Embed Size (px)
Citation preview

DEacuteTERMINATION DU MODE DrsquoACTION ET DE LA CIBLE CELLULAIRE DE LA TOMATIDINE CHEZ Staphylococcus aureus
Par
Isabelle Guay
meacutemoire preacutesenteacute au deacutepartement de biologie en vue de lrsquoobtention du grade de maicirctre egraves science (M Sc)
FACULTEacute DES SCIENCES UNIVERSITEacute DE SHERBROOKE
Sherbrooke Queacutebec Canada 17 Juillet 2014
Le 17 juillet 2014
le jury a accepteacute le meacutemoire de Madame Isabelle Guay
dans sa version finale
Membres du jury
Professeur Franccedilois Malouin
Directeur de recherche
Deacutepartement de Biologie
Professeur Eacuteric Marsault
Codirecteur de recherche
Deacutepartement de Pharmacologie
Professeur Seacutebastien Rodrigue
Membre interne
Deacutepartement de Biologie
Professeur Kamal Bouarab
Preacutesident-rapporteur
Deacutepartement de Biologie
ii
SOMMAIRE
Dans le but de mieux comprendre le mode drsquoaction et de nous permettre de
deacuteterminer la cible de la tomatidine nous avons dans un premier temps tenteacute de
mieux circonscrire le spectre drsquoactiviteacute de la tomatidine Gracircce agrave ces travaux nous
sommes en effet maintenant en mesure de dire que la tomatidine possegravede une
activiteacute antibacteacuterienne contre les espegraveces de la divison des Firmicutes et plus
preacuteciseacutement contre les bacteacuteries de lrsquoordre des Bacillales dont font partie les genres
Bacillus Staphylococcus et Listeria Nous avons eacutegalement deacutecouvert gracircce agrave des
expeacuteriences en collaboration avec le laboratoire drsquoEacuteric Marsault qursquoun analogue de
la tomatidine (FC04-100) avait non seulement des proprieacuteteacutes similaires agrave la
moleacutecule naturelle mais deacutemontrait une activiteacute par lui-mecircme contre S aureus agrave
pheacutenotype normal alors que la tomatidine possegravede uniquement une activiteacute contre
les laquo small colony variants raquo De plus alors que la tomatidine possegravede plutocirct une
activiteacute bacteacuteriostatique contre la forme SCV de L monocytogenes le nouveau
composeacute (FC04-100) deacutemontre quant agrave lui une forte activiteacute bacteacutericide contre
cette souche tout comme contre la forme SCV des autres Bacillales
Parallegravelement et toujours dans le but de rechercher le mode drsquoaction et la cible de
la tomatidine nous avons obtenu par passages successifs dans un milieu avec
antibiotiques des mutants de S aureus agrave pheacutenotype normal et des SCV reacutesistants
agrave la tomatidine ou agrave la combinaison tomatidine et gentamicine Apregraves le
seacutequenccedilage de ces mutants lrsquoeacutetude de la position de ces mutations agrave lrsquoaide de
diffeacuterents logiciels de bio-informatique nous a permis drsquoeacutemettre un modegravele-
hypothegravese quant au mode drsquoaction et agrave la cible de la tomatidine Selon les reacutesultats
que nous avons agrave ce stade-ci la cible de la tomatidine chez S aureus serait la
sous-uniteacute c de lrsquoATP synthase Cependant son mode drsquoaction serait eacutegalement
deacutependant de la fonctionnaliteacute de la chaine de transport des eacutelectrons et donc de la
iii
polarisation membranaire et de la production de ROS intracellulaire ce qui
expliquerait la diffeacuterence drsquoactiviteacute entre les souches agrave pheacutenotype normal et les
SCV
Mots cleacutes tomatidine ATP synthase chaine de transport des eacutelectrons synergie laquo small-colony variants raquo ROS S aureus analogues
iv
REMERCIEMENTS
Je voudrais remercier mon directeur de recherche Pr Franccedilois Malouin qui a cru
en moi pour mrsquoavoir proposeacute ce beau projet de maitrise Merci pour sa grande
disponibiliteacute son eacutecoute et ses critiques toujours constructives et jamais
condescendantes
Je voudrais eacutegalement remercier mes conseilleacutes Kamal Bouarab Seacutebastien
Rodrigue et Eacuteric Marsault leur implication dans mon projet leur occasionnant un
surplus de travail Merci pour leur support technique et leur disponibiliteacute agrave reacutepondre
agrave mes questions
Un gros merci agrave Gabriel Mitchell qui a eacuteteacute mon mentor au deacutebut de ma maitrise et
qui mrsquoa introduit au projet laquo Tomatidine raquo Un merci particulier eacutegalement agrave
Maxime-Lamontagne-Boulet pour son aide en tant que stagiaire lors de mes
derniegraveres expeacuteriences Je voudrais eacutegalement remercier tous les membres du
laboratoire preacutesents et passeacutes qui ont toujours eacuteteacute disponibles pour partager avec
moi leurs connaissances theacuteoriques et techniques
Je ne peux passer sous silence lrsquoimplication de lrsquoUniversiteacute de Sherbrooke et du
FQRNT comme organismes subventionnaires au projet Merci eacutegalement agrave Fibrose
Kystique Canada qui a participeacute eacutegalement agrave la subvention drsquoune partie du projet
En terminant je voudrais remercier ma famille qui a crue en moi et qui mrsquoencourage
agrave perseacuteveacuterer depuis toujours Un merci tout particulier agrave mon conjoint et ma belle-
fille pour qui je nrsquoai pas toujours eacuteteacute aussi disponible que jrsquoaurais voulu lrsquoecirctre et qui
mrsquoont supporteacutee au quotidien et ce dans tous les sens du terme
v
TABLE DES MATIEgraveRES
SOMMAIRE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ii
REMERCIEMENTS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip iv
TABLE DES MATIEgraveRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip v
LISTE DES ABBREacuteVIATIONS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip vii
LISTE DES TABLEAUX helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ix
LISTE DES FIGURES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip x
CHAPITRE 1 INTRODUCTION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
11 Staphylococcus aureus helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
111 Importance cliniquehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)helliphellip 2
1121 Diffeacuterences et similariteacutes helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 2
1122 La chaine de transport des eacutelectrons helliphelliphelliphelliphellip 4
12 Antibiotiques Modes drsquoaction cibles cellulaire et
meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles helliphelliphelliphellip 25
122 Les aminoglycosides helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1221 Mode drsquoaction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1222 Meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphellip 31
123 La tomatidine helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 33
13 Projet de maitrise helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
CHAPITRE 2 ARTICLE SCIENTIFIQUE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 37
ldquoTomatidine and derivative FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Firmicutes Including Those of Listeria monocytogenes
and Are Bactericidal in Combination With Aminoglycosides Against their Normal
Phenotyperdquo
vi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE helliphelliphelliphellip 76
31 Obtention des mutants helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
32 Seacutequenccedilage et analyses bio-informatiques helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 79
321 Identification des mutations en lien avec la tomatidine helliphellip 89
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct helliphellip 90
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine helliphelliphelliphelliphellip 99
CHAPITRE 4 DISCUSSION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 113
CHAPITRE 5 CONCLUSION ET PERSPECTIVES helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 124
CONTRIBUTIONS SUPPLEacuteMENTAIRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 1 Application de brevet helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 2 Article scientifique helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 129
laquo Unraveling the structure-activity relationship of tomatidine
a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureus raquo
BIBLIOGRAPHIE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 131
vii
LISTE DES ABBREacuteVIATIONS
Adh Alcool deacuteshydrogeacutenase
ADN Acide deacutesoxyribo-nucleacuteique
ADP Adeacutenosine diphosphate
ARN Acide ribo-nucleacuteique
ATP Adeacutenosine triphosphate
CA-MRSA Community-associated MRSA
CMI concentration minimale inhibitrice
CoQ Coenzyme Q
Cyt Cytochrome
DMQ laquo demethylmenaquinone raquo
dTMP Deacutesoxythymidine monophosphate
dUMP Deacutesoxyuridine monophosphate
FADH2 Flavine adenine dinucleacuteotide
GEN gentamicine
HA-MRSA Healthcare-associated MRSA
Ldh Lactate deacuteshydrogeacutenase
ml millilitre
MQ meacutenaquinone
MRSA methycilin resistant Staphylococcus aureus
Mqo2 Quninol oxidoreacuteductase
NADH Nicotinamide adenine dinucleacuteotide (forme reacuteduite)
Pdh Pyruvate deacuteshydrogeacutenase
Pfl Pyruvate formate lyase
PyrD Dihydroorotate deacuteshydrogeacutenase
Q Quinone
Rif rifampicine
ROS laquo reactive oxygen species raquo espegravece reacuteactive de lrsquooxygegravene
viii
[S] Polysulfide
S aureus Staphylococcus aureus
SARM Staphylococcus aureus resistant agrave la meacutethycilline
SCV laquo small colony variants raquo
SHCHC 2-succinyl-6-hydroxy-24-cyclohexadiene-1-carboxylate syntase
TCA Tricarboxylic acid cycle
TCD tableau croiseacute dynamique
TO tomatidine UFC Uniteacute formatrice de colonies
UQ Ubiquinone
microg microgramme
ix
LISTE DES TABLEAUX
CHAPITRE 1 INTRODUCTION
1 Cibles et modes drsquoaction des principales classes drsquoantibiotique 27
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants (B) chez les mutants NewbouldΔhemB reacutesistants 95-96
CHAPITRE 4 DISCUSSION
1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 123
x
LISTE DES FIGURES
CHAPITRE 1 INTRODUCTION
1 Diffeacuterence entre les pheacutenotypes normaux et SCV de S aureus 3
2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes 6
3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale 8
4 Chaines respiratoires bacteacuteriennes contenant des cytochromes 9
5 Flexibiliteacute respiratoire (A) chez Paracoccus denitrificans (B) chez
Escherichia coli 10
6 Chaine de transport des eacutelectrons proposeacutee chez les staphylocoques 12
7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies 17
8 Modegravele de reacutegulation du cycle TCA 20
9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS 23
10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de O2-bull 24
11 Voies de formation et de deacutetoxification des ROS 25
12 Cineacutetique drsquoentreacutee des aminoglycosides 30
13 Sites de mutations de certains reacutesistants aux aminoglycosides 32
14 Structures chimiques de (A) tomatine (B) tomatidine et (C) Analogue
de tomatidine FC04-100 34
xi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Graphiques des CMI pour chacun des passages pour (A) la
tomatidine sur NewbouldΔhemB (B) la tomatidine en preacutesence de
gentamicine sur NewbouldΔhemB (C) la gentamicine sur S aureus
ATCC 29213 (D) la gentamicine en preacutesence de tomatidine sur S
aureus ATCC 29213 77-79
2 Organigramme des mutants reacutesistants seacutequenceacutes 80-81
3 Eacutetapes du seacutequenccedilage Illumina 83
4 Meacutethode drsquoamplification par la meacutethode Illumina 85
5 Correction du programme RATT sur les annotations transfeacutereacutees 88
6 Exemple drsquoune partie du tableau compilation des mutations 89
7 Seacutelection des gegravenes drsquointeacuterecirct chez ATCC 29213 dans le TCD par
lrsquoutilisation de diffeacuterents filtres 91-92
8 Seacutelection des gegravenes drsquointeacuterecirct chez NewbouldΔhemB dans le TCD par
lrsquoutilisation de diffeacuterents filtres 93-94
9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct 97-98
10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants 100
11 Courbe de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison gentamicine (+TO) du premier et troisiegraveme niveau de
reacutesistance 101
12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B)
NewbouldΔhemB 102
xii
13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213 104
14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicinelorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC
29213 105
15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants
ATCC 29213 reacutesistants agrave la combinaison gentamicine (+TO) 106
16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la
tomatidine 107
17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la
tomatidine 108
18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB
reacutesistants agrave la combinaison tomatidine (+gentamicine) 109
19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane
Potential Kit (Invitrogen) 111
20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du
gegravene atpE de S aureus Newman 112
xiii
CHAPITRE 4 DISCUSSION
1 Site drsquoinhibition de lrsquoATP synthase par diffeacuterentes moleacutecules 114
2 Structure chimique de (A) TMC207 et (B) lrsquoanalogue anti Gram
positifs 115
3 Seacutequence proteacuteique de la proteacuteine AtpE chez diffeacuterentes espegraveces 118
4 Modeacutelisation de la structure tridimentionnelle de lrsquoATP synthase de S
aureus (atpE) montrant les acides amineacutes muteacutes chez les mutants
reacutesistants 121
- 1 -
CHAPITRE 1
INTRODUCTION
11 Staphylococcus aureus
111 Importance clinique
Staphylococcus aureus est une bacteacuterie agrave Gram positif anaeacuterobie facultative Crsquoest un
microorganisme opportuniste pouvant infecter diffeacuterents hocirctes diffeacuterents organes ou
sites drsquoinfection et ayant la capaciteacute de causer tant des infections aiguumles que
chroniques (Archer 1998 Goerke et Wolz 2004)
Staphylococcus aureus reacutesistant agrave la meacutethilline le SARM (ou laquo MRSA raquo) est une
souche de S aureus devenue reacutesistante aux β-Lactamines par lrsquoacquisition du gegravene
mecA dans son chromosome (Hiramatsu 1995) Le SARM a eacuteteacute isoleacute pour la
premiegravere fois en 1960 en Angleterre (Jevons 1961) et est devenu eacutepideacutemique agrave
travers le monde depuis les anneacutees 1970 Au moins trois geacutenotypes distincts eacutetaient
recenseacutes dans les anneacutees 1980 (Hiramatsu 1995) dont deux sont toujours preacutevalents
agrave travers le monde comme eacutetant des souches HA-MRSA (Healthcare-associated
MRSA) des souches acquises agrave lrsquohocircpital (Enright et al 2002) Ces souches ont
acquis des gegravenes de reacutesistance agrave pratiquement tous les antibiotiques introduits dans
lrsquoutilisation clinique depuis les 50 derniegraveres anneacutees mecircme agrave lrsquoantibiotique de dernier
ressort la vancomycine(Hiramitsu 1997 Chen et al 2003) Nous sommes
maintenant en manque drsquoagents theacuterapeutiques efficaces contre les SARM touchant
les personnes immunodeacuteprimeacutees qui sont dans les hocircpitaux En plus depuis le deacutebut
des anneacutees 1990 des clones de SARM dont plusieurs deacutemontraient un potentiel
pathogeacutenique supeacuterieur aux souches classiques de HA-MRSA retrouveacutees agrave lrsquohocircpital
- 2 -
sont maintenant retrouveacutes dans la communauteacute (CA-MRSA) agrave travers le monde (Udo
et al 1993 Riley et al 1995 Moreno et al 1995 Aubry-Damon et al 1997) et ont
la capaciteacute drsquoaffecter des personnes en santeacute en causant des infections aiguumles
(Hiramatsu et al 2013) Il apparaicirct donc urgent de trouver des drogues alternatives
afin de surmonter le nombre grandissant de souches de SARM
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)
1121 Diffeacuterences et similariteacutes
Les laquo small colony variants raquo (SCV) sont des bacteacuteries souvent isoleacutees lors des
infections chroniques comme crsquoest le cas des infections pulmonaires chez les
patients atteints de fibrose kystique mais aussi dans des cas drsquoosteacuteomyeacutelite
drsquoarthrite septique drsquoinfections de prothegraveses orthopeacutediques et lors de la mammite
bovine (Alexender et Hudson 2001 Moisan et al 2006 Proctor et al 2006) Les
SCV sont caracteacuteriseacutes par soit un meacutetabolisme oxydatif dysfonctionnel ou un manque
dans la biosynthegravese de la thymidine les deux amenant une alteacuteration dans
lrsquoexpression des facteurs de virulence une croissance plus lente et une diminution
de la pigmentation des colonies (Figure 1) (Proctor et al 2006) Cependant alors
que les souches normales reacutepriment lrsquoexpression des proteacuteines de surface et
expriment les exoproteacuteines lors de la phase stationnaire (Novick 2003) les SCV
expriment de faccedilon stable les gegravenes Sig-B deacutependants codant pour des proteacuteines de
surface telle les adheacutesines au lieu drsquoactiver le systegraveme agr deacutependant du quorum-
sensing et produire des exoproteacuteines (Moisan et al 2006) Ces diffeacuterences dans
lrsquoexpression des facteurs de virulence reacutesultent chez les SCV en une meilleure
habileteacute agrave adheacuterer aux composantes de lrsquohocircte (Mitchell et al 2008) et agrave former du
biofilm (Mitchell et al 2010a Mitchell et al 20010b) et seraient eacutegalement lieacutees agrave
lrsquohabileteacute des SCV agrave lrsquoinvasion et la persistance des cellules de lrsquohocircte (Proctor et al
2006 Sendi et Proctor 2009 Mitchell et al 2011b)
- 3 -
S aureus est donc un pathogegravene qui peut se preacutesenter sous deux pheacutenotypes qui
sont souvent retrouveacutes simultaneacutement lors drsquoune mecircme infection et qui ont le pouvoir
de reacuteverter drsquoun pheacutenotype agrave lrsquoautre selon les conditions du milieu (Figure 1)
(Tuchscherr et al 2011)
Le pheacutenotype dit rdquonormalrdquo est associeacute aux infections aiguumles agrave une forte production
de facteurs de virulence (ex heacutemolysine) et est associeacute agrave la phase de disseacutemination
de la bacteacuterie Lrsquoautre pheacutenotype le laquo small-colony variants raquo ou SCV est affecteacute
dans sa chaine respiratoire ce qui amegravene une croissance beaucoup plus lente des
colonies de 8 -10 fois plus petites que les prototypes et une faible susceptibiliteacute aux
aminoglycosides Les SCV sont plutocirct associeacutes aux infections chroniques agrave S aureus
de par leur capaciteacute agrave srsquointernaliser dans les cellules de lrsquohocircte leur forte capaciteacute agrave
produire du biofilm et leur capaciteacute agrave produire des facteurs de colonisation
(Tuchscherr et al 2011 Proctor et al 2006) (Figure 1)
Figure 1 Diffeacuterences entre les pheacutenotypes normaux et SCV de S aureus
S aureus agrave
pheacutenotype normal
Disseacutemination
amp
Infections aiguumles
Colonisation
amp
Infections persistantes
SCVs
(laquo small-colony
variants raquo)
Reacutevers
ion
pheacutenoty
piq
ue
Norm
al
SC
V
Croissance
Biofilm
Internalisationheacutemolyse
Mitchell G et Malouin F 2012
- 4 -
1122 La chaine de transport des eacutelectrons
Chaine respiratoire tableau geacuteneacuteral
Les chaines respiratoires bacteacuteriennes bien que tregraves variables dans leurs
composantes parmi les diffeacuterentes espegraveces sont toutes constitueacutees sur le mecircme
modegravele de base Elles sont composeacutees de trois composantes majeures soit
- les deacuteshydrogeacutenases primaires qui sont chargeacutees drsquooxyder des substrats
principalement les coenzymes NADH et FADH2 (issues du cycle de Krebs) (mais
aussi selon le cas le succinate le lactate lrsquohydrogegravene hellip) et de pomper deux
protons du cocircteacute externe de la membrane en plus de transfeacuterer deux eacutelectrons agrave un
groupement de quinones
- Les quinones sont des moleacutecules liposolubles non proteacuteiques mobiles dans la
membrane Elles servent drsquointermeacutediaires entre les deacuteshydrogeacutenases primaires et
les oxydases terminales pour le transport des eacutelectrons Il existe deux sortes de
quinones les benzoquinones dont font partie les ubiquinones et les
naphtoquinones dont font partie les meacutenaquinones Les bacteacuteries agrave Gram positif
ne possegravedent que des meacutenaquinones alors que les deux sont preacutesentes chez les
bacteacuteries agrave Gram neacutegatif (Hederstedt 1993 Pratique et Theraulaz 2007)
- Les oxidoreacuteductases terminales sont chargeacutees de reacuteduire lrsquoaccepteur final
drsquoeacutelectron lrsquooxygegravene eacutetant lrsquoaccepteur final privileacutegieacute
De faccedilon plus deacutetailleacutee la chaine respiratoire est composeacutee de cinq
complexes proteacuteiques principaux (Figure 2)
- complexe I NADH deacuteshydrogeacutenase ou NADH reacuteductase ou NADHCoQ
oxidoreacuteductase ou NADH-ubiquinone oxidoreacuteductase
- complexe II Succinate deacuteshydrogeacutenase
- complexe III Cytochrome bc1 complexe ou ubiquinol cytochrome c
reacuteductaseoxydoreacuteductase
- complexe IV Cytochrome c oxydase
- 5 -
- complexe V ATP synthase (Creacutepin-Gilbert et al 2006)
La phosphorylation oxydative est deacuteclencheacutee dans la chaine de complexes
proteacuteiques par la preacutesence drsquoO2 accepteur terminal privileacutegieacute drsquoeacutelectrons (ou drsquoautres
accepteurs finaux tel le nitrate) dans le milieu ainsi que par les eacutelectrons ameneacutes par
le NADH et le FADH2 au complexe I et II respectivement (Creacutepin-Gilbert et al 2006)
Les eacutelectrons vont ensuite transiter par le complexe III jusqursquoau complexe IV ou les
eacutelectrons seront utiliseacutes afin de reacuteduire lrsquooxygegravene en eau le complexe IV est
lrsquooxydase terminale consideacutereacutee comme eacutetant lrsquoaccepteur final drsquoeacutelectrons Le transfert
drsquoeacutelectrons est possible gracircce aux reacuteactions drsquooxydoreacuteduction en cascade dans les
diffeacuterents complexes proteacuteiques Ces reacuteactions fournissent de lrsquoeacutenergie libre utiliseacutee
par les complexes I III et IV afin de transfeacuterer des protons agrave lrsquoexteacuterieur de la cellule
(Creacutepin-Gilbert et al 2006) Le complexe V (ATP synthase) quant agrave lui utilise les
eacutelectrons pompeacutes agrave lrsquoexteacuterieur par les autres complexes comme force proton-motrice
afin de phosphoryler trois moleacutecules drsquoADP et ainsi produire trois ATP Le transport
des eacutelectrons entre les diffeacuterents complexes proteacuteiques est possible gracircce agrave la
preacutesence dans la membrane de deux types de moleacutecules liposolubles non proteacuteiques
mobiles les meacutenaquinones et le cytochrome c Les meacutenaquinones seules quinones
preacutesentes chez les bacteacuteries agrave Gram positif assurent le transport des eacutelectrons entre
le complexe I et le complexe III et entre le complexe II et III Le cytochrome c pour sa
part veacutehicule les eacutelectrons entre le complexe III et le complexe IV Le transport des
eacutelectrons srsquoeffectue de la composante ayant le plus faible potentiel drsquooxydoreacuteduction
vers celle ayant le plus fort potentiel
- 6 -
La chaine respiratoire bacteacuterienne
La mitochondrie montre assez peu de flexibiliteacute respiratoire Il y a une certaine
flexibiliteacute au niveau de lrsquoentreacutee des eacutelectrons mais aucune au niveau de la sortie des
eacutelectrons ougrave la cytochrome aa3 oxydase reacuteduit lrsquooxygegravene en eau (Richardson 2000)
(Figure 3) Chez les bacteacuteries alors que le complexe II fonctionne sensiblement de
faccedilon identique agrave celui des eucaryotes les autres complexes peuvent se composer
de diffeacuterents complexes enzymatiques (reacuteductases) selon les espegraveces et les
conditions environnementales (Richardson 2000 Thoumlny-Meyer 1997) En outre
chez les bacteacuteries contrairement aux eucaryotes la chaine respiratoire comporte
souvent des voies de transport des eacutelectrons alternatives on dit alors qursquoelles ont une
chaine respiratoire agrave branches ou brancheacutee ou ramifieacutee (Artzatbanov et Petrov 1990)
Plusieurs espegraveces bacteacuteriennes possegravedent une chaine respiratoire ramifieacutee dont la
ramification se situe habituellement suite agrave la reacuteduction des quinones en quinols
(Lauraeus et Wikstrom 1993 Thoumlny-Meyer 1997) (Figure 4) Une des deux
Complexe I NADH
deacuteshydrogeacutenase
Complexe II Succinate
deacuteshydrogeacutenase
Complexe III
bc 1
Complexe IV Cytochrome c
oxydase
Complexe V
ATP synthase
Modifieacute de wikipedia httpfrwikipediaorgwikiChaine_respiratoire
Figure 2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes
Leurs rocircles dans le transport des eacutelectrons et dans lrsquoeacutelaboration du gradient de
protons et le lien entre la chaine respiratoire et le cycle de Krebs Q Quinones
- 7 -
branches connue comme la branche cytochrome c oxyde les quinols (meacutenaquinol
QH2) via la voie complexe bc1-cytochrome c- cytochrome c oxydase Lrsquoautre
branche-type consiste en une seule uniteacute enzymatique (cytochrome bd) laquelle
transfegravere directement les eacutelectrons des quinols agrave lrsquooxygegravene Cette voie est connue
comme la branche quinol oxydase il srsquoagit drsquoune oxydase terminale transfeacuterant
directement les eacutelectrons agrave lrsquooxygegravene eacutevitant de passer par lrsquointermeacutediaire du
complexe III (bc1) et du cytochrome c (Artzatbanov et Petrov 1990 Lauraeus et
Wikstrom 1993) Alors que la voie du cytochrome c permet le pompage de protons agrave
lrsquoexteacuterieur par lrsquoaction du complexe bc1 (ou complexe III) et du cytochrome c oxydase
le cytochrome bd ne permet aucun pompage de protons (Lauraeus et Wikstrom
1993) La voie passant par le complexe bc1-cytochrome c-cytochrome c oxydase est
plus efficace dans la production drsquoeacutenergie Alors que les 2 voies peuvent ecirctre
exprimeacutees simultaneacutement chez quelques espegraveces il semble que lrsquoexpression de ces
voies soit inductible selon les conditions environnementales chez Bacillus subtilis et
les Staphylocoques (Lauraeus et Wikstrom 1993) On pourrait dire eacutegalement que la
chaine respiratoire est seacutepareacutee en deux eacutetapes la premiegravere partie est la phase
quinone-reacuteductase (Q-QH2) et la seconde la phase quinol-oxydase (QH2-Q) menant
agrave lrsquooxydase terminale La phase de reacuteduction est responsable du transfert des
eacutelectrons drsquoune varieacuteteacute de substrats tels le NADH le succinate le lactate le formate
et lrsquohydrogegravene vers les quinones et inclut diffeacuterentes reacuteductases (Thoumlny-Meyer 1997)
(Figure 4)
- 8 -
Figure 3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale UQ ubiquinone UQH2 ubiquinol Cyt cytochrome Il semble y avoir presque autant de chaines de transport des eacutelectrons diffeacuterentes
qursquoil y a drsquoespegraveces bacteacuteriennes (par exemple Paracoccus denitrificans et Escherichia
coli (Figure 5)) De plus il ne semble pas y avoir consensus dans la litteacuterature sur
les noms attribueacutes aux oxydases terminales Les noms pour les oxydases terminales
deacutependent des types de cytochromes dont elles sont composeacutees Il ne semble donc
pas eacutevident de connaicirctre de faccedilon preacutecise les types de cytochromes qui font partie
des oxydases terminales Pour rajouter agrave la confusion les gegravenes et opeacuterons ne sont
pas toujours associeacutes agrave leur oxydase terminale dans la litteacuterature ce qui fait qursquoon ne
peut savoir de faccedilon certaine de quelle oxydase on parle Le Cytochrome aa3 peut
eacutegalement porter le nom de Cyt ba3 et le cytochrome bd peux eacutegalement porter les
noms de Cyt bo
Tireacute de Richardson DJ 2000
- 9 -
Succinate
Formate
H2
SDH
FDH
HYD
QQH 2 bc1 c
Reacuteductase alternatives
b
c
cd1c
cb
c
b
c
d
Siroheme
Siroheme
b
NO3-
NO2-
NO
TMAO
DMSOS2O3
2-
[S]
SO32-
Fumarate
Quinol oxidases
ba3bb3bo3d
bd
aa3
O2
Cyt c oxidases
aa3ba3baa3caa3cbb3cao
co
O2
Modifieacute de Thony-Mayer L 1997
NADH
NDH
Figure 4 Chaines respiratoires bacteacuteriennes contenant des cytochromes
Du cocircteacute gauche on retrouve la partie quinone (ubiquinone ou meacutenaquinone)-
reacuteductase Du cocircteacute droit on retrouve la partie quinol-oxydase avec les oxydases
terminales reacuteduisant lrsquoO2 ou lrsquoaccepteur terminal alternatif tel que montreacute Les
oxydoreacuteductases sont surligneacutees QQH2 repreacutesente le pool de quinonesquinols
dans la membrane Les polysulfides sont repreacutesenteacutes par [S] Les chaines
respiratoires ne contenant pas de cytochromes ne sont pas montreacutees NDH
NADH deacuteshydrogeacutenase
Modifieacute de Thoumlny-Mayer L 1997
- 10 -
Figure 5 Flexibiliteacute respiratoire chez Paracoccus denitrificans (a) et chez
Escherichia coli (b) MQ menaquinone UQ ubiquinone DMQ
demethylmenoquinone
Chez S aureus
Comme plusieurs autres espegraveces bacteacuteriennes anaeacuterobies facultatives S aureus a
la possibiliteacute drsquoexprimer diffeacuterentes voies de synthegravese eacutenergeacutetique dont lrsquoexpression
est finement reacuteguleacutee selon des conditions environnementales du moment (Hammer et
Tireacute de Richardson DJ 2000
- 11 -
al 2013 Lauraeus et Wikstrom 1993) Mecircme si S aureus peut utiliser la
fermentation la respiration (aeacuterobie et anaeacuterobie) sera privileacutegieacutee par la bacteacuterie
puisque celle-ci amegravene une bien meilleure production drsquoeacutenergie (Goumltz et al 2006
Hammer et al 2013 ) La respiration aeacuterobie avec lrsquooxygegravene comme accepteur final
drsquoeacutelectron sera utiliseacutee de faccedilon preacutefeacuterentielle en preacutesence drsquooxygegravene (Hammer et al
2013) Malgreacute sa versatiliteacute dans ses voies de synthegravese eacutenergeacutetique S aureus et les
autres bacteacuteries agrave Gram positif ne produisent qursquoun seul type de quinone les
meacutenaquinones lesquelles sont utiliseacutees pour la croissance degraves qursquoil y a de lrsquooxygegravene
de preacutesent dans le milieu (Somerville et Proctor 2009) La reacuteduction de lrsquooxygegravene en
eau est meacutedieacutee par des oxydases terminales hegraveme deacutependant (Hammer et al 2013)
Selon certains auteurs S aureus possegravederait deux oxydases terminales la
cytochrome oxydase aa3 (ou ba3) (QoxABCD) et la cytochrome bd quinol oxydase
(CydAB) (Goumltz et al 2006 Hammer et al 2013 Kohler et al 2008) Cette derniegravere
est exclusive aux procaryotes et est une voie de contournement du complexe III qui
transmet directement les eacutelectrons du laquo pool raquo de quinol (QH2) agrave lrsquooxygegravene Selon
certains auteurs il y aurait eacutegalement chez S aureus une troisiegraveme oxydase
terminale le cytochrome o (CyoABCDE) (Thoumlny-Meyer 1997 Voggu et al 2006)
qui selon certain serait un cytochrome oxydase terminal alternatif au cytochrome aa3
et au cytochrome bd (Voggu et al 2006 Goumltz et al 2006 Taber et Morrison 1964)
(Figure 6) Ces voies alternatives permettent agrave S aureus de srsquoadapter rapidement agrave
un changement drsquoenvironnement (Hammer et al 2013) In vivo chaque organe est
un environnement diffeacuterent preacutesentant des deacutefis respiratoires distincts dus agrave la
variation dans 1- la quantiteacute drsquooxygegravene disponible 2- la disponibiliteacute drsquoaccepteurs
drsquoeacutelectrons alternatifs 3- la concentration drsquoinhibiteurs de la respiration dans le milieu
tels les oxydes nitriques (NO) produits par les neutrophiles et 4- la disponibiliteacute des
cofacteurs neacutecessaires agrave la production drsquoeacutenergie (Hammer et al 2013) Le
cytochrome est exprimeacute de faccedilon modeacutereacutee dans un milieu riche en oxygegravene mais est
fortement exprimeacute dans un milieu pauvre en oxygegravene mais riche en oxyde nitrique
De plus le type drsquooxydoreacuteductase terminale exprimeacute est influenceacute eacutegalement par la
- 12 -
source de carbone disponible la phase de croissance le pH extracellulaire et la
tension en oxygegravene (Lauraeus et Wikstrom 1993 Tynecka et al 1999)
La chaine respiratoire chez les SCV
S aureus est particuliegraverement doueacute agrave surpasser les deacutefis respiratoires en acqueacuterant
une reacutesistance agrave lrsquoacide nitrique produit par les neutrophiles et une habiliteacute agrave persister
dans les cellules de lrsquohocircte en se preacutesentant comme un variant agrave petites colonies
ayant une chaine respiratoire deacuteficiente (SCV) (Hammer et al 2013)
Deux principaux types de SCV sont retrouveacutes dans les isolats cliniques 1- les SCV
ayant une chaine de transport des eacutelectrons deacutefectueuse et 2- les SCV auxotrophes
pour la thymidine (Proctor et Peters 1998 Samuelsen et al 2005) Ces derniers
Figure 6 Chaine de transport des eacutelectrons proposeacutee chez les
staphylocoques Les informations disponibles sur les composantes de la chaine
respiratoire des staphylocoques suggegraverent une chaine ramifieacutee consistant en 2
oxydases terminales meacutenaquinol-deacutependante un cytochrome o et un cytochrome
aa3 oxydase NADH deacuteshydrogeacutenase (DHase) transmet les eacutelectrons au laquo pool raquo
de meacutenaquinones De lagrave les eacutelectrons sont transfeacutereacutes soit au cytochrome b ou agrave
lrsquoaccepteur alternatif drsquoeacutelectrons le cytochrome bd quinol oxydase Le cytochrome
b transfegravere les eacutelectrons agrave une des deux oxydases terminales Ces oxydases
terminales transfegraverent les eacutelectrons agrave lrsquooxygegravene
Modifieacute de Voggu L et al 2006
- 13 -
arborent une ou plusieurs mutations dans le gegravene thyA codant pour la thymidylate
synthase laquelle catalyse la derniegravere eacutetape dans la voie de biosynthegravese de la
thymidine crsquoest-agrave-dire la formation de dTMP agrave partir du dUMP (Samuelsen et al
2005) Le dTMP est essentiel pour la synthegravese de lrsquoADN (Besier et al 2007) Les
mutants thymidine-auxotrophes sont souvent la conseacutequence drsquoune pression
seacutelective par lrsquoutilisation prolongeacutee de trimethoprim-sulfamethoxazole (TMP-SMX)
(souvent utiliseacute pour le traitement des patients fibroses kystiques) un antibiotique qui
interfegravere dans la biosynthegravese de lrsquoacide tetrahydrofolique qui agit comme cofacteur de
la thymidylate synthase (Besier et al 2007 Lannergard et al 2008 Samuelsen et al
2005) Mecircme si de premiers abords les SCV thymidine-auxotrophes ne semblent pas
avoir de lien avec la chaine transport des eacutelectrons il se pourrait que celle-ci ait sont
importance chez ces SCV puisque lrsquoentreacutee de la thymidine passe par NupC dont le
fonctionnement neacutecessite un gradient eacutelectrochimique creacuteeacute par la chaine de transport
des eacutelectrons (Proctor et Peters 1998 ) Les caracteacuteristiques pheacutenotypiques des
SCV deacutecrites ci-apregraves sont toutes lieacutees agrave lrsquoalteacuteration de la chaine de transport des
eacutelectrons 1- la croissance plus lente parce que la synthegravese de la paroi cellulaire
neacutecessite une grande quantiteacute drsquoATP 2-la diminution de la formation de pigment
parce que la biosynthegravese des caroteacutenoiumldes requiert le transport des eacutelectrons 3- une
reacutesistance accrue agrave la gentamicine et autres aminoglycosides parce que lrsquoentreacutee de
ces composeacutes neacutecessite un grand potentiel membranaire geacuteneacutereacute par le transport des
eacutelectrons (Lannergard et al 2008) et 4- une perte partielle ou complegravete de la
fermentation du mannitol parce que lrsquoutilisation des sucres-alcool tels que le
mannitol est diminueacutee lorsque la chaine de transport des eacutelectrons ne peut oxyder le
NADH produit lorsque le mannitol est meacutetaboliseacute en fructose-6-phosphate (Kohler et
al 2003 Proctor et Peters 1998 Samuelsen et al 2005)
Les SCV posseacutedant une chaine de transport des eacutelectrons deacutefectueuse ont une voie
de biosynthegravese de lrsquoheacutemine ou de la meacutenadione deacuteficiente et le pheacutenotype est
reacuteversible lorsque compleacutementeacute avec lrsquoheacutemine ou la meacutenadione (Hammer et al
- 14 -
2013 Kohler et al 2008 Lannergard et al 2008 Proctor et Peters 1998) La
meacutenadione est essentielle agrave la biosynthegravese des meacutenaquinones et lrsquoheacutemine est
essentielle agrave la biosynthegravese de lrsquohegraveme qui entre dans formation des cytochromes
(Hammer et al 2013 Kohler et al 2008 Lannergard et al 2008 Thoumlny-Meyer
1997) Les meacutenaquinones et les cytochromes sont essentiels au fonctionnement de
la chaine de transport des eacutelectrons (vers lrsquoO2 autant que vers le nitrate) (Kohler et al
2008) Il se pourrait qursquoil existe eacutegalement des SCV causeacutes par drsquoautres deacuteficiences
telles que lrsquoalteacuteration de lrsquoATP synthase qui ne reacutesulterait pas drsquoune auxotrophie pour
lrsquoheacutemine ou la meacutenadione mais reacutesulterait tout de mecircme en une deacuteficience dans la
chaine de transport des eacutelectrons (Proctor et Peters 1998 )
Comme lrsquohegraveme est un cofacteur essentiel pour les oxydases terminales et que la
meacutenaquinone est un transporteur drsquoeacutelectrons utiliseacute par S aureus on comprend que
la faible croissance des mutants hemB et menD repreacutesente un pheacutenotype mimant la
perte des oxydases terminales (Hammer et al 2013) Les deux oxydases terminales
de S aureus QoxABCD (cyt aa3) et CydAB (cyt bd) semblent ecirctre diffeacuteremment
reacuteguleacutees chez les mutants menD et la souche sauvage alors que lrsquoopeacuteron cydAB est
fortement sous exprimeacute chez les mutants aucune diffeacuterence de niveau de
transcription de qoxABCD nrsquoa eacuteteacute observeacutee entre les mutants menD et leur souche
parente (Kohler et al 2008)
Des mutations dans les gegravenes menD hemB et ctaA peuvent reacutesulter en un
pheacutenotype SCV posseacutedant une chaine de transport des eacutelectrons deacuteficiente
(Clements et al 1999 Proctor et Peters 1998 ) Les gegravenes codant pour les eacutetapes
de formation de lrsquohegraveme se preacutesentent sous la forme drsquoun opeacuteron lrsquoopeacuteron
hemAXCDBL Chacun des gegravenes est essentiel (sauf hemL) et la deacuteleacutetion drsquoun seul
drsquoentre eux empecircchera la formation de lrsquohegraveme moleacutecule essentielle agrave la formation
des cytochromes (Taber 1993) hemB code pour la porphobilinogene synthase qui
catalyse une eacutetape dans la voie de biosynthegravese de lrsquohegraveme (Kohler et al 2008 Taber
- 15 -
1993 Thoumlny-Meyer 1997) Une mutation dans le gegravene ctaA (laquo hegraveme A synthase raquo)
empecircche la biosynthegravese de lrsquohegraveme A lequel est aussi impliqueacute dans la biosynthegravese
des cytochromes et donc altegraverera le fonctionnement de la chaine de transport des
eacutelectrons (Clements et al 1999 Proctor et Peters 1998) La capaciteacute de la bacteacuterie
agrave biosyntheacutetiser de lrsquohegraveme de faccedilon endogegravene est essentielle afin drsquoinitier une
infection systeacutemique Le gegravene menD code pour la 2-succinyl-6-hydroxy-24-
cyclohexadiene-1-carboxylate syntase (SHCHC) qui est une enzyme essentielle agrave la
voie de biosynthegravese des meacutenaquinones (Kohler et al 2008) De plus les
meacutenaquinones sont eacutegalement requises pour le fonctionnement de plusieurs
enzymes impliqueacutees dans des voies meacutetaboliques essentielles telles la malate quinol
oxidoreacuteductase (Mqo2) et la dihydroorotate deacuteshydrogeacutenase (PyrD) (Kohler et al
2008) Les mutants menD seront donc incapables de convertir lrsquooxaloaceacutetate en
malate ducirc agrave la perte de lrsquoactiviteacute du Mqo2 (Kohler et al 2008 Proctor et Peters
1998)
Les bacteacuteries agrave pheacutenotype SCV deacutemontreront eacutegalement un gradient eacutelectrochimique
nettement plus faible que les bacteacuteries agrave pheacutenotype normal de par lrsquoincapaciteacute de la
chaine de transport des eacutelectrons agrave y pourvoir
De par leur chaine de transport des eacutelectrons interrompue les SCV sont incapables
drsquoutiliser lrsquoO2 et le nitrate comme accepteur final drsquoeacutelectrons (Kohler et al 2008) Par
ce fait les proteacuteines impliqueacutees dans la voie de la glycolyse ainsi que dans la voie de
la fermentation sont surexprimeacutees (Pfl Ldh Adh) alors que les enzymes faisant
partie du cycle du citrate (TCA) sont significativement reacuteguleacutees agrave la baisse (Kohler et
al 2003 Kohler et al 2008 Kriegeskorte et al 2011) Ceci indique clairement que
les SCV (ex hemB) geacutenegraverent leur ATP agrave partir du glucose ou fructose seulement par
la phosphorylation des substrats Lrsquoanalyse de la reacuteaction de fermentation indique
que le produit principal est le lactate Les SCV reacutegeacutenegraverent leur NADH en reacuteduisant le
pyruvate en lactate par la voie fermentaire (Kohler et al 2003 Kohler et al 2008)
- 16 -
emmenant ainsi une acidification du milieu (Kriegeskorte et al 2011) La conversion
du pyruvate en aceacutetate via lrsquoacetyl-CoA est restreinte chez les mutants hemB mecircme
si la pyruvate formate lyase (Pfl) et les composantes du pyruvate deacuteshydrogeacutenase
sont preacutesentes (Figure 7) Lrsquoactiviteacute de la pyruvate deacuteshydrogeacutenase est tregraves faible
probablement en lien avec le haut niveau de NADH chez les SCV et il est preacutesumeacute
que la Pfl nrsquoest pas activeacutee en preacutesence drsquooxygegravene (La pyruvate deacuteshydrogeacutenase
converti le pyruvate en Co-A en condition anaeacuterobie) (Figure 7) Chez les mutants
hemB la voie de lrsquoarginine deacuteiminase est aussi induite et pourrait eacutegalement
participer agrave la production drsquoATP (Kohler et al 2003) Les gegravenes exprimeacutes chez les
mutants hemB sont similaires agrave ceux exprimeacutes chez les mutants menD ainsi que
chez les souches agrave pheacutenotypes normaux en condition anaeacuterobie (Kohler et al 2008)
- 17 -
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Figure 7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies Les enzymes suivantes sont retrouveacutees agrave une forte
concentration chez les mutants hemb phosphoglucomutase (Pgi) fructose
bisphosphate aldolase (FbaFda) glyceacuteraldeacutehyde deacuteshydrogeacutenase (Gap)
phosphoglyceacuterate kinase (Pgk) eacutenolase (Eno) pyruvate kinase (Pyk) L-lactate
deacuteshydrogeacutenase (L-Ldh) pyruvate formate lyase (PflB) alcool deacuteshydrogeacutenase
(Adh) La formation drsquoaceacutetate et de formate et tregraves limiteacutee (X rouges)
Modifieacute de Kolher C et al 2003
- 18 -
La chaine respiratoire en anaeacuterobie (S aureus)
Il existe 3 voies distinctes de geacuteneacuteration drsquoeacutenergie chez les bacteacuteries 1- en preacutesence
drsquooxygegravene la bacteacuterie respire de faccedilon aeacuterobique et lrsquooxygegravene est lrsquoaccepteur final
drsquoeacutelectron 2- la bacteacuterie peut aussi respirer de maniegravere anaeacuterobique lorsqursquoun
accepteur terminal drsquoeacutelectron autre que lrsquooxygegravene est preacutesent 3- en absence
drsquooxygegravene et drsquoun accepteur drsquoeacutelectrons alternatif la bacteacuterie peut produire de
lrsquoeacutenergie de faccedilon limiteacutee par fermentation (Hammer et al 2013)
Chez S aureus le nitrate (NO3-) et dans un troisiegraveme temps le nitrite (NO2
-)
peuvent ecirctre utiliseacutes comme accepteur final drsquoeacutelectron alternatif lorsque lrsquooxygegravene
nrsquoest pas disponible (respiration anaeacuterobie) Les enzymes de cette voie meacutetabolique
ne sont retrouveacutees que chez les bacteacuteries (Goumltz et al 2006) La croissance
bacteacuterienne est consideacuterablement reacuteduite lorsque la bacteacuterie passe drsquoune respiration
aeacuterobie agrave une respiration anaeacuterobie La nitrate reacuteductase fonctionne probablement in
vivo gracircce au L-lactate comme principal donneur drsquohydrogegravene (Burke et Lasccelles
1975) La deacuteshydrogeacutenase primaire pour ce substrat est preacutesumeacutement la L-lactate
deacuteshydrogeacutenase associeacutee agrave la membrane laquelle meacutedie lrsquooxydation du lactate par la
chaine respiratoire aeacuterobique (Burke et Lasccelles 1975) La preacutedominance du
lactate parmi les produits finaux de la fermentation par S aureus est correacuteleacutee avec
son efficaciteacute comme principal donneur drsquohydrogegravene lorsque le nitrate est lrsquoaccepteur
final drsquoeacutelectrons Le transfert des eacutelectrons du Lactate vers le nitrate implique des
composantes qui sont eacutegalement preacutesentes dans la chaine respiratoire La nitrate
reacuteductase du type respiratoire est habituellement associeacutee agrave la membrane
cytoplasmique (Burke et Lasccelles 1975) Les reacuteductases sont inhibeacutees par
lrsquoazoture et le cyanure (Burke et Lascelles 1976)
Lrsquoenzyme membranaire nitrate reacuteductase NarGHI geacutenegravere des nitrites lesquels seront
reacuteduits en ammonium (NH4+) par lrsquoenzyme cytoplasmique NADH-deacutependant nitrite
reacuteductase encodeacutee par les gegravenes nirBD (Goumltz et al 2006) La nitrate reacuteductase 1-
- 19 -
est reacuteprimeacutee et inhibeacutee par la preacutesence drsquooxygegravene 2- est induite par lrsquoanaeacuterobiose et
la preacutesence de nitrate ou nitrite 3- nrsquoest pas inhibeacutee par lrsquoammoniac 4- permet la
production drsquoeacutenergie dans des conditions anaeacuterobies et 5- est localiseacutee dans la
membrane (Goumltz et al 2006) La preacutesence drsquoun systegraveme de senseur pour lrsquooxygegravene
est cruciale pour le controcircle transcriptionnel des gegravenes facilitant lrsquoadaptation de la
bacteacuterie agrave lrsquoanaeacuterobiose ou aux conditions microaeacuterophiles Chez S aureus il existe
deux systegravemes de senseur 1- le systegraveme SrrAB (pour staphylococcal respiratory
response AB synonyme SrhSR) (Figure 8) homologue agrave ResDE chez Bacillus
subtilis (Yan et al 2011) et 2- le systegraveme NreABC (pour nitrogen regulation chez S
carnosus correspondant agrave SA2181 (NreA) SA2180 (NreB) SA2179 (NreC) chez S
aureus) NreBC est un systegraveme agrave deux composantes qui mesure la concentration
drsquooxygegravene directement par un groupement Fe-S de type Fnr lequel est instable en
preacutesence drsquooxygegravene et par lequel est controcircleacute le systegraveme membranaire de reacuteduction
du nitratenitrite (Yan et al 2011) Srr est aussi un systegraveme agrave deux composantes qui
reacutegule positivement la lactate deacuteshydrogeacutenase et lrsquoalcool deacuteshydrogeacutenase et semble
reacuteguler neacutegativement plusieurs enzymes du cycle du citrate (TCA) (Figure 8)
Lrsquoexpression des enzymes fermentaires comme la lactate deacuteshydrogeacutenase (LDH)
pyruvate formate lyase (PFL) et lrsquoalcool deacuteshydrogeacutenase sont aussi fortement induite
dans des conditions aeacuterobies lorsque la chaine de transport des eacutelectrons est
interrompue (Kohler et al 2003) Donc il se peut que lrsquoeacutetat reacuteduit des composantes
de la chaine respiratoire le potentiel membranaire etou le niveau de NADH soit un
signal pour la reacutegulation des gegravenes laquo anaeacuterobies raquo chez S aureus Dans des
conditions aeacuterobies la conversion du pyruvate en aceacutetyl-coA est catalyseacutee par le
complexe pyruvate deacuteshydrogeacutenase (PDH) et caracteacuteriseacutee par la production de
NADH (Somerville et Proctor 2009) En condition anaeacuterobie lrsquoexpression de la
pyruvate deacuteshydrogeacutenase (PDH) est fortement reacuteprimeacutee alors que lrsquoexpression de la
pyruvate formate lyase (PFL) est fortement induite Lrsquoactiviteacute de la PDH est
remplaceacutee en anaeacuterobiose par lrsquoactiviteacute de la PFL qui geacutenegravere lrsquoacetyl-CoA et le
formate agrave partir du pyruvate Le NADH nrsquoeacutetant pas reacuteoxydeacute par une chaine de
- 20 -
transport des eacutelectrons fonctionnelle celui-ci est recycleacute par la reacuteduction du pyruvate
en lactate (Kohler et al 2003 Somerville et Proctor 2009)
La meacutenaquinone et lrsquohegraveme sont esssentiels agrave la respiration anaeacuterobie (Kohler et al
2008) Le pheacutenotype de S aureus en condition anaeacuterobie est tregraves similaire au
pheacutenotype SCV (Somerville et Proctor 2009)
Figure 8 Modegravele de reacutegulation du cycle TCA (Pour simplifier les reacutegulateurs de
facteurs de virulence bien caracteacuteriseacutes [ie RNAIII Rot SarA et σB] ont eacuteteacute omis
dans ce modegravele)
Les sous-produits de la phosphorylation oxydative les ROS
La reacuteduction de lrsquooxygegravene en eau se fait un eacutelectron agrave la fois (Turrens 2003) Le
complexe IV retient tous les intermeacutediaires partiellement reacuteduits jusqursquoagrave ce que la
Tireacute de Somerville GA Proctor RA 2009
- 21 -
reacuteduction soit complegravetement termineacutee (Turrens 2003) Cependant certains autres
centres redox de la chaine respiratoire laissent passer des eacutelectrons agrave lrsquooxygegravene
reacuteduisant partiellement cette moleacutecule en anion superoxyde (O2-bull) (Liu et al 2002
Turrens 2003) Mecircme si O2-bull nrsquoest pas un agent oxydant tregraves puissant il est le
preacutecurseur de plusieurs autres laquo reactive oxygen species raquo (ROS) et il est aussi
impliqueacute dans la propagation de la chaine de reacuteaction oxydative (Turrens 2003)
LrsquoO2-bull est un intermeacutediaire instable qui est rapidement converti en H2O2 par les
superoxide dismutase (SOD) (S aureus en possegravede deux SodA (le principal) et
SodM) (10 18) ou se convertit de faccedilon spontaneacutee lorsqursquoen concentration eacuteleveacutee
(Starkov 2010 Turrens 2003) Le H2O2 peut ensuite ecirctre complegravetement reacuteduit en
H2O ou partiellement reacuteduit en radical hydroxyle (bullOH) un des plus puissants
oxydants (Starkov 2010) LrsquoO2-bull peut eacutegalement reacuteagir avec drsquoautres radicaux
incluant lrsquooxyde nitrique (NObull) selon le taux de diffusion des 2 radicaux (Turrens
2003) Le peroxynitrite est eacutegalement un oxydant puissant (Turrens 2003) Les
oxydants deacuterivant du NObull sont appeleacutes des laquo reactive nitrogen species raquo (RNS)
(Turrens 2003) mais seront consideacutereacutes comme faisant partie des ROS dans ce
travail Les ROS peuvent endommager lrsquoADN lrsquoARN les proteacuteines et les lipides
reacutesultants agrave lrsquoapoptose de la cellule lorsque le niveau de ROS produit excegravede la
capaciteacute de deacutetoxifications et de reacuteparation de la cellule (Brynildsen et al 2013
Wells et al 2005) Malgreacute ce danger il srsquoagit drsquoun risque compenseacute par
lrsquoaugmentation du rendement eacutenergeacutetique (Brynildsen et al 2013) Le complexe I (et
II) et III sont les principaux sites de production de ROS (Batandier et al 2006 Chen
et al 2003 Liu et al 2002) Lrsquoinhibition du complexe I par la rotenone provoque une
augmentation de la production de ROS Lrsquooxydation des substrats du complexe I ou
du complexe II en preacutesence du complexe III inhibeacute par lrsquoantimicine A amegravene
eacutegalement une augmentation de la production de ROS (Batandier et al 2006 Chen
et al 2003) (Figure 9) De plus mecircme si le complexe cytochrome oxydase nrsquoest pas
un site de production de ROS son inhibition par le KCN (cyanure de potassium)
facilite la production de ROS par les complexes I et II (Chen et al 2003) Le
- 22 -
complexe ATP synthase nrsquoest pas reconnu pour produire directement des ROS
Cependant la liaison drsquoun inhibiteur tel lrsquooligomycine bloque le passage des protons
agrave travers le complexe et inhibe ainsi la production drsquoATP De plus lrsquoATP synthase ne
fonctionnant plus les pompes agrave protons de la chaine de transport des eacutelectrons sont
incapables de fonctionner car le gradient de protons est beaucoup trop eacuteleveacute ce
provoque une surproduction de ROS par les complexes I et III Conseacutequemment le
NADH nrsquoest plus oxydeacute et le cycle du TCA cesse drsquoopeacuterer car la concentration de
NAD nrsquoest plus suffisante pour le fonctionnement des enzymes du TCA (Joshi et
Huang 1991)
Une production significative de O2-bull principalement par le complexe I peut ecirctre le
reacutesultat de deux modes drsquoopeacuteration 1- lorsque la cellule ne produit pas drsquoATP et
conseacutequemment possegravede une grande force proton-motrice et possegravede un laquo pool raquo de
quinone agrave lrsquoeacutetat reacuteduit (QH2) 2- lorsque la cellule possegravede un haut ratio de
NADHNAD (Figure 10) Lorsque la cellule produit activement de lrsquoATP elle possegravede
alors une moins grande force proton-motrice son ratio NADHNAD est plus faible et
la formation de O2-bull est beaucoup plus faible (Murphy 2009)
Le H2O2 est le substrat pour deux enzymes la catalase et la glutathione (GSH)
peroxydase qui convertissent le H2O2 en H2O+O2 le H2O2 est ainsi deacutetoxifieacute (Figure
9 Figure 11) La reacuteaction catalyseacutee par la glutathione peroxydase neacutecessite de la
glutathione (GSH) comme substrats et deacutepend en partie du ratio de GSSG GSH et
de la concentration des reacuteactants laquelle est le reflet de lrsquoeacutetat reacutedox de la cellule De
faccedilon similaire la concentration de meacutetaux sous leur forme reacuteduite tel le Fer catalyse
la formation de certains ROS (Figure 11) Ceci peut ecirctre minimiseacute en tenant cette
concentration ions meacutetallique tregraves basse entre autres par leur emmagasinage et leur
liaison aux proteacuteines de transport (ei ferritine transferrine lactoferrine) et ainsi
minimiser la formation de bullOH Finalement certains antioxydants (eg vitamine E)
peuvent interrompre la chaine de reacuteactions par la capture des radicaux le radical-
vitamine E est relativement stable et peut ecirctre enzymatiquement reconverti agrave sa
- 23 -
forme non-radical (Figure 11) Il est maintenant accepteacute que certaines enzymes du
cycle de Krebs (ie αKGDH) soient non seulement des cibles des ROS mais aussi
des producteurs non neacutegligeables de ROS (Brynildsen et al 2013)
Figure 9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS Le complexe I ainsi que le site Qi du complexe III relacircchent des
ROS directement vers les deacutefenses drsquoantioxydants intracellulaires Les antioxidants
intracellulaire tel la superoxide dismutase (MnSOD) la catalase la glutathione (GSH)
la glutathione peroxydase (GPX) et la glutathione reacuteductase (GRase) sont montreacute Le
site Qo du complexe III quant agrave lui relacircche les ROS dans le milieu extra cellulaire
Modifieacute de Chen Q et al 2003
Oligomycine
- 24 -
Figure 10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de
O2-bull 1- Le pool de NADH est dans un eacutetat reacuteduit par exemple par des dommages agrave
la chaine respiratoire la perte de cytochrome c durant lrsquoapoptose ou agrave un faible
besoin en ATP Ceci megravene agrave une production de O2-bull au complexe I qui deacutecoule de la
reacuteduction du FMN laquelle est reacuteguleacutee par le ratio NADHNAD+ Drsquoautres sites
autres que le complexe I tel lrsquoα-ceacutetoglutatrate deacuteshydrogeacutenase (αKGDH) peut
aussi contribuer agrave la production de O2-bull 2- Il nrsquoy a pas (peu) de production drsquoATP il y
a un fort potentiel membranaire et un pool de CoQ (meacutenaqinone) agrave lrsquoeacutetat reacuteduit
important qui provoque une reacuteversion de la chaine de transport des eacutelectrons agrave
travers le complexe I amenant une forte production de O2-bull 3- Il y a une production
active drsquoATP et conseacutequemment un potentiel membranaire plus bas que dans la
situation 2 et une plus grande quantiteacute de NADH oxydeacute que dans la situation 1
Dans ces conditions la quantiteacute de O2-bull produit est plus faible que dans la situation
1 et 2 et la provenance des O2-bull est moins certaine
Tireacute de Murphy MP 2009
- 25 -
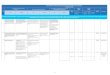
12 Antibiotiques Mode drsquoaction cible cellulaire et meacutecanismes de reacutesistance
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles
Notre compreacutehension de la faccedilon que les antibiotiques induisent la mort cellulaire est
principalement baseacutee sur la fonction cellulaire essentielle inhibeacutee par lrsquointeraction de
lrsquoantibiotique avec sa cible Les antibiotiques peuvent ecirctre classeacutes selon la
composante ou le systegraveme cellulaire qursquoils affectent ainsi que par le fait qursquoils
induisent la mort cellulaire (antibiotiques bacteacutericides) ou srsquoils inhibent la croissance
Figure 11 Voies de formation et de deacutetoxification des ROS Tireacute de Wikipeacutedia
httpfrwikipediaorgwikiDC3A9rivC3A9_rC3A9actif_de_l27oxygC3A8ne
Consulteacute le 11 deacutec 2013
- 26 -
bacteacuterienne (antibiotique bacteacuteriostatique) La plupart des antibiotiques bacteacutericides
inhibent la synthegravese de lrsquoADN ou de lrsquoARN la synthegravese de la paroi bacteacuterienne ou la
synthegravese proteacuteique (Kohanski et al 2010) Le tableau 1 est une synthegravese des
principales classes drsquoantibiotiques ainsi que leur mode drsquoaction et leurs cibles
cellulaires Ce tableau nrsquoest cependant pas exhaustif et il existe plusieurs autres
classes drsquoantibiotiques et plusieurs autres exemples drsquoantibiotiques dans les classes
identifieacutees dans ce tableau
Depuis la deacutecouverte de la peacutenicilline en 1929 plusieurs autres antibiotiques ont eacuteteacute
deacutecouverts et deacuteveloppeacutes suite agrave lrsquoeacutelucidation des interactions entre lrsquoantibiotique et
sa cible et en apportant des modifications structurales et chimiques aux moleacutecules
deacutejagrave connues Ces efforts ont significativement augmenteacute le nombre drsquoantibiotiques
disponibles dans le milieu clinique Le processus de mort cellulaire induit par lrsquoaction
drsquoun antibiotique est complexe et deacutebute avec une interaction physique entre la
moleacutecule antibiotique et sa cible bacteacuterienne speacutecifique et implique des modifications
qui affectent la bacteacuterie tant au niveau biochimique que moleacuteculaire et structural
27
Acid
e n
alid
ixiq
ue (
1 iegravere
geacuteneacutera
tion)
Cip
rofloxacin
e (
2 iegravem
e g
eacuteneacutera
tion)
Levofloxacin
e (
3 iegravem
e g
eacuteneacutera
tion)
Gem
iflo
xacin
e (
4 iegravem
e g
eacuteneacutera
tion)
Flu
oro
quin
olo
ne (
Norf
loxacin
e)
Peacutenic
iline
Ceacutephalo
sporine (
ceacutefa
lexin
e)
Carb
apenem
Vancom
ycin
e
Teacutela
vancin
e
Fosfo
mycin
Dapto
mycin
Poly
mix
ine B
Bacitra
cin
Valin
om
ycin
e
Nom
encin
e
Genta
mic
in
Kanam
ycin
e
Tobra
mycin
e
Str
epto
mycin
e
Teacutetr
acyclin
e
Chlo
rteacutetr
acyclin
e
Macro
lides
Ery
thro
mycin
eLie
sous-u
niteacute 23S
du r
ibosom
e 5
0S
et em
pecircche
leacutelo
ngation d
u p
eptide (
apregrave
s 6
-8 a
a)
sta
tique
Lin
cosam
ides
Clin
dam
ycin
Inhib
e d
irecte
ment la
ctiviteacute p
eptidyl
transfeacute
rase d
u
ribosom
esta
tique
Str
epto
gra
min
es
DalfopristinQ
uin
upristin
Com
bin
ais
on d
e 2
antibio
tiques B
loque la
transla
tion p
eptidiq
ue e
n lia
nt le
site P
du r
ibosom
e
et en b
loquant au p
eptide la
ccegraves a
u tunnel de s
ort
ie
cid
al
Am
phenic
ol
Chlo
ram
phenic
ol
Em
pecircche la lia
ison d
u a
a-
AR
Nt au s
ite A
du
ribosom
e e
t in
terf
egravere
dans la f
orm
ation d
u lie
n
peptidiq
ue
sta
tique
Oxazolid
inones
Lin
ezolid
Blo
que initia
tion d
e la tra
nsla
tion p
roteacute
ique lors
de la
reacuteaction d
eacutelo
ngation p
ar
la p
eptidyl
transfeacute
rase
sta
tique
Trim
eth
oprim
Sulfam
eth
oxazole
sta
tique
cid
al
Ionophore
Alteacutera
tion d
e lin
teacutegriteacute
mem
bra
naire
Se
nchasse d
ans la m
em
bra
ne fo
rme u
n p
ore
ce
qui am
egravene u
ne p
ert
urb
ation d
e leacute
quili
bre
ioniq
ue
cid
al
Inhib
itio
n d
e la s
ynth
egravese d
e
la p
aro
i cellu
laire
(peptidogly
can)
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
cid
al
cid
al
sta
tique
AR
Nr
16S
(S
ite A
)(fo
rmation d
e p
roteacute
ines n
on-
fonctionnelle
s)
cid
al
Inhib
itio
n d
e la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 3
0S
)
Inhib
iteur
de la s
ynth
egravese d
e
lacid
e f
oliq
ue
Inhib
itio
n d
e c
ert
ain
es r
eacuteactions e
nzym
atiques
essentielle
s agrave
la f
orm
ation d
e la
cid
e f
oliq
ue
Sulfonam
ides
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
AR
Nr
16S
(preacute
vie
nt la
ttachem
ent de la
min
oacyl-
AR
Nt au r
ibosom
e)
Teacutetr
acyclin
e
Poly
peptides
β-L
acta
ms
Gly
copeptides
Lip
opeptides
Inhib
iteur
de la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 5
0S
)
Am
inogly
cosid
es
Cib
le c
ell
ula
ire
Sta
tiq
ue
Cid
al
Rifam
ycin
eR
ifam
pic
ine
Cla
ss
es
Ex
d
an
tib
ioti
qu
eM
od
e d
ac
tio
n
Inhib
itio
n d
e la s
ynth
egravese d
e
lAR
N (
AD
N d
eacutependant)
AR
N p
oly
meacutera
se s
ous-u
niteacute β
(codeacute p
ar
rpoB
)
Quin
olo
nes
cid
al
Inhib
itio
n d
e la r
eacuteplic
ation d
e
lAD
N
Com
ple
xe A
DN
-topois
om
eacutera
se (
AD
N
gyr
ase(t
opois
om
eacutera
se II)
et to
pois
om
eacutera
se IV
)cid
al
Tra
nspeptidase (
penic
illin
bin
din
g p
rote
in
(PB
P))
cid
al
Tab
leau
1
Cib
les e
t m
od
e d
rsquoacti
on
des p
rin
cip
ale
s c
lasses d
rsquoan
tib
ioti
qu
es
28
122 Les aminoglycosides
1221 Mode drsquoaction
Les aminoglycosides peacutenegravetrent la paroi cellulaire en suivant un processus
comprenant 3 eacutetapes une premiegravere eacutetape eacutenergie- indeacutependante suivie de deux
eacutetapes eacutenergie deacutependantes Deux composantes sont essentielles pour
lrsquoaccumulation intracellulaire des aminoglycosides les ribosomes et la chaine de
transport des eacutelectrons (Ramirez et Tolmasky 2010)
Eacutetape 1- Lorsque les aminoglycosides entrent en contact avec la paroi
cellulaire la moleacutecule polycationique se lie aux composantes de surface anioniques
tels les lipopolysaccharides les phospholipides et autres proteacuteines membranaires
chez les bacteacuteries agrave Gram neacutegatif et les moleacutecules drsquoacides teacuteichoiumlques et les
phospholipides des bacteacuteries agrave Gram positif (Ramirez et Tolmasky 2010) Apregraves
cette liaison aux moleacutecules polaires les aminoglycosides (telle la gentamicine)
srsquoassocient aux transporteurs membranaires sur la base de leur charge positive et
sont transporteacutes agrave travers la membrane cytoplasmique gracircce au potentiel
membranaire (intra cellulaire neacutegatif) (Bryan et al 1980 Proctor et al 2006) Cette
liaison agrave la membrane entraine une augmentation de la permeacuteabiliteacute membranaire
menant agrave lrsquoentreacutee des aminoglycosides dans lrsquoespace peacuteriplasmique (Gram neacutegatif)
ou dans le cytoplasme (Gram positif) (Ramirez et Tolmasky 2010) (Figure 12A)
Eacutetape 2- Lrsquoeacutetape suivante connue comme la laquo phase eacutenergie-deacutependante
1 raquo (EDP-I) est bloqueacutee par les inhibiteurs de la chaine de transport des eacutelectrons et
de la phosphorylation oxydative ainsi que par les ions divalents lrsquohyperosmolariteacute le
faible pH et lrsquoanaeacuterobiose dans cette phase la dureacutee et le taux drsquoentreacutee de
lrsquoantibiotique sont en fonction de la concentration (Miller et al 1980 Mingeot-
Leclercq et al 1999 Ramirez et Tolmasky 2010) Agrave cette eacutetape le transport
transmembranaire des aminoglycosides et plus speacutecifiquement de la gentamicine
neacutecessite 2 composantes majeures a) un transporteur anionique non speacutecifique dont
la charge est directement lieacutee agrave lrsquoactiviteacute des cytochromes oxydases terminales et des
29
nitrates reacuteductases de la chaine de transport des eacutelectrons Selon certaines eacutetudes
les transporteurs pourraient ecirctre les quinones reacuteduites de la chaine de transport des
eacutelectrons (Bryan et al 1981 Miller et al 1980) et donc le cycle drsquooxydation-reacuteduction
des quinones est essentiel aux transports des aminoglycosides (Bryan et al 1981)
mais selon drsquoautres eacutetudes les transporteurs pourraient ecirctre toute autre proteacuteine non
deacutefinie impliqueacutee dans la polarisation membranaire lieacutee agrave lrsquoactiviteacute de la chaine de
transport des eacutelectrons (Miller et al 1980) La croissance anaeacuterobie ainsi que le
sodium azide (inhibiteur de la chaine respiratoire) inhibent lrsquoentreacutee de la gentamicine
et lrsquoeffet bacteacutericide indiquant que le transport actif de cet antibiotique neacutecessite la
respiration b) un potentiel eacutelectrique avec un milieu intra cellulaire neacutegatif par rapport
agrave lrsquoexteacuterieur En effet lrsquoinhibition de lrsquoentreacutee dans la bacteacuterie des aminoglycosides
suite agrave lrsquoajout de CCCP (agent deacutepolarisant) chez des souches de S aureus laquo wild-
type raquo deacutemontre la neacutecessiteacute de la force proton-motrice pour lrsquoentreacutee des
aminoglycosides dans les cellules (Miller et al 1980)
Eacutetape 3- La petite quantiteacute drsquoaminoglycosides srsquoeacutetant rendu au cytoplasme
durant cette premiegravere phase eacutenergie-deacutependante induisent apregraves liaison aux
ribosomes des erreurs dans la synthegravese proteacuteique Les erreurs dans la traduction
des proteacuteines membranaires produites lors de cette eacutetape connue comme la laquo phase
eacutenergie-deacutependante 2 raquo (EDP-II) provoquent des dommages dans lrsquointeacutegriteacute de la
membrane cytoplasmique (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky
2010) Les proteacuteines membranaires aberrantes dans la membrane facilitent le
transport drsquoune plus grande quantiteacute drsquoaminoglycosides lesquels augmenteront
encore le niveau drsquointerfeacuterence avec la synthegravese proteacuteique normale amenant encore
plus de dommage agrave la membrane provoquant ainsi une acceacuteleacuteration importante du
taux drsquoentreacutee des moleacutecules antibiotique et qui ultimement reacutesultera en la mort de la
cellule (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky 2010) (Figure 12B)
Des eacutetudes ont deacutemontreacute que la susceptibiliteacute agrave la gentamicine streptomycine
amikacine kanamycine et tobramycine est supeacuterieure lorsque lrsquooxygegravene est
30
lrsquoaccepteur terminal drsquoeacutelectrons (Bryan et Kwan 1981) Une diminution significative
de la susceptibiliteacute est remarqueacutee lorsque le nitrate est lrsquoaccepteur final drsquoeacutelectrons et
une diminution encore plus marqueacutee est observeacutee lorsque le fumarate est lrsquoaccepteur
final drsquoeacutelectron ou lorsque les conditions sont fermentaires Lrsquoassociation entre le
transport de la gentamicine et le complexe terminal de la chaine de transport des
eacutelectrons particuliegraverement lrsquoactiviteacute cytochrome oxydase
Le ribosome possegravede trois sites de liaison importants de lrsquoARNt deacutesigneacute comme site
A (pour aminoacyl) site P (pour peptidyl) et E (pour exit) (Vakulenko et Mobashery
2003) Ils sont situeacutes sur lrsquoARN16S (lequel avec plus de 21 proteacuteines compose la
sous-uniteacute 30S du ribosome bacteacuterien) (Ramirez et Tolmasky 2010 Vakulenko et
Mobashery 2003) Durant la synthegravese proteacuteique le ribosome deacutecode lrsquoinformation de
lrsquoARNm et catalyse lrsquoincorporation seacutequentielle de chaque acide amineacute faisant partie
de la chaine polypeptidique en formation La haute fideacuteliteacute de la traduction (les
ribosomes incorporent un mauvais acide amineacute par approximativement 3000 liaisons
peptidiques effectueacutees) est possible gracircce agrave la capaciteacute de discriminer entre des
changements conformationels dans le ribosome induit par la liaison drsquoun ARNt correct
Modifieacute de Davis BD 1987
Figure 12 Cineacutetique drsquoentreacutee des aminoglycosides
A
B
31
ou incorrect au site A La reconnaissance drsquoune liaison correcte anticodoncodon par
le ribosome se passe en deux temps la reconnaissance initiale de lrsquoappariement du
codon sur lrsquoARNm avec lrsquoanticodon de lrsquoARNt et subseacutequemment la correction
(Vakulenko et Mobashery 2003)
La plupart des aminoglycosides se lient agrave des oligonucleacuteotides faisant partie du site
A Les diffeacuterents aminoglycosides ne lient pas tous le mecircme site de lrsquoARN 16S mais
lrsquoeffet commun agrave tous est que cette liaison amegravene un changement de conformation du
site A vers une conformation mimant la conformation fermeacutee de la sous uniteacute 30S qui
est induite par une interaction entre lrsquoARNm et son ARNt apparenteacute eacuteliminant la
possibiliteacute de correction du ribosome ce qui promeut les erreurs de traduction
(Ramirez et Tolmasky 2010 Vakulenko et Mobashery 2003) La grande majoriteacute des
aminoglycosides sont bacteacutericides et cette leacutetaliteacute est la conseacutequence de cette
mauvaise traduction (Vakulenko et Mobashery 2003)
1222 Meacutecanismes de reacutesistance
La reacutesistance contre les aminoglycosides se produit par diffeacuterents meacutecanismes qui
peuvent coexister simultaneacutement dans la mecircme bacteacuterie (Vakulenko et Mobashery
2003) Ces meacutecanismes incluent 1- la modification de la cible par des mutations
dans lrsquoARN 16S ou autres proteacuteines ribosomales associeacutees (Figure 13) 2- la
meacutethylation de lrsquoARN 16S un meacutecanisme retrouveacute chez plusieurs organismes
producteurs drsquoaminoglycosides et chez des souches cliniques Cette meacutethylation
empecircche les aminoglycosides drsquoavoir accegraves agrave leur site de liaison (Doi et Arakawa
2007) 3- la reacuteduction de la permeacuteabiliteacute par une alteacuteration de la permeacuteabiliteacute de la
membrane externe ou une diminution du transport agrave travers la membrane interne 4-
lrsquoexportation des aminoglycosides agrave lrsquoexteacuterieur de la cellule par les pompes agrave efflux
Eacutetant des moleacutecules hydrophiliques les aminoglycosides ont naturellement peu
drsquoaffiniteacute avec les pompes agrave efflux 5- le swarming actif probablement un meacutecanisme
32
non speacutecifique deacutemontreacute chez Pseudomonas aeruginosa lequel deacutemontre des
reacutesistances agrave plusieurs antibiotiques 6- la seacutequestration de la drogue par des liaisons
agrave un aceacutetyltransfeacuterase agrave tregraves faible activiteacute 7- une inactivation enzymatique de la
moleacutecule antibiotique le meacutecanisme de reacutesistance le plus preacutevalent au niveau
clinique (Vakulenko et Mobashery 2003) Lrsquoinactivation enzymatique des
aminoglycosides est le plus souvent due agrave des aceacutetyltransfeacuterases des
nucleacuteotidyltransfeacuterases ou des phosphotransfeacuterases (Dickgiesser et Kreiswirth 1986)
Ces enzymes sont souvent encodeacutees sur des plasmides mais sont eacutegalement
associeacutees agrave des eacuteleacutements transposables Lrsquoeacutechange de plasmides et la disseacutemination
de transposons facilitent lrsquoacquisition rapide de pheacutenotypes reacutesistants aux
antibiotiques non seulement au sein drsquoune mecircme espegravece mais parmi un large
eacuteventail drsquoespegraveces bacteacuteriennes (Mingeot-Leclercq et al 1999)
La reacutesistance naturelle des S aureus laquo small colony variants raquo agrave la majoriteacute des
aminoglycosides est la conseacutequence de lrsquoimpossibiliteacute agrave lrsquoantibiotique de peacuteneacutetrer une
cellule posseacutedant une faible polariteacute membranaire qui est due chez les SCV agrave leur
Tireacute de Doi Y Arakawa Y 2007
Figure 13 Sites de mutation de certains reacutesistants aux aminoglycosides
33
incapaciteacute agrave activer le cycle de Krebs (Miller et al 1980 Proctor et al 2006 Tubby
et al 2013) Les SCV ont non seulement une polariteacute membranaire faible mais (ceci
eacutetant la conseacutequence de cela) aussi une force proton motrice inhibeacutee et une chaine
des eacutelectrons deacuteficiente De ce fait le transport transmembranaire des
aminoglycosides chez les SCV est grandement reacuteduit et conseacutequemment la
susceptibiliteacute des SCVs pour cette classe drsquoantibiotique est 110-113 de celle des
souches parentes (Proctor et al 2006)
123 La tomatidine
Plusieurs espegraveces veacutegeacutetales accumulent des steacuterols et des triterpegravenes antimicrobiens
appeleacutes saponines (Osborn 1996) Ces moleacutecules sont produites de faccedilon
constitutive par la plante et jouent un important rocircle de deacutefense contre diffeacuterents
pathogegravenes comme des insectes ou des microbes Parmi les saponines la tomatine
(Figure 14A) est un glycoalcaloiumlde steacuteroiumldien preacutesentant une action antimicrobienne
contre des levures et une quantiteacute drsquoautres microorganismes (Bednarek et Osboum
2009 Friedman 2002 Osborn 1996 Sandrock et Vanetten 1998) Plusieurs
espegraveces de champignons pathogegravenes de la tomate produisent des enzymes
extracellulaires communeacutement appeleacutes tomatinase qui sont capables de deacutetoxifier
lrsquoα-tomatine (Martin-Hernandez et al 2000 Roddick 1974 Ruiz-Rubbio et al 2001)
Par exemple Fusarium oxysporum f sp lycopersici est reconnu pour cliver lrsquoα-
tomatine en ses sous-produits soit un tetrasaccharide (lycotreacutetraose) et un alcaloiumlde
steacuteroiumldien non glycosyleacute la tomatidine (Figure 14B) Ces sous-produits nrsquoont pas ou
peu drsquoactiviteacute antifongique (Ruiz-Rubbio et al 2001 Simon et al 2006)
34
Figure 14 Structures chimiques de la (A) tomatine de (B) tomatidine et de (C)
lrsquoanalogue de tomatidine FC04-100
Il a eacuteteacute deacutecouvert il a quelques anneacutees que la tomatidine avait un effet antibacteacuterien
sur les S aureus de pheacutenotype SCV mais aucune activiteacute contre les S aureus agrave
pheacutenotype normal Par contre lorsqursquoutiliseacute en combinaison avec les
aminoglycosides lrsquoajout de la tomatidine permettait drsquoaugmenter de 8 agrave 16 fois
lrsquoactiviteacute des aminoglycosides contre S aureus agrave pheacutenotype normal (Mitchell et al
2011a Mitchell et al 2012)
Donc on savait que la tomatidine avait une activiteacute antibacteacuterienne tregraves inteacuteressante
mais on ne connaissait ni sa cible ni son mode drsquoaction De plus si la tomatidine a
une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait donc avoir un
spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Par ailleurs de faccedilon parallegravele la production drsquoanalogues structuraux de la tomatidine
permettrait de mieux comprendre la relation structure-activiteacute (SAR) de la moleacutecule et
donc de preacuteciser que lrsquoactiviteacute biologique de la tomatidine neacutecessite une orientation
3
A B
C
35
de groupements fonctionnels bien speacutecifique De plus lrsquoeacutetude de lrsquoactiviteacute de
plusieurs analogues permettrait de confirmer si la tomatidine possegravede un meacutecanisme
drsquoaction deacutependant drsquoune interaction speacutecifique avec une ou des cibles biologiques
preacutecises
Le laboratoire du Pr Eacuteric Marsault du deacutepartement de pharmacologie de lrsquoUniversiteacute
de Sherbrooke et lrsquoeacutetudiant agrave la maitrise Feacutelix Chagnon ont produit plus de 33
analogues de la tomatidine (voir Annexe 2 article laquo Unraveling the structure-activity
relationship of tomatidine a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureusraquo dans la section laquo Contributions
Supleacutementaires raquo du meacutemoire) dont lrsquoactiviteacute antibacteacuterienne a eacuteteacute eacutevalueacutee par des
tests de susceptibiliteacute agrave la moleacutecule (CMI) sur S aureus SCV et sur S aureus agrave
pheacutenotype normal en combinaison avec la gentamicine Plusieurs de ces moleacutecules
font lrsquoobjet drsquoune application de brevet (voir annexe 1) dont la prometteuse moleacutecule
FC04-100 (Figure 14C) (voir laquo Annexe 1 Application de brevet raquo dans la section
laquo Contributions suppleacutementaires raquo du meacutemoire)
13 Projet de maitrise
Premiegravere hypothegravese
Si la tomatidine a une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait
donc avoir un spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Objectif Provoquer lrsquoapparition de pheacutenotypes SCV chez diffeacuterentes espegraveces
bacteacuterienne afin de pouvoir veacuterifier leur niveau de sensibiliteacute agrave la tomatidine
Deuxiegraveme hypothegravese
Lrsquoidentification des gegravenes preacutesentant des mutations parmi les souches de S aureus
reacutesistantes agrave la tomatidine pourrait reacuteveacuteler la ou les cibles cellulaires de la moleacutecule
et nous aider agrave comprendre son mode drsquoaction
36
Objectif Obtenir des mutants reacutesistants avant de les envoyer agrave sequencer
faire les analyses comparatives des geacutenomes et finalement identifier les mutations
37
CHAPITRE 2
ARTICLE SCIENTIFIQUE
TITRE Tomatidine and Analog FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Some Firmicutes Including Those of Listeria
monocytogenes and Are Bactericidal in Combination With Aminoglycosides Against
their Normal Phenotype
AUTEURS ET CONTRIBUTIONS
Isabelle Guay1 Expeacuteriences biologiques avec le produit naturel tomatidine et son
analogue FC04-100 sur diffeacuterentes espegraveces bacteacuteriennes afin de comparer les deux
activiteacutes
Simon Boulanger1 Expeacuterience biologique permettant de quantifier lrsquoactiviteacute de
lrsquoanalogue FC04-100 sur la production de biofilm et son activiteacute intracellulaire
Feacutelix Chagnon2 Production de lrsquoanalogue chimique de la tomatidine FC04-100
Kamal Bouarab1 Initiateur du projet gracircce agrave lrsquoisolement du produit naturel tomatidine
Eacuteric Marsault2 Directeur de recherche de Feacutelix Chagnon
Franccedilois Malouin1 Directeur de recherche drsquoIsabelle Guay et Simon Boulanger et
auteur de correspondance
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
38
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
Corresponding author Mailing address Universiteacute de Sherbrooke
Deacutepartement de Biologie Faculteacute des Sciences 2500 Boul
Universiteacute Sherbrooke Quebec Canada J1K2R1 Phone (819) 821-
8000 ext 61202 Fax (819) 821-8049 E-mail FrancoisMalouin
USherbrookeca
39
Tomatidine and Analog FC04-100 Possess Bactericidal Activities
Against the Small Colony Variants of Some Firmicutes Including
Those of Listeria monocytogenes and Are Bactericidal in Combination
With Aminoglycosides Against their Normal Phenotype
Running Title Tomatidine is bactericidal against several Firmicutes
Isabelle Guay1
Simon Boulanger1
Feacutelix Chagnon2 Kamal Bouarab
1 Eacuteric Marsault
2 and
Franccedilois Malouin1
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM) Deacutepartement de
Biologie Faculteacute des Sciences Universiteacute de Sherbrooke Sherbrooke Queacutebec Canada J1K
2R1
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute Universiteacute
de Sherbrooke Queacutebec Canada J1H 5N4
Have contributed equally to the work
Corresponding author
Mailing address Universiteacute de Sherbrooke Deacutepartement de Biologie Faculteacute des Sciences
2500 Boul Universiteacute Sherbrooke Quebec Canada J1K2R1
Phone (819) 821-8000 ext 61202 Fax (819) 821-8049
E-mail FrancoisMalouinUSherbrookeca
40
ABSTRACT
Tomatidine (TO) is a plant steroidal alkaloid that possesses an antibacterial activity against the
small colony variants (SCVs) of Staphylococcus aureus We report here the spectrum of
activity of TO against species of the Firmicutes and the improved antibacterial activity of a
chemically-modified TO derivative (FC04-100) In addition to that seen on S aureus TO
showed a bactericidal activity against SCVs of Bacillus cereus B subtilis and Listeria
monocytogenes The combination of an aminoglycoside antibiotic and TO also generated an
antibacterial synergy against strains of the normal phenotype in these species In contrast to
TO which has no relevant activity by itself against S aureus of the normal phenotype (MIC
gt64 mgL) the TO analog FC04-100 showed a MIC of 8-16 mgL Furthermore FC04-100
showed a strong bactericidal activity against L monocytogenes SCVs in kill kinetic
experiments while TO did not FC04-100 also improved the bactericidal activity of
gentamicin against L monocytogenes of the normal phenotype The addition of FC04-100 (4
mgL) to a cephalexinkanamycin (32) combination improved the activity of the combination
by 32 fold against cephalexin and kanamycin-resistant MRSA strains In combination with
gentamicin FC04-100 also showed a strong bactericidal activity against biofilm-embedded S
aureus and when used alone was superior to TO for its biofilm-preventing activity Finally
FC04-100 and TO showed comparable intracellular killing of S aureus SCVs in a polarized
Calu-3 cell culture model In conclusion chemical modifications of TO allowed improvement
of its antibacterial activity against L monocytogenes SCVs and prototypical S aureus
41
Keywords Tomatidine aminoglycoside synergy SCV Firmicutes S aureus L
monocytogenes
Introduction
The Firmicutes are Gram-positive bacteria with genomes presenting less than 50 of G+C
The Firmicutes constitutes one of the main divisions within the Bacteria and is divided into
three classes Bacilli Mollicutes and Clostridia The class of Bacilli is further divided into the
orders of the Lactobacillales and the Bacillales The Bacillales are divided into the genus of
Staphylococcus Listeria and Bacillus A number of bacterial species such as Listeria spp and
Bacillus spp can contaminate food and cause infections in humans (Magalhaes 2013) To
name a few Listeria monocytogenes L ivanovii and Bacillus cereus can cause listeriosis
(Guillet et al 2010) and food poisoining (Bad Bug Book FDA) Bacillus subtilis B
coagulans B licheniformis and B sphaericus are also known to cause illnesses Bacillus
anthracis causes anthrax and can often be acquired by contact with food producing animals
and cattle (beef cattle sheeps etc) and this bacterium is also well-known for its endospores
that have been used as biological weapons (Beierlein and Anderson 2011) Staphylococci are
divided in coagulase-positive species Staphylococcus aureus being the most clinically
relevant of this group and coagulase-negative species such as S epidermidis the most
prevalent pathogen associated with infections of implanted medical devices (Vuong and Otto
2002) The emergence and spread of resistance to multiple antibiotics in staphylococci is now
considered a real health treat and impaired therapeutic endeavor to combat these bacteria
42
(Witte et al 2007) Notably the prevalence of methicillin-resistant S aureus (MRSA) has
steadily increased over the recent years not only in hospitals but also in the community
(Hiramatsu et al 2013)
Since their first description in 1910 for Salmonella enterica serovar Typhi (S typhi) small-
colony variants (SCVs) have been described for a wide range of bacterial genera and species
and have been isolated from both laboratory and clinical environments (Proctor 2006)
Staphylococcus aureus SCVs have attracted a great deal of interest over the past recent years
S aureus SCVs often present a dysfunctional oxidative metabolism causing an alteration in
the expression of virulence factors a slow growth and a loss of colony pigmentation (Proctor
2006) This dysfunctional oxidative metabolism is also responsible for a decreased
susceptibility to aminoglycoside antibiotics because this class of molecules requires the
proton-motive force in order to penetrate the bacterium (Bryan LE 1981) This respiratory
deficiency is often caused by mutations affecting the electron transport system and several
SCV isolates can recover normal growth with supplemental hemin or menadione which are
needed to synthesize electron transport system components S aureus SCVs often are isolated
from chronic infections such as lung infections in cystic fibrosis (CF) patients osteomyelitis
septic arthritis bovine mastitis and from infections associated with orthopedic devices (Atalla
2008 Moisan 2006 Proctor 2006) The ability of S aureus to switch back and forth from the
prototypic to the SCV phenotypes in vivo is an integral part of the pathogenesis of S aureus
and may be responsible for the establishment of chronic infections (Tuchscherr 2011
Mitchell 2011 inf imm)
43
Tomatidine (TO) is a steroidal alkaloid produced by the Solanaceae plant family such as the
tomato (Ruiz-Rubio 2001 Simons 2006) We showed previously the antibacterial activity of
TO against S aureus SCVs and also documented a strong synergic activity of TO in
combination with aminoglycoside antibiotics against prototypic S aureus (Mitchell G AAC
2011 Mitchell G 2012 JAC) Recently we synthesized a variety of TO analogs in order to
explore the structure-activity relationship of this new class of antibiotics able to act on SCVs
(Chagnon et al 2014) One analog FC04-100 showed the same steroidal backbone as the
natural molecule but with an additional carbon chain on the A cycle (Fig 1) Preliminary
characterization of FC04-100 revealed an antibacterial activity that was similar to that of TO
against S aureus SCVs and also preserved the synergic activity with aminoglycosides against
prototypic S aureus (Chagnon et al 2014) However contrary to TO FC04-100 showed
notable antibiotic activity by itself against prototypic S aureus with a MIC of 8-16 gL
whereas that of TO was gt64 gL It would be of great interest to see if this property of FC04-
100 modifies or improves its spectrum of antibacterial activity
The aim of this study was to further describe the spectrum activity of TO and FC04-100 The
susceptibility of several species among the Firmicutes to TO and FC04-100 alone or in
combination with other antibiotics was investigated The improved antibacterial activity of
FC04-100 was demonstrated against Listeria monocytogenes and methicillin-resistant S
aureus (MRSA)
44
Materials and methods
Chemical reagents and antibiotics
Tomatidine (TO) (Sigma Aldrich Oakville Ontario Canada) (Fig 1A) was solubilised in
dimethylsulfoxide (DMSO) at a concentration of 2 mgmL The TO analog FC04-100 (Fig
1B) was synthesized (Chagnon et al 2014) and solubilised in DMSO at 20 gL Cefalexin
kanamycin gentamicine (GEN) (all from Sigma Aldrich) were solubilised in water at a
concentration of 10 gL Menadione was solubilized in DMSO hemin in 14 M NH4OH and
thymidine in water (all from Sigma Aldrich) and were prepared at a concentration of 10 gL
45
3
Fig 1 Tomatidine (A) is a steroid alkaloid structurally characterized by 6 rings 12
stereogenic centers a 3 β-hydroxyl group and spiro-fused E F rings in the form of an
aminoketal The tomatidine analog FC04-100 (B) contains a diamine in position 3 of ring A
Bacterial strains and growth conditions
Staphylococcus aureus ATCC 29213 S aureus Newbould (ATCC 29740) Listeria
monocytogenes ATCC 13932 Bacillus subtilis ATCC 6333 and Bacillus cereus ATCC 11778
were used as prototypic strains Methicillin-resistant S aureus (MRSA) strains COL and
USA100 (hospital acquired-MRSA) and strain USA300 (community acquired-MRSA) were
also used in this study The reference laboratory strain S aureus SH-1000 an isogenic mutant
strain derived from S aureus 8523-4 but with a functional rsbU allele (HorsburghMJ 2002)
was used for its good yield of biofilm production The S aureus strains CF07-L (prototypic)
A
B
A B
C D
E
F
46
and CF07-S (SCV phenotype) are genetically related clinical strains originally isolated from a
cystic fibrosis patient (MitchellG 2011) Strain CF07-S was used in the intracellular
infection model Staphylococcus and Bacillus were maintained on Tryptic Soy agar (TSA BD
Mississauga Ontario Canada) whereas L monocytogenes was grown on brain-heart infusion
(BHI) agar (BD Mississauga Ontario Canada)
Small colony variants (SCVs)
For S aureus strain NewbouldhemB was used as the reference SCV NewbouldΔhemB was
generated from strain Newbould (ATCC 29740) by disrupting the hemB gene with the ermA
cassette by homologous recombination (Brouillette 2004) SCVs from B cereus B subtillis
and L monocytogenes were generated by growth in presence of a subinhibitory concentration
of GEN Briefly overnight broth cultures (18ndash20 h) were used to inoculate BHI broth at a
dilution of 1100 supplemented or not with 025 to 1X the MIC of GEN Cultures were
incubated 18 h at 35degC with shaking (225 rpm) and then adjusted to an A595 nm of 20 in PBS at
4degC Determination of CFU and SCV colonies was done by serial dilution plating SCV were
obtained by plating on TSA (BHI agar for L monocytogenes) containing gentamicin at a
concentration of 8 to 16 times the induction concentration followed by an incubation of 48 h at
35degC The pinpoint colonies selected by this method were confirmed to be SCVs by streaking
several of them on agar plates without antibiotic SCV that conserved their phenotype after
two passages were considered to have a stable phenotype and were used in subsequent
experiments Selected SCVs had a colony size a tenth of the size of a normal colony and
showed no pigmentation or hemolytic activity on blood agar plates The SCV species were
47
confirmed by rDNA 16S sequencing after PCR amplification using universal primers (Gomaa
2007)
Auxotrophy assays
For SCVs auxotrophism is defined as the requirement of specific compounds in order to
regain a normal growth phenotype [Sendie 2009] An agar diffusion method was used to
characterize the auxotrophism of SCVs using hemin or menadione (1-10 μg eachwell) or
thymidine (100 microgwell) on an inoculated Mueller-Hinton agar (MHA) or Brain-heart infusion
agar (BHIA) plate [Besier 2007] Auxotrophy for specific supplements was detected by a
zone of normal growth surrounding the well after 18 h of incubation at 35degC The results were
confirmed by two indepedent experiments
Antibiotic susceptibility testing
MICs were determined by a broth microdilution technique following the recommendations of
the Clinical and Laboratory Standards Institute (CLSI) except that the incubation period was
extended to 48 h and the medium used was BHI to allow SCVs to reach maximal growth as
previously described (Mitchell 2010) As recommended by the CLSI L monocytogenes (and
its SCVs) were grown in cation-adjusted Muller-Hinton broth (CAMHB BD) containing 3
lysed horse blood (LHB Remel Hartford CT USA) The reported results were obtained from
three independent experiments
48
Time-kill experiments
Kill kinetics were performed to determine whether the effect of compounds alone or in
combination was bacteriostatic or bactericidal Bacteria were inoculated at 105-10
6 CFUmL in
appropriate medium in the absence or presence of the different antibiotic compounds at
concentrations specified in figure legends At several points in time at 35degC (225 rpm)
bacteria were sampled serially diluted and plated on TSA for CFU determinations Plates
were incubated for 24 or 48 h at 35degC for normal and SCV strains respectively The data were
collected from a minimum of three independents experiments
Antibiotic activity in biofilms
The viability of bacteria in biofilms was evaluated based on previously described protocols
(CeriH 2001 MoskowitzSM 2004 HarrisonJJ 2010) For these assays 96-peg lids and
corresponding 96-well plates were used (Nunc CITY STATE) Wells were inoculated with a
suspension of S aureus SH-1000 adjusted to a 05 McFarland standard in TSB (150 microL per
wells) and plates with 96-well lids were incubated at 35degC with an agitation of 120 rpm for 24
h The biofilms on pegs were then washed three times with 200 microL PBS Biofilms on pegs
were further incubated in fresh 96-well plates containing 200 microL of TSB containing or not
antibiotic compounds or a combination of molecules at 35degC 120 rpm for 24 h The treated
biofilms on pegs were washed three times in PBS and bacteria in biofilms were recovered by
sonication in a new microtiter plate containing 200 microL of PBS per wells using an
ultrasonicator bath for 10 min followed by centrifugation for five min at 1000 RPM The
49
bacteria recovered in each well were resuspended serially diluted plated on TSA and
incubated at 35degC for 24 h before CFU determination
For an equivalent series of wells instead of disrupting the biofilm by sonication biofilm
formation on pegs was evaluated through crystal violet staining using a procedure adapted
from Mitchell et al(2010) Following exposure to antibiotics as described above biofilms on
pegs were washed three times using 200 microL of PBS before biofilms were air-dried for 30
minutes Biofilm on pegs were then stained by dipping in 200 microL of a crystal violet solution
(01) for 30 minutes rinsed in 200 microL of distilled water and air-dried for 30 min Finally the
crystal violet from stained biofilms was solubilized in 200 microL of ethanol 95 at room
temperature and the absorbance was measured at 562 nm using a plate reader (nom de
lrsquoappareil)
Cell invasion and measurement of intracellular antibiotic activity
The Calu-3 cell line (ATCC HTB 55 Homo sapiens lung adenocarcinoma) was cultured in
Eaglersquos Minimum Essential Medium (EMEM) supplemented with 01 mM MEM nonessential
amino acids 1 mM of sodium pyruvate 100 UmL penicillin 100 gL streptomycin 25 gL of
Fungizone and 10 fetal bovine serum (FBS) at 37oC in 5 CO2 For routine culture 4 gL of
puromycin was added to culture media All cell culture reagents were purchased from Wisent
(St-Bruno QC Canada)
50
The cell invasion assay was performed with the Calu-3 cells in an air-liquid interface as
previously described with few modifications (Mitchell TO I paperAAC 2011) Briefly cells
were seeded at 15 x 105 cells per insert in a 12-well Transwell plate (Corning CITY STATE)
and cultured for 10 days with the apical medium replaced each day The complete medium in
the basal compartment was replaced by the invasion medium (1 FBS and no antibiotics) 18
h before invasion assays Inocula were prepared by suspending bacteria grown 20 h on BHI
agar plates in ice-cold PBS Bacteria were then washed three times in ice-cold PBS and
suspended in the invasion medium supplemented with 05 BSA at a density of
approximately 4 x 108 CFUmL Cells were washed twice with PBS and 250 μL of bacterial
suspension were apically added to each insert Invasion of cells by bacteria was allowed for 3
h inserts were emptied and washed three times with PBS The basal medium was changed for
the invasion medium with 20 gL of lysostaphin (Sigma) to kill extracellular bacteria and with
or without the tested antibiotic compounds The infected cells were incubated for a total of 48
h with a change of medium at 23 h (invasion medium with lysostaphin with or without the
tested antibiotic compounds) The invasion medium with lysostaphin but without tested
compound was also apically added 1 h before cell lysis to ensure that only intracellular
bacteria were counted At 48 h following three washes with PBS the apical and basal media
were removed and cells were detached with 100 μL of trypsin 025 and lysed for 10 min by
the addition of 400 μL of water containing 005 of Triton X-100 Then 50 μL of PBS (50X)
was added and mixed Lysates were serially diluted 10-fold and plated on TSA for CFU
determination Plates were incubated at 35oC for 48 h
51
RESULTS
Emergence the SCV phenotype
The SCV phenotype is characterized by a slow growth yielding small colony size on agar
plates (15 to 110 of the parent phenotype) Figure 2 illustrates the ability of a subinhibitory
concentration of GEN to promote the emergence of the SCV phenotype for B cereus B
subtilis and L monocytogenes B cereus SCVs were obtained at an inducing concentration of
1 X MIC of GEN (Fig 2C) B subtilis SCVs at 05 X MIC (Fig 2D) and L monocytogenes
SCVs at 025 X MIC (Fig 2E) Two prototypic S aureus Newbould and CF07-L and their
SCV counterpart NewbouldhemB and CF07-S respectively were used as control strains (Fig
2A and 2B) The SCV isolates selected for the rest of the study add a low reversion rate and
kept their phenotype without a GEN selection pressure The B cereus SCVs showed
auxotrophy for menadione (Fig 1F Table 1) some of the L monocytogenes SCVs showed
auxotrophy for hemin (Table 1) while the B subtilis SCVs showed no apparent auxotrophy to
hemin menadione or thymidine (Table 1) These results show that SCVs with defects in their
respiratory chain (ie hemin or menadione auxotrophy) can indeed emerge from a selective
pressure with aminoglycosides
52
A B
Fig 2 Typical colony sizes for SCV and protocipic strains (left and right sides of the plates
respectively) for (A) S aureus CF07-S and CF07-L (B) S aureus NewbouldhemB and
Newbould (C) B cereus ATCC 11778 (SCV1 and prototypic) (D) B subtilis ATCC 6333
(SCV2 and prototypic) and (E) L monocytogenes ATCC 13932 (SCV1 and prototypic) In
(F) a TSA plate was inoculated with a B cereus SCV1 and wells were filled with (i) 10 microg
hemin (ii) 10 microg menadione (iii) 100microg thymidine and diluents (iv) DMSO and (v) NH4OH
14N
C D
E
i ii
iii iv
v
F
53
Antibacterial activity of TO against the various species of SCVs and synergy of
the TO and GEN combination against some prototypic Firmicutes
All the SCVs tested showed a MIC for TO of 006 mgL against S aureus and lt003 mgL
against L monocytogenes B cereus and B subtilis respectivly whereas TO showed no
activity (MIC gt64 microgmL) against their prototypic counterparts (Table 1) A synergic effect of
TO with GEN against all the prototypic strains tested was also observed (Table 1) Indeed the
addition of TO provoked a 4-8 fold decrease of GEN MIC against all species tested (Table 1)
These results are consistent with our previous studies on the activity of TO on S aureus
((Mitchell 2011 AAC 2012 JAC)) No change in the MIC of GEN was observed against the
streptococci (S pyogenes S suis S agalactiae S mitis and S pneumoniae) when tested in
combination with TO (data not shown) These results show that TO is active against some but
not all Firmicutes
54
Table 1 Susceptibility of prototypic and SCV strains and species to TO andor GEN
Ratio of activity of GEN+TO vs GEN alone
Bactericidal activity of TO against SCVs and bactericidal synergy of the TO
and GEN combination against prototypic strains
As shown above the addition of 8 mgL of TO to GEN reduced the MIC of GEN against some
Firmicutes (Table 1) but also time-kill kinetics showed that TO improved the bactericidal
effect of GEN against prototypic B subtilis and B cereus (Fig 3A and 3B respectively) This
bactericidal synergy was also previously described for S aureus (Mitchell 2012 JAC)
However although the MIC of GEN was improved when in combination with TO against L
monocytogenes (Table 1) time-kill experiments failed to demonstrate a bactericidal synergy
TO GENGEN
(+TO)
Saureus ATCC 29213 gt64 05 006 8
Saureus NewbouldΔhemB 006 8 Hemin
B cereus ATCC 11778 gt64 1 025 4
B cereus ATCC 11778 (SCV1) lt003 16 Menadione
B cereus ATCC 11778 (SCV2) lt003 16 Menadione
B subtilis ATCC 6333 gt64 0125 003 4
B subtilis ATCC 6333 (SCV1) lt003 2 unknown
B subtilis ATCC 6333 (SCV2) lt003 4 unknown
B subtilis ATCC 9372 gt64 0125 lt0015 gt8
B subtilis ATCC 9372 (SCV1) lt003 1 unknown
B subtilis ATCC 9372 (SCV2) lt003 1 unknown
L monocytogenes gt64 05 0125 4
L monocytogenes (SCV1) lt003 gt64 unknown
L monocytogenes (SCV2) lt003 gt64 Hemin
Strains
MIC (gL)
Synergy
foldAuxotrophy
55
with an improvement of at most 1 log10 in killing compared to that achieved with GEN alone
(Fig 3C) Similarly the effect of TO used alone against SCVs was strongly bactericidal
against S aureus B subtilis and B cereus but not against the L monocytogenes SCV (Fig
3D) and this even if the MIC of TO against L monocytogenes SCVs was inferior to 003
mgL (Table 1) These results show that the activity of TO against L monocytogenes needs to
be improved
56
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 0 6 m g L G E N
0 0 6 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 5 m g L G E N
0 5 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
2 m g L G E N
2 m g L G E N + T O
8 m g L T O
C T R L
Lo
g1
0 C
FU
ml
0
2
4
6
8
1 0
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
CT
RL
CT
RL
CT
RL
CT
RL
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
S a u re u s
h e m B
S C V
B c e r e u s
A T C C 1 1 7 7 8
S C V 2
B s u b t i l i s
A T C C 6 3 3 3
S C V 2
L m o n o c y to g e n e s
A T C C 1 3 9 3 2
S C V 1
Fig 3 Time-kill kinetics of TO andor GEN against (A) prototypic B subtilis ATCC 6333(B)
B cereus ATCC 11778 and (C) L monocytogenes ATCC 13932 The bactericidal effect of
TO against SCVs is also shown at 8 and 24 h (D) The results were obtained from at least
three independent experiments
A B
C
D
Detection
threshold
57
Bactericidal activiy of the TO analog FC04-100 against L monocytogenes
SCVs and synergy with aminoglycosides against prototypic strains and MRSA
The addition of 8 mgL of FC04-100 to GEN (2 mgL) decreased by an average of 5 log10
prototypic L monocytogenes CFUmL after 24 h of treatment in comparison to GEN alone
(Fig 4A) Also the bactericidal activity of FC04-100 used alone against the L monocytogenes
SCV was shown to be better than that observed for TO (Fig 3D) To facilitate comparison
between TO and FC04-100 Figure 4B shows the residual CFU of the L monocytogenes SCV
strain after treatment with TO or FC04-100 and calculated as the percentage of live CFU in
comparison with untreated L monocytogenes SCVs after 24 h of incubation These results
show that TO can be chemically modified (eg derivative FC04-100) and improve its activity
against L monocytogenes and thus the antibacterial activity of FC04-100 against S aureus
was further investigated Interestingly the MIC of FC04-100 against the SCV strain S aureus
NewbouldhemB (MIC of 05 -1 mgL Table 2) was higher than that found for TO (MIC of
006 mgL Table 1) On the other hand the MIC of FC04-100 against the prototypic S
aureus ATCC 29213 is lower than that of TO (8-16 mgL vs gt 64 mgL respectively) (Tables
1 and 2) This newly detected antibacterial activity of FC04-100 was also noted against
prototypic MRSA strains (Table 2) Furthermore the ability of FC04-100 to potentiate the
activity of GEN was similar to that of TO with a 4 to 8 fold improvement of the
aminoglycoside MIC (Table 2)
Although the multi-resistant S aureus MRSA USA100 USA300 and COL are resistant to
cefalexin (beta-lactam antibiotic MIC of 256 mgL) and to the aminoglycoside kanamycin
(MIC of 256 mgL and gt 1024 mgL for USA 100 and USA300 respectively Table 3) the
58
combination of both antibiotics decreased their respective MICs (Table 3) Indeed the
combination of cefalexin and kanamycin in a proportion of 32 (commercialized under the
name UbrolexinTM
) decreased the MIC of one or both antibiotics This is as expected for the
combination of a beta-lactam with an aminoglycoside (Davis 1982) However the synergy is
weak with MRSA strains that are resistant to one or the other or both antibiotics and the
addition of 4 mgL of FC04-100 further improved by 8 to 32 fold the MIC of the cephalexin-
kanamycin combination against these strains (Table 3)
59
te m p s (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
F C 0 4 -1 0 0 8 m g L
2 m g L G E N +
F C 0 4 -1 0 0 8 m g L
C T R L
G E N 2 m g L (B H I)
Re
sid
ua
l C
FU
(
)
CT
RL
TO
8 g
L
FC
04-1
00 8
gL
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
F C 0 4 -1 0 0 8 g L
T O 8 g L
C TR L
T=
24
h
T=
8h
T=
8h
T=
24
h
T=
24
h
Fig 4 Time-kill kinetics of FC04-100 andor GEN against (A) prototypic L monocytogenes
ATCC 13932 The bactericidal effect of FC04-100 used alone against L monocytogenes
SCV1 is also shown at 8 and 24 h (B) In (B) the residual CFU were calculated as the
percentage of live CFU in each condition in comparison with the untreated L monocytogenes
SCV1 after 24h of incubation The results were obtained from at least three independent
experiments
A
B
60
Table 2 Susceptibility of prototypic and SCV S aureus strains to FC04-100 with or without
GEN
a FC04-100 w as used at 4 microgmL
b Ratio of activity of GEN + FC04-100 vs activity of GEN alone
Table 3 Synergistic activity of FC04-100 with the combinaison cefalexin-kanamycin
aFC04-100 was used at 4 microgmL
bRatio of the MIC of cefalexin+kanamycin with FC04-100 vs the MIC of
cefalexin+kanamycin alone
Staphylococcus aureus ATCC 29213 8-16 05 006-0125 4-8
S aureus SH1000 8-16 05 006-0125 4-8
S aureus NewbouldΔhemB 1 8
MRSA USA100 16 05 0125 4
MRSA USA300 16 05 006 8
MRSA Col 16 025 006 4
Strains FC04-100 GENGEN
(+FC04-100 )fold
MIC (gL)
MRSA USA100 256 256 12886 43 32
MRSA USA300 256 gt1024 256171 86 32
MRSA Col 256 4 43 05035 8
MIC (gL)
Strains cefalexin kanamycin cef+kana (32)cef+kana (32)
(+FC04-100 )fold
a b
a b
61
FC04-100 potentiates GEN against S aureus embedded in biofilm and inhibits
intracellular replication of SCVs
Figure 5 demonstrates that compound FC04-100 efficiently kills S aureus embedded in
biofilms when in combination with GEN ie there was an important decrease of bacterial
CFUs compared to the no drug control or to either FC04-100 used alone at 4 ugmL (ie 025-
05MIC Table 2) or GEN used alone at 025 05 or 1 X MIC Besides we previously
demonstrated that TO was able to inhibit intracellular replication of SCVs (Mitchell AAC)
which is an important aspect of virulence for S aureus Here (Fig 6) we show that FC04-100
like TO can significantly decrease the infection of polarized airway epithelial cells by SCVs
62
05
mgL
GE
N
05
mgL
GE
N +
FC
04
02
5 m
gL
GE
N
02
5 m
gL
GE
N +
FC
04
01
2 m
gL
GE
N
01
2 m
gL
GE
N +
FC
04
No d
rug
FC
04
0
2
4
6
8
log
10
CF
Up
eg
= 3 2
= 3 8
= 3 5
= 2
Fig 5 Bactericidal effect of FC04-100 alone or in combination with GEN against prototypic
S aureus SH100 embedded in biofilm FC04-100 (FC04) was used at 4 mgL alone or in
combination with GEN Data are from three independent experiments performed in triplicate
Significant differences in comparison to the no drug control are shown ( Plt 0001 and
Plt 00001 one-way ANOVA with a Dunnettrsquos post test) Data are presented as means with
standard deviations
63
No d
rug
TO
8 m
gL
FC
04 8
mgL
0
2
4
6
log
10
CF
Ui
ns
er
t
Fig 6 Effect of TO and FC04-100 on the intracellular load of CF07-S 48 h post-
internalization Data are from three independent experiments performed in duplicate
Significant differences among groups are shown ( Plt 00001One-way ANOVA with
Turkeyrsquos post test) Data are presented as means with standard deviations
64
DISCUSSION
In previous studies we have demonstrated that TO has a strong inhibitory activity against S
aureus SCVs and improves the bactericidal activity of aminoglycoside antibiotics against
prototypic S aureus and also more broadly against staphylococci This effect was documented
for several strains of diverse clinical origins including aminoglycoside-resistant S aureus
carrying aminoglycoside modifying enzymes TO showed however no synergistic effect on the
activity of aminoglycosides against P aeruginosa E coli or Enterococcus spp (Mitchell
2012) In the present study we showed that the antibacterial spectrum of TO can be extended
to species of the Bacillales order with very low MICs against L monocytogenes B cereus and
B subtillis SCVs (lt003 microgmL) Time-kill kinetics showed that the combination of TO and
GEN creates a bactericidal synergy against prototypic of B subtilis and B cereus similarly to
that previously demonstrated against S aureus However the mixture of TO and GEN did not
demonstrated much improvement over the activity of GEN alone against prototypic L
monocytogenes in time-kill experiments Likewise investigations with Bacillales SCVs
showed that TO efficiently killed S aureus B cereus and B subtillis but not L
monocytogenes SCVs despite its very low MIC and was thus bacteriostatic against that species
Therefore improvement of the antibacterial activity against this important pathogen seems
inevitable for future development of steroidal alkaloids such as TO
TO is a steroid alkaloid structurally characterized by 6 rings 12 stereogenic centers a 3-
hydroxyl group and spiro-fused E F rings in the form of an aminoketal (Fig 1) It is generally
65
prepared by hydrolysis of its glycosylated analog tomatine while both molecules can be found
in the Solanaceae plants (Ruiz-Rubio 2001 Simons 2006) Although we previously
documented some interesting antibacterial activity for TO the absence of an identified cellular
target makes difficult the establishment of a structure-activity relationship (SAR) Our initial
attempt to elucidate SAR was therefore based on the hypothesis that the steroidal A-D rings
act as a rigid scaffold to orient pharmacophores defined by the 3-hydroxyl group on ring A
and the spiroaminoketal group on rings E F Noteworthy we prepared analogs bearing
modifications on ring A of TO and schegravemes for chemistry synthesis were recently published
(Chagnon et al 2014) From seven compounds keeping rings E and F of TO and possessing
structural variations on position 3 of ring A we were able to ascertain some structure activity
relationships Results suggest that the hydrogen bond donor effect of the 3-hydroxyl group
was not important for activity although the stereochemistry of the 3 position substitution
moderately affected activity against both S aureus SCVs and prototypic strains (in
combination with an aminoglycoside) In addition although the presence of one or two
ammonium groups in position C3 reduced the activity against S aureus SCVs (from a MIC of
006 to 05-1 microgmL) this change was highly beneficial for antibiotic activity against normal
strains (from a MIC of gt64 to 8-16 microgmL) without the presence of an aminoglycoside This
change was unexpected and one of such analogs FC04-100 increased its solubility by more
than ten times (from 2 mgmL to 26 mgmL in DMSO) The biological properties of FC04-
100 were therefore further investigated in the present study especially that TO showed some
weak bactericidal activity against Listeria and that the solubility of TO may limit eventual
clinical applications
66
Here we demonstrated that FC04-100 was able to kill both L monocytogenes SCVs as well as
the normal phenotype alone and in the presence of GEN respectively (Fig 4) Furthermore
FC04-100 showed a noticeable antibacterial activity on its own (MIC of 16 microgmL) against the
MRSA strains USA100 USA300 and COL (Table 2) FC04-100 was also synergistic in
combination with an aminoglycoside against such MRSA strains with a 4 to 8 fold gain in the
MIC of GEN Because MRSA strains are often multiresistant to antibiotics and often carry
AMEs we examined the possibility of using a triple antibiotic combination Indeed the
combination of kanamycin and cefalexin in a proportion 32 is already approved to treat
bovine mastitis pathogens under the brand name of UbrolexinTM
(Ganiegravere 2009) This
antibiotic combination offers an extended spectrum of activity compared to each individual
drug and is expected to cover both S aureus Streptococcus uberis and E coli (Ganiere
2009 Maneke et al 2011 Silley 2012) Unfortunately the increased frequency of livestock-
associated MRSA (Garcia-Alvarez et al 2011 Graveland et al 2011 Leonard and Markey
2008) and frequent incidence of strains and species carrying AMEs (Ramirez and Tolmasky
2010) may limit the spectrum of activity of the combination of a beta-lactam and an
aminoglycoside With this in mind we showed that the use of FC04-100 in combination with
kanamycin and cefalexin improves by 32 fold the activity of this mixture against MRSA
strains carrying AMEs (Table 3) Overall the addition of FC04-100 to the aminoglycoside-
beta-lactam combination decreased the MICs of kanamycin and cefalexin below their
resistance breakpoints and this for all the MRSA strains tested
67
L monocytogenes is a highly specialized intracellular pathogen (Hamon et al 2006) causing
life-threatening infections especially in pregnant women (Janakiraman 2008) L
monocytogenes in a foodborne pathogen and because of its intracellular life cycle and
propagation it is able to hide from host defenses and antibiotic treatment Similarly S aureus
although not primarily recognized as a typical intracellular pathogen is also able to enter and
survive in host cells (Lowy 2000) Moreover the ability of S aureus to convert to the SCV
phenotype showing increased biofilm production improved adherence to host cells and tissues
as well as increased intracellular persistence allows this pathogen to cause chronic and
difficult to treat infections (Garcia et al 2013 Mitchell and Malouin 2012) Discovering
antibacterial drugs able to act in the intracellular compartment is generally recognized as a
difficult challenge Previously we demonstrated that steroidal alkaloids such as TO had such
an ability and could act on intracellular S aureus SCVs (Mitchell et al 2011) and here we
demonstrated that this was also the case for the TO analog FC04-100 (Fig 6) In addition we
also show in the present study that FC04-100 greatly improves the bactericidal activity of
aminoglycosides such as GEN against bacteria embedded in biofilms Biofilms are
recognized to be a major hindrance to antibiotic action and are responsible for persistent
colonization in many diseases (Costerton et al 1999) and in the food industry (Srey et al
2013) The intracellular and intrabiofilm antibacterial properties of FC04-100 warrant further
investigation of this compound and this class of molecules in general
In conclusion we showed in this study that the spectrum of activity of TO is related to the
Firmicutes division and more precisely to the order of the Bacillales TO possesses an
68
antibacterial activity against SCVs and a synergistic activity with animoglycoside antibiotics
against prototypic strains The novel TO analog FC04-100 showed very promising new
characteristics that include a much improved bactericidal effect on L monocytogenes and
killing of bacteria in biofilms
ACKNOWLEDGEMENTS
This study was supported by an operation grant from Cystic Fibrosis Canada to FM and by a
team grant from the Fonds Queacutebeacutecois de la Recherche sur la Nature et les Technologies
(FQRNT) to FM KB and EM
REFERENCES
Atalla H Gyles C Jacob CL Moisan H Malouin F Mallard B 2008 Characterization of
a Staphylococcus aureus small colony variant (SCV) associated with persistent bovine mastitis
Foodborne Pathog Dis 5785ndash799
Bad Bug Book Foodborne pathogenic microorganisms and natural toxins 2012 Second
edition Lampel KA Al-Khaldi S Cahill SM (Ed) Published by the Center for Food Safety and
Applied Nutrition of the Food and Drug Administration (FDA) US Department of Health and
Human Services pp 87-103
Beierlein JM Anderson AC 2011 New developments in vaccines inhibitors of anthrax
toxins and antibiotic therapeutics for Bacillus anthracis Curr Med Chem 185083-5094
69
Brouillette E A Martinez B J Boyll N E Allen and F Malouin 2004 Persistence of
a Staphylococcus aureus small-colony variant under antibiotic pressure in vivo FEMS
Immunol Med Microbiol 4135ndash41
Bryan L E and S Kwan 1981 Aminoglycoside-resistant mutants of Pseudomonas
aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob
Agents Chemother 19958ndash964
Ceri H Olson M Morck D Storey D Read R Buret A Olson B 2001 The MBEC Assay
System multiple equivalent biofilms for antibiotic and biocide susceptibility testing Methods
Enzymol 337377-385
Chagnon F I Guay M-A Bonin G Mitchell K Bouarab F Malouin E Marsault
2014 Unraveling the structure-activity relationship of tomatidine a steroid alkaloid with
antibiotic properties against persistent forms of Staphylococcus aureus Eur J Med Chem (In
Press EJMECH-S-13-01625-3)
Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial
Susceptibility Testing Twenty-First Informational Supplement M100-S21 CLSI Wayne PA
USA 2011
70
Costerton JW Stewart PS Greenberg EP 1999 Bacterial biofilms a common cause of
persistent infections Science 284 1318-1322
Davis BD 1982 Bactericidal synergism between beta-lactams and aminoglycosides
mechanism and possible therapeutic implications Rev Infect Dis 4237-245
Ganiegravere JP Denuault L 2009 Synergetic interaction between cefalexin and kanamycin in
Muller-Hinton broth medium and in milk J appl microbiol 107 117-125
Garcia LG S Lemaire B C Kahl K Becker R A Proctor O Denis P M Tulkens
F Van Bambeke 2013 Antibiotic activity against small-colony variants of Staphylococcus
aureus review of in vitro animal and clinical data J Antimicrob Chemother 68 1455ndash1464
Garcia-Alvarez L M T Holden H Lindsay C R Webb D F Brown M D Curran
E Walpole K Brooks D J Pickard C Teale J Parkhill S D Bentley G F Edwards
E K Girvan A M Kearns B Pichon R L Hill A R Larsen R L Skov S J Peacock
D J Maskell and M A Holmes 2011 Methicillin-resistant Staphylococcus aureus with a
novel mecA homologue in human and bovine populations in the UK and Denmark a
descriptive study Lancet Infect Dis 11595-603
Gomaa OM Momtaz OA 2007 16 rRNA characterization of Bacillus isolate and its
tolerance profile after subsequent subculturing Arab J Biotech 10 107-116
71
Graveland H Duim B van Duijkeren E Heederik D Wagenaar JA 2011 Livestock-
associated methicillin-resistant Staphylococcus aureus in animals and humans Int J Med
Microbiol 301630-634
Guillet C Join-Lambert O Le Monnier A Leclercq A Mechaiuml F Mamzer-Bruneel MF
Bielecka MK Scortti M Disson O Berche P Vazquez-Boland J Lortholary O Lecuit M
2010 Human listeriosis caused by Listeria ivanovii Emerg Infect Dis 16136-138
Hamon M H Bierne P Cossart 2006 Listeria monocytogenes a multifaceted model Nat
Rev Microbiol 4 423-434
Harrison JJ Stremick CA Turner RJ Allan ND Olson ME Ceri H 2010 Microtiter
susceptibility testing of microbes growing on peg lids a miniaturized biofilm model for high-
throughput screening Nat Protoc 51236-1254
Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y
Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T Baba T 2013 Genomic
Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
Horsburgh MJ Aish JL White IJ Shaw L Lithgow JK Foster SJ 2002 SigmaB
modulates virulence determinant expression and stress resistance characterization of a
72
functional rsbU strain derived from Staphylococcus aureus 8325-4 J Bacteriol 1845457-
5467
Janakiraman V 2008 Listeriosis in Pregnancy Diagnosis Treatment and Prevention Rev
Obstet Gynecol 1 179ndash185
Leonard FC Markey BK 2008 Methicillin-resistant Staphylococcus aureus in animals a
review Vet J 175 27-36
Lowy FD 2000 Is S aureus an intracellular pathogen Trends Microbiol 8341-342
Magalhatildees L Nitschke M 2013 Antimicrobial activity of rhamnolipids against Listeria
monocytogenes and their synergistic interaction with nisin Food Control 29 138-142
Maneke E Pridmore A Goby L Lang I 2011 Kill rate of mastitis pathogens by a
combination of cefalexin and kanamycin J Appl Microbiol 110184-190
Mitchell G Brouillette Eacute Lalonde-Seacuteguin D Asselin AE Lebeau Jacob C Malouin F
2009 A role for sigma factor B in the emergence of Staphylococcus aureus small-colony
variants and elevated biofilm production resulting from an exposure to aminoglycosides
Microb Pathog 4818-27
73
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K Malouin F 2011
Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic
fibrosis airway epithelial cells Antimicrob Agents and Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM Malouin F 2011 Infection of Polarized
airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is
increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function
and is influenced by NF- κ B Infect Immun 793541-3551
Mitchell G Lafrance M Boulanger S Lalonde-Seacuteguin D Guay I Gattuso M Marsault
Eacute Bourab K Malouin F 2012 Tomatidine acts in synergy with aminoglycoside antibiotics
against multiresistant Staphylococcus aureus and prevents virulence gene expression J
Antimicrob Chemother 67 559-568
Mitchell G Lalonde-Seacuteguin D Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S
Malouin F 2010 Staphylococcus aureus sigma B-dependent emergence of small-colony
variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-
heptylquinoline-N-oxide BMC Microbiology 10 33-48
Mitchell G and F Malouin 2012 Outcome and prevention of Pseudomonas aeruginosa-
Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic
Fibrosis InTech Open Access Publisher ISBN 979-953-307-059-8
74
Moisan H Brouillette E Jacob CL Langlois-Beacutegin P Michaud S Malouin F 2006
Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated
from cystic fibrosis patients is influ- enced by SigB J Bacteriol 18864ndash76
Moskowitz SM Foster JM Emerson J Burns JL 2004 Clinically feasible biofilm
susceptibility assay for isolates of Pseudomonas aeruginosa from patients with cystic fibrosis
J Clin Microbiol 421915-1922
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M Peters G 2006
Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent
infections Nat Rev Micro- biol 4295ndash305
Ramirez MS M E Tolmasky 2010 Aminoglycoside modifying enzymes Drug Res
Updates 13 151ndash171
Ruiz-Rubio M Peacuterez-Espinosa A Lairini K Roldagraven-Arjona T Dipietro A Anaya N
2001 Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi p 293ndash326 In
A Rahman (ed) Studies in natural products chemistry vol 25 Elsevier Oxford United
Kingdom
75
Sendi P Proctor RA 2009 Staphylococcus aureus as an intracellular pathogen the role of
small colony variants Trends Microbiol 1754-58
Silley P Goby L Pillar CM 2012 Susceptibility of coagulase-negative staphylococci to a
kanamycin and cefalexin combination J Dairy Sci 953448-3453
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C Osbourn A
2006 Dual effects of plant steroidal alkaloids on Saccharomyces cerevisiae Antimicrob
Agents Chemother 502732ndash2740
Srey S Jahid IK Ha S-D 2013 Biofilm formation in food industries A food safety concern
Food control 31572-585
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D
Roth J Proctor RA Becker K Peters G Loumlffler B 2011 Staphylococcus aureus
phenotype switching an effective bacterial strategy to escape host immune response and
establish a chronic infection EMBO Mol Med 3129ndash141
Vuong C Otto M 2002 Staphylococcus epidermidis infections Microbes Infect 4481-489
Witte W Strommenger B Stanek C Cuny C 2007 Methicillin-resistant Staphylococcus
aureus ST398 in Humans and Animals Central Europe Emerg Infect Dis 13 255ndash258
76
CHAPITRE 3
MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
31 Obtention des mutants reacutesistants
Afin de nous aider agrave mieux deacutefinir la cible et le mode drsquoaction de la tomatidine (TO)
nous avons forceacute lrsquoapparition de reacutesistances chez Staphylococcus aureus Comme
souche controcircle prototype nous avons choisi ATCC 29213 puisque crsquoest une souche
tregraves freacutequemment utiliseacutee pour des expeacuteriences avec des antibiotiques et nous avons
choisi la souche NewbouldΔhemB comme S aureus laquo small colony variant raquo puisque
crsquoest une souche SCV qui est stable et bien connue nrsquoayant pas de capaciteacute de
reacuteversion Par passages successifs dans une concentration sous inhibitrice
drsquoantibiotique (technique de CMI (concentration minimale inhibitrice)) (CLSI 2011)
nous avons obtenu des mutants deacutemontrant une reacutesistance croissante agrave lrsquoantibiotique
au fil des passages (Figure 1) Nous avons obtenu des mutants ATCC 29213
reacutesistants agrave la gentamicine (GEN) et agrave la GEN lorsque combineacutee agrave la TO ainsi que
des mutants reacutesistants NewbouldΔhemB reacutesistants agrave la GEN seule agrave la TO seule et
agrave la TO lorsque combineacute agrave la GEN
Certains passages ont ensuite eacuteteacute seacutelectionneacutes parce qursquoils deacutemontraient une hausse
plus marqueacutee de leur CMI Ces passages ont eacuteteacute choisis car nous supposions que
ces hausses marqueacutees de CMI correspondraient donc agrave des hausses de la
reacutesistance de cette souche agrave lrsquoantibiotique testeacute et que cette reacutesistance pouvait ecirctre la
conseacutequence drsquoune ou plusieurs mutations en lien avec la cible ou le mode drsquoaction
de lrsquoantibiotique preacutesent Trois agrave quatre passages (ou paliers de reacutesistance) par
condition ont eacuteteacute seacutelectionneacutes Nous avons ensuite reacuteensemenceacute le laquo pool raquo
bacteacuterien de chacun des passages choisis afin de pouvoir isoler 3 isolats ayant la
mecircme CMI (ou tregraves similaire (CMI vraies)) que celui du laquo pool raquo bacteacuterien des
77
passages drsquoougrave ils provenaient Soixante quatre mutants reacutesistants ont ainsi eacuteteacute
seacutelectionneacutes en vue drsquoecirctre seacutequenceacutes (Figure 2)
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt 1
6
CM
I v
raie
s
02
5
CM
I v
raie
s
02
5-1
A
78
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
en
pr
eacutes
en
ce
de
ge
n (
4 micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
01
25
1
6
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
8
CM
I v
raie
s
2
4
CM
I v
raie
s
2
B
C
79
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
en
pr
eacutes
en
ce
de
TO
(8
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
1
2
3
4
5
CM
I v
ra
ies
4
CM
I v
ra
ies
2
CM
I v
ra
ies
0
5
1
CM
I v
ra
ies
2
4
Figure 1 Graphiques des CMI pour chacun des passages pour (A) la tomatidine sur
NewbouldΔhemB (B) pour la tomatidine en preacutesence de gentamicine sur
NewbouldΔhemB (C) pour la gentamicine sur S aureus ATCC 29213 (D) pour la
gentamicine en preacutesence de tomatidine sur S aureus ATCC 29213
32 Seacutequenccedilage et analyses bioinformatiques
Une extraction drsquoADN geacutenomique a ensuite eacuteteacute effectueacutee sur chacun des isolats en
utilisant le Kit drsquoextraction drsquoADN geacutenomique miniKit de Qiagen (QIAamp DNA minikit)
(Li et al 2003) Puisque leurs seacutequences geacutenomiques nrsquoeacutetaient pas connues nous
avons eacutegalement extrait lrsquoADN geacutenomique de nos deux souches de reacutefeacuterences
ATCC29213 et NewbouldΔhemB (provenant de quatre colonies isoleacutees chacune)
afin drsquoavoir les seacutequences de comparaison pour nos mutants (Figure 2)
D
80
Ne
wm
an
(g
en
om
ede r
ef h
ttpw
ww
ncbinlm
nih
govn
uccore
150373012)
AT
CC
29
213
(A p
art
ir d
e 4
AT
CC
29
21
3 r
eco
nstr
uire
AT
CC
29
21
3 c
om
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
5
10
7
10
8 1
09
)
Ne
wbo
uld
Δhe
mB
(A p
art
ir d
e 4
Ne
wb
ou
ldh
em
Bre
co
nstr
uire
Ne
wb
ou
ldh
em
Bcom
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
1
agraveG
EN
2
5 (
CM
I2
)
2
7 (
CM
I2
)
2
8 (
CM
I2
)
AT
CC
29
21
3
Pa
lier
de
reacutesis
tance
1
agravecom
bo
29
(C
MI
05
)
3
0 (
CM
I 0
5
3
1 (
CM
I 0
5)
Suite
fe
uill
e s
uiv
ante
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agraveG
EN
3
(C
MI
4)
4
(C
MI
4)
3
9 (
CM
I4
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agraveG
EN
4
6 (
CM
I8
)
4
7 (
CM
I8
)
4
8 (
CM
I8
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agravecom
bo
1
1 (
CM
I 2
)
1
2 (
CM
I 2
)
2
1 (
CM
I 2
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agravecom
bo
5 (
CM
I 2
-4)
3
6 (
CM
I 2
-4)
3
7 (
CM
I 2
-4)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
4
agravecom
bo
3
8 (
CM
I4
)
6
7 (
CM
I 4
)
81
Ne
wb
ould
Δhem
B(A
part
ir d
e 4
Ne
wb
ou
ldΔ
he
mB
reco
nstr
uire
Ne
wb
ou
ldΔ
hem
Bco
mp
let e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveG
EN
24
(C
MI
64
)
4
1 (
CM
I 1
6)
4
2 (
CM
I 6
4)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveT
O
95
(C
MI
02
5)
9
6 (
CM
I 0
25)
9
7 (
CM
I 0
25)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agravecom
bo
33
34
35
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveG
EN
49
(C
MI
12
8)
5
1 (
CM
I 1
28)
5
2 (
CM
I 1
28)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agraveG
EN
7
(C
MI
128
)
4
3 (
CM
I 1
28)
4
5 (
CM
I1
28
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agraveT
O
6
4 (
CM
I gt
16)
6
5 (
CM
I gt
16)
6
6 (
CM
I gt
16)
75
76
77
79
80
81
82
83
84
85
86
87
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agravecom
bo
15
16
18
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agravecom
bo
59
60
61)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agravecom
bo
54
56
58)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveT
O
90
(C
MI
02
5)
9
1 (
CM
I 0
25)
9
3 (
CM
I 0
5)
Ne
wb
ou
ldh
em
B
palie
r d
e r
eacutesis
tance
3
agraveT
O
11
0 (
CM
I 1
)
1
12 (
CM
I 0
25
-05
)
1
13 (
CM
I 0
5)
Fig
ure
2
Org
an
igra
mm
e d
es
mu
tan
ts r
eacutes
ista
nts
seacute
qu
en
ceacutes
se
lon
leu
r pa
lier
de
reacute
sis
tan
ce e
t a
vec leu
r n
um
eacutero
drsquoeacute
chan
tillo
n
co
rre
spo
ndan
t
82
Un minimum de 20 ngmicrol drsquoADN geacutenomique de chacun des isolats a ensuite eacuteteacute
achemineacute au laboratoire du Pr Seacutebastien Rodrigue (Deacutepartement de biologie Faculteacute
des sciences Universiteacute de Sherbrooke) qui a effectueacute la librairie de fragments Pour
se faire il y a eu dans un premier temps digestion enzymatique de lrsquoADN afin
drsquoobtenir des fragments drsquoADN double brin Ensuite tous les fragments ont eacuteteacute
purifieacutes suivant la meacutethode SPRI (Solid-Phase Reversible Immobilization) Il srsquoagit
drsquoune meacutethode simple et tregraves efficace drsquoisolation drsquoacides nucleacuteiques sur microplaque
(Rodrigue S et al 2010) Les acides nucleacuteiques cibles sont immobiliseacutes sur des
microparticules paramagneacutetiques dans des conditions tampons-speacutecifiques La
flexibiliteacute du systegraveme se fait par lrsquoutilisation de diffeacuterents tampons de liaison (laquo binding
buffer raquo) qui modifient le type et la grandeur des acides nucleacuteiques immobiliseacutes Les
contaminants peuvent ainsi ecirctre retireacutes de lrsquoeacutechantillon sans besoin de filtration ou
centrifugation (Beckman Coulter website 2013 DeAngelis et al 1995) Suite agrave la
purification par meacutethode SPRI lrsquoeacutequipe du Pr Rodrigue a obtenu des fragments
drsquoADN de ~250pb auxquels ont eacuteteacute ajouteacutes par ligation des adapteurs aux deux
extreacutemiteacutes des fragments Dans un deuxiegraveme temps des codes-bars speacutecifiques agrave
chaque eacutechantillon drsquoADN geacutenomique ont eacutegalement eacuteteacute lieacutes agrave chacun des fragments
Alors que les adapteurs permettent aux fragments de se lier agrave une matrice solide afin
de pouvoir ecirctre amplifieacutes les codes-bars permettront de classer chacun des
fragments selon leur appartenance agrave un eacutechantillon drsquoADN geacutenomique speacutecifique
83
Figure 3 Eacutetape du seacutequenccedilage Illumina
La librairie de fragments a ensuite eacuteteacute envoyeacutee chez Geacutenome-Queacutebec afin que
chacun des geacutenomes puisse ecirctre seacutequenceacute Geacutenome ndashQueacutebec utilise la meacutethode
Illumina comme outil de seacutequenccedilage (Figure 3) La grande innovation de la meacutethode
Illumina est lrsquoamplification des fragments par liaison de ceux-ci agrave une surface solide
(Flowcell) ce qui permet par un processus de ponts drsquoamplification de geacuteneacuterer plus
drsquoun million de laquo clusters raquo comprenant eux-mecircme au plus un millier de copies de la
moleacutecule originale de deacutepart (solid-phase PCR) Chaque regroupement de fragments
provenant de lrsquoamplification drsquoun mecircme fragment original est tregraves pregraves les uns des
Modifieacute de Quail MA et al 2008
84
autres formant une zone distincte drsquoun regroupement de fragments agrave un autre
Lrsquoincorporation des nucleacuteotides eacutetiqueteacutes preacutealablement avec des moleacutecules
fluorescentes speacutecifiques est deacutetecteacutee suite agrave leur excitation par un laser et les
images sont compileacutees pour produire la seacutequence de bases pour chaque fragment
ADN (Figure 4) Les nouvelles seacuteries de bases appeleacutees laquo readsraquo seront ensuite
reacuteassembleacutees (aligneacutes) en se basant sur un geacutenome de reacutefeacuterence connu (appeleacute
laquo resequencing raquo) (crsquoest notre cas ici) ou en absence de geacutenome de reacutefeacuterence
(appeleacute laquo De novo sequencing raquo) Au final lrsquoensemble des laquo reads raquo aligneacutes
correspondra agrave la seacutequence de chaque chromosome de lrsquoeacutechantillon drsquoADN
geacutenomique de deacutepart
85
Figure 4 Meacutethode drsquoamplification par la meacutethode Illumina
Suite au seacutequenccedilage chez Geacutenome ndashQueacutebec nous avions en main les seacutequences
geacutenomiques de tous nos mutants et de nos souches de reacutefeacuterence sous forme de
Tireacute de BiteSizeBio httpbitesizebiocom13546sequencing-by-synthesis-
explaining-the-illumina-sequencing-technology consulteacute le 3 deacutec 2013
86
contigs Nous avions donc le casse-tecircte mais en piegraveces deacutetacheacutees pecircle-mecircle et
sans avoir la photo de lrsquoimage de reacutefeacuterence sur laquelle se baser pour pouvoir
compleacuteter le casse-tecircte
1- Lrsquoassemblage
Les diffeacuterents geacutenomes avaient eacuteteacute preacutealablement fractionneacutes aleacuteatoirement
en petits fragments de 250 pb et amplifieacutes en vue du seacutequenccedilage Suite au
seacutequenccedilage les seacutequences de chaque fragment (appeleacute laquo reads raquo) sont ensuite
assembleacutees en contigs crsquoest-agrave-dire des seacutequences plus ou mois longues continues
et ordonneacutees geacuteneacutereacutees par lassemblage des clones dune bibliothegraveque geacutenomique
qui se chevauche La qualiteacute de lrsquoassemblage en contigs deacutepend de la capaciteacute de
superposition des laquo reads raquo Parce que la digestion enzymatique de lrsquoADN se fait de
faccedilon aleacuteatoire et se fait sur plusieurs copies drsquoADN chaque portion du geacutenome est
repreacutesenteacutee plusieurs fois dans diffeacuterents modegraveles de fragments Donc les
seacutequences identiques des diffeacuterents fragments (ou laquo reads raquo) peuvent ecirctre
superposeacutees par chevauchegravement et les laquo reads raquo superposeacutes peuvent ecirctre
assembleacutes en contigs par un logiciel drsquoassemblage En ce qui nous concerne cette
eacutetape a eacuteteacute faite agrave lrsquoaide du programme de bio-informatique Newbler (Zhang et al
2012) Lrsquoassemblage geacutenomique peut ecirctre probleacutematique pour les informaticiens car
les geacutenomes contiennent des seacutequences qui se reacutepegravetent et ces reacutepeacutetitions peuvent
ecirctre assez longues en fonction des organismes Suite agrave lrsquoassemblage les geacutenomes
sont donc sous forme de contigs
2- Lrsquoalignement
En se basant sur une seacutequence geacutenomique de reacutefeacuterence (notre reacutefeacuterence est
Staphylococcus aureus str Newman (~ 288Mb)
( httpwwwncbinlmnihgovgenome154project_id=58839) ) le programme de bio-
informatique ABACAS permet de rapidement aligner ordonner et orienter les contigs
Selon les besoins ce qui nrsquoa pas eacuteteacute notre cas ABACAS permet eacutegalement de
87
visualiser et designer les amorces permettant de fermer les gaps (espaces) entre les
contigs en faisant des PCR (Assefa et al 2009)
3- La fermeture des gaps
La fermeture des gaps nrsquoa pas eacuteteacute faite avec le programme ABACAS mais
avec lrsquoaide du programme de bio-informatique GapFiller Lrsquoutilisation de GapFiller
pour la fermeture des gaps possegravede 3 principaux avantages Premiegraverement le
programme prend en compte lrsquoestimation de la grandeur du gap agrave fermer donc il
exclut automatiquement les fermetures de gaps erroneacutes qui sont plus courts ou plus
longs que la grandeur estimeacutee Deuxiegravemement il nrsquoessaie pas de remplir un gap
avec un assemblage de laquo reads raquo en contigs mais cherche plutocirct agrave allonger les
contigs agrave partir de leurs extreacutemiteacutes jusqursquoagrave obtenir des seacutequences superposeacutees en
passant par la creacuteation de K-mer (seacuteparation des reads en plus petits fragments) Et
troisiegravemement il prend en compte que les extreacutemiteacutes des contigs sont souvent
source de mauvais assemblage et reacute eacutevalue ces extreacutemiteacutes pendant la fermeture des
gaps (Boetzer et Pirovano 2012)
4- Lrsquoannotation des geacutenomes
Suite agrave la fermeture des gaps il a fallu annoter les geacutenomes en se basant sur
lrsquoannotation du geacutenome de reacutefeacuterence Staphylococcus aureus str Newman RATT est
un outil de transfert rapide drsquoannotation (Rapid Annotation Transfert Tool) qui a eacuteteacute
deacuteveloppeacute afin de fournir rapidement une annotation preacutecise pour des geacutenomes
inconnus en se basant sur des geacutenomes deacutejagrave annoteacutes comme reacutefeacuterences (Figure 5)
La meacutethode transfert les annotations drsquoune reacutefeacuterence de haute qualiteacute agrave un nouveau
geacutenome sur la base de la conservation des synteny (ordre des gegravenes) (Otto et al
2011) Suite agrave lrsquoannotation les gegravenes tant chez les mutants que chez les souches
parentes portent les noms auxquels ils correspondent chez Staphylococcus aureus
str Newman
88
Figure 5 Corrections du programme RATT sur les annotations transfeacutereacutees Les
annotations de H37Rv ont eacuteteacute transfeacutereacutees sur la seacutequence F11 (bleu pacircle) ont eacuteteacute corrigeacutees
(vert) et ensuite compareacutees agrave lrsquoannotation de la souche existante F11 EMBL (jaune) (A et B)
La correction des codons ldquostartrdquo et ldquostoprdquo respectivement Dans une situation de laquo mapping raquo
plus complexe (C) ougrave les cadres de lecture sont montreacutes pour plus de clarteacute RATT
cartographie une large seacutequence codante (CDS) de H37Rv vers un locus dans F11 qui inclue
plusieurs codons laquo stop raquo laquo in-frame raquo En inseacuterant un deacutecalage du cadre de lecture (ie un
pseudogegravene) la traduction conceptuelle est conserveacutee Ceci diffegravere avec les deux gegravenes se
superposant preacutedis comme faisant partie du geacutenome F11
Tireacute de Otto TD et al 2011
89
321 Identification des mutations en lien avec la tomatidine
Apregraves avoir obtenu une seacutequence consensus agrave partir des seacutequences geacutenomiques
des 4 copies de chacune des souches parente S aureus ATCC 29213 et S aureus
NewbouldΔhemB les diffeacuterences geacutenomiques entre les souches parentes et le
geacutenome de reacutefeacuterence S aureus str Newman ont eacuteteacute eacutelimineacutees afin que ces
diffeacuterences nrsquoapparaissent pas comme des mutations lors de la comparaison des
seacutequences geacutenomiques des mutants avec celles de leur souche parente ces
diffeacuterences eacutetant plutocirct attribuables agrave la diffeacuterence qursquoil y a entre S aureus str
Newman et la souche parente et non pas agrave des mutations en lien avec la reacutesistance
des mutants
Le service bio-informatique du deacutepartement de biologie (Faculteacute des sciences
Universiteacute de Sherbrooke) a ensuite produit un tableau contenant pour chacun des
mutants tous les gegravenes muteacutes apregraves comparaison avec leur souche parentes Agrave cette
eacutetape on pouvait donc savoir dans quel gegravene se situaient les mutations quelle eacutetait
lrsquoeffet de cette mutation (mutation synonyme deacuteleacutetion insertion SNP) et en principe
srsquoil srsquoagissait pour un mecircme gegravene muteacute de la mecircme mutation puisque dans ce
tableau pour un mecircme gegravene chaque ligne correspond agrave des mutations diffeacuterentes
(Figure 6)
Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4
Figure 6 Exemple drsquoune partie du tableau compilation des mutations chez
les mutants ATCC 29213 reacutesistants agrave la GEN seule et agrave la GEN (+TO)
90
Agrave lrsquoaide de ce tableau (Figure 6) jrsquoai chercheacute les mutations qui pouvaient ecirctre
attribuables agrave la TO Chez les mutants S aureus ATCC 29213 reacutesistants jrsquoai
chercheacute les gegravenes qui eacutetaient muteacutes chez tous les mutants reacutesistants agrave la
combinaison GEN (+TO) ou dont la mutation apparaissait graduellement au fil des
paliers de reacutesistance mais dont ce mecircme gegravene nrsquoeacutetait pas muteacute chez les mutants
reacutesistants agrave la GEN seule Je pouvais donc estimer que cette mutation chez les
reacutesistants agrave la combinaison eacutetait en lien avec la preacutesence de la TO dans le milieu
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct
Afin de mrsquoaider dans ma recherche jrsquoai eu lrsquoaide du Pr Pierre-Eacutetienne Jacques
(Deacutepartement de biologie Faculteacute des sciences Universiteacute de Sherbrooke)qui a
mis les donneacutees de ce tableau dans un tableau croiseacute dynamique (TCD) ce qui a
permis de filtrer les donneacutees selon des critegraveres bien preacutecis me permettant drsquoarriver agrave
mes fins plus facilement Dans un premier temps un TCD a eacuteteacute creacuteeacute ougrave pour
chacun des mutants chaque gegravene ayant au moins une mutation eacutetait indiqueacute par un
laquo 1 raquo alors qursquoun laquo 0 raquo eacutetait marqueacute si ce gegravene nrsquoeacutetait pas muteacute Dans un second
temps la quantiteacute de laquo 1 raquo par palier de reacutesistance (correspondant au nombre de
mutants de ce palier posseacutedant une mutation dans ce gegravene) eacutetait indiqueacutee par un
chiffre allant de 0 pour laquo aucun mutant de ce palier de reacutesistance ne posseacutedait de
mutation dans ce gegravene raquo agrave 1 2 ou 3 selon le nombre de mutants de ce palier ayant
ce gegravene muteacute Jrsquoai ensuite pu soumettre le TCD agrave des filtres approprieacutes afin de faire
ressortir les gegravenes drsquointeacuterecircts selon des critegraveres de seacutelection preacutecis (Figures 7-8)
91
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
1
00
33
32
1
1
11
4
5
0
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
10
03
33
21
11
14
50
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_04
12
lpl9
nm
20
12
32
23
92
44
1
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
NW
MN
_00
46
_2
10
13
21
37
24
31
NW
MN
_09
55
_2
01
23
11
37
24
21
NW
MN
_13
83
gpsA
21
02
31
13
72
42
1
NW
MN
_03
39
_1
11
21
31
37
34
42
NW
MN
_15
97
dn
aE2
10
22
30
37
23
31
NW
MN
_23
30
_2
10
23
20
37
23
21
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_2
07
0_
10
01
23
21
81
45
0
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_10
28
_1
22
12
32
58
34
54
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
A B C
92
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
39
8h
sdM
31
33
33
27
11
34
54
NW
MN
_04
26
_3
31
33
32
71
13
45
4
NW
MN
_05
24
sdrD
32
33
33
28
11
34
55
NW
MN
_07
39
_3
33
33
32
91
13
45
6
NW
MN
_11
64
hsl
U3
33
33
32
91
13
45
6
NW
MN
_05
23
sdrC
22
33
32
27
10
34
45
NW
MN
_20
61
fmtB
23
23
32
27
10
34
45
NW
MN
_04
03
lpl1
nm
33
22
33
28
10
34
55
NW
MN
_13
45
ebh
13
32
33
31
81
03
44
5
NW
MN
_16
49
_3
32
33
31
81
03
44
5
NW
MN
_17
00
hsd
M3
23
33
22
81
03
44
5
NW
MN
_00
42
_3
33
33
22
91
03
44
6
NW
MN
_05
80
_3
33
33
31
91
03
44
6
NW
MN
_13
44
ebh
23
31
32
22
79
34
44
NW
MN
_13
75
aro
A3
13
23
31
79
34
44
NW
MN
_23
97
fnb
B2
32
23
22
79
34
45
NW
MN
_04
01
_3
32
23
31
89
34
45
NW
MN
_07
33
_3
32
32
31
89
34
45
NW
MN
_17
45
_2
33
22
32
89
34
56
NW
MN
_01
66
coa
33
32
32
29
93
44
6
NW
MN
_05
25
sdrE
33
32
33
19
93
44
6
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D Fig
ure
7
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez A
TC
C 2
921
3 d
an
s le
TC
D p
ar
dif
feacutere
nts
fil
tre
s
(A)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts la
ge
nta
mic
ine
ge2
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(C
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts d
an
s le
pa
lier
1
de
reacute
sis
tan
ce
agravela
ge
nta
mic
ine
le1
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
et
nom
bre
de m
uta
nts
reacute
sis
tan
ts agrave
la la
gen
tam
icin
e (
+T
O)
de
s 2
de
rnie
rs p
alie
rs d
e r
eacutesis
tan
ce
ge5
(D
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
7 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
ge
nta
mic
ine
(+
TO
) ge
9
93
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_16
29
ccp
A0
01
33
31
50
00
01
24
0
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_21
28
rpsM
11
12
21
12
22
00
31
74
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
NW
MN
_25
29
clfB
33
03
33
80
11
26
17
4
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e reacute
sist
ance
agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
On
b m
uta
nts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
16
29
ccp
A0
01
33
31
50
00
01
24
0N
WM
N_
21
28
rpsM
11
12
21
12
22
00
31
74
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
A B C
94
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
04
6_
1
01
00
28
22
32
2
10
9
NW
MN
_17
33
prs
A0
11
12
01
12
12
22
14
7
NW
MN
_19
23
_0
11
31
15
13
22
21
08
NW
MN
_20
37
_0
02
32
07
23
11
21
27
NW
MN
_22
77
_1
02
21
11
02
12
33
14
8
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
01
43
_0
03
00
04
23
33
34
11
NW
MN
_0
40
6lp
l3n
m2
01
00
03
12
22
33
7
NW
MN
_1
65
2_
00
10
00
62
12
31
68
NW
MN
_2
01
2_
00
00
00
32
33
30
31
1N
WM
N_
20
15
mn
aA0
00
00
01
23
33
01
11
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
o
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D E Fig
ure
8
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez N
ew
bo
uld
Δh
em
Bd
an
s l
e T
CD
par
dif
feacutere
nts
fil
tres
(A)
Nom
bre
de
mu
tan
ts
reacutesis
tan
ts agrave
la tom
atid
ine
ge15
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atid
ine
ge15
et no
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
(C)
No
mbre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la tom
atid
ine
ge15
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3(
D)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atidin
ege
10
et no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
tom
atid
ine
(+G
EN
) ge
7
(E)
No
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la to
ma
tid
ine
le6
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
95
Le TCD mrsquoa permis de rapidement identifier les gegravenes drsquointeacuterecircts Agrave lrsquoaide du site CMR
(Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) 2013) jrsquoai
ensuite veacuterifieacute la fonction des gegravenes identifieacutes afin de confirmer leur inteacuterecirct (Tableau
1)
Tableau 1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants et (B) chez les mutants NewbouldΔhemB reacutesistants
Gegravene Alias Fonction
NWMN_0811 NADH dh complexe 1 chaine de transport des eacutelectrons
NWMN_2070 _membrane prot Putative- Toxin production+resistance drug transporteur-
similar to multidrug transporteur 85
NWMN_0635 _membrane prot Putative- Bordeacute par AraC(reacutegulateur opeacuteron arabinose) ou
SsaA-likeprot(lieacute au quorum sensing RNAIII(secretory antigen)
NWMN_2084 _truncated transposase for Is1272- mobile+extrachromosomal element
fonctiontransposon fonction
NWMN_0928 qoxC quinol oxidase polypeptide IIIcomplexe 4 chaine de transport des eacutelectrons
NWMN_0739 _ conserved hypothetical prot fonction inconnues
NWMN_1164 hslU ATP-dependent protease ATP-binding subunit
NWMN_0524 sdrD Ser-Asp rich fibrinogenbone sialoprotein-binding protein sdrD
NWMN_0042 _ lipoprotein putativecell enveloppe- lipoprot en tendem
NWMN_0580 _conserved hypothetical prot fonction inconnuesDNA metabolism DNA
replication recombination and repair
NWMN_1745 _
NWMN_0166 coa coagulase precursor
NWMN_0525 sdrE Ser-Asp rich fibrinogenbone sialoprotein-binding protein SdrE
A
96
Apregraves avoir aligneacute speacutecifiquement une par une chacune des seacutequences (Fasta) des
gegravenes drsquointeacuterecirct de chaque mutant avec leur souche parente jrsquoai utiliseacute le programme
de visualisation et de comparaison de seacutequences ClustalW2 (ClustalW2 2013) afin
de confirmer que la mutation eacutetait une vraie mutation et non une erreur de
seacutequenccedilage ou drsquoalignement et que la mutation eacutetait identique chez tous les mutants
ayant le gegravene drsquointeacuterecirct muteacute Cette eacutetape a permis de restreindre encore la liste des
gegravenes pouvant ecirctre en lien avec la cible ou le mode drsquoaction de la TO (Figure 9)
Gene Alias Fonction
NWMN_0896 _conserved hypothetical prot fonction inconnue- comK family prot-
regulation DNA interaction- ORFIDresssemble agrave facteur de transcription
NWMN_0996 _ conserved hypothetical prot fonction inconnue- prophage fonction
NWMN_1025 _ phage major tail protprophage fonction
NWMN_1629 ccpAcatabollite control protA virulence determinant production and antibiotique
resistance
NWMN_1916 _ conserved hypothetical prot-fonction inconnue ou prophage prot
NWMN_2314 _drug resistance transporteur- EmrBQacA subfamily prot- toxin
production+resistance- transport and binding prot
NWMN_0046 _ hypothetical prot
NWMN_2015 mnaA UDP-Glc-Nac 2 epimerase
NWMN_2012 atpE ATP synthase subunit C
NWMN_1652 _ multidrug resistance transporter protein B
NWMN_0143 _ oligopeptide ABC transporter ATP-binding protein
B
97
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
A
B
98
Jrsquoai ensuite chercheacute le rocircle de ces gegravenes sur le site CMR afin de veacuterifier si leur
mutation pouvait expliquer la reacutesistance agrave lrsquoantibiotique (Comprehensive Microbial
Ressouce (J Craig Venter Institute (JCVI) 2013) Dans une premiegravere eacutetape le rocircle
drsquoun gegravene a eacuteteacute deacutetermineacute en allant tout simplement voir directement le rocircle de ce
gegravene chez Staphylococcus aureus str Newman Dans le cas ougrave le gegravene nrsquoavait pas
de fonction connue chez Saureus str Newman je suis alleacutee voir quelle eacutetait la
fonction de ce mecircme gegravene (et des gegravenes flanquant celui-ci) chez diffeacuterentes souches
de Staphylococcus aureus Advenant le cas ougrave malgreacute tout je nrsquoeacutetais toujours pas
capable de deacuteterminer le rocircle de ce gegravene je suis alleacutee veacuterifier si sa seacutequence pouvait
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 hellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Mutants resistants agrave GEN (4 microgml)+TO
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip 185-AATTGCATTCA-197helliphelliphellip495-ATAGCAATGG-506hellip
hellipnwmn_1629helliphelliphelliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_0143hellip
ccpAATP synthase c
ABC transp
Niveau 1
Niveau 2
Niveau 3
Niveau 4
TO +GEN (4microgml)
Figure 9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct (A) nwmn_0811 et
nwmn_0928 chez les mutants ATCC 29213 reacutesistants agrave la combinaison gentamicine
(+TO) leur souche parente ATCC 29213 (B) nwmn_1629 et nwmn_2012 chez les
mutants hemB reacutesistants agrave la TO et leur souche parente NewbouldΔhemB (C)
nwmn_1629 nwmn_2012 et nwmn_0143 chez les mutants hemB reacutesistant agrave la
tomatidine (+genta) et leur souche parente hemB
C
99
avoir une forte homologie avec un gegravene drsquoune autre espegravece Malgreacute tout certains
gegravenes portent toujours la mention laquo de fonction inconnue raquo
La meacutethodologie que nous avons employeacutee nous a permis drsquoidentifier plusieurs
gegravenes possiblement impliqueacutes dans reacutesistance de nos souches aux antibiotiques
testeacutes
1- chez Staphylococcus aureus ATCC 29213 reacutesistant agrave la gentamicine (+TO)
(Figure 9A)
- NWMN_0811 codant possiblement pour la NADH deacuteshydrogeacutenase
- qoxC (NWMN_0928) codant pour la quinol oxidase polypeptide III
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
2- chez NewbouldΔhemB reacutesistant agrave la tomatidine (Figure 9B)
- ccpA (NWMN_1629) codant pour la laquo catabolite control protein A raquo
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
3- chez NewbouldΔhemB reacutesistant agrave la tomatidine (+gentamicine) (Figure 9C)
- NWMN_0143 codant pour un ABC transporteur
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
Lrsquoanalyse de ces diffeacuterents gegravenes nous amegravene agrave suspecter la sous-uniteacute c de lrsquoATP
synthase comme eacutetant la cible potentielle de la tomatidine chez Staphylococcus
aureus
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine
Les gegravenes muteacutes pointent en grande partie vers la chaine respiratoire qursquoon appelle
aussi chaine de transport des eacutelectrons
100
La NADH deacuteshydrogeacutenase (NWMN_0811) fait partie du complexes I de la chaine de
transport de eacutelectrons alors que la quinol oxidase polypeptide III (NWMN_0928 ou
QoxC) est une composante du complex III et la sous-uniteacute c de lrsquoATP synthase (AtpE)
fait partie eacutevidement du complexe de lrsquoATP synthase ou complexe V de la chaine
respiratoire (Figure 10)
Figure 10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants
Les courbes de croissance des mutants ATCC29213 reacutesistants agrave la combinaison
GEN(+TO) appuient le fait que ces mutations affectent la chaine de transport des
eacutelectrons ce qui affecte le niveau de production drsquoATP et donc la croissance
bacteacuterienne En effet les mutants du troisiegraveme niveau de reacutesistance (qui possegravede
deux gegravenes de la chaine de transport des eacutelectrons de muteacutes) ont une croissance
plus lente que les mutants du premier niveau de reacutesistance qui eux ont un seul gegravene
dans la chaine de transport des eacutelectrons de muteacute (Figure 11)
Quinoloxidase
NADHdehydrogenase
ATPsynthetase
Proton gradient
XXX
101
Figure 11 Courbes de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison GEN+TO du premier niveau de reacutesistance (bleu) et du troisiegraveme niveau
de reacutesistance (vert)
Voici plus preacuteciseacutement comment fonctionne la chaine de transport des eacutelectrons chez
les 2 pheacutenotypes-parents selon le modegravele-hypothegravese eacutetabli avec nos reacutesultats et
analyses
Tout drsquoabord chez le prototype ou souche de pheacutenotype normal (ex ATCC 29213) la
chaine de transport des eacutelectrons fonctionne normalement Les complexes de la
chaine de transport des eacutelectrons sont chargeacutes de transfeacuterer des protons vers
lrsquoexteacuterieur de la cellule en transfeacuterant en mecircme temps des eacutelectrons drsquoun complexe agrave
un autre jusqursquoagrave un accepteur final qui est lrsquoO2 lorsque crsquoest possible Se faisant il se
creacutee un gradient eacutelectrochimique de part et drsquoautre de la membrane et les protons
accumuleacutes du cocircteacute externe sont utiliseacutes par la suite par lrsquoATP synthase afin de
pouvoir produire de lrsquoATP dans la cellule
0 4 8 1 2 1 6 2 0 2 4
0 0 1
0 1
1
C o u r b e d e c r o is s a n c e d e s m u ta n ts A T C C 2 9 2 1 3
reacute s is ta n ts agrave G E N + T O (8 m g L )
te m p s (h )
OD
60
0n
m
Niveau 3
Niveau 1
102
Le transfert drsquoeacutelectrons ne se fait pas sans coucirct et des produits toxiques pour la
cellule sont produits en mecircme temps ce sont les espegraveces reacuteactives de lrsquooxygegravene ou
ROS (Reactive Oxygen Species) Cependant les prototypes par leur reacuteponse au
stress produisent des enzymes capables de deacutetoxifier le milieu (Figure 12A)
Chez les souches SCV ici la souche controcircle NewbouldΔhemB la chaine de
transport des eacutelectrons est deacuteficiente parce que le gegravene permettant la production
drsquoheacutemine qui entre dans la composition des cytochromes est supprimeacute Les
cytochromes entrent dans la composition de plusieurs composantes de la chaine
respiratoire
Lorsque les complexes de la chaine sont alteacutereacutes il y a diminution du transfert de
protons vers lrsquoexteacuterieur une diminution de la production drsquoATP et une plus grande
production de ROS (Figure 12B)
Figure 12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B) NewbouldΔhemB
I III IV V I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
eacute eacute
eacute
H+ H+
ATPO2 H2O
ROS +ROS
Prototype SCV (hemB)
H+
0811 0928 2012 0811 0928
hegraveme
2012
A B
103
La gentamicine a besoin drsquoune bonne polarisation membranaire donc drsquoune chaine de
transport des eacutelectrons fonctionnelle pour pouvoir peacuteneacutetrer dans la cellule (Bryan et
Kwan 1983 (Bryan and van den Elzen 1977 Davis 1987 Taber et al 1987) Une
fois entreacutee elle inhibe la synthegravese proteacuteique ce qui provoque la mort cellulaire La
CMI de la gentamicine contre ATCC 29213 est de 05-1 microgml (Figure 13A)
La tomatidine elle irait se lier agrave lrsquoATP synthase pour en diminuer la fonctionnaliteacute et
ce tant chez ATCC 29213 que chez hemB Cette liaison amegravenerait une diminution
de la production drsquoATP et conseacutequemment provoquerait agrave plus ou moins long terme
une augmentation de production de ROS par lrsquoalteacuteration de la chaine de transport des
eacutelectrons Cependant comme la cellule prototype produit efficacement les enzymes
de deacutetoxification lrsquoimpact de la tomatidine est pratiquement nulle sur les souches
prototypes La CMI de la tomatidine est gt16microml contre ATCC 29213 (Figure 13B)
104
Figure 13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213
Chez ATCC 29213 lrsquoajout de 8microgml de tomatidine agrave la Gentamicine amegravene une
diminution de 8X agrave 16X la CMI de la gentamicine En effet la CMI de la gentamicine
passe de 05-1microgml agrave 006microgml (Figure 14AB)
Lors de lrsquoajout de la tomatidine agrave la gentamicine crsquoest la tomatidine qui se lie dans
un premier temps agrave lrsquoATP synthase reacuteduisant ainsi sont niveau drsquoactiviteacute Il y a alors
de faccedilon transitoire une augmentation de la concentration de protons agrave lrsquoexteacuterieur de
la cellule ce qui permet une entreacutee massive de gentamicine ainsi qursquoune rapide et
forte inhibition de la synthegravese proteacuteique Dans un second temps il y aura eacutegalement
une alteacuteration de la chaine de transport des eacutelectrons conduisant agrave une
augmentation de la production de ROS Crsquoest cependant lrsquoentreacutee massive de
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI TO gt16 microgmlCMI GEN 05-1 microgml
+ROS
ROS mais deacutetoxification efficace
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
105
gentamicine entrainant une forte inhibition de la synthegravese proteacuteique qui provoquera
dans ce cas-ci la mort cellulaire (Figure 14B)
Figure 14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicine lorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC 29213
Chez les mutants ATCC29213 reacutesistants agrave la combinaison GEN+TO les complexes I
et III sont muteacutes et donc ont une fonctionnaliteacute alteacutereacutee ce qui amegravene une
deacutepolarisation membranaire Une membrane moins polariseacutee amegravene une plus faible
entreacutee de gentamicine et donc une plus faible inhibition proteacuteique La CMI de la
gentamicine passe en effet de 05-1 microgml chez la souche parente agrave 2 microgml chez les
mutants reacutesistants
La tomatidine en se liant agrave lrsquoATP synthase accentuera la deacutepolarisation
membranaire provoquant une augmentation de la production des ROS et une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN (TO 8 microgml) 006 microgmlCMI GEN 05-1 microgml
+ROS
polarisation entreacutee massive de GEN
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
GENH+
BA
106
diminution de la production drsquoATP De fait les mutants reacutesistants ont une croissance
plus lente (Figure 15)(Figure 11)
Figure 15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants ATCC 29213
reacutesistants agrave la combinaison gentamicine (+TO)
Chez les SCV la tomatidine se lie aussi agrave lrsquoATP synthase et ce faisant amegravene une
diminution de la production drsquoATP Cette diminution de la production drsquoATP megravenera
ultimement agrave une alteacuteration de la chaine de transport des eacutelectrons et une
augmentation des ROS (Murphy 2009) tout comme chez les souches prototypes
Cependant comme le SCV a deacutejagrave une chaine de transport des eacutelectrons deacuteficiente
une faible production drsquoATP par lrsquoATP synthase et une forte production de ROS
lrsquoajout drsquoune faible concentration de tomatidine (la CMI est de 006microgml) amegravene une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype R GEN (+TO [8 microgml])
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN 2 microgmlCMI GEN 05-1 microgml
+ROS
R agrave GEN (+TO) mais mutants ont une
croissance diminueacutee (+ROS et -ATP)
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
107
augmentation de chacun de ces pheacutenomegravenes ce qui est suffisant pour provoquer la
mort cellulaire (Figure 16A)
Dans les premiers niveaux de reacutesistance agrave la tomatidine chez les mutants hemB la
reacutesistance (la CMI passe de 006 agrave 025-1microgml) est ameneacutee par la mutation du gegravene
ccpA (Catabolite controcircle protein A) ce qui megravenerait agrave une augmentation de la
production drsquoATP par lrsquoutilisation drsquoune autre voie que lrsquoATP synthase etou agrave une
diminution de la production de ROS (Figure 16B) Le rocircle de la CcpA en lien avec la
reacutesistance agrave la tomatidine est encore neacutebuleux Toutefois on sait que normalement
ce gegravene agit comme reacutegulateur transcriptionnel des gegravenes codant pour des enzymes
du cycle de Krebs et qursquoil a un rocircle de reacutegulation dans le meacutetabolisme du carbone et
lrsquoutilisation des substrats (Somerville et Proctor 2009 Goumltz et al 2006)
Figure 16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO 006 microgml
H+
SCV
I III IVeacute eacute
eacuteO2 H2O
+ROS
H+TO
Niveau 1
CcpA (CMI TO 025-1 microgml)
ROS
ATP
V
H+
ATP
TO0811 0928
2012
1629
0811 0928
2012
BA
108
Chez les SCV du deuxiegraveme palier de reacutesistance agrave la tomatidine non seulement le
gegravene ccpA est toujours muteacute mais le gegravene de la sous-uniteacute c de lrsquoATP synthase est
eacutegalement muteacute La mutation dans le site de liaison mecircme de la tomatidine empecircche
sa liaison et donc empecircche son effet sa CMI passant de 006 agrave gt16microgml (Figure
17B)
Malgreacute la mutation dans le gegravene lrsquoATP synthase resterait quand mecircme fonctionnel
puisque la mutation nrsquoamegravene pas la mort cellulaire comme crsquoest le cas lorsque la
tomatidine lie lrsquoATP synthase chez le SCV parent (Figure 17A)
Figure 17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la tomatidine
Tout comme chez les mutants hemB reacutesistant agrave la tomatidine seule les mutants
hemB reacutesistant agrave la tomatidine lorsqursquoen preacutesence de gentamicine possegravedent
eacutegalement une mutation dans le gegravene de lrsquoATP synthase ce qui rend la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO006 microgml
H+
SCV Niveau 2CMI TO gt16 microgml
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+TO
TO
Niveaux 1 et 2
CcpA (CMI TO 025-1 microgml)
ROS
ATP
0811 0928
2012
1629
0811 0928
2012
A B
109
inactive puisqursquoincapable de srsquoy lier (CMI gt16microgml) (Figure 18B) Chez les SCV non
muteacutes (hemB) la gentamicine nrsquoentre que faiblement ducirc agrave la deacutepolarisation de sa
membrane (CMI 4-8 microgml) (Figure 18A) La mutation dans le gegravene codant pour une
pompe (NWMN_0143 codant pour un ABC transporteur) chez les mutants hemB
reacutesistant agrave la tomatidine (+gentamicine) amegravenerait une augmentation de lrsquoactiviteacute de
cette pompe permettant de rejeter hors de la cellule le peu de gentamicine eacutetant
entreacutee La mutation de cette pompe fait passer la CMI de la gentamicine de 4-8
microgml pour hemB agrave 16 microgml pour les mutants hemB reacutesistant agrave la combinaison
gentamicine (+TO) ayant cette mutation (Figure 18B)
Figure 18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB reacutesistants
agrave la combinaison tomatidine (+gentamicine)
Donc selon notre modegravele-hypothegravese lrsquoaction de la tomatidine et de la gentamicine
sur S aureus deacutepend de la liaison agrave leur cible mais a aussi un lien avec la
SCV R agrave TO (+GEN [4microgml])
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI GEN 4-8 microgml
H+
SCV
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+ TO
GEN
CMI GEN 16 microgml
CMI TO gt16 microgml
0811 0928
2012
0811 0928 20120143
GEN
Synthegravese proteacuteique
BA
110
polarisation membranaire et la production de ROS La mesure de la polarisation et
concentration cellulaire en ROS chez nos mutants et nos souches parents en
preacutesence ou non drsquoantibiotique nous permettra de valider notre modegravele-hypothegravese
Chez une bacteacuterie meacutetaboliquement active et posseacutedant une membrane
cytoplasmique intacte il y a normalement une diffeacuterence de potentiel eacutelectrique entre
le cocircteacute inteacuterieur et le cocircteacute exteacuterieur de la membrane lrsquointeacuterieur eacutetant plus neacutegatif de
100 agrave 200 mV par rapport agrave lrsquoexteacuterieur Une diminution de la magnitude de ce
potentiel membranaire est appeleacutee deacutepolarisation alors qursquoune augmentation de ce
potentiel est appeleacutee hyperpolarisation Le potentiel membranaire sera reacuteduit agrave zeacutero
srsquoil y a rupture de la membrane cest-agrave-dire srsquoil y a formation de trous assez grands
permettant aux ions inorganiques de passer librement Ce pheacutenomegravene peut arriver
lorsque les cellules sont tueacutees par la chaleur le froid ou les antibiotiques β-Lactams
Dans ces circonstances la membrane devient permeacuteable agrave diffeacuterents colorants
comme le propidium iodide (PI) et le TO-PRO3 et le changement est irreacuteversible
Plusieurs classes de composeacutes chimiques notamment les ionophores peuvent
eacutegalement alteacuterer le potentiel membranaire sans pour autant permettre la
permeacuteabiliteacute au PI TO-PRO3 et autres colorants similaires chez les bacteacuteries
lrsquoadministration drsquoun proton-ionophore tel le carbonyl cyanide 3-
chlorophenylhydrazone (CCCP) reacuteduira le potentiel membranaire agrave zeacutero
Des tests preacuteliminaires sur la mesure du potentiel membranaire (BacLighttrade Bacterial
Membrane Potential Kit (B34950) drsquoInvitrogen et lecture au cytomegravetre de flux (FACS))
(Novo et al 2000) chez nos deux souches parentes mets en eacutevidence que ATCC
29213 est plus polariseacute que NewbouldΔhemB (Figure 19A) Toutefois malgreacute
qursquoayant une plus faible polarisation membranaire NewbouldΔhemB nrsquoest pas
complegravetement deacutepolariseacute puisqursquoil est possible de le deacutepolariser davantage agrave lrsquoaide
drsquoun agent deacutepolarisant tel que le CCCP (carbonyl cyanide 3-chlorophenylhydrazone)
(Figure 19B) Le TO-PRO3 a eacuteteacute utiliseacute comme controcircle afin de confirmer la viabiliteacute
de nos cellules (donneacutees non montreacutees)
111
A B
Figure 19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane Potential Kit
(Invitrogen) (Lecture au cytomegravetre de flux agrave lrsquoaide du programme FCS Express) (A)
Ratio FL3-H (axe des Y)FL1-H (axe des X) de ATCC 29213 (rouge) et de
NewbouldΔhemB (noir) (B) Lecture des rouges (FL3-H) sur NewbouldΔhemB traiteacute
avec le DiOC2(3) (33prime-diethyloxacarbocyanine iodide) (noir) ou avec le CCCP (bleu)
ou sur les cellules chauffeacutees (rouge)
La sous-uniteacute c de lrsquoATP synthase (AtpE) semble donc la cible la plus probable de la
tomatidine Afin de veacuterifier si les mutations identifieacutees pouvaient avoir un effet
veacuteritable sur la proteacuteine nous avons fait un alignement de seacutequences noteacute les bases
muteacutees et veacuterifieacute si la mutation amenait un changement drsquoacide amineacute et si ce
changement drsquoacide amineacute pouvait avoir un impact sur la proteacuteine finale De fait
nous avons trouveacute que les deux diffeacuterentes mutations dans lrsquoAtpE amenaient un
changement significatif drsquoacides amineacutes pouvant amener une variation fonctionnelle
FL3-H
Count
100
101
102
103
104
0
18
36
53
71
Chauffeacute
+CCCP+DiOC2
Chauffeacute
FL1-H
FL3-H
100
101
102
103
104
100
101
102
103
104
hemB
29213
VERT
RO
UG
E
ROUGE
ΔhemB
112
de la proteacuteine La mutation de la base 49 change un G pour un T il srsquoagit drsquoun SNP
nrsquoamenant pas de changement de cadre de lecture La mutation fait passer lrsquoacide
amineacute alanine (A) qui est apolaire agrave une seacuterine (S) qui est polaire La mutation de la
base 190 change un G pour un A et est aussi un SNP nrsquoamenant pas de changement
de cadre de lecture Cette mutation fait passer lrsquoacide amineacute alanine (A) qui est
apolaire agrave une threacuteonine (T) qui est polaire (Figure 20)
gtlcl||NWMN_2012|atpE|72493880 F0F1 ATP synthase subunit C
1 10 19 31 43 49 58
ATG AAT TTA ATC GCA GCA GCA ATC GCA ATT GGT TTA TCA GCA TTA GGA GCA GGT ATC GGT
M N L I A A A I A I G L S A L G A G I G
(S)
17 61 73 85 97 109 118
AAC GGT TTA ATC GTT TCA AGA ACA GTT GAA GGT GTA GCA CGT CAA CCA GAA GCA CGT GGT
N G L I V S R T V E G V A R Q P E A R G
21 40
121 133 145 157 169 178
CAA TTA ATG GGT ATC ATG TTC ATT GGT GTA GGT TTA GTT GAG GCA TTA CCT ATC ATC GGT
Q L M G I M F I G V G L V E A L P I I G
41 60
181 190 199 211
GTA GTA ATT GCA TTC ATG ACA TTT GCT GGA TAA
V V I A F M T F A G
(T)
61 64 70
Figure 20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du gegravene
atpE de S aureus Newman avec mise en eacutevidence des mutations (rose) en lien avec
la cible de la tomatidine Lrsquoalignement a eacuteteacute effectueacute agrave lrsquoaide du site CMR de JCVI
113
CHAPITRE 4
DISCUSSION
LrsquoATP synthase est eacutevidemment un complexe proteacuteique ubiquitaire tant chez les
procaryotes que les eucaryotes Malgreacute ce fait il semble que la cible de la tomatidine
soit speacutecifique aux bacillales celle-ci ne deacutemontrant aucune activiteacute contre les
bacteacuteries agrave Gram neacutegatif et les lactobacillales On suppose donc que lrsquoATP synthase
de ces espegraveces doit ecirctre speacutecifique au niveau du site de liaison de la tomatidine
Lrsquoecirctre humain en tant qursquoespegravece eucaryote est eacutegalement pourvu de ce complexe
proteacuteique et on peut naturellement se demander si lrsquoutilisation clinique de cette
moleacutecule pourrait ecirctre cytotoxique De fait plusieurs composeacutes naturels tel
lrsquooligomycine et la venturicidine sont connus pour inhiber lrsquoactiviteacute de lrsquoATP synthase
par leur interaction avec la sous-uniteacute c de ce complexe Cependant ces moleacutecules
ne sont pas seacutelectives et nrsquoinhibent pas seulement lrsquoATP synthase bacteacuterien mais
aussi lrsquoATP synthase mitochondrial (Haagsma et al 2009 Matsuno-Yagi et Hatefi
1993) (Figure 1) Neacuteanmoins il existe maintenant une moleacutecule qui a pour cible
lrsquoATP synthase bacteacuterien et qui est en processus en vue drsquoune utilisation clinique
chez lrsquohomme le TMC207 (Haagsma et al 2009))
114
Figure 1 Sites drsquoinhibition de lrsquoATP synthase Les sites de liaison dans lrsquoATP
synthase des inhibiteurs reacuteveacuteleacutes par des eacutetudes biochimiques et structurales sont
indiqueacutes par des cercles rouges et les sites de liaisons qui nrsquoont pas eacuteteacute totalement
clarifieacutes sont indiqueacutes par des cercles verts (Hong et Pedersen 2008)
Le TMC207 (Figure 2A) est un composeacute de la classe des diarylquinoline posseacutedant
un nouveau meacutecanisme drsquoaction lrsquoinhibition de lrsquoATP synthase bacteacuterien De fait la
cible et le meacutecanisme drsquoaction du TMC207 sont diffeacuterents des autres agents
antituberculeux Lrsquoinhibition de lrsquoATP synthase peut mener agrave une baisse de la
production drsquoATP et un deacutebalancement dans lrsquohomeacuteostasie du pH ces 2 effets
pouvant contribuer agrave la diminution de la survie de la bacteacuterie (Rao et al 2001
Andries et al 2005)
Le TMC207 est une moleacutecule ayant une forte activiteacute bacteacutericide contre
Mycobacterium tuberculosis et autres espegraveces mycobacteacuteriennes mais peu drsquoactiviteacute
115
contres les autres espegraveces bacteacuteriennes Lrsquoindex de seacutelectiviteacute de TMC207 pour
lrsquoATP synthase mycobacteacuterienne (CMI50 gt200 microml) compareacute agrave lrsquoATP synthase
mitochondriale (CMI50 001 microgml) est de plus de 20000 Un indice de seacutelectiviteacute au
dessus de 1000 est consideacutereacute dans le deacuteveloppement drsquoune drogue comme un
preacutealable pour une moleacutecule candidate (Haagsma et al 2009) La diversiteacute dans les
gegravenes atpE explique la reacutesistance au TMC207 et justifie la faible affiniteacute de cette
drogue agrave lrsquoATP synthase mitochondriale (Matteelli et al 2010) (Figure 3)
Figure 2 Structure chimique de (A) TMC207 et (B) son analogue anti bacteacuteries
agrave Gram positifs (Balemans et al 2012)
Lorsqursquoon compare la seacutequence proteacuteique de la sous-uniteacute c de lrsquoATP synthase
(atpE) de quelques espegraveces de lrsquoordre des Bacillales (tel Saureus B cereus B
subtilis et L monocytogenes) avec des espegraveces appartenant agrave drsquoautres ordres
bacteacuteriens on constate que les seacutequences divergent ce qui est en accord avec la
speacutecificiteacute des moleacutecules Les points de mutations amenant une reacutesistance au
TMC207 chez Mycobacterium smegmatis sont speacutecifiques aux mycobacteacuteries et les
deux principaux points de mutations D32V(ouP) et A67P chez M smegmatis sont
situeacutes tous les deux dans les heacutelices alpha qui entourent le site catalytique de la
proteacuteine (E65) qui est impliqueacute dans le transport des protons (Andries et al 2005)
Lrsquoacide amineacute aspartate (D) agrave la position 32 et lrsquoalanine (A) agrave la position 67 sont tregraves
speacutecifiques agrave M smegmatis
A B
116
Reacutecemment un groupe de chercheurs (Balemans et al 2012) on mit en lumiegravere une
autre moleacutecule (Figure 2B) de la classe des diarylquinolines dont le meacutecanisme
drsquoaction eacutetait aussi lrsquoinhibition de la sous-uniteacute c de lrsquoATP synthase mais chez les
bacteacuteries agrave Gram positif tels Streptococcus pneumoniae et S aureus Cette moleacutecule
irait lier les acides amineacutes valines (V) situeacutes agrave la position 50 et 62 de lrsquoAtpE de S
aureus (Donneacutees non montreacutees chez S pneumoniae(Balemans et al 2012)) En
effet une mutation amenant le changement de la valine en position 50 par une
isoleucine (I) etou une mutation amenant le changement de la valine en position 62
par une alanine (A) amegravene la reacutesistance de la bacteacuterie agrave la moleacutecule Cependant L
monocytogenes et Bacillus subtilis ne possegravedent pas de valine agrave la position 50 alors
qursquoils font eacutegalement partie des bacteacuteries agrave Gram positif (Il faut noter que chez
Bacillus sp PS3 (Swiss-Prot assession Ndeg P 00845) possegravede une valine agrave la position
50 (Balemans et al 2012)) Par contre tout comme pour les autres bacteacuteries agrave Gram
positif testeacutees les deux espegraveces possegravedent une valine agrave la position 62 (Figure 3)
Chez B cereus B subtilis B anthracis S aureus et L monocytogenes les points de
divergence sont plutocirct A17S et A64T Cependant B pseudofirmus qui fait eacutegalement
partie de lrsquoordre des Bacillales possegravede une glycine (G) agrave la position 17 plutocirct qursquoune
alanine (A) et une seacuterine (S) plutocirct qursquoune alanine (A) agrave la position 64 comme les
espegraveces preacuteceacutedentes Suivant ces reacutesultats et si la cible de la tomatidine est bien
ces deux points de mutation la tomatidine ne devrait pas pouvoir se lier agrave lrsquoAtpE de B
pseudofirmus et devrait donc nrsquoavoir aucune activiteacute contre cette souche alors que
nous avions extrapoleacute que le spectre drsquoactiviteacute de la tomatidine eacutetait tous les
Bacillales De plus E coli et M smegmatis possegravedent eacutegalement une alanine (A) agrave la
position 17 alors que la tomatidine ne possegravede aucune activiteacute contre ces espegraveces (M
smegmatis nrsquoa pas eacuteteacute eacutevalueacute il srsquoagit ici drsquoune extrapolation des reacutesultats de CMI)
par contre ces deux espegraveces possegravedent des acides amineacutes diffeacuterents drsquoune alanine
(A) agrave la position 64 ce qui pourrait leur confeacuterer une reacutesistance agrave la tomatidine telle
qursquoobserveacutee On remarque eacutegalement que les acides amineacutes agrave ces deux positions
117
sont totalement diffeacuterents chez lrsquohomme (Homo sapiens) ce qui suggeacutererait que la
tomatidine ne possegravederait pas drsquoactiviteacute antimitochondriale (Figure 3)
Selon ces reacutesultats on peut se demander si les deux mutations sont essentielles agrave la
liaison de la moleacutecule ou si seulement lrsquoalanine (A) agrave la position 64 est neacutecessaire
On doit se rappeler que la reacutesistance apporteacutee par la mutation faisant en sorte que
lrsquoalanine (A) en position 17 devient une seacuterine (S) apparaicirct chez les mutants
NewbouldΔhemb reacutesistant agrave la tomatidine et que cette mutation nrsquoapparaicirct qursquoau
quatriegraveme et dernier niveau de reacutesistance On se rappelle eacutegalement que la mutation
faisant en sorte que lrsquoalanine (A) agrave la position 64 devient une threacuteonine (T) apparaicirct
chez les mutants NewbouldΔhemb reacutesistants agrave la tomatidine en combinaison avec la
gentamicine et ce degraves le premier niveau de reacutesistance
118
Bcereus G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Banthracis G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Lmonocytogenes G---------VIAAAIAVG---LGALGAGIGNGLIVSKTVEGVARQPEARSMLQTIMFV- 50
Saureus_NWMN_2012 -----------IAAAIAIG---LSALGAGIGNGLIVSRTVEGVARQPEARGQLMGIMFIG 49
Bsubtilis -----------IAAAIAIG---LGALGAGIGNGLIVSRTVEGIARQPEAGKELRTLMFMG 49
Bpseudofirmus -----------LGAAIAAG---LAAVAGAIAVAIIVKATIEGTTRQPELRGTLQTLMFIG 49
Ecoli L---------YMAAAVMMG---LAAIGAAIGIGILGGKFLEGAARQPDLIPLLRTQFFIV 56
Msmegmatis DPNAIITAGALIGGGLIMG---GGAIGAGIGDGIAGNALISGIARQPEAQGRLFTPFFIT 60
Homo sapiens QTSAISRDIDTAAKFIGAGAATVGVAGSGAGIGTVFGSLIIGYARNPSLKQQLFSYAILG 120
MyR V
MyR P
SaR S
Bcereus VALVEALPIIGVVIAFIVMNK----- 72
Banthracis VALVEALPIIGVVIAFIVMNK----- 72
Lmonocytogenes ----EALPIIAVVIAFMVLNK----- 67
Saureus_NWMN_2012 VGLVEALPIIGVVIAFMTFAG----- 70
Bsubtilis IALVEALPIIAVVIAFLAFFG----- 70
Bpseudofirmus VPLAEAVPIIAIVISLLILF------ 69
Ecoli MGLVDAIPMIAVGLGLYVMFAVA--- 79
Msmegmatis VGLVEAAYFINLAFMALFVFATPGLQ 86
Homo sapiens FALSEAMGLFCLMVAFLILFAM---- 142
MyR D P M
SpR I A
SaR T
Fig
ure
3
Seacuteq
uen
ce
pro
teacuteiq
ue d
e la p
roteacute
ine
Atp
Ec
hez d
iffeacute
ren
tes
es
pegravec
es e
t m
ise e
n eacute
vid
en
ce
de
s s
ite
s d
e
mu
tati
on
pe
rmett
an
t la
reacutesis
tan
ce agrave
dif
feacutere
nte
s m
oleacute
cu
les
M
yR M
tu
berc
ulo
sis
ou
M sm
eg
matis
reacutesis
tan
t a
u
TM
C2
07 S
pR
S
p
neu
mon
iae
reacutesis
tan
t agrave
un a
na
log
ue d
u T
MC
20
7
SaR
S
a
ure
us
SC
V r
eacutesis
tant agrave
la to
matid
ine
En
cad
reacutero
uge
S
ite d
e m
uta
tion a
men
ant la
reacutesis
tance
agravela
to
matid
ine E
ncad
reacuteno
ir
Site
cata
lytiq
ue
im
pliq
ueacute
da
ns le
tra
nspo
rt d
es p
roto
ns
Su
rlig
nag
etu
rqu
ois
e S
ite d
e m
uta
tion a
me
nant u
ne
reacutesis
tan
ce a
u T
MC
20
7
119
Les seacutequences proteacuteiques des souches bacteacuteriennes de la figure 3 ont eacuteteacute trouveacutees
sur pubmedproteacuteine httpwwwncbinlmnihgovpubmed
laquo ATP synthase subunit c raquo Assession number
Streptococcus pneumoniae D39 YP_8168031 (NCBI)
S aureus Newman BAF682841 (GenBank)
E coli AAA247321 (GenBank)
Bacillus cereus ISO75 EJP900901 (GenBank)
Bacillus anthracis str H9401 YP_0062123491 (NCBI)
Mycobacterium smegmatis MC2 155 YP_0065695541 (NCBI)
Bacillus subtilis str 168 CAA822551 (GenBank)
Listeria monocytogenes YP_0076049931 (NCBI)
Homo sapiens P482011 (UniProtKBSwiss-Prot)
Lrsquoalignement comparatif des seacutequences drsquoacides amineacutes a eacuteteacute fait agrave lrsquoaide du
programme ClustalW2 sur le web
httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2
Le modegravele 3D de lrsquoAtpE a eacuteteacute obtenu gracircce agrave lrsquoaide du Pr Ryszard Brzezinski du
Deacutepartement de biologie de lrsquoUniversiteacute de Sherbrooke (Figure 4) Une recherche
dans le Protein Data Bank a drsquoabord eacuteteacute effectueacutee afin de veacuterifier parmi les proteacuteines
AtpE (sous-uniteacute C de lrsquoATP synthase) pour lesquelles il y avait des structures 3D
reacutesolues laquelle ressemblait le plus agrave celle de Staphylococcus aureus Les
alignements ont montreacute que cest la proteacuteine de Bacillus pseudofirmus qui avait la
plus grande similitude de seacutequence La structure est stockeacutee dans la banque PDB
sous le symbole 3ZO6 Il a eacuteteacute confirmeacute par quelques alignements que cette sous-
uniteacute C de lrsquoATP synthase eacutetait plus proche par sa seacutequence de celle de
Staphylococcus aureus que de celle de Mycobacterium pour laquelle la structure 3D
avait eacuteteacute deacutetermineacutee plus tocirct Ce reacutesultat sexplique par le fait que Staphylococcus et
Bacillus font tous deux partie de la branche A+T-riche des bacteacuteries agrave Gram+ alors
que Mycobacterium appartient agrave la branche G+C-riche La seacutequence de lAtpE de
120
Staphylococcus aureus a ensuite eacuteteacute envoyeacutee au serveur internet de modeacutelisation
3D-Jigsaw (httpbmmcancerresearchukorg~3djigsaw) en indiquant que le modegravele
devait ecirctre bacircti en prenant comme matrice le fichier PDB 3ZO6 Le modegravele nous a
eacuteteacute retourneacute sous format PDB ndash comme les laquo vrais raquo cristaux dans la Protein Data
Bank Les fichiers au format PDB peuvent ecirctre visualiseacutes par le logiciel RasMol ndash
gratuit sur internet Cest avec RasMol que limage (Figure 4) a eacuteteacute obtenue
121
Figure 4 Modeacutelisation de la structure tridimensionelle de la sous-uniteacute c de
lrsquoATP synthase de S aureus (AtpE) montrant les acides amineacutes muteacutes chez les
mutants reacutesistantsEn blanc et vert les deux acides amineacutes muteacutes chez S aureus
reacutesistant agrave la tomatidine En rose les sites de mutations chez Mycobacterium
smegmatis reacutesistant au TMC207 et des sites de mutations chez Streptoccocus
pneumoniae reacutesistants agrave un analogue de TMC207 En bleu le site actif de la proteacuteine
Agrave la vue de ces reacutesultats certains pourront mettre en doute lrsquointeacuterecirct de la tomatidine
eacutetant donneacute lrsquoapparition relativement rapide de reacutesistance chez S aureus Cependant
122
nos reacutesultats deacutemontrent clairement que lrsquoajout de tomatidine ralentit le
deacuteveloppement de reacutesistance agrave la gentamicine chez S aureus ATCC29213 (Tableau
1) De plus cette augmentation de la CMI de la gentamicine en preacutesence de
tomatidine au fil des passages suit lrsquoaugmentation de la CMI de la ciprofloxacine lors
de ces mecircmes passages alors que la ciprofloxacine est consideacutereacutee comme la drogue
de reacutefeacuterences dans les bonnes drogues (Tableau 1) Tout comme pour la
ciprofloxacine pour laquelle la reacutesistance chez S aureus peut ecirctre la conseacutequence
de deux mutations dans le gegravene cible gyrA la reacutesistance agrave la tomatidine peut ecirctre la
conseacutequence de deux mutations dans le gegravene cible atpE Par comparaison la
rifampicine est consideacutereacutee comme une mauvaise drogue de par lrsquoapparition rapide
drsquoune forte reacutesistance chez S aureus conseacutequence de ses 17 mutations possibles
dans le gegravene cible rpoB (Tableau 1)
De plus il sera important de relativiser lrsquoapparition de reacutesistance agrave la tomatidine en
calculant le taux drsquoapparition de ces colonies reacutesistantes ceci nrsquoayant pas eacuteteacute fait
jusqursquoici De fait la freacutequence de mutation chez S aureus amenant une reacutesistance agrave
