Embed Size (px)
Citation preview

DNA Structure, Replication, Repair and RNA Transcription
Presented by
Dr. Mohammad Saadeh
The requirements for the Pharmaceutical Biochemistry I
Philadelphia University
Faculty of pharmacy
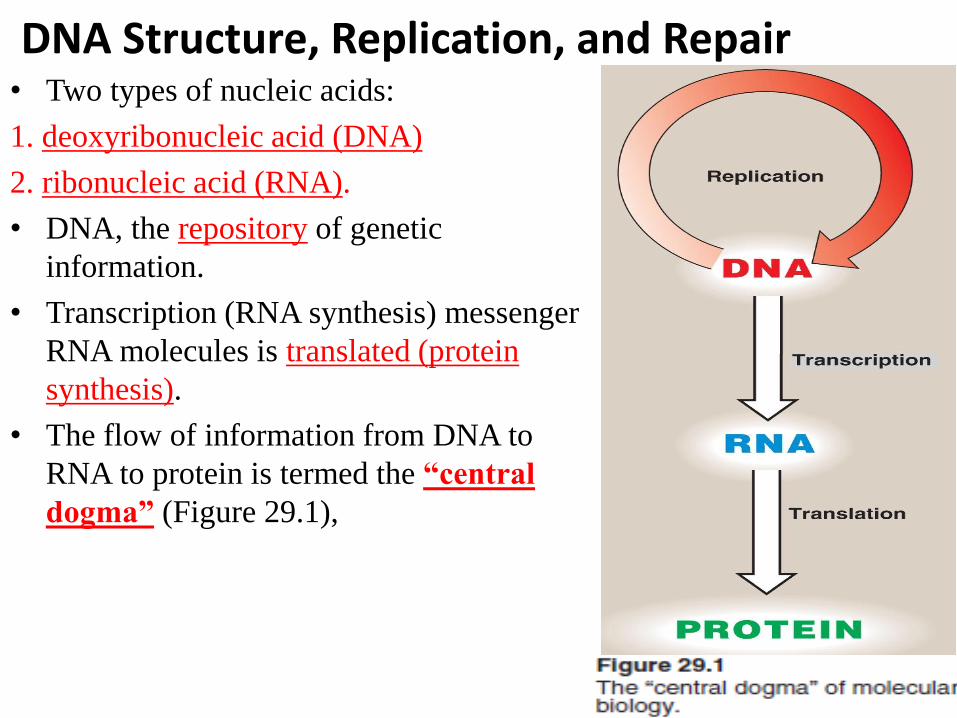
DNA Structure, Replication, and Repair • Two types of nucleic acids:
1. deoxyribonucleic acid (DNA)
2. ribonucleic acid (RNA).
• DNA, the repository of genetic
information.
• Transcription (RNA synthesis) messenger
RNA molecules is translated (protein
synthesis).
• The flow of information from DNA to
RNA to protein is termed the “central
dogma” (Figure 29.1),
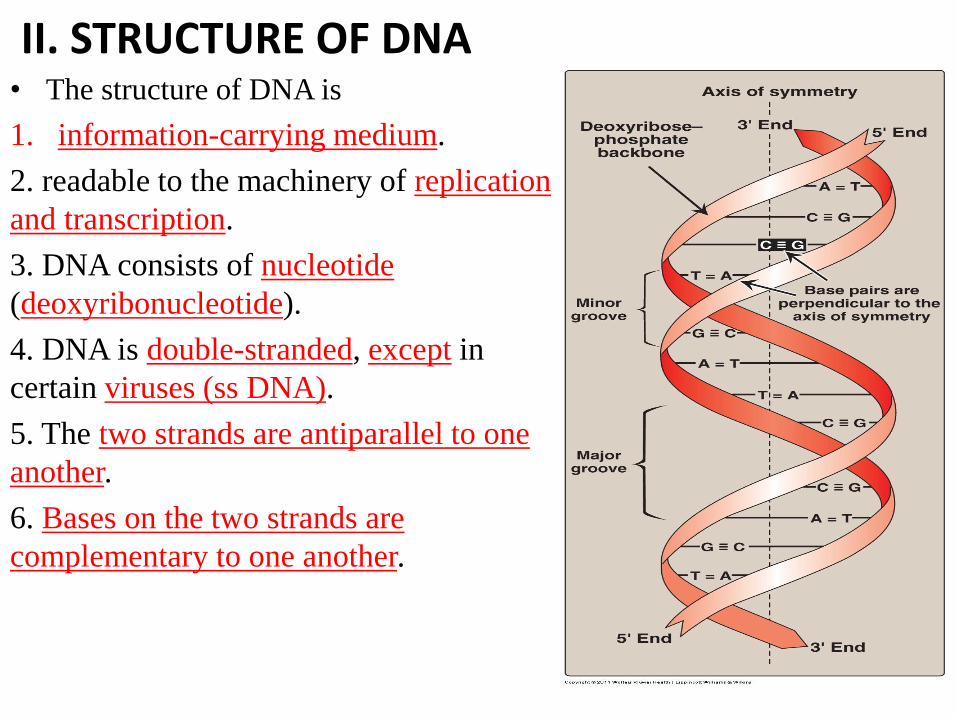
II. STRUCTURE OF DNA • The structure of DNA is
1. information-carrying medium.
2. readable to the machinery of replication
and transcription.
3. DNA consists of nucleotide
(deoxyribonucleotide).
4. DNA is double-stranded, except in
certain viruses (ss DNA).
5. The two strands are antiparallel to one
another.
6. Bases on the two strands are
complementary to one another.
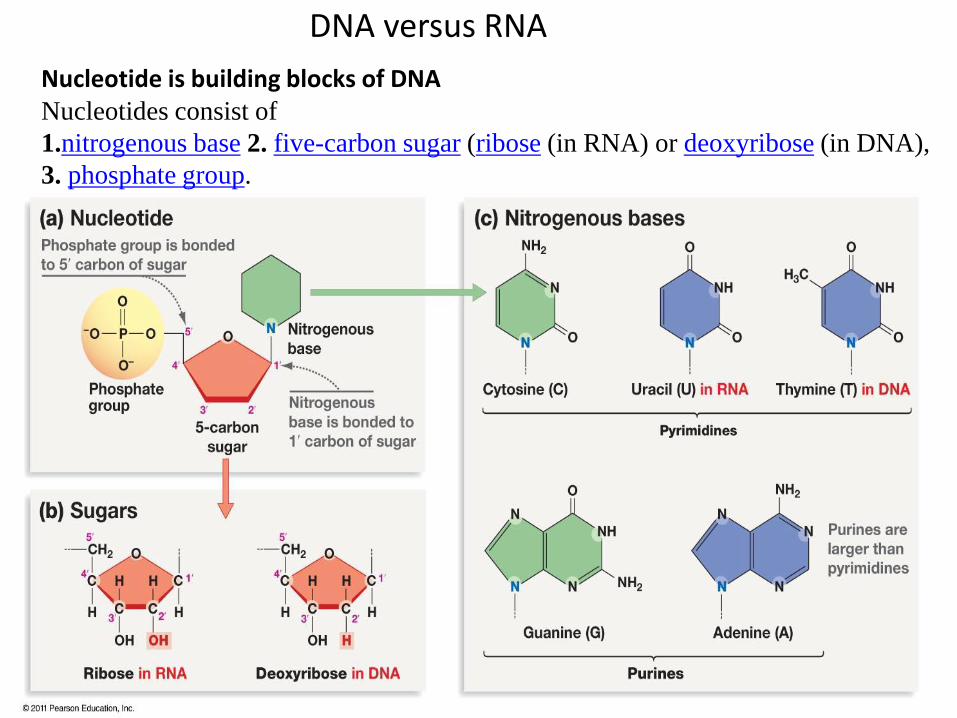
DNA versus RNA
Nucleotide is building blocks of DNA Nucleotides consist of
1.nitrogenous base 2. five-carbon sugar (ribose (in RNA) or deoxyribose (in DNA),
3. phosphate group.
