Embed Size (px)
Citation preview

ENZİMLER

Jon BerzeliusPatateste katalitik görev yapan maddeler olduğunu,
Louis PasteurKatalitik aktivitenin hücre içinde olduğunu, canlı hücreden ayrılamayacağını,
Hans ve Eduard Büchner (1897)Canlı hücreden ayırdığı maya ekstraktı ile glukozu parçaladı.
Duclaux (19. yy sonu)Enzim substratının sonuna “az” eki getirilmesini önermiştir. (Örn: proteaz, üreaz, lipaz, amilaz, vb.)
James Sumner (1926)İlk kez bir enzimin (üreaz) kristalizasyonunu gerçekleştirdi ve bunun bir protein olduğunu gösterdi.
Thomas Ceck ve Sidney Altman (1980)Belli tip RNA moleküllerinin enzim özelliği gösterdiğini tespit ettiler (ribozimler: RNA enzimleri)

International Union of Biochemistry (IUB) adlandırma sisteminin özellikleri (1965)
1. Reaksiyonlar ve onları katalizleyen enzimler 6sınıfa ayrılırlar. Her birinin 4-13 alt sınıfı vardır.
2. Enzimin adı 2 kısımdan oluşmaktadır. (substrat veya substratların adı)+ (katalize edilen reaksiyon tipi)-az
Örn: L-malat= NADP+ oksiredüktaz
3. Söz konusu reaksiyonu aydınlatmak için ilave bilgi gerekirse parantez içinde verilebilir.
Örn: L-malat + NADP+ Piruvat + CO2 + NADPH + H+
L-malat : NADP+ oksiredüktaz (dekarboksile eden)

4. Her enzimin bir kod numarası vardır.Reaksiyon tipi - Alt sınıf - Alt alt sınıf - Enzim için spesifik
EC 2. 7. 1. 1
1. sayı 2. sayı 3. sayı 4. sayı
2: Transferaz
7: Fosfat transferi
1: Fosfat transferi görevini yapan alkoldür
1: Glukozun 6. karbonuna fosfat transferi yapan hegzokinaz

ÖRNEK: 4. Liyazlar
4.1. Karbon- karbon liyazlar (C-C bağının parçalanması)
4.1.1. Karboksilazlar (C-COO- bağının parç.)
4.1.1.1 ……………………..
4.1.1.2. Histidin karboksilaz
(histidindeki C-COO- bağının parç.)
…………………………..
4.1.2. Aldehit liyazlar (C-CHO bağını parçalar)
4.2. Karbon-Oksijen liyazlar (C-O)
4.3. Karbon-Azot liyazlar (C-N)
4.4. Karbon-Sülfür liyazlar (C-S)

ENZİMLERİN SINIFLANDIRILMASI� 1. Oksiredüktazlar
� Dehidrogenazlar
� Peroksidazlar
� Redüktazlar
� 2. Transferazlar� Transaminazlar
� Transmetilazlar
� 3. Hidrolazlar� Esterazlar
� Glikozidazlar
� Peptidazlar
� Fosfatazlar
Oksidasyon-redüksiyon
Bir fonksiyonel grubun bir molekülden diğerine taşınması
Hidrolitik parçalanma
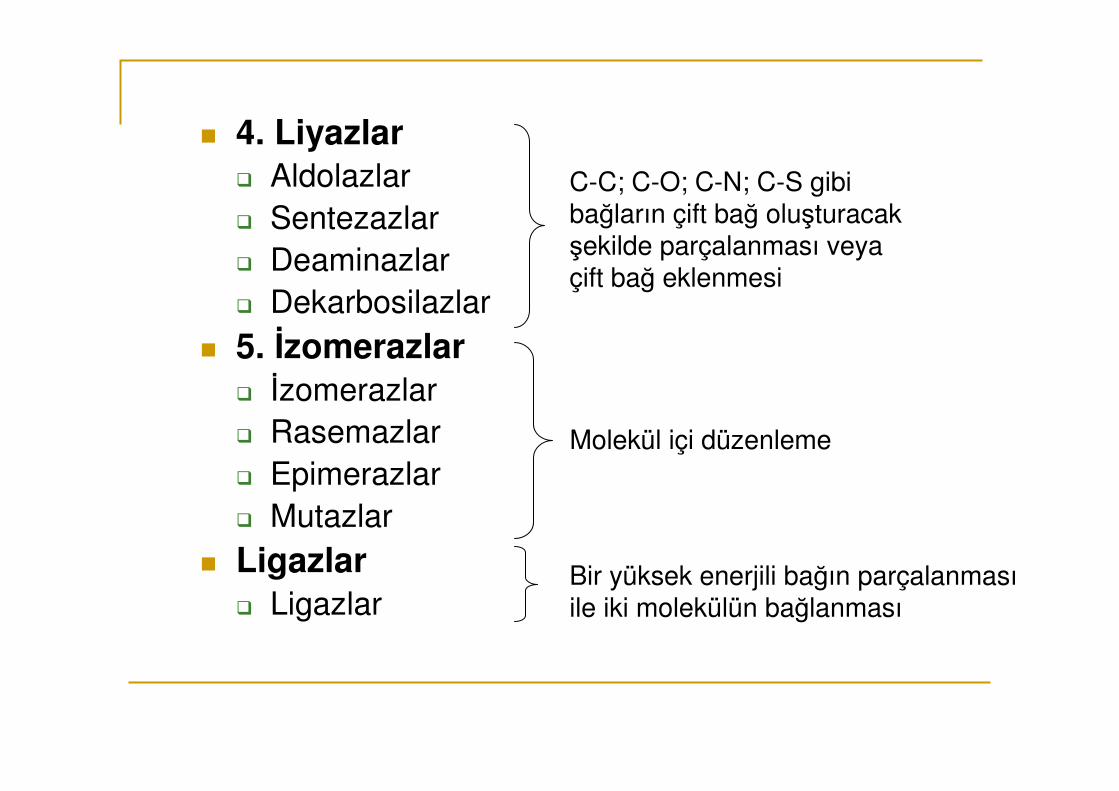
� 4. Liyazlar� Aldolazlar� Sentezazlar� Deaminazlar� Dekarbosilazlar
� 5. İzomerazlar� İzomerazlar� Rasemazlar� Epimerazlar� Mutazlar
� Ligazlar� Ligazlar
C-C; C-O; C-N; C-S gibi bağların çift bağ oluşturacak şekilde parçalanması veya çift bağ eklenmesi
Molekül içi düzenleme
Bir yüksek enerjili bağın parçalanması ile iki molekülün bağlanması

