual desta plantação com 87,99% de toda sua
cobertura.
As áreas Nativa, Figueira e Delta apresentam maiores faixas de reflorestamento,
tendo, respectivamente, 16,68%, 16,76% e 17,25% de suas áreas cobertas por matas
somadas aos fragmentos florestais. Em Santa Bárbara e Noboro, apresentam apenas
10,96% e 4,02% de cobertura de matas.
A porção de área urbana (estradas, casas, indústrias) em Delta foi a maior dentre
as áreas, apresentando 9,41%, em contraste com Santa Bárbara com 0,17% destas
áreas. A área Nativa apresenta uma porcentagem um pouco menor de áreas de
agricultura, tais como pasto (0,7%) e canavial (65,07 %), quando comparada às áreas
de Delta e Figueira, o que é compensado por maiores valores para outras ocupações
como 12,97% de áreas de Cerrado e 14,29 % de fragmentos de mata.
Nas matrizes das cinco áreas ocorrem pequenos corpos d’água, tendo Delta o
maior percentual destes em sua paisagem, com 4%. Santa Bárbara apresenta a menor
cobertura com 0,89%.
Noboro é a única área que apresenta matriz arbórea no entorno do fragmento
reflorestado, caracterizada por um seringal que corresponde a 18,25% da paisagem
desta área.
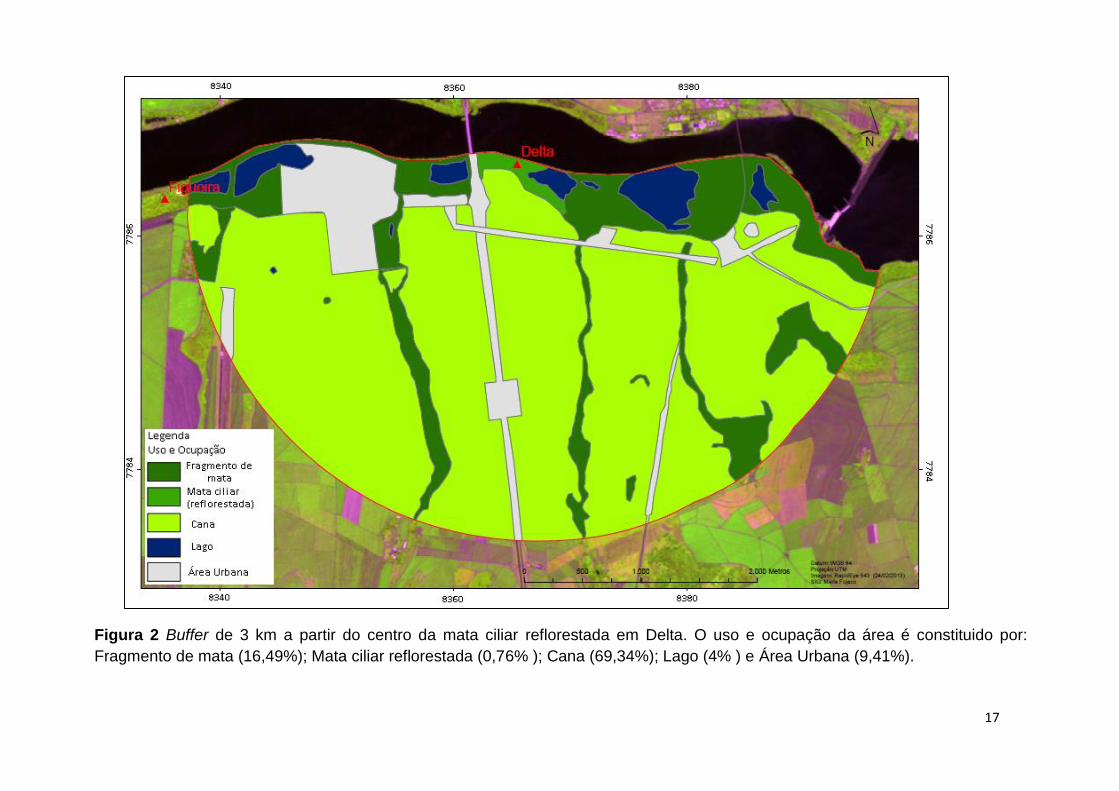
17
Figura 2 Buffer de 3 km a partir do centro da mata ciliar reflorestada em Delta. O uso e ocupação da área é constituido por:
Fragmento de mata (16,49%); Mata ciliar reflorestada (0,76% ); Cana (69,34%); Lago (4% ) e Área Urbana (9,41%).
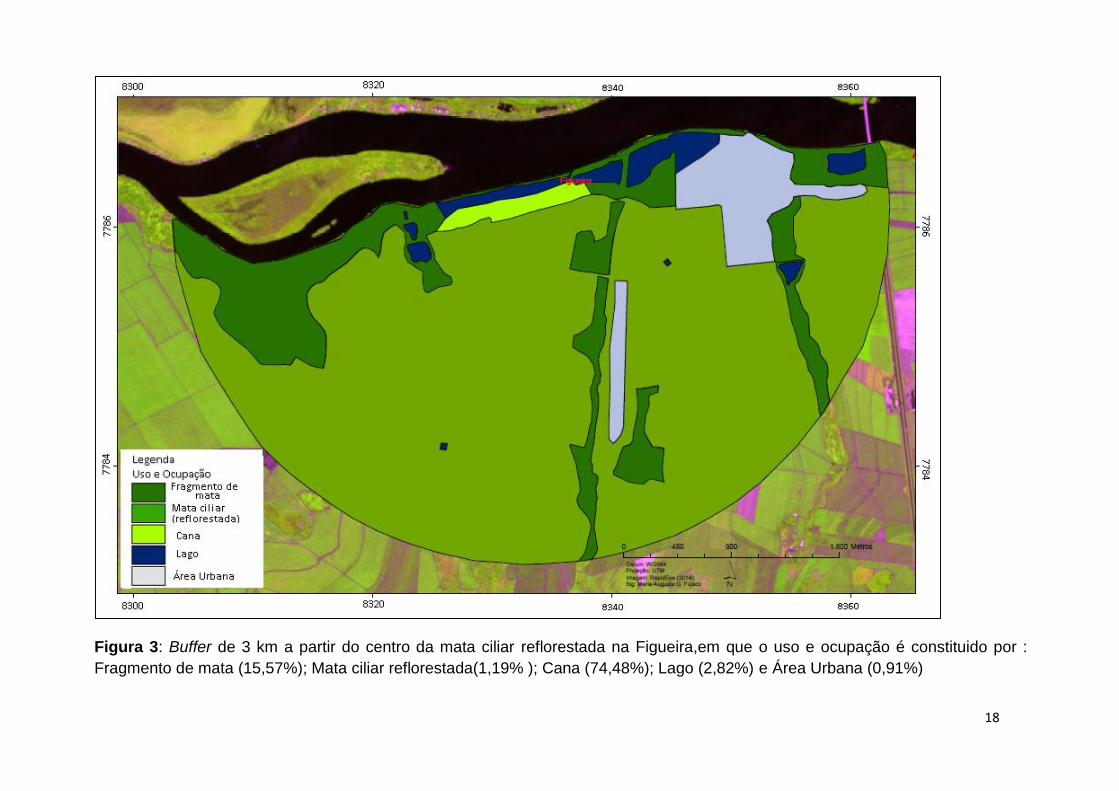
18
Figura 3: Buffer de 3 km a partir do centro da mata ciliar reflorestada na Figueira,em que o uso e ocupação é constituido por :
Fragmento de mata (15,57%); Mata ciliar reflorestada(1,19% ); Cana (74,48%); Lago (2,82%) e Área Urbana (0,91%)
19
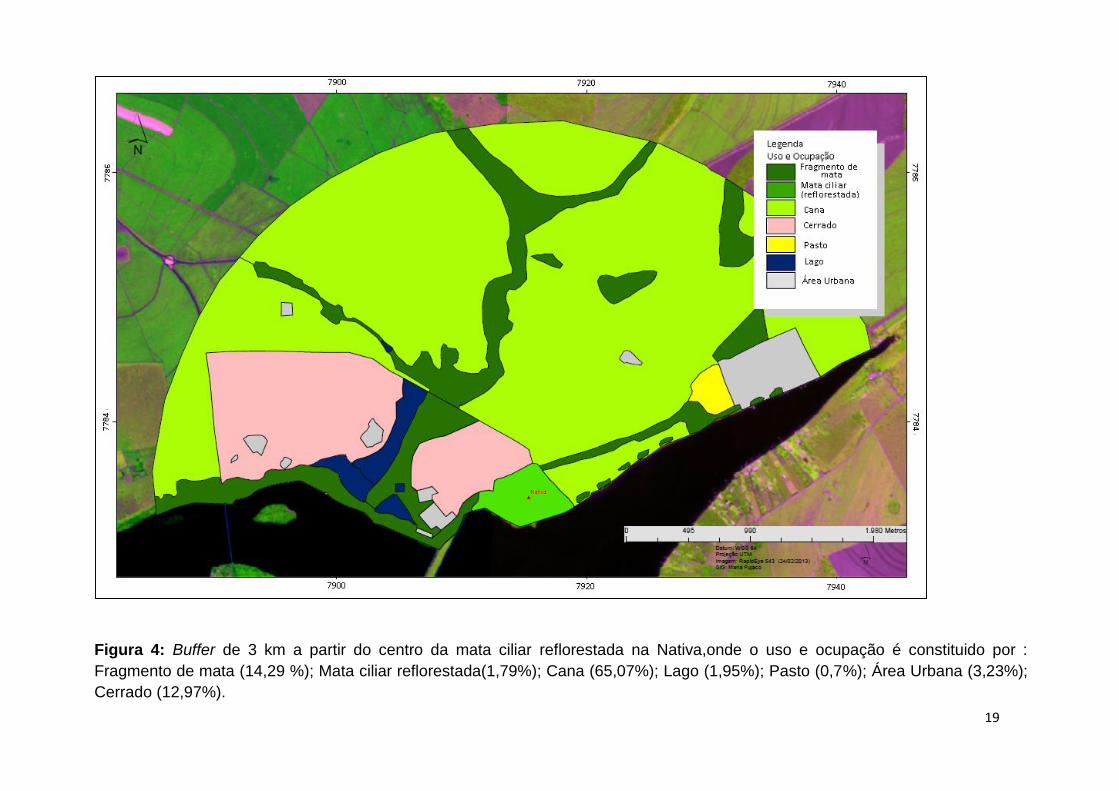
Figura 4: Buffer de 3 km a partir do centro da mata ciliar reflorestada na Nativa,onde o uso e ocupação é constituido por :
Fragmento de mata (14,29 %); Mata ciliar reflorestada(1,79%); Cana (65,07%); Lago (1,95%); Pasto (0,7%); Área Urbana (3,23%);
Cerrado (12,97%).
20
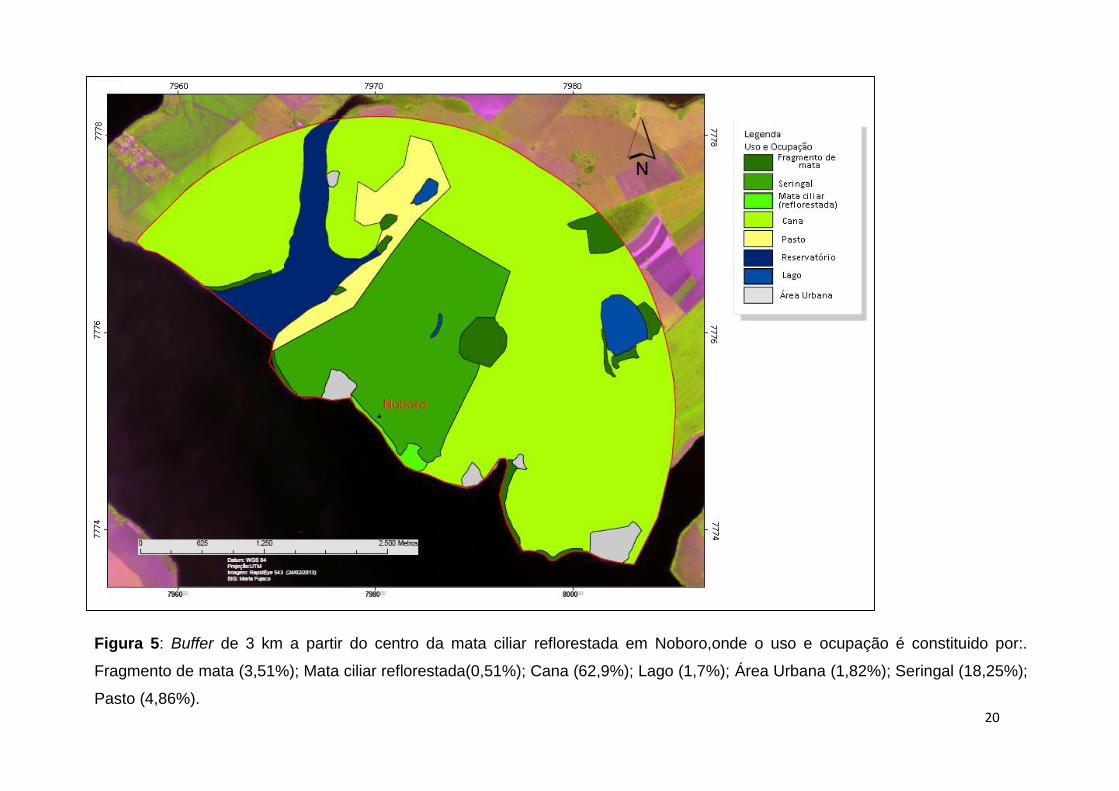
Figura 5: Buffer de 3 km a partir do centro da mata ciliar reflorestada em Noboro,onde o uso e ocupação é constituido por:.
Fragmento de mata (3,51%); Mata ciliar reflorestada(0,51%); Cana (62,9%); Lago (1,7%); Área Urbana (1,82%); Seringal (18,25%);
Pasto (4,86%).
21
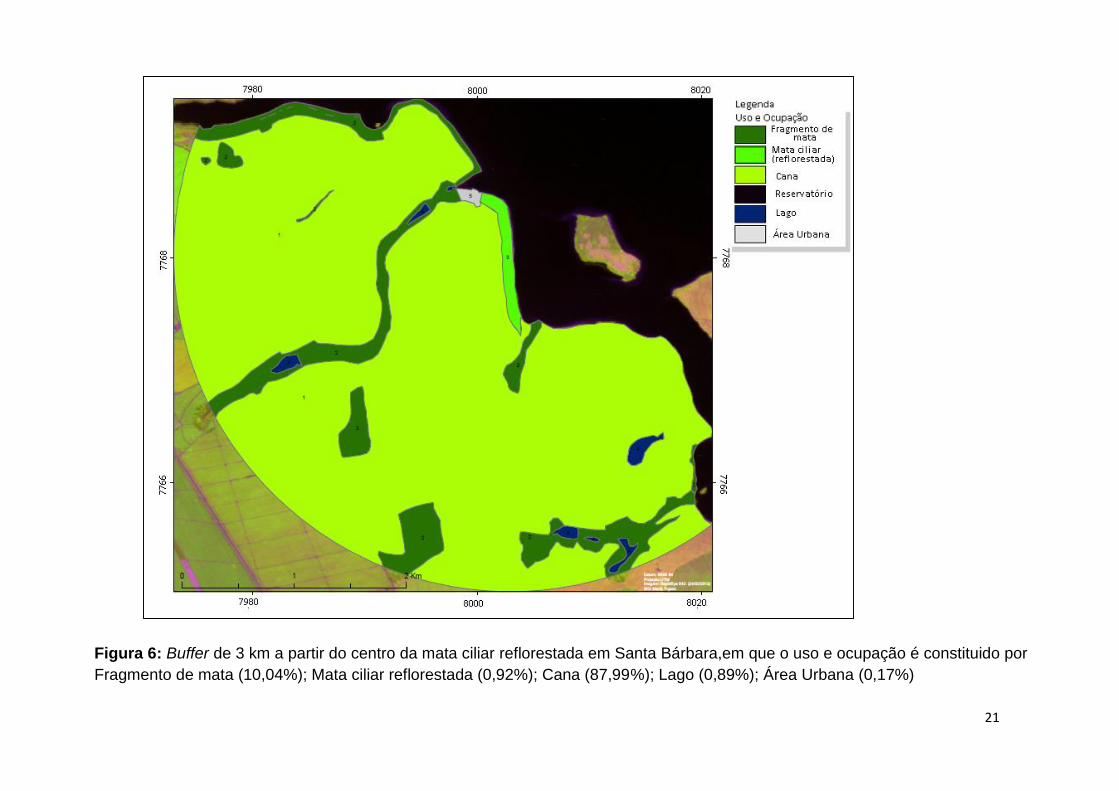
Figura 6: Buffer de 3 km a partir do centro da mata ciliar reflorestada em Santa Bárbara,em que o uso e ocupação é constituido por
Fragmento de mata (10,04%); Mata ciliar reflorestada (0,92%); Cana (87,99%); Lago (0,89%); Área Urbana (0,17%)

