-
7/30/2019 Elasmo GG Gilnet
1/6
Preliminary data on elasmobranch gillnet fishery in the Gulf of
Gabe` s, Tunisia
By K. Echwikhi1,2, B. Saidi1, M. N. Bradai1 and A. Bouain2
1National Institute of Sea Sciences and Technologies, Sfax,
Tunisia; 2Sfax Faculty of Sciences, Sfax, Tunisia
Summary
Elasmobranchs are an important catch component in Tuni-
sian artisanal fisheries. Generally, species-specific
information
is largely unavailable for artisanal fisheries; an increase
in
knowledge thereof is essential to ensure proper management
of these species. The study analyzed the gillnet fishery
elas-
mobranch catches in the Gulf of Gabes, whereby 45 fishing
trips were conducted from April to June 2007 and 2008 for
the capture of a total of six species: three Rajiformes andthree
Carchariniformes. Rhinobatos cemiculus was the most
important species (52%; 4.588 ind km2 net per day).
Among shark species, Mustelus mustelus was the most impor-
tant (66.8%; 2.21 ind km2 net per day). Discards repre-
sented 6.87% of total catch in weight and 14.28% in number
of specimens caught. Size composition of captures varied by
species, but usually mature, mainly gravid females were
abundant. Further investigations are needed to obtain more
information on such fragile species for the development of
protective measures.
Introduction
Due to their typically vulnerable life cycle characterized
by
slow growth rates, late maturity, and low fecundity (Corte
s,
2000), elasmobranch fishes are highly susceptible to fishing
mortality. In these animals, overexploitation can occur even
with low levels of fishing mortality (Stevens et al., 2000);
once they begin to decline, populations may need decades to
recover (Anderson, 1990).
Industrial fisheries have been highlighted as a major source
of bycatch and mortality for elasmobranchs (Baum et al.,
2003); however, artisanal fisheries represent a considerable
portion of global shark and ray landings (Bonfil, 1994;
Vannuccini, 1999). Artisanal fishery exploitation can
greatly
affect the abundance and size composition of long-lived spe-
cies such as elasmobranchs (Pinnegar and Engelhard,
2008).Although essential for the monitoring of exploited
popula-
tions and the development of effective management plans,
species-specific information is unavailable for artisanal
elas-
mobranch fisheries (White et al., 2006; Henderson et al.,
2007).
In the Mediterranean Sea, few elasmobranchs are subject
to direct fishery; local fisheries land elasmobranchs mostly
as
by-catch (Fowler et al., 2005). Elasmobranchs currently rep-
resent 0.78% of the total landings in the Mediterranean Sea
(FAO-FIGIS, 2006). Between 1970 and 1985, landings of
elasmobranch fishes in the Mediterranean increased from
10 000 tonnes (t) to 25 000 t. Subsequently, reported land-
ings declined to 1000 t by 2004 (FAO-FIGIS, 2006).
In Tunisia, the annual catch of elasmobranchs is about
2000 t, of which over 62% (over 1200 t/year) were from the
Gulf of Gabe` s where elasmobranchs are landed as either
bycatch or target species. Artisanal fisheries targeting
elasmo-
branchs are seasonal; they operate in springsummer when
species move to shallow waters. The target species are
Carcharhinus plumbeus, Mustelus punctulatus, M. Mustelus,
Rhinobathos cemiculus and R. rhinobathos (Bradai et al.,
2005a). Although elasmobranchs are a significant component
of the Gulf of Gabe` s fisheries, there is no quantification
of
the retained or discarded portion, which is never accountedfor
official fishery statistics.
Gillnets are well represented in the Tunisian artisanal
fish-
eries and are highly selective for those species captured
mainly by gilling (i.e. captured behind the gill cover) or
wedged (being held by a mesh around their maximum body
girth). Because of the unregulated nature of most gillnet
fish-
eries, the information describing their operational
character-
istics and associated bycatch in the Mediterranean Sea is
limited. This study is the first field assessment of the
ecosys-
tem impact of the small scale fleet using gillnets and
operat-
ing in the Gulf of Gabe` s (southern Tunisia; central
Mediterranean Sea) that currently counts among the infre-
quent fisheries targeting elasmobranchs in the entire
Mediter-
ranean basin. Specific project objectives were to:(i) determine
the species composition and catch per unit
effort (CPUE); and (ii) provide biological information (size
distribution and sex ratios).
