Embed Size (px)
Citation preview

Aus dem Physiologischen Institut der Tierärztlichen Hochschule Hannover
___________________________________________________________________________
Expression der Ca2+-Kanäle ECaC1 und ECaC2 im
Dünndarm von Saug- und Absetzferkeln
INAUGURAL-DISSERTATION Zur Erlangung des Grades eines
Doktors der Veterinärmedizin (Dr. med. vet.)
durch die Tierärztliche Hochschule Hannover
Vorgelegt von Thomas Hinterding
aus Krefeld
Hannover 2002

Wissenschaftliche Betreuung: Prof. Dr. B. Schröder 1. Gutachter: Prof. Dr. B. Schröder 2. Gutachter: Prof. Dr. U. Ebert Tag der mündlichen Prüfung: 22. November 2002 Gefördert durch die H. WILHELM SCHAUMANN STIFTUNG

Für meine Eltern
Karl & Maria
Hinterding


Inhaltsverzeichnis Inhaltsverzeichnis
1 Einleitung 1
2 Literaturübersicht 2
2.1 Biologische Bedeutung von Calcium im Säugetierorganismus 2
2.2 Regulation der zellulären Ca2+-Homöostase 2
2.3 Regulation der systemischen Ca2+-Homöostase 3
2.4 Alimentäre Ca2+-Aufnahme 5
2.5 Mechanismen des intestinalen Ca2+-Transports 6
2.5.1 Parazellulärer Ca2+-Transport 6
2.5.2 Transzellulärer Ca2+-Transport 7
2.6 Neonatale Entwicklung des Gastrointestinaltrakts 12
2.7 Besonderheiten des transzellulären Ca2+-Transports bei Saugferkeln 14
3 Material und Methoden 15
3.1 Versuchstiere 15
3.2 Probenentnahmen 16
3.2.1 Probenentnahme für die Präparation von Bürstensaummembranvesikeln 16
3.2.2 Probenentnahme für die RNA-Präparation 16

Inhaltsverzeichnis
3.3 Präparation der Bürstensaummembranvesikel 17
3.3.1 Prinzip 17
3.3.2 Durchführung 17
3.3.3 Präparationspuffer 19
3.4 Messung der Aufnahmeraten in die Bürstensaummembranvesikel 21
3.4.1 Puffer für die Aufnahmestudien 21
3.4.2 Kalkulation von [Ca2+]f 22
3.4.3 Studien zur zeitabhängigen Glucose-Aufnahme 23
3.4.4 Studien zur zeitabhängigen Calcium-Aufnahme 23
3.4.5 Studien zur konzentrationsabhängigen Calcium-Aufnahme 25
3.4.6 Kalkulation der kinetischen Kenngrößen der Ca2+-Aufnahme 26
3.4.7 Ermittlung des Vesikelvolumens 27
3.5 Protein- und Enzym-Bestimmungen im Mukosahomogenat
und in der Vesikelsuspension 28
3.5.1 Protein 28
3.5.1.1 Prinzip 28
3.5.1.2 Durchführung 28
3.5.2 Aktivität der alkalische Phosphatase (AP) 29
3.5.2.1 Prinzip 29
3.5.2.2 Durchführung 29
3.5.3 Aktivität der Na+/K+-ATPase 29
3.5.3.1 Prinzip 29
3.5.3.2 Durchführung 30

Inhaltsverzeichnis
3.6 Gesamt-RNA-Isolierung und DNase-Verdau 30
3.6.1 Prinzip 31
3.6.2 Durchführung 31
3.7 cDNA-Synthese 32
3.7.1 Prinzip 32
3.7.2 Durchführung 32
3.8 Polymerasekettenreaktion (PCR) 33
3.8.1 Prinzip 33
3.8.2 Durchführung 35
3.9 Darstellung der DNA auf einem Agarosegel 36
3.9.1 Prinzip 36
3.9.2 Durchführung 36
3.10 Isolierung der PCR-Produkte 37
3.10.1 Prinzip 37
3.10.2 Durchführung 37
3.11 Ligation der PCR-Produkte 38
3.11.1 Prinzip 38
3.11.2 Durchführung 38

Inhaltsverzeichnis
3.12 Transformation der Bakterien 39
3.12.1 Prinzip 39
3.12.2 Durchführung 39
3.13 Plasmidpräparation 40
3.13.1 Prinzip 40
3.13.2 Durchführung 40
3.14 Restriktionsverdau 41
3.14.1 Prinzip 41
3.14.2 Durchführung 41
3.15 Sequenzierung 41
3.16 mRNA-Isolierung 42
3.16.1 Prinzip 42
3.16.2 Durchführung 42
3.17 Northern-Blot 43
3.17.1 Prinzip 43
3.17.2 Größenfraktionierung der RNA in einem denaturierenden Agarosegel 43
3.17.3 Übertragen der RNA auf eine Nitrocellulosemembran 44
3.17.4 Erstellung von spezifischen, radioaktiv markierten Sonden 46
3.17.5 Hybridisierung der RNA mit den radioaktiv markierten Sonden 47
3.17.6 Darstellung der RNA 48
3.17.7 Auswertung der Northern-Blots 48

Inhaltsverzeichnis
3.18 Real-Time-PCR 49
3.18.1 Prinzip 49
3.18.2 Durchführung 50
3.18.3 Auswertung der Real-Time-PCR 52
3.19 Chemikalien 53
3.20 Statistik 54
4 Ergebnisse 55
4.1 Effekte des Alters der Versuchstiere auf die Ca2+-Aufnahme
in BSMV 55
4.1.1 Charakterisierung der BSMV 55
4.1.2 Ca2+-Aufnahmeraten in die BSMV 60
4.2 Ergebnisse der PCR 65
4.3 Teilsequenz des ECaC im konservierten Bereich 66
4.4 Teilsequenz des ECaC1 im hypervariablen Bereich 67
4.5 Teilsequenz des ECaC2 im hypervariablen Bereich 69
4.6 Northern-Blots mit verschiedenen Sonden entlang der Darmachse 72
4.6.1 Northern-Blot mit der „ECaC-Sonde“ 72
4.6.2 Northern-Blot mit der „ECaC1-Sonde“ 73
4.6.3 Northern-Blot mit der „ECaC2-Sonde“ 73

Inhaltsverzeichnis
4.7 Ergebnisse der Real-Time-PCR bei Saug- und Absetzferkeln 74
5 Diskussion 79
5.1 Untersuchungen zur Funktion der Ca2+-Kanäle 79
5.2 Untersuchungen zur Struktur der Ca2+-Kanäle 84
5.3 Untersuchungen zur Quantifizierung der Ca2+-Kanäle 85
5.4 Schlussbetrachtung und Ausblick 86
6 Zusammenfassung 88
7 Summary 90
8 Literaturverzeichnis 92

Abkürzungsverzeichnis ABKÜRZUNGSVERZEICHNIS
ATP Adenosintriphosphat
as antisense
ave average
bp Basenpaare
BSMV Bürstensaummembranvesikel
Ca2+ Calcium in ionisierter Form
[Ca2+]ges Gesamt- Ca2+-Konzentration
[Ca2+]f Konzentration freier Ca2+-Ionen
cDNA komplementäre DNA (complementary DNA)
cpm Impulse pro Minute (counts per minute)
Ct-Wert der Ct-Wert gibt an, wie viele Zyklen in der PCR für den jeweiligen Ansatz
gebraucht werden, bis das Fluoreszenzsignal einen definierten Schwellenwert
überschritten hat (cycle; threshold)
dCTP 2`Desoxycytidin-5`triphosphat
DMSO Dimethylsulphoxid
DNA Desoxyribonukleinsäure (desoxyribonucleic acid)
dNTP 2`Desoxyribonucleosid-5`triphosphat
DTT Dithiothreitol
EDTA Ethylen-Diamin-tetra-Essigsäure (-Acetic-Acid)
EGTA Ethylen-Glykol-bis (β-Aminoethylether)-N,N´-tetra-Essigsäure (-Acetic-Acid)
g Zentrifugalbeschleunigung
GM Größenmarker
HEPES 4-(2-Hydroxy-Ethyl)-1-Piperazin-Ethan-Sulfonsäure
Kap. Kapitel
Km Michaelis-Menten-Konstante; gibt die Menge an Substrat an, die für die
halbmaximale Transportrate notwendig ist
LB-Medium Luria-Bertani-Medium

Abkürzungsverzeichnis MOPS 3-(N-Morpholino)Propansulfonsäure
µCi mikro Curie (1 Ci = 37 x 109 Bq)
mRNA Boten-RNA (messenger-RNA)
p Irrtumswahrscheinlichkeit
PCR Polymerase Kettenreaktion (polymerase chain reaction)
PMCA Plasmamembran-Calciumpumpe
PTH Parathormon
RNA Ribonukleinsäure (ribonucleic acid)
rpm Umdrehungen pro min (rounds per minute)
RT Raumtemperatur
RT-PCR Reverse Transcriptase-PCR
s sense
SEM Standardfehler des Mittelwertes (standard error of the mean)
SDS Sodiumdodecylsulfat
spez. A. spezifische Aktivität
Tris Tris (Hydroxymethyl) Aminomethan
VDR Vitamin D-Rezeptor
Vmax maximale Transportkapazität
v/v Volumen zu Volumen
w/v Gewicht zu Volumen

Einleitung 1 EINLEITUNG Beim Schwein ist das Duodenum wichtigster Ort der aktiven Ca2+-Absorption, der damit
wesentlich an der Aufrechterhaltung der Calcium-Homöostase beteiligt ist. Dieser Prozess ist
zunächst in der frühen postnatalen Lebensphase bis etwa zur 4. Lebenswoche unabhängig von
Calcitriol, dem klassischen Vitamin D-Hormon, kommt aber anschließend mehr und mehr
unter Kontrolle dieses Hormons. Über den physiologischen Sinn dieser postnatalen
Anpassung kann bislang nur diskutiert werden.
Für viele Spezies wurde gezeigt, dass Ca2+ durch einen aus mindestens drei Einzelschritten
bestehenden Mechanismus vom Darmlumen durch die Enterozyten zur Blutseite transportiert
wird. Im ersten Teilschritt, der für den transzellulären Ca2+-Transport wahrscheinlich
geschwindigkeitsbestimmend ist, tritt Ca2+ entlang eines chemischen Gradienten über die
Bürstensaummembran durch Ca2+-Kanäle in die Zelle ein. Im zweiten Teilschritt wird Ca2+
auf bisher nicht genau geklärte Weise unter Beteiligung von Calbindin-D9k durch das Cytosol
transportiert. Die Ausschleusung aus der Zelle durch die basolaterale Membran wird durch
die ATP-abhängige Plasmamembran-Calciumpumpe vermittelt und stellt den dritten
Teilschritt dar.
Beim Menschen und bei der Ratte konnten vor kurzem jeweils zwei verschiedene Ca2+-
Kanäle kloniert werden, die eine 75%ige Aminosäurenidentität zueinander aufwiesen. Es gibt
Hinweise darauf, dass ECaC1 und ECaC2 im Dünndarm möglicherweise in
unterschiedlichem Maße durch Calcitriol reguliert werden können.
Der intestinale Ca2+-Transport beim Schwein ist bislang noch nicht mit
molekularbiologischen Methoden untersucht worden. Die Erkenntnisse solcher
Untersuchungen zum Ca2+-Transport berechtigen jedoch zu der Annahme, dass auch beim
Schwein Ca2+-Kanäle an der duodenalen Ca2+-Absorption beteiligt sind. Daher war es das
primäre Ziel der vorliegenden Untersuchungen, dieser Hypothese nachzugehen. Dazu sollten
zunächst die für Human- und Rattenproben beschriebenen molekularbiologischen Methoden
zur Darstellung von Ca2+-Kanälen auf mRNA-Ebene für den Schweinedarm adaptiert werden.
Anschließend sollte der Frage nachgegangen werden, ob eine veränderte Expression von
ECaC1 und/oder ECaC2 mit dem Wechsel vom calcitriol-unabhängigen zum calcitriol-
abhängigen Ca2+-Transport korreliert werden kann.
1

Literaturübersicht 2 LITERATURÜBERSICHT
2.1 Biologische Bedeutung von Calcium im Säugetierorganismus
Das Erdalkalielement Calcium ist aufgrund seiner multiplen Aufgaben eines der wichtigsten
kationischen Mengenelemente im Säugetierorganismus. Als essenzielles Strukturelement für
die Knochen und Zähne ist es genauso von Bedeutung, wie als Second messenger innerhalb
von Zellen. In der Funktion als Second messenger beeinflusst Calcium in ionisierter Form
(Ca2+) zum Beispiel die präsynaptische Ausschüttung von Neurotransmittern, die molekularen
Mechanismen der Muskelkontraktion (VAN BREEMEN u. SAIDA 1989), sowie die
Sekretionstätigkeit bestimmter exo- und endokriner Drüsen (MUALLEM 1989). Eine weitere
wichtige Rolle spielt Ca2+ in der Blutgerinnung als Aktivator der Thrombokinase (MORIN
1980) und auch beim epithelialen Transport von z. B. Natrium, Kalium und Chlorid wirkt
Ca2+ regulatorisch mit (DONOWITZ u. WELSH 1986).
2.2 Regulation der zellulären Ca2+-Homöostase
Die Konzentration an freien Ca2+-Ionen [Ca2+]f im Cytosol der ruhenden Zelle liegt bei vielen
Zelltypen in etwa bei 10-7 bis 10-8 mol.l-1 und steigt im stimulierten Zustand um bis zu 2
Zehnerpotenzen an (CAMPBELL 1988), was als funktionelles Signal interpretiert werden
kann (NEMERE und NORMAN 1991). Zur Aufrechterhaltung der Second messenger-
Funktion muss [Ca2+]f wieder auf die Ausgangskonzentration zurückgeführt werden. Dies
geschieht durch primär aktiven Transport mit Hilfe der Adenosintriphosphat (ATP)-
abhängigen Ca2+-Pumpen oder durch erleichterte Diffusion mittels des 3Na+/Ca2+-
Austauschers. Dabei kann Ca2+ entweder in Zellorganellen, wie die Mitochondrien und das
endoplasmatische Retikulum oder in den Extrazelluärraum transportiert werden (CARAFOLI
1987). Zur Regulation des [Ca2+]f kann dieses an Ca2+-bindende Proteine gebunden werden.
Ein ubiquitär vorkommendes Protein mit hoher Ca2+-Affinität ist das Calmodulin, das z. B. in
2

Literaturübersicht glatten Muskelzellen oder in den Mikrovilli von Enterozyten gefunden wird (GRAND et al.
1979, GLENNEY u. WEBER 1980). Enterozyten sind täglich mit relativ großen Mengen an
Ca2+ konfrontiert, das das Cytosol passieren muss, ohne dass die Second messenger-Funktion
eingeschränkt werden darf. Um dies zu gewährleisten, stehen der Darmschleimhautzelle
spezifische Ca2+-bindende Proteine zur Verfügung, die auf Grund ihrer Abhängigkeit vom
Vitamin D-Hormon Calbindin-D genannt werden (TAYLOR u. WASSERMAN 1967,
CORRADINO u. WASSERMAN 1968, WAREMBOURG et al. 1986). Dabei induziert
Calcitriol nach Bindung an den kernständigen Vitamin D-Rezeptor (VDR) die Bildung von
Calbindin-D, wobei sich auf Grund des Molekulargewichtes 2 Typen unterscheiden lassen,
nämlich Calbindin-D9k und –D28k (NORMAN et al. 1999). Im Vogelintestinum wurde das
Calbindin-D28k nachgewiesen, das je Molekül 4 Moleküle Ca2+ binden kann (WASSERMAN
u. TAYLOR 1966). Das Calbindin-D9k, das nur 2 Moleküle Ca2+ binden kann, wurde dagegen
im Säugetierintestinum gefunden (JOHNSON u. KUMAR 1994). Man nimmt an, dass durch
die Bindung an das Calbindin die Second messenger-Funktion des Ca2+ trotz des
gleichzeitigen Ca2+-Transports weitestgehend unbeeinflusst bleibt (FEHER et al. 1992,
SCHRÖDER et al. 1996).
2.3 Regulation der systemischen Ca2+-Homöostase
Die Ca2+-Konzentrationen im Blutplasma bei Schweinen variieren unter physiologischen
Bedingungen in Abhängigkeit vom Alter, von der Rasse und vom Geschlecht, wobei bei
weiblichen Tieren der Reproduktionszustand mit entscheidend ist. Auch individuelle
Schwankungen und Beeinflussungen durch den Ca2+-Gehalt des Futters sind beschrieben
worden (SCHRÖDER 1996). So liegt der Plasmacalciumspiegel beim adulten Schwein bei
2,3 bis 2,8 mmol.l-1 und beim Ferkel während der Saugperiode zwischen 2,7 und 3,1 mmol.l-1
(KOLB 1989). Calcium wird im Blutplasma zu etwa 40 % in Komplexverbindungen oder an
Proteine (z. B. Albumine, Globuline) gebunden. Für die Ca2+-Homöostase im Organismus ist
nur das ionisiert vorliegende Calcium (etwa 60 %) von Relevanz (BROWN 1991).
Für die hormonelle Regulation des Plasmacalciumspiegels sind das Parathormon (PTH),
Calcitonin und Calcitriol, der biologisch aktive Metabolit des Vitamins D3, von
entscheidender Bedeutung. Die an der Regulation relevant beteiligten Organe sind die
3

Literaturübersicht Knochen, der Darm, die Nieren und die Nebenschilddrüsen (AUDRAN u. KUMAR 1985,
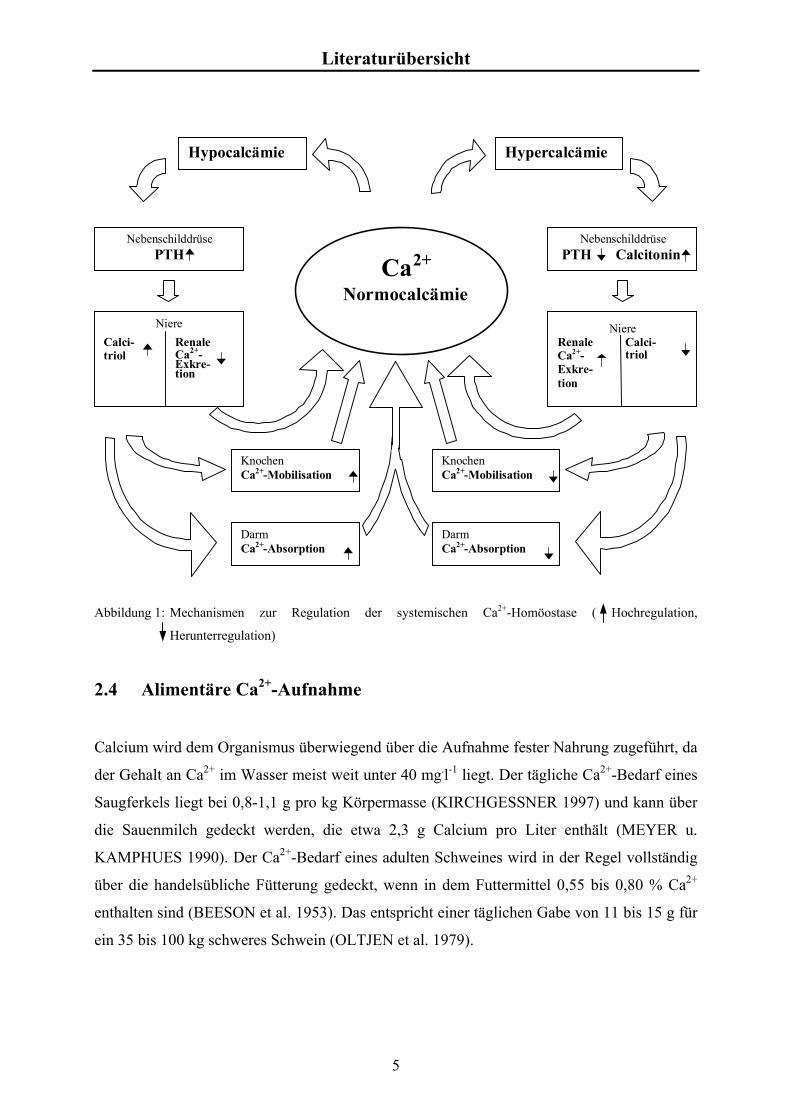
NORMAN 1987, DELUCA 1986, BROWN 1991). Abbildung 1 beschreibt in einer Übersicht
die Beeinflussungen und Anpassungsmechanismen zur Regulation der Ca2+-Homöostase.
PTH ist ein Peptidhormon, das bei niedrigen Plasmacalciumspiegel aus der Nebenschilddrüse
ausgeschüttet wird, und eine erhöhte renal-tubuläre Absorption von Ca2+ (BINDELS et al.
1991), eine erhöhte Mobilisation von Ca2+ aus dem Knochen und eine verstärkte Biosynthese
von Calcitriol in den proximalen Tubulusepithelzellen der Niere bewirkt (KOVARIK 1983).
Calcitonin, ein Peptidhormon, wird bei einem erhöhten Plasmacalciumspiegel überwiegend
aus den C-Zellen der Nebenschilddrüse freigesetzt. Calcitonin vermindert die Mobilisation
von Ca2+ aus dem Knochen (PECHET et al. 1967) und die renale Absorption (ZUO et al.
1997) und wirkt somit der PTH-Wirkung entgegen. Ein Einfluss von Calcitonin auf die
intestinale Ca2+-Absorption konnte bis jetzt nicht nachgewiesen werden.
Vitamin D ist ein Steroidhormon und wird entweder als Ergocalciferol (Vitamin D2) aus der
pflanzlichen Nahrung, als Cholecalciferol (Vitamin D3) aus tierischer Nahrung aufgenommen
oder im Körper selbst unter Einfluss von ultraviolettem Licht in der Haut aus Provitamin D3
synthetisiert. Vitamin D2 und D3 bzw. deren Metaboliten wirken im Säugetierorganismus über
den gleichen Mechanismus (NORMAN u. ROSS 1979). Vom Vitamin D3 sind mehr als 37
Metaboliten unter physiologischen Bedingungen bekannt (HENRY u. NORMAN 1984,
1991), wobei die biologisch aktivste Form für den intestinalen Ca2+-Transport das Calcitriol
ist (FAVUS 1985, NEMERE u. NORMAN 1990). Calcitriol (1α,25-Dihydroxyvitamin D3
oder 1α,25-Dihydroxychole-calciferol) entsteht aus Vitamin D3, das in der Leber am C25-
Atom und in der Niere am C1-Atom hydroxyliert wird. Calcitriol stimuliert die aktive
Absorption von Ca2+ aus dem Darm und fördert in Anwesenheit von PTH sowohl die
Absorption von Ca2+ in der Niere, als auch die Auflösung von Hydroxylapatit aus der
Knochenmatrix (HARMEYER u. KAUNE 1988).
Es sind weitere Substanzen, die die Ca2+-Homöostase beeinflussen, beschrieben worden, doch
sind deren Bedeutung sowie deren Wirkmechanismen bislang unklar. Zu diesen Substanzen
gehören das PTHrP (parathyroid hormone-related protein) (BLIND et al. 1993), das Prolaktin
(PAHUJA und DELUCA 1981) und das Stanniocalcin (MADSEN et al. 1998).
4
Literaturübersicht
Abbildung 1: Mechanismen zur Regulation der systemischen Ca2+-Homöostase ( Hochregulation,
Herunterregulation)
HypercalcämieHypocalcämie
NebenschilddrüsePTH
NebenschilddrüsePTH Calcitonin
NiereCalci-triol Ca2+-
Renale
Exkre-tion
NiereRenale Calci-Ca2+- triolExkre-tion
KnochenCa2+-Mobilisation
KnochenCa2+-Mobilisation
Ca2+
Normocalcämie
DarmCa2+-Absorption
DarmCa2+-Absorption
2.4 Alimentäre Ca2+-Aufnahme
Calcium wird dem Organismus überwiegend über die Aufnahme fester Nahrung zugeführt, da
der Gehalt an Ca2+ im Wasser meist weit unter 40 mg.l-1 liegt. Der tägliche Ca2+-Bedarf eines
Saugferkels liegt bei 0,8-1,1 g pro kg Körpermasse (KIRCHGESSNER 1997) und kann über
die Sauenmilch gedeckt werden, die etwa 2,3 g Calcium pro Liter enthält (MEYER u.
KAMPHUES 1990). Der Ca2+-Bedarf eines adulten Schweines wird in der Regel vollständig
über die handelsübliche Fütterung gedeckt, wenn in dem Futtermittel 0,55 bis 0,80 % Ca2+
enthalten sind (BEESON et al. 1953). Das entspricht einer täglichen Gabe von 11 bis 15 g für
ein 35 bis 100 kg schweres Schwein (OLTJEN et al. 1979).
5

Literaturübersicht Die Ca2+-Absorption im Magen des Schweines ist vernachlässigbar. Die Nettoabsorption
findet fast vollständig (über 90 %) im Dünndarm statt, wobei dem proximalen Teil die größte
Bedeutung zukommt (PARTRIDGE 1978). Die Befunde bezüglich der Ca2+-Absorption im
Dickdarm sind sehr widersprüchlich und reichen von einer täglichen Ca2+-Nettosekretion von
1,9 g bis zu einer gleichhohen Nettoabsorption (SAUER et al. 1982, LARSEN u.
SANDSTRÖM 1993). Die Ca2+-Aufnahme über den gesamten Darm variiert je nach Autor
von 3,0 bis 5,3 g pro Tag bei einer Aufnahme von 9,2 bis 15,1 g (HENNING et al. 1988,
LARSEN u. SANDSTRÖM 1993). Dies führt zu einer scheinbaren Verdaulichkeit des
Calciums von etwa 34 %.
2.5 Mechanismen des intestinalen Ca2+-Transports
Der intestinale Ca2+-Transport kann prinzipiell parazellulär und/oder transzellulär verlaufen.
Die quantitativen Anteile dieser Transportwege an der Ca2+-Nettoabsorption sind unter In-
vivo-Bedingungen nicht geklärt.
2.5.1 Parazellulärer Ca2+-Transport
Der parazelluläre Transport verläuft passiv und ist nicht sättigbar. Er nimmt mit der luminalen
Ca2+-Konzentration linear zu und ist außerdem abhängig vom elektrischen Gradienten
zwischen der luminalen und basolateralen Seite des Epithels (PANSU et al. 1983a). Diese
Transportform kann über den gesamten Darmtrakt nachgewiesen werden (PANSU et al.
1981), findet sich aber vor allem im distalen Dünndarm (NELLANS 1990, BRONNER 1992).
Sie kann aber auch im proximalen Dünndarm in gleichem Ausmaß auftreten, wenn
entsprechend hohe Ca2+-Konzentrationen vorliegen (PANSU et al. 1983b). Eine besondere
Bedeutung hat der parazelluläre Transport anscheinend bei jungen Ratten. Bei In-situ-
Untersuchungen mit ligierten Darmschlingen wurde festgestellt, dass neugeborene Ratten bis
zum 3. Lebenstag fast ausschließlich auf parazellulärem Wege transportieren. Während der
Zeit bis zum 35. Lebenstag verliert der passive Transport an Bedeutung und stagniert auf
niedrigem Niveau (PANSU 1983a, b).
6

Literaturübersicht 2.5.2 Transzellulärer Ca2+-Transport
Der transzelluläre Transport ist sättigbar und stellt einen aktiven Vorgang dar. Er erfolgt im
proximalen Dünndarm, vor allem in Duodenum (PANSU et al. 1981, FAVUS 1985) und ist
auch beim Saugferkel nachzuweisen (SCHRÖDER et al. 1993). Dabei wird Ca2+ durch einen
aus mindestens drei Einzelschritten bestehenden Mechanismus vom Darmlumen durch die
Enterozyten zur Blutseite transportiert (Abbildung 2). In dem ersten Teilschritt tritt Ca2+
durch die Bürstensaummembran in die Zelle ein, im zweiten Teilschritt wird Ca2+ durch das
Cytosol transportiert. Die Ausschleusung aus der Zelle durch die basolaterale Membran stellt
den dritten Teilschritt dar (KAUNE et al. 1992, SCHRÖDER et al. 1998a, VAN OS 1987,
TIMMERMANS et al. 1991).
?ECaC1
ECaC2
Lumen Enterozyt Blut
PMCA
Ca2+Ca2+
Calcitriol
VDR
?
?
Ca2+
CAL
+
+ -
Abbildung 2: Modell der aktiven Ca2+-Absorption im Dünndarm (CAL = Calbindin-D9k, PMCA =
Plasmamembran-Calciumpumpe, VDR = Vitamin D-Rezeptor).
7

