Embed Size (px)
Citation preview

Evaluation of Lovastatin Effects on Expressionof Anti-apoptotic Nrf2 and PGC-1α Genes in NeuralStem Cells Treated with Hydrogen Peroxide
Alireza Abdanipour & Taki Tiraihi & Ali Noori-Zadeh &
Arezo Majdi & Ramin Gosaili
Received: 29 August 2013 /Accepted: 8 December 2013# Springer Science+Business Media New York 2014
Abstract Reactive oxygen species and oxidative stress areassociated with various cell processes, including cell survivaland apoptosis. Oxidative stress has been implicated in thepathogenesis of several neurological disorders includingAlzheimer's disease (AD), Parkinson's disease (PD), amyotro-phic lateral sclerosis (ALS), Huntington's disease (HD), andmultiple sclerosis (MS). In the present study, we evaluated theeffects of lovastatin chemoprotection against hydrogenperoxide-induced oxidative stress in bone marrow stromalcell-derived neural stem cells (BMSC-derived NSCs) andwhether it has protective effects. BMSC-derived NSCs werepretreated with different doses of lovastatin for 48 h and thenexposed to 125 μM H2O2 for 30 min. Using MTT, TUNELassay, and real-time RT-PCR, we evaluated the effects oflovastatin on cell survival, apoptosis, and PGC-1α and Nrf2expression rates in pretreated BMSC-derived NSCs comparedto control groups. Results showed that apoptosis rate in thelovastatin-pretreated BMSC-derived NSCs was significantlydecreased compared to the control group. Our findings sug-gest that lovastatin protects NSCs against oxidative stress-induced cell death, and therefore, it may be used to promotethe survival rate of NSCs and can be a candidate for treatmentof oxidative stress-mediated neurological diseases.
Keywords Lovastatin . Oxidative stress . Nrf2 . PGC-1α .
Neural stem cells
Introduction
Oxidative stress has been implicated in the etiology of neuro-degenerative diseases, aging, and cancer. Oxidative stress andoverproduction of reactive oxygen species can lead to DNAdamage and degradation of proteins and lipids, causing neu-rodegenerative diseases including Alzheimer's disease (AD),Parkinson's disease (PD), amyotrophic lateral sclerosis (ALS),multiple sclerosis (MS), and Huntington's disease (HD) [1–3].Although the precise pathophysiological mechanisms remainunknown, oxidative stress may be the major triggering factorin the apoptosis events of the aforementioned diseases [4].The 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA)reductase inhibitors or statins are the most widely prescribeddrugs in the world for the primary and secondary preventionof cardiovascular disease. Indeed, decreases in LDL choles-terol with statin therapy correlate almost linearly with reduc-tions in cardiovascular events [5, 6]. In recent years, accumu-lating evidence indicates that statins suppressed H2O2-in-duced oxidative stress and attenuated its induced cell injuryvia expression induction of the Nrf2 (nuclear factor(erythroid-derived 2)-like 2) [7]. According to the solubilityproperties, statins can be categorized into two major groups:lipophilic and hydrophilic. The former group which can read-ily cross the blood–brain barrier (BBB) includes simvastatin,lovastatin, atorvastatin, and cerivastatin, while hydrophilicstatins include rosuvastatin and pravastatin [6]. In the presentstudy, we investigated the in vitro therapeutic effects of lova-statin on apoptosis inhibition and cell survival of bonemarrowstromal cell-derived neural stem cells (BMSC-derived NSCs)treated with hydrogen peroxide. Our results showed that PGC-1α (peroxisome proliferator-activated receptor γ coactivator-1α) and Nrf2 expression rates increased more significantly incomparison with control groups and that H2O2-induced cellinjury was attenuated markedly.
A. Abdanipour (*) :A. Majdi : R. GosailiStem Cells Research Laboratory, Department of Medical Sciences,Islamic Azad University, Ardabil Branch, Ardabil, Irane-mail: [email protected]
T. Tiraihi :A. Noori-ZadehShefa Neuroscience Research Center, Khatam Al-Anbia Hospital,Tehran, Iran
Mol NeurobiolDOI 10.1007/s12035-013-8613-5

Materials and Methods
BMSC Culture
Wistar rat (200–300 g) BMSCs were initially cultivated inDulbecco's Modified Eagle Medium (DMEM), supplementedwith 15 % fetal bovine serum (FBS, Gibco), 100 U/ml peni-cillin, and 100 mg/ml streptomycin. The cells were incubatedat 37 °C and 5 % CO2 in a 25-cm
2 plastic flask for 2 days, andnon-adherent cells were removed by replacement of the me-dium. The medium was changed every other day until cellsbecame confluent. The cells were harvested with trypsin–EDTA and passaged up to three times.
Neurosphere Formation and Expansion
Rat BMSCs were dissociated with trypsin–EDTA and platedon a 25-cm2 non-adherent plastic flask (105 cells/ml) inDMEM/F12 medium supplemented with 2 % B27 (Gibco),20 ng/ml basic fibroblast growth factor (bFGF, Invitrogen),20 ng/ml epidermal growth factor (EGF, Invitrogen), 100 U/ml penicillin, and 100 mg/ml streptomycin. The medium andgrowth factors were added every 2 days [8]. After severaldays, neurosphere-like structures were observed. The floatingneurospheres were collected by centrifuging at 300g for 5 min.Then, they were dissociated enzymatically and then mechan-ically (pipetting up and down) to single cells and suspended inDMEM/F12 supplemented with 2 % B27 (Gibco), 20 ng/mlbFGF, 20 ng/ml EGF, and 5 % FBS for 1 week in six-welladherent plates coated with poly-L-lysine (Sigma-Aldrich) andpassaged up to three times [9]. For these cells, immunocyto-chemical evaluation was performed for nestin marker.