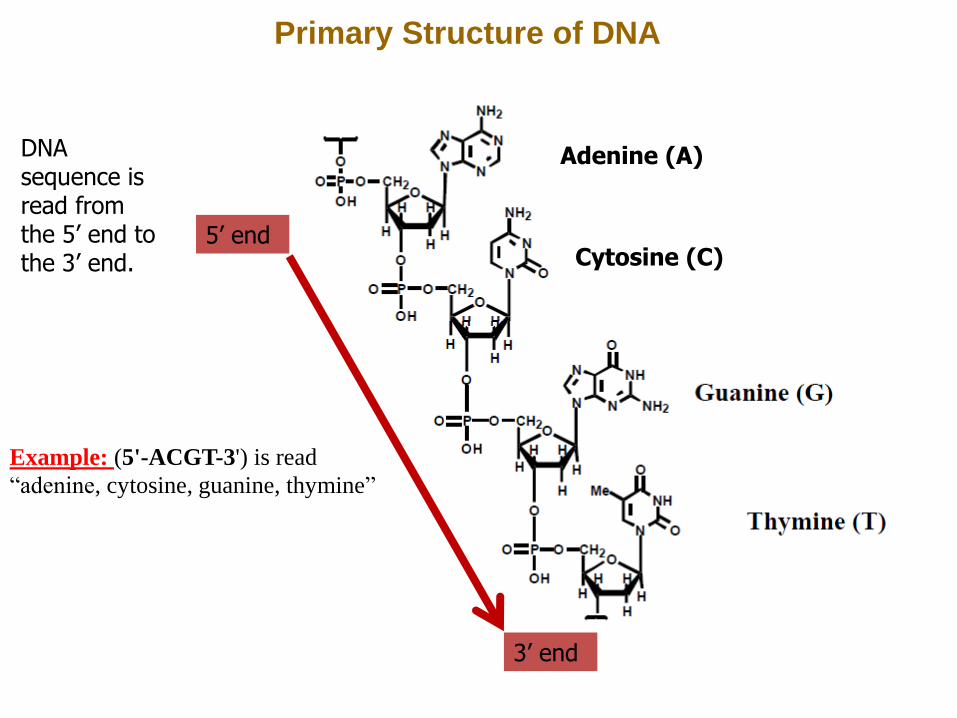
Primary Structure of DNA
Adenine (A)
Cytosine (C) 5’ end
3’ end
DNA sequence is read from the 5’ end to the 3’ end.
Example: (5'-ACGT-3') is read
“adenine, cytosine, guanine, thymine”
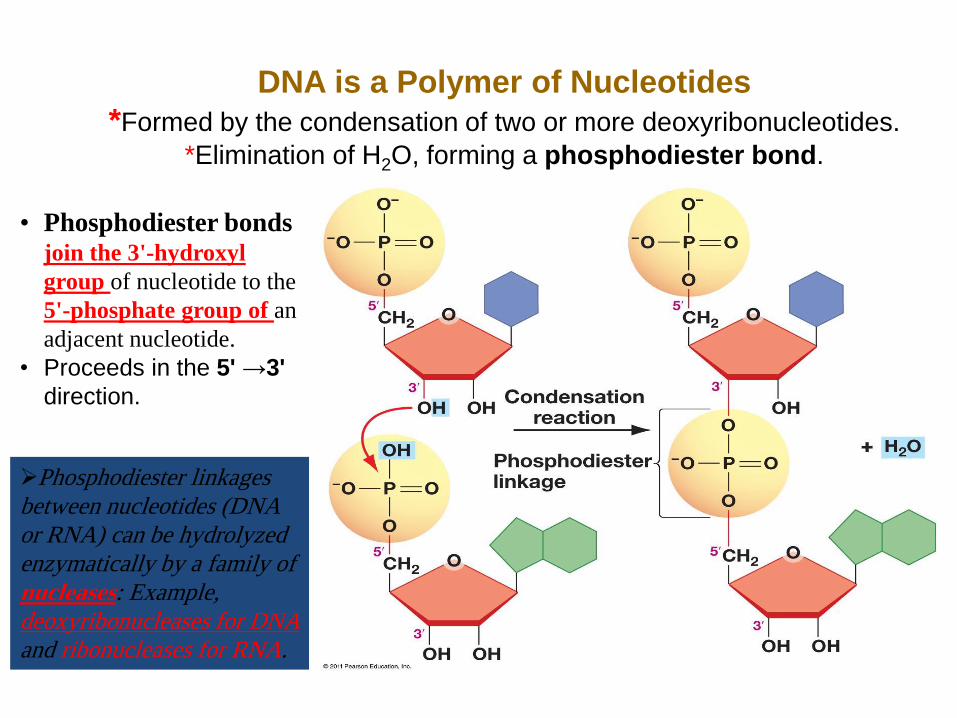
DNA is a Polymer of Nucleotides
*Formed by the condensation of two or more deoxyribonucleotides.
*Elimination of H2O, forming a phosphodiester bond.
• Phosphodiester bonds join the 3'-hydroxyl
group of nucleotide to the
5'-phosphate group of an
adjacent nucleotide.
• Proceeds in the 5' →3'
direction.
Phosphodiester linkages
between nucleotides (DNA
or RNA) can be hydrolyzed
enzymatically by a family of
nucleases: Example,
deoxyribonucleases for DNA
and ribonucleases for RNA.

II. STRUCTURE OF DNA B. Double helix
• d.S DNA are paired in an antiparallel manner, and wind around each other, forming a double helix (Figure 29.3).
• (the 5'-end of one strand is paired with the 3'-end of the other strand) Figure 29.3.
• The spatial relationship between the two strands in the helix creates a major (wide) groove and a minor (narrow) groove.
*These grooves provide access for (VVI): (Figure 29.3)
1. the binding of regulatory proteins to their specific recognition sequences along the DNA chain.
2. Certain anticancer drugs, such as actinomycin D, exert their cytotoxic effect by intercalating into the narrow groove of the DNA double helix, thus interfering with DNA and RNA synthesis.
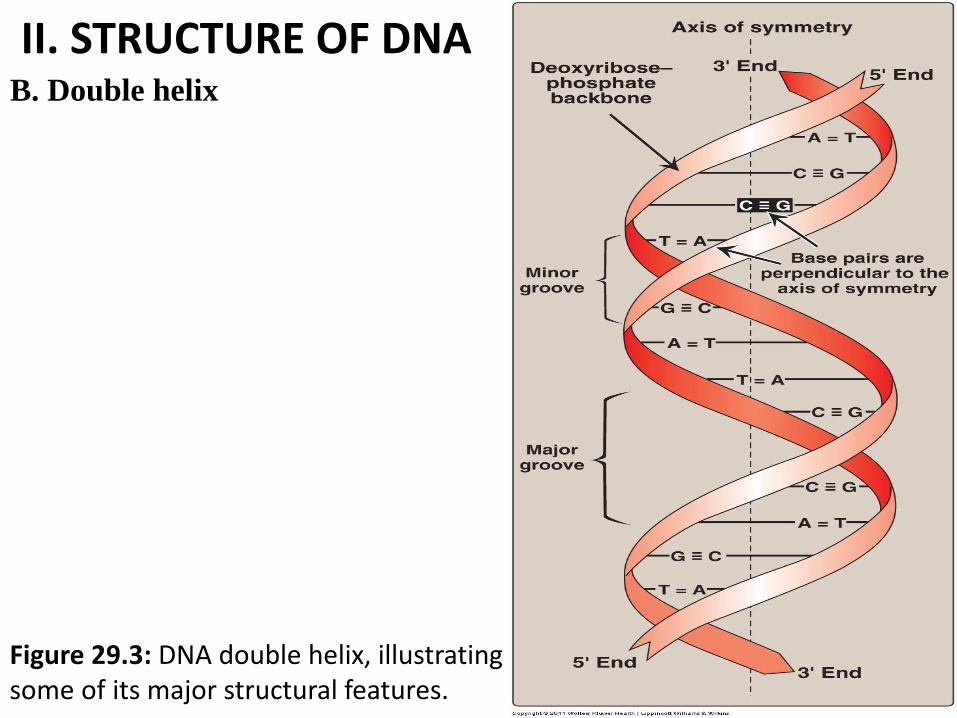
II. STRUCTURE OF DNA B. Double helix
Figure 29.3: DNA double helix, illustrating some of its major structural features.