Literaturübersicht Der Eintritt von Ca2+ in die Zelle erfolgt sowohl entlang eines chemischen Gradienten mit
einer Ca2+-Konzentration im Darmlumen im millimolaren Bereich (FULLMER 1992) und
einer Ca2+-Konzentration im Zellinneren, die um den Faktor 1000 bis 10000 niedriger ist
(Kap. 2.2), als auch entlang eines elektrischen Gradienten mit einer intra/extrazellulären
Potentialdifferenz von etwa 50 mV (FULLMER 1992). Aufgrund des elektrochemischen
Gradienten erfordert dieser Schritt keine Energie, doch kann Ca2+ aufgrund der Ladung nicht
in ausreichendem Maße ohne spezifische Transportsysteme wie z. B. Kanäle, durch die
Lipiddoppelschicht gelangen. Es gibt sichere Hinweise darauf, dass dieser Schritt für die
Ca2+-Aufnahme geschwindigkeitslimitierend ist (HOENDEROP et al. 1999b, 2000c).
Untersuchungen von KAUNE et al. (1992) haben gezeigt, dass sich die Ca2+-Aufnahmen in
Bürstensaummembranvesikel durch Verapamil, einem Antagonisten der
spannungsgesteuerten Ca2+-Kanäle vom L-Typ, die aus erregbaren Zellen bekannt sind,
hemmen ließen. Bei Ratten wurde der Effekt des Verapamils ebenfalls beschrieben (MILLER
u. BRONNER 1981), doch muss aufgrund der hohen eingesetzten Konzentration des
Kanalblockers auch ein unspezifischer Effekt auf das Epithel diskutiert werden. Beim
Kaninchen konnte ein Ca2+-Kanal (ECaC), der Verapamil-insensitiv ist, kloniert werden.
Dieser Kanal wurde im proximalen Dünndarm, in der Plazenta und in den distalen
Abschnitten des Nephrons nachgewiesen (HOENDEROP et al. 1999a, b). Bei der Ratte
wurde ein Ca2+-Transportprotein (CaT1) mit einer 75%igen Homologie zum ECaC aus dem
Dünndarm kloniert. Eine Northern-Blot Analyse zeigte seine Präsenz auf mRNA-Ebene im
Duodenum, proximalen Jejunum, Caecum und Colon (PENG et al. 1999).
Da bei der Ratte das Homologon zum ECaC (bezeichnet als ECaC1, ISHIBASHI et al. 2000
bzw. CaT2, PENG et al. 2000b), bei der Maus das Homologon zum CaT1 (bezeichnet als
CaT) (SUZUKI et al. 2000) und beim Menschen sowohl das Homologon zum ECaC
(MÜLLER et al. 2000b), als auch zum CaT1 (PENG et al. 2000a) kloniert wurden und beide
Proteine als Ca2+-selektive Ionenkanäle funktionieren (HOENDEROP et al 2001), werden im
folgenden die homologen Transportproteine zum ECaC als ECaC1 und die homologen
Transportproteine zum CaT1 als ECaC2 bezeichnet. Diese Transportproteine, ECaC1 und
ECaC2, werden den „TRP cation channels“ (transient receptor potential cation channels)
zugeordnet und sind Bestandteil der Unterfamilie TRPV. Der ECaC1 wird auch als TRPV5
und der ECaC2 als TRPV6 bezeichnet (MONTELL et al. 2002).
8

Literaturübersicht Der ECaC1 hat bei den verschiedenen Spezies eine primäre Aminosäurensequenz von 724
(Ratte) bis 730 (Mensch und Kaninchen) Aminosäuren bei einer mRNA-Größe von 2190
(Mensch) bis 4860 (Ratte) Basenpaaren (HOENDEROP et al. 1999a, MÜLLER et al. 2000b,
ISHIBASHI et al. 2000). Der ECaC2 hat bei den verschiedenen Spezies eine primäre
Aminosäurensequenz von 726 (Mensch) bis 730 (Maus) Aminosäuren bei einer mRNA-
Größe von 2850 (Maus) bis 2955 (Ratte) Basenpaaren (PENG et al. 1999, 2000b, SUZUKI et
al. 2000). Beide Transporter haben 6 transmembranale Domänen und eine Porenregion
zwischen den Membrandurchgängen 5 und 6 (Abbildung 3). In diesen Bereichen besteht die
höchste Homologie zwischen den verschiedenen ECaC. Sie unterscheiden sich aber in den für
die Regulation wichtigen Bereichen mutmaßlicher Glycosylierungsstellen und in den
„Ankyrin-Repeats“, die für die Verankerung am Cytoskelett in der apikalen Membran wichtig
sind. In besonderem Maße lassen sich Unterschiede im Bereich des N- und C-terminalen
Ende finden (HOENDEROP et al. 1999a, 2000a, b, PENG et al. 1999).
Lumen
Zytosol
1 2 3 4 5 6
P
C
N
AA
A
Abbildung 3: Modell des ECaC (Epithelial Calcium Channel) in der Zellmembran (1-6 = transmembranale
Domänen, A = Ankyrin-Repeats, C = C-terminales Ende, N = N-terminales Ende, P =
Porenregion; MÜLLER et al. 2000b, modifiziert).
Die Km-Werte als Maß für die Spezifität des Ca2+-Transports durch ECaC unterscheiden sich
deutlich sowohl bei den verschiedenen Transportproteinen als auch zwischen den Spezies.
Die bekannten Km-Werte für die einzelnen Transportproteine, die bisher alle über die Ca2+-
Aufnahmen in ECaC-exprimierende Oozyten von Xenopus laevis bestimmt wurden, variieren
9

Literaturübersicht beim ECaC1 von 0,2 mmol.l-1 (HOENDEROP et al. 1999a) bis 0,66 mmol.l-1 (PENG et al.
2000b) und beim ECaC2 von 0,25 mmol.l-1 (PENG et al. 2000a) bis 0,44 mmol.l-1 (PENG et
al. 1999). In Studien zur Ca2+-Aufnahme mit BSMV wurden Km-Werte für den Gesamt-Ca2+-
Transport ermittelt. Dabei hat KAUNE 1992 für das Schwein eine Km von 0,03 mmol.l-1
ermittelt. Diese liegt in der gleichen Größenordnung wie die Km-Werte für den Menschen
(GHISHAN et al. 1989), Ratten und Hamster (SCHEDL u. WILSON 1985, WILSON et al.
1989). MILLER u. BRONNER haben allerdings 1981 eine apparente Km für die Ratten von
0,27 mmol.l-1 beschrieben und für Hühner lag der Km-Wert bei 0,54 mmol.l-1 (LIANG et al.
1986).
Wie die einzelnen Transportproteine reguliert werden, ist noch weitgehend unklar. RT-PCR
Analysen mit unterschiedlichen Humangeweben haben erbracht, dass beide Kanaltypen in
Calbindin-D enthaltenden Geweben coexprimiert werden, wie z. B. im Dünndarm, Plazenta
und Pankreas (HOENDEROP et al. 2001). Dieser Befund könnte auf eine Regulation von
ECaC1 und/oder ECaC2 durch das Calcitriol/VDR-System hindeuten. Eine ausschließliche
Expression von ECaC1 wurde für die Niere, das Colon und das Gehirn festgestellt, während
sich im Magen nur ECaC2 nachweisen ließ. In der Promotorregion des ECaC1 wurde eine
Struktur ermittelt, die vier Vitamin D-abhängige Elemente sowohl nach CARLBERG 1995
als auch nach CHRISTAKOS et al. 1996 enthält (MÜLLER et al. 2000a). Der ECaC2 ist im
Darm insensitiv gegenüber Vitamin D (PENG et al. 1999, BARLEY et al. 2001), jedoch
reagiert die ECaC2-Expression in Caco-2-Zellen signifikant auf den Zusatz von Vitamin D3
(WOOD et al. 2001). Im Gegensatz dazu schließen WEBER et al. (2001) für beide
Transporter eine Beeinflussung durch Vitamin D aus und postulieren eine Regulation über
extrazelluläres Calcium. Andererseits weisen Studien an zwei verschiedenen VDR-Knockout-
Mäusemodellen auf eine Beeinflussung von ECaC1 und ECaC2 durch Calcitriol hin (VAN
CROMPHAUT et al. 2001), jedoch konnten WEBER et al. 2001 dies nicht durch Studien an
Mäusen mit einem mutierten und dadurch funktionsunfähigen VDR bestätigen.
Die Funktion von ECaC1 und ECaC2 wird durch die intrazelluläre Ca2+-Konzentration
insofern reguliert (HOENDEROP et al. 1999b, SUZUKI et al. 2000, NILIUS et al. 2001a), als
eine Reduktion der intrazellulären Ca2+-Konzentration den Ca2+-Einstrom in die Zelle fördert,
während ein Anstieg der intrazellulären Ca2+-Konzentration die ECaC-Aktivität inhibiert
(YUE et al. 2001). Extrazelluläres Ca2+ kann die Expression der Transportproteine
10

Literaturübersicht beeinflussen (WEBER et al. 2001) und in HEK293-Zellen führte extrazelluläres Ca2+ in
physiologischen Konzentrationen zu einer schnellen und reversiblen Abnahme der ECaC-
Aktivität (NILIUS et al. 2000, 2001b, VENNEKENS et al. 2000). Beide Transportproteine
lassen sich durch Rutheniumrot hemmen, wobei der ECaC1 eine 100fach höhere Sensitivität
für den Blocker als der ECaC2 hat (HOENDEROP et al. 2001). ECaC2 lässt sich bis zu 15 %
durch Verapamil hemmen (PENG et al. 1999). In exokrinen Geweben wurde ein Protein
beschrieben, dass eine sehr hohe Homologie zu den bekannten Sequenzen des ECaC1 bzw.
des ECaC2 hat. Es wurde daher als CaT-like bezeichnet und scheint aber ausschließlich eine
Bedeutung im Zusammenhang mit der Malignität von Prostataentartungen zu haben
(WISSENBACH et al. 2001, NIEMEYER et al. 2001). Dieses Protein kann durch Calmodulin
und Proteinkinase C reguliert werden, indem Calmodulin an das Protein bindet bzw. diese
Bindungsstelle durch die Proteinkinase C phosphoryliert wird (NIEMEYER et al. 2001).
Diese potentiellen Phosphorylierungsstellen wurden auch im ECaC1 bzw. ECaC2 identifiziert
(HOENDEROP et al. 1999a, PENG et al. 1999).
Beim 2. Schritt des transzellulären Ca2+-Transports, der Durchschleusung durch das Cytosol,
muss simultan die Second messenger-Funktion des freien Ca2+ erhalten bleiben. Es wird
angenommen, dass dies überwiegend dadurch erreicht wird, dass Ca2+ an Calbindin-D
gebunden wird (SCHRÖDER et al. 1996). In dieser Form kann Ca2+ etwa 70-mal schneller
durch die Zelle diffundieren als bei freier Diffusion (BRONNER et al. 1986). Die genauen
Vorgänge sind zur Zeit nicht geklärt.
Nach einem weiteren Modell werden Ca2+-Ionen in Vesikel verpackt oder an diese gebunden
durch das Cytosol transportiert. Dieser als Transcaltachia bezeichnete Effekt wurde als „Fast
Response“ auf Calcitriol beobachtet und wird wahrscheinlich unter Einbeziehung des
Cytoskeletts über spezifische Membranrezeptoren vermittelt. Die physiologische Bedeutung
dieses Vorganges ist allerdings noch umstritten (NEMERE et al. 1986, NEMERE u.
NORMAN 1988, 1990, NEMERE u. FARACH-CARSON 1998, NEMERE et al. 2000,
NORMAN et al. 1999, SCHWARTZ et al. 2002).
Die Ausschleusung des Ca2+ durch die basolaterale Membran als Finalschritt der Ca2+-
Absorption verläuft entgegen des elektrochemischen Gradienten und benötigt daher Energie.
11

Literaturübersicht Plasmamembranale Ca2+-Pumpen (PMCA) transportieren Ca2+ unter ATP-Verbrauch. Bei mit
Vitamin D behandelten Küken, die vorher Vitamin D freie Nahrung erhielten, konnte dieser
Mechanismus stimuliert werden (WASSERMAN et al. 1992a, b), während Untersuchungen
an duodenalen Enterozyten von Ratte und Schwein einen Vitamin D-Effekt nicht bestätigen
konnten (VAN CORVEN et al. 1987a,b, KAUNE et al. 1990). In den basolateralen
Membranen von Enterozyten wurde außerdem ein 3Na+/Ca2+-Austauscher nachgewiesen, der
jedoch in quantitativer Hinsicht von untergeordneter Relevanz zu sein scheint (VAN OS
1987, KAUNE et al. 1992).
2.6 Neonatale Entwicklung des Gastrointestinaltrakts
Während der postnatalen Entwicklung des Magen-Darm-Traktes zeigen sich zahlreiche
morphologische und funktionelle Veränderungen, die einerseits altersabhängig, andererseits
aber auch alimentär bedingt sein können. Diese Vorgänge können sich prinzipiell auch auf
den Ca2+-Transport auswirken.
Bis zum 3. Lebenstag nimmt die Masse des Dünndarms um 72 % zu, was in einer Zunahme
der Dünndarmlänge und Schleimhautmasse begründet ist (XU et al. 1992), ohne das
Verhältnis der Schleimhautoberfläche zum Körpergewicht wesentlich zu verändern
(BUDDINGTON et al. 2001). In den ersten Lebenstagen nehmen die einzelnen Zotten sowohl
in ihrer Länge als auch in ihrem Umfang zu, wobei die Zottenlänge ab der 2. bis 5. Woche
wieder abnimmt und in dieser Zeit die Krypten etwas tiefer werden, was auf eine vermehrte
Zellproliferation hinweist (XU et al. 1992, CERA et al. 1988). Durch die Reduktion der
Zottenlänge und die gleichzeitige stärkere Ausprägung der Krypten verringert sich das
Längenverhältnis von Zotte zu Krypte in den ersten drei Lebenswochen um den Faktor 3
(MOON 1971). Da die Zellen der Darmschleimhaut von den Stammzellen der Krypten
gebildet werden, erklärt dies die Altersabhängigkeit der Lebensdauer der Enterozyten. Die
mittlere Lebensdauer der Enterozyten beträgt bei neugeborenen Ferkeln etwa 7 bis 10 Tage
(MARTINSSON u. JÖNSSON 1976, MOON 1971) und bei 4 Wochen alten Ferkeln etwa 2
bis 4 Tage (MOON 1971, KOLDOVSKY et al. 1966). Im apikalen Bereich der Enterozyten
des gesamten Dünndarms zeigen sich beim Saugferkel bis zur 4. Lebenswoche
charakteristische Vakuolisierungen (MOON 1972), in denen größere Mengen an
12

Literaturübersicht Immunglobulinen nachgewiesen wurden (KÖMÜVES u. HEATH 1992). Während sich die
morphologischen Veränderungen im Dünndarm schnell entwickeln und in besonderem Maße
vom Alter des Tieres abhängig sind (CERA et al. 1988), entwickelt sich der Dickdarm
langsam und hängt vor allem von der Fütterung insbesondere der Umstellung der reinen
Milchfütterung in der Saugperiode auf die faserreiche Fütterung ab (MCCANCE 1974).
Dennoch beeinflusst die Fütterung auch die Morphologie des Dünndarm insofern, dass nach
dem Absetzen eine deutliche Abnahme der Zottenlänge und Zunahme der Kryptentiefe
einsetzt (HAMPSON 1986). Die Zottenform ändert sich von fingerförmig beim Saugferkel
nach zungen- oder blattförmig beim abgesetzten Tier (HALL u. BYRNE 1989) und kann
ebenfalls durch verschiedene Futterbestandteile bzw. Futterzubereitungen modifiziert werden
(DUNSFORD et al. 1989, MILLER et al. 1984, KELLY et al. 1990).
Die in der postnatalen Phase ablaufenden Veränderungen auf funktioneller Ebene
insbesondere auf die Anpassung der Transportvorgänge für die Hauptnährstoffe und
Mineralstoffe sind mindestens so komplex wie die morphologischen Veränderungen. Es soll
an dieser Stelle nicht näher darauf eingegangen werden, stattdessen wird auf die
umfangreichen Arbeiten zur postnatalen Entwicklung des Schweinedarms der Arbeitsgruppe
um R.K. Buddington hingewiesen (BUDDINGTON u. DIAMOND 1989, PUCHAL u.
BUDDINGTON 1992, BUDDINGTON 1994, BUDDINGTON u. MALO 1996, ZHANG et
al. 1997, 1998, BUDDINGTON et al. 2001).
13

Literaturübersicht 2.7 Besonderheiten des Ca2+-Transports bei Saugferkeln
Bei neugeborenen Ferkeln wurde im Gegensatz zu den Absetzferkeln, bei denen die
intestinale Ca2+-Absorption deutlich durch Calcitriol stimuliert wurde, gezeigt, dass die aktive
Ca2+-Absorption im vorderen Dünndarm in den ersten 2 bis 4 Lebenswochen nicht oder nur
partiell durch das Calcitriol/VDR-System kontrolliert wird (KAUNE et al. 1990,
SCHRÖDER et al. 1990, 1993). In den ersten 10 Lebenstagen ist nicht nur der vordere
Dünndarm, sondern auch das Ileum zur aktiven Ca2+-Resorpion befähigt (RADDE et al.
1980). Bei milchfrei ernährten Saugferkeln ist der aktive Ca2+-Transport signifikant
erniedrigt, was auf eine calcitrophische Wirkung bestimmter Milchinhaltsstoffe hinweist
(KLEIN 1999). Sowohl im Duodenum als auch im Jejunum wurde eine Abnahme der aktiven
Ca2+-Absorption bei 15 bis 35 Tage alten Ferkeln nach dem Absetzen im Vergleich zu 1 bis
14 Tage alten Saugferkeln nachgewiesen, jedoch war dieser Alterseffekt nicht signifikant
(RADDE et al. 1980).
Über den genauen Mechanismus der Ca2+-Absorption und die einzelnen Teilschritte beim
Saugferkel ist relativ wenig bekannt. Ferkel mit unphysiologisch niedrigem Plasma-
Calcitriolspiegel, wie z. B. bei Neugeborenen mit angeborenem Calcitriolmangel, können
Ca2+ in der gleichen Größenordnung resorbieren wie gesunde Tier mit physiologischen
Plasma-Calcitriolspiegel (LACHENMAIER-CURRLE u. HARMEYER 1988, SCHRÖDER
et al. 1998a). Ob die Durchschleusung von Ca2+ durch die Zelle bei Schweinen dieses Alters
weniger von calcitriol-induziertem Calbindin abhängig ist, oder ob noch andere Mechanismen
eine Rolle spielen ist nicht geklärt. Untersuchungen an Küken und jungen Ratten haben
Hinweise auf eine Beteiligung von Mikrotubuli und Actinfilamenten gegeben (NEMERE et
al. 1984, NASSAR et al. 1988), die möglicherweise auch beim Saugferkel eine Rolle spielen
(SCHRÖDER et al. 1998b).
14

Material und Methoden 3. MATERIAL UND METHODEN
3.1 Versuchstiere
Bei den verwendeten Schweinen handelte es sich um Kreuzungen der Rassen Deutsches
Edelschwein und Pietrain aus dem Lehr- und Forschungsgut Ruthe der Tierärztlichen
Hochschule Hannover. Dabei standen drei Altersgruppen zur Verfügung (Tabelle 1). Die
Saugferkel waren zwischen 5 und 6 Tage alt und wogen zwischen 2,1 und 2,8 kg. Bis zum
Zeitpunkt der Schlachtung wurden sie bei der Muttersau gehalten und ausschließlich mit
Muttermilch ernährt. Die Absetzferkel waren 60 Tage alt und wogen zwischen 8,1 und
11,7 kg. Sie wurden nach der dritten Woche abgesetzt und mit handelsüblichem Ferkel-Starter
Futter ernährt. Die Mastferkel waren zwischen 88 und 117 Tage alt und wogen zwischen 16,8
und 25,5 kg. Sie wurden mit handelsüblichem Anfangsmastfutter ernährt.
Tabelle 1: Lebendgewicht und Alter der Versuchstiere zum Zeitpunkt der Schlachtung.
( x ± SEM, n = Anzahl der Tiere)
Gruppe n Alter [Tage] Gewicht [kg]
Saugferkel 5 5,2 ± 0,2 2,4 ± 0,1
Absetzferkel 5 60 ± 0 10,0 ± 0,6
Mastferkel 5 102,4 ± 4,7 19,2 ± 1,6
15

Material und Methoden 3.2 Probenentnahmen
Die Tiere wurden durch einen Bolzenschuss betäubt und anschließend durch Eröffnen der
Arteriae carotides communes, der Venae jugulares externae und internae entblutet. Danach
wurde die Bauchhöhle eröffnet und das gesamte Darmkonvolut entnommen.
3.2.1 Probenentnahme für die Präparation von Bürstensaummembran-
vesikeln
Für die Herstellung der Bürstensaummembranvesikel (BSMV) wurde das restliche
Duodenum, je nach Größe des Tieres, mit einer Länge von 15 bis 80 cm, caudal der
Entnahmestelle für die RNA-Präparation (Kap. 3.2.2) entnommen. Der Darmabschnitt wurde
mit eiskalter physiologischer Kochsalzlösung (NaCl 0,9 % w/v) gespült, am Mesenterial-
ansatz eröffnet und in etwa 10 cm lange Stücke geschnitten, die in flüssigem Stickstoff
tiefgefroren und bis zur Präparation der Vesikel bei mindestens –70 °C gelagert wurden. Das
Gewebe der Saugferkel wurde maximal drei Tage bis zur Präparation aufbewahrt. Das
Gewebe der übrigen Ferkel wurde nicht länger als 5 Wochen eingefroren.
3.2.2 Probenentnahme für die RNA-Präparation
Es wurden jeweils die ersten 10 cm des Duodenums, des Jejunums, des Ileums, des
proximalen Colons sowie des distalen Colons entnommen, am Mesenterialansatz
aufgeschnitten und in eiskalter physiologischer Kochsalzlösung partikelfrei gespült. Danach
wurde das Darmstück mit der serosalen Seite auf eine Glasplatte gelegt und mittels zweier
Objektträger wurde die Darmwand im Bereich der Tunica mucosa, bestehend aus Lamina
epithelialis mucosae und Lamina propria mucosae, von der Lamina muscularis mucosae, der
Tela submucosa, der Tunica muscularis und von der Lamina serosa (soweit noch vorhanden)
getrennt. Die Mucosa wurde in flüssigem Stickstoff tiefgefroren und bis zur weiteren
Verwendung bei mindestens –70 °C gelagert. Die restlichen Darmschichten wurden
verworfen.
16

Material und Methoden 3.3 Präparation der Bürstensaummembranvesikel
3.3.1 Prinzip
Die verschiedenen Membranen der Zelle, die nach mechanischem und osmotischem
Aufbrechen der Zellen bzw. Abtrennen der schweren Organellen zurückbleiben, so dass sie in
unterschiedlichem Maße mit Magnesiumionen aggregieren. Dieser Umstand kann bei
geeigneter Zentrifugation dazu verwendet werden, verschiedene Membranfraktionen
voneinander zu trennen.
3.3.2 Durchführung
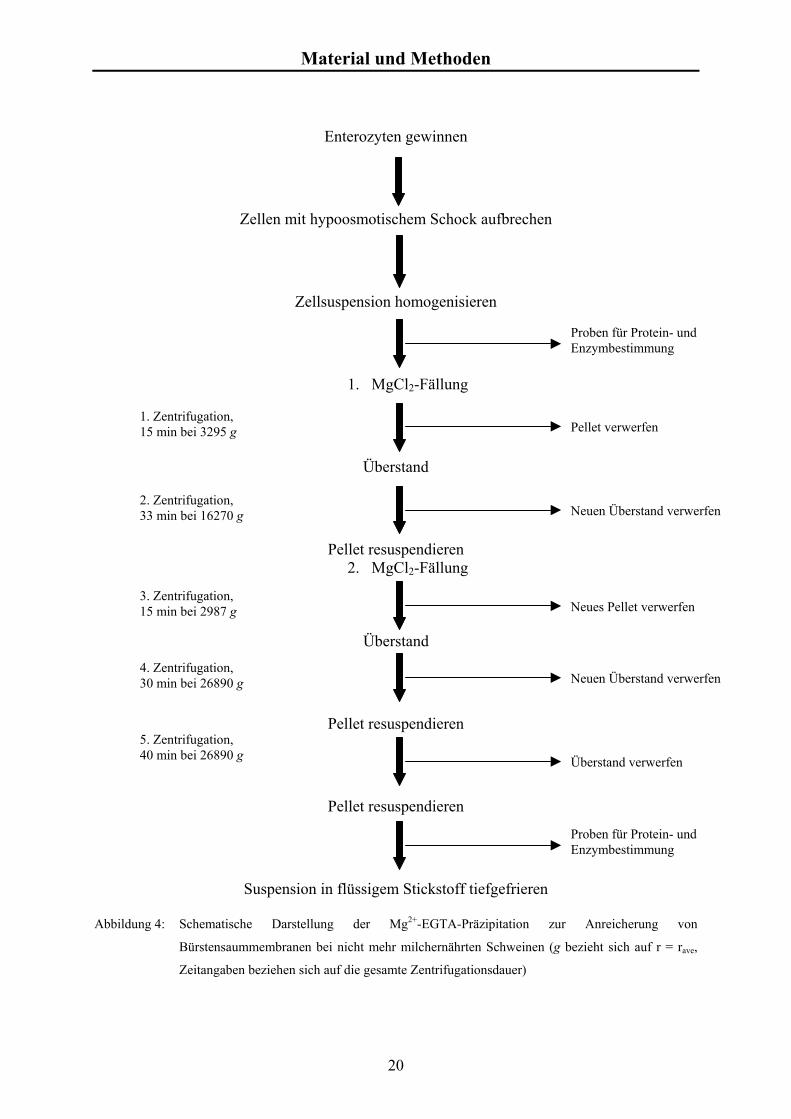
Die Präparation der BSMV erfolgte nach einer modifizierten Mg2+-EGTA-
Präzipitationsmethode mit Differentialzentrifugation (KAUNE el al. 1992, SCHRÖDER et al.
1998c). Abbildung 4 gibt die Präparationsschritte in einer Übersicht wieder.
Das Gewebe wurde 24 Stunden vor der Präparation bei –20 °C gelagert, um den
Auftauvorgang zu beschleunigen. Die gesamte Präparation erfolgte anschließend bei 0-4 °C
auf Eis. Zwischen 3,5 und 40 g Gewebe wurden mit der gleichen Menge an
Präparationspuffer I aufgetaut. Anschließend wurden die Enterozyten mit einem Vibro Mixer
(E1; Chemap AG, CH-8604 Volketswil) 10 min bei höchster Stufe von der Darmwand gelöst.
Die Flüssigkeit wurde durch ein herkömmliches Haushaltssieb gegeben und mit der 4fachen
Menge (bezogen auf das Volumen des Präparationspuffers I) H2O nachgespült. Dadurch
konnten die Enterozyten osmotisch aufgebrochen werden. Die Darmreste im Sieb wurden
verworfen und die Zellsuspension in einem Küchenmixer (Turboblender, Modell D70,
Moulinex) dreimal je 1 min auf der 4. Stufe homogenisiert. Bei dem Material der Saugferkeln
wurde der letzte Schritt durch eine zweimalige Homogenisierung mit dem Ultraturrax
(Heidolph DIAX 900, Stufe 5, Heidolph Instruments, D-91126 Schwabach) von je 30 s
ersetzt. Nach jedem Homogenisierungsschritt wurde der entstandene Schaum, nachdem er
sich 1 min abgesetzt hatte, mit einer Wasserstrahlpumpe abgesaugt. Nach Entnahme von bis
zu 1 ml Homogenat, zur Bestimmung des anfänglichen Proteingehaltes und der
17

