Embed Size (px)
Citation preview

2013; doi: 10.1101/cshperspect.a013847Cold Spring Harb Perspect Med Thomas Brade, Luna S. Pane, Alessandra Moretti, Kenneth R. Chien and Karl-Ludwig Laugwitz Embryonic Heart Progenitors and Cardiogenesis
Subject Collection The Biology of Heart Disease
The Genetic Basis of Aortic AneurysmMark E. Lindsay and Harry C. Dietz
Cardiac Cell Lineages that Form the Heart
Blanpain, et al.Sigolène M. Meilhac, Fabienne Lescroart, Cédric
Discovery and Development ParadigmRegenerative Medicine: Transforming the Drug
Sotirios K. KarathanasisCongenital Heart DiseaseComplex Genetics and the Etiology of Human
Bruce D. Gelb and Wendy K. Chung
from a Research-Based Pharmaceutical CompanyCardiovascular Drug Discovery: A Perspective
G. Gromo, J. Mann and J.D. Fitzgerald
Myocardial Tissue Engineering: In Vitro Models
and Christine MummeryGordana Vunjak Novakovic, Thomas Eschenhagen
Genetics and Disease of Ventricular Muscle
G. SeidmanDiane Fatkin, Christine E. Seidman and Jonathan Disease
Pluripotent Stem Cell Models of Human Heart
Dorn, et al.Alessandra Moretti, Karl-Ludwig Laugwitz, Tatjana
Embryonic Heart Progenitors and Cardiogenesis
et al.Thomas Brade, Luna S. Pane, Alessandra Moretti,
MedicineDirection for Cardiovascular Regenerative Direct Cardiomyocyte Reprogramming: A New
Kenneth R. ChienB. Alexander Yi, Christine L. Mummery and
BiologyModel Systems for Cardiovascular Regenerative
Richard T. LeeJessica C. Garbern, Christine L. Mummery and
http://perspectivesinmedicine.cshlp.org/cgi/collection/ For additional articles in this collection, see
Copyright © 2013 Cold Spring Harbor Laboratory Press; all rights reserved
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

Embryonic Heart Progenitors and Cardiogenesis
Thomas Brade1,5, Luna S. Pane1,5, Alessandra Moretti1,2, Kenneth R. Chien3,4,and Karl-Ludwig Laugwitz1,2
1Klinikum rechts der Isar, Technische Universitat Munchen, I. Medical Department, Cardiology, 81675 Munich,Germany
2DZHK (German Centre for Cardiovascular Research), Munich Heart Alliance, Munich, Germany3Karolinska Institutet, Department of Cell and Molecular Biology and Medicine, Berzelius Vag, Stockholm,Sweden
4Harvard University, Department of Stem Cell and Regenerative Biology, Cambridge, Massachusetts 02138
Correspondence: [email protected]
The mammalian heart is a highly specialized organ, comprised of many different cell typesarising from distinct embryonic progenitor populations during cardiogenesis. Three precur-sor populations have been identified to contribute to different myocytic and nonmyocyticcell lineages of the heart: cardiogenic mesoderm cells (CMC), the proepicardium (PE), andcardiac neural crest cells (CNCCs). This review will focus on molecular cues necessary forproper induction, expansion, and lineage-specific differentiation of these progenitor popu-lations during cardiac development in vivo. Moreover, we will briefly discuss how theknowledge gained on embryonic heart progenitor biology can be used to develop noveltherapeutic strategies for the management of congenital heart disease as well as for improve-ment of cardiac function in ischemic heart disease.
The functional heart is comprised of diversemuscle and nonmuscle cell lineages: atrial
and ventricular cardiomyocytes, endocardialcells, valvular components, and connective tis-sues, conduction system cells, as well as smoothmuscle and endothelial cells of the coronary ar-teries and veins. Understanding the molecularmechanisms that drive the specification of thesecell lineages from naıve progenitor populationsinto terminally differentiated cell types withinthe four-chambered embryonic heart is of fun-damental importance to elucidate the patholog-ical mechanisms underlying congenital heart
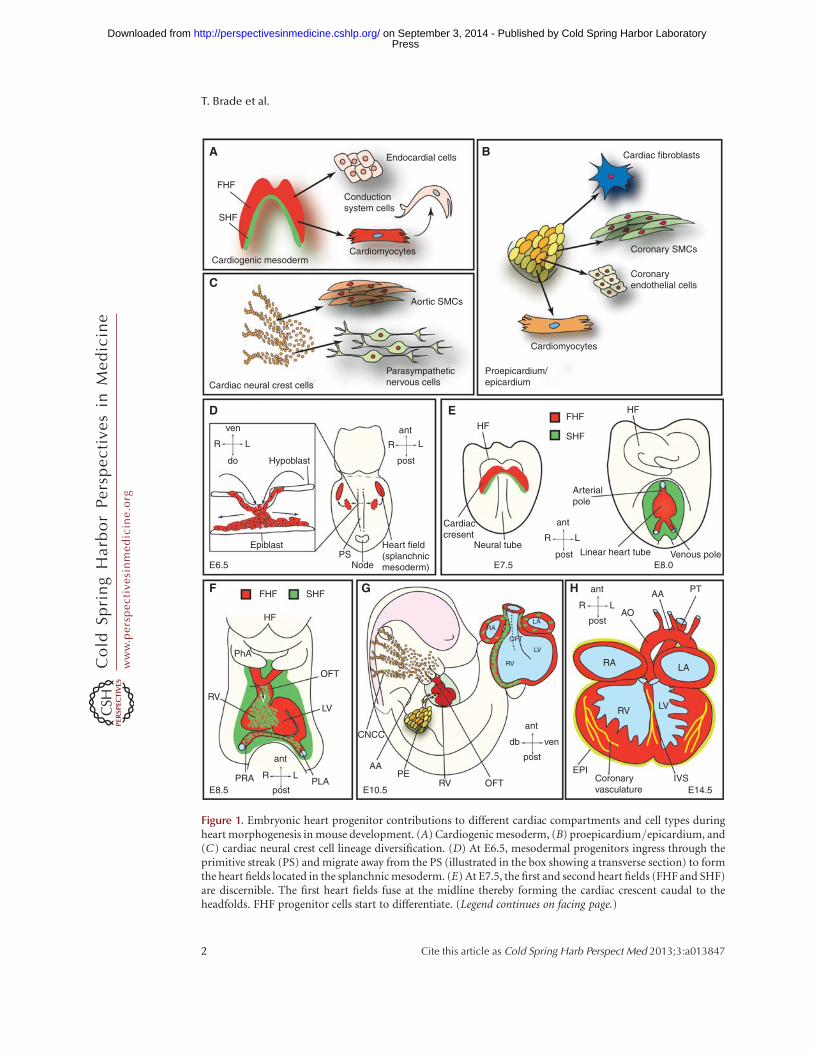
disease and to improve management of ische-mic heart disease (Olson 2004; Srivastava 2006).Three spatiallyandtemporallydistinct sourcesofheart cell precursors have been identified in theembryo: cardiogenic mesoderm cells (CMCs),the proepicardium (PE), and cardiac neural crestcells (CNCCs) (Fig. 1A–C).
The cardiogenic mesoderm, which harborsthe so-called first and second heart fields (FHFand SHF), forms the major proportion of theventricular, atrial, and outflow tract (OFT) myo-cardium. Additionally, these progenitors con-tribute cells to the endocardium, the conduction
5These authors contributed equally to this work.
Editors: Margaret Buckingham, Christine L. Mummery, and Kenneth R. Chien
Additional Perspectives on The Biology of Heart Disease available at www.perspectivesinmedicine.org
Copyright # 2013 Cold Spring Harbor Laboratory Press; all rights reserved; doi: 10.1101/cshperspect.a013847
Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847
1
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from
Conductionsystem cells
Cardiomyocytes
Aortic SMCs
Parasympatheticnervous cellsCardiac neural crest cells
Endocardial cells Cardiac fibroblasts
Coronary SMCs
Coronaryendothelial cells
Cardiomyocytes
ven
L
ant HFFHF
Proepicardium/epicardium
FHF SHF
HF
PhA
RV
OFT
CNCC
RA
RV
LV
OFT
LA
AAPE
RV OFT
LV
PRA PLAE8.5 E10.5
HF
Arterialpole
ant
L
Linear heart tubeE8.0
AO
AA PT
RA LA
RV
EPICoronaryvasculature
IVS
LV
E7.5
Neural tube
Cardiaccresent
Venous pole
SHFR
do
LR
R
post
post
ant
LR
post
ant
LR
post
ant
vendb
post
PSNode
Heart field(splanchnicmesoderm)
Hypoblast
Epiblast
E6.5
A
FHF
SHF
Cardiogenic mesoderm
C
D
F G H
E
B
E14.5
Figure 1. Embryonic heart progenitor contributions to different cardiac compartments and cell types duringheart morphogenesis in mouse development. (A) Cardiogenic mesoderm, (B) proepicardium/epicardium, and(C) cardiac neural crest cell lineage diversification. (D) At E6.5, mesodermal progenitors ingress through theprimitive streak (PS) and migrate away from the PS (illustrated in the box showing a transverse section) to formthe heart fields located in the splanchnic mesoderm. (E) At E7.5, the first and second heart fields (FHF and SHF)are discernible. The first heart fields fuse at the midline thereby forming the cardiac crescent caudal to theheadfolds. FHF progenitor cells start to differentiate. (Legend continues on facing page.)
T. Brade et al.
2 Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

system, and the aortic and pulmonary cushions(Fig. 1A).
Progenitors originating from the PE com-prise the epicardium and differentiate into inter-stitial fibroblasts embedded in the myocardium,vascular smooth muscle cells and endothelialcells of the coronary vessels, and some myocytes,mainly in the atrioventricular septum. Addi-tionally, interaction of the epicardium withthe underlying myocardium is crucial for cham-ber maturation and ventricular muscle growth(Fig. 1B).
Finally, the CNC progenitors give rise to thedistal smooth muscle cells of the OFT and theaorticopulmonary ridge as well as to the auton-omous nervous system of the heart. Cardiacvalve development and septation of the heart isalso critically dependent on proper CNC devel-opment (Fig. 1C).
Here we will describe “the lives” of these dif-ferent cardiac progenitors, from their embryon-ic specification to their differentiation into ma-ture cells of the heart, and we will discuss howthis knowledge could inform the future devel-opment of novel therapies for cardiovascularheart disease.
