Embed Size (px)
Citation preview

POTENTIAL CLINICAL RELEVANCE
Nanomedicine: Nanotechnology, Biology, and Medicine7 (2011) 541–550
Original Article
Effect of curcumin-associated and lipid ligand-functionalizednanoliposomes on aggregation of the Alzheimer's Aβ peptide
Mark Taylor, PhDa, Susan Moore, PhDa, Spyridon Mourtas, PhDb,Anna Niarakis, PhDb, Francesca Re, PhDc, Cristiano Zona, PhDd,
Barbara La Ferla, PhDd, Francesco Nicotra, PhDd, Massimo Masserini, PhDc,Sophia G. Antimisiaris, PhDb,e, Maria Gregori, PhDc, David Allsop, PhDa,⁎
aDivision of Biomedical and Life Sciences, School of Health and Medicine, University of Lancaster, Lancaster, UKbLaboratory of Pharmaceutical Technology, Department of Pharmacy, School of Health Sciences, University of Patras, Patras, Greece
cDepartment of Experimental Medicine, University of Milano-Bicocca, Monza (MBI), ItalydDepartment of Biotechnology and Bioscience, University of Milano-Bicocca, Milano, Italy
eInstitute of Chemical Engineering and High Temperature Processes, FORTH-ICEHT, Patras, Greece
Received 25 January 2011; accepted 18 June 2011
nanomedjournal.com
Abstract
The effect of various types of nanoliposomes (associated with curcumin, phosphatidic acid, cardiolipin, or GM1 ganglioside) on theaggregation of the amyloid-β1-42 (Aβ1-42) peptide was investigated. Nanoliposomes incorporating curcumin (curcumin-liposomes) wereprepared by adding curcumin in the lipid phase during liposome preparation, whereas curcumin surface-decorated liposomes were preparedby using a curcumin-lipid conjugate (lipid-S-curcumin liposomes) or by attaching a curcumin derivative on preformed liposomes by clickchemistry (click-curcumin liposomes). The lipid ligands (phosphatidic acid, cardiolipin, or GM1) were also incorporated into nanoliposomesduring their formation. All nanoliposomes with curcumin, or the curcumin derivative, were able to inhibit the formation of fibrillar and/oroligomeric Aβ in vitro. Of the three forms of curcumin liposomes tested, the click-curcumin type was by far the most effective. Liposomeswith lipid ligands only inhibited Aβ fibril and oligomer formation at a very high ratio of liposome to peptide. Curcumin-based liposomescould be further developed as a novel treatment for Alzheimer's disease.
From the Clinical Editor: In this paper, the potential clinical applicability of curcumin-based nanpoarticles is investigated on the aggregationof amyloid-β1-42, a key protein in the pathogenesis of Alzheimer’s disease. Future extensions of this work may pave the way to a noveltherapeutic approach to AD.© 2011 Elsevier Inc. All rights reserved.
Key words: Liposomes; Curcumin; Amyloid-β; Oligomer; Fibril
Alzheimer's disease (AD) is the most common cause ofdementia in the elderly population, and the fourth most commoncause of death in western countries after heart disease, cancer,and stroke.1-3 The pathogenic mechanisms underlying AD arenot yet completely clear; however, in recent years, severalapproaches aimed at inhibiting disease progression haveadvanced to clinical trials. Among these, strategies targeting
The research leading to these results has received funding from theEuropean Community's Seventh Framework Programme (FP7/2007-2013)under grant agreement no. 212043.
⁎Corresponding author: Division of Biomedical and Life Sciences,School of Health and Medicine, University of Lancaster, Lancaster, UK.
E-mail address: [email protected] (D. Allsop).
Please cite this article as: M., Taylor, et al, Effect of curcumin-associated and lipAβ peptide. Nanomedicine: NBM 2011;7:541-550, doi:10.1016/j.nano.2011.06
1549-9634/$ – see front matter © 2011 Elsevier Inc. All rights reserved.doi:10.1016/j.nano.2011.06.015
the production and clearance of the amyloid-β peptide (Aβ) arethe most advanced.4 The production and progressive aggregationof Aβ play a central role in AD pathogenesis.5-7 The predominantand initial peptide deposited in the brain parenchyma is Aβ1-42,which is highly fibrillogenic.8,9
We have been investigating a number of different ligands fortheir ability to bind to Aβ with high affinity, and so, potentially,to interfere with the aggregation process. These includecurcumin and its derivatives, and also certain lipid ligands.
Curcumin is a naturally occurring phytochemical withantioxidant and anti-inflammatory activity and a favorabletoxicity profile10 that protects the brain from lipid peroxidation.11
Curcumin inhibits amyloid Aβ1-42 oligomer formation and celltoxicity at micromolar concentrations in vitro12-16 and binds tosenile plaques, reducing amyloid levels in vivo.14
id ligand-functionalized nanoliposomes on aggregation of the Alzheimer's.015

542 M. Taylor et al / Nanomedicine: Nanotechnology, Biology, and Medicine 7 (2011) 541–550
Currently under development are curcumin analogues withsimilar biological activity to curcumin itself, but with improvedpharmacokinetics, water solubility, and stability.17-20 A furtherstrategy for application of the beneficial effects of curcumincould be attachment of the molecule, in a stable way and inan active structural conformation, onto the surface ofbiocompatible/biodegradable and stealth nanoparticles (NPs).Such attachment could help to increase the bioavailability ofthe drug and, perhaps, further increase the binding affinity ofcurcumin or its derivatives for Aβ peptides, due to the effectsof multivalency.21-23 Among all the different NPs, liposomeshave many advantages for drug delivery applications becauseof their nontoxic and nonimmunogenic, fully biodegradable,and structurally versatile nature.24 Previous experiments bymembers of our group, using the surface plasmon resonance(SPR) technique, have shown that nanoliposomes exposing acurcumin derivative on their surface demonstrated a very highaffinity (1–5 nM) for binding to Aβ1-42 fibrils.25
Aβ is also known to interact with membrane lipids,26 par-ticularly ionic lipids27 and gangliosides28 that have been dem-onstrated to act as a seeds for Aβ aggregation.29-31 Thus, NPsdecorated with these lipid ligands might also be expected to bindto Aβ and affect the aggregation process. Members of our grouphave previously reported that nanoliposomes containing phos-phatidic acid (PA) and cardiolipin (CL) targeted aggregatedforms of Aβ1-42 with high binding affinity (22-60 nM).32
In the present study we designed and formulated differenttypes of nanosized liposomes incorporating (in their lipidmembrane) or decorated (on their surface) with curcumin, thecurcumin derivative, or lipid ligands (PA, CL, or GM1ganglioside), and then evaluated their ability to influence Aβ1-42
peptide aggregation in vitro. Two different techniques wereused to monitor Aβ aggregation: an assay based on thefluorescent dye thioflavin T (ThT), which binds to fibrillarforms of Aβ, and a sandwich immunoassay that detects smalloligomeric forms of Aβ with high sensitivity.33
Methods
Cholesterol (Chol), Sephadex G50 (coarse), and all chemicalsfor the curcumin derivative and lipid synthesis were purchasedfrom Sigma-Aldrich (St Louis, Missouri). All reagents requiringanhydrous solvents were dried with molecular sieves for at least24 hours before use. Phosphatidyl choline (PC), sphingomyelin(SM), 1,2-distearoyl-sn-glycerol-3-phosphatidylcholine (DSPC),1,2-dipalmitoyl-sn-glycerol-3-phosphatidylcholine (DPPC), andphosphatidylglycerol (PG) were purchased from Avanti PolarLipids Inc. (Alabaster, Alabama). Spectrapore dialysis tubing(MW cutoff 10,000) was from Serva Electrophoresis (Heidel-berg, Germany). Recombinant Aβ1-42, Ultrapure, was purchasedfrom rPeptide (Bogart, Georgia). All other chemicals werereagent grade. Ultrapure water was produced with a MilliporeDirect-Q system (Millipore, Billerica, Massachusetts).