Material und Methoden Ausgangsenzymaktivitäten, wurde dem restlichen Homogenat MgCl2 bis zu einer
Endkonzentration von 10 mmol.l-1 unter langsamem Rühren hinzugefügt und 15 min
inkubiert. Nach der Inkubation wurde das Homogenat 15 min lang bei 3295 g und 4 °C
zentrifugiert (Zentrifuge RC5C, Sorvall Instruments, Du Pont Company, Wilmington Delware
19898, USA; Rotor GSA), das Pellet verworfen und der Überstand 33 min lang bei 16270 g
und 4 °C zentrifugiert. Bei den Saugferkeln wurde das Homogenat 15 min bei 2987 g und
4 °C (Rotor SS34) und der entstandene Überstand 30 min bei 26890 g zentrifugiert. Nach
dieser Zentrifugation wurde der Überstand verworfen und das Pellet mit der dreifachen
Menge Präparationspuffer II, in Bezug auf die eingesetzte Gewebemenge, jedoch maximal
35 ml, in einem Elvehjem-Potter (Potter S 30; B. Braun, D-34212 Melsungen) mit
10 Schüben bei 1500 rpm homogenisiert. Diesem Homogenat wurde MgCl2 bis zu einer
Endkonzentration von 10 mmol.l-1 unter langsamem Rühren hinzugefügt und 15 min
inkubiert. Nach der Inkubation wurde das Homogenat 15 min bei 2987 g und 4 °C
zentrifugiert (Rotor SS34), das Pellet verworfen und der Überstand 30 min bei 26890 g und
4 °C zentrifugiert. Diesmal wurde der Überstand verworfen, das Pellet mit 30 ml
Vesikelpuffer in einem Elvehjem-Potter mit 10 Schüben bei 1500 rpm homogenisiert und das
Homogenat 40 min bei 26890 g und 4 °C zentrifugiert. Das entstandene Pellet wurde unter
Zugabe von 0,7 bis 3 ml Vesikelpuffer mit einer 1 ml-Spritze (Kanüle 0,45 x 0,23 mm)
homogenisiert. Es wurden bis zu 500 µl Vesikelsuspension zur Bestimmung der
Endproteinkonzentration und Enzymaktivitäten entnommen und bei –20 °C gelagert. Der Rest
der Vesikelsuspension wurde in flüssigem Stickstoff tiefgefroren und bis zur weiteren
Verwendung bei mindestens –70 °C gelagert. Die Vesikelsuspension der Saugferkel wurde
nicht eingefroren, sondern unmittelbar bei den Aufnahmestudien eingesetzt.
18

Material und Methoden 3.3.3 Präparationspuffer
Präparationspuffer I 300 mmol.l-1 D-Mannit
12 mmol.l-1 Tris, basisch
5 mmol.l-1 EGTA
HCl (1 mol.l-1) pH 7,4 bei 4 °C
Präparationspuffer II 60 mmol.l-1 D-Mannit
2,4 mmol.l-1 Tris, basisch
1 mmol.l-1 EGTA
HCl (1 mol.l-1) pH 7,4 bei 4 °C
Vesikelpuffer 100 mmol.l-1 D-Mannit
100 mmol.l-1 KCl
10 mmol.l-1 HEPES
Tris, basisch (1 mol.l-1) pH 7,4 bei 20 °C
19
Material und Methoden
Enterozyten gewinnen
Zellen mit hypoosmotischem Schock aufbrechen
Zellsuspension homogenisieren
Proben für Protein- und Enzymbestimmung
1. MgCl2-Fällung
Pellet verwerfen
1. Zentrifugation, 15 min bei 3295 g
Überstand
2. Zentrifugation, 33 min bei 16270 g
Pellet resuspendieren 2. MgCl2-Fällung
3. Zentrifugation, 15 min bei 2987 g
Überstand
4. Zentrifugation, 30 min bei 26890 g
Pellet resuspendieren 5. Zentrifugation, 40 min bei 26890 g
Pellet resuspendieren
Suspension in flüssigem Stickstoff tiefgefrieren
Abbildung 4: Schematische Darstellung der Mg2+-EGTA-Präzipitation zur
Bürstensaummembranen bei nicht mehr milchernährten Schweinen (g
Zeitangaben beziehen sich auf die gesamte Zentrifugationsdauer)
20
Neuen Überstand verwerfen
Neues Pellet verwerfen
Neuen Überstand verwerfen Überstand verwerfenProben für Protein- und Enzymbestimmung
Anreicherung von
bezieht sich auf r = rave,

Material und Methoden 3.4 Messung der Aufnahmeraten in die Bürstensaummembranvesikel
Die BSMV der Absetz- und der Mastferkel wurden bei RT aufgetaut. Die Aufnahmestudien
wurden bei RT durchgeführt. Vor dem Einsatz der Vesikelsuspension wurde diese nochmals
mittels 1 ml-Spritze (Kanüle 0,45 x 0,23 mm) homogenisiert. Je Messpunkt wurde der
Inkubationsansatz mit der Vesikelsuspension im Verhältnis 60 µl zu 20 µl versetzt und auf
einem Schüttler (Heidolph Reax 2000, Heidolph Instruments, D-91126 Schwabach)
durchmischt. Nach der Inkubation wurden bis zu 100 µl entnommen und die Ca2+-Aufnahme
durch Zugabe von 1 ml eiskalter Stopp-Lösung (Kap. 3.4.1) beendet. Anschließend wurden
die Vesikel durch Schnellfiltration (Cellulose-Nitrat-Filter, Porengröße 0,65 µm, Sartorius, D-
55555 Göttingen) von dem Reaktionsmedium getrennt, das Reaktionsgefäß mit 1 ml eiskalter
Stopp-Lösung gespült und der Filter nochmals zweimal mit je 5 ml eiskalter Stopp-Lösung
nachgespült. Die Filter wurden mit 4,3 ml Szintillationsflüssigkeit gemischt und die Aktivität
in einem Flüssigkeitsszintillationsmessgerät (Tri-Carb 2500; Canberra Packard GmbH, D-
63266 Dreieich) bestimmt. Die Zähldauer betrug stets 10 min. Der statistische Zählfehler
betrug 2-3 %. Alle Proben wurden in Doppelbestimmung gemessen.
3.4.1 Puffer für die Aufnahmestudien
Transportpuffer-NaCl 200 mmol.l-1 D-Mannit
(zweifach konzentriert) 200 mmol.l-1 NaCl
20 mmol.l-1 HEPES
Tris, basisch (1 mol.l-1) pH 7,4 bei 20 °C
Transportpuffer-KCl 200 mmol.l-1 D-Mannit
(zweifach konzentriert) 200 mmol.l-1 KCl
20 mmol.l-1 HEPES
Tris, basisch (1 mol.l-1) pH 7,4 bei 20 °C
21

Material und Methoden Stopp-Lösung 5 mmol.l-1 D-Mannit
150 mmol.l-1 NaCl/KCl
10 mmol.l-1 HEPES
Tris, basisch ( 1 mol.l-1) pH 7,4 bei 4 °C
CaCl2-Stammlösung 10 mmol.l-1
Valinomycin in Methanol 500 µmol.l-1
A23187 in DMSO 1 mg.ml-1
EGTA in H2O 10 mmol.l-1
Glucose in H2O 0,5 mmol.l-1
3.4.2 Kalkulation von [Ca2+]f
[Ca2+]f wurde mit Hilfe des Computerprogrammes Chelator ermittelt (SCHOENMAKERS et
al. 1992). Die Gesamt-Ca2+-Konzentrationen [Ca2+]ges in Bezug auf [Ca2+]f sind unter den
Bedingungen der Inkubationsansätze in Tabelle 2 wiedergegeben. Der Anteil an Ca2+-Ionen,
der durch die Applikation des Radiotracers 45Ca zusätzlich in den Inkubationsansatz verbracht
wurde, musste berücksichtigt werden und wurde deshalb über die spezifische Aktivität
(spez. A.) der Tracer-Stammlösung ermittelt:
X [mCi] . 1000 Ca-Konzentration [mmol.l-1]: . durch Zugabe von 45Ca 40,08 [g.mol-1] . spez. A. [mCi.mg-1] . Volumen [ml] Tabelle 2: Vergleich der Gesamt-Ca2+-Konzentrationen [Ca2+]ges mit den freien Ca2+-
Konzentrationen [Ca2+]f unter den Bedingungen im Inkubationsansatz mit 0,5 mmol.l-1 EGTA.
22
[Ca2+]ges [mmol.l-1] 0,5107 0,5278 0,5893 0,8998 1,2499 2,5 4,5
[Ca2+]f [mmol.l-1] 0,015 0,03 0,09 0,4 0,75 2,0 4,0

Material und Methoden 3.4.3 Studien zur zeitabhängigen Glucose-Aufnahme
Für die zeitabhängigen Messungen der Glucose-Aufnahmen wurde zum Zeitpunkt t = 0 min
570 µl Inkubationsansatz mit 190 µl Vesikelsuspension versetzt und durchmischt. Nach 0,5,
1, 2, 3, 5 60 und 180 min wurden je 100 µl entnommen und wie in Kap. 3.4 beschrieben
weiter bearbeitet.
Es wurden Aufnahmestudien in An- und Abwesenheit von Na+-Ionen im extravesikulären
Kompartiment durchgeführt.
Aus zweifach konzentriertem Transportpuffer (KCl bzw. NaCl), Glucose-Stammlösung und
radioaktiv markierter 3H-Glucose (~10 µCi bzw. 0,37 x 106 Bq) wurde der Inkubationsansatz
mit einem Gesamtvolumen von 1,5 ml hergestellt. Die Endkonzentration für Glucose betrug
10 µmol.l-1.
Zur Bestimmung der Gesamtaktivität je Probe wurden 75 µl Inkubationsansatz direkt mit
Szintillationsflüssigkeit gemischt und gemessen.
Zur Bestimmung des Leerwertes wurden 75 µl Inkubationsansatz analog zu den Proben
filtriert und gemessen.
Die Glucose-Aufnahmen in die BSMV als Funktion der Zeit werden in nmol Glucose je mg
Protein angegeben. Die Berechnung der Glucoseaufnahmen erfolgte analog zu der
Berechnung der Calciumaufnahmen (siehe Kap. 3.4.4).
3.4.4 Studien zur zeitabhängigen Calcium-Aufnahme
Für die zeitabhängigen Messungen der Ca2+-Aufnahmen wurde zum Zeitpunkt t = 0 min
570 µl Inkubationsansatz mit 190 µl Vesikelsuspension versetzt und durchmischt. Nach 0,5,
1, 2, 6, 10 und 60 min wurden je 100 µl entnommen und wie in Kap. 3.4 beschrieben weiter
bearbeitet.
Es wurden Aufnahmestudien in An- und Abwesenheit des Calcium-Ionophors A23187
durchgeführt.
Aus zweifach konzentriertem Transportpuffer-KCl, EGTA-Stammlösung, Valinomycin-
Stammlösung, A23187-Stammlösung bzw. DMSO, Ca2+-Stammlösung und radioaktivem
23

Material und Methoden 45Ca (~50 µCi bzw. 2,05 x 106 Bq) wurde der Inkubationsansatz mit einem Gesamtvolumen
von 1,5 ml hergestellt. Die Endkonzentrationen betrugen für EGTA 0,5 mmol.l-1, für
Valinomycin 5 µmol.l-1, für A23187 10 µg.ml-1 und für [Ca2+]f 0,03 mmol.l-1.
Zur Bestimmung der Gesamtaktivität je Probe wurden 75 µl Inkubationsansatz direkt mit
Szintillationsflüssigkeit gemischt und gemessen.
Zur Bestimmung des Leerwertes wurden 75 µl Inkubationsansatz analog zu den Proben
filtriert und gemessen.
Die Ca2+-Aufnahme (CA) in die BSMV als Funktion der Zeit werden in nmol Ca2+ je mg
Protein angegeben und lassen sich nach folgender Formel berechnen:
(cpmP-cpmL) . [Ca2+]f CA = cpmT . Pr
mit
cpmP = gezählte Zerfälle pro Minute in der Probe [cpm],
cpmL = gezählte Zerfälle pro Minute im Leerwert [cpm],
[Ca2+]f = Konzentration an ionisiertem, freiem Ca2+ je Probe [µmol.l-1],
cpmT = Gesamtaktivität je Probe [cpm],
Pr = Proteinkonzentration im Inkubationsansatz [mg.l-1].
Die Ca2+-Aufnahmen in Abhängigkeit der Inkubationsdauer können durch eine exponentielle
Sättigungskurve der Gleichung y = a . (1-e-kt) annähernd beschrieben und damit die Parameter
a und k sowie der Standardfehler ermittelt werden. Dazu wurde das Programm „GraphPad
Prism“ (ISI, Philadelphia, USA; www.graphpad.com) benutzt.
24

Material und Methoden 3.4.5 Studien zur konzentrationsabhängigen Calcium-Aufnahme
Für die Messungen der Ca2+-Aufnahme als Funktion der [Ca2+]f wurde zum Zeitpunkt t = 0
min 60 µl Inkubationsansatz mit 20 µl Vesikelsuspension versetzt und durchmischt. Nach 0,5
min wurde die Reaktion gestoppt und wie in Kap. 3.4 beschrieben weiter bearbeitet.
Es wurden Aufnahmestudien in An- und Abwesenheit des Calcium-Ionophors A23187
durchgeführt.
Aus zweifach konzentriertem Transportpuffer (KCl), EGTA-Stammlösung, A23187-
Stammlösung bzw. DMSO, Ca2+-Stammlösung und radioaktivem 45Ca (~40 µCi bzw.
1,48 x 106 Bq) wurden die Inkubationsansätze mit einem Gesamtvolumen von je 1,2 ml
hergestellt. Die Endkonzentrationen betrugen für EGTA 0,5 mmol.l-1, für A23187 10 µg.ml-1
und für [Ca2+]f 0,015; 0,03; 0,09; 0,4; 0,75; 2 und 4 mmol.l-1 (Tabelle 2). In Anwesenheit von
A23187 wurde ein Ansatz mit 0,03 mmol.l-1 an [Ca2+]f untersucht.
Zur Bestimmung der Gesamtaktivität je Probe wurden je 60 µl Inkubationsansatz direkt mit
Szintillationsflüssigkeit gemischt und gemessen.
Zur Bestimmung des Leerwertes wurden je 60 µl Inkubationsansatz analog zu den Proben
filtriert und gemessen.
25

Material und Methoden 3.4.6 Kalkulation der kinetischen Kenngrößen der Ca2+-Aufnahme
Die Ca2+-Aufnahmeraten in Abhängigkeit von [Ca2+]f wurden mit Hilfe eines Algorithmus
des Programmpakets „GraphPad Prism“ (ISI, Philadelphia, USA) ermittelt. Dabei wurde
folgende Formel zu Grunde gelegt:
Vmax . [Ca2+]f Ca2+-Aufnahme = + U . [Ca2+]f Km + [Ca2+]f mit
Vmax = Maximum der sättigbaren Aufnahme [nmol.mg-1],
Km = Michaelis-Menten-Konstante [mmol.l-1],
[Ca2+]f = Konzentration des freien Calciums [mmol.l-1],
U = nicht sättigbarer Anteil der Ca2+-Aufnahme. Dies ermöglicht die Kalkulation von Vmax und Km im Sinne einer typischen Michaelis-
Menten-Kinetik.
26

Material und Methoden 3.4.7 Ermittlung des Vesikelvolumens
Aus der Glucoseaufnahme zum Equilibriumszeitpunkt nach 180 min Inkubation wurde das
Vesikelvolumen berechnet. Das Vesikelvolumen wird in µl.mg-1 Protein angegeben und
errechnet sich durch Anwendung folgender Formeln:
VA cpmp - cpmL VV = ; VA = . AV ; P = Pr . AV P cpmT mit
VV = Vesikelvolumen [µl.mg-1],
VA = Vesikelvolumen im Ansatz [µl],
cpmp = gezählte Zerfälle pro Minute in der Probe beim Ausgleichswert [cpm],
cpmL = gezählte Zerfälle pro Minute im Leerwert [cpm],
cpmT = gezählte Zerfälle pro Minute im Gesamtansatz je Probe [cpm],
P = Proteinmenge im Ansatz [mg],
Pr = Proteinkonzentration im Ansatz [mg.l-1],
AV = Ansatzvolumen [ml].
27

Material und Methoden 3.5 Protein- und Enzym-Bestimmungen im Mukosahomogenat und in der
Vesikelsuspension
3.5.1 Protein
3.5.1.1 Prinzip
Der Farbstoff Coomassie Brilliant Blue G 250 hat die Eigenschaft, sich im sauren Milieu an
Protein zu binden. Das ursprünglich rote Kation wird durch die Bindung als blaues Anion
stabilisiert, was zu einer Verschiebung des Absorptionsmaximums von 465 nm nach 595 nm
führt. Somit kann die Menge des gebildeten Farbstoffes bei 595 nm photometrisch bestimmt
werden. Die Menge des gebildeten Farbstoffes ist dabei proportional zur Proteinmenge. Vor
der Farbreaktion wird den Proben oberflächenaktives Saponin zugegeben, um möglichst alle
Proteinbindungsstellen zugänglich zu machen.
3.5.1.2 Durchführung
Die Proteinbestimmung erfolgte nach der von BRADFORD (1976) beschriebenen Methode.
Das Farbstoffkonzentrat wurde 1:5 mit H2O verdünnt. Fünfzig µl der Probe wurden mit 50 µl
1%igem Saponin versetzt und 20 min bei 20 °C inkubiert. Nach Zugabe von 2,5 ml
verdünntem Farbkonzentrat wurde die Probe erneut 20 min bei 20 °C inkubiert. In gleicher
Weise wurde mit einer Standardreihe aus bovinem γ-Globulin aus Plasma sowie H2O als
Leerwert verfahren. Danach wurden die Proben im Photometer (DU®-8 Spectrophotometer;
Beckman Instruments GmbH, D-80807 München) bei 595 nm gegen den Leerwert gemessen.
Schließlich wurden den Proben Proteinwerte auf der linearen Regressionsgeraden der
Standardreihe zugeordnet.
28

Material und Methoden 3.5.2 Aktivität der alkalische Phosphatase (AP)
3.5.2.1 Prinzip
Die AP hat die Eigenschaft p-Nitrophenylphosphat hydrolytisch in p-Nitrophenyl und
Phosphat zu spalten. Die entstandene Menge an gelbem p-Nitrophenyl ist proportional zur
Aktivität der AP und kann photometrisch bei 405 nm gemessen werden.
3.5.2.2 Durchführung
Die Bestimmung der Aktivität der AP erfolgte nach der von KAWADE (1964) beschriebenen
Methode. Je 50 µl Probe wurden mit 3 ml AP-Puffer (1,02 mol.l-1 Diethanolamin,
0,51 mmol.l-1 MgCl2, 10 mmol.l-1 Na-Nitrophenylphosphat, mit HCl auf pH 9,8 eingestellt)
versetzt und gemischt. Im Photometer (Kap. 3.5.1.2) wurde nach 1, 2 und 3 min die
Extinktion bei 405 nm gegen Luft gemessen. Zur Berechnung wurde die Differenz der
Extinktion pro Minute mit dem versuchsinternen Faktor 3300 bei 1 cm Schichtdicke
entsprechend dem Lambert-Beerschen Gesetz multipliziert. Daraus ergab sich die Einheit
U.l-1, die anschließend in U.g-1 Protein umgerechnet wurde.
3.5.3 Aktivität der Na+/K+-ATPase
3.5.3.1 Prinzip
Ouabain hat die Eigenschaft, spezifisch die Aktivität der Na+/K+-ATPase zu hemmen.
Üblicherweise wird durch die intakte ATPase-Aktivität anorganisches Phosphat frei gesetzt,
das in saurer Lösung mit Molybdat Phosphormolybdänsäure bildet, die zu Molybdänblau
reduziert werden kann. Die Extinktion dieses blauen Farbkomplexes wird bei 609 nm
photometrisch gemessen. Die Aktivität der Na+/K+-ATPase entspricht der Differenz der
ATPase-Aktivität mit und ohne Ouabainzusatz.
29

Material und Methoden 3.5.3.2 Durchführung
Die Bestimmung der Na+/K+-ATPase-Aktivität erfolgte nach der von FUJITA et al. (1971)
beschriebenen Methode. Von der Probe wurden 20 µl mit 0,5 ml Assaymix-Puffer ohne
Ouabain (5 mmol.l-1 MgCl2; 100 mmol.l-1 NaCl; 10 mmol.l-1 KCl; 100 mmol.l-1 Tris, basisch;
2 mmol.l-1 ATP; 3 mmol.l-1 EDTA; 32%ige HCl pH 7,4 bei 37°C) bzw. 0,5 ml Assaymix-
Puffer mit Ouabain (5 mmol.l-1 MgCl2; 100 mmol.l-1 NaCl; 10 mmol.l-1 KCl; 100 mmol.l-1
Tris, basisch; 2 mmol.l-1 ATP; 3 mmol.l-1 EDTA; 5 mg.ml-1 Ouabain, 32%ige HCl pH 7,4
bei 37°C) 30 min bei 37 °C inkubiert. Als Leerwert wurden 20 µl H2O als Probe eingesetzt.
Nach der Inkubation wurde die Reaktion mit 1,5 ml eiskaltem Farbreagenz (760 mmol.l-1
H2SO4; 730 mg.ml-1 Ammoniumheptamolybdat; 2,667 g.ml-1 FeSO4.7H2O; 16,7 mg.ml-1
Trichloressigsäure) gestoppt und 30 min bei Raumtemperatur inkubiert. Danach wurde die
Extinktion in einem Photometer (Kap. 3.5.1.2) bei 690 nm gegen den Leerwert gemessen. Zur
Berechnung wurde die Extinktion der Probe mit Ouabain von der Extinktion der Probe ohne
Ouabain subtrahiert und die Differenz mit dem Faktor 862 bei 1 cm Schichtdicke gemäß einer
Standardreihe multipliziert. Daraus ergab sich die Einheit U.l-1, die anschließend in U.g-1
Protein umgerechnet wurde.
3.6 Gesamt-RNA-Isolierung und DNase-Verdau
Zur Gesamt-RNA-Isolierung wurde ein kommerziell erhältliches System (RNeasy Mini Kit
(50)) der Firma Quiagen (Quiagen GmbH, D-40724 Hilden) verwendet, das mit dem DNase-
Set (RNase-Free DNase Set (50)) derselben Firma kombiniert wurde
(www.quiagen.com/literature/malit.asp).
30

Material und Methoden 3.6.1 Prinzip
Die Zellmembranen des Gewebes werden mechanisch und chemisch desintegriert und damit
die RNA freigesetzt. Die freigesetzte RNA wird bei hohen Salzkonzentrationen reversibel an
eine Kieselgel-Membran gebunden. Diese Membran wird gewaschen, getrocknet und
anschließend wird die RNA mit H2O eluiert. Um eine Kontamination der RNA mit
genomischer DNA auszuschließen, wird die an die Kieselgel-Membran gebundene RNA vor
der Elution mit DNase behandelt.
3.6.2 Durchführung
Ein ca. 1 g schweres Stück Gewebe wurde mit Hilfe eines Mörsers und eines Pistills in
flüssigem Stickstoff zu einem Pulver verrieben. Von diesem Pulver wurden 30 mg in einem
vorgekühlten Polyethylen-Gefäß (Eppendorf-Gefäß) mit 600 µl RLT-Puffer, dem 1 %
Mercapto-Ethanol zugesetzt worden war, versetzt und mit einer 1-ml Spritze und einer 20 G-
Kanüle durch fünfmaliges Aufziehen homogenisiert. Das Homogenat wurde 3 min bei
13000 rpm und RT in einer Zentrifuge (Biofuge pico; Heraeus Instruments GmbH, D-63450
Hanau) zentrifugiert. Der separierte und klare Überstand wurde mit 600 µl Ethanol (70 % v/v)
versetzt und gemischt. Dieses Gemisch wurde in zwei Partien auf eine Kieselgel-Membran-
Säule gegeben und jeweils 15 s bei 10000 rpm und RT zentrifugiert. Die Säule wurde mit
350 µl RW1-Puffer gewaschen, indem der Puffer auf die Säule gegeben wurde und diese
anschließend 15 s bei 10000 rpm und RT zentrifugiert wurde. Daraufhin wurde die Säule
15 min bei RT mit 10 µl DNase und 70 µl RDD-Puffer inkubiert und noch einmal mit 350 µl
RW1-Puffer wie oben angegeben gewaschen. In analoger Weise wurde die Säule mit 500 µl
RPE-Puffer gewaschen. Anschließend wurde die Säule mit 500 µl RPE-Puffer gewaschen,
wobei die Säule 2 min bei 13000 rpm und RT zentrifugiert wurde, um diese zu trocknen. Zur
Eluierung der RNA wurden 50 µl H2O auf die Säule gegeben und 1 min bei 10000 rpm und
RT zentrifugiert. Die Gesamt-RNA wurde photometrisch quantifiziert (Eppendorf
Biophotometer; Eppendorf-Netheler-Hinz GmbH, D-22331 Hamburg) und bis zur weiteren
Verwendung bei –20 °C gelagert.
31

Material und Methoden 3.7 cDNA-Synthese
3.7.1 Prinzip
Das Enzym Reverse Transkriptase vermittelt die Umsetzung einer RNA-Sequenz in eine
entsprechende cDNA. Es benötigt dazu geeignete Temperatur-Bedingungen und
Salzkonzentrationen sowie die einzelnen Bestandteile der DNA und unspezifische Primer.
3.7.2 Durchführung
In einem 0,5 ml fassenden Eppendorf-Gefäß wurde 1 µg in H2O gelöster Gesamt-RNA mit
1 µl Oligo-dT-Primer (500 µg.ml-1) und 1 µl dNTP-Mix (je 10 mmol.l-1) versetzt, auf 13 µl
mit H2O aufgefüllt, gemischt, 4 s in der Zentrifuge (Kap. 3.6.2) angeschleudert und 5 min bei
65 °C in einem Thermomixer (Thermomixer comfort, 1,5 ml; Eppendorf-Netheler-Hinz
GmbH, D-22331 Hamburg) inkubiert. Anschließend wurde dieses Gemisch 2 min in Eis
(0-4 °C) inkubiert. Nach der Inkubation wurde dem Gemisch 4 µl Puffer (250 mmol.l-1 Tris-
HCl, pH 8,3; 375 mmol.l-1 KCl; 15 mmol.l-1 MgCl2; 50 mmol.l-1 Dithiothreitol (DTT)) und
2 µl DTT (0,1 mol.l-1) hinzugefügt, angeschleudert und 2 min bei 42 °C im Thermocycler
(Mastercycler gradient; Eppendorf-Netheler-Hinz GmbH, D-22331 Hamburg) inkubiert.
Daraufhin wurde 1 µl Superscript II RNase H-RT (200 U.µl-1) zu dem Gemisch hinzugegeben,
wiederum angeschleudert und 50 min bei 42 °C im Thermocycler inkubiert. Hiernach erfolgte
eine Inkubation von 15 min bei 70 °C im Thermomixer. Bis zur weiteren Verwendung wurde
die cDNA bei –20 °C gelagert.
32