THE CARDIOGENIC MESODERMPROGENITORS
Cardiogenic Mesoderm Progenitors in EarlyVertebrate Cardiogenesis
The embryonic heart is the first organ to func-tion during development. Fate mapping studiesin mouse and chicken embryos have shown thattime and location of cell ingression through theprimitive streak (PS) (Fig. 1D section detail inbox) determines the fate of gastrulating cells.
Consequently, cardiac mesoderm progenitorsare among the first cells to ingress through thePS during gastrulation. These naıve cardiogenicmesoderm cells migrate away from the PS andcoalesce in an anterior lateral region relative tothe streak, the so-called splanchnic mesoderm(Fig. 1D), where they form the cardiac crescent(at mouse embryonic day E7.5, correspondingroughly to week 2 of human gestation); at thisstage, their commitment to a cardiac fate re-mains plastic (Fig. 1E) (Buckingham et al.2005). At E8, or 3 weeks in human gestation,the cardiac crescent fuses at the midline andgives rise to the FHF-derived linear heart tube(Fig. 1E), which subsequently commences beat-ing and undergoes rightward looping and rapidgrowth (mouse E8.5, 4 weeks in human devel-opment) (Fig. 1F) (Zaffran et al. 2004). As de-velopment proceeds, the linear heart tube ex-pands, essentially by two mechanisms: (i) cellproliferation, and (ii) recruitment of additionalcells. These additional cells, which are added tothe arterial and venous poles of the linear hearttube, originate from the pharyngeal mesodermlocated caudal to the pharynx, and represent theSHF (Mjaatvedt et al. 2001; Waldo et al. 2001;Buckingham et al. 2005; Kelly 2012; Moormanet al. 2013). SHF progenitors mainly contributeto the OFT, the right ventricle, and a large por-tion of the inflow region (atria), whereas the leftventricle mainly derives from the FHF (Fig. 1F).The septated four-chambered fetal heart is sub-sequently generated involving intricate interac-tions of cardiomyocytes with epicardial and en-docardial cells as well as CCNCs. At day E10.5 inmice (day 32 in human) the developing heartshows well-defined chambers (Fig. 1G), whichare fully septated and connected to the pulmo-
Figure 1. (Continued) At E8.0, the cardiac crescent forms the beating, linear heart tube. SHF progenitors willgradually migrate into the linear heart tube and differentiate then. (F) At E8.5, the linear heart tube undergoesrightward looping. (G) At E10.5, cardiac neural crest and proepicardial cells contribute to the heart, whichalready shows a defined four-chamber morphology. (H ) At E14.5, the heart shows four fully septated chambersand a septated outflow tract connected to the pulmonary trunk and the dorsal aorta. AA, Aortic arch; ant,anterior; AO, dorsal aorta; CNCC, cardiac neural crest cells; do, dorsal; EPI, epicardium; FHF, first heart field;HF, headfolds; IVS, interventricular septum; L, left; LA, left atrium; LV, left ventricle; OFT, outflow tract; PE,proepicardium; PhA, pharyngeal arch; PLA, primitive left atrium; post, posterior; PRA, primitive right atrium;PS, primitive streak; PT, pulmonary trunk; R, right; RA, right atrium; RV, right ventricle; SHF, second heart field;SMCs, smooth muscle cells; ven, ventral.
Embryonic Heart Progenitors and Cardiogenesis
Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847 3
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

nary trunk and aorta at E14.5 (�7 weeks in hu-man gestation) (Fig. 1H).
From Mesoderm Induction to CardiogenicMesoderm Progenitors
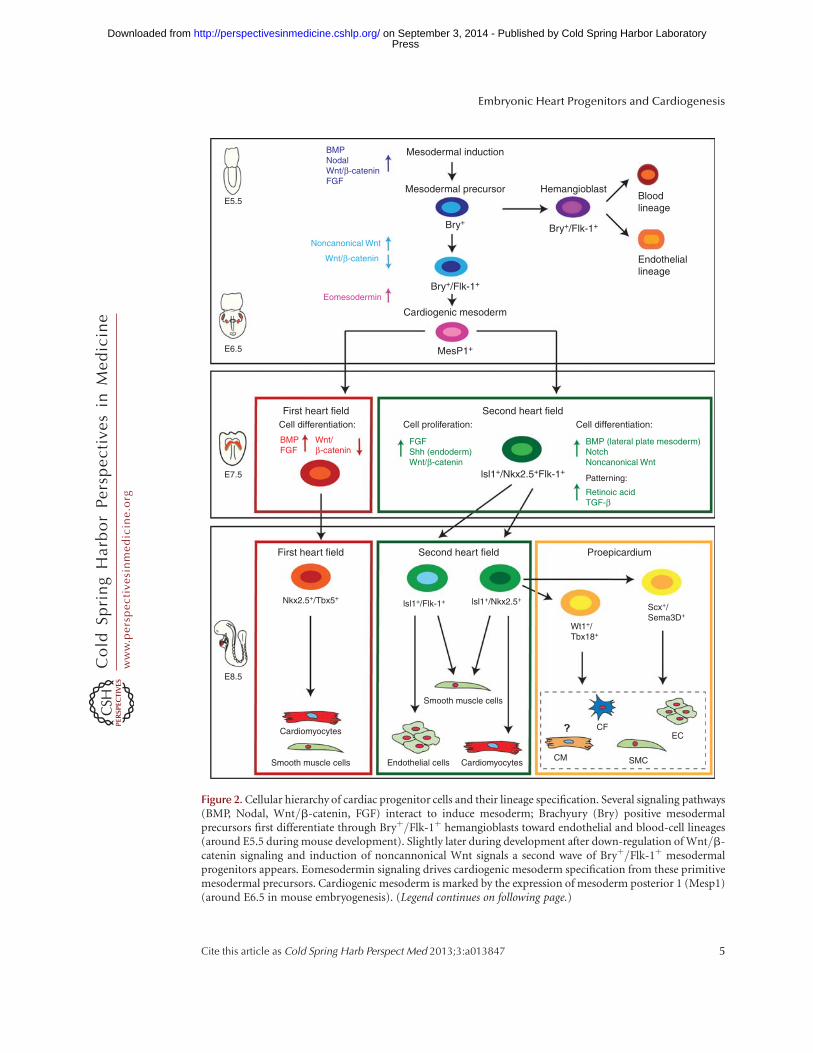
Mesoderm induction, a quintessential prerequi-site for heart development, is evolutionarilyconserved and regulated by numerous signalingpathways. Key players are Nodal and bone mor-phogenetic protein (BMP) signals as well as Wntand fibroblast growth factors (FGF) (Kimelman2006; Noseda et al. 2011). Expression of theT-box transcription factor Brachyury/T (Bry),a direct target gene of Wnt/b-catenin signal-ing, marks mesodermal cells ingressing throughthe PS (Showell et al. 2004). Commitment ofnascent Bryþ mesodermal progenitors towarda cardiogenic fate requires inhibition of canoni-cal Wnt/b-catenin signaling and activation ofnoncanonical Wnt signaling (Gessert and Kuhl2010). Moreover, intricate interactions betweenthe mesodermal germ layer and the endodermare additionally necessary for the induction of acommon primordial cardiovascular progenitorpopulation from Bryþ mesodermal precursors,which latergiverise tobothfirstandsecondheartfield progenitors (Figs. 1 and 2) (Abu-Issa et al.2004; Buckingham et al. 2005). Tracing cardio-genic fate back to a Bryþ population sparked theidea of a clonal model of heart lineage diversifi-cation analogous to hematopoiesis, in which asingle hematopoietic stem cell can generate allblood-cell lineages (Weissman 2000). Accordingto this hierarchy model, diverse muscle andnonmuscle lineages arise from a single-cell-level decision of multipotent, primordial car-diovascular stem cells, which in turn give riseto downstream intermediates representing tis-sue-restricted precursors of fully differentiatedheart cells (Fig. 2). First, data supporting thishypothesis have been gained by lineage-trac-ing studies, and marker gene expression analysisin mice showing that the endocardial endotheli-um and the myocytic lineage develop from anintermediate mesodermal population that ex-presses vascularendothelial growth factor recep-tor 2 (VEGFR2, Flk-1, KDR) (Motoike et al.2003; Ema et al. 2006). These initial findings
were corroborated by additional in vivo and invitro studies. During development as well as dur-ing in vitro differentiation of mouse and humanembryonic stem cells, two distinct Bryþ cell pop-ulations arise (Fig. 2). First, a Bryþ/Flk-1þ
hemangioblast precursor population contrib-utes cells to the hematopoietic system and thedeveloping vasculature. During subsequent de-velopment, Bryþ/Flk-12 mesodermal progeni-tors initiate a new wave of Flk-1 expression andconstitute cardiovascular progenitors (Fig. 2)(Kouskoff et al. 2005; Kattman et al. 2006; Yanget al. 2008). Later, this primitive Bryþ/Flk-1þ
cardiogenic mesoderm population down-regu-lates Bry and activates, upon T-box transcrip-tion factor Eomesodermin action, expressionof the mesoderm posterior 1 (Mesp1) gene(Fig. 2) (Bondue and Blanpain 2010; Costelloet al. 2011). Mesp1þ mesodermal progenitorscontribute to paraxial mesoderm and skeletalmuscle of the head as well as cardiac muscle(Saga et al. 2000; Scott 2012). By turning onthe core cardiogenic transcriptional network,Mesp1 has been shown to further restrict theseprimitive mesodermal precursors toward a car-diovascular and anterior mesodermal fate (Bon-due et al. 2008; Martin-Puig et al. 2008; Bondueand Blanpain 2010).
Differentiation of Early CardiogenicMesoderm Progenitors—Formation of theCardiac Crescent and the Linear Heart Tube
After ingression through the PS, cardiogenicprogenitor cells migrate to an anterior lateralposition caudal to the headfolds and form theso-called cardiac crescent. At this time in devel-opment, the first and second heart fields canbe distinguished (Moorman et al. 2013). Al-though the FHF progenitors already differenti-ate at this stage, cells within the SHF remain in aproliferative, undifferentiated progenitor stateuntil they enter the heart tube at a later timepoint (Kelly 2012). In fact, because of their po-sition in the crescent (more anterior and lateralin respect to SHF progenitors) (Fig. 1E), FHFprogenitor cells are exposed to cytokines of theBMP (Schultheiss et al. 1997) and FGF (Reiferset al. 2000) families as well as to inhibitors of the
T. Brade et al.
4 Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from
BMPNodalWnt/β-cateninFGF
E5.5
E6.5
Mesodermal induction
Mesodermal precursor
Cardiogenic mesoderm
First heart field
First heart field Second heart field Proepicardium
Second heart field
FGFShh (endoderm)Wnt/β-catenin
Cell differentiation: Cell differentiation:Cell proliferation:
BMPFGF
E7.5
E8.5
Wnt/β-catenin
MesP1+
Eomesodermin
Hemangioblast
Endotheliallineage
Bry+
Noncanonical Wnt
Wnt/β-catenin
Bry+/Flk-1+
Bloodlineage
Bry+/Flk-1+
lsl1+/Nkx2.5+Flk-1+
Nkx2.5+/Tbx5+lsl1+/Flk-1+ lsl1+/Nkx2.5+
Scx+/Sema3D+
Wt1+/Tbx18+
CM SMC
ECCF
Smooth muscle cells
Endothelial cells Cardiomyocytes
Cardiomyocytes
Smooth muscle cells
BMP (lateral plate mesoderm)NotchNoncanonical Wnt
Patterning:
Retinoic acidTGF-β
Figure 2. Cellular hierarchy of cardiac progenitor cells and their lineage specification. Several signaling pathways(BMP, Nodal, Wnt/b-catenin, FGF) interact to induce mesoderm; Brachyury (Bry) positive mesodermalprecursors first differentiate through Bryþ/Flk-1þ hemangioblasts toward endothelial and blood-cell lineages(around E5.5 during mouse development). Slightly later during development after down-regulation of Wnt/b-catenin signaling and induction of noncannonical Wnt signals a second wave of Bryþ/Flk-1þ mesodermalprogenitors appears. Eomesodermin signaling drives cardiogenic mesoderm specification from these primitivemesodermal precursors. Cardiogenic mesoderm is marked by the expression of mesoderm posterior 1 (Mesp1)(around E6.5 in mouse embryogenesis). (Legend continues on following page.)
Embryonic Heart Progenitors and Cardiogenesis
Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847 5
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

Wnt pathway (Marvin et al. 2001; Schneider andMercola 2001; Tzahor and Lassar 2001; Nosedaet al. 2011), resulting in the onset of cardiacdifferentiation marked by the expression of keyregulators of the lineage, namely, Nkx2.5 (Lintset al. 1993), Gata-4 (Arceci et al. 1993; Kelley etal. 1993; Heikinheimo et al. 1994; Zeisberg et al.2005), and Tbx5 (Harvey 2002). Myocytic line-age commitment is associated with expression ofcontractile proteins including myosin lightchain-2a (MLC2a) (Kubalaket al. 1994) and sar-comeric myosin heavy chain (MHC) first in thecardiac crescent and then throughout the linearheart tube.
Although no genes uniquely expressed inearly FHF progenitors have been identified yet,SHF precursors are marked by the LIM-homeo-domain transcription factor Isl-1. Although sev-eral studies suggest Isl-1 to be a very early pan-cardiac marker, after formation of the linearheart tube, Isl-1 clearly demarcates the SHFand its expression is absent in differentiatedFHF derivatives (Prall et al. 2007; Kelly 2012).Isl-1 expression is dependent on canonical Wntsignaling (Tzahor 2007; Cohen et al. 2008), andits function is required for survival, prolifera-tion, and migration of into the primitive hearttube of the SHF progenitors. Isl-1 expression isextinguished as progenitor cells reach the devel-oping heart and differentiate (Cai et al. 2003).Several studies have shown that the molecularsignature Isl-1þ/Nkx2.5þ/Flk-1þ marks a spe-cific pool of primitive SHF progenitors that aremultipotent and give rise, by further lineage
restriction, to downstream tissue-specific inter-mediates generating both myocytic and vascularcells (Fig. 2) (Kattman et al. 2006; Moretti et al.2006; Wu et al. 2006; Bu et al. 2009). In partic-ular, Isl-1þ/Nkx2.5þdescendants (that have lostFlk-1 expression) are committed to form cardio-myocytes and smooth muscle cells (SMCs),extensively contributing to the proepicardium/epicardium (Cai et al. 2008; Zhou et al. 2008a,b),whereas the Isl-1þ/Flk-1þ subset (in whichNkx2.5 expression has ceased) differentiates toform endothelial cells and SMCs (Fig. 2) (Mo-retti et al. 2006). The fate of SHF progenitors iscontrolled by many different signaling pathways.Among them, FGF signaling within the SHFpromotes progenitor cell proliferation; Shh-me-diated signals from the endoderm and canonicalWnt signaling from the midline (neural tube)are important for the maintenance of a prolifer-ative state and inhibition of differentiation (Kel-ly 2012). On the contrary, BMPs secreted fromlateral plate mesoderm as well as Notch and non-canonical Wnt signals promote cardiac differen-tiation of SHF progenitors (Vincent and Buck-ingham 2010). Furthermore, mechanisms ofepigenetic control (e.g., miRNA, lncRNA) arealso important regulators of the progression ofprogenitors to terminally differentiated cardiacmuscle and nonmuscle lineages (Liu and Olson2010).
Taking together two decades’ worth of re-search data, we can now construct a core tran-scriptional and signaling network regulatingearly cardiac development. This highly com-
Figure 2. (Continued) Early mesoderm-derived cardiac precursors undergo further lineage restriction anddifferentiate into progenitor pools that populate the FHF and SHF, respectively. At this stage (E7.5 mousedevelopment) FHF progenitors start to differentiate upon BMP and FGF action toward cardiomyocytes andsmooth muscle cells, whereas Wnt/b-catenin, FGF, and endodermal Shh signaling keeps SHF progenitors in aproliferative state. These SHF progenitors are defined by the molecular signature Isl-1þ/Nkx2.5þ/Flk-1þ. SHFprogenitors are now gradually added to the looping heart tube and get further restricted in their differentiationpotential (E8.5). Two subpopulations of SHF progenitors can be distinguished. One population marked by theexpression of Isl-1 and Flk-1 differentiates into endothelial cells and smooth muscle cells, whereas a second poolof Isl-1þ/Nkx2.5þ SHF precursors provides smooth muscle cells and cardiomyocytes as well as contributing tothe proepicardial lineages (Wt1þ/Tbx18þ and Scxþ/Sema3Dþ populations), which later form cardiac fibro-blasts (CF), smooth muscle cells (SMCs), endothelial cells (EC), and cardiomyocytes (CM), with the lattercontribution being still unclear. These distinct SHF progenitor populations differentiate upon BMP signals fromthe lateral plate mesoderm as well as Notch and noncanonical Wnt signals. SHF patterning is governed by RAand TGF-b signals.
T. Brade et al.
6 Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

plex network of spatiotemporal interactions ofgrowth factors and transcriptional regulators(e.g., BMP, Wnt, FGF, Nkx2.5, and Gata4) gov-erns both FHF and SHF development (Kelly2012; Harvey 2013; Moorman et al. 2013). Im-portantly, ourcurrent knowledge supports a newparadigm for cardiovascular development in-volving the divergence of myocardial, smoothmuscle and endothelial cell lineages from a com-mon precursor.
THE PROEPICARDIUM
The early embryonic heart tube developingfrom the FHF and SHF progenitors consistsonly of two cell layers, the endocardium andthe myocardium. The epicardium, the outer-most layer of the heart, and the epicardium-de-rived cells (EPDCs), which are found in the ma-ture heart, such as cardiac fibroblasts and cells ofthe coronary vasculature, are not present at thetubular heart stage. These cell lineages arise later(between E9.5 and E11.5 in mouse develop-ment) (Fig. 1) and are derived from a cluster ofcells, which is known as the proepicardium (PE).
Induction of the Proepicardium andEpicardium Formation
The transitory structure of the PE arises fromthe coelomic mesenchyme of the septum trans-versum in close proximity to the venous pole ofthe linear heart tube at E 8.5 (d21 in humangestation) (Manner et al. 2001). PE induction,growth, and maintenance depends, among oth-ers, on opposing interaction between FGF sig-naling, which induces a proepicardial fate in theposterior splanchnic mesoderm, and BMP sig-naling, driving myocardial differentiation ofthis cell population (Kruithof et al. 2006; vanWijk et al. 2009; Torlopp et al. 2010; Schlueterand Brand 2012). PE progenitors are marked bythe expression of Tbx18 and Wt1 (Kraus etal. 2001; Cai et al. 2008; Zhou et al. 2008b) aswell as by FHF and SHF markers Nkx2-5 andIsl-1 (Zhou et al. 2008b). One important ques-tion concerns heterogeneity of PE progenitors,which could represent early-segregated progen-itor pools. Earlier studies in chicken embryos
showed at least two morphologically differentproepicardial cell populations: (1) an externalmesothelial epithelium, and (2) an internalextracellular matrix (ECM)-rich mesenchymalcore (Nahirney et al. 2003). Although no mor-phological differences have been described inmammalian PE cells, a very recent lineage-trac-ing study in mouse analyzing semaphorin3Dþ
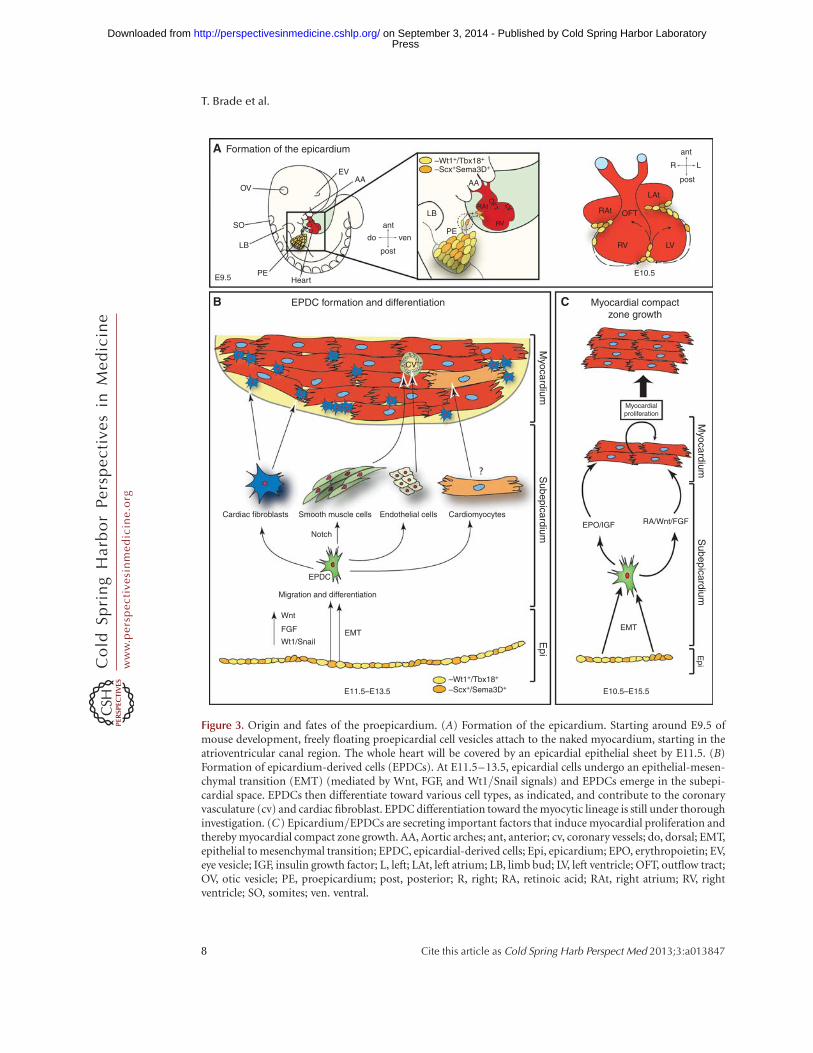
(Sema3Dþ) and scleraxisþ (Scxþ) PE progeni-tors revealed that these cells show a distinct dif-ferentiation potential from earlier describedTbx18þ and Wt1þ proepicardial cells, confirm-ing the data on PE heterogeneity evident fromstudying the avian system (Katz et al. 2012). Inmammals, freely floating PE cell vesicles are re-leased from the PE anlage, which flatten andspread out on contact to the naked myocardium,thereby forming the epicardium between E9.5and E11.5 in mouse development (Fig. 3A). Ge-netic evidence from mouse embryos suggests acrucial role for cell adhesion molecules (vascularcell adhesion molecule [VCAM] and b4-a1-in-tegrin) in this process (Kwee et al. 1995; Yanget al. 1995).
The Epicardium- and Epicardial-Derived CellsGovern Chamber Maturation and CoronaryVessel Formation
After the epicardium has been formed, a wholearray of interacting signaling pathways drive (1)epicardial epithelial to mesenchymal transition(EMT) and formation of EPDCs, (2) differenti-ation of EPDCs into different cell lineages, (3)induction of compact zone growth, as well as (4)establishment of coronary vessel formation (Fig.3B,C). Once the epithelial cell sheet of the epi-cardium is formed, directed proliferation paral-lel or perpendicular to the cellular basementmembrane leads to the expansion of the epicar-dium in the formercase or the induction of EMTand subsequent migration and differentiation ofEPDCs in the latter. Wt1 signaling is a key factorfor normal progression of epicardial EMT aswell as EPDC differentiation and myocardialgrowth (Martinez-Estrada et al. 2010; von Giseet al. 2011; Lim and Thiery 2012).
EPDC differentiation and myocardial com-pact zonegrowthdependona networkofrecipro-
Embryonic Heart Progenitors and Cardiogenesis
Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847 7
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from
A
B C
Formation of the epicardium
EPDC formation and differentiation
CV
Myocardial compactzone growth
Myocardialproliferation
EPO/IGF RA/Wnt/FGF
EMT
Epi
Myocardium
Subepicardium
Cardiac fibroblasts Smooth muscle cells Endothelial cells Cardiomyocytes
Myocardium
Subepicardium
Epi
EPDC
Notch
Migration and differentiation
Wnt
FGF
Wt1/SnailEMT
E11.5–E13.5 E10.5–E15.5
–Wt1+/Tbx18+
–Scx+/Sema3D+
OV
SO
LB
PEHeartE9.5
EV–Wt1+/Tbx18+
–Scx+Sema3D+
LB
PE
LV
RV
AA
RAt RAt OFT
LAt
RV LV
ant
R L
post
E10.5
OFT
AA
ant
do ven
post
Figure 3. Origin and fates of the proepicardium. (A) Formation of the epicardium. Starting around E9.5 ofmouse development, freely floating proepicardial cell vesicles attach to the naked myocardium, starting in theatrioventricular canal region. The whole heart will be covered by an epicardial epithelial sheet by E11.5. (B)Formation of epicardium-derived cells (EPDCs). At E11.5–13.5, epicardial cells undergo an epithelial-mesen-chymal transition (EMT) (mediated by Wnt, FGF, and Wt1/Snail signals) and EPDCs emerge in the subepi-cardial space. EPDCs then differentiate toward various cell types, as indicated, and contribute to the coronaryvasculature (cv) and cardiac fibroblast. EPDC differentiation toward the myocytic lineage is still under thoroughinvestigation. (C) Epicardium/EPDCs are secreting important factors that induce myocardial proliferation andthereby myocardial compact zone growth. AA, Aortic arches; ant, anterior; cv, coronary vessels; do, dorsal; EMT,epithelial to mesenchymal transition; EPDC, epicardial-derived cells; Epi, epicardium; EPO, erythropoietin; EV,eye vesicle; IGF, insulin growth factor; L, left; LAt, left atrium; LB, limb bud; LV, left ventricle; OFT, outflow tract;OV, otic vesicle; PE, proepicardium; post, posterior; R, right; RA, retinoic acid; RAt, right atrium; RV, rightventricle; SO, somites; ven. ventral.
T. Brade et al.
8 Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

cal epicardium-myocardium signaling of largelyunknown nature (Fig. 3C) (Perez-Pomares andde la Pompa 2011). There is still extensive dis-cussion about the potential contribution ofEPDCs to different cell lineages (Gittenberger-de Groot et al. 2012). Although it is consensus inthe field that EPDCs can differentiate toward acardiac fibroblast and vascular smooth musclecell fate, formation of coronary endothelial cellsand cardiomyocytes by EPDCs is less clear (Fig.3B). Tabin andcolleagues have shownfor thefirsttime that the mammalian epicardium consists ofdistinct cell populations that can give rise to en-dothelial cells (Katz et al. 2012). The vast major-ityofcardiomyocytes,on theotherhand, seem tobe derived from other sources (Gittenberger-deGroot et al. 2012). Signaling molecules with aprominent role in EPDC differentiation andcompact zone growth are FGFs, Notch and reti-noic acid (Sucov et al. 2009). Two different RAsignaling networks have been proposed. Onestudy showed that an RA stimulus is necessaryto induce hepatic erythropoietin (Epo) produc-tion. Epo in turn induces epicardial IGF-2 secre-tion, which then acts as a proliferative factor oncompact zone myocytes (Brade et al. 2011; Liet al. 2011). Other studies placed Wt1 upstreamof epicardial Raldh2 (a key molecule in RA me-tabolism) expression (Guadix et al. 2011). RAsignaling in turn has been described earlier tospearhead a signaling network that includescross talk with Wnt and FGF signaling molecules(Fig. 3C) (Merki et al. 2005; Lavine et al. 2006;Zamora et al. 2007).
Formation of the coronary vessels is tightlylinked to compact zone growth and relies onoverlapping signaling molecules (Olivey andSvensson 2010; Perez-Pomares and de la Pompa2011). Mesenchymal cells in the subepicardialspace and in the myocardial wall (most ofthem if not all are EPDCs) will coalesce anddifferentiate, thereby forming the primitivecoronary plexus around E11.5 during mousedevelopment (Reese et al. 2002). Primary coro-nary vessels spread over the entire ventricle untilE13.5; afterward, intricate remodeling and ar-terial and venous differentiation takes place.Important signaling molecules driving theseprocesses include the aforementioned FGFs and
RA signaling as well as ligands belonging to theHedgehog and VEGF families and the TGF-bsuperfamily (Lavine and Ornitz 2008; Oliveyand Svensson 2010; Perez-Pomares and de laPompa 2011). Moreover, epicardial Notch sig-naling is required for vSMC differentiation,whereas it is dispensable for EMT or migrationand myocardial growth (del Monte et al. 2011;Grieskamp et al. 2011).
CARDIAC NEURAL CREST PROGENITORS
The third distinct embryonic heart progenitorpopulation that is involved in cardiogenesisconsists of the CNCCs. CNCCs comprise a non-cardiac cell type important for proper pattern-ing of the aortic arteries, for OFT developmentand septation, and normal myocardial function.Although CNCCs are differentiating into ecto-mesenchyme and aortic smooth muscle cells aswell as forming the aorticopulmonary septum,which divides the arterial outlet of the heart, itseems that the main role of CNCCs in the above-mentioned developmental processes is to pro-vide signals rather than actively contribute dif-ferentiated cells to the specific cardiac structures.That said, there is a direct pivotal cellular con-tribution of the crest to the heart valves and theyprovide all parasympathetic innervation of theheart (Fig. 1C).
Signaling Pathways Governing Cardiac NeuralCrest Cell Induction and Migration
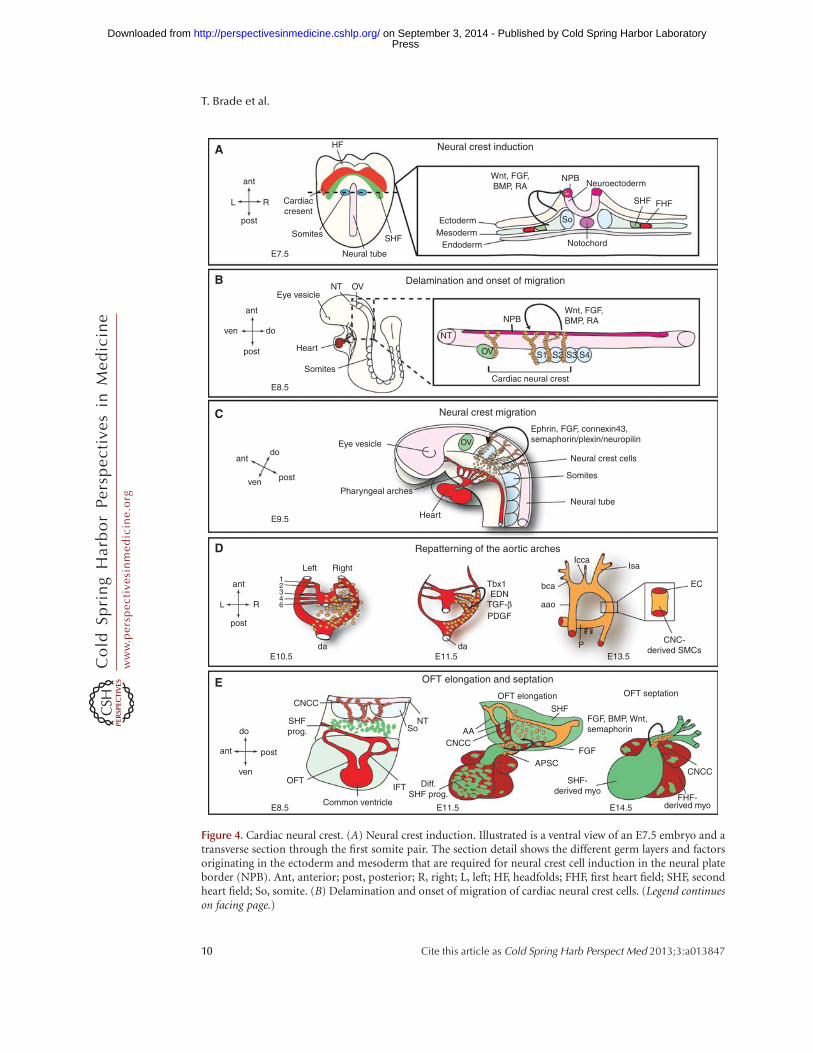
CNCCs are a subpopulation of the cranial neu-ral crest cells and arise from the dorsal neuraltube between the mid-otic placode and the pos-terior border of somite 3 (Fig. 4A) (Keyte andHutson 2012). The crest cells delaminate fromthe neural tube and migrate on preset routes to-ward the heart, reaching the pharyngeal arches 3,4, and 6 (Fig. 4B). Signals important for the in-duction of the CNC progenitors belong to theBMP/TGF-b superfamily of growth factors aswell as to the FGF, Wnt/b-catenin, and retinoicacid signaling pathways. These different familiesof secreted molecules control key aspects of neu-ral crest biology, starting from the establishmentof the neural plate border up to the cytosekeletal
Embryonic Heart Progenitors and Cardiogenesis
Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847 9
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from
A
B
C
D
E
Neural crest induction
Delamination and onset of migration
Neural crest migration
OFT elongation and septation
Repatterning of the aortic arches
CNCC
SHFprog.
ant
venOFT
E8.5 E11.5 E14.5
APSCFGF
SHF
OFT elongation
SHF-derived myo
FGF, BMP, Wnt,semaphorin
OFT septation
CNCC
FHF-derived myoCommon ventricle
IFT
SoNT
AACNCC
Diff.SHF prog.
Left Right
Tbx1EDN
TGF-βPDGF
da
bca
aao
P
IccaIsa
EC
CNC-derived SMCs
12346
da
ant
RL
post
post
NT OVEye vesicle
Eye vesicle
Heart
Heart
Pharyngeal arches
Ephrin, FGF, connexin43,semaphorin/plexin/neuropilin
Neural crest cells
Neural tube
Somites
Somites
E8.5
E9.5
E10.5 E11.5 E13.5
NT
NPBWnt, FGF,BMP, RA
S1OV
Cardiac neural crest
S2 S3 S4
HF
SHF
Wnt, FGF,BMP, RA
NPBNeuroectoderm
SHF FHF
So
Notochord
EctodermMesoderm
Endoderm
ant
ant
ant
ven
ven
do
do
do
post
post
OV
L R
post
Cardiaccresent
Somites
E7.5 Neural tube
Figure 4. Cardiac neural crest. (A) Neural crest induction. Illustrated is a ventral view of an E7.5 embryo and atransverse section through the first somite pair. The section detail shows the different germ layers and factorsoriginating in the ectoderm and mesoderm that are required for neural crest cell induction in the neural plateborder (NPB). Ant, anterior; post, posterior; R, right; L, left; HF, headfolds; FHF, first heart field; SHF, secondheart field; So, somite. (B) Delamination and onset of migration of cardiac neural crest cells. (Legend continueson facing page.)
T. Brade et al.
10 Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

rearrangements required for delamination ofneural crest cells from the neural tube and on-set of migration (Fig. 4A,B) (Scholl and Kirby2009).
After induction and delamination the crestcells migrate along specific routes guided bychemical attractant and repellent cues towardthe heart (Fig. 4C). Important molecules forthis process belong to the ephrin family of li-gands and receptors that ensure specific migra-tion patterns alongside FGF signaling molecules,semaphorins and connexin 43 (Kuriyama andMayor 2008). Semaphorin/plexin/neuropilinsignaling is suggested as a targeting cue to theOFT cushions and Cx43 seems to facilitate prop-er migration and survival. Intricate interactionsalong their path between the pharyngeal ecto-derm and endoderm and continuous prolifera-tion ensure that the CNC progenitors reach theirtarget areas in the heart in sufficient numbers(Xu et al. 2006; Keyte and Hutson 2012).
Signaling Pathways Instructing CNC Cellsduring Aortic Arch Artery Patterning andOFT Septation
Once the crest cells reached the pharyngeal arch-es, endothelin, TGF-b, and PDGF signalingpathways are crucial for the CNCCs to exert theirrole in patterning the aortic arch arteries (Fig.4D). Endothelin (END) signals are important
for the maintenance of the instructive cues ofCNCCsduring this processand, althoughdown-stream effectors are not well understood yet,Hand2 seems to process endothelin signals inthe aortic arches. TGF-b signals are thought toprovide important antiapoptotic effects, andPDGF signals ensure proliferation and mainte-nance (Hutson and Kirby 2007). T-box tran-scription factors (namely, Tbx1) also play a keyrole in neural crest biology. Although not ex-pressed in CNCCs themselves, Tbx1 regulatesthe expression of the guidance molecule Slit inthe pharyngeal endoderm. Loss of Slit functionhas been shown to prevent CNCC migration intoaortic arch 4 and thereby leading to interruptedaortic arch. Besides that, Tbx1 also interacts withFGF-8, whose levels need to be regulated in averyconfined manner for CNCC survival as well asfor timing and targeting of the migrating pro-genitors to the pharynx (Abu-Issa et al. 2002;Sato et al. 2011).
Additional to their role in aortic arch pat-terning CNCCs exert another important func-tion in OFT development and septation (Fig.4E). One of the earliest detectable defects afterCNCC ablation is defective looping because of ashortened OFT. This phenotype is caused by al-tered SHF progenitor addition to the developingOFT. In CNCC ablated chick embryos alteredFGF-8 levels keep the SHF progenitors in a pro-liferative state, precluding proper migration and
Figure 4. (Continued) At E8.5, cardiac neural crest cells delaminate from the NPB between the mid-otic vesicle(ov) and the posterior border of somite 3 (S3) and start on their migratory path toward the heart. Theseprocesses are mediated by Wnt, FGF, BMP, and RA signals. Please see text for more details. ant, anterior;post, posterior; ven, ventral; do, dorsal; S1,2,3,4, somite 1,2,3,4. (C) Neural crest migration. By E9.5, the CNCCsreach the aortic arches and are proceeding toward the OFT. Signals targeting the CNCCs to the aortic arches areindicated. ov, otic vesicle. (D) The CNCCs are vital for the repatterning of the aortic arches. There are initially sixsymmetrical arteries attached to the parallel dorsal aorta (da). Sequentially these arteries are remodeled and giverise to a separate ascending aorta (aao) and pulmonary trunk (P) with two pulmonary arteries attached.Additionally, the cardiac neural crest cells are differentiating into aortic smooth muscle cells and envelopethe nascent endothelial cells forming the aortic arches. Required factors for both processes are indicated(END, endothelin); see text for details. bca, Brachiochepalic artery; lcca, left common carotid artery; lsa, leftsubclavian artery. (E) Interaction of CNCCs and SHF progenitors leads to outflow tract (OFT) elongation andseptation. At E11.5, cardiac neural crest cells condense and form the U-shaped aorticopulmonary septationcomplex (APSC); see text for details. CNCC, Cardiac neural crest cells; SHF prog., second heart field progenitors;NT, neural tube; So, somites; IFT, inflow tract; diff. SHF prog., differentiated second heart field progenitors; AA,aortic arches; FHF derived myo., first heart field-derived myocardium; SHF derived myo, second heart field-derived myocardium.
Embryonic Heart Progenitors and Cardiogenesis
Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847 11
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

eventually initiation of myocardial differentia-tion (Waldo et al. 2005; Ilagan et al. 2006; Scholland Kirby 2009). Furthermore, upon CNCC ab-lation myocardial function is deranged, which isindicated by disrupted Ca2þ handling and de-pressed ejection fraction (Hutson et al. 2006).Further down the developmental road, CNCCsgovern OFT septation. In the avian model sys-tem, the CNCCs reach the OFT cushions sub-endocardially and submyocardially where theyform together with mesenchymal cells the so-called aorticopulmonary septation complex,which will initiate the division of the systemicand pulmonary blood flow (Keyte and Hutson2012). Multiple signaling pathways are implicat-ed to instruct CNCCs during this process, themajor players being BMP, Wnt, and semaphorinsignaling (Fig. 4E). The cellular contribution oftheCNCCsto thecardiacconduction system andthe cardiac valves remains an ongoing debate butdevelopment of both structures is obscuredupon loss of neural crest function, indicatingthat at least signals from CNCCs are requiredfor their normal development (Hutson andKirby 2007).