Materials and methods
In the Gulf of Gabe` s, gillnets targeting sharks and
guitarfish
are used in the spring and at the beginning of summer
(March to July). Gillnets are constructed of polyamide
monofilament netting and consist of several pieces of net
(panels) tied together to form a single curtain positioned
upright in the water. These nets are fixed to the bottom by
means of anchors or ballast sufficiently heavy to neutralizethe
buoyancy of the floats. During the fishery season, gillnets
are checked daily and cleared of catch, or pulled and reset.
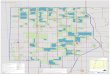
The investigation was carried out from April to June 2007
and 2008, onboard four artisanal boats associated with the
ports of Zarzis and Jerba (Fig. 1). Although mesh sizes from
14 to 16 cm were observed in the study area, bottom gillnets
consisted of 14 cm mesh and varied between 2500 and
3000 m in length and 2.5 to 3 m in height. Soak time could
reach from two to five days, depending on weather condi-
tions. For each set, the length and height of the gillnet
and
the station data (time of set and retrieval, position, depth
and course) were recorded. After retrieval and untangling of
specimens from the nets, fishermen decided whether the fish
were retained or discarded. All individuals were identified
following Compagno (1984) and Fischer et al. (1987) and
J. Appl. Ichthyol. (2012), 16 2012 Blackwell Verlag, BerlinISSN
01758659
Received: February 28, 2012
Accepted: March 19, 2012
doi: 10.1111/jai.12022
Applied IchthyologyJournal of
-
7/30/2019 Elasmo GG Gilnet
2/6
total length (TL), disc width of batoids (DW) (to the
nearest
mm) and weight (to the nearest 5 g) were recorded. Dis-
carded individuals were processed onboard and returned tothe
sea. The maturity status and stages (juveniles and
mature) of specimens were determined based on previous
studies conducted in the area (Sadi et al., 2005; 2008,
2009;
Enajjar et al., 2008, 2012).
There is no generally recognized standard measure of catch
per unit effort (CPUE) for nets. Given this, the CPUE was
chosen taking into account as many parameters as possible.
To assess the relative importance of details such as total
net
area and soak time for quantifying the interaction of
differ-
ent species with gillnets, CPUE 1 was calculated as the num-
ber of specimens captured per km2 of nets per day:
CPUE 1 n=S d
Where n is the total number of specimens caught, S is the
total surface area of gillnets used in the fishing
operations
expressed in km2 and d is the total duration of soak time
expressed in number of days.
In addition, the number of each species captured per set,
CPUE 2, was calculated:
CPUE 2 n/N
Where N is the total number of sets. Then the total cap-
ture was estimated using CPUE 2 and the total number of
fishing sets conducted by the gillnet fishery in the study
area
(ET) obtained from the DGPA (General Directorate of Fish-
ing and Aquaculture).
CT CPUE 2 ET
All measured specimens were utilized to determine size
composition and sex ratio of capture. Size data were evalu-
ated using two-tailed non-parametric MannWhitney U tests
(Zar, 1996). Additionally, the assumption of equal sex
ratios
(1:1) was tested using chi-square analysis with Yates
correc-
tion for continuity (Zar, 1996).
Results
During the study period, 45 fishing sets were analyzed
corre-
sponding to a soak time of 108.8 days and a total gillnet
sur-
face of 0.627 km2 (Table 1). A total of 602 specimens
belonging to six elasmobranch species were caught: three
Rajiformes and three Carchariniformes (Table 2). All speci-
mens captured were identified to the species level, except
for
three individuals that could only be identified as Raja
spp.,
due to advanced decomposition. Batoids dominated gillnet
capture, accounting for 62.46% of the overall recorded catch
number.
The blackchin guitarfish, Rhinobatos cemiculus, was the
most abundant species, comprising 52% of catches (4.588
ind km2 net per day), followed by the common smooth-
hound Mustelus mustelus (25.08%; 2.213 ind km2 net per
day), as described in Table 2. Considering values of biomass
caught, R. cemiculus was the most important species com-
prising 62.24% of total weight, followed by M. mustelus
(16.15%) (Table 2). Total captures were estimated (Table
3)taking into account the annual total fishing effort (554.5
sets)
and catch rates (CPUE 2) in the study area.
The main elasmobranch portions caught were retained
(85.72% in number; 93.12% in weight) (Table 2). Among
retained captures, a small portion was for own consumption
(7.22% in number; 0.923% in weight) of fishermen, with the
remainder landed. Discarded elasmobranchs represented
14.28% in number of individuals caught (Table 2). Main rea-
sons for target species discards were damaged conditions
when disentangled (63.95%) and size (10.46%); species with
low or no commercial value, such as Raja radula, were
always discarded (25.58%) (Table 2). Captures in poor con-
dition, e.g. entangled individuals showing advanced decom-
position, were found especially after stormy days (which led
to higher soak times since fishermen failed to retrieve nets
during these periods). Although less important, fish size
also
seemed to contribute to discards in that discarded specimens
were generally smaller than retained individuals.
Species-specific size and sex composition were available for
all recorded elasmobranchs. Specimens were often examined
prior to a vessels arrival in the harbor. The observed sex
ratio of R. cemiculus (n = 313) differed significantly from
a
1 : 1 female-to-male ratio (v2 = 54.827, d.f. = 1, P <
0.05)
(Fig. 2; Table 4). Size composition of females and males did
not differ significantly (U = 9821.500, P = 0.700). More
than
40% of females and 70% of males captured were mature
(Table 4).Of the 41 R. rhinobathos examined, females were
signifi-
cantly more common than males (v2 = 4.12, d.f. = 1,
P = 0.04) (Fig. 2; Table 4). Size composition of females and
males did not differ significantly (U = 172.500, P = 0.64).
Among captured individuals, 48% and 57% of females and
males, respectively, were mature (Table 4).
The examined size range of M. mustelus (n = 151) was 620
1580 mm TL (Fig. 2; Table 4). Females were significantly
more abundant, comprising 60.20% of the total number of
individuals documented (v2 = 6.364, d.f. = 1, P = 0.011).
Females were significantly larger than males (U = 1369.500,
P < 0.0001). All females and 90% of males captured were
mature (Fig. 2; Table 4).
The recorded size composition of M. punctulatus ranged
from 530 to 1190 cm TL (Fig. 2; Table 4). The number of
Fig. 1. Gulf of Gabe` s, Tunisia
Table 1Fishing effort in the study area
2007 2008 Total
Number of sets 17 28 45Total surface of gillnets (km2) 0.181
0.445 0.627Total duration of soak time (days) 35.94 72.9 108.8
2 K. Echwikhi et al.
-
7/30/2019 Elasmo GG Gilnet
3/6
females and males did not depart significantly from a 1 : 1
ratio (v2 = 0.75, d.f. = 1, P = 0.38). Females were signifi-
cantly larger than males (U = 69.500, P < 0.0001). Mature
females (70%) dominated captures, while males were mainly
juveniles (71.43%) (Fig. 2; Table 4).
Carcharhinus plumbeus individuals ranged from 1310 to
1980 mm TL; size composition of females and males did not
differ significantly (U = 61.500, P = 0.237) (Fig. 2; Table
4).
The overall male to female ratio was not significantly
differ-
ent from 1:1 (v2 = 1.81, d.f. = 1, P = 0.178). Most captured
specimens were mature: 70% of males and 60% of females
(Fig. 2; Table 4).
Female R. radula outnumbered males within sampled
catches, which consisted primarily of individuals >220 mm
DW (Fig. 2; Table 4). The sex ratio was not significantly
dif-
ferent from 1 : 1 (v2 = 2.909, d.f. = 1, P = 0.08). Size
com-
position of females and males differed significantly
(U = 21.000, P = 0.011). The majority of specimens, i.e. 80%
of the females and 57% of the males, were mature.
Discussion
This study represents to date the most detailed evaluation
of
elasmobranch capture in the Gulf of Gabe` s. Previous
investi-
gations were either limited in scope or generalized,
preclud-
ing a detailed understanding of fishery characteristics and
species-specific data (Bradai et al., 2005a).
Results show that R. cemiculus is the common component
of gillnet elasmobranch fisheries in the Gulf of Gabe` s.
Previ-
ous information reported that this species was the most
abundant ray species landed in the area, where it was caught
between April and August (Bradai et al., 2005a). According
to catch statistics from the Zarzis fish market, which is
the
largest landing port for shark fishery in Tunisia, annual
land-
ings by target fisheries of this species are about 250 t
(Enajjar
et al., 2012). Because of its shape and schooling behavior,
R. cemiculus appears to be especially susceptible to capture
in bottom gillnets.
Although C. plumbeus is considered to be a target species,
its low catch rate compared to others species could be
related
to the fishing sites. In fact, the fishing gear types operate
in
diverse habitats (at different depths), where the relative
abun-
dances of species may be different. The importance in num-bers
and catch rates of Mustelus species reflect their
abundance in the fishing area. These species appear to be
more abundant in the study area, compared to the northern
coasts of the Mediterranean where they are considered as
extirpated locally (Aldebert, 1997; Jukic-Peladic et al.,
2001;
Ferretti et al., 2005). Indeed, specific elasmobranch
fisheries
already subsist, showing an important exploitation
potential.
In addition, their abundance suggests that Mustelus spp. in
the Gulf of Gabe` s find the conditions to reproduce and
develop for consideration as a nursery area (Bradai et al.,
2005a). Diversity of catch composition of gillnet fishery
probably reflects the highly opportunistic tendency of
fisher-
men and the mixed-species nature of the fishery. Fishermen
tended to target whatever demersal elasmobranchs were
locally present and retained all, or nearly all, of the
species.
In the present study, discards accounting for 14.28% of
the elasmobranch catch were higher than those recorded in
southern Portugal (5.4%) (Coelho et al., 2005) and lower
than on the Portuguese west coast (24.8%) (Baeta et al.,
2010). Differences between discard rates could be related to
local species diversity, environmental constrains,
social-eco-
nomic aspects or simply due to random factors (Batista
et al., 2009). Nevertheless, the choice of fishing grounds,
reduction in the soak time as well as the use of larger mesh
sizes may reduce bycatch and discards (Goncalves et al.,
2007). A decrease in soak time during the warmer months
could also contribute to a higher quality of landings, sincefish
and invertebrates degrade faster at higher water tempera-
tures and the small fleet does not have refrigerated storage
onboard.
In Tunisia, catch statistics lack information on species
composition, even for the most important elasmobranch spe-
cies in the capture, and only catch trends can be assessed.
Annual elasmobranch production from the Gulf of Gabe` s,
considered as the largest in Tunisia, showed a steady
increase
during the late 1990s. The highest annual catch of 1650 t
was attained in 2002. From 2003, landings decreased, falling
to 950 t in 2007 (Fig. 3). The same trend was reported for
shark and batoid production in Zarzis, where gillnet activi-
ties were the most developed (Fig. 3). In fact, more than
70% of elasmobranchs were landed by artisanal fisheries.
The concentration of fishing effort on selected species
(and/
Table 2Elasmobranch catches in gillnet fishery. Total catches,
discards and relative importance of each species captured
(percentages in brackets).CPUE 1 (per km2 of net per day)
Number Weight
N (%) CPUE 1Retained(%)
Discarded(%)
W (kg)(%)
Retained(%)
Discarded(%)
Rhinobatos cemiculus 313 (52.00) 4.588 287 (91.69) 26 (8.30)
2850 (62.24) 2715 (95.26) 135 (4.73)Rhinobatos rhinobathos 41
(06.81) 0.601 34 (82.92) 7 (17.07) 320 (6.98) 275 (85.93) 45
(14.06)Mustelus mustelus 151 (25.08) 2.213 134 (88.74) 17 (11.25)
740 (16.160) 676 (91.35) 64 (8.64)Mustelus puctulatus 48 (07.97)
0.703 39 (81.25) 9 (18.75) 285 (6.22) 253 (88.77) 32
(11.22)Carcharhinus plumbeus 27 (04.49) 0.395 22 (81.48) 5 (18.51)
375 (8.187) 345 (92) 30 (8)Raja radula 22 (03.65) 0.322 00 (00) 22
(100) 9 (0.19) 00 (0) 9 (100)Total 602 8.824 516 (85.71) 86 (14.28)
4580 4265 (93.13) 315 (6.87)
Table 3Catch rate (CPUE 2) and estimated total capture of
different species
Species
Numberofspecimens
CPUE 2(Individualsper set)
Totalcapture(Individuals/year)
R. cemiculus 313 6.95 3856.85R. rhinobathos 41 0.91 505.21M.
mustelus 151 3.35 1860.65M. puctulatus 48 1.06 591.46C. plumbeus 27
0.6 332.70R. radula 22 0.48 271.08
Elasmobranch captures in Tunisian waters 3
-
7/30/2019 Elasmo GG Gilnet
4/6
or specific maturity stages, mainly adults) could have
adverse
consequences, such the decreasing trend observed in elasmo-
branch landings in the Gulf of Gabes.
A visual check of histograms of the size distribution for
females, except Rhinobatids, showed that mature specimens
dominated captures. For males, with the exception of
M. punctulatus, mature individuals also constituted the
greater proportion of the catches. Neonates and small size
classes were absent; possibly because of gillnet selectivity
for
larger specimens. Size composition of Rhinobatids consisted
of a mixture of mature and medium-sized individuals. The
two Rhinobatid species show significantly different sex
ratios:
females dominate, undergo seasonal movements, and are
especially abundant during warm water periods, often for
breeding and nursery functions (Enajjar et al., 2008, 2012).
In the Gulf of Gabes, fishermen target aggregations of
gravid
females when they migrate to shallow coastal and insular
waters to pup and breed (Bradai et al., 2005a; Enajjar et
al.,
Fig. 2. Length-frequencies distribution of elasmobranch species
caught by gillnet, Gulf of Gabe` s, 20072008. [Size at maturity of
males ( )and females ( )]
Table 4Size composition and importance of mature individuals
caught by gillnet fishery, Gulf of Gabe` s, 20072008: (a) Enajjar
et al., 2012; (b) Enaj-jar et al., 2008; (c) Sadi et al., 2008; (d)
Sadi et al., 2009; (e) Sadi et al., 2005; (f) Capape , 1974.
Species Sex Number
Size range(mm)
Mean SD Size at maturity
Percentage in catch
Min Max Juveniles Mature
R. cemiculus F 222 520 1730 1278.42 323.17 1381 (a) 127 (57.20)
95 (42.80)M 91 550 1720 1265.82 320.14 1118 (a) 28 (30.77) 63
(69.23)
R. rhinobathos F 27 530 1180 817.40 204.91 790 (b) 14 (51.85) 13
(48.15)M 14 550 1120 846.42 211.42 700 (b) 6 (42.86) 8 (57.14)
M. mustelus F 91 1220 1550 1364.40 103.65 1172 (c) 0 (0) 91
(100)M 60 620 1340 1208.50 179.90 971 (c) 6 (10) 54 (90)
M. puncutulatus F 27 640 1160 1011.11 176.90 956 (d) 8 (29.62)
19 (70.38)
M 21 580 950 739.52 113.68 814 (d) 15 (71.43) 6 (28.57)C.
plumbeus F 17 1310 1960 1781.76 159.14 1600 (e) 5 (29.42) 12
(70.58)
M 10 1420 1920 1656.00 254.61 1720 (e) 4 (40) 6 (60)R. radula F
15 220 420 350.00 60.47 340 (f) 3 (20) 12 (80)
M 7 220 370 295.71 57.40 320 (f) 3 (42.85) 4 (57.15)
4 K. Echwikhi et al.
-
7/30/2019 Elasmo GG Gilnet
5/6
2008, 2012). Captured M. mustelus and M. punctulatus
included juvenile and mature individuals of both sexes. As
indicated by the size frequency distribution of both
species,mature females were an important component of the
elasmo-
branch gillnets capture, whereas juveniles were largely com-
posed of males. Mustelus spp. dominated shark landings in
this region in the spring, when females moved to near shore
waters to reproduce (Sadi et al., 2008, 2009). The capture
of
sandbar sharks consisted especially of mature, usually near-
term pregnant females that apparently approached the coast
to give birth in a nursery area with advantageous environ-
mental conditions (Bradai et al., 2005b; Sadi et al., 2005).
Records of sandbar sharks were reported throughout the
Mediterranean, but the species seemed to be more abundant
in the southern coasts, where it found favorable conditions
for reproduction (Bradai et al., 2005b; Sadi et al., 2005).
In the Gulf of Gabe` s, gillnet fisheries are mostly
restricted
to shallow waters. This activity is related to the timing
migration of gravid females to nearshore waters to pup and
breed (Bradai et al., 2005a). Investigations conducted along
Tunisian coasts suggested that the Gulf of Gabe` s could
con-
stitute a nursery for some elasmobranch species (Bradai
et al., 2005a,b). The targeting of gravid females of
exploited
species reduces the productivity, resilience, and
sustainability
of such populations (Kokko et al., 2001; Smith et al.,
2008).
In addition, Simpfendorfer (1999) suggested that, for some
elasmobranch species, sustainable exploitation might be pos-
sible if the youngest age-classes were targeted and older
ages
left unfished.
This study provides the first detailed, quantitative
informa-tion on the artisanal elasmobranch fisheries in the Gulf
of
Gabe` s, one of the most productive areas of Tunisia in
terms
of elasmobranch landings. Captures of elasmobranchs were
substantial during the survey years and probably represent a
considerable source of mortality for exploited populations.
The increasing domestic demand for elasmobranch fishing
permits coupled with the lack of regulatory actions poses a
threat to the elasmobranch stocks harvested in Tunisian
waters.
Successful management of elasmobranch fisheries is com-
plicated by a general lack of fishery information and life
his-
tory characteristics of this k-selected group of fishes
(Musick,
1999). Conservation of these populations is further con-
founded by the delayed responses in the implementation of
fishery restrictions, typically resulting in the
implementation
of management strategies following the overexploitation of
targeted elasmobranch populations (Stevens et al., 2000).
In the study area, for gillnet fisheries an effective
manage-
ment plan might include the closure of nearshore and inshore
waters to gillnetting during the migration of gravid
females,
or the establishment of mesh-size requirements to target
spe-
cific population segments. Successful conservation and sus-
tainable use of this fishery resource requires action:(i)
continuance of the collection and survey of species-specific
information on catches and landings, by number and weight;
(ii) improvement in statistical data collection at a
specific
level; (iii) delineation of the nursery area in the Gulf of
Gabe` s; (iv) assessment of the impact of fishing mortality
on
juveniles and gravid females in elasmobranchs/nursery areas;
and (v) socio-economic studies of the coastal artisanal
fisheries.
Acknowledgements
We wish to thank all fishermen who allowed us to operate
on their boats, all of the crews, as well as the scientists
and
onboard observers who participated in this work. We
thankanonymous reviewers for their insightful comments, which
led to a much-improved manuscript.
References
Aldebert, Y., 1997: Demersal resources of the Gulf of Lions
(NWMediterranean). Impact of exploitation on fish diversity.
VieMilieu 47, 275284.
Anderson, E. D., 1990: Fishery models as applied to
elasmobranchfisheries. In: Elasmobranchs as living resources:
advancesin the biology, ecology, systematics, and the status of
fisheries.H. L. Pratt, S. H. Gruber, T. S. Taniuchi (Eds). NOAA
Techni-cal Report NMFS, East-West Center, Honolulu, Hawaii
(USDepartment of Commerce), pp. 473484.
Baeta, F.; Batista, M.; Maiab, A.; Costaa, M. J.; Cabrala, H.,
2010:Elasmobranch bycatch in a trammel net fishery in the
Portu-guese west coast. Fish. Res. 102, 123129.
Batista, M. I.; Teixeira, C. M.; Cabral, H. N., 2009: Catches of
tar-get species and bycatches of an artisanal fishery: the case
studyof a trammel net fishery in the Portuguese coast. Fish. Res.
100,167177.
Baum, J. K.; Myers, R. A.; Kehler, D. G.; Worm, B.; Harley, S.
J.;Doherty, P. A., 2003: Collapse and conservation of shark
popu-lations in the northwest Atlantic. Science 299, 389392.
Bonfil, R., 1994: Overview of world elasmobranch fisheries.
FAOFisheries Techn Pap. No 341, Rome, Italy, pp. 119.
Bradai, M. N.; Saidi, B.; Enajjar, S.; Bouain, A., 2005a: The
Gulf ofGabe` s: a spot for the Mediterranean Elasmobranchs.
In:Proceedings of the International workshop on
Mediterraneancartilaginous fish with emphasis on southeastern
Mediterranean.N. Bausta, C . Keskin, F. Serena, B. Seret (Eds.).
Turkish
Marine Research Foundation, Istanbul, Turkey, pp. 107
117.Bradai, M. N.; Saidi, B.; Bouain, A.; Gue lorget, O.; Capape
, C.,
2005b: The Gulf of Gabe` s (Central Mediterranean): nurseryarea
for the sandbar shark, Carcharhinus plumbeus (Nardo,1827)
(Chondrichthyes: Cacharhinidae). Ann. Ser. Hist. Nat.
15,187194.
Capape, C., 1974: Contribution a la biologie des Rajides des
cotestunisiennes. II. Raja radula Delaroche 1809. Re partition ge
o-graphique et bathyme trique, sexualite , reproduction et fe
condite .Arch. Inst. Pasteur Tunis 51, 211228.
Coelho, R.; Erzini, K.; Bentes, L.; Correia, C.; Lino, P.
G.;Monteiro, P.; Ribeiro, J.; Goncalves, J. M. S., 2005:
Semi-pela-gic longline and trammel net elasmobranch catches in
southernPortugal: catch composition, catch rates and discards.J.
Northw. Atl. Fish. Sci. 35, 531537.
Compagno, L. J. V., 1984: FAO species catalogue. Vol. 4. Sharks
ofthe World. An annotated and illustrated catalogue of shark
spe-cies known to date. Part 2 Carcharhiniformes. FAO Fish.Synop.
125, 251655.
Fig. 3. Elasmobranch production in the Gulf of Gabe` s vs shark
andbatoid landings in the Zarzis zone, 19952008
Elasmobranch captures in Tunisian waters 5
-
7/30/2019 Elasmo GG Gilnet
6/6
Corte s, E., 2000: Life history patterns and correlations in
sharks.Rev. Fish. Sci. 8, 299344.
Enajjar, S.; Brada, M. N.; Bouan, A., 2008: New data on the
repro-ductive biology of the common guitarfish of the Gulf of Gabe`
s(southern Tunisia, central Mediterranean). J. Mar. Biol.
Assoc.U.K. 88, 10631068.
Enajjar, S.; Brada, M. N.; Bouan, A., 2012: Age, growth and
sexualmaturity of the blackchin guitarfish Rhinobatos cemiculus in
theGulf of Gabe` s (southern Tunisia, central Mediterranean).
Cah.
Biol. Mar. 88, 1063
1068.FAO-FIGIS, 2006: FAO-FIGIS (fisheries global information
sys-
tem). Available at: http://www.fao.org (accessed on 19
May2006).
Ferretti, F.; Myers, R. A.; Serena, F.; Sartor, P., 2005:
Long-termdynamics of chondrichthyan fish community in the upper
Tyr-rhenian Sea. Theme session on Elasmobranch fisheries
science.ICES, 2005, ICES Document CM. 25, pp. 156165.
Fischer, W.; Bauchot, M. L.; Schneider, M., 1987: Fiches
FAOdidentification des espe` ces pour les besoins de la peche. Me
dit-errane e et Mer Noire, Zone de peche 37, II: Verte bre s. Rev.
1.FAO, Rome, pp. 7611530.
Fowler, S.; Cavanagh, R.; Camhi, M.; Burgess, G.; Cailliet,
G.;Fordham, S.; Simpfendorfer, C.; Musick, J., 2005: Sharks,
Raysand Chimaeras: the status of the Chondrichthyan fishes.
IUCNPublication Services Unit, Cambridge, UK. pp. x+461.
Goncalves, J. M. S.; Stergiou, K. I.; Hernando, J. A.; Puente,
E.;Moutopoulos, D. K.; Arregi, L.; Soriguer, M. C.; Vilas,
C.;Coelho, R.; Erzini, K., 2007: Discards from experimental
tram-mel nets in southern European small-scale fisheries. Fish.