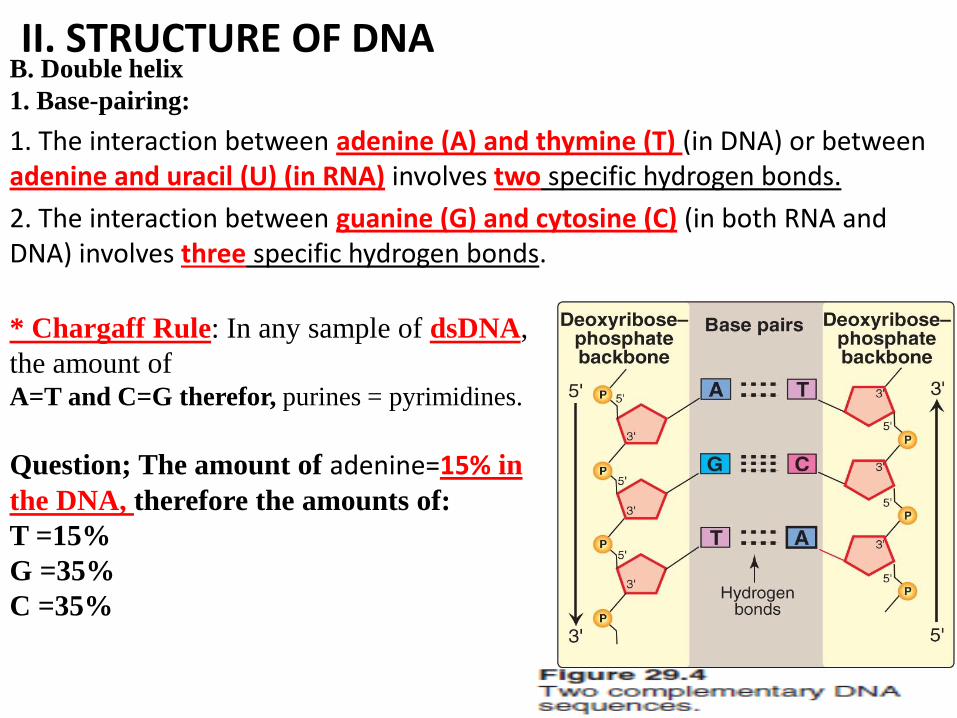
II. STRUCTURE OF DNA B. Double helix
1. Base-pairing:
1. The interaction between adenine (A) and thymine (T) (in DNA) or between adenine and uracil (U) (in RNA) involves two specific hydrogen bonds.
2. The interaction between guanine (G) and cytosine (C) (in both RNA and DNA) involves three specific hydrogen bonds.
* Chargaff Rule: In any sample of dsDNA,
the amount of A=T and C=G therefor, purines = pyrimidines.
Question; The amount of adenine=15% in
the DNA, therefore the amounts of:
T =15%
G =35%
C =35%

II. STRUCTURE OF DNA B. Double helix
2. Separation of the two DNA strands in the double helix; when
hydrogen bonds between the paired bases are disrupted.
Disruption of DNA in the laboratory:
1. pH of the DNA solution is altered so that the nucleotide bases ionize,
2. When DNA is heated, Cause loss of helical structure in DNA, called
denaturation, can be monitored by measuring its absorbance at 260
nm.
3. DNA that contains high concentrations of G and C denatures at high
temperature, Because there are three hydrogen bonds between G
and C (Figure 29.6).
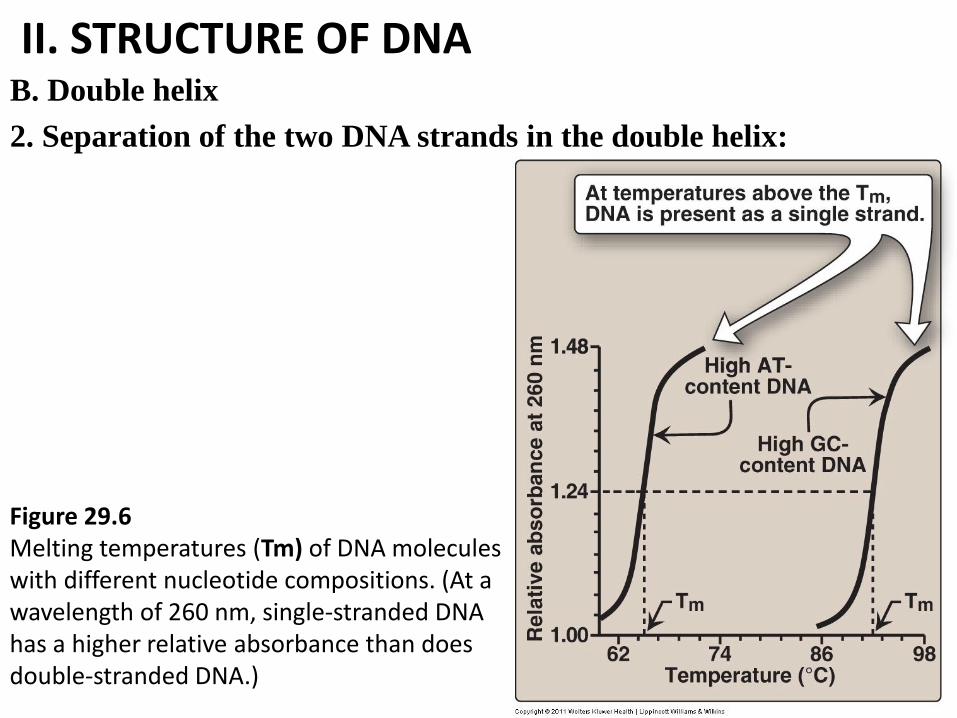
II. STRUCTURE OF DNA B. Double helix
2. Separation of the two DNA strands in the double helix:
Figure 29.6 Melting temperatures (Tm) of DNA molecules with different nucleotide compositions. (At a wavelength of 260 nm, single-stranded DNA has a higher relative absorbance than does double-stranded DNA.)
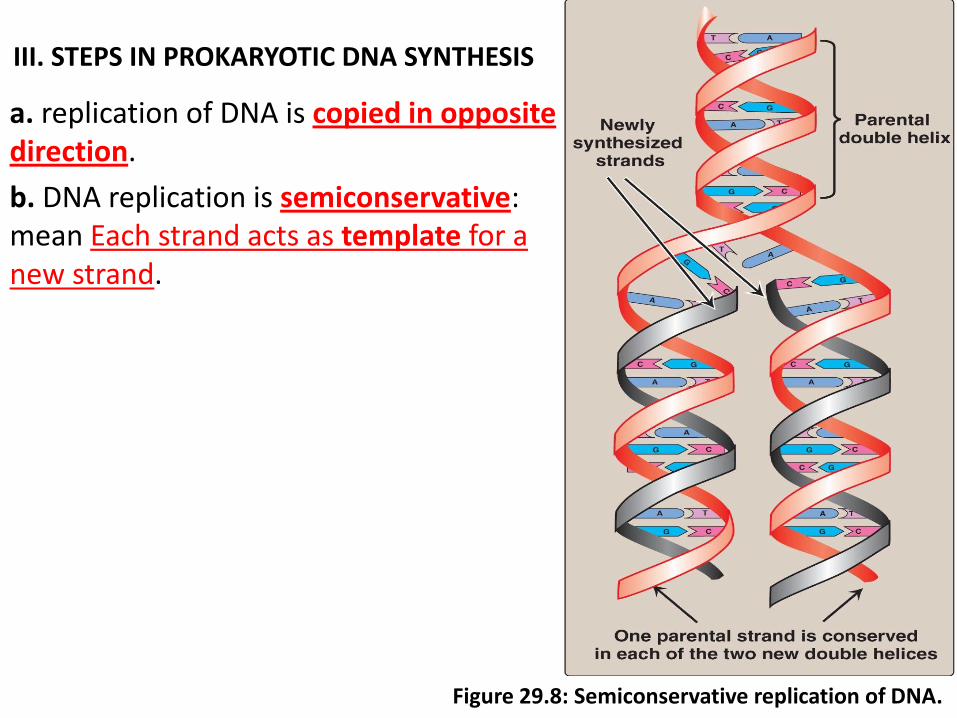
III. STEPS IN PROKARYOTIC DNA SYNTHESIS
a. replication of DNA is copied in opposite direction.
b. DNA replication is semiconservative: mean Each strand acts as template for a new strand.