Curcumin-associating liposome preparation
For all types of curcumin-associating liposomes, the thin-film hydration method was initially applied.24,34 In brief, the
lipids used were solvated in a chloroform/methanol (2:1 v/v)solution, and the appropriate amount of each lipid was placed ina 50-mL round-bottomed flask. The lipid mixture solution wasdried by connection to a rotary evaporator, until a thin film oflipid was developed on the flask. Residual organic solventswere removed by flushing with nitrogen for 5 minutes, and thefilms were hydrated by adding the required amount ofphosphate buffered saline (PBS) (pH 7.40) at a temperatureabove the transition temperature of the lipid used in each case.The lipid dispersion was then sonicated by a probe (Sonics andMaterials, Suffolk, United kingdom) using a microtip for two10-minute cycles, or until the dispersion became completelytransparent. In all cases, the liposome dispersions were left forannealing of structural defects at a temperature above the lipidtransition temperature for 1-2 hours.
Liposomes incorporating curcumin (curcumin-liposomes)In the case of these liposomes (hereafter named
“curcumin-liposomes”), curcumin was added in the lipid mixture(as a solution in methanol) before the thin-film formation(procedure above). Separation of liposome-incorporated and freecurcumin was accomplished by size exclusion chromatographyon a Sephadex G-50 column and eluted with PBS, pH 7.40. Thelipid-containing fractions were identified (by phospholipiddetermination) and pooled; if needed the liposome dispersionswere concentrated by ultrafiltration using Amicon ultra-15 tubes(Millipore). SM/Chol (2:1) lipid composition was used.
Liposomes decorated by click chemistry(click-curcumin liposomes)
These liposomes were functionalized with curcuminderivative X [N-propargyl 2-(3′,5′-di(4-hydroxy-3-metoxys-tyryl)-1H-pyrazol-1′-yl)-acetamide] (Figure 1; B. La Ferla,unpublished data) by using the click-chemistry method (herafterreferred to as “click-curcumin liposomes”). In more detail, weperformed copper(I)-catalyzed azide-alkyne cycloaddition with aliposomal dispersion consisted of DPPC/DPPG/Chol/Y (8:2:10:1or 2 molar ratio), which was prepared by thin-film hydration (seeabove) in PBS, pH 6.50, and this was reacted with the curcuminderivative X (Figure 1), under click-reaction conditions.34,35 Y isa lipid-polyethylene glycol-N3-3-deoxy-1,2-dipalmitoyl-3-(4′-methyl (O-(2-azidoethyl)-heptaethylenglycol-2-yl)-ethylcarba-moylmethoxy ethylcarbamoyl-1H-1′,2′,3′-triazol-1′-yl)-sn-glyc-erol (Figure 1) that was incorporated into the liposomalmembrane during liposome preparation.
For the click reaction, 8 mM CuSO4·5H2O (in water) and 145mM sodium ascorbate (in buffer) were mixed, and the solutionwas stirred until the formation of an orange suspension. Freshlyprepared bathophenanthrolinedisulfonate (28 mM, in buffer)catalyst was added to this, and the resulting mixture was addedto the preformed liposomes. Finally curcumin-alkyne derivativeX dissolved in dimethyl sulfoxide was added, and the reactionmixture was left for 6 hours at 25°C under a continuousflow of N2. The liposomal dispersion was purified by dialysisand size exclusion chromatography on Sephadex G-50. High-performance liquid chromatography analysis of the finalliposomal dispersion proved quantitative reaction of theliposome surface azide groups with the curcumin derivative.

Figure 1. Chemical structures of the curcumin (X) and phospholipid derivatives (Y) used for the preparation of click-curcumin liposomes, and the phospholipidderivative (lipid-S-curcumin) used for the lipid-S-curcumin liposomes.
543M. Taylor et al / Nanomedicine: Nanotechnology, Biology, and Medicine 7 (2011) 541–550
Liposomes decorated by curcumin-phospholipid conjugate(lipid-S-curcumin liposomes)
A phospholipid conjugate was synthesized via Michaeladdition of a thiol derivative to curcumin.36 For this, 1,2-dipalmitoyl-sn-glycero-3-phosphothioethanol (sodium salt)[lipid-SH] reacted with curcumin and 1,2-dipalmitoyl-3-(2-(1,7-bis(4-hydroxy-3-methoxyphenyl)-3,5-dioxohept-6-enylthio)ethylphospho)–sn-glycerol (lipid-S-curcumin) was obtained. Usinglipid-S-curcumin (Figure 1), we prepared small unilamellarliposomes comprising DPPC/Chol/lipid-S-curcumin (2:1:0.2or 0.4 molar ratio), by the thin-film hydration technique(see above). These liposomes are named “lipid-S-curcuminliposomes” and are characterized as containing 10% or 20%lipid-S-curcumin in the following sections (as the approximatepercentage of curcumin per phospholipid).