Material und Methoden 3.8 Polymerasekettenreaktion (PCR)
3.8.1 Prinzip
Die Polymerase bildet mit Hilfe von zielsequenzspezifischen Primern Kopien von DNA-
Einzelsträngen. Dieser Vorgang wird bis zu 40-mal wiederholt.
Jede PCR stellt eine mehrfache Abfolge von Zyklen dar, wobei jeder Zyklus aus drei Schritten
besteht. Bei dem ersten Schritt, der „Denaturierung“, werden durch Erhitzen auf eine
Temperatur von 95 °C die beiden Stränge der DNA bzw. cDNA (Kap. 3.7) voneinander
getrennt, indem die Wasserstoffbrückenbindungen zwischen den komplementären Basen
durch Hitze zerstört werden. Bei dem zweiten Schritt, dem „Annealing“, lagern sich zwei
Primer über Wasserstoffbrückenbindungen an die DNA-Einzelstränge. Primer sind kleine
synthetisch hergestellte Abschnitte von einzelsträngiger DNA mit einer Länge von 15 bis 35
Nukleotiden (Oligonukleotide). Sie werden so ausgewählt, dass sie komplementär zu den
Zielsträngen sind, wobei sie die zu vermehrende Region begrenzen und ein Primer der
Sequenz des einen DNA-Stranges entspricht (sense) und der andere der Sequenz des
komplementären Stranges (antisense). Die Primer werden im relativen Überschuß zur Menge
an Ziel-DNA zum Reaktionsansatz gegeben, so dass sich statistisch gesehen die beiden
komplementären Zielstränge der denaturierten Matrize häufiger mit den Primern als
untereinander verbinden. Die Annealing-Temperatur hängt von der Basenzusammensetzung
der Primer ab und liegt üblicherweise zwischen 50 °C und 60 °C. Sie sollte möglichst hoch
gewählt werden, um unspezifische Produkte zu vermeiden (RYCHLIK et al. 1990). Bei dem
dritten Schritt der PCR, der „Elongation“, synthetisiert eine DNA-Polymerase ausgehend von
dem Primer als Startpunkt matrizenabhängig den komplementären Strang der DNA. Die
optimale Arbeitstemperatur der für die PCR verwendeten, hitzestabilen Enzyme liegt bei 72
°C. Die Zeitdauer der einzelnen Schritte ist von der Länge des PCR-Produktes und den
verwendeten Reagenzien und Geräten abhängig und liegt zwischen 15 s und 2 min. Diese
Zyklen werden etwa 25 bis 40-mal wiederholt. Nach jedem Zyklus verdoppelt sich theoretisch
die Anzahl der amplifizierten Fragmente (1, 2, 4, 8, 32, 64, ... ) (NEDELMAN et al. 1992).
33

Material und Methoden Von BELL u. DEMARINI (1991) werden kurze Reaktionszeiten und möglichst wenige Zyklen
zur Vermeidung von unspezifischen Amplifikaten empfohlen.
Ein Reaktionsansatz für die PCR besteht aus der DNA, Puffer-Systemen (Salzen),
Magnesiumchlorid, Primern, Nukleotiden und der thermostabilen DNA-Polymerase
(REMICK et al. 1990). Die eingesetzten thermostabilen Polymerasen haben ihr
Aktivitätsmaximum bei einem pH-Wert oberhalb von 8,0. Zur Pufferung wird am häufigsten
ein Tris-Puffer mit einem pH-Wert von 8,5 bis 9,0 verwendet. Kaliumchlorid und
Ammoniumsulfat werden zur Steigerung der Ausbeute dem PCR-Ansatz zugesetzt.
Natriumchlorid kann die Amplifikation hemmen (MUELHARDT 2002). Magnesiumionen sind
der wichtigste Kofaktor der DNA-Polymerase und stimulieren ihre Aktivität. Außerdem bilden
sie zusammen mit den Nukleotiden einen löslichen Komplex, der für den Nukleotid-Einbau
entscheidend ist. Auch beeinflussen sie die Bindung der Primer und die Annealing-
Temperatur. Die Magnesiumchlorid-Konzentration ist somit für die Spezifität und Ausbeute
der PCR von wesentlicher Bedeutung. Sie variiert in einem Bereich zwischen 0,5 mmol.l-1 und
5 mmol.l-1. Je niedriger die Magnesiumchlorid-Konzentration ist, desto weniger unspezifische
Amplifikate treten auf (LYONS 1992). Entscheidend für den Erfolg der PCR sind die Primer.
Neben ihrer Länge ist auch die Basenzusammensetzung von Bedeutung. Der G/C-Gehalt
sollte bei ca. 50 % liegen. Des Weiteren sollten komplementäre Bereiche zwischen den
Primern, die zur Bildung von Primer-Dimeren führen können und auch signifikante
Sekundärstrukturen der Primer-Sequenzen vermieden werden. Die optimale
Primerkonzentration liegt zwischen 0,1 µmol.l-1 und 1 µmol.l-1. Die Nukleotide stellen die
Grundbausteine der neu zu bildenden DNA dar. Ihre optimale Konzentration hängt von der
Länge des zu amplifizierenden Produktes, der Primerkonzentration und der
Magnesiumchlorid-Konzentration ab. Die normalerweise verwendete Konzentration der
Nukleotide liegt bei etwa 100 µmol.l-l. Bei niedrigen Konzentrationen arbeitet die DNA-
Polymerase präziser (FINCKH et al. 1991). Eine Voraussetzung zur automatischen
Durchführung der PCR, ohne die nach jedem Denaturierungsschritt neue DNA-Polymerase
zum Reaktionsansatz hinzugegeben werden müsste, ist der Einsatz von thermostabilen
Polymerasen (SAIKI et al. 1988). Thermostabile Polymerasen ergänzen, ausgehend vom 3‘-
Ende der an die Ziel-DNA gebundenen Primer, die einzelsträngige Matrize durch Einbau der
vier Nukleotide im Sinne einer komplementären Basenpaarung zu einem Doppelstrang. Sie
34

Material und Methoden wurden ursprünglich aus thermostabilen Mikroorganismen isoliert (z. B. Thermus aquaticus:
Taq-Polymerase, CHIEN et al. 1976; Thermus thermophilus: Tth-Polymerase, MYERS u.
GELFAND 1991) und werden inzwischen gentechnisch hergestellt. Sie unterscheiden sich
bezüglich der Thermostabilität, der Aktivität und der 3‘-5‘-Exonuklease-Funktion, die einer
Korrekturaktivität entspricht und damit deutlich verringerte Fehlerraten ermöglicht (CLINE
et al. 1996, TINDALL u. KUNKEL 1988).
3.8.2 Durchführung
In den Reaktionsansatz wurden folgende Substanzen gegeben: 2 µl cDNA (Kap. 3.7), 5 µl
Puffer (0,1 mol.l-1 Tris HCl, pH 8,3; 0,5 mol.l-1 KCl; 1 mg.ml-1 Gelatine; 25 mmol.l-1 MgCl2),
0,5 µl Primer1 (10 pmol.l-1), 0,5 µl Primer2 (10 pmol.l-1), 0,5 µl dNTPs (10 mmol.l-1), 41 µl
H2O und 0,2 µl Taq-Polymerase (5 U.µl-1). Bei mehr als 4 Ansätze wurden die Substanzen
erst als Mix gemischt und dann auf die Reaktionsansätze verteilt.
Soweit nichts anderes beschrieben ist, lief folgendes Programm in dem Thermocycler
(Kap. 3.7.2, MOSSA et al. 1991) ab:
1. 94 °C 2 min lang,
2. 94 °C 30 s lang,
3. 60 °C 1 min lang,
4. 72 °C 1 min lang,
5. 39-mal Wiederholung der Schritte 2 bis 4,
6. 72 °C 15 min lang,
Die verwendeten Primer wurden aus bekannten Sequenzen anderer Spezies hergeleitet und
wurden über die GenBank (National Center for Biotechnology Information [NCBI], Bethesda,
MD, USA) ermittelt.
Für den konservierten ECaC-Bereich, also für den Bereich in dem sich ECaC1 und ECaC2
nur geringfügig unterscheiden, wurden folgende Primer gewählt:
5`>CAGATGTACAACCTGCTGCT<3`, s, ECaC1 (Kaninchen, AJ133128, Basen: 738ff),
5`>CACATGGTGAGCAGATGATG<3`, as, ECaC2 (Mensch, AF365927, Basen: 1233ff).
35

Material und Methoden Für den hypervariablen Bereich des ECaC1 und ECaC2 wurden folgende Primer gewählt:
ECaC1
5`>GAATCCGTATCGAGTACTT<3`, s, ECaC1 (Ratte, AB032019, Basen: 2061ff),
5`>GGAGAATTTCCCATCCACGG<3`, as, ECaC1 (Ratte, AB0322019, Basen: 2241ff),
ECaC2
5`>GGCCCTGCGAACTATGACGTGGATCTGCCCTT<3`, s, ECaC2 (Maus, AB037373,
Basen: 1826ff),
5`>TGGAAGGCCTGTGCGTAGCG<3`, as, ECaC2 (Mensch, AF365927, Basen: 2157ff).
3.9 Darstellung der DNA auf einem Agarosegel
3.9.1 Prinzip
Die DNA wird im Agarosegel durch Anlegen eines elektrischen Feldes größenfraktioniert und
anschließend mit Ethidiumbromid angefärbt. Ethidiumbromid bindet an DNA und kann im
ultravioletten Licht sichtbar gemacht werden (KEMP et al. 1989). Durch einen parallel
aufgetragenen Größenmarker (GM) lässt sich die Größe des DNA-Stückes bestimmen
(MULLIS u. FALOONA 1987).
3.9.2 Durchführung
500 mg Agarose wurden mit TAE-Puffer (Tris, basisch, 242,0 g.l-1; Eisessig 57,1 ml.l-1;
EDTA-Na.2 H2O 100 ml.l-1, pH 8,0) versetzt, in einer Mikrowelle zweimal bis zum Kochen
erhitzt und in einen Gelgießstand verbracht. Anschließend wurde ein Kamm in den
Gelgießstand gehängt, so dass im Gel Taschen von etwa 15 µl Fassungsvermögen entstanden.
Nachdem das Gel erstarrt war, wurde es in die Elektrophorese-Kammer (wide mini Sub® Cell
GT; Bio-Rad-Laboratories GmbH, D-80939 München) überführt und mit TAE-Puffer
bedeckt. Je 5 µl des PCR-Gemisches wurden mit 2 µl Loading-dye (DNA) gemischt und in
die Taschen des Agarosegels gegeben. Die DNA wurde bei 60 Volt (Spannungsgeber: Power
Pac 3000; Bio-Rad-Laboratories GmbH, D-80939 München) über 25 min größenfraktioniert,
36

Material und Methoden anschließend 10 min in einer Ethidiumbromidlösung geschwenkt und nochmals 10 min in
H2O geschwenkt. Schließlich wurde das Gel über einer ultravioletten Lampe (Biometra TI 1;
Biometra, D-37079 Göttingen) betrachtet und in dieser Form abgelichtet.
3.10 Isolierung der PCR-Produkte
3.10.1 Prinzip
Die Agarosegel-Struktur wird zerstört und die DNA in Lösung gebracht. DNA bindet bei
hohen Salzkonzentrationen reversibel an Glas und kann nach Separierung wieder mit H2O
abgelöst werden.
3.10.2 Durchführung
Es wurde der QIAquick PCR Purification Kit (Quiagen GmbH, D-40724 Hilden) nach
Anleitung des Herstellers verwendet, wenn im Agarosegel nur ein PCR-Produkt nachweisbar
war (www.quiagen.com/literature/cleanlit.asp):
Zu 45 µl PCR-Mix wurden 225 µl PB-Puffer hinzugegeben und gemischt. Dieses Gemisch
wurde auf eine QIAquick-Säule gegeben und bei 10000 g und RT 1 min lang zentrifugiert
(Zentrifuge: Kap. 3.6.2). Dann wurden 0,75 ml PE-Puffer auf die Säule gegeben und zweimal
je 1 min bei 10000 g und RT zentrifugiert. Schließlich wurden 30 µl EB-Puffer auf die Säule
gegeben, 1 min bei RT inkubiert und bei 10000 g und RT 1 min zentrifugiert. Das Eluat
enthielt die DNA und wurde bis zum weiteren Gebrauch bei –20 °C gelagert. Zur Kontrolle
wurden 5 µl des Eluates auf ein Agarosegel aufgetragen (Kap. 3.9).
Waren zwei oder mehr PCR-Produkte im Agarosegel sichtbar, wurde der QIAEX II Agarose
Gel Extractions-Kit (Quiagen GmbH, D-40724 Hilden) verwendet (www.quiagen.com/
literature/cleanlit.asp):
Die DNA wurde im Agarosegel sichtbar gemacht, das gewünschte PCR-Produkt mit einem
sterilen Skalpell herausgeschnitten und gewogen. Zu 100 mg Gel wurden in einem
Eppendorf-Gefäß 300 µl QX1-Puffer hinzugefügt und mindestens 30 s geschüttelt. Dem
37

Material und Methoden Gemisch wurden 30 µl QIAEX II hinzugefügt, gemischt und bei 50 °C 10 min im Wasserbad
inkubiert, wobei die Suspension alle 2 min geschüttelt wurde. Es folgte ein
Zentrifugationsschritt bei 10000 g und RT für eine Dauer von 30 s. Der Überstand wurde
sorgfältig entfernt und das Pellet in 500 µl QX1-Puffer durch Schütteln resuspendiert.
Wiederum wurde 30 s bei 10000 g und RT zentrifugiert und der Überstand sorgfältig entfernt.
Diesmal wurde das Pellet in 500 µl PE-Puffer durch Schütteln resuspendiert, bei 10000 g und
RT 30 s lang zentrifugiert und der Überstand sorgfältig entfernt. Das Pellet wurde ein zweites
Mal in 500 µl PE-Puffer durch Schütteln resuspendiert, bei 10000 g und RT 30 s lang
zentrifugiert, der Überstand sorgfältig entfernt, und schließlich wurde das Pellet 10 min bei
RT getrocknet. Dem getrocknetem Pellet wurden 20 µl H2O hinzugefügt und geschüttelt.
Nach einer Inkubation von 5 min bei RT wurde bei 10000 g und RT 30 s lang zentrifugiert.
Der DNA-haltige Überstand wurde separiert und bis zum weiteren Gebrauch bei –20 °C
gelagert. Zur Kontrolle wurden 5 µl des Eluates auf ein Agarosegel aufgetragen (Kap. 3.9).
3.11 Ligation der PCR-Produkte
3.11.1 Prinzip
Die PCR-Produkte werden zur späteren Vermehrung in Bakterien zuerst durch eine Ligase in
einen spezifischen Vektor ligiert. Auf diesem Vektor befindet sich zur Selektion der Bakterien
eine Ampicillin-Resistenz.
3.11.2 Durchführung
Zu 5 µl PCR-Produkt wurden 1 µl Vektor (pGEMEX®-2 Vector; Promega GmbH, D-68199
Mannheim), 1 µl Ligase und 7 µl Ligations-Puffer (60 mmol.l-1 Tris-HCl, pH 7,8; 20 mmol.l-1
MgCl2; 20 mmol.l-1 DTT, 2 mmol.l-1 ATP) hinzugegeben, gemischt und 5½ Stunden bei RT
inkubiert.
38

Material und Methoden 3.12 Transformation der Bakterien
3.12.1 Prinzip Bei der Transformation werden z. B. Bakterien, sogenannte Kompetente Zellen, verwendet,
die die Eigenschaft haben, zugefügte Plasmide unter bestimmten Salzkonzentrationen und
Temperaturbedingungen aufzunehmen.
3.12.2 Durchführung
Zu dem Ligationsansatz (Kap. 3.11) wurden 100 µl CaCl2 (0,1 mol.l-1) hinzugefügt, gemischt
und auf Eis 10 min inkubiert. Danach wurden 10 µl Bakteriensuspension (Select 96™
Competent Cells) hinzugefügt, gemischt und 20 min auf Eis inkubiert. Es folgte ein
Hitzeschock bei 42 °C von exakt 90 s im Wasserbad mit anschließender Kühlung der
Suspension auf Eis über eine Dauer von 2 min. Nach Zugabe von 800 µl Luria-Bertani-
Medium (LB-Medium, WANG u. KOCH 1978) wurde die Suspension 35 min im Wasserbad
bei 37 °C inkubiert. Nach dieser Inkubation wurde die Suspension bei 9000 g (Zentrifuge,
Kap. 3.6.2) und RT 1 min lang zentrifugiert und der Überstand bis auf etwa 100 µl reduziert.
Anschließend wurde das Pellet in dem Rest des Überstandes resuspendiert und dieses
Gemisch auf LB-Medium-Agarplatten mit Ampicillin (0,1 mg.ml-1) ausgestrichen und 12 bis
14 Stunden bei 37 °C im Brutschrank (BBD 6220; Heraeus Instruments, D-63450 Hanau)
inkubiert. Die gewachsenen Bakterienkolonien wurden bis zum weiteren Gebrauch auf den
Platten im Kühlschrank aufbewahrt. Einzelne Kolonien wurden je in 2,5 ml LB-Medium mit
Ampicillin (0,1 mg.ml-1) versetzt und 12-14 Stunden bei 37°C und 180 rpm im
Schüttelinkubator (Typ 3031; GFL, Gesellschaft für Labortechnik mbH, D-30938 Burgwedel)
inkubiert.
39

Material und Methoden 3.13 Plasmidpräparation
Bei der Plasmidpräparation fand ein kommerziell erhältliches System (GenElute™ Plasmid
Purification Kit Miniprep; Sigma-Aldrich Chemicals, D-82041 Deisenhofen) Anwendung
(www.sigmaaldrich.com).
3.13.1 Prinzip
Nach Lyse der Bakterien mit alkalischem SDS und anschließender Neutralisation werden die
Plasmide reversibel an eine glasbeschichtete Säule gebunden. Die Säule wird gewaschen und
danach werden die Plasmide wieder eluiert.
3.13.2 Durchführung
1,5 ml der Bakterienkultur (Kap. 3.12) wurden bei 12000 g und RT 1 min lang zentrifugiert
(Zentrifuge, Kap. 3.6.2) und der Überstand sorgfältig entfernt. Das Pellet wurde in 200 µl
Resuspensionspuffer mit der Pipette resuspendiert. Der Suspension wurden 200 µl
Lysispuffer zugefügt, vorsichtig geschwenkt und 10 min bei 12000 g und RT zentrifugiert.
Der Überstand wurde auf eine Säule gegeben und diese bei 12000 g und RT 1 min lang
zentrifugiert. Die Säule wurde mit 500 µl Optional-Wasch-Lösung versetzt und 1 min bei
12000 g und RT zentrifugiert. Anschließend wurden 750 µl Wasch-Lösung auf die Säule
gegeben und diese 1 min bei 12000 g und RT zentrifugiert. Um die Säule zu trocknen wurde
ein erneuter Zentrifugationsschritt bei 12000 g und RT für eine Dauer von 1 min angehängt.
Die DNA wurde nun mittels 50 µl Elutions-Puffer, der auf die Säule gegeben wurde welche
hiernach bei 12000 g und RT 1 min lang zentrifugiert wurde, eluiert.
40

Material und Methoden 3.14 Restriktionsverdau
3.14.1 Prinzip
In unmittelbarer Nähe der Ligationsstellen im Plasmid liegen spezifische Enzymschnittstellen.
Mit dem entsprechenden Enzym ist es möglich, das spezifische PCR-Produkt mit einem
geringen Überhang wieder aus dem Plasmid zu schneiden, auf ein Agarosegel aufzutragen
und die Größe mit dem erwarteten PCR-Produkt zu vergleichen.
3.14.2 Durchführung
Zu 2 µl Plasmidlösung wurden 15 µl H2O, 2 µl Puffer (10 mmol.l-1 MgCl2, 100 mmol.l-1
NaCl, 0,1 mg.ml-1 bovines Serum-Albumin, Tris/HCl pH 7,5 bei 37 °C) und 1 µl Enzym
(EcoRI) hinzugegeben, gemischt und bei 37 °C 1,5 Stunden im Wasserbad inkubiert.
Anschließend wurden 5 µl auf ein Agarosegel aufgetragen (Kap. 3.9).
3.15 Sequenzierung
Zur Sequenzierung wurden die Plasmide zum Sequenzierservice der „Faculty of Medical
Sciences“ der Universität Nijmwegen (Niederlande) gegeben. Um auszuschließen, dass eine
fehlerhafte Sequenz sequenziert wurde, die dadurch entsteht, dass die Polymerase einen
Einbaufehler hat (CLINE et al. 1996), wurden stets mehrere Plasmide sequenziert.
41

Material und Methoden 3.16 mRNA-Isolierung
3.16.1 Prinzip
Die Poly (A)+-Überhänge der mRNA binden bei hohen Salzkonzentrationen reversibel in einer
Säule an Oligo-dT-Zellulose (AVIV u. LEDER 1972). Diese wird gewaschen und
anschließend wird die mRNA aus dem Säulenmaterial eluiert. Bevor die Gesamt-RNA auf die
Säule gegeben wird, wird sie hitzedenaturiert.
3.16.2 Durchführung
Die Gesamt-RNA wurde auf Eis aufgetaut und mit 14000 rpm bei 4 °C 20 min lang
zentrifugiert (Megafuge 1.0R, Rotor-Nummer: 3041; Heraeus Instruments, D-63450 Hanau).
Das entstandene Pellet wurde zweimal mit je 500 µl Ethanol (70 % v/v) überschichtet,
gemischt, mit 14000 rpm bei 4 °C 3 min lang zentrifugiert und der Überstand anschließend
sorgfältig entfernt. Nach einer zweiminütigen Trocknung in der Vakuum-Zentrifuge (Speed
VacR Plus SC 110A; Savant Instruments Inc., Holbrook, NY, USA) wurde das Pellet in
800 µl H2O gelöst und nach einer fünfminütigen Inkubation bei 65 °C im Thermomixer
(Kap. 3.7), 3 min auf Eis inkubiert. Die Probe wurde mit 100 µl einer NaCl-Lösung
(5 mol.l-1), 100 µl einer Sodiumdodecylsulfat (SDS)-Lösung (1% w/v) und 10 µl einer
Tris/HCl-Lösung (1 mol.l-1) versetzt, gemischt und mit 13000 rpm bei RT 2 min lang
zentrifugiert (Kap. 3.6.2). Dieses Gemisch wurde auf eine Oligo-dT-Zellulose-Säule, die
zuvor 5-mal mit je 1 ml Loading-Puffer (10 mmol.l-1 Tris/HCl; 0,5 mol.l-1 NaCl; 0,1% SDS)
gespült worden war, zusammen mit 500 µl Loading-Puffer gegeben, aufgefangen und noch
insgesamt 4-mal über die Säule gegeben. Anschließend wurde die Säule 5-mal mit je 1 ml
Loading-Puffer und 5-mal mit je 1 ml Washing-Puffer (10 mmol.l-1 Tris/HCl; 0,1 mol.l-1
NaCl) gespült. Die mRNA wurde schließlich mit 2,5 ml Elution-Puffer (10 mmol.l-1 Tris/HCl)
wieder aus der Säule eluiert. Je 0,5 ml Eluat wurden mit 10 µl Glycogen (Typ IX, 2 mg.ml-1),
50 µl Natriumacetat (2 mol.l-1) und 500 µl Isopropanol versetzt, gemischt und in mindestens 8
Stunden bei –20 °C ausgefällt.
42

Material und Methoden 3.17 Northern-Blot
3.17.1 Prinzip
Die mRNA wird in einem denaturierendem Agarosegel der Größe nach elektrophoretisch
aufgetrennt. Durch Kapillarkräfte wird die RNA aus dem Gel auf eine Nitrozellulosemembran
transferiert. Die spezifische RNA wird mit Hilfe einer radioaktiv markierten DNA-Sonde, die
aus einem Stück komplementärer Basensequenz zur gesuchten RNA besteht, hybridisiert und
anschließend auf einem Film sichtbar gemacht (modifizierte Methode von SOUTHERN 1975).
3.17.2 Größenfraktionierung der RNA in einem denaturierenden
Agarosegel Zur Auftrennung der mRNA nach dem Molekulargewicht wurden intramolekulare
Basenpaarungen geschmolzen und die denaturierte RNA-Struktur durch Formaldehyd und
Formamid fixiert. Zuvor musste die mRNA (Kap. 3.16) in H2O gelöst werden. Dazu wurde
sie bei 13000 rpm (Zentrifuge, Kap. 3.16.2) und 4 °C 20 min lang zentrifugiert und der
Überstand sorgfältig entfernt. Das entstandene Pellet wurde mit 500 µl Ethanol (70 % v/v)
überschichtet und nach einer 3 min langen Zentrifugation bei 13000 rpm und 4 °C wurde der
Überstand sorgfältig entfernt. Dieser Schritt wurde einmal wiederholt. Um das Ethanol zu
entfernen, wurde die RNA in einer Vakuum-Zentrifuge (Kap. 3.16.2) in 2 min auf kleiner
Stufe getrocknet. Danach wurde die pelletierte RNA zum Lösen in 800 µl H2O je Probe
15 min auf Eis inkubiert. Die gelöste RNA wurde in einem Photometer (Kap. 3.6.2)
quantifiziert. Etwa 10 µg RNA wurden mit 10 µl Loading-Puffer (48 % Formamid, 17,5 %
Formaldehyd (37%ig), 35 % Loading dye (RNA); v/v) versetzt, gemischt und 3 min bei 95 °C
denaturiert. Anschließend wurde das Gemisch 3 min auf Eis inkubiert, um eine Renaturierung
zu verhindern. Vor dem Auftragen der Proben auf das Gel wurden diese in der Zentrifuge
angeschleudert.
43

Material und Methoden Zur Anfertigung des Gels wurden 1 g Agarose mit 85 ml H2O zweimal in der Mikrowelle
aufgekocht. Nachdem sich das Gel auf etwa 50 °C abgekühlt hatte, wurden diesem 10 ml
sterilfiltrierter 10x MOPS-Puffer (0,2 mol.l-1 MOPS; 0,05 mol.l-1 EDTA-Natriumsalz; pH 5,5
bei RT) und 5,4 ml Formaldehydlösung (37 % v/v) zugesetzt und das Gel in einem
Gelgießstand gegossen. Anschließend wurde ein Kamm in den Gelgießstand gehängt, so dass
im Gel Taschen von etwa 25 µl Fassungsvermögen entstanden. Nachdem das Gel erstarrt war,
wurde es in die Elektrophorese-Kammer (Kap. 3.9.2) überführt und mit 1x MOPS-Puffer (10x
MOPS-Puffer, 1 zu 10 verdünnt) bedeckt.
Neben den Proben wurde ein kommerziell erhältlicher RNA-Größenmarker mit auf das Gel
aufgetragen. Die RNA wurde bei 40 V (Spannungsgeber, Kap. 3.9.2) über 4 Stunden
größenfraktioniert. Anschließend wurde der Größenmarker abgeschnitten, 10 min in
Ethidiumbromidlösung und 15 min in H2O geschwenkt und über einer ultravioletten Lampe
(Kap. 3.9.2) betrachtet. Zur späteren Analyse wurde der Größenmarker zusammen mit der
mm-Einteilung eines Lineals abgelichtet.
3.17.3 Übertragen der RNA auf eine Nitrocellulosemembran
Das Agarosegel (Kap. 3.17.2) wurde 15 min in H2O und 15 min in 20x SSC-Puffer (3 mol.l-1
NaCl, 0,3 mol.l-1 Natriumcitrat; pH 7,0 bei RT) geschwenkt, um es von dem MOPS-Puffer zu
reinigen und bereits mit dem Transfer-Puffer vorzuinkubieren. Danach wurde der Anteil,
welcher die mRNA-Proben enthielt, aus dem Gesamtgel herausgeschnitten.
Eine Glaswanne wurde mit 20x SSC-Puffer gefüllt und mit einer Glasplatte abgedeckt, so
dass zwei gegenüberliegende Seiten der Wanne nicht durch sie bedeckt waren. Aus Whatman-
Papier (Biometra, D-37079 Göttingen) wurde eine, zuvor in den Puffer eingeweichte, Brücke
luftblasenfrei über die Glasplatte gelegt, wobei die Enden der Brücke in den 20x SSC-Puffer
tauchten. Das Gel wurde mit den Taschenöffnungen nach unten ebenfalls luftblasenfrei auf
die Whatman-Papierbrücke gelegt und die Ränder des Gels mit Parafilm (American National
CanTM, USA) abgedeckt, um zu verhindern, dass der Transfer-Puffer an dem Gel vorbei
gelangen konnte. Auf das Gel wurde eine Nitrocellulosemembran (Hybond N+; Amersham
Pharmacia Biotech, Uppsala, Schweden), welche zuvor auf die exakte Größe des Gels
geschnitten worden und nach Anfeuchtung mit H2O 10 min in 20x SSC-Puffer inkubiert
44

