Embed Size (px)
Citation preview

Doktori (Ph.D.) értekezés
Fehér Balázs
Biológia Doktori Iskola
Szeged
2011

Az UV-B fény és a növényi cirkadián óra
kapcsolatának vizsgálata
Doktori (Ph.D.) értekezés
Készítette: Fehér Balázs
Témavezető: Dr. Nagy Ferenc
Magyar Tudományos Akadémia
Szegedi Biológiai Kutatóközpont
Növénybiológiai Intézet
Foto- és Kronobiológiai Csoport
Szegedi Tudományegyetem
Természettudományi és Informatikai Kar
Biológia Doktori Iskola
Szeged
2011

TARTALOMJEGYZÉK II _________________________________________________________________________________________________
TARTALOMJEGYZÉK
RÖVIDÍTÉSEK JEGYZÉKE ......................................................................................................................................2
BEVEZETÉS................................................................................................................................................................3
SZAKIRODALMI ÁTTEKINTÉS ............................................................................................................................5
Ritmikus folyamatok ......................................................................................................................................................... .5
Cirkadián ritmusok ............................................................................................................................................................ .5
Alapismeretek, alapfogalmak .............................................................................................................................. 5
Mérési módszerek ..................................................................................................................................................... 8
A cirkadián óra................................................................................................................................................................... 10
A központi oszcillátor felépítése .......................................................................................................................11
Az óra kapcsolata a külvilággal (bemenet, input).....................................................................................11
Az óra kimeneti elemei által szabályozott folyamatok (output) .........................................................12
Az Arabidopsis cirkadián órájának modellje ......................................................................................................... 12
A központi óramű felépítése ...............................................................................................................................12
A bemeneti oldal elemei .......................................................................................................................................16
UV-B érzékelés és jelátvitel .......................................................................................................................................... 23
UV-B: stresszfaktor és környezeti jel ...............................................................................................................23
Az UV-B fotomorfogenikus jellemzői...............................................................................................................24
UV-B fotoreceptor? .................................................................................................................................................25
A COP1 új funkciója ................................................................................................................................................27
Az UVR8 jellemzői ...................................................................................................................................................28
UVR8 a lehetséges UV-B fotoreceptor .............................................................................................................30
CÉLKITŰZÉSEK...................................................................................................................................................... 32
ANYAGOK ÉS MÓDSZEREK................................................................................................................................ 33
Kísérleti anyagok és élőlények.................................................................................................................................... 33
Tápoldatok, táptalajok, antibiotikumok, szelekció ...................................................................................33
Baktériumok .............................................................................................................................................................34
Növények ....................................................................................................................................................................34
Molekuláris biológiai módszerek............................................................................................................................... 35
Riportergén konstrukciók készítése, transzgénikus növények előállítása .......................................35
Agrobaktérium konjugáció .................................................................................................................................37
Növényi DNS tisztítása..........................................................................................................................................38

TARTALOMJEGYZÉK II _________________________________________________________________________________________________
Növényi RNS tisztítása..........................................................................................................................................38
mRNS szint meghatározása RT-PCR-rel ........................................................................................................38
Növényi összfehérje tisztítása ............................................................................................................................40
Fehérjeszint meghatározása Western-blot módszerrel ..........................................................................40
Növényeken alkalmazott technikák ......................................................................................................................... 41
Magsterilizálás, növénynevelés .........................................................................................................................41
Fénykezelések ...........................................................................................................................................................41
Alkalmazott UV szűrők .........................................................................................................................................42
Transzgénkus növények előállítása .................................................................................................................43
In vivo lumineszcencia mérés luminométerrel............................................................................................43
Periódus adatok számolása ................................................................................................................................44
A fázis igazíthatóságának mérése (PRC).......................................................................................................44
A kapuzás (gating) vizsgálata ...........................................................................................................................45
UV-B tolerancia vizsgálata .................................................................................................................................45
Klorofilltartalom meghatározása ....................................................................................................................46
EREDMÉNYEK........................................................................................................................................................ 47
A cirkadián óra beállíthatósága UV-B fénnyel...................................................................................................... 47
Az UV-B befolyásolja a cirkadián óra sebességét .......................................................................................47
UV-B pulzusokkal a cirkadián óra késleltetése idézhető elő..................................................................49
UV-B kiváltotta fényindukció cirkadián szabályozása ..................................................................................... 51
Optimális indukciós és mintaszedési körülmények megállapítása .....................................................51
Az UV-B képes indukálni a központi óraelemeket .....................................................................................53
A cirkadián óra a nem ritmikus gének UV-B indukcióját is kapuzza .................................................56
A HY5 és a HYH nem szükségesek sem az óragének UV-B indukciójához, sem az UV-B
indukálta génexpresszió kapuzásához .......................................................................................................... 61
Aritmiás mutánsokban hiányzik az UV-B indukált génkifejeződés kapuzása................................63
Az óra által szabályozott UV-B génindukció fiziológiai hatása ...........................................................67
AZ EREDMÉNYEK ÉRTELMEZÉSE ................................................................................................................... 69
Az UV-B szerepet játszik a növényi cirkadián óra beállításában................................................................. 69
Az UV-B indukció ritmikus szabályozása ............................................................................................................... 71
Az óramodulált UV-B válaszok fiziológiai jelentősége ..................................................................................... 73
KÖVETKEZTETÉSEK............................................................................................................................................ 75
SAJÁT KÖZLEMÉNYEK ........................................................................................................................................ 77
IDÉZETT KÖZLEMÉNYEK................................................................................................................................... 78
ÖSSZEFOGLALÁS................................................................................................................................................... 86
SUMMARY .............................................................................................................................................................. 90
KÖSZÖNETNYILVÁNÍTÁS .................................................................................................................................. 99

RÖVIDÍTÉSEK JEGYZÉKE
AhR Aril-hidrokarbon receptor AM Arabidopsis Medium (növényi táptalaj) BCIP 5-bróm-4-klór-3-indolil foszfát bHLH Bázikus Helix-Loop-Helix fehérjeelem BiFC Bimolecular Fluorescent Complementation BRASS Biological Rhythms Analysis Software System BSA Bovine serum albumin CBS CCA1 binding site A CCA1 transzkripciós faktor által felismert szekvenciamotívum CCD Charge coupled device Col-0 Arabidopsis thaliana (lúdfű), Columbia természetes ökotípusa CPD Ciklobután pirimidin dimer cps Counts per second (másodpercenként beérkező fotonok száma) CTAB Cetil-trimetil-ammónium bromid EDTA Etilén-diamin-tetraacetát EE Esti kifejeződésű promóterekben feldúsult szekvenciamotívum FFT-NLLS Fast Fourier Transformation Non Linear Least Squares FICZ 6-formilindol[3,2-b]carbazol FMN Flavin-mononukleotid FRC Fluence rate curve (egy jelleg függése a fényerősségtől) FRET Fluorescent Resonance Energy Transfer GEF Guanine nucleotide exchange factor IPTG Izopropil-β-D-galakto-piranozid LB Luria-Bertani baktérium táptalaj LOV (light/oxigen/voltage) fehérjeelem MS Murashige-Skoog növényi táptalaj NBT Nitroblue tetrazólium NES Nuclear export signal NLS Nuclear localization signal PAS (Per/Arnt/Sim) fehérjeelem Pfr A fitokróm távoli vörös fényt elnyelő alakja pPCV Plant Cloning Vector (növényi kétgazdás plazmid) Pr A fitokróm vörös fényt elnyelő alakja PRC Phase response curve (az óra beállítható-ságát az idő függvényében leíró görbe) SCF SKP-Cullin-F-box SDS Nátrium-dodecil-szulfát SEM Standard error of the means (a középértékek standard hibája) TBS TCP transzkripciós faktorok által felismert szekvenciamotívum TBST Tris-Buffered Saline Tween-20 TCP (TB1, CYC, PCFs) fehérjeelem Tris·HCl Trisz-(hidroximetil)-amino-metán hidroklorid sója UV-A/B Ibolyántúli-A/B sugárzás Ws Arabidopsis thaliana (lúdfű), Wassilewskija természetes változata X-gal 5-bróm-4-klór-3-indolil-β-D-galakto-piranozid YEB Yeast Beef (Agrobacterium táptalaj)

RÖVIDÍTÉSEK JEGYZÉKE 2 __________________________________________________________________________________________
Gén szimbólumok
ARR ARABIDOPSIS RESPONSE REGULATOR
ASK ARABIDOPSIS SKP
β-TUB β-TUBULIN
BRI1 BRASSINOSTEROID INSENSITIVE1
CAB2 CHLOROPHYLL A/B-BINDING 2
CCA1 CIRCADIAN CLOCK ASSOCIATED 1
CCR2 COLD CIRCADIAN CLOCK REGULATED 2
CDF1 CYCLING DOF-FACTOR 1
CFP CYAN FLUORESCENT PROTEIN
CHE CCA1 HIKING EXPEDITION
CHS CHALCONE SYNTHASE
CO CONSTANS
COP1 CONSTITUTIVE PHOTOMORPHOGENIC 1
CRY CRYPTOCHROME = kriptokróm
CUL1 CULLIN 1
DET1 DE-ETIOLATED 1
ELF EARLY FLOWERING
ELIP EARLY LIGHT INDUCIBLE PROTEIN
ETR1 ETHYLENE RESPONSE 1
FAH1 FERULIC ACID 5-HYDROXILASE 1
FHL FH1-LIKE
FHY FAR RED ELONGATED HYPOCOTYL
FKF1 FLAVIN-BINDING, KELCH-REPEAT, F-BOX 1
FLC FLOWERING LOCUS C
FT FLORAL TRANSITION
GFP GREEN FLUORESCENT PROTEIN
GI GIGANTEA
GPX7 GLUTATION PEROXIDASE 7
HFR1 LONG HYPOCOTYL IN FAR-RED
HPT HYGROMYCIN PHOSPHOTRANSFERASE
HY5 LONG HYPOCOTYL 5
HYH HY5 HOMOLOGUE
JAR1 JASMONATE RESISTANT1
LAF1 LONG AFTER FAR-RED LIGHT 1 LHY LATE ELONGATED HYPOCOTYL
LKP2 LOV KELCH PROTEIN 2
LUC Photinus pyralis (szentjánosbogár) luciferáz LUX LUX ARRHYTHMO
PHOT PHOTOTROPIN = fototropin
PHY PHYTOCHROME = fitokróm
PIF PHYTOCHROME-INTERACTING FACTOR
PRR PSEUDO RESPONSE REGULATOR
RAN RAS RELATED NUCLEAR PROTEIN
RCC1 REGULATOR OF CHROMOSOME CONDENSATION 1
SRR1 SENSITIVITY TO RED LIGHT REDUCED 1
TIC TIME FOR COFFEE
TOC1 TIMING OF CAB 1
TT TRANSPARENT TESTA
UVH1 ULTRAVIOLET HYPERSENSITIVE 1
UVR UV RESISTANCE LOCUS
UVS ULTRAVIOLET SENSITIVE
WC WHITE COLLAR
YFP YELLOW FLUORESCENT PROTEIN
ZTL ZEITLUPE

BEVEZETÉS
A cirkadián órák az élővilág egészében elterjedt biokémiai időmérők. Működésükkel
nagymértékben segítik az élőlényeket a Föld tengely körüli forgása következtében
kialakult, ritmikusan változó környezethez való alkalmazkodásban. Különösen fontos ez
az alkalmazkodás a növények számára, hiszen egyfelől, mint helyhez kötött élőlények
különösen kiszolgáltatottak a kedvezőtlen tényezőknek; másfelől, a fotoszintézishez
szükséges energia minél hatákonyabb felhasználásához szükséges, hogy az
életfolyamataikat a fény napi eloszlásának megfelelően időzítsék. A cirkadián óra
segítségével képesek ezeket a folyamatokat a megfelelő időpontokra időzíteni, illetve a
kedvezőtlenekben gátolni. Ennek az „órarendnek” segítségével energiát takarítanak meg
és hatákonyabbá válnak (Dodd és mtsai, 2005).
A cirkadián óra hatékonysága függ attól, hogy mennyire van szinkronban az óra által
mért belső idő a külső környezet valós idejével. A szubjektív idő pontosításának
érdekében a cirkadián órát a külső környezet periodikus változásai – melyek közül
növényekben a fény a legfontosabb - képesek beállítani. A fény így kettős szerepet
játszik a növények életében, egyrészt a fotoszintézisen keresztül a fő energiaforrás,
másrészt a környezet állapotáról információt hordozó jel. A növények a fényt speciális
fehérjék, úgynevezett fotoreceptorok segítségével fogják fel. Az impulzus a jelátviteli
láncon keresztül eléri a cirkadián órát, ahol megváltoztatja a szintjét és/vagy aktivitását
egy vagy több órakomponensnek, ezáltal másik fázisba állítva azt (Kozma-Bognár és
Káldi, 2008). A vörös/távoli vörös fényelnyelésű fitokrómok, illetve a kék fényelnyelésű
kriptokrómok szerepe a cirkadián óra fotoreceptoraként már bizonyított Arabidopsisban
(Somers és mtsai, 1998a; Devlin és Kay, 2000).
Az ultraibolya-B (UV-B, 280-315 nm) sugárzás természetes része a Föld felszínét
elérő napfénynek. A felszínt elérő UV-B sugárzás igen változatos fiziológiai folyamatokat
vált ki a növényekből. A legismertebbek azok, amiket összefoglalóan stresszhatásoknak
nevezünk (DNS-, és fehérjekárosodás, reaktív oxigéngyökök felhalmozódása, lipid

BEVEZETÉS 4 __________________________________________________________________________________________
peroxidáció) (Jansen és mtsai, 1998). Azonban van egy másik, kevésbé ismert hatása is,
az alacsony intenzitású UV-B fotomorfogenikus hatással is rendelkezik (gátolja a
hipokotil megnyúlását, serkenti a sziklevél növekedését, illetve a flavonoid
felhalmozódást (Ulm és mtsai, 2004; Brown és mtsai, 2005; Oravecz és mtsai, 2006;
Stracke és mtsai, 2010). Vagyis a növények ugyanúgy környezeti jelként érzékelik ezt a
sugárzást, mint a fényspektrum látható részét.
Munkánk során az alacsony intenzitású UV-B kapcsolatát vizsgáltuk a cirkadián
órával. Eddig semilyen adat nem volt arról, hogy a fénynek ez a természetes
komponense befolyásolja-e a cirkadián óra működését, illetve, hogy a cirkadián óra
szabályozza-e az UV-B által kiváltott élettani folyamatokat.

SZAKIRODALMI ÁTTEKINTÉS
Ritmikus folyamatok
Az élővilágban általánosan megfigyelhetőek olyan folyamatok, melyek időben
ismétlődnek, ritmikusak. Ezeket az élő rendszerekben megjelenő periodikus
folyamatokat csoportosíthatjuk az ismétlődésük gyakorisága szerint. Ultradián
ritmusoknak nevezzük a latin ultra diem (egy napon túli) kifejezés alapján azokat,
melyek naponta többször ismétlődnek, Ilyenek például az agyhullámok
(tizedmásodperc), a szívverés (~ egy másodperc), a légzés, a madárdal, a tücsökciripelés
(néhány másodperc), az ember alvásszakaszainak ismétlődése (90 perc), vagy a
csíranövények hipokotiljának köröző mozgásában egy kör megtétele (néhány óra). Más
folyamatok sokkal ritkábban, csak több nap alatt ismétlődnek. Ilyenek például a
gerincesek menstruációs ciklusai (rágcsálók: 3-5 nap, ember: 25-35 nap), vagy azok a
leghosszabb biológiai ritmusok, melyek évente csak egyszer ismétlődnek, mint a
vándormadarak költözése, vagy a téli álmot alvó állatok nyugalmi időszaka. Ezeket
infradián ritmusoknak nevezzük (latin infra diem, egy nap alatti). A kettő között
találhatóak azok a folyamatok, melyek megközelítőleg naponta ismétlődnek, amelyeket
ezért cirkadián ritmusoknak hívunk (latin circa diem egy nap körüli). A cirkadián
ritmusok nagyon elterjedtek, szinte minden a fény érzékelésére képes élőlényben
megfigyelhetők.
Cirkadián ritmusok
Alapismeretek, alapfogalmak
Összefoglalóan napszakos ritmusról beszélünk, ha egy jelleg természetes viszonyok
között 24 órás gyakorisággal ismétlődik. A cirkadián ritmusoknak azonban van néhány

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
6
olyan jellemvonása, ami megkülönbözteti őket az egyszerű, a környezeti jelek által
vezérelt napszakos ritmusoktól.
A cirkadián ritmusok nemcsak ciklusosan változó körülmények (pl. fény/sötét vagy
hőmérséklet ciklusok) hanem állandó körülmények között is több cikluson keresztül
fennmaradnak. Ilyenkor kialakul a cirkadián ritmusokra jellemző saját periódus, mely
közelítőleg, bár soha nem pontosan 24 óra. Az ilyen ritmust szabadon futó ritmusnak
nevezzük.
A cirkadián ritmusok ugyan külső környezeti jelek nélkül is fennmaradnak, de
azokkal beállíthatóak. A beállítás során a ritmus belső időzítése (szubjektív idő) a külső
környezet idejéhez (objektív idő) igazodik. A cirkadián órát beállító két legfontosabb
tényező a fény/sötét átmenet, illetve a meleg/hideg szakaszok periodikus váltakozása.
Az óra érzékenysége a beállító jelekre azonban nem egyforma a nap folyamán, hanem
hullámzó mintázatot mutat. A belső óra ugyanis a bemeneti elemek cirkadián
szabályzásával bizonyos napszakokra korlátozza a saját beállíthatóságát. Ezt a jelenséget
hívjuk kapuzásnak (gating).
A cirkadián ritmusok hőmérséklet-kompenzáltak, azaz képesek ellensúlyozni a
hőmérséklet kémiai reakciókat befolyásoló hatását. A biokémiai folyamatok
sebességének hőmérsékletfüggését az ún. Q10 érték mutatja. Ez azt fejezi ki, hogy egy
adott folyamat sebessége hányszorosára növekszik, ha a hőmérsékletet 10 ⁰C-al emeljük.
Az átlagos biokémiai folyamatok Q10 értéke 2-4 közötti, a cirkadián periódusoké
azonban csak 0,8-1,2. Ennek köszönhetően a szabadon futó ritmusuk tág hőmérsékleti
határok között is csak kis mértékben változik.
A természetben ugyan soha nincsenek állandó fény- vagy hőmérsékleti viszonyok,
mégis a kutatásban sokszor alkalmazzuk ezt a kísérleti elrendezést. Ilyen körülmények
között ugyanis a környezet hatása elhanyagolható, a ritmus jellegét kizárólag a cirkadián
rendszer belső törvényszerűségei alakítják. Ebben az egyszerűsített vizsgálati
rendszerben a szabadon futó ritmusok tanulmányozásával könnyebben megérthetjük a
cirkadián szabályozásban résztvevő mechanizmusokat.
A cirkadián ritmusokat az alábbi fogalmakkal tudjuk leírni, amelyeket
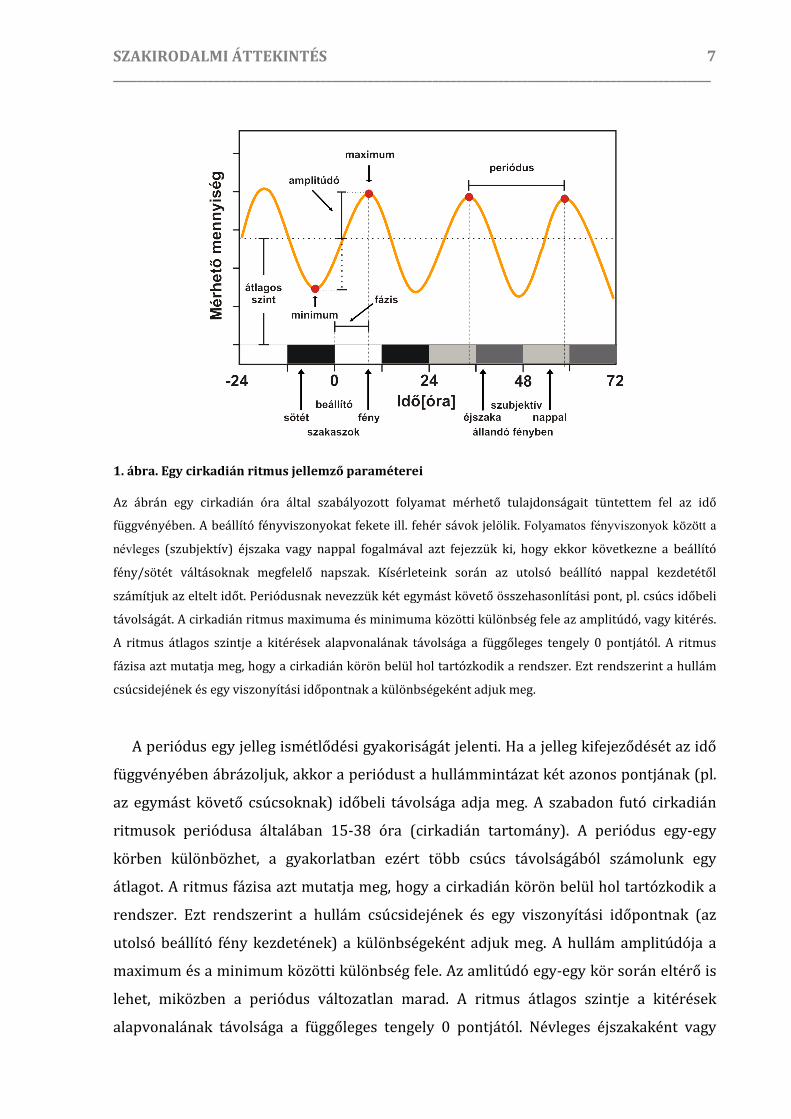
legegyszerűbben egy vázlatos ábrán lehet szemléltetni (1. ábra).
SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
7
1. ábra. Egy cirkadián ritmus jellemző paraméterei
Az ábrán egy cirkadián óra által szabályozott folyamat mérhető tulajdonságait tüntettem fel az idő
függvényében. A beállító fényviszonyokat fekete ill. fehér sávok jelölik. Folyamatos fényviszonyok között a
névleges (szubjektív) éjszaka vagy nappal fogalmával azt fejezzük ki, hogy ekkor következne a beállító
fény/sötét váltásoknak megfelelő napszak. Kísérleteink során az utolsó beállító nappal kezdetétől
számítjuk az eltelt időt. Periódusnak nevezzük két egymást követő összehasonlítási pont, pl. csúcs időbeli
távolságát. A cirkadián ritmus maximuma és minimuma közötti különbség fele az amplitúdó, vagy kitérés.
A ritmus átlagos szintje a kitérések alapvonalának távolsága a függőleges tengely 0 pontjától. A ritmus
fázisa azt mutatja meg, hogy a cirkadián körön belül hol tartózkodik a rendszer. Ezt rendszerint a hullám
csúcsidejének és egy viszonyítási időpontnak a különbségeként adjuk meg.
A periódus egy jelleg ismétlődési gyakoriságát jelenti. Ha a jelleg kifejeződését az idő
függvényében ábrázoljuk, akkor a periódust a hullámmintázat két azonos pontjának (pl.
az egymást követő csúcsoknak) időbeli távolsága adja meg. A szabadon futó cirkadián
ritmusok periódusa általában 15-38 óra (cirkadián tartomány). A periódus egy-egy
körben különbözhet, a gyakorlatban ezért több csúcs távolságából számolunk egy
átlagot. A ritmus fázisa azt mutatja meg, hogy a cirkadián körön belül hol tartózkodik a
rendszer. Ezt rendszerint a hullám csúcsidejének és egy viszonyítási időpontnak (az
utolsó beállító fény kezdetének) a különbségeként adjuk meg. A hullám amplitúdója a
maximum és a minimum közötti különbség fele. Az amlitúdó egy-egy kör során eltérő is
lehet, miközben a periódus változatlan marad. A ritmus átlagos szintje a kitérések
alapvonalának távolsága a függőleges tengely 0 pontjától. Névleges éjszakaként vagy

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
8
nappalként tüntetjük fel folyamatos körülmények alatt a beállító fény/sötét váltásoknak
megfelelő időszakokat. Ezzel azt fejezzük ki, hogy ekkor következne a megfelelő
napszak. Kísérleteink során az utolsó beállító nappal kezdetétől számítjuk az eltelt időt.
Mérési módszerek
A szabadon futó körülmények között mérhető periódus a legtöbb élőlényben függ az
alkalmazott fény erősségétől (Aschoff, 1979). Növényekben ez a viszony fordított: minél
erősebb a fény, annál rövidebb a periódus. A periódus fényerősségtől való függését úgy
vizsgáljuk, hogy az azonos módon beállított növényeket eltérő erősségű állandó fénybe
helyezzük néhány napra, majd a kialakuló periódust az alkalmazott fényerősség
függvényében ábrázoljuk. Az így kialakuló fényerősség-függés görbe (FRC, Fluence Rate
Curve) segítségével szerezhetünk ismereteket a különböző hullámhosszú és erősségű
fényben működő fényérzékelő rendszerek érintettségéről.
A cirkadián órát be lehet állítani egyetlen fényvillanással is. Ez lényegében azt jelenti,
hogy a központi óra valamely kulcselemének működését a fény közvetlenül vagy
közvetve befolyásolja, például ideiglenesen megemeli annak szintjét. Az óra fázisától
függően ez késést vagy sietést okozhat. Ez a kettős válasz az alapja a beállíthatóságnak
(2. ábra, A mező), amelynek során a túl gyorsan vagy túl lassan futó belső óra a külső
környezet idejéhez igazodik.
Korábban már említettem, hogy az óra saját beállíthatóságát cirkadián szabályozás
alatt tartja. Ennek köszönhetően az óra a nap nagy részében szinte teljesen érzéketlen a
beállító fényjelekre, míg a legnagyobb fáziscsúszásokat a névleges éjszaka alatt lehet
kiváltani. A természetben ilyenkor nincs fény, ezért valójában az óra a napfelkelte
és/vagy a naplemente idején a legérzékenyebb a beállításra. A természetben e két
időpontban változik legnagyobb mértékben a fény erőssége (és színösszetétele), ezért ez
a két legbiztosabb pont a nap folyamán, amelyekhez igazodni lehet. Az egyéb
időpontokban fellépő ideiglenes, nem kiszámíthatóan bekövetkező környezeti
változások (pl. egy felhő eltakarja a napot) cirkadián szempontból nem hasznosak, így
érthető, hogy az óra a nappal során érzéketlen a beállító jelekre.
Ha a szabadon futó óra fázisát különböző időpontokban adott, azonos erősségű és
időtartamú fénypulzussal állítjuk be, és a tapasztalt fáziscsúszást az adott fényvillanás
idejének függvényében ábrázoljuk, megkapjuk az ún. fázisválasz görbét (PRC, Phase
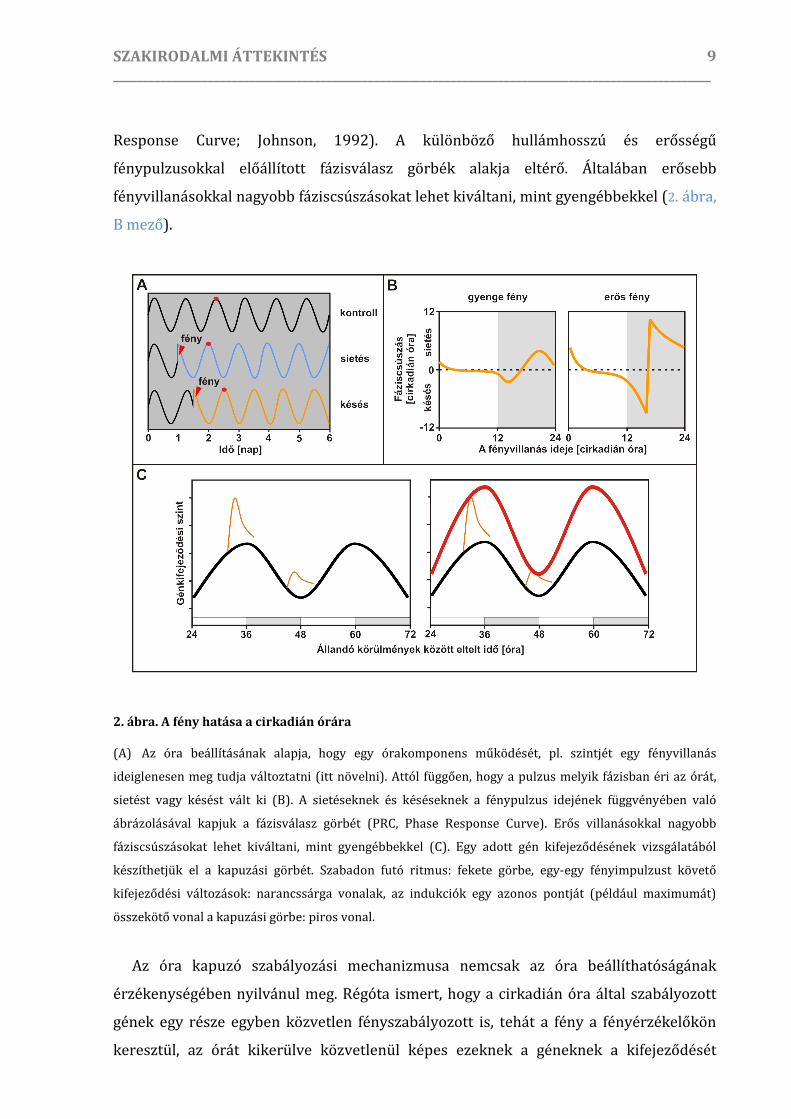
SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
9
Response Curve; Johnson, 1992). A különböző hullámhosszú és erősségű
fénypulzusokkal előállított fázisválasz görbék alakja eltérő. Általában erősebb
fényvillanásokkal nagyobb fáziscsúszásokat lehet kiváltani, mint gyengébbekkel (2. ábra,
B mező).
2. ábra. A fény hatása a cirkadián órára
(A) Az óra beállításának alapja, hogy egy órakomponens működését, pl. szintjét egy fényvillanás
ideiglenesen meg tudja változtatni (itt növelni). Attól függően, hogy a pulzus melyik fázisban éri az órát,
sietést vagy késést vált ki (B). A sietéseknek és késéseknek a fénypulzus idejének függvényében való
ábrázolásával kapjuk a fázisválasz görbét (PRC, Phase Response Curve). Erős villanásokkal nagyobb
fáziscsúszásokat lehet kiváltani, mint gyengébbekkel (C). Egy adott gén kifejeződésének vizsgálatából
készíthetjük el a kapuzási görbét. Szabadon futó ritmus: fekete görbe, egy-egy fényimpulzust követő
kifejeződési változások: narancssárga vonalak, az indukciók egy azonos pontját (például maximumát)
összekötő vonal a kapuzási görbe: piros vonal.
Az óra kapuzó szabályozási mechanizmusa nemcsak az óra beállíthatóságának
érzékenységében nyilvánul meg. Régóta ismert, hogy a cirkadián óra által szabályozott
gének egy része egyben közvetlen fényszabályozott is, tehát a fény a fényérzékelőkön
keresztül, az órát kikerülve közvetlenül képes ezeknek a géneknek a kifejeződését

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
10
szabályozni. A fénynek ezt a gyors, közvetlen hatását akut válasznak nevezzük. A
cirkadián óra a kapuzási mechanizmus révén képes a fázisának megfelelően gátolni a
fényre adott akut válasz erősségét, ami az óra fázisának megfelelően csökkenti, vagy
növeli az adott folyamat fényérzékenységét. A gátló hatás tehát maga is cirkadián
ritmust mutat. A szabadon futó periódusú növényt különböző időpontokban azonos
erősségű és időtartamú fényfelpulzusokkal kezelve, és a kezelés után egy-egy ilyen
kettős szabályzású gén megemelkedett szintjeit az idő függvényében ábrázolva kapjuk
az adott gén kapuzási görbéjét (2. ábra, C panel).
A cirkadián óra
Cirkadián órának nevezzük azt a belső időmérő mechanizmust, amely a külső
környezettől függetlenül is képes a cirkadián ritmusok létrehozására, fenntartására és
irányítására. A cirkadián óra három fő részből épül fel: főrezgőkör, vagy központi
oszcillátor, bemenet (input) és kimenet (output). Vázlatos felépítését a 3. ábra
szemlélteti.
3. ábra. A cirkadián óra vázlatos felépítése
Működés szempontjából a cirkadián órát három egységre bonthatjuk. A főrezgőkör, amely a környezettől
függetlenül képes ismétlődő mintázat létrehozására, legalább egy serkentő és egy gátló folyamatból álló
negatív visszacsatoláson alapul. A hullámmintázat létrejöttének feltétele a visszacsatolás késleltetése. A
főrezgőkör ugyan képes a környezettől függetlenül is működni, de az alkalmazkodás szempontjából az is
fontos, hogy a belső ritmus a külső, természetes ritmusra hangolódjék. A naponkénti órabeállítást a
környezeti jeleket érzékelő molekulák, és a hozzájuk kapcsolódó jelátviteli elemek főrezgőkörhöz való
csatolása teszi lehetővé (bemenet). Az órát sokféle környezeti jellel be lehet állítani, de a két
legjelentősebb a fény és a hőmérséklet. A létrehozott ismétlődő mintázat a kimeneti jelátviteli utakon
keresztül nyilvánul meg a különféle élettani folyamatokban, amelyek a génkifejeződéstől kezdve a
szervmozgásokig minden szinten megfigyelhetők.