22
2. Coleta dos dados
Em cada uma das cinco áreas de amostragem, foram instalados três transectos
lineares de armadilhas de queda (pitfall traps, figura 7) (Cechin & Martins 2000). Esses
transectos foram posicionados paralelamente à represa e organizados em sequência,
mantendo intervalos de 50m entre eles. Cada transecto consistiu de 10 baldes de 60
litros enterrados até a boca, respeitando um distanciamento de 5m entre os baldes.
Uma cerca-guia de 1m de altura acompanhava a linha de baldes, incidindo no centro
destes e tendo a toda à extensão de sua base enterrada no solo, no espaço entre os
baldes. Essa cerca foi confeccionada com tela tipo mosquiteiro e sustentada por
estacas de madeira.
Figura 7: Desenho esquemático mostrando a disposição dos transectos de pitfall (traços em cinza) em cada área amostral de mata ciliar. As faixas azuis onduladas representam a represa.
Entre março de 2013 e janeiro de 2014, foram realizadas nove campanhas
mensais de coleta, totalizando 36 dias de amostragem. Não houve coleta em agosto e
setembro. Em cada campanha, as amostragens por meio das armadilhas de pitfall
ocorreram ao longo de quatro dias consecutivos. No primeiro dia de cada campanha,

23
os baldes eram abertos, sendo fechados após as coletas do último dia e assim
permaneciam no intervalo entre as campanhas. Os baldes eram vistoriados no período
da manhã.
Os anfíbios anuros foram também amostrados por meio de busca ativa associada
à zoofonia (Crump & Scott 1994). Para tanto, cada área foi amostrada por uma noite a
cada mês. As buscas foram realizadas entre 19 e 21h, totalizando esforço amostral de
12 horas/pessoa/área. O período de procura noturna foi distribuído em três meses na
estação seca em junho, julho e outubro, e três meses na chuvosa em novembro,
dezembro de 2013 e janeiro de 2014.
Os animais capturados foram marcados com elastômero e soltos no local de
captura. Esta metodologia de marcação foi utilizada para que os indivíduos
recapturados nas armadilhas não fossem recontados nas amostragens subsequentes.
O elastômero é um polímero pastoso fluorescente, disponível em diversas cores. Esse
material deve ser aplicado subcutaneamente, criando marcas únicas de acordo com a
cor e local de aplicação no corpo do animal. Após a aplicação o polímero se solidifica,
mantendo-se flexível e visível sob luz UV (Freitas et al. 2013).
Os exemplares coletados foram fixados com formalina 10%, transferidos para
álcool 70% e tombados na coleção Herpetológica do Laboratório de Zoologia dos
Vertebrados da Universidade Federal de Ouro Preto (LZV/UFOP). Todos os
procedimentos de coleta, transporte, marcação, tombamento em coleção científica
foram autorizados pelo comitê de Ética da UFOP n°. 142/2013 e pela licença do
ICMBio n° 36206-1. Para a identificação, os espécimes foram comparados com os
indivíduos da coleção Herpetológica do Laboratório de Zoologia dos Vertebrados da
Universidade Federal de Ouro Preto e foi utilizada a chave dicotômica de Provete et al.
(2011).
Foram obtidos dados da literatura sobre a ecologia e história natural das espécies
encontradas nas áreas amostrais (tabela 2). Para isto, utilizamos os trabalhos de
Brandão & Araújo 2001, Bastos 2007, Uetanabaro et al. 2008, Brassaloti et al. 2010,
Valdujo et al. 2011 e IUCN 2014.