ENZİMLERİN SAFLAŞTIRILMASI
� Amaç:
Enzimin içinde bulunduğu çok sayıda farklı madde içeren hücre ekstraktından ayrılmasıdır.Örnek:
Kaba homojenat Berrak sıvı Çökeltisantrifüj Amonyum sülfat
Kolon kromatografisi
Kristalizasyon

ENZİMLERİN ÖZELLİKLERİENZİMLER BİYOLOJİK KATALİZÖRLERDİR.
� Katalizörler reaksiyonun hızını arttıran maddelerdir.� Katalizörler reaksiyon sonunda kaybolmazlar.� Katalizörlere sadece çok küçük miktarlarda
gereksinim duyulur.� Reaksiyonun denge noktasını değiştirmezler.
Sadece dengeye ulaşmak için gerekli süreyi kısaltırlar.
� Katalizörlerin genel özelliklerinden farklı olarak enzimler bazı farklılıklara sahiptir.
� Enzim etkinliği� Enzim spesifikliği� Enzim kontrolü

� 1. Enzim etkinliği: � Enzimler diğer katalizörlerden daha etkilidirler. Örn.,
2H2O 2H2O2 + O2
� Katalizör yoksa reaksiyon çok yavaş gerçekleşir.Ferrik iyon (Fe+3) reaksiyon hızını 30.000 kat arttırırken,Katalaz enzimi reaksiyon hızını 108 kat arttırır.
� 2. Enzim spesifikliği� Organik ve inorganik katalizörler çok sayıda farklı substratlara
etkilidirler.� Enzimler ise, hem substrat yapısı hem de reaksiyon tipi açısından
son derece seçicidirler.� Mutlak spesifiklik� Grup spesifikliği� Reaksiyon veya bağ spesifikliği� Stereo-kimyasal spesifiklik
� 3. Enzim kontrolü:� Enzimlerin katalitik etkinliği kontrol edilebilir, enzim aktivitesi
arttırılabilir, azaltılabilir veya tamamen durdurulabilir.