Material und Methoden worden war, gelegt. Diese wurde wiederum mit 3 Lagen exakt geschnittenem Whatman-
Papier, das in 20x SSC-Puffer vorgeweicht worden war, abgedeckt. Auf das Whatman-Papier
wurde ein mindestens 8 cm hoher Turm aus Saugpapier (Handy Einzeltücher; Migros-
Genossenschaftsbund, CH-8031 Zürich) gelegt, der mit einem Gewicht von etwa 300 g
beschwert wurde. In 12 Stunden wurde die negativ geladene RNA durch Kapillarkräfte auf
die positiv geladene Nitrocellulosemembran transferiert und dort kovalent gebunden. Auf der
Membran wurde die Höhe der Taschen im Gel markiert, um den Startpunkt der
Elektrophorese auch auf der Membran nachvollziehen zu können. Daraufhin wurde sie
10 min in 2x SSC-Puffer inkubiert. Die im Anschluß luftgetrocknete Membran wurde
zwischen 2 Whatman-Papierstücke gelegt und unter Vakuum (Slab Gel Dryer SGD 2000;
Savant Instruments Inc., Holbrook, New York) 2 Stunden lang bei 80 °C getrocknet. Die
Nitrocellulosemembran wurde bis zum weiteren Gebrauch bei –20 °C gelagert.
Abbildung 5: Schematische Darstellung des Kapillar-Blot-Verfahrens
Nitrocellulosemembran
Whatman-Papierbrücke Glasplatte
3x Whatman-Papier Agarosegel
20x SSC
45

Material und Methoden 3.17.4 Erstellung von spezifischen, radioaktiv markierten Sonden
Die radioaktiv markierten Sonden wurden nach der „Random priming“-Methode
(FEINBERG u. VOGELSTEIN 1984) unter Verwendung des „Rediprime II“-Kits
(Amersham Pharmacia Biotech, Uppsala, Schweden), eines kommerziell erhältlichen
Systems, hergestellt. Hierfür wurden 5 µl des PCR-Produktes der Primer, die auch für die
Real-Time-PCR (Kap. 3.18) verwendet wurden, mit TE-Puffer (10 mmol.l-1 Tris/HCl,
1 mmol.l-1 EDTA, pH 8,0) auf 40 µl aufgefüllt und 5 min bei 95 °C im Wasserbad denaturiert.
Um den denaturierten Zustand der DNA zu erhalten, wurde sie anschließend sofort 5 min lang
auf Eis inkubiert. Diese 40 µl wurden kurz angeschleudert (Zentrifuge, Kap. 3.6.2),
zusammen mit 50 µCi (2,05 x 106 Bq) α32P-dCTP in das „Rediprime“-Reaktionsgefäß
gegeben und mit TE-Puffer auf ein Gesamtvolumen von 50 µl aufgefüllt. In dem
Reaktionsgefäß befand sich das sogenannte Klenow-Fragment, das große Fragment der DNA-
Polymerase I aus Escherichia coli. Dieses Enzym synthetisiert, nach Anlagerung von
Random-Primern, unter gleichzeitigem Einbau von α32P-dCTP den komplementären DNA-
Strang. Dem Klenow-Fragment fehlt die 5` 3`-Exonukleaseaktivität. Dadurch ist es nicht in
der Lage, das 5`-Ende der an die Matrizen-DNA gebundenen Primer zu entfernen. Da
mehrere Primer an einen Strang binden, wodurch Teilsequenzen erstellt werden, entsteht ein
Gemisch aus unterschiedlich langen komplementären DNA-Stücken. Das Gemisch in dem
„Rediprime“-Reaktionsgefäß wurde durch 12-maliges auf- und abpipettieren gemischt und
1 Stunde bei 37 °C im Thermomixer (Kap. 3.7.2) inkubiert. Die Reaktion wurde mit 5 µl
EDTA-Lösung (0,2 mol.l-1) gestoppt. Zur Kontrolle der Einbaurate wurden 1 µl der Sonde mit
49 µl H2O versetzt. Fünf µl der verdünnten Sonde wurden mit 2,5 µl „Herring Sperm“ DNA
(10 mg.ml-1), 22,5 µl H2O und 1 ml eisgekühlter Trichloressigsäure (10 % w/v) versetzt und
10 min auf Eis inkubiert, um die DNA zu fällen. Diese Probe wurde auf einen Glasfaserfilter
(APFCO2500, Millipore, Irland) gegeben und zunächst 10-mal mit je 1 ml Trichloressigsäure
und anschließend 10-mal mit je 1 ml Ethanol (100%ig) gewaschen. Der Filter wurde
luftgetrocknet, in einem Glasszintillationsgefäß mit 12 ml Szintillationsflüssigkeit versetzt
und nach sorgfältigem Schütteln im Szintillationsmessgerät (Kap. 3.4) gemessen. Die
Zähldauer betrug stets 10 min. Der statistische Zählfehler betrug 2-3 %.
46

Material und Methoden 3.17.5 Hybridisierung der RNA mit den spezifischen, radioaktiv markierten
Sonden
Um alle unspezifischen Bindungsstellen der Nitrocellulosemembran (Kap. 3.17.3) zu
besetzen, wurde diese mit 200 µl „Herring Sperm“ DNA (10 mg.ml-1) im Prähybridisierungs-
puffer (5x Denhardt`s-Reagenz, 0,1 % w/v SDS, 5x SSC, 40 % v/v Formamid) versetzt und
7 Stunden bei 42 °C im Hybridisierofen (Rolleninkubator 4020; Gesellschaft für
Labortechnik mbH, D-30938 Burgwedel) prähybridisiert. Dazu wurde die „Herring Sperm“
DNA zuerst bei 95 °C 3 min lang denaturiert, dann 3 min auf Eis inkubiert und anschließend
zusammen mit 15 ml Prähybridisierungspuffer auf die Membran in die Prähybridisierröhren
gegeben. Die Membran wurde zuvor in der Waschlösung I (2x SSC, 0,1 % w/v SDS)
vorgeweicht und mit der RNA-behafteten Seite zum Lumen der Prähybridisierröhre zeigend,
luftblasenfrei an die Wand der Röhre angelegt.
Nach der Prähybridisierung wurden 5 ml Prähybridisierungspuffer mit „Herring Sperm“ DNA
aus der Röhre entfernt und eine spezifische, radioaktiv markierte Sonde (Kap. 3.17.4) in den
verbliebenen Puffer gegeben. Die Sonde wurde zuvor bei 95 °C 3 min lang denaturiert, 3 min
auf Eis inkubiert und anschließend kurz angeschleudert (Zentrifuge: Kap. 3.6.2). Die
Nitrocellulosemembran wurde 16 Stunden bei 42 °C hybridisiert.
Nach der Hybridisierung mussten eventuell unspezifisch gebundene Sondenanteile, nicht
gebundene Sondenanteile und freie radioaktive Nukleotide entfernt werden. Dies wurde durch
wiederholte Waschschritte mit abnehmender Salzkonzentration und zunehmender Temperatur
durchgeführt, wobei sich die Stringenz in jedem Waschvorgang erhöhte. Die Stringenz kann
auch über die Waschdauer beeinflusst werden. Durch eine höhere Spezifität können über
einen größeren Bereich mehr Wasserstoffbrückenbindungen zwischen den komplementären
Basenpaaren ausgebildet werden, wodurch eine höhere Stringenz beim Waschvorgang
gewählt werden kann, was zu einer höheren Spezifität und geringerem Hintergrund des
resultierenden Signals führt (LOTTSPEICH und ZORBAS 1998).
Der erste Waschschritt wurde nach Abgießen der Sonde bei RT durchgeführt. Die Membran
wurde im Hybridisierofen 10 min lang mit je 15 ml Waschpuffer 1 gewaschen, wobei der
Puffer zweimal gewechselt wurde.
47

Material und Methoden Der zweite Waschschritt wurde bei 37 °C mit dem Waschpuffer 2 (0,5x SSC, 0,1 % w/v SDS)
im Hybridisierofen für eine Dauer von 10 min mit je 15 ml Puffer durchgeführt, wobei der
Puffer zweimal gewechselt wurde.
Der dritte Waschschritt erfolgte analog der ersten beiden Waschschritte, jedoch mit dem
Waschpuffer 3 (0,2x SSC, 0,1 % w/v SDS) bei 42 °C.
Nach diesen Waschschritten wurde die Membran zum Trocknen auf ein Whatman-Papier
(Kap. 3.17.3) gelegt und in eine „Saran Wrap“ Folie eingeschlagen.
3.17.6 Darstellung der RNA
Mit Hilfe eines Phosphor-Imagers (Personal molecular imager FX, Bio-Rad Laboratories
GmbH, D-80939 München) wurden die Banden auf der Nitrocellulosemembran (Kap. 3.17.5)
detektiert. Dafür wurde ein mit BaFBr:Eu-Kristallen beschichteter „Film“ (Imaging Screen K
20x25 cm; Bio-Rad Laboratories GmbH, D-80939 München) auf die Membran in einer
Photofilmkassette (Exposure Cassette-K, 20x25 cm; Bio-Rad Laboratories GmbH, D-80939
München) gelegt und 4 Stunden exponiert. In dieser Zeit strahlte die radioaktiv markierte
Sonde ionisierende Energie ab und überführte die BaFBr:Eu-Kristalle in einen stabilen,
angeregten Zustand (Eu Eu2+). Der Screen wurde anschließend in den Phosphor-Imager
verbracht, wo er von einem Laser mit einer Wellenlänge von 635 nm abgetastet wurde. Durch
das Abtasten wurden die Kristalle weiter angeregt und somit in einen instabilen Zustand
(Eu2+ Eu3+) überführt. Die Elektronen fallen in ihren Grundzustand zurück (Eu3+ Eu) und
geben dabei Energie in Form von Photonen ab, die im Phosphor-Imager gemessen wird
(MUELHARDT 2002).
3.17.7 Auswertung der Northern-Blots
Die Auswertung der Daten aus Kap. 3.17.6 erfolgte mit der Quantifizierungs-Software
„Quantity One“ für Windows (Bio-Rad-Laboratories GmbH, D-80939 München). Diese
Software stellt die Energie, die von den BaFBr:Eu-Kristallen freigesetzt wurde, graphisch dar.
So entstehen Banden auf den RNA-Spuren, die spezifisch für die gewählte Sonde
48

Material und Methoden (Kap. 3.17.4) sind. Jede dieser Banden wurde vermessen, wobei sowohl Fläche als auch die
Pixeldichte (Intensität) berücksichtigt wurden. Da diese Quantifizierung der RNA abhängig
von der aufgetragenen RNA-Menge ist, wurde ein interner Standard gewählt, um die Werte
zu relativieren und vergleichbar zu machen. Dies führt zu einer semiquantitativen
Bestimmung der spezifischen mRNA-Menge. Als interner Standard wurde das in jeder
Körperzelle vorkommende β-Aktin gewählt. Jeder Blot wurde mit einer spezifischen Sonde
für das β-Aktin hybridisiert und die RNA-Menge des β-Aktins ermittelt. Bei verschiedenen
Tieren kann so der Quotient aus der Menge an gesuchter RNA und der Menge an β-Aktin-
RNA verglichen werden.
3.18 Real-Time-PCR
3.18.1 Prinzip
Die sogenannte Real-Time-PCR funktioniert im Prinzip wie eine standardmäßige PCR
(Kap. 3.8), jedoch wird dem PCR-Ansatz ein Farbstoff zugegeben, der die Eigenschaft hat,
sich an Doppelstrang-DNA zu binden und der in diesem Zustand detektiert werden kann.
Eingeführt wurde diese Technik von HIGUCHI et al. 1993. Dabei wurde mit einer
Videokamera die in der PCR entstandene DNA mit Hilfe von Ethidiumbromid detektiert.
Diese Technik wurde weiterentwickelt, indem eine digitale Aufnahmetechnik eingeführt wurde
und ein fluoreszierender Farbstoff zur Verfügung stand, der sich durch eine erhöhte
Sensitivität auszeichnete (BECKER et al. 1996). Die modernste Version der Real-Time-PCR
erlaubt die Echtzeit-Messung der ansteigenden Fluoreszenz mit jedem Zyklus. Der Anstieg
der Fluoreszenz wird, um die Hintergrundfluoreszenz korrigiert, für jeden einzelnen Zyklus
dargestellt und lässt sich im linearen Abschnitt der aufsteigenden Kurve prinzipiell direkt mit
der Menge an vorliegender DNA korrelieren (WOO et al. 1998, MORRISON et al. 1998).
49

Material und Methoden 3.18.2 Durchführung
Bei der Durchführung der Real-Time-PCR wurde der BrilliantTM Quantitative PCR Core
Reagent Kit der Firma Stratagene (La Jolla, Ca 92037, USA) nach Anweisung des Herstellers
verwendet. Zum Nachweis der DNA diente der fluoreszierende Farbstoff SYBR® Green I
nucleic acid gel stain der Firma Molecular Probes (Molecular Probes Europe BV, Nl-2333
AA Leiden). Die Reaktion lief in einer Real-Time-PCR-Maschine (Mx4000TM Multiplex
Quantitative PCR System; Stratagene, La Jolla, Ca 92037, USA) ab.
Zunächst wurden die optimalen Reaktionsbedingungen für jedes einzelne PCR-Produkt
ermittelt. In jeden Reaktionsansatz wurden folgende Substanzen gegeben: cDNA (Kap. 3.7),
Puffer (10fach konzentriert), Primer1 (10 pmol.l-1), Primer2 (10 pmol.l-1), dNTPs
(20 mmol.l-1), H2O, SYBR® Green I (1/3000), ROX (1mmol.l-1) und Sure StartTM Taq DNA
polymerase (5 U.µl-1). Dabei wurden immer 1 µl dNTPs, 2,5 µl SYBR® Green I, 0,375 µl
ROX (1/200) und 0,25 µl Sure StartTM Taq DNA Polymerase eingesetzt. Der Reaktionsansatz
wurde mit H20 auf 25 µl aufgefüllt. Die Menge an eingesetztem MgCl2, Primern und cDNA
wurde über Konzentrationsreihen ermittelt, wobei sich für die verschiedenen zu
untersuchenden Sequenzen folgende Mengen als optimal erwiesen:
ECaC1: 1,5 µl MgCl2, 0,25 µl je Primer und 4 µl cDNA,
ECaC2: 2,0 µl MgCl2, 0,1 µl je Primer und 1 µl cDNA,
β-Aktin: 1,5 µl MgCl2, 1,5 µl je Primer und 0,1 µl cDNA.
Bei jedem Durchgang wurde ein Doppelansatz untersucht.
Die verwendeten Primer waren spezifische Primer für die bekannten bzw. ermittelten
Teilsequenzen der entsprechenden RNAs. Die Teilsequenz für das β-Aktin ist unter
AF054837 in der GenBank (National Center for Biotechnology Information [NCBI],
Bethesda, MD, USA) zu finden, die Teilsequenzen von ECaC1 und ECaC2 wurden wie in
Kap. 3.8 beschrieben ermittelt. Die Primer wurden so gewählt, dass bei den verschiedenen
Produkten Bereiche von 110 bis 139 Basenpaaren amplifiziert wurden. Ihre Spezifität wurde
durch das Programm BLAST der Genbank überprüft. Für das β-Aktin sind die Lokalisationen
der Primer in Abbildung 6 dargestellt. Diese wurden so gewählt, dass sie aus zwei
verschiedenen Exons stammen, um eine Kontamination der cDNA mit genomischer DNA
50

Material und Methoden auszuschließen. Diese Primer bilden mit der cDNA-Matrize ein kurzes Produkt und würden
mit genomischer DNA ein zweites, größeres Amplifikat erzeugen. Die Lokalisationen der
Primer für ECaC1 und ECaC2 sind in Kap. 4.4 und 4.5 dargestellt.
1 10 20 30 40 50 60 70 ATGTTTGAGACCTTCAACACGCCGGCCATGTACGTGGCCATCCAGGCTGTGCTGTCCCTGTACGCCTCTG 80 90 100 110 120 130 140 GCCGCACCACTGGCATTGTCATGGACTCTGGGGATGGGGTCACCCACACGGTGCCCATCTACGAGGGGTA 150 160 170 180 190 200 210 CGCCCTGCCCCACGCCATCCTGCGTCTGGACCTGGCTGGCCGGGACCTGACCGACTACCTCATGAAGATC 220 230 240 250 260 270 280 CTCACGGAGCGGGGCTACAGCTTCACCACCACGGCCGAGCGGGAGATCGTGCGGGACATCAAGGAGAAGC 290 300 310 320 330 340 350 TCTGCTACGTCGCCCTGGACTTCGAGCAGGAGATGGCCACGGCCGCCTCCTCCTCCTCCCTGGAGAAGAG 360 370 380 390 400 410 420 CTACGAGCTGCCCGACGGGCAGGTCATCACCATCGGCAACGAGCGCTTCCGGTGTCCAGAGGCGCTCTTC 430 440 450 460 470 480 490 CAGCCCTCCTTCCTGGGCATGGAGTCCTGCGGCATCCACGAGACCACCTTCAACTCGATCATGAAGTGCG 500 510 520 530 540 550 560 ACGTGGACATCAGGAAGGACCTCTACGCCAACACGGTGCTGTCTGGCGGGACCACCATGTACCCCGGCAT 570 580 590 600 610 620 630 CGCCGACAGGATGCAGAAGGAGATCACGGCCCTGGCGCCCAGCACCATGAAGATCAAGATCATCGCCCCT
Abbildung 6: Sus scrofa, β-Aktin, Teilsequenz; die ausgewählten Primer sind mit grauem Hintergrund abgesetzt,
der schwarze Pfeil markiert den Übergang von zwei verschiedenen Exons.
In der Real-Time-PCR-Maschine lief folgendes Programm ab:
1. 95 °C 10 min lang,
2. 95 °C 15 s lang,
3. 60 °C 1 min lang,
4. 39-mal Wiederholung der Schritte 2 und 3,
5. schrittweise Temperaturerhöhung zur Erstellung einer
Schmelzkurve.
Die Verwendung eines modifizierten Programms erzielte keine besseren Ergebnisse.
51

Material und Methoden 3.18.3 Auswertung der Real-Time-PCR
Bei der Auswertung der Real-Time-PCR wurde die Software des Mx4000TM Multiplex
Quantitative PCR System (Kap. 3.19.2) verwendet. Stellt man die Zunahme an Fluoreszenz
abzüglich der Hintergrundfluoreszenz (dRn) gegenüber der Zykluszahl graphisch dar und legt
eine Basislinie oberhalb der Hintergrundfluoreszenz, so ergibt sich ein gemeinsamer
Schnittpunkt im Bereich der linearen Zunahme. Es wurden immer die Mittelwerte des
Doppelansatzes betrachtet. Der Schnittpunkt kann durch die entsprechende Zyklusanzahl
charakterisiert werden (Ct-Wert). Um alle Versuchsproben zu vergleichen, wurden sie mit
dem internen Standard, β-Aktin, gleichgesetzt. Dies bedeutet, dass zur weiteren Auswertung
die Differenzen der Ct-Werte der Ziel-DNA und dem dazugehörigen β-Aktin betrachtet
wurden. Die so ermittelten Differenzen der Ct-Werte verschiedener Tiere wurden nun
miteinander verglichen: eine um 1 Ct-Wert größere Differenz des einen Tieres gegenüber
eines anderen lässt den Schluss zu, dass in der cDNA nur die Hälfte der Matrize der Ziel-
DNA vorhanden war. Dabei wurde vorausgesetzt, dass sich die Anzahl der amplifizierten
Fragmente nach jedem Zyklus verdoppelt (NEDELMANN et al. 1992), jedoch muss bei der
Interpretation der Ergebnisse kritisch betrachtet werden, dass die Effizienz eines jeden Zyklus
niedriger ist (WANG et al. 1989). MUEHLHARDT (2002) gibt die Effizienz mit 1,6 bis 1,7
pro Zyklus an, allerdings gibt es Untersuchungen unter Verwendung der Real-Time-PCR, die
eine Effizienz von annähernd 2 zeigten (PFAFFL 2001).
Setzt man die größte vorhandene Matrizenanzahl (Tier1) mit 100 % gleich, so ergibt sich für
die anderen Tiere:
M = . 100 %, 1 D2
mit
M = relative Anzahl der Matrizen [%],
D = Ct-Wert-Differenz Tier2 – Ct-Wert-Differenz Tier1.
52

Material und Methoden 3.19 Chemikalien
Soweit die Chemikalien nicht im Folgenden aufgeführt werden, wurden sie von Sigma-
Aldrich Chemie GmbH, D-89552 Steinheim bezogen und hatten mindestens den
Qualitätsstandard pro Analysis.
α32P-dCTP; Hartmann Analytic, D-38124 Braunschweig
Agarose; Qualex Gold™ Agarose, Hybaid GmbH, D-69123 Heidelberg
Bovines Plasma γ-Globulin; Bio-Rad Laboratories GmbH, D-80939 München 45Ca, Calcium 45 (NEZ-013); Perkin Elmer™, B-1930 Zaventem
DNA-Größenmarker 100 bp (0,5 mg.ml-1); MBI Fermentas GmbH, D-68789 St. Leon-Rot
dNTP-Mix (10 mmol.l-1); MBI Fermentas GmbH, D-68789 St. Leon-Rot
DTT (0,1 mol.l-1); Gibco™ Invitrogen GmbH, D-76131 Karlsruhe
EcoRI; MBI Fermentas GmbH, D-68789 St. Leon-Rot
Farbstoff Coomassie Brilliant Blue G 250; Bio-Rad Laboratories GmbH, D-80939 München
„Herring Sperm“ DNA (10 mg.ml-1); Gibco™ Invitrogen GmbH, D-76131 Karlsruhe 3H-Glucose, Tritium-markierte Glucose (NET-238C); Perkin Elmer™, B-1930 Zaventem
Oligo-dT-Primer (500 µg.ml-1); Promega GmbH, D-68199 Mannheim
Primer; Carl Roth GmbH + Co, D-76185 Karlsruhe
Select 96™ Competent Cells; Promega GmbH, D-68199 Mannheim
Superscript II RNase H-RT (200 U.l-1); Gibco™ Invitrogen GmbH, D-76131 Karlsruhe
Szintillationsflüssigkeit, Lumasafe™ Plus; Canberra Packard GmbH, D-63266 Dreieich
Taq Polymerase; Gibco™ Invitrogen GmbH, D-76131 Karlsruhe
53

Material und Methoden 3.20 Statistik
Aus den Versuchsdaten wurden arithmetische Mittelwerte ( x ) und Standardfehler der
Mittelwerte (SEM) berechnet. Aus früheren Untersuchungen mit vergleichbaren Tiergruppen
und Messparametern war bekannt, dass die zu erhaltenen Werte bei den Aufnahmestudien
normalverteilt sind (SCHRÖDER et al. 1993, SCHRÖDER et al. 1998a). Die statistische
Prüfung der vorliegenden Werte erfolgte daher mit parametrischen Testverfahren („One way
ANOVA“). Ergaben sich daraus signifikante Unterschiede, erfolgte der Gruppenvergleich
über einen nachfolgenden Tukey-Test. Wurden nur zwei Gruppen miteinander verglichen, so
erfolgte dies über den Student´s t-Test.
Insgesamt wurden folgende statistische Verfahren angewandt:
-„One way ANOVA“ mit nachfolgendem Tukey-Test
-Student´s t-Test für verbundene oder unverbundene Stichproben.
Die statistischen Berechnungen wurden mit GraphPad Prism (GraphPad Software Inc.,
Philadelphia, USA) durchgeführt.
Für die Signifikanzangaben gilt:
n.s. = nicht signifikant,
p<0,05 = *, schwach signifikant,
p<0,01 = **, signifikant,
p<0,001 = ***, hoch signifikant.
Bei der Ermittlung der linearen Regressionsgeraden für die Proteinbestimmung und bei der
Berechnung der Aufnahmeraten in die BSMV wurde die Software „GraphPad Prism“
(Kap. 3.4.9) genutzt.
Die Auswertung der Northern Blots erfolgte mit der Software „Quantity One“ für Windows
(Kap. 3.17.7).
Die Auswertung der Real-Time-PCR erfolgte mit der Software des Mx4000™ Multiplex
Quantitative PCR Systems (Kap. 3.18.3).
54