24
Análise dos dados
Para avaliar se o esforço de coleta empregado foi suficiente para amostrar a
anurofauna local, foi construída a curva do coletor e a curva de riqueza estimada,
utilizando o programa Estimates versão 6.0b1 (Colwel 2000). A riqueza esperada foi
calculada pelo estimador Jacknife de primeira ordem com mil aleatorizações, utilizando
dados de presença e ausência de cada espécie por cada dia de procura (Burnham &
Overton 1979).
Os métodos de coleta, procura ativa e pitffall, não seguiram o mesmo padrão,
sendo assim, as analises estatísticas foram realizadas utilizando somente os dados das
armadilhas de interceptação e queda.
As espécies foram classificadas como dependentes ou independentes de matas
ciliares, conforme proposto por Brandão & Araújo (2001), que se basearam nas
características reprodutivas e especificidade de habitat da anurofauna.
Devido à dificuldade na diferenciação taxonômica dos juvenis das espécies
Physalaemus centralis e P. cuvieri (Giaretta et al. 2008; Araújo et al. 2013), os
espécimes foram considerados conjuntamente para a determinação da riqueza de
espécies e contagem de indivíduos.
O modelo linear generalizado (GLM) foi empregado para verificar a influência das
variáveis explicativas (tipo de matriz, idade, e largura do fragmento) sobre as variáveis
respostas (riqueza e abundância). Por meio dessas análises, foram estabelecidas
comparações entre as áreas amostrais. Após a análise utilizando o GLM, foi feita uma
análise de contraste, visando apontar qual variável explicativa teve maior influência
sobre a riqueza e abundância.
A variação da composição da anurofauna entre as áreas foi testada por meio de
uma análise multivariada de variância permutacional (PERMANOVA, Anderson 2001,
2006). Utilizamos dados de abundância e riqueza agrupados em cinco categorias de
acordo com as unidades amostrais. Medidas de dissimilaridade de Jaccard foram
utilizadas e 1000 permutações foram geradas. Após a PERMANOVA, uma análise
multivariada de distância permutacional (PERMDISP) foi feita com a matriz dos valores
de dissimilaridade de Jaccard gerados por meio da PERMANOVA, para testar a

25
homogeneidade da anurofauna entre as áreas (Anderson 2001, 2006). Um
escalonamento multidimencional não métrico (NMDS) foi utilizado para representar
graficamente a variação na composição (Clarke 1993). Todas as análises foram
realizadas utilizando o software R (R Development Core Team 2014). Foram utilizados
os pacotes GLM e Vegan para as analises de GLM e PERMANOVA respectivamente.
A significância para p foi <0,05.
Para determinar a diversidade total entre as áreas utilizamos o índice de Simpson,
pois captura bem a variação das distribuições de abundância e estabiliza com menores
tamanhos de amostras (Gimaret-Carpentier et al. 1998). Junto ao índice de
diversidade, foram gerados dados de dominância e equitabilidade das espécies. Todos
calculados pelo programa Past 2.08 (Hammer et al. 2001).
RESULTADOS
Foram registradas na RVG 21 espécies de anfíbios anuros pertencentes a 10
gêneros, distribuídos em 4 famílias: Bufonidae, Hylidae, Leptodactylidae e Microhylidae
(tabela 2). Leptodactylidae foi a família com maior riqueza (nove espécies), seguida por
Hylidae (oito espécies).
A curva de rarefação de espécies não se estabilizou, ou seja, não atingiu um
platô, indicando que não foram encontradas todas as espécies que ocorrem nesses
ambientes (figura 8). No entanto, a busca ativa foi uma forma de complementar o
número de espécies nas áreas.
26
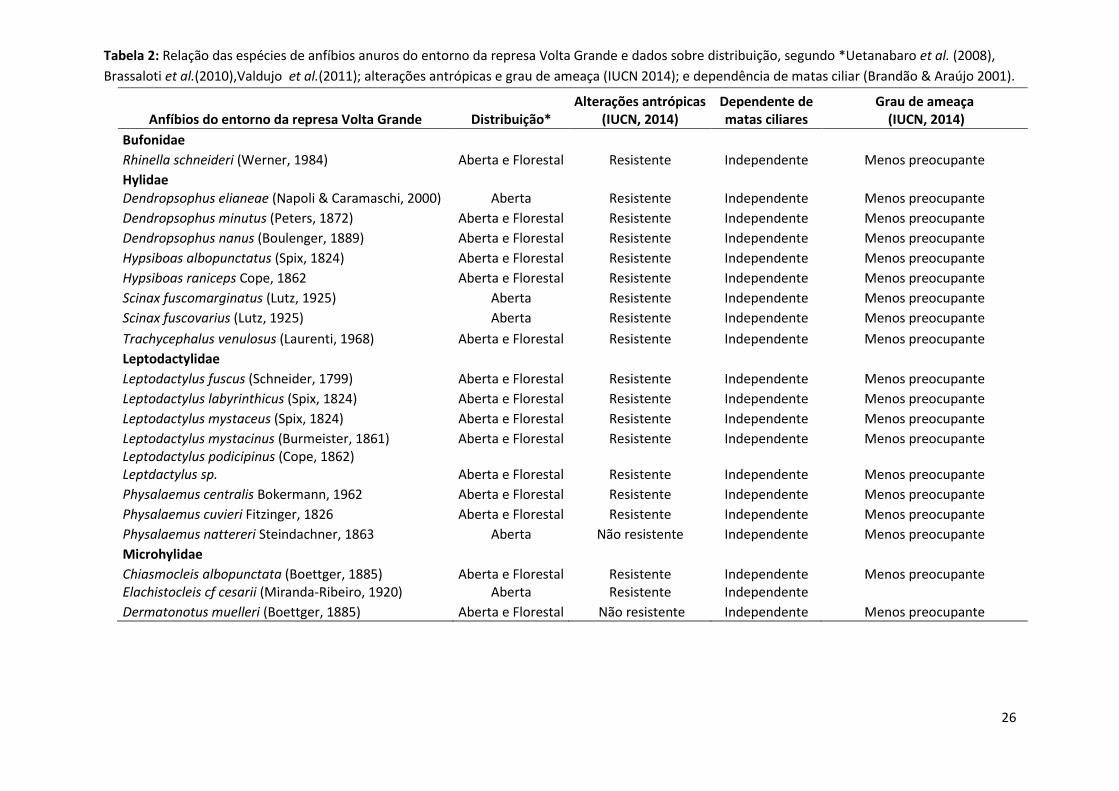
Tabela 2: Relação das espécies de anfíbios anuros do entorno da represa Volta Grande e dados sobre distribuição, segundo *Uetanabaro et al. (2008),
Brassaloti et al.(2010),Valdujo et al.(2011); alterações antrópicas e grau de ameaça (IUCN 2014); e dependência de matas ciliar (Brandão & Araújo 2001).
Anfíbios do entorno da represa Volta Grande Distribuição* Alterações antrópicas
(IUCN, 2014) Dependente de matas ciliares
Grau de ameaça (IUCN, 2014)
Bufonidae Rhinella schneideri (Werner, 1984) Aberta e Florestal Resistente Independente Menos preocupante
Hylidae Dendropsophus elianeae (Napoli & Caramaschi, 2000) Aberta Resistente Independente Menos preocupante
Dendropsophus minutus (Peters, 1872) Aberta e Florestal Resistente Independente Menos preocupante
Dendropsophus nanus (Boulenger, 1889) Aberta e Florestal Resistente Independente Menos preocupante
Hypsiboas albopunctatus (Spix, 1824) Aberta e Florestal Resistente Independente Menos preocupante
Hypsiboas raniceps Cope, 1862 Aberta e Florestal Resistente Independente Menos preocupante
Scinax fuscomarginatus (Lutz, 1925) Aberta Resistente Independente Menos preocupante
Scinax fuscovarius (Lutz, 1925) Aberta Resistente Independente Menos preocupante
Trachycephalus venulosus (Laurenti, 1968) Aberta e Florestal Resistente Independente Menos preocupante
Leptodactylidae Leptodactylus fuscus (Schneider, 1799) Aberta e Florestal Resistente Independente Menos preocupante
Leptodactylus labyrinthicus (Spix, 1824) Aberta e Florestal Resistente Independente Menos preocupante
Leptodactylus mystaceus (Spix, 1824) Aberta e Florestal Resistente Independente Menos preocupante
Leptodactylus mystacinus (Burmeister, 1861) Aberta e Florestal Resistente Independente Menos preocupante Leptodactylus podicipinus (Cope, 1862) Leptdactylus sp. Aberta e Florestal Resistente Independente Menos preocupante
Physalaemus centralis Bokermann, 1962 Aberta e Florestal Resistente Independente Menos preocupante
Physalaemus cuvieri Fitzinger, 1826 Aberta e Florestal Resistente Independente Menos preocupante
Physalaemus nattereri Steindachner, 1863 Aberta Não resistente Independente Menos preocupante
Microhylidae Chiasmocleis albopunctata (Boettger, 1885) Aberta e Florestal Resistente Independente Menos preocupante
Elachistocleis cf cesarii (Miranda-Ribeiro, 1920) Aberta Resistente Independente Dermatonotus muelleri (Boettger, 1885) Aberta e Florestal Não resistente Independente Menos preocupante
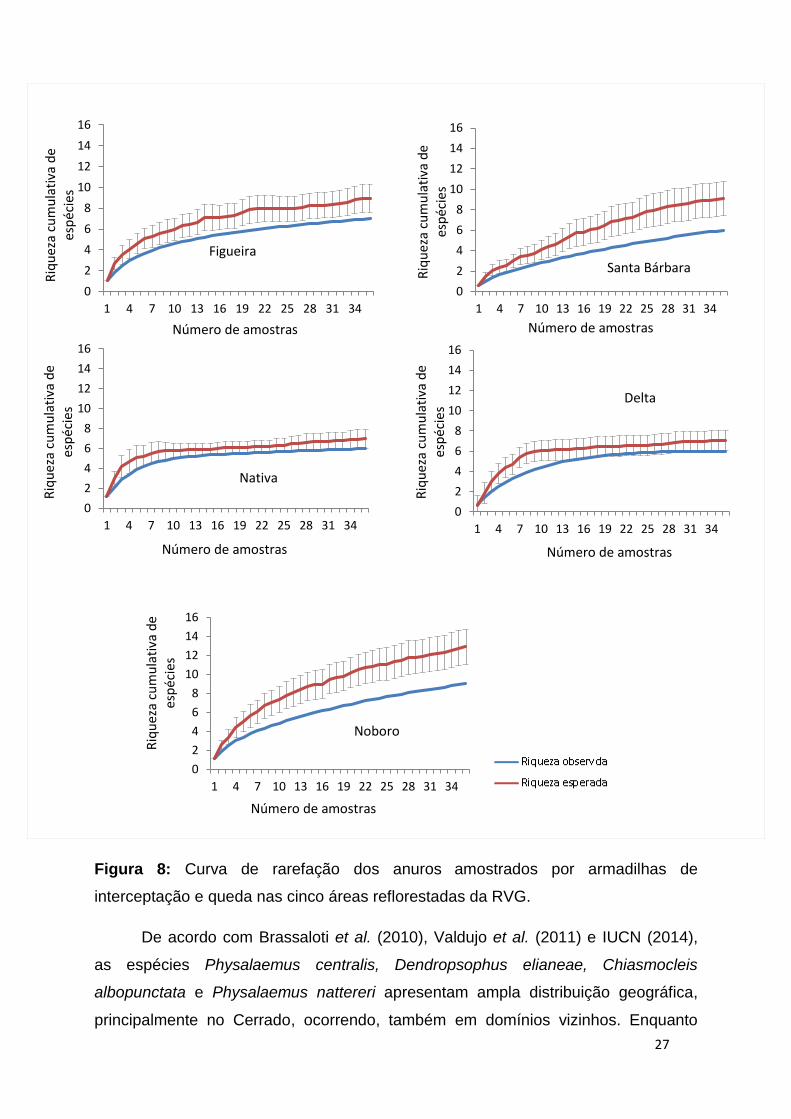
27
Figura 8: Curva de rarefação dos anuros amostrados por armadilhas de
interceptação e queda nas cinco áreas reflorestadas da RVG.
De acordo com Brassaloti et al. (2010), Valdujo et al. (2011) e IUCN (2014),
as espécies Physalaemus centralis, Dendropsophus elianeae, Chiasmocleis
albopunctata e Physalaemus nattereri apresentam ampla distribuição geográfica,
principalmente no Cerrado, ocorrendo, também em domínios vizinhos. Enquanto
0
2
4
6
8
10
12
14
16
1 4 7 10 13 16 19 22 25 28 31 34
Figueira
0
2
4
6
8
10
12
14
16
1 4 7 10 13 16 19 22 25 28 31 34
Santa Bárbara
0
2
4
6
8
10
12
14
16
1 4 7 10 13 16 19 22 25 28 31 34
Nativa
0
2
4
6
8
10
12
14
16
1 4 7 10 13 16 19 22 25 28 31 34
Delta
0
2
4
6
8
10
12
14
16
1 4 7 10 13 16 19 22 25 28 31 34
Noboro
Riq
uez
a cu
mu
lati
va d
e es
pé
cies
Riq
uez
a cu
mu
lati
va d
e es
péc
ies
Riq
uez
a cu
mu
lati
va d
e es
pé
cies
Riq
uez
a cu
mu
lati
va d
e es
péc
ies
Riq
uez
a cu
mu
lati
va d
e es
péc
ies
Número de amostras Número de amostras
Número de amostras Número de amostras
Número de amostras

