Embed Size (px)
Citation preview

http://msj.sagepub.com/Multiple Sclerosis
http://msj.sagepub.com/content/11/3/310The online version of this article can be found at:
DOI: 10.1191/1352458505ms1184oa
2005 11: 310Mult SclerMinagar
Joaquin J Jimenez, Wenche Jy, Lucia M Mauro, Lawrence L Horstman, Eugene R Ahn, Yeon S Ahn and Alirezaand transendothelial migration of monocyte-endothelial microparticle complexes
1b on release of endothelial microparticles, formationβand the inhibitory effects of interferon-monocyte complexes induced by multiple sclerosis plasma−−Elevated endothelial microparticle
Published by:
http://www.sagepublications.com
On behalf of:
European Committee for Treatment and Research in Multiple Sclerosis
Americas Committee for Treatment and Research in Multiple Sclerosis
Pan-Asian Committee for Treatment and Research in Multiple Sclerosis
Latin American Committee on Treatment and Research of Multiple Sclerosis
can be found at:Multiple SclerosisAdditional services and information for
http://msj.sagepub.com/cgi/alertsEmail Alerts:
at UNIV TORONTO on August 12, 2014msj.sagepub.comDownloaded from at UNIV TORONTO on August 12, 2014msj.sagepub.comDownloaded from

http://msj.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://msj.sagepub.com/content/11/3/310.refs.htmlCitations:
What is This?
- Jun 1, 2005Version of Record >>
at UNIV TORONTO on August 12, 2014msj.sagepub.comDownloaded from at UNIV TORONTO on August 12, 2014msj.sagepub.comDownloaded from

Elevated endothelial microparticle�/monocyte complexes induced bymultiple sclerosis plasma and the inhibitory effects of interferon-b1bon release of endothelial microparticles, formation andtransendothelial migration of monocyte�/endothelial microparticlecomplexes
Joaquin J Jimenez*,1, Wenche Jy1, Lucia M Mauro1, Lawrence L Horstman1, Eugene R Ahn1,Yeon S Ahn1 and Alireza Minagar2
1Wallace H Coulter Platelet Laboratory, Division of Hematology/Oncology, University of Miami School of Medicine,Miami, FL 33136, USA; 2Department of Neurology, Louisiana State University Health Sciences Center, Shreveport, LA71130, USA
Monocyte migration through the disrupted cerebral endothelial cell (EC) junctions plays an essential role in formation of multiple sclerosis(MS) demyelinating lesions. During pathogenesis of MS, activated ECs release endothelial microparticles (EMP), which possibly facilitatetransendothelial migration (TEMIG) of monocytes. To assess functional roles of EMP in MS, specifically, their (i) interaction withmonocytes, (ii) effect on monocyte TEMIG in an in vitro model of the brain microvascular endothelial cells (BMVEC), (iii) phenotypicprofiles of EMP elicited by MS plasma and (iv) the effects of IFN-b1b on release of EMP and on TEMIG of monocytes (mono) andmonocytes:EMP complexes (mono:EMP) through the BMVEC. The effect of IFN-b1b on the release of EMP and the TEMIG of mono andmono:EMP was assessed by preincubating BMVEC cultures of IFN-b1b prior to addition of plasma. Three EMP phenotypes, CD54, CD62Eand CD31 were assayed. Plasma specimens from 20 patients with relapsing�/remitting MS (11 in exacerbation, MS-E, and 9 in remission,ME-R) and 10 healthy controls were studied. Incubation of BMVEC with MS-E plasma yielded elevated levels of EMPCD54, EMP62E andEMPCD31 relative to MS-R and control plasmas. MS-E but not MS-R or control plasma also augmented TEMIG of monocytes, respectively.Mono:EMP complexes further augmented TEMIG relative to mono alone, but only in the presence of MS-E plasma; there was nosignificant effect with MS-R or control plasmas. The presence of IFN-b1b inhibited TEMIG of mono and mono:EMP by 20% and 30%,respectively. MS-E but not MS-R plasma elicited release of activation-derived EMP and enhanced TEMIG of mono and mono:EMP. IFN-b1binhibited TEMIG and release of EMP, suggesting a role of EMP and a novel therapeutic mechanism for IFN-b1b in MS.Multiple Sclerosis (2005) 11, 310�/315
Key words: endothelial microparticle; exacerbation; interferon-b1b; monocyte; multiple sclerosis; remission
Multiple sclerosis (MS) is an immune-mediated neurolo-
gical disease characterized by the formation of multiple
foci of inflammatory demyelination within the white and
grey matter of the central nervous system (CNS).1 Adhe-
sion of activated leukocytes �/ chiefly myelin basic
protein-specific CD4�/ T lymphocytes and monocytes �/
to cerebral endothelial cells (ECs) and their transendothe-
lial migration (TEMIG) through a disrupted blood�/brain
barrier (BBB) into the CNS contributes to formation of
demyelinating lesions.2 �5 The sequence of events in
TEMIG are known to include activation of leukocytes
and ECs, chemoattraction, leukocyte-EC adhesion, proteo-
lysis of the basal membrane surrounding the BBB by
matrix metalloproteinases (MMP),6 and finally, extravasa-
tion of activated leukocytes.IFN-b1b has been approved by the FDA for treatment of
relapsing�/remitting MS (RRMS),7 although the mechan-
isms of its beneficial effects on leukocyte trafficking
through the BBB have only been partially explored.8,9
EC dysfunction appears to be centrally involved in the
disruption of the BBB. In this regard, several studies have
documented elevated circulating markers of EC distur-
bance in MS patients, especially during exacerbations.10
We demonstrated that MS patients in exacerbation but not
remission exhibit significantly elevated plasma endothe-
lial microparticles (EMP).11
EMP are small (�/B/1 mm) membranous vesicles bearing
many of the adhesion molecules and other antigens of the
parent EC, notably including platelet-EC adhesion mole-
cule-1 (PECAM-1/CD31).12,13 Plasma levels of CD31�/
EMP (EMPCD31) were elevated during exacerbations of
MS and were associated with disease progression as
judged by clinical activity and presence of contrast-
*Correspondence: Alireza Minagar, Department ofNeurology, Louisiana State University Health SciencesCenter, 1501 Kings Highway, Shreveport, LA 71130, USA.E-mail: [email protected] 11 June 2004; revised 5 October 2004;accepted 21 December 2004
Multiple Sclerosis 2005; 11: 310�/315www.multiplesclerosisjournal.com
# 2005 Edward Arnold (Publishers) Ltd 10.1191/1352458505ms1184oa at UNIV TORONTO on August 12, 2014msj.sagepub.comDownloaded from

enhancing lesions on brain magnetic resonance imaging(MRI).11 However, while that study documented thepresence of CD31�/ and CD51�/ EMP (EMPCD51), it didnot address their possible functional role in MS. Previouswork has shown that some EMP, particularly thosecarrying CD54 (EMPCD54), preferentially interact withmonocytes to form leukocyte�/EMP complexes. Sabatieret al. have also demonstrated that EMP bind to monocytes,eliciting the expression of tissue factor.14 Jy et al. demon-strated that EMPCD54 bind to monocytes and form EMP-monocyte conjugates in MS.15
We therefore hypothesized that EMP may interact withleukocytes and influence their TEMIG through the en-dothelial barrier. Thus, the present report evaluates theeffect of MS-E plasma on the release of phenotypicallydistinct EMP as well as TEMIG of monocytes and mono:-EMP complexes. Additionally, the effect of IFN-b1b on therelease of EMP and TEMIG of monocytes and mono:EMPcomplexes was assessed.
Patients and methods
PatientsA total of 20 patients who met the criteria for MS16 werestudied and compared to 10 normal controls. The studywas approved by the Institutional Review Board and allpatients signed informed consent forms. Prior to enrol-ment, all patients underwent neurological examination.Eleven of them had exacerbation documented by theexamining neurologist within 24 hours of onset of theattack. The other nine MS patients were confirmed to be inremission. Exacerbation was defined as a worsening ofneurologic impairment or the appearance of a newsymptom or abnormality attributable to MS andpersisting�/24 hours, preceded by stability of at leastone month. No patient had received any immunosuppres-sive treatment for at least three months prior to enrolment;i.e., no corticosteroids or immunomodulators such as IFN-b1a, IFN-b1b or glatiramer acetate. None of the 20 MSpatients had any infections at the time of presentation.The control group consisted of 10 healthy volunteers.After neurologic evaluation, peripheral blood was drawnin citrate Vacutainers and plasma was separated asdetailed below.
NeuroimagingBrain and spinal cord MRI were performed in all patientson a 1.5 T machine with standard head coils. The imagingprotocol included sagittal T1-, axial T1-, T2- and FLAIRimages. All MRI scans were performed after infusion ofgadolinium diethylenetriamine pentaacetic acid (Gd).Axial T2- and T1-weighted postcontrast images wereused for assessment of MS plaques. The images wereindependently interpreted by neuroradiologists blinded tothe study, using visual inspection.
Antibodies and other reagentsThe following fluorescent-tagged anti-human monoclonalantibodies (mAbs) were obtained from commercial sup-
pliers: from Pharmingen, anti-CD31 (Cat. #555446, PElabel), anti-CD54 (Cat. #F-0549, FITC label) and anti-CD62E (Cat. #F-0674, FITC label). The lectin, Ulex euro-paeus FITC-labelled, was obtained from Sigma (Cat.#L9006).
BMVEC culturesMVEC of brain origin (BMVEC) were obtained from CellSystems (Kirkland, WA, USA; Cat. no ACBRI 128) andcultured as previously described.17 To assess the effect ofMS plasma on the release of EMP, BMVEC were plated on12-well multiwell clusters at a concentration of 5�/105/well. After 48 hours, cultures were confluent and ready foraddition of plasmas. For TEMIG studies, BMVEC wereresuspended in CS-C medium and replated in 6.5-mm poreTranswell multiwell clusters precoated with collagen.Cultures were closely monitored until confluency wasachieved, then MS-E, MS-R or control plasma was added(20% v/v) and incubated for 18 h. To assess effects of IFN-b1b on TEMIG of monocyte:EMP complexes and mono-cytes alone, BMVEC was preincubated with 1000 IU/wellof IFN-b1b for 6 hours prior to addition of plasmas. MS-E,MS-R and normal plasma specimens were added andincubated for 18 hours.
Monocyte and mono:EMP complexes for TEMIGThe monocytic cell line U937, here abbreviated to mono,was obtained from ATCC (Cat. #CRL-1593.2) and wascultured in RPMI 1640 medium supplemented with 10%fetal bovine serum (FBS). Mono:EMP binding was per-formed as follows. Briefly, EMP were generated by treatingBMVEC cultures for 24 hours with TNF-a (10 ng/mL), thensupernatants were collected by centrifugation for 15minutes at 42 000�/g and the resulting pellets wereconcentrated 10-fold relative to original volume by resus-pending in culture medium devoid of growth factors orserum. EMP preparations were then labelled with FITC-tagged U. europaeus for 15 minutes, washed 2�/ withmedium (RPMI 1640) and resuspended in medium with-out FBS. They were then incubated with 1�/105 mono/mLin a ratio of 1:100 mono/EMP for 30 minutes. As EMP aremuch smaller and less dense than mono, centrifugation at100�/g allowed separation of the EMP:mono complexesfrom unbound EMP. Aliquots of EMP:mono conjugateswere assayed by flow cytometry as before. More than 75%of the mono formed complexes (bound at least 1 EMP) asjudged by fluorescent intensity of FITC-Ulex label on themono.
Transendothelial migration (TEM) assayThese were done using U937 cells (mono), untreated orpretreated by exposure to EMP as above (mono:EMP).Cells (1 mL) at 1�/105 cells/mL were added to a confluentmonolayer of BMVEC and the percentage of cells passedthrough after six hours was evaluated by flow cytometry.
Assay of endothelial microparticles (EMP)
Anti-CD31-PE (4 mL) or anti-CD62E-FITC (4 mL) was addedto 50 mL of supernatant, then incubated for 20 min withgentle (100 rpm) orbital shaking. Diluting buffer (PBS)
Interferon-b1b and endothelial microparticlesJJ Jimenez et al.
311
Multiple Sclerosis at UNIV TORONTO on August 12, 2014msj.sagepub.comDownloaded from
(1 mL) was then added for flow cytometry on a Coulter
EPICS XL flow cytometer. Detection of particles was by
triggering on the CD31 or CD62E fluorescence signal
exceeding noise, and fluorescent particles were further
separated on a histogram based on size. EMP are defined
as particles of size B/1.5 mm expressing either marker.
Statistical analysis
One-way ANOVA (analysis of variance) was used to
analyse differences among three or more groups. If there
were significant differences (P B/0.05), then Student’s t test
was used to evaluate significance between pairs of groups.
If the data failed the normality test, then the Mann�/
Whitney rank sum test was used. The Chi-square test
was used to compare frequency data between groups. Data
analyses were performed using Statmost and Sigmaplot v.
4.0.
Results
Patient demographics and MRI findingsThe mean age of the 20 MS patients was 33 years and 80%
were female. The 10 healthy controls had a mean age of 35
years and 80% were female. Eleven of the MS patients
were in exacerbation confirmed by the examining neurol-
ogist and contrast-enhancing lesions on their T1-weighetd
postcontrast MRIs of brain. Nine MS patients in exacer-
bation had one and two had two contract-enhancing
lesions on their T1-weighted postcontrast brain MRI.
The other nine MS patients were clinically in remission
and exhibited no contrast-enhancing lesions on their brain
MRI.
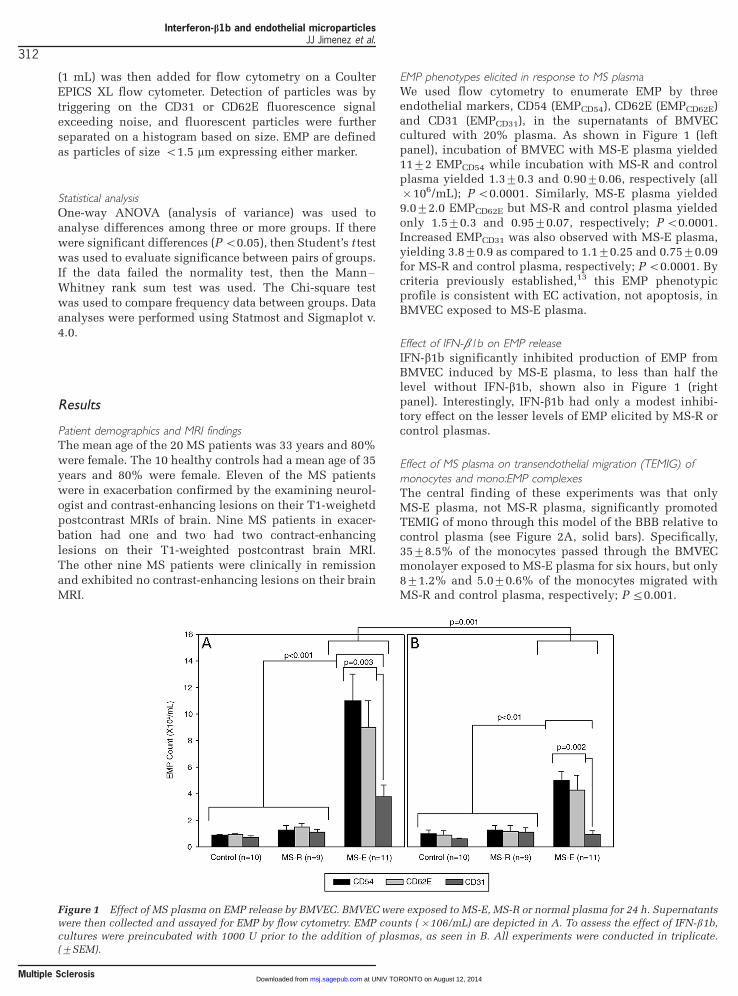
EMP phenotypes elicited in response to MS plasmaWe used flow cytometry to enumerate EMP by threeendothelial markers, CD54 (EMPCD54), CD62E (EMPCD62E)and CD31 (EMPCD31), in the supernatants of BMVECcultured with 20% plasma. As shown in Figure 1 (leftpanel), incubation of BMVEC with MS-E plasma yielded119/2 EMPCD54 while incubation with MS-R and controlplasma yielded 1.39/0.3 and 0.909/0.06, respectively (all�/106/mL); P B/0.0001. Similarly, MS-E plasma yielded9.09/2.0 EMPCD62E but MS-R and control plasma yieldedonly 1.59/0.3 and 0.959/0.07, respectively; P B/0.0001.Increased EMPCD31 was also observed with MS-E plasma,yielding 3.89/0.9 as compared to 1.19/0.25 and 0.759/0.09for MS-R and control plasma, respectively; P B/0.0001. Bycriteria previously established,13 this EMP phenotypicprofile is consistent with EC activation, not apoptosis, inBMVEC exposed to MS-E plasma.
Effect of IFN-b1b on EMP releaseIFN-b1b significantly inhibited production of EMP fromBMVEC induced by MS-E plasma, to less than half thelevel without IFN-b1b, shown also in Figure 1 (rightpanel). Interestingly, IFN-b1b had only a modest inhibi-tory effect on the lesser levels of EMP elicited by MS-R orcontrol plasmas.
Effect of MS plasma on transendothelial migration (TEMIG) of
monocytes and mono:EMP complexesThe central finding of these experiments was that onlyMS-E plasma, not MS-R plasma, significantly promotedTEMIG of mono through this model of the BBB relative tocontrol plasma (see Figure 2A, solid bars). Specifically,359/8.5% of the monocytes passed through the BMVECmonolayer exposed to MS-E plasma for six hours, but only89/1.2% and 5.09/0.6% of the monocytes migrated withMS-R and control plasma, respectively; P 5/0.001.
Figure 1 Effect of MS plasma on EMP release by BMVEC. BMVEC were exposed to MS-E, MS-R or normal plasma for 24 h. Supernatants
were then collected and assayed for EMP by flow cytometry. EMP counts (�/106/mL) are depicted in A. To assess the effect of IFN-b1b,cultures were preincubated with 1000 U prior to the addition of plasmas, as seen in B. All experiments were conducted in triplicate.
(9/SEM).
Interferon-b1b and endothelial microparticlesJJ Jimenez et al.
312
Multiple Sclerosis at UNIV TORONTO on August 12, 2014msj.sagepub.comDownloaded from
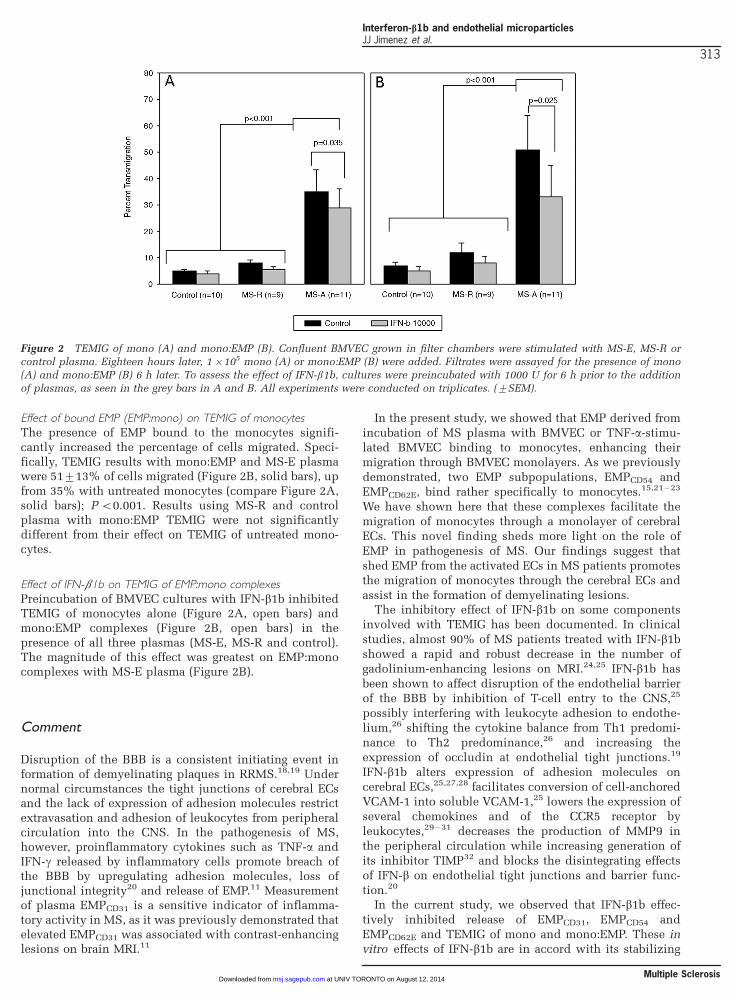
Effect of bound EMP (EMP:mono) on TEMIG of monocytesThe presence of EMP bound to the monocytes signifi-cantly increased the percentage of cells migrated. Speci-fically, TEMIG results with mono:EMP and MS-E plasmawere 519/13% of cells migrated (Figure 2B, solid bars), upfrom 35% with untreated monocytes (compare Figure 2A,solid bars); P B/0.001. Results using MS-R and controlplasma with mono:EMP TEMIG were not significantlydifferent from their effect on TEMIG of untreated mono-cytes.
Effect of IFN-b1b on TEMIG of EMP:mono complexes
Preincubation of BMVEC cultures with IFN-b1b inhibitedTEMIG of monocytes alone (Figure 2A, open bars) andmono:EMP complexes (Figure 2B, open bars) in thepresence of all three plasmas (MS-E, MS-R and control).The magnitude of this effect was greatest on EMP:monocomplexes with MS-E plasma (Figure 2B).
Comment
Disruption of the BBB is a consistent initiating event information of demyelinating plaques in RRMS.18,19 Undernormal circumstances the tight junctions of cerebral ECsand the lack of expression of adhesion molecules restrictextravasation and adhesion of leukocytes from peripheralcirculation into the CNS. In the pathogenesis of MS,however, proinflammatory cytokines such as TNF-a andIFN-g released by inflammatory cells promote breach ofthe BBB by upregulating adhesion molecules, loss ofjunctional integrity20 and release of EMP.11 Measurementof plasma EMPCD31 is a sensitive indicator of inflamma-tory activity in MS, as it was previously demonstrated thatelevated EMPCD31 was associated with contrast-enhancinglesions on brain MRI.11
In the present study, we showed that EMP derived from
incubation of MS plasma with BMVEC or TNF-a-stimu-lated BMVEC binding to monocytes, enhancing their
migration through BMVEC monolayers. As we previously
demonstrated, two EMP subpopulations, EMPCD54 and
EMPCD62E, bind rather specifically to monocytes.15,21 �23
We have shown here that these complexes facilitate themigration of monocytes through a monolayer of cerebral
ECs. This novel finding sheds more light on the role of
EMP in pathogenesis of MS. Our findings suggest that
shed EMP from the activated ECs in MS patients promotesthe migration of monocytes through the cerebral ECs and
assist in the formation of demyelinating lesions.The inhibitory effect of IFN-b1b on some components
involved with TEMIG has been documented. In clinical
studies, almost 90% of MS patients treated with IFN-b1b
showed a rapid and robust decrease in the number ofgadolinium-enhancing lesions on MRI.24,25 IFN-b1b has
been shown to affect disruption of the endothelial barrier
of the BBB by inhibition of T-cell entry to the CNS,25
possibly interfering with leukocyte adhesion to endothe-lium,26 shifting the cytokine balance from Th1 predomi-
nance to Th2 predominance,26 and increasing the
expression of occludin at endothelial tight junctions.19
IFN-b1b alters expression of adhesion molecules oncerebral ECs,25,27,28 facilitates conversion of cell-anchored
VCAM-1 into soluble VCAM-1,25 lowers the expression of
several chemokines and of the CCR5 receptor by
leukocytes,29 �31 decreases the production of MMP9 inthe peripheral circulation while increasing generation of
its inhibitor TIMP32 and blocks the disintegrating effects
of IFN-b on endothelial tight junctions and barrier func-
tion.20
In the current study, we observed that IFN-b1b effec-
tively inhibited release of EMPCD31, EMPCD54 and
EMPCD62E and TEMIG of mono and mono:EMP. These invitro effects of IFN-b1b are in accord with its stabilizing
Figure 2 TEMIG of mono (A) and mono:EMP (B). Confluent BMVEC grown in filter chambers were stimulated with MS-E, MS-R orcontrol plasma. Eighteen hours later, 1�/105 mono (A) or mono:EMP (B) were added. Filtrates were assayed for the presence of mono
(A) and mono:EMP (B) 6 h later. To assess the effect of IFN-b1b, cultures were preincubated with 1000 U for 6 h prior to the addition
of plasmas, as seen in the grey bars in A and B. All experiments were conducted on triplicates. (9/SEM).
Interferon-b1b and endothelial microparticlesJJ Jimenez et al.
313
Multiple Sclerosis at UNIV TORONTO on August 12, 2014msj.sagepub.comDownloaded from

effects20,25,28 on endothelial integrity and activity. Inaddition, Corsini et al ., using an in vitro model of humanumbilical vein endothelial cell monolayers, demonstratedthat treatment of MS patients with IFN-b1b induced areduction in TEMIG of monocytes through ECs.9 Theinvestigators attributed their finding to a decrease in theserum levels of MMP-9. It is known that under stress,expression of MMP-9 by cerebral ECs is increased33 and itis possible that in our in vitro model, the expression ofMMP-9 by BMVEC was inhibited by the IFN-b1b. Alter-natively, IFN-b1b may directly interact with monocytesand impair the formation of monocyte:EMP complexes. Ithas been shown that increased migration of monocytesacross a fibronectin matrix in response to the chemokinesRANTES, MIP-1 and MCP-1 is antagonized by IFN-b1b.34
MCP-1 treatment of monocytes elevates their mRNA leveland secretion of a matrix degrading enzyme, MMP-9,which is abrogated by IFN-b1b.
However, our findings suggest a novel protective me-chanism for IFN-b1b in patients with MS. This inhibitoryeffect of IFN-b1b on TEMIG of monocytes and monocy-te:EMP complexes may be explained by our findings. Wepreviously demonstrated that treatment of MS patientswith IFN-b1a reduces plasma concentrations ofEMPCD31.35 It is possible that as demonstrated in vitro ,IFN-b1b reduces EMP release, thereby leading to de-creased formation of EMP�/monocyte complexes and alowered rate of TEMIG of these cells in vivo .
In summary, our findings suggest a novel effect ofplasma EMP in MS patients, namely that these particlesmay enhance monocyte migration through the endothelialmonolayer in vivo . The enhancing effect is furtherinhibited by IFN-b1b; our TEMIG model demonstrates anovel mechanism for its beneficial effects in patients withRRMS. Further studies of EMP in the context of MS canhighlight more details of TEMIG of activated leukocytesthrough the cerebral EC into the CNS.
AcknowledgementsThis work was supported by the Wallace H CoulterFoundation and Funds from Mary Beth Weiss, Jane andCharles Bosco, and gifts from Frank Smather.
References
1 Noseworthy JH, Lucchinetti C, Rodriguez M, Weinshenker BG.
Multiple sclerosis. N Engl J Med 2000; 343: 938�/52.