Lovastatin Cell Treatments
To induce oxidative stress, H2O2 was freshly prepared from30% stock solution. The third passage of NSCswas cultured in96-well plates (105 cells/well) in DMEM/F12 medium supple-mented with 2 % B27, 20 ng/ml bFGF, and 20 ng/ml EGF, andthen pretreated NSCs were incubated with 125 μM H2O2 for30 min. To evaluate the protective effects of lovastatin, cellswere treated with different concentrations of lovastatin (0, 2, 4,6, 8, and 10 μM) 48 h before the H2O2 treatment.
Assessment of Cell Proliferation
Cell viability and proliferation were evaluated using the3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium (MTT)bromide assay, which is based on the conversion of MTT toformazan crystals [10]. The cells were treated with 125 μMH2O2 and different concentrations of lovastatin (0, 2, 4, 6, 8,and 10 μM) 48 h before the H2O2 treatment. Then, the cellswere incubated with 1mg/mlMTT for 4 h. The culture medium
was removed, and 100μl dimethyl sulfoxide was added to eachwell to dissolve the formazan crystals. The amount of formazanwas quantified at 570 nm using amicroplate ELISA reader. Therelative cell viability in percentage was calculated as (A570 oftreated samples/A570 of untreated samples)×100.
Detection of Apoptotic Cells (TUNEL Assay)
After treatment, the cells were fixed with 4 % paraformalde-hyde in phosphate-buffered saline (PBS, Gibco) for 30 min atroom temperature (RT). The cells were subjected to a terminaldeoxynucleotidyl transferase-mediated deoxyuridine triphos-phate nick end labeling (TUNEL) assay using the In Situ CellDeath Detection Kit (Roche, Germany) according to the man-ufacturer's instructions. TUNEL-positive cells were coloredusing diaminobenzidine (DAB) as the chromogen and coun-terstained with hematoxylin. The percentage of TUNEL-positive cells was assessed in five randomly selected fieldsfor each well.
Immunostaining
BMSC-derived NSCs were cultured on cover slides and fixedin 3 % paraformaldehyde for 20 min at RT, followed by apermeabilization step in 100%methanol for 30min at RT. Forimmunofluorescence, cells were incubated with anti-CD90(for BMSCs) and anti-nestin (for BMSC-derived NSCs)monoclonal antibodies (both from Abcam), followed by incu-bation with a fluorescein isothiocyanate (FITC)-conjugatedrabbit anti-mouse antibody (Millipore). Nuclei were counter-s t a i n ed w i t h e t h i d i um b rom id e . Fo r i nd i r e c timmunoperoxidase labeling, 6 μM-treated BMSC-derivedNSCs were permeabilized with 0.4 % Triton X-100, followedby immersion in 3 % H2O2/methanol for 15 min to blockendogenous peroxidase. They were incubated with anti-PGC-1α and Nrf2 antibodies (both from Abcam) overnight at 4 °C.Then, BMSC-derived NSCs were processed with avidin–bio-tin complex (ABC kit) according to the manufacturer's in-structions. Primary antibodies were visualized using the di-aminobenzidine method.
Real-Time RT-PCR
Real-time RT-PCR was carried out with cDNA from N (un-treated BMSC-derived NSCs), NL (BMSC-derived NSCstreated with 6 μM lovastatin), NH (BMSC-derived NSCstreated with 125 μMH2O2), and NLH (BMSC-derived NSCstreated with 6 μM lovastatin and 125 μM H2O2) groups aftertreatments. In all groups, 1,000 ng purified RNA from cul-tured cells was used to synthesize 20 μl cDNA, using Revertaid™ first-strand cDNA synthesis kit (Fermentas, Germany)according to the manufacturer's instructions. cDNA (500 ng)was used to quantify PGC-1α and Nrf2 mRNA levels. As an
Mol Neurobiol

internal control for normalization, beta-2-microglobulin(B2M) was used. All primers have been listed in Table 1.The PCR reaction was performed in a 25-μl final volume(sense and anti-sense primers, cDNA and SYBR® Green I)and carried out for 40 cycles (Applied Biosystems ThermalCycler). For analyzing relative changes in mRNA levels, weused delta CT method [11].
Statistics
Statistical analysis was performed using SPSS 15 software.All data are presented as means±SEM. To compare multiplemeans in groups, one-way ANOVA followed by Tukey's posthoc comparison was used. Values of P<0.05 were consideredstatistically significant.