la rifampicine est de lrsquoordre de 10-7 10-8 alors qursquoil est de 10-11 pour lrsquoacide fusidique
(Canfield et al 2013 Orsquoneill et al 2001 Price et Gustafson 2001) Pour ce qui est
de la nouvelle moleacutecule anti-bacteacuteries agrave Gram positif de la classe des diarylquinoline
une freacutequence de mutation sur S pneumoniae ATCC49619 de 81- 83 x 10-7 a eacuteteacute
observeacutee agrave 5x la CMI et une freacutequence de mutation de 46 x 10-7 agrave 17 x 10-6 agrave 50x la
CMI (Balemans et al 2012)
123
Tableau 1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 par la meacutethode de CMI lors de passages successifs dans un milieu avec
lrsquoantibiotique
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
Typiqueldquomauvaiserdquodrogue (17 mutations possibles dansrpoB)
Typique ldquobonnerdquo drogue (2 mutations possibles dans gyrA)
Preacutesence de TO ralenti le deacuteveloppement de reacutesistances vs GEN seul
124
CHAPITRE 5
CONCLUSION ET PERSPECTIVES
Mon projet de maitrise a permis de preacuteciser le spectre drsquoactiviteacute de la tomatidine
Nous savons maintenant que la tomatidine possegravede une activiteacute antibacteacuterienne
contre la plupart des Firmicutes et plus speacutecifiquement contre les Bacillales Plus
preacuteciseacutement la tomatidine est bacteacutericide contre les pheacutenotypes SCV et agit en
synergie avec les aminoglycosides augmentant lrsquoactiviteacute bacteacutericide de ces derniers
de 8-16 fois contre les souches agrave pheacutenotype normal
De plus parmi tous les analogues de la tomatidine produits par le laboratoire du Pr
Eacuteric Marsault (voir article en annexe 2) le composeacute FC04-100 a eacuteteacute plus
speacutecifiquement mis en lumiegravere Celui-ci conserve non seulement une aussi bonne
activiteacute synergique avec les aminoglicosides contre les Saureus au pheacutenotype
normal et une activiteacute modeacutereacutee contre les S aureus SCV mais montre eacutegalement de
faccedilon inespeacutereacutee une activiteacute par lui-mecircme contre les S aureus de pheacutenotype normal
incluant plusieurs MRSA une proprieacuteteacute qui nrsquoeacutetait pas observeacutee pour la tomatidine
Une autre activiteacute inteacuteressante de FC04-100 est sa capaciteacute drsquoameacuteliorer lrsquoactiviteacute
bacteacutericide des aminoglicosides contre L monocytogenes agrave pheacutenotype normal apregraves
un traitement de 24h alors que la tomatidine ne deacutemontrait qursquoune faible synergie
avec les aminoglycosides contre cette souche De plus FC04-100 deacutemontre une
activiteacute bacteacutericide contre les SCV de L monocytogenes alors que la tomatidine ne
deacutemontrait qursquoune activiteacute bacteacuteriostatique FC04-100 montre eacutegalement un effet
potentiateur de lrsquoUbrolexinTM (Ceacutefalexin Kanamycin (3 2)) une combinaison
drsquoantibiotique utiliseacute dans le traitement de mammites bovines en abaissant de 8 agrave 16
fois leur CMI amenant celles-ci sous le seuil de reacutesistance Finalement FC04-100
possegravede une meilleure solubiliteacute que la tomatidine une proprieacuteteacute qui facilitera son
utilisation in vitro
125
Nos reacutesultats deacutemontrent que FC04-100 conserve une certaine activiteacute contre les
SCV reacutesistant agrave la tomatidine ce qui suggegravere possiblement que FC04-100 a plus
daffiniteacute pour la cible ou plus de sites dinteraction avec AtpE En effet les mutants
SCV reacutesistants agrave la tomatidine deviennent gt512 fois plus reacutesistants alors que FC04-
100 ne perd que 8 agrave 16 fois son activiteacute contre ces mecircmes souches Ceci deacutemontre
quil est possible dameacuteliorer significativement lactiviteacute de ces alkaloiumldes steacuteroidiens
ce qui est tregraves prometteur pour lavenir de cette moleacutecule
Dans un second temps lrsquoanalyse des mutations chez les mutants reacutesistants agrave la
tomatidine a permis de non seulement de mieux comprendre le mode drsquoaction de la
tomatidine mais eacutegalement de preacuteciser la cible potentielle de ce nouvel antibiotique
De fait son mode drsquoaction serait en lien direct avec la fonctionnaliteacute de la chaine de
transport des eacutelectrons chez la bacteacuterie lrsquoactiviteacute de la tomatidine serait inversement
proportionnelle agrave la fonctionnaliteacute de la chaine de transport des eacutelectrons et donc en
lien eacutegalement avec la polarisation membranaire et la production de ROS
intracellulaire Afin de confirmer le modegravele-hypothegravese la mesure de la polarisation
membranaire et la concentration de ROS intracellulaire chez les mutants ainsi que
chez les souches parentes suite agrave un traitement antibiotique devront ecirctre effectueacutees
Les reacutesultats de mes recherches suggegraverent que la cible bacteacuterienne speacutecifique de la
tomatidine serait la sous-uniteacute C de lrsquoATP synthase Afin de confirmer la cible
Maxime Lamontagne-Boulet tente au moment drsquoeacutecrire ces lignes drsquoobtenir des
mutants de S aureus Newman reacutesistants au composeacute FC04-100 et dont la reacutesistance
serait attribuable agrave une mutation dans le gegravene atpE Par la suite on devra confirmer
la fonctionnaliteacute de la cible ATP synthase par quantification de lrsquoATP produit agrave lrsquoaide
de la lucifeacuterase tel que lrsquoon fait Balemans et al en 2012 Lrsquoattachegravement agrave la cible
devra eacutegalement ecirctre confirmeacute et la meacutethode de SPR (Surface Plasmon Resonance)
pourrait ecirctre tout indiqueacutee pour ce faire (Balemans et al 2012) On devra eacutegalement
126
deacutemontrer la seacutelectiviteacute des moleacutecules tomatidine et FC04-100 pour lrsquoATP synthase
des Bacillales
Les infections agrave bacteacuteries multireacutesistantes deviennent de plus en plus preacutevalentes et
sont un des problegravemes de santeacute majeurs auxquels nous devons faire face aujourdrsquohui
Cette augmentation de la reacutesistance limite le reacutepertoire drsquoantibiotiques efficace creacuteant
une situation probleacutematique laquelle est exacerbeacutee par le trop petit nombre de
nouveaux antibiotiques introduits sur le marcheacute ces derniegraveres anneacutees Il est donc
drsquoune grande importance de travailler agrave la mise en marcheacute de nouveaux antibiotiques
et plus particuliegraverement de nouvelles classes drsquoantibiotiques espeacuterant ainsi deacutejouer la
multireacutesistance bacteacuterienne du moins pour un temps La tomatidine de par sa
structure sa cible et son mode drsquoaction speacutecifique est une moleacutecule antibiotique
dans une classe agrave part et de ce fait une moleacutecule plus qursquointeacuteressante pour le
traitement drsquoinfection agrave S aureus De plus comme elle deacutemontre une activiteacute
bacteacutericide contre les deux pheacutenotypes de S aureus (et autres Bacillales) lesquels
sont retrouveacutes simultaneacutement dans les cas drsquoinfections chroniques son pouvoir
antibiotique devient inestimables drsquoun point de vue clinique
127
CONTRIBUTIONS SUPPLEacuteMENTAIRES
ANNEXE 1 Application de brevet Internationnal publication number WO
2012109752 A1
INVENTEURS Franccedilois Malouin2 Gabriel Mitchell2 Kamal Bouarab2 Eacuteric Marsault1
Feacutelix Chagnon1 Simon Boulanger 2 Isabelle Guay2
1Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
2Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
128
129
ANNEXE 2 Article scientifique dans laquo Europeen Journal of medicinal chemistry raquo
2014 80 605-620
TITRE Unraveling the structure-activity relationship of tomatidine a steroid alkaloid
with unique antibiotic properties against persistent forms of
Staphylococcus aureus
ABSTRACT Staphylococcus aureus is responsible for difficult-to-treat and relapsing
infections and constitutes one of the most problematic pathogens due to its multiple
resistances to clinically available antibiotics Additionally this pathogenrsquos ability to
develop small-colony variants is associated with a reduced susceptibility to
aminoglycoside antibiotics and in vivo persistence We have recently demonstrated
that tomatidine a steroid alkaloid isolated from tomato plants possesses anti-
virulence activity against normal strains of S aureus as well as the ability to
potentiate the effect of aminoglycosides against this pathogen In addition tomatidine
has shown antibiotic activity against small-colony variants We herein report the first
study of the structure-activity relationship of tomatidine against S aureus
AUTEURS Feacutelix Chagnon1 Isabelle Guay2 Marc-Andreacute Bonin1 Gabriel Mitchell2
Kamal Bouarab2Franccedilois Malouin2 Eacuteric Marsault1
MA CONTRIBUTION Responsable des tests biologiques de tous les composeacutes
autant envers lrsquoeffet potentiateur de la tomatidine sur lrsquoactiviteacute de la gentamicine
contre la souche ATCC29213 de S aureus que pour lrsquoactiviteacute bacteacuteriostatique sur la
souche NewbouldΔhem imitant le pheacutenotype SCV Ces travaux furent effectueacutes sous
la tutelle du professeur Franccedilois Malouin
130
1Institut de Pharmacologie de Sherbrooke Deacutepartement de pharmacologie
Universiteacute de Sherbrooke 3001 12e av nord Sherbrooke Queacutebec Canada J1H
5N4
2Centre drsquoeacutetude et de valorisation de la diversiteacute microbienne (CEVDM) Deacutepartement
de biologie Universiteacute de Sherbrooke Sherbrooke QC Canada J1K 2R1
131
BIBLIOGRAPHIE Alexander E H and Hudson MC (2001) Factors influencing the internal- ization of Staphylococcus aureus and impacts on the course of infections in humans Appl Microbiol Biotechnol 56 361ndash366
Andries K Verhasselt P Guillemont J Goumlhlmann HW Neefs JM Winkler H Van Gestel J Timmerman P Zhu M Lee E et al (2005) A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis Science 307 223-227
Artzatbanov VY and Petrov VV (1990) Branched respiratory chain in aerobically grown Staphylococcus aureus--oxidation of ethanol by cells and protoplasts Arch Microbiol 153 580- 584 Archer GL (1998) Staphylococcus aureus a well-armed pathogen Clin Infect Dis 26 1179ndash1181 Assefa S Keane TM Otto TD Newbold C and Berriman M (2009) ABACAS algorithm-based automatic contiguation of assembled sequences Bioinformatics 25 1968-1969 Aubry-Damon H Legrand P Brun-Buisson C Astier A Soussy CJ and Leclercq R (1997) Reemergence of gentamicin-sus- ceptible strains of methicillin-resistant Staphylococcus aureus roles of an infection control program and changes in aminoglycoside use Clin Infect Dis 25 647-653
Balemans W Vranckx L Lounis N Pop O Guillemont J Vergauwen K Mol S Gilissen R Motte M Lanccedilois et al (2012) Novel antibiotics targeting respiratory ATP synthesis in Gram-positive pathogenic bacteria Antimicrob Agents Chemother 56 4131-4139
Batandier C Guigas B Detaille D El-Mir MY Fontaine E Rigoulet M and Leverve XM (2006) The ROS production induced by a reverse-electron flux at respiratory-chain complex 1 is hampered by metformin J Bioenerg Biomembr 38 33ndash 42
132
Beckman Coulter website httpswwwbeckmancoultercomwsrportalwsrresearch-and-discoveryproducts-and-servicesnucleic-acid-sample-preparationspri-laboratory-automationindexhtm (consulteacute le 3 Deacutec 2013) Bednarek P and Osbourn A (2009) Plant-microbe interactions chemical diversity in plant defense Science 324 746ndash748 Besier S Ludwig A Ohlsen K Brade V and Wichelhaus TA (2007) Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus Int J Med Microbiol 297 217-225 BiteSizeBio Brain food for biologists httpnxseqbitesizebiocomarticlessequencing-by-synthesis-explaining-the-illumina-sequencing-technology (consulteacute le 3 deacutec 2013) Boetzer M and Pirovano W (2012) Toward almost closed genomes with GapFiller Genome Biology 13 R56 Bouarab K Ordi E Gattuso M Moisan H and Malouin F (2007) Plant stress response agents affect Staphylococcus aureus virulence genes Abstr 47th Intersci Conf Antimicrob Agents Chemother abstr C1-1483 Bryan LE and Kwan S (1981) Aminoglycoside-resistant mutants of Pseudomonas aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob Agents Chemother 19 958- 964 Bryan LE and Kwan S (1983) Roles of ribosomal binding membrane potential and electron transport in bacterial uptake of streptomycin and gentamicin Antimicrob Agents Chemother 23 835ndash845
Bryan LE Nicas T Holloway BW and Crowther C (1980) Aminoglycoside-resistant mutation of Pseudomonas aeruginosa defective in cytochrome C552 and nitrate reductase Antimicrob Agents Chemother 17 71- 79
133
Bryan LE and van den Elzen HM (1977) Effects of membrane-energy mutations and cations on streptomycin and gentamicin accumulation by bacteria a model for entry of streptomycin and gentamicin in susceptible and resistant bacteria Antimicrob Agents Chemother 12 163-177 Brynildsen MP Winkler JA Spina CS MacDonald IC and Collins JJ (2013) Potentiating antibacterial activity by predictably enhancing endogenous microbial ROS production Nat Biotechnol 31 160ndash 165 Burke KA and Lasccelles J (1975) Nitrate reductase system in Staphylococcus aureus wild type and mutants J Bacteriol 123 308- 316 Burke KA and Lascelles J (1976) Nitrate reductase activity in hegraveme-deficient mutants of Staphylococcus aureus J Bacteriol 126 225- 231 Canfield GS Schwingel JM Foley MH Vore KL Boonanantanasarn K Gill AL Sutton MD and Gill SR (2013) Evolution in fast forward a potential role for mutators in accelerating Staphylococcus aureus pathoadaptation J Bacteriol 195 615-628 Chang S Sievert DM Hageman JC Boulton ML Tenover FC Downes FP Shah S Rudrik JT Pupp GR Brown WJ et al (2003) Infection with vancomycin- resistant Staphylococcus aureus containing the vanA resistance gene N Engl J Med 348 1342-1347 Chen Q Vazquez EJ Moghaddas S Hoppel CL and Lesnefsky EJ (2003) Production of reactive oxygen species by mitochondria central role of complex III J Biol Chem 278 36027-36031 Clements MO Watson SP Poole RK and Foster SJ (1999) CtaA of Staphylococcus aureus is required for starvation survival recovery and cytochrome biosynthesis J Bacteriol 181 501- 507
134
Clinical and Laboratory Standards Institute (2011) Performance standards for antimicrobial susceptibility testing Twenty-first informational supplement M100-S21 CLSI Wayne PA USA ClustalW2 httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2 (consulteacute le 3 deacutec 2013) Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) httpcmrjcviorgtigr-scriptsCMRCmrHomePagecgi (consulteacute le 3 deacutec 2013) Creacutepin-Gilbert C (2006) Transfert de ligand dans la cytochrome c oxydase observeacute par des expeacuteriences femtosecondes infrarouge inteacutegreacutees et reacutesolues spectralement Thegravese de doctorat de l rsquoEacutecole Polytechnique Paris Pastel-00002200 version 1 29 jul 2010 Davis BD (1987) Mechanism of bactericidal action of aminoglycosides Microbiol Rev 51 341-350 DeAngelis MM Wang DG and Hawkins TL (1995) Solid-phase reversible immobilization for the isolation of PCR products Nucl Acids Res 22 4742- 4743 Dickgiesser N and Kreiswirth BN (1986) Determination of aminoglycoside resistance in Staphylococcus aureus by DNA hybridization Antimicrob Agents Chemother 29 930-932 Doi Y and Arakawa Y (2007) 16S ribosomal RNA methylation Emerging resistance mechanism against aminoglycosides Clin Infect Dis 45 88ndash94 Enright MC Robinson DA Randle G Feil EJ Grundmann H and Spratt BG (2002) The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA) Proc Natl Acad Sci USA 99 7687-7692 Friedman M (2002) Tomato glycoalkaloids role in the plant and in the diet J Agric Food Chem 50 5751ndash5780
135
Goerke C and Wolz C (2004) Regulatory and genomic plasticity of Staph- ylococcus aureus during persistent colonization and infection Int J Med Microbiol 294 195ndash202 Goumltz F Bannerman T and Schleifer K-H (2006) The genera Staphylococcus and Macrococcus In The Prokaryotes Vol 4 Bacteria Firmicutes Cyanobacteria M Dworkin (ed) (Springer New-York USA) pp5- 75 Haagsma AC Abdillahi-Ibrahim R Wagner MJ Krab K Vergauwen K Guillemont J AndriesK Lill H Koul A and Bald D (2009) Selectivity of TMC207 towards mycobacterial ATP synthase compared with that towards the eukaryotic homologue Antimicrob Agents Chemother 53 1290-1292 Hammer ND Reniere ML Cassat JE Zhang Y Hirsch AO Indriati Hood M and Skaar EP (2013) Two hegraveme-dependent terminal oxydases power Staphylococcus aureus organ-specific colonization of the vertebrate host MBio 4 e00241- 13 Hederstedt L (1993) The Krebs citric acid cycle In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of Microbiology Washington DC) pp 181-198 Hiramatsu K (1995) Molecular evolution of MRSA Microbiol Immunol 39 531-543 Hiramatsu K Hanaki H Ino T Yabuta K Oguri T and Tenover FC (1997) Methicillin-resistant Staphylococcus aureus clinical strain with reduced vancomycin susceptibility J Antimicrob Chemother 40 135-136 Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T et al (2013) Genomic Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
136
Hong S and Pedersen PL (2008) ATP synthase and the actions of inhibitors utilized to study its roles in human health disease and other scientific areas Microbiol Mol Biol Rev 72 590-641
Jevons MP (1961) lsquoCelbeninrsquo-resistant staphylococci BMJ 1 124-125
Joshi S and Huang YG (1991) ATP synthase complex from bovine heart mitochondria the oligomycin sensitivity conferring protein is essential for dicyclohexyl carbodiimidesensitive ATPase Biochim Biophys Acta 1067 255ndash258
Kohanski MA Dwyer DJ and Collins JJ (2010) How antibiotics kill bacteria from targets to networks Nat Rev Microbiol 8 423-435 Kohler C von Eiff C Liebeke M McNamara PJ Lalk M Proctor RA Hecker M and Engelmann S (2008) A defect in menadione biosynthesis induces global changes in gene expression in Staphylococcus aureus J Bacteriol 190 6351- 6364 Kohler C von Eiff C Peters G Proctor RA Hecker M and Engelmann S (2003) Physiological characterization of a hegraveme-deficient mutant of Staphylococcus aureus by a proteomic approach J Bacteriol 185 6928- 6937 Kriegeskorte A Koumlnig S Sander G Pirkl A Mahabir E Proctor RA von Eiff C Peters G and Becker K (2011) Small colony variants of Staphylococcus aureus reveal distinct protein profiles Proteomics 11 2476-2490 Lannergard J von Eiff C Sander G Cordes T Seggewiss J Peters G Proctor RA Becker K and Hughes D (2008) Identification of the genetic basis for clinical menadione-auxotrophic small-colony variant isolates of Staphylococcus aureus Antimicrob Agents Chemother 52 4017-4022
Lauraeus M and Wikstrom M (1993) The terminal quinol oxydases of Bacillus subtilis have different energy conservation properties J Biol Chem 268 11470-11473
137
Li M Gong J Cottrill M Yu H de Lange C Burton J and Topp E (2003) Evaluation of QIAamp DNA Stool Mini Kit for ecological studies of gut microbiota J Microbiol Methods 54 13-20
Liu Y Fiskum G and Schubert D (2002) Generation of reactive oxygen species by the mitochondrial electron transport chain J Neurochem 80 780-787
Martin-Hernandez A M Dufresne M Hugouvieux V Melton R and Osbourn A (2000) Effects of targeted replacement of the tomatinase gene on the interaction of Septoria lycopersici with tomato plants Mol Plant Microbe Interact 13 1301ndash1311
Matteelli A Carvalho AC Dooley KE and Kritski A (2010) TMC207 the first compound of a new class of potent anti-tuberculosis drugs Future Microbiol 5 849-858
Matsuno-Yagi A and Hatefi Y (1993) Studies on the mechanism of oxidative phosphorylation ATP synthesis by submitochondrial particles inhibited at F0 by venturicidin and organotin compounds J Biol Chem 268 6168ndash6173
Miller MH Edberg SC Mandel LJ Behar CF and Steigbigel NH (1980) Gentamicin uptake in wild-type and aminoglycoside-resistant small-colony mutants of Staphylococcus aureus Antimicrob Agents Chemother 18 722- 729
Mingeot-Leclercq M-P Glupczynski Y and Tulkens PM (1999) Aminoglycosides Activity and Resistance Antimicrob Agents Chemother 43 727- 737
Mitchell G Brouillette Eacute Seacuteguin DL Asselin AE Jacob CL and Malouin F (2010) A role for sigma factor B in the emergence of Staphylococcus aureus small-colony variants and elevated biofilm production resulting from an exposure to aminoglycosides Microb Pathog 48 18-27
Mitchell G Gattuso M Bouarab K and Malouin F (2009) Tomatidine (TO) affects virulence regulators of prototypical Staphylococcus aureus (SA) and small colony variants (SCV) of cystic fibrosis (CF) patients Abstr 49th Intersci Conf Antimicrob Agents Chemother abstr C1-1341
138
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K and Malouin F (2011) Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic fibrosis airway epithelial cells Antimicrob Agents Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM and Malouin F (2011) Infection of polarized airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function and is influenced by NF-κB Infect Immun 79 3541-3551
Mitchell G Lafrance M Boulanger S Seacuteguin DL Guay I Gattuso M Marsault Eacute Bouarab K and Malouin F (2012) Tomatidine acts in synergy with aminoglycoside antibiotics against multiresistant Staphylococcus aureus and prevents virulence gene expression J Antimicrob Chemother 67 559-568
Mitchell G Lamontagne CA Brouillette Eacute Grondin G Talbot BG Grandbois M and Malouin F (2008) Staphylococcus aureus SigB activity promotes a strong fibronectin-bacterium interaction which may sustain host tissue colonization by small-colony variants isolated from cystic fibrosis patients Mol Microbiol 70 1540-1555
Mitchell G Malouin F 2012 Outcome and prevention of Pseudomonas aeruginosa-Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic Fibrosis Chap 8 pp 181-206 Ed D Sriramulu InTech Open Access Publisher ISBN 979-953-307-059-8
Mitchell G Seacuteguin DL Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S and Malouin F (2010) Staphylococcus aureus sigma B-dependent emergence of small-colony variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-heptylquinoline-N-oxide BMC Microbiol 30 10-33
Moisan H Brouillette Eacute Jacob CL Langlois-Beacutegin P Michaud S and Malouin F (2006) Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated from cystic fibrosis patients is influenced by SigB J Bacteriol 188 64-76
139
Moreno F Crisp C Jorgensen JH and Patterson JE (1995) Methicillin- resistant Staphylococcus aureus as a community organism Clin Infect Dis 21 1308-1312
Murphy MP (2009) How mitochondria produce reactive oxygen species Biochem J 417 1- 13
Novick RP (2003) Autoinduction and signal transduction in the regulation of staphylococcal virulence Mol Microbiol 48 1429-1449
Novo DJ Perlmutter NG Hunt RH and Shapiro HM (2000) Multiparameter flow cytometric analysis of antibiotic effects on membrane potential membrane permeability and bacterial counts of Staphylococcus aureus and Micrococcus luteus Antimicrob Agents Chemother 44 827-834
ONeill AJ Cove JH and Chopra I (2001) Mutation frequencies for resistance to fusidic acid and rifampicin in Staphylococcus aureus J Antimicrob Chemother 47 647-650
Osbourn A E (1996) Preformed antimicrobial compounds and plant defense against fungal attack Plant Cell 8 1821ndash1831
Otto TD Dillon GP Degrave WS and Berriman M (2011) RATT Rapid annotation transfert tool Nucl Acids Res 39 doi101093 Pratique E Theraulaz L (2007) Eacutetude de la reacutegulation du systegraveme respiratoire TMAO reacuteductase chez Escherichia coli Meacutemoire de maitrise de lrsquoeacutecole pratique des hautes eacutetudes Paris
Price CT and Gustafson JE (2001) Increases in the mutation frequency at which fusidic acid-resistant Staphylococcus aureus arise with salicylate J Med Microbiol 50 104-106
Proctor RA and Peters G (1998) Small colony variants in staphylococcal infections Diagnostic and therapeutic implications Clin Infect Dis 27 419- 423
140
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M and Peters G (2006) Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent infections Nat Rev Microbiol 4 295ndash305
Quail MA Kozarewa I Smith F Scally A Stephens PJ Durbin R Swerdlow H and Turner DJ (2008) A large genome centers improvements to the Illumina sequencing system Nat methods 5 1005-1010
Ramirez MS and Tolmasky ME (2010) Aminoglycoside modifying enzymes Drug Resist Updat 13 151ndash171
Rao M Streur TL Aldwell FE and Cook GM (2001) Intracellular pH regulation by Mycobacterium smegmatis and Mycobacterium bovis BCG Microbiology 147 1017-1024
Richardson DJ (2000) Bacterial respiration a flexible process for a changing environment Microbiology 146 551- 571
Riley TV Pearman JW and Rouse IL (1995) Changing epidemiology of methicillin-resistant Staphylococcus aureus in Western Australia Med J Aust 163 412-414
Roddick J G (1974) The steroidal glycoalkaloid α-tomatine Phytochemistry 13 25-36
Rodrigue S Materna AC Timberlake SC Blackburn MC Malmstrom RR et al(2010) Unlocking Short Read Sequencing for Metagenomics PLoS ONE 5(7) e11840doi101371journalpone0011840 Ruiz-Rubio M Alonso Peacuterez-Espinosa A Lairini K RoldAacuten-Arjona T Dipietro A and Anaya N (2001) Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi In Studies in natural products chemistry vol 25 A Rahman (ed) (Elsevier Oxford United Kingdom) pp 293ndash326
141
Samuelsen oslash Haukland HH Kahl BC von Eiff C Proctor RA Ulvatne H Sandvik K and Vorland LH (2005) Staphylococcus aureus small colony variants are resistant to the antimicrobial peptide lactoferricin B J Antimicrob Chemother 56 1126- 1129
Sandrock R W and Vanetten HD (1998) Fungal sensitivity to and enzymatic degradation of the phytoanticipin α-tomatine Phytopathology 88 137ndash143
Sendi P and Proctor RA (2009) Staphylococcus aureus as an intracellular pathogen the role of small colony variants Trends Microbiol 17 54ndash58
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C and Osbourn A (2006) Dual effects of plant steroidal alkaloids on Saccha- romyces cerevisiae Antimicrob Agents Chemother 50 2732ndash2740
Somerville GA and Proctor RA (2009) At the crossroads of bacterial metabolism and virulence factor synthesis in Staphylococci Microbiol Mol Biol Rev 73 233- 248
Starkov AA (2010) Measurement of mitochondrial ROS production Methods Mol Biol 648 245-255
Taber HW (1993) Respiratory chain In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of MicrobiologyWashington DC) pp 199-212
Taber HW and Morrison M (1964) Electron transport in Staphylococci Properties of a particle preparation from exponential phase Staphylococcus aureus Arch Biochem Biophys 105 367- 379
Taber HW et al(1987) Bacterial uptake of aminoglycoside antibiotics Microbiol Rev 51 439-457
Thoumlny-Meyer L (1997) Biogenesis of respiratory cytochromes in bacteria Microbiol Mol Biol Rev 61 337- 376
142
Tubby S Wilson M Wright JA Zhang P and Nair SP (2013) Staphylococcus aureus small colony variants are susceptible to light activated antimicrobial agents BMC Microbiol 13 201- 204
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D Roth J Proctor RA Becker K et al (2011) Staphylococcus aureus phenotype switching an effective bacterial strategy to escape host immune response and establish a chronic infection EMBO Mol Med 3 129ndash41
Turrens JF (2003) Mitochondrial formation of reactive oxygen species J Physiol 552 335ndash 344
Tynecka Z Szcześniak Z Malm A and Los R (1999) Energy conservation in aerobically grown Staphylococcus aureus Res Microbiol 150 555- 566
Udo EE Pearman JW and Grubb WB(1993) Genetic analysis of community isolates of methicillin-resistant Staphylococcus aureus in Western Australia J Hosp Infect 25 97-108
Vakulenko SB and Mobashery S (2003) Versatility of Aminoglycosides and Prospects for Their Future Clin Microbiol Rev 16 430-450
Voggu L Schlag S Biswas R Rosenstein R Rausch C and Goumltz F (2006) Microevolution of cytochrome bd oxydase in Staphylococci and its implication in resistance to respiratory toxins released by Pseudomonas J Bacteriol 188 8079- 8086
Wells PG Bhuller Y Chen CS Jeng W Kasapinovic S Kennedy JC Kim PM Laposa RR McCallum GP Nicol CJ et al (2005) Molecular and biochemical mechanisms in teratogenesis involving reactive oxygen species Toxicol Appl Pharmacol 207 354-366
143
Yan M Yu C Yang J and Ji Y (2011) The essential two-component system YhcSR is involved in regulation of the nitrate respiratory pathway of Staphylococcus aureus J Bacteriol 193 1799- 1805
Zhang T Luo Y Chen Y Li X and Yu J (2012) BIGrat a repeat resolver for pyrosequencing-based re-sequencing with Newbler BMC Res Notes 5 567
144

Le 17 juillet 2014
le jury a accepteacute le meacutemoire de Madame Isabelle Guay
dans sa version finale
Membres du jury
Professeur Franccedilois Malouin
Directeur de recherche
Deacutepartement de Biologie
Professeur Eacuteric Marsault
Codirecteur de recherche
Deacutepartement de Pharmacologie
Professeur Seacutebastien Rodrigue
Membre interne
Deacutepartement de Biologie
Professeur Kamal Bouarab
Preacutesident-rapporteur
Deacutepartement de Biologie
ii
SOMMAIRE
Dans le but de mieux comprendre le mode drsquoaction et de nous permettre de
deacuteterminer la cible de la tomatidine nous avons dans un premier temps tenteacute de
mieux circonscrire le spectre drsquoactiviteacute de la tomatidine Gracircce agrave ces travaux nous
sommes en effet maintenant en mesure de dire que la tomatidine possegravede une
activiteacute antibacteacuterienne contre les espegraveces de la divison des Firmicutes et plus
preacuteciseacutement contre les bacteacuteries de lrsquoordre des Bacillales dont font partie les genres
Bacillus Staphylococcus et Listeria Nous avons eacutegalement deacutecouvert gracircce agrave des
expeacuteriences en collaboration avec le laboratoire drsquoEacuteric Marsault qursquoun analogue de
la tomatidine (FC04-100) avait non seulement des proprieacuteteacutes similaires agrave la
moleacutecule naturelle mais deacutemontrait une activiteacute par lui-mecircme contre S aureus agrave
pheacutenotype normal alors que la tomatidine possegravede uniquement une activiteacute contre
les laquo small colony variants raquo De plus alors que la tomatidine possegravede plutocirct une
activiteacute bacteacuteriostatique contre la forme SCV de L monocytogenes le nouveau
composeacute (FC04-100) deacutemontre quant agrave lui une forte activiteacute bacteacutericide contre
cette souche tout comme contre la forme SCV des autres Bacillales
Parallegravelement et toujours dans le but de rechercher le mode drsquoaction et la cible de
la tomatidine nous avons obtenu par passages successifs dans un milieu avec
antibiotiques des mutants de S aureus agrave pheacutenotype normal et des SCV reacutesistants
agrave la tomatidine ou agrave la combinaison tomatidine et gentamicine Apregraves le
seacutequenccedilage de ces mutants lrsquoeacutetude de la position de ces mutations agrave lrsquoaide de
diffeacuterents logiciels de bio-informatique nous a permis drsquoeacutemettre un modegravele-
hypothegravese quant au mode drsquoaction et agrave la cible de la tomatidine Selon les reacutesultats
que nous avons agrave ce stade-ci la cible de la tomatidine chez S aureus serait la
sous-uniteacute c de lrsquoATP synthase Cependant son mode drsquoaction serait eacutegalement
deacutependant de la fonctionnaliteacute de la chaine de transport des eacutelectrons et donc de la
iii
polarisation membranaire et de la production de ROS intracellulaire ce qui
expliquerait la diffeacuterence drsquoactiviteacute entre les souches agrave pheacutenotype normal et les
SCV
Mots cleacutes tomatidine ATP synthase chaine de transport des eacutelectrons synergie laquo small-colony variants raquo ROS S aureus analogues
iv
REMERCIEMENTS
Je voudrais remercier mon directeur de recherche Pr Franccedilois Malouin qui a cru
en moi pour mrsquoavoir proposeacute ce beau projet de maitrise Merci pour sa grande
disponibiliteacute son eacutecoute et ses critiques toujours constructives et jamais
condescendantes
Je voudrais eacutegalement remercier mes conseilleacutes Kamal Bouarab Seacutebastien
Rodrigue et Eacuteric Marsault leur implication dans mon projet leur occasionnant un
surplus de travail Merci pour leur support technique et leur disponibiliteacute agrave reacutepondre
agrave mes questions
Un gros merci agrave Gabriel Mitchell qui a eacuteteacute mon mentor au deacutebut de ma maitrise et
qui mrsquoa introduit au projet laquo Tomatidine raquo Un merci particulier eacutegalement agrave
Maxime-Lamontagne-Boulet pour son aide en tant que stagiaire lors de mes
derniegraveres expeacuteriences Je voudrais eacutegalement remercier tous les membres du
laboratoire preacutesents et passeacutes qui ont toujours eacuteteacute disponibles pour partager avec
moi leurs connaissances theacuteoriques et techniques
Je ne peux passer sous silence lrsquoimplication de lrsquoUniversiteacute de Sherbrooke et du
FQRNT comme organismes subventionnaires au projet Merci eacutegalement agrave Fibrose
Kystique Canada qui a participeacute eacutegalement agrave la subvention drsquoune partie du projet
En terminant je voudrais remercier ma famille qui a crue en moi et qui mrsquoencourage
agrave perseacuteveacuterer depuis toujours Un merci tout particulier agrave mon conjoint et ma belle-
fille pour qui je nrsquoai pas toujours eacuteteacute aussi disponible que jrsquoaurais voulu lrsquoecirctre et qui
mrsquoont supporteacutee au quotidien et ce dans tous les sens du terme
v
TABLE DES MATIEgraveRES
SOMMAIRE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ii
REMERCIEMENTS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip iv
TABLE DES MATIEgraveRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip v
LISTE DES ABBREacuteVIATIONS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip vii
LISTE DES TABLEAUX helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ix
LISTE DES FIGURES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip x
CHAPITRE 1 INTRODUCTION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
11 Staphylococcus aureus helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
111 Importance cliniquehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)helliphellip 2
1121 Diffeacuterences et similariteacutes helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 2
1122 La chaine de transport des eacutelectrons helliphelliphelliphelliphellip 4
12 Antibiotiques Modes drsquoaction cibles cellulaire et
meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles helliphelliphelliphellip 25
122 Les aminoglycosides helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1221 Mode drsquoaction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1222 Meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphellip 31
123 La tomatidine helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 33
13 Projet de maitrise helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
CHAPITRE 2 ARTICLE SCIENTIFIQUE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 37
ldquoTomatidine and derivative FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Firmicutes Including Those of Listeria monocytogenes
and Are Bactericidal in Combination With Aminoglycosides Against their Normal
Phenotyperdquo
vi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE helliphelliphelliphellip 76
31 Obtention des mutants helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
32 Seacutequenccedilage et analyses bio-informatiques helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 79
321 Identification des mutations en lien avec la tomatidine helliphellip 89
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct helliphellip 90
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine helliphelliphelliphelliphellip 99
CHAPITRE 4 DISCUSSION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 113
CHAPITRE 5 CONCLUSION ET PERSPECTIVES helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 124
CONTRIBUTIONS SUPPLEacuteMENTAIRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 1 Application de brevet helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 2 Article scientifique helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 129
laquo Unraveling the structure-activity relationship of tomatidine
a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureus raquo
BIBLIOGRAPHIE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 131
vii
LISTE DES ABBREacuteVIATIONS
Adh Alcool deacuteshydrogeacutenase
ADN Acide deacutesoxyribo-nucleacuteique
ADP Adeacutenosine diphosphate
ARN Acide ribo-nucleacuteique
ATP Adeacutenosine triphosphate
CA-MRSA Community-associated MRSA
CMI concentration minimale inhibitrice
CoQ Coenzyme Q
Cyt Cytochrome
DMQ laquo demethylmenaquinone raquo
dTMP Deacutesoxythymidine monophosphate
dUMP Deacutesoxyuridine monophosphate
FADH2 Flavine adenine dinucleacuteotide
GEN gentamicine
HA-MRSA Healthcare-associated MRSA
Ldh Lactate deacuteshydrogeacutenase
ml millilitre
MQ meacutenaquinone
MRSA methycilin resistant Staphylococcus aureus
Mqo2 Quninol oxidoreacuteductase
NADH Nicotinamide adenine dinucleacuteotide (forme reacuteduite)
Pdh Pyruvate deacuteshydrogeacutenase
Pfl Pyruvate formate lyase
PyrD Dihydroorotate deacuteshydrogeacutenase
Q Quinone
Rif rifampicine
ROS laquo reactive oxygen species raquo espegravece reacuteactive de lrsquooxygegravene
viii
[S] Polysulfide
S aureus Staphylococcus aureus
SARM Staphylococcus aureus resistant agrave la meacutethycilline
SCV laquo small colony variants raquo
SHCHC 2-succinyl-6-hydroxy-24-cyclohexadiene-1-carboxylate syntase
TCA Tricarboxylic acid cycle
TCD tableau croiseacute dynamique
TO tomatidine UFC Uniteacute formatrice de colonies
UQ Ubiquinone
microg microgramme
ix
LISTE DES TABLEAUX
CHAPITRE 1 INTRODUCTION
1 Cibles et modes drsquoaction des principales classes drsquoantibiotique 27
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants (B) chez les mutants NewbouldΔhemB reacutesistants 95-96
CHAPITRE 4 DISCUSSION
1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 123
x
LISTE DES FIGURES
CHAPITRE 1 INTRODUCTION
1 Diffeacuterence entre les pheacutenotypes normaux et SCV de S aureus 3
2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes 6
3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale 8
4 Chaines respiratoires bacteacuteriennes contenant des cytochromes 9
5 Flexibiliteacute respiratoire (A) chez Paracoccus denitrificans (B) chez
Escherichia coli 10
6 Chaine de transport des eacutelectrons proposeacutee chez les staphylocoques 12
7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies 17
8 Modegravele de reacutegulation du cycle TCA 20
9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS 23
10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de O2-bull 24
11 Voies de formation et de deacutetoxification des ROS 25
12 Cineacutetique drsquoentreacutee des aminoglycosides 30
13 Sites de mutations de certains reacutesistants aux aminoglycosides 32
14 Structures chimiques de (A) tomatine (B) tomatidine et (C) Analogue
de tomatidine FC04-100 34
xi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Graphiques des CMI pour chacun des passages pour (A) la
tomatidine sur NewbouldΔhemB (B) la tomatidine en preacutesence de
gentamicine sur NewbouldΔhemB (C) la gentamicine sur S aureus
ATCC 29213 (D) la gentamicine en preacutesence de tomatidine sur S
aureus ATCC 29213 77-79
2 Organigramme des mutants reacutesistants seacutequenceacutes 80-81
3 Eacutetapes du seacutequenccedilage Illumina 83
4 Meacutethode drsquoamplification par la meacutethode Illumina 85
5 Correction du programme RATT sur les annotations transfeacutereacutees 88
6 Exemple drsquoune partie du tableau compilation des mutations 89
7 Seacutelection des gegravenes drsquointeacuterecirct chez ATCC 29213 dans le TCD par
lrsquoutilisation de diffeacuterents filtres 91-92
8 Seacutelection des gegravenes drsquointeacuterecirct chez NewbouldΔhemB dans le TCD par
lrsquoutilisation de diffeacuterents filtres 93-94
9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct 97-98
10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants 100
11 Courbe de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison gentamicine (+TO) du premier et troisiegraveme niveau de
reacutesistance 101
12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B)
NewbouldΔhemB 102
xii
13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213 104
14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicinelorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC
29213 105
15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants
ATCC 29213 reacutesistants agrave la combinaison gentamicine (+TO) 106
16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la
tomatidine 107
17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la
tomatidine 108
18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB
reacutesistants agrave la combinaison tomatidine (+gentamicine) 109
19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane
Potential Kit (Invitrogen) 111
20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du
gegravene atpE de S aureus Newman 112
xiii
CHAPITRE 4 DISCUSSION
1 Site drsquoinhibition de lrsquoATP synthase par diffeacuterentes moleacutecules 114
2 Structure chimique de (A) TMC207 et (B) lrsquoanalogue anti Gram
positifs 115
3 Seacutequence proteacuteique de la proteacuteine AtpE chez diffeacuterentes espegraveces 118
4 Modeacutelisation de la structure tridimentionnelle de lrsquoATP synthase de S
aureus (atpE) montrant les acides amineacutes muteacutes chez les mutants
reacutesistants 121
- 1 -
CHAPITRE 1
INTRODUCTION
11 Staphylococcus aureus
111 Importance clinique
Staphylococcus aureus est une bacteacuterie agrave Gram positif anaeacuterobie facultative Crsquoest un
microorganisme opportuniste pouvant infecter diffeacuterents hocirctes diffeacuterents organes ou
sites drsquoinfection et ayant la capaciteacute de causer tant des infections aiguumles que
chroniques (Archer 1998 Goerke et Wolz 2004)
Staphylococcus aureus reacutesistant agrave la meacutethilline le SARM (ou laquo MRSA raquo) est une
souche de S aureus devenue reacutesistante aux β-Lactamines par lrsquoacquisition du gegravene
mecA dans son chromosome (Hiramatsu 1995) Le SARM a eacuteteacute isoleacute pour la
premiegravere fois en 1960 en Angleterre (Jevons 1961) et est devenu eacutepideacutemique agrave
travers le monde depuis les anneacutees 1970 Au moins trois geacutenotypes distincts eacutetaient
recenseacutes dans les anneacutees 1980 (Hiramatsu 1995) dont deux sont toujours preacutevalents
agrave travers le monde comme eacutetant des souches HA-MRSA (Healthcare-associated
MRSA) des souches acquises agrave lrsquohocircpital (Enright et al 2002) Ces souches ont
acquis des gegravenes de reacutesistance agrave pratiquement tous les antibiotiques introduits dans
lrsquoutilisation clinique depuis les 50 derniegraveres anneacutees mecircme agrave lrsquoantibiotique de dernier
ressort la vancomycine(Hiramitsu 1997 Chen et al 2003) Nous sommes
maintenant en manque drsquoagents theacuterapeutiques efficaces contre les SARM touchant
les personnes immunodeacuteprimeacutees qui sont dans les hocircpitaux En plus depuis le deacutebut
des anneacutees 1990 des clones de SARM dont plusieurs deacutemontraient un potentiel
pathogeacutenique supeacuterieur aux souches classiques de HA-MRSA retrouveacutees agrave lrsquohocircpital
- 2 -
sont maintenant retrouveacutes dans la communauteacute (CA-MRSA) agrave travers le monde (Udo
et al 1993 Riley et al 1995 Moreno et al 1995 Aubry-Damon et al 1997) et ont
la capaciteacute drsquoaffecter des personnes en santeacute en causant des infections aiguumles
(Hiramatsu et al 2013) Il apparaicirct donc urgent de trouver des drogues alternatives
afin de surmonter le nombre grandissant de souches de SARM
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)
1121 Diffeacuterences et similariteacutes
Les laquo small colony variants raquo (SCV) sont des bacteacuteries souvent isoleacutees lors des
infections chroniques comme crsquoest le cas des infections pulmonaires chez les
patients atteints de fibrose kystique mais aussi dans des cas drsquoosteacuteomyeacutelite
drsquoarthrite septique drsquoinfections de prothegraveses orthopeacutediques et lors de la mammite
bovine (Alexender et Hudson 2001 Moisan et al 2006 Proctor et al 2006) Les
SCV sont caracteacuteriseacutes par soit un meacutetabolisme oxydatif dysfonctionnel ou un manque
dans la biosynthegravese de la thymidine les deux amenant une alteacuteration dans
lrsquoexpression des facteurs de virulence une croissance plus lente et une diminution
de la pigmentation des colonies (Figure 1) (Proctor et al 2006) Cependant alors
que les souches normales reacutepriment lrsquoexpression des proteacuteines de surface et
expriment les exoproteacuteines lors de la phase stationnaire (Novick 2003) les SCV
expriment de faccedilon stable les gegravenes Sig-B deacutependants codant pour des proteacuteines de
surface telle les adheacutesines au lieu drsquoactiver le systegraveme agr deacutependant du quorum-
sensing et produire des exoproteacuteines (Moisan et al 2006) Ces diffeacuterences dans
lrsquoexpression des facteurs de virulence reacutesultent chez les SCV en une meilleure
habileteacute agrave adheacuterer aux composantes de lrsquohocircte (Mitchell et al 2008) et agrave former du
biofilm (Mitchell et al 2010a Mitchell et al 20010b) et seraient eacutegalement lieacutees agrave
lrsquohabileteacute des SCV agrave lrsquoinvasion et la persistance des cellules de lrsquohocircte (Proctor et al
2006 Sendi et Proctor 2009 Mitchell et al 2011b)
- 3 -
S aureus est donc un pathogegravene qui peut se preacutesenter sous deux pheacutenotypes qui
sont souvent retrouveacutes simultaneacutement lors drsquoune mecircme infection et qui ont le pouvoir
de reacuteverter drsquoun pheacutenotype agrave lrsquoautre selon les conditions du milieu (Figure 1)
(Tuchscherr et al 2011)
Le pheacutenotype dit rdquonormalrdquo est associeacute aux infections aiguumles agrave une forte production
de facteurs de virulence (ex heacutemolysine) et est associeacute agrave la phase de disseacutemination
de la bacteacuterie Lrsquoautre pheacutenotype le laquo small-colony variants raquo ou SCV est affecteacute
dans sa chaine respiratoire ce qui amegravene une croissance beaucoup plus lente des
colonies de 8 -10 fois plus petites que les prototypes et une faible susceptibiliteacute aux
aminoglycosides Les SCV sont plutocirct associeacutes aux infections chroniques agrave S aureus
de par leur capaciteacute agrave srsquointernaliser dans les cellules de lrsquohocircte leur forte capaciteacute agrave
produire du biofilm et leur capaciteacute agrave produire des facteurs de colonisation
(Tuchscherr et al 2011 Proctor et al 2006) (Figure 1)
Figure 1 Diffeacuterences entre les pheacutenotypes normaux et SCV de S aureus
S aureus agrave
pheacutenotype normal
Disseacutemination
amp
Infections aiguumles
Colonisation
amp
Infections persistantes
SCVs
(laquo small-colony
variants raquo)
Reacutevers
ion
pheacutenoty
piq
ue
Norm
al
SC
V
Croissance
Biofilm
Internalisationheacutemolyse
Mitchell G et Malouin F 2012
- 4 -
1122 La chaine de transport des eacutelectrons
Chaine respiratoire tableau geacuteneacuteral
Les chaines respiratoires bacteacuteriennes bien que tregraves variables dans leurs
composantes parmi les diffeacuterentes espegraveces sont toutes constitueacutees sur le mecircme
modegravele de base Elles sont composeacutees de trois composantes majeures soit
- les deacuteshydrogeacutenases primaires qui sont chargeacutees drsquooxyder des substrats
principalement les coenzymes NADH et FADH2 (issues du cycle de Krebs) (mais
aussi selon le cas le succinate le lactate lrsquohydrogegravene hellip) et de pomper deux
protons du cocircteacute externe de la membrane en plus de transfeacuterer deux eacutelectrons agrave un
groupement de quinones
- Les quinones sont des moleacutecules liposolubles non proteacuteiques mobiles dans la
membrane Elles servent drsquointermeacutediaires entre les deacuteshydrogeacutenases primaires et
les oxydases terminales pour le transport des eacutelectrons Il existe deux sortes de
quinones les benzoquinones dont font partie les ubiquinones et les
naphtoquinones dont font partie les meacutenaquinones Les bacteacuteries agrave Gram positif
ne possegravedent que des meacutenaquinones alors que les deux sont preacutesentes chez les
bacteacuteries agrave Gram neacutegatif (Hederstedt 1993 Pratique et Theraulaz 2007)
- Les oxidoreacuteductases terminales sont chargeacutees de reacuteduire lrsquoaccepteur final
drsquoeacutelectron lrsquooxygegravene eacutetant lrsquoaccepteur final privileacutegieacute
De faccedilon plus deacutetailleacutee la chaine respiratoire est composeacutee de cinq
complexes proteacuteiques principaux (Figure 2)
- complexe I NADH deacuteshydrogeacutenase ou NADH reacuteductase ou NADHCoQ
oxidoreacuteductase ou NADH-ubiquinone oxidoreacuteductase
- complexe II Succinate deacuteshydrogeacutenase
- complexe III Cytochrome bc1 complexe ou ubiquinol cytochrome c
reacuteductaseoxydoreacuteductase
- complexe IV Cytochrome c oxydase
- 5 -
- complexe V ATP synthase (Creacutepin-Gilbert et al 2006)
La phosphorylation oxydative est deacuteclencheacutee dans la chaine de complexes
proteacuteiques par la preacutesence drsquoO2 accepteur terminal privileacutegieacute drsquoeacutelectrons (ou drsquoautres
accepteurs finaux tel le nitrate) dans le milieu ainsi que par les eacutelectrons ameneacutes par
le NADH et le FADH2 au complexe I et II respectivement (Creacutepin-Gilbert et al 2006)
Les eacutelectrons vont ensuite transiter par le complexe III jusqursquoau complexe IV ou les
eacutelectrons seront utiliseacutes afin de reacuteduire lrsquooxygegravene en eau le complexe IV est
lrsquooxydase terminale consideacutereacutee comme eacutetant lrsquoaccepteur final drsquoeacutelectrons Le transfert
drsquoeacutelectrons est possible gracircce aux reacuteactions drsquooxydoreacuteduction en cascade dans les
diffeacuterents complexes proteacuteiques Ces reacuteactions fournissent de lrsquoeacutenergie libre utiliseacutee
par les complexes I III et IV afin de transfeacuterer des protons agrave lrsquoexteacuterieur de la cellule
(Creacutepin-Gilbert et al 2006) Le complexe V (ATP synthase) quant agrave lui utilise les
eacutelectrons pompeacutes agrave lrsquoexteacuterieur par les autres complexes comme force proton-motrice
afin de phosphoryler trois moleacutecules drsquoADP et ainsi produire trois ATP Le transport
des eacutelectrons entre les diffeacuterents complexes proteacuteiques est possible gracircce agrave la
preacutesence dans la membrane de deux types de moleacutecules liposolubles non proteacuteiques
mobiles les meacutenaquinones et le cytochrome c Les meacutenaquinones seules quinones
preacutesentes chez les bacteacuteries agrave Gram positif assurent le transport des eacutelectrons entre
le complexe I et le complexe III et entre le complexe II et III Le cytochrome c pour sa
part veacutehicule les eacutelectrons entre le complexe III et le complexe IV Le transport des
eacutelectrons srsquoeffectue de la composante ayant le plus faible potentiel drsquooxydoreacuteduction
vers celle ayant le plus fort potentiel
- 6 -
La chaine respiratoire bacteacuterienne
La mitochondrie montre assez peu de flexibiliteacute respiratoire Il y a une certaine
flexibiliteacute au niveau de lrsquoentreacutee des eacutelectrons mais aucune au niveau de la sortie des
eacutelectrons ougrave la cytochrome aa3 oxydase reacuteduit lrsquooxygegravene en eau (Richardson 2000)
(Figure 3) Chez les bacteacuteries alors que le complexe II fonctionne sensiblement de
faccedilon identique agrave celui des eucaryotes les autres complexes peuvent se composer
de diffeacuterents complexes enzymatiques (reacuteductases) selon les espegraveces et les
conditions environnementales (Richardson 2000 Thoumlny-Meyer 1997) En outre
chez les bacteacuteries contrairement aux eucaryotes la chaine respiratoire comporte
souvent des voies de transport des eacutelectrons alternatives on dit alors qursquoelles ont une
chaine respiratoire agrave branches ou brancheacutee ou ramifieacutee (Artzatbanov et Petrov 1990)
Plusieurs espegraveces bacteacuteriennes possegravedent une chaine respiratoire ramifieacutee dont la
ramification se situe habituellement suite agrave la reacuteduction des quinones en quinols
(Lauraeus et Wikstrom 1993 Thoumlny-Meyer 1997) (Figure 4) Une des deux
Complexe I NADH
deacuteshydrogeacutenase
Complexe II Succinate
deacuteshydrogeacutenase
Complexe III
bc 1
Complexe IV Cytochrome c
oxydase
Complexe V
ATP synthase
Modifieacute de wikipedia httpfrwikipediaorgwikiChaine_respiratoire
Figure 2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes
Leurs rocircles dans le transport des eacutelectrons et dans lrsquoeacutelaboration du gradient de
protons et le lien entre la chaine respiratoire et le cycle de Krebs Q Quinones
- 7 -
branches connue comme la branche cytochrome c oxyde les quinols (meacutenaquinol
QH2) via la voie complexe bc1-cytochrome c- cytochrome c oxydase Lrsquoautre
branche-type consiste en une seule uniteacute enzymatique (cytochrome bd) laquelle
transfegravere directement les eacutelectrons des quinols agrave lrsquooxygegravene Cette voie est connue
comme la branche quinol oxydase il srsquoagit drsquoune oxydase terminale transfeacuterant
directement les eacutelectrons agrave lrsquooxygegravene eacutevitant de passer par lrsquointermeacutediaire du
complexe III (bc1) et du cytochrome c (Artzatbanov et Petrov 1990 Lauraeus et
Wikstrom 1993) Alors que la voie du cytochrome c permet le pompage de protons agrave
lrsquoexteacuterieur par lrsquoaction du complexe bc1 (ou complexe III) et du cytochrome c oxydase
le cytochrome bd ne permet aucun pompage de protons (Lauraeus et Wikstrom
1993) La voie passant par le complexe bc1-cytochrome c-cytochrome c oxydase est
plus efficace dans la production drsquoeacutenergie Alors que les 2 voies peuvent ecirctre
exprimeacutees simultaneacutement chez quelques espegraveces il semble que lrsquoexpression de ces
voies soit inductible selon les conditions environnementales chez Bacillus subtilis et
les Staphylocoques (Lauraeus et Wikstrom 1993) On pourrait dire eacutegalement que la
chaine respiratoire est seacutepareacutee en deux eacutetapes la premiegravere partie est la phase
quinone-reacuteductase (Q-QH2) et la seconde la phase quinol-oxydase (QH2-Q) menant
agrave lrsquooxydase terminale La phase de reacuteduction est responsable du transfert des
eacutelectrons drsquoune varieacuteteacute de substrats tels le NADH le succinate le lactate le formate
et lrsquohydrogegravene vers les quinones et inclut diffeacuterentes reacuteductases (Thoumlny-Meyer 1997)
(Figure 4)
- 8 -
Figure 3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale UQ ubiquinone UQH2 ubiquinol Cyt cytochrome Il semble y avoir presque autant de chaines de transport des eacutelectrons diffeacuterentes
qursquoil y a drsquoespegraveces bacteacuteriennes (par exemple Paracoccus denitrificans et Escherichia
coli (Figure 5)) De plus il ne semble pas y avoir consensus dans la litteacuterature sur
les noms attribueacutes aux oxydases terminales Les noms pour les oxydases terminales
deacutependent des types de cytochromes dont elles sont composeacutees Il ne semble donc
pas eacutevident de connaicirctre de faccedilon preacutecise les types de cytochromes qui font partie
des oxydases terminales Pour rajouter agrave la confusion les gegravenes et opeacuterons ne sont
pas toujours associeacutes agrave leur oxydase terminale dans la litteacuterature ce qui fait qursquoon ne
peut savoir de faccedilon certaine de quelle oxydase on parle Le Cytochrome aa3 peut
eacutegalement porter le nom de Cyt ba3 et le cytochrome bd peux eacutegalement porter les
noms de Cyt bo
Tireacute de Richardson DJ 2000
- 9 -
Succinate
Formate
H2
SDH
FDH
HYD
QQH 2 bc1 c
Reacuteductase alternatives
b
c
cd1c
cb
c
b
c
d
Siroheme
Siroheme
b
NO3-
NO2-
NO
TMAO
DMSOS2O3
2-
[S]
SO32-
Fumarate
Quinol oxidases
ba3bb3bo3d
bd
aa3
O2
Cyt c oxidases
aa3ba3baa3caa3cbb3cao
co
O2
Modifieacute de Thony-Mayer L 1997
NADH
NDH
Figure 4 Chaines respiratoires bacteacuteriennes contenant des cytochromes
Du cocircteacute gauche on retrouve la partie quinone (ubiquinone ou meacutenaquinone)-
reacuteductase Du cocircteacute droit on retrouve la partie quinol-oxydase avec les oxydases
terminales reacuteduisant lrsquoO2 ou lrsquoaccepteur terminal alternatif tel que montreacute Les
oxydoreacuteductases sont surligneacutees QQH2 repreacutesente le pool de quinonesquinols
dans la membrane Les polysulfides sont repreacutesenteacutes par [S] Les chaines
respiratoires ne contenant pas de cytochromes ne sont pas montreacutees NDH
NADH deacuteshydrogeacutenase
Modifieacute de Thoumlny-Mayer L 1997
- 10 -
Figure 5 Flexibiliteacute respiratoire chez Paracoccus denitrificans (a) et chez
Escherichia coli (b) MQ menaquinone UQ ubiquinone DMQ
demethylmenoquinone
Chez S aureus
Comme plusieurs autres espegraveces bacteacuteriennes anaeacuterobies facultatives S aureus a
la possibiliteacute drsquoexprimer diffeacuterentes voies de synthegravese eacutenergeacutetique dont lrsquoexpression
est finement reacuteguleacutee selon des conditions environnementales du moment (Hammer et
Tireacute de Richardson DJ 2000
- 11 -
al 2013 Lauraeus et Wikstrom 1993) Mecircme si S aureus peut utiliser la
fermentation la respiration (aeacuterobie et anaeacuterobie) sera privileacutegieacutee par la bacteacuterie
puisque celle-ci amegravene une bien meilleure production drsquoeacutenergie (Goumltz et al 2006
Hammer et al 2013 ) La respiration aeacuterobie avec lrsquooxygegravene comme accepteur final
drsquoeacutelectron sera utiliseacutee de faccedilon preacutefeacuterentielle en preacutesence drsquooxygegravene (Hammer et al
2013) Malgreacute sa versatiliteacute dans ses voies de synthegravese eacutenergeacutetique S aureus et les
autres bacteacuteries agrave Gram positif ne produisent qursquoun seul type de quinone les
meacutenaquinones lesquelles sont utiliseacutees pour la croissance degraves qursquoil y a de lrsquooxygegravene
de preacutesent dans le milieu (Somerville et Proctor 2009) La reacuteduction de lrsquooxygegravene en
eau est meacutedieacutee par des oxydases terminales hegraveme deacutependant (Hammer et al 2013)
Selon certains auteurs S aureus possegravederait deux oxydases terminales la
cytochrome oxydase aa3 (ou ba3) (QoxABCD) et la cytochrome bd quinol oxydase
(CydAB) (Goumltz et al 2006 Hammer et al 2013 Kohler et al 2008) Cette derniegravere
est exclusive aux procaryotes et est une voie de contournement du complexe III qui
transmet directement les eacutelectrons du laquo pool raquo de quinol (QH2) agrave lrsquooxygegravene Selon
certains auteurs il y aurait eacutegalement chez S aureus une troisiegraveme oxydase
terminale le cytochrome o (CyoABCDE) (Thoumlny-Meyer 1997 Voggu et al 2006)
qui selon certain serait un cytochrome oxydase terminal alternatif au cytochrome aa3
et au cytochrome bd (Voggu et al 2006 Goumltz et al 2006 Taber et Morrison 1964)
(Figure 6) Ces voies alternatives permettent agrave S aureus de srsquoadapter rapidement agrave
un changement drsquoenvironnement (Hammer et al 2013) In vivo chaque organe est
un environnement diffeacuterent preacutesentant des deacutefis respiratoires distincts dus agrave la
variation dans 1- la quantiteacute drsquooxygegravene disponible 2- la disponibiliteacute drsquoaccepteurs
drsquoeacutelectrons alternatifs 3- la concentration drsquoinhibiteurs de la respiration dans le milieu
tels les oxydes nitriques (NO) produits par les neutrophiles et 4- la disponibiliteacute des
cofacteurs neacutecessaires agrave la production drsquoeacutenergie (Hammer et al 2013) Le
cytochrome est exprimeacute de faccedilon modeacutereacutee dans un milieu riche en oxygegravene mais est
fortement exprimeacute dans un milieu pauvre en oxygegravene mais riche en oxyde nitrique
De plus le type drsquooxydoreacuteductase terminale exprimeacute est influenceacute eacutegalement par la
- 12 -
source de carbone disponible la phase de croissance le pH extracellulaire et la
tension en oxygegravene (Lauraeus et Wikstrom 1993 Tynecka et al 1999)
La chaine respiratoire chez les SCV
S aureus est particuliegraverement doueacute agrave surpasser les deacutefis respiratoires en acqueacuterant
une reacutesistance agrave lrsquoacide nitrique produit par les neutrophiles et une habiliteacute agrave persister
dans les cellules de lrsquohocircte en se preacutesentant comme un variant agrave petites colonies
ayant une chaine respiratoire deacuteficiente (SCV) (Hammer et al 2013)
Deux principaux types de SCV sont retrouveacutes dans les isolats cliniques 1- les SCV
ayant une chaine de transport des eacutelectrons deacutefectueuse et 2- les SCV auxotrophes
pour la thymidine (Proctor et Peters 1998 Samuelsen et al 2005) Ces derniers
Figure 6 Chaine de transport des eacutelectrons proposeacutee chez les
staphylocoques Les informations disponibles sur les composantes de la chaine
respiratoire des staphylocoques suggegraverent une chaine ramifieacutee consistant en 2
oxydases terminales meacutenaquinol-deacutependante un cytochrome o et un cytochrome
aa3 oxydase NADH deacuteshydrogeacutenase (DHase) transmet les eacutelectrons au laquo pool raquo
de meacutenaquinones De lagrave les eacutelectrons sont transfeacutereacutes soit au cytochrome b ou agrave
lrsquoaccepteur alternatif drsquoeacutelectrons le cytochrome bd quinol oxydase Le cytochrome
b transfegravere les eacutelectrons agrave une des deux oxydases terminales Ces oxydases
terminales transfegraverent les eacutelectrons agrave lrsquooxygegravene
Modifieacute de Voggu L et al 2006
- 13 -
arborent une ou plusieurs mutations dans le gegravene thyA codant pour la thymidylate
synthase laquelle catalyse la derniegravere eacutetape dans la voie de biosynthegravese de la
thymidine crsquoest-agrave-dire la formation de dTMP agrave partir du dUMP (Samuelsen et al
2005) Le dTMP est essentiel pour la synthegravese de lrsquoADN (Besier et al 2007) Les
mutants thymidine-auxotrophes sont souvent la conseacutequence drsquoune pression
seacutelective par lrsquoutilisation prolongeacutee de trimethoprim-sulfamethoxazole (TMP-SMX)
(souvent utiliseacute pour le traitement des patients fibroses kystiques) un antibiotique qui
interfegravere dans la biosynthegravese de lrsquoacide tetrahydrofolique qui agit comme cofacteur de
la thymidylate synthase (Besier et al 2007 Lannergard et al 2008 Samuelsen et al
2005) Mecircme si de premiers abords les SCV thymidine-auxotrophes ne semblent pas
avoir de lien avec la chaine transport des eacutelectrons il se pourrait que celle-ci ait sont
importance chez ces SCV puisque lrsquoentreacutee de la thymidine passe par NupC dont le
fonctionnement neacutecessite un gradient eacutelectrochimique creacuteeacute par la chaine de transport
des eacutelectrons (Proctor et Peters 1998 ) Les caracteacuteristiques pheacutenotypiques des
SCV deacutecrites ci-apregraves sont toutes lieacutees agrave lrsquoalteacuteration de la chaine de transport des
eacutelectrons 1- la croissance plus lente parce que la synthegravese de la paroi cellulaire
neacutecessite une grande quantiteacute drsquoATP 2-la diminution de la formation de pigment
parce que la biosynthegravese des caroteacutenoiumldes requiert le transport des eacutelectrons 3- une
reacutesistance accrue agrave la gentamicine et autres aminoglycosides parce que lrsquoentreacutee de
ces composeacutes neacutecessite un grand potentiel membranaire geacuteneacutereacute par le transport des
eacutelectrons (Lannergard et al 2008) et 4- une perte partielle ou complegravete de la
fermentation du mannitol parce que lrsquoutilisation des sucres-alcool tels que le
mannitol est diminueacutee lorsque la chaine de transport des eacutelectrons ne peut oxyder le
NADH produit lorsque le mannitol est meacutetaboliseacute en fructose-6-phosphate (Kohler et
al 2003 Proctor et Peters 1998 Samuelsen et al 2005)
Les SCV posseacutedant une chaine de transport des eacutelectrons deacutefectueuse ont une voie
de biosynthegravese de lrsquoheacutemine ou de la meacutenadione deacuteficiente et le pheacutenotype est
reacuteversible lorsque compleacutementeacute avec lrsquoheacutemine ou la meacutenadione (Hammer et al
- 14 -
2013 Kohler et al 2008 Lannergard et al 2008 Proctor et Peters 1998) La
meacutenadione est essentielle agrave la biosynthegravese des meacutenaquinones et lrsquoheacutemine est
essentielle agrave la biosynthegravese de lrsquohegraveme qui entre dans formation des cytochromes
(Hammer et al 2013 Kohler et al 2008 Lannergard et al 2008 Thoumlny-Meyer
1997) Les meacutenaquinones et les cytochromes sont essentiels au fonctionnement de
la chaine de transport des eacutelectrons (vers lrsquoO2 autant que vers le nitrate) (Kohler et al
2008) Il se pourrait qursquoil existe eacutegalement des SCV causeacutes par drsquoautres deacuteficiences
telles que lrsquoalteacuteration de lrsquoATP synthase qui ne reacutesulterait pas drsquoune auxotrophie pour
lrsquoheacutemine ou la meacutenadione mais reacutesulterait tout de mecircme en une deacuteficience dans la
chaine de transport des eacutelectrons (Proctor et Peters 1998 )
Comme lrsquohegraveme est un cofacteur essentiel pour les oxydases terminales et que la
meacutenaquinone est un transporteur drsquoeacutelectrons utiliseacute par S aureus on comprend que
la faible croissance des mutants hemB et menD repreacutesente un pheacutenotype mimant la
perte des oxydases terminales (Hammer et al 2013) Les deux oxydases terminales
de S aureus QoxABCD (cyt aa3) et CydAB (cyt bd) semblent ecirctre diffeacuteremment
reacuteguleacutees chez les mutants menD et la souche sauvage alors que lrsquoopeacuteron cydAB est
fortement sous exprimeacute chez les mutants aucune diffeacuterence de niveau de
transcription de qoxABCD nrsquoa eacuteteacute observeacutee entre les mutants menD et leur souche
parente (Kohler et al 2008)
Des mutations dans les gegravenes menD hemB et ctaA peuvent reacutesulter en un
pheacutenotype SCV posseacutedant une chaine de transport des eacutelectrons deacuteficiente
(Clements et al 1999 Proctor et Peters 1998 ) Les gegravenes codant pour les eacutetapes
de formation de lrsquohegraveme se preacutesentent sous la forme drsquoun opeacuteron lrsquoopeacuteron
hemAXCDBL Chacun des gegravenes est essentiel (sauf hemL) et la deacuteleacutetion drsquoun seul
drsquoentre eux empecircchera la formation de lrsquohegraveme moleacutecule essentielle agrave la formation
des cytochromes (Taber 1993) hemB code pour la porphobilinogene synthase qui
catalyse une eacutetape dans la voie de biosynthegravese de lrsquohegraveme (Kohler et al 2008 Taber
- 15 -
1993 Thoumlny-Meyer 1997) Une mutation dans le gegravene ctaA (laquo hegraveme A synthase raquo)
empecircche la biosynthegravese de lrsquohegraveme A lequel est aussi impliqueacute dans la biosynthegravese
des cytochromes et donc altegraverera le fonctionnement de la chaine de transport des
eacutelectrons (Clements et al 1999 Proctor et Peters 1998) La capaciteacute de la bacteacuterie
agrave biosyntheacutetiser de lrsquohegraveme de faccedilon endogegravene est essentielle afin drsquoinitier une
infection systeacutemique Le gegravene menD code pour la 2-succinyl-6-hydroxy-24-
cyclohexadiene-1-carboxylate syntase (SHCHC) qui est une enzyme essentielle agrave la
voie de biosynthegravese des meacutenaquinones (Kohler et al 2008) De plus les
meacutenaquinones sont eacutegalement requises pour le fonctionnement de plusieurs
enzymes impliqueacutees dans des voies meacutetaboliques essentielles telles la malate quinol
oxidoreacuteductase (Mqo2) et la dihydroorotate deacuteshydrogeacutenase (PyrD) (Kohler et al
2008) Les mutants menD seront donc incapables de convertir lrsquooxaloaceacutetate en
malate ducirc agrave la perte de lrsquoactiviteacute du Mqo2 (Kohler et al 2008 Proctor et Peters
1998)
Les bacteacuteries agrave pheacutenotype SCV deacutemontreront eacutegalement un gradient eacutelectrochimique
nettement plus faible que les bacteacuteries agrave pheacutenotype normal de par lrsquoincapaciteacute de la
chaine de transport des eacutelectrons agrave y pourvoir
De par leur chaine de transport des eacutelectrons interrompue les SCV sont incapables
drsquoutiliser lrsquoO2 et le nitrate comme accepteur final drsquoeacutelectrons (Kohler et al 2008) Par
ce fait les proteacuteines impliqueacutees dans la voie de la glycolyse ainsi que dans la voie de
la fermentation sont surexprimeacutees (Pfl Ldh Adh) alors que les enzymes faisant
partie du cycle du citrate (TCA) sont significativement reacuteguleacutees agrave la baisse (Kohler et
al 2003 Kohler et al 2008 Kriegeskorte et al 2011) Ceci indique clairement que
les SCV (ex hemB) geacutenegraverent leur ATP agrave partir du glucose ou fructose seulement par
la phosphorylation des substrats Lrsquoanalyse de la reacuteaction de fermentation indique
que le produit principal est le lactate Les SCV reacutegeacutenegraverent leur NADH en reacuteduisant le
pyruvate en lactate par la voie fermentaire (Kohler et al 2003 Kohler et al 2008)
- 16 -
emmenant ainsi une acidification du milieu (Kriegeskorte et al 2011) La conversion
du pyruvate en aceacutetate via lrsquoacetyl-CoA est restreinte chez les mutants hemB mecircme
si la pyruvate formate lyase (Pfl) et les composantes du pyruvate deacuteshydrogeacutenase
sont preacutesentes (Figure 7) Lrsquoactiviteacute de la pyruvate deacuteshydrogeacutenase est tregraves faible
probablement en lien avec le haut niveau de NADH chez les SCV et il est preacutesumeacute
que la Pfl nrsquoest pas activeacutee en preacutesence drsquooxygegravene (La pyruvate deacuteshydrogeacutenase
converti le pyruvate en Co-A en condition anaeacuterobie) (Figure 7) Chez les mutants
hemB la voie de lrsquoarginine deacuteiminase est aussi induite et pourrait eacutegalement
participer agrave la production drsquoATP (Kohler et al 2003) Les gegravenes exprimeacutes chez les
mutants hemB sont similaires agrave ceux exprimeacutes chez les mutants menD ainsi que
chez les souches agrave pheacutenotypes normaux en condition anaeacuterobie (Kohler et al 2008)
- 17 -
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Figure 7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies Les enzymes suivantes sont retrouveacutees agrave une forte
concentration chez les mutants hemb phosphoglucomutase (Pgi) fructose
bisphosphate aldolase (FbaFda) glyceacuteraldeacutehyde deacuteshydrogeacutenase (Gap)
phosphoglyceacuterate kinase (Pgk) eacutenolase (Eno) pyruvate kinase (Pyk) L-lactate
deacuteshydrogeacutenase (L-Ldh) pyruvate formate lyase (PflB) alcool deacuteshydrogeacutenase
(Adh) La formation drsquoaceacutetate et de formate et tregraves limiteacutee (X rouges)
Modifieacute de Kolher C et al 2003
- 18 -
La chaine respiratoire en anaeacuterobie (S aureus)
Il existe 3 voies distinctes de geacuteneacuteration drsquoeacutenergie chez les bacteacuteries 1- en preacutesence
drsquooxygegravene la bacteacuterie respire de faccedilon aeacuterobique et lrsquooxygegravene est lrsquoaccepteur final
drsquoeacutelectron 2- la bacteacuterie peut aussi respirer de maniegravere anaeacuterobique lorsqursquoun
accepteur terminal drsquoeacutelectron autre que lrsquooxygegravene est preacutesent 3- en absence
drsquooxygegravene et drsquoun accepteur drsquoeacutelectrons alternatif la bacteacuterie peut produire de
lrsquoeacutenergie de faccedilon limiteacutee par fermentation (Hammer et al 2013)
Chez S aureus le nitrate (NO3-) et dans un troisiegraveme temps le nitrite (NO2
-)
peuvent ecirctre utiliseacutes comme accepteur final drsquoeacutelectron alternatif lorsque lrsquooxygegravene
nrsquoest pas disponible (respiration anaeacuterobie) Les enzymes de cette voie meacutetabolique
ne sont retrouveacutees que chez les bacteacuteries (Goumltz et al 2006) La croissance
bacteacuterienne est consideacuterablement reacuteduite lorsque la bacteacuterie passe drsquoune respiration
aeacuterobie agrave une respiration anaeacuterobie La nitrate reacuteductase fonctionne probablement in
vivo gracircce au L-lactate comme principal donneur drsquohydrogegravene (Burke et Lasccelles
1975) La deacuteshydrogeacutenase primaire pour ce substrat est preacutesumeacutement la L-lactate
deacuteshydrogeacutenase associeacutee agrave la membrane laquelle meacutedie lrsquooxydation du lactate par la
chaine respiratoire aeacuterobique (Burke et Lasccelles 1975) La preacutedominance du
lactate parmi les produits finaux de la fermentation par S aureus est correacuteleacutee avec
son efficaciteacute comme principal donneur drsquohydrogegravene lorsque le nitrate est lrsquoaccepteur
final drsquoeacutelectrons Le transfert des eacutelectrons du Lactate vers le nitrate implique des
composantes qui sont eacutegalement preacutesentes dans la chaine respiratoire La nitrate
reacuteductase du type respiratoire est habituellement associeacutee agrave la membrane
cytoplasmique (Burke et Lasccelles 1975) Les reacuteductases sont inhibeacutees par
lrsquoazoture et le cyanure (Burke et Lascelles 1976)
Lrsquoenzyme membranaire nitrate reacuteductase NarGHI geacutenegravere des nitrites lesquels seront
reacuteduits en ammonium (NH4+) par lrsquoenzyme cytoplasmique NADH-deacutependant nitrite
reacuteductase encodeacutee par les gegravenes nirBD (Goumltz et al 2006) La nitrate reacuteductase 1-
- 19 -
est reacuteprimeacutee et inhibeacutee par la preacutesence drsquooxygegravene 2- est induite par lrsquoanaeacuterobiose et
la preacutesence de nitrate ou nitrite 3- nrsquoest pas inhibeacutee par lrsquoammoniac 4- permet la
production drsquoeacutenergie dans des conditions anaeacuterobies et 5- est localiseacutee dans la
membrane (Goumltz et al 2006) La preacutesence drsquoun systegraveme de senseur pour lrsquooxygegravene
est cruciale pour le controcircle transcriptionnel des gegravenes facilitant lrsquoadaptation de la
bacteacuterie agrave lrsquoanaeacuterobiose ou aux conditions microaeacuterophiles Chez S aureus il existe
deux systegravemes de senseur 1- le systegraveme SrrAB (pour staphylococcal respiratory
response AB synonyme SrhSR) (Figure 8) homologue agrave ResDE chez Bacillus
subtilis (Yan et al 2011) et 2- le systegraveme NreABC (pour nitrogen regulation chez S
carnosus correspondant agrave SA2181 (NreA) SA2180 (NreB) SA2179 (NreC) chez S
aureus) NreBC est un systegraveme agrave deux composantes qui mesure la concentration
drsquooxygegravene directement par un groupement Fe-S de type Fnr lequel est instable en
preacutesence drsquooxygegravene et par lequel est controcircleacute le systegraveme membranaire de reacuteduction
du nitratenitrite (Yan et al 2011) Srr est aussi un systegraveme agrave deux composantes qui
reacutegule positivement la lactate deacuteshydrogeacutenase et lrsquoalcool deacuteshydrogeacutenase et semble
reacuteguler neacutegativement plusieurs enzymes du cycle du citrate (TCA) (Figure 8)
Lrsquoexpression des enzymes fermentaires comme la lactate deacuteshydrogeacutenase (LDH)
pyruvate formate lyase (PFL) et lrsquoalcool deacuteshydrogeacutenase sont aussi fortement induite
dans des conditions aeacuterobies lorsque la chaine de transport des eacutelectrons est
interrompue (Kohler et al 2003) Donc il se peut que lrsquoeacutetat reacuteduit des composantes
de la chaine respiratoire le potentiel membranaire etou le niveau de NADH soit un
signal pour la reacutegulation des gegravenes laquo anaeacuterobies raquo chez S aureus Dans des
conditions aeacuterobies la conversion du pyruvate en aceacutetyl-coA est catalyseacutee par le
complexe pyruvate deacuteshydrogeacutenase (PDH) et caracteacuteriseacutee par la production de
NADH (Somerville et Proctor 2009) En condition anaeacuterobie lrsquoexpression de la
pyruvate deacuteshydrogeacutenase (PDH) est fortement reacuteprimeacutee alors que lrsquoexpression de la
pyruvate formate lyase (PFL) est fortement induite Lrsquoactiviteacute de la PDH est
remplaceacutee en anaeacuterobiose par lrsquoactiviteacute de la PFL qui geacutenegravere lrsquoacetyl-CoA et le
formate agrave partir du pyruvate Le NADH nrsquoeacutetant pas reacuteoxydeacute par une chaine de
- 20 -
transport des eacutelectrons fonctionnelle celui-ci est recycleacute par la reacuteduction du pyruvate
en lactate (Kohler et al 2003 Somerville et Proctor 2009)
La meacutenaquinone et lrsquohegraveme sont esssentiels agrave la respiration anaeacuterobie (Kohler et al
2008) Le pheacutenotype de S aureus en condition anaeacuterobie est tregraves similaire au
pheacutenotype SCV (Somerville et Proctor 2009)
Figure 8 Modegravele de reacutegulation du cycle TCA (Pour simplifier les reacutegulateurs de
facteurs de virulence bien caracteacuteriseacutes [ie RNAIII Rot SarA et σB] ont eacuteteacute omis
dans ce modegravele)
Les sous-produits de la phosphorylation oxydative les ROS
La reacuteduction de lrsquooxygegravene en eau se fait un eacutelectron agrave la fois (Turrens 2003) Le
complexe IV retient tous les intermeacutediaires partiellement reacuteduits jusqursquoagrave ce que la
Tireacute de Somerville GA Proctor RA 2009
- 21 -
reacuteduction soit complegravetement termineacutee (Turrens 2003) Cependant certains autres
centres redox de la chaine respiratoire laissent passer des eacutelectrons agrave lrsquooxygegravene
reacuteduisant partiellement cette moleacutecule en anion superoxyde (O2-bull) (Liu et al 2002
Turrens 2003) Mecircme si O2-bull nrsquoest pas un agent oxydant tregraves puissant il est le
preacutecurseur de plusieurs autres laquo reactive oxygen species raquo (ROS) et il est aussi
impliqueacute dans la propagation de la chaine de reacuteaction oxydative (Turrens 2003)
LrsquoO2-bull est un intermeacutediaire instable qui est rapidement converti en H2O2 par les
superoxide dismutase (SOD) (S aureus en possegravede deux SodA (le principal) et
SodM) (10 18) ou se convertit de faccedilon spontaneacutee lorsqursquoen concentration eacuteleveacutee
(Starkov 2010 Turrens 2003) Le H2O2 peut ensuite ecirctre complegravetement reacuteduit en
H2O ou partiellement reacuteduit en radical hydroxyle (bullOH) un des plus puissants
oxydants (Starkov 2010) LrsquoO2-bull peut eacutegalement reacuteagir avec drsquoautres radicaux
incluant lrsquooxyde nitrique (NObull) selon le taux de diffusion des 2 radicaux (Turrens
2003) Le peroxynitrite est eacutegalement un oxydant puissant (Turrens 2003) Les
oxydants deacuterivant du NObull sont appeleacutes des laquo reactive nitrogen species raquo (RNS)
(Turrens 2003) mais seront consideacutereacutes comme faisant partie des ROS dans ce
travail Les ROS peuvent endommager lrsquoADN lrsquoARN les proteacuteines et les lipides
reacutesultants agrave lrsquoapoptose de la cellule lorsque le niveau de ROS produit excegravede la
capaciteacute de deacutetoxifications et de reacuteparation de la cellule (Brynildsen et al 2013
Wells et al 2005) Malgreacute ce danger il srsquoagit drsquoun risque compenseacute par
lrsquoaugmentation du rendement eacutenergeacutetique (Brynildsen et al 2013) Le complexe I (et
II) et III sont les principaux sites de production de ROS (Batandier et al 2006 Chen
et al 2003 Liu et al 2002) Lrsquoinhibition du complexe I par la rotenone provoque une
augmentation de la production de ROS Lrsquooxydation des substrats du complexe I ou
du complexe II en preacutesence du complexe III inhibeacute par lrsquoantimicine A amegravene
eacutegalement une augmentation de la production de ROS (Batandier et al 2006 Chen
et al 2003) (Figure 9) De plus mecircme si le complexe cytochrome oxydase nrsquoest pas
un site de production de ROS son inhibition par le KCN (cyanure de potassium)
facilite la production de ROS par les complexes I et II (Chen et al 2003) Le
- 22 -
complexe ATP synthase nrsquoest pas reconnu pour produire directement des ROS
Cependant la liaison drsquoun inhibiteur tel lrsquooligomycine bloque le passage des protons
agrave travers le complexe et inhibe ainsi la production drsquoATP De plus lrsquoATP synthase ne
fonctionnant plus les pompes agrave protons de la chaine de transport des eacutelectrons sont
incapables de fonctionner car le gradient de protons est beaucoup trop eacuteleveacute ce
provoque une surproduction de ROS par les complexes I et III Conseacutequemment le
NADH nrsquoest plus oxydeacute et le cycle du TCA cesse drsquoopeacuterer car la concentration de
NAD nrsquoest plus suffisante pour le fonctionnement des enzymes du TCA (Joshi et
Huang 1991)
Une production significative de O2-bull principalement par le complexe I peut ecirctre le
reacutesultat de deux modes drsquoopeacuteration 1- lorsque la cellule ne produit pas drsquoATP et
conseacutequemment possegravede une grande force proton-motrice et possegravede un laquo pool raquo de
quinone agrave lrsquoeacutetat reacuteduit (QH2) 2- lorsque la cellule possegravede un haut ratio de
NADHNAD (Figure 10) Lorsque la cellule produit activement de lrsquoATP elle possegravede
alors une moins grande force proton-motrice son ratio NADHNAD est plus faible et
la formation de O2-bull est beaucoup plus faible (Murphy 2009)
Le H2O2 est le substrat pour deux enzymes la catalase et la glutathione (GSH)
peroxydase qui convertissent le H2O2 en H2O+O2 le H2O2 est ainsi deacutetoxifieacute (Figure
9 Figure 11) La reacuteaction catalyseacutee par la glutathione peroxydase neacutecessite de la
glutathione (GSH) comme substrats et deacutepend en partie du ratio de GSSG GSH et
de la concentration des reacuteactants laquelle est le reflet de lrsquoeacutetat reacutedox de la cellule De
faccedilon similaire la concentration de meacutetaux sous leur forme reacuteduite tel le Fer catalyse
la formation de certains ROS (Figure 11) Ceci peut ecirctre minimiseacute en tenant cette
concentration ions meacutetallique tregraves basse entre autres par leur emmagasinage et leur
liaison aux proteacuteines de transport (ei ferritine transferrine lactoferrine) et ainsi
minimiser la formation de bullOH Finalement certains antioxydants (eg vitamine E)
peuvent interrompre la chaine de reacuteactions par la capture des radicaux le radical-
vitamine E est relativement stable et peut ecirctre enzymatiquement reconverti agrave sa
- 23 -
forme non-radical (Figure 11) Il est maintenant accepteacute que certaines enzymes du
cycle de Krebs (ie αKGDH) soient non seulement des cibles des ROS mais aussi
des producteurs non neacutegligeables de ROS (Brynildsen et al 2013)
Figure 9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS Le complexe I ainsi que le site Qi du complexe III relacircchent des
ROS directement vers les deacutefenses drsquoantioxydants intracellulaires Les antioxidants
intracellulaire tel la superoxide dismutase (MnSOD) la catalase la glutathione (GSH)
la glutathione peroxydase (GPX) et la glutathione reacuteductase (GRase) sont montreacute Le
site Qo du complexe III quant agrave lui relacircche les ROS dans le milieu extra cellulaire
Modifieacute de Chen Q et al 2003
Oligomycine
- 24 -
Figure 10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de
O2-bull 1- Le pool de NADH est dans un eacutetat reacuteduit par exemple par des dommages agrave
la chaine respiratoire la perte de cytochrome c durant lrsquoapoptose ou agrave un faible
besoin en ATP Ceci megravene agrave une production de O2-bull au complexe I qui deacutecoule de la
reacuteduction du FMN laquelle est reacuteguleacutee par le ratio NADHNAD+ Drsquoautres sites
autres que le complexe I tel lrsquoα-ceacutetoglutatrate deacuteshydrogeacutenase (αKGDH) peut
aussi contribuer agrave la production de O2-bull 2- Il nrsquoy a pas (peu) de production drsquoATP il y
a un fort potentiel membranaire et un pool de CoQ (meacutenaqinone) agrave lrsquoeacutetat reacuteduit
important qui provoque une reacuteversion de la chaine de transport des eacutelectrons agrave
travers le complexe I amenant une forte production de O2-bull 3- Il y a une production
active drsquoATP et conseacutequemment un potentiel membranaire plus bas que dans la
situation 2 et une plus grande quantiteacute de NADH oxydeacute que dans la situation 1
Dans ces conditions la quantiteacute de O2-bull produit est plus faible que dans la situation
1 et 2 et la provenance des O2-bull est moins certaine
Tireacute de Murphy MP 2009
- 25 -
12 Antibiotiques Mode drsquoaction cible cellulaire et meacutecanismes de reacutesistance
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles
Notre compreacutehension de la faccedilon que les antibiotiques induisent la mort cellulaire est
principalement baseacutee sur la fonction cellulaire essentielle inhibeacutee par lrsquointeraction de
lrsquoantibiotique avec sa cible Les antibiotiques peuvent ecirctre classeacutes selon la
composante ou le systegraveme cellulaire qursquoils affectent ainsi que par le fait qursquoils
induisent la mort cellulaire (antibiotiques bacteacutericides) ou srsquoils inhibent la croissance
Figure 11 Voies de formation et de deacutetoxification des ROS Tireacute de Wikipeacutedia
httpfrwikipediaorgwikiDC3A9rivC3A9_rC3A9actif_de_l27oxygC3A8ne
Consulteacute le 11 deacutec 2013
- 26 -
bacteacuterienne (antibiotique bacteacuteriostatique) La plupart des antibiotiques bacteacutericides
inhibent la synthegravese de lrsquoADN ou de lrsquoARN la synthegravese de la paroi bacteacuterienne ou la
synthegravese proteacuteique (Kohanski et al 2010) Le tableau 1 est une synthegravese des
principales classes drsquoantibiotiques ainsi que leur mode drsquoaction et leurs cibles
cellulaires Ce tableau nrsquoest cependant pas exhaustif et il existe plusieurs autres
classes drsquoantibiotiques et plusieurs autres exemples drsquoantibiotiques dans les classes
identifieacutees dans ce tableau
Depuis la deacutecouverte de la peacutenicilline en 1929 plusieurs autres antibiotiques ont eacuteteacute
deacutecouverts et deacuteveloppeacutes suite agrave lrsquoeacutelucidation des interactions entre lrsquoantibiotique et
sa cible et en apportant des modifications structurales et chimiques aux moleacutecules
deacutejagrave connues Ces efforts ont significativement augmenteacute le nombre drsquoantibiotiques
disponibles dans le milieu clinique Le processus de mort cellulaire induit par lrsquoaction
drsquoun antibiotique est complexe et deacutebute avec une interaction physique entre la
moleacutecule antibiotique et sa cible bacteacuterienne speacutecifique et implique des modifications
qui affectent la bacteacuterie tant au niveau biochimique que moleacuteculaire et structural
27
Acid
e n
alid
ixiq
ue (
1 iegravere
geacuteneacutera
tion)
Cip
rofloxacin
e (
2 iegravem
e g
eacuteneacutera
tion)
Levofloxacin
e (
3 iegravem
e g
eacuteneacutera
tion)
Gem
iflo
xacin
e (
4 iegravem
e g
eacuteneacutera
tion)
Flu
oro
quin
olo
ne (
Norf
loxacin
e)
Peacutenic
iline
Ceacutephalo
sporine (
ceacutefa
lexin
e)
Carb
apenem
Vancom
ycin
e
Teacutela
vancin
e
Fosfo
mycin
Dapto
mycin
Poly
mix
ine B
Bacitra
cin
Valin
om
ycin
e
Nom
encin
e
Genta
mic
in
Kanam
ycin
e
Tobra
mycin
e
Str
epto
mycin
e
Teacutetr
acyclin
e
Chlo
rteacutetr
acyclin
e
Macro
lides
Ery
thro
mycin
eLie
sous-u
niteacute 23S
du r
ibosom
e 5
0S
et em
pecircche
leacutelo
ngation d
u p
eptide (
apregrave
s 6
-8 a
a)
sta
tique
Lin
cosam
ides
Clin
dam
ycin
Inhib
e d
irecte
ment la
ctiviteacute p
eptidyl
transfeacute
rase d
u
ribosom
esta
tique
Str
epto
gra
min
es
DalfopristinQ
uin
upristin
Com
bin
ais
on d
e 2
antibio
tiques B
loque la
transla
tion p
eptidiq
ue e
n lia
nt le
site P
du r
ibosom
e
et en b
loquant au p
eptide la
ccegraves a
u tunnel de s
ort
ie
cid
al
Am
phenic
ol
Chlo
ram
phenic
ol
Em
pecircche la lia
ison d
u a
a-
AR
Nt au s
ite A
du
ribosom
e e
t in
terf
egravere
dans la f
orm
ation d
u lie
n
peptidiq
ue
sta
tique
Oxazolid
inones
Lin
ezolid
Blo
que initia
tion d
e la tra
nsla
tion p
roteacute
ique lors
de la
reacuteaction d
eacutelo
ngation p
ar
la p
eptidyl
transfeacute
rase
sta
tique
Trim
eth
oprim
Sulfam
eth
oxazole
sta
tique
cid
al
Ionophore
Alteacutera
tion d
e lin
teacutegriteacute
mem
bra
naire
Se
nchasse d
ans la m
em
bra
ne fo
rme u
n p
ore
ce
qui am
egravene u
ne p
ert
urb
ation d
e leacute
quili
bre
ioniq
ue
cid
al
Inhib
itio
n d
e la s
ynth
egravese d
e
la p
aro
i cellu
laire
(peptidogly
can)
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
cid
al
cid
al
sta
tique
AR
Nr
16S
(S
ite A
)(fo
rmation d
e p
roteacute
ines n
on-
fonctionnelle
s)
cid
al
Inhib
itio
n d
e la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 3
0S
)
Inhib
iteur
de la s
ynth
egravese d
e
lacid
e f
oliq
ue
Inhib
itio
n d
e c
ert
ain
es r
eacuteactions e
nzym
atiques
essentielle
s agrave
la f
orm
ation d
e la
cid
e f
oliq
ue
Sulfonam
ides
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
AR
Nr
16S
(preacute
vie
nt la
ttachem
ent de la
min
oacyl-
AR
Nt au r
ibosom
e)
Teacutetr
acyclin
e
Poly
peptides
β-L
acta
ms
Gly
copeptides
Lip
opeptides
Inhib
iteur
de la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 5
0S
)
Am
inogly
cosid
es
Cib
le c
ell
ula
ire
Sta
tiq
ue
Cid
al
Rifam
ycin
eR
ifam
pic
ine
Cla
ss
es
Ex
d
an
tib
ioti
qu
eM
od
e d
ac
tio
n
Inhib
itio
n d
e la s
ynth
egravese d
e
lAR
N (
AD
N d
eacutependant)
AR
N p
oly
meacutera
se s
ous-u
niteacute β
(codeacute p
ar
rpoB
)
Quin
olo
nes
cid
al
Inhib
itio
n d
e la r
eacuteplic
ation d
e
lAD
N
Com
ple
xe A
DN
-topois
om
eacutera
se (
AD
N
gyr
ase(t
opois
om
eacutera
se II)
et to
pois
om
eacutera
se IV
)cid
al
Tra
nspeptidase (
penic
illin
bin
din
g p
rote
in
(PB
P))
cid
al
Tab
leau
1
Cib
les e
t m
od
e d
rsquoacti
on
des p
rin
cip
ale
s c
lasses d
rsquoan
tib
ioti
qu
es
28
122 Les aminoglycosides
1221 Mode drsquoaction
Les aminoglycosides peacutenegravetrent la paroi cellulaire en suivant un processus
comprenant 3 eacutetapes une premiegravere eacutetape eacutenergie- indeacutependante suivie de deux
eacutetapes eacutenergie deacutependantes Deux composantes sont essentielles pour
lrsquoaccumulation intracellulaire des aminoglycosides les ribosomes et la chaine de
transport des eacutelectrons (Ramirez et Tolmasky 2010)
Eacutetape 1- Lorsque les aminoglycosides entrent en contact avec la paroi
cellulaire la moleacutecule polycationique se lie aux composantes de surface anioniques
tels les lipopolysaccharides les phospholipides et autres proteacuteines membranaires
chez les bacteacuteries agrave Gram neacutegatif et les moleacutecules drsquoacides teacuteichoiumlques et les
phospholipides des bacteacuteries agrave Gram positif (Ramirez et Tolmasky 2010) Apregraves
cette liaison aux moleacutecules polaires les aminoglycosides (telle la gentamicine)
srsquoassocient aux transporteurs membranaires sur la base de leur charge positive et
sont transporteacutes agrave travers la membrane cytoplasmique gracircce au potentiel
membranaire (intra cellulaire neacutegatif) (Bryan et al 1980 Proctor et al 2006) Cette
liaison agrave la membrane entraine une augmentation de la permeacuteabiliteacute membranaire
menant agrave lrsquoentreacutee des aminoglycosides dans lrsquoespace peacuteriplasmique (Gram neacutegatif)
ou dans le cytoplasme (Gram positif) (Ramirez et Tolmasky 2010) (Figure 12A)
Eacutetape 2- Lrsquoeacutetape suivante connue comme la laquo phase eacutenergie-deacutependante
1 raquo (EDP-I) est bloqueacutee par les inhibiteurs de la chaine de transport des eacutelectrons et
de la phosphorylation oxydative ainsi que par les ions divalents lrsquohyperosmolariteacute le
faible pH et lrsquoanaeacuterobiose dans cette phase la dureacutee et le taux drsquoentreacutee de
lrsquoantibiotique sont en fonction de la concentration (Miller et al 1980 Mingeot-
Leclercq et al 1999 Ramirez et Tolmasky 2010) Agrave cette eacutetape le transport
transmembranaire des aminoglycosides et plus speacutecifiquement de la gentamicine
neacutecessite 2 composantes majeures a) un transporteur anionique non speacutecifique dont
la charge est directement lieacutee agrave lrsquoactiviteacute des cytochromes oxydases terminales et des
29
nitrates reacuteductases de la chaine de transport des eacutelectrons Selon certaines eacutetudes
les transporteurs pourraient ecirctre les quinones reacuteduites de la chaine de transport des
eacutelectrons (Bryan et al 1981 Miller et al 1980) et donc le cycle drsquooxydation-reacuteduction
des quinones est essentiel aux transports des aminoglycosides (Bryan et al 1981)
mais selon drsquoautres eacutetudes les transporteurs pourraient ecirctre toute autre proteacuteine non
deacutefinie impliqueacutee dans la polarisation membranaire lieacutee agrave lrsquoactiviteacute de la chaine de
transport des eacutelectrons (Miller et al 1980) La croissance anaeacuterobie ainsi que le
sodium azide (inhibiteur de la chaine respiratoire) inhibent lrsquoentreacutee de la gentamicine
et lrsquoeffet bacteacutericide indiquant que le transport actif de cet antibiotique neacutecessite la
respiration b) un potentiel eacutelectrique avec un milieu intra cellulaire neacutegatif par rapport
agrave lrsquoexteacuterieur En effet lrsquoinhibition de lrsquoentreacutee dans la bacteacuterie des aminoglycosides
suite agrave lrsquoajout de CCCP (agent deacutepolarisant) chez des souches de S aureus laquo wild-
type raquo deacutemontre la neacutecessiteacute de la force proton-motrice pour lrsquoentreacutee des
aminoglycosides dans les cellules (Miller et al 1980)
Eacutetape 3- La petite quantiteacute drsquoaminoglycosides srsquoeacutetant rendu au cytoplasme
durant cette premiegravere phase eacutenergie-deacutependante induisent apregraves liaison aux
ribosomes des erreurs dans la synthegravese proteacuteique Les erreurs dans la traduction
des proteacuteines membranaires produites lors de cette eacutetape connue comme la laquo phase
eacutenergie-deacutependante 2 raquo (EDP-II) provoquent des dommages dans lrsquointeacutegriteacute de la
membrane cytoplasmique (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky
2010) Les proteacuteines membranaires aberrantes dans la membrane facilitent le
transport drsquoune plus grande quantiteacute drsquoaminoglycosides lesquels augmenteront
encore le niveau drsquointerfeacuterence avec la synthegravese proteacuteique normale amenant encore
plus de dommage agrave la membrane provoquant ainsi une acceacuteleacuteration importante du
taux drsquoentreacutee des moleacutecules antibiotique et qui ultimement reacutesultera en la mort de la
cellule (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky 2010) (Figure 12B)
Des eacutetudes ont deacutemontreacute que la susceptibiliteacute agrave la gentamicine streptomycine
amikacine kanamycine et tobramycine est supeacuterieure lorsque lrsquooxygegravene est
30
lrsquoaccepteur terminal drsquoeacutelectrons (Bryan et Kwan 1981) Une diminution significative
de la susceptibiliteacute est remarqueacutee lorsque le nitrate est lrsquoaccepteur final drsquoeacutelectrons et
une diminution encore plus marqueacutee est observeacutee lorsque le fumarate est lrsquoaccepteur
final drsquoeacutelectron ou lorsque les conditions sont fermentaires Lrsquoassociation entre le
transport de la gentamicine et le complexe terminal de la chaine de transport des
eacutelectrons particuliegraverement lrsquoactiviteacute cytochrome oxydase
Le ribosome possegravede trois sites de liaison importants de lrsquoARNt deacutesigneacute comme site
A (pour aminoacyl) site P (pour peptidyl) et E (pour exit) (Vakulenko et Mobashery
2003) Ils sont situeacutes sur lrsquoARN16S (lequel avec plus de 21 proteacuteines compose la
sous-uniteacute 30S du ribosome bacteacuterien) (Ramirez et Tolmasky 2010 Vakulenko et
Mobashery 2003) Durant la synthegravese proteacuteique le ribosome deacutecode lrsquoinformation de
lrsquoARNm et catalyse lrsquoincorporation seacutequentielle de chaque acide amineacute faisant partie
de la chaine polypeptidique en formation La haute fideacuteliteacute de la traduction (les
ribosomes incorporent un mauvais acide amineacute par approximativement 3000 liaisons
peptidiques effectueacutees) est possible gracircce agrave la capaciteacute de discriminer entre des
changements conformationels dans le ribosome induit par la liaison drsquoun ARNt correct
Modifieacute de Davis BD 1987
Figure 12 Cineacutetique drsquoentreacutee des aminoglycosides
A
B
31
ou incorrect au site A La reconnaissance drsquoune liaison correcte anticodoncodon par
le ribosome se passe en deux temps la reconnaissance initiale de lrsquoappariement du
codon sur lrsquoARNm avec lrsquoanticodon de lrsquoARNt et subseacutequemment la correction
(Vakulenko et Mobashery 2003)
La plupart des aminoglycosides se lient agrave des oligonucleacuteotides faisant partie du site
A Les diffeacuterents aminoglycosides ne lient pas tous le mecircme site de lrsquoARN 16S mais
lrsquoeffet commun agrave tous est que cette liaison amegravene un changement de conformation du
site A vers une conformation mimant la conformation fermeacutee de la sous uniteacute 30S qui
est induite par une interaction entre lrsquoARNm et son ARNt apparenteacute eacuteliminant la
possibiliteacute de correction du ribosome ce qui promeut les erreurs de traduction
(Ramirez et Tolmasky 2010 Vakulenko et Mobashery 2003) La grande majoriteacute des
aminoglycosides sont bacteacutericides et cette leacutetaliteacute est la conseacutequence de cette
mauvaise traduction (Vakulenko et Mobashery 2003)
1222 Meacutecanismes de reacutesistance
La reacutesistance contre les aminoglycosides se produit par diffeacuterents meacutecanismes qui
peuvent coexister simultaneacutement dans la mecircme bacteacuterie (Vakulenko et Mobashery
2003) Ces meacutecanismes incluent 1- la modification de la cible par des mutations
dans lrsquoARN 16S ou autres proteacuteines ribosomales associeacutees (Figure 13) 2- la
meacutethylation de lrsquoARN 16S un meacutecanisme retrouveacute chez plusieurs organismes
producteurs drsquoaminoglycosides et chez des souches cliniques Cette meacutethylation
empecircche les aminoglycosides drsquoavoir accegraves agrave leur site de liaison (Doi et Arakawa
2007) 3- la reacuteduction de la permeacuteabiliteacute par une alteacuteration de la permeacuteabiliteacute de la
membrane externe ou une diminution du transport agrave travers la membrane interne 4-
lrsquoexportation des aminoglycosides agrave lrsquoexteacuterieur de la cellule par les pompes agrave efflux
Eacutetant des moleacutecules hydrophiliques les aminoglycosides ont naturellement peu
drsquoaffiniteacute avec les pompes agrave efflux 5- le swarming actif probablement un meacutecanisme
32
non speacutecifique deacutemontreacute chez Pseudomonas aeruginosa lequel deacutemontre des
reacutesistances agrave plusieurs antibiotiques 6- la seacutequestration de la drogue par des liaisons
agrave un aceacutetyltransfeacuterase agrave tregraves faible activiteacute 7- une inactivation enzymatique de la
moleacutecule antibiotique le meacutecanisme de reacutesistance le plus preacutevalent au niveau
clinique (Vakulenko et Mobashery 2003) Lrsquoinactivation enzymatique des
aminoglycosides est le plus souvent due agrave des aceacutetyltransfeacuterases des
nucleacuteotidyltransfeacuterases ou des phosphotransfeacuterases (Dickgiesser et Kreiswirth 1986)
Ces enzymes sont souvent encodeacutees sur des plasmides mais sont eacutegalement
associeacutees agrave des eacuteleacutements transposables Lrsquoeacutechange de plasmides et la disseacutemination
de transposons facilitent lrsquoacquisition rapide de pheacutenotypes reacutesistants aux
antibiotiques non seulement au sein drsquoune mecircme espegravece mais parmi un large
eacuteventail drsquoespegraveces bacteacuteriennes (Mingeot-Leclercq et al 1999)
La reacutesistance naturelle des S aureus laquo small colony variants raquo agrave la majoriteacute des
aminoglycosides est la conseacutequence de lrsquoimpossibiliteacute agrave lrsquoantibiotique de peacuteneacutetrer une
cellule posseacutedant une faible polariteacute membranaire qui est due chez les SCV agrave leur
Tireacute de Doi Y Arakawa Y 2007
Figure 13 Sites de mutation de certains reacutesistants aux aminoglycosides
33
incapaciteacute agrave activer le cycle de Krebs (Miller et al 1980 Proctor et al 2006 Tubby
et al 2013) Les SCV ont non seulement une polariteacute membranaire faible mais (ceci
eacutetant la conseacutequence de cela) aussi une force proton motrice inhibeacutee et une chaine
des eacutelectrons deacuteficiente De ce fait le transport transmembranaire des
aminoglycosides chez les SCV est grandement reacuteduit et conseacutequemment la
susceptibiliteacute des SCVs pour cette classe drsquoantibiotique est 110-113 de celle des
souches parentes (Proctor et al 2006)
123 La tomatidine
Plusieurs espegraveces veacutegeacutetales accumulent des steacuterols et des triterpegravenes antimicrobiens
appeleacutes saponines (Osborn 1996) Ces moleacutecules sont produites de faccedilon
constitutive par la plante et jouent un important rocircle de deacutefense contre diffeacuterents
pathogegravenes comme des insectes ou des microbes Parmi les saponines la tomatine
(Figure 14A) est un glycoalcaloiumlde steacuteroiumldien preacutesentant une action antimicrobienne
contre des levures et une quantiteacute drsquoautres microorganismes (Bednarek et Osboum
2009 Friedman 2002 Osborn 1996 Sandrock et Vanetten 1998) Plusieurs
espegraveces de champignons pathogegravenes de la tomate produisent des enzymes
extracellulaires communeacutement appeleacutes tomatinase qui sont capables de deacutetoxifier
lrsquoα-tomatine (Martin-Hernandez et al 2000 Roddick 1974 Ruiz-Rubbio et al 2001)
Par exemple Fusarium oxysporum f sp lycopersici est reconnu pour cliver lrsquoα-
tomatine en ses sous-produits soit un tetrasaccharide (lycotreacutetraose) et un alcaloiumlde
steacuteroiumldien non glycosyleacute la tomatidine (Figure 14B) Ces sous-produits nrsquoont pas ou
peu drsquoactiviteacute antifongique (Ruiz-Rubbio et al 2001 Simon et al 2006)
34
Figure 14 Structures chimiques de la (A) tomatine de (B) tomatidine et de (C)
lrsquoanalogue de tomatidine FC04-100
Il a eacuteteacute deacutecouvert il a quelques anneacutees que la tomatidine avait un effet antibacteacuterien
sur les S aureus de pheacutenotype SCV mais aucune activiteacute contre les S aureus agrave
pheacutenotype normal Par contre lorsqursquoutiliseacute en combinaison avec les
aminoglycosides lrsquoajout de la tomatidine permettait drsquoaugmenter de 8 agrave 16 fois
lrsquoactiviteacute des aminoglycosides contre S aureus agrave pheacutenotype normal (Mitchell et al
2011a Mitchell et al 2012)
Donc on savait que la tomatidine avait une activiteacute antibacteacuterienne tregraves inteacuteressante
mais on ne connaissait ni sa cible ni son mode drsquoaction De plus si la tomatidine a
une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait donc avoir un
spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Par ailleurs de faccedilon parallegravele la production drsquoanalogues structuraux de la tomatidine
permettrait de mieux comprendre la relation structure-activiteacute (SAR) de la moleacutecule et
donc de preacuteciser que lrsquoactiviteacute biologique de la tomatidine neacutecessite une orientation
3
A B
C
35
de groupements fonctionnels bien speacutecifique De plus lrsquoeacutetude de lrsquoactiviteacute de
plusieurs analogues permettrait de confirmer si la tomatidine possegravede un meacutecanisme
drsquoaction deacutependant drsquoune interaction speacutecifique avec une ou des cibles biologiques
preacutecises
Le laboratoire du Pr Eacuteric Marsault du deacutepartement de pharmacologie de lrsquoUniversiteacute
de Sherbrooke et lrsquoeacutetudiant agrave la maitrise Feacutelix Chagnon ont produit plus de 33
analogues de la tomatidine (voir Annexe 2 article laquo Unraveling the structure-activity
relationship of tomatidine a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureusraquo dans la section laquo Contributions
Supleacutementaires raquo du meacutemoire) dont lrsquoactiviteacute antibacteacuterienne a eacuteteacute eacutevalueacutee par des
tests de susceptibiliteacute agrave la moleacutecule (CMI) sur S aureus SCV et sur S aureus agrave
pheacutenotype normal en combinaison avec la gentamicine Plusieurs de ces moleacutecules
font lrsquoobjet drsquoune application de brevet (voir annexe 1) dont la prometteuse moleacutecule
FC04-100 (Figure 14C) (voir laquo Annexe 1 Application de brevet raquo dans la section
laquo Contributions suppleacutementaires raquo du meacutemoire)
13 Projet de maitrise
Premiegravere hypothegravese
Si la tomatidine a une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait
donc avoir un spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Objectif Provoquer lrsquoapparition de pheacutenotypes SCV chez diffeacuterentes espegraveces
bacteacuterienne afin de pouvoir veacuterifier leur niveau de sensibiliteacute agrave la tomatidine
Deuxiegraveme hypothegravese
Lrsquoidentification des gegravenes preacutesentant des mutations parmi les souches de S aureus
reacutesistantes agrave la tomatidine pourrait reacuteveacuteler la ou les cibles cellulaires de la moleacutecule
et nous aider agrave comprendre son mode drsquoaction
36
Objectif Obtenir des mutants reacutesistants avant de les envoyer agrave sequencer
faire les analyses comparatives des geacutenomes et finalement identifier les mutations
37
CHAPITRE 2
ARTICLE SCIENTIFIQUE
TITRE Tomatidine and Analog FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Some Firmicutes Including Those of Listeria
monocytogenes and Are Bactericidal in Combination With Aminoglycosides Against
their Normal Phenotype
AUTEURS ET CONTRIBUTIONS
Isabelle Guay1 Expeacuteriences biologiques avec le produit naturel tomatidine et son
analogue FC04-100 sur diffeacuterentes espegraveces bacteacuteriennes afin de comparer les deux
activiteacutes
Simon Boulanger1 Expeacuterience biologique permettant de quantifier lrsquoactiviteacute de
lrsquoanalogue FC04-100 sur la production de biofilm et son activiteacute intracellulaire
Feacutelix Chagnon2 Production de lrsquoanalogue chimique de la tomatidine FC04-100
Kamal Bouarab1 Initiateur du projet gracircce agrave lrsquoisolement du produit naturel tomatidine
Eacuteric Marsault2 Directeur de recherche de Feacutelix Chagnon
Franccedilois Malouin1 Directeur de recherche drsquoIsabelle Guay et Simon Boulanger et
auteur de correspondance
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
38
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
Corresponding author Mailing address Universiteacute de Sherbrooke
Deacutepartement de Biologie Faculteacute des Sciences 2500 Boul
Universiteacute Sherbrooke Quebec Canada J1K2R1 Phone (819) 821-
8000 ext 61202 Fax (819) 821-8049 E-mail FrancoisMalouin
USherbrookeca
39
Tomatidine and Analog FC04-100 Possess Bactericidal Activities
Against the Small Colony Variants of Some Firmicutes Including
Those of Listeria monocytogenes and Are Bactericidal in Combination
With Aminoglycosides Against their Normal Phenotype
Running Title Tomatidine is bactericidal against several Firmicutes
Isabelle Guay1
Simon Boulanger1
Feacutelix Chagnon2 Kamal Bouarab
1 Eacuteric Marsault
2 and
Franccedilois Malouin1
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM) Deacutepartement de
Biologie Faculteacute des Sciences Universiteacute de Sherbrooke Sherbrooke Queacutebec Canada J1K
2R1
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute Universiteacute
de Sherbrooke Queacutebec Canada J1H 5N4
Have contributed equally to the work
Corresponding author
Mailing address Universiteacute de Sherbrooke Deacutepartement de Biologie Faculteacute des Sciences
2500 Boul Universiteacute Sherbrooke Quebec Canada J1K2R1
Phone (819) 821-8000 ext 61202 Fax (819) 821-8049
E-mail FrancoisMalouinUSherbrookeca
40
ABSTRACT
Tomatidine (TO) is a plant steroidal alkaloid that possesses an antibacterial activity against the
small colony variants (SCVs) of Staphylococcus aureus We report here the spectrum of
activity of TO against species of the Firmicutes and the improved antibacterial activity of a
chemically-modified TO derivative (FC04-100) In addition to that seen on S aureus TO
showed a bactericidal activity against SCVs of Bacillus cereus B subtilis and Listeria
monocytogenes The combination of an aminoglycoside antibiotic and TO also generated an
antibacterial synergy against strains of the normal phenotype in these species In contrast to
TO which has no relevant activity by itself against S aureus of the normal phenotype (MIC
gt64 mgL) the TO analog FC04-100 showed a MIC of 8-16 mgL Furthermore FC04-100
showed a strong bactericidal activity against L monocytogenes SCVs in kill kinetic
experiments while TO did not FC04-100 also improved the bactericidal activity of
gentamicin against L monocytogenes of the normal phenotype The addition of FC04-100 (4
mgL) to a cephalexinkanamycin (32) combination improved the activity of the combination
by 32 fold against cephalexin and kanamycin-resistant MRSA strains In combination with
gentamicin FC04-100 also showed a strong bactericidal activity against biofilm-embedded S
aureus and when used alone was superior to TO for its biofilm-preventing activity Finally
FC04-100 and TO showed comparable intracellular killing of S aureus SCVs in a polarized
Calu-3 cell culture model In conclusion chemical modifications of TO allowed improvement
of its antibacterial activity against L monocytogenes SCVs and prototypical S aureus
41
Keywords Tomatidine aminoglycoside synergy SCV Firmicutes S aureus L
monocytogenes
Introduction
The Firmicutes are Gram-positive bacteria with genomes presenting less than 50 of G+C
The Firmicutes constitutes one of the main divisions within the Bacteria and is divided into
three classes Bacilli Mollicutes and Clostridia The class of Bacilli is further divided into the
orders of the Lactobacillales and the Bacillales The Bacillales are divided into the genus of
Staphylococcus Listeria and Bacillus A number of bacterial species such as Listeria spp and
Bacillus spp can contaminate food and cause infections in humans (Magalhaes 2013) To
name a few Listeria monocytogenes L ivanovii and Bacillus cereus can cause listeriosis
(Guillet et al 2010) and food poisoining (Bad Bug Book FDA) Bacillus subtilis B
coagulans B licheniformis and B sphaericus are also known to cause illnesses Bacillus
anthracis causes anthrax and can often be acquired by contact with food producing animals
and cattle (beef cattle sheeps etc) and this bacterium is also well-known for its endospores
that have been used as biological weapons (Beierlein and Anderson 2011) Staphylococci are
divided in coagulase-positive species Staphylococcus aureus being the most clinically
relevant of this group and coagulase-negative species such as S epidermidis the most
prevalent pathogen associated with infections of implanted medical devices (Vuong and Otto
2002) The emergence and spread of resistance to multiple antibiotics in staphylococci is now
considered a real health treat and impaired therapeutic endeavor to combat these bacteria
42
(Witte et al 2007) Notably the prevalence of methicillin-resistant S aureus (MRSA) has
steadily increased over the recent years not only in hospitals but also in the community
(Hiramatsu et al 2013)
Since their first description in 1910 for Salmonella enterica serovar Typhi (S typhi) small-
colony variants (SCVs) have been described for a wide range of bacterial genera and species
and have been isolated from both laboratory and clinical environments (Proctor 2006)
Staphylococcus aureus SCVs have attracted a great deal of interest over the past recent years
S aureus SCVs often present a dysfunctional oxidative metabolism causing an alteration in
the expression of virulence factors a slow growth and a loss of colony pigmentation (Proctor
2006) This dysfunctional oxidative metabolism is also responsible for a decreased
susceptibility to aminoglycoside antibiotics because this class of molecules requires the
proton-motive force in order to penetrate the bacterium (Bryan LE 1981) This respiratory
deficiency is often caused by mutations affecting the electron transport system and several
SCV isolates can recover normal growth with supplemental hemin or menadione which are
needed to synthesize electron transport system components S aureus SCVs often are isolated
from chronic infections such as lung infections in cystic fibrosis (CF) patients osteomyelitis
septic arthritis bovine mastitis and from infections associated with orthopedic devices (Atalla
2008 Moisan 2006 Proctor 2006) The ability of S aureus to switch back and forth from the
prototypic to the SCV phenotypes in vivo is an integral part of the pathogenesis of S aureus
and may be responsible for the establishment of chronic infections (Tuchscherr 2011
Mitchell 2011 inf imm)
43
Tomatidine (TO) is a steroidal alkaloid produced by the Solanaceae plant family such as the
tomato (Ruiz-Rubio 2001 Simons 2006) We showed previously the antibacterial activity of
TO against S aureus SCVs and also documented a strong synergic activity of TO in
combination with aminoglycoside antibiotics against prototypic S aureus (Mitchell G AAC
2011 Mitchell G 2012 JAC) Recently we synthesized a variety of TO analogs in order to
explore the structure-activity relationship of this new class of antibiotics able to act on SCVs
(Chagnon et al 2014) One analog FC04-100 showed the same steroidal backbone as the
natural molecule but with an additional carbon chain on the A cycle (Fig 1) Preliminary
characterization of FC04-100 revealed an antibacterial activity that was similar to that of TO
against S aureus SCVs and also preserved the synergic activity with aminoglycosides against
prototypic S aureus (Chagnon et al 2014) However contrary to TO FC04-100 showed
notable antibiotic activity by itself against prototypic S aureus with a MIC of 8-16 gL
whereas that of TO was gt64 gL It would be of great interest to see if this property of FC04-
100 modifies or improves its spectrum of antibacterial activity
The aim of this study was to further describe the spectrum activity of TO and FC04-100 The
susceptibility of several species among the Firmicutes to TO and FC04-100 alone or in
combination with other antibiotics was investigated The improved antibacterial activity of
FC04-100 was demonstrated against Listeria monocytogenes and methicillin-resistant S
aureus (MRSA)
44
Materials and methods
Chemical reagents and antibiotics
Tomatidine (TO) (Sigma Aldrich Oakville Ontario Canada) (Fig 1A) was solubilised in
dimethylsulfoxide (DMSO) at a concentration of 2 mgmL The TO analog FC04-100 (Fig
1B) was synthesized (Chagnon et al 2014) and solubilised in DMSO at 20 gL Cefalexin
kanamycin gentamicine (GEN) (all from Sigma Aldrich) were solubilised in water at a
concentration of 10 gL Menadione was solubilized in DMSO hemin in 14 M NH4OH and
thymidine in water (all from Sigma Aldrich) and were prepared at a concentration of 10 gL
45
3
Fig 1 Tomatidine (A) is a steroid alkaloid structurally characterized by 6 rings 12
stereogenic centers a 3 β-hydroxyl group and spiro-fused E F rings in the form of an
aminoketal The tomatidine analog FC04-100 (B) contains a diamine in position 3 of ring A
Bacterial strains and growth conditions
Staphylococcus aureus ATCC 29213 S aureus Newbould (ATCC 29740) Listeria
monocytogenes ATCC 13932 Bacillus subtilis ATCC 6333 and Bacillus cereus ATCC 11778
were used as prototypic strains Methicillin-resistant S aureus (MRSA) strains COL and
USA100 (hospital acquired-MRSA) and strain USA300 (community acquired-MRSA) were
also used in this study The reference laboratory strain S aureus SH-1000 an isogenic mutant
strain derived from S aureus 8523-4 but with a functional rsbU allele (HorsburghMJ 2002)
was used for its good yield of biofilm production The S aureus strains CF07-L (prototypic)
A
B
A B
C D
E
F
46
and CF07-S (SCV phenotype) are genetically related clinical strains originally isolated from a
cystic fibrosis patient (MitchellG 2011) Strain CF07-S was used in the intracellular
infection model Staphylococcus and Bacillus were maintained on Tryptic Soy agar (TSA BD
Mississauga Ontario Canada) whereas L monocytogenes was grown on brain-heart infusion
(BHI) agar (BD Mississauga Ontario Canada)
Small colony variants (SCVs)
For S aureus strain NewbouldhemB was used as the reference SCV NewbouldΔhemB was
generated from strain Newbould (ATCC 29740) by disrupting the hemB gene with the ermA
cassette by homologous recombination (Brouillette 2004) SCVs from B cereus B subtillis
and L monocytogenes were generated by growth in presence of a subinhibitory concentration
of GEN Briefly overnight broth cultures (18ndash20 h) were used to inoculate BHI broth at a
dilution of 1100 supplemented or not with 025 to 1X the MIC of GEN Cultures were
incubated 18 h at 35degC with shaking (225 rpm) and then adjusted to an A595 nm of 20 in PBS at
4degC Determination of CFU and SCV colonies was done by serial dilution plating SCV were
obtained by plating on TSA (BHI agar for L monocytogenes) containing gentamicin at a
concentration of 8 to 16 times the induction concentration followed by an incubation of 48 h at
35degC The pinpoint colonies selected by this method were confirmed to be SCVs by streaking
several of them on agar plates without antibiotic SCV that conserved their phenotype after
two passages were considered to have a stable phenotype and were used in subsequent
experiments Selected SCVs had a colony size a tenth of the size of a normal colony and
showed no pigmentation or hemolytic activity on blood agar plates The SCV species were
47
confirmed by rDNA 16S sequencing after PCR amplification using universal primers (Gomaa
2007)
Auxotrophy assays
For SCVs auxotrophism is defined as the requirement of specific compounds in order to
regain a normal growth phenotype [Sendie 2009] An agar diffusion method was used to
characterize the auxotrophism of SCVs using hemin or menadione (1-10 μg eachwell) or
thymidine (100 microgwell) on an inoculated Mueller-Hinton agar (MHA) or Brain-heart infusion
agar (BHIA) plate [Besier 2007] Auxotrophy for specific supplements was detected by a
zone of normal growth surrounding the well after 18 h of incubation at 35degC The results were
confirmed by two indepedent experiments
Antibiotic susceptibility testing
MICs were determined by a broth microdilution technique following the recommendations of
the Clinical and Laboratory Standards Institute (CLSI) except that the incubation period was
extended to 48 h and the medium used was BHI to allow SCVs to reach maximal growth as
previously described (Mitchell 2010) As recommended by the CLSI L monocytogenes (and
its SCVs) were grown in cation-adjusted Muller-Hinton broth (CAMHB BD) containing 3
lysed horse blood (LHB Remel Hartford CT USA) The reported results were obtained from
three independent experiments
48
Time-kill experiments
Kill kinetics were performed to determine whether the effect of compounds alone or in
combination was bacteriostatic or bactericidal Bacteria were inoculated at 105-10
6 CFUmL in
appropriate medium in the absence or presence of the different antibiotic compounds at
concentrations specified in figure legends At several points in time at 35degC (225 rpm)
bacteria were sampled serially diluted and plated on TSA for CFU determinations Plates
were incubated for 24 or 48 h at 35degC for normal and SCV strains respectively The data were
collected from a minimum of three independents experiments
Antibiotic activity in biofilms
The viability of bacteria in biofilms was evaluated based on previously described protocols
(CeriH 2001 MoskowitzSM 2004 HarrisonJJ 2010) For these assays 96-peg lids and
corresponding 96-well plates were used (Nunc CITY STATE) Wells were inoculated with a
suspension of S aureus SH-1000 adjusted to a 05 McFarland standard in TSB (150 microL per
wells) and plates with 96-well lids were incubated at 35degC with an agitation of 120 rpm for 24
h The biofilms on pegs were then washed three times with 200 microL PBS Biofilms on pegs
were further incubated in fresh 96-well plates containing 200 microL of TSB containing or not
antibiotic compounds or a combination of molecules at 35degC 120 rpm for 24 h The treated
biofilms on pegs were washed three times in PBS and bacteria in biofilms were recovered by
sonication in a new microtiter plate containing 200 microL of PBS per wells using an
ultrasonicator bath for 10 min followed by centrifugation for five min at 1000 RPM The
49
bacteria recovered in each well were resuspended serially diluted plated on TSA and
incubated at 35degC for 24 h before CFU determination
For an equivalent series of wells instead of disrupting the biofilm by sonication biofilm
formation on pegs was evaluated through crystal violet staining using a procedure adapted
from Mitchell et al(2010) Following exposure to antibiotics as described above biofilms on
pegs were washed three times using 200 microL of PBS before biofilms were air-dried for 30
minutes Biofilm on pegs were then stained by dipping in 200 microL of a crystal violet solution
(01) for 30 minutes rinsed in 200 microL of distilled water and air-dried for 30 min Finally the
crystal violet from stained biofilms was solubilized in 200 microL of ethanol 95 at room
temperature and the absorbance was measured at 562 nm using a plate reader (nom de
lrsquoappareil)
Cell invasion and measurement of intracellular antibiotic activity
The Calu-3 cell line (ATCC HTB 55 Homo sapiens lung adenocarcinoma) was cultured in
Eaglersquos Minimum Essential Medium (EMEM) supplemented with 01 mM MEM nonessential
amino acids 1 mM of sodium pyruvate 100 UmL penicillin 100 gL streptomycin 25 gL of
Fungizone and 10 fetal bovine serum (FBS) at 37oC in 5 CO2 For routine culture 4 gL of
puromycin was added to culture media All cell culture reagents were purchased from Wisent
(St-Bruno QC Canada)
50
The cell invasion assay was performed with the Calu-3 cells in an air-liquid interface as
previously described with few modifications (Mitchell TO I paperAAC 2011) Briefly cells
were seeded at 15 x 105 cells per insert in a 12-well Transwell plate (Corning CITY STATE)
and cultured for 10 days with the apical medium replaced each day The complete medium in
the basal compartment was replaced by the invasion medium (1 FBS and no antibiotics) 18
h before invasion assays Inocula were prepared by suspending bacteria grown 20 h on BHI
agar plates in ice-cold PBS Bacteria were then washed three times in ice-cold PBS and
suspended in the invasion medium supplemented with 05 BSA at a density of
approximately 4 x 108 CFUmL Cells were washed twice with PBS and 250 μL of bacterial
suspension were apically added to each insert Invasion of cells by bacteria was allowed for 3
h inserts were emptied and washed three times with PBS The basal medium was changed for
the invasion medium with 20 gL of lysostaphin (Sigma) to kill extracellular bacteria and with
or without the tested antibiotic compounds The infected cells were incubated for a total of 48
h with a change of medium at 23 h (invasion medium with lysostaphin with or without the
tested antibiotic compounds) The invasion medium with lysostaphin but without tested
compound was also apically added 1 h before cell lysis to ensure that only intracellular
bacteria were counted At 48 h following three washes with PBS the apical and basal media
were removed and cells were detached with 100 μL of trypsin 025 and lysed for 10 min by
the addition of 400 μL of water containing 005 of Triton X-100 Then 50 μL of PBS (50X)
was added and mixed Lysates were serially diluted 10-fold and plated on TSA for CFU
determination Plates were incubated at 35oC for 48 h
51
RESULTS
Emergence the SCV phenotype
The SCV phenotype is characterized by a slow growth yielding small colony size on agar
plates (15 to 110 of the parent phenotype) Figure 2 illustrates the ability of a subinhibitory
concentration of GEN to promote the emergence of the SCV phenotype for B cereus B
subtilis and L monocytogenes B cereus SCVs were obtained at an inducing concentration of
1 X MIC of GEN (Fig 2C) B subtilis SCVs at 05 X MIC (Fig 2D) and L monocytogenes
SCVs at 025 X MIC (Fig 2E) Two prototypic S aureus Newbould and CF07-L and their
SCV counterpart NewbouldhemB and CF07-S respectively were used as control strains (Fig
2A and 2B) The SCV isolates selected for the rest of the study add a low reversion rate and
kept their phenotype without a GEN selection pressure The B cereus SCVs showed
auxotrophy for menadione (Fig 1F Table 1) some of the L monocytogenes SCVs showed
auxotrophy for hemin (Table 1) while the B subtilis SCVs showed no apparent auxotrophy to
hemin menadione or thymidine (Table 1) These results show that SCVs with defects in their
respiratory chain (ie hemin or menadione auxotrophy) can indeed emerge from a selective
pressure with aminoglycosides
52
A B
Fig 2 Typical colony sizes for SCV and protocipic strains (left and right sides of the plates
respectively) for (A) S aureus CF07-S and CF07-L (B) S aureus NewbouldhemB and
Newbould (C) B cereus ATCC 11778 (SCV1 and prototypic) (D) B subtilis ATCC 6333
(SCV2 and prototypic) and (E) L monocytogenes ATCC 13932 (SCV1 and prototypic) In
(F) a TSA plate was inoculated with a B cereus SCV1 and wells were filled with (i) 10 microg
hemin (ii) 10 microg menadione (iii) 100microg thymidine and diluents (iv) DMSO and (v) NH4OH
14N
C D
E
i ii
iii iv
v
F
53
Antibacterial activity of TO against the various species of SCVs and synergy of
the TO and GEN combination against some prototypic Firmicutes
All the SCVs tested showed a MIC for TO of 006 mgL against S aureus and lt003 mgL
against L monocytogenes B cereus and B subtilis respectivly whereas TO showed no
activity (MIC gt64 microgmL) against their prototypic counterparts (Table 1) A synergic effect of
TO with GEN against all the prototypic strains tested was also observed (Table 1) Indeed the
addition of TO provoked a 4-8 fold decrease of GEN MIC against all species tested (Table 1)
These results are consistent with our previous studies on the activity of TO on S aureus
((Mitchell 2011 AAC 2012 JAC)) No change in the MIC of GEN was observed against the
streptococci (S pyogenes S suis S agalactiae S mitis and S pneumoniae) when tested in
combination with TO (data not shown) These results show that TO is active against some but
not all Firmicutes
54
Table 1 Susceptibility of prototypic and SCV strains and species to TO andor GEN
Ratio of activity of GEN+TO vs GEN alone
Bactericidal activity of TO against SCVs and bactericidal synergy of the TO
and GEN combination against prototypic strains
As shown above the addition of 8 mgL of TO to GEN reduced the MIC of GEN against some
Firmicutes (Table 1) but also time-kill kinetics showed that TO improved the bactericidal
effect of GEN against prototypic B subtilis and B cereus (Fig 3A and 3B respectively) This
bactericidal synergy was also previously described for S aureus (Mitchell 2012 JAC)
However although the MIC of GEN was improved when in combination with TO against L
monocytogenes (Table 1) time-kill experiments failed to demonstrate a bactericidal synergy
TO GENGEN
(+TO)
Saureus ATCC 29213 gt64 05 006 8
Saureus NewbouldΔhemB 006 8 Hemin
B cereus ATCC 11778 gt64 1 025 4
B cereus ATCC 11778 (SCV1) lt003 16 Menadione
B cereus ATCC 11778 (SCV2) lt003 16 Menadione
B subtilis ATCC 6333 gt64 0125 003 4
B subtilis ATCC 6333 (SCV1) lt003 2 unknown
B subtilis ATCC 6333 (SCV2) lt003 4 unknown
B subtilis ATCC 9372 gt64 0125 lt0015 gt8
B subtilis ATCC 9372 (SCV1) lt003 1 unknown
B subtilis ATCC 9372 (SCV2) lt003 1 unknown
L monocytogenes gt64 05 0125 4
L monocytogenes (SCV1) lt003 gt64 unknown
L monocytogenes (SCV2) lt003 gt64 Hemin
Strains
MIC (gL)
Synergy
foldAuxotrophy
55
with an improvement of at most 1 log10 in killing compared to that achieved with GEN alone
(Fig 3C) Similarly the effect of TO used alone against SCVs was strongly bactericidal
against S aureus B subtilis and B cereus but not against the L monocytogenes SCV (Fig
3D) and this even if the MIC of TO against L monocytogenes SCVs was inferior to 003
mgL (Table 1) These results show that the activity of TO against L monocytogenes needs to
be improved
56
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 0 6 m g L G E N
0 0 6 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 5 m g L G E N
0 5 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
2 m g L G E N
2 m g L G E N + T O
8 m g L T O
C T R L
Lo
g1
0 C
FU
ml
0
2
4
6
8
1 0
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
CT
RL
CT
RL
CT
RL
CT
RL
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
S a u re u s
h e m B
S C V
B c e r e u s
A T C C 1 1 7 7 8
S C V 2
B s u b t i l i s
A T C C 6 3 3 3
S C V 2
L m o n o c y to g e n e s
A T C C 1 3 9 3 2
S C V 1
Fig 3 Time-kill kinetics of TO andor GEN against (A) prototypic B subtilis ATCC 6333(B)
B cereus ATCC 11778 and (C) L monocytogenes ATCC 13932 The bactericidal effect of
TO against SCVs is also shown at 8 and 24 h (D) The results were obtained from at least
three independent experiments
A B
C
D
Detection
threshold
57
Bactericidal activiy of the TO analog FC04-100 against L monocytogenes
SCVs and synergy with aminoglycosides against prototypic strains and MRSA
The addition of 8 mgL of FC04-100 to GEN (2 mgL) decreased by an average of 5 log10
prototypic L monocytogenes CFUmL after 24 h of treatment in comparison to GEN alone
(Fig 4A) Also the bactericidal activity of FC04-100 used alone against the L monocytogenes
SCV was shown to be better than that observed for TO (Fig 3D) To facilitate comparison
between TO and FC04-100 Figure 4B shows the residual CFU of the L monocytogenes SCV
strain after treatment with TO or FC04-100 and calculated as the percentage of live CFU in
comparison with untreated L monocytogenes SCVs after 24 h of incubation These results
show that TO can be chemically modified (eg derivative FC04-100) and improve its activity
against L monocytogenes and thus the antibacterial activity of FC04-100 against S aureus
was further investigated Interestingly the MIC of FC04-100 against the SCV strain S aureus
NewbouldhemB (MIC of 05 -1 mgL Table 2) was higher than that found for TO (MIC of
006 mgL Table 1) On the other hand the MIC of FC04-100 against the prototypic S
aureus ATCC 29213 is lower than that of TO (8-16 mgL vs gt 64 mgL respectively) (Tables
1 and 2) This newly detected antibacterial activity of FC04-100 was also noted against
prototypic MRSA strains (Table 2) Furthermore the ability of FC04-100 to potentiate the
activity of GEN was similar to that of TO with a 4 to 8 fold improvement of the
aminoglycoside MIC (Table 2)
Although the multi-resistant S aureus MRSA USA100 USA300 and COL are resistant to
cefalexin (beta-lactam antibiotic MIC of 256 mgL) and to the aminoglycoside kanamycin
(MIC of 256 mgL and gt 1024 mgL for USA 100 and USA300 respectively Table 3) the
58
combination of both antibiotics decreased their respective MICs (Table 3) Indeed the
combination of cefalexin and kanamycin in a proportion of 32 (commercialized under the
name UbrolexinTM
) decreased the MIC of one or both antibiotics This is as expected for the
combination of a beta-lactam with an aminoglycoside (Davis 1982) However the synergy is
weak with MRSA strains that are resistant to one or the other or both antibiotics and the
addition of 4 mgL of FC04-100 further improved by 8 to 32 fold the MIC of the cephalexin-
kanamycin combination against these strains (Table 3)
59
te m p s (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
F C 0 4 -1 0 0 8 m g L
2 m g L G E N +
F C 0 4 -1 0 0 8 m g L
C T R L
G E N 2 m g L (B H I)
Re
sid
ua
l C
FU
(
)
CT
RL
TO
8 g
L
FC
04-1
00 8
gL
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
F C 0 4 -1 0 0 8 g L
T O 8 g L
C TR L
T=
24
h
T=
8h
T=
8h
T=
24
h
T=
24
h
Fig 4 Time-kill kinetics of FC04-100 andor GEN against (A) prototypic L monocytogenes
ATCC 13932 The bactericidal effect of FC04-100 used alone against L monocytogenes
SCV1 is also shown at 8 and 24 h (B) In (B) the residual CFU were calculated as the
percentage of live CFU in each condition in comparison with the untreated L monocytogenes
SCV1 after 24h of incubation The results were obtained from at least three independent
experiments
A
B
60
Table 2 Susceptibility of prototypic and SCV S aureus strains to FC04-100 with or without
GEN
a FC04-100 w as used at 4 microgmL
b Ratio of activity of GEN + FC04-100 vs activity of GEN alone
Table 3 Synergistic activity of FC04-100 with the combinaison cefalexin-kanamycin
aFC04-100 was used at 4 microgmL
bRatio of the MIC of cefalexin+kanamycin with FC04-100 vs the MIC of
cefalexin+kanamycin alone
Staphylococcus aureus ATCC 29213 8-16 05 006-0125 4-8
S aureus SH1000 8-16 05 006-0125 4-8
S aureus NewbouldΔhemB 1 8
MRSA USA100 16 05 0125 4
MRSA USA300 16 05 006 8
MRSA Col 16 025 006 4
Strains FC04-100 GENGEN
(+FC04-100 )fold
MIC (gL)
MRSA USA100 256 256 12886 43 32
MRSA USA300 256 gt1024 256171 86 32
MRSA Col 256 4 43 05035 8
MIC (gL)
Strains cefalexin kanamycin cef+kana (32)cef+kana (32)
(+FC04-100 )fold
a b
a b
61
FC04-100 potentiates GEN against S aureus embedded in biofilm and inhibits
intracellular replication of SCVs
Figure 5 demonstrates that compound FC04-100 efficiently kills S aureus embedded in
biofilms when in combination with GEN ie there was an important decrease of bacterial
CFUs compared to the no drug control or to either FC04-100 used alone at 4 ugmL (ie 025-
05MIC Table 2) or GEN used alone at 025 05 or 1 X MIC Besides we previously
demonstrated that TO was able to inhibit intracellular replication of SCVs (Mitchell AAC)
which is an important aspect of virulence for S aureus Here (Fig 6) we show that FC04-100
like TO can significantly decrease the infection of polarized airway epithelial cells by SCVs
62
05
mgL
GE
N
05
mgL
GE
N +
FC
04
02
5 m
gL
GE
N
02
5 m
gL
GE
N +
FC
04
01
2 m
gL
GE
N
01
2 m
gL
GE
N +
FC
04
No d
rug
FC
04
0
2
4
6
8
log
10
CF
Up
eg
= 3 2
= 3 8
= 3 5
= 2
Fig 5 Bactericidal effect of FC04-100 alone or in combination with GEN against prototypic
S aureus SH100 embedded in biofilm FC04-100 (FC04) was used at 4 mgL alone or in
combination with GEN Data are from three independent experiments performed in triplicate
Significant differences in comparison to the no drug control are shown ( Plt 0001 and
Plt 00001 one-way ANOVA with a Dunnettrsquos post test) Data are presented as means with
standard deviations
63
No d
rug
TO
8 m
gL
FC
04 8
mgL
0
2
4
6
log
10
CF
Ui
ns
er
t
Fig 6 Effect of TO and FC04-100 on the intracellular load of CF07-S 48 h post-
internalization Data are from three independent experiments performed in duplicate
Significant differences among groups are shown ( Plt 00001One-way ANOVA with
Turkeyrsquos post test) Data are presented as means with standard deviations
64
DISCUSSION
In previous studies we have demonstrated that TO has a strong inhibitory activity against S
aureus SCVs and improves the bactericidal activity of aminoglycoside antibiotics against
prototypic S aureus and also more broadly against staphylococci This effect was documented
for several strains of diverse clinical origins including aminoglycoside-resistant S aureus
carrying aminoglycoside modifying enzymes TO showed however no synergistic effect on the
activity of aminoglycosides against P aeruginosa E coli or Enterococcus spp (Mitchell
2012) In the present study we showed that the antibacterial spectrum of TO can be extended
to species of the Bacillales order with very low MICs against L monocytogenes B cereus and
B subtillis SCVs (lt003 microgmL) Time-kill kinetics showed that the combination of TO and
GEN creates a bactericidal synergy against prototypic of B subtilis and B cereus similarly to
that previously demonstrated against S aureus However the mixture of TO and GEN did not
demonstrated much improvement over the activity of GEN alone against prototypic L
monocytogenes in time-kill experiments Likewise investigations with Bacillales SCVs
showed that TO efficiently killed S aureus B cereus and B subtillis but not L
monocytogenes SCVs despite its very low MIC and was thus bacteriostatic against that species
Therefore improvement of the antibacterial activity against this important pathogen seems
inevitable for future development of steroidal alkaloids such as TO
TO is a steroid alkaloid structurally characterized by 6 rings 12 stereogenic centers a 3-
hydroxyl group and spiro-fused E F rings in the form of an aminoketal (Fig 1) It is generally
65
prepared by hydrolysis of its glycosylated analog tomatine while both molecules can be found
in the Solanaceae plants (Ruiz-Rubio 2001 Simons 2006) Although we previously
documented some interesting antibacterial activity for TO the absence of an identified cellular
target makes difficult the establishment of a structure-activity relationship (SAR) Our initial
attempt to elucidate SAR was therefore based on the hypothesis that the steroidal A-D rings
act as a rigid scaffold to orient pharmacophores defined by the 3-hydroxyl group on ring A
and the spiroaminoketal group on rings E F Noteworthy we prepared analogs bearing
modifications on ring A of TO and schegravemes for chemistry synthesis were recently published
(Chagnon et al 2014) From seven compounds keeping rings E and F of TO and possessing
structural variations on position 3 of ring A we were able to ascertain some structure activity
relationships Results suggest that the hydrogen bond donor effect of the 3-hydroxyl group
was not important for activity although the stereochemistry of the 3 position substitution
moderately affected activity against both S aureus SCVs and prototypic strains (in
combination with an aminoglycoside) In addition although the presence of one or two
ammonium groups in position C3 reduced the activity against S aureus SCVs (from a MIC of
006 to 05-1 microgmL) this change was highly beneficial for antibiotic activity against normal
strains (from a MIC of gt64 to 8-16 microgmL) without the presence of an aminoglycoside This
change was unexpected and one of such analogs FC04-100 increased its solubility by more
than ten times (from 2 mgmL to 26 mgmL in DMSO) The biological properties of FC04-
100 were therefore further investigated in the present study especially that TO showed some
weak bactericidal activity against Listeria and that the solubility of TO may limit eventual
clinical applications
66
Here we demonstrated that FC04-100 was able to kill both L monocytogenes SCVs as well as
the normal phenotype alone and in the presence of GEN respectively (Fig 4) Furthermore
FC04-100 showed a noticeable antibacterial activity on its own (MIC of 16 microgmL) against the
MRSA strains USA100 USA300 and COL (Table 2) FC04-100 was also synergistic in
combination with an aminoglycoside against such MRSA strains with a 4 to 8 fold gain in the
MIC of GEN Because MRSA strains are often multiresistant to antibiotics and often carry
AMEs we examined the possibility of using a triple antibiotic combination Indeed the
combination of kanamycin and cefalexin in a proportion 32 is already approved to treat
bovine mastitis pathogens under the brand name of UbrolexinTM
(Ganiegravere 2009) This
antibiotic combination offers an extended spectrum of activity compared to each individual
drug and is expected to cover both S aureus Streptococcus uberis and E coli (Ganiere
2009 Maneke et al 2011 Silley 2012) Unfortunately the increased frequency of livestock-
associated MRSA (Garcia-Alvarez et al 2011 Graveland et al 2011 Leonard and Markey
2008) and frequent incidence of strains and species carrying AMEs (Ramirez and Tolmasky
2010) may limit the spectrum of activity of the combination of a beta-lactam and an
aminoglycoside With this in mind we showed that the use of FC04-100 in combination with
kanamycin and cefalexin improves by 32 fold the activity of this mixture against MRSA
strains carrying AMEs (Table 3) Overall the addition of FC04-100 to the aminoglycoside-
beta-lactam combination decreased the MICs of kanamycin and cefalexin below their
resistance breakpoints and this for all the MRSA strains tested
67
L monocytogenes is a highly specialized intracellular pathogen (Hamon et al 2006) causing
life-threatening infections especially in pregnant women (Janakiraman 2008) L
monocytogenes in a foodborne pathogen and because of its intracellular life cycle and
propagation it is able to hide from host defenses and antibiotic treatment Similarly S aureus
although not primarily recognized as a typical intracellular pathogen is also able to enter and
survive in host cells (Lowy 2000) Moreover the ability of S aureus to convert to the SCV
phenotype showing increased biofilm production improved adherence to host cells and tissues
as well as increased intracellular persistence allows this pathogen to cause chronic and
difficult to treat infections (Garcia et al 2013 Mitchell and Malouin 2012) Discovering
antibacterial drugs able to act in the intracellular compartment is generally recognized as a
difficult challenge Previously we demonstrated that steroidal alkaloids such as TO had such
an ability and could act on intracellular S aureus SCVs (Mitchell et al 2011) and here we
demonstrated that this was also the case for the TO analog FC04-100 (Fig 6) In addition we
also show in the present study that FC04-100 greatly improves the bactericidal activity of
aminoglycosides such as GEN against bacteria embedded in biofilms Biofilms are
recognized to be a major hindrance to antibiotic action and are responsible for persistent
colonization in many diseases (Costerton et al 1999) and in the food industry (Srey et al
2013) The intracellular and intrabiofilm antibacterial properties of FC04-100 warrant further
investigation of this compound and this class of molecules in general
In conclusion we showed in this study that the spectrum of activity of TO is related to the
Firmicutes division and more precisely to the order of the Bacillales TO possesses an
68
antibacterial activity against SCVs and a synergistic activity with animoglycoside antibiotics
against prototypic strains The novel TO analog FC04-100 showed very promising new
characteristics that include a much improved bactericidal effect on L monocytogenes and
killing of bacteria in biofilms
ACKNOWLEDGEMENTS
This study was supported by an operation grant from Cystic Fibrosis Canada to FM and by a
team grant from the Fonds Queacutebeacutecois de la Recherche sur la Nature et les Technologies
(FQRNT) to FM KB and EM
REFERENCES
Atalla H Gyles C Jacob CL Moisan H Malouin F Mallard B 2008 Characterization of
a Staphylococcus aureus small colony variant (SCV) associated with persistent bovine mastitis
Foodborne Pathog Dis 5785ndash799
Bad Bug Book Foodborne pathogenic microorganisms and natural toxins 2012 Second
edition Lampel KA Al-Khaldi S Cahill SM (Ed) Published by the Center for Food Safety and
Applied Nutrition of the Food and Drug Administration (FDA) US Department of Health and
Human Services pp 87-103
Beierlein JM Anderson AC 2011 New developments in vaccines inhibitors of anthrax
toxins and antibiotic therapeutics for Bacillus anthracis Curr Med Chem 185083-5094
69
Brouillette E A Martinez B J Boyll N E Allen and F Malouin 2004 Persistence of
a Staphylococcus aureus small-colony variant under antibiotic pressure in vivo FEMS
Immunol Med Microbiol 4135ndash41
Bryan L E and S Kwan 1981 Aminoglycoside-resistant mutants of Pseudomonas
aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob
Agents Chemother 19958ndash964
Ceri H Olson M Morck D Storey D Read R Buret A Olson B 2001 The MBEC Assay
System multiple equivalent biofilms for antibiotic and biocide susceptibility testing Methods
Enzymol 337377-385
Chagnon F I Guay M-A Bonin G Mitchell K Bouarab F Malouin E Marsault
2014 Unraveling the structure-activity relationship of tomatidine a steroid alkaloid with
antibiotic properties against persistent forms of Staphylococcus aureus Eur J Med Chem (In
Press EJMECH-S-13-01625-3)
Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial
Susceptibility Testing Twenty-First Informational Supplement M100-S21 CLSI Wayne PA
USA 2011
70
Costerton JW Stewart PS Greenberg EP 1999 Bacterial biofilms a common cause of
persistent infections Science 284 1318-1322
Davis BD 1982 Bactericidal synergism between beta-lactams and aminoglycosides
mechanism and possible therapeutic implications Rev Infect Dis 4237-245
Ganiegravere JP Denuault L 2009 Synergetic interaction between cefalexin and kanamycin in
Muller-Hinton broth medium and in milk J appl microbiol 107 117-125
Garcia LG S Lemaire B C Kahl K Becker R A Proctor O Denis P M Tulkens
F Van Bambeke 2013 Antibiotic activity against small-colony variants of Staphylococcus
aureus review of in vitro animal and clinical data J Antimicrob Chemother 68 1455ndash1464
Garcia-Alvarez L M T Holden H Lindsay C R Webb D F Brown M D Curran
E Walpole K Brooks D J Pickard C Teale J Parkhill S D Bentley G F Edwards
E K Girvan A M Kearns B Pichon R L Hill A R Larsen R L Skov S J Peacock
D J Maskell and M A Holmes 2011 Methicillin-resistant Staphylococcus aureus with a
novel mecA homologue in human and bovine populations in the UK and Denmark a
descriptive study Lancet Infect Dis 11595-603
Gomaa OM Momtaz OA 2007 16 rRNA characterization of Bacillus isolate and its
tolerance profile after subsequent subculturing Arab J Biotech 10 107-116
71
Graveland H Duim B van Duijkeren E Heederik D Wagenaar JA 2011 Livestock-
associated methicillin-resistant Staphylococcus aureus in animals and humans Int J Med
Microbiol 301630-634
Guillet C Join-Lambert O Le Monnier A Leclercq A Mechaiuml F Mamzer-Bruneel MF
Bielecka MK Scortti M Disson O Berche P Vazquez-Boland J Lortholary O Lecuit M
2010 Human listeriosis caused by Listeria ivanovii Emerg Infect Dis 16136-138
Hamon M H Bierne P Cossart 2006 Listeria monocytogenes a multifaceted model Nat
Rev Microbiol 4 423-434
Harrison JJ Stremick CA Turner RJ Allan ND Olson ME Ceri H 2010 Microtiter
susceptibility testing of microbes growing on peg lids a miniaturized biofilm model for high-
throughput screening Nat Protoc 51236-1254
Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y
Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T Baba T 2013 Genomic
Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
Horsburgh MJ Aish JL White IJ Shaw L Lithgow JK Foster SJ 2002 SigmaB
modulates virulence determinant expression and stress resistance characterization of a
72
functional rsbU strain derived from Staphylococcus aureus 8325-4 J Bacteriol 1845457-
5467
Janakiraman V 2008 Listeriosis in Pregnancy Diagnosis Treatment and Prevention Rev
Obstet Gynecol 1 179ndash185
Leonard FC Markey BK 2008 Methicillin-resistant Staphylococcus aureus in animals a
review Vet J 175 27-36
Lowy FD 2000 Is S aureus an intracellular pathogen Trends Microbiol 8341-342
Magalhatildees L Nitschke M 2013 Antimicrobial activity of rhamnolipids against Listeria
monocytogenes and their synergistic interaction with nisin Food Control 29 138-142
Maneke E Pridmore A Goby L Lang I 2011 Kill rate of mastitis pathogens by a
combination of cefalexin and kanamycin J Appl Microbiol 110184-190
Mitchell G Brouillette Eacute Lalonde-Seacuteguin D Asselin AE Lebeau Jacob C Malouin F
2009 A role for sigma factor B in the emergence of Staphylococcus aureus small-colony
variants and elevated biofilm production resulting from an exposure to aminoglycosides
Microb Pathog 4818-27
73
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K Malouin F 2011
Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic
fibrosis airway epithelial cells Antimicrob Agents and Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM Malouin F 2011 Infection of Polarized
airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is
increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function
and is influenced by NF- κ B Infect Immun 793541-3551
Mitchell G Lafrance M Boulanger S Lalonde-Seacuteguin D Guay I Gattuso M Marsault
Eacute Bourab K Malouin F 2012 Tomatidine acts in synergy with aminoglycoside antibiotics
against multiresistant Staphylococcus aureus and prevents virulence gene expression J
Antimicrob Chemother 67 559-568
Mitchell G Lalonde-Seacuteguin D Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S
Malouin F 2010 Staphylococcus aureus sigma B-dependent emergence of small-colony
variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-
heptylquinoline-N-oxide BMC Microbiology 10 33-48
Mitchell G and F Malouin 2012 Outcome and prevention of Pseudomonas aeruginosa-
Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic
Fibrosis InTech Open Access Publisher ISBN 979-953-307-059-8
74
Moisan H Brouillette E Jacob CL Langlois-Beacutegin P Michaud S Malouin F 2006
Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated
from cystic fibrosis patients is influ- enced by SigB J Bacteriol 18864ndash76
Moskowitz SM Foster JM Emerson J Burns JL 2004 Clinically feasible biofilm
susceptibility assay for isolates of Pseudomonas aeruginosa from patients with cystic fibrosis
J Clin Microbiol 421915-1922
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M Peters G 2006
Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent
infections Nat Rev Micro- biol 4295ndash305
Ramirez MS M E Tolmasky 2010 Aminoglycoside modifying enzymes Drug Res
Updates 13 151ndash171
Ruiz-Rubio M Peacuterez-Espinosa A Lairini K Roldagraven-Arjona T Dipietro A Anaya N
2001 Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi p 293ndash326 In
A Rahman (ed) Studies in natural products chemistry vol 25 Elsevier Oxford United
Kingdom
75
Sendi P Proctor RA 2009 Staphylococcus aureus as an intracellular pathogen the role of
small colony variants Trends Microbiol 1754-58
Silley P Goby L Pillar CM 2012 Susceptibility of coagulase-negative staphylococci to a
kanamycin and cefalexin combination J Dairy Sci 953448-3453
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C Osbourn A
2006 Dual effects of plant steroidal alkaloids on Saccharomyces cerevisiae Antimicrob
Agents Chemother 502732ndash2740
Srey S Jahid IK Ha S-D 2013 Biofilm formation in food industries A food safety concern
Food control 31572-585
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D
Roth J Proctor RA Becker K Peters G Loumlffler B 2011 Staphylococcus aureus
phenotype switching an effective bacterial strategy to escape host immune response and
establish a chronic infection EMBO Mol Med 3129ndash141
Vuong C Otto M 2002 Staphylococcus epidermidis infections Microbes Infect 4481-489
Witte W Strommenger B Stanek C Cuny C 2007 Methicillin-resistant Staphylococcus
aureus ST398 in Humans and Animals Central Europe Emerg Infect Dis 13 255ndash258
76
CHAPITRE 3
MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
31 Obtention des mutants reacutesistants
Afin de nous aider agrave mieux deacutefinir la cible et le mode drsquoaction de la tomatidine (TO)
nous avons forceacute lrsquoapparition de reacutesistances chez Staphylococcus aureus Comme
souche controcircle prototype nous avons choisi ATCC 29213 puisque crsquoest une souche
tregraves freacutequemment utiliseacutee pour des expeacuteriences avec des antibiotiques et nous avons
choisi la souche NewbouldΔhemB comme S aureus laquo small colony variant raquo puisque
crsquoest une souche SCV qui est stable et bien connue nrsquoayant pas de capaciteacute de
reacuteversion Par passages successifs dans une concentration sous inhibitrice
drsquoantibiotique (technique de CMI (concentration minimale inhibitrice)) (CLSI 2011)
nous avons obtenu des mutants deacutemontrant une reacutesistance croissante agrave lrsquoantibiotique
au fil des passages (Figure 1) Nous avons obtenu des mutants ATCC 29213
reacutesistants agrave la gentamicine (GEN) et agrave la GEN lorsque combineacutee agrave la TO ainsi que
des mutants reacutesistants NewbouldΔhemB reacutesistants agrave la GEN seule agrave la TO seule et
agrave la TO lorsque combineacute agrave la GEN
Certains passages ont ensuite eacuteteacute seacutelectionneacutes parce qursquoils deacutemontraient une hausse
plus marqueacutee de leur CMI Ces passages ont eacuteteacute choisis car nous supposions que
ces hausses marqueacutees de CMI correspondraient donc agrave des hausses de la
reacutesistance de cette souche agrave lrsquoantibiotique testeacute et que cette reacutesistance pouvait ecirctre la
conseacutequence drsquoune ou plusieurs mutations en lien avec la cible ou le mode drsquoaction
de lrsquoantibiotique preacutesent Trois agrave quatre passages (ou paliers de reacutesistance) par
condition ont eacuteteacute seacutelectionneacutes Nous avons ensuite reacuteensemenceacute le laquo pool raquo
bacteacuterien de chacun des passages choisis afin de pouvoir isoler 3 isolats ayant la
mecircme CMI (ou tregraves similaire (CMI vraies)) que celui du laquo pool raquo bacteacuterien des
77
passages drsquoougrave ils provenaient Soixante quatre mutants reacutesistants ont ainsi eacuteteacute
seacutelectionneacutes en vue drsquoecirctre seacutequenceacutes (Figure 2)
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt 1
6
CM
I v
raie
s
02
5
CM
I v
raie
s
02
5-1
A
78
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
en
pr
eacutes
en
ce
de
ge
n (
4 micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
01
25
1
6
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
8
CM
I v
raie
s
2
4
CM
I v
raie
s
2
B
C
79
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
en
pr
eacutes
en
ce
de
TO
(8
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
1
2
3
4
5
CM
I v
ra
ies
4
CM
I v
ra
ies
2
CM
I v
ra
ies
0
5
1
CM
I v
ra
ies
2
4
Figure 1 Graphiques des CMI pour chacun des passages pour (A) la tomatidine sur
NewbouldΔhemB (B) pour la tomatidine en preacutesence de gentamicine sur
NewbouldΔhemB (C) pour la gentamicine sur S aureus ATCC 29213 (D) pour la
gentamicine en preacutesence de tomatidine sur S aureus ATCC 29213
32 Seacutequenccedilage et analyses bioinformatiques
Une extraction drsquoADN geacutenomique a ensuite eacuteteacute effectueacutee sur chacun des isolats en
utilisant le Kit drsquoextraction drsquoADN geacutenomique miniKit de Qiagen (QIAamp DNA minikit)
(Li et al 2003) Puisque leurs seacutequences geacutenomiques nrsquoeacutetaient pas connues nous
avons eacutegalement extrait lrsquoADN geacutenomique de nos deux souches de reacutefeacuterences
ATCC29213 et NewbouldΔhemB (provenant de quatre colonies isoleacutees chacune)
afin drsquoavoir les seacutequences de comparaison pour nos mutants (Figure 2)
D
80
Ne
wm
an
(g
en
om
ede r
ef h
ttpw
ww
ncbinlm
nih
govn
uccore
150373012)
AT
CC
29
213
(A p
art
ir d
e 4
AT
CC
29
21
3 r
eco
nstr
uire
AT
CC
29
21
3 c
om
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
5
10
7
10
8 1
09
)
Ne
wbo
uld
Δhe
mB
(A p
art
ir d
e 4
Ne
wb
ou
ldh
em
Bre
co
nstr
uire
Ne
wb
ou
ldh
em
Bcom
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
1
agraveG
EN
2
5 (
CM
I2
)
2
7 (
CM
I2
)
2
8 (
CM
I2
)
AT
CC
29
21
3
Pa
lier
de
reacutesis
tance
1
agravecom
bo
29
(C
MI
05
)
3
0 (
CM
I 0
5
3
1 (
CM
I 0
5)
Suite
fe
uill
e s
uiv
ante
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agraveG
EN
3
(C
MI
4)
4
(C
MI
4)
3
9 (
CM
I4
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agraveG
EN
4
6 (
CM
I8
)
4
7 (
CM
I8
)
4
8 (
CM
I8
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agravecom
bo
1
1 (
CM
I 2
)
1
2 (
CM
I 2
)
2
1 (
CM
I 2
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agravecom
bo
5 (
CM
I 2
-4)
3
6 (
CM
I 2
-4)
3
7 (
CM
I 2
-4)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
4
agravecom
bo
3
8 (
CM
I4
)
6
7 (
CM
I 4
)
81
Ne
wb
ould
Δhem
B(A
part
ir d
e 4
Ne
wb
ou
ldΔ
he
mB
reco
nstr
uire
Ne
wb
ou
ldΔ
hem
Bco
mp
let e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveG
EN
24
(C
MI
64
)
4
1 (
CM
I 1
6)
4
2 (
CM
I 6
4)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveT
O
95
(C
MI
02
5)
9
6 (
CM
I 0
25)
9
7 (
CM
I 0
25)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agravecom
bo
33
34
35
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveG
EN
49
(C
MI
12
8)
5
1 (
CM
I 1
28)
5
2 (
CM
I 1
28)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agraveG
EN
7
(C
MI
128
)
4
3 (
CM
I 1
28)
4
5 (
CM
I1
28
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agraveT
O
6
4 (
CM
I gt
16)
6
5 (
CM
I gt
16)
6
6 (
CM
I gt
16)
75
76
77
79
80
81
82
83
84
85
86
87
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agravecom
bo
15
16
18
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agravecom
bo
59
60
61)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agravecom
bo
54
56
58)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveT
O
90
(C
MI
02
5)
9
1 (
CM
I 0
25)
9
3 (
CM
I 0
5)
Ne
wb
ou
ldh
em
B
palie
r d
e r
eacutesis
tance
3
agraveT
O
11
0 (
CM
I 1
)
1
12 (
CM
I 0
25
-05
)
1
13 (
CM
I 0
5)
Fig
ure
2
Org
an
igra
mm
e d
es
mu
tan
ts r
eacutes
ista
nts
seacute
qu
en
ceacutes
se
lon
leu
r pa
lier
de
reacute
sis
tan
ce e
t a
vec leu
r n
um
eacutero
drsquoeacute
chan
tillo
n
co
rre
spo
ndan
t
82
Un minimum de 20 ngmicrol drsquoADN geacutenomique de chacun des isolats a ensuite eacuteteacute
achemineacute au laboratoire du Pr Seacutebastien Rodrigue (Deacutepartement de biologie Faculteacute
des sciences Universiteacute de Sherbrooke) qui a effectueacute la librairie de fragments Pour
se faire il y a eu dans un premier temps digestion enzymatique de lrsquoADN afin
drsquoobtenir des fragments drsquoADN double brin Ensuite tous les fragments ont eacuteteacute
purifieacutes suivant la meacutethode SPRI (Solid-Phase Reversible Immobilization) Il srsquoagit
drsquoune meacutethode simple et tregraves efficace drsquoisolation drsquoacides nucleacuteiques sur microplaque
(Rodrigue S et al 2010) Les acides nucleacuteiques cibles sont immobiliseacutes sur des
microparticules paramagneacutetiques dans des conditions tampons-speacutecifiques La
flexibiliteacute du systegraveme se fait par lrsquoutilisation de diffeacuterents tampons de liaison (laquo binding
buffer raquo) qui modifient le type et la grandeur des acides nucleacuteiques immobiliseacutes Les
contaminants peuvent ainsi ecirctre retireacutes de lrsquoeacutechantillon sans besoin de filtration ou
centrifugation (Beckman Coulter website 2013 DeAngelis et al 1995) Suite agrave la
purification par meacutethode SPRI lrsquoeacutequipe du Pr Rodrigue a obtenu des fragments
drsquoADN de ~250pb auxquels ont eacuteteacute ajouteacutes par ligation des adapteurs aux deux
extreacutemiteacutes des fragments Dans un deuxiegraveme temps des codes-bars speacutecifiques agrave
chaque eacutechantillon drsquoADN geacutenomique ont eacutegalement eacuteteacute lieacutes agrave chacun des fragments
Alors que les adapteurs permettent aux fragments de se lier agrave une matrice solide afin
de pouvoir ecirctre amplifieacutes les codes-bars permettront de classer chacun des
fragments selon leur appartenance agrave un eacutechantillon drsquoADN geacutenomique speacutecifique
83
Figure 3 Eacutetape du seacutequenccedilage Illumina
La librairie de fragments a ensuite eacuteteacute envoyeacutee chez Geacutenome-Queacutebec afin que
chacun des geacutenomes puisse ecirctre seacutequenceacute Geacutenome ndashQueacutebec utilise la meacutethode
Illumina comme outil de seacutequenccedilage (Figure 3) La grande innovation de la meacutethode
Illumina est lrsquoamplification des fragments par liaison de ceux-ci agrave une surface solide
(Flowcell) ce qui permet par un processus de ponts drsquoamplification de geacuteneacuterer plus
drsquoun million de laquo clusters raquo comprenant eux-mecircme au plus un millier de copies de la
moleacutecule originale de deacutepart (solid-phase PCR) Chaque regroupement de fragments
provenant de lrsquoamplification drsquoun mecircme fragment original est tregraves pregraves les uns des
Modifieacute de Quail MA et al 2008
84
autres formant une zone distincte drsquoun regroupement de fragments agrave un autre
Lrsquoincorporation des nucleacuteotides eacutetiqueteacutes preacutealablement avec des moleacutecules
fluorescentes speacutecifiques est deacutetecteacutee suite agrave leur excitation par un laser et les
images sont compileacutees pour produire la seacutequence de bases pour chaque fragment
ADN (Figure 4) Les nouvelles seacuteries de bases appeleacutees laquo readsraquo seront ensuite
reacuteassembleacutees (aligneacutes) en se basant sur un geacutenome de reacutefeacuterence connu (appeleacute
laquo resequencing raquo) (crsquoest notre cas ici) ou en absence de geacutenome de reacutefeacuterence
(appeleacute laquo De novo sequencing raquo) Au final lrsquoensemble des laquo reads raquo aligneacutes
correspondra agrave la seacutequence de chaque chromosome de lrsquoeacutechantillon drsquoADN
geacutenomique de deacutepart
85
Figure 4 Meacutethode drsquoamplification par la meacutethode Illumina
Suite au seacutequenccedilage chez Geacutenome ndashQueacutebec nous avions en main les seacutequences
geacutenomiques de tous nos mutants et de nos souches de reacutefeacuterence sous forme de
Tireacute de BiteSizeBio httpbitesizebiocom13546sequencing-by-synthesis-
explaining-the-illumina-sequencing-technology consulteacute le 3 deacutec 2013
86
contigs Nous avions donc le casse-tecircte mais en piegraveces deacutetacheacutees pecircle-mecircle et
sans avoir la photo de lrsquoimage de reacutefeacuterence sur laquelle se baser pour pouvoir
compleacuteter le casse-tecircte
1- Lrsquoassemblage
Les diffeacuterents geacutenomes avaient eacuteteacute preacutealablement fractionneacutes aleacuteatoirement
en petits fragments de 250 pb et amplifieacutes en vue du seacutequenccedilage Suite au
seacutequenccedilage les seacutequences de chaque fragment (appeleacute laquo reads raquo) sont ensuite
assembleacutees en contigs crsquoest-agrave-dire des seacutequences plus ou mois longues continues
et ordonneacutees geacuteneacutereacutees par lassemblage des clones dune bibliothegraveque geacutenomique
qui se chevauche La qualiteacute de lrsquoassemblage en contigs deacutepend de la capaciteacute de
superposition des laquo reads raquo Parce que la digestion enzymatique de lrsquoADN se fait de
faccedilon aleacuteatoire et se fait sur plusieurs copies drsquoADN chaque portion du geacutenome est
repreacutesenteacutee plusieurs fois dans diffeacuterents modegraveles de fragments Donc les
seacutequences identiques des diffeacuterents fragments (ou laquo reads raquo) peuvent ecirctre
superposeacutees par chevauchegravement et les laquo reads raquo superposeacutes peuvent ecirctre
assembleacutes en contigs par un logiciel drsquoassemblage En ce qui nous concerne cette
eacutetape a eacuteteacute faite agrave lrsquoaide du programme de bio-informatique Newbler (Zhang et al
2012) Lrsquoassemblage geacutenomique peut ecirctre probleacutematique pour les informaticiens car
les geacutenomes contiennent des seacutequences qui se reacutepegravetent et ces reacutepeacutetitions peuvent
ecirctre assez longues en fonction des organismes Suite agrave lrsquoassemblage les geacutenomes
sont donc sous forme de contigs
2- Lrsquoalignement
En se basant sur une seacutequence geacutenomique de reacutefeacuterence (notre reacutefeacuterence est
Staphylococcus aureus str Newman (~ 288Mb)
( httpwwwncbinlmnihgovgenome154project_id=58839) ) le programme de bio-
informatique ABACAS permet de rapidement aligner ordonner et orienter les contigs
Selon les besoins ce qui nrsquoa pas eacuteteacute notre cas ABACAS permet eacutegalement de
87
visualiser et designer les amorces permettant de fermer les gaps (espaces) entre les
contigs en faisant des PCR (Assefa et al 2009)
3- La fermeture des gaps
La fermeture des gaps nrsquoa pas eacuteteacute faite avec le programme ABACAS mais
avec lrsquoaide du programme de bio-informatique GapFiller Lrsquoutilisation de GapFiller
pour la fermeture des gaps possegravede 3 principaux avantages Premiegraverement le
programme prend en compte lrsquoestimation de la grandeur du gap agrave fermer donc il
exclut automatiquement les fermetures de gaps erroneacutes qui sont plus courts ou plus
longs que la grandeur estimeacutee Deuxiegravemement il nrsquoessaie pas de remplir un gap
avec un assemblage de laquo reads raquo en contigs mais cherche plutocirct agrave allonger les
contigs agrave partir de leurs extreacutemiteacutes jusqursquoagrave obtenir des seacutequences superposeacutees en
passant par la creacuteation de K-mer (seacuteparation des reads en plus petits fragments) Et
troisiegravemement il prend en compte que les extreacutemiteacutes des contigs sont souvent
source de mauvais assemblage et reacute eacutevalue ces extreacutemiteacutes pendant la fermeture des
gaps (Boetzer et Pirovano 2012)
4- Lrsquoannotation des geacutenomes
Suite agrave la fermeture des gaps il a fallu annoter les geacutenomes en se basant sur
lrsquoannotation du geacutenome de reacutefeacuterence Staphylococcus aureus str Newman RATT est
un outil de transfert rapide drsquoannotation (Rapid Annotation Transfert Tool) qui a eacuteteacute
deacuteveloppeacute afin de fournir rapidement une annotation preacutecise pour des geacutenomes
inconnus en se basant sur des geacutenomes deacutejagrave annoteacutes comme reacutefeacuterences (Figure 5)
La meacutethode transfert les annotations drsquoune reacutefeacuterence de haute qualiteacute agrave un nouveau
geacutenome sur la base de la conservation des synteny (ordre des gegravenes) (Otto et al
2011) Suite agrave lrsquoannotation les gegravenes tant chez les mutants que chez les souches
parentes portent les noms auxquels ils correspondent chez Staphylococcus aureus
str Newman
88
Figure 5 Corrections du programme RATT sur les annotations transfeacutereacutees Les
annotations de H37Rv ont eacuteteacute transfeacutereacutees sur la seacutequence F11 (bleu pacircle) ont eacuteteacute corrigeacutees
(vert) et ensuite compareacutees agrave lrsquoannotation de la souche existante F11 EMBL (jaune) (A et B)
La correction des codons ldquostartrdquo et ldquostoprdquo respectivement Dans une situation de laquo mapping raquo
plus complexe (C) ougrave les cadres de lecture sont montreacutes pour plus de clarteacute RATT
cartographie une large seacutequence codante (CDS) de H37Rv vers un locus dans F11 qui inclue
plusieurs codons laquo stop raquo laquo in-frame raquo En inseacuterant un deacutecalage du cadre de lecture (ie un
pseudogegravene) la traduction conceptuelle est conserveacutee Ceci diffegravere avec les deux gegravenes se
superposant preacutedis comme faisant partie du geacutenome F11
Tireacute de Otto TD et al 2011
89
321 Identification des mutations en lien avec la tomatidine
Apregraves avoir obtenu une seacutequence consensus agrave partir des seacutequences geacutenomiques
des 4 copies de chacune des souches parente S aureus ATCC 29213 et S aureus
NewbouldΔhemB les diffeacuterences geacutenomiques entre les souches parentes et le
geacutenome de reacutefeacuterence S aureus str Newman ont eacuteteacute eacutelimineacutees afin que ces
diffeacuterences nrsquoapparaissent pas comme des mutations lors de la comparaison des
seacutequences geacutenomiques des mutants avec celles de leur souche parente ces
diffeacuterences eacutetant plutocirct attribuables agrave la diffeacuterence qursquoil y a entre S aureus str
Newman et la souche parente et non pas agrave des mutations en lien avec la reacutesistance
des mutants
Le service bio-informatique du deacutepartement de biologie (Faculteacute des sciences
Universiteacute de Sherbrooke) a ensuite produit un tableau contenant pour chacun des
mutants tous les gegravenes muteacutes apregraves comparaison avec leur souche parentes Agrave cette
eacutetape on pouvait donc savoir dans quel gegravene se situaient les mutations quelle eacutetait
lrsquoeffet de cette mutation (mutation synonyme deacuteleacutetion insertion SNP) et en principe
srsquoil srsquoagissait pour un mecircme gegravene muteacute de la mecircme mutation puisque dans ce
tableau pour un mecircme gegravene chaque ligne correspond agrave des mutations diffeacuterentes
(Figure 6)
Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4
Figure 6 Exemple drsquoune partie du tableau compilation des mutations chez
les mutants ATCC 29213 reacutesistants agrave la GEN seule et agrave la GEN (+TO)
90
Agrave lrsquoaide de ce tableau (Figure 6) jrsquoai chercheacute les mutations qui pouvaient ecirctre
attribuables agrave la TO Chez les mutants S aureus ATCC 29213 reacutesistants jrsquoai
chercheacute les gegravenes qui eacutetaient muteacutes chez tous les mutants reacutesistants agrave la
combinaison GEN (+TO) ou dont la mutation apparaissait graduellement au fil des
paliers de reacutesistance mais dont ce mecircme gegravene nrsquoeacutetait pas muteacute chez les mutants
reacutesistants agrave la GEN seule Je pouvais donc estimer que cette mutation chez les
reacutesistants agrave la combinaison eacutetait en lien avec la preacutesence de la TO dans le milieu
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct
Afin de mrsquoaider dans ma recherche jrsquoai eu lrsquoaide du Pr Pierre-Eacutetienne Jacques
(Deacutepartement de biologie Faculteacute des sciences Universiteacute de Sherbrooke)qui a
mis les donneacutees de ce tableau dans un tableau croiseacute dynamique (TCD) ce qui a
permis de filtrer les donneacutees selon des critegraveres bien preacutecis me permettant drsquoarriver agrave
mes fins plus facilement Dans un premier temps un TCD a eacuteteacute creacuteeacute ougrave pour
chacun des mutants chaque gegravene ayant au moins une mutation eacutetait indiqueacute par un
laquo 1 raquo alors qursquoun laquo 0 raquo eacutetait marqueacute si ce gegravene nrsquoeacutetait pas muteacute Dans un second
temps la quantiteacute de laquo 1 raquo par palier de reacutesistance (correspondant au nombre de
mutants de ce palier posseacutedant une mutation dans ce gegravene) eacutetait indiqueacutee par un
chiffre allant de 0 pour laquo aucun mutant de ce palier de reacutesistance ne posseacutedait de
mutation dans ce gegravene raquo agrave 1 2 ou 3 selon le nombre de mutants de ce palier ayant
ce gegravene muteacute Jrsquoai ensuite pu soumettre le TCD agrave des filtres approprieacutes afin de faire
ressortir les gegravenes drsquointeacuterecircts selon des critegraveres de seacutelection preacutecis (Figures 7-8)
91
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
1
00
33
32
1
1
11
4
5
0
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
10
03
33
21
11
14
50
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_04
12
lpl9
nm
20
12
32
23
92
44
1
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
NW
MN
_00
46
_2
10
13
21
37
24
31
NW
MN
_09
55
_2
01
23
11
37
24
21
NW
MN
_13
83
gpsA
21
02
31
13
72
42
1
NW
MN
_03
39
_1
11
21
31
37
34
42
NW
MN
_15
97
dn
aE2
10
22
30
37
23
31
NW
MN
_23
30
_2
10
23
20
37
23
21
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_2
07
0_
10
01
23
21
81
45
0
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_10
28
_1
22
12
32
58
34
54
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
A B C
92
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
39
8h
sdM
31
33
33
27
11
34
54
NW
MN
_04
26
_3
31
33
32
71
13
45
4
NW
MN
_05
24
sdrD
32
33
33
28
11
34
55
NW
MN
_07
39
_3
33
33
32
91
13
45
6
NW
MN
_11
64
hsl
U3
33
33
32
91
13
45
6
NW
MN
_05
23
sdrC
22
33
32
27
10
34
45
NW
MN
_20
61
fmtB
23
23
32
27
10
34
45
NW
MN
_04
03
lpl1
nm
33
22
33
28
10
34
55
NW
MN
_13
45
ebh
13
32
33
31
81
03
44
5
NW
MN
_16
49
_3
32
33
31
81
03
44
5
NW
MN
_17
00
hsd
M3
23
33
22
81
03
44
5
NW
MN
_00
42
_3
33
33
22
91
03
44
6
NW
MN
_05
80
_3
33
33
31
91
03
44
6
NW
MN
_13
44
ebh
23
31
32
22
79
34
44
NW
MN
_13
75
aro
A3
13
23
31
79
34
44
NW
MN
_23
97
fnb
B2
32
23
22
79
34
45
NW
MN
_04
01
_3
32
23
31
89
34
45
NW
MN
_07
33
_3
32
32
31
89
34
45
NW
MN
_17
45
_2
33
22
32
89
34
56
NW
MN
_01
66
coa
33
32
32
29
93
44
6
NW
MN
_05
25
sdrE
33
32
33
19
93
44
6
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D Fig
ure
7
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez A
TC
C 2
921
3 d
an
s le
TC
D p
ar
dif
feacutere
nts
fil
tre
s
(A)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts la
ge
nta
mic
ine
ge2
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(C
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts d
an
s le
pa
lier
1
de
reacute
sis
tan
ce
agravela
ge
nta
mic
ine
le1
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
et
nom
bre
de m
uta
nts
reacute
sis
tan
ts agrave
la la
gen
tam
icin
e (
+T
O)
de
s 2
de
rnie
rs p
alie
rs d
e r
eacutesis
tan
ce
ge5
(D
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
7 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
ge
nta
mic
ine
(+
TO
) ge
9
93
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_16
29
ccp
A0
01
33
31
50
00
01
24
0
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_21
28
rpsM
11
12
21
12
22
00
31
74
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
NW
MN
_25
29
clfB
33
03
33
80
11
26
17
4
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e reacute
sist
ance
agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
On
b m
uta
nts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
16
29
ccp
A0
01
33
31
50
00
01
24
0N
WM
N_
21
28
rpsM
11
12
21
12
22
00
31
74
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
A B C
94
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
04
6_
1
01
00
28
22
32
2
10
9
NW
MN
_17
33
prs
A0
11
12
01
12
12
22
14
7
NW
MN
_19
23
_0
11
31
15
13
22
21
08
NW
MN
_20
37
_0
02
32
07
23
11
21
27
NW
MN
_22
77
_1
02
21
11
02
12
33
14
8
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
01
43
_0
03
00
04
23
33
34
11
NW
MN
_0
40
6lp
l3n
m2
01
00
03
12
22
33
7
NW
MN
_1
65
2_
00
10
00
62
12
31
68
NW
MN
_2
01
2_
00
00
00
32
33
30
31
1N
WM
N_
20
15
mn
aA0
00
00
01
23
33
01
11
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
o
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D E Fig
ure
8
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez N
ew
bo
uld
Δh
em
Bd
an
s l
e T
CD
par
dif
feacutere
nts
fil
tres
(A)
Nom
bre
de
mu
tan
ts
reacutesis
tan
ts agrave
la tom
atid
ine
ge15
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atid
ine
ge15
et no
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
(C)
No
mbre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la tom
atid
ine
ge15
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3(
D)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atidin
ege
10
et no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
tom
atid
ine
(+G
EN
) ge
7
(E)
No
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la to
ma
tid
ine
le6
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
95
Le TCD mrsquoa permis de rapidement identifier les gegravenes drsquointeacuterecircts Agrave lrsquoaide du site CMR
(Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) 2013) jrsquoai
ensuite veacuterifieacute la fonction des gegravenes identifieacutes afin de confirmer leur inteacuterecirct (Tableau
1)
Tableau 1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants et (B) chez les mutants NewbouldΔhemB reacutesistants
Gegravene Alias Fonction
NWMN_0811 NADH dh complexe 1 chaine de transport des eacutelectrons
NWMN_2070 _membrane prot Putative- Toxin production+resistance drug transporteur-
similar to multidrug transporteur 85
NWMN_0635 _membrane prot Putative- Bordeacute par AraC(reacutegulateur opeacuteron arabinose) ou
SsaA-likeprot(lieacute au quorum sensing RNAIII(secretory antigen)
NWMN_2084 _truncated transposase for Is1272- mobile+extrachromosomal element
fonctiontransposon fonction
NWMN_0928 qoxC quinol oxidase polypeptide IIIcomplexe 4 chaine de transport des eacutelectrons
NWMN_0739 _ conserved hypothetical prot fonction inconnues
NWMN_1164 hslU ATP-dependent protease ATP-binding subunit
NWMN_0524 sdrD Ser-Asp rich fibrinogenbone sialoprotein-binding protein sdrD
NWMN_0042 _ lipoprotein putativecell enveloppe- lipoprot en tendem
NWMN_0580 _conserved hypothetical prot fonction inconnuesDNA metabolism DNA
replication recombination and repair
NWMN_1745 _
NWMN_0166 coa coagulase precursor
NWMN_0525 sdrE Ser-Asp rich fibrinogenbone sialoprotein-binding protein SdrE
A
96
Apregraves avoir aligneacute speacutecifiquement une par une chacune des seacutequences (Fasta) des
gegravenes drsquointeacuterecirct de chaque mutant avec leur souche parente jrsquoai utiliseacute le programme
de visualisation et de comparaison de seacutequences ClustalW2 (ClustalW2 2013) afin
de confirmer que la mutation eacutetait une vraie mutation et non une erreur de
seacutequenccedilage ou drsquoalignement et que la mutation eacutetait identique chez tous les mutants
ayant le gegravene drsquointeacuterecirct muteacute Cette eacutetape a permis de restreindre encore la liste des
gegravenes pouvant ecirctre en lien avec la cible ou le mode drsquoaction de la TO (Figure 9)
Gene Alias Fonction
NWMN_0896 _conserved hypothetical prot fonction inconnue- comK family prot-
regulation DNA interaction- ORFIDresssemble agrave facteur de transcription
NWMN_0996 _ conserved hypothetical prot fonction inconnue- prophage fonction
NWMN_1025 _ phage major tail protprophage fonction
NWMN_1629 ccpAcatabollite control protA virulence determinant production and antibiotique
resistance
NWMN_1916 _ conserved hypothetical prot-fonction inconnue ou prophage prot
NWMN_2314 _drug resistance transporteur- EmrBQacA subfamily prot- toxin
production+resistance- transport and binding prot
NWMN_0046 _ hypothetical prot
NWMN_2015 mnaA UDP-Glc-Nac 2 epimerase
NWMN_2012 atpE ATP synthase subunit C
NWMN_1652 _ multidrug resistance transporter protein B
NWMN_0143 _ oligopeptide ABC transporter ATP-binding protein
B
97
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
A
B
98
Jrsquoai ensuite chercheacute le rocircle de ces gegravenes sur le site CMR afin de veacuterifier si leur
mutation pouvait expliquer la reacutesistance agrave lrsquoantibiotique (Comprehensive Microbial
Ressouce (J Craig Venter Institute (JCVI) 2013) Dans une premiegravere eacutetape le rocircle
drsquoun gegravene a eacuteteacute deacutetermineacute en allant tout simplement voir directement le rocircle de ce
gegravene chez Staphylococcus aureus str Newman Dans le cas ougrave le gegravene nrsquoavait pas
de fonction connue chez Saureus str Newman je suis alleacutee voir quelle eacutetait la
fonction de ce mecircme gegravene (et des gegravenes flanquant celui-ci) chez diffeacuterentes souches
de Staphylococcus aureus Advenant le cas ougrave malgreacute tout je nrsquoeacutetais toujours pas
capable de deacuteterminer le rocircle de ce gegravene je suis alleacutee veacuterifier si sa seacutequence pouvait
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 hellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Mutants resistants agrave GEN (4 microgml)+TO
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip 185-AATTGCATTCA-197helliphelliphellip495-ATAGCAATGG-506hellip
hellipnwmn_1629helliphelliphelliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_0143hellip
ccpAATP synthase c
ABC transp
Niveau 1
Niveau 2
Niveau 3
Niveau 4
TO +GEN (4microgml)
Figure 9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct (A) nwmn_0811 et
nwmn_0928 chez les mutants ATCC 29213 reacutesistants agrave la combinaison gentamicine
(+TO) leur souche parente ATCC 29213 (B) nwmn_1629 et nwmn_2012 chez les
mutants hemB reacutesistants agrave la TO et leur souche parente NewbouldΔhemB (C)
nwmn_1629 nwmn_2012 et nwmn_0143 chez les mutants hemB reacutesistant agrave la
tomatidine (+genta) et leur souche parente hemB
C
99
avoir une forte homologie avec un gegravene drsquoune autre espegravece Malgreacute tout certains
gegravenes portent toujours la mention laquo de fonction inconnue raquo
La meacutethodologie que nous avons employeacutee nous a permis drsquoidentifier plusieurs
gegravenes possiblement impliqueacutes dans reacutesistance de nos souches aux antibiotiques
testeacutes
1- chez Staphylococcus aureus ATCC 29213 reacutesistant agrave la gentamicine (+TO)
(Figure 9A)
- NWMN_0811 codant possiblement pour la NADH deacuteshydrogeacutenase
- qoxC (NWMN_0928) codant pour la quinol oxidase polypeptide III
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
2- chez NewbouldΔhemB reacutesistant agrave la tomatidine (Figure 9B)
- ccpA (NWMN_1629) codant pour la laquo catabolite control protein A raquo
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
3- chez NewbouldΔhemB reacutesistant agrave la tomatidine (+gentamicine) (Figure 9C)
- NWMN_0143 codant pour un ABC transporteur
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
Lrsquoanalyse de ces diffeacuterents gegravenes nous amegravene agrave suspecter la sous-uniteacute c de lrsquoATP
synthase comme eacutetant la cible potentielle de la tomatidine chez Staphylococcus
aureus
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine
Les gegravenes muteacutes pointent en grande partie vers la chaine respiratoire qursquoon appelle
aussi chaine de transport des eacutelectrons
100
La NADH deacuteshydrogeacutenase (NWMN_0811) fait partie du complexes I de la chaine de
transport de eacutelectrons alors que la quinol oxidase polypeptide III (NWMN_0928 ou
QoxC) est une composante du complex III et la sous-uniteacute c de lrsquoATP synthase (AtpE)
fait partie eacutevidement du complexe de lrsquoATP synthase ou complexe V de la chaine
respiratoire (Figure 10)
Figure 10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants
Les courbes de croissance des mutants ATCC29213 reacutesistants agrave la combinaison
GEN(+TO) appuient le fait que ces mutations affectent la chaine de transport des
eacutelectrons ce qui affecte le niveau de production drsquoATP et donc la croissance
bacteacuterienne En effet les mutants du troisiegraveme niveau de reacutesistance (qui possegravede
deux gegravenes de la chaine de transport des eacutelectrons de muteacutes) ont une croissance
plus lente que les mutants du premier niveau de reacutesistance qui eux ont un seul gegravene
dans la chaine de transport des eacutelectrons de muteacute (Figure 11)
Quinoloxidase
NADHdehydrogenase
ATPsynthetase
Proton gradient
XXX
101
Figure 11 Courbes de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison GEN+TO du premier niveau de reacutesistance (bleu) et du troisiegraveme niveau
de reacutesistance (vert)
Voici plus preacuteciseacutement comment fonctionne la chaine de transport des eacutelectrons chez
les 2 pheacutenotypes-parents selon le modegravele-hypothegravese eacutetabli avec nos reacutesultats et
analyses
Tout drsquoabord chez le prototype ou souche de pheacutenotype normal (ex ATCC 29213) la
chaine de transport des eacutelectrons fonctionne normalement Les complexes de la
chaine de transport des eacutelectrons sont chargeacutes de transfeacuterer des protons vers
lrsquoexteacuterieur de la cellule en transfeacuterant en mecircme temps des eacutelectrons drsquoun complexe agrave
un autre jusqursquoagrave un accepteur final qui est lrsquoO2 lorsque crsquoest possible Se faisant il se
creacutee un gradient eacutelectrochimique de part et drsquoautre de la membrane et les protons
accumuleacutes du cocircteacute externe sont utiliseacutes par la suite par lrsquoATP synthase afin de
pouvoir produire de lrsquoATP dans la cellule
0 4 8 1 2 1 6 2 0 2 4
0 0 1
0 1
1
C o u r b e d e c r o is s a n c e d e s m u ta n ts A T C C 2 9 2 1 3
reacute s is ta n ts agrave G E N + T O (8 m g L )
te m p s (h )
OD
60
0n
m
Niveau 3
Niveau 1
102
Le transfert drsquoeacutelectrons ne se fait pas sans coucirct et des produits toxiques pour la
cellule sont produits en mecircme temps ce sont les espegraveces reacuteactives de lrsquooxygegravene ou
ROS (Reactive Oxygen Species) Cependant les prototypes par leur reacuteponse au
stress produisent des enzymes capables de deacutetoxifier le milieu (Figure 12A)
Chez les souches SCV ici la souche controcircle NewbouldΔhemB la chaine de
transport des eacutelectrons est deacuteficiente parce que le gegravene permettant la production
drsquoheacutemine qui entre dans la composition des cytochromes est supprimeacute Les
cytochromes entrent dans la composition de plusieurs composantes de la chaine
respiratoire
Lorsque les complexes de la chaine sont alteacutereacutes il y a diminution du transfert de
protons vers lrsquoexteacuterieur une diminution de la production drsquoATP et une plus grande
production de ROS (Figure 12B)
Figure 12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B) NewbouldΔhemB
I III IV V I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
eacute eacute
eacute
H+ H+
ATPO2 H2O
ROS +ROS
Prototype SCV (hemB)
H+
0811 0928 2012 0811 0928
hegraveme
2012
A B
103
La gentamicine a besoin drsquoune bonne polarisation membranaire donc drsquoune chaine de
transport des eacutelectrons fonctionnelle pour pouvoir peacuteneacutetrer dans la cellule (Bryan et
Kwan 1983 (Bryan and van den Elzen 1977 Davis 1987 Taber et al 1987) Une