EMBRYONIC CARDIAC PROGENITORSAND CONGENITAL HEART DISEASE
Congenital heart disease (CHD) is one of themost prevalent forms of human birth defects,affecting 0.4%–5% of live births, dependingon which level of severity of CHD is includedin the statistics. Moreover, around 30% of pre-natal loss is owing to heart malformations (Hoff-man 1995; Bruneau 2008). CHD most common-ly arises from defective cardiac morphogenesis.In the last few decades, with our growing knowl-edge of normal cardiac development and of car-diac precursor origin and regulation, the role ofthe different embryonic cardiac progenitor pop-ulations (cardiac mesoderm progenitors, epi-cardial progenitors, and CNCCs) in the etiologyof CHD has begun to gain increasing interest.Although disruption of embryonic cardiac pro-genitor biology (specification, proliferation, mi-gration, lineage-specific differentiation) has notyet been assigned to be causative for any humanCHD, identification of transcriptional programs
that are impaired in CHD patients and controlkey aspects of cardiac progenitor lineage diversi-fication and fate strongly suggests compro-mised cardiac progenitor biology at the root ofat least some human CHDs. Mutations inNkx2.5 (e.g., atrial septal defects [ASD] and te-tralogy of Fallot), Tbx5 (e.g., Holt-Oram syn-drome, ASD, and ventricular septal defects[VSD]), and GATA-4 (e.g., ASD and VSD) pointto cardiogenic mesoderm progenitor involve-ment, whereas mutations in the Tbx1 gene (Di-George syndrome, 22q11 deletions) implicatesCNCCs. The involvement of epicardial pro-genitors in CHD is not as straightforward, buta role in hypoplastic left heart syndrome hasbeen suggested along with contributions to theleft ventricular noncompaction phenotype andvalve malformations (Bruneau 2008; Musunuruet al. 2010; Gittenberger-de Groot et al. 2012;Keyte and Hutson 2012).
The possibility of derailed embryonic cardi-ac progenitors contributing to the developingheart as a causative factor for human CHD en-ables intriguing explanations for some unre-solved questions in CHD etiology. It is, for ex-ample, unclear why most CHDs are polygenicand havevariable penetranceand disease severityevenwithin one family (Fahed et al. 2013). Giventhe complex networks of cytokines, transcrip-tion factors as well as physical stimuli (e.g.,cell–cell interactions, hemodynamic flow, cellmigration) that are regulating cardiac progenitordevelopment, it becomes feasible that a plethoraof variables dictates the phenotypic characteris-tics of a certain CHD, which undoubtedly canvary even between siblings. Another puzzlingquestion is how the same morphological pheno-type is caused by seemingly unrelated geneticcauses (Lage et al. 2012). This could be explainedif we consider progenitor development goneawry as a basis for the cardiac malformation athand.
CARDIAC STEM AND PROGENITOR CELLSIN REGENERATIVE MEDICINE
Ischemic heart disease and heart failure are theleading causes of death in the western world.Even if patients survive an ischemic event such
T. Brade et al.
12 Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

as acute myocardial infarction (MI) their prog-nosis can be very poor. Diminished cardiac func-tion owing to a massive loss of cardiomyocytesinduces hypertrophic remodeling and eventual-ly heart failure. Novel therapeutic avenues aim-ing at disrupting this vicious circle by renewal oflost cardiac muscle are greatly needed to improveour means to manage ischemic heart disease.Unfortunately, the regenerative capacity of theheart is extremely limited. Until recently, it wasbelieved that the mammalian heart cannot re-generate at all owing to the inabilityof terminallydifferentiated cardiomyocytes to reenter the cellcycle and proliferate. Studies of the last few yearshave now provided evidence for cardiomyocyteturnover in the adult human heart. Althoughthis process occurs at a low rate during normalhomeostasis and aging of the heart, it seems to beelevated after injury (Bergmann et al. 2009; Par-macek and Epstein 2009; Kajstura et al. 2010;Senyo et al. 2013). It has been shown that micecan regenerate lost heart muscle within their firstweek of life, similarly to adult zebrafish, and thatmost of the new cardiomyocytes are mainly de-rived by cell division from preexisting cardio-myocytes (Jopling et al. 2010; Kikuchi et al.2010; Porrello et al. 2011). Recently, a role ofthe epicardium and EPDCs in cardiac regenera-tion has been suggested. The epicardium seemsto contribute mainly paracrine factors for myo-cardial proliferation and neovascularization aswell as nonmyocytic lineages, therefore reca-pitulating embryonic development (Perez-Po-mares and de la Pompa 2011; Kikuchi and Poss2012).
Alternative options for improving cardiacfunction after MI arise from the purification ofcardiac progenitors or cardiomyocytes differen-tiated from human embryonic or induced plu-ripotent stem cells (Burridge et al. 2012; Aguirreet al. 2013; Chien and Mummery 2013; Mum-mery 2013) and from direct in vivo reprogram-ming approaches. In recent years, a plethora ofdifferentiation protocols driving hPSCs towarda cardiogenic fate have been established (Bur-ridge et al. 2012; Mummery et al. 2012). How-ever, clinical applications of hPSC-derived cellshave so far been hampered by safety issues andlow engraftment after injection in the injured
heart (Aguirre et al. 2013). An exciting hope inthe field of cardiac repair has come from therecent success in direct in vivo reprogrammingof cardiac fibroblasts into cardiomyocytes usingoverexpression of Tbx5, Mef2c, and Gata4 incombination with Hand2 or only using miRNAs(miRs-1/133/208/499) (Song et al. 2010; Jaya-wardena et al. 2012; Qian et al. 2012). Thesestudies have shown improvements of cardiacfunction after direct reprogramming in experi-mental rodent models of MI, suggesting the po-tential of this new technology as a viable strategyfor cardiac regeneration.
Although a lot of problems still need to betackled, activation of endogenous regenerativeprocesses, along with stem cell-based technolo-gies, is holding great promise for possible futureclinical applications. In the past 20 years we im-proved the path to be traveled to reach this goaland further efforts of many scientists and clini-cians around the world will provide a silver lin-ing in the battle against the world’s biggest lifethreat (see also Chien and Mummery 2013 andMummery 2013 for more details).
CONCLUDING REMARKS
The different embryonic heart progenitor line-ages all rely on a great variety of spatiotemporalsignals that tightly regulate their terminal differ-entiation. But it is noteworthy that recurring ge-netic regulatory networks are now emerging. Forinduction/specification of progenitor popula-tions EMT is indispensable. Here factors of theTGF-b superfamily play a vital role. Once themultipotent progenitor cell state is reached,this population needs to proliferate and main-tain an undifferentiated state to ensure enoughterminally differentiated cells for the developingorgan. Here canonical Wnt and FGF signalingfactor as well as RA are used, among others. Dif-ferentiation toward specialized cell types is thenachieved by BMP growth factors (myocardium)and Notch signaling (vascular cell types). Thesegenetic loops are fine-tuned by transcription fac-tors and noncoding RNAs, gradually restrictingthe differentiation potential of the specific line-age. To transfer our knowledge of cardiac devel-opment to the bedside, the field faces new chal-
Embryonic Heart Progenitors and Cardiogenesis
Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847 13
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

lenges within the coming years. One task wouldbe to decipher definitive molecular signaturesthat mark cardiac precursors in a spatiotempor-ally defined manner and enables researchers tospecifically isolate and analyze distinct progeni-tor populations in greater detail. Although wehave elucidated some marker genes for embry-onic heart progenitors, the segregation of dis-tinct progenitor populations with defined dif-ferentiation potential remains incomplete, andknown marker genes are so far not exclusive for agiven population. Additionally, molecular sig-natures comprised by cell surface moleculesrather than transcription factors would greatlyhelp to better analyze precursor biology in vitro,which would give us tools at hand to furtherdelineate factors needed for survival and differ-entiation. Great progress has been made in ourunderstanding of the embryonic developmentof the heart. But so far we have only understoodthe broad concept of induction, proliferation/maintenance, and differentiation. It will requirefurther efforts in filling the gaps to fully compre-hend complex etiologies of human CHDs and tomodulate embryonic developmental processesin vitro and in vivo to restore myocardial func-tion in ischemic heart diseases.
ACKNOWLEDGMENTS
K.-L.L. acknowledges and thanks the GermanResearch Foundation and the European Re-search Council for their ongoing support of re-search in the Laugwitz laboratory. A.M. is sup-ported by grants of the German ResearchFoundation and the German Ministry for Edu-cation and Research. K.-L.L. and A.M. also ac-knowledge the Munich Heart Alliance, a mem-ber of the German Centre for CardiovascularResearch.
REFERENCES�Reference is also in this collection.
Abu-Issa R, Smyth G, Smoak I, Yamamura K, Meyers EN.2002. Fgf8 is required for pharyngeal arch and cardio-vascular development in the mouse. Development 129:4613–4625.
Abu-Issa R, Waldo K, Kirby ML. 2004. Heart fields: One,two or more? Dev Biol 272: 281–285.
Aguirre A, Sancho-Martinez I, Izpisua Belmonte JC. 2013.Reprogramming toward heart regeneration: Stem cellsand beyond. Cell Stem Cell 12: 275–284.
Arceci RJ, King AA, Simon MC, Orkin SH, Wilson DB. 1993.Mouse GATA-4: A retinoic acid-inducible GATA-bindingtranscription factor expressed in endodermally derivedtissues and heart. Mol Cell Biol 13: 2235–2246.
Bergmann O, Bhardwaj RD, Bernard S, Zdunek S, Barnabe-Heider F, Walsh S, Zupicich J, Alkass K, Buchholz BA,Druid H, et al. 2009. Evidence for cardiomyocyte renewalin humans. Science 324: 98–102.
Bondue A, Blanpain C. 2010. Mesp1: A key regulator of car-diovascular lineage commitment. Circ Res 107: 1414–1427.
Bondue A, Lapouge G, Paulissen C, Semeraro C, IacovinoM, Kyba M, Blanpain C. 2008. Mesp1 acts as a masterregulator of multipotent cardiovascular progenitor spec-ification. Cell Stem Cell 3: 69–84.
Brade T, Kumar S, Cunningham TJ, Chatzi C, Zhao X, Cav-allero S, Li P, Sucov HM, Ruiz-Lozano P, Duester G. 2011.Retinoic acid stimulates myocardial expansion by induc-tion of hepatic erythropoietin which activates epicardialIgf2. Development 138: 139–148.
Bruneau BG. 2008. The developmental genetics of congen-ital heart disease. Nature 451: 943–948.
Bu L, Jiang X, Martin-Puig S, Caron L, Zhu S, Shao Y,Roberts DJ, Huang PL, Domian IJ, Chien KR. 2009. Hu-man ISL1 heart progenitors generate diverse multipotentcardiovascular cell lineages. Nature 460: 113–117.
Buckingham M, Meilhac S, Zaffran S. 2005. Building themammalian heart from two sources of myocardial cells.Nat Rev Genet 6: 826–835.
Burridge PW, Keller G, Gold JD, Wu JC. 2012. Production ofde novo cardiomyocytes: Human pluripotent stem celldifferentiation and direct reprogramming. Cell Stem Cell10: 16–28.
Cai CL, Liang X, Shi Y, Chu PH, Pfaff SL, Chen J, Evans S.2003. Isl1 identifies a cardiac progenitor population thatproliferates prior to differentiation and contributes a ma-jority of cells to the heart. Dev Cell 5: 877–889.
Cai CL, Martin JC, Sun Y, Cui L, Wang L, Ouyang K, Yang L,Bu L, Liang X, Zhang X, et al. 2008. A myocardial lineagederives from Tbx18 epicardial cells. Nature 454: 104–108.
� Chien K. 2013. Pluripotent stem cell technology and regen-erative cardiovascular medicine. Cold Spring Harb Per-spect Med doi: 10.1101/cshperspect.a013870.
Cohen ED, Tian Y, Morrisey EE. 2008. Wnt signaling:An essential regulator of cardiovascular differentiation,morphogenesis and progenitor self-renewal. Develop-ment 135: 789–798.
Costello I, Pimeisl IM, Drager S, Bikoff EK, Robertson EJ,Arnold SJ. 2011. The T-box transcription factor Eome-sodermin acts upstream of Mesp1 to specify cardiac me-soderm during mouse gastrulation. Nat Cell Biol 13:1084–1091.
del Monte G, Casanova JC, Guadix JA, MacGrogan D, BurchJB, Perez-Pomares JM, de la Pompa JL. 2011. DifferentialNotch signaling in the epicardium is required for cardiacinflow development and coronary vessel morphogenesis.Circ Res 108: 824–836.
T. Brade et al.
14 Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

Ema M, Takahashi S, Rossant J. 2006. Deletion of the selec-tion cassette, but not cis-acting elements, in targetedFlk1-lacZ allele reveals Flk1 expression in multipotentmesodermal progenitors. Blood 107: 111–117.
Fahed AC, Gelb BD, Seidman JG, Seidman CE. 2013. Ge-netics of congenital heart disease: The glass half empty.Circ Res 112: 707–720.
Gessert S, Kuhl M. 2010. The multiple phases and faces ofwnt signaling during cardiac differentiation and develop-ment. Circ Res 107: 186–199.
Gittenberger-de Groot AC, Winter EM, Bartelings MM,Goumans MJ, DeRuiter MC, Poelmann RE. 2012. Thearterial and cardiac epicardium in development, diseaseand repair. Differentiation 84: 41–53.
Grieskamp T, Rudat C, Ludtke TH, Norden J, Kispert A.2011. Notch signaling regulates smooth muscle differen-tiation of epicardium-derived cells. Circ Res 108: 813–823.
Guadix JA, Ruiz-Villalba A, Lettice L, Velecela V, Munoz-Chapuli R, Hastie ND, Perez-Pomares JM, Martinez-Es-trada OM. 2011. Wt1 controls retinoic acid signalling inembryonic epicardium through transcriptional activa-tion of Raldh2. Development 138: 1093–1097.
Harvey RP. 2002. Patterning the vertebrate heart. Nat RevGenet 3: 544–556.
� Harvey R. 2013. Genetic networks. Cold Spring Harb Per-spect Med doi: 10.1101/cshperspect.a013839.
Heikinheimo M, Scandrett JM, Wilson DB. 1994. Localiza-tion of transcription factor GATA-4 to regions of themouse embryo involved in cardiac development. DevBiol 164: 361–373.
Hoffman JI. 1995. Incidence of congenital heart disease.I: Postnatal incidence. Pediatr Cardiol 16: 103–113.
Hutson MR, Kirby ML. 2007. Model systems for the studyof heart development and disease. Cardiac neural crestand conotruncal malformations. Semin Cell Dev Biol 18:101–110.
Hutson MR, Zhang P, Stadt HA, Sato AK, Li YX, Burch J,Creazzo TL, Kirby ML. 2006. Cardiac arterial pole align-ment is sensitive to FGF8 signaling in the pharynx. DevBiol 295: 486–497.
Ilagan R, Abu-Issa R, Brown D, Yang YP, Jiao K, SchwartzRJ, Klingensmith J, Meyers EN. 2006. Fgf8 is required foranterior heart field development. Development 133:2435–2445.
Jayawardena TM, Egemnazarov B, Finch EA, Zhang L, PayneJA, Pandya K, Zhang Z, Rosenberg P, Mirotsou M, DzauVJ. 2012. MicroRNA-mediated in vitro and in vivo directreprogramming of cardiac fibroblasts to cardiomyocytes.Circ Res 110: 1465–1473.
Jopling C, Sleep E, Raya M, Marti M, Raya A, Izpisua Bel-monte JC. 2010. Zebrafish heart regeneration occurs bycardiomyocyte dedifferentiation and proliferation. Na-ture 464: 606–609.
Kajstura J, Gurusamy N, Ogorek B, Goichberg P, Clavo-Rondon C, Hosoda T, D’Amario D, Bardelli S, BeltramiAP, Cesselli D, et al. 2010. Myocyte turnover in the aginghuman heart. Circ Res 107: 1374–1386.
Kattman SJ, Huber TL, Keller GM. 2006. Multipotent flk-1þ
cardiovascular progenitor cells give rise to the cardio-
myocyte, endothelial, and vascular smooth muscle line-ages. Dev Cell 11: 723–732.
Katz TC, Singh MK, Degenhardt K, Rivera-Feliciano J, John-son RL, Epstein JA, Tabin CJ. 2012. Distinct compart-ments of the proepicardial organ give rise to coronaryvascular endothelial cells. Dev Cell 22: 639–650.
Kelley C, Blumberg H, Zon LI, Evans T. 1993. GATA-4 is anovel transcription factor expressed in endocardium ofthe developing heart. Development 118: 817–827.
Kelly RG. 2012. The second heart field. Curr Top Dev Biol100: 33–65.
Keyte A, Hutson MR. 2012. The neural crest in cardiac con-genital anomalies. Differentiation 84: 25–40.
Kikuchi K, Poss KD. 2012. Cardiac regenerative capacity andmechanisms. Annu Rev Cell Dev Biol 28: 719–741.
Kikuchi K, Holdway JE, Werdich AA, Anderson RM, Fang Y,Egnaczyk GF, Evans T, Macrae CA, Stainier DY, Poss KD.2010. Primary contribution to zebrafish heart regenera-tion by gata4þ cardiomyocytes. Nature 464: 601–605.
Kimelman D. 2006. Mesoderm induction: From caps tochips. Nat Rev Genet 7: 360–372.
Kouskoff V, Lacaud G, Schwantz S, Fehling HJ, Keller G.2005. Sequential development of hematopoietic and car-diac mesoderm during embryonic stem cell differentia-tion. Proc Natl Acad Sci 102: 13170–13175.
Kraus F, Haenig B, Kispert A. 2001. Cloning and expressionanalysis of the mouse T-box gene Tbx18. Mech Dev 100:83–86.
Kruithof BP, van Wijk B, Somi S, Kruithof-de Julio M, PerezPomares JM, Weesie F, Wessels A, Moorman AF, van denHoff MJ. 2006. BMP and FGF regulate the differentiationof multipotential pericardial mesoderm into the myocar-dial or epicardial lineage. Dev Biol 295: 507–522.
Kubalak SW, Miller-Hance WC, O’Brien TX, Dyson E,Chien KR. 1994. Chamber specification of atrial myosinlight chain-2 expression precedes septation during mu-rine cardiogenesis. J Biol Chem 269: 16961–16970.
Kuriyama S, Mayor R. 2008. Molecular analysis of neuralcrest migration. Philos Trans R Soc Lond B Biol Sci 363:1349–1362.
Kwee L, Baldwin HS, Shen HM, Stewart CL, Buck C, BuckCA, Labow MA. 1995. Defective development of the em-bryonic and extraembryonic circulatory systems in vas-cular cell adhesion molecule (VCAM-1) deficient mice.Development 121: 489–503.
Lage K, Greenway SC, Rosenfeld JA, Wakimoto H, GorhamJM, Segre AV, Roberts AE, Smoot LB, Pu WT, Pereira AC,et al. 2012. Genetic and environmental risk factors incongenital heart disease functionally converge in proteinnetworks driving heart development. Proc Natl Acad Sci109: 14035–14040.
Lavine KJ, Ornitz DM. 2008. Fibroblast growth factors andHedgehogs: At the heart of the epicardial signaling cen-ter. Trends Genet 24: 33–40.
Lavine KJ, White AC, Park C, Smith CS, Choi K, Long F, HuiCC, Ornitz DM. 2006. Fibroblast growth factor signalsregulate a wave of Hedgehog activation that is essentialfor coronary vascular development. Genes Dev 20: 1651–1666.
Li P, Cavallero S, Gu Y, Chen TH, Hughes J, Hassan AB,Bruning JC, Pashmforoush M, Sucov HM. 2011. IGF
Embryonic Heart Progenitors and Cardiogenesis
Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847 15
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

signaling directs ventricular cardiomyocyte proliferationduring embryonic heart development. Development 138:1795–1805.
Lim J, Thiery JP. 2012. Epithelial-mesenchymal transitions:Insights from development. Development 139: 3471–3486.
Lints TJ, Parsons LM, Hartley L, Lyons I, Harvey RP. 1993.Nkx-2.5: A novel murine homeobox gene expressed inearly heart progenitor cells and their myogenic descen-dants. Development 119: 969.
Liu N, Olson EN. 2010. MicroRNA regulatory networks incardiovascular development. Dev Cell 18: 510–525.
Manner J, Perez-Pomares JM, Macias D, Munoz-Chapuli R.2001. The origin, formation and developmental signifi-cance of the epicardium: A review. Cells Tissues Organs169: 89–103.
Martinez-Estrada OM, Lettice LA, Essafi A, Guadix JA,Slight J, Velecela V, Hall E, Reichmann J, Devenney PS,Hohenstein P, et al. 2010. Wt1 is required for cardiovas-cular progenitor cell formation through transcriptionalcontrol of Snail and E-cadherin. Nat Genet 42: 89–93.
Martin-Puig S, Wang Z, Chien KR. 2008. Lives of a heartcell: Tracing the origins of cardiac progenitors. Cell StemCell 2: 320–331.
Marvin MJ, Di Rocco G, Gardiner A, Bush SM, Lassar AB.2001. Inhibition of Wnt activity induces heart formationfrom posterior mesoderm. Genes Dev 15: 316–327.
Merki E, Zamora M, Raya A, Kawakami Y, Wang J, Zhang X,Burch J, Kubalak SW, Kaliman P, Izpisua Belmonte JC, etal. 2005. Epicardial retinoid X receptor a is required formyocardial growth and coronary artery formation. ProcNatl Acad Sci 102: 18455–18460.