Results
BMSC Culture and Neurosphere and NSC Formation
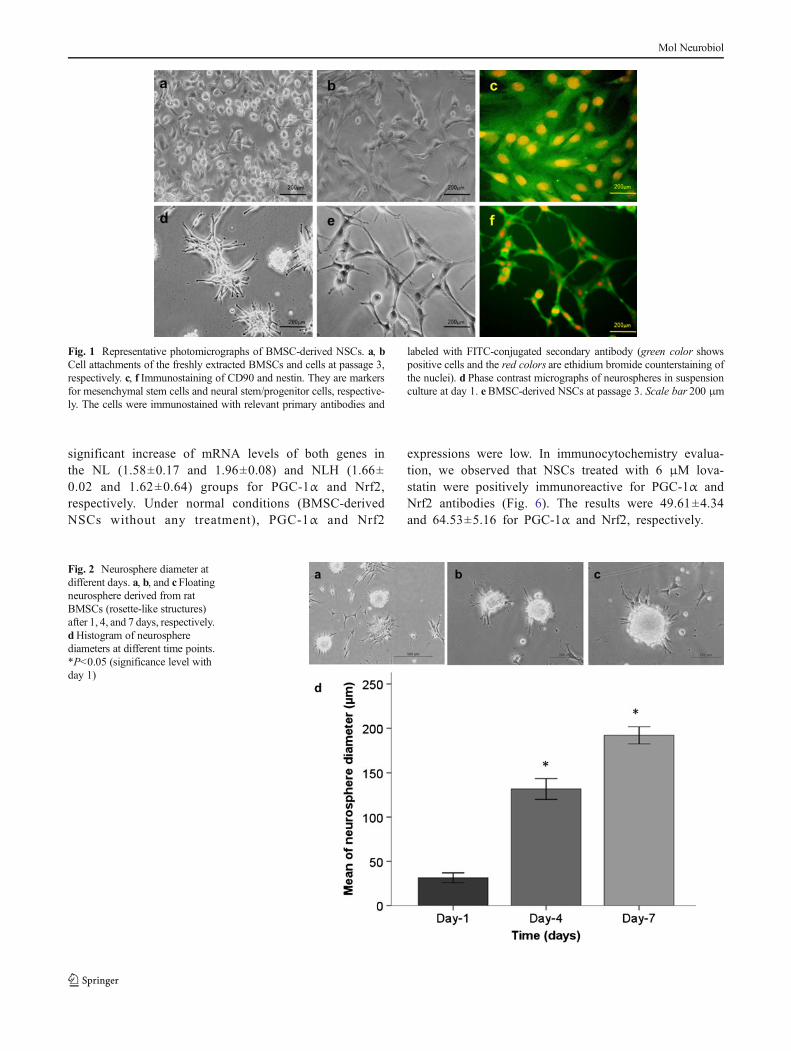
The results of BMSC culture showed that, at earlier hours, thecells were floating and at 6–8 h began to attach to the dishsurface. The adherent cells initially showed an irregular ap-pearance and then grew rapidly and became fibroblast-likeshapes (Fig. 1a, b). At 7–10 days, the adherent cells becameconfluent and could be passaged once every 3–4 days.BMSCs were found to express CD90 (98.7 %±3.2)(Fig. 1c). Then, BMSCs were induced to be differentiated inculture with neuronal induction media. After a few hours,neural-like cells with multipolar processes and growth cone-like features were identified. Small spheroids were observed atseveral days (Fig. 1d). We observed that spheroid diameters atday 4 (131.75±5.85 μm) and day 7 (192.02±4.83 μm) sig-nificantly increase compared to day 1 (31.49±3.7 μm). Itseems that some cells separate from the primary spheroidand proliferate and form secondary spheroid (Fig. 2a–d). Also,we cultured the intact neurospheres without dissociation in thesix-well adherent plates coated with poly-L-lysine. In thiscontext, the cells formed rosette-like structures (Fig. 2c). The
BMSC-derived NSCs from spheroids strongly were positivefor nestin immunostaining (99.23 %±0.17) (Fig. 1f).
Cell Viability
To examine the protective effects of lovastatin, BMSC-derivedNSCs were treated with various concentrations of lovastatinbefore the H2O2 exposure. As seen in Fig. 3a, b, H2O2-inducedcytotoxicity in BMSC-derived NSCs was significantly reversedby pretreatment with lovastatin. But there is no significantdifference between N and NL groups. Lovastatin promoted cellproliferation compared to untreated cells. Thus, the effects oflovastatin was dose dependent, and we chose 6 μM lovastatin(71.95 %±1.56) as the optimum dose which was selected forfurther neuroprotection studies because higher concentration oflovastatin had neurotoxic effects (Fig. 3a).
Evaluation Anti-apoptotic Effect of Lovastatin
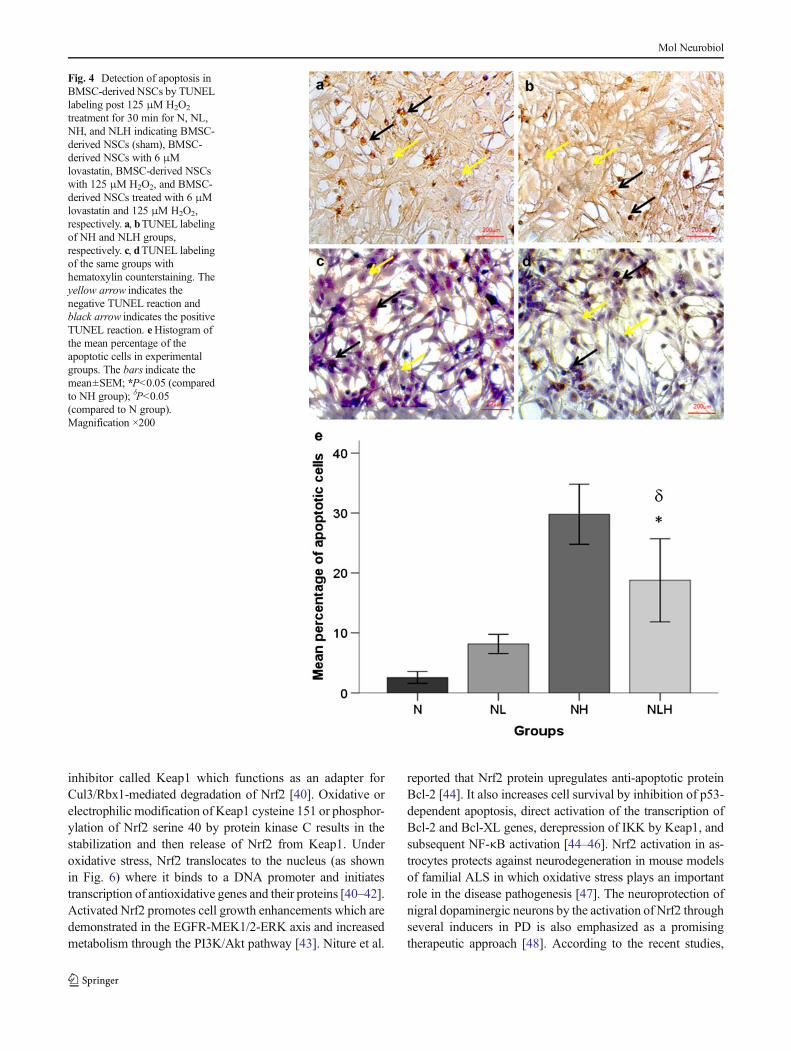
To determine anti-apoptotic effects of lovastatin in BMSC-derived NSCs treated with H2O2, TUNEL assay was used. Asshown in Fig. 4a–d, TUNEL-positive nuclei staining demon-strates that H2O2 induces apoptotic cell death. The percentageof TUNEL-positive cells decreased in the NLH group com-pared to the NH group (18.77±3.46 % versus 29.78±2.5 %,respectively; P<0.05) (Fig. 4e).
PGC-1α and Nrf2 Expression Evaluation
Dose-dependent increasing of PGC-1α and Nrf2 mRNAexpressions in BMSC-derived NSCs treated with variousconcentrations of lovastatin and various groups (N, NL,NH, and NLH) at 48 h was confirmed by quantitativereal-time RT-PCR. The results of mRNA expressionpattern have been shown in Fig. 5a, b. Our data showedthat mRNA expressions of both genes increased when arelatively higher concentration of lovastatin was used.Also, in the 6 μM lovastatin-treated group, mRNAexpressions were significantly upregulated compared toNH and N groups at 48 h. The results demonstrated a
Table 1 Primer sequences and PCR parameters. Primers for amplification of target sequences, their GenBank accession number, and size of thefragment amplified are presented
Gene Gene accession no. Sense 5→3 Anti-sense 5→3 Size (bp)
B2M NM_012512.2 CTTGCCATTCAGAAAACTCC CATCGGTCTCGGTGGGTG 235
PGC-1α NM_031347.1 GACAGAACTGAGAGACCGCT CTGAGTTTGAATCTAGGTCTGC 236
Nrf2 NM_031789.2 ATTCCCGAGTTACAGTGTCT CGTGGAGAGGATGCTGCT 204
Primers were designed by Gene Runner 3.05 software (product by [email protected])
B2M beta-2-microglobulin, PGC-1α peroxisome proliferator-activated receptor gamma coactivator-1 alpha, Nrf2 nuclear factor (erythroid-derived 2)-like 2 also known as NFE2L2
Mol Neurobiol
significant increase of mRNA levels of both genes inthe NL (1.58±0.17 and 1.96±0.08) and NLH (1.66±0.02 and 1.62±0.64) groups for PGC-1α and Nrf2,respectively. Under normal conditions (BMSC-derivedNSCs without any treatment), PGC-1α and Nrf2
expressions were low. In immunocytochemistry evalua-tion, we observed that NSCs treated with 6 μM lova-statin were positively immunoreactive for PGC-1α andNrf2 antibodies (Fig. 6). The results were 49.61±4.34and 64.53±5.16 for PGC-1α and Nrf2, respectively.
Fig. 1 Representative photomicrographs of BMSC-derived NSCs. a, bCell attachments of the freshly extracted BMSCs and cells at passage 3,respectively. c, f Immunostaining of CD90 and nestin. They are markersfor mesenchymal stem cells and neural stem/progenitor cells, respective-ly. The cells were immunostained with relevant primary antibodies and
labeled with FITC-conjugated secondary antibody (green color showspositive cells and the red colors are ethidium bromide counterstaining ofthe nuclei). d Phase contrast micrographs of neurospheres in suspensionculture at day 1. eBMSC-derived NSCs at passage 3. Scale bar 200 μm
Fig. 2 Neurosphere diameter atdifferent days. a, b, and cFloatingneurosphere derived from ratBMSCs (rosette-like structures)after 1, 4, and 7 days, respectively.dHistogram of neurospherediameters at different time points.*P<0.05 (significance level withday 1)
Mol Neurobiol

Discussion
In the present study, we demonstrated that lovastatin increasesPGC-1α and Nrf2 mRNA expressions in BMSC-derivedNSCs and protects them against hydrogen peroxide-inducedapoptosis. Outside of their conventional use in cholesterollowering, research on statins is ongoing for many new clinicalapplications. The favorable effects of statins on ischemicstroke have been well documented, and various mechanismsare involved [12]. Studies showed that statins completelyprevent the progression of PD [13, 14] and can be used forearly treatment of MS [15, 16]. By a variety of mechanisms,statins display favorable effects in AD as well [17, 18]. In vivo