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
11
A központi oszcillátor felépítése
A központi oszcillátor egy olyan mechanizmus, amely a külső környezettől
függetlenül képes a ritmus létrehozására és fenntartására. Az oszcillátor elemei (az ún.
óragének ill. órafehérjék) negatív visszacsatolás útján szabályozzák önmaguk és egymás
kifejeződését, működését. A negatív visszacsatolás lényege, hogy egy elem több lépcsőn
keresztül végül a saját működésének gátlását idézi elő. A cirkadián órák működésének és
a 24 órás periódus kialakításának alapvető feltétele a késleltetés, amelynek révén az
egyes órafehérjék mennyisége csak a megfelelő óragén indukciója után több órával éri el
a maximumát. Ennek hiányában a szabályozó ciklus jóval rövidebb idő alatt futna körbe,
mint egy nap.
Hagyományosan azokat az elemeket tekintjük a főrezgőkör részeinek, amelyek
törlése vagy folyamatos működése minden megfigyelhető ritmus megszűnéséhez vezet,
szintjük vagy működésük cirkadián módon hullámzik, továbbá amelyek szintje vagy
működése a fény és egyéb beállítójelek által pillanatszerűen megváltoztatható, és ez
egyben minden megfigyelhető ritmus fázisának átállítódásához is vezet.
Az óra kapcsolata a külvilággal (bemenet, input)
A cirkadián óra létezésére a napszakos változások előrejelzése miatt van szükség. A
belső ritmus külsőhöz való igazítását a környezeti jeleket érzékelő jelátviteli rendszerek,
és a központi oszcillátor kapcsolata teszi lehetővé. Ezt az összeköttetést biztosító
jelátviteli rendszert hívjuk a cirkadián óra bemenetének. A legfontosabb beállító jel a
fény és a hőmérséklet, amelyek meghatározott pontoknál befolyásolják az óraelemek
működését.
Számos esetben kimutatták, hogy saját bemeneti elemeinek működését a cirkadián
óra szabályozza. Modellek tanúsága szerint ez a fajta szabályozás erőteljesebbé teszi a
rezgést (Roenneberg és Merrow, 1998), ezen kívül pedig biztosítja az óra
beállíthatóságának bizonyos napszakra való korlátozását (kapuzás).
Azokat a komponenseket tekintjük a cirkadián óra bemeneti elemeinek, amelyek
elrontása csak az óra működésének környezettől (pl. fénytől) való függését változtatja
meg, de nem szünteti meg a ritmust. Így például megváltozhat a periódus, a hullám
tartóssága (lecsengés), de a ritmus nem szűnik meg.

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
12
Az óra kimeneti elemei által szabályozott folyamatok (output)
Azokat a jelátviteli utakat, amelyeken keresztül az óra a különféle életfolyamatokat
vezérli az óra kimeneteinek nevezzük. Cirkadián szabályozás a génkifejeződéstől a
viselkedésig minden szinten megfigyelhető. Cianobaktériumokban az összes promóter
működése cirkadián szabályozás alatt áll (Liu és mtsai, 1995), és növényekben is a gének
89%-ának kifejeződése változik valamilyen napszakos mintázat szerint (Michael és
mtsai, 2008). A különböző gének kifejeződési csúcsai (fázisai) ugyan lefedik az egész
napot, mégis a hasonló szerepet betöltő gének időzítése ugyanazon napszakra,
legtöbbjük vagy a napfelkelte vagy a naplemente környékére esik (Harmer és mtsai,
2000; Michael és mtsai, 2008). A génkifejeződéshez hasonlóan az összes többi cirkadián-
szabályozott folyamat is a számára legmegfelelőbb napszakra időzítődik. A központi
oszcillátor értelemszerűen csak egyféle fázissal rezeg, az ettől eltérő fázisokat módosító
kimeneti elemek közbeiktatásával lehet létrehozni (fázisátalakítók). Az eltérő
hullámformák kialakulásáért is módosító kimeneti elemek felelősek.
Azokat az elemeket tekintjük az óra kimenetének, amelyek elrontása nem szünteti
meg a ritmusképzést, és nem változtatja meg a periódust sem, hanem csak egyes
ritmusok kifejeződésére van hatással.
Az Arabidopsis cirkadián órájának modellje
A központi óramű felépítése
A közelmúltban végzett kísérletek és a matematikai modellek szerint az Arabidopsis
központi oszcillátora három – óragének és termékeik által alkotott – összekapcsolt,
negatív visszacsatoláson alapuló szabályozóhurokból épül fel (4. ábra). Az elsőként
azonosított hurok három összetevőből áll. A TIMING OF CAB 1 (TOC1), vagy más néven
PSEUDO RESPONSE REGULATOR 1 (PRR1) egy esti kifejeződésű, cirkadián-szabályozott
gén (Somers és mtsai, 1998b). A sejtmagban elhelyezkedő TOC1 fehérje közvetett
módon serkenti a két reggeli MYB-szerű transzkripciós faktor, a CIRCADIAN CLOCK
ASSOCIATED 1 (CCA1) és a LATE ELONGATED HYPOCOTYL (LHY) génjének kifejeződését
(Alabadi és mtsai, 2001). A CCA1 és LHY olyan szokatlan felépítésű, MYB-elem tartalmú
DNS-kötő fehérjék, amelyek a DNS-en aszimmetrikus nukleotidsorrendet, az ún. esti
elemet (EE), (AAATATCT) ismerik fel (Harmer és mtsai, 2000). Az mRNS-ek és fehérjék

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
13
cirkadián módon hullámzanak, reggeli csúcsokkal. Kifejeződésük fénnyel serkenhető.
Szerepük részben átfed: a két gén bármelyikének túltermelése minden vizsgált ritmus
megszűnését (Schaffer és mtsai, 1998; Wang és Tobin, 1998), hiánya a ritmusok
periódusrövidülését okozza (Green és Tobin, 1999; Mizoguchi és mtsai, 2002); együttes
hiányuk csaknem teljes ritmustalansággal jár (Ding és mtsai, 2007a). Közvetlenül
kötődnek a TOC1 promóteréhez és gátolják annak működését (Alabadi és mtsai, 2001).
Ennek a kölcsönös szabályozásnak köszönhetően a TOC1 és a CCA1/LHY gének cirkadián
kifejeződése ellentétes fázisú. A matematikai modell egy ismeretlen X elem jelenlétét is
feltételezi, amely a TOC1 fehérje és a CCA1/LHY gének között helyezkedik el, feladata az
utóbbi gének serkentése. Ez a lépés magyarázná a jelentős időbeni eltérést a TOC1
fehérje maximális mennyisége, és a CCA1/LHY gének maximális átíródása között (Locke
és mtsai, 2005). Elképzelhető, hogy ezt a szerepkört az EARLY FLOWERING 3 (ELF3)
és/vagy az ELF4 molekulák töltik be (Locke és mtsai, 2005), mivel hiányukban a
CCA1/LHY kifejeződése drámaian lecsökken (Doyle és mtsai, 2002; Kikis és mtsai, 2005).
Az ELF3 és ELF4 gének kifejeződése cirkadián mintázatú, az ELF4-é ráadásul
fényérzékeny is. A fehérjék sejtmagiak, és nem tartalmaznak egyetlen ismert
fehérjeelemet sem.
Az újabb kutatások szerint a CCA1 HIKING EXPEDITION (CHE) fehérje lehet egy
összekötő elem a CCA1 és a TOC1 között. A CHE egy TCP transzkripciós faktor ami in
vivo képes kötődni a CCA1 promóterében lévő TBS (TCP binding site) szekvenciához
(GGNCCCAC). A CHE fehérje gátolja a CCA1 átíródását, míg a CCA1/LHY fehérjék a CHE
promóterében lévő CBS (CCA1 binding site) elemhez (AAAAATCT) kötődve gátolják a
CHE átíródását, ezzel egy kis szabályzókört alkotva. A CHE in vivo képes kölcsönhatásba
lépni a TOC1 fehérjével, így azt a CCA1 promóter egy adott területére irányítani
(Pruneda-Paz és mtsai, 2009).
Az eredeti egyhurkos modell azonban olyan fontos kísérleti eredményekre nem
tudott magyarázatot adni mint az, hogy a cca1 lhy dupla mutáns nem veszti el teljesen a
ritmikusságát. A továbbfejlesztett modellben már egy esti időzítésű hurok is szerepel.
Ennek lényeges összetevője egy ismeretlen Y elem, amelynek szerepe a TOC1 gén
kifejeződésének serkentése. Az Y gént a modell alapján mind a TOC1, mind pedig a
CCA1/LHY gátolja (Locke és mtsai, 2005). Egyes adatok szerint az Y tényező egy részéért
a GIGANTEA (GI) lehet a felelős (Locke és mtsai, 2006; Locke és mtsai, 2005), mások

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
14
azonban csak áttételes szerepet tulajdonítanak a GI fehérjének ebben a tekintetben (Kim
és mtsai, 2007; Martin-Tryon és mtsai, 2007; Sawa és mtsai, 2007). A GI (Fowler és
mtsai, 1999; Park és mtsai, 1999) egy nagy méretű sejtmagi fehérje (Huq és mtsai,
2000), amely nem tartalmaz ismert fehérjeelemet. A GI mRNS és fehérje mennyisége
cirkadián módon hullámzik, kora esti csúcsokkal (David és mtsai, 2006). A gi növényben
az LHY és CCA1 gének kifejeződése erősen csökkent, ami arra utal, hogy a GI a megfelelő
LHY és CCA1 szint kialakításában játszik szerepet.
4. ábra. A növényi cirkadián óra modellje
A jelenlegi elképzelés szerint a központi rezgőkör három egymáshoz kapcsolt, negatív visszacsatoláson
alapuló szabályozó hurokból épül fel. A „reggeli” hurkot a CIRCADIAN CLOCK REGULATED 1 (CCA1) és a
LATE ELONGATED HYPOCOTYL (LHY), valamint a PSEUDO RESPONSE REGULATOR 7 és 9 (PRR7,9)
fehérjék és génjeik alkotják. A CCA1/LHY fehérjék közvetlenül serkentik a PRR7/9 gének átíródását, míg a
PRR-ek közvetett módon gátolják a CCA1/LHY géneket. Az „esti” hurok tagjai a PRR-családba tartozó
TIMING OF CAB 1 (TOC1) és a GIGANTEA (GI). Az GI ismeretlen módon serkenti a TOC1 gén kifejeződését,
míg a TOC1 fehérje szintén ismeretlen módon gátolja a GI gént. A két hurok több ponton is egymáshoz
kapcsolódik: a TOC1 egy feltételezett X elemen keresztül serkenti a CCA1/LHY géneket, míg a CCA1/LHY
fehérjék közvetlenül a TOC1 promóteréhez kapcsolódva gátolják annak működését. A CCA1/LHY nem csak
a TOC1, hanem a GI, valamint saját génjeik kifejeződésére is gátlólag hat. A sárga villámok jelölik az adott
gének fényindukált átíródását. Az ábra eredeti verziója a Harmer által 2009-ben közölt összefoglaló
cikkből származik (Harmer, 2009).

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
15
Az Arabidopsis génállománya négy TOC1-hez hasonló gént tartalmaz, cirkadián
mintázatú kifejeződésük csúcsa különböző napszakokra esik (Matsushika és mtsai,
2000). Legkorábbi a PRR9, majd sorban következik a PRR7, PRR5, PRR3, végül az esti
TOC1/PRR1. A TOC1 és PRR5 hiánya rövid, a PRR7 és 9-é hosszú periódust okoz
(Eriksson és mtsai, 2003; Somers és mtsai, 1998b; Yamamoto és mtsai, 2003). A PRR3, 5,
7, 9 fehérjék, valamint a TOC1 az amino-végükön a baktériumok kételemű jeltovábbító
rendszerének jelfogó tagjára jellemző fehérjeelemet tartalmaznak. Az ilyen molekulák
nem közvetlenül szabályozzák az átírást, hanem feltételezhetően egyéb segédfehérjéken
keresztül (Ben-Naim és mtsai, 2006). A TOC1-re hasonlító gének mindegyikéről
kimutatták, hogy közük van a cirkadián órához, noha az egyszeres mutánsaik cirkadián
fenotípusa nem túl feltűnő (Eriksson és mtsai, 2003; Farré és mtsai, 2005; Kaczorowski
és Quail, 2003; Nakamichi és mtsai, 2005a). A többszörös mutánsok azonban már
súlyosabban sérültek (Farré és mtsai, 2005; Nakamichi és mtsai, 2005a). A
legszélsőségesebb a prr5/prr7/prr9 mutáns, amelyben megszűnik mindenféle ritmus
(Nakamichi és mtsai, 2005b).
A toc1 null mutáns növényekben viszont határozott cirkadián ritmus mérhető, és
mivel a TOC1 közös komponense mindkét korábban leírt szabályozó körnek, ezért
mutációja esetén aritmia várható. Ezek az eredmények ismét arra utaltak, hogy egy
további szabályozó hiányzik. Ennek leírása 2006-ban következett be, mikor többszörös
mutánsok, ill. túltermelő vonalak vizsgálatával kimutatták, hogy a harmadik szabályozó
kört a CCA1/LHY valamint a PRR7 és PRR9 gének/fehérjék alkotják. A CCA1/LHY
fehérje serkenti a két reggeli kifejeződésű gén, a PRR7 és 9 kifejeződését, a PRR7 és 9
fehérje viszont (a TOC1-hez hasonlóan közvetetten) gátolja a CCA1/LHY géneket. Ez a
negatív visszacsatolás egy reggeli szabályozó hurkot alkot (Locke és mtsai, 2006;
Zeilinger és mtsai, 2006).
Mutánsok vizsgálatával igazolták, hogy a reggeli és esti szabályozó körök
önmagukban is képesek oszcilláció létrehozására, de ezen ritmusok periódusa
jelentősen rövidebb a vad típusú óráénál. Ez azt jelenti, hogy a vad típusú növényekben
mérhető kb. 24 órás periódus azáltal alakul ki, hogy a két rövid periódusú hurok
összekapcsolódik egymással a központi szabályozó körön keresztül. Mivel a CCA1/LHY,
PRR9 és GI gének átíródása fényérzékeny, gyakorlatilag mindhárom szabályozó hurok

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
16
rendelkezik fénybemenettel, amelyen keresztül a különböző fotoreceptorok által
továbbított fényjelek hatással lehetnek az egyes szabályozó körök működésére.
Több olyan óraelem is ismert, amelynek hatása az órára jelentős, mégsincs benne a
modellben. Ilyenek például a LUX ARRHYTHMO (LUX), amelynek hiányában teljes
ritmustalanság tapasztalható (Hazen és mtsai, 2005; Onai és Ishiura, 2005). A LUX egy
háromtagú géncsalád egyik eleme, egy MYB-szerű DNS-kötő elemmel rendelkező
transzkripciós faktor, amely szükséges a CCA1/LHY megfelelő szintjének kialakításához
és a TOC1 ritmusának kialakulásához. A LUX a TOC1-hez hasonlóan szabályozódik, mivel
a CCA1/LHY a promóterében levő esti elemhez kötődve gátolja a kifejeződést. A
BROTHER OF LUX ARHHYTMO (BOA) a géncsalád egy másik tagja, túltermelése
jelentősen megnyújtja a periódust. In vivo képes serkenteni a CCA1 átíródását, míg a
CCA1/LHY fehérjék a LUX-hoz hasonlóan gátolják a BOA kifejeződését, így egy alhurkot
kialakítva (Dai és mtsai, 2011).
A bemeneti oldal elemei
Ahogy azt már korábban kifejtettem, a cirkadián óra nyújtotta előnyök
érvényesüléséhez a belső időnek azonos fázisban kell lennie a valós idővel. Mivel a
szabadon futó cirkadián ritmusok periódusa nem pontosan 24 óra, ezért a szétcsúszás
elkerülése érdekében az órát naponta hozzá kell igazítani a külső időhöz. A beállítást
különböző környezeti tényezők (leginkább a fény és a hőmérséklet) változása váltja ki,
amelyet az erre érzékeny receptormolekulák fordítanak le biokémiai jelekre. Az
érzékelők a jelet ezután a hozzájuk kapcsolt jelátviteli hálózaton keresztül juttatják el a
központi oszcillátorhoz. A jel az órafehérjék szintjét vagy működését oly módon
változtatja meg, hogy az a cirkadián kör egy másik fázisára lesz jellemző. Az óra ezután
ettől a ponttól folytatja tovább működését. Ez a beállítás molekulaszintű magyarázata.
A főrezgőkör egyes elemei fényérzékenyek. Ezek azok a belépési pontok, amelyeken
keresztül a beállító fényjelek elérik az órát. Így például a fény serkenti a CCA1/LHY (Kim
és mtsai, 2003a; Wang és Tobin, 1998) és a PRR9 (Makino és mtsai, 2001), valamint az
ELF4 (Kikis és mtsai, 2005) és a GI (Locke és mtsai, 2005) gén kifejeződését, ugyanakkor
a CCA1 mRNS lebomlását (Yakir és mtsai, 2007) és az LHY mRNS lefordítását is (Kim és
mtsai, 2003a). A fény az átírás utáni szinteken is befolyásolja az óra működését. A GI és a

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
17
ZTL fehérje például csak sötétben bomlik, a kék fény meggátolja ezt a folyamatot (Kim és
mtsai, 2007; Kim és mtsai, 2003b; Yu és mtsai, 2008).
Fényérzékelők
A cirkadián órát a fény nem a fotoszintézis fényelnyelő és energia-átalakító
rendszerén, hanem speciális fényérzékelő-fehérjéken keresztül szabályozza. Ezek a
fehérjék a fényt a hozzájuk kötött jellegzetes (kromofor) molekulák segítségével nyelik
el. A kromofor molekulák fényelnyelés által kiváltott szerkezetváltozása torzulásra
kényszeríti az őket megkötő fehérjéket is, ami elindítja a jelátvitelt. A fényérzékelőket az
általuk elnyelt fény hullámhossza alapján három nagy csoportba oszthatjuk (Chen és
mtsai, 2004).
1. vörös/távoli vörös fényt érzékelő fitokrómok (Bae és Choi, 2008; Kevei és mtsai,
2007)
2. UV-A és kék tartományban elnyelő kriptokrómok, fototropinok és a ZTL-család
tagjai (Briggs, 2007; Briggs és Christie, 2002; Briggs és Huala, 1999; Cashmore
és mtsai, 1999; Christie, 2007; Kim és mtsai, 2007; Lin és Shalitin, 2003)
3. UV-B fényérzékelő molekulák (Jenkins, 2009; Rizzini et al, 2011)
Jelenlegi tudásunk szerint a fototropinok nem vesznek részt a cirkadián óra
szabályozásában. Az UV-B érzékeléssel pedig a - dolgozat témája miatt - külön fejezetben
foglalkozom.
A fitokrómok olyan, ~125 kDa molekulatömegű hisztidin kináz-szerű fehérjék,
amelyek amino-végi fehérjeeleméhez kovalensen kötődik egy nyílt láncú tetrapirrol
természetű fitokromobilin molekula. Ezen kromofor fényelnyelési tulajdonságainak
köszönhetően a fitokrómoknak két alakjuk van: a vörös fényt elnyelő, élettanilag
hatástalan Pr (λmax=660 nm), valamint a távoli vöröst elnyelő, hatásos Pfr (λmax=730 nm).
A fitokrómok a sejtben Pr alakban képződnek, majd vörös fény elnyelésekor átalakulnak
Pfr alakba. Ez az alak távoli vörös fény hatására képes visszakerülni az eredeti Pr
állapotba, amelybe egyébként idővel sötétben magától is visszaalakul. A fitokrómok
dimer alakban működnek, a párbaállásért felelős elem a molekula közepén helyezkedik
el. A fitokrómok sötétben a sejtplazmában helyezkednek el, fény hatására pedig a
sejtmagba vándorolnak, ahol közvetlenül transzkripciós faktorokhoz kapcsolódva

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
18
fényfüggő gének kifejeződését szabályozzák (Kevei és mtsai, 2007). A fitokrómokat
Arabidopsisban öt gén kódolja, a PHYA, -B, -C, -D és -E. Az egyes gének által kódolt
fehérjék szekvenciája ugyan nagymértékben hasonlít egymásra, a köztük lévő kis
különbségek azonban eltérő működéssel ruházzák fel őket. A fitokrómok működésük
alapján két csoportba sorolhatók: az I-es csoport tagjai (Arabidopsisban csak egy van, a
PHYA) fényben gyorsan lebomlanak, így legnagyobb mennyiségben a sötétben csírázó
(föld alatti) és sötétben nőtt növényekben találhatók meg (Furuya, 1989; Furuya és
Schäfer, 1996). E tulajdonságok egyértelműen meghatározzák a PHYA szerepét:
elsősorban a csírázás megindításában és nagyon gyenge fényben a zöldülés
szabályozásában vesz részt. A II-es csoportot a többi (PHYB–E) gén terméke alkotja.
Ezek a fehérjék fényben nehezebben bomlanak, ezért a zöld növények uralkodó
fitokrómjai. Többek között a zöldülés folyamatát, a szár megnyúlását, az
árnyékelkerülést és a virágzást szabályozzák. A II-csoport tagjai képesek egymással
heterodimereket képezni. (Sharrock és Clack, 2004).
A kriptokrómokat elsőként növényekben azonosították, majd állatokban is leírták
szerepüket a cirkadián óra működésében (Cashmore és mtsai, 1999). Növényekben
elsősorban a kék/UV-A-függő élettani folyamatok (pl. zöldülés, virágzás, fényfüggő
génkifejeződés) szabályozásában játszanak szerepet. Amino-végükön egy FAD és MTHF
kromofort kötő fotoliáz-szerű, karboxi-végükön pedig egy szerin/treonin kináz
fehérjeelemet tartalmaznak (Lin és Shalitin, 2003). Sötétben a molekula csukott
állapotban van, a karboxi-végi elem elfedi az amino-végen található jelátvivő elemet. Kék
fény hatására a kromoforkötő elem szerkezetváltozása következtében a molekula saját
magát foszforilálja. Ennek hatására az amino-végi elem szabaddá válik, lehetővé téve a
jelátviteli molekulákkal való kölcsönhatást (Shalitin és mtsai, 2003; Yu és mtsai, 2007b).
A kriptokrómok sejtmagi elhelyezkedésűek. Arabidopsisban két eltérően viselkedő
kriptokrómot azonosítottak, amelyeket a CRY1 és a CRY2 gének kódolnak. A két fehérje
fényérzékenysége különbözik: kék fény hatására a CRY2 (foszforilálódás után)
ubikvitinálódik és lebomlik, míg a CRY1 fehérje szintje nem változik (Yu és mtsai,
2007a). Ennek megfelelően a CRY2 szerepe gyenge, míg a CRY1-é inkább erősebb
fényben fontosabb.
Az egyedi fitokróm- és kriptokrómhiányos (phyA, phyB, cry1, cry2), illetve ezek kettős,
hármas és négyes mutánsainak vizsgálata során sikerült meghatározni, hogy az egyes

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
19
fényérzékelők milyen mértékben vesznek részt az óra fénybemenetének
szabályozásában (Yanovsky és mtsai, 2001). A kísérletek során a beállított növényeket
folyamatos gyenge fénybe helyezésük után különböző színű fénypulzusokkal kezelték és
a levélmozgás ritmusában bekövetkező fáziscsúszást mérték. A kísérletek kimutatták,
hogy a PHYA a távoli vörös- és kék, a CRY1 és CRY2 a kék, a PHYB pedig a vörös fény
érzékelésében vesz részt. A phyA/phyB kétszeres mutánsban csökkent, de nem szűnt
meg a vörös fényre adott fázisválasz, ami további fényérzékelők (PHYC–E) szerepére
utal.
Növényekben a szabadon futó cirkadián ritmus periódusa a fény erősségének
növelésével csökken (Aschoff-szabály) (Aschoff, 1979). A fényérzékelők hiánya a
fényerősség csökkenéséhez hasonlóan a folyamatos fényben megfigyelhető periódus
megnyúlásával jár. Az egyes érzékelők hiánya csak az általuk elnyelt hullámhossz-
tartományban és jellegzetes fényerősségnél mutatkozik meg. Az egyes fotoreceptorok
periódusra gyakorolt hatását Somers és munkatársai a különböző mutánsokban olyan
jelzőgén segítségével vizsgálták, amelyben a cirkadián óra által vezérelt CAB2 gén
promótere a szentjánosbogár luciferáz génjének kifejeződését szabályozta (Somers és
mtsai, 1998a). A kutatók kimutatták, hogy a phyA növényekben a CAB2:LUC jelzőgén
kifejeződésének periódusa gyenge vörös és kék fényben a vad típushoz képest hosszabb.
A phyB növényekben viszont erős vörös fényben figyelhető meg ugyanez a jelenség. A
PHYB mellett minden bizonnyal további fitokrómok is részt vesznek a vörös fény
érzékelésében, mivel a phyB növényekben továbbra is észlelhető a fény periódusrövidítő
hatása. Hasonló kísérletben igazolták a CRY1 és CRY2 fehérjék szerepét a kék fény
periódus hangolásában. Érdekes, hogy a phyA/phyB/cry1/cry2 mutáns továbbra is képes
a fényjelek továbbítására a cirkadián óra felé, miközben fotomorfogenikus képessége
gyakorlatilag nincs (Yanovsky és mtsai, 2000a). Eszerint az említett fényérzékelőkön
kívül mások (PHYC–E, ZTL-család) is szabályozzák az óra fénybemenetét.
A ZEITLUPE (ZTL), a LOV KELCH PROTEIN 2 (LKP2) és FLAVIN-BINDING, KELCH
REPEAT, F-BOX 1 (FKF1) egy újabb fényérzékelő fehérjecsalád tagjai. E fehérjék három
jellegzetes fehérjedomént tartalmaznak: LOV (Light, Oxigen, Voltage), F-box és Kelch.
Ezek együttes előfordulása egy molekulán belül sejteti a fehérjék működésének
mikéntjét. A LOV elemet eredetileg olyan baktériumfehérjékben azonosították, amelyek
különféle környezeti tényezők, pl. fény, redox állapot ill. feszültség érzékelésében

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
20
vesznek részt. Ilyen elemet tartalmaznak a fototropinok, valamint a fonalasgomba
cirkadián órafehérjéi, a WHITE COLLAR (WC) 1 és 2 is. A LOV-elem FMN redoxfestéket
(kromofort) köt, amelynek külső hatásra bekövetkező elektronszerkezeti változása teszi
lehetővé a fent említett környezeti jelek érzékelését (Crosson és Moffat, 2001). A
fototropinok, a WC fehérjék és az FKF1 LOV-elemeinek vizsgálatával sikerült kimutatni,
hogy ez a fehérjeelem általános (kék) fényérzékelőként működik (Cheng és mtsai, 2003;
Imaizumi és mtsai, 2003; Zikihara és mtsai, 2006). Az F-box elem olyan fehérjékre
jellemző, amelyek az ún. SCF (Skp1-Cul1-F-box) ubikvitin-ligáz együttes részeként más
fehérjék irányított lebomlásában működnek közre (Cardozo és Pagano, 2004; Ho és
mtsai, 2008; Kipreos és Pagano, 2000). Az F-box fehérjék felismerik a lebontandó
célfehérjéket és az ubikvitin-ligázhoz kapcsolják őket, ezzel biztosítva az ubikvitinnel
történő jelölés célzottságát. Arabidopsisban igen sok, mintegy 700 F-box tartalmú fehérje
található, amelyhez változatos fehérjefelismerő elemek társulnak (Xiao és Jang, 2000). A
Kelch viszont egy általánosan elterjedt, β-propeller szerkezetű fehérjefelismerő elem,
amelynek változékonysága lehetővé teszi a célfehérjék egyedi felismerését (Adams és
mtsai, 2000). Ez felépítés azt sugallja, hogy a ZTL család tagjai célfehérjék
fényszabályozott lebomlásában vesznek részt.
A ZTL gén elrontása a cirkadián ritmusok periódusának hosszabbodását okozza
(Somers és mtsai, 2000). A periódus a fényerősség függvényében erőteljesebben
változik, mint a vad hátterű növényben. Ez arra utal, hogy a ZTL a fénybemenet
gátlóeleme. A hosszú periódus sötétben is megfigyelhető. Az FKF1 a virágzás
nappalhosszúságtól függő szabályozásában kulcsszerepet játszó fehérje. A GIGANTEA
(GI) fehérjével együtt a CONSTANS (CO) gén kifejeződését gátló transzkripciós faktor, a
CYCLING DOF FACTOR1 (CDF1) lebomlását irányítja (Imaizumi és mtsai, 2005; Sawa és
mtsai, 2007). Az fkf1 mutánsban emellett a cirkadián ritmusok hullámalakja is
megváltozik, tehát ez a fehérje is hozzájárul a megfelelő óraműködéshez (Nelson és
mtsai, 2000). Az LKP2 túltermelése a ritmusok megszűnését okozza, míg mutációjának
nincs cirkadián fenotípusa. (Schultz és mtsai, 2001). Az LKP2 fehérjéről kimutatták, hogy
élesztő kéthibrid rendszerben (a ZTL-hez hasonlóan) összekapcsolódik a TOC1 és PRR5
fehérjével is, bár ezen kölcsönhatások élettani jelentőségéről még nem sokat tudunk
(Yasuhara és mtsai, 2004). A ZTL-család egyes fehérjéi önmagukkal illetve egymással
dimert képezve működnek (Yasuhara és mtsai, 2004). A fénybemenet és a főrezgőkör
közti kölcsönhatást szemlélteti az a tény, hogy a fitokróm és kriptokróm gének (PHYA, B,

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
21
D, E; CRY1, 2) kifejeződése cirkadián mintázatú (Tóth és mtsai, 2001). Ez lehet az alapja
annak a korábban már említett jelenségnek, hogy az óra a saját fényérzékenységét csak
bizonyos időszakokban kapuzza (lásd: fázisválasz görbe).
Az eddig leírt kísérleti eredményeket összefoglalva megállapíthatjuk, hogy a növényi
cirkadián óra fénybemenetében az egyes fényérzékelők a következő módon jutnak
szerephez: a PHYA, CRY1, CRY2, ZTL egyaránt részt vesznek a kék fénybemenetben, míg
a távoli vörös fény érzékelésére egyedül a PHYA képes. A PHYA, PHYB, PHYD és PHYE
egymást kiegészítve juttatja el a vörös fény hatásait az órához (Devlin és Kay, 2000). A
fényérzékelők egymás működését is befolyásolják. Leírták például, hogy a PHYA
megfelelő működéséhez vörös és kék fényben a CRY1 (Devlin és Kay, 2000), a CRY2
fehér fényben való működéséhez pedig a PHYB jelenléte szükséges (Más és mtsai, 2000).
Jelátviteli utak
Génkölcsönhatásokat feltérképező kísérletekből sikerült néhány olyan elemet
azonosítani, amelyek a fényérzékelők és a központi óraelemek közé helyezhetők. A
fotomorfogenezisben sérült constitutive photomorphogenic 1 (cop1) és de-etiolated 1
(det1) mutánsokban a vizsgált ritmusok periódusa állandó sötétben rövidebb (Millar és
mtsai, 1995b). Hatásuk a fényfejlődésre és a génkifejeződés genomszintű mintázatára
nagyon hasonló, ezért feltételezhetően ugyanabban a szabályozási egységben (E3
ubikvitin-ligáz együttes) működnek. Nemrég mutatták ki, hogy a COP1 az ELF3
fehérjével együtt felelős a GI fehérje ubikvitinálásáért és lebomlásáért, és ezt a
folyamatot a kék fény a CRY2-n keresztül gátolja (Yu és mtsai, 2008). A DET1 pontos
szerepét az órában nem ismerjük, csak annyit figyeltek meg, hogy hiányában az LHY
fehérje lebomlása felgyorsul (Song és Carré, 2005). Ez a hatás fényfüggetlen, így a DET1
nem is a fénybemenet, sokkal inkább a központi óramű része lehet. A FAR RED
ELONGATED HYPOCOTYL 1 és 3 (FHY1 és FHY3) hiányában távoli vörös fénnyel nem
állítható át megfelelően a fázis (Yanovsky és mtsai, 2001; Yanovsky és mtsai, 2000b), így
ezen fehérjék is nélkülözhetetlenek az óra megfelelő működéséhez. Az FHY3 ezen túl
részt vesz a beállító fényjelek kapuzásában is (Allen és mtsai, 2006).
A kriptokrómok eredendően a sejtmagban találhatók, a fitokrómok pedig fényfüggő
módon vándorolnak a sejtmagba. Ez arra enged következtetni, hogy a fénybemeneti
folyamatok korai lépései a sejtmagban játszódnak le, ahol a fényérzékelők közvetlenül,
vagy jelátvivő molekulákon keresztül szabályozzák a génkifejeződést. A fitokrómok

SZAKIRODALMI ÁTTEKINTÉS _____________________________________________________________________________________________________
22
esetében a közvetlen kapcsolat különféle transzkripciós faktorokkal régóta ismert. Az
elsőként azonosított PHYB-vel kölcsönható fehérje a PHYTOCHROME INTERACTING
FACTOR 3 (PIF3) volt (Ni és mtsai, 1998). Később számos rokon fehérjét azonosítottak,
amelyek mindegyike kölcsönhat a fitokrómokkal, élettani szerepük azonban sokféle
(Castillon és mtsai, 2007; Josse és mtsai, 2008; Khanna és mtsai, 2004). A PIF3 egy bHLH
transzkripciós faktor, amely a bekapcsolt fitokrómhoz tapadva kötődik számos
fényszabályozott gén promóterében található ún. G-box mintázathoz (Martínez-García
és mtsai, 2000). Ilyen G-box mintázat található a CCA1 és LHY óragének promóterében is,
amelyekhez a PIF3 szintén képes kapcsolódni. PIF3-hiányos növényekben az LHY és
CCA1 gének fényválasza erősen csökken, ami arra utal, hogy a PIF3 fehérje szerepet
játszik a két óragén kifejeződésének fényszabályozásában. Ez a megfigyelés azt sugallta,
hogy az óra fázisának beállítása ezen az úton, a PIF3 közreműködésével történik.
Kiderült azonban, hogy a PIF3 egyáltalán nem vesz részt a cirkadián óra beállításában,
szerepe a fotomorfogenikus folyamatok szabályozására korlátozódik (Viczián és mtsai,
2005).
A TIME FOR COFFEE (TIC) szerepe elsősorban a cirkadián ritmusok erőteljességének
és periódusának fenntartásában van (Hall és mtsai, 2003). A TIC elrontása nem
egyformán hat minden kimenetre: egyeseknél periódus hosszabbodást, másoknál
rövidülést okoz. A TIC fényben és sötétben is szükséges a cirkadián óra megfelelő
működéséhez, valamint részt vesz a kapuzásban is. A TIC egy igen nagy méretű
növényjellegzetes sejtmagi molekula, ismert fehérjeelem (egy P-hurkon kívül) nem
található benne (Ding és mtsai, 2007b).
A SENSITIVITY TO RED LIGHT REDUCED 1 (SRR1) fehérjét egy olyan munka során
azonosították, amelynek célja fényszabályozott fejlődési folyamatokban résztvevő
elemek azonosítása volt (Staiger és mtsai, 2003). Az srr1 mutáns kevésbé érzékeny a
vörös fényre, és számos egyéb, PHYB-függő fényválasza is sérült. Igen valószínű, hogy az
SRR1 a fényszabályozásban a PHYB jelátviteli lánc részeként működik. Állandó
körülmények között az srr1 szabadon futó ritmusainak periódusa a vad típuséhoz képest
rövidebb. Az SRR1 kifejeződése vörös fény hatására megemelkedik, ami arra utal, hogy
az SRR1 az óra fényérzékenységének beállításában vesz részt. A fehérjében nem
található semmilyen ismert fehérjeelem, és a molekula működési módját sem ismerjük.

SZAKIRODALMI ÁTTEKINTÉS ______________________________________________________________________________________________________
23
UV-B érzékelés és jelátvitel
UV-B: stresszfaktor és környezeti jel
Az ultraibolya-B (UV-B, 280-315 nm) sugárzás természetes összetevője a napfénynek.
Ugyan az ózonpajzs szűrő hatásaként számottevő mennyiségben csak a 295 nm–es
hullámhossz feletti része éri el a felszínt, azonban még ez a tartomány is képes
károsítani a DNS-t, a fehérjéket, és a lipideket; elősegíti a reaktív oxigén származékok
(ROS) keletkeződését (Brosché és Strid, 2003; Frohnmeyer és Staiger, 2003). Az
élőlényeknek ezért ki kellett fejleszteniük a védelmüket a káros UV-B sugárzással
szemben, és a javító mechanizmusaikat az általa okozott károk enyhítésére. A
magasabbrendű növények UV-B elnyelő fenolszármazékokat, és a ROS-ok
közömbösítésére antioxidánsokat halmoznak fel az epidermisz sejtjeikben (Bornmann
és mtsai, 1997), a DNS károk kijavításában pedig a többi élőlényhez hasonló,
fénystimulált DNS fotoliázaik játszanak szerepet (Britt, 2004).
Az UV-B által kiváltott válaszok nagyban függenek az intenzitástól, az időtartamtól és
a hullámhossztól (Brosché és Strid, 2003; Jenkins és Brown, 2007). Általánosságban, a
nagy intenzitású és rövid hullámhosszú UV-B stresszválaszokat indukál, míg az alacsony
intenzitású UV-B képes szabályzó folyamatokat kiváltani. Ilyenek például a
hipokotilmegnyúlás gátlása, a sziklevelek kiterülésének serkentése, illetve a
génkifejeződés szabályzásán keresztül többek között a flavonoid felhalmozódás
beindítása (Boccalandro és mtsai, 2001; Brown és mtsai, 2005; Frohnmeyer és Staiger,
2003; Kim és mtsai, 1998). Az alacsony intenzitású UV-B hatására stimulálódnak az UV
védelemben és az UV okozta hibák kijavításában közreműködő gének, ami így
akklimációt, adaptációt biztosít elsősorban az UV-B, de a stresszjelátviteli utak
kapcsoltsága miatt a kártevők, patogének ellen is (Brosché és Strid, 2003; Jenkins és
mtsai, 2001; Jenkins és Brown, 2007; Jansen és mtsai, 1998; Stratmann, 2003). Ezek a
szabályzó folyamatok nyilvánvalóan nem stresszválaszok, inkább hasonlóak az általános
fotoreceptorok (fitokrómok, kriptokrómok, fototropinok) által szabályozott
fotomorfogenikus folyamatokhoz.

SZAKIRODALMI ÁTTEKINTÉS ______________________________________________________________________________________________________
24
Az UV-B fotomorfogenikus jellemzői
A két legjobban jellemzett, alacsony intenzitású UV-B által kiváltott fotomorfogenikus
folyamat a hipokotilmegyúlás gátlása (Ballaré és mtsai, 1995; Ballaré és mtsai, 1996;
Boccalandro és mtsai, 2001; Kim és mtsai, 1998) és a flavonoid bioszintézisben szerepet
játszó gének (elsősorban a kalkon–szintáz, CHS) indukciójának vizsgálata (Frohnmeyer
és mtsai, 1999; Jenkins és mtsai, 1997; Jenkins és mtsai, 2001). A kísérletek eredményei
azt mutatják, hogy ezek az UV-B által kiváltott folyamatok nem stresszválaszok és
nincsenek egyik, már korábban leírt fotoreceptor közvetlen szabályozása alatt sem:
Ezek a fotomorfogenikus folyamatok már egészen alacsony UV-B intenzitással
(kevesebb mint 0,1 μmol/m-2 s-1, kb. a napsugárzás UV-B tartalmának 1/40-ed része), és
igen rövid pulzussal (kevesebb, mint öt másodperc) kiválthatóak. Ez az intenzitás nem
váltja ki a stresszválaszokért felelős gének mérhető indukcióját.
A UV-B által kiváltott fotomorfogenikus génindukció mértéke nem arányos az UV-B
fény által képzett ciklobután pirimidin dimérek (CPD) mennyiségével (Frohnmeyer és
mtsai, 1999; Kalbin és mtsai, 2001).
A DNS hibajavításban mutáns növények, melyeket az UV-B hatására megemelkedett
stresszválaszok jellemeznek nem térnek el a vad típustól a hipokotilmegnyúlás
gátlásában, vagy a szíklevél kiterülés serkentésében (Boccalandro és mtsai, 2001; Kim és
mtsai, 1998). UV-B-vel együtt alkalmazott látható fényű megvilágítás a fotoliáz
aktiválásán keresztül csökkenti a DNS károsodást azonban a CHS gén UV-B indukcióját
megnöveli (Jenkins, 2009).
A nagy intenzitású UV-B által kiváltott stressz jelátvitelében a jázmonsav, az etilén és
a ROS-ok játszanak döntő szerepet (A-H-Mackerness és mtsai, 1999). Míg azonban a
JASMONATE RESISTANT 1, és az ETHYLENE RESPONSE 1 hiányos jelátviteli
mutánsokban UV-B besugárzás hatására a védelemben fontos gének indukciója valóban
alacsonyabb, a CHS-é nem változik (Jenkins, 2009).
Hasonlóan, míg az UV-B hatására bekövetkező védelmi gén indukció gátolható
antioxidáns adásával (Green és Fluhr, 1995; Surplus és mtsai, 1998), a CHS gén
indukciója nem (Jenkins és mtsai, 2001).

SZAKIRODALMI ÁTTEKINTÉS ______________________________________________________________________________________________________
25
A CHS kifejeződés nem stimulálható reaktív oxigéngyökökkel sem Arabidopsis
sejtkultúrában (Jenkins és mtsai, 2001), sem növényekben (Desikan és mtsai, 2001);
(Gadjev és mtsai, 2006).
A már ismert fotoreceptorok mindegyike képes ugyan elnyelni UV-B-t és ezért
lehetséges receptorai annak, azonban a fotoreceptor mutánsok nem mutatnak csökkent
CHS indukciót alacsony intenzitású UV-B megvilágításra és a hipokotilmegnyúlás gátlása
is megmarad fitokróm, és kriptokróm mutánsokban (Ballaré és mtsai, 1991;
Boccalandro és mtsai, 2001; Suesslin és Frohnmeyer, 2003).
Mindazonáltal a fotomorfogenikus UV-B jelátvitel nem teljesen független a fény
jelátviteli utaktól. Az UV-B és a fitokróm B jelátvitel átfedően szabályozza a sziklevelek
kinyílását (Boccalandro és mtsai, 2001). Az UV-B által kiváltott CHS indukciót a PHYB
gátolja, míg az UV-A és a kék fény ismeretlen fotoreceptoron keresztül serkenti azt
(Wade és mtsai, 2001).
UV-B fotoreceptor?
A fenti kísérletek már jóideje felvetették annak lehetőségét, hogy a növényekben
megtalálható egy specifikus UV-B fotoreceptor. Az azonosítását megnehezíti, hogy
nagyon sok sejtben található anyag (pl. a nukleinsavak, a fehérjék, a fenolszármaszékok)
képes elnyelni UV-B-t. Tovább nehezíti a keresést, hogy az UV-B által kiváltott
folyamatoknak nincs olyan karakterisztikus jellemzője, mint például a fotoreverzió a
fitokrómoknál. A különböző UV-B által kiváltott folyamatok akciós spektruma is sok
esetben eltér egymástól, így ma sem tudni biztosan, hogy egyetlen érzékelő felelős az
UV-B válaszokért, vagy különböző rendszerek váltják ki őket. Az utóbbi mellett szól,
hogy Arabidopsisban egyes gének másként reagálnak a 280-290, illetve a 300-310 nm-es
hullámhossz-tartományra (Ulm és mtsai, 2004; Kalbina és mtsai, 2008).
Az ismert fotoreceptorok mind tartalmaznak egy nem fehérje jellegű kromofórt, ami
kulcsfontosságú a fényérzékelésben. Egyes festékmolekulák, mint például a redukált
pterinek, flavinok, vagy a p-kumarinsav képesek elnyelni UV-B-t, ezáltal lehetséges UV-B
érzékelő kromofórok (Galland, 1988a; Galland, 1988b; Imamoto és Kataoka, 2007).
Kísérletekkel sikerült azt is bebizonyítani, hogy hiányukban alacsonyabb az UV-B által
kiváltott antocianin termelés kukoricában (Khare és Guruprasad, 1993), csakúgy, mint a
hipokotilmegnyúlás gátlása paradicsomban (Ballaré és mtsai, 1995). Külsőleg bevitt

SZAKIRODALMI ÁTTEKINTÉS ______________________________________________________________________________________________________
26
riboflavin hatására pedig több CHS fehérje, és nagyobb flavonoid felhalmozódás
tapasztalható petrezselyem szövetkultúrában UV-B fény hatására, míg a kék fény hatása
azonos marad. Azonban olyan Arabidopsis mutáns is mutat HY5 génindukciót melyben a
teljes fenilpropanoid-bioszintézis sérült, és az előbbi festékmolekulák hiányoznak
(Jenkins, 2009). Ezért továbbra sincs bizonyíték egy, az ismert fotoreceptorokhoz
hasonló, festékanyagot tartalmazó UV-B receptor létére.
Emlős sejtekben bebizonyosodott, hogy UV-B sugárzás következtében termelődött
ROS-ok képesek ligand független módon aktiválni a membránkötött tirozin kináz
receptorokat (Bender és mtsai, 1997; Herrlich és Bohmer, 2000). Ugyan a növényekben
nincsenek tirozin kináz receptorok, de vannak más receptor kinázaik (Johnson és
Ingram, 2005). Az elvi lehetőség tehát fennáll, azonban alacsony UV-B intenzitásnál
aligha termelődik elegendő ROS a fotomorfogenikus reakciókat kiváltó receptor
aktivációhoz, és a folyamat nem is lenne UV-B secifikus. A brasszinoszteroid receptor a
BRASSINOSTEROID INSENSETIVE 1 (BRI1) egy receptor-szerű kináz. Érdekesen a bri1
mutánsban néhány gén UV-B indukciója lecsökkent (Savenstrand és mtsai, 2004). Az
azonban vitatott, hogy ez mennyire UV-B specifikus; továbbá amennyiben a BRI1 lenne a
felelős az UV-B érzékeléséért azt várhatnánk, hogy az indukciók nemcsak lecsökkennek,
hanem teljesen el is tűnnek. Természetesen elképzelhető, hogy több receptor kináz
egymást kiegészítve játszik szerepet az UV-B érzékelésben.
Egy másik, szintén az emlős sejtekben vizsgált lehetőség az aril-hidrokarbon receptor
(AhR) lehetséges részvétele az UV-B jelátvitelben. Az aril-hidrokarbon receptor egy
bHLH/PAS transzkripciós faktor, inaktív formában a citoplazmában helyezkedik el,
szubsztrátkötés hatására a sejtmagba helyeződik, ahol a DNS-hez kötődve elősegíti
bizonyos gének kifejeződését. Az AhR egyik lehetséges szubsztrátja a FICZ
(6-formilindol[3,2-b]carbazol), ami triptofánból keletkezik UV-B sugárzás hatására
(Fritsche és mtsai, 2007). A sejten belüli triptofánszint csökkenése valóban az UV-B
indukálta génkifejeződés csökkenését eredményezi, azonban jelenleg nincs bizonyíték
az AhR meglétére növényekben.
Az aromás aminósavak –elsősorban a triptofán– azon tulajdonsága, hogy képesek
elnyelni UV-B-t, vetette fel a lehetőségét annak, hogy egy fehérje közvetlen UV-B
elnyelése válthatja ki az UV-B választ (Ballaré és mtsai, 1995; Ensminger, 1993;
Gerhardt és mtsai, 2005). Minden fehérje képes elnyelni UV-B-t, tehát potenciálisan

SZAKIRODALMI ÁTTEKINTÉS ______________________________________________________________________________________________________
27
bármelyik lehet UV-B receptor, amennyiben az elnyelés elindít benne egy egyedi fizikai
vagy kémiai változást, ami az UV-B érzékelés kiindulása.
A triptofán UV-B elnyelési maximuma 280 nm, vagyis vagy egy különleges
fehérjekörnyezet szükséges az akár 300 nm feletti elnyeléshez, vagy egy UV-B elnyelő
kromofor lehet az, ami képes a gerjesztett állapotát átadni neki (Fluorescent Resonance
Energy Transfer, FRET).
A COP1 új funkciója
A fitokróm és kriptokróm fotoreceptorok kutatásához hasonlóan az áttörést az UV-B
fotoreceptor megtalálásához a molekuláris biológiai kísérletek, a mutáns izolálások,
genetikai szűrővizsgálatok hozták. Segítségükkel sikerült azonosítani több UV-B
érzékenységet okozó mutációt. Ezek legtöbbjében vagy a DNS hibajavítás sérült: UV
resistance 1 (uvr1) (Britt és mtsai, 1993), UV resistance 2 (uvr2) (Jiang és mtsai, 1997;
Landry és mtsai, 1997), UV resistance3 (uvr3) (Jiang és mtsai, 1997; Nakajima és mtsai,
1998), ultraviolet hypersensitive 1 (uvh1) (Harlow és mtsai, 1994), vagy az UV-B
védelemben fontos színanyagok forrása, a fenilpropanoid bioszintézis: transparent testa
4 és 5 (tt4, tt5) (Li és mtsai, 1993), ultraviolet sensitive (uvs) (Lois és Buchanan, 1994),
ferulic acid hidroxilase (fah1) (Landry és mtsai, 1995), UV tolerant 1 (uvt1) (Bieza és
Lois, 2001). Kliebenstein és munkatársai 2002-ben egy olyan mutánst izoláltak, (uv
resistance locus8, uvr8) mely érzékenyebb volt UV-B vel szemben, mint a flavonoid
bioszintézis mutánsok, és UV-B hatására nem mutatott CHS génindukciót (Kliebenstein
és mtsai, 2002). A további mutáns izolálási szűrővizsgálatok, melyeket CHS, illetve HY5
promóter-szentjánosbogár luciferáz jelzőgént hordozó növényeken végeztek további
uvr8 allélokat, illetve egy CONSTITUTIVELY PHOTOMORPHOGENIC1 (COP1) hiányos
mutánst eredményeztek.
A COP1 az egyik fő negatív szabályzója a fotomorfogenezisnek (Chen és mtsai, 2004;
Yi és Deng, 2005). A COP1 gátolja a fotomorfogenikus gének kifejeződését sötétben,
ezért a cop1 mutáns sok tekintetben a fényen nőtt növények jellemzőit mutatja. A COP1
fehérje három szerkezeti egységből, egy RING finger motívumból, egy coiled-coil
doménből és WD40 ismétlődésekből felépülő E3 ubikvitin ligáz. A COP9
szignaloszómával kölcsönhatva irányítja olyan pozitív faktorok mint a LONG
HYPOCOTYL 5 (HY5), HY5 HOMOLOGUE (HYH), LONG HYPOCOTYL IN FAR-RED (HFR1),
LONG AFTER FAR RED LIGHT 1 (LAF1) lebomlását a 35S proteoszómán keresztül (Chen

SZAKIRODALMI ÁTTEKINTÉS ______________________________________________________________________________________________________
28
és mtsai, 2004; Yi és Deng, 2005). Megvilágítást követően a COP1 inaktiválódik, lassan
elhagyja a sejtmagot hagyva, hogy a HY5 és a többi transzkripciós faktor felhalmozódjon,
és elősegítse a fotomorfogenezist.
A fotomorfogenezisben betöltött negatív szabályzó funkciójával szöges ellentétben a
COP1 az UV-B válaszok pozitív szabályzója. Génchip kisérletekből kiderült, hogy sok, az
alacsony intenzitású UV-B által indukált gén kifejeződése jelentősen alacsonyabb cop1-4
mutánsban, és az egyik közülük éppen a HY5. A COP1 által szabályzott funkciók
majdnem felét a HY5 is szabályozza, vagyis a HY5 a COP1 jelátvitel kulcsszabályozója.
Tehát a COP1 és a HY5 a sejtmagban együttműködnek az UV-B válaszok kialakításában,
míg sötétben a COP1 hatására lebomlik a HY5. A cop1-4 mutáns UV-B stresszre
érzékenyebb a vad típusú növénynél, összhangban a védelemben szerepet játszó gének
csökkent UV-B indukciójával. Viszont kevésbé érzékeny mint a hy5, jóllehet ezt
magyarázhatja a az UV-B védelemben szerepet játszó gének magasabb alapaktivitása.
Fehér fény hatásával szemben UV-B (fény) elősegíti a COP1 sejtmagi akkumulációját
(Oravecz és mtsai, 2006).
Az UVR8 jellemzői
A kutatások szerint az Arabidopsis UVR8 kifejezetten UV-B válasz specifikus. Az uvr8
mutánsban hiányzik a HY5 és CHS gén UV-B indukciója, azonban megmarad az indukció
látható fényre, alacsony hőmérsékletre, és szaharózra is (Brown és mtsai, 2005).
Transzkriptóma analízis szerint alacsony intenzitású UV-B hatására vad típusú
növényben egy órán belül 377 gén kifejeződése növekedett, illetve 102 géné csökkent
legalább kétszeres mértékben. Az indukálódó gének 99.5%-a volt COP1 és 98.6%-a UVR8
függő, míg a csökkenést mutatók kivétel nélkül mind a két gén szabályozása alatt álltak
(Favory és mtsai, 2009). A növekedést mutató gének közül soknak szerepe van az UV
védelemben, mint például a flavonoid bioszintézis egyes enzimeinek, DNS fotoliázoknak,
oxidatív stresszt enyhítő, és fotooxidatív károsodást javító fehérjéknek. Ez jól
magyarázza, hogy miért mutat az uvr8 mutáns komoly nekrózist napfényen, noha UV-B
hiányában megkülönböztethetetlen a vad típustól. Az UVR8 az alacsony, (akár
0.1 μmol m-2 s-1) intenzitású UV-B közvetítője, egyetlen más komponens sem ismert, ami
UV-B specifikus lenne a fotomorfogenezisben.

SZAKIRODALMI ÁTTEKINTÉS ______________________________________________________________________________________________________
29
Az UVR8 35%-os azonosságot mutat fehérje szinten az emberi REGULATOR OF
CHROMATIN CONDENSATION 1 (RCC1) fehérjével. Az RCC1 egy nukleotidcserélő
segédfehérjéje (guanin nucleotide exchange factor, GEF) a RAN kis GTP-kötő
fehérjéknek, amik például a sejtmagi transzportban, a sejtciklus szabályzásában, vagy a
mitózisban játszanak szerepet (Moore, 2001). Az RCC1-nek egy speciális hétlapátú
hajócsavart mintázó alakja van és ezt fehérje struktúrát kialakító aminósavak
konzerváltak UVR8-ban (Renault és mtsai, 1998). Ennek fontossága azonban vitatott,
mert a formai hasonlóság ellenére úgy tűnik, az RCC1 és az UVR8 különbözik funkcióban
és aktivitásban. Az UVR8-nak igen kicsi a nukleotidcserélő aktivitása, és míg az élesző
vagy emlős sejtek életképtelenek RCC1 hiányában (Moore, 2001), az Arabidopsis uvr8
mutáns UV-B hiányában megkülönböztethetetlen a vad típustól.
Az egyetlen funkcionális hasonlóság az RCC1, és az UVR8 között az, hogy mind a kettő
képes a kromatinnal (elsősorban a H2B hisztonnal) kölcsönhatni (Cloix és Jenkins,
2008). A kromatin immunoprecipitációs (ChIP) kísérletek alapján, úgy tűnik, az UVR8
kötődik egyes UV-B által indukált gének (HY5, MYB12) kromatinjához, míg másokéhoz
(HYH, CHS) nem. A kötődés a HY5 esetében nem csak a promóterrégióban (Brown és
mtsai, 2005), hanem a kódoló, illetve az 5’ és 3’ nem kódoló szakaszokon is megvalósul
(Cloix és Jenkins, 2008). A kölcsönhatáshoz UV-B nem szükséges.
A zöld fluoreszcens fehérjével (GFP) jelölt UVR8 fehérje UV-B nélküli, fehér fényen
nőtt növényekben mind a citoplazmában, mind a sejtmagban megtalálható (Brown és
mtsai, 2005). öt perc alacsony intenzitású UV-B megvilágítás hatására jelentős sejtmagi
felhalmozódás indul meg (Kaiserli és Jenkins, 2007), melyhez elegendő az N-terminális
23 aminósav. Ez a szakasz nem tartalmaz nukleáris lokalizációs szignált (NLS), így
valószínűleg az UVR8 sejtmagi transzportjában szerepet játszó, NLS tartalmú fehérjével
való kapcsolódásért felelős, hasonlóan a PHYA és FAR-RED ELONGATED HYPOCOTYL
(FHY1), illetve FHY1 LIKE (FHL) által közvetített áthelyeződéséhez. A nukleáris export
szignált (NES) hordozó GFP-UVR8 konstrukció a citoplazmában helyezkedik el, azonban
rövid UV-B megvilágítás hatására ez is a sejtmagba vándorol (Kaiserli és Jenkins, 2007).
Az NLS-t hordozó pedig ugyan folyamatosan a sejtmagban található, azonban
funkcióképessége ellenére UV-B hiányában nem vált ki HY5 átíródást. Vagyis az UV-B
fény két lépcsőben hat az UVR8-ra, egyrészt a sejtmagi felhalmozódásán, másrészt az
aktivációján keresztül (Jenkins, 2009).

SZAKIRODALMI ÁTTEKINTÉS ______________________________________________________________________________________________________
30
UVR8, a lehetséges UV-B fotoreceptor
Favory és munkatársai (Favory és mtsai, 2009) fluoreszcens fehérjék alkalmazásával
vizsgálták a COP1 és az UVR8 sejten belüli elhelyezkedését. Megállapították, hogy a
sárga fluoreszcens fehérjével (YFP) jelölt COP1 molekulák az UV-B megvilágítástól
függetlenül a sejtmagon belül, egyenlőtlenül helyezkednek el, csoportokban, sejtmagi
foltokat (nuclear body, specle) alkotva. A kék fluoreszcens fehérjével (CFP) jelölt UVR8
molekulák UV-B hiányában egyenletes sejtmagi elhelyezkedést mutattak, míg UV-B
megvilágítás hatására szintén foltokat alkottak, melyek helyzete jórészt egybeesett a
jelölt COP1 fehérjék pozíciójával. A közvetlen kölcsönhatás bizonyítására a kutatók egy
olyan módszert választottak, melyben a “kettévágott” fluoreszcens jelzőfehérje N-,
illetve C-terminális részeit illesztették külön-külön a vizsgálandó fehérjékhez. Ebben az
esetben csak akkor jön létre fluoreszcencia, ha a vizsgálandó fehérjék kölcsönhatásának
következtében a fluoreszcens fehérje két része kellő közelségbe kerül egymáshoz
(bimolecular fluorescent complementation, BiFC) (Kerppola, 2006). A kölcsönhatás ez
esetben szintén UV-B függő volt, megvilágítás hiányában alig volt észlelhető
fluoreszcencia. Mutáns alléleket használva kiderült, hogy a vizsgált UVR8 mutációk
mindegyike, és a legtöbb COP1 mutáció is megakadályozza a kölcsönhatást. A cop1eid6
mutánsban továbbra is létrehozható volt a kölcsönhatás, ami nem meglepő annak
fényében, hogy ebben a mutánsban nem sérült az UV-B kiváltotta flavonoid
felhalmozódás (Oravecz és mtsai, 2006). Ez arra utal, hogy a kölcsönhatás
szempontjából a COP1 WD40 ismétlődéseinek van nagyobb jelentősége, és nem a RING
finger doménnek. Tovább vizsgálva a kölcsönhatást sikerült immunoprecipitálni az
UVR8 fehérjét az YFP-vel jelölt COP1 molekula segítségével, pár percig UV-B-vel
megvilágított növényekből. A kísérletek során kiderült az is, hogy az UVR8 képes saját
magával is kölcsönhatásba lépni UV-B megvilágítástól függetlenül (Favory és mtsai,
2009).
A COP1-UVR8 kölcsönhatás létrejött in vitro úgy is, ha a jelölt fehérjéket hordozó
növényekből fehérjekivonatot készítettek, és azt világították meg UV-B-vel. Pontosabban
csak akkor, ha azt a kivonatot világították meg, amelyik az UVR8-at tartalmazta. Ez
bizonyította, hogy az UVR8 fehérjét tartalmazó kivonat UV-B megvilágítása szükséges és
elégséges feltétel az UVR8-COP1 kölcsönhatáshoz, jelezve, hogy ez az UVR8 elsődleges
válasza UV-B jelátvitelben.

SZAKIRODALMI ÁTTEKINTÉS ______________________________________________________________________________________________________
31
A kölcsönhatás UV-B megvilágítás hatására élesztőben is létrejön. A különböző uvr8
és cop1 mutánsok a cop1eid6 kivételével szintén nem hatnak kölcsön, illetve a COP1
C-terminális 340 aminósavát (a WD40 ismétlődést) tartalmazó csonka fehérje UV-B
függő módon kölcsönhat az UVR8-al. Az UVR8 homodimereket nem denaturált tisztítás
után vizsgálva kiderült, hogy az UVR8 UV-B megvilágítás nélkül dimer formában
található, és UV-B megvilágítás hatására monomerizálódik. A monomerizáció igen gyors,
már öt másodperc megvilágítás után észlelhető és intenzitásfüggő, valamint nem
szükséges hozzá a COP1 fehérje. A dimer-monomer átalakulás élesztőben is megvalósul.
A mutáns, COP1-el nem kölcsönható UVR8 fehérjék nem formálnak dimereket sem.
Az UVR8 fehérje az RCC1-el összehasonlítva feltünően sok triptofánt tartalmaz, 14-et
a 4-el szemben. Ezek a triptofánok ráadásul valamennyien a feltételezett β-propeller
struktúra felszinén helyezkednek el. A fehérjestruktúra közepén a 233., 285. és 337.
pozícióban lévő triptofánok különösen érdekes fehérjekörnyezetben vannak.
Bármelyikük kicserélése funkcióvesztést okoz, a 285. triptofán fenilalaninra cserélése
pedig folyamatos dimer állapotot. Ez különösen érdekes, hiszen a dimerizációra való
képesség azt mutatja, hogy a molekula szerkezetét alig érinti az aminósavcsere, UV-B
érzékelésre mégis képtelen (Rizzini és mtsai, 2011).
Ezek a legújabb vizsgálatok azt mutatják, hogy az UVR8 a specifikus UV-B
fotoreceptor. Alighanem a molekulában található triptofánok, elsősorban a 285.
triptofán UV-B elnyelésének következtében jön létre az a molekulán belüli változás,
aminek eredményeként az UVR8 monomerizálódik, aktiválódik, majd kölcsönhatásba
lép a COP1 fehérjével, elindítva a jelátvitelt.

CÉLKITŰZÉSEK
A növényi cirkadián óra legfontosabb beállító tényezője a fény. A szabadon futó
ritmus periódusa fordított arányban áll fény erősségével, a fényimpulzusok pedig
képesek az óra fázisát átállítani. Intenzíven kutatott terület azon jelátviteli utak
vizsgálata amelyeken keresztül a fény kifejti a hatását a cirkadián órára. Ennek
köszönhetően igen sok információnk van arról, hogy a látható fény egyes komponensei
milyen fotoreceptorokon és jelátviteli elemeken keresztül befolyásolják az órát. Azonban
mostanáig nem volt adat arról, hogy a fiziológiai mennyiségben jelenlévő UV-B milyen
hatással van a növényi cirkadián órára.
Mostanáig még nem sikerült minden kétséget kizáróan egyetlen molekulához, vagy
molekulacsaládhoz kötni az UV-B érzékelést. Azt azonban már sikerült igazolni, hogy az
alacsony fényintenzitású UV-B által kiváltott fotomorfogenikus folyamatok
fotoreceptora az UVR8 fehérje. Annak vizsgálatára, hogy az UV-B fénynek az UVR8 által
szabályozott alacsony intenzitású része hogyan befolyásolja a cirkadián órát. Illetve
fordítva, a cirkadián óra hogyan szabályozza az UV-B által kiváltott folyamatokat az
alábbi célokat tűztük ki:
1. A cirkadián óra UV-B fénnyel való beállíthatóságának a vizsgálata
2. A cirkadián óra hatásának jellemzése az UV-B által kiváltott fényindukciókra
3. Az UV-B jelátvitel hatásának vizsgálata a kapuzásra
4. Cirkadián óra hiányában történő UV-B indukciók jellemzése
5. A Cirkadián óra szerepé nek vizsgálata az UV-B védelemben

ANYAGOK ÉS MÓDSZEREK
Kísérleti anyagok és élőlények
Tápoldatok, táptalajok, antibiotikumok, szelekció
Bakteriális táptalajok
LB (Luria-Bertani Medium) (pH=7.0): 1% tripton (Reanal), 0.5% élesztő kivonat
(Reanal), 1% NaCl (Reanal); táptalajhoz: 1.5% agar (Reanal)
YEB (pH=7.0): 0.5% marhahús kivonat (Difco) 0.1% élesztő kivonat, 0.5% Bacto® -
pepton (Difco), 0.5% szaharóz (Reanal), 2 mM MgCl2 (sterilre szűrve, autoklávozás után
hozzáadva; Reanal); táptalajhoz:1.5% Bacto® -agar (Difco).
Növényi táptalajok
MS3 (Murashige-Skoog Medium) (pH=5.6): 4.3 g/l MS (Murashige-Skoog sókeverék;
Sigma), 3% szaharóz, 1% agar (Reanal)
MS1 (Murashige-Skoog Medium) (pH=5.6): 4.3 g/l MS (Murashige-Skoog sókeverék;
Sigma), 1% szaharóz, 0,8% agar (Reanal)
MS0 (Murashige-Skoog Medium) (pH=5.6): 4.3 g/l MS (Murashige-Skoog sókeverék,
Sigma), 0,8% agar (Reanal)
AM (Arabidopsis Medium) (pH=5.6): 2.16 g/l MS (Murashige-Skoog sókeverék, Sigma)
1% szaharóz, 0.2% phytagel (Sigma)
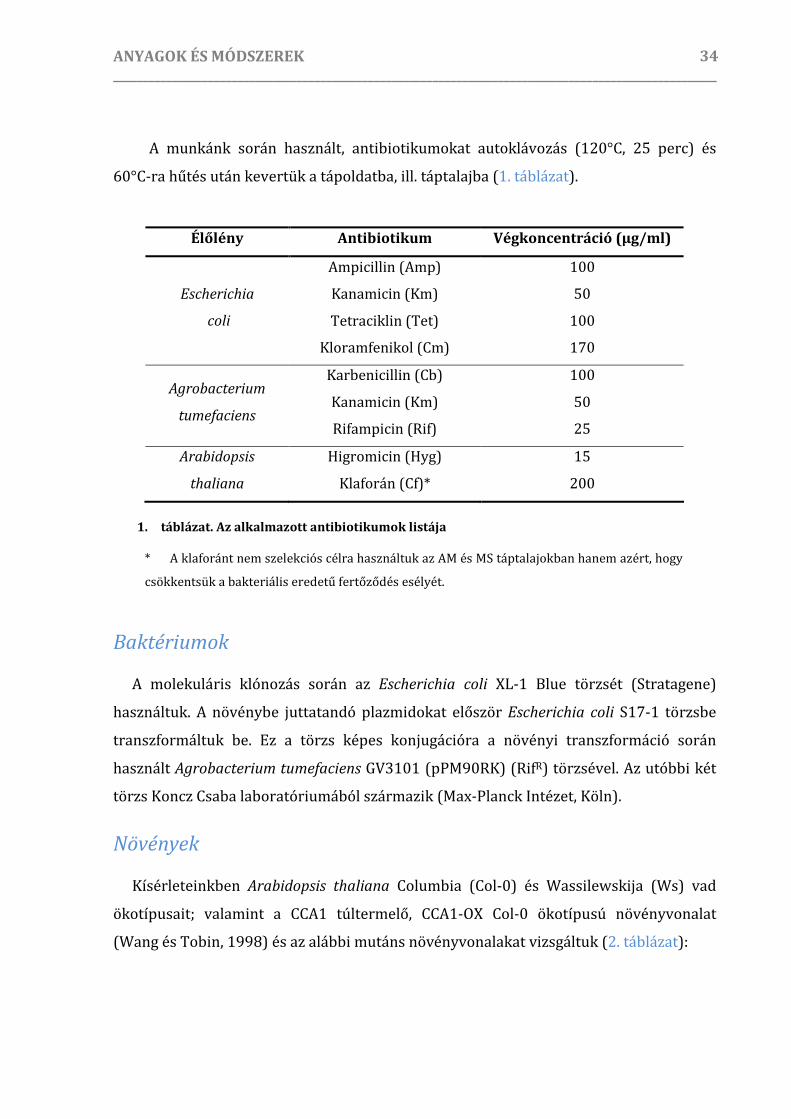
ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
34
A munkánk során használt, antibiotikumokat autoklávozás (120°C, 25 perc) és
60°C-ra hűtés után kevertük a tápoldatba, ill. táptalajba (1. táblázat).
Élőlény Antibiotikum Végkoncentráció (µg/ml)
Escherichia
coli
Ampicillin (Amp)
Kanamicin (Km)
Tetraciklin (Tet)
Kloramfenikol (Cm)
100
50
100
170
Agrobacterium
tumefaciens
Karbenicillin (Cb)
Kanamicin (Km)
Rifampicin (Rif)
100
50
25
Arabidopsis
thaliana
Higromicin (Hyg)
Klaforán (Cf)*
15
200
1. táblázat. Az alkalmazott antibiotikumok listája
* A klaforánt nem szelekciós célra használtuk az AM és MS táptalajokban hanem azért, hogy
csökkentsük a bakteriális eredetű fertőződés esélyét.
Baktériumok
A molekuláris klónozás során az Escherichia coli XL-1 Blue törzsét (Stratagene)
használtuk. A növénybe juttatandó plazmidokat először Escherichia coli S17-1 törzsbe
transzformáltuk be. Ez a törzs képes konjugációra a növényi transzformáció során
használt Agrobacterium tumefaciens GV3101 (pPM90RK) (RifR) törzsével. Az utóbbi két
törzs Koncz Csaba laboratóriumából származik (Max-Planck Intézet, Köln).
Növények
Kísérleteinkben Arabidopsis thaliana Columbia (Col-0) és Wassilewskija (Ws) vad
ökotípusait; valamint a CCA1 túltermelő, CCA1-OX Col-0 ökotípusú növényvonalat
(Wang és Tobin, 1998) és az alábbi mutáns növényvonalakat vizsgáltuk (2. táblázat):

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
35
Vonal neve
Hivatkozás Mutáció típusa
Mutáció helye
Fnkcióvesztés foka
Háttér
uvr8-6 Favory és
mtsai, 2009 T-DNS inszerció 9. intron nullmutáns Col-0
cop1-4 McNellis és mtsai, 1994
pontmutáció Gln283STOP részleges mutáns Col-0
hy5-ks50 hyh Brown és
Jenkins, 2008 T-DNS
inszerció/deléció
HY5: promóter, első exon
HYH: első exon
kétszres nullmutáns
Ws
elf3-4 Hicks és mtsai,
1996 T-DNS
inszerció/deléció első exon nullmutáns Ws
2. táblázat. A használt mutáns növényvonalak jellemzői
Molekuláris biológiai módszerek
Riportergén konstrukciók készítése, transzgenikus növények
előállítása
A növények vizsgálatakor alkalmazott riportergén konstrukciókat (CCR2:LUC+,
HY5:LUC+ és UVR8:LUC+) Agrobacterium-közvetített géntranszfer (Clough és Bent,
1998) segítségével juttattuk be a mutáns és a megfelelő vad típusú növényekbe. A
riportergének expresszióját mindegyik háttér esetében (mutáns és vad típus) legalább
öt független transzformáns növényi vonalban vizsgáltuk meg. A vizsgált riportergén
kifejeződése minden azonos genotípusba tartozó transzgenikus növényi vonalban
hasonló mintázatot mutatott.
A vizsgált gének (CCR2, HY5, UVR8) promótereit Arabidopsis Col-0 változatának
genomi DNS-éből (a növényi genomi DNS tisztítását lásd: 39. oldal) sokszorosítottuk
PCR-rel, a kívánt restrikciós enzim hasítóhelyeket tartalmazó oligonukleotidok
segítségével. A PCR-ben hibajavító enzimkeveréket (Stratagene PfuUltra™ II Fusion HS)
használtunk a gyártó útmutatása szerint.
A restrikciós endonukleáz emésztéseket és egyéb DNS módosító kezeléseket a gyártó
(Fermentas) által megadott körülmények között végeztük, majd a termékeket agaróz

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
36
gélen (SeaKem® LE, Cambrex), elektroforézissel választottuk el. A megfelelő DNS
darabokat a gélből kivágás után fenolozással tisztítottuk. A darabok ligálását a T4 DNS
ligáz gyártójának (Fermentas) útmutatásai alapján végeztük. Az összeillesztett DNS-eket
Inoue módszere (Inoue és mtsai, 1990) szerint előkészített XL-1 Blue sejtekbe egy gyors
és hatékony módszerrel (Pope és Kent, 1996) juttattuk be. A plazmidot hordozó
baktériumsejtek kiválogatását a megfelelő antibiotikumot tartalmazó LB lemezeken
37 °C-on 16 órán át végeztük. A plazmidokat Escherichia coli sejtekből a hagyományos
lúgos feltárás módosított változatával tisztítottuk (Ahn és mtsai, 2000), amelyeket
azután restrikciós enzimekkel, majd szekvenálással ellenőriztünk.
A riportergén-konstrukciókban használt szentjánosbogár LUCIFERÁZ (LUC)
módosított (erősebb fénykibocsátású) változatának (LUC+) cDNS-ét a Promega cégtől
vásároltuk.
A CCR2:LUC+, a HY5:LUC+ és az UVR8:LUC+ jelzőgének létrehozásához a CCR2 kódoló
része előtti 2233 bp-nyi DNS darabot, a HY5 kódoló része előtti 565 bp-nyi DNS darabot,
illetve az UVR8 kódoló része előtti 2568 bp-nyi szakaszt sokszorosítottuk Arabidopsis
Col-0 változatából a következő primerek felhasználásával:
CCR2 pro F: 5’-CGCAAGCTTAAATCTTCTCCTTCATCACCGCATA-3’
CCR2 pro R: 5’-CGCGGATCCTGAAATTTGAAAAGAAGATCTAAGGGA-3’
HY5 pro F: 5’-AAAGAATTCTCTAATGTTAACGTTGAGATGGCAATG-3’
HY5 pro R: 5’-TTTGTCGACTTTTCTTACTCTTTGAAGATCGATCAGG-3’
UVR8 pro F: 5’-ATAGTCGAC TAGTTCTTAGTTTCTTGGTTCC-3’
UVR8 pro R: 5’-ATAGGATCC GATCACAGTTGCAGTTTTC-3’
A termékeket közvetlenül a növényi kétgazdás pPCVH LUC+ plazmidba klónoztuk
HindIII–BamHI (CCR2), EcoRI-SalI (HY5), illetve SalI–BamHI (UVR8) helyekre.
Az Arabidopsis transzformálására használt pPCV (Plant Cloning Vector) bináris
plazmidot Koncz Csaba csoportja fejlesztette ki (Koncz és mtsai, 1994). A pPCV bináris
vektor vázlata az 5. ábrán látható.

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
37
5. ábra. A pPCV növényi klónozó vektor szerkezete
A 7.5 kb méretű, Agrobacterium Ti-plazmid alapú vektor tartalmazza a jobb- és baloldali integrációs
határszekvenciákat (LB ill. RB), ezek közé építve a bakteriális replikációs origót (ORIColE1) és az
ampicillin/karbenicillin rezisztenciát okozó β-laktamáz kazettát (ApR), valamint a növényi szelekciót
lehetővé tevő – a higromicin rezisztenciáért felelős – higromicin-foszfotranszferáz (HPT) fehérjét
konstitutívan kifejező kazettát is. A plazmidba építettek egy, a molekuláris klónozást megkönnyítő, egyedi
DNS endonukleáz hasítóhelyeket hordozó régiót, amelyet 3’ irányban a nopalin szintáz gén
3’ poliadenilációs szignálja (Tnos) követ. A NOS szekvencia az expresszált mRNS megfelelő processzálását
irányítja növényben.
Agrobacterium konjugáció
Az elkészült bináris plazmid vektort Escherichia coli S17-1 törzsbe transzformáltuk,
majd 2 ml-es, ampicillinnel kiegészített LB folyadékkultúrában 16 órán át növesztettük.
Agrobacterium tumefaciens GV3101 törzséből YEB tápoldatban, 24 óra alatt szintén friss
folyadékkultúrát neveltünk. A két baktériumszuszpenzióból 1-1 ml-t összekevertünk és
YEB táplemezen együtt növesztettünk. 24 óra után a növekedésnek indult
baktériumtenyészetből kis mennyiséget rifampicin és karbenicillin tartalmú YEB
lemezre szélesztettünk, majd a felnövekvő rezisztens Agrobacterium-telepeket legalább

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
38
három alkalommal passzáltuk. Az E. colitól ilyen módon megtisztított Agrobacteriumot
használtuk Arabidopsis virág-infiltrációs transzformálásra (lásd:43. oldal).
Növényi DNS tisztítás
Körülbelül egy cm2 felületű levelet/levéldarabot folyékony nitrogénben
lefagyasztottunk, majd 1.5 ml-es Eppendorf csőben elporítottunk. A mintára 500 μl
60°C-ra előmelegített 2×CTAB puffert (2% CTAB, 100 mM Tris·HCl pH=8.0, 20 mM EDTA
pH=8.0, 1.4 M NaCl, 1% PVP 40, 0.5% β-merkaptoetanol) mértünk rá, majd többször
összerázva a csövet, 30 percig 65°C-on tartottuk. öt percnyi ülepítés után a
törmelékmentes felülúszót új csőbe pipettáztuk át és azonos térfogatú kloroformmal
ráztuk össze. A vizes és a szerves fázist 10 perc centrifugálással választottuk szét. A felső
vizes fázisból a nukleinsavakat 0.75 térfogatnyi 2-propanollal szobahőmérsékleten
10 perc alatt csaptuk ki, majd 10 perces centrifugálással ülepítettük. A csapadékot
70%-os etanollal mostuk, légszivattyúval megszárítottuk és 100 μl csíramentes TE
pufferben (10 mM Tris·HCl pH=7.5, 1 mM EDTA) feloldottuk. A mintát 10 μg RNáz-A
enzimmel kezeltük 37°C-on egy órán át, majd fenol-kloroform (1:1) elegyével ráztuk
össze. Centrifugálás után a vizes fázist kloroformmal is tisztítottuk, majd a vizes fázisból
a DNS-t 0.2 térfogatnyi 3 M nátrium-acetát pH=5.2 és 0.75 térfogatnyi 2-propanol
hozzáadásával szobahőmérsékleten 10 perc alatt csaptuk ki. Centrifugálásos ülepítés és
70%-os etanollal történő mosás után a csapadékot megszárítottuk és 100 μl csíramentes
TE-ben oldottuk fel. A mintából 1–2 μl-t használtunk egy PCR-hez.
Növényi RNS tisztítása
Össz-RNS-t ~100 mg, folyékony nitrogénben porított növényi anyagból a Qiagen
RNeasy® Plant Mini Kit-jével tisztítottuk (oszlopon történő RNáz-mentes DNáz
kezeléssel kiegészítve) a gyártó leírása alapján. A kapott RNS mennyiségét és minőségét
a minta 260 és 280 nm-en mért fényelnyeléséből határoztuk meg.
mRNS szint meghatározása RT-PCR-rel
Mérési pontonként ~20 db csíranövényből tisztítottunk össz-RNS-t. 0.5–1 µg RNS-ből
cDNS-t a Fermentas által gyártott RevertAid™ First-Strand cDNA Synthesis Kit
felhasználásával készítettünk a gyártó útmutatásainak megfelelően. A termékeket RNáz-

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
39
mentes vízzel ötszörösére hígítottuk. Az mRNS-ek szintjét valós idejű PCR-rel
ABI PRISM® 7700 (Applied Biosystems) gépen, ABI SYBR® Green PCR Master Mix
(Applied Biosystems) alkalmazásával 10 µl térfogatban határoztuk meg.
Minden kísérletnél készítettünk egy cDNS hígítási sort: UV-B-fénnyel kezelt vad
típusú növények a fénykezelés után 90 perccel gyűjtött mintáiból cDNS keveréket
készítettünk, amelyet 1-, 10-, 100-, 1000-szeresre hígítottunk. Ezekben megmértük a
kérdéses mRNS-ek szintjét. Az egyes primerpárokkal kapott értékekre egyenest
illesztettünk, amelynek meghatároztuk az egyenletét. Az ismeretlen mintákban az mRNS
szinteket ennek a hitelesítő egyenesnek a segítségével számítottuk ki. A kapott értékeket
a minták TUB2 és TUB3 össz mRNS-szintjére vonatkoztattuk. Minden mintát háromszor
mértünk meg. A sokszorozáshoz használt primerek az alábbiak voltak, amelyeket 0.3 µM
töménységben alkalmaztunk (3. táblázat).
Gén neve Forward primer (5’→3’) Reverse primer (5’→3’)
CCA1 (At2g46830) CTGTGTCTGACGAGGGTCGAA ATATGTAAAACTTTGCGGCAATACCT
PRR9 (At2g46790) GCCTTCTCAAGATTTGAGGAAAGC TTTGGCTCACCTGAAGTACTCTC
GI (At1g22770) AATTCAGCACGCGCCTATTG GTTGCTTCTGCTGCAGGAACTT
ELF4 (At2g40080) CGACAATCACCAATCGAGAATG AATGTTTCCGTTGAGTTCTTGAATC
CHS (At5g13930) TGAGAACCATGTGCTTCAGGCGGAG TGTCGACTTGTCGCACATGCGCT
GPX7 (At4g31870) GACAAGGTTGATGTGAATGGA GATGATATCACCAAGGAAACCA
ELIP2 (At4g14690) CTCCTATGAAACCTAAGGTGAG GCTAAACGTCCGTTAATCCT
HY5 (At5g11260) CAGGCGACTGTCGGAGAAAGTCAAAGG TCAACAACCTCTTCAGCCGCTTGTTCTC
HYH (At3g17609) GCCTCAAGAGATTATTGAGGA CTCTTGATTCCAAATCACTCAC
At3g16350 ATTCGACATACTAGTTCAAGCC CGATGAATCGGTTACCATCTC
At3g57830 TTACTATGCTGAGGACGAGAG GTGAAGGCAGAGTATTCGAG
TUB2/TUB3
(At5g62690/At5g62700) CCAGCTTTGGTGATTTGAAC CAAGCTTTCGGAGGTCAGAG
3. Táblázat Az RT-PCR reakciókban használt oligonukleotidok listája
A PCR körülményei az alábbiak voltak: 94°C 2.5 perc, 40×(95°C 15 mp, 60°C 1 perc),
95°C 15 mp, 60°C 1 perc, 95°C 15 mp, 60°C 15 mp.

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
40
Növényi összfehérje tisztítása
50–100 mg növényi anyagot folyékony nitrogénben lefagyasztottunk és 1.5 ml-es
csőben porrá őröltük. Az elporított mintára 300 μl/100 mg 60°C-ra előmelegített
feltárópuffert mértünk (4 M urea, 5% SDS, 5% β-merkaptoetanol, 16.66% glicerin). A
mintákat tovább őröltük addig, amíg nagyobb darabokat már nem láttunk az oldatban,
majd 60°C-os vízfürdőbe helyeztük őket 10 percre. Időnként erőteljesen összeráztuk a
csöveket. A törmeléket ezután centrifugálással ülepítettük 30 percig. A tiszta felülúszót
új csövekbe mértük át. Az így elkészített minták azonnal felvihetők poliakrilamid gélre.
Fehérjeszint meghatározása Western blot módszerrel
Mintánként 10μl fehérjekivonatot futtattunk 10%-os SDS-poliakrilamid gélen
Tris/glicin/SDS futattópufferben. Futtatás után a gélből 2×15 percen keresztül
átvivőpufferrel (12mM Tris·HCl pH=8.3, 96mM glicin, 20% metanol) mostuk ki az SDS és
urea maradványokat. A gélből az elválasztott fehérjéket elektroblot készülékkel (Bio-
Rad; 25V, 1h) átvittük Immobilon®-P PVDF (polivinilidén-difluorid) membránra
(Millipore). Az átvitel után a membránról levágtuk a belső kontrollt (aktin) tartalmazó
részt, majd a membránokat TBST pufferrel (20mM Tris·HCl pH=7.5, 150mM NaCl, 0.05%
Tween® 20) öblítettük át. A szabadon maradt kötőhelyeket kioltópufferrel (5% sovány
tejpor, 1% szérum albumin (Bovine Serum Albumin, BSA) TBST pufferben)
semlegesítettük 4°C-on egy éjszakán át. Ezután a membránokat műanyag zacskóba
helyeztük, amelyet (Dr Roman Ulm csoportja által készített) UVR8-, illetve az aktin elleni
elsődleges ellenanyag hozzáadása után lehegesztettünk. Az ellenanyagokat
kioltópufferben egy órán keresztül hagytuk a membránon, majd a membránt kivéve
3×5 perc TBST pufferben történő mosással távolítottuk el róla a felesleges ellenanyag
maradványokat. Ezután kioltópufferben 10000-szeresre hígított, alkalikus-foszfatázhoz
kapcsolt nyúl-IgG elleni másodlagos ellenanyagot (BioRad, 170-6518) adtunk hozzá, és
így tartottuk szobahőmérsékleten egy óráig. Ezt követően 3×10 percig TBST pufferben
mostuk. A felesleges folyadék leitatása után előhívó oldatban (100mM Tris·HCl pH=9.5,
100mM NaCl, 5mM MgCl2, 0.02% 5-Bróm-4-klór-3-indolil foszfát (BCIP),
0.03% Nitroblue-tetrazólium (NBT) tartottuk szobahőmérsékleten, míg a fehérjesávok
láthatóvá nem váltak. A reakciót vizes öblítéssel leállítottuk, majd a lapot megszárítottuk
és képet készítettünk róla.

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
41
Növényeken alkalmazott technikák
Magsterilizálás, növénynevelés
A sterilizálandó magokat vízbe áztatás után két napig 4°C-on tároltuk, majd 10 percig
30%-os Domestos oldatban áztattuk. Ezután steril desztillált vízzel ötször átmostuk,
majd a steril magokat táptalajra szélesztettük. Ezt követően a növényeket 12 h fehér
fény (50 μmol m-2 s-1)/12 h sötétben, 22°C-on neveltük steril körülmények között,
növénynevelő kamrában (MLR-350, Sanyo) hét napig.
Fénykezelések
A méréseket 22°C-on végeztük. A kísérletekben használt fehér fényt fluoreszcens
fénycsövek (PHILIPS TL-D 18W/33-640 biztosították, a megvilágítás intenzitása
10 μmol m-2 s-1 volt. A monokromatikus vörös (λmax=667nm), és kék (λmax=470nm)
fénypulzusokhoz Quantum Devices Inc (USA) Snaplite™ LED-eket alkalmaztunk. Az
alkalmazott fényintenzitások 40 μmol m-2 s-1 (vörös), és 25 μmol m-2 s-1 (kék) volt. Az
kísérleteinkbhez használt alacsony intenzitású (1.5 μmol m-2 s-1) UV-B-t PHILIPS
ULTRAVIOLET-B TL 20W/01 RS; a tolerancia teszthez alkalmazott nagy intenzitású
(14 μmol m-2 s-1) UV-B-t pedig PHILIPS ULTRAVIOLET-B TL 40w/12 RS fénycsövek
segítségével állítottuk elő ezen fénycsövek fénykibocsátását a 6. ábrán ábrázoltuk.
Kísérleteinkben az UV-B megvilágítást minden esetben fehér fénnyel együtt alkalmaztuk,
ezzel közelítve modellrendszerünket a külső környezetben tapasztalható
körülményekhez, hiszen a természetben sincsen UV-B látható fény nélkül.
A fényintenzitás méréséhez látható tartományban a Gigahertz-Optik cég X11
fénymérőjét használtuk XD-9504 mérőfejjel, míg az UV-B fényt a Cole-Palmer cég
fénymérőjével (katalógus szám: 97503-00), 312 nm-es szenzorral mértük.

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
42
Alkalmazott UV szűrők
A kísérletekben „alulvágó” (cut-off) filtereket alkalmaztunk. UV-B kezelésnek
alávetett mintáknál alkalmazott LP305 filter nem engedi át a fényforrásból származó
UV-C fényt, viszont átengedi az UV-B jelentős részét. A kontroll növények esetében
használt LP385 filter pedig kiszűri a teljes UV-B tartományt. A szűrőket a Rapp
Optoelectronic GmbH (Hamburg, Németország)-tól vásároltuk, jellemzőik a 7. ábrán
láthatóak.
6. Ábra. Az alkalmazott UV-B
fényforrások fénykibocsátási
spektruma
(A) A cirkadián és génindukciós
kísérletekhez alkalmazott PHILIPS
ULTRAVIOLET-B TL 20W/01 RS, illetve
(B) az UV-B stressztolerancia tesztben
használt PHILIPS ULTRAVIOLET-B TL
40w/12 RS fénycsövek hullámhossz
tartománya.
7. Ábra. A használt UV-B szűrők jellemzői
Fekete vonal:LP305, piros vonal: LP385.

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
43
Transzgenikus növények előállítása
A megfelelő pPCV alapú vektorkonstrukciót hordozó Agrobacterium törzset 300 ml,
100 µg/ml karbenicillint tartalmazó YEB tápoldatban növesztettük 25°C-on két napig
(OD600= 2.5-3.0), majd az Agrobacteriumot centrifugálással (4000 g, 20 perc, 25°C)
összegyűjtöttük és 3 ml YEB oldatban felszuszpendáltuk. Ezután folyamatos kevergetés
mellett 300 ml 3%-os (w/v) szaharózoldatot, végül 40 µl Silwet L-77-et (Lehle Seeds)
adtunk a szuszpenzióhoz. A transzformálandó, üvegházban nevelt Arabidopsis növények
teljes virágzatát 15 másodpercre a szuszpenzióba merítettük, majd a növényeket egy
napig letakarva, magas páratartalmat biztosítva, tartottuk. A felnevelt növényekről
magot gyűjtöttünk és a megfelelő szelekciós ágenssel és klaforánnal kiegészített AM
táptalajon steril körülmények között szelektáltuk a transzformánsokat (T1 nemzedék),
amelyeket, miután megfelelő erősségű gyökérzetet fejlesztettek (10-12 nap), földbe
ültettünk és üvegházban felneveltünk. Minden konstrukció esetében 20-30 független T1
transzformáns vonalat állítottunk elő. Vizsgálatainkat a T3 nemzedék transzgénre nézve
homozigóta, a transzgént egy kópiában hordozó egyedein végeztük.
In vivo lumineszcencia mérés luminométerrel
A luciferáz jelzőgéneket kifejező csíranövényeket 12 óra fény (50 µmol m-2 s-1) /12
óra sötét fénycikluson, 22°C-on neveltük MS3 táptalajon, hét napig. A mérés kezdete
előtt min. hat órával a csíranövényeket steril körülmények között egyenként egy 96
zsebes mikrotiter lemezbe helyeztük, amelynek minden zsebe 0.25 ml MS3 táptalajt
tartalmazott. Minden növényre 20 µl 2 mM-os luciferin oldatot cseppentettünk. A
méréseket TopCount® NXT™ automatizált luminométerrel (Perkin-Elmer) végeztük,
amely minden egyes - a mikrotiter lemez zsebeiben elhelyezett - csíranövény által
kibocsátott lumineszcenciát 1-2 óránként rögzítette, a mérési idő végéig (2-7 nap). A
mérés a csírázást követő 7. napon indult, a nevelés során alkalmazott fényciklusoknak
megfelelően a fény-sötét átmenet pillanatában. A mérés módjától függetlenül a
növények lumineszcenciáját az első 12 óra során (a nevelési körülményeknek megfelelő
éjszakai fázisban) sötétben mértük. Ezt követően a mintákat a mérés módjától függően
sötétben, illetve folyamatos fehér fényben tartottuk. A fényben mért minták csupán a
mérések alkalmával (egy-két óránként öt percre) kerültek a mérőtér sötétjébe, ami nem
befolyásolja a cirkadián óra működését (Millar és mtsai, 1995a). A mérési eredmények

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
44
feldolgozása a TopTempII nevű Microsoft Excel makro programmal (Dr. Andrew Millar,
University of Edinburgh, UK ajándéka) történt. A luminométer által mért lumineszcencia
értékek az egy másodperc alatt beérkezett fotonok számát (cps = count per seconds)
mutatják. Egy csíranövény által adott időpontban kibocsátott lumineszcencia értékét az
adott növény által a teljes mérés során kibocsátott lumineszcencia átlagával osztottuk el,
így kaptuk meg az adott időponthoz tartozó normalizált lumineszcencia értéket. Az
ábrákon –ha másképpen nem tüntettük fel– legalább 16 csíranövény (normalizált)
lumineszcencia értékéből számolt átlagértéket ábrázoltunk. az idő függvényében. Az
időtengely 0 pontja az utolsó sötét/fehér fény átmenet időpontja.
Periódus adatok számolása
A periódusok becslését a Biological Rhythms Analysis Software System (BRASS)
(Locke és mtsai, 2005; Southern és Millar, 2005) segítségével végeztük el. A Microsoft
Excel alapú makró képes a TopCount®NXT™ luminométer használata során nyert adatok
kiértékelésére. A program a „Fast Fourier Transform-Non-Linear Least Squares (FFT-
NLLS)” módszerrel (Plautz és mtsai, 1997) minden egyes növény adatsorára illesztett
görbe alapján ad becslést az egyedek periódusára. Minden kísérletben legalább 80,
legfeljebb 120 órás mérési időtartamból származó adatokat használtunk fel.
Csoportonként legalább 20 növény periódusának statisztikai elemzését BRASS
programmal végeztük el. A periódus középértékének becslése során csak azokat az
értékeket vettük figyelembe, amelyek 15 és 38 óra közé estek (cirkadián tartomány).
A fázis igazíthatóságának mérése (PRC)
A CCR2:LUC+ jelzőgént hordozó növényeket 12 óra sötét/12 óra fehér fény
(50 µmol m-2 s-1) váltásokkal állítottuk be hét napig, majd folyamatos sötétbe helyeztük
őket. Ezután háromóránként egyórás, ugyanolyan erősségű fénypulzusokat kaptak. A
pulzusok által kiváltott fáziscsúszást a pulzus időpontjának függvényében ábrázoltuk. A
különböző periódusú növények fázisainak összehasonlíthatósága érdekében a
fáziscsúszásokat a következő módon számítottuk ki: a kezelt növények új fázisát (a
folyamatos sötétbe helyezés után jelentkező harmadik cirkadián csúcs idejét) a
kezeletlen, folyamatosan sötétben mérődő növények fázisából kivontuk. A negatív
értékek jelzik ezért az óra késését, mert a csúcs későbbi időpontban van, mint a
kezeletlenben. A pozitív értékek a hamarabb érkező, siető csúcsot jelölik. Megmértük

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
45
továbbá az adott növényvonal folyamatos sötétben mérhető periódusát is. Ezt az értéket
elosztottuk 24-gyel, majd a kapott arányszámmal szoroztuk be az összes
fáziskülönbséget, valamint a pulzusok időpontjait is. Így minden értéket egy névleges
24 órás napon belül tudtunk feltüntetni (cirkadián idő). A fáziscsúszás hibáját a kezelt és
a kezeletlen növényekben mért fázisok középértékhibáiból (SEM) számoltuk ki az alábbi
képlet szerint:
A kapuzás (gating) vizsgálata
A csíranövényeket 12 óra fény (50 µmol m-2 s-1) /12 óra sötét fénycikluson, 22°C-on
neveltük MS3 táptalajon, hét napig, majd az utolsó sötét fázis után alacsony intenzitású
(10 μmol m-2 s-1) folymatos fehér fénybe helyeztük őket (ZT0 időpont). Harminchat
órával később másfélnapos időszakon (ZT36-ZT72) keresztül háromóránként
növényanyagot szedtünk az indukció nélküli mRNS szintek megállapításához. Minden
mintaszedés alkalmával egy-egy elkülönített növénycsoportot 30 perc, 1.5 μmol m-2 s-1
UV-B kezelésnek vetettünk alá. A kezelések után a növényeket visszahelyeztük a fehér
fénybe és a kezelés kezdetétől számított 90. percben leszedett mintáikból határoztuk
meg az indukált mRNS szinteket. A vörös fénnyel végzett kísérletnél a növények
folyamatos sötétbe kerültek, és oda lettek visszahelyezve a fényindukció
(60 perc, 40 μmol m-2 s-1) után.
UV-B tolerancia vizsgálata
A csíranövényeket 12 óra fény (50 µmol m-2 s-1) /12 óra sötét fénycikluson, 22°C-on
neveltük MS3 táptalajon hét napig, majd 12 óra alacsony intenzitású (10 μmol m-2 s-1)
fehér fénnyel kiegészített 1.5 μmol m-2 s-1 UV-B fény / 12 óra sötét fényciklusra
helyeztük át két napra. Az áthelyezés utáni harmadik napon, háromóránként egy-egy
elkülönített növénycsoportot nagy intenzitású (14 μmol m-2 s-1) UV-B besugárzásnak
vetettünk alá, míg a kontroll növényeket LP385-ös filterrel megszűrt fénnyel kezeltük. A
kezelések után a növényeket visszahelyeztük a kísérlet elején is alkalmazott
körülmények közé, majd hét nap után készítettük el a fotókat és szedtünk növényi
mintákat a klorofilltartalom meghatározásához.

ANYAGOK ÉS MÓDSZEREK ______________________________________________________________________________________________________
46
Klorofilltartalom meghatározása
A klorofill méréshez mintánként 10-10 csíranövényt Eppendorf csőbe helyeztünk.
Lemértük a nedvestömegüket, majd 1-1 ml N,N-dimetilformamidot mértünk rájuk. A
mintákat időnkénti összerázással 24 órán keresztül sötétben, szobahőmérsékleten
tartottuk, majd lecentrifugáltuk. A növényi anyagtól mentes felülúszó abszorbanciáját
Shimadzu spektrofotométerrel 664 nm-en lemértük, és a klorofill mennyiséget ebből
számítottuk ki (Inskeep és Bloom, 1985) A klorofill mennyiségeket a nedves tömegek
mennyiségeivel arányosítottuk, és a kontrollhoz viszonyítva ábrázoltuk.

EREDMÉNYEK
Cirkadián óra beállíthatósága UV-B fénnyel
Az UV-B befolyásolja a cirkadián óra sebességét
Ismert biológiai tény, hogy a cirkadián rendszerek beállításában a (látható) fény
elsőrendű szerepű. Munkánk során arra kerestük a választ, hogy a növények képesek-e
életfolyamataik időzítéséhez felhasználni a fotoszintézisben nem hasznosuló UV-B fényt.
Folyamatos fényben a mért periódus és az alkalmazott fény erőssége között fordított
összefüggés van (parametrikus beállítás; Devlin és Kay, 2000), emiatt a fényválasz görbe
(FRC, a periódus ábrázolása az alkalmazott fény erősségének függvényében)
meredeksége növényekben negatív.
A fényválasz görbe előállításához CCR2:LUC+ jelzőgént kifejező vad hátterű és uvr8-6
csíranövényeket állítottunk be 12 óra fény/12 óra sötét váltásokkal egy héten keresztül,
majd olyan azonos fényintenzitású fehér fénybe helyeztük őket, melyet két különböző
UV-B intenzitással egészítettünk ki. A csíranövények által kibocsátott fényt
luminométerrel rögzítettük, a kialakuló periódusokat a BRASS alkalmazással
számszerűsítettük, majd a fényerősség függvényében ábrázoltuk (8. ábra, A mező). A
kapott eredmények alapján megállapíthatjuk, hogy vad típusban a periódus UV-B fény
intenzitásának emelkedésével csökken akárcsak látható fényben, míg uvr8-6
növényekben nem tapasztalható periódus változás.

EREDMÉNYEK ______________________________________________________________________________________________________
48
Annak eldöntésére, hogy az uvr8-6 növények fényválasza eltérést mutat-e a vad
típushoz képest a kísérletet elvégeztük különböző intenzitású fehér fénnyel is UV-B
hiányában (8. ábra, B mező). Megfigyelhető, hogy az uvr8-6 periódusa UV-B hiányában
nem tér el a vad típusétól sem sötétben, sem állandó fényben.
Hasonló kísérleti elrendezésben megvizsgáltuk az UV-B jelátvitel más elemei
hiányának a hatását is. Amint az a 4. táblázatban látható, a hy5 hyh dupla mutáns
periódusa nem tér el a vad típusétól fehér fényben és ahhoz hasonlóan körülbelül egy
órával rövidül a periódus az UV-B hatására. A cop1-4 periódusa a szakirodalomban
(Yu és mtsai, 2008) leírtaknak megfelelően nagyjából két órával rövidebb mint a vad
típusé és az UV-B periódus csökkentő hatására ugyanúgy érzéketlen, mint az uvr8-6. A
HY5:LUC+ jelzőgén periódusának UV-B hatására történő csökkenése pedig azt mutatja,
8. ábra. A CCR2:LUC+ jelzőgén
periódusának fényerősségtől való
függése
A vad típusú Col-0 (fekete négyzet) és uvr8-6 mutáns (piros háromszög) háttérben CCR2:LUC+ jelzőgént kifejező hét napos csíranövényeket azonos fényerősségű fehér (10µmol m-2 s-1), de eltérő intenzitású UV-B fénybe (A), illetve különbözű intenzitású folyamatos fehér fénybe (B) helyeztük. A jelzőgén működését luminométerrel 5 napig követtük nyomon, az egyedi növények periódusait BRASS alkalmazással becsültük meg.

EREDMÉNYEK ______________________________________________________________________________________________________
49
hogy az UV-B a hatását a központi órán fejti ki, nem csak a CCR2 kimeneti elemet
befolyásolja.
4. táblázat. CCR2:LUC+ és HY5:LUC+ periódusa UV-B jelátviteli mutáns és vad hátterű
növényekben
A jelzőgének vizsgálatakor a csíranövények cirkadián óráját 12 óra fény/12 óra sötét váltásokkal
állítottuk be hét napon keresztül, majd folyamatos fehér (10 μmol m-2 s-1), illetve UV-B-vel
(1.5 μmol m-2 s-1) kiegészített fehér fénybe helyeztük őket. A luciferáz működését a szubsztrát
hozzáadása után luminométerben mértük öt napon keresztül. (A periódusbecslést a BRASS
alkalmazással végeztük.)
UV-B pulzusokkal a cirkadián óra késleltetése idézhető elő
A fény szükséges a periódus finomhangolása mellett a naponkénti órabeállításhoz is.
A cirkadián óra a nap egy bizonyos szakára korlátozza a saját beállíthatóságát,
legfőképpen a napfelkelte vagy a naplemente környékére. Ekkor legnagyobb ugyanis a
fényerősség változása, ami egy jó tájékozódási pont a beállításhoz. Érzékenységének
időzítését az óra saját bemeneti elemeinek cirkadián szabályozásával oldja meg (Lakin-
Thomas, 2000; Roenneberg és Merrow, 1998; Tóth és mtsai, 2001). A beállíthatóság
vizsgálatánál az állandó körülmények között, szabadon futó cirkadián órát érő
fénypulzusok által kiváltott perióduscsúszást mérjük. Vad típusú növényekben kék vagy
vörös pulzusok hatására jelentős fáziscsúszások történnek, leginkább a szubjektív
éjszaka során, amikor az egyre nagyobb mértékű késések sietésbe csapnak át, igen
jellegzetes fázisválasz görbét mutatva (Covington és mtsai, 2001).
Annak vizsgálatára, hogy a jelenség kiváltható-e UV-B fénnyel is, a CCR2:LUC+
jelzőgént hordozó vad típusú és uvr8-6 növényeket egy hétig 12 óra fény/12 óra sötét
cikluson tartottuk, majd alacsony intenzitású folyamatos fehér fénybe helyeztük át. Az
áthelyezés után 36 órával háromóránként 30 perces 1.5 μmol m-2 s-1 intenzitású UV-B
pulzussal kezeltük őket. Az egyes pulzusok által kiváltott CCR2:LUC+ fáziscsúszások
Periódus (óra) ±SEM
CCR2:LUC+ HY5:LUC+
Megvilágítás
Col-0 uvr8-6 cop1-4 Ws hy5 hyh Ws
fehér 25.36±0.18 25.21±0.13 22.07±0.14 25.97±0.13 25.83±0.17 27.25±0.26
fehér és UV-B 24.45±0.22 25.35±0.25 22.0±0.13 24.82±0.08 24.61±0.2 24.95±0.46

EREDMÉNYEK ______________________________________________________________________________________________________
50
irányát és erősségét nem kezelt kontroll növényekhez viszonyítva kaptuk meg az
aktuális sietés/késés értékét, amit a fénypulzusok idejének cirkadián időbe történt
átszámításának a függvényében ábrázoltunk (9.ábra, A mező). UV-B pulzussal készült
PRC-ből kiderült, hogy vad típusú növényben UV-B fénypulzusok gyenge késést okoznak
a szubjektív éjszaka során felében, viszont nem okoznak periódusváltozást a szubjektív
nappal alatt. uvr8-6 és cop1-4 növényekben pedig nem volt tapasztalható egyáltalán
fázisváltozás.
9. ábra. A cirkadián óra beállíthatóságának vizsgálata fénypulzusokkal
A CCR2:LUC+ jelzőgént hordozó vad típusú Col-0 (fekete négyzet) uvr8-6 (piros háromszög), és cop1-4
mutáns (kék kör) hét napos csíranövényeket folyamatos, alacsony intenzitású (10 μmol m-2 s-1) fehér
fénybe (A) vagy sötétbe (B, C, D) helyeztük. Ezután háromóránként 30 perces (A), illetve egyórás (B, C, D),
ugyanolyan erősségű fénypulzusokat kaptak. A pulusok által kiváltott fáziscsúszást a pulzus időpontjának
függvényében ábrázoltuk. Az értékek számolásának és feltüntetésének módját lásd a 44-45. oldalon. A
növényeket a következő színű és erősségű fénypulzusokkal kezeltük: (A) UV-B (1,5 μmol m-2 s-1); (B)
vörös (40 μmol m-2 s-1); (C) kék (25 μmol m-2 s-1); (D) fehér (10 μmol m-2 s-1). A fáziscsúszásokat és a
fénypulzusok idejét az összehasonlíthatóság miatt 24 órára viszonyítva adtuk meg.

EREDMÉNYEK ______________________________________________________________________________________________________
51
A kísérletet megismételtük látható fényt alkalmazva. A növényeket egyhetes beállítás
után folyamatos sötétbe helyeztük, majd 36 órával később 1-1 órás vörös
(40 µmol m-2 s-1) kék (25 µmol m-2 s-1), illetve fehér (10 µmol m-2 s-1) fénypulzusokat
alkalmaztunk (9. ábra, B, C, D mező)). Ezen körülmények között az uvr8-6 növények
fázisválasza megkülönböztethetetlen volt a vad típusétól. Megállapíthatjuk, hogy UV-B
fény képes beállítani a cirkadián órát és ebben a folyamatban az UVR8 és a COP1
kulcsfontosságú.
UV-B általi fényindukció cirkadián szabályozása
Optimális indukciós és mintaszedési körülmények megállapítása
Mielőtt megvizsgáltuk, hogy a cirkadián óra szempontjából fontos gének a látható
fényhez hasonlóan indukálódnak-e UV-B hatására, szükséges volt elvégeznünk még két
kísérletet. Az előzményekből az feltételezhető volt, hogy az UV-B indukálja legalább az
egyik óragént, viszont nem tudhattuk mekkora az optimális UV-B dózis, illetve, hogy az
indukció hatására milyen a célgének mRNS-einek átíródási kinetikája, vagyis az
indukciót után mennyi idővel érdemes mintát szedni ahhoz, hogy maximális, vagy
legalábbis jelentős mértékű legyen a szintnövekedés.
Az mRNS szint felfutásának a vizsgálatához alacsony intenzitású, állandó fehér fénybe
helyezett vad típusú csíranövényeket 51 órával az áthelyezés után 10 perces
1,5 µmol m-2 s-1 erősségű UV-B pulzussal kezeltük. A kezelés előtt közvetlenül, illetve
utána 20 perccel, majd ezt követően 10 percenként mintát szedtünk, amiből össz RNS-t
tisztítottunk. Az mRNS szinteket valós idejű PCR segítségével határoztuk meg. Az
értékeket a TUBULIN mRNS szintjéhez viszonyítottuk, majd a kezdeti indukálatlan minta
növekményeként ábrázoltuk (10. ábra).

EREDMÉNYEK ______________________________________________________________________________________________________
52
10. ábra. Néhány UV-B indukált gén mRNS szintje az indukciót követően
A vad típusú (Col-0) hét napos csíranövényeket alacsony intenzitású (10 µmol m-2 s-1) fehér fénybe helyeztük. 51 órával később 10 perces 1,5 µmol m-2 s-1 erősségű UV-B pulzussal kezeltük őket. A kezelést közvetlenül megelőzően (0 időpont) illetve utána 10 percenként mintát vettünk, amelyekből össz-RNS-t tisztítottunk. Az mRNS-szinteket valós idejű PCR segítségével határoztuk meg, az értékeket a TUBULIN mRNS szintjéhez viszonyítottuk és a kezeletlen minta növekményeként ábrázoltuk. (A) Óragének: PRR9 (fekete), GI,
(piros), CCA1 (narancs) (B) UV-B indukálódó nem óragének:
HY5 (zöld), glutation peroxidáz (kék). A glutation peroxidáz mRNS mennyiségeket a bal- a HY5 mRNS mennyiségeket pedig a jobboldali skála mutatja.
A kísérletből megállapíthattuk, hogy mindhárom fényindukciót mutató óragén
indukálható UV-B-vel is. A három gén közül az egyébként legnagyobb növekedést
produkáló PRR9 mRNS-e érte el legelőször a maximumát, az indukció kezdete után
90 perccel. Ezt az időpontot választottuk indukció utáni mintavételi pontnak, amit az is
alátámasztott, hogy más UV-B indukciót mutató gének mRNS-ei (10. ábra, B panel) még
később érik el a csúcsértéküket.
A következő kísérletben az előbbiekkel azonos módon nevelt vad típusú növényeket
különböző idejű és erőségű UV-B pulzusokkal kezeltük. A kezelés megkezdése előtt majd
a kezelések kezdetétől számított 90. percben gyűjtött mintákból tisztítottunk össz RNS-t.
Ezekből a PRR9 mRNS szinteket valós idejű PCR-segítségével határoztuk meg, az
értékeket az előző kísérlethez hasonlóan ábrázoltuk (11. ábra).

EREDMÉNYEK ______________________________________________________________________________________________________
53
11. ábra. A PRR9 mRNS szint növekedése
különböző erősségű UV-B sugárzás
hatására
A vad típusú (Col-0) hét napos csíranövényeket alacsony intenzitású (10 µmol m-2 s-1) fehér fénybe helyeztük. 51 órával később különböző idejű (10-40 perc) és erősségű (1.5-4 µmol m-2 s-1) UV-B pulzussal kezeltük őket. A kezelést megelőzően illetve utána 90 perccel mintát vettünk. A mintákat az előző kísérlethez hasonlóan kezeltük illetve az mRNS értékeket ugyanúgy ábrázoltuk. Az alkalmazott fényintenzitásokat különböző
szinekkel jelöltük. LP 385 mutatja a kezeletlen kontroll növekményét a vizsgált időszakban.
Megállapítható volt, hogy a 1,5 µmol m-2 s-1 UV-B intenzitás 30 percen keresztül elég a
maximális válasz kiváltásához, valamint a 385 nm-es filterrel megszűrt fényforrás nem
okoz mRNS szint emelkedést. (A tapasztalható gyenge növekedés a kontroll mintában a
PRR9 mRNS napi ciklusa miatt van.)
Az UV-B képes indukálni a központi óraelemeket
Jelenleg még nem teljesen ismert az a pontos jelátviteli mechanizmus, amelyen
keresztül a fény képes beállítani a cirkadián órát. Az aktuális óramodell szerint a
növényi cirkadián óra lelke az óragének átírásán és mRNS-ének lefordításán alapuló
visszacsatolási kör. Az elmélet szerint az óragének mRNS szintjének változása szükséges
a hullámmintázat létrehozásához és a fázisbeállításhoz egyaránt. Egyes óragének (CCA1,
LHY, PRR9, GI), illetve a központi órával kölcsönható ELF4 átírása közvetlen
fényindukciót mutat (Wang és mtsai, 1997; Tepperman és mtsai, 2001; Locke és mtsai,
2005; Ito és mtsai, 2003; Kikis és mtsai, 2005). Jelenlegi ismereteink szerint ez az mRNS
szint növekedés a kiváltója a fázisátállításoknak.
Ezek a gének nem csak fényindukálhatóak, hanem a cirkadián óra szigorú kontrollja
alatt is állnak, azaz a kifejeződésük állandó körülmények között (akár fény hiányában) is
ritmikus, a fényindukciójuk pedig kapuzott Ez úgy értendő, hogy egy azonos erősségű
fénypulzus más és más mértékű indukciót vált ki a cirkadián nap során. Maximálisat a
gén saját (indukció nélküli) cirkadián csúcsa körül, és egészen kicsit a cirkadián
minimum környékén.

EREDMÉNYEK ______________________________________________________________________________________________________
54
Annak megállapítására, hogy mindez igaz-e UV-B indukcióra is, megvizsgáltuk a
fényindukált óragének (CCA1, PRR9, GI, ELF4) akut indukcióját valós idejű PCR
segítségével vad típusú, illetve uvr8-6 és cop1-4 mutáns hátterekben. A növényeket az
előzőekhez hasonlóan állítottuk be, fény-sötét ciklusokkal, majd folyamatos, alacsony
intenzitású (10 µmol m-2 s-1) fehér fénybe helyeztük őket. Az egyes növénycsoportokat
1,5 µmol m-2 s-1 intenzitású 30 perces UV-B pulzusokkal kezeltük háromóránként az
áthelyezés utáni 36. órától kezdődően. A kezelés után a növényeket visszahelyeztük
folyamatos fehér fénybe, majd egy órával később gyűjtöttünk róluk mintát és ebből
határoztuk meg az indukált génkifejeződés szintjét. Közvetlenül a kezelés előtt vett
mintákból pedig a vizsgált gének alapaktivitásait állapítottuk meg (12. ábra).

EREDMÉNYEK ______________________________________________________________________________________________________
55
12.ábra. Központi óragének UV-B indukciója uvr8-6 és cop1-4 háttérben
A vad típusú Col-0 (fekete négyzet), uvr8-6 (piros háromszög) és cop1-4 mutáns (kék kör) hét napos csíranövényeket alacsony intenzitású (10 µmol m-2 s-1) folyamatos fehér fénybe helyeztük és háromóránként 30 perces 1.5 µmol m-2 s-1 erősségű UV-B pulzussal kezeltük őket. Közvetlenül a kezelést megelőzően szedett mintákból készítettük az alapaktivitások görbéit (telt szimbólumok) és a 90 perccel későbbiekből az indukált görbéket (üres szimbólumok). A mintákból össz RNS-t tisztítottunk. A központi óragének mRNS-szintjét valós idejű PCR segítségével határoztuk meg, az értékeket a TUBULIN mRNS szintjéhez viszonyítva ábrázoltuk. A grafikonok három független kísérlet átlageredményét mutatják. A szürke sávok az időtengelyen a névleges éjszakát jelölik. A vizsgált gének: CCA1 (A, B), PRR9 (C, D), GI (E, F), ELF4 (G, H).
Megfigyelhettük, hogy mind a négy gén indukálható UV-B fénnyel és az indukció
kapuzott. Az indukció a cirkadián maximum értékénél a CCA1 esetében majdnem
kétszeres, míg a másik három gén esetében 2-4 szeres mRNS szint adódott az indukció
után. Az uvr8-6 mutáció nem érintette az alap kifejeződési szintet, az indukciót viszont
teljes mértékben eltörölte. Csakúgy, mint a cop1-4 mutáció. Ennek az utóbbinak már
korábban leírt korai fázisú és rövid periódusú fenotípusa van, ami itt is látható volt
minden vizsgált génen. A cop1-4 mutáció sem volt hatással a PRR9, GI, ELF4
alapkifejeződésre, azonban a CCA1 mRNS amplitúdója jelentősen megemelkedett ebben
a mutánsban.

EREDMÉNYEK ______________________________________________________________________________________________________
56
13. ábra. TOC1 gén indukciója
A vad típusú (Col-0) növények beállítása, kísérleti körülményeik, mintafeldolgozásuk, illetve a kapott adatok ábrázolása megegyezett az előző kísérletben alkalmazottakkal. Az alapaktivitások görbéit telt-, míg az indukált görbéket üres szimbólumokkal ábrázoltuk. A szürke sávok az időtengelyen a névleges éjszakát jelölik.
A TOC1 óragén mRNS-ének kifejeződésére nincs hatással a (látható) fény (Makino és
mtsai, 2001). Megvizsgálva úgy találtuk, az UV-B szintén nem változtat a szintjén
(13. ábra).
Az adatok alapján úgy tűnik, az UV-B hatása az
óragének indukciójára nagyon hasonló a látható fényhez. Az uvr8-6 és a cop1-4 mutáció
pedig teljes mértékben eltörli ezt a indukciót. Azonban látható fényben az uvr8-6
mutációnak nincs semmilyen hatása az órára nézve.
A cirkadián óra a nem ritmikus gének UV-B indukcióját is kapuzza
A cirkadián óra központi génjein kivül is sok olyan gén van, mely kettős szabályozás
alatt áll. A fény képes közvetlenlül indukálni őket, de ez az indukció nem mindig azonos
mértékű a cirkadián óra kapuzási szabályozása miatt. Annak vizsgálatára, hogy ez az
UV-B-re is igaz-e megnéztük néhány olyan gén kifejeződését és indukcióját, melyek nem
részei a központi órának. A szakirodalomban közölt, cirkadián kifejeződést vizsgáló
(Covington és mtsai, 2008), illetve az alacsony intenzitású UV-B génindukciókat
feltérképező (Favory és mtsai, 2009) génchip adatbázisok összevetésével az alábbi
géneket választottuk ki: CHALCONE SYNTHASE (CHS, At5g13930, UV-B védelemben
fontos flavonoidok termelődésének kulcsenzimét kódolja), GLUTATHIONE PEROXIDASE7
(GPX7, At4g31870, antioxidáns enzim génje, a GPX7 a hidrogén peroxidot redukálja
glutation segítségével), EARLY LIGHT INDUCED PROTEIN 2 (ELIP2, At4g14690,
fotoprotektív gén), ELONGATED HYPOCOTYL 5 (HY5, At5g11260 Bzip transzkripciós
faktor génje a fotomorfogenezis fontos pozitív faktora) valamint a HY5-HOMOLOG (HYH,

EREDMÉNYEK ______________________________________________________________________________________________________
57
At3g17609) melyek mind UV-B indukciót, mind cirkadián mintázatot mutattak. Az
indukciós tesztet az óragénekével teljesen megegyezően vizsgáltuk vad típusú uvr8-6 és
cop1-4 mutáns növényeken (14. ábra).

EREDMÉNYEK ______________________________________________________________________________________________________
58
14. ábra. UV-B indukált és cirkadián mintázatot mutató gének UV-B indukciója
A vad típusú Col-0 (fekete négyzet), uvr8-6 (piros háromszög) és cop1-4 mutáns (kék kör) növények
beállítása, kísérleti körülményeik, mintafeldolgozásuk, illetve a kapott adatok ábrázolása megegyezett a
központi óraelemek vizsgálatában alkalmazottakkal. Az alapaktivitások görbéit telt-, míg az indukált
görbéket üres szimbólumokkal ábrázoltuk. A szürke sávok az időtengelyen a névleges éjszakát jelölik.
A megvizsgált gének: CHS (A, B), GPX7 (C, D), ELIP2 (E, F), HY5 (G, H), HYH (I, J). Második (jobboldali) Y
tengelyen ábrázoltuk a következő mRNS adatokat: CHS cop1-4 háttérben (B), indukált GPX7 vad típusban
(C, D), GPX7 cop1-4 háttérben (D), indukált ELIP2 vad típusban (E, F), ELIP2 cop1-4 háttérben (F), és
indukált HY5 vad típusban (G, H).

EREDMÉNYEK ______________________________________________________________________________________________________
59
A CHS, az ELIP2 a GPX7 és a HYH mRNS-ek kifejeződése cirkadián szabályzást követ,
az alapszintjük az óragénekkel összevethető kitéréseket mutatott és a legnagyobb
indukció időben egybeesett a cirkadián csúcsokkal. Azonban a cirkadián alapvonal és a
nagymértékű indukciók ellenére a HY5 mRNS-en nem látható kapuzottság. Annak
eldöntésére, hogy ez általános hiány-e minden fénykörülmény esetén, elvégeztük a
„klasszikus” kapuzási kísérletet vörös fénnyel. A növényeket hét napos beállítás után
folyamatos sötétbe helyeztük, ahol háromóránként egyórás 40 µmol m-2 s-1 erősségű
vörös pulzust kaptak. A pulzus előtt, illetve után 90 perccel gyűjtött mintákból össz RNS-
t tisztítottunk, amiből valós idejű PCR segítségével határoztuk meg a HY5, és HYH mRNS
szinteket, melyeket a TUBULIN mRNS szintjéhez viszonyítva ábrázoltunk (15. ábra).
15. ábra. A HY5 és a HYH gének vörös indukciója
A vad típusú Col-0 (fekete négyzet) és uvr8-6 mutáns (piros háromszög) hét napos csíranövényeket
folyamatos sötétbe helyeztük és háromóránként egyórás 40 µmol m-2 s-1 intenzitású vörös pulzussalokkal
kezeltük őket. Mintafeldolgozásuk, illetve a kapott adatok ábrázolása megegyezett a korábbi UV-B
kapuzást vizsgáló kísérletekben alkalmazottakkal. Az alapaktivitások görbéit telt-, míg az indukált
görbéket üres szimbólumokkal ábrázoltuk. A szürke sávok az időtengelyen a névleges nappalt jelölik.
HY5 (A), HYH (B).
Amint látható, nem általános jelenségről van szó, mert a HY5 és a HYH vörös
indukcióját az óra kapuzza. Az uvr8-6 -csakúgy, mint korábban- UV-B hiányában
megkülönböztethetetlen a vad típustól.
Vizsgáltuk továbbá az At3g16350, At3g57830 géneket, melyek nem cirkadián
kifejeződésűek, de jelentős UV-B indukciót mutatnak. Az előbbi egy ismeretlen szerepű
Myb transzkripciós faktort (Ikeda és Ohme-Takagi, 2009), míg az utóbbi egy
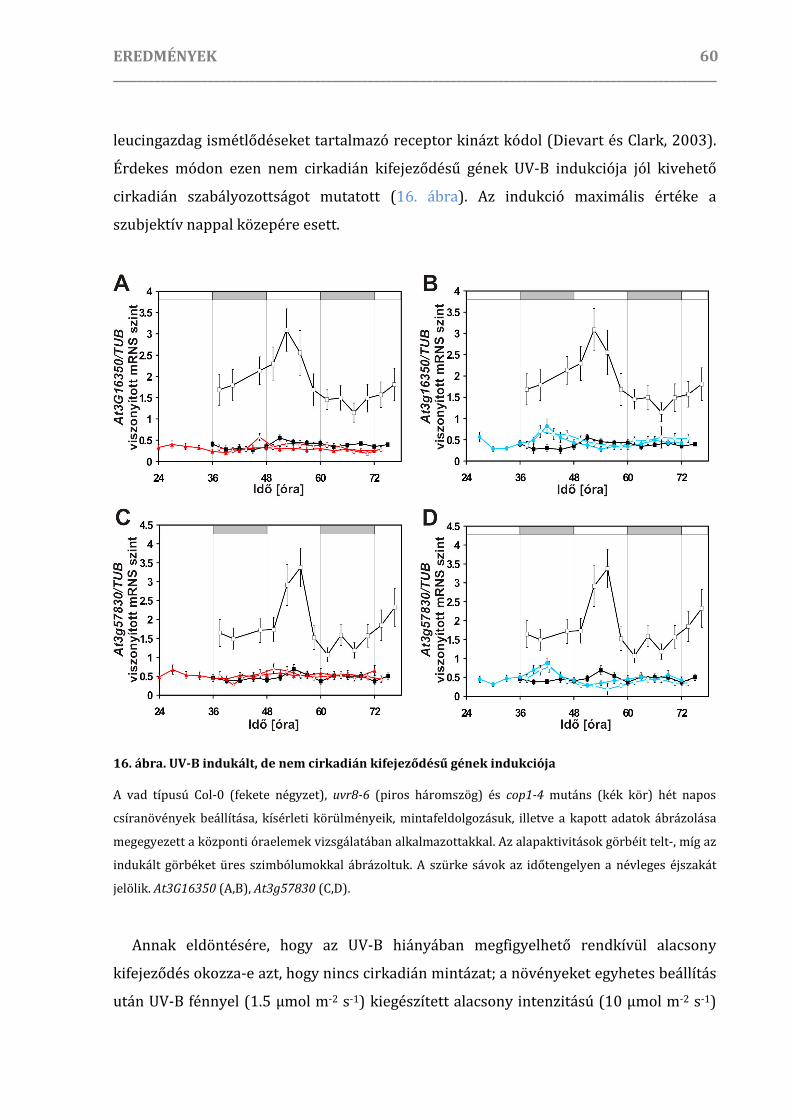
EREDMÉNYEK ______________________________________________________________________________________________________
60
leucingazdag ismétlődéseket tartalmazó receptor kinázt kódol (Dievart és Clark, 2003).
Érdekes módon ezen nem cirkadián kifejeződésű gének UV-B indukciója jól kivehető
cirkadián szabályozottságot mutatott (16. ábra). Az indukció maximális értéke a
szubjektív nappal közepére esett.
16. ábra. UV-B indukált, de nem cirkadián kifejeződésű gének indukciója
A vad típusú Col-0 (fekete négyzet), uvr8-6 (piros háromszög) és cop1-4 mutáns (kék kör) hét napos
csíranövények beállítása, kísérleti körülményeik, mintafeldolgozásuk, illetve a kapott adatok ábrázolása
megegyezett a központi óraelemek vizsgálatában alkalmazottakkal. Az alapaktivitások görbéit telt-, míg az
indukált görbéket üres szimbólumokkal ábrázoltuk. A szürke sávok az időtengelyen a névleges éjszakát
jelölik. At3G16350 (A,B), At3g57830 (C,D).
Annak eldöntésére, hogy az UV-B hiányában megfigyelhető rendkívül alacsony
kifejeződés okozza-e azt, hogy nincs cirkadián mintázat; a növényeket egyhetes beállítás
után UV-B fénnyel (1.5 µmol m-2 s-1) kiegészített alacsony intenzitású (10 µmol m-2 s-1)

EREDMÉNYEK ______________________________________________________________________________________________________
61
folyamatos fehér fénybe helyeztük (17.ábra). Ahogy az ábrák mutatják, a magasabb
mRNS szint ellenére a gének nem cirkadián kifejeződésűek.
17. ábra. Az At3g16350, és At3g57830 gének kifelyeződése UV-B vel kiegészített fehér fényben
A vad típusú (Col-0) hét napos csíranövényeket UV-B fénnyel (1.5 µmol m-2 s-1) kiegészített alacsony
intenzitású (10 µmol m-2 s-1) folyamatos fehér fénybe helyeztük. háromóránként mintát vettünk belőlük,
amiből össz RNS-t risztítottunk. A gének mRNS-szintjét valós idejű PCR segítségével határoztuk meg, az
értékeket a TUBULIN mRNS szintjéhez viszonyítva ábrázoltuk. A grafikonok három független kísérlet
átlageredményét mutatják, a szürke sávok az időtengelyen a névleges éjszakát jelölik.
Az uvr8-6 és a cop1-4 mutáció az általunk vizsgált összes gén UV-B indukcióját
eltüntette. Az uvr8-6 mutánsnak nincsen semmilyen hatása az alap kifejeződési
mintázatra, viszont a cop1-4 a már leírt korai fázis és rövid periódus mellett megemelte
az alapszintjét a CHS, ELIP2 és HY5 mRNS-eknek. Mindent egybevéve, ezek az adatok azt
sugallják, hogy legalábbis a génindukció szinjén másként kontrollált a kapuzás az UV-B,
mint a látható fény esetében.
A HY5 és a HYH nem szükségesek sem az óragének UV-B
indukciójához, sem az UV-B indukálta génexpresszió kapuzásához
Korábbi, génchipekkel megerősített kutatások azt mutatták, hogy a HY5 és a HYH a
legtöbb UV-B indukálta gént kontrolállják (Brown és Jenkins, 2008). Hogy megvizsgáljuk
ezeknek a transzkripciós faktoroknak a hatását a mi kísérleti körülményeink között;
megnéztük a hy5 hyh kettős mutáció hatásait a fentebb már leírt kapuzást vizsgáló

EREDMÉNYEK ______________________________________________________________________________________________________
62
kísérleti rendszerben. A növesztési és kísérleti körülmények azonosak voltak az
előzőekkel (18.ábra).
18. ábra. Néhány gén UV-B indukciója hy5 hyh dupla mutáns háttérben
A vad típusú Col-0 (fekete négyzet) és hy5-ks50 hyh mutáns (narancs rombusz) hét napos csíranövények
beállítása, kísérleti körülményeik, mintafeldolgozásuk, illetve a kapott adatok ábrázolása megegyezett a
korábbi, UV-B kapuzást vizsgáló kísérletekben alkalmazottakkal. Az alapaktivitások görbéit telt-, míg az
indukált görbéket üres szimbólumokkal ábrázoltuk. A szürke sávok az időtengelyen a névleges éjszakát
jelölik. A vizsgált gének: CCA1 (A), PRR9, (B), CHS (C), At3g16350 (D). Az indukált CHS mRNS szinteket, vad
típusú háttérben másodlagos Y tengelyen ábrázoltuk (C)
A kétszeres mutáns háttérben a CCA1 és a PRR9 mRNS-ek alapszintje csak egészen
kicsit csökkent le, míg indukciójuk gyakorlatilag nem változott. Ezzel szemben a CHS
alapszintje drasztikusan lecsökkent, indukciója teljesen eltűnt a mutánsban, ahogy már
korábban leírták (Brown és Jenkins, 2008; Stracke és mtsai, 2010). Az At3g16350

EREDMÉNYEK ______________________________________________________________________________________________________
63
indukciója erőteljesen lecsökkent, de nem tűnt el teljesen és a megmaradt indukció
cirkadián kapuzást mutatott.
Mindezek az adatok azt mutatják, hogy az óragének UV-B indukciója nem függ a HY5,
HYH transzkripciós faktoroktól és az UV-B indukált génkifejeződés cirkadián
szabályzása sem rajtuk keresztül történik.
Aritmiás mutánsokban hiányzik az UV-B indukált génkifejeződés
kapuzása
Az UV-B kiváltotta génkifejeződési mintázat napi különbségeit a cirkadián óra
szabályzása biztosítja. Vagyis az elmélet szerint, ha az óra szabályzása kimarad, a
kapuzás megszűnik, viszont az UV-B indukálhatóság meg kell maradjon egy többé-
kevésbé állandó szinten. Ezért megvizsgáltuk az UV-B indukált génkifejeződést olyan
növényekben, melyek nem rendelkeznek működő cirkadián órával (19. ábra).

EREDMÉNYEK ______________________________________________________________________________________________________
64
19. ábra. Néhány gén UV-B indukciója elf3-4 mutáns háttérben
A vad típusú Ws (fekete négyzet) és elf3-4 mutáns (magenta háromszög) hét napos csíranövények
beállítása, kísérleti körülményeik, mintafeldolgozásuk, illetve a kapott adatok ábrázolása megegyezett a
korábbi, UV-B kapuzást vizsgáló kísérletekben alkalmazottakkal. Az alapaktivitások görbéit telt-, míg az
indukált görbéket üres szimbólumokkal ábrázoltuk. A szürke sávok az időtengelyen a névleges éjszakát
jelölik. A vizsgált gének: CCA1 (A), PRR9, (B), CHS (C), At3g16350 (D). Az indukált At3g16350 mRNS
szinteket, elf3-4 háttérben másodlagos Y tengelyen ábrázoltuk (D)
Az elf3-4 növényben a CCA1 mRNS indukció nélküli szintje folyamatosan alacsony
értékű volt, míg a PRR9 mRNS magas szinten mutatta a folyamatos kifejeződést. Ezek az
adatok egybecsengtek a korábbi megfigyelésekkel (Alabadi és mtsai, 2001; Kim és mtsai,
2005; Thines és Harmon, 2010). Az óravezérelt CHS és a nem ritmikus At3g16350 a
PRR9-hez hasonlóan, magas alapszintet mutatott.
Minden általunk tesztelt gén UV-B indukciója azonos szintű, nem kapuzott jelleget
mutatott az elf3-4 mutánsban. A CCA1 kifejeződés minden időpontban kétszeres mRNS
szintbeli különbséget mutatott az elf3-4 mutánsban, ami egyezik a maximális CCA1
mRNS indukcióval a vad típusban. Azonban az alacsony alapszint miatt, az UV-B indukált

EREDMÉNYEK ______________________________________________________________________________________________________
65
CCA1 mRNS felhalmozódása alacsonyabb az elf3-4 mutánsban, mint a vad típusban.
Szemben a CCA1-gyel a többi gén indukált mRNS szintje azonos vagy magasabb szintű
volt, mint a vad típus maximális értéke.
A kísérleteket elvégeztük egy másik ritmus nélküli növényvonalon is, a CCA1-et
folyamatosan magas szinten kifejező CCA1-OX 38-as vonalon (Wang és Tobin, 1998).
A kísérletek az elf3-4 mutánshoz hasonló eredményt adtak. UV-B megvilágítás magas
szintű génindukciót okozott függetlenül a kezelés időpontjától. Érdekes módon
kétszeres CCA1 mRNS szint növekedést láttunk ebben a CCA1-et folyamatosan
túltermelő vonalban, ami tekintve hogy a 35S promóter nem indukálódik UV-B sugárzás
hatására (Boyko és mtsai, 2010) azt sugallaja, hogy az UV-B stabilizálja a CCA1 mRNS-t
(20. ábra).

EREDMÉNYEK ______________________________________________________________________________________________________
66
20. ábra. Néhány gén UV-B indukciója CCA1 túltermelő háttérben
A vad típusú Ws (fekete négyzet) és CCA1-OX 38 (zöld kör) hét napos csíranövények beállítása, kísérleti
körülményeik, mintafeldolgozásuk, illetve a kapott adatok ábrázolása megegyezett a korábbi, UV-B
kapuzást vizsgáló kísérletekben alkalmazottakkal. Az alapaktivitások görbéit telt-, míg az indukált
görbéket üres szimbólumokkal ábrázoltuk. A szürke sávok az időtengelyen a névleges éjszakát jelölik.
A vizsgált gének: CCA1 (A), PRR9, (B), CHS (C), At3g16350 (D).
Ezek az adatok azt mutatják, hogy a ritmikus kapuzás elvész, de az UV-B indukált
génkifejeződés növekedés megmarad ezekben a ritmustalan vonalakban. Az indukciós
szintek arra utalnak, hogy az óra megreked egy olyan sajátos állapotban, amikor az UV-B
válaszok nincsenek gátolva.

EREDMÉNYEK ______________________________________________________________________________________________________
67
Az óra által szabályozott UV-B génindukció fiziológiai hatása
Az a megfigyelés, hogy az elf3-4 mutánsban a vad típushoz képest a legtöbb UV-B
indukálta gén erősebb választ ad; felvetette annak a lehetőségét, hogy az elf3-4 növény
ellenállóbb lehet UV-B okozta stresszel szemben. Ennek a feltételezésnek a
kipróbálására a növényeket a szokásos egyhetes beállítás után UV-B vel
(1.5 µmol m-2 s-1) kiegészített alacsony intenzitású (10 µmol m-2 s-1) fehér fénybe
helyeztük. 24 illetve 35 órával később (szubjektív reggel és este), 20 perces
(14 µmol m-2 s-1) UV-B pulzust adtunk nekik. A növényeket további egy hétre
visszahelyeztük a nevelés körülményei közé (12 h fehér fény (50 μmol m-2 s-1)/12 h
sötét, 22°C), majd analizáltuk.
Ahogyan a 21. ábra mutatja, a mutáns és a vad típusú növények között nem volt
különbség, jelezve, hogy ez a részleges mRNS szintemelkedés nem biztosított nagyobb
ellenállást a nagy intenzitású UV-val szemben az elf3-4 növényeknek. Az UV-B dózis nem
volt halálos, de lelassult a fejlődés és az elsődleges levelek növekedése, valamint
csökkent a klorofilltartalom. Az elf3-4 növények eleve kevesebb klorofillt tartalmaztak,
de a csökkenés (körülbelül 40% a kezeletlen növényekhez képest) egyforma volt a két
genotípusnál. Továbbá, az UV-B stressz mértéke független volt attól, hogy mely
napszakban kapták azt a növények.

EREDMÉNYEK ______________________________________________________________________________________________________
68
21. ábra UV-B ellenállóképesség
vizsgálata
A vad típusú és elf3-4 mutáns hét
napos csíranövényeket alacsony
intenzitású (1.5µmol m-2 s-1) UV-B-
vel kiegészített fehér fényben
(10µmol m-2 s-1) helyeztük
(aklimáltuk). 24, illetve 35 órával
később (szubjektív reggel és este)
20 perces 14 µmol m-2 s-1 UV-B
pulzussal kezeltük őket. A
növényeket ezután további egy
hétre visszahelyeztük fény-sötét
ciklusokra. A növényeket lefotóztuk
(A), majd mintát szedtünk. A
mintákból kivontuk a klorofillt,
aminek mennyiségét fotométerrel
határoztuk meg és a nedves
tömegre (NT) vonatkoztatva (B),
illetve a kontrollhoz viszonyítva (C)
adtuk meg.

AZ EREDMÉNYEK ÉRTELMEZÉSE
Az UV-B fény szerepet játszik a növényi cirkadián óra beállításában
A fényimpulzusok az időtartalmuktól függően különböző paramétereit módosíthatják
a szabadon futó cirkadián órának. Növényekben a folyamatos megvilágítás az
intenzitásával arányosan rövidíti a szabadon futó óra periódusát, míg a fénypulzusok
fáziscsúszást okoznak.
Kísérleteink elsődleges célja az volt, hogy megvizsgáljuk az UV-B egy olyan
részletének a hatását a cirkadián órára, melynek szerepe már bizonyított a
fotomorfogenezisben és melynek az UVR8 a receptora. Ennek érdekében olyan alacsony
intenzitású (1.5 μmol m-2 s-1) UV-B fényt használtunk beállító fénynek, amelynek nincsen
káros hatása a növényekre.
Adataink megmutatták, hogy az UV-B képes a cirkadián óra beállítására. A 8. ábra,
illetve a 4. táblázat bemutatja, hogy vad típusú növényekben mindkét általunk használt
kimenet jelzőgén (CCR2:LUC+, HY5:LUC+) szabadon futó ritmusa szignifikánsan, 1-2
órával rövidebb periódusú volt, amennyiben a folyamatos fehér fényt (10 μmol m-2 s-1)
kiegészítetük UV-B-vel.
A 9/A ábra mutatja, hogy UV-B pulzus a folyamatos fehér fényben tartott
növényeknél fáziskéséseket okozott a szubjektív éjszaka során, azonban azonos mértékű
pulzus nem okozott eltérést a szubjektív nappal alatt. Érdekes, hogy egy korábbi
kísérletben leírt, DNS károsodást okozó, erős UV-B pulzusok újraindították az órát és
csakis fázissietéseket okoztak fibroblaszt kultúrában. (Oklejewicz és mtsai, 2008) Ugyan
kísérleteinknek nem képezte célját megvizsgálni, hogy UV-B-stressz hogyan hat a
növényi circadian órára, egy kísérletünk azonban azt mutatta, hogy az erős UV-B pulzus
drámaian csökkenti a cirkadián ritmusok kitérését, ami lehetetlenné teszi a beállítás
vizsgálatát (22. ábra).

AZ EREDMÉNYEK ÉRTELMEZÉSE ______________________________________________________________________________________________________
70
UV-B által kiváltott órabeállítás lehetséges molekuláris működésének vizsgálatához
több mint egy cirkadián cikluson keresztül követtük a központi óragének (CCA1,PRR9,
GI) illetve az óraközeli ELF4 indukálhatóságát. A 10. ábra mutatja, hogy minden gén
ritmikusan kapuzott UV-B indukciót mutatott vad típusú növényekben, míg az indukciók
megszűntek uvr8-6 illetve cop1-4 mutáns hátterekben. Érdekes módon a HY5 illetve
HYH transzkripciós faktorok hiánya nem érintette az óra periódusát (4. táblázat), illetve
az óragének UV-B indukálhatóságát sem fehér fényben, sem UV-B-ben (18. ábra). Ezek a
megfigyelések meglehetősen váratlanok voltak, hiszen a HY5:
• In vivo kötődik több óragén (CCA1, LHY, ELF4, TOC1) promóteréhez fehér
fényben (Lee és mtsai, 2007).
• A hy5 és a hy5 hyh mutáns, hasonlóan az uvr8-hoz, sérült az UV-B függő
fotomorfogenezisben (Ulm és mtsai, 2004; Oravecz és mtsai, 2006; Brown és
Jenkins, 2008; Stracke és mtsai, 2010)
• A HY5 befolyásolja a legtöbb UVR8, illetve COP1 szabályozott gén UV-B
indukcióját (Brown és mtsai, 2005; Oravecz és mtsai, 2006).
Adataink alapján az összefüggések a fiziológiai (a periódus módosítása) és a
molekuláris (óragén indukciók) adatok között a vad típusú és a mutáns növényekben azt
mutatják, hogy az UV-B órabeállító hatása az óragének átírási szabályozásán alapul. Az
általunk tesztelt óragének közül a PRR9 mutatta a legjelentősebb indukciót UV-B-re.
A megemelkedett PRR9 szint a folyamatos UV-B kezelés alatt egy lehetséges magyarázat
22. ábra Az erős UV-B hatása a
cirkadián órára
CCR2:LUC+ jelzőgént hordozó vad
típusú Col-0 (kék rombusz) és
cop1-4 mutáns (magenta négyzet)
hét napos csíranövényeket
folyamatos sötétbe helyeztük és a
fénykibocsátást luminométerrel
mértük. ZT47 időpontban (fekete
nyilak) 60 μmol m-2 s-1 UV-B
pulzussal kezeltük őket.

AZ EREDMÉNYEK ÉRTELMEZÉSE ______________________________________________________________________________________________________
71
az UV-B periódus módosítására, mivel a PRR9 túltermelése hasonló mértékű periódus
rövidülést eredményez (Matsushika és mtsai, 2002).
Az UV-B indukció ritmikus szabályozása
A cirkadián kapuzás az a folyamat, amelyben az óra ritmikusan gátolja az akut hatását
egy külső környezeti, vagy akár egy belső jelnek a megfelelő célfolyamaton. A
növényekben a cirkadián kapuzás első leírása a CHLOROPHYLL A/B-BINDING PROTEIN 2
(CAB2) gén napi ritmusú fényindukciója volt (Millar és Kay, 1996). A 12-14. ábrák
bemutatják, hogy az összes általunk vizsgált gén UV-B indukciója -egy kivétellel- az óra
által kapuzott volt; hasonló módon, ahogy a látható fény által kiváltott indukciók. Az
egyetlen, ám annál fontosabb kivétel a HY5 volt. Ez a gén az általunk vizsgált
időtartományban a vörös fényre kapuzott indukciót mutatott, UV-B-re viszont nem. Ezek
az adatok pedig arra utalnak, hogy a HY5 UV-B indukcióját irányító jelátviteli lánc nincs
az óra hatásköre alatt és különbözik a látható fényszignál által bejárt útvonaltól.
Szintén váratlanul az At3g16350, és At3g57830 nem ritmikus gének
(http://diurnal.cgrb.oregonstate.edu) UV-B indukciója szép cirkadián kapuzást
mutatott, maximális indukcióval a szubjektív nap közepén (16. ábra).
A kapuzás jelensége nem korlátozódik a nap egy adott pontjára, illetve az érintett
gének sincsenek egy fázisban. Ezért a működése olyan UV-B jelátviteli elemekkel
magyarázható, amik párhuzamos szignál utakon hatnak és az óra különböző fázisban
szabályozza őket.
23. ábra. Az UVR8:LUC jelzőgén
kifejeződésének mintázata
folyamatos fényben
A jelzőgént hordozó vad típusú Col-
0 hét napos csíranövényeke
fénykibocsátását folyamatos fehér
fényben (10µmol m-2 s-1) mértük
luminométerrel.

AZ EREDMÉNYEK ÉRTELMEZÉSE ______________________________________________________________________________________________________
72
Az UVR8 átírása ritmikus, naplemente körüli maximummal (23. ábra). Azonban az
UVR8 fehérje nem mutat napi oszcillációt (24. ábra), vagyis valószínűtlen, hogy
közvetíthetné az UV-B kapuzás mechanizmusát. Nem zárható ki azonban az UVR8
fehérje óravezérelt átfordítás utáni módosításának és ezáltali aktiválásának a
lehetősége.
24. ábra. UVR8 fehérje szintjének
változása fény-sötét cikluson
A vad típusú Col-0 hét napos
csíranövényeket folyamatos fehér fénybe
(10µmol m-2 s-1) helyeztük. háromóránként
mintát szedtünk, amelyekből fehérjét tisztítottunk. Az UVR8, és a belső kontrollként használt aktin
fehérjeszinteket Western-blot módszerrel vizsgáltuk.
A HY5 és a HYH alacsony amplitúdójú ritmust mutatnak fehér fényben, viszont a két
transzkripciós faktor közül csak a HYH UV-B indukciója kapuzott. Elméletileg
elképzelhető, hogy aktív HY5/HYH heterodimert alkotva a felhalmozódásuk közvetíteni
képes az UV-B válasz kapuzását a reggel kifejeződő géneknek. Azonban adataink azt
mutatják, hogy az óragének UV-B indukciója érintetlen a hy5 hyh kétszeres mutánsban
reggel (CCA1) illetve a nap folyamán is (PRR9) (18. ábra). A CHS indukciója hy5 hyh
háttérben teljesen hiányzik, míg némi kapuzott indukciója fennmarad az At3g16350
génnek. (18. ábra) Ez azt jelzi, hogy a HY5/HYH fehérjék fontosak egy sor gén akut UV-B
indukciójában, de nélkülözhetőek a válasz időbeni módosításában.
Korábbi adatok bizonyították, hogy a cirkadián órának befolyása van a hormon
jelátvitelben, és több stresszválasz szabályzásában. A folyamatok ezen összehangolása
emeli a fitnesszt (Covington és mtsai, 2008). Kísérleteink megmutatták, hogy a gének
UV-B indukciója szintén jelentősen szabályozott az óra által. Ezért az UVR8-függő
génszabályozás egy újabb jelátvitel, ami az órához köthető. Kísérleteink alapján az alábbi
modellt vázoltuk fel, ami bemutatja az UV-B és a cirkadián óra által szabályozott
folyamatok közötti kölcsönhatásokat (25. ábra).

AZ EREDMÉNYEK ÉRTELMEZÉSE ______________________________________________________________________________________________________
73
25. ábra. UV-B jelátvitel és a cirkadián óra közötti kapcsolatokat leíró modell
Kísérleteink alapján az UV-B fény az UVR8 és a COP1 fehérjéken keresztül, a HY5 és HYH transzkripciós
faktroktól függetlenül szabályozza a cirkadián órát. A cirkadián óra kapuzza mind a saját UV-B
beállíthatóságát mind a HYH fehérje transzkripcióját; valamint befolyással van a HY5, HYH transzkripciós
faktorok által szabályozott gének napszakos kifejeződésére is. Azonban a HY5 fehérje UV indukciójára
nincsen hatással.
Az óramodulált UV-B válaszok fiziológiai jelentősége
Az alacsony intenzitású UV-B sugárzásra válaszul kialakuló génindukciónak igen
fontos szerepe van a fotoprotektív pigmentek (flavonoidok, antocianinok)
termelődésének serkentésében, melyek védelmet nyújtanak a nagy intenzitású, káros
UV-B-vel szemben (aklimáció) (Favory és mtsai, 2009). Eredményeink azt mutatják,
hogy az összes általunk vizsgált nem óragén indukciója a reggeli, illetve délelőtti
időszakban volt a legnagyobb. Ez arra utal, hogy az alacsony intenzitású UV-B jel
érzékelése és feldolgozása a nap korai részében fontosabb, azért hogy a növény felépítse
a flavonoid védelmet és túlélje a nap további részében várható UV-B terhelést.
Az alacsony intenzitású UV-B sugárzás hatására kialakuló, óra által szabályozott
génindukció fiziológiai fontosságának a felderítésére megvizsgáltuk az aritmikus elf3-4
mutáns molekuláris és fiziológiai válaszait. Adataink azt mutatták, hogy majdnem
minden általunk vizsgált gén erős indukciót mutatott az alacsony intenzitású UV-B–re az
egész nap során ebben a mutánsban.

AZ EREDMÉNYEK ÉRTELMEZÉSE ______________________________________________________________________________________________________
74
Az indukált mRNS szintek a CCA1 kivételével többnyire magasabbak, mint a
maximális szintek a vad típusban, ami arra utalt, hogy az elf3-4 növény nagyobb UV-B-
stressz toleranciával rendelkezik, mint a vad típus. Azonban ahogyan 21. ábra mutatja,
az aklimált elf3-4 mutánsok nem voltak ellenállóbbak a nap különböző részein adott
pulzusokkal szemben, mint vad típusú növények. Egy másik aritmiás növényvonal, a
CCA1 óragént túltermelő CCA1-OX azonos eredményeket adott. A génkifejeződésekben
megfigyelhető UV-B okozta indukció magasabb volt mint a vad típusban, azonban
UV-B-stressz tűrőképessége nem haladta meg a vad típusét. Mindez arra utal, hogy egy
rövid idejű, alacsony intenzitású UV-B-függő génexpresszió és az azt követő flavonoid
felhalmozás a nap egy rövid szakaszán elégséges a normális védelemhez a nagy
intenzitású UV-B káros hatásaival szemben. Azt az erőforrás-igényt, ami egy
folyamatosan aktív UV-B jelátvithez szükséges, jól illusztrálja az UVR8 gént túltermelő
növények törpe fenotípusa alacsony intenziású UV-B fényben (Favory és mtsai, 2009).
Eredményeink arra utalnak, hogy a növény számára előnyösebb az UV-B válaszokat a
cirkadián óra segítségével a megfelelő napszakra időzíteni, mint folyamatosan nagyobb
toleranciát biztosítani. Így minimalizálható a ráfordított energia, azonban az
ellenállóképesség sem lesz kisebb.

KÖVETKEZTETÉSEK
A dolgozatomban ismertetett kísérletek eredményét és a belőlük levonható
következtetéseket az alábbiakban foglalhatjuk össze:
1. Vad típusú növényekben a cirkadián óra periódusa alacsony intenzitású UV-B
hatására fényintenzitás függő módon rövidül. Míg az uvr8 és cop1 mutáns
növényekben a periódus nem rövidül alacsony intenzitású UV-B hatására.
2. A hy5 hyh mutáns növények periódusa nem tér el a vad típusétól, sem fehér, sem
UV-B-vel kiegészített fehér fényben. Mivel a HY5-ről korábban leírták, hogy in vivo
kötődik több óragén (CCA1, LHY, TOC1, ELF4) promóteréhez, felvetődik, hogy ennek
ellenére sem vesz részt a cirkadián szabályozásban.
3. Alacsony intenzitású UV-B pulzusok hatására vad típusú növényekben fáziskésés
figyelhető meg a szubjektív éjszaka során és nem tapasztalható fázisváltozás a
szubjektív nappal alatt. Az uvr8 és a cop1 mutáns növények nem mutatnak
fázisváltozást alacsony intenzitású UV-B pulzusok hatására, vagyis az egyes ponttal
összegezve: a cirkadián óra beállítható UV-B fénnyel, az UVR8 és a COP1 pedig
kulcsfontosságú ebben a beállításában.
4. Minden általunk vizsgált fényérzékeny óragén (CCA1, PRR9, GI, ELF4) indukálható
volt alacsony intenzitású UV-B vel is. Az indukciójuk kapuzott mintázatot mutatott.
A legnagyobb mértékű indukciót a PRR9 mutatta, a megemelkedett PRR9 szint
magában magyarázhatja a periódus rövidülését
5. Az uvr8 mutánsban a központi óra általunk vizsgált minden eleme a vad típusénak
megfelelő szinten volt UV-B hiányában és egyik sem volt indukálható UV-B
pulzusokkal. A cop1 mutáció az uvr8 hoz hasonlóan eltörölte az óragének UV-B
indukálhatóságát.
6. Az általunk vizsgált kettős (cirkadián és közvetlen fény) szabályozású gének nagy
része kapuzott indukciót mutatott alacsony intenzitású UV-B fénypulzusok hatására.

KÖVETKEZTETÉSEK ______________________________________________________________________________________________________
76
7. A HY5 UV-B indukcióját nem szabályozza a cirkadián óra. A HYH átíródását viszont
igen. Elképzelhető, hogy a két fehérje heterodimerként összekapcsolódva működik
és ez esetben a HYH cirkadián átíródási szabályzása kihat a komplex működésére.
8. Az általunk vizsgált nem cirkadián kifejeződésű, de UV-B indukálható gének szintén
kapuzott indukciót mutattak alacsony intenzitású UV-B pulzusok hatására. A
maximális indukciók –akár a többi vizsgált nem óragén esetében- a reggeli/délelőtti
órákban voltak, ami feltételezi azt, hogy a növény számára ekkor a legfontosabb az
alacsony intenzitású UV-B jel érzékelése. Ennek hatására építi fel UV-B elleni
védelmét a később érkező nagyobb intenzitású sugárzással szemben.
9. A megvizsgált aritmiás növényekben (elf3, CCA1-OX) nem volt tapasztalható
kapuzás, azonban majdnem minden megvizsgált gén magasabb indukciót mutatott,
mint a vad típusban. A minden időpontban magasabb indukció azonban nem tette
védettebbé ezeket a növényeket a nagy intenzitású UV-B sugárzással szemben. Ez
arra utal, hogy egy rövid idejű, alacsony intenzitású UV-B függő génexpresszió és az
azt követő flavonoid felhalmozás a nap egy rövid szakaszán elégséges a normális
védelemhez a nagy intenzitású UV-B káros hatásaival szemben.

SAJÁT KÖZLEMÉNYEK
Kiss I., Kesserű P., Feher B., Bihari Z. és Polyák B.: Co-immobilization of symbiotic green algae and Saccharomyces unispora. Symbiosis 2002, 32(3): 247-256.
Kevei, E., Gyula, P., Hall, A., Kozma-Bognar, L., Kim, W.-Y., Erikson, M.E., Toth, R.,
Hanano, S., Feher, B., Southern, M.M., Bastow, R.M., Viczian, A., Hibberd, V., Davis, S.J., Somers., D.E., Nagy,F. and Millar, A.J: Forward genetic analysis of the circadian clock
separates the multiple functions of ZEITLUPE. Plant Physiology 2006, 140(3): 933-945.
James C.W. Locke, Laszlo Kozma-Bognar, Peter Gould, Balazs Feher, Eva Kevei, Ferenc Nagy, Matthew S. Turner, Anthony Hall, Andrew J. Millar: Experimental validation of a
predicted feedback loop in the multi-oscillator clock of Arabidopsis. Molecular Systems
Biology 2006, 2:59.
Kevei E, Gyula P, Fehér B, Tóth R, Viczián A, Kircher S, Rea D, Dorjgotov D, Schäfer E, Millar AJ, Kozma-Bognár L, Nagy F.: Arabidopsis thaliana circadian clock is Regulated by
the small GTPase LIP1. Current Biology 2007, 17(17):1456-1464.
Safrany J, Haasz V, Mate Z, Ciolfi A, Feher B, Oravecz A, Stec A, Dallmann G, Morelli G, Ulm R, Nagy F.: Identification of a novel cis-regulatory element for UV-B induced
transcription in Arabidopsis. Plant Journal. 2008, 54(3):402-414.
Fehér B*, Kozma-Bognár L
*, Kevei E, Hajdu A, Binkert M, Davis SJ, Schäfer E, Ulm R,
Nagy F.: Functional interaction of the circadian clock and UV RESISTANCE LOCUS 8-
controlled UV-B signaling pathways in Arabidopsis thaliana. Plant Journal. 2011 (In Press).†
* Megosztott első szerzők † A dolgozat alapjául szolgáló közlemény

IDÉZETT KÖZLEMÉNYEK
Adams J, Kelso R, Cooley L (2000) The kelch repeat superfamily of proteins: propellers of cell function. Trends Cell Biol 10: 17-24
A-H-Mackerness S, Surplus SL, Blake P, John CF, Buchanan-Wollaston V. (1999) Ultraviolet-B-induced stress and changes in gene expression in Arabidopsis thaliana: role of signalling pathways controlled by jasmonic acid, ethylene and reactive oxygen species. Plant Cell Environ. 22: 1413–23
Ahn SC, Baek BS, Oh T, Song CS, Chatterjee B (2000) Rapid mini-scale plasmid isolation for DNA sequencing and restriction mapping. BioTechniques 29: 466-468
Alabadi D, Oyama T, Yanovsky MJ, Harmon FG, Mas P, Kay SA (2001) Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293: 880-883
Allen T, Koustenis A, Theodorou G, Somers DE, Kay SA, Whitelam GC, Devlin PF (2006) Arabidopsis FHY3 specifically gates phytochrome signaling to the circadian clock. Plant Cell 18: 2506-2516
Aschoff J (1979) Circadian rhythms: influences of internal and external factors on the period measured in constant conditions. Z Tierpsychol 49: 225-249
Bae G, Choi G (2008) Decoding of light signals by plant phytochromes and their interacting proteins. Annu Rev Plant Biol 59: 281-311
Ballaré CL, Scopel AL, Stapleton AE, Yanovsky MJ (1996) Solar Ultraviolet-B Radiation Affects Seedling Emergence, DNA Integrity, Plant Morphology, Growth Rate, and Attractiveness to Herbivore Insects in Datura ferox. Plant Physiol 112: 161-170
Ballaré CL, Barnes PW, Flint SD. (1995) Inhibition of hypocotyl elongation by UV-B radiation in deetiolating tomato seedlings. 1. The photoreceptor. Physiol. Plant. 93: 584–92
Ballaré CL, Barnes PW, Kendrick RE. (1991) Photomorphogenic effects of UV-B radiation on hypocotyl elongation in wild-type and stable-phytochrome-deficient mutant seedlings of cucumber. Physiol. Plant. 83: 652–58
Ben-Naim O, Eshed R, Parnis A, Teper-Bamnolker P, Shalit A, Coupland G, Samach A, Lifschitz E (2006) The CCAAT binding factor can mediate interactions between CONSTANS-like proteins and DNA. Plant J 46: 462-476
Bender K, Blattner C, Knebel A, Iordanov M, Herrlich P, Rahmsdorf HJ (1997) UV-induced signal transduction. J Photochem Photobiol B 37: 1-17
Bieza K, Lois R (2001) An Arabidopsis mutant tolerant to lethal ultraviolet-B levels shows constitutively elevated accumulation of flavonoids and other phenolics. Plant Physiol 126: 1105-1115
Boccalandro HE, Mazza CA, Mazzella MA, Casal JJ, Ballare CL (2001) Ultraviolet B radiation enhances a phytochrome-B-mediated photomorphogenic response in Arabidopsis. Plant Physiol 126: 780-788
Bornman JF, Reuber S, Cen Y-P, Weissenböck G. (1997) Ultraviolet radiation as a stress factor and the role of protective pigments. In Plants and UV-B: Responses to Environmental Change, ed. P Lumsden, pp. 157–68. Cambridge: Cambridge Univ. Press
Boyko A, Molinier J, Chatter W, Laroche A, Kovalchuk I (2010) Acute but not chronic exposure to abiotic stress results in transient reduction of expression levels of the transgene driven by the 35S promoter. N Biotechnol 27: 70-77
Briggs WR (2007) The LOV domain: a chromophore module servicing multiple photoreceptors. J Biomed Sci 14: 499-504
Briggs WR, Christie JM (2002) Phototropins 1 and 2: versatile plant blue-light receptors. Trends Plant Sci 7: 204-210
Briggs WR, Huala E (1999) Blue-light photoreceptors in higher plants. Annu Rev Cell Dev Biol 15: 33-62

IDÉZETT KÖZLEMÉNYEK ______________________________________________________________________________________________________
79
Britt AB, Chen JJ, Wykoff D, Mitchell D (1993) A UV-sensitive mutant of Arabidopsis defective in the repair of pyrimidine-pyrimidinone(6-4) dimers. Science 261: 1571-1574
Britt AB. (2004) Repair of DNA damage induced by solar UV. Photosynth. Res. 81:105–12
Brosché N, Strid A. (2003) Molecular events following perception of UV-B radiation by plants. Physiol. Plant. 117: 1–10
Brown BA, Cloix C, Jiang GH, Kaiserli E, Herzyk P, Kliebenstein DJ, Jenkins GI (2005) A UV-B-specific signaling component orchestrates plant UV protection. Proc Natl Acad Sci U S A 102: 18225-18230
Brown BA, Jenkins GI (2008) UV-B signaling pathways with different fluence-rate response profiles are distinguished in mature Arabidopsis leaf tissue by requirement for UVR8, HY5, and HYH. Plant Physiol 146: 576-588
Cardozo T, Pagano M (2004) The SCF ubiquitin ligase: insights into a molecular machine. Nat Rev Mol Cell Biol 5: 739-751
Cashmore AR, Jarillo JA, Wu YJ, Liu D (1999) Cryptochromes: blue light receptors for plants and animals. Science 284: 760-765
Castillon A, Shen H, Huq E (2007) Phytochrome Interacting Factors: central players in phytochrome-mediated light signaling networks. Trends Plant Sci 12: 514-521
Chen M, Chory J, Fankhauser C (2004) Light signal transduction in higher plants. Annu Rev Genet 38: 87-117
Cheng P, He Q, Yang Y, Wang L, Liu Y (2003) Functional conservation of light, oxygen, or voltage domains in light sensing. Proc Natl Acad Sci U S A 100: 5938-5943
Christie JM (2007) Phototropin blue-light receptors. Annu Rev Plant Biol 58: 21-45
Cloix C, Jenkins GI (2008) Interaction of the Arabidopsis UV-B-specific signaling component UVR8 with chromatin. Mol Plant 1: 118-128
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16: 735-743
Covington MF, Maloof JN, Straume M, Kay SA, Harmer SL (2008) Global transcriptome analysis reveals circadian regulation of key pathways in plant growth and development. Genome Biol 9: R130
Covington MF, Panda S, Liu XL, Strayer CA, Wagner DR, Kay SA (2001) ELF3 modulates resetting of the circadian clock in Arabidopsis. Plant Cell 13: 1305-1315
Crosson S, Moffat K (2001) Structure of a flavin-binding plant photoreceptor domain: insights into light-mediated signal transduction. Proc Natl Acad Sci U S A 98: 2995-3000
Dai S, Wei X, Pei L, Thompson RL, Liu Y, Heard JE, Ruff TG, Beachy RN (2011) BROTHER OF LUX ARRHYTHMO Is a Component of the Arabidopsis Circadian Clock. Plant Cell 23: 961-972
David KM, Armbruster U, Tama N, Putterill J (2006) Arabidopsis GIGANTEA protein is post-transcriptionally regulated by light and dark. FEBS Lett 580: 1193-1197
Desikan R, S AH-M, Hancock JT, Neill SJ (2001) Regulation of the Arabidopsis transcriptome by oxidative stress. Plant Physiol 127: 159-172
Devlin PF, Kay SA (2000) Cryptochromes are required for phytochrome signaling to the circadian clock but not for rhythmicity. Plant Cell 12: 2499-2510
Dievart A, Clark SE (2003) Using mutant alleles to determine the structure and function of leucine-rich repeat receptor-like kinases. Curr Opin Plant Biol 6: 507-516
Ding Z, Doyle MR, Amasino RM, Davis SJ (2007a) A complex genetic interaction between Arabidopsis thaliana TOC1 and CCA1/LHY in driving the circadian clock and in output regulation. Genetics 176: 1501-1510
Ding Z, Millar AJ, Davis AM, Davis SJ (2007b) TIME FOR COFFEE encodes a nuclear regulator in the Arabidopsis
thaliana circadian clock. Plant Cell 19: 1522-1536
Dodd AN, Salathia N, Hall A, Kevei E, Toth R, Nagy F, Hibberd JM, Millar AJ, Webb AA (2005) Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage. Science 309: 630-633
Doyle MR, Davis SJ, Bastow RM, McWatters HG, Kozma-Bognár L, Nagy F, Millar AJ, Amasino RM (2002) The ELF4 gene controls circadian rhythms and flowering time in Arabidopsis thaliana. Nature 419: 74-77
Ensminger PA. (1993) Control of development in plants and fungi by far-UV radiation. Physiol. Plant. 88: 501–8
Eriksson ME, Hanano S, Southern MM, Hall A, Millar AJ (2003) Response regulator homologues have complementary, light-dependent functions in the Arabidopsis circadian clock. Planta 218: 159-162

IDÉZETT KÖZLEMÉNYEK ______________________________________________________________________________________________________
80
Farré EM, Harmer SL, Harmon FG, Yanovsky MJ, Kay SA (2005) Overlapping and distinct roles of PRR7 and PRR9 in the Arabidopsis circadian clock. Curr Biol 15: 47-54
Favory JJ, Stec A, Gruber H, Rizzini L, Oravecz A, Funk M, Albert A, Cloix C, Jenkins GI, Oakeley EJ, Seidlitz HK, Nagy F, Ulm R (2009) Interaction of COP1 and UVR8 regulates UV-B-induced photomorphogenesis and stress acclimation in Arabidopsis. Embo J 28: 591-601
Fowler S, Lee K, Onouchi H, Samach A, Richardson K, Morris B, Coupland G, Putterill J (1999) GIGANTEA: a circadian clock-controlled gene that regulates photoperiodic flowering in Arabidopsis and encodes a protein with several possible membrane-spanning domains. Embo J 18: 4679-4688
Fritsche E, Schafer C, Calles C, Bernsmann T, Bernshausen T, Wurm M, Hubenthal U, Cline JE, Hajimiragha H, Schroeder P, Klotz LO, Rannug A, Furst P, Hanenberg H, Abel J, Krutmann J (2007) Lightening up the UV response by identification of the arylhydrocarbon receptor as a cytoplasmatic target for ultraviolet B radiation. Proc Natl Acad Sci U S A 104: 8851-8856
Frohnmeyer H, Loyall L, Blatt MR, Grabov A (1999) Millisecond UV-B irradiation evokes prolonged elevation of cytosolic-free Ca2+ and stimulates gene expression in transgenic parsley cell cultures. Plant J 20: 109-117
Frohnmeyer H, Staiger D (2003) Ultraviolet-B radiation-mediated responses in plants. Balancing damage and protection. Plant Physiol 133: 1420-1428
Furuya M (1989) Molecular properties and biogenesis of phytochrome I and II. Adv Biophys 25: 133-167
Furuya M, Schäfer E (1996) Photoperception and signalling of induction reactions by different phytochromes. Trends Plant Sci 1: 301-307
Galland P, Senger H. (1988) The role of flavins as photoreceptors. J. Photochem. Photobiol. B Biol. 1: 277–94
Galland P, Senger H. (1988) The role of pterins in the photoreception and metabolism of plants. Photochem. Photobiol. 48: 811–20
Gadjev I, Vanderauwera S, Gechev TS, Laloi C, Minkov IN, Shulaev V, Apel K, Inze D, Mittler R, Van Breusegem F (2006) Transcriptomic footprints disclose specificity of reactive oxygen species signaling in Arabidopsis. Plant Physiol 141: 436-445
Gerhardt KE, Wilson MI, Greenberg BM (2005) Ultraviolet wavelength dependence of photomorphological and photosynthetic responses in Brassica napus and Arabidopsis thaliana. Photochem Photobiol 81: 1061-1068
Green R, Fluhr R (1995) UV-B-Induced PR-1 Accumulation Is Mediated by Active Oxygen Species. Plant Cell 7: 203-212
Green RM, Tobin EM (1999) Loss of the circadian clock-associated protein 1 in Arabidopsis results in altered clock-regulated gene expression. Proc Natl Acad Sci U S A 96: 4176-4179
Hall A, Bastow RM, Davis SJ, Hanano S, McWatters HG, Hibberd V, Doyle MR, Sung S, Halliday KJ, Amasino RM, Millar AJ (2003) The TIME FOR COFFEE gene maintains the amplitude and timing of Arabidopsis circadian clocks. Plant Cell 15: 2719-2729
Harlow GR, Jenkins ME, Pittalwala TS, Mount DW (1994) Isolation of uvh1, an Arabidopsis mutant hypersensitive to ultraviolet light and ionizing radiation. Plant Cell 6: 227-235
Harmer SL (2009) The circadian system in higher plants. Annu Rev Plant Biol 60: 357-377
Harmer SL, Hogenesch JB, Straume M, Chang HS, Han B, Zhu T, Wang X, Kreps JA, Kay SA (2000) Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science 290: 2110-2113
Hazen SP, Schultz TF, Pruneda-Paz JL, Borevitz JO, Ecker JR, Kay SA (2005) LUX ARRHYTHMO encodes a Myb domain protein essential for circadian rhythms. Proc Natl Acad Sci U S A 102: 10387-10392
Herrlich P, Bohmer FD (2000) Redox regulation of signal transduction in mammalian cells. Biochem Pharmacol 59: 35-41
Hicks KA, Millar AJ, Carré IA, Somers DE, Straume M, Meeks-Wagner DR, Kay SA (1996) Conditional circadian dysfunction of the Arabidopsis early-flowering 3 mutant. Science 274: 790-792
Ho MS, Ou C, Chan YR, Chien CT, Pi H (2008) The utility F-box for protein destruction. Cell Mol Life Sci 65: 1977-2000
Huq E, Tepperman JM, Quail PH (2000) GIGANTEA is a nuclear protein involved in phytochrome signaling in Arabidopsis. Proc Natl Acad Sci U S A 97: 9789-9794
Ikeda M, Ohme-Takagi M (2009) A novel group of transcriptional repressors in Arabidopsis. Plant Cell Physiol 50: 970-975

IDÉZETT KÖZLEMÉNYEK ______________________________________________________________________________________________________
81
Imaizumi T, Schultz TF, Harmon FG, Ho LA, Kay SA (2005) FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis. Science 309: 293-297
Imaizumi T, Tran HG, Swartz TE, Briggs WR, Kay SA (2003) FKF1 is essential for photoperiodic-specific light signalling in Arabidopsis. Nature 426: 302-306
Imamoto Y, Kataoka M (2007) Structure and photoreaction of photoactive yellow protein, a structural prototype of the PAS domain superfamily. Photochem Photobiol 83: 40-49
Inoue H, Nojima H, Okayama H (1990) High efficiency transformation of Escherichia coli with plasmids. Gene 96: 23-28
Inskeep WP, Bloom PR (1985) Extinction coefficients of chlorophyll a and B in n,n-dimethylformamide and 80% acetone. Plant Physiol 77: 483-485
Ito S, Matsushika A, Yamada H, Sato S, Kato T, Tabata S, Yamashino T, Mizuno T (2003) Characterization of the APRR9 pseudo-response regulator belonging to the APRR1/TOC1 quintet in Arabidopsis thaliana. Plant Cell Physiol 44: 1237-1245
Jansen, M.A.K., Gaba, V., and Greenberg, B.M. (1998). Higher plants and UV-B radiation: balancing damage, repair and acclimation (vol 3, pg 131, 1998) Trends Plant Sci. 3: 243-243.
Jenkins GI (2009) Signal transduction in responses to UV-B radiation. Annu Rev Plant Biol 60: 407-431
Jenkins GI, Brown BA. (2007) UV-B perception and signal transduction. In Light and Plant Development, ed. GC Whitlam, KJ Halliday, pp. 155–182. Oxford: Blackwell
Jenkins GI, Long JC, Wade HK, Shenton MR, Bibikova TN. (2001) UV and blue light signalling: pathways regulating chalcone synthase gene expression in Arabidopsis. New Phytol. 151: 121–31
Jenkins GI, Fuglevand G, Christie JM. (1997) UV-B perception and signal transduction. In Plants and UV-B:
Responses to Environmental Change, ed. P Lumsden, pp. 135–56. Cambridge: Cambridge Univ. Press
Jiang CZ, Yee J, Mitchell DL, Britt AB (1997) Photorepair mutants of Arabidopsis. Proc Natl Acad Sci U S A 94: 7441-7445
Johnson CH (1992) Phase Response Curves: what can they tell us about circadian clocks? In Circadian clocks from
cell to human, Hiroshige T, Honma K (eds), 1 edn, pp 209-249. Sapporo: Hokkaido University Press
Johnson KL, Ingram GC (2005) Sending the right signals: regulating receptor kinase activity. Curr Opin Plant Biol 8: 648-656
Josse EM, Foreman J, Halliday KJ (2008) Paths through the phytochrome network. Plant Cell Environ 31: 667-678
Kaczorowski KA, Quail PH (2003) Arabidopsis PSEUDO-RESPONSE REGULATOR7 is a signaling intermediate in phytochrome-regulated seedling deetiolation and phasing of the circadian clock. Plant Cell 15: 2654-2665
Kaiserli E, Jenkins GI (2007) UV-B promotes rapid nuclear translocation of the Arabidopsis UV-B specific signaling component UVR8 and activates its function in the nucleus. Plant Cell 19: 2662-2673
Kalbin G, Hidema J, Brosché M, Kumagai T, Bornman JF, Strid A. (2001) UV-B-induced DNA damage and expression of defence genes under UV-B stress: tissue-specific molecular marker analysis in leaves. Plant Cell Environ. 24: 983–90
Kalbina I, Li S, Kalbin G, Björn LO, Strid A. (2008) Two separate UV-B radiation wavelength regions control expression of different molecular markers in Arabidopsis thaliana. Funct. Plant Biol. 35: 222–27
Kerppola TK (2006) Design and implementation of bimolecular fluorescence complementation (BiFC) assays for the visualization of protein interactions in living cells. Nat Protoc 1: 1278-1286
Kevei É, Schäfer E, Nagy F (2007) Light-regulated nucleo-cytoplasmic partitioning of phytochromes. J Exp Bot 58: 3113-3124
Khanna R, Huq E, Kikis EA, Al-Sady B, Lanzatella C, Quail PH (2004) A novel molecular recognition motif necessary for targeting photoactivated phytochrome signaling to specific basic helix-loop-helix transcription factors. Plant Cell 16: 3033-3044
Khare M, Guruprasad KN. (1993) UV-B-induced anthocyanin synthesis in maize regulated by FMN and inhibitors of FMN photoreactions. Plant Sci. 91: 1–5
Kikis EA, Khanna R, Quail PH (2005) ELF4 is a phytochrome-regulated component of a negative-feedback loop involving the central oscillator components CCA1 and LHY. Plant J 44: 300-313
Kim BC, Tennessen DJ, Last RL (1998) UV-B-induced photomorphogenesis in Arabidopsis thaliana. Plant J 15: 667-674
Kim JY, Song HR, Taylor BL, Carré IA (2003a) Light-regulated translation mediates gated induction of the Arabidopsis clock protein LHY. Embo J 22: 935-944

IDÉZETT KÖZLEMÉNYEK ______________________________________________________________________________________________________
82
Kim WY, Fujiwara S, Suh SS, Kim J, Kim Y, Han L, David K, Putterill J, Nam HG, Somers DE (2007) ZEITLUPE is a circadian photoreceptor stabilized by GIGANTEA in blue light. Nature 449: 356-360
Kim WY, Geng R, Somers DE (2003b) Circadian phase-specific degradation of the F-box protein ZTL is mediated by the proteasome. Proc Natl Acad Sci U S A 100: 4933-4938
Kim WY, Hicks KA, Somers DE (2005) Independent roles for EARLY FLOWERING 3 and ZEITLUPE in the control of circadian timing, hypocotyl length, and flowering time. Plant Physiol 139: 1557-1569
Kipreos ET, Pagano M (2000) The F-box protein family. Genome Biol 1: REVIEWS3002
Kliebenstein DJ, Lim JE, Landry LG, Last RL (2002) Arabidopsis UVR8 regulates ultraviolet-B signal transduction and tolerance and contains sequence similarity to human regulator of chromatin condensation 1. Plant Physiol 130: 234-243
Koncz, C., Martini, N., Szabados, L., Hrouda, M., Bachmair, A., Schell, J. (1994) Specialized vectors for gene tagging and expression studies. In Plant Molecular Biology Manual, Gelvin, B.S., Schilperoot, R.A. (eds), Kluwer Academic Publishers, Dordrecht, The Netherlands, pp. 1-22.
Kozma-Bognár L, Káldi K (2008) Synchronization of the fungal and the plant circadian clock by light. Chembiochem 9: 2565-2573
Lakin-Thomas PL (2000) Circadian rhythms: new functions for old clock genes. Trends Genet 16: 135-142
Landry LG, Chapple CC, Last RL (1995) Arabidopsis mutants lacking phenolic sunscreens exhibit enhanced ultraviolet-B injury and oxidative damage. Plant Physiol 109: 1159-1166
Landry LG, Stapleton AE, Lim J, Hoffman P, Hays JB, Walbot V, Last RL (1997) An Arabidopsis photolyase mutant is hypersensitive to ultraviolet-B radiation. Proc Natl Acad Sci U S A 94: 328-332
Lee J, He K, Stolc V, Lee H, Figueroa P, Gao Y, Tongprasit W, Zhao H, Lee I, Deng XW (2007) Analysis of transcription factor HY5 genomic binding sites revealed its hierarchical role in light regulation of development. Plant Cell 19: 731-749
Li J, Ou-Lee TM, Raba R, Amundson RG, Last RL (1993) Arabidopsis Flavonoid Mutants Are Hypersensitive to UV-B Irradiation. Plant Cell 5: 171-179
Lin C, Shalitin D (2003) Cryptochrome structure and signal transduction. Annu Rev Plant Biol 54: 469-496
Liu Y, Tsinoremas NF, Johnson CH, Lebedeva NV, Golden SS, Ishiura M, Kondo T (1995) Circadian orchestration of gene expression in cyanobacteria. Genes Dev 9: 1469-1478
Locke JC, Kozma-Bognar L, Gould PD, Feher B, Kevei E, Nagy F, Turner MS, Hall A, Millar AJ (2006) Experimental validation of a predicted feedback loop in the multi-oscillator clock of Arabidopsis thaliana. Mol Syst Biol 2: 59
Locke JC, Southern MM, Kozma-Bognar L, Hibberd V, Brown PE, Turner MS, Millar AJ (2005) Extension of a genetic network model by iterative experimentation and mathematical analysis. Mol Syst Biol 1: 2005 0013
Lois R, Buchanan BB (1994) Severe sensitivity to ultraviolet radiation in an Arabidopsis mutant deficient in flavonoid accumulation. Planta 194: 504–509
Makino S, Matsushika A, Kojima M, Oda Y, Mizuno T (2001) Light response of the circadian waves of the APRR1/TOC1 quintet: when does the quintet start singing rhythmically in Arabidopsis? Plant Cell Physiol 42: 334-339
Martin-Tryon EL, Kreps JA, Harmer SL (2007) GIGANTEA acts in blue light signaling and has biochemically separable roles in circadian clock and flowering time regulation. Plant Physiol 143: 473-486
Martínez-García JF, Huq E, Quail PH (2000) Direct targeting of light signals to a promoter element-bound transcription factor. Science 288: 859-863
Más P, Devlin PF, Panda S, Kay SA (2000) Functional interaction of phytochrome B and cryptochrome 2. Nature 408: 207-211
Matsushika A, Imamura A, Yamashino T, Mizuno T (2002) Aberrant expression of the light-inducible and circadian-regulated APRR9 gene belonging to the circadian-associated APRR1/TOC1 quintet results in the phenotype of early flowering in Arabidopsis thaliana. Plant Cell Physiol 43: 833-843
Matsushika A, Makino S, Kojima M, Mizuno T (2000) Circadian waves of expression of the APRR1/TOC1 family of pseudo-response regulators in Arabidopsis thaliana: insight into the plant circadian clock. Plant Cell Physiol 41: 1002-1012

IDÉZETT KÖZLEMÉNYEK ______________________________________________________________________________________________________
83
McNellis TW, von Arnim AG, Araki T, Komeda Y, Miséra S, Deng XW (1994) Genetic and molecular analysis of an allelic series of cop1 mutants suggests functional roles for the multiple protein domains. Plant Cell 6: 487-500
Michael TP, Mockler TC, Breton G, McEntee C, Byer A, Trout JD, Hazen SP, Shen R, Priest HD, Sullivan CM, Givan SA, Yanovsky M, Hong F, Kay SA, Chory J (2008) Network discovery pipeline elucidates conserved time-of-day-specific cis-regulatory modules. PLoS genetics 4: e14
Millar AJ, Carre IA, Strayer CA, Chua NH, Kay SA (1995a) Circadian clock mutants in Arabidopsis identified by luciferase imaging. Science 267: 1161-1163
Millar AJ, Kay SA (1996) Integration of circadian and phototransduction pathways in the network controlling CAB gene transcription in Arabidopsis. Proc Natl Acad Sci U S A 93: 15491-15496
Millar AJ, Straume M, Chory J, Chua NH, Kay SA (1995b) The regulation of circadian period by phototransduction pathways in Arabidopsis. Science 267: 1163-1166
Mizoguchi T, Wheatley K, Hanzawa Y, Wright L, Mizoguchi M, Song HR, Carré IA, Coupland G (2002) LHY and CCA1 are partially redundant genes required to maintain circadian rhythms in Arabidopsis. Dev Cell 2: 629-641
Moore JD (2001) The Ran-GTPase and cell-cycle control. Bioessays 23: 77-85
Nakajima S, Sugiyama M, Iwai S, Hitomi K, Otoshi E, Kim ST, Jiang CZ, Todo T, Britt AB, Yamamoto K (1998) Cloning and characterization of a gene (UVR3) required for photorepair of 6-4 photoproducts in Arabidopsis thaliana. Nucleic Acids Res 26: 638-644
Nakamichi N, Kita M, Ito S, Sato E, Yamashino T, Mizuno T (2005a) The Arabidopsis pseudo-response regulators, PRR5 and PRR7, coordinately play essential roles for circadian clock function. Plant Cell Physiol 46: 609-619
Nakamichi N, Kita M, Ito S, Yamashino T, Mizuno T (2005b) PSEUDO-RESPONSE REGULATORS, PRR9, PRR7 and PRR5, together play essential roles close to the circadian clock of Arabidopsis thaliana. Plant Cell Physiol 46: 686-698
Nelson DC, Lasswell J, Rogg LE, Cohen MA, Bartel B (2000) FKF1, a clock-controlled gene that regulates the transition to flowering in Arabidopsis. Cell 101: 331-340
Ni M, Tepperman JM, Quail PH (1998) PIF3, a phytochrome-interacting factor necessary for normal photoinduced signal transduction, is a novel basic helix-loop-helix protein. Cell 95: 657-667
Oklejewicz M, Destici E, Tamanini F, Hut RA, Janssens R, van der Horst GT (2008) Phase resetting of the mammalian circadian clock by DNA damage. Curr Biol 18: 286-291
Onai K, Ishiura M (2005) PHYTOCLOCK 1 encoding a novel GARP protein essential for the Arabidopsis circadian clock. Genes Cells 10: 963-972
Oravecz A, Baumann A, Mate Z, Brzezinska A, Molinier J, Oakeley EJ, Adam E, Schafer E, Nagy F, Ulm R (2006) CONSTITUTIVELY PHOTOMORPHOGENIC1 is required for the UV-B response in Arabidopsis. Plant Cell 18: 1975-1990
Park DH, Somers DE, Kim YS, Choy YH, Lim HK, Soh MS, Kim HJ, Kay SA, Nam HG (1999) Control of circadian rhythms and photoperiodic flowering by the Arabidopsis GIGANTEA gene. Science 285: 1579-1582
Plautz JD, Straume M, Stanewsky R, Jamison CF, Brandes C, Dowse HB, Hall JC, Kay SA (1997) Quantitative analysis of Drosophila period gene transcription in living animals. J Biol Rhythms 12: 204-217
Pope B, Kent HM (1996) High efficiency 5 min transformation of Escherichia coli. Nucleic Acids Res 24: 536-537
Pruneda-Paz JL, Breton G, Para A, Kay SA (2009) A functional genomics approach reveals CHE as a component of the Arabidopsis circadian clock. Science 323: 1481-1485
Renault L, Nassar N, Vetter I, Becker J, Klebe C, Roth M, Wittinghofer A (1998) The 1.7 A crystal structure of the regulator of chromosome condensation (RCC1) reveals a seven-bladed propeller. Nature 392: 97-101
Rizzini L, Favory JJ, Cloix C, Faggionato D, O'Hara A, Kaiserli E, Baumeister R, Schafer E, Nagy F, Jenkins GI, Ulm R (2011) Perception of UV-B by the Arabidopsis UVR8 protein. Science 332: 103-106
Roenneberg T, Merrow M (1998) Molecular circadian oscillators: an alternative hypothesis. J Biol Rhythms 13: 167-179
Savenstrand H, Brosche M, Strid A (2004) Ultraviolet-B signalling: Arabidopsis brassinosteroid mutants are defective in UV-B regulated defence gene expression. Plant Physiol Biochem 42: 687-694
Sawa M, Nusinow DA, Kay SA, Imaizumi T (2007) FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science 318: 261-265

IDÉZETT KÖZLEMÉNYEK ______________________________________________________________________________________________________
84
Schaffer R, Ramsay N, Samach A, Corden S, Putterill J, Carré IA, Coupland G (1998) The late elongated hypocotyl mutation of Arabidopsis disrupts circadian rhythms and the photoperiodic control of flowering. Cell 93: 1219-1229
Schultz TF, Kiyosue T, Yanovsky M, Wada M, Kay SA (2001) A role for LKP2 in the circadian clock of Arabidopsis. Plant Cell 13: 2659-2670
Shalitin D, Yu X, Maymon M, Mockler T, Lin C (2003) Blue light-dependent in vivo and in vitro phosphorylation of Arabidopsis cryptochrome 1. Plant Cell 15: 2421-2429
Sharrock RA, Clack T (2004) Heterodimerization of type II phytochromes in Arabidopsis. Proc Natl Acad Sci U S A 101: 11500-11505
Somers DE, Devlin PF, Kay SA (1998a) Phytochromes and cryptochromes in the entrainment of the Arabidopsis circadian clock. Science 282: 1488-1490
Somers DE, Schultz TF, Milnamow M, Kay SA (2000) ZEITLUPE encodes a novel clock-associated PAS protein from Arabidopsis. Cell 101: 319-329
Somers DE, Webb AA, Pearson M, Kay SA (1998b) The short-period mutant, toc1-1, alters circadian clock regulation of multiple outputs throughout development in Arabidopsis thaliana. Development 125: 485-494
Song HR, Carré IA (2005) DET1 regulates the proteasomal degradation of LHY, a component of the Arabidopsis circadian clock. Plant Mol Biol 57: 761-771
Southern MM, Millar AJ (2005) Circadian genetics in the model higher plant, Arabidopsis thaliana. Methods
Enzymol 393: 23-35
Staiger D, Allenbach L, Salathia N, Fiechter V, Davis SJ, Millar AJ, Chory J, Fankhauser C (2003) The Arabidopsis
SRR1 gene mediates phyB signaling and is required for normal circadian clock function. Genes Dev 17: 256-268
Stracke R, Favory JJ, Gruber H, Bartelniewoehner L, Bartels S, Binkert M, Funk M, Weisshaar B, Ulm R (2010) The Arabidopsis bZIP transcription factor HY5 regulates expression of the PFG1/MYB12 gene in response to light and ultraviolet-B radiation. Plant Cell Environ 33: 88-103
Stratmann J (2003) Ultraviolet-B radiation co-opts defense signaling pathways. Trends Plant Sci 8: 526-533
Suesslin C, Frohnmeyer H (2003) An Arabidopsis mutant defective in UV-B light-mediated responses. Plant J 33: 591-601
Surplus SL, Jordan BR, Murphy AM, Carr JP, Thomas B, A-H-Mackerness S. (1998) Ultraviolet-Binduced responses in Arabidopsis thaliana: role of salicylic acid and reactive oxygen species in the regulation of transcripts encoding photosynthetic and acidic pathogenesis-related proteins. Plant Cell Environ. 21: 685–94
Tepperman JM, Zhu T, Chang HS, Wang X, Quail PH (2001) Multiple transcription-factor genes are early targets of phytochrome A signaling. Proc Natl Acad Sci U S A 98: 9437-9442
Thines B, Harmon FG (2010) Ambient temperature response establishes ELF3 as a required component of the core Arabidopsis circadian clock. Proc Natl Acad Sci U S A 107: 3257-3262
Tóth R, Kevei É, Hall A, Millar AJ, Nagy F, Kozma-Bognár L (2001) Circadian clock-regulated expression of phytochrome and cryptochrome genes in Arabidopsis. Plant Physiol 127: 1607-1616
Ulm R, Baumann A, Oravecz A, Mate Z, Adam E, Oakeley EJ, Schafer E, Nagy F (2004) Genome-wide analysis of gene expression reveals function of the bZIP transcription factor HY5 in the UV-B response of Arabidopsis. Proc Natl Acad Sci U S A 101: 1397-1402
Viczián A, Kircher S, Fejes E, Millar AJ, Schäfer E, Kozma-Bognár L, Nagy F (2005) Functional characterization of phytochrome interacting factor 3 for the Arabidopsis thaliana circadian clockwork. Plant Cell Physiol 46: 1591-1602
Wade HK, Bibikova TN, Valentine WJ, Jenkins GI (2001) Interactions within a network of phytochrome, cryptochrome and UV-B phototransduction pathways regulate chalcone synthase gene expression in Arabidopsis leaf tissue. Plant J 25: 675-685
Wang ZY, Kenigsbuch D, Sun L, Harel E, Ong MS, Tobin EM (1997) A Myb-related transcription factor is involved in the phytochrome regulation of an Arabidopsis Lhcb gene. Plant Cell 9: 491-507
Wang ZY, Tobin EM (1998) Constitutive expression of the CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) gene disrupts circadian rhythms and suppresses its own expression. Cell 93: 1207-1217
Xiao W, Jang J (2000) F-box proteins in Arabidopsis. Trends Plant Sci 5: 454-457

IDÉZETT KÖZLEMÉNYEK ______________________________________________________________________________________________________
85
Yakir E, Hilman D, Hassidim M, Green RM (2007) CIRCADIAN CLOCK ASSOCIATED1 transcript stability and the entrainment of the circadian clock in Arabidopsis. Plant Physiol 145: 925-932
Yamamoto Y, Sato E, Shimizu T, Nakamich N, Sato S, Kato T, Tabata S, Nagatani A, Yamashino T, Mizuno T (2003) Comparative genetic studies on the APRR5 and APRR7 genes belonging to the APRR1/TOC1 quintet implicated in circadian rhythm, control of flowering time, and early photomorphogenesis. Plant Cell Physiol 44: 1119-1130
Yanovsky MJ, Mazzella MA, Casal JJ (2000a) A quadruple photoreceptor mutant still keeps track of time. Curr Biol 10: 1013-1015
Yanovsky MJ, Mazzella MA, Whitelam GC, Casal JJ (2001) Resetting of the circadian clock by phytochromes and cryptochromes in Arabidopsis. J Biol Rhythms 16: 523-530
Yanovsky MJ, Whitelam GC, Casal JJ (2000b) fhy3-1 retains inductive responses of phytochrome A. Plant Physiol 123: 235-242
Yasuhara M, Mitsui S, Hirano H, Takanabe R, Tokioka Y, Ihara N, Komatsu A, Seki M, Shinozaki K, Kiyosue T (2004) Identification of ASK and clock-associated proteins as molecular partners of LKP2 (LOV kelch protein 2) in Arabidopsis. J Exp Bot 55: 2015-2027
Yi C, Deng XW (2005) COP1 - from plant photomorphogenesis to mammalian tumorigenesis. Trends Cell Biol 15: 618-625
Yu JW, Rubio V, Lee NY, Bai S, Lee SY, Kim SS, Liu L, Zhang Y, Irigoyen ML, Sullivan JA, Lee I, Xie Q, Paek NC, Deng XW (2008) COP1 and ELF3 control circadian function and photoperiodic flowering by regulating GI stability. Mol Cell 32: 617-630
Yu X, Klejnot J, Zhao X, Shalitin D, Maymon M, Yang H, Lee J, Liu X, Lopez J, Lin C (2007a) Arabidopsis cryptochrome 2 completes its posttranslational life cycle in the nucleus. Plant Cell 19: 3146-3156
Yu X, Shalitin D, Liu X, Maymon M, Klejnot J, Yang H, Lopez J, Zhao X, Bendehakkalu KT, Lin C (2007b) Derepression of the NC80 motif is critical for the photoactivation of Arabidopsis CRY2. Proc Natl Acad Sci U S A 104: 7289-7294
Zeilinger MN, Farré EM, Taylor SR, Kay SA, Doyle FJ, 3rd (2006) A novel computational model of the circadian clock in Arabidopsis that incorporates PRR7 and PRR9. Mol Syst Biol 2: 58
Zikihara K, Iwata T, Matsuoka D, Kandori H, Todo T, Tokutomi S (2006) Photoreaction cycle of the light, oxygen, and voltage domain in FKF1 determined by low-temperature absorption spectroscopy. Biochemistry 45: 10828-10837

ÖSSZEFOGLALÁS
A Földön nem minden környezeti tényező változása véletlenszerű. Bolygónk tengely
körüli forgása következtében a felszínen a fény erőssége, összetétele és a hőmérséklet
24 óránként ismétlődő mintázat szerint változik. Ehhez a kiszámítható jelenséghez
szinte minden földi élőlény alkalmazkodott. Kifejleszettek egy általánosan elterjedt belső
időmérő mechanizmust, a cirkadián órát, melynek segítségével életfolyamataikat
képesek bizonyos napszakokhoz igazítani, valamint a környezet napszakos változásaira
felkészülni. A fotoszintetizáló élőlények számára a fény az elsőrendű energiaforrás, ezért
számukra különösen fontos, hogy ezt az erőforrást minél hatékonyabban tudják
hasznosítani. Ennek feltétele, hogy már napfelkelte előtt felkészítsék fényelnyelő és
hasznosító, valamint a fénykárosodástól védő apparátusaikat. A cirkadián rendszer
megfelelő működéséhez a belső biológiai óra által mért időnek azonos fázisban kell
lennie az abszolút, környezetben mérhető idővel. A cirkadián óra és a környezetben
mért idő szinkronizációja a biológiai óra újra és újra történő beállításával valósul meg,
amely folyamatban ritmikusan változó környezeti faktorok vesznek részt. A növényi
cirkadián óra legfontosabb beállító tényezője a fény. A vörös/távoli vörös fényelnyelésű
fitokrómok, illetve a kék fényelnyelésű kriptokrómok szerepe a cirkadián óra
fotoreceptoraként már bizonyított Arabidopsisban (Somers és mtsai, 1998a; Devlin és
Kay, 2000).
Munkánk során az alacsony intenzitású UV-B kapcsolatát vizsgáltuk a cirkadián
órával. Eddig semilyen adat nem volt arról, hogy a fénynek ez a természetes
komponense befolyásolja-e a cirkadián óra működését, illetve az alacsony intenzitású
UV-B érzékelésében kulcsfontosságú UVR8, COP1, HY5 illetve HYH fehérjék milyen
szerepet játszanak az UV-B jelek közvetítésében a cirkadián órához. Vizsgáltuk továbbá,
hogy a cirkadián óra szabályozza-e az UV-B által kiváltott élettani folyamatokat, szerepet
játszik-e az UV-B védelemben.