Figure 29.8: Semiconservative replication of DNA.

III. STEPS IN PROKARYOTIC DNA SYNTHESIS
Steps of DNA synthesis (replication):
1. Separation of ds DNA (or “melt”) at origin of replication (rich in A=T), because the polymerases use only ssDNA as a template by DNA helicase (Figure 29.9).
An origin of replication (rich in A=T) is a site in a DNA molecules at which helicase unwinds the double helix. Allowing DNA replication to commence.
A prokaryotic chromosome has a single origin (figure 29.9A) of replication.
A eukaryotic chromosome has many origins of replication (figure 29.9B).
DNA replication occurs in the S phase of the cell cycle
The strands are separated locally, forming two replication forks. Replication of dsDNA is bidirectional.
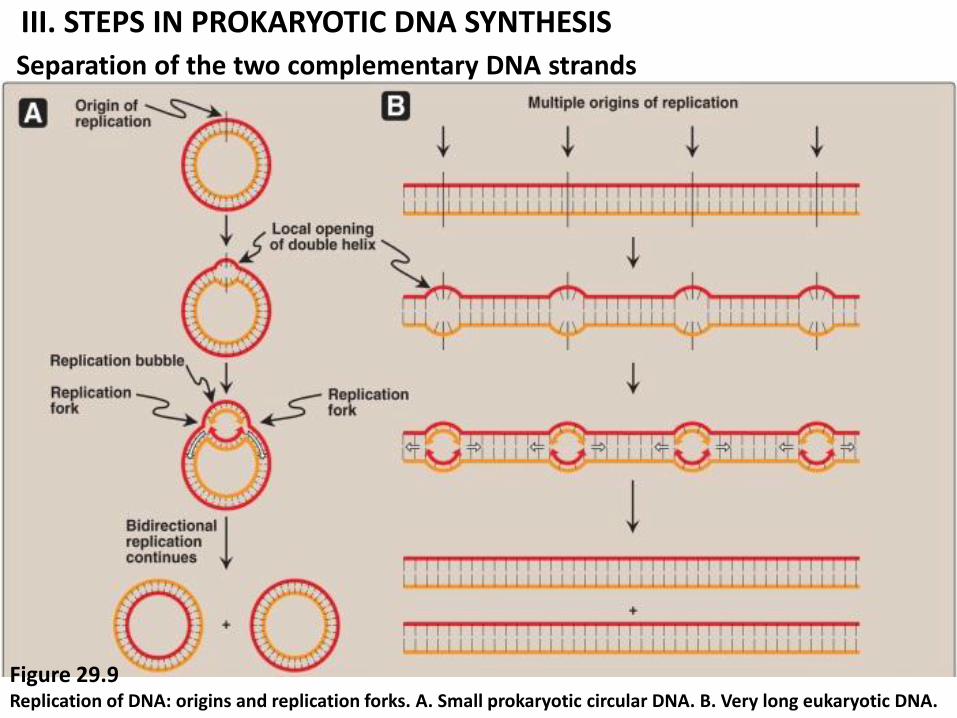
III. STEPS IN PROKARYOTIC DNA SYNTHESIS
Separation of the two complementary DNA strands
Figure 29.9 Replication of DNA: origins and replication forks. A. Small prokaryotic circular DNA. B. Very long eukaryotic DNA.
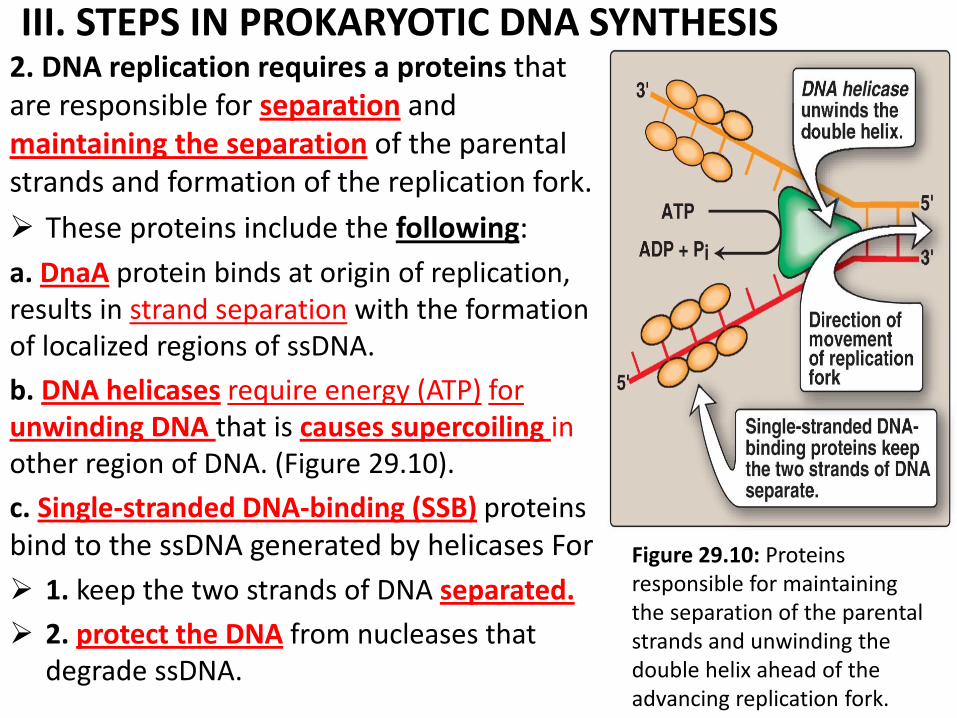
III. STEPS IN PROKARYOTIC DNA SYNTHESIS 2. DNA replication requires a proteins that are responsible for separation and maintaining the separation of the parental strands and formation of the replication fork.
These proteins include the following:
a. DnaA protein binds at origin of replication, results in strand separation with the formation of localized regions of ssDNA.
b. DNA helicases require energy (ATP) for unwinding DNA that is causes supercoiling in other region of DNA. (Figure 29.10).
c. Single-stranded DNA-binding (SSB) proteins bind to the ssDNA generated by helicases For
1. keep the two strands of DNA separated.
2. protect the DNA from nucleases that degrade ssDNA.
.
Figure 29.10: Proteins responsible for maintaining the separation of the parental strands and unwinding the double helix ahead of the advancing replication fork.

III. STEPS IN PROKARYOTIC DNA SYNTHESIS
3. Solving the problem of supercoils: when ds DNA are separated, a problem is encountered are positive supercoils in the region of DNA ahead of the replication fork as result of overwinding (Figure 29.11), and negative supercoils in the region behind the fork.
To solve this problem,
a. Type I DNA topoisomerases, cut one strand of the DNA duplex, type I
topoisomerases are able to relax negative supercoiled DNA (Figure
29.12).
b. Type II DNA topoisomerases, cut both strands of a DNA duplex, can
relax either negatively or positively supercoild DNA molecules (Figure
29.13).
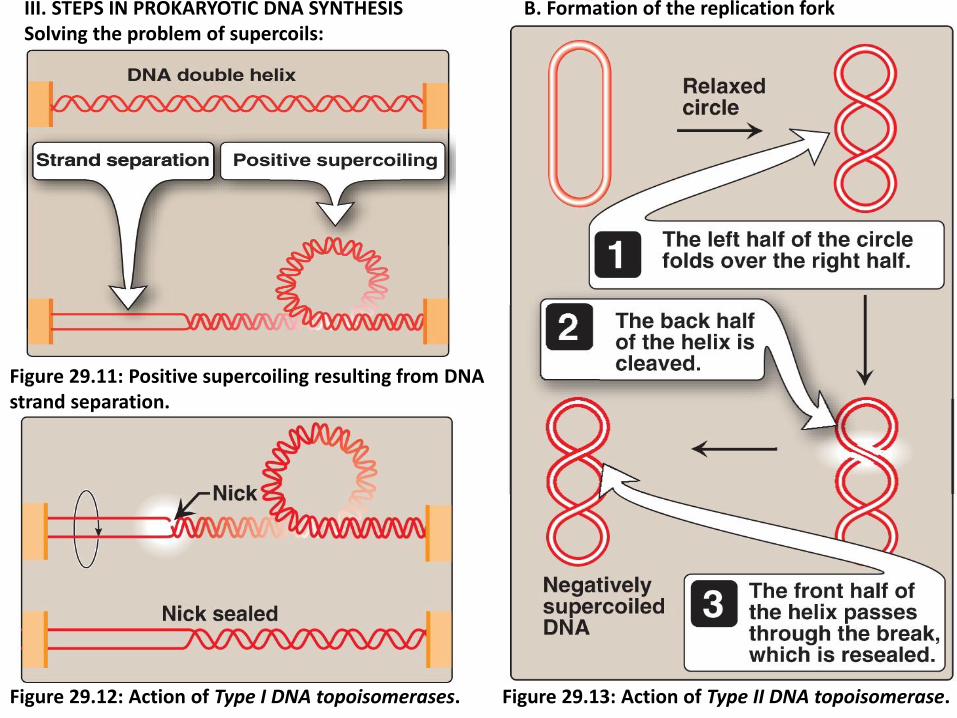
III. STEPS IN PROKARYOTIC DNA SYNTHESIS B. Formation of the replication fork Solving the problem of supercoils:
Figure 29.11: Positive supercoiling resulting from DNA strand separation.
Figure 29.12: Action of Type I DNA topoisomerases. Figure 29.13: Action of Type II DNA topoisomerase.
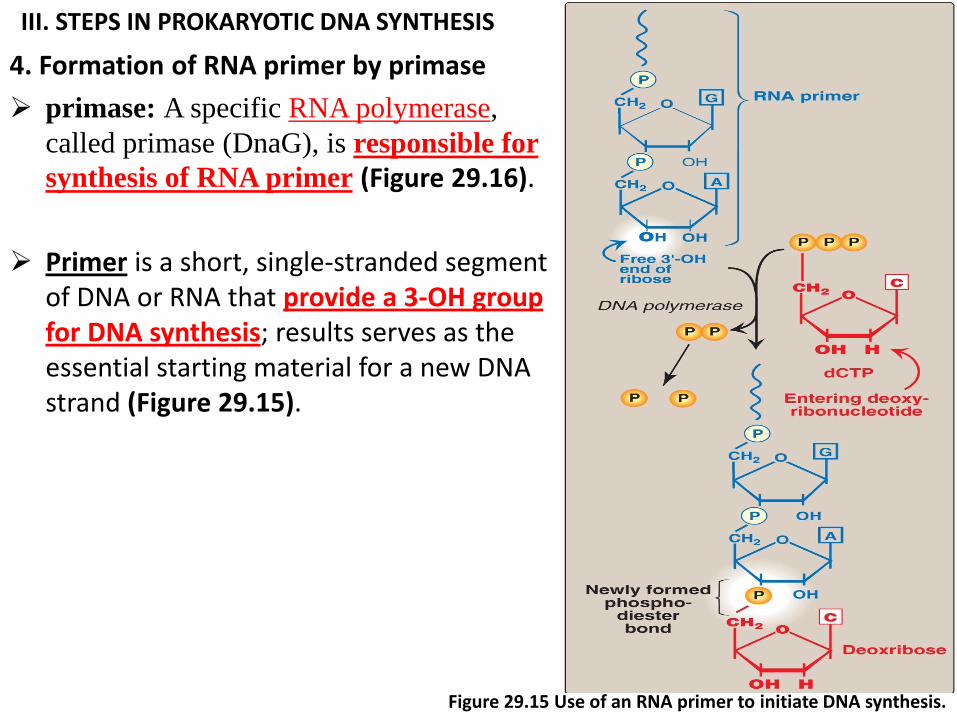
III. STEPS IN PROKARYOTIC DNA SYNTHESIS
4. Formation of RNA primer by primase
primase: A specific RNA polymerase,
called primase (DnaG), is responsible for
synthesis of RNA primer (Figure 29.16).
Primer is a short, single-stranded segment of DNA or RNA that provide a 3-OH group for DNA synthesis; results serves as the essential starting material for a new DNA strand (Figure 29.15).
Figure 29.15 Use of an RNA primer to initiate DNA synthesis.

III. STEPS IN PROKARYOTIC DNA SYNTHESIS
5. Elongation of the leading and lagging strands.
DNA elongation (DNA pol III) is catalyzed by DNA polymerase III.
Properties of DNA polymerase III:
a. synthesis of DNA in 5'→3' direction using templet (ssDNA), antiparallel to the
parental strand (Figure 29.15 and Figure 29.16).
b. DNA polymerase III has a “proofreading” and erorr correction activity (3'→5'
exonuclease, Figure 29.17).
*3'→5' exonuclease remove the misplaced nucleotide The 5' →3„ polymerase activity then replaces it with the correct nucleotide (Figure 29.17).
1. Leading strand: one strand is leading strand, grows continuously and
uninterruptedly from its 5 ˋ→3ˋ in the same direction of movement of the
replication fork. (Figure 29.14).
2. Lagging strand: direction of DNA on away from the replication fork is
synthesized discontinuously from their 5 ˋ→3 until it is blocked by proximity to
an RNA primer (The strand growing in opposite direction to unwinding = lagging strand.
3. Many short DNA fragments newly synthesized from lagging strand = Okazaki fragments (joined together by DNA ligase) Figure 29.14.
Note: Elongation and polymerization of lagging strand proceeds via generation of okazzki fragments
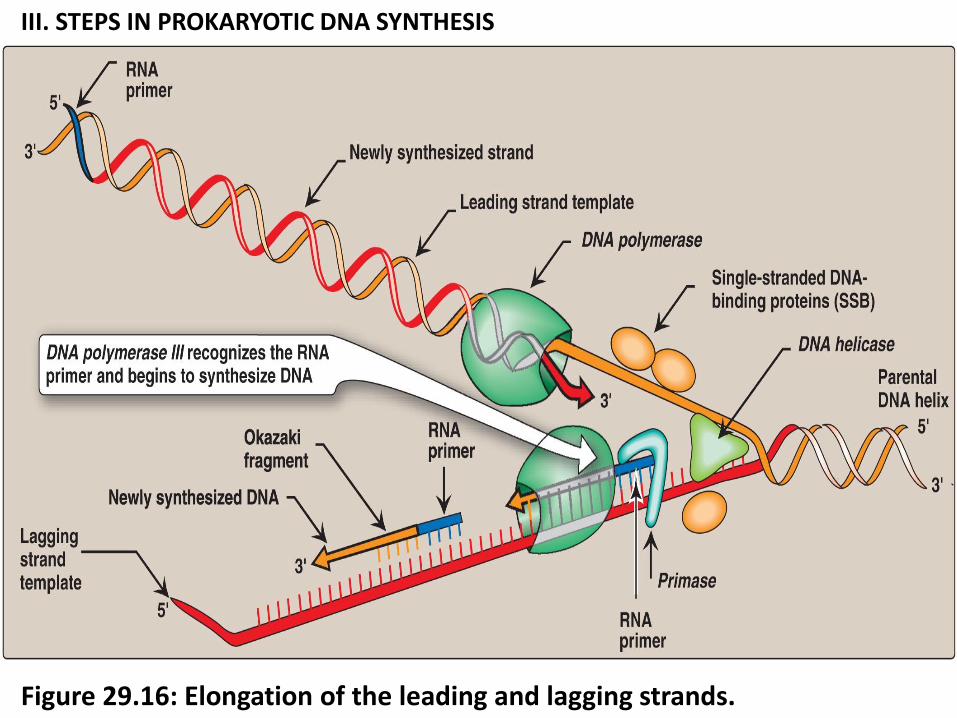
III. STEPS IN PROKARYOTIC DNA SYNTHESIS
Figure 29.16: Elongation of the leading and lagging strands.