experiments showed that statin treatment after brain damageincreases synaptogenesis and neurogenesis without alteringserum cholesterol levels. Also, it is suggested that statinsinduce neuroprotection by promoting the release of neuro-trophic factors and induction of gene expression such as thebrain-derived neurotrophic factor [19, 20]. Some studies pro-vide evidence that lovastatin activates the neuroprotectiveprotein kinase B (Akt), and it is attested in both lovastatin-treated cultured neurons [21] and central nervous systemlovastatin-treated animals. Also, it has been shown that pre-treatment of cultured cortical neurons with lovastatin canprotect them against excitotoxic damage [22]. Interestingly,it has been reported that lovastatin has neuroprotective im-pacts on the amyloid-β challenge through the Wnt signalingpathway [23]. Statins reduce axonal injury, increase neuriteoutgrowth, and help to recover neurological functions. It isbelieved that neurite outgrowth may be mediated throughmanipulation of the PI3K/Akt/mTOR and PI3K/GSK-3β/APC pathways [24, 25]. Also, it seems that statins can inducecell death in neurons and glia [26, 27]. Analysis of the cho-lesterol biosynthetic pathway revealed that the lack ofmevalonate and of its downstream metabolites, mainlygeranylgeranyl-pyrophosphate, is responsible for the statin-induced apoptosis of neurons and astrocytes [26]. The toxiceffect on oligodendrocytes can be reversed by farnesyl pyro-phosphate [27]. These results suggest that the inactivation ofprenylated proteins can cause cell death, and factors like celllinage properties and experimental techniques can influencethe outcomes. However, the effects of statins in proliferationand survival of oligodendrocyte precursors under in vivo andin vitro conditions are controversial [28, 29]. Overproductionof free radicals can cause oxidative damage to biomoleculeslike lipids, proteins, and DNA [30]. Numerous studies con-firmed that free radicals play a casual role in the neuronal celldeath [19, 31, 32]. In the neurodegenerative disorders such asPD, AD, and MS, reduction of oxidative stress can lead toinhibition of apoptotic cell death and disease progression[33–36]. In this study, to evaluate the protective effects oflovastatin, we first treated BMSC-derived NSCs with variousconcentrations of lovastatin and observed a dose dependentincrease in Nrf2 and PGC-1αmRNA expressions. Our resultsshowed increases of both gene mRNA levels in the 6 μMlovastatin-treated group after 48 h of treatment. PGC-1α is themaster regulator of mitochondrial function and serves as aninducible coactivator for a number of transcription factors tocontrol energy metabolism. It is expressed at high levels inoxidative tissues such as the brain [28, 37, 38]. IncreasedPGC-1α induces the transcription of nuclear respiratory factor(Nrf1) and Nrf2, leading to the increased expression of mito-chondrial transcription factor A (mtTFA) [39]. Recent studieshave shown that Nrf1, Nrf2, and PGC-1 family coactivatorscontrol mitochondrial transcription specificity factors [29].Studies show that Nrf2 is retained in the cytoplasm by an
Fig. 3 Protective effects of lovastatin treatment on cell viability. a Thedose–response BMSC-derived NSC viability at different concentrationsof lovastatin detected by MTT assay. Results show the mean percentviability (relative to untreated cells). Each bar represents the averagemeasurement from five replicates. bMean of absorbance at 570 nm usinga microplate reader for N, NL, NH, and NLH indicating BMSC-derivedNSCs (sham), BMSC-derived NSCs treated with 6 μM lovastatin,BMSC-derived NSCs with 125 μM H2O2 (untreatment), and BMSC-derived NSCs treated with 6 μM lovastatin and 125 μM H2O2, respec-tively. Asterisk denotes statistically significant difference in cell viabilitybetween 6 μM and 0, 2, and 10 μM concentrations. Double asteriskdenotes statistically significant difference between NLH and NH. Bargraphs indicate the mean±SEM
Mol Neurobiol
inhibitor called Keap1 which functions as an adapter forCul3/Rbx1-mediated degradation of Nrf2 [40]. Oxidative orelectrophilic modification of Keap1 cysteine 151 or phosphor-ylation of Nrf2 serine 40 by protein kinase C results in thestabilization and then release of Nrf2 from Keap1. Underoxidative stress, Nrf2 translocates to the nucleus (as shownin Fig. 6) where it binds to a DNA promoter and initiatestranscription of antioxidative genes and their proteins [40–42].Activated Nrf2 promotes cell growth enhancements which aredemonstrated in the EGFR-MEK1/2-ERK axis and increasedmetabolism through the PI3K/Akt pathway [43]. Niture et al.