fois entreacutee elle inhibe la synthegravese proteacuteique ce qui provoque la mort cellulaire La
CMI de la gentamicine contre ATCC 29213 est de 05-1 microgml (Figure 13A)
La tomatidine elle irait se lier agrave lrsquoATP synthase pour en diminuer la fonctionnaliteacute et
ce tant chez ATCC 29213 que chez hemB Cette liaison amegravenerait une diminution
de la production drsquoATP et conseacutequemment provoquerait agrave plus ou moins long terme
une augmentation de production de ROS par lrsquoalteacuteration de la chaine de transport des
eacutelectrons Cependant comme la cellule prototype produit efficacement les enzymes
de deacutetoxification lrsquoimpact de la tomatidine est pratiquement nulle sur les souches
prototypes La CMI de la tomatidine est gt16microml contre ATCC 29213 (Figure 13B)
104
Figure 13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213
Chez ATCC 29213 lrsquoajout de 8microgml de tomatidine agrave la Gentamicine amegravene une
diminution de 8X agrave 16X la CMI de la gentamicine En effet la CMI de la gentamicine
passe de 05-1microgml agrave 006microgml (Figure 14AB)
Lors de lrsquoajout de la tomatidine agrave la gentamicine crsquoest la tomatidine qui se lie dans
un premier temps agrave lrsquoATP synthase reacuteduisant ainsi sont niveau drsquoactiviteacute Il y a alors
de faccedilon transitoire une augmentation de la concentration de protons agrave lrsquoexteacuterieur de
la cellule ce qui permet une entreacutee massive de gentamicine ainsi qursquoune rapide et
forte inhibition de la synthegravese proteacuteique Dans un second temps il y aura eacutegalement
une alteacuteration de la chaine de transport des eacutelectrons conduisant agrave une
augmentation de la production de ROS Crsquoest cependant lrsquoentreacutee massive de
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI TO gt16 microgmlCMI GEN 05-1 microgml
+ROS
ROS mais deacutetoxification efficace
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
105
gentamicine entrainant une forte inhibition de la synthegravese proteacuteique qui provoquera
dans ce cas-ci la mort cellulaire (Figure 14B)
Figure 14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicine lorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC 29213
Chez les mutants ATCC29213 reacutesistants agrave la combinaison GEN+TO les complexes I
et III sont muteacutes et donc ont une fonctionnaliteacute alteacutereacutee ce qui amegravene une
deacutepolarisation membranaire Une membrane moins polariseacutee amegravene une plus faible
entreacutee de gentamicine et donc une plus faible inhibition proteacuteique La CMI de la
gentamicine passe en effet de 05-1 microgml chez la souche parente agrave 2 microgml chez les
mutants reacutesistants
La tomatidine en se liant agrave lrsquoATP synthase accentuera la deacutepolarisation
membranaire provoquant une augmentation de la production des ROS et une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN (TO 8 microgml) 006 microgmlCMI GEN 05-1 microgml
+ROS
polarisation entreacutee massive de GEN
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
GENH+
BA
106
diminution de la production drsquoATP De fait les mutants reacutesistants ont une croissance
plus lente (Figure 15)(Figure 11)
Figure 15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants ATCC 29213
reacutesistants agrave la combinaison gentamicine (+TO)
Chez les SCV la tomatidine se lie aussi agrave lrsquoATP synthase et ce faisant amegravene une
diminution de la production drsquoATP Cette diminution de la production drsquoATP megravenera
ultimement agrave une alteacuteration de la chaine de transport des eacutelectrons et une
augmentation des ROS (Murphy 2009) tout comme chez les souches prototypes
Cependant comme le SCV a deacutejagrave une chaine de transport des eacutelectrons deacuteficiente
une faible production drsquoATP par lrsquoATP synthase et une forte production de ROS
lrsquoajout drsquoune faible concentration de tomatidine (la CMI est de 006microgml) amegravene une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype R GEN (+TO [8 microgml])
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN 2 microgmlCMI GEN 05-1 microgml
+ROS
R agrave GEN (+TO) mais mutants ont une
croissance diminueacutee (+ROS et -ATP)
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
107
augmentation de chacun de ces pheacutenomegravenes ce qui est suffisant pour provoquer la
mort cellulaire (Figure 16A)
Dans les premiers niveaux de reacutesistance agrave la tomatidine chez les mutants hemB la
reacutesistance (la CMI passe de 006 agrave 025-1microgml) est ameneacutee par la mutation du gegravene
ccpA (Catabolite controcircle protein A) ce qui megravenerait agrave une augmentation de la
production drsquoATP par lrsquoutilisation drsquoune autre voie que lrsquoATP synthase etou agrave une
diminution de la production de ROS (Figure 16B) Le rocircle de la CcpA en lien avec la
reacutesistance agrave la tomatidine est encore neacutebuleux Toutefois on sait que normalement
ce gegravene agit comme reacutegulateur transcriptionnel des gegravenes codant pour des enzymes
du cycle de Krebs et qursquoil a un rocircle de reacutegulation dans le meacutetabolisme du carbone et
lrsquoutilisation des substrats (Somerville et Proctor 2009 Goumltz et al 2006)
Figure 16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO 006 microgml
H+
SCV
I III IVeacute eacute
eacuteO2 H2O
+ROS
H+TO
Niveau 1
CcpA (CMI TO 025-1 microgml)
ROS
ATP
V
H+
ATP
TO0811 0928
2012
1629
0811 0928
2012
BA
108
Chez les SCV du deuxiegraveme palier de reacutesistance agrave la tomatidine non seulement le
gegravene ccpA est toujours muteacute mais le gegravene de la sous-uniteacute c de lrsquoATP synthase est
eacutegalement muteacute La mutation dans le site de liaison mecircme de la tomatidine empecircche
sa liaison et donc empecircche son effet sa CMI passant de 006 agrave gt16microgml (Figure
17B)
Malgreacute la mutation dans le gegravene lrsquoATP synthase resterait quand mecircme fonctionnel
puisque la mutation nrsquoamegravene pas la mort cellulaire comme crsquoest le cas lorsque la
tomatidine lie lrsquoATP synthase chez le SCV parent (Figure 17A)
Figure 17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la tomatidine
Tout comme chez les mutants hemB reacutesistant agrave la tomatidine seule les mutants
hemB reacutesistant agrave la tomatidine lorsqursquoen preacutesence de gentamicine possegravedent
eacutegalement une mutation dans le gegravene de lrsquoATP synthase ce qui rend la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO006 microgml
H+
SCV Niveau 2CMI TO gt16 microgml
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+TO
TO
Niveaux 1 et 2
CcpA (CMI TO 025-1 microgml)
ROS
ATP
0811 0928
2012
1629
0811 0928
2012
A B
109
inactive puisqursquoincapable de srsquoy lier (CMI gt16microgml) (Figure 18B) Chez les SCV non
muteacutes (hemB) la gentamicine nrsquoentre que faiblement ducirc agrave la deacutepolarisation de sa
membrane (CMI 4-8 microgml) (Figure 18A) La mutation dans le gegravene codant pour une
pompe (NWMN_0143 codant pour un ABC transporteur) chez les mutants hemB
reacutesistant agrave la tomatidine (+gentamicine) amegravenerait une augmentation de lrsquoactiviteacute de
cette pompe permettant de rejeter hors de la cellule le peu de gentamicine eacutetant
entreacutee La mutation de cette pompe fait passer la CMI de la gentamicine de 4-8
microgml pour hemB agrave 16 microgml pour les mutants hemB reacutesistant agrave la combinaison
gentamicine (+TO) ayant cette mutation (Figure 18B)
Figure 18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB reacutesistants
agrave la combinaison tomatidine (+gentamicine)
Donc selon notre modegravele-hypothegravese lrsquoaction de la tomatidine et de la gentamicine
sur S aureus deacutepend de la liaison agrave leur cible mais a aussi un lien avec la
SCV R agrave TO (+GEN [4microgml])
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI GEN 4-8 microgml
H+
SCV
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+ TO
GEN
CMI GEN 16 microgml
CMI TO gt16 microgml
0811 0928
2012
0811 0928 20120143
GEN
Synthegravese proteacuteique
BA
110
polarisation membranaire et la production de ROS La mesure de la polarisation et
concentration cellulaire en ROS chez nos mutants et nos souches parents en
preacutesence ou non drsquoantibiotique nous permettra de valider notre modegravele-hypothegravese
Chez une bacteacuterie meacutetaboliquement active et posseacutedant une membrane
cytoplasmique intacte il y a normalement une diffeacuterence de potentiel eacutelectrique entre
le cocircteacute inteacuterieur et le cocircteacute exteacuterieur de la membrane lrsquointeacuterieur eacutetant plus neacutegatif de
100 agrave 200 mV par rapport agrave lrsquoexteacuterieur Une diminution de la magnitude de ce
potentiel membranaire est appeleacutee deacutepolarisation alors qursquoune augmentation de ce
potentiel est appeleacutee hyperpolarisation Le potentiel membranaire sera reacuteduit agrave zeacutero
srsquoil y a rupture de la membrane cest-agrave-dire srsquoil y a formation de trous assez grands
permettant aux ions inorganiques de passer librement Ce pheacutenomegravene peut arriver
lorsque les cellules sont tueacutees par la chaleur le froid ou les antibiotiques β-Lactams
Dans ces circonstances la membrane devient permeacuteable agrave diffeacuterents colorants
comme le propidium iodide (PI) et le TO-PRO3 et le changement est irreacuteversible
Plusieurs classes de composeacutes chimiques notamment les ionophores peuvent
eacutegalement alteacuterer le potentiel membranaire sans pour autant permettre la
permeacuteabiliteacute au PI TO-PRO3 et autres colorants similaires chez les bacteacuteries
lrsquoadministration drsquoun proton-ionophore tel le carbonyl cyanide 3-
chlorophenylhydrazone (CCCP) reacuteduira le potentiel membranaire agrave zeacutero
Des tests preacuteliminaires sur la mesure du potentiel membranaire (BacLighttrade Bacterial
Membrane Potential Kit (B34950) drsquoInvitrogen et lecture au cytomegravetre de flux (FACS))
(Novo et al 2000) chez nos deux souches parentes mets en eacutevidence que ATCC
29213 est plus polariseacute que NewbouldΔhemB (Figure 19A) Toutefois malgreacute
qursquoayant une plus faible polarisation membranaire NewbouldΔhemB nrsquoest pas
complegravetement deacutepolariseacute puisqursquoil est possible de le deacutepolariser davantage agrave lrsquoaide
drsquoun agent deacutepolarisant tel que le CCCP (carbonyl cyanide 3-chlorophenylhydrazone)
(Figure 19B) Le TO-PRO3 a eacuteteacute utiliseacute comme controcircle afin de confirmer la viabiliteacute
de nos cellules (donneacutees non montreacutees)
111
A B
Figure 19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane Potential Kit
(Invitrogen) (Lecture au cytomegravetre de flux agrave lrsquoaide du programme FCS Express) (A)
Ratio FL3-H (axe des Y)FL1-H (axe des X) de ATCC 29213 (rouge) et de
NewbouldΔhemB (noir) (B) Lecture des rouges (FL3-H) sur NewbouldΔhemB traiteacute
avec le DiOC2(3) (33prime-diethyloxacarbocyanine iodide) (noir) ou avec le CCCP (bleu)
ou sur les cellules chauffeacutees (rouge)
La sous-uniteacute c de lrsquoATP synthase (AtpE) semble donc la cible la plus probable de la
tomatidine Afin de veacuterifier si les mutations identifieacutees pouvaient avoir un effet
veacuteritable sur la proteacuteine nous avons fait un alignement de seacutequences noteacute les bases
muteacutees et veacuterifieacute si la mutation amenait un changement drsquoacide amineacute et si ce
changement drsquoacide amineacute pouvait avoir un impact sur la proteacuteine finale De fait
nous avons trouveacute que les deux diffeacuterentes mutations dans lrsquoAtpE amenaient un
changement significatif drsquoacides amineacutes pouvant amener une variation fonctionnelle
FL3-H
Count
100
101
102
103
104
0
18
36
53
71
Chauffeacute
+CCCP+DiOC2
Chauffeacute
FL1-H
FL3-H
100
101
102
103
104
100
101
102
103
104
hemB
29213
VERT
RO
UG
E
ROUGE
ΔhemB
112
de la proteacuteine La mutation de la base 49 change un G pour un T il srsquoagit drsquoun SNP
nrsquoamenant pas de changement de cadre de lecture La mutation fait passer lrsquoacide
amineacute alanine (A) qui est apolaire agrave une seacuterine (S) qui est polaire La mutation de la
base 190 change un G pour un A et est aussi un SNP nrsquoamenant pas de changement
de cadre de lecture Cette mutation fait passer lrsquoacide amineacute alanine (A) qui est
apolaire agrave une threacuteonine (T) qui est polaire (Figure 20)
gtlcl||NWMN_2012|atpE|72493880 F0F1 ATP synthase subunit C
1 10 19 31 43 49 58
ATG AAT TTA ATC GCA GCA GCA ATC GCA ATT GGT TTA TCA GCA TTA GGA GCA GGT ATC GGT
M N L I A A A I A I G L S A L G A G I G
(S)
17 61 73 85 97 109 118
AAC GGT TTA ATC GTT TCA AGA ACA GTT GAA GGT GTA GCA CGT CAA CCA GAA GCA CGT GGT
N G L I V S R T V E G V A R Q P E A R G
21 40
121 133 145 157 169 178
CAA TTA ATG GGT ATC ATG TTC ATT GGT GTA GGT TTA GTT GAG GCA TTA CCT ATC ATC GGT
Q L M G I M F I G V G L V E A L P I I G
41 60
181 190 199 211
GTA GTA ATT GCA TTC ATG ACA TTT GCT GGA TAA
V V I A F M T F A G
(T)
61 64 70
Figure 20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du gegravene
atpE de S aureus Newman avec mise en eacutevidence des mutations (rose) en lien avec
la cible de la tomatidine Lrsquoalignement a eacuteteacute effectueacute agrave lrsquoaide du site CMR de JCVI
113
CHAPITRE 4
DISCUSSION
LrsquoATP synthase est eacutevidemment un complexe proteacuteique ubiquitaire tant chez les
procaryotes que les eucaryotes Malgreacute ce fait il semble que la cible de la tomatidine
soit speacutecifique aux bacillales celle-ci ne deacutemontrant aucune activiteacute contre les
bacteacuteries agrave Gram neacutegatif et les lactobacillales On suppose donc que lrsquoATP synthase
de ces espegraveces doit ecirctre speacutecifique au niveau du site de liaison de la tomatidine
Lrsquoecirctre humain en tant qursquoespegravece eucaryote est eacutegalement pourvu de ce complexe
proteacuteique et on peut naturellement se demander si lrsquoutilisation clinique de cette
moleacutecule pourrait ecirctre cytotoxique De fait plusieurs composeacutes naturels tel
lrsquooligomycine et la venturicidine sont connus pour inhiber lrsquoactiviteacute de lrsquoATP synthase
par leur interaction avec la sous-uniteacute c de ce complexe Cependant ces moleacutecules
ne sont pas seacutelectives et nrsquoinhibent pas seulement lrsquoATP synthase bacteacuterien mais
aussi lrsquoATP synthase mitochondrial (Haagsma et al 2009 Matsuno-Yagi et Hatefi
1993) (Figure 1) Neacuteanmoins il existe maintenant une moleacutecule qui a pour cible
lrsquoATP synthase bacteacuterien et qui est en processus en vue drsquoune utilisation clinique
chez lrsquohomme le TMC207 (Haagsma et al 2009))
114
Figure 1 Sites drsquoinhibition de lrsquoATP synthase Les sites de liaison dans lrsquoATP
synthase des inhibiteurs reacuteveacuteleacutes par des eacutetudes biochimiques et structurales sont
indiqueacutes par des cercles rouges et les sites de liaisons qui nrsquoont pas eacuteteacute totalement
clarifieacutes sont indiqueacutes par des cercles verts (Hong et Pedersen 2008)
Le TMC207 (Figure 2A) est un composeacute de la classe des diarylquinoline posseacutedant
un nouveau meacutecanisme drsquoaction lrsquoinhibition de lrsquoATP synthase bacteacuterien De fait la
cible et le meacutecanisme drsquoaction du TMC207 sont diffeacuterents des autres agents
antituberculeux Lrsquoinhibition de lrsquoATP synthase peut mener agrave une baisse de la
production drsquoATP et un deacutebalancement dans lrsquohomeacuteostasie du pH ces 2 effets
pouvant contribuer agrave la diminution de la survie de la bacteacuterie (Rao et al 2001
Andries et al 2005)
Le TMC207 est une moleacutecule ayant une forte activiteacute bacteacutericide contre
Mycobacterium tuberculosis et autres espegraveces mycobacteacuteriennes mais peu drsquoactiviteacute
115
contres les autres espegraveces bacteacuteriennes Lrsquoindex de seacutelectiviteacute de TMC207 pour
lrsquoATP synthase mycobacteacuterienne (CMI50 gt200 microml) compareacute agrave lrsquoATP synthase
mitochondriale (CMI50 001 microgml) est de plus de 20000 Un indice de seacutelectiviteacute au
dessus de 1000 est consideacutereacute dans le deacuteveloppement drsquoune drogue comme un
preacutealable pour une moleacutecule candidate (Haagsma et al 2009) La diversiteacute dans les
gegravenes atpE explique la reacutesistance au TMC207 et justifie la faible affiniteacute de cette
drogue agrave lrsquoATP synthase mitochondriale (Matteelli et al 2010) (Figure 3)
Figure 2 Structure chimique de (A) TMC207 et (B) son analogue anti bacteacuteries
agrave Gram positifs (Balemans et al 2012)
Lorsqursquoon compare la seacutequence proteacuteique de la sous-uniteacute c de lrsquoATP synthase
(atpE) de quelques espegraveces de lrsquoordre des Bacillales (tel Saureus B cereus B
subtilis et L monocytogenes) avec des espegraveces appartenant agrave drsquoautres ordres
bacteacuteriens on constate que les seacutequences divergent ce qui est en accord avec la
speacutecificiteacute des moleacutecules Les points de mutations amenant une reacutesistance au
TMC207 chez Mycobacterium smegmatis sont speacutecifiques aux mycobacteacuteries et les
deux principaux points de mutations D32V(ouP) et A67P chez M smegmatis sont
situeacutes tous les deux dans les heacutelices alpha qui entourent le site catalytique de la
proteacuteine (E65) qui est impliqueacute dans le transport des protons (Andries et al 2005)
Lrsquoacide amineacute aspartate (D) agrave la position 32 et lrsquoalanine (A) agrave la position 67 sont tregraves
speacutecifiques agrave M smegmatis
A B
116
Reacutecemment un groupe de chercheurs (Balemans et al 2012) on mit en lumiegravere une
autre moleacutecule (Figure 2B) de la classe des diarylquinolines dont le meacutecanisme
drsquoaction eacutetait aussi lrsquoinhibition de la sous-uniteacute c de lrsquoATP synthase mais chez les
bacteacuteries agrave Gram positif tels Streptococcus pneumoniae et S aureus Cette moleacutecule
irait lier les acides amineacutes valines (V) situeacutes agrave la position 50 et 62 de lrsquoAtpE de S
aureus (Donneacutees non montreacutees chez S pneumoniae(Balemans et al 2012)) En
effet une mutation amenant le changement de la valine en position 50 par une
isoleucine (I) etou une mutation amenant le changement de la valine en position 62
par une alanine (A) amegravene la reacutesistance de la bacteacuterie agrave la moleacutecule Cependant L
monocytogenes et Bacillus subtilis ne possegravedent pas de valine agrave la position 50 alors
qursquoils font eacutegalement partie des bacteacuteries agrave Gram positif (Il faut noter que chez
Bacillus sp PS3 (Swiss-Prot assession Ndeg P 00845) possegravede une valine agrave la position
50 (Balemans et al 2012)) Par contre tout comme pour les autres bacteacuteries agrave Gram
positif testeacutees les deux espegraveces possegravedent une valine agrave la position 62 (Figure 3)
Chez B cereus B subtilis B anthracis S aureus et L monocytogenes les points de
divergence sont plutocirct A17S et A64T Cependant B pseudofirmus qui fait eacutegalement
partie de lrsquoordre des Bacillales possegravede une glycine (G) agrave la position 17 plutocirct qursquoune
alanine (A) et une seacuterine (S) plutocirct qursquoune alanine (A) agrave la position 64 comme les
espegraveces preacuteceacutedentes Suivant ces reacutesultats et si la cible de la tomatidine est bien
ces deux points de mutation la tomatidine ne devrait pas pouvoir se lier agrave lrsquoAtpE de B
pseudofirmus et devrait donc nrsquoavoir aucune activiteacute contre cette souche alors que
nous avions extrapoleacute que le spectre drsquoactiviteacute de la tomatidine eacutetait tous les
Bacillales De plus E coli et M smegmatis possegravedent eacutegalement une alanine (A) agrave la
position 17 alors que la tomatidine ne possegravede aucune activiteacute contre ces espegraveces (M
smegmatis nrsquoa pas eacuteteacute eacutevalueacute il srsquoagit ici drsquoune extrapolation des reacutesultats de CMI)
par contre ces deux espegraveces possegravedent des acides amineacutes diffeacuterents drsquoune alanine
(A) agrave la position 64 ce qui pourrait leur confeacuterer une reacutesistance agrave la tomatidine telle
qursquoobserveacutee On remarque eacutegalement que les acides amineacutes agrave ces deux positions
117
sont totalement diffeacuterents chez lrsquohomme (Homo sapiens) ce qui suggeacutererait que la
tomatidine ne possegravederait pas drsquoactiviteacute antimitochondriale (Figure 3)
Selon ces reacutesultats on peut se demander si les deux mutations sont essentielles agrave la
liaison de la moleacutecule ou si seulement lrsquoalanine (A) agrave la position 64 est neacutecessaire
On doit se rappeler que la reacutesistance apporteacutee par la mutation faisant en sorte que
lrsquoalanine (A) en position 17 devient une seacuterine (S) apparaicirct chez les mutants
NewbouldΔhemb reacutesistant agrave la tomatidine et que cette mutation nrsquoapparaicirct qursquoau
quatriegraveme et dernier niveau de reacutesistance On se rappelle eacutegalement que la mutation
faisant en sorte que lrsquoalanine (A) agrave la position 64 devient une threacuteonine (T) apparaicirct
chez les mutants NewbouldΔhemb reacutesistants agrave la tomatidine en combinaison avec la
gentamicine et ce degraves le premier niveau de reacutesistance
118
Bcereus G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Banthracis G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Lmonocytogenes G---------VIAAAIAVG---LGALGAGIGNGLIVSKTVEGVARQPEARSMLQTIMFV- 50
Saureus_NWMN_2012 -----------IAAAIAIG---LSALGAGIGNGLIVSRTVEGVARQPEARGQLMGIMFIG 49
Bsubtilis -----------IAAAIAIG---LGALGAGIGNGLIVSRTVEGIARQPEAGKELRTLMFMG 49
Bpseudofirmus -----------LGAAIAAG---LAAVAGAIAVAIIVKATIEGTTRQPELRGTLQTLMFIG 49
Ecoli L---------YMAAAVMMG---LAAIGAAIGIGILGGKFLEGAARQPDLIPLLRTQFFIV 56
Msmegmatis DPNAIITAGALIGGGLIMG---GGAIGAGIGDGIAGNALISGIARQPEAQGRLFTPFFIT 60
Homo sapiens QTSAISRDIDTAAKFIGAGAATVGVAGSGAGIGTVFGSLIIGYARNPSLKQQLFSYAILG 120
MyR V
MyR P
SaR S
Bcereus VALVEALPIIGVVIAFIVMNK----- 72
Banthracis VALVEALPIIGVVIAFIVMNK----- 72
Lmonocytogenes ----EALPIIAVVIAFMVLNK----- 67
Saureus_NWMN_2012 VGLVEALPIIGVVIAFMTFAG----- 70
Bsubtilis IALVEALPIIAVVIAFLAFFG----- 70
Bpseudofirmus VPLAEAVPIIAIVISLLILF------ 69
Ecoli MGLVDAIPMIAVGLGLYVMFAVA--- 79
Msmegmatis VGLVEAAYFINLAFMALFVFATPGLQ 86
Homo sapiens FALSEAMGLFCLMVAFLILFAM---- 142
MyR D P M
SpR I A
SaR T
Fig
ure
3
Seacuteq
uen
ce
pro
teacuteiq
ue d
e la p
roteacute
ine
Atp
Ec
hez d
iffeacute
ren
tes
es
pegravec
es e
t m
ise e
n eacute
vid
en
ce
de
s s
ite
s d
e
mu
tati
on
pe
rmett
an
t la
reacutesis
tan
ce agrave
dif
feacutere
nte
s m
oleacute
cu
les
M
yR M
tu
berc
ulo
sis
ou
M sm
eg
matis
reacutesis
tan
t a
u
TM
C2
07 S
pR
S
p
neu
mon
iae
reacutesis
tan
t agrave
un a
na
log
ue d
u T
MC
20
7
SaR
S
a
ure
us
SC
V r
eacutesis
tant agrave
la to
matid
ine
En
cad
reacutero
uge
S
ite d
e m
uta
tion a
men
ant la
reacutesis
tance
agravela
to
matid
ine E
ncad
reacuteno
ir
Site
cata
lytiq
ue
im
pliq
ueacute
da
ns le
tra
nspo
rt d
es p
roto
ns
Su
rlig
nag
etu
rqu
ois
e S
ite d
e m
uta
tion a
me
nant u
ne
reacutesis
tan
ce a
u T
MC
20
7
119
Les seacutequences proteacuteiques des souches bacteacuteriennes de la figure 3 ont eacuteteacute trouveacutees
sur pubmedproteacuteine httpwwwncbinlmnihgovpubmed
laquo ATP synthase subunit c raquo Assession number
Streptococcus pneumoniae D39 YP_8168031 (NCBI)
S aureus Newman BAF682841 (GenBank)
E coli AAA247321 (GenBank)
Bacillus cereus ISO75 EJP900901 (GenBank)
Bacillus anthracis str H9401 YP_0062123491 (NCBI)
Mycobacterium smegmatis MC2 155 YP_0065695541 (NCBI)
Bacillus subtilis str 168 CAA822551 (GenBank)
Listeria monocytogenes YP_0076049931 (NCBI)
Homo sapiens P482011 (UniProtKBSwiss-Prot)
Lrsquoalignement comparatif des seacutequences drsquoacides amineacutes a eacuteteacute fait agrave lrsquoaide du
programme ClustalW2 sur le web
httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2
Le modegravele 3D de lrsquoAtpE a eacuteteacute obtenu gracircce agrave lrsquoaide du Pr Ryszard Brzezinski du
Deacutepartement de biologie de lrsquoUniversiteacute de Sherbrooke (Figure 4) Une recherche
dans le Protein Data Bank a drsquoabord eacuteteacute effectueacutee afin de veacuterifier parmi les proteacuteines
AtpE (sous-uniteacute C de lrsquoATP synthase) pour lesquelles il y avait des structures 3D
reacutesolues laquelle ressemblait le plus agrave celle de Staphylococcus aureus Les
alignements ont montreacute que cest la proteacuteine de Bacillus pseudofirmus qui avait la
plus grande similitude de seacutequence La structure est stockeacutee dans la banque PDB
sous le symbole 3ZO6 Il a eacuteteacute confirmeacute par quelques alignements que cette sous-
uniteacute C de lrsquoATP synthase eacutetait plus proche par sa seacutequence de celle de
Staphylococcus aureus que de celle de Mycobacterium pour laquelle la structure 3D
avait eacuteteacute deacutetermineacutee plus tocirct Ce reacutesultat sexplique par le fait que Staphylococcus et
Bacillus font tous deux partie de la branche A+T-riche des bacteacuteries agrave Gram+ alors
que Mycobacterium appartient agrave la branche G+C-riche La seacutequence de lAtpE de
120
Staphylococcus aureus a ensuite eacuteteacute envoyeacutee au serveur internet de modeacutelisation
3D-Jigsaw (httpbmmcancerresearchukorg~3djigsaw) en indiquant que le modegravele
devait ecirctre bacircti en prenant comme matrice le fichier PDB 3ZO6 Le modegravele nous a
eacuteteacute retourneacute sous format PDB ndash comme les laquo vrais raquo cristaux dans la Protein Data
Bank Les fichiers au format PDB peuvent ecirctre visualiseacutes par le logiciel RasMol ndash
gratuit sur internet Cest avec RasMol que limage (Figure 4) a eacuteteacute obtenue
121
Figure 4 Modeacutelisation de la structure tridimensionelle de la sous-uniteacute c de
lrsquoATP synthase de S aureus (AtpE) montrant les acides amineacutes muteacutes chez les
mutants reacutesistantsEn blanc et vert les deux acides amineacutes muteacutes chez S aureus
reacutesistant agrave la tomatidine En rose les sites de mutations chez Mycobacterium
smegmatis reacutesistant au TMC207 et des sites de mutations chez Streptoccocus
pneumoniae reacutesistants agrave un analogue de TMC207 En bleu le site actif de la proteacuteine
Agrave la vue de ces reacutesultats certains pourront mettre en doute lrsquointeacuterecirct de la tomatidine
eacutetant donneacute lrsquoapparition relativement rapide de reacutesistance chez S aureus Cependant
122
nos reacutesultats deacutemontrent clairement que lrsquoajout de tomatidine ralentit le
deacuteveloppement de reacutesistance agrave la gentamicine chez S aureus ATCC29213 (Tableau
1) De plus cette augmentation de la CMI de la gentamicine en preacutesence de
tomatidine au fil des passages suit lrsquoaugmentation de la CMI de la ciprofloxacine lors
de ces mecircmes passages alors que la ciprofloxacine est consideacutereacutee comme la drogue
de reacutefeacuterences dans les bonnes drogues (Tableau 1) Tout comme pour la
ciprofloxacine pour laquelle la reacutesistance chez S aureus peut ecirctre la conseacutequence
de deux mutations dans le gegravene cible gyrA la reacutesistance agrave la tomatidine peut ecirctre la
conseacutequence de deux mutations dans le gegravene cible atpE Par comparaison la
rifampicine est consideacutereacutee comme une mauvaise drogue de par lrsquoapparition rapide
drsquoune forte reacutesistance chez S aureus conseacutequence de ses 17 mutations possibles
dans le gegravene cible rpoB (Tableau 1)
De plus il sera important de relativiser lrsquoapparition de reacutesistance agrave la tomatidine en
calculant le taux drsquoapparition de ces colonies reacutesistantes ceci nrsquoayant pas eacuteteacute fait
jusqursquoici De fait la freacutequence de mutation chez S aureus amenant une reacutesistance agrave
la rifampicine est de lrsquoordre de 10-7 10-8 alors qursquoil est de 10-11 pour lrsquoacide fusidique
(Canfield et al 2013 Orsquoneill et al 2001 Price et Gustafson 2001) Pour ce qui est
de la nouvelle moleacutecule anti-bacteacuteries agrave Gram positif de la classe des diarylquinoline
une freacutequence de mutation sur S pneumoniae ATCC49619 de 81- 83 x 10-7 a eacuteteacute
observeacutee agrave 5x la CMI et une freacutequence de mutation de 46 x 10-7 agrave 17 x 10-6 agrave 50x la
CMI (Balemans et al 2012)
123
Tableau 1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 par la meacutethode de CMI lors de passages successifs dans un milieu avec
lrsquoantibiotique
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
Typiqueldquomauvaiserdquodrogue (17 mutations possibles dansrpoB)
Typique ldquobonnerdquo drogue (2 mutations possibles dans gyrA)
Preacutesence de TO ralenti le deacuteveloppement de reacutesistances vs GEN seul
124
CHAPITRE 5
CONCLUSION ET PERSPECTIVES
Mon projet de maitrise a permis de preacuteciser le spectre drsquoactiviteacute de la tomatidine
Nous savons maintenant que la tomatidine possegravede une activiteacute antibacteacuterienne
contre la plupart des Firmicutes et plus speacutecifiquement contre les Bacillales Plus
preacuteciseacutement la tomatidine est bacteacutericide contre les pheacutenotypes SCV et agit en
synergie avec les aminoglycosides augmentant lrsquoactiviteacute bacteacutericide de ces derniers
de 8-16 fois contre les souches agrave pheacutenotype normal
De plus parmi tous les analogues de la tomatidine produits par le laboratoire du Pr
Eacuteric Marsault (voir article en annexe 2) le composeacute FC04-100 a eacuteteacute plus
speacutecifiquement mis en lumiegravere Celui-ci conserve non seulement une aussi bonne
activiteacute synergique avec les aminoglicosides contre les Saureus au pheacutenotype
normal et une activiteacute modeacutereacutee contre les S aureus SCV mais montre eacutegalement de
faccedilon inespeacutereacutee une activiteacute par lui-mecircme contre les S aureus de pheacutenotype normal
incluant plusieurs MRSA une proprieacuteteacute qui nrsquoeacutetait pas observeacutee pour la tomatidine
Une autre activiteacute inteacuteressante de FC04-100 est sa capaciteacute drsquoameacuteliorer lrsquoactiviteacute
bacteacutericide des aminoglicosides contre L monocytogenes agrave pheacutenotype normal apregraves
un traitement de 24h alors que la tomatidine ne deacutemontrait qursquoune faible synergie
avec les aminoglycosides contre cette souche De plus FC04-100 deacutemontre une
activiteacute bacteacutericide contre les SCV de L monocytogenes alors que la tomatidine ne
deacutemontrait qursquoune activiteacute bacteacuteriostatique FC04-100 montre eacutegalement un effet
potentiateur de lrsquoUbrolexinTM (Ceacutefalexin Kanamycin (3 2)) une combinaison
drsquoantibiotique utiliseacute dans le traitement de mammites bovines en abaissant de 8 agrave 16
fois leur CMI amenant celles-ci sous le seuil de reacutesistance Finalement FC04-100
possegravede une meilleure solubiliteacute que la tomatidine une proprieacuteteacute qui facilitera son
utilisation in vitro
125
Nos reacutesultats deacutemontrent que FC04-100 conserve une certaine activiteacute contre les
SCV reacutesistant agrave la tomatidine ce qui suggegravere possiblement que FC04-100 a plus
daffiniteacute pour la cible ou plus de sites dinteraction avec AtpE En effet les mutants
SCV reacutesistants agrave la tomatidine deviennent gt512 fois plus reacutesistants alors que FC04-
100 ne perd que 8 agrave 16 fois son activiteacute contre ces mecircmes souches Ceci deacutemontre
quil est possible dameacuteliorer significativement lactiviteacute de ces alkaloiumldes steacuteroidiens
ce qui est tregraves prometteur pour lavenir de cette moleacutecule
Dans un second temps lrsquoanalyse des mutations chez les mutants reacutesistants agrave la
tomatidine a permis de non seulement de mieux comprendre le mode drsquoaction de la
tomatidine mais eacutegalement de preacuteciser la cible potentielle de ce nouvel antibiotique
De fait son mode drsquoaction serait en lien direct avec la fonctionnaliteacute de la chaine de
transport des eacutelectrons chez la bacteacuterie lrsquoactiviteacute de la tomatidine serait inversement
proportionnelle agrave la fonctionnaliteacute de la chaine de transport des eacutelectrons et donc en
lien eacutegalement avec la polarisation membranaire et la production de ROS
intracellulaire Afin de confirmer le modegravele-hypothegravese la mesure de la polarisation
membranaire et la concentration de ROS intracellulaire chez les mutants ainsi que
chez les souches parentes suite agrave un traitement antibiotique devront ecirctre effectueacutees
Les reacutesultats de mes recherches suggegraverent que la cible bacteacuterienne speacutecifique de la
tomatidine serait la sous-uniteacute C de lrsquoATP synthase Afin de confirmer la cible
Maxime Lamontagne-Boulet tente au moment drsquoeacutecrire ces lignes drsquoobtenir des
mutants de S aureus Newman reacutesistants au composeacute FC04-100 et dont la reacutesistance
serait attribuable agrave une mutation dans le gegravene atpE Par la suite on devra confirmer
la fonctionnaliteacute de la cible ATP synthase par quantification de lrsquoATP produit agrave lrsquoaide
de la lucifeacuterase tel que lrsquoon fait Balemans et al en 2012 Lrsquoattachegravement agrave la cible
devra eacutegalement ecirctre confirmeacute et la meacutethode de SPR (Surface Plasmon Resonance)
pourrait ecirctre tout indiqueacutee pour ce faire (Balemans et al 2012) On devra eacutegalement
126
deacutemontrer la seacutelectiviteacute des moleacutecules tomatidine et FC04-100 pour lrsquoATP synthase
des Bacillales
Les infections agrave bacteacuteries multireacutesistantes deviennent de plus en plus preacutevalentes et
sont un des problegravemes de santeacute majeurs auxquels nous devons faire face aujourdrsquohui
Cette augmentation de la reacutesistance limite le reacutepertoire drsquoantibiotiques efficace creacuteant
une situation probleacutematique laquelle est exacerbeacutee par le trop petit nombre de
nouveaux antibiotiques introduits sur le marcheacute ces derniegraveres anneacutees Il est donc
drsquoune grande importance de travailler agrave la mise en marcheacute de nouveaux antibiotiques
et plus particuliegraverement de nouvelles classes drsquoantibiotiques espeacuterant ainsi deacutejouer la
multireacutesistance bacteacuterienne du moins pour un temps La tomatidine de par sa
structure sa cible et son mode drsquoaction speacutecifique est une moleacutecule antibiotique
dans une classe agrave part et de ce fait une moleacutecule plus qursquointeacuteressante pour le
traitement drsquoinfection agrave S aureus De plus comme elle deacutemontre une activiteacute
bacteacutericide contre les deux pheacutenotypes de S aureus (et autres Bacillales) lesquels
sont retrouveacutes simultaneacutement dans les cas drsquoinfections chroniques son pouvoir
antibiotique devient inestimables drsquoun point de vue clinique
127
CONTRIBUTIONS SUPPLEacuteMENTAIRES
ANNEXE 1 Application de brevet Internationnal publication number WO
2012109752 A1
INVENTEURS Franccedilois Malouin2 Gabriel Mitchell2 Kamal Bouarab2 Eacuteric Marsault1
Feacutelix Chagnon1 Simon Boulanger 2 Isabelle Guay2
1Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
2Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
128
129
ANNEXE 2 Article scientifique dans laquo Europeen Journal of medicinal chemistry raquo
2014 80 605-620
TITRE Unraveling the structure-activity relationship of tomatidine a steroid alkaloid
with unique antibiotic properties against persistent forms of
Staphylococcus aureus
ABSTRACT Staphylococcus aureus is responsible for difficult-to-treat and relapsing
infections and constitutes one of the most problematic pathogens due to its multiple
resistances to clinically available antibiotics Additionally this pathogenrsquos ability to
develop small-colony variants is associated with a reduced susceptibility to
aminoglycoside antibiotics and in vivo persistence We have recently demonstrated
that tomatidine a steroid alkaloid isolated from tomato plants possesses anti-
virulence activity against normal strains of S aureus as well as the ability to
potentiate the effect of aminoglycosides against this pathogen In addition tomatidine
has shown antibiotic activity against small-colony variants We herein report the first
study of the structure-activity relationship of tomatidine against S aureus
AUTEURS Feacutelix Chagnon1 Isabelle Guay2 Marc-Andreacute Bonin1 Gabriel Mitchell2
Kamal Bouarab2Franccedilois Malouin2 Eacuteric Marsault1
MA CONTRIBUTION Responsable des tests biologiques de tous les composeacutes
autant envers lrsquoeffet potentiateur de la tomatidine sur lrsquoactiviteacute de la gentamicine
contre la souche ATCC29213 de S aureus que pour lrsquoactiviteacute bacteacuteriostatique sur la
souche NewbouldΔhem imitant le pheacutenotype SCV Ces travaux furent effectueacutes sous
la tutelle du professeur Franccedilois Malouin
130
1Institut de Pharmacologie de Sherbrooke Deacutepartement de pharmacologie
Universiteacute de Sherbrooke 3001 12e av nord Sherbrooke Queacutebec Canada J1H
5N4
2Centre drsquoeacutetude et de valorisation de la diversiteacute microbienne (CEVDM) Deacutepartement
de biologie Universiteacute de Sherbrooke Sherbrooke QC Canada J1K 2R1
131
BIBLIOGRAPHIE Alexander E H and Hudson MC (2001) Factors influencing the internal- ization of Staphylococcus aureus and impacts on the course of infections in humans Appl Microbiol Biotechnol 56 361ndash366
Andries K Verhasselt P Guillemont J Goumlhlmann HW Neefs JM Winkler H Van Gestel J Timmerman P Zhu M Lee E et al (2005) A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis Science 307 223-227
Artzatbanov VY and Petrov VV (1990) Branched respiratory chain in aerobically grown Staphylococcus aureus--oxidation of ethanol by cells and protoplasts Arch Microbiol 153 580- 584 Archer GL (1998) Staphylococcus aureus a well-armed pathogen Clin Infect Dis 26 1179ndash1181 Assefa S Keane TM Otto TD Newbold C and Berriman M (2009) ABACAS algorithm-based automatic contiguation of assembled sequences Bioinformatics 25 1968-1969 Aubry-Damon H Legrand P Brun-Buisson C Astier A Soussy CJ and Leclercq R (1997) Reemergence of gentamicin-sus- ceptible strains of methicillin-resistant Staphylococcus aureus roles of an infection control program and changes in aminoglycoside use Clin Infect Dis 25 647-653
Balemans W Vranckx L Lounis N Pop O Guillemont J Vergauwen K Mol S Gilissen R Motte M Lanccedilois et al (2012) Novel antibiotics targeting respiratory ATP synthesis in Gram-positive pathogenic bacteria Antimicrob Agents Chemother 56 4131-4139
Batandier C Guigas B Detaille D El-Mir MY Fontaine E Rigoulet M and Leverve XM (2006) The ROS production induced by a reverse-electron flux at respiratory-chain complex 1 is hampered by metformin J Bioenerg Biomembr 38 33ndash 42
132
Beckman Coulter website httpswwwbeckmancoultercomwsrportalwsrresearch-and-discoveryproducts-and-servicesnucleic-acid-sample-preparationspri-laboratory-automationindexhtm (consulteacute le 3 Deacutec 2013) Bednarek P and Osbourn A (2009) Plant-microbe interactions chemical diversity in plant defense Science 324 746ndash748 Besier S Ludwig A Ohlsen K Brade V and Wichelhaus TA (2007) Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus Int J Med Microbiol 297 217-225 BiteSizeBio Brain food for biologists httpnxseqbitesizebiocomarticlessequencing-by-synthesis-explaining-the-illumina-sequencing-technology (consulteacute le 3 deacutec 2013) Boetzer M and Pirovano W (2012) Toward almost closed genomes with GapFiller Genome Biology 13 R56 Bouarab K Ordi E Gattuso M Moisan H and Malouin F (2007) Plant stress response agents affect Staphylococcus aureus virulence genes Abstr 47th Intersci Conf Antimicrob Agents Chemother abstr C1-1483 Bryan LE and Kwan S (1981) Aminoglycoside-resistant mutants of Pseudomonas aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob Agents Chemother 19 958- 964 Bryan LE and Kwan S (1983) Roles of ribosomal binding membrane potential and electron transport in bacterial uptake of streptomycin and gentamicin Antimicrob Agents Chemother 23 835ndash845
Bryan LE Nicas T Holloway BW and Crowther C (1980) Aminoglycoside-resistant mutation of Pseudomonas aeruginosa defective in cytochrome C552 and nitrate reductase Antimicrob Agents Chemother 17 71- 79
133
Bryan LE and van den Elzen HM (1977) Effects of membrane-energy mutations and cations on streptomycin and gentamicin accumulation by bacteria a model for entry of streptomycin and gentamicin in susceptible and resistant bacteria Antimicrob Agents Chemother 12 163-177 Brynildsen MP Winkler JA Spina CS MacDonald IC and Collins JJ (2013) Potentiating antibacterial activity by predictably enhancing endogenous microbial ROS production Nat Biotechnol 31 160ndash 165 Burke KA and Lasccelles J (1975) Nitrate reductase system in Staphylococcus aureus wild type and mutants J Bacteriol 123 308- 316 Burke KA and Lascelles J (1976) Nitrate reductase activity in hegraveme-deficient mutants of Staphylococcus aureus J Bacteriol 126 225- 231 Canfield GS Schwingel JM Foley MH Vore KL Boonanantanasarn K Gill AL Sutton MD and Gill SR (2013) Evolution in fast forward a potential role for mutators in accelerating Staphylococcus aureus pathoadaptation J Bacteriol 195 615-628 Chang S Sievert DM Hageman JC Boulton ML Tenover FC Downes FP Shah S Rudrik JT Pupp GR Brown WJ et al (2003) Infection with vancomycin- resistant Staphylococcus aureus containing the vanA resistance gene N Engl J Med 348 1342-1347 Chen Q Vazquez EJ Moghaddas S Hoppel CL and Lesnefsky EJ (2003) Production of reactive oxygen species by mitochondria central role of complex III J Biol Chem 278 36027-36031 Clements MO Watson SP Poole RK and Foster SJ (1999) CtaA of Staphylococcus aureus is required for starvation survival recovery and cytochrome biosynthesis J Bacteriol 181 501- 507
134
Clinical and Laboratory Standards Institute (2011) Performance standards for antimicrobial susceptibility testing Twenty-first informational supplement M100-S21 CLSI Wayne PA USA ClustalW2 httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2 (consulteacute le 3 deacutec 2013) Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) httpcmrjcviorgtigr-scriptsCMRCmrHomePagecgi (consulteacute le 3 deacutec 2013) Creacutepin-Gilbert C (2006) Transfert de ligand dans la cytochrome c oxydase observeacute par des expeacuteriences femtosecondes infrarouge inteacutegreacutees et reacutesolues spectralement Thegravese de doctorat de l rsquoEacutecole Polytechnique Paris Pastel-00002200 version 1 29 jul 2010 Davis BD (1987) Mechanism of bactericidal action of aminoglycosides Microbiol Rev 51 341-350 DeAngelis MM Wang DG and Hawkins TL (1995) Solid-phase reversible immobilization for the isolation of PCR products Nucl Acids Res 22 4742- 4743 Dickgiesser N and Kreiswirth BN (1986) Determination of aminoglycoside resistance in Staphylococcus aureus by DNA hybridization Antimicrob Agents Chemother 29 930-932 Doi Y and Arakawa Y (2007) 16S ribosomal RNA methylation Emerging resistance mechanism against aminoglycosides Clin Infect Dis 45 88ndash94 Enright MC Robinson DA Randle G Feil EJ Grundmann H and Spratt BG (2002) The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA) Proc Natl Acad Sci USA 99 7687-7692 Friedman M (2002) Tomato glycoalkaloids role in the plant and in the diet J Agric Food Chem 50 5751ndash5780
135
Goerke C and Wolz C (2004) Regulatory and genomic plasticity of Staph- ylococcus aureus during persistent colonization and infection Int J Med Microbiol 294 195ndash202 Goumltz F Bannerman T and Schleifer K-H (2006) The genera Staphylococcus and Macrococcus In The Prokaryotes Vol 4 Bacteria Firmicutes Cyanobacteria M Dworkin (ed) (Springer New-York USA) pp5- 75 Haagsma AC Abdillahi-Ibrahim R Wagner MJ Krab K Vergauwen K Guillemont J AndriesK Lill H Koul A and Bald D (2009) Selectivity of TMC207 towards mycobacterial ATP synthase compared with that towards the eukaryotic homologue Antimicrob Agents Chemother 53 1290-1292 Hammer ND Reniere ML Cassat JE Zhang Y Hirsch AO Indriati Hood M and Skaar EP (2013) Two hegraveme-dependent terminal oxydases power Staphylococcus aureus organ-specific colonization of the vertebrate host MBio 4 e00241- 13 Hederstedt L (1993) The Krebs citric acid cycle In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of Microbiology Washington DC) pp 181-198 Hiramatsu K (1995) Molecular evolution of MRSA Microbiol Immunol 39 531-543 Hiramatsu K Hanaki H Ino T Yabuta K Oguri T and Tenover FC (1997) Methicillin-resistant Staphylococcus aureus clinical strain with reduced vancomycin susceptibility J Antimicrob Chemother 40 135-136 Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T et al (2013) Genomic Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
136
Hong S and Pedersen PL (2008) ATP synthase and the actions of inhibitors utilized to study its roles in human health disease and other scientific areas Microbiol Mol Biol Rev 72 590-641
Jevons MP (1961) lsquoCelbeninrsquo-resistant staphylococci BMJ 1 124-125
Joshi S and Huang YG (1991) ATP synthase complex from bovine heart mitochondria the oligomycin sensitivity conferring protein is essential for dicyclohexyl carbodiimidesensitive ATPase Biochim Biophys Acta 1067 255ndash258
Kohanski MA Dwyer DJ and Collins JJ (2010) How antibiotics kill bacteria from targets to networks Nat Rev Microbiol 8 423-435 Kohler C von Eiff C Liebeke M McNamara PJ Lalk M Proctor RA Hecker M and Engelmann S (2008) A defect in menadione biosynthesis induces global changes in gene expression in Staphylococcus aureus J Bacteriol 190 6351- 6364 Kohler C von Eiff C Peters G Proctor RA Hecker M and Engelmann S (2003) Physiological characterization of a hegraveme-deficient mutant of Staphylococcus aureus by a proteomic approach J Bacteriol 185 6928- 6937 Kriegeskorte A Koumlnig S Sander G Pirkl A Mahabir E Proctor RA von Eiff C Peters G and Becker K (2011) Small colony variants of Staphylococcus aureus reveal distinct protein profiles Proteomics 11 2476-2490 Lannergard J von Eiff C Sander G Cordes T Seggewiss J Peters G Proctor RA Becker K and Hughes D (2008) Identification of the genetic basis for clinical menadione-auxotrophic small-colony variant isolates of Staphylococcus aureus Antimicrob Agents Chemother 52 4017-4022
Lauraeus M and Wikstrom M (1993) The terminal quinol oxydases of Bacillus subtilis have different energy conservation properties J Biol Chem 268 11470-11473
137
Li M Gong J Cottrill M Yu H de Lange C Burton J and Topp E (2003) Evaluation of QIAamp DNA Stool Mini Kit for ecological studies of gut microbiota J Microbiol Methods 54 13-20
Liu Y Fiskum G and Schubert D (2002) Generation of reactive oxygen species by the mitochondrial electron transport chain J Neurochem 80 780-787
Martin-Hernandez A M Dufresne M Hugouvieux V Melton R and Osbourn A (2000) Effects of targeted replacement of the tomatinase gene on the interaction of Septoria lycopersici with tomato plants Mol Plant Microbe Interact 13 1301ndash1311
Matteelli A Carvalho AC Dooley KE and Kritski A (2010) TMC207 the first compound of a new class of potent anti-tuberculosis drugs Future Microbiol 5 849-858
Matsuno-Yagi A and Hatefi Y (1993) Studies on the mechanism of oxidative phosphorylation ATP synthesis by submitochondrial particles inhibited at F0 by venturicidin and organotin compounds J Biol Chem 268 6168ndash6173
Miller MH Edberg SC Mandel LJ Behar CF and Steigbigel NH (1980) Gentamicin uptake in wild-type and aminoglycoside-resistant small-colony mutants of Staphylococcus aureus Antimicrob Agents Chemother 18 722- 729
Mingeot-Leclercq M-P Glupczynski Y and Tulkens PM (1999) Aminoglycosides Activity and Resistance Antimicrob Agents Chemother 43 727- 737
Mitchell G Brouillette Eacute Seacuteguin DL Asselin AE Jacob CL and Malouin F (2010) A role for sigma factor B in the emergence of Staphylococcus aureus small-colony variants and elevated biofilm production resulting from an exposure to aminoglycosides Microb Pathog 48 18-27
Mitchell G Gattuso M Bouarab K and Malouin F (2009) Tomatidine (TO) affects virulence regulators of prototypical Staphylococcus aureus (SA) and small colony variants (SCV) of cystic fibrosis (CF) patients Abstr 49th Intersci Conf Antimicrob Agents Chemother abstr C1-1341
138
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K and Malouin F (2011) Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic fibrosis airway epithelial cells Antimicrob Agents Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM and Malouin F (2011) Infection of polarized airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function and is influenced by NF-κB Infect Immun 79 3541-3551
Mitchell G Lafrance M Boulanger S Seacuteguin DL Guay I Gattuso M Marsault Eacute Bouarab K and Malouin F (2012) Tomatidine acts in synergy with aminoglycoside antibiotics against multiresistant Staphylococcus aureus and prevents virulence gene expression J Antimicrob Chemother 67 559-568
Mitchell G Lamontagne CA Brouillette Eacute Grondin G Talbot BG Grandbois M and Malouin F (2008) Staphylococcus aureus SigB activity promotes a strong fibronectin-bacterium interaction which may sustain host tissue colonization by small-colony variants isolated from cystic fibrosis patients Mol Microbiol 70 1540-1555
Mitchell G Malouin F 2012 Outcome and prevention of Pseudomonas aeruginosa-Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic Fibrosis Chap 8 pp 181-206 Ed D Sriramulu InTech Open Access Publisher ISBN 979-953-307-059-8
Mitchell G Seacuteguin DL Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S and Malouin F (2010) Staphylococcus aureus sigma B-dependent emergence of small-colony variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-heptylquinoline-N-oxide BMC Microbiol 30 10-33
Moisan H Brouillette Eacute Jacob CL Langlois-Beacutegin P Michaud S and Malouin F (2006) Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated from cystic fibrosis patients is influenced by SigB J Bacteriol 188 64-76
139
Moreno F Crisp C Jorgensen JH and Patterson JE (1995) Methicillin- resistant Staphylococcus aureus as a community organism Clin Infect Dis 21 1308-1312
Murphy MP (2009) How mitochondria produce reactive oxygen species Biochem J 417 1- 13
Novick RP (2003) Autoinduction and signal transduction in the regulation of staphylococcal virulence Mol Microbiol 48 1429-1449
Novo DJ Perlmutter NG Hunt RH and Shapiro HM (2000) Multiparameter flow cytometric analysis of antibiotic effects on membrane potential membrane permeability and bacterial counts of Staphylococcus aureus and Micrococcus luteus Antimicrob Agents Chemother 44 827-834
ONeill AJ Cove JH and Chopra I (2001) Mutation frequencies for resistance to fusidic acid and rifampicin in Staphylococcus aureus J Antimicrob Chemother 47 647-650
Osbourn A E (1996) Preformed antimicrobial compounds and plant defense against fungal attack Plant Cell 8 1821ndash1831
Otto TD Dillon GP Degrave WS and Berriman M (2011) RATT Rapid annotation transfert tool Nucl Acids Res 39 doi101093 Pratique E Theraulaz L (2007) Eacutetude de la reacutegulation du systegraveme respiratoire TMAO reacuteductase chez Escherichia coli Meacutemoire de maitrise de lrsquoeacutecole pratique des hautes eacutetudes Paris
Price CT and Gustafson JE (2001) Increases in the mutation frequency at which fusidic acid-resistant Staphylococcus aureus arise with salicylate J Med Microbiol 50 104-106
Proctor RA and Peters G (1998) Small colony variants in staphylococcal infections Diagnostic and therapeutic implications Clin Infect Dis 27 419- 423
140
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M and Peters G (2006) Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent infections Nat Rev Microbiol 4 295ndash305
Quail MA Kozarewa I Smith F Scally A Stephens PJ Durbin R Swerdlow H and Turner DJ (2008) A large genome centers improvements to the Illumina sequencing system Nat methods 5 1005-1010
Ramirez MS and Tolmasky ME (2010) Aminoglycoside modifying enzymes Drug Resist Updat 13 151ndash171
Rao M Streur TL Aldwell FE and Cook GM (2001) Intracellular pH regulation by Mycobacterium smegmatis and Mycobacterium bovis BCG Microbiology 147 1017-1024
Richardson DJ (2000) Bacterial respiration a flexible process for a changing environment Microbiology 146 551- 571
Riley TV Pearman JW and Rouse IL (1995) Changing epidemiology of methicillin-resistant Staphylococcus aureus in Western Australia Med J Aust 163 412-414
Roddick J G (1974) The steroidal glycoalkaloid α-tomatine Phytochemistry 13 25-36
Rodrigue S Materna AC Timberlake SC Blackburn MC Malmstrom RR et al(2010) Unlocking Short Read Sequencing for Metagenomics PLoS ONE 5(7) e11840doi101371journalpone0011840 Ruiz-Rubio M Alonso Peacuterez-Espinosa A Lairini K RoldAacuten-Arjona T Dipietro A and Anaya N (2001) Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi In Studies in natural products chemistry vol 25 A Rahman (ed) (Elsevier Oxford United Kingdom) pp 293ndash326
141
Samuelsen oslash Haukland HH Kahl BC von Eiff C Proctor RA Ulvatne H Sandvik K and Vorland LH (2005) Staphylococcus aureus small colony variants are resistant to the antimicrobial peptide lactoferricin B J Antimicrob Chemother 56 1126- 1129
Sandrock R W and Vanetten HD (1998) Fungal sensitivity to and enzymatic degradation of the phytoanticipin α-tomatine Phytopathology 88 137ndash143
Sendi P and Proctor RA (2009) Staphylococcus aureus as an intracellular pathogen the role of small colony variants Trends Microbiol 17 54ndash58
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C and Osbourn A (2006) Dual effects of plant steroidal alkaloids on Saccha- romyces cerevisiae Antimicrob Agents Chemother 50 2732ndash2740
Somerville GA and Proctor RA (2009) At the crossroads of bacterial metabolism and virulence factor synthesis in Staphylococci Microbiol Mol Biol Rev 73 233- 248
Starkov AA (2010) Measurement of mitochondrial ROS production Methods Mol Biol 648 245-255
Taber HW (1993) Respiratory chain In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of MicrobiologyWashington DC) pp 199-212
Taber HW and Morrison M (1964) Electron transport in Staphylococci Properties of a particle preparation from exponential phase Staphylococcus aureus Arch Biochem Biophys 105 367- 379
Taber HW et al(1987) Bacterial uptake of aminoglycoside antibiotics Microbiol Rev 51 439-457
Thoumlny-Meyer L (1997) Biogenesis of respiratory cytochromes in bacteria Microbiol Mol Biol Rev 61 337- 376
142
Tubby S Wilson M Wright JA Zhang P and Nair SP (2013) Staphylococcus aureus small colony variants are susceptible to light activated antimicrobial agents BMC Microbiol 13 201- 204
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D Roth J Proctor RA Becker K et al (2011) Staphylococcus aureus phenotype switching an effective bacterial strategy to escape host immune response and establish a chronic infection EMBO Mol Med 3 129ndash41
Turrens JF (2003) Mitochondrial formation of reactive oxygen species J Physiol 552 335ndash 344
Tynecka Z Szcześniak Z Malm A and Los R (1999) Energy conservation in aerobically grown Staphylococcus aureus Res Microbiol 150 555- 566
Udo EE Pearman JW and Grubb WB(1993) Genetic analysis of community isolates of methicillin-resistant Staphylococcus aureus in Western Australia J Hosp Infect 25 97-108
Vakulenko SB and Mobashery S (2003) Versatility of Aminoglycosides and Prospects for Their Future Clin Microbiol Rev 16 430-450
Voggu L Schlag S Biswas R Rosenstein R Rausch C and Goumltz F (2006) Microevolution of cytochrome bd oxydase in Staphylococci and its implication in resistance to respiratory toxins released by Pseudomonas J Bacteriol 188 8079- 8086
Wells PG Bhuller Y Chen CS Jeng W Kasapinovic S Kennedy JC Kim PM Laposa RR McCallum GP Nicol CJ et al (2005) Molecular and biochemical mechanisms in teratogenesis involving reactive oxygen species Toxicol Appl Pharmacol 207 354-366
143
Yan M Yu C Yang J and Ji Y (2011) The essential two-component system YhcSR is involved in regulation of the nitrate respiratory pathway of Staphylococcus aureus J Bacteriol 193 1799- 1805
Zhang T Luo Y Chen Y Li X and Yu J (2012) BIGrat a repeat resolver for pyrosequencing-based re-sequencing with Newbler BMC Res Notes 5 567
144

ii
SOMMAIRE
Dans le but de mieux comprendre le mode drsquoaction et de nous permettre de
deacuteterminer la cible de la tomatidine nous avons dans un premier temps tenteacute de
mieux circonscrire le spectre drsquoactiviteacute de la tomatidine Gracircce agrave ces travaux nous
sommes en effet maintenant en mesure de dire que la tomatidine possegravede une
activiteacute antibacteacuterienne contre les espegraveces de la divison des Firmicutes et plus
preacuteciseacutement contre les bacteacuteries de lrsquoordre des Bacillales dont font partie les genres
Bacillus Staphylococcus et Listeria Nous avons eacutegalement deacutecouvert gracircce agrave des
expeacuteriences en collaboration avec le laboratoire drsquoEacuteric Marsault qursquoun analogue de
la tomatidine (FC04-100) avait non seulement des proprieacuteteacutes similaires agrave la
moleacutecule naturelle mais deacutemontrait une activiteacute par lui-mecircme contre S aureus agrave
pheacutenotype normal alors que la tomatidine possegravede uniquement une activiteacute contre
les laquo small colony variants raquo De plus alors que la tomatidine possegravede plutocirct une
activiteacute bacteacuteriostatique contre la forme SCV de L monocytogenes le nouveau
composeacute (FC04-100) deacutemontre quant agrave lui une forte activiteacute bacteacutericide contre
cette souche tout comme contre la forme SCV des autres Bacillales
Parallegravelement et toujours dans le but de rechercher le mode drsquoaction et la cible de
la tomatidine nous avons obtenu par passages successifs dans un milieu avec
antibiotiques des mutants de S aureus agrave pheacutenotype normal et des SCV reacutesistants
agrave la tomatidine ou agrave la combinaison tomatidine et gentamicine Apregraves le
seacutequenccedilage de ces mutants lrsquoeacutetude de la position de ces mutations agrave lrsquoaide de
diffeacuterents logiciels de bio-informatique nous a permis drsquoeacutemettre un modegravele-
hypothegravese quant au mode drsquoaction et agrave la cible de la tomatidine Selon les reacutesultats
que nous avons agrave ce stade-ci la cible de la tomatidine chez S aureus serait la
sous-uniteacute c de lrsquoATP synthase Cependant son mode drsquoaction serait eacutegalement
deacutependant de la fonctionnaliteacute de la chaine de transport des eacutelectrons et donc de la
iii
polarisation membranaire et de la production de ROS intracellulaire ce qui
expliquerait la diffeacuterence drsquoactiviteacute entre les souches agrave pheacutenotype normal et les
SCV
Mots cleacutes tomatidine ATP synthase chaine de transport des eacutelectrons synergie laquo small-colony variants raquo ROS S aureus analogues
iv
REMERCIEMENTS
Je voudrais remercier mon directeur de recherche Pr Franccedilois Malouin qui a cru
en moi pour mrsquoavoir proposeacute ce beau projet de maitrise Merci pour sa grande
disponibiliteacute son eacutecoute et ses critiques toujours constructives et jamais
condescendantes
Je voudrais eacutegalement remercier mes conseilleacutes Kamal Bouarab Seacutebastien
Rodrigue et Eacuteric Marsault leur implication dans mon projet leur occasionnant un
surplus de travail Merci pour leur support technique et leur disponibiliteacute agrave reacutepondre
agrave mes questions
Un gros merci agrave Gabriel Mitchell qui a eacuteteacute mon mentor au deacutebut de ma maitrise et
qui mrsquoa introduit au projet laquo Tomatidine raquo Un merci particulier eacutegalement agrave
Maxime-Lamontagne-Boulet pour son aide en tant que stagiaire lors de mes
derniegraveres expeacuteriences Je voudrais eacutegalement remercier tous les membres du
laboratoire preacutesents et passeacutes qui ont toujours eacuteteacute disponibles pour partager avec
moi leurs connaissances theacuteoriques et techniques
Je ne peux passer sous silence lrsquoimplication de lrsquoUniversiteacute de Sherbrooke et du
FQRNT comme organismes subventionnaires au projet Merci eacutegalement agrave Fibrose
Kystique Canada qui a participeacute eacutegalement agrave la subvention drsquoune partie du projet
En terminant je voudrais remercier ma famille qui a crue en moi et qui mrsquoencourage
agrave perseacuteveacuterer depuis toujours Un merci tout particulier agrave mon conjoint et ma belle-
fille pour qui je nrsquoai pas toujours eacuteteacute aussi disponible que jrsquoaurais voulu lrsquoecirctre et qui
mrsquoont supporteacutee au quotidien et ce dans tous les sens du terme
v
TABLE DES MATIEgraveRES
SOMMAIRE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ii
REMERCIEMENTS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip iv
TABLE DES MATIEgraveRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip v
LISTE DES ABBREacuteVIATIONS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip vii
LISTE DES TABLEAUX helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ix
LISTE DES FIGURES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip x
CHAPITRE 1 INTRODUCTION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
11 Staphylococcus aureus helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
111 Importance cliniquehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)helliphellip 2
1121 Diffeacuterences et similariteacutes helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 2
1122 La chaine de transport des eacutelectrons helliphelliphelliphelliphellip 4
12 Antibiotiques Modes drsquoaction cibles cellulaire et
meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles helliphelliphelliphellip 25
122 Les aminoglycosides helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1221 Mode drsquoaction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1222 Meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphellip 31
123 La tomatidine helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 33
13 Projet de maitrise helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
CHAPITRE 2 ARTICLE SCIENTIFIQUE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 37
ldquoTomatidine and derivative FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Firmicutes Including Those of Listeria monocytogenes
and Are Bactericidal in Combination With Aminoglycosides Against their Normal
Phenotyperdquo
vi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE helliphelliphelliphellip 76
31 Obtention des mutants helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
32 Seacutequenccedilage et analyses bio-informatiques helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 79
321 Identification des mutations en lien avec la tomatidine helliphellip 89
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct helliphellip 90
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine helliphelliphelliphelliphellip 99
CHAPITRE 4 DISCUSSION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 113
CHAPITRE 5 CONCLUSION ET PERSPECTIVES helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 124
CONTRIBUTIONS SUPPLEacuteMENTAIRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 1 Application de brevet helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 2 Article scientifique helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 129
laquo Unraveling the structure-activity relationship of tomatidine
a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureus raquo
BIBLIOGRAPHIE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 131
vii
LISTE DES ABBREacuteVIATIONS
Adh Alcool deacuteshydrogeacutenase
ADN Acide deacutesoxyribo-nucleacuteique
ADP Adeacutenosine diphosphate
ARN Acide ribo-nucleacuteique
ATP Adeacutenosine triphosphate
CA-MRSA Community-associated MRSA
CMI concentration minimale inhibitrice
CoQ Coenzyme Q
Cyt Cytochrome
DMQ laquo demethylmenaquinone raquo
dTMP Deacutesoxythymidine monophosphate
dUMP Deacutesoxyuridine monophosphate
FADH2 Flavine adenine dinucleacuteotide
GEN gentamicine
HA-MRSA Healthcare-associated MRSA
Ldh Lactate deacuteshydrogeacutenase
ml millilitre
MQ meacutenaquinone
MRSA methycilin resistant Staphylococcus aureus
Mqo2 Quninol oxidoreacuteductase
NADH Nicotinamide adenine dinucleacuteotide (forme reacuteduite)
Pdh Pyruvate deacuteshydrogeacutenase
Pfl Pyruvate formate lyase
PyrD Dihydroorotate deacuteshydrogeacutenase
Q Quinone
Rif rifampicine
ROS laquo reactive oxygen species raquo espegravece reacuteactive de lrsquooxygegravene
viii
[S] Polysulfide
S aureus Staphylococcus aureus
SARM Staphylococcus aureus resistant agrave la meacutethycilline
SCV laquo small colony variants raquo
SHCHC 2-succinyl-6-hydroxy-24-cyclohexadiene-1-carboxylate syntase
TCA Tricarboxylic acid cycle
TCD tableau croiseacute dynamique
TO tomatidine UFC Uniteacute formatrice de colonies
UQ Ubiquinone
microg microgramme
ix
LISTE DES TABLEAUX
CHAPITRE 1 INTRODUCTION
1 Cibles et modes drsquoaction des principales classes drsquoantibiotique 27
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants (B) chez les mutants NewbouldΔhemB reacutesistants 95-96
CHAPITRE 4 DISCUSSION
1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 123
x
LISTE DES FIGURES
CHAPITRE 1 INTRODUCTION
1 Diffeacuterence entre les pheacutenotypes normaux et SCV de S aureus 3
2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes 6
3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale 8
4 Chaines respiratoires bacteacuteriennes contenant des cytochromes 9
5 Flexibiliteacute respiratoire (A) chez Paracoccus denitrificans (B) chez
Escherichia coli 10
6 Chaine de transport des eacutelectrons proposeacutee chez les staphylocoques 12
7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies 17
8 Modegravele de reacutegulation du cycle TCA 20
9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS 23
10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de O2-bull 24
11 Voies de formation et de deacutetoxification des ROS 25
12 Cineacutetique drsquoentreacutee des aminoglycosides 30
13 Sites de mutations de certains reacutesistants aux aminoglycosides 32
14 Structures chimiques de (A) tomatine (B) tomatidine et (C) Analogue
de tomatidine FC04-100 34
xi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Graphiques des CMI pour chacun des passages pour (A) la
tomatidine sur NewbouldΔhemB (B) la tomatidine en preacutesence de
gentamicine sur NewbouldΔhemB (C) la gentamicine sur S aureus
ATCC 29213 (D) la gentamicine en preacutesence de tomatidine sur S
aureus ATCC 29213 77-79
2 Organigramme des mutants reacutesistants seacutequenceacutes 80-81
3 Eacutetapes du seacutequenccedilage Illumina 83
4 Meacutethode drsquoamplification par la meacutethode Illumina 85
5 Correction du programme RATT sur les annotations transfeacutereacutees 88
6 Exemple drsquoune partie du tableau compilation des mutations 89
7 Seacutelection des gegravenes drsquointeacuterecirct chez ATCC 29213 dans le TCD par
lrsquoutilisation de diffeacuterents filtres 91-92
8 Seacutelection des gegravenes drsquointeacuterecirct chez NewbouldΔhemB dans le TCD par
lrsquoutilisation de diffeacuterents filtres 93-94
9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct 97-98
10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants 100
11 Courbe de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison gentamicine (+TO) du premier et troisiegraveme niveau de
reacutesistance 101
12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B)
NewbouldΔhemB 102
xii
13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213 104
14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicinelorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC
29213 105
15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants
ATCC 29213 reacutesistants agrave la combinaison gentamicine (+TO) 106
16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la
tomatidine 107
17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la
tomatidine 108
18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB
reacutesistants agrave la combinaison tomatidine (+gentamicine) 109
19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane
Potential Kit (Invitrogen) 111
20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du
gegravene atpE de S aureus Newman 112
xiii
CHAPITRE 4 DISCUSSION
1 Site drsquoinhibition de lrsquoATP synthase par diffeacuterentes moleacutecules 114
2 Structure chimique de (A) TMC207 et (B) lrsquoanalogue anti Gram
positifs 115
3 Seacutequence proteacuteique de la proteacuteine AtpE chez diffeacuterentes espegraveces 118
4 Modeacutelisation de la structure tridimentionnelle de lrsquoATP synthase de S
aureus (atpE) montrant les acides amineacutes muteacutes chez les mutants
reacutesistants 121
- 1 -
CHAPITRE 1
INTRODUCTION
11 Staphylococcus aureus
111 Importance clinique
Staphylococcus aureus est une bacteacuterie agrave Gram positif anaeacuterobie facultative Crsquoest un
microorganisme opportuniste pouvant infecter diffeacuterents hocirctes diffeacuterents organes ou
sites drsquoinfection et ayant la capaciteacute de causer tant des infections aiguumles que
chroniques (Archer 1998 Goerke et Wolz 2004)
Staphylococcus aureus reacutesistant agrave la meacutethilline le SARM (ou laquo MRSA raquo) est une
souche de S aureus devenue reacutesistante aux β-Lactamines par lrsquoacquisition du gegravene
mecA dans son chromosome (Hiramatsu 1995) Le SARM a eacuteteacute isoleacute pour la
premiegravere fois en 1960 en Angleterre (Jevons 1961) et est devenu eacutepideacutemique agrave
travers le monde depuis les anneacutees 1970 Au moins trois geacutenotypes distincts eacutetaient
recenseacutes dans les anneacutees 1980 (Hiramatsu 1995) dont deux sont toujours preacutevalents
agrave travers le monde comme eacutetant des souches HA-MRSA (Healthcare-associated
MRSA) des souches acquises agrave lrsquohocircpital (Enright et al 2002) Ces souches ont
acquis des gegravenes de reacutesistance agrave pratiquement tous les antibiotiques introduits dans
lrsquoutilisation clinique depuis les 50 derniegraveres anneacutees mecircme agrave lrsquoantibiotique de dernier
ressort la vancomycine(Hiramitsu 1997 Chen et al 2003) Nous sommes
maintenant en manque drsquoagents theacuterapeutiques efficaces contre les SARM touchant
les personnes immunodeacuteprimeacutees qui sont dans les hocircpitaux En plus depuis le deacutebut
des anneacutees 1990 des clones de SARM dont plusieurs deacutemontraient un potentiel
pathogeacutenique supeacuterieur aux souches classiques de HA-MRSA retrouveacutees agrave lrsquohocircpital
- 2 -
sont maintenant retrouveacutes dans la communauteacute (CA-MRSA) agrave travers le monde (Udo
et al 1993 Riley et al 1995 Moreno et al 1995 Aubry-Damon et al 1997) et ont
la capaciteacute drsquoaffecter des personnes en santeacute en causant des infections aiguumles
(Hiramatsu et al 2013) Il apparaicirct donc urgent de trouver des drogues alternatives
afin de surmonter le nombre grandissant de souches de SARM
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)
1121 Diffeacuterences et similariteacutes
Les laquo small colony variants raquo (SCV) sont des bacteacuteries souvent isoleacutees lors des
infections chroniques comme crsquoest le cas des infections pulmonaires chez les
patients atteints de fibrose kystique mais aussi dans des cas drsquoosteacuteomyeacutelite
drsquoarthrite septique drsquoinfections de prothegraveses orthopeacutediques et lors de la mammite
bovine (Alexender et Hudson 2001 Moisan et al 2006 Proctor et al 2006) Les
SCV sont caracteacuteriseacutes par soit un meacutetabolisme oxydatif dysfonctionnel ou un manque
dans la biosynthegravese de la thymidine les deux amenant une alteacuteration dans
lrsquoexpression des facteurs de virulence une croissance plus lente et une diminution
de la pigmentation des colonies (Figure 1) (Proctor et al 2006) Cependant alors
que les souches normales reacutepriment lrsquoexpression des proteacuteines de surface et
expriment les exoproteacuteines lors de la phase stationnaire (Novick 2003) les SCV
expriment de faccedilon stable les gegravenes Sig-B deacutependants codant pour des proteacuteines de
surface telle les adheacutesines au lieu drsquoactiver le systegraveme agr deacutependant du quorum-
sensing et produire des exoproteacuteines (Moisan et al 2006) Ces diffeacuterences dans
lrsquoexpression des facteurs de virulence reacutesultent chez les SCV en une meilleure
habileteacute agrave adheacuterer aux composantes de lrsquohocircte (Mitchell et al 2008) et agrave former du
biofilm (Mitchell et al 2010a Mitchell et al 20010b) et seraient eacutegalement lieacutees agrave
lrsquohabileteacute des SCV agrave lrsquoinvasion et la persistance des cellules de lrsquohocircte (Proctor et al
2006 Sendi et Proctor 2009 Mitchell et al 2011b)
- 3 -
S aureus est donc un pathogegravene qui peut se preacutesenter sous deux pheacutenotypes qui
sont souvent retrouveacutes simultaneacutement lors drsquoune mecircme infection et qui ont le pouvoir
de reacuteverter drsquoun pheacutenotype agrave lrsquoautre selon les conditions du milieu (Figure 1)
(Tuchscherr et al 2011)
Le pheacutenotype dit rdquonormalrdquo est associeacute aux infections aiguumles agrave une forte production
de facteurs de virulence (ex heacutemolysine) et est associeacute agrave la phase de disseacutemination
de la bacteacuterie Lrsquoautre pheacutenotype le laquo small-colony variants raquo ou SCV est affecteacute
dans sa chaine respiratoire ce qui amegravene une croissance beaucoup plus lente des
colonies de 8 -10 fois plus petites que les prototypes et une faible susceptibiliteacute aux
aminoglycosides Les SCV sont plutocirct associeacutes aux infections chroniques agrave S aureus
de par leur capaciteacute agrave srsquointernaliser dans les cellules de lrsquohocircte leur forte capaciteacute agrave
produire du biofilm et leur capaciteacute agrave produire des facteurs de colonisation
(Tuchscherr et al 2011 Proctor et al 2006) (Figure 1)
Figure 1 Diffeacuterences entre les pheacutenotypes normaux et SCV de S aureus
S aureus agrave
pheacutenotype normal
Disseacutemination
amp
Infections aiguumles
Colonisation
amp
Infections persistantes
SCVs
(laquo small-colony
variants raquo)
Reacutevers
ion
pheacutenoty
piq
ue
Norm
al
SC
V
Croissance
Biofilm
Internalisationheacutemolyse
Mitchell G et Malouin F 2012
- 4 -
1122 La chaine de transport des eacutelectrons
Chaine respiratoire tableau geacuteneacuteral
Les chaines respiratoires bacteacuteriennes bien que tregraves variables dans leurs
composantes parmi les diffeacuterentes espegraveces sont toutes constitueacutees sur le mecircme
modegravele de base Elles sont composeacutees de trois composantes majeures soit
- les deacuteshydrogeacutenases primaires qui sont chargeacutees drsquooxyder des substrats
principalement les coenzymes NADH et FADH2 (issues du cycle de Krebs) (mais
aussi selon le cas le succinate le lactate lrsquohydrogegravene hellip) et de pomper deux
protons du cocircteacute externe de la membrane en plus de transfeacuterer deux eacutelectrons agrave un
groupement de quinones
- Les quinones sont des moleacutecules liposolubles non proteacuteiques mobiles dans la
membrane Elles servent drsquointermeacutediaires entre les deacuteshydrogeacutenases primaires et
les oxydases terminales pour le transport des eacutelectrons Il existe deux sortes de
quinones les benzoquinones dont font partie les ubiquinones et les
naphtoquinones dont font partie les meacutenaquinones Les bacteacuteries agrave Gram positif
ne possegravedent que des meacutenaquinones alors que les deux sont preacutesentes chez les
bacteacuteries agrave Gram neacutegatif (Hederstedt 1993 Pratique et Theraulaz 2007)
- Les oxidoreacuteductases terminales sont chargeacutees de reacuteduire lrsquoaccepteur final
drsquoeacutelectron lrsquooxygegravene eacutetant lrsquoaccepteur final privileacutegieacute
De faccedilon plus deacutetailleacutee la chaine respiratoire est composeacutee de cinq
complexes proteacuteiques principaux (Figure 2)
- complexe I NADH deacuteshydrogeacutenase ou NADH reacuteductase ou NADHCoQ
oxidoreacuteductase ou NADH-ubiquinone oxidoreacuteductase
- complexe II Succinate deacuteshydrogeacutenase
- complexe III Cytochrome bc1 complexe ou ubiquinol cytochrome c
reacuteductaseoxydoreacuteductase
- complexe IV Cytochrome c oxydase
- 5 -
- complexe V ATP synthase (Creacutepin-Gilbert et al 2006)
La phosphorylation oxydative est deacuteclencheacutee dans la chaine de complexes
proteacuteiques par la preacutesence drsquoO2 accepteur terminal privileacutegieacute drsquoeacutelectrons (ou drsquoautres
accepteurs finaux tel le nitrate) dans le milieu ainsi que par les eacutelectrons ameneacutes par
le NADH et le FADH2 au complexe I et II respectivement (Creacutepin-Gilbert et al 2006)
Les eacutelectrons vont ensuite transiter par le complexe III jusqursquoau complexe IV ou les
eacutelectrons seront utiliseacutes afin de reacuteduire lrsquooxygegravene en eau le complexe IV est
lrsquooxydase terminale consideacutereacutee comme eacutetant lrsquoaccepteur final drsquoeacutelectrons Le transfert
drsquoeacutelectrons est possible gracircce aux reacuteactions drsquooxydoreacuteduction en cascade dans les
diffeacuterents complexes proteacuteiques Ces reacuteactions fournissent de lrsquoeacutenergie libre utiliseacutee
par les complexes I III et IV afin de transfeacuterer des protons agrave lrsquoexteacuterieur de la cellule
(Creacutepin-Gilbert et al 2006) Le complexe V (ATP synthase) quant agrave lui utilise les
eacutelectrons pompeacutes agrave lrsquoexteacuterieur par les autres complexes comme force proton-motrice
afin de phosphoryler trois moleacutecules drsquoADP et ainsi produire trois ATP Le transport
des eacutelectrons entre les diffeacuterents complexes proteacuteiques est possible gracircce agrave la
preacutesence dans la membrane de deux types de moleacutecules liposolubles non proteacuteiques
mobiles les meacutenaquinones et le cytochrome c Les meacutenaquinones seules quinones
preacutesentes chez les bacteacuteries agrave Gram positif assurent le transport des eacutelectrons entre
le complexe I et le complexe III et entre le complexe II et III Le cytochrome c pour sa
part veacutehicule les eacutelectrons entre le complexe III et le complexe IV Le transport des
eacutelectrons srsquoeffectue de la composante ayant le plus faible potentiel drsquooxydoreacuteduction
vers celle ayant le plus fort potentiel
- 6 -
La chaine respiratoire bacteacuterienne
La mitochondrie montre assez peu de flexibiliteacute respiratoire Il y a une certaine
flexibiliteacute au niveau de lrsquoentreacutee des eacutelectrons mais aucune au niveau de la sortie des
eacutelectrons ougrave la cytochrome aa3 oxydase reacuteduit lrsquooxygegravene en eau (Richardson 2000)
(Figure 3) Chez les bacteacuteries alors que le complexe II fonctionne sensiblement de
faccedilon identique agrave celui des eucaryotes les autres complexes peuvent se composer
de diffeacuterents complexes enzymatiques (reacuteductases) selon les espegraveces et les
conditions environnementales (Richardson 2000 Thoumlny-Meyer 1997) En outre
chez les bacteacuteries contrairement aux eucaryotes la chaine respiratoire comporte
souvent des voies de transport des eacutelectrons alternatives on dit alors qursquoelles ont une
chaine respiratoire agrave branches ou brancheacutee ou ramifieacutee (Artzatbanov et Petrov 1990)
Plusieurs espegraveces bacteacuteriennes possegravedent une chaine respiratoire ramifieacutee dont la
ramification se situe habituellement suite agrave la reacuteduction des quinones en quinols
(Lauraeus et Wikstrom 1993 Thoumlny-Meyer 1997) (Figure 4) Une des deux
Complexe I NADH
deacuteshydrogeacutenase
Complexe II Succinate
deacuteshydrogeacutenase
Complexe III
bc 1
Complexe IV Cytochrome c
oxydase
Complexe V
ATP synthase
Modifieacute de wikipedia httpfrwikipediaorgwikiChaine_respiratoire
Figure 2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes
Leurs rocircles dans le transport des eacutelectrons et dans lrsquoeacutelaboration du gradient de
protons et le lien entre la chaine respiratoire et le cycle de Krebs Q Quinones
- 7 -
branches connue comme la branche cytochrome c oxyde les quinols (meacutenaquinol
QH2) via la voie complexe bc1-cytochrome c- cytochrome c oxydase Lrsquoautre
branche-type consiste en une seule uniteacute enzymatique (cytochrome bd) laquelle
transfegravere directement les eacutelectrons des quinols agrave lrsquooxygegravene Cette voie est connue
comme la branche quinol oxydase il srsquoagit drsquoune oxydase terminale transfeacuterant
directement les eacutelectrons agrave lrsquooxygegravene eacutevitant de passer par lrsquointermeacutediaire du
complexe III (bc1) et du cytochrome c (Artzatbanov et Petrov 1990 Lauraeus et
Wikstrom 1993) Alors que la voie du cytochrome c permet le pompage de protons agrave
lrsquoexteacuterieur par lrsquoaction du complexe bc1 (ou complexe III) et du cytochrome c oxydase
le cytochrome bd ne permet aucun pompage de protons (Lauraeus et Wikstrom
1993) La voie passant par le complexe bc1-cytochrome c-cytochrome c oxydase est
plus efficace dans la production drsquoeacutenergie Alors que les 2 voies peuvent ecirctre
exprimeacutees simultaneacutement chez quelques espegraveces il semble que lrsquoexpression de ces
voies soit inductible selon les conditions environnementales chez Bacillus subtilis et
les Staphylocoques (Lauraeus et Wikstrom 1993) On pourrait dire eacutegalement que la
chaine respiratoire est seacutepareacutee en deux eacutetapes la premiegravere partie est la phase
quinone-reacuteductase (Q-QH2) et la seconde la phase quinol-oxydase (QH2-Q) menant
agrave lrsquooxydase terminale La phase de reacuteduction est responsable du transfert des
eacutelectrons drsquoune varieacuteteacute de substrats tels le NADH le succinate le lactate le formate
et lrsquohydrogegravene vers les quinones et inclut diffeacuterentes reacuteductases (Thoumlny-Meyer 1997)
(Figure 4)
- 8 -
Figure 3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale UQ ubiquinone UQH2 ubiquinol Cyt cytochrome Il semble y avoir presque autant de chaines de transport des eacutelectrons diffeacuterentes
qursquoil y a drsquoespegraveces bacteacuteriennes (par exemple Paracoccus denitrificans et Escherichia
coli (Figure 5)) De plus il ne semble pas y avoir consensus dans la litteacuterature sur
les noms attribueacutes aux oxydases terminales Les noms pour les oxydases terminales
deacutependent des types de cytochromes dont elles sont composeacutees Il ne semble donc
pas eacutevident de connaicirctre de faccedilon preacutecise les types de cytochromes qui font partie
des oxydases terminales Pour rajouter agrave la confusion les gegravenes et opeacuterons ne sont
pas toujours associeacutes agrave leur oxydase terminale dans la litteacuterature ce qui fait qursquoon ne
peut savoir de faccedilon certaine de quelle oxydase on parle Le Cytochrome aa3 peut
eacutegalement porter le nom de Cyt ba3 et le cytochrome bd peux eacutegalement porter les
noms de Cyt bo
Tireacute de Richardson DJ 2000
- 9 -
Succinate
Formate
H2
SDH
FDH
HYD
QQH 2 bc1 c
Reacuteductase alternatives
b
c
cd1c
cb
c
b
c
d
Siroheme
Siroheme
b
NO3-
NO2-
NO
TMAO
DMSOS2O3
2-
[S]
SO32-
Fumarate
Quinol oxidases
ba3bb3bo3d
bd
aa3
O2
Cyt c oxidases
aa3ba3baa3caa3cbb3cao
co
O2
Modifieacute de Thony-Mayer L 1997
NADH
NDH
Figure 4 Chaines respiratoires bacteacuteriennes contenant des cytochromes
Du cocircteacute gauche on retrouve la partie quinone (ubiquinone ou meacutenaquinone)-
reacuteductase Du cocircteacute droit on retrouve la partie quinol-oxydase avec les oxydases
terminales reacuteduisant lrsquoO2 ou lrsquoaccepteur terminal alternatif tel que montreacute Les
oxydoreacuteductases sont surligneacutees QQH2 repreacutesente le pool de quinonesquinols
dans la membrane Les polysulfides sont repreacutesenteacutes par [S] Les chaines
respiratoires ne contenant pas de cytochromes ne sont pas montreacutees NDH
NADH deacuteshydrogeacutenase
Modifieacute de Thoumlny-Mayer L 1997
- 10 -
Figure 5 Flexibiliteacute respiratoire chez Paracoccus denitrificans (a) et chez
Escherichia coli (b) MQ menaquinone UQ ubiquinone DMQ
demethylmenoquinone
Chez S aureus
Comme plusieurs autres espegraveces bacteacuteriennes anaeacuterobies facultatives S aureus a
la possibiliteacute drsquoexprimer diffeacuterentes voies de synthegravese eacutenergeacutetique dont lrsquoexpression
est finement reacuteguleacutee selon des conditions environnementales du moment (Hammer et
Tireacute de Richardson DJ 2000
- 11 -
al 2013 Lauraeus et Wikstrom 1993) Mecircme si S aureus peut utiliser la
fermentation la respiration (aeacuterobie et anaeacuterobie) sera privileacutegieacutee par la bacteacuterie
puisque celle-ci amegravene une bien meilleure production drsquoeacutenergie (Goumltz et al 2006
Hammer et al 2013 ) La respiration aeacuterobie avec lrsquooxygegravene comme accepteur final
drsquoeacutelectron sera utiliseacutee de faccedilon preacutefeacuterentielle en preacutesence drsquooxygegravene (Hammer et al
2013) Malgreacute sa versatiliteacute dans ses voies de synthegravese eacutenergeacutetique S aureus et les
autres bacteacuteries agrave Gram positif ne produisent qursquoun seul type de quinone les
meacutenaquinones lesquelles sont utiliseacutees pour la croissance degraves qursquoil y a de lrsquooxygegravene
de preacutesent dans le milieu (Somerville et Proctor 2009) La reacuteduction de lrsquooxygegravene en
eau est meacutedieacutee par des oxydases terminales hegraveme deacutependant (Hammer et al 2013)
Selon certains auteurs S aureus possegravederait deux oxydases terminales la
cytochrome oxydase aa3 (ou ba3) (QoxABCD) et la cytochrome bd quinol oxydase
(CydAB) (Goumltz et al 2006 Hammer et al 2013 Kohler et al 2008) Cette derniegravere
est exclusive aux procaryotes et est une voie de contournement du complexe III qui
transmet directement les eacutelectrons du laquo pool raquo de quinol (QH2) agrave lrsquooxygegravene Selon
certains auteurs il y aurait eacutegalement chez S aureus une troisiegraveme oxydase
terminale le cytochrome o (CyoABCDE) (Thoumlny-Meyer 1997 Voggu et al 2006)
qui selon certain serait un cytochrome oxydase terminal alternatif au cytochrome aa3
et au cytochrome bd (Voggu et al 2006 Goumltz et al 2006 Taber et Morrison 1964)
(Figure 6) Ces voies alternatives permettent agrave S aureus de srsquoadapter rapidement agrave
un changement drsquoenvironnement (Hammer et al 2013) In vivo chaque organe est
un environnement diffeacuterent preacutesentant des deacutefis respiratoires distincts dus agrave la
variation dans 1- la quantiteacute drsquooxygegravene disponible 2- la disponibiliteacute drsquoaccepteurs
drsquoeacutelectrons alternatifs 3- la concentration drsquoinhibiteurs de la respiration dans le milieu
tels les oxydes nitriques (NO) produits par les neutrophiles et 4- la disponibiliteacute des
cofacteurs neacutecessaires agrave la production drsquoeacutenergie (Hammer et al 2013) Le
cytochrome est exprimeacute de faccedilon modeacutereacutee dans un milieu riche en oxygegravene mais est
fortement exprimeacute dans un milieu pauvre en oxygegravene mais riche en oxyde nitrique
De plus le type drsquooxydoreacuteductase terminale exprimeacute est influenceacute eacutegalement par la
- 12 -
source de carbone disponible la phase de croissance le pH extracellulaire et la
tension en oxygegravene (Lauraeus et Wikstrom 1993 Tynecka et al 1999)
La chaine respiratoire chez les SCV
S aureus est particuliegraverement doueacute agrave surpasser les deacutefis respiratoires en acqueacuterant
une reacutesistance agrave lrsquoacide nitrique produit par les neutrophiles et une habiliteacute agrave persister
dans les cellules de lrsquohocircte en se preacutesentant comme un variant agrave petites colonies
ayant une chaine respiratoire deacuteficiente (SCV) (Hammer et al 2013)
Deux principaux types de SCV sont retrouveacutes dans les isolats cliniques 1- les SCV
ayant une chaine de transport des eacutelectrons deacutefectueuse et 2- les SCV auxotrophes
pour la thymidine (Proctor et Peters 1998 Samuelsen et al 2005) Ces derniers
Figure 6 Chaine de transport des eacutelectrons proposeacutee chez les
staphylocoques Les informations disponibles sur les composantes de la chaine
respiratoire des staphylocoques suggegraverent une chaine ramifieacutee consistant en 2
oxydases terminales meacutenaquinol-deacutependante un cytochrome o et un cytochrome
aa3 oxydase NADH deacuteshydrogeacutenase (DHase) transmet les eacutelectrons au laquo pool raquo
de meacutenaquinones De lagrave les eacutelectrons sont transfeacutereacutes soit au cytochrome b ou agrave
lrsquoaccepteur alternatif drsquoeacutelectrons le cytochrome bd quinol oxydase Le cytochrome
b transfegravere les eacutelectrons agrave une des deux oxydases terminales Ces oxydases
terminales transfegraverent les eacutelectrons agrave lrsquooxygegravene
Modifieacute de Voggu L et al 2006
- 13 -
arborent une ou plusieurs mutations dans le gegravene thyA codant pour la thymidylate
synthase laquelle catalyse la derniegravere eacutetape dans la voie de biosynthegravese de la
thymidine crsquoest-agrave-dire la formation de dTMP agrave partir du dUMP (Samuelsen et al
2005) Le dTMP est essentiel pour la synthegravese de lrsquoADN (Besier et al 2007) Les
mutants thymidine-auxotrophes sont souvent la conseacutequence drsquoune pression
seacutelective par lrsquoutilisation prolongeacutee de trimethoprim-sulfamethoxazole (TMP-SMX)
(souvent utiliseacute pour le traitement des patients fibroses kystiques) un antibiotique qui
interfegravere dans la biosynthegravese de lrsquoacide tetrahydrofolique qui agit comme cofacteur de
la thymidylate synthase (Besier et al 2007 Lannergard et al 2008 Samuelsen et al
2005) Mecircme si de premiers abords les SCV thymidine-auxotrophes ne semblent pas
avoir de lien avec la chaine transport des eacutelectrons il se pourrait que celle-ci ait sont
importance chez ces SCV puisque lrsquoentreacutee de la thymidine passe par NupC dont le
fonctionnement neacutecessite un gradient eacutelectrochimique creacuteeacute par la chaine de transport
des eacutelectrons (Proctor et Peters 1998 ) Les caracteacuteristiques pheacutenotypiques des
SCV deacutecrites ci-apregraves sont toutes lieacutees agrave lrsquoalteacuteration de la chaine de transport des
eacutelectrons 1- la croissance plus lente parce que la synthegravese de la paroi cellulaire
neacutecessite une grande quantiteacute drsquoATP 2-la diminution de la formation de pigment
parce que la biosynthegravese des caroteacutenoiumldes requiert le transport des eacutelectrons 3- une
reacutesistance accrue agrave la gentamicine et autres aminoglycosides parce que lrsquoentreacutee de
ces composeacutes neacutecessite un grand potentiel membranaire geacuteneacutereacute par le transport des
eacutelectrons (Lannergard et al 2008) et 4- une perte partielle ou complegravete de la
fermentation du mannitol parce que lrsquoutilisation des sucres-alcool tels que le
mannitol est diminueacutee lorsque la chaine de transport des eacutelectrons ne peut oxyder le
NADH produit lorsque le mannitol est meacutetaboliseacute en fructose-6-phosphate (Kohler et
al 2003 Proctor et Peters 1998 Samuelsen et al 2005)
Les SCV posseacutedant une chaine de transport des eacutelectrons deacutefectueuse ont une voie
de biosynthegravese de lrsquoheacutemine ou de la meacutenadione deacuteficiente et le pheacutenotype est
reacuteversible lorsque compleacutementeacute avec lrsquoheacutemine ou la meacutenadione (Hammer et al
- 14 -
2013 Kohler et al 2008 Lannergard et al 2008 Proctor et Peters 1998) La
meacutenadione est essentielle agrave la biosynthegravese des meacutenaquinones et lrsquoheacutemine est
essentielle agrave la biosynthegravese de lrsquohegraveme qui entre dans formation des cytochromes
(Hammer et al 2013 Kohler et al 2008 Lannergard et al 2008 Thoumlny-Meyer
1997) Les meacutenaquinones et les cytochromes sont essentiels au fonctionnement de
la chaine de transport des eacutelectrons (vers lrsquoO2 autant que vers le nitrate) (Kohler et al
2008) Il se pourrait qursquoil existe eacutegalement des SCV causeacutes par drsquoautres deacuteficiences
telles que lrsquoalteacuteration de lrsquoATP synthase qui ne reacutesulterait pas drsquoune auxotrophie pour
lrsquoheacutemine ou la meacutenadione mais reacutesulterait tout de mecircme en une deacuteficience dans la
chaine de transport des eacutelectrons (Proctor et Peters 1998 )
Comme lrsquohegraveme est un cofacteur essentiel pour les oxydases terminales et que la
meacutenaquinone est un transporteur drsquoeacutelectrons utiliseacute par S aureus on comprend que
la faible croissance des mutants hemB et menD repreacutesente un pheacutenotype mimant la
perte des oxydases terminales (Hammer et al 2013) Les deux oxydases terminales
de S aureus QoxABCD (cyt aa3) et CydAB (cyt bd) semblent ecirctre diffeacuteremment
reacuteguleacutees chez les mutants menD et la souche sauvage alors que lrsquoopeacuteron cydAB est
fortement sous exprimeacute chez les mutants aucune diffeacuterence de niveau de
transcription de qoxABCD nrsquoa eacuteteacute observeacutee entre les mutants menD et leur souche
parente (Kohler et al 2008)
Des mutations dans les gegravenes menD hemB et ctaA peuvent reacutesulter en un
pheacutenotype SCV posseacutedant une chaine de transport des eacutelectrons deacuteficiente
(Clements et al 1999 Proctor et Peters 1998 ) Les gegravenes codant pour les eacutetapes
de formation de lrsquohegraveme se preacutesentent sous la forme drsquoun opeacuteron lrsquoopeacuteron
hemAXCDBL Chacun des gegravenes est essentiel (sauf hemL) et la deacuteleacutetion drsquoun seul
drsquoentre eux empecircchera la formation de lrsquohegraveme moleacutecule essentielle agrave la formation
des cytochromes (Taber 1993) hemB code pour la porphobilinogene synthase qui
catalyse une eacutetape dans la voie de biosynthegravese de lrsquohegraveme (Kohler et al 2008 Taber
- 15 -
1993 Thoumlny-Meyer 1997) Une mutation dans le gegravene ctaA (laquo hegraveme A synthase raquo)
empecircche la biosynthegravese de lrsquohegraveme A lequel est aussi impliqueacute dans la biosynthegravese
des cytochromes et donc altegraverera le fonctionnement de la chaine de transport des
eacutelectrons (Clements et al 1999 Proctor et Peters 1998) La capaciteacute de la bacteacuterie
agrave biosyntheacutetiser de lrsquohegraveme de faccedilon endogegravene est essentielle afin drsquoinitier une
infection systeacutemique Le gegravene menD code pour la 2-succinyl-6-hydroxy-24-
cyclohexadiene-1-carboxylate syntase (SHCHC) qui est une enzyme essentielle agrave la
voie de biosynthegravese des meacutenaquinones (Kohler et al 2008) De plus les
meacutenaquinones sont eacutegalement requises pour le fonctionnement de plusieurs
enzymes impliqueacutees dans des voies meacutetaboliques essentielles telles la malate quinol
oxidoreacuteductase (Mqo2) et la dihydroorotate deacuteshydrogeacutenase (PyrD) (Kohler et al
2008) Les mutants menD seront donc incapables de convertir lrsquooxaloaceacutetate en
malate ducirc agrave la perte de lrsquoactiviteacute du Mqo2 (Kohler et al 2008 Proctor et Peters
1998)
Les bacteacuteries agrave pheacutenotype SCV deacutemontreront eacutegalement un gradient eacutelectrochimique
nettement plus faible que les bacteacuteries agrave pheacutenotype normal de par lrsquoincapaciteacute de la
chaine de transport des eacutelectrons agrave y pourvoir
De par leur chaine de transport des eacutelectrons interrompue les SCV sont incapables
drsquoutiliser lrsquoO2 et le nitrate comme accepteur final drsquoeacutelectrons (Kohler et al 2008) Par
ce fait les proteacuteines impliqueacutees dans la voie de la glycolyse ainsi que dans la voie de
la fermentation sont surexprimeacutees (Pfl Ldh Adh) alors que les enzymes faisant
partie du cycle du citrate (TCA) sont significativement reacuteguleacutees agrave la baisse (Kohler et
al 2003 Kohler et al 2008 Kriegeskorte et al 2011) Ceci indique clairement que
les SCV (ex hemB) geacutenegraverent leur ATP agrave partir du glucose ou fructose seulement par
la phosphorylation des substrats Lrsquoanalyse de la reacuteaction de fermentation indique
que le produit principal est le lactate Les SCV reacutegeacutenegraverent leur NADH en reacuteduisant le
pyruvate en lactate par la voie fermentaire (Kohler et al 2003 Kohler et al 2008)
- 16 -
emmenant ainsi une acidification du milieu (Kriegeskorte et al 2011) La conversion
du pyruvate en aceacutetate via lrsquoacetyl-CoA est restreinte chez les mutants hemB mecircme
si la pyruvate formate lyase (Pfl) et les composantes du pyruvate deacuteshydrogeacutenase
sont preacutesentes (Figure 7) Lrsquoactiviteacute de la pyruvate deacuteshydrogeacutenase est tregraves faible
probablement en lien avec le haut niveau de NADH chez les SCV et il est preacutesumeacute
que la Pfl nrsquoest pas activeacutee en preacutesence drsquooxygegravene (La pyruvate deacuteshydrogeacutenase
converti le pyruvate en Co-A en condition anaeacuterobie) (Figure 7) Chez les mutants
hemB la voie de lrsquoarginine deacuteiminase est aussi induite et pourrait eacutegalement
participer agrave la production drsquoATP (Kohler et al 2003) Les gegravenes exprimeacutes chez les
mutants hemB sont similaires agrave ceux exprimeacutes chez les mutants menD ainsi que
chez les souches agrave pheacutenotypes normaux en condition anaeacuterobie (Kohler et al 2008)
- 17 -
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Figure 7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies Les enzymes suivantes sont retrouveacutees agrave une forte
concentration chez les mutants hemb phosphoglucomutase (Pgi) fructose
bisphosphate aldolase (FbaFda) glyceacuteraldeacutehyde deacuteshydrogeacutenase (Gap)
phosphoglyceacuterate kinase (Pgk) eacutenolase (Eno) pyruvate kinase (Pyk) L-lactate
deacuteshydrogeacutenase (L-Ldh) pyruvate formate lyase (PflB) alcool deacuteshydrogeacutenase
(Adh) La formation drsquoaceacutetate et de formate et tregraves limiteacutee (X rouges)
Modifieacute de Kolher C et al 2003
- 18 -
La chaine respiratoire en anaeacuterobie (S aureus)
Il existe 3 voies distinctes de geacuteneacuteration drsquoeacutenergie chez les bacteacuteries 1- en preacutesence
drsquooxygegravene la bacteacuterie respire de faccedilon aeacuterobique et lrsquooxygegravene est lrsquoaccepteur final
drsquoeacutelectron 2- la bacteacuterie peut aussi respirer de maniegravere anaeacuterobique lorsqursquoun
accepteur terminal drsquoeacutelectron autre que lrsquooxygegravene est preacutesent 3- en absence
drsquooxygegravene et drsquoun accepteur drsquoeacutelectrons alternatif la bacteacuterie peut produire de
lrsquoeacutenergie de faccedilon limiteacutee par fermentation (Hammer et al 2013)
Chez S aureus le nitrate (NO3-) et dans un troisiegraveme temps le nitrite (NO2
-)
peuvent ecirctre utiliseacutes comme accepteur final drsquoeacutelectron alternatif lorsque lrsquooxygegravene
nrsquoest pas disponible (respiration anaeacuterobie) Les enzymes de cette voie meacutetabolique
ne sont retrouveacutees que chez les bacteacuteries (Goumltz et al 2006) La croissance
bacteacuterienne est consideacuterablement reacuteduite lorsque la bacteacuterie passe drsquoune respiration
aeacuterobie agrave une respiration anaeacuterobie La nitrate reacuteductase fonctionne probablement in
vivo gracircce au L-lactate comme principal donneur drsquohydrogegravene (Burke et Lasccelles
1975) La deacuteshydrogeacutenase primaire pour ce substrat est preacutesumeacutement la L-lactate
deacuteshydrogeacutenase associeacutee agrave la membrane laquelle meacutedie lrsquooxydation du lactate par la
chaine respiratoire aeacuterobique (Burke et Lasccelles 1975) La preacutedominance du
lactate parmi les produits finaux de la fermentation par S aureus est correacuteleacutee avec
son efficaciteacute comme principal donneur drsquohydrogegravene lorsque le nitrate est lrsquoaccepteur
final drsquoeacutelectrons Le transfert des eacutelectrons du Lactate vers le nitrate implique des
composantes qui sont eacutegalement preacutesentes dans la chaine respiratoire La nitrate
reacuteductase du type respiratoire est habituellement associeacutee agrave la membrane
cytoplasmique (Burke et Lasccelles 1975) Les reacuteductases sont inhibeacutees par
lrsquoazoture et le cyanure (Burke et Lascelles 1976)
Lrsquoenzyme membranaire nitrate reacuteductase NarGHI geacutenegravere des nitrites lesquels seront
reacuteduits en ammonium (NH4+) par lrsquoenzyme cytoplasmique NADH-deacutependant nitrite
reacuteductase encodeacutee par les gegravenes nirBD (Goumltz et al 2006) La nitrate reacuteductase 1-
- 19 -
est reacuteprimeacutee et inhibeacutee par la preacutesence drsquooxygegravene 2- est induite par lrsquoanaeacuterobiose et
la preacutesence de nitrate ou nitrite 3- nrsquoest pas inhibeacutee par lrsquoammoniac 4- permet la
production drsquoeacutenergie dans des conditions anaeacuterobies et 5- est localiseacutee dans la
membrane (Goumltz et al 2006) La preacutesence drsquoun systegraveme de senseur pour lrsquooxygegravene
est cruciale pour le controcircle transcriptionnel des gegravenes facilitant lrsquoadaptation de la
bacteacuterie agrave lrsquoanaeacuterobiose ou aux conditions microaeacuterophiles Chez S aureus il existe
deux systegravemes de senseur 1- le systegraveme SrrAB (pour staphylococcal respiratory
response AB synonyme SrhSR) (Figure 8) homologue agrave ResDE chez Bacillus
subtilis (Yan et al 2011) et 2- le systegraveme NreABC (pour nitrogen regulation chez S
carnosus correspondant agrave SA2181 (NreA) SA2180 (NreB) SA2179 (NreC) chez S
aureus) NreBC est un systegraveme agrave deux composantes qui mesure la concentration
drsquooxygegravene directement par un groupement Fe-S de type Fnr lequel est instable en
preacutesence drsquooxygegravene et par lequel est controcircleacute le systegraveme membranaire de reacuteduction
du nitratenitrite (Yan et al 2011) Srr est aussi un systegraveme agrave deux composantes qui
reacutegule positivement la lactate deacuteshydrogeacutenase et lrsquoalcool deacuteshydrogeacutenase et semble
reacuteguler neacutegativement plusieurs enzymes du cycle du citrate (TCA) (Figure 8)
Lrsquoexpression des enzymes fermentaires comme la lactate deacuteshydrogeacutenase (LDH)
pyruvate formate lyase (PFL) et lrsquoalcool deacuteshydrogeacutenase sont aussi fortement induite
dans des conditions aeacuterobies lorsque la chaine de transport des eacutelectrons est
interrompue (Kohler et al 2003) Donc il se peut que lrsquoeacutetat reacuteduit des composantes
de la chaine respiratoire le potentiel membranaire etou le niveau de NADH soit un
signal pour la reacutegulation des gegravenes laquo anaeacuterobies raquo chez S aureus Dans des
conditions aeacuterobies la conversion du pyruvate en aceacutetyl-coA est catalyseacutee par le
complexe pyruvate deacuteshydrogeacutenase (PDH) et caracteacuteriseacutee par la production de
NADH (Somerville et Proctor 2009) En condition anaeacuterobie lrsquoexpression de la
pyruvate deacuteshydrogeacutenase (PDH) est fortement reacuteprimeacutee alors que lrsquoexpression de la
pyruvate formate lyase (PFL) est fortement induite Lrsquoactiviteacute de la PDH est
remplaceacutee en anaeacuterobiose par lrsquoactiviteacute de la PFL qui geacutenegravere lrsquoacetyl-CoA et le
formate agrave partir du pyruvate Le NADH nrsquoeacutetant pas reacuteoxydeacute par une chaine de
- 20 -
transport des eacutelectrons fonctionnelle celui-ci est recycleacute par la reacuteduction du pyruvate
en lactate (Kohler et al 2003 Somerville et Proctor 2009)
La meacutenaquinone et lrsquohegraveme sont esssentiels agrave la respiration anaeacuterobie (Kohler et al
2008) Le pheacutenotype de S aureus en condition anaeacuterobie est tregraves similaire au
pheacutenotype SCV (Somerville et Proctor 2009)
Figure 8 Modegravele de reacutegulation du cycle TCA (Pour simplifier les reacutegulateurs de
facteurs de virulence bien caracteacuteriseacutes [ie RNAIII Rot SarA et σB] ont eacuteteacute omis
dans ce modegravele)
Les sous-produits de la phosphorylation oxydative les ROS
La reacuteduction de lrsquooxygegravene en eau se fait un eacutelectron agrave la fois (Turrens 2003) Le
complexe IV retient tous les intermeacutediaires partiellement reacuteduits jusqursquoagrave ce que la
Tireacute de Somerville GA Proctor RA 2009
- 21 -
reacuteduction soit complegravetement termineacutee (Turrens 2003) Cependant certains autres
centres redox de la chaine respiratoire laissent passer des eacutelectrons agrave lrsquooxygegravene
reacuteduisant partiellement cette moleacutecule en anion superoxyde (O2-bull) (Liu et al 2002
Turrens 2003) Mecircme si O2-bull nrsquoest pas un agent oxydant tregraves puissant il est le
preacutecurseur de plusieurs autres laquo reactive oxygen species raquo (ROS) et il est aussi
impliqueacute dans la propagation de la chaine de reacuteaction oxydative (Turrens 2003)
LrsquoO2-bull est un intermeacutediaire instable qui est rapidement converti en H2O2 par les
superoxide dismutase (SOD) (S aureus en possegravede deux SodA (le principal) et
SodM) (10 18) ou se convertit de faccedilon spontaneacutee lorsqursquoen concentration eacuteleveacutee
(Starkov 2010 Turrens 2003) Le H2O2 peut ensuite ecirctre complegravetement reacuteduit en
H2O ou partiellement reacuteduit en radical hydroxyle (bullOH) un des plus puissants
oxydants (Starkov 2010) LrsquoO2-bull peut eacutegalement reacuteagir avec drsquoautres radicaux
incluant lrsquooxyde nitrique (NObull) selon le taux de diffusion des 2 radicaux (Turrens
2003) Le peroxynitrite est eacutegalement un oxydant puissant (Turrens 2003) Les
oxydants deacuterivant du NObull sont appeleacutes des laquo reactive nitrogen species raquo (RNS)
(Turrens 2003) mais seront consideacutereacutes comme faisant partie des ROS dans ce
travail Les ROS peuvent endommager lrsquoADN lrsquoARN les proteacuteines et les lipides
reacutesultants agrave lrsquoapoptose de la cellule lorsque le niveau de ROS produit excegravede la
capaciteacute de deacutetoxifications et de reacuteparation de la cellule (Brynildsen et al 2013
Wells et al 2005) Malgreacute ce danger il srsquoagit drsquoun risque compenseacute par
lrsquoaugmentation du rendement eacutenergeacutetique (Brynildsen et al 2013) Le complexe I (et
II) et III sont les principaux sites de production de ROS (Batandier et al 2006 Chen
et al 2003 Liu et al 2002) Lrsquoinhibition du complexe I par la rotenone provoque une
augmentation de la production de ROS Lrsquooxydation des substrats du complexe I ou
du complexe II en preacutesence du complexe III inhibeacute par lrsquoantimicine A amegravene
eacutegalement une augmentation de la production de ROS (Batandier et al 2006 Chen
et al 2003) (Figure 9) De plus mecircme si le complexe cytochrome oxydase nrsquoest pas
un site de production de ROS son inhibition par le KCN (cyanure de potassium)
facilite la production de ROS par les complexes I et II (Chen et al 2003) Le
- 22 -
complexe ATP synthase nrsquoest pas reconnu pour produire directement des ROS
Cependant la liaison drsquoun inhibiteur tel lrsquooligomycine bloque le passage des protons
agrave travers le complexe et inhibe ainsi la production drsquoATP De plus lrsquoATP synthase ne
fonctionnant plus les pompes agrave protons de la chaine de transport des eacutelectrons sont
incapables de fonctionner car le gradient de protons est beaucoup trop eacuteleveacute ce
provoque une surproduction de ROS par les complexes I et III Conseacutequemment le
NADH nrsquoest plus oxydeacute et le cycle du TCA cesse drsquoopeacuterer car la concentration de
NAD nrsquoest plus suffisante pour le fonctionnement des enzymes du TCA (Joshi et
Huang 1991)
Une production significative de O2-bull principalement par le complexe I peut ecirctre le
reacutesultat de deux modes drsquoopeacuteration 1- lorsque la cellule ne produit pas drsquoATP et
conseacutequemment possegravede une grande force proton-motrice et possegravede un laquo pool raquo de
quinone agrave lrsquoeacutetat reacuteduit (QH2) 2- lorsque la cellule possegravede un haut ratio de
NADHNAD (Figure 10) Lorsque la cellule produit activement de lrsquoATP elle possegravede
alors une moins grande force proton-motrice son ratio NADHNAD est plus faible et
la formation de O2-bull est beaucoup plus faible (Murphy 2009)
Le H2O2 est le substrat pour deux enzymes la catalase et la glutathione (GSH)
peroxydase qui convertissent le H2O2 en H2O+O2 le H2O2 est ainsi deacutetoxifieacute (Figure
9 Figure 11) La reacuteaction catalyseacutee par la glutathione peroxydase neacutecessite de la
glutathione (GSH) comme substrats et deacutepend en partie du ratio de GSSG GSH et
de la concentration des reacuteactants laquelle est le reflet de lrsquoeacutetat reacutedox de la cellule De
faccedilon similaire la concentration de meacutetaux sous leur forme reacuteduite tel le Fer catalyse
la formation de certains ROS (Figure 11) Ceci peut ecirctre minimiseacute en tenant cette
concentration ions meacutetallique tregraves basse entre autres par leur emmagasinage et leur
liaison aux proteacuteines de transport (ei ferritine transferrine lactoferrine) et ainsi
minimiser la formation de bullOH Finalement certains antioxydants (eg vitamine E)
peuvent interrompre la chaine de reacuteactions par la capture des radicaux le radical-
vitamine E est relativement stable et peut ecirctre enzymatiquement reconverti agrave sa
- 23 -
forme non-radical (Figure 11) Il est maintenant accepteacute que certaines enzymes du
cycle de Krebs (ie αKGDH) soient non seulement des cibles des ROS mais aussi
des producteurs non neacutegligeables de ROS (Brynildsen et al 2013)
Figure 9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS Le complexe I ainsi que le site Qi du complexe III relacircchent des
ROS directement vers les deacutefenses drsquoantioxydants intracellulaires Les antioxidants
intracellulaire tel la superoxide dismutase (MnSOD) la catalase la glutathione (GSH)
la glutathione peroxydase (GPX) et la glutathione reacuteductase (GRase) sont montreacute Le
site Qo du complexe III quant agrave lui relacircche les ROS dans le milieu extra cellulaire
Modifieacute de Chen Q et al 2003
Oligomycine
- 24 -
Figure 10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de
O2-bull 1- Le pool de NADH est dans un eacutetat reacuteduit par exemple par des dommages agrave
la chaine respiratoire la perte de cytochrome c durant lrsquoapoptose ou agrave un faible
besoin en ATP Ceci megravene agrave une production de O2-bull au complexe I qui deacutecoule de la
reacuteduction du FMN laquelle est reacuteguleacutee par le ratio NADHNAD+ Drsquoautres sites
autres que le complexe I tel lrsquoα-ceacutetoglutatrate deacuteshydrogeacutenase (αKGDH) peut
aussi contribuer agrave la production de O2-bull 2- Il nrsquoy a pas (peu) de production drsquoATP il y
a un fort potentiel membranaire et un pool de CoQ (meacutenaqinone) agrave lrsquoeacutetat reacuteduit
important qui provoque une reacuteversion de la chaine de transport des eacutelectrons agrave
travers le complexe I amenant une forte production de O2-bull 3- Il y a une production
active drsquoATP et conseacutequemment un potentiel membranaire plus bas que dans la
situation 2 et une plus grande quantiteacute de NADH oxydeacute que dans la situation 1
Dans ces conditions la quantiteacute de O2-bull produit est plus faible que dans la situation
1 et 2 et la provenance des O2-bull est moins certaine
Tireacute de Murphy MP 2009
- 25 -
12 Antibiotiques Mode drsquoaction cible cellulaire et meacutecanismes de reacutesistance
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles
Notre compreacutehension de la faccedilon que les antibiotiques induisent la mort cellulaire est
principalement baseacutee sur la fonction cellulaire essentielle inhibeacutee par lrsquointeraction de
lrsquoantibiotique avec sa cible Les antibiotiques peuvent ecirctre classeacutes selon la
composante ou le systegraveme cellulaire qursquoils affectent ainsi que par le fait qursquoils
induisent la mort cellulaire (antibiotiques bacteacutericides) ou srsquoils inhibent la croissance
Figure 11 Voies de formation et de deacutetoxification des ROS Tireacute de Wikipeacutedia
httpfrwikipediaorgwikiDC3A9rivC3A9_rC3A9actif_de_l27oxygC3A8ne
Consulteacute le 11 deacutec 2013
- 26 -
bacteacuterienne (antibiotique bacteacuteriostatique) La plupart des antibiotiques bacteacutericides
inhibent la synthegravese de lrsquoADN ou de lrsquoARN la synthegravese de la paroi bacteacuterienne ou la
synthegravese proteacuteique (Kohanski et al 2010) Le tableau 1 est une synthegravese des
principales classes drsquoantibiotiques ainsi que leur mode drsquoaction et leurs cibles
cellulaires Ce tableau nrsquoest cependant pas exhaustif et il existe plusieurs autres
classes drsquoantibiotiques et plusieurs autres exemples drsquoantibiotiques dans les classes
identifieacutees dans ce tableau
Depuis la deacutecouverte de la peacutenicilline en 1929 plusieurs autres antibiotiques ont eacuteteacute
deacutecouverts et deacuteveloppeacutes suite agrave lrsquoeacutelucidation des interactions entre lrsquoantibiotique et
sa cible et en apportant des modifications structurales et chimiques aux moleacutecules
deacutejagrave connues Ces efforts ont significativement augmenteacute le nombre drsquoantibiotiques
disponibles dans le milieu clinique Le processus de mort cellulaire induit par lrsquoaction
drsquoun antibiotique est complexe et deacutebute avec une interaction physique entre la
moleacutecule antibiotique et sa cible bacteacuterienne speacutecifique et implique des modifications
qui affectent la bacteacuterie tant au niveau biochimique que moleacuteculaire et structural
27
Acid
e n
alid
ixiq
ue (
1 iegravere
geacuteneacutera
tion)
Cip
rofloxacin
e (
2 iegravem
e g
eacuteneacutera
tion)
Levofloxacin
e (
3 iegravem
e g
eacuteneacutera
tion)
Gem
iflo
xacin
e (
4 iegravem
e g
eacuteneacutera
tion)
Flu
oro
quin
olo
ne (
Norf
loxacin
e)
Peacutenic
iline
Ceacutephalo
sporine (
ceacutefa
lexin
e)
Carb
apenem
Vancom
ycin
e
Teacutela
vancin
e
Fosfo
mycin
Dapto
mycin
Poly
mix
ine B
Bacitra
cin
Valin
om
ycin
e
Nom
encin
e
Genta
mic
in
Kanam
ycin
e
Tobra
mycin
e
Str
epto
mycin
e
Teacutetr
acyclin
e
Chlo
rteacutetr
acyclin
e
Macro
lides
Ery
thro
mycin
eLie
sous-u
niteacute 23S
du r
ibosom
e 5
0S
et em
pecircche
leacutelo
ngation d
u p
eptide (
apregrave
s 6
-8 a
a)
sta
tique
Lin
cosam
ides
Clin
dam
ycin
Inhib
e d
irecte
ment la
ctiviteacute p
eptidyl
transfeacute
rase d
u
ribosom
esta
tique
Str
epto
gra
min
es
DalfopristinQ
uin
upristin
Com
bin
ais
on d
e 2
antibio
tiques B
loque la
transla
tion p
eptidiq
ue e
n lia
nt le
site P
du r
ibosom
e
et en b
loquant au p
eptide la
ccegraves a
u tunnel de s
ort
ie
cid
al
Am
phenic
ol
Chlo
ram
phenic
ol
Em
pecircche la lia
ison d
u a
a-
AR
Nt au s
ite A
du
ribosom
e e
t in
terf
egravere
dans la f
orm
ation d
u lie
n
peptidiq
ue
sta
tique
Oxazolid
inones
Lin
ezolid
Blo
que initia
tion d
e la tra
nsla
tion p
roteacute
ique lors
de la
reacuteaction d
eacutelo
ngation p
ar
la p
eptidyl
transfeacute
rase
sta
tique
Trim
eth
oprim
Sulfam
eth
oxazole
sta
tique
cid
al
Ionophore
Alteacutera
tion d
e lin
teacutegriteacute
mem
bra
naire
Se
nchasse d
ans la m
em
bra
ne fo
rme u
n p
ore
ce
qui am
egravene u
ne p
ert
urb
ation d
e leacute
quili
bre
ioniq
ue
cid
al
Inhib
itio
n d
e la s
ynth
egravese d
e
la p
aro
i cellu
laire
(peptidogly
can)
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
cid
al
cid
al
sta
tique
AR
Nr
16S
(S
ite A
)(fo
rmation d
e p
roteacute
ines n
on-
fonctionnelle
s)
cid
al
Inhib
itio
n d
e la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 3
0S
)
Inhib
iteur
de la s
ynth
egravese d
e
lacid
e f
oliq
ue
Inhib
itio
n d
e c
ert
ain
es r
eacuteactions e
nzym
atiques
essentielle
s agrave
la f
orm
ation d
e la
cid
e f
oliq
ue
Sulfonam
ides
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
AR
Nr
16S
(preacute
vie
nt la
ttachem
ent de la
min
oacyl-
AR
Nt au r
ibosom
e)
Teacutetr
acyclin
e
Poly
peptides
β-L
acta
ms
Gly
copeptides
Lip
opeptides
Inhib
iteur
de la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 5
0S
)
Am
inogly
cosid
es
Cib
le c
ell
ula
ire
Sta
tiq
ue
Cid
al
Rifam
ycin
eR
ifam
pic
ine
Cla
ss
es
Ex
d
an
tib
ioti
qu
eM
od
e d
ac
tio
n
Inhib
itio
n d
e la s
ynth
egravese d
e
lAR
N (
AD
N d
eacutependant)
AR
N p
oly
meacutera
se s
ous-u
niteacute β
(codeacute p
ar
rpoB
)
Quin
olo
nes
cid
al
Inhib
itio
n d
e la r
eacuteplic
ation d
e
lAD
N
Com
ple
xe A
DN
-topois
om
eacutera
se (
AD
N
gyr
ase(t
opois
om
eacutera
se II)
et to
pois
om
eacutera
se IV
)cid
al
Tra
nspeptidase (
penic
illin
bin
din
g p
rote
in
(PB
P))
cid
al
Tab
leau
1
Cib
les e
t m
od
e d
rsquoacti
on
des p
rin
cip
ale
s c
lasses d
rsquoan
tib
ioti
qu
es
28
122 Les aminoglycosides
1221 Mode drsquoaction
Les aminoglycosides peacutenegravetrent la paroi cellulaire en suivant un processus
comprenant 3 eacutetapes une premiegravere eacutetape eacutenergie- indeacutependante suivie de deux
eacutetapes eacutenergie deacutependantes Deux composantes sont essentielles pour
lrsquoaccumulation intracellulaire des aminoglycosides les ribosomes et la chaine de
transport des eacutelectrons (Ramirez et Tolmasky 2010)
Eacutetape 1- Lorsque les aminoglycosides entrent en contact avec la paroi
cellulaire la moleacutecule polycationique se lie aux composantes de surface anioniques
tels les lipopolysaccharides les phospholipides et autres proteacuteines membranaires
chez les bacteacuteries agrave Gram neacutegatif et les moleacutecules drsquoacides teacuteichoiumlques et les
phospholipides des bacteacuteries agrave Gram positif (Ramirez et Tolmasky 2010) Apregraves
cette liaison aux moleacutecules polaires les aminoglycosides (telle la gentamicine)
srsquoassocient aux transporteurs membranaires sur la base de leur charge positive et
sont transporteacutes agrave travers la membrane cytoplasmique gracircce au potentiel
membranaire (intra cellulaire neacutegatif) (Bryan et al 1980 Proctor et al 2006) Cette
liaison agrave la membrane entraine une augmentation de la permeacuteabiliteacute membranaire
menant agrave lrsquoentreacutee des aminoglycosides dans lrsquoespace peacuteriplasmique (Gram neacutegatif)
ou dans le cytoplasme (Gram positif) (Ramirez et Tolmasky 2010) (Figure 12A)
Eacutetape 2- Lrsquoeacutetape suivante connue comme la laquo phase eacutenergie-deacutependante
1 raquo (EDP-I) est bloqueacutee par les inhibiteurs de la chaine de transport des eacutelectrons et
de la phosphorylation oxydative ainsi que par les ions divalents lrsquohyperosmolariteacute le
faible pH et lrsquoanaeacuterobiose dans cette phase la dureacutee et le taux drsquoentreacutee de
lrsquoantibiotique sont en fonction de la concentration (Miller et al 1980 Mingeot-
Leclercq et al 1999 Ramirez et Tolmasky 2010) Agrave cette eacutetape le transport
transmembranaire des aminoglycosides et plus speacutecifiquement de la gentamicine
neacutecessite 2 composantes majeures a) un transporteur anionique non speacutecifique dont
la charge est directement lieacutee agrave lrsquoactiviteacute des cytochromes oxydases terminales et des
29
nitrates reacuteductases de la chaine de transport des eacutelectrons Selon certaines eacutetudes
les transporteurs pourraient ecirctre les quinones reacuteduites de la chaine de transport des
eacutelectrons (Bryan et al 1981 Miller et al 1980) et donc le cycle drsquooxydation-reacuteduction
des quinones est essentiel aux transports des aminoglycosides (Bryan et al 1981)
mais selon drsquoautres eacutetudes les transporteurs pourraient ecirctre toute autre proteacuteine non
deacutefinie impliqueacutee dans la polarisation membranaire lieacutee agrave lrsquoactiviteacute de la chaine de
transport des eacutelectrons (Miller et al 1980) La croissance anaeacuterobie ainsi que le
sodium azide (inhibiteur de la chaine respiratoire) inhibent lrsquoentreacutee de la gentamicine
et lrsquoeffet bacteacutericide indiquant que le transport actif de cet antibiotique neacutecessite la
respiration b) un potentiel eacutelectrique avec un milieu intra cellulaire neacutegatif par rapport
agrave lrsquoexteacuterieur En effet lrsquoinhibition de lrsquoentreacutee dans la bacteacuterie des aminoglycosides
suite agrave lrsquoajout de CCCP (agent deacutepolarisant) chez des souches de S aureus laquo wild-
type raquo deacutemontre la neacutecessiteacute de la force proton-motrice pour lrsquoentreacutee des
aminoglycosides dans les cellules (Miller et al 1980)
Eacutetape 3- La petite quantiteacute drsquoaminoglycosides srsquoeacutetant rendu au cytoplasme
durant cette premiegravere phase eacutenergie-deacutependante induisent apregraves liaison aux
ribosomes des erreurs dans la synthegravese proteacuteique Les erreurs dans la traduction
des proteacuteines membranaires produites lors de cette eacutetape connue comme la laquo phase
eacutenergie-deacutependante 2 raquo (EDP-II) provoquent des dommages dans lrsquointeacutegriteacute de la
membrane cytoplasmique (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky
2010) Les proteacuteines membranaires aberrantes dans la membrane facilitent le
transport drsquoune plus grande quantiteacute drsquoaminoglycosides lesquels augmenteront
encore le niveau drsquointerfeacuterence avec la synthegravese proteacuteique normale amenant encore
plus de dommage agrave la membrane provoquant ainsi une acceacuteleacuteration importante du
taux drsquoentreacutee des moleacutecules antibiotique et qui ultimement reacutesultera en la mort de la
cellule (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky 2010) (Figure 12B)
Des eacutetudes ont deacutemontreacute que la susceptibiliteacute agrave la gentamicine streptomycine
amikacine kanamycine et tobramycine est supeacuterieure lorsque lrsquooxygegravene est
30
lrsquoaccepteur terminal drsquoeacutelectrons (Bryan et Kwan 1981) Une diminution significative
de la susceptibiliteacute est remarqueacutee lorsque le nitrate est lrsquoaccepteur final drsquoeacutelectrons et
une diminution encore plus marqueacutee est observeacutee lorsque le fumarate est lrsquoaccepteur
final drsquoeacutelectron ou lorsque les conditions sont fermentaires Lrsquoassociation entre le
transport de la gentamicine et le complexe terminal de la chaine de transport des
eacutelectrons particuliegraverement lrsquoactiviteacute cytochrome oxydase
Le ribosome possegravede trois sites de liaison importants de lrsquoARNt deacutesigneacute comme site
A (pour aminoacyl) site P (pour peptidyl) et E (pour exit) (Vakulenko et Mobashery
2003) Ils sont situeacutes sur lrsquoARN16S (lequel avec plus de 21 proteacuteines compose la
sous-uniteacute 30S du ribosome bacteacuterien) (Ramirez et Tolmasky 2010 Vakulenko et
Mobashery 2003) Durant la synthegravese proteacuteique le ribosome deacutecode lrsquoinformation de
lrsquoARNm et catalyse lrsquoincorporation seacutequentielle de chaque acide amineacute faisant partie
de la chaine polypeptidique en formation La haute fideacuteliteacute de la traduction (les
ribosomes incorporent un mauvais acide amineacute par approximativement 3000 liaisons
peptidiques effectueacutees) est possible gracircce agrave la capaciteacute de discriminer entre des
changements conformationels dans le ribosome induit par la liaison drsquoun ARNt correct
Modifieacute de Davis BD 1987
Figure 12 Cineacutetique drsquoentreacutee des aminoglycosides
A
B
31
ou incorrect au site A La reconnaissance drsquoune liaison correcte anticodoncodon par
le ribosome se passe en deux temps la reconnaissance initiale de lrsquoappariement du
codon sur lrsquoARNm avec lrsquoanticodon de lrsquoARNt et subseacutequemment la correction
(Vakulenko et Mobashery 2003)
La plupart des aminoglycosides se lient agrave des oligonucleacuteotides faisant partie du site
A Les diffeacuterents aminoglycosides ne lient pas tous le mecircme site de lrsquoARN 16S mais
lrsquoeffet commun agrave tous est que cette liaison amegravene un changement de conformation du
site A vers une conformation mimant la conformation fermeacutee de la sous uniteacute 30S qui
est induite par une interaction entre lrsquoARNm et son ARNt apparenteacute eacuteliminant la
possibiliteacute de correction du ribosome ce qui promeut les erreurs de traduction
(Ramirez et Tolmasky 2010 Vakulenko et Mobashery 2003) La grande majoriteacute des
aminoglycosides sont bacteacutericides et cette leacutetaliteacute est la conseacutequence de cette
mauvaise traduction (Vakulenko et Mobashery 2003)
1222 Meacutecanismes de reacutesistance
La reacutesistance contre les aminoglycosides se produit par diffeacuterents meacutecanismes qui
peuvent coexister simultaneacutement dans la mecircme bacteacuterie (Vakulenko et Mobashery
2003) Ces meacutecanismes incluent 1- la modification de la cible par des mutations
dans lrsquoARN 16S ou autres proteacuteines ribosomales associeacutees (Figure 13) 2- la
meacutethylation de lrsquoARN 16S un meacutecanisme retrouveacute chez plusieurs organismes
producteurs drsquoaminoglycosides et chez des souches cliniques Cette meacutethylation
empecircche les aminoglycosides drsquoavoir accegraves agrave leur site de liaison (Doi et Arakawa
2007) 3- la reacuteduction de la permeacuteabiliteacute par une alteacuteration de la permeacuteabiliteacute de la
membrane externe ou une diminution du transport agrave travers la membrane interne 4-
lrsquoexportation des aminoglycosides agrave lrsquoexteacuterieur de la cellule par les pompes agrave efflux
Eacutetant des moleacutecules hydrophiliques les aminoglycosides ont naturellement peu
drsquoaffiniteacute avec les pompes agrave efflux 5- le swarming actif probablement un meacutecanisme
32
non speacutecifique deacutemontreacute chez Pseudomonas aeruginosa lequel deacutemontre des
reacutesistances agrave plusieurs antibiotiques 6- la seacutequestration de la drogue par des liaisons
agrave un aceacutetyltransfeacuterase agrave tregraves faible activiteacute 7- une inactivation enzymatique de la
moleacutecule antibiotique le meacutecanisme de reacutesistance le plus preacutevalent au niveau
clinique (Vakulenko et Mobashery 2003) Lrsquoinactivation enzymatique des
aminoglycosides est le plus souvent due agrave des aceacutetyltransfeacuterases des
nucleacuteotidyltransfeacuterases ou des phosphotransfeacuterases (Dickgiesser et Kreiswirth 1986)
Ces enzymes sont souvent encodeacutees sur des plasmides mais sont eacutegalement
associeacutees agrave des eacuteleacutements transposables Lrsquoeacutechange de plasmides et la disseacutemination
de transposons facilitent lrsquoacquisition rapide de pheacutenotypes reacutesistants aux
antibiotiques non seulement au sein drsquoune mecircme espegravece mais parmi un large
eacuteventail drsquoespegraveces bacteacuteriennes (Mingeot-Leclercq et al 1999)
La reacutesistance naturelle des S aureus laquo small colony variants raquo agrave la majoriteacute des
aminoglycosides est la conseacutequence de lrsquoimpossibiliteacute agrave lrsquoantibiotique de peacuteneacutetrer une
cellule posseacutedant une faible polariteacute membranaire qui est due chez les SCV agrave leur
Tireacute de Doi Y Arakawa Y 2007
Figure 13 Sites de mutation de certains reacutesistants aux aminoglycosides
33
incapaciteacute agrave activer le cycle de Krebs (Miller et al 1980 Proctor et al 2006 Tubby
et al 2013) Les SCV ont non seulement une polariteacute membranaire faible mais (ceci
eacutetant la conseacutequence de cela) aussi une force proton motrice inhibeacutee et une chaine
des eacutelectrons deacuteficiente De ce fait le transport transmembranaire des
aminoglycosides chez les SCV est grandement reacuteduit et conseacutequemment la
susceptibiliteacute des SCVs pour cette classe drsquoantibiotique est 110-113 de celle des
souches parentes (Proctor et al 2006)
123 La tomatidine
Plusieurs espegraveces veacutegeacutetales accumulent des steacuterols et des triterpegravenes antimicrobiens
appeleacutes saponines (Osborn 1996) Ces moleacutecules sont produites de faccedilon
constitutive par la plante et jouent un important rocircle de deacutefense contre diffeacuterents
pathogegravenes comme des insectes ou des microbes Parmi les saponines la tomatine
(Figure 14A) est un glycoalcaloiumlde steacuteroiumldien preacutesentant une action antimicrobienne
contre des levures et une quantiteacute drsquoautres microorganismes (Bednarek et Osboum
2009 Friedman 2002 Osborn 1996 Sandrock et Vanetten 1998) Plusieurs
espegraveces de champignons pathogegravenes de la tomate produisent des enzymes
extracellulaires communeacutement appeleacutes tomatinase qui sont capables de deacutetoxifier
lrsquoα-tomatine (Martin-Hernandez et al 2000 Roddick 1974 Ruiz-Rubbio et al 2001)
Par exemple Fusarium oxysporum f sp lycopersici est reconnu pour cliver lrsquoα-
tomatine en ses sous-produits soit un tetrasaccharide (lycotreacutetraose) et un alcaloiumlde
steacuteroiumldien non glycosyleacute la tomatidine (Figure 14B) Ces sous-produits nrsquoont pas ou
peu drsquoactiviteacute antifongique (Ruiz-Rubbio et al 2001 Simon et al 2006)
34
Figure 14 Structures chimiques de la (A) tomatine de (B) tomatidine et de (C)
lrsquoanalogue de tomatidine FC04-100
Il a eacuteteacute deacutecouvert il a quelques anneacutees que la tomatidine avait un effet antibacteacuterien
sur les S aureus de pheacutenotype SCV mais aucune activiteacute contre les S aureus agrave
pheacutenotype normal Par contre lorsqursquoutiliseacute en combinaison avec les
aminoglycosides lrsquoajout de la tomatidine permettait drsquoaugmenter de 8 agrave 16 fois
lrsquoactiviteacute des aminoglycosides contre S aureus agrave pheacutenotype normal (Mitchell et al
2011a Mitchell et al 2012)
Donc on savait que la tomatidine avait une activiteacute antibacteacuterienne tregraves inteacuteressante
mais on ne connaissait ni sa cible ni son mode drsquoaction De plus si la tomatidine a
une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait donc avoir un
spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Par ailleurs de faccedilon parallegravele la production drsquoanalogues structuraux de la tomatidine
permettrait de mieux comprendre la relation structure-activiteacute (SAR) de la moleacutecule et
donc de preacuteciser que lrsquoactiviteacute biologique de la tomatidine neacutecessite une orientation
3
A B
C
35
de groupements fonctionnels bien speacutecifique De plus lrsquoeacutetude de lrsquoactiviteacute de
plusieurs analogues permettrait de confirmer si la tomatidine possegravede un meacutecanisme
drsquoaction deacutependant drsquoune interaction speacutecifique avec une ou des cibles biologiques
preacutecises
Le laboratoire du Pr Eacuteric Marsault du deacutepartement de pharmacologie de lrsquoUniversiteacute
de Sherbrooke et lrsquoeacutetudiant agrave la maitrise Feacutelix Chagnon ont produit plus de 33
analogues de la tomatidine (voir Annexe 2 article laquo Unraveling the structure-activity
relationship of tomatidine a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureusraquo dans la section laquo Contributions
Supleacutementaires raquo du meacutemoire) dont lrsquoactiviteacute antibacteacuterienne a eacuteteacute eacutevalueacutee par des
tests de susceptibiliteacute agrave la moleacutecule (CMI) sur S aureus SCV et sur S aureus agrave
pheacutenotype normal en combinaison avec la gentamicine Plusieurs de ces moleacutecules
font lrsquoobjet drsquoune application de brevet (voir annexe 1) dont la prometteuse moleacutecule
FC04-100 (Figure 14C) (voir laquo Annexe 1 Application de brevet raquo dans la section
laquo Contributions suppleacutementaires raquo du meacutemoire)
13 Projet de maitrise
Premiegravere hypothegravese
Si la tomatidine a une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait
donc avoir un spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Objectif Provoquer lrsquoapparition de pheacutenotypes SCV chez diffeacuterentes espegraveces
bacteacuterienne afin de pouvoir veacuterifier leur niveau de sensibiliteacute agrave la tomatidine
Deuxiegraveme hypothegravese
Lrsquoidentification des gegravenes preacutesentant des mutations parmi les souches de S aureus
reacutesistantes agrave la tomatidine pourrait reacuteveacuteler la ou les cibles cellulaires de la moleacutecule
et nous aider agrave comprendre son mode drsquoaction
36
Objectif Obtenir des mutants reacutesistants avant de les envoyer agrave sequencer
faire les analyses comparatives des geacutenomes et finalement identifier les mutations
37
CHAPITRE 2
ARTICLE SCIENTIFIQUE
TITRE Tomatidine and Analog FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Some Firmicutes Including Those of Listeria
monocytogenes and Are Bactericidal in Combination With Aminoglycosides Against
their Normal Phenotype
AUTEURS ET CONTRIBUTIONS
Isabelle Guay1 Expeacuteriences biologiques avec le produit naturel tomatidine et son
analogue FC04-100 sur diffeacuterentes espegraveces bacteacuteriennes afin de comparer les deux
activiteacutes
Simon Boulanger1 Expeacuterience biologique permettant de quantifier lrsquoactiviteacute de
lrsquoanalogue FC04-100 sur la production de biofilm et son activiteacute intracellulaire
Feacutelix Chagnon2 Production de lrsquoanalogue chimique de la tomatidine FC04-100
Kamal Bouarab1 Initiateur du projet gracircce agrave lrsquoisolement du produit naturel tomatidine
Eacuteric Marsault2 Directeur de recherche de Feacutelix Chagnon
Franccedilois Malouin1 Directeur de recherche drsquoIsabelle Guay et Simon Boulanger et
auteur de correspondance
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
38
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
Corresponding author Mailing address Universiteacute de Sherbrooke
Deacutepartement de Biologie Faculteacute des Sciences 2500 Boul
Universiteacute Sherbrooke Quebec Canada J1K2R1 Phone (819) 821-
8000 ext 61202 Fax (819) 821-8049 E-mail FrancoisMalouin
USherbrookeca
39
Tomatidine and Analog FC04-100 Possess Bactericidal Activities
Against the Small Colony Variants of Some Firmicutes Including
Those of Listeria monocytogenes and Are Bactericidal in Combination
With Aminoglycosides Against their Normal Phenotype
Running Title Tomatidine is bactericidal against several Firmicutes
Isabelle Guay1
Simon Boulanger1
Feacutelix Chagnon2 Kamal Bouarab
1 Eacuteric Marsault
2 and
Franccedilois Malouin1
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM) Deacutepartement de
Biologie Faculteacute des Sciences Universiteacute de Sherbrooke Sherbrooke Queacutebec Canada J1K
2R1
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute Universiteacute
de Sherbrooke Queacutebec Canada J1H 5N4
Have contributed equally to the work
Corresponding author
Mailing address Universiteacute de Sherbrooke Deacutepartement de Biologie Faculteacute des Sciences
2500 Boul Universiteacute Sherbrooke Quebec Canada J1K2R1
Phone (819) 821-8000 ext 61202 Fax (819) 821-8049
E-mail FrancoisMalouinUSherbrookeca
40
ABSTRACT
Tomatidine (TO) is a plant steroidal alkaloid that possesses an antibacterial activity against the
small colony variants (SCVs) of Staphylococcus aureus We report here the spectrum of
activity of TO against species of the Firmicutes and the improved antibacterial activity of a
chemically-modified TO derivative (FC04-100) In addition to that seen on S aureus TO
showed a bactericidal activity against SCVs of Bacillus cereus B subtilis and Listeria
monocytogenes The combination of an aminoglycoside antibiotic and TO also generated an
antibacterial synergy against strains of the normal phenotype in these species In contrast to
TO which has no relevant activity by itself against S aureus of the normal phenotype (MIC
gt64 mgL) the TO analog FC04-100 showed a MIC of 8-16 mgL Furthermore FC04-100
showed a strong bactericidal activity against L monocytogenes SCVs in kill kinetic
experiments while TO did not FC04-100 also improved the bactericidal activity of
gentamicin against L monocytogenes of the normal phenotype The addition of FC04-100 (4
mgL) to a cephalexinkanamycin (32) combination improved the activity of the combination
by 32 fold against cephalexin and kanamycin-resistant MRSA strains In combination with
gentamicin FC04-100 also showed a strong bactericidal activity against biofilm-embedded S
aureus and when used alone was superior to TO for its biofilm-preventing activity Finally
FC04-100 and TO showed comparable intracellular killing of S aureus SCVs in a polarized
Calu-3 cell culture model In conclusion chemical modifications of TO allowed improvement
of its antibacterial activity against L monocytogenes SCVs and prototypical S aureus
41
Keywords Tomatidine aminoglycoside synergy SCV Firmicutes S aureus L
monocytogenes
Introduction
The Firmicutes are Gram-positive bacteria with genomes presenting less than 50 of G+C
The Firmicutes constitutes one of the main divisions within the Bacteria and is divided into
three classes Bacilli Mollicutes and Clostridia The class of Bacilli is further divided into the
orders of the Lactobacillales and the Bacillales The Bacillales are divided into the genus of
Staphylococcus Listeria and Bacillus A number of bacterial species such as Listeria spp and
Bacillus spp can contaminate food and cause infections in humans (Magalhaes 2013) To
name a few Listeria monocytogenes L ivanovii and Bacillus cereus can cause listeriosis
(Guillet et al 2010) and food poisoining (Bad Bug Book FDA) Bacillus subtilis B
coagulans B licheniformis and B sphaericus are also known to cause illnesses Bacillus
anthracis causes anthrax and can often be acquired by contact with food producing animals
and cattle (beef cattle sheeps etc) and this bacterium is also well-known for its endospores
that have been used as biological weapons (Beierlein and Anderson 2011) Staphylococci are
divided in coagulase-positive species Staphylococcus aureus being the most clinically
relevant of this group and coagulase-negative species such as S epidermidis the most
prevalent pathogen associated with infections of implanted medical devices (Vuong and Otto
2002) The emergence and spread of resistance to multiple antibiotics in staphylococci is now
considered a real health treat and impaired therapeutic endeavor to combat these bacteria
42
(Witte et al 2007) Notably the prevalence of methicillin-resistant S aureus (MRSA) has
steadily increased over the recent years not only in hospitals but also in the community
(Hiramatsu et al 2013)
Since their first description in 1910 for Salmonella enterica serovar Typhi (S typhi) small-
colony variants (SCVs) have been described for a wide range of bacterial genera and species
and have been isolated from both laboratory and clinical environments (Proctor 2006)
Staphylococcus aureus SCVs have attracted a great deal of interest over the past recent years
S aureus SCVs often present a dysfunctional oxidative metabolism causing an alteration in
the expression of virulence factors a slow growth and a loss of colony pigmentation (Proctor
2006) This dysfunctional oxidative metabolism is also responsible for a decreased
susceptibility to aminoglycoside antibiotics because this class of molecules requires the
proton-motive force in order to penetrate the bacterium (Bryan LE 1981) This respiratory
deficiency is often caused by mutations affecting the electron transport system and several
SCV isolates can recover normal growth with supplemental hemin or menadione which are
needed to synthesize electron transport system components S aureus SCVs often are isolated
from chronic infections such as lung infections in cystic fibrosis (CF) patients osteomyelitis
septic arthritis bovine mastitis and from infections associated with orthopedic devices (Atalla
2008 Moisan 2006 Proctor 2006) The ability of S aureus to switch back and forth from the
prototypic to the SCV phenotypes in vivo is an integral part of the pathogenesis of S aureus
and may be responsible for the establishment of chronic infections (Tuchscherr 2011
Mitchell 2011 inf imm)
43
Tomatidine (TO) is a steroidal alkaloid produced by the Solanaceae plant family such as the
tomato (Ruiz-Rubio 2001 Simons 2006) We showed previously the antibacterial activity of
TO against S aureus SCVs and also documented a strong synergic activity of TO in
combination with aminoglycoside antibiotics against prototypic S aureus (Mitchell G AAC
2011 Mitchell G 2012 JAC) Recently we synthesized a variety of TO analogs in order to
explore the structure-activity relationship of this new class of antibiotics able to act on SCVs
(Chagnon et al 2014) One analog FC04-100 showed the same steroidal backbone as the
natural molecule but with an additional carbon chain on the A cycle (Fig 1) Preliminary
characterization of FC04-100 revealed an antibacterial activity that was similar to that of TO
against S aureus SCVs and also preserved the synergic activity with aminoglycosides against
prototypic S aureus (Chagnon et al 2014) However contrary to TO FC04-100 showed
notable antibiotic activity by itself against prototypic S aureus with a MIC of 8-16 gL
whereas that of TO was gt64 gL It would be of great interest to see if this property of FC04-
100 modifies or improves its spectrum of antibacterial activity
The aim of this study was to further describe the spectrum activity of TO and FC04-100 The
susceptibility of several species among the Firmicutes to TO and FC04-100 alone or in
combination with other antibiotics was investigated The improved antibacterial activity of
FC04-100 was demonstrated against Listeria monocytogenes and methicillin-resistant S
aureus (MRSA)
44
Materials and methods
Chemical reagents and antibiotics
Tomatidine (TO) (Sigma Aldrich Oakville Ontario Canada) (Fig 1A) was solubilised in
dimethylsulfoxide (DMSO) at a concentration of 2 mgmL The TO analog FC04-100 (Fig
1B) was synthesized (Chagnon et al 2014) and solubilised in DMSO at 20 gL Cefalexin
kanamycin gentamicine (GEN) (all from Sigma Aldrich) were solubilised in water at a
concentration of 10 gL Menadione was solubilized in DMSO hemin in 14 M NH4OH and
thymidine in water (all from Sigma Aldrich) and were prepared at a concentration of 10 gL
45
3
Fig 1 Tomatidine (A) is a steroid alkaloid structurally characterized by 6 rings 12
stereogenic centers a 3 β-hydroxyl group and spiro-fused E F rings in the form of an
aminoketal The tomatidine analog FC04-100 (B) contains a diamine in position 3 of ring A
Bacterial strains and growth conditions
Staphylococcus aureus ATCC 29213 S aureus Newbould (ATCC 29740) Listeria
monocytogenes ATCC 13932 Bacillus subtilis ATCC 6333 and Bacillus cereus ATCC 11778
were used as prototypic strains Methicillin-resistant S aureus (MRSA) strains COL and
USA100 (hospital acquired-MRSA) and strain USA300 (community acquired-MRSA) were
also used in this study The reference laboratory strain S aureus SH-1000 an isogenic mutant
strain derived from S aureus 8523-4 but with a functional rsbU allele (HorsburghMJ 2002)
was used for its good yield of biofilm production The S aureus strains CF07-L (prototypic)
A
B
A B
C D
E
F
46
and CF07-S (SCV phenotype) are genetically related clinical strains originally isolated from a
cystic fibrosis patient (MitchellG 2011) Strain CF07-S was used in the intracellular
infection model Staphylococcus and Bacillus were maintained on Tryptic Soy agar (TSA BD
Mississauga Ontario Canada) whereas L monocytogenes was grown on brain-heart infusion
(BHI) agar (BD Mississauga Ontario Canada)
Small colony variants (SCVs)
For S aureus strain NewbouldhemB was used as the reference SCV NewbouldΔhemB was
generated from strain Newbould (ATCC 29740) by disrupting the hemB gene with the ermA
cassette by homologous recombination (Brouillette 2004) SCVs from B cereus B subtillis
and L monocytogenes were generated by growth in presence of a subinhibitory concentration
of GEN Briefly overnight broth cultures (18ndash20 h) were used to inoculate BHI broth at a
dilution of 1100 supplemented or not with 025 to 1X the MIC of GEN Cultures were
incubated 18 h at 35degC with shaking (225 rpm) and then adjusted to an A595 nm of 20 in PBS at
4degC Determination of CFU and SCV colonies was done by serial dilution plating SCV were
obtained by plating on TSA (BHI agar for L monocytogenes) containing gentamicin at a
concentration of 8 to 16 times the induction concentration followed by an incubation of 48 h at
35degC The pinpoint colonies selected by this method were confirmed to be SCVs by streaking
several of them on agar plates without antibiotic SCV that conserved their phenotype after
two passages were considered to have a stable phenotype and were used in subsequent
experiments Selected SCVs had a colony size a tenth of the size of a normal colony and
showed no pigmentation or hemolytic activity on blood agar plates The SCV species were
47
confirmed by rDNA 16S sequencing after PCR amplification using universal primers (Gomaa
2007)
Auxotrophy assays
For SCVs auxotrophism is defined as the requirement of specific compounds in order to
regain a normal growth phenotype [Sendie 2009] An agar diffusion method was used to
characterize the auxotrophism of SCVs using hemin or menadione (1-10 μg eachwell) or
thymidine (100 microgwell) on an inoculated Mueller-Hinton agar (MHA) or Brain-heart infusion
agar (BHIA) plate [Besier 2007] Auxotrophy for specific supplements was detected by a
zone of normal growth surrounding the well after 18 h of incubation at 35degC The results were
confirmed by two indepedent experiments
Antibiotic susceptibility testing
MICs were determined by a broth microdilution technique following the recommendations of
the Clinical and Laboratory Standards Institute (CLSI) except that the incubation period was
extended to 48 h and the medium used was BHI to allow SCVs to reach maximal growth as
previously described (Mitchell 2010) As recommended by the CLSI L monocytogenes (and
its SCVs) were grown in cation-adjusted Muller-Hinton broth (CAMHB BD) containing 3
lysed horse blood (LHB Remel Hartford CT USA) The reported results were obtained from
three independent experiments
48
Time-kill experiments
Kill kinetics were performed to determine whether the effect of compounds alone or in
combination was bacteriostatic or bactericidal Bacteria were inoculated at 105-10
6 CFUmL in
appropriate medium in the absence or presence of the different antibiotic compounds at
concentrations specified in figure legends At several points in time at 35degC (225 rpm)
bacteria were sampled serially diluted and plated on TSA for CFU determinations Plates
were incubated for 24 or 48 h at 35degC for normal and SCV strains respectively The data were
collected from a minimum of three independents experiments
Antibiotic activity in biofilms
The viability of bacteria in biofilms was evaluated based on previously described protocols
(CeriH 2001 MoskowitzSM 2004 HarrisonJJ 2010) For these assays 96-peg lids and
corresponding 96-well plates were used (Nunc CITY STATE) Wells were inoculated with a
suspension of S aureus SH-1000 adjusted to a 05 McFarland standard in TSB (150 microL per
wells) and plates with 96-well lids were incubated at 35degC with an agitation of 120 rpm for 24
h The biofilms on pegs were then washed three times with 200 microL PBS Biofilms on pegs
were further incubated in fresh 96-well plates containing 200 microL of TSB containing or not
antibiotic compounds or a combination of molecules at 35degC 120 rpm for 24 h The treated
biofilms on pegs were washed three times in PBS and bacteria in biofilms were recovered by
sonication in a new microtiter plate containing 200 microL of PBS per wells using an
ultrasonicator bath for 10 min followed by centrifugation for five min at 1000 RPM The
49
bacteria recovered in each well were resuspended serially diluted plated on TSA and
incubated at 35degC for 24 h before CFU determination
For an equivalent series of wells instead of disrupting the biofilm by sonication biofilm
formation on pegs was evaluated through crystal violet staining using a procedure adapted
from Mitchell et al(2010) Following exposure to antibiotics as described above biofilms on
pegs were washed three times using 200 microL of PBS before biofilms were air-dried for 30
minutes Biofilm on pegs were then stained by dipping in 200 microL of a crystal violet solution
(01) for 30 minutes rinsed in 200 microL of distilled water and air-dried for 30 min Finally the
crystal violet from stained biofilms was solubilized in 200 microL of ethanol 95 at room
temperature and the absorbance was measured at 562 nm using a plate reader (nom de
lrsquoappareil)
Cell invasion and measurement of intracellular antibiotic activity
The Calu-3 cell line (ATCC HTB 55 Homo sapiens lung adenocarcinoma) was cultured in
Eaglersquos Minimum Essential Medium (EMEM) supplemented with 01 mM MEM nonessential
amino acids 1 mM of sodium pyruvate 100 UmL penicillin 100 gL streptomycin 25 gL of
Fungizone and 10 fetal bovine serum (FBS) at 37oC in 5 CO2 For routine culture 4 gL of
puromycin was added to culture media All cell culture reagents were purchased from Wisent
(St-Bruno QC Canada)
50
The cell invasion assay was performed with the Calu-3 cells in an air-liquid interface as
previously described with few modifications (Mitchell TO I paperAAC 2011) Briefly cells
were seeded at 15 x 105 cells per insert in a 12-well Transwell plate (Corning CITY STATE)
and cultured for 10 days with the apical medium replaced each day The complete medium in
the basal compartment was replaced by the invasion medium (1 FBS and no antibiotics) 18
h before invasion assays Inocula were prepared by suspending bacteria grown 20 h on BHI
agar plates in ice-cold PBS Bacteria were then washed three times in ice-cold PBS and
suspended in the invasion medium supplemented with 05 BSA at a density of
approximately 4 x 108 CFUmL Cells were washed twice with PBS and 250 μL of bacterial
suspension were apically added to each insert Invasion of cells by bacteria was allowed for 3
h inserts were emptied and washed three times with PBS The basal medium was changed for
the invasion medium with 20 gL of lysostaphin (Sigma) to kill extracellular bacteria and with
or without the tested antibiotic compounds The infected cells were incubated for a total of 48
h with a change of medium at 23 h (invasion medium with lysostaphin with or without the
tested antibiotic compounds) The invasion medium with lysostaphin but without tested
compound was also apically added 1 h before cell lysis to ensure that only intracellular
bacteria were counted At 48 h following three washes with PBS the apical and basal media
were removed and cells were detached with 100 μL of trypsin 025 and lysed for 10 min by
the addition of 400 μL of water containing 005 of Triton X-100 Then 50 μL of PBS (50X)
was added and mixed Lysates were serially diluted 10-fold and plated on TSA for CFU
determination Plates were incubated at 35oC for 48 h
51
RESULTS
Emergence the SCV phenotype
The SCV phenotype is characterized by a slow growth yielding small colony size on agar
plates (15 to 110 of the parent phenotype) Figure 2 illustrates the ability of a subinhibitory
concentration of GEN to promote the emergence of the SCV phenotype for B cereus B
subtilis and L monocytogenes B cereus SCVs were obtained at an inducing concentration of
1 X MIC of GEN (Fig 2C) B subtilis SCVs at 05 X MIC (Fig 2D) and L monocytogenes
SCVs at 025 X MIC (Fig 2E) Two prototypic S aureus Newbould and CF07-L and their
SCV counterpart NewbouldhemB and CF07-S respectively were used as control strains (Fig
2A and 2B) The SCV isolates selected for the rest of the study add a low reversion rate and
kept their phenotype without a GEN selection pressure The B cereus SCVs showed
auxotrophy for menadione (Fig 1F Table 1) some of the L monocytogenes SCVs showed
auxotrophy for hemin (Table 1) while the B subtilis SCVs showed no apparent auxotrophy to
hemin menadione or thymidine (Table 1) These results show that SCVs with defects in their
respiratory chain (ie hemin or menadione auxotrophy) can indeed emerge from a selective
pressure with aminoglycosides
52
A B
Fig 2 Typical colony sizes for SCV and protocipic strains (left and right sides of the plates
respectively) for (A) S aureus CF07-S and CF07-L (B) S aureus NewbouldhemB and
Newbould (C) B cereus ATCC 11778 (SCV1 and prototypic) (D) B subtilis ATCC 6333
(SCV2 and prototypic) and (E) L monocytogenes ATCC 13932 (SCV1 and prototypic) In
(F) a TSA plate was inoculated with a B cereus SCV1 and wells were filled with (i) 10 microg
hemin (ii) 10 microg menadione (iii) 100microg thymidine and diluents (iv) DMSO and (v) NH4OH
14N
C D
E
i ii
iii iv
v
F
53
Antibacterial activity of TO against the various species of SCVs and synergy of
the TO and GEN combination against some prototypic Firmicutes
All the SCVs tested showed a MIC for TO of 006 mgL against S aureus and lt003 mgL
against L monocytogenes B cereus and B subtilis respectivly whereas TO showed no
activity (MIC gt64 microgmL) against their prototypic counterparts (Table 1) A synergic effect of
TO with GEN against all the prototypic strains tested was also observed (Table 1) Indeed the
addition of TO provoked a 4-8 fold decrease of GEN MIC against all species tested (Table 1)
These results are consistent with our previous studies on the activity of TO on S aureus
((Mitchell 2011 AAC 2012 JAC)) No change in the MIC of GEN was observed against the
streptococci (S pyogenes S suis S agalactiae S mitis and S pneumoniae) when tested in
combination with TO (data not shown) These results show that TO is active against some but
not all Firmicutes
54
Table 1 Susceptibility of prototypic and SCV strains and species to TO andor GEN
Ratio of activity of GEN+TO vs GEN alone
Bactericidal activity of TO against SCVs and bactericidal synergy of the TO
and GEN combination against prototypic strains
As shown above the addition of 8 mgL of TO to GEN reduced the MIC of GEN against some
Firmicutes (Table 1) but also time-kill kinetics showed that TO improved the bactericidal
effect of GEN against prototypic B subtilis and B cereus (Fig 3A and 3B respectively) This
bactericidal synergy was also previously described for S aureus (Mitchell 2012 JAC)
However although the MIC of GEN was improved when in combination with TO against L
monocytogenes (Table 1) time-kill experiments failed to demonstrate a bactericidal synergy
TO GENGEN
(+TO)
Saureus ATCC 29213 gt64 05 006 8
Saureus NewbouldΔhemB 006 8 Hemin
B cereus ATCC 11778 gt64 1 025 4
B cereus ATCC 11778 (SCV1) lt003 16 Menadione
B cereus ATCC 11778 (SCV2) lt003 16 Menadione
B subtilis ATCC 6333 gt64 0125 003 4
B subtilis ATCC 6333 (SCV1) lt003 2 unknown
B subtilis ATCC 6333 (SCV2) lt003 4 unknown
B subtilis ATCC 9372 gt64 0125 lt0015 gt8
B subtilis ATCC 9372 (SCV1) lt003 1 unknown
B subtilis ATCC 9372 (SCV2) lt003 1 unknown
L monocytogenes gt64 05 0125 4
L monocytogenes (SCV1) lt003 gt64 unknown
L monocytogenes (SCV2) lt003 gt64 Hemin
Strains
MIC (gL)
Synergy
foldAuxotrophy
55
with an improvement of at most 1 log10 in killing compared to that achieved with GEN alone
(Fig 3C) Similarly the effect of TO used alone against SCVs was strongly bactericidal
against S aureus B subtilis and B cereus but not against the L monocytogenes SCV (Fig
3D) and this even if the MIC of TO against L monocytogenes SCVs was inferior to 003
mgL (Table 1) These results show that the activity of TO against L monocytogenes needs to
be improved
56
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 0 6 m g L G E N
0 0 6 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 5 m g L G E N
0 5 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
2 m g L G E N
2 m g L G E N + T O
8 m g L T O
C T R L
Lo
g1
0 C
FU
ml
0
2
4
6
8
1 0
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
CT
RL
CT
RL
CT
RL
CT
RL
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
S a u re u s
h e m B
S C V
B c e r e u s
A T C C 1 1 7 7 8
S C V 2
B s u b t i l i s
A T C C 6 3 3 3
S C V 2
L m o n o c y to g e n e s
A T C C 1 3 9 3 2
S C V 1
Fig 3 Time-kill kinetics of TO andor GEN against (A) prototypic B subtilis ATCC 6333(B)
B cereus ATCC 11778 and (C) L monocytogenes ATCC 13932 The bactericidal effect of
TO against SCVs is also shown at 8 and 24 h (D) The results were obtained from at least
three independent experiments
A B
C
D
Detection
threshold
57
Bactericidal activiy of the TO analog FC04-100 against L monocytogenes
SCVs and synergy with aminoglycosides against prototypic strains and MRSA
The addition of 8 mgL of FC04-100 to GEN (2 mgL) decreased by an average of 5 log10
prototypic L monocytogenes CFUmL after 24 h of treatment in comparison to GEN alone
(Fig 4A) Also the bactericidal activity of FC04-100 used alone against the L monocytogenes
SCV was shown to be better than that observed for TO (Fig 3D) To facilitate comparison
between TO and FC04-100 Figure 4B shows the residual CFU of the L monocytogenes SCV
strain after treatment with TO or FC04-100 and calculated as the percentage of live CFU in
comparison with untreated L monocytogenes SCVs after 24 h of incubation These results
show that TO can be chemically modified (eg derivative FC04-100) and improve its activity
against L monocytogenes and thus the antibacterial activity of FC04-100 against S aureus
was further investigated Interestingly the MIC of FC04-100 against the SCV strain S aureus
NewbouldhemB (MIC of 05 -1 mgL Table 2) was higher than that found for TO (MIC of
006 mgL Table 1) On the other hand the MIC of FC04-100 against the prototypic S
aureus ATCC 29213 is lower than that of TO (8-16 mgL vs gt 64 mgL respectively) (Tables
1 and 2) This newly detected antibacterial activity of FC04-100 was also noted against
prototypic MRSA strains (Table 2) Furthermore the ability of FC04-100 to potentiate the
activity of GEN was similar to that of TO with a 4 to 8 fold improvement of the
aminoglycoside MIC (Table 2)
Although the multi-resistant S aureus MRSA USA100 USA300 and COL are resistant to
cefalexin (beta-lactam antibiotic MIC of 256 mgL) and to the aminoglycoside kanamycin
(MIC of 256 mgL and gt 1024 mgL for USA 100 and USA300 respectively Table 3) the
58
combination of both antibiotics decreased their respective MICs (Table 3) Indeed the
combination of cefalexin and kanamycin in a proportion of 32 (commercialized under the
name UbrolexinTM
) decreased the MIC of one or both antibiotics This is as expected for the
combination of a beta-lactam with an aminoglycoside (Davis 1982) However the synergy is
weak with MRSA strains that are resistant to one or the other or both antibiotics and the
addition of 4 mgL of FC04-100 further improved by 8 to 32 fold the MIC of the cephalexin-
kanamycin combination against these strains (Table 3)
59
te m p s (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
F C 0 4 -1 0 0 8 m g L
2 m g L G E N +
F C 0 4 -1 0 0 8 m g L
C T R L
G E N 2 m g L (B H I)
Re
sid
ua
l C
FU
(
)
CT
RL
TO
8 g
L
FC
04-1
00 8
gL
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
F C 0 4 -1 0 0 8 g L
T O 8 g L
C TR L
T=
24
h
T=
8h
T=
8h
T=
24
h
T=
24
h
Fig 4 Time-kill kinetics of FC04-100 andor GEN against (A) prototypic L monocytogenes
ATCC 13932 The bactericidal effect of FC04-100 used alone against L monocytogenes
SCV1 is also shown at 8 and 24 h (B) In (B) the residual CFU were calculated as the
percentage of live CFU in each condition in comparison with the untreated L monocytogenes
SCV1 after 24h of incubation The results were obtained from at least three independent
experiments
A
B
60
Table 2 Susceptibility of prototypic and SCV S aureus strains to FC04-100 with or without
GEN
a FC04-100 w as used at 4 microgmL
b Ratio of activity of GEN + FC04-100 vs activity of GEN alone
Table 3 Synergistic activity of FC04-100 with the combinaison cefalexin-kanamycin
aFC04-100 was used at 4 microgmL
bRatio of the MIC of cefalexin+kanamycin with FC04-100 vs the MIC of
cefalexin+kanamycin alone
Staphylococcus aureus ATCC 29213 8-16 05 006-0125 4-8
S aureus SH1000 8-16 05 006-0125 4-8
S aureus NewbouldΔhemB 1 8
MRSA USA100 16 05 0125 4
MRSA USA300 16 05 006 8
MRSA Col 16 025 006 4
Strains FC04-100 GENGEN
(+FC04-100 )fold
MIC (gL)
MRSA USA100 256 256 12886 43 32
MRSA USA300 256 gt1024 256171 86 32
MRSA Col 256 4 43 05035 8
MIC (gL)
Strains cefalexin kanamycin cef+kana (32)cef+kana (32)
(+FC04-100 )fold
a b
a b
61
FC04-100 potentiates GEN against S aureus embedded in biofilm and inhibits
intracellular replication of SCVs
Figure 5 demonstrates that compound FC04-100 efficiently kills S aureus embedded in
biofilms when in combination with GEN ie there was an important decrease of bacterial
CFUs compared to the no drug control or to either FC04-100 used alone at 4 ugmL (ie 025-
05MIC Table 2) or GEN used alone at 025 05 or 1 X MIC Besides we previously
demonstrated that TO was able to inhibit intracellular replication of SCVs (Mitchell AAC)
which is an important aspect of virulence for S aureus Here (Fig 6) we show that FC04-100
like TO can significantly decrease the infection of polarized airway epithelial cells by SCVs
62
05
mgL
GE
N
05
mgL
GE
N +
FC
04
02
5 m
gL
GE
N
02
5 m
gL
GE
N +
FC
04
01
2 m
gL
GE
N
01
2 m
gL
GE
N +
FC
04
No d
rug
FC
04
0
2
4
6
8
log
10
CF
Up
eg
= 3 2
= 3 8
= 3 5
= 2
Fig 5 Bactericidal effect of FC04-100 alone or in combination with GEN against prototypic
S aureus SH100 embedded in biofilm FC04-100 (FC04) was used at 4 mgL alone or in
combination with GEN Data are from three independent experiments performed in triplicate
Significant differences in comparison to the no drug control are shown ( Plt 0001 and
Plt 00001 one-way ANOVA with a Dunnettrsquos post test) Data are presented as means with
standard deviations
63
No d
rug
TO
8 m
gL
FC
04 8
mgL
0
2
4
6
log
10
CF
Ui
ns
er
t
Fig 6 Effect of TO and FC04-100 on the intracellular load of CF07-S 48 h post-
internalization Data are from three independent experiments performed in duplicate
Significant differences among groups are shown ( Plt 00001One-way ANOVA with
Turkeyrsquos post test) Data are presented as means with standard deviations
64
DISCUSSION
In previous studies we have demonstrated that TO has a strong inhibitory activity against S
aureus SCVs and improves the bactericidal activity of aminoglycoside antibiotics against
prototypic S aureus and also more broadly against staphylococci This effect was documented
for several strains of diverse clinical origins including aminoglycoside-resistant S aureus
carrying aminoglycoside modifying enzymes TO showed however no synergistic effect on the
activity of aminoglycosides against P aeruginosa E coli or Enterococcus spp (Mitchell
2012) In the present study we showed that the antibacterial spectrum of TO can be extended
to species of the Bacillales order with very low MICs against L monocytogenes B cereus and
B subtillis SCVs (lt003 microgmL) Time-kill kinetics showed that the combination of TO and
GEN creates a bactericidal synergy against prototypic of B subtilis and B cereus similarly to
that previously demonstrated against S aureus However the mixture of TO and GEN did not
demonstrated much improvement over the activity of GEN alone against prototypic L
monocytogenes in time-kill experiments Likewise investigations with Bacillales SCVs
showed that TO efficiently killed S aureus B cereus and B subtillis but not L
monocytogenes SCVs despite its very low MIC and was thus bacteriostatic against that species
Therefore improvement of the antibacterial activity against this important pathogen seems
inevitable for future development of steroidal alkaloids such as TO
TO is a steroid alkaloid structurally characterized by 6 rings 12 stereogenic centers a 3-
hydroxyl group and spiro-fused E F rings in the form of an aminoketal (Fig 1) It is generally
65
prepared by hydrolysis of its glycosylated analog tomatine while both molecules can be found
in the Solanaceae plants (Ruiz-Rubio 2001 Simons 2006) Although we previously
documented some interesting antibacterial activity for TO the absence of an identified cellular
target makes difficult the establishment of a structure-activity relationship (SAR) Our initial
attempt to elucidate SAR was therefore based on the hypothesis that the steroidal A-D rings
act as a rigid scaffold to orient pharmacophores defined by the 3-hydroxyl group on ring A
and the spiroaminoketal group on rings E F Noteworthy we prepared analogs bearing
modifications on ring A of TO and schegravemes for chemistry synthesis were recently published
(Chagnon et al 2014) From seven compounds keeping rings E and F of TO and possessing
structural variations on position 3 of ring A we were able to ascertain some structure activity
relationships Results suggest that the hydrogen bond donor effect of the 3-hydroxyl group
was not important for activity although the stereochemistry of the 3 position substitution
moderately affected activity against both S aureus SCVs and prototypic strains (in
combination with an aminoglycoside) In addition although the presence of one or two
ammonium groups in position C3 reduced the activity against S aureus SCVs (from a MIC of
006 to 05-1 microgmL) this change was highly beneficial for antibiotic activity against normal
strains (from a MIC of gt64 to 8-16 microgmL) without the presence of an aminoglycoside This
change was unexpected and one of such analogs FC04-100 increased its solubility by more
than ten times (from 2 mgmL to 26 mgmL in DMSO) The biological properties of FC04-
100 were therefore further investigated in the present study especially that TO showed some
weak bactericidal activity against Listeria and that the solubility of TO may limit eventual
clinical applications
66
Here we demonstrated that FC04-100 was able to kill both L monocytogenes SCVs as well as
the normal phenotype alone and in the presence of GEN respectively (Fig 4) Furthermore
FC04-100 showed a noticeable antibacterial activity on its own (MIC of 16 microgmL) against the
MRSA strains USA100 USA300 and COL (Table 2) FC04-100 was also synergistic in
combination with an aminoglycoside against such MRSA strains with a 4 to 8 fold gain in the
MIC of GEN Because MRSA strains are often multiresistant to antibiotics and often carry
AMEs we examined the possibility of using a triple antibiotic combination Indeed the
combination of kanamycin and cefalexin in a proportion 32 is already approved to treat
bovine mastitis pathogens under the brand name of UbrolexinTM
(Ganiegravere 2009) This
antibiotic combination offers an extended spectrum of activity compared to each individual
drug and is expected to cover both S aureus Streptococcus uberis and E coli (Ganiere
2009 Maneke et al 2011 Silley 2012) Unfortunately the increased frequency of livestock-
associated MRSA (Garcia-Alvarez et al 2011 Graveland et al 2011 Leonard and Markey
2008) and frequent incidence of strains and species carrying AMEs (Ramirez and Tolmasky
2010) may limit the spectrum of activity of the combination of a beta-lactam and an
aminoglycoside With this in mind we showed that the use of FC04-100 in combination with
kanamycin and cefalexin improves by 32 fold the activity of this mixture against MRSA
strains carrying AMEs (Table 3) Overall the addition of FC04-100 to the aminoglycoside-
beta-lactam combination decreased the MICs of kanamycin and cefalexin below their
resistance breakpoints and this for all the MRSA strains tested
67
L monocytogenes is a highly specialized intracellular pathogen (Hamon et al 2006) causing
life-threatening infections especially in pregnant women (Janakiraman 2008) L
monocytogenes in a foodborne pathogen and because of its intracellular life cycle and
propagation it is able to hide from host defenses and antibiotic treatment Similarly S aureus
although not primarily recognized as a typical intracellular pathogen is also able to enter and
survive in host cells (Lowy 2000) Moreover the ability of S aureus to convert to the SCV
phenotype showing increased biofilm production improved adherence to host cells and tissues
as well as increased intracellular persistence allows this pathogen to cause chronic and
difficult to treat infections (Garcia et al 2013 Mitchell and Malouin 2012) Discovering
antibacterial drugs able to act in the intracellular compartment is generally recognized as a
difficult challenge Previously we demonstrated that steroidal alkaloids such as TO had such
an ability and could act on intracellular S aureus SCVs (Mitchell et al 2011) and here we
demonstrated that this was also the case for the TO analog FC04-100 (Fig 6) In addition we
also show in the present study that FC04-100 greatly improves the bactericidal activity of
aminoglycosides such as GEN against bacteria embedded in biofilms Biofilms are
recognized to be a major hindrance to antibiotic action and are responsible for persistent
colonization in many diseases (Costerton et al 1999) and in the food industry (Srey et al
2013) The intracellular and intrabiofilm antibacterial properties of FC04-100 warrant further
investigation of this compound and this class of molecules in general
In conclusion we showed in this study that the spectrum of activity of TO is related to the
Firmicutes division and more precisely to the order of the Bacillales TO possesses an
68
antibacterial activity against SCVs and a synergistic activity with animoglycoside antibiotics
against prototypic strains The novel TO analog FC04-100 showed very promising new
characteristics that include a much improved bactericidal effect on L monocytogenes and
killing of bacteria in biofilms
ACKNOWLEDGEMENTS
This study was supported by an operation grant from Cystic Fibrosis Canada to FM and by a
team grant from the Fonds Queacutebeacutecois de la Recherche sur la Nature et les Technologies
(FQRNT) to FM KB and EM
REFERENCES
Atalla H Gyles C Jacob CL Moisan H Malouin F Mallard B 2008 Characterization of
a Staphylococcus aureus small colony variant (SCV) associated with persistent bovine mastitis
Foodborne Pathog Dis 5785ndash799
Bad Bug Book Foodborne pathogenic microorganisms and natural toxins 2012 Second
edition Lampel KA Al-Khaldi S Cahill SM (Ed) Published by the Center for Food Safety and
Applied Nutrition of the Food and Drug Administration (FDA) US Department of Health and
Human Services pp 87-103
Beierlein JM Anderson AC 2011 New developments in vaccines inhibitors of anthrax
toxins and antibiotic therapeutics for Bacillus anthracis Curr Med Chem 185083-5094
69
Brouillette E A Martinez B J Boyll N E Allen and F Malouin 2004 Persistence of
a Staphylococcus aureus small-colony variant under antibiotic pressure in vivo FEMS
Immunol Med Microbiol 4135ndash41
Bryan L E and S Kwan 1981 Aminoglycoside-resistant mutants of Pseudomonas
aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob
Agents Chemother 19958ndash964
Ceri H Olson M Morck D Storey D Read R Buret A Olson B 2001 The MBEC Assay
System multiple equivalent biofilms for antibiotic and biocide susceptibility testing Methods
Enzymol 337377-385
Chagnon F I Guay M-A Bonin G Mitchell K Bouarab F Malouin E Marsault
2014 Unraveling the structure-activity relationship of tomatidine a steroid alkaloid with
antibiotic properties against persistent forms of Staphylococcus aureus Eur J Med Chem (In
Press EJMECH-S-13-01625-3)
Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial
Susceptibility Testing Twenty-First Informational Supplement M100-S21 CLSI Wayne PA
USA 2011
70
Costerton JW Stewart PS Greenberg EP 1999 Bacterial biofilms a common cause of
persistent infections Science 284 1318-1322
Davis BD 1982 Bactericidal synergism between beta-lactams and aminoglycosides
mechanism and possible therapeutic implications Rev Infect Dis 4237-245
Ganiegravere JP Denuault L 2009 Synergetic interaction between cefalexin and kanamycin in
Muller-Hinton broth medium and in milk J appl microbiol 107 117-125
Garcia LG S Lemaire B C Kahl K Becker R A Proctor O Denis P M Tulkens
F Van Bambeke 2013 Antibiotic activity against small-colony variants of Staphylococcus
aureus review of in vitro animal and clinical data J Antimicrob Chemother 68 1455ndash1464
Garcia-Alvarez L M T Holden H Lindsay C R Webb D F Brown M D Curran
E Walpole K Brooks D J Pickard C Teale J Parkhill S D Bentley G F Edwards
E K Girvan A M Kearns B Pichon R L Hill A R Larsen R L Skov S J Peacock
D J Maskell and M A Holmes 2011 Methicillin-resistant Staphylococcus aureus with a
novel mecA homologue in human and bovine populations in the UK and Denmark a
descriptive study Lancet Infect Dis 11595-603
Gomaa OM Momtaz OA 2007 16 rRNA characterization of Bacillus isolate and its
tolerance profile after subsequent subculturing Arab J Biotech 10 107-116
71
Graveland H Duim B van Duijkeren E Heederik D Wagenaar JA 2011 Livestock-
associated methicillin-resistant Staphylococcus aureus in animals and humans Int J Med
Microbiol 301630-634
Guillet C Join-Lambert O Le Monnier A Leclercq A Mechaiuml F Mamzer-Bruneel MF
Bielecka MK Scortti M Disson O Berche P Vazquez-Boland J Lortholary O Lecuit M
2010 Human listeriosis caused by Listeria ivanovii Emerg Infect Dis 16136-138
Hamon M H Bierne P Cossart 2006 Listeria monocytogenes a multifaceted model Nat
Rev Microbiol 4 423-434
Harrison JJ Stremick CA Turner RJ Allan ND Olson ME Ceri H 2010 Microtiter
susceptibility testing of microbes growing on peg lids a miniaturized biofilm model for high-
throughput screening Nat Protoc 51236-1254
Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y
Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T Baba T 2013 Genomic
Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
Horsburgh MJ Aish JL White IJ Shaw L Lithgow JK Foster SJ 2002 SigmaB
modulates virulence determinant expression and stress resistance characterization of a
72
functional rsbU strain derived from Staphylococcus aureus 8325-4 J Bacteriol 1845457-
5467
Janakiraman V 2008 Listeriosis in Pregnancy Diagnosis Treatment and Prevention Rev
Obstet Gynecol 1 179ndash185
Leonard FC Markey BK 2008 Methicillin-resistant Staphylococcus aureus in animals a
review Vet J 175 27-36
Lowy FD 2000 Is S aureus an intracellular pathogen Trends Microbiol 8341-342
Magalhatildees L Nitschke M 2013 Antimicrobial activity of rhamnolipids against Listeria
monocytogenes and their synergistic interaction with nisin Food Control 29 138-142
Maneke E Pridmore A Goby L Lang I 2011 Kill rate of mastitis pathogens by a
combination of cefalexin and kanamycin J Appl Microbiol 110184-190
Mitchell G Brouillette Eacute Lalonde-Seacuteguin D Asselin AE Lebeau Jacob C Malouin F
2009 A role for sigma factor B in the emergence of Staphylococcus aureus small-colony
variants and elevated biofilm production resulting from an exposure to aminoglycosides
Microb Pathog 4818-27
73
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K Malouin F 2011
Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic
fibrosis airway epithelial cells Antimicrob Agents and Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM Malouin F 2011 Infection of Polarized
airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is
increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function
and is influenced by NF- κ B Infect Immun 793541-3551
Mitchell G Lafrance M Boulanger S Lalonde-Seacuteguin D Guay I Gattuso M Marsault
Eacute Bourab K Malouin F 2012 Tomatidine acts in synergy with aminoglycoside antibiotics
against multiresistant Staphylococcus aureus and prevents virulence gene expression J
Antimicrob Chemother 67 559-568
Mitchell G Lalonde-Seacuteguin D Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S
Malouin F 2010 Staphylococcus aureus sigma B-dependent emergence of small-colony
variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-
heptylquinoline-N-oxide BMC Microbiology 10 33-48
Mitchell G and F Malouin 2012 Outcome and prevention of Pseudomonas aeruginosa-
Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic
Fibrosis InTech Open Access Publisher ISBN 979-953-307-059-8
74
Moisan H Brouillette E Jacob CL Langlois-Beacutegin P Michaud S Malouin F 2006
Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated
from cystic fibrosis patients is influ- enced by SigB J Bacteriol 18864ndash76
Moskowitz SM Foster JM Emerson J Burns JL 2004 Clinically feasible biofilm
susceptibility assay for isolates of Pseudomonas aeruginosa from patients with cystic fibrosis
J Clin Microbiol 421915-1922
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M Peters G 2006
Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent
infections Nat Rev Micro- biol 4295ndash305
Ramirez MS M E Tolmasky 2010 Aminoglycoside modifying enzymes Drug Res
Updates 13 151ndash171
Ruiz-Rubio M Peacuterez-Espinosa A Lairini K Roldagraven-Arjona T Dipietro A Anaya N
2001 Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi p 293ndash326 In
A Rahman (ed) Studies in natural products chemistry vol 25 Elsevier Oxford United
Kingdom
75
Sendi P Proctor RA 2009 Staphylococcus aureus as an intracellular pathogen the role of
small colony variants Trends Microbiol 1754-58
Silley P Goby L Pillar CM 2012 Susceptibility of coagulase-negative staphylococci to a
kanamycin and cefalexin combination J Dairy Sci 953448-3453
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C Osbourn A
2006 Dual effects of plant steroidal alkaloids on Saccharomyces cerevisiae Antimicrob
Agents Chemother 502732ndash2740
Srey S Jahid IK Ha S-D 2013 Biofilm formation in food industries A food safety concern
Food control 31572-585
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D
Roth J Proctor RA Becker K Peters G Loumlffler B 2011 Staphylococcus aureus
phenotype switching an effective bacterial strategy to escape host immune response and
establish a chronic infection EMBO Mol Med 3129ndash141
Vuong C Otto M 2002 Staphylococcus epidermidis infections Microbes Infect 4481-489
Witte W Strommenger B Stanek C Cuny C 2007 Methicillin-resistant Staphylococcus
aureus ST398 in Humans and Animals Central Europe Emerg Infect Dis 13 255ndash258
76
CHAPITRE 3
MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
31 Obtention des mutants reacutesistants
Afin de nous aider agrave mieux deacutefinir la cible et le mode drsquoaction de la tomatidine (TO)
nous avons forceacute lrsquoapparition de reacutesistances chez Staphylococcus aureus Comme
souche controcircle prototype nous avons choisi ATCC 29213 puisque crsquoest une souche
tregraves freacutequemment utiliseacutee pour des expeacuteriences avec des antibiotiques et nous avons
choisi la souche NewbouldΔhemB comme S aureus laquo small colony variant raquo puisque
crsquoest une souche SCV qui est stable et bien connue nrsquoayant pas de capaciteacute de
reacuteversion Par passages successifs dans une concentration sous inhibitrice
drsquoantibiotique (technique de CMI (concentration minimale inhibitrice)) (CLSI 2011)
nous avons obtenu des mutants deacutemontrant une reacutesistance croissante agrave lrsquoantibiotique
au fil des passages (Figure 1) Nous avons obtenu des mutants ATCC 29213
reacutesistants agrave la gentamicine (GEN) et agrave la GEN lorsque combineacutee agrave la TO ainsi que
des mutants reacutesistants NewbouldΔhemB reacutesistants agrave la GEN seule agrave la TO seule et
agrave la TO lorsque combineacute agrave la GEN
Certains passages ont ensuite eacuteteacute seacutelectionneacutes parce qursquoils deacutemontraient une hausse
plus marqueacutee de leur CMI Ces passages ont eacuteteacute choisis car nous supposions que
ces hausses marqueacutees de CMI correspondraient donc agrave des hausses de la
reacutesistance de cette souche agrave lrsquoantibiotique testeacute et que cette reacutesistance pouvait ecirctre la
conseacutequence drsquoune ou plusieurs mutations en lien avec la cible ou le mode drsquoaction
de lrsquoantibiotique preacutesent Trois agrave quatre passages (ou paliers de reacutesistance) par
condition ont eacuteteacute seacutelectionneacutes Nous avons ensuite reacuteensemenceacute le laquo pool raquo
bacteacuterien de chacun des passages choisis afin de pouvoir isoler 3 isolats ayant la
mecircme CMI (ou tregraves similaire (CMI vraies)) que celui du laquo pool raquo bacteacuterien des
77
passages drsquoougrave ils provenaient Soixante quatre mutants reacutesistants ont ainsi eacuteteacute
seacutelectionneacutes en vue drsquoecirctre seacutequenceacutes (Figure 2)
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt 1
6
CM
I v
raie
s
02
5
CM
I v
raie
s
02
5-1
A
78
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
en
pr
eacutes
en
ce
de
ge
n (
4 micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
01
25
1
6
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
8
CM
I v
raie
s
2
4
CM
I v
raie
s
2
B
C
79
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
en
pr
eacutes
en
ce
de
TO
(8
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
1
2
3
4
5
CM
I v
ra
ies
4
CM
I v
ra
ies
2
CM
I v
ra
ies
0
5
1
CM
I v
ra
ies
2
4
Figure 1 Graphiques des CMI pour chacun des passages pour (A) la tomatidine sur
NewbouldΔhemB (B) pour la tomatidine en preacutesence de gentamicine sur
NewbouldΔhemB (C) pour la gentamicine sur S aureus ATCC 29213 (D) pour la
gentamicine en preacutesence de tomatidine sur S aureus ATCC 29213
32 Seacutequenccedilage et analyses bioinformatiques
Une extraction drsquoADN geacutenomique a ensuite eacuteteacute effectueacutee sur chacun des isolats en
utilisant le Kit drsquoextraction drsquoADN geacutenomique miniKit de Qiagen (QIAamp DNA minikit)
(Li et al 2003) Puisque leurs seacutequences geacutenomiques nrsquoeacutetaient pas connues nous
avons eacutegalement extrait lrsquoADN geacutenomique de nos deux souches de reacutefeacuterences
ATCC29213 et NewbouldΔhemB (provenant de quatre colonies isoleacutees chacune)
afin drsquoavoir les seacutequences de comparaison pour nos mutants (Figure 2)
D
80
Ne
wm
an
(g
en
om
ede r
ef h
ttpw
ww
ncbinlm
nih
govn
uccore
150373012)
AT
CC
29
213
(A p
art
ir d
e 4
AT
CC
29
21
3 r
eco
nstr
uire
AT
CC
29
21
3 c
om
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
5
10
7
10
8 1
09
)
Ne
wbo
uld
Δhe
mB
(A p
art
ir d
e 4
Ne
wb
ou
ldh
em
Bre
co
nstr
uire
Ne
wb
ou
ldh
em
Bcom
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
1
agraveG
EN
2
5 (
CM
I2
)
2
7 (
CM
I2
)
2
8 (
CM
I2
)
AT
CC
29
21
3
Pa
lier
de
reacutesis
tance
1
agravecom
bo
29
(C
MI
05
)
3
0 (
CM
I 0
5
3
1 (
CM
I 0
5)
Suite
fe
uill
e s
uiv
ante
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agraveG
EN
3
(C
MI
4)
4
(C
MI
4)
3
9 (
CM
I4
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agraveG
EN
4
6 (
CM
I8
)
4
7 (
CM
I8
)
4
8 (
CM
I8
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agravecom
bo
1
1 (
CM
I 2
)
1
2 (
CM
I 2
)
2
1 (
CM
I 2
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agravecom
bo
5 (
CM
I 2
-4)
3
6 (
CM
I 2
-4)
3
7 (
CM
I 2
-4)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
4
agravecom
bo
3
8 (
CM
I4
)
6
7 (
CM
I 4
)
81
Ne
wb
ould
Δhem
B(A
part
ir d
e 4
Ne
wb
ou
ldΔ
he
mB
reco
nstr
uire
Ne
wb
ou
ldΔ
hem
Bco
mp
let e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveG
EN
24
(C
MI
64
)
4
1 (
CM
I 1
6)
4
2 (
CM
I 6
4)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveT
O
95
(C
MI
02
5)
9
6 (
CM
I 0
25)
9
7 (
CM
I 0
25)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agravecom
bo
33
34
35
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveG
EN
49
(C
MI
12
8)
5
1 (
CM
I 1
28)
5
2 (
CM
I 1
28)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agraveG
EN
7
(C
MI
128
)
4
3 (
CM
I 1
28)
4
5 (
CM
I1
28
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agraveT
O
6
4 (
CM
I gt
16)
6
5 (
CM
I gt
16)
6
6 (
CM
I gt
16)
75
76
77
79
80
81
82
83
84
85
86
87
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agravecom
bo
15
16
18
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agravecom
bo
59
60
61)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agravecom
bo
54
56
58)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveT
O
90
(C
MI
02
5)
9
1 (
CM
I 0
25)
9
3 (
CM
I 0
5)
Ne
wb
ou
ldh
em
B
palie
r d
e r
eacutesis
tance
3
agraveT
O
11
0 (
CM
I 1
)
1
12 (
CM
I 0
25
-05
)
1
13 (
CM
I 0
5)
Fig
ure
2
Org
an
igra
mm
e d
es
mu
tan
ts r
eacutes
ista
nts
seacute
qu
en
ceacutes
se
lon
leu
r pa
lier
de
reacute
sis
tan
ce e
t a
vec leu
r n
um
eacutero
drsquoeacute
chan
tillo
n
co
rre
spo
ndan
t
82
Un minimum de 20 ngmicrol drsquoADN geacutenomique de chacun des isolats a ensuite eacuteteacute
achemineacute au laboratoire du Pr Seacutebastien Rodrigue (Deacutepartement de biologie Faculteacute
des sciences Universiteacute de Sherbrooke) qui a effectueacute la librairie de fragments Pour
se faire il y a eu dans un premier temps digestion enzymatique de lrsquoADN afin
drsquoobtenir des fragments drsquoADN double brin Ensuite tous les fragments ont eacuteteacute
purifieacutes suivant la meacutethode SPRI (Solid-Phase Reversible Immobilization) Il srsquoagit
drsquoune meacutethode simple et tregraves efficace drsquoisolation drsquoacides nucleacuteiques sur microplaque
(Rodrigue S et al 2010) Les acides nucleacuteiques cibles sont immobiliseacutes sur des
microparticules paramagneacutetiques dans des conditions tampons-speacutecifiques La
flexibiliteacute du systegraveme se fait par lrsquoutilisation de diffeacuterents tampons de liaison (laquo binding
buffer raquo) qui modifient le type et la grandeur des acides nucleacuteiques immobiliseacutes Les
contaminants peuvent ainsi ecirctre retireacutes de lrsquoeacutechantillon sans besoin de filtration ou
centrifugation (Beckman Coulter website 2013 DeAngelis et al 1995) Suite agrave la
purification par meacutethode SPRI lrsquoeacutequipe du Pr Rodrigue a obtenu des fragments
drsquoADN de ~250pb auxquels ont eacuteteacute ajouteacutes par ligation des adapteurs aux deux
extreacutemiteacutes des fragments Dans un deuxiegraveme temps des codes-bars speacutecifiques agrave
chaque eacutechantillon drsquoADN geacutenomique ont eacutegalement eacuteteacute lieacutes agrave chacun des fragments
Alors que les adapteurs permettent aux fragments de se lier agrave une matrice solide afin
de pouvoir ecirctre amplifieacutes les codes-bars permettront de classer chacun des
fragments selon leur appartenance agrave un eacutechantillon drsquoADN geacutenomique speacutecifique
83
Figure 3 Eacutetape du seacutequenccedilage Illumina
La librairie de fragments a ensuite eacuteteacute envoyeacutee chez Geacutenome-Queacutebec afin que
chacun des geacutenomes puisse ecirctre seacutequenceacute Geacutenome ndashQueacutebec utilise la meacutethode
Illumina comme outil de seacutequenccedilage (Figure 3) La grande innovation de la meacutethode
Illumina est lrsquoamplification des fragments par liaison de ceux-ci agrave une surface solide
(Flowcell) ce qui permet par un processus de ponts drsquoamplification de geacuteneacuterer plus
drsquoun million de laquo clusters raquo comprenant eux-mecircme au plus un millier de copies de la
moleacutecule originale de deacutepart (solid-phase PCR) Chaque regroupement de fragments
provenant de lrsquoamplification drsquoun mecircme fragment original est tregraves pregraves les uns des
Modifieacute de Quail MA et al 2008
84
autres formant une zone distincte drsquoun regroupement de fragments agrave un autre
Lrsquoincorporation des nucleacuteotides eacutetiqueteacutes preacutealablement avec des moleacutecules
fluorescentes speacutecifiques est deacutetecteacutee suite agrave leur excitation par un laser et les
images sont compileacutees pour produire la seacutequence de bases pour chaque fragment
ADN (Figure 4) Les nouvelles seacuteries de bases appeleacutees laquo readsraquo seront ensuite
reacuteassembleacutees (aligneacutes) en se basant sur un geacutenome de reacutefeacuterence connu (appeleacute
laquo resequencing raquo) (crsquoest notre cas ici) ou en absence de geacutenome de reacutefeacuterence
(appeleacute laquo De novo sequencing raquo) Au final lrsquoensemble des laquo reads raquo aligneacutes
correspondra agrave la seacutequence de chaque chromosome de lrsquoeacutechantillon drsquoADN
geacutenomique de deacutepart
85
Figure 4 Meacutethode drsquoamplification par la meacutethode Illumina
Suite au seacutequenccedilage chez Geacutenome ndashQueacutebec nous avions en main les seacutequences
geacutenomiques de tous nos mutants et de nos souches de reacutefeacuterence sous forme de
Tireacute de BiteSizeBio httpbitesizebiocom13546sequencing-by-synthesis-
explaining-the-illumina-sequencing-technology consulteacute le 3 deacutec 2013
86
contigs Nous avions donc le casse-tecircte mais en piegraveces deacutetacheacutees pecircle-mecircle et
sans avoir la photo de lrsquoimage de reacutefeacuterence sur laquelle se baser pour pouvoir
compleacuteter le casse-tecircte
1- Lrsquoassemblage
Les diffeacuterents geacutenomes avaient eacuteteacute preacutealablement fractionneacutes aleacuteatoirement
en petits fragments de 250 pb et amplifieacutes en vue du seacutequenccedilage Suite au
seacutequenccedilage les seacutequences de chaque fragment (appeleacute laquo reads raquo) sont ensuite
assembleacutees en contigs crsquoest-agrave-dire des seacutequences plus ou mois longues continues
et ordonneacutees geacuteneacutereacutees par lassemblage des clones dune bibliothegraveque geacutenomique
qui se chevauche La qualiteacute de lrsquoassemblage en contigs deacutepend de la capaciteacute de
superposition des laquo reads raquo Parce que la digestion enzymatique de lrsquoADN se fait de
faccedilon aleacuteatoire et se fait sur plusieurs copies drsquoADN chaque portion du geacutenome est
repreacutesenteacutee plusieurs fois dans diffeacuterents modegraveles de fragments Donc les
seacutequences identiques des diffeacuterents fragments (ou laquo reads raquo) peuvent ecirctre
superposeacutees par chevauchegravement et les laquo reads raquo superposeacutes peuvent ecirctre
assembleacutes en contigs par un logiciel drsquoassemblage En ce qui nous concerne cette
eacutetape a eacuteteacute faite agrave lrsquoaide du programme de bio-informatique Newbler (Zhang et al
2012) Lrsquoassemblage geacutenomique peut ecirctre probleacutematique pour les informaticiens car
les geacutenomes contiennent des seacutequences qui se reacutepegravetent et ces reacutepeacutetitions peuvent
ecirctre assez longues en fonction des organismes Suite agrave lrsquoassemblage les geacutenomes
sont donc sous forme de contigs
2- Lrsquoalignement
En se basant sur une seacutequence geacutenomique de reacutefeacuterence (notre reacutefeacuterence est
Staphylococcus aureus str Newman (~ 288Mb)
( httpwwwncbinlmnihgovgenome154project_id=58839) ) le programme de bio-
informatique ABACAS permet de rapidement aligner ordonner et orienter les contigs
Selon les besoins ce qui nrsquoa pas eacuteteacute notre cas ABACAS permet eacutegalement de
87
visualiser et designer les amorces permettant de fermer les gaps (espaces) entre les
contigs en faisant des PCR (Assefa et al 2009)
3- La fermeture des gaps
La fermeture des gaps nrsquoa pas eacuteteacute faite avec le programme ABACAS mais
avec lrsquoaide du programme de bio-informatique GapFiller Lrsquoutilisation de GapFiller
pour la fermeture des gaps possegravede 3 principaux avantages Premiegraverement le
programme prend en compte lrsquoestimation de la grandeur du gap agrave fermer donc il
exclut automatiquement les fermetures de gaps erroneacutes qui sont plus courts ou plus
longs que la grandeur estimeacutee Deuxiegravemement il nrsquoessaie pas de remplir un gap
avec un assemblage de laquo reads raquo en contigs mais cherche plutocirct agrave allonger les
contigs agrave partir de leurs extreacutemiteacutes jusqursquoagrave obtenir des seacutequences superposeacutees en
passant par la creacuteation de K-mer (seacuteparation des reads en plus petits fragments) Et
troisiegravemement il prend en compte que les extreacutemiteacutes des contigs sont souvent
source de mauvais assemblage et reacute eacutevalue ces extreacutemiteacutes pendant la fermeture des
gaps (Boetzer et Pirovano 2012)
4- Lrsquoannotation des geacutenomes
Suite agrave la fermeture des gaps il a fallu annoter les geacutenomes en se basant sur
lrsquoannotation du geacutenome de reacutefeacuterence Staphylococcus aureus str Newman RATT est
un outil de transfert rapide drsquoannotation (Rapid Annotation Transfert Tool) qui a eacuteteacute
deacuteveloppeacute afin de fournir rapidement une annotation preacutecise pour des geacutenomes
inconnus en se basant sur des geacutenomes deacutejagrave annoteacutes comme reacutefeacuterences (Figure 5)
La meacutethode transfert les annotations drsquoune reacutefeacuterence de haute qualiteacute agrave un nouveau
geacutenome sur la base de la conservation des synteny (ordre des gegravenes) (Otto et al
2011) Suite agrave lrsquoannotation les gegravenes tant chez les mutants que chez les souches
parentes portent les noms auxquels ils correspondent chez Staphylococcus aureus
str Newman
88
Figure 5 Corrections du programme RATT sur les annotations transfeacutereacutees Les
annotations de H37Rv ont eacuteteacute transfeacutereacutees sur la seacutequence F11 (bleu pacircle) ont eacuteteacute corrigeacutees
(vert) et ensuite compareacutees agrave lrsquoannotation de la souche existante F11 EMBL (jaune) (A et B)
La correction des codons ldquostartrdquo et ldquostoprdquo respectivement Dans une situation de laquo mapping raquo
plus complexe (C) ougrave les cadres de lecture sont montreacutes pour plus de clarteacute RATT
cartographie une large seacutequence codante (CDS) de H37Rv vers un locus dans F11 qui inclue
plusieurs codons laquo stop raquo laquo in-frame raquo En inseacuterant un deacutecalage du cadre de lecture (ie un
pseudogegravene) la traduction conceptuelle est conserveacutee Ceci diffegravere avec les deux gegravenes se
superposant preacutedis comme faisant partie du geacutenome F11
Tireacute de Otto TD et al 2011
89
321 Identification des mutations en lien avec la tomatidine
Apregraves avoir obtenu une seacutequence consensus agrave partir des seacutequences geacutenomiques
des 4 copies de chacune des souches parente S aureus ATCC 29213 et S aureus
NewbouldΔhemB les diffeacuterences geacutenomiques entre les souches parentes et le
geacutenome de reacutefeacuterence S aureus str Newman ont eacuteteacute eacutelimineacutees afin que ces
diffeacuterences nrsquoapparaissent pas comme des mutations lors de la comparaison des
seacutequences geacutenomiques des mutants avec celles de leur souche parente ces
diffeacuterences eacutetant plutocirct attribuables agrave la diffeacuterence qursquoil y a entre S aureus str
Newman et la souche parente et non pas agrave des mutations en lien avec la reacutesistance
des mutants
Le service bio-informatique du deacutepartement de biologie (Faculteacute des sciences
Universiteacute de Sherbrooke) a ensuite produit un tableau contenant pour chacun des
mutants tous les gegravenes muteacutes apregraves comparaison avec leur souche parentes Agrave cette
eacutetape on pouvait donc savoir dans quel gegravene se situaient les mutations quelle eacutetait
lrsquoeffet de cette mutation (mutation synonyme deacuteleacutetion insertion SNP) et en principe
srsquoil srsquoagissait pour un mecircme gegravene muteacute de la mecircme mutation puisque dans ce
tableau pour un mecircme gegravene chaque ligne correspond agrave des mutations diffeacuterentes
(Figure 6)
Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4
Figure 6 Exemple drsquoune partie du tableau compilation des mutations chez
les mutants ATCC 29213 reacutesistants agrave la GEN seule et agrave la GEN (+TO)
90
Agrave lrsquoaide de ce tableau (Figure 6) jrsquoai chercheacute les mutations qui pouvaient ecirctre
attribuables agrave la TO Chez les mutants S aureus ATCC 29213 reacutesistants jrsquoai
chercheacute les gegravenes qui eacutetaient muteacutes chez tous les mutants reacutesistants agrave la
combinaison GEN (+TO) ou dont la mutation apparaissait graduellement au fil des
paliers de reacutesistance mais dont ce mecircme gegravene nrsquoeacutetait pas muteacute chez les mutants
reacutesistants agrave la GEN seule Je pouvais donc estimer que cette mutation chez les
reacutesistants agrave la combinaison eacutetait en lien avec la preacutesence de la TO dans le milieu
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct
Afin de mrsquoaider dans ma recherche jrsquoai eu lrsquoaide du Pr Pierre-Eacutetienne Jacques
(Deacutepartement de biologie Faculteacute des sciences Universiteacute de Sherbrooke)qui a
mis les donneacutees de ce tableau dans un tableau croiseacute dynamique (TCD) ce qui a
permis de filtrer les donneacutees selon des critegraveres bien preacutecis me permettant drsquoarriver agrave
mes fins plus facilement Dans un premier temps un TCD a eacuteteacute creacuteeacute ougrave pour
chacun des mutants chaque gegravene ayant au moins une mutation eacutetait indiqueacute par un
laquo 1 raquo alors qursquoun laquo 0 raquo eacutetait marqueacute si ce gegravene nrsquoeacutetait pas muteacute Dans un second
temps la quantiteacute de laquo 1 raquo par palier de reacutesistance (correspondant au nombre de
mutants de ce palier posseacutedant une mutation dans ce gegravene) eacutetait indiqueacutee par un
chiffre allant de 0 pour laquo aucun mutant de ce palier de reacutesistance ne posseacutedait de
mutation dans ce gegravene raquo agrave 1 2 ou 3 selon le nombre de mutants de ce palier ayant
ce gegravene muteacute Jrsquoai ensuite pu soumettre le TCD agrave des filtres approprieacutes afin de faire
ressortir les gegravenes drsquointeacuterecircts selon des critegraveres de seacutelection preacutecis (Figures 7-8)
91
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
1
00
33
32
1
1
11
4
5
0
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
10
03
33
21
11
14
50
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_04
12
lpl9
nm
20
12
32
23
92
44
1
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
NW
MN
_00
46
_2
10
13
21
37
24
31
NW
MN
_09
55
_2
01
23
11
37
24
21
NW
MN
_13
83
gpsA
21
02
31
13
72
42
1
NW
MN
_03
39
_1
11
21
31
37
34
42
NW
MN
_15
97
dn
aE2
10
22
30
37
23
31
NW
MN
_23
30
_2
10
23
20
37
23
21
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_2
07
0_
10
01
23
21
81
45
0
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_10
28
_1
22
12
32
58
34
54
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
A B C
92
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
39
8h
sdM
31
33
33
27
11
34
54
NW
MN
_04
26
_3
31
33
32
71
13
45
4
NW
MN
_05
24
sdrD
32
33
33
28
11
34
55
NW
MN
_07
39
_3
33
33
32
91
13
45
6
NW
MN
_11
64
hsl
U3
33
33
32
91
13
45
6
NW
MN
_05
23
sdrC
22
33
32
27
10
34
45
NW
MN
_20
61
fmtB
23
23
32
27
10
34
45
NW
MN
_04
03
lpl1
nm
33
22
33
28
10
34
55
NW
MN
_13
45
ebh
13
32
33
31
81
03
44
5
NW
MN
_16
49
_3
32
33
31
81
03
44
5
NW
MN
_17
00
hsd
M3
23
33
22
81
03
44
5
NW
MN
_00
42
_3
33
33
22
91
03
44
6
NW
MN
_05
80
_3
33
33
31
91
03
44
6
NW
MN
_13
44
ebh
23
31
32
22
79
34
44
NW
MN
_13
75
aro
A3
13
23
31
79
34
44
NW
MN
_23
97
fnb
B2
32
23
22
79
34
45
NW
MN
_04
01
_3
32
23
31
89
34
45
NW
MN
_07
33
_3
32
32
31
89
34
45
NW
MN
_17
45
_2
33
22
32
89
34
56
NW
MN
_01
66
coa
33
32
32
29
93
44
6
NW
MN
_05
25
sdrE
33
32
33
19
93
44
6
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D Fig
ure
7
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez A
TC
C 2
921
3 d
an
s le
TC
D p
ar
dif
feacutere
nts
fil
tre
s
(A)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts la
ge
nta
mic
ine
ge2
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(C
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts d
an
s le
pa
lier
1
de
reacute
sis
tan
ce
agravela
ge
nta
mic
ine
le1
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
et
nom
bre
de m
uta
nts
reacute
sis
tan
ts agrave
la la
gen
tam
icin
e (
+T
O)
de
s 2
de
rnie
rs p
alie
rs d
e r
eacutesis
tan
ce
ge5
(D
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
7 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
ge
nta
mic
ine
(+
TO
) ge
9
93
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_16
29
ccp
A0
01
33
31
50
00
01
24
0
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_21
28
rpsM
11
12
21
12
22
00
31
74
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
NW
MN
_25
29
clfB
33
03
33
80
11
26
17
4
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e reacute
sist
ance
agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
On
b m
uta
nts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
16
29
ccp
A0
01
33
31
50
00
01
24
0N
WM
N_
21
28
rpsM
11
12
21
12
22
00
31
74
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
A B C
94
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
04
6_
1
01
00
28
22
32
2
10
9
NW
MN
_17
33
prs
A0
11
12
01
12
12
22
14
7
NW
MN
_19
23
_0
11
31
15
13
22
21
08
NW
MN
_20
37
_0
02
32
07
23
11
21
27
NW
MN
_22
77
_1
02
21
11
02
12
33
14
8
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
01
43
_0
03
00
04
23
33
34
11
NW
MN
_0
40
6lp
l3n
m2
01
00
03
12
22
33
7
NW
MN
_1
65
2_
00
10
00
62
12
31
68
NW
MN
_2
01
2_
00
00
00
32
33
30
31
1N
WM
N_
20
15
mn
aA0
00
00
01
23
33
01
11
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
o
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D E Fig
ure
8
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez N
ew
bo
uld
Δh
em
Bd
an
s l
e T
CD
par
dif
feacutere
nts
fil
tres
(A)
Nom
bre
de
mu
tan
ts
reacutesis
tan
ts agrave
la tom
atid
ine
ge15
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atid
ine
ge15
et no
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
(C)
No
mbre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la tom
atid
ine
ge15
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3(
D)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atidin
ege
10
et no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
tom
atid
ine
(+G
EN
) ge
7
(E)
No
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la to
ma
tid
ine
le6
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
95
Le TCD mrsquoa permis de rapidement identifier les gegravenes drsquointeacuterecircts Agrave lrsquoaide du site CMR
(Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) 2013) jrsquoai
ensuite veacuterifieacute la fonction des gegravenes identifieacutes afin de confirmer leur inteacuterecirct (Tableau
1)
Tableau 1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants et (B) chez les mutants NewbouldΔhemB reacutesistants
Gegravene Alias Fonction
NWMN_0811 NADH dh complexe 1 chaine de transport des eacutelectrons
NWMN_2070 _membrane prot Putative- Toxin production+resistance drug transporteur-
similar to multidrug transporteur 85
NWMN_0635 _membrane prot Putative- Bordeacute par AraC(reacutegulateur opeacuteron arabinose) ou
SsaA-likeprot(lieacute au quorum sensing RNAIII(secretory antigen)
NWMN_2084 _truncated transposase for Is1272- mobile+extrachromosomal element
fonctiontransposon fonction
NWMN_0928 qoxC quinol oxidase polypeptide IIIcomplexe 4 chaine de transport des eacutelectrons
NWMN_0739 _ conserved hypothetical prot fonction inconnues
NWMN_1164 hslU ATP-dependent protease ATP-binding subunit
NWMN_0524 sdrD Ser-Asp rich fibrinogenbone sialoprotein-binding protein sdrD
NWMN_0042 _ lipoprotein putativecell enveloppe- lipoprot en tendem
NWMN_0580 _conserved hypothetical prot fonction inconnuesDNA metabolism DNA
replication recombination and repair
NWMN_1745 _
NWMN_0166 coa coagulase precursor
NWMN_0525 sdrE Ser-Asp rich fibrinogenbone sialoprotein-binding protein SdrE
A
96
Apregraves avoir aligneacute speacutecifiquement une par une chacune des seacutequences (Fasta) des
gegravenes drsquointeacuterecirct de chaque mutant avec leur souche parente jrsquoai utiliseacute le programme
de visualisation et de comparaison de seacutequences ClustalW2 (ClustalW2 2013) afin
de confirmer que la mutation eacutetait une vraie mutation et non une erreur de
seacutequenccedilage ou drsquoalignement et que la mutation eacutetait identique chez tous les mutants
ayant le gegravene drsquointeacuterecirct muteacute Cette eacutetape a permis de restreindre encore la liste des
gegravenes pouvant ecirctre en lien avec la cible ou le mode drsquoaction de la TO (Figure 9)
Gene Alias Fonction
NWMN_0896 _conserved hypothetical prot fonction inconnue- comK family prot-
regulation DNA interaction- ORFIDresssemble agrave facteur de transcription
NWMN_0996 _ conserved hypothetical prot fonction inconnue- prophage fonction
NWMN_1025 _ phage major tail protprophage fonction
NWMN_1629 ccpAcatabollite control protA virulence determinant production and antibiotique
resistance
NWMN_1916 _ conserved hypothetical prot-fonction inconnue ou prophage prot
NWMN_2314 _drug resistance transporteur- EmrBQacA subfamily prot- toxin
production+resistance- transport and binding prot
NWMN_0046 _ hypothetical prot
NWMN_2015 mnaA UDP-Glc-Nac 2 epimerase
NWMN_2012 atpE ATP synthase subunit C
NWMN_1652 _ multidrug resistance transporter protein B
NWMN_0143 _ oligopeptide ABC transporter ATP-binding protein
B
97
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
A
B
98
Jrsquoai ensuite chercheacute le rocircle de ces gegravenes sur le site CMR afin de veacuterifier si leur
mutation pouvait expliquer la reacutesistance agrave lrsquoantibiotique (Comprehensive Microbial
Ressouce (J Craig Venter Institute (JCVI) 2013) Dans une premiegravere eacutetape le rocircle
drsquoun gegravene a eacuteteacute deacutetermineacute en allant tout simplement voir directement le rocircle de ce
gegravene chez Staphylococcus aureus str Newman Dans le cas ougrave le gegravene nrsquoavait pas
de fonction connue chez Saureus str Newman je suis alleacutee voir quelle eacutetait la
fonction de ce mecircme gegravene (et des gegravenes flanquant celui-ci) chez diffeacuterentes souches
de Staphylococcus aureus Advenant le cas ougrave malgreacute tout je nrsquoeacutetais toujours pas
capable de deacuteterminer le rocircle de ce gegravene je suis alleacutee veacuterifier si sa seacutequence pouvait
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 hellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Mutants resistants agrave GEN (4 microgml)+TO
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip 185-AATTGCATTCA-197helliphelliphellip495-ATAGCAATGG-506hellip
hellipnwmn_1629helliphelliphelliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_0143hellip
ccpAATP synthase c
ABC transp
Niveau 1
Niveau 2
Niveau 3
Niveau 4
TO +GEN (4microgml)
Figure 9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct (A) nwmn_0811 et
nwmn_0928 chez les mutants ATCC 29213 reacutesistants agrave la combinaison gentamicine
(+TO) leur souche parente ATCC 29213 (B) nwmn_1629 et nwmn_2012 chez les
mutants hemB reacutesistants agrave la TO et leur souche parente NewbouldΔhemB (C)
nwmn_1629 nwmn_2012 et nwmn_0143 chez les mutants hemB reacutesistant agrave la
tomatidine (+genta) et leur souche parente hemB
C
99
avoir une forte homologie avec un gegravene drsquoune autre espegravece Malgreacute tout certains
gegravenes portent toujours la mention laquo de fonction inconnue raquo
La meacutethodologie que nous avons employeacutee nous a permis drsquoidentifier plusieurs
gegravenes possiblement impliqueacutes dans reacutesistance de nos souches aux antibiotiques
testeacutes
1- chez Staphylococcus aureus ATCC 29213 reacutesistant agrave la gentamicine (+TO)
(Figure 9A)
- NWMN_0811 codant possiblement pour la NADH deacuteshydrogeacutenase
- qoxC (NWMN_0928) codant pour la quinol oxidase polypeptide III
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
2- chez NewbouldΔhemB reacutesistant agrave la tomatidine (Figure 9B)
- ccpA (NWMN_1629) codant pour la laquo catabolite control protein A raquo
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
3- chez NewbouldΔhemB reacutesistant agrave la tomatidine (+gentamicine) (Figure 9C)
- NWMN_0143 codant pour un ABC transporteur
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
Lrsquoanalyse de ces diffeacuterents gegravenes nous amegravene agrave suspecter la sous-uniteacute c de lrsquoATP
synthase comme eacutetant la cible potentielle de la tomatidine chez Staphylococcus
aureus
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine
Les gegravenes muteacutes pointent en grande partie vers la chaine respiratoire qursquoon appelle
aussi chaine de transport des eacutelectrons
100
La NADH deacuteshydrogeacutenase (NWMN_0811) fait partie du complexes I de la chaine de
transport de eacutelectrons alors que la quinol oxidase polypeptide III (NWMN_0928 ou
QoxC) est une composante du complex III et la sous-uniteacute c de lrsquoATP synthase (AtpE)
fait partie eacutevidement du complexe de lrsquoATP synthase ou complexe V de la chaine
respiratoire (Figure 10)
Figure 10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants
Les courbes de croissance des mutants ATCC29213 reacutesistants agrave la combinaison
GEN(+TO) appuient le fait que ces mutations affectent la chaine de transport des
eacutelectrons ce qui affecte le niveau de production drsquoATP et donc la croissance
bacteacuterienne En effet les mutants du troisiegraveme niveau de reacutesistance (qui possegravede
deux gegravenes de la chaine de transport des eacutelectrons de muteacutes) ont une croissance
plus lente que les mutants du premier niveau de reacutesistance qui eux ont un seul gegravene
dans la chaine de transport des eacutelectrons de muteacute (Figure 11)
Quinoloxidase
NADHdehydrogenase
ATPsynthetase
Proton gradient
XXX
101
Figure 11 Courbes de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison GEN+TO du premier niveau de reacutesistance (bleu) et du troisiegraveme niveau
de reacutesistance (vert)
Voici plus preacuteciseacutement comment fonctionne la chaine de transport des eacutelectrons chez
les 2 pheacutenotypes-parents selon le modegravele-hypothegravese eacutetabli avec nos reacutesultats et
analyses
Tout drsquoabord chez le prototype ou souche de pheacutenotype normal (ex ATCC 29213) la
chaine de transport des eacutelectrons fonctionne normalement Les complexes de la
chaine de transport des eacutelectrons sont chargeacutes de transfeacuterer des protons vers
lrsquoexteacuterieur de la cellule en transfeacuterant en mecircme temps des eacutelectrons drsquoun complexe agrave
un autre jusqursquoagrave un accepteur final qui est lrsquoO2 lorsque crsquoest possible Se faisant il se
creacutee un gradient eacutelectrochimique de part et drsquoautre de la membrane et les protons
accumuleacutes du cocircteacute externe sont utiliseacutes par la suite par lrsquoATP synthase afin de
pouvoir produire de lrsquoATP dans la cellule
0 4 8 1 2 1 6 2 0 2 4
0 0 1
0 1
1
C o u r b e d e c r o is s a n c e d e s m u ta n ts A T C C 2 9 2 1 3
reacute s is ta n ts agrave G E N + T O (8 m g L )
te m p s (h )
OD
60
0n
m
Niveau 3
Niveau 1
102
Le transfert drsquoeacutelectrons ne se fait pas sans coucirct et des produits toxiques pour la
cellule sont produits en mecircme temps ce sont les espegraveces reacuteactives de lrsquooxygegravene ou
ROS (Reactive Oxygen Species) Cependant les prototypes par leur reacuteponse au
stress produisent des enzymes capables de deacutetoxifier le milieu (Figure 12A)
Chez les souches SCV ici la souche controcircle NewbouldΔhemB la chaine de
transport des eacutelectrons est deacuteficiente parce que le gegravene permettant la production
drsquoheacutemine qui entre dans la composition des cytochromes est supprimeacute Les
cytochromes entrent dans la composition de plusieurs composantes de la chaine
respiratoire
Lorsque les complexes de la chaine sont alteacutereacutes il y a diminution du transfert de
protons vers lrsquoexteacuterieur une diminution de la production drsquoATP et une plus grande
production de ROS (Figure 12B)
Figure 12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B) NewbouldΔhemB
I III IV V I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
eacute eacute
eacute
H+ H+
ATPO2 H2O
ROS +ROS
Prototype SCV (hemB)
H+
0811 0928 2012 0811 0928
hegraveme
2012
A B
103
La gentamicine a besoin drsquoune bonne polarisation membranaire donc drsquoune chaine de
transport des eacutelectrons fonctionnelle pour pouvoir peacuteneacutetrer dans la cellule (Bryan et
Kwan 1983 (Bryan and van den Elzen 1977 Davis 1987 Taber et al 1987) Une
fois entreacutee elle inhibe la synthegravese proteacuteique ce qui provoque la mort cellulaire La
CMI de la gentamicine contre ATCC 29213 est de 05-1 microgml (Figure 13A)
La tomatidine elle irait se lier agrave lrsquoATP synthase pour en diminuer la fonctionnaliteacute et
ce tant chez ATCC 29213 que chez hemB Cette liaison amegravenerait une diminution
de la production drsquoATP et conseacutequemment provoquerait agrave plus ou moins long terme
une augmentation de production de ROS par lrsquoalteacuteration de la chaine de transport des
eacutelectrons Cependant comme la cellule prototype produit efficacement les enzymes
de deacutetoxification lrsquoimpact de la tomatidine est pratiquement nulle sur les souches
prototypes La CMI de la tomatidine est gt16microml contre ATCC 29213 (Figure 13B)
104
Figure 13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213
Chez ATCC 29213 lrsquoajout de 8microgml de tomatidine agrave la Gentamicine amegravene une
diminution de 8X agrave 16X la CMI de la gentamicine En effet la CMI de la gentamicine
passe de 05-1microgml agrave 006microgml (Figure 14AB)
Lors de lrsquoajout de la tomatidine agrave la gentamicine crsquoest la tomatidine qui se lie dans
un premier temps agrave lrsquoATP synthase reacuteduisant ainsi sont niveau drsquoactiviteacute Il y a alors
de faccedilon transitoire une augmentation de la concentration de protons agrave lrsquoexteacuterieur de
la cellule ce qui permet une entreacutee massive de gentamicine ainsi qursquoune rapide et
forte inhibition de la synthegravese proteacuteique Dans un second temps il y aura eacutegalement
une alteacuteration de la chaine de transport des eacutelectrons conduisant agrave une
augmentation de la production de ROS Crsquoest cependant lrsquoentreacutee massive de
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI TO gt16 microgmlCMI GEN 05-1 microgml
+ROS
ROS mais deacutetoxification efficace
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
105
gentamicine entrainant une forte inhibition de la synthegravese proteacuteique qui provoquera
dans ce cas-ci la mort cellulaire (Figure 14B)
Figure 14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicine lorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC 29213
Chez les mutants ATCC29213 reacutesistants agrave la combinaison GEN+TO les complexes I
et III sont muteacutes et donc ont une fonctionnaliteacute alteacutereacutee ce qui amegravene une
deacutepolarisation membranaire Une membrane moins polariseacutee amegravene une plus faible
entreacutee de gentamicine et donc une plus faible inhibition proteacuteique La CMI de la
gentamicine passe en effet de 05-1 microgml chez la souche parente agrave 2 microgml chez les
mutants reacutesistants
La tomatidine en se liant agrave lrsquoATP synthase accentuera la deacutepolarisation
membranaire provoquant une augmentation de la production des ROS et une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN (TO 8 microgml) 006 microgmlCMI GEN 05-1 microgml
+ROS
polarisation entreacutee massive de GEN
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
GENH+
BA
106
diminution de la production drsquoATP De fait les mutants reacutesistants ont une croissance
plus lente (Figure 15)(Figure 11)
Figure 15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants ATCC 29213
reacutesistants agrave la combinaison gentamicine (+TO)
Chez les SCV la tomatidine se lie aussi agrave lrsquoATP synthase et ce faisant amegravene une
diminution de la production drsquoATP Cette diminution de la production drsquoATP megravenera
ultimement agrave une alteacuteration de la chaine de transport des eacutelectrons et une
augmentation des ROS (Murphy 2009) tout comme chez les souches prototypes
Cependant comme le SCV a deacutejagrave une chaine de transport des eacutelectrons deacuteficiente
une faible production drsquoATP par lrsquoATP synthase et une forte production de ROS
lrsquoajout drsquoune faible concentration de tomatidine (la CMI est de 006microgml) amegravene une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype R GEN (+TO [8 microgml])
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN 2 microgmlCMI GEN 05-1 microgml
+ROS
R agrave GEN (+TO) mais mutants ont une
croissance diminueacutee (+ROS et -ATP)
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
107
augmentation de chacun de ces pheacutenomegravenes ce qui est suffisant pour provoquer la
mort cellulaire (Figure 16A)
Dans les premiers niveaux de reacutesistance agrave la tomatidine chez les mutants hemB la
reacutesistance (la CMI passe de 006 agrave 025-1microgml) est ameneacutee par la mutation du gegravene
ccpA (Catabolite controcircle protein A) ce qui megravenerait agrave une augmentation de la
production drsquoATP par lrsquoutilisation drsquoune autre voie que lrsquoATP synthase etou agrave une
diminution de la production de ROS (Figure 16B) Le rocircle de la CcpA en lien avec la
reacutesistance agrave la tomatidine est encore neacutebuleux Toutefois on sait que normalement
ce gegravene agit comme reacutegulateur transcriptionnel des gegravenes codant pour des enzymes
du cycle de Krebs et qursquoil a un rocircle de reacutegulation dans le meacutetabolisme du carbone et
lrsquoutilisation des substrats (Somerville et Proctor 2009 Goumltz et al 2006)
Figure 16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO 006 microgml
H+
SCV
I III IVeacute eacute
eacuteO2 H2O
+ROS
H+TO
Niveau 1
CcpA (CMI TO 025-1 microgml)
ROS
ATP
V
H+
ATP
TO0811 0928
2012
1629
0811 0928
2012
BA
108
Chez les SCV du deuxiegraveme palier de reacutesistance agrave la tomatidine non seulement le
gegravene ccpA est toujours muteacute mais le gegravene de la sous-uniteacute c de lrsquoATP synthase est
eacutegalement muteacute La mutation dans le site de liaison mecircme de la tomatidine empecircche
sa liaison et donc empecircche son effet sa CMI passant de 006 agrave gt16microgml (Figure
17B)
Malgreacute la mutation dans le gegravene lrsquoATP synthase resterait quand mecircme fonctionnel
puisque la mutation nrsquoamegravene pas la mort cellulaire comme crsquoest le cas lorsque la
tomatidine lie lrsquoATP synthase chez le SCV parent (Figure 17A)
Figure 17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la tomatidine
Tout comme chez les mutants hemB reacutesistant agrave la tomatidine seule les mutants
hemB reacutesistant agrave la tomatidine lorsqursquoen preacutesence de gentamicine possegravedent
eacutegalement une mutation dans le gegravene de lrsquoATP synthase ce qui rend la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO006 microgml
H+
SCV Niveau 2CMI TO gt16 microgml
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+TO
TO
Niveaux 1 et 2
CcpA (CMI TO 025-1 microgml)
ROS
ATP
0811 0928
2012
1629
0811 0928
2012
A B
109
inactive puisqursquoincapable de srsquoy lier (CMI gt16microgml) (Figure 18B) Chez les SCV non
muteacutes (hemB) la gentamicine nrsquoentre que faiblement ducirc agrave la deacutepolarisation de sa
membrane (CMI 4-8 microgml) (Figure 18A) La mutation dans le gegravene codant pour une
pompe (NWMN_0143 codant pour un ABC transporteur) chez les mutants hemB
reacutesistant agrave la tomatidine (+gentamicine) amegravenerait une augmentation de lrsquoactiviteacute de
cette pompe permettant de rejeter hors de la cellule le peu de gentamicine eacutetant
entreacutee La mutation de cette pompe fait passer la CMI de la gentamicine de 4-8
microgml pour hemB agrave 16 microgml pour les mutants hemB reacutesistant agrave la combinaison
gentamicine (+TO) ayant cette mutation (Figure 18B)
Figure 18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB reacutesistants
agrave la combinaison tomatidine (+gentamicine)
Donc selon notre modegravele-hypothegravese lrsquoaction de la tomatidine et de la gentamicine
sur S aureus deacutepend de la liaison agrave leur cible mais a aussi un lien avec la
SCV R agrave TO (+GEN [4microgml])
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI GEN 4-8 microgml
H+
SCV
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+ TO
GEN
CMI GEN 16 microgml
CMI TO gt16 microgml
0811 0928
2012
0811 0928 20120143
GEN
Synthegravese proteacuteique
BA
110
polarisation membranaire et la production de ROS La mesure de la polarisation et
concentration cellulaire en ROS chez nos mutants et nos souches parents en
preacutesence ou non drsquoantibiotique nous permettra de valider notre modegravele-hypothegravese
Chez une bacteacuterie meacutetaboliquement active et posseacutedant une membrane
cytoplasmique intacte il y a normalement une diffeacuterence de potentiel eacutelectrique entre
le cocircteacute inteacuterieur et le cocircteacute exteacuterieur de la membrane lrsquointeacuterieur eacutetant plus neacutegatif de
100 agrave 200 mV par rapport agrave lrsquoexteacuterieur Une diminution de la magnitude de ce
potentiel membranaire est appeleacutee deacutepolarisation alors qursquoune augmentation de ce
potentiel est appeleacutee hyperpolarisation Le potentiel membranaire sera reacuteduit agrave zeacutero
srsquoil y a rupture de la membrane cest-agrave-dire srsquoil y a formation de trous assez grands
permettant aux ions inorganiques de passer librement Ce pheacutenomegravene peut arriver
lorsque les cellules sont tueacutees par la chaleur le froid ou les antibiotiques β-Lactams
Dans ces circonstances la membrane devient permeacuteable agrave diffeacuterents colorants
comme le propidium iodide (PI) et le TO-PRO3 et le changement est irreacuteversible
Plusieurs classes de composeacutes chimiques notamment les ionophores peuvent
eacutegalement alteacuterer le potentiel membranaire sans pour autant permettre la
permeacuteabiliteacute au PI TO-PRO3 et autres colorants similaires chez les bacteacuteries
lrsquoadministration drsquoun proton-ionophore tel le carbonyl cyanide 3-
chlorophenylhydrazone (CCCP) reacuteduira le potentiel membranaire agrave zeacutero
Des tests preacuteliminaires sur la mesure du potentiel membranaire (BacLighttrade Bacterial
Membrane Potential Kit (B34950) drsquoInvitrogen et lecture au cytomegravetre de flux (FACS))
(Novo et al 2000) chez nos deux souches parentes mets en eacutevidence que ATCC
29213 est plus polariseacute que NewbouldΔhemB (Figure 19A) Toutefois malgreacute
qursquoayant une plus faible polarisation membranaire NewbouldΔhemB nrsquoest pas
complegravetement deacutepolariseacute puisqursquoil est possible de le deacutepolariser davantage agrave lrsquoaide
drsquoun agent deacutepolarisant tel que le CCCP (carbonyl cyanide 3-chlorophenylhydrazone)
(Figure 19B) Le TO-PRO3 a eacuteteacute utiliseacute comme controcircle afin de confirmer la viabiliteacute
de nos cellules (donneacutees non montreacutees)
111
A B
Figure 19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane Potential Kit
(Invitrogen) (Lecture au cytomegravetre de flux agrave lrsquoaide du programme FCS Express) (A)
Ratio FL3-H (axe des Y)FL1-H (axe des X) de ATCC 29213 (rouge) et de
NewbouldΔhemB (noir) (B) Lecture des rouges (FL3-H) sur NewbouldΔhemB traiteacute
avec le DiOC2(3) (33prime-diethyloxacarbocyanine iodide) (noir) ou avec le CCCP (bleu)
ou sur les cellules chauffeacutees (rouge)
La sous-uniteacute c de lrsquoATP synthase (AtpE) semble donc la cible la plus probable de la
tomatidine Afin de veacuterifier si les mutations identifieacutees pouvaient avoir un effet
veacuteritable sur la proteacuteine nous avons fait un alignement de seacutequences noteacute les bases
muteacutees et veacuterifieacute si la mutation amenait un changement drsquoacide amineacute et si ce
changement drsquoacide amineacute pouvait avoir un impact sur la proteacuteine finale De fait
nous avons trouveacute que les deux diffeacuterentes mutations dans lrsquoAtpE amenaient un
changement significatif drsquoacides amineacutes pouvant amener une variation fonctionnelle
FL3-H
Count
100
101
102
103
104
0
18
36
53
71
Chauffeacute
+CCCP+DiOC2
Chauffeacute
FL1-H
FL3-H
100
101
102
103
104
100
101
102
103
104
hemB
29213
VERT
RO
UG
E
ROUGE
ΔhemB
112
de la proteacuteine La mutation de la base 49 change un G pour un T il srsquoagit drsquoun SNP
nrsquoamenant pas de changement de cadre de lecture La mutation fait passer lrsquoacide
amineacute alanine (A) qui est apolaire agrave une seacuterine (S) qui est polaire La mutation de la
base 190 change un G pour un A et est aussi un SNP nrsquoamenant pas de changement
de cadre de lecture Cette mutation fait passer lrsquoacide amineacute alanine (A) qui est
apolaire agrave une threacuteonine (T) qui est polaire (Figure 20)
gtlcl||NWMN_2012|atpE|72493880 F0F1 ATP synthase subunit C
1 10 19 31 43 49 58
ATG AAT TTA ATC GCA GCA GCA ATC GCA ATT GGT TTA TCA GCA TTA GGA GCA GGT ATC GGT
M N L I A A A I A I G L S A L G A G I G
(S)
17 61 73 85 97 109 118
AAC GGT TTA ATC GTT TCA AGA ACA GTT GAA GGT GTA GCA CGT CAA CCA GAA GCA CGT GGT
N G L I V S R T V E G V A R Q P E A R G
21 40
121 133 145 157 169 178
CAA TTA ATG GGT ATC ATG TTC ATT GGT GTA GGT TTA GTT GAG GCA TTA CCT ATC ATC GGT
Q L M G I M F I G V G L V E A L P I I G
41 60
181 190 199 211
GTA GTA ATT GCA TTC ATG ACA TTT GCT GGA TAA
V V I A F M T F A G
(T)
61 64 70
Figure 20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du gegravene
atpE de S aureus Newman avec mise en eacutevidence des mutations (rose) en lien avec
la cible de la tomatidine Lrsquoalignement a eacuteteacute effectueacute agrave lrsquoaide du site CMR de JCVI
113
CHAPITRE 4
DISCUSSION
LrsquoATP synthase est eacutevidemment un complexe proteacuteique ubiquitaire tant chez les
procaryotes que les eucaryotes Malgreacute ce fait il semble que la cible de la tomatidine
soit speacutecifique aux bacillales celle-ci ne deacutemontrant aucune activiteacute contre les
bacteacuteries agrave Gram neacutegatif et les lactobacillales On suppose donc que lrsquoATP synthase
de ces espegraveces doit ecirctre speacutecifique au niveau du site de liaison de la tomatidine
Lrsquoecirctre humain en tant qursquoespegravece eucaryote est eacutegalement pourvu de ce complexe
proteacuteique et on peut naturellement se demander si lrsquoutilisation clinique de cette
moleacutecule pourrait ecirctre cytotoxique De fait plusieurs composeacutes naturels tel
lrsquooligomycine et la venturicidine sont connus pour inhiber lrsquoactiviteacute de lrsquoATP synthase
par leur interaction avec la sous-uniteacute c de ce complexe Cependant ces moleacutecules
ne sont pas seacutelectives et nrsquoinhibent pas seulement lrsquoATP synthase bacteacuterien mais
aussi lrsquoATP synthase mitochondrial (Haagsma et al 2009 Matsuno-Yagi et Hatefi
1993) (Figure 1) Neacuteanmoins il existe maintenant une moleacutecule qui a pour cible
lrsquoATP synthase bacteacuterien et qui est en processus en vue drsquoune utilisation clinique
chez lrsquohomme le TMC207 (Haagsma et al 2009))
114
Figure 1 Sites drsquoinhibition de lrsquoATP synthase Les sites de liaison dans lrsquoATP
synthase des inhibiteurs reacuteveacuteleacutes par des eacutetudes biochimiques et structurales sont
indiqueacutes par des cercles rouges et les sites de liaisons qui nrsquoont pas eacuteteacute totalement
clarifieacutes sont indiqueacutes par des cercles verts (Hong et Pedersen 2008)
Le TMC207 (Figure 2A) est un composeacute de la classe des diarylquinoline posseacutedant
un nouveau meacutecanisme drsquoaction lrsquoinhibition de lrsquoATP synthase bacteacuterien De fait la
cible et le meacutecanisme drsquoaction du TMC207 sont diffeacuterents des autres agents
antituberculeux Lrsquoinhibition de lrsquoATP synthase peut mener agrave une baisse de la
production drsquoATP et un deacutebalancement dans lrsquohomeacuteostasie du pH ces 2 effets
pouvant contribuer agrave la diminution de la survie de la bacteacuterie (Rao et al 2001
Andries et al 2005)
Le TMC207 est une moleacutecule ayant une forte activiteacute bacteacutericide contre
Mycobacterium tuberculosis et autres espegraveces mycobacteacuteriennes mais peu drsquoactiviteacute
115
contres les autres espegraveces bacteacuteriennes Lrsquoindex de seacutelectiviteacute de TMC207 pour
lrsquoATP synthase mycobacteacuterienne (CMI50 gt200 microml) compareacute agrave lrsquoATP synthase
mitochondriale (CMI50 001 microgml) est de plus de 20000 Un indice de seacutelectiviteacute au
dessus de 1000 est consideacutereacute dans le deacuteveloppement drsquoune drogue comme un
preacutealable pour une moleacutecule candidate (Haagsma et al 2009) La diversiteacute dans les
gegravenes atpE explique la reacutesistance au TMC207 et justifie la faible affiniteacute de cette
drogue agrave lrsquoATP synthase mitochondriale (Matteelli et al 2010) (Figure 3)
Figure 2 Structure chimique de (A) TMC207 et (B) son analogue anti bacteacuteries
agrave Gram positifs (Balemans et al 2012)
Lorsqursquoon compare la seacutequence proteacuteique de la sous-uniteacute c de lrsquoATP synthase
(atpE) de quelques espegraveces de lrsquoordre des Bacillales (tel Saureus B cereus B
subtilis et L monocytogenes) avec des espegraveces appartenant agrave drsquoautres ordres
bacteacuteriens on constate que les seacutequences divergent ce qui est en accord avec la
speacutecificiteacute des moleacutecules Les points de mutations amenant une reacutesistance au
TMC207 chez Mycobacterium smegmatis sont speacutecifiques aux mycobacteacuteries et les
deux principaux points de mutations D32V(ouP) et A67P chez M smegmatis sont
situeacutes tous les deux dans les heacutelices alpha qui entourent le site catalytique de la
proteacuteine (E65) qui est impliqueacute dans le transport des protons (Andries et al 2005)
Lrsquoacide amineacute aspartate (D) agrave la position 32 et lrsquoalanine (A) agrave la position 67 sont tregraves
speacutecifiques agrave M smegmatis
A B
116
Reacutecemment un groupe de chercheurs (Balemans et al 2012) on mit en lumiegravere une
autre moleacutecule (Figure 2B) de la classe des diarylquinolines dont le meacutecanisme
drsquoaction eacutetait aussi lrsquoinhibition de la sous-uniteacute c de lrsquoATP synthase mais chez les
bacteacuteries agrave Gram positif tels Streptococcus pneumoniae et S aureus Cette moleacutecule
irait lier les acides amineacutes valines (V) situeacutes agrave la position 50 et 62 de lrsquoAtpE de S
aureus (Donneacutees non montreacutees chez S pneumoniae(Balemans et al 2012)) En
effet une mutation amenant le changement de la valine en position 50 par une
isoleucine (I) etou une mutation amenant le changement de la valine en position 62
par une alanine (A) amegravene la reacutesistance de la bacteacuterie agrave la moleacutecule Cependant L
monocytogenes et Bacillus subtilis ne possegravedent pas de valine agrave la position 50 alors
qursquoils font eacutegalement partie des bacteacuteries agrave Gram positif (Il faut noter que chez
Bacillus sp PS3 (Swiss-Prot assession Ndeg P 00845) possegravede une valine agrave la position
50 (Balemans et al 2012)) Par contre tout comme pour les autres bacteacuteries agrave Gram
positif testeacutees les deux espegraveces possegravedent une valine agrave la position 62 (Figure 3)
Chez B cereus B subtilis B anthracis S aureus et L monocytogenes les points de
divergence sont plutocirct A17S et A64T Cependant B pseudofirmus qui fait eacutegalement
partie de lrsquoordre des Bacillales possegravede une glycine (G) agrave la position 17 plutocirct qursquoune
alanine (A) et une seacuterine (S) plutocirct qursquoune alanine (A) agrave la position 64 comme les
espegraveces preacuteceacutedentes Suivant ces reacutesultats et si la cible de la tomatidine est bien
ces deux points de mutation la tomatidine ne devrait pas pouvoir se lier agrave lrsquoAtpE de B
pseudofirmus et devrait donc nrsquoavoir aucune activiteacute contre cette souche alors que
nous avions extrapoleacute que le spectre drsquoactiviteacute de la tomatidine eacutetait tous les
Bacillales De plus E coli et M smegmatis possegravedent eacutegalement une alanine (A) agrave la
position 17 alors que la tomatidine ne possegravede aucune activiteacute contre ces espegraveces (M
smegmatis nrsquoa pas eacuteteacute eacutevalueacute il srsquoagit ici drsquoune extrapolation des reacutesultats de CMI)
par contre ces deux espegraveces possegravedent des acides amineacutes diffeacuterents drsquoune alanine
(A) agrave la position 64 ce qui pourrait leur confeacuterer une reacutesistance agrave la tomatidine telle
qursquoobserveacutee On remarque eacutegalement que les acides amineacutes agrave ces deux positions
117
sont totalement diffeacuterents chez lrsquohomme (Homo sapiens) ce qui suggeacutererait que la
tomatidine ne possegravederait pas drsquoactiviteacute antimitochondriale (Figure 3)
Selon ces reacutesultats on peut se demander si les deux mutations sont essentielles agrave la
liaison de la moleacutecule ou si seulement lrsquoalanine (A) agrave la position 64 est neacutecessaire
On doit se rappeler que la reacutesistance apporteacutee par la mutation faisant en sorte que
lrsquoalanine (A) en position 17 devient une seacuterine (S) apparaicirct chez les mutants
NewbouldΔhemb reacutesistant agrave la tomatidine et que cette mutation nrsquoapparaicirct qursquoau
quatriegraveme et dernier niveau de reacutesistance On se rappelle eacutegalement que la mutation
faisant en sorte que lrsquoalanine (A) agrave la position 64 devient une threacuteonine (T) apparaicirct
chez les mutants NewbouldΔhemb reacutesistants agrave la tomatidine en combinaison avec la
gentamicine et ce degraves le premier niveau de reacutesistance
118
Bcereus G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Banthracis G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Lmonocytogenes G---------VIAAAIAVG---LGALGAGIGNGLIVSKTVEGVARQPEARSMLQTIMFV- 50
Saureus_NWMN_2012 -----------IAAAIAIG---LSALGAGIGNGLIVSRTVEGVARQPEARGQLMGIMFIG 49
Bsubtilis -----------IAAAIAIG---LGALGAGIGNGLIVSRTVEGIARQPEAGKELRTLMFMG 49
Bpseudofirmus -----------LGAAIAAG---LAAVAGAIAVAIIVKATIEGTTRQPELRGTLQTLMFIG 49
Ecoli L---------YMAAAVMMG---LAAIGAAIGIGILGGKFLEGAARQPDLIPLLRTQFFIV 56
Msmegmatis DPNAIITAGALIGGGLIMG---GGAIGAGIGDGIAGNALISGIARQPEAQGRLFTPFFIT 60
Homo sapiens QTSAISRDIDTAAKFIGAGAATVGVAGSGAGIGTVFGSLIIGYARNPSLKQQLFSYAILG 120
MyR V
MyR P
SaR S
Bcereus VALVEALPIIGVVIAFIVMNK----- 72
Banthracis VALVEALPIIGVVIAFIVMNK----- 72
Lmonocytogenes ----EALPIIAVVIAFMVLNK----- 67
Saureus_NWMN_2012 VGLVEALPIIGVVIAFMTFAG----- 70
Bsubtilis IALVEALPIIAVVIAFLAFFG----- 70
Bpseudofirmus VPLAEAVPIIAIVISLLILF------ 69
Ecoli MGLVDAIPMIAVGLGLYVMFAVA--- 79
Msmegmatis VGLVEAAYFINLAFMALFVFATPGLQ 86
Homo sapiens FALSEAMGLFCLMVAFLILFAM---- 142
MyR D P M
SpR I A
SaR T
Fig
ure
3
Seacuteq
uen
ce
pro
teacuteiq
ue d
e la p
roteacute
ine
Atp
Ec
hez d
iffeacute
ren
tes
es
pegravec
es e
t m
ise e
n eacute
vid
en
ce
de
s s
ite
s d
e
mu
tati
on
pe
rmett
an
t la
reacutesis
tan
ce agrave
dif
feacutere
nte
s m
oleacute
cu
les
M
yR M
tu
berc
ulo
sis
ou
M sm
eg
matis
reacutesis
tan
t a
u
TM
C2
07 S
pR
S
p
neu
mon
iae
reacutesis
tan
t agrave
un a
na
log
ue d
u T
MC
20
7
SaR
S
a
ure
us
SC
V r
eacutesis
tant agrave
la to
matid
ine
En
cad
reacutero
uge
S
ite d
e m
uta
tion a
men
ant la
reacutesis
tance
agravela
to
matid
ine E
ncad
reacuteno
ir
Site
cata
lytiq
ue
im
pliq
ueacute
da
ns le
tra
nspo
rt d
es p
roto
ns
Su
rlig
nag
etu
rqu
ois
e S
ite d
e m
uta
tion a
me
nant u
ne
reacutesis
tan
ce a
u T
MC
20
7
119
Les seacutequences proteacuteiques des souches bacteacuteriennes de la figure 3 ont eacuteteacute trouveacutees
sur pubmedproteacuteine httpwwwncbinlmnihgovpubmed
laquo ATP synthase subunit c raquo Assession number
Streptococcus pneumoniae D39 YP_8168031 (NCBI)
S aureus Newman BAF682841 (GenBank)
E coli AAA247321 (GenBank)
Bacillus cereus ISO75 EJP900901 (GenBank)
Bacillus anthracis str H9401 YP_0062123491 (NCBI)
Mycobacterium smegmatis MC2 155 YP_0065695541 (NCBI)
Bacillus subtilis str 168 CAA822551 (GenBank)
Listeria monocytogenes YP_0076049931 (NCBI)
Homo sapiens P482011 (UniProtKBSwiss-Prot)
Lrsquoalignement comparatif des seacutequences drsquoacides amineacutes a eacuteteacute fait agrave lrsquoaide du
programme ClustalW2 sur le web
httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2
Le modegravele 3D de lrsquoAtpE a eacuteteacute obtenu gracircce agrave lrsquoaide du Pr Ryszard Brzezinski du
Deacutepartement de biologie de lrsquoUniversiteacute de Sherbrooke (Figure 4) Une recherche
dans le Protein Data Bank a drsquoabord eacuteteacute effectueacutee afin de veacuterifier parmi les proteacuteines
AtpE (sous-uniteacute C de lrsquoATP synthase) pour lesquelles il y avait des structures 3D
reacutesolues laquelle ressemblait le plus agrave celle de Staphylococcus aureus Les
alignements ont montreacute que cest la proteacuteine de Bacillus pseudofirmus qui avait la
plus grande similitude de seacutequence La structure est stockeacutee dans la banque PDB
sous le symbole 3ZO6 Il a eacuteteacute confirmeacute par quelques alignements que cette sous-
uniteacute C de lrsquoATP synthase eacutetait plus proche par sa seacutequence de celle de
Staphylococcus aureus que de celle de Mycobacterium pour laquelle la structure 3D
avait eacuteteacute deacutetermineacutee plus tocirct Ce reacutesultat sexplique par le fait que Staphylococcus et
Bacillus font tous deux partie de la branche A+T-riche des bacteacuteries agrave Gram+ alors
que Mycobacterium appartient agrave la branche G+C-riche La seacutequence de lAtpE de
120
Staphylococcus aureus a ensuite eacuteteacute envoyeacutee au serveur internet de modeacutelisation
3D-Jigsaw (httpbmmcancerresearchukorg~3djigsaw) en indiquant que le modegravele
devait ecirctre bacircti en prenant comme matrice le fichier PDB 3ZO6 Le modegravele nous a
eacuteteacute retourneacute sous format PDB ndash comme les laquo vrais raquo cristaux dans la Protein Data
Bank Les fichiers au format PDB peuvent ecirctre visualiseacutes par le logiciel RasMol ndash
gratuit sur internet Cest avec RasMol que limage (Figure 4) a eacuteteacute obtenue
121
Figure 4 Modeacutelisation de la structure tridimensionelle de la sous-uniteacute c de
lrsquoATP synthase de S aureus (AtpE) montrant les acides amineacutes muteacutes chez les
mutants reacutesistantsEn blanc et vert les deux acides amineacutes muteacutes chez S aureus
reacutesistant agrave la tomatidine En rose les sites de mutations chez Mycobacterium
smegmatis reacutesistant au TMC207 et des sites de mutations chez Streptoccocus
pneumoniae reacutesistants agrave un analogue de TMC207 En bleu le site actif de la proteacuteine
Agrave la vue de ces reacutesultats certains pourront mettre en doute lrsquointeacuterecirct de la tomatidine
eacutetant donneacute lrsquoapparition relativement rapide de reacutesistance chez S aureus Cependant
122
nos reacutesultats deacutemontrent clairement que lrsquoajout de tomatidine ralentit le
deacuteveloppement de reacutesistance agrave la gentamicine chez S aureus ATCC29213 (Tableau
1) De plus cette augmentation de la CMI de la gentamicine en preacutesence de
tomatidine au fil des passages suit lrsquoaugmentation de la CMI de la ciprofloxacine lors
de ces mecircmes passages alors que la ciprofloxacine est consideacutereacutee comme la drogue
de reacutefeacuterences dans les bonnes drogues (Tableau 1) Tout comme pour la
ciprofloxacine pour laquelle la reacutesistance chez S aureus peut ecirctre la conseacutequence
de deux mutations dans le gegravene cible gyrA la reacutesistance agrave la tomatidine peut ecirctre la
conseacutequence de deux mutations dans le gegravene cible atpE Par comparaison la
rifampicine est consideacutereacutee comme une mauvaise drogue de par lrsquoapparition rapide
drsquoune forte reacutesistance chez S aureus conseacutequence de ses 17 mutations possibles
dans le gegravene cible rpoB (Tableau 1)
De plus il sera important de relativiser lrsquoapparition de reacutesistance agrave la tomatidine en
calculant le taux drsquoapparition de ces colonies reacutesistantes ceci nrsquoayant pas eacuteteacute fait
jusqursquoici De fait la freacutequence de mutation chez S aureus amenant une reacutesistance agrave
la rifampicine est de lrsquoordre de 10-7 10-8 alors qursquoil est de 10-11 pour lrsquoacide fusidique
(Canfield et al 2013 Orsquoneill et al 2001 Price et Gustafson 2001) Pour ce qui est
de la nouvelle moleacutecule anti-bacteacuteries agrave Gram positif de la classe des diarylquinoline
une freacutequence de mutation sur S pneumoniae ATCC49619 de 81- 83 x 10-7 a eacuteteacute
observeacutee agrave 5x la CMI et une freacutequence de mutation de 46 x 10-7 agrave 17 x 10-6 agrave 50x la
CMI (Balemans et al 2012)
123
Tableau 1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 par la meacutethode de CMI lors de passages successifs dans un milieu avec
lrsquoantibiotique
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
Typiqueldquomauvaiserdquodrogue (17 mutations possibles dansrpoB)
Typique ldquobonnerdquo drogue (2 mutations possibles dans gyrA)
Preacutesence de TO ralenti le deacuteveloppement de reacutesistances vs GEN seul
124
CHAPITRE 5
CONCLUSION ET PERSPECTIVES
Mon projet de maitrise a permis de preacuteciser le spectre drsquoactiviteacute de la tomatidine
Nous savons maintenant que la tomatidine possegravede une activiteacute antibacteacuterienne
contre la plupart des Firmicutes et plus speacutecifiquement contre les Bacillales Plus
preacuteciseacutement la tomatidine est bacteacutericide contre les pheacutenotypes SCV et agit en
synergie avec les aminoglycosides augmentant lrsquoactiviteacute bacteacutericide de ces derniers
de 8-16 fois contre les souches agrave pheacutenotype normal
De plus parmi tous les analogues de la tomatidine produits par le laboratoire du Pr
Eacuteric Marsault (voir article en annexe 2) le composeacute FC04-100 a eacuteteacute plus
speacutecifiquement mis en lumiegravere Celui-ci conserve non seulement une aussi bonne
activiteacute synergique avec les aminoglicosides contre les Saureus au pheacutenotype
normal et une activiteacute modeacutereacutee contre les S aureus SCV mais montre eacutegalement de
faccedilon inespeacutereacutee une activiteacute par lui-mecircme contre les S aureus de pheacutenotype normal
incluant plusieurs MRSA une proprieacuteteacute qui nrsquoeacutetait pas observeacutee pour la tomatidine
Une autre activiteacute inteacuteressante de FC04-100 est sa capaciteacute drsquoameacuteliorer lrsquoactiviteacute
bacteacutericide des aminoglicosides contre L monocytogenes agrave pheacutenotype normal apregraves
un traitement de 24h alors que la tomatidine ne deacutemontrait qursquoune faible synergie
avec les aminoglycosides contre cette souche De plus FC04-100 deacutemontre une
activiteacute bacteacutericide contre les SCV de L monocytogenes alors que la tomatidine ne
deacutemontrait qursquoune activiteacute bacteacuteriostatique FC04-100 montre eacutegalement un effet
potentiateur de lrsquoUbrolexinTM (Ceacutefalexin Kanamycin (3 2)) une combinaison
drsquoantibiotique utiliseacute dans le traitement de mammites bovines en abaissant de 8 agrave 16
fois leur CMI amenant celles-ci sous le seuil de reacutesistance Finalement FC04-100
possegravede une meilleure solubiliteacute que la tomatidine une proprieacuteteacute qui facilitera son
utilisation in vitro
125
Nos reacutesultats deacutemontrent que FC04-100 conserve une certaine activiteacute contre les
SCV reacutesistant agrave la tomatidine ce qui suggegravere possiblement que FC04-100 a plus
daffiniteacute pour la cible ou plus de sites dinteraction avec AtpE En effet les mutants
SCV reacutesistants agrave la tomatidine deviennent gt512 fois plus reacutesistants alors que FC04-
100 ne perd que 8 agrave 16 fois son activiteacute contre ces mecircmes souches Ceci deacutemontre
quil est possible dameacuteliorer significativement lactiviteacute de ces alkaloiumldes steacuteroidiens
ce qui est tregraves prometteur pour lavenir de cette moleacutecule
Dans un second temps lrsquoanalyse des mutations chez les mutants reacutesistants agrave la
tomatidine a permis de non seulement de mieux comprendre le mode drsquoaction de la
tomatidine mais eacutegalement de preacuteciser la cible potentielle de ce nouvel antibiotique
De fait son mode drsquoaction serait en lien direct avec la fonctionnaliteacute de la chaine de
transport des eacutelectrons chez la bacteacuterie lrsquoactiviteacute de la tomatidine serait inversement
proportionnelle agrave la fonctionnaliteacute de la chaine de transport des eacutelectrons et donc en
lien eacutegalement avec la polarisation membranaire et la production de ROS
intracellulaire Afin de confirmer le modegravele-hypothegravese la mesure de la polarisation
membranaire et la concentration de ROS intracellulaire chez les mutants ainsi que
chez les souches parentes suite agrave un traitement antibiotique devront ecirctre effectueacutees
Les reacutesultats de mes recherches suggegraverent que la cible bacteacuterienne speacutecifique de la
tomatidine serait la sous-uniteacute C de lrsquoATP synthase Afin de confirmer la cible
Maxime Lamontagne-Boulet tente au moment drsquoeacutecrire ces lignes drsquoobtenir des
mutants de S aureus Newman reacutesistants au composeacute FC04-100 et dont la reacutesistance
serait attribuable agrave une mutation dans le gegravene atpE Par la suite on devra confirmer
la fonctionnaliteacute de la cible ATP synthase par quantification de lrsquoATP produit agrave lrsquoaide
de la lucifeacuterase tel que lrsquoon fait Balemans et al en 2012 Lrsquoattachegravement agrave la cible
devra eacutegalement ecirctre confirmeacute et la meacutethode de SPR (Surface Plasmon Resonance)
pourrait ecirctre tout indiqueacutee pour ce faire (Balemans et al 2012) On devra eacutegalement
126
deacutemontrer la seacutelectiviteacute des moleacutecules tomatidine et FC04-100 pour lrsquoATP synthase
des Bacillales
Les infections agrave bacteacuteries multireacutesistantes deviennent de plus en plus preacutevalentes et
sont un des problegravemes de santeacute majeurs auxquels nous devons faire face aujourdrsquohui
Cette augmentation de la reacutesistance limite le reacutepertoire drsquoantibiotiques efficace creacuteant
une situation probleacutematique laquelle est exacerbeacutee par le trop petit nombre de
nouveaux antibiotiques introduits sur le marcheacute ces derniegraveres anneacutees Il est donc
drsquoune grande importance de travailler agrave la mise en marcheacute de nouveaux antibiotiques
et plus particuliegraverement de nouvelles classes drsquoantibiotiques espeacuterant ainsi deacutejouer la
multireacutesistance bacteacuterienne du moins pour un temps La tomatidine de par sa
structure sa cible et son mode drsquoaction speacutecifique est une moleacutecule antibiotique
dans une classe agrave part et de ce fait une moleacutecule plus qursquointeacuteressante pour le
traitement drsquoinfection agrave S aureus De plus comme elle deacutemontre une activiteacute
bacteacutericide contre les deux pheacutenotypes de S aureus (et autres Bacillales) lesquels
sont retrouveacutes simultaneacutement dans les cas drsquoinfections chroniques son pouvoir
antibiotique devient inestimables drsquoun point de vue clinique
127
CONTRIBUTIONS SUPPLEacuteMENTAIRES
ANNEXE 1 Application de brevet Internationnal publication number WO
2012109752 A1
INVENTEURS Franccedilois Malouin2 Gabriel Mitchell2 Kamal Bouarab2 Eacuteric Marsault1
Feacutelix Chagnon1 Simon Boulanger 2 Isabelle Guay2
1Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
2Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
128
129
ANNEXE 2 Article scientifique dans laquo Europeen Journal of medicinal chemistry raquo
2014 80 605-620
TITRE Unraveling the structure-activity relationship of tomatidine a steroid alkaloid
with unique antibiotic properties against persistent forms of
Staphylococcus aureus
ABSTRACT Staphylococcus aureus is responsible for difficult-to-treat and relapsing
infections and constitutes one of the most problematic pathogens due to its multiple
resistances to clinically available antibiotics Additionally this pathogenrsquos ability to
develop small-colony variants is associated with a reduced susceptibility to
aminoglycoside antibiotics and in vivo persistence We have recently demonstrated
that tomatidine a steroid alkaloid isolated from tomato plants possesses anti-
virulence activity against normal strains of S aureus as well as the ability to
potentiate the effect of aminoglycosides against this pathogen In addition tomatidine
has shown antibiotic activity against small-colony variants We herein report the first
study of the structure-activity relationship of tomatidine against S aureus
AUTEURS Feacutelix Chagnon1 Isabelle Guay2 Marc-Andreacute Bonin1 Gabriel Mitchell2
Kamal Bouarab2Franccedilois Malouin2 Eacuteric Marsault1
MA CONTRIBUTION Responsable des tests biologiques de tous les composeacutes
autant envers lrsquoeffet potentiateur de la tomatidine sur lrsquoactiviteacute de la gentamicine
contre la souche ATCC29213 de S aureus que pour lrsquoactiviteacute bacteacuteriostatique sur la
souche NewbouldΔhem imitant le pheacutenotype SCV Ces travaux furent effectueacutes sous
la tutelle du professeur Franccedilois Malouin
130
1Institut de Pharmacologie de Sherbrooke Deacutepartement de pharmacologie
Universiteacute de Sherbrooke 3001 12e av nord Sherbrooke Queacutebec Canada J1H
5N4
2Centre drsquoeacutetude et de valorisation de la diversiteacute microbienne (CEVDM) Deacutepartement
de biologie Universiteacute de Sherbrooke Sherbrooke QC Canada J1K 2R1
131
BIBLIOGRAPHIE Alexander E H and Hudson MC (2001) Factors influencing the internal- ization of Staphylococcus aureus and impacts on the course of infections in humans Appl Microbiol Biotechnol 56 361ndash366
Andries K Verhasselt P Guillemont J Goumlhlmann HW Neefs JM Winkler H Van Gestel J Timmerman P Zhu M Lee E et al (2005) A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis Science 307 223-227
Artzatbanov VY and Petrov VV (1990) Branched respiratory chain in aerobically grown Staphylococcus aureus--oxidation of ethanol by cells and protoplasts Arch Microbiol 153 580- 584 Archer GL (1998) Staphylococcus aureus a well-armed pathogen Clin Infect Dis 26 1179ndash1181 Assefa S Keane TM Otto TD Newbold C and Berriman M (2009) ABACAS algorithm-based automatic contiguation of assembled sequences Bioinformatics 25 1968-1969 Aubry-Damon H Legrand P Brun-Buisson C Astier A Soussy CJ and Leclercq R (1997) Reemergence of gentamicin-sus- ceptible strains of methicillin-resistant Staphylococcus aureus roles of an infection control program and changes in aminoglycoside use Clin Infect Dis 25 647-653
Balemans W Vranckx L Lounis N Pop O Guillemont J Vergauwen K Mol S Gilissen R Motte M Lanccedilois et al (2012) Novel antibiotics targeting respiratory ATP synthesis in Gram-positive pathogenic bacteria Antimicrob Agents Chemother 56 4131-4139
Batandier C Guigas B Detaille D El-Mir MY Fontaine E Rigoulet M and Leverve XM (2006) The ROS production induced by a reverse-electron flux at respiratory-chain complex 1 is hampered by metformin J Bioenerg Biomembr 38 33ndash 42
132
Beckman Coulter website httpswwwbeckmancoultercomwsrportalwsrresearch-and-discoveryproducts-and-servicesnucleic-acid-sample-preparationspri-laboratory-automationindexhtm (consulteacute le 3 Deacutec 2013) Bednarek P and Osbourn A (2009) Plant-microbe interactions chemical diversity in plant defense Science 324 746ndash748 Besier S Ludwig A Ohlsen K Brade V and Wichelhaus TA (2007) Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus Int J Med Microbiol 297 217-225 BiteSizeBio Brain food for biologists httpnxseqbitesizebiocomarticlessequencing-by-synthesis-explaining-the-illumina-sequencing-technology (consulteacute le 3 deacutec 2013) Boetzer M and Pirovano W (2012) Toward almost closed genomes with GapFiller Genome Biology 13 R56 Bouarab K Ordi E Gattuso M Moisan H and Malouin F (2007) Plant stress response agents affect Staphylococcus aureus virulence genes Abstr 47th Intersci Conf Antimicrob Agents Chemother abstr C1-1483 Bryan LE and Kwan S (1981) Aminoglycoside-resistant mutants of Pseudomonas aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob Agents Chemother 19 958- 964 Bryan LE and Kwan S (1983) Roles of ribosomal binding membrane potential and electron transport in bacterial uptake of streptomycin and gentamicin Antimicrob Agents Chemother 23 835ndash845
Bryan LE Nicas T Holloway BW and Crowther C (1980) Aminoglycoside-resistant mutation of Pseudomonas aeruginosa defective in cytochrome C552 and nitrate reductase Antimicrob Agents Chemother 17 71- 79
133
Bryan LE and van den Elzen HM (1977) Effects of membrane-energy mutations and cations on streptomycin and gentamicin accumulation by bacteria a model for entry of streptomycin and gentamicin in susceptible and resistant bacteria Antimicrob Agents Chemother 12 163-177 Brynildsen MP Winkler JA Spina CS MacDonald IC and Collins JJ (2013) Potentiating antibacterial activity by predictably enhancing endogenous microbial ROS production Nat Biotechnol 31 160ndash 165 Burke KA and Lasccelles J (1975) Nitrate reductase system in Staphylococcus aureus wild type and mutants J Bacteriol 123 308- 316 Burke KA and Lascelles J (1976) Nitrate reductase activity in hegraveme-deficient mutants of Staphylococcus aureus J Bacteriol 126 225- 231 Canfield GS Schwingel JM Foley MH Vore KL Boonanantanasarn K Gill AL Sutton MD and Gill SR (2013) Evolution in fast forward a potential role for mutators in accelerating Staphylococcus aureus pathoadaptation J Bacteriol 195 615-628 Chang S Sievert DM Hageman JC Boulton ML Tenover FC Downes FP Shah S Rudrik JT Pupp GR Brown WJ et al (2003) Infection with vancomycin- resistant Staphylococcus aureus containing the vanA resistance gene N Engl J Med 348 1342-1347 Chen Q Vazquez EJ Moghaddas S Hoppel CL and Lesnefsky EJ (2003) Production of reactive oxygen species by mitochondria central role of complex III J Biol Chem 278 36027-36031 Clements MO Watson SP Poole RK and Foster SJ (1999) CtaA of Staphylococcus aureus is required for starvation survival recovery and cytochrome biosynthesis J Bacteriol 181 501- 507
134
Clinical and Laboratory Standards Institute (2011) Performance standards for antimicrobial susceptibility testing Twenty-first informational supplement M100-S21 CLSI Wayne PA USA ClustalW2 httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2 (consulteacute le 3 deacutec 2013) Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) httpcmrjcviorgtigr-scriptsCMRCmrHomePagecgi (consulteacute le 3 deacutec 2013) Creacutepin-Gilbert C (2006) Transfert de ligand dans la cytochrome c oxydase observeacute par des expeacuteriences femtosecondes infrarouge inteacutegreacutees et reacutesolues spectralement Thegravese de doctorat de l rsquoEacutecole Polytechnique Paris Pastel-00002200 version 1 29 jul 2010 Davis BD (1987) Mechanism of bactericidal action of aminoglycosides Microbiol Rev 51 341-350 DeAngelis MM Wang DG and Hawkins TL (1995) Solid-phase reversible immobilization for the isolation of PCR products Nucl Acids Res 22 4742- 4743 Dickgiesser N and Kreiswirth BN (1986) Determination of aminoglycoside resistance in Staphylococcus aureus by DNA hybridization Antimicrob Agents Chemother 29 930-932 Doi Y and Arakawa Y (2007) 16S ribosomal RNA methylation Emerging resistance mechanism against aminoglycosides Clin Infect Dis 45 88ndash94 Enright MC Robinson DA Randle G Feil EJ Grundmann H and Spratt BG (2002) The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA) Proc Natl Acad Sci USA 99 7687-7692 Friedman M (2002) Tomato glycoalkaloids role in the plant and in the diet J Agric Food Chem 50 5751ndash5780
135
Goerke C and Wolz C (2004) Regulatory and genomic plasticity of Staph- ylococcus aureus during persistent colonization and infection Int J Med Microbiol 294 195ndash202 Goumltz F Bannerman T and Schleifer K-H (2006) The genera Staphylococcus and Macrococcus In The Prokaryotes Vol 4 Bacteria Firmicutes Cyanobacteria M Dworkin (ed) (Springer New-York USA) pp5- 75 Haagsma AC Abdillahi-Ibrahim R Wagner MJ Krab K Vergauwen K Guillemont J AndriesK Lill H Koul A and Bald D (2009) Selectivity of TMC207 towards mycobacterial ATP synthase compared with that towards the eukaryotic homologue Antimicrob Agents Chemother 53 1290-1292 Hammer ND Reniere ML Cassat JE Zhang Y Hirsch AO Indriati Hood M and Skaar EP (2013) Two hegraveme-dependent terminal oxydases power Staphylococcus aureus organ-specific colonization of the vertebrate host MBio 4 e00241- 13 Hederstedt L (1993) The Krebs citric acid cycle In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of Microbiology Washington DC) pp 181-198 Hiramatsu K (1995) Molecular evolution of MRSA Microbiol Immunol 39 531-543 Hiramatsu K Hanaki H Ino T Yabuta K Oguri T and Tenover FC (1997) Methicillin-resistant Staphylococcus aureus clinical strain with reduced vancomycin susceptibility J Antimicrob Chemother 40 135-136 Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T et al (2013) Genomic Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
136
Hong S and Pedersen PL (2008) ATP synthase and the actions of inhibitors utilized to study its roles in human health disease and other scientific areas Microbiol Mol Biol Rev 72 590-641
Jevons MP (1961) lsquoCelbeninrsquo-resistant staphylococci BMJ 1 124-125
Joshi S and Huang YG (1991) ATP synthase complex from bovine heart mitochondria the oligomycin sensitivity conferring protein is essential for dicyclohexyl carbodiimidesensitive ATPase Biochim Biophys Acta 1067 255ndash258
Kohanski MA Dwyer DJ and Collins JJ (2010) How antibiotics kill bacteria from targets to networks Nat Rev Microbiol 8 423-435 Kohler C von Eiff C Liebeke M McNamara PJ Lalk M Proctor RA Hecker M and Engelmann S (2008) A defect in menadione biosynthesis induces global changes in gene expression in Staphylococcus aureus J Bacteriol 190 6351- 6364 Kohler C von Eiff C Peters G Proctor RA Hecker M and Engelmann S (2003) Physiological characterization of a hegraveme-deficient mutant of Staphylococcus aureus by a proteomic approach J Bacteriol 185 6928- 6937 Kriegeskorte A Koumlnig S Sander G Pirkl A Mahabir E Proctor RA von Eiff C Peters G and Becker K (2011) Small colony variants of Staphylococcus aureus reveal distinct protein profiles Proteomics 11 2476-2490 Lannergard J von Eiff C Sander G Cordes T Seggewiss J Peters G Proctor RA Becker K and Hughes D (2008) Identification of the genetic basis for clinical menadione-auxotrophic small-colony variant isolates of Staphylococcus aureus Antimicrob Agents Chemother 52 4017-4022
Lauraeus M and Wikstrom M (1993) The terminal quinol oxydases of Bacillus subtilis have different energy conservation properties J Biol Chem 268 11470-11473
137
Li M Gong J Cottrill M Yu H de Lange C Burton J and Topp E (2003) Evaluation of QIAamp DNA Stool Mini Kit for ecological studies of gut microbiota J Microbiol Methods 54 13-20
Liu Y Fiskum G and Schubert D (2002) Generation of reactive oxygen species by the mitochondrial electron transport chain J Neurochem 80 780-787
Martin-Hernandez A M Dufresne M Hugouvieux V Melton R and Osbourn A (2000) Effects of targeted replacement of the tomatinase gene on the interaction of Septoria lycopersici with tomato plants Mol Plant Microbe Interact 13 1301ndash1311
Matteelli A Carvalho AC Dooley KE and Kritski A (2010) TMC207 the first compound of a new class of potent anti-tuberculosis drugs Future Microbiol 5 849-858
Matsuno-Yagi A and Hatefi Y (1993) Studies on the mechanism of oxidative phosphorylation ATP synthesis by submitochondrial particles inhibited at F0 by venturicidin and organotin compounds J Biol Chem 268 6168ndash6173
Miller MH Edberg SC Mandel LJ Behar CF and Steigbigel NH (1980) Gentamicin uptake in wild-type and aminoglycoside-resistant small-colony mutants of Staphylococcus aureus Antimicrob Agents Chemother 18 722- 729
Mingeot-Leclercq M-P Glupczynski Y and Tulkens PM (1999) Aminoglycosides Activity and Resistance Antimicrob Agents Chemother 43 727- 737
Mitchell G Brouillette Eacute Seacuteguin DL Asselin AE Jacob CL and Malouin F (2010) A role for sigma factor B in the emergence of Staphylococcus aureus small-colony variants and elevated biofilm production resulting from an exposure to aminoglycosides Microb Pathog 48 18-27
Mitchell G Gattuso M Bouarab K and Malouin F (2009) Tomatidine (TO) affects virulence regulators of prototypical Staphylococcus aureus (SA) and small colony variants (SCV) of cystic fibrosis (CF) patients Abstr 49th Intersci Conf Antimicrob Agents Chemother abstr C1-1341
138
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K and Malouin F (2011) Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic fibrosis airway epithelial cells Antimicrob Agents Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM and Malouin F (2011) Infection of polarized airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function and is influenced by NF-κB Infect Immun 79 3541-3551
Mitchell G Lafrance M Boulanger S Seacuteguin DL Guay I Gattuso M Marsault Eacute Bouarab K and Malouin F (2012) Tomatidine acts in synergy with aminoglycoside antibiotics against multiresistant Staphylococcus aureus and prevents virulence gene expression J Antimicrob Chemother 67 559-568
Mitchell G Lamontagne CA Brouillette Eacute Grondin G Talbot BG Grandbois M and Malouin F (2008) Staphylococcus aureus SigB activity promotes a strong fibronectin-bacterium interaction which may sustain host tissue colonization by small-colony variants isolated from cystic fibrosis patients Mol Microbiol 70 1540-1555
Mitchell G Malouin F 2012 Outcome and prevention of Pseudomonas aeruginosa-Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic Fibrosis Chap 8 pp 181-206 Ed D Sriramulu InTech Open Access Publisher ISBN 979-953-307-059-8
Mitchell G Seacuteguin DL Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S and Malouin F (2010) Staphylococcus aureus sigma B-dependent emergence of small-colony variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-heptylquinoline-N-oxide BMC Microbiol 30 10-33
Moisan H Brouillette Eacute Jacob CL Langlois-Beacutegin P Michaud S and Malouin F (2006) Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated from cystic fibrosis patients is influenced by SigB J Bacteriol 188 64-76
139
Moreno F Crisp C Jorgensen JH and Patterson JE (1995) Methicillin- resistant Staphylococcus aureus as a community organism Clin Infect Dis 21 1308-1312
Murphy MP (2009) How mitochondria produce reactive oxygen species Biochem J 417 1- 13
Novick RP (2003) Autoinduction and signal transduction in the regulation of staphylococcal virulence Mol Microbiol 48 1429-1449
Novo DJ Perlmutter NG Hunt RH and Shapiro HM (2000) Multiparameter flow cytometric analysis of antibiotic effects on membrane potential membrane permeability and bacterial counts of Staphylococcus aureus and Micrococcus luteus Antimicrob Agents Chemother 44 827-834
ONeill AJ Cove JH and Chopra I (2001) Mutation frequencies for resistance to fusidic acid and rifampicin in Staphylococcus aureus J Antimicrob Chemother 47 647-650
Osbourn A E (1996) Preformed antimicrobial compounds and plant defense against fungal attack Plant Cell 8 1821ndash1831
Otto TD Dillon GP Degrave WS and Berriman M (2011) RATT Rapid annotation transfert tool Nucl Acids Res 39 doi101093 Pratique E Theraulaz L (2007) Eacutetude de la reacutegulation du systegraveme respiratoire TMAO reacuteductase chez Escherichia coli Meacutemoire de maitrise de lrsquoeacutecole pratique des hautes eacutetudes Paris
Price CT and Gustafson JE (2001) Increases in the mutation frequency at which fusidic acid-resistant Staphylococcus aureus arise with salicylate J Med Microbiol 50 104-106
Proctor RA and Peters G (1998) Small colony variants in staphylococcal infections Diagnostic and therapeutic implications Clin Infect Dis 27 419- 423
140
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M and Peters G (2006) Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent infections Nat Rev Microbiol 4 295ndash305
Quail MA Kozarewa I Smith F Scally A Stephens PJ Durbin R Swerdlow H and Turner DJ (2008) A large genome centers improvements to the Illumina sequencing system Nat methods 5 1005-1010
Ramirez MS and Tolmasky ME (2010) Aminoglycoside modifying enzymes Drug Resist Updat 13 151ndash171
Rao M Streur TL Aldwell FE and Cook GM (2001) Intracellular pH regulation by Mycobacterium smegmatis and Mycobacterium bovis BCG Microbiology 147 1017-1024
Richardson DJ (2000) Bacterial respiration a flexible process for a changing environment Microbiology 146 551- 571
Riley TV Pearman JW and Rouse IL (1995) Changing epidemiology of methicillin-resistant Staphylococcus aureus in Western Australia Med J Aust 163 412-414
Roddick J G (1974) The steroidal glycoalkaloid α-tomatine Phytochemistry 13 25-36
Rodrigue S Materna AC Timberlake SC Blackburn MC Malmstrom RR et al(2010) Unlocking Short Read Sequencing for Metagenomics PLoS ONE 5(7) e11840doi101371journalpone0011840 Ruiz-Rubio M Alonso Peacuterez-Espinosa A Lairini K RoldAacuten-Arjona T Dipietro A and Anaya N (2001) Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi In Studies in natural products chemistry vol 25 A Rahman (ed) (Elsevier Oxford United Kingdom) pp 293ndash326
141
Samuelsen oslash Haukland HH Kahl BC von Eiff C Proctor RA Ulvatne H Sandvik K and Vorland LH (2005) Staphylococcus aureus small colony variants are resistant to the antimicrobial peptide lactoferricin B J Antimicrob Chemother 56 1126- 1129
Sandrock R W and Vanetten HD (1998) Fungal sensitivity to and enzymatic degradation of the phytoanticipin α-tomatine Phytopathology 88 137ndash143
Sendi P and Proctor RA (2009) Staphylococcus aureus as an intracellular pathogen the role of small colony variants Trends Microbiol 17 54ndash58
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C and Osbourn A (2006) Dual effects of plant steroidal alkaloids on Saccha- romyces cerevisiae Antimicrob Agents Chemother 50 2732ndash2740
Somerville GA and Proctor RA (2009) At the crossroads of bacterial metabolism and virulence factor synthesis in Staphylococci Microbiol Mol Biol Rev 73 233- 248
Starkov AA (2010) Measurement of mitochondrial ROS production Methods Mol Biol 648 245-255
Taber HW (1993) Respiratory chain In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of MicrobiologyWashington DC) pp 199-212
Taber HW and Morrison M (1964) Electron transport in Staphylococci Properties of a particle preparation from exponential phase Staphylococcus aureus Arch Biochem Biophys 105 367- 379
Taber HW et al(1987) Bacterial uptake of aminoglycoside antibiotics Microbiol Rev 51 439-457
Thoumlny-Meyer L (1997) Biogenesis of respiratory cytochromes in bacteria Microbiol Mol Biol Rev 61 337- 376
142
Tubby S Wilson M Wright JA Zhang P and Nair SP (2013) Staphylococcus aureus small colony variants are susceptible to light activated antimicrobial agents BMC Microbiol 13 201- 204
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D Roth J Proctor RA Becker K et al (2011) Staphylococcus aureus phenotype switching an effective bacterial strategy to escape host immune response and establish a chronic infection EMBO Mol Med 3 129ndash41
Turrens JF (2003) Mitochondrial formation of reactive oxygen species J Physiol 552 335ndash 344
Tynecka Z Szcześniak Z Malm A and Los R (1999) Energy conservation in aerobically grown Staphylococcus aureus Res Microbiol 150 555- 566
Udo EE Pearman JW and Grubb WB(1993) Genetic analysis of community isolates of methicillin-resistant Staphylococcus aureus in Western Australia J Hosp Infect 25 97-108
Vakulenko SB and Mobashery S (2003) Versatility of Aminoglycosides and Prospects for Their Future Clin Microbiol Rev 16 430-450
Voggu L Schlag S Biswas R Rosenstein R Rausch C and Goumltz F (2006) Microevolution of cytochrome bd oxydase in Staphylococci and its implication in resistance to respiratory toxins released by Pseudomonas J Bacteriol 188 8079- 8086
Wells PG Bhuller Y Chen CS Jeng W Kasapinovic S Kennedy JC Kim PM Laposa RR McCallum GP Nicol CJ et al (2005) Molecular and biochemical mechanisms in teratogenesis involving reactive oxygen species Toxicol Appl Pharmacol 207 354-366
143
Yan M Yu C Yang J and Ji Y (2011) The essential two-component system YhcSR is involved in regulation of the nitrate respiratory pathway of Staphylococcus aureus J Bacteriol 193 1799- 1805
Zhang T Luo Y Chen Y Li X and Yu J (2012) BIGrat a repeat resolver for pyrosequencing-based re-sequencing with Newbler BMC Res Notes 5 567
144

iii
polarisation membranaire et de la production de ROS intracellulaire ce qui
expliquerait la diffeacuterence drsquoactiviteacute entre les souches agrave pheacutenotype normal et les
SCV
Mots cleacutes tomatidine ATP synthase chaine de transport des eacutelectrons synergie laquo small-colony variants raquo ROS S aureus analogues
iv
REMERCIEMENTS
Je voudrais remercier mon directeur de recherche Pr Franccedilois Malouin qui a cru
en moi pour mrsquoavoir proposeacute ce beau projet de maitrise Merci pour sa grande
disponibiliteacute son eacutecoute et ses critiques toujours constructives et jamais
condescendantes
Je voudrais eacutegalement remercier mes conseilleacutes Kamal Bouarab Seacutebastien
Rodrigue et Eacuteric Marsault leur implication dans mon projet leur occasionnant un
surplus de travail Merci pour leur support technique et leur disponibiliteacute agrave reacutepondre
agrave mes questions
Un gros merci agrave Gabriel Mitchell qui a eacuteteacute mon mentor au deacutebut de ma maitrise et
qui mrsquoa introduit au projet laquo Tomatidine raquo Un merci particulier eacutegalement agrave
Maxime-Lamontagne-Boulet pour son aide en tant que stagiaire lors de mes
derniegraveres expeacuteriences Je voudrais eacutegalement remercier tous les membres du
laboratoire preacutesents et passeacutes qui ont toujours eacuteteacute disponibles pour partager avec
moi leurs connaissances theacuteoriques et techniques
Je ne peux passer sous silence lrsquoimplication de lrsquoUniversiteacute de Sherbrooke et du
FQRNT comme organismes subventionnaires au projet Merci eacutegalement agrave Fibrose
Kystique Canada qui a participeacute eacutegalement agrave la subvention drsquoune partie du projet
En terminant je voudrais remercier ma famille qui a crue en moi et qui mrsquoencourage
agrave perseacuteveacuterer depuis toujours Un merci tout particulier agrave mon conjoint et ma belle-
fille pour qui je nrsquoai pas toujours eacuteteacute aussi disponible que jrsquoaurais voulu lrsquoecirctre et qui
mrsquoont supporteacutee au quotidien et ce dans tous les sens du terme
v
TABLE DES MATIEgraveRES
SOMMAIRE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ii
REMERCIEMENTS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip iv
TABLE DES MATIEgraveRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip v
LISTE DES ABBREacuteVIATIONS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip vii
LISTE DES TABLEAUX helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ix
LISTE DES FIGURES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip x
CHAPITRE 1 INTRODUCTION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
11 Staphylococcus aureus helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
111 Importance cliniquehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)helliphellip 2
1121 Diffeacuterences et similariteacutes helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 2
1122 La chaine de transport des eacutelectrons helliphelliphelliphelliphellip 4
12 Antibiotiques Modes drsquoaction cibles cellulaire et
meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles helliphelliphelliphellip 25
122 Les aminoglycosides helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1221 Mode drsquoaction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1222 Meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphellip 31
123 La tomatidine helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 33
13 Projet de maitrise helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
CHAPITRE 2 ARTICLE SCIENTIFIQUE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 37
ldquoTomatidine and derivative FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Firmicutes Including Those of Listeria monocytogenes
and Are Bactericidal in Combination With Aminoglycosides Against their Normal
Phenotyperdquo
vi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE helliphelliphelliphellip 76
31 Obtention des mutants helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
32 Seacutequenccedilage et analyses bio-informatiques helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 79
321 Identification des mutations en lien avec la tomatidine helliphellip 89
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct helliphellip 90
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine helliphelliphelliphelliphellip 99
CHAPITRE 4 DISCUSSION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 113
CHAPITRE 5 CONCLUSION ET PERSPECTIVES helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 124
CONTRIBUTIONS SUPPLEacuteMENTAIRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 1 Application de brevet helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 2 Article scientifique helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 129
laquo Unraveling the structure-activity relationship of tomatidine
a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureus raquo
BIBLIOGRAPHIE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 131
vii
LISTE DES ABBREacuteVIATIONS
Adh Alcool deacuteshydrogeacutenase
ADN Acide deacutesoxyribo-nucleacuteique
ADP Adeacutenosine diphosphate
ARN Acide ribo-nucleacuteique
ATP Adeacutenosine triphosphate
CA-MRSA Community-associated MRSA
CMI concentration minimale inhibitrice
CoQ Coenzyme Q
Cyt Cytochrome
DMQ laquo demethylmenaquinone raquo
dTMP Deacutesoxythymidine monophosphate
dUMP Deacutesoxyuridine monophosphate
FADH2 Flavine adenine dinucleacuteotide
GEN gentamicine
HA-MRSA Healthcare-associated MRSA
Ldh Lactate deacuteshydrogeacutenase
ml millilitre
MQ meacutenaquinone
MRSA methycilin resistant Staphylococcus aureus
Mqo2 Quninol oxidoreacuteductase
NADH Nicotinamide adenine dinucleacuteotide (forme reacuteduite)
Pdh Pyruvate deacuteshydrogeacutenase
Pfl Pyruvate formate lyase
PyrD Dihydroorotate deacuteshydrogeacutenase
Q Quinone
Rif rifampicine
ROS laquo reactive oxygen species raquo espegravece reacuteactive de lrsquooxygegravene
viii
[S] Polysulfide
S aureus Staphylococcus aureus
SARM Staphylococcus aureus resistant agrave la meacutethycilline
SCV laquo small colony variants raquo
SHCHC 2-succinyl-6-hydroxy-24-cyclohexadiene-1-carboxylate syntase
TCA Tricarboxylic acid cycle
TCD tableau croiseacute dynamique
TO tomatidine UFC Uniteacute formatrice de colonies
UQ Ubiquinone
microg microgramme
ix
LISTE DES TABLEAUX
CHAPITRE 1 INTRODUCTION
1 Cibles et modes drsquoaction des principales classes drsquoantibiotique 27
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants (B) chez les mutants NewbouldΔhemB reacutesistants 95-96
CHAPITRE 4 DISCUSSION
1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 123
x
LISTE DES FIGURES
CHAPITRE 1 INTRODUCTION
1 Diffeacuterence entre les pheacutenotypes normaux et SCV de S aureus 3
2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes 6
3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale 8
4 Chaines respiratoires bacteacuteriennes contenant des cytochromes 9
5 Flexibiliteacute respiratoire (A) chez Paracoccus denitrificans (B) chez
Escherichia coli 10
6 Chaine de transport des eacutelectrons proposeacutee chez les staphylocoques 12
7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies 17
8 Modegravele de reacutegulation du cycle TCA 20
9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS 23
10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de O2-bull 24
11 Voies de formation et de deacutetoxification des ROS 25
12 Cineacutetique drsquoentreacutee des aminoglycosides 30
13 Sites de mutations de certains reacutesistants aux aminoglycosides 32
14 Structures chimiques de (A) tomatine (B) tomatidine et (C) Analogue
de tomatidine FC04-100 34
xi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Graphiques des CMI pour chacun des passages pour (A) la
tomatidine sur NewbouldΔhemB (B) la tomatidine en preacutesence de
gentamicine sur NewbouldΔhemB (C) la gentamicine sur S aureus
ATCC 29213 (D) la gentamicine en preacutesence de tomatidine sur S
aureus ATCC 29213 77-79
2 Organigramme des mutants reacutesistants seacutequenceacutes 80-81
3 Eacutetapes du seacutequenccedilage Illumina 83
4 Meacutethode drsquoamplification par la meacutethode Illumina 85
5 Correction du programme RATT sur les annotations transfeacutereacutees 88
6 Exemple drsquoune partie du tableau compilation des mutations 89
7 Seacutelection des gegravenes drsquointeacuterecirct chez ATCC 29213 dans le TCD par
lrsquoutilisation de diffeacuterents filtres 91-92
8 Seacutelection des gegravenes drsquointeacuterecirct chez NewbouldΔhemB dans le TCD par
lrsquoutilisation de diffeacuterents filtres 93-94
9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct 97-98
10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants 100
11 Courbe de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison gentamicine (+TO) du premier et troisiegraveme niveau de
reacutesistance 101
12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B)
NewbouldΔhemB 102
xii
13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213 104
14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicinelorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC
29213 105
15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants
ATCC 29213 reacutesistants agrave la combinaison gentamicine (+TO) 106
16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la
tomatidine 107
17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la
tomatidine 108
18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB
reacutesistants agrave la combinaison tomatidine (+gentamicine) 109
19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane
Potential Kit (Invitrogen) 111
20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du
gegravene atpE de S aureus Newman 112
xiii
CHAPITRE 4 DISCUSSION
1 Site drsquoinhibition de lrsquoATP synthase par diffeacuterentes moleacutecules 114
2 Structure chimique de (A) TMC207 et (B) lrsquoanalogue anti Gram
positifs 115
3 Seacutequence proteacuteique de la proteacuteine AtpE chez diffeacuterentes espegraveces 118
4 Modeacutelisation de la structure tridimentionnelle de lrsquoATP synthase de S
aureus (atpE) montrant les acides amineacutes muteacutes chez les mutants
reacutesistants 121
- 1 -
CHAPITRE 1
INTRODUCTION
11 Staphylococcus aureus
111 Importance clinique
Staphylococcus aureus est une bacteacuterie agrave Gram positif anaeacuterobie facultative Crsquoest un
microorganisme opportuniste pouvant infecter diffeacuterents hocirctes diffeacuterents organes ou
sites drsquoinfection et ayant la capaciteacute de causer tant des infections aiguumles que
chroniques (Archer 1998 Goerke et Wolz 2004)
Staphylococcus aureus reacutesistant agrave la meacutethilline le SARM (ou laquo MRSA raquo) est une
souche de S aureus devenue reacutesistante aux β-Lactamines par lrsquoacquisition du gegravene
mecA dans son chromosome (Hiramatsu 1995) Le SARM a eacuteteacute isoleacute pour la
premiegravere fois en 1960 en Angleterre (Jevons 1961) et est devenu eacutepideacutemique agrave
travers le monde depuis les anneacutees 1970 Au moins trois geacutenotypes distincts eacutetaient
recenseacutes dans les anneacutees 1980 (Hiramatsu 1995) dont deux sont toujours preacutevalents
agrave travers le monde comme eacutetant des souches HA-MRSA (Healthcare-associated
MRSA) des souches acquises agrave lrsquohocircpital (Enright et al 2002) Ces souches ont
acquis des gegravenes de reacutesistance agrave pratiquement tous les antibiotiques introduits dans
lrsquoutilisation clinique depuis les 50 derniegraveres anneacutees mecircme agrave lrsquoantibiotique de dernier
ressort la vancomycine(Hiramitsu 1997 Chen et al 2003) Nous sommes
maintenant en manque drsquoagents theacuterapeutiques efficaces contre les SARM touchant
les personnes immunodeacuteprimeacutees qui sont dans les hocircpitaux En plus depuis le deacutebut
des anneacutees 1990 des clones de SARM dont plusieurs deacutemontraient un potentiel
pathogeacutenique supeacuterieur aux souches classiques de HA-MRSA retrouveacutees agrave lrsquohocircpital
- 2 -
sont maintenant retrouveacutes dans la communauteacute (CA-MRSA) agrave travers le monde (Udo
et al 1993 Riley et al 1995 Moreno et al 1995 Aubry-Damon et al 1997) et ont
la capaciteacute drsquoaffecter des personnes en santeacute en causant des infections aiguumles
(Hiramatsu et al 2013) Il apparaicirct donc urgent de trouver des drogues alternatives
afin de surmonter le nombre grandissant de souches de SARM
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)
1121 Diffeacuterences et similariteacutes
Les laquo small colony variants raquo (SCV) sont des bacteacuteries souvent isoleacutees lors des
infections chroniques comme crsquoest le cas des infections pulmonaires chez les
patients atteints de fibrose kystique mais aussi dans des cas drsquoosteacuteomyeacutelite
drsquoarthrite septique drsquoinfections de prothegraveses orthopeacutediques et lors de la mammite
bovine (Alexender et Hudson 2001 Moisan et al 2006 Proctor et al 2006) Les
SCV sont caracteacuteriseacutes par soit un meacutetabolisme oxydatif dysfonctionnel ou un manque
dans la biosynthegravese de la thymidine les deux amenant une alteacuteration dans
lrsquoexpression des facteurs de virulence une croissance plus lente et une diminution
de la pigmentation des colonies (Figure 1) (Proctor et al 2006) Cependant alors
que les souches normales reacutepriment lrsquoexpression des proteacuteines de surface et
expriment les exoproteacuteines lors de la phase stationnaire (Novick 2003) les SCV
expriment de faccedilon stable les gegravenes Sig-B deacutependants codant pour des proteacuteines de
surface telle les adheacutesines au lieu drsquoactiver le systegraveme agr deacutependant du quorum-
sensing et produire des exoproteacuteines (Moisan et al 2006) Ces diffeacuterences dans
lrsquoexpression des facteurs de virulence reacutesultent chez les SCV en une meilleure
habileteacute agrave adheacuterer aux composantes de lrsquohocircte (Mitchell et al 2008) et agrave former du
biofilm (Mitchell et al 2010a Mitchell et al 20010b) et seraient eacutegalement lieacutees agrave
lrsquohabileteacute des SCV agrave lrsquoinvasion et la persistance des cellules de lrsquohocircte (Proctor et al
2006 Sendi et Proctor 2009 Mitchell et al 2011b)
- 3 -
S aureus est donc un pathogegravene qui peut se preacutesenter sous deux pheacutenotypes qui
sont souvent retrouveacutes simultaneacutement lors drsquoune mecircme infection et qui ont le pouvoir
de reacuteverter drsquoun pheacutenotype agrave lrsquoautre selon les conditions du milieu (Figure 1)
(Tuchscherr et al 2011)
Le pheacutenotype dit rdquonormalrdquo est associeacute aux infections aiguumles agrave une forte production
de facteurs de virulence (ex heacutemolysine) et est associeacute agrave la phase de disseacutemination
de la bacteacuterie Lrsquoautre pheacutenotype le laquo small-colony variants raquo ou SCV est affecteacute
dans sa chaine respiratoire ce qui amegravene une croissance beaucoup plus lente des
colonies de 8 -10 fois plus petites que les prototypes et une faible susceptibiliteacute aux
aminoglycosides Les SCV sont plutocirct associeacutes aux infections chroniques agrave S aureus
de par leur capaciteacute agrave srsquointernaliser dans les cellules de lrsquohocircte leur forte capaciteacute agrave
produire du biofilm et leur capaciteacute agrave produire des facteurs de colonisation
(Tuchscherr et al 2011 Proctor et al 2006) (Figure 1)
Figure 1 Diffeacuterences entre les pheacutenotypes normaux et SCV de S aureus
S aureus agrave
pheacutenotype normal
Disseacutemination
amp
Infections aiguumles
Colonisation
amp
Infections persistantes
SCVs
(laquo small-colony
variants raquo)
Reacutevers
ion
pheacutenoty
piq
ue
Norm
al
SC
V
Croissance
Biofilm
Internalisationheacutemolyse
Mitchell G et Malouin F 2012
- 4 -
1122 La chaine de transport des eacutelectrons
Chaine respiratoire tableau geacuteneacuteral
Les chaines respiratoires bacteacuteriennes bien que tregraves variables dans leurs
composantes parmi les diffeacuterentes espegraveces sont toutes constitueacutees sur le mecircme
modegravele de base Elles sont composeacutees de trois composantes majeures soit
- les deacuteshydrogeacutenases primaires qui sont chargeacutees drsquooxyder des substrats
principalement les coenzymes NADH et FADH2 (issues du cycle de Krebs) (mais
aussi selon le cas le succinate le lactate lrsquohydrogegravene hellip) et de pomper deux
protons du cocircteacute externe de la membrane en plus de transfeacuterer deux eacutelectrons agrave un
groupement de quinones
- Les quinones sont des moleacutecules liposolubles non proteacuteiques mobiles dans la
membrane Elles servent drsquointermeacutediaires entre les deacuteshydrogeacutenases primaires et
les oxydases terminales pour le transport des eacutelectrons Il existe deux sortes de
quinones les benzoquinones dont font partie les ubiquinones et les
naphtoquinones dont font partie les meacutenaquinones Les bacteacuteries agrave Gram positif
ne possegravedent que des meacutenaquinones alors que les deux sont preacutesentes chez les
bacteacuteries agrave Gram neacutegatif (Hederstedt 1993 Pratique et Theraulaz 2007)
- Les oxidoreacuteductases terminales sont chargeacutees de reacuteduire lrsquoaccepteur final
drsquoeacutelectron lrsquooxygegravene eacutetant lrsquoaccepteur final privileacutegieacute
De faccedilon plus deacutetailleacutee la chaine respiratoire est composeacutee de cinq
complexes proteacuteiques principaux (Figure 2)
- complexe I NADH deacuteshydrogeacutenase ou NADH reacuteductase ou NADHCoQ
oxidoreacuteductase ou NADH-ubiquinone oxidoreacuteductase
- complexe II Succinate deacuteshydrogeacutenase
- complexe III Cytochrome bc1 complexe ou ubiquinol cytochrome c
reacuteductaseoxydoreacuteductase
- complexe IV Cytochrome c oxydase
- 5 -
- complexe V ATP synthase (Creacutepin-Gilbert et al 2006)
La phosphorylation oxydative est deacuteclencheacutee dans la chaine de complexes
proteacuteiques par la preacutesence drsquoO2 accepteur terminal privileacutegieacute drsquoeacutelectrons (ou drsquoautres
accepteurs finaux tel le nitrate) dans le milieu ainsi que par les eacutelectrons ameneacutes par
le NADH et le FADH2 au complexe I et II respectivement (Creacutepin-Gilbert et al 2006)
Les eacutelectrons vont ensuite transiter par le complexe III jusqursquoau complexe IV ou les
eacutelectrons seront utiliseacutes afin de reacuteduire lrsquooxygegravene en eau le complexe IV est
lrsquooxydase terminale consideacutereacutee comme eacutetant lrsquoaccepteur final drsquoeacutelectrons Le transfert
drsquoeacutelectrons est possible gracircce aux reacuteactions drsquooxydoreacuteduction en cascade dans les
diffeacuterents complexes proteacuteiques Ces reacuteactions fournissent de lrsquoeacutenergie libre utiliseacutee
par les complexes I III et IV afin de transfeacuterer des protons agrave lrsquoexteacuterieur de la cellule
(Creacutepin-Gilbert et al 2006) Le complexe V (ATP synthase) quant agrave lui utilise les
eacutelectrons pompeacutes agrave lrsquoexteacuterieur par les autres complexes comme force proton-motrice
afin de phosphoryler trois moleacutecules drsquoADP et ainsi produire trois ATP Le transport
des eacutelectrons entre les diffeacuterents complexes proteacuteiques est possible gracircce agrave la
preacutesence dans la membrane de deux types de moleacutecules liposolubles non proteacuteiques
mobiles les meacutenaquinones et le cytochrome c Les meacutenaquinones seules quinones
preacutesentes chez les bacteacuteries agrave Gram positif assurent le transport des eacutelectrons entre
le complexe I et le complexe III et entre le complexe II et III Le cytochrome c pour sa
part veacutehicule les eacutelectrons entre le complexe III et le complexe IV Le transport des
eacutelectrons srsquoeffectue de la composante ayant le plus faible potentiel drsquooxydoreacuteduction
vers celle ayant le plus fort potentiel
- 6 -
La chaine respiratoire bacteacuterienne
La mitochondrie montre assez peu de flexibiliteacute respiratoire Il y a une certaine
flexibiliteacute au niveau de lrsquoentreacutee des eacutelectrons mais aucune au niveau de la sortie des
eacutelectrons ougrave la cytochrome aa3 oxydase reacuteduit lrsquooxygegravene en eau (Richardson 2000)
(Figure 3) Chez les bacteacuteries alors que le complexe II fonctionne sensiblement de
faccedilon identique agrave celui des eucaryotes les autres complexes peuvent se composer
de diffeacuterents complexes enzymatiques (reacuteductases) selon les espegraveces et les
conditions environnementales (Richardson 2000 Thoumlny-Meyer 1997) En outre
chez les bacteacuteries contrairement aux eucaryotes la chaine respiratoire comporte
souvent des voies de transport des eacutelectrons alternatives on dit alors qursquoelles ont une
chaine respiratoire agrave branches ou brancheacutee ou ramifieacutee (Artzatbanov et Petrov 1990)
Plusieurs espegraveces bacteacuteriennes possegravedent une chaine respiratoire ramifieacutee dont la
ramification se situe habituellement suite agrave la reacuteduction des quinones en quinols
(Lauraeus et Wikstrom 1993 Thoumlny-Meyer 1997) (Figure 4) Une des deux
Complexe I NADH
deacuteshydrogeacutenase
Complexe II Succinate
deacuteshydrogeacutenase
Complexe III
bc 1
Complexe IV Cytochrome c
oxydase
Complexe V
ATP synthase
Modifieacute de wikipedia httpfrwikipediaorgwikiChaine_respiratoire
Figure 2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes
Leurs rocircles dans le transport des eacutelectrons et dans lrsquoeacutelaboration du gradient de
protons et le lien entre la chaine respiratoire et le cycle de Krebs Q Quinones
- 7 -
branches connue comme la branche cytochrome c oxyde les quinols (meacutenaquinol
QH2) via la voie complexe bc1-cytochrome c- cytochrome c oxydase Lrsquoautre
branche-type consiste en une seule uniteacute enzymatique (cytochrome bd) laquelle
transfegravere directement les eacutelectrons des quinols agrave lrsquooxygegravene Cette voie est connue
comme la branche quinol oxydase il srsquoagit drsquoune oxydase terminale transfeacuterant
directement les eacutelectrons agrave lrsquooxygegravene eacutevitant de passer par lrsquointermeacutediaire du
complexe III (bc1) et du cytochrome c (Artzatbanov et Petrov 1990 Lauraeus et
Wikstrom 1993) Alors que la voie du cytochrome c permet le pompage de protons agrave
lrsquoexteacuterieur par lrsquoaction du complexe bc1 (ou complexe III) et du cytochrome c oxydase
le cytochrome bd ne permet aucun pompage de protons (Lauraeus et Wikstrom
1993) La voie passant par le complexe bc1-cytochrome c-cytochrome c oxydase est
plus efficace dans la production drsquoeacutenergie Alors que les 2 voies peuvent ecirctre
exprimeacutees simultaneacutement chez quelques espegraveces il semble que lrsquoexpression de ces
voies soit inductible selon les conditions environnementales chez Bacillus subtilis et
les Staphylocoques (Lauraeus et Wikstrom 1993) On pourrait dire eacutegalement que la
chaine respiratoire est seacutepareacutee en deux eacutetapes la premiegravere partie est la phase
quinone-reacuteductase (Q-QH2) et la seconde la phase quinol-oxydase (QH2-Q) menant
agrave lrsquooxydase terminale La phase de reacuteduction est responsable du transfert des
eacutelectrons drsquoune varieacuteteacute de substrats tels le NADH le succinate le lactate le formate
et lrsquohydrogegravene vers les quinones et inclut diffeacuterentes reacuteductases (Thoumlny-Meyer 1997)
(Figure 4)
- 8 -
Figure 3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale UQ ubiquinone UQH2 ubiquinol Cyt cytochrome Il semble y avoir presque autant de chaines de transport des eacutelectrons diffeacuterentes
qursquoil y a drsquoespegraveces bacteacuteriennes (par exemple Paracoccus denitrificans et Escherichia
coli (Figure 5)) De plus il ne semble pas y avoir consensus dans la litteacuterature sur
les noms attribueacutes aux oxydases terminales Les noms pour les oxydases terminales
deacutependent des types de cytochromes dont elles sont composeacutees Il ne semble donc
pas eacutevident de connaicirctre de faccedilon preacutecise les types de cytochromes qui font partie
des oxydases terminales Pour rajouter agrave la confusion les gegravenes et opeacuterons ne sont
pas toujours associeacutes agrave leur oxydase terminale dans la litteacuterature ce qui fait qursquoon ne
peut savoir de faccedilon certaine de quelle oxydase on parle Le Cytochrome aa3 peut
eacutegalement porter le nom de Cyt ba3 et le cytochrome bd peux eacutegalement porter les
noms de Cyt bo
Tireacute de Richardson DJ 2000
- 9 -
Succinate
Formate
H2
SDH
FDH
HYD
QQH 2 bc1 c
Reacuteductase alternatives
b
c
cd1c
cb
c
b
c
d
Siroheme
Siroheme
b
NO3-
NO2-
NO
TMAO
DMSOS2O3
2-
[S]
SO32-
Fumarate
Quinol oxidases
ba3bb3bo3d
bd
aa3
O2
Cyt c oxidases
aa3ba3baa3caa3cbb3cao
co
O2
Modifieacute de Thony-Mayer L 1997
NADH
NDH
Figure 4 Chaines respiratoires bacteacuteriennes contenant des cytochromes
Du cocircteacute gauche on retrouve la partie quinone (ubiquinone ou meacutenaquinone)-
reacuteductase Du cocircteacute droit on retrouve la partie quinol-oxydase avec les oxydases
terminales reacuteduisant lrsquoO2 ou lrsquoaccepteur terminal alternatif tel que montreacute Les
oxydoreacuteductases sont surligneacutees QQH2 repreacutesente le pool de quinonesquinols
dans la membrane Les polysulfides sont repreacutesenteacutes par [S] Les chaines
respiratoires ne contenant pas de cytochromes ne sont pas montreacutees NDH
NADH deacuteshydrogeacutenase
Modifieacute de Thoumlny-Mayer L 1997
- 10 -
Figure 5 Flexibiliteacute respiratoire chez Paracoccus denitrificans (a) et chez
Escherichia coli (b) MQ menaquinone UQ ubiquinone DMQ
demethylmenoquinone
Chez S aureus
Comme plusieurs autres espegraveces bacteacuteriennes anaeacuterobies facultatives S aureus a
la possibiliteacute drsquoexprimer diffeacuterentes voies de synthegravese eacutenergeacutetique dont lrsquoexpression
est finement reacuteguleacutee selon des conditions environnementales du moment (Hammer et
Tireacute de Richardson DJ 2000
- 11 -
al 2013 Lauraeus et Wikstrom 1993) Mecircme si S aureus peut utiliser la
fermentation la respiration (aeacuterobie et anaeacuterobie) sera privileacutegieacutee par la bacteacuterie
puisque celle-ci amegravene une bien meilleure production drsquoeacutenergie (Goumltz et al 2006
Hammer et al 2013 ) La respiration aeacuterobie avec lrsquooxygegravene comme accepteur final
drsquoeacutelectron sera utiliseacutee de faccedilon preacutefeacuterentielle en preacutesence drsquooxygegravene (Hammer et al
2013) Malgreacute sa versatiliteacute dans ses voies de synthegravese eacutenergeacutetique S aureus et les
autres bacteacuteries agrave Gram positif ne produisent qursquoun seul type de quinone les
meacutenaquinones lesquelles sont utiliseacutees pour la croissance degraves qursquoil y a de lrsquooxygegravene
de preacutesent dans le milieu (Somerville et Proctor 2009) La reacuteduction de lrsquooxygegravene en
eau est meacutedieacutee par des oxydases terminales hegraveme deacutependant (Hammer et al 2013)
Selon certains auteurs S aureus possegravederait deux oxydases terminales la
cytochrome oxydase aa3 (ou ba3) (QoxABCD) et la cytochrome bd quinol oxydase
(CydAB) (Goumltz et al 2006 Hammer et al 2013 Kohler et al 2008) Cette derniegravere
est exclusive aux procaryotes et est une voie de contournement du complexe III qui
transmet directement les eacutelectrons du laquo pool raquo de quinol (QH2) agrave lrsquooxygegravene Selon
certains auteurs il y aurait eacutegalement chez S aureus une troisiegraveme oxydase
terminale le cytochrome o (CyoABCDE) (Thoumlny-Meyer 1997 Voggu et al 2006)
qui selon certain serait un cytochrome oxydase terminal alternatif au cytochrome aa3
et au cytochrome bd (Voggu et al 2006 Goumltz et al 2006 Taber et Morrison 1964)
(Figure 6) Ces voies alternatives permettent agrave S aureus de srsquoadapter rapidement agrave
un changement drsquoenvironnement (Hammer et al 2013) In vivo chaque organe est
un environnement diffeacuterent preacutesentant des deacutefis respiratoires distincts dus agrave la
variation dans 1- la quantiteacute drsquooxygegravene disponible 2- la disponibiliteacute drsquoaccepteurs
drsquoeacutelectrons alternatifs 3- la concentration drsquoinhibiteurs de la respiration dans le milieu
tels les oxydes nitriques (NO) produits par les neutrophiles et 4- la disponibiliteacute des
cofacteurs neacutecessaires agrave la production drsquoeacutenergie (Hammer et al 2013) Le
cytochrome est exprimeacute de faccedilon modeacutereacutee dans un milieu riche en oxygegravene mais est
fortement exprimeacute dans un milieu pauvre en oxygegravene mais riche en oxyde nitrique
De plus le type drsquooxydoreacuteductase terminale exprimeacute est influenceacute eacutegalement par la
- 12 -
source de carbone disponible la phase de croissance le pH extracellulaire et la
tension en oxygegravene (Lauraeus et Wikstrom 1993 Tynecka et al 1999)
La chaine respiratoire chez les SCV
S aureus est particuliegraverement doueacute agrave surpasser les deacutefis respiratoires en acqueacuterant
une reacutesistance agrave lrsquoacide nitrique produit par les neutrophiles et une habiliteacute agrave persister
dans les cellules de lrsquohocircte en se preacutesentant comme un variant agrave petites colonies
ayant une chaine respiratoire deacuteficiente (SCV) (Hammer et al 2013)
Deux principaux types de SCV sont retrouveacutes dans les isolats cliniques 1- les SCV
ayant une chaine de transport des eacutelectrons deacutefectueuse et 2- les SCV auxotrophes
pour la thymidine (Proctor et Peters 1998 Samuelsen et al 2005) Ces derniers
Figure 6 Chaine de transport des eacutelectrons proposeacutee chez les
staphylocoques Les informations disponibles sur les composantes de la chaine
respiratoire des staphylocoques suggegraverent une chaine ramifieacutee consistant en 2
oxydases terminales meacutenaquinol-deacutependante un cytochrome o et un cytochrome
aa3 oxydase NADH deacuteshydrogeacutenase (DHase) transmet les eacutelectrons au laquo pool raquo
de meacutenaquinones De lagrave les eacutelectrons sont transfeacutereacutes soit au cytochrome b ou agrave
lrsquoaccepteur alternatif drsquoeacutelectrons le cytochrome bd quinol oxydase Le cytochrome
b transfegravere les eacutelectrons agrave une des deux oxydases terminales Ces oxydases
terminales transfegraverent les eacutelectrons agrave lrsquooxygegravene
Modifieacute de Voggu L et al 2006
- 13 -
arborent une ou plusieurs mutations dans le gegravene thyA codant pour la thymidylate
synthase laquelle catalyse la derniegravere eacutetape dans la voie de biosynthegravese de la
thymidine crsquoest-agrave-dire la formation de dTMP agrave partir du dUMP (Samuelsen et al
2005) Le dTMP est essentiel pour la synthegravese de lrsquoADN (Besier et al 2007) Les
mutants thymidine-auxotrophes sont souvent la conseacutequence drsquoune pression
seacutelective par lrsquoutilisation prolongeacutee de trimethoprim-sulfamethoxazole (TMP-SMX)
(souvent utiliseacute pour le traitement des patients fibroses kystiques) un antibiotique qui
interfegravere dans la biosynthegravese de lrsquoacide tetrahydrofolique qui agit comme cofacteur de
la thymidylate synthase (Besier et al 2007 Lannergard et al 2008 Samuelsen et al
2005) Mecircme si de premiers abords les SCV thymidine-auxotrophes ne semblent pas
avoir de lien avec la chaine transport des eacutelectrons il se pourrait que celle-ci ait sont
importance chez ces SCV puisque lrsquoentreacutee de la thymidine passe par NupC dont le
fonctionnement neacutecessite un gradient eacutelectrochimique creacuteeacute par la chaine de transport
des eacutelectrons (Proctor et Peters 1998 ) Les caracteacuteristiques pheacutenotypiques des
SCV deacutecrites ci-apregraves sont toutes lieacutees agrave lrsquoalteacuteration de la chaine de transport des
eacutelectrons 1- la croissance plus lente parce que la synthegravese de la paroi cellulaire
neacutecessite une grande quantiteacute drsquoATP 2-la diminution de la formation de pigment
parce que la biosynthegravese des caroteacutenoiumldes requiert le transport des eacutelectrons 3- une
reacutesistance accrue agrave la gentamicine et autres aminoglycosides parce que lrsquoentreacutee de
ces composeacutes neacutecessite un grand potentiel membranaire geacuteneacutereacute par le transport des
eacutelectrons (Lannergard et al 2008) et 4- une perte partielle ou complegravete de la
fermentation du mannitol parce que lrsquoutilisation des sucres-alcool tels que le
mannitol est diminueacutee lorsque la chaine de transport des eacutelectrons ne peut oxyder le
NADH produit lorsque le mannitol est meacutetaboliseacute en fructose-6-phosphate (Kohler et
al 2003 Proctor et Peters 1998 Samuelsen et al 2005)
Les SCV posseacutedant une chaine de transport des eacutelectrons deacutefectueuse ont une voie
de biosynthegravese de lrsquoheacutemine ou de la meacutenadione deacuteficiente et le pheacutenotype est
reacuteversible lorsque compleacutementeacute avec lrsquoheacutemine ou la meacutenadione (Hammer et al
- 14 -
2013 Kohler et al 2008 Lannergard et al 2008 Proctor et Peters 1998) La
meacutenadione est essentielle agrave la biosynthegravese des meacutenaquinones et lrsquoheacutemine est
essentielle agrave la biosynthegravese de lrsquohegraveme qui entre dans formation des cytochromes
(Hammer et al 2013 Kohler et al 2008 Lannergard et al 2008 Thoumlny-Meyer
1997) Les meacutenaquinones et les cytochromes sont essentiels au fonctionnement de
la chaine de transport des eacutelectrons (vers lrsquoO2 autant que vers le nitrate) (Kohler et al
2008) Il se pourrait qursquoil existe eacutegalement des SCV causeacutes par drsquoautres deacuteficiences
telles que lrsquoalteacuteration de lrsquoATP synthase qui ne reacutesulterait pas drsquoune auxotrophie pour
lrsquoheacutemine ou la meacutenadione mais reacutesulterait tout de mecircme en une deacuteficience dans la
chaine de transport des eacutelectrons (Proctor et Peters 1998 )
Comme lrsquohegraveme est un cofacteur essentiel pour les oxydases terminales et que la
meacutenaquinone est un transporteur drsquoeacutelectrons utiliseacute par S aureus on comprend que
la faible croissance des mutants hemB et menD repreacutesente un pheacutenotype mimant la
perte des oxydases terminales (Hammer et al 2013) Les deux oxydases terminales
de S aureus QoxABCD (cyt aa3) et CydAB (cyt bd) semblent ecirctre diffeacuteremment
reacuteguleacutees chez les mutants menD et la souche sauvage alors que lrsquoopeacuteron cydAB est
fortement sous exprimeacute chez les mutants aucune diffeacuterence de niveau de
transcription de qoxABCD nrsquoa eacuteteacute observeacutee entre les mutants menD et leur souche
parente (Kohler et al 2008)
Des mutations dans les gegravenes menD hemB et ctaA peuvent reacutesulter en un
pheacutenotype SCV posseacutedant une chaine de transport des eacutelectrons deacuteficiente
(Clements et al 1999 Proctor et Peters 1998 ) Les gegravenes codant pour les eacutetapes
de formation de lrsquohegraveme se preacutesentent sous la forme drsquoun opeacuteron lrsquoopeacuteron
hemAXCDBL Chacun des gegravenes est essentiel (sauf hemL) et la deacuteleacutetion drsquoun seul
drsquoentre eux empecircchera la formation de lrsquohegraveme moleacutecule essentielle agrave la formation
des cytochromes (Taber 1993) hemB code pour la porphobilinogene synthase qui
catalyse une eacutetape dans la voie de biosynthegravese de lrsquohegraveme (Kohler et al 2008 Taber
- 15 -
1993 Thoumlny-Meyer 1997) Une mutation dans le gegravene ctaA (laquo hegraveme A synthase raquo)
empecircche la biosynthegravese de lrsquohegraveme A lequel est aussi impliqueacute dans la biosynthegravese
des cytochromes et donc altegraverera le fonctionnement de la chaine de transport des
eacutelectrons (Clements et al 1999 Proctor et Peters 1998) La capaciteacute de la bacteacuterie
agrave biosyntheacutetiser de lrsquohegraveme de faccedilon endogegravene est essentielle afin drsquoinitier une
infection systeacutemique Le gegravene menD code pour la 2-succinyl-6-hydroxy-24-
cyclohexadiene-1-carboxylate syntase (SHCHC) qui est une enzyme essentielle agrave la
voie de biosynthegravese des meacutenaquinones (Kohler et al 2008) De plus les
meacutenaquinones sont eacutegalement requises pour le fonctionnement de plusieurs
enzymes impliqueacutees dans des voies meacutetaboliques essentielles telles la malate quinol
oxidoreacuteductase (Mqo2) et la dihydroorotate deacuteshydrogeacutenase (PyrD) (Kohler et al
2008) Les mutants menD seront donc incapables de convertir lrsquooxaloaceacutetate en
malate ducirc agrave la perte de lrsquoactiviteacute du Mqo2 (Kohler et al 2008 Proctor et Peters
1998)
Les bacteacuteries agrave pheacutenotype SCV deacutemontreront eacutegalement un gradient eacutelectrochimique
nettement plus faible que les bacteacuteries agrave pheacutenotype normal de par lrsquoincapaciteacute de la
chaine de transport des eacutelectrons agrave y pourvoir
De par leur chaine de transport des eacutelectrons interrompue les SCV sont incapables
drsquoutiliser lrsquoO2 et le nitrate comme accepteur final drsquoeacutelectrons (Kohler et al 2008) Par
ce fait les proteacuteines impliqueacutees dans la voie de la glycolyse ainsi que dans la voie de
la fermentation sont surexprimeacutees (Pfl Ldh Adh) alors que les enzymes faisant
partie du cycle du citrate (TCA) sont significativement reacuteguleacutees agrave la baisse (Kohler et
al 2003 Kohler et al 2008 Kriegeskorte et al 2011) Ceci indique clairement que
les SCV (ex hemB) geacutenegraverent leur ATP agrave partir du glucose ou fructose seulement par
la phosphorylation des substrats Lrsquoanalyse de la reacuteaction de fermentation indique
que le produit principal est le lactate Les SCV reacutegeacutenegraverent leur NADH en reacuteduisant le
pyruvate en lactate par la voie fermentaire (Kohler et al 2003 Kohler et al 2008)
- 16 -
emmenant ainsi une acidification du milieu (Kriegeskorte et al 2011) La conversion
du pyruvate en aceacutetate via lrsquoacetyl-CoA est restreinte chez les mutants hemB mecircme
si la pyruvate formate lyase (Pfl) et les composantes du pyruvate deacuteshydrogeacutenase
sont preacutesentes (Figure 7) Lrsquoactiviteacute de la pyruvate deacuteshydrogeacutenase est tregraves faible
probablement en lien avec le haut niveau de NADH chez les SCV et il est preacutesumeacute
que la Pfl nrsquoest pas activeacutee en preacutesence drsquooxygegravene (La pyruvate deacuteshydrogeacutenase
converti le pyruvate en Co-A en condition anaeacuterobie) (Figure 7) Chez les mutants
hemB la voie de lrsquoarginine deacuteiminase est aussi induite et pourrait eacutegalement
participer agrave la production drsquoATP (Kohler et al 2003) Les gegravenes exprimeacutes chez les
mutants hemB sont similaires agrave ceux exprimeacutes chez les mutants menD ainsi que
chez les souches agrave pheacutenotypes normaux en condition anaeacuterobie (Kohler et al 2008)
- 17 -
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Figure 7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies Les enzymes suivantes sont retrouveacutees agrave une forte
concentration chez les mutants hemb phosphoglucomutase (Pgi) fructose
bisphosphate aldolase (FbaFda) glyceacuteraldeacutehyde deacuteshydrogeacutenase (Gap)
phosphoglyceacuterate kinase (Pgk) eacutenolase (Eno) pyruvate kinase (Pyk) L-lactate
deacuteshydrogeacutenase (L-Ldh) pyruvate formate lyase (PflB) alcool deacuteshydrogeacutenase
(Adh) La formation drsquoaceacutetate et de formate et tregraves limiteacutee (X rouges)
Modifieacute de Kolher C et al 2003
- 18 -
La chaine respiratoire en anaeacuterobie (S aureus)
Il existe 3 voies distinctes de geacuteneacuteration drsquoeacutenergie chez les bacteacuteries 1- en preacutesence
drsquooxygegravene la bacteacuterie respire de faccedilon aeacuterobique et lrsquooxygegravene est lrsquoaccepteur final
drsquoeacutelectron 2- la bacteacuterie peut aussi respirer de maniegravere anaeacuterobique lorsqursquoun
accepteur terminal drsquoeacutelectron autre que lrsquooxygegravene est preacutesent 3- en absence
drsquooxygegravene et drsquoun accepteur drsquoeacutelectrons alternatif la bacteacuterie peut produire de
lrsquoeacutenergie de faccedilon limiteacutee par fermentation (Hammer et al 2013)
Chez S aureus le nitrate (NO3-) et dans un troisiegraveme temps le nitrite (NO2
-)
peuvent ecirctre utiliseacutes comme accepteur final drsquoeacutelectron alternatif lorsque lrsquooxygegravene
nrsquoest pas disponible (respiration anaeacuterobie) Les enzymes de cette voie meacutetabolique
ne sont retrouveacutees que chez les bacteacuteries (Goumltz et al 2006) La croissance
bacteacuterienne est consideacuterablement reacuteduite lorsque la bacteacuterie passe drsquoune respiration
aeacuterobie agrave une respiration anaeacuterobie La nitrate reacuteductase fonctionne probablement in
vivo gracircce au L-lactate comme principal donneur drsquohydrogegravene (Burke et Lasccelles
1975) La deacuteshydrogeacutenase primaire pour ce substrat est preacutesumeacutement la L-lactate
deacuteshydrogeacutenase associeacutee agrave la membrane laquelle meacutedie lrsquooxydation du lactate par la
chaine respiratoire aeacuterobique (Burke et Lasccelles 1975) La preacutedominance du
lactate parmi les produits finaux de la fermentation par S aureus est correacuteleacutee avec
son efficaciteacute comme principal donneur drsquohydrogegravene lorsque le nitrate est lrsquoaccepteur
final drsquoeacutelectrons Le transfert des eacutelectrons du Lactate vers le nitrate implique des
composantes qui sont eacutegalement preacutesentes dans la chaine respiratoire La nitrate
reacuteductase du type respiratoire est habituellement associeacutee agrave la membrane
cytoplasmique (Burke et Lasccelles 1975) Les reacuteductases sont inhibeacutees par
lrsquoazoture et le cyanure (Burke et Lascelles 1976)
Lrsquoenzyme membranaire nitrate reacuteductase NarGHI geacutenegravere des nitrites lesquels seront
reacuteduits en ammonium (NH4+) par lrsquoenzyme cytoplasmique NADH-deacutependant nitrite
reacuteductase encodeacutee par les gegravenes nirBD (Goumltz et al 2006) La nitrate reacuteductase 1-
- 19 -
est reacuteprimeacutee et inhibeacutee par la preacutesence drsquooxygegravene 2- est induite par lrsquoanaeacuterobiose et
la preacutesence de nitrate ou nitrite 3- nrsquoest pas inhibeacutee par lrsquoammoniac 4- permet la
production drsquoeacutenergie dans des conditions anaeacuterobies et 5- est localiseacutee dans la
membrane (Goumltz et al 2006) La preacutesence drsquoun systegraveme de senseur pour lrsquooxygegravene
est cruciale pour le controcircle transcriptionnel des gegravenes facilitant lrsquoadaptation de la
bacteacuterie agrave lrsquoanaeacuterobiose ou aux conditions microaeacuterophiles Chez S aureus il existe
deux systegravemes de senseur 1- le systegraveme SrrAB (pour staphylococcal respiratory
response AB synonyme SrhSR) (Figure 8) homologue agrave ResDE chez Bacillus
subtilis (Yan et al 2011) et 2- le systegraveme NreABC (pour nitrogen regulation chez S
carnosus correspondant agrave SA2181 (NreA) SA2180 (NreB) SA2179 (NreC) chez S
aureus) NreBC est un systegraveme agrave deux composantes qui mesure la concentration
drsquooxygegravene directement par un groupement Fe-S de type Fnr lequel est instable en
preacutesence drsquooxygegravene et par lequel est controcircleacute le systegraveme membranaire de reacuteduction
du nitratenitrite (Yan et al 2011) Srr est aussi un systegraveme agrave deux composantes qui
reacutegule positivement la lactate deacuteshydrogeacutenase et lrsquoalcool deacuteshydrogeacutenase et semble
reacuteguler neacutegativement plusieurs enzymes du cycle du citrate (TCA) (Figure 8)
Lrsquoexpression des enzymes fermentaires comme la lactate deacuteshydrogeacutenase (LDH)
pyruvate formate lyase (PFL) et lrsquoalcool deacuteshydrogeacutenase sont aussi fortement induite
dans des conditions aeacuterobies lorsque la chaine de transport des eacutelectrons est
interrompue (Kohler et al 2003) Donc il se peut que lrsquoeacutetat reacuteduit des composantes
de la chaine respiratoire le potentiel membranaire etou le niveau de NADH soit un
signal pour la reacutegulation des gegravenes laquo anaeacuterobies raquo chez S aureus Dans des
conditions aeacuterobies la conversion du pyruvate en aceacutetyl-coA est catalyseacutee par le
complexe pyruvate deacuteshydrogeacutenase (PDH) et caracteacuteriseacutee par la production de
NADH (Somerville et Proctor 2009) En condition anaeacuterobie lrsquoexpression de la
pyruvate deacuteshydrogeacutenase (PDH) est fortement reacuteprimeacutee alors que lrsquoexpression de la
pyruvate formate lyase (PFL) est fortement induite Lrsquoactiviteacute de la PDH est
remplaceacutee en anaeacuterobiose par lrsquoactiviteacute de la PFL qui geacutenegravere lrsquoacetyl-CoA et le
formate agrave partir du pyruvate Le NADH nrsquoeacutetant pas reacuteoxydeacute par une chaine de
- 20 -
transport des eacutelectrons fonctionnelle celui-ci est recycleacute par la reacuteduction du pyruvate
en lactate (Kohler et al 2003 Somerville et Proctor 2009)
La meacutenaquinone et lrsquohegraveme sont esssentiels agrave la respiration anaeacuterobie (Kohler et al
2008) Le pheacutenotype de S aureus en condition anaeacuterobie est tregraves similaire au
pheacutenotype SCV (Somerville et Proctor 2009)
Figure 8 Modegravele de reacutegulation du cycle TCA (Pour simplifier les reacutegulateurs de
facteurs de virulence bien caracteacuteriseacutes [ie RNAIII Rot SarA et σB] ont eacuteteacute omis
dans ce modegravele)
Les sous-produits de la phosphorylation oxydative les ROS
La reacuteduction de lrsquooxygegravene en eau se fait un eacutelectron agrave la fois (Turrens 2003) Le
complexe IV retient tous les intermeacutediaires partiellement reacuteduits jusqursquoagrave ce que la
Tireacute de Somerville GA Proctor RA 2009
- 21 -
reacuteduction soit complegravetement termineacutee (Turrens 2003) Cependant certains autres
centres redox de la chaine respiratoire laissent passer des eacutelectrons agrave lrsquooxygegravene
reacuteduisant partiellement cette moleacutecule en anion superoxyde (O2-bull) (Liu et al 2002
Turrens 2003) Mecircme si O2-bull nrsquoest pas un agent oxydant tregraves puissant il est le
preacutecurseur de plusieurs autres laquo reactive oxygen species raquo (ROS) et il est aussi
impliqueacute dans la propagation de la chaine de reacuteaction oxydative (Turrens 2003)
LrsquoO2-bull est un intermeacutediaire instable qui est rapidement converti en H2O2 par les
superoxide dismutase (SOD) (S aureus en possegravede deux SodA (le principal) et
SodM) (10 18) ou se convertit de faccedilon spontaneacutee lorsqursquoen concentration eacuteleveacutee
(Starkov 2010 Turrens 2003) Le H2O2 peut ensuite ecirctre complegravetement reacuteduit en
H2O ou partiellement reacuteduit en radical hydroxyle (bullOH) un des plus puissants
oxydants (Starkov 2010) LrsquoO2-bull peut eacutegalement reacuteagir avec drsquoautres radicaux
incluant lrsquooxyde nitrique (NObull) selon le taux de diffusion des 2 radicaux (Turrens
2003) Le peroxynitrite est eacutegalement un oxydant puissant (Turrens 2003) Les
oxydants deacuterivant du NObull sont appeleacutes des laquo reactive nitrogen species raquo (RNS)
(Turrens 2003) mais seront consideacutereacutes comme faisant partie des ROS dans ce
travail Les ROS peuvent endommager lrsquoADN lrsquoARN les proteacuteines et les lipides
reacutesultants agrave lrsquoapoptose de la cellule lorsque le niveau de ROS produit excegravede la
capaciteacute de deacutetoxifications et de reacuteparation de la cellule (Brynildsen et al 2013
Wells et al 2005) Malgreacute ce danger il srsquoagit drsquoun risque compenseacute par
lrsquoaugmentation du rendement eacutenergeacutetique (Brynildsen et al 2013) Le complexe I (et
II) et III sont les principaux sites de production de ROS (Batandier et al 2006 Chen
et al 2003 Liu et al 2002) Lrsquoinhibition du complexe I par la rotenone provoque une
augmentation de la production de ROS Lrsquooxydation des substrats du complexe I ou
du complexe II en preacutesence du complexe III inhibeacute par lrsquoantimicine A amegravene
eacutegalement une augmentation de la production de ROS (Batandier et al 2006 Chen
et al 2003) (Figure 9) De plus mecircme si le complexe cytochrome oxydase nrsquoest pas
un site de production de ROS son inhibition par le KCN (cyanure de potassium)
facilite la production de ROS par les complexes I et II (Chen et al 2003) Le
- 22 -
complexe ATP synthase nrsquoest pas reconnu pour produire directement des ROS
Cependant la liaison drsquoun inhibiteur tel lrsquooligomycine bloque le passage des protons
agrave travers le complexe et inhibe ainsi la production drsquoATP De plus lrsquoATP synthase ne
fonctionnant plus les pompes agrave protons de la chaine de transport des eacutelectrons sont
incapables de fonctionner car le gradient de protons est beaucoup trop eacuteleveacute ce
provoque une surproduction de ROS par les complexes I et III Conseacutequemment le
NADH nrsquoest plus oxydeacute et le cycle du TCA cesse drsquoopeacuterer car la concentration de
NAD nrsquoest plus suffisante pour le fonctionnement des enzymes du TCA (Joshi et
Huang 1991)
Une production significative de O2-bull principalement par le complexe I peut ecirctre le
reacutesultat de deux modes drsquoopeacuteration 1- lorsque la cellule ne produit pas drsquoATP et
conseacutequemment possegravede une grande force proton-motrice et possegravede un laquo pool raquo de
quinone agrave lrsquoeacutetat reacuteduit (QH2) 2- lorsque la cellule possegravede un haut ratio de
NADHNAD (Figure 10) Lorsque la cellule produit activement de lrsquoATP elle possegravede
alors une moins grande force proton-motrice son ratio NADHNAD est plus faible et
la formation de O2-bull est beaucoup plus faible (Murphy 2009)
Le H2O2 est le substrat pour deux enzymes la catalase et la glutathione (GSH)
peroxydase qui convertissent le H2O2 en H2O+O2 le H2O2 est ainsi deacutetoxifieacute (Figure
9 Figure 11) La reacuteaction catalyseacutee par la glutathione peroxydase neacutecessite de la
glutathione (GSH) comme substrats et deacutepend en partie du ratio de GSSG GSH et
de la concentration des reacuteactants laquelle est le reflet de lrsquoeacutetat reacutedox de la cellule De
faccedilon similaire la concentration de meacutetaux sous leur forme reacuteduite tel le Fer catalyse
la formation de certains ROS (Figure 11) Ceci peut ecirctre minimiseacute en tenant cette
concentration ions meacutetallique tregraves basse entre autres par leur emmagasinage et leur
liaison aux proteacuteines de transport (ei ferritine transferrine lactoferrine) et ainsi
minimiser la formation de bullOH Finalement certains antioxydants (eg vitamine E)
peuvent interrompre la chaine de reacuteactions par la capture des radicaux le radical-
vitamine E est relativement stable et peut ecirctre enzymatiquement reconverti agrave sa
- 23 -
forme non-radical (Figure 11) Il est maintenant accepteacute que certaines enzymes du
cycle de Krebs (ie αKGDH) soient non seulement des cibles des ROS mais aussi
des producteurs non neacutegligeables de ROS (Brynildsen et al 2013)
Figure 9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS Le complexe I ainsi que le site Qi du complexe III relacircchent des
ROS directement vers les deacutefenses drsquoantioxydants intracellulaires Les antioxidants
intracellulaire tel la superoxide dismutase (MnSOD) la catalase la glutathione (GSH)
la glutathione peroxydase (GPX) et la glutathione reacuteductase (GRase) sont montreacute Le
site Qo du complexe III quant agrave lui relacircche les ROS dans le milieu extra cellulaire
Modifieacute de Chen Q et al 2003
Oligomycine
- 24 -
Figure 10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de
O2-bull 1- Le pool de NADH est dans un eacutetat reacuteduit par exemple par des dommages agrave
la chaine respiratoire la perte de cytochrome c durant lrsquoapoptose ou agrave un faible
besoin en ATP Ceci megravene agrave une production de O2-bull au complexe I qui deacutecoule de la
reacuteduction du FMN laquelle est reacuteguleacutee par le ratio NADHNAD+ Drsquoautres sites
autres que le complexe I tel lrsquoα-ceacutetoglutatrate deacuteshydrogeacutenase (αKGDH) peut
aussi contribuer agrave la production de O2-bull 2- Il nrsquoy a pas (peu) de production drsquoATP il y
a un fort potentiel membranaire et un pool de CoQ (meacutenaqinone) agrave lrsquoeacutetat reacuteduit
important qui provoque une reacuteversion de la chaine de transport des eacutelectrons agrave
travers le complexe I amenant une forte production de O2-bull 3- Il y a une production
active drsquoATP et conseacutequemment un potentiel membranaire plus bas que dans la
situation 2 et une plus grande quantiteacute de NADH oxydeacute que dans la situation 1
Dans ces conditions la quantiteacute de O2-bull produit est plus faible que dans la situation
1 et 2 et la provenance des O2-bull est moins certaine
Tireacute de Murphy MP 2009
- 25 -
12 Antibiotiques Mode drsquoaction cible cellulaire et meacutecanismes de reacutesistance
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles
Notre compreacutehension de la faccedilon que les antibiotiques induisent la mort cellulaire est
principalement baseacutee sur la fonction cellulaire essentielle inhibeacutee par lrsquointeraction de
lrsquoantibiotique avec sa cible Les antibiotiques peuvent ecirctre classeacutes selon la
composante ou le systegraveme cellulaire qursquoils affectent ainsi que par le fait qursquoils
induisent la mort cellulaire (antibiotiques bacteacutericides) ou srsquoils inhibent la croissance
Figure 11 Voies de formation et de deacutetoxification des ROS Tireacute de Wikipeacutedia
httpfrwikipediaorgwikiDC3A9rivC3A9_rC3A9actif_de_l27oxygC3A8ne
Consulteacute le 11 deacutec 2013
- 26 -
bacteacuterienne (antibiotique bacteacuteriostatique) La plupart des antibiotiques bacteacutericides
inhibent la synthegravese de lrsquoADN ou de lrsquoARN la synthegravese de la paroi bacteacuterienne ou la
synthegravese proteacuteique (Kohanski et al 2010) Le tableau 1 est une synthegravese des
principales classes drsquoantibiotiques ainsi que leur mode drsquoaction et leurs cibles
cellulaires Ce tableau nrsquoest cependant pas exhaustif et il existe plusieurs autres
classes drsquoantibiotiques et plusieurs autres exemples drsquoantibiotiques dans les classes
identifieacutees dans ce tableau
Depuis la deacutecouverte de la peacutenicilline en 1929 plusieurs autres antibiotiques ont eacuteteacute
deacutecouverts et deacuteveloppeacutes suite agrave lrsquoeacutelucidation des interactions entre lrsquoantibiotique et
sa cible et en apportant des modifications structurales et chimiques aux moleacutecules
deacutejagrave connues Ces efforts ont significativement augmenteacute le nombre drsquoantibiotiques
disponibles dans le milieu clinique Le processus de mort cellulaire induit par lrsquoaction
drsquoun antibiotique est complexe et deacutebute avec une interaction physique entre la
moleacutecule antibiotique et sa cible bacteacuterienne speacutecifique et implique des modifications
qui affectent la bacteacuterie tant au niveau biochimique que moleacuteculaire et structural
27
Acid
e n
alid
ixiq
ue (
1 iegravere
geacuteneacutera
tion)
Cip
rofloxacin
e (
2 iegravem
e g
eacuteneacutera
tion)
Levofloxacin
e (
3 iegravem
e g
eacuteneacutera
tion)
Gem
iflo
xacin
e (
4 iegravem
e g
eacuteneacutera
tion)
Flu
oro
quin
olo
ne (
Norf
loxacin
e)
Peacutenic
iline
Ceacutephalo
sporine (
ceacutefa
lexin
e)
Carb
apenem
Vancom
ycin
e
Teacutela
vancin
e
Fosfo
mycin
Dapto
mycin
Poly
mix
ine B
Bacitra
cin
Valin
om
ycin
e
Nom
encin
e
Genta
mic
in
Kanam
ycin
e
Tobra
mycin
e
Str
epto
mycin
e
Teacutetr
acyclin
e
Chlo
rteacutetr
acyclin
e
Macro
lides
Ery
thro
mycin
eLie
sous-u
niteacute 23S
du r
ibosom
e 5
0S
et em
pecircche
leacutelo
ngation d
u p
eptide (
apregrave
s 6
-8 a
a)
sta
tique
Lin
cosam
ides
Clin
dam
ycin
Inhib
e d
irecte
ment la
ctiviteacute p
eptidyl
transfeacute
rase d
u
ribosom
esta
tique
Str
epto
gra
min
es
DalfopristinQ
uin
upristin
Com
bin
ais
on d
e 2
antibio
tiques B
loque la
transla
tion p
eptidiq
ue e
n lia
nt le
site P
du r
ibosom
e
et en b
loquant au p
eptide la
ccegraves a
u tunnel de s
ort
ie
cid
al
Am
phenic
ol
Chlo
ram
phenic
ol
Em
pecircche la lia
ison d
u a
a-
AR
Nt au s
ite A
du
ribosom
e e
t in
terf
egravere
dans la f
orm
ation d
u lie
n
peptidiq
ue
sta
tique
Oxazolid
inones
Lin
ezolid
Blo
que initia
tion d
e la tra
nsla
tion p
roteacute
ique lors
de la
reacuteaction d
eacutelo
ngation p
ar
la p
eptidyl
transfeacute
rase
sta
tique
Trim
eth
oprim
Sulfam
eth
oxazole
sta
tique
cid
al
Ionophore
Alteacutera
tion d
e lin
teacutegriteacute
mem
bra
naire
Se
nchasse d
ans la m
em
bra
ne fo
rme u
n p
ore
ce
qui am
egravene u
ne p
ert
urb
ation d
e leacute
quili
bre
ioniq
ue
cid
al
Inhib
itio
n d
e la s
ynth
egravese d
e
la p
aro
i cellu
laire
(peptidogly
can)
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
cid
al
cid
al
sta
tique
AR
Nr
16S
(S
ite A
)(fo
rmation d
e p
roteacute
ines n
on-
fonctionnelle
s)
cid
al
Inhib
itio
n d
e la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 3
0S
)
Inhib
iteur
de la s
ynth
egravese d
e
lacid
e f
oliq
ue
Inhib
itio
n d
e c
ert
ain
es r
eacuteactions e
nzym
atiques
essentielle
s agrave
la f
orm
ation d
e la
cid
e f
oliq
ue
Sulfonam
ides
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
AR
Nr
16S
(preacute
vie
nt la
ttachem
ent de la
min
oacyl-
AR
Nt au r
ibosom
e)
Teacutetr
acyclin
e
Poly
peptides
β-L
acta
ms
Gly
copeptides
Lip
opeptides
Inhib
iteur
de la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 5
0S
)
Am
inogly
cosid
es
Cib
le c
ell
ula
ire
Sta
tiq
ue
Cid
al
Rifam
ycin
eR
ifam
pic
ine
Cla
ss
es
Ex
d
an
tib
ioti
qu
eM
od
e d
ac
tio
n
Inhib
itio
n d
e la s
ynth
egravese d
e
lAR
N (
AD
N d
eacutependant)
AR
N p
oly
meacutera
se s
ous-u
niteacute β
(codeacute p
ar
rpoB
)
Quin
olo
nes
cid
al
Inhib
itio
n d
e la r
eacuteplic
ation d
e
lAD
N
Com
ple
xe A
DN
-topois
om
eacutera
se (
AD
N
gyr
ase(t
opois
om
eacutera
se II)
et to
pois
om
eacutera
se IV
)cid
al
Tra
nspeptidase (
penic
illin
bin
din
g p
rote
in
(PB
P))
cid
al
Tab
leau
1
Cib
les e
t m
od
e d
rsquoacti
on
des p
rin
cip
ale
s c
lasses d
rsquoan
tib
ioti
qu
es
28
122 Les aminoglycosides
1221 Mode drsquoaction
Les aminoglycosides peacutenegravetrent la paroi cellulaire en suivant un processus
comprenant 3 eacutetapes une premiegravere eacutetape eacutenergie- indeacutependante suivie de deux
eacutetapes eacutenergie deacutependantes Deux composantes sont essentielles pour
lrsquoaccumulation intracellulaire des aminoglycosides les ribosomes et la chaine de
transport des eacutelectrons (Ramirez et Tolmasky 2010)
Eacutetape 1- Lorsque les aminoglycosides entrent en contact avec la paroi
cellulaire la moleacutecule polycationique se lie aux composantes de surface anioniques
tels les lipopolysaccharides les phospholipides et autres proteacuteines membranaires
chez les bacteacuteries agrave Gram neacutegatif et les moleacutecules drsquoacides teacuteichoiumlques et les
phospholipides des bacteacuteries agrave Gram positif (Ramirez et Tolmasky 2010) Apregraves
cette liaison aux moleacutecules polaires les aminoglycosides (telle la gentamicine)
srsquoassocient aux transporteurs membranaires sur la base de leur charge positive et
sont transporteacutes agrave travers la membrane cytoplasmique gracircce au potentiel
membranaire (intra cellulaire neacutegatif) (Bryan et al 1980 Proctor et al 2006) Cette
liaison agrave la membrane entraine une augmentation de la permeacuteabiliteacute membranaire
menant agrave lrsquoentreacutee des aminoglycosides dans lrsquoespace peacuteriplasmique (Gram neacutegatif)
ou dans le cytoplasme (Gram positif) (Ramirez et Tolmasky 2010) (Figure 12A)
Eacutetape 2- Lrsquoeacutetape suivante connue comme la laquo phase eacutenergie-deacutependante
1 raquo (EDP-I) est bloqueacutee par les inhibiteurs de la chaine de transport des eacutelectrons et
de la phosphorylation oxydative ainsi que par les ions divalents lrsquohyperosmolariteacute le
faible pH et lrsquoanaeacuterobiose dans cette phase la dureacutee et le taux drsquoentreacutee de
lrsquoantibiotique sont en fonction de la concentration (Miller et al 1980 Mingeot-
Leclercq et al 1999 Ramirez et Tolmasky 2010) Agrave cette eacutetape le transport
transmembranaire des aminoglycosides et plus speacutecifiquement de la gentamicine
neacutecessite 2 composantes majeures a) un transporteur anionique non speacutecifique dont
la charge est directement lieacutee agrave lrsquoactiviteacute des cytochromes oxydases terminales et des
29
nitrates reacuteductases de la chaine de transport des eacutelectrons Selon certaines eacutetudes
les transporteurs pourraient ecirctre les quinones reacuteduites de la chaine de transport des
eacutelectrons (Bryan et al 1981 Miller et al 1980) et donc le cycle drsquooxydation-reacuteduction
des quinones est essentiel aux transports des aminoglycosides (Bryan et al 1981)
mais selon drsquoautres eacutetudes les transporteurs pourraient ecirctre toute autre proteacuteine non
deacutefinie impliqueacutee dans la polarisation membranaire lieacutee agrave lrsquoactiviteacute de la chaine de
transport des eacutelectrons (Miller et al 1980) La croissance anaeacuterobie ainsi que le
sodium azide (inhibiteur de la chaine respiratoire) inhibent lrsquoentreacutee de la gentamicine
et lrsquoeffet bacteacutericide indiquant que le transport actif de cet antibiotique neacutecessite la
respiration b) un potentiel eacutelectrique avec un milieu intra cellulaire neacutegatif par rapport
agrave lrsquoexteacuterieur En effet lrsquoinhibition de lrsquoentreacutee dans la bacteacuterie des aminoglycosides
suite agrave lrsquoajout de CCCP (agent deacutepolarisant) chez des souches de S aureus laquo wild-
type raquo deacutemontre la neacutecessiteacute de la force proton-motrice pour lrsquoentreacutee des
aminoglycosides dans les cellules (Miller et al 1980)
Eacutetape 3- La petite quantiteacute drsquoaminoglycosides srsquoeacutetant rendu au cytoplasme
durant cette premiegravere phase eacutenergie-deacutependante induisent apregraves liaison aux
ribosomes des erreurs dans la synthegravese proteacuteique Les erreurs dans la traduction
des proteacuteines membranaires produites lors de cette eacutetape connue comme la laquo phase
eacutenergie-deacutependante 2 raquo (EDP-II) provoquent des dommages dans lrsquointeacutegriteacute de la
membrane cytoplasmique (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky
2010) Les proteacuteines membranaires aberrantes dans la membrane facilitent le
transport drsquoune plus grande quantiteacute drsquoaminoglycosides lesquels augmenteront
encore le niveau drsquointerfeacuterence avec la synthegravese proteacuteique normale amenant encore
plus de dommage agrave la membrane provoquant ainsi une acceacuteleacuteration importante du
taux drsquoentreacutee des moleacutecules antibiotique et qui ultimement reacutesultera en la mort de la
cellule (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky 2010) (Figure 12B)
Des eacutetudes ont deacutemontreacute que la susceptibiliteacute agrave la gentamicine streptomycine
amikacine kanamycine et tobramycine est supeacuterieure lorsque lrsquooxygegravene est
30
lrsquoaccepteur terminal drsquoeacutelectrons (Bryan et Kwan 1981) Une diminution significative
de la susceptibiliteacute est remarqueacutee lorsque le nitrate est lrsquoaccepteur final drsquoeacutelectrons et
une diminution encore plus marqueacutee est observeacutee lorsque le fumarate est lrsquoaccepteur
final drsquoeacutelectron ou lorsque les conditions sont fermentaires Lrsquoassociation entre le
transport de la gentamicine et le complexe terminal de la chaine de transport des
eacutelectrons particuliegraverement lrsquoactiviteacute cytochrome oxydase
Le ribosome possegravede trois sites de liaison importants de lrsquoARNt deacutesigneacute comme site
A (pour aminoacyl) site P (pour peptidyl) et E (pour exit) (Vakulenko et Mobashery
2003) Ils sont situeacutes sur lrsquoARN16S (lequel avec plus de 21 proteacuteines compose la
sous-uniteacute 30S du ribosome bacteacuterien) (Ramirez et Tolmasky 2010 Vakulenko et
Mobashery 2003) Durant la synthegravese proteacuteique le ribosome deacutecode lrsquoinformation de
lrsquoARNm et catalyse lrsquoincorporation seacutequentielle de chaque acide amineacute faisant partie
de la chaine polypeptidique en formation La haute fideacuteliteacute de la traduction (les
ribosomes incorporent un mauvais acide amineacute par approximativement 3000 liaisons
peptidiques effectueacutees) est possible gracircce agrave la capaciteacute de discriminer entre des
changements conformationels dans le ribosome induit par la liaison drsquoun ARNt correct
Modifieacute de Davis BD 1987
Figure 12 Cineacutetique drsquoentreacutee des aminoglycosides
A
B
31
ou incorrect au site A La reconnaissance drsquoune liaison correcte anticodoncodon par
le ribosome se passe en deux temps la reconnaissance initiale de lrsquoappariement du
codon sur lrsquoARNm avec lrsquoanticodon de lrsquoARNt et subseacutequemment la correction
(Vakulenko et Mobashery 2003)
La plupart des aminoglycosides se lient agrave des oligonucleacuteotides faisant partie du site
A Les diffeacuterents aminoglycosides ne lient pas tous le mecircme site de lrsquoARN 16S mais
lrsquoeffet commun agrave tous est que cette liaison amegravene un changement de conformation du
site A vers une conformation mimant la conformation fermeacutee de la sous uniteacute 30S qui
est induite par une interaction entre lrsquoARNm et son ARNt apparenteacute eacuteliminant la
possibiliteacute de correction du ribosome ce qui promeut les erreurs de traduction
(Ramirez et Tolmasky 2010 Vakulenko et Mobashery 2003) La grande majoriteacute des
aminoglycosides sont bacteacutericides et cette leacutetaliteacute est la conseacutequence de cette
mauvaise traduction (Vakulenko et Mobashery 2003)
1222 Meacutecanismes de reacutesistance
La reacutesistance contre les aminoglycosides se produit par diffeacuterents meacutecanismes qui
peuvent coexister simultaneacutement dans la mecircme bacteacuterie (Vakulenko et Mobashery
2003) Ces meacutecanismes incluent 1- la modification de la cible par des mutations
dans lrsquoARN 16S ou autres proteacuteines ribosomales associeacutees (Figure 13) 2- la
meacutethylation de lrsquoARN 16S un meacutecanisme retrouveacute chez plusieurs organismes
producteurs drsquoaminoglycosides et chez des souches cliniques Cette meacutethylation
empecircche les aminoglycosides drsquoavoir accegraves agrave leur site de liaison (Doi et Arakawa
2007) 3- la reacuteduction de la permeacuteabiliteacute par une alteacuteration de la permeacuteabiliteacute de la
membrane externe ou une diminution du transport agrave travers la membrane interne 4-
lrsquoexportation des aminoglycosides agrave lrsquoexteacuterieur de la cellule par les pompes agrave efflux
Eacutetant des moleacutecules hydrophiliques les aminoglycosides ont naturellement peu
drsquoaffiniteacute avec les pompes agrave efflux 5- le swarming actif probablement un meacutecanisme
32
non speacutecifique deacutemontreacute chez Pseudomonas aeruginosa lequel deacutemontre des
reacutesistances agrave plusieurs antibiotiques 6- la seacutequestration de la drogue par des liaisons
agrave un aceacutetyltransfeacuterase agrave tregraves faible activiteacute 7- une inactivation enzymatique de la
moleacutecule antibiotique le meacutecanisme de reacutesistance le plus preacutevalent au niveau
clinique (Vakulenko et Mobashery 2003) Lrsquoinactivation enzymatique des
aminoglycosides est le plus souvent due agrave des aceacutetyltransfeacuterases des
nucleacuteotidyltransfeacuterases ou des phosphotransfeacuterases (Dickgiesser et Kreiswirth 1986)
Ces enzymes sont souvent encodeacutees sur des plasmides mais sont eacutegalement
associeacutees agrave des eacuteleacutements transposables Lrsquoeacutechange de plasmides et la disseacutemination
de transposons facilitent lrsquoacquisition rapide de pheacutenotypes reacutesistants aux
antibiotiques non seulement au sein drsquoune mecircme espegravece mais parmi un large
eacuteventail drsquoespegraveces bacteacuteriennes (Mingeot-Leclercq et al 1999)
La reacutesistance naturelle des S aureus laquo small colony variants raquo agrave la majoriteacute des
aminoglycosides est la conseacutequence de lrsquoimpossibiliteacute agrave lrsquoantibiotique de peacuteneacutetrer une
cellule posseacutedant une faible polariteacute membranaire qui est due chez les SCV agrave leur
Tireacute de Doi Y Arakawa Y 2007
Figure 13 Sites de mutation de certains reacutesistants aux aminoglycosides
33
incapaciteacute agrave activer le cycle de Krebs (Miller et al 1980 Proctor et al 2006 Tubby
et al 2013) Les SCV ont non seulement une polariteacute membranaire faible mais (ceci
eacutetant la conseacutequence de cela) aussi une force proton motrice inhibeacutee et une chaine
des eacutelectrons deacuteficiente De ce fait le transport transmembranaire des
aminoglycosides chez les SCV est grandement reacuteduit et conseacutequemment la
susceptibiliteacute des SCVs pour cette classe drsquoantibiotique est 110-113 de celle des
souches parentes (Proctor et al 2006)
123 La tomatidine
Plusieurs espegraveces veacutegeacutetales accumulent des steacuterols et des triterpegravenes antimicrobiens
appeleacutes saponines (Osborn 1996) Ces moleacutecules sont produites de faccedilon
constitutive par la plante et jouent un important rocircle de deacutefense contre diffeacuterents
pathogegravenes comme des insectes ou des microbes Parmi les saponines la tomatine
(Figure 14A) est un glycoalcaloiumlde steacuteroiumldien preacutesentant une action antimicrobienne
contre des levures et une quantiteacute drsquoautres microorganismes (Bednarek et Osboum
2009 Friedman 2002 Osborn 1996 Sandrock et Vanetten 1998) Plusieurs
espegraveces de champignons pathogegravenes de la tomate produisent des enzymes
extracellulaires communeacutement appeleacutes tomatinase qui sont capables de deacutetoxifier
lrsquoα-tomatine (Martin-Hernandez et al 2000 Roddick 1974 Ruiz-Rubbio et al 2001)
Par exemple Fusarium oxysporum f sp lycopersici est reconnu pour cliver lrsquoα-
tomatine en ses sous-produits soit un tetrasaccharide (lycotreacutetraose) et un alcaloiumlde
steacuteroiumldien non glycosyleacute la tomatidine (Figure 14B) Ces sous-produits nrsquoont pas ou
peu drsquoactiviteacute antifongique (Ruiz-Rubbio et al 2001 Simon et al 2006)
34
Figure 14 Structures chimiques de la (A) tomatine de (B) tomatidine et de (C)
lrsquoanalogue de tomatidine FC04-100
Il a eacuteteacute deacutecouvert il a quelques anneacutees que la tomatidine avait un effet antibacteacuterien
sur les S aureus de pheacutenotype SCV mais aucune activiteacute contre les S aureus agrave
pheacutenotype normal Par contre lorsqursquoutiliseacute en combinaison avec les
aminoglycosides lrsquoajout de la tomatidine permettait drsquoaugmenter de 8 agrave 16 fois
lrsquoactiviteacute des aminoglycosides contre S aureus agrave pheacutenotype normal (Mitchell et al
2011a Mitchell et al 2012)
Donc on savait que la tomatidine avait une activiteacute antibacteacuterienne tregraves inteacuteressante
mais on ne connaissait ni sa cible ni son mode drsquoaction De plus si la tomatidine a
une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait donc avoir un
spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Par ailleurs de faccedilon parallegravele la production drsquoanalogues structuraux de la tomatidine
permettrait de mieux comprendre la relation structure-activiteacute (SAR) de la moleacutecule et
donc de preacuteciser que lrsquoactiviteacute biologique de la tomatidine neacutecessite une orientation
3
A B
C
35
de groupements fonctionnels bien speacutecifique De plus lrsquoeacutetude de lrsquoactiviteacute de
plusieurs analogues permettrait de confirmer si la tomatidine possegravede un meacutecanisme
drsquoaction deacutependant drsquoune interaction speacutecifique avec une ou des cibles biologiques
preacutecises
Le laboratoire du Pr Eacuteric Marsault du deacutepartement de pharmacologie de lrsquoUniversiteacute
de Sherbrooke et lrsquoeacutetudiant agrave la maitrise Feacutelix Chagnon ont produit plus de 33
analogues de la tomatidine (voir Annexe 2 article laquo Unraveling the structure-activity
relationship of tomatidine a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureusraquo dans la section laquo Contributions
Supleacutementaires raquo du meacutemoire) dont lrsquoactiviteacute antibacteacuterienne a eacuteteacute eacutevalueacutee par des
tests de susceptibiliteacute agrave la moleacutecule (CMI) sur S aureus SCV et sur S aureus agrave
pheacutenotype normal en combinaison avec la gentamicine Plusieurs de ces moleacutecules
font lrsquoobjet drsquoune application de brevet (voir annexe 1) dont la prometteuse moleacutecule
FC04-100 (Figure 14C) (voir laquo Annexe 1 Application de brevet raquo dans la section
laquo Contributions suppleacutementaires raquo du meacutemoire)
13 Projet de maitrise
Premiegravere hypothegravese
Si la tomatidine a une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait
donc avoir un spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Objectif Provoquer lrsquoapparition de pheacutenotypes SCV chez diffeacuterentes espegraveces
bacteacuterienne afin de pouvoir veacuterifier leur niveau de sensibiliteacute agrave la tomatidine
Deuxiegraveme hypothegravese
Lrsquoidentification des gegravenes preacutesentant des mutations parmi les souches de S aureus
reacutesistantes agrave la tomatidine pourrait reacuteveacuteler la ou les cibles cellulaires de la moleacutecule
et nous aider agrave comprendre son mode drsquoaction
36
Objectif Obtenir des mutants reacutesistants avant de les envoyer agrave sequencer
faire les analyses comparatives des geacutenomes et finalement identifier les mutations
37
CHAPITRE 2
ARTICLE SCIENTIFIQUE
TITRE Tomatidine and Analog FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Some Firmicutes Including Those of Listeria
monocytogenes and Are Bactericidal in Combination With Aminoglycosides Against
their Normal Phenotype
AUTEURS ET CONTRIBUTIONS
Isabelle Guay1 Expeacuteriences biologiques avec le produit naturel tomatidine et son
analogue FC04-100 sur diffeacuterentes espegraveces bacteacuteriennes afin de comparer les deux
activiteacutes
Simon Boulanger1 Expeacuterience biologique permettant de quantifier lrsquoactiviteacute de
lrsquoanalogue FC04-100 sur la production de biofilm et son activiteacute intracellulaire
Feacutelix Chagnon2 Production de lrsquoanalogue chimique de la tomatidine FC04-100
Kamal Bouarab1 Initiateur du projet gracircce agrave lrsquoisolement du produit naturel tomatidine
Eacuteric Marsault2 Directeur de recherche de Feacutelix Chagnon
Franccedilois Malouin1 Directeur de recherche drsquoIsabelle Guay et Simon Boulanger et
auteur de correspondance
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
38
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
Corresponding author Mailing address Universiteacute de Sherbrooke
Deacutepartement de Biologie Faculteacute des Sciences 2500 Boul
Universiteacute Sherbrooke Quebec Canada J1K2R1 Phone (819) 821-
8000 ext 61202 Fax (819) 821-8049 E-mail FrancoisMalouin
USherbrookeca
39
Tomatidine and Analog FC04-100 Possess Bactericidal Activities
Against the Small Colony Variants of Some Firmicutes Including
Those of Listeria monocytogenes and Are Bactericidal in Combination
With Aminoglycosides Against their Normal Phenotype
Running Title Tomatidine is bactericidal against several Firmicutes
Isabelle Guay1
Simon Boulanger1
Feacutelix Chagnon2 Kamal Bouarab
1 Eacuteric Marsault
2 and
Franccedilois Malouin1
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM) Deacutepartement de
Biologie Faculteacute des Sciences Universiteacute de Sherbrooke Sherbrooke Queacutebec Canada J1K
2R1
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute Universiteacute
de Sherbrooke Queacutebec Canada J1H 5N4
Have contributed equally to the work
Corresponding author
Mailing address Universiteacute de Sherbrooke Deacutepartement de Biologie Faculteacute des Sciences
2500 Boul Universiteacute Sherbrooke Quebec Canada J1K2R1
Phone (819) 821-8000 ext 61202 Fax (819) 821-8049
E-mail FrancoisMalouinUSherbrookeca
40
ABSTRACT
Tomatidine (TO) is a plant steroidal alkaloid that possesses an antibacterial activity against the
small colony variants (SCVs) of Staphylococcus aureus We report here the spectrum of
activity of TO against species of the Firmicutes and the improved antibacterial activity of a
chemically-modified TO derivative (FC04-100) In addition to that seen on S aureus TO
showed a bactericidal activity against SCVs of Bacillus cereus B subtilis and Listeria
monocytogenes The combination of an aminoglycoside antibiotic and TO also generated an
antibacterial synergy against strains of the normal phenotype in these species In contrast to
TO which has no relevant activity by itself against S aureus of the normal phenotype (MIC
gt64 mgL) the TO analog FC04-100 showed a MIC of 8-16 mgL Furthermore FC04-100
showed a strong bactericidal activity against L monocytogenes SCVs in kill kinetic
experiments while TO did not FC04-100 also improved the bactericidal activity of
gentamicin against L monocytogenes of the normal phenotype The addition of FC04-100 (4
mgL) to a cephalexinkanamycin (32) combination improved the activity of the combination
by 32 fold against cephalexin and kanamycin-resistant MRSA strains In combination with
gentamicin FC04-100 also showed a strong bactericidal activity against biofilm-embedded S
aureus and when used alone was superior to TO for its biofilm-preventing activity Finally
FC04-100 and TO showed comparable intracellular killing of S aureus SCVs in a polarized
Calu-3 cell culture model In conclusion chemical modifications of TO allowed improvement
of its antibacterial activity against L monocytogenes SCVs and prototypical S aureus
41
Keywords Tomatidine aminoglycoside synergy SCV Firmicutes S aureus L
monocytogenes
Introduction
The Firmicutes are Gram-positive bacteria with genomes presenting less than 50 of G+C
The Firmicutes constitutes one of the main divisions within the Bacteria and is divided into
three classes Bacilli Mollicutes and Clostridia The class of Bacilli is further divided into the
orders of the Lactobacillales and the Bacillales The Bacillales are divided into the genus of
Staphylococcus Listeria and Bacillus A number of bacterial species such as Listeria spp and
Bacillus spp can contaminate food and cause infections in humans (Magalhaes 2013) To
name a few Listeria monocytogenes L ivanovii and Bacillus cereus can cause listeriosis
(Guillet et al 2010) and food poisoining (Bad Bug Book FDA) Bacillus subtilis B
coagulans B licheniformis and B sphaericus are also known to cause illnesses Bacillus
anthracis causes anthrax and can often be acquired by contact with food producing animals
and cattle (beef cattle sheeps etc) and this bacterium is also well-known for its endospores
that have been used as biological weapons (Beierlein and Anderson 2011) Staphylococci are
divided in coagulase-positive species Staphylococcus aureus being the most clinically
relevant of this group and coagulase-negative species such as S epidermidis the most
prevalent pathogen associated with infections of implanted medical devices (Vuong and Otto
2002) The emergence and spread of resistance to multiple antibiotics in staphylococci is now
considered a real health treat and impaired therapeutic endeavor to combat these bacteria
42
(Witte et al 2007) Notably the prevalence of methicillin-resistant S aureus (MRSA) has
steadily increased over the recent years not only in hospitals but also in the community
(Hiramatsu et al 2013)
Since their first description in 1910 for Salmonella enterica serovar Typhi (S typhi) small-
colony variants (SCVs) have been described for a wide range of bacterial genera and species
and have been isolated from both laboratory and clinical environments (Proctor 2006)
Staphylococcus aureus SCVs have attracted a great deal of interest over the past recent years
S aureus SCVs often present a dysfunctional oxidative metabolism causing an alteration in
the expression of virulence factors a slow growth and a loss of colony pigmentation (Proctor
2006) This dysfunctional oxidative metabolism is also responsible for a decreased
susceptibility to aminoglycoside antibiotics because this class of molecules requires the
proton-motive force in order to penetrate the bacterium (Bryan LE 1981) This respiratory
deficiency is often caused by mutations affecting the electron transport system and several
SCV isolates can recover normal growth with supplemental hemin or menadione which are
needed to synthesize electron transport system components S aureus SCVs often are isolated
from chronic infections such as lung infections in cystic fibrosis (CF) patients osteomyelitis
septic arthritis bovine mastitis and from infections associated with orthopedic devices (Atalla
2008 Moisan 2006 Proctor 2006) The ability of S aureus to switch back and forth from the
prototypic to the SCV phenotypes in vivo is an integral part of the pathogenesis of S aureus
and may be responsible for the establishment of chronic infections (Tuchscherr 2011
Mitchell 2011 inf imm)
43
Tomatidine (TO) is a steroidal alkaloid produced by the Solanaceae plant family such as the
tomato (Ruiz-Rubio 2001 Simons 2006) We showed previously the antibacterial activity of
TO against S aureus SCVs and also documented a strong synergic activity of TO in
combination with aminoglycoside antibiotics against prototypic S aureus (Mitchell G AAC
2011 Mitchell G 2012 JAC) Recently we synthesized a variety of TO analogs in order to
explore the structure-activity relationship of this new class of antibiotics able to act on SCVs
(Chagnon et al 2014) One analog FC04-100 showed the same steroidal backbone as the
natural molecule but with an additional carbon chain on the A cycle (Fig 1) Preliminary
characterization of FC04-100 revealed an antibacterial activity that was similar to that of TO
against S aureus SCVs and also preserved the synergic activity with aminoglycosides against
prototypic S aureus (Chagnon et al 2014) However contrary to TO FC04-100 showed
notable antibiotic activity by itself against prototypic S aureus with a MIC of 8-16 gL
whereas that of TO was gt64 gL It would be of great interest to see if this property of FC04-
100 modifies or improves its spectrum of antibacterial activity
The aim of this study was to further describe the spectrum activity of TO and FC04-100 The
susceptibility of several species among the Firmicutes to TO and FC04-100 alone or in
combination with other antibiotics was investigated The improved antibacterial activity of
FC04-100 was demonstrated against Listeria monocytogenes and methicillin-resistant S
aureus (MRSA)
44
Materials and methods
Chemical reagents and antibiotics
Tomatidine (TO) (Sigma Aldrich Oakville Ontario Canada) (Fig 1A) was solubilised in
dimethylsulfoxide (DMSO) at a concentration of 2 mgmL The TO analog FC04-100 (Fig
1B) was synthesized (Chagnon et al 2014) and solubilised in DMSO at 20 gL Cefalexin
kanamycin gentamicine (GEN) (all from Sigma Aldrich) were solubilised in water at a
concentration of 10 gL Menadione was solubilized in DMSO hemin in 14 M NH4OH and
thymidine in water (all from Sigma Aldrich) and were prepared at a concentration of 10 gL
45
3
Fig 1 Tomatidine (A) is a steroid alkaloid structurally characterized by 6 rings 12
stereogenic centers a 3 β-hydroxyl group and spiro-fused E F rings in the form of an
aminoketal The tomatidine analog FC04-100 (B) contains a diamine in position 3 of ring A
Bacterial strains and growth conditions
Staphylococcus aureus ATCC 29213 S aureus Newbould (ATCC 29740) Listeria
monocytogenes ATCC 13932 Bacillus subtilis ATCC 6333 and Bacillus cereus ATCC 11778
were used as prototypic strains Methicillin-resistant S aureus (MRSA) strains COL and
USA100 (hospital acquired-MRSA) and strain USA300 (community acquired-MRSA) were
also used in this study The reference laboratory strain S aureus SH-1000 an isogenic mutant
strain derived from S aureus 8523-4 but with a functional rsbU allele (HorsburghMJ 2002)
was used for its good yield of biofilm production The S aureus strains CF07-L (prototypic)
A
B
A B
C D
E
F
46
and CF07-S (SCV phenotype) are genetically related clinical strains originally isolated from a
cystic fibrosis patient (MitchellG 2011) Strain CF07-S was used in the intracellular
infection model Staphylococcus and Bacillus were maintained on Tryptic Soy agar (TSA BD
Mississauga Ontario Canada) whereas L monocytogenes was grown on brain-heart infusion
(BHI) agar (BD Mississauga Ontario Canada)
Small colony variants (SCVs)
For S aureus strain NewbouldhemB was used as the reference SCV NewbouldΔhemB was
generated from strain Newbould (ATCC 29740) by disrupting the hemB gene with the ermA
cassette by homologous recombination (Brouillette 2004) SCVs from B cereus B subtillis
and L monocytogenes were generated by growth in presence of a subinhibitory concentration
of GEN Briefly overnight broth cultures (18ndash20 h) were used to inoculate BHI broth at a
dilution of 1100 supplemented or not with 025 to 1X the MIC of GEN Cultures were
incubated 18 h at 35degC with shaking (225 rpm) and then adjusted to an A595 nm of 20 in PBS at
4degC Determination of CFU and SCV colonies was done by serial dilution plating SCV were
obtained by plating on TSA (BHI agar for L monocytogenes) containing gentamicin at a
concentration of 8 to 16 times the induction concentration followed by an incubation of 48 h at
35degC The pinpoint colonies selected by this method were confirmed to be SCVs by streaking
several of them on agar plates without antibiotic SCV that conserved their phenotype after
two passages were considered to have a stable phenotype and were used in subsequent
experiments Selected SCVs had a colony size a tenth of the size of a normal colony and
showed no pigmentation or hemolytic activity on blood agar plates The SCV species were
47
confirmed by rDNA 16S sequencing after PCR amplification using universal primers (Gomaa
2007)
Auxotrophy assays
For SCVs auxotrophism is defined as the requirement of specific compounds in order to
regain a normal growth phenotype [Sendie 2009] An agar diffusion method was used to
characterize the auxotrophism of SCVs using hemin or menadione (1-10 μg eachwell) or
thymidine (100 microgwell) on an inoculated Mueller-Hinton agar (MHA) or Brain-heart infusion
agar (BHIA) plate [Besier 2007] Auxotrophy for specific supplements was detected by a
zone of normal growth surrounding the well after 18 h of incubation at 35degC The results were
confirmed by two indepedent experiments
Antibiotic susceptibility testing
MICs were determined by a broth microdilution technique following the recommendations of
the Clinical and Laboratory Standards Institute (CLSI) except that the incubation period was
extended to 48 h and the medium used was BHI to allow SCVs to reach maximal growth as
previously described (Mitchell 2010) As recommended by the CLSI L monocytogenes (and
its SCVs) were grown in cation-adjusted Muller-Hinton broth (CAMHB BD) containing 3
lysed horse blood (LHB Remel Hartford CT USA) The reported results were obtained from
three independent experiments
48
Time-kill experiments
Kill kinetics were performed to determine whether the effect of compounds alone or in
combination was bacteriostatic or bactericidal Bacteria were inoculated at 105-10
6 CFUmL in
appropriate medium in the absence or presence of the different antibiotic compounds at
concentrations specified in figure legends At several points in time at 35degC (225 rpm)
bacteria were sampled serially diluted and plated on TSA for CFU determinations Plates
were incubated for 24 or 48 h at 35degC for normal and SCV strains respectively The data were
collected from a minimum of three independents experiments
Antibiotic activity in biofilms
The viability of bacteria in biofilms was evaluated based on previously described protocols
(CeriH 2001 MoskowitzSM 2004 HarrisonJJ 2010) For these assays 96-peg lids and
corresponding 96-well plates were used (Nunc CITY STATE) Wells were inoculated with a
suspension of S aureus SH-1000 adjusted to a 05 McFarland standard in TSB (150 microL per
wells) and plates with 96-well lids were incubated at 35degC with an agitation of 120 rpm for 24
h The biofilms on pegs were then washed three times with 200 microL PBS Biofilms on pegs
were further incubated in fresh 96-well plates containing 200 microL of TSB containing or not
antibiotic compounds or a combination of molecules at 35degC 120 rpm for 24 h The treated
biofilms on pegs were washed three times in PBS and bacteria in biofilms were recovered by
sonication in a new microtiter plate containing 200 microL of PBS per wells using an
ultrasonicator bath for 10 min followed by centrifugation for five min at 1000 RPM The
49
bacteria recovered in each well were resuspended serially diluted plated on TSA and
incubated at 35degC for 24 h before CFU determination
For an equivalent series of wells instead of disrupting the biofilm by sonication biofilm
formation on pegs was evaluated through crystal violet staining using a procedure adapted
from Mitchell et al(2010) Following exposure to antibiotics as described above biofilms on
pegs were washed three times using 200 microL of PBS before biofilms were air-dried for 30
minutes Biofilm on pegs were then stained by dipping in 200 microL of a crystal violet solution
(01) for 30 minutes rinsed in 200 microL of distilled water and air-dried for 30 min Finally the
crystal violet from stained biofilms was solubilized in 200 microL of ethanol 95 at room
temperature and the absorbance was measured at 562 nm using a plate reader (nom de
lrsquoappareil)
Cell invasion and measurement of intracellular antibiotic activity
The Calu-3 cell line (ATCC HTB 55 Homo sapiens lung adenocarcinoma) was cultured in
Eaglersquos Minimum Essential Medium (EMEM) supplemented with 01 mM MEM nonessential
amino acids 1 mM of sodium pyruvate 100 UmL penicillin 100 gL streptomycin 25 gL of
Fungizone and 10 fetal bovine serum (FBS) at 37oC in 5 CO2 For routine culture 4 gL of
puromycin was added to culture media All cell culture reagents were purchased from Wisent
(St-Bruno QC Canada)
50
The cell invasion assay was performed with the Calu-3 cells in an air-liquid interface as
previously described with few modifications (Mitchell TO I paperAAC 2011) Briefly cells
were seeded at 15 x 105 cells per insert in a 12-well Transwell plate (Corning CITY STATE)
and cultured for 10 days with the apical medium replaced each day The complete medium in
the basal compartment was replaced by the invasion medium (1 FBS and no antibiotics) 18
h before invasion assays Inocula were prepared by suspending bacteria grown 20 h on BHI
agar plates in ice-cold PBS Bacteria were then washed three times in ice-cold PBS and
suspended in the invasion medium supplemented with 05 BSA at a density of
approximately 4 x 108 CFUmL Cells were washed twice with PBS and 250 μL of bacterial
suspension were apically added to each insert Invasion of cells by bacteria was allowed for 3
h inserts were emptied and washed three times with PBS The basal medium was changed for
the invasion medium with 20 gL of lysostaphin (Sigma) to kill extracellular bacteria and with
or without the tested antibiotic compounds The infected cells were incubated for a total of 48
h with a change of medium at 23 h (invasion medium with lysostaphin with or without the
tested antibiotic compounds) The invasion medium with lysostaphin but without tested
compound was also apically added 1 h before cell lysis to ensure that only intracellular
bacteria were counted At 48 h following three washes with PBS the apical and basal media
were removed and cells were detached with 100 μL of trypsin 025 and lysed for 10 min by
the addition of 400 μL of water containing 005 of Triton X-100 Then 50 μL of PBS (50X)
was added and mixed Lysates were serially diluted 10-fold and plated on TSA for CFU
determination Plates were incubated at 35oC for 48 h
51
RESULTS
Emergence the SCV phenotype
The SCV phenotype is characterized by a slow growth yielding small colony size on agar
plates (15 to 110 of the parent phenotype) Figure 2 illustrates the ability of a subinhibitory
concentration of GEN to promote the emergence of the SCV phenotype for B cereus B
subtilis and L monocytogenes B cereus SCVs were obtained at an inducing concentration of
1 X MIC of GEN (Fig 2C) B subtilis SCVs at 05 X MIC (Fig 2D) and L monocytogenes
SCVs at 025 X MIC (Fig 2E) Two prototypic S aureus Newbould and CF07-L and their
SCV counterpart NewbouldhemB and CF07-S respectively were used as control strains (Fig
2A and 2B) The SCV isolates selected for the rest of the study add a low reversion rate and
kept their phenotype without a GEN selection pressure The B cereus SCVs showed
auxotrophy for menadione (Fig 1F Table 1) some of the L monocytogenes SCVs showed
auxotrophy for hemin (Table 1) while the B subtilis SCVs showed no apparent auxotrophy to
hemin menadione or thymidine (Table 1) These results show that SCVs with defects in their
respiratory chain (ie hemin or menadione auxotrophy) can indeed emerge from a selective
pressure with aminoglycosides
52
A B
Fig 2 Typical colony sizes for SCV and protocipic strains (left and right sides of the plates
respectively) for (A) S aureus CF07-S and CF07-L (B) S aureus NewbouldhemB and
Newbould (C) B cereus ATCC 11778 (SCV1 and prototypic) (D) B subtilis ATCC 6333
(SCV2 and prototypic) and (E) L monocytogenes ATCC 13932 (SCV1 and prototypic) In
(F) a TSA plate was inoculated with a B cereus SCV1 and wells were filled with (i) 10 microg
hemin (ii) 10 microg menadione (iii) 100microg thymidine and diluents (iv) DMSO and (v) NH4OH
14N
C D
E
i ii
iii iv
v
F
53
Antibacterial activity of TO against the various species of SCVs and synergy of
the TO and GEN combination against some prototypic Firmicutes
All the SCVs tested showed a MIC for TO of 006 mgL against S aureus and lt003 mgL
against L monocytogenes B cereus and B subtilis respectivly whereas TO showed no
activity (MIC gt64 microgmL) against their prototypic counterparts (Table 1) A synergic effect of
TO with GEN against all the prototypic strains tested was also observed (Table 1) Indeed the
addition of TO provoked a 4-8 fold decrease of GEN MIC against all species tested (Table 1)
These results are consistent with our previous studies on the activity of TO on S aureus
((Mitchell 2011 AAC 2012 JAC)) No change in the MIC of GEN was observed against the
streptococci (S pyogenes S suis S agalactiae S mitis and S pneumoniae) when tested in
combination with TO (data not shown) These results show that TO is active against some but
not all Firmicutes
54
Table 1 Susceptibility of prototypic and SCV strains and species to TO andor GEN
Ratio of activity of GEN+TO vs GEN alone
Bactericidal activity of TO against SCVs and bactericidal synergy of the TO
and GEN combination against prototypic strains
As shown above the addition of 8 mgL of TO to GEN reduced the MIC of GEN against some
Firmicutes (Table 1) but also time-kill kinetics showed that TO improved the bactericidal
effect of GEN against prototypic B subtilis and B cereus (Fig 3A and 3B respectively) This
bactericidal synergy was also previously described for S aureus (Mitchell 2012 JAC)
However although the MIC of GEN was improved when in combination with TO against L
monocytogenes (Table 1) time-kill experiments failed to demonstrate a bactericidal synergy
TO GENGEN
(+TO)
Saureus ATCC 29213 gt64 05 006 8
Saureus NewbouldΔhemB 006 8 Hemin
B cereus ATCC 11778 gt64 1 025 4
B cereus ATCC 11778 (SCV1) lt003 16 Menadione
B cereus ATCC 11778 (SCV2) lt003 16 Menadione
B subtilis ATCC 6333 gt64 0125 003 4
B subtilis ATCC 6333 (SCV1) lt003 2 unknown
B subtilis ATCC 6333 (SCV2) lt003 4 unknown
B subtilis ATCC 9372 gt64 0125 lt0015 gt8
B subtilis ATCC 9372 (SCV1) lt003 1 unknown
B subtilis ATCC 9372 (SCV2) lt003 1 unknown
L monocytogenes gt64 05 0125 4
L monocytogenes (SCV1) lt003 gt64 unknown
L monocytogenes (SCV2) lt003 gt64 Hemin
Strains
MIC (gL)
Synergy
foldAuxotrophy
55
with an improvement of at most 1 log10 in killing compared to that achieved with GEN alone
(Fig 3C) Similarly the effect of TO used alone against SCVs was strongly bactericidal
against S aureus B subtilis and B cereus but not against the L monocytogenes SCV (Fig
3D) and this even if the MIC of TO against L monocytogenes SCVs was inferior to 003
mgL (Table 1) These results show that the activity of TO against L monocytogenes needs to
be improved
56
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 0 6 m g L G E N
0 0 6 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 5 m g L G E N
0 5 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
2 m g L G E N
2 m g L G E N + T O
8 m g L T O
C T R L
Lo
g1
0 C
FU
ml
0
2
4
6
8
1 0
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
CT
RL
CT
RL
CT
RL
CT
RL
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
S a u re u s
h e m B
S C V
B c e r e u s
A T C C 1 1 7 7 8
S C V 2
B s u b t i l i s
A T C C 6 3 3 3
S C V 2
L m o n o c y to g e n e s
A T C C 1 3 9 3 2
S C V 1
Fig 3 Time-kill kinetics of TO andor GEN against (A) prototypic B subtilis ATCC 6333(B)
B cereus ATCC 11778 and (C) L monocytogenes ATCC 13932 The bactericidal effect of
TO against SCVs is also shown at 8 and 24 h (D) The results were obtained from at least
three independent experiments
A B
C
D
Detection
threshold
57
Bactericidal activiy of the TO analog FC04-100 against L monocytogenes
SCVs and synergy with aminoglycosides against prototypic strains and MRSA
The addition of 8 mgL of FC04-100 to GEN (2 mgL) decreased by an average of 5 log10
prototypic L monocytogenes CFUmL after 24 h of treatment in comparison to GEN alone
(Fig 4A) Also the bactericidal activity of FC04-100 used alone against the L monocytogenes
SCV was shown to be better than that observed for TO (Fig 3D) To facilitate comparison
between TO and FC04-100 Figure 4B shows the residual CFU of the L monocytogenes SCV
strain after treatment with TO or FC04-100 and calculated as the percentage of live CFU in
comparison with untreated L monocytogenes SCVs after 24 h of incubation These results
show that TO can be chemically modified (eg derivative FC04-100) and improve its activity
against L monocytogenes and thus the antibacterial activity of FC04-100 against S aureus
was further investigated Interestingly the MIC of FC04-100 against the SCV strain S aureus
NewbouldhemB (MIC of 05 -1 mgL Table 2) was higher than that found for TO (MIC of
006 mgL Table 1) On the other hand the MIC of FC04-100 against the prototypic S
aureus ATCC 29213 is lower than that of TO (8-16 mgL vs gt 64 mgL respectively) (Tables
1 and 2) This newly detected antibacterial activity of FC04-100 was also noted against
prototypic MRSA strains (Table 2) Furthermore the ability of FC04-100 to potentiate the
activity of GEN was similar to that of TO with a 4 to 8 fold improvement of the
aminoglycoside MIC (Table 2)
Although the multi-resistant S aureus MRSA USA100 USA300 and COL are resistant to
cefalexin (beta-lactam antibiotic MIC of 256 mgL) and to the aminoglycoside kanamycin
(MIC of 256 mgL and gt 1024 mgL for USA 100 and USA300 respectively Table 3) the
58
combination of both antibiotics decreased their respective MICs (Table 3) Indeed the
combination of cefalexin and kanamycin in a proportion of 32 (commercialized under the
name UbrolexinTM
) decreased the MIC of one or both antibiotics This is as expected for the
combination of a beta-lactam with an aminoglycoside (Davis 1982) However the synergy is
weak with MRSA strains that are resistant to one or the other or both antibiotics and the
addition of 4 mgL of FC04-100 further improved by 8 to 32 fold the MIC of the cephalexin-
kanamycin combination against these strains (Table 3)
59
te m p s (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
F C 0 4 -1 0 0 8 m g L
2 m g L G E N +
F C 0 4 -1 0 0 8 m g L
C T R L
G E N 2 m g L (B H I)
Re
sid
ua
l C
FU
(
)
CT
RL
TO
8 g
L
FC
04-1
00 8
gL
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
F C 0 4 -1 0 0 8 g L
T O 8 g L
C TR L
T=
24
h
T=
8h
T=
8h
T=
24
h
T=
24
h
Fig 4 Time-kill kinetics of FC04-100 andor GEN against (A) prototypic L monocytogenes
ATCC 13932 The bactericidal effect of FC04-100 used alone against L monocytogenes
SCV1 is also shown at 8 and 24 h (B) In (B) the residual CFU were calculated as the
percentage of live CFU in each condition in comparison with the untreated L monocytogenes
SCV1 after 24h of incubation The results were obtained from at least three independent
experiments
A
B
60
Table 2 Susceptibility of prototypic and SCV S aureus strains to FC04-100 with or without
GEN
a FC04-100 w as used at 4 microgmL
b Ratio of activity of GEN + FC04-100 vs activity of GEN alone
Table 3 Synergistic activity of FC04-100 with the combinaison cefalexin-kanamycin
aFC04-100 was used at 4 microgmL
bRatio of the MIC of cefalexin+kanamycin with FC04-100 vs the MIC of
cefalexin+kanamycin alone
Staphylococcus aureus ATCC 29213 8-16 05 006-0125 4-8
S aureus SH1000 8-16 05 006-0125 4-8
S aureus NewbouldΔhemB 1 8
MRSA USA100 16 05 0125 4
MRSA USA300 16 05 006 8
MRSA Col 16 025 006 4
Strains FC04-100 GENGEN
(+FC04-100 )fold
MIC (gL)
MRSA USA100 256 256 12886 43 32
MRSA USA300 256 gt1024 256171 86 32
MRSA Col 256 4 43 05035 8
MIC (gL)
Strains cefalexin kanamycin cef+kana (32)cef+kana (32)
(+FC04-100 )fold
a b
a b
61
FC04-100 potentiates GEN against S aureus embedded in biofilm and inhibits
intracellular replication of SCVs
Figure 5 demonstrates that compound FC04-100 efficiently kills S aureus embedded in
biofilms when in combination with GEN ie there was an important decrease of bacterial
CFUs compared to the no drug control or to either FC04-100 used alone at 4 ugmL (ie 025-
05MIC Table 2) or GEN used alone at 025 05 or 1 X MIC Besides we previously
demonstrated that TO was able to inhibit intracellular replication of SCVs (Mitchell AAC)
which is an important aspect of virulence for S aureus Here (Fig 6) we show that FC04-100
like TO can significantly decrease the infection of polarized airway epithelial cells by SCVs
62
05
mgL
GE
N
05
mgL
GE
N +
FC
04
02
5 m
gL
GE
N
02
5 m
gL
GE
N +
FC
04
01
2 m
gL
GE
N
01
2 m
gL
GE
N +
FC
04
No d
rug
FC
04
0
2
4
6
8
log
10
CF
Up
eg
= 3 2
= 3 8
= 3 5
= 2
Fig 5 Bactericidal effect of FC04-100 alone or in combination with GEN against prototypic
S aureus SH100 embedded in biofilm FC04-100 (FC04) was used at 4 mgL alone or in
combination with GEN Data are from three independent experiments performed in triplicate
Significant differences in comparison to the no drug control are shown ( Plt 0001 and
Plt 00001 one-way ANOVA with a Dunnettrsquos post test) Data are presented as means with
standard deviations
63
No d
rug
TO
8 m
gL
FC
04 8
mgL
0
2
4
6
log
10
CF
Ui
ns
er
t
Fig 6 Effect of TO and FC04-100 on the intracellular load of CF07-S 48 h post-
internalization Data are from three independent experiments performed in duplicate
Significant differences among groups are shown ( Plt 00001One-way ANOVA with
Turkeyrsquos post test) Data are presented as means with standard deviations
64
DISCUSSION
In previous studies we have demonstrated that TO has a strong inhibitory activity against S
aureus SCVs and improves the bactericidal activity of aminoglycoside antibiotics against
prototypic S aureus and also more broadly against staphylococci This effect was documented
for several strains of diverse clinical origins including aminoglycoside-resistant S aureus
carrying aminoglycoside modifying enzymes TO showed however no synergistic effect on the
activity of aminoglycosides against P aeruginosa E coli or Enterococcus spp (Mitchell
2012) In the present study we showed that the antibacterial spectrum of TO can be extended
to species of the Bacillales order with very low MICs against L monocytogenes B cereus and
B subtillis SCVs (lt003 microgmL) Time-kill kinetics showed that the combination of TO and
GEN creates a bactericidal synergy against prototypic of B subtilis and B cereus similarly to
that previously demonstrated against S aureus However the mixture of TO and GEN did not
demonstrated much improvement over the activity of GEN alone against prototypic L
monocytogenes in time-kill experiments Likewise investigations with Bacillales SCVs
showed that TO efficiently killed S aureus B cereus and B subtillis but not L
monocytogenes SCVs despite its very low MIC and was thus bacteriostatic against that species
Therefore improvement of the antibacterial activity against this important pathogen seems
inevitable for future development of steroidal alkaloids such as TO
TO is a steroid alkaloid structurally characterized by 6 rings 12 stereogenic centers a 3-
hydroxyl group and spiro-fused E F rings in the form of an aminoketal (Fig 1) It is generally
65
prepared by hydrolysis of its glycosylated analog tomatine while both molecules can be found
in the Solanaceae plants (Ruiz-Rubio 2001 Simons 2006) Although we previously
documented some interesting antibacterial activity for TO the absence of an identified cellular
target makes difficult the establishment of a structure-activity relationship (SAR) Our initial
attempt to elucidate SAR was therefore based on the hypothesis that the steroidal A-D rings
act as a rigid scaffold to orient pharmacophores defined by the 3-hydroxyl group on ring A
and the spiroaminoketal group on rings E F Noteworthy we prepared analogs bearing
modifications on ring A of TO and schegravemes for chemistry synthesis were recently published
(Chagnon et al 2014) From seven compounds keeping rings E and F of TO and possessing
structural variations on position 3 of ring A we were able to ascertain some structure activity
relationships Results suggest that the hydrogen bond donor effect of the 3-hydroxyl group
was not important for activity although the stereochemistry of the 3 position substitution
moderately affected activity against both S aureus SCVs and prototypic strains (in
combination with an aminoglycoside) In addition although the presence of one or two
ammonium groups in position C3 reduced the activity against S aureus SCVs (from a MIC of
006 to 05-1 microgmL) this change was highly beneficial for antibiotic activity against normal
strains (from a MIC of gt64 to 8-16 microgmL) without the presence of an aminoglycoside This
change was unexpected and one of such analogs FC04-100 increased its solubility by more
than ten times (from 2 mgmL to 26 mgmL in DMSO) The biological properties of FC04-
100 were therefore further investigated in the present study especially that TO showed some
weak bactericidal activity against Listeria and that the solubility of TO may limit eventual
clinical applications
66
Here we demonstrated that FC04-100 was able to kill both L monocytogenes SCVs as well as
the normal phenotype alone and in the presence of GEN respectively (Fig 4) Furthermore
FC04-100 showed a noticeable antibacterial activity on its own (MIC of 16 microgmL) against the
MRSA strains USA100 USA300 and COL (Table 2) FC04-100 was also synergistic in
combination with an aminoglycoside against such MRSA strains with a 4 to 8 fold gain in the
MIC of GEN Because MRSA strains are often multiresistant to antibiotics and often carry
AMEs we examined the possibility of using a triple antibiotic combination Indeed the
combination of kanamycin and cefalexin in a proportion 32 is already approved to treat
bovine mastitis pathogens under the brand name of UbrolexinTM
(Ganiegravere 2009) This
antibiotic combination offers an extended spectrum of activity compared to each individual
drug and is expected to cover both S aureus Streptococcus uberis and E coli (Ganiere
2009 Maneke et al 2011 Silley 2012) Unfortunately the increased frequency of livestock-
associated MRSA (Garcia-Alvarez et al 2011 Graveland et al 2011 Leonard and Markey
2008) and frequent incidence of strains and species carrying AMEs (Ramirez and Tolmasky
2010) may limit the spectrum of activity of the combination of a beta-lactam and an
aminoglycoside With this in mind we showed that the use of FC04-100 in combination with
kanamycin and cefalexin improves by 32 fold the activity of this mixture against MRSA
strains carrying AMEs (Table 3) Overall the addition of FC04-100 to the aminoglycoside-
beta-lactam combination decreased the MICs of kanamycin and cefalexin below their
resistance breakpoints and this for all the MRSA strains tested
67
L monocytogenes is a highly specialized intracellular pathogen (Hamon et al 2006) causing
life-threatening infections especially in pregnant women (Janakiraman 2008) L
monocytogenes in a foodborne pathogen and because of its intracellular life cycle and
propagation it is able to hide from host defenses and antibiotic treatment Similarly S aureus
although not primarily recognized as a typical intracellular pathogen is also able to enter and
survive in host cells (Lowy 2000) Moreover the ability of S aureus to convert to the SCV
phenotype showing increased biofilm production improved adherence to host cells and tissues
as well as increased intracellular persistence allows this pathogen to cause chronic and
difficult to treat infections (Garcia et al 2013 Mitchell and Malouin 2012) Discovering
antibacterial drugs able to act in the intracellular compartment is generally recognized as a
difficult challenge Previously we demonstrated that steroidal alkaloids such as TO had such
an ability and could act on intracellular S aureus SCVs (Mitchell et al 2011) and here we
demonstrated that this was also the case for the TO analog FC04-100 (Fig 6) In addition we
also show in the present study that FC04-100 greatly improves the bactericidal activity of
aminoglycosides such as GEN against bacteria embedded in biofilms Biofilms are
recognized to be a major hindrance to antibiotic action and are responsible for persistent
colonization in many diseases (Costerton et al 1999) and in the food industry (Srey et al
2013) The intracellular and intrabiofilm antibacterial properties of FC04-100 warrant further
investigation of this compound and this class of molecules in general
In conclusion we showed in this study that the spectrum of activity of TO is related to the
Firmicutes division and more precisely to the order of the Bacillales TO possesses an
68
antibacterial activity against SCVs and a synergistic activity with animoglycoside antibiotics
against prototypic strains The novel TO analog FC04-100 showed very promising new
characteristics that include a much improved bactericidal effect on L monocytogenes and
killing of bacteria in biofilms
ACKNOWLEDGEMENTS
This study was supported by an operation grant from Cystic Fibrosis Canada to FM and by a
team grant from the Fonds Queacutebeacutecois de la Recherche sur la Nature et les Technologies
(FQRNT) to FM KB and EM
REFERENCES
Atalla H Gyles C Jacob CL Moisan H Malouin F Mallard B 2008 Characterization of
a Staphylococcus aureus small colony variant (SCV) associated with persistent bovine mastitis
Foodborne Pathog Dis 5785ndash799
Bad Bug Book Foodborne pathogenic microorganisms and natural toxins 2012 Second
edition Lampel KA Al-Khaldi S Cahill SM (Ed) Published by the Center for Food Safety and
Applied Nutrition of the Food and Drug Administration (FDA) US Department of Health and
Human Services pp 87-103
Beierlein JM Anderson AC 2011 New developments in vaccines inhibitors of anthrax
toxins and antibiotic therapeutics for Bacillus anthracis Curr Med Chem 185083-5094
69
Brouillette E A Martinez B J Boyll N E Allen and F Malouin 2004 Persistence of
a Staphylococcus aureus small-colony variant under antibiotic pressure in vivo FEMS
Immunol Med Microbiol 4135ndash41
Bryan L E and S Kwan 1981 Aminoglycoside-resistant mutants of Pseudomonas
aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob
Agents Chemother 19958ndash964
Ceri H Olson M Morck D Storey D Read R Buret A Olson B 2001 The MBEC Assay
System multiple equivalent biofilms for antibiotic and biocide susceptibility testing Methods
Enzymol 337377-385
Chagnon F I Guay M-A Bonin G Mitchell K Bouarab F Malouin E Marsault
2014 Unraveling the structure-activity relationship of tomatidine a steroid alkaloid with
antibiotic properties against persistent forms of Staphylococcus aureus Eur J Med Chem (In
Press EJMECH-S-13-01625-3)
Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial
Susceptibility Testing Twenty-First Informational Supplement M100-S21 CLSI Wayne PA
USA 2011
70
Costerton JW Stewart PS Greenberg EP 1999 Bacterial biofilms a common cause of
persistent infections Science 284 1318-1322
Davis BD 1982 Bactericidal synergism between beta-lactams and aminoglycosides
mechanism and possible therapeutic implications Rev Infect Dis 4237-245
Ganiegravere JP Denuault L 2009 Synergetic interaction between cefalexin and kanamycin in
Muller-Hinton broth medium and in milk J appl microbiol 107 117-125
Garcia LG S Lemaire B C Kahl K Becker R A Proctor O Denis P M Tulkens
F Van Bambeke 2013 Antibiotic activity against small-colony variants of Staphylococcus
aureus review of in vitro animal and clinical data J Antimicrob Chemother 68 1455ndash1464
Garcia-Alvarez L M T Holden H Lindsay C R Webb D F Brown M D Curran
E Walpole K Brooks D J Pickard C Teale J Parkhill S D Bentley G F Edwards
E K Girvan A M Kearns B Pichon R L Hill A R Larsen R L Skov S J Peacock
D J Maskell and M A Holmes 2011 Methicillin-resistant Staphylococcus aureus with a
novel mecA homologue in human and bovine populations in the UK and Denmark a
descriptive study Lancet Infect Dis 11595-603
Gomaa OM Momtaz OA 2007 16 rRNA characterization of Bacillus isolate and its
tolerance profile after subsequent subculturing Arab J Biotech 10 107-116
71
Graveland H Duim B van Duijkeren E Heederik D Wagenaar JA 2011 Livestock-
associated methicillin-resistant Staphylococcus aureus in animals and humans Int J Med
Microbiol 301630-634
Guillet C Join-Lambert O Le Monnier A Leclercq A Mechaiuml F Mamzer-Bruneel MF
Bielecka MK Scortti M Disson O Berche P Vazquez-Boland J Lortholary O Lecuit M
2010 Human listeriosis caused by Listeria ivanovii Emerg Infect Dis 16136-138
Hamon M H Bierne P Cossart 2006 Listeria monocytogenes a multifaceted model Nat
Rev Microbiol 4 423-434
Harrison JJ Stremick CA Turner RJ Allan ND Olson ME Ceri H 2010 Microtiter
susceptibility testing of microbes growing on peg lids a miniaturized biofilm model for high-
throughput screening Nat Protoc 51236-1254
Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y
Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T Baba T 2013 Genomic
Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
Horsburgh MJ Aish JL White IJ Shaw L Lithgow JK Foster SJ 2002 SigmaB
modulates virulence determinant expression and stress resistance characterization of a
72
functional rsbU strain derived from Staphylococcus aureus 8325-4 J Bacteriol 1845457-
5467
Janakiraman V 2008 Listeriosis in Pregnancy Diagnosis Treatment and Prevention Rev
Obstet Gynecol 1 179ndash185
Leonard FC Markey BK 2008 Methicillin-resistant Staphylococcus aureus in animals a
review Vet J 175 27-36
Lowy FD 2000 Is S aureus an intracellular pathogen Trends Microbiol 8341-342
Magalhatildees L Nitschke M 2013 Antimicrobial activity of rhamnolipids against Listeria
monocytogenes and their synergistic interaction with nisin Food Control 29 138-142
Maneke E Pridmore A Goby L Lang I 2011 Kill rate of mastitis pathogens by a
combination of cefalexin and kanamycin J Appl Microbiol 110184-190
Mitchell G Brouillette Eacute Lalonde-Seacuteguin D Asselin AE Lebeau Jacob C Malouin F
2009 A role for sigma factor B in the emergence of Staphylococcus aureus small-colony
variants and elevated biofilm production resulting from an exposure to aminoglycosides
Microb Pathog 4818-27
73
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K Malouin F 2011
Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic
fibrosis airway epithelial cells Antimicrob Agents and Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM Malouin F 2011 Infection of Polarized
airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is
increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function
and is influenced by NF- κ B Infect Immun 793541-3551
Mitchell G Lafrance M Boulanger S Lalonde-Seacuteguin D Guay I Gattuso M Marsault
Eacute Bourab K Malouin F 2012 Tomatidine acts in synergy with aminoglycoside antibiotics
against multiresistant Staphylococcus aureus and prevents virulence gene expression J
Antimicrob Chemother 67 559-568
Mitchell G Lalonde-Seacuteguin D Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S
Malouin F 2010 Staphylococcus aureus sigma B-dependent emergence of small-colony
variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-
heptylquinoline-N-oxide BMC Microbiology 10 33-48
Mitchell G and F Malouin 2012 Outcome and prevention of Pseudomonas aeruginosa-
Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic
Fibrosis InTech Open Access Publisher ISBN 979-953-307-059-8
74
Moisan H Brouillette E Jacob CL Langlois-Beacutegin P Michaud S Malouin F 2006
Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated
from cystic fibrosis patients is influ- enced by SigB J Bacteriol 18864ndash76
Moskowitz SM Foster JM Emerson J Burns JL 2004 Clinically feasible biofilm
susceptibility assay for isolates of Pseudomonas aeruginosa from patients with cystic fibrosis
J Clin Microbiol 421915-1922
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M Peters G 2006
Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent
infections Nat Rev Micro- biol 4295ndash305
Ramirez MS M E Tolmasky 2010 Aminoglycoside modifying enzymes Drug Res
Updates 13 151ndash171
Ruiz-Rubio M Peacuterez-Espinosa A Lairini K Roldagraven-Arjona T Dipietro A Anaya N
2001 Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi p 293ndash326 In
A Rahman (ed) Studies in natural products chemistry vol 25 Elsevier Oxford United
Kingdom
75
Sendi P Proctor RA 2009 Staphylococcus aureus as an intracellular pathogen the role of
small colony variants Trends Microbiol 1754-58
Silley P Goby L Pillar CM 2012 Susceptibility of coagulase-negative staphylococci to a
kanamycin and cefalexin combination J Dairy Sci 953448-3453
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C Osbourn A
2006 Dual effects of plant steroidal alkaloids on Saccharomyces cerevisiae Antimicrob
Agents Chemother 502732ndash2740
Srey S Jahid IK Ha S-D 2013 Biofilm formation in food industries A food safety concern
Food control 31572-585
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D
Roth J Proctor RA Becker K Peters G Loumlffler B 2011 Staphylococcus aureus
phenotype switching an effective bacterial strategy to escape host immune response and
establish a chronic infection EMBO Mol Med 3129ndash141
Vuong C Otto M 2002 Staphylococcus epidermidis infections Microbes Infect 4481-489
Witte W Strommenger B Stanek C Cuny C 2007 Methicillin-resistant Staphylococcus
aureus ST398 in Humans and Animals Central Europe Emerg Infect Dis 13 255ndash258
76
CHAPITRE 3
MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
31 Obtention des mutants reacutesistants
Afin de nous aider agrave mieux deacutefinir la cible et le mode drsquoaction de la tomatidine (TO)
nous avons forceacute lrsquoapparition de reacutesistances chez Staphylococcus aureus Comme
souche controcircle prototype nous avons choisi ATCC 29213 puisque crsquoest une souche
tregraves freacutequemment utiliseacutee pour des expeacuteriences avec des antibiotiques et nous avons
choisi la souche NewbouldΔhemB comme S aureus laquo small colony variant raquo puisque
crsquoest une souche SCV qui est stable et bien connue nrsquoayant pas de capaciteacute de
reacuteversion Par passages successifs dans une concentration sous inhibitrice
drsquoantibiotique (technique de CMI (concentration minimale inhibitrice)) (CLSI 2011)
nous avons obtenu des mutants deacutemontrant une reacutesistance croissante agrave lrsquoantibiotique
au fil des passages (Figure 1) Nous avons obtenu des mutants ATCC 29213
reacutesistants agrave la gentamicine (GEN) et agrave la GEN lorsque combineacutee agrave la TO ainsi que
des mutants reacutesistants NewbouldΔhemB reacutesistants agrave la GEN seule agrave la TO seule et
agrave la TO lorsque combineacute agrave la GEN
Certains passages ont ensuite eacuteteacute seacutelectionneacutes parce qursquoils deacutemontraient une hausse
plus marqueacutee de leur CMI Ces passages ont eacuteteacute choisis car nous supposions que
ces hausses marqueacutees de CMI correspondraient donc agrave des hausses de la
reacutesistance de cette souche agrave lrsquoantibiotique testeacute et que cette reacutesistance pouvait ecirctre la
conseacutequence drsquoune ou plusieurs mutations en lien avec la cible ou le mode drsquoaction
de lrsquoantibiotique preacutesent Trois agrave quatre passages (ou paliers de reacutesistance) par
condition ont eacuteteacute seacutelectionneacutes Nous avons ensuite reacuteensemenceacute le laquo pool raquo
bacteacuterien de chacun des passages choisis afin de pouvoir isoler 3 isolats ayant la
mecircme CMI (ou tregraves similaire (CMI vraies)) que celui du laquo pool raquo bacteacuterien des
77
passages drsquoougrave ils provenaient Soixante quatre mutants reacutesistants ont ainsi eacuteteacute
seacutelectionneacutes en vue drsquoecirctre seacutequenceacutes (Figure 2)
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt 1
6
CM
I v
raie
s
02
5
CM
I v
raie
s
02
5-1
A
78
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
en
pr
eacutes
en
ce
de
ge
n (
4 micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
01
25
1
6
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
8
CM
I v
raie
s
2
4
CM
I v
raie
s
2
B
C
79
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
en
pr
eacutes
en
ce
de
TO
(8
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
1
2
3
4
5
CM
I v
ra
ies
4
CM
I v
ra
ies
2
CM
I v
ra
ies
0
5
1
CM
I v
ra
ies
2
4
Figure 1 Graphiques des CMI pour chacun des passages pour (A) la tomatidine sur
NewbouldΔhemB (B) pour la tomatidine en preacutesence de gentamicine sur
NewbouldΔhemB (C) pour la gentamicine sur S aureus ATCC 29213 (D) pour la
gentamicine en preacutesence de tomatidine sur S aureus ATCC 29213
32 Seacutequenccedilage et analyses bioinformatiques
Une extraction drsquoADN geacutenomique a ensuite eacuteteacute effectueacutee sur chacun des isolats en
utilisant le Kit drsquoextraction drsquoADN geacutenomique miniKit de Qiagen (QIAamp DNA minikit)
(Li et al 2003) Puisque leurs seacutequences geacutenomiques nrsquoeacutetaient pas connues nous
avons eacutegalement extrait lrsquoADN geacutenomique de nos deux souches de reacutefeacuterences
ATCC29213 et NewbouldΔhemB (provenant de quatre colonies isoleacutees chacune)
afin drsquoavoir les seacutequences de comparaison pour nos mutants (Figure 2)
D
80
Ne
wm
an
(g
en
om
ede r
ef h
ttpw
ww
ncbinlm
nih
govn
uccore
150373012)
AT
CC
29
213
(A p
art
ir d
e 4
AT
CC
29
21
3 r
eco
nstr
uire
AT
CC
29
21
3 c
om
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
5
10
7
10
8 1
09
)
Ne
wbo
uld
Δhe
mB
(A p
art
ir d
e 4
Ne
wb
ou
ldh
em
Bre
co
nstr
uire
Ne
wb
ou
ldh
em
Bcom
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
1
agraveG
EN
2
5 (
CM
I2
)
2
7 (
CM
I2
)
2
8 (
CM
I2
)
AT
CC
29
21
3
Pa
lier
de
reacutesis
tance
1
agravecom
bo
29
(C
MI
05
)
3
0 (
CM
I 0
5
3
1 (
CM
I 0
5)
Suite
fe
uill
e s
uiv
ante
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agraveG
EN
3
(C
MI
4)
4
(C
MI
4)
3
9 (
CM
I4
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agraveG
EN
4
6 (
CM
I8
)
4
7 (
CM
I8
)
4
8 (
CM
I8
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agravecom
bo
1
1 (
CM
I 2
)
1
2 (
CM
I 2
)
2
1 (
CM
I 2
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agravecom
bo
5 (
CM
I 2
-4)
3
6 (
CM
I 2
-4)
3
7 (
CM
I 2
-4)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
4
agravecom
bo
3
8 (
CM
I4
)
6
7 (
CM
I 4
)
81
Ne
wb
ould
Δhem
B(A
part
ir d
e 4
Ne
wb
ou
ldΔ
he
mB
reco
nstr
uire
Ne
wb
ou
ldΔ
hem
Bco
mp
let e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveG
EN
24
(C
MI
64
)
4
1 (
CM
I 1
6)
4
2 (
CM
I 6
4)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveT
O
95
(C
MI
02
5)
9
6 (
CM
I 0
25)
9
7 (
CM
I 0
25)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agravecom
bo
33
34
35
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveG
EN
49
(C
MI
12
8)
5
1 (
CM
I 1
28)
5
2 (
CM
I 1
28)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agraveG
EN
7
(C
MI
128
)
4
3 (
CM
I 1
28)
4
5 (
CM
I1
28
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agraveT
O
6
4 (
CM
I gt
16)
6
5 (
CM
I gt
16)
6
6 (
CM
I gt
16)
75
76
77
79
80
81
82
83
84
85
86
87
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agravecom
bo
15
16
18
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agravecom
bo
59
60
61)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agravecom
bo
54
56
58)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveT
O
90
(C
MI
02
5)
9
1 (
CM
I 0
25)
9
3 (
CM
I 0
5)
Ne
wb
ou
ldh
em
B
palie
r d
e r
eacutesis
tance
3
agraveT
O
11
0 (
CM
I 1
)
1
12 (
CM
I 0
25
-05
)
1
13 (
CM
I 0
5)
Fig
ure
2
Org
an
igra
mm
e d
es
mu
tan
ts r
eacutes
ista
nts
seacute
qu
en
ceacutes
se
lon
leu
r pa
lier
de
reacute
sis
tan
ce e
t a
vec leu
r n
um
eacutero
drsquoeacute
chan
tillo
n
co
rre
spo
ndan
t
82
Un minimum de 20 ngmicrol drsquoADN geacutenomique de chacun des isolats a ensuite eacuteteacute
achemineacute au laboratoire du Pr Seacutebastien Rodrigue (Deacutepartement de biologie Faculteacute
des sciences Universiteacute de Sherbrooke) qui a effectueacute la librairie de fragments Pour
se faire il y a eu dans un premier temps digestion enzymatique de lrsquoADN afin
drsquoobtenir des fragments drsquoADN double brin Ensuite tous les fragments ont eacuteteacute
purifieacutes suivant la meacutethode SPRI (Solid-Phase Reversible Immobilization) Il srsquoagit
drsquoune meacutethode simple et tregraves efficace drsquoisolation drsquoacides nucleacuteiques sur microplaque
(Rodrigue S et al 2010) Les acides nucleacuteiques cibles sont immobiliseacutes sur des
microparticules paramagneacutetiques dans des conditions tampons-speacutecifiques La
flexibiliteacute du systegraveme se fait par lrsquoutilisation de diffeacuterents tampons de liaison (laquo binding
buffer raquo) qui modifient le type et la grandeur des acides nucleacuteiques immobiliseacutes Les
contaminants peuvent ainsi ecirctre retireacutes de lrsquoeacutechantillon sans besoin de filtration ou
centrifugation (Beckman Coulter website 2013 DeAngelis et al 1995) Suite agrave la
purification par meacutethode SPRI lrsquoeacutequipe du Pr Rodrigue a obtenu des fragments
drsquoADN de ~250pb auxquels ont eacuteteacute ajouteacutes par ligation des adapteurs aux deux
extreacutemiteacutes des fragments Dans un deuxiegraveme temps des codes-bars speacutecifiques agrave
chaque eacutechantillon drsquoADN geacutenomique ont eacutegalement eacuteteacute lieacutes agrave chacun des fragments
Alors que les adapteurs permettent aux fragments de se lier agrave une matrice solide afin
de pouvoir ecirctre amplifieacutes les codes-bars permettront de classer chacun des
fragments selon leur appartenance agrave un eacutechantillon drsquoADN geacutenomique speacutecifique
83
Figure 3 Eacutetape du seacutequenccedilage Illumina
La librairie de fragments a ensuite eacuteteacute envoyeacutee chez Geacutenome-Queacutebec afin que
chacun des geacutenomes puisse ecirctre seacutequenceacute Geacutenome ndashQueacutebec utilise la meacutethode
Illumina comme outil de seacutequenccedilage (Figure 3) La grande innovation de la meacutethode
Illumina est lrsquoamplification des fragments par liaison de ceux-ci agrave une surface solide
(Flowcell) ce qui permet par un processus de ponts drsquoamplification de geacuteneacuterer plus
drsquoun million de laquo clusters raquo comprenant eux-mecircme au plus un millier de copies de la
moleacutecule originale de deacutepart (solid-phase PCR) Chaque regroupement de fragments
provenant de lrsquoamplification drsquoun mecircme fragment original est tregraves pregraves les uns des
Modifieacute de Quail MA et al 2008
84
autres formant une zone distincte drsquoun regroupement de fragments agrave un autre
Lrsquoincorporation des nucleacuteotides eacutetiqueteacutes preacutealablement avec des moleacutecules
fluorescentes speacutecifiques est deacutetecteacutee suite agrave leur excitation par un laser et les
images sont compileacutees pour produire la seacutequence de bases pour chaque fragment
ADN (Figure 4) Les nouvelles seacuteries de bases appeleacutees laquo readsraquo seront ensuite
reacuteassembleacutees (aligneacutes) en se basant sur un geacutenome de reacutefeacuterence connu (appeleacute
laquo resequencing raquo) (crsquoest notre cas ici) ou en absence de geacutenome de reacutefeacuterence
(appeleacute laquo De novo sequencing raquo) Au final lrsquoensemble des laquo reads raquo aligneacutes
correspondra agrave la seacutequence de chaque chromosome de lrsquoeacutechantillon drsquoADN
geacutenomique de deacutepart
85
Figure 4 Meacutethode drsquoamplification par la meacutethode Illumina
Suite au seacutequenccedilage chez Geacutenome ndashQueacutebec nous avions en main les seacutequences
geacutenomiques de tous nos mutants et de nos souches de reacutefeacuterence sous forme de
Tireacute de BiteSizeBio httpbitesizebiocom13546sequencing-by-synthesis-
explaining-the-illumina-sequencing-technology consulteacute le 3 deacutec 2013
86
contigs Nous avions donc le casse-tecircte mais en piegraveces deacutetacheacutees pecircle-mecircle et
sans avoir la photo de lrsquoimage de reacutefeacuterence sur laquelle se baser pour pouvoir
compleacuteter le casse-tecircte
1- Lrsquoassemblage
Les diffeacuterents geacutenomes avaient eacuteteacute preacutealablement fractionneacutes aleacuteatoirement
en petits fragments de 250 pb et amplifieacutes en vue du seacutequenccedilage Suite au
seacutequenccedilage les seacutequences de chaque fragment (appeleacute laquo reads raquo) sont ensuite
assembleacutees en contigs crsquoest-agrave-dire des seacutequences plus ou mois longues continues
et ordonneacutees geacuteneacutereacutees par lassemblage des clones dune bibliothegraveque geacutenomique
qui se chevauche La qualiteacute de lrsquoassemblage en contigs deacutepend de la capaciteacute de
superposition des laquo reads raquo Parce que la digestion enzymatique de lrsquoADN se fait de
faccedilon aleacuteatoire et se fait sur plusieurs copies drsquoADN chaque portion du geacutenome est
repreacutesenteacutee plusieurs fois dans diffeacuterents modegraveles de fragments Donc les
seacutequences identiques des diffeacuterents fragments (ou laquo reads raquo) peuvent ecirctre
superposeacutees par chevauchegravement et les laquo reads raquo superposeacutes peuvent ecirctre
assembleacutes en contigs par un logiciel drsquoassemblage En ce qui nous concerne cette
eacutetape a eacuteteacute faite agrave lrsquoaide du programme de bio-informatique Newbler (Zhang et al
2012) Lrsquoassemblage geacutenomique peut ecirctre probleacutematique pour les informaticiens car
les geacutenomes contiennent des seacutequences qui se reacutepegravetent et ces reacutepeacutetitions peuvent
ecirctre assez longues en fonction des organismes Suite agrave lrsquoassemblage les geacutenomes
sont donc sous forme de contigs
2- Lrsquoalignement
En se basant sur une seacutequence geacutenomique de reacutefeacuterence (notre reacutefeacuterence est
Staphylococcus aureus str Newman (~ 288Mb)
( httpwwwncbinlmnihgovgenome154project_id=58839) ) le programme de bio-
informatique ABACAS permet de rapidement aligner ordonner et orienter les contigs
Selon les besoins ce qui nrsquoa pas eacuteteacute notre cas ABACAS permet eacutegalement de
87
visualiser et designer les amorces permettant de fermer les gaps (espaces) entre les
contigs en faisant des PCR (Assefa et al 2009)
3- La fermeture des gaps
La fermeture des gaps nrsquoa pas eacuteteacute faite avec le programme ABACAS mais
avec lrsquoaide du programme de bio-informatique GapFiller Lrsquoutilisation de GapFiller
pour la fermeture des gaps possegravede 3 principaux avantages Premiegraverement le
programme prend en compte lrsquoestimation de la grandeur du gap agrave fermer donc il
exclut automatiquement les fermetures de gaps erroneacutes qui sont plus courts ou plus
longs que la grandeur estimeacutee Deuxiegravemement il nrsquoessaie pas de remplir un gap
avec un assemblage de laquo reads raquo en contigs mais cherche plutocirct agrave allonger les
contigs agrave partir de leurs extreacutemiteacutes jusqursquoagrave obtenir des seacutequences superposeacutees en
passant par la creacuteation de K-mer (seacuteparation des reads en plus petits fragments) Et
troisiegravemement il prend en compte que les extreacutemiteacutes des contigs sont souvent
source de mauvais assemblage et reacute eacutevalue ces extreacutemiteacutes pendant la fermeture des
gaps (Boetzer et Pirovano 2012)
4- Lrsquoannotation des geacutenomes
Suite agrave la fermeture des gaps il a fallu annoter les geacutenomes en se basant sur
lrsquoannotation du geacutenome de reacutefeacuterence Staphylococcus aureus str Newman RATT est
un outil de transfert rapide drsquoannotation (Rapid Annotation Transfert Tool) qui a eacuteteacute
deacuteveloppeacute afin de fournir rapidement une annotation preacutecise pour des geacutenomes
inconnus en se basant sur des geacutenomes deacutejagrave annoteacutes comme reacutefeacuterences (Figure 5)
La meacutethode transfert les annotations drsquoune reacutefeacuterence de haute qualiteacute agrave un nouveau
geacutenome sur la base de la conservation des synteny (ordre des gegravenes) (Otto et al
2011) Suite agrave lrsquoannotation les gegravenes tant chez les mutants que chez les souches
parentes portent les noms auxquels ils correspondent chez Staphylococcus aureus
str Newman
88
Figure 5 Corrections du programme RATT sur les annotations transfeacutereacutees Les
annotations de H37Rv ont eacuteteacute transfeacutereacutees sur la seacutequence F11 (bleu pacircle) ont eacuteteacute corrigeacutees
(vert) et ensuite compareacutees agrave lrsquoannotation de la souche existante F11 EMBL (jaune) (A et B)
La correction des codons ldquostartrdquo et ldquostoprdquo respectivement Dans une situation de laquo mapping raquo
plus complexe (C) ougrave les cadres de lecture sont montreacutes pour plus de clarteacute RATT
cartographie une large seacutequence codante (CDS) de H37Rv vers un locus dans F11 qui inclue
plusieurs codons laquo stop raquo laquo in-frame raquo En inseacuterant un deacutecalage du cadre de lecture (ie un
pseudogegravene) la traduction conceptuelle est conserveacutee Ceci diffegravere avec les deux gegravenes se
superposant preacutedis comme faisant partie du geacutenome F11
Tireacute de Otto TD et al 2011
89
321 Identification des mutations en lien avec la tomatidine
Apregraves avoir obtenu une seacutequence consensus agrave partir des seacutequences geacutenomiques
des 4 copies de chacune des souches parente S aureus ATCC 29213 et S aureus
NewbouldΔhemB les diffeacuterences geacutenomiques entre les souches parentes et le
geacutenome de reacutefeacuterence S aureus str Newman ont eacuteteacute eacutelimineacutees afin que ces
diffeacuterences nrsquoapparaissent pas comme des mutations lors de la comparaison des
seacutequences geacutenomiques des mutants avec celles de leur souche parente ces
diffeacuterences eacutetant plutocirct attribuables agrave la diffeacuterence qursquoil y a entre S aureus str
Newman et la souche parente et non pas agrave des mutations en lien avec la reacutesistance
des mutants
Le service bio-informatique du deacutepartement de biologie (Faculteacute des sciences
Universiteacute de Sherbrooke) a ensuite produit un tableau contenant pour chacun des
mutants tous les gegravenes muteacutes apregraves comparaison avec leur souche parentes Agrave cette
eacutetape on pouvait donc savoir dans quel gegravene se situaient les mutations quelle eacutetait
lrsquoeffet de cette mutation (mutation synonyme deacuteleacutetion insertion SNP) et en principe
srsquoil srsquoagissait pour un mecircme gegravene muteacute de la mecircme mutation puisque dans ce
tableau pour un mecircme gegravene chaque ligne correspond agrave des mutations diffeacuterentes
(Figure 6)
Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4
Figure 6 Exemple drsquoune partie du tableau compilation des mutations chez
les mutants ATCC 29213 reacutesistants agrave la GEN seule et agrave la GEN (+TO)
90
Agrave lrsquoaide de ce tableau (Figure 6) jrsquoai chercheacute les mutations qui pouvaient ecirctre
attribuables agrave la TO Chez les mutants S aureus ATCC 29213 reacutesistants jrsquoai
chercheacute les gegravenes qui eacutetaient muteacutes chez tous les mutants reacutesistants agrave la
combinaison GEN (+TO) ou dont la mutation apparaissait graduellement au fil des
paliers de reacutesistance mais dont ce mecircme gegravene nrsquoeacutetait pas muteacute chez les mutants
reacutesistants agrave la GEN seule Je pouvais donc estimer que cette mutation chez les
reacutesistants agrave la combinaison eacutetait en lien avec la preacutesence de la TO dans le milieu
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct
Afin de mrsquoaider dans ma recherche jrsquoai eu lrsquoaide du Pr Pierre-Eacutetienne Jacques
(Deacutepartement de biologie Faculteacute des sciences Universiteacute de Sherbrooke)qui a
mis les donneacutees de ce tableau dans un tableau croiseacute dynamique (TCD) ce qui a
permis de filtrer les donneacutees selon des critegraveres bien preacutecis me permettant drsquoarriver agrave
mes fins plus facilement Dans un premier temps un TCD a eacuteteacute creacuteeacute ougrave pour
chacun des mutants chaque gegravene ayant au moins une mutation eacutetait indiqueacute par un
laquo 1 raquo alors qursquoun laquo 0 raquo eacutetait marqueacute si ce gegravene nrsquoeacutetait pas muteacute Dans un second
temps la quantiteacute de laquo 1 raquo par palier de reacutesistance (correspondant au nombre de
mutants de ce palier posseacutedant une mutation dans ce gegravene) eacutetait indiqueacutee par un
chiffre allant de 0 pour laquo aucun mutant de ce palier de reacutesistance ne posseacutedait de
mutation dans ce gegravene raquo agrave 1 2 ou 3 selon le nombre de mutants de ce palier ayant
ce gegravene muteacute Jrsquoai ensuite pu soumettre le TCD agrave des filtres approprieacutes afin de faire
ressortir les gegravenes drsquointeacuterecircts selon des critegraveres de seacutelection preacutecis (Figures 7-8)
91
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
1
00
33
32
1
1
11
4
5
0
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
10
03
33
21
11
14
50
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_04
12
lpl9
nm
20
12
32
23
92
44
1
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
NW
MN
_00
46
_2
10
13
21
37
24
31
NW
MN
_09
55
_2
01
23
11
37
24
21
NW
MN
_13
83
gpsA
21
02
31
13
72
42
1
NW
MN
_03
39
_1
11
21
31
37
34
42
NW
MN
_15
97
dn
aE2
10
22
30
37
23
31
NW
MN
_23
30
_2
10
23
20
37
23
21
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_2
07
0_
10
01
23
21
81
45
0
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_10
28
_1
22
12
32
58
34
54
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
A B C
92
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
39
8h
sdM
31
33
33
27
11
34
54
NW
MN
_04
26
_3
31
33
32
71
13
45
4
NW
MN
_05
24
sdrD
32
33
33
28
11
34
55
NW
MN
_07
39
_3
33
33
32
91
13
45
6
NW
MN
_11
64
hsl
U3
33
33
32
91
13
45
6
NW
MN
_05
23
sdrC
22
33
32
27
10
34
45
NW
MN
_20
61
fmtB
23
23
32
27
10
34
45
NW
MN
_04
03
lpl1
nm
33
22
33
28
10
34
55
NW
MN
_13
45
ebh
13
32
33
31
81
03
44
5
NW
MN
_16
49
_3
32
33
31
81
03
44
5
NW
MN
_17
00
hsd
M3
23
33
22
81
03
44
5
NW
MN
_00
42
_3
33
33
22
91
03
44
6
NW
MN
_05
80
_3
33
33
31
91
03
44
6
NW
MN
_13
44
ebh
23
31
32
22
79
34
44
NW
MN
_13
75
aro
A3
13
23
31
79
34
44
NW
MN
_23
97
fnb
B2
32
23
22
79
34
45
NW
MN
_04
01
_3
32
23
31
89
34
45
NW
MN
_07
33
_3
32
32
31
89
34
45
NW
MN
_17
45
_2
33
22
32
89
34
56
NW
MN
_01
66
coa
33
32
32
29
93
44
6
NW
MN
_05
25
sdrE
33
32
33
19
93
44
6
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D Fig
ure
7
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez A
TC
C 2
921
3 d
an
s le
TC
D p
ar
dif
feacutere
nts
fil
tre
s
(A)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts la
ge
nta
mic
ine
ge2
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(C
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts d
an
s le
pa
lier
1
de
reacute
sis
tan
ce
agravela
ge
nta
mic
ine
le1
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
et
nom
bre
de m
uta
nts
reacute
sis
tan
ts agrave
la la
gen
tam
icin
e (
+T
O)
de
s 2
de
rnie
rs p
alie
rs d
e r
eacutesis
tan
ce
ge5
(D
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
7 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
ge
nta
mic
ine
(+
TO
) ge
9
93
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_16
29
ccp
A0
01
33
31
50
00
01
24
0
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_21
28
rpsM
11
12
21
12
22
00
31
74
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
NW
MN
_25
29
clfB
33
03
33
80
11
26
17
4
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e reacute
sist
ance
agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
On
b m
uta
nts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
16
29
ccp
A0
01
33
31
50
00
01
24
0N
WM
N_
21
28
rpsM
11
12
21
12
22
00
31
74
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
A B C
94
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
04
6_
1
01
00
28
22
32
2
10
9
NW
MN
_17
33
prs
A0
11
12
01
12
12
22
14
7
NW
MN
_19
23
_0
11
31
15
13
22
21
08
NW
MN
_20
37
_0
02
32
07
23
11
21
27
NW
MN
_22
77
_1
02
21
11
02
12
33
14
8
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
01
43
_0
03
00
04
23
33
34
11
NW
MN
_0
40
6lp
l3n
m2
01
00
03
12
22
33
7
NW
MN
_1
65
2_
00
10
00
62
12
31
68
NW
MN
_2
01
2_
00
00
00
32
33
30
31
1N
WM
N_
20
15
mn
aA0
00
00
01
23
33
01
11
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
o
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D E Fig
ure
8
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez N
ew
bo
uld
Δh
em
Bd
an
s l
e T
CD
par
dif
feacutere
nts
fil
tres
(A)
Nom
bre
de
mu
tan
ts
reacutesis
tan
ts agrave
la tom
atid
ine
ge15
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atid
ine
ge15
et no
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
(C)
No
mbre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la tom
atid
ine
ge15
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3(
D)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atidin
ege
10
et no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
tom
atid
ine
(+G
EN
) ge
7
(E)
No
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la to
ma
tid
ine
le6
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
95
Le TCD mrsquoa permis de rapidement identifier les gegravenes drsquointeacuterecircts Agrave lrsquoaide du site CMR
(Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) 2013) jrsquoai
ensuite veacuterifieacute la fonction des gegravenes identifieacutes afin de confirmer leur inteacuterecirct (Tableau
1)
Tableau 1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants et (B) chez les mutants NewbouldΔhemB reacutesistants
Gegravene Alias Fonction
NWMN_0811 NADH dh complexe 1 chaine de transport des eacutelectrons
NWMN_2070 _membrane prot Putative- Toxin production+resistance drug transporteur-
similar to multidrug transporteur 85
NWMN_0635 _membrane prot Putative- Bordeacute par AraC(reacutegulateur opeacuteron arabinose) ou
SsaA-likeprot(lieacute au quorum sensing RNAIII(secretory antigen)
NWMN_2084 _truncated transposase for Is1272- mobile+extrachromosomal element
fonctiontransposon fonction
NWMN_0928 qoxC quinol oxidase polypeptide IIIcomplexe 4 chaine de transport des eacutelectrons
NWMN_0739 _ conserved hypothetical prot fonction inconnues
NWMN_1164 hslU ATP-dependent protease ATP-binding subunit
NWMN_0524 sdrD Ser-Asp rich fibrinogenbone sialoprotein-binding protein sdrD
NWMN_0042 _ lipoprotein putativecell enveloppe- lipoprot en tendem
NWMN_0580 _conserved hypothetical prot fonction inconnuesDNA metabolism DNA
replication recombination and repair
NWMN_1745 _
NWMN_0166 coa coagulase precursor
NWMN_0525 sdrE Ser-Asp rich fibrinogenbone sialoprotein-binding protein SdrE
A
96
Apregraves avoir aligneacute speacutecifiquement une par une chacune des seacutequences (Fasta) des
gegravenes drsquointeacuterecirct de chaque mutant avec leur souche parente jrsquoai utiliseacute le programme
de visualisation et de comparaison de seacutequences ClustalW2 (ClustalW2 2013) afin
de confirmer que la mutation eacutetait une vraie mutation et non une erreur de
seacutequenccedilage ou drsquoalignement et que la mutation eacutetait identique chez tous les mutants
ayant le gegravene drsquointeacuterecirct muteacute Cette eacutetape a permis de restreindre encore la liste des
gegravenes pouvant ecirctre en lien avec la cible ou le mode drsquoaction de la TO (Figure 9)
Gene Alias Fonction
NWMN_0896 _conserved hypothetical prot fonction inconnue- comK family prot-
regulation DNA interaction- ORFIDresssemble agrave facteur de transcription
NWMN_0996 _ conserved hypothetical prot fonction inconnue- prophage fonction
NWMN_1025 _ phage major tail protprophage fonction
NWMN_1629 ccpAcatabollite control protA virulence determinant production and antibiotique
resistance
NWMN_1916 _ conserved hypothetical prot-fonction inconnue ou prophage prot
NWMN_2314 _drug resistance transporteur- EmrBQacA subfamily prot- toxin
production+resistance- transport and binding prot
NWMN_0046 _ hypothetical prot
NWMN_2015 mnaA UDP-Glc-Nac 2 epimerase
NWMN_2012 atpE ATP synthase subunit C
NWMN_1652 _ multidrug resistance transporter protein B
NWMN_0143 _ oligopeptide ABC transporter ATP-binding protein
B
97
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
A
B
98
Jrsquoai ensuite chercheacute le rocircle de ces gegravenes sur le site CMR afin de veacuterifier si leur
mutation pouvait expliquer la reacutesistance agrave lrsquoantibiotique (Comprehensive Microbial
Ressouce (J Craig Venter Institute (JCVI) 2013) Dans une premiegravere eacutetape le rocircle
drsquoun gegravene a eacuteteacute deacutetermineacute en allant tout simplement voir directement le rocircle de ce
gegravene chez Staphylococcus aureus str Newman Dans le cas ougrave le gegravene nrsquoavait pas
de fonction connue chez Saureus str Newman je suis alleacutee voir quelle eacutetait la
fonction de ce mecircme gegravene (et des gegravenes flanquant celui-ci) chez diffeacuterentes souches
de Staphylococcus aureus Advenant le cas ougrave malgreacute tout je nrsquoeacutetais toujours pas
capable de deacuteterminer le rocircle de ce gegravene je suis alleacutee veacuterifier si sa seacutequence pouvait
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 hellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Mutants resistants agrave GEN (4 microgml)+TO
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip 185-AATTGCATTCA-197helliphelliphellip495-ATAGCAATGG-506hellip
hellipnwmn_1629helliphelliphelliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_0143hellip
ccpAATP synthase c
ABC transp
Niveau 1
Niveau 2
Niveau 3
Niveau 4
TO +GEN (4microgml)
Figure 9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct (A) nwmn_0811 et
nwmn_0928 chez les mutants ATCC 29213 reacutesistants agrave la combinaison gentamicine
(+TO) leur souche parente ATCC 29213 (B) nwmn_1629 et nwmn_2012 chez les
mutants hemB reacutesistants agrave la TO et leur souche parente NewbouldΔhemB (C)
nwmn_1629 nwmn_2012 et nwmn_0143 chez les mutants hemB reacutesistant agrave la
tomatidine (+genta) et leur souche parente hemB
C
99
avoir une forte homologie avec un gegravene drsquoune autre espegravece Malgreacute tout certains
gegravenes portent toujours la mention laquo de fonction inconnue raquo
La meacutethodologie que nous avons employeacutee nous a permis drsquoidentifier plusieurs
gegravenes possiblement impliqueacutes dans reacutesistance de nos souches aux antibiotiques
testeacutes
1- chez Staphylococcus aureus ATCC 29213 reacutesistant agrave la gentamicine (+TO)
(Figure 9A)
- NWMN_0811 codant possiblement pour la NADH deacuteshydrogeacutenase
- qoxC (NWMN_0928) codant pour la quinol oxidase polypeptide III
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
2- chez NewbouldΔhemB reacutesistant agrave la tomatidine (Figure 9B)
- ccpA (NWMN_1629) codant pour la laquo catabolite control protein A raquo
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
3- chez NewbouldΔhemB reacutesistant agrave la tomatidine (+gentamicine) (Figure 9C)
- NWMN_0143 codant pour un ABC transporteur
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
Lrsquoanalyse de ces diffeacuterents gegravenes nous amegravene agrave suspecter la sous-uniteacute c de lrsquoATP
synthase comme eacutetant la cible potentielle de la tomatidine chez Staphylococcus
aureus
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine
Les gegravenes muteacutes pointent en grande partie vers la chaine respiratoire qursquoon appelle
aussi chaine de transport des eacutelectrons
100
La NADH deacuteshydrogeacutenase (NWMN_0811) fait partie du complexes I de la chaine de
transport de eacutelectrons alors que la quinol oxidase polypeptide III (NWMN_0928 ou
QoxC) est une composante du complex III et la sous-uniteacute c de lrsquoATP synthase (AtpE)
fait partie eacutevidement du complexe de lrsquoATP synthase ou complexe V de la chaine
respiratoire (Figure 10)
Figure 10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants
Les courbes de croissance des mutants ATCC29213 reacutesistants agrave la combinaison
GEN(+TO) appuient le fait que ces mutations affectent la chaine de transport des
eacutelectrons ce qui affecte le niveau de production drsquoATP et donc la croissance
bacteacuterienne En effet les mutants du troisiegraveme niveau de reacutesistance (qui possegravede
deux gegravenes de la chaine de transport des eacutelectrons de muteacutes) ont une croissance
plus lente que les mutants du premier niveau de reacutesistance qui eux ont un seul gegravene
dans la chaine de transport des eacutelectrons de muteacute (Figure 11)
Quinoloxidase
NADHdehydrogenase
ATPsynthetase
Proton gradient
XXX
101
Figure 11 Courbes de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison GEN+TO du premier niveau de reacutesistance (bleu) et du troisiegraveme niveau
de reacutesistance (vert)
Voici plus preacuteciseacutement comment fonctionne la chaine de transport des eacutelectrons chez
les 2 pheacutenotypes-parents selon le modegravele-hypothegravese eacutetabli avec nos reacutesultats et
analyses
Tout drsquoabord chez le prototype ou souche de pheacutenotype normal (ex ATCC 29213) la
chaine de transport des eacutelectrons fonctionne normalement Les complexes de la
chaine de transport des eacutelectrons sont chargeacutes de transfeacuterer des protons vers
lrsquoexteacuterieur de la cellule en transfeacuterant en mecircme temps des eacutelectrons drsquoun complexe agrave
un autre jusqursquoagrave un accepteur final qui est lrsquoO2 lorsque crsquoest possible Se faisant il se
creacutee un gradient eacutelectrochimique de part et drsquoautre de la membrane et les protons
accumuleacutes du cocircteacute externe sont utiliseacutes par la suite par lrsquoATP synthase afin de
pouvoir produire de lrsquoATP dans la cellule
0 4 8 1 2 1 6 2 0 2 4
0 0 1
0 1
1
C o u r b e d e c r o is s a n c e d e s m u ta n ts A T C C 2 9 2 1 3
reacute s is ta n ts agrave G E N + T O (8 m g L )
te m p s (h )
OD
60
0n
m
Niveau 3
Niveau 1
102
Le transfert drsquoeacutelectrons ne se fait pas sans coucirct et des produits toxiques pour la
cellule sont produits en mecircme temps ce sont les espegraveces reacuteactives de lrsquooxygegravene ou
ROS (Reactive Oxygen Species) Cependant les prototypes par leur reacuteponse au
stress produisent des enzymes capables de deacutetoxifier le milieu (Figure 12A)
Chez les souches SCV ici la souche controcircle NewbouldΔhemB la chaine de
transport des eacutelectrons est deacuteficiente parce que le gegravene permettant la production
drsquoheacutemine qui entre dans la composition des cytochromes est supprimeacute Les
cytochromes entrent dans la composition de plusieurs composantes de la chaine
respiratoire
Lorsque les complexes de la chaine sont alteacutereacutes il y a diminution du transfert de
protons vers lrsquoexteacuterieur une diminution de la production drsquoATP et une plus grande
production de ROS (Figure 12B)
Figure 12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B) NewbouldΔhemB
I III IV V I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
eacute eacute
eacute
H+ H+
ATPO2 H2O
ROS +ROS
Prototype SCV (hemB)
H+
0811 0928 2012 0811 0928
hegraveme
2012
A B
103
La gentamicine a besoin drsquoune bonne polarisation membranaire donc drsquoune chaine de
transport des eacutelectrons fonctionnelle pour pouvoir peacuteneacutetrer dans la cellule (Bryan et
Kwan 1983 (Bryan and van den Elzen 1977 Davis 1987 Taber et al 1987) Une
fois entreacutee elle inhibe la synthegravese proteacuteique ce qui provoque la mort cellulaire La
CMI de la gentamicine contre ATCC 29213 est de 05-1 microgml (Figure 13A)
La tomatidine elle irait se lier agrave lrsquoATP synthase pour en diminuer la fonctionnaliteacute et
ce tant chez ATCC 29213 que chez hemB Cette liaison amegravenerait une diminution
de la production drsquoATP et conseacutequemment provoquerait agrave plus ou moins long terme
une augmentation de production de ROS par lrsquoalteacuteration de la chaine de transport des
eacutelectrons Cependant comme la cellule prototype produit efficacement les enzymes
de deacutetoxification lrsquoimpact de la tomatidine est pratiquement nulle sur les souches
prototypes La CMI de la tomatidine est gt16microml contre ATCC 29213 (Figure 13B)
104
Figure 13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213
Chez ATCC 29213 lrsquoajout de 8microgml de tomatidine agrave la Gentamicine amegravene une
diminution de 8X agrave 16X la CMI de la gentamicine En effet la CMI de la gentamicine
passe de 05-1microgml agrave 006microgml (Figure 14AB)
Lors de lrsquoajout de la tomatidine agrave la gentamicine crsquoest la tomatidine qui se lie dans
un premier temps agrave lrsquoATP synthase reacuteduisant ainsi sont niveau drsquoactiviteacute Il y a alors
de faccedilon transitoire une augmentation de la concentration de protons agrave lrsquoexteacuterieur de
la cellule ce qui permet une entreacutee massive de gentamicine ainsi qursquoune rapide et
forte inhibition de la synthegravese proteacuteique Dans un second temps il y aura eacutegalement
une alteacuteration de la chaine de transport des eacutelectrons conduisant agrave une
augmentation de la production de ROS Crsquoest cependant lrsquoentreacutee massive de
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI TO gt16 microgmlCMI GEN 05-1 microgml
+ROS
ROS mais deacutetoxification efficace
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
105
gentamicine entrainant une forte inhibition de la synthegravese proteacuteique qui provoquera
dans ce cas-ci la mort cellulaire (Figure 14B)
Figure 14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicine lorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC 29213
Chez les mutants ATCC29213 reacutesistants agrave la combinaison GEN+TO les complexes I
et III sont muteacutes et donc ont une fonctionnaliteacute alteacutereacutee ce qui amegravene une
deacutepolarisation membranaire Une membrane moins polariseacutee amegravene une plus faible
entreacutee de gentamicine et donc une plus faible inhibition proteacuteique La CMI de la
gentamicine passe en effet de 05-1 microgml chez la souche parente agrave 2 microgml chez les
mutants reacutesistants
La tomatidine en se liant agrave lrsquoATP synthase accentuera la deacutepolarisation
membranaire provoquant une augmentation de la production des ROS et une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN (TO 8 microgml) 006 microgmlCMI GEN 05-1 microgml
+ROS
polarisation entreacutee massive de GEN
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
GENH+
BA
106
diminution de la production drsquoATP De fait les mutants reacutesistants ont une croissance
plus lente (Figure 15)(Figure 11)
Figure 15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants ATCC 29213
reacutesistants agrave la combinaison gentamicine (+TO)
Chez les SCV la tomatidine se lie aussi agrave lrsquoATP synthase et ce faisant amegravene une
diminution de la production drsquoATP Cette diminution de la production drsquoATP megravenera
ultimement agrave une alteacuteration de la chaine de transport des eacutelectrons et une
augmentation des ROS (Murphy 2009) tout comme chez les souches prototypes
Cependant comme le SCV a deacutejagrave une chaine de transport des eacutelectrons deacuteficiente
une faible production drsquoATP par lrsquoATP synthase et une forte production de ROS
lrsquoajout drsquoune faible concentration de tomatidine (la CMI est de 006microgml) amegravene une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype R GEN (+TO [8 microgml])
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN 2 microgmlCMI GEN 05-1 microgml
+ROS
R agrave GEN (+TO) mais mutants ont une
croissance diminueacutee (+ROS et -ATP)
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
107
augmentation de chacun de ces pheacutenomegravenes ce qui est suffisant pour provoquer la
mort cellulaire (Figure 16A)
Dans les premiers niveaux de reacutesistance agrave la tomatidine chez les mutants hemB la
reacutesistance (la CMI passe de 006 agrave 025-1microgml) est ameneacutee par la mutation du gegravene
ccpA (Catabolite controcircle protein A) ce qui megravenerait agrave une augmentation de la
production drsquoATP par lrsquoutilisation drsquoune autre voie que lrsquoATP synthase etou agrave une
diminution de la production de ROS (Figure 16B) Le rocircle de la CcpA en lien avec la
reacutesistance agrave la tomatidine est encore neacutebuleux Toutefois on sait que normalement
ce gegravene agit comme reacutegulateur transcriptionnel des gegravenes codant pour des enzymes
du cycle de Krebs et qursquoil a un rocircle de reacutegulation dans le meacutetabolisme du carbone et
lrsquoutilisation des substrats (Somerville et Proctor 2009 Goumltz et al 2006)
Figure 16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO 006 microgml
H+
SCV
I III IVeacute eacute
eacuteO2 H2O
+ROS
H+TO
Niveau 1
CcpA (CMI TO 025-1 microgml)
ROS
ATP
V
H+
ATP
TO0811 0928
2012
1629
0811 0928
2012
BA
108
Chez les SCV du deuxiegraveme palier de reacutesistance agrave la tomatidine non seulement le
gegravene ccpA est toujours muteacute mais le gegravene de la sous-uniteacute c de lrsquoATP synthase est
eacutegalement muteacute La mutation dans le site de liaison mecircme de la tomatidine empecircche
sa liaison et donc empecircche son effet sa CMI passant de 006 agrave gt16microgml (Figure
17B)
Malgreacute la mutation dans le gegravene lrsquoATP synthase resterait quand mecircme fonctionnel
puisque la mutation nrsquoamegravene pas la mort cellulaire comme crsquoest le cas lorsque la
tomatidine lie lrsquoATP synthase chez le SCV parent (Figure 17A)
Figure 17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la tomatidine
Tout comme chez les mutants hemB reacutesistant agrave la tomatidine seule les mutants
hemB reacutesistant agrave la tomatidine lorsqursquoen preacutesence de gentamicine possegravedent
eacutegalement une mutation dans le gegravene de lrsquoATP synthase ce qui rend la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO006 microgml
H+
SCV Niveau 2CMI TO gt16 microgml
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+TO
TO
Niveaux 1 et 2
CcpA (CMI TO 025-1 microgml)
ROS
ATP
0811 0928
2012
1629
0811 0928
2012
A B
109
inactive puisqursquoincapable de srsquoy lier (CMI gt16microgml) (Figure 18B) Chez les SCV non
muteacutes (hemB) la gentamicine nrsquoentre que faiblement ducirc agrave la deacutepolarisation de sa
membrane (CMI 4-8 microgml) (Figure 18A) La mutation dans le gegravene codant pour une
pompe (NWMN_0143 codant pour un ABC transporteur) chez les mutants hemB
reacutesistant agrave la tomatidine (+gentamicine) amegravenerait une augmentation de lrsquoactiviteacute de
cette pompe permettant de rejeter hors de la cellule le peu de gentamicine eacutetant
entreacutee La mutation de cette pompe fait passer la CMI de la gentamicine de 4-8
microgml pour hemB agrave 16 microgml pour les mutants hemB reacutesistant agrave la combinaison
gentamicine (+TO) ayant cette mutation (Figure 18B)
Figure 18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB reacutesistants
agrave la combinaison tomatidine (+gentamicine)
Donc selon notre modegravele-hypothegravese lrsquoaction de la tomatidine et de la gentamicine
sur S aureus deacutepend de la liaison agrave leur cible mais a aussi un lien avec la
SCV R agrave TO (+GEN [4microgml])
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI GEN 4-8 microgml
H+
SCV
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+ TO
GEN
CMI GEN 16 microgml
CMI TO gt16 microgml
0811 0928
2012
0811 0928 20120143
GEN
Synthegravese proteacuteique
BA
110
polarisation membranaire et la production de ROS La mesure de la polarisation et
concentration cellulaire en ROS chez nos mutants et nos souches parents en
preacutesence ou non drsquoantibiotique nous permettra de valider notre modegravele-hypothegravese
Chez une bacteacuterie meacutetaboliquement active et posseacutedant une membrane
cytoplasmique intacte il y a normalement une diffeacuterence de potentiel eacutelectrique entre
le cocircteacute inteacuterieur et le cocircteacute exteacuterieur de la membrane lrsquointeacuterieur eacutetant plus neacutegatif de
100 agrave 200 mV par rapport agrave lrsquoexteacuterieur Une diminution de la magnitude de ce
potentiel membranaire est appeleacutee deacutepolarisation alors qursquoune augmentation de ce
potentiel est appeleacutee hyperpolarisation Le potentiel membranaire sera reacuteduit agrave zeacutero
srsquoil y a rupture de la membrane cest-agrave-dire srsquoil y a formation de trous assez grands
permettant aux ions inorganiques de passer librement Ce pheacutenomegravene peut arriver
lorsque les cellules sont tueacutees par la chaleur le froid ou les antibiotiques β-Lactams
Dans ces circonstances la membrane devient permeacuteable agrave diffeacuterents colorants
comme le propidium iodide (PI) et le TO-PRO3 et le changement est irreacuteversible
Plusieurs classes de composeacutes chimiques notamment les ionophores peuvent
eacutegalement alteacuterer le potentiel membranaire sans pour autant permettre la
permeacuteabiliteacute au PI TO-PRO3 et autres colorants similaires chez les bacteacuteries
lrsquoadministration drsquoun proton-ionophore tel le carbonyl cyanide 3-
chlorophenylhydrazone (CCCP) reacuteduira le potentiel membranaire agrave zeacutero
Des tests preacuteliminaires sur la mesure du potentiel membranaire (BacLighttrade Bacterial
Membrane Potential Kit (B34950) drsquoInvitrogen et lecture au cytomegravetre de flux (FACS))
(Novo et al 2000) chez nos deux souches parentes mets en eacutevidence que ATCC
29213 est plus polariseacute que NewbouldΔhemB (Figure 19A) Toutefois malgreacute
qursquoayant une plus faible polarisation membranaire NewbouldΔhemB nrsquoest pas
complegravetement deacutepolariseacute puisqursquoil est possible de le deacutepolariser davantage agrave lrsquoaide
drsquoun agent deacutepolarisant tel que le CCCP (carbonyl cyanide 3-chlorophenylhydrazone)
(Figure 19B) Le TO-PRO3 a eacuteteacute utiliseacute comme controcircle afin de confirmer la viabiliteacute
de nos cellules (donneacutees non montreacutees)
111
A B
Figure 19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane Potential Kit
(Invitrogen) (Lecture au cytomegravetre de flux agrave lrsquoaide du programme FCS Express) (A)
Ratio FL3-H (axe des Y)FL1-H (axe des X) de ATCC 29213 (rouge) et de
NewbouldΔhemB (noir) (B) Lecture des rouges (FL3-H) sur NewbouldΔhemB traiteacute
avec le DiOC2(3) (33prime-diethyloxacarbocyanine iodide) (noir) ou avec le CCCP (bleu)
ou sur les cellules chauffeacutees (rouge)
La sous-uniteacute c de lrsquoATP synthase (AtpE) semble donc la cible la plus probable de la
tomatidine Afin de veacuterifier si les mutations identifieacutees pouvaient avoir un effet
veacuteritable sur la proteacuteine nous avons fait un alignement de seacutequences noteacute les bases
muteacutees et veacuterifieacute si la mutation amenait un changement drsquoacide amineacute et si ce
changement drsquoacide amineacute pouvait avoir un impact sur la proteacuteine finale De fait
nous avons trouveacute que les deux diffeacuterentes mutations dans lrsquoAtpE amenaient un
changement significatif drsquoacides amineacutes pouvant amener une variation fonctionnelle
FL3-H
Count
100
101
102
103
104
0
18
36
53
71
Chauffeacute
+CCCP+DiOC2
Chauffeacute
FL1-H
FL3-H
100
101
102
103
104
100
101
102
103
104
hemB
29213
VERT
RO
UG
E
ROUGE
ΔhemB
112
de la proteacuteine La mutation de la base 49 change un G pour un T il srsquoagit drsquoun SNP
nrsquoamenant pas de changement de cadre de lecture La mutation fait passer lrsquoacide
amineacute alanine (A) qui est apolaire agrave une seacuterine (S) qui est polaire La mutation de la
base 190 change un G pour un A et est aussi un SNP nrsquoamenant pas de changement
de cadre de lecture Cette mutation fait passer lrsquoacide amineacute alanine (A) qui est
apolaire agrave une threacuteonine (T) qui est polaire (Figure 20)
gtlcl||NWMN_2012|atpE|72493880 F0F1 ATP synthase subunit C
1 10 19 31 43 49 58
ATG AAT TTA ATC GCA GCA GCA ATC GCA ATT GGT TTA TCA GCA TTA GGA GCA GGT ATC GGT
M N L I A A A I A I G L S A L G A G I G
(S)
17 61 73 85 97 109 118
AAC GGT TTA ATC GTT TCA AGA ACA GTT GAA GGT GTA GCA CGT CAA CCA GAA GCA CGT GGT
N G L I V S R T V E G V A R Q P E A R G
21 40
121 133 145 157 169 178
CAA TTA ATG GGT ATC ATG TTC ATT GGT GTA GGT TTA GTT GAG GCA TTA CCT ATC ATC GGT
Q L M G I M F I G V G L V E A L P I I G
41 60
181 190 199 211
GTA GTA ATT GCA TTC ATG ACA TTT GCT GGA TAA
V V I A F M T F A G
(T)
61 64 70
Figure 20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du gegravene
atpE de S aureus Newman avec mise en eacutevidence des mutations (rose) en lien avec
la cible de la tomatidine Lrsquoalignement a eacuteteacute effectueacute agrave lrsquoaide du site CMR de JCVI
113
CHAPITRE 4
DISCUSSION
LrsquoATP synthase est eacutevidemment un complexe proteacuteique ubiquitaire tant chez les
procaryotes que les eucaryotes Malgreacute ce fait il semble que la cible de la tomatidine
soit speacutecifique aux bacillales celle-ci ne deacutemontrant aucune activiteacute contre les
bacteacuteries agrave Gram neacutegatif et les lactobacillales On suppose donc que lrsquoATP synthase
de ces espegraveces doit ecirctre speacutecifique au niveau du site de liaison de la tomatidine
Lrsquoecirctre humain en tant qursquoespegravece eucaryote est eacutegalement pourvu de ce complexe
proteacuteique et on peut naturellement se demander si lrsquoutilisation clinique de cette
moleacutecule pourrait ecirctre cytotoxique De fait plusieurs composeacutes naturels tel
lrsquooligomycine et la venturicidine sont connus pour inhiber lrsquoactiviteacute de lrsquoATP synthase
par leur interaction avec la sous-uniteacute c de ce complexe Cependant ces moleacutecules
ne sont pas seacutelectives et nrsquoinhibent pas seulement lrsquoATP synthase bacteacuterien mais
aussi lrsquoATP synthase mitochondrial (Haagsma et al 2009 Matsuno-Yagi et Hatefi
1993) (Figure 1) Neacuteanmoins il existe maintenant une moleacutecule qui a pour cible
lrsquoATP synthase bacteacuterien et qui est en processus en vue drsquoune utilisation clinique
chez lrsquohomme le TMC207 (Haagsma et al 2009))
114
Figure 1 Sites drsquoinhibition de lrsquoATP synthase Les sites de liaison dans lrsquoATP
synthase des inhibiteurs reacuteveacuteleacutes par des eacutetudes biochimiques et structurales sont
indiqueacutes par des cercles rouges et les sites de liaisons qui nrsquoont pas eacuteteacute totalement
clarifieacutes sont indiqueacutes par des cercles verts (Hong et Pedersen 2008)
Le TMC207 (Figure 2A) est un composeacute de la classe des diarylquinoline posseacutedant
un nouveau meacutecanisme drsquoaction lrsquoinhibition de lrsquoATP synthase bacteacuterien De fait la
cible et le meacutecanisme drsquoaction du TMC207 sont diffeacuterents des autres agents
antituberculeux Lrsquoinhibition de lrsquoATP synthase peut mener agrave une baisse de la
production drsquoATP et un deacutebalancement dans lrsquohomeacuteostasie du pH ces 2 effets
pouvant contribuer agrave la diminution de la survie de la bacteacuterie (Rao et al 2001
Andries et al 2005)
Le TMC207 est une moleacutecule ayant une forte activiteacute bacteacutericide contre
Mycobacterium tuberculosis et autres espegraveces mycobacteacuteriennes mais peu drsquoactiviteacute
115
contres les autres espegraveces bacteacuteriennes Lrsquoindex de seacutelectiviteacute de TMC207 pour
lrsquoATP synthase mycobacteacuterienne (CMI50 gt200 microml) compareacute agrave lrsquoATP synthase
mitochondriale (CMI50 001 microgml) est de plus de 20000 Un indice de seacutelectiviteacute au
dessus de 1000 est consideacutereacute dans le deacuteveloppement drsquoune drogue comme un
preacutealable pour une moleacutecule candidate (Haagsma et al 2009) La diversiteacute dans les
gegravenes atpE explique la reacutesistance au TMC207 et justifie la faible affiniteacute de cette
drogue agrave lrsquoATP synthase mitochondriale (Matteelli et al 2010) (Figure 3)
Figure 2 Structure chimique de (A) TMC207 et (B) son analogue anti bacteacuteries
agrave Gram positifs (Balemans et al 2012)
Lorsqursquoon compare la seacutequence proteacuteique de la sous-uniteacute c de lrsquoATP synthase
(atpE) de quelques espegraveces de lrsquoordre des Bacillales (tel Saureus B cereus B
subtilis et L monocytogenes) avec des espegraveces appartenant agrave drsquoautres ordres
bacteacuteriens on constate que les seacutequences divergent ce qui est en accord avec la
speacutecificiteacute des moleacutecules Les points de mutations amenant une reacutesistance au
TMC207 chez Mycobacterium smegmatis sont speacutecifiques aux mycobacteacuteries et les
deux principaux points de mutations D32V(ouP) et A67P chez M smegmatis sont
situeacutes tous les deux dans les heacutelices alpha qui entourent le site catalytique de la
proteacuteine (E65) qui est impliqueacute dans le transport des protons (Andries et al 2005)
Lrsquoacide amineacute aspartate (D) agrave la position 32 et lrsquoalanine (A) agrave la position 67 sont tregraves
speacutecifiques agrave M smegmatis
A B
116
Reacutecemment un groupe de chercheurs (Balemans et al 2012) on mit en lumiegravere une
autre moleacutecule (Figure 2B) de la classe des diarylquinolines dont le meacutecanisme
drsquoaction eacutetait aussi lrsquoinhibition de la sous-uniteacute c de lrsquoATP synthase mais chez les
bacteacuteries agrave Gram positif tels Streptococcus pneumoniae et S aureus Cette moleacutecule
irait lier les acides amineacutes valines (V) situeacutes agrave la position 50 et 62 de lrsquoAtpE de S
aureus (Donneacutees non montreacutees chez S pneumoniae(Balemans et al 2012)) En
effet une mutation amenant le changement de la valine en position 50 par une
isoleucine (I) etou une mutation amenant le changement de la valine en position 62
par une alanine (A) amegravene la reacutesistance de la bacteacuterie agrave la moleacutecule Cependant L
monocytogenes et Bacillus subtilis ne possegravedent pas de valine agrave la position 50 alors
qursquoils font eacutegalement partie des bacteacuteries agrave Gram positif (Il faut noter que chez
Bacillus sp PS3 (Swiss-Prot assession Ndeg P 00845) possegravede une valine agrave la position
50 (Balemans et al 2012)) Par contre tout comme pour les autres bacteacuteries agrave Gram
positif testeacutees les deux espegraveces possegravedent une valine agrave la position 62 (Figure 3)
Chez B cereus B subtilis B anthracis S aureus et L monocytogenes les points de
divergence sont plutocirct A17S et A64T Cependant B pseudofirmus qui fait eacutegalement
partie de lrsquoordre des Bacillales possegravede une glycine (G) agrave la position 17 plutocirct qursquoune
alanine (A) et une seacuterine (S) plutocirct qursquoune alanine (A) agrave la position 64 comme les
espegraveces preacuteceacutedentes Suivant ces reacutesultats et si la cible de la tomatidine est bien
ces deux points de mutation la tomatidine ne devrait pas pouvoir se lier agrave lrsquoAtpE de B
pseudofirmus et devrait donc nrsquoavoir aucune activiteacute contre cette souche alors que
nous avions extrapoleacute que le spectre drsquoactiviteacute de la tomatidine eacutetait tous les
Bacillales De plus E coli et M smegmatis possegravedent eacutegalement une alanine (A) agrave la
position 17 alors que la tomatidine ne possegravede aucune activiteacute contre ces espegraveces (M
smegmatis nrsquoa pas eacuteteacute eacutevalueacute il srsquoagit ici drsquoune extrapolation des reacutesultats de CMI)
par contre ces deux espegraveces possegravedent des acides amineacutes diffeacuterents drsquoune alanine
(A) agrave la position 64 ce qui pourrait leur confeacuterer une reacutesistance agrave la tomatidine telle
qursquoobserveacutee On remarque eacutegalement que les acides amineacutes agrave ces deux positions
117
sont totalement diffeacuterents chez lrsquohomme (Homo sapiens) ce qui suggeacutererait que la
tomatidine ne possegravederait pas drsquoactiviteacute antimitochondriale (Figure 3)
Selon ces reacutesultats on peut se demander si les deux mutations sont essentielles agrave la
liaison de la moleacutecule ou si seulement lrsquoalanine (A) agrave la position 64 est neacutecessaire
On doit se rappeler que la reacutesistance apporteacutee par la mutation faisant en sorte que
lrsquoalanine (A) en position 17 devient une seacuterine (S) apparaicirct chez les mutants
NewbouldΔhemb reacutesistant agrave la tomatidine et que cette mutation nrsquoapparaicirct qursquoau
quatriegraveme et dernier niveau de reacutesistance On se rappelle eacutegalement que la mutation
faisant en sorte que lrsquoalanine (A) agrave la position 64 devient une threacuteonine (T) apparaicirct
chez les mutants NewbouldΔhemb reacutesistants agrave la tomatidine en combinaison avec la
gentamicine et ce degraves le premier niveau de reacutesistance
118
Bcereus G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Banthracis G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Lmonocytogenes G---------VIAAAIAVG---LGALGAGIGNGLIVSKTVEGVARQPEARSMLQTIMFV- 50
Saureus_NWMN_2012 -----------IAAAIAIG---LSALGAGIGNGLIVSRTVEGVARQPEARGQLMGIMFIG 49
Bsubtilis -----------IAAAIAIG---LGALGAGIGNGLIVSRTVEGIARQPEAGKELRTLMFMG 49
Bpseudofirmus -----------LGAAIAAG---LAAVAGAIAVAIIVKATIEGTTRQPELRGTLQTLMFIG 49
Ecoli L---------YMAAAVMMG---LAAIGAAIGIGILGGKFLEGAARQPDLIPLLRTQFFIV 56
Msmegmatis DPNAIITAGALIGGGLIMG---GGAIGAGIGDGIAGNALISGIARQPEAQGRLFTPFFIT 60
Homo sapiens QTSAISRDIDTAAKFIGAGAATVGVAGSGAGIGTVFGSLIIGYARNPSLKQQLFSYAILG 120
MyR V
MyR P
SaR S
Bcereus VALVEALPIIGVVIAFIVMNK----- 72
Banthracis VALVEALPIIGVVIAFIVMNK----- 72
Lmonocytogenes ----EALPIIAVVIAFMVLNK----- 67
Saureus_NWMN_2012 VGLVEALPIIGVVIAFMTFAG----- 70
Bsubtilis IALVEALPIIAVVIAFLAFFG----- 70
Bpseudofirmus VPLAEAVPIIAIVISLLILF------ 69
Ecoli MGLVDAIPMIAVGLGLYVMFAVA--- 79
Msmegmatis VGLVEAAYFINLAFMALFVFATPGLQ 86
Homo sapiens FALSEAMGLFCLMVAFLILFAM---- 142
MyR D P M
SpR I A
SaR T
Fig
ure
3
Seacuteq
uen
ce
pro
teacuteiq
ue d
e la p
roteacute
ine
Atp
Ec
hez d
iffeacute
ren
tes
es
pegravec
es e
t m
ise e
n eacute
vid
en
ce
de
s s
ite
s d
e
mu
tati
on
pe
rmett
an
t la
reacutesis
tan
ce agrave
dif
feacutere
nte
s m
oleacute
cu
les
M
yR M
tu
berc
ulo
sis
ou
M sm
eg
matis
reacutesis
tan
t a
u
TM
C2
07 S
pR
S
p
neu
mon
iae
reacutesis
tan
t agrave
un a
na
log
ue d
u T
MC
20
7
SaR
S
a
ure
us
SC
V r
eacutesis
tant agrave
la to
matid
ine
En
cad
reacutero
uge
S
ite d
e m
uta
tion a
men
ant la
reacutesis
tance
agravela
to
matid
ine E
ncad
reacuteno
ir
Site
cata
lytiq
ue
im
pliq
ueacute
da
ns le
tra
nspo
rt d
es p
roto
ns
Su
rlig
nag
etu
rqu
ois
e S
ite d
e m
uta
tion a
me
nant u
ne
reacutesis
tan
ce a
u T
MC
20
7
119
Les seacutequences proteacuteiques des souches bacteacuteriennes de la figure 3 ont eacuteteacute trouveacutees
sur pubmedproteacuteine httpwwwncbinlmnihgovpubmed
laquo ATP synthase subunit c raquo Assession number
Streptococcus pneumoniae D39 YP_8168031 (NCBI)
S aureus Newman BAF682841 (GenBank)
E coli AAA247321 (GenBank)
Bacillus cereus ISO75 EJP900901 (GenBank)
Bacillus anthracis str H9401 YP_0062123491 (NCBI)
Mycobacterium smegmatis MC2 155 YP_0065695541 (NCBI)
Bacillus subtilis str 168 CAA822551 (GenBank)
Listeria monocytogenes YP_0076049931 (NCBI)
Homo sapiens P482011 (UniProtKBSwiss-Prot)
Lrsquoalignement comparatif des seacutequences drsquoacides amineacutes a eacuteteacute fait agrave lrsquoaide du
programme ClustalW2 sur le web
httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2
Le modegravele 3D de lrsquoAtpE a eacuteteacute obtenu gracircce agrave lrsquoaide du Pr Ryszard Brzezinski du
Deacutepartement de biologie de lrsquoUniversiteacute de Sherbrooke (Figure 4) Une recherche
dans le Protein Data Bank a drsquoabord eacuteteacute effectueacutee afin de veacuterifier parmi les proteacuteines
AtpE (sous-uniteacute C de lrsquoATP synthase) pour lesquelles il y avait des structures 3D
reacutesolues laquelle ressemblait le plus agrave celle de Staphylococcus aureus Les
alignements ont montreacute que cest la proteacuteine de Bacillus pseudofirmus qui avait la
plus grande similitude de seacutequence La structure est stockeacutee dans la banque PDB
sous le symbole 3ZO6 Il a eacuteteacute confirmeacute par quelques alignements que cette sous-
uniteacute C de lrsquoATP synthase eacutetait plus proche par sa seacutequence de celle de
Staphylococcus aureus que de celle de Mycobacterium pour laquelle la structure 3D
avait eacuteteacute deacutetermineacutee plus tocirct Ce reacutesultat sexplique par le fait que Staphylococcus et
Bacillus font tous deux partie de la branche A+T-riche des bacteacuteries agrave Gram+ alors
que Mycobacterium appartient agrave la branche G+C-riche La seacutequence de lAtpE de
120
Staphylococcus aureus a ensuite eacuteteacute envoyeacutee au serveur internet de modeacutelisation
3D-Jigsaw (httpbmmcancerresearchukorg~3djigsaw) en indiquant que le modegravele
devait ecirctre bacircti en prenant comme matrice le fichier PDB 3ZO6 Le modegravele nous a
eacuteteacute retourneacute sous format PDB ndash comme les laquo vrais raquo cristaux dans la Protein Data
Bank Les fichiers au format PDB peuvent ecirctre visualiseacutes par le logiciel RasMol ndash
gratuit sur internet Cest avec RasMol que limage (Figure 4) a eacuteteacute obtenue
121
Figure 4 Modeacutelisation de la structure tridimensionelle de la sous-uniteacute c de
lrsquoATP synthase de S aureus (AtpE) montrant les acides amineacutes muteacutes chez les
mutants reacutesistantsEn blanc et vert les deux acides amineacutes muteacutes chez S aureus
reacutesistant agrave la tomatidine En rose les sites de mutations chez Mycobacterium
smegmatis reacutesistant au TMC207 et des sites de mutations chez Streptoccocus
pneumoniae reacutesistants agrave un analogue de TMC207 En bleu le site actif de la proteacuteine
Agrave la vue de ces reacutesultats certains pourront mettre en doute lrsquointeacuterecirct de la tomatidine
eacutetant donneacute lrsquoapparition relativement rapide de reacutesistance chez S aureus Cependant
122
nos reacutesultats deacutemontrent clairement que lrsquoajout de tomatidine ralentit le
deacuteveloppement de reacutesistance agrave la gentamicine chez S aureus ATCC29213 (Tableau
1) De plus cette augmentation de la CMI de la gentamicine en preacutesence de
tomatidine au fil des passages suit lrsquoaugmentation de la CMI de la ciprofloxacine lors
de ces mecircmes passages alors que la ciprofloxacine est consideacutereacutee comme la drogue
de reacutefeacuterences dans les bonnes drogues (Tableau 1) Tout comme pour la
ciprofloxacine pour laquelle la reacutesistance chez S aureus peut ecirctre la conseacutequence
de deux mutations dans le gegravene cible gyrA la reacutesistance agrave la tomatidine peut ecirctre la
conseacutequence de deux mutations dans le gegravene cible atpE Par comparaison la
rifampicine est consideacutereacutee comme une mauvaise drogue de par lrsquoapparition rapide
drsquoune forte reacutesistance chez S aureus conseacutequence de ses 17 mutations possibles
dans le gegravene cible rpoB (Tableau 1)
De plus il sera important de relativiser lrsquoapparition de reacutesistance agrave la tomatidine en
calculant le taux drsquoapparition de ces colonies reacutesistantes ceci nrsquoayant pas eacuteteacute fait
jusqursquoici De fait la freacutequence de mutation chez S aureus amenant une reacutesistance agrave
la rifampicine est de lrsquoordre de 10-7 10-8 alors qursquoil est de 10-11 pour lrsquoacide fusidique
(Canfield et al 2013 Orsquoneill et al 2001 Price et Gustafson 2001) Pour ce qui est
de la nouvelle moleacutecule anti-bacteacuteries agrave Gram positif de la classe des diarylquinoline
une freacutequence de mutation sur S pneumoniae ATCC49619 de 81- 83 x 10-7 a eacuteteacute
observeacutee agrave 5x la CMI et une freacutequence de mutation de 46 x 10-7 agrave 17 x 10-6 agrave 50x la
CMI (Balemans et al 2012)
123
Tableau 1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 par la meacutethode de CMI lors de passages successifs dans un milieu avec
lrsquoantibiotique
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
Typiqueldquomauvaiserdquodrogue (17 mutations possibles dansrpoB)
Typique ldquobonnerdquo drogue (2 mutations possibles dans gyrA)
Preacutesence de TO ralenti le deacuteveloppement de reacutesistances vs GEN seul
124
CHAPITRE 5
CONCLUSION ET PERSPECTIVES
Mon projet de maitrise a permis de preacuteciser le spectre drsquoactiviteacute de la tomatidine
Nous savons maintenant que la tomatidine possegravede une activiteacute antibacteacuterienne
contre la plupart des Firmicutes et plus speacutecifiquement contre les Bacillales Plus
preacuteciseacutement la tomatidine est bacteacutericide contre les pheacutenotypes SCV et agit en
synergie avec les aminoglycosides augmentant lrsquoactiviteacute bacteacutericide de ces derniers
de 8-16 fois contre les souches agrave pheacutenotype normal
De plus parmi tous les analogues de la tomatidine produits par le laboratoire du Pr
Eacuteric Marsault (voir article en annexe 2) le composeacute FC04-100 a eacuteteacute plus
speacutecifiquement mis en lumiegravere Celui-ci conserve non seulement une aussi bonne
activiteacute synergique avec les aminoglicosides contre les Saureus au pheacutenotype
normal et une activiteacute modeacutereacutee contre les S aureus SCV mais montre eacutegalement de
faccedilon inespeacutereacutee une activiteacute par lui-mecircme contre les S aureus de pheacutenotype normal
incluant plusieurs MRSA une proprieacuteteacute qui nrsquoeacutetait pas observeacutee pour la tomatidine
Une autre activiteacute inteacuteressante de FC04-100 est sa capaciteacute drsquoameacuteliorer lrsquoactiviteacute
bacteacutericide des aminoglicosides contre L monocytogenes agrave pheacutenotype normal apregraves
un traitement de 24h alors que la tomatidine ne deacutemontrait qursquoune faible synergie
avec les aminoglycosides contre cette souche De plus FC04-100 deacutemontre une
activiteacute bacteacutericide contre les SCV de L monocytogenes alors que la tomatidine ne
deacutemontrait qursquoune activiteacute bacteacuteriostatique FC04-100 montre eacutegalement un effet
potentiateur de lrsquoUbrolexinTM (Ceacutefalexin Kanamycin (3 2)) une combinaison
drsquoantibiotique utiliseacute dans le traitement de mammites bovines en abaissant de 8 agrave 16
fois leur CMI amenant celles-ci sous le seuil de reacutesistance Finalement FC04-100
possegravede une meilleure solubiliteacute que la tomatidine une proprieacuteteacute qui facilitera son
utilisation in vitro
125
Nos reacutesultats deacutemontrent que FC04-100 conserve une certaine activiteacute contre les
SCV reacutesistant agrave la tomatidine ce qui suggegravere possiblement que FC04-100 a plus
daffiniteacute pour la cible ou plus de sites dinteraction avec AtpE En effet les mutants
SCV reacutesistants agrave la tomatidine deviennent gt512 fois plus reacutesistants alors que FC04-
100 ne perd que 8 agrave 16 fois son activiteacute contre ces mecircmes souches Ceci deacutemontre
quil est possible dameacuteliorer significativement lactiviteacute de ces alkaloiumldes steacuteroidiens
ce qui est tregraves prometteur pour lavenir de cette moleacutecule
Dans un second temps lrsquoanalyse des mutations chez les mutants reacutesistants agrave la
tomatidine a permis de non seulement de mieux comprendre le mode drsquoaction de la
tomatidine mais eacutegalement de preacuteciser la cible potentielle de ce nouvel antibiotique
De fait son mode drsquoaction serait en lien direct avec la fonctionnaliteacute de la chaine de
transport des eacutelectrons chez la bacteacuterie lrsquoactiviteacute de la tomatidine serait inversement
proportionnelle agrave la fonctionnaliteacute de la chaine de transport des eacutelectrons et donc en
lien eacutegalement avec la polarisation membranaire et la production de ROS
intracellulaire Afin de confirmer le modegravele-hypothegravese la mesure de la polarisation
membranaire et la concentration de ROS intracellulaire chez les mutants ainsi que
chez les souches parentes suite agrave un traitement antibiotique devront ecirctre effectueacutees
Les reacutesultats de mes recherches suggegraverent que la cible bacteacuterienne speacutecifique de la
tomatidine serait la sous-uniteacute C de lrsquoATP synthase Afin de confirmer la cible
Maxime Lamontagne-Boulet tente au moment drsquoeacutecrire ces lignes drsquoobtenir des
mutants de S aureus Newman reacutesistants au composeacute FC04-100 et dont la reacutesistance
serait attribuable agrave une mutation dans le gegravene atpE Par la suite on devra confirmer
la fonctionnaliteacute de la cible ATP synthase par quantification de lrsquoATP produit agrave lrsquoaide
de la lucifeacuterase tel que lrsquoon fait Balemans et al en 2012 Lrsquoattachegravement agrave la cible
devra eacutegalement ecirctre confirmeacute et la meacutethode de SPR (Surface Plasmon Resonance)
pourrait ecirctre tout indiqueacutee pour ce faire (Balemans et al 2012) On devra eacutegalement
126
deacutemontrer la seacutelectiviteacute des moleacutecules tomatidine et FC04-100 pour lrsquoATP synthase
des Bacillales
Les infections agrave bacteacuteries multireacutesistantes deviennent de plus en plus preacutevalentes et
sont un des problegravemes de santeacute majeurs auxquels nous devons faire face aujourdrsquohui
Cette augmentation de la reacutesistance limite le reacutepertoire drsquoantibiotiques efficace creacuteant
une situation probleacutematique laquelle est exacerbeacutee par le trop petit nombre de
nouveaux antibiotiques introduits sur le marcheacute ces derniegraveres anneacutees Il est donc
drsquoune grande importance de travailler agrave la mise en marcheacute de nouveaux antibiotiques
et plus particuliegraverement de nouvelles classes drsquoantibiotiques espeacuterant ainsi deacutejouer la
multireacutesistance bacteacuterienne du moins pour un temps La tomatidine de par sa
structure sa cible et son mode drsquoaction speacutecifique est une moleacutecule antibiotique
dans une classe agrave part et de ce fait une moleacutecule plus qursquointeacuteressante pour le
traitement drsquoinfection agrave S aureus De plus comme elle deacutemontre une activiteacute
bacteacutericide contre les deux pheacutenotypes de S aureus (et autres Bacillales) lesquels
sont retrouveacutes simultaneacutement dans les cas drsquoinfections chroniques son pouvoir
antibiotique devient inestimables drsquoun point de vue clinique
127
CONTRIBUTIONS SUPPLEacuteMENTAIRES
ANNEXE 1 Application de brevet Internationnal publication number WO
2012109752 A1
INVENTEURS Franccedilois Malouin2 Gabriel Mitchell2 Kamal Bouarab2 Eacuteric Marsault1
Feacutelix Chagnon1 Simon Boulanger 2 Isabelle Guay2
1Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
2Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
128
129
ANNEXE 2 Article scientifique dans laquo Europeen Journal of medicinal chemistry raquo
2014 80 605-620
TITRE Unraveling the structure-activity relationship of tomatidine a steroid alkaloid
with unique antibiotic properties against persistent forms of
Staphylococcus aureus
ABSTRACT Staphylococcus aureus is responsible for difficult-to-treat and relapsing
infections and constitutes one of the most problematic pathogens due to its multiple
resistances to clinically available antibiotics Additionally this pathogenrsquos ability to
develop small-colony variants is associated with a reduced susceptibility to
aminoglycoside antibiotics and in vivo persistence We have recently demonstrated
that tomatidine a steroid alkaloid isolated from tomato plants possesses anti-
virulence activity against normal strains of S aureus as well as the ability to
potentiate the effect of aminoglycosides against this pathogen In addition tomatidine
has shown antibiotic activity against small-colony variants We herein report the first
study of the structure-activity relationship of tomatidine against S aureus
AUTEURS Feacutelix Chagnon1 Isabelle Guay2 Marc-Andreacute Bonin1 Gabriel Mitchell2
Kamal Bouarab2Franccedilois Malouin2 Eacuteric Marsault1
MA CONTRIBUTION Responsable des tests biologiques de tous les composeacutes
autant envers lrsquoeffet potentiateur de la tomatidine sur lrsquoactiviteacute de la gentamicine
contre la souche ATCC29213 de S aureus que pour lrsquoactiviteacute bacteacuteriostatique sur la
souche NewbouldΔhem imitant le pheacutenotype SCV Ces travaux furent effectueacutes sous
la tutelle du professeur Franccedilois Malouin
130
1Institut de Pharmacologie de Sherbrooke Deacutepartement de pharmacologie
Universiteacute de Sherbrooke 3001 12e av nord Sherbrooke Queacutebec Canada J1H
5N4
2Centre drsquoeacutetude et de valorisation de la diversiteacute microbienne (CEVDM) Deacutepartement
de biologie Universiteacute de Sherbrooke Sherbrooke QC Canada J1K 2R1
131
BIBLIOGRAPHIE Alexander E H and Hudson MC (2001) Factors influencing the internal- ization of Staphylococcus aureus and impacts on the course of infections in humans Appl Microbiol Biotechnol 56 361ndash366
Andries K Verhasselt P Guillemont J Goumlhlmann HW Neefs JM Winkler H Van Gestel J Timmerman P Zhu M Lee E et al (2005) A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis Science 307 223-227
Artzatbanov VY and Petrov VV (1990) Branched respiratory chain in aerobically grown Staphylococcus aureus--oxidation of ethanol by cells and protoplasts Arch Microbiol 153 580- 584 Archer GL (1998) Staphylococcus aureus a well-armed pathogen Clin Infect Dis 26 1179ndash1181 Assefa S Keane TM Otto TD Newbold C and Berriman M (2009) ABACAS algorithm-based automatic contiguation of assembled sequences Bioinformatics 25 1968-1969 Aubry-Damon H Legrand P Brun-Buisson C Astier A Soussy CJ and Leclercq R (1997) Reemergence of gentamicin-sus- ceptible strains of methicillin-resistant Staphylococcus aureus roles of an infection control program and changes in aminoglycoside use Clin Infect Dis 25 647-653
Balemans W Vranckx L Lounis N Pop O Guillemont J Vergauwen K Mol S Gilissen R Motte M Lanccedilois et al (2012) Novel antibiotics targeting respiratory ATP synthesis in Gram-positive pathogenic bacteria Antimicrob Agents Chemother 56 4131-4139
Batandier C Guigas B Detaille D El-Mir MY Fontaine E Rigoulet M and Leverve XM (2006) The ROS production induced by a reverse-electron flux at respiratory-chain complex 1 is hampered by metformin J Bioenerg Biomembr 38 33ndash 42
132
Beckman Coulter website httpswwwbeckmancoultercomwsrportalwsrresearch-and-discoveryproducts-and-servicesnucleic-acid-sample-preparationspri-laboratory-automationindexhtm (consulteacute le 3 Deacutec 2013) Bednarek P and Osbourn A (2009) Plant-microbe interactions chemical diversity in plant defense Science 324 746ndash748 Besier S Ludwig A Ohlsen K Brade V and Wichelhaus TA (2007) Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus Int J Med Microbiol 297 217-225 BiteSizeBio Brain food for biologists httpnxseqbitesizebiocomarticlessequencing-by-synthesis-explaining-the-illumina-sequencing-technology (consulteacute le 3 deacutec 2013) Boetzer M and Pirovano W (2012) Toward almost closed genomes with GapFiller Genome Biology 13 R56 Bouarab K Ordi E Gattuso M Moisan H and Malouin F (2007) Plant stress response agents affect Staphylococcus aureus virulence genes Abstr 47th Intersci Conf Antimicrob Agents Chemother abstr C1-1483 Bryan LE and Kwan S (1981) Aminoglycoside-resistant mutants of Pseudomonas aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob Agents Chemother 19 958- 964 Bryan LE and Kwan S (1983) Roles of ribosomal binding membrane potential and electron transport in bacterial uptake of streptomycin and gentamicin Antimicrob Agents Chemother 23 835ndash845
Bryan LE Nicas T Holloway BW and Crowther C (1980) Aminoglycoside-resistant mutation of Pseudomonas aeruginosa defective in cytochrome C552 and nitrate reductase Antimicrob Agents Chemother 17 71- 79
133
Bryan LE and van den Elzen HM (1977) Effects of membrane-energy mutations and cations on streptomycin and gentamicin accumulation by bacteria a model for entry of streptomycin and gentamicin in susceptible and resistant bacteria Antimicrob Agents Chemother 12 163-177 Brynildsen MP Winkler JA Spina CS MacDonald IC and Collins JJ (2013) Potentiating antibacterial activity by predictably enhancing endogenous microbial ROS production Nat Biotechnol 31 160ndash 165 Burke KA and Lasccelles J (1975) Nitrate reductase system in Staphylococcus aureus wild type and mutants J Bacteriol 123 308- 316 Burke KA and Lascelles J (1976) Nitrate reductase activity in hegraveme-deficient mutants of Staphylococcus aureus J Bacteriol 126 225- 231 Canfield GS Schwingel JM Foley MH Vore KL Boonanantanasarn K Gill AL Sutton MD and Gill SR (2013) Evolution in fast forward a potential role for mutators in accelerating Staphylococcus aureus pathoadaptation J Bacteriol 195 615-628 Chang S Sievert DM Hageman JC Boulton ML Tenover FC Downes FP Shah S Rudrik JT Pupp GR Brown WJ et al (2003) Infection with vancomycin- resistant Staphylococcus aureus containing the vanA resistance gene N Engl J Med 348 1342-1347 Chen Q Vazquez EJ Moghaddas S Hoppel CL and Lesnefsky EJ (2003) Production of reactive oxygen species by mitochondria central role of complex III J Biol Chem 278 36027-36031 Clements MO Watson SP Poole RK and Foster SJ (1999) CtaA of Staphylococcus aureus is required for starvation survival recovery and cytochrome biosynthesis J Bacteriol 181 501- 507
134
Clinical and Laboratory Standards Institute (2011) Performance standards for antimicrobial susceptibility testing Twenty-first informational supplement M100-S21 CLSI Wayne PA USA ClustalW2 httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2 (consulteacute le 3 deacutec 2013) Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) httpcmrjcviorgtigr-scriptsCMRCmrHomePagecgi (consulteacute le 3 deacutec 2013) Creacutepin-Gilbert C (2006) Transfert de ligand dans la cytochrome c oxydase observeacute par des expeacuteriences femtosecondes infrarouge inteacutegreacutees et reacutesolues spectralement Thegravese de doctorat de l rsquoEacutecole Polytechnique Paris Pastel-00002200 version 1 29 jul 2010 Davis BD (1987) Mechanism of bactericidal action of aminoglycosides Microbiol Rev 51 341-350 DeAngelis MM Wang DG and Hawkins TL (1995) Solid-phase reversible immobilization for the isolation of PCR products Nucl Acids Res 22 4742- 4743 Dickgiesser N and Kreiswirth BN (1986) Determination of aminoglycoside resistance in Staphylococcus aureus by DNA hybridization Antimicrob Agents Chemother 29 930-932 Doi Y and Arakawa Y (2007) 16S ribosomal RNA methylation Emerging resistance mechanism against aminoglycosides Clin Infect Dis 45 88ndash94 Enright MC Robinson DA Randle G Feil EJ Grundmann H and Spratt BG (2002) The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA) Proc Natl Acad Sci USA 99 7687-7692 Friedman M (2002) Tomato glycoalkaloids role in the plant and in the diet J Agric Food Chem 50 5751ndash5780
135
Goerke C and Wolz C (2004) Regulatory and genomic plasticity of Staph- ylococcus aureus during persistent colonization and infection Int J Med Microbiol 294 195ndash202 Goumltz F Bannerman T and Schleifer K-H (2006) The genera Staphylococcus and Macrococcus In The Prokaryotes Vol 4 Bacteria Firmicutes Cyanobacteria M Dworkin (ed) (Springer New-York USA) pp5- 75 Haagsma AC Abdillahi-Ibrahim R Wagner MJ Krab K Vergauwen K Guillemont J AndriesK Lill H Koul A and Bald D (2009) Selectivity of TMC207 towards mycobacterial ATP synthase compared with that towards the eukaryotic homologue Antimicrob Agents Chemother 53 1290-1292 Hammer ND Reniere ML Cassat JE Zhang Y Hirsch AO Indriati Hood M and Skaar EP (2013) Two hegraveme-dependent terminal oxydases power Staphylococcus aureus organ-specific colonization of the vertebrate host MBio 4 e00241- 13 Hederstedt L (1993) The Krebs citric acid cycle In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of Microbiology Washington DC) pp 181-198 Hiramatsu K (1995) Molecular evolution of MRSA Microbiol Immunol 39 531-543 Hiramatsu K Hanaki H Ino T Yabuta K Oguri T and Tenover FC (1997) Methicillin-resistant Staphylococcus aureus clinical strain with reduced vancomycin susceptibility J Antimicrob Chemother 40 135-136 Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T et al (2013) Genomic Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
136
Hong S and Pedersen PL (2008) ATP synthase and the actions of inhibitors utilized to study its roles in human health disease and other scientific areas Microbiol Mol Biol Rev 72 590-641
Jevons MP (1961) lsquoCelbeninrsquo-resistant staphylococci BMJ 1 124-125
Joshi S and Huang YG (1991) ATP synthase complex from bovine heart mitochondria the oligomycin sensitivity conferring protein is essential for dicyclohexyl carbodiimidesensitive ATPase Biochim Biophys Acta 1067 255ndash258
Kohanski MA Dwyer DJ and Collins JJ (2010) How antibiotics kill bacteria from targets to networks Nat Rev Microbiol 8 423-435 Kohler C von Eiff C Liebeke M McNamara PJ Lalk M Proctor RA Hecker M and Engelmann S (2008) A defect in menadione biosynthesis induces global changes in gene expression in Staphylococcus aureus J Bacteriol 190 6351- 6364 Kohler C von Eiff C Peters G Proctor RA Hecker M and Engelmann S (2003) Physiological characterization of a hegraveme-deficient mutant of Staphylococcus aureus by a proteomic approach J Bacteriol 185 6928- 6937 Kriegeskorte A Koumlnig S Sander G Pirkl A Mahabir E Proctor RA von Eiff C Peters G and Becker K (2011) Small colony variants of Staphylococcus aureus reveal distinct protein profiles Proteomics 11 2476-2490 Lannergard J von Eiff C Sander G Cordes T Seggewiss J Peters G Proctor RA Becker K and Hughes D (2008) Identification of the genetic basis for clinical menadione-auxotrophic small-colony variant isolates of Staphylococcus aureus Antimicrob Agents Chemother 52 4017-4022
Lauraeus M and Wikstrom M (1993) The terminal quinol oxydases of Bacillus subtilis have different energy conservation properties J Biol Chem 268 11470-11473
137
Li M Gong J Cottrill M Yu H de Lange C Burton J and Topp E (2003) Evaluation of QIAamp DNA Stool Mini Kit for ecological studies of gut microbiota J Microbiol Methods 54 13-20
Liu Y Fiskum G and Schubert D (2002) Generation of reactive oxygen species by the mitochondrial electron transport chain J Neurochem 80 780-787
Martin-Hernandez A M Dufresne M Hugouvieux V Melton R and Osbourn A (2000) Effects of targeted replacement of the tomatinase gene on the interaction of Septoria lycopersici with tomato plants Mol Plant Microbe Interact 13 1301ndash1311
Matteelli A Carvalho AC Dooley KE and Kritski A (2010) TMC207 the first compound of a new class of potent anti-tuberculosis drugs Future Microbiol 5 849-858
Matsuno-Yagi A and Hatefi Y (1993) Studies on the mechanism of oxidative phosphorylation ATP synthesis by submitochondrial particles inhibited at F0 by venturicidin and organotin compounds J Biol Chem 268 6168ndash6173
Miller MH Edberg SC Mandel LJ Behar CF and Steigbigel NH (1980) Gentamicin uptake in wild-type and aminoglycoside-resistant small-colony mutants of Staphylococcus aureus Antimicrob Agents Chemother 18 722- 729
Mingeot-Leclercq M-P Glupczynski Y and Tulkens PM (1999) Aminoglycosides Activity and Resistance Antimicrob Agents Chemother 43 727- 737
Mitchell G Brouillette Eacute Seacuteguin DL Asselin AE Jacob CL and Malouin F (2010) A role for sigma factor B in the emergence of Staphylococcus aureus small-colony variants and elevated biofilm production resulting from an exposure to aminoglycosides Microb Pathog 48 18-27
Mitchell G Gattuso M Bouarab K and Malouin F (2009) Tomatidine (TO) affects virulence regulators of prototypical Staphylococcus aureus (SA) and small colony variants (SCV) of cystic fibrosis (CF) patients Abstr 49th Intersci Conf Antimicrob Agents Chemother abstr C1-1341
138
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K and Malouin F (2011) Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic fibrosis airway epithelial cells Antimicrob Agents Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM and Malouin F (2011) Infection of polarized airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function and is influenced by NF-κB Infect Immun 79 3541-3551
Mitchell G Lafrance M Boulanger S Seacuteguin DL Guay I Gattuso M Marsault Eacute Bouarab K and Malouin F (2012) Tomatidine acts in synergy with aminoglycoside antibiotics against multiresistant Staphylococcus aureus and prevents virulence gene expression J Antimicrob Chemother 67 559-568
Mitchell G Lamontagne CA Brouillette Eacute Grondin G Talbot BG Grandbois M and Malouin F (2008) Staphylococcus aureus SigB activity promotes a strong fibronectin-bacterium interaction which may sustain host tissue colonization by small-colony variants isolated from cystic fibrosis patients Mol Microbiol 70 1540-1555
Mitchell G Malouin F 2012 Outcome and prevention of Pseudomonas aeruginosa-Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic Fibrosis Chap 8 pp 181-206 Ed D Sriramulu InTech Open Access Publisher ISBN 979-953-307-059-8
Mitchell G Seacuteguin DL Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S and Malouin F (2010) Staphylococcus aureus sigma B-dependent emergence of small-colony variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-heptylquinoline-N-oxide BMC Microbiol 30 10-33
Moisan H Brouillette Eacute Jacob CL Langlois-Beacutegin P Michaud S and Malouin F (2006) Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated from cystic fibrosis patients is influenced by SigB J Bacteriol 188 64-76
139
Moreno F Crisp C Jorgensen JH and Patterson JE (1995) Methicillin- resistant Staphylococcus aureus as a community organism Clin Infect Dis 21 1308-1312
Murphy MP (2009) How mitochondria produce reactive oxygen species Biochem J 417 1- 13
Novick RP (2003) Autoinduction and signal transduction in the regulation of staphylococcal virulence Mol Microbiol 48 1429-1449
Novo DJ Perlmutter NG Hunt RH and Shapiro HM (2000) Multiparameter flow cytometric analysis of antibiotic effects on membrane potential membrane permeability and bacterial counts of Staphylococcus aureus and Micrococcus luteus Antimicrob Agents Chemother 44 827-834
ONeill AJ Cove JH and Chopra I (2001) Mutation frequencies for resistance to fusidic acid and rifampicin in Staphylococcus aureus J Antimicrob Chemother 47 647-650
Osbourn A E (1996) Preformed antimicrobial compounds and plant defense against fungal attack Plant Cell 8 1821ndash1831
Otto TD Dillon GP Degrave WS and Berriman M (2011) RATT Rapid annotation transfert tool Nucl Acids Res 39 doi101093 Pratique E Theraulaz L (2007) Eacutetude de la reacutegulation du systegraveme respiratoire TMAO reacuteductase chez Escherichia coli Meacutemoire de maitrise de lrsquoeacutecole pratique des hautes eacutetudes Paris
Price CT and Gustafson JE (2001) Increases in the mutation frequency at which fusidic acid-resistant Staphylococcus aureus arise with salicylate J Med Microbiol 50 104-106
Proctor RA and Peters G (1998) Small colony variants in staphylococcal infections Diagnostic and therapeutic implications Clin Infect Dis 27 419- 423
140
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M and Peters G (2006) Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent infections Nat Rev Microbiol 4 295ndash305
Quail MA Kozarewa I Smith F Scally A Stephens PJ Durbin R Swerdlow H and Turner DJ (2008) A large genome centers improvements to the Illumina sequencing system Nat methods 5 1005-1010
Ramirez MS and Tolmasky ME (2010) Aminoglycoside modifying enzymes Drug Resist Updat 13 151ndash171
Rao M Streur TL Aldwell FE and Cook GM (2001) Intracellular pH regulation by Mycobacterium smegmatis and Mycobacterium bovis BCG Microbiology 147 1017-1024
Richardson DJ (2000) Bacterial respiration a flexible process for a changing environment Microbiology 146 551- 571
Riley TV Pearman JW and Rouse IL (1995) Changing epidemiology of methicillin-resistant Staphylococcus aureus in Western Australia Med J Aust 163 412-414
Roddick J G (1974) The steroidal glycoalkaloid α-tomatine Phytochemistry 13 25-36
Rodrigue S Materna AC Timberlake SC Blackburn MC Malmstrom RR et al(2010) Unlocking Short Read Sequencing for Metagenomics PLoS ONE 5(7) e11840doi101371journalpone0011840 Ruiz-Rubio M Alonso Peacuterez-Espinosa A Lairini K RoldAacuten-Arjona T Dipietro A and Anaya N (2001) Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi In Studies in natural products chemistry vol 25 A Rahman (ed) (Elsevier Oxford United Kingdom) pp 293ndash326
141
Samuelsen oslash Haukland HH Kahl BC von Eiff C Proctor RA Ulvatne H Sandvik K and Vorland LH (2005) Staphylococcus aureus small colony variants are resistant to the antimicrobial peptide lactoferricin B J Antimicrob Chemother 56 1126- 1129
Sandrock R W and Vanetten HD (1998) Fungal sensitivity to and enzymatic degradation of the phytoanticipin α-tomatine Phytopathology 88 137ndash143
Sendi P and Proctor RA (2009) Staphylococcus aureus as an intracellular pathogen the role of small colony variants Trends Microbiol 17 54ndash58
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C and Osbourn A (2006) Dual effects of plant steroidal alkaloids on Saccha- romyces cerevisiae Antimicrob Agents Chemother 50 2732ndash2740
Somerville GA and Proctor RA (2009) At the crossroads of bacterial metabolism and virulence factor synthesis in Staphylococci Microbiol Mol Biol Rev 73 233- 248
Starkov AA (2010) Measurement of mitochondrial ROS production Methods Mol Biol 648 245-255
Taber HW (1993) Respiratory chain In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of MicrobiologyWashington DC) pp 199-212
Taber HW and Morrison M (1964) Electron transport in Staphylococci Properties of a particle preparation from exponential phase Staphylococcus aureus Arch Biochem Biophys 105 367- 379
Taber HW et al(1987) Bacterial uptake of aminoglycoside antibiotics Microbiol Rev 51 439-457
Thoumlny-Meyer L (1997) Biogenesis of respiratory cytochromes in bacteria Microbiol Mol Biol Rev 61 337- 376
142
Tubby S Wilson M Wright JA Zhang P and Nair SP (2013) Staphylococcus aureus small colony variants are susceptible to light activated antimicrobial agents BMC Microbiol 13 201- 204
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D Roth J Proctor RA Becker K et al (2011) Staphylococcus aureus phenotype switching an effective bacterial strategy to escape host immune response and establish a chronic infection EMBO Mol Med 3 129ndash41
Turrens JF (2003) Mitochondrial formation of reactive oxygen species J Physiol 552 335ndash 344
Tynecka Z Szcześniak Z Malm A and Los R (1999) Energy conservation in aerobically grown Staphylococcus aureus Res Microbiol 150 555- 566
Udo EE Pearman JW and Grubb WB(1993) Genetic analysis of community isolates of methicillin-resistant Staphylococcus aureus in Western Australia J Hosp Infect 25 97-108
Vakulenko SB and Mobashery S (2003) Versatility of Aminoglycosides and Prospects for Their Future Clin Microbiol Rev 16 430-450
Voggu L Schlag S Biswas R Rosenstein R Rausch C and Goumltz F (2006) Microevolution of cytochrome bd oxydase in Staphylococci and its implication in resistance to respiratory toxins released by Pseudomonas J Bacteriol 188 8079- 8086
Wells PG Bhuller Y Chen CS Jeng W Kasapinovic S Kennedy JC Kim PM Laposa RR McCallum GP Nicol CJ et al (2005) Molecular and biochemical mechanisms in teratogenesis involving reactive oxygen species Toxicol Appl Pharmacol 207 354-366
143
Yan M Yu C Yang J and Ji Y (2011) The essential two-component system YhcSR is involved in regulation of the nitrate respiratory pathway of Staphylococcus aureus J Bacteriol 193 1799- 1805
Zhang T Luo Y Chen Y Li X and Yu J (2012) BIGrat a repeat resolver for pyrosequencing-based re-sequencing with Newbler BMC Res Notes 5 567
144

iv
REMERCIEMENTS
Je voudrais remercier mon directeur de recherche Pr Franccedilois Malouin qui a cru
en moi pour mrsquoavoir proposeacute ce beau projet de maitrise Merci pour sa grande
disponibiliteacute son eacutecoute et ses critiques toujours constructives et jamais
condescendantes
Je voudrais eacutegalement remercier mes conseilleacutes Kamal Bouarab Seacutebastien
Rodrigue et Eacuteric Marsault leur implication dans mon projet leur occasionnant un
surplus de travail Merci pour leur support technique et leur disponibiliteacute agrave reacutepondre
agrave mes questions
Un gros merci agrave Gabriel Mitchell qui a eacuteteacute mon mentor au deacutebut de ma maitrise et
qui mrsquoa introduit au projet laquo Tomatidine raquo Un merci particulier eacutegalement agrave
Maxime-Lamontagne-Boulet pour son aide en tant que stagiaire lors de mes
derniegraveres expeacuteriences Je voudrais eacutegalement remercier tous les membres du
laboratoire preacutesents et passeacutes qui ont toujours eacuteteacute disponibles pour partager avec
moi leurs connaissances theacuteoriques et techniques
Je ne peux passer sous silence lrsquoimplication de lrsquoUniversiteacute de Sherbrooke et du
FQRNT comme organismes subventionnaires au projet Merci eacutegalement agrave Fibrose
Kystique Canada qui a participeacute eacutegalement agrave la subvention drsquoune partie du projet
En terminant je voudrais remercier ma famille qui a crue en moi et qui mrsquoencourage
agrave perseacuteveacuterer depuis toujours Un merci tout particulier agrave mon conjoint et ma belle-
fille pour qui je nrsquoai pas toujours eacuteteacute aussi disponible que jrsquoaurais voulu lrsquoecirctre et qui
mrsquoont supporteacutee au quotidien et ce dans tous les sens du terme
v
TABLE DES MATIEgraveRES
SOMMAIRE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ii
REMERCIEMENTS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip iv
TABLE DES MATIEgraveRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip v
LISTE DES ABBREacuteVIATIONS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip vii
LISTE DES TABLEAUX helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ix
LISTE DES FIGURES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip x
CHAPITRE 1 INTRODUCTION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
11 Staphylococcus aureus helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
111 Importance cliniquehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)helliphellip 2
1121 Diffeacuterences et similariteacutes helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 2
1122 La chaine de transport des eacutelectrons helliphelliphelliphelliphellip 4
12 Antibiotiques Modes drsquoaction cibles cellulaire et
meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles helliphelliphelliphellip 25
122 Les aminoglycosides helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1221 Mode drsquoaction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1222 Meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphellip 31
123 La tomatidine helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 33
13 Projet de maitrise helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
CHAPITRE 2 ARTICLE SCIENTIFIQUE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 37
ldquoTomatidine and derivative FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Firmicutes Including Those of Listeria monocytogenes
and Are Bactericidal in Combination With Aminoglycosides Against their Normal
Phenotyperdquo
vi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE helliphelliphelliphellip 76
31 Obtention des mutants helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
32 Seacutequenccedilage et analyses bio-informatiques helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 79
321 Identification des mutations en lien avec la tomatidine helliphellip 89
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct helliphellip 90
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine helliphelliphelliphelliphellip 99
CHAPITRE 4 DISCUSSION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 113
CHAPITRE 5 CONCLUSION ET PERSPECTIVES helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 124
CONTRIBUTIONS SUPPLEacuteMENTAIRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 1 Application de brevet helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 2 Article scientifique helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 129
laquo Unraveling the structure-activity relationship of tomatidine
a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureus raquo
BIBLIOGRAPHIE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 131
vii
LISTE DES ABBREacuteVIATIONS
Adh Alcool deacuteshydrogeacutenase
ADN Acide deacutesoxyribo-nucleacuteique
ADP Adeacutenosine diphosphate
ARN Acide ribo-nucleacuteique
ATP Adeacutenosine triphosphate
CA-MRSA Community-associated MRSA
CMI concentration minimale inhibitrice
CoQ Coenzyme Q
Cyt Cytochrome
DMQ laquo demethylmenaquinone raquo
dTMP Deacutesoxythymidine monophosphate
dUMP Deacutesoxyuridine monophosphate
FADH2 Flavine adenine dinucleacuteotide
GEN gentamicine
HA-MRSA Healthcare-associated MRSA
Ldh Lactate deacuteshydrogeacutenase
ml millilitre
MQ meacutenaquinone
MRSA methycilin resistant Staphylococcus aureus
Mqo2 Quninol oxidoreacuteductase
NADH Nicotinamide adenine dinucleacuteotide (forme reacuteduite)
Pdh Pyruvate deacuteshydrogeacutenase
Pfl Pyruvate formate lyase
PyrD Dihydroorotate deacuteshydrogeacutenase
Q Quinone
Rif rifampicine
ROS laquo reactive oxygen species raquo espegravece reacuteactive de lrsquooxygegravene
viii
[S] Polysulfide
S aureus Staphylococcus aureus
SARM Staphylococcus aureus resistant agrave la meacutethycilline
SCV laquo small colony variants raquo
SHCHC 2-succinyl-6-hydroxy-24-cyclohexadiene-1-carboxylate syntase
TCA Tricarboxylic acid cycle
TCD tableau croiseacute dynamique
TO tomatidine UFC Uniteacute formatrice de colonies
UQ Ubiquinone
microg microgramme
ix
LISTE DES TABLEAUX
CHAPITRE 1 INTRODUCTION
1 Cibles et modes drsquoaction des principales classes drsquoantibiotique 27
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants (B) chez les mutants NewbouldΔhemB reacutesistants 95-96
CHAPITRE 4 DISCUSSION
1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 123
x
LISTE DES FIGURES
CHAPITRE 1 INTRODUCTION
1 Diffeacuterence entre les pheacutenotypes normaux et SCV de S aureus 3
2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes 6
3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale 8
4 Chaines respiratoires bacteacuteriennes contenant des cytochromes 9
5 Flexibiliteacute respiratoire (A) chez Paracoccus denitrificans (B) chez
Escherichia coli 10
6 Chaine de transport des eacutelectrons proposeacutee chez les staphylocoques 12
7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies 17
8 Modegravele de reacutegulation du cycle TCA 20
9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS 23
10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de O2-bull 24
11 Voies de formation et de deacutetoxification des ROS 25
12 Cineacutetique drsquoentreacutee des aminoglycosides 30
13 Sites de mutations de certains reacutesistants aux aminoglycosides 32
14 Structures chimiques de (A) tomatine (B) tomatidine et (C) Analogue
de tomatidine FC04-100 34
xi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Graphiques des CMI pour chacun des passages pour (A) la
tomatidine sur NewbouldΔhemB (B) la tomatidine en preacutesence de
gentamicine sur NewbouldΔhemB (C) la gentamicine sur S aureus
ATCC 29213 (D) la gentamicine en preacutesence de tomatidine sur S
aureus ATCC 29213 77-79
2 Organigramme des mutants reacutesistants seacutequenceacutes 80-81
3 Eacutetapes du seacutequenccedilage Illumina 83
4 Meacutethode drsquoamplification par la meacutethode Illumina 85
5 Correction du programme RATT sur les annotations transfeacutereacutees 88
6 Exemple drsquoune partie du tableau compilation des mutations 89
7 Seacutelection des gegravenes drsquointeacuterecirct chez ATCC 29213 dans le TCD par
lrsquoutilisation de diffeacuterents filtres 91-92
8 Seacutelection des gegravenes drsquointeacuterecirct chez NewbouldΔhemB dans le TCD par
lrsquoutilisation de diffeacuterents filtres 93-94
9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct 97-98
10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants 100
11 Courbe de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison gentamicine (+TO) du premier et troisiegraveme niveau de
reacutesistance 101
12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B)
NewbouldΔhemB 102
xii
13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213 104
14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicinelorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC
29213 105
15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants
ATCC 29213 reacutesistants agrave la combinaison gentamicine (+TO) 106
16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la
tomatidine 107
17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la
tomatidine 108
18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB
reacutesistants agrave la combinaison tomatidine (+gentamicine) 109
19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane
Potential Kit (Invitrogen) 111
20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du
gegravene atpE de S aureus Newman 112
xiii
CHAPITRE 4 DISCUSSION
1 Site drsquoinhibition de lrsquoATP synthase par diffeacuterentes moleacutecules 114
2 Structure chimique de (A) TMC207 et (B) lrsquoanalogue anti Gram
positifs 115
3 Seacutequence proteacuteique de la proteacuteine AtpE chez diffeacuterentes espegraveces 118
4 Modeacutelisation de la structure tridimentionnelle de lrsquoATP synthase de S
aureus (atpE) montrant les acides amineacutes muteacutes chez les mutants
reacutesistants 121
- 1 -
CHAPITRE 1
INTRODUCTION
11 Staphylococcus aureus
111 Importance clinique
Staphylococcus aureus est une bacteacuterie agrave Gram positif anaeacuterobie facultative Crsquoest un
microorganisme opportuniste pouvant infecter diffeacuterents hocirctes diffeacuterents organes ou
sites drsquoinfection et ayant la capaciteacute de causer tant des infections aiguumles que
chroniques (Archer 1998 Goerke et Wolz 2004)
Staphylococcus aureus reacutesistant agrave la meacutethilline le SARM (ou laquo MRSA raquo) est une
souche de S aureus devenue reacutesistante aux β-Lactamines par lrsquoacquisition du gegravene
mecA dans son chromosome (Hiramatsu 1995) Le SARM a eacuteteacute isoleacute pour la
premiegravere fois en 1960 en Angleterre (Jevons 1961) et est devenu eacutepideacutemique agrave
travers le monde depuis les anneacutees 1970 Au moins trois geacutenotypes distincts eacutetaient
recenseacutes dans les anneacutees 1980 (Hiramatsu 1995) dont deux sont toujours preacutevalents
agrave travers le monde comme eacutetant des souches HA-MRSA (Healthcare-associated
MRSA) des souches acquises agrave lrsquohocircpital (Enright et al 2002) Ces souches ont
acquis des gegravenes de reacutesistance agrave pratiquement tous les antibiotiques introduits dans
lrsquoutilisation clinique depuis les 50 derniegraveres anneacutees mecircme agrave lrsquoantibiotique de dernier
ressort la vancomycine(Hiramitsu 1997 Chen et al 2003) Nous sommes
maintenant en manque drsquoagents theacuterapeutiques efficaces contre les SARM touchant
les personnes immunodeacuteprimeacutees qui sont dans les hocircpitaux En plus depuis le deacutebut
des anneacutees 1990 des clones de SARM dont plusieurs deacutemontraient un potentiel
pathogeacutenique supeacuterieur aux souches classiques de HA-MRSA retrouveacutees agrave lrsquohocircpital
- 2 -
sont maintenant retrouveacutes dans la communauteacute (CA-MRSA) agrave travers le monde (Udo
et al 1993 Riley et al 1995 Moreno et al 1995 Aubry-Damon et al 1997) et ont
la capaciteacute drsquoaffecter des personnes en santeacute en causant des infections aiguumles
(Hiramatsu et al 2013) Il apparaicirct donc urgent de trouver des drogues alternatives
afin de surmonter le nombre grandissant de souches de SARM
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)
1121 Diffeacuterences et similariteacutes
Les laquo small colony variants raquo (SCV) sont des bacteacuteries souvent isoleacutees lors des
infections chroniques comme crsquoest le cas des infections pulmonaires chez les
patients atteints de fibrose kystique mais aussi dans des cas drsquoosteacuteomyeacutelite
drsquoarthrite septique drsquoinfections de prothegraveses orthopeacutediques et lors de la mammite
bovine (Alexender et Hudson 2001 Moisan et al 2006 Proctor et al 2006) Les
SCV sont caracteacuteriseacutes par soit un meacutetabolisme oxydatif dysfonctionnel ou un manque
dans la biosynthegravese de la thymidine les deux amenant une alteacuteration dans
lrsquoexpression des facteurs de virulence une croissance plus lente et une diminution
de la pigmentation des colonies (Figure 1) (Proctor et al 2006) Cependant alors
que les souches normales reacutepriment lrsquoexpression des proteacuteines de surface et
expriment les exoproteacuteines lors de la phase stationnaire (Novick 2003) les SCV
expriment de faccedilon stable les gegravenes Sig-B deacutependants codant pour des proteacuteines de
surface telle les adheacutesines au lieu drsquoactiver le systegraveme agr deacutependant du quorum-
sensing et produire des exoproteacuteines (Moisan et al 2006) Ces diffeacuterences dans
lrsquoexpression des facteurs de virulence reacutesultent chez les SCV en une meilleure
habileteacute agrave adheacuterer aux composantes de lrsquohocircte (Mitchell et al 2008) et agrave former du
biofilm (Mitchell et al 2010a Mitchell et al 20010b) et seraient eacutegalement lieacutees agrave
lrsquohabileteacute des SCV agrave lrsquoinvasion et la persistance des cellules de lrsquohocircte (Proctor et al
2006 Sendi et Proctor 2009 Mitchell et al 2011b)
- 3 -
S aureus est donc un pathogegravene qui peut se preacutesenter sous deux pheacutenotypes qui
sont souvent retrouveacutes simultaneacutement lors drsquoune mecircme infection et qui ont le pouvoir
de reacuteverter drsquoun pheacutenotype agrave lrsquoautre selon les conditions du milieu (Figure 1)
(Tuchscherr et al 2011)
Le pheacutenotype dit rdquonormalrdquo est associeacute aux infections aiguumles agrave une forte production
de facteurs de virulence (ex heacutemolysine) et est associeacute agrave la phase de disseacutemination
de la bacteacuterie Lrsquoautre pheacutenotype le laquo small-colony variants raquo ou SCV est affecteacute
dans sa chaine respiratoire ce qui amegravene une croissance beaucoup plus lente des
colonies de 8 -10 fois plus petites que les prototypes et une faible susceptibiliteacute aux
aminoglycosides Les SCV sont plutocirct associeacutes aux infections chroniques agrave S aureus
de par leur capaciteacute agrave srsquointernaliser dans les cellules de lrsquohocircte leur forte capaciteacute agrave
produire du biofilm et leur capaciteacute agrave produire des facteurs de colonisation
(Tuchscherr et al 2011 Proctor et al 2006) (Figure 1)
Figure 1 Diffeacuterences entre les pheacutenotypes normaux et SCV de S aureus
S aureus agrave
pheacutenotype normal
Disseacutemination
amp
Infections aiguumles
Colonisation
amp
Infections persistantes
SCVs
(laquo small-colony
variants raquo)
Reacutevers
ion
pheacutenoty
piq
ue
Norm
al
SC
V
Croissance
Biofilm
Internalisationheacutemolyse
Mitchell G et Malouin F 2012
- 4 -
1122 La chaine de transport des eacutelectrons
Chaine respiratoire tableau geacuteneacuteral
Les chaines respiratoires bacteacuteriennes bien que tregraves variables dans leurs
composantes parmi les diffeacuterentes espegraveces sont toutes constitueacutees sur le mecircme
modegravele de base Elles sont composeacutees de trois composantes majeures soit
- les deacuteshydrogeacutenases primaires qui sont chargeacutees drsquooxyder des substrats
principalement les coenzymes NADH et FADH2 (issues du cycle de Krebs) (mais
aussi selon le cas le succinate le lactate lrsquohydrogegravene hellip) et de pomper deux
protons du cocircteacute externe de la membrane en plus de transfeacuterer deux eacutelectrons agrave un
groupement de quinones
- Les quinones sont des moleacutecules liposolubles non proteacuteiques mobiles dans la
membrane Elles servent drsquointermeacutediaires entre les deacuteshydrogeacutenases primaires et
les oxydases terminales pour le transport des eacutelectrons Il existe deux sortes de
quinones les benzoquinones dont font partie les ubiquinones et les
naphtoquinones dont font partie les meacutenaquinones Les bacteacuteries agrave Gram positif
ne possegravedent que des meacutenaquinones alors que les deux sont preacutesentes chez les
bacteacuteries agrave Gram neacutegatif (Hederstedt 1993 Pratique et Theraulaz 2007)
- Les oxidoreacuteductases terminales sont chargeacutees de reacuteduire lrsquoaccepteur final
drsquoeacutelectron lrsquooxygegravene eacutetant lrsquoaccepteur final privileacutegieacute
De faccedilon plus deacutetailleacutee la chaine respiratoire est composeacutee de cinq
complexes proteacuteiques principaux (Figure 2)
- complexe I NADH deacuteshydrogeacutenase ou NADH reacuteductase ou NADHCoQ
oxidoreacuteductase ou NADH-ubiquinone oxidoreacuteductase
- complexe II Succinate deacuteshydrogeacutenase
- complexe III Cytochrome bc1 complexe ou ubiquinol cytochrome c
reacuteductaseoxydoreacuteductase
- complexe IV Cytochrome c oxydase
- 5 -
- complexe V ATP synthase (Creacutepin-Gilbert et al 2006)
La phosphorylation oxydative est deacuteclencheacutee dans la chaine de complexes
proteacuteiques par la preacutesence drsquoO2 accepteur terminal privileacutegieacute drsquoeacutelectrons (ou drsquoautres
accepteurs finaux tel le nitrate) dans le milieu ainsi que par les eacutelectrons ameneacutes par
le NADH et le FADH2 au complexe I et II respectivement (Creacutepin-Gilbert et al 2006)
Les eacutelectrons vont ensuite transiter par le complexe III jusqursquoau complexe IV ou les
eacutelectrons seront utiliseacutes afin de reacuteduire lrsquooxygegravene en eau le complexe IV est
lrsquooxydase terminale consideacutereacutee comme eacutetant lrsquoaccepteur final drsquoeacutelectrons Le transfert
drsquoeacutelectrons est possible gracircce aux reacuteactions drsquooxydoreacuteduction en cascade dans les
diffeacuterents complexes proteacuteiques Ces reacuteactions fournissent de lrsquoeacutenergie libre utiliseacutee
par les complexes I III et IV afin de transfeacuterer des protons agrave lrsquoexteacuterieur de la cellule
(Creacutepin-Gilbert et al 2006) Le complexe V (ATP synthase) quant agrave lui utilise les
eacutelectrons pompeacutes agrave lrsquoexteacuterieur par les autres complexes comme force proton-motrice
afin de phosphoryler trois moleacutecules drsquoADP et ainsi produire trois ATP Le transport
des eacutelectrons entre les diffeacuterents complexes proteacuteiques est possible gracircce agrave la
preacutesence dans la membrane de deux types de moleacutecules liposolubles non proteacuteiques
mobiles les meacutenaquinones et le cytochrome c Les meacutenaquinones seules quinones
preacutesentes chez les bacteacuteries agrave Gram positif assurent le transport des eacutelectrons entre
le complexe I et le complexe III et entre le complexe II et III Le cytochrome c pour sa
part veacutehicule les eacutelectrons entre le complexe III et le complexe IV Le transport des
eacutelectrons srsquoeffectue de la composante ayant le plus faible potentiel drsquooxydoreacuteduction
vers celle ayant le plus fort potentiel
- 6 -
La chaine respiratoire bacteacuterienne
La mitochondrie montre assez peu de flexibiliteacute respiratoire Il y a une certaine
flexibiliteacute au niveau de lrsquoentreacutee des eacutelectrons mais aucune au niveau de la sortie des
eacutelectrons ougrave la cytochrome aa3 oxydase reacuteduit lrsquooxygegravene en eau (Richardson 2000)
(Figure 3) Chez les bacteacuteries alors que le complexe II fonctionne sensiblement de
faccedilon identique agrave celui des eucaryotes les autres complexes peuvent se composer
de diffeacuterents complexes enzymatiques (reacuteductases) selon les espegraveces et les
conditions environnementales (Richardson 2000 Thoumlny-Meyer 1997) En outre
chez les bacteacuteries contrairement aux eucaryotes la chaine respiratoire comporte
souvent des voies de transport des eacutelectrons alternatives on dit alors qursquoelles ont une
chaine respiratoire agrave branches ou brancheacutee ou ramifieacutee (Artzatbanov et Petrov 1990)
Plusieurs espegraveces bacteacuteriennes possegravedent une chaine respiratoire ramifieacutee dont la
ramification se situe habituellement suite agrave la reacuteduction des quinones en quinols
(Lauraeus et Wikstrom 1993 Thoumlny-Meyer 1997) (Figure 4) Une des deux
Complexe I NADH
deacuteshydrogeacutenase
Complexe II Succinate
deacuteshydrogeacutenase
Complexe III
bc 1
Complexe IV Cytochrome c
oxydase
Complexe V
ATP synthase
Modifieacute de wikipedia httpfrwikipediaorgwikiChaine_respiratoire
Figure 2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes
Leurs rocircles dans le transport des eacutelectrons et dans lrsquoeacutelaboration du gradient de
protons et le lien entre la chaine respiratoire et le cycle de Krebs Q Quinones
- 7 -
branches connue comme la branche cytochrome c oxyde les quinols (meacutenaquinol
QH2) via la voie complexe bc1-cytochrome c- cytochrome c oxydase Lrsquoautre
branche-type consiste en une seule uniteacute enzymatique (cytochrome bd) laquelle
transfegravere directement les eacutelectrons des quinols agrave lrsquooxygegravene Cette voie est connue
comme la branche quinol oxydase il srsquoagit drsquoune oxydase terminale transfeacuterant
directement les eacutelectrons agrave lrsquooxygegravene eacutevitant de passer par lrsquointermeacutediaire du
complexe III (bc1) et du cytochrome c (Artzatbanov et Petrov 1990 Lauraeus et
Wikstrom 1993) Alors que la voie du cytochrome c permet le pompage de protons agrave
lrsquoexteacuterieur par lrsquoaction du complexe bc1 (ou complexe III) et du cytochrome c oxydase
le cytochrome bd ne permet aucun pompage de protons (Lauraeus et Wikstrom
1993) La voie passant par le complexe bc1-cytochrome c-cytochrome c oxydase est
plus efficace dans la production drsquoeacutenergie Alors que les 2 voies peuvent ecirctre
exprimeacutees simultaneacutement chez quelques espegraveces il semble que lrsquoexpression de ces
voies soit inductible selon les conditions environnementales chez Bacillus subtilis et
les Staphylocoques (Lauraeus et Wikstrom 1993) On pourrait dire eacutegalement que la
chaine respiratoire est seacutepareacutee en deux eacutetapes la premiegravere partie est la phase
quinone-reacuteductase (Q-QH2) et la seconde la phase quinol-oxydase (QH2-Q) menant
agrave lrsquooxydase terminale La phase de reacuteduction est responsable du transfert des
eacutelectrons drsquoune varieacuteteacute de substrats tels le NADH le succinate le lactate le formate
et lrsquohydrogegravene vers les quinones et inclut diffeacuterentes reacuteductases (Thoumlny-Meyer 1997)
(Figure 4)
- 8 -
Figure 3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale UQ ubiquinone UQH2 ubiquinol Cyt cytochrome Il semble y avoir presque autant de chaines de transport des eacutelectrons diffeacuterentes
qursquoil y a drsquoespegraveces bacteacuteriennes (par exemple Paracoccus denitrificans et Escherichia
coli (Figure 5)) De plus il ne semble pas y avoir consensus dans la litteacuterature sur
les noms attribueacutes aux oxydases terminales Les noms pour les oxydases terminales
deacutependent des types de cytochromes dont elles sont composeacutees Il ne semble donc
pas eacutevident de connaicirctre de faccedilon preacutecise les types de cytochromes qui font partie
des oxydases terminales Pour rajouter agrave la confusion les gegravenes et opeacuterons ne sont
pas toujours associeacutes agrave leur oxydase terminale dans la litteacuterature ce qui fait qursquoon ne
peut savoir de faccedilon certaine de quelle oxydase on parle Le Cytochrome aa3 peut
eacutegalement porter le nom de Cyt ba3 et le cytochrome bd peux eacutegalement porter les
noms de Cyt bo
Tireacute de Richardson DJ 2000
- 9 -
Succinate
Formate
H2
SDH
FDH
HYD
QQH 2 bc1 c
Reacuteductase alternatives
b
c
cd1c
cb
c
b
c
d
Siroheme
Siroheme
b
NO3-
NO2-
NO
TMAO
DMSOS2O3
2-
[S]
SO32-
Fumarate
Quinol oxidases
ba3bb3bo3d
bd
aa3
O2
Cyt c oxidases
aa3ba3baa3caa3cbb3cao
co
O2
Modifieacute de Thony-Mayer L 1997
NADH
NDH
Figure 4 Chaines respiratoires bacteacuteriennes contenant des cytochromes
Du cocircteacute gauche on retrouve la partie quinone (ubiquinone ou meacutenaquinone)-
reacuteductase Du cocircteacute droit on retrouve la partie quinol-oxydase avec les oxydases
terminales reacuteduisant lrsquoO2 ou lrsquoaccepteur terminal alternatif tel que montreacute Les
oxydoreacuteductases sont surligneacutees QQH2 repreacutesente le pool de quinonesquinols
dans la membrane Les polysulfides sont repreacutesenteacutes par [S] Les chaines
respiratoires ne contenant pas de cytochromes ne sont pas montreacutees NDH
NADH deacuteshydrogeacutenase
Modifieacute de Thoumlny-Mayer L 1997
- 10 -
Figure 5 Flexibiliteacute respiratoire chez Paracoccus denitrificans (a) et chez
Escherichia coli (b) MQ menaquinone UQ ubiquinone DMQ
demethylmenoquinone
Chez S aureus
Comme plusieurs autres espegraveces bacteacuteriennes anaeacuterobies facultatives S aureus a
la possibiliteacute drsquoexprimer diffeacuterentes voies de synthegravese eacutenergeacutetique dont lrsquoexpression
est finement reacuteguleacutee selon des conditions environnementales du moment (Hammer et
Tireacute de Richardson DJ 2000
- 11 -
al 2013 Lauraeus et Wikstrom 1993) Mecircme si S aureus peut utiliser la
fermentation la respiration (aeacuterobie et anaeacuterobie) sera privileacutegieacutee par la bacteacuterie
puisque celle-ci amegravene une bien meilleure production drsquoeacutenergie (Goumltz et al 2006
Hammer et al 2013 ) La respiration aeacuterobie avec lrsquooxygegravene comme accepteur final
drsquoeacutelectron sera utiliseacutee de faccedilon preacutefeacuterentielle en preacutesence drsquooxygegravene (Hammer et al
2013) Malgreacute sa versatiliteacute dans ses voies de synthegravese eacutenergeacutetique S aureus et les
autres bacteacuteries agrave Gram positif ne produisent qursquoun seul type de quinone les
meacutenaquinones lesquelles sont utiliseacutees pour la croissance degraves qursquoil y a de lrsquooxygegravene
de preacutesent dans le milieu (Somerville et Proctor 2009) La reacuteduction de lrsquooxygegravene en
eau est meacutedieacutee par des oxydases terminales hegraveme deacutependant (Hammer et al 2013)
Selon certains auteurs S aureus possegravederait deux oxydases terminales la
cytochrome oxydase aa3 (ou ba3) (QoxABCD) et la cytochrome bd quinol oxydase
(CydAB) (Goumltz et al 2006 Hammer et al 2013 Kohler et al 2008) Cette derniegravere
est exclusive aux procaryotes et est une voie de contournement du complexe III qui
transmet directement les eacutelectrons du laquo pool raquo de quinol (QH2) agrave lrsquooxygegravene Selon
certains auteurs il y aurait eacutegalement chez S aureus une troisiegraveme oxydase
terminale le cytochrome o (CyoABCDE) (Thoumlny-Meyer 1997 Voggu et al 2006)
qui selon certain serait un cytochrome oxydase terminal alternatif au cytochrome aa3
et au cytochrome bd (Voggu et al 2006 Goumltz et al 2006 Taber et Morrison 1964)
(Figure 6) Ces voies alternatives permettent agrave S aureus de srsquoadapter rapidement agrave
un changement drsquoenvironnement (Hammer et al 2013) In vivo chaque organe est
un environnement diffeacuterent preacutesentant des deacutefis respiratoires distincts dus agrave la
variation dans 1- la quantiteacute drsquooxygegravene disponible 2- la disponibiliteacute drsquoaccepteurs
drsquoeacutelectrons alternatifs 3- la concentration drsquoinhibiteurs de la respiration dans le milieu
tels les oxydes nitriques (NO) produits par les neutrophiles et 4- la disponibiliteacute des
cofacteurs neacutecessaires agrave la production drsquoeacutenergie (Hammer et al 2013) Le
cytochrome est exprimeacute de faccedilon modeacutereacutee dans un milieu riche en oxygegravene mais est
fortement exprimeacute dans un milieu pauvre en oxygegravene mais riche en oxyde nitrique
De plus le type drsquooxydoreacuteductase terminale exprimeacute est influenceacute eacutegalement par la
- 12 -
source de carbone disponible la phase de croissance le pH extracellulaire et la
tension en oxygegravene (Lauraeus et Wikstrom 1993 Tynecka et al 1999)
La chaine respiratoire chez les SCV
S aureus est particuliegraverement doueacute agrave surpasser les deacutefis respiratoires en acqueacuterant
une reacutesistance agrave lrsquoacide nitrique produit par les neutrophiles et une habiliteacute agrave persister
dans les cellules de lrsquohocircte en se preacutesentant comme un variant agrave petites colonies
ayant une chaine respiratoire deacuteficiente (SCV) (Hammer et al 2013)
Deux principaux types de SCV sont retrouveacutes dans les isolats cliniques 1- les SCV
ayant une chaine de transport des eacutelectrons deacutefectueuse et 2- les SCV auxotrophes
pour la thymidine (Proctor et Peters 1998 Samuelsen et al 2005) Ces derniers
Figure 6 Chaine de transport des eacutelectrons proposeacutee chez les
staphylocoques Les informations disponibles sur les composantes de la chaine
respiratoire des staphylocoques suggegraverent une chaine ramifieacutee consistant en 2
oxydases terminales meacutenaquinol-deacutependante un cytochrome o et un cytochrome
aa3 oxydase NADH deacuteshydrogeacutenase (DHase) transmet les eacutelectrons au laquo pool raquo
de meacutenaquinones De lagrave les eacutelectrons sont transfeacutereacutes soit au cytochrome b ou agrave
lrsquoaccepteur alternatif drsquoeacutelectrons le cytochrome bd quinol oxydase Le cytochrome
b transfegravere les eacutelectrons agrave une des deux oxydases terminales Ces oxydases
terminales transfegraverent les eacutelectrons agrave lrsquooxygegravene
Modifieacute de Voggu L et al 2006
- 13 -
arborent une ou plusieurs mutations dans le gegravene thyA codant pour la thymidylate
synthase laquelle catalyse la derniegravere eacutetape dans la voie de biosynthegravese de la
thymidine crsquoest-agrave-dire la formation de dTMP agrave partir du dUMP (Samuelsen et al
2005) Le dTMP est essentiel pour la synthegravese de lrsquoADN (Besier et al 2007) Les
mutants thymidine-auxotrophes sont souvent la conseacutequence drsquoune pression
seacutelective par lrsquoutilisation prolongeacutee de trimethoprim-sulfamethoxazole (TMP-SMX)
(souvent utiliseacute pour le traitement des patients fibroses kystiques) un antibiotique qui
interfegravere dans la biosynthegravese de lrsquoacide tetrahydrofolique qui agit comme cofacteur de
la thymidylate synthase (Besier et al 2007 Lannergard et al 2008 Samuelsen et al
2005) Mecircme si de premiers abords les SCV thymidine-auxotrophes ne semblent pas
avoir de lien avec la chaine transport des eacutelectrons il se pourrait que celle-ci ait sont
importance chez ces SCV puisque lrsquoentreacutee de la thymidine passe par NupC dont le
fonctionnement neacutecessite un gradient eacutelectrochimique creacuteeacute par la chaine de transport
des eacutelectrons (Proctor et Peters 1998 ) Les caracteacuteristiques pheacutenotypiques des
SCV deacutecrites ci-apregraves sont toutes lieacutees agrave lrsquoalteacuteration de la chaine de transport des
eacutelectrons 1- la croissance plus lente parce que la synthegravese de la paroi cellulaire
neacutecessite une grande quantiteacute drsquoATP 2-la diminution de la formation de pigment
parce que la biosynthegravese des caroteacutenoiumldes requiert le transport des eacutelectrons 3- une
reacutesistance accrue agrave la gentamicine et autres aminoglycosides parce que lrsquoentreacutee de
ces composeacutes neacutecessite un grand potentiel membranaire geacuteneacutereacute par le transport des
eacutelectrons (Lannergard et al 2008) et 4- une perte partielle ou complegravete de la
fermentation du mannitol parce que lrsquoutilisation des sucres-alcool tels que le
mannitol est diminueacutee lorsque la chaine de transport des eacutelectrons ne peut oxyder le
NADH produit lorsque le mannitol est meacutetaboliseacute en fructose-6-phosphate (Kohler et
al 2003 Proctor et Peters 1998 Samuelsen et al 2005)
Les SCV posseacutedant une chaine de transport des eacutelectrons deacutefectueuse ont une voie
de biosynthegravese de lrsquoheacutemine ou de la meacutenadione deacuteficiente et le pheacutenotype est
reacuteversible lorsque compleacutementeacute avec lrsquoheacutemine ou la meacutenadione (Hammer et al
- 14 -
2013 Kohler et al 2008 Lannergard et al 2008 Proctor et Peters 1998) La
meacutenadione est essentielle agrave la biosynthegravese des meacutenaquinones et lrsquoheacutemine est
essentielle agrave la biosynthegravese de lrsquohegraveme qui entre dans formation des cytochromes
(Hammer et al 2013 Kohler et al 2008 Lannergard et al 2008 Thoumlny-Meyer
1997) Les meacutenaquinones et les cytochromes sont essentiels au fonctionnement de
la chaine de transport des eacutelectrons (vers lrsquoO2 autant que vers le nitrate) (Kohler et al
2008) Il se pourrait qursquoil existe eacutegalement des SCV causeacutes par drsquoautres deacuteficiences
telles que lrsquoalteacuteration de lrsquoATP synthase qui ne reacutesulterait pas drsquoune auxotrophie pour
lrsquoheacutemine ou la meacutenadione mais reacutesulterait tout de mecircme en une deacuteficience dans la
chaine de transport des eacutelectrons (Proctor et Peters 1998 )
Comme lrsquohegraveme est un cofacteur essentiel pour les oxydases terminales et que la
meacutenaquinone est un transporteur drsquoeacutelectrons utiliseacute par S aureus on comprend que
la faible croissance des mutants hemB et menD repreacutesente un pheacutenotype mimant la
perte des oxydases terminales (Hammer et al 2013) Les deux oxydases terminales
de S aureus QoxABCD (cyt aa3) et CydAB (cyt bd) semblent ecirctre diffeacuteremment
reacuteguleacutees chez les mutants menD et la souche sauvage alors que lrsquoopeacuteron cydAB est
fortement sous exprimeacute chez les mutants aucune diffeacuterence de niveau de
transcription de qoxABCD nrsquoa eacuteteacute observeacutee entre les mutants menD et leur souche
parente (Kohler et al 2008)
Des mutations dans les gegravenes menD hemB et ctaA peuvent reacutesulter en un
pheacutenotype SCV posseacutedant une chaine de transport des eacutelectrons deacuteficiente
(Clements et al 1999 Proctor et Peters 1998 ) Les gegravenes codant pour les eacutetapes
de formation de lrsquohegraveme se preacutesentent sous la forme drsquoun opeacuteron lrsquoopeacuteron
hemAXCDBL Chacun des gegravenes est essentiel (sauf hemL) et la deacuteleacutetion drsquoun seul
drsquoentre eux empecircchera la formation de lrsquohegraveme moleacutecule essentielle agrave la formation
des cytochromes (Taber 1993) hemB code pour la porphobilinogene synthase qui
catalyse une eacutetape dans la voie de biosynthegravese de lrsquohegraveme (Kohler et al 2008 Taber
- 15 -
1993 Thoumlny-Meyer 1997) Une mutation dans le gegravene ctaA (laquo hegraveme A synthase raquo)
empecircche la biosynthegravese de lrsquohegraveme A lequel est aussi impliqueacute dans la biosynthegravese
des cytochromes et donc altegraverera le fonctionnement de la chaine de transport des
eacutelectrons (Clements et al 1999 Proctor et Peters 1998) La capaciteacute de la bacteacuterie
agrave biosyntheacutetiser de lrsquohegraveme de faccedilon endogegravene est essentielle afin drsquoinitier une
infection systeacutemique Le gegravene menD code pour la 2-succinyl-6-hydroxy-24-
cyclohexadiene-1-carboxylate syntase (SHCHC) qui est une enzyme essentielle agrave la
voie de biosynthegravese des meacutenaquinones (Kohler et al 2008) De plus les
meacutenaquinones sont eacutegalement requises pour le fonctionnement de plusieurs
enzymes impliqueacutees dans des voies meacutetaboliques essentielles telles la malate quinol
oxidoreacuteductase (Mqo2) et la dihydroorotate deacuteshydrogeacutenase (PyrD) (Kohler et al
2008) Les mutants menD seront donc incapables de convertir lrsquooxaloaceacutetate en
malate ducirc agrave la perte de lrsquoactiviteacute du Mqo2 (Kohler et al 2008 Proctor et Peters
1998)
Les bacteacuteries agrave pheacutenotype SCV deacutemontreront eacutegalement un gradient eacutelectrochimique
nettement plus faible que les bacteacuteries agrave pheacutenotype normal de par lrsquoincapaciteacute de la
chaine de transport des eacutelectrons agrave y pourvoir
De par leur chaine de transport des eacutelectrons interrompue les SCV sont incapables
drsquoutiliser lrsquoO2 et le nitrate comme accepteur final drsquoeacutelectrons (Kohler et al 2008) Par
ce fait les proteacuteines impliqueacutees dans la voie de la glycolyse ainsi que dans la voie de
la fermentation sont surexprimeacutees (Pfl Ldh Adh) alors que les enzymes faisant
partie du cycle du citrate (TCA) sont significativement reacuteguleacutees agrave la baisse (Kohler et
al 2003 Kohler et al 2008 Kriegeskorte et al 2011) Ceci indique clairement que
les SCV (ex hemB) geacutenegraverent leur ATP agrave partir du glucose ou fructose seulement par
la phosphorylation des substrats Lrsquoanalyse de la reacuteaction de fermentation indique
que le produit principal est le lactate Les SCV reacutegeacutenegraverent leur NADH en reacuteduisant le
pyruvate en lactate par la voie fermentaire (Kohler et al 2003 Kohler et al 2008)
- 16 -
emmenant ainsi une acidification du milieu (Kriegeskorte et al 2011) La conversion
du pyruvate en aceacutetate via lrsquoacetyl-CoA est restreinte chez les mutants hemB mecircme
si la pyruvate formate lyase (Pfl) et les composantes du pyruvate deacuteshydrogeacutenase
sont preacutesentes (Figure 7) Lrsquoactiviteacute de la pyruvate deacuteshydrogeacutenase est tregraves faible
probablement en lien avec le haut niveau de NADH chez les SCV et il est preacutesumeacute
que la Pfl nrsquoest pas activeacutee en preacutesence drsquooxygegravene (La pyruvate deacuteshydrogeacutenase
converti le pyruvate en Co-A en condition anaeacuterobie) (Figure 7) Chez les mutants
hemB la voie de lrsquoarginine deacuteiminase est aussi induite et pourrait eacutegalement
participer agrave la production drsquoATP (Kohler et al 2003) Les gegravenes exprimeacutes chez les
mutants hemB sont similaires agrave ceux exprimeacutes chez les mutants menD ainsi que
chez les souches agrave pheacutenotypes normaux en condition anaeacuterobie (Kohler et al 2008)
- 17 -
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Figure 7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies Les enzymes suivantes sont retrouveacutees agrave une forte
concentration chez les mutants hemb phosphoglucomutase (Pgi) fructose
bisphosphate aldolase (FbaFda) glyceacuteraldeacutehyde deacuteshydrogeacutenase (Gap)
phosphoglyceacuterate kinase (Pgk) eacutenolase (Eno) pyruvate kinase (Pyk) L-lactate
deacuteshydrogeacutenase (L-Ldh) pyruvate formate lyase (PflB) alcool deacuteshydrogeacutenase
(Adh) La formation drsquoaceacutetate et de formate et tregraves limiteacutee (X rouges)
Modifieacute de Kolher C et al 2003
- 18 -
La chaine respiratoire en anaeacuterobie (S aureus)
Il existe 3 voies distinctes de geacuteneacuteration drsquoeacutenergie chez les bacteacuteries 1- en preacutesence
drsquooxygegravene la bacteacuterie respire de faccedilon aeacuterobique et lrsquooxygegravene est lrsquoaccepteur final
drsquoeacutelectron 2- la bacteacuterie peut aussi respirer de maniegravere anaeacuterobique lorsqursquoun
accepteur terminal drsquoeacutelectron autre que lrsquooxygegravene est preacutesent 3- en absence
drsquooxygegravene et drsquoun accepteur drsquoeacutelectrons alternatif la bacteacuterie peut produire de
lrsquoeacutenergie de faccedilon limiteacutee par fermentation (Hammer et al 2013)
Chez S aureus le nitrate (NO3-) et dans un troisiegraveme temps le nitrite (NO2
-)
peuvent ecirctre utiliseacutes comme accepteur final drsquoeacutelectron alternatif lorsque lrsquooxygegravene
nrsquoest pas disponible (respiration anaeacuterobie) Les enzymes de cette voie meacutetabolique
ne sont retrouveacutees que chez les bacteacuteries (Goumltz et al 2006) La croissance
bacteacuterienne est consideacuterablement reacuteduite lorsque la bacteacuterie passe drsquoune respiration
aeacuterobie agrave une respiration anaeacuterobie La nitrate reacuteductase fonctionne probablement in
vivo gracircce au L-lactate comme principal donneur drsquohydrogegravene (Burke et Lasccelles
1975) La deacuteshydrogeacutenase primaire pour ce substrat est preacutesumeacutement la L-lactate
deacuteshydrogeacutenase associeacutee agrave la membrane laquelle meacutedie lrsquooxydation du lactate par la
chaine respiratoire aeacuterobique (Burke et Lasccelles 1975) La preacutedominance du
lactate parmi les produits finaux de la fermentation par S aureus est correacuteleacutee avec
son efficaciteacute comme principal donneur drsquohydrogegravene lorsque le nitrate est lrsquoaccepteur
final drsquoeacutelectrons Le transfert des eacutelectrons du Lactate vers le nitrate implique des
composantes qui sont eacutegalement preacutesentes dans la chaine respiratoire La nitrate
reacuteductase du type respiratoire est habituellement associeacutee agrave la membrane
cytoplasmique (Burke et Lasccelles 1975) Les reacuteductases sont inhibeacutees par
lrsquoazoture et le cyanure (Burke et Lascelles 1976)
Lrsquoenzyme membranaire nitrate reacuteductase NarGHI geacutenegravere des nitrites lesquels seront
reacuteduits en ammonium (NH4+) par lrsquoenzyme cytoplasmique NADH-deacutependant nitrite
reacuteductase encodeacutee par les gegravenes nirBD (Goumltz et al 2006) La nitrate reacuteductase 1-
- 19 -
est reacuteprimeacutee et inhibeacutee par la preacutesence drsquooxygegravene 2- est induite par lrsquoanaeacuterobiose et
la preacutesence de nitrate ou nitrite 3- nrsquoest pas inhibeacutee par lrsquoammoniac 4- permet la
production drsquoeacutenergie dans des conditions anaeacuterobies et 5- est localiseacutee dans la
membrane (Goumltz et al 2006) La preacutesence drsquoun systegraveme de senseur pour lrsquooxygegravene
est cruciale pour le controcircle transcriptionnel des gegravenes facilitant lrsquoadaptation de la
bacteacuterie agrave lrsquoanaeacuterobiose ou aux conditions microaeacuterophiles Chez S aureus il existe
deux systegravemes de senseur 1- le systegraveme SrrAB (pour staphylococcal respiratory
response AB synonyme SrhSR) (Figure 8) homologue agrave ResDE chez Bacillus
subtilis (Yan et al 2011) et 2- le systegraveme NreABC (pour nitrogen regulation chez S
carnosus correspondant agrave SA2181 (NreA) SA2180 (NreB) SA2179 (NreC) chez S
aureus) NreBC est un systegraveme agrave deux composantes qui mesure la concentration
drsquooxygegravene directement par un groupement Fe-S de type Fnr lequel est instable en
preacutesence drsquooxygegravene et par lequel est controcircleacute le systegraveme membranaire de reacuteduction
du nitratenitrite (Yan et al 2011) Srr est aussi un systegraveme agrave deux composantes qui
reacutegule positivement la lactate deacuteshydrogeacutenase et lrsquoalcool deacuteshydrogeacutenase et semble
reacuteguler neacutegativement plusieurs enzymes du cycle du citrate (TCA) (Figure 8)
Lrsquoexpression des enzymes fermentaires comme la lactate deacuteshydrogeacutenase (LDH)
pyruvate formate lyase (PFL) et lrsquoalcool deacuteshydrogeacutenase sont aussi fortement induite
dans des conditions aeacuterobies lorsque la chaine de transport des eacutelectrons est
interrompue (Kohler et al 2003) Donc il se peut que lrsquoeacutetat reacuteduit des composantes
de la chaine respiratoire le potentiel membranaire etou le niveau de NADH soit un
signal pour la reacutegulation des gegravenes laquo anaeacuterobies raquo chez S aureus Dans des
conditions aeacuterobies la conversion du pyruvate en aceacutetyl-coA est catalyseacutee par le
complexe pyruvate deacuteshydrogeacutenase (PDH) et caracteacuteriseacutee par la production de
NADH (Somerville et Proctor 2009) En condition anaeacuterobie lrsquoexpression de la
pyruvate deacuteshydrogeacutenase (PDH) est fortement reacuteprimeacutee alors que lrsquoexpression de la
pyruvate formate lyase (PFL) est fortement induite Lrsquoactiviteacute de la PDH est
remplaceacutee en anaeacuterobiose par lrsquoactiviteacute de la PFL qui geacutenegravere lrsquoacetyl-CoA et le
formate agrave partir du pyruvate Le NADH nrsquoeacutetant pas reacuteoxydeacute par une chaine de
- 20 -
transport des eacutelectrons fonctionnelle celui-ci est recycleacute par la reacuteduction du pyruvate
en lactate (Kohler et al 2003 Somerville et Proctor 2009)
La meacutenaquinone et lrsquohegraveme sont esssentiels agrave la respiration anaeacuterobie (Kohler et al
2008) Le pheacutenotype de S aureus en condition anaeacuterobie est tregraves similaire au
pheacutenotype SCV (Somerville et Proctor 2009)
Figure 8 Modegravele de reacutegulation du cycle TCA (Pour simplifier les reacutegulateurs de
facteurs de virulence bien caracteacuteriseacutes [ie RNAIII Rot SarA et σB] ont eacuteteacute omis
dans ce modegravele)
Les sous-produits de la phosphorylation oxydative les ROS
La reacuteduction de lrsquooxygegravene en eau se fait un eacutelectron agrave la fois (Turrens 2003) Le
complexe IV retient tous les intermeacutediaires partiellement reacuteduits jusqursquoagrave ce que la
Tireacute de Somerville GA Proctor RA 2009
- 21 -
reacuteduction soit complegravetement termineacutee (Turrens 2003) Cependant certains autres
centres redox de la chaine respiratoire laissent passer des eacutelectrons agrave lrsquooxygegravene
reacuteduisant partiellement cette moleacutecule en anion superoxyde (O2-bull) (Liu et al 2002
Turrens 2003) Mecircme si O2-bull nrsquoest pas un agent oxydant tregraves puissant il est le
preacutecurseur de plusieurs autres laquo reactive oxygen species raquo (ROS) et il est aussi
impliqueacute dans la propagation de la chaine de reacuteaction oxydative (Turrens 2003)
LrsquoO2-bull est un intermeacutediaire instable qui est rapidement converti en H2O2 par les
superoxide dismutase (SOD) (S aureus en possegravede deux SodA (le principal) et
SodM) (10 18) ou se convertit de faccedilon spontaneacutee lorsqursquoen concentration eacuteleveacutee
(Starkov 2010 Turrens 2003) Le H2O2 peut ensuite ecirctre complegravetement reacuteduit en
H2O ou partiellement reacuteduit en radical hydroxyle (bullOH) un des plus puissants
oxydants (Starkov 2010) LrsquoO2-bull peut eacutegalement reacuteagir avec drsquoautres radicaux
incluant lrsquooxyde nitrique (NObull) selon le taux de diffusion des 2 radicaux (Turrens
2003) Le peroxynitrite est eacutegalement un oxydant puissant (Turrens 2003) Les
oxydants deacuterivant du NObull sont appeleacutes des laquo reactive nitrogen species raquo (RNS)
(Turrens 2003) mais seront consideacutereacutes comme faisant partie des ROS dans ce
travail Les ROS peuvent endommager lrsquoADN lrsquoARN les proteacuteines et les lipides
reacutesultants agrave lrsquoapoptose de la cellule lorsque le niveau de ROS produit excegravede la
capaciteacute de deacutetoxifications et de reacuteparation de la cellule (Brynildsen et al 2013
Wells et al 2005) Malgreacute ce danger il srsquoagit drsquoun risque compenseacute par
lrsquoaugmentation du rendement eacutenergeacutetique (Brynildsen et al 2013) Le complexe I (et
II) et III sont les principaux sites de production de ROS (Batandier et al 2006 Chen
et al 2003 Liu et al 2002) Lrsquoinhibition du complexe I par la rotenone provoque une
augmentation de la production de ROS Lrsquooxydation des substrats du complexe I ou
du complexe II en preacutesence du complexe III inhibeacute par lrsquoantimicine A amegravene
eacutegalement une augmentation de la production de ROS (Batandier et al 2006 Chen
et al 2003) (Figure 9) De plus mecircme si le complexe cytochrome oxydase nrsquoest pas
un site de production de ROS son inhibition par le KCN (cyanure de potassium)
facilite la production de ROS par les complexes I et II (Chen et al 2003) Le
- 22 -
complexe ATP synthase nrsquoest pas reconnu pour produire directement des ROS
Cependant la liaison drsquoun inhibiteur tel lrsquooligomycine bloque le passage des protons
agrave travers le complexe et inhibe ainsi la production drsquoATP De plus lrsquoATP synthase ne
fonctionnant plus les pompes agrave protons de la chaine de transport des eacutelectrons sont
incapables de fonctionner car le gradient de protons est beaucoup trop eacuteleveacute ce
provoque une surproduction de ROS par les complexes I et III Conseacutequemment le
NADH nrsquoest plus oxydeacute et le cycle du TCA cesse drsquoopeacuterer car la concentration de
NAD nrsquoest plus suffisante pour le fonctionnement des enzymes du TCA (Joshi et
Huang 1991)
Une production significative de O2-bull principalement par le complexe I peut ecirctre le
reacutesultat de deux modes drsquoopeacuteration 1- lorsque la cellule ne produit pas drsquoATP et
conseacutequemment possegravede une grande force proton-motrice et possegravede un laquo pool raquo de
quinone agrave lrsquoeacutetat reacuteduit (QH2) 2- lorsque la cellule possegravede un haut ratio de
NADHNAD (Figure 10) Lorsque la cellule produit activement de lrsquoATP elle possegravede
alors une moins grande force proton-motrice son ratio NADHNAD est plus faible et
la formation de O2-bull est beaucoup plus faible (Murphy 2009)
Le H2O2 est le substrat pour deux enzymes la catalase et la glutathione (GSH)
peroxydase qui convertissent le H2O2 en H2O+O2 le H2O2 est ainsi deacutetoxifieacute (Figure
9 Figure 11) La reacuteaction catalyseacutee par la glutathione peroxydase neacutecessite de la
glutathione (GSH) comme substrats et deacutepend en partie du ratio de GSSG GSH et
de la concentration des reacuteactants laquelle est le reflet de lrsquoeacutetat reacutedox de la cellule De
faccedilon similaire la concentration de meacutetaux sous leur forme reacuteduite tel le Fer catalyse
la formation de certains ROS (Figure 11) Ceci peut ecirctre minimiseacute en tenant cette
concentration ions meacutetallique tregraves basse entre autres par leur emmagasinage et leur
liaison aux proteacuteines de transport (ei ferritine transferrine lactoferrine) et ainsi
minimiser la formation de bullOH Finalement certains antioxydants (eg vitamine E)
peuvent interrompre la chaine de reacuteactions par la capture des radicaux le radical-
vitamine E est relativement stable et peut ecirctre enzymatiquement reconverti agrave sa
- 23 -
forme non-radical (Figure 11) Il est maintenant accepteacute que certaines enzymes du
cycle de Krebs (ie αKGDH) soient non seulement des cibles des ROS mais aussi
des producteurs non neacutegligeables de ROS (Brynildsen et al 2013)
Figure 9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS Le complexe I ainsi que le site Qi du complexe III relacircchent des
ROS directement vers les deacutefenses drsquoantioxydants intracellulaires Les antioxidants
intracellulaire tel la superoxide dismutase (MnSOD) la catalase la glutathione (GSH)
la glutathione peroxydase (GPX) et la glutathione reacuteductase (GRase) sont montreacute Le
site Qo du complexe III quant agrave lui relacircche les ROS dans le milieu extra cellulaire
Modifieacute de Chen Q et al 2003
Oligomycine
- 24 -
Figure 10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de
O2-bull 1- Le pool de NADH est dans un eacutetat reacuteduit par exemple par des dommages agrave
la chaine respiratoire la perte de cytochrome c durant lrsquoapoptose ou agrave un faible
besoin en ATP Ceci megravene agrave une production de O2-bull au complexe I qui deacutecoule de la
reacuteduction du FMN laquelle est reacuteguleacutee par le ratio NADHNAD+ Drsquoautres sites
autres que le complexe I tel lrsquoα-ceacutetoglutatrate deacuteshydrogeacutenase (αKGDH) peut
aussi contribuer agrave la production de O2-bull 2- Il nrsquoy a pas (peu) de production drsquoATP il y
a un fort potentiel membranaire et un pool de CoQ (meacutenaqinone) agrave lrsquoeacutetat reacuteduit
important qui provoque une reacuteversion de la chaine de transport des eacutelectrons agrave
travers le complexe I amenant une forte production de O2-bull 3- Il y a une production
active drsquoATP et conseacutequemment un potentiel membranaire plus bas que dans la
situation 2 et une plus grande quantiteacute de NADH oxydeacute que dans la situation 1
Dans ces conditions la quantiteacute de O2-bull produit est plus faible que dans la situation
1 et 2 et la provenance des O2-bull est moins certaine
Tireacute de Murphy MP 2009
- 25 -
12 Antibiotiques Mode drsquoaction cible cellulaire et meacutecanismes de reacutesistance
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles
Notre compreacutehension de la faccedilon que les antibiotiques induisent la mort cellulaire est
principalement baseacutee sur la fonction cellulaire essentielle inhibeacutee par lrsquointeraction de
lrsquoantibiotique avec sa cible Les antibiotiques peuvent ecirctre classeacutes selon la
composante ou le systegraveme cellulaire qursquoils affectent ainsi que par le fait qursquoils
induisent la mort cellulaire (antibiotiques bacteacutericides) ou srsquoils inhibent la croissance
Figure 11 Voies de formation et de deacutetoxification des ROS Tireacute de Wikipeacutedia
httpfrwikipediaorgwikiDC3A9rivC3A9_rC3A9actif_de_l27oxygC3A8ne
Consulteacute le 11 deacutec 2013
- 26 -
bacteacuterienne (antibiotique bacteacuteriostatique) La plupart des antibiotiques bacteacutericides
inhibent la synthegravese de lrsquoADN ou de lrsquoARN la synthegravese de la paroi bacteacuterienne ou la
synthegravese proteacuteique (Kohanski et al 2010) Le tableau 1 est une synthegravese des
principales classes drsquoantibiotiques ainsi que leur mode drsquoaction et leurs cibles
cellulaires Ce tableau nrsquoest cependant pas exhaustif et il existe plusieurs autres
classes drsquoantibiotiques et plusieurs autres exemples drsquoantibiotiques dans les classes
identifieacutees dans ce tableau
Depuis la deacutecouverte de la peacutenicilline en 1929 plusieurs autres antibiotiques ont eacuteteacute
deacutecouverts et deacuteveloppeacutes suite agrave lrsquoeacutelucidation des interactions entre lrsquoantibiotique et
sa cible et en apportant des modifications structurales et chimiques aux moleacutecules
deacutejagrave connues Ces efforts ont significativement augmenteacute le nombre drsquoantibiotiques
disponibles dans le milieu clinique Le processus de mort cellulaire induit par lrsquoaction
drsquoun antibiotique est complexe et deacutebute avec une interaction physique entre la
moleacutecule antibiotique et sa cible bacteacuterienne speacutecifique et implique des modifications
qui affectent la bacteacuterie tant au niveau biochimique que moleacuteculaire et structural
27
Acid
e n
alid
ixiq
ue (
1 iegravere
geacuteneacutera
tion)
Cip
rofloxacin
e (
2 iegravem
e g
eacuteneacutera
tion)
Levofloxacin
e (
3 iegravem
e g
eacuteneacutera
tion)
Gem
iflo
xacin
e (
4 iegravem
e g
eacuteneacutera
tion)
Flu
oro
quin
olo
ne (
Norf
loxacin
e)
Peacutenic
iline
Ceacutephalo
sporine (
ceacutefa
lexin
e)
Carb
apenem
Vancom
ycin
e
Teacutela
vancin
e
Fosfo
mycin
Dapto
mycin
Poly
mix
ine B
Bacitra
cin
Valin
om
ycin
e
Nom
encin
e
Genta
mic
in
Kanam
ycin
e
Tobra
mycin
e
Str
epto
mycin
e
Teacutetr
acyclin
e
Chlo
rteacutetr
acyclin
e
Macro
lides
Ery
thro
mycin
eLie
sous-u
niteacute 23S
du r
ibosom
e 5
0S
et em
pecircche
leacutelo
ngation d
u p
eptide (
apregrave
s 6
-8 a
a)
sta
tique
Lin
cosam
ides
Clin
dam
ycin
Inhib
e d
irecte
ment la
ctiviteacute p
eptidyl
transfeacute
rase d
u
ribosom
esta
tique
Str
epto
gra
min
es
DalfopristinQ
uin
upristin
Com
bin
ais
on d
e 2
antibio
tiques B
loque la
transla
tion p
eptidiq
ue e
n lia
nt le
site P
du r
ibosom
e
et en b
loquant au p
eptide la
ccegraves a
u tunnel de s
ort
ie
cid
al
Am
phenic
ol
Chlo
ram
phenic
ol
Em
pecircche la lia
ison d
u a
a-
AR
Nt au s
ite A
du
ribosom
e e
t in
terf
egravere
dans la f
orm
ation d
u lie
n
peptidiq
ue
sta
tique
Oxazolid
inones
Lin
ezolid
Blo
que initia
tion d
e la tra
nsla
tion p
roteacute
ique lors
de la
reacuteaction d
eacutelo
ngation p
ar
la p
eptidyl
transfeacute
rase
sta
tique
Trim
eth
oprim
Sulfam
eth
oxazole
sta
tique
cid
al
Ionophore
Alteacutera
tion d
e lin
teacutegriteacute
mem
bra
naire
Se
nchasse d
ans la m
em
bra
ne fo
rme u
n p
ore
ce
qui am
egravene u
ne p
ert
urb
ation d
e leacute
quili
bre
ioniq
ue
cid
al
Inhib
itio
n d
e la s
ynth
egravese d
e
la p
aro
i cellu
laire
(peptidogly
can)
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
cid
al
cid
al
sta
tique
AR
Nr
16S
(S
ite A
)(fo
rmation d
e p
roteacute
ines n
on-
fonctionnelle
s)
cid
al
Inhib
itio
n d
e la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 3
0S
)
Inhib
iteur
de la s
ynth
egravese d
e
lacid
e f
oliq
ue
Inhib
itio
n d
e c
ert
ain
es r
eacuteactions e
nzym
atiques
essentielle
s agrave
la f
orm
ation d
e la
cid
e f
oliq
ue
Sulfonam
ides
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
AR
Nr
16S
(preacute
vie
nt la
ttachem
ent de la
min
oacyl-
AR
Nt au r
ibosom
e)
Teacutetr
acyclin
e
Poly
peptides
β-L
acta
ms
Gly
copeptides
Lip
opeptides
Inhib
iteur
de la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 5
0S
)
Am
inogly
cosid
es
Cib
le c
ell
ula
ire
Sta
tiq
ue
Cid
al
Rifam
ycin
eR
ifam
pic
ine
Cla
ss
es
Ex
d
an
tib
ioti
qu
eM
od
e d
ac
tio
n
Inhib
itio
n d
e la s
ynth
egravese d
e
lAR
N (
AD
N d
eacutependant)
AR
N p
oly
meacutera
se s
ous-u
niteacute β
(codeacute p
ar
rpoB
)
Quin
olo
nes
cid
al
Inhib
itio
n d
e la r
eacuteplic
ation d
e
lAD
N
Com
ple
xe A
DN
-topois
om
eacutera
se (
AD
N
gyr
ase(t
opois
om
eacutera
se II)
et to
pois
om
eacutera
se IV
)cid
al
Tra
nspeptidase (
penic
illin
bin
din
g p
rote
in
(PB
P))
cid
al
Tab
leau
1
Cib
les e
t m
od
e d
rsquoacti
on
des p
rin
cip
ale
s c
lasses d
rsquoan
tib
ioti
qu
es
28
122 Les aminoglycosides
1221 Mode drsquoaction
Les aminoglycosides peacutenegravetrent la paroi cellulaire en suivant un processus
comprenant 3 eacutetapes une premiegravere eacutetape eacutenergie- indeacutependante suivie de deux
eacutetapes eacutenergie deacutependantes Deux composantes sont essentielles pour
lrsquoaccumulation intracellulaire des aminoglycosides les ribosomes et la chaine de
transport des eacutelectrons (Ramirez et Tolmasky 2010)
Eacutetape 1- Lorsque les aminoglycosides entrent en contact avec la paroi
cellulaire la moleacutecule polycationique se lie aux composantes de surface anioniques
tels les lipopolysaccharides les phospholipides et autres proteacuteines membranaires
chez les bacteacuteries agrave Gram neacutegatif et les moleacutecules drsquoacides teacuteichoiumlques et les
phospholipides des bacteacuteries agrave Gram positif (Ramirez et Tolmasky 2010) Apregraves
cette liaison aux moleacutecules polaires les aminoglycosides (telle la gentamicine)
srsquoassocient aux transporteurs membranaires sur la base de leur charge positive et
sont transporteacutes agrave travers la membrane cytoplasmique gracircce au potentiel
membranaire (intra cellulaire neacutegatif) (Bryan et al 1980 Proctor et al 2006) Cette
liaison agrave la membrane entraine une augmentation de la permeacuteabiliteacute membranaire
menant agrave lrsquoentreacutee des aminoglycosides dans lrsquoespace peacuteriplasmique (Gram neacutegatif)
ou dans le cytoplasme (Gram positif) (Ramirez et Tolmasky 2010) (Figure 12A)
Eacutetape 2- Lrsquoeacutetape suivante connue comme la laquo phase eacutenergie-deacutependante
1 raquo (EDP-I) est bloqueacutee par les inhibiteurs de la chaine de transport des eacutelectrons et
de la phosphorylation oxydative ainsi que par les ions divalents lrsquohyperosmolariteacute le
faible pH et lrsquoanaeacuterobiose dans cette phase la dureacutee et le taux drsquoentreacutee de
lrsquoantibiotique sont en fonction de la concentration (Miller et al 1980 Mingeot-
Leclercq et al 1999 Ramirez et Tolmasky 2010) Agrave cette eacutetape le transport
transmembranaire des aminoglycosides et plus speacutecifiquement de la gentamicine
neacutecessite 2 composantes majeures a) un transporteur anionique non speacutecifique dont
la charge est directement lieacutee agrave lrsquoactiviteacute des cytochromes oxydases terminales et des
29
nitrates reacuteductases de la chaine de transport des eacutelectrons Selon certaines eacutetudes
les transporteurs pourraient ecirctre les quinones reacuteduites de la chaine de transport des
eacutelectrons (Bryan et al 1981 Miller et al 1980) et donc le cycle drsquooxydation-reacuteduction
des quinones est essentiel aux transports des aminoglycosides (Bryan et al 1981)
mais selon drsquoautres eacutetudes les transporteurs pourraient ecirctre toute autre proteacuteine non
deacutefinie impliqueacutee dans la polarisation membranaire lieacutee agrave lrsquoactiviteacute de la chaine de
transport des eacutelectrons (Miller et al 1980) La croissance anaeacuterobie ainsi que le
sodium azide (inhibiteur de la chaine respiratoire) inhibent lrsquoentreacutee de la gentamicine
et lrsquoeffet bacteacutericide indiquant que le transport actif de cet antibiotique neacutecessite la
respiration b) un potentiel eacutelectrique avec un milieu intra cellulaire neacutegatif par rapport
agrave lrsquoexteacuterieur En effet lrsquoinhibition de lrsquoentreacutee dans la bacteacuterie des aminoglycosides
suite agrave lrsquoajout de CCCP (agent deacutepolarisant) chez des souches de S aureus laquo wild-
type raquo deacutemontre la neacutecessiteacute de la force proton-motrice pour lrsquoentreacutee des
aminoglycosides dans les cellules (Miller et al 1980)
Eacutetape 3- La petite quantiteacute drsquoaminoglycosides srsquoeacutetant rendu au cytoplasme
durant cette premiegravere phase eacutenergie-deacutependante induisent apregraves liaison aux
ribosomes des erreurs dans la synthegravese proteacuteique Les erreurs dans la traduction
des proteacuteines membranaires produites lors de cette eacutetape connue comme la laquo phase
eacutenergie-deacutependante 2 raquo (EDP-II) provoquent des dommages dans lrsquointeacutegriteacute de la
membrane cytoplasmique (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky
2010) Les proteacuteines membranaires aberrantes dans la membrane facilitent le
transport drsquoune plus grande quantiteacute drsquoaminoglycosides lesquels augmenteront
encore le niveau drsquointerfeacuterence avec la synthegravese proteacuteique normale amenant encore
plus de dommage agrave la membrane provoquant ainsi une acceacuteleacuteration importante du
taux drsquoentreacutee des moleacutecules antibiotique et qui ultimement reacutesultera en la mort de la
cellule (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky 2010) (Figure 12B)
Des eacutetudes ont deacutemontreacute que la susceptibiliteacute agrave la gentamicine streptomycine
amikacine kanamycine et tobramycine est supeacuterieure lorsque lrsquooxygegravene est
30
lrsquoaccepteur terminal drsquoeacutelectrons (Bryan et Kwan 1981) Une diminution significative
de la susceptibiliteacute est remarqueacutee lorsque le nitrate est lrsquoaccepteur final drsquoeacutelectrons et
une diminution encore plus marqueacutee est observeacutee lorsque le fumarate est lrsquoaccepteur
final drsquoeacutelectron ou lorsque les conditions sont fermentaires Lrsquoassociation entre le
transport de la gentamicine et le complexe terminal de la chaine de transport des
eacutelectrons particuliegraverement lrsquoactiviteacute cytochrome oxydase
Le ribosome possegravede trois sites de liaison importants de lrsquoARNt deacutesigneacute comme site
A (pour aminoacyl) site P (pour peptidyl) et E (pour exit) (Vakulenko et Mobashery
2003) Ils sont situeacutes sur lrsquoARN16S (lequel avec plus de 21 proteacuteines compose la
sous-uniteacute 30S du ribosome bacteacuterien) (Ramirez et Tolmasky 2010 Vakulenko et
Mobashery 2003) Durant la synthegravese proteacuteique le ribosome deacutecode lrsquoinformation de
lrsquoARNm et catalyse lrsquoincorporation seacutequentielle de chaque acide amineacute faisant partie
de la chaine polypeptidique en formation La haute fideacuteliteacute de la traduction (les
ribosomes incorporent un mauvais acide amineacute par approximativement 3000 liaisons
peptidiques effectueacutees) est possible gracircce agrave la capaciteacute de discriminer entre des
changements conformationels dans le ribosome induit par la liaison drsquoun ARNt correct
Modifieacute de Davis BD 1987
Figure 12 Cineacutetique drsquoentreacutee des aminoglycosides
A
B
31
ou incorrect au site A La reconnaissance drsquoune liaison correcte anticodoncodon par
le ribosome se passe en deux temps la reconnaissance initiale de lrsquoappariement du
codon sur lrsquoARNm avec lrsquoanticodon de lrsquoARNt et subseacutequemment la correction
(Vakulenko et Mobashery 2003)
La plupart des aminoglycosides se lient agrave des oligonucleacuteotides faisant partie du site
A Les diffeacuterents aminoglycosides ne lient pas tous le mecircme site de lrsquoARN 16S mais
lrsquoeffet commun agrave tous est que cette liaison amegravene un changement de conformation du
site A vers une conformation mimant la conformation fermeacutee de la sous uniteacute 30S qui
est induite par une interaction entre lrsquoARNm et son ARNt apparenteacute eacuteliminant la
possibiliteacute de correction du ribosome ce qui promeut les erreurs de traduction
(Ramirez et Tolmasky 2010 Vakulenko et Mobashery 2003) La grande majoriteacute des
aminoglycosides sont bacteacutericides et cette leacutetaliteacute est la conseacutequence de cette
mauvaise traduction (Vakulenko et Mobashery 2003)
1222 Meacutecanismes de reacutesistance
La reacutesistance contre les aminoglycosides se produit par diffeacuterents meacutecanismes qui
peuvent coexister simultaneacutement dans la mecircme bacteacuterie (Vakulenko et Mobashery
2003) Ces meacutecanismes incluent 1- la modification de la cible par des mutations
dans lrsquoARN 16S ou autres proteacuteines ribosomales associeacutees (Figure 13) 2- la
meacutethylation de lrsquoARN 16S un meacutecanisme retrouveacute chez plusieurs organismes
producteurs drsquoaminoglycosides et chez des souches cliniques Cette meacutethylation
empecircche les aminoglycosides drsquoavoir accegraves agrave leur site de liaison (Doi et Arakawa
2007) 3- la reacuteduction de la permeacuteabiliteacute par une alteacuteration de la permeacuteabiliteacute de la
membrane externe ou une diminution du transport agrave travers la membrane interne 4-
lrsquoexportation des aminoglycosides agrave lrsquoexteacuterieur de la cellule par les pompes agrave efflux
Eacutetant des moleacutecules hydrophiliques les aminoglycosides ont naturellement peu
drsquoaffiniteacute avec les pompes agrave efflux 5- le swarming actif probablement un meacutecanisme
32
non speacutecifique deacutemontreacute chez Pseudomonas aeruginosa lequel deacutemontre des
reacutesistances agrave plusieurs antibiotiques 6- la seacutequestration de la drogue par des liaisons
agrave un aceacutetyltransfeacuterase agrave tregraves faible activiteacute 7- une inactivation enzymatique de la
moleacutecule antibiotique le meacutecanisme de reacutesistance le plus preacutevalent au niveau
clinique (Vakulenko et Mobashery 2003) Lrsquoinactivation enzymatique des
aminoglycosides est le plus souvent due agrave des aceacutetyltransfeacuterases des
nucleacuteotidyltransfeacuterases ou des phosphotransfeacuterases (Dickgiesser et Kreiswirth 1986)
Ces enzymes sont souvent encodeacutees sur des plasmides mais sont eacutegalement
associeacutees agrave des eacuteleacutements transposables Lrsquoeacutechange de plasmides et la disseacutemination
de transposons facilitent lrsquoacquisition rapide de pheacutenotypes reacutesistants aux
antibiotiques non seulement au sein drsquoune mecircme espegravece mais parmi un large
eacuteventail drsquoespegraveces bacteacuteriennes (Mingeot-Leclercq et al 1999)
La reacutesistance naturelle des S aureus laquo small colony variants raquo agrave la majoriteacute des
aminoglycosides est la conseacutequence de lrsquoimpossibiliteacute agrave lrsquoantibiotique de peacuteneacutetrer une
cellule posseacutedant une faible polariteacute membranaire qui est due chez les SCV agrave leur
Tireacute de Doi Y Arakawa Y 2007
Figure 13 Sites de mutation de certains reacutesistants aux aminoglycosides
33
incapaciteacute agrave activer le cycle de Krebs (Miller et al 1980 Proctor et al 2006 Tubby
et al 2013) Les SCV ont non seulement une polariteacute membranaire faible mais (ceci
eacutetant la conseacutequence de cela) aussi une force proton motrice inhibeacutee et une chaine
des eacutelectrons deacuteficiente De ce fait le transport transmembranaire des
aminoglycosides chez les SCV est grandement reacuteduit et conseacutequemment la
susceptibiliteacute des SCVs pour cette classe drsquoantibiotique est 110-113 de celle des
souches parentes (Proctor et al 2006)
123 La tomatidine
Plusieurs espegraveces veacutegeacutetales accumulent des steacuterols et des triterpegravenes antimicrobiens
appeleacutes saponines (Osborn 1996) Ces moleacutecules sont produites de faccedilon
constitutive par la plante et jouent un important rocircle de deacutefense contre diffeacuterents
pathogegravenes comme des insectes ou des microbes Parmi les saponines la tomatine
(Figure 14A) est un glycoalcaloiumlde steacuteroiumldien preacutesentant une action antimicrobienne
contre des levures et une quantiteacute drsquoautres microorganismes (Bednarek et Osboum
2009 Friedman 2002 Osborn 1996 Sandrock et Vanetten 1998) Plusieurs
espegraveces de champignons pathogegravenes de la tomate produisent des enzymes
extracellulaires communeacutement appeleacutes tomatinase qui sont capables de deacutetoxifier
lrsquoα-tomatine (Martin-Hernandez et al 2000 Roddick 1974 Ruiz-Rubbio et al 2001)
Par exemple Fusarium oxysporum f sp lycopersici est reconnu pour cliver lrsquoα-
tomatine en ses sous-produits soit un tetrasaccharide (lycotreacutetraose) et un alcaloiumlde
steacuteroiumldien non glycosyleacute la tomatidine (Figure 14B) Ces sous-produits nrsquoont pas ou
peu drsquoactiviteacute antifongique (Ruiz-Rubbio et al 2001 Simon et al 2006)
34
Figure 14 Structures chimiques de la (A) tomatine de (B) tomatidine et de (C)
lrsquoanalogue de tomatidine FC04-100
Il a eacuteteacute deacutecouvert il a quelques anneacutees que la tomatidine avait un effet antibacteacuterien
sur les S aureus de pheacutenotype SCV mais aucune activiteacute contre les S aureus agrave
pheacutenotype normal Par contre lorsqursquoutiliseacute en combinaison avec les
aminoglycosides lrsquoajout de la tomatidine permettait drsquoaugmenter de 8 agrave 16 fois
lrsquoactiviteacute des aminoglycosides contre S aureus agrave pheacutenotype normal (Mitchell et al
2011a Mitchell et al 2012)
Donc on savait que la tomatidine avait une activiteacute antibacteacuterienne tregraves inteacuteressante
mais on ne connaissait ni sa cible ni son mode drsquoaction De plus si la tomatidine a
une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait donc avoir un
spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Par ailleurs de faccedilon parallegravele la production drsquoanalogues structuraux de la tomatidine
permettrait de mieux comprendre la relation structure-activiteacute (SAR) de la moleacutecule et
donc de preacuteciser que lrsquoactiviteacute biologique de la tomatidine neacutecessite une orientation
3
A B
C
35
de groupements fonctionnels bien speacutecifique De plus lrsquoeacutetude de lrsquoactiviteacute de
plusieurs analogues permettrait de confirmer si la tomatidine possegravede un meacutecanisme
drsquoaction deacutependant drsquoune interaction speacutecifique avec une ou des cibles biologiques
preacutecises
Le laboratoire du Pr Eacuteric Marsault du deacutepartement de pharmacologie de lrsquoUniversiteacute
de Sherbrooke et lrsquoeacutetudiant agrave la maitrise Feacutelix Chagnon ont produit plus de 33
analogues de la tomatidine (voir Annexe 2 article laquo Unraveling the structure-activity
relationship of tomatidine a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureusraquo dans la section laquo Contributions
Supleacutementaires raquo du meacutemoire) dont lrsquoactiviteacute antibacteacuterienne a eacuteteacute eacutevalueacutee par des
tests de susceptibiliteacute agrave la moleacutecule (CMI) sur S aureus SCV et sur S aureus agrave
pheacutenotype normal en combinaison avec la gentamicine Plusieurs de ces moleacutecules
font lrsquoobjet drsquoune application de brevet (voir annexe 1) dont la prometteuse moleacutecule
FC04-100 (Figure 14C) (voir laquo Annexe 1 Application de brevet raquo dans la section
laquo Contributions suppleacutementaires raquo du meacutemoire)
13 Projet de maitrise
Premiegravere hypothegravese
Si la tomatidine a une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait
donc avoir un spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Objectif Provoquer lrsquoapparition de pheacutenotypes SCV chez diffeacuterentes espegraveces
bacteacuterienne afin de pouvoir veacuterifier leur niveau de sensibiliteacute agrave la tomatidine
Deuxiegraveme hypothegravese
Lrsquoidentification des gegravenes preacutesentant des mutations parmi les souches de S aureus
reacutesistantes agrave la tomatidine pourrait reacuteveacuteler la ou les cibles cellulaires de la moleacutecule
et nous aider agrave comprendre son mode drsquoaction
36
Objectif Obtenir des mutants reacutesistants avant de les envoyer agrave sequencer
faire les analyses comparatives des geacutenomes et finalement identifier les mutations
37
CHAPITRE 2
ARTICLE SCIENTIFIQUE
TITRE Tomatidine and Analog FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Some Firmicutes Including Those of Listeria
monocytogenes and Are Bactericidal in Combination With Aminoglycosides Against
their Normal Phenotype
AUTEURS ET CONTRIBUTIONS
Isabelle Guay1 Expeacuteriences biologiques avec le produit naturel tomatidine et son
analogue FC04-100 sur diffeacuterentes espegraveces bacteacuteriennes afin de comparer les deux
activiteacutes
Simon Boulanger1 Expeacuterience biologique permettant de quantifier lrsquoactiviteacute de
lrsquoanalogue FC04-100 sur la production de biofilm et son activiteacute intracellulaire
Feacutelix Chagnon2 Production de lrsquoanalogue chimique de la tomatidine FC04-100
Kamal Bouarab1 Initiateur du projet gracircce agrave lrsquoisolement du produit naturel tomatidine
Eacuteric Marsault2 Directeur de recherche de Feacutelix Chagnon
Franccedilois Malouin1 Directeur de recherche drsquoIsabelle Guay et Simon Boulanger et
auteur de correspondance
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
38
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
Corresponding author Mailing address Universiteacute de Sherbrooke
Deacutepartement de Biologie Faculteacute des Sciences 2500 Boul
Universiteacute Sherbrooke Quebec Canada J1K2R1 Phone (819) 821-
8000 ext 61202 Fax (819) 821-8049 E-mail FrancoisMalouin
USherbrookeca
39
Tomatidine and Analog FC04-100 Possess Bactericidal Activities
Against the Small Colony Variants of Some Firmicutes Including
Those of Listeria monocytogenes and Are Bactericidal in Combination
With Aminoglycosides Against their Normal Phenotype
Running Title Tomatidine is bactericidal against several Firmicutes
Isabelle Guay1
Simon Boulanger1
Feacutelix Chagnon2 Kamal Bouarab
1 Eacuteric Marsault
2 and
Franccedilois Malouin1
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM) Deacutepartement de
Biologie Faculteacute des Sciences Universiteacute de Sherbrooke Sherbrooke Queacutebec Canada J1K
2R1
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute Universiteacute
de Sherbrooke Queacutebec Canada J1H 5N4
Have contributed equally to the work
Corresponding author
Mailing address Universiteacute de Sherbrooke Deacutepartement de Biologie Faculteacute des Sciences
2500 Boul Universiteacute Sherbrooke Quebec Canada J1K2R1
Phone (819) 821-8000 ext 61202 Fax (819) 821-8049
E-mail FrancoisMalouinUSherbrookeca
40
ABSTRACT
Tomatidine (TO) is a plant steroidal alkaloid that possesses an antibacterial activity against the
small colony variants (SCVs) of Staphylococcus aureus We report here the spectrum of
activity of TO against species of the Firmicutes and the improved antibacterial activity of a
chemically-modified TO derivative (FC04-100) In addition to that seen on S aureus TO
showed a bactericidal activity against SCVs of Bacillus cereus B subtilis and Listeria
monocytogenes The combination of an aminoglycoside antibiotic and TO also generated an
antibacterial synergy against strains of the normal phenotype in these species In contrast to
TO which has no relevant activity by itself against S aureus of the normal phenotype (MIC
gt64 mgL) the TO analog FC04-100 showed a MIC of 8-16 mgL Furthermore FC04-100
showed a strong bactericidal activity against L monocytogenes SCVs in kill kinetic
experiments while TO did not FC04-100 also improved the bactericidal activity of
gentamicin against L monocytogenes of the normal phenotype The addition of FC04-100 (4
mgL) to a cephalexinkanamycin (32) combination improved the activity of the combination
by 32 fold against cephalexin and kanamycin-resistant MRSA strains In combination with
gentamicin FC04-100 also showed a strong bactericidal activity against biofilm-embedded S
aureus and when used alone was superior to TO for its biofilm-preventing activity Finally
FC04-100 and TO showed comparable intracellular killing of S aureus SCVs in a polarized
Calu-3 cell culture model In conclusion chemical modifications of TO allowed improvement
of its antibacterial activity against L monocytogenes SCVs and prototypical S aureus
41
Keywords Tomatidine aminoglycoside synergy SCV Firmicutes S aureus L
monocytogenes
Introduction
The Firmicutes are Gram-positive bacteria with genomes presenting less than 50 of G+C
The Firmicutes constitutes one of the main divisions within the Bacteria and is divided into
three classes Bacilli Mollicutes and Clostridia The class of Bacilli is further divided into the
orders of the Lactobacillales and the Bacillales The Bacillales are divided into the genus of
Staphylococcus Listeria and Bacillus A number of bacterial species such as Listeria spp and
Bacillus spp can contaminate food and cause infections in humans (Magalhaes 2013) To
name a few Listeria monocytogenes L ivanovii and Bacillus cereus can cause listeriosis
(Guillet et al 2010) and food poisoining (Bad Bug Book FDA) Bacillus subtilis B
coagulans B licheniformis and B sphaericus are also known to cause illnesses Bacillus
anthracis causes anthrax and can often be acquired by contact with food producing animals
and cattle (beef cattle sheeps etc) and this bacterium is also well-known for its endospores
that have been used as biological weapons (Beierlein and Anderson 2011) Staphylococci are
divided in coagulase-positive species Staphylococcus aureus being the most clinically
relevant of this group and coagulase-negative species such as S epidermidis the most
prevalent pathogen associated with infections of implanted medical devices (Vuong and Otto
2002) The emergence and spread of resistance to multiple antibiotics in staphylococci is now
considered a real health treat and impaired therapeutic endeavor to combat these bacteria
42
(Witte et al 2007) Notably the prevalence of methicillin-resistant S aureus (MRSA) has
steadily increased over the recent years not only in hospitals but also in the community
(Hiramatsu et al 2013)
Since their first description in 1910 for Salmonella enterica serovar Typhi (S typhi) small-
colony variants (SCVs) have been described for a wide range of bacterial genera and species
and have been isolated from both laboratory and clinical environments (Proctor 2006)
Staphylococcus aureus SCVs have attracted a great deal of interest over the past recent years
S aureus SCVs often present a dysfunctional oxidative metabolism causing an alteration in
the expression of virulence factors a slow growth and a loss of colony pigmentation (Proctor
2006) This dysfunctional oxidative metabolism is also responsible for a decreased
susceptibility to aminoglycoside antibiotics because this class of molecules requires the
proton-motive force in order to penetrate the bacterium (Bryan LE 1981) This respiratory
deficiency is often caused by mutations affecting the electron transport system and several
SCV isolates can recover normal growth with supplemental hemin or menadione which are
needed to synthesize electron transport system components S aureus SCVs often are isolated
from chronic infections such as lung infections in cystic fibrosis (CF) patients osteomyelitis
septic arthritis bovine mastitis and from infections associated with orthopedic devices (Atalla
2008 Moisan 2006 Proctor 2006) The ability of S aureus to switch back and forth from the
prototypic to the SCV phenotypes in vivo is an integral part of the pathogenesis of S aureus
and may be responsible for the establishment of chronic infections (Tuchscherr 2011
Mitchell 2011 inf imm)
43
Tomatidine (TO) is a steroidal alkaloid produced by the Solanaceae plant family such as the
tomato (Ruiz-Rubio 2001 Simons 2006) We showed previously the antibacterial activity of
TO against S aureus SCVs and also documented a strong synergic activity of TO in
combination with aminoglycoside antibiotics against prototypic S aureus (Mitchell G AAC
2011 Mitchell G 2012 JAC) Recently we synthesized a variety of TO analogs in order to
explore the structure-activity relationship of this new class of antibiotics able to act on SCVs
(Chagnon et al 2014) One analog FC04-100 showed the same steroidal backbone as the
natural molecule but with an additional carbon chain on the A cycle (Fig 1) Preliminary
characterization of FC04-100 revealed an antibacterial activity that was similar to that of TO
against S aureus SCVs and also preserved the synergic activity with aminoglycosides against
prototypic S aureus (Chagnon et al 2014) However contrary to TO FC04-100 showed
notable antibiotic activity by itself against prototypic S aureus with a MIC of 8-16 gL
whereas that of TO was gt64 gL It would be of great interest to see if this property of FC04-
100 modifies or improves its spectrum of antibacterial activity
The aim of this study was to further describe the spectrum activity of TO and FC04-100 The
susceptibility of several species among the Firmicutes to TO and FC04-100 alone or in
combination with other antibiotics was investigated The improved antibacterial activity of
FC04-100 was demonstrated against Listeria monocytogenes and methicillin-resistant S
aureus (MRSA)
44
Materials and methods
Chemical reagents and antibiotics
Tomatidine (TO) (Sigma Aldrich Oakville Ontario Canada) (Fig 1A) was solubilised in
dimethylsulfoxide (DMSO) at a concentration of 2 mgmL The TO analog FC04-100 (Fig
1B) was synthesized (Chagnon et al 2014) and solubilised in DMSO at 20 gL Cefalexin
kanamycin gentamicine (GEN) (all from Sigma Aldrich) were solubilised in water at a
concentration of 10 gL Menadione was solubilized in DMSO hemin in 14 M NH4OH and
thymidine in water (all from Sigma Aldrich) and were prepared at a concentration of 10 gL
45
3
Fig 1 Tomatidine (A) is a steroid alkaloid structurally characterized by 6 rings 12
stereogenic centers a 3 β-hydroxyl group and spiro-fused E F rings in the form of an
aminoketal The tomatidine analog FC04-100 (B) contains a diamine in position 3 of ring A
Bacterial strains and growth conditions
Staphylococcus aureus ATCC 29213 S aureus Newbould (ATCC 29740) Listeria
monocytogenes ATCC 13932 Bacillus subtilis ATCC 6333 and Bacillus cereus ATCC 11778
were used as prototypic strains Methicillin-resistant S aureus (MRSA) strains COL and
USA100 (hospital acquired-MRSA) and strain USA300 (community acquired-MRSA) were
also used in this study The reference laboratory strain S aureus SH-1000 an isogenic mutant
strain derived from S aureus 8523-4 but with a functional rsbU allele (HorsburghMJ 2002)
was used for its good yield of biofilm production The S aureus strains CF07-L (prototypic)
A
B
A B
C D
E
F
46
and CF07-S (SCV phenotype) are genetically related clinical strains originally isolated from a
cystic fibrosis patient (MitchellG 2011) Strain CF07-S was used in the intracellular
infection model Staphylococcus and Bacillus were maintained on Tryptic Soy agar (TSA BD
Mississauga Ontario Canada) whereas L monocytogenes was grown on brain-heart infusion
(BHI) agar (BD Mississauga Ontario Canada)
Small colony variants (SCVs)
For S aureus strain NewbouldhemB was used as the reference SCV NewbouldΔhemB was
generated from strain Newbould (ATCC 29740) by disrupting the hemB gene with the ermA
cassette by homologous recombination (Brouillette 2004) SCVs from B cereus B subtillis
and L monocytogenes were generated by growth in presence of a subinhibitory concentration
of GEN Briefly overnight broth cultures (18ndash20 h) were used to inoculate BHI broth at a
dilution of 1100 supplemented or not with 025 to 1X the MIC of GEN Cultures were
incubated 18 h at 35degC with shaking (225 rpm) and then adjusted to an A595 nm of 20 in PBS at
4degC Determination of CFU and SCV colonies was done by serial dilution plating SCV were
obtained by plating on TSA (BHI agar for L monocytogenes) containing gentamicin at a
concentration of 8 to 16 times the induction concentration followed by an incubation of 48 h at
35degC The pinpoint colonies selected by this method were confirmed to be SCVs by streaking
several of them on agar plates without antibiotic SCV that conserved their phenotype after
two passages were considered to have a stable phenotype and were used in subsequent
experiments Selected SCVs had a colony size a tenth of the size of a normal colony and
showed no pigmentation or hemolytic activity on blood agar plates The SCV species were
47
confirmed by rDNA 16S sequencing after PCR amplification using universal primers (Gomaa
2007)
Auxotrophy assays
For SCVs auxotrophism is defined as the requirement of specific compounds in order to
regain a normal growth phenotype [Sendie 2009] An agar diffusion method was used to
characterize the auxotrophism of SCVs using hemin or menadione (1-10 μg eachwell) or
thymidine (100 microgwell) on an inoculated Mueller-Hinton agar (MHA) or Brain-heart infusion
agar (BHIA) plate [Besier 2007] Auxotrophy for specific supplements was detected by a
zone of normal growth surrounding the well after 18 h of incubation at 35degC The results were
confirmed by two indepedent experiments
Antibiotic susceptibility testing
MICs were determined by a broth microdilution technique following the recommendations of
the Clinical and Laboratory Standards Institute (CLSI) except that the incubation period was
extended to 48 h and the medium used was BHI to allow SCVs to reach maximal growth as
previously described (Mitchell 2010) As recommended by the CLSI L monocytogenes (and
its SCVs) were grown in cation-adjusted Muller-Hinton broth (CAMHB BD) containing 3
lysed horse blood (LHB Remel Hartford CT USA) The reported results were obtained from
three independent experiments
48
Time-kill experiments
Kill kinetics were performed to determine whether the effect of compounds alone or in
combination was bacteriostatic or bactericidal Bacteria were inoculated at 105-10
6 CFUmL in
appropriate medium in the absence or presence of the different antibiotic compounds at
concentrations specified in figure legends At several points in time at 35degC (225 rpm)
bacteria were sampled serially diluted and plated on TSA for CFU determinations Plates
were incubated for 24 or 48 h at 35degC for normal and SCV strains respectively The data were
collected from a minimum of three independents experiments
Antibiotic activity in biofilms
The viability of bacteria in biofilms was evaluated based on previously described protocols
(CeriH 2001 MoskowitzSM 2004 HarrisonJJ 2010) For these assays 96-peg lids and
corresponding 96-well plates were used (Nunc CITY STATE) Wells were inoculated with a
suspension of S aureus SH-1000 adjusted to a 05 McFarland standard in TSB (150 microL per
wells) and plates with 96-well lids were incubated at 35degC with an agitation of 120 rpm for 24
h The biofilms on pegs were then washed three times with 200 microL PBS Biofilms on pegs
were further incubated in fresh 96-well plates containing 200 microL of TSB containing or not
antibiotic compounds or a combination of molecules at 35degC 120 rpm for 24 h The treated
biofilms on pegs were washed three times in PBS and bacteria in biofilms were recovered by
sonication in a new microtiter plate containing 200 microL of PBS per wells using an
ultrasonicator bath for 10 min followed by centrifugation for five min at 1000 RPM The
49
bacteria recovered in each well were resuspended serially diluted plated on TSA and
incubated at 35degC for 24 h before CFU determination
For an equivalent series of wells instead of disrupting the biofilm by sonication biofilm
formation on pegs was evaluated through crystal violet staining using a procedure adapted
from Mitchell et al(2010) Following exposure to antibiotics as described above biofilms on
pegs were washed three times using 200 microL of PBS before biofilms were air-dried for 30
minutes Biofilm on pegs were then stained by dipping in 200 microL of a crystal violet solution
(01) for 30 minutes rinsed in 200 microL of distilled water and air-dried for 30 min Finally the
crystal violet from stained biofilms was solubilized in 200 microL of ethanol 95 at room
temperature and the absorbance was measured at 562 nm using a plate reader (nom de
lrsquoappareil)
Cell invasion and measurement of intracellular antibiotic activity
The Calu-3 cell line (ATCC HTB 55 Homo sapiens lung adenocarcinoma) was cultured in
Eaglersquos Minimum Essential Medium (EMEM) supplemented with 01 mM MEM nonessential
amino acids 1 mM of sodium pyruvate 100 UmL penicillin 100 gL streptomycin 25 gL of
Fungizone and 10 fetal bovine serum (FBS) at 37oC in 5 CO2 For routine culture 4 gL of
puromycin was added to culture media All cell culture reagents were purchased from Wisent
(St-Bruno QC Canada)
50
The cell invasion assay was performed with the Calu-3 cells in an air-liquid interface as
previously described with few modifications (Mitchell TO I paperAAC 2011) Briefly cells
were seeded at 15 x 105 cells per insert in a 12-well Transwell plate (Corning CITY STATE)
and cultured for 10 days with the apical medium replaced each day The complete medium in
the basal compartment was replaced by the invasion medium (1 FBS and no antibiotics) 18
h before invasion assays Inocula were prepared by suspending bacteria grown 20 h on BHI
agar plates in ice-cold PBS Bacteria were then washed three times in ice-cold PBS and
suspended in the invasion medium supplemented with 05 BSA at a density of
approximately 4 x 108 CFUmL Cells were washed twice with PBS and 250 μL of bacterial
suspension were apically added to each insert Invasion of cells by bacteria was allowed for 3
h inserts were emptied and washed three times with PBS The basal medium was changed for
the invasion medium with 20 gL of lysostaphin (Sigma) to kill extracellular bacteria and with
or without the tested antibiotic compounds The infected cells were incubated for a total of 48
h with a change of medium at 23 h (invasion medium with lysostaphin with or without the
tested antibiotic compounds) The invasion medium with lysostaphin but without tested
compound was also apically added 1 h before cell lysis to ensure that only intracellular
bacteria were counted At 48 h following three washes with PBS the apical and basal media
were removed and cells were detached with 100 μL of trypsin 025 and lysed for 10 min by
the addition of 400 μL of water containing 005 of Triton X-100 Then 50 μL of PBS (50X)
was added and mixed Lysates were serially diluted 10-fold and plated on TSA for CFU
determination Plates were incubated at 35oC for 48 h
51
RESULTS
Emergence the SCV phenotype
The SCV phenotype is characterized by a slow growth yielding small colony size on agar
plates (15 to 110 of the parent phenotype) Figure 2 illustrates the ability of a subinhibitory
concentration of GEN to promote the emergence of the SCV phenotype for B cereus B
subtilis and L monocytogenes B cereus SCVs were obtained at an inducing concentration of
1 X MIC of GEN (Fig 2C) B subtilis SCVs at 05 X MIC (Fig 2D) and L monocytogenes
SCVs at 025 X MIC (Fig 2E) Two prototypic S aureus Newbould and CF07-L and their
SCV counterpart NewbouldhemB and CF07-S respectively were used as control strains (Fig
2A and 2B) The SCV isolates selected for the rest of the study add a low reversion rate and
kept their phenotype without a GEN selection pressure The B cereus SCVs showed
auxotrophy for menadione (Fig 1F Table 1) some of the L monocytogenes SCVs showed
auxotrophy for hemin (Table 1) while the B subtilis SCVs showed no apparent auxotrophy to
hemin menadione or thymidine (Table 1) These results show that SCVs with defects in their
respiratory chain (ie hemin or menadione auxotrophy) can indeed emerge from a selective
pressure with aminoglycosides
52
A B
Fig 2 Typical colony sizes for SCV and protocipic strains (left and right sides of the plates
respectively) for (A) S aureus CF07-S and CF07-L (B) S aureus NewbouldhemB and
Newbould (C) B cereus ATCC 11778 (SCV1 and prototypic) (D) B subtilis ATCC 6333
(SCV2 and prototypic) and (E) L monocytogenes ATCC 13932 (SCV1 and prototypic) In
(F) a TSA plate was inoculated with a B cereus SCV1 and wells were filled with (i) 10 microg
hemin (ii) 10 microg menadione (iii) 100microg thymidine and diluents (iv) DMSO and (v) NH4OH
14N
C D
E
i ii
iii iv
v
F
53
Antibacterial activity of TO against the various species of SCVs and synergy of
the TO and GEN combination against some prototypic Firmicutes
All the SCVs tested showed a MIC for TO of 006 mgL against S aureus and lt003 mgL
against L monocytogenes B cereus and B subtilis respectivly whereas TO showed no
activity (MIC gt64 microgmL) against their prototypic counterparts (Table 1) A synergic effect of
TO with GEN against all the prototypic strains tested was also observed (Table 1) Indeed the
addition of TO provoked a 4-8 fold decrease of GEN MIC against all species tested (Table 1)
These results are consistent with our previous studies on the activity of TO on S aureus
((Mitchell 2011 AAC 2012 JAC)) No change in the MIC of GEN was observed against the
streptococci (S pyogenes S suis S agalactiae S mitis and S pneumoniae) when tested in
combination with TO (data not shown) These results show that TO is active against some but
not all Firmicutes
54
Table 1 Susceptibility of prototypic and SCV strains and species to TO andor GEN
Ratio of activity of GEN+TO vs GEN alone
Bactericidal activity of TO against SCVs and bactericidal synergy of the TO
and GEN combination against prototypic strains
As shown above the addition of 8 mgL of TO to GEN reduced the MIC of GEN against some
Firmicutes (Table 1) but also time-kill kinetics showed that TO improved the bactericidal
effect of GEN against prototypic B subtilis and B cereus (Fig 3A and 3B respectively) This
bactericidal synergy was also previously described for S aureus (Mitchell 2012 JAC)
However although the MIC of GEN was improved when in combination with TO against L
monocytogenes (Table 1) time-kill experiments failed to demonstrate a bactericidal synergy
TO GENGEN
(+TO)
Saureus ATCC 29213 gt64 05 006 8
Saureus NewbouldΔhemB 006 8 Hemin
B cereus ATCC 11778 gt64 1 025 4
B cereus ATCC 11778 (SCV1) lt003 16 Menadione
B cereus ATCC 11778 (SCV2) lt003 16 Menadione
B subtilis ATCC 6333 gt64 0125 003 4
B subtilis ATCC 6333 (SCV1) lt003 2 unknown
B subtilis ATCC 6333 (SCV2) lt003 4 unknown
B subtilis ATCC 9372 gt64 0125 lt0015 gt8
B subtilis ATCC 9372 (SCV1) lt003 1 unknown
B subtilis ATCC 9372 (SCV2) lt003 1 unknown
L monocytogenes gt64 05 0125 4
L monocytogenes (SCV1) lt003 gt64 unknown
L monocytogenes (SCV2) lt003 gt64 Hemin
Strains
MIC (gL)
Synergy
foldAuxotrophy
55
with an improvement of at most 1 log10 in killing compared to that achieved with GEN alone
(Fig 3C) Similarly the effect of TO used alone against SCVs was strongly bactericidal
against S aureus B subtilis and B cereus but not against the L monocytogenes SCV (Fig
3D) and this even if the MIC of TO against L monocytogenes SCVs was inferior to 003
mgL (Table 1) These results show that the activity of TO against L monocytogenes needs to
be improved
56
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 0 6 m g L G E N
0 0 6 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 5 m g L G E N
0 5 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
2 m g L G E N
2 m g L G E N + T O
8 m g L T O
C T R L
Lo
g1
0 C
FU
ml
0
2
4
6
8
1 0
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
CT
RL
CT
RL
CT
RL
CT
RL
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
S a u re u s
h e m B
S C V
B c e r e u s
A T C C 1 1 7 7 8
S C V 2
B s u b t i l i s
A T C C 6 3 3 3
S C V 2
L m o n o c y to g e n e s
A T C C 1 3 9 3 2
S C V 1
Fig 3 Time-kill kinetics of TO andor GEN against (A) prototypic B subtilis ATCC 6333(B)
B cereus ATCC 11778 and (C) L monocytogenes ATCC 13932 The bactericidal effect of
TO against SCVs is also shown at 8 and 24 h (D) The results were obtained from at least
three independent experiments
A B
C
D
Detection
threshold
57
Bactericidal activiy of the TO analog FC04-100 against L monocytogenes
SCVs and synergy with aminoglycosides against prototypic strains and MRSA
The addition of 8 mgL of FC04-100 to GEN (2 mgL) decreased by an average of 5 log10
prototypic L monocytogenes CFUmL after 24 h of treatment in comparison to GEN alone
(Fig 4A) Also the bactericidal activity of FC04-100 used alone against the L monocytogenes
SCV was shown to be better than that observed for TO (Fig 3D) To facilitate comparison
between TO and FC04-100 Figure 4B shows the residual CFU of the L monocytogenes SCV
strain after treatment with TO or FC04-100 and calculated as the percentage of live CFU in
comparison with untreated L monocytogenes SCVs after 24 h of incubation These results
show that TO can be chemically modified (eg derivative FC04-100) and improve its activity
against L monocytogenes and thus the antibacterial activity of FC04-100 against S aureus
was further investigated Interestingly the MIC of FC04-100 against the SCV strain S aureus
NewbouldhemB (MIC of 05 -1 mgL Table 2) was higher than that found for TO (MIC of
006 mgL Table 1) On the other hand the MIC of FC04-100 against the prototypic S
aureus ATCC 29213 is lower than that of TO (8-16 mgL vs gt 64 mgL respectively) (Tables
1 and 2) This newly detected antibacterial activity of FC04-100 was also noted against
prototypic MRSA strains (Table 2) Furthermore the ability of FC04-100 to potentiate the
activity of GEN was similar to that of TO with a 4 to 8 fold improvement of the
aminoglycoside MIC (Table 2)
Although the multi-resistant S aureus MRSA USA100 USA300 and COL are resistant to
cefalexin (beta-lactam antibiotic MIC of 256 mgL) and to the aminoglycoside kanamycin
(MIC of 256 mgL and gt 1024 mgL for USA 100 and USA300 respectively Table 3) the
58
combination of both antibiotics decreased their respective MICs (Table 3) Indeed the
combination of cefalexin and kanamycin in a proportion of 32 (commercialized under the
name UbrolexinTM
) decreased the MIC of one or both antibiotics This is as expected for the
combination of a beta-lactam with an aminoglycoside (Davis 1982) However the synergy is
weak with MRSA strains that are resistant to one or the other or both antibiotics and the
addition of 4 mgL of FC04-100 further improved by 8 to 32 fold the MIC of the cephalexin-
kanamycin combination against these strains (Table 3)
59
te m p s (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
F C 0 4 -1 0 0 8 m g L
2 m g L G E N +
F C 0 4 -1 0 0 8 m g L
C T R L
G E N 2 m g L (B H I)
Re
sid
ua
l C
FU
(
)
CT
RL
TO
8 g
L
FC
04-1
00 8
gL
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
F C 0 4 -1 0 0 8 g L
T O 8 g L
C TR L
T=
24
h
T=
8h
T=
8h
T=
24
h
T=
24
h
Fig 4 Time-kill kinetics of FC04-100 andor GEN against (A) prototypic L monocytogenes
ATCC 13932 The bactericidal effect of FC04-100 used alone against L monocytogenes
SCV1 is also shown at 8 and 24 h (B) In (B) the residual CFU were calculated as the
percentage of live CFU in each condition in comparison with the untreated L monocytogenes
SCV1 after 24h of incubation The results were obtained from at least three independent
experiments
A
B
60
Table 2 Susceptibility of prototypic and SCV S aureus strains to FC04-100 with or without
GEN
a FC04-100 w as used at 4 microgmL
b Ratio of activity of GEN + FC04-100 vs activity of GEN alone
Table 3 Synergistic activity of FC04-100 with the combinaison cefalexin-kanamycin
aFC04-100 was used at 4 microgmL
bRatio of the MIC of cefalexin+kanamycin with FC04-100 vs the MIC of
cefalexin+kanamycin alone
Staphylococcus aureus ATCC 29213 8-16 05 006-0125 4-8
S aureus SH1000 8-16 05 006-0125 4-8
S aureus NewbouldΔhemB 1 8
MRSA USA100 16 05 0125 4
MRSA USA300 16 05 006 8
MRSA Col 16 025 006 4
Strains FC04-100 GENGEN
(+FC04-100 )fold
MIC (gL)
MRSA USA100 256 256 12886 43 32
MRSA USA300 256 gt1024 256171 86 32
MRSA Col 256 4 43 05035 8
MIC (gL)
Strains cefalexin kanamycin cef+kana (32)cef+kana (32)
(+FC04-100 )fold
a b
a b
61
FC04-100 potentiates GEN against S aureus embedded in biofilm and inhibits
intracellular replication of SCVs
Figure 5 demonstrates that compound FC04-100 efficiently kills S aureus embedded in
biofilms when in combination with GEN ie there was an important decrease of bacterial
CFUs compared to the no drug control or to either FC04-100 used alone at 4 ugmL (ie 025-
05MIC Table 2) or GEN used alone at 025 05 or 1 X MIC Besides we previously
demonstrated that TO was able to inhibit intracellular replication of SCVs (Mitchell AAC)
which is an important aspect of virulence for S aureus Here (Fig 6) we show that FC04-100
like TO can significantly decrease the infection of polarized airway epithelial cells by SCVs
62
05
mgL
GE
N
05
mgL
GE
N +
FC
04
02
5 m
gL
GE
N
02
5 m
gL
GE
N +
FC
04
01
2 m
gL
GE
N
01
2 m
gL
GE
N +
FC
04
No d
rug
FC
04
0
2
4
6
8
log
10
CF
Up
eg
= 3 2
= 3 8
= 3 5
= 2
Fig 5 Bactericidal effect of FC04-100 alone or in combination with GEN against prototypic
S aureus SH100 embedded in biofilm FC04-100 (FC04) was used at 4 mgL alone or in
combination with GEN Data are from three independent experiments performed in triplicate
Significant differences in comparison to the no drug control are shown ( Plt 0001 and
Plt 00001 one-way ANOVA with a Dunnettrsquos post test) Data are presented as means with
standard deviations
63
No d
rug
TO
8 m
gL
FC
04 8
mgL
0
2
4
6
log
10
CF
Ui
ns
er
t
Fig 6 Effect of TO and FC04-100 on the intracellular load of CF07-S 48 h post-
internalization Data are from three independent experiments performed in duplicate
Significant differences among groups are shown ( Plt 00001One-way ANOVA with
Turkeyrsquos post test) Data are presented as means with standard deviations
64
DISCUSSION
In previous studies we have demonstrated that TO has a strong inhibitory activity against S
aureus SCVs and improves the bactericidal activity of aminoglycoside antibiotics against
prototypic S aureus and also more broadly against staphylococci This effect was documented
for several strains of diverse clinical origins including aminoglycoside-resistant S aureus
carrying aminoglycoside modifying enzymes TO showed however no synergistic effect on the
activity of aminoglycosides against P aeruginosa E coli or Enterococcus spp (Mitchell
2012) In the present study we showed that the antibacterial spectrum of TO can be extended
to species of the Bacillales order with very low MICs against L monocytogenes B cereus and
B subtillis SCVs (lt003 microgmL) Time-kill kinetics showed that the combination of TO and
GEN creates a bactericidal synergy against prototypic of B subtilis and B cereus similarly to
that previously demonstrated against S aureus However the mixture of TO and GEN did not
demonstrated much improvement over the activity of GEN alone against prototypic L
monocytogenes in time-kill experiments Likewise investigations with Bacillales SCVs
showed that TO efficiently killed S aureus B cereus and B subtillis but not L
monocytogenes SCVs despite its very low MIC and was thus bacteriostatic against that species
Therefore improvement of the antibacterial activity against this important pathogen seems
inevitable for future development of steroidal alkaloids such as TO
TO is a steroid alkaloid structurally characterized by 6 rings 12 stereogenic centers a 3-
hydroxyl group and spiro-fused E F rings in the form of an aminoketal (Fig 1) It is generally
65
prepared by hydrolysis of its glycosylated analog tomatine while both molecules can be found
in the Solanaceae plants (Ruiz-Rubio 2001 Simons 2006) Although we previously
documented some interesting antibacterial activity for TO the absence of an identified cellular
target makes difficult the establishment of a structure-activity relationship (SAR) Our initial
attempt to elucidate SAR was therefore based on the hypothesis that the steroidal A-D rings
act as a rigid scaffold to orient pharmacophores defined by the 3-hydroxyl group on ring A
and the spiroaminoketal group on rings E F Noteworthy we prepared analogs bearing
modifications on ring A of TO and schegravemes for chemistry synthesis were recently published
(Chagnon et al 2014) From seven compounds keeping rings E and F of TO and possessing
structural variations on position 3 of ring A we were able to ascertain some structure activity
relationships Results suggest that the hydrogen bond donor effect of the 3-hydroxyl group
was not important for activity although the stereochemistry of the 3 position substitution
moderately affected activity against both S aureus SCVs and prototypic strains (in
combination with an aminoglycoside) In addition although the presence of one or two
ammonium groups in position C3 reduced the activity against S aureus SCVs (from a MIC of
006 to 05-1 microgmL) this change was highly beneficial for antibiotic activity against normal
strains (from a MIC of gt64 to 8-16 microgmL) without the presence of an aminoglycoside This
change was unexpected and one of such analogs FC04-100 increased its solubility by more
than ten times (from 2 mgmL to 26 mgmL in DMSO) The biological properties of FC04-
100 were therefore further investigated in the present study especially that TO showed some
weak bactericidal activity against Listeria and that the solubility of TO may limit eventual
clinical applications
66
Here we demonstrated that FC04-100 was able to kill both L monocytogenes SCVs as well as
the normal phenotype alone and in the presence of GEN respectively (Fig 4) Furthermore
FC04-100 showed a noticeable antibacterial activity on its own (MIC of 16 microgmL) against the
MRSA strains USA100 USA300 and COL (Table 2) FC04-100 was also synergistic in
combination with an aminoglycoside against such MRSA strains with a 4 to 8 fold gain in the
MIC of GEN Because MRSA strains are often multiresistant to antibiotics and often carry
AMEs we examined the possibility of using a triple antibiotic combination Indeed the
combination of kanamycin and cefalexin in a proportion 32 is already approved to treat
bovine mastitis pathogens under the brand name of UbrolexinTM
(Ganiegravere 2009) This
antibiotic combination offers an extended spectrum of activity compared to each individual
drug and is expected to cover both S aureus Streptococcus uberis and E coli (Ganiere
2009 Maneke et al 2011 Silley 2012) Unfortunately the increased frequency of livestock-
associated MRSA (Garcia-Alvarez et al 2011 Graveland et al 2011 Leonard and Markey
2008) and frequent incidence of strains and species carrying AMEs (Ramirez and Tolmasky
2010) may limit the spectrum of activity of the combination of a beta-lactam and an
aminoglycoside With this in mind we showed that the use of FC04-100 in combination with
kanamycin and cefalexin improves by 32 fold the activity of this mixture against MRSA
strains carrying AMEs (Table 3) Overall the addition of FC04-100 to the aminoglycoside-
beta-lactam combination decreased the MICs of kanamycin and cefalexin below their
resistance breakpoints and this for all the MRSA strains tested
67
L monocytogenes is a highly specialized intracellular pathogen (Hamon et al 2006) causing
life-threatening infections especially in pregnant women (Janakiraman 2008) L
monocytogenes in a foodborne pathogen and because of its intracellular life cycle and
propagation it is able to hide from host defenses and antibiotic treatment Similarly S aureus
although not primarily recognized as a typical intracellular pathogen is also able to enter and
survive in host cells (Lowy 2000) Moreover the ability of S aureus to convert to the SCV
phenotype showing increased biofilm production improved adherence to host cells and tissues
as well as increased intracellular persistence allows this pathogen to cause chronic and
difficult to treat infections (Garcia et al 2013 Mitchell and Malouin 2012) Discovering
antibacterial drugs able to act in the intracellular compartment is generally recognized as a
difficult challenge Previously we demonstrated that steroidal alkaloids such as TO had such
an ability and could act on intracellular S aureus SCVs (Mitchell et al 2011) and here we
demonstrated that this was also the case for the TO analog FC04-100 (Fig 6) In addition we
also show in the present study that FC04-100 greatly improves the bactericidal activity of
aminoglycosides such as GEN against bacteria embedded in biofilms Biofilms are
recognized to be a major hindrance to antibiotic action and are responsible for persistent
colonization in many diseases (Costerton et al 1999) and in the food industry (Srey et al
2013) The intracellular and intrabiofilm antibacterial properties of FC04-100 warrant further
investigation of this compound and this class of molecules in general
In conclusion we showed in this study that the spectrum of activity of TO is related to the
Firmicutes division and more precisely to the order of the Bacillales TO possesses an
68
antibacterial activity against SCVs and a synergistic activity with animoglycoside antibiotics
against prototypic strains The novel TO analog FC04-100 showed very promising new
characteristics that include a much improved bactericidal effect on L monocytogenes and
killing of bacteria in biofilms
ACKNOWLEDGEMENTS
This study was supported by an operation grant from Cystic Fibrosis Canada to FM and by a
team grant from the Fonds Queacutebeacutecois de la Recherche sur la Nature et les Technologies
(FQRNT) to FM KB and EM
REFERENCES
Atalla H Gyles C Jacob CL Moisan H Malouin F Mallard B 2008 Characterization of
a Staphylococcus aureus small colony variant (SCV) associated with persistent bovine mastitis
Foodborne Pathog Dis 5785ndash799
Bad Bug Book Foodborne pathogenic microorganisms and natural toxins 2012 Second
edition Lampel KA Al-Khaldi S Cahill SM (Ed) Published by the Center for Food Safety and
Applied Nutrition of the Food and Drug Administration (FDA) US Department of Health and
Human Services pp 87-103
Beierlein JM Anderson AC 2011 New developments in vaccines inhibitors of anthrax
toxins and antibiotic therapeutics for Bacillus anthracis Curr Med Chem 185083-5094
69
Brouillette E A Martinez B J Boyll N E Allen and F Malouin 2004 Persistence of
a Staphylococcus aureus small-colony variant under antibiotic pressure in vivo FEMS
Immunol Med Microbiol 4135ndash41
Bryan L E and S Kwan 1981 Aminoglycoside-resistant mutants of Pseudomonas
aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob
Agents Chemother 19958ndash964
Ceri H Olson M Morck D Storey D Read R Buret A Olson B 2001 The MBEC Assay
System multiple equivalent biofilms for antibiotic and biocide susceptibility testing Methods
Enzymol 337377-385
Chagnon F I Guay M-A Bonin G Mitchell K Bouarab F Malouin E Marsault
2014 Unraveling the structure-activity relationship of tomatidine a steroid alkaloid with
antibiotic properties against persistent forms of Staphylococcus aureus Eur J Med Chem (In
Press EJMECH-S-13-01625-3)
Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial
Susceptibility Testing Twenty-First Informational Supplement M100-S21 CLSI Wayne PA
USA 2011
70
Costerton JW Stewart PS Greenberg EP 1999 Bacterial biofilms a common cause of
persistent infections Science 284 1318-1322
Davis BD 1982 Bactericidal synergism between beta-lactams and aminoglycosides
mechanism and possible therapeutic implications Rev Infect Dis 4237-245
Ganiegravere JP Denuault L 2009 Synergetic interaction between cefalexin and kanamycin in
Muller-Hinton broth medium and in milk J appl microbiol 107 117-125
Garcia LG S Lemaire B C Kahl K Becker R A Proctor O Denis P M Tulkens
F Van Bambeke 2013 Antibiotic activity against small-colony variants of Staphylococcus
aureus review of in vitro animal and clinical data J Antimicrob Chemother 68 1455ndash1464
Garcia-Alvarez L M T Holden H Lindsay C R Webb D F Brown M D Curran
E Walpole K Brooks D J Pickard C Teale J Parkhill S D Bentley G F Edwards
E K Girvan A M Kearns B Pichon R L Hill A R Larsen R L Skov S J Peacock
D J Maskell and M A Holmes 2011 Methicillin-resistant Staphylococcus aureus with a
novel mecA homologue in human and bovine populations in the UK and Denmark a
descriptive study Lancet Infect Dis 11595-603
Gomaa OM Momtaz OA 2007 16 rRNA characterization of Bacillus isolate and its
tolerance profile after subsequent subculturing Arab J Biotech 10 107-116
71
Graveland H Duim B van Duijkeren E Heederik D Wagenaar JA 2011 Livestock-
associated methicillin-resistant Staphylococcus aureus in animals and humans Int J Med
Microbiol 301630-634
Guillet C Join-Lambert O Le Monnier A Leclercq A Mechaiuml F Mamzer-Bruneel MF
Bielecka MK Scortti M Disson O Berche P Vazquez-Boland J Lortholary O Lecuit M
2010 Human listeriosis caused by Listeria ivanovii Emerg Infect Dis 16136-138
Hamon M H Bierne P Cossart 2006 Listeria monocytogenes a multifaceted model Nat
Rev Microbiol 4 423-434
Harrison JJ Stremick CA Turner RJ Allan ND Olson ME Ceri H 2010 Microtiter
susceptibility testing of microbes growing on peg lids a miniaturized biofilm model for high-
throughput screening Nat Protoc 51236-1254
Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y
Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T Baba T 2013 Genomic
Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
Horsburgh MJ Aish JL White IJ Shaw L Lithgow JK Foster SJ 2002 SigmaB
modulates virulence determinant expression and stress resistance characterization of a
72
functional rsbU strain derived from Staphylococcus aureus 8325-4 J Bacteriol 1845457-
5467
Janakiraman V 2008 Listeriosis in Pregnancy Diagnosis Treatment and Prevention Rev
Obstet Gynecol 1 179ndash185
Leonard FC Markey BK 2008 Methicillin-resistant Staphylococcus aureus in animals a
review Vet J 175 27-36
Lowy FD 2000 Is S aureus an intracellular pathogen Trends Microbiol 8341-342
Magalhatildees L Nitschke M 2013 Antimicrobial activity of rhamnolipids against Listeria
monocytogenes and their synergistic interaction with nisin Food Control 29 138-142
Maneke E Pridmore A Goby L Lang I 2011 Kill rate of mastitis pathogens by a
combination of cefalexin and kanamycin J Appl Microbiol 110184-190
Mitchell G Brouillette Eacute Lalonde-Seacuteguin D Asselin AE Lebeau Jacob C Malouin F
2009 A role for sigma factor B in the emergence of Staphylococcus aureus small-colony
variants and elevated biofilm production resulting from an exposure to aminoglycosides
Microb Pathog 4818-27
73
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K Malouin F 2011
Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic
fibrosis airway epithelial cells Antimicrob Agents and Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM Malouin F 2011 Infection of Polarized
airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is
increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function
and is influenced by NF- κ B Infect Immun 793541-3551
Mitchell G Lafrance M Boulanger S Lalonde-Seacuteguin D Guay I Gattuso M Marsault
Eacute Bourab K Malouin F 2012 Tomatidine acts in synergy with aminoglycoside antibiotics
against multiresistant Staphylococcus aureus and prevents virulence gene expression J
Antimicrob Chemother 67 559-568
Mitchell G Lalonde-Seacuteguin D Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S
Malouin F 2010 Staphylococcus aureus sigma B-dependent emergence of small-colony
variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-
heptylquinoline-N-oxide BMC Microbiology 10 33-48
Mitchell G and F Malouin 2012 Outcome and prevention of Pseudomonas aeruginosa-
Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic
Fibrosis InTech Open Access Publisher ISBN 979-953-307-059-8
74
Moisan H Brouillette E Jacob CL Langlois-Beacutegin P Michaud S Malouin F 2006
Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated
from cystic fibrosis patients is influ- enced by SigB J Bacteriol 18864ndash76
Moskowitz SM Foster JM Emerson J Burns JL 2004 Clinically feasible biofilm
susceptibility assay for isolates of Pseudomonas aeruginosa from patients with cystic fibrosis
J Clin Microbiol 421915-1922
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M Peters G 2006
Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent
infections Nat Rev Micro- biol 4295ndash305
Ramirez MS M E Tolmasky 2010 Aminoglycoside modifying enzymes Drug Res
Updates 13 151ndash171
Ruiz-Rubio M Peacuterez-Espinosa A Lairini K Roldagraven-Arjona T Dipietro A Anaya N
2001 Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi p 293ndash326 In
A Rahman (ed) Studies in natural products chemistry vol 25 Elsevier Oxford United
Kingdom
75
Sendi P Proctor RA 2009 Staphylococcus aureus as an intracellular pathogen the role of
small colony variants Trends Microbiol 1754-58
Silley P Goby L Pillar CM 2012 Susceptibility of coagulase-negative staphylococci to a
kanamycin and cefalexin combination J Dairy Sci 953448-3453
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C Osbourn A
2006 Dual effects of plant steroidal alkaloids on Saccharomyces cerevisiae Antimicrob
Agents Chemother 502732ndash2740
Srey S Jahid IK Ha S-D 2013 Biofilm formation in food industries A food safety concern
Food control 31572-585
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D
Roth J Proctor RA Becker K Peters G Loumlffler B 2011 Staphylococcus aureus
phenotype switching an effective bacterial strategy to escape host immune response and
establish a chronic infection EMBO Mol Med 3129ndash141
Vuong C Otto M 2002 Staphylococcus epidermidis infections Microbes Infect 4481-489
Witte W Strommenger B Stanek C Cuny C 2007 Methicillin-resistant Staphylococcus
aureus ST398 in Humans and Animals Central Europe Emerg Infect Dis 13 255ndash258
76
CHAPITRE 3
MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
31 Obtention des mutants reacutesistants
Afin de nous aider agrave mieux deacutefinir la cible et le mode drsquoaction de la tomatidine (TO)
nous avons forceacute lrsquoapparition de reacutesistances chez Staphylococcus aureus Comme
souche controcircle prototype nous avons choisi ATCC 29213 puisque crsquoest une souche
tregraves freacutequemment utiliseacutee pour des expeacuteriences avec des antibiotiques et nous avons
choisi la souche NewbouldΔhemB comme S aureus laquo small colony variant raquo puisque
crsquoest une souche SCV qui est stable et bien connue nrsquoayant pas de capaciteacute de
reacuteversion Par passages successifs dans une concentration sous inhibitrice
drsquoantibiotique (technique de CMI (concentration minimale inhibitrice)) (CLSI 2011)
nous avons obtenu des mutants deacutemontrant une reacutesistance croissante agrave lrsquoantibiotique
au fil des passages (Figure 1) Nous avons obtenu des mutants ATCC 29213
reacutesistants agrave la gentamicine (GEN) et agrave la GEN lorsque combineacutee agrave la TO ainsi que
des mutants reacutesistants NewbouldΔhemB reacutesistants agrave la GEN seule agrave la TO seule et
agrave la TO lorsque combineacute agrave la GEN
Certains passages ont ensuite eacuteteacute seacutelectionneacutes parce qursquoils deacutemontraient une hausse
plus marqueacutee de leur CMI Ces passages ont eacuteteacute choisis car nous supposions que
ces hausses marqueacutees de CMI correspondraient donc agrave des hausses de la
reacutesistance de cette souche agrave lrsquoantibiotique testeacute et que cette reacutesistance pouvait ecirctre la
conseacutequence drsquoune ou plusieurs mutations en lien avec la cible ou le mode drsquoaction
de lrsquoantibiotique preacutesent Trois agrave quatre passages (ou paliers de reacutesistance) par
condition ont eacuteteacute seacutelectionneacutes Nous avons ensuite reacuteensemenceacute le laquo pool raquo
bacteacuterien de chacun des passages choisis afin de pouvoir isoler 3 isolats ayant la
mecircme CMI (ou tregraves similaire (CMI vraies)) que celui du laquo pool raquo bacteacuterien des
77
passages drsquoougrave ils provenaient Soixante quatre mutants reacutesistants ont ainsi eacuteteacute
seacutelectionneacutes en vue drsquoecirctre seacutequenceacutes (Figure 2)
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt 1
6
CM
I v
raie
s
02
5
CM
I v
raie
s
02
5-1
A
78
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
en
pr
eacutes
en
ce
de
ge
n (
4 micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
01
25
1
6
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
8
CM
I v
raie
s
2
4
CM
I v
raie
s
2
B
C
79
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
en
pr
eacutes
en
ce
de
TO
(8
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
1
2
3
4
5
CM
I v
ra
ies
4
CM
I v
ra
ies
2
CM
I v
ra
ies
0
5
1
CM
I v
ra
ies
2
4
Figure 1 Graphiques des CMI pour chacun des passages pour (A) la tomatidine sur
NewbouldΔhemB (B) pour la tomatidine en preacutesence de gentamicine sur
NewbouldΔhemB (C) pour la gentamicine sur S aureus ATCC 29213 (D) pour la
gentamicine en preacutesence de tomatidine sur S aureus ATCC 29213
32 Seacutequenccedilage et analyses bioinformatiques
Une extraction drsquoADN geacutenomique a ensuite eacuteteacute effectueacutee sur chacun des isolats en
utilisant le Kit drsquoextraction drsquoADN geacutenomique miniKit de Qiagen (QIAamp DNA minikit)
(Li et al 2003) Puisque leurs seacutequences geacutenomiques nrsquoeacutetaient pas connues nous
avons eacutegalement extrait lrsquoADN geacutenomique de nos deux souches de reacutefeacuterences
ATCC29213 et NewbouldΔhemB (provenant de quatre colonies isoleacutees chacune)
afin drsquoavoir les seacutequences de comparaison pour nos mutants (Figure 2)
D
80
Ne
wm
an
(g
en
om
ede r
ef h
ttpw
ww
ncbinlm
nih
govn
uccore
150373012)
AT
CC
29
213
(A p
art
ir d
e 4
AT
CC
29
21
3 r
eco
nstr
uire
AT
CC
29
21
3 c
om
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
5
10
7
10
8 1
09
)
Ne
wbo
uld
Δhe
mB
(A p
art
ir d
e 4
Ne
wb
ou
ldh
em
Bre
co
nstr
uire
Ne
wb
ou
ldh
em
Bcom
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
1
agraveG
EN
2
5 (
CM
I2
)
2
7 (
CM
I2
)
2
8 (
CM
I2
)
AT
CC
29
21
3
Pa
lier
de
reacutesis
tance
1
agravecom
bo
29
(C
MI
05
)
3
0 (
CM
I 0
5
3
1 (
CM
I 0
5)
Suite
fe
uill
e s
uiv
ante
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agraveG
EN
3
(C
MI
4)
4
(C
MI
4)
3
9 (
CM
I4
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agraveG
EN
4
6 (
CM
I8
)
4
7 (
CM
I8
)
4
8 (
CM
I8
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agravecom
bo
1
1 (
CM
I 2
)
1
2 (
CM
I 2
)
2
1 (
CM
I 2
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agravecom
bo
5 (
CM
I 2
-4)
3
6 (
CM
I 2
-4)
3
7 (
CM
I 2
-4)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
4
agravecom
bo
3
8 (
CM
I4
)
6
7 (
CM
I 4
)
81
Ne
wb
ould
Δhem
B(A
part
ir d
e 4
Ne
wb
ou
ldΔ
he
mB
reco
nstr
uire
Ne
wb
ou
ldΔ
hem
Bco
mp
let e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveG
EN
24
(C
MI
64
)
4
1 (
CM
I 1
6)
4
2 (
CM
I 6
4)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveT
O
95
(C
MI
02
5)
9
6 (
CM
I 0
25)
9
7 (
CM
I 0
25)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agravecom
bo
33
34
35
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveG
EN
49
(C
MI
12
8)
5
1 (
CM
I 1
28)
5
2 (
CM
I 1
28)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agraveG
EN
7
(C
MI
128
)
4
3 (
CM
I 1
28)
4
5 (
CM
I1
28
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agraveT
O
6
4 (
CM
I gt
16)
6
5 (
CM
I gt
16)
6
6 (
CM
I gt
16)
75
76
77
79
80
81
82
83
84
85
86
87
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agravecom
bo
15
16
18
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agravecom
bo
59
60
61)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agravecom
bo
54
56
58)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveT
O
90
(C
MI
02
5)
9
1 (
CM
I 0
25)
9
3 (
CM
I 0
5)
Ne
wb
ou
ldh
em
B
palie
r d
e r
eacutesis
tance
3
agraveT
O
11
0 (
CM
I 1
)
1
12 (
CM
I 0
25
-05
)
1
13 (
CM
I 0
5)
Fig
ure
2
Org
an
igra
mm
e d
es
mu
tan
ts r
eacutes
ista
nts
seacute
qu
en
ceacutes
se
lon
leu
r pa
lier
de
reacute
sis
tan
ce e
t a
vec leu
r n
um
eacutero
drsquoeacute
chan
tillo
n
co
rre
spo
ndan
t
82
Un minimum de 20 ngmicrol drsquoADN geacutenomique de chacun des isolats a ensuite eacuteteacute
achemineacute au laboratoire du Pr Seacutebastien Rodrigue (Deacutepartement de biologie Faculteacute
des sciences Universiteacute de Sherbrooke) qui a effectueacute la librairie de fragments Pour
se faire il y a eu dans un premier temps digestion enzymatique de lrsquoADN afin
drsquoobtenir des fragments drsquoADN double brin Ensuite tous les fragments ont eacuteteacute
purifieacutes suivant la meacutethode SPRI (Solid-Phase Reversible Immobilization) Il srsquoagit
drsquoune meacutethode simple et tregraves efficace drsquoisolation drsquoacides nucleacuteiques sur microplaque
(Rodrigue S et al 2010) Les acides nucleacuteiques cibles sont immobiliseacutes sur des
microparticules paramagneacutetiques dans des conditions tampons-speacutecifiques La
flexibiliteacute du systegraveme se fait par lrsquoutilisation de diffeacuterents tampons de liaison (laquo binding
buffer raquo) qui modifient le type et la grandeur des acides nucleacuteiques immobiliseacutes Les
contaminants peuvent ainsi ecirctre retireacutes de lrsquoeacutechantillon sans besoin de filtration ou
centrifugation (Beckman Coulter website 2013 DeAngelis et al 1995) Suite agrave la
purification par meacutethode SPRI lrsquoeacutequipe du Pr Rodrigue a obtenu des fragments
drsquoADN de ~250pb auxquels ont eacuteteacute ajouteacutes par ligation des adapteurs aux deux
extreacutemiteacutes des fragments Dans un deuxiegraveme temps des codes-bars speacutecifiques agrave
chaque eacutechantillon drsquoADN geacutenomique ont eacutegalement eacuteteacute lieacutes agrave chacun des fragments
Alors que les adapteurs permettent aux fragments de se lier agrave une matrice solide afin
de pouvoir ecirctre amplifieacutes les codes-bars permettront de classer chacun des
fragments selon leur appartenance agrave un eacutechantillon drsquoADN geacutenomique speacutecifique
83
Figure 3 Eacutetape du seacutequenccedilage Illumina
La librairie de fragments a ensuite eacuteteacute envoyeacutee chez Geacutenome-Queacutebec afin que
chacun des geacutenomes puisse ecirctre seacutequenceacute Geacutenome ndashQueacutebec utilise la meacutethode
Illumina comme outil de seacutequenccedilage (Figure 3) La grande innovation de la meacutethode
Illumina est lrsquoamplification des fragments par liaison de ceux-ci agrave une surface solide
(Flowcell) ce qui permet par un processus de ponts drsquoamplification de geacuteneacuterer plus
drsquoun million de laquo clusters raquo comprenant eux-mecircme au plus un millier de copies de la
moleacutecule originale de deacutepart (solid-phase PCR) Chaque regroupement de fragments
provenant de lrsquoamplification drsquoun mecircme fragment original est tregraves pregraves les uns des
Modifieacute de Quail MA et al 2008
84
autres formant une zone distincte drsquoun regroupement de fragments agrave un autre
Lrsquoincorporation des nucleacuteotides eacutetiqueteacutes preacutealablement avec des moleacutecules
fluorescentes speacutecifiques est deacutetecteacutee suite agrave leur excitation par un laser et les
images sont compileacutees pour produire la seacutequence de bases pour chaque fragment
ADN (Figure 4) Les nouvelles seacuteries de bases appeleacutees laquo readsraquo seront ensuite
reacuteassembleacutees (aligneacutes) en se basant sur un geacutenome de reacutefeacuterence connu (appeleacute
laquo resequencing raquo) (crsquoest notre cas ici) ou en absence de geacutenome de reacutefeacuterence
(appeleacute laquo De novo sequencing raquo) Au final lrsquoensemble des laquo reads raquo aligneacutes
correspondra agrave la seacutequence de chaque chromosome de lrsquoeacutechantillon drsquoADN
geacutenomique de deacutepart
85
Figure 4 Meacutethode drsquoamplification par la meacutethode Illumina
Suite au seacutequenccedilage chez Geacutenome ndashQueacutebec nous avions en main les seacutequences
geacutenomiques de tous nos mutants et de nos souches de reacutefeacuterence sous forme de
Tireacute de BiteSizeBio httpbitesizebiocom13546sequencing-by-synthesis-
explaining-the-illumina-sequencing-technology consulteacute le 3 deacutec 2013
86
contigs Nous avions donc le casse-tecircte mais en piegraveces deacutetacheacutees pecircle-mecircle et
sans avoir la photo de lrsquoimage de reacutefeacuterence sur laquelle se baser pour pouvoir
compleacuteter le casse-tecircte
1- Lrsquoassemblage
Les diffeacuterents geacutenomes avaient eacuteteacute preacutealablement fractionneacutes aleacuteatoirement
en petits fragments de 250 pb et amplifieacutes en vue du seacutequenccedilage Suite au
seacutequenccedilage les seacutequences de chaque fragment (appeleacute laquo reads raquo) sont ensuite
assembleacutees en contigs crsquoest-agrave-dire des seacutequences plus ou mois longues continues
et ordonneacutees geacuteneacutereacutees par lassemblage des clones dune bibliothegraveque geacutenomique
qui se chevauche La qualiteacute de lrsquoassemblage en contigs deacutepend de la capaciteacute de
superposition des laquo reads raquo Parce que la digestion enzymatique de lrsquoADN se fait de
faccedilon aleacuteatoire et se fait sur plusieurs copies drsquoADN chaque portion du geacutenome est
repreacutesenteacutee plusieurs fois dans diffeacuterents modegraveles de fragments Donc les
seacutequences identiques des diffeacuterents fragments (ou laquo reads raquo) peuvent ecirctre
superposeacutees par chevauchegravement et les laquo reads raquo superposeacutes peuvent ecirctre
assembleacutes en contigs par un logiciel drsquoassemblage En ce qui nous concerne cette
eacutetape a eacuteteacute faite agrave lrsquoaide du programme de bio-informatique Newbler (Zhang et al
2012) Lrsquoassemblage geacutenomique peut ecirctre probleacutematique pour les informaticiens car
les geacutenomes contiennent des seacutequences qui se reacutepegravetent et ces reacutepeacutetitions peuvent
ecirctre assez longues en fonction des organismes Suite agrave lrsquoassemblage les geacutenomes
sont donc sous forme de contigs
2- Lrsquoalignement
En se basant sur une seacutequence geacutenomique de reacutefeacuterence (notre reacutefeacuterence est
Staphylococcus aureus str Newman (~ 288Mb)
( httpwwwncbinlmnihgovgenome154project_id=58839) ) le programme de bio-
informatique ABACAS permet de rapidement aligner ordonner et orienter les contigs
Selon les besoins ce qui nrsquoa pas eacuteteacute notre cas ABACAS permet eacutegalement de
87
visualiser et designer les amorces permettant de fermer les gaps (espaces) entre les
contigs en faisant des PCR (Assefa et al 2009)
3- La fermeture des gaps
La fermeture des gaps nrsquoa pas eacuteteacute faite avec le programme ABACAS mais
avec lrsquoaide du programme de bio-informatique GapFiller Lrsquoutilisation de GapFiller
pour la fermeture des gaps possegravede 3 principaux avantages Premiegraverement le
programme prend en compte lrsquoestimation de la grandeur du gap agrave fermer donc il
exclut automatiquement les fermetures de gaps erroneacutes qui sont plus courts ou plus
longs que la grandeur estimeacutee Deuxiegravemement il nrsquoessaie pas de remplir un gap
avec un assemblage de laquo reads raquo en contigs mais cherche plutocirct agrave allonger les
contigs agrave partir de leurs extreacutemiteacutes jusqursquoagrave obtenir des seacutequences superposeacutees en
passant par la creacuteation de K-mer (seacuteparation des reads en plus petits fragments) Et
troisiegravemement il prend en compte que les extreacutemiteacutes des contigs sont souvent
source de mauvais assemblage et reacute eacutevalue ces extreacutemiteacutes pendant la fermeture des
gaps (Boetzer et Pirovano 2012)
4- Lrsquoannotation des geacutenomes
Suite agrave la fermeture des gaps il a fallu annoter les geacutenomes en se basant sur
lrsquoannotation du geacutenome de reacutefeacuterence Staphylococcus aureus str Newman RATT est
un outil de transfert rapide drsquoannotation (Rapid Annotation Transfert Tool) qui a eacuteteacute
deacuteveloppeacute afin de fournir rapidement une annotation preacutecise pour des geacutenomes
inconnus en se basant sur des geacutenomes deacutejagrave annoteacutes comme reacutefeacuterences (Figure 5)
La meacutethode transfert les annotations drsquoune reacutefeacuterence de haute qualiteacute agrave un nouveau
geacutenome sur la base de la conservation des synteny (ordre des gegravenes) (Otto et al
2011) Suite agrave lrsquoannotation les gegravenes tant chez les mutants que chez les souches
parentes portent les noms auxquels ils correspondent chez Staphylococcus aureus
str Newman
88
Figure 5 Corrections du programme RATT sur les annotations transfeacutereacutees Les
annotations de H37Rv ont eacuteteacute transfeacutereacutees sur la seacutequence F11 (bleu pacircle) ont eacuteteacute corrigeacutees
(vert) et ensuite compareacutees agrave lrsquoannotation de la souche existante F11 EMBL (jaune) (A et B)
La correction des codons ldquostartrdquo et ldquostoprdquo respectivement Dans une situation de laquo mapping raquo
plus complexe (C) ougrave les cadres de lecture sont montreacutes pour plus de clarteacute RATT
cartographie une large seacutequence codante (CDS) de H37Rv vers un locus dans F11 qui inclue
plusieurs codons laquo stop raquo laquo in-frame raquo En inseacuterant un deacutecalage du cadre de lecture (ie un
pseudogegravene) la traduction conceptuelle est conserveacutee Ceci diffegravere avec les deux gegravenes se
superposant preacutedis comme faisant partie du geacutenome F11
Tireacute de Otto TD et al 2011
89
321 Identification des mutations en lien avec la tomatidine
Apregraves avoir obtenu une seacutequence consensus agrave partir des seacutequences geacutenomiques
des 4 copies de chacune des souches parente S aureus ATCC 29213 et S aureus
NewbouldΔhemB les diffeacuterences geacutenomiques entre les souches parentes et le
geacutenome de reacutefeacuterence S aureus str Newman ont eacuteteacute eacutelimineacutees afin que ces
diffeacuterences nrsquoapparaissent pas comme des mutations lors de la comparaison des
seacutequences geacutenomiques des mutants avec celles de leur souche parente ces
diffeacuterences eacutetant plutocirct attribuables agrave la diffeacuterence qursquoil y a entre S aureus str
Newman et la souche parente et non pas agrave des mutations en lien avec la reacutesistance
des mutants
Le service bio-informatique du deacutepartement de biologie (Faculteacute des sciences
Universiteacute de Sherbrooke) a ensuite produit un tableau contenant pour chacun des
mutants tous les gegravenes muteacutes apregraves comparaison avec leur souche parentes Agrave cette
eacutetape on pouvait donc savoir dans quel gegravene se situaient les mutations quelle eacutetait
lrsquoeffet de cette mutation (mutation synonyme deacuteleacutetion insertion SNP) et en principe
srsquoil srsquoagissait pour un mecircme gegravene muteacute de la mecircme mutation puisque dans ce
tableau pour un mecircme gegravene chaque ligne correspond agrave des mutations diffeacuterentes
(Figure 6)
Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4
Figure 6 Exemple drsquoune partie du tableau compilation des mutations chez
les mutants ATCC 29213 reacutesistants agrave la GEN seule et agrave la GEN (+TO)
90
Agrave lrsquoaide de ce tableau (Figure 6) jrsquoai chercheacute les mutations qui pouvaient ecirctre
attribuables agrave la TO Chez les mutants S aureus ATCC 29213 reacutesistants jrsquoai
chercheacute les gegravenes qui eacutetaient muteacutes chez tous les mutants reacutesistants agrave la
combinaison GEN (+TO) ou dont la mutation apparaissait graduellement au fil des
paliers de reacutesistance mais dont ce mecircme gegravene nrsquoeacutetait pas muteacute chez les mutants
reacutesistants agrave la GEN seule Je pouvais donc estimer que cette mutation chez les
reacutesistants agrave la combinaison eacutetait en lien avec la preacutesence de la TO dans le milieu
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct
Afin de mrsquoaider dans ma recherche jrsquoai eu lrsquoaide du Pr Pierre-Eacutetienne Jacques
(Deacutepartement de biologie Faculteacute des sciences Universiteacute de Sherbrooke)qui a
mis les donneacutees de ce tableau dans un tableau croiseacute dynamique (TCD) ce qui a
permis de filtrer les donneacutees selon des critegraveres bien preacutecis me permettant drsquoarriver agrave
mes fins plus facilement Dans un premier temps un TCD a eacuteteacute creacuteeacute ougrave pour
chacun des mutants chaque gegravene ayant au moins une mutation eacutetait indiqueacute par un
laquo 1 raquo alors qursquoun laquo 0 raquo eacutetait marqueacute si ce gegravene nrsquoeacutetait pas muteacute Dans un second
temps la quantiteacute de laquo 1 raquo par palier de reacutesistance (correspondant au nombre de
mutants de ce palier posseacutedant une mutation dans ce gegravene) eacutetait indiqueacutee par un
chiffre allant de 0 pour laquo aucun mutant de ce palier de reacutesistance ne posseacutedait de
mutation dans ce gegravene raquo agrave 1 2 ou 3 selon le nombre de mutants de ce palier ayant
ce gegravene muteacute Jrsquoai ensuite pu soumettre le TCD agrave des filtres approprieacutes afin de faire
ressortir les gegravenes drsquointeacuterecircts selon des critegraveres de seacutelection preacutecis (Figures 7-8)
91
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
1
00
33
32
1
1
11
4
5
0
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
10
03
33
21
11
14
50
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_04
12
lpl9
nm
20
12
32
23
92
44
1
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
NW
MN
_00
46
_2
10
13
21
37
24
31
NW
MN
_09
55
_2
01
23
11
37
24
21
NW
MN
_13
83
gpsA
21
02
31
13
72
42
1
NW
MN
_03
39
_1
11
21
31
37
34
42
NW
MN
_15
97
dn
aE2
10
22
30
37
23
31
NW
MN
_23
30
_2
10
23
20
37
23
21
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_2
07
0_
10
01
23
21
81
45
0
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_10
28
_1
22
12
32
58
34
54
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
A B C
92
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
39
8h
sdM
31
33
33
27
11
34
54
NW
MN
_04
26
_3
31
33
32
71
13
45
4
NW
MN
_05
24
sdrD
32
33
33
28
11
34
55
NW
MN
_07
39
_3
33
33
32
91
13
45
6
NW
MN
_11
64
hsl
U3
33
33
32
91
13
45
6
NW
MN
_05
23
sdrC
22
33
32
27
10
34
45
NW
MN
_20
61
fmtB
23
23
32
27
10
34
45
NW
MN
_04
03
lpl1
nm
33
22
33
28
10
34
55
NW
MN
_13
45
ebh
13
32
33
31
81
03
44
5
NW
MN
_16
49
_3
32
33
31
81
03
44
5
NW
MN
_17
00
hsd
M3
23
33
22
81
03
44
5
NW
MN
_00
42
_3
33
33
22
91
03
44
6
NW
MN
_05
80
_3
33
33
31
91
03
44
6
NW
MN
_13
44
ebh
23
31
32
22
79
34
44
NW
MN
_13
75
aro
A3
13
23
31
79
34
44
NW
MN
_23
97
fnb
B2
32
23
22
79
34
45
NW
MN
_04
01
_3
32
23
31
89
34
45
NW
MN
_07
33
_3
32
32
31
89
34
45
NW
MN
_17
45
_2
33
22
32
89
34
56
NW
MN
_01
66
coa
33
32
32
29
93
44
6
NW
MN
_05
25
sdrE
33
32
33
19
93
44
6
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D Fig
ure
7
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez A
TC
C 2
921
3 d
an
s le
TC
D p
ar
dif
feacutere
nts
fil
tre
s
(A)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts la
ge
nta
mic
ine
ge2
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(C
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts d
an
s le
pa
lier
1
de
reacute
sis
tan
ce
agravela
ge
nta
mic
ine
le1
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
et
nom
bre
de m
uta
nts
reacute
sis
tan
ts agrave
la la
gen
tam
icin
e (
+T
O)
de
s 2
de
rnie
rs p
alie
rs d
e r
eacutesis
tan
ce
ge5
(D
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
7 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
ge
nta
mic
ine
(+
TO
) ge
9
93
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_16
29
ccp
A0
01
33
31
50
00
01
24
0
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_21
28
rpsM
11
12
21
12
22
00
31
74
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
NW
MN
_25
29
clfB
33
03
33
80
11
26
17
4
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e reacute
sist
ance
agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
On
b m
uta
nts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
16
29
ccp
A0
01
33
31
50
00
01
24
0N
WM
N_
21
28
rpsM
11
12
21
12
22
00
31
74
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
A B C
94
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
04
6_
1
01
00
28
22
32
2
10
9
NW
MN
_17
33
prs
A0
11
12
01
12
12
22
14
7
NW
MN
_19
23
_0
11
31
15
13
22
21
08
NW
MN
_20
37
_0
02
32
07
23
11
21
27
NW
MN
_22
77
_1
02
21
11
02
12
33
14
8
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
01
43
_0
03
00
04
23
33
34
11
NW
MN
_0
40
6lp
l3n
m2
01
00
03
12
22
33
7
NW
MN
_1
65
2_
00
10
00
62
12
31
68
NW
MN
_2
01
2_
00
00
00
32
33
30
31
1N
WM
N_
20
15
mn
aA0
00
00
01
23
33
01
11
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
o
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D E Fig
ure
8
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez N
ew
bo
uld
Δh
em
Bd
an
s l
e T
CD
par
dif
feacutere
nts
fil
tres
(A)
Nom
bre
de
mu
tan
ts
reacutesis
tan
ts agrave
la tom
atid
ine
ge15
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atid
ine
ge15
et no
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
(C)
No
mbre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la tom
atid
ine
ge15
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3(
D)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atidin
ege
10
et no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
tom
atid
ine
(+G
EN
) ge
7
(E)
No
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la to
ma
tid
ine
le6
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
95
Le TCD mrsquoa permis de rapidement identifier les gegravenes drsquointeacuterecircts Agrave lrsquoaide du site CMR
(Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) 2013) jrsquoai
ensuite veacuterifieacute la fonction des gegravenes identifieacutes afin de confirmer leur inteacuterecirct (Tableau
1)
Tableau 1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants et (B) chez les mutants NewbouldΔhemB reacutesistants
Gegravene Alias Fonction
NWMN_0811 NADH dh complexe 1 chaine de transport des eacutelectrons
NWMN_2070 _membrane prot Putative- Toxin production+resistance drug transporteur-
similar to multidrug transporteur 85
NWMN_0635 _membrane prot Putative- Bordeacute par AraC(reacutegulateur opeacuteron arabinose) ou
SsaA-likeprot(lieacute au quorum sensing RNAIII(secretory antigen)
NWMN_2084 _truncated transposase for Is1272- mobile+extrachromosomal element
fonctiontransposon fonction
NWMN_0928 qoxC quinol oxidase polypeptide IIIcomplexe 4 chaine de transport des eacutelectrons
NWMN_0739 _ conserved hypothetical prot fonction inconnues
NWMN_1164 hslU ATP-dependent protease ATP-binding subunit
NWMN_0524 sdrD Ser-Asp rich fibrinogenbone sialoprotein-binding protein sdrD
NWMN_0042 _ lipoprotein putativecell enveloppe- lipoprot en tendem
NWMN_0580 _conserved hypothetical prot fonction inconnuesDNA metabolism DNA
replication recombination and repair
NWMN_1745 _
NWMN_0166 coa coagulase precursor
NWMN_0525 sdrE Ser-Asp rich fibrinogenbone sialoprotein-binding protein SdrE
A
96
Apregraves avoir aligneacute speacutecifiquement une par une chacune des seacutequences (Fasta) des
gegravenes drsquointeacuterecirct de chaque mutant avec leur souche parente jrsquoai utiliseacute le programme
de visualisation et de comparaison de seacutequences ClustalW2 (ClustalW2 2013) afin
de confirmer que la mutation eacutetait une vraie mutation et non une erreur de
seacutequenccedilage ou drsquoalignement et que la mutation eacutetait identique chez tous les mutants
ayant le gegravene drsquointeacuterecirct muteacute Cette eacutetape a permis de restreindre encore la liste des
gegravenes pouvant ecirctre en lien avec la cible ou le mode drsquoaction de la TO (Figure 9)
Gene Alias Fonction
NWMN_0896 _conserved hypothetical prot fonction inconnue- comK family prot-
regulation DNA interaction- ORFIDresssemble agrave facteur de transcription
NWMN_0996 _ conserved hypothetical prot fonction inconnue- prophage fonction
NWMN_1025 _ phage major tail protprophage fonction
NWMN_1629 ccpAcatabollite control protA virulence determinant production and antibiotique
resistance
NWMN_1916 _ conserved hypothetical prot-fonction inconnue ou prophage prot
NWMN_2314 _drug resistance transporteur- EmrBQacA subfamily prot- toxin
production+resistance- transport and binding prot
NWMN_0046 _ hypothetical prot
NWMN_2015 mnaA UDP-Glc-Nac 2 epimerase
NWMN_2012 atpE ATP synthase subunit C
NWMN_1652 _ multidrug resistance transporter protein B
NWMN_0143 _ oligopeptide ABC transporter ATP-binding protein
B
97
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
A
B
98
Jrsquoai ensuite chercheacute le rocircle de ces gegravenes sur le site CMR afin de veacuterifier si leur
mutation pouvait expliquer la reacutesistance agrave lrsquoantibiotique (Comprehensive Microbial
Ressouce (J Craig Venter Institute (JCVI) 2013) Dans une premiegravere eacutetape le rocircle
drsquoun gegravene a eacuteteacute deacutetermineacute en allant tout simplement voir directement le rocircle de ce
gegravene chez Staphylococcus aureus str Newman Dans le cas ougrave le gegravene nrsquoavait pas
de fonction connue chez Saureus str Newman je suis alleacutee voir quelle eacutetait la
fonction de ce mecircme gegravene (et des gegravenes flanquant celui-ci) chez diffeacuterentes souches
de Staphylococcus aureus Advenant le cas ougrave malgreacute tout je nrsquoeacutetais toujours pas
capable de deacuteterminer le rocircle de ce gegravene je suis alleacutee veacuterifier si sa seacutequence pouvait
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 hellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Mutants resistants agrave GEN (4 microgml)+TO
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip 185-AATTGCATTCA-197helliphelliphellip495-ATAGCAATGG-506hellip
hellipnwmn_1629helliphelliphelliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_0143hellip
ccpAATP synthase c
ABC transp
Niveau 1
Niveau 2
Niveau 3
Niveau 4
TO +GEN (4microgml)
Figure 9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct (A) nwmn_0811 et
nwmn_0928 chez les mutants ATCC 29213 reacutesistants agrave la combinaison gentamicine
(+TO) leur souche parente ATCC 29213 (B) nwmn_1629 et nwmn_2012 chez les
mutants hemB reacutesistants agrave la TO et leur souche parente NewbouldΔhemB (C)
nwmn_1629 nwmn_2012 et nwmn_0143 chez les mutants hemB reacutesistant agrave la
tomatidine (+genta) et leur souche parente hemB
C
99
avoir une forte homologie avec un gegravene drsquoune autre espegravece Malgreacute tout certains
gegravenes portent toujours la mention laquo de fonction inconnue raquo
La meacutethodologie que nous avons employeacutee nous a permis drsquoidentifier plusieurs
gegravenes possiblement impliqueacutes dans reacutesistance de nos souches aux antibiotiques
testeacutes
1- chez Staphylococcus aureus ATCC 29213 reacutesistant agrave la gentamicine (+TO)
(Figure 9A)
- NWMN_0811 codant possiblement pour la NADH deacuteshydrogeacutenase
- qoxC (NWMN_0928) codant pour la quinol oxidase polypeptide III
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
2- chez NewbouldΔhemB reacutesistant agrave la tomatidine (Figure 9B)
- ccpA (NWMN_1629) codant pour la laquo catabolite control protein A raquo
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
3- chez NewbouldΔhemB reacutesistant agrave la tomatidine (+gentamicine) (Figure 9C)
- NWMN_0143 codant pour un ABC transporteur
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
Lrsquoanalyse de ces diffeacuterents gegravenes nous amegravene agrave suspecter la sous-uniteacute c de lrsquoATP
synthase comme eacutetant la cible potentielle de la tomatidine chez Staphylococcus
aureus
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine
Les gegravenes muteacutes pointent en grande partie vers la chaine respiratoire qursquoon appelle
aussi chaine de transport des eacutelectrons
100
La NADH deacuteshydrogeacutenase (NWMN_0811) fait partie du complexes I de la chaine de
transport de eacutelectrons alors que la quinol oxidase polypeptide III (NWMN_0928 ou
QoxC) est une composante du complex III et la sous-uniteacute c de lrsquoATP synthase (AtpE)
fait partie eacutevidement du complexe de lrsquoATP synthase ou complexe V de la chaine
respiratoire (Figure 10)
Figure 10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants
Les courbes de croissance des mutants ATCC29213 reacutesistants agrave la combinaison
GEN(+TO) appuient le fait que ces mutations affectent la chaine de transport des
eacutelectrons ce qui affecte le niveau de production drsquoATP et donc la croissance
bacteacuterienne En effet les mutants du troisiegraveme niveau de reacutesistance (qui possegravede
deux gegravenes de la chaine de transport des eacutelectrons de muteacutes) ont une croissance
plus lente que les mutants du premier niveau de reacutesistance qui eux ont un seul gegravene
dans la chaine de transport des eacutelectrons de muteacute (Figure 11)
Quinoloxidase
NADHdehydrogenase
ATPsynthetase
Proton gradient
XXX
101
Figure 11 Courbes de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison GEN+TO du premier niveau de reacutesistance (bleu) et du troisiegraveme niveau
de reacutesistance (vert)
Voici plus preacuteciseacutement comment fonctionne la chaine de transport des eacutelectrons chez
les 2 pheacutenotypes-parents selon le modegravele-hypothegravese eacutetabli avec nos reacutesultats et
analyses
Tout drsquoabord chez le prototype ou souche de pheacutenotype normal (ex ATCC 29213) la
chaine de transport des eacutelectrons fonctionne normalement Les complexes de la
chaine de transport des eacutelectrons sont chargeacutes de transfeacuterer des protons vers
lrsquoexteacuterieur de la cellule en transfeacuterant en mecircme temps des eacutelectrons drsquoun complexe agrave
un autre jusqursquoagrave un accepteur final qui est lrsquoO2 lorsque crsquoest possible Se faisant il se
creacutee un gradient eacutelectrochimique de part et drsquoautre de la membrane et les protons
accumuleacutes du cocircteacute externe sont utiliseacutes par la suite par lrsquoATP synthase afin de
pouvoir produire de lrsquoATP dans la cellule
0 4 8 1 2 1 6 2 0 2 4
0 0 1
0 1
1
C o u r b e d e c r o is s a n c e d e s m u ta n ts A T C C 2 9 2 1 3
reacute s is ta n ts agrave G E N + T O (8 m g L )
te m p s (h )
OD
60
0n
m
Niveau 3
Niveau 1
102
Le transfert drsquoeacutelectrons ne se fait pas sans coucirct et des produits toxiques pour la
cellule sont produits en mecircme temps ce sont les espegraveces reacuteactives de lrsquooxygegravene ou
ROS (Reactive Oxygen Species) Cependant les prototypes par leur reacuteponse au
stress produisent des enzymes capables de deacutetoxifier le milieu (Figure 12A)
Chez les souches SCV ici la souche controcircle NewbouldΔhemB la chaine de
transport des eacutelectrons est deacuteficiente parce que le gegravene permettant la production
drsquoheacutemine qui entre dans la composition des cytochromes est supprimeacute Les
cytochromes entrent dans la composition de plusieurs composantes de la chaine
respiratoire
Lorsque les complexes de la chaine sont alteacutereacutes il y a diminution du transfert de
protons vers lrsquoexteacuterieur une diminution de la production drsquoATP et une plus grande
production de ROS (Figure 12B)
Figure 12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B) NewbouldΔhemB
I III IV V I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
eacute eacute
eacute
H+ H+
ATPO2 H2O
ROS +ROS
Prototype SCV (hemB)
H+
0811 0928 2012 0811 0928
hegraveme
2012
A B
103
La gentamicine a besoin drsquoune bonne polarisation membranaire donc drsquoune chaine de
transport des eacutelectrons fonctionnelle pour pouvoir peacuteneacutetrer dans la cellule (Bryan et
Kwan 1983 (Bryan and van den Elzen 1977 Davis 1987 Taber et al 1987) Une
fois entreacutee elle inhibe la synthegravese proteacuteique ce qui provoque la mort cellulaire La
CMI de la gentamicine contre ATCC 29213 est de 05-1 microgml (Figure 13A)
La tomatidine elle irait se lier agrave lrsquoATP synthase pour en diminuer la fonctionnaliteacute et
ce tant chez ATCC 29213 que chez hemB Cette liaison amegravenerait une diminution
de la production drsquoATP et conseacutequemment provoquerait agrave plus ou moins long terme
une augmentation de production de ROS par lrsquoalteacuteration de la chaine de transport des
eacutelectrons Cependant comme la cellule prototype produit efficacement les enzymes
de deacutetoxification lrsquoimpact de la tomatidine est pratiquement nulle sur les souches
prototypes La CMI de la tomatidine est gt16microml contre ATCC 29213 (Figure 13B)
104
Figure 13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213
Chez ATCC 29213 lrsquoajout de 8microgml de tomatidine agrave la Gentamicine amegravene une
diminution de 8X agrave 16X la CMI de la gentamicine En effet la CMI de la gentamicine
passe de 05-1microgml agrave 006microgml (Figure 14AB)
Lors de lrsquoajout de la tomatidine agrave la gentamicine crsquoest la tomatidine qui se lie dans
un premier temps agrave lrsquoATP synthase reacuteduisant ainsi sont niveau drsquoactiviteacute Il y a alors
de faccedilon transitoire une augmentation de la concentration de protons agrave lrsquoexteacuterieur de
la cellule ce qui permet une entreacutee massive de gentamicine ainsi qursquoune rapide et
forte inhibition de la synthegravese proteacuteique Dans un second temps il y aura eacutegalement
une alteacuteration de la chaine de transport des eacutelectrons conduisant agrave une
augmentation de la production de ROS Crsquoest cependant lrsquoentreacutee massive de
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI TO gt16 microgmlCMI GEN 05-1 microgml
+ROS
ROS mais deacutetoxification efficace
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
105
gentamicine entrainant une forte inhibition de la synthegravese proteacuteique qui provoquera
dans ce cas-ci la mort cellulaire (Figure 14B)
Figure 14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicine lorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC 29213
Chez les mutants ATCC29213 reacutesistants agrave la combinaison GEN+TO les complexes I
et III sont muteacutes et donc ont une fonctionnaliteacute alteacutereacutee ce qui amegravene une
deacutepolarisation membranaire Une membrane moins polariseacutee amegravene une plus faible
entreacutee de gentamicine et donc une plus faible inhibition proteacuteique La CMI de la
gentamicine passe en effet de 05-1 microgml chez la souche parente agrave 2 microgml chez les
mutants reacutesistants
La tomatidine en se liant agrave lrsquoATP synthase accentuera la deacutepolarisation
membranaire provoquant une augmentation de la production des ROS et une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN (TO 8 microgml) 006 microgmlCMI GEN 05-1 microgml
+ROS
polarisation entreacutee massive de GEN
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
GENH+
BA
106
diminution de la production drsquoATP De fait les mutants reacutesistants ont une croissance
plus lente (Figure 15)(Figure 11)
Figure 15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants ATCC 29213
reacutesistants agrave la combinaison gentamicine (+TO)
Chez les SCV la tomatidine se lie aussi agrave lrsquoATP synthase et ce faisant amegravene une
diminution de la production drsquoATP Cette diminution de la production drsquoATP megravenera
ultimement agrave une alteacuteration de la chaine de transport des eacutelectrons et une
augmentation des ROS (Murphy 2009) tout comme chez les souches prototypes
Cependant comme le SCV a deacutejagrave une chaine de transport des eacutelectrons deacuteficiente
une faible production drsquoATP par lrsquoATP synthase et une forte production de ROS
lrsquoajout drsquoune faible concentration de tomatidine (la CMI est de 006microgml) amegravene une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype R GEN (+TO [8 microgml])
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN 2 microgmlCMI GEN 05-1 microgml
+ROS
R agrave GEN (+TO) mais mutants ont une
croissance diminueacutee (+ROS et -ATP)
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
107
augmentation de chacun de ces pheacutenomegravenes ce qui est suffisant pour provoquer la
mort cellulaire (Figure 16A)
Dans les premiers niveaux de reacutesistance agrave la tomatidine chez les mutants hemB la
reacutesistance (la CMI passe de 006 agrave 025-1microgml) est ameneacutee par la mutation du gegravene
ccpA (Catabolite controcircle protein A) ce qui megravenerait agrave une augmentation de la
production drsquoATP par lrsquoutilisation drsquoune autre voie que lrsquoATP synthase etou agrave une
diminution de la production de ROS (Figure 16B) Le rocircle de la CcpA en lien avec la
reacutesistance agrave la tomatidine est encore neacutebuleux Toutefois on sait que normalement
ce gegravene agit comme reacutegulateur transcriptionnel des gegravenes codant pour des enzymes
du cycle de Krebs et qursquoil a un rocircle de reacutegulation dans le meacutetabolisme du carbone et
lrsquoutilisation des substrats (Somerville et Proctor 2009 Goumltz et al 2006)
Figure 16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO 006 microgml
H+
SCV
I III IVeacute eacute
eacuteO2 H2O
+ROS
H+TO
Niveau 1
CcpA (CMI TO 025-1 microgml)
ROS
ATP
V
H+
ATP
TO0811 0928
2012
1629
0811 0928
2012
BA
108
Chez les SCV du deuxiegraveme palier de reacutesistance agrave la tomatidine non seulement le
gegravene ccpA est toujours muteacute mais le gegravene de la sous-uniteacute c de lrsquoATP synthase est
eacutegalement muteacute La mutation dans le site de liaison mecircme de la tomatidine empecircche
sa liaison et donc empecircche son effet sa CMI passant de 006 agrave gt16microgml (Figure
17B)
Malgreacute la mutation dans le gegravene lrsquoATP synthase resterait quand mecircme fonctionnel
puisque la mutation nrsquoamegravene pas la mort cellulaire comme crsquoest le cas lorsque la
tomatidine lie lrsquoATP synthase chez le SCV parent (Figure 17A)
Figure 17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la tomatidine
Tout comme chez les mutants hemB reacutesistant agrave la tomatidine seule les mutants
hemB reacutesistant agrave la tomatidine lorsqursquoen preacutesence de gentamicine possegravedent
eacutegalement une mutation dans le gegravene de lrsquoATP synthase ce qui rend la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO006 microgml
H+
SCV Niveau 2CMI TO gt16 microgml
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+TO
TO
Niveaux 1 et 2
CcpA (CMI TO 025-1 microgml)
ROS
ATP
0811 0928
2012
1629
0811 0928
2012
A B
109
inactive puisqursquoincapable de srsquoy lier (CMI gt16microgml) (Figure 18B) Chez les SCV non
muteacutes (hemB) la gentamicine nrsquoentre que faiblement ducirc agrave la deacutepolarisation de sa
membrane (CMI 4-8 microgml) (Figure 18A) La mutation dans le gegravene codant pour une
pompe (NWMN_0143 codant pour un ABC transporteur) chez les mutants hemB
reacutesistant agrave la tomatidine (+gentamicine) amegravenerait une augmentation de lrsquoactiviteacute de
cette pompe permettant de rejeter hors de la cellule le peu de gentamicine eacutetant
entreacutee La mutation de cette pompe fait passer la CMI de la gentamicine de 4-8
microgml pour hemB agrave 16 microgml pour les mutants hemB reacutesistant agrave la combinaison
gentamicine (+TO) ayant cette mutation (Figure 18B)
Figure 18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB reacutesistants
agrave la combinaison tomatidine (+gentamicine)
Donc selon notre modegravele-hypothegravese lrsquoaction de la tomatidine et de la gentamicine
sur S aureus deacutepend de la liaison agrave leur cible mais a aussi un lien avec la
SCV R agrave TO (+GEN [4microgml])
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI GEN 4-8 microgml
H+
SCV
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+ TO
GEN
CMI GEN 16 microgml
CMI TO gt16 microgml
0811 0928
2012
0811 0928 20120143
GEN
Synthegravese proteacuteique
BA
110
polarisation membranaire et la production de ROS La mesure de la polarisation et
concentration cellulaire en ROS chez nos mutants et nos souches parents en
preacutesence ou non drsquoantibiotique nous permettra de valider notre modegravele-hypothegravese
Chez une bacteacuterie meacutetaboliquement active et posseacutedant une membrane
cytoplasmique intacte il y a normalement une diffeacuterence de potentiel eacutelectrique entre
le cocircteacute inteacuterieur et le cocircteacute exteacuterieur de la membrane lrsquointeacuterieur eacutetant plus neacutegatif de
100 agrave 200 mV par rapport agrave lrsquoexteacuterieur Une diminution de la magnitude de ce
potentiel membranaire est appeleacutee deacutepolarisation alors qursquoune augmentation de ce
potentiel est appeleacutee hyperpolarisation Le potentiel membranaire sera reacuteduit agrave zeacutero
srsquoil y a rupture de la membrane cest-agrave-dire srsquoil y a formation de trous assez grands
permettant aux ions inorganiques de passer librement Ce pheacutenomegravene peut arriver
lorsque les cellules sont tueacutees par la chaleur le froid ou les antibiotiques β-Lactams
Dans ces circonstances la membrane devient permeacuteable agrave diffeacuterents colorants
comme le propidium iodide (PI) et le TO-PRO3 et le changement est irreacuteversible
Plusieurs classes de composeacutes chimiques notamment les ionophores peuvent
eacutegalement alteacuterer le potentiel membranaire sans pour autant permettre la
permeacuteabiliteacute au PI TO-PRO3 et autres colorants similaires chez les bacteacuteries
lrsquoadministration drsquoun proton-ionophore tel le carbonyl cyanide 3-
chlorophenylhydrazone (CCCP) reacuteduira le potentiel membranaire agrave zeacutero
Des tests preacuteliminaires sur la mesure du potentiel membranaire (BacLighttrade Bacterial
Membrane Potential Kit (B34950) drsquoInvitrogen et lecture au cytomegravetre de flux (FACS))
(Novo et al 2000) chez nos deux souches parentes mets en eacutevidence que ATCC
29213 est plus polariseacute que NewbouldΔhemB (Figure 19A) Toutefois malgreacute
qursquoayant une plus faible polarisation membranaire NewbouldΔhemB nrsquoest pas
complegravetement deacutepolariseacute puisqursquoil est possible de le deacutepolariser davantage agrave lrsquoaide
drsquoun agent deacutepolarisant tel que le CCCP (carbonyl cyanide 3-chlorophenylhydrazone)
(Figure 19B) Le TO-PRO3 a eacuteteacute utiliseacute comme controcircle afin de confirmer la viabiliteacute
de nos cellules (donneacutees non montreacutees)
111
A B
Figure 19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane Potential Kit
(Invitrogen) (Lecture au cytomegravetre de flux agrave lrsquoaide du programme FCS Express) (A)
Ratio FL3-H (axe des Y)FL1-H (axe des X) de ATCC 29213 (rouge) et de
NewbouldΔhemB (noir) (B) Lecture des rouges (FL3-H) sur NewbouldΔhemB traiteacute
avec le DiOC2(3) (33prime-diethyloxacarbocyanine iodide) (noir) ou avec le CCCP (bleu)
ou sur les cellules chauffeacutees (rouge)
La sous-uniteacute c de lrsquoATP synthase (AtpE) semble donc la cible la plus probable de la
tomatidine Afin de veacuterifier si les mutations identifieacutees pouvaient avoir un effet
veacuteritable sur la proteacuteine nous avons fait un alignement de seacutequences noteacute les bases
muteacutees et veacuterifieacute si la mutation amenait un changement drsquoacide amineacute et si ce
changement drsquoacide amineacute pouvait avoir un impact sur la proteacuteine finale De fait
nous avons trouveacute que les deux diffeacuterentes mutations dans lrsquoAtpE amenaient un
changement significatif drsquoacides amineacutes pouvant amener une variation fonctionnelle
FL3-H
Count
100
101
102
103
104
0
18
36
53
71
Chauffeacute
+CCCP+DiOC2
Chauffeacute
FL1-H
FL3-H
100
101
102
103
104
100
101
102
103
104
hemB
29213
VERT
RO
UG
E
ROUGE
ΔhemB
112
de la proteacuteine La mutation de la base 49 change un G pour un T il srsquoagit drsquoun SNP
nrsquoamenant pas de changement de cadre de lecture La mutation fait passer lrsquoacide
amineacute alanine (A) qui est apolaire agrave une seacuterine (S) qui est polaire La mutation de la
base 190 change un G pour un A et est aussi un SNP nrsquoamenant pas de changement
de cadre de lecture Cette mutation fait passer lrsquoacide amineacute alanine (A) qui est
apolaire agrave une threacuteonine (T) qui est polaire (Figure 20)
gtlcl||NWMN_2012|atpE|72493880 F0F1 ATP synthase subunit C
1 10 19 31 43 49 58
ATG AAT TTA ATC GCA GCA GCA ATC GCA ATT GGT TTA TCA GCA TTA GGA GCA GGT ATC GGT
M N L I A A A I A I G L S A L G A G I G
(S)
17 61 73 85 97 109 118
AAC GGT TTA ATC GTT TCA AGA ACA GTT GAA GGT GTA GCA CGT CAA CCA GAA GCA CGT GGT
N G L I V S R T V E G V A R Q P E A R G
21 40
121 133 145 157 169 178
CAA TTA ATG GGT ATC ATG TTC ATT GGT GTA GGT TTA GTT GAG GCA TTA CCT ATC ATC GGT
Q L M G I M F I G V G L V E A L P I I G
41 60
181 190 199 211
GTA GTA ATT GCA TTC ATG ACA TTT GCT GGA TAA
V V I A F M T F A G
(T)
61 64 70
Figure 20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du gegravene
atpE de S aureus Newman avec mise en eacutevidence des mutations (rose) en lien avec
la cible de la tomatidine Lrsquoalignement a eacuteteacute effectueacute agrave lrsquoaide du site CMR de JCVI
113
CHAPITRE 4
DISCUSSION
LrsquoATP synthase est eacutevidemment un complexe proteacuteique ubiquitaire tant chez les
procaryotes que les eucaryotes Malgreacute ce fait il semble que la cible de la tomatidine
soit speacutecifique aux bacillales celle-ci ne deacutemontrant aucune activiteacute contre les
bacteacuteries agrave Gram neacutegatif et les lactobacillales On suppose donc que lrsquoATP synthase
de ces espegraveces doit ecirctre speacutecifique au niveau du site de liaison de la tomatidine
Lrsquoecirctre humain en tant qursquoespegravece eucaryote est eacutegalement pourvu de ce complexe
proteacuteique et on peut naturellement se demander si lrsquoutilisation clinique de cette
moleacutecule pourrait ecirctre cytotoxique De fait plusieurs composeacutes naturels tel
lrsquooligomycine et la venturicidine sont connus pour inhiber lrsquoactiviteacute de lrsquoATP synthase
par leur interaction avec la sous-uniteacute c de ce complexe Cependant ces moleacutecules
ne sont pas seacutelectives et nrsquoinhibent pas seulement lrsquoATP synthase bacteacuterien mais
aussi lrsquoATP synthase mitochondrial (Haagsma et al 2009 Matsuno-Yagi et Hatefi
1993) (Figure 1) Neacuteanmoins il existe maintenant une moleacutecule qui a pour cible
lrsquoATP synthase bacteacuterien et qui est en processus en vue drsquoune utilisation clinique
chez lrsquohomme le TMC207 (Haagsma et al 2009))
114
Figure 1 Sites drsquoinhibition de lrsquoATP synthase Les sites de liaison dans lrsquoATP
synthase des inhibiteurs reacuteveacuteleacutes par des eacutetudes biochimiques et structurales sont
indiqueacutes par des cercles rouges et les sites de liaisons qui nrsquoont pas eacuteteacute totalement
clarifieacutes sont indiqueacutes par des cercles verts (Hong et Pedersen 2008)
Le TMC207 (Figure 2A) est un composeacute de la classe des diarylquinoline posseacutedant
un nouveau meacutecanisme drsquoaction lrsquoinhibition de lrsquoATP synthase bacteacuterien De fait la
cible et le meacutecanisme drsquoaction du TMC207 sont diffeacuterents des autres agents
antituberculeux Lrsquoinhibition de lrsquoATP synthase peut mener agrave une baisse de la
production drsquoATP et un deacutebalancement dans lrsquohomeacuteostasie du pH ces 2 effets
pouvant contribuer agrave la diminution de la survie de la bacteacuterie (Rao et al 2001
Andries et al 2005)
Le TMC207 est une moleacutecule ayant une forte activiteacute bacteacutericide contre
Mycobacterium tuberculosis et autres espegraveces mycobacteacuteriennes mais peu drsquoactiviteacute
115
contres les autres espegraveces bacteacuteriennes Lrsquoindex de seacutelectiviteacute de TMC207 pour
lrsquoATP synthase mycobacteacuterienne (CMI50 gt200 microml) compareacute agrave lrsquoATP synthase
mitochondriale (CMI50 001 microgml) est de plus de 20000 Un indice de seacutelectiviteacute au
dessus de 1000 est consideacutereacute dans le deacuteveloppement drsquoune drogue comme un
preacutealable pour une moleacutecule candidate (Haagsma et al 2009) La diversiteacute dans les
gegravenes atpE explique la reacutesistance au TMC207 et justifie la faible affiniteacute de cette
drogue agrave lrsquoATP synthase mitochondriale (Matteelli et al 2010) (Figure 3)
Figure 2 Structure chimique de (A) TMC207 et (B) son analogue anti bacteacuteries
agrave Gram positifs (Balemans et al 2012)
Lorsqursquoon compare la seacutequence proteacuteique de la sous-uniteacute c de lrsquoATP synthase
(atpE) de quelques espegraveces de lrsquoordre des Bacillales (tel Saureus B cereus B
subtilis et L monocytogenes) avec des espegraveces appartenant agrave drsquoautres ordres
bacteacuteriens on constate que les seacutequences divergent ce qui est en accord avec la
speacutecificiteacute des moleacutecules Les points de mutations amenant une reacutesistance au
TMC207 chez Mycobacterium smegmatis sont speacutecifiques aux mycobacteacuteries et les
deux principaux points de mutations D32V(ouP) et A67P chez M smegmatis sont
situeacutes tous les deux dans les heacutelices alpha qui entourent le site catalytique de la
proteacuteine (E65) qui est impliqueacute dans le transport des protons (Andries et al 2005)
Lrsquoacide amineacute aspartate (D) agrave la position 32 et lrsquoalanine (A) agrave la position 67 sont tregraves
speacutecifiques agrave M smegmatis
A B
116
Reacutecemment un groupe de chercheurs (Balemans et al 2012) on mit en lumiegravere une
autre moleacutecule (Figure 2B) de la classe des diarylquinolines dont le meacutecanisme
drsquoaction eacutetait aussi lrsquoinhibition de la sous-uniteacute c de lrsquoATP synthase mais chez les
bacteacuteries agrave Gram positif tels Streptococcus pneumoniae et S aureus Cette moleacutecule
irait lier les acides amineacutes valines (V) situeacutes agrave la position 50 et 62 de lrsquoAtpE de S
aureus (Donneacutees non montreacutees chez S pneumoniae(Balemans et al 2012)) En
effet une mutation amenant le changement de la valine en position 50 par une
isoleucine (I) etou une mutation amenant le changement de la valine en position 62
par une alanine (A) amegravene la reacutesistance de la bacteacuterie agrave la moleacutecule Cependant L
monocytogenes et Bacillus subtilis ne possegravedent pas de valine agrave la position 50 alors
qursquoils font eacutegalement partie des bacteacuteries agrave Gram positif (Il faut noter que chez
Bacillus sp PS3 (Swiss-Prot assession Ndeg P 00845) possegravede une valine agrave la position
50 (Balemans et al 2012)) Par contre tout comme pour les autres bacteacuteries agrave Gram
positif testeacutees les deux espegraveces possegravedent une valine agrave la position 62 (Figure 3)
Chez B cereus B subtilis B anthracis S aureus et L monocytogenes les points de
divergence sont plutocirct A17S et A64T Cependant B pseudofirmus qui fait eacutegalement
partie de lrsquoordre des Bacillales possegravede une glycine (G) agrave la position 17 plutocirct qursquoune
alanine (A) et une seacuterine (S) plutocirct qursquoune alanine (A) agrave la position 64 comme les
espegraveces preacuteceacutedentes Suivant ces reacutesultats et si la cible de la tomatidine est bien
ces deux points de mutation la tomatidine ne devrait pas pouvoir se lier agrave lrsquoAtpE de B
pseudofirmus et devrait donc nrsquoavoir aucune activiteacute contre cette souche alors que
nous avions extrapoleacute que le spectre drsquoactiviteacute de la tomatidine eacutetait tous les
Bacillales De plus E coli et M smegmatis possegravedent eacutegalement une alanine (A) agrave la
position 17 alors que la tomatidine ne possegravede aucune activiteacute contre ces espegraveces (M
smegmatis nrsquoa pas eacuteteacute eacutevalueacute il srsquoagit ici drsquoune extrapolation des reacutesultats de CMI)
par contre ces deux espegraveces possegravedent des acides amineacutes diffeacuterents drsquoune alanine
(A) agrave la position 64 ce qui pourrait leur confeacuterer une reacutesistance agrave la tomatidine telle
qursquoobserveacutee On remarque eacutegalement que les acides amineacutes agrave ces deux positions
117
sont totalement diffeacuterents chez lrsquohomme (Homo sapiens) ce qui suggeacutererait que la
tomatidine ne possegravederait pas drsquoactiviteacute antimitochondriale (Figure 3)
Selon ces reacutesultats on peut se demander si les deux mutations sont essentielles agrave la
liaison de la moleacutecule ou si seulement lrsquoalanine (A) agrave la position 64 est neacutecessaire
On doit se rappeler que la reacutesistance apporteacutee par la mutation faisant en sorte que
lrsquoalanine (A) en position 17 devient une seacuterine (S) apparaicirct chez les mutants
NewbouldΔhemb reacutesistant agrave la tomatidine et que cette mutation nrsquoapparaicirct qursquoau
quatriegraveme et dernier niveau de reacutesistance On se rappelle eacutegalement que la mutation
faisant en sorte que lrsquoalanine (A) agrave la position 64 devient une threacuteonine (T) apparaicirct
chez les mutants NewbouldΔhemb reacutesistants agrave la tomatidine en combinaison avec la
gentamicine et ce degraves le premier niveau de reacutesistance
118
Bcereus G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Banthracis G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Lmonocytogenes G---------VIAAAIAVG---LGALGAGIGNGLIVSKTVEGVARQPEARSMLQTIMFV- 50
Saureus_NWMN_2012 -----------IAAAIAIG---LSALGAGIGNGLIVSRTVEGVARQPEARGQLMGIMFIG 49
Bsubtilis -----------IAAAIAIG---LGALGAGIGNGLIVSRTVEGIARQPEAGKELRTLMFMG 49
Bpseudofirmus -----------LGAAIAAG---LAAVAGAIAVAIIVKATIEGTTRQPELRGTLQTLMFIG 49
Ecoli L---------YMAAAVMMG---LAAIGAAIGIGILGGKFLEGAARQPDLIPLLRTQFFIV 56
Msmegmatis DPNAIITAGALIGGGLIMG---GGAIGAGIGDGIAGNALISGIARQPEAQGRLFTPFFIT 60
Homo sapiens QTSAISRDIDTAAKFIGAGAATVGVAGSGAGIGTVFGSLIIGYARNPSLKQQLFSYAILG 120
MyR V
MyR P
SaR S
Bcereus VALVEALPIIGVVIAFIVMNK----- 72
Banthracis VALVEALPIIGVVIAFIVMNK----- 72
Lmonocytogenes ----EALPIIAVVIAFMVLNK----- 67
Saureus_NWMN_2012 VGLVEALPIIGVVIAFMTFAG----- 70
Bsubtilis IALVEALPIIAVVIAFLAFFG----- 70
Bpseudofirmus VPLAEAVPIIAIVISLLILF------ 69
Ecoli MGLVDAIPMIAVGLGLYVMFAVA--- 79
Msmegmatis VGLVEAAYFINLAFMALFVFATPGLQ 86
Homo sapiens FALSEAMGLFCLMVAFLILFAM---- 142
MyR D P M
SpR I A
SaR T
Fig
ure
3
Seacuteq
uen
ce
pro
teacuteiq
ue d
e la p
roteacute
ine
Atp
Ec
hez d
iffeacute
ren
tes
es
pegravec
es e
t m
ise e
n eacute
vid
en
ce
de
s s
ite
s d
e
mu
tati
on
pe
rmett
an
t la
reacutesis
tan
ce agrave
dif
feacutere
nte
s m
oleacute
cu
les
M
yR M
tu
berc
ulo
sis
ou
M sm
eg
matis
reacutesis
tan
t a
u
TM
C2
07 S
pR
S
p
neu
mon
iae
reacutesis
tan
t agrave
un a
na
log
ue d
u T
MC
20
7
SaR
S
a
ure
us
SC
V r
eacutesis
tant agrave
la to
matid
ine
En
cad
reacutero
uge
S
ite d
e m
uta
tion a
men
ant la
reacutesis
tance
agravela
to
matid
ine E
ncad
reacuteno
ir
Site
cata
lytiq
ue
im
pliq
ueacute
da
ns le
tra
nspo
rt d
es p
roto
ns
Su
rlig
nag
etu
rqu
ois
e S
ite d
e m
uta
tion a
me
nant u
ne
reacutesis
tan
ce a
u T
MC
20
7
119
Les seacutequences proteacuteiques des souches bacteacuteriennes de la figure 3 ont eacuteteacute trouveacutees
sur pubmedproteacuteine httpwwwncbinlmnihgovpubmed
laquo ATP synthase subunit c raquo Assession number
Streptococcus pneumoniae D39 YP_8168031 (NCBI)
S aureus Newman BAF682841 (GenBank)
E coli AAA247321 (GenBank)
Bacillus cereus ISO75 EJP900901 (GenBank)
Bacillus anthracis str H9401 YP_0062123491 (NCBI)
Mycobacterium smegmatis MC2 155 YP_0065695541 (NCBI)
Bacillus subtilis str 168 CAA822551 (GenBank)
Listeria monocytogenes YP_0076049931 (NCBI)
Homo sapiens P482011 (UniProtKBSwiss-Prot)
Lrsquoalignement comparatif des seacutequences drsquoacides amineacutes a eacuteteacute fait agrave lrsquoaide du
programme ClustalW2 sur le web
httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2
Le modegravele 3D de lrsquoAtpE a eacuteteacute obtenu gracircce agrave lrsquoaide du Pr Ryszard Brzezinski du
Deacutepartement de biologie de lrsquoUniversiteacute de Sherbrooke (Figure 4) Une recherche
dans le Protein Data Bank a drsquoabord eacuteteacute effectueacutee afin de veacuterifier parmi les proteacuteines
AtpE (sous-uniteacute C de lrsquoATP synthase) pour lesquelles il y avait des structures 3D
reacutesolues laquelle ressemblait le plus agrave celle de Staphylococcus aureus Les
alignements ont montreacute que cest la proteacuteine de Bacillus pseudofirmus qui avait la
plus grande similitude de seacutequence La structure est stockeacutee dans la banque PDB
sous le symbole 3ZO6 Il a eacuteteacute confirmeacute par quelques alignements que cette sous-
uniteacute C de lrsquoATP synthase eacutetait plus proche par sa seacutequence de celle de
Staphylococcus aureus que de celle de Mycobacterium pour laquelle la structure 3D
avait eacuteteacute deacutetermineacutee plus tocirct Ce reacutesultat sexplique par le fait que Staphylococcus et
Bacillus font tous deux partie de la branche A+T-riche des bacteacuteries agrave Gram+ alors
que Mycobacterium appartient agrave la branche G+C-riche La seacutequence de lAtpE de
120
Staphylococcus aureus a ensuite eacuteteacute envoyeacutee au serveur internet de modeacutelisation
3D-Jigsaw (httpbmmcancerresearchukorg~3djigsaw) en indiquant que le modegravele
devait ecirctre bacircti en prenant comme matrice le fichier PDB 3ZO6 Le modegravele nous a
eacuteteacute retourneacute sous format PDB ndash comme les laquo vrais raquo cristaux dans la Protein Data
Bank Les fichiers au format PDB peuvent ecirctre visualiseacutes par le logiciel RasMol ndash
gratuit sur internet Cest avec RasMol que limage (Figure 4) a eacuteteacute obtenue
121
Figure 4 Modeacutelisation de la structure tridimensionelle de la sous-uniteacute c de
lrsquoATP synthase de S aureus (AtpE) montrant les acides amineacutes muteacutes chez les
mutants reacutesistantsEn blanc et vert les deux acides amineacutes muteacutes chez S aureus
reacutesistant agrave la tomatidine En rose les sites de mutations chez Mycobacterium
smegmatis reacutesistant au TMC207 et des sites de mutations chez Streptoccocus
pneumoniae reacutesistants agrave un analogue de TMC207 En bleu le site actif de la proteacuteine
Agrave la vue de ces reacutesultats certains pourront mettre en doute lrsquointeacuterecirct de la tomatidine
eacutetant donneacute lrsquoapparition relativement rapide de reacutesistance chez S aureus Cependant
122
nos reacutesultats deacutemontrent clairement que lrsquoajout de tomatidine ralentit le
deacuteveloppement de reacutesistance agrave la gentamicine chez S aureus ATCC29213 (Tableau
1) De plus cette augmentation de la CMI de la gentamicine en preacutesence de
tomatidine au fil des passages suit lrsquoaugmentation de la CMI de la ciprofloxacine lors
de ces mecircmes passages alors que la ciprofloxacine est consideacutereacutee comme la drogue
de reacutefeacuterences dans les bonnes drogues (Tableau 1) Tout comme pour la
ciprofloxacine pour laquelle la reacutesistance chez S aureus peut ecirctre la conseacutequence
de deux mutations dans le gegravene cible gyrA la reacutesistance agrave la tomatidine peut ecirctre la
conseacutequence de deux mutations dans le gegravene cible atpE Par comparaison la
rifampicine est consideacutereacutee comme une mauvaise drogue de par lrsquoapparition rapide
drsquoune forte reacutesistance chez S aureus conseacutequence de ses 17 mutations possibles
dans le gegravene cible rpoB (Tableau 1)
De plus il sera important de relativiser lrsquoapparition de reacutesistance agrave la tomatidine en
calculant le taux drsquoapparition de ces colonies reacutesistantes ceci nrsquoayant pas eacuteteacute fait
jusqursquoici De fait la freacutequence de mutation chez S aureus amenant une reacutesistance agrave
la rifampicine est de lrsquoordre de 10-7 10-8 alors qursquoil est de 10-11 pour lrsquoacide fusidique
(Canfield et al 2013 Orsquoneill et al 2001 Price et Gustafson 2001) Pour ce qui est
de la nouvelle moleacutecule anti-bacteacuteries agrave Gram positif de la classe des diarylquinoline
une freacutequence de mutation sur S pneumoniae ATCC49619 de 81- 83 x 10-7 a eacuteteacute
observeacutee agrave 5x la CMI et une freacutequence de mutation de 46 x 10-7 agrave 17 x 10-6 agrave 50x la
CMI (Balemans et al 2012)
123
Tableau 1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 par la meacutethode de CMI lors de passages successifs dans un milieu avec
lrsquoantibiotique
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
Typiqueldquomauvaiserdquodrogue (17 mutations possibles dansrpoB)
Typique ldquobonnerdquo drogue (2 mutations possibles dans gyrA)
Preacutesence de TO ralenti le deacuteveloppement de reacutesistances vs GEN seul
124
CHAPITRE 5
CONCLUSION ET PERSPECTIVES
Mon projet de maitrise a permis de preacuteciser le spectre drsquoactiviteacute de la tomatidine
Nous savons maintenant que la tomatidine possegravede une activiteacute antibacteacuterienne
contre la plupart des Firmicutes et plus speacutecifiquement contre les Bacillales Plus
preacuteciseacutement la tomatidine est bacteacutericide contre les pheacutenotypes SCV et agit en
synergie avec les aminoglycosides augmentant lrsquoactiviteacute bacteacutericide de ces derniers
de 8-16 fois contre les souches agrave pheacutenotype normal
De plus parmi tous les analogues de la tomatidine produits par le laboratoire du Pr
Eacuteric Marsault (voir article en annexe 2) le composeacute FC04-100 a eacuteteacute plus
speacutecifiquement mis en lumiegravere Celui-ci conserve non seulement une aussi bonne
activiteacute synergique avec les aminoglicosides contre les Saureus au pheacutenotype
normal et une activiteacute modeacutereacutee contre les S aureus SCV mais montre eacutegalement de
faccedilon inespeacutereacutee une activiteacute par lui-mecircme contre les S aureus de pheacutenotype normal
incluant plusieurs MRSA une proprieacuteteacute qui nrsquoeacutetait pas observeacutee pour la tomatidine
Une autre activiteacute inteacuteressante de FC04-100 est sa capaciteacute drsquoameacuteliorer lrsquoactiviteacute
bacteacutericide des aminoglicosides contre L monocytogenes agrave pheacutenotype normal apregraves
un traitement de 24h alors que la tomatidine ne deacutemontrait qursquoune faible synergie
avec les aminoglycosides contre cette souche De plus FC04-100 deacutemontre une
activiteacute bacteacutericide contre les SCV de L monocytogenes alors que la tomatidine ne
deacutemontrait qursquoune activiteacute bacteacuteriostatique FC04-100 montre eacutegalement un effet
potentiateur de lrsquoUbrolexinTM (Ceacutefalexin Kanamycin (3 2)) une combinaison
drsquoantibiotique utiliseacute dans le traitement de mammites bovines en abaissant de 8 agrave 16
fois leur CMI amenant celles-ci sous le seuil de reacutesistance Finalement FC04-100
possegravede une meilleure solubiliteacute que la tomatidine une proprieacuteteacute qui facilitera son
utilisation in vitro
125
Nos reacutesultats deacutemontrent que FC04-100 conserve une certaine activiteacute contre les
SCV reacutesistant agrave la tomatidine ce qui suggegravere possiblement que FC04-100 a plus
daffiniteacute pour la cible ou plus de sites dinteraction avec AtpE En effet les mutants
SCV reacutesistants agrave la tomatidine deviennent gt512 fois plus reacutesistants alors que FC04-
100 ne perd que 8 agrave 16 fois son activiteacute contre ces mecircmes souches Ceci deacutemontre
quil est possible dameacuteliorer significativement lactiviteacute de ces alkaloiumldes steacuteroidiens
ce qui est tregraves prometteur pour lavenir de cette moleacutecule
Dans un second temps lrsquoanalyse des mutations chez les mutants reacutesistants agrave la
tomatidine a permis de non seulement de mieux comprendre le mode drsquoaction de la
tomatidine mais eacutegalement de preacuteciser la cible potentielle de ce nouvel antibiotique
De fait son mode drsquoaction serait en lien direct avec la fonctionnaliteacute de la chaine de
transport des eacutelectrons chez la bacteacuterie lrsquoactiviteacute de la tomatidine serait inversement
proportionnelle agrave la fonctionnaliteacute de la chaine de transport des eacutelectrons et donc en
lien eacutegalement avec la polarisation membranaire et la production de ROS
intracellulaire Afin de confirmer le modegravele-hypothegravese la mesure de la polarisation
membranaire et la concentration de ROS intracellulaire chez les mutants ainsi que
chez les souches parentes suite agrave un traitement antibiotique devront ecirctre effectueacutees
Les reacutesultats de mes recherches suggegraverent que la cible bacteacuterienne speacutecifique de la
tomatidine serait la sous-uniteacute C de lrsquoATP synthase Afin de confirmer la cible
Maxime Lamontagne-Boulet tente au moment drsquoeacutecrire ces lignes drsquoobtenir des
mutants de S aureus Newman reacutesistants au composeacute FC04-100 et dont la reacutesistance
serait attribuable agrave une mutation dans le gegravene atpE Par la suite on devra confirmer
la fonctionnaliteacute de la cible ATP synthase par quantification de lrsquoATP produit agrave lrsquoaide
de la lucifeacuterase tel que lrsquoon fait Balemans et al en 2012 Lrsquoattachegravement agrave la cible
devra eacutegalement ecirctre confirmeacute et la meacutethode de SPR (Surface Plasmon Resonance)
pourrait ecirctre tout indiqueacutee pour ce faire (Balemans et al 2012) On devra eacutegalement
126
deacutemontrer la seacutelectiviteacute des moleacutecules tomatidine et FC04-100 pour lrsquoATP synthase
des Bacillales
Les infections agrave bacteacuteries multireacutesistantes deviennent de plus en plus preacutevalentes et
sont un des problegravemes de santeacute majeurs auxquels nous devons faire face aujourdrsquohui
Cette augmentation de la reacutesistance limite le reacutepertoire drsquoantibiotiques efficace creacuteant
une situation probleacutematique laquelle est exacerbeacutee par le trop petit nombre de
nouveaux antibiotiques introduits sur le marcheacute ces derniegraveres anneacutees Il est donc
drsquoune grande importance de travailler agrave la mise en marcheacute de nouveaux antibiotiques
et plus particuliegraverement de nouvelles classes drsquoantibiotiques espeacuterant ainsi deacutejouer la
multireacutesistance bacteacuterienne du moins pour un temps La tomatidine de par sa
structure sa cible et son mode drsquoaction speacutecifique est une moleacutecule antibiotique
dans une classe agrave part et de ce fait une moleacutecule plus qursquointeacuteressante pour le
traitement drsquoinfection agrave S aureus De plus comme elle deacutemontre une activiteacute
bacteacutericide contre les deux pheacutenotypes de S aureus (et autres Bacillales) lesquels
sont retrouveacutes simultaneacutement dans les cas drsquoinfections chroniques son pouvoir
antibiotique devient inestimables drsquoun point de vue clinique
127
CONTRIBUTIONS SUPPLEacuteMENTAIRES
ANNEXE 1 Application de brevet Internationnal publication number WO
2012109752 A1
INVENTEURS Franccedilois Malouin2 Gabriel Mitchell2 Kamal Bouarab2 Eacuteric Marsault1
Feacutelix Chagnon1 Simon Boulanger 2 Isabelle Guay2
1Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
2Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
128
129
ANNEXE 2 Article scientifique dans laquo Europeen Journal of medicinal chemistry raquo
2014 80 605-620
TITRE Unraveling the structure-activity relationship of tomatidine a steroid alkaloid
with unique antibiotic properties against persistent forms of
Staphylococcus aureus
ABSTRACT Staphylococcus aureus is responsible for difficult-to-treat and relapsing
infections and constitutes one of the most problematic pathogens due to its multiple
resistances to clinically available antibiotics Additionally this pathogenrsquos ability to
develop small-colony variants is associated with a reduced susceptibility to
aminoglycoside antibiotics and in vivo persistence We have recently demonstrated
that tomatidine a steroid alkaloid isolated from tomato plants possesses anti-
virulence activity against normal strains of S aureus as well as the ability to
potentiate the effect of aminoglycosides against this pathogen In addition tomatidine
has shown antibiotic activity against small-colony variants We herein report the first
study of the structure-activity relationship of tomatidine against S aureus
AUTEURS Feacutelix Chagnon1 Isabelle Guay2 Marc-Andreacute Bonin1 Gabriel Mitchell2
Kamal Bouarab2Franccedilois Malouin2 Eacuteric Marsault1
MA CONTRIBUTION Responsable des tests biologiques de tous les composeacutes
autant envers lrsquoeffet potentiateur de la tomatidine sur lrsquoactiviteacute de la gentamicine
contre la souche ATCC29213 de S aureus que pour lrsquoactiviteacute bacteacuteriostatique sur la
souche NewbouldΔhem imitant le pheacutenotype SCV Ces travaux furent effectueacutes sous
la tutelle du professeur Franccedilois Malouin
130
1Institut de Pharmacologie de Sherbrooke Deacutepartement de pharmacologie
Universiteacute de Sherbrooke 3001 12e av nord Sherbrooke Queacutebec Canada J1H
5N4
2Centre drsquoeacutetude et de valorisation de la diversiteacute microbienne (CEVDM) Deacutepartement
de biologie Universiteacute de Sherbrooke Sherbrooke QC Canada J1K 2R1
131
BIBLIOGRAPHIE Alexander E H and Hudson MC (2001) Factors influencing the internal- ization of Staphylococcus aureus and impacts on the course of infections in humans Appl Microbiol Biotechnol 56 361ndash366
Andries K Verhasselt P Guillemont J Goumlhlmann HW Neefs JM Winkler H Van Gestel J Timmerman P Zhu M Lee E et al (2005) A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis Science 307 223-227
Artzatbanov VY and Petrov VV (1990) Branched respiratory chain in aerobically grown Staphylococcus aureus--oxidation of ethanol by cells and protoplasts Arch Microbiol 153 580- 584 Archer GL (1998) Staphylococcus aureus a well-armed pathogen Clin Infect Dis 26 1179ndash1181 Assefa S Keane TM Otto TD Newbold C and Berriman M (2009) ABACAS algorithm-based automatic contiguation of assembled sequences Bioinformatics 25 1968-1969 Aubry-Damon H Legrand P Brun-Buisson C Astier A Soussy CJ and Leclercq R (1997) Reemergence of gentamicin-sus- ceptible strains of methicillin-resistant Staphylococcus aureus roles of an infection control program and changes in aminoglycoside use Clin Infect Dis 25 647-653
Balemans W Vranckx L Lounis N Pop O Guillemont J Vergauwen K Mol S Gilissen R Motte M Lanccedilois et al (2012) Novel antibiotics targeting respiratory ATP synthesis in Gram-positive pathogenic bacteria Antimicrob Agents Chemother 56 4131-4139
Batandier C Guigas B Detaille D El-Mir MY Fontaine E Rigoulet M and Leverve XM (2006) The ROS production induced by a reverse-electron flux at respiratory-chain complex 1 is hampered by metformin J Bioenerg Biomembr 38 33ndash 42
132
Beckman Coulter website httpswwwbeckmancoultercomwsrportalwsrresearch-and-discoveryproducts-and-servicesnucleic-acid-sample-preparationspri-laboratory-automationindexhtm (consulteacute le 3 Deacutec 2013) Bednarek P and Osbourn A (2009) Plant-microbe interactions chemical diversity in plant defense Science 324 746ndash748 Besier S Ludwig A Ohlsen K Brade V and Wichelhaus TA (2007) Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus Int J Med Microbiol 297 217-225 BiteSizeBio Brain food for biologists httpnxseqbitesizebiocomarticlessequencing-by-synthesis-explaining-the-illumina-sequencing-technology (consulteacute le 3 deacutec 2013) Boetzer M and Pirovano W (2012) Toward almost closed genomes with GapFiller Genome Biology 13 R56 Bouarab K Ordi E Gattuso M Moisan H and Malouin F (2007) Plant stress response agents affect Staphylococcus aureus virulence genes Abstr 47th Intersci Conf Antimicrob Agents Chemother abstr C1-1483 Bryan LE and Kwan S (1981) Aminoglycoside-resistant mutants of Pseudomonas aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob Agents Chemother 19 958- 964 Bryan LE and Kwan S (1983) Roles of ribosomal binding membrane potential and electron transport in bacterial uptake of streptomycin and gentamicin Antimicrob Agents Chemother 23 835ndash845
Bryan LE Nicas T Holloway BW and Crowther C (1980) Aminoglycoside-resistant mutation of Pseudomonas aeruginosa defective in cytochrome C552 and nitrate reductase Antimicrob Agents Chemother 17 71- 79
133
Bryan LE and van den Elzen HM (1977) Effects of membrane-energy mutations and cations on streptomycin and gentamicin accumulation by bacteria a model for entry of streptomycin and gentamicin in susceptible and resistant bacteria Antimicrob Agents Chemother 12 163-177 Brynildsen MP Winkler JA Spina CS MacDonald IC and Collins JJ (2013) Potentiating antibacterial activity by predictably enhancing endogenous microbial ROS production Nat Biotechnol 31 160ndash 165 Burke KA and Lasccelles J (1975) Nitrate reductase system in Staphylococcus aureus wild type and mutants J Bacteriol 123 308- 316 Burke KA and Lascelles J (1976) Nitrate reductase activity in hegraveme-deficient mutants of Staphylococcus aureus J Bacteriol 126 225- 231 Canfield GS Schwingel JM Foley MH Vore KL Boonanantanasarn K Gill AL Sutton MD and Gill SR (2013) Evolution in fast forward a potential role for mutators in accelerating Staphylococcus aureus pathoadaptation J Bacteriol 195 615-628 Chang S Sievert DM Hageman JC Boulton ML Tenover FC Downes FP Shah S Rudrik JT Pupp GR Brown WJ et al (2003) Infection with vancomycin- resistant Staphylococcus aureus containing the vanA resistance gene N Engl J Med 348 1342-1347 Chen Q Vazquez EJ Moghaddas S Hoppel CL and Lesnefsky EJ (2003) Production of reactive oxygen species by mitochondria central role of complex III J Biol Chem 278 36027-36031 Clements MO Watson SP Poole RK and Foster SJ (1999) CtaA of Staphylococcus aureus is required for starvation survival recovery and cytochrome biosynthesis J Bacteriol 181 501- 507
134
Clinical and Laboratory Standards Institute (2011) Performance standards for antimicrobial susceptibility testing Twenty-first informational supplement M100-S21 CLSI Wayne PA USA ClustalW2 httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2 (consulteacute le 3 deacutec 2013) Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) httpcmrjcviorgtigr-scriptsCMRCmrHomePagecgi (consulteacute le 3 deacutec 2013) Creacutepin-Gilbert C (2006) Transfert de ligand dans la cytochrome c oxydase observeacute par des expeacuteriences femtosecondes infrarouge inteacutegreacutees et reacutesolues spectralement Thegravese de doctorat de l rsquoEacutecole Polytechnique Paris Pastel-00002200 version 1 29 jul 2010 Davis BD (1987) Mechanism of bactericidal action of aminoglycosides Microbiol Rev 51 341-350 DeAngelis MM Wang DG and Hawkins TL (1995) Solid-phase reversible immobilization for the isolation of PCR products Nucl Acids Res 22 4742- 4743 Dickgiesser N and Kreiswirth BN (1986) Determination of aminoglycoside resistance in Staphylococcus aureus by DNA hybridization Antimicrob Agents Chemother 29 930-932 Doi Y and Arakawa Y (2007) 16S ribosomal RNA methylation Emerging resistance mechanism against aminoglycosides Clin Infect Dis 45 88ndash94 Enright MC Robinson DA Randle G Feil EJ Grundmann H and Spratt BG (2002) The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA) Proc Natl Acad Sci USA 99 7687-7692 Friedman M (2002) Tomato glycoalkaloids role in the plant and in the diet J Agric Food Chem 50 5751ndash5780
135
Goerke C and Wolz C (2004) Regulatory and genomic plasticity of Staph- ylococcus aureus during persistent colonization and infection Int J Med Microbiol 294 195ndash202 Goumltz F Bannerman T and Schleifer K-H (2006) The genera Staphylococcus and Macrococcus In The Prokaryotes Vol 4 Bacteria Firmicutes Cyanobacteria M Dworkin (ed) (Springer New-York USA) pp5- 75 Haagsma AC Abdillahi-Ibrahim R Wagner MJ Krab K Vergauwen K Guillemont J AndriesK Lill H Koul A and Bald D (2009) Selectivity of TMC207 towards mycobacterial ATP synthase compared with that towards the eukaryotic homologue Antimicrob Agents Chemother 53 1290-1292 Hammer ND Reniere ML Cassat JE Zhang Y Hirsch AO Indriati Hood M and Skaar EP (2013) Two hegraveme-dependent terminal oxydases power Staphylococcus aureus organ-specific colonization of the vertebrate host MBio 4 e00241- 13 Hederstedt L (1993) The Krebs citric acid cycle In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of Microbiology Washington DC) pp 181-198 Hiramatsu K (1995) Molecular evolution of MRSA Microbiol Immunol 39 531-543 Hiramatsu K Hanaki H Ino T Yabuta K Oguri T and Tenover FC (1997) Methicillin-resistant Staphylococcus aureus clinical strain with reduced vancomycin susceptibility J Antimicrob Chemother 40 135-136 Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T et al (2013) Genomic Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
136
Hong S and Pedersen PL (2008) ATP synthase and the actions of inhibitors utilized to study its roles in human health disease and other scientific areas Microbiol Mol Biol Rev 72 590-641
Jevons MP (1961) lsquoCelbeninrsquo-resistant staphylococci BMJ 1 124-125
Joshi S and Huang YG (1991) ATP synthase complex from bovine heart mitochondria the oligomycin sensitivity conferring protein is essential for dicyclohexyl carbodiimidesensitive ATPase Biochim Biophys Acta 1067 255ndash258
Kohanski MA Dwyer DJ and Collins JJ (2010) How antibiotics kill bacteria from targets to networks Nat Rev Microbiol 8 423-435 Kohler C von Eiff C Liebeke M McNamara PJ Lalk M Proctor RA Hecker M and Engelmann S (2008) A defect in menadione biosynthesis induces global changes in gene expression in Staphylococcus aureus J Bacteriol 190 6351- 6364 Kohler C von Eiff C Peters G Proctor RA Hecker M and Engelmann S (2003) Physiological characterization of a hegraveme-deficient mutant of Staphylococcus aureus by a proteomic approach J Bacteriol 185 6928- 6937 Kriegeskorte A Koumlnig S Sander G Pirkl A Mahabir E Proctor RA von Eiff C Peters G and Becker K (2011) Small colony variants of Staphylococcus aureus reveal distinct protein profiles Proteomics 11 2476-2490 Lannergard J von Eiff C Sander G Cordes T Seggewiss J Peters G Proctor RA Becker K and Hughes D (2008) Identification of the genetic basis for clinical menadione-auxotrophic small-colony variant isolates of Staphylococcus aureus Antimicrob Agents Chemother 52 4017-4022
Lauraeus M and Wikstrom M (1993) The terminal quinol oxydases of Bacillus subtilis have different energy conservation properties J Biol Chem 268 11470-11473
137
Li M Gong J Cottrill M Yu H de Lange C Burton J and Topp E (2003) Evaluation of QIAamp DNA Stool Mini Kit for ecological studies of gut microbiota J Microbiol Methods 54 13-20
Liu Y Fiskum G and Schubert D (2002) Generation of reactive oxygen species by the mitochondrial electron transport chain J Neurochem 80 780-787
Martin-Hernandez A M Dufresne M Hugouvieux V Melton R and Osbourn A (2000) Effects of targeted replacement of the tomatinase gene on the interaction of Septoria lycopersici with tomato plants Mol Plant Microbe Interact 13 1301ndash1311
Matteelli A Carvalho AC Dooley KE and Kritski A (2010) TMC207 the first compound of a new class of potent anti-tuberculosis drugs Future Microbiol 5 849-858
Matsuno-Yagi A and Hatefi Y (1993) Studies on the mechanism of oxidative phosphorylation ATP synthesis by submitochondrial particles inhibited at F0 by venturicidin and organotin compounds J Biol Chem 268 6168ndash6173
Miller MH Edberg SC Mandel LJ Behar CF and Steigbigel NH (1980) Gentamicin uptake in wild-type and aminoglycoside-resistant small-colony mutants of Staphylococcus aureus Antimicrob Agents Chemother 18 722- 729
Mingeot-Leclercq M-P Glupczynski Y and Tulkens PM (1999) Aminoglycosides Activity and Resistance Antimicrob Agents Chemother 43 727- 737
Mitchell G Brouillette Eacute Seacuteguin DL Asselin AE Jacob CL and Malouin F (2010) A role for sigma factor B in the emergence of Staphylococcus aureus small-colony variants and elevated biofilm production resulting from an exposure to aminoglycosides Microb Pathog 48 18-27
Mitchell G Gattuso M Bouarab K and Malouin F (2009) Tomatidine (TO) affects virulence regulators of prototypical Staphylococcus aureus (SA) and small colony variants (SCV) of cystic fibrosis (CF) patients Abstr 49th Intersci Conf Antimicrob Agents Chemother abstr C1-1341
138
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K and Malouin F (2011) Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic fibrosis airway epithelial cells Antimicrob Agents Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM and Malouin F (2011) Infection of polarized airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function and is influenced by NF-κB Infect Immun 79 3541-3551
Mitchell G Lafrance M Boulanger S Seacuteguin DL Guay I Gattuso M Marsault Eacute Bouarab K and Malouin F (2012) Tomatidine acts in synergy with aminoglycoside antibiotics against multiresistant Staphylococcus aureus and prevents virulence gene expression J Antimicrob Chemother 67 559-568
Mitchell G Lamontagne CA Brouillette Eacute Grondin G Talbot BG Grandbois M and Malouin F (2008) Staphylococcus aureus SigB activity promotes a strong fibronectin-bacterium interaction which may sustain host tissue colonization by small-colony variants isolated from cystic fibrosis patients Mol Microbiol 70 1540-1555
Mitchell G Malouin F 2012 Outcome and prevention of Pseudomonas aeruginosa-Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic Fibrosis Chap 8 pp 181-206 Ed D Sriramulu InTech Open Access Publisher ISBN 979-953-307-059-8
Mitchell G Seacuteguin DL Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S and Malouin F (2010) Staphylococcus aureus sigma B-dependent emergence of small-colony variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-heptylquinoline-N-oxide BMC Microbiol 30 10-33
Moisan H Brouillette Eacute Jacob CL Langlois-Beacutegin P Michaud S and Malouin F (2006) Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated from cystic fibrosis patients is influenced by SigB J Bacteriol 188 64-76
139
Moreno F Crisp C Jorgensen JH and Patterson JE (1995) Methicillin- resistant Staphylococcus aureus as a community organism Clin Infect Dis 21 1308-1312
Murphy MP (2009) How mitochondria produce reactive oxygen species Biochem J 417 1- 13
Novick RP (2003) Autoinduction and signal transduction in the regulation of staphylococcal virulence Mol Microbiol 48 1429-1449
Novo DJ Perlmutter NG Hunt RH and Shapiro HM (2000) Multiparameter flow cytometric analysis of antibiotic effects on membrane potential membrane permeability and bacterial counts of Staphylococcus aureus and Micrococcus luteus Antimicrob Agents Chemother 44 827-834
ONeill AJ Cove JH and Chopra I (2001) Mutation frequencies for resistance to fusidic acid and rifampicin in Staphylococcus aureus J Antimicrob Chemother 47 647-650
Osbourn A E (1996) Preformed antimicrobial compounds and plant defense against fungal attack Plant Cell 8 1821ndash1831
Otto TD Dillon GP Degrave WS and Berriman M (2011) RATT Rapid annotation transfert tool Nucl Acids Res 39 doi101093 Pratique E Theraulaz L (2007) Eacutetude de la reacutegulation du systegraveme respiratoire TMAO reacuteductase chez Escherichia coli Meacutemoire de maitrise de lrsquoeacutecole pratique des hautes eacutetudes Paris
Price CT and Gustafson JE (2001) Increases in the mutation frequency at which fusidic acid-resistant Staphylococcus aureus arise with salicylate J Med Microbiol 50 104-106
Proctor RA and Peters G (1998) Small colony variants in staphylococcal infections Diagnostic and therapeutic implications Clin Infect Dis 27 419- 423
140
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M and Peters G (2006) Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent infections Nat Rev Microbiol 4 295ndash305
Quail MA Kozarewa I Smith F Scally A Stephens PJ Durbin R Swerdlow H and Turner DJ (2008) A large genome centers improvements to the Illumina sequencing system Nat methods 5 1005-1010
Ramirez MS and Tolmasky ME (2010) Aminoglycoside modifying enzymes Drug Resist Updat 13 151ndash171
Rao M Streur TL Aldwell FE and Cook GM (2001) Intracellular pH regulation by Mycobacterium smegmatis and Mycobacterium bovis BCG Microbiology 147 1017-1024
Richardson DJ (2000) Bacterial respiration a flexible process for a changing environment Microbiology 146 551- 571
Riley TV Pearman JW and Rouse IL (1995) Changing epidemiology of methicillin-resistant Staphylococcus aureus in Western Australia Med J Aust 163 412-414
Roddick J G (1974) The steroidal glycoalkaloid α-tomatine Phytochemistry 13 25-36
Rodrigue S Materna AC Timberlake SC Blackburn MC Malmstrom RR et al(2010) Unlocking Short Read Sequencing for Metagenomics PLoS ONE 5(7) e11840doi101371journalpone0011840 Ruiz-Rubio M Alonso Peacuterez-Espinosa A Lairini K RoldAacuten-Arjona T Dipietro A and Anaya N (2001) Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi In Studies in natural products chemistry vol 25 A Rahman (ed) (Elsevier Oxford United Kingdom) pp 293ndash326
141
Samuelsen oslash Haukland HH Kahl BC von Eiff C Proctor RA Ulvatne H Sandvik K and Vorland LH (2005) Staphylococcus aureus small colony variants are resistant to the antimicrobial peptide lactoferricin B J Antimicrob Chemother 56 1126- 1129
Sandrock R W and Vanetten HD (1998) Fungal sensitivity to and enzymatic degradation of the phytoanticipin α-tomatine Phytopathology 88 137ndash143
Sendi P and Proctor RA (2009) Staphylococcus aureus as an intracellular pathogen the role of small colony variants Trends Microbiol 17 54ndash58
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C and Osbourn A (2006) Dual effects of plant steroidal alkaloids on Saccha- romyces cerevisiae Antimicrob Agents Chemother 50 2732ndash2740
Somerville GA and Proctor RA (2009) At the crossroads of bacterial metabolism and virulence factor synthesis in Staphylococci Microbiol Mol Biol Rev 73 233- 248
Starkov AA (2010) Measurement of mitochondrial ROS production Methods Mol Biol 648 245-255
Taber HW (1993) Respiratory chain In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of MicrobiologyWashington DC) pp 199-212
Taber HW and Morrison M (1964) Electron transport in Staphylococci Properties of a particle preparation from exponential phase Staphylococcus aureus Arch Biochem Biophys 105 367- 379
Taber HW et al(1987) Bacterial uptake of aminoglycoside antibiotics Microbiol Rev 51 439-457
Thoumlny-Meyer L (1997) Biogenesis of respiratory cytochromes in bacteria Microbiol Mol Biol Rev 61 337- 376
142
Tubby S Wilson M Wright JA Zhang P and Nair SP (2013) Staphylococcus aureus small colony variants are susceptible to light activated antimicrobial agents BMC Microbiol 13 201- 204
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D Roth J Proctor RA Becker K et al (2011) Staphylococcus aureus phenotype switching an effective bacterial strategy to escape host immune response and establish a chronic infection EMBO Mol Med 3 129ndash41
Turrens JF (2003) Mitochondrial formation of reactive oxygen species J Physiol 552 335ndash 344
Tynecka Z Szcześniak Z Malm A and Los R (1999) Energy conservation in aerobically grown Staphylococcus aureus Res Microbiol 150 555- 566
Udo EE Pearman JW and Grubb WB(1993) Genetic analysis of community isolates of methicillin-resistant Staphylococcus aureus in Western Australia J Hosp Infect 25 97-108
Vakulenko SB and Mobashery S (2003) Versatility of Aminoglycosides and Prospects for Their Future Clin Microbiol Rev 16 430-450
Voggu L Schlag S Biswas R Rosenstein R Rausch C and Goumltz F (2006) Microevolution of cytochrome bd oxydase in Staphylococci and its implication in resistance to respiratory toxins released by Pseudomonas J Bacteriol 188 8079- 8086
Wells PG Bhuller Y Chen CS Jeng W Kasapinovic S Kennedy JC Kim PM Laposa RR McCallum GP Nicol CJ et al (2005) Molecular and biochemical mechanisms in teratogenesis involving reactive oxygen species Toxicol Appl Pharmacol 207 354-366
143
Yan M Yu C Yang J and Ji Y (2011) The essential two-component system YhcSR is involved in regulation of the nitrate respiratory pathway of Staphylococcus aureus J Bacteriol 193 1799- 1805
Zhang T Luo Y Chen Y Li X and Yu J (2012) BIGrat a repeat resolver for pyrosequencing-based re-sequencing with Newbler BMC Res Notes 5 567
144

v
TABLE DES MATIEgraveRES
SOMMAIRE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ii
REMERCIEMENTS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip iv
TABLE DES MATIEgraveRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip v
LISTE DES ABBREacuteVIATIONS helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip vii
LISTE DES TABLEAUX helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip ix
LISTE DES FIGURES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip x
CHAPITRE 1 INTRODUCTION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
11 Staphylococcus aureus helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
111 Importance cliniquehelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)helliphellip 2
1121 Diffeacuterences et similariteacutes helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 2
1122 La chaine de transport des eacutelectrons helliphelliphelliphelliphellip 4
12 Antibiotiques Modes drsquoaction cibles cellulaire et
meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles helliphelliphelliphellip 25
122 Les aminoglycosides helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1221 Mode drsquoaction helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 28
1222 Meacutecanismes de reacutesistance helliphelliphelliphelliphelliphelliphelliphelliphellip 31
123 La tomatidine helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 33
13 Projet de maitrise helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
CHAPITRE 2 ARTICLE SCIENTIFIQUE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 37
ldquoTomatidine and derivative FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Firmicutes Including Those of Listeria monocytogenes
and Are Bactericidal in Combination With Aminoglycosides Against their Normal
Phenotyperdquo
vi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE helliphelliphelliphellip 76
31 Obtention des mutants helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
32 Seacutequenccedilage et analyses bio-informatiques helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 79
321 Identification des mutations en lien avec la tomatidine helliphellip 89
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct helliphellip 90
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine helliphelliphelliphelliphellip 99
CHAPITRE 4 DISCUSSION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 113
CHAPITRE 5 CONCLUSION ET PERSPECTIVES helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 124
CONTRIBUTIONS SUPPLEacuteMENTAIRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 1 Application de brevet helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 2 Article scientifique helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 129
laquo Unraveling the structure-activity relationship of tomatidine
a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureus raquo
BIBLIOGRAPHIE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 131
vii
LISTE DES ABBREacuteVIATIONS
Adh Alcool deacuteshydrogeacutenase
ADN Acide deacutesoxyribo-nucleacuteique
ADP Adeacutenosine diphosphate
ARN Acide ribo-nucleacuteique
ATP Adeacutenosine triphosphate
CA-MRSA Community-associated MRSA
CMI concentration minimale inhibitrice
CoQ Coenzyme Q
Cyt Cytochrome
DMQ laquo demethylmenaquinone raquo
dTMP Deacutesoxythymidine monophosphate
dUMP Deacutesoxyuridine monophosphate
FADH2 Flavine adenine dinucleacuteotide
GEN gentamicine
HA-MRSA Healthcare-associated MRSA
Ldh Lactate deacuteshydrogeacutenase
ml millilitre
MQ meacutenaquinone
MRSA methycilin resistant Staphylococcus aureus
Mqo2 Quninol oxidoreacuteductase
NADH Nicotinamide adenine dinucleacuteotide (forme reacuteduite)
Pdh Pyruvate deacuteshydrogeacutenase
Pfl Pyruvate formate lyase
PyrD Dihydroorotate deacuteshydrogeacutenase
Q Quinone
Rif rifampicine
ROS laquo reactive oxygen species raquo espegravece reacuteactive de lrsquooxygegravene
viii
[S] Polysulfide
S aureus Staphylococcus aureus
SARM Staphylococcus aureus resistant agrave la meacutethycilline
SCV laquo small colony variants raquo
SHCHC 2-succinyl-6-hydroxy-24-cyclohexadiene-1-carboxylate syntase
TCA Tricarboxylic acid cycle
TCD tableau croiseacute dynamique
TO tomatidine UFC Uniteacute formatrice de colonies
UQ Ubiquinone
microg microgramme
ix
LISTE DES TABLEAUX
CHAPITRE 1 INTRODUCTION
1 Cibles et modes drsquoaction des principales classes drsquoantibiotique 27
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants (B) chez les mutants NewbouldΔhemB reacutesistants 95-96
CHAPITRE 4 DISCUSSION
1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 123
x
LISTE DES FIGURES
CHAPITRE 1 INTRODUCTION
1 Diffeacuterence entre les pheacutenotypes normaux et SCV de S aureus 3
2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes 6
3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale 8
4 Chaines respiratoires bacteacuteriennes contenant des cytochromes 9
5 Flexibiliteacute respiratoire (A) chez Paracoccus denitrificans (B) chez
Escherichia coli 10
6 Chaine de transport des eacutelectrons proposeacutee chez les staphylocoques 12
7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies 17
8 Modegravele de reacutegulation du cycle TCA 20
9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS 23
10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de O2-bull 24
11 Voies de formation et de deacutetoxification des ROS 25
12 Cineacutetique drsquoentreacutee des aminoglycosides 30
13 Sites de mutations de certains reacutesistants aux aminoglycosides 32
14 Structures chimiques de (A) tomatine (B) tomatidine et (C) Analogue
de tomatidine FC04-100 34
xi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Graphiques des CMI pour chacun des passages pour (A) la
tomatidine sur NewbouldΔhemB (B) la tomatidine en preacutesence de
gentamicine sur NewbouldΔhemB (C) la gentamicine sur S aureus
ATCC 29213 (D) la gentamicine en preacutesence de tomatidine sur S
aureus ATCC 29213 77-79
2 Organigramme des mutants reacutesistants seacutequenceacutes 80-81
3 Eacutetapes du seacutequenccedilage Illumina 83
4 Meacutethode drsquoamplification par la meacutethode Illumina 85
5 Correction du programme RATT sur les annotations transfeacutereacutees 88
6 Exemple drsquoune partie du tableau compilation des mutations 89
7 Seacutelection des gegravenes drsquointeacuterecirct chez ATCC 29213 dans le TCD par
lrsquoutilisation de diffeacuterents filtres 91-92
8 Seacutelection des gegravenes drsquointeacuterecirct chez NewbouldΔhemB dans le TCD par
lrsquoutilisation de diffeacuterents filtres 93-94
9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct 97-98
10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants 100
11 Courbe de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison gentamicine (+TO) du premier et troisiegraveme niveau de
reacutesistance 101
12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B)
NewbouldΔhemB 102
xii
13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213 104
14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicinelorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC
29213 105
15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants
ATCC 29213 reacutesistants agrave la combinaison gentamicine (+TO) 106
16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la
tomatidine 107
17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la
tomatidine 108
18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB
reacutesistants agrave la combinaison tomatidine (+gentamicine) 109
19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane
Potential Kit (Invitrogen) 111
20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du
gegravene atpE de S aureus Newman 112
xiii
CHAPITRE 4 DISCUSSION
1 Site drsquoinhibition de lrsquoATP synthase par diffeacuterentes moleacutecules 114
2 Structure chimique de (A) TMC207 et (B) lrsquoanalogue anti Gram
positifs 115
3 Seacutequence proteacuteique de la proteacuteine AtpE chez diffeacuterentes espegraveces 118
4 Modeacutelisation de la structure tridimentionnelle de lrsquoATP synthase de S
aureus (atpE) montrant les acides amineacutes muteacutes chez les mutants
reacutesistants 121
- 1 -
CHAPITRE 1
INTRODUCTION
11 Staphylococcus aureus
111 Importance clinique
Staphylococcus aureus est une bacteacuterie agrave Gram positif anaeacuterobie facultative Crsquoest un
microorganisme opportuniste pouvant infecter diffeacuterents hocirctes diffeacuterents organes ou
sites drsquoinfection et ayant la capaciteacute de causer tant des infections aiguumles que
chroniques (Archer 1998 Goerke et Wolz 2004)
Staphylococcus aureus reacutesistant agrave la meacutethilline le SARM (ou laquo MRSA raquo) est une
souche de S aureus devenue reacutesistante aux β-Lactamines par lrsquoacquisition du gegravene
mecA dans son chromosome (Hiramatsu 1995) Le SARM a eacuteteacute isoleacute pour la
premiegravere fois en 1960 en Angleterre (Jevons 1961) et est devenu eacutepideacutemique agrave
travers le monde depuis les anneacutees 1970 Au moins trois geacutenotypes distincts eacutetaient
recenseacutes dans les anneacutees 1980 (Hiramatsu 1995) dont deux sont toujours preacutevalents
agrave travers le monde comme eacutetant des souches HA-MRSA (Healthcare-associated
MRSA) des souches acquises agrave lrsquohocircpital (Enright et al 2002) Ces souches ont
acquis des gegravenes de reacutesistance agrave pratiquement tous les antibiotiques introduits dans
lrsquoutilisation clinique depuis les 50 derniegraveres anneacutees mecircme agrave lrsquoantibiotique de dernier
ressort la vancomycine(Hiramitsu 1997 Chen et al 2003) Nous sommes
maintenant en manque drsquoagents theacuterapeutiques efficaces contre les SARM touchant
les personnes immunodeacuteprimeacutees qui sont dans les hocircpitaux En plus depuis le deacutebut
des anneacutees 1990 des clones de SARM dont plusieurs deacutemontraient un potentiel
pathogeacutenique supeacuterieur aux souches classiques de HA-MRSA retrouveacutees agrave lrsquohocircpital
- 2 -
sont maintenant retrouveacutes dans la communauteacute (CA-MRSA) agrave travers le monde (Udo
et al 1993 Riley et al 1995 Moreno et al 1995 Aubry-Damon et al 1997) et ont
la capaciteacute drsquoaffecter des personnes en santeacute en causant des infections aiguumles
(Hiramatsu et al 2013) Il apparaicirct donc urgent de trouver des drogues alternatives
afin de surmonter le nombre grandissant de souches de SARM
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)
1121 Diffeacuterences et similariteacutes
Les laquo small colony variants raquo (SCV) sont des bacteacuteries souvent isoleacutees lors des
infections chroniques comme crsquoest le cas des infections pulmonaires chez les
patients atteints de fibrose kystique mais aussi dans des cas drsquoosteacuteomyeacutelite
drsquoarthrite septique drsquoinfections de prothegraveses orthopeacutediques et lors de la mammite
bovine (Alexender et Hudson 2001 Moisan et al 2006 Proctor et al 2006) Les
SCV sont caracteacuteriseacutes par soit un meacutetabolisme oxydatif dysfonctionnel ou un manque
dans la biosynthegravese de la thymidine les deux amenant une alteacuteration dans
lrsquoexpression des facteurs de virulence une croissance plus lente et une diminution
de la pigmentation des colonies (Figure 1) (Proctor et al 2006) Cependant alors
que les souches normales reacutepriment lrsquoexpression des proteacuteines de surface et
expriment les exoproteacuteines lors de la phase stationnaire (Novick 2003) les SCV
expriment de faccedilon stable les gegravenes Sig-B deacutependants codant pour des proteacuteines de
surface telle les adheacutesines au lieu drsquoactiver le systegraveme agr deacutependant du quorum-
sensing et produire des exoproteacuteines (Moisan et al 2006) Ces diffeacuterences dans
lrsquoexpression des facteurs de virulence reacutesultent chez les SCV en une meilleure
habileteacute agrave adheacuterer aux composantes de lrsquohocircte (Mitchell et al 2008) et agrave former du
biofilm (Mitchell et al 2010a Mitchell et al 20010b) et seraient eacutegalement lieacutees agrave
lrsquohabileteacute des SCV agrave lrsquoinvasion et la persistance des cellules de lrsquohocircte (Proctor et al
2006 Sendi et Proctor 2009 Mitchell et al 2011b)
- 3 -
S aureus est donc un pathogegravene qui peut se preacutesenter sous deux pheacutenotypes qui
sont souvent retrouveacutes simultaneacutement lors drsquoune mecircme infection et qui ont le pouvoir
de reacuteverter drsquoun pheacutenotype agrave lrsquoautre selon les conditions du milieu (Figure 1)
(Tuchscherr et al 2011)
Le pheacutenotype dit rdquonormalrdquo est associeacute aux infections aiguumles agrave une forte production
de facteurs de virulence (ex heacutemolysine) et est associeacute agrave la phase de disseacutemination
de la bacteacuterie Lrsquoautre pheacutenotype le laquo small-colony variants raquo ou SCV est affecteacute
dans sa chaine respiratoire ce qui amegravene une croissance beaucoup plus lente des
colonies de 8 -10 fois plus petites que les prototypes et une faible susceptibiliteacute aux
aminoglycosides Les SCV sont plutocirct associeacutes aux infections chroniques agrave S aureus
de par leur capaciteacute agrave srsquointernaliser dans les cellules de lrsquohocircte leur forte capaciteacute agrave
produire du biofilm et leur capaciteacute agrave produire des facteurs de colonisation
(Tuchscherr et al 2011 Proctor et al 2006) (Figure 1)
Figure 1 Diffeacuterences entre les pheacutenotypes normaux et SCV de S aureus
S aureus agrave
pheacutenotype normal
Disseacutemination
amp
Infections aiguumles
Colonisation
amp
Infections persistantes
SCVs
(laquo small-colony
variants raquo)
Reacutevers
ion
pheacutenoty
piq
ue
Norm
al
SC
V
Croissance
Biofilm
Internalisationheacutemolyse
Mitchell G et Malouin F 2012
- 4 -
1122 La chaine de transport des eacutelectrons
Chaine respiratoire tableau geacuteneacuteral
Les chaines respiratoires bacteacuteriennes bien que tregraves variables dans leurs
composantes parmi les diffeacuterentes espegraveces sont toutes constitueacutees sur le mecircme
modegravele de base Elles sont composeacutees de trois composantes majeures soit
- les deacuteshydrogeacutenases primaires qui sont chargeacutees drsquooxyder des substrats
principalement les coenzymes NADH et FADH2 (issues du cycle de Krebs) (mais
aussi selon le cas le succinate le lactate lrsquohydrogegravene hellip) et de pomper deux
protons du cocircteacute externe de la membrane en plus de transfeacuterer deux eacutelectrons agrave un
groupement de quinones
- Les quinones sont des moleacutecules liposolubles non proteacuteiques mobiles dans la
membrane Elles servent drsquointermeacutediaires entre les deacuteshydrogeacutenases primaires et
les oxydases terminales pour le transport des eacutelectrons Il existe deux sortes de
quinones les benzoquinones dont font partie les ubiquinones et les
naphtoquinones dont font partie les meacutenaquinones Les bacteacuteries agrave Gram positif
ne possegravedent que des meacutenaquinones alors que les deux sont preacutesentes chez les
bacteacuteries agrave Gram neacutegatif (Hederstedt 1993 Pratique et Theraulaz 2007)
- Les oxidoreacuteductases terminales sont chargeacutees de reacuteduire lrsquoaccepteur final
drsquoeacutelectron lrsquooxygegravene eacutetant lrsquoaccepteur final privileacutegieacute
De faccedilon plus deacutetailleacutee la chaine respiratoire est composeacutee de cinq
complexes proteacuteiques principaux (Figure 2)
- complexe I NADH deacuteshydrogeacutenase ou NADH reacuteductase ou NADHCoQ
oxidoreacuteductase ou NADH-ubiquinone oxidoreacuteductase
- complexe II Succinate deacuteshydrogeacutenase
- complexe III Cytochrome bc1 complexe ou ubiquinol cytochrome c
reacuteductaseoxydoreacuteductase
- complexe IV Cytochrome c oxydase
- 5 -
- complexe V ATP synthase (Creacutepin-Gilbert et al 2006)
La phosphorylation oxydative est deacuteclencheacutee dans la chaine de complexes
proteacuteiques par la preacutesence drsquoO2 accepteur terminal privileacutegieacute drsquoeacutelectrons (ou drsquoautres
accepteurs finaux tel le nitrate) dans le milieu ainsi que par les eacutelectrons ameneacutes par
le NADH et le FADH2 au complexe I et II respectivement (Creacutepin-Gilbert et al 2006)
Les eacutelectrons vont ensuite transiter par le complexe III jusqursquoau complexe IV ou les
eacutelectrons seront utiliseacutes afin de reacuteduire lrsquooxygegravene en eau le complexe IV est
lrsquooxydase terminale consideacutereacutee comme eacutetant lrsquoaccepteur final drsquoeacutelectrons Le transfert
drsquoeacutelectrons est possible gracircce aux reacuteactions drsquooxydoreacuteduction en cascade dans les
diffeacuterents complexes proteacuteiques Ces reacuteactions fournissent de lrsquoeacutenergie libre utiliseacutee
par les complexes I III et IV afin de transfeacuterer des protons agrave lrsquoexteacuterieur de la cellule
(Creacutepin-Gilbert et al 2006) Le complexe V (ATP synthase) quant agrave lui utilise les
eacutelectrons pompeacutes agrave lrsquoexteacuterieur par les autres complexes comme force proton-motrice
afin de phosphoryler trois moleacutecules drsquoADP et ainsi produire trois ATP Le transport
des eacutelectrons entre les diffeacuterents complexes proteacuteiques est possible gracircce agrave la
preacutesence dans la membrane de deux types de moleacutecules liposolubles non proteacuteiques
mobiles les meacutenaquinones et le cytochrome c Les meacutenaquinones seules quinones
preacutesentes chez les bacteacuteries agrave Gram positif assurent le transport des eacutelectrons entre
le complexe I et le complexe III et entre le complexe II et III Le cytochrome c pour sa
part veacutehicule les eacutelectrons entre le complexe III et le complexe IV Le transport des
eacutelectrons srsquoeffectue de la composante ayant le plus faible potentiel drsquooxydoreacuteduction
vers celle ayant le plus fort potentiel
- 6 -
La chaine respiratoire bacteacuterienne
La mitochondrie montre assez peu de flexibiliteacute respiratoire Il y a une certaine
flexibiliteacute au niveau de lrsquoentreacutee des eacutelectrons mais aucune au niveau de la sortie des
eacutelectrons ougrave la cytochrome aa3 oxydase reacuteduit lrsquooxygegravene en eau (Richardson 2000)
(Figure 3) Chez les bacteacuteries alors que le complexe II fonctionne sensiblement de
faccedilon identique agrave celui des eucaryotes les autres complexes peuvent se composer
de diffeacuterents complexes enzymatiques (reacuteductases) selon les espegraveces et les
conditions environnementales (Richardson 2000 Thoumlny-Meyer 1997) En outre
chez les bacteacuteries contrairement aux eucaryotes la chaine respiratoire comporte
souvent des voies de transport des eacutelectrons alternatives on dit alors qursquoelles ont une
chaine respiratoire agrave branches ou brancheacutee ou ramifieacutee (Artzatbanov et Petrov 1990)
Plusieurs espegraveces bacteacuteriennes possegravedent une chaine respiratoire ramifieacutee dont la
ramification se situe habituellement suite agrave la reacuteduction des quinones en quinols
(Lauraeus et Wikstrom 1993 Thoumlny-Meyer 1997) (Figure 4) Une des deux
Complexe I NADH
deacuteshydrogeacutenase
Complexe II Succinate
deacuteshydrogeacutenase
Complexe III
bc 1
Complexe IV Cytochrome c
oxydase
Complexe V
ATP synthase
Modifieacute de wikipedia httpfrwikipediaorgwikiChaine_respiratoire
Figure 2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes
Leurs rocircles dans le transport des eacutelectrons et dans lrsquoeacutelaboration du gradient de
protons et le lien entre la chaine respiratoire et le cycle de Krebs Q Quinones
- 7 -
branches connue comme la branche cytochrome c oxyde les quinols (meacutenaquinol
QH2) via la voie complexe bc1-cytochrome c- cytochrome c oxydase Lrsquoautre
branche-type consiste en une seule uniteacute enzymatique (cytochrome bd) laquelle
transfegravere directement les eacutelectrons des quinols agrave lrsquooxygegravene Cette voie est connue
comme la branche quinol oxydase il srsquoagit drsquoune oxydase terminale transfeacuterant
directement les eacutelectrons agrave lrsquooxygegravene eacutevitant de passer par lrsquointermeacutediaire du
complexe III (bc1) et du cytochrome c (Artzatbanov et Petrov 1990 Lauraeus et
Wikstrom 1993) Alors que la voie du cytochrome c permet le pompage de protons agrave
lrsquoexteacuterieur par lrsquoaction du complexe bc1 (ou complexe III) et du cytochrome c oxydase
le cytochrome bd ne permet aucun pompage de protons (Lauraeus et Wikstrom
1993) La voie passant par le complexe bc1-cytochrome c-cytochrome c oxydase est
plus efficace dans la production drsquoeacutenergie Alors que les 2 voies peuvent ecirctre
exprimeacutees simultaneacutement chez quelques espegraveces il semble que lrsquoexpression de ces
voies soit inductible selon les conditions environnementales chez Bacillus subtilis et
les Staphylocoques (Lauraeus et Wikstrom 1993) On pourrait dire eacutegalement que la
chaine respiratoire est seacutepareacutee en deux eacutetapes la premiegravere partie est la phase
quinone-reacuteductase (Q-QH2) et la seconde la phase quinol-oxydase (QH2-Q) menant
agrave lrsquooxydase terminale La phase de reacuteduction est responsable du transfert des
eacutelectrons drsquoune varieacuteteacute de substrats tels le NADH le succinate le lactate le formate
et lrsquohydrogegravene vers les quinones et inclut diffeacuterentes reacuteductases (Thoumlny-Meyer 1997)
(Figure 4)
- 8 -
Figure 3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale UQ ubiquinone UQH2 ubiquinol Cyt cytochrome Il semble y avoir presque autant de chaines de transport des eacutelectrons diffeacuterentes
qursquoil y a drsquoespegraveces bacteacuteriennes (par exemple Paracoccus denitrificans et Escherichia
coli (Figure 5)) De plus il ne semble pas y avoir consensus dans la litteacuterature sur
les noms attribueacutes aux oxydases terminales Les noms pour les oxydases terminales
deacutependent des types de cytochromes dont elles sont composeacutees Il ne semble donc
pas eacutevident de connaicirctre de faccedilon preacutecise les types de cytochromes qui font partie
des oxydases terminales Pour rajouter agrave la confusion les gegravenes et opeacuterons ne sont
pas toujours associeacutes agrave leur oxydase terminale dans la litteacuterature ce qui fait qursquoon ne
peut savoir de faccedilon certaine de quelle oxydase on parle Le Cytochrome aa3 peut
eacutegalement porter le nom de Cyt ba3 et le cytochrome bd peux eacutegalement porter les
noms de Cyt bo
Tireacute de Richardson DJ 2000
- 9 -
Succinate
Formate
H2
SDH
FDH
HYD
QQH 2 bc1 c
Reacuteductase alternatives
b
c
cd1c
cb
c
b
c
d
Siroheme
Siroheme
b
NO3-
NO2-
NO
TMAO
DMSOS2O3
2-
[S]
SO32-
Fumarate
Quinol oxidases
ba3bb3bo3d
bd
aa3
O2
Cyt c oxidases
aa3ba3baa3caa3cbb3cao
co
O2
Modifieacute de Thony-Mayer L 1997
NADH
NDH
Figure 4 Chaines respiratoires bacteacuteriennes contenant des cytochromes
Du cocircteacute gauche on retrouve la partie quinone (ubiquinone ou meacutenaquinone)-
reacuteductase Du cocircteacute droit on retrouve la partie quinol-oxydase avec les oxydases
terminales reacuteduisant lrsquoO2 ou lrsquoaccepteur terminal alternatif tel que montreacute Les
oxydoreacuteductases sont surligneacutees QQH2 repreacutesente le pool de quinonesquinols
dans la membrane Les polysulfides sont repreacutesenteacutes par [S] Les chaines
respiratoires ne contenant pas de cytochromes ne sont pas montreacutees NDH
NADH deacuteshydrogeacutenase
Modifieacute de Thoumlny-Mayer L 1997
- 10 -
Figure 5 Flexibiliteacute respiratoire chez Paracoccus denitrificans (a) et chez
Escherichia coli (b) MQ menaquinone UQ ubiquinone DMQ
demethylmenoquinone
Chez S aureus
Comme plusieurs autres espegraveces bacteacuteriennes anaeacuterobies facultatives S aureus a
la possibiliteacute drsquoexprimer diffeacuterentes voies de synthegravese eacutenergeacutetique dont lrsquoexpression
est finement reacuteguleacutee selon des conditions environnementales du moment (Hammer et
Tireacute de Richardson DJ 2000
- 11 -
al 2013 Lauraeus et Wikstrom 1993) Mecircme si S aureus peut utiliser la
fermentation la respiration (aeacuterobie et anaeacuterobie) sera privileacutegieacutee par la bacteacuterie
puisque celle-ci amegravene une bien meilleure production drsquoeacutenergie (Goumltz et al 2006
Hammer et al 2013 ) La respiration aeacuterobie avec lrsquooxygegravene comme accepteur final
drsquoeacutelectron sera utiliseacutee de faccedilon preacutefeacuterentielle en preacutesence drsquooxygegravene (Hammer et al
2013) Malgreacute sa versatiliteacute dans ses voies de synthegravese eacutenergeacutetique S aureus et les
autres bacteacuteries agrave Gram positif ne produisent qursquoun seul type de quinone les
meacutenaquinones lesquelles sont utiliseacutees pour la croissance degraves qursquoil y a de lrsquooxygegravene
de preacutesent dans le milieu (Somerville et Proctor 2009) La reacuteduction de lrsquooxygegravene en
eau est meacutedieacutee par des oxydases terminales hegraveme deacutependant (Hammer et al 2013)
Selon certains auteurs S aureus possegravederait deux oxydases terminales la
cytochrome oxydase aa3 (ou ba3) (QoxABCD) et la cytochrome bd quinol oxydase
(CydAB) (Goumltz et al 2006 Hammer et al 2013 Kohler et al 2008) Cette derniegravere
est exclusive aux procaryotes et est une voie de contournement du complexe III qui
transmet directement les eacutelectrons du laquo pool raquo de quinol (QH2) agrave lrsquooxygegravene Selon
certains auteurs il y aurait eacutegalement chez S aureus une troisiegraveme oxydase
terminale le cytochrome o (CyoABCDE) (Thoumlny-Meyer 1997 Voggu et al 2006)
qui selon certain serait un cytochrome oxydase terminal alternatif au cytochrome aa3
et au cytochrome bd (Voggu et al 2006 Goumltz et al 2006 Taber et Morrison 1964)
(Figure 6) Ces voies alternatives permettent agrave S aureus de srsquoadapter rapidement agrave
un changement drsquoenvironnement (Hammer et al 2013) In vivo chaque organe est
un environnement diffeacuterent preacutesentant des deacutefis respiratoires distincts dus agrave la
variation dans 1- la quantiteacute drsquooxygegravene disponible 2- la disponibiliteacute drsquoaccepteurs
drsquoeacutelectrons alternatifs 3- la concentration drsquoinhibiteurs de la respiration dans le milieu
tels les oxydes nitriques (NO) produits par les neutrophiles et 4- la disponibiliteacute des
cofacteurs neacutecessaires agrave la production drsquoeacutenergie (Hammer et al 2013) Le
cytochrome est exprimeacute de faccedilon modeacutereacutee dans un milieu riche en oxygegravene mais est
fortement exprimeacute dans un milieu pauvre en oxygegravene mais riche en oxyde nitrique
De plus le type drsquooxydoreacuteductase terminale exprimeacute est influenceacute eacutegalement par la
- 12 -
source de carbone disponible la phase de croissance le pH extracellulaire et la
tension en oxygegravene (Lauraeus et Wikstrom 1993 Tynecka et al 1999)
La chaine respiratoire chez les SCV
S aureus est particuliegraverement doueacute agrave surpasser les deacutefis respiratoires en acqueacuterant
une reacutesistance agrave lrsquoacide nitrique produit par les neutrophiles et une habiliteacute agrave persister
dans les cellules de lrsquohocircte en se preacutesentant comme un variant agrave petites colonies
ayant une chaine respiratoire deacuteficiente (SCV) (Hammer et al 2013)
Deux principaux types de SCV sont retrouveacutes dans les isolats cliniques 1- les SCV
ayant une chaine de transport des eacutelectrons deacutefectueuse et 2- les SCV auxotrophes
pour la thymidine (Proctor et Peters 1998 Samuelsen et al 2005) Ces derniers
Figure 6 Chaine de transport des eacutelectrons proposeacutee chez les
staphylocoques Les informations disponibles sur les composantes de la chaine
respiratoire des staphylocoques suggegraverent une chaine ramifieacutee consistant en 2
oxydases terminales meacutenaquinol-deacutependante un cytochrome o et un cytochrome
aa3 oxydase NADH deacuteshydrogeacutenase (DHase) transmet les eacutelectrons au laquo pool raquo
de meacutenaquinones De lagrave les eacutelectrons sont transfeacutereacutes soit au cytochrome b ou agrave
lrsquoaccepteur alternatif drsquoeacutelectrons le cytochrome bd quinol oxydase Le cytochrome
b transfegravere les eacutelectrons agrave une des deux oxydases terminales Ces oxydases
terminales transfegraverent les eacutelectrons agrave lrsquooxygegravene
Modifieacute de Voggu L et al 2006
- 13 -
arborent une ou plusieurs mutations dans le gegravene thyA codant pour la thymidylate
synthase laquelle catalyse la derniegravere eacutetape dans la voie de biosynthegravese de la
thymidine crsquoest-agrave-dire la formation de dTMP agrave partir du dUMP (Samuelsen et al
2005) Le dTMP est essentiel pour la synthegravese de lrsquoADN (Besier et al 2007) Les
mutants thymidine-auxotrophes sont souvent la conseacutequence drsquoune pression
seacutelective par lrsquoutilisation prolongeacutee de trimethoprim-sulfamethoxazole (TMP-SMX)
(souvent utiliseacute pour le traitement des patients fibroses kystiques) un antibiotique qui
interfegravere dans la biosynthegravese de lrsquoacide tetrahydrofolique qui agit comme cofacteur de
la thymidylate synthase (Besier et al 2007 Lannergard et al 2008 Samuelsen et al
2005) Mecircme si de premiers abords les SCV thymidine-auxotrophes ne semblent pas
avoir de lien avec la chaine transport des eacutelectrons il se pourrait que celle-ci ait sont
importance chez ces SCV puisque lrsquoentreacutee de la thymidine passe par NupC dont le
fonctionnement neacutecessite un gradient eacutelectrochimique creacuteeacute par la chaine de transport
des eacutelectrons (Proctor et Peters 1998 ) Les caracteacuteristiques pheacutenotypiques des
SCV deacutecrites ci-apregraves sont toutes lieacutees agrave lrsquoalteacuteration de la chaine de transport des
eacutelectrons 1- la croissance plus lente parce que la synthegravese de la paroi cellulaire
neacutecessite une grande quantiteacute drsquoATP 2-la diminution de la formation de pigment
parce que la biosynthegravese des caroteacutenoiumldes requiert le transport des eacutelectrons 3- une
reacutesistance accrue agrave la gentamicine et autres aminoglycosides parce que lrsquoentreacutee de
ces composeacutes neacutecessite un grand potentiel membranaire geacuteneacutereacute par le transport des
eacutelectrons (Lannergard et al 2008) et 4- une perte partielle ou complegravete de la
fermentation du mannitol parce que lrsquoutilisation des sucres-alcool tels que le
mannitol est diminueacutee lorsque la chaine de transport des eacutelectrons ne peut oxyder le
NADH produit lorsque le mannitol est meacutetaboliseacute en fructose-6-phosphate (Kohler et
al 2003 Proctor et Peters 1998 Samuelsen et al 2005)
Les SCV posseacutedant une chaine de transport des eacutelectrons deacutefectueuse ont une voie
de biosynthegravese de lrsquoheacutemine ou de la meacutenadione deacuteficiente et le pheacutenotype est
reacuteversible lorsque compleacutementeacute avec lrsquoheacutemine ou la meacutenadione (Hammer et al
- 14 -
2013 Kohler et al 2008 Lannergard et al 2008 Proctor et Peters 1998) La
meacutenadione est essentielle agrave la biosynthegravese des meacutenaquinones et lrsquoheacutemine est
essentielle agrave la biosynthegravese de lrsquohegraveme qui entre dans formation des cytochromes
(Hammer et al 2013 Kohler et al 2008 Lannergard et al 2008 Thoumlny-Meyer
1997) Les meacutenaquinones et les cytochromes sont essentiels au fonctionnement de
la chaine de transport des eacutelectrons (vers lrsquoO2 autant que vers le nitrate) (Kohler et al
2008) Il se pourrait qursquoil existe eacutegalement des SCV causeacutes par drsquoautres deacuteficiences
telles que lrsquoalteacuteration de lrsquoATP synthase qui ne reacutesulterait pas drsquoune auxotrophie pour
lrsquoheacutemine ou la meacutenadione mais reacutesulterait tout de mecircme en une deacuteficience dans la
chaine de transport des eacutelectrons (Proctor et Peters 1998 )
Comme lrsquohegraveme est un cofacteur essentiel pour les oxydases terminales et que la
meacutenaquinone est un transporteur drsquoeacutelectrons utiliseacute par S aureus on comprend que
la faible croissance des mutants hemB et menD repreacutesente un pheacutenotype mimant la
perte des oxydases terminales (Hammer et al 2013) Les deux oxydases terminales
de S aureus QoxABCD (cyt aa3) et CydAB (cyt bd) semblent ecirctre diffeacuteremment
reacuteguleacutees chez les mutants menD et la souche sauvage alors que lrsquoopeacuteron cydAB est
fortement sous exprimeacute chez les mutants aucune diffeacuterence de niveau de
transcription de qoxABCD nrsquoa eacuteteacute observeacutee entre les mutants menD et leur souche
parente (Kohler et al 2008)
Des mutations dans les gegravenes menD hemB et ctaA peuvent reacutesulter en un
pheacutenotype SCV posseacutedant une chaine de transport des eacutelectrons deacuteficiente
(Clements et al 1999 Proctor et Peters 1998 ) Les gegravenes codant pour les eacutetapes
de formation de lrsquohegraveme se preacutesentent sous la forme drsquoun opeacuteron lrsquoopeacuteron
hemAXCDBL Chacun des gegravenes est essentiel (sauf hemL) et la deacuteleacutetion drsquoun seul
drsquoentre eux empecircchera la formation de lrsquohegraveme moleacutecule essentielle agrave la formation
des cytochromes (Taber 1993) hemB code pour la porphobilinogene synthase qui
catalyse une eacutetape dans la voie de biosynthegravese de lrsquohegraveme (Kohler et al 2008 Taber
- 15 -
1993 Thoumlny-Meyer 1997) Une mutation dans le gegravene ctaA (laquo hegraveme A synthase raquo)
empecircche la biosynthegravese de lrsquohegraveme A lequel est aussi impliqueacute dans la biosynthegravese
des cytochromes et donc altegraverera le fonctionnement de la chaine de transport des
eacutelectrons (Clements et al 1999 Proctor et Peters 1998) La capaciteacute de la bacteacuterie
agrave biosyntheacutetiser de lrsquohegraveme de faccedilon endogegravene est essentielle afin drsquoinitier une
infection systeacutemique Le gegravene menD code pour la 2-succinyl-6-hydroxy-24-
cyclohexadiene-1-carboxylate syntase (SHCHC) qui est une enzyme essentielle agrave la
voie de biosynthegravese des meacutenaquinones (Kohler et al 2008) De plus les
meacutenaquinones sont eacutegalement requises pour le fonctionnement de plusieurs
enzymes impliqueacutees dans des voies meacutetaboliques essentielles telles la malate quinol
oxidoreacuteductase (Mqo2) et la dihydroorotate deacuteshydrogeacutenase (PyrD) (Kohler et al
2008) Les mutants menD seront donc incapables de convertir lrsquooxaloaceacutetate en
malate ducirc agrave la perte de lrsquoactiviteacute du Mqo2 (Kohler et al 2008 Proctor et Peters
1998)
Les bacteacuteries agrave pheacutenotype SCV deacutemontreront eacutegalement un gradient eacutelectrochimique
nettement plus faible que les bacteacuteries agrave pheacutenotype normal de par lrsquoincapaciteacute de la
chaine de transport des eacutelectrons agrave y pourvoir
De par leur chaine de transport des eacutelectrons interrompue les SCV sont incapables
drsquoutiliser lrsquoO2 et le nitrate comme accepteur final drsquoeacutelectrons (Kohler et al 2008) Par
ce fait les proteacuteines impliqueacutees dans la voie de la glycolyse ainsi que dans la voie de
la fermentation sont surexprimeacutees (Pfl Ldh Adh) alors que les enzymes faisant
partie du cycle du citrate (TCA) sont significativement reacuteguleacutees agrave la baisse (Kohler et
al 2003 Kohler et al 2008 Kriegeskorte et al 2011) Ceci indique clairement que
les SCV (ex hemB) geacutenegraverent leur ATP agrave partir du glucose ou fructose seulement par
la phosphorylation des substrats Lrsquoanalyse de la reacuteaction de fermentation indique
que le produit principal est le lactate Les SCV reacutegeacutenegraverent leur NADH en reacuteduisant le
pyruvate en lactate par la voie fermentaire (Kohler et al 2003 Kohler et al 2008)
- 16 -
emmenant ainsi une acidification du milieu (Kriegeskorte et al 2011) La conversion
du pyruvate en aceacutetate via lrsquoacetyl-CoA est restreinte chez les mutants hemB mecircme
si la pyruvate formate lyase (Pfl) et les composantes du pyruvate deacuteshydrogeacutenase
sont preacutesentes (Figure 7) Lrsquoactiviteacute de la pyruvate deacuteshydrogeacutenase est tregraves faible
probablement en lien avec le haut niveau de NADH chez les SCV et il est preacutesumeacute
que la Pfl nrsquoest pas activeacutee en preacutesence drsquooxygegravene (La pyruvate deacuteshydrogeacutenase
converti le pyruvate en Co-A en condition anaeacuterobie) (Figure 7) Chez les mutants
hemB la voie de lrsquoarginine deacuteiminase est aussi induite et pourrait eacutegalement
participer agrave la production drsquoATP (Kohler et al 2003) Les gegravenes exprimeacutes chez les
mutants hemB sont similaires agrave ceux exprimeacutes chez les mutants menD ainsi que
chez les souches agrave pheacutenotypes normaux en condition anaeacuterobie (Kohler et al 2008)
- 17 -
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Figure 7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies Les enzymes suivantes sont retrouveacutees agrave une forte
concentration chez les mutants hemb phosphoglucomutase (Pgi) fructose
bisphosphate aldolase (FbaFda) glyceacuteraldeacutehyde deacuteshydrogeacutenase (Gap)
phosphoglyceacuterate kinase (Pgk) eacutenolase (Eno) pyruvate kinase (Pyk) L-lactate
deacuteshydrogeacutenase (L-Ldh) pyruvate formate lyase (PflB) alcool deacuteshydrogeacutenase
(Adh) La formation drsquoaceacutetate et de formate et tregraves limiteacutee (X rouges)
Modifieacute de Kolher C et al 2003
- 18 -
La chaine respiratoire en anaeacuterobie (S aureus)
Il existe 3 voies distinctes de geacuteneacuteration drsquoeacutenergie chez les bacteacuteries 1- en preacutesence
drsquooxygegravene la bacteacuterie respire de faccedilon aeacuterobique et lrsquooxygegravene est lrsquoaccepteur final
drsquoeacutelectron 2- la bacteacuterie peut aussi respirer de maniegravere anaeacuterobique lorsqursquoun
accepteur terminal drsquoeacutelectron autre que lrsquooxygegravene est preacutesent 3- en absence
drsquooxygegravene et drsquoun accepteur drsquoeacutelectrons alternatif la bacteacuterie peut produire de
lrsquoeacutenergie de faccedilon limiteacutee par fermentation (Hammer et al 2013)
Chez S aureus le nitrate (NO3-) et dans un troisiegraveme temps le nitrite (NO2
-)
peuvent ecirctre utiliseacutes comme accepteur final drsquoeacutelectron alternatif lorsque lrsquooxygegravene
nrsquoest pas disponible (respiration anaeacuterobie) Les enzymes de cette voie meacutetabolique
ne sont retrouveacutees que chez les bacteacuteries (Goumltz et al 2006) La croissance
bacteacuterienne est consideacuterablement reacuteduite lorsque la bacteacuterie passe drsquoune respiration
aeacuterobie agrave une respiration anaeacuterobie La nitrate reacuteductase fonctionne probablement in
vivo gracircce au L-lactate comme principal donneur drsquohydrogegravene (Burke et Lasccelles
1975) La deacuteshydrogeacutenase primaire pour ce substrat est preacutesumeacutement la L-lactate
deacuteshydrogeacutenase associeacutee agrave la membrane laquelle meacutedie lrsquooxydation du lactate par la
chaine respiratoire aeacuterobique (Burke et Lasccelles 1975) La preacutedominance du
lactate parmi les produits finaux de la fermentation par S aureus est correacuteleacutee avec
son efficaciteacute comme principal donneur drsquohydrogegravene lorsque le nitrate est lrsquoaccepteur
final drsquoeacutelectrons Le transfert des eacutelectrons du Lactate vers le nitrate implique des
composantes qui sont eacutegalement preacutesentes dans la chaine respiratoire La nitrate
reacuteductase du type respiratoire est habituellement associeacutee agrave la membrane
cytoplasmique (Burke et Lasccelles 1975) Les reacuteductases sont inhibeacutees par
lrsquoazoture et le cyanure (Burke et Lascelles 1976)
Lrsquoenzyme membranaire nitrate reacuteductase NarGHI geacutenegravere des nitrites lesquels seront
reacuteduits en ammonium (NH4+) par lrsquoenzyme cytoplasmique NADH-deacutependant nitrite
reacuteductase encodeacutee par les gegravenes nirBD (Goumltz et al 2006) La nitrate reacuteductase 1-
- 19 -
est reacuteprimeacutee et inhibeacutee par la preacutesence drsquooxygegravene 2- est induite par lrsquoanaeacuterobiose et
la preacutesence de nitrate ou nitrite 3- nrsquoest pas inhibeacutee par lrsquoammoniac 4- permet la
production drsquoeacutenergie dans des conditions anaeacuterobies et 5- est localiseacutee dans la
membrane (Goumltz et al 2006) La preacutesence drsquoun systegraveme de senseur pour lrsquooxygegravene
est cruciale pour le controcircle transcriptionnel des gegravenes facilitant lrsquoadaptation de la
bacteacuterie agrave lrsquoanaeacuterobiose ou aux conditions microaeacuterophiles Chez S aureus il existe
deux systegravemes de senseur 1- le systegraveme SrrAB (pour staphylococcal respiratory
response AB synonyme SrhSR) (Figure 8) homologue agrave ResDE chez Bacillus
subtilis (Yan et al 2011) et 2- le systegraveme NreABC (pour nitrogen regulation chez S
carnosus correspondant agrave SA2181 (NreA) SA2180 (NreB) SA2179 (NreC) chez S
aureus) NreBC est un systegraveme agrave deux composantes qui mesure la concentration
drsquooxygegravene directement par un groupement Fe-S de type Fnr lequel est instable en
preacutesence drsquooxygegravene et par lequel est controcircleacute le systegraveme membranaire de reacuteduction
du nitratenitrite (Yan et al 2011) Srr est aussi un systegraveme agrave deux composantes qui
reacutegule positivement la lactate deacuteshydrogeacutenase et lrsquoalcool deacuteshydrogeacutenase et semble
reacuteguler neacutegativement plusieurs enzymes du cycle du citrate (TCA) (Figure 8)
Lrsquoexpression des enzymes fermentaires comme la lactate deacuteshydrogeacutenase (LDH)
pyruvate formate lyase (PFL) et lrsquoalcool deacuteshydrogeacutenase sont aussi fortement induite
dans des conditions aeacuterobies lorsque la chaine de transport des eacutelectrons est
interrompue (Kohler et al 2003) Donc il se peut que lrsquoeacutetat reacuteduit des composantes
de la chaine respiratoire le potentiel membranaire etou le niveau de NADH soit un
signal pour la reacutegulation des gegravenes laquo anaeacuterobies raquo chez S aureus Dans des
conditions aeacuterobies la conversion du pyruvate en aceacutetyl-coA est catalyseacutee par le
complexe pyruvate deacuteshydrogeacutenase (PDH) et caracteacuteriseacutee par la production de
NADH (Somerville et Proctor 2009) En condition anaeacuterobie lrsquoexpression de la
pyruvate deacuteshydrogeacutenase (PDH) est fortement reacuteprimeacutee alors que lrsquoexpression de la
pyruvate formate lyase (PFL) est fortement induite Lrsquoactiviteacute de la PDH est
remplaceacutee en anaeacuterobiose par lrsquoactiviteacute de la PFL qui geacutenegravere lrsquoacetyl-CoA et le
formate agrave partir du pyruvate Le NADH nrsquoeacutetant pas reacuteoxydeacute par une chaine de
- 20 -
transport des eacutelectrons fonctionnelle celui-ci est recycleacute par la reacuteduction du pyruvate
en lactate (Kohler et al 2003 Somerville et Proctor 2009)
La meacutenaquinone et lrsquohegraveme sont esssentiels agrave la respiration anaeacuterobie (Kohler et al
2008) Le pheacutenotype de S aureus en condition anaeacuterobie est tregraves similaire au
pheacutenotype SCV (Somerville et Proctor 2009)
Figure 8 Modegravele de reacutegulation du cycle TCA (Pour simplifier les reacutegulateurs de
facteurs de virulence bien caracteacuteriseacutes [ie RNAIII Rot SarA et σB] ont eacuteteacute omis
dans ce modegravele)
Les sous-produits de la phosphorylation oxydative les ROS
La reacuteduction de lrsquooxygegravene en eau se fait un eacutelectron agrave la fois (Turrens 2003) Le
complexe IV retient tous les intermeacutediaires partiellement reacuteduits jusqursquoagrave ce que la
Tireacute de Somerville GA Proctor RA 2009
- 21 -
reacuteduction soit complegravetement termineacutee (Turrens 2003) Cependant certains autres
centres redox de la chaine respiratoire laissent passer des eacutelectrons agrave lrsquooxygegravene
reacuteduisant partiellement cette moleacutecule en anion superoxyde (O2-bull) (Liu et al 2002
Turrens 2003) Mecircme si O2-bull nrsquoest pas un agent oxydant tregraves puissant il est le
preacutecurseur de plusieurs autres laquo reactive oxygen species raquo (ROS) et il est aussi
impliqueacute dans la propagation de la chaine de reacuteaction oxydative (Turrens 2003)
LrsquoO2-bull est un intermeacutediaire instable qui est rapidement converti en H2O2 par les
superoxide dismutase (SOD) (S aureus en possegravede deux SodA (le principal) et
SodM) (10 18) ou se convertit de faccedilon spontaneacutee lorsqursquoen concentration eacuteleveacutee
(Starkov 2010 Turrens 2003) Le H2O2 peut ensuite ecirctre complegravetement reacuteduit en
H2O ou partiellement reacuteduit en radical hydroxyle (bullOH) un des plus puissants
oxydants (Starkov 2010) LrsquoO2-bull peut eacutegalement reacuteagir avec drsquoautres radicaux
incluant lrsquooxyde nitrique (NObull) selon le taux de diffusion des 2 radicaux (Turrens
2003) Le peroxynitrite est eacutegalement un oxydant puissant (Turrens 2003) Les
oxydants deacuterivant du NObull sont appeleacutes des laquo reactive nitrogen species raquo (RNS)
(Turrens 2003) mais seront consideacutereacutes comme faisant partie des ROS dans ce
travail Les ROS peuvent endommager lrsquoADN lrsquoARN les proteacuteines et les lipides
reacutesultants agrave lrsquoapoptose de la cellule lorsque le niveau de ROS produit excegravede la
capaciteacute de deacutetoxifications et de reacuteparation de la cellule (Brynildsen et al 2013
Wells et al 2005) Malgreacute ce danger il srsquoagit drsquoun risque compenseacute par
lrsquoaugmentation du rendement eacutenergeacutetique (Brynildsen et al 2013) Le complexe I (et
II) et III sont les principaux sites de production de ROS (Batandier et al 2006 Chen
et al 2003 Liu et al 2002) Lrsquoinhibition du complexe I par la rotenone provoque une
augmentation de la production de ROS Lrsquooxydation des substrats du complexe I ou
du complexe II en preacutesence du complexe III inhibeacute par lrsquoantimicine A amegravene
eacutegalement une augmentation de la production de ROS (Batandier et al 2006 Chen
et al 2003) (Figure 9) De plus mecircme si le complexe cytochrome oxydase nrsquoest pas
un site de production de ROS son inhibition par le KCN (cyanure de potassium)
facilite la production de ROS par les complexes I et II (Chen et al 2003) Le
- 22 -
complexe ATP synthase nrsquoest pas reconnu pour produire directement des ROS
Cependant la liaison drsquoun inhibiteur tel lrsquooligomycine bloque le passage des protons
agrave travers le complexe et inhibe ainsi la production drsquoATP De plus lrsquoATP synthase ne
fonctionnant plus les pompes agrave protons de la chaine de transport des eacutelectrons sont
incapables de fonctionner car le gradient de protons est beaucoup trop eacuteleveacute ce
provoque une surproduction de ROS par les complexes I et III Conseacutequemment le
NADH nrsquoest plus oxydeacute et le cycle du TCA cesse drsquoopeacuterer car la concentration de
NAD nrsquoest plus suffisante pour le fonctionnement des enzymes du TCA (Joshi et
Huang 1991)
Une production significative de O2-bull principalement par le complexe I peut ecirctre le
reacutesultat de deux modes drsquoopeacuteration 1- lorsque la cellule ne produit pas drsquoATP et
conseacutequemment possegravede une grande force proton-motrice et possegravede un laquo pool raquo de
quinone agrave lrsquoeacutetat reacuteduit (QH2) 2- lorsque la cellule possegravede un haut ratio de
NADHNAD (Figure 10) Lorsque la cellule produit activement de lrsquoATP elle possegravede
alors une moins grande force proton-motrice son ratio NADHNAD est plus faible et
la formation de O2-bull est beaucoup plus faible (Murphy 2009)
Le H2O2 est le substrat pour deux enzymes la catalase et la glutathione (GSH)
peroxydase qui convertissent le H2O2 en H2O+O2 le H2O2 est ainsi deacutetoxifieacute (Figure
9 Figure 11) La reacuteaction catalyseacutee par la glutathione peroxydase neacutecessite de la
glutathione (GSH) comme substrats et deacutepend en partie du ratio de GSSG GSH et
de la concentration des reacuteactants laquelle est le reflet de lrsquoeacutetat reacutedox de la cellule De
faccedilon similaire la concentration de meacutetaux sous leur forme reacuteduite tel le Fer catalyse
la formation de certains ROS (Figure 11) Ceci peut ecirctre minimiseacute en tenant cette
concentration ions meacutetallique tregraves basse entre autres par leur emmagasinage et leur
liaison aux proteacuteines de transport (ei ferritine transferrine lactoferrine) et ainsi
minimiser la formation de bullOH Finalement certains antioxydants (eg vitamine E)
peuvent interrompre la chaine de reacuteactions par la capture des radicaux le radical-
vitamine E est relativement stable et peut ecirctre enzymatiquement reconverti agrave sa
- 23 -
forme non-radical (Figure 11) Il est maintenant accepteacute que certaines enzymes du
cycle de Krebs (ie αKGDH) soient non seulement des cibles des ROS mais aussi
des producteurs non neacutegligeables de ROS (Brynildsen et al 2013)
Figure 9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS Le complexe I ainsi que le site Qi du complexe III relacircchent des
ROS directement vers les deacutefenses drsquoantioxydants intracellulaires Les antioxidants
intracellulaire tel la superoxide dismutase (MnSOD) la catalase la glutathione (GSH)
la glutathione peroxydase (GPX) et la glutathione reacuteductase (GRase) sont montreacute Le
site Qo du complexe III quant agrave lui relacircche les ROS dans le milieu extra cellulaire
Modifieacute de Chen Q et al 2003
Oligomycine
- 24 -
Figure 10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de
O2-bull 1- Le pool de NADH est dans un eacutetat reacuteduit par exemple par des dommages agrave
la chaine respiratoire la perte de cytochrome c durant lrsquoapoptose ou agrave un faible
besoin en ATP Ceci megravene agrave une production de O2-bull au complexe I qui deacutecoule de la
reacuteduction du FMN laquelle est reacuteguleacutee par le ratio NADHNAD+ Drsquoautres sites
autres que le complexe I tel lrsquoα-ceacutetoglutatrate deacuteshydrogeacutenase (αKGDH) peut
aussi contribuer agrave la production de O2-bull 2- Il nrsquoy a pas (peu) de production drsquoATP il y
a un fort potentiel membranaire et un pool de CoQ (meacutenaqinone) agrave lrsquoeacutetat reacuteduit
important qui provoque une reacuteversion de la chaine de transport des eacutelectrons agrave
travers le complexe I amenant une forte production de O2-bull 3- Il y a une production
active drsquoATP et conseacutequemment un potentiel membranaire plus bas que dans la
situation 2 et une plus grande quantiteacute de NADH oxydeacute que dans la situation 1
Dans ces conditions la quantiteacute de O2-bull produit est plus faible que dans la situation
1 et 2 et la provenance des O2-bull est moins certaine
Tireacute de Murphy MP 2009
- 25 -
12 Antibiotiques Mode drsquoaction cible cellulaire et meacutecanismes de reacutesistance
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles
Notre compreacutehension de la faccedilon que les antibiotiques induisent la mort cellulaire est
principalement baseacutee sur la fonction cellulaire essentielle inhibeacutee par lrsquointeraction de
lrsquoantibiotique avec sa cible Les antibiotiques peuvent ecirctre classeacutes selon la
composante ou le systegraveme cellulaire qursquoils affectent ainsi que par le fait qursquoils
induisent la mort cellulaire (antibiotiques bacteacutericides) ou srsquoils inhibent la croissance
Figure 11 Voies de formation et de deacutetoxification des ROS Tireacute de Wikipeacutedia
httpfrwikipediaorgwikiDC3A9rivC3A9_rC3A9actif_de_l27oxygC3A8ne
Consulteacute le 11 deacutec 2013
- 26 -
bacteacuterienne (antibiotique bacteacuteriostatique) La plupart des antibiotiques bacteacutericides
inhibent la synthegravese de lrsquoADN ou de lrsquoARN la synthegravese de la paroi bacteacuterienne ou la
synthegravese proteacuteique (Kohanski et al 2010) Le tableau 1 est une synthegravese des
principales classes drsquoantibiotiques ainsi que leur mode drsquoaction et leurs cibles
cellulaires Ce tableau nrsquoest cependant pas exhaustif et il existe plusieurs autres
classes drsquoantibiotiques et plusieurs autres exemples drsquoantibiotiques dans les classes
identifieacutees dans ce tableau
Depuis la deacutecouverte de la peacutenicilline en 1929 plusieurs autres antibiotiques ont eacuteteacute
deacutecouverts et deacuteveloppeacutes suite agrave lrsquoeacutelucidation des interactions entre lrsquoantibiotique et
sa cible et en apportant des modifications structurales et chimiques aux moleacutecules
deacutejagrave connues Ces efforts ont significativement augmenteacute le nombre drsquoantibiotiques
disponibles dans le milieu clinique Le processus de mort cellulaire induit par lrsquoaction
drsquoun antibiotique est complexe et deacutebute avec une interaction physique entre la
moleacutecule antibiotique et sa cible bacteacuterienne speacutecifique et implique des modifications
qui affectent la bacteacuterie tant au niveau biochimique que moleacuteculaire et structural
27
Acid
e n
alid
ixiq
ue (
1 iegravere
geacuteneacutera
tion)
Cip
rofloxacin
e (
2 iegravem
e g
eacuteneacutera
tion)
Levofloxacin
e (
3 iegravem
e g
eacuteneacutera
tion)
Gem
iflo
xacin
e (
4 iegravem
e g
eacuteneacutera
tion)
Flu
oro
quin
olo
ne (
Norf
loxacin
e)
Peacutenic
iline
Ceacutephalo
sporine (
ceacutefa
lexin
e)
Carb
apenem
Vancom
ycin
e
Teacutela
vancin
e
Fosfo
mycin
Dapto
mycin
Poly
mix
ine B
Bacitra
cin
Valin
om
ycin
e
Nom
encin
e
Genta
mic
in
Kanam
ycin
e
Tobra
mycin
e
Str
epto
mycin
e
Teacutetr
acyclin
e
Chlo
rteacutetr
acyclin
e
Macro
lides
Ery
thro
mycin
eLie
sous-u
niteacute 23S
du r
ibosom
e 5
0S
et em
pecircche
leacutelo
ngation d
u p
eptide (
apregrave
s 6
-8 a
a)
sta
tique
Lin
cosam
ides
Clin
dam
ycin
Inhib
e d
irecte
ment la
ctiviteacute p
eptidyl
transfeacute
rase d
u
ribosom
esta
tique
Str
epto
gra
min
es
DalfopristinQ
uin
upristin
Com
bin
ais
on d
e 2
antibio
tiques B
loque la
transla
tion p
eptidiq
ue e
n lia
nt le
site P
du r
ibosom
e
et en b
loquant au p
eptide la
ccegraves a
u tunnel de s
ort
ie
cid
al
Am
phenic
ol
Chlo
ram
phenic
ol
Em
pecircche la lia
ison d
u a
a-
AR
Nt au s
ite A
du
ribosom
e e
t in
terf
egravere
dans la f
orm
ation d
u lie
n
peptidiq
ue
sta
tique
Oxazolid
inones
Lin
ezolid
Blo
que initia
tion d
e la tra
nsla
tion p
roteacute
ique lors
de la
reacuteaction d
eacutelo
ngation p
ar
la p
eptidyl
transfeacute
rase
sta
tique
Trim
eth
oprim
Sulfam
eth
oxazole
sta
tique
cid
al
Ionophore
Alteacutera
tion d
e lin
teacutegriteacute
mem
bra
naire
Se
nchasse d
ans la m
em
bra
ne fo
rme u
n p
ore
ce
qui am
egravene u
ne p
ert
urb
ation d
e leacute
quili
bre
ioniq
ue
cid
al
Inhib
itio
n d
e la s
ynth
egravese d
e
la p
aro
i cellu
laire
(peptidogly
can)
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
cid
al
cid
al
sta
tique
AR
Nr
16S
(S
ite A
)(fo
rmation d
e p
roteacute
ines n
on-
fonctionnelle
s)
cid
al
Inhib
itio
n d
e la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 3
0S
)
Inhib
iteur
de la s
ynth
egravese d
e
lacid
e f
oliq
ue
Inhib
itio
n d
e c
ert
ain
es r
eacuteactions e
nzym
atiques
essentielle
s agrave
la f
orm
ation d
e la
cid
e f
oliq
ue
Sulfonam
ides
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
AR
Nr
16S
(preacute
vie
nt la
ttachem
ent de la
min
oacyl-
AR
Nt au r
ibosom
e)
Teacutetr
acyclin
e
Poly
peptides
β-L
acta
ms
Gly
copeptides
Lip
opeptides
Inhib
iteur
de la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 5
0S
)
Am
inogly
cosid
es
Cib
le c
ell
ula
ire
Sta
tiq
ue
Cid
al
Rifam
ycin
eR
ifam
pic
ine
Cla
ss
es
Ex
d
an
tib
ioti
qu
eM
od
e d
ac
tio
n
Inhib
itio
n d
e la s
ynth
egravese d
e
lAR
N (
AD
N d
eacutependant)
AR
N p
oly
meacutera
se s
ous-u
niteacute β
(codeacute p
ar
rpoB
)
Quin
olo
nes
cid
al
Inhib
itio
n d
e la r
eacuteplic
ation d
e
lAD
N
Com
ple
xe A
DN
-topois
om
eacutera
se (
AD
N
gyr
ase(t
opois
om
eacutera
se II)
et to
pois
om
eacutera
se IV
)cid
al
Tra
nspeptidase (
penic
illin
bin
din
g p
rote
in
(PB
P))
cid
al
Tab
leau
1
Cib
les e
t m
od
e d
rsquoacti
on
des p
rin
cip
ale
s c
lasses d
rsquoan
tib
ioti
qu
es
28
122 Les aminoglycosides
1221 Mode drsquoaction
Les aminoglycosides peacutenegravetrent la paroi cellulaire en suivant un processus
comprenant 3 eacutetapes une premiegravere eacutetape eacutenergie- indeacutependante suivie de deux
eacutetapes eacutenergie deacutependantes Deux composantes sont essentielles pour
lrsquoaccumulation intracellulaire des aminoglycosides les ribosomes et la chaine de
transport des eacutelectrons (Ramirez et Tolmasky 2010)
Eacutetape 1- Lorsque les aminoglycosides entrent en contact avec la paroi
cellulaire la moleacutecule polycationique se lie aux composantes de surface anioniques
tels les lipopolysaccharides les phospholipides et autres proteacuteines membranaires
chez les bacteacuteries agrave Gram neacutegatif et les moleacutecules drsquoacides teacuteichoiumlques et les
phospholipides des bacteacuteries agrave Gram positif (Ramirez et Tolmasky 2010) Apregraves
cette liaison aux moleacutecules polaires les aminoglycosides (telle la gentamicine)
srsquoassocient aux transporteurs membranaires sur la base de leur charge positive et
sont transporteacutes agrave travers la membrane cytoplasmique gracircce au potentiel
membranaire (intra cellulaire neacutegatif) (Bryan et al 1980 Proctor et al 2006) Cette
liaison agrave la membrane entraine une augmentation de la permeacuteabiliteacute membranaire
menant agrave lrsquoentreacutee des aminoglycosides dans lrsquoespace peacuteriplasmique (Gram neacutegatif)
ou dans le cytoplasme (Gram positif) (Ramirez et Tolmasky 2010) (Figure 12A)
Eacutetape 2- Lrsquoeacutetape suivante connue comme la laquo phase eacutenergie-deacutependante
1 raquo (EDP-I) est bloqueacutee par les inhibiteurs de la chaine de transport des eacutelectrons et
de la phosphorylation oxydative ainsi que par les ions divalents lrsquohyperosmolariteacute le
faible pH et lrsquoanaeacuterobiose dans cette phase la dureacutee et le taux drsquoentreacutee de
lrsquoantibiotique sont en fonction de la concentration (Miller et al 1980 Mingeot-
Leclercq et al 1999 Ramirez et Tolmasky 2010) Agrave cette eacutetape le transport
transmembranaire des aminoglycosides et plus speacutecifiquement de la gentamicine
neacutecessite 2 composantes majeures a) un transporteur anionique non speacutecifique dont
la charge est directement lieacutee agrave lrsquoactiviteacute des cytochromes oxydases terminales et des
29
nitrates reacuteductases de la chaine de transport des eacutelectrons Selon certaines eacutetudes
les transporteurs pourraient ecirctre les quinones reacuteduites de la chaine de transport des
eacutelectrons (Bryan et al 1981 Miller et al 1980) et donc le cycle drsquooxydation-reacuteduction
des quinones est essentiel aux transports des aminoglycosides (Bryan et al 1981)
mais selon drsquoautres eacutetudes les transporteurs pourraient ecirctre toute autre proteacuteine non
deacutefinie impliqueacutee dans la polarisation membranaire lieacutee agrave lrsquoactiviteacute de la chaine de
transport des eacutelectrons (Miller et al 1980) La croissance anaeacuterobie ainsi que le
sodium azide (inhibiteur de la chaine respiratoire) inhibent lrsquoentreacutee de la gentamicine
et lrsquoeffet bacteacutericide indiquant que le transport actif de cet antibiotique neacutecessite la
respiration b) un potentiel eacutelectrique avec un milieu intra cellulaire neacutegatif par rapport
agrave lrsquoexteacuterieur En effet lrsquoinhibition de lrsquoentreacutee dans la bacteacuterie des aminoglycosides
suite agrave lrsquoajout de CCCP (agent deacutepolarisant) chez des souches de S aureus laquo wild-
type raquo deacutemontre la neacutecessiteacute de la force proton-motrice pour lrsquoentreacutee des
aminoglycosides dans les cellules (Miller et al 1980)
Eacutetape 3- La petite quantiteacute drsquoaminoglycosides srsquoeacutetant rendu au cytoplasme
durant cette premiegravere phase eacutenergie-deacutependante induisent apregraves liaison aux
ribosomes des erreurs dans la synthegravese proteacuteique Les erreurs dans la traduction
des proteacuteines membranaires produites lors de cette eacutetape connue comme la laquo phase
eacutenergie-deacutependante 2 raquo (EDP-II) provoquent des dommages dans lrsquointeacutegriteacute de la
membrane cytoplasmique (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky
2010) Les proteacuteines membranaires aberrantes dans la membrane facilitent le
transport drsquoune plus grande quantiteacute drsquoaminoglycosides lesquels augmenteront
encore le niveau drsquointerfeacuterence avec la synthegravese proteacuteique normale amenant encore
plus de dommage agrave la membrane provoquant ainsi une acceacuteleacuteration importante du
taux drsquoentreacutee des moleacutecules antibiotique et qui ultimement reacutesultera en la mort de la
cellule (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky 2010) (Figure 12B)
Des eacutetudes ont deacutemontreacute que la susceptibiliteacute agrave la gentamicine streptomycine
amikacine kanamycine et tobramycine est supeacuterieure lorsque lrsquooxygegravene est
30
lrsquoaccepteur terminal drsquoeacutelectrons (Bryan et Kwan 1981) Une diminution significative
de la susceptibiliteacute est remarqueacutee lorsque le nitrate est lrsquoaccepteur final drsquoeacutelectrons et
une diminution encore plus marqueacutee est observeacutee lorsque le fumarate est lrsquoaccepteur
final drsquoeacutelectron ou lorsque les conditions sont fermentaires Lrsquoassociation entre le
transport de la gentamicine et le complexe terminal de la chaine de transport des
eacutelectrons particuliegraverement lrsquoactiviteacute cytochrome oxydase
Le ribosome possegravede trois sites de liaison importants de lrsquoARNt deacutesigneacute comme site
A (pour aminoacyl) site P (pour peptidyl) et E (pour exit) (Vakulenko et Mobashery
2003) Ils sont situeacutes sur lrsquoARN16S (lequel avec plus de 21 proteacuteines compose la
sous-uniteacute 30S du ribosome bacteacuterien) (Ramirez et Tolmasky 2010 Vakulenko et
Mobashery 2003) Durant la synthegravese proteacuteique le ribosome deacutecode lrsquoinformation de
lrsquoARNm et catalyse lrsquoincorporation seacutequentielle de chaque acide amineacute faisant partie
de la chaine polypeptidique en formation La haute fideacuteliteacute de la traduction (les
ribosomes incorporent un mauvais acide amineacute par approximativement 3000 liaisons
peptidiques effectueacutees) est possible gracircce agrave la capaciteacute de discriminer entre des
changements conformationels dans le ribosome induit par la liaison drsquoun ARNt correct
Modifieacute de Davis BD 1987
Figure 12 Cineacutetique drsquoentreacutee des aminoglycosides
A
B
31
ou incorrect au site A La reconnaissance drsquoune liaison correcte anticodoncodon par
le ribosome se passe en deux temps la reconnaissance initiale de lrsquoappariement du
codon sur lrsquoARNm avec lrsquoanticodon de lrsquoARNt et subseacutequemment la correction
(Vakulenko et Mobashery 2003)
La plupart des aminoglycosides se lient agrave des oligonucleacuteotides faisant partie du site
A Les diffeacuterents aminoglycosides ne lient pas tous le mecircme site de lrsquoARN 16S mais
lrsquoeffet commun agrave tous est que cette liaison amegravene un changement de conformation du
site A vers une conformation mimant la conformation fermeacutee de la sous uniteacute 30S qui
est induite par une interaction entre lrsquoARNm et son ARNt apparenteacute eacuteliminant la
possibiliteacute de correction du ribosome ce qui promeut les erreurs de traduction
(Ramirez et Tolmasky 2010 Vakulenko et Mobashery 2003) La grande majoriteacute des
aminoglycosides sont bacteacutericides et cette leacutetaliteacute est la conseacutequence de cette
mauvaise traduction (Vakulenko et Mobashery 2003)
1222 Meacutecanismes de reacutesistance
La reacutesistance contre les aminoglycosides se produit par diffeacuterents meacutecanismes qui
peuvent coexister simultaneacutement dans la mecircme bacteacuterie (Vakulenko et Mobashery
2003) Ces meacutecanismes incluent 1- la modification de la cible par des mutations
dans lrsquoARN 16S ou autres proteacuteines ribosomales associeacutees (Figure 13) 2- la
meacutethylation de lrsquoARN 16S un meacutecanisme retrouveacute chez plusieurs organismes
producteurs drsquoaminoglycosides et chez des souches cliniques Cette meacutethylation
empecircche les aminoglycosides drsquoavoir accegraves agrave leur site de liaison (Doi et Arakawa
2007) 3- la reacuteduction de la permeacuteabiliteacute par une alteacuteration de la permeacuteabiliteacute de la
membrane externe ou une diminution du transport agrave travers la membrane interne 4-
lrsquoexportation des aminoglycosides agrave lrsquoexteacuterieur de la cellule par les pompes agrave efflux
Eacutetant des moleacutecules hydrophiliques les aminoglycosides ont naturellement peu
drsquoaffiniteacute avec les pompes agrave efflux 5- le swarming actif probablement un meacutecanisme
32
non speacutecifique deacutemontreacute chez Pseudomonas aeruginosa lequel deacutemontre des
reacutesistances agrave plusieurs antibiotiques 6- la seacutequestration de la drogue par des liaisons
agrave un aceacutetyltransfeacuterase agrave tregraves faible activiteacute 7- une inactivation enzymatique de la
moleacutecule antibiotique le meacutecanisme de reacutesistance le plus preacutevalent au niveau
clinique (Vakulenko et Mobashery 2003) Lrsquoinactivation enzymatique des
aminoglycosides est le plus souvent due agrave des aceacutetyltransfeacuterases des
nucleacuteotidyltransfeacuterases ou des phosphotransfeacuterases (Dickgiesser et Kreiswirth 1986)
Ces enzymes sont souvent encodeacutees sur des plasmides mais sont eacutegalement
associeacutees agrave des eacuteleacutements transposables Lrsquoeacutechange de plasmides et la disseacutemination
de transposons facilitent lrsquoacquisition rapide de pheacutenotypes reacutesistants aux
antibiotiques non seulement au sein drsquoune mecircme espegravece mais parmi un large
eacuteventail drsquoespegraveces bacteacuteriennes (Mingeot-Leclercq et al 1999)
La reacutesistance naturelle des S aureus laquo small colony variants raquo agrave la majoriteacute des
aminoglycosides est la conseacutequence de lrsquoimpossibiliteacute agrave lrsquoantibiotique de peacuteneacutetrer une
cellule posseacutedant une faible polariteacute membranaire qui est due chez les SCV agrave leur
Tireacute de Doi Y Arakawa Y 2007
Figure 13 Sites de mutation de certains reacutesistants aux aminoglycosides
33
incapaciteacute agrave activer le cycle de Krebs (Miller et al 1980 Proctor et al 2006 Tubby
et al 2013) Les SCV ont non seulement une polariteacute membranaire faible mais (ceci
eacutetant la conseacutequence de cela) aussi une force proton motrice inhibeacutee et une chaine
des eacutelectrons deacuteficiente De ce fait le transport transmembranaire des
aminoglycosides chez les SCV est grandement reacuteduit et conseacutequemment la
susceptibiliteacute des SCVs pour cette classe drsquoantibiotique est 110-113 de celle des
souches parentes (Proctor et al 2006)
123 La tomatidine
Plusieurs espegraveces veacutegeacutetales accumulent des steacuterols et des triterpegravenes antimicrobiens
appeleacutes saponines (Osborn 1996) Ces moleacutecules sont produites de faccedilon
constitutive par la plante et jouent un important rocircle de deacutefense contre diffeacuterents
pathogegravenes comme des insectes ou des microbes Parmi les saponines la tomatine
(Figure 14A) est un glycoalcaloiumlde steacuteroiumldien preacutesentant une action antimicrobienne
contre des levures et une quantiteacute drsquoautres microorganismes (Bednarek et Osboum
2009 Friedman 2002 Osborn 1996 Sandrock et Vanetten 1998) Plusieurs
espegraveces de champignons pathogegravenes de la tomate produisent des enzymes
extracellulaires communeacutement appeleacutes tomatinase qui sont capables de deacutetoxifier
lrsquoα-tomatine (Martin-Hernandez et al 2000 Roddick 1974 Ruiz-Rubbio et al 2001)
Par exemple Fusarium oxysporum f sp lycopersici est reconnu pour cliver lrsquoα-
tomatine en ses sous-produits soit un tetrasaccharide (lycotreacutetraose) et un alcaloiumlde
steacuteroiumldien non glycosyleacute la tomatidine (Figure 14B) Ces sous-produits nrsquoont pas ou
peu drsquoactiviteacute antifongique (Ruiz-Rubbio et al 2001 Simon et al 2006)
34
Figure 14 Structures chimiques de la (A) tomatine de (B) tomatidine et de (C)
lrsquoanalogue de tomatidine FC04-100
Il a eacuteteacute deacutecouvert il a quelques anneacutees que la tomatidine avait un effet antibacteacuterien
sur les S aureus de pheacutenotype SCV mais aucune activiteacute contre les S aureus agrave
pheacutenotype normal Par contre lorsqursquoutiliseacute en combinaison avec les
aminoglycosides lrsquoajout de la tomatidine permettait drsquoaugmenter de 8 agrave 16 fois
lrsquoactiviteacute des aminoglycosides contre S aureus agrave pheacutenotype normal (Mitchell et al
2011a Mitchell et al 2012)
Donc on savait que la tomatidine avait une activiteacute antibacteacuterienne tregraves inteacuteressante
mais on ne connaissait ni sa cible ni son mode drsquoaction De plus si la tomatidine a
une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait donc avoir un
spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Par ailleurs de faccedilon parallegravele la production drsquoanalogues structuraux de la tomatidine
permettrait de mieux comprendre la relation structure-activiteacute (SAR) de la moleacutecule et
donc de preacuteciser que lrsquoactiviteacute biologique de la tomatidine neacutecessite une orientation
3
A B
C
35
de groupements fonctionnels bien speacutecifique De plus lrsquoeacutetude de lrsquoactiviteacute de
plusieurs analogues permettrait de confirmer si la tomatidine possegravede un meacutecanisme
drsquoaction deacutependant drsquoune interaction speacutecifique avec une ou des cibles biologiques
preacutecises
Le laboratoire du Pr Eacuteric Marsault du deacutepartement de pharmacologie de lrsquoUniversiteacute
de Sherbrooke et lrsquoeacutetudiant agrave la maitrise Feacutelix Chagnon ont produit plus de 33
analogues de la tomatidine (voir Annexe 2 article laquo Unraveling the structure-activity
relationship of tomatidine a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureusraquo dans la section laquo Contributions
Supleacutementaires raquo du meacutemoire) dont lrsquoactiviteacute antibacteacuterienne a eacuteteacute eacutevalueacutee par des
tests de susceptibiliteacute agrave la moleacutecule (CMI) sur S aureus SCV et sur S aureus agrave
pheacutenotype normal en combinaison avec la gentamicine Plusieurs de ces moleacutecules
font lrsquoobjet drsquoune application de brevet (voir annexe 1) dont la prometteuse moleacutecule
FC04-100 (Figure 14C) (voir laquo Annexe 1 Application de brevet raquo dans la section
laquo Contributions suppleacutementaires raquo du meacutemoire)
13 Projet de maitrise
Premiegravere hypothegravese
Si la tomatidine a une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait
donc avoir un spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Objectif Provoquer lrsquoapparition de pheacutenotypes SCV chez diffeacuterentes espegraveces
bacteacuterienne afin de pouvoir veacuterifier leur niveau de sensibiliteacute agrave la tomatidine
Deuxiegraveme hypothegravese
Lrsquoidentification des gegravenes preacutesentant des mutations parmi les souches de S aureus
reacutesistantes agrave la tomatidine pourrait reacuteveacuteler la ou les cibles cellulaires de la moleacutecule
et nous aider agrave comprendre son mode drsquoaction
36
Objectif Obtenir des mutants reacutesistants avant de les envoyer agrave sequencer
faire les analyses comparatives des geacutenomes et finalement identifier les mutations
37
CHAPITRE 2
ARTICLE SCIENTIFIQUE
TITRE Tomatidine and Analog FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Some Firmicutes Including Those of Listeria
monocytogenes and Are Bactericidal in Combination With Aminoglycosides Against
their Normal Phenotype
AUTEURS ET CONTRIBUTIONS
Isabelle Guay1 Expeacuteriences biologiques avec le produit naturel tomatidine et son
analogue FC04-100 sur diffeacuterentes espegraveces bacteacuteriennes afin de comparer les deux
activiteacutes
Simon Boulanger1 Expeacuterience biologique permettant de quantifier lrsquoactiviteacute de
lrsquoanalogue FC04-100 sur la production de biofilm et son activiteacute intracellulaire
Feacutelix Chagnon2 Production de lrsquoanalogue chimique de la tomatidine FC04-100
Kamal Bouarab1 Initiateur du projet gracircce agrave lrsquoisolement du produit naturel tomatidine
Eacuteric Marsault2 Directeur de recherche de Feacutelix Chagnon
Franccedilois Malouin1 Directeur de recherche drsquoIsabelle Guay et Simon Boulanger et
auteur de correspondance
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
38
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
Corresponding author Mailing address Universiteacute de Sherbrooke
Deacutepartement de Biologie Faculteacute des Sciences 2500 Boul
Universiteacute Sherbrooke Quebec Canada J1K2R1 Phone (819) 821-
8000 ext 61202 Fax (819) 821-8049 E-mail FrancoisMalouin
USherbrookeca
39
Tomatidine and Analog FC04-100 Possess Bactericidal Activities
Against the Small Colony Variants of Some Firmicutes Including
Those of Listeria monocytogenes and Are Bactericidal in Combination
With Aminoglycosides Against their Normal Phenotype
Running Title Tomatidine is bactericidal against several Firmicutes
Isabelle Guay1
Simon Boulanger1
Feacutelix Chagnon2 Kamal Bouarab
1 Eacuteric Marsault
2 and
Franccedilois Malouin1
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM) Deacutepartement de
Biologie Faculteacute des Sciences Universiteacute de Sherbrooke Sherbrooke Queacutebec Canada J1K
2R1
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute Universiteacute
de Sherbrooke Queacutebec Canada J1H 5N4
Have contributed equally to the work
Corresponding author
Mailing address Universiteacute de Sherbrooke Deacutepartement de Biologie Faculteacute des Sciences
2500 Boul Universiteacute Sherbrooke Quebec Canada J1K2R1
Phone (819) 821-8000 ext 61202 Fax (819) 821-8049
E-mail FrancoisMalouinUSherbrookeca
40
ABSTRACT
Tomatidine (TO) is a plant steroidal alkaloid that possesses an antibacterial activity against the
small colony variants (SCVs) of Staphylococcus aureus We report here the spectrum of
activity of TO against species of the Firmicutes and the improved antibacterial activity of a
chemically-modified TO derivative (FC04-100) In addition to that seen on S aureus TO
showed a bactericidal activity against SCVs of Bacillus cereus B subtilis and Listeria
monocytogenes The combination of an aminoglycoside antibiotic and TO also generated an
antibacterial synergy against strains of the normal phenotype in these species In contrast to
TO which has no relevant activity by itself against S aureus of the normal phenotype (MIC
gt64 mgL) the TO analog FC04-100 showed a MIC of 8-16 mgL Furthermore FC04-100
showed a strong bactericidal activity against L monocytogenes SCVs in kill kinetic
experiments while TO did not FC04-100 also improved the bactericidal activity of
gentamicin against L monocytogenes of the normal phenotype The addition of FC04-100 (4
mgL) to a cephalexinkanamycin (32) combination improved the activity of the combination
by 32 fold against cephalexin and kanamycin-resistant MRSA strains In combination with
gentamicin FC04-100 also showed a strong bactericidal activity against biofilm-embedded S
aureus and when used alone was superior to TO for its biofilm-preventing activity Finally
FC04-100 and TO showed comparable intracellular killing of S aureus SCVs in a polarized
Calu-3 cell culture model In conclusion chemical modifications of TO allowed improvement
of its antibacterial activity against L monocytogenes SCVs and prototypical S aureus
41
Keywords Tomatidine aminoglycoside synergy SCV Firmicutes S aureus L
monocytogenes
Introduction
The Firmicutes are Gram-positive bacteria with genomes presenting less than 50 of G+C
The Firmicutes constitutes one of the main divisions within the Bacteria and is divided into
three classes Bacilli Mollicutes and Clostridia The class of Bacilli is further divided into the
orders of the Lactobacillales and the Bacillales The Bacillales are divided into the genus of
Staphylococcus Listeria and Bacillus A number of bacterial species such as Listeria spp and
Bacillus spp can contaminate food and cause infections in humans (Magalhaes 2013) To
name a few Listeria monocytogenes L ivanovii and Bacillus cereus can cause listeriosis
(Guillet et al 2010) and food poisoining (Bad Bug Book FDA) Bacillus subtilis B
coagulans B licheniformis and B sphaericus are also known to cause illnesses Bacillus
anthracis causes anthrax and can often be acquired by contact with food producing animals
and cattle (beef cattle sheeps etc) and this bacterium is also well-known for its endospores
that have been used as biological weapons (Beierlein and Anderson 2011) Staphylococci are
divided in coagulase-positive species Staphylococcus aureus being the most clinically
relevant of this group and coagulase-negative species such as S epidermidis the most
prevalent pathogen associated with infections of implanted medical devices (Vuong and Otto
2002) The emergence and spread of resistance to multiple antibiotics in staphylococci is now
considered a real health treat and impaired therapeutic endeavor to combat these bacteria
42
(Witte et al 2007) Notably the prevalence of methicillin-resistant S aureus (MRSA) has
steadily increased over the recent years not only in hospitals but also in the community
(Hiramatsu et al 2013)
Since their first description in 1910 for Salmonella enterica serovar Typhi (S typhi) small-
colony variants (SCVs) have been described for a wide range of bacterial genera and species
and have been isolated from both laboratory and clinical environments (Proctor 2006)
Staphylococcus aureus SCVs have attracted a great deal of interest over the past recent years
S aureus SCVs often present a dysfunctional oxidative metabolism causing an alteration in
the expression of virulence factors a slow growth and a loss of colony pigmentation (Proctor
2006) This dysfunctional oxidative metabolism is also responsible for a decreased
susceptibility to aminoglycoside antibiotics because this class of molecules requires the
proton-motive force in order to penetrate the bacterium (Bryan LE 1981) This respiratory
deficiency is often caused by mutations affecting the electron transport system and several
SCV isolates can recover normal growth with supplemental hemin or menadione which are
needed to synthesize electron transport system components S aureus SCVs often are isolated
from chronic infections such as lung infections in cystic fibrosis (CF) patients osteomyelitis
septic arthritis bovine mastitis and from infections associated with orthopedic devices (Atalla
2008 Moisan 2006 Proctor 2006) The ability of S aureus to switch back and forth from the
prototypic to the SCV phenotypes in vivo is an integral part of the pathogenesis of S aureus
and may be responsible for the establishment of chronic infections (Tuchscherr 2011
Mitchell 2011 inf imm)
43
Tomatidine (TO) is a steroidal alkaloid produced by the Solanaceae plant family such as the
tomato (Ruiz-Rubio 2001 Simons 2006) We showed previously the antibacterial activity of
TO against S aureus SCVs and also documented a strong synergic activity of TO in
combination with aminoglycoside antibiotics against prototypic S aureus (Mitchell G AAC
2011 Mitchell G 2012 JAC) Recently we synthesized a variety of TO analogs in order to
explore the structure-activity relationship of this new class of antibiotics able to act on SCVs
(Chagnon et al 2014) One analog FC04-100 showed the same steroidal backbone as the
natural molecule but with an additional carbon chain on the A cycle (Fig 1) Preliminary
characterization of FC04-100 revealed an antibacterial activity that was similar to that of TO
against S aureus SCVs and also preserved the synergic activity with aminoglycosides against
prototypic S aureus (Chagnon et al 2014) However contrary to TO FC04-100 showed
notable antibiotic activity by itself against prototypic S aureus with a MIC of 8-16 gL
whereas that of TO was gt64 gL It would be of great interest to see if this property of FC04-
100 modifies or improves its spectrum of antibacterial activity
The aim of this study was to further describe the spectrum activity of TO and FC04-100 The
susceptibility of several species among the Firmicutes to TO and FC04-100 alone or in
combination with other antibiotics was investigated The improved antibacterial activity of
FC04-100 was demonstrated against Listeria monocytogenes and methicillin-resistant S
aureus (MRSA)
44
Materials and methods
Chemical reagents and antibiotics
Tomatidine (TO) (Sigma Aldrich Oakville Ontario Canada) (Fig 1A) was solubilised in
dimethylsulfoxide (DMSO) at a concentration of 2 mgmL The TO analog FC04-100 (Fig
1B) was synthesized (Chagnon et al 2014) and solubilised in DMSO at 20 gL Cefalexin
kanamycin gentamicine (GEN) (all from Sigma Aldrich) were solubilised in water at a
concentration of 10 gL Menadione was solubilized in DMSO hemin in 14 M NH4OH and
thymidine in water (all from Sigma Aldrich) and were prepared at a concentration of 10 gL
45
3
Fig 1 Tomatidine (A) is a steroid alkaloid structurally characterized by 6 rings 12
stereogenic centers a 3 β-hydroxyl group and spiro-fused E F rings in the form of an
aminoketal The tomatidine analog FC04-100 (B) contains a diamine in position 3 of ring A
Bacterial strains and growth conditions
Staphylococcus aureus ATCC 29213 S aureus Newbould (ATCC 29740) Listeria
monocytogenes ATCC 13932 Bacillus subtilis ATCC 6333 and Bacillus cereus ATCC 11778
were used as prototypic strains Methicillin-resistant S aureus (MRSA) strains COL and
USA100 (hospital acquired-MRSA) and strain USA300 (community acquired-MRSA) were
also used in this study The reference laboratory strain S aureus SH-1000 an isogenic mutant
strain derived from S aureus 8523-4 but with a functional rsbU allele (HorsburghMJ 2002)
was used for its good yield of biofilm production The S aureus strains CF07-L (prototypic)
A
B
A B
C D
E
F
46
and CF07-S (SCV phenotype) are genetically related clinical strains originally isolated from a
cystic fibrosis patient (MitchellG 2011) Strain CF07-S was used in the intracellular
infection model Staphylococcus and Bacillus were maintained on Tryptic Soy agar (TSA BD
Mississauga Ontario Canada) whereas L monocytogenes was grown on brain-heart infusion
(BHI) agar (BD Mississauga Ontario Canada)
Small colony variants (SCVs)
For S aureus strain NewbouldhemB was used as the reference SCV NewbouldΔhemB was
generated from strain Newbould (ATCC 29740) by disrupting the hemB gene with the ermA
cassette by homologous recombination (Brouillette 2004) SCVs from B cereus B subtillis
and L monocytogenes were generated by growth in presence of a subinhibitory concentration
of GEN Briefly overnight broth cultures (18ndash20 h) were used to inoculate BHI broth at a
dilution of 1100 supplemented or not with 025 to 1X the MIC of GEN Cultures were
incubated 18 h at 35degC with shaking (225 rpm) and then adjusted to an A595 nm of 20 in PBS at
4degC Determination of CFU and SCV colonies was done by serial dilution plating SCV were
obtained by plating on TSA (BHI agar for L monocytogenes) containing gentamicin at a
concentration of 8 to 16 times the induction concentration followed by an incubation of 48 h at
35degC The pinpoint colonies selected by this method were confirmed to be SCVs by streaking
several of them on agar plates without antibiotic SCV that conserved their phenotype after
two passages were considered to have a stable phenotype and were used in subsequent
experiments Selected SCVs had a colony size a tenth of the size of a normal colony and
showed no pigmentation or hemolytic activity on blood agar plates The SCV species were
47
confirmed by rDNA 16S sequencing after PCR amplification using universal primers (Gomaa
2007)
Auxotrophy assays
For SCVs auxotrophism is defined as the requirement of specific compounds in order to
regain a normal growth phenotype [Sendie 2009] An agar diffusion method was used to
characterize the auxotrophism of SCVs using hemin or menadione (1-10 μg eachwell) or
thymidine (100 microgwell) on an inoculated Mueller-Hinton agar (MHA) or Brain-heart infusion
agar (BHIA) plate [Besier 2007] Auxotrophy for specific supplements was detected by a
zone of normal growth surrounding the well after 18 h of incubation at 35degC The results were
confirmed by two indepedent experiments
Antibiotic susceptibility testing
MICs were determined by a broth microdilution technique following the recommendations of
the Clinical and Laboratory Standards Institute (CLSI) except that the incubation period was
extended to 48 h and the medium used was BHI to allow SCVs to reach maximal growth as
previously described (Mitchell 2010) As recommended by the CLSI L monocytogenes (and
its SCVs) were grown in cation-adjusted Muller-Hinton broth (CAMHB BD) containing 3
lysed horse blood (LHB Remel Hartford CT USA) The reported results were obtained from
three independent experiments
48
Time-kill experiments
Kill kinetics were performed to determine whether the effect of compounds alone or in
combination was bacteriostatic or bactericidal Bacteria were inoculated at 105-10
6 CFUmL in
appropriate medium in the absence or presence of the different antibiotic compounds at
concentrations specified in figure legends At several points in time at 35degC (225 rpm)
bacteria were sampled serially diluted and plated on TSA for CFU determinations Plates
were incubated for 24 or 48 h at 35degC for normal and SCV strains respectively The data were
collected from a minimum of three independents experiments
Antibiotic activity in biofilms
The viability of bacteria in biofilms was evaluated based on previously described protocols
(CeriH 2001 MoskowitzSM 2004 HarrisonJJ 2010) For these assays 96-peg lids and
corresponding 96-well plates were used (Nunc CITY STATE) Wells were inoculated with a
suspension of S aureus SH-1000 adjusted to a 05 McFarland standard in TSB (150 microL per
wells) and plates with 96-well lids were incubated at 35degC with an agitation of 120 rpm for 24
h The biofilms on pegs were then washed three times with 200 microL PBS Biofilms on pegs
were further incubated in fresh 96-well plates containing 200 microL of TSB containing or not
antibiotic compounds or a combination of molecules at 35degC 120 rpm for 24 h The treated
biofilms on pegs were washed three times in PBS and bacteria in biofilms were recovered by
sonication in a new microtiter plate containing 200 microL of PBS per wells using an
ultrasonicator bath for 10 min followed by centrifugation for five min at 1000 RPM The
49
bacteria recovered in each well were resuspended serially diluted plated on TSA and
incubated at 35degC for 24 h before CFU determination
For an equivalent series of wells instead of disrupting the biofilm by sonication biofilm
formation on pegs was evaluated through crystal violet staining using a procedure adapted
from Mitchell et al(2010) Following exposure to antibiotics as described above biofilms on
pegs were washed three times using 200 microL of PBS before biofilms were air-dried for 30
minutes Biofilm on pegs were then stained by dipping in 200 microL of a crystal violet solution
(01) for 30 minutes rinsed in 200 microL of distilled water and air-dried for 30 min Finally the
crystal violet from stained biofilms was solubilized in 200 microL of ethanol 95 at room
temperature and the absorbance was measured at 562 nm using a plate reader (nom de
lrsquoappareil)
Cell invasion and measurement of intracellular antibiotic activity
The Calu-3 cell line (ATCC HTB 55 Homo sapiens lung adenocarcinoma) was cultured in
Eaglersquos Minimum Essential Medium (EMEM) supplemented with 01 mM MEM nonessential
amino acids 1 mM of sodium pyruvate 100 UmL penicillin 100 gL streptomycin 25 gL of
Fungizone and 10 fetal bovine serum (FBS) at 37oC in 5 CO2 For routine culture 4 gL of
puromycin was added to culture media All cell culture reagents were purchased from Wisent
(St-Bruno QC Canada)
50
The cell invasion assay was performed with the Calu-3 cells in an air-liquid interface as
previously described with few modifications (Mitchell TO I paperAAC 2011) Briefly cells
were seeded at 15 x 105 cells per insert in a 12-well Transwell plate (Corning CITY STATE)
and cultured for 10 days with the apical medium replaced each day The complete medium in
the basal compartment was replaced by the invasion medium (1 FBS and no antibiotics) 18
h before invasion assays Inocula were prepared by suspending bacteria grown 20 h on BHI
agar plates in ice-cold PBS Bacteria were then washed three times in ice-cold PBS and
suspended in the invasion medium supplemented with 05 BSA at a density of
approximately 4 x 108 CFUmL Cells were washed twice with PBS and 250 μL of bacterial
suspension were apically added to each insert Invasion of cells by bacteria was allowed for 3
h inserts were emptied and washed three times with PBS The basal medium was changed for
the invasion medium with 20 gL of lysostaphin (Sigma) to kill extracellular bacteria and with
or without the tested antibiotic compounds The infected cells were incubated for a total of 48
h with a change of medium at 23 h (invasion medium with lysostaphin with or without the
tested antibiotic compounds) The invasion medium with lysostaphin but without tested
compound was also apically added 1 h before cell lysis to ensure that only intracellular
bacteria were counted At 48 h following three washes with PBS the apical and basal media
were removed and cells were detached with 100 μL of trypsin 025 and lysed for 10 min by
the addition of 400 μL of water containing 005 of Triton X-100 Then 50 μL of PBS (50X)
was added and mixed Lysates were serially diluted 10-fold and plated on TSA for CFU
determination Plates were incubated at 35oC for 48 h
51
RESULTS
Emergence the SCV phenotype
The SCV phenotype is characterized by a slow growth yielding small colony size on agar
plates (15 to 110 of the parent phenotype) Figure 2 illustrates the ability of a subinhibitory
concentration of GEN to promote the emergence of the SCV phenotype for B cereus B
subtilis and L monocytogenes B cereus SCVs were obtained at an inducing concentration of
1 X MIC of GEN (Fig 2C) B subtilis SCVs at 05 X MIC (Fig 2D) and L monocytogenes
SCVs at 025 X MIC (Fig 2E) Two prototypic S aureus Newbould and CF07-L and their
SCV counterpart NewbouldhemB and CF07-S respectively were used as control strains (Fig
2A and 2B) The SCV isolates selected for the rest of the study add a low reversion rate and
kept their phenotype without a GEN selection pressure The B cereus SCVs showed
auxotrophy for menadione (Fig 1F Table 1) some of the L monocytogenes SCVs showed
auxotrophy for hemin (Table 1) while the B subtilis SCVs showed no apparent auxotrophy to
hemin menadione or thymidine (Table 1) These results show that SCVs with defects in their
respiratory chain (ie hemin or menadione auxotrophy) can indeed emerge from a selective
pressure with aminoglycosides
52
A B
Fig 2 Typical colony sizes for SCV and protocipic strains (left and right sides of the plates
respectively) for (A) S aureus CF07-S and CF07-L (B) S aureus NewbouldhemB and
Newbould (C) B cereus ATCC 11778 (SCV1 and prototypic) (D) B subtilis ATCC 6333
(SCV2 and prototypic) and (E) L monocytogenes ATCC 13932 (SCV1 and prototypic) In
(F) a TSA plate was inoculated with a B cereus SCV1 and wells were filled with (i) 10 microg
hemin (ii) 10 microg menadione (iii) 100microg thymidine and diluents (iv) DMSO and (v) NH4OH
14N
C D
E
i ii
iii iv
v
F
53
Antibacterial activity of TO against the various species of SCVs and synergy of
the TO and GEN combination against some prototypic Firmicutes
All the SCVs tested showed a MIC for TO of 006 mgL against S aureus and lt003 mgL
against L monocytogenes B cereus and B subtilis respectivly whereas TO showed no
activity (MIC gt64 microgmL) against their prototypic counterparts (Table 1) A synergic effect of
TO with GEN against all the prototypic strains tested was also observed (Table 1) Indeed the
addition of TO provoked a 4-8 fold decrease of GEN MIC against all species tested (Table 1)
These results are consistent with our previous studies on the activity of TO on S aureus
((Mitchell 2011 AAC 2012 JAC)) No change in the MIC of GEN was observed against the
streptococci (S pyogenes S suis S agalactiae S mitis and S pneumoniae) when tested in
combination with TO (data not shown) These results show that TO is active against some but
not all Firmicutes
54
Table 1 Susceptibility of prototypic and SCV strains and species to TO andor GEN
Ratio of activity of GEN+TO vs GEN alone
Bactericidal activity of TO against SCVs and bactericidal synergy of the TO
and GEN combination against prototypic strains
As shown above the addition of 8 mgL of TO to GEN reduced the MIC of GEN against some
Firmicutes (Table 1) but also time-kill kinetics showed that TO improved the bactericidal
effect of GEN against prototypic B subtilis and B cereus (Fig 3A and 3B respectively) This
bactericidal synergy was also previously described for S aureus (Mitchell 2012 JAC)
However although the MIC of GEN was improved when in combination with TO against L
monocytogenes (Table 1) time-kill experiments failed to demonstrate a bactericidal synergy
TO GENGEN
(+TO)
Saureus ATCC 29213 gt64 05 006 8
Saureus NewbouldΔhemB 006 8 Hemin
B cereus ATCC 11778 gt64 1 025 4
B cereus ATCC 11778 (SCV1) lt003 16 Menadione
B cereus ATCC 11778 (SCV2) lt003 16 Menadione
B subtilis ATCC 6333 gt64 0125 003 4
B subtilis ATCC 6333 (SCV1) lt003 2 unknown
B subtilis ATCC 6333 (SCV2) lt003 4 unknown
B subtilis ATCC 9372 gt64 0125 lt0015 gt8
B subtilis ATCC 9372 (SCV1) lt003 1 unknown
B subtilis ATCC 9372 (SCV2) lt003 1 unknown
L monocytogenes gt64 05 0125 4
L monocytogenes (SCV1) lt003 gt64 unknown
L monocytogenes (SCV2) lt003 gt64 Hemin
Strains
MIC (gL)
Synergy
foldAuxotrophy
55
with an improvement of at most 1 log10 in killing compared to that achieved with GEN alone
(Fig 3C) Similarly the effect of TO used alone against SCVs was strongly bactericidal
against S aureus B subtilis and B cereus but not against the L monocytogenes SCV (Fig
3D) and this even if the MIC of TO against L monocytogenes SCVs was inferior to 003
mgL (Table 1) These results show that the activity of TO against L monocytogenes needs to
be improved
56
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 0 6 m g L G E N
0 0 6 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 5 m g L G E N
0 5 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
2 m g L G E N
2 m g L G E N + T O
8 m g L T O
C T R L
Lo
g1
0 C
FU
ml
0
2
4
6
8
1 0
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
CT
RL
CT
RL
CT
RL
CT
RL
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
S a u re u s
h e m B
S C V
B c e r e u s
A T C C 1 1 7 7 8
S C V 2
B s u b t i l i s
A T C C 6 3 3 3
S C V 2
L m o n o c y to g e n e s
A T C C 1 3 9 3 2
S C V 1
Fig 3 Time-kill kinetics of TO andor GEN against (A) prototypic B subtilis ATCC 6333(B)
B cereus ATCC 11778 and (C) L monocytogenes ATCC 13932 The bactericidal effect of
TO against SCVs is also shown at 8 and 24 h (D) The results were obtained from at least
three independent experiments
A B
C
D
Detection
threshold
57
Bactericidal activiy of the TO analog FC04-100 against L monocytogenes
SCVs and synergy with aminoglycosides against prototypic strains and MRSA
The addition of 8 mgL of FC04-100 to GEN (2 mgL) decreased by an average of 5 log10
prototypic L monocytogenes CFUmL after 24 h of treatment in comparison to GEN alone
(Fig 4A) Also the bactericidal activity of FC04-100 used alone against the L monocytogenes
SCV was shown to be better than that observed for TO (Fig 3D) To facilitate comparison
between TO and FC04-100 Figure 4B shows the residual CFU of the L monocytogenes SCV
strain after treatment with TO or FC04-100 and calculated as the percentage of live CFU in
comparison with untreated L monocytogenes SCVs after 24 h of incubation These results
show that TO can be chemically modified (eg derivative FC04-100) and improve its activity
against L monocytogenes and thus the antibacterial activity of FC04-100 against S aureus
was further investigated Interestingly the MIC of FC04-100 against the SCV strain S aureus
NewbouldhemB (MIC of 05 -1 mgL Table 2) was higher than that found for TO (MIC of
006 mgL Table 1) On the other hand the MIC of FC04-100 against the prototypic S
aureus ATCC 29213 is lower than that of TO (8-16 mgL vs gt 64 mgL respectively) (Tables
1 and 2) This newly detected antibacterial activity of FC04-100 was also noted against
prototypic MRSA strains (Table 2) Furthermore the ability of FC04-100 to potentiate the
activity of GEN was similar to that of TO with a 4 to 8 fold improvement of the
aminoglycoside MIC (Table 2)
Although the multi-resistant S aureus MRSA USA100 USA300 and COL are resistant to
cefalexin (beta-lactam antibiotic MIC of 256 mgL) and to the aminoglycoside kanamycin
(MIC of 256 mgL and gt 1024 mgL for USA 100 and USA300 respectively Table 3) the
58
combination of both antibiotics decreased their respective MICs (Table 3) Indeed the
combination of cefalexin and kanamycin in a proportion of 32 (commercialized under the
name UbrolexinTM
) decreased the MIC of one or both antibiotics This is as expected for the
combination of a beta-lactam with an aminoglycoside (Davis 1982) However the synergy is
weak with MRSA strains that are resistant to one or the other or both antibiotics and the
addition of 4 mgL of FC04-100 further improved by 8 to 32 fold the MIC of the cephalexin-
kanamycin combination against these strains (Table 3)
59
te m p s (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
F C 0 4 -1 0 0 8 m g L
2 m g L G E N +
F C 0 4 -1 0 0 8 m g L
C T R L
G E N 2 m g L (B H I)
Re
sid
ua
l C
FU
(
)
CT
RL
TO
8 g
L
FC
04-1
00 8
gL
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
F C 0 4 -1 0 0 8 g L
T O 8 g L
C TR L
T=
24
h
T=
8h
T=
8h
T=
24
h
T=
24
h
Fig 4 Time-kill kinetics of FC04-100 andor GEN against (A) prototypic L monocytogenes
ATCC 13932 The bactericidal effect of FC04-100 used alone against L monocytogenes
SCV1 is also shown at 8 and 24 h (B) In (B) the residual CFU were calculated as the
percentage of live CFU in each condition in comparison with the untreated L monocytogenes
SCV1 after 24h of incubation The results were obtained from at least three independent
experiments
A
B
60
Table 2 Susceptibility of prototypic and SCV S aureus strains to FC04-100 with or without
GEN
a FC04-100 w as used at 4 microgmL
b Ratio of activity of GEN + FC04-100 vs activity of GEN alone
Table 3 Synergistic activity of FC04-100 with the combinaison cefalexin-kanamycin
aFC04-100 was used at 4 microgmL
bRatio of the MIC of cefalexin+kanamycin with FC04-100 vs the MIC of
cefalexin+kanamycin alone
Staphylococcus aureus ATCC 29213 8-16 05 006-0125 4-8
S aureus SH1000 8-16 05 006-0125 4-8
S aureus NewbouldΔhemB 1 8
MRSA USA100 16 05 0125 4
MRSA USA300 16 05 006 8
MRSA Col 16 025 006 4
Strains FC04-100 GENGEN
(+FC04-100 )fold
MIC (gL)
MRSA USA100 256 256 12886 43 32
MRSA USA300 256 gt1024 256171 86 32
MRSA Col 256 4 43 05035 8
MIC (gL)
Strains cefalexin kanamycin cef+kana (32)cef+kana (32)
(+FC04-100 )fold
a b
a b
61
FC04-100 potentiates GEN against S aureus embedded in biofilm and inhibits
intracellular replication of SCVs
Figure 5 demonstrates that compound FC04-100 efficiently kills S aureus embedded in
biofilms when in combination with GEN ie there was an important decrease of bacterial
CFUs compared to the no drug control or to either FC04-100 used alone at 4 ugmL (ie 025-
05MIC Table 2) or GEN used alone at 025 05 or 1 X MIC Besides we previously
demonstrated that TO was able to inhibit intracellular replication of SCVs (Mitchell AAC)
which is an important aspect of virulence for S aureus Here (Fig 6) we show that FC04-100
like TO can significantly decrease the infection of polarized airway epithelial cells by SCVs
62
05
mgL
GE
N
05
mgL
GE
N +
FC
04
02
5 m
gL
GE
N
02
5 m
gL
GE
N +
FC
04
01
2 m
gL
GE
N
01
2 m
gL
GE
N +
FC
04
No d
rug
FC
04
0
2
4
6
8
log
10
CF
Up
eg
= 3 2
= 3 8
= 3 5
= 2
Fig 5 Bactericidal effect of FC04-100 alone or in combination with GEN against prototypic
S aureus SH100 embedded in biofilm FC04-100 (FC04) was used at 4 mgL alone or in
combination with GEN Data are from three independent experiments performed in triplicate
Significant differences in comparison to the no drug control are shown ( Plt 0001 and
Plt 00001 one-way ANOVA with a Dunnettrsquos post test) Data are presented as means with
standard deviations
63
No d
rug
TO
8 m
gL
FC
04 8
mgL
0
2
4
6
log
10
CF
Ui
ns
er
t
Fig 6 Effect of TO and FC04-100 on the intracellular load of CF07-S 48 h post-
internalization Data are from three independent experiments performed in duplicate
Significant differences among groups are shown ( Plt 00001One-way ANOVA with
Turkeyrsquos post test) Data are presented as means with standard deviations
64
DISCUSSION
In previous studies we have demonstrated that TO has a strong inhibitory activity against S
aureus SCVs and improves the bactericidal activity of aminoglycoside antibiotics against
prototypic S aureus and also more broadly against staphylococci This effect was documented
for several strains of diverse clinical origins including aminoglycoside-resistant S aureus
carrying aminoglycoside modifying enzymes TO showed however no synergistic effect on the
activity of aminoglycosides against P aeruginosa E coli or Enterococcus spp (Mitchell
2012) In the present study we showed that the antibacterial spectrum of TO can be extended
to species of the Bacillales order with very low MICs against L monocytogenes B cereus and
B subtillis SCVs (lt003 microgmL) Time-kill kinetics showed that the combination of TO and
GEN creates a bactericidal synergy against prototypic of B subtilis and B cereus similarly to
that previously demonstrated against S aureus However the mixture of TO and GEN did not
demonstrated much improvement over the activity of GEN alone against prototypic L
monocytogenes in time-kill experiments Likewise investigations with Bacillales SCVs
showed that TO efficiently killed S aureus B cereus and B subtillis but not L
monocytogenes SCVs despite its very low MIC and was thus bacteriostatic against that species
Therefore improvement of the antibacterial activity against this important pathogen seems
inevitable for future development of steroidal alkaloids such as TO
TO is a steroid alkaloid structurally characterized by 6 rings 12 stereogenic centers a 3-
hydroxyl group and spiro-fused E F rings in the form of an aminoketal (Fig 1) It is generally
65
prepared by hydrolysis of its glycosylated analog tomatine while both molecules can be found
in the Solanaceae plants (Ruiz-Rubio 2001 Simons 2006) Although we previously
documented some interesting antibacterial activity for TO the absence of an identified cellular
target makes difficult the establishment of a structure-activity relationship (SAR) Our initial
attempt to elucidate SAR was therefore based on the hypothesis that the steroidal A-D rings
act as a rigid scaffold to orient pharmacophores defined by the 3-hydroxyl group on ring A
and the spiroaminoketal group on rings E F Noteworthy we prepared analogs bearing
modifications on ring A of TO and schegravemes for chemistry synthesis were recently published
(Chagnon et al 2014) From seven compounds keeping rings E and F of TO and possessing
structural variations on position 3 of ring A we were able to ascertain some structure activity
relationships Results suggest that the hydrogen bond donor effect of the 3-hydroxyl group
was not important for activity although the stereochemistry of the 3 position substitution
moderately affected activity against both S aureus SCVs and prototypic strains (in
combination with an aminoglycoside) In addition although the presence of one or two
ammonium groups in position C3 reduced the activity against S aureus SCVs (from a MIC of
006 to 05-1 microgmL) this change was highly beneficial for antibiotic activity against normal
strains (from a MIC of gt64 to 8-16 microgmL) without the presence of an aminoglycoside This
change was unexpected and one of such analogs FC04-100 increased its solubility by more
than ten times (from 2 mgmL to 26 mgmL in DMSO) The biological properties of FC04-
100 were therefore further investigated in the present study especially that TO showed some
weak bactericidal activity against Listeria and that the solubility of TO may limit eventual
clinical applications
66
Here we demonstrated that FC04-100 was able to kill both L monocytogenes SCVs as well as
the normal phenotype alone and in the presence of GEN respectively (Fig 4) Furthermore
FC04-100 showed a noticeable antibacterial activity on its own (MIC of 16 microgmL) against the
MRSA strains USA100 USA300 and COL (Table 2) FC04-100 was also synergistic in
combination with an aminoglycoside against such MRSA strains with a 4 to 8 fold gain in the
MIC of GEN Because MRSA strains are often multiresistant to antibiotics and often carry
AMEs we examined the possibility of using a triple antibiotic combination Indeed the
combination of kanamycin and cefalexin in a proportion 32 is already approved to treat
bovine mastitis pathogens under the brand name of UbrolexinTM
(Ganiegravere 2009) This
antibiotic combination offers an extended spectrum of activity compared to each individual
drug and is expected to cover both S aureus Streptococcus uberis and E coli (Ganiere
2009 Maneke et al 2011 Silley 2012) Unfortunately the increased frequency of livestock-
associated MRSA (Garcia-Alvarez et al 2011 Graveland et al 2011 Leonard and Markey
2008) and frequent incidence of strains and species carrying AMEs (Ramirez and Tolmasky
2010) may limit the spectrum of activity of the combination of a beta-lactam and an
aminoglycoside With this in mind we showed that the use of FC04-100 in combination with
kanamycin and cefalexin improves by 32 fold the activity of this mixture against MRSA
strains carrying AMEs (Table 3) Overall the addition of FC04-100 to the aminoglycoside-
beta-lactam combination decreased the MICs of kanamycin and cefalexin below their
resistance breakpoints and this for all the MRSA strains tested
67
L monocytogenes is a highly specialized intracellular pathogen (Hamon et al 2006) causing
life-threatening infections especially in pregnant women (Janakiraman 2008) L
monocytogenes in a foodborne pathogen and because of its intracellular life cycle and
propagation it is able to hide from host defenses and antibiotic treatment Similarly S aureus
although not primarily recognized as a typical intracellular pathogen is also able to enter and
survive in host cells (Lowy 2000) Moreover the ability of S aureus to convert to the SCV
phenotype showing increased biofilm production improved adherence to host cells and tissues
as well as increased intracellular persistence allows this pathogen to cause chronic and
difficult to treat infections (Garcia et al 2013 Mitchell and Malouin 2012) Discovering
antibacterial drugs able to act in the intracellular compartment is generally recognized as a
difficult challenge Previously we demonstrated that steroidal alkaloids such as TO had such
an ability and could act on intracellular S aureus SCVs (Mitchell et al 2011) and here we
demonstrated that this was also the case for the TO analog FC04-100 (Fig 6) In addition we
also show in the present study that FC04-100 greatly improves the bactericidal activity of
aminoglycosides such as GEN against bacteria embedded in biofilms Biofilms are
recognized to be a major hindrance to antibiotic action and are responsible for persistent
colonization in many diseases (Costerton et al 1999) and in the food industry (Srey et al
2013) The intracellular and intrabiofilm antibacterial properties of FC04-100 warrant further
investigation of this compound and this class of molecules in general
In conclusion we showed in this study that the spectrum of activity of TO is related to the
Firmicutes division and more precisely to the order of the Bacillales TO possesses an
68
antibacterial activity against SCVs and a synergistic activity with animoglycoside antibiotics
against prototypic strains The novel TO analog FC04-100 showed very promising new
characteristics that include a much improved bactericidal effect on L monocytogenes and
killing of bacteria in biofilms
ACKNOWLEDGEMENTS
This study was supported by an operation grant from Cystic Fibrosis Canada to FM and by a
team grant from the Fonds Queacutebeacutecois de la Recherche sur la Nature et les Technologies
(FQRNT) to FM KB and EM
REFERENCES
Atalla H Gyles C Jacob CL Moisan H Malouin F Mallard B 2008 Characterization of
a Staphylococcus aureus small colony variant (SCV) associated with persistent bovine mastitis
Foodborne Pathog Dis 5785ndash799
Bad Bug Book Foodborne pathogenic microorganisms and natural toxins 2012 Second
edition Lampel KA Al-Khaldi S Cahill SM (Ed) Published by the Center for Food Safety and
Applied Nutrition of the Food and Drug Administration (FDA) US Department of Health and
Human Services pp 87-103
Beierlein JM Anderson AC 2011 New developments in vaccines inhibitors of anthrax
toxins and antibiotic therapeutics for Bacillus anthracis Curr Med Chem 185083-5094
69
Brouillette E A Martinez B J Boyll N E Allen and F Malouin 2004 Persistence of
a Staphylococcus aureus small-colony variant under antibiotic pressure in vivo FEMS
Immunol Med Microbiol 4135ndash41
Bryan L E and S Kwan 1981 Aminoglycoside-resistant mutants of Pseudomonas
aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob
Agents Chemother 19958ndash964
Ceri H Olson M Morck D Storey D Read R Buret A Olson B 2001 The MBEC Assay
System multiple equivalent biofilms for antibiotic and biocide susceptibility testing Methods
Enzymol 337377-385
Chagnon F I Guay M-A Bonin G Mitchell K Bouarab F Malouin E Marsault
2014 Unraveling the structure-activity relationship of tomatidine a steroid alkaloid with
antibiotic properties against persistent forms of Staphylococcus aureus Eur J Med Chem (In
Press EJMECH-S-13-01625-3)
Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial
Susceptibility Testing Twenty-First Informational Supplement M100-S21 CLSI Wayne PA
USA 2011
70
Costerton JW Stewart PS Greenberg EP 1999 Bacterial biofilms a common cause of
persistent infections Science 284 1318-1322
Davis BD 1982 Bactericidal synergism between beta-lactams and aminoglycosides
mechanism and possible therapeutic implications Rev Infect Dis 4237-245
Ganiegravere JP Denuault L 2009 Synergetic interaction between cefalexin and kanamycin in
Muller-Hinton broth medium and in milk J appl microbiol 107 117-125
Garcia LG S Lemaire B C Kahl K Becker R A Proctor O Denis P M Tulkens
F Van Bambeke 2013 Antibiotic activity against small-colony variants of Staphylococcus
aureus review of in vitro animal and clinical data J Antimicrob Chemother 68 1455ndash1464
Garcia-Alvarez L M T Holden H Lindsay C R Webb D F Brown M D Curran
E Walpole K Brooks D J Pickard C Teale J Parkhill S D Bentley G F Edwards
E K Girvan A M Kearns B Pichon R L Hill A R Larsen R L Skov S J Peacock
D J Maskell and M A Holmes 2011 Methicillin-resistant Staphylococcus aureus with a
novel mecA homologue in human and bovine populations in the UK and Denmark a
descriptive study Lancet Infect Dis 11595-603
Gomaa OM Momtaz OA 2007 16 rRNA characterization of Bacillus isolate and its
tolerance profile after subsequent subculturing Arab J Biotech 10 107-116
71
Graveland H Duim B van Duijkeren E Heederik D Wagenaar JA 2011 Livestock-
associated methicillin-resistant Staphylococcus aureus in animals and humans Int J Med
Microbiol 301630-634
Guillet C Join-Lambert O Le Monnier A Leclercq A Mechaiuml F Mamzer-Bruneel MF
Bielecka MK Scortti M Disson O Berche P Vazquez-Boland J Lortholary O Lecuit M
2010 Human listeriosis caused by Listeria ivanovii Emerg Infect Dis 16136-138
Hamon M H Bierne P Cossart 2006 Listeria monocytogenes a multifaceted model Nat
Rev Microbiol 4 423-434
Harrison JJ Stremick CA Turner RJ Allan ND Olson ME Ceri H 2010 Microtiter
susceptibility testing of microbes growing on peg lids a miniaturized biofilm model for high-
throughput screening Nat Protoc 51236-1254
Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y
Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T Baba T 2013 Genomic
Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
Horsburgh MJ Aish JL White IJ Shaw L Lithgow JK Foster SJ 2002 SigmaB
modulates virulence determinant expression and stress resistance characterization of a
72
functional rsbU strain derived from Staphylococcus aureus 8325-4 J Bacteriol 1845457-
5467
Janakiraman V 2008 Listeriosis in Pregnancy Diagnosis Treatment and Prevention Rev
Obstet Gynecol 1 179ndash185
Leonard FC Markey BK 2008 Methicillin-resistant Staphylococcus aureus in animals a
review Vet J 175 27-36
Lowy FD 2000 Is S aureus an intracellular pathogen Trends Microbiol 8341-342
Magalhatildees L Nitschke M 2013 Antimicrobial activity of rhamnolipids against Listeria
monocytogenes and their synergistic interaction with nisin Food Control 29 138-142
Maneke E Pridmore A Goby L Lang I 2011 Kill rate of mastitis pathogens by a
combination of cefalexin and kanamycin J Appl Microbiol 110184-190
Mitchell G Brouillette Eacute Lalonde-Seacuteguin D Asselin AE Lebeau Jacob C Malouin F
2009 A role for sigma factor B in the emergence of Staphylococcus aureus small-colony
variants and elevated biofilm production resulting from an exposure to aminoglycosides
Microb Pathog 4818-27
73
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K Malouin F 2011
Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic
fibrosis airway epithelial cells Antimicrob Agents and Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM Malouin F 2011 Infection of Polarized
airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is
increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function
and is influenced by NF- κ B Infect Immun 793541-3551
Mitchell G Lafrance M Boulanger S Lalonde-Seacuteguin D Guay I Gattuso M Marsault
Eacute Bourab K Malouin F 2012 Tomatidine acts in synergy with aminoglycoside antibiotics
against multiresistant Staphylococcus aureus and prevents virulence gene expression J
Antimicrob Chemother 67 559-568
Mitchell G Lalonde-Seacuteguin D Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S
Malouin F 2010 Staphylococcus aureus sigma B-dependent emergence of small-colony
variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-
heptylquinoline-N-oxide BMC Microbiology 10 33-48
Mitchell G and F Malouin 2012 Outcome and prevention of Pseudomonas aeruginosa-
Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic
Fibrosis InTech Open Access Publisher ISBN 979-953-307-059-8
74
Moisan H Brouillette E Jacob CL Langlois-Beacutegin P Michaud S Malouin F 2006
Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated
from cystic fibrosis patients is influ- enced by SigB J Bacteriol 18864ndash76
Moskowitz SM Foster JM Emerson J Burns JL 2004 Clinically feasible biofilm
susceptibility assay for isolates of Pseudomonas aeruginosa from patients with cystic fibrosis
J Clin Microbiol 421915-1922
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M Peters G 2006
Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent
infections Nat Rev Micro- biol 4295ndash305
Ramirez MS M E Tolmasky 2010 Aminoglycoside modifying enzymes Drug Res
Updates 13 151ndash171
Ruiz-Rubio M Peacuterez-Espinosa A Lairini K Roldagraven-Arjona T Dipietro A Anaya N
2001 Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi p 293ndash326 In
A Rahman (ed) Studies in natural products chemistry vol 25 Elsevier Oxford United
Kingdom
75
Sendi P Proctor RA 2009 Staphylococcus aureus as an intracellular pathogen the role of
small colony variants Trends Microbiol 1754-58
Silley P Goby L Pillar CM 2012 Susceptibility of coagulase-negative staphylococci to a
kanamycin and cefalexin combination J Dairy Sci 953448-3453
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C Osbourn A
2006 Dual effects of plant steroidal alkaloids on Saccharomyces cerevisiae Antimicrob
Agents Chemother 502732ndash2740
Srey S Jahid IK Ha S-D 2013 Biofilm formation in food industries A food safety concern
Food control 31572-585
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D
Roth J Proctor RA Becker K Peters G Loumlffler B 2011 Staphylococcus aureus
phenotype switching an effective bacterial strategy to escape host immune response and
establish a chronic infection EMBO Mol Med 3129ndash141
Vuong C Otto M 2002 Staphylococcus epidermidis infections Microbes Infect 4481-489
Witte W Strommenger B Stanek C Cuny C 2007 Methicillin-resistant Staphylococcus
aureus ST398 in Humans and Animals Central Europe Emerg Infect Dis 13 255ndash258
76
CHAPITRE 3
MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
31 Obtention des mutants reacutesistants
Afin de nous aider agrave mieux deacutefinir la cible et le mode drsquoaction de la tomatidine (TO)
nous avons forceacute lrsquoapparition de reacutesistances chez Staphylococcus aureus Comme
souche controcircle prototype nous avons choisi ATCC 29213 puisque crsquoest une souche
tregraves freacutequemment utiliseacutee pour des expeacuteriences avec des antibiotiques et nous avons
choisi la souche NewbouldΔhemB comme S aureus laquo small colony variant raquo puisque
crsquoest une souche SCV qui est stable et bien connue nrsquoayant pas de capaciteacute de
reacuteversion Par passages successifs dans une concentration sous inhibitrice
drsquoantibiotique (technique de CMI (concentration minimale inhibitrice)) (CLSI 2011)
nous avons obtenu des mutants deacutemontrant une reacutesistance croissante agrave lrsquoantibiotique
au fil des passages (Figure 1) Nous avons obtenu des mutants ATCC 29213
reacutesistants agrave la gentamicine (GEN) et agrave la GEN lorsque combineacutee agrave la TO ainsi que
des mutants reacutesistants NewbouldΔhemB reacutesistants agrave la GEN seule agrave la TO seule et
agrave la TO lorsque combineacute agrave la GEN
Certains passages ont ensuite eacuteteacute seacutelectionneacutes parce qursquoils deacutemontraient une hausse
plus marqueacutee de leur CMI Ces passages ont eacuteteacute choisis car nous supposions que
ces hausses marqueacutees de CMI correspondraient donc agrave des hausses de la
reacutesistance de cette souche agrave lrsquoantibiotique testeacute et que cette reacutesistance pouvait ecirctre la
conseacutequence drsquoune ou plusieurs mutations en lien avec la cible ou le mode drsquoaction
de lrsquoantibiotique preacutesent Trois agrave quatre passages (ou paliers de reacutesistance) par
condition ont eacuteteacute seacutelectionneacutes Nous avons ensuite reacuteensemenceacute le laquo pool raquo
bacteacuterien de chacun des passages choisis afin de pouvoir isoler 3 isolats ayant la
mecircme CMI (ou tregraves similaire (CMI vraies)) que celui du laquo pool raquo bacteacuterien des
77
passages drsquoougrave ils provenaient Soixante quatre mutants reacutesistants ont ainsi eacuteteacute
seacutelectionneacutes en vue drsquoecirctre seacutequenceacutes (Figure 2)
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt 1
6
CM
I v
raie
s
02
5
CM
I v
raie
s
02
5-1
A
78
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
en
pr
eacutes
en
ce
de
ge
n (
4 micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
01
25
1
6
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
8
CM
I v
raie
s
2
4
CM
I v
raie
s
2
B
C
79
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
en
pr
eacutes
en
ce
de
TO
(8
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
1
2
3
4
5
CM
I v
ra
ies
4
CM
I v
ra
ies
2
CM
I v
ra
ies
0
5
1
CM
I v
ra
ies
2
4
Figure 1 Graphiques des CMI pour chacun des passages pour (A) la tomatidine sur
NewbouldΔhemB (B) pour la tomatidine en preacutesence de gentamicine sur
NewbouldΔhemB (C) pour la gentamicine sur S aureus ATCC 29213 (D) pour la
gentamicine en preacutesence de tomatidine sur S aureus ATCC 29213
32 Seacutequenccedilage et analyses bioinformatiques
Une extraction drsquoADN geacutenomique a ensuite eacuteteacute effectueacutee sur chacun des isolats en
utilisant le Kit drsquoextraction drsquoADN geacutenomique miniKit de Qiagen (QIAamp DNA minikit)
(Li et al 2003) Puisque leurs seacutequences geacutenomiques nrsquoeacutetaient pas connues nous
avons eacutegalement extrait lrsquoADN geacutenomique de nos deux souches de reacutefeacuterences
ATCC29213 et NewbouldΔhemB (provenant de quatre colonies isoleacutees chacune)
afin drsquoavoir les seacutequences de comparaison pour nos mutants (Figure 2)
D
80
Ne
wm
an
(g
en
om
ede r
ef h
ttpw
ww
ncbinlm
nih
govn
uccore
150373012)
AT
CC
29
213
(A p
art
ir d
e 4
AT
CC
29
21
3 r
eco
nstr
uire
AT
CC
29
21
3 c
om
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
5
10
7
10
8 1
09
)
Ne
wbo
uld
Δhe
mB
(A p
art
ir d
e 4
Ne
wb
ou
ldh
em
Bre
co
nstr
uire
Ne
wb
ou
ldh
em
Bcom
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
1
agraveG
EN
2
5 (
CM
I2
)
2
7 (
CM
I2
)
2
8 (
CM
I2
)
AT
CC
29
21
3
Pa
lier
de
reacutesis
tance
1
agravecom
bo
29
(C
MI
05
)
3
0 (
CM
I 0
5
3
1 (
CM
I 0
5)
Suite
fe
uill
e s
uiv
ante
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agraveG
EN
3
(C
MI
4)
4
(C
MI
4)
3
9 (
CM
I4
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agraveG
EN
4
6 (
CM
I8
)
4
7 (
CM
I8
)
4
8 (
CM
I8
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agravecom
bo
1
1 (
CM
I 2
)
1
2 (
CM
I 2
)
2
1 (
CM
I 2
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agravecom
bo
5 (
CM
I 2
-4)
3
6 (
CM
I 2
-4)
3
7 (
CM
I 2
-4)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
4
agravecom
bo
3
8 (
CM
I4
)
6
7 (
CM
I 4
)
81
Ne
wb
ould
Δhem
B(A
part
ir d
e 4
Ne
wb
ou
ldΔ
he
mB
reco
nstr
uire
Ne
wb
ou
ldΔ
hem
Bco
mp
let e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveG
EN
24
(C
MI
64
)
4
1 (
CM
I 1
6)
4
2 (
CM
I 6
4)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveT
O
95
(C
MI
02
5)
9
6 (
CM
I 0
25)
9
7 (
CM
I 0
25)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agravecom
bo
33
34
35
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveG
EN
49
(C
MI
12
8)
5
1 (
CM
I 1
28)
5
2 (
CM
I 1
28)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agraveG
EN
7
(C
MI
128
)
4
3 (
CM
I 1
28)
4
5 (
CM
I1
28
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agraveT
O
6
4 (
CM
I gt
16)
6
5 (
CM
I gt
16)
6
6 (
CM
I gt
16)
75
76
77
79
80
81
82
83
84
85
86
87
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agravecom
bo
15
16
18
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agravecom
bo
59
60
61)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agravecom
bo
54
56
58)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveT
O
90
(C
MI
02
5)
9
1 (
CM
I 0
25)
9
3 (
CM
I 0
5)
Ne
wb
ou
ldh
em
B
palie
r d
e r
eacutesis
tance
3
agraveT
O
11
0 (
CM
I 1
)
1
12 (
CM
I 0
25
-05
)
1
13 (
CM
I 0
5)
Fig
ure
2
Org
an
igra
mm
e d
es
mu
tan
ts r
eacutes
ista
nts
seacute
qu
en
ceacutes
se
lon
leu
r pa
lier
de
reacute
sis
tan
ce e
t a
vec leu
r n
um
eacutero
drsquoeacute
chan
tillo
n
co
rre
spo
ndan
t
82
Un minimum de 20 ngmicrol drsquoADN geacutenomique de chacun des isolats a ensuite eacuteteacute
achemineacute au laboratoire du Pr Seacutebastien Rodrigue (Deacutepartement de biologie Faculteacute
des sciences Universiteacute de Sherbrooke) qui a effectueacute la librairie de fragments Pour
se faire il y a eu dans un premier temps digestion enzymatique de lrsquoADN afin
drsquoobtenir des fragments drsquoADN double brin Ensuite tous les fragments ont eacuteteacute
purifieacutes suivant la meacutethode SPRI (Solid-Phase Reversible Immobilization) Il srsquoagit
drsquoune meacutethode simple et tregraves efficace drsquoisolation drsquoacides nucleacuteiques sur microplaque
(Rodrigue S et al 2010) Les acides nucleacuteiques cibles sont immobiliseacutes sur des
microparticules paramagneacutetiques dans des conditions tampons-speacutecifiques La
flexibiliteacute du systegraveme se fait par lrsquoutilisation de diffeacuterents tampons de liaison (laquo binding
buffer raquo) qui modifient le type et la grandeur des acides nucleacuteiques immobiliseacutes Les
contaminants peuvent ainsi ecirctre retireacutes de lrsquoeacutechantillon sans besoin de filtration ou
centrifugation (Beckman Coulter website 2013 DeAngelis et al 1995) Suite agrave la
purification par meacutethode SPRI lrsquoeacutequipe du Pr Rodrigue a obtenu des fragments
drsquoADN de ~250pb auxquels ont eacuteteacute ajouteacutes par ligation des adapteurs aux deux
extreacutemiteacutes des fragments Dans un deuxiegraveme temps des codes-bars speacutecifiques agrave
chaque eacutechantillon drsquoADN geacutenomique ont eacutegalement eacuteteacute lieacutes agrave chacun des fragments
Alors que les adapteurs permettent aux fragments de se lier agrave une matrice solide afin
de pouvoir ecirctre amplifieacutes les codes-bars permettront de classer chacun des
fragments selon leur appartenance agrave un eacutechantillon drsquoADN geacutenomique speacutecifique
83
Figure 3 Eacutetape du seacutequenccedilage Illumina
La librairie de fragments a ensuite eacuteteacute envoyeacutee chez Geacutenome-Queacutebec afin que
chacun des geacutenomes puisse ecirctre seacutequenceacute Geacutenome ndashQueacutebec utilise la meacutethode
Illumina comme outil de seacutequenccedilage (Figure 3) La grande innovation de la meacutethode
Illumina est lrsquoamplification des fragments par liaison de ceux-ci agrave une surface solide
(Flowcell) ce qui permet par un processus de ponts drsquoamplification de geacuteneacuterer plus
drsquoun million de laquo clusters raquo comprenant eux-mecircme au plus un millier de copies de la
moleacutecule originale de deacutepart (solid-phase PCR) Chaque regroupement de fragments
provenant de lrsquoamplification drsquoun mecircme fragment original est tregraves pregraves les uns des
Modifieacute de Quail MA et al 2008
84
autres formant une zone distincte drsquoun regroupement de fragments agrave un autre
Lrsquoincorporation des nucleacuteotides eacutetiqueteacutes preacutealablement avec des moleacutecules
fluorescentes speacutecifiques est deacutetecteacutee suite agrave leur excitation par un laser et les
images sont compileacutees pour produire la seacutequence de bases pour chaque fragment
ADN (Figure 4) Les nouvelles seacuteries de bases appeleacutees laquo readsraquo seront ensuite
reacuteassembleacutees (aligneacutes) en se basant sur un geacutenome de reacutefeacuterence connu (appeleacute
laquo resequencing raquo) (crsquoest notre cas ici) ou en absence de geacutenome de reacutefeacuterence
(appeleacute laquo De novo sequencing raquo) Au final lrsquoensemble des laquo reads raquo aligneacutes
correspondra agrave la seacutequence de chaque chromosome de lrsquoeacutechantillon drsquoADN
geacutenomique de deacutepart
85
Figure 4 Meacutethode drsquoamplification par la meacutethode Illumina
Suite au seacutequenccedilage chez Geacutenome ndashQueacutebec nous avions en main les seacutequences
geacutenomiques de tous nos mutants et de nos souches de reacutefeacuterence sous forme de
Tireacute de BiteSizeBio httpbitesizebiocom13546sequencing-by-synthesis-
explaining-the-illumina-sequencing-technology consulteacute le 3 deacutec 2013
86
contigs Nous avions donc le casse-tecircte mais en piegraveces deacutetacheacutees pecircle-mecircle et
sans avoir la photo de lrsquoimage de reacutefeacuterence sur laquelle se baser pour pouvoir
compleacuteter le casse-tecircte
1- Lrsquoassemblage
Les diffeacuterents geacutenomes avaient eacuteteacute preacutealablement fractionneacutes aleacuteatoirement
en petits fragments de 250 pb et amplifieacutes en vue du seacutequenccedilage Suite au
seacutequenccedilage les seacutequences de chaque fragment (appeleacute laquo reads raquo) sont ensuite
assembleacutees en contigs crsquoest-agrave-dire des seacutequences plus ou mois longues continues
et ordonneacutees geacuteneacutereacutees par lassemblage des clones dune bibliothegraveque geacutenomique
qui se chevauche La qualiteacute de lrsquoassemblage en contigs deacutepend de la capaciteacute de
superposition des laquo reads raquo Parce que la digestion enzymatique de lrsquoADN se fait de
faccedilon aleacuteatoire et se fait sur plusieurs copies drsquoADN chaque portion du geacutenome est
repreacutesenteacutee plusieurs fois dans diffeacuterents modegraveles de fragments Donc les
seacutequences identiques des diffeacuterents fragments (ou laquo reads raquo) peuvent ecirctre
superposeacutees par chevauchegravement et les laquo reads raquo superposeacutes peuvent ecirctre
assembleacutes en contigs par un logiciel drsquoassemblage En ce qui nous concerne cette
eacutetape a eacuteteacute faite agrave lrsquoaide du programme de bio-informatique Newbler (Zhang et al
2012) Lrsquoassemblage geacutenomique peut ecirctre probleacutematique pour les informaticiens car
les geacutenomes contiennent des seacutequences qui se reacutepegravetent et ces reacutepeacutetitions peuvent
ecirctre assez longues en fonction des organismes Suite agrave lrsquoassemblage les geacutenomes
sont donc sous forme de contigs
2- Lrsquoalignement
En se basant sur une seacutequence geacutenomique de reacutefeacuterence (notre reacutefeacuterence est
Staphylococcus aureus str Newman (~ 288Mb)
( httpwwwncbinlmnihgovgenome154project_id=58839) ) le programme de bio-
informatique ABACAS permet de rapidement aligner ordonner et orienter les contigs
Selon les besoins ce qui nrsquoa pas eacuteteacute notre cas ABACAS permet eacutegalement de
87
visualiser et designer les amorces permettant de fermer les gaps (espaces) entre les
contigs en faisant des PCR (Assefa et al 2009)
3- La fermeture des gaps
La fermeture des gaps nrsquoa pas eacuteteacute faite avec le programme ABACAS mais
avec lrsquoaide du programme de bio-informatique GapFiller Lrsquoutilisation de GapFiller
pour la fermeture des gaps possegravede 3 principaux avantages Premiegraverement le
programme prend en compte lrsquoestimation de la grandeur du gap agrave fermer donc il
exclut automatiquement les fermetures de gaps erroneacutes qui sont plus courts ou plus
longs que la grandeur estimeacutee Deuxiegravemement il nrsquoessaie pas de remplir un gap
avec un assemblage de laquo reads raquo en contigs mais cherche plutocirct agrave allonger les
contigs agrave partir de leurs extreacutemiteacutes jusqursquoagrave obtenir des seacutequences superposeacutees en
passant par la creacuteation de K-mer (seacuteparation des reads en plus petits fragments) Et
troisiegravemement il prend en compte que les extreacutemiteacutes des contigs sont souvent
source de mauvais assemblage et reacute eacutevalue ces extreacutemiteacutes pendant la fermeture des
gaps (Boetzer et Pirovano 2012)
4- Lrsquoannotation des geacutenomes
Suite agrave la fermeture des gaps il a fallu annoter les geacutenomes en se basant sur
lrsquoannotation du geacutenome de reacutefeacuterence Staphylococcus aureus str Newman RATT est
un outil de transfert rapide drsquoannotation (Rapid Annotation Transfert Tool) qui a eacuteteacute
deacuteveloppeacute afin de fournir rapidement une annotation preacutecise pour des geacutenomes
inconnus en se basant sur des geacutenomes deacutejagrave annoteacutes comme reacutefeacuterences (Figure 5)
La meacutethode transfert les annotations drsquoune reacutefeacuterence de haute qualiteacute agrave un nouveau
geacutenome sur la base de la conservation des synteny (ordre des gegravenes) (Otto et al
2011) Suite agrave lrsquoannotation les gegravenes tant chez les mutants que chez les souches
parentes portent les noms auxquels ils correspondent chez Staphylococcus aureus
str Newman
88
Figure 5 Corrections du programme RATT sur les annotations transfeacutereacutees Les
annotations de H37Rv ont eacuteteacute transfeacutereacutees sur la seacutequence F11 (bleu pacircle) ont eacuteteacute corrigeacutees
(vert) et ensuite compareacutees agrave lrsquoannotation de la souche existante F11 EMBL (jaune) (A et B)
La correction des codons ldquostartrdquo et ldquostoprdquo respectivement Dans une situation de laquo mapping raquo
plus complexe (C) ougrave les cadres de lecture sont montreacutes pour plus de clarteacute RATT
cartographie une large seacutequence codante (CDS) de H37Rv vers un locus dans F11 qui inclue
plusieurs codons laquo stop raquo laquo in-frame raquo En inseacuterant un deacutecalage du cadre de lecture (ie un
pseudogegravene) la traduction conceptuelle est conserveacutee Ceci diffegravere avec les deux gegravenes se
superposant preacutedis comme faisant partie du geacutenome F11
Tireacute de Otto TD et al 2011
89
321 Identification des mutations en lien avec la tomatidine
Apregraves avoir obtenu une seacutequence consensus agrave partir des seacutequences geacutenomiques
des 4 copies de chacune des souches parente S aureus ATCC 29213 et S aureus
NewbouldΔhemB les diffeacuterences geacutenomiques entre les souches parentes et le
geacutenome de reacutefeacuterence S aureus str Newman ont eacuteteacute eacutelimineacutees afin que ces
diffeacuterences nrsquoapparaissent pas comme des mutations lors de la comparaison des
seacutequences geacutenomiques des mutants avec celles de leur souche parente ces
diffeacuterences eacutetant plutocirct attribuables agrave la diffeacuterence qursquoil y a entre S aureus str
Newman et la souche parente et non pas agrave des mutations en lien avec la reacutesistance
des mutants
Le service bio-informatique du deacutepartement de biologie (Faculteacute des sciences
Universiteacute de Sherbrooke) a ensuite produit un tableau contenant pour chacun des
mutants tous les gegravenes muteacutes apregraves comparaison avec leur souche parentes Agrave cette
eacutetape on pouvait donc savoir dans quel gegravene se situaient les mutations quelle eacutetait
lrsquoeffet de cette mutation (mutation synonyme deacuteleacutetion insertion SNP) et en principe
srsquoil srsquoagissait pour un mecircme gegravene muteacute de la mecircme mutation puisque dans ce
tableau pour un mecircme gegravene chaque ligne correspond agrave des mutations diffeacuterentes
(Figure 6)
Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4
Figure 6 Exemple drsquoune partie du tableau compilation des mutations chez
les mutants ATCC 29213 reacutesistants agrave la GEN seule et agrave la GEN (+TO)
90
Agrave lrsquoaide de ce tableau (Figure 6) jrsquoai chercheacute les mutations qui pouvaient ecirctre
attribuables agrave la TO Chez les mutants S aureus ATCC 29213 reacutesistants jrsquoai
chercheacute les gegravenes qui eacutetaient muteacutes chez tous les mutants reacutesistants agrave la
combinaison GEN (+TO) ou dont la mutation apparaissait graduellement au fil des
paliers de reacutesistance mais dont ce mecircme gegravene nrsquoeacutetait pas muteacute chez les mutants
reacutesistants agrave la GEN seule Je pouvais donc estimer que cette mutation chez les
reacutesistants agrave la combinaison eacutetait en lien avec la preacutesence de la TO dans le milieu
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct
Afin de mrsquoaider dans ma recherche jrsquoai eu lrsquoaide du Pr Pierre-Eacutetienne Jacques
(Deacutepartement de biologie Faculteacute des sciences Universiteacute de Sherbrooke)qui a
mis les donneacutees de ce tableau dans un tableau croiseacute dynamique (TCD) ce qui a
permis de filtrer les donneacutees selon des critegraveres bien preacutecis me permettant drsquoarriver agrave
mes fins plus facilement Dans un premier temps un TCD a eacuteteacute creacuteeacute ougrave pour
chacun des mutants chaque gegravene ayant au moins une mutation eacutetait indiqueacute par un
laquo 1 raquo alors qursquoun laquo 0 raquo eacutetait marqueacute si ce gegravene nrsquoeacutetait pas muteacute Dans un second
temps la quantiteacute de laquo 1 raquo par palier de reacutesistance (correspondant au nombre de
mutants de ce palier posseacutedant une mutation dans ce gegravene) eacutetait indiqueacutee par un
chiffre allant de 0 pour laquo aucun mutant de ce palier de reacutesistance ne posseacutedait de
mutation dans ce gegravene raquo agrave 1 2 ou 3 selon le nombre de mutants de ce palier ayant
ce gegravene muteacute Jrsquoai ensuite pu soumettre le TCD agrave des filtres approprieacutes afin de faire
ressortir les gegravenes drsquointeacuterecircts selon des critegraveres de seacutelection preacutecis (Figures 7-8)
91
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
1
00
33
32
1
1
11
4
5
0
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
10
03
33
21
11
14
50
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_04
12
lpl9
nm
20
12
32
23
92
44
1
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
NW
MN
_00
46
_2
10
13
21
37
24
31
NW
MN
_09
55
_2
01
23
11
37
24
21
NW
MN
_13
83
gpsA
21
02
31
13
72
42
1
NW
MN
_03
39
_1
11
21
31
37
34
42
NW
MN
_15
97
dn
aE2
10
22
30
37
23
31
NW
MN
_23
30
_2
10
23
20
37
23
21
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_2
07
0_
10
01
23
21
81
45
0
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_10
28
_1
22
12
32
58
34
54
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
A B C
92
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
39
8h
sdM
31
33
33
27
11
34
54
NW
MN
_04
26
_3
31
33
32
71
13
45
4
NW
MN
_05
24
sdrD
32
33
33
28
11
34
55
NW
MN
_07
39
_3
33
33
32
91
13
45
6
NW
MN
_11
64
hsl
U3
33
33
32
91
13
45
6
NW
MN
_05
23
sdrC
22
33
32
27
10
34
45
NW
MN
_20
61
fmtB
23
23
32
27
10
34
45
NW
MN
_04
03
lpl1
nm
33
22
33
28
10
34
55
NW
MN
_13
45
ebh
13
32
33
31
81
03
44
5
NW
MN
_16
49
_3
32
33
31
81
03
44
5
NW
MN
_17
00
hsd
M3
23
33
22
81
03
44
5
NW
MN
_00
42
_3
33
33
22
91
03
44
6
NW
MN
_05
80
_3
33
33
31
91
03
44
6
NW
MN
_13
44
ebh
23
31
32
22
79
34
44
NW
MN
_13
75
aro
A3
13
23
31
79
34
44
NW
MN
_23
97
fnb
B2
32
23
22
79
34
45
NW
MN
_04
01
_3
32
23
31
89
34
45
NW
MN
_07
33
_3
32
32
31
89
34
45
NW
MN
_17
45
_2
33
22
32
89
34
56
NW
MN
_01
66
coa
33
32
32
29
93
44
6
NW
MN
_05
25
sdrE
33
32
33
19
93
44
6
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D Fig
ure
7
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez A
TC
C 2
921
3 d
an
s le
TC
D p
ar
dif
feacutere
nts
fil
tre
s
(A)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts la
ge
nta
mic
ine
ge2
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(C
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts d
an
s le
pa
lier
1
de
reacute
sis
tan
ce
agravela
ge
nta
mic
ine
le1
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
et
nom
bre
de m
uta
nts
reacute
sis
tan
ts agrave
la la
gen
tam
icin
e (
+T
O)
de
s 2
de
rnie
rs p
alie
rs d
e r
eacutesis
tan
ce
ge5
(D
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
7 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
ge
nta
mic
ine
(+
TO
) ge
9
93
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_16
29
ccp
A0
01
33
31
50
00
01
24
0
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_21
28
rpsM
11
12
21
12
22
00
31
74
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
NW
MN
_25
29
clfB
33
03
33
80
11
26
17
4
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e reacute
sist
ance
agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
On
b m
uta
nts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
16
29
ccp
A0
01
33
31
50
00
01
24
0N
WM
N_
21
28
rpsM
11
12
21
12
22
00
31
74
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
A B C
94
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
04
6_
1
01
00
28
22
32
2
10
9
NW
MN
_17
33
prs
A0
11
12
01
12
12
22
14
7
NW
MN
_19
23
_0
11
31
15
13
22
21
08
NW
MN
_20
37
_0
02
32
07
23
11
21
27
NW
MN
_22
77
_1
02
21
11
02
12
33
14
8
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
01
43
_0
03
00
04
23
33
34
11
NW
MN
_0
40
6lp
l3n
m2
01
00
03
12
22
33
7
NW
MN
_1
65
2_
00
10
00
62
12
31
68
NW
MN
_2
01
2_
00
00
00
32
33
30
31
1N
WM
N_
20
15
mn
aA0
00
00
01
23
33
01
11
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
o
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D E Fig
ure
8
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez N
ew
bo
uld
Δh
em
Bd
an
s l
e T
CD
par
dif
feacutere
nts
fil
tres
(A)
Nom
bre
de
mu
tan
ts
reacutesis
tan
ts agrave
la tom
atid
ine
ge15
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atid
ine
ge15
et no
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
(C)
No
mbre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la tom
atid
ine
ge15
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3(
D)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atidin
ege
10
et no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
tom
atid
ine
(+G
EN
) ge
7
(E)
No
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la to
ma
tid
ine
le6
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
95
Le TCD mrsquoa permis de rapidement identifier les gegravenes drsquointeacuterecircts Agrave lrsquoaide du site CMR
(Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) 2013) jrsquoai
ensuite veacuterifieacute la fonction des gegravenes identifieacutes afin de confirmer leur inteacuterecirct (Tableau
1)
Tableau 1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants et (B) chez les mutants NewbouldΔhemB reacutesistants
Gegravene Alias Fonction
NWMN_0811 NADH dh complexe 1 chaine de transport des eacutelectrons
NWMN_2070 _membrane prot Putative- Toxin production+resistance drug transporteur-
similar to multidrug transporteur 85
NWMN_0635 _membrane prot Putative- Bordeacute par AraC(reacutegulateur opeacuteron arabinose) ou
SsaA-likeprot(lieacute au quorum sensing RNAIII(secretory antigen)
NWMN_2084 _truncated transposase for Is1272- mobile+extrachromosomal element
fonctiontransposon fonction
NWMN_0928 qoxC quinol oxidase polypeptide IIIcomplexe 4 chaine de transport des eacutelectrons
NWMN_0739 _ conserved hypothetical prot fonction inconnues
NWMN_1164 hslU ATP-dependent protease ATP-binding subunit
NWMN_0524 sdrD Ser-Asp rich fibrinogenbone sialoprotein-binding protein sdrD
NWMN_0042 _ lipoprotein putativecell enveloppe- lipoprot en tendem
NWMN_0580 _conserved hypothetical prot fonction inconnuesDNA metabolism DNA
replication recombination and repair
NWMN_1745 _
NWMN_0166 coa coagulase precursor
NWMN_0525 sdrE Ser-Asp rich fibrinogenbone sialoprotein-binding protein SdrE
A
96
Apregraves avoir aligneacute speacutecifiquement une par une chacune des seacutequences (Fasta) des
gegravenes drsquointeacuterecirct de chaque mutant avec leur souche parente jrsquoai utiliseacute le programme
de visualisation et de comparaison de seacutequences ClustalW2 (ClustalW2 2013) afin
de confirmer que la mutation eacutetait une vraie mutation et non une erreur de
seacutequenccedilage ou drsquoalignement et que la mutation eacutetait identique chez tous les mutants
ayant le gegravene drsquointeacuterecirct muteacute Cette eacutetape a permis de restreindre encore la liste des
gegravenes pouvant ecirctre en lien avec la cible ou le mode drsquoaction de la TO (Figure 9)
Gene Alias Fonction
NWMN_0896 _conserved hypothetical prot fonction inconnue- comK family prot-
regulation DNA interaction- ORFIDresssemble agrave facteur de transcription
NWMN_0996 _ conserved hypothetical prot fonction inconnue- prophage fonction
NWMN_1025 _ phage major tail protprophage fonction
NWMN_1629 ccpAcatabollite control protA virulence determinant production and antibiotique
resistance
NWMN_1916 _ conserved hypothetical prot-fonction inconnue ou prophage prot
NWMN_2314 _drug resistance transporteur- EmrBQacA subfamily prot- toxin
production+resistance- transport and binding prot
NWMN_0046 _ hypothetical prot
NWMN_2015 mnaA UDP-Glc-Nac 2 epimerase
NWMN_2012 atpE ATP synthase subunit C
NWMN_1652 _ multidrug resistance transporter protein B
NWMN_0143 _ oligopeptide ABC transporter ATP-binding protein
B
97
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
A
B
98
Jrsquoai ensuite chercheacute le rocircle de ces gegravenes sur le site CMR afin de veacuterifier si leur
mutation pouvait expliquer la reacutesistance agrave lrsquoantibiotique (Comprehensive Microbial
Ressouce (J Craig Venter Institute (JCVI) 2013) Dans une premiegravere eacutetape le rocircle
drsquoun gegravene a eacuteteacute deacutetermineacute en allant tout simplement voir directement le rocircle de ce
gegravene chez Staphylococcus aureus str Newman Dans le cas ougrave le gegravene nrsquoavait pas
de fonction connue chez Saureus str Newman je suis alleacutee voir quelle eacutetait la
fonction de ce mecircme gegravene (et des gegravenes flanquant celui-ci) chez diffeacuterentes souches
de Staphylococcus aureus Advenant le cas ougrave malgreacute tout je nrsquoeacutetais toujours pas
capable de deacuteterminer le rocircle de ce gegravene je suis alleacutee veacuterifier si sa seacutequence pouvait
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 hellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Mutants resistants agrave GEN (4 microgml)+TO
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip 185-AATTGCATTCA-197helliphelliphellip495-ATAGCAATGG-506hellip
hellipnwmn_1629helliphelliphelliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_0143hellip
ccpAATP synthase c
ABC transp
Niveau 1
Niveau 2
Niveau 3
Niveau 4
TO +GEN (4microgml)
Figure 9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct (A) nwmn_0811 et
nwmn_0928 chez les mutants ATCC 29213 reacutesistants agrave la combinaison gentamicine
(+TO) leur souche parente ATCC 29213 (B) nwmn_1629 et nwmn_2012 chez les
mutants hemB reacutesistants agrave la TO et leur souche parente NewbouldΔhemB (C)
nwmn_1629 nwmn_2012 et nwmn_0143 chez les mutants hemB reacutesistant agrave la
tomatidine (+genta) et leur souche parente hemB
C
99
avoir une forte homologie avec un gegravene drsquoune autre espegravece Malgreacute tout certains
gegravenes portent toujours la mention laquo de fonction inconnue raquo
La meacutethodologie que nous avons employeacutee nous a permis drsquoidentifier plusieurs
gegravenes possiblement impliqueacutes dans reacutesistance de nos souches aux antibiotiques
testeacutes
1- chez Staphylococcus aureus ATCC 29213 reacutesistant agrave la gentamicine (+TO)
(Figure 9A)
- NWMN_0811 codant possiblement pour la NADH deacuteshydrogeacutenase
- qoxC (NWMN_0928) codant pour la quinol oxidase polypeptide III
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
2- chez NewbouldΔhemB reacutesistant agrave la tomatidine (Figure 9B)
- ccpA (NWMN_1629) codant pour la laquo catabolite control protein A raquo
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
3- chez NewbouldΔhemB reacutesistant agrave la tomatidine (+gentamicine) (Figure 9C)
- NWMN_0143 codant pour un ABC transporteur
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
Lrsquoanalyse de ces diffeacuterents gegravenes nous amegravene agrave suspecter la sous-uniteacute c de lrsquoATP
synthase comme eacutetant la cible potentielle de la tomatidine chez Staphylococcus
aureus
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine
Les gegravenes muteacutes pointent en grande partie vers la chaine respiratoire qursquoon appelle
aussi chaine de transport des eacutelectrons
100
La NADH deacuteshydrogeacutenase (NWMN_0811) fait partie du complexes I de la chaine de
transport de eacutelectrons alors que la quinol oxidase polypeptide III (NWMN_0928 ou
QoxC) est une composante du complex III et la sous-uniteacute c de lrsquoATP synthase (AtpE)
fait partie eacutevidement du complexe de lrsquoATP synthase ou complexe V de la chaine
respiratoire (Figure 10)
Figure 10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants
Les courbes de croissance des mutants ATCC29213 reacutesistants agrave la combinaison
GEN(+TO) appuient le fait que ces mutations affectent la chaine de transport des
eacutelectrons ce qui affecte le niveau de production drsquoATP et donc la croissance
bacteacuterienne En effet les mutants du troisiegraveme niveau de reacutesistance (qui possegravede
deux gegravenes de la chaine de transport des eacutelectrons de muteacutes) ont une croissance
plus lente que les mutants du premier niveau de reacutesistance qui eux ont un seul gegravene
dans la chaine de transport des eacutelectrons de muteacute (Figure 11)
Quinoloxidase
NADHdehydrogenase
ATPsynthetase
Proton gradient
XXX
101
Figure 11 Courbes de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison GEN+TO du premier niveau de reacutesistance (bleu) et du troisiegraveme niveau
de reacutesistance (vert)
Voici plus preacuteciseacutement comment fonctionne la chaine de transport des eacutelectrons chez
les 2 pheacutenotypes-parents selon le modegravele-hypothegravese eacutetabli avec nos reacutesultats et
analyses
Tout drsquoabord chez le prototype ou souche de pheacutenotype normal (ex ATCC 29213) la
chaine de transport des eacutelectrons fonctionne normalement Les complexes de la
chaine de transport des eacutelectrons sont chargeacutes de transfeacuterer des protons vers
lrsquoexteacuterieur de la cellule en transfeacuterant en mecircme temps des eacutelectrons drsquoun complexe agrave
un autre jusqursquoagrave un accepteur final qui est lrsquoO2 lorsque crsquoest possible Se faisant il se
creacutee un gradient eacutelectrochimique de part et drsquoautre de la membrane et les protons
accumuleacutes du cocircteacute externe sont utiliseacutes par la suite par lrsquoATP synthase afin de
pouvoir produire de lrsquoATP dans la cellule
0 4 8 1 2 1 6 2 0 2 4
0 0 1
0 1
1
C o u r b e d e c r o is s a n c e d e s m u ta n ts A T C C 2 9 2 1 3
reacute s is ta n ts agrave G E N + T O (8 m g L )
te m p s (h )
OD
60
0n
m
Niveau 3
Niveau 1
102
Le transfert drsquoeacutelectrons ne se fait pas sans coucirct et des produits toxiques pour la
cellule sont produits en mecircme temps ce sont les espegraveces reacuteactives de lrsquooxygegravene ou
ROS (Reactive Oxygen Species) Cependant les prototypes par leur reacuteponse au
stress produisent des enzymes capables de deacutetoxifier le milieu (Figure 12A)
Chez les souches SCV ici la souche controcircle NewbouldΔhemB la chaine de
transport des eacutelectrons est deacuteficiente parce que le gegravene permettant la production
drsquoheacutemine qui entre dans la composition des cytochromes est supprimeacute Les
cytochromes entrent dans la composition de plusieurs composantes de la chaine
respiratoire
Lorsque les complexes de la chaine sont alteacutereacutes il y a diminution du transfert de
protons vers lrsquoexteacuterieur une diminution de la production drsquoATP et une plus grande
production de ROS (Figure 12B)
Figure 12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B) NewbouldΔhemB
I III IV V I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
eacute eacute
eacute
H+ H+
ATPO2 H2O
ROS +ROS
Prototype SCV (hemB)
H+
0811 0928 2012 0811 0928
hegraveme
2012
A B
103
La gentamicine a besoin drsquoune bonne polarisation membranaire donc drsquoune chaine de
transport des eacutelectrons fonctionnelle pour pouvoir peacuteneacutetrer dans la cellule (Bryan et
Kwan 1983 (Bryan and van den Elzen 1977 Davis 1987 Taber et al 1987) Une
fois entreacutee elle inhibe la synthegravese proteacuteique ce qui provoque la mort cellulaire La
CMI de la gentamicine contre ATCC 29213 est de 05-1 microgml (Figure 13A)
La tomatidine elle irait se lier agrave lrsquoATP synthase pour en diminuer la fonctionnaliteacute et
ce tant chez ATCC 29213 que chez hemB Cette liaison amegravenerait une diminution
de la production drsquoATP et conseacutequemment provoquerait agrave plus ou moins long terme
une augmentation de production de ROS par lrsquoalteacuteration de la chaine de transport des
eacutelectrons Cependant comme la cellule prototype produit efficacement les enzymes
de deacutetoxification lrsquoimpact de la tomatidine est pratiquement nulle sur les souches
prototypes La CMI de la tomatidine est gt16microml contre ATCC 29213 (Figure 13B)
104
Figure 13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213
Chez ATCC 29213 lrsquoajout de 8microgml de tomatidine agrave la Gentamicine amegravene une
diminution de 8X agrave 16X la CMI de la gentamicine En effet la CMI de la gentamicine
passe de 05-1microgml agrave 006microgml (Figure 14AB)
Lors de lrsquoajout de la tomatidine agrave la gentamicine crsquoest la tomatidine qui se lie dans
un premier temps agrave lrsquoATP synthase reacuteduisant ainsi sont niveau drsquoactiviteacute Il y a alors
de faccedilon transitoire une augmentation de la concentration de protons agrave lrsquoexteacuterieur de
la cellule ce qui permet une entreacutee massive de gentamicine ainsi qursquoune rapide et
forte inhibition de la synthegravese proteacuteique Dans un second temps il y aura eacutegalement
une alteacuteration de la chaine de transport des eacutelectrons conduisant agrave une
augmentation de la production de ROS Crsquoest cependant lrsquoentreacutee massive de
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI TO gt16 microgmlCMI GEN 05-1 microgml
+ROS
ROS mais deacutetoxification efficace
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
105
gentamicine entrainant une forte inhibition de la synthegravese proteacuteique qui provoquera
dans ce cas-ci la mort cellulaire (Figure 14B)
Figure 14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicine lorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC 29213
Chez les mutants ATCC29213 reacutesistants agrave la combinaison GEN+TO les complexes I
et III sont muteacutes et donc ont une fonctionnaliteacute alteacutereacutee ce qui amegravene une
deacutepolarisation membranaire Une membrane moins polariseacutee amegravene une plus faible
entreacutee de gentamicine et donc une plus faible inhibition proteacuteique La CMI de la
gentamicine passe en effet de 05-1 microgml chez la souche parente agrave 2 microgml chez les
mutants reacutesistants
La tomatidine en se liant agrave lrsquoATP synthase accentuera la deacutepolarisation
membranaire provoquant une augmentation de la production des ROS et une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN (TO 8 microgml) 006 microgmlCMI GEN 05-1 microgml
+ROS
polarisation entreacutee massive de GEN
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
GENH+
BA
106
diminution de la production drsquoATP De fait les mutants reacutesistants ont une croissance
plus lente (Figure 15)(Figure 11)
Figure 15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants ATCC 29213
reacutesistants agrave la combinaison gentamicine (+TO)
Chez les SCV la tomatidine se lie aussi agrave lrsquoATP synthase et ce faisant amegravene une
diminution de la production drsquoATP Cette diminution de la production drsquoATP megravenera
ultimement agrave une alteacuteration de la chaine de transport des eacutelectrons et une
augmentation des ROS (Murphy 2009) tout comme chez les souches prototypes
Cependant comme le SCV a deacutejagrave une chaine de transport des eacutelectrons deacuteficiente
une faible production drsquoATP par lrsquoATP synthase et une forte production de ROS
lrsquoajout drsquoune faible concentration de tomatidine (la CMI est de 006microgml) amegravene une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype R GEN (+TO [8 microgml])
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN 2 microgmlCMI GEN 05-1 microgml
+ROS
R agrave GEN (+TO) mais mutants ont une
croissance diminueacutee (+ROS et -ATP)
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
107
augmentation de chacun de ces pheacutenomegravenes ce qui est suffisant pour provoquer la
mort cellulaire (Figure 16A)
Dans les premiers niveaux de reacutesistance agrave la tomatidine chez les mutants hemB la
reacutesistance (la CMI passe de 006 agrave 025-1microgml) est ameneacutee par la mutation du gegravene
ccpA (Catabolite controcircle protein A) ce qui megravenerait agrave une augmentation de la
production drsquoATP par lrsquoutilisation drsquoune autre voie que lrsquoATP synthase etou agrave une
diminution de la production de ROS (Figure 16B) Le rocircle de la CcpA en lien avec la
reacutesistance agrave la tomatidine est encore neacutebuleux Toutefois on sait que normalement
ce gegravene agit comme reacutegulateur transcriptionnel des gegravenes codant pour des enzymes
du cycle de Krebs et qursquoil a un rocircle de reacutegulation dans le meacutetabolisme du carbone et
lrsquoutilisation des substrats (Somerville et Proctor 2009 Goumltz et al 2006)
Figure 16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO 006 microgml
H+
SCV
I III IVeacute eacute
eacuteO2 H2O
+ROS
H+TO
Niveau 1
CcpA (CMI TO 025-1 microgml)
ROS
ATP
V
H+
ATP
TO0811 0928
2012
1629
0811 0928
2012
BA
108
Chez les SCV du deuxiegraveme palier de reacutesistance agrave la tomatidine non seulement le
gegravene ccpA est toujours muteacute mais le gegravene de la sous-uniteacute c de lrsquoATP synthase est
eacutegalement muteacute La mutation dans le site de liaison mecircme de la tomatidine empecircche
sa liaison et donc empecircche son effet sa CMI passant de 006 agrave gt16microgml (Figure
17B)
Malgreacute la mutation dans le gegravene lrsquoATP synthase resterait quand mecircme fonctionnel
puisque la mutation nrsquoamegravene pas la mort cellulaire comme crsquoest le cas lorsque la
tomatidine lie lrsquoATP synthase chez le SCV parent (Figure 17A)
Figure 17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la tomatidine
Tout comme chez les mutants hemB reacutesistant agrave la tomatidine seule les mutants
hemB reacutesistant agrave la tomatidine lorsqursquoen preacutesence de gentamicine possegravedent
eacutegalement une mutation dans le gegravene de lrsquoATP synthase ce qui rend la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO006 microgml
H+
SCV Niveau 2CMI TO gt16 microgml
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+TO
TO
Niveaux 1 et 2
CcpA (CMI TO 025-1 microgml)
ROS
ATP
0811 0928
2012
1629
0811 0928
2012
A B
109
inactive puisqursquoincapable de srsquoy lier (CMI gt16microgml) (Figure 18B) Chez les SCV non
muteacutes (hemB) la gentamicine nrsquoentre que faiblement ducirc agrave la deacutepolarisation de sa
membrane (CMI 4-8 microgml) (Figure 18A) La mutation dans le gegravene codant pour une
pompe (NWMN_0143 codant pour un ABC transporteur) chez les mutants hemB
reacutesistant agrave la tomatidine (+gentamicine) amegravenerait une augmentation de lrsquoactiviteacute de
cette pompe permettant de rejeter hors de la cellule le peu de gentamicine eacutetant
entreacutee La mutation de cette pompe fait passer la CMI de la gentamicine de 4-8
microgml pour hemB agrave 16 microgml pour les mutants hemB reacutesistant agrave la combinaison
gentamicine (+TO) ayant cette mutation (Figure 18B)
Figure 18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB reacutesistants
agrave la combinaison tomatidine (+gentamicine)
Donc selon notre modegravele-hypothegravese lrsquoaction de la tomatidine et de la gentamicine
sur S aureus deacutepend de la liaison agrave leur cible mais a aussi un lien avec la
SCV R agrave TO (+GEN [4microgml])
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI GEN 4-8 microgml
H+
SCV
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+ TO
GEN
CMI GEN 16 microgml
CMI TO gt16 microgml
0811 0928
2012
0811 0928 20120143
GEN
Synthegravese proteacuteique
BA
110
polarisation membranaire et la production de ROS La mesure de la polarisation et
concentration cellulaire en ROS chez nos mutants et nos souches parents en
preacutesence ou non drsquoantibiotique nous permettra de valider notre modegravele-hypothegravese
Chez une bacteacuterie meacutetaboliquement active et posseacutedant une membrane
cytoplasmique intacte il y a normalement une diffeacuterence de potentiel eacutelectrique entre
le cocircteacute inteacuterieur et le cocircteacute exteacuterieur de la membrane lrsquointeacuterieur eacutetant plus neacutegatif de
100 agrave 200 mV par rapport agrave lrsquoexteacuterieur Une diminution de la magnitude de ce
potentiel membranaire est appeleacutee deacutepolarisation alors qursquoune augmentation de ce
potentiel est appeleacutee hyperpolarisation Le potentiel membranaire sera reacuteduit agrave zeacutero
srsquoil y a rupture de la membrane cest-agrave-dire srsquoil y a formation de trous assez grands
permettant aux ions inorganiques de passer librement Ce pheacutenomegravene peut arriver
lorsque les cellules sont tueacutees par la chaleur le froid ou les antibiotiques β-Lactams
Dans ces circonstances la membrane devient permeacuteable agrave diffeacuterents colorants
comme le propidium iodide (PI) et le TO-PRO3 et le changement est irreacuteversible
Plusieurs classes de composeacutes chimiques notamment les ionophores peuvent
eacutegalement alteacuterer le potentiel membranaire sans pour autant permettre la
permeacuteabiliteacute au PI TO-PRO3 et autres colorants similaires chez les bacteacuteries
lrsquoadministration drsquoun proton-ionophore tel le carbonyl cyanide 3-
chlorophenylhydrazone (CCCP) reacuteduira le potentiel membranaire agrave zeacutero
Des tests preacuteliminaires sur la mesure du potentiel membranaire (BacLighttrade Bacterial
Membrane Potential Kit (B34950) drsquoInvitrogen et lecture au cytomegravetre de flux (FACS))
(Novo et al 2000) chez nos deux souches parentes mets en eacutevidence que ATCC
29213 est plus polariseacute que NewbouldΔhemB (Figure 19A) Toutefois malgreacute
qursquoayant une plus faible polarisation membranaire NewbouldΔhemB nrsquoest pas
complegravetement deacutepolariseacute puisqursquoil est possible de le deacutepolariser davantage agrave lrsquoaide
drsquoun agent deacutepolarisant tel que le CCCP (carbonyl cyanide 3-chlorophenylhydrazone)
(Figure 19B) Le TO-PRO3 a eacuteteacute utiliseacute comme controcircle afin de confirmer la viabiliteacute
de nos cellules (donneacutees non montreacutees)
111
A B
Figure 19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane Potential Kit
(Invitrogen) (Lecture au cytomegravetre de flux agrave lrsquoaide du programme FCS Express) (A)
Ratio FL3-H (axe des Y)FL1-H (axe des X) de ATCC 29213 (rouge) et de
NewbouldΔhemB (noir) (B) Lecture des rouges (FL3-H) sur NewbouldΔhemB traiteacute
avec le DiOC2(3) (33prime-diethyloxacarbocyanine iodide) (noir) ou avec le CCCP (bleu)
ou sur les cellules chauffeacutees (rouge)
La sous-uniteacute c de lrsquoATP synthase (AtpE) semble donc la cible la plus probable de la
tomatidine Afin de veacuterifier si les mutations identifieacutees pouvaient avoir un effet
veacuteritable sur la proteacuteine nous avons fait un alignement de seacutequences noteacute les bases
muteacutees et veacuterifieacute si la mutation amenait un changement drsquoacide amineacute et si ce
changement drsquoacide amineacute pouvait avoir un impact sur la proteacuteine finale De fait
nous avons trouveacute que les deux diffeacuterentes mutations dans lrsquoAtpE amenaient un
changement significatif drsquoacides amineacutes pouvant amener une variation fonctionnelle
FL3-H
Count
100
101
102
103
104
0
18
36
53
71
Chauffeacute
+CCCP+DiOC2
Chauffeacute
FL1-H
FL3-H
100
101
102
103
104
100
101
102
103
104
hemB
29213
VERT
RO
UG
E
ROUGE
ΔhemB
112
de la proteacuteine La mutation de la base 49 change un G pour un T il srsquoagit drsquoun SNP
nrsquoamenant pas de changement de cadre de lecture La mutation fait passer lrsquoacide
amineacute alanine (A) qui est apolaire agrave une seacuterine (S) qui est polaire La mutation de la
base 190 change un G pour un A et est aussi un SNP nrsquoamenant pas de changement
de cadre de lecture Cette mutation fait passer lrsquoacide amineacute alanine (A) qui est
apolaire agrave une threacuteonine (T) qui est polaire (Figure 20)
gtlcl||NWMN_2012|atpE|72493880 F0F1 ATP synthase subunit C
1 10 19 31 43 49 58
ATG AAT TTA ATC GCA GCA GCA ATC GCA ATT GGT TTA TCA GCA TTA GGA GCA GGT ATC GGT
M N L I A A A I A I G L S A L G A G I G
(S)
17 61 73 85 97 109 118
AAC GGT TTA ATC GTT TCA AGA ACA GTT GAA GGT GTA GCA CGT CAA CCA GAA GCA CGT GGT
N G L I V S R T V E G V A R Q P E A R G
21 40
121 133 145 157 169 178
CAA TTA ATG GGT ATC ATG TTC ATT GGT GTA GGT TTA GTT GAG GCA TTA CCT ATC ATC GGT
Q L M G I M F I G V G L V E A L P I I G
41 60
181 190 199 211
GTA GTA ATT GCA TTC ATG ACA TTT GCT GGA TAA
V V I A F M T F A G
(T)
61 64 70
Figure 20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du gegravene
atpE de S aureus Newman avec mise en eacutevidence des mutations (rose) en lien avec
la cible de la tomatidine Lrsquoalignement a eacuteteacute effectueacute agrave lrsquoaide du site CMR de JCVI
113
CHAPITRE 4
DISCUSSION
LrsquoATP synthase est eacutevidemment un complexe proteacuteique ubiquitaire tant chez les
procaryotes que les eucaryotes Malgreacute ce fait il semble que la cible de la tomatidine
soit speacutecifique aux bacillales celle-ci ne deacutemontrant aucune activiteacute contre les
bacteacuteries agrave Gram neacutegatif et les lactobacillales On suppose donc que lrsquoATP synthase
de ces espegraveces doit ecirctre speacutecifique au niveau du site de liaison de la tomatidine
Lrsquoecirctre humain en tant qursquoespegravece eucaryote est eacutegalement pourvu de ce complexe
proteacuteique et on peut naturellement se demander si lrsquoutilisation clinique de cette
moleacutecule pourrait ecirctre cytotoxique De fait plusieurs composeacutes naturels tel
lrsquooligomycine et la venturicidine sont connus pour inhiber lrsquoactiviteacute de lrsquoATP synthase
par leur interaction avec la sous-uniteacute c de ce complexe Cependant ces moleacutecules
ne sont pas seacutelectives et nrsquoinhibent pas seulement lrsquoATP synthase bacteacuterien mais
aussi lrsquoATP synthase mitochondrial (Haagsma et al 2009 Matsuno-Yagi et Hatefi
1993) (Figure 1) Neacuteanmoins il existe maintenant une moleacutecule qui a pour cible
lrsquoATP synthase bacteacuterien et qui est en processus en vue drsquoune utilisation clinique
chez lrsquohomme le TMC207 (Haagsma et al 2009))
114
Figure 1 Sites drsquoinhibition de lrsquoATP synthase Les sites de liaison dans lrsquoATP
synthase des inhibiteurs reacuteveacuteleacutes par des eacutetudes biochimiques et structurales sont
indiqueacutes par des cercles rouges et les sites de liaisons qui nrsquoont pas eacuteteacute totalement
clarifieacutes sont indiqueacutes par des cercles verts (Hong et Pedersen 2008)
Le TMC207 (Figure 2A) est un composeacute de la classe des diarylquinoline posseacutedant
un nouveau meacutecanisme drsquoaction lrsquoinhibition de lrsquoATP synthase bacteacuterien De fait la
cible et le meacutecanisme drsquoaction du TMC207 sont diffeacuterents des autres agents
antituberculeux Lrsquoinhibition de lrsquoATP synthase peut mener agrave une baisse de la
production drsquoATP et un deacutebalancement dans lrsquohomeacuteostasie du pH ces 2 effets
pouvant contribuer agrave la diminution de la survie de la bacteacuterie (Rao et al 2001
Andries et al 2005)
Le TMC207 est une moleacutecule ayant une forte activiteacute bacteacutericide contre
Mycobacterium tuberculosis et autres espegraveces mycobacteacuteriennes mais peu drsquoactiviteacute
115
contres les autres espegraveces bacteacuteriennes Lrsquoindex de seacutelectiviteacute de TMC207 pour
lrsquoATP synthase mycobacteacuterienne (CMI50 gt200 microml) compareacute agrave lrsquoATP synthase
mitochondriale (CMI50 001 microgml) est de plus de 20000 Un indice de seacutelectiviteacute au
dessus de 1000 est consideacutereacute dans le deacuteveloppement drsquoune drogue comme un
preacutealable pour une moleacutecule candidate (Haagsma et al 2009) La diversiteacute dans les
gegravenes atpE explique la reacutesistance au TMC207 et justifie la faible affiniteacute de cette
drogue agrave lrsquoATP synthase mitochondriale (Matteelli et al 2010) (Figure 3)
Figure 2 Structure chimique de (A) TMC207 et (B) son analogue anti bacteacuteries
agrave Gram positifs (Balemans et al 2012)
Lorsqursquoon compare la seacutequence proteacuteique de la sous-uniteacute c de lrsquoATP synthase
(atpE) de quelques espegraveces de lrsquoordre des Bacillales (tel Saureus B cereus B
subtilis et L monocytogenes) avec des espegraveces appartenant agrave drsquoautres ordres
bacteacuteriens on constate que les seacutequences divergent ce qui est en accord avec la
speacutecificiteacute des moleacutecules Les points de mutations amenant une reacutesistance au
TMC207 chez Mycobacterium smegmatis sont speacutecifiques aux mycobacteacuteries et les
deux principaux points de mutations D32V(ouP) et A67P chez M smegmatis sont
situeacutes tous les deux dans les heacutelices alpha qui entourent le site catalytique de la
proteacuteine (E65) qui est impliqueacute dans le transport des protons (Andries et al 2005)
Lrsquoacide amineacute aspartate (D) agrave la position 32 et lrsquoalanine (A) agrave la position 67 sont tregraves
speacutecifiques agrave M smegmatis
A B
116
Reacutecemment un groupe de chercheurs (Balemans et al 2012) on mit en lumiegravere une
autre moleacutecule (Figure 2B) de la classe des diarylquinolines dont le meacutecanisme
drsquoaction eacutetait aussi lrsquoinhibition de la sous-uniteacute c de lrsquoATP synthase mais chez les
bacteacuteries agrave Gram positif tels Streptococcus pneumoniae et S aureus Cette moleacutecule
irait lier les acides amineacutes valines (V) situeacutes agrave la position 50 et 62 de lrsquoAtpE de S
aureus (Donneacutees non montreacutees chez S pneumoniae(Balemans et al 2012)) En
effet une mutation amenant le changement de la valine en position 50 par une
isoleucine (I) etou une mutation amenant le changement de la valine en position 62
par une alanine (A) amegravene la reacutesistance de la bacteacuterie agrave la moleacutecule Cependant L
monocytogenes et Bacillus subtilis ne possegravedent pas de valine agrave la position 50 alors
qursquoils font eacutegalement partie des bacteacuteries agrave Gram positif (Il faut noter que chez
Bacillus sp PS3 (Swiss-Prot assession Ndeg P 00845) possegravede une valine agrave la position
50 (Balemans et al 2012)) Par contre tout comme pour les autres bacteacuteries agrave Gram
positif testeacutees les deux espegraveces possegravedent une valine agrave la position 62 (Figure 3)
Chez B cereus B subtilis B anthracis S aureus et L monocytogenes les points de
divergence sont plutocirct A17S et A64T Cependant B pseudofirmus qui fait eacutegalement
partie de lrsquoordre des Bacillales possegravede une glycine (G) agrave la position 17 plutocirct qursquoune
alanine (A) et une seacuterine (S) plutocirct qursquoune alanine (A) agrave la position 64 comme les
espegraveces preacuteceacutedentes Suivant ces reacutesultats et si la cible de la tomatidine est bien
ces deux points de mutation la tomatidine ne devrait pas pouvoir se lier agrave lrsquoAtpE de B
pseudofirmus et devrait donc nrsquoavoir aucune activiteacute contre cette souche alors que
nous avions extrapoleacute que le spectre drsquoactiviteacute de la tomatidine eacutetait tous les
Bacillales De plus E coli et M smegmatis possegravedent eacutegalement une alanine (A) agrave la
position 17 alors que la tomatidine ne possegravede aucune activiteacute contre ces espegraveces (M
smegmatis nrsquoa pas eacuteteacute eacutevalueacute il srsquoagit ici drsquoune extrapolation des reacutesultats de CMI)
par contre ces deux espegraveces possegravedent des acides amineacutes diffeacuterents drsquoune alanine
(A) agrave la position 64 ce qui pourrait leur confeacuterer une reacutesistance agrave la tomatidine telle
qursquoobserveacutee On remarque eacutegalement que les acides amineacutes agrave ces deux positions
117
sont totalement diffeacuterents chez lrsquohomme (Homo sapiens) ce qui suggeacutererait que la
tomatidine ne possegravederait pas drsquoactiviteacute antimitochondriale (Figure 3)
Selon ces reacutesultats on peut se demander si les deux mutations sont essentielles agrave la
liaison de la moleacutecule ou si seulement lrsquoalanine (A) agrave la position 64 est neacutecessaire
On doit se rappeler que la reacutesistance apporteacutee par la mutation faisant en sorte que
lrsquoalanine (A) en position 17 devient une seacuterine (S) apparaicirct chez les mutants
NewbouldΔhemb reacutesistant agrave la tomatidine et que cette mutation nrsquoapparaicirct qursquoau
quatriegraveme et dernier niveau de reacutesistance On se rappelle eacutegalement que la mutation
faisant en sorte que lrsquoalanine (A) agrave la position 64 devient une threacuteonine (T) apparaicirct
chez les mutants NewbouldΔhemb reacutesistants agrave la tomatidine en combinaison avec la
gentamicine et ce degraves le premier niveau de reacutesistance
118
Bcereus G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Banthracis G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Lmonocytogenes G---------VIAAAIAVG---LGALGAGIGNGLIVSKTVEGVARQPEARSMLQTIMFV- 50
Saureus_NWMN_2012 -----------IAAAIAIG---LSALGAGIGNGLIVSRTVEGVARQPEARGQLMGIMFIG 49
Bsubtilis -----------IAAAIAIG---LGALGAGIGNGLIVSRTVEGIARQPEAGKELRTLMFMG 49
Bpseudofirmus -----------LGAAIAAG---LAAVAGAIAVAIIVKATIEGTTRQPELRGTLQTLMFIG 49
Ecoli L---------YMAAAVMMG---LAAIGAAIGIGILGGKFLEGAARQPDLIPLLRTQFFIV 56
Msmegmatis DPNAIITAGALIGGGLIMG---GGAIGAGIGDGIAGNALISGIARQPEAQGRLFTPFFIT 60
Homo sapiens QTSAISRDIDTAAKFIGAGAATVGVAGSGAGIGTVFGSLIIGYARNPSLKQQLFSYAILG 120
MyR V
MyR P
SaR S
Bcereus VALVEALPIIGVVIAFIVMNK----- 72
Banthracis VALVEALPIIGVVIAFIVMNK----- 72
Lmonocytogenes ----EALPIIAVVIAFMVLNK----- 67
Saureus_NWMN_2012 VGLVEALPIIGVVIAFMTFAG----- 70
Bsubtilis IALVEALPIIAVVIAFLAFFG----- 70
Bpseudofirmus VPLAEAVPIIAIVISLLILF------ 69
Ecoli MGLVDAIPMIAVGLGLYVMFAVA--- 79
Msmegmatis VGLVEAAYFINLAFMALFVFATPGLQ 86
Homo sapiens FALSEAMGLFCLMVAFLILFAM---- 142
MyR D P M
SpR I A
SaR T
Fig
ure
3
Seacuteq
uen
ce
pro
teacuteiq
ue d
e la p
roteacute
ine
Atp
Ec
hez d
iffeacute
ren
tes
es
pegravec
es e
t m
ise e
n eacute
vid
en
ce
de
s s
ite
s d
e
mu
tati
on
pe
rmett
an
t la
reacutesis
tan
ce agrave
dif
feacutere
nte
s m
oleacute
cu
les
M
yR M
tu
berc
ulo
sis
ou
M sm
eg
matis
reacutesis
tan
t a
u
TM
C2
07 S
pR
S
p
neu
mon
iae
reacutesis
tan
t agrave
un a
na
log
ue d
u T
MC
20
7
SaR
S
a
ure
us
SC
V r
eacutesis
tant agrave
la to
matid
ine
En
cad
reacutero
uge
S
ite d
e m
uta
tion a
men
ant la
reacutesis
tance
agravela
to
matid
ine E
ncad
reacuteno
ir
Site
cata
lytiq
ue
im
pliq
ueacute
da
ns le
tra
nspo
rt d
es p
roto
ns
Su
rlig
nag
etu
rqu
ois
e S
ite d
e m
uta
tion a
me
nant u
ne
reacutesis
tan
ce a
u T
MC
20
7
119
Les seacutequences proteacuteiques des souches bacteacuteriennes de la figure 3 ont eacuteteacute trouveacutees
sur pubmedproteacuteine httpwwwncbinlmnihgovpubmed
laquo ATP synthase subunit c raquo Assession number
Streptococcus pneumoniae D39 YP_8168031 (NCBI)
S aureus Newman BAF682841 (GenBank)
E coli AAA247321 (GenBank)
Bacillus cereus ISO75 EJP900901 (GenBank)
Bacillus anthracis str H9401 YP_0062123491 (NCBI)
Mycobacterium smegmatis MC2 155 YP_0065695541 (NCBI)
Bacillus subtilis str 168 CAA822551 (GenBank)
Listeria monocytogenes YP_0076049931 (NCBI)
Homo sapiens P482011 (UniProtKBSwiss-Prot)
Lrsquoalignement comparatif des seacutequences drsquoacides amineacutes a eacuteteacute fait agrave lrsquoaide du
programme ClustalW2 sur le web
httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2
Le modegravele 3D de lrsquoAtpE a eacuteteacute obtenu gracircce agrave lrsquoaide du Pr Ryszard Brzezinski du
Deacutepartement de biologie de lrsquoUniversiteacute de Sherbrooke (Figure 4) Une recherche
dans le Protein Data Bank a drsquoabord eacuteteacute effectueacutee afin de veacuterifier parmi les proteacuteines
AtpE (sous-uniteacute C de lrsquoATP synthase) pour lesquelles il y avait des structures 3D
reacutesolues laquelle ressemblait le plus agrave celle de Staphylococcus aureus Les
alignements ont montreacute que cest la proteacuteine de Bacillus pseudofirmus qui avait la
plus grande similitude de seacutequence La structure est stockeacutee dans la banque PDB
sous le symbole 3ZO6 Il a eacuteteacute confirmeacute par quelques alignements que cette sous-
uniteacute C de lrsquoATP synthase eacutetait plus proche par sa seacutequence de celle de
Staphylococcus aureus que de celle de Mycobacterium pour laquelle la structure 3D
avait eacuteteacute deacutetermineacutee plus tocirct Ce reacutesultat sexplique par le fait que Staphylococcus et
Bacillus font tous deux partie de la branche A+T-riche des bacteacuteries agrave Gram+ alors
que Mycobacterium appartient agrave la branche G+C-riche La seacutequence de lAtpE de
120
Staphylococcus aureus a ensuite eacuteteacute envoyeacutee au serveur internet de modeacutelisation
3D-Jigsaw (httpbmmcancerresearchukorg~3djigsaw) en indiquant que le modegravele
devait ecirctre bacircti en prenant comme matrice le fichier PDB 3ZO6 Le modegravele nous a
eacuteteacute retourneacute sous format PDB ndash comme les laquo vrais raquo cristaux dans la Protein Data
Bank Les fichiers au format PDB peuvent ecirctre visualiseacutes par le logiciel RasMol ndash
gratuit sur internet Cest avec RasMol que limage (Figure 4) a eacuteteacute obtenue
121
Figure 4 Modeacutelisation de la structure tridimensionelle de la sous-uniteacute c de
lrsquoATP synthase de S aureus (AtpE) montrant les acides amineacutes muteacutes chez les
mutants reacutesistantsEn blanc et vert les deux acides amineacutes muteacutes chez S aureus
reacutesistant agrave la tomatidine En rose les sites de mutations chez Mycobacterium
smegmatis reacutesistant au TMC207 et des sites de mutations chez Streptoccocus
pneumoniae reacutesistants agrave un analogue de TMC207 En bleu le site actif de la proteacuteine
Agrave la vue de ces reacutesultats certains pourront mettre en doute lrsquointeacuterecirct de la tomatidine
eacutetant donneacute lrsquoapparition relativement rapide de reacutesistance chez S aureus Cependant
122
nos reacutesultats deacutemontrent clairement que lrsquoajout de tomatidine ralentit le
deacuteveloppement de reacutesistance agrave la gentamicine chez S aureus ATCC29213 (Tableau
1) De plus cette augmentation de la CMI de la gentamicine en preacutesence de
tomatidine au fil des passages suit lrsquoaugmentation de la CMI de la ciprofloxacine lors
de ces mecircmes passages alors que la ciprofloxacine est consideacutereacutee comme la drogue
de reacutefeacuterences dans les bonnes drogues (Tableau 1) Tout comme pour la
ciprofloxacine pour laquelle la reacutesistance chez S aureus peut ecirctre la conseacutequence
de deux mutations dans le gegravene cible gyrA la reacutesistance agrave la tomatidine peut ecirctre la
conseacutequence de deux mutations dans le gegravene cible atpE Par comparaison la
rifampicine est consideacutereacutee comme une mauvaise drogue de par lrsquoapparition rapide
drsquoune forte reacutesistance chez S aureus conseacutequence de ses 17 mutations possibles
dans le gegravene cible rpoB (Tableau 1)
De plus il sera important de relativiser lrsquoapparition de reacutesistance agrave la tomatidine en
calculant le taux drsquoapparition de ces colonies reacutesistantes ceci nrsquoayant pas eacuteteacute fait
jusqursquoici De fait la freacutequence de mutation chez S aureus amenant une reacutesistance agrave
la rifampicine est de lrsquoordre de 10-7 10-8 alors qursquoil est de 10-11 pour lrsquoacide fusidique
(Canfield et al 2013 Orsquoneill et al 2001 Price et Gustafson 2001) Pour ce qui est
de la nouvelle moleacutecule anti-bacteacuteries agrave Gram positif de la classe des diarylquinoline
une freacutequence de mutation sur S pneumoniae ATCC49619 de 81- 83 x 10-7 a eacuteteacute
observeacutee agrave 5x la CMI et une freacutequence de mutation de 46 x 10-7 agrave 17 x 10-6 agrave 50x la
CMI (Balemans et al 2012)
123
Tableau 1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 par la meacutethode de CMI lors de passages successifs dans un milieu avec
lrsquoantibiotique
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
Typiqueldquomauvaiserdquodrogue (17 mutations possibles dansrpoB)
Typique ldquobonnerdquo drogue (2 mutations possibles dans gyrA)
Preacutesence de TO ralenti le deacuteveloppement de reacutesistances vs GEN seul
124
CHAPITRE 5
CONCLUSION ET PERSPECTIVES
Mon projet de maitrise a permis de preacuteciser le spectre drsquoactiviteacute de la tomatidine
Nous savons maintenant que la tomatidine possegravede une activiteacute antibacteacuterienne
contre la plupart des Firmicutes et plus speacutecifiquement contre les Bacillales Plus
preacuteciseacutement la tomatidine est bacteacutericide contre les pheacutenotypes SCV et agit en
synergie avec les aminoglycosides augmentant lrsquoactiviteacute bacteacutericide de ces derniers
de 8-16 fois contre les souches agrave pheacutenotype normal
De plus parmi tous les analogues de la tomatidine produits par le laboratoire du Pr
Eacuteric Marsault (voir article en annexe 2) le composeacute FC04-100 a eacuteteacute plus
speacutecifiquement mis en lumiegravere Celui-ci conserve non seulement une aussi bonne
activiteacute synergique avec les aminoglicosides contre les Saureus au pheacutenotype
normal et une activiteacute modeacutereacutee contre les S aureus SCV mais montre eacutegalement de
faccedilon inespeacutereacutee une activiteacute par lui-mecircme contre les S aureus de pheacutenotype normal
incluant plusieurs MRSA une proprieacuteteacute qui nrsquoeacutetait pas observeacutee pour la tomatidine
Une autre activiteacute inteacuteressante de FC04-100 est sa capaciteacute drsquoameacuteliorer lrsquoactiviteacute
bacteacutericide des aminoglicosides contre L monocytogenes agrave pheacutenotype normal apregraves
un traitement de 24h alors que la tomatidine ne deacutemontrait qursquoune faible synergie
avec les aminoglycosides contre cette souche De plus FC04-100 deacutemontre une
activiteacute bacteacutericide contre les SCV de L monocytogenes alors que la tomatidine ne
deacutemontrait qursquoune activiteacute bacteacuteriostatique FC04-100 montre eacutegalement un effet
potentiateur de lrsquoUbrolexinTM (Ceacutefalexin Kanamycin (3 2)) une combinaison
drsquoantibiotique utiliseacute dans le traitement de mammites bovines en abaissant de 8 agrave 16
fois leur CMI amenant celles-ci sous le seuil de reacutesistance Finalement FC04-100
possegravede une meilleure solubiliteacute que la tomatidine une proprieacuteteacute qui facilitera son
utilisation in vitro
125
Nos reacutesultats deacutemontrent que FC04-100 conserve une certaine activiteacute contre les
SCV reacutesistant agrave la tomatidine ce qui suggegravere possiblement que FC04-100 a plus
daffiniteacute pour la cible ou plus de sites dinteraction avec AtpE En effet les mutants
SCV reacutesistants agrave la tomatidine deviennent gt512 fois plus reacutesistants alors que FC04-
100 ne perd que 8 agrave 16 fois son activiteacute contre ces mecircmes souches Ceci deacutemontre
quil est possible dameacuteliorer significativement lactiviteacute de ces alkaloiumldes steacuteroidiens
ce qui est tregraves prometteur pour lavenir de cette moleacutecule
Dans un second temps lrsquoanalyse des mutations chez les mutants reacutesistants agrave la
tomatidine a permis de non seulement de mieux comprendre le mode drsquoaction de la
tomatidine mais eacutegalement de preacuteciser la cible potentielle de ce nouvel antibiotique
De fait son mode drsquoaction serait en lien direct avec la fonctionnaliteacute de la chaine de
transport des eacutelectrons chez la bacteacuterie lrsquoactiviteacute de la tomatidine serait inversement
proportionnelle agrave la fonctionnaliteacute de la chaine de transport des eacutelectrons et donc en
lien eacutegalement avec la polarisation membranaire et la production de ROS
intracellulaire Afin de confirmer le modegravele-hypothegravese la mesure de la polarisation
membranaire et la concentration de ROS intracellulaire chez les mutants ainsi que
chez les souches parentes suite agrave un traitement antibiotique devront ecirctre effectueacutees
Les reacutesultats de mes recherches suggegraverent que la cible bacteacuterienne speacutecifique de la
tomatidine serait la sous-uniteacute C de lrsquoATP synthase Afin de confirmer la cible
Maxime Lamontagne-Boulet tente au moment drsquoeacutecrire ces lignes drsquoobtenir des
mutants de S aureus Newman reacutesistants au composeacute FC04-100 et dont la reacutesistance
serait attribuable agrave une mutation dans le gegravene atpE Par la suite on devra confirmer
la fonctionnaliteacute de la cible ATP synthase par quantification de lrsquoATP produit agrave lrsquoaide
de la lucifeacuterase tel que lrsquoon fait Balemans et al en 2012 Lrsquoattachegravement agrave la cible
devra eacutegalement ecirctre confirmeacute et la meacutethode de SPR (Surface Plasmon Resonance)
pourrait ecirctre tout indiqueacutee pour ce faire (Balemans et al 2012) On devra eacutegalement
126
deacutemontrer la seacutelectiviteacute des moleacutecules tomatidine et FC04-100 pour lrsquoATP synthase
des Bacillales
Les infections agrave bacteacuteries multireacutesistantes deviennent de plus en plus preacutevalentes et
sont un des problegravemes de santeacute majeurs auxquels nous devons faire face aujourdrsquohui
Cette augmentation de la reacutesistance limite le reacutepertoire drsquoantibiotiques efficace creacuteant
une situation probleacutematique laquelle est exacerbeacutee par le trop petit nombre de
nouveaux antibiotiques introduits sur le marcheacute ces derniegraveres anneacutees Il est donc
drsquoune grande importance de travailler agrave la mise en marcheacute de nouveaux antibiotiques
et plus particuliegraverement de nouvelles classes drsquoantibiotiques espeacuterant ainsi deacutejouer la
multireacutesistance bacteacuterienne du moins pour un temps La tomatidine de par sa
structure sa cible et son mode drsquoaction speacutecifique est une moleacutecule antibiotique
dans une classe agrave part et de ce fait une moleacutecule plus qursquointeacuteressante pour le
traitement drsquoinfection agrave S aureus De plus comme elle deacutemontre une activiteacute
bacteacutericide contre les deux pheacutenotypes de S aureus (et autres Bacillales) lesquels
sont retrouveacutes simultaneacutement dans les cas drsquoinfections chroniques son pouvoir
antibiotique devient inestimables drsquoun point de vue clinique
127
CONTRIBUTIONS SUPPLEacuteMENTAIRES
ANNEXE 1 Application de brevet Internationnal publication number WO
2012109752 A1
INVENTEURS Franccedilois Malouin2 Gabriel Mitchell2 Kamal Bouarab2 Eacuteric Marsault1
Feacutelix Chagnon1 Simon Boulanger 2 Isabelle Guay2
1Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
2Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
128
129
ANNEXE 2 Article scientifique dans laquo Europeen Journal of medicinal chemistry raquo
2014 80 605-620
TITRE Unraveling the structure-activity relationship of tomatidine a steroid alkaloid
with unique antibiotic properties against persistent forms of
Staphylococcus aureus
ABSTRACT Staphylococcus aureus is responsible for difficult-to-treat and relapsing
infections and constitutes one of the most problematic pathogens due to its multiple
resistances to clinically available antibiotics Additionally this pathogenrsquos ability to
develop small-colony variants is associated with a reduced susceptibility to
aminoglycoside antibiotics and in vivo persistence We have recently demonstrated
that tomatidine a steroid alkaloid isolated from tomato plants possesses anti-
virulence activity against normal strains of S aureus as well as the ability to
potentiate the effect of aminoglycosides against this pathogen In addition tomatidine
has shown antibiotic activity against small-colony variants We herein report the first
study of the structure-activity relationship of tomatidine against S aureus
AUTEURS Feacutelix Chagnon1 Isabelle Guay2 Marc-Andreacute Bonin1 Gabriel Mitchell2
Kamal Bouarab2Franccedilois Malouin2 Eacuteric Marsault1
MA CONTRIBUTION Responsable des tests biologiques de tous les composeacutes
autant envers lrsquoeffet potentiateur de la tomatidine sur lrsquoactiviteacute de la gentamicine
contre la souche ATCC29213 de S aureus que pour lrsquoactiviteacute bacteacuteriostatique sur la
souche NewbouldΔhem imitant le pheacutenotype SCV Ces travaux furent effectueacutes sous
la tutelle du professeur Franccedilois Malouin
130
1Institut de Pharmacologie de Sherbrooke Deacutepartement de pharmacologie
Universiteacute de Sherbrooke 3001 12e av nord Sherbrooke Queacutebec Canada J1H
5N4
2Centre drsquoeacutetude et de valorisation de la diversiteacute microbienne (CEVDM) Deacutepartement
de biologie Universiteacute de Sherbrooke Sherbrooke QC Canada J1K 2R1
131
BIBLIOGRAPHIE Alexander E H and Hudson MC (2001) Factors influencing the internal- ization of Staphylococcus aureus and impacts on the course of infections in humans Appl Microbiol Biotechnol 56 361ndash366
Andries K Verhasselt P Guillemont J Goumlhlmann HW Neefs JM Winkler H Van Gestel J Timmerman P Zhu M Lee E et al (2005) A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis Science 307 223-227
Artzatbanov VY and Petrov VV (1990) Branched respiratory chain in aerobically grown Staphylococcus aureus--oxidation of ethanol by cells and protoplasts Arch Microbiol 153 580- 584 Archer GL (1998) Staphylococcus aureus a well-armed pathogen Clin Infect Dis 26 1179ndash1181 Assefa S Keane TM Otto TD Newbold C and Berriman M (2009) ABACAS algorithm-based automatic contiguation of assembled sequences Bioinformatics 25 1968-1969 Aubry-Damon H Legrand P Brun-Buisson C Astier A Soussy CJ and Leclercq R (1997) Reemergence of gentamicin-sus- ceptible strains of methicillin-resistant Staphylococcus aureus roles of an infection control program and changes in aminoglycoside use Clin Infect Dis 25 647-653
Balemans W Vranckx L Lounis N Pop O Guillemont J Vergauwen K Mol S Gilissen R Motte M Lanccedilois et al (2012) Novel antibiotics targeting respiratory ATP synthesis in Gram-positive pathogenic bacteria Antimicrob Agents Chemother 56 4131-4139
Batandier C Guigas B Detaille D El-Mir MY Fontaine E Rigoulet M and Leverve XM (2006) The ROS production induced by a reverse-electron flux at respiratory-chain complex 1 is hampered by metformin J Bioenerg Biomembr 38 33ndash 42
132
Beckman Coulter website httpswwwbeckmancoultercomwsrportalwsrresearch-and-discoveryproducts-and-servicesnucleic-acid-sample-preparationspri-laboratory-automationindexhtm (consulteacute le 3 Deacutec 2013) Bednarek P and Osbourn A (2009) Plant-microbe interactions chemical diversity in plant defense Science 324 746ndash748 Besier S Ludwig A Ohlsen K Brade V and Wichelhaus TA (2007) Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus Int J Med Microbiol 297 217-225 BiteSizeBio Brain food for biologists httpnxseqbitesizebiocomarticlessequencing-by-synthesis-explaining-the-illumina-sequencing-technology (consulteacute le 3 deacutec 2013) Boetzer M and Pirovano W (2012) Toward almost closed genomes with GapFiller Genome Biology 13 R56 Bouarab K Ordi E Gattuso M Moisan H and Malouin F (2007) Plant stress response agents affect Staphylococcus aureus virulence genes Abstr 47th Intersci Conf Antimicrob Agents Chemother abstr C1-1483 Bryan LE and Kwan S (1981) Aminoglycoside-resistant mutants of Pseudomonas aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob Agents Chemother 19 958- 964 Bryan LE and Kwan S (1983) Roles of ribosomal binding membrane potential and electron transport in bacterial uptake of streptomycin and gentamicin Antimicrob Agents Chemother 23 835ndash845
Bryan LE Nicas T Holloway BW and Crowther C (1980) Aminoglycoside-resistant mutation of Pseudomonas aeruginosa defective in cytochrome C552 and nitrate reductase Antimicrob Agents Chemother 17 71- 79
133
Bryan LE and van den Elzen HM (1977) Effects of membrane-energy mutations and cations on streptomycin and gentamicin accumulation by bacteria a model for entry of streptomycin and gentamicin in susceptible and resistant bacteria Antimicrob Agents Chemother 12 163-177 Brynildsen MP Winkler JA Spina CS MacDonald IC and Collins JJ (2013) Potentiating antibacterial activity by predictably enhancing endogenous microbial ROS production Nat Biotechnol 31 160ndash 165 Burke KA and Lasccelles J (1975) Nitrate reductase system in Staphylococcus aureus wild type and mutants J Bacteriol 123 308- 316 Burke KA and Lascelles J (1976) Nitrate reductase activity in hegraveme-deficient mutants of Staphylococcus aureus J Bacteriol 126 225- 231 Canfield GS Schwingel JM Foley MH Vore KL Boonanantanasarn K Gill AL Sutton MD and Gill SR (2013) Evolution in fast forward a potential role for mutators in accelerating Staphylococcus aureus pathoadaptation J Bacteriol 195 615-628 Chang S Sievert DM Hageman JC Boulton ML Tenover FC Downes FP Shah S Rudrik JT Pupp GR Brown WJ et al (2003) Infection with vancomycin- resistant Staphylococcus aureus containing the vanA resistance gene N Engl J Med 348 1342-1347 Chen Q Vazquez EJ Moghaddas S Hoppel CL and Lesnefsky EJ (2003) Production of reactive oxygen species by mitochondria central role of complex III J Biol Chem 278 36027-36031 Clements MO Watson SP Poole RK and Foster SJ (1999) CtaA of Staphylococcus aureus is required for starvation survival recovery and cytochrome biosynthesis J Bacteriol 181 501- 507
134
Clinical and Laboratory Standards Institute (2011) Performance standards for antimicrobial susceptibility testing Twenty-first informational supplement M100-S21 CLSI Wayne PA USA ClustalW2 httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2 (consulteacute le 3 deacutec 2013) Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) httpcmrjcviorgtigr-scriptsCMRCmrHomePagecgi (consulteacute le 3 deacutec 2013) Creacutepin-Gilbert C (2006) Transfert de ligand dans la cytochrome c oxydase observeacute par des expeacuteriences femtosecondes infrarouge inteacutegreacutees et reacutesolues spectralement Thegravese de doctorat de l rsquoEacutecole Polytechnique Paris Pastel-00002200 version 1 29 jul 2010 Davis BD (1987) Mechanism of bactericidal action of aminoglycosides Microbiol Rev 51 341-350 DeAngelis MM Wang DG and Hawkins TL (1995) Solid-phase reversible immobilization for the isolation of PCR products Nucl Acids Res 22 4742- 4743 Dickgiesser N and Kreiswirth BN (1986) Determination of aminoglycoside resistance in Staphylococcus aureus by DNA hybridization Antimicrob Agents Chemother 29 930-932 Doi Y and Arakawa Y (2007) 16S ribosomal RNA methylation Emerging resistance mechanism against aminoglycosides Clin Infect Dis 45 88ndash94 Enright MC Robinson DA Randle G Feil EJ Grundmann H and Spratt BG (2002) The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA) Proc Natl Acad Sci USA 99 7687-7692 Friedman M (2002) Tomato glycoalkaloids role in the plant and in the diet J Agric Food Chem 50 5751ndash5780
135
Goerke C and Wolz C (2004) Regulatory and genomic plasticity of Staph- ylococcus aureus during persistent colonization and infection Int J Med Microbiol 294 195ndash202 Goumltz F Bannerman T and Schleifer K-H (2006) The genera Staphylococcus and Macrococcus In The Prokaryotes Vol 4 Bacteria Firmicutes Cyanobacteria M Dworkin (ed) (Springer New-York USA) pp5- 75 Haagsma AC Abdillahi-Ibrahim R Wagner MJ Krab K Vergauwen K Guillemont J AndriesK Lill H Koul A and Bald D (2009) Selectivity of TMC207 towards mycobacterial ATP synthase compared with that towards the eukaryotic homologue Antimicrob Agents Chemother 53 1290-1292 Hammer ND Reniere ML Cassat JE Zhang Y Hirsch AO Indriati Hood M and Skaar EP (2013) Two hegraveme-dependent terminal oxydases power Staphylococcus aureus organ-specific colonization of the vertebrate host MBio 4 e00241- 13 Hederstedt L (1993) The Krebs citric acid cycle In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of Microbiology Washington DC) pp 181-198 Hiramatsu K (1995) Molecular evolution of MRSA Microbiol Immunol 39 531-543 Hiramatsu K Hanaki H Ino T Yabuta K Oguri T and Tenover FC (1997) Methicillin-resistant Staphylococcus aureus clinical strain with reduced vancomycin susceptibility J Antimicrob Chemother 40 135-136 Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T et al (2013) Genomic Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
136
Hong S and Pedersen PL (2008) ATP synthase and the actions of inhibitors utilized to study its roles in human health disease and other scientific areas Microbiol Mol Biol Rev 72 590-641
Jevons MP (1961) lsquoCelbeninrsquo-resistant staphylococci BMJ 1 124-125
Joshi S and Huang YG (1991) ATP synthase complex from bovine heart mitochondria the oligomycin sensitivity conferring protein is essential for dicyclohexyl carbodiimidesensitive ATPase Biochim Biophys Acta 1067 255ndash258
Kohanski MA Dwyer DJ and Collins JJ (2010) How antibiotics kill bacteria from targets to networks Nat Rev Microbiol 8 423-435 Kohler C von Eiff C Liebeke M McNamara PJ Lalk M Proctor RA Hecker M and Engelmann S (2008) A defect in menadione biosynthesis induces global changes in gene expression in Staphylococcus aureus J Bacteriol 190 6351- 6364 Kohler C von Eiff C Peters G Proctor RA Hecker M and Engelmann S (2003) Physiological characterization of a hegraveme-deficient mutant of Staphylococcus aureus by a proteomic approach J Bacteriol 185 6928- 6937 Kriegeskorte A Koumlnig S Sander G Pirkl A Mahabir E Proctor RA von Eiff C Peters G and Becker K (2011) Small colony variants of Staphylococcus aureus reveal distinct protein profiles Proteomics 11 2476-2490 Lannergard J von Eiff C Sander G Cordes T Seggewiss J Peters G Proctor RA Becker K and Hughes D (2008) Identification of the genetic basis for clinical menadione-auxotrophic small-colony variant isolates of Staphylococcus aureus Antimicrob Agents Chemother 52 4017-4022
Lauraeus M and Wikstrom M (1993) The terminal quinol oxydases of Bacillus subtilis have different energy conservation properties J Biol Chem 268 11470-11473
137
Li M Gong J Cottrill M Yu H de Lange C Burton J and Topp E (2003) Evaluation of QIAamp DNA Stool Mini Kit for ecological studies of gut microbiota J Microbiol Methods 54 13-20
Liu Y Fiskum G and Schubert D (2002) Generation of reactive oxygen species by the mitochondrial electron transport chain J Neurochem 80 780-787
Martin-Hernandez A M Dufresne M Hugouvieux V Melton R and Osbourn A (2000) Effects of targeted replacement of the tomatinase gene on the interaction of Septoria lycopersici with tomato plants Mol Plant Microbe Interact 13 1301ndash1311
Matteelli A Carvalho AC Dooley KE and Kritski A (2010) TMC207 the first compound of a new class of potent anti-tuberculosis drugs Future Microbiol 5 849-858
Matsuno-Yagi A and Hatefi Y (1993) Studies on the mechanism of oxidative phosphorylation ATP synthesis by submitochondrial particles inhibited at F0 by venturicidin and organotin compounds J Biol Chem 268 6168ndash6173
Miller MH Edberg SC Mandel LJ Behar CF and Steigbigel NH (1980) Gentamicin uptake in wild-type and aminoglycoside-resistant small-colony mutants of Staphylococcus aureus Antimicrob Agents Chemother 18 722- 729
Mingeot-Leclercq M-P Glupczynski Y and Tulkens PM (1999) Aminoglycosides Activity and Resistance Antimicrob Agents Chemother 43 727- 737
Mitchell G Brouillette Eacute Seacuteguin DL Asselin AE Jacob CL and Malouin F (2010) A role for sigma factor B in the emergence of Staphylococcus aureus small-colony variants and elevated biofilm production resulting from an exposure to aminoglycosides Microb Pathog 48 18-27
Mitchell G Gattuso M Bouarab K and Malouin F (2009) Tomatidine (TO) affects virulence regulators of prototypical Staphylococcus aureus (SA) and small colony variants (SCV) of cystic fibrosis (CF) patients Abstr 49th Intersci Conf Antimicrob Agents Chemother abstr C1-1341
138
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K and Malouin F (2011) Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic fibrosis airway epithelial cells Antimicrob Agents Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM and Malouin F (2011) Infection of polarized airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function and is influenced by NF-κB Infect Immun 79 3541-3551
Mitchell G Lafrance M Boulanger S Seacuteguin DL Guay I Gattuso M Marsault Eacute Bouarab K and Malouin F (2012) Tomatidine acts in synergy with aminoglycoside antibiotics against multiresistant Staphylococcus aureus and prevents virulence gene expression J Antimicrob Chemother 67 559-568
Mitchell G Lamontagne CA Brouillette Eacute Grondin G Talbot BG Grandbois M and Malouin F (2008) Staphylococcus aureus SigB activity promotes a strong fibronectin-bacterium interaction which may sustain host tissue colonization by small-colony variants isolated from cystic fibrosis patients Mol Microbiol 70 1540-1555
Mitchell G Malouin F 2012 Outcome and prevention of Pseudomonas aeruginosa-Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic Fibrosis Chap 8 pp 181-206 Ed D Sriramulu InTech Open Access Publisher ISBN 979-953-307-059-8
Mitchell G Seacuteguin DL Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S and Malouin F (2010) Staphylococcus aureus sigma B-dependent emergence of small-colony variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-heptylquinoline-N-oxide BMC Microbiol 30 10-33
Moisan H Brouillette Eacute Jacob CL Langlois-Beacutegin P Michaud S and Malouin F (2006) Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated from cystic fibrosis patients is influenced by SigB J Bacteriol 188 64-76
139
Moreno F Crisp C Jorgensen JH and Patterson JE (1995) Methicillin- resistant Staphylococcus aureus as a community organism Clin Infect Dis 21 1308-1312
Murphy MP (2009) How mitochondria produce reactive oxygen species Biochem J 417 1- 13
Novick RP (2003) Autoinduction and signal transduction in the regulation of staphylococcal virulence Mol Microbiol 48 1429-1449
Novo DJ Perlmutter NG Hunt RH and Shapiro HM (2000) Multiparameter flow cytometric analysis of antibiotic effects on membrane potential membrane permeability and bacterial counts of Staphylococcus aureus and Micrococcus luteus Antimicrob Agents Chemother 44 827-834
ONeill AJ Cove JH and Chopra I (2001) Mutation frequencies for resistance to fusidic acid and rifampicin in Staphylococcus aureus J Antimicrob Chemother 47 647-650
Osbourn A E (1996) Preformed antimicrobial compounds and plant defense against fungal attack Plant Cell 8 1821ndash1831
Otto TD Dillon GP Degrave WS and Berriman M (2011) RATT Rapid annotation transfert tool Nucl Acids Res 39 doi101093 Pratique E Theraulaz L (2007) Eacutetude de la reacutegulation du systegraveme respiratoire TMAO reacuteductase chez Escherichia coli Meacutemoire de maitrise de lrsquoeacutecole pratique des hautes eacutetudes Paris
Price CT and Gustafson JE (2001) Increases in the mutation frequency at which fusidic acid-resistant Staphylococcus aureus arise with salicylate J Med Microbiol 50 104-106
Proctor RA and Peters G (1998) Small colony variants in staphylococcal infections Diagnostic and therapeutic implications Clin Infect Dis 27 419- 423
140
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M and Peters G (2006) Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent infections Nat Rev Microbiol 4 295ndash305
Quail MA Kozarewa I Smith F Scally A Stephens PJ Durbin R Swerdlow H and Turner DJ (2008) A large genome centers improvements to the Illumina sequencing system Nat methods 5 1005-1010
Ramirez MS and Tolmasky ME (2010) Aminoglycoside modifying enzymes Drug Resist Updat 13 151ndash171
Rao M Streur TL Aldwell FE and Cook GM (2001) Intracellular pH regulation by Mycobacterium smegmatis and Mycobacterium bovis BCG Microbiology 147 1017-1024
Richardson DJ (2000) Bacterial respiration a flexible process for a changing environment Microbiology 146 551- 571
Riley TV Pearman JW and Rouse IL (1995) Changing epidemiology of methicillin-resistant Staphylococcus aureus in Western Australia Med J Aust 163 412-414
Roddick J G (1974) The steroidal glycoalkaloid α-tomatine Phytochemistry 13 25-36
Rodrigue S Materna AC Timberlake SC Blackburn MC Malmstrom RR et al(2010) Unlocking Short Read Sequencing for Metagenomics PLoS ONE 5(7) e11840doi101371journalpone0011840 Ruiz-Rubio M Alonso Peacuterez-Espinosa A Lairini K RoldAacuten-Arjona T Dipietro A and Anaya N (2001) Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi In Studies in natural products chemistry vol 25 A Rahman (ed) (Elsevier Oxford United Kingdom) pp 293ndash326
141
Samuelsen oslash Haukland HH Kahl BC von Eiff C Proctor RA Ulvatne H Sandvik K and Vorland LH (2005) Staphylococcus aureus small colony variants are resistant to the antimicrobial peptide lactoferricin B J Antimicrob Chemother 56 1126- 1129
Sandrock R W and Vanetten HD (1998) Fungal sensitivity to and enzymatic degradation of the phytoanticipin α-tomatine Phytopathology 88 137ndash143
Sendi P and Proctor RA (2009) Staphylococcus aureus as an intracellular pathogen the role of small colony variants Trends Microbiol 17 54ndash58
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C and Osbourn A (2006) Dual effects of plant steroidal alkaloids on Saccha- romyces cerevisiae Antimicrob Agents Chemother 50 2732ndash2740
Somerville GA and Proctor RA (2009) At the crossroads of bacterial metabolism and virulence factor synthesis in Staphylococci Microbiol Mol Biol Rev 73 233- 248
Starkov AA (2010) Measurement of mitochondrial ROS production Methods Mol Biol 648 245-255
Taber HW (1993) Respiratory chain In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of MicrobiologyWashington DC) pp 199-212
Taber HW and Morrison M (1964) Electron transport in Staphylococci Properties of a particle preparation from exponential phase Staphylococcus aureus Arch Biochem Biophys 105 367- 379
Taber HW et al(1987) Bacterial uptake of aminoglycoside antibiotics Microbiol Rev 51 439-457
Thoumlny-Meyer L (1997) Biogenesis of respiratory cytochromes in bacteria Microbiol Mol Biol Rev 61 337- 376
142
Tubby S Wilson M Wright JA Zhang P and Nair SP (2013) Staphylococcus aureus small colony variants are susceptible to light activated antimicrobial agents BMC Microbiol 13 201- 204
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D Roth J Proctor RA Becker K et al (2011) Staphylococcus aureus phenotype switching an effective bacterial strategy to escape host immune response and establish a chronic infection EMBO Mol Med 3 129ndash41
Turrens JF (2003) Mitochondrial formation of reactive oxygen species J Physiol 552 335ndash 344
Tynecka Z Szcześniak Z Malm A and Los R (1999) Energy conservation in aerobically grown Staphylococcus aureus Res Microbiol 150 555- 566
Udo EE Pearman JW and Grubb WB(1993) Genetic analysis of community isolates of methicillin-resistant Staphylococcus aureus in Western Australia J Hosp Infect 25 97-108
Vakulenko SB and Mobashery S (2003) Versatility of Aminoglycosides and Prospects for Their Future Clin Microbiol Rev 16 430-450
Voggu L Schlag S Biswas R Rosenstein R Rausch C and Goumltz F (2006) Microevolution of cytochrome bd oxydase in Staphylococci and its implication in resistance to respiratory toxins released by Pseudomonas J Bacteriol 188 8079- 8086
Wells PG Bhuller Y Chen CS Jeng W Kasapinovic S Kennedy JC Kim PM Laposa RR McCallum GP Nicol CJ et al (2005) Molecular and biochemical mechanisms in teratogenesis involving reactive oxygen species Toxicol Appl Pharmacol 207 354-366
143
Yan M Yu C Yang J and Ji Y (2011) The essential two-component system YhcSR is involved in regulation of the nitrate respiratory pathway of Staphylococcus aureus J Bacteriol 193 1799- 1805
Zhang T Luo Y Chen Y Li X and Yu J (2012) BIGrat a repeat resolver for pyrosequencing-based re-sequencing with Newbler BMC Res Notes 5 567
144

vi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE helliphelliphelliphellip 76
31 Obtention des mutants helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
32 Seacutequenccedilage et analyses bio-informatiques helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 79
321 Identification des mutations en lien avec la tomatidine helliphellip 89
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct helliphellip 90
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine helliphelliphelliphelliphellip 99
CHAPITRE 4 DISCUSSION helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 113
CHAPITRE 5 CONCLUSION ET PERSPECTIVES helliphelliphelliphelliphelliphelliphelliphelliphelliphellip 124
CONTRIBUTIONS SUPPLEacuteMENTAIRES helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 1 Application de brevet helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 127
Annexe 2 Article scientifique helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 129
laquo Unraveling the structure-activity relationship of tomatidine
a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureus raquo
BIBLIOGRAPHIE helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 131
vii
LISTE DES ABBREacuteVIATIONS
Adh Alcool deacuteshydrogeacutenase
ADN Acide deacutesoxyribo-nucleacuteique
ADP Adeacutenosine diphosphate
ARN Acide ribo-nucleacuteique
ATP Adeacutenosine triphosphate
CA-MRSA Community-associated MRSA
CMI concentration minimale inhibitrice
CoQ Coenzyme Q
Cyt Cytochrome
DMQ laquo demethylmenaquinone raquo
dTMP Deacutesoxythymidine monophosphate
dUMP Deacutesoxyuridine monophosphate
FADH2 Flavine adenine dinucleacuteotide
GEN gentamicine
HA-MRSA Healthcare-associated MRSA
Ldh Lactate deacuteshydrogeacutenase
ml millilitre
MQ meacutenaquinone
MRSA methycilin resistant Staphylococcus aureus
Mqo2 Quninol oxidoreacuteductase
NADH Nicotinamide adenine dinucleacuteotide (forme reacuteduite)
Pdh Pyruvate deacuteshydrogeacutenase
Pfl Pyruvate formate lyase
PyrD Dihydroorotate deacuteshydrogeacutenase
Q Quinone
Rif rifampicine
ROS laquo reactive oxygen species raquo espegravece reacuteactive de lrsquooxygegravene
viii
[S] Polysulfide
S aureus Staphylococcus aureus
SARM Staphylococcus aureus resistant agrave la meacutethycilline
SCV laquo small colony variants raquo
SHCHC 2-succinyl-6-hydroxy-24-cyclohexadiene-1-carboxylate syntase
TCA Tricarboxylic acid cycle
TCD tableau croiseacute dynamique
TO tomatidine UFC Uniteacute formatrice de colonies
UQ Ubiquinone
microg microgramme
ix
LISTE DES TABLEAUX
CHAPITRE 1 INTRODUCTION
1 Cibles et modes drsquoaction des principales classes drsquoantibiotique 27
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants (B) chez les mutants NewbouldΔhemB reacutesistants 95-96
CHAPITRE 4 DISCUSSION
1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 123
x
LISTE DES FIGURES
CHAPITRE 1 INTRODUCTION
1 Diffeacuterence entre les pheacutenotypes normaux et SCV de S aureus 3
2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes 6
3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale 8
4 Chaines respiratoires bacteacuteriennes contenant des cytochromes 9
5 Flexibiliteacute respiratoire (A) chez Paracoccus denitrificans (B) chez
Escherichia coli 10
6 Chaine de transport des eacutelectrons proposeacutee chez les staphylocoques 12
7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies 17
8 Modegravele de reacutegulation du cycle TCA 20
9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS 23
10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de O2-bull 24
11 Voies de formation et de deacutetoxification des ROS 25
12 Cineacutetique drsquoentreacutee des aminoglycosides 30
13 Sites de mutations de certains reacutesistants aux aminoglycosides 32
14 Structures chimiques de (A) tomatine (B) tomatidine et (C) Analogue
de tomatidine FC04-100 34
xi
CHAPITRE 3 MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
1 Graphiques des CMI pour chacun des passages pour (A) la
tomatidine sur NewbouldΔhemB (B) la tomatidine en preacutesence de
gentamicine sur NewbouldΔhemB (C) la gentamicine sur S aureus
ATCC 29213 (D) la gentamicine en preacutesence de tomatidine sur S
aureus ATCC 29213 77-79
2 Organigramme des mutants reacutesistants seacutequenceacutes 80-81
3 Eacutetapes du seacutequenccedilage Illumina 83
4 Meacutethode drsquoamplification par la meacutethode Illumina 85
5 Correction du programme RATT sur les annotations transfeacutereacutees 88
6 Exemple drsquoune partie du tableau compilation des mutations 89
7 Seacutelection des gegravenes drsquointeacuterecirct chez ATCC 29213 dans le TCD par
lrsquoutilisation de diffeacuterents filtres 91-92
8 Seacutelection des gegravenes drsquointeacuterecirct chez NewbouldΔhemB dans le TCD par
lrsquoutilisation de diffeacuterents filtres 93-94
9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct 97-98
10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants 100
11 Courbe de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison gentamicine (+TO) du premier et troisiegraveme niveau de
reacutesistance 101
12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B)
NewbouldΔhemB 102
xii
13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213 104
14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicinelorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC
29213 105
15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants
ATCC 29213 reacutesistants agrave la combinaison gentamicine (+TO) 106
16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la
tomatidine 107
17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la
tomatidine 108
18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB
reacutesistants agrave la combinaison tomatidine (+gentamicine) 109
19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane
Potential Kit (Invitrogen) 111
20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du
gegravene atpE de S aureus Newman 112
xiii
CHAPITRE 4 DISCUSSION
1 Site drsquoinhibition de lrsquoATP synthase par diffeacuterentes moleacutecules 114
2 Structure chimique de (A) TMC207 et (B) lrsquoanalogue anti Gram
positifs 115
3 Seacutequence proteacuteique de la proteacuteine AtpE chez diffeacuterentes espegraveces 118
4 Modeacutelisation de la structure tridimentionnelle de lrsquoATP synthase de S
aureus (atpE) montrant les acides amineacutes muteacutes chez les mutants
reacutesistants 121
- 1 -
CHAPITRE 1
INTRODUCTION
11 Staphylococcus aureus
111 Importance clinique
Staphylococcus aureus est une bacteacuterie agrave Gram positif anaeacuterobie facultative Crsquoest un
microorganisme opportuniste pouvant infecter diffeacuterents hocirctes diffeacuterents organes ou
sites drsquoinfection et ayant la capaciteacute de causer tant des infections aiguumles que
chroniques (Archer 1998 Goerke et Wolz 2004)
Staphylococcus aureus reacutesistant agrave la meacutethilline le SARM (ou laquo MRSA raquo) est une
souche de S aureus devenue reacutesistante aux β-Lactamines par lrsquoacquisition du gegravene
mecA dans son chromosome (Hiramatsu 1995) Le SARM a eacuteteacute isoleacute pour la
premiegravere fois en 1960 en Angleterre (Jevons 1961) et est devenu eacutepideacutemique agrave
travers le monde depuis les anneacutees 1970 Au moins trois geacutenotypes distincts eacutetaient
recenseacutes dans les anneacutees 1980 (Hiramatsu 1995) dont deux sont toujours preacutevalents
agrave travers le monde comme eacutetant des souches HA-MRSA (Healthcare-associated
MRSA) des souches acquises agrave lrsquohocircpital (Enright et al 2002) Ces souches ont
acquis des gegravenes de reacutesistance agrave pratiquement tous les antibiotiques introduits dans
lrsquoutilisation clinique depuis les 50 derniegraveres anneacutees mecircme agrave lrsquoantibiotique de dernier
ressort la vancomycine(Hiramitsu 1997 Chen et al 2003) Nous sommes
maintenant en manque drsquoagents theacuterapeutiques efficaces contre les SARM touchant
les personnes immunodeacuteprimeacutees qui sont dans les hocircpitaux En plus depuis le deacutebut
des anneacutees 1990 des clones de SARM dont plusieurs deacutemontraient un potentiel
pathogeacutenique supeacuterieur aux souches classiques de HA-MRSA retrouveacutees agrave lrsquohocircpital
- 2 -
sont maintenant retrouveacutes dans la communauteacute (CA-MRSA) agrave travers le monde (Udo
et al 1993 Riley et al 1995 Moreno et al 1995 Aubry-Damon et al 1997) et ont
la capaciteacute drsquoaffecter des personnes en santeacute en causant des infections aiguumles
(Hiramatsu et al 2013) Il apparaicirct donc urgent de trouver des drogues alternatives
afin de surmonter le nombre grandissant de souches de SARM
112 Pheacutenotypes normal et laquo small colony variants raquo (SCV)
1121 Diffeacuterences et similariteacutes
Les laquo small colony variants raquo (SCV) sont des bacteacuteries souvent isoleacutees lors des
infections chroniques comme crsquoest le cas des infections pulmonaires chez les
patients atteints de fibrose kystique mais aussi dans des cas drsquoosteacuteomyeacutelite
drsquoarthrite septique drsquoinfections de prothegraveses orthopeacutediques et lors de la mammite
bovine (Alexender et Hudson 2001 Moisan et al 2006 Proctor et al 2006) Les
SCV sont caracteacuteriseacutes par soit un meacutetabolisme oxydatif dysfonctionnel ou un manque
dans la biosynthegravese de la thymidine les deux amenant une alteacuteration dans
lrsquoexpression des facteurs de virulence une croissance plus lente et une diminution
de la pigmentation des colonies (Figure 1) (Proctor et al 2006) Cependant alors
que les souches normales reacutepriment lrsquoexpression des proteacuteines de surface et
expriment les exoproteacuteines lors de la phase stationnaire (Novick 2003) les SCV
expriment de faccedilon stable les gegravenes Sig-B deacutependants codant pour des proteacuteines de
surface telle les adheacutesines au lieu drsquoactiver le systegraveme agr deacutependant du quorum-
sensing et produire des exoproteacuteines (Moisan et al 2006) Ces diffeacuterences dans
lrsquoexpression des facteurs de virulence reacutesultent chez les SCV en une meilleure
habileteacute agrave adheacuterer aux composantes de lrsquohocircte (Mitchell et al 2008) et agrave former du
biofilm (Mitchell et al 2010a Mitchell et al 20010b) et seraient eacutegalement lieacutees agrave
lrsquohabileteacute des SCV agrave lrsquoinvasion et la persistance des cellules de lrsquohocircte (Proctor et al
2006 Sendi et Proctor 2009 Mitchell et al 2011b)
- 3 -
S aureus est donc un pathogegravene qui peut se preacutesenter sous deux pheacutenotypes qui
sont souvent retrouveacutes simultaneacutement lors drsquoune mecircme infection et qui ont le pouvoir
de reacuteverter drsquoun pheacutenotype agrave lrsquoautre selon les conditions du milieu (Figure 1)
(Tuchscherr et al 2011)
Le pheacutenotype dit rdquonormalrdquo est associeacute aux infections aiguumles agrave une forte production
de facteurs de virulence (ex heacutemolysine) et est associeacute agrave la phase de disseacutemination
de la bacteacuterie Lrsquoautre pheacutenotype le laquo small-colony variants raquo ou SCV est affecteacute
dans sa chaine respiratoire ce qui amegravene une croissance beaucoup plus lente des
colonies de 8 -10 fois plus petites que les prototypes et une faible susceptibiliteacute aux
aminoglycosides Les SCV sont plutocirct associeacutes aux infections chroniques agrave S aureus
de par leur capaciteacute agrave srsquointernaliser dans les cellules de lrsquohocircte leur forte capaciteacute agrave
produire du biofilm et leur capaciteacute agrave produire des facteurs de colonisation
(Tuchscherr et al 2011 Proctor et al 2006) (Figure 1)
Figure 1 Diffeacuterences entre les pheacutenotypes normaux et SCV de S aureus
S aureus agrave
pheacutenotype normal
Disseacutemination
amp
Infections aiguumles
Colonisation
amp
Infections persistantes
SCVs
(laquo small-colony
variants raquo)
Reacutevers
ion
pheacutenoty
piq
ue
Norm
al
SC
V
Croissance
Biofilm
Internalisationheacutemolyse
Mitchell G et Malouin F 2012
- 4 -
1122 La chaine de transport des eacutelectrons
Chaine respiratoire tableau geacuteneacuteral
Les chaines respiratoires bacteacuteriennes bien que tregraves variables dans leurs
composantes parmi les diffeacuterentes espegraveces sont toutes constitueacutees sur le mecircme
modegravele de base Elles sont composeacutees de trois composantes majeures soit
- les deacuteshydrogeacutenases primaires qui sont chargeacutees drsquooxyder des substrats
principalement les coenzymes NADH et FADH2 (issues du cycle de Krebs) (mais
aussi selon le cas le succinate le lactate lrsquohydrogegravene hellip) et de pomper deux
protons du cocircteacute externe de la membrane en plus de transfeacuterer deux eacutelectrons agrave un
groupement de quinones
- Les quinones sont des moleacutecules liposolubles non proteacuteiques mobiles dans la
membrane Elles servent drsquointermeacutediaires entre les deacuteshydrogeacutenases primaires et
les oxydases terminales pour le transport des eacutelectrons Il existe deux sortes de
quinones les benzoquinones dont font partie les ubiquinones et les
naphtoquinones dont font partie les meacutenaquinones Les bacteacuteries agrave Gram positif
ne possegravedent que des meacutenaquinones alors que les deux sont preacutesentes chez les
bacteacuteries agrave Gram neacutegatif (Hederstedt 1993 Pratique et Theraulaz 2007)
- Les oxidoreacuteductases terminales sont chargeacutees de reacuteduire lrsquoaccepteur final
drsquoeacutelectron lrsquooxygegravene eacutetant lrsquoaccepteur final privileacutegieacute
De faccedilon plus deacutetailleacutee la chaine respiratoire est composeacutee de cinq
complexes proteacuteiques principaux (Figure 2)
- complexe I NADH deacuteshydrogeacutenase ou NADH reacuteductase ou NADHCoQ
oxidoreacuteductase ou NADH-ubiquinone oxidoreacuteductase
- complexe II Succinate deacuteshydrogeacutenase
- complexe III Cytochrome bc1 complexe ou ubiquinol cytochrome c
reacuteductaseoxydoreacuteductase
- complexe IV Cytochrome c oxydase
- 5 -
- complexe V ATP synthase (Creacutepin-Gilbert et al 2006)
La phosphorylation oxydative est deacuteclencheacutee dans la chaine de complexes
proteacuteiques par la preacutesence drsquoO2 accepteur terminal privileacutegieacute drsquoeacutelectrons (ou drsquoautres
accepteurs finaux tel le nitrate) dans le milieu ainsi que par les eacutelectrons ameneacutes par
le NADH et le FADH2 au complexe I et II respectivement (Creacutepin-Gilbert et al 2006)
Les eacutelectrons vont ensuite transiter par le complexe III jusqursquoau complexe IV ou les
eacutelectrons seront utiliseacutes afin de reacuteduire lrsquooxygegravene en eau le complexe IV est
lrsquooxydase terminale consideacutereacutee comme eacutetant lrsquoaccepteur final drsquoeacutelectrons Le transfert
drsquoeacutelectrons est possible gracircce aux reacuteactions drsquooxydoreacuteduction en cascade dans les
diffeacuterents complexes proteacuteiques Ces reacuteactions fournissent de lrsquoeacutenergie libre utiliseacutee
par les complexes I III et IV afin de transfeacuterer des protons agrave lrsquoexteacuterieur de la cellule
(Creacutepin-Gilbert et al 2006) Le complexe V (ATP synthase) quant agrave lui utilise les
eacutelectrons pompeacutes agrave lrsquoexteacuterieur par les autres complexes comme force proton-motrice
afin de phosphoryler trois moleacutecules drsquoADP et ainsi produire trois ATP Le transport
des eacutelectrons entre les diffeacuterents complexes proteacuteiques est possible gracircce agrave la
preacutesence dans la membrane de deux types de moleacutecules liposolubles non proteacuteiques
mobiles les meacutenaquinones et le cytochrome c Les meacutenaquinones seules quinones
preacutesentes chez les bacteacuteries agrave Gram positif assurent le transport des eacutelectrons entre
le complexe I et le complexe III et entre le complexe II et III Le cytochrome c pour sa
part veacutehicule les eacutelectrons entre le complexe III et le complexe IV Le transport des
eacutelectrons srsquoeffectue de la composante ayant le plus faible potentiel drsquooxydoreacuteduction
vers celle ayant le plus fort potentiel
- 6 -
La chaine respiratoire bacteacuterienne
La mitochondrie montre assez peu de flexibiliteacute respiratoire Il y a une certaine
flexibiliteacute au niveau de lrsquoentreacutee des eacutelectrons mais aucune au niveau de la sortie des
eacutelectrons ougrave la cytochrome aa3 oxydase reacuteduit lrsquooxygegravene en eau (Richardson 2000)
(Figure 3) Chez les bacteacuteries alors que le complexe II fonctionne sensiblement de
faccedilon identique agrave celui des eucaryotes les autres complexes peuvent se composer
de diffeacuterents complexes enzymatiques (reacuteductases) selon les espegraveces et les
conditions environnementales (Richardson 2000 Thoumlny-Meyer 1997) En outre
chez les bacteacuteries contrairement aux eucaryotes la chaine respiratoire comporte
souvent des voies de transport des eacutelectrons alternatives on dit alors qursquoelles ont une
chaine respiratoire agrave branches ou brancheacutee ou ramifieacutee (Artzatbanov et Petrov 1990)
Plusieurs espegraveces bacteacuteriennes possegravedent une chaine respiratoire ramifieacutee dont la
ramification se situe habituellement suite agrave la reacuteduction des quinones en quinols
(Lauraeus et Wikstrom 1993 Thoumlny-Meyer 1997) (Figure 4) Une des deux
Complexe I NADH
deacuteshydrogeacutenase
Complexe II Succinate
deacuteshydrogeacutenase
Complexe III
bc 1
Complexe IV Cytochrome c
oxydase
Complexe V
ATP synthase
Modifieacute de wikipedia httpfrwikipediaorgwikiChaine_respiratoire
Figure 2 Chaine de transport des eacutelectrons et ses diffeacuterents complexes
Leurs rocircles dans le transport des eacutelectrons et dans lrsquoeacutelaboration du gradient de
protons et le lien entre la chaine respiratoire et le cycle de Krebs Q Quinones
- 7 -
branches connue comme la branche cytochrome c oxyde les quinols (meacutenaquinol
QH2) via la voie complexe bc1-cytochrome c- cytochrome c oxydase Lrsquoautre
branche-type consiste en une seule uniteacute enzymatique (cytochrome bd) laquelle
transfegravere directement les eacutelectrons des quinols agrave lrsquooxygegravene Cette voie est connue
comme la branche quinol oxydase il srsquoagit drsquoune oxydase terminale transfeacuterant
directement les eacutelectrons agrave lrsquooxygegravene eacutevitant de passer par lrsquointermeacutediaire du
complexe III (bc1) et du cytochrome c (Artzatbanov et Petrov 1990 Lauraeus et
Wikstrom 1993) Alors que la voie du cytochrome c permet le pompage de protons agrave
lrsquoexteacuterieur par lrsquoaction du complexe bc1 (ou complexe III) et du cytochrome c oxydase
le cytochrome bd ne permet aucun pompage de protons (Lauraeus et Wikstrom
1993) La voie passant par le complexe bc1-cytochrome c-cytochrome c oxydase est
plus efficace dans la production drsquoeacutenergie Alors que les 2 voies peuvent ecirctre
exprimeacutees simultaneacutement chez quelques espegraveces il semble que lrsquoexpression de ces
voies soit inductible selon les conditions environnementales chez Bacillus subtilis et
les Staphylocoques (Lauraeus et Wikstrom 1993) On pourrait dire eacutegalement que la
chaine respiratoire est seacutepareacutee en deux eacutetapes la premiegravere partie est la phase
quinone-reacuteductase (Q-QH2) et la seconde la phase quinol-oxydase (QH2-Q) menant
agrave lrsquooxydase terminale La phase de reacuteduction est responsable du transfert des
eacutelectrons drsquoune varieacuteteacute de substrats tels le NADH le succinate le lactate le formate
et lrsquohydrogegravene vers les quinones et inclut diffeacuterentes reacuteductases (Thoumlny-Meyer 1997)
(Figure 4)
- 8 -
Figure 3 Reacutesumeacute de la chaine de transport des eacutelectrons mitochondriale UQ ubiquinone UQH2 ubiquinol Cyt cytochrome Il semble y avoir presque autant de chaines de transport des eacutelectrons diffeacuterentes
qursquoil y a drsquoespegraveces bacteacuteriennes (par exemple Paracoccus denitrificans et Escherichia
coli (Figure 5)) De plus il ne semble pas y avoir consensus dans la litteacuterature sur
les noms attribueacutes aux oxydases terminales Les noms pour les oxydases terminales
deacutependent des types de cytochromes dont elles sont composeacutees Il ne semble donc
pas eacutevident de connaicirctre de faccedilon preacutecise les types de cytochromes qui font partie
des oxydases terminales Pour rajouter agrave la confusion les gegravenes et opeacuterons ne sont
pas toujours associeacutes agrave leur oxydase terminale dans la litteacuterature ce qui fait qursquoon ne
peut savoir de faccedilon certaine de quelle oxydase on parle Le Cytochrome aa3 peut
eacutegalement porter le nom de Cyt ba3 et le cytochrome bd peux eacutegalement porter les
noms de Cyt bo
Tireacute de Richardson DJ 2000
- 9 -
Succinate
Formate
H2
SDH
FDH
HYD
QQH 2 bc1 c
Reacuteductase alternatives
b
c
cd1c
cb
c
b
c
d
Siroheme
Siroheme
b
NO3-
NO2-
NO
TMAO
DMSOS2O3
2-
[S]
SO32-
Fumarate
Quinol oxidases
ba3bb3bo3d
bd
aa3
O2
Cyt c oxidases
aa3ba3baa3caa3cbb3cao
co
O2
Modifieacute de Thony-Mayer L 1997
NADH
NDH
Figure 4 Chaines respiratoires bacteacuteriennes contenant des cytochromes
Du cocircteacute gauche on retrouve la partie quinone (ubiquinone ou meacutenaquinone)-
reacuteductase Du cocircteacute droit on retrouve la partie quinol-oxydase avec les oxydases
terminales reacuteduisant lrsquoO2 ou lrsquoaccepteur terminal alternatif tel que montreacute Les
oxydoreacuteductases sont surligneacutees QQH2 repreacutesente le pool de quinonesquinols
dans la membrane Les polysulfides sont repreacutesenteacutes par [S] Les chaines
respiratoires ne contenant pas de cytochromes ne sont pas montreacutees NDH
NADH deacuteshydrogeacutenase
Modifieacute de Thoumlny-Mayer L 1997
- 10 -
Figure 5 Flexibiliteacute respiratoire chez Paracoccus denitrificans (a) et chez
Escherichia coli (b) MQ menaquinone UQ ubiquinone DMQ
demethylmenoquinone
Chez S aureus
Comme plusieurs autres espegraveces bacteacuteriennes anaeacuterobies facultatives S aureus a
la possibiliteacute drsquoexprimer diffeacuterentes voies de synthegravese eacutenergeacutetique dont lrsquoexpression
est finement reacuteguleacutee selon des conditions environnementales du moment (Hammer et
Tireacute de Richardson DJ 2000
- 11 -
al 2013 Lauraeus et Wikstrom 1993) Mecircme si S aureus peut utiliser la
fermentation la respiration (aeacuterobie et anaeacuterobie) sera privileacutegieacutee par la bacteacuterie
puisque celle-ci amegravene une bien meilleure production drsquoeacutenergie (Goumltz et al 2006
Hammer et al 2013 ) La respiration aeacuterobie avec lrsquooxygegravene comme accepteur final
drsquoeacutelectron sera utiliseacutee de faccedilon preacutefeacuterentielle en preacutesence drsquooxygegravene (Hammer et al
2013) Malgreacute sa versatiliteacute dans ses voies de synthegravese eacutenergeacutetique S aureus et les
autres bacteacuteries agrave Gram positif ne produisent qursquoun seul type de quinone les
meacutenaquinones lesquelles sont utiliseacutees pour la croissance degraves qursquoil y a de lrsquooxygegravene
de preacutesent dans le milieu (Somerville et Proctor 2009) La reacuteduction de lrsquooxygegravene en
eau est meacutedieacutee par des oxydases terminales hegraveme deacutependant (Hammer et al 2013)
Selon certains auteurs S aureus possegravederait deux oxydases terminales la
cytochrome oxydase aa3 (ou ba3) (QoxABCD) et la cytochrome bd quinol oxydase
(CydAB) (Goumltz et al 2006 Hammer et al 2013 Kohler et al 2008) Cette derniegravere
est exclusive aux procaryotes et est une voie de contournement du complexe III qui
transmet directement les eacutelectrons du laquo pool raquo de quinol (QH2) agrave lrsquooxygegravene Selon
certains auteurs il y aurait eacutegalement chez S aureus une troisiegraveme oxydase
terminale le cytochrome o (CyoABCDE) (Thoumlny-Meyer 1997 Voggu et al 2006)
qui selon certain serait un cytochrome oxydase terminal alternatif au cytochrome aa3
et au cytochrome bd (Voggu et al 2006 Goumltz et al 2006 Taber et Morrison 1964)
(Figure 6) Ces voies alternatives permettent agrave S aureus de srsquoadapter rapidement agrave
un changement drsquoenvironnement (Hammer et al 2013) In vivo chaque organe est
un environnement diffeacuterent preacutesentant des deacutefis respiratoires distincts dus agrave la
variation dans 1- la quantiteacute drsquooxygegravene disponible 2- la disponibiliteacute drsquoaccepteurs
drsquoeacutelectrons alternatifs 3- la concentration drsquoinhibiteurs de la respiration dans le milieu
tels les oxydes nitriques (NO) produits par les neutrophiles et 4- la disponibiliteacute des
cofacteurs neacutecessaires agrave la production drsquoeacutenergie (Hammer et al 2013) Le
cytochrome est exprimeacute de faccedilon modeacutereacutee dans un milieu riche en oxygegravene mais est
fortement exprimeacute dans un milieu pauvre en oxygegravene mais riche en oxyde nitrique
De plus le type drsquooxydoreacuteductase terminale exprimeacute est influenceacute eacutegalement par la
- 12 -
source de carbone disponible la phase de croissance le pH extracellulaire et la
tension en oxygegravene (Lauraeus et Wikstrom 1993 Tynecka et al 1999)
La chaine respiratoire chez les SCV
S aureus est particuliegraverement doueacute agrave surpasser les deacutefis respiratoires en acqueacuterant
une reacutesistance agrave lrsquoacide nitrique produit par les neutrophiles et une habiliteacute agrave persister
dans les cellules de lrsquohocircte en se preacutesentant comme un variant agrave petites colonies
ayant une chaine respiratoire deacuteficiente (SCV) (Hammer et al 2013)
Deux principaux types de SCV sont retrouveacutes dans les isolats cliniques 1- les SCV
ayant une chaine de transport des eacutelectrons deacutefectueuse et 2- les SCV auxotrophes
pour la thymidine (Proctor et Peters 1998 Samuelsen et al 2005) Ces derniers
Figure 6 Chaine de transport des eacutelectrons proposeacutee chez les
staphylocoques Les informations disponibles sur les composantes de la chaine
respiratoire des staphylocoques suggegraverent une chaine ramifieacutee consistant en 2
oxydases terminales meacutenaquinol-deacutependante un cytochrome o et un cytochrome
aa3 oxydase NADH deacuteshydrogeacutenase (DHase) transmet les eacutelectrons au laquo pool raquo
de meacutenaquinones De lagrave les eacutelectrons sont transfeacutereacutes soit au cytochrome b ou agrave
lrsquoaccepteur alternatif drsquoeacutelectrons le cytochrome bd quinol oxydase Le cytochrome
b transfegravere les eacutelectrons agrave une des deux oxydases terminales Ces oxydases
terminales transfegraverent les eacutelectrons agrave lrsquooxygegravene
Modifieacute de Voggu L et al 2006
- 13 -
arborent une ou plusieurs mutations dans le gegravene thyA codant pour la thymidylate
synthase laquelle catalyse la derniegravere eacutetape dans la voie de biosynthegravese de la
thymidine crsquoest-agrave-dire la formation de dTMP agrave partir du dUMP (Samuelsen et al
2005) Le dTMP est essentiel pour la synthegravese de lrsquoADN (Besier et al 2007) Les
mutants thymidine-auxotrophes sont souvent la conseacutequence drsquoune pression
seacutelective par lrsquoutilisation prolongeacutee de trimethoprim-sulfamethoxazole (TMP-SMX)
(souvent utiliseacute pour le traitement des patients fibroses kystiques) un antibiotique qui
interfegravere dans la biosynthegravese de lrsquoacide tetrahydrofolique qui agit comme cofacteur de
la thymidylate synthase (Besier et al 2007 Lannergard et al 2008 Samuelsen et al
2005) Mecircme si de premiers abords les SCV thymidine-auxotrophes ne semblent pas
avoir de lien avec la chaine transport des eacutelectrons il se pourrait que celle-ci ait sont