Res.88, 514.
Henderson, A. C.; McIlwain, J. L.; Al-Oufi, H. S.; Al-Shelii,
S.,2007: The Sultanate of Oman shark fishery: species
composition,seasonality, and diversity. Fish. Res. 86, 159168.
Jukic-Peladic, S.; Vrgoc, N.; Krstulovic-Sifner, S.; Piccinetti,
C.;Piccinetti-Manfrin, G.; Marano, G.; Ungaro, N., 2001: Long-term
changes in demersal resources of the Adriatic Sea: compar-ison
between trawl surveys carried out in 1948 and 1998. Fish.Res. 53,
95104.
Kokko, H.; Linsdstrom, J.; Ranta, E., 2001: Life histories and
sus-tainable harvesting. In: Conservation of exploited species. J.
D.
Reynolds, K. H. Redford, G. M. Mace, J. G. Robinson.
(Eds)Cambridge University Press, Cambridge, pp. 301322.
Pinnegar, J. K.; Engelhard, G. H., 2008: The shifting
baselinephenomenon: a global perspective. Rev. Fish Biol. Fisher.
18,116.
Sadi, B.; Brada, M. N.; Bouan, A.; Gue orget, O.; Capape ,
C.,2005: Reproductive biology of the sandbar shark,
Carcharhinusplumbeus (Chondrichthyes: Carcharhinidae) from the Gulf
ofGabe` s (southern Tunisia, Central Mediterranean). Acta
Adriat.
46, 47
62.Sadi, B.; Brada, M. N.; Bouan, A., 2008: Reproductive biology
of
the smooth-hound shark Mustelus mustelus in the Gulf of Gabe`
s(south-central Mediterranean Sea). J. Fish Biol. 72, 13431354.
Sadi, B.; Brada, M. N.; Bouan, A., 2009: Reproductive biologyand
diet of Mustelus punctulatus (Risso, 1826)
(Chondrichthyes:Triakidae) from the Gulf of Gabe` s, central
Mediterranean Sea.Sci. Mar. 73, 249258.
Simpfendorfer, C. A., 1999: Demographic analysis of the
duskyshark fishery in southwestern Australia. Amer. Fish. Soc.
Sym.23, 149160.
Smith, W. D.; Cailliet, G. M.; Corte s, E., 2008: Demography
andelasticity of the diamond stingray, Dasyatis dipterura:
parameteruncertainty and resilience to fishing pressure. Mar.
Freshwat.Res. 59, 575586.
Stevens, J. D.; Bonfil, R.; Dulvy, N. K.; Walker, P. A., 2000:
Theeffects of fishing on sharks, rays, and chimaeras
(chondrichth-yans), and the implications for marine ecosystems.
ICES J. Mar.Sci. 57, 476494.
Vannuccini, S., 1999: Shark utilization, marketing, and trade.
FAOTech, Rome, Italy. Paper No. 389.
White, W. T.; Giles, J.; Dharmadi, Potter, I. C., 2006: Data on
thebycatch fishery and reproductive biology of mobulid
rays(Myliobatiformes) in Indonesia. Fish. Res. 82, 6573.
Zar, J. H., 1996: Biostatistical analysis, 4th edn.
Prentice-Hall,Upper Saddle River, NJ, pp. 663.
Authors address: K. Echwikhi, National Institute of Sea
Sciencesand Technologies, P. O. Box 1035, 3018 Sfax,Tunisia.E-mail:
[email protected]
6 K. Echwikhi et al.












![· 2020-04-15 · ˚ ˘ [f /] -˚2 5j- 48k 5; ˝˚ p ˝k ,. ˘ gg jp ˆ ˜˝ gg / gg 4˘ ggj 5gg6 5gg ,gg 2 3 / ˆ7 gg 5gg6 ˝ gg˚ q gg :˝ ˜ ˘ ˝ w l; 6 p„ t 6 .˝- ˘ ˘ -](https://img.pdfslide.tips/doc/110x75/5ea9c71468ab2b044305bbac/2020-04-15-f-2-5j-48k-5-p-k-gg-jp-oe-gg-gg.jpg)