reported that Nrf2 protein upregulates anti-apoptotic proteinBcl-2 [44]. It also increases cell survival by inhibition of p53-dependent apoptosis, direct activation of the transcription ofBcl-2 and Bcl-XL genes, derepression of IKK by Keap1, andsubsequent NF-κB activation [44–46]. Nrf2 activation in as-trocytes protects against neurodegeneration in mouse modelsof familial ALS in which oxidative stress plays an importantrole in the disease pathogenesis [47]. The neuroprotection ofnigral dopaminergic neurons by the activation of Nrf2 throughseveral inducers in PD is also emphasized as a promisingtherapeutic approach [48]. According to the recent studies,
Fig. 4 Detection of apoptosis inBMSC-derived NSCs by TUNELlabeling post 125 μM H2O2
treatment for 30 min for N, NL,NH, and NLH indicating BMSC-derived NSCs (sham), BMSC-derived NSCs with 6 μMlovastatin, BMSC-derived NSCswith 125 μM H2O2, and BMSC-derived NSCs treated with 6 μMlovastatin and 125 μM H2O2,respectively. a, bTUNEL labelingof NH and NLH groups,respectively. c, dTUNEL labelingof the same groups withhematoxylin counterstaining. Theyellow arrow indicates thenegative TUNEL reaction andblack arrow indicates the positiveTUNEL reaction. eHistogram ofthe mean percentage of theapoptotic cells in experimentalgroups. The bars indicate themean±SEM; *P<0.05 (comparedto NH group); δP<0.05(compared to N group).Magnification ×200
Mol Neurobiol

Nrf2 can autoregulate its own expression through an antioxi-dant response element-like sequence located in the proximalregion of its promoter, leading to persistent nuclear accumu-lation of Nrf2, and thus, in response to chemopreventiveagents [49], it can guard the cells against oxidative stress moreproperly. Among other effects, Nrf2 increases the expressionof several antioxidant enzymes. The Nrf2 transcription factorcontrols the expression of a number of protective genes inresponse to oxidative stress [50, 52]. Protective effects oflovastatin are still not clearly understood, but we suggest thatNrf2 and PGC-1α are responsible for the anti-apoptotic effectsof lovastatin. Other studies support this opinion that HMG-CoA reductase inhibitors can confer antioxidant properties inpart via the activation of Nrf2 expression [51, 52].Meanwhile,a recent study shows that lovastatin induces apoptosis throughthe mitochondrial pathway in an undifferentiated SH-SY5Yneuroblastoma cell line. However, these different results canbe attributed to different doses (10 μM versus 6 μM oflovastatin), cell type (SH-SY5Y neuroblastoma cell line ver-sus BMSC-derived NSCs or normal versus cancerous cell),treatment time methods, and different gene expression detec-tion time courses (24 h versus 48 h). Totally, our results fromtreatment of the BMSC-derived NSCs with lovastatin are alsocompatible with mechanisms in which increases in the Nrf2PGC-1α expression enhance cell survival. Also, TUNEL-positive cell percentage decreased in the NLH, compared toNH group. Habeos et al. reported that activation of the Keap1/Nrf2 signaling pathway by simvastatin might provide effec-tive protection of the cell from the deleterious effects ofoxidative stress [52]. Our data provide preliminary evidencesuggesting that using lovastatin may be a safe and effectiveway to protect NSCs against oxidative stress. Moreover, lov-astatin can pass through the BBB and thus acts as a pharma-cological support in the survival promotion of endogenous ortransplanted NSCs, and this phenomenon is a great opportu-nity in the brain repair challenge. As a treatment option,
Fig. 5 PGC-1α and Nrf2mRNA expressions. Fold change ratio of PGC-1α and Nrf2 mRNA of BMSC-derived NSCs treated with various con-centrations of lovastatin (a) and various groups (b). Real-time PCR resultshave been presented as relative expression normalized to B2M mRNAamplification. Amplification of the PGC-1α and Nrf2 mRNA derivedfrom N, NL, NH, and NL groups shows increases in the level of bothmRNA after 6 μM lovastatin treatment. The bars indicate the mean±SEM. *P<0.05 (compared to N group), ΩP<0.05 (compared to N group)
Fig. 6 PGC-1α and Nrf2 proteinexpressions. Representative DABstaining photomicrographsshowing Nrf2 (upper panel) andPGC-1α (lower panel)immunoreactivity in N, NL, andNLH groups after 48 h oftreatments. Black arrows indicateimmunopositive cells.Magnification ×200
Mol Neurobiol

lovastatin can be utilized as an adjuvant in combination withimplanted BMSC-derived NSCs. Therefore, it is suggestedthat lovastatin might be used as a drug candidate in theneurological disorders in which oxidative stress plays animportant role in their pathogenesis.
Acknowledgments The project was funded byArdabil Branch, IslamicAzad University, Ardabil, Iran. Also, we are grateful to Dr. SaeidHashemin for his help in carrying out our project.
Conflict of Interest The authors declare that they have no conflict ofinterest.
References
1. Di Carlo M, Giacomazza D, Picone P, Nuzzo D, San Biagio PL(2012) Are oxidative stress and mitochondrial dysfunction the keyplayers in the neurodegenerative diseases? Free Radic Res 46:1327–1338
2. Lin MT, Beal MF (2006) Mitochondrial dysfunction and oxidativestress in neurodegenerative diseases. Nature 443(7113):787–795
3. Gilgun-Sherki Y, Melamed E, Offen D (2001) Oxidative stressinduced-neurodegenerative diseases: the need for antioxidants thatpenetrate the blood brain barrier. Neuropharmacology 40(8):959–975
4. Crunkhorn S (2011) Neurodegenerative disorders: restoring the bal-ance. Nat Rev Drug Discov 10(8):576
5. Grundy SM, Cleeman JI, Merz CN, Brewer HB Jr, Clark LT,Hunninghake DB, Pasternak RC, Smith SC Jr, Stone NJ (2004)Cholesterol Education Program. Implications of recent clinical trialsfor the National Cholesterol Education Program Adult TreatmentPanel III guidelines. Arterioscler ThrombVasc Biol 24(8):e149–e161
6. Schachter M (2005) Chemical, pharmacokinetic and pharmacody-namic properties of statins: an update. Fundam Clin Pharmacol19:117–125
7. Huang W, Shang WL, Li DH, WuWW, Hou SX (2012) Simvastatinprotects osteoblast against H2O2-induced oxidative damage viainhibiting the upregulation of Nox4. Mol Cell Biochem360(1–2):71–77
8. Laks DR, Masterman-Smith M, Visnyei K, Angenieux B, OrozcoNM, Foran I, YongWH, Vinters HV, Liau LM, Lazareff JA, MischelPS, Cloughesy TF, Horvath S, Kornblum HI (2009) Neurosphereformation is an independent predictor of clinical outcome in malig-nant glioma. Stem Cells 27(4):980–987
9. Yang Q, Mu J, Li Q, Li A, Zeng Z, Yang J, Zhang X, Tang J, Xie P(2008) A simple and efficient method for deriving neurospheres frombone marrow stromal cells. Biochem Biophys Res Commun372(4):520–524
10. Radad K, RauschWD, Gille G (2006) Rotenone induces cell death inprimary dopaminergic culture by increasing ROS production andinhibiting mitochondrial respiration. Neurochem Int 49:379–386
11. Yuan JS, Reed A, Chen F, Stewart CN Jr (2006) Statistical analysis ofreal-time PCR data. BMC Bioinformatics 7:85
12. Chen PS, Cheng CL, Kao YangYH, Yeh PS, Li YH (2013) Impact ofearly statin therapy in patients with ischemic stroke or transientischemic attack. Acta Neurol Scand 129:41–48
13. Ghosh A, Roy A, Matras J, Brahmachari S, Gendelman HE, Pahan K(2009) Simvastatin inhibits the activation of p21ras and prevents theloss of dopaminergic neurons in a mouse model of Parkinson'sdisease. J Neurosci 29:13543–13556
14. Roy A, Pahan K (2011) Prospects of statins in Parkinson disease.Neuroscientist 17:244–255
15. Neuhaus O, Strasser-Fuchs S, Fazekas F, Kieseier BC, Niederwieser G,Hartung HP, Archelos JJ (2002) Statins as immunomodulators: com-parison with interferon-beta 1b in MS. Neurology 59(7):990–997
16. Darlington CL (2005) The potential role for statins in the treatment ofmultiple sclerosis. Curr Opin Investig Drugs 6:667–671
17. Silva T, Teixeira J, Remiao F, Borges F (2013) Alzheimer's disease,cholesterol, and statins: the junctions of important metabolic path-ways. Angew Chem Int Ed Engl 52:1110–1121
18. Sparks DL (2011) Alzheimer disease: statins in the treatment ofAlzheimer disease. Nat Rev Neurol 7:662–663
19. Chen J, Zhang C, Jiang H, Li Y, Zhang L, Robin A, Katakowski M,Lu M, Chopp M (2005) Atorvastatin induction of VEGF and BDNFpromotes brain plasticity after stroke in mice. J Cereb Blood FlowMetab 25(2):281–290
20. Chen J, Zhang ZG, Li Y, Wang Y, Wang L, Jiang H, Zhang C, Lu M,Katakowski M, Feldkamp CS, Chopp M (2003) Statins induceangiogenesis, neurogenesis, and synaptogenesis after stroke. AnnNeurol 53(6):743–751
21. Dolga AM, Nijholt IM, Ostroveanu A, Ten Bosch Q, Luiten PG, EiselUL (2008) Lovastatin induces neuroprotection through tumor necrosisfactor receptor 2 signaling pathways. J Alzheimers Dis 13:111–122
22. Dolga AM, Granic I, Nijholt IM, Nyakas C, van der Zee EA, LuitenPG, Eisel UL (2009) Pretreatment with lovastatin prevents N-methyl-D-aspartate-induced neurodegeneration in the magnocellular nucleusbasalis and behavioral dysfunction. J Alzheimers Dis 17(2):327–336
23. Salins P, Shawesh S, He Y, Dibrov A, Kashour T, Arthur G, Amara F(2007) Lovastatin protects human neurons against Abeta-inducedtoxicity and causes activation of beta-catenin-TCF/LEF signaling.Neurosci Lett 412(3):211–216
24. Jin Y, Sui HJ, Dong Y, Ding Q, Qu WH, Yu SX, Jin YX (2012)Atorvastatin enhances neurite outgrowth in cortical neurons in vitrovia up-regulating the Akt/mTOR and Akt/GSK-3β signaling path-ways. Acta Pharmacol Sin 33(7):861–872
25. Wu H, Mahmood A, Qu C, Xiong Y, Chopp M (2012) Simvastatinattenuates axonal injury after experimental traumatic brain injury andpromotes neurite outgrowth of primary cortical neurons. Brain Res1486:121–130
26. Marz P, Otten U, Miserez AR (2007) Statins induce differentiationand cell death in neurons and astroglia. Glia 55:1–12
27. Xiang Z, Reeves SA (2009) Simvastatin induces cell death in amouse cerebellar slice culture (CSC) model of developmentalmyelination. Exp Neurol 215:41–47
28. Chen SD, Yang DI, Lin TK, Shaw FZ, Liou CW, Chuang YC (2011)Roles of oxidative stress, apoptosis, PGC-1alpha and mitochondrialbiogenesis in cerebral ischemia. Int J Mol Sci 12:7199–7215
29. Gleyzer N, Vercauteren K, Scarpulla RC (2005) Control of mito-chondrial transcription specificity factors (TFB1M and TFB2M) bynuclear respiratory factors (NRF-1 and NRF-2) and PGC-1 familycoactivators. Mol Cell Biol 25:1354–1366
30. Uttara B, Singh AV, Zamboni P, Mahajan RT (2009) Oxidative stressand neurodegenerative diseases: a review of upstream and down-stream antioxidant therapeutic options. Curr Neuropharmacol7(1):65–74
31. Emerit J, EdeasM, Bricaire F (2004) Neurodegenerative diseases andoxidative stress. Biomed Pharmacother 58(1):39–46
32. Hernández-Saavedra D, McCord JM (2007) Evolution and free rad-icals. Importance of oxidative stress in human pathology. Rev MedInst Mex Seguro Soc 45(5):477–484
33. Ischiropoulos H, Beckman JS (2003) Oxidative stress and nitration inneurodegeneration: cause, effect, or association? J Clin Invest 111:163–169
34. Yokota T, Igarashi K, Uchihara T, Jishage K, Tomita H, Inaba A, LiY, Arita M, Suzuki H, Mizusawa H, Arai H (2001) Delayed-onsetataxia in mice lacking alpha-tocopherol transfer protein: model forneuronal degeneration caused by chronic oxidative stress. Proc NatlAcad Sci U S A 98(26):15185–15190
Mol Neurobiol

35. Sayre LM, Smith MA, Perry G (2001) Chemistry and biochemistryof oxidative stress in neurodegenerative disease. Curr Med Chem8(7):721–738
36. Butterfield DA, Kanski J (2001) Brain protein oxidation in age-related neurodegenerative disorders that are associated with aggre-gated proteins. Mech Ageing Dev 122(9):945–962
37. WangW,Wong CW (2010) Statins enhance peroxisome proliferator-activated receptor gamma coactivator-1alpha activity to regulateenergy metabolism. J Mol Med (Berl) 88(3):309–317
38. Lin J, Puigserver P, Donovan J, Tarr P, Spiegelman BM (2002)Peroxisome proliferator-activated receptor gamma coactivator 1beta(PGC-1beta), a novel PGC-1-related transcription coactivator associ-ated with host cell factor. J Biol Chem 277(3):1645–1648
39. Wu Z, Puigserver P, Andersson U, Zhang C, Adelmant G, Mootha V,Troy A, Cinti S, Lowell B, Scarpulla RC, Spiegelman BM (1999)Mechanisms controlling mitochondrial biogenesis and respirationthrough the thermogenic coactivator PGC-1. Cell 98:115–124
40. Kobayashi A, Kang MI, Okawa H, Ohtsuji M, Zenke Y, Chiba T,Igarashi K, Yamamoto M (2004) Oxidative stress sensor Keap1functions as an adaptor for Cul3-based E3 ligase to regulateproteasomal degradation of Nrf2. Mol Cell Biol 24(16):7130–7139
41. Itoh K, Wakabayashi N, Katoh Y, Ishii T, Igarashi K, Engel JD,Yamamoto M (1999) Keap1 represses nuclear activation of antioxi-dant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev 13(1):76–86
42. Kaspar JW, Niture SK, Jaiswal AK (2009) Nrf2:INrf2 (Keap1)signaling in oxidative stress. Free Radic Biol Med 47:1304–1309
43. Xu Y, Duan C, Kuang Z, Hao Y, Jeffries JL, Lau GW (2013)Pseudomonas aeruginosa pyocyanin activates NRF2-ARE-mediated transcriptional response via the ROS-EGFR-PI3K-AKT/MEK-ERK MAP kinase signaling in pulmonary epithelial cells.PLoS One 8:e72528
44. Niture SK, Jaiswal AK (2012) Nrf2 protein up-regulatesantiapoptotic protein Bcl-2 and prevents cellular apoptosis. J BiolChem 287(13):9873–9886
45. Chen W, Jiang T, Wang H, Tao S, Lau A, Fang D, Zhang DD (2012)Does Nrf2 contribute to p53-mediated control of cell survival anddeath? Antioxid Redox Signal 17(12):1670–1675
46. Niture SK, Jaiswal AK (2013) Nrf2-induced antiapoptotic Bcl-xLprotein enhances cell survival and drug resistance. Free Radic BiolMed 57:119–131
47. Vargas MR, Johnson DA, Sirkis DW,Messing A, Johnson JA (2008)Nrf2 activation in astrocytes protects against neurodegeneration inmouse models of familial amyotrophic lateral sclerosis. J Neurosci28:13574–13581
48. Tufekci KU, Civi Bayin E, Genc S, Genc K (2011) The Nrf2/AREpathway: a promising target to counteract mitochondrial dysfunctionin Parkinson's disease. Park Dis 2011:314082
49. Kwak MK, Itoh K, Yamamoto M, Kensler TW (2002) Enhancedexpression of the transcription factor Nrf2 by cancer chemopreven-tive agents: role of antioxidant response element-like sequences in thenrf2 promoter. Mol Cell Biol 22:2883–2892
50. Moi P, Chan K,Asunis I, Cao A,KanYW (1994) Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcrip-tional activator that binds to the tandem NF-E2/AP1 repeat of thebeta-globin locus control region. Proc Natl Acad Sci U S A 91(21):9926–9930
51. Macari ER, Schaeffer EK, West RJ, Lowrey CH (2013) Simvastatinand t-butylhydroquinone suppress KLF1 and BCL11A gene expres-sion and additively increase fetal hemoglobin in primary humanerythroid cells. Blood 121(5):830–839
52. Habeos IG, Ziros PG, Chartoumpekis D, Psyrogiannis A,Kyriazopoulou V, Papavassiliou AG (2008) Simvastatin activatesKeap1/Nrf2 signaling in rat liver. JMolMed (Berl) 86(11):1279–1285
Mol Neurobiol





