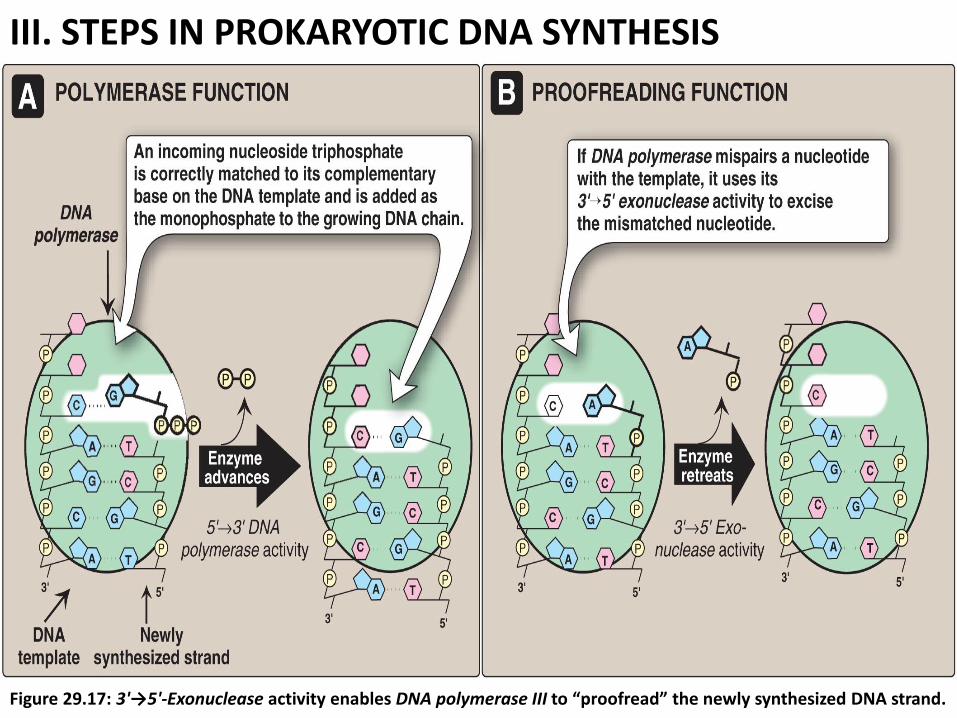
III. STEPS IN PROKARYOTIC DNA SYNTHESIS
Figure 29.17: 3'→5'-Exonuclease activity enables DNA polymerase III to “proofread” the newly synthesized DNA strand.
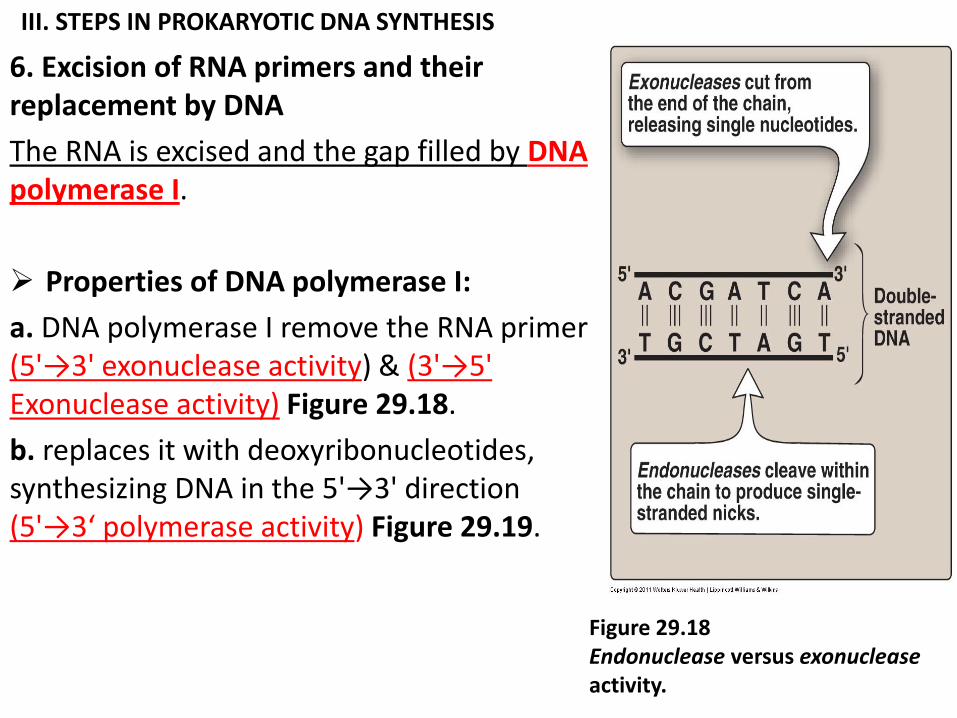
III. STEPS IN PROKARYOTIC DNA SYNTHESIS
6. Excision of RNA primers and their replacement by DNA
The RNA is excised and the gap filled by DNA polymerase I.
Properties of DNA polymerase I:
a. DNA polymerase I remove the RNA primer (5'→3' exonuclease activity) & (3'→5' Exonuclease activity) Figure 29.18.
b. replaces it with deoxyribonucleotides, synthesizing DNA in the 5'→3' direction (5'→3‘ polymerase activity) Figure 29.19.
Figure 29.18 Endonuclease versus exonuclease activity.
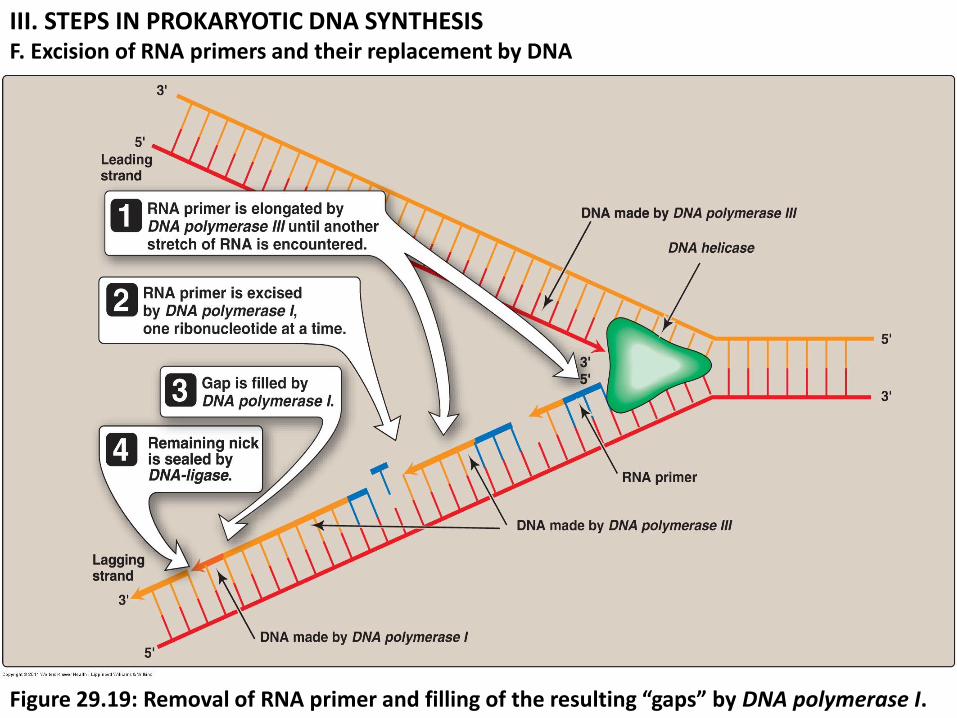
III. STEPS IN PROKARYOTIC DNA SYNTHESIS F. Excision of RNA primers and their replacement by DNA
Figure 29.19: Removal of RNA primer and filling of the resulting “gaps” by DNA polymerase I.
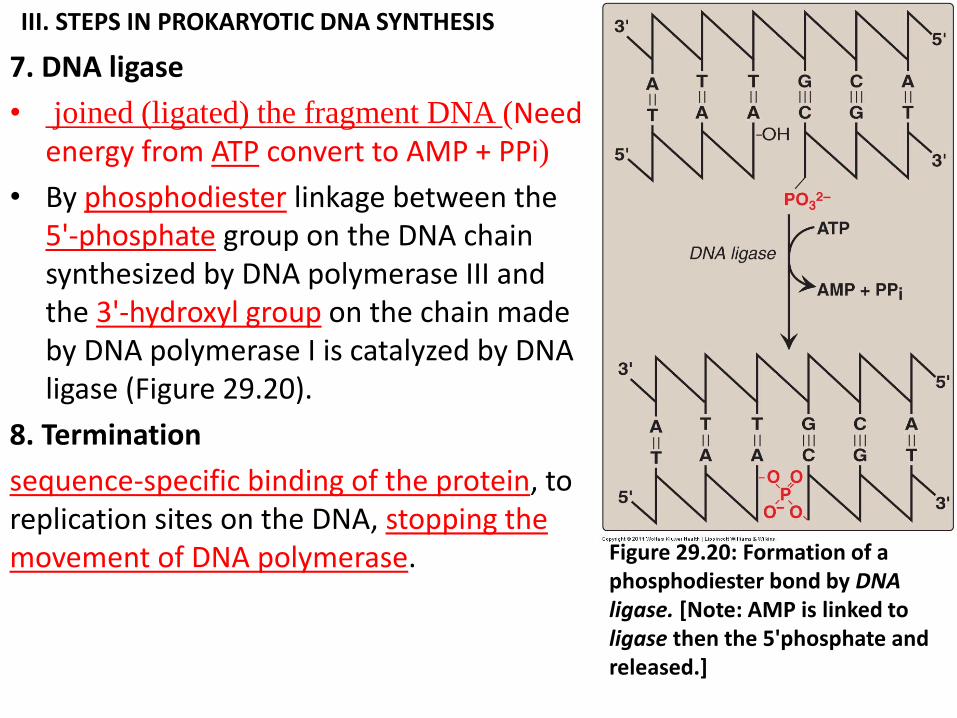
III. STEPS IN PROKARYOTIC DNA SYNTHESIS
7. DNA ligase
• joined (ligated) the fragment DNA (Need energy from ATP convert to AMP + PPi)
• By phosphodiester linkage between the 5'-phosphate group on the DNA chain synthesized by DNA polymerase III and the 3'-hydroxyl group on the chain made by DNA polymerase I is catalyzed by DNA ligase (Figure 29.20).
8. Termination
sequence-specific binding of the protein, to replication sites on the DNA, stopping the movement of DNA polymerase. Figure 29.20: Formation of a
phosphodiester bond by DNA ligase. [Note: AMP is linked to ligase then the 5'phosphate and released.]

IV. EUKARYOTIC DNA REPLICATION
eukaryotic DNA replication closely of prokaryotic DNA synthesis. Some differences, such as
What is the different between DNA replication in EUKARYOTIC and prokaryotic?
1. multiple origins of replication in eukaryotic cells versus single origins of replication in prokaryotes.
2. RNA primers are removed by RNase H rather than by a DNA polymerase.
3. Some of the enzymes involved in eukaryotic replication are different.
These include 5 enzymes
Pol α and pol γ
(see figure 29.23):
Figure 29.22: Activities of eukaryotic

IV. EUKARYOTIC DNA REPLICATION 4. Telomeres are repetitive non coding DNA sequences plus protein at the end
of linear chromosomes.
Function of telomeres:
1. preventing attack by nucleases.
2. As most cells divide and age, these sequences are shortened, contributing
to senescence. In cells that do not senesce (for example, germ line and
cancer cells).
5. Telomerase : are complex contains (1. protein + 2. short RNA)
1. protein that acts as a reverse transcriptase, and
2. Short piece of RNA that acts as a template. The CA-rich RNA template
base-pairs with the GT-rich, single-stranded 3'-end of telomeric DNA
(Figure 29.23). employs its enzyme component reverse transcriptase,
RNA as a template for the 5'→3„synthesis 5'→3„of DNA.
6. Nucleoside analogs containing modified sugars can be used to block DNA
chain growth. They are useful in anticancer and antiviral chemotherapy.
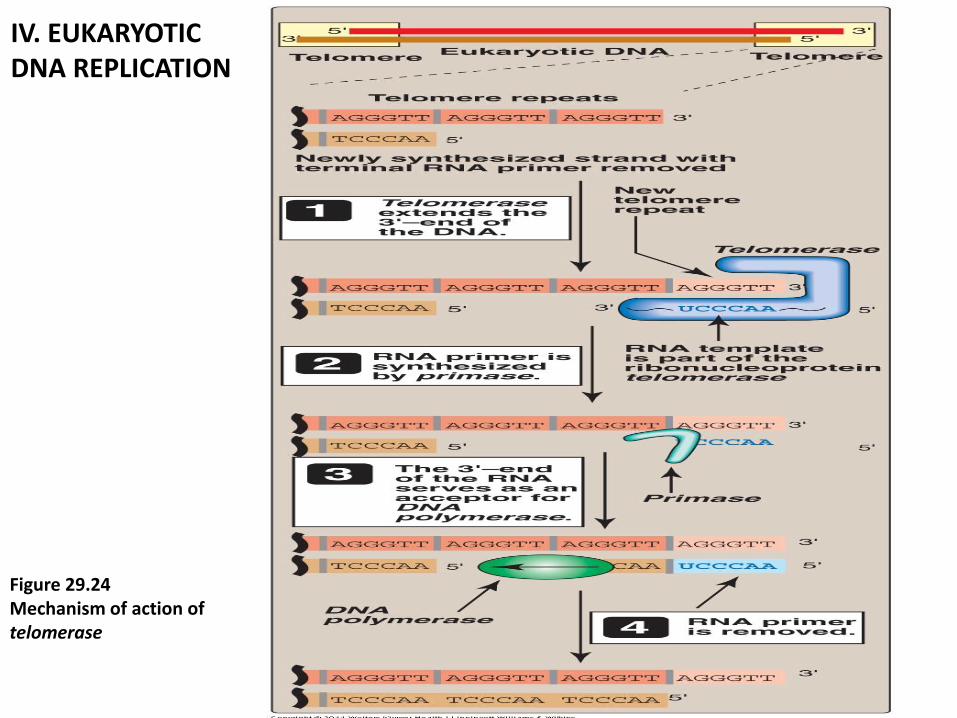
IV. EUKARYOTIC DNA REPLICATION
Figure 29.24 Mechanism of action of telomerase
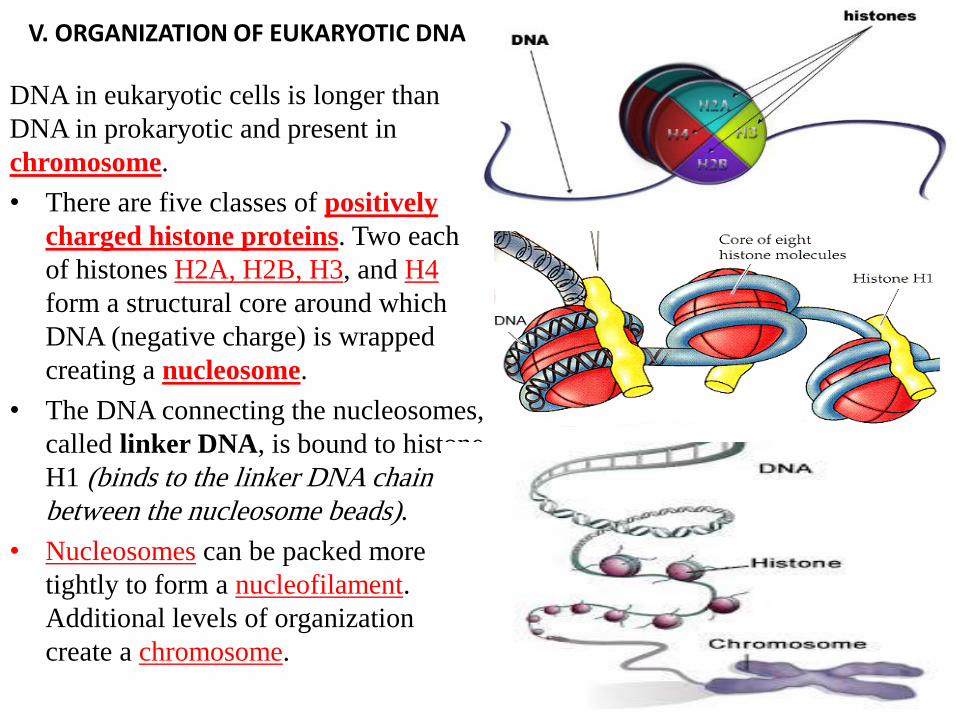
V. ORGANIZATION OF EUKARYOTIC DNA
DNA in eukaryotic cells is longer than
DNA in prokaryotic and present in
chromosome.