ENZİM AKTİVİTESİ� Enzimler protein yapısında oldukları için 20 a.a’in
sayısız kombinasyonu sonucu üretilebilirler ve çok yüksek yapısal ve fonksiyonel farklılıklar gösterirler.
Enzimlerin çalışmasını anlayabilmek için öncelikle:� Kimyasal Reaksiyonlar İçin Kinetik (Çarpışma) Teorisi
A. Bir reaksiyonun olması için moleküller çarpışmalıdır. (Yani birbirlerine bağ yapabilecek uzaklıkta bulunmalıdır.)
B. Çarpışmanın bir reaksiyona sebep olabilmesi için, moleküller o reaksiyona ait enerji engelini yenecek enerjiye sahip olmalıdırlar.
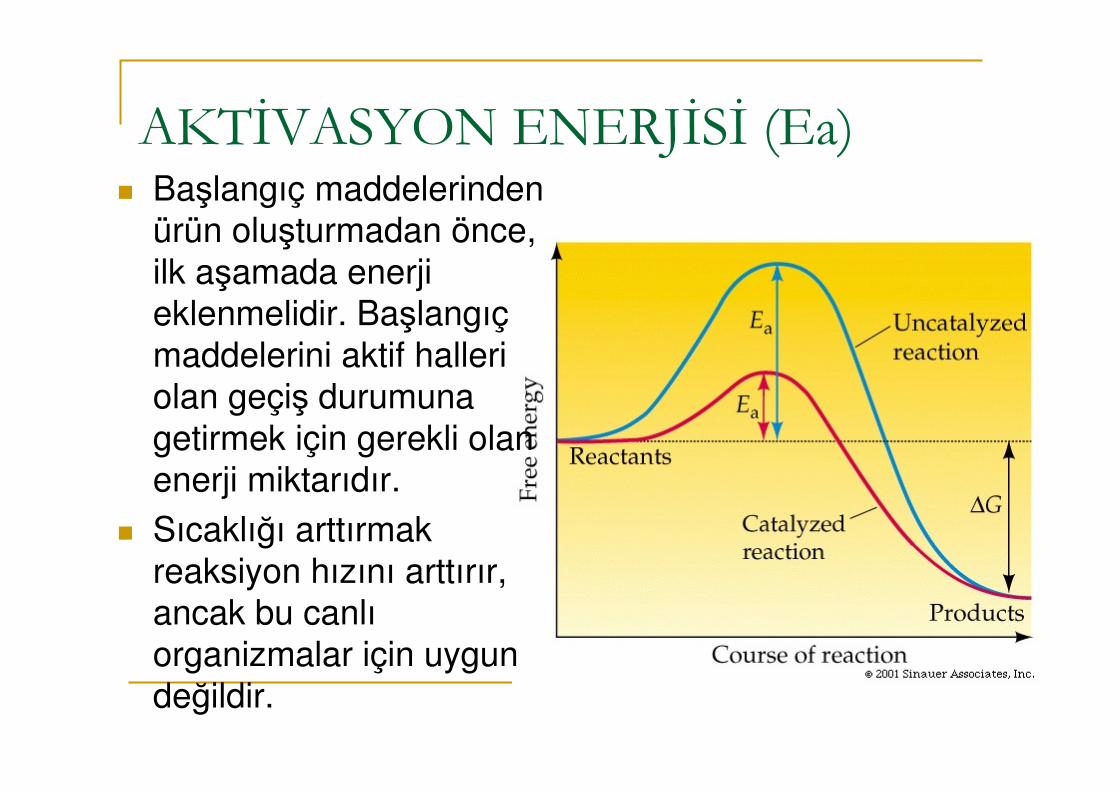
AKTİVASYON ENERJİSİ (Ea)� Başlangıç maddelerinden
ürün oluşturmadan önce, ilk aşamada enerji eklenmelidir. Başlangıç maddelerini aktif halleri olan geçiş durumuna getirmek için gerekli olan enerji miktarıdır.
� Sıcaklığı arttırmak reaksiyon hızını arttırır, ancak bu canlı organizmalar için uygun değildir.
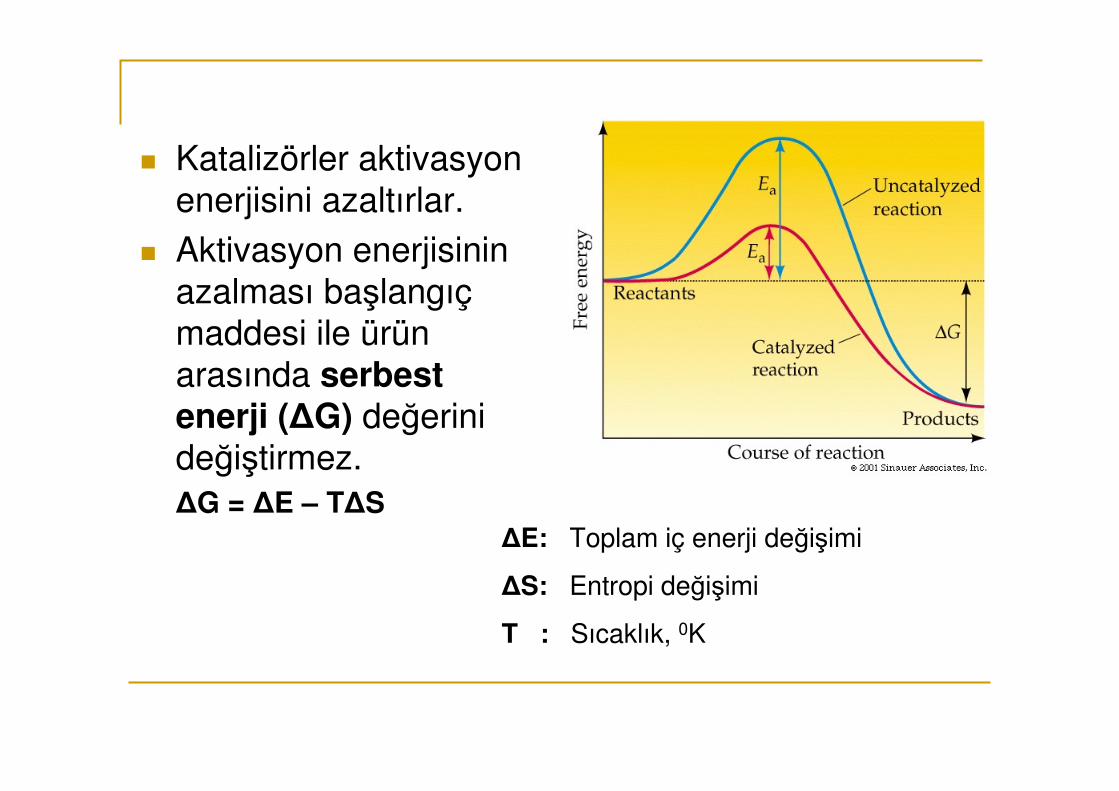
� Katalizörler aktivasyon enerjisini azaltırlar.
� Aktivasyon enerjisinin azalması başlangıç maddesi ile ürün arasında serbest enerji (∆G) değerini değiştirmez.∆G = ∆E – T∆S
∆E: Toplam iç enerji değişimi
∆S: Entropi değişimi
T : Sıcaklık, 0K
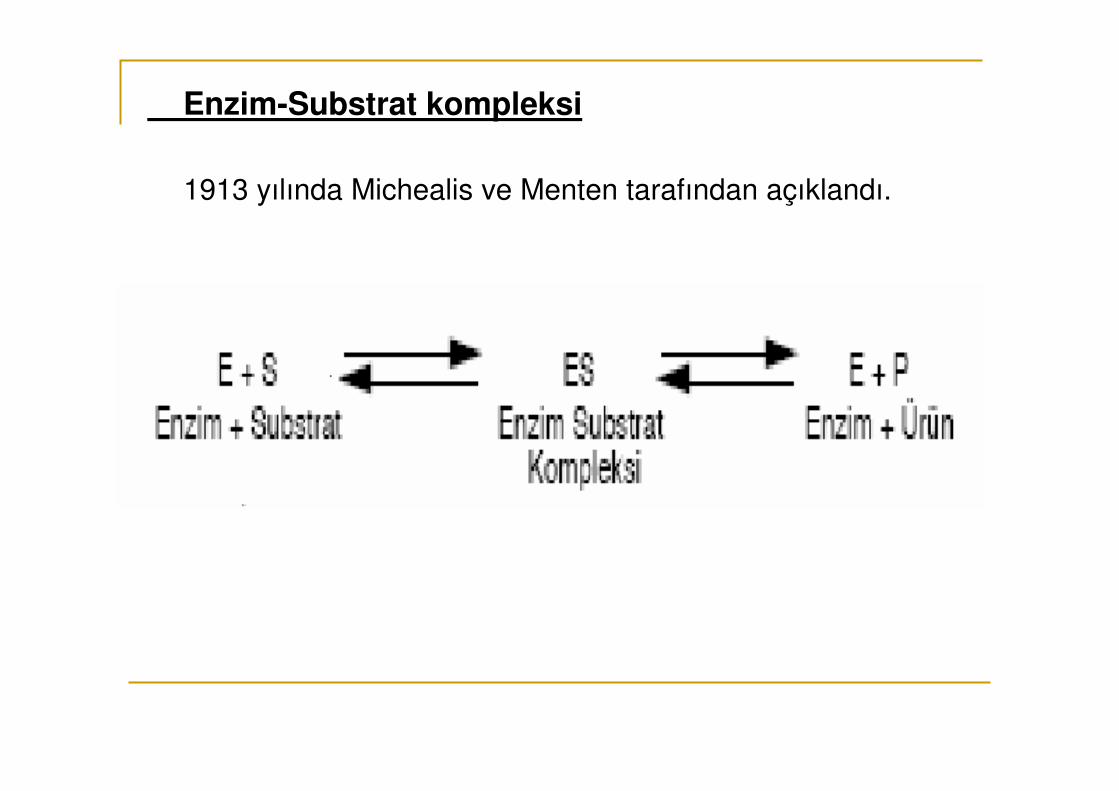
Enzim-Substrat kompleksi
1913 yılında Michealis ve Menten tarafından açıklandı.
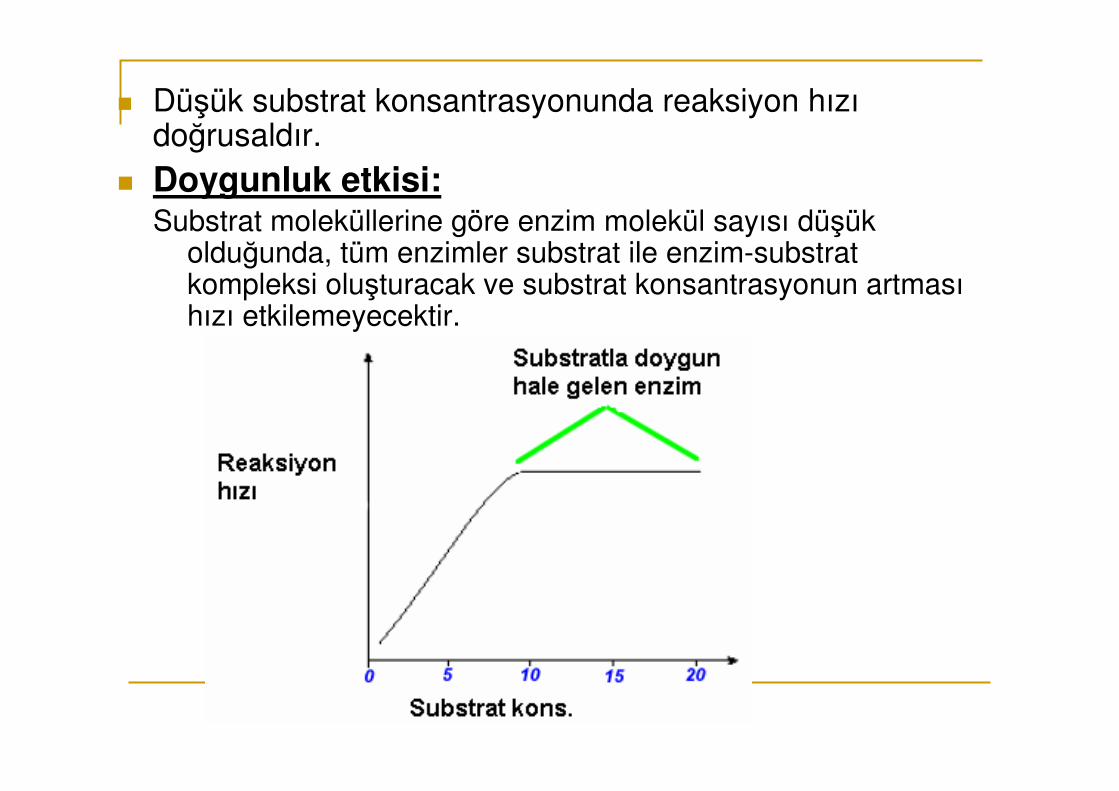
� Düşük substrat konsantrasyonunda reaksiyon hızı doğrusaldır.
� Doygunluk etkisi:Substrat moleküllerine göre enzim molekül sayısı düşük
olduğunda, tüm enzimler substrat ile enzim-substrat kompleksi oluşturacak ve substrat konsantrasyonun artması hızı etkilemeyecektir.