Characterization of curcumin-associating liposomes
Vesicle entrapment efficiencyIn the curcumin-liposomes, the curcumin-to-lipid (mol/mol)
ratio was calculated. For this, the curcumin concentration wasmeasured in the liposomes after vesicle disruption by methanol;in brief, 100 μL of the liposome dispersion was mixed with 2 mLof methanol, and the curcumin concentration was calculated bymeasuring the optical density at 420 nm, from a calibration curvethat was created from curcumin solutions of known concentra-tion. The lipid concentration of all the liposome dispersions wasmeasured by the Stewart colorimetric assay, as previouslydescribed.37 In this assay, phospholipids form a colored complexwith ammonium ferrothiocyanate (optical density 485 nm) that is
extracted with chloroform. Lipid concentration in samples iscalculated on the basis of a standard curve prepared with aknown amount of each phospholipid. After measurement, thelipid concentrations of the liposome dispersions were adjusted atthe desired value, for each study conducted. In the followingparts, the curcumin content of the various types of curcumin-associated liposomes is always calculated as a percentage of thephospholipid content of the liposomes (which is the lipidmeasured by the Stewart assay).
Vesicle size distribution, polydispersity index (PI), and zetapotential measurements
Particle size of diluted (with PBS pH 7.40) vesicle dispersions(0.4 mg/mL) was measured by dynamic light scattering (DLS)technique (Malvern Nano-ZS; Malvern Instruments, Worcester-shire, United kingdom) at 25°C at a 173-degree angle. Zetapotential was measured for the same samples (dispersed in 10mM PBS, pH 7.40) at 25°C, by the same instrument (utilizing theDoppler electrophoresis technique).
PA, CL, and GM1 liposome preparation
Liposomes were composed of a matrix of Sm/Chol (1:1molar ratio) mixed or not with 5 or 20 mol % of PA, CL, orGM1. We have previously documented the preparation proce-dure for these liposomes.38 Briefly, lipids were resuspended inchloroform-methanol (2:1, v:v) and dried under a gentle stream ofnitrogen. The resulting lipid film was resuspended in Tris buffer(10mMTris, 150mMNaCl, pH 7.40) vortexed, and then extruded(using a Lipex Biomembranes extruder) 10 times through a 100-

Table 1Physicochemical properties of curcumin-liposomes and liposomes decorated with different curcumin derivatives (click-curcumin liposomes and lipid-S-curcumin liposomes), PA, CL, and GM1
Lipid composition Mean diameter (nm) PI Zeta potential (mV)
Curcumin-liposomesSM/Chol 2:1 (control) 57.7 ± 1.4 0.215 3.9 ± 1.7SM/Chol/CURC 1:0.5:0.013 97.6 ± 3.1 0.223 3.88 ± 0.25Click-curcumin liposomesControl (without Y) 52.8 ± 5.5 0.097 –7.6 ± 1.7DPPC/DPPG/Chol/Y 8:2:10:2 (before reaction) 159.5 ± 3.94 0.097 –DPPC/DPPG/Chol/Y 8:2:10:1 188 ± 16 0.183 –19.4 ± 3.6DPPC/DPPG/Chol/Y 8:2:10:2 218 ± 11 0.237 –19.2 ± 4.5Lipid-S-curcumin liposomesControl (no Lipid-S-CURC) 63.1 ± 6.2 0.195 –6.44 ± 0.49DPPC/Chol/Lipid-S-CURC 2:1:0.4 135.30 ± 0.17 0.170 –DPPC/Chol/Lipid-S-CURC 2:1:0.2 180 ± 13 0.193 –14.5 ± 2.4DPPC/Chol/Lipid-S-CURC 2:1:0.4 207.2 ± 8.0 0.255 –10.5 ± 1.2PA, CL, and GM1 liposomesSM/Chol 1:1 (control) 140 ± 6.4 0.255 –23.6 ± 1.5SM/Chol/PA 47.5:47.5:5 151 ± 6.2 0.132 –39.5 ± 1.9SM/Chol/PA 40:40:20 151 ± 6.0 0.143 –49.36 ± 1.2SM/Chol/CL 47.5:47.5:5 147 ± 8.2 0.129 –36.25 ± 2.4SM/Chol/CL 40:40:20 143 ± 8.3 0.098 –48.51 ± 1.4SM/Chol/GM1 40:40:20 141 ± 12.5 0.235 –29.01 ± 1.8
In all cases the lipid compositions appear as molar ratios, whereas each measurement is the mean of at least three different preparations. PI is polydispersityIndex. Y and lipid-S-CURC are the lipid derivatives presented in Figure 1.
544 M. Taylor et al / Nanomedicine: Nanotechnology, Biology, and Medicine 7 (2011) 541–550
nm pore polycarbonate filter (Millipore) under 20 bar nitrogenpressure, at room temperature (22–25°C). Lipid recovery afterextrusion was assessed by assaying the individual components, asdescribed.38 The total lipid recovery was about 90% for all thesamples. The presence of an aqueous core was established byincorporating calcein, using the procedure described.38
Characterization of PA, CL, and GM1 liposomes
The size, PI, and zeta potential of the liposomes weredetermined using a ZetaPlus particle sizer and zeta-potentialanalyzer (Brookhaven Instruments Corp., Holtsville, New York).The size and PI measurements were performed at 25°C.Liposomes, prepared in 10 mM Tris, 150 mM NaCl, pH 7.4,were diluted at 0.25 mM total lipid concentration. The particle sizewas assessed by DLS with a 652-nm laser beam. Particle size andPI were obtained from the intensity autocorrelation function of thelight scattered at a fixed angle of 90 degrees. The correlationfunction was analyzed by means of a two-cumulant expansion.The zeta potential was measured at 25°C. Each measurement wasperformed on freshly prepared liposomes samples diluted 1:25with deionized water. The liposome measurements wereperformed with a ZetaPALS device (Brookhaven), which isbasically an interferometic Doppler velocimetry. The reporteddata are the mean of at least five different measurements. Stabilitywas verified by DLS by following size and PI, which remainedconstant in PBS for up to 3 days (data not shown). Size and zetapotential distributions of liposomal preparations were measuredby DLS and are reported in Table 1.
Aβ aggregation assays
The ThT assay and the immunoassay for oligomeric Aβ wereperformed using Aβ1-42 that was de-seeded using a method based
on the report of Manzoni et al.39 This was achieved by dissolvinga 1-mg aliquot of recombinant Ultrapure Aβ1-42 (rPeptide) in 1mL trifluoroacetic acid containing 45 μL of thioanisol. Thepeptide was left for 1 hour, and then all liquid was removed usinga stream of N2. The peptide was further treated by dissolving inhexafluoro-2-propanol and then evaporating the liquid, with thiswhole process being repeated three times. Solid peptide wasstored in aliquots at –20°C until use.
ThT assays were conducted in 384-well, clear-bottomedmicrotiter plates, with 25 μmol Aβ1-42, 15 μmol ThT, 40 μLliposome (total lipids) (for all curcumin and curcumin derivativeliposomes), or 1 mM liposome (total lipids) (for PA, CL, or GM1liposomes), in 10 mM PBS pH 7.4, with a total reaction volumeof 60 μL. Aggregation was monitored using a BioTek Synergyplate reader (BioTek, Winooski, Vermont) (λex = 442 nm, λem =483 nm) over 48 hours at 30°C, with the plate being shaken andthen read every 10 minutes. Control assays were also performedinvolving the incubation of pre-aggregated Aβ peptide with ThTin the presence of each inhibitor, at the same concentration asbefore, with the peptide being pre-incubated at 25 C for 24 hours.These “pre-aggregation controls” were designed to rule out thepossibility that the inhibitors might interfere with the binding ofThT to Aβ fibrils. If these inhibitors can compete with ThT forbinding to fibrils, this would result in clear inhibition of thefluorescent signal.
A dose-dependency study, involving the click-curcuminliposomes, followed the same method as before, except for theconcentration of liposomes, which was adjusted to 5:1, 2:1, 1:1,1:2, 1:5 molar ratio of liposome to Aβ. A similar experiment wasalso carried out with “empty” control liposomes.
The sandwich immunoassay followed the method of Tayloret al.33 Here, Aβ oligomers are captured by mouse monoclonalantibody 6E10 and detected by a biotinylated form of the same

545M. Taylor et al / Nanomedicine: Nanotechnology, Biology, and Medicine 7 (2011) 541–550
antibody. Monomeric Aβ does not produce a signal in thisassay, because the capture antibody occupies the only 6E10epitope available, and so the biotinylated 6E10 antibody cannotsubsequently bind. However, in the case of oligomers, multiple6E10 epitopes are available, permitting both capture anddetection. We have found that this is a very sensitive techniquefor detecting early-stage oligomers, and, in aggregation time-course experiments, produces a clear signal well before the ThTmethod.33 Briefly, 96-well microtiter plates (Maxisorp; Nunc,Roskilde, Denmark) were coated with mouse monoclonalantibody 6E10, diluted 1:1000 in assay buffer [Tris-bufferedsaline (TBS) pH 7.4, containing 0.05% γ-globulins and 0.005%v/v Tween 20). The incubated samples of peptide, with orwithout liposome (12.5 μM Aβ and 20 μg/mL liposome (totallipids) (for all curcumin and curcumin-derivative liposomes) or500 μM liposome (total lipid) (for PA, CL, or GM1 liposomes)in PBS, pH 7.4, at 25°C), were diluted to 1 μM Aβ andincubated, in triplicate, in the 96-well plates for 1 hour at 37°C.The plates were washed with 10 mM PBS, containing 0.05%v/v Tween 20. Following this, 100 μL of TBS containing a1:1000 dilution of biotinylated 6E10 was added to each of the96 wells and incubated for 1 hour at 37°C. After the sampleshad been washed further, europium-linked streptavidin wasadded at 1:500 dilution in StrepE buffer (TBS containing20 μM DTPA, 0.5% bovine serum albumin, and 0.05% γ-globulins), again incubated for 1 hour, and washed. Enhancersolution was added, and the plates were read using the time-resolved fluorescence setting for europium on a Wallac Victor 2plate reader (Perkin-Elmer, Boston, Massachusetts). Pre-aggregated peptide controls were also performed (see explana-tion above) to rule out the possibility of the liposome inhibitorsblocking the binding of 6E10 to Aβ.
The ThT and immunoassay results shown are averagedata from one example of two or more repeat experiments,each of which was performed in triplicate (error bars, ± SD).Although only one representative example is shown, therepeats were similar in terms of the percentage inhibitions ofaggregation seen.
Results
Liposome physicochemical properties
Curcumin incorporation in the curcumin-liposomes wascalculated as 1.3 mol% (in respect to total phospholipidcontent). The mean diameters, PIs, and zeta potential values ofall the curcumin-associated liposomes prepared are presented inTable 1. As can be seen, when curcumin is associated to theliposomes a gradual increase in the mean diameter is noticed asthe amount of curcumin or curcumin-lipid incorporated in theliposome membranes increases. The highest increase is noticedin the case of the click-curcumin liposomes, which is logicalbecause in this method of ligation a polyethylene glycol chainis also included between the vesicle surface and the curcuminderivative attached on the vesicles (as seen in Figure 1; Yderivative). Nevertheless, in all three cases of curcumin-associating liposomes the mean diameters are below or justaround 200 nm, which is the requirement for targeting
applications. Furthermore, the PIs of all liposome preparationsare low (below or around 0.25), which indicates that theliposome dispersions are monodisperse. In fact, the two typesof liposomes that are decorated with curcumin (the lipid-S-curcumin and the click-curcumin liposomes) have demonstratedgood colloidal stability in size stability studies performedbefore.40 As also noticed before, the surface charge of thevesicles decreases when curcumin is immobilized on thevesicle surface (lipid-S-curcumin and click-curcumin lipo-somes) but not when curcumin is incorporated in the membrane(curcumin-liposomes) which may in fact correlate with thelower amount of curcumin associated with the liposomes in thiscase (compared to the two other cases).
Aβ peptide aggregation studies
PA-, CL-, and GM1-decorated liposomesThe results of the ThT assay (Figure 2, A) for 1 mM
liposomes indicate that there was little or no inhibitory effect onthe formation of amyloid fibrils by liposomes containing thelipid ligands PA, CL, and GM1 relative to Aβ alone, or to Aβplus unadorned liposomes. This is evidenced by the observationthat the final ThT signal (plateau level) was similar in all cases. Ifanything, liposomes with GM1, PA, and CL appeared to slightlyincrease aggregation at early time points (up to 18 hours). Theimmunoassay data (Figure 2, B) show that there was noinhibitory effect on oligomer formation with the PA-, CL-, orGM1 decorated liposomes at a 2:1 molar ratio of liposome to Aβ.Some inhibition was seen at 10:1 and/or 40:1 molar ratios, butthis was also seen with the “liposome only” control. The resultsshown here refer to liposomes with 20% lipid ligand, but similarresults were seen for 5% lipid ligand (data not shown). Thecontrols involving addition of each of these liposomes to pre-aggregated Aβ showed that they did not interfere with either theThT assay or the immunoassay.
Curcumin-liposomesThe curcumin-liposomes, at 40 μM, were found to inhibit the
formation of Aβ1-42 fibrils (Figure 3, A), as indicated by areduction in the final ThT signal. Some inhibition of late-stageaggregation was also found with the liposomes alone, but theamount of inhibition was clearly greater when the liposomescontained curcumin. There was no depression of the ThT signalwhen the curcumin-liposomes were added to pre-aggregatedAβ1-42, showing that the inhibition seen in the assay is not due tothe liposomes interfering with either the binding of ThT to fibrils,or its fluorescence. Over the first 24 hours, the presence ofcurcumin-liposomes did not appear to inhibit formation ofoligomeric Aβ, as monitored by the immunoassay technique(Figure 3, B). However, by 48 hours the presence ofthese liposomes did reduce the amount of aggregated Aβ,and this reduction was enhanced (and only reached statisticalsignificance) when the liposomes contained curcumin. The pre-incubated control data indicate that there was no interference ofthe liposomes with the immunoassay method.
Lipid-S-curcumin liposomesIn this case, the presence of 40 μM unadorned liposomes
appeared to decrease the time at which aggregation was detected,
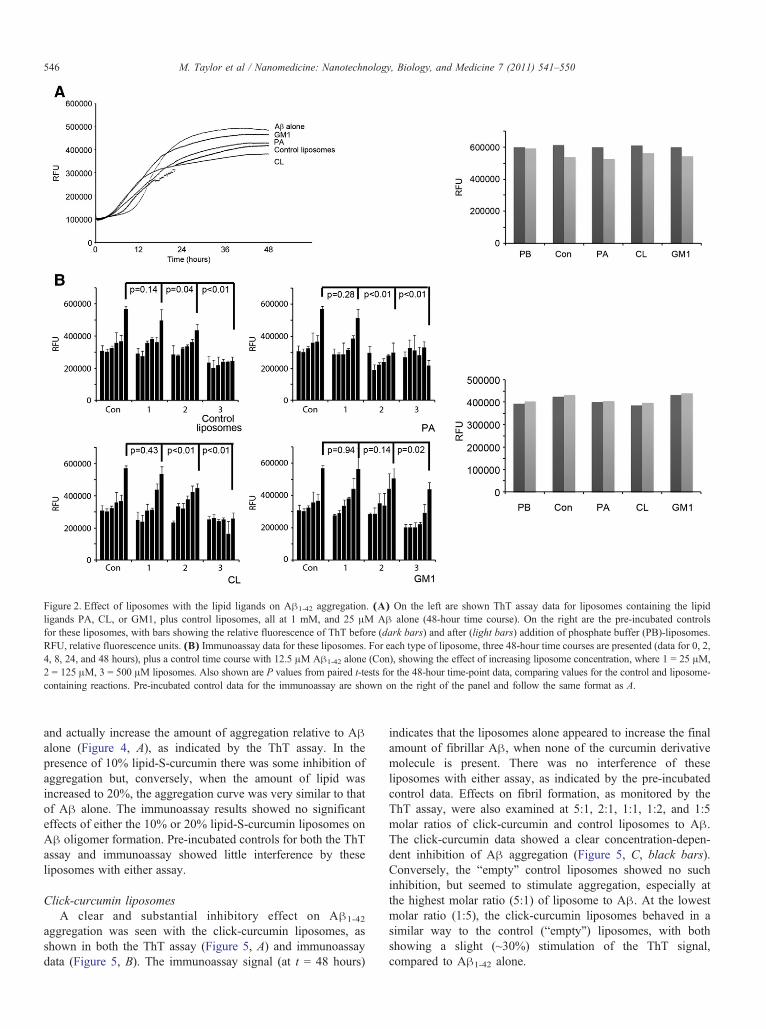
Figure 2. Effect of liposomes with the lipid ligands on Aβ1-42 aggregation. (A) On the left are shown ThT assay data for liposomes containing the lipidligands PA, CL, or GM1, plus control liposomes, all at 1 mM, and 25 μM Aβ alone (48-hour time course). On the right are the pre-incubated controlsfor these liposomes, with bars showing the relative fluorescence of ThT before (dark bars) and after (light bars) addition of phosphate buffer (PB)-liposomes.RFU, relative fluorescence units. (B) Immunoassay data for these liposomes. For each type of liposome, three 48-hour time courses are presented (data for 0, 2,4, 8, 24, and 48 hours), plus a control time course with 12.5 μM Aβ1-42 alone (Con), showing the effect of increasing liposome concentration, where 1 = 25 μM,2 = 125 μM, 3 = 500 μM liposomes. Also shown are P values from paired t-tests for the 48-hour time-point data, comparing values for the control and liposome-containing reactions. Pre-incubated control data for the immunoassay are shown on the right of the panel and follow the same format as A.
546 M. Taylor et al / Nanomedicine: Nanotechnology, Biology, and Medicine 7 (2011) 541–550
and actually increase the amount of aggregation relative to Aβalone (Figure 4, A), as indicated by the ThT assay. In thepresence of 10% lipid-S-curcumin there was some inhibition ofaggregation but, conversely, when the amount of lipid wasincreased to 20%, the aggregation curve was very similar to thatof Aβ alone. The immunoassay results showed no significanteffects of either the 10% or 20% lipid-S-curcumin liposomes onAβ oligomer formation. Pre-incubated controls for both the ThTassay and immunoassay showed little interference by theseliposomes with either assay.
Click-curcumin liposomesA clear and substantial inhibitory effect on Aβ1-42
aggregation was seen with the click-curcumin liposomes, asshown in both the ThT assay (Figure 5, A) and immunoassaydata (Figure 5, B). The immunoassay signal (at t = 48 hours)
indicates that the liposomes alone appeared to increase the finalamount of fibrillar Aβ, when none of the curcumin derivativemolecule is present. There was no interference of theseliposomes with either assay, as indicated by the pre-incubatedcontrol data. Effects on fibril formation, as monitored by theThT assay, were also examined at 5:1, 2:1, 1:1, 1:2, and 1:5molar ratios of click-curcumin and control liposomes to Aβ.The click-curcumin data showed a clear concentration-depen-dent inhibition of Aβ aggregation (Figure 5, C, black bars).Conversely, the “empty” control liposomes showed no suchinhibition, but seemed to stimulate aggregation, especially atthe highest molar ratio (5:1) of liposome to Aβ. At the lowestmolar ratio (1:5), the click-curcumin liposomes behaved in asimilar way to the control (“empty”) liposomes, with bothshowing a slight (~30%) stimulation of the ThT signal,compared to Aβ1-42 alone.

Figure 3. Effect of curcumin-liposomes on Aβ1-42 aggregation. (A) Data for the ThT assay of Aβ fibril formation in the presence and absence of 40 μMliposomes (total phospholipids). On the left is a time course of aggregation, and on the right are data for the pre-incubated controls, with columns showingrelative fluorescence units (RFU) before (dark columns) and after (light columns) addition of liposomes. 1, control (phosphate buffer); 2, liposomes alone;3, curcumin-liposomes. (B) Data for the immunoassay for Aβ oligomers in the presence and absence of 20 μM liposomes (total phospholipids). On theleft, three time courses are shown with samples taken at 0, 2, 4, 8, 24, and 48 hours. The results of paired t-tests are shown, comparing data from 48-hourtime points as indicated by the lines. The right side shows the pre-incubated controls for these samples.
547M. Taylor et al / Nanomedicine: Nanotechnology, Biology, and Medicine 7 (2011) 541–550
Discussion
The Aβ peptide is the major component of senile plaque (andcerebrovascular) amyloid in AD, and data from moleculargenetics, together with biochemical and animal model studies, allsuggest that the self-association and aggregation of Aβ is aseminal event in the early stages of AD pathogenesis.5,7 Solubleoligomers that precede amyloid fibril formation are now thoughtto be partly responsible for neurodegeneration and dementia.4
Not surprisingly, therefore, there has been a considerable effortto try and develop new drugs that block the formation oraggregation of Aβ, or ameliorate its toxic effects. One recentpromising development has been the discovery that curcumin, apolyphenolic compound derived from the dietary spice turmeric,is an effective inhibitor of Aβ oligomer formation.12-14
Curcumin has many other beneficial properties, including anti-inflammatory, antioxidant, antiproliferative, and antiangiogenicactivities, and so has been widely studied as a potential drug.However, even though curcumin is safe at high doses, it exhibitspoor bioavailability, due in part to its rapid metabolism andsystemic elimination. We have therefore incorporated thismolecule, and a derivative of it, into liposomes, with the aimof improving its bioavailability profile.
Curcumin-associating nanoliposomes were prepared in threedifferent ways: (1) by adding curcumin in the lipid phase duringliposome formation (curcumin-liposomes); (2) by using a
curcumin-lipid conjugate to make surface-decorated liposomes(lipid-S-curcumin liposomes); and (3) by attaching the curcuminderivative on preformed liposomes by click chemistry (click-curcumin liposomes). These liposomes were then tested for theirability to inhibit the formation of Aβ fibrils (by ThT assay) andAβ oligomers (by immunoassay). We have shown previouslythat the latter assay detects early Aβ oligomers (and notmonomers) with high sensitivity.33 The curcumin-liposomesshowed some inhibition of the final ThT assay signal (Figure 3)but appeared to stimulate aggregation at earlier time points, andgave no significant inhibition until the last incubation time point(48 hours) in the immunoassay. The lipid-S-curcumin liposomesshowed some inhibition in the ThT assay (for 10% lipid-S-curcumin only), but failed to demonstrate any significantinhibition in the immunoassay (Figure 4). However, the click-curcumin liposomes showed convincing and substantial inhibi-tion of Aβ1-42 aggregation in both of these assays (Figure 5). Inthe presence of the click-curcumin liposomes, a clear decrease insignal relative to control (with Aβ alone) was seen at all timepoints in the ThT assay, and from 4 hours' incubation onward inthe immunoassay. This suggests good inhibition of both fibriland early oligomer formation. Previous results have shown high-affinity (1-5 nM) binding of these same click-curcuminliposomes to immobilized Aβ1-42 fibrils by using the SPRtechnique.25 This high-affinity binding may be due to theoccurrence of multivalent interactions between the click-

Figure 4. Effect of lipid-S-curcumin liposomes on Aβ1-42 aggregation. (A) The ThT assay data for Aβ aggregation over a 48-hour time course in the presenceand absence of 40 μM liposomes (total phospholipids). The right side of the panel shows the pre-incubated controls for this assay (the darker bar shows thesignal before and the lighter bar after addition of liposome to pre-aggregated Aβ1-42): 1, PB; 2, control liposomes without lipid-S-curcumin; 3, liposomes with10% lipid-S-curcumin (DPPC/Chol/lipid-S-curcumin 2:1:0.2; curcumin is referred to as percent of DPPC lipid); 4, liposome with 20% lipid-S-curcumin (DPPC/Chol/lipid-S-curcumin 2:1:0.4; curcumin is referred to as percent of DPPC lipid). (B) Data for the immunoassay with the same liposome samples, 20 μMliposomes. Data for 0, 2, 4, 8, 24, and 48 hours are shown, for four different conditions. The results of paired t-tests are shown, comparing data from 48-hour timepoints as indicated by the lines. To the right of the panel are data for the pre-incubated controls for these reactions.
548 M. Taylor et al / Nanomedicine: Nanotechnology, Biology, and Medicine 7 (2011) 541–550
curcumin liposomes and Aβ, which could also make them veryeffective as aggregation inhibitors.25 We calculate that theconcentration of exposed curcumin for the 40 μM click-liposomes used in the Aβ aggregation studies (at 8:2:10:2composition) is 4 μM, which is approximately 13 times higherthan the concentration of 300 nM of exposed curcumin thatshowed a clear binding to Aβ fibrils in the study by Mourtas etal.25 We suppose that click-curcumin liposomes are an effectiveaggregation inhibitor because the curcumin-derivative moleculesprotrude from the liposome surface rather than entering theliposome bilayer and so they are more amenable to the interactionwith Aβ species. The click-curcumin liposomes also showedclear concentration-dependent inhibition of Aβ1-42 fibril forma-tion in the ThT assay (Figure 5, C), although at a lowconcentration relative to Aβ (1:5 molar ratio of liposome toAβ) they appeared to show a slight (approximately 30%)stimulation of the ThT signal. We have also noted that the“empty” control liposomes, especially at higher concentrations,show a similar effect (Figure 5, C). Further investigation isrequired to understand this phenomenon. This potential stimu-lation with “empty” liposomes could lead to problems in vivo ifthe click-curcumin liposomes are not stable in biological fluids.However, our data so far indicate that these liposomes are stable
when incubated in the presence of blood plasma proteins.25 Otherresearchers have developed a polymeric NP-encapsulated formof curcumin (NanoCurc; SignPath Pharma, Inc., Quakertown,Pennsylvania), which was shown to protect neuronally differen-tiated human SK-N-SH cells from reactive oxygen species-mediated insults, but the effects of these NPs on Aβ aggregationwere not tested.40 Moreover, apolipoprotein E3-mediated poly(butyl) cyanoacrylate NPs containing curcumin have shownenhanced protection against Aβ-induced cytotoxicity in SH-SY5Y neuroblastoma cells, compared to plain curcumin solution,but, again, any effects on Aβ aggregation were not determined.41
We also tried an alternative strategy of attaching certain lipidligands (PA, CL, or GM1) to liposomes, because previous workbased on ultracentrifugation and SPR had indicated that theselipid-based NPs can also bind to Aβ with high (22-60 nM)affinity.32 However, these lipid-based NPs proved to be poorinhibitors of Aβ aggregation, with no convincing inhibition overand above that seen with “empty liposome” controls, even atvery high ratios of liposome to peptide (Figure 2). This is inaccord with the fact that none of these lipids have been shownpreviously to inhibit Aβ aggregation. However, the lack of anydirect effect on aggregation does not rule out the possible futureuse of these lipid-based NPs as “sinks” to remove Aβ from

Figure 5. Click-curcumin liposomes inhibit the aggregation of Aβ1-42 in both the ThT assay (40 μM liposomes) and immunoassay (20 μM liposomes). (A) Theeffect of nanoliposomes on the aggregation of Aβ as monitored by the ThT assay. On the left of the panel there is a 48-hour incubation time course, and the pre-incubated controls for these liposomes are on the right: 1, PB; 2, control liposomes; 3, click-curcumin liposomes (DPPC/DPPG/Chol/Y; 8:2:10:2). The dark-gray bar shows the signal before addition of PB/liposome and the light-gray bar after. (B) Data from the immunoassay. Three 48-hour time courses (data for 0,2-, 4-, 8-, 24-, and 48-hour time points) are shown for Aβ alone, Aβ plus click-curcumin liposomes, and Aβ plus control (“empty”) liposomes. The results fromt-tests comparing 48-hour time-point data paired as indicated by the lines are also shown. Pre-incubated controls presented on the right: 1, PB; 2, click-curcuminderivative liposomes; 3, control liposomes. (C) ThT data for a Aβ alone (gray bar) and Aβ incubated with decreasing molar ratios of click-curcumin liposomesto peptide (black bars) or control liposomes to peptide (white bars). Data are presented as the percentage of RFU seen for Aβ alone, at the 48-hour time point.