28
que, Rhinella schneideri, D. minutus, D. nanus, Hypsiboas albopunctatus, H.
raniceps, Scinax fuscomarginatus, S. fuscovarius, Trachycephalus venulosus, P.
cuvieri, Lepstodactylus fuscus, L. labyrinthicus, L. mystaceus, L. mystacinus, L.
podicipinus, C. albopunctata, Elachistocleis cf cesarii e D. muelleri encontram-se
amplamente distribuídas pelo Brasil, ocorrendo em vários biomas (Brassaloti et al.
2010; IUCN 2014).
Uma parcela significativa, ou seja, 80% dos anuros encontrados nas áreas de
estudo ocorrem principalmente em áreas abertas e, eventualmente, são encontrados
em áreas florestais. Apenas Dendropsophus elianeae, Scinax fuscomarginatus, S
fuscovarius e Physalaemus nattereri são exclusivas de formações abertas. Como
nenhuma das espécies utilizam apenas ambientes florestais, foram classificadas
como independentes de mata de ciliar (tabela 2). Apenas P. nattereri e D. muelleri
são descritas como não resistentes a impactos antrópicos (IUCN 2014). Quanto a
questão do grau de ameaça de extinção das espécies do estudo, todas são
descritas como pouco preocupante.
Dentre os 39 modos reprodutivos registrados para anfíbios anuros (Haddad &
Prado 2005), ocorrem entre as espécies do entorno da represa os modos 1, 2, 11,
13 e 30 (tabela 3). O modo 1 envolve ovos e girinos exotróficos que se desenvolvem
em corpos d’agua lênticos (Haddad & Prado 2005). Esse modo reprodutivo foi o
mais comum entre as famílias, ocorrendo em 12 espécies. Foi o único modo
reprodutivo encontrado entre os Hilídeos e Microhilideos. A espécie Rhinella
schneideri a única representante da família Bufonidae, pode apresentar o modo
reprodutivo 1 e 2, ou seja, ovos e girinos exotróficos que se desenvolvem tanto em
corpos d’água lênticos, quanto em lóticos.
A família Leptodactylidae apresentou maior número de modos reprodutivos
(três) e estes não são descritos para as espécies que ocorreram nas outras famílias.
O modo reprodutivo 11, que ocorreu apenas para os Physalaemus, é descrito pela
postura de ovos em ninhos de espuma flutuante em corpo d’agua lêntico e girinos
exotróficos que se desenvolvem nesses corpos d’água (Haddad & Prado 2005). Os
outros dois modos reprodutivos ocorreram apenas para o gênero Leptodactylus,
sendo o modo 13 descrito pela postura de ovos em ninhos de espuma flutuante em
água acumulada em piscinas construídas e girinos exotróficos em corpos d’água
lênticos (Haddad & Prado 2005). O modo 30 é caracterizado por apresentar ninho de
29
espuma com ovos e estágios larvais iniciais em ninhos subterrâneos construídos;
após inundação, se desenvolvem os girinos exotróficos em corpos d’ água lênticos
(Haddad & Prado 2005).
Tabela 3: Modos reprodutivos das espécies encontradas no entorno da represa Volta Grande, segundo Bastos (2007), Uetanabaro et al. (2008) e Haddad et al. (2013), habitat onde foram coletadas: mata ciliar (MC), corpos d água próximos à mata ciliar (EG) e métodos de coleta: pitfall traps (PT) e procura ativa (PA).
Anfíbios do entorno da represa Volta Grande
Modo Reprodutivo Habitat Método
de coleta
Bufonidae
Rhinella schneideri Modo 1 e 2 MC e EG PT e PA
Hylidae
Dendropsophus elianeae Modo 1 EG PA
Dendropsophus minutus Modo 1 EG PA
Dendropsophus nanus Modo 1 EG PA
Hypsiboas albopunctatus Modo 1 EG PA
Hypsiboas raniceps Modo 1 EG PA
Scinax fuscomarginatus Modo 1 EG PA
Scinax fuscovarius Modo 1 MC e EG PT e PA
Trachycephalus venulosus Modo 1 EG PA
Leptodactylidae
Leptodactylus fuscus Modo 30 MC e EG PT e PA
Leptodactylus labyrinthicus Modo 13 EG PA
Leptodactylus mystaceus Modo 30 MC e EG PT e PA
Leptodactylus mystacinus Modo 30 MC PT
Leptodactylus podicipinus Modo 13 MC e EG PT e PA
Leptodactylus sp. MC PT
Physalaemus centralis Modo 11 EG PA
Physalaemus cuvieri Modo 11 MC e EG PT e PA
Physalaemus nattereri Modo 11 MC PT
Microhylidae
Chiasmocleis albopunctata Modo 1 MC PT Elachistocleis cf cesarii Modo 1 MC PT
Dermatonotus muelleri Modo 1 MC e EG PT e PA
Das 21 espécies registradas, cinco foram coletadas apenas por pitfall; nove
por procura ativa e sete pelos dois métodos. Todos os Hilídeos foram encontrados
por busca ativa em lagoas, somente S. fuscovarius foi também capturada nas
armadilhas de pitfall (tabela 3). As espécies coletadas pelos dois métodos são
encontradas nos dois tipos de ambiente, mata ciliar e corpos d’água em áreas
abertas do entorno.
30
Physalaemus cf. cuvieri e Rhinella schneideri se destacaram, pois foram
presentes em todas as áreas e também foram as espécies mais abundantes,
totalizando mais que 70% do total de indivíduos da RVG. Em contraste, somada a
abundância de indivíduos das espécies Scinax fuscovarius, Leptodactylus fuscus,
Leptodactylus sp. e Elachistocleis cf. cesarii representam 1,5% da abundância total.
Foram encontrados apenas um individuo das espécies Elachistocleis cf cesarii
(Nativa) e Leptodactylus fuscus (Noboro) e três indivíduos das espécies S.
fuscovarius e Leptodactylus sp. (tabela 4).
Tabela 4: Distribuição e abundância das espécies de anfíbios em cada área amostrada da represa Volta Grande entre março de 2013 e janeiro de 2014.
Abundância de anfíbios Total Geral
Família/Espécie Delta Figueira Nativa Noboro Santa
Bárbara
Bufonidae Rhinella schneideri 5 28 14 34 115 196
Hylidae Scinax fuscovarius 0 2 0 0 1 3
Leiuperidae Physalaemus cuvieri 11 9 25 135 1 181
Physalaemus nattereri 0 26 0 4 2 32 Leptodactylidae
Leptodactylus fuscus 0 0 0 1 0 1 Leptodactylus mystaceus 22 4 0 2 0 28 Leptodactylus mystacinus 3 4 5 3 0 15 Leptodactylus podicipinus 11 1 0 0 8 20 Leptodactylus sp. 0 0 0 3 0 3 Microhylidae
Chiasmocleis albopunctata 6 0 13 1 1 21 Elachistocleis cf cesarii 0 0 1 0 0 1 Dermatonotus muelleri 0 0 25 2 0 27 Abundância Total 58 75 83 185 128 529 Riqueza Total 6 8 6 9 6 13
As espécies Physalaemus cf. cuvieri, em Noboro, e Rhinella schneideri, em
Santa Bárbara, representaram 73% e 90% do total de indivíduos para as respectivas
áreas, onde foram consideradas dominantes (tabela 5). Na Nativa, as espécies mais
abundantes foram P. cf. cuvieri e D. muelleri, e seus valores individuais de
abundância representaram 30% do total de espécies para esta área. Em Figueira e
Delta, as espécies mais abundantes foram R. schneideri e L. mystaceus,
respectivamente. Estas espécies mais abundantes nas áreas Nativa, Figueira e
Delta não se configuram como espécies dominantes. Estas áreas apresentaram
31
maior valor de equitabilidade, com a abundância melhor distribuída entre as
espécies.
Tabela 5: Riqueza de espécies, número de indivíduos encontrados, índices de dominância, diversidade e equitabilidade das cinco áreas reflorestadas no entorno da RVG.
Delta Figueira Nativa Noboro Santa
Bárbara Geral: RVG
Riqueza de espécies 6 8 6 9 6 13
Número de indivíduos 58 75 83 185 129 529
Dominância (D) 0,23 0,28 0,23 0,56 0,81 0,26
Simpson (1-D) 0,76 0,71 0,76 0,43 0,18 0,73
Equitabilidade (J) 0,89 0,72 0,85 0,41 0,25 0,64
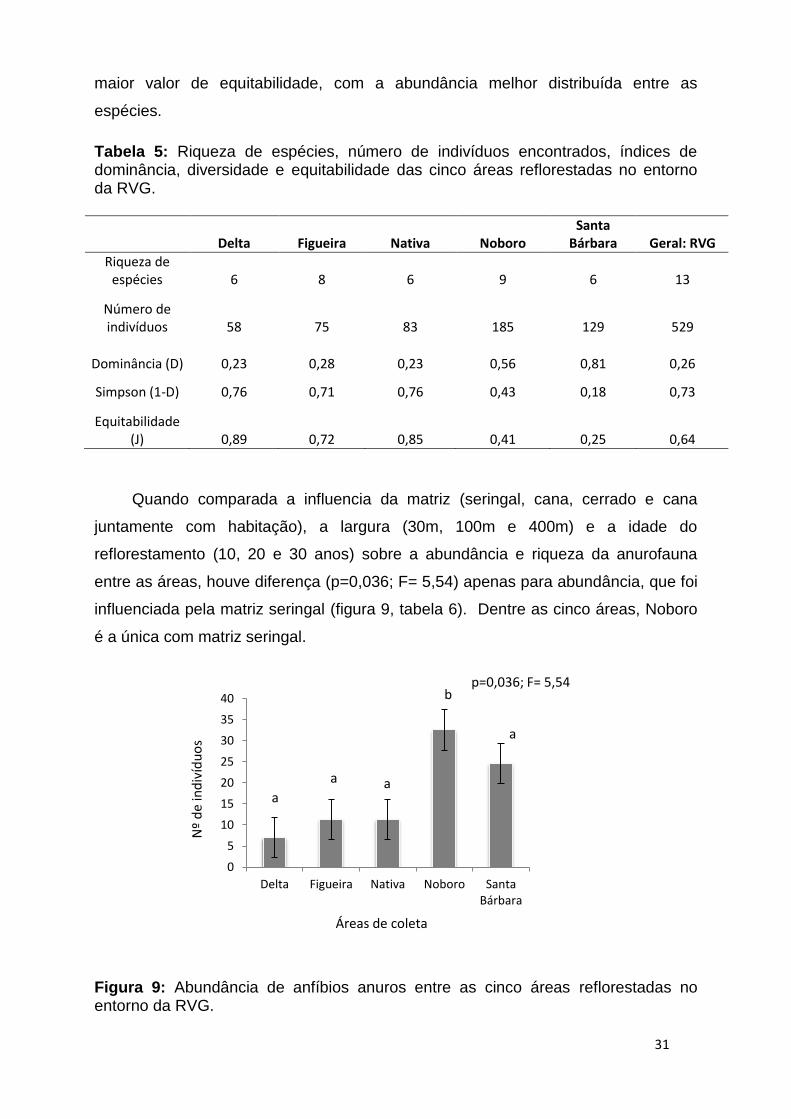
Quando comparada a influencia da matriz (seringal, cana, cerrado e cana
juntamente com habitação), a largura (30m, 100m e 400m) e a idade do
reflorestamento (10, 20 e 30 anos) sobre a abundância e riqueza da anurofauna
entre as áreas, houve diferença (p=0,036; F= 5,54) apenas para abundância, que foi
influenciada pela matriz seringal (figura 9, tabela 6). Dentre as cinco áreas, Noboro
é a única com matriz seringal.
Figura 9: Abundância de anfíbios anuros entre as cinco áreas reflorestadas no entorno da RVG.
0
5
10
15
20
25
30
35
40
Delta Figueira Nativa Noboro SantaBárbara
Nº
de
ind
ivíd
uo
s
a
a a
a
bp=0,036; F= 5,54
Áreas de coleta
32
Tabela 6: Modelos Lineares Generalizados (GLM) e Modelos Generalizados Mistos
(GLMM) construídos com abundância de anfíbios em áreas de mata ciliar do
reservatório de Volta Grande e as variáveis explicativas testadas.
Variável resposta
Variável explicativa
Deviance Distribuição
de erro (família)
Grau de Liberdade
F-valor p-valor
Abundância de anuros
Matriz 39,49 Quasi-poisson 13 5,54 0,03*
Abundância de anuros
Largura 23,916 Quasi-poisson 12 3,36 0,09
Abundância de anuros
Idade 0,53 Quasi-poisson 11 0,07 0,78
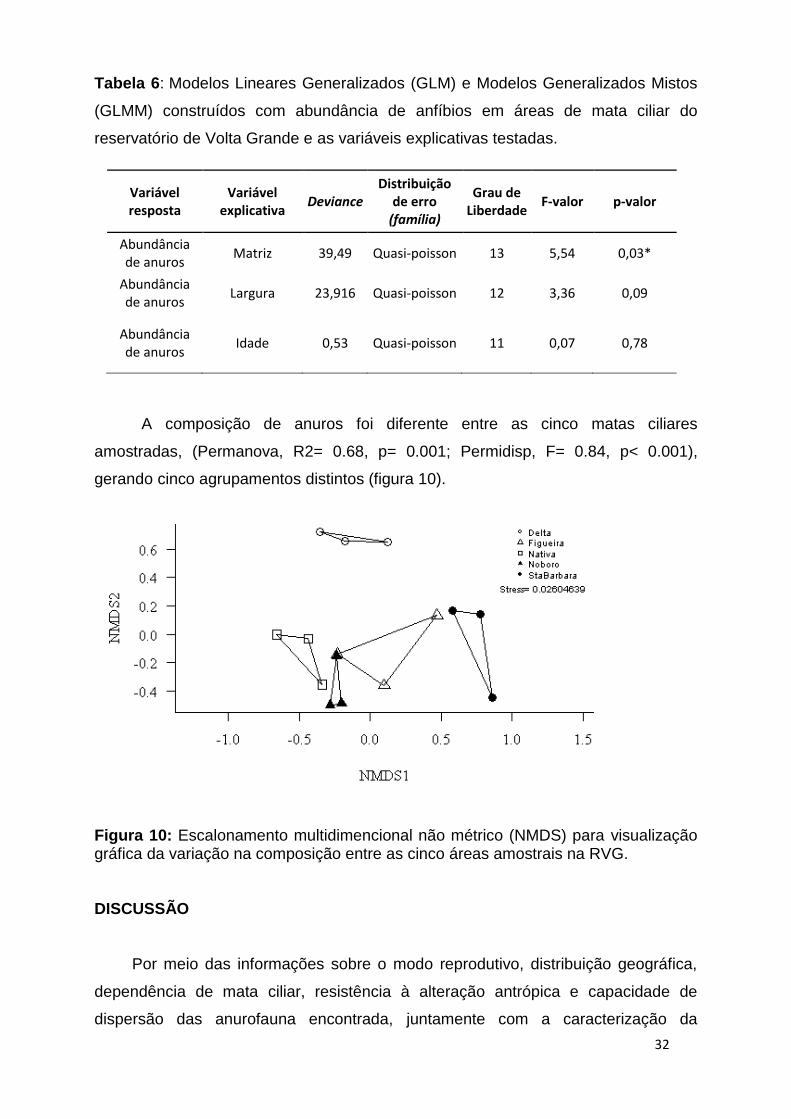
A composição de anuros foi diferente entre as cinco matas ciliares
amostradas, (Permanova, R2= 0.68, p= 0.001; Permidisp, F= 0.84, p< 0.001),
gerando cinco agrupamentos distintos (figura 10).
Figura 10: Escalonamento multidimencional não métrico (NMDS) para visualização gráfica da variação na composição entre as cinco áreas amostrais na RVG. DISCUSSÃO
Por meio das informações sobre o modo reprodutivo, distribuição geográfica,
dependência de mata ciliar, resistência à alteração antrópica e capacidade de
dispersão das anurofauna encontrada, juntamente com a caracterização da

33
paisagem, foi possível avaliar colonização das áreas de mata ciliar reflorestas no
cerrado. O reflorestamento do entorno da RVG, utilizado como medida de
recuperação de áreas modificadas pela construção da hidrelétrica, não se mostrou
eficaz em manter variabilidade espécies de anuros, de modo que selecionou apenas
espécies com ecologia e história natural semelhante.
A região do estudo parece ter sido modificada a tal ponto que as espécies
endêmicas foram extintas localmente. De acordo com Ricklefs (1993), mudanças
locais no habitat ou clima podem levar espécies endêmicas à extinção. Estas
espécies não foram encontradas, sendo esperadas para as áreas de estudo mesmo
que em baixa proporção, uma vez que no Cerrado 50% das espécies de anuros são
endêmicas (Valdujo, 2011).
Outro grupo ausente nas áreas de estudo, é referente as espécies
dependentes de mata ciliar, o que aponta para a eliminação das mesmas nas áreas,
sendo a construção do reservatório uma das possíveis causas. O mesmo foi
observado por Brandão & Araújo (2008), após a construção da Usina Hidrelétrica de
Serra da Mesa. Era esperado o encontro de espécies hábitat- especialistas dado
que as matas ciliares normalmente abrigam estas espécies (Brandão & Araújo
2001), no entanto, pode ser que as áreas antes da construção da hidrelétrica não
apresentavam estas espécies.
A classificação quanto a dependência por mata ciliar foi baseada no conceito
de Brandão & Araújo (2001), contudo, esta classificação ainda não é definitiva,
devido à escassez de informações sobre a anurofauna nestes ambientes. Estes dois
autores classificaram Leptodactylus podicipinus e L. mystacinus como dependentes
de mata de galeria no Distrito Federal. Contudo foram aqui classificadas como
independentes, pois são espécies que ocorrem tanto em áreas abertas, quanto
florestais (Uetanabaro et al. 2008; Brassaloti et al. 2010; IUCN 2014).
Nenhuma das áreas de mata de galeria no entorno da represa é nativa. Essas
matas foram constituídas por reflorestamentos (Noboro, Santa Bárbara, Delta e
Figueira) e processos de sucessão natural (Nativa) e se encontram envolvidas por
extensas matrizes agrícolas, principalmente canaviais. Em algumas áreas, ocorrem
estreitos corredores ecológicos, os quais são interrompidos por estradas ou outras
barreiras antrópicas como moradias. Tudo isto quando analisado em função da
composição da anurofauna da RVG, nos leva a supor que as espécies dependentes

34
de mata ciliar e outras mais sensíveis não conseguiriam atravessar as matrizes
agrícolas, nem os estreitos corredores ecológicos, de modo a alcançar os
fragmentos ciliares.
Outra explicação para a não ocorrência de anuros especialistas de matas de
ciliares no entorno da RVG pode estar relacionada ao fato de que as áreas
reflorestadas não estão sendo eficazes para evitar a dessecação dos ovos. Dessa
forma, as áreas são dominadas por espécies com modo reprodutivo característico
de áreas abertas. Espécies de ambientes não florestados apresentam pequena
diversidade de modos reprodutivos (Duellman & Trueb 1994), nas áreas de estudo
identificamos cinco modos reprodutivos.
Espécies de áreas abertas são mais resistentes à dessecação e apresentam
modos reprodutivos mais generalizados (modos 1 e 2) ou modos adaptados a
insolação (por exemplo, modos de 11, 13 e 30) (Haddad & Prado 2005). Os modos
reprodutivos 11, 13 e 30 incluem ninhos de espuma, um modo reprodutivo
especializado que impede a dessecação dos ovos, sendo considerada como uma
adaptação a ambientes sazonais (Heyer 1969). O ninho de espuma é característico
da maioria das espécies das famílias Leptodactilidae (Frost et al. 2006), que reúnem
40% das espécies encontradas nas áreas de estudo. Os modos reprodutivos dos
anfíbios nas áreas do estudo se caracterizam como sendo resistentes à insolação.
Desta forma, mesmo após decorridos 10 ou 20 anos de reflorestamento, as áreas
não foram colonizadas por espécies com modo reprodutivo característico de um
ambiente mais ameno.
Informações quanto à capacidade de dispersão dos anfíbios são praticamente
inexistentes, sendo assim discutiram aqui a possibilidade de classificação de alguns
anuros das áreas de estudo através de informações da paisagem local e
informações da literatura. As áreas de mata ciliar próximas a Noboro e Santa
Barbara são descontínuas e toda a região é cercada por extensas matrizes
agrícolas. Mesmo assim, estas duas áreas foram colonizadas por 11 espécies de
anfíbios. As espécies encontradas para estas duas áreas foram Rhinella schneideri,
Scinax fuscovarius, Physalaemus cuvieri, Physalaemus nattereri, Leptodactylus
fuscus, L. mystaceus, L. mystacinus, L. podicipinus, Leptodactylus sp. e
Dermatonotus. muelleri. A capacidade de atravessar a matriz é um fator
determinante para a persistência de algumas espécies em áreas fragmentadas

35
(Laurance 1991 e Gascon et al. 1999), sendo assim, supomos que as espécies
presentes nas duas áreas reflorestadas apresentam grande capacidade de
dispersão.
Existe uma relação positiva entre a capacidade de deslocamento de uma
espécie e a sua habilidade para estabelecer populações em áreas fragmentadas
(Otani 2011). Em poças temporárias inseridas em canavial no cerrado, algumas das
espécies foram as mesmas que encontramos para Noboro e Santa Barbara: Scinax
fuscovarius, Physalaemus nattereri, Physalaemus centralis, P. cuvieri, Leptodactylus
fuscus, L. mystaceus e L. mystacinus (Zina et al. 2007), o que sustenta nossa
suspeita destas espécies apresentarem grande capacidade de dispersão e
resistência às condições da matriz.
Diante das características predominantes das espécies encontradas
concluímos que está ocorrendo nas áreas um processo conhecido como
homogeneização biótica, considerado um dos mais acentuados formas de
empobrecimento da biodiversidade em todo mundo (Olden, 2004). Ela consiste na
substituição das espécies nativas (endêmicas ou não) por uma biota de ampla
distribuição geográfica e elevada plasticidade adaptativa (Vitule e Pozenato, 2012).
Estas espécies se beneficiam das atividades humanas e são também conhecidas
como “vencedoras” (McKinney & Lockwood, 1999), no entanto, não se tem
informação se este sucesso frente a homogeneização, persistirá a longo prazo
(Olden, 2004).
As alterações antrópicas vem dificultando a heterogeneidade biótica no local de
estudo. Estas pressões atuam continuamente no interior e nas proximidades das
áreas reflorestadas, de forma a não permitir o estabelecimento de condições para
que a “próxima” espécie venha colonizá-la. As moradias ao longo das matas ciliares
interferem em seu papel de conectividade (Crema 2008); o uso de agrotóxicos pode
acarretar em castração química de anfíbios (Sparling et al. 2000; Hayes et al. 2010)
e as áreas de agricultura representam barreiras para algumas espécies (Gascon et
al. 1999; Ricketts 2001). Esse efeito pode variar ao longo do ano, devido ao corte
periódico da cana.
Cada evento de corte de cana pode estar atuando como um novo evento de
fragmentação, gerando efeitos de borda que oscilam em função da altura das
plantas, as quais vão regular incidência de luz, vento e temperatura sobre as áreas
reflorestadas. Além disso, o despejo de contaminantes químicos pode também ser

36
considerado como uma pressão antrópica constante nessas áreas. Alguns tipos de
contaminantes químicos são apontados como causas do declínio de anfíbios no
mundo (Young 2004). Somente por meio de um monitoramento em longo prazo,
seria possível mostrar se essas espécies continuarão sendo as únicas capazes de
colonizar o local.
A maior abundância de anuros influenciada pela matriz seringal na área
Noboro, pode significar que este tipo de matriz arbórea seja como uma zona de
amortecimento, em que o efeito de borda seja menor. No entanto, esta área teve
grande dominância da espécie Physalaemus cf. cuvieri, seguida por Rhinella
schneideri, uma das áreas com menor equitabilidade, podendo ser apenas um
reflexo da reprodução da espécie na área, ou seja, uma alta abundância ao acaso.
Quanto a composição da anurofauna, houve diferença entre as cinco áreas.
Este fato se deu pelo conjunto de alguns fatores. As espécies dominantes foram
diferentes para cada área, sendo que em Figueira, Delta e Nativa houve maior
equitabilidade e nas áreas Santa Bárbara e Noboro, as espécies Physalaemus cf.
cuvieri e Rhinella schneideri foram as espécies dominantes, respectivamente. As
áreas também diferiram quanto ao padrão de distribuição de abundância das
espécies comuns, como exemplo: Physalaemus. nattereri, Dermatonotus. muelleri e
Leptodactylus mystaceus, que foram as mais abundantes em Figueira, Nativa e
Delta, respectivamente.
A variação na composição também pode ser analisada a partir de espécies que
foram encontrados exclusivamente em uma das áreas, como Elachistocleis cf.
cesarii, encontrada apenas na área nativa. Apesar de Leptodactylus fuscus ter sido
encontrada no entorno de algumas áreas reflorestadas por meio da busca ativa, só
foi encontrado nas armadilhas da área Noboro. Embora exista esta diferença na
composição entre as áreas, a característica predominante da anurofauna em cada
uma delas leva ao mesmo diagnóstico, ou seja, espécies generalistas, resistentes a
impactos antrópicos, ampla distribuição e com modos reprodutivos propícios para
evitar insolação e dessecação.
Conclusão
Para a anurofauna, apenas o reflorestamento não é suficiente. Está ocorrendo nas
áreas do entorno da RVG o processo de homogeneização biótica da anurofauna.

37
Devido aos constantes impactos antrópicos, próximos e no interior das matas, este
processo tende a continuar. O tipo de matriz e a idade e largura do reflorestamento,
não mostraram ter influência sobre a riqueza de espécies, sendo assim a estrutura
da paisagem que envolve as matas reflorestadas, principalmente a dimensão dos
canaviais, devem ser os grandes selecionadores das espécies colonizadoras das
áreas de estudo.
REFERÊNCIA BIBLIOGRÁFICA
Anderson MJ (2001). A new method for non-parametric multivariate analysis of
variance. Austral Ecology 26:32-46.
Anderson MJ (2006) Distance-based tests for homogeneity of multivariate
dispersions. Biometrics 62:245-253.
ANEEL (Agência Nacional De Energia Elétrica). Atlas de Energia Elétrica do Brasil /
Agência Nacional de Energia Elétrica: ANEEL. (2002)
http://www.aneel.gov.br/arquivos/pdf/livro_atlas.pdf. Acesso em: 02 set. 2014.
Araújo CO, Corrêa DT, Almeida-Santos SM (2013) Anurans of the Estação Ecológica
de Santa Bárbara, a remant of open Cerrado formations in the state of São
Paulo. Biota Neotropica 13(3): 230 -240.
Azevedo-Ramos C, Carvalho Jr., Nasi R (2005) Animais como indicadores: uma
ferramenta para acessar a integridade biológica após a exploração madeireira
em florestas tropicais. CIFOR: IPAM, Belém, Pará.
Bastos RP (2007) Anfíbios do Cerrado. Páginas 87-100 In: Nascimento LB, Oliveira
ME (eds) Herpetologia no Brasil II. Sociedade Brasileira de Herpetologia, Belo
Horizonte.
Braga FMS (2001) Crescimento e mortalidade de Leporinus friderici (Ostariophysi,
Anostomidae) na represa de Volta Grande, Rio Grande, localizada entre os
Estados de Minas Gerais e São Paulo, Brasil. Acta Scientiarum 23 (2): 415-
420.
Brandão RA, Araújo AFB (2001) A herpetofauna associada às matas de galeria do
Distrito Federal. Páginas 561-604 In: Ribeiro JF, Fonseca CEL, Sousa-Silva JC

38
(eds) Cerrado: Caracterização e Recuperação de Matas de Galeria. Embrapa
Cerrados, Planaltina.
Brandão RA, Araújo AFB (2008) Changes in anuran species richness and
abundance resulting from hydroelectric dam flooding in Central Brazil.
Biotropica 40(2): 263–266.
Brasil. Decreto N° 97.632, de 10 de abril de 1989 (1989) Dispõe sobre a
regulamentação do artigo 2°, Inciso VIII, da Lei n° 6.938, de 31 de Agosto de
1981, e dá outras providências
http://www.planalto.gov.br/ccivil_03/decreto/1980-1989/D97632.htm. (Acesso
em: 28 Ago. 2014).
Brasil. Lei n. 9.985, de 18 de julho de 2000 (2000) Regulamenta o art. 225, § 1o,
incisos I, II, III e VII da Constituição Federal, institui o Sistema Nacional de
Unidades de Conservação da Natureza e dá outras providências
http://www.planalto.gov.br/ccivil_03/leis/l9985.htm. (Acesso em: 28 Ago. 2014).
Brasil. Resolução CONAMA Nº 302, de 20 de março de 2002 (2002) Dispõe sobre
os parâmetros, definições e limites de Áreas de Preservação Permanente de
reservatórios artificiais e o regime de uso do entorno.
http://www.mma.gov.br/port/conama/res/res02/res30202.html (Acesso em: 28
Ago. 2014).
Brasil. Ministério do Meio Ambiente – MMA Instituto Brasileiro do Meio Ambiente e
dos Recursos Naturais Renováveis – IBAMA (2011) Monitoramento do
desmatamento nos biomas brasileiros por satélite: acordo de cooperação
técnica MMA/IBAMA monitoramento do bioma Cerrado 2009-2010.
http://www.mma.gov.br/estruturas/sbf_chm_rbbio/_arquivos/relatoriofinal_cerra
do_2010_final_72_1.pdf (Acesso em: 28 Ago. 2014).
Brassaloti RA, Rossa-Feres DC, Bertoluci J (2010) Anurofauna da Floresta
Estacional Semidecidual da Estação Ecológica dos Caetetus, sudeste do
Brasil. Biota Neotropica 10(1): 275-292.
Burnham KP, Overton WS (1979) Robust estimation of population size when capture
probabilities vary among animals. Ecology 60: 927-936.

39
Cechin SZ, Martins M (2000) Eficiência das armadilhas de queda (pitfall traps) em
amostragens de anfíbios e répteis no Brasil. Revista Brasileira de Zoologia. 17
(3): 729-740.
Clarke KR (1993) Non-parametric multivariate analysis of changes in community
structure. Austral Ecology 18:117-143.
Colli GR, Bastos RP, Araújo AB (2002) The character and dynamics of the Cerrado
herpetofauna. Páginas 223-241 In: Oliveira PS, Marquis RJ (eds) The Cerrados
of Brazil. Ecology and Natural History of a Neotropical Savanna. Columbia
University Press, New York.
Cosson JF, Ringuet S, Claessens O, Massary JC, Dalecky A, Villiers JF, Granjon L,
Pons JM (1999) Ecological changes in recent land-bridge island in French
Guiana, with emphasis on vertebrate communities. Biological Conservation 91:
213-222.
Colwell RK (2000) EstimateS 6.0b1: Statistical Estimation of Species Richness and
Shared Species from Samples. University of Connecticut.
Crema A (2008) Diversidade e distribuição de anfíbios anuros associados a matas
de galeria dentro e fora de Unidades de Conservação do Distrito Federal.
Dissertação Mestrado. Universidade de Brasília, Brasília-DF.
Crump ML, Scott NJJ (1994) Visual encounter surveys. Páginas 84-92 In: Heyer,
WR, Donnelly MA, McDiarmid RW, Hayek LAC, Foster MS (eds) Measuring and
Monitoring Biological Diversity: Standard Methods for Amphibians. Smithsonian
Institution Press, Washington.
Cullen WR, Wheater CP, Dunleavy PJ (1998). Establishment of species-rich
vegetation on reclaimed limestone quarry faces in Derbyshire, UK. Biological
Conservation 84: 25-33.
Dias BFS (1994) A conservação da natureza. Páginas 607-663 In: Pinto MN (eds)
Cerrado: Caracterização, Ocupação e Perspectivas. Universidade de Brasília,
Brasília-DF.
Duellman WE, Trueb L, (1994) Biology of Amphibians. The Johns Hopkins University
Press, Baltimore.

40
Dynesius M, Nilsson C (1994) Fragmentation and flow regulation of river systems in
the northern third of the world. Science 266: 753–762.
Feio RN, Braga UML, Wiederhecker H, Santos PA (1998) Anfíbios do Parque
Estadual do Rio Doce (Minas Gerais). Universidade Federal de Viçosa. Instituto
Estadual de Florestas. Imprensa Universitária, Viçosa.
Ferreira WC (2009) Estoque biomassa e carbono e parâmetros indicadores de
recuperação de mata ciliar. Tese Doutorado. Universidade Federal De Lavras.
Lavras.
Filardi FLR, Garcia FCP, Carvalho-Okano RM (2007) Espécies lenhosas de
Papilionoideae (Leguminosae) na Estação Ambiental de Volta Grande, Minas
Gerais, Brasil. Rodriguesia 58:363-378.
Fonseca CEL, Ribeiro JF, Souza CC, Rezende RP, Balbino VK (2001) Recuperação
da vegetação de matas de Galeria: estudos de caso no Distrito Federal e
Entorno. Páginas 815-870 In: Ribeiro JF, Fonseca CEL, Sousa-Silva JC (eds)
Cerrado: Caracterização e recuperação de Matas de Galeria. Embrapa
Cerrados, Planaltina.
Forman RTT, GODRON M (1986) Landspace Ecology. John Wiley & Sons, New
York.
Fox BJ (1982) Fire and mammalian secondary succession in an australian coastal
heath. Ecology 5: 1332-1341.
Freitas PRS, Mesquita DO, França FGR (2013) Uso do implante visível de
elastômero fluorescente (IVE) para marcação de lagartos Phyllopezus pollicaris
(Squamata: Phyllodactylidae). Biotemas 26 (4): 271-276.
Frost DRT, Grant T, Faivovich J, Bain RH, Haas A, Haddad CFB, De Sá RO,
Channing A, Wilkinson M, Donnellan S, Raxworthy CJ, Campbell JA, Blotto BL,
Moler P, Drenes RC, Nussbaum RA, Lynch JD, Green DM, Wheeler WC
(2006). The Amphibian Tree of Life. Bulletin of the American Museum of Natural
Hystory, 297:1-370.
Gardner RH, O’neil RV (1991) Pattern, process and predictability: The use of neutral
models for landspace analysis. Páginas 289-308 In: Turner GM, Gardner RH

41
(eds) Quantitative Methods in Landscape Ecology: The Analyses and
Interpretation of Landspace Heterogeneity. SPB Academic Publishing, The
Hague. New York.
Gimaret-Carpentier C, Pélissier R, Pascal JP, Houllier F (1998) Sampling strategies
for the assessment of tree species diversity. Journal of Vegetation Science 9:
161-172.
Giaretta AA, Menin M, Facure KG, Kokubum MNC, Filho JCO (2008) Species
richness, relative abundance, and habitat of reproduction of terrestrial frogs in
the Triângulo Mineiro region, Cerrado biome, southeastern Brazil. Iheringia,
Série Zoológica, Porto Alegre 98(2):181-188.
Gascon C, Lovejoy TE, Bierregaard jr. RO, Malcolm JR, Stouer PC, Vasconcelos HL,
Laurance WL, Zimmerman B, Tocher M, Borges S (1999) Matrix habitat and
species richness in tropical forest remnants. Biological Conservation 91: 223-
229.
Haddad CFB, Prado CPA (2005) Reproductive modes in frogs and their unexpected
diversity in the Atlantic Forest of Brazil. BioScience 55:207-217.
Haddad CFB, Toledo LF, Prado CPA, Loebmann D, Gasparini JL, Sazima I (2013)
Guia dos Anfíbios da Mata Atlântica: Diversidade e biologia. Anolis books, São
Paulo.
Hayes TB, Anderson LL, Beasley VR, Solla SR, Igushi T, Ingraham H (2011)
Demasculinization and feminization of male gonads by atrazine: Consistent
effects across vertebrate classes. Journal of Steroid Biochemistry & Molecular
Biology, 127(2):64-73
Heyer WR (1969) The adaptive ecology of the species groups of the genus
Leptodactylus (Amphibia, Leptodactylidae). Evolution 23: 421-428.
Hammer O, Harper DAT, Ryan PD (2001) PAST: Paleontological Statistics Software
Package for Education and Data Analysis. Paleontologia Electronica 4(1):9.
IUCN (2014) IUCN Red List of Threatened Species http://www.iucnredlist.org.
(Acesso em: 28 Ago. 2014).