Ergebnisse 4 ERGEBNISSE
4.1 Effekte des Alters der Versuchstiere auf die Ca2+-Aufnahme in BSMV
4.1.1 Charakterisierung der BSMV
Zur Abschätzung der Anreicherungen der BSMV wurden charakteristische
Membranleitenzyme für das Ausgangshomogenat und die Vesikelsuspension untersucht. Als
spezifische Markerenzyme wurden für die Bürstensaummembranen die Aktivität der
alkalischen Phosphatase und für die basolateralen Membranen die Aktivität der Na+/K+-
ATPase bestimmt.
Tabelle 3: Spezifische Aktivität der alkalischen Phosphatase als Leitenzym für die
Bürstensaummembranen und der Na+/K+-ATPase als Leitenzym der basolateralen
Membranen im Homogenat und der Vesikelsuspension und deren Anreicherungen
in den verschiedenen Altersgruppen. Der Anreicherungsfaktor ergab sich aus dem
Vergleich der entsprechenden Aktivitäten im Homogenat und der
Vesikelsuspension. ( x ± SEM, n = Anzahl der Tiere).
Aktivität der alkalischen Phosphatase Aktivität der Na+/K+-ATPase
Gruppe Homogenat U.g-1 Protein
Vesikel-suspension
U.g-1 Protein
Anreicher-ungsfaktor
Homogenat U.g-1 Protein
Vesikel-suspension
U.g-1 Protein
Anreicher-ungsfaktor
Saugferkel (n = 5)
197 ± 10
1971 ± 152
10,1 ± 0,8
33,6 ± 3,8
73,0 ± 13,0
2,3 ± 0,5
Absetzferkel (n = 5)
174 ± 14
2476 ± 359
15,1 ± 1,3
34,7 ± 3,7
107,7 ± 23,8
3,3 ± 0,8
Mastferkel (n = 5)
165 ± 18
2155 ± 347
12,5 ± 0,9
36,8 ± 4,1
116,4 ± 14,5
3,2 ± 0,3
55
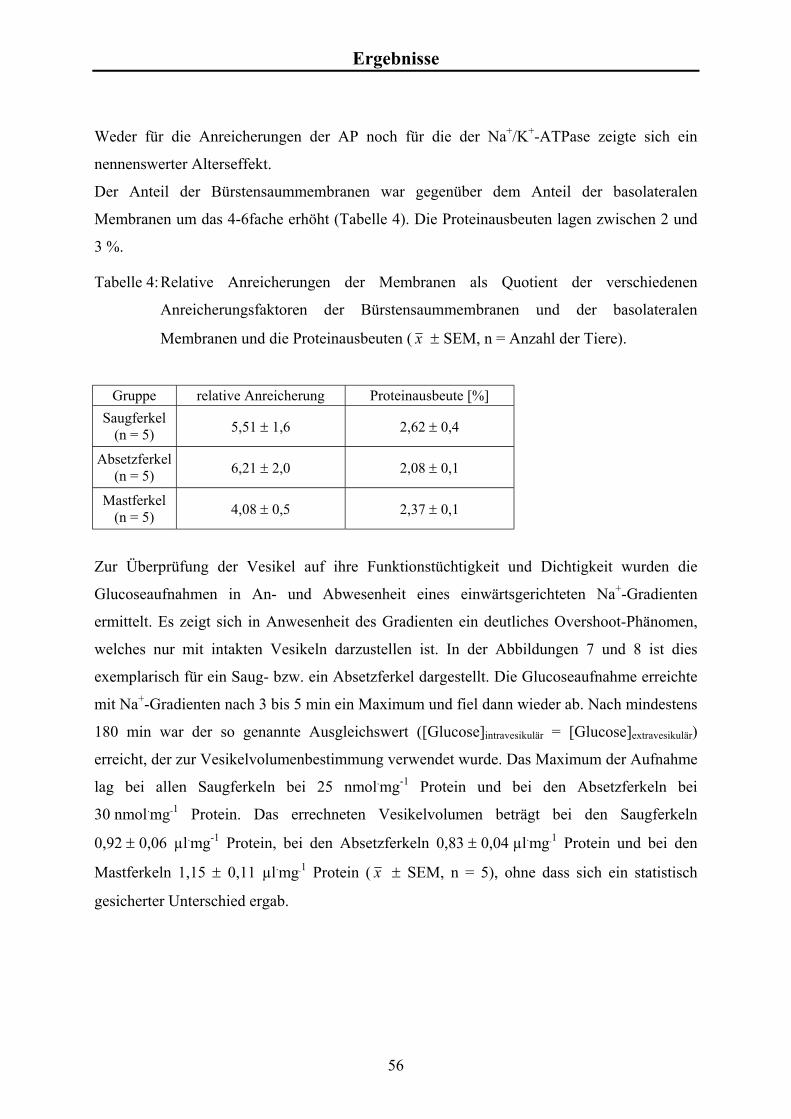
Ergebnisse Weder für die Anreicherungen der AP noch für die der Na+/K+-ATPase zeigte sich ein
nennenswerter Alterseffekt.
Der Anteil der Bürstensaummembranen war gegenüber dem Anteil der basolateralen
Membranen um das 4-6fache erhöht (Tabelle 4). Die Proteinausbeuten lagen zwischen 2 und
3 %.
Tabelle 4: Relative Anreicherungen der Membranen als Quotient der verschiedenen
Anreicherungsfaktoren der Bürstensaummembranen und der basolateralen
Membranen und die Proteinausbeuten ( x ± SEM, n = Anzahl der Tiere).
Gruppe relative Anreicherung Proteinausbeute [%] Saugferkel
(n = 5) 5,51 ± 1,6 2,62 ± 0,4
Absetzferkel (n = 5) 6,21 ± 2,0 2,08 ± 0,1
Mastferkel (n = 5) 4,08 ± 0,5 2,37 ± 0,1
Zur Überprüfung der Vesikel auf ihre Funktionstüchtigkeit und Dichtigkeit wurden die
Glucoseaufnahmen in An- und Abwesenheit eines einwärtsgerichteten Na+-Gradienten
ermittelt. Es zeigt sich in Anwesenheit des Gradienten ein deutliches Overshoot-Phänomen,
welches nur mit intakten Vesikeln darzustellen ist. In der Abbildungen 7 und 8 ist dies
exemplarisch für ein Saug- bzw. ein Absetzferkel dargestellt. Die Glucoseaufnahme erreichte
mit Na+-Gradienten nach 3 bis 5 min ein Maximum und fiel dann wieder ab. Nach mindestens
180 min war der so genannte Ausgleichswert ([Glucose]intravesikulär = [Glucose]extravesikulär)
erreicht, der zur Vesikelvolumenbestimmung verwendet wurde. Das Maximum der Aufnahme
lag bei allen Saugferkeln bei 25 nmol.mg-1 Protein und bei den Absetzferkeln bei
30 nmol.mg-1 Protein. Das errechneten Vesikelvolumen beträgt bei den Saugferkeln
0,92 ± 0,06 µl.mg-1 Protein, bei den Absetzferkeln 0,83 ± 0,04 µl.mg-1 Protein und bei den
Mastferkeln 1,15 ± 0,11 µl.mg-1 Protein ( x ± SEM, n = 5), ohne dass sich ein statistisch
gesicherter Unterschied ergab.
56
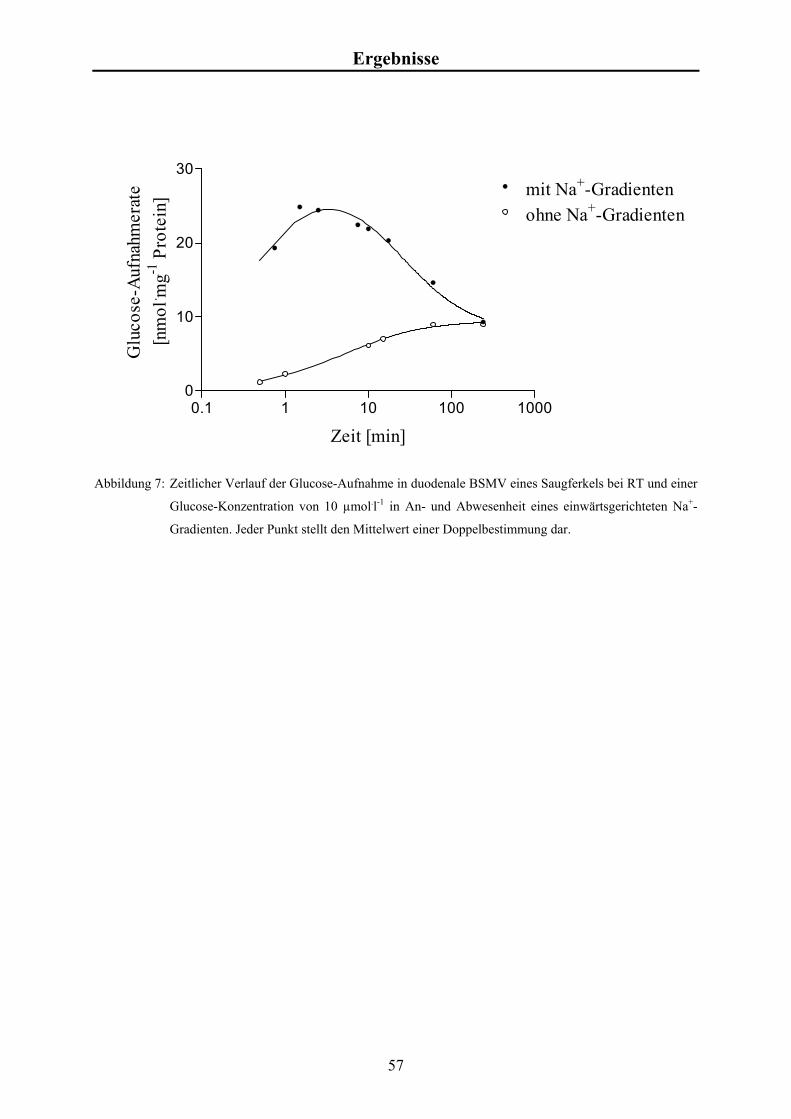
Ergebnisse
0.1 1 10 100 10000
10
20
30mit Na+-Gradientenohne Na+-Gradienten
Zeit [min]
Glu
cose
-Auf
nahm
erat
e[n
mol
. mg-1
Pro
tein
]
Abbildung 7: Zeitlicher Verlauf der Glucose-Aufnahme in duodenale BSMV eines Saugferkels bei RT und einer
Glucose-Konzentration von 10 µmol.l-1 in An- und Abwesenheit eines einwärtsgerichteten Na+-
Gradienten. Jeder Punkt stellt den Mittelwert einer Doppelbestimmung dar.
57
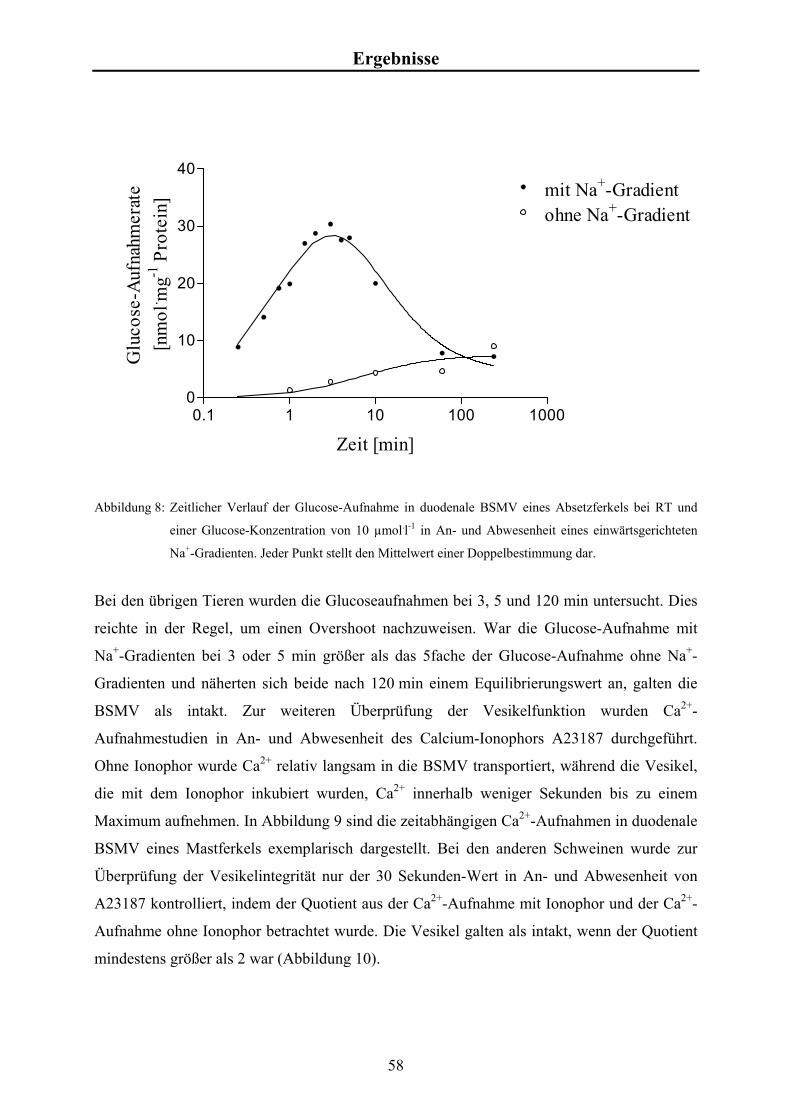
Ergebnisse
0.1 1 10 100 10000
10
20
30
40mit Na+-Gradientohne Na+-Gradient
Zeit [min]
Glu
cose
-Auf
nahm
erat
e[n
mol
. mg-1
Pro
tein
]
Abbildung 8: Zeitlicher Verlauf der Glucose-Aufnahme in duodenale BSMV eines Absetzferkels bei RT und
einer Glucose-Konzentration von 10 µmol.l-1 in An- und Abwesenheit eines einwärtsgerichteten
Na+-Gradienten. Jeder Punkt stellt den Mittelwert einer Doppelbestimmung dar.
Bei den übrigen Tieren wurden die Glucoseaufnahmen bei 3, 5 und 120 min untersucht. Dies
reichte in der Regel, um einen Overshoot nachzuweisen. War die Glucose-Aufnahme mit
Na+-Gradienten bei 3 oder 5 min größer als das 5fache der Glucose-Aufnahme ohne Na+-
Gradienten und näherten sich beide nach 120 min einem Equilibrierungswert an, galten die
BSMV als intakt. Zur weiteren Überprüfung der Vesikelfunktion wurden Ca2+-
Aufnahmestudien in An- und Abwesenheit des Calcium-Ionophors A23187 durchgeführt.
Ohne Ionophor wurde Ca2+ relativ langsam in die BSMV transportiert, während die Vesikel,
die mit dem Ionophor inkubiert wurden, Ca2+ innerhalb weniger Sekunden bis zu einem
Maximum aufnehmen. In Abbildung 9 sind die zeitabhängigen Ca2+-Aufnahmen in duodenale
BSMV eines Mastferkels exemplarisch dargestellt. Bei den anderen Schweinen wurde zur
Überprüfung der Vesikelintegrität nur der 30 Sekunden-Wert in An- und Abwesenheit von
A23187 kontrolliert, indem der Quotient aus der Ca2+-Aufnahme mit Ionophor und der Ca2+-
Aufnahme ohne Ionophor betrachtet wurde. Die Vesikel galten als intakt, wenn der Quotient
mindestens größer als 2 war (Abbildung 10).
58
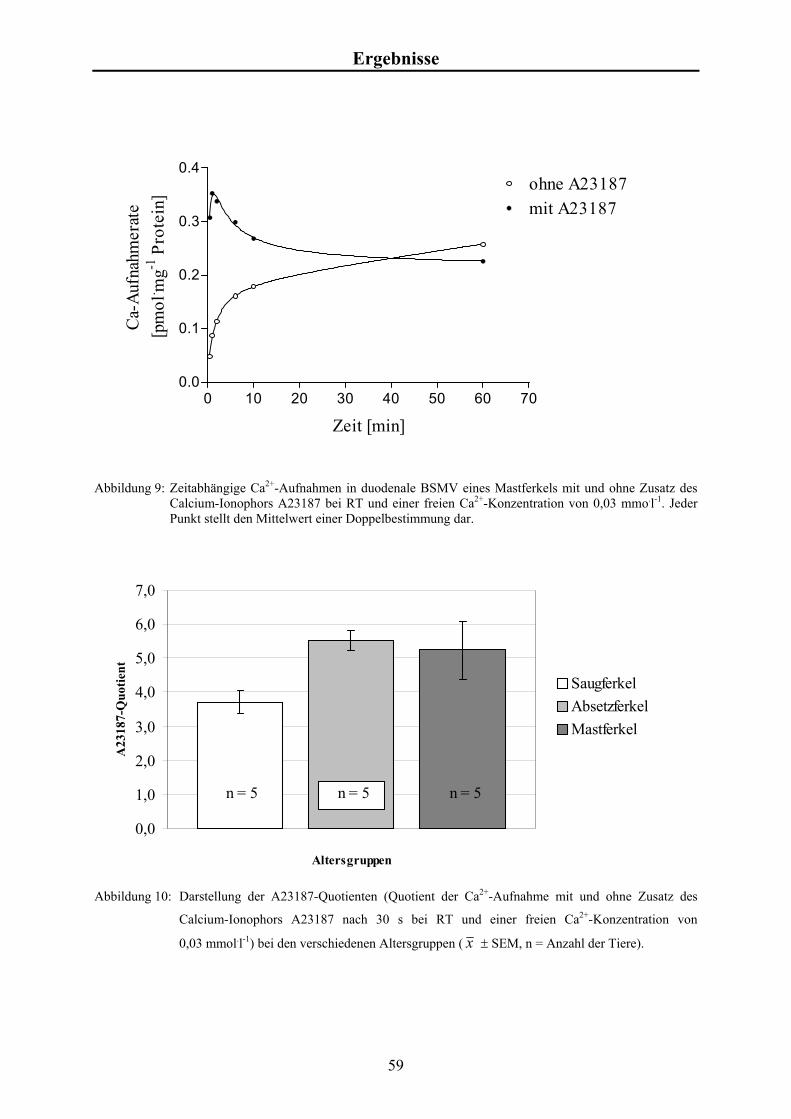
Ergebnisse
0 10 20 30 40 50 60 700.0
0.1
0.2
0.3
0.4ohne A23187mit A23187
Zeit [min]
Ca-
Auf
nahm
erat
e[p
mol
. mg-1
Pro
tein
]
Abbildung 9: Zeitabhängige Ca2+-Aufnahmen in duodenale BSMV eines Mastferkels mit und ohne Zusatz des Calcium-Ionophors A23187 bei RT und einer freien Ca2+-Konzentration von 0,03 mmo.l-1. Jeder Punkt stellt den Mittelwert einer Doppelbestimmung dar.
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
Altersgruppen
A23
187-
Quo
tient
SaugferkelAbsetzferkelMastferkel
n = 5 n = 5 n = 5
Abbildung 10: Darstellung der A23187-Quotienten (Quotient der Ca2+-Aufnahme mit und ohne Zusatz des
Calcium-Ionophors A23187 nach 30 s bei RT und einer freien Ca2+-Konzentration von
0,03 mmol.l-1) bei den verschiedenen Altersgruppen ( x ± SEM, n = Anzahl der Tiere).
59

Ergebnisse 4.1.2 Ca2+-Aufnahmeraten in die BSMV
Die Gesamt-Ca2+-Aufnahmeraten in duodenale BSMV der verschiedenen Altersgruppen sind
in Abbildung 11 dargestellt. Dabei waren die Ca2+-Aufnahmeraten in BSMV der Saugferkel
jeweils höher als die in BSMV der Absetz- und Mastferkel, die sich ihrerseits nicht
wesentlich unterschieden. Die Gesamt-Aufnahmerate setzt sich aus einem unspezifischen,
nicht sättigbarem Anteil und einem sättigbarem Anteil zusammen, dessen Kurvenverlauf
einer typischen Michaelis-Menten-Kinetik entspricht und aus dem sich die Parameter Km und
Vmax bestimmen lassen. Die Michaelis-Menten-Konstante (Km), die die Menge an Substrat
angibt, die für die halbmaximale Transportrate notwendig ist, ist ein Maß für die Ca2+-
Affinität des Transportsystems und die maximale Transportrate (Vmax) gibt einen Hinweis auf
die Transportkapazität bzw. Anzahl der Transporter. Berechnet man die Parameter Km, Vmax
und den unspezifischen Anteil mit den Mittelwerten der gemessenen Aufnahmen für die
jeweiligen Tiergruppen, so ergeben sich für die Saugferkel eine Vmax von
3,39 nmol.mg-1 Protein.30s-1, eine apparente Km von 1,41 mmol.l-1 bei einem
Bestimmtheitsmaß (r2) von 0,89 , für die Absetzferkel eine Vmax von
2,04 nmol.mg-1 Protein.30s-1, eine apparente Km von 1,19 mmol.l-1 bei einem r2 von 0,97 und
für die Mastferkel eine Vmax von 2,08 nmol.mg-1 Protein.30s-1 und eine apparente Km von
1,11 mmol.l-1 bei einem r2 von 0,98.
60
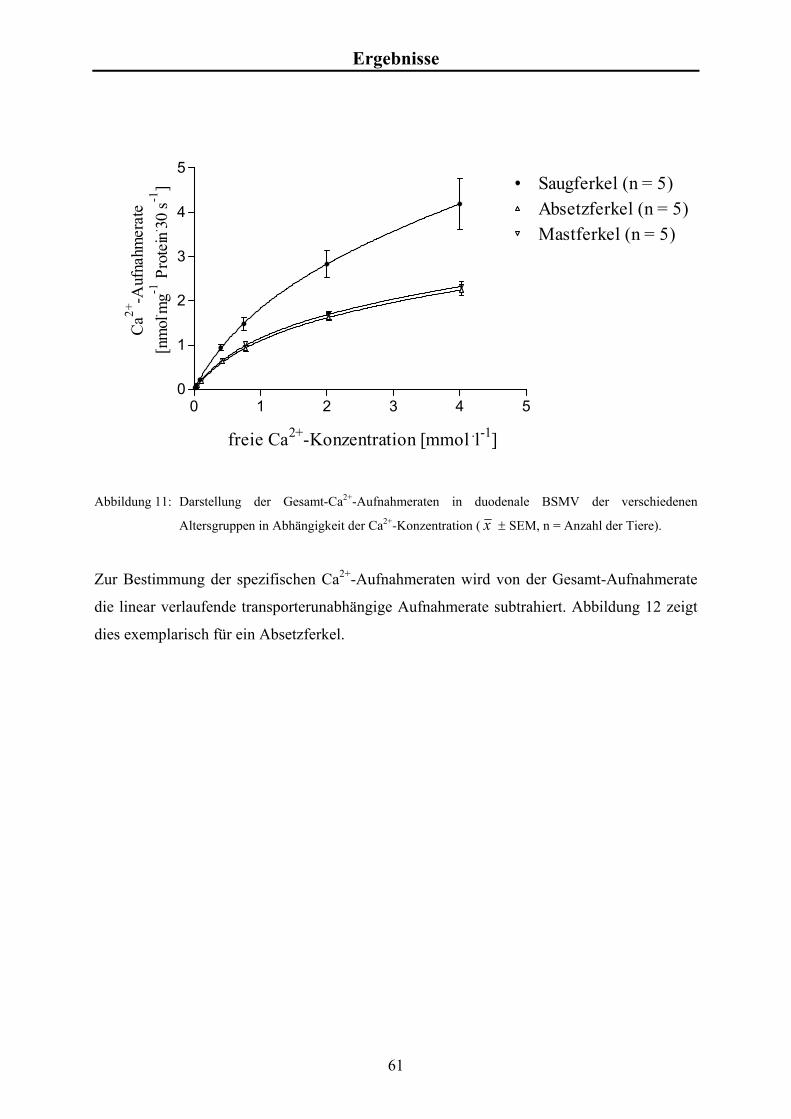
Ergebnisse
0 1 2 3 4 50
1
2
3
4
5Saugferkel (n = 5)Absetzferkel (n = 5)Mastferkel (n = 5)
freie Ca2+-Konzentration [mmol .l-1]
Ca2+
-Auf
nahm
erat
e[n
mol
. mg-1
Pro
tein
. 30 s
-1]
Abbildung 11: Darstellung der Gesamt-Ca2+-Aufnahmeraten in duodenale BSMV der verschiedenen
Altersgruppen in Abhängigkeit der Ca2+-Konzentration ( ± SEM, n = Anzahl der Tiere). x
Zur Bestimmung der spezifischen Ca2+-Aufnahmeraten wird von der Gesamt-Aufnahmerate
die linear verlaufende transporterunabhängige Aufnahmerate subtrahiert. Abbildung 12 zeigt
dies exemplarisch für ein Absetzferkel.
61
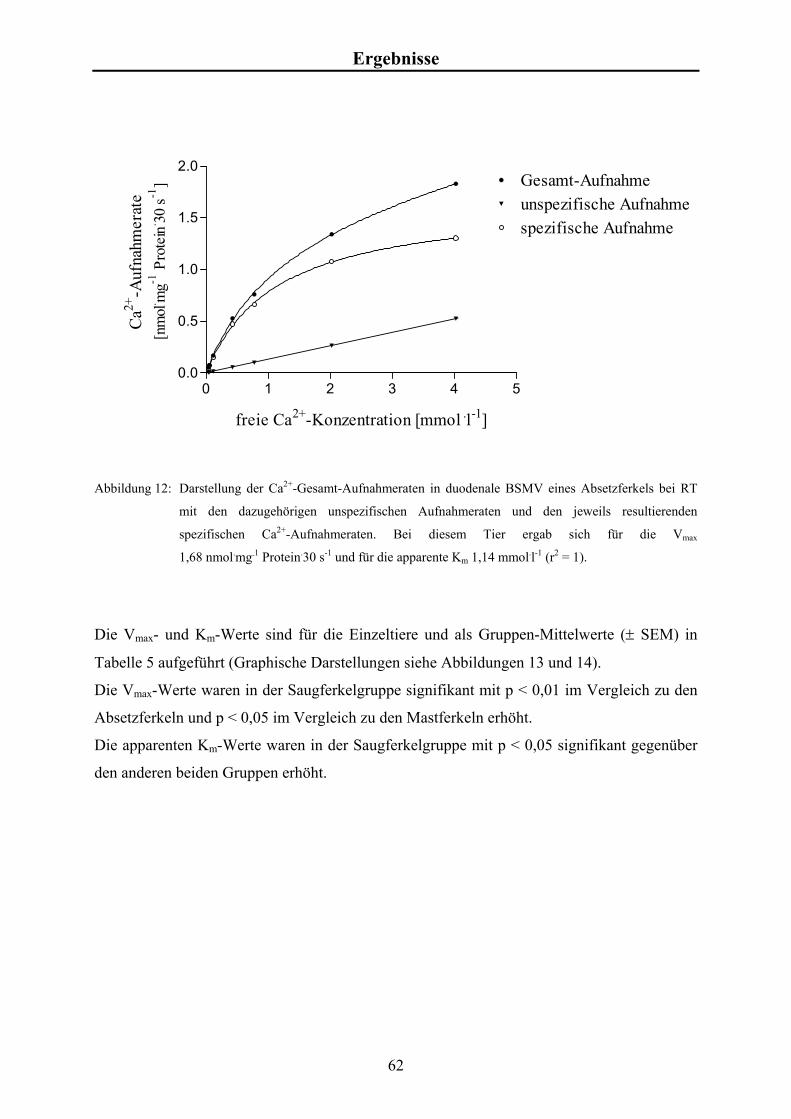
Ergebnisse
0 1 2 3 4 50.0
0.5
1.0
1.5
2.0Gesamt-Aufnahmeunspezifische Aufnahmespezifische Aufnahme
freie Ca2+-Konzentration [mmol .l-1]
Ca2+
-Auf
nahm
erat
e[n
mol
. mg-1
Pro
tein
. 30 s
-1]
Abbildung 12: Darstellung der Ca2+-Gesamt-Aufnahmeraten in duodenale BSMV eines Absetzferkels bei RT
mit den dazugehörigen unspezifischen Aufnahmeraten und den jeweils resultierenden
spezifischen Ca2+-Aufnahmeraten. Bei diesem Tier ergab sich für die Vmax
1,68 nmol.mg-1 Protein.30 s-1 und für die apparente Km 1,14 mmol.l-1 (r2 = 1).
Die Vmax- und Km-Werte sind für die Einzeltiere und als Gruppen-Mittelwerte (± SEM) in
Tabelle 5 aufgeführt (Graphische Darstellungen siehe Abbildungen 13 und 14).
Die Vmax-Werte waren in der Saugferkelgruppe signifikant mit p < 0,01 im Vergleich zu den
Absetzferkeln und p < 0,05 im Vergleich zu den Mastferkeln erhöht.
Die apparenten Km-Werte waren in der Saugferkelgruppe mit p < 0,05 signifikant gegenüber
den anderen beiden Gruppen erhöht.
62
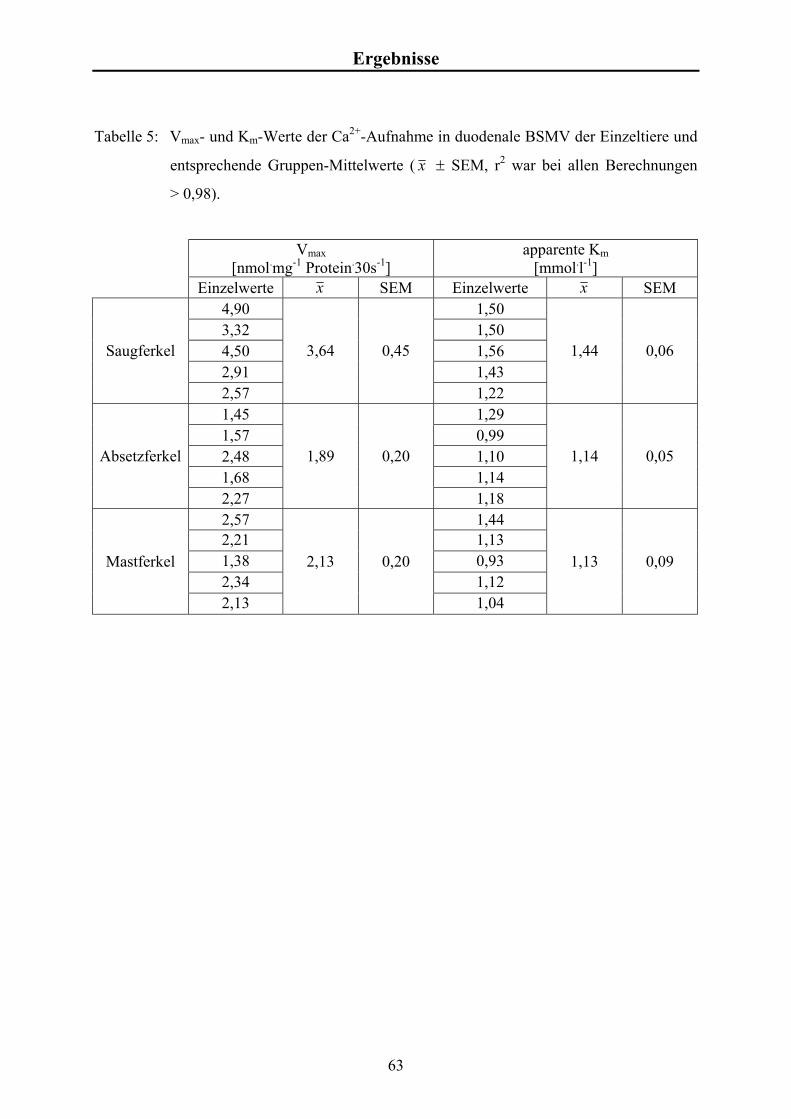
Ergebnisse Tabelle 5: Vmax- und Km-Werte der Ca2+-Aufnahme in duodenale BSMV der Einzeltiere und
entsprechende Gruppen-Mittelwerte ( x ± SEM, r2 war bei allen Berechnungen
> 0,98).
Vmax [nmol.mg-1 Protein.30s-1]
apparente Km [mmol.l-1]
Einzelwerte x SEM Einzelwerte x SEM 4,90 1,50 3,32 1,50 4,50 1,56 2,91 1,43
Saugferkel
2,57
3,64 0,45
1,22
1,44 0,06
1,45 1,29 1,57 0,99 2,48 1,10 1,68 1,14
Absetzferkel
2,27
1,89 0,20
1,18
1,14 0,05
2,57 1,44 2,21 1,13 1,38 0,93 2,34 1,12
Mastferkel
2,13
2,13 0,20
1,04
1,13 0,09
63
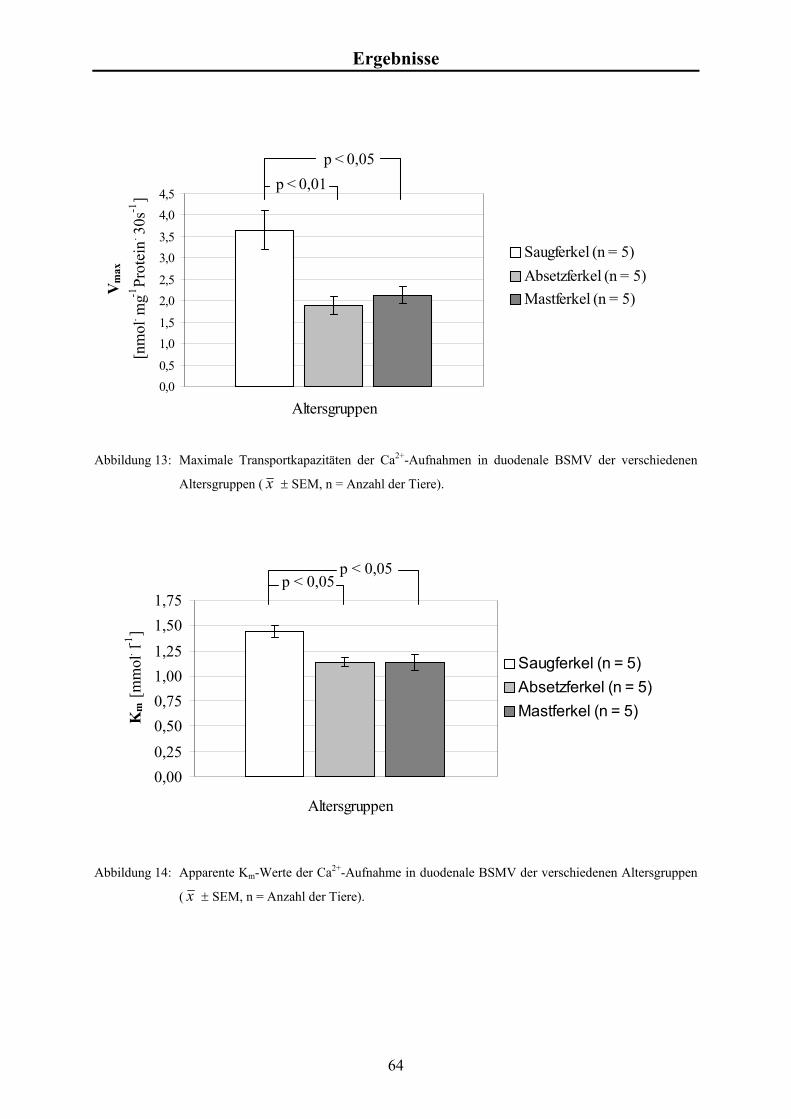
Ergebnisse
0,00,5
1,01,5
2,02,5
3,03,5
4,04,5
Altersgruppen
Vm
ax
[nm
ol . m
g-1Pr
otei
n . 30s-1
]
Saugferkel (n = 5)Absetzferkel (n = 5)Mastferkel (n = 5)
p < 0,01p < 0,05
Abbildung 13: Maximale Transportkapazitäten der Ca2+-Aufnahmen in duodenale BSMV der verschiedenen
Altersgruppen ( ± SEM, n = Anzahl der Tiere). x
0,000,250,500,751,001,251,501,75
Altersgruppen
Km
[mm
ol . l-1
]
Saugferkel (n = 5)Absetzferkel (n = 5)Mastferkel (n = 5)
p < 0,05p < 0,05
Abbildung 14: Apparente Km-Werte der Ca2+-Aufnahme in duodenale BSMV der verschiedenen Altersgruppen
( x ± SEM, n = Anzahl der Tiere).
64
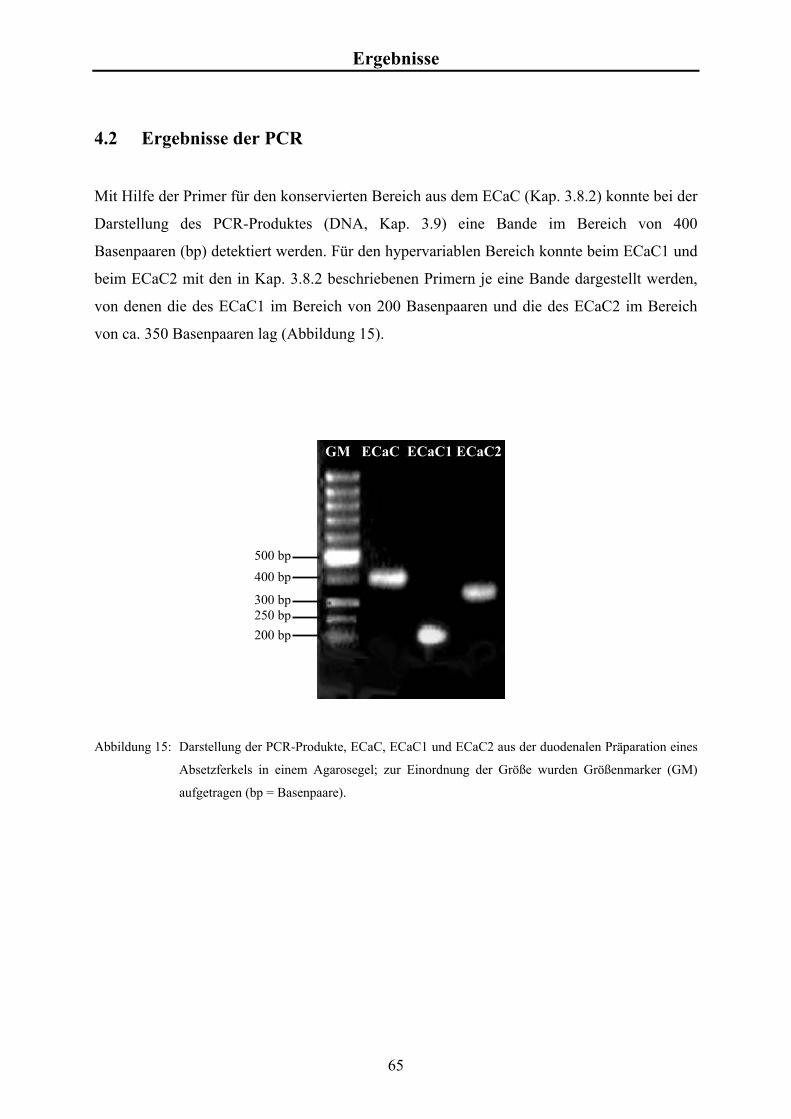
Ergebnisse 4.2 Ergebnisse der PCR
Mit Hilfe der Primer für den konservierten Bereich aus dem ECaC (Kap. 3.8.2) konnte bei der
Darstellung des PCR-Produktes (DNA, Kap. 3.9) eine Bande im Bereich von 400
Basenpaaren (bp) detektiert werden. Für den hypervariablen Bereich konnte beim ECaC1 und
beim ECaC2 mit den in Kap. 3.8.2 beschriebenen Primern je eine Bande dargestellt werden,
von denen die des ECaC1 im Bereich von 200 Basenpaaren und die des ECaC2 im Bereich
von ca. 350 Basenpaaren lag (Abbildung 15).
500 bp 400 bp
300 bp 250 bp 200 bp
GM ECaC ECaC1 ECaC2
Abbildung 15: Darstellung der PCR-Produkte, ECaC, ECaC1 und ECaC2 aus der duodenalen Präparation eines
Absetzferkels in einem Agarosegel; zur Einordnung der Größe wurden Größenmarker (GM)
aufgetragen (bp = Basenpaare).
65

Ergebnisse 4.3 Teilsequenz des ECaC im konservierten Bereich
Die ermittelte Basensequenz für den ECaC im konservierten Bereich besteht aus 398
Basenpaaren und wird im Folgenden mit der dazugehörigen Aminosäurensequenz dargestellt.
9 18 27 36 45 54 5' CAG ATG TAC AAC CTG CTG CTG TCC TAT GAC GGT CGT GGG GAC CAC CTA CAG TCC --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- Q M Y N L L L S Y D G R G D H L Q S 63 72 81 90 99 108 CTG GAG CTC GTG CCC AAT CAC GAG GGT CTC ACC CCG TTC AAG CTG GCC GGA GTG --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- L E L V P N H E G L T P F K L A G V 117 126 135 144 153 162 GAG GGC AAC ACC GTG ATG TTC CAG CAC CTG ATG CAG AAG CGG AAG CAC ATC CAG --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- E G N T V M F Q H L M Q K R K H I Q 171 180 189 198 207 216 TGG GTC CTC GGG CCC CTG ACC TCC ACT CTC TAC GAC CTG ACG GAG ATC GAC TCC --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- W V L G P L T S T L Y D L T E I D S 225 234 243 252 261 270 TCG GGG GAT GAG CAG TCC CTG CTG GAA CTT CTT GTC ACC TCC AAG AAG CGG GAG --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- S G D E Q S L L E L L V T S K K R E 279 288 297 306 315 324 GCT CGC CAG ATC CTG GAC CAG ACG CCA GTG AAG GAG CTG GTG AGC CTC AAG TGG --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- A R Q I L D Q T P V K E L V S L K W 333 342 351 360 369 378 AGG AGA TAC GGG CGG CGG TAC TTC TGG GGG CTG GCT GCT GGG TAC CTG CTG TAC --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- R R Y G R R Y F W G L A A G Y L L Y 387 396 ATC ATC TGC TTC ACC ATG TG 3' --- --- --- --- --- --- -- I I C F T M
Abbildung 16: Darstellung der Basensequenz mit der dazugehörigen Aminosäurensequenz des ECaC im
konservierten Bereich aus dem Duodenum eines Absetzferkels ( = Primer siehe Kap. 3.8.2).
66

Ergebnisse Die Basensequenz hat eine 84%ige Homologie zu der des ECaC1 des Kaninchen und eine
88%ige Homologie zu der des ECaC2 des Menschen.
Die Aminosäurensequenz hat eine 81%ige Homologie zu der des ECaC1 des Kaninchen und
eine 86%ige Homologie zu der des ECaC2 der Ratte (Abbildung 19).
Die gesamte Basensequenz mit ihrer Länge von 398 Basenpaaren wurde als „ECaC-Sonde“
beim Northern-Blot (Kap. 3.17) eingesetzt.
4.4 Teilsequenz des ECaC1 im hypervariablen Bereich
Die ermittelte Basensequenz für den ECaC1 im hypervariablen Bereich besteht aus 201
Basenpaaren und wird im Folgenden mit der dazugehörigen Aminosäurensequenz dargestellt.
10 19 28 37 46 55 5'G AAT CCG TAT CGA GTA CTT CGA TAT GTG GAA GCT TTC AAA TCT TCA GAC AAA GAG --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- N P Y R V L R Y V E A F K S S D K E 64 73 82 91 100 109 GAA GTA CAA GAG CAG CTA TCT GAG AAA CAA CCT TCT GGG ACT GAG ACT GGG ACT --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- E V Q E Q L S E K Q P S G T E T G T 118 127 136 145 154 163 TTA GCT AGA GGC TCT GTG GTT CTC CAA ACA CCT CCT TTG TCC AGG ACT ACA TCC --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- L A R G S V V L Q T P P L S R T T S 172 181 190 199 CTA AGC AGC AAT AGC CAC CGT GGA TGG GAA ATT CTC C 3' --- --- --- --- --- --- --- --- --- --- --- --- - L S S N S H R G W E I L Abbildung 17: Darstellung der Basensequenz mit der dazugehörigen Aminosäurensequenz des ECaC1 im
hypervariablen Bereich aus dem Duodenum eines Absetzferkels ( = Primer siehe Kap. 3.8.2, = schweinespezifische Primer für die Real-Time-PCR und die „ECaC1-Sonde“ beim Northern-Blot).
67

Ergebnisse Die Basensequenz hat eine 95%ige Homologie zu der des ECaC1 der Ratte und eine 33%ige
Homologie zu der des ECaC2 des Menschen.
Die Aminosäurensequenz hat eine 97%ige Homologie zu der des ECaC1 der Ratte und eine
33%ige Homologie zu der des ECaC2 der Ratte (Abbildung 19).
Von dieser Sequenz wurden für die Real-Time-PCR schweinespezifische Primer abgeleitet:
5`>GGAAGTACAAGAGCAGCTATCTGAGA<3`, s, ECaC1 (Schwein, Basen: 55ff),
5`>GGGATGTAGTCCTGGACAAAGG<3`, as, ECaC1 (Schwein, Basen: 143ff).
Die Basensequenz zwischen und mit diesen Primern wurde als „ECaC1-Sonde“ mit einer
Länge von 109 Basenpaaren beim Northern-Blot (Kap. 3.17) eingesetzt.
68
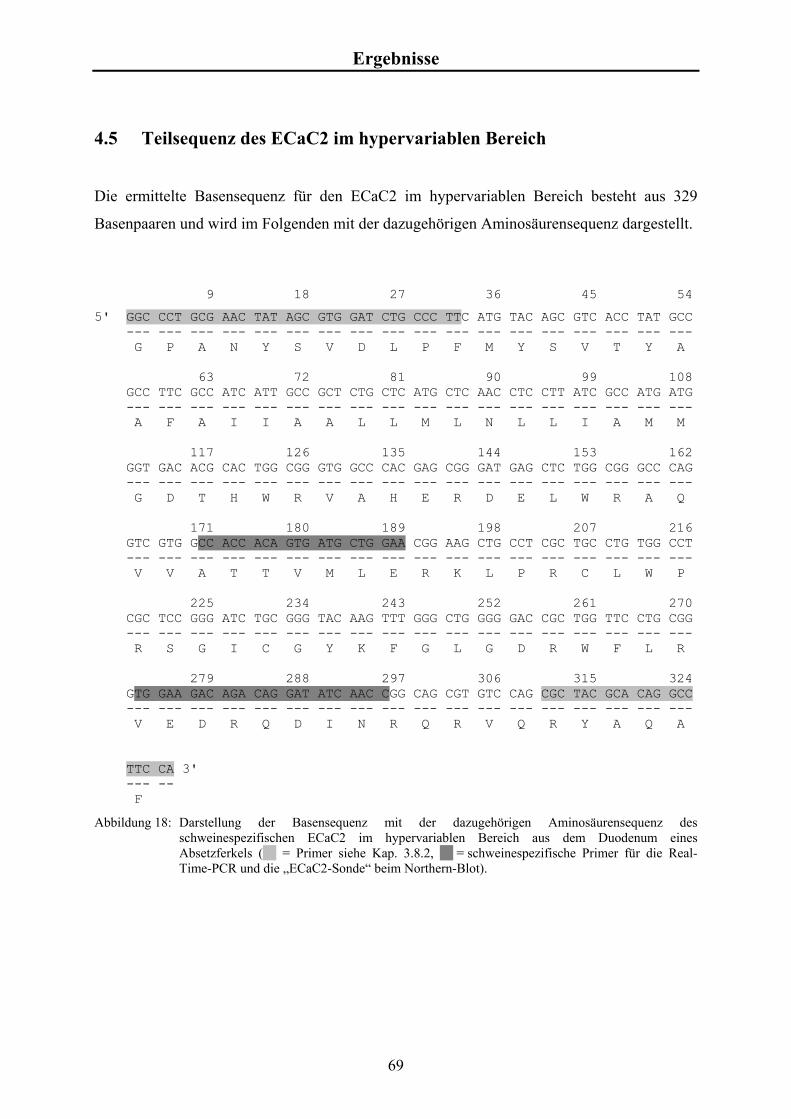
Ergebnisse 4.5 Teilsequenz des ECaC2 im hypervariablen Bereich
Die ermittelte Basensequenz für den ECaC2 im hypervariablen Bereich besteht aus 329
Basenpaaren und wird im Folgenden mit der dazugehörigen Aminosäurensequenz dargestellt.
9 18 27 36 45 54
5' GGC CCT GCG AAC TAT AGC GTG GAT CTG CCC TTC ATG TAC AGC GTC ACC TAT GCC --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- G P A N Y S V D L P F M Y S V T Y A 63 72 81 90 99 108 GCC TTC GCC ATC ATT GCC GCT CTG CTC ATG CTC AAC CTC CTT ATC GCC ATG ATG --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- A F A I I A A L L M L N L L I A M M 117 126 135 144 153 162 GGT GAC ACG CAC TGG CGG GTG GCC CAC GAG CGG GAT GAG CTC TGG CGG GCC CAG --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- G D T H W R V A H E R D E L W R A Q 171 180 189 198 207 216 GTC GTG GCC ACC ACA GTG ATG CTG GAA CGG AAG CTG CCT CGC TGC CTG TGG CCT --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- V V A T T V M L E R K L P R C L W P 225 234 243 252 261 270 CGC TCC GGG ATC TGC GGG TAC AAG TTT GGG CTG GGG GAC CGC TGG TTC CTG CGG --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- R S G I C G Y K F G L G D R W F L R 279 288 297 306 315 324 GTG GAA GAC AGA CAG GAT ATC AAC CGG CAG CGT GTC CAG CGC TAC GCA CAG GCC --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- --- V E D R Q D I N R Q R V Q R Y A Q A TTC CA 3' --- -- F
Abbildung 18: Darstellung der Basensequenz mit der dazugehörigen Aminosäurensequenz des schweinespezifischen ECaC2 im hypervariablen Bereich aus dem Duodenum eines Absetzferkels ( = Primer siehe Kap. 3.8.2, = schweinespezifische Primer für die Real-Time-PCR und die „ECaC2-Sonde“ beim Northern-Blot).
69

Ergebnisse Die Basensequenz hat eine 81%ige Homologie zu der des ECaC1 der Ratte und eine 88%ige
Homologie zu der des ECaC2 des Menschen.
Die Aminosäurensequenz hat eine 84%ige Homologie zu der des ECaC1 des Kaninchen und
eine 93%ige Homologie zu der des ECaC2 der Ratte (Abbildung 19).
Von dieser Sequenz wurden für die Real-Time-PCR schweinespezifische Primer abgeleitet:
5`>CCACCACAGTGATGCTGGAA<3`, s, ECaC1 (Schwein, Basen: 170ff),
5`>GGTTGATATCCTGTCTGTCTTCCA<3`, as, ECaC1 (Schwein, Basen: 272ff).
Die Basensequenz zwischen und mit diesen Primern wurde als „ECaC2-Sonde“ mit ihrer
Länge von 126 Basenpaaren beim Northern-Blot (Kap. 3.17) eingesetzt.
70

Ergebnisse
71
ensch
MGGFLPKAEGPGSQLQKLLPSFLVREQDWDQHLDKLHMLQQKRILESPLLRASKENDLSVLRQLLLDCTCDVRQRGALGETALHIAALYDNLEAAL
aninchen
MGACPPKAKGPWAQLQKLLISWPVGEQDWEQYRDRVNMLQQERIRDSPLLQAAKENDLRLLKILLLNQSCDFQQRGAVGETALHVAALYDNLEAAT
atte
MGWSLPKEKGLILCLWNKFCRWFHRRESWAQSRDEQNLLQQKRIWESPLLLAAKENNVQALIKLLKFEGCEVHQKGAMGETALHIAALYDNLEAAM
ensch
VLMEAAPELVFEPTTCEAFAGQTALHIAVVNQNVNLVRALLARRASVSARATGTAFRRSPCNLIYFGEHPLSFAACVNSEEIVRLLIEHGADIRAQ
aninchen
LLMEAAPELAKEPALCEPFVGQTALHIAVMNQNLNLVRALLARGASVSARATGAAFRRSPHNLIYYGEHPLSFAACVGSEEIVRLLIEHGADIRAQ
atte
VLMEAAPELVFEPMTSELYEGQTALHIAVINQNVNLVRALLARGASVSARATGSVFHYRPHNLIYYGEHPLSFAACVGSEEIVRLLIEHGADIRAQ
ensch
DSLGNTVLHILILQPNKTFACQMYNLLLTYDRHGDHLQPLDLWPNHQGLTPFKLAGVEGNTVMFQHLMQKRKHVQWTCGPLTSTLYDLTEIDSWGE
aninchen
DSLGNTVLHILILQPNKTFACQMYNLLLSYDEHSDHLQSLELVPNHQGLTPFKLAGVEGNTVMFQHLMQKRKHVQWTCGPLTSTLYDLTEIDSWGE
atte
DSLGNTVLHILILQPNKTFACQMYNLLLSYD-GGDHLKSLELVPNNQGLTPFKLAGVEGNIVMFQHLMQKRKHIQWTYGPLTSTLYDLTEIDSSGD
Schwein
QMYNLLLSYDGRGDHLQSLELVPNHEGLTPFKLAGVEGNTVMFQHLMQKRKHIQWVLGPLTSTLYDLTEIDSSGD
ensch
ELSFLELVVSSDKREARQILEQTPVKEPVSFKWNKYGRPYFCILAALYLLYMICFTTCCVYRPLKFRGGNRTHSRDITILQQKLLQEAYETREDII
aninche
ELSFLELVVSSKKREARQILEQTPVKELVSFKWKKYGRPYFCVLASLYILYMICFTTCCIYRPLKLRDDNRTDPRDITILQQKLLQEAYVTHQDNI
atte
DQSLLELIVTTKKREARQILDQTPVKELVSLKWKRYGRPYFCVLGAIYVLYIICFTMCCVYRPLKPRITNRTNPRDNTLLQQKLLQEAYVTPKDDL
Schwein
EQSLLELLVTSKKREARQILDQTPVKELVSLKWRRYGRRYFWGLAAGYLLYIICFTM
ensch
RLVGELVSIVGAVIILLLEIPDIFRVGASRYFGKTILGGPFHVIIIT
VLVTMVMRLTNTNGEVVPMSFALVLGWCSVMYFTRGFQMLGPFTD
aninchen
RLVGELVTVTGAVIILLLEIPDIFRVGASRYFGQTILGGPFHVIIITYASLVLLTMVMRLTNMNGEVVPLSFALVLGWCSVMYFARGFQMLGPFTD
atte
RLVGELVSIVGAVIILLVEIPDIFRLGVTRFFGQTILGGPFHVIIVTYAFMVLVTMVMRLTNSDGEVVPMSFALVLGWCNVMYFARGFQMLGPFTD
ensch
MIQKMIFGD
ITYAAFAIIATLLMLNLFIA
aninchen
MIQKMIFGDLMRFCWLMAVVILGFASAFHITFQTEDPNNLGEFSDYPTALFSTFELFLTIIDGPANYSVDLPFMYCITYAAFAIIATLLMLNLFIA
atte
MIQKMIFGDLMRFCWLMAVVILGFASAFYIIFQTEDPDELGHFYDYPMALFSTFELFLTIIDGPANYDVDLPFMYSITYAAFAIIATLLMLNLLIA
Schwein
GPANYSVDLPFMYSVTYAAFAIIAALLMLNLLIA
ensch
MMGDTHWRVAHERDELWR
KQPSGAE
aninchen
MMGDTHWRVAQERDELWRAQVVATTVMLERKMPRFLWPRSGICGYEYGLGDRWFLRVENHHDQNPLRVLRYVEAFKCSDKEDGQEQLSEKRPSTVE
atte
MMGDTHWRVAHERDELWRAQVVATTVMLERKLPRCLWPRSGICGREYGLGDRWFLRVEDRQDLNRQRIRRYAQAFQQQD-DLYSEDLEKDSGEKLE
Schwein
MMGDTHWRVAHERDELWRAQVVATTVMLERKLPRCLWPRSGICGYKFGLGDRWFLRVEDRQDINRQRVQRYAQAF
Schwein
NPYRVLRYVEAFKSSDKEEVQEQLSEKQPSGTE
ensch
SGTLARASLALPTSSLSRTASQSS-SHRGWEILRQNTL-GHLN--LGLNLSEGDGEEVYHF
aninchen
SGMLSRASVAFQTPSLSRTTSQSSNSHRGWEILRRNTL-GHLN--LGLDLGEGDGEEVYHF
atte
MARPFGAYLSFPTPSVSRSTSRSSTN---WDRLRQGALRKDLQGIINRGLEDGEGWE-YQI
Schw
ein
TGTLARGSVVLQTPPLSRTTSLSSNSHRGWEILQ
***
**
***
AN
K
AN
K
A
NK
M K R
Schwein
**
AQVVATTVMLERKLPRCLWPRSGICGCEFGLGDRWFLRVENHNDQNPLRVLRYVEVFKNSDKEDDQEHPSE
TM
5
P
OR
E
T
M6
M K R
LMRLCWLMAVVILGFASAFYIIFQTEDPTSLGQFYDYPMALFSTFELFLTIIDGPANYNVDLPFMYS
TM
2
TM
3
T
M4
M K R
YASL
TM
1 M K R
n * *
M K RM K RM K RM K R Abb
ildun
g 19
: D
arst
ellu
ng d
er A
min
osäu
rese
quen
zen
des E
CaC
1 de
s Men
sche
n un
d de
s Kan
inch
ens,
des E
CaC
2 de
r Rat
te u
nd d
es E
CaC
(*) b
zw. E
CaC
1(*
*) u
nd E
CaC
2 (*
**)
des
Schw
eins
im d
irekt
en V
ergl
eich
zum
EC
aC1
(Men
sch
und
Kan
inch
en)
und
ECaC
2 (R
atte
) (A
NK
= A
nkyr
in-
Rep
eats
, TM
1-6
= tra
nsm
embr
anal
e D
omän
e, P
OR
E =
Pore
nreg
ion)
.