549M. Taylor et al / Nanomedicine: Nanotechnology, Biology, and Medicine 7 (2011) 541–550
human blood plasma, and so indirectly affect the accumulation ofamyloid in the brain.
Other researchers have taken several very different ap-proaches to the development of NPs for the treatment of AD.Fluorinated NPs that promote an increase in the α-helical contentof Aβ have been shown to inhibit the formation of Aβ oligomers,whereas this was not the case with hydrogenated NPs.42 NPsconjugated to iron chelators may have the potential to inhibit Aβaggregation and/or protect human cortical neurons from Aβ-associated oxidative toxicity.43 NPs synthesized by sulfonationand sulfation of polystyrene have been shown to affect theconformation of Aβ by inducing an unordered state and reducingAβ oligomerization and cytotoxicity.44 Liposomes derivatizedwith fragments of the Aβ peptide are also under development asa potential vaccine.45 However, based on the results presentedhere, in conjunction with previous binding data,25 curcumin-
based liposomes, particularly the ones characterized as click-curcumin liposomes, should be considered as a seriouspossibility for further development as a novel AD therapeutic.
References
1. Brookmeyer R, Gray S, Kawas C. Projections of Alzheimer's disease inthe United States and the public health impact of delaying disease onset.Am J Public Health 1998;88:1337-42.
2. American Health Assistance Foundation. Alzheimer disease research:about Alzheimer. Available from: http://www.ahaf.org/alzheimers/about/.
3. Brookmeyer R, Corrada MM, Curriero FC, Kawas C. Survival followinga diagnosis of Alzheimer disease. Arch Neurol 2002;59:1764-7.
4. Citron M. Alzheimer's disease: strategies for disease modification. NatRev Drug Discov 2010;9:387-98.
5. Hardy J, Allsop D. Amyloid deposition as the central event in theaetiology of Alzheimer's disease. Trends Pharmacol Sci 1991;12:383-8.

550 M. Taylor et al / Nanomedicine: Nanotechnology, Biology, and Medicine 7 (2011) 541–550
6. Mann DM, Iwatsubo T, Ihara Y, Cairns NJ, Lantos PL, Bogdanovic N,et al. Predominant deposition of amyloid-β 42(43) in plaques in casesof Alzheimer's disease and hereditary cerebral hemorrhage associatedwith mutations in the amyloid precursor protein gene. Am J Pathol1996;148:1257-66.
7. Selkoe DJ. Alzheimer's disease: genes, proteins, and therapy. PhysiolRev 2001;81:741-66.
8. Siemers E, DeMattos RB, May PC, Dean RA. Role of biochemicalAlzheimer's disease biomarkers as end points in clinical trials. BiomarkMed 2010;4:81-9.
9. Yin YI, Bassit B, Zhu L, Yang X, Wang C. Li YM. γ-Secretase substrateconcentration modulates the Aβ42/Aβ40 ratio: implications forAlzheimer disease. J Biol Chem 2007;282:23639-44.
10. Shen Y, Yu LC. Potential protection of curcumin against hypoxia-induced decreases in β-III tubulin content in rat prefrontal corticalneurons. Neurochem Res 2008;33:2112-7.
11. Thomas T, Nadackal TG, Thomas K. Aspirin and non-steroidal anti-inflammatory drugs inhibit amyloid-β aggregation. Neuroreport 2001;12:3263-7.
12. Ono K, Hasegawa K, Naiki H, Yamada M. Curcumin has potentanti-amyloidogenic effects for Alzheimer's β-amyloid fibrils in vitro.J Neurosci Res 2004;M75:742-50.
13. Kim H, Park BS, Lee KG, Choi CY, Jang SS, Kim YH, et al. Effects ofnaturally occurring compounds on fibril formation and oxidative stressof β-amyloid. J Agric Food Chem 2005;53:8537-41.
14. Yang F, Lim GP, Begum AN, Ubeda OJ, Simmons MR, AmbegaokarSS, et al. Curcumin inhibits formation of amyloid β oligomers andfibrils, binds plaques, and reduces amyloid in vivo. J Biol Chem 2005;280:5892-901.
15. Kim DS, Park SY, Kim JK. Curcuminoids from Curcuma longa L.(Zingiberaceae) that protect PC12 rat pheochromocytoma and normalhuman umbilical vein endothelial cells from βA(1-42) insult. NeurosciLett 2001;303:57-61.
16. Re F, Airoldi C, Zona C, Masserini M, La Ferla B, Quattrocchi N, et al.β-Amyloid aggregation inhibitors: small molecules as candidate drugsfor therapy of Alzheimer's disease. Curr Med Chem 2010;17:2990-3006.
17. Anand P, Thomas SG, Kunnumakkara AB, Sundaram C, Harikumar KB,Sung B, et al. Biological activities of curcumin and its analogues(congeners) made by man and mother nature. Biochem Pharmacol 2008;76:1590-611.