2 Steinman L. Multiple sclerosis: a coordinated immunological
attack against myelin in the central nervous system. Cell 1996;
85: 299�/302.
3 Brosnan CF, Raine CS. Mechanisms of immune injury in
multiple sclerosis. Brain Pathol 1996; 6: 243�/57.
4 Prat A, Biernacki K, Lavoie JF, Poireir J, Duquette P, Antel JP.
Migration of multiple sclerosis lymphocytes through brain
endothelium. Arch Neurol 2002; 59: 391�/97.
5 Hartung HP, Reiners K, Archelos JJ, Michels M, Seeldrayers P,
Heidenreich F et al . Circulating adhesion molecules and
tumor necrosis factor receptor in multiple sclerosis: correla-
tion with magnetic resonance imaging. Ann Neurol 1995; 38:
186�/93.
6 Leppert D, Lindberg RL, Kappos L, Leib SL. Matrix metallo-
proteinases: multifunctional effectors of inflammation in
multiple sclerosis and bacterial meningitis. Brain Res Brain
Res Rev 2001; 36: 249�/57.
7 The IFNb Multiple Sclerosis Study Group. Interferon beta-1b
is effective in relapsing�/remitting multiple sclerosis. I. Clin-
ical results of a multicenter, randomized, double-blind,
placebo-controlled trial. The IFNB Multiple Sclerosis Study
Group. Neurology 1993; 43: 655�/61.
8 Uhm JH, Dooley NP, Stuve O, Francis GS, Duquette P, Antel JP
et al . Migratory behavior of lymphocytes isolated from multi-
ple sclerosis patients: effects of interferon beta-1b therapy.
Ann Neurol 1999; 46: 319�/24.
9 Corsini E, Gelati M, Dufour A, Ciusani E, La Mantia L, Massa G
et al . Reduction of transendothelial migration of mononuclear
cells in interferon-beta1b-treated multiple sclerosis patients.
Ann Neurol 1999; 46: 435.
10 Losy J, Niezgoda A, Wender M. Increased serum levels of
soluble PECAM-1 in multiple sclerosis patients with brain
gadolinium-enhancing lesions. J Neuroimmunol 1999; 99:
169�/72.
11 Minagar A, Jy W, Jimenez JJ, Sheremata WA, Mauro LM, Mao
WW et al . Elevated plasma endothelial microparticles in
multiple sclerosis. Neurology 2001; 56: 1319�/24.
12 Combes V, Simon AC, Grau GE, Arnoux D, Camoin L, Sabatier
F et al . In vitro generation of endothelial microparticles and
possible prothrombotic activity in patients with lupus antic-
oagulant. J Clin Invest 1999; 104: 93�/102.
13 Jimenez JJ, Jy W, Mauro LM, Soderland C, Horstman LL, Ahn
YS. Endothelial cells release phenotypically and quantita-
tively distinct microparticles in activation and apoptosis.
Thromb Res 2003; 109: 175�/80.
14 Sabatier F, Roux V, Anfosso F, Camoin L, Sampol J, Dignat-
George F. Interaction of endothelial microparticles with
monocytic cells in vitro induces tissue factor-dependent
procoagulant activity. Blood 2002; 99: 3962�/70.
15 Jy W, Minagar A, Jimenez JJ, Sheremata WA, Mauro LM,
Horstman LL et al . Endothelial microparticles (EMP) bind and
activate monocytes: elevated EMP�/monocyte conjugates in
multiple sclerosis. Front Biosci 2004; 9: 3137�/44.
16 Poser CM, Paty DW, Scheinberg L, McDonald WI, Davis FA,
Ebers GC et al . New diagnostic criteria for multiple sclerosis:
guidelines for research protocols. Ann Neurol 1983; 13: 227�/
31.
17 Jimenez JJ, Jy W, Mauro LM, Horstman LL, Ahn YS. Elevated
endothelial microparticles in thrombotic thrombocytopenic
purpura: findings from brain and renal microvascular cell
culture and patients with active disease. Br J Haematol 2001;
112: 81�/90.
18 Kermode AG, Thompson AJ, Tofts P, MacManus DG, Kendall
BE, Kingsley DP et al. Breakdown of the blood�/brain barrier
precedes symptoms and other MRI signs of new lesions in
multiple sclerosis. Pathogenetic and clinical implications.
Brain 1990; 113: 1477�/89.
19 Minagar A, Ostanin D, Long AC, Jennings M, Kelley RE, Sasaki
M et al . Serum from patients with multiple sclerosis down-
regulates occludin and VE-cadherin expression in cultured
endothelial cells. Mult Scler 2003; 9: 235�/38.
20 Minagar A, Long A, Ma T, Jackson TH, Kelley RE, Ostanin DV
et al . Interferon (IFN)-beta 1a and IFN-beta 1b block IFN-
gamma-induced disintegration of endothelial junction integ-
rity and barrier. Endothelium 2003; 10: 299�/307.
21 Jy W, Jimenez JJ, Minagar A, Mauro L, Horstmann LL, Bidot CJ
et al . Endothelial microparticles (EMP) enhance adhesion and
Interferon-b1b and endothelial microparticlesJJ Jimenez et al.
314
Multiple Sclerosis at UNIV TORONTO on August 12, 2014msj.sagepub.comDownloaded from

transmigration of monocytes: EMP�/monocyte conjugates as a
marker of disease activity in multiple sclerosis (MS). Blood
2002; 100: 460a.
22 Jy W, Jimenez JJ, Mauro L, Horstmann LL, Bernal-Mizrachi LL,
Minagar A et al . Interaction of endothelial microparticles
(EMP) with leukocytes: potential roles of EMP in thrombosis
and inflammation. Blood 2001; 98: 226a.
23 Horstman LL, Jy W, Jimenez JJ, Ahn YS. Endothelial micro-
particles as markers of endothelial dysfunction. Front Biosci
2004; 9: 1118�/35.
24 Stone LA, Frank JA, Albert PS, Bash C, Smith ME, Maloni H et
al . The effect of interferon-beta on blood�/brain barrier
disruptions demonstrated by contrast-enhanced magnetic
resonance imaging in relapsing�/remitting multiple sclerosis.
Ann Neurol 1995; 37: 611�/19.
25 Calabresi PA, Tranquill LR, Dambrosia JM, Stone LA, Maloni
H, Bash CN et al . Increases in soluble VCAM-1 correlate with
a decrease in MRI lesions in multiple sclerosis treated with
interferon beta-1b. Ann Neurol 1997; 41: 669�/74.
26 Dhib-Jalbut S, Jiang H, Williams GJ. The effect of interferon
beta-1b on lymphocyte-endothelial cell adhesion. J Neuroim-
munol 1996; 71: 215�/22.
27 Floris S, Ruuls SR, Wierinckx A, van der Pol SM, Dopp E, van
der Meide PH et al . Interferon-beta directly influences mono-
cyte infiltration into the central nervous system. J Neuroim-
munol 2002; 127: 69�/79.
28 Corsini E, Gelati M, Dufour A, Massa G, Nespolo A, Ciusani E
et al . Effects of beta-IFN-1b treatment in MS patients on
adhesion between PBMNCs, HUVECs and MS-HBECs: an in
vivo and in vitro study. J Neuroimmunol 1997; 79: 76�/83.
29 Sorensen TL, Roed H, Sellebjerg F. Chemokine receptor
expression on B cells and effect of interferon-beta in multiple
sclerosis. J Neuroimmunol 2002; 122: 125�/31.
30 Iarlori C, Reale M, De Luca G, Di Iorio A, Feliciani C, Tulli A et
al . Interferon beta-1b modulates MCP-1 expression and
production in relapsing�/remitting multiple sclerosis. J Neu-
roimmunol 2002; 123: 170�/79.
31 Zang YC, Halder JB, Samanta AK, Samanta AK, Hong J, Rivera
VM, Zhang JZ. Regulation of chemokine receptor CCR5 and
production of RANTES and MIP-1alpha by interferon-beta. JNeuroimmunol 2001; 112: 174�/80.
32 Waubant E, Goodkin D, Bostrom A, Bacchetti P, Hietpas J,
Lindberg R et al IFNbeta lowers MMP-9/TIMP-1 ratio, which
predicts new enhancing lesions in patients with SPMS.
Neurology 2003; 60: 52�/57.
33 Romanic AM, White RF, Arleth AJ, Ohlstein EH, Barone FC.
Matrix metalloproteinase expression increases after cerebral
focal ischemia in rats: inhibition of matrix metalloproteinase-9
reduces infarct size. Stroke 1998; 29: 1020�/30.
34 Stuve O, Chabot S, Jung SS, Williams G, Yong VW. Chemo-
kine-enhanced migration of human peripheral blood mono-
nuclear cells is antagonized by interferon beta-1b through an
effect on matrix metalloproteinase-9. J Neuroimmunol 1997;
80: 38�/46.
35 Jy W, Delgado S, Minagar A, Ahn Y, Sheremata W. Interferon-
b1a reduces plasma CD31�/ endothelial microparticles in MS.
Ann Neurol 2003; 54: S60.
Interferon-b1b and endothelial microparticlesJJ Jimenez et al.
315
Multiple Sclerosis





