ENZİMLERİN AKTİF BÖLGELERİ� Enzim spesifikliği aktif bölge şekli ile açılanabilir.
� Her proteinin özgün 3 boyutlu bir yapısı vardır.
� Emil Fischer, enzimin aktif bölgesinin, etki ettiği substrat ile uygun olduğunu bildirmiştir.
� Aktif Bölge, bir enzimin substrata bağlanma noktasıdır.
� Enzimler dinamik bir yapıya sahiptir. Yapıları substrat bağlanması ile değişime uğrayabilir.

Kimotripsin Enziminin Bir Peptid Bağını Hidrolize Etmesi
� Bir substrat molekülü kimotripsine bağlandığı zaman enzimin aktif bölgesinde bulunan Ser-195 hidrolize edilecek peptid bağının karbon atomu ile kovalent bir bağ yapar. (2)
� Bu olay proton alıcısı rolü oynayan His-57 tarafından kolaylaştırılır. (4) Sonra His-57 aldığı protonu, hidroliz olan peptid bağının azotuna geri vererek peptid bağının parçalanmasını kolaylaştırır. (2)-(3)
� Reaksiyonun son aşamasında His-57 su molekülünden proton alır(4) ve Ser-195’e iletir(5) ve enzime kovalent olarak bağlı bulunan peptid parçaları enzimden ayrılırlar.(6)

Aktif Bölgenin Katalitik Etkinliği� Substrat aktif bölgeye bağlandıktan sonra reaksiyon
hızı bazı olaylara bağlıdır.1. Birden fazla substrat gerektiren reaksiyonlarda, aktif bölge
reaksiyona giren maddeleri birbirine yaklaştırır. Uygun bir konuma getirerek reaksiyon oluşma şansını optimize eder.
2. Bazı enzimler substrat ile stabil olmayan bir kovalent ara yapı oluşturur. Bu son ürün oluşumunu hızlandırır.
3. Çoğu enzimin aktif bölgesinde elektron/proton alabilen veya verebilen fonksiyonel gruplar bulunur. Bu e/p transferi ürün oluşumunu hızlandıran geçiş durumu yaratarak reaksiyon hızını arttırır.
4. Substratın aktif bölgeye bağlanmasının substrat molekülün aktivitesini arttıran yapısal gerilime veya bozulmaya neden olduğu düşünülmektedir.