Ergebnisse 4.6 Northern-Blots mit verschiedenen Sonden entlang der Darmachse
4.6.1 Northern-Blot mit der „ECaC-Sonde“
Für den Northern-Blot mit der „ECaC-Sonde“ wurde die mRNA von 5 verschiedenen
Darmabschnitten eines Absetzferkels im Gel aufgetrennt und auf eine Nitrocellulosemembran
transferiert. Diese Membran wurde mit der „ECaC-Sonde“, die eine Aktivität von 70267 cpm
pro in einem µl Sonde enthaltener DNA aufwies, hybridisiert. Die Sonde hatte eine Länge von
398 Basenpaaren.
D = Duodenum J = Jejunum I = Ileum pC = proximales Colon dC = distales Colon
-- β-Aktin
-- ECaC
D J I pC dC
9,0 3,0 0,0 0,5 0,7 -- ECaC/β-Aktin-Verhältnis Abbildung 20: Darstellung der ECaC-Expression entlang der cranial-caudalen Darmachse eines Absetzferkels
mittels Northern-Blot und der „ECaC-Sonde“. Das ECaC/β-Aktin-Verhältnis lässt einen
relativen Vergleich der ECaC-Expression der einzelnen Darmabschnitte zu.
Mit der „ECaC-Sonde“ wurde maximal eine Bande je Darmabschnitt erzeugt. Das ECaC/β-
Aktin-Verhältnis zeigt eine deutlich höhere ECaC-Expression im Duodenum als in den
anderen Darmabschnitten. Im Jejunum war im Gegensatz zum Ileum, wo das ECaC/β-Aktin-
Verhältnis Null war noch eine relativ hohe Expression nachzuweisen. Gering, aber deutlich
nachweisbar war die Expression in beiden Abschnitten des Colons.
72

Ergebnisse 4.6.2 Northern-Blot mit der „ECaC1-Sonde“
Für den Northern-Blot mit der „ECaC1-Sonde“ wurde die mRNA von 5 verschiedenen
Darmabschnitten eines Absetzferkels und eines Saugferkels im Gel aufgetrennt und auf eine
Nitrocellulosemembran transferiert. Diese Membran wurde mit der „ECaC1-Sonde“, die eine
Aktivität von 57143 cpm pro in einem µl Sonde enthaltener DNA aufwies, hybridisiert. Die
Sonde hat eine Länge von 109 Basenpaaren.
-- β-Aktin
-- Bande 2
-- Bande 1
Saugferkel Absetzferkel D J I pC dC D J I pC dC D = Duodenum
J = Jejunum I = Ileum pC = proximales Colon dC = distales Colon
Abbildung 21: Darstellung der ECaC-Expression entlang der cranial-caudalen Darmachse eines Saugferkels
und eines Absetzferkels mittels Northern-Blot und der „ECaC1-Sonde“.
Mit der „ECaC1-Sonde“ wurden zwei Banden dargestellt. Eine Zuordnung der Banden zu
ECaC1 oder ECaC2 war nicht möglich.
4.6.3 Northern-Blot mit der „ECaC2-Sonde“
Für den Northern-Blot mit der „ECaC2-Sonde“ wurde die mRNA von 5 verschiedenen
Darmabschnitten eines Absetzferkels und eines Saugferkels im Gel aufgetrennt und auf eine
Nitrocellulosemembran transferiert. Diese Membran wurde mit der „ECaC2-Sonde“, die eine
Aktivität von 79604 cpm pro in einem µl Sonde enthaltener DNA aufwies, hybridisiert. Die
Sonde hat eine Länge von 126 Basenpaaren.
73

Ergebnisse
-- β-Aktin
-- Bande 2
-- Bande 1
Saugferkel Absetzferkel D J I pC dC D J I pC dC D = Duodenum
J = Jejunum I = Ileum pC = proximales Colon dC = distales Colon
Abbildung 22: Darstellung der ECaC-Expression entlang der cranial-caudalen Darmachse eines Saugferkels
und eines Absetzferkels mittels Northern-Blot und der „ECaC2-Sonde“.
Auch mit der „ECaC2-Sonde“ wurden zwei Banden dargestellt. Erneut war eine Zuordnung
der Banden zu entweder ECaC1 oder ECaC2 nicht möglich.
4.7 Ergebnisse der Real-Time-PCR bei Saug- und Absetzferkeln
Mit Hilfe der Real-Time-PCR wurden die mRNA-Gehalte der Saug- und Absetzferkel an
ECaC1 und ECaC2 relativ zum β-Aktin verglichen. Dies stellt eine semiquantitative
Bestimmung der mRNA-Gehalte dar. Jede Tierprobe wurde im Doppelansatz gemessen und
jeder Doppelansatz wurde wiederholt. Bei den Absetzferkeln konnte für 5 Tiere der ECaC1,
der ECaC2 und das β-Aktin quantifiziert werden. Für 5 Saugferkel konnte der quantitative
Nachweis des ECaC2 und des β-Aktins geführt werden, der quantitative Nachweis für den
ECaC1 gelang bei 4 Tieren, bei einem Tier lagen die Werte unterhalb der Nachweisgrenze.
Die Abbildung 23 zeigt die hintergrundkorrigierte Fluoreszenz-Zunahme (dRn) exemplarisch
für ein Saug- und ein Absetzferkel bei ECaC1, ECaC2 und β-Aktin. Für jede
Primerkombination wurde ein Leerwert, dem H2O anstelle der cDNA zugegeben wurde,
quantifiziert.
74
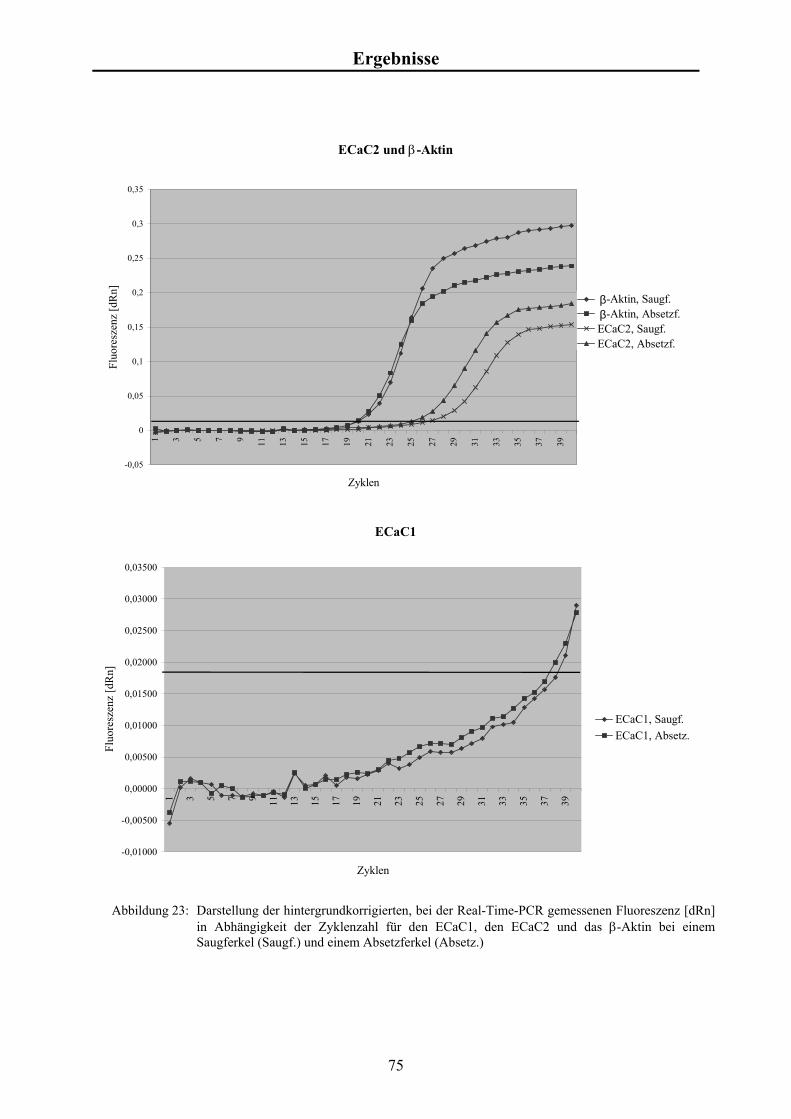
Ergebnisse
ECaC2 und β-Aktin
-0,05
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39
Zyklen
Fluo
resz
enz
[dR
n]
-Aktin, Saugf. -Aktin, Absetzf.ECaC2, Saugf.ECaC2, Absetzf.
ββ
ECaC1
-0,01000
-0,00500
0,00000
0,00500
0,01000
0,01500
0,02000
0,02500
0,03000
0,03500
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39
Zyklen
Fluo
resz
enz
[dR
n]
ECaC1, Saugf.ECaC1, Absetz.
Abbildung 23: Darstellung der hintergrundkorrigierten, bei der Real-Time-PCR gemessenen Fluoreszenz [dRn]in Abhängigkeit der Zyklenzahl für den ECaC1, den ECaC2 und das β-Aktin bei einemSaugferkel (Saugf.) und einem Absetzferkel (Absetz.)
75
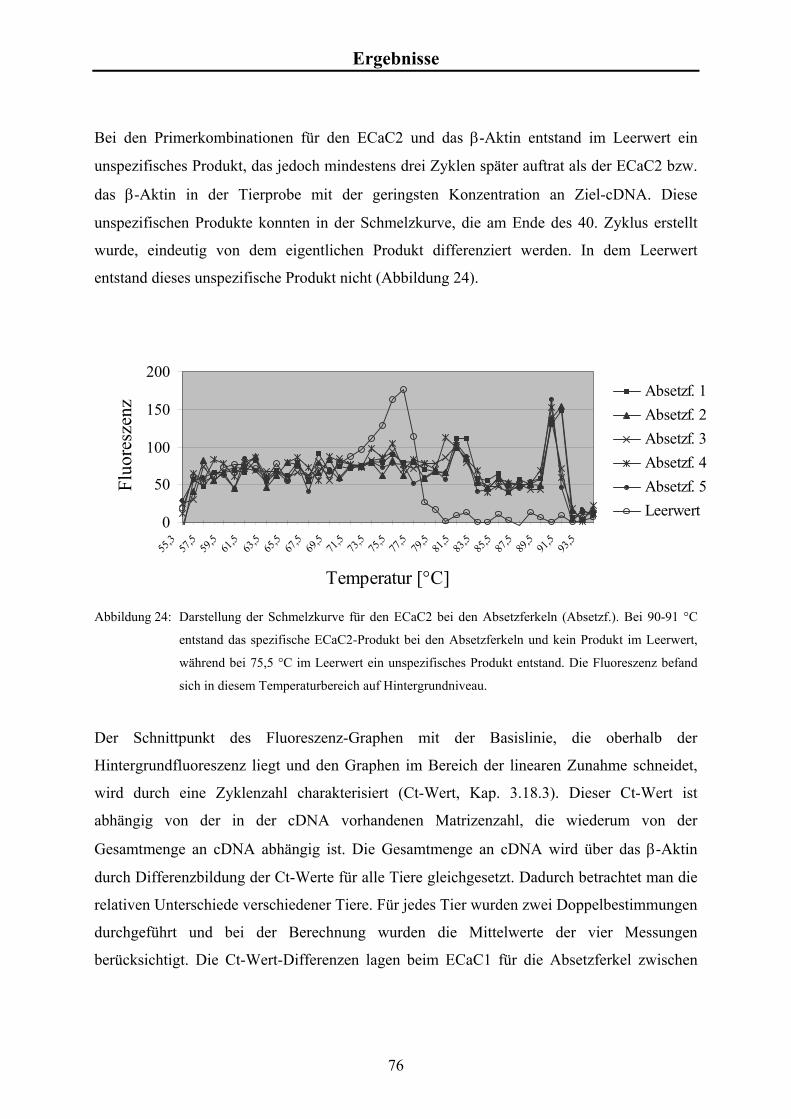
Ergebnisse Bei den Primerkombinationen für den ECaC2 und das β-Aktin entstand im Leerwert ein
unspezifisches Produkt, das jedoch mindestens drei Zyklen später auftrat als der ECaC2 bzw.
das β-Aktin in der Tierprobe mit der geringsten Konzentration an Ziel-cDNA. Diese
unspezifischen Produkte konnten in der Schmelzkurve, die am Ende des 40. Zyklus erstellt
wurde, eindeutig von dem eigentlichen Produkt differenziert werden. In dem Leerwert
entstand dieses unspezifische Produkt nicht (Abbildung 24).
0
50
100
150
200
55,3
57,5
59,5
61,5
63,5
65,5
67,5
69,5
71,5
73,5
75,5
77,5
79,5
81,5
83,5
85,5
87,5
89,5
91,5
93,5
Temperatur [°C]
Fluo
resz
enz Absetzf. 1
Absetzf. 2Absetzf. 3Absetzf. 4Absetzf. 5Leerwert
Abbildung 24: Darstellung der Schmelzkurve für den ECaC2 bei den Absetzferkeln (Absetzf.). Bei 90-91 °C
entstand das spezifische ECaC2-Produkt bei den Absetzferkeln und kein Produkt im Leerwert,
während bei 75,5 °C im Leerwert ein unspezifisches Produkt entstand. Die Fluoreszenz befand
sich in diesem Temperaturbereich auf Hintergrundniveau.
Der Schnittpunkt des Fluoreszenz-Graphen mit der Basislinie, die oberhalb der
Hintergrundfluoreszenz liegt und den Graphen im Bereich der linearen Zunahme schneidet,
wird durch eine Zyklenzahl charakterisiert (Ct-Wert, Kap. 3.18.3). Dieser Ct-Wert ist
abhängig von der in der cDNA vorhandenen Matrizenzahl, die wiederum von der
Gesamtmenge an cDNA abhängig ist. Die Gesamtmenge an cDNA wird über das β-Aktin
durch Differenzbildung der Ct-Werte für alle Tiere gleichgesetzt. Dadurch betrachtet man die
relativen Unterschiede verschiedener Tiere. Für jedes Tier wurden zwei Doppelbestimmungen
durchgeführt und bei der Berechnung wurden die Mittelwerte der vier Messungen
berücksichtigt. Die Ct-Wert-Differenzen lagen beim ECaC1 für die Absetzferkel zwischen
76
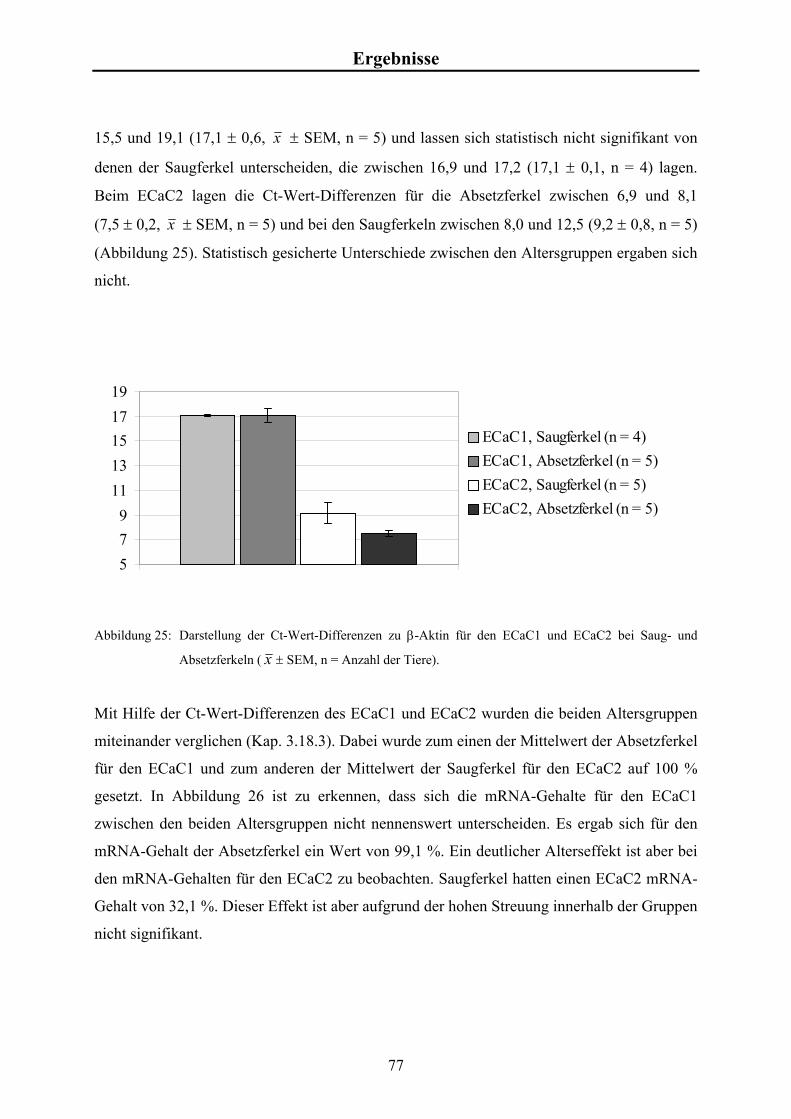
Ergebnisse 15,5 und 19,1 (17,1 ± 0,6, x ± SEM, n = 5) und lassen sich statistisch nicht signifikant von
denen der Saugferkel unterscheiden, die zwischen 16,9 und 17,2 (17,1 ± 0,1, n = 4) lagen.
Beim ECaC2 lagen die Ct-Wert-Differenzen für die Absetzferkel zwischen 6,9 und 8,1
(7,5 ± 0,2, x ± SEM, n = 5) und bei den Saugferkeln zwischen 8,0 und 12,5 (9,2 ± 0,8, n = 5)
(Abbildung 25). Statistisch gesicherte Unterschiede zwischen den Altersgruppen ergaben sich
nicht.
579
1113151719
1
ECaC1, Saugferkel (n = 4)ECaC1, Absetzferkel (n = 5)ECaC2, Saugferkel (n = 5)ECaC2, Absetzferkel (n = 5)
Abbildung 25: Darstellung der Ct-Wert-Differenzen zu β-Aktin für den ECaC1 und ECaC2 bei Saug- und
Absetzferkeln ( x ± SEM, n = Anzahl der Tiere).
Mit Hilfe der Ct-Wert-Differenzen des ECaC1 und ECaC2 wurden die beiden Altersgruppen
miteinander verglichen (Kap. 3.18.3). Dabei wurde zum einen der Mittelwert der Absetzferkel
für den ECaC1 und zum anderen der Mittelwert der Saugferkel für den ECaC2 auf 100 %
gesetzt. In Abbildung 26 ist zu erkennen, dass sich die mRNA-Gehalte für den ECaC1
zwischen den beiden Altersgruppen nicht nennenswert unterscheiden. Es ergab sich für den
mRNA-Gehalt der Absetzferkel ein Wert von 99,1 %. Ein deutlicher Alterseffekt ist aber bei
den mRNA-Gehalten für den ECaC2 zu beobachten. Saugferkel hatten einen ECaC2 mRNA-
Gehalt von 32,1 %. Dieser Effekt ist aber aufgrund der hohen Streuung innerhalb der Gruppen
nicht signifikant.
77
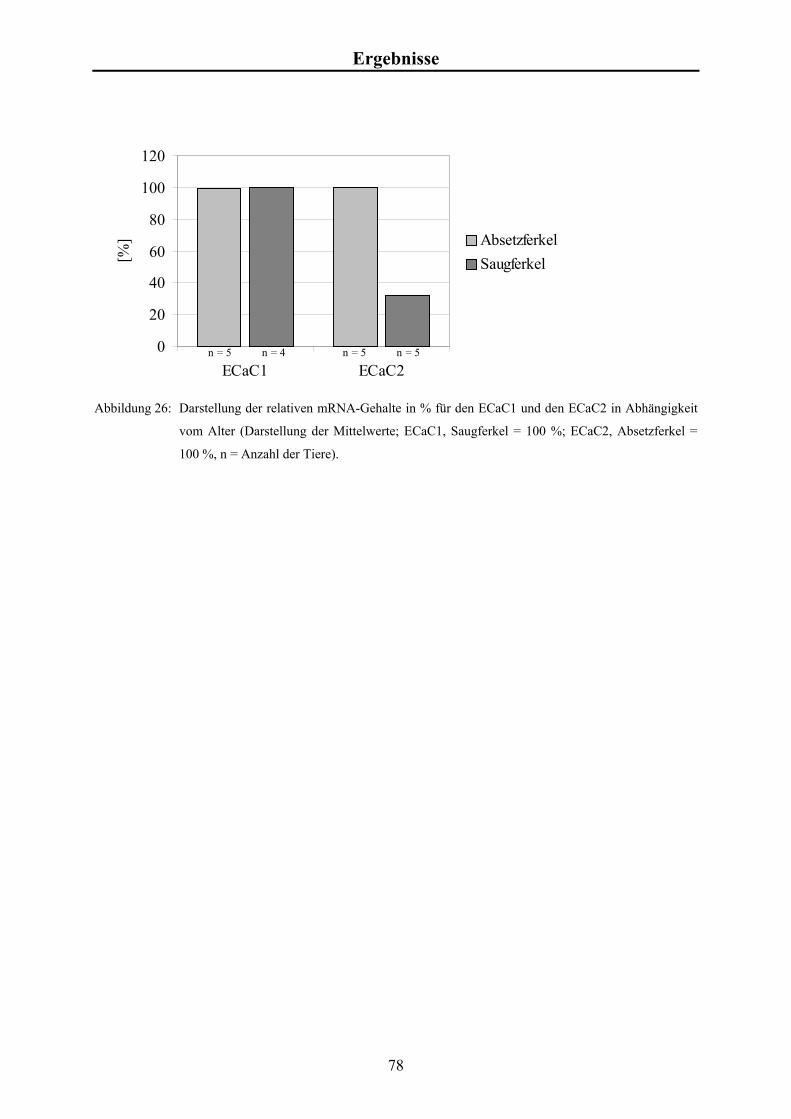
Ergebnisse
0
20
40
60
80
100
120
ECaC1 ECaC2
[%] Absetzferkel
Saugferkel
n = 5 n = 4 n = 5 n = 5
Abbildung 26: Darstellung der relativen mRNA-Gehalte in % für den ECaC1 und den ECaC2 in Abhängigkeit
vom Alter (Darstellung der Mittelwerte; ECaC1, Saugferkel = 100 %; ECaC2, Absetzferkel =
100 %, n = Anzahl der Tiere).
78

Diskussion 5 DISKUSSION
In der vorliegenden Arbeit wurde die Ca2+-Aufnahme in duodenale BSMV während der
postnatalen Entwicklung untersucht. Außerdem wurde mit der RT-PCR gezeigt, dass beim
Schwein wie bei verschiedenen anderen Säugerspezies mindestens zwei verschiedene Ca2+-
Kanäle an der aktiven intestinalen Ca2+-Absorption beteiligt sind (WEBER et al. 2001,
SLEPCHENKO u. BRONNER 2001), die möglicherweise in unterschiedlichem Ausmaß
unter Kontrolle von Calcitriol stehen. Da es aus Untersuchungen auf funktioneller Ebene
sichere Hinweise gab, dass der transepitheliale Ca2+-Transport in der frühen postnatalen Phase
nicht wie später im Absetzalter durch Calcitriol reguliert wird (SCHRÖDER et al. 1990,
1993, 1998a), lag es nahe, die jeweils anteilige Expression der Ca2+-Kanäle während der
Umstellung von flüssiger auf feste Nahrung zu untersuchen, um damit zum Verständnis der
Rolle des Vitamin D-Hormonsystems bei diesen Vorgängen erweiternd beizutragen.
5.1 Untersuchungen zur Funktion der Ca2+-Kanäle
Die BSMV wurden in Anlehnung an eine für Absetzferkel etablierte Methode von
Enterozyten des Duodenums mittels Mg2+-Präzipitation und Differentialzentrifugation
hergestellt (KAUNE et al. 1992, SCHRÖDER et al. 1998c). In zahlreichen Arbeiten wurde
schon zuvor der Ca2+-Transport in BSMV sowohl bei Schweinen (KASSIANOFF 1989,
KAUNE 1992, SCHRÖDER et al. 1998a, DAHL 1999) als auch bei anderen Spezies
untersucht (RASMUSSEN et al. 1979, WILSON u. LAWSON 1980, MILLER u. BRONNER
1981, SHULTZ et al. 1982, BIKLE et al. 1983, KREUTTER et al. 1984, SCHEDL u.
WILSON 1985, LIANG et al. 1986, GHISHAN et al. 1989, WILSON et al. 1989). Generell
hat sich dabei ergeben, dass die BSMV eine große Oberfläche für den Stoff- und
Flüssigkeitstransport bieten und dass sie eine für die Versuchsdauer ausreichende
mechanische Stabilität aufweisen. Durch die Isolierung der Bürstensaummembranen unter
weitgehendem Ausschluss bzw. Abreicherung anderer Zellkomponenten können die
79

Diskussion Vorgänge an der apikalen Zellmembran solitär betrachtet und beschrieben werden. Durch die
Trennung zwischen intra- und extravesikulärem Kompartiment können die
Transportbedingungen variiert und die Wirkungen verschiedener Substanzen auf den
betrachteten Transportvorgang untersucht werden. Dazu müssen in den Präparationsschritten
die Bürstensaummembranen gegenüber anderen Membranfragmenten wie z. B. denen der
basolateralen Zellseite ausreichend angereichert werden.
Die Anreicherung der Bürstensaummembranen als Qualitätsmerkmal der BSMV-Präparation
lässt sich mit Hilfe von membrantypischen Leitenzymen bestimmen, die in dem
Ausgangshomogenat und der Vesikelsuspension bestimmt und miteinander verglichen werden
(MIRCHEFF u. WRIGHT 1976). Als Leitenzym für die Bürstensaummembranen wurde die
Aktivität der alkalischen Phosphatase gemessen, die im Mittel um das 12fache angereichert
werden konnte, wobei kein Unterschied zwischen den unterschiedlich alten Ferkeln auftrat.
Ähnliche Anreicherungen wurden auch in anderen Arbeiten über den Schweinedarm ermittelt
(KELJO et al. 1985, BRANDIS et al. 1987, KASSINOFF 1989, KAUNE et al. 1992,
HATTENHAUER 1998, DAHL 1999). Auch für weitere Spezies wurden Werte in dieser
Größenordnung angegeben (Hühner: BIKLE et al. 1983, Kaninchen: DANISI et al 1984,
AHEARN u. MURER 1984, Schaf: SHIRAZI-BEECHEY et al. 1991, Ziege: RÜBELT
1995). Als Leitenzym für die basolateralen Membranen wurde die Aktivität der Na+/K+-
ATPase bestimmt. Diese war im Mittel nur um das 3fache angereichert und lag somit in
einem Bereich, wie er bei anderen Spezies gefunden wurde (MILLER u. BRONNER 1981,
AHEARN u. MURER 1984, KELJO et al. 1985, SCHEDL u. WILSON 1985, KASSIANOFF
1989, KAUNE 1992, RÜBELT 1995, HATTENHAUER 1998). Eine gewisse Kontamination
der apikalen Vesikelsuspension mit basolateralen Membranen ist bei der Präzipitation mit
divalenten Kationen (Ca2+ oder Mg2+) und Differentialzentrifugation nicht zu vermeiden
(MURER et al. 1989), aber entscheidend ist die deutlich höhere Anreicherung der
Bürstensaummembranen gegenüber den basolateralen Membranen.
Mit Hilfe der zeitabhängigen Glucose-Aufnahme in Anwesenheit eines einwärts gerichteten
Na+-Gradienten wurde wie von HOPFER et al. (1973, 1976) vorgeschlagen die funktionelle
Integrität der BSMV bestimmt. Durch die Triebkraft des Na+-Gradienten wird initial mehr
Glucose in die Vesikel aufgenommen als bei einem Konzentrationsausgleich zwischen extra-
und intravesikulärem Kompartiment zu erwarten wäre (Abbildung 6). Diese übermäßige
80

Diskussion Aufnahme wird als „Overshoot“ bezeichnet, dessen Maximalwert unter den gewählten
Versuchsbedingungen bei den Saugferkeln nach 2 min und bei den Absetzferkeln nach 3 min
erreicht war. Zu diesem Zeitpunkt war die Glucose-Aufnahme in Gegenwart des Na+-
Gradienten 10-mal höher als ohne Na+-Gradient. Der Maximalwert ist u. a. abhängig von der
Inkubationstemperatur, der Lokalisation der zur Präparation genutzten Dünndarmabschnitte
und dem Alter des jeweiligen Versuchstieres (BUDDINGTON u. MALO 1996). Das
„Overshoot“-Phänomen kann ausschließlich bei einem geschlossenen Kompartiment
beobachtet werden. In Abwesenheit von Na+ gelangt die Glucose entlang ihres chemischen
Gradienten per Diffusion in die Vesikel und erreicht den Maximalwert unter den vorliegenden
Bedingungen nach 120 min, wenn die extra- und intravesikuläre Glucosekonzentration im
Equilibrium vorliegt. Auch in Anwesenheit des Na+-Gradienten erfolgt während der 120-
minütigen Inkubation ein Konzentrationsausgleich (Abbildung 6). Der Ausgleichswert wurde
zur Bestimmung der Vesikelvolumina herangezogen, die altersunabhängig bei ca. 1 µl.mg-1
Protein lagen und damit eine ähnliche Größenordnung aufwiesen wie in anderen
Untersuchungen mit Enterozyten beschrieben (BERNER et al. 1976, BOROWITZ u.
GHISHAN 1985, RÜBELT 1995, WALTER 1999, HATTENHAUER 1998).
Die Untersuchung der zeitabhängigen Ca2+-Aufnahme in An- und Abwesenheit des Ca2+-
Ionophors A23187 stellt prinzipiell eine weitere Möglichkeit dar, die Vesikelintegrität zu
überprüfen. Werden intakte BSMV mit dem Ionophor vermischt, so werden sie permeabel für
Ca2+ und füllen sich ungehindert. Dies führt zu einer „Ad hoc-Aufnahme“ von Ca2+ in die
BSMV, was sich graphisch durch einen steileren Kurvenverlauf als bei der erleichterten
Diffusion durch die Bürstensaummembran darstellt (Abbildung 7). Um die Integrität der
Vesikel zu beurteilen, wurden regelmäßig die Ca2+-Aufnahmewerte nach 30 s in An- und
Abwesenheit des Ca2+-Ionophors verglichen. Dabei ergab sich eine um das 3,3 bis 6,3fache
initial höhere Ca2+-Aufnahme, was nur bei einem geschlossenen Kