Embed Size (px)
Citation preview

Universidade Federal do Pará Campus Universitário de Bragança
Instituto de Estudos Costeiros Faculdade de Ciências Biológicas
JUÇARA DA SILVA CORDEIRO
DIVERSIDADE DE ARANHAS (ARACHNIDA, ARANEAE) EM
DUAS ÁREAS DE MANGUEZAL DA PENÍNSULA DE
AJURUTEUA, BRAGANÇA, PARÁ.
BRAGANÇA
2008

JUÇARA DA SILVA CORDEIRO
DIVERSIDADE DE ARANHAS (ARACHNIDA, ARANEAE) EM
DUAS ÁREAS DE MANGUEZAL DA PENÍNSULA DE
AJURUTEUA, BRAGANÇA, PARÁ.
Trabalho de Conclusão de Curso apresentado ao
Colegiado da Faculdade de Ciências Biológicas, do
Instituto de Estudos Costeiros da Universidade
Federal do Pará, Campus de Bragança, como
requisito parcial para obtenção de Grau de
Licenciado em Ciências Biológicas.
Orientador: Prof Dr. Marcus Emanuel Barroncas
Fernandes. UFPA, Campus de Bragança.
BRAGANÇA
2008

JUÇARA DA SILVA CORDEIRO
DIVERSIDADE DE ARANHAS (ARACHNIDA, ARANEAE) EM
DUAS ÁREAS DE MANGUEZAL DA PENÍNSULA DE
AJURUTEUA, BRAGANÇA, PARÁ.
Este trabalho foi julgado para obtenção do Grau de
Licenciado em Ciências Biológicas da Faculdade de
Ciências Biológicas do Instituto de estudos Costeiros da
Universidade Federal do Pará no Campus de Bragança.
BANCA EXAMINADORA
_____________________________________ Prof. Dr. Marcus E. B. Fernandes (Orientador) Campus de Bragança, UFPA ______________________________________ Profª M.Sc. Rita de Cássia Oliveira Santos (Titular) Campus de Bragança, UFPA ______________________________________ M.Sc. Sidclay Calaça Dias (Titular) UFPA/Museu Paraense Emílio Goeldi ______________________________________ M.Sc. Darlan Simith (Suplente) Campus de Bragança, UFPA
BRAGANÇA
2008

i
“O sucesso nasce do querer, da determinação e persistência em se chegar a um objetivo. Mesmo não atingindo o alvo,
quem busca e vence obstáculos, no mínimo fará coisas admiráveis."
José de Alencar

ii
Dedico este trabalho:
Ao meu pai Olavo e minha mãe Jurema, vocês são as pessoas mais importante da minha vida!

iii
AGRADECIMENTOS
A Universidade Federal do Pará, pela vaga no curso e oportunidade de ensino
gratuita.
Ao Laboratório de Ecologia de Manguezal e ao Prof. Dr. Marcus E. B. Fernandes
pela amizade, dedicação, compreensão, paciência e pelo apoio durante todo o
desenvolvimento e orientação deste trabalho, dentro e fora do laboratório.
Ao Laboratório de Aracnologia do Museu Paraense Emílio Goeldi por ter me
concedido oportunidade de estágio e ajuda na identificação das aranhas, em
especial ao Sidclay Dias, Nancy França, Laura Miglio, David Candiani e Danni
Roberto que me auxiliaram muito durante o estágio e sempre que precisei de ajuda.
A José Augusto Barreiros (in memoriam) pelo incentivo e orientação neste trabalho
em quanto esteve entre nós. Foi ele quem me apresentou o mundo das aranhas,
obrigado!
A minha mãe, que apesar de nossas diferenças, sei que me ama e nunca deixa de
orar e cuidar de mim mesmo que estejamos distantes.
Ao meu pai, com muito carinho, pois sei que apesar de todas as dificuldades e com
todas as limitações financeiras, apostou e confiou em mim.
A minha tia Júlia, ao meu avô Acácio Joaquim (in memoriam) pelo carinho, meus
irmãos, simplesmente por existirem! E a toda a minha família que é maravilhosa.
Ao meu namorado Christóvam Pamplona Neto, por todo auxílio e companheirismo.
Você depois de mim foi o maior responsável por este trabalho, venci muita coisa e
muita dificuldade ao seu lado, você com seu jeito estranho que muitas vezes não
compreendi me mostrou a principal motivação de chegar ao fim, te amo e admiro
muito.
Aos meus amigos da turma de Biologia 2003 (Galera da Irmandade) que foram
muito importantes durante ao longo do curso pela união e companheirismo.
As minhas amigas, Aline Carvalho, Elia Lindoso e Kely Melo pelo apoio e auxílio
sempre em todas as horas.
Aos professores(as) em especial, Fernando Abrunhosa, Colin Beasley, Claudia
Tagliaro, Nils Asp, Rita Santos, Moirah Menezes e Marivana Borges, pela amizade,
atenção, compreensão e prestatividade em todos os momentos que precisei, dentro
e fora da sala de aula.
Por fim, agradeço a Deus o mestre supremo, que me deu o dom da vida e me
ampara com seu amor e sabedoria fazendo com que eu me sinta cada vez mais feliz
a cada dia que se renova! Sem ele nada disso seria possível.

iv
SUMÁRIO
1. INTRODUÇÃO .................................................................................................. 1
1.1. Objetivo Geral ........................................................................................... 3
1.1.1. Objetivos específicos .....................................................................
3
2. MATERIAIS E MÉTODOS ................................................................................ 4
2.1. Área de estudo .......................................................................................... 4
2.2. Sítios de trabalho ..................................................................................... 5
2.3. Procedimento ............................................................................................ 6
2.4. Guildas de aranhas .................................................................................. 6
2.5. Análise de dados ......................................................................................
8
3. RESULTADOS .................................................................................................. 9
3.1. Composição e abundância ...................................................................... 9
3.2. Diversidade e densidade ......................................................................... 13
3.3. Guildas Comportamentais .......................................................................
13
4. DISCUSSÃO E CONCLUSÃO .........................................................................
15
5. REFERÊNCIAS BIBLIOGRÁFICAS ................................................................ 17

v
LISTA DE TABELAS E FIGURAS
Figura 1. Localização da área de estudo e dos sítios de trabalho (Furo do Taici e
Furo Grande) na península de Ajuruteua, Bragança - PA. Modificado de Krause
et al. (2001). ........................................................................................................... 4
Figura 2. Sítios de trabalho na península de Ajuruteua, Bragança - PA. A = Furo
do Taici e B = Furo Grande..................................................................................... 5
Figura 3. Representação esquemática: arranjo das parcelas perpendiculares à
rodovia PA–458, que liga a cidade de Bragança à praia de Ajuruteua, Bragança -
PA............................................................................................................................. 7
Figura 4. Curva de acumulação representada pelo número de espécies e pelo
número de parcelas (representando o número de indivíduos coletados) nos dois
sítios de trabalho, Furo do Taici e Furo Grande, Bragança - PA. ........................... 10
Figura 5. Número total de indivíduos por família nas duas áreas de estudo, Furo
do Taici e Furo Grande, península de Ajuruteua, Bragança - Pará. ....................... 12
Figura 6. Densidade (ind.m2) das famílias de Araneae no Furo do Taici e Furo
Grande, península de Ajuruteua, Bragança-PA. ..................................................... 13
Tabela 1. Lista de táxons, número de indivíduos e abundância relativa nos dois
sítios de trabalho, Furo do Taici e Furo Grande, Bragança - PA. ........................... 11
Tabela 2. Guildas e o número de indivíduos por famílias nas duas áreas de
estudo. ..................................................................................................................... 14

vi
RESUMO
As aranhas são predadoras generalistas que habitam a maioria dos
ambientes terrestres. No entanto, nenhum trabalho sistematizado sobre a
araneofauna de manguezal foi publicado até o momento. Assim, o presente trabalho
objetivou caracterizar a assembléia de aranhas quanto a sua composição e
diversidade em dois sítios de manguezal na península de Ajuruteua, Bragança-PA.
O trabalho foi realizado no Furo do Taici e no Furo Grande, onde espécimes de
aranhas foram coletados entre dezembro de 2007 e janeiro de 2008, com guarda-
chuva entomológico em parcelas de 10x10 m, totalizando 20 parcelas para cada
sítio de trabalho. Foram coletadas 247 aranhas pertencentes a 12 famílias.
Salticidae foi a família mais abundante para o Furo do Taici, enquanto
Tetragnathidae foi a mais abundante para o Furo Grande. Não houve diferença
significativa entre os valores de abundância por família dos dois sítios de trabalho (t
=0,13; gl=1; p>0,05). A análise de similaridade ANOSIM, mostrou que houve
diferença significativa entre o número de indivíduos das duas áreas de estudo
(R=0,045; p<0,01). Os valores estimados do índice de Shannon e sua respectiva
equitabilidade para os dois sítios de trabalhos foram os seguintes: Furo do Taici (
H’=2,27; E=0,84) e o Furo Grande (H’=2,65; E=0,98), havendo diferença significativa
entre os sítios (M=0,4; 2=6,53; gl=1; p<0,05). O valor de densidade variou de
0,0005 a 0,008 ind.m2 para o Furo do Taici e 0,0005 a 0,0035 para o Furo Grande.
Os coeficientes de Jaccard (CJaccard= 0,04) e Sorenson (CSorensen= 0,13) mostraram
uma baixa similaridade entre os sítios de trabalho. Sendo assim, foi registrada uma
diferença na composição de espécies dos dois sítios de trabalho, o que pode estar
relacionada com a localização de cada sítio e a estrutura dos bosques de mangue. A
inundação também pode ser um fator importante e decisivo para a composição da
araneofauna no manguezal, influenciando diretamente na diversidade e na
abundância desse grupo nas florestas alagáveis e de terra firme.

1
1. INTRODUÇÃO
De acordo com Rupert & Barnes (2005), Araneae é a maior ordem de
aracnídeos perdendo somente para a ordem Acari. Essa ordem, segundo Brescovit
(1999), diferencia-se das outras ordens por apresentar fiandeiras na região posterior
do abdômen, glândulas de veneno conectadas às quelíceras e os pedipalpos dos
machos modificados em órgãos copuladores. A ordem Araneae está dividida nas
subordens Mesothelae e Opistothele, a qual inclui as infraordens Mygalomorphae e
Araneomorphae (Barreiros et al., 2001). Foram descritas 40.462 espécies de
aranhas, distribuídas em 109 famílias e 3.694 gêneros, sendo estimadas, pelo
menos, 170.000 espécies no mundo inteiro (Platinick, 2008).
As aranhas são caracterizadas por possuírem um hábito exclusivamente
predador, além de serem importantes reguladoras de populações de insetos e outros
invertebrados (Flórez, 2000). São encontradas na maioria dos ambientes terrestres,
mas estão ausentes no mar (Rupert & Barnes, 2005). Possuem grande plasticidade
adaptativa, colonizando inclusive os ambientes urbanos e áreas de cultivo
(Brescovit, 1999), tornando-se importantes objetos de estudo para o monitoramento
da diversidade de espécies em ecossistemas terrestres (Marusik & Koponen, 2000).
Apesar da alta diversidade nas regiões tropicais e subtropicais, estas áreas
são pouco estudadas no que se refere à fauna araneológica. Estima-se que apenas
um terço das espécies dessa fauna seja conhecido, atualmente (Coddington & Levi,
1991), sendo que para a região Neotropical essa estimativa é de apenas 50%
(Platinick, 1999). Apenas recentemente os países da região Neotropical dirigiram
seus estudos para a investigação da diversidade da classe Arachnida. No Brasil,
esses trabalhos ainda não são representativos e estima-se que as áreas com maior
diversidade seriam as da Mata Atlântica e da Amazônia. O território brasileiro
também é uma das áreas com maior diversidade de espécies do mundo, mas
acredita-se que sejam conhecidos apenas 30% da sua fauna de aranhas (Brescovit,
1999).
Durante os últimos 20 anos foram realizados alguns inventários
sistematizados de aranhas brasileiras, incluindo aqueles relacionados à região
amazônica (Brescovit & Francesconi, 2002). Estes inventários incluem o estado do
Amazonas (Höfer 1990, Höfer & Beck 1996, Borges & Brescovit 1996) e do Pará
(Martins & Lise 1997). No Pará, a lista mais recente da fauna aranealógica é aquela

2
produzida por Ricetti & Bonaldo (2008), produto do inventário realizado na Serra do
Caximbo. Contudo, nenhum dos trabalhos realizados sobre a araneofauna na
Amazônia e no restante do Brasil refere-se àquela presente no ecossistema
manguezal.
Na costa norte brasileira são encontradas as florestas de mangue mais
exuberantes do país (Mendes, 2005), as quais representam cerca de 90% desse
ecossistema ao longo da costa brasileira (Herz, 1991), sendo considerado por
Fernandes (2003) um dos principais ecossistemas costeiros da Amazônia brasileira.
Só o Estado do Pará possui cerca de 600 km de costa (Mendes, 2005), onde os
manguezais ocupam uma área que corresponde aproximadamente 2.177 Km2
(Souza Filho, 2005). De acordo com Schaeffer-Novelli (1995), os manguezais são
ecossistemas costeiros de transição entre os ambientes terrestre e marinho. São
caracterizados pela ocorrência de espécies vegetais lenhosas, adaptadas ao
ambiente salino, periodicamente inundado pelas marés.
A fauna desse ecossistema é caracterizada por animais que habitam zonas
permanentemente inundadas pelas marés, com maior ou menor grau de
dependência da água e por espécies arborícolas locais ou provenientes de sistemas
adjacentes (Hogarth, 1999). A maioria dos grupos animais associado aos
manguezais apresentam diferentes status de residência no sistema (Fernandes,
2000). Os grupos animais são representados por espécies de mamíferos, aves,
répteis, anfíbios, peixes, crustáceos, anelídeos, insetos e outros invertebrados
incluindo os aracnídeos, cuja interação com o manguezal é resultado da busca,
principalmente, por abrigo e alimento (Fernandes, 2000).
Segundo Hogarth (1999), entre os invertebrados de origem terrestre no
manguezal, as aranhas são as mais conhecidas, ou talvez, as mais evidentes entre
os artrópodes. Esse autor cita a presença de aranhas construtoras de teias, como as
mais abundantes no manguezal, bem como a presença de aranhas caçadoras, as
quais descem das árvores para forragear no chão da floresta de mangue por
ocasião da baixa-mar. Essas aranhas caçadoras, geralmente, alimentam-se de
insetos como formigas, no entanto uma espécie do gênero Pardosa, da Família
Lycosidae, foi observada alimentando-se de caranguejos juvenis. As aranhas
encontradas no ecossistema de manguezal estão também presentes nas florestas
adjacentes, não sendo, portanto, próprias deste ambiente, ou mesmo adaptadas a
este sistema.

3
Até o presente, nenhum trabalho sistematizado a respeito da fauna
araneológica presente em ecossistema de manguezal foi registrado na literatura
nacional, dessa forma é importante que estudos pioneiros sobre a caracterização
dessas assembléias nas florestas de mangue sejam realizados, para que possam
servir de base a futuros estudos.
1.1. Objetivo geral
Descrever a composição e conhecer a diversidade da assembléia de aranhas
em duas florestas de mangue na península de Ajuruteua, Bragança, Pará.
1.1.1. Objetivos específicos
i. Produzir uma lista de espécies de aranhas associadas às florestas de
mangue;
ii. Citar a composição das guildas da assembléia de aranhas presentes no
manguezal;
iii. Calcular e comparar os valores de diversidade, abundância e densidade da
araneofauna entre dois sítios de manguezal, na península de Ajuruteua.

4
2. MATERIAIS E MÉTODOS
2.1. Área de estudo
Este trabalho foi desenvolvido na península de Ajuruteua, Bragança-PA, entre
as coordenadas geográficas 46º50’– 46º38’W e 00º05’–01º00’S (Figura 1). A região
é entrecortada por canais de maré que ligam o manguezal ao estuário (Fernandes,
2003).
Figura 1 – Localização da área de estudo e dos sítios de trabalho (Furo do Taici e
Furo Grande) na península de Ajuruteua, Bragança - PA. Modificado de Krause et al.
(2001).
Km

5
Dois sítios de trabalho foram escolhidos para a realização das coletas.
Segundo Matni et al. (2006), essas duas localidades apresentam manguezais sob
diferentes condições, desde alta salinidade e baixa inundação até manguezais de
estuário sob a influência da água doce do rio Caeté.
As espécies arbóreas encontradas nos dois sítios estudados são compostas
por três espécies típicas de manguezal: Rhizophora mangle L. (Rhizophoraceae),
Avicennia germinans (L.) Stearn (Aviceniaceae) e Laguncularia racemosa L. Gaertn
f. (Combretaceae) (Matni, 2007).
2.2. Sítios de trabalho
i. Furo do Taici - localizado no início da península (Km 10), próximo às áreas de
terra firme e com maior influência de água doce. Apresenta salinidade média
da água em torno de16 (Figura 2A);
ii. Furo Grande - localizado na porção norte da península. Apresenta salinidade
média da água de aproximadamente 33 (Figura 2B).
Figura 2. Sítios de trabalho na península de Ajuruteua, Bragança - PA. A =
Furo do Taici e B = Furo Grande.
Foto: J. Cordeiro Foto: C. Pamplona
A B

6
2.3. Procedimento
As coletas foram feitas entre dezembro de 2007 e janeiro de 2008. Cada sítio
foi amostrado através da abertura de 20 parcelas medindo 10x10 m (Figura 3), e em
cada parcela coletou-se no tempo de uma hora, totalizando 40 horas para os dois
sítios de estudo. As aranhas foram coletadas, dentro de cada parcela, com auxílio
de um guarda-chuva entomológico, sendo os indivíduos coletados acondicionados
em álcool 70% e, posteriormente, levados para triagem no laboratório. O método de
coleta utilizado no presente trabalho é uma adaptação daqueles utilizados
comumente em trabalhos com aranhas, onde se aplica, normalmente, um protocolo
adaptado de Coddington & Levi. (1991), com o uso de transecções de 30 m como
unidades amostrais, para a utilização do guarda-chuva entomológico.
A identificação do material foi feita, primeiramente, em nível de família,
através da chave dicotômica proposta por Brescovit et al. (2002). Uma análise
posterior tentou alcançar o nível taxonômico específico, na impossibilidade os
exemplares foram registrados como morfoespécies.
Os espécimes jovens foram identificados em nível de família, não sendo
incluídos nas análises estatísticas. O material testemunho ficou depositado na
Coleção de Aracnídeos do Museu Paraense Emílio Goeldi (MPEG), Belém-PA e
uma coleção de referência foi depositada na Coleção de Zoologia de Bragança
(CZB), do Instituto de Estudos Costeiros (IECOS), da Universidade Federal do Pará
(UFPA), Campus de Bragança, Bragança-PA.
2.4. Guildas
Conforme Simberloff & Dayan (1991), guilda é definido como um grupo que
explora as mesmas classes de recursos ambientais de um modo semelhante. No
caso das aranhas, elas são classificadas a partir de suas estratégias de caças. Os
grupos funcionais (guildas) das aranhas foram separados com base no trabalho de
Uetz et al. (1999), onde todas as aranhas capturadas, separadas por famílias, foram
agrupadas em uma das seguintes guildas: a) entre as construtoras de teias: 1.
construtoras de teias orbiculares (constroem teias bidimensionais) e 2. construtoras
de teias irregulares (constroem teias tridimensionais) e b) entre as caçadoras: 1.

7
caçadoras corredoras (buscam suas presas ativamente) e 2. caçadoras de
emboscada (não constroem teias e aguardam suas presas por espreita).
Figura 3 – Representação esquemática: arranjo das
parcelas perpendiculares à rodovia PA–458, que liga a
cidade de Bragança à praia de Ajuruteua, Bragança - PA.
Bragança Ajuruteua

8
2.5. Análise de dados
A abundância relativa (%) e a densidade (ind./m2) foram calculadas de acordo
com as fórmulas abaixo:
(Equação 1)
(Equação 2)
Para calcular o índice de diversidade foi usado o Índice de Shannon (H’), que
é baseado na abundância proporcional das espécies, sendo estimado através da
seguinte fórmula:
(Equação 3)
onde: ni = no total de indivíduos da espécie;
N = no total de indivíduos na amostra;
ni /N = pi abundância proporcional de cada espécie na amostra total.
A equitabilidade, utilizada para verificar o grau de uniformidade, distribuição
dos indivíduos entre as espécies das assembléias de aranhas em cada sítio de
trabalho, foi estimada pela fórmula:
(Equação 4)
onde: S = é o número total de espécies na amostra
H’ = valor obtido do índice de Shannon
Para averiguar se houve diferença significativa na composição das espécies
entre os pontos amostrados aplicou-se a Análise de Similaridade (ANOSIM). Esta
análise multivariada foi baseada nos dados de abundância dos espécimes de cada
sítio, operando sobre uma matriz de similaridade, gerada com os dados

9
transformados para raiz-quarta e o uso do índice de similaridade de Bray-Curtis,
sendo realizada no programa estatístico PRIMER 6.0 (Clarke & Gorley, 2006).
Para comparar o número de indivíduos por famílias entre as duas áreas de
trabalho usou-se o teste t. Os valores do índice de diversidade (H’) foram
comparados com o Teste da Mediana, este teste é um procedimento destinado a
comprovar se duas amostras independentes, do mesmo tamanho ou desiguais, são
provenientes da mesma população com o mesmo valor médio. Para ambos os
testes, o valor de significância (p) foi de 0,05. Para a execução dessas análises
utilizou-se o software Bioestat 5.0 (Ayres et al., 2007).
Os coeficientes de similaridade de Jacard e Sorenson foram utilizados para
avaliar o grau de semelhança da composição de espécies entre os dois sítios de
trabalho. Esses índices foram estimados a partir das seguintes fórmulas:
(Equação 5)
(Equação 6)
3. RESULTADOS
3.1. Composição e Abundância
Foi coletado um total de 247 aranhas pertencentes a 12 famílias, sendo que
135 indivíduos de 10 famílias foram capturados no Furo do Taici e 112 indivíduos,
distribuídos também em 10 famílias, no Furo Grande. Analisando a curva de
acumulação das espécies, observou-se que os valores encontrados estão distantes
de alcançar a assíntota da curva (Figura 4).
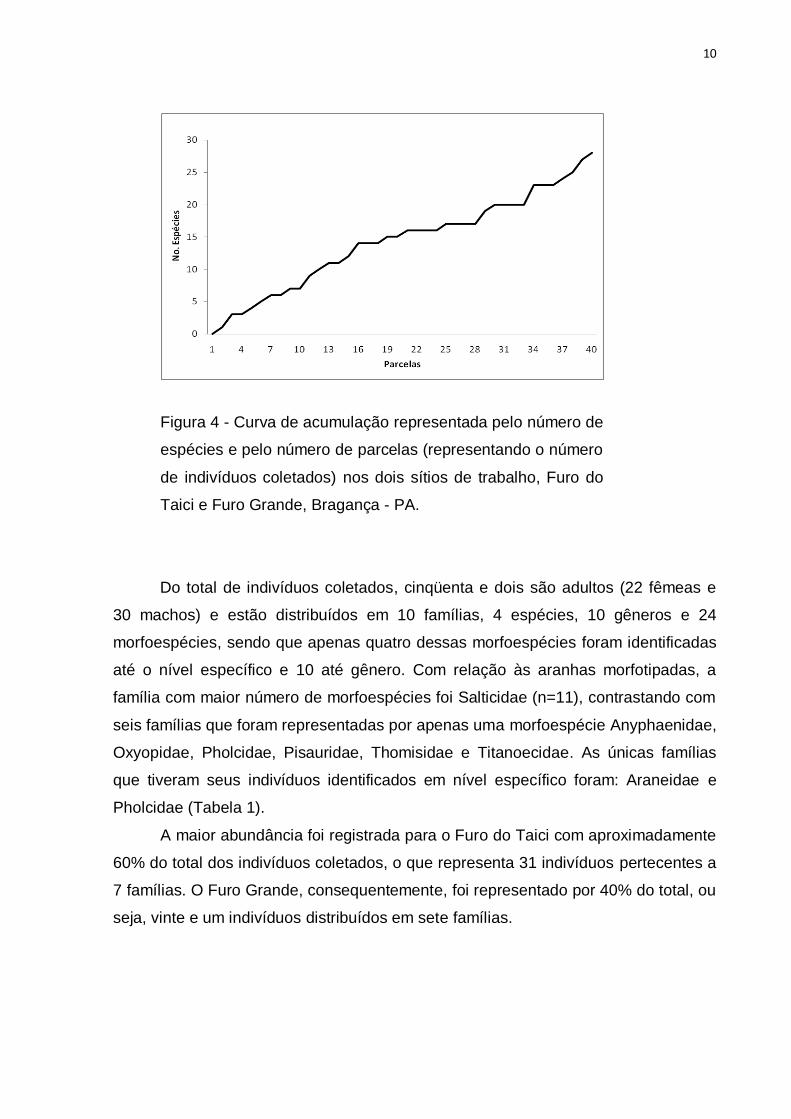
10
Figura 4 - Curva de acumulação representada pelo número de
espécies e pelo número de parcelas (representando o número
de indivíduos coletados) nos dois sítios de trabalho, Furo do
Taici e Furo Grande, Bragança - PA.
Do total de indivíduos coletados, cinqüenta e dois são adultos (22 fêmeas e
30 machos) e estão distribuídos em 10 famílias, 4 espécies, 10 gêneros e 24
morfoespécies, sendo que apenas quatro dessas morfoespécies foram identificadas
até o nível específico e 10 até gênero. Com relação às aranhas morfotipadas, a
família com maior número de morfoespécies foi Salticidae (n=11), contrastando com
seis famílias que foram representadas por apenas uma morfoespécie Anyphaenidae,
Oxyopidae, Pholcidae, Pisauridae, Thomisidae e Titanoecidae. As únicas famílias
que tiveram seus indivíduos identificados em nível específico foram: Araneidae e
Pholcidae (Tabela 1).
A maior abundância foi registrada para o Furo do Taici com aproximadamente
60% do total dos indivíduos coletados, o que representa 31 indivíduos pertecentes a
7 famílias. O Furo Grande, consequentemente, foi representado por 40% do total, ou
seja, vinte e um indivíduos distribuídos em sete famílias.

11
Tabela 1- Lista de táxons (morfoespécies), número de indivíduos e abundância relativa nos dois sítios de trabalho, Furo
do Taici e Furo Grande, Bragança - PA.
Furo do Taici Furo Grande
Família Morfoespécies Nº de indivíduos Abundância Relativa (%) Nº de indivíduos Abundância Relativa (%)
Anyphaenidae Anyphaenidae sp1 1 3,22 - -
Araneidae Gasterachanta cancriformes - - 1 4,76
Manogea porracea - - 1 4,76
Eustala sp1 - - 1 4,76
Araneus guttatus - - 1 4,76
Oxyopidae Oxyopes sp1 2 6,45 2 9, 52
Pholcidae Mesobolivar aurantiaus 8 25,81 - -
Pisauridae Pisauridae sp1 1 3,22 1 4,76
Salticidae Lyssomanes sp1 8 25,81 - -
Lyssomanes sp2 1 3,22 - -
Lyssomanes sp3 1 3,22 - -
Salticidae sp1 - - 1 4,76
Salticidae sp2 1 3,22 - -
Salticidae sp3 1 3,22 - -
Salticidae sp4 1 3,22 - -
Salticidae sp5 - - 1 4,76
Salticidae sp6 1 3,22 - -
Salticidae sp7 1 3,22 - -
Salticidae sp8 1 3,22 - -
Tetragnathidae Azilia sp1 - - 2 9, 52
Tetragnathidae sp1 - - 2 9, 52
Tetragnathidae sp2 - - 2 9, 52
Tetragnathidae sp3 - - 2 9, 52
Theridiidae Coleossoma sp1 - - 1 4,76
Chrisso sp1 2 6,45 - -
Faiditus sp1 - - 2 9, 52
Thomisidae Thomisidae sp1 1 3,22 - -
Titanoecidae Goeldia sp1 - - 1 4,76
Total 31 100 21 100
11

12
Considerando-se o número total de indivíduos coletados por família no Furo
do Taici, constatou-se que Salticidae foi a família mais abundante (n=55), ao passo
que Theridiidae (n=3) e Corinidae (n=1) apresentaram o menor número de
indivíduos. No Furo Grande, a família mais abundante foi Tetragnathidae com 56
indivíduos, enquanto que Thomisidae, Titanoecidae e Nephilidae apresentaram
apenas um indivíduo (Figura 5).
O teste t mostrou que não houve diferença significativa entre os valores de
abundância por família das duas áreas (t =0,13; gl=1; p>0,05). Contudo, a análise de
similaridade ANOSIM, com base nos dados de abundância por unidade amostral,
mostrou que houve diferença significativa entre o número de indivíduos das duas
áreas de estudo (R=0,045; p<0,01). Para tanto, se aceita o resultado de diferença na
composição da assembléia de aranhas entre os dois sítios de trabalho, baseado na
hipótese de que a proximidade com a terra firme possa ter influenciado na
abundancia e distribuição das famílias.
Os coeficientes de similaridade de Jaccard (CJaccard= 0,04) e Sorenson
(CSorensen= 0,13) mostraram uma baixa similaridade entre os sítios de trabalho.
Figura 5. Número total de indivíduos por família nas duas
áreas de estudo, Furo do Taici e Furo Grande, península de
Ajuruteua, Bragança - Pará.
0
10
20
30
40
50
60
Tetra
gnath
idae
Saltic
idae
Pho
lcidae
Ara
neida
e
Pisau
ridae
Thom
isidae
Therid
iidae
Oxy
opidae
Any
phaen
idae
Corin
idae
Neph
ilida
e
Titano
ecidae
Famílias
nº
de i
nd
ivíd
uo
s
Furo do Taici
Furo Grande

13
3.2. Diversidade e densidade
O valor estimado através do índice de Shannon para o Furo do Taici foi H’=
2,27 e para o Furo Grande H’= 2,65, ao passo que os valores da equitabilidade
foram E=0,84 e E=0,98, respectivamente. O teste da Mediana mostrou que houve
diferença significativa entre os valores de diversidade de Shannon (M=0,4; 2=6,53;
gl=1; p<0,05).
Os valores de densidade apresentaram-se muito baixos, variando de 0,0005 a
0,008 ind.m2 para o Furo do Taici e 0,0005 a 0,0035 para o Furo Grande (Figura 6).
Figura 6 - Densidade (ind.m2) das famílias de Araneae no Furo
do Taici e Furo Grande, península de Ajuruteua, Bragança-PA.
3.3. Guildas
A assembléia de aranhas registrada para os dois sítios de trabalho foi
representada por quatro guildas entre caçadoras e tecelãs (Tabela 2).
0
0,001
0,002
0,003
0,004
0,005
0,006
0,007
0,008
0,009
Saltic
idae
Pho
lcidae
Tetra
gnath
idae
Ara
neida
e
Oxy
opidae
Therid
iidae
Any
phaen
idae
Pisau
ridae
Thom
isidae
Titano
ecidae
Famílias
Den
sid
ad
e (
ind
./m
2)
Furo do Taici
Furo Grande

14

15
Tabela 2 – Guildas comportamentais e número de indivíduos por famílias nas duas áreas de estudo.
Furo Taici Furo Grande
Guildas Famílias Indivíduos
(N)
Abundância Relativa
(%)
Indivíduos (N)
Abundância Relativa
(%) Total
Construtoras de teias Σ (%)
Construtoras de teias orbiculares
Araneidae 0 0 4 19,0 4 7,7
Tetragnathidae 0 0 8 38,0 8 15,4
Construtoras de teias irregulares
Theridiidae 2 6,5 3 14,0 5 9,7
Pholcidae 8 25,8 0 0 8 15,4
Titanoecidae 0 0 1 4,8 1 1,9
Caçadoras
Corredoras Anyphaenidae 1 3,2 0 0 1 1,9
Emboscada
Oxyopidae 2 6,5 2 9,5 4 7,7
Pisauridae 1 3,2 1 4,8 2 3,8
Salticidae 16 51,6 2 9,5 18 34,6
Thomisidae 1 3,2 0 0 1 1,9
Total 31 100 21 100 52 100
14

16
4. DISCUSSÃO E CONCLUSÃO
Devido à falta de trabalhos sobre a araneofauna de manguezais, os
resultados aqui obtidos foram comparados com alguns trabalhos realizados em
outros ecossistemas.
Oliveira-Alves (2005), em um estudo das assembléias de aranhas na Mata
Atlântica, no Parque do Pituaçu, Salvador, BA, também registrou a família Salticidae
como a mais abundante, seguida de Thomisidae e Araneidae. Outro estudo
realizado no Parque Estadual do Turvo, localizado às margens do rio Uruguai no Rio
Grande do Sul também registrou Salticidae, Araneidae e Thomisidae como as mais
abundantes (Podgaiski et al., 2007). Borges e Brescovit (1996) fizeram um inventário
da araneofauna de duas áreas localizadas na Amazônia Ocidental. Uma delas foi a
Reserva de Desenvolvimento Sustentável Mamirauá, com florestas inundáveis de
várzea. Em ambiente de chavascal, caracterizado por apresentar um sub-bosque
bastante emaranhado e sujeito às inundações de água doce sazonais, os autores
registraram com maior abundância as famílias Araneidae e Theridiidae. Raizer et al.
(2005), em um trabalho realizado em vários hábitats no Pantanal, coletaram próximo
aos córregos, rios, cachoeiras e nascentes, obtendo na seguinte ordem de
abundância as famílias Araneidae, Theridiidae e Salticidae. Essa mesma ordem de
abundância das famílias foi registrada no estudo sobre a riqueza e diversidade de
formigas e aranhas numa floresta de terra firme na Amazônia Central, com o uso do
método de parcelas (1m2) em ambientes de platô e baixio (Almeida, 2005).
No presente estudo, a família Salticidae foi registrada como a mais abundante
e bem representada dos dois sítios de coleta. Considerando os trabalhos
supracitados pode-se observar que nenhum deles apresenta a mesma ordem de
abundância das famílias aqui registradas, isto é, Salticidae, Pholcidae e
Tetragnathidae. É importante também ressaltar que as famílias Pholcidae e
Tetragnathidae, ao contrário do presente trabalho, aparecem com valores baixos de
abundância. Raizer et al. (2005) registrou Goeldia sp. (Titanoecidae) e Mesobolivar
aurantiacus (Pholcidae) com poucos indivíduos em 52 unidades amostrais,
realizadas com guarda-chuva entomológico. Resultado similar foi obtido no presente
estudo, onde as duas espécies foram pouco registradas, sugerindo que elas talvez
não sejam comuns em assembléias arbóreas.

17
Brescovit et al. (2002) cita que a família Salticidae é encontrada em todas as
áreas amostradas, habitando da serapilheira à copa das árvores, sendo considerada
a mais abundante da Amazônia Central. Estes resultados corroboram com aqueles
encontrados nos trabalhos supracitados e no presente trabalho, sugerindo que esta
família ocorre nos habitas mais distintos, independente das condições locais.
Assim como Goeldia sp. outras espécies coletadas no manguezal também
aparecem com um único registro. Isto pode ter origem no fato de serem espécies
raras no manguezal ou, simplesmente ser resultado do pequeno esforço amostral.
Entretanto, Borges e Brescovit (1996) sugerem que esta alta representatividade de
indivíduos com baixa freqüência parece ser característica das assembléias de
aranhas tropicais.
Os resultados do presente estudo registraram uma diferença na composição
de espécies dos dois sítios de trabalho (Furo do Taici e Furo Grande). Tal diferença
pode estar relacionada com a localização de cada sítio, ou seja, maior ou menor
proximidade da terra firme. A menor proximidade da terra firme, por exemplo, parece
influenciar a presença de espécies de aranhas de hábito arborícola, como é o caso
da assembléia de espécies registrada para o Furo Grande, certamente em função da
inundação diária ocasionada pelo regime de marés.
No geral, os bosques de mangue aqui inventariados apresentaram uma
composição faunística diferenciada. Tal diferença também pode ser registrada
quando a araneofauna do manguezal é comparada com a de outros sistemas. Isto
pode ocorrer devido à estrutura dos bosques de mangue, cujas características
diferem bastante dos outros sistemas florestais. As florestas de mangue não
apresentam, em geral, estratos inferiores característicos (ex. estrato arbustivo e sub-
bosque), o que pode ter interferido na baixa diversidade e abundância de aranhas.
Segundo Almeida (2005), a alta densidade de plantas nos estratos inferiores parece
aumentar a riqueza e abundância das aranhas, pois as plantas servem de suporte
para teias e ao mesmo tempo de área para o forrageamento. Além do mais, a
inundação também pode ser um fator importante e decisivo para a composição da
araneofauna no manguezal. Borges & Brescovit (1996), por exemplo, comparando a
composição das assembléias de aranhas da várzea e da terra firme, mostraram que
a várzea apresentou menor riqueza de espécies. Dessa forma, espera-se que a
inundação, tanto por água doce quanto por água salobra, possa exercer uma

18
seleção das espécies de aranhas, influenciando diretamente na diversidade e na
abundância desse grupo nas florestas alagáveis e de terra firme.
5. REFERÊNCIAS BIBLIOGRÁFICAS
Almeida, W. R. (2005). Riqueza e abundância de formigas e aranhas de vegetação
em baixios e platôs em uma área de floresta de terra firme na Amazônia
central. Disponível em: <www.inpa.gov.br/~pdbff/download/efa/livro/2005/
pdfs/rlfwal.pdf> [Acesso em 20/10/2008]
Ayres, M., Ayres Jr. M., Ayres, D. L. & Santos, A. S. (2007). BioEstat 5.0: aplicações
estatísticas nas áreas das Ciências Biológicas e médicas. Belém: Sociedade
Civil Mamirauá: Brasília CNPq.
Barreiros, J. A. P., Miglio L. T., Caxias, F. C., Araújo, C. O., Rocha, R. P. & Bonaldo,
A. B. (2001). Composição e Riqueza de aranhas e escorpiões (Arachinida:
Araneae, Scorpiones) cursoriais de serrapilheira na Estaçõa Ciêntífica
Ferreira Penna (ECFPn) Melgaço, Pará. Estação Ciêntífica Ferreira Penna -
10 anos de pesquisa na Amazônia.
Borges, S. H. & Brescovit, A. D. (1996). Inventário preliminar da aracnofauna
(Araneae) de duas localidades na Amazônia Ocidental. Boletim Museu
Paraense Emílo Goeldi, série Zool. , 12, 9-21.
Brescovit, A. D. (1999). Invertebrados Terrestres. Em: Biodiversidade do estado de
São Paulo Síntese do conhecimento ao final do século XX . C. B. Roberto.
São Paulo: Museu de Zoologia da Universidade de São Paulo.
Brescovit, A. D., Bonaldo, A. B., Bertani, R., Rheims, C. A. (2002). Araneae. Em:
Amazonian Arachnida and Myriapoda: Identification keys to all classes,
orders, families, some genera. Adis, J., Sofia-Moscow: Pensoft Publishers p.
303-343..
Brescovit, A. D., Francesconi, P. (2002). Implementação de um banco de dados da
Araneofauna Neotropical (Araneae) com ênfase na diversidade de espécies

19
brasileiras. Resumo apresentado no III Simpósio do Programa Biota/Fadesp
de 26 a 28 de Novembro , 21. Universidade Federal de São Carlos, São
Paulo, Brasil.
Clarke, K. R., Gorley, R. N. (2006). PRIMER-E ver. 6— manual: Plymouth Routines
in Multivariate Ecological Research, PRIMER-E, Ltd. p 190.
Coddington, J. A., Levi, H. W. (1991). Systematics and evolutions of spiders
(Araneae). Annual Review of Ecology and Systematics , 22, p 565-592.
Fernandes, M. E. (2003). Prefácio. Em: Os Manguezais da Costa Norte Brasileira.
Fernandes M. E. São Luiz, MA: Fundação Rio Bacanga.
Fernandes, B. E. (2000). Association of mammals with mangrove forest: a world wide
review. Boletim do Laboratório de Hidrobiologia , 13, 83-108.
Flórez, E. (2000). Comunidade de Arañas de La Región Pacífica Del departamento
Del Valle Del Cauca, Colômbia. Revista Colombiana de Entomologia , 26, p
77-81.
Herz, R. (1991). Manguezais do Brasil. São Paulo: Universidade de São Paulo.
Höfer, H. (1990). The spider community (Araneae) of Central Amazonian blackwater
inundation forest (Igapó). Acta Zoologia Fennica , 190, p 173-179.
Höfer, H. & Beck, L. (1996). Die spinnentienrfauna des regenwaldreservats "Reserva
Duke" in zentralamazonien II. Natur und Museum , 126, 69-107.
Hogarth, J. P. (1999). Sipders. In: The Biology of Mangroves (p. 228). New York:
Oxford University press.
krause, G. (2001). Spatial patters of mangroves ecossystems: the bragantinian
mangrove of Northern Brazil (Bragabça, Pará). Ecotropica , 7, 93-107.
Martins, M. & Lise. (1997). As aranhas. Em: Caxiuanã, Belém, Pará: Museu
Paraense Emílio Goeldi Museu Paraense Emílio Goeldi, Lisboa P. L., p. 381-
388.
Marusik, Y. M., Koponen, S. (2000). Diversity of spiders in boreal and artic zones.
The Journal of Arachnology , 30, p 205-210.

20
Matni, S. A., Menezes, M. P. M., Mehlig, U. (2006). Estrutura dos bosques de
mangue da península de Bragança, Pará, Brasil. Boetim do Museu Paraense
Emílio Goeldi, Ciências Naturais. , 1(3), 3-52.
Matni, S. A. (2007). Estudo Comparativo da Fenologia Reprodutiva de Rhiziphora
mangle, Avicennia germinans e Lagunculária racemosa ao longo da
Península de Ajuruteua, Bragança, Pará. Dissertação de mestrado
apresentada ao Programa de Pós-Graduação em Biologia Ambiental,
Mestrado em Ecologia de Ecossistemas Costeiros e Estuarinos da
Universidade Federal do Pará .
Mendes, C. A. (2005). Geomorfologia e Sedimentologia. In: B. E. Fernandes, Os
Manguezais da Costa Nort Brasileira (Vol. 2, p. 165). Belém, Pará: Fundação
Rio Bacanga.
Oliveira-Alves, A., Peres, M. C. L., Dias, M. A., Cazais-Ferreira, G. S. & Souto, L. R.
A. (2005). Estudo das Comunidades de Aranhas (Arachnida: Araneae) em
Ambiente de Mata Atlântica no Parque Metropolitano de Pituaçu- PMP,
Salvador, Bahia. Biota Neotropica , 5(1), p 1-8.
Platnick, N. I. The world spider catalog, version 9.0. 2008. American Museum of
Natural History, online at http://research.amnh.org/entomology/spiders/
catalog/index.html. Acessado em: jun. 2008.
Platinick, N. I. (1999). Dimensions of bodiversity: targeting megadiverse groups. Em:,
The living planet in crisis: biodiversity science and policy. Cracraft J. A.
Columbia University Press, p. 33-52.
Podgaiski, L. R., Ott, R., Rodrigues, E. N. L., Buckup, E. H. & Marques, M. A. (2007).
Araneofauna (Arachnida; Araneae) do Parque Estadual do Turvo, Rio Grande
do Sul, Brasil. Biota Neotropica , 7(2), p 2-15.
Raizer, J., Japyassú, H. F., Indicatti, R. P. & Brescovit, A. D. (2005). Comunidade de
Aranhas (Arachnida, Araneae) do Pantanal Norte (Mato Grosso, Brasil) e Sua
Similaridade com a Araneofauna Amazônica. Biota Neotropica , 5(1), p 1-16.

21
Ricetti, J., Bonaldo, A. B. (2008). Diversidade e estimativas de riqueza de aranhas
em quatro fitofisionomias na Serra do Cachimbo, Pará, Brasil. Iheringia, Sér.
Zool , 98(1), p 88-99.
Rupert, E. E.; Barnes, R. D. (2005). Ordem Aranae. Em: E. E. Rupert, & R. D.
Barnes, Zoologia dos Invertebrados (7ª ed.). Roca.
Schaeffer-Novelli, Y. (1995). Introdução. Em: Manguezal ecossistema entre a terra e
o mar. Y. Schaeffer-Novelli. São Paulo: Caribbean Ecological Research, p 64.
Simberloff, D., Dayan, T. (1991). The Guild Concept and the Structure of Ecological
Communities. Annual Reviews Ecol. Syst. , 22, p 115-143.
Souza Filho, P. W. (2005). Costa de manguezais de macromarés da Amazônia:
cenários morfológicos, mapeamento e quantificação de áreas usando dados
de sensores remotos. Revista Brasileira de Geofíısica , 23(4), p 427-435.
Szlafsztein, C., Lara, R. J., Cohen, M. C. L. (2000). Coastal management: some
studies of the past and present of the Bragança region (Pará, Brasil). The
MADAM project. Journal of International Environmental Creation .
Uetz, G. W., Halaj, J., Cady, A. B. (1999). Guild Structure of Spiders in Major Crops.
The Journal of Arachnology , 27, p 270–280.





