Mjaatvedt CH, Nakaoka T, Moreno-Rodriguez R, NorrisRA, Kern MJ, Eisenberg CA, Turner D, Markwald RR.2001. The outflow tract of the heart is recruited from anovel heart-forming field. Dev Biol 238: 97–109.
� Moorman A. 2013. Heart fields and cardiac morphogenesis.Cold Spring Harb Perspect Med doi: 10.1101/cshper-spect.a015750.
Moretti A, Caron L, Nakano A, Lam JT, Bernshausen A,Chen Y, Qyang Y, Bu L, Sasaki M, Martin-Puig S, et al.2006. Multipotent embryonic isl1þ progenitor cells leadto cardiac, smooth muscle, and endothelial cell diversifi-cation. Cell 127: 1151–1165.
Motoike T, Markham DW, Rossant J, Sato TN. 2003. Evi-dence for novel fate of Flk1þ progenitor: Contribution tomuscle lineage. Genesis 35: 153–159.
� Mummery C. 2013. Paradigms for heart cell therapeutics.Cold Spring Harb Perspect Med doi: 10.1101/cshper-spect.a013862.
Mummery CL, Zhang J, Ng ES, Elliott DA, Elefanty AG,Kamp TJ. 2012. Differentiation of human embryonicstem cells and induced pluripotent stem cells to cardio-myocytes: A methods overview. Circ Res 111: 344–358.
Musunuru K, Domian IJ, Chien KR. 2010. Stem cell modelsof cardiac development and disease. Annu Rev Cell DevBiol 26: 667–687.
Nahirney PC, Mikawa T, Fischman DA. 2003. Evidence foran extracellular matrix bridge guiding proepicardial cellmigration to the myocardium of chick embryos. Dev Dyn227: 511–523.
Noseda M, Peterkin T, Simoes FC, Patient R, Schneider MD.2011. Cardiopoietic factors: Extracellular signals for car-diac lineage commitment. Circ Res 108: 129–152.
Olivey HE, Svensson EC. 2010. Epicardial-myocardial sig-naling directing coronary vasculogenesis. Circ Res 106:818–832.
Olson EN. 2004. A decade of discoveries in cardiac biology.Nat Med 10: 467–474.
Parmacek MS, Epstein JA. 2009. Cardiomyocyte renewal. NEngl J Med 361: 86–88.
Perez-Pomares JM, de la Pompa JL. 2011. Signaling duringepicardium and coronary vessel development. Circ Res109: 1429–1442.
Porrello ER, Mahmoud AI, Simpson E, Hill JA, RichardsonJA, Olson EN, Sadek HA. 2011. Transient regenerativepotential of the neonatal mouse heart. Science 331:1078–1080.
Prall OW, Menon MK, Solloway MJ, Watanabe Y, Zaffran S,Bajolle F, Biben C, McBride JJ, Robertson BR, Chaulet H,et al. 2007. An Nkx2-5/Bmp2/Smad1 negative feedbackloop controls heart progenitor specification and prolif-eration. Cell 128: 947–959.
Qian L, Huang Y, Spencer CI, Foley A, Vedantham V, Liu L,Conway SJ, Fu JD, Srivastava D. 2012. In vivo reprogram-ming of murine cardiac fibroblasts into induced cardio-myocytes. Nature 485: 593–598.
Reese DE, Mikawa T, Bader DM. 2002. Development of thecoronary vessel system. Circ Res 91: 761–768.
Reifers F, Walsh EC, Leger S, Stainier DY, Brand M. 2000.Induction and differentiation of the zebrafish heart re-quires fibroblast growth factor 8 (fgf8/acerebellar). De-velopment 127: 225–235.
Saga Y, Kitajima S, Miyagawa-Tomita S. 2000. Mesp1 expres-sion is the earliest sign of cardiovascular development.Trends Cardiovasc Med 10: 345–352.
Sato A, Scholl AM, Kuhn EN, Stadt HA, Decker JR, PegramK, Hutson MR, Kirby ML. 2011. FGF8 signaling is che-motactic for cardiac neural crest cells. Dev Biol 354: 18–30.
Schlueter J, Brand T. 2012. Epicardial progenitor cells incardiacdevelopmentandregeneration. J CardiovascTranslRes 5: 641–653.
Schneider VA, Mercola M. 2001. Wnt antagonism initiatescardiogenesis in Xenopus laevis. Genes Dev 15: 304–315.
Scholl AM, Kirby ML. 2009. Signals controlling neural crestcontributions to the heart. Wiley Interdiscip Rev Syst BiolMed 1: 220–227.
Schultheiss TM, Burch JB, Lassar AB. 1997. A role for bonemorphogenetic proteins in the induction of cardiac myo-genesis. Genes Dev 11: 451–462.
Scott IC. 2012. Life before Nkx2.5: Cardiovascular progen-itor cells: Embryonic origins and development. Curr TopDev Biol 100: 1–31.
Senyo SE, Steinhauser ML, Pizzimenti CL, Yang VK, Cai L,Wang M, Wu TD, Guerquin-Kern JL, Lechene CP, Lee RT.2013. Mammalian heart renewal by pre-existing cardio-myocytes. Nature 493: 433–436.
Showell C, Binder O, Conlon FL. 2004. T-box genes in earlyembryogenesis. Dev Dyn 229: 201–218.
T. Brade et al.
16 Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from

Song H, Chung SK, Xu Y. 2010. Modeling disease in humanESCs using an efficient BAC-based homologous recom-bination system. Cell Stem Cell 6: 80–89.
Srivastava D. 2006. Genetic regulation of cardiogenesis andcongenital heart disease. Annu Rev Pathol 1: 199–213.
Sucov HM, Gu Y, Thomas S, Li P, Pashmforoush M. 2009.Epicardial control of myocardial proliferation and mor-phogenesis. Pediatr Cardiol 30: 617–625.
Torlopp A, Schlueter J, Brand T. 2010. Role of fibroblastgrowth factor signaling during proepicardium formationin the chick embryo. Dev Dyn 239: 2393–2403.
Tzahor E. 2007. Wnt/b-catenin signaling and cardiogene-sis: Timing does matter. Dev Cell 13: 10–13.
Tzahor E, Lassar AB. 2001. Wnt signals from the neural tubeblock ectopic cardiogenesis. Genes Dev 15: 255–260.
van Wijk B, van den Berg G, Abu-Issa R, Barnett P, van derVelden S, Schmidt M, Ruijter JM, Kirby ML, MoormanAF, van den Hoff MJ. 2009. Epicardium and myocardi-um separate from a common precursor pool by crosstalkbetween bone morphogenetic protein- and fibroblastgrowth factor-signaling pathways. Circ Res 105: 431–441.
Vincent SD, Buckingham ME. 2010. How to make a heart:The origin and regulation of cardiac progenitor cells.Curr Top Dev Biol 90: 1–41.
von Gise A, Zhou B, Honor LB, Ma Q, Petryk A, Pu WT.2011. WT1 regulates epicardial epithelial to mesenchy-mal transition through b-catenin and retinoic acid sig-naling pathways. Dev Biol 356: 421–431.
Waldo KL, Kumiski DH, Wallis KT, Stadt HA, Hutson MR,Platt DH, Kirby ML. 2001. Conotruncal myocardiumarises from a secondary heart field. Development 128:3179–3188.
Waldo KL, Hutson MR, Ward CC, Zdanowicz M, Stadt HA,Kumiski D, Abu-Issa R, Kirby ML. 2005. Secondary heartfield contributes myocardium and smooth muscle tothe arterial pole of the developing heart. Dev Biol 281:78–90.
Weissman IL. 2000. Stem cells: Units of development, unitsof regeneration, and units in evolution. Cell 100: 157–168.
Wu SM, Fujiwara Y, Cibulsky SM, Clapham DE, Lien CL,Schultheiss TM, Orkin SH. 2006. Developmental originof a bipotential myocardial and smooth muscle cell pre-cursor in the mammalian heart. Cell 127: 1137–1150.
Xu X, Francis R, Wei CJ, Linask KL, Lo CW. 2006. Connexin43-mediated modulation of polarized cell movement andthe directional migration of cardiac neural crest cells.Development 133: 3629–3639.
Yang JT, Rayburn H, Hynes RO. 1995. Cell adhesion eventsmediated by a4 integrins are essential in placental andcardiac development. Development 121: 549–560.
Yang L, Soonpaa MH, Adler ED, Roepke TK, Kattman SJ,Kennedy M, Henckaerts E, Bonham K, Abbott GW, Lin-den RM, et al. 2008. Human cardiovascular progenitorcells develop from a KDRþ embryonic-stem-cell-derivedpopulation. Nature 453: 524–528.
Zaffran S, Kelly RG, Meilhac SM, Buckingham ME, BrownNA. 2004. Right ventricular myocardium derives fromthe anterior heart field. Circ Res 95: 261–268.
Zamora M, Manner J, Ruiz-Lozano P. 2007. Epicardium-derived progenitor cells require b-catenin for coronaryartery formation. Proc Natl Acad Sci 104: 18109–18114.
Zeisberg EM, Ma Q, Juraszek AL, Moses K, Schwartz RJ,Izumo S, Pu WT. 2005. Morphogenesis of the right ven-tricle requires myocardial expression of Gata4. J ClinInvest 115: 1522–1531.
Zhou B, Ma Q, Rajagopal S, Wu SM, Domian I, Rivera-Feliciano J, Jiang D, von Gise A, Ikeda S, Chien KR, etal. 2008a. Epicardial progenitors contribute to the cardi-omyocyte lineage in the developing heart. Nature 454:109–113.
Zhou B, von Gise A, Ma Q, Rivera-Feliciano J, Pu WT.2008b. Nkx2–5- and Isl1-expressing cardiac progenitorscontribute to proepicardium. Biochem Biophys Res Com-mun 375: 450–453.
Embryonic Heart Progenitors and Cardiogenesis
Cite this article as Cold Spring Harb Perspect Med 2013;3:a013847 17
ww
w.p
ersp
ecti
vesi
nm
edic
ine.
org
Press on September 3, 2014 - Published by Cold Spring Harbor Laboratoryhttp://perspectivesinmedicine.cshlp.org/Downloaded from





