ENZİM KİNETİĞİ� Vmax = Belli bir miktar enzim varlığında reaksiyonun
ulaşabileceği maksimum hız (Doygunluk hızı)
� Km = Michaelis-Menten sabiti= Reaksiyon hızının maksimum hızın yarısına eşit olduğu andaki substrat konsantrasyonu. (~10-7-10-1M)
� Vi= Tepkime hızıV i =
Vmax
Km + [S]
Enzim konsantrasyonu iki katına çıkınca max. hız iki kat artar ancak Km sabit kalır.
Eğer Km = [S] olursa;
V i =Vmax
2

ENZİM AKTİVİTESİNİ ETKİLEYEN FAKTÖRLER
1. Süre:Başlangıçta hız yüksektir. Oluşan ürünler enzimi inhibe ettiği için reax. hızı azalır.
2. Enzim Konsantrasyonu: Hız ile konsantrasyon doğrusaldır. 3. Substrat Konsantrasyonu:
Düşük substrat kons. da reaksiyon hızı substrat konsantrasyonu ile doğrusaldır.

4. Sıcaklık ve pH� Enzim aktivitesi sıcaklık ve pH’daki
küçük değişikliklere duyarlıdır.� Aşırı sıcaklık ve pH değişmelerinde ise
enzim inaktive olur. � Optimum enzim aktivitesi pH 7
civarındadır. Memeli canlıların enzimleri optimum 370C’de aktivite gösterir.
� Sıcaklığın artması ile enzim aktivitesi artar.
� Sıcaklık daha da artarsa enzim denatüre olur ve aktivite düşer.
� Pepsin ve arginaz gibi asidik ve bazik pH’da görev yapan enzimlerin optimum pH’sı görev yaptıkları çevrenin pH’sınayakındır.

KOFAKTÖRLER
1. Koenzimler
2. Prostetik gruplar
3. Metal iyonları
Apoenzim
(protein kısmı)
Kofaktör
(protein olmayan)
Haloenzim
(aktif enzim)
+

1. Koenzimler
Enzime geri dönüşümlü olarak bağlanırlar ve prosese doğrudan katılırlar. Apoenzime gevşek bağlandıkları için, ürün oluşumundan hemen sonra ayrılır. Genellikle vitamin yapısındadırlar.
Vitamin Koenzim Fonksiyonu
� Nikotinik asit NAH+ oksidasyon-redüksiyon
� Riboflavin FMN oksidasyon-redüksiyon
� Pantotenik asit koenzim A açil grubu transreri
� Biyotin biyositin karbondioksit transferi
� Tiamin tiaminpirofosfat amino grubu transferi

2. Prostetik gruplarKoenzimlere benzemekle birlikte enzime daha sıkı (kovalent bağ) bağlıdırlar. Örn: Katalaza hem grubu bağlanmadan H2O2’i parçalayamaz.
3. Metal iyonlarıFe, Cu, Mn, Mg, Ca, Zn… enzimlerin katalitik mekanizmasında görev yaparlar.Enzim veya substratın uygun konformasyonunu stabilize eder veya katalize edilen reaksiyona doğrudan katılırlar. Metal iyonları prostetik gruba bağlı da olabilirler.Örn: katalaz enzimi: protein + hem(Fe) (Katalazdaki Fe atomu (e-) alma ve verme görevi yapar)

ENZİM İNHİBİTÖRLERİ
� Pek çok doğal ve sentetik kimyasallar enzim aktivitesini engellemektedir. Etki mekanizmalarına göre inhibitörler;1. Geri dönüşümsüz (irreversible) inhibitörler,
2. Yarışmalı (competitive) inhibitörler,
3. Yarışmasız (non-competitive) inhibitörler.

1. Geri Dönüşümsüz İnhibitörler:Enzime kovalent bağ ile bağlanarak aktiviteyi tamamen durduran toksik maddelerdir. Bu maddeler toksik olup, çevre için sorun yaratırlar.
Örn: DİPF (diisopropil fosfofloridat) insektisitler ve sinir gazlarının temel bileşenidir. Serin amino asidinin OH grubuna kovalent olarak bağlanırlar.
Serin pek çok enzimin aktif bölgelerinde rol aldığı için enzimler inaktif hale gelir.
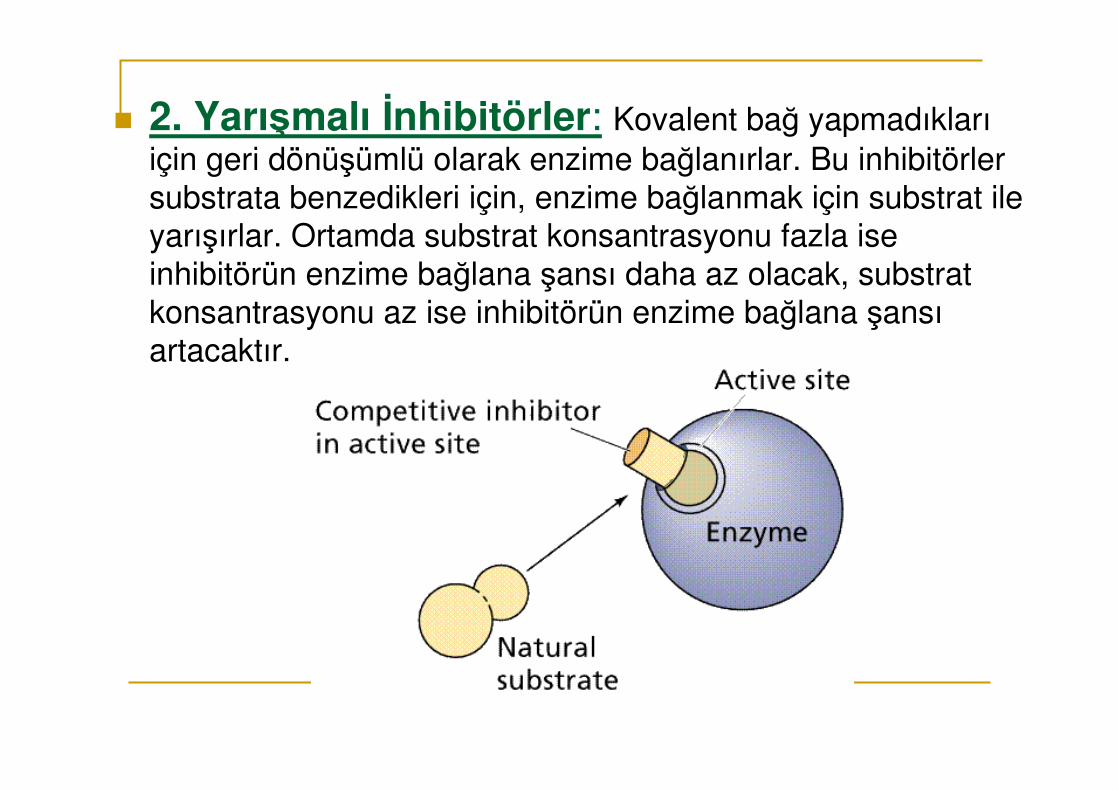
� 2. Yarışmalı İnhibitörler: Kovalent bağ yapmadıkları için geri dönüşümlü olarak enzime bağlanırlar. Bu inhibitörler substrata benzedikleri için, enzime bağlanmak için substrat ile yarışırlar. Ortamda substrat konsantrasyonu fazla ise inhibitörün enzime bağlana şansı daha az olacak, substrat konsantrasyonu az ise inhibitörün enzime bağlana şansı artacaktır.

3. Yarışmasız İnhibitörler:Kovalent bağ yapmadan, enzime geri dönüşümlü
olarak bağlanırlar. Yarışmalı inhibitörlerden farkı;fazla substrat ilavesi yarışmasız inhibitörlerin enzimden ayrılmasını sağlamaz. Bunlar substrata benzemez ve aktif bölgeden başka bir yere bağlanarak enzim aktivitesini azaltırlar.

ENZİM KONTROLÜ
� Diğer katalizörlerden farklı olarak enzimlerin aktiviteleri kontrol edilebilmektedir.
� Enzim aktivitesini düzenleyen mekanizmalar� Enzim miktarını değiştirmek,
� Substrat konsantrasyonunu değiştirmek,
� Enzimin katalitik etkinliğini değiştirmek.

FEED-BACK İNHİBİSYONU
� Metabolik enzimlerin kontrolü, genellikle metabolik olay sonucu üretilen ürün tarafından gerçekleşir. Böylece bir ürünün aşırı miktarda üretimi engellenmiş olur.
� 1963’de ortaya atılan allosterik kontrol teorisine göre;� Feedback inhibitörler inhibe ettikleri enzimin aktif bölgesine
değil allosterik bölge denen özel bir kontrol bölgesine bağlanır.
� Allosterik madde de aktif bölge gibi sadece belli moleküllere özgüdür.
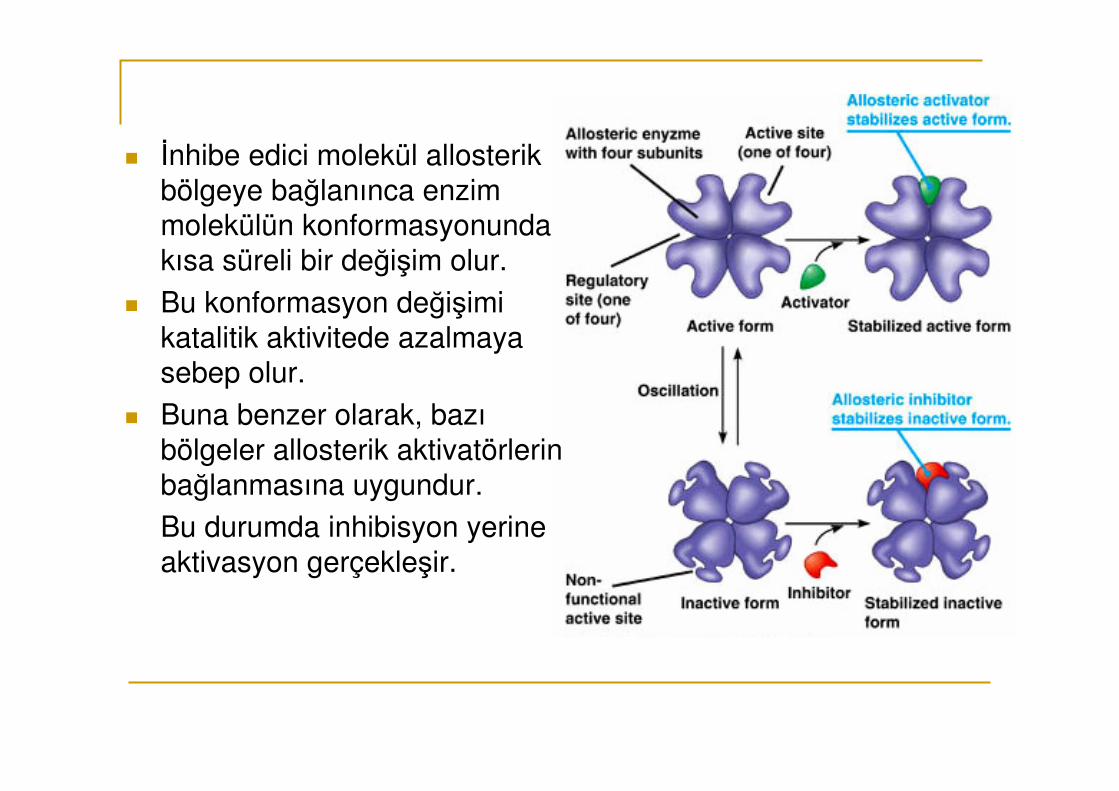
� İnhibe edici molekül allosterik bölgeye bağlanınca enzim molekülün konformasyonunda kısa süreli bir değişim olur.
� Bu konformasyon değişimi katalitik aktivitede azalmaya sebep olur.
� Buna benzer olarak, bazı bölgeler allosterik aktivatörlerin bağlanmasına uygundur.
Bu durumda inhibisyon yerine aktivasyon gerçekleşir.

ENZİMLERİN EŞLEŞMİŞ REAKSİYONLARI
� Kontrolör ve katalizör görevlerine ek olarak enzimler, termodinamik olarak kendiliğinden oluşmayan (enerji gerektiren) reaksiyonları, kendiliğinden oluşan reaksiyonlarla eşleştirerek, oluşmasını sağlar.
A B ∆G= +4 kcal/mol (endojen)
C D ∆G= -7 kcal/mol (eksojen) A + C B + D ∆G= -3 kcal/mol (eksojen)
� Eşleşme reaksiyonları canlılar için hayati öneme sahiptir. Canlı hücrede termodinamik açıdan mümkün olmayan reaksiyonlar da gerçekleşmiş olur.

� 1 Enzim Ünitesi (IUB’ye göre)� Bir dakika içinde 1 µmol substratın belli koşullar
altında ürüne dönüşümünü sağlayan enzim miktarıdır. Sıcaklık genellikle 300C, pH ise enzimin optimum pH’sıdır.
� 1 Katal (SI’ a göre)� Saniyede 1 mol substratın ürüne dönüşümünü
sağlayan enzim miktarıdır.

ENZİM AKTİVİTESİNİN REJENERASYONU
� Bazı enzimler ısıl işlemle inaktive edildikten bir süre sonra aktivitelerini yeniden kazanabilirler.
� Sütteki peroksidaz 750C’da inaktive olur. Ancak 24 saat sonra yeniden aktif hale geçer.
� Bezelyedeki peroksidaz 60sn haşlama ile inaktive olur. Ancak -180C’da 2 ay depolama sonunda bile enzim aktivitesi devam eder.

ENDÜSTRİYEL ENZİM ÜRETİMİ
� Hayvansal veya bitkisel kaynaklardan üretilebilirler.
Üretilen enzimler aşağıdaki özellikleri taşımalıdır:� Hammadde ucuz ve kolay bulunur olmalı,
� Enzimin ekstraksiyonu kolay olmalı,
� Kısmen saflaştırma ile kullanılabilmeli,
� Çevreyi kirletmemeli.

� Yukarıdaki kriterler göz önüne alındığında mikroorganizmalardan enzim üretiminde yararlanmak daha avantajlıdır.
� Mikroorganizmalardan enzim üretiminde, öncelikle;
� Uygun mikroorganizma seçilir,
� En yüksek verimle en aktif enzimin üretilebileceği besiyeri ve fermantasyon koşulları saptanır.

� Besiyerlerinde karbon kaynağı olarak;� nişasta hidrolizatı,
� melas,
� mısır,
� arpa,
� buğday;
� protein kaynağı olarak;� soya unu,
� pamuk küspesi,
� yer fıstığı unu,
� maya hidrolizatı,
� peyniraltı suyu,
� gluten kullanılabilir.

BAZI ENZİMLERİN ELDE EDİLDİKLERİ KAYNAKLAR
Fisinİncir
BromelinAnanas
PapainPapaya
ß-amilazTatlı patates
α ve ß-amilazArpa maltı
Bitkisel Kaynaklar
KatalazSığır ciğeri
RenninBuzağıların 4. işkembesi
PepsinDomuz mide mukozası
Pankreatin, tripsin,
imotripsin, lipaz, α-amilaz
Sığır ve domuz pankreası
EnzimHayvansal Kaynaklar

BAZI ENZİMLERİN ELDE EDİLDİKLERİ KAYNAKLAR
LaktazKluyveromyces fragilis
İnvertazSaccharomyces carlsbergensis
İnvertazSaccharomyces cerevisiae
KatalazMicrococcus lysodeikticus
α-amilaz, proteazBacillus subtilis
Fungal rennetEndothia parasitica
α-amilaz, glikoamilaz, pektinaz,
lipaz
Rhizopus spp.
α-amilaz, glukoamilaz, selülaz,
pektinaz, katalaz, glikozoksidaz
Aspergillus niger
α-amilaz, proteazAspergillus oryzae
EnzimMikroorganizma

MİKROORGANİZMALARDAN ENZİM ÜRETİMİNDE KULLANILAN YÖNTEMLER
1. Yüzey Yöntemi:Mik.org. lar sığ tavalar içindeki besiyerlerinde geliştirilirler (ör: küfler)2. Daldırma Yöntemi:Mikroorganizmalar havalandırılan tüp, şişe veya fermantörlerde
geliştirilir.Mikroorganizmaların gelişme aşamaları;
� Lag fazı� Log fazı� Durgun faz� Ölüm fazı
Bazı enzimler logaritmik fazda (örn:bakteriyel proteazlar) veya durgun fazda (örn: bakteriyel amilazlar) daha fazla enzim üretirler.
� Her mikroorganizmanın gelişme eğrisi bilinmelidir.

DALDIRMA VE YÜZEY YÖNTEMLERİNİN KARŞILAŞTIRILMASI
Enerji ihtiyacı fazlaEnerji ihtiyacı az
Kontaminasyon kaybı fazlaKontaminasyon kaybı az
Kontrolü zorKontrolü kolay
İşçilik azFazla iş gücü
Yüksek basınçlı havalandırmaDüşük basınçlı havalandırma
Kapalı fermantöre gerek varDaha geniş alan
Daldırma YöntemiYüzey Yöntemi

MİKROORGANİZMALARDAN ENZİMLERİN İZOLE EDİLMESİ
� Mikroorganizmalar üretildikten sonra gelişme sıvısından uzaklaştırılır. Bunun için santrifüj, filtrasyon vb. yöntemler uygulanır.
� Üretilen enzim extraselüler (hücre dışı) veya intraselüler (hücre içi) olabilir.
� Hücre içi enzimler hücre duvarı parçalanarak dışarı çıkarılır.
� Hayvansal hücrelerde izolasyon daha basittir.
� Saflaştırma işlemlerinin her aşamasında pH ve sıcaklık sürekli kontrol edilmelidir.

SAFLAŞTIRMA YÖNTEMLERİa. Tuzla Saflaştırma:
b. Adsorbsiyon:Adsorban madde enzim proteinini veya diğer maddeleri adsorbe
ederek saflaştırma yapılır. En çok kullanılan adsorbanlar: Ca-fosfat, aktif kömür, çinko hidroksit
c. Kolon Kromatografisi:Adsorbsiyon, iyon değiştirme ve moleküler ağırlığa göre ayırma
yapan kolonlarda saflaştırma yapılır.
Ham ekstrakt +Amonyum sülfat veya sodyum sülfat
15 dk bekleme Santrifüj(10000rpm)
Filtrasyon
Tortu

d. Elektroforez:� Protein molekülleri elektriksel alanda yüklerine göre
anoda yada katoda giderek saflaştırılabilirler. Laboratuar çalışmasına uygundur. Endüstriyel olarak kullanılamaz.
e. Kristalizasyon:� Amonyum sülfat ilavesi ile yapılır.
f. Diğer Yöntemler:� Ağır metal iyonları ile çöktürme,
� nükleik asitlerle ayırma

DEPOLAMA� Enzim aktivitesi, sterilite ve kuruluk kontrolleri
yapılmalıdır.
� Et tenderizasyonunda (yumuşatma) kullanılan proteazlar propilen glikol, ve NaCl içinde %3.5’lik çözelti halinde pazarlanır.
� Toz şeklinde enzimler ise kapsüllenerek pazara sunulur.

ENDÜSTRİYEL ENZİMLER� α- Amilaz: α-1,4 glikozit bağını parçalar. Hammadde olarak
arpa maltından elde edilir. Tükürük, pankreas ve bazıbitkilerde de bulunur. Nişastanın viskozitesini azaltır. Nişasta veya glikoz şurubu eldesinde diğer enzimlerle beraber kullanılır.
� ß-Amilaz: Polisakkaritlerdeki α-1,4 bağlarını indirgen olmayan uçlardan başlayarak maltoz birimlerine parçalar. Arpa, buğday ve pirinçte bulunur.
� Amiloglukozidaz: α-1,4 ve α-1,6 bağlarını parçalar. Nişastayı glikoz birimlerine kadar parçalar. Aspergillussuşlarından elde edilir.
� Maltaz: (glukozidaz) Maltozu glukoza parçalar. Nişasta sanayinde kullanılır.

� Selülaz: 3 enzimin karışımından oluşur. Selülozdaki β-1,4 bağlarına etki eder. Ticari olarak Tichaderma suşlarından elde edilir. Narenciye sularındaki bulanıklığı gidermek, birada filtrasyonu kolaylaştırmak, bazı ilaçlarda sindirim yardımcısıolarak kullanılır.
� Endoglukonaz: Selülozun β-1,4 bağlarını rasgele parçalar.
� Eksoglukonaz: β-1,4 glukan zincirinden bir glukozu ayırır.
� Sellobiyohidrolaz: Selülozu sellobioz(glu+glu)’a parçalar.
� Sellobiaz: Sellobiyozu glukoza parçalar.
� Hemiselülaz: Hemiselülozu parçalayan enzimlerdir. Örn: Galaktanazlar, ksilanazlar, mannazlar, ksiloglukanazlar…

İMMOBİLİZE ENZİMLER
Enzimlerin katalizör olarak pek çok avantajları olmasına rağmen, çoğu enzimin endüstriyel uygulaması,
� Enzimlerin izolasyon ve saflaştırma masraflarının yüksekliği� Bazı enzimlerin canlı hücreden ayrıldığı zaman aktivitelerini veya
stabilitelerini kaybedebilmeleri,� Enzimatik işlem bittikten sonra, reaksiyon karışımından enzimi
ayırmanın zor ve pahalı olması
� Bu sorunlar, enzimlerin suda çözünmeyen taşıyıcılar üzerinde immobilize edilmeleri ile çözülmüştür.
� Reaksiyon sonunda ortamdan kolaylıkla uzaklaştırılabildiği için, daha ekonomik bir uygulamadır.

İmmobilize enzim kullanmanın avantajları:� Çevre koşullarına daha dayanıklıdırlar� Reaksiyon sonunda ortamdan kolaylıkla
uzaklaştırılabildiği için ürünle kirlenme sorunu yoktur.� Pek çok kez ve uzun süre kullanılabilir.� Sürekli işlemlerde kullanılabilirler� Ürün oluşumu daha kolay kontrol edilebilir� Bazı durumlarda, serbest enzimlerden daha yüksek
aktivite gösterebilirler.� Enzimin kendi kendini parçalama olasılığı daha azdır.

İMMOBİLİZASYON YÖNTEMLERİ1. Tutuklama� Enzim bir polimer matriks veya yarı-geçirgen membran içinde
tutuklanmıştır. 2. Çapraz bağlama� Küçük moleküllü iki veya çok fonksiyonlu polimerleştirici maddeler
yardımı ile enzim molekülleri arasında bağlar oluşturarak suda çözünmeyen kompleksler meydana getirilir.
3. Taşıyıcıya bağlanmaEnzimler kendilerine uygun bir taşıyıcıya,
� Fiziksel adsorbsiyon� Kovalent bağlama� İyonik bağlama ile bağlanabilir.
Doğal taşıyıcılar: Selüloz, nişasta, kollagen, kitin Yapay taşıyıcılar: Polistiren, poliakrilamid, polivinil alkol, vb





