importance chez ces SCV puisque lrsquoentreacutee de la thymidine passe par NupC dont le
fonctionnement neacutecessite un gradient eacutelectrochimique creacuteeacute par la chaine de transport
des eacutelectrons (Proctor et Peters 1998 ) Les caracteacuteristiques pheacutenotypiques des
SCV deacutecrites ci-apregraves sont toutes lieacutees agrave lrsquoalteacuteration de la chaine de transport des
eacutelectrons 1- la croissance plus lente parce que la synthegravese de la paroi cellulaire
neacutecessite une grande quantiteacute drsquoATP 2-la diminution de la formation de pigment
parce que la biosynthegravese des caroteacutenoiumldes requiert le transport des eacutelectrons 3- une
reacutesistance accrue agrave la gentamicine et autres aminoglycosides parce que lrsquoentreacutee de
ces composeacutes neacutecessite un grand potentiel membranaire geacuteneacutereacute par le transport des
eacutelectrons (Lannergard et al 2008) et 4- une perte partielle ou complegravete de la
fermentation du mannitol parce que lrsquoutilisation des sucres-alcool tels que le
mannitol est diminueacutee lorsque la chaine de transport des eacutelectrons ne peut oxyder le
NADH produit lorsque le mannitol est meacutetaboliseacute en fructose-6-phosphate (Kohler et
al 2003 Proctor et Peters 1998 Samuelsen et al 2005)
Les SCV posseacutedant une chaine de transport des eacutelectrons deacutefectueuse ont une voie
de biosynthegravese de lrsquoheacutemine ou de la meacutenadione deacuteficiente et le pheacutenotype est
reacuteversible lorsque compleacutementeacute avec lrsquoheacutemine ou la meacutenadione (Hammer et al
- 14 -
2013 Kohler et al 2008 Lannergard et al 2008 Proctor et Peters 1998) La
meacutenadione est essentielle agrave la biosynthegravese des meacutenaquinones et lrsquoheacutemine est
essentielle agrave la biosynthegravese de lrsquohegraveme qui entre dans formation des cytochromes
(Hammer et al 2013 Kohler et al 2008 Lannergard et al 2008 Thoumlny-Meyer
1997) Les meacutenaquinones et les cytochromes sont essentiels au fonctionnement de
la chaine de transport des eacutelectrons (vers lrsquoO2 autant que vers le nitrate) (Kohler et al
2008) Il se pourrait qursquoil existe eacutegalement des SCV causeacutes par drsquoautres deacuteficiences
telles que lrsquoalteacuteration de lrsquoATP synthase qui ne reacutesulterait pas drsquoune auxotrophie pour
lrsquoheacutemine ou la meacutenadione mais reacutesulterait tout de mecircme en une deacuteficience dans la
chaine de transport des eacutelectrons (Proctor et Peters 1998 )
Comme lrsquohegraveme est un cofacteur essentiel pour les oxydases terminales et que la
meacutenaquinone est un transporteur drsquoeacutelectrons utiliseacute par S aureus on comprend que
la faible croissance des mutants hemB et menD repreacutesente un pheacutenotype mimant la
perte des oxydases terminales (Hammer et al 2013) Les deux oxydases terminales
de S aureus QoxABCD (cyt aa3) et CydAB (cyt bd) semblent ecirctre diffeacuteremment
reacuteguleacutees chez les mutants menD et la souche sauvage alors que lrsquoopeacuteron cydAB est
fortement sous exprimeacute chez les mutants aucune diffeacuterence de niveau de
transcription de qoxABCD nrsquoa eacuteteacute observeacutee entre les mutants menD et leur souche
parente (Kohler et al 2008)
Des mutations dans les gegravenes menD hemB et ctaA peuvent reacutesulter en un
pheacutenotype SCV posseacutedant une chaine de transport des eacutelectrons deacuteficiente
(Clements et al 1999 Proctor et Peters 1998 ) Les gegravenes codant pour les eacutetapes
de formation de lrsquohegraveme se preacutesentent sous la forme drsquoun opeacuteron lrsquoopeacuteron
hemAXCDBL Chacun des gegravenes est essentiel (sauf hemL) et la deacuteleacutetion drsquoun seul
drsquoentre eux empecircchera la formation de lrsquohegraveme moleacutecule essentielle agrave la formation
des cytochromes (Taber 1993) hemB code pour la porphobilinogene synthase qui
catalyse une eacutetape dans la voie de biosynthegravese de lrsquohegraveme (Kohler et al 2008 Taber
- 15 -
1993 Thoumlny-Meyer 1997) Une mutation dans le gegravene ctaA (laquo hegraveme A synthase raquo)
empecircche la biosynthegravese de lrsquohegraveme A lequel est aussi impliqueacute dans la biosynthegravese
des cytochromes et donc altegraverera le fonctionnement de la chaine de transport des
eacutelectrons (Clements et al 1999 Proctor et Peters 1998) La capaciteacute de la bacteacuterie
agrave biosyntheacutetiser de lrsquohegraveme de faccedilon endogegravene est essentielle afin drsquoinitier une
infection systeacutemique Le gegravene menD code pour la 2-succinyl-6-hydroxy-24-
cyclohexadiene-1-carboxylate syntase (SHCHC) qui est une enzyme essentielle agrave la
voie de biosynthegravese des meacutenaquinones (Kohler et al 2008) De plus les
meacutenaquinones sont eacutegalement requises pour le fonctionnement de plusieurs
enzymes impliqueacutees dans des voies meacutetaboliques essentielles telles la malate quinol
oxidoreacuteductase (Mqo2) et la dihydroorotate deacuteshydrogeacutenase (PyrD) (Kohler et al
2008) Les mutants menD seront donc incapables de convertir lrsquooxaloaceacutetate en
malate ducirc agrave la perte de lrsquoactiviteacute du Mqo2 (Kohler et al 2008 Proctor et Peters
1998)
Les bacteacuteries agrave pheacutenotype SCV deacutemontreront eacutegalement un gradient eacutelectrochimique
nettement plus faible que les bacteacuteries agrave pheacutenotype normal de par lrsquoincapaciteacute de la
chaine de transport des eacutelectrons agrave y pourvoir
De par leur chaine de transport des eacutelectrons interrompue les SCV sont incapables
drsquoutiliser lrsquoO2 et le nitrate comme accepteur final drsquoeacutelectrons (Kohler et al 2008) Par
ce fait les proteacuteines impliqueacutees dans la voie de la glycolyse ainsi que dans la voie de
la fermentation sont surexprimeacutees (Pfl Ldh Adh) alors que les enzymes faisant
partie du cycle du citrate (TCA) sont significativement reacuteguleacutees agrave la baisse (Kohler et
al 2003 Kohler et al 2008 Kriegeskorte et al 2011) Ceci indique clairement que
les SCV (ex hemB) geacutenegraverent leur ATP agrave partir du glucose ou fructose seulement par
la phosphorylation des substrats Lrsquoanalyse de la reacuteaction de fermentation indique
que le produit principal est le lactate Les SCV reacutegeacutenegraverent leur NADH en reacuteduisant le
pyruvate en lactate par la voie fermentaire (Kohler et al 2003 Kohler et al 2008)
- 16 -
emmenant ainsi une acidification du milieu (Kriegeskorte et al 2011) La conversion
du pyruvate en aceacutetate via lrsquoacetyl-CoA est restreinte chez les mutants hemB mecircme
si la pyruvate formate lyase (Pfl) et les composantes du pyruvate deacuteshydrogeacutenase
sont preacutesentes (Figure 7) Lrsquoactiviteacute de la pyruvate deacuteshydrogeacutenase est tregraves faible
probablement en lien avec le haut niveau de NADH chez les SCV et il est preacutesumeacute
que la Pfl nrsquoest pas activeacutee en preacutesence drsquooxygegravene (La pyruvate deacuteshydrogeacutenase
converti le pyruvate en Co-A en condition anaeacuterobie) (Figure 7) Chez les mutants
hemB la voie de lrsquoarginine deacuteiminase est aussi induite et pourrait eacutegalement
participer agrave la production drsquoATP (Kohler et al 2003) Les gegravenes exprimeacutes chez les
mutants hemB sont similaires agrave ceux exprimeacutes chez les mutants menD ainsi que
chez les souches agrave pheacutenotypes normaux en condition anaeacuterobie (Kohler et al 2008)
- 17 -
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Glucose
Glucose-6-P
Fructose-6-P
Fructose-16-P
ATP
ATP
ADP
ADP
DHA-P Glyceraldehyde-3-P
Glycerate-13-P
Glycerate-3-P
Glycerate-2-P
Phosphoenolpyruvate
Pyruvate
Acetyl-CoA
TCA
cycle
Formate
AcetaldehydeEtOH
Lactate
Acetate
ATP
ADP
ATP
ADP
ATP
ADP
NADH2
NAD+Tpi
Pgi
FbaFda
Gap
PgK
Eno
Pyk
PflB
O2
Adh
NADH2NAD+
NADH2
NAD+
NADH2 NAD+
L-Ldh
Figure 7 Voie meacutetabolique des sucres chez les mutants hemB dans des
conditions aeacuterobies Les enzymes suivantes sont retrouveacutees agrave une forte
concentration chez les mutants hemb phosphoglucomutase (Pgi) fructose
bisphosphate aldolase (FbaFda) glyceacuteraldeacutehyde deacuteshydrogeacutenase (Gap)
phosphoglyceacuterate kinase (Pgk) eacutenolase (Eno) pyruvate kinase (Pyk) L-lactate
deacuteshydrogeacutenase (L-Ldh) pyruvate formate lyase (PflB) alcool deacuteshydrogeacutenase
(Adh) La formation drsquoaceacutetate et de formate et tregraves limiteacutee (X rouges)
Modifieacute de Kolher C et al 2003
- 18 -
La chaine respiratoire en anaeacuterobie (S aureus)
Il existe 3 voies distinctes de geacuteneacuteration drsquoeacutenergie chez les bacteacuteries 1- en preacutesence
drsquooxygegravene la bacteacuterie respire de faccedilon aeacuterobique et lrsquooxygegravene est lrsquoaccepteur final
drsquoeacutelectron 2- la bacteacuterie peut aussi respirer de maniegravere anaeacuterobique lorsqursquoun
accepteur terminal drsquoeacutelectron autre que lrsquooxygegravene est preacutesent 3- en absence
drsquooxygegravene et drsquoun accepteur drsquoeacutelectrons alternatif la bacteacuterie peut produire de
lrsquoeacutenergie de faccedilon limiteacutee par fermentation (Hammer et al 2013)
Chez S aureus le nitrate (NO3-) et dans un troisiegraveme temps le nitrite (NO2
-)
peuvent ecirctre utiliseacutes comme accepteur final drsquoeacutelectron alternatif lorsque lrsquooxygegravene
nrsquoest pas disponible (respiration anaeacuterobie) Les enzymes de cette voie meacutetabolique
ne sont retrouveacutees que chez les bacteacuteries (Goumltz et al 2006) La croissance
bacteacuterienne est consideacuterablement reacuteduite lorsque la bacteacuterie passe drsquoune respiration
aeacuterobie agrave une respiration anaeacuterobie La nitrate reacuteductase fonctionne probablement in
vivo gracircce au L-lactate comme principal donneur drsquohydrogegravene (Burke et Lasccelles
1975) La deacuteshydrogeacutenase primaire pour ce substrat est preacutesumeacutement la L-lactate
deacuteshydrogeacutenase associeacutee agrave la membrane laquelle meacutedie lrsquooxydation du lactate par la
chaine respiratoire aeacuterobique (Burke et Lasccelles 1975) La preacutedominance du
lactate parmi les produits finaux de la fermentation par S aureus est correacuteleacutee avec
son efficaciteacute comme principal donneur drsquohydrogegravene lorsque le nitrate est lrsquoaccepteur
final drsquoeacutelectrons Le transfert des eacutelectrons du Lactate vers le nitrate implique des
composantes qui sont eacutegalement preacutesentes dans la chaine respiratoire La nitrate
reacuteductase du type respiratoire est habituellement associeacutee agrave la membrane
cytoplasmique (Burke et Lasccelles 1975) Les reacuteductases sont inhibeacutees par
lrsquoazoture et le cyanure (Burke et Lascelles 1976)
Lrsquoenzyme membranaire nitrate reacuteductase NarGHI geacutenegravere des nitrites lesquels seront
reacuteduits en ammonium (NH4+) par lrsquoenzyme cytoplasmique NADH-deacutependant nitrite
reacuteductase encodeacutee par les gegravenes nirBD (Goumltz et al 2006) La nitrate reacuteductase 1-
- 19 -
est reacuteprimeacutee et inhibeacutee par la preacutesence drsquooxygegravene 2- est induite par lrsquoanaeacuterobiose et
la preacutesence de nitrate ou nitrite 3- nrsquoest pas inhibeacutee par lrsquoammoniac 4- permet la
production drsquoeacutenergie dans des conditions anaeacuterobies et 5- est localiseacutee dans la
membrane (Goumltz et al 2006) La preacutesence drsquoun systegraveme de senseur pour lrsquooxygegravene
est cruciale pour le controcircle transcriptionnel des gegravenes facilitant lrsquoadaptation de la
bacteacuterie agrave lrsquoanaeacuterobiose ou aux conditions microaeacuterophiles Chez S aureus il existe
deux systegravemes de senseur 1- le systegraveme SrrAB (pour staphylococcal respiratory
response AB synonyme SrhSR) (Figure 8) homologue agrave ResDE chez Bacillus
subtilis (Yan et al 2011) et 2- le systegraveme NreABC (pour nitrogen regulation chez S
carnosus correspondant agrave SA2181 (NreA) SA2180 (NreB) SA2179 (NreC) chez S
aureus) NreBC est un systegraveme agrave deux composantes qui mesure la concentration
drsquooxygegravene directement par un groupement Fe-S de type Fnr lequel est instable en
preacutesence drsquooxygegravene et par lequel est controcircleacute le systegraveme membranaire de reacuteduction
du nitratenitrite (Yan et al 2011) Srr est aussi un systegraveme agrave deux composantes qui
reacutegule positivement la lactate deacuteshydrogeacutenase et lrsquoalcool deacuteshydrogeacutenase et semble
reacuteguler neacutegativement plusieurs enzymes du cycle du citrate (TCA) (Figure 8)
Lrsquoexpression des enzymes fermentaires comme la lactate deacuteshydrogeacutenase (LDH)
pyruvate formate lyase (PFL) et lrsquoalcool deacuteshydrogeacutenase sont aussi fortement induite
dans des conditions aeacuterobies lorsque la chaine de transport des eacutelectrons est
interrompue (Kohler et al 2003) Donc il se peut que lrsquoeacutetat reacuteduit des composantes
de la chaine respiratoire le potentiel membranaire etou le niveau de NADH soit un
signal pour la reacutegulation des gegravenes laquo anaeacuterobies raquo chez S aureus Dans des
conditions aeacuterobies la conversion du pyruvate en aceacutetyl-coA est catalyseacutee par le
complexe pyruvate deacuteshydrogeacutenase (PDH) et caracteacuteriseacutee par la production de
NADH (Somerville et Proctor 2009) En condition anaeacuterobie lrsquoexpression de la
pyruvate deacuteshydrogeacutenase (PDH) est fortement reacuteprimeacutee alors que lrsquoexpression de la
pyruvate formate lyase (PFL) est fortement induite Lrsquoactiviteacute de la PDH est
remplaceacutee en anaeacuterobiose par lrsquoactiviteacute de la PFL qui geacutenegravere lrsquoacetyl-CoA et le
formate agrave partir du pyruvate Le NADH nrsquoeacutetant pas reacuteoxydeacute par une chaine de
- 20 -
transport des eacutelectrons fonctionnelle celui-ci est recycleacute par la reacuteduction du pyruvate
en lactate (Kohler et al 2003 Somerville et Proctor 2009)
La meacutenaquinone et lrsquohegraveme sont esssentiels agrave la respiration anaeacuterobie (Kohler et al
2008) Le pheacutenotype de S aureus en condition anaeacuterobie est tregraves similaire au
pheacutenotype SCV (Somerville et Proctor 2009)
Figure 8 Modegravele de reacutegulation du cycle TCA (Pour simplifier les reacutegulateurs de
facteurs de virulence bien caracteacuteriseacutes [ie RNAIII Rot SarA et σB] ont eacuteteacute omis
dans ce modegravele)
Les sous-produits de la phosphorylation oxydative les ROS
La reacuteduction de lrsquooxygegravene en eau se fait un eacutelectron agrave la fois (Turrens 2003) Le
complexe IV retient tous les intermeacutediaires partiellement reacuteduits jusqursquoagrave ce que la
Tireacute de Somerville GA Proctor RA 2009
- 21 -
reacuteduction soit complegravetement termineacutee (Turrens 2003) Cependant certains autres
centres redox de la chaine respiratoire laissent passer des eacutelectrons agrave lrsquooxygegravene
reacuteduisant partiellement cette moleacutecule en anion superoxyde (O2-bull) (Liu et al 2002
Turrens 2003) Mecircme si O2-bull nrsquoest pas un agent oxydant tregraves puissant il est le
preacutecurseur de plusieurs autres laquo reactive oxygen species raquo (ROS) et il est aussi
impliqueacute dans la propagation de la chaine de reacuteaction oxydative (Turrens 2003)
LrsquoO2-bull est un intermeacutediaire instable qui est rapidement converti en H2O2 par les
superoxide dismutase (SOD) (S aureus en possegravede deux SodA (le principal) et
SodM) (10 18) ou se convertit de faccedilon spontaneacutee lorsqursquoen concentration eacuteleveacutee
(Starkov 2010 Turrens 2003) Le H2O2 peut ensuite ecirctre complegravetement reacuteduit en
H2O ou partiellement reacuteduit en radical hydroxyle (bullOH) un des plus puissants
oxydants (Starkov 2010) LrsquoO2-bull peut eacutegalement reacuteagir avec drsquoautres radicaux
incluant lrsquooxyde nitrique (NObull) selon le taux de diffusion des 2 radicaux (Turrens
2003) Le peroxynitrite est eacutegalement un oxydant puissant (Turrens 2003) Les
oxydants deacuterivant du NObull sont appeleacutes des laquo reactive nitrogen species raquo (RNS)
(Turrens 2003) mais seront consideacutereacutes comme faisant partie des ROS dans ce
travail Les ROS peuvent endommager lrsquoADN lrsquoARN les proteacuteines et les lipides
reacutesultants agrave lrsquoapoptose de la cellule lorsque le niveau de ROS produit excegravede la
capaciteacute de deacutetoxifications et de reacuteparation de la cellule (Brynildsen et al 2013
Wells et al 2005) Malgreacute ce danger il srsquoagit drsquoun risque compenseacute par
lrsquoaugmentation du rendement eacutenergeacutetique (Brynildsen et al 2013) Le complexe I (et
II) et III sont les principaux sites de production de ROS (Batandier et al 2006 Chen
et al 2003 Liu et al 2002) Lrsquoinhibition du complexe I par la rotenone provoque une
augmentation de la production de ROS Lrsquooxydation des substrats du complexe I ou
du complexe II en preacutesence du complexe III inhibeacute par lrsquoantimicine A amegravene
eacutegalement une augmentation de la production de ROS (Batandier et al 2006 Chen
et al 2003) (Figure 9) De plus mecircme si le complexe cytochrome oxydase nrsquoest pas
un site de production de ROS son inhibition par le KCN (cyanure de potassium)
facilite la production de ROS par les complexes I et II (Chen et al 2003) Le
- 22 -
complexe ATP synthase nrsquoest pas reconnu pour produire directement des ROS
Cependant la liaison drsquoun inhibiteur tel lrsquooligomycine bloque le passage des protons
agrave travers le complexe et inhibe ainsi la production drsquoATP De plus lrsquoATP synthase ne
fonctionnant plus les pompes agrave protons de la chaine de transport des eacutelectrons sont
incapables de fonctionner car le gradient de protons est beaucoup trop eacuteleveacute ce
provoque une surproduction de ROS par les complexes I et III Conseacutequemment le
NADH nrsquoest plus oxydeacute et le cycle du TCA cesse drsquoopeacuterer car la concentration de
NAD nrsquoest plus suffisante pour le fonctionnement des enzymes du TCA (Joshi et
Huang 1991)
Une production significative de O2-bull principalement par le complexe I peut ecirctre le
reacutesultat de deux modes drsquoopeacuteration 1- lorsque la cellule ne produit pas drsquoATP et
conseacutequemment possegravede une grande force proton-motrice et possegravede un laquo pool raquo de
quinone agrave lrsquoeacutetat reacuteduit (QH2) 2- lorsque la cellule possegravede un haut ratio de
NADHNAD (Figure 10) Lorsque la cellule produit activement de lrsquoATP elle possegravede
alors une moins grande force proton-motrice son ratio NADHNAD est plus faible et
la formation de O2-bull est beaucoup plus faible (Murphy 2009)
Le H2O2 est le substrat pour deux enzymes la catalase et la glutathione (GSH)
peroxydase qui convertissent le H2O2 en H2O+O2 le H2O2 est ainsi deacutetoxifieacute (Figure
9 Figure 11) La reacuteaction catalyseacutee par la glutathione peroxydase neacutecessite de la
glutathione (GSH) comme substrats et deacutepend en partie du ratio de GSSG GSH et
de la concentration des reacuteactants laquelle est le reflet de lrsquoeacutetat reacutedox de la cellule De
faccedilon similaire la concentration de meacutetaux sous leur forme reacuteduite tel le Fer catalyse
la formation de certains ROS (Figure 11) Ceci peut ecirctre minimiseacute en tenant cette
concentration ions meacutetallique tregraves basse entre autres par leur emmagasinage et leur
liaison aux proteacuteines de transport (ei ferritine transferrine lactoferrine) et ainsi
minimiser la formation de bullOH Finalement certains antioxydants (eg vitamine E)
peuvent interrompre la chaine de reacuteactions par la capture des radicaux le radical-
vitamine E est relativement stable et peut ecirctre enzymatiquement reconverti agrave sa
- 23 -
forme non-radical (Figure 11) Il est maintenant accepteacute que certaines enzymes du
cycle de Krebs (ie αKGDH) soient non seulement des cibles des ROS mais aussi
des producteurs non neacutegligeables de ROS (Brynildsen et al 2013)
Figure 9 Portrait de la chaine de transport des eacutelectrons avec les sites de
production de ROS Le complexe I ainsi que le site Qi du complexe III relacircchent des
ROS directement vers les deacutefenses drsquoantioxydants intracellulaires Les antioxidants
intracellulaire tel la superoxide dismutase (MnSOD) la catalase la glutathione (GSH)
la glutathione peroxydase (GPX) et la glutathione reacuteductase (GRase) sont montreacute Le
site Qo du complexe III quant agrave lui relacircche les ROS dans le milieu extra cellulaire
Modifieacute de Chen Q et al 2003
Oligomycine
- 24 -
Figure 10 Meacutecanismes chez la mitochondrie qui megravenent agrave la production de
O2-bull 1- Le pool de NADH est dans un eacutetat reacuteduit par exemple par des dommages agrave
la chaine respiratoire la perte de cytochrome c durant lrsquoapoptose ou agrave un faible
besoin en ATP Ceci megravene agrave une production de O2-bull au complexe I qui deacutecoule de la
reacuteduction du FMN laquelle est reacuteguleacutee par le ratio NADHNAD+ Drsquoautres sites
autres que le complexe I tel lrsquoα-ceacutetoglutatrate deacuteshydrogeacutenase (αKGDH) peut
aussi contribuer agrave la production de O2-bull 2- Il nrsquoy a pas (peu) de production drsquoATP il y
a un fort potentiel membranaire et un pool de CoQ (meacutenaqinone) agrave lrsquoeacutetat reacuteduit
important qui provoque une reacuteversion de la chaine de transport des eacutelectrons agrave
travers le complexe I amenant une forte production de O2-bull 3- Il y a une production
active drsquoATP et conseacutequemment un potentiel membranaire plus bas que dans la
situation 2 et une plus grande quantiteacute de NADH oxydeacute que dans la situation 1
Dans ces conditions la quantiteacute de O2-bull produit est plus faible que dans la situation
1 et 2 et la provenance des O2-bull est moins certaine
Tireacute de Murphy MP 2009
- 25 -
12 Antibiotiques Mode drsquoaction cible cellulaire et meacutecanismes de reacutesistance
121 Diffeacuterentes classes drsquoantibiotiques et leurs cibles
Notre compreacutehension de la faccedilon que les antibiotiques induisent la mort cellulaire est
principalement baseacutee sur la fonction cellulaire essentielle inhibeacutee par lrsquointeraction de
lrsquoantibiotique avec sa cible Les antibiotiques peuvent ecirctre classeacutes selon la
composante ou le systegraveme cellulaire qursquoils affectent ainsi que par le fait qursquoils
induisent la mort cellulaire (antibiotiques bacteacutericides) ou srsquoils inhibent la croissance
Figure 11 Voies de formation et de deacutetoxification des ROS Tireacute de Wikipeacutedia
httpfrwikipediaorgwikiDC3A9rivC3A9_rC3A9actif_de_l27oxygC3A8ne
Consulteacute le 11 deacutec 2013
- 26 -
bacteacuterienne (antibiotique bacteacuteriostatique) La plupart des antibiotiques bacteacutericides
inhibent la synthegravese de lrsquoADN ou de lrsquoARN la synthegravese de la paroi bacteacuterienne ou la
synthegravese proteacuteique (Kohanski et al 2010) Le tableau 1 est une synthegravese des
principales classes drsquoantibiotiques ainsi que leur mode drsquoaction et leurs cibles
cellulaires Ce tableau nrsquoest cependant pas exhaustif et il existe plusieurs autres
classes drsquoantibiotiques et plusieurs autres exemples drsquoantibiotiques dans les classes
identifieacutees dans ce tableau
Depuis la deacutecouverte de la peacutenicilline en 1929 plusieurs autres antibiotiques ont eacuteteacute
deacutecouverts et deacuteveloppeacutes suite agrave lrsquoeacutelucidation des interactions entre lrsquoantibiotique et
sa cible et en apportant des modifications structurales et chimiques aux moleacutecules
deacutejagrave connues Ces efforts ont significativement augmenteacute le nombre drsquoantibiotiques
disponibles dans le milieu clinique Le processus de mort cellulaire induit par lrsquoaction
drsquoun antibiotique est complexe et deacutebute avec une interaction physique entre la
moleacutecule antibiotique et sa cible bacteacuterienne speacutecifique et implique des modifications
qui affectent la bacteacuterie tant au niveau biochimique que moleacuteculaire et structural
27
Acid
e n
alid
ixiq
ue (
1 iegravere
geacuteneacutera
tion)
Cip
rofloxacin
e (
2 iegravem
e g
eacuteneacutera
tion)
Levofloxacin
e (
3 iegravem
e g
eacuteneacutera
tion)
Gem
iflo
xacin
e (
4 iegravem
e g
eacuteneacutera
tion)
Flu
oro
quin
olo
ne (
Norf
loxacin
e)
Peacutenic
iline
Ceacutephalo
sporine (
ceacutefa
lexin
e)
Carb
apenem
Vancom
ycin
e
Teacutela
vancin
e
Fosfo
mycin
Dapto
mycin
Poly
mix
ine B
Bacitra
cin
Valin
om
ycin
e
Nom
encin
e
Genta
mic
in
Kanam
ycin
e
Tobra
mycin
e
Str
epto
mycin
e
Teacutetr
acyclin
e
Chlo
rteacutetr
acyclin
e
Macro
lides
Ery
thro
mycin
eLie
sous-u
niteacute 23S
du r
ibosom
e 5
0S
et em
pecircche
leacutelo
ngation d
u p
eptide (
apregrave
s 6
-8 a
a)
sta
tique
Lin
cosam
ides
Clin
dam
ycin
Inhib
e d
irecte
ment la
ctiviteacute p
eptidyl
transfeacute
rase d
u
ribosom
esta
tique
Str
epto
gra
min
es
DalfopristinQ
uin
upristin
Com
bin
ais
on d
e 2
antibio
tiques B
loque la
transla
tion p
eptidiq
ue e
n lia
nt le
site P
du r
ibosom
e
et en b
loquant au p
eptide la
ccegraves a
u tunnel de s
ort
ie
cid
al
Am
phenic
ol
Chlo
ram
phenic
ol
Em
pecircche la lia
ison d
u a
a-
AR
Nt au s
ite A
du
ribosom
e e
t in
terf
egravere
dans la f
orm
ation d
u lie
n
peptidiq
ue
sta
tique
Oxazolid
inones
Lin
ezolid
Blo
que initia
tion d
e la tra
nsla
tion p
roteacute
ique lors
de la
reacuteaction d
eacutelo
ngation p
ar
la p
eptidyl
transfeacute
rase
sta
tique
Trim
eth
oprim
Sulfam
eth
oxazole
sta
tique
cid
al
Ionophore
Alteacutera
tion d
e lin
teacutegriteacute
mem
bra
naire
Se
nchasse d
ans la m
em
bra
ne fo
rme u
n p
ore
ce
qui am
egravene u
ne p
ert
urb
ation d
e leacute
quili
bre
ioniq
ue
cid
al
Inhib
itio
n d
e la s
ynth
egravese d
e
la p
aro
i cellu
laire
(peptidogly
can)
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
cid
al
cid
al
sta
tique
AR
Nr
16S
(S
ite A
)(fo
rmation d
e p
roteacute
ines n
on-
fonctionnelle
s)
cid
al
Inhib
itio
n d
e la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 3
0S
)
Inhib
iteur
de la s
ynth
egravese d
e
lacid
e f
oliq
ue
Inhib
itio
n d
e c
ert
ain
es r
eacuteactions e
nzym
atiques
essentielle
s agrave
la f
orm
ation d
e la
cid
e f
oliq
ue
Sulfonam
ides
Inhib
itio
n d
e la tra
nslo
cation d
u p
reacutecurs
eur
et de s
on
utilis
ation
AR
Nr
16S
(preacute
vie
nt la
ttachem
ent de la
min
oacyl-
AR
Nt au r
ibosom
e)
Teacutetr
acyclin
e
Poly
peptides
β-L
acta
ms
Gly
copeptides
Lip
opeptides
Inhib
iteur
de la s
ynth
egravese
pro
teacuteiq
ue (
ribosom
e 5
0S
)
Am
inogly
cosid
es
Cib
le c
ell
ula
ire
Sta
tiq
ue
Cid
al
Rifam
ycin
eR
ifam
pic
ine
Cla
ss
es
Ex
d
an
tib
ioti
qu
eM
od
e d
ac
tio
n
Inhib
itio
n d
e la s
ynth
egravese d
e
lAR
N (
AD
N d
eacutependant)
AR
N p
oly
meacutera
se s
ous-u
niteacute β
(codeacute p
ar
rpoB
)
Quin
olo
nes
cid
al
Inhib
itio
n d
e la r
eacuteplic
ation d
e
lAD
N
Com
ple
xe A
DN
-topois
om
eacutera
se (
AD
N
gyr
ase(t
opois
om
eacutera
se II)
et to
pois
om
eacutera
se IV
)cid
al
Tra
nspeptidase (
penic
illin
bin
din
g p
rote
in
(PB
P))
cid
al
Tab
leau
1
Cib
les e
t m
od
e d
rsquoacti
on
des p
rin
cip
ale
s c
lasses d
rsquoan
tib
ioti
qu
es
28
122 Les aminoglycosides
1221 Mode drsquoaction
Les aminoglycosides peacutenegravetrent la paroi cellulaire en suivant un processus
comprenant 3 eacutetapes une premiegravere eacutetape eacutenergie- indeacutependante suivie de deux
eacutetapes eacutenergie deacutependantes Deux composantes sont essentielles pour
lrsquoaccumulation intracellulaire des aminoglycosides les ribosomes et la chaine de
transport des eacutelectrons (Ramirez et Tolmasky 2010)
Eacutetape 1- Lorsque les aminoglycosides entrent en contact avec la paroi
cellulaire la moleacutecule polycationique se lie aux composantes de surface anioniques
tels les lipopolysaccharides les phospholipides et autres proteacuteines membranaires
chez les bacteacuteries agrave Gram neacutegatif et les moleacutecules drsquoacides teacuteichoiumlques et les
phospholipides des bacteacuteries agrave Gram positif (Ramirez et Tolmasky 2010) Apregraves
cette liaison aux moleacutecules polaires les aminoglycosides (telle la gentamicine)
srsquoassocient aux transporteurs membranaires sur la base de leur charge positive et
sont transporteacutes agrave travers la membrane cytoplasmique gracircce au potentiel
membranaire (intra cellulaire neacutegatif) (Bryan et al 1980 Proctor et al 2006) Cette
liaison agrave la membrane entraine une augmentation de la permeacuteabiliteacute membranaire
menant agrave lrsquoentreacutee des aminoglycosides dans lrsquoespace peacuteriplasmique (Gram neacutegatif)
ou dans le cytoplasme (Gram positif) (Ramirez et Tolmasky 2010) (Figure 12A)
Eacutetape 2- Lrsquoeacutetape suivante connue comme la laquo phase eacutenergie-deacutependante
1 raquo (EDP-I) est bloqueacutee par les inhibiteurs de la chaine de transport des eacutelectrons et
de la phosphorylation oxydative ainsi que par les ions divalents lrsquohyperosmolariteacute le
faible pH et lrsquoanaeacuterobiose dans cette phase la dureacutee et le taux drsquoentreacutee de
lrsquoantibiotique sont en fonction de la concentration (Miller et al 1980 Mingeot-
Leclercq et al 1999 Ramirez et Tolmasky 2010) Agrave cette eacutetape le transport
transmembranaire des aminoglycosides et plus speacutecifiquement de la gentamicine
neacutecessite 2 composantes majeures a) un transporteur anionique non speacutecifique dont
la charge est directement lieacutee agrave lrsquoactiviteacute des cytochromes oxydases terminales et des
29
nitrates reacuteductases de la chaine de transport des eacutelectrons Selon certaines eacutetudes
les transporteurs pourraient ecirctre les quinones reacuteduites de la chaine de transport des
eacutelectrons (Bryan et al 1981 Miller et al 1980) et donc le cycle drsquooxydation-reacuteduction
des quinones est essentiel aux transports des aminoglycosides (Bryan et al 1981)
mais selon drsquoautres eacutetudes les transporteurs pourraient ecirctre toute autre proteacuteine non
deacutefinie impliqueacutee dans la polarisation membranaire lieacutee agrave lrsquoactiviteacute de la chaine de
transport des eacutelectrons (Miller et al 1980) La croissance anaeacuterobie ainsi que le
sodium azide (inhibiteur de la chaine respiratoire) inhibent lrsquoentreacutee de la gentamicine
et lrsquoeffet bacteacutericide indiquant que le transport actif de cet antibiotique neacutecessite la
respiration b) un potentiel eacutelectrique avec un milieu intra cellulaire neacutegatif par rapport
agrave lrsquoexteacuterieur En effet lrsquoinhibition de lrsquoentreacutee dans la bacteacuterie des aminoglycosides
suite agrave lrsquoajout de CCCP (agent deacutepolarisant) chez des souches de S aureus laquo wild-
type raquo deacutemontre la neacutecessiteacute de la force proton-motrice pour lrsquoentreacutee des
aminoglycosides dans les cellules (Miller et al 1980)
Eacutetape 3- La petite quantiteacute drsquoaminoglycosides srsquoeacutetant rendu au cytoplasme
durant cette premiegravere phase eacutenergie-deacutependante induisent apregraves liaison aux
ribosomes des erreurs dans la synthegravese proteacuteique Les erreurs dans la traduction
des proteacuteines membranaires produites lors de cette eacutetape connue comme la laquo phase
eacutenergie-deacutependante 2 raquo (EDP-II) provoquent des dommages dans lrsquointeacutegriteacute de la
membrane cytoplasmique (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky
2010) Les proteacuteines membranaires aberrantes dans la membrane facilitent le
transport drsquoune plus grande quantiteacute drsquoaminoglycosides lesquels augmenteront
encore le niveau drsquointerfeacuterence avec la synthegravese proteacuteique normale amenant encore
plus de dommage agrave la membrane provoquant ainsi une acceacuteleacuteration importante du
taux drsquoentreacutee des moleacutecules antibiotique et qui ultimement reacutesultera en la mort de la
cellule (Mingeot-Leclercq et al 1999 Ramirez et Tolmasky 2010) (Figure 12B)
Des eacutetudes ont deacutemontreacute que la susceptibiliteacute agrave la gentamicine streptomycine
amikacine kanamycine et tobramycine est supeacuterieure lorsque lrsquooxygegravene est
30
lrsquoaccepteur terminal drsquoeacutelectrons (Bryan et Kwan 1981) Une diminution significative
de la susceptibiliteacute est remarqueacutee lorsque le nitrate est lrsquoaccepteur final drsquoeacutelectrons et
une diminution encore plus marqueacutee est observeacutee lorsque le fumarate est lrsquoaccepteur
final drsquoeacutelectron ou lorsque les conditions sont fermentaires Lrsquoassociation entre le
transport de la gentamicine et le complexe terminal de la chaine de transport des
eacutelectrons particuliegraverement lrsquoactiviteacute cytochrome oxydase
Le ribosome possegravede trois sites de liaison importants de lrsquoARNt deacutesigneacute comme site
A (pour aminoacyl) site P (pour peptidyl) et E (pour exit) (Vakulenko et Mobashery
2003) Ils sont situeacutes sur lrsquoARN16S (lequel avec plus de 21 proteacuteines compose la
sous-uniteacute 30S du ribosome bacteacuterien) (Ramirez et Tolmasky 2010 Vakulenko et
Mobashery 2003) Durant la synthegravese proteacuteique le ribosome deacutecode lrsquoinformation de
lrsquoARNm et catalyse lrsquoincorporation seacutequentielle de chaque acide amineacute faisant partie
de la chaine polypeptidique en formation La haute fideacuteliteacute de la traduction (les
ribosomes incorporent un mauvais acide amineacute par approximativement 3000 liaisons
peptidiques effectueacutees) est possible gracircce agrave la capaciteacute de discriminer entre des
changements conformationels dans le ribosome induit par la liaison drsquoun ARNt correct
Modifieacute de Davis BD 1987
Figure 12 Cineacutetique drsquoentreacutee des aminoglycosides
A
B
31
ou incorrect au site A La reconnaissance drsquoune liaison correcte anticodoncodon par
le ribosome se passe en deux temps la reconnaissance initiale de lrsquoappariement du
codon sur lrsquoARNm avec lrsquoanticodon de lrsquoARNt et subseacutequemment la correction
(Vakulenko et Mobashery 2003)
La plupart des aminoglycosides se lient agrave des oligonucleacuteotides faisant partie du site
A Les diffeacuterents aminoglycosides ne lient pas tous le mecircme site de lrsquoARN 16S mais
lrsquoeffet commun agrave tous est que cette liaison amegravene un changement de conformation du
site A vers une conformation mimant la conformation fermeacutee de la sous uniteacute 30S qui
est induite par une interaction entre lrsquoARNm et son ARNt apparenteacute eacuteliminant la
possibiliteacute de correction du ribosome ce qui promeut les erreurs de traduction
(Ramirez et Tolmasky 2010 Vakulenko et Mobashery 2003) La grande majoriteacute des
aminoglycosides sont bacteacutericides et cette leacutetaliteacute est la conseacutequence de cette
mauvaise traduction (Vakulenko et Mobashery 2003)
1222 Meacutecanismes de reacutesistance
La reacutesistance contre les aminoglycosides se produit par diffeacuterents meacutecanismes qui
peuvent coexister simultaneacutement dans la mecircme bacteacuterie (Vakulenko et Mobashery
2003) Ces meacutecanismes incluent 1- la modification de la cible par des mutations
dans lrsquoARN 16S ou autres proteacuteines ribosomales associeacutees (Figure 13) 2- la
meacutethylation de lrsquoARN 16S un meacutecanisme retrouveacute chez plusieurs organismes
producteurs drsquoaminoglycosides et chez des souches cliniques Cette meacutethylation
empecircche les aminoglycosides drsquoavoir accegraves agrave leur site de liaison (Doi et Arakawa
2007) 3- la reacuteduction de la permeacuteabiliteacute par une alteacuteration de la permeacuteabiliteacute de la
membrane externe ou une diminution du transport agrave travers la membrane interne 4-
lrsquoexportation des aminoglycosides agrave lrsquoexteacuterieur de la cellule par les pompes agrave efflux
Eacutetant des moleacutecules hydrophiliques les aminoglycosides ont naturellement peu
drsquoaffiniteacute avec les pompes agrave efflux 5- le swarming actif probablement un meacutecanisme
32
non speacutecifique deacutemontreacute chez Pseudomonas aeruginosa lequel deacutemontre des
reacutesistances agrave plusieurs antibiotiques 6- la seacutequestration de la drogue par des liaisons
agrave un aceacutetyltransfeacuterase agrave tregraves faible activiteacute 7- une inactivation enzymatique de la
moleacutecule antibiotique le meacutecanisme de reacutesistance le plus preacutevalent au niveau
clinique (Vakulenko et Mobashery 2003) Lrsquoinactivation enzymatique des
aminoglycosides est le plus souvent due agrave des aceacutetyltransfeacuterases des
nucleacuteotidyltransfeacuterases ou des phosphotransfeacuterases (Dickgiesser et Kreiswirth 1986)
Ces enzymes sont souvent encodeacutees sur des plasmides mais sont eacutegalement
associeacutees agrave des eacuteleacutements transposables Lrsquoeacutechange de plasmides et la disseacutemination
de transposons facilitent lrsquoacquisition rapide de pheacutenotypes reacutesistants aux
antibiotiques non seulement au sein drsquoune mecircme espegravece mais parmi un large
eacuteventail drsquoespegraveces bacteacuteriennes (Mingeot-Leclercq et al 1999)
La reacutesistance naturelle des S aureus laquo small colony variants raquo agrave la majoriteacute des
aminoglycosides est la conseacutequence de lrsquoimpossibiliteacute agrave lrsquoantibiotique de peacuteneacutetrer une
cellule posseacutedant une faible polariteacute membranaire qui est due chez les SCV agrave leur
Tireacute de Doi Y Arakawa Y 2007
Figure 13 Sites de mutation de certains reacutesistants aux aminoglycosides
33
incapaciteacute agrave activer le cycle de Krebs (Miller et al 1980 Proctor et al 2006 Tubby
et al 2013) Les SCV ont non seulement une polariteacute membranaire faible mais (ceci
eacutetant la conseacutequence de cela) aussi une force proton motrice inhibeacutee et une chaine
des eacutelectrons deacuteficiente De ce fait le transport transmembranaire des
aminoglycosides chez les SCV est grandement reacuteduit et conseacutequemment la
susceptibiliteacute des SCVs pour cette classe drsquoantibiotique est 110-113 de celle des
souches parentes (Proctor et al 2006)
123 La tomatidine
Plusieurs espegraveces veacutegeacutetales accumulent des steacuterols et des triterpegravenes antimicrobiens
appeleacutes saponines (Osborn 1996) Ces moleacutecules sont produites de faccedilon
constitutive par la plante et jouent un important rocircle de deacutefense contre diffeacuterents
pathogegravenes comme des insectes ou des microbes Parmi les saponines la tomatine
(Figure 14A) est un glycoalcaloiumlde steacuteroiumldien preacutesentant une action antimicrobienne
contre des levures et une quantiteacute drsquoautres microorganismes (Bednarek et Osboum
2009 Friedman 2002 Osborn 1996 Sandrock et Vanetten 1998) Plusieurs
espegraveces de champignons pathogegravenes de la tomate produisent des enzymes
extracellulaires communeacutement appeleacutes tomatinase qui sont capables de deacutetoxifier
lrsquoα-tomatine (Martin-Hernandez et al 2000 Roddick 1974 Ruiz-Rubbio et al 2001)
Par exemple Fusarium oxysporum f sp lycopersici est reconnu pour cliver lrsquoα-
tomatine en ses sous-produits soit un tetrasaccharide (lycotreacutetraose) et un alcaloiumlde
steacuteroiumldien non glycosyleacute la tomatidine (Figure 14B) Ces sous-produits nrsquoont pas ou
peu drsquoactiviteacute antifongique (Ruiz-Rubbio et al 2001 Simon et al 2006)
34
Figure 14 Structures chimiques de la (A) tomatine de (B) tomatidine et de (C)
lrsquoanalogue de tomatidine FC04-100
Il a eacuteteacute deacutecouvert il a quelques anneacutees que la tomatidine avait un effet antibacteacuterien
sur les S aureus de pheacutenotype SCV mais aucune activiteacute contre les S aureus agrave
pheacutenotype normal Par contre lorsqursquoutiliseacute en combinaison avec les
aminoglycosides lrsquoajout de la tomatidine permettait drsquoaugmenter de 8 agrave 16 fois
lrsquoactiviteacute des aminoglycosides contre S aureus agrave pheacutenotype normal (Mitchell et al
2011a Mitchell et al 2012)
Donc on savait que la tomatidine avait une activiteacute antibacteacuterienne tregraves inteacuteressante
mais on ne connaissait ni sa cible ni son mode drsquoaction De plus si la tomatidine a
une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait donc avoir un
spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Par ailleurs de faccedilon parallegravele la production drsquoanalogues structuraux de la tomatidine
permettrait de mieux comprendre la relation structure-activiteacute (SAR) de la moleacutecule et
donc de preacuteciser que lrsquoactiviteacute biologique de la tomatidine neacutecessite une orientation
3
A B
C
35
de groupements fonctionnels bien speacutecifique De plus lrsquoeacutetude de lrsquoactiviteacute de
plusieurs analogues permettrait de confirmer si la tomatidine possegravede un meacutecanisme
drsquoaction deacutependant drsquoune interaction speacutecifique avec une ou des cibles biologiques
preacutecises
Le laboratoire du Pr Eacuteric Marsault du deacutepartement de pharmacologie de lrsquoUniversiteacute
de Sherbrooke et lrsquoeacutetudiant agrave la maitrise Feacutelix Chagnon ont produit plus de 33
analogues de la tomatidine (voir Annexe 2 article laquo Unraveling the structure-activity
relationship of tomatidine a steroid alkaloid with unique antibiotic properties against
persistent forms of Staphylococcus aureusraquo dans la section laquo Contributions
Supleacutementaires raquo du meacutemoire) dont lrsquoactiviteacute antibacteacuterienne a eacuteteacute eacutevalueacutee par des
tests de susceptibiliteacute agrave la moleacutecule (CMI) sur S aureus SCV et sur S aureus agrave
pheacutenotype normal en combinaison avec la gentamicine Plusieurs de ces moleacutecules
font lrsquoobjet drsquoune application de brevet (voir annexe 1) dont la prometteuse moleacutecule
FC04-100 (Figure 14C) (voir laquo Annexe 1 Application de brevet raquo dans la section
laquo Contributions suppleacutementaires raquo du meacutemoire)
13 Projet de maitrise
Premiegravere hypothegravese
Si la tomatidine a une activiteacute antibacteacuterienne et un meacutecanisme speacutecifique on devrait
donc avoir un spectre drsquoactiviteacute seacutelectif en lien avec le mode drsquoaction
Objectif Provoquer lrsquoapparition de pheacutenotypes SCV chez diffeacuterentes espegraveces
bacteacuterienne afin de pouvoir veacuterifier leur niveau de sensibiliteacute agrave la tomatidine
Deuxiegraveme hypothegravese
Lrsquoidentification des gegravenes preacutesentant des mutations parmi les souches de S aureus
reacutesistantes agrave la tomatidine pourrait reacuteveacuteler la ou les cibles cellulaires de la moleacutecule
et nous aider agrave comprendre son mode drsquoaction
36
Objectif Obtenir des mutants reacutesistants avant de les envoyer agrave sequencer
faire les analyses comparatives des geacutenomes et finalement identifier les mutations
37
CHAPITRE 2
ARTICLE SCIENTIFIQUE
TITRE Tomatidine and Analog FC04-100 Possess Bactericidal Activities Against the
Small Colony Variants of Some Firmicutes Including Those of Listeria
monocytogenes and Are Bactericidal in Combination With Aminoglycosides Against
their Normal Phenotype
AUTEURS ET CONTRIBUTIONS
Isabelle Guay1 Expeacuteriences biologiques avec le produit naturel tomatidine et son
analogue FC04-100 sur diffeacuterentes espegraveces bacteacuteriennes afin de comparer les deux
activiteacutes
Simon Boulanger1 Expeacuterience biologique permettant de quantifier lrsquoactiviteacute de
lrsquoanalogue FC04-100 sur la production de biofilm et son activiteacute intracellulaire
Feacutelix Chagnon2 Production de lrsquoanalogue chimique de la tomatidine FC04-100
Kamal Bouarab1 Initiateur du projet gracircce agrave lrsquoisolement du produit naturel tomatidine
Eacuteric Marsault2 Directeur de recherche de Feacutelix Chagnon
Franccedilois Malouin1 Directeur de recherche drsquoIsabelle Guay et Simon Boulanger et
auteur de correspondance
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
38
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
Corresponding author Mailing address Universiteacute de Sherbrooke
Deacutepartement de Biologie Faculteacute des Sciences 2500 Boul
Universiteacute Sherbrooke Quebec Canada J1K2R1 Phone (819) 821-
8000 ext 61202 Fax (819) 821-8049 E-mail FrancoisMalouin
USherbrookeca
39
Tomatidine and Analog FC04-100 Possess Bactericidal Activities
Against the Small Colony Variants of Some Firmicutes Including
Those of Listeria monocytogenes and Are Bactericidal in Combination
With Aminoglycosides Against their Normal Phenotype
Running Title Tomatidine is bactericidal against several Firmicutes
Isabelle Guay1
Simon Boulanger1
Feacutelix Chagnon2 Kamal Bouarab
1 Eacuteric Marsault
2 and
Franccedilois Malouin1
1Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM) Deacutepartement de
Biologie Faculteacute des Sciences Universiteacute de Sherbrooke Sherbrooke Queacutebec Canada J1K
2R1
2Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute Universiteacute
de Sherbrooke Queacutebec Canada J1H 5N4
Have contributed equally to the work
Corresponding author
Mailing address Universiteacute de Sherbrooke Deacutepartement de Biologie Faculteacute des Sciences
2500 Boul Universiteacute Sherbrooke Quebec Canada J1K2R1
Phone (819) 821-8000 ext 61202 Fax (819) 821-8049
E-mail FrancoisMalouinUSherbrookeca
40
ABSTRACT
Tomatidine (TO) is a plant steroidal alkaloid that possesses an antibacterial activity against the
small colony variants (SCVs) of Staphylococcus aureus We report here the spectrum of
activity of TO against species of the Firmicutes and the improved antibacterial activity of a
chemically-modified TO derivative (FC04-100) In addition to that seen on S aureus TO
showed a bactericidal activity against SCVs of Bacillus cereus B subtilis and Listeria
monocytogenes The combination of an aminoglycoside antibiotic and TO also generated an
antibacterial synergy against strains of the normal phenotype in these species In contrast to
TO which has no relevant activity by itself against S aureus of the normal phenotype (MIC
gt64 mgL) the TO analog FC04-100 showed a MIC of 8-16 mgL Furthermore FC04-100
showed a strong bactericidal activity against L monocytogenes SCVs in kill kinetic
experiments while TO did not FC04-100 also improved the bactericidal activity of
gentamicin against L monocytogenes of the normal phenotype The addition of FC04-100 (4
mgL) to a cephalexinkanamycin (32) combination improved the activity of the combination
by 32 fold against cephalexin and kanamycin-resistant MRSA strains In combination with
gentamicin FC04-100 also showed a strong bactericidal activity against biofilm-embedded S
aureus and when used alone was superior to TO for its biofilm-preventing activity Finally
FC04-100 and TO showed comparable intracellular killing of S aureus SCVs in a polarized
Calu-3 cell culture model In conclusion chemical modifications of TO allowed improvement
of its antibacterial activity against L monocytogenes SCVs and prototypical S aureus
41
Keywords Tomatidine aminoglycoside synergy SCV Firmicutes S aureus L
monocytogenes
Introduction
The Firmicutes are Gram-positive bacteria with genomes presenting less than 50 of G+C
The Firmicutes constitutes one of the main divisions within the Bacteria and is divided into
three classes Bacilli Mollicutes and Clostridia The class of Bacilli is further divided into the
orders of the Lactobacillales and the Bacillales The Bacillales are divided into the genus of
Staphylococcus Listeria and Bacillus A number of bacterial species such as Listeria spp and
Bacillus spp can contaminate food and cause infections in humans (Magalhaes 2013) To
name a few Listeria monocytogenes L ivanovii and Bacillus cereus can cause listeriosis
(Guillet et al 2010) and food poisoining (Bad Bug Book FDA) Bacillus subtilis B
coagulans B licheniformis and B sphaericus are also known to cause illnesses Bacillus
anthracis causes anthrax and can often be acquired by contact with food producing animals
and cattle (beef cattle sheeps etc) and this bacterium is also well-known for its endospores
that have been used as biological weapons (Beierlein and Anderson 2011) Staphylococci are
divided in coagulase-positive species Staphylococcus aureus being the most clinically
relevant of this group and coagulase-negative species such as S epidermidis the most
prevalent pathogen associated with infections of implanted medical devices (Vuong and Otto
2002) The emergence and spread of resistance to multiple antibiotics in staphylococci is now
considered a real health treat and impaired therapeutic endeavor to combat these bacteria
42
(Witte et al 2007) Notably the prevalence of methicillin-resistant S aureus (MRSA) has
steadily increased over the recent years not only in hospitals but also in the community
(Hiramatsu et al 2013)
Since their first description in 1910 for Salmonella enterica serovar Typhi (S typhi) small-
colony variants (SCVs) have been described for a wide range of bacterial genera and species
and have been isolated from both laboratory and clinical environments (Proctor 2006)
Staphylococcus aureus SCVs have attracted a great deal of interest over the past recent years
S aureus SCVs often present a dysfunctional oxidative metabolism causing an alteration in
the expression of virulence factors a slow growth and a loss of colony pigmentation (Proctor
2006) This dysfunctional oxidative metabolism is also responsible for a decreased
susceptibility to aminoglycoside antibiotics because this class of molecules requires the
proton-motive force in order to penetrate the bacterium (Bryan LE 1981) This respiratory
deficiency is often caused by mutations affecting the electron transport system and several
SCV isolates can recover normal growth with supplemental hemin or menadione which are
needed to synthesize electron transport system components S aureus SCVs often are isolated
from chronic infections such as lung infections in cystic fibrosis (CF) patients osteomyelitis
septic arthritis bovine mastitis and from infections associated with orthopedic devices (Atalla
2008 Moisan 2006 Proctor 2006) The ability of S aureus to switch back and forth from the
prototypic to the SCV phenotypes in vivo is an integral part of the pathogenesis of S aureus
and may be responsible for the establishment of chronic infections (Tuchscherr 2011
Mitchell 2011 inf imm)
43
Tomatidine (TO) is a steroidal alkaloid produced by the Solanaceae plant family such as the
tomato (Ruiz-Rubio 2001 Simons 2006) We showed previously the antibacterial activity of
TO against S aureus SCVs and also documented a strong synergic activity of TO in
combination with aminoglycoside antibiotics against prototypic S aureus (Mitchell G AAC
2011 Mitchell G 2012 JAC) Recently we synthesized a variety of TO analogs in order to
explore the structure-activity relationship of this new class of antibiotics able to act on SCVs
(Chagnon et al 2014) One analog FC04-100 showed the same steroidal backbone as the
natural molecule but with an additional carbon chain on the A cycle (Fig 1) Preliminary
characterization of FC04-100 revealed an antibacterial activity that was similar to that of TO
against S aureus SCVs and also preserved the synergic activity with aminoglycosides against
prototypic S aureus (Chagnon et al 2014) However contrary to TO FC04-100 showed
notable antibiotic activity by itself against prototypic S aureus with a MIC of 8-16 gL
whereas that of TO was gt64 gL It would be of great interest to see if this property of FC04-
100 modifies or improves its spectrum of antibacterial activity
The aim of this study was to further describe the spectrum activity of TO and FC04-100 The
susceptibility of several species among the Firmicutes to TO and FC04-100 alone or in
combination with other antibiotics was investigated The improved antibacterial activity of
FC04-100 was demonstrated against Listeria monocytogenes and methicillin-resistant S
aureus (MRSA)
44
Materials and methods
Chemical reagents and antibiotics
Tomatidine (TO) (Sigma Aldrich Oakville Ontario Canada) (Fig 1A) was solubilised in
dimethylsulfoxide (DMSO) at a concentration of 2 mgmL The TO analog FC04-100 (Fig
1B) was synthesized (Chagnon et al 2014) and solubilised in DMSO at 20 gL Cefalexin
kanamycin gentamicine (GEN) (all from Sigma Aldrich) were solubilised in water at a
concentration of 10 gL Menadione was solubilized in DMSO hemin in 14 M NH4OH and
thymidine in water (all from Sigma Aldrich) and were prepared at a concentration of 10 gL
45
3
Fig 1 Tomatidine (A) is a steroid alkaloid structurally characterized by 6 rings 12
stereogenic centers a 3 β-hydroxyl group and spiro-fused E F rings in the form of an
aminoketal The tomatidine analog FC04-100 (B) contains a diamine in position 3 of ring A
Bacterial strains and growth conditions
Staphylococcus aureus ATCC 29213 S aureus Newbould (ATCC 29740) Listeria
monocytogenes ATCC 13932 Bacillus subtilis ATCC 6333 and Bacillus cereus ATCC 11778
were used as prototypic strains Methicillin-resistant S aureus (MRSA) strains COL and
USA100 (hospital acquired-MRSA) and strain USA300 (community acquired-MRSA) were
also used in this study The reference laboratory strain S aureus SH-1000 an isogenic mutant
strain derived from S aureus 8523-4 but with a functional rsbU allele (HorsburghMJ 2002)
was used for its good yield of biofilm production The S aureus strains CF07-L (prototypic)
A
B
A B
C D
E
F
46
and CF07-S (SCV phenotype) are genetically related clinical strains originally isolated from a
cystic fibrosis patient (MitchellG 2011) Strain CF07-S was used in the intracellular
infection model Staphylococcus and Bacillus were maintained on Tryptic Soy agar (TSA BD
Mississauga Ontario Canada) whereas L monocytogenes was grown on brain-heart infusion
(BHI) agar (BD Mississauga Ontario Canada)
Small colony variants (SCVs)
For S aureus strain NewbouldhemB was used as the reference SCV NewbouldΔhemB was
generated from strain Newbould (ATCC 29740) by disrupting the hemB gene with the ermA
cassette by homologous recombination (Brouillette 2004) SCVs from B cereus B subtillis
and L monocytogenes were generated by growth in presence of a subinhibitory concentration
of GEN Briefly overnight broth cultures (18ndash20 h) were used to inoculate BHI broth at a
dilution of 1100 supplemented or not with 025 to 1X the MIC of GEN Cultures were
incubated 18 h at 35degC with shaking (225 rpm) and then adjusted to an A595 nm of 20 in PBS at
4degC Determination of CFU and SCV colonies was done by serial dilution plating SCV were
obtained by plating on TSA (BHI agar for L monocytogenes) containing gentamicin at a
concentration of 8 to 16 times the induction concentration followed by an incubation of 48 h at
35degC The pinpoint colonies selected by this method were confirmed to be SCVs by streaking
several of them on agar plates without antibiotic SCV that conserved their phenotype after
two passages were considered to have a stable phenotype and were used in subsequent
experiments Selected SCVs had a colony size a tenth of the size of a normal colony and
showed no pigmentation or hemolytic activity on blood agar plates The SCV species were
47
confirmed by rDNA 16S sequencing after PCR amplification using universal primers (Gomaa
2007)
Auxotrophy assays
For SCVs auxotrophism is defined as the requirement of specific compounds in order to
regain a normal growth phenotype [Sendie 2009] An agar diffusion method was used to
characterize the auxotrophism of SCVs using hemin or menadione (1-10 μg eachwell) or
thymidine (100 microgwell) on an inoculated Mueller-Hinton agar (MHA) or Brain-heart infusion
agar (BHIA) plate [Besier 2007] Auxotrophy for specific supplements was detected by a
zone of normal growth surrounding the well after 18 h of incubation at 35degC The results were
confirmed by two indepedent experiments
Antibiotic susceptibility testing
MICs were determined by a broth microdilution technique following the recommendations of
the Clinical and Laboratory Standards Institute (CLSI) except that the incubation period was
extended to 48 h and the medium used was BHI to allow SCVs to reach maximal growth as
previously described (Mitchell 2010) As recommended by the CLSI L monocytogenes (and
its SCVs) were grown in cation-adjusted Muller-Hinton broth (CAMHB BD) containing 3
lysed horse blood (LHB Remel Hartford CT USA) The reported results were obtained from
three independent experiments
48
Time-kill experiments
Kill kinetics were performed to determine whether the effect of compounds alone or in
combination was bacteriostatic or bactericidal Bacteria were inoculated at 105-10
6 CFUmL in
appropriate medium in the absence or presence of the different antibiotic compounds at
concentrations specified in figure legends At several points in time at 35degC (225 rpm)
bacteria were sampled serially diluted and plated on TSA for CFU determinations Plates
were incubated for 24 or 48 h at 35degC for normal and SCV strains respectively The data were
collected from a minimum of three independents experiments
Antibiotic activity in biofilms
The viability of bacteria in biofilms was evaluated based on previously described protocols
(CeriH 2001 MoskowitzSM 2004 HarrisonJJ 2010) For these assays 96-peg lids and
corresponding 96-well plates were used (Nunc CITY STATE) Wells were inoculated with a
suspension of S aureus SH-1000 adjusted to a 05 McFarland standard in TSB (150 microL per
wells) and plates with 96-well lids were incubated at 35degC with an agitation of 120 rpm for 24
h The biofilms on pegs were then washed three times with 200 microL PBS Biofilms on pegs
were further incubated in fresh 96-well plates containing 200 microL of TSB containing or not
antibiotic compounds or a combination of molecules at 35degC 120 rpm for 24 h The treated
biofilms on pegs were washed three times in PBS and bacteria in biofilms were recovered by
sonication in a new microtiter plate containing 200 microL of PBS per wells using an
ultrasonicator bath for 10 min followed by centrifugation for five min at 1000 RPM The
49
bacteria recovered in each well were resuspended serially diluted plated on TSA and
incubated at 35degC for 24 h before CFU determination
For an equivalent series of wells instead of disrupting the biofilm by sonication biofilm
formation on pegs was evaluated through crystal violet staining using a procedure adapted
from Mitchell et al(2010) Following exposure to antibiotics as described above biofilms on
pegs were washed three times using 200 microL of PBS before biofilms were air-dried for 30
minutes Biofilm on pegs were then stained by dipping in 200 microL of a crystal violet solution
(01) for 30 minutes rinsed in 200 microL of distilled water and air-dried for 30 min Finally the
crystal violet from stained biofilms was solubilized in 200 microL of ethanol 95 at room
temperature and the absorbance was measured at 562 nm using a plate reader (nom de
lrsquoappareil)
Cell invasion and measurement of intracellular antibiotic activity
The Calu-3 cell line (ATCC HTB 55 Homo sapiens lung adenocarcinoma) was cultured in
Eaglersquos Minimum Essential Medium (EMEM) supplemented with 01 mM MEM nonessential
amino acids 1 mM of sodium pyruvate 100 UmL penicillin 100 gL streptomycin 25 gL of
Fungizone and 10 fetal bovine serum (FBS) at 37oC in 5 CO2 For routine culture 4 gL of
puromycin was added to culture media All cell culture reagents were purchased from Wisent
(St-Bruno QC Canada)
50
The cell invasion assay was performed with the Calu-3 cells in an air-liquid interface as
previously described with few modifications (Mitchell TO I paperAAC 2011) Briefly cells
were seeded at 15 x 105 cells per insert in a 12-well Transwell plate (Corning CITY STATE)
and cultured for 10 days with the apical medium replaced each day The complete medium in
the basal compartment was replaced by the invasion medium (1 FBS and no antibiotics) 18
h before invasion assays Inocula were prepared by suspending bacteria grown 20 h on BHI
agar plates in ice-cold PBS Bacteria were then washed three times in ice-cold PBS and
suspended in the invasion medium supplemented with 05 BSA at a density of
approximately 4 x 108 CFUmL Cells were washed twice with PBS and 250 μL of bacterial
suspension were apically added to each insert Invasion of cells by bacteria was allowed for 3
h inserts were emptied and washed three times with PBS The basal medium was changed for
the invasion medium with 20 gL of lysostaphin (Sigma) to kill extracellular bacteria and with
or without the tested antibiotic compounds The infected cells were incubated for a total of 48
h with a change of medium at 23 h (invasion medium with lysostaphin with or without the
tested antibiotic compounds) The invasion medium with lysostaphin but without tested
compound was also apically added 1 h before cell lysis to ensure that only intracellular
bacteria were counted At 48 h following three washes with PBS the apical and basal media
were removed and cells were detached with 100 μL of trypsin 025 and lysed for 10 min by
the addition of 400 μL of water containing 005 of Triton X-100 Then 50 μL of PBS (50X)
was added and mixed Lysates were serially diluted 10-fold and plated on TSA for CFU
determination Plates were incubated at 35oC for 48 h
51
RESULTS
Emergence the SCV phenotype
The SCV phenotype is characterized by a slow growth yielding small colony size on agar
plates (15 to 110 of the parent phenotype) Figure 2 illustrates the ability of a subinhibitory
concentration of GEN to promote the emergence of the SCV phenotype for B cereus B
subtilis and L monocytogenes B cereus SCVs were obtained at an inducing concentration of
1 X MIC of GEN (Fig 2C) B subtilis SCVs at 05 X MIC (Fig 2D) and L monocytogenes
SCVs at 025 X MIC (Fig 2E) Two prototypic S aureus Newbould and CF07-L and their
SCV counterpart NewbouldhemB and CF07-S respectively were used as control strains (Fig
2A and 2B) The SCV isolates selected for the rest of the study add a low reversion rate and
kept their phenotype without a GEN selection pressure The B cereus SCVs showed
auxotrophy for menadione (Fig 1F Table 1) some of the L monocytogenes SCVs showed
auxotrophy for hemin (Table 1) while the B subtilis SCVs showed no apparent auxotrophy to
hemin menadione or thymidine (Table 1) These results show that SCVs with defects in their
respiratory chain (ie hemin or menadione auxotrophy) can indeed emerge from a selective
pressure with aminoglycosides
52
A B
Fig 2 Typical colony sizes for SCV and protocipic strains (left and right sides of the plates
respectively) for (A) S aureus CF07-S and CF07-L (B) S aureus NewbouldhemB and
Newbould (C) B cereus ATCC 11778 (SCV1 and prototypic) (D) B subtilis ATCC 6333
(SCV2 and prototypic) and (E) L monocytogenes ATCC 13932 (SCV1 and prototypic) In
(F) a TSA plate was inoculated with a B cereus SCV1 and wells were filled with (i) 10 microg
hemin (ii) 10 microg menadione (iii) 100microg thymidine and diluents (iv) DMSO and (v) NH4OH
14N
C D
E
i ii
iii iv
v
F
53
Antibacterial activity of TO against the various species of SCVs and synergy of
the TO and GEN combination against some prototypic Firmicutes
All the SCVs tested showed a MIC for TO of 006 mgL against S aureus and lt003 mgL
against L monocytogenes B cereus and B subtilis respectivly whereas TO showed no
activity (MIC gt64 microgmL) against their prototypic counterparts (Table 1) A synergic effect of
TO with GEN against all the prototypic strains tested was also observed (Table 1) Indeed the
addition of TO provoked a 4-8 fold decrease of GEN MIC against all species tested (Table 1)
These results are consistent with our previous studies on the activity of TO on S aureus
((Mitchell 2011 AAC 2012 JAC)) No change in the MIC of GEN was observed against the
streptococci (S pyogenes S suis S agalactiae S mitis and S pneumoniae) when tested in
combination with TO (data not shown) These results show that TO is active against some but
not all Firmicutes
54
Table 1 Susceptibility of prototypic and SCV strains and species to TO andor GEN
Ratio of activity of GEN+TO vs GEN alone
Bactericidal activity of TO against SCVs and bactericidal synergy of the TO
and GEN combination against prototypic strains
As shown above the addition of 8 mgL of TO to GEN reduced the MIC of GEN against some
Firmicutes (Table 1) but also time-kill kinetics showed that TO improved the bactericidal
effect of GEN against prototypic B subtilis and B cereus (Fig 3A and 3B respectively) This
bactericidal synergy was also previously described for S aureus (Mitchell 2012 JAC)
However although the MIC of GEN was improved when in combination with TO against L
monocytogenes (Table 1) time-kill experiments failed to demonstrate a bactericidal synergy
TO GENGEN
(+TO)
Saureus ATCC 29213 gt64 05 006 8
Saureus NewbouldΔhemB 006 8 Hemin
B cereus ATCC 11778 gt64 1 025 4
B cereus ATCC 11778 (SCV1) lt003 16 Menadione
B cereus ATCC 11778 (SCV2) lt003 16 Menadione
B subtilis ATCC 6333 gt64 0125 003 4
B subtilis ATCC 6333 (SCV1) lt003 2 unknown
B subtilis ATCC 6333 (SCV2) lt003 4 unknown
B subtilis ATCC 9372 gt64 0125 lt0015 gt8
B subtilis ATCC 9372 (SCV1) lt003 1 unknown
B subtilis ATCC 9372 (SCV2) lt003 1 unknown
L monocytogenes gt64 05 0125 4
L monocytogenes (SCV1) lt003 gt64 unknown
L monocytogenes (SCV2) lt003 gt64 Hemin
Strains
MIC (gL)
Synergy
foldAuxotrophy
55
with an improvement of at most 1 log10 in killing compared to that achieved with GEN alone
(Fig 3C) Similarly the effect of TO used alone against SCVs was strongly bactericidal
against S aureus B subtilis and B cereus but not against the L monocytogenes SCV (Fig
3D) and this even if the MIC of TO against L monocytogenes SCVs was inferior to 003
mgL (Table 1) These results show that the activity of TO against L monocytogenes needs to
be improved
56
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 0 6 m g L G E N
0 0 6 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
0 5 m g L G E N
0 5 m g L G E N + T O
8 m g L T O
C T R L
tim e (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
2 m g L G E N
2 m g L G E N + T O
8 m g L T O
C T R L
Lo
g1
0 C
FU
ml
0
2
4
6
8
1 0
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
T=
24
h
T=
8h
T=
24
h
CT
RL
CT
RL
CT
RL
CT
RL
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
TO
(8 m
gL
)
S a u re u s
h e m B
S C V
B c e r e u s
A T C C 1 1 7 7 8
S C V 2
B s u b t i l i s
A T C C 6 3 3 3
S C V 2
L m o n o c y to g e n e s
A T C C 1 3 9 3 2
S C V 1
Fig 3 Time-kill kinetics of TO andor GEN against (A) prototypic B subtilis ATCC 6333(B)
B cereus ATCC 11778 and (C) L monocytogenes ATCC 13932 The bactericidal effect of
TO against SCVs is also shown at 8 and 24 h (D) The results were obtained from at least
three independent experiments
A B
C
D
Detection
threshold
57
Bactericidal activiy of the TO analog FC04-100 against L monocytogenes
SCVs and synergy with aminoglycosides against prototypic strains and MRSA
The addition of 8 mgL of FC04-100 to GEN (2 mgL) decreased by an average of 5 log10
prototypic L monocytogenes CFUmL after 24 h of treatment in comparison to GEN alone
(Fig 4A) Also the bactericidal activity of FC04-100 used alone against the L monocytogenes
SCV was shown to be better than that observed for TO (Fig 3D) To facilitate comparison
between TO and FC04-100 Figure 4B shows the residual CFU of the L monocytogenes SCV
strain after treatment with TO or FC04-100 and calculated as the percentage of live CFU in
comparison with untreated L monocytogenes SCVs after 24 h of incubation These results
show that TO can be chemically modified (eg derivative FC04-100) and improve its activity
against L monocytogenes and thus the antibacterial activity of FC04-100 against S aureus
was further investigated Interestingly the MIC of FC04-100 against the SCV strain S aureus
NewbouldhemB (MIC of 05 -1 mgL Table 2) was higher than that found for TO (MIC of
006 mgL Table 1) On the other hand the MIC of FC04-100 against the prototypic S
aureus ATCC 29213 is lower than that of TO (8-16 mgL vs gt 64 mgL respectively) (Tables
1 and 2) This newly detected antibacterial activity of FC04-100 was also noted against
prototypic MRSA strains (Table 2) Furthermore the ability of FC04-100 to potentiate the
activity of GEN was similar to that of TO with a 4 to 8 fold improvement of the
aminoglycoside MIC (Table 2)
Although the multi-resistant S aureus MRSA USA100 USA300 and COL are resistant to
cefalexin (beta-lactam antibiotic MIC of 256 mgL) and to the aminoglycoside kanamycin
(MIC of 256 mgL and gt 1024 mgL for USA 100 and USA300 respectively Table 3) the
58
combination of both antibiotics decreased their respective MICs (Table 3) Indeed the
combination of cefalexin and kanamycin in a proportion of 32 (commercialized under the
name UbrolexinTM
) decreased the MIC of one or both antibiotics This is as expected for the
combination of a beta-lactam with an aminoglycoside (Davis 1982) However the synergy is
weak with MRSA strains that are resistant to one or the other or both antibiotics and the
addition of 4 mgL of FC04-100 further improved by 8 to 32 fold the MIC of the cephalexin-
kanamycin combination against these strains (Table 3)
59
te m p s (h )
Lo
g1
0 C
FU
ml
0 4 8 1 2 1 6 2 0 2 4
0
1
2
3
4
5
6
7
8
9
1 0
F C 0 4 -1 0 0 8 m g L
2 m g L G E N +
F C 0 4 -1 0 0 8 m g L
C T R L
G E N 2 m g L (B H I)
Re
sid
ua
l C
FU
(
)
CT
RL
TO
8 g
L
FC
04-1
00 8
gL
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
F C 0 4 -1 0 0 8 g L
T O 8 g L
C TR L
T=
24
h
T=
8h
T=
8h
T=
24
h
T=
24
h
Fig 4 Time-kill kinetics of FC04-100 andor GEN against (A) prototypic L monocytogenes
ATCC 13932 The bactericidal effect of FC04-100 used alone against L monocytogenes
SCV1 is also shown at 8 and 24 h (B) In (B) the residual CFU were calculated as the
percentage of live CFU in each condition in comparison with the untreated L monocytogenes
SCV1 after 24h of incubation The results were obtained from at least three independent
experiments
A
B
60
Table 2 Susceptibility of prototypic and SCV S aureus strains to FC04-100 with or without
GEN
a FC04-100 w as used at 4 microgmL
b Ratio of activity of GEN + FC04-100 vs activity of GEN alone
Table 3 Synergistic activity of FC04-100 with the combinaison cefalexin-kanamycin
aFC04-100 was used at 4 microgmL
bRatio of the MIC of cefalexin+kanamycin with FC04-100 vs the MIC of
cefalexin+kanamycin alone
Staphylococcus aureus ATCC 29213 8-16 05 006-0125 4-8
S aureus SH1000 8-16 05 006-0125 4-8
S aureus NewbouldΔhemB 1 8
MRSA USA100 16 05 0125 4
MRSA USA300 16 05 006 8
MRSA Col 16 025 006 4
Strains FC04-100 GENGEN
(+FC04-100 )fold
MIC (gL)
MRSA USA100 256 256 12886 43 32
MRSA USA300 256 gt1024 256171 86 32
MRSA Col 256 4 43 05035 8
MIC (gL)
Strains cefalexin kanamycin cef+kana (32)cef+kana (32)
(+FC04-100 )fold
a b
a b
61
FC04-100 potentiates GEN against S aureus embedded in biofilm and inhibits
intracellular replication of SCVs
Figure 5 demonstrates that compound FC04-100 efficiently kills S aureus embedded in
biofilms when in combination with GEN ie there was an important decrease of bacterial
CFUs compared to the no drug control or to either FC04-100 used alone at 4 ugmL (ie 025-
05MIC Table 2) or GEN used alone at 025 05 or 1 X MIC Besides we previously
demonstrated that TO was able to inhibit intracellular replication of SCVs (Mitchell AAC)
which is an important aspect of virulence for S aureus Here (Fig 6) we show that FC04-100
like TO can significantly decrease the infection of polarized airway epithelial cells by SCVs
62
05
mgL
GE
N
05
mgL
GE
N +
FC
04
02
5 m
gL
GE
N
02
5 m
gL
GE
N +
FC
04
01
2 m
gL
GE
N
01
2 m
gL
GE
N +
FC
04
No d
rug
FC
04
0
2
4
6
8
log
10
CF
Up
eg
= 3 2
= 3 8
= 3 5
= 2
Fig 5 Bactericidal effect of FC04-100 alone or in combination with GEN against prototypic
S aureus SH100 embedded in biofilm FC04-100 (FC04) was used at 4 mgL alone or in
combination with GEN Data are from three independent experiments performed in triplicate
Significant differences in comparison to the no drug control are shown ( Plt 0001 and
Plt 00001 one-way ANOVA with a Dunnettrsquos post test) Data are presented as means with
standard deviations
63
No d
rug
TO
8 m
gL
FC
04 8
mgL
0
2
4
6
log
10
CF
Ui
ns
er
t
Fig 6 Effect of TO and FC04-100 on the intracellular load of CF07-S 48 h post-
internalization Data are from three independent experiments performed in duplicate
Significant differences among groups are shown ( Plt 00001One-way ANOVA with
Turkeyrsquos post test) Data are presented as means with standard deviations
64
DISCUSSION
In previous studies we have demonstrated that TO has a strong inhibitory activity against S
aureus SCVs and improves the bactericidal activity of aminoglycoside antibiotics against
prototypic S aureus and also more broadly against staphylococci This effect was documented
for several strains of diverse clinical origins including aminoglycoside-resistant S aureus
carrying aminoglycoside modifying enzymes TO showed however no synergistic effect on the
activity of aminoglycosides against P aeruginosa E coli or Enterococcus spp (Mitchell
2012) In the present study we showed that the antibacterial spectrum of TO can be extended
to species of the Bacillales order with very low MICs against L monocytogenes B cereus and
B subtillis SCVs (lt003 microgmL) Time-kill kinetics showed that the combination of TO and
GEN creates a bactericidal synergy against prototypic of B subtilis and B cereus similarly to
that previously demonstrated against S aureus However the mixture of TO and GEN did not
demonstrated much improvement over the activity of GEN alone against prototypic L
monocytogenes in time-kill experiments Likewise investigations with Bacillales SCVs
showed that TO efficiently killed S aureus B cereus and B subtillis but not L
monocytogenes SCVs despite its very low MIC and was thus bacteriostatic against that species
Therefore improvement of the antibacterial activity against this important pathogen seems
inevitable for future development of steroidal alkaloids such as TO
TO is a steroid alkaloid structurally characterized by 6 rings 12 stereogenic centers a 3-
hydroxyl group and spiro-fused E F rings in the form of an aminoketal (Fig 1) It is generally
65
prepared by hydrolysis of its glycosylated analog tomatine while both molecules can be found
in the Solanaceae plants (Ruiz-Rubio 2001 Simons 2006) Although we previously
documented some interesting antibacterial activity for TO the absence of an identified cellular
target makes difficult the establishment of a structure-activity relationship (SAR) Our initial
attempt to elucidate SAR was therefore based on the hypothesis that the steroidal A-D rings
act as a rigid scaffold to orient pharmacophores defined by the 3-hydroxyl group on ring A
and the spiroaminoketal group on rings E F Noteworthy we prepared analogs bearing
modifications on ring A of TO and schegravemes for chemistry synthesis were recently published
(Chagnon et al 2014) From seven compounds keeping rings E and F of TO and possessing
structural variations on position 3 of ring A we were able to ascertain some structure activity
relationships Results suggest that the hydrogen bond donor effect of the 3-hydroxyl group
was not important for activity although the stereochemistry of the 3 position substitution
moderately affected activity against both S aureus SCVs and prototypic strains (in
combination with an aminoglycoside) In addition although the presence of one or two
ammonium groups in position C3 reduced the activity against S aureus SCVs (from a MIC of
006 to 05-1 microgmL) this change was highly beneficial for antibiotic activity against normal
strains (from a MIC of gt64 to 8-16 microgmL) without the presence of an aminoglycoside This
change was unexpected and one of such analogs FC04-100 increased its solubility by more
than ten times (from 2 mgmL to 26 mgmL in DMSO) The biological properties of FC04-
100 were therefore further investigated in the present study especially that TO showed some
weak bactericidal activity against Listeria and that the solubility of TO may limit eventual
clinical applications
66
Here we demonstrated that FC04-100 was able to kill both L monocytogenes SCVs as well as
the normal phenotype alone and in the presence of GEN respectively (Fig 4) Furthermore
FC04-100 showed a noticeable antibacterial activity on its own (MIC of 16 microgmL) against the
MRSA strains USA100 USA300 and COL (Table 2) FC04-100 was also synergistic in
combination with an aminoglycoside against such MRSA strains with a 4 to 8 fold gain in the
MIC of GEN Because MRSA strains are often multiresistant to antibiotics and often carry
AMEs we examined the possibility of using a triple antibiotic combination Indeed the
combination of kanamycin and cefalexin in a proportion 32 is already approved to treat
bovine mastitis pathogens under the brand name of UbrolexinTM
(Ganiegravere 2009) This
antibiotic combination offers an extended spectrum of activity compared to each individual
drug and is expected to cover both S aureus Streptococcus uberis and E coli (Ganiere
2009 Maneke et al 2011 Silley 2012) Unfortunately the increased frequency of livestock-
associated MRSA (Garcia-Alvarez et al 2011 Graveland et al 2011 Leonard and Markey
2008) and frequent incidence of strains and species carrying AMEs (Ramirez and Tolmasky
2010) may limit the spectrum of activity of the combination of a beta-lactam and an
aminoglycoside With this in mind we showed that the use of FC04-100 in combination with
kanamycin and cefalexin improves by 32 fold the activity of this mixture against MRSA
strains carrying AMEs (Table 3) Overall the addition of FC04-100 to the aminoglycoside-
beta-lactam combination decreased the MICs of kanamycin and cefalexin below their
resistance breakpoints and this for all the MRSA strains tested
67
L monocytogenes is a highly specialized intracellular pathogen (Hamon et al 2006) causing
life-threatening infections especially in pregnant women (Janakiraman 2008) L
monocytogenes in a foodborne pathogen and because of its intracellular life cycle and
propagation it is able to hide from host defenses and antibiotic treatment Similarly S aureus
although not primarily recognized as a typical intracellular pathogen is also able to enter and
survive in host cells (Lowy 2000) Moreover the ability of S aureus to convert to the SCV
phenotype showing increased biofilm production improved adherence to host cells and tissues
as well as increased intracellular persistence allows this pathogen to cause chronic and
difficult to treat infections (Garcia et al 2013 Mitchell and Malouin 2012) Discovering
antibacterial drugs able to act in the intracellular compartment is generally recognized as a
difficult challenge Previously we demonstrated that steroidal alkaloids such as TO had such
an ability and could act on intracellular S aureus SCVs (Mitchell et al 2011) and here we
demonstrated that this was also the case for the TO analog FC04-100 (Fig 6) In addition we
also show in the present study that FC04-100 greatly improves the bactericidal activity of
aminoglycosides such as GEN against bacteria embedded in biofilms Biofilms are
recognized to be a major hindrance to antibiotic action and are responsible for persistent
colonization in many diseases (Costerton et al 1999) and in the food industry (Srey et al
2013) The intracellular and intrabiofilm antibacterial properties of FC04-100 warrant further
investigation of this compound and this class of molecules in general
In conclusion we showed in this study that the spectrum of activity of TO is related to the
Firmicutes division and more precisely to the order of the Bacillales TO possesses an
68
antibacterial activity against SCVs and a synergistic activity with animoglycoside antibiotics
against prototypic strains The novel TO analog FC04-100 showed very promising new
characteristics that include a much improved bactericidal effect on L monocytogenes and
killing of bacteria in biofilms
ACKNOWLEDGEMENTS
This study was supported by an operation grant from Cystic Fibrosis Canada to FM and by a
team grant from the Fonds Queacutebeacutecois de la Recherche sur la Nature et les Technologies
(FQRNT) to FM KB and EM
REFERENCES
Atalla H Gyles C Jacob CL Moisan H Malouin F Mallard B 2008 Characterization of
a Staphylococcus aureus small colony variant (SCV) associated with persistent bovine mastitis
Foodborne Pathog Dis 5785ndash799
Bad Bug Book Foodborne pathogenic microorganisms and natural toxins 2012 Second
edition Lampel KA Al-Khaldi S Cahill SM (Ed) Published by the Center for Food Safety and
Applied Nutrition of the Food and Drug Administration (FDA) US Department of Health and
Human Services pp 87-103
Beierlein JM Anderson AC 2011 New developments in vaccines inhibitors of anthrax
toxins and antibiotic therapeutics for Bacillus anthracis Curr Med Chem 185083-5094
69
Brouillette E A Martinez B J Boyll N E Allen and F Malouin 2004 Persistence of
a Staphylococcus aureus small-colony variant under antibiotic pressure in vivo FEMS
Immunol Med Microbiol 4135ndash41
Bryan L E and S Kwan 1981 Aminoglycoside-resistant mutants of Pseudomonas
aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob
Agents Chemother 19958ndash964
Ceri H Olson M Morck D Storey D Read R Buret A Olson B 2001 The MBEC Assay
System multiple equivalent biofilms for antibiotic and biocide susceptibility testing Methods
Enzymol 337377-385
Chagnon F I Guay M-A Bonin G Mitchell K Bouarab F Malouin E Marsault
2014 Unraveling the structure-activity relationship of tomatidine a steroid alkaloid with
antibiotic properties against persistent forms of Staphylococcus aureus Eur J Med Chem (In
Press EJMECH-S-13-01625-3)
Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial
Susceptibility Testing Twenty-First Informational Supplement M100-S21 CLSI Wayne PA
USA 2011
70
Costerton JW Stewart PS Greenberg EP 1999 Bacterial biofilms a common cause of
persistent infections Science 284 1318-1322
Davis BD 1982 Bactericidal synergism between beta-lactams and aminoglycosides
mechanism and possible therapeutic implications Rev Infect Dis 4237-245
Ganiegravere JP Denuault L 2009 Synergetic interaction between cefalexin and kanamycin in
Muller-Hinton broth medium and in milk J appl microbiol 107 117-125
Garcia LG S Lemaire B C Kahl K Becker R A Proctor O Denis P M Tulkens
F Van Bambeke 2013 Antibiotic activity against small-colony variants of Staphylococcus
aureus review of in vitro animal and clinical data J Antimicrob Chemother 68 1455ndash1464
Garcia-Alvarez L M T Holden H Lindsay C R Webb D F Brown M D Curran
E Walpole K Brooks D J Pickard C Teale J Parkhill S D Bentley G F Edwards
E K Girvan A M Kearns B Pichon R L Hill A R Larsen R L Skov S J Peacock
D J Maskell and M A Holmes 2011 Methicillin-resistant Staphylococcus aureus with a
novel mecA homologue in human and bovine populations in the UK and Denmark a
descriptive study Lancet Infect Dis 11595-603
Gomaa OM Momtaz OA 2007 16 rRNA characterization of Bacillus isolate and its
tolerance profile after subsequent subculturing Arab J Biotech 10 107-116
71
Graveland H Duim B van Duijkeren E Heederik D Wagenaar JA 2011 Livestock-
associated methicillin-resistant Staphylococcus aureus in animals and humans Int J Med
Microbiol 301630-634
Guillet C Join-Lambert O Le Monnier A Leclercq A Mechaiuml F Mamzer-Bruneel MF
Bielecka MK Scortti M Disson O Berche P Vazquez-Boland J Lortholary O Lecuit M
2010 Human listeriosis caused by Listeria ivanovii Emerg Infect Dis 16136-138
Hamon M H Bierne P Cossart 2006 Listeria monocytogenes a multifaceted model Nat
Rev Microbiol 4 423-434
Harrison JJ Stremick CA Turner RJ Allan ND Olson ME Ceri H 2010 Microtiter
susceptibility testing of microbes growing on peg lids a miniaturized biofilm model for high-
throughput screening Nat Protoc 51236-1254
Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y
Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T Baba T 2013 Genomic
Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
Horsburgh MJ Aish JL White IJ Shaw L Lithgow JK Foster SJ 2002 SigmaB
modulates virulence determinant expression and stress resistance characterization of a
72
functional rsbU strain derived from Staphylococcus aureus 8325-4 J Bacteriol 1845457-
5467
Janakiraman V 2008 Listeriosis in Pregnancy Diagnosis Treatment and Prevention Rev
Obstet Gynecol 1 179ndash185
Leonard FC Markey BK 2008 Methicillin-resistant Staphylococcus aureus in animals a
review Vet J 175 27-36
Lowy FD 2000 Is S aureus an intracellular pathogen Trends Microbiol 8341-342
Magalhatildees L Nitschke M 2013 Antimicrobial activity of rhamnolipids against Listeria
monocytogenes and their synergistic interaction with nisin Food Control 29 138-142
Maneke E Pridmore A Goby L Lang I 2011 Kill rate of mastitis pathogens by a
combination of cefalexin and kanamycin J Appl Microbiol 110184-190
Mitchell G Brouillette Eacute Lalonde-Seacuteguin D Asselin AE Lebeau Jacob C Malouin F
2009 A role for sigma factor B in the emergence of Staphylococcus aureus small-colony
variants and elevated biofilm production resulting from an exposure to aminoglycosides
Microb Pathog 4818-27
73
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K Malouin F 2011
Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic
fibrosis airway epithelial cells Antimicrob Agents and Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM Malouin F 2011 Infection of Polarized
airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is
increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function
and is influenced by NF- κ B Infect Immun 793541-3551
Mitchell G Lafrance M Boulanger S Lalonde-Seacuteguin D Guay I Gattuso M Marsault
Eacute Bourab K Malouin F 2012 Tomatidine acts in synergy with aminoglycoside antibiotics
against multiresistant Staphylococcus aureus and prevents virulence gene expression J
Antimicrob Chemother 67 559-568
Mitchell G Lalonde-Seacuteguin D Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S
Malouin F 2010 Staphylococcus aureus sigma B-dependent emergence of small-colony
variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-
heptylquinoline-N-oxide BMC Microbiology 10 33-48
Mitchell G and F Malouin 2012 Outcome and prevention of Pseudomonas aeruginosa-
Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic
Fibrosis InTech Open Access Publisher ISBN 979-953-307-059-8
74
Moisan H Brouillette E Jacob CL Langlois-Beacutegin P Michaud S Malouin F 2006
Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated
from cystic fibrosis patients is influ- enced by SigB J Bacteriol 18864ndash76
Moskowitz SM Foster JM Emerson J Burns JL 2004 Clinically feasible biofilm
susceptibility assay for isolates of Pseudomonas aeruginosa from patients with cystic fibrosis
J Clin Microbiol 421915-1922
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M Peters G 2006
Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent
infections Nat Rev Micro- biol 4295ndash305
Ramirez MS M E Tolmasky 2010 Aminoglycoside modifying enzymes Drug Res
Updates 13 151ndash171
Ruiz-Rubio M Peacuterez-Espinosa A Lairini K Roldagraven-Arjona T Dipietro A Anaya N
2001 Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi p 293ndash326 In
A Rahman (ed) Studies in natural products chemistry vol 25 Elsevier Oxford United
Kingdom
75
Sendi P Proctor RA 2009 Staphylococcus aureus as an intracellular pathogen the role of
small colony variants Trends Microbiol 1754-58
Silley P Goby L Pillar CM 2012 Susceptibility of coagulase-negative staphylococci to a
kanamycin and cefalexin combination J Dairy Sci 953448-3453
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C Osbourn A
2006 Dual effects of plant steroidal alkaloids on Saccharomyces cerevisiae Antimicrob
Agents Chemother 502732ndash2740
Srey S Jahid IK Ha S-D 2013 Biofilm formation in food industries A food safety concern
Food control 31572-585
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D
Roth J Proctor RA Becker K Peters G Loumlffler B 2011 Staphylococcus aureus
phenotype switching an effective bacterial strategy to escape host immune response and
establish a chronic infection EMBO Mol Med 3129ndash141
Vuong C Otto M 2002 Staphylococcus epidermidis infections Microbes Infect 4481-489
Witte W Strommenger B Stanek C Cuny C 2007 Methicillin-resistant Staphylococcus
aureus ST398 in Humans and Animals Central Europe Emerg Infect Dis 13 255ndash258
76
CHAPITRE 3
MODE DrsquoACTION ET CIBLE DE LA TOMATIDINE
31 Obtention des mutants reacutesistants
Afin de nous aider agrave mieux deacutefinir la cible et le mode drsquoaction de la tomatidine (TO)
nous avons forceacute lrsquoapparition de reacutesistances chez Staphylococcus aureus Comme
souche controcircle prototype nous avons choisi ATCC 29213 puisque crsquoest une souche
tregraves freacutequemment utiliseacutee pour des expeacuteriences avec des antibiotiques et nous avons
choisi la souche NewbouldΔhemB comme S aureus laquo small colony variant raquo puisque
crsquoest une souche SCV qui est stable et bien connue nrsquoayant pas de capaciteacute de
reacuteversion Par passages successifs dans une concentration sous inhibitrice
drsquoantibiotique (technique de CMI (concentration minimale inhibitrice)) (CLSI 2011)
nous avons obtenu des mutants deacutemontrant une reacutesistance croissante agrave lrsquoantibiotique
au fil des passages (Figure 1) Nous avons obtenu des mutants ATCC 29213
reacutesistants agrave la gentamicine (GEN) et agrave la GEN lorsque combineacutee agrave la TO ainsi que
des mutants reacutesistants NewbouldΔhemB reacutesistants agrave la GEN seule agrave la TO seule et
agrave la TO lorsque combineacute agrave la GEN
Certains passages ont ensuite eacuteteacute seacutelectionneacutes parce qursquoils deacutemontraient une hausse
plus marqueacutee de leur CMI Ces passages ont eacuteteacute choisis car nous supposions que
ces hausses marqueacutees de CMI correspondraient donc agrave des hausses de la
reacutesistance de cette souche agrave lrsquoantibiotique testeacute et que cette reacutesistance pouvait ecirctre la
conseacutequence drsquoune ou plusieurs mutations en lien avec la cible ou le mode drsquoaction
de lrsquoantibiotique preacutesent Trois agrave quatre passages (ou paliers de reacutesistance) par
condition ont eacuteteacute seacutelectionneacutes Nous avons ensuite reacuteensemenceacute le laquo pool raquo
bacteacuterien de chacun des passages choisis afin de pouvoir isoler 3 isolats ayant la
mecircme CMI (ou tregraves similaire (CMI vraies)) que celui du laquo pool raquo bacteacuterien des
77
passages drsquoougrave ils provenaient Soixante quatre mutants reacutesistants ont ainsi eacuteteacute
seacutelectionneacutes en vue drsquoecirctre seacutequenceacutes (Figure 2)
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt 1
6
CM
I v
raie
s
02
5
CM
I v
raie
s
02
5-1
A
78
n o m b r e d e p a s s a g e s
CM
I d
e T
O (
microg
ml)
en
pr
eacutes
en
ce
de
ge
n (
4 micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
gt1
6
CM
I v
raie
s
01
25
1
6
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
2
4
6
8
1 0
1 2
1 4
1 6
CM
I v
raie
s
8
CM
I v
raie
s
2
4
CM
I v
raie
s
2
B
C
79
n o m b r e d e p a s s a g e s
CM
I d
e G
EN
(micro
gm
l)
en
pr
eacutes
en
ce
de
TO
(8
microg
ml)
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0
0
1
2
3
4
5
CM
I v
ra
ies
4
CM
I v
ra
ies
2
CM
I v
ra
ies
0
5
1
CM
I v
ra
ies
2
4
Figure 1 Graphiques des CMI pour chacun des passages pour (A) la tomatidine sur
NewbouldΔhemB (B) pour la tomatidine en preacutesence de gentamicine sur
NewbouldΔhemB (C) pour la gentamicine sur S aureus ATCC 29213 (D) pour la
gentamicine en preacutesence de tomatidine sur S aureus ATCC 29213
32 Seacutequenccedilage et analyses bioinformatiques
Une extraction drsquoADN geacutenomique a ensuite eacuteteacute effectueacutee sur chacun des isolats en
utilisant le Kit drsquoextraction drsquoADN geacutenomique miniKit de Qiagen (QIAamp DNA minikit)
(Li et al 2003) Puisque leurs seacutequences geacutenomiques nrsquoeacutetaient pas connues nous
avons eacutegalement extrait lrsquoADN geacutenomique de nos deux souches de reacutefeacuterences
ATCC29213 et NewbouldΔhemB (provenant de quatre colonies isoleacutees chacune)
afin drsquoavoir les seacutequences de comparaison pour nos mutants (Figure 2)
D
80
Ne
wm
an
(g
en
om
ede r
ef h
ttpw
ww
ncbinlm
nih
govn
uccore
150373012)
AT
CC
29
213
(A p
art
ir d
e 4
AT
CC
29
21
3 r
eco
nstr
uire
AT
CC
29
21
3 c
om
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
5
10
7
10
8 1
09
)
Ne
wbo
uld
Δhe
mB
(A p
art
ir d
e 4
Ne
wb
ou
ldh
em
Bre
co
nstr
uire
Ne
wb
ou
ldh
em
Bcom
ple
t e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
1
agraveG
EN
2
5 (
CM
I2
)
2
7 (
CM
I2
)
2
8 (
CM
I2
)
AT
CC
29
21
3
Pa
lier
de
reacutesis
tance
1
agravecom
bo
29
(C
MI
05
)
3
0 (
CM
I 0
5
3
1 (
CM
I 0
5)
Suite
fe
uill
e s
uiv
ante
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agraveG
EN
3
(C
MI
4)
4
(C
MI
4)
3
9 (
CM
I4
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agraveG
EN
4
6 (
CM
I8
)
4
7 (
CM
I8
)
4
8 (
CM
I8
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
2
agravecom
bo
1
1 (
CM
I 2
)
1
2 (
CM
I 2
)
2
1 (
CM
I 2
)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
3
agravecom
bo
5 (
CM
I 2
-4)
3
6 (
CM
I 2
-4)
3
7 (
CM
I 2
-4)
AT
CC
29
21
3
palie
r d
e r
eacutesis
tance
4
agravecom
bo
3
8 (
CM
I4
)
6
7 (
CM
I 4
)
81
Ne
wb
ould
Δhem
B(A
part
ir d
e 4
Ne
wb
ou
ldΔ
he
mB
reco
nstr
uire
Ne
wb
ou
ldΔ
hem
Bco
mp
let e
t
sa
ns in
ce
rtitu
de
en
utilis
ant
la s
ouch
e d
e r
eacutefeacute
rence
New
ma
n)
(eacutech
an
tillo
n
10
0
10
1
10
2 1
03
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveG
EN
24
(C
MI
64
)
4
1 (
CM
I 1
6)
4
2 (
CM
I 6
4)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agraveT
O
95
(C
MI
02
5)
9
6 (
CM
I 0
25)
9
7 (
CM
I 0
25)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
1
agravecom
bo
33
34
35
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveG
EN
49
(C
MI
12
8)
5
1 (
CM
I 1
28)
5
2 (
CM
I 1
28)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agraveG
EN
7
(C
MI
128
)
4
3 (
CM
I 1
28)
4
5 (
CM
I1
28
)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agraveT
O
6
4 (
CM
I gt
16)
6
5 (
CM
I gt
16)
6
6 (
CM
I gt
16)
75
76
77
79
80
81
82
83
84
85
86
87
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agravecom
bo
15
16
18
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
3
agravecom
bo
59
60
61)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
4
agravecom
bo
54
56
58)
Ne
wb
ou
ldΔ
hem
B
palie
r d
e r
eacutesis
tance
2
agraveT
O
90
(C
MI
02
5)
9
1 (
CM
I 0
25)
9
3 (
CM
I 0
5)
Ne
wb
ou
ldh
em
B
palie
r d
e r
eacutesis
tance
3
agraveT
O
11
0 (
CM
I 1
)
1
12 (
CM
I 0
25
-05
)
1
13 (
CM
I 0
5)
Fig
ure
2
Org
an
igra
mm
e d
es
mu
tan
ts r
eacutes
ista
nts
seacute
qu
en
ceacutes
se
lon
leu
r pa
lier
de
reacute
sis
tan
ce e
t a
vec leu
r n
um
eacutero
drsquoeacute
chan
tillo
n
co
rre
spo
ndan
t
82
Un minimum de 20 ngmicrol drsquoADN geacutenomique de chacun des isolats a ensuite eacuteteacute
achemineacute au laboratoire du Pr Seacutebastien Rodrigue (Deacutepartement de biologie Faculteacute
des sciences Universiteacute de Sherbrooke) qui a effectueacute la librairie de fragments Pour
se faire il y a eu dans un premier temps digestion enzymatique de lrsquoADN afin
drsquoobtenir des fragments drsquoADN double brin Ensuite tous les fragments ont eacuteteacute
purifieacutes suivant la meacutethode SPRI (Solid-Phase Reversible Immobilization) Il srsquoagit
drsquoune meacutethode simple et tregraves efficace drsquoisolation drsquoacides nucleacuteiques sur microplaque
(Rodrigue S et al 2010) Les acides nucleacuteiques cibles sont immobiliseacutes sur des
microparticules paramagneacutetiques dans des conditions tampons-speacutecifiques La
flexibiliteacute du systegraveme se fait par lrsquoutilisation de diffeacuterents tampons de liaison (laquo binding
buffer raquo) qui modifient le type et la grandeur des acides nucleacuteiques immobiliseacutes Les
contaminants peuvent ainsi ecirctre retireacutes de lrsquoeacutechantillon sans besoin de filtration ou
centrifugation (Beckman Coulter website 2013 DeAngelis et al 1995) Suite agrave la
purification par meacutethode SPRI lrsquoeacutequipe du Pr Rodrigue a obtenu des fragments
drsquoADN de ~250pb auxquels ont eacuteteacute ajouteacutes par ligation des adapteurs aux deux
extreacutemiteacutes des fragments Dans un deuxiegraveme temps des codes-bars speacutecifiques agrave
chaque eacutechantillon drsquoADN geacutenomique ont eacutegalement eacuteteacute lieacutes agrave chacun des fragments
Alors que les adapteurs permettent aux fragments de se lier agrave une matrice solide afin
de pouvoir ecirctre amplifieacutes les codes-bars permettront de classer chacun des
fragments selon leur appartenance agrave un eacutechantillon drsquoADN geacutenomique speacutecifique
83
Figure 3 Eacutetape du seacutequenccedilage Illumina
La librairie de fragments a ensuite eacuteteacute envoyeacutee chez Geacutenome-Queacutebec afin que
chacun des geacutenomes puisse ecirctre seacutequenceacute Geacutenome ndashQueacutebec utilise la meacutethode
Illumina comme outil de seacutequenccedilage (Figure 3) La grande innovation de la meacutethode
Illumina est lrsquoamplification des fragments par liaison de ceux-ci agrave une surface solide
(Flowcell) ce qui permet par un processus de ponts drsquoamplification de geacuteneacuterer plus
drsquoun million de laquo clusters raquo comprenant eux-mecircme au plus un millier de copies de la
moleacutecule originale de deacutepart (solid-phase PCR) Chaque regroupement de fragments
provenant de lrsquoamplification drsquoun mecircme fragment original est tregraves pregraves les uns des
Modifieacute de Quail MA et al 2008
84
autres formant une zone distincte drsquoun regroupement de fragments agrave un autre
Lrsquoincorporation des nucleacuteotides eacutetiqueteacutes preacutealablement avec des moleacutecules
fluorescentes speacutecifiques est deacutetecteacutee suite agrave leur excitation par un laser et les
images sont compileacutees pour produire la seacutequence de bases pour chaque fragment
ADN (Figure 4) Les nouvelles seacuteries de bases appeleacutees laquo readsraquo seront ensuite
reacuteassembleacutees (aligneacutes) en se basant sur un geacutenome de reacutefeacuterence connu (appeleacute
laquo resequencing raquo) (crsquoest notre cas ici) ou en absence de geacutenome de reacutefeacuterence
(appeleacute laquo De novo sequencing raquo) Au final lrsquoensemble des laquo reads raquo aligneacutes
correspondra agrave la seacutequence de chaque chromosome de lrsquoeacutechantillon drsquoADN
geacutenomique de deacutepart
85
Figure 4 Meacutethode drsquoamplification par la meacutethode Illumina
Suite au seacutequenccedilage chez Geacutenome ndashQueacutebec nous avions en main les seacutequences
geacutenomiques de tous nos mutants et de nos souches de reacutefeacuterence sous forme de
Tireacute de BiteSizeBio httpbitesizebiocom13546sequencing-by-synthesis-
explaining-the-illumina-sequencing-technology consulteacute le 3 deacutec 2013
86
contigs Nous avions donc le casse-tecircte mais en piegraveces deacutetacheacutees pecircle-mecircle et
sans avoir la photo de lrsquoimage de reacutefeacuterence sur laquelle se baser pour pouvoir
compleacuteter le casse-tecircte
1- Lrsquoassemblage
Les diffeacuterents geacutenomes avaient eacuteteacute preacutealablement fractionneacutes aleacuteatoirement
en petits fragments de 250 pb et amplifieacutes en vue du seacutequenccedilage Suite au
seacutequenccedilage les seacutequences de chaque fragment (appeleacute laquo reads raquo) sont ensuite
assembleacutees en contigs crsquoest-agrave-dire des seacutequences plus ou mois longues continues
et ordonneacutees geacuteneacutereacutees par lassemblage des clones dune bibliothegraveque geacutenomique
qui se chevauche La qualiteacute de lrsquoassemblage en contigs deacutepend de la capaciteacute de
superposition des laquo reads raquo Parce que la digestion enzymatique de lrsquoADN se fait de
faccedilon aleacuteatoire et se fait sur plusieurs copies drsquoADN chaque portion du geacutenome est
repreacutesenteacutee plusieurs fois dans diffeacuterents modegraveles de fragments Donc les
seacutequences identiques des diffeacuterents fragments (ou laquo reads raquo) peuvent ecirctre
superposeacutees par chevauchegravement et les laquo reads raquo superposeacutes peuvent ecirctre
assembleacutes en contigs par un logiciel drsquoassemblage En ce qui nous concerne cette
eacutetape a eacuteteacute faite agrave lrsquoaide du programme de bio-informatique Newbler (Zhang et al
2012) Lrsquoassemblage geacutenomique peut ecirctre probleacutematique pour les informaticiens car
les geacutenomes contiennent des seacutequences qui se reacutepegravetent et ces reacutepeacutetitions peuvent
ecirctre assez longues en fonction des organismes Suite agrave lrsquoassemblage les geacutenomes
sont donc sous forme de contigs
2- Lrsquoalignement
En se basant sur une seacutequence geacutenomique de reacutefeacuterence (notre reacutefeacuterence est
Staphylococcus aureus str Newman (~ 288Mb)
( httpwwwncbinlmnihgovgenome154project_id=58839) ) le programme de bio-
informatique ABACAS permet de rapidement aligner ordonner et orienter les contigs
Selon les besoins ce qui nrsquoa pas eacuteteacute notre cas ABACAS permet eacutegalement de
87
visualiser et designer les amorces permettant de fermer les gaps (espaces) entre les
contigs en faisant des PCR (Assefa et al 2009)
3- La fermeture des gaps
La fermeture des gaps nrsquoa pas eacuteteacute faite avec le programme ABACAS mais
avec lrsquoaide du programme de bio-informatique GapFiller Lrsquoutilisation de GapFiller
pour la fermeture des gaps possegravede 3 principaux avantages Premiegraverement le
programme prend en compte lrsquoestimation de la grandeur du gap agrave fermer donc il
exclut automatiquement les fermetures de gaps erroneacutes qui sont plus courts ou plus
longs que la grandeur estimeacutee Deuxiegravemement il nrsquoessaie pas de remplir un gap
avec un assemblage de laquo reads raquo en contigs mais cherche plutocirct agrave allonger les
contigs agrave partir de leurs extreacutemiteacutes jusqursquoagrave obtenir des seacutequences superposeacutees en
passant par la creacuteation de K-mer (seacuteparation des reads en plus petits fragments) Et
troisiegravemement il prend en compte que les extreacutemiteacutes des contigs sont souvent
source de mauvais assemblage et reacute eacutevalue ces extreacutemiteacutes pendant la fermeture des
gaps (Boetzer et Pirovano 2012)
4- Lrsquoannotation des geacutenomes
Suite agrave la fermeture des gaps il a fallu annoter les geacutenomes en se basant sur
lrsquoannotation du geacutenome de reacutefeacuterence Staphylococcus aureus str Newman RATT est
un outil de transfert rapide drsquoannotation (Rapid Annotation Transfert Tool) qui a eacuteteacute
deacuteveloppeacute afin de fournir rapidement une annotation preacutecise pour des geacutenomes
inconnus en se basant sur des geacutenomes deacutejagrave annoteacutes comme reacutefeacuterences (Figure 5)
La meacutethode transfert les annotations drsquoune reacutefeacuterence de haute qualiteacute agrave un nouveau
geacutenome sur la base de la conservation des synteny (ordre des gegravenes) (Otto et al
2011) Suite agrave lrsquoannotation les gegravenes tant chez les mutants que chez les souches
parentes portent les noms auxquels ils correspondent chez Staphylococcus aureus
str Newman
88
Figure 5 Corrections du programme RATT sur les annotations transfeacutereacutees Les
annotations de H37Rv ont eacuteteacute transfeacutereacutees sur la seacutequence F11 (bleu pacircle) ont eacuteteacute corrigeacutees
(vert) et ensuite compareacutees agrave lrsquoannotation de la souche existante F11 EMBL (jaune) (A et B)
La correction des codons ldquostartrdquo et ldquostoprdquo respectivement Dans une situation de laquo mapping raquo
plus complexe (C) ougrave les cadres de lecture sont montreacutes pour plus de clarteacute RATT
cartographie une large seacutequence codante (CDS) de H37Rv vers un locus dans F11 qui inclue
plusieurs codons laquo stop raquo laquo in-frame raquo En inseacuterant un deacutecalage du cadre de lecture (ie un
pseudogegravene) la traduction conceptuelle est conserveacutee Ceci diffegravere avec les deux gegravenes se
superposant preacutedis comme faisant partie du geacutenome F11
Tireacute de Otto TD et al 2011
89
321 Identification des mutations en lien avec la tomatidine
Apregraves avoir obtenu une seacutequence consensus agrave partir des seacutequences geacutenomiques
des 4 copies de chacune des souches parente S aureus ATCC 29213 et S aureus
NewbouldΔhemB les diffeacuterences geacutenomiques entre les souches parentes et le
geacutenome de reacutefeacuterence S aureus str Newman ont eacuteteacute eacutelimineacutees afin que ces
diffeacuterences nrsquoapparaissent pas comme des mutations lors de la comparaison des
seacutequences geacutenomiques des mutants avec celles de leur souche parente ces
diffeacuterences eacutetant plutocirct attribuables agrave la diffeacuterence qursquoil y a entre S aureus str
Newman et la souche parente et non pas agrave des mutations en lien avec la reacutesistance
des mutants
Le service bio-informatique du deacutepartement de biologie (Faculteacute des sciences
Universiteacute de Sherbrooke) a ensuite produit un tableau contenant pour chacun des
mutants tous les gegravenes muteacutes apregraves comparaison avec leur souche parentes Agrave cette
eacutetape on pouvait donc savoir dans quel gegravene se situaient les mutations quelle eacutetait
lrsquoeffet de cette mutation (mutation synonyme deacuteleacutetion insertion SNP) et en principe
srsquoil srsquoagissait pour un mecircme gegravene muteacute de la mecircme mutation puisque dans ce
tableau pour un mecircme gegravene chaque ligne correspond agrave des mutations diffeacuterentes
(Figure 6)
Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4Reacutesistance P1 Reacutesistance P2 Reacutesistance P3
1 2 3 1 2 3 1 2 3
Reacutesistance P2Reacutesistance P1 Reacutesistance P3
1 2 3 1 2 3 1 2 3 1 2
Reacutes P4
Figure 6 Exemple drsquoune partie du tableau compilation des mutations chez
les mutants ATCC 29213 reacutesistants agrave la GEN seule et agrave la GEN (+TO)
90
Agrave lrsquoaide de ce tableau (Figure 6) jrsquoai chercheacute les mutations qui pouvaient ecirctre
attribuables agrave la TO Chez les mutants S aureus ATCC 29213 reacutesistants jrsquoai
chercheacute les gegravenes qui eacutetaient muteacutes chez tous les mutants reacutesistants agrave la
combinaison GEN (+TO) ou dont la mutation apparaissait graduellement au fil des
paliers de reacutesistance mais dont ce mecircme gegravene nrsquoeacutetait pas muteacute chez les mutants
reacutesistants agrave la GEN seule Je pouvais donc estimer que cette mutation chez les
reacutesistants agrave la combinaison eacutetait en lien avec la preacutesence de la TO dans le milieu
322 Cheminement pour lrsquoidentification des gegravenes drsquointeacuterecirct
Afin de mrsquoaider dans ma recherche jrsquoai eu lrsquoaide du Pr Pierre-Eacutetienne Jacques
(Deacutepartement de biologie Faculteacute des sciences Universiteacute de Sherbrooke)qui a
mis les donneacutees de ce tableau dans un tableau croiseacute dynamique (TCD) ce qui a
permis de filtrer les donneacutees selon des critegraveres bien preacutecis me permettant drsquoarriver agrave
mes fins plus facilement Dans un premier temps un TCD a eacuteteacute creacuteeacute ougrave pour
chacun des mutants chaque gegravene ayant au moins une mutation eacutetait indiqueacute par un
laquo 1 raquo alors qursquoun laquo 0 raquo eacutetait marqueacute si ce gegravene nrsquoeacutetait pas muteacute Dans un second
temps la quantiteacute de laquo 1 raquo par palier de reacutesistance (correspondant au nombre de
mutants de ce palier posseacutedant une mutation dans ce gegravene) eacutetait indiqueacutee par un
chiffre allant de 0 pour laquo aucun mutant de ce palier de reacutesistance ne posseacutedait de
mutation dans ce gegravene raquo agrave 1 2 ou 3 selon le nombre de mutants de ce palier ayant
ce gegravene muteacute Jrsquoai ensuite pu soumettre le TCD agrave des filtres approprieacutes afin de faire
ressortir les gegravenes drsquointeacuterecircts selon des critegraveres de seacutelection preacutecis (Figures 7-8)
91
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
1
00
33
32
1
1
11
4
5
0
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
81
1_
10
03
33
21
11
14
50
NW
MN
_20
70
_1
00
12
32
18
14
50
NW
MN
_06
35
_0
01
13
21
17
14
31
NW
MN
_20
84
_0
02
12
22
27
14
42
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_04
12
lpl9
nm
20
12
32
23
92
44
1
NW
MN
_11
79
rbfA
11
02
12
22
72
44
1
NW
MN
_rR
NA
02
_0
11
21
22
27
24
42
NW
MN
_00
46
_2
10
13
21
37
24
31
NW
MN
_09
55
_2
01
23
11
37
24
21
NW
MN
_13
83
gpsA
21
02
31
13
72
42
1
NW
MN
_03
39
_1
11
21
31
37
34
42
NW
MN
_15
97
dn
aE2
10
22
30
37
23
31
NW
MN
_23
30
_2
10
23
20
37
23
21
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_2
07
0_
10
01
23
21
81
45
0
NW
MN
_09
28
qo
xC0
00
03
32
08
03
50
NW
MN
_10
28
_1
22
12
32
58
34
54
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
A B C
92
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENco
mb
oG
ENco
mb
oco
mb
oG
ENN
WM
N_0
39
8h
sdM
31
33
33
27
11
34
54
NW
MN
_04
26
_3
31
33
32
71
13
45
4
NW
MN
_05
24
sdrD
32
33
33
28
11
34
55
NW
MN
_07
39
_3
33
33
32
91
13
45
6
NW
MN
_11
64
hsl
U3
33
33
32
91
13
45
6
NW
MN
_05
23
sdrC
22
33
32
27
10
34
45
NW
MN
_20
61
fmtB
23
23
32
27
10
34
45
NW
MN
_04
03
lpl1
nm
33
22
33
28
10
34
55
NW
MN
_13
45
ebh
13
32
33
31
81
03
44
5
NW
MN
_16
49
_3
32
33
31
81
03
44
5
NW
MN
_17
00
hsd
M3
23
33
22
81
03
44
5
NW
MN
_00
42
_3
33
33
22
91
03
44
6
NW
MN
_05
80
_3
33
33
31
91
03
44
6
NW
MN
_13
44
ebh
23
31
32
22
79
34
44
NW
MN
_13
75
aro
A3
13
23
31
79
34
44
NW
MN
_23
97
fnb
B2
32
23
22
79
34
45
NW
MN
_04
01
_3
32
23
31
89
34
45
NW
MN
_07
33
_3
32
32
31
89
34
45
NW
MN
_17
45
_2
33
22
32
89
34
56
NW
MN
_01
66
coa
33
32
32
29
93
44
6
NW
MN
_05
25
sdrE
33
32
33
19
93
44
6
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
pal
iers
nb
pal
iers
2 d
ern
iers
pal
iers
2 d
ern
iers
pal
iers
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D Fig
ure
7
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez A
TC
C 2
921
3 d
an
s le
TC
D p
ar
dif
feacutere
nts
fil
tre
s
(A)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts la
ge
nta
mic
ine
ge2
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
(C
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts d
an
s le
pa
lier
1
de
reacute
sis
tan
ce
agravela
ge
nta
mic
ine
le1
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e (
+T
O)
ge7
et
nom
bre
de m
uta
nts
reacute
sis
tan
ts agrave
la la
gen
tam
icin
e (
+T
O)
de
s 2
de
rnie
rs p
alie
rs d
e r
eacutesis
tan
ce
ge5
(D
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e ge
7 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
ge
nta
mic
ine
(+
TO
) ge
9
93
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_16
29
ccp
A0
01
33
31
50
00
01
24
0
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_21
28
rpsM
11
12
21
12
22
00
31
74
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
NW
MN
_25
29
clfB
33
03
33
80
11
26
17
4
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e reacute
sist
ance
agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
89
6_
33
33
23
15
23
33
92
31
1
NW
MN
_09
96
_3
33
33
31
12
33
39
20
11
NW
MN
_10
25
_3
33
32
31
32
33
39
21
11
NW
MN
_10
28
_3
22
13
39
22
22
71
68
NW
MN
_13
45
ebh
12
23
22
11
12
32
27
16
9
NW
MN
_17
80
_3
22
13
29
21
22
71
57
NW
MN
_19
16
_3
32
23
31
32
33
38
21
11
NW
MN
_23
14
_3
33
32
31
52
33
39
23
11
NW
MN
_23
92
_2
33
32
38
22
32
81
69
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
On
b m
uta
nts
mu
teacutes
Pal
ier
de
reacute
sist
ance
agrave c
om
bo
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
16
29
ccp
A0
01
33
31
50
00
01
24
0N
WM
N_
21
28
rpsM
11
12
21
12
22
00
31
74
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
A B C
94
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_0
04
6_
1
01
00
28
22
32
2
10
9
NW
MN
_17
33
prs
A0
11
12
01
12
12
22
14
7
NW
MN
_19
23
_0
11
31
15
13
22
21
08
NW
MN
_20
37
_0
02
32
07
23
11
21
27
NW
MN
_22
77
_1
02
21
11
02
12
33
14
8
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
on
b m
uta
nts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Gen
eA
lias
Pal
ier
1P
alie
r 2
Pal
ier
3P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4P
alie
r 1
Pal
ier
2P
alie
r 3
Pal
ier
4G
ENTO
Co
mb
oN
WM
N_
01
43
_0
03
00
04
23
33
34
11
NW
MN
_0
40
6lp
l3n
m2
01
00
03
12
22
33
7
NW
MN
_1
65
2_
00
10
00
62
12
31
68
NW
MN
_2
01
2_
00
00
00
32
33
30
31
1N
WM
N_
20
15
mn
aA0
00
00
01
23
33
01
11
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
nb
mu
tan
ts
mu
teacutes
Pal
ier
de
reacutesi
stan
ce agrave
GEN
Pal
ier
de
reacute
sist
ance
agrave T
OP
alie
r d
e r
eacutesi
stan
ce agrave
co
mb
o
nom
bre
de m
uta
nts
de c
e p
alie
r ayant au m
oin
s 1
muta
tion d
ans c
e g
egravene
n
om
bre
de m
uta
nts
de c
ette c
ateacute
gorie a
yant au m
oin
s 1
muta
tion d
ans c
e g
egravene to
us p
alie
rs c
onfo
ndus
nom
bre
de p
alie
rs p
osseacutedant au m
oin
s 1
muta
nts
pour
ce g
egravenes
nom
bre
de m
uta
nts
muteacute
s p
our
ce g
egravenes d
ans les 2
dern
iers
palie
rs d
e r
eacutesis
tance d
e c
ette c
ateacute
gorie
D E Fig
ure
8
Seacute
lec
tio
n d
es
gegrave
nes
drsquoi
nteacute
recirct
ch
ez N
ew
bo
uld
Δh
em
Bd
an
s l
e T
CD
par
dif
feacutere
nts
fil
tres
(A)
Nom
bre
de
mu
tan
ts
reacutesis
tan
ts agrave
la tom
atid
ine
ge15
(B
) N
om
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atid
ine
ge15
et no
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
(C)
No
mbre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la tom
atid
ine
ge15
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3(
D)
Nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la t
om
atidin
ege
10
et no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la
tom
atid
ine
(+G
EN
) ge
7
(E)
No
mb
re d
e m
uta
nts
reacute
sis
tan
ts agrave
la to
ma
tid
ine
le6
et
nom
bre
de
mu
tan
ts r
eacutesis
tan
ts agrave
la g
en
tam
icin
e le
3 e
t
no
mb
re d
e m
uta
nts
reacutesis
tan
ts agrave
la t
om
atid
ine
(+G
EN
) ge
7
95
Le TCD mrsquoa permis de rapidement identifier les gegravenes drsquointeacuterecircts Agrave lrsquoaide du site CMR
(Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) 2013) jrsquoai
ensuite veacuterifieacute la fonction des gegravenes identifieacutes afin de confirmer leur inteacuterecirct (Tableau
1)
Tableau 1 Fonction des gegravenes drsquointeacuterecirct identifieacutes (A) chez les mutants ATCC
29213 reacutesistants et (B) chez les mutants NewbouldΔhemB reacutesistants
Gegravene Alias Fonction
NWMN_0811 NADH dh complexe 1 chaine de transport des eacutelectrons
NWMN_2070 _membrane prot Putative- Toxin production+resistance drug transporteur-
similar to multidrug transporteur 85
NWMN_0635 _membrane prot Putative- Bordeacute par AraC(reacutegulateur opeacuteron arabinose) ou
SsaA-likeprot(lieacute au quorum sensing RNAIII(secretory antigen)
NWMN_2084 _truncated transposase for Is1272- mobile+extrachromosomal element
fonctiontransposon fonction
NWMN_0928 qoxC quinol oxidase polypeptide IIIcomplexe 4 chaine de transport des eacutelectrons
NWMN_0739 _ conserved hypothetical prot fonction inconnues
NWMN_1164 hslU ATP-dependent protease ATP-binding subunit
NWMN_0524 sdrD Ser-Asp rich fibrinogenbone sialoprotein-binding protein sdrD
NWMN_0042 _ lipoprotein putativecell enveloppe- lipoprot en tendem
NWMN_0580 _conserved hypothetical prot fonction inconnuesDNA metabolism DNA
replication recombination and repair
NWMN_1745 _
NWMN_0166 coa coagulase precursor
NWMN_0525 sdrE Ser-Asp rich fibrinogenbone sialoprotein-binding protein SdrE
A
96
Apregraves avoir aligneacute speacutecifiquement une par une chacune des seacutequences (Fasta) des
gegravenes drsquointeacuterecirct de chaque mutant avec leur souche parente jrsquoai utiliseacute le programme
de visualisation et de comparaison de seacutequences ClustalW2 (ClustalW2 2013) afin
de confirmer que la mutation eacutetait une vraie mutation et non une erreur de
seacutequenccedilage ou drsquoalignement et que la mutation eacutetait identique chez tous les mutants
ayant le gegravene drsquointeacuterecirct muteacute Cette eacutetape a permis de restreindre encore la liste des
gegravenes pouvant ecirctre en lien avec la cible ou le mode drsquoaction de la TO (Figure 9)
Gene Alias Fonction
NWMN_0896 _conserved hypothetical prot fonction inconnue- comK family prot-
regulation DNA interaction- ORFIDresssemble agrave facteur de transcription
NWMN_0996 _ conserved hypothetical prot fonction inconnue- prophage fonction
NWMN_1025 _ phage major tail protprophage fonction
NWMN_1629 ccpAcatabollite control protA virulence determinant production and antibiotique
resistance
NWMN_1916 _ conserved hypothetical prot-fonction inconnue ou prophage prot
NWMN_2314 _drug resistance transporteur- EmrBQacA subfamily prot- toxin
production+resistance- transport and binding prot
NWMN_0046 _ hypothetical prot
NWMN_2015 mnaA UDP-Glc-Nac 2 epimerase
NWMN_2012 atpE ATP synthase subunit C
NWMN_1652 _ multidrug resistance transporter protein B
NWMN_0143 _ oligopeptide ABC transporter ATP-binding protein
B
97
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
GegravenehelliphelliphelliphelliphellipNWMN_811helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellipNWMN_928helliphelliphelliphelliphellip
hellip 25-TTGTACTTGGTGCTGGTTA-45hellip 409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGCATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
hellip 25-TTGTACTTGGT - CTGGTTA-45hellip409-TAGGAACGAATGGTTG-426hellip
ATCC29213
Isolat 1
Isolat 2
Isolat 3
Isolat 1
Isolat 2
Isolat 3
Niveau reacutesistance 3
Niveau reacutesistance 1
NADH dh Quinol ox
Niveau 1
Niveau 3
ATCC 29213 reacutesistants
agrave TO (+GEN)
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
hellipnwmn_1629helliphelliphelliphelliphellipnwmn_2012helliphellip
Mutants reacutesistants agrave TO
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellipIsolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGAGCAGGT-55hellip
Isolat 1 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 2 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Isolat 3 hellip440-AAATGTTAAGG-452helliphelliphelliphellip43-TAGGATCAGGT-55hellip
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55hellip
ccpA ATP synthase c
Niveau 1
Niveau 2
Niveau 3
Niveau 4
A
B
98
Jrsquoai ensuite chercheacute le rocircle de ces gegravenes sur le site CMR afin de veacuterifier si leur
mutation pouvait expliquer la reacutesistance agrave lrsquoantibiotique (Comprehensive Microbial
Ressouce (J Craig Venter Institute (JCVI) 2013) Dans une premiegravere eacutetape le rocircle
drsquoun gegravene a eacuteteacute deacutetermineacute en allant tout simplement voir directement le rocircle de ce
gegravene chez Staphylococcus aureus str Newman Dans le cas ougrave le gegravene nrsquoavait pas
de fonction connue chez Saureus str Newman je suis alleacutee voir quelle eacutetait la
fonction de ce mecircme gegravene (et des gegravenes flanquant celui-ci) chez diffeacuterentes souches
de Staphylococcus aureus Advenant le cas ougrave malgreacute tout je nrsquoeacutetais toujours pas
capable de deacuteterminer le rocircle de ce gegravene je suis alleacutee veacuterifier si sa seacutequence pouvait
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 hellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55 helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellipIsolat 3 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 1 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Isolat 2 hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip185-AATTACATTCA-197helliphelliphellip495-ATAACAATGG-506hellip
Mutants resistants agrave GEN (4 microgml)+TO
Newbould_hemB hellip440-AAATGGTAAGG-452helliphelliphellip43-TAGGAGCAGGT-55helliphellip 185-AATTGCATTCA-197helliphelliphellip495-ATAGCAATGG-506hellip
hellipnwmn_1629helliphelliphelliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_2012helliphelliphelliphellipnwmn_0143hellip
ccpAATP synthase c
ABC transp
Niveau 1
Niveau 2
Niveau 3
Niveau 4
TO +GEN (4microgml)
Figure 9 Comparaison de seacutequences des gegravenes drsquointeacuterecirct (A) nwmn_0811 et
nwmn_0928 chez les mutants ATCC 29213 reacutesistants agrave la combinaison gentamicine
(+TO) leur souche parente ATCC 29213 (B) nwmn_1629 et nwmn_2012 chez les
mutants hemB reacutesistants agrave la TO et leur souche parente NewbouldΔhemB (C)
nwmn_1629 nwmn_2012 et nwmn_0143 chez les mutants hemB reacutesistant agrave la
tomatidine (+genta) et leur souche parente hemB
C
99
avoir une forte homologie avec un gegravene drsquoune autre espegravece Malgreacute tout certains
gegravenes portent toujours la mention laquo de fonction inconnue raquo
La meacutethodologie que nous avons employeacutee nous a permis drsquoidentifier plusieurs
gegravenes possiblement impliqueacutes dans reacutesistance de nos souches aux antibiotiques
testeacutes
1- chez Staphylococcus aureus ATCC 29213 reacutesistant agrave la gentamicine (+TO)
(Figure 9A)
- NWMN_0811 codant possiblement pour la NADH deacuteshydrogeacutenase
- qoxC (NWMN_0928) codant pour la quinol oxidase polypeptide III
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
2- chez NewbouldΔhemB reacutesistant agrave la tomatidine (Figure 9B)
- ccpA (NWMN_1629) codant pour la laquo catabolite control protein A raquo
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
3- chez NewbouldΔhemB reacutesistant agrave la tomatidine (+gentamicine) (Figure 9C)
- NWMN_0143 codant pour un ABC transporteur
- atpE (NWMN_2012) codant pour la sous-uniteacute C de lrsquoATP synthase
Lrsquoanalyse de ces diffeacuterents gegravenes nous amegravene agrave suspecter la sous-uniteacute c de lrsquoATP
synthase comme eacutetant la cible potentielle de la tomatidine chez Staphylococcus
aureus
33 Modegravele-hypothegravese du mode drsquoaction de la tomatidine
Les gegravenes muteacutes pointent en grande partie vers la chaine respiratoire qursquoon appelle
aussi chaine de transport des eacutelectrons
100
La NADH deacuteshydrogeacutenase (NWMN_0811) fait partie du complexes I de la chaine de
transport de eacutelectrons alors que la quinol oxidase polypeptide III (NWMN_0928 ou
QoxC) est une composante du complex III et la sous-uniteacute c de lrsquoATP synthase (AtpE)
fait partie eacutevidement du complexe de lrsquoATP synthase ou complexe V de la chaine
respiratoire (Figure 10)
Figure 10 Chaine de transport des eacutelectrons et identification des complexes
muteacutes chez les mutants reacutesistants
Les courbes de croissance des mutants ATCC29213 reacutesistants agrave la combinaison
GEN(+TO) appuient le fait que ces mutations affectent la chaine de transport des
eacutelectrons ce qui affecte le niveau de production drsquoATP et donc la croissance
bacteacuterienne En effet les mutants du troisiegraveme niveau de reacutesistance (qui possegravede
deux gegravenes de la chaine de transport des eacutelectrons de muteacutes) ont une croissance
plus lente que les mutants du premier niveau de reacutesistance qui eux ont un seul gegravene
dans la chaine de transport des eacutelectrons de muteacute (Figure 11)
Quinoloxidase
NADHdehydrogenase
ATPsynthetase
Proton gradient
XXX
101
Figure 11 Courbes de croissance des mutants ATCC 29213 reacutesistants agrave la
combinaison GEN+TO du premier niveau de reacutesistance (bleu) et du troisiegraveme niveau
de reacutesistance (vert)
Voici plus preacuteciseacutement comment fonctionne la chaine de transport des eacutelectrons chez
les 2 pheacutenotypes-parents selon le modegravele-hypothegravese eacutetabli avec nos reacutesultats et
analyses
Tout drsquoabord chez le prototype ou souche de pheacutenotype normal (ex ATCC 29213) la
chaine de transport des eacutelectrons fonctionne normalement Les complexes de la
chaine de transport des eacutelectrons sont chargeacutes de transfeacuterer des protons vers
lrsquoexteacuterieur de la cellule en transfeacuterant en mecircme temps des eacutelectrons drsquoun complexe agrave
un autre jusqursquoagrave un accepteur final qui est lrsquoO2 lorsque crsquoest possible Se faisant il se
creacutee un gradient eacutelectrochimique de part et drsquoautre de la membrane et les protons
accumuleacutes du cocircteacute externe sont utiliseacutes par la suite par lrsquoATP synthase afin de
pouvoir produire de lrsquoATP dans la cellule
0 4 8 1 2 1 6 2 0 2 4
0 0 1
0 1
1
C o u r b e d e c r o is s a n c e d e s m u ta n ts A T C C 2 9 2 1 3
reacute s is ta n ts agrave G E N + T O (8 m g L )
te m p s (h )
OD
60
0n
m
Niveau 3
Niveau 1
102
Le transfert drsquoeacutelectrons ne se fait pas sans coucirct et des produits toxiques pour la
cellule sont produits en mecircme temps ce sont les espegraveces reacuteactives de lrsquooxygegravene ou
ROS (Reactive Oxygen Species) Cependant les prototypes par leur reacuteponse au
stress produisent des enzymes capables de deacutetoxifier le milieu (Figure 12A)
Chez les souches SCV ici la souche controcircle NewbouldΔhemB la chaine de
transport des eacutelectrons est deacuteficiente parce que le gegravene permettant la production
drsquoheacutemine qui entre dans la composition des cytochromes est supprimeacute Les
cytochromes entrent dans la composition de plusieurs composantes de la chaine
respiratoire
Lorsque les complexes de la chaine sont alteacutereacutes il y a diminution du transfert de
protons vers lrsquoexteacuterieur une diminution de la production drsquoATP et une plus grande
production de ROS (Figure 12B)
Figure 12 Scheacutematisation du fonctionnement de la chaine de transport des
eacutelectrons chez les souches parentes (A) ATCC 29213 et (B) NewbouldΔhemB
I III IV V I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
eacute eacute
eacute
H+ H+
ATPO2 H2O
ROS +ROS
Prototype SCV (hemB)
H+
0811 0928 2012 0811 0928
hegraveme
2012
A B
103
La gentamicine a besoin drsquoune bonne polarisation membranaire donc drsquoune chaine de
transport des eacutelectrons fonctionnelle pour pouvoir peacuteneacutetrer dans la cellule (Bryan et
Kwan 1983 (Bryan and van den Elzen 1977 Davis 1987 Taber et al 1987) Une
fois entreacutee elle inhibe la synthegravese proteacuteique ce qui provoque la mort cellulaire La
CMI de la gentamicine contre ATCC 29213 est de 05-1 microgml (Figure 13A)
La tomatidine elle irait se lier agrave lrsquoATP synthase pour en diminuer la fonctionnaliteacute et
ce tant chez ATCC 29213 que chez hemB Cette liaison amegravenerait une diminution
de la production drsquoATP et conseacutequemment provoquerait agrave plus ou moins long terme
une augmentation de production de ROS par lrsquoalteacuteration de la chaine de transport des
eacutelectrons Cependant comme la cellule prototype produit efficacement les enzymes
de deacutetoxification lrsquoimpact de la tomatidine est pratiquement nulle sur les souches
prototypes La CMI de la tomatidine est gt16microml contre ATCC 29213 (Figure 13B)
104
Figure 13 Scheacutematisation du mode drsquoaction de (A) la gentamicine et (B) la
tomatidine sur la souche ATCC 29213
Chez ATCC 29213 lrsquoajout de 8microgml de tomatidine agrave la Gentamicine amegravene une
diminution de 8X agrave 16X la CMI de la gentamicine En effet la CMI de la gentamicine
passe de 05-1microgml agrave 006microgml (Figure 14AB)
Lors de lrsquoajout de la tomatidine agrave la gentamicine crsquoest la tomatidine qui se lie dans
un premier temps agrave lrsquoATP synthase reacuteduisant ainsi sont niveau drsquoactiviteacute Il y a alors
de faccedilon transitoire une augmentation de la concentration de protons agrave lrsquoexteacuterieur de
la cellule ce qui permet une entreacutee massive de gentamicine ainsi qursquoune rapide et
forte inhibition de la synthegravese proteacuteique Dans un second temps il y aura eacutegalement
une alteacuteration de la chaine de transport des eacutelectrons conduisant agrave une
augmentation de la production de ROS Crsquoest cependant lrsquoentreacutee massive de
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI TO gt16 microgmlCMI GEN 05-1 microgml
+ROS
ROS mais deacutetoxification efficace
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
105
gentamicine entrainant une forte inhibition de la synthegravese proteacuteique qui provoquera
dans ce cas-ci la mort cellulaire (Figure 14B)
Figure 14 Scheacutematisation de lrsquoaction de (A) la gentamicine seule et de (B) la
gentamicine lorsqursquoon y ajoute 8 microgml de tomatidine contre ATCC 29213
Chez les mutants ATCC29213 reacutesistants agrave la combinaison GEN+TO les complexes I
et III sont muteacutes et donc ont une fonctionnaliteacute alteacutereacutee ce qui amegravene une
deacutepolarisation membranaire Une membrane moins polariseacutee amegravene une plus faible
entreacutee de gentamicine et donc une plus faible inhibition proteacuteique La CMI de la
gentamicine passe en effet de 05-1 microgml chez la souche parente agrave 2 microgml chez les
mutants reacutesistants
La tomatidine en se liant agrave lrsquoATP synthase accentuera la deacutepolarisation
membranaire provoquant une augmentation de la production des ROS et une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN (TO 8 microgml) 006 microgmlCMI GEN 05-1 microgml
+ROS
polarisation entreacutee massive de GEN
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
GENH+
BA
106
diminution de la production drsquoATP De fait les mutants reacutesistants ont une croissance
plus lente (Figure 15)(Figure 11)
Figure 15 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur ATCC
29213 et de (B) la gentamicine et la tomatidine sur les mutants ATCC 29213
reacutesistants agrave la combinaison gentamicine (+TO)
Chez les SCV la tomatidine se lie aussi agrave lrsquoATP synthase et ce faisant amegravene une
diminution de la production drsquoATP Cette diminution de la production drsquoATP megravenera
ultimement agrave une alteacuteration de la chaine de transport des eacutelectrons et une
augmentation des ROS (Murphy 2009) tout comme chez les souches prototypes
Cependant comme le SCV a deacutejagrave une chaine de transport des eacutelectrons deacuteficiente
une faible production drsquoATP par lrsquoATP synthase et une forte production de ROS
lrsquoajout drsquoune faible concentration de tomatidine (la CMI est de 006microgml) amegravene une
I III IV Veacute eacute
eacute
H+ H+
ATPO2 H2O
H+ H+
ROS
Prototype Prototype R GEN (+TO [8 microgml])
H+
I III IVeacute eacute
eacuteO2 H2O
H+
GEN GEN
Synthegravese proteacuteique Synthegravese proteacuteique
CMI GEN 2 microgmlCMI GEN 05-1 microgml
+ROS
R agrave GEN (+TO) mais mutants ont une
croissance diminueacutee (+ROS et -ATP)
V
H+
ATP
TO
H+
0811 0928 2012 0811 0928
A B
107
augmentation de chacun de ces pheacutenomegravenes ce qui est suffisant pour provoquer la
mort cellulaire (Figure 16A)
Dans les premiers niveaux de reacutesistance agrave la tomatidine chez les mutants hemB la
reacutesistance (la CMI passe de 006 agrave 025-1microgml) est ameneacutee par la mutation du gegravene
ccpA (Catabolite controcircle protein A) ce qui megravenerait agrave une augmentation de la
production drsquoATP par lrsquoutilisation drsquoune autre voie que lrsquoATP synthase etou agrave une
diminution de la production de ROS (Figure 16B) Le rocircle de la CcpA en lien avec la
reacutesistance agrave la tomatidine est encore neacutebuleux Toutefois on sait que normalement
ce gegravene agit comme reacutegulateur transcriptionnel des gegravenes codant pour des enzymes
du cycle de Krebs et qursquoil a un rocircle de reacutegulation dans le meacutetabolisme du carbone et
lrsquoutilisation des substrats (Somerville et Proctor 2009 Goumltz et al 2006)
Figure 16 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du premier palier de reacutesistance agrave la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO 006 microgml
H+
SCV
I III IVeacute eacute
eacuteO2 H2O
+ROS
H+TO
Niveau 1
CcpA (CMI TO 025-1 microgml)
ROS
ATP
V
H+
ATP
TO0811 0928
2012
1629
0811 0928
2012
BA
108
Chez les SCV du deuxiegraveme palier de reacutesistance agrave la tomatidine non seulement le
gegravene ccpA est toujours muteacute mais le gegravene de la sous-uniteacute c de lrsquoATP synthase est
eacutegalement muteacute La mutation dans le site de liaison mecircme de la tomatidine empecircche
sa liaison et donc empecircche son effet sa CMI passant de 006 agrave gt16microgml (Figure
17B)
Malgreacute la mutation dans le gegravene lrsquoATP synthase resterait quand mecircme fonctionnel
puisque la mutation nrsquoamegravene pas la mort cellulaire comme crsquoest le cas lorsque la
tomatidine lie lrsquoATP synthase chez le SCV parent (Figure 17A)
Figure 17 Scheacutematisation de lrsquoaction de la tomatidine chez (A) les SCV (hemB)
et (B) chez les SCV (hemB) du deuxiegraveme palier de reacutesistance agrave la tomatidine
Tout comme chez les mutants hemB reacutesistant agrave la tomatidine seule les mutants
hemB reacutesistant agrave la tomatidine lorsqursquoen preacutesence de gentamicine possegravedent
eacutegalement une mutation dans le gegravene de lrsquoATP synthase ce qui rend la tomatidine
SCV R agrave TO
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI TO006 microgml
H+
SCV Niveau 2CMI TO gt16 microgml
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+TO
TO
Niveaux 1 et 2
CcpA (CMI TO 025-1 microgml)
ROS
ATP
0811 0928
2012
1629
0811 0928
2012
A B
109
inactive puisqursquoincapable de srsquoy lier (CMI gt16microgml) (Figure 18B) Chez les SCV non
muteacutes (hemB) la gentamicine nrsquoentre que faiblement ducirc agrave la deacutepolarisation de sa
membrane (CMI 4-8 microgml) (Figure 18A) La mutation dans le gegravene codant pour une
pompe (NWMN_0143 codant pour un ABC transporteur) chez les mutants hemB
reacutesistant agrave la tomatidine (+gentamicine) amegravenerait une augmentation de lrsquoactiviteacute de
cette pompe permettant de rejeter hors de la cellule le peu de gentamicine eacutetant
entreacutee La mutation de cette pompe fait passer la CMI de la gentamicine de 4-8
microgml pour hemB agrave 16 microgml pour les mutants hemB reacutesistant agrave la combinaison
gentamicine (+TO) ayant cette mutation (Figure 18B)
Figure 18 Scheacutematisation de lrsquoaction de (A) la gentamicine seule sur le SCV
hemB et de (B) la gentamicine et la tomatidine sur les mutants hemB reacutesistants
agrave la combinaison tomatidine (+gentamicine)
Donc selon notre modegravele-hypothegravese lrsquoaction de la tomatidine et de la gentamicine
sur S aureus deacutepend de la liaison agrave leur cible mais a aussi un lien avec la
SCV R agrave TO (+GEN [4microgml])
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
++ROS
CMI GEN 4-8 microgml
H+
SCV
I III IV Veacute eacute
eacute
H+
ATPO2 H2O
+ROS
H+ TO
GEN
CMI GEN 16 microgml
CMI TO gt16 microgml
0811 0928
2012
0811 0928 20120143
GEN
Synthegravese proteacuteique
BA
110
polarisation membranaire et la production de ROS La mesure de la polarisation et
concentration cellulaire en ROS chez nos mutants et nos souches parents en
preacutesence ou non drsquoantibiotique nous permettra de valider notre modegravele-hypothegravese
Chez une bacteacuterie meacutetaboliquement active et posseacutedant une membrane
cytoplasmique intacte il y a normalement une diffeacuterence de potentiel eacutelectrique entre
le cocircteacute inteacuterieur et le cocircteacute exteacuterieur de la membrane lrsquointeacuterieur eacutetant plus neacutegatif de
100 agrave 200 mV par rapport agrave lrsquoexteacuterieur Une diminution de la magnitude de ce
potentiel membranaire est appeleacutee deacutepolarisation alors qursquoune augmentation de ce
potentiel est appeleacutee hyperpolarisation Le potentiel membranaire sera reacuteduit agrave zeacutero
srsquoil y a rupture de la membrane cest-agrave-dire srsquoil y a formation de trous assez grands
permettant aux ions inorganiques de passer librement Ce pheacutenomegravene peut arriver
lorsque les cellules sont tueacutees par la chaleur le froid ou les antibiotiques β-Lactams
Dans ces circonstances la membrane devient permeacuteable agrave diffeacuterents colorants
comme le propidium iodide (PI) et le TO-PRO3 et le changement est irreacuteversible
Plusieurs classes de composeacutes chimiques notamment les ionophores peuvent
eacutegalement alteacuterer le potentiel membranaire sans pour autant permettre la
permeacuteabiliteacute au PI TO-PRO3 et autres colorants similaires chez les bacteacuteries
lrsquoadministration drsquoun proton-ionophore tel le carbonyl cyanide 3-
chlorophenylhydrazone (CCCP) reacuteduira le potentiel membranaire agrave zeacutero
Des tests preacuteliminaires sur la mesure du potentiel membranaire (BacLighttrade Bacterial
Membrane Potential Kit (B34950) drsquoInvitrogen et lecture au cytomegravetre de flux (FACS))
(Novo et al 2000) chez nos deux souches parentes mets en eacutevidence que ATCC
29213 est plus polariseacute que NewbouldΔhemB (Figure 19A) Toutefois malgreacute
qursquoayant une plus faible polarisation membranaire NewbouldΔhemB nrsquoest pas
complegravetement deacutepolariseacute puisqursquoil est possible de le deacutepolariser davantage agrave lrsquoaide
drsquoun agent deacutepolarisant tel que le CCCP (carbonyl cyanide 3-chlorophenylhydrazone)
(Figure 19B) Le TO-PRO3 a eacuteteacute utiliseacute comme controcircle afin de confirmer la viabiliteacute
de nos cellules (donneacutees non montreacutees)
111
A B
Figure 19 Mesure de la polarisation membranaire de S aureus ATCC 29213 et
de NewbouldΔhemB agrave lrsquoaide du BacLighttrade Bacterial Membrane Potential Kit
(Invitrogen) (Lecture au cytomegravetre de flux agrave lrsquoaide du programme FCS Express) (A)
Ratio FL3-H (axe des Y)FL1-H (axe des X) de ATCC 29213 (rouge) et de
NewbouldΔhemB (noir) (B) Lecture des rouges (FL3-H) sur NewbouldΔhemB traiteacute
avec le DiOC2(3) (33prime-diethyloxacarbocyanine iodide) (noir) ou avec le CCCP (bleu)
ou sur les cellules chauffeacutees (rouge)
La sous-uniteacute c de lrsquoATP synthase (AtpE) semble donc la cible la plus probable de la
tomatidine Afin de veacuterifier si les mutations identifieacutees pouvaient avoir un effet
veacuteritable sur la proteacuteine nous avons fait un alignement de seacutequences noteacute les bases
muteacutees et veacuterifieacute si la mutation amenait un changement drsquoacide amineacute et si ce
changement drsquoacide amineacute pouvait avoir un impact sur la proteacuteine finale De fait
nous avons trouveacute que les deux diffeacuterentes mutations dans lrsquoAtpE amenaient un
changement significatif drsquoacides amineacutes pouvant amener une variation fonctionnelle
FL3-H
Count
100
101
102
103
104
0
18
36
53
71
Chauffeacute
+CCCP+DiOC2
Chauffeacute
FL1-H
FL3-H
100
101
102
103
104
100
101
102
103
104
hemB
29213
VERT
RO
UG
E
ROUGE
ΔhemB
112
de la proteacuteine La mutation de la base 49 change un G pour un T il srsquoagit drsquoun SNP
nrsquoamenant pas de changement de cadre de lecture La mutation fait passer lrsquoacide
amineacute alanine (A) qui est apolaire agrave une seacuterine (S) qui est polaire La mutation de la
base 190 change un G pour un A et est aussi un SNP nrsquoamenant pas de changement
de cadre de lecture Cette mutation fait passer lrsquoacide amineacute alanine (A) qui est
apolaire agrave une threacuteonine (T) qui est polaire (Figure 20)
gtlcl||NWMN_2012|atpE|72493880 F0F1 ATP synthase subunit C
1 10 19 31 43 49 58
ATG AAT TTA ATC GCA GCA GCA ATC GCA ATT GGT TTA TCA GCA TTA GGA GCA GGT ATC GGT
M N L I A A A I A I G L S A L G A G I G
(S)
17 61 73 85 97 109 118
AAC GGT TTA ATC GTT TCA AGA ACA GTT GAA GGT GTA GCA CGT CAA CCA GAA GCA CGT GGT
N G L I V S R T V E G V A R Q P E A R G
21 40
121 133 145 157 169 178
CAA TTA ATG GGT ATC ATG TTC ATT GGT GTA GGT TTA GTT GAG GCA TTA CCT ATC ATC GGT
Q L M G I M F I G V G L V E A L P I I G
41 60
181 190 199 211
GTA GTA ATT GCA TTC ATG ACA TTT GCT GGA TAA
V V I A F M T F A G
(T)
61 64 70
Figure 20 Alignement de la seacutequence geacutenique et de la seacutequence proteacuteique du gegravene
atpE de S aureus Newman avec mise en eacutevidence des mutations (rose) en lien avec
la cible de la tomatidine Lrsquoalignement a eacuteteacute effectueacute agrave lrsquoaide du site CMR de JCVI
113
CHAPITRE 4
DISCUSSION
LrsquoATP synthase est eacutevidemment un complexe proteacuteique ubiquitaire tant chez les
procaryotes que les eucaryotes Malgreacute ce fait il semble que la cible de la tomatidine
soit speacutecifique aux bacillales celle-ci ne deacutemontrant aucune activiteacute contre les
bacteacuteries agrave Gram neacutegatif et les lactobacillales On suppose donc que lrsquoATP synthase
de ces espegraveces doit ecirctre speacutecifique au niveau du site de liaison de la tomatidine
Lrsquoecirctre humain en tant qursquoespegravece eucaryote est eacutegalement pourvu de ce complexe
proteacuteique et on peut naturellement se demander si lrsquoutilisation clinique de cette
moleacutecule pourrait ecirctre cytotoxique De fait plusieurs composeacutes naturels tel
lrsquooligomycine et la venturicidine sont connus pour inhiber lrsquoactiviteacute de lrsquoATP synthase
par leur interaction avec la sous-uniteacute c de ce complexe Cependant ces moleacutecules
ne sont pas seacutelectives et nrsquoinhibent pas seulement lrsquoATP synthase bacteacuterien mais
aussi lrsquoATP synthase mitochondrial (Haagsma et al 2009 Matsuno-Yagi et Hatefi
1993) (Figure 1) Neacuteanmoins il existe maintenant une moleacutecule qui a pour cible
lrsquoATP synthase bacteacuterien et qui est en processus en vue drsquoune utilisation clinique
chez lrsquohomme le TMC207 (Haagsma et al 2009))
114
Figure 1 Sites drsquoinhibition de lrsquoATP synthase Les sites de liaison dans lrsquoATP
synthase des inhibiteurs reacuteveacuteleacutes par des eacutetudes biochimiques et structurales sont
indiqueacutes par des cercles rouges et les sites de liaisons qui nrsquoont pas eacuteteacute totalement
clarifieacutes sont indiqueacutes par des cercles verts (Hong et Pedersen 2008)
Le TMC207 (Figure 2A) est un composeacute de la classe des diarylquinoline posseacutedant
un nouveau meacutecanisme drsquoaction lrsquoinhibition de lrsquoATP synthase bacteacuterien De fait la
cible et le meacutecanisme drsquoaction du TMC207 sont diffeacuterents des autres agents
antituberculeux Lrsquoinhibition de lrsquoATP synthase peut mener agrave une baisse de la
production drsquoATP et un deacutebalancement dans lrsquohomeacuteostasie du pH ces 2 effets
pouvant contribuer agrave la diminution de la survie de la bacteacuterie (Rao et al 2001
Andries et al 2005)
Le TMC207 est une moleacutecule ayant une forte activiteacute bacteacutericide contre
Mycobacterium tuberculosis et autres espegraveces mycobacteacuteriennes mais peu drsquoactiviteacute
115
contres les autres espegraveces bacteacuteriennes Lrsquoindex de seacutelectiviteacute de TMC207 pour
lrsquoATP synthase mycobacteacuterienne (CMI50 gt200 microml) compareacute agrave lrsquoATP synthase
mitochondriale (CMI50 001 microgml) est de plus de 20000 Un indice de seacutelectiviteacute au
dessus de 1000 est consideacutereacute dans le deacuteveloppement drsquoune drogue comme un
preacutealable pour une moleacutecule candidate (Haagsma et al 2009) La diversiteacute dans les
gegravenes atpE explique la reacutesistance au TMC207 et justifie la faible affiniteacute de cette
drogue agrave lrsquoATP synthase mitochondriale (Matteelli et al 2010) (Figure 3)
Figure 2 Structure chimique de (A) TMC207 et (B) son analogue anti bacteacuteries
agrave Gram positifs (Balemans et al 2012)
Lorsqursquoon compare la seacutequence proteacuteique de la sous-uniteacute c de lrsquoATP synthase
(atpE) de quelques espegraveces de lrsquoordre des Bacillales (tel Saureus B cereus B
subtilis et L monocytogenes) avec des espegraveces appartenant agrave drsquoautres ordres
bacteacuteriens on constate que les seacutequences divergent ce qui est en accord avec la
speacutecificiteacute des moleacutecules Les points de mutations amenant une reacutesistance au
TMC207 chez Mycobacterium smegmatis sont speacutecifiques aux mycobacteacuteries et les
deux principaux points de mutations D32V(ouP) et A67P chez M smegmatis sont
situeacutes tous les deux dans les heacutelices alpha qui entourent le site catalytique de la
proteacuteine (E65) qui est impliqueacute dans le transport des protons (Andries et al 2005)
Lrsquoacide amineacute aspartate (D) agrave la position 32 et lrsquoalanine (A) agrave la position 67 sont tregraves
speacutecifiques agrave M smegmatis
A B
116
Reacutecemment un groupe de chercheurs (Balemans et al 2012) on mit en lumiegravere une
autre moleacutecule (Figure 2B) de la classe des diarylquinolines dont le meacutecanisme
drsquoaction eacutetait aussi lrsquoinhibition de la sous-uniteacute c de lrsquoATP synthase mais chez les
bacteacuteries agrave Gram positif tels Streptococcus pneumoniae et S aureus Cette moleacutecule
irait lier les acides amineacutes valines (V) situeacutes agrave la position 50 et 62 de lrsquoAtpE de S
aureus (Donneacutees non montreacutees chez S pneumoniae(Balemans et al 2012)) En
effet une mutation amenant le changement de la valine en position 50 par une
isoleucine (I) etou une mutation amenant le changement de la valine en position 62
par une alanine (A) amegravene la reacutesistance de la bacteacuterie agrave la moleacutecule Cependant L
monocytogenes et Bacillus subtilis ne possegravedent pas de valine agrave la position 50 alors
qursquoils font eacutegalement partie des bacteacuteries agrave Gram positif (Il faut noter que chez
Bacillus sp PS3 (Swiss-Prot assession Ndeg P 00845) possegravede une valine agrave la position
50 (Balemans et al 2012)) Par contre tout comme pour les autres bacteacuteries agrave Gram
positif testeacutees les deux espegraveces possegravedent une valine agrave la position 62 (Figure 3)
Chez B cereus B subtilis B anthracis S aureus et L monocytogenes les points de
divergence sont plutocirct A17S et A64T Cependant B pseudofirmus qui fait eacutegalement
partie de lrsquoordre des Bacillales possegravede une glycine (G) agrave la position 17 plutocirct qursquoune
alanine (A) et une seacuterine (S) plutocirct qursquoune alanine (A) agrave la position 64 comme les
espegraveces preacuteceacutedentes Suivant ces reacutesultats et si la cible de la tomatidine est bien
ces deux points de mutation la tomatidine ne devrait pas pouvoir se lier agrave lrsquoAtpE de B
pseudofirmus et devrait donc nrsquoavoir aucune activiteacute contre cette souche alors que
nous avions extrapoleacute que le spectre drsquoactiviteacute de la tomatidine eacutetait tous les
Bacillales De plus E coli et M smegmatis possegravedent eacutegalement une alanine (A) agrave la
position 17 alors que la tomatidine ne possegravede aucune activiteacute contre ces espegraveces (M
smegmatis nrsquoa pas eacuteteacute eacutevalueacute il srsquoagit ici drsquoune extrapolation des reacutesultats de CMI)
par contre ces deux espegraveces possegravedent des acides amineacutes diffeacuterents drsquoune alanine
(A) agrave la position 64 ce qui pourrait leur confeacuterer une reacutesistance agrave la tomatidine telle
qursquoobserveacutee On remarque eacutegalement que les acides amineacutes agrave ces deux positions
117
sont totalement diffeacuterents chez lrsquohomme (Homo sapiens) ce qui suggeacutererait que la
tomatidine ne possegravederait pas drsquoactiviteacute antimitochondriale (Figure 3)
Selon ces reacutesultats on peut se demander si les deux mutations sont essentielles agrave la
liaison de la moleacutecule ou si seulement lrsquoalanine (A) agrave la position 64 est neacutecessaire
On doit se rappeler que la reacutesistance apporteacutee par la mutation faisant en sorte que
lrsquoalanine (A) en position 17 devient une seacuterine (S) apparaicirct chez les mutants
NewbouldΔhemb reacutesistant agrave la tomatidine et que cette mutation nrsquoapparaicirct qursquoau
quatriegraveme et dernier niveau de reacutesistance On se rappelle eacutegalement que la mutation
faisant en sorte que lrsquoalanine (A) agrave la position 64 devient une threacuteonine (T) apparaicirct
chez les mutants NewbouldΔhemb reacutesistants agrave la tomatidine en combinaison avec la
gentamicine et ce degraves le premier niveau de reacutesistance
118
Bcereus G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Banthracis G---------VIAAAIAIG---LSALGAGIGNGLIVSRTIEGVARQPELKGALQTIMFIG 51
Lmonocytogenes G---------VIAAAIAVG---LGALGAGIGNGLIVSKTVEGVARQPEARSMLQTIMFV- 50
Saureus_NWMN_2012 -----------IAAAIAIG---LSALGAGIGNGLIVSRTVEGVARQPEARGQLMGIMFIG 49
Bsubtilis -----------IAAAIAIG---LGALGAGIGNGLIVSRTVEGIARQPEAGKELRTLMFMG 49
Bpseudofirmus -----------LGAAIAAG---LAAVAGAIAVAIIVKATIEGTTRQPELRGTLQTLMFIG 49
Ecoli L---------YMAAAVMMG---LAAIGAAIGIGILGGKFLEGAARQPDLIPLLRTQFFIV 56
Msmegmatis DPNAIITAGALIGGGLIMG---GGAIGAGIGDGIAGNALISGIARQPEAQGRLFTPFFIT 60
Homo sapiens QTSAISRDIDTAAKFIGAGAATVGVAGSGAGIGTVFGSLIIGYARNPSLKQQLFSYAILG 120
MyR V
MyR P
SaR S
Bcereus VALVEALPIIGVVIAFIVMNK----- 72
Banthracis VALVEALPIIGVVIAFIVMNK----- 72
Lmonocytogenes ----EALPIIAVVIAFMVLNK----- 67
Saureus_NWMN_2012 VGLVEALPIIGVVIAFMTFAG----- 70
Bsubtilis IALVEALPIIAVVIAFLAFFG----- 70
Bpseudofirmus VPLAEAVPIIAIVISLLILF------ 69
Ecoli MGLVDAIPMIAVGLGLYVMFAVA--- 79
Msmegmatis VGLVEAAYFINLAFMALFVFATPGLQ 86
Homo sapiens FALSEAMGLFCLMVAFLILFAM---- 142
MyR D P M
SpR I A
SaR T
Fig
ure
3
Seacuteq
uen
ce
pro
teacuteiq
ue d
e la p
roteacute
ine
Atp
Ec
hez d
iffeacute
ren
tes
es
pegravec
es e
t m
ise e
n eacute
vid
en
ce
de
s s
ite
s d
e
mu
tati
on
pe
rmett
an
t la
reacutesis
tan
ce agrave
dif
feacutere
nte
s m
oleacute
cu
les
M
yR M
tu
berc
ulo
sis
ou
M sm
eg
matis
reacutesis
tan
t a
u
TM
C2
07 S
pR
S
p
neu
mon
iae
reacutesis
tan
t agrave
un a
na
log
ue d
u T
MC
20
7
SaR
S
a
ure
us
SC
V r
eacutesis
tant agrave
la to
matid
ine
En
cad
reacutero
uge
S
ite d
e m
uta
tion a
men
ant la
reacutesis
tance
agravela
to
matid
ine E
ncad
reacuteno
ir
Site
cata
lytiq
ue
im
pliq
ueacute
da
ns le
tra
nspo
rt d
es p
roto
ns
Su
rlig
nag
etu
rqu
ois
e S
ite d
e m
uta
tion a
me
nant u
ne
reacutesis
tan
ce a
u T
MC
20
7
119
Les seacutequences proteacuteiques des souches bacteacuteriennes de la figure 3 ont eacuteteacute trouveacutees
sur pubmedproteacuteine httpwwwncbinlmnihgovpubmed
laquo ATP synthase subunit c raquo Assession number
Streptococcus pneumoniae D39 YP_8168031 (NCBI)
S aureus Newman BAF682841 (GenBank)
E coli AAA247321 (GenBank)
Bacillus cereus ISO75 EJP900901 (GenBank)
Bacillus anthracis str H9401 YP_0062123491 (NCBI)
Mycobacterium smegmatis MC2 155 YP_0065695541 (NCBI)
Bacillus subtilis str 168 CAA822551 (GenBank)
Listeria monocytogenes YP_0076049931 (NCBI)
Homo sapiens P482011 (UniProtKBSwiss-Prot)
Lrsquoalignement comparatif des seacutequences drsquoacides amineacutes a eacuteteacute fait agrave lrsquoaide du
programme ClustalW2 sur le web
httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2
Le modegravele 3D de lrsquoAtpE a eacuteteacute obtenu gracircce agrave lrsquoaide du Pr Ryszard Brzezinski du
Deacutepartement de biologie de lrsquoUniversiteacute de Sherbrooke (Figure 4) Une recherche
dans le Protein Data Bank a drsquoabord eacuteteacute effectueacutee afin de veacuterifier parmi les proteacuteines
AtpE (sous-uniteacute C de lrsquoATP synthase) pour lesquelles il y avait des structures 3D
reacutesolues laquelle ressemblait le plus agrave celle de Staphylococcus aureus Les
alignements ont montreacute que cest la proteacuteine de Bacillus pseudofirmus qui avait la
plus grande similitude de seacutequence La structure est stockeacutee dans la banque PDB
sous le symbole 3ZO6 Il a eacuteteacute confirmeacute par quelques alignements que cette sous-
uniteacute C de lrsquoATP synthase eacutetait plus proche par sa seacutequence de celle de
Staphylococcus aureus que de celle de Mycobacterium pour laquelle la structure 3D
avait eacuteteacute deacutetermineacutee plus tocirct Ce reacutesultat sexplique par le fait que Staphylococcus et
Bacillus font tous deux partie de la branche A+T-riche des bacteacuteries agrave Gram+ alors
que Mycobacterium appartient agrave la branche G+C-riche La seacutequence de lAtpE de
120
Staphylococcus aureus a ensuite eacuteteacute envoyeacutee au serveur internet de modeacutelisation
3D-Jigsaw (httpbmmcancerresearchukorg~3djigsaw) en indiquant que le modegravele
devait ecirctre bacircti en prenant comme matrice le fichier PDB 3ZO6 Le modegravele nous a
eacuteteacute retourneacute sous format PDB ndash comme les laquo vrais raquo cristaux dans la Protein Data
Bank Les fichiers au format PDB peuvent ecirctre visualiseacutes par le logiciel RasMol ndash
gratuit sur internet Cest avec RasMol que limage (Figure 4) a eacuteteacute obtenue
121
Figure 4 Modeacutelisation de la structure tridimensionelle de la sous-uniteacute c de
lrsquoATP synthase de S aureus (AtpE) montrant les acides amineacutes muteacutes chez les
mutants reacutesistantsEn blanc et vert les deux acides amineacutes muteacutes chez S aureus
reacutesistant agrave la tomatidine En rose les sites de mutations chez Mycobacterium
smegmatis reacutesistant au TMC207 et des sites de mutations chez Streptoccocus
pneumoniae reacutesistants agrave un analogue de TMC207 En bleu le site actif de la proteacuteine
Agrave la vue de ces reacutesultats certains pourront mettre en doute lrsquointeacuterecirct de la tomatidine
eacutetant donneacute lrsquoapparition relativement rapide de reacutesistance chez S aureus Cependant
122
nos reacutesultats deacutemontrent clairement que lrsquoajout de tomatidine ralentit le
deacuteveloppement de reacutesistance agrave la gentamicine chez S aureus ATCC29213 (Tableau
1) De plus cette augmentation de la CMI de la gentamicine en preacutesence de
tomatidine au fil des passages suit lrsquoaugmentation de la CMI de la ciprofloxacine lors
de ces mecircmes passages alors que la ciprofloxacine est consideacutereacutee comme la drogue
de reacutefeacuterences dans les bonnes drogues (Tableau 1) Tout comme pour la
ciprofloxacine pour laquelle la reacutesistance chez S aureus peut ecirctre la conseacutequence
de deux mutations dans le gegravene cible gyrA la reacutesistance agrave la tomatidine peut ecirctre la
conseacutequence de deux mutations dans le gegravene cible atpE Par comparaison la
rifampicine est consideacutereacutee comme une mauvaise drogue de par lrsquoapparition rapide
drsquoune forte reacutesistance chez S aureus conseacutequence de ses 17 mutations possibles
dans le gegravene cible rpoB (Tableau 1)
De plus il sera important de relativiser lrsquoapparition de reacutesistance agrave la tomatidine en
calculant le taux drsquoapparition de ces colonies reacutesistantes ceci nrsquoayant pas eacuteteacute fait
jusqursquoici De fait la freacutequence de mutation chez S aureus amenant une reacutesistance agrave
la rifampicine est de lrsquoordre de 10-7 10-8 alors qursquoil est de 10-11 pour lrsquoacide fusidique
(Canfield et al 2013 Orsquoneill et al 2001 Price et Gustafson 2001) Pour ce qui est
de la nouvelle moleacutecule anti-bacteacuteries agrave Gram positif de la classe des diarylquinoline
une freacutequence de mutation sur S pneumoniae ATCC49619 de 81- 83 x 10-7 a eacuteteacute
observeacutee agrave 5x la CMI et une freacutequence de mutation de 46 x 10-7 agrave 17 x 10-6 agrave 50x la
CMI (Balemans et al 2012)
123
Tableau 1 Mesure de la susceptibiliteacute agrave diffeacuterentes drogues de S aureus ATCC
29213 par la meacutethode de CMI lors de passages successifs dans un milieu avec
lrsquoantibiotique
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
CMI des antibiotiques agrave chacun des passages (microgmL)
passage GEN GEN+TO CIPRO RIF
0 05 006 025 006
~6 1 025 05 512
~12 2 1-2 1 512
~24 4 2 2 1024
~30 8 2-4 4 1024
Typiqueldquomauvaiserdquodrogue (17 mutations possibles dansrpoB)
Typique ldquobonnerdquo drogue (2 mutations possibles dans gyrA)
Preacutesence de TO ralenti le deacuteveloppement de reacutesistances vs GEN seul
124
CHAPITRE 5
CONCLUSION ET PERSPECTIVES
Mon projet de maitrise a permis de preacuteciser le spectre drsquoactiviteacute de la tomatidine
Nous savons maintenant que la tomatidine possegravede une activiteacute antibacteacuterienne
contre la plupart des Firmicutes et plus speacutecifiquement contre les Bacillales Plus
preacuteciseacutement la tomatidine est bacteacutericide contre les pheacutenotypes SCV et agit en
synergie avec les aminoglycosides augmentant lrsquoactiviteacute bacteacutericide de ces derniers
de 8-16 fois contre les souches agrave pheacutenotype normal
De plus parmi tous les analogues de la tomatidine produits par le laboratoire du Pr
Eacuteric Marsault (voir article en annexe 2) le composeacute FC04-100 a eacuteteacute plus
speacutecifiquement mis en lumiegravere Celui-ci conserve non seulement une aussi bonne
activiteacute synergique avec les aminoglicosides contre les Saureus au pheacutenotype
normal et une activiteacute modeacutereacutee contre les S aureus SCV mais montre eacutegalement de
faccedilon inespeacutereacutee une activiteacute par lui-mecircme contre les S aureus de pheacutenotype normal
incluant plusieurs MRSA une proprieacuteteacute qui nrsquoeacutetait pas observeacutee pour la tomatidine
Une autre activiteacute inteacuteressante de FC04-100 est sa capaciteacute drsquoameacuteliorer lrsquoactiviteacute
bacteacutericide des aminoglicosides contre L monocytogenes agrave pheacutenotype normal apregraves
un traitement de 24h alors que la tomatidine ne deacutemontrait qursquoune faible synergie
avec les aminoglycosides contre cette souche De plus FC04-100 deacutemontre une
activiteacute bacteacutericide contre les SCV de L monocytogenes alors que la tomatidine ne
deacutemontrait qursquoune activiteacute bacteacuteriostatique FC04-100 montre eacutegalement un effet
potentiateur de lrsquoUbrolexinTM (Ceacutefalexin Kanamycin (3 2)) une combinaison
drsquoantibiotique utiliseacute dans le traitement de mammites bovines en abaissant de 8 agrave 16
fois leur CMI amenant celles-ci sous le seuil de reacutesistance Finalement FC04-100
possegravede une meilleure solubiliteacute que la tomatidine une proprieacuteteacute qui facilitera son
utilisation in vitro
125
Nos reacutesultats deacutemontrent que FC04-100 conserve une certaine activiteacute contre les
SCV reacutesistant agrave la tomatidine ce qui suggegravere possiblement que FC04-100 a plus
daffiniteacute pour la cible ou plus de sites dinteraction avec AtpE En effet les mutants
SCV reacutesistants agrave la tomatidine deviennent gt512 fois plus reacutesistants alors que FC04-
100 ne perd que 8 agrave 16 fois son activiteacute contre ces mecircmes souches Ceci deacutemontre
quil est possible dameacuteliorer significativement lactiviteacute de ces alkaloiumldes steacuteroidiens
ce qui est tregraves prometteur pour lavenir de cette moleacutecule
Dans un second temps lrsquoanalyse des mutations chez les mutants reacutesistants agrave la
tomatidine a permis de non seulement de mieux comprendre le mode drsquoaction de la
tomatidine mais eacutegalement de preacuteciser la cible potentielle de ce nouvel antibiotique
De fait son mode drsquoaction serait en lien direct avec la fonctionnaliteacute de la chaine de
transport des eacutelectrons chez la bacteacuterie lrsquoactiviteacute de la tomatidine serait inversement
proportionnelle agrave la fonctionnaliteacute de la chaine de transport des eacutelectrons et donc en
lien eacutegalement avec la polarisation membranaire et la production de ROS
intracellulaire Afin de confirmer le modegravele-hypothegravese la mesure de la polarisation
membranaire et la concentration de ROS intracellulaire chez les mutants ainsi que
chez les souches parentes suite agrave un traitement antibiotique devront ecirctre effectueacutees
Les reacutesultats de mes recherches suggegraverent que la cible bacteacuterienne speacutecifique de la
tomatidine serait la sous-uniteacute C de lrsquoATP synthase Afin de confirmer la cible
Maxime Lamontagne-Boulet tente au moment drsquoeacutecrire ces lignes drsquoobtenir des
mutants de S aureus Newman reacutesistants au composeacute FC04-100 et dont la reacutesistance
serait attribuable agrave une mutation dans le gegravene atpE Par la suite on devra confirmer
la fonctionnaliteacute de la cible ATP synthase par quantification de lrsquoATP produit agrave lrsquoaide
de la lucifeacuterase tel que lrsquoon fait Balemans et al en 2012 Lrsquoattachegravement agrave la cible
devra eacutegalement ecirctre confirmeacute et la meacutethode de SPR (Surface Plasmon Resonance)
pourrait ecirctre tout indiqueacutee pour ce faire (Balemans et al 2012) On devra eacutegalement
126
deacutemontrer la seacutelectiviteacute des moleacutecules tomatidine et FC04-100 pour lrsquoATP synthase
des Bacillales
Les infections agrave bacteacuteries multireacutesistantes deviennent de plus en plus preacutevalentes et
sont un des problegravemes de santeacute majeurs auxquels nous devons faire face aujourdrsquohui
Cette augmentation de la reacutesistance limite le reacutepertoire drsquoantibiotiques efficace creacuteant
une situation probleacutematique laquelle est exacerbeacutee par le trop petit nombre de
nouveaux antibiotiques introduits sur le marcheacute ces derniegraveres anneacutees Il est donc
drsquoune grande importance de travailler agrave la mise en marcheacute de nouveaux antibiotiques
et plus particuliegraverement de nouvelles classes drsquoantibiotiques espeacuterant ainsi deacutejouer la
multireacutesistance bacteacuterienne du moins pour un temps La tomatidine de par sa
structure sa cible et son mode drsquoaction speacutecifique est une moleacutecule antibiotique
dans une classe agrave part et de ce fait une moleacutecule plus qursquointeacuteressante pour le
traitement drsquoinfection agrave S aureus De plus comme elle deacutemontre une activiteacute
bacteacutericide contre les deux pheacutenotypes de S aureus (et autres Bacillales) lesquels
sont retrouveacutes simultaneacutement dans les cas drsquoinfections chroniques son pouvoir
antibiotique devient inestimables drsquoun point de vue clinique
127
CONTRIBUTIONS SUPPLEacuteMENTAIRES
ANNEXE 1 Application de brevet Internationnal publication number WO
2012109752 A1
INVENTEURS Franccedilois Malouin2 Gabriel Mitchell2 Kamal Bouarab2 Eacuteric Marsault1
Feacutelix Chagnon1 Simon Boulanger 2 Isabelle Guay2
1Deacutepartement de Pharmacologie Faculteacute de Meacutedecine et des Sciences de la Santeacute
Universiteacute de Sherbrooke Queacutebec Canada J1H 5N4
2Centre drsquoEacutetude et de Valorisation de la Diversiteacute Microbienne (CEVDM)
Deacutepartement de Biologie Faculteacute des Sciences Universiteacute de Sherbrooke
Sherbrooke Queacutebec Canada J1K 2R1
128
129
ANNEXE 2 Article scientifique dans laquo Europeen Journal of medicinal chemistry raquo
2014 80 605-620
TITRE Unraveling the structure-activity relationship of tomatidine a steroid alkaloid
with unique antibiotic properties against persistent forms of
Staphylococcus aureus
ABSTRACT Staphylococcus aureus is responsible for difficult-to-treat and relapsing
infections and constitutes one of the most problematic pathogens due to its multiple
resistances to clinically available antibiotics Additionally this pathogenrsquos ability to
develop small-colony variants is associated with a reduced susceptibility to
aminoglycoside antibiotics and in vivo persistence We have recently demonstrated
that tomatidine a steroid alkaloid isolated from tomato plants possesses anti-
virulence activity against normal strains of S aureus as well as the ability to
potentiate the effect of aminoglycosides against this pathogen In addition tomatidine
has shown antibiotic activity against small-colony variants We herein report the first
study of the structure-activity relationship of tomatidine against S aureus
AUTEURS Feacutelix Chagnon1 Isabelle Guay2 Marc-Andreacute Bonin1 Gabriel Mitchell2
Kamal Bouarab2Franccedilois Malouin2 Eacuteric Marsault1
MA CONTRIBUTION Responsable des tests biologiques de tous les composeacutes
autant envers lrsquoeffet potentiateur de la tomatidine sur lrsquoactiviteacute de la gentamicine
contre la souche ATCC29213 de S aureus que pour lrsquoactiviteacute bacteacuteriostatique sur la
souche NewbouldΔhem imitant le pheacutenotype SCV Ces travaux furent effectueacutes sous
la tutelle du professeur Franccedilois Malouin
130
1Institut de Pharmacologie de Sherbrooke Deacutepartement de pharmacologie
Universiteacute de Sherbrooke 3001 12e av nord Sherbrooke Queacutebec Canada J1H
5N4
2Centre drsquoeacutetude et de valorisation de la diversiteacute microbienne (CEVDM) Deacutepartement
de biologie Universiteacute de Sherbrooke Sherbrooke QC Canada J1K 2R1
131
BIBLIOGRAPHIE Alexander E H and Hudson MC (2001) Factors influencing the internal- ization of Staphylococcus aureus and impacts on the course of infections in humans Appl Microbiol Biotechnol 56 361ndash366
Andries K Verhasselt P Guillemont J Goumlhlmann HW Neefs JM Winkler H Van Gestel J Timmerman P Zhu M Lee E et al (2005) A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis Science 307 223-227
Artzatbanov VY and Petrov VV (1990) Branched respiratory chain in aerobically grown Staphylococcus aureus--oxidation of ethanol by cells and protoplasts Arch Microbiol 153 580- 584 Archer GL (1998) Staphylococcus aureus a well-armed pathogen Clin Infect Dis 26 1179ndash1181 Assefa S Keane TM Otto TD Newbold C and Berriman M (2009) ABACAS algorithm-based automatic contiguation of assembled sequences Bioinformatics 25 1968-1969 Aubry-Damon H Legrand P Brun-Buisson C Astier A Soussy CJ and Leclercq R (1997) Reemergence of gentamicin-sus- ceptible strains of methicillin-resistant Staphylococcus aureus roles of an infection control program and changes in aminoglycoside use Clin Infect Dis 25 647-653
Balemans W Vranckx L Lounis N Pop O Guillemont J Vergauwen K Mol S Gilissen R Motte M Lanccedilois et al (2012) Novel antibiotics targeting respiratory ATP synthesis in Gram-positive pathogenic bacteria Antimicrob Agents Chemother 56 4131-4139
Batandier C Guigas B Detaille D El-Mir MY Fontaine E Rigoulet M and Leverve XM (2006) The ROS production induced by a reverse-electron flux at respiratory-chain complex 1 is hampered by metformin J Bioenerg Biomembr 38 33ndash 42
132
Beckman Coulter website httpswwwbeckmancoultercomwsrportalwsrresearch-and-discoveryproducts-and-servicesnucleic-acid-sample-preparationspri-laboratory-automationindexhtm (consulteacute le 3 Deacutec 2013) Bednarek P and Osbourn A (2009) Plant-microbe interactions chemical diversity in plant defense Science 324 746ndash748 Besier S Ludwig A Ohlsen K Brade V and Wichelhaus TA (2007) Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus Int J Med Microbiol 297 217-225 BiteSizeBio Brain food for biologists httpnxseqbitesizebiocomarticlessequencing-by-synthesis-explaining-the-illumina-sequencing-technology (consulteacute le 3 deacutec 2013) Boetzer M and Pirovano W (2012) Toward almost closed genomes with GapFiller Genome Biology 13 R56 Bouarab K Ordi E Gattuso M Moisan H and Malouin F (2007) Plant stress response agents affect Staphylococcus aureus virulence genes Abstr 47th Intersci Conf Antimicrob Agents Chemother abstr C1-1483 Bryan LE and Kwan S (1981) Aminoglycoside-resistant mutants of Pseudomonas aeruginosa deficient in cytochrome d nitrite reductase and aerobic transport Antimicrob Agents Chemother 19 958- 964 Bryan LE and Kwan S (1983) Roles of ribosomal binding membrane potential and electron transport in bacterial uptake of streptomycin and gentamicin Antimicrob Agents Chemother 23 835ndash845
Bryan LE Nicas T Holloway BW and Crowther C (1980) Aminoglycoside-resistant mutation of Pseudomonas aeruginosa defective in cytochrome C552 and nitrate reductase Antimicrob Agents Chemother 17 71- 79
133
Bryan LE and van den Elzen HM (1977) Effects of membrane-energy mutations and cations on streptomycin and gentamicin accumulation by bacteria a model for entry of streptomycin and gentamicin in susceptible and resistant bacteria Antimicrob Agents Chemother 12 163-177 Brynildsen MP Winkler JA Spina CS MacDonald IC and Collins JJ (2013) Potentiating antibacterial activity by predictably enhancing endogenous microbial ROS production Nat Biotechnol 31 160ndash 165 Burke KA and Lasccelles J (1975) Nitrate reductase system in Staphylococcus aureus wild type and mutants J Bacteriol 123 308- 316 Burke KA and Lascelles J (1976) Nitrate reductase activity in hegraveme-deficient mutants of Staphylococcus aureus J Bacteriol 126 225- 231 Canfield GS Schwingel JM Foley MH Vore KL Boonanantanasarn K Gill AL Sutton MD and Gill SR (2013) Evolution in fast forward a potential role for mutators in accelerating Staphylococcus aureus pathoadaptation J Bacteriol 195 615-628 Chang S Sievert DM Hageman JC Boulton ML Tenover FC Downes FP Shah S Rudrik JT Pupp GR Brown WJ et al (2003) Infection with vancomycin- resistant Staphylococcus aureus containing the vanA resistance gene N Engl J Med 348 1342-1347 Chen Q Vazquez EJ Moghaddas S Hoppel CL and Lesnefsky EJ (2003) Production of reactive oxygen species by mitochondria central role of complex III J Biol Chem 278 36027-36031 Clements MO Watson SP Poole RK and Foster SJ (1999) CtaA of Staphylococcus aureus is required for starvation survival recovery and cytochrome biosynthesis J Bacteriol 181 501- 507
134
Clinical and Laboratory Standards Institute (2011) Performance standards for antimicrobial susceptibility testing Twenty-first informational supplement M100-S21 CLSI Wayne PA USA ClustalW2 httpwwwebiacukToolsserviceswebtoolformebitool=clustalw2 (consulteacute le 3 deacutec 2013) Comprehensive Microbial Ressouce (J Craig Venter Institute (JCVI) httpcmrjcviorgtigr-scriptsCMRCmrHomePagecgi (consulteacute le 3 deacutec 2013) Creacutepin-Gilbert C (2006) Transfert de ligand dans la cytochrome c oxydase observeacute par des expeacuteriences femtosecondes infrarouge inteacutegreacutees et reacutesolues spectralement Thegravese de doctorat de l rsquoEacutecole Polytechnique Paris Pastel-00002200 version 1 29 jul 2010 Davis BD (1987) Mechanism of bactericidal action of aminoglycosides Microbiol Rev 51 341-350 DeAngelis MM Wang DG and Hawkins TL (1995) Solid-phase reversible immobilization for the isolation of PCR products Nucl Acids Res 22 4742- 4743 Dickgiesser N and Kreiswirth BN (1986) Determination of aminoglycoside resistance in Staphylococcus aureus by DNA hybridization Antimicrob Agents Chemother 29 930-932 Doi Y and Arakawa Y (2007) 16S ribosomal RNA methylation Emerging resistance mechanism against aminoglycosides Clin Infect Dis 45 88ndash94 Enright MC Robinson DA Randle G Feil EJ Grundmann H and Spratt BG (2002) The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA) Proc Natl Acad Sci USA 99 7687-7692 Friedman M (2002) Tomato glycoalkaloids role in the plant and in the diet J Agric Food Chem 50 5751ndash5780
135
Goerke C and Wolz C (2004) Regulatory and genomic plasticity of Staph- ylococcus aureus during persistent colonization and infection Int J Med Microbiol 294 195ndash202 Goumltz F Bannerman T and Schleifer K-H (2006) The genera Staphylococcus and Macrococcus In The Prokaryotes Vol 4 Bacteria Firmicutes Cyanobacteria M Dworkin (ed) (Springer New-York USA) pp5- 75 Haagsma AC Abdillahi-Ibrahim R Wagner MJ Krab K Vergauwen K Guillemont J AndriesK Lill H Koul A and Bald D (2009) Selectivity of TMC207 towards mycobacterial ATP synthase compared with that towards the eukaryotic homologue Antimicrob Agents Chemother 53 1290-1292 Hammer ND Reniere ML Cassat JE Zhang Y Hirsch AO Indriati Hood M and Skaar EP (2013) Two hegraveme-dependent terminal oxydases power Staphylococcus aureus organ-specific colonization of the vertebrate host MBio 4 e00241- 13 Hederstedt L (1993) The Krebs citric acid cycle In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of Microbiology Washington DC) pp 181-198 Hiramatsu K (1995) Molecular evolution of MRSA Microbiol Immunol 39 531-543 Hiramatsu K Hanaki H Ino T Yabuta K Oguri T and Tenover FC (1997) Methicillin-resistant Staphylococcus aureus clinical strain with reduced vancomycin susceptibility J Antimicrob Chemother 40 135-136 Hiramatsu K Ito T Tsubakishita S Sasaki T Takeuchi F Morimoto Y Katayama Y Matsuo M Kuwahara-Arai K Hishinuma T et al (2013) Genomic Basis for Methicillin Resistance in Staphylococcus aureus Infect Chemother 45 117- 136
136
Hong S and Pedersen PL (2008) ATP synthase and the actions of inhibitors utilized to study its roles in human health disease and other scientific areas Microbiol Mol Biol Rev 72 590-641
Jevons MP (1961) lsquoCelbeninrsquo-resistant staphylococci BMJ 1 124-125
Joshi S and Huang YG (1991) ATP synthase complex from bovine heart mitochondria the oligomycin sensitivity conferring protein is essential for dicyclohexyl carbodiimidesensitive ATPase Biochim Biophys Acta 1067 255ndash258
Kohanski MA Dwyer DJ and Collins JJ (2010) How antibiotics kill bacteria from targets to networks Nat Rev Microbiol 8 423-435 Kohler C von Eiff C Liebeke M McNamara PJ Lalk M Proctor RA Hecker M and Engelmann S (2008) A defect in menadione biosynthesis induces global changes in gene expression in Staphylococcus aureus J Bacteriol 190 6351- 6364 Kohler C von Eiff C Peters G Proctor RA Hecker M and Engelmann S (2003) Physiological characterization of a hegraveme-deficient mutant of Staphylococcus aureus by a proteomic approach J Bacteriol 185 6928- 6937 Kriegeskorte A Koumlnig S Sander G Pirkl A Mahabir E Proctor RA von Eiff C Peters G and Becker K (2011) Small colony variants of Staphylococcus aureus reveal distinct protein profiles Proteomics 11 2476-2490 Lannergard J von Eiff C Sander G Cordes T Seggewiss J Peters G Proctor RA Becker K and Hughes D (2008) Identification of the genetic basis for clinical menadione-auxotrophic small-colony variant isolates of Staphylococcus aureus Antimicrob Agents Chemother 52 4017-4022
Lauraeus M and Wikstrom M (1993) The terminal quinol oxydases of Bacillus subtilis have different energy conservation properties J Biol Chem 268 11470-11473
137
Li M Gong J Cottrill M Yu H de Lange C Burton J and Topp E (2003) Evaluation of QIAamp DNA Stool Mini Kit for ecological studies of gut microbiota J Microbiol Methods 54 13-20
Liu Y Fiskum G and Schubert D (2002) Generation of reactive oxygen species by the mitochondrial electron transport chain J Neurochem 80 780-787
Martin-Hernandez A M Dufresne M Hugouvieux V Melton R and Osbourn A (2000) Effects of targeted replacement of the tomatinase gene on the interaction of Septoria lycopersici with tomato plants Mol Plant Microbe Interact 13 1301ndash1311
Matteelli A Carvalho AC Dooley KE and Kritski A (2010) TMC207 the first compound of a new class of potent anti-tuberculosis drugs Future Microbiol 5 849-858
Matsuno-Yagi A and Hatefi Y (1993) Studies on the mechanism of oxidative phosphorylation ATP synthesis by submitochondrial particles inhibited at F0 by venturicidin and organotin compounds J Biol Chem 268 6168ndash6173
Miller MH Edberg SC Mandel LJ Behar CF and Steigbigel NH (1980) Gentamicin uptake in wild-type and aminoglycoside-resistant small-colony mutants of Staphylococcus aureus Antimicrob Agents Chemother 18 722- 729
Mingeot-Leclercq M-P Glupczynski Y and Tulkens PM (1999) Aminoglycosides Activity and Resistance Antimicrob Agents Chemother 43 727- 737
Mitchell G Brouillette Eacute Seacuteguin DL Asselin AE Jacob CL and Malouin F (2010) A role for sigma factor B in the emergence of Staphylococcus aureus small-colony variants and elevated biofilm production resulting from an exposure to aminoglycosides Microb Pathog 48 18-27
Mitchell G Gattuso M Bouarab K and Malouin F (2009) Tomatidine (TO) affects virulence regulators of prototypical Staphylococcus aureus (SA) and small colony variants (SCV) of cystic fibrosis (CF) patients Abstr 49th Intersci Conf Antimicrob Agents Chemother abstr C1-1341
138
Mitchell G Gattuso M Grondin G Marsault Eacute Bouarab K and Malouin F (2011) Tomatidine inhibits replication of Staphylococcus aureus small-colony variants in cystic fibrosis airway epithelial cells Antimicrob Agents Chemother 55 1937-1945
Mitchell G Grondin G Bilodeau G Cantin AM and Malouin F (2011) Infection of polarized airway epithelial cells by normal and small-colony variant strains of Staphylococcus aureus is increased in cells with abnormal cystic fibrosis transmembrane conductance regulator function and is influenced by NF-κB Infect Immun 79 3541-3551
Mitchell G Lafrance M Boulanger S Seacuteguin DL Guay I Gattuso M Marsault Eacute Bouarab K and Malouin F (2012) Tomatidine acts in synergy with aminoglycoside antibiotics against multiresistant Staphylococcus aureus and prevents virulence gene expression J Antimicrob Chemother 67 559-568
Mitchell G Lamontagne CA Brouillette Eacute Grondin G Talbot BG Grandbois M and Malouin F (2008) Staphylococcus aureus SigB activity promotes a strong fibronectin-bacterium interaction which may sustain host tissue colonization by small-colony variants isolated from cystic fibrosis patients Mol Microbiol 70 1540-1555
Mitchell G Malouin F 2012 Outcome and prevention of Pseudomonas aeruginosa-Staphylococcus aureus interactions during pulmonary infections in cystic fibrosis In Cystic Fibrosis Chap 8 pp 181-206 Ed D Sriramulu InTech Open Access Publisher ISBN 979-953-307-059-8
Mitchell G Seacuteguin DL Asselin AE Deacuteziel Eacute Cantin AM Frost EH Michaud S and Malouin F (2010) Staphylococcus aureus sigma B-dependent emergence of small-colony variants and biofilm production following exposure to Pseudomonas aeruginosa 4-hydroxy-2-heptylquinoline-N-oxide BMC Microbiol 30 10-33
Moisan H Brouillette Eacute Jacob CL Langlois-Beacutegin P Michaud S and Malouin F (2006) Transcription of virulence factors in Staphylococcus aureus small-colony variants isolated from cystic fibrosis patients is influenced by SigB J Bacteriol 188 64-76
139
Moreno F Crisp C Jorgensen JH and Patterson JE (1995) Methicillin- resistant Staphylococcus aureus as a community organism Clin Infect Dis 21 1308-1312
Murphy MP (2009) How mitochondria produce reactive oxygen species Biochem J 417 1- 13
Novick RP (2003) Autoinduction and signal transduction in the regulation of staphylococcal virulence Mol Microbiol 48 1429-1449
Novo DJ Perlmutter NG Hunt RH and Shapiro HM (2000) Multiparameter flow cytometric analysis of antibiotic effects on membrane potential membrane permeability and bacterial counts of Staphylococcus aureus and Micrococcus luteus Antimicrob Agents Chemother 44 827-834
ONeill AJ Cove JH and Chopra I (2001) Mutation frequencies for resistance to fusidic acid and rifampicin in Staphylococcus aureus J Antimicrob Chemother 47 647-650
Osbourn A E (1996) Preformed antimicrobial compounds and plant defense against fungal attack Plant Cell 8 1821ndash1831
Otto TD Dillon GP Degrave WS and Berriman M (2011) RATT Rapid annotation transfert tool Nucl Acids Res 39 doi101093 Pratique E Theraulaz L (2007) Eacutetude de la reacutegulation du systegraveme respiratoire TMAO reacuteductase chez Escherichia coli Meacutemoire de maitrise de lrsquoeacutecole pratique des hautes eacutetudes Paris
Price CT and Gustafson JE (2001) Increases in the mutation frequency at which fusidic acid-resistant Staphylococcus aureus arise with salicylate J Med Microbiol 50 104-106
Proctor RA and Peters G (1998) Small colony variants in staphylococcal infections Diagnostic and therapeutic implications Clin Infect Dis 27 419- 423
140
Proctor RA von Eiff C Kahl BC Becker K McNamara P Herrmann M and Peters G (2006) Small colony variants a pathogenic form of bacteria that facilitates persistent and recurrent infections Nat Rev Microbiol 4 295ndash305
Quail MA Kozarewa I Smith F Scally A Stephens PJ Durbin R Swerdlow H and Turner DJ (2008) A large genome centers improvements to the Illumina sequencing system Nat methods 5 1005-1010
Ramirez MS and Tolmasky ME (2010) Aminoglycoside modifying enzymes Drug Resist Updat 13 151ndash171
Rao M Streur TL Aldwell FE and Cook GM (2001) Intracellular pH regulation by Mycobacterium smegmatis and Mycobacterium bovis BCG Microbiology 147 1017-1024
Richardson DJ (2000) Bacterial respiration a flexible process for a changing environment Microbiology 146 551- 571
Riley TV Pearman JW and Rouse IL (1995) Changing epidemiology of methicillin-resistant Staphylococcus aureus in Western Australia Med J Aust 163 412-414
Roddick J G (1974) The steroidal glycoalkaloid α-tomatine Phytochemistry 13 25-36
Rodrigue S Materna AC Timberlake SC Blackburn MC Malmstrom RR et al(2010) Unlocking Short Read Sequencing for Metagenomics PLoS ONE 5(7) e11840doi101371journalpone0011840 Ruiz-Rubio M Alonso Peacuterez-Espinosa A Lairini K RoldAacuten-Arjona T Dipietro A and Anaya N (2001) Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi In Studies in natural products chemistry vol 25 A Rahman (ed) (Elsevier Oxford United Kingdom) pp 293ndash326
141
Samuelsen oslash Haukland HH Kahl BC von Eiff C Proctor RA Ulvatne H Sandvik K and Vorland LH (2005) Staphylococcus aureus small colony variants are resistant to the antimicrobial peptide lactoferricin B J Antimicrob Chemother 56 1126- 1129
Sandrock R W and Vanetten HD (1998) Fungal sensitivity to and enzymatic degradation of the phytoanticipin α-tomatine Phytopathology 88 137ndash143
Sendi P and Proctor RA (2009) Staphylococcus aureus as an intracellular pathogen the role of small colony variants Trends Microbiol 17 54ndash58
Simons V Morrissey JP Latijnhouwers M Csukai M Cleaver A Yarrow C and Osbourn A (2006) Dual effects of plant steroidal alkaloids on Saccha- romyces cerevisiae Antimicrob Agents Chemother 50 2732ndash2740
Somerville GA and Proctor RA (2009) At the crossroads of bacterial metabolism and virulence factor synthesis in Staphylococci Microbiol Mol Biol Rev 73 233- 248
Starkov AA (2010) Measurement of mitochondrial ROS production Methods Mol Biol 648 245-255
Taber HW (1993) Respiratory chain In Bacillus subtilis and other Gram-positive bacteria Sonenshein AL Hoch JA Losick R (ed) (American society of MicrobiologyWashington DC) pp 199-212
Taber HW and Morrison M (1964) Electron transport in Staphylococci Properties of a particle preparation from exponential phase Staphylococcus aureus Arch Biochem Biophys 105 367- 379
Taber HW et al(1987) Bacterial uptake of aminoglycoside antibiotics Microbiol Rev 51 439-457
Thoumlny-Meyer L (1997) Biogenesis of respiratory cytochromes in bacteria Microbiol Mol Biol Rev 61 337- 376
142
Tubby S Wilson M Wright JA Zhang P and Nair SP (2013) Staphylococcus aureus small colony variants are susceptible to light activated antimicrobial agents BMC Microbiol 13 201- 204
Tuchscherr L Medina E Hussain M Voumllker W Heitmann V Niemann S Holzinger D Roth J Proctor RA Becker K et al (2011) Staphylococcus aureus phenotype switching an effective bacterial strategy to escape host immune response and establish a chronic infection EMBO Mol Med 3 129ndash41
Turrens JF (2003) Mitochondrial formation of reactive oxygen species J Physiol 552 335ndash 344
Tynecka Z Szcześniak Z Malm A and Los R (1999) Energy conservation in aerobically grown Staphylococcus aureus Res Microbiol 150 555- 566
Udo EE Pearman JW and Grubb WB(1993) Genetic analysis of community isolates of methicillin-resistant Staphylococcus aureus in Western Australia J Hosp Infect 25 97-108
Vakulenko SB and Mobashery S (2003) Versatility of Aminoglycosides and Prospects for Their Future Clin Microbiol Rev 16 430-450
Voggu L Schlag S Biswas R Rosenstein R Rausch C and Goumltz F (2006) Microevolution of cytochrome bd oxydase in Staphylococci and its implication in resistance to respiratory toxins released by Pseudomonas J Bacteriol 188 8079- 8086
Wells PG Bhuller Y Chen CS Jeng W Kasapinovic S Kennedy JC Kim PM Laposa RR McCallum GP Nicol CJ et al (2005) Molecular and biochemical mechanisms in teratogenesis involving reactive oxygen species Toxicol Appl Pharmacol 207 354-366
143
Yan M Yu C Yang J and Ji Y (2011) The essential two-component system YhcSR is involved in regulation of the nitrate respiratory pathway of Staphylococcus aureus J Bacteriol 193 1799- 1805
Zhang T Luo Y Chen Y Li X and Yu J (2012) BIGrat a repeat resolver for pyrosequencing-based re-sequencing with Newbler BMC Res Notes 5 567
144