Embed Size (px)
Citation preview

식물 세포벽 대사 관련 효소의
연구동향
이상필․손은수

머 리 말
21세기는 지식과 정보가 그 국가의 경쟁력을 좌우하는 지식
기반 산업사회로 나아가고 있으며, 최고가 아니면 살아남을
수 없는 무한경쟁시 가 되어가고 있습니다. 이러한 변화 속
에서 각 국가에서는 미래 유망기술(Emerging Technology)을
선정하여 국가 역량을 집 함으로써 차세 국가경쟁력을 확
보하려는 여러 가지 노력을 기울이고 있습니다.
최근 우리나라에서도 미래 유망기술에 한 심이 어느 때
보다도 증 되고 있는 가운데, 한국과학기술정보연구원에서는
과학계량학 인 방법으로 미래 국가 유망기술을 측하기
한 일련의 연구를 수행하고 있습니다.
본 보고서는 과학기술정보데이터베이스(SCIE)에서 최근 6
년간 분야별 피인용도가 높은 핵심논문들을 가지고 정보계량
학 인 분석을 행하여 선정된 핵심 유망 연구 역에 해
련 국내 문가들의 자문을 토 로 작성된 R&D 동향보고서입
니다. 본 보고서가 련 과학기술정보를 국내에 확산시키고,
미래 국가유망기술의 략 육성을 한 연구개발 활동에 작
으나마 도움이 되었으면 합니다.
마지막으로 본 보고서를 집필한 자들의 노고에 감사드리
며, 본고의 내용은 한국과학기술정보연구원의 공식의견이 아
님을 밝 둡니다.
2005년 12월
한국과학기술정보연구원
원 장

ⅰ
목 차
제1장 서 론 ······················································································1
1. 연구의 배경 ····························································································1
2. 연구의 방법 ····························································································2
제2장 기술의 개요 ·············································································3
1. 기술의 정의와 범 ················································································3
2. 기술의 요성 ·························································································4
3. 기술의 배경 ····························································································5
4. 기술의 특성 ····························································································6
5. 기술의 안문제 ·····················································································7
제3장 연구개발동향 ···········································································9
1. 개요 ·········································································································9
2. 식물에서 세포벽의 요성 ···································································10
3. 식물 세포벽의 구조 ··············································································11
4. 식물 세포벽의 기능 ··············································································13
가. 세포의 신장 ·························································································13
나. 식물의 발생 ·························································································14
5. 식물 세포벽의 형성 ··············································································15
가. 1차 세포벽의 구조 ·················································································15
나. Cellulose Microfibrils의 합성 ··································································17
다. Matrix Polymers 합성 ············································································18
라. 1차세포벽의 형성 - 세포분열 ·································································21
마. 2차세포벽의 형성 ··················································································22
6. Cellulose synthase ················································································23
가. 개요 ···································································································23
나. Cellulose synthase와 관련 유전자 ···························································24
다. 물관 발생과 cellulose 합성 ·····································································25
라. 제초제 내성 돌연변이를 사용한 연구 ························································27

ⅱ
마. Microtubule 배치와의 관계 ·····································································28
7. Expansins ······························································································29
8. Glucanases 등 기타 가수분해효소 ······················································30
9. 세포벽 분해 효소에 의한 생리 작용 ···············································31
가. 과일의 성숙 ·························································································31
나. 병원성 생명체에 대한 반응 ·····································································32
제4장 결론 제언 ·········································································33
참고문헌 ····························································································35

ⅲ
그림 목차
<그림 2-1> 세포벽의 구조가 형성되는 과정 ···············································15
<그림 2-2> Cellulose Microfibrils의 합성 ·····················································17

1
제1장
서 론
1. 연구의 배경
○ 21세기 지식기반사회에서 과학기술경쟁력은 국가경쟁력
의 원천이며, 이에 세계 각국들은 미래의 경쟁에 살아남
기 해 핵심기술과제를 선정하여 연구개발에 박차를 가
하고 있다.
○ 우리나라 과학기술부도 2005년 6월 ‘미래국가유망기술
원회’를 구성하여 ‘과학기술 측조사(2005-2030)’ 결과
(2005년 5월, 국가과학기술 원회 보고)에서 도출된 기
술후보군을 바탕으로 『미래 국가유망기술 21』을 선정
하여 발표한 바 있다.
○ 한 한국과학기술정보연구원(KISTI)에서는 2005년 SCIE
논문데이터베이스를 이용한 정보계량학 분석을 통해
『미래 유망연구 역 선정연구』를 시도하 으며, 본 보
고서는 그 결과에 기 하여 최근 2~3년간 논문의 인용도
가 속히 높아지고 있는 유망 연구 역을 심으로 기술
논평 형식으로 풀이한 심층 Expert Review이다.

2 식물 세포벽 대사 관련 효소의 연구동향
2. 연구의 방법
○ 한국과학기술정보연구원에서는 SCIE 데이터베이스에 등
록된 논문(1999~2005년 상반기까지 발표된 논문) 에
서, 각 연도 각 분야별( 분류 22분야)로 피인용수
가 상 1%인 고인용 논문(HCP; Highly cited papers)
을 추출하고 공인용분석(Co-citation analysis) 동시단
어분석(Co-word analysis) 등의 과학계량학 방법들과
문가 평가(Expert evaluation)를 통해 ‘미래 유망연구
역’을 도출하 다.
○ 상기 도출된 미래 유망연구 역 에서 통계학 방법으
로 최근 논문의 인용도가 격히 상승하는 연구 역을
과학기술 분야별로 추출하여 본 테크이슈 보고서의 주제
로 삼았다.
○ 본 보고서는 식물 세포벽 사 련 효소의 연구 분야에
있어서 최근 많이 발표되고 있는 논문들을 종합하여
련 분야 연구에 한 기 지식과 함께 세계 인 연구동
향을 개 으로 살펴보고, 미래 핵심기술로 자리잡기
한 연구개발 략을 제시하 다.

3
제2장
기술의 개요
1. 기술의 정의와 범위
○ 본 주제에서 다루는 세포벽은 세균과 균류를 제외한 식
물의 세포벽이다. 세포벽의 사란 세포벽 성분의 합성
과 분해, 구성 성분 사이의 상호작용의 변화 등을 포함
한다.
○ 세포벽은 크게 1차 세포벽, 2차 세포벽, 엽 (middle
lamella)으로 나뉘는데, 엽과 1차 세포벽은 세포가 새
롭게 형성되면서 합성되고 2차 세포벽은 세포가 생성된
후 발생이 진행되면서 1차 세포벽 안쪽에 비교 두텁게
형성된다. 개의 경우, 1차 세포벽을 지닌 세포는 발생
는 생장 인 세포로 세포벽의 변화가 크게 일어나는
데에 반해 2차 세포벽의 경우에는 세포벽의 변화가 비교
크지 않다. 따라서 구체 으로 세포벽의 사란 1, 2
차 세포벽의 형성 과정과 형성된 1차 세포벽에서 일어나
는 변화를 가리킨다.
○ 세포벽은 에 해당하는 microfibril과 pectin, 이 두 가
지를 연결하는 hemicellulose로 구성된다. 이러한 각 성분

4 식물 세포벽 대사 관련 효소의 연구동향
의 변화는 세포벽 사 과정에서 일어날 수밖에 없다.
그 에서 본 주제는 주로 microfibril의 주성분인
cellulose의 합성을 다루게 될 것이다.
2. 기술의 중요성
○ 동물은 내골격 는 외골격 (곤충의 경우) 등을 지니고
있어 형태를 유지하고 운동을 수행할 수 있다. 식물은
이러한 골격을 지니고 있지 않은 신 세포벽을 지니고
있어 개체의 모양을 유지할 수 있고 굴 성, 굴 성, 굴
성 등의 운동 (movement)을 수행하게 된다.
○ 다른 생물과 마찬가지로 식물 역시, 생장과 발생을 하게
되는데 이 과정에 일어날 수밖에 없는 세포의 신장과 분
열 동안 세포벽의 변화는 필수 이다. 따라서 식물에서
세포벽 변화를 알기 한 세포벽 사에 한 연구는 식
물의 생장과 발생을 이해하는 데에 요한 치를 차지
한다고 볼 수 있다. 더 나아가 경제 으로도 식물의 세
포벽은 여러 가지로 활용되고 있다. 식물 세포벽이 상
으로 얇고 강한 성질을 지니고 있어 종이, 옷감, 섬유,
활성탄(charcoal), 목재, 목제품 등에 사용되어 왔다.
한 세포벽에 추출된 다당류는 라스틱, 필름, 코 제,
착제, 젤 등을 제작하는 원료로 사용되어 왔다. 한
과일이나 야채의 세포벽은 직장암, 심장병, 당뇨 등을
방하는 데에 여하는 것으로 알려졌다. 세포벽 성분 ,

제2장 기술의 개요 5
β-glucan은 청내의 콜 스테롤의 수치를 감소시키고
당뇨환자의 insulin 요구량을 감소시키다. 따라서 세포벽
의 사에 한 연구는 비교 경제성이 큰 부가가치를
창출할 수 있다.
3. 기술의 배경
○ 에서 기술한 로 세포벽은 식물의 생장과 발생에 있
어서 요한 역할을 한다. 그 cellulose는 세포벽의
를 형성하는 것으로 생장과 발생에 따른 세포의 변화
에 따라 그 함량과 구조가 변화할 수밖에 없다.
○ 세포벽과 련하여 살펴볼 수 있는 발생과정은 물 의
형성과정이다. 식물은 동물의 순환계와 같은 구조로 물
과 체 을 지니고 있다. 이들 기 은 식물의 세포질에
변화가 생긴 결과 형성되는 구조인데 물 의 경우 세포
질이 제거된, 즉, 세포가 죽고 세포벽만이 남는 발생 과
정을 통해 그 구조가 완성된다. 물 의 경우, 물과 뿌리
에서 흡수된 양분 (nutrients)을 수송하게 되는데 증산
작용에 의해 형성되는 물의 음압을 견딜 정도로 강해야
한다. 이런 강도는 세포질이 제거된 상태에서 남아있는
2차 세포벽에 의해 유지된다. 한 세포질에 제거된 상
태에서 서로 연결된 세포벽은 세포들의 연결체와 비교하
여 훨씬 효과 인 수송 수단이 될 수 있다. 게다가 2차
세포벽이 형성되면서 suberin이나 cutin 등이 침착되어

6 식물 세포벽 대사 관련 효소의 연구동향
식물이 물을 구획화 하는 (새지 않게 만드는) 요한 기
능을 한다.
○ 이러한 물 의 발생, 분화의 처럼 식물의 세포벽은 식
물의 생존에 요한 구조물로 역할을 하고 이 구조물을
형성하는 는 cellulose로 이루어져 있다. 따라서 유럽
이나 미국의 경우, 농학, 식물학, 조경학 등의 분야에서
는 이미 오래 부터 cellulose 합성을 심으로 하여 세
포벽의 생성과 분해 즉, 사에 한 연구가 진행되어
왔다.
4. 기술의 특성
○ 세포벽의 주성분이 탄수화물이라는 은 통 인 생화
학 연구에 있어서 하나의 장애로 간주되어 왔다. 탄수
화물을 구성하는 5탄당, 6탄당 등 단당류는 다양한 구조
, 기하학 enantiomer 등을 지니고 있어 이들을 재료
로 하여 형성된 다당류의 구조는 매우 다양하여 세포벽
을 이루는 다당류들의 구조와 기능을 연구하는 것이 쉽
지 않다. 더구나 식물의 경우, 녹말, sucrose, cellulose를
심으로 하여 진행되는 탄수화물의 사 과정이 동물
등 기타 생물에 비해 상당히 복잡하다. 따라서 통 인
생화학 근과 함께 새로운 근 방법이 요구된다.
○ 근래에 이르러 생물학은 비약 인 발 을 이루어내고 있

제2장 기술의 개요 7
다. 1990년 에 시작한 Human Genome Project가 진
행되면서 식물의 유 체도 함께 밝 져 왔다. 재 애기
장 (Arabidopsis thaliana)와 벼 (Oryza sativa)의 유
체가 해독되고 다양한 종류의 돌연변이가 제작되어 일정
유 자의 구조와 기능에 한 연구가 한층 수월해졌다.
세포벽 사의 경우도 외는 아니라고 단된다.
Cellulose 사의 경우, cellulose 합성 효소가 돌연변이
된 식물을 제작하거나 골라내 이들의 생리학 , 발생학
인 특징을 연구할 수 있었고 1990년 부터 이러한 경
향의 연구는 뚜렷하다. 제 제에 한 내성을 지닌 돌연
변이나 물 의 발생에 문제가 생긴 돌연변이들이 그
이다. 이외의 다양한 돌연변이를 포함하여 돌연변이에
한 연구는 cellulose 합성 효소 (다른 세포벽 사
련 효소도 마찬가지임)를 분자 차원에서 그 구조와 기능
을 구명할 수 있게 하 다. 본고는 이러한 최근의 연구
동향을 기술하고자 하 다.
5. 기술의 현안문제
○ 국내의 경우, 세포벽에 한 본격 인 연구가 진행되지
않아왔고 재도 활발하다고 볼 수 없다. 세포벽 사
연구에 한한 국내의 안문제는 이 연구가 거의 수행
되지 않는다는 것이다. 따라서 외국에서 진행되는 연구
에 심을 기울임은 물론 국내에서도 이 분야에 한 연
구를 장려하는 정책이 만들어져야 할 것으로 단되다.

8 식물 세포벽 대사 관련 효소의 연구동향
다른 분야와 마찬가지로 이 분야 역시 기 연구를 튼튼
하게 만들 수 있는 정부와 학계의 노력이 아쉽다.

9
제3장
연구개발동향
1. 개요
○ 세포벽 사에 한 기 인 연구를 바탕으로 유럽과
미국에서는 이미 상당한 수 의 경제 인 가치를 얻어내
고 있다. 그러나 국내의 경우 정부 는 기업이 주도하
여 세포벽 연구의 경제 가치를 인식하여 연구를 진행
한 사례는 찾아볼 수 없어 정책 심이 필요한 분야라
할 수 있다. 따라서 세포벽을 주요한 주제로 삼아 연구
가 진행된 사례는 흔하지 않다. 몇몇 학의 연구
를 들어, 서울38, 포항공
49, 충남
13, 14, 15, 16, 연세
56
가 식물의 생리, 발생학 상에 따라 세포벽 사에
여하는 효소를 연구한 사례 정도가 있을 뿐이다. 요컨
cellulose synthase를 비롯한 세포벽의 합성을 심으
로 세포벽의 사를 본격 으로 연구하는 집단은 국내에
서 무한 실정이다.
○ 이러한 사실을 근거로 하여 아래에서는 외국의 연구개발
동향이 기술될 것이다. 이 은 세포벽의 요성과 세포
벽의 사과정에 해 설명한 후, 이 사과정에 여하
는 각 효소에 한 내용을 담을 것이다

10 식물 세포벽 대사 관련 효소의 연구동향
2. 식물에서 세포벽의 중요성
○ 동물과 달리 식물은 식물만의 구조를 지니고 있다. 식물
은 원천 으로 양을 얻는 방법이 달라, 즉 합성을
하여 유기 분자를 얻게 되므로 엽록체라는 세포내 소기
을 지니고 있다. 이와 함께 식물만이 지니는 독특한
구조가 세포를 감싸고 있는 세포벽이다.
○ 다음과 같은 면에서 세포벽은 식물에게 필수 이다.
- 우선 세포벽은 식물의 생장이 가능하도록 물리 인 지
지 를 제공한다. 동물에 비교하자면 내골격을 이루는
와 같은 작용을 한다고 볼 수 있다.
- 둘째로 세포벽은 세포와 세포를 고착시킨다. 이는 동
물 세포의 경우, 단백질에 의해 세포간의 고착과 이동
이 이루어진 것과는 다른 양식이다. 따라서 발생 과정
에서 세포의 움직임이 용이한 동물과 달리 세포의 움
직임이 발견되지 않는다.
- 셋째로 세포벽은 세포의 모양을 결정하고 팽압이 생기
는 원인을 제공함으로써 식물의 외골격으로 기능한다.
따라서 원천 으로 세포벽이 팽창할 수 있는 능력에
따라 식물의 형태형성이 진행된다. 이는 팽압과 세포
의 부피 사이의 계를 결정하여 정상 인 물의 출입
을 가능 함으로써 가능하게 된다.
- 넷째로 세포의 분화 결과 형성된 물 의 경우 세포질
은 제거된 세포벽끼리의 연결구조이다. 따라서 세포벽
은 물을 량으로 통과시키는 물 을 구성하여 동물의

제3장 연구개발동향 11
순환계에 비견되는 기능을 수행한다.
- 다섯번째로 세포벽은 고분자가 확산에 의해 세포에
근하는 것을 막고 pathogen (병원성 생명체, 바이러
스, 세균, 균류 등)의 요한 장벽으로 기능한다. 즉
세포내외로의 물질 이동과 식물의 방어에 여하게 된
다.
- 이외에도 세포벽의 분해산물은 씨의 경우, 양분으로
쓰이고 이 oligosaccharides는 신호로 세포에 자극
을 달하는 역할을 할 수 있다. 요약하면 식물의 생
존, 발생, 성장에 세포벽은 필수불가결한 요소로서 작
용하고 모든 식물의 모든 부 에 있는 만큼 그 기능은
물론 구조도 매우 다양하다 (참조 Brett and Waldron,
1996; Carpita and McCann, 2000).
3. 식물 세포벽의 구조
○ 식물의 세포벽은 존재하는 치에 따라 다양한 구조를
지닐 수밖에 없다. 피층에 있는 유조직 (parenchyma)
세포벽은 얇고 구조 특징이 뚜렷하지 않다. 표피세포,
후각세포 (collenchyma), 체 섬유 (phloem fibers), 헛
물 요소 (xylem tracheary elements), 후벽조직
(sclerenchyma) 등의 세포벽은 두껍고 여러 층으로 이루
어져 있으며 lignin, cutin, suberin, waxes, silica, 구조 단
백질 등이 내재되어 있다. 따라서 세포벽의 두께, 침착된
물질, plasmodesmata의 수 등에 따라 측면성(sidedness)이

12 식물 세포벽 대사 관련 효소의 연구동향
생길 수 있고 이에 따라 세포의 극성, 세포 기능의 분화,
세포벽 성분의 세포막으로의 표 화 (targeting) 등의 기
능을 나타낼 수 있다. 표피세포벽의 바깥쪽은 비교 두
텁고 plasmodesmata가 으며 cutin, waxes 등이 침
되어 있는 상이나 기공세포의 경우, 기공 쪽의 세포벽
이 더 두터운 것 등을 로 들 수 있다.
○ 이러한 다양한 구조가 존재함에도 불구하고 세포벽의 구
조는 기본 으로 주요한 2가지로 나 어진다. 첫째는 1
차 세포벽 (primary wall)으로 생장 인 세포와 비교
덜 분화되어 모든 세포에서 발견되는 것으로 공통 인
구조를 띤다. 를 들어 양 유조직 세포에는 매우 얇
은 (두께가 약 100nm) 세포벽이 발견된다. 둘째로 2차
세포벽 (secondary wall)은 물 (xylem) 세포와 같은
분화된 세포에서 나타나는데 lignin이 있어 두텁고 매우
강하다. 이외에도 세포벽과 세포벽을 부착시키는 역할을
담당하는 엽 (middle lamella)과 세포질과 세포질을
연결하여 단백질, 핵산을 비롯한 다양한 분자의 이동 경
로로서 기능하는 plasmodesmata 등의 구조가 있다 (참
조 Brett and Waldron, 1996; Carpita and McCann,
2000).

제3장 연구개발동향 13
4. 식물 세포벽의 기능
가. 세포의 신장
○ 식물 세포벽의 가장 큰 기능은 세포생장에 여한다는
것이다. 세포생장은 극성을 띠거나 (tip growth: 뿌리털,
pollen tubes) 골고루 일어난다. 그러나 많은 경우, 를
들어 섬유, tichomes, sclereids 등에는 두 가지 양상
의 생장이 모두 발견된다. 이러한 생장은 세포벽에 가해
진 압력, 효소의 활성 차이, 구조 차이에 의해 세포마
다 생장 양상이 달라져 독특한 모양을 나타낸다. 세포
차원에서 보면 세포벽 체에 고루 힘이 가해지는 팽압
을 이용, microfibril의 배열 상태에 따라 생장 방향이 결
정된다. 즉 세포의 장축에 직각으로 microfibril이 배열되
고 세포신장 과정에서 외부의 세포벽은 얇아짐으로 인해
microfibril의 배열이 자유로워진다. 이러한 세포생장의
방향은 가장 안쪽 1/4의 세포벽에 의해 결정된다. 한
세포질의 microtubules가 세포벽의 microfibrils와 같은
방향으로 배열되어 있어 microtubule이 microfibril의 배
열을 유도하는 것으로 알려져 있다.
○ 요컨 식물 세포벽은 식물 세포 생장의 두 가지 구성
요소인 팽압과 세포벽의 신장도 (extensibility) 한 가
지를 담당하고 있는 것이다. 이러한 세포생장에 의해 식
물세포는 세포벽이 얇아지지 않고 강도는 유지하면서 성
숙하기까지 10~100배 정도 부피가 증가한다.

14 식물 세포벽 대사 관련 효소의 연구동향
나. 식물의 발생
○ 이외에도 세포벽은 물리 인 장벽으로서 작용하고 세포
막 단백질과 상호작용을 하여 세포발생에 있어서 치와
발생에 한 정보를 제공한다. 한 많은 효소와 활성이
있는 분자를 지녀 세포벽의 물리 성질을 빠르게 변
화시키고 다양한 환경조건에 응하여 신호를 생산한다.
이러한 특징에 의해 세포생장이 끝난 다음에도 세포벽은
실질 인 변형을 일으킨다. 를 들어, 과일과 발아 인
씨의 배젖에서 분해가 매우 활발하다든지 잎과 과일의
낙엽층에서 엽만 분해되어 세포가 분리되는 것 등이
그 이다.
○ 한 발아 인 씨에서 뿌리가 자라면서 세포 간 공기로
채워진 공간이 형성되는 동안 세포가 분리되고 pathogen
의 공격에 의해 세포벽이 분해된다 (참조 Brett and
Waldron, 1996; Carpita and McCann, 2000).

제3장 연구개발동향 15
5. 식물 세포벽의 형성
가. 1차 세포벽의 구조
○ 1차 세포벽은 다당류 기질 (matrix)에 묻 있는
cellulose microfibrils로 구성되어 있다. 콘크리트로 비유
하자면 microfibril은 철근에, 기질은 시멘트에 해당한다
고 볼 수 있다. Glucose의 polymer인 cellulose의 집합체
인 microfibril은 체 세포벽의 25%를 차지하는데
<그림 2-1> 세포벽의 구조가 형성되는 과정
Microfibril을 구성하는 cellulose는 원형질막에 치한 cellulose synthase에 의해 합성되
는데, 이 때 세포의 원형질막 안쪽에 배치된 microtubule의 방향에 의해 새롭게 합성
되는 microfibril의 방향이 조 된다 (출처: Taiz and Zeiger, Plant Physiolgy, 3rd ed.,
Sinauer Associates Inc, 2002).

16 식물 세포벽 대사 관련 효소의 연구동향
crystalline ribbon 구조로서 효소의 근이 어렵고 주변
에 물이 별로 없는 단단한 구조이다. 구조 인 특징으로
인해 microfibril은 세포벽에 유연성 (flexibility)과 강도
(strength)를 제공한다.
○ 기질 (matrix)은 pectin, hemicellulose, 구조 단백질로 구
성된다. Pectin은 glucan, xylan, galactan, glucomannan
등 매우 다양한 다당류로 구성되어 있다. Pectin은
hydrated gel로 체 세포벽의 35%를 차지한다. 이외에
도 pectin은 glucan 와 xylose side chain으로 구성된
xyloglucan과 glucuronoarabinoxylan도 포함한다.
○ 결국 pectin은 microfibril과 후술하는 hemicellulose가 박힌
구조로 세포벽의 구멍을 형성하게 된다. Hemicellulose는
세포벽의 25% ( 본의 자엽 의 경우는 60~70%까지 차
지)를 차지하고 유연한 성질을 지니는데 이들의 역할은
microfibril 사이를 연결하는 역할을 한다.
○ 구조단백질은 세포벽의 1~8%를 차지하며 세포벽의 물
리 인 강도를 유지하는 데에 여하는 것으로 알려져
있다 (참조 Brett and Waldron, 1996; Carpita and
McCann, 2000).
제3장 연구개발동향 17
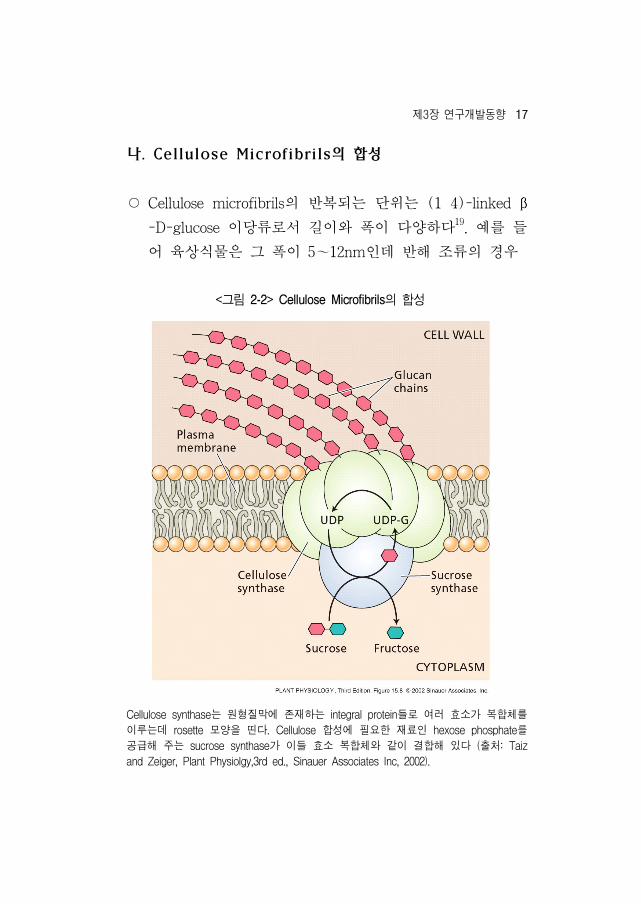
나. Cellulose Microfibrils의 합성
○ Cellulose microfibrils의 반복되는 단 는 (1 4)-linked β
-D-glucose 이당류로서 길이와 폭이 다양하다19. 를 들
어 육상식물은 그 폭이 5~12nm인데 반해 조류의 경우
<그림 2-2> Cellulose Microfibrils의 합성
Cellulose synthase는 원형질막에 존재하는 integral protein들로 여러 효소가 복합체를
이루는데 rosette 모양을 띤다. Cellulose 합성에 필요한 재료인 hexose phosphate를
공 해 주는 sucrose synthase가 이들 효소 복합체와 같이 결합해 있다 (출처: Taiz
and Zeiger, Plant Physiolgy,3rd ed., Sinauer Associates Inc, 2002).

18 식물 세포벽 대사 관련 효소의 연구동향
에는 그 폭이 30nm에 달한다. Cellulose microfibril은
crystalline domain과 amorphous domain으로 구성되어
있다. 이 crystalline domain은 매우 질서정연한 구조
로 수소, 소수성 결합으로 이루어져 있고 2,000~25,000
glucose로 구성되어 있는데 철 정도의 강도를 유지한다.
한 소수성으로 효소나 화학물질에 의한 분해정도가 약
하다19. 따라서 fungal cellulases는 amorphous regions부
터 분해를 시작한다.
○ Cellulose microfibrils의 합성은 원형질막에 cellulose
synthase들로 이루어진 particle rosettes와 terminal
complex에서 이루어진다. 세포내의 cellulose synthase는
sterol-glucosides를 primer로 glucose를 연결시킨 후 세포
밖으로 수송하는데, 이 때 sterol은 endoglucanase에 의해
잘린다. 세포 밖에서 glucan chain들은 crystalline을 형성
하고 xyloglucan과 결합한다.
○ In vitro 실험에서는 xyloglucan과의 결합이 찰되었다.
이 과정에서 cellulose synthase와 같이 붙어서 작용하는
sucrose synthase는 sucrose를 분해하여 glucose를 공 하
는데, 이 glucose는 UDP-glucose로 환되어 cellulose 합
성 재료로 쓰인다37, 46, 46
.
다. Matrix Polymers 합성
○ 이들은 Golgi에서 합성되어 세포 밖으로 수송된다. 이 과정
에서 sugar-nucleotide polysaccharide glycosyltransferases

제3장 연구개발동향 19
는 다당류 chain에 sugar nucleotide에서 얻은 다당류를 더
해 다. 이러한 합성 과정은 부분 무정형으로 진행되는
데 cellulose 장축과 연 되어 분포하는 부분 질서를 나타
낸다.
○ Hemicellulose는 cellulose에 결합되어 존재한다.
Hemicellulose는 다양한 종류의 다당류로 구성된다. 이
hemicellulose는 pectin을 제거한 후 1-4 M NaOH로 분리
할 수 있다. 떡잎식물의 1차 세포벽에서는 xyloglucan이
가장 풍부한데 (14)-linked β-D-glucose 에 xylose,
galactose, terminal fucose 등이 연결되어 있다. 이러한
side chains에 의해 crystalline 구조를 이루기 어렵다 (참
조 Brett and Waldron, 1996; Carpita and McCann,
2000). 게다가 microfibril 사이의 간격 (20~40nm)보다
길어 (50~500nm) 여러 개의 microfibrils를 연결할 수 있
다. 이들의 종류는 glucuronoarabinoxylan, glucomannans
등으로 발생에 따라 hemicellulose의 주요 성분이 바 게
된다. 한 곡류의 2차 세포벽에는 xyloglucan 신
glucuronoarabinoxylans과 (13,14) β-D-glucan이 많다9.
○ Pectins는 matrix의 gel 상태를 구성한다.
Pectins는 다양한 pectic polysaccharides로 구성된 매우
크고 복잡한 분자이다. 이들은 수용성이라 calcium
chelator가 포함된 뜨거운 물로 추출할 수 있고 산성 당
인 galacturonic acid와 성 당인 rhamnose, galactose,
arabinose 등으로 구성되어 있다. 이들 당으로 구성된 구
조는 간단한 homogalacturonan (polygalacturonic acid)과

20 식물 세포벽 대사 관련 효소의 연구동향
가장 풍부한 구성 다당류로 (14)-linked β-D-glucuronic
acid의 합체인 rhamnogalacturonan I (RG I)를 들 수
있다. RG I은 한 가장 풍부한 pectin으로 구조에
가지를 지닌 부 와 그 지 않은 부 를 지니고 있다.
Rhamnogalacturonan II (RG II)는 10가지 이상의 당이
복잡한 형태로 결합되어 있고 borate diesters에 의해
cross-link 된다41. 이들 구조는 gel을 형성하거나 딱딱해
지는데 물과 결합하거나 calcium과 결합하여 calcium-
bridged network를 형성한다. 한 산성기가 methyl,
acetyl 등으로 ester 구조가 형성되면 calcium과의 결합을
억제하여 gel 상태가 되기 어렵다. 만약 이 구조들이 세
포 밖으로 분비되면 esterase에 의해 ester group이 제거
되어 단단한 gel이 형성되는데 이 때에는 ion 상태의 분
자가 증가하고 세포벽 효소 활성이 증가하며 ferulic acid
등 phenolic residues와 pectin 분자가 공유 결합을 하게
된다35.
○ Structural Proteins는 세포벽을 횡 으로 연결한다.
구조 단백질은 hydroxyproline-rich protein (HRGP),
glycine-rich protein (GRP), proline-rich protein (PRP)
등의 주요한 3가지로 나뉘어진다. 많은 경우, 이들은 매
우 반복 인 구조를 지니거나 매우 높은 비율로
glycosylation 되어 있다. 합성되어 분비된 단백질은 친수
성이지만 세포가 성숙되거나 상처에 의해 소수성으로 바
다. 세포의 종류, 성숙 정도, 상처, elicitor, pathogen
등에 의한 양 변화가 다양하고 세포 는 조직에 따른
분포 역시 다양하다. 를 들어 HRGPs는 cambium, 체

제3장 연구개발동향 21
유조직, 후각조직 (sclerenchyma)에서, GRPs와
PRPs는 물 등 분화된 세포벽에서 발견된다.
Aarabinogalactan proteins (AGPs)는 세포벽의 건량비
로 1%를 차지한다. 이 단백질의 90% 이상이 galactose
와 arabinose 등으로 glycosylation되어 있다. 이들은 원
형질막에 붙어있고 조직, 세포에 따라 유 자의 발 양
상이 다르다. 한 세포의 고착, 세포 분화동안의
signaling에 여한다. 따라서 탁 배양 인 세포에
AGPs나 AGPs 결합단백질을 처리하면 세포의 증식, 배
의 발생 (embryogenesis)이 진되어, 이 단백질이 발생
과정, 생장, 양, pollen tube의 생장 방향 결정 등에
여함을 알 수 있다. 이 단백질들은 분비 소낭 (secretory
vesicle)내에서 chaperone으로 작용하여 새로 합성된 다
당류가 세포벽으로 분비되기 까지 다른 부 에 결합하
는 것을 억제한다 (참조 Gasper et al., 2001).
라. 1차세포벽의 형성 - 세포분열
○ 1차 세포벽은 세포가 분열되면서 주로 형성된다. 세포분
열 후기에 생긴 phragmoplast에는 microtubules와 막, 그
리고 소낭 (vesicles) 등이 모여 새롭게 세포벽을 형성하
게 된다. 이들은 유사분열이 끝나면서 세포 (cell
plate)을 형성하게 되고, 세포벽이 형성되는데 이 때 세
포벽 구성 물질이 분비된 후, 한 배열을 갖기 해
조립 (assemble)되어야 한다. 이러한 조립 과정은 자동
으로 이루어지기도 하지만 효소의 도움을 받는 경우가
많다. 를 들어 cellulose는 추출한 뒤 강한 유기용매로

22 식물 세포벽 대사 관련 효소의 연구동향
녹이면 rayon이라는 섬유를 형성하고 hemicellulose는 강
알칼리로 녹이면 세포벽과 비슷한 배열로 모이게 된다.
한 pectins는 매우 수용성으로 추출 후 모이지 않고
퍼지게 된다. 이 과정에 여하는 효소는 xyloglucan을
잘라 다른 xyloglucan과 연결하는 xyloglucan
endotransglycosylase, hemicellulose의 side chains를 잘라
microfibril 표면에 잘 붙게 하는 glycosidase 종류,
methyl ester를 잘라 carboxyl기를 노출, Ca2+와 붙게 하
는 pectin methyl esterase, 세포벽 단백질의 tyrosine,
phenylalanine, ferulic acid 등 phenolic group, pectins,
다른 세포벽 구성원과의 cross-linking을 매하고 lignin
cross-links 등에 요한 peroxidase 등이 보고되었다 (참
조 Brett and Waldron, 1996; Carpita and McCann,
2000).
마. 2차세포벽의 형성
○ 세포신장이 끝난 세포 안쪽으로 2차 세포벽이 생긴다.
이 게 생긴 세포벽은 매우 두껍고 물리 강도가 커서
물 을 형성하게 된다. 2차 세포벽은 세포벽이 덜 약해
지는 신 조성 성분 사이의 결합이 늘게 되면서 여러
층을 이루고 1차 세포벽과 구성성분이 다르다. 즉,
cellulose 는 다른 polymer와 단단한 결합을 하는
matrix 다당류가 분비되고 β-D-glucans가 제거되며
xyloglucan 신 xylan이 많게 된다. Pectin을
de-esterification시켜 더 단단한 pectin gels를 형성하고
HRGPs의 phenolic groups의 cross-linking을 증가시킨

제3장 연구개발동향 23
결과 microfibril의 배열이 더 평행으로 이루어지게 된다.
한 lignin이 많이 들어 있고 matrix에서 물을 제거하고
cellulose 사이에 소수성 network를 형성하여 세포 신장
을 억제한다. 이로 인해 세포벽의 물리 강도를 높이고
pathogens에 한 감수성을 감소시킨다. 따라서 동물이
세포벽을 분해, 소화시키기 어렵게 된다 (참조 Brett
and Waldron, 1996; Carpita and McCann, 2000).
6. Cellulose synthase
가. 개요
○ 식물은 동물과 달리 세포벽을 지니고 있고 이 세포벽을
통해 병원균에 응하고 생장과 발생 동안 필요한 양
분의 흡수, 수송, 분비 등의 기능을 수행하는데, 이 세포
벽의 역할을 하는 구조가 cellulose로 구성된
microfibril이다. 이 microfibril의 생성은 cellulose의 합성
을 기 로 한다.
○ 세포벽의 주요 성분은 β-(1,4)-linked glucose 분자의 연
결체인 선구조의 cellulose이다. 이 리본 모양의 glucan
연결체는 결정화하여 microfibril을 구성하는데 이 구조로
인해 세포벽은 탄성과 강한 성질을 지니게 된다26.
○ 식물에서 cellulose는 세포의 원형질막에 치한 rosette

24 식물 세포벽 대사 관련 효소의 연구동향
모양의 단백질 복합체 즉, cellulose synthase 복합체에
의해 합성된다37, 40, 46. 그리고 합성된 cellulose는 주변의
다른 단백질에 의해 결합되어 microfibril를 형성하게 된
다23.
○ 이러한 잘 알려진 결과에도 불구하고 식물의 cellulose
synthase는 정제되거나 그 특징이 밝 지기 어려웠다19.
최근에 이르러 돌연변이를 제작하거나 유 자의 기능을
억제하는 분자유 학 인 방법과 제 제 연구를 통해,
rosette 복합체의 subunits가 CesA (cellulose synthase)
유 자에 의해 암호화되는 것이 밝 졌다1, 8, 42, 48. 밝 진
유 체의 염기 서열과 많은 양의 expressed sequence
tag (EST) database에 따르면, 식물의 CesA 유 자는
multigene family의 일종이다. CesA 유 자가 애기장
에서는 10개, 벼 (Oryza sativa)에서는 12개45 (
http://cellwall.stanford.edu/), Populus trichocarpa에서
는 18개22, 보리 (Hordeum vulgare)에서는 어도 8개7,
옥수수 (Zea mays)에서는 어도 9개 정도가 있는 것24,
34으로 밝 졌다.
나. Cellulose synthase와 관련 유전자
○ 애기장 에서 각각의 CesA 유 자를 돌연변이 시켜 표
형을 찰한 보고1, 2, 6, 9, 11, 28, 31, 48, 52, 53, 54
에 따르면, 이
유 자의 발 이 조직에 따라 다르고 세포벽의 종류 (1
차 는 2차)에 따라 달라 특화된 기능을 지니는 방향으
로 진화해 왔음을 알 수 있다. 이 몇몇 CesA 단백질

제3장 연구개발동향 25
은 애기장 에서 rosette 모양이 제 로 형성되는 데에
필수 임23, 52이 보고되었다. CesA 유 자들의 다른
기능은 cellulose가 아닌 다당류를 합성하는 것이다.
○ Cellulose가 아닌 세포벽 구성 성분 에서 를 구성
하는 heteroxylans, xyloglucans, mannans, (13,14) β
--D-glucans 등은 화학 으로 cellulose와 비슷하기 때문
에9, 29, CesA 유 자들이나 Csl (cellulose synthase-like)
유 자들이9, 23, 24, 55
이러한 다당류를 합성하는 것은 있을
수 있는 일이다. 를 들어 최근의 Liepman 등39의 연구
에 따르면 Csl gene families (CslA, CslC, CslD, CslE,
CslH) 몇가지는 β-mannan synthases로서 작용하는데
GDP-mannose와 GDP-glucose를 재료로 사용하여 β
-linked mannan 합체 는 β-linked glucan 합체를
각각 합성하고 두 재료를 섞어 공 하면 β-linked
glucomannan 합체를 형성하는 것으로 알려졌다.
한 애기장 와 벼의 씨의 배젖에 존재하는 CsI의 일종인
mannan synthase는 장 기능을 지닌 hemicellulose의
일종인 galactomannan의 β 1,4-mannan 를 형성한
다고 보고되었다24.
다. 물관 발생과 cellulose 합성
○ Cellulose synthase의 각 유 자에 한 연구는 주로 애
기장 를 사용하여 수행되어 왔다. 애기장 의 물 의
형태 형성에 문제가 있는 돌연변이를 사용한 연구에서 2
차 세포벽에서 cellulose가 심각하게 부족한 결과를 발견

26 식물 세포벽 대사 관련 효소의 연구동향
하여 이에 련된 유 자가 irx1과 irx3로 밝 졌고 이들
이 cellulose 합성에 필수 임이 알려졌다53.
○ 이러한 물 의 형성에 문제가 있는 돌연변이 식물은 일
감치 알려져 있었는데 구체 으로 어떤 cellulose
synthase에 문제가 생긴 것인지는 이후에 밝 졌다. 클
론된 irx1은 CesA8과 립인자이고 irx3은 CesA7, irx5
는 CesA4와 각각 서로에 해 립인자임이 보고되었다52.
○ 병원성 균류에 한 반응에 cellulose 합성이 여되어
있는데 이 때 여하는 효소가 CesA3임이 보고되어
cellulose 합성이 stress에 응하는 식물의 신호 달 과
정에 여됨이 보고되었다27. 이러한 IRX1과 IRX3의 역
할에 한 결과들은 최근에 microarray data에 의해 다
시 확인되었다5.
○ 이와 함께, Persson 등43 역시 Affymetrix Arabidopsis
microarray 방법을 사용하여 물 의 발생에 irx8과 irx13
등의 유 자가 2차 세포벽의 합성에 여함을 보고하여
기존에 알려진 것보다 더 많은 유 자가 물 의 형성에
여하는 것으로 단된다.
○ Szyjanowicz 등51은 irx2 분자가 KORRIGAN과 마찬가
지로 2차 세포벽 형성의 후기에 작용하여 신장하는
microfibril의 가공(processing)과 cellulose synthase 복합
체의 방출에 여하는 결과를 발표하 다.

제3장 연구개발동향 27
라. 제초제 내성 돌연변이를 사용한 연구
○ 애기장 에서 두 가지의 유 자 CesA1과 CesA6의 돌연
변이를 사용한 연구에서 세포의 신장이 일어나지 않는
결과를 얻어 이 유 자들이 세포의 신장에 필요하고 뿌
리와 하배축에서 동 으로 발 하여 세포 신장에 여
함이 알려졌다28. CesA1은 한 부피 생장 돌연변이인
rsw1에서 발 이 해됨이 밝 져 rsw1 유 자와 립
인자임이 보고되었다1. 돌연변이 ixr1라는 이름은 제 제
인 isoxaben에 항성을 띠어 isoxaben-resistant (ixr)에
서 유래하 다. 이 제 제는 다른 제 제, 즉
thiazolidinone과 함께 cellulose의 합성을 억제하는 것으
로 알려져 있었는데 이를 근거로 ixr1은 CesA3와 ixr2는
CesA6와 각각 립인자이고 이들은 1차 세포벽 합성에
필수 임이 알려졌다20, 48
.
○ 이러한 연구 결과를 종합해 보면 CesA1, CesA3, CesA6
는 cellulose synthase 복합체를 이루어 1차 세포벽의 합
성에 여하고 CesA4, CesA7, CesA8 역시 복합체를 이
루어 2차 세포벽을 생성한다는 것을 알 수 있다. 그러나
1차 세포벽과 비교하여 2차 세포벽의 경우, microfibril
당 β-glucan의 함량과 cellulose의 함량이 더 많은 사실을
감안한다면 2차 세포벽의 생성에 여하는 각 CesA의
기능은 1차 세포벽 합성에 여하는 CesA와 다를 것으
로 보인다.

28 식물 세포벽 대사 관련 효소의 연구동향
마. Microtubule 배치와의 관계
○ Gardiner 등31은 물 조직이 발생하는 에 IRX1/
CesA8, IRX3/CesA7, IRX5/CesA4 등의 효소들이 기능
인 cellulose synthase 복합체를 형성하는 데에 필요하
고 항체를 이용하여 이 단백질들이 microtubule에 의해
2차 세포벽에 형성하는 데에 향을 받는다는 것을 보여
주었다. 그러나 이 과정에 actin filament는 주요한 역할
을 하지 않는 것으로 보인다. 한 3가지의 단백질이
모두 있어야 CesA 복합체가 제 기능을 발휘할 수 있음
을 나타내 이들 3가지 단백질이 상호 작용한다는 것을
나타냈다.
○ Fisher와 Cyr30는 microtubule의 배치와 안정화를 해
cellulose의 생합성이 필요하다고 주장하 다. 이러한 주
장은 2차 세포벽에서 microtubule의 배치에 의해
cellulose의 침착과 배치가 결정된다는 기존의 학설을
microtubule과 microfibril이 상호간의 배치에 향을 끼
치는 학설로 체하게 되는 것이다. 그러나 2차 세포벽
에서 CesA 복합체가 cellulose 합성을 못하게 된 돌연변
이에서 microtubule의 배치가 변하지 않는 결과50로 보아
microfibril이 microtubule의 배치에 향을 미치는 것으
로 보이지는 않는다.
○ Sugimoto 등50은 한 RSW1/CesA1와 and MOR1가 독
립 인 경로를 통해 작용할 것이라 제안하 는데, 그 이
유는 mor1 rsw1 이 돌연변이가 각각의 단일 돌연변이

제3장 연구개발동향 29
와 비교하여 부가 인 표 형을 보 기 때문이다. 이 이
돌연변이와 rsw1 돌연변이는 모두 온도에 민감하고,
이 두 가지 돌연변이 사이에는 cellulose의 함량에 차이
를 발견할 수 없었는데, 이 결과를 토 로 MOR1에 의
한 microtubule의 배치가 cellulose의 합성에 향을 미친
다는 기존의 다양한 보고들을 지지한다고 볼 수 있다.
○ 식물 세포의 신장에는 세포벽의 변화가 따를 수밖에 없
다. 암조건에서 애기장 하배축의 길이생장은 처음에는
천천히 그리고 이후에는 빨리 일어나는 것이 찰되었는
데, cesA6(prc1-1) 유 자의 돌연변이 식물이나
isoxaben을 처리한 야생형 식물은 빠른 생장이 찰되지
않았다. 이러한 결과와 자 미경을 사용한 결과에 따
르면 빠른 생장이 일어나는 단계에서 microfibril의 배치
가 요한 역할을 하는 것으로 제안되었다44.
○ 이외에도 가뭄과 삼투 stress에 응하여 AtCesA8/IRX1
유 자의 발 이 증가하고 이 유 자의 돌연변이에서
abscisic acid, proline 수용성 당 등의 양 변화가 수반
되는 것이 보고되었다12.
7. Expansins
○ 산성 pH에서 세포벽의 생장은 더 빠르게 일어난다. 이러
한 사실은 산성 완충액이나 곰팡이 독소인 fusicoccin

30 식물 세포벽 대사 관련 효소의 연구동향
는 auxin 등을 처리한 식물의 길이 생장이 증가하는 결과
로부터 확인되기 시작하 다. 분자 차원에서는 원형질막
에 있는 H+-ATPase가 세포질의 H
+를 세포벽으로 수송
하는 능력이 fusicoccin이나 auxin에 의해 증가해서 세포
벽의 pH를 낮추는 결과, 세포 신장이 일어나는 것으로 알
려져 있다. 이러한 상은 육상식물에서 공통 으로 나타
난다. 기와 뿌리의 길이 신장에서도 발견되는 상이지
만 뿌리털이 생기면서 표피세포 부 의 pH가 4.5로 감소
하는 상을 로 들 수 있다. 세포벽에 형성된 산성 pH
는 세포벽을 비가역 으로 신장시켜 세포의 신장을 가능
하게 한다. 이러한 조건에서 세포의 신장에 여하는 단백
질이 expansin이다. 세포벽을 분리하여 산성 조건에서
extensometer를 사용하여 신장이 일어나는 상을 찰하
는 가운데 열과 protease에 민감한 요소가 여하는 것을
발견하고 생장 인 세포벽에서 추출한 단백질이
expansin이다. 이 게 발견된 expansin은 cellulose 사이
는 hemicellulose에 결합하여 작용한다. 식물 세포의 생장
에 여하는 만큼 그 요성을 감안한다면 이 단백질이
애기장 와 벼를 비롯한 식물에서 multigene family로 존
재한다는 것은 이상한 결과가 아닐 것이다 (참조,
http://www. bio.psu.edu/expansins/).
8. Glucanases 등 기타 가수분해효소
○ (1, 4)β-D-glucanase는 hemicellulose를 구성하는

제3장 연구개발동향 31
xyloglucan 분해에 여한다33. 이 효소는 auxin에 의해 세
포가 생장할 때 발 이 증가하는 것으로 알려져 있다. In
vitro 상태에서 세포에 더해주면 식물의 생장이 진된다.
이 효소는 곡류의 세포벽에서는 (1,3)(1,4)β-D-glucans를
분해하는 기능을 나타내고 expansin에 의한 세포벽의 변
화를 간 으로 조 하게 된다9. 이외에도 Xyloglucan
endotransglycosylase, pectinase 등이 세포벽의 사에
여하는 것으로 알려져 있다18.
9. 세포벽 분해 효소에 의한 생리적 작용
가. 과일의 성숙
○ 과일은 성숙되면서 세포벽의 viscosity가 감소한다. 그 이
유는 세포벽의 성분 , hemicellulose와 cellulose의 결합
양상이 바 기 때문이다. 이 과정에 여하는 효소는 β
-D-glucan을 분해하는 glucanase 종류와 xyloglucan의
side chain들을 제거하는 xylosidase, transglycosylases 등
이다. 한 expansin, methyl esterase 등이 발 되어 이
들 효소가 여할 가능성도 제안되었는데 이들의 역할은
pectin의 분해를 담당하는 pectinase의 근이 용이해지
기 때문일 것으로 알려졌다47.

32 식물 세포벽 대사 관련 효소의 연구동향
나. 병원성 생명체에 대한 반응
○ 병원성 균류나 세균의 공격, 는 상처 등에 식물은 활
성산소종의 생성을 증폭시켜 방어 mechanism을 유도하
는 것으로 알려져 있다. 활성산소종은 병원성 생물을 직
공격하거나 간 으로는 phenolic compounds를
cross-link시켜 병원성 생물의 공격을 막는다. 한,
proline을 많이 함유하는 구조단백질은 소수성 정도를 증
가시켜 세포벽을 단단하게 한다4.
○ 병원성 생물의 공격에 의해 세포벽의 사가 일어나게
되는데 이 사로 인해 10~15개의 당으로 구성된
oligasaccharin이 생성된다. 이 oligosaccharin은 세포 내의
신호 달 과정을 일으켜 세균을 공격하는 phytoalexin의 합
성을 진하고, 활성산소종의 증폭, ethylene의 합성, 막
, 세포 내 calcium의 농도, chitinase, glucanase 등의 병원
성 생물과 련된 세포벽 효소 단백질의 합성, 다른 병원성
련 신호, 일정 조직의 생장과 발생 등에 변화를 일으킨다17. 일례로 Phytophthora는 endopolygalacturonase를 분비하
여 분해된 pectin이 분비되면서 oligogalacturonans가 생성되
면서 방어 mechanism을 작동시키는 것으로 알려졌다. 한
균류의 공격은 세포벽에서 특정 β-D-glucanase를 분비하게
되어 β-D-glucans가 분해되면서 정상 인 세포 생장과 분화
에 변화를 유도하게 된다. 이외에도 2,4-D에 의한 생장 억
제의 과정에서는 xyloglucan으로부터 nanosaccharide가 만들
어져 auxin에 의해 생장에 feedback inhibitor로 작용하는
것으로 알려졌다 (참조 John et al., 1997).

33
제4장
결론 및 제언
○ 동물과 달리 식물은 식물만의 구조를 지니고 있다. 식물
은 원천 으로 양을 얻는 방법이 달라, 즉 합성을
하여 유기 분자를 얻게 되므로 엽록체라는 세포내 소기
을 지니고 있다. 이와 함께 식물만이 지니는 독특한
구조가 세포를 감싸고 있는 세포벽이다. 세포벽은 식물
의 발생, 생장, 병원성 생명체에 한 항 등에 필수불
가결한 요소로서 작용하며, 모든 식물의 모든 부 에 있
는 만큼 그 기능과 함께 구조도 매우 다양하다. 그러나
세포벽은 공통 으로 다당류 기질 (matrix)에 묻 있는
cellulose microfibrils로 구성되어 있다. 따라서 cellulose로
구성된 microfibril은 로서 작용하는 요한 역할을
수행한다. Cellulose microfibrils의 합성은 원형질막에
cellulose synthase들로 이루어진 particle rosettes에서 이
루어지고 matrix polymers는 Golgi에서 합성되어 세포
밖으로 수송된다.
○ 식물에서 cellulose는 세포의 원형질막에 치한 rosette
모양의 단백질 복합체 즉, cellulose synthase 복합체에 의
해 합성된다. 이 게 합성된 cellulose는 주변의 다른 단백
질에 의해 결합되어 microfibril를 형성하게 된다. 최근의

34 식물 세포벽 대사 관련 효소의 연구동향
분자유 학 방법에 의해 cellulose synthase 의 유 자
가 애기장 , 벼 (Oryza sativa), Populus trichocarpa, 보
리 (Hordeum vulgare), 옥수수 (Zea mays) 등에서
multigene family로 존재함이 밝 졌다. 이들 다양한
cellulose synthase 유 자 종류는 물 형성 과정, 세포신
장 과정, microtubule 형성과 련되어 그 구조와 기능이
밝 졌다. 세포벽의 다른 요한 성분인 matrix의
사에 여하는 단백질로 expansin, (1, 4)β-D-glucanase,
xyloglucan endotransglycosylase, pectinase, xylosidase,
methyl esterase 등의 구조와 기능에 한 연구 역시 활
발히 진행되고 있다.
○ 세포벽 사에 한 기 인 연구를 바탕으로 유럽과
미국에서는 이미 상당한 수 의 경제 인 가치를 얻어내
고 있다. 그러나 국내의 경우 정부 는 기업이 주도하
여 세포벽 연구의 경제 가치를 인식하여 연구를 진행
한 사례는 찾아볼 수 없어 정책 심이 필요한 분야라
할 수 있다.

35
참고 문헌
1. Arioli T, Peng L, Betzner AS, Burn J, Wittke W, Herth W,
Camilleri C, Hofte H, Plazinski J, Birch R et al., Molecular
analysis of cellulose biosynthesis in Arabidopsis. Science 279:
717-720, 1998
2. Beeckman T, Przemeck GHK, Stamatiou G, Lau R, Terryn
N, de Rycke R, Inze D, Berleth T, Genetic complexity of
cellulose synthase A gene function in Arabidopsis
embryogenesis. Plant Physiol 130: 1883-1893, 2002
3. Brett CT, Waldron KW, Physiology and Biochemistry of
Plant Cell Walls, 2nd ed., Chapman and Hall, London, 1996
4. Brisson LF, Tenhaken R, Lamb C, Function of oxidative
cross-linking of cell wall structural proteins in plant disease
resistance. Plant Cell 6: 1703-1712, 1994
5. Brown DM, Zeef LA, Ellis J, Goodacre R, Turner SR,
Identification of novel genes in Arabidopsis involved in
secondary cell wall formation using expression profiling and
reverse genetics. Plant Cell 17: 2281-2295, 2005
6. Burn JE, Hocart CH, Birch RJ, Cork AC, Williamson RE,
Functional analysis of the cellulose synthase genes CesA1,
CesA2, and CesA3 in Arabidopsis. Plant Physiol 129:
797-807, 2002
7. Burton RA, Shirley NJ, King BJ, Harvey AJ, Fincher GB,
The CesA gene family of barley. Quantitative analysis of
transcripts reveals two groups of co-expressed genes. Plant
Physiol 134:224-236, 2004
8. Burton RA, Gibeaut DM, Bacic A, Findlay K, Roberts K,
Hamilton A, Baulcombe DC, Fincher GB, Virus-induced

36 식물 세포벽 대사 관련 효소의 연구동향
silencing of a plant cellulose synthase gene. Plant Cell 12:
691-705, 2000
9. Carpita NC, Structure and biogenesis of the cell walls of
grasses. Annu Rev Plant Physiol Plant Mol Biol 47:
445-476, 1996
10. Carpita NC, McCann M, The cell wall. In Biochemistry and
Molecular Biology of Plants, BB Buchanan, W Cruissem,
and RL Jones, eds., American Society of Plant Biologists,
Rockville, MD, pp52-108, 2000
11. Caño-Delgado A, Penfield S, Smith C, Catley M, Bevan M,
Reduced cellulose synthesis invokes lignification and defense
responses in Arabidopsis thaliana. Plant J 34: 351-362, 2003
12. Chen Z, Hong X, Zhang H, Wang Y, Li X, Zhu JK, Gong
Z, Disruption of the cellulose synthase gene, AtCesA8/IRX1,
enhances drought and osmotic stress tolerance in
Arabidopsis. Plant J 43:273-83, 2005
13. Cho H-T, Kende H, Expansin in deepwater rice internodes.
Plant Physiol 113: 1137-1143, 1997a.
14. Cho H-T, Kende H, Expansins and internodal growth of
deepwater rice. Plant Physiol 113: 1145-1151, 1997b.
15. Cho H-T, Kende H, Expression of expansin genes is
correlated with growth in deepwater rice. Plant Cell 9:
1661-1671, 1997c
16. Cho H-T, Kende H, Tissue localization of expansins in
deepwater rice. Plant J 15: 805-812, 1998.
17. Creelman RA, Mullet JE, Oligosaccharins, brassinolides, and
jasmonates: Nontraditional regulators of plant growth,
development, and gene expression. Plant Cell 9: 1211-1223,
1997

참고문헌 37
18. Darley CP, Forrester AM, McQueen-Mason SJ, The
molecular basis of plant cell wall extension. Plant Mol Biol
47: 179-195, 2001
19. Delmer DP, Cellulose synthesis: exciting times for a difficult
field. Annu Rev Plant Physiol Plant Mol Biol 50: 245-276,
1999
20. Desprez T, Vernhettes S, Fagard M, Refrégier G, Desnos T,
Aletti E, Py N, Pelletier S, Höfte H, Resistance against
herbicide isoxaben and cellulose deficiency caused by
distinct mutations in same cellulose synthase isoform
CESA6. Plant Physiol 128: 482–490, 2002
22. Djerbi S, Lindskog M, Arvestad L, Sterky F, Teeri TT,
The genome sequence of black cottonwood (Populus
trichocarpa) reveals 18 conserved cellulose synthase (CesA)
genes. Planta 221:739-46, 2005
23. Doblin MS, Kurek I, Jacob-Wilk D, Delmer DP, Cellulose
biosynthesis in plants: from genes to rosettes. Plant Cell
Physiol 43: 1407-1420, 2002
24. Dhugga KS, Building the wall: genes and enzyme
complexes for polysaccharide synthases. Curr Opin Plant
Biol 4: 488-493, 2001
25. Dhugga KS, Barreiro R, Whitten B, Stecca K, Hazebroek J,
Randhawa GS, Dolan M, Kinney AJ, Tomes D, Nichols S,
Anderson P, Guar seed beta-mannan synthase is a member
of the cellulose synthase super gene family. Science
303:363-366, 2004
26. Eckardt NA, Cellulose Synthesis Takes the CesA Train.
Plant Cell 15: 1685-1687, 2003
27. Ellis C, Karafyllidis I, Wasternack C, Turner JG, The

38 식물 세포벽 대사 관련 효소의 연구동향
Arabidopsis Mutant cev1 Links Cell Wall Signaling to
Jasmonate and Ethylene Responses. Plant Cell 14:
1557-1566, 2002
28. Fagard M, Desnos T, Desprez T, Goubet F, Refregier G,
Mouille G, McCann M, Rayon C, Vernhettes S, Hofte H,
PROCUSTE1 encodes a cellulose synthase required for
normal cell elongation specifically in roots and dark-grown
hypocotyls of Arabidopsis. Plant Cell 12: 2409-2423, 2000
29. Fincher GB, Stone BA, The physiology and biochemistry of
germination in barley. In AW MacGregor, R Bhatty, S
Barley, eds, Chemistry and Technology. American
Association of Cereal Chemists, St. Paul, MN pp 247-295,
1993
30. Fisher DD, Cyr RJ, Extending the microtubule/microfibril
paradigm: Cellulose synthesis is required for normal cortical
microtubule alignment in elongating cells. Plant Physiol. 116,
1043–1051, 1998
31. Gardiner JC, Taylor NG, Turner SR, Control of cellulose
synthase complex localization in developing xylem. Plant
Cell 15: 1740-1748, 2003
32. Gasper Y, Johnson KL, McKenna JA, Bacic A, Schultz CJ,
The complex structures of arabinogalactan-proteins and the
journey towards understanding function. Plant Mol Biol 47:
161-176, 2001
33. Hayashi T, Xylogulcans in the primary cell wall. Annu Rev
Plant Physiol Plant Biol 40: 139-168, 1989
34. Holland N, Holland D, Helentjaris T, Dhugga KS,
Xoconostle-Cazares B, Delmer DP, A comparative analysis
of the plant cellulose synthase (CesA) gene family. Plant

참고문헌 39
Physiol 123: 1313-1323, 2000
35. Ishii T, Matsunaga T, Pellerin P, O'Neill MA, Darvill A,
Albersheim P, the plant cell wall polysaccharide
rhamnogalacturonan II self-assembles into a covalently
cross-linked dimer. J Biol Chem 274: 13098-13104, 1999
36. John M, Roehrig H, Schmidt J, Walden R, Schell J, Cell
signaling by oligosaccharides. Trends Plant Sci 2: 111-115,
1997
37. Kimura S, Laosinchai W, Itoh T, Cui X, Linder CR, Brown
RM Jr, Immunogold labeling of rosette terminal
cellulose-synthesizing complexes in the vascular plant Vigna
angularis. Plant Cell 11: 2075-2085, 1999
38. Lee D-K, Ahn JH, Song D-K, Choi YD, Lee JS, Expression
of an expansin gene is correlated with root elongation in
soybean, Plant Physiol 131: 985-997, 2003
39. Liepman AH, Wilkerson CG, Keegstra K, Expression of
cellulose synthase-like (Csl) genes in insect cells reveals
that CslA family members encode mannan synthases. Proc
Natl Acad Sci USA 102:2221-2226, 2005
40. Mueller SC, Brown RM Jr, Evidence for an
intramembranous component associated with a cellulose
microfibril synthesizing complex in higher plants. J Cell Biol
84: 315-326, 1980
41. O'Neill MA, Eberhard S, Albersheim P, Darvill AG,
Requirement of borate cross-linking of cell wall
rhamnogalacturonan II for Arabidopsis growth. Science 294:
846-849, 2001
42. Pear JR, Kawagoe Y, Schreckengost WE, Delmer DP,
Stalker DM, Higher plants contain homologs of the bacterial

40 식물 세포벽 대사 관련 효소의 연구동향
celA genes encoding the catalytic subunit of cellulose
synthase. Proc Natl Acad Sci USA 93: 12637-12642, 1996
43. Persson S, Wei H, Milne J, Page GP, Somerville CR,
Identification of genes required for cellulose synthesis by
regression analysis of public microarray data sets. Proc Natl
Acad Sci USA 102:8633-8638, 2005
44. Refregier G, Pelletier S, Jaillard D, Hofte H, Interaction
between wall deposition and cell elongation in dark-grown
hypocotyl cells in Arabidopsis. Plant Physiol 135:959-968,
2004
45. Richmond TA, Somerville CR, The cellulose synthase
superfamily. Plant Physiol 124: 495–498, 2000
46. Roelofsen A, Cell wall structure as related to surface
growth. Acta Bot Neerl 7: 77-89, 1958
47. Rose JKC, Bennett AB, Cooperative disassembly of the
cellulose-xyloglucan network of plant cell walls: Parallels
between cell expansion and fruit ripening. Trends Plant Sci
4: 176-183, 1999
48. Scheible W-R, Eshed R, Richmond T, Delmer D, Somerville
C, Modifications of cellulose synthase confer resistance to
isoxaben and thiazolidinone herbicides in Arabidopsis ixr1
mutants. Proc Natl Acad Sci USA 98: 10079-10084, 2001
49. Shin J-H, Jeong D-H, Park MC, An G, Characterization
and Transcriptional Expression of the a-Expansin Gene
Family in Rice. Mol Cells 20: 210-218, 2005
50. Sugimoto K, Williamson RE, Wasteneys GO, Wall
architecture in the cellulose-deficient rsw1 mutant of
Arabidopsis thaliana: Microfibrils but not microtubules lose
their transverse alignment before microfibrils become

참고문헌 41
unrecognizable in the mitotic and elongation zones of roots.
Protoplasma 215, 172–183, 2001
51. Szyjanowicz PM, McKinnon I, Taylor NG, Gardiner J, Jarvis
MC, Turner SR, The irregular xylem 2 mutant is an allele
of korrigan that affects the secondary cell wall of
Arabidopsis thaliana. Plant J 37:730-740, 2004
52. Taylor NG, Howells RM, Huttly AK, Vickers K, Turner
SR, Interactions among three distinct CesA proteins
essential for cellulose synthesis. Proc Natl Acad Sci USA
100: 1450-1455, 2003
53. Taylor NG, Laurie S, Turner SR, Multiple cellulose
synthesis catalytic subunits are required for cellulose
synthesis in Arabidopsis. Plant Cell 12: 2529-2539, 2000
54. Taylor NG, Scheible WR, Cutler S, Somerville CR, Turner
SR, The irregular xylem3 locus of Arabidopsis encodes a
cellulose synthase required for secondary cell wall synthesis.
Plant Cell 11: 769-780, 1999
55. Vergara CE, Carpita NC, β-D-Glycan synthases and the
CesA gene family: lessons to be learned from the
mixed-linkage (13),(14)-d-glucan synthase. Plant Mol Biol
47: 145-160, 2001
56. Yun HS, Kwon C, Kang BG, Lee JS, Han T-J, Chang SC,
Kim S-K, A xyloglucan endotransglycosylase/hydrolase1,
VrXTH1, is associated with cell elongation in mungbean
hypocotyls. Physiol Plant 125: 106-113, 2005.

저자소개 자문위원
이 상 필
․공학박사
․ , 한국과학기술정보연구원 책임연구원
․ 서: 기능성화장품, 바이오매스 인공 등
손 은 수
․약사
․ , 한국과학기술정보연구원 선임연구원
․ 서: 바이오의약품의 개발 특허동향 등
장 수 철
․이학박사
․ , 연세 학교 학부 학 교수
BB077 이상필․손은수
식물 세포벽 대사 관련 효소의 연구동향
2005년 12월 19일 인쇄
2005년 12월 23일 발행
발행처
서울특별시 동 문구 청량리동 206-9
ꂕ 130-742
화 : 3299-6114
등록: 1991년 2월 12일 제5-258호
발행인
조 화
인쇄처
신기획









![[정보보호] 모바일 보안 표준화 동향weekly.tta.or.kr/weekly/files/2005/213/2005-213-490.pdf · 련 보안 요소 집합, 그리고 이를 위한 정보 요소를 정의하고](https://img.pdfslide.tips/doc/110x75/5fc0536e473dd732042fe1ad/ee-ee-e-oe-e-e-e-oe-eee.jpg)



