18. Bernabe-Pineda M, Ramirez-Silva MT, Romero-Romo M, Gonzalez-Vergara E, Rojas-Hernandez A. Determination of acidity constants ofcurcumin in aqueous solution and apparent rate constant of its decompo-sition. Spectrochim Acta A Mol Biomol Spectrosc 2004;60:1091-7.
19. Wang YJ, Pan MH, Cheng AL, Lin LI, Ho YS, Hsieh CY, et al. Stabilityof curcumin in buffer solutions and characterization of its degradationproducts. J Pharm Biomed Anal 1997;15:1867-76.
20. Tonnesen HH, Karlsen J. Studies on curcumin and curcuminoids. VI.Kinetics of curcumin degradation in aqueous solution. Z LebensmUnters Forsch 1985;180:402-4.
21. Montet X, Funovics M, Montet-Abou K, Weissleder R, Josephson L.Multivalent effects of RGD peptides obtained by nanoparticle display. JMed Chem 2006;49:6087-93.
22. Hong S, Leroueil PR, Majoros IJ, Orr BG, Baker Jr JR, Banaszak HollMM. The binding avidity of a nanoparticle-based multivalent targeteddrug delivery platform. Chem Biol 2007;14:107-15.
23. Tassa C, Duffner JL, Lewis TA, Weissleder R, Schreiber SL, KoehlerAN, et al. Binding affinity and kinetic analysis of targeted smallmolecule-modified nanoparticles. Bioconjug Chem 2010;21:14-9.
24. Antimisiaris SG, Kallinteri P, Fatouros D. Liposomes and drug delivery.In: Gad SC, editor. Pharmaceutical manufacturing handbook productionand processes. New York: John Wiley & Sons; 2008. p. 443-533.
25. Mourtas S, Canovi M, Zona C, Aurilia D, Niarakis A, La Ferla B, et al.Curcumin-decorated nanoliposomes with very high affinity for amyloid-β1-42 peptide. Biomaterials 2011;32:1635-45.
26. Matsuzaki K. Physicochemical interactions of amyloid β-peptide withlipid bilayers. Biochim Biophys Acta 2007;1768:1935-42.
27. Chauhan A, Ray I, Chauhan VP. Interaction of amyloid β-protein withanionic phospholipids: possible involvement of Lys28 and C-terminusaliphatic amino acids. Neurochem Res 2000;25:423-9.
28. McLaurin J, Chakrabartty A. Membrane disruption by Alzheimerβ-amyloid peptides mediated through specific binding to either phospho-lipids or gangliosides. Implications for neurotoxicity. J Biol Chem 1996;271:26482-9.
29. Kakio A, Nishimoto SI, Yanagisawa K, Kozutsumii Y, Matsuzaki K.Cholesterol-dependent formation of GM1 ganglioside-bound amyloidβ-protein, an endogenous seed for Alzheimer amyloid. J Biol Chem2001;276:24985-90.
30. Aisenbrey C, Bechinger B, Gröbner G. Macromolecular crowding atmembrane interfaces: adsorption and alignment of membrane peptides.J Mol Biol 2008;375:376-85.
31. Yanagisawa K. Role of gangliosides in Alzheimer's disease. BiochimBiophys Acta 2007;1768:1943-51.
32. Gobbi M, Re F, Canovi M, Beeg M, Gregori M, Sesana S, et al. Lipid-based nanoparticles with high binding affinity for amyloid-β1-42peptide. Biomaterials 2010;31:6519-29.
33. Taylor M, Moore S, Mayes J, Parkin E, Beeg M, Canovi M, et al.Development of a proteolytically stable retro-inverso peptide inhibitor ofβ-amyloid oligomerization as a potential novel treatment for Alzhei-mer's disease. Biochemistry 2010;49:3261-72.
34. Kokona M, Kallinteri P, Fatouros D, Antimisiaris SG. Stability of SUVliposomes in the presence of cholate salts and pancreatic lipases: effect oflipid composition. Eur J Pharm Sci 2000;9:245-52.
35. Hassane F, Frisch B. Schuber F. Targeted liposomes: convenientcoupling of ligands to preformed vesicles using “click chemistry”.Bioconjug Chem 2006;17:849-54.
36. Usta M, Wortelboer HM, Vervoort J, Boersma MG, Rietjens IM, vanBladeren PJ, et al. Human glutathione S-transferase-mediated glutathi-one conjugation of curcumin and efflux of these conjugates in Caco-2cells. Chem Res Toxicol 2007;20:1895-902.
37. Stewart JCM. Colorimetric determination of phospholipids withammonium ferrothiocyanate. Anal Biochem 1980;104:10-4.
38. Zhang L, Rozek A, Hancock RE. Interaction of cationic antimicrobialpeptides with model membranes. J Biol Chem 2001;276:35714-22.
39. Manzoni C, Colombo L, Messa M, Cagnotto A, Cantu L, Del Favero E,et al. Overcoming synthetic Aβ peptide aging: a new approach to an age-old problem. Amyloid 2009;16:71-80.
40. Ray B, Bisht S, Maitra A, Maitra A, Lahiri DK. Neuroprotective andneurorescue effects of a novel polymeric nanoparticle formulation ofcurcumin (NanoCurc) in the neuronal cell culture and animal model:implications for Alzheimer's disease. J Alzheimer's Dis 2011;23:61-77.
41. Mulik RS, Mönkkönen J, Juvonen RO, Mahadik KR, Paradkar AR.ApoE3 mediated poly(butyl) cyanoacrylate nanoparticles containingcurcumin: study of enhanced activity of curcumin against β amyloidinduced cytotoxicity using in vitro cell culture model. Mol Pharm 2010;7:815-25.
42. Saraiva AM, Cardoso I, Pereira MC, Coelho MA, Saraiva MJ, MöhwaldH, et al. Controlling amyloid-β peptide(1-42) oligomerization andtoxicity by fluorinated nanoparticles. Chembiochem 2010;11:1905-13.
43. Liu G, Men P, Kudo W, Perry G, Smith MA. Nanoparticle-chelatorconjugates as inhibitors of amyloid-β aggregation and neurotoxicity: anovel therapeutic approach for Alzheimer disease. Neurosci Lett 2009;455:187-90.
44. Saraiva AM, Cardoso I, Saraiva MJ, Tauer K, Pereira MC, CoelhoMA, et al. Randomization of amyloid-β-peptide(1-42) conformationby sulfonated and sulfated nanoparticles reduces aggregation andcytotoxicity. Macromol Biosci 2010;10:1152-63.
45. Muhs A, Hickman DT, Pihlgren M, Chuard N, Giriens V, MeerschmanC, et al. Liposomal vaccines with conformation-specific amyloid peptideantigens define immune response and efficacy in APP transgenic mice.Proc Natl Acad Sci USA 2007;104:9810-5.