42
Klink CA, Machado RB (2005) A conservação do Cerrado brasileiro.
Megadiversidade 1:147-155.
Laurance WF, Ecological Correlates of Extinction Proneness in Australian Tropical
Rain Forest Mammals (1991). Conservation Biology 5 (1): 79-89.
Laurance WF, Vasconcelos HL (2009) Consequências ecológicas da fragmentação
florestal na Amazônia. Oecologia Brasiliensis 13(3): 434-451.
Magalhães AP (2012) Composição e diversidade da anurofauna de serapilheira da
Serra do Ouro Branco, Minas Gerais. Dissertação Mestrado. Universidade
Federal de Ouro Preto, Ouro Preto.
Majer JD (1989) Long-term colonization of fauna in reclaimed land. Páginas143-174
In Majer JD (eds) Animals in Primary Succession - The Role of Fauna in
Reclaimed Lands. Cambridge University Press, Cambridge.
Martins R (2014) Síndromes de polinização em diferentes fisionomias de uma área
de mata ciliar no reservatório de Volta Grande, Minas Gerais, Brasil.
Dissertação Mestrado. Universidade Federal de Ouro Preto. Ouro Preto.
Myers N, Mittermeier RA, Mittermeier CG, Fonseca GAB, Kent J (2000) Biodiversity
hotspots for conservation priorities. Nature 403:853-858.
Mexia TMM (2008) Revegetação e seus efeitos na sucessão ecológica em pedreiras
calcárias após exploração: a pedreira da secil como caso-estudo. Dissertação
Mestrado. Universidade De Lisboa.Lisboa.
Moraes RA, Sawaya RJ, Barrela W (2007). Composição e diversidade de anfíbios
anuros em dois ambientes de Mata Atlântica no Parque Estadual Carlos
Botelho, São Paulo, sudeste do Brasil. Biota Neotropica. 7 (2):27-36.
Olden JD, Poff NL, Douglas MR. Douglas ME, Fausch KD (2004) Ecological and
evolutionary consequences of biotic homogenization. Trends in Ecology &
Evolution 19:18–24.
Otani L (2011). Aspectos da fisiologia metabólica e do desempenho locomotor em
anfíbios anuros: implicações da fragmentação ambiental. Tese Doutorado.
Universidade de São Paulo. São Paulo.

43
Pavan D (2007) Assembleias de répteis e anfíbios do Cerrado ao longo da bacia do
rio Tocantins e o impacto do aproveitamento hidrelétrico da região na sua
conservação. Dissertação Mestrado, Universidade de São Paulo. São Paulo.
Power ME, Dietrich WE, Finlay JC (1996) Dams and downstream aquatic
biodiversity: potential food web consequences of hydrologic and geomorphic
change. Environmental Management 20:887–895.
Provete DB, Garey MV, Silva FR, Rossa-Feres DC (2011) Anurofauna do noroeste
paulista: lista de espécies e chave de identificação para adultos. Biota
Neotropica 11 (2):000-000.
R Development Core Team (2014) R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria.
http://www.R-project.org/ (Acesso em: 02 set. 2014).
Ribeiro JF, Walter BMT (2001). As Matas de Galeria no contexto do bioma Cerrado.
Páginas 29-47 In: Ribeiro JF, Fonseca CEL, Sousa-Silva JC (eds) Cerrado:
Caracterização e Recuperação de Matas de Galeria. Embrapa Cerrados,
Planaltina.
Ricklefs RE (1993) A Economia da Natureza. Guanabara: Koogan. Rio de Janeiro.
Ricketts TH (2001) The matrix matters: effective isolation in fragmented landscapes.
American Naturalist 158:87-99.
Rosenberg DM, Berkes F, Bodaly RA, Hecky RE, Kelly CA, Rudd JWM (1997) Large-
scale impacts of hydroelectric development. Environmental Reviews. 5:27 – 54.
Sparling DW, Linder G, Bishop CA (2000) Ecotoxicology of amphibians and reptiles.
SETAC Press, Pensacola.
TOLEDO, L.F., CARVALHO-E-SILVA, S.P., SÁNCHEZ, C., ALMEIDA, M.A. &
HADDAD, C.F.B. The review of the Brazilian Forest Act: harmful effects on
amphibian conservation. Biota Neotrop. 10(4):
http://www.biotaneotropica.org.br/v10n4/en/abstract?article+bn00410042010.
Uetanabaro M, Prado CPA, Rodrigues DJ, Gordo M, Campos Z (2008) Guia de
Campo dos Anuros do Pantanal e Planaltos de Entorno. Universidade Federal
Mato Grosso do Sul, Universidade Federal Mato Grosso. Cuiabá, Mato Grosso.

44
Valdujo PH, Camacho A, Recoder RS, Teixeira Jr M, Ghellere JMB, Mott T, Nunes
PMS, Nogueira C, Rodrigues MT (2011). Anfíbios da estação Ecológica Serra
Geral do Tocantins, região do Jalapão, Estados do Tocantins e Bahia Biota
Neotropica 11(1): 251-262.
Young BE, Stuart SN, Chanson JS, Cox NA, Boucher TM (2004) Disappearing
Jewels: The Status of New World Amphibians. Nature Serve, Arlington.
Zhong Y, Power G (1996) Environmental impacts of hydroelectric projects on fish
resources in China. Regulated Rivers: Research & Management 12: 81–98.
Zina J, Ennser J, Pinheiro SCP, Haddad CFB, Toledo LF (2007) Taxocenose de
anuros de uma mata semidecídua do interior do Estado de São Paulo e
comparações com outras taxocenoses do Estado, sudeste do Brasil. Biota
Neotropica 7(2):000-000.

45
ANEXO 1
Lista das espécies de anfíbios coletados no entorno da represa de Volta Grande,
depositados na coleção herpetológica do Laboratório de Zoologia dos Vertebrados
(LZV) da Universidade Federal de Ouro Preto.
Chiasmocleis albopunctata : (LZV 2361 A; 2384 A; 2394 A; 2395 A; 2626 A; 2627 A;
2628 A; 2629 A; 2630 A).
Elachistocleis cf cesarii: (LZV 2748 A).
Dendropsophus elianeae: (LZV 2633 A; 2634 A; 2635 A; 2636 A; 2649 A).
Dendropsophus nanus: (LZV 2638 A; 2639 A; 2640 A; 2641 A; 2642 A; 2643 A;2644 A;
2645 A; 2646 A; 2647 A; 2845 A; 2648 A; 2649 A; 2650 A).
Dermatonotus muelleri: (LZV 2340 A; 2341 A; 2353 A; 2356 A; 2357 A; 2358 A; 2359 A;
2370 A; 2371 A; 2372 A; 2373 A; 2374 A; 2396 A; 2567 A; 2568 A; 2569 A; 2570 A;
2571 A; 2572 A; 2573 A; 2574 A; 2575 A; 2576 A;2577 A; 2578 A; 2579 A; 2580 A;
2581 A).
Physalaemus nattereri (LZV 2319 A; 2363 A; 2364 A; 2368 A; 2369 A; 2378 A; 2379 A;
2380 A; 2381 A; 2552 A; 2553 A; 2554 A; 2555 A; 2556 A; 2557 A; 2558A; 2559 A;
2560 A; 2561 A; 2562 A; 2563 A; 2564 A; 2565 A).
Hypsiboas raniceps (LZV 2582 A; 2583 A; 2584 A; 2585 A).
Leptodactylus sp. (LZV 2330 A; 2385 A; 2653 A; 2652 A).
Leptodactylus fuscus (LZV 2651 A; 2586 A; 2587 A; 2588 A; 2589 A; 2590 A; 2591 A;
2654 A).
Leptodactylus mystaceus (LXV 2592 A; 2593 A; 2594 A; 2595 A; 2596 A; 2597 A; 2598
A; 2599 A; 2600 A; 2601 A; 2602 A; 2603 A; 2604 A; 2605 A; 2606 A).
Leptodactylus mystacinus (LZV 2362 A; 2397 A; 2607 A; 2608 A; 2609 A; 2610 A; 2611
A).
Leptodactylus podicipinus (LZV 2328 A; 2329 A; 2365 A; 2366 A; 2612 A; 2613 A; 2614
A; 2615 A; 2616 A; 2617 A; 2618 A; 2834 A; 2619 A; 2620 A; 2621 A; 2622 A; 2623 A;
2624 A; 2625 A).
Physalaemus centralis (LZV 2749 A).

46
Physalaemus cf. cuvieri (LZV 2308 A; 2309 A; 2310 A; 2311 A; 2312 A; 2313 A; 2314
A; 2315 A; 2316 A; 2317 A; 2318 A; 2334 A; 2335 A; 2336 A; 2337 A; 2346 A; 2347 A;
2348 A; 2349 A; 2350 A; 2351 A; 2352 A; 2354 A; 2355 A; 2360 A; 2367 A; 2375 A;
2376 A; 2377 A; 2382 A; 2383 A; 2387 A; 2390 A; 2391 A; 2392 A; 2393 A; 2504 A;
2505 A; 2506 A; 2507 A; 2508 A; 2509 A; 2510 A; 2511 A; 2512 A; 2513 A; 2514 A;
2515 A; 2516 A; 2517 A; 2518 A; 2519 A; 2520 A; 2521 A; 2522 A; 2523 A; 2524 A;
2525 A; 2526 A; 2527 A; 2528 A; 2529 A; 2530 A; 2531 A; 2532 A; 2533 A; 2534 A;
2535 A; 2536 A; 2537 A; 2538 A; 2539 A; 2540 A; 2541 A; 2542 A; 2543 A; 2544 A;
2545 A; 2546 A; 2547 A; 2548 A; 2846 A; 2549 A; 2751 A; 2752 A; 2753 A; 2754 A;
2755 A; 2756 A; 2757 A; 2758 A; 2759 A; 2760 A; 2761 A; 2762 A; 2763 A; 2764 A;
2765 A; 2766 A; 2767 A; 2768 A; 2769 A; 2770 A; 2771 A; 2772 A; 2773 A; 2607 A;
2844 A; 2837 A; 2838 A; 2839 A; 2840 A; 2841 A; 2842 A; 2843 A; 2775).

47
ANEXO 2
Fotos de algumas das espécies de anuros coletados na Represa de Volta Grande-MC/SP
A) Chiasmocleis albopunctata; B) Dermatonotus mulleri; C) Physalaemus centralis; D) Dendropsophus nanus;
E)Physalaemus nattereri; F) Rhinella schneideri; G) Hypsiboas raniceps; H) Leptodactylus mystacinus; I) Elachistocleis cf.
cesarii; J) Leptodactylus podicipinus; K) Leptodactylus mystaceus; L) Trachycephalus venulosus.
A B C D
E F G H
I J K L
M N