ÖSSZEFOGLALÁS ______________________________________________________________________________________________________
87
A cirkadián óra UV-B fénnyel való beállíthatóságának vizsgálata során kapott
eredmények alapján az alábbi következtetéseket tehetjük:
• Vad típusú növényekben a cirkadián óra periódusa alacsony intenzitású UV-B
hatására fényintenzitás függő módon rövidül.
• Az uvr8 mutáns növényben a periódus nem rövidül alacsony intenzitású UV-B
hatására, viszont a látható fény a vad típushoz hasonlóan rövidíti a periódust. A
kísérlet megerősíti azt a korábbi megfigyelést, miszerint az uvr8 mutáns UV-B
hiányában megkülönböztethetetlen a vad típustól.
• A cop1 mutáns növények periódusa sem rövidül alacsony intenzitású UV-B
hatására.
• A hy5 hyh mutáns növények periódusa nem tér el a vad típusétól, sem fehér, sem
UV-B-vel kiegészített fehér fényben. Mivel a HY5-ről korábban leírták, hogy in
vivo kötődik több óragén (CCA1, LHY, TOC1, ELF4) promóteréhez, felvetődik, hogy
ennek ellenére sem vesz részt a cirkadián szabályozásban.
• Alacsony intenzitású UV-B pulzusok hatására vad típusú növényekben fáziskésés
figyelhető meg a szubjektív éjszaka során és nem tapasztalható fázisváltozás a
szubjektív nappal alatt.
• Sem az uvr8, sem a cop1 mutáns növények nem mutatnak fázisváltozást alacsony
intenzitású UV-B pulzusok hatására.
• Az uvr8 mutáns növények látható fénypulzusokra adott fázisváltozásaikban vad
típusú válaszokat adnak minden általunk vizsgált körülményben. Látható fényben
szintén nem okoz fenotípust a mutáció.

ÖSSZEFOGLALÁS ______________________________________________________________________________________________________
88
A cirkadián óra UV-B által kiváltott fényindukciókra gyakorolt hatásának vizsgálata
alapján az alábbi megállapítások tehetőek:
• Minden általunk vizsgált fényérzékeny óragén (CCA1, PRR9, GI, ELF4) indukálható
volt alacsony intenzitású UV-B vel is. Az indukciójuk kapuzott mintázatot
mutatott. A legnagyobb mértékű indukciót a PRR9 mutatta, a megemelkedett
PRR9 szint magában magyarázhatja a periódus rövidülését
• Az uvr8 mutánsban a központi óra általunk vizsgált minden eleme a vad típusnak
megfelelő szinten volt UV-B hiányában és egyik sem volt indukálható UV-B
pulzusokkal.
• A cop1 mutáció az uvr8 hoz hasonlóan eltörölte az óragének UV-B
indukálhatóságát.
• Az általunk vizsgált kettős (cirkadián és közvetlen fény) szabályozású gének nagy
része kapuzott indukciót mutatott alacsony intenzitású UV-B fénypulzusok
hatására.
• A HY5 UV-B indukcióját nem szabályozza a cirkadián óra. A HYH átíródását
viszont igen. Elképzelhető, hogy a két fehérje heterodimerként összekapcsolódva
működik és ez esetben a HYH cirkadián átíródási szabályzása kihat a komplex
működésére.
• Az általunk vizsgált nem cirkadián kifejeződésű, de UV-B indukálható gének
szintén kapuzott indukciót mutattak alacsony intenzitású UV-B pulzusok
hatására. A maximális indukciók – akár a többi vizsgált nem óragén esetében - a
reggeli/délelőtti órákban voltak, ami feltételezi azt, hogy a növény számára ekkor
a legfontosabb az alacsony intenzitású UV-B jel érzékelése. Ennek hatására építi
fel UV-B elleni védelmét a később érkező nagyobb intenzitású sugárzással
szemben.

ÖSSZEFOGLALÁS ______________________________________________________________________________________________________
89
A megvizsgált aritmiás növényekben (elf3, CCA1-OX) nem volt tapasztalható kapuzás,
azonban majdnem minden megvizsgált gén magasabb indukciót mutatott, mint a vad
típusban. A minden időpontban magasabb indukció azonban nem tette védettebbé
ezeket a növényeket a nagy intenzitású UV-B sugárzással szemben. Ez arra utal, hogy
egy rövid idejű, alacsony intenzitású UV-B függő génexpresszió és az azt követő
flavonoid felhalmozás a nap egy rövid szakaszán elégséges a normális védelemhez a
nagy intenzitású UV-B káros hatásaival szemben.
Összegezve az eredményeket megállapítható, hogy a növényi cirkadián óra
beállítható UV-B fénnyel, és ebben az UVR8 és COP1 fehérjék kulcsfontosságúak. A HY5
és HYH transzkripciós faktorok nem komponensei a cirkadián óra fénybemenetének
sem UV-B-ben sem látható fényben. A központi óra fényindukálható génjei UV-B
indukálhatóak is, a cirkadián óra pedig kapuzza a legtöbb UV-B indukálható gén UV-B
indukcióját, a nem cirkadián kifejeződésű génekét is beleértve. Az egyetlen általunk
talált kivétel a HY5 UV-B indukciója volt, mely nincsen cirkadián szabályozás alatt,
szemben a gén vörös indukciójával, mely kapuzott. Az uvr8 és cop1 mutánsokban nem
figyelhető meg egyáltalán UV-B génindukció, míg a hy5 hyh kétszeres mutánsban a
központi óra génjeinek UV-B indukciója nem változott a vad típushoz képest. A vizsgált
aritmiás mutánsokban nincs kapuzása az UV-B indukcióknak. A vizsgált gének indukciós
szintje általánosan magasabb a mutáns növényekben a vad típushoz képest. A nagyobb
indukció viszont nem ad számukra nagyobb védettséget a nagy intenzitású
UV-B sugárzással szemben.

SUMMARY
Introduction Circadian clocks are ubiquitous biochemical timing mechanisms that rhythmically
regulate a wide range of molecular and physiological processes in most of the living
organisms exposed to the daily succession of days and nights (Harmer, 2009). Due to the
function of the clock, certain processes are timed to the most appropriate time of the day
and are attenuated, when they are not needed. This temporal organization results in
significant saving of chemical energy and contributes to the fitness of the organism
(Dodd et al., 2005).
In Arabidopsis the central oscillator relies on the function of three regulatory circuits
(Locke et al., 2006). The “morning loop” consists of the Myb-related CIRCADIAN CLOCK
ASSOCIATED 1 (CCA1) and the LATE ELONGATED HYPOCOTYL (LHY) transcription
factors, which facilitate the transcription of the PSEUDO RESPONSE REGULATOR 7/9
(PRR7/9) genes in the morning. PRR7/9 proteins inhibit transcription of CCA1/LHY
during the day (Farré et al., 2005). The evening loop is formed by TIMING OF CAB
EXPRESSION 1 (TOC1) and the hypothetical component Y. Y positively regulates TOC1
expression in the evening, whereas TOC1 attenuates Y expression during the night.
GIGANTEA (GI) has been proposed to contribute to the function of Y (Locke et al., 2005).
The morning and evening loops are coupled by a third circuit, where TOC1 indirectly
induces transcription of CCA1/LHY during the late night and CCA1/LHY inhibit TOC1
expression in the morning (Alabadi et al., 2001). The coordinated function of these three
circuits is required to generate the basic oscillation in the level of the clock.
The biological impact of the clock depends on the synchrony between the subjective
time provided by the oscillator and the real time of the environment. To achieve this, the
phase of the oscillator is reset to the day/night cycles by periodic environmental signals
among which light is the most significant. Light is perceived by photoreceptors and

SUMMARY ______________________________________________________________________________________________________
91
signals are transduced to the oscillator via the light input pathway, where they affect the
level and/or activity the clock components (Kozma-Bognar and Kaldi, 2008).
UV-B light has long been known as a harmful component of the sunlight causing
damage to DNA, protein and other macromolecules capable of direct absorption of these
wavelengths of light (Jansen et al., 1998). However, UV-B at lower fluence rates has been
shown to act as an environmental signal to control development, promote
photomorphogenesis and drive the expression of genes required for the production of
flavonoids (Ulm et al., 2004; Brown et al., 2005). Although the photoreceptor, which is
thought to mediate all of the UV-B effects, is still unknown, several components of the
signaling cascade have been identified recently. UV RESISTANCE LOCUS 8 (UVR8) is a
protein required for virtually all physiological UV-B responses, indicating that it is
located very close to the putative receptor in the signaling cascade (Favory et al., 2009)
or it is the low fluence UV-B photoreceptor itself (Rizzini et al., 2011). UVR8 is highly
specialized for UV-B signal transduction since uvr8 mutants show UV-B-dependent
phenotypes only (Brown et al., 2005; Favory et al., 2009). The E3 ubiquitin ligase
CONSTITUTIVELY PHOTOMORPHOGENIC 1 (COP1) physically interacts with UVR8 in a
UV-B dependent manner and this interaction is suggested to be required for signaling
(Favory et al., 2009). Under supplementary UV-B, COP1 promotes transcription of the
ELONGATED HYPOCOTYL 5 (HY5) gene (Favory et al., 2009). HY5 and HY5 HOMOLOG
(HYH) act as transcription factors and play crucial roles in mediating molecular and
physiological processes in response to photomorphogenic UV-B light (Ulm et al., 2004;
Brown and Jenkins, 2008; Stracke et al., 2010). Low intensity UV-B light induces the
expression of genes implicated in flavonoid biosynthesis. In turn, accumulation of UV-B
absorbing flavonoids confers protection to plants against the damaging effects of
highintensity UVB, which is apparent under natural conditions (Stracke et al., 2010).
Research objectives
Entrainment is required to set and maintain the correct and stable phase-relationship
between the endogenous circadian clock and environmental light/dark cycle. Since UV-B
is an important component of sunlight, we ought to test if low intensity, non-damaging
UV-B contributes to the light-mediated entrainment of the circadian clock in Arabidopsis.

SUMMARY ______________________________________________________________________________________________________
92
Most biochemical processes are organized by the circadian clock. This timing results
in saving of resources and improves fitness. Therefore we want to know, how does the
circadian clock regulate the UV-B responsive processes.
Results
UV-B light mediates entrainment of the plant circadian clock
The function of the light input pathway affects the pace or the phase of the oscillator,
depending on the light conditions. In plants, continuous irradiation shortens the
reerunning period length in a fluence rate dependent manner. To test the effect of UV-B
in this process, WT, uvr8-6, cop1-4, and hy5 hyh mutant plants expressing the CCR2:LUC+
reporter were grown in 12/12 LD cycles for a week and transferred to continuous white
light (WLL) or WLL supplemented by UVB of different fluence rates. Rhythmic
CCR2:LUC+ expression was monitored and freerunning periods were calculated. UV-B
shortens the period in WT and in hy5 hyh mutant a fluence rate dependent manner, but
not in uvr8-6 and in cop1-4 mutants. The effect of the uvr8-6 mutation is specific for UV-
B light, since it does not affect period length in plants free-running in WLL or in
darkness.
In contrast to the effect of continuous irradiation, discrete light pulses elicit
characteristic phase shifts of the free-running oscillator. Phase response curves (PRCs)
are constructed by plotting the magnitude of the phase change against the circadian time
when the light pulse was administered. It has been demonstrated that red or blue light
pulses given during the early or late subjective night trigger phase delays (negative
phase changes) or phase advances (positive phase changes), respectively (Covington et
al., 2001). To test the effect of UV-B in phase re-setting, plants described above were
grown in 12/12 LD cycles for a week and transferred to WLL. Short UV-B pulses
(1.5 μmol m-2 s-1 for 30 min) were given to parallel sets of plants in every 3 hr starting at
36 hr after the transfer to WLL. The magnitude and direction of phase shifts of
CCR2:LUC+ expression were determined relative to the non-pulsed control plants. UV-B
pulses cause phase delays during the subjective night, but apparently have no effect
during the subjective day in WT plants. In contrast, no significant phase shifts were
detected in uvr8-6 or in cop1-4 mutant plants at any time of the circadian cycle. On the

SUMMARY ______________________________________________________________________________________________________
93
other hand, when red, blue or white light pulses were used to induce phase shifts in
plants free-running in DD, WT and uvr8-6 plants displayed identical PRCs with the shape
consistent with published data (Covington et al., 2001). Collectively, these data
demonstrate that UV-B light does entrain the plant circadian clock, and UVR8 and COP1
play essential roles in this process.
UV-B acutely induces transcription of core clock genes
The precise molecular mechanism by which light signals entrain the plant circadian
oscillator is not fully understood yet, but light-modulated transcription of core oscillator
genes is thought to contribute to this process. In Arabidopsis, transcription of clock
genes CCA1, LHY, PRR9 and GI and the clock-associated gene ELF4 is acutely induced by
visible light (Wang et al., 1997; Tepperman et al., 2001; Locke et al., 2005; Ito et al.,
2003; Kikis et al., 2005). Induction of these rhythmically expressed genes is gated by the
clock, which means that the same light treatment causes maximal induction at the time
around the circadian peak of a given gene, but only a small increase is detected at the
time of the circadian trough.
To see if UV-B affects the transcription of clock genes in Arabidopsis, the acute
induction of CCA1, PRR9, GI and ELF4 genes was tested by quantitative RT-PCR. WT,
uvr8-6 and cop1-4 plants were grown in 12/12 LD cycles for a week and transferred to
WLL. Plants were irradiated by UV-B (1.5 μmol m-2 s-1 for 30 min) at 3 hr intervals
starting from 36 hr after the transfer, returned to WLL and harvested 1 hr later. The
control, non-pulsed samples were harvested at 3 hr intervals and provided data on the
basal expression of the corresponding genes. CCA1, PRR9, GI and ELF4 showed acute and
gated induction in response to UV-B pulses. The uvr8-6 mutation had no effect on the
basal circadian expression of these genes, but eliminated the response to UV-B pulses. In
contrast, the early phase and short period phenotype of the cop1-4 mutation was clearly
seen in the case of all clock genes tested and UV-B induction of all tested was
diminished. Expression of the clock gene TOC1 is not affected by visible light, and TOC1
is not affected by UV-B light either. These data suggest that UV-B induces the expression
of a very similar set of clock genes as visible light and demonstrate that UVR8 regulates
clock gene expression in response to UV-B, but not visible light.

SUMMARY ______________________________________________________________________________________________________
94
Circadian gated induction of rhythmic and non-rhythmic genes
The circadian clock rhythmically attenuates visible light responses (Hotta et al.,
2007). To see if UV-B responses are influenced similarly, UV-B induced circadian genes
CHALCONE SYNTHASE (CHS, At5g13930), GLUTATHIONE PEROXIDASE 7 (GPX7,
At4g31870), EARLY LIGHT INDUCED PROTEIN 2 (ELIP2, At4g14690), ELONGATED
HYPOCOTYL 5 (HY5, At5g11260) and HY5-HOMOLOG (HYH, At3g17609) and non-
circadian genes (At3g16350 and At3g57830) were selected and their UV-B inducibility
was tested in WT, uvr8-6 and cop1-4 plants. Induction of CHS, GPX7, ELIP2 and HYH
showed clear circadian control with amplitude comparable to that of the clock genes. In
contrast, inducibility of HY5 did not display any circadian patterns. To see if the lack of
gating is general for any light responses of HY5, the effect of red light pulses was tested
in a “classical” gating experiment. Plants were grown in 12/12 LD cycles for a week and
transferred to DD. Red light pulses (40 μmol m-2 s-1 for 60 min) were given at 3 hr
intervals and HY5 and HYH mRNA levels were determined in pulsed and nonpulsed
samples. It seems, red light induction of HY5 and HYH is gated by the clock.
Interestingly, UV-B induction of the non-rhythmic At3g16350 and At3g57830 genes
showed clear circadian modulation, and the maximal induction was detected around
mid-day. To see if the low expression level of these two genes in the absence of UV-B
masks circadian rhythmicity, we measured transcript levels in plants transferred to WLL
supplemented with low intensity UV-B. We conclude, despite the elevated expression
levels under these conditions, these genes are expressed non-rhythmically.
UV-B induction of all genes tested was completely eliminated in uvr8-6 or cop1-4
plants. The uvr8-6 mutation had no effects on basal expression levels/patterns, but
cop1-4 caused an early phase/short period phenotype as expected. Moreover, the basal
expression level of CHS, ELIP2 and HY5 was elevated in cop1-4 plants. Taken together,
these data suggest that -at least regarding gene expression- gating of UV-B and visible
light induction is differently regulated.
HY5 and HYH are not essential for UV-B induction or for gating
The HY5 and HYH transcription factors were suggested to control the UV-B induction
of most responsive genes in Arabidopsis (Brown and Jenkins, 2008). To test their
function in our conditions and for the particular target genes used in this study, a UV-B

SUMMARY ______________________________________________________________________________________________________
95
gating experiment was performed using hy5 hyh double mutant plants. The growth and
induction conditions were essentially the same as described in the previous sections. In
double mutant plants the basal expression level and UV-B induction of CCA1 and PRR9
was only slightly affected, whereas gating was clearly retained. In contrast, expression of
CHS was severely reduced and UV-B induction was completely lost in hy5 hyh, as
described before (Brown and Jenkins, 2008; Stracke et al., 2010). In the case of
At3g16350 UV-B induction was diminished, but not completely lost. Importantly, the
retained inducibility showed clear circadian gating. Collectively, these results
demonstrate that UV-B induction of clock genes does not rely on HY5/HYH and
circadian modulation of UV-B induced gene expression is not mediated by these
transcription factors.
Circadian gating of UV-B induced gene expression is lost in the
arrhythmic elf3-4 mutant and CCA1-OX plants
Circadian modulation of UV-B induction is provided by the circadian clock. In
principle, if clock regulation is eliminated, gating is lost, but UV-B inducibility should be
retained at a certain constant level. To test this, UV-B induced gene expression was
investigated in plants with non-functional circadian clocks. CCA1 showed low and
constitutive basal expression level, whereas PRR9 was expressed at high and
constitutive level in non-induced elf3-4 plants, in agreement with published data
(Alabadi et al., 2001; Kim et al., 2005; Thines and Harmon, 2010). The clock-controlled
CHS and the non-rhythmic At3g16350 showed increased basal expression levels in
elf3-4, similarly to PRR9. UV-B induction of all tested genes displayed constitutive, non-
gated pattern in elf3-4. The induced mRNA levels of PRR9, GI, CHS and At3g16350 in
elf3-4 were similar to or higher than the maximal induced levels in WT.
Analysis of the arrhythmic line constitutively over-expressing CCA1 (CCA1-OX, line 38
(Wang and Tobin, 1998) yielded similar results. UV-B irradiation resulted in high levels
of gene induction independent of the time of the treatment. Interestingly, two-fold
induction of CCA1 was observed in the CCA1-OX line. Since the 35S promoter is not up-
regulated by UV-B (Boyko et al., 2010), this result suggests an effect of UV-B on the
stability of CCA1 mRNA. These data indicate that rhythmic gating is lost, but UV-B
induction of gene expression is retained in these arrhythmic lines.

SUMMARY ______________________________________________________________________________________________________
96
Physiological impact of clock-modulated UV-B induction
The observed hypersensitivity of UV-B dependent gene expression has suggested that
elf3-4 mutant plants could be more tolerant to UV-B stress compared to WT. To test this
hypothesis, plants were grown in 12/12 LD cycles for a week, transferred to continuous
white light supplemented with UV-B (1.5 μmol m-2 s-1) and were given high-intensity
(14 μmol m-2 s-1) UV-B pulse for 20 min in the subjective morning (24h after the
transfer) and in the subjective evening (35h after the transfer). Plants were returned to
normal growth conditions and analyzed one week later. WT and elf3-4 plants were
similarly affected by the UV-B pulse, indicating that the particular high induced levels of
genes dependent on low intensity UV-B do not confer resistance to elf3-4 plants against
high intensity UV-B. The elf3-4 mutant accumulates less chlorophyll than WT, but the
UV-B induced decrease is the same in both genotypes. Moreover, the effect of stressing
UV-B pulses was independent of the time of the day in both genotypes.
Discussion
UV-B light mediates the entrainment of the plant circadian clock
Entraining light signals affect different parameters of the free-running clock,
depending on the duration of irradiation. In plants, continuous irradiation shortens the
free-running period length in a fluence rate dependent manner, whereas light pulses
elicit discrete phase shifts of the oscillator. Our data demonstrate that UV-B acts as a
signal for both modes of entrainment. In WT seedlings the clock output markers
exhibited significantly (1-2 h) shorter free-running periods if continuous white light was
supplemented by low fluences of UV-B. UV-B pulses applied to plants free-running in
WLL were effective in causing phase delays during the night, but no phase shifts were
elicited during the day.
To provide a possible molecular mechanism for UV-B mediated entrainment, the
inducibility of the core clock genes CCA1, PRR9, GI and the clock-associated gene ELF4
was tested over one circadian cycle. All genes showed rhythmically gated UV-B
induction in WT seedlings and that their induction was eliminated in uvr8-6 and cop1-4
plants. Surprisingly, the loss of HY5/HYH did not affect the period length of the clock in
WL or in UV-B and the UV-B inducibility of clock genes. These findings are somewhat

SUMMARY ______________________________________________________________________________________________________
97
unexpected as HY5 was shown (i) to bind in vivo to the promoters of several clock genes,
including CCA1, LHY, ELF4 and TOC1 in seedlings grown in WL (Lee et al., 2007), (ii) the
hy5 and hy5 hyh double mutant, similarly to uvr8, is impaired in UV-B dependent
photomorphogenesis, (Ulm et al., 2004; Oravecz et al., 2006) and (iii) hy5 was thought to
mediate UV-B inducibility of the majority of genes regulated by UVR8 and COP1 (Brown
et al., 2005; Oravecz et al., 2006).
Taken together, the correlation of physiological (modulation of period length) and
molecular (clock gene induction) data from UV-B signaling mutants suggests that this
effect of UV-B is mediated through the transcriptional regulation of clock genes. Among
the clock genes tested, PRR9 showed the most significant induction to UV-B. The
elevated level of PRR9 in continuous UV-B light could explain the effect of UV-B on
period length, since over-expression of PRR9 resulted in a similar degree of period
shortening (Matsushika et al., 2002).
The circadian clock rhythmically modulates UV-B induction of
responsive genes
Circadian gating is the process by which the clock rhythmically inhibits the acute
effect of environmental or endogenous signals on the corresponding target processes.
UV-B induction of all but one tested clock and clock-controlled genes, including HYH,
was gated by the clock in a manner similar to gating visible light responses. The only, but
significant exception was HY5, which showed a flat, non-gated induction by UV-B but a
gated induction by red light over the time course tested. These results demonstrate that
the signaling pathway mediating the UV-B induction of HY5 is not targeted by the clock
and it is different from the cascades transducing visible light signals to HY5.
Unexpectedly, UV-B induction of the non-rhythmic At3g16350 and At3g57830 showed
clear circadian gating with peak levels of induction at the middle of the subjective day.
Gating is not restricted to a single time of day, but works for a range of genes with
markedly different phases of expression. Therefore the mechanism underlying gating
could be explained by UV-B signaling components acting in parallel routes and
controlled by the clock in different phases. UVR8 is transcribed rhythmically and shows
the peak of expression close to dusk. However, UVR8 protein levels show no daily
oscillations, so it is unlikely that this mechanism mediates gating of UV-B induction,

SUMMARY ______________________________________________________________________________________________________
98
although the possibility of clock-regulated post-translational modification of UVR8
cannot be excluded. HY5 and HYH show a low amplitude rhythm in WL but UV-B
induction of HY5 lacks clock control whereas the UV-B induction of HYH is gated;
therefore, accumulation of HY5/HYH heterodimers in the morning could mediate gating
of UV-B responses of morning expressed genes. However, our data demonstrate that
clock gene induction by UV-B is unaffected in the hy5 hyh double mutant in the morning
(CCA1) or during the day (PRR9). UV-B induction of CHS was completely absent, but a
residual and gated induction of At3g16350 was retained in hy5 hyh, indicating that
HY5/HYH are important for the acute UV-B induction of a set of genes, but they are
dispensable for the temporal modulation of the response.
Physiological significance of clock-modulated UV-B responses
Induction of gene expression in response to low fluences of UV-B light (acclimation)
has an important function in triggering the production of pigments (flavonoids,
anthocyanin) that are protective against high intensity, damaging UV-B (Favory et al.,
2009). Our results showed that induction of all tested non-clock genes was gated to the
subjective morning or the first part of the subjective day. This indicated that perception
and processing of low intensity UV-B signals in the early part of the day is important to
set up the flavonoid “shield” and survive UV-B stress expected during the subsequent
hours of the day. In order to test the physiological relevance of clock modulated
induction of gene expression in response to low intensity UV-B, we characterised
molecular and physiological UV-B responses in the arrhythmic elf3-4 mutant. Our data
demonstrated that almost all of the genes tested here showed a high induction level to
low fluences of UV-B in elf3-4 at all times of the day. The induced mRNA levels, except
for CCA1, were usually higher than the maximal induction level achieved in WT which in
turn suggested that elf3-4 plants may display increased stress tolerance to UV-B
compared to WT plants. However, acclimated elf3-4 mutants were as resistant to UV-B
stress pulses applied at different times of the day as were the acclimated WT controls.
Analysis of another arrhythmic mutant, CCA1-OX, has provided similar results.. This
indicates that limiting low intensity UV-B dependent gene expression and the
subsequent accumulation of flavonoids to a relatively short interval of the day is
sufficient to reach normal protection against the damaging effect of high intensityUVB.

KÖSZÖNETNYILVÁNÍTÁS
A dolgozatban szereplő kísérleteket a Magyar Tudományos Akadémia Szegedi
Biológiai Központjának Növénybiológiai Intézetében, a Foto- és Kronobiológiai Csoport
tagjaként végezhettem el. Köszönettel tartozom téma- és csoportvezetőmnek, Dr. Nagy
Ferencnek jelentős anyagi és szellemi támogatásáért, amellyel mindvégig segítette
munkámat és akinek nem kis szerepe volt abban, hogy eredményeinket megfelelő
színvonalon tudtuk közzétenni. Köszönöm Dr. Dudits Dénes volt-, Dr Ormos Pál jelenlegi
főigazgató és Dr. Vass Imre igazgató uraknak, hogy munkámat lehetővé tették.
Köszönettel tartozom a Foto- és Kronobiológiai csoport minden egyes tagjának a
munkám során nyújtott segítségükért és a közvetlen munkahelyi légkör
megteremtéséért. Köszönöm Dr. Kozma-Bognár Lászlónak a laboratóriumban eltöltött
éveim alatt köztünk lezajlott rendkívül értékes és izgalmas tudományos eszmecseréket,
valamint tekintélyes mértékű hozzájárulását a dolgozatom alapját képező közlemény
megírásához. Köszönöm Dr. Gyula Péternek az önzetlen szakmai segítségét a munkám
kezdeti időszakában. Köszönöm Dr. Kevei Évának az UV-B fénnyel végzett munka első
lépéseiben nyújtott szakmai tanácsait.
Külön köszönöm Dr. Szekeres Miklósnak az értekezésem tudományos bírálatát,
értékes észrevételeit, javaslatait.
Köszönöm Jószai Katalin, Majzik Hedvig és Veres Gabriella asszisztenseknek a
kísérletek előkészítésében és a növények gondozásában nyújtott pótolhatatlan
segítségét és özv. Hajó Róbertnének az Arabidopsis magok tisztításában végzett
munkáját.
Köszönöm Terecskei Katának a figyelmét és türelmét a munkám során, valamint a
dolgozat javításában tett felbecsülhetetlen segítségét.
Végül köszönöm családomnak a kitartó érdeklődést és támogatást, amelyet munkám
és tudományos előmenetelem iránt mindvégig tanúsítottak.