• There are five classes of positively
charged histone proteins. Two each
of histones H2A, H2B, H3, and H4
form a structural core around which
DNA (negative charge) is wrapped
creating a nucleosome.
• The DNA connecting the nucleosomes,
called linker DNA, is bound to histone
H1 (binds to the linker DNA chain
between the nucleosome beads).
• Nucleosomes can be packed more
tightly to form a nucleofilament.
Additional levels of organization
create a chromosome.

VI. DNA REPAIR
• Most DNA damage can be corrected by excision repair involving
recognition and removal of the damage by repair proteins, followed by
replacement by DNA polymerase and joining by ligase.
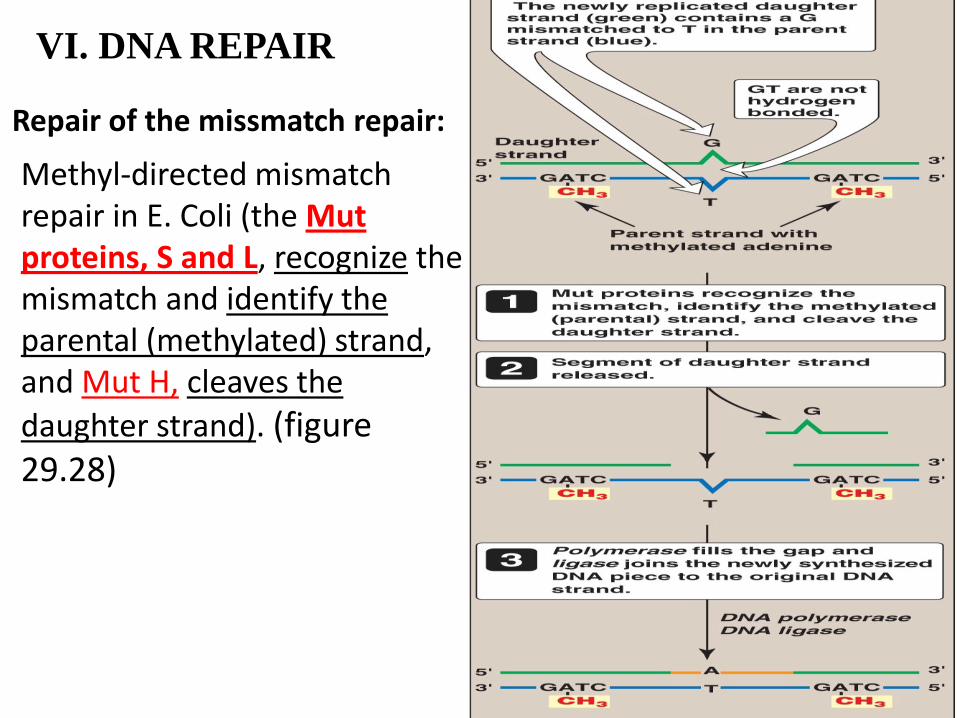
VI. DNA REPAIR
Repair of the missmatch repair:
Methyl-directed mismatch repair in E. Coli (the Mut proteins, S and L, recognize the mismatch and identify the parental (methylated) strand, and Mut H, cleaves the
daughter strand). (figure 29.28)
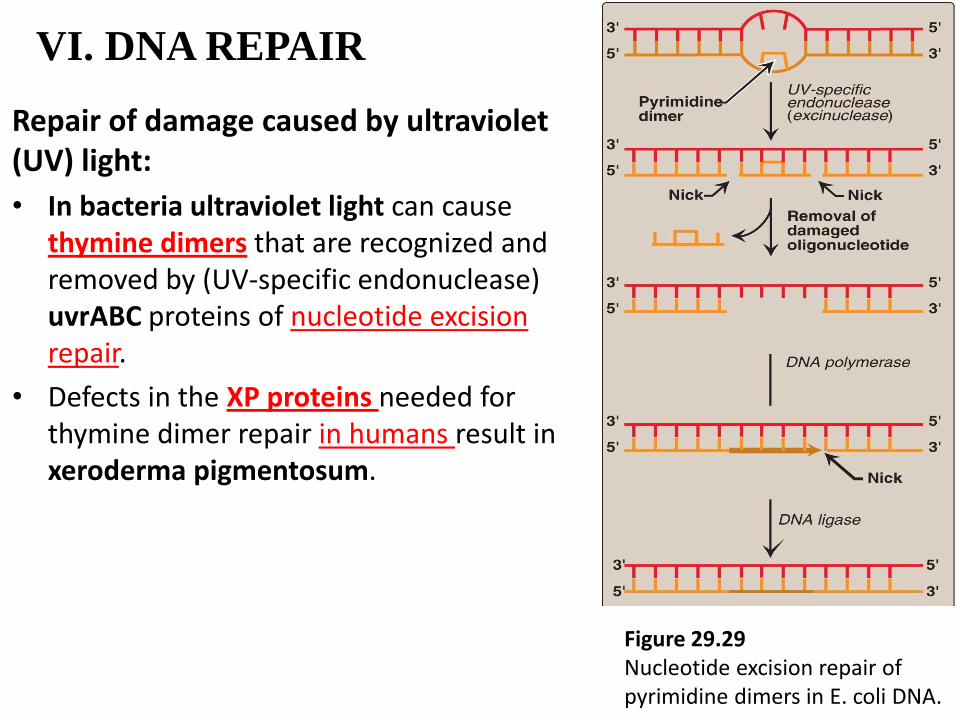
VI. DNA REPAIR
Repair of damage caused by ultraviolet (UV) light:
• In bacteria ultraviolet light can cause thymine dimers that are recognized and removed by (UV-specific endonuclease) uvrABC proteins of nucleotide excision repair.
• Defects in the XP proteins needed for thymine dimer repair in humans result in xeroderma pigmentosum.
Figure 29.29 Nucleotide excision repair of pyrimidine dimers in E. coli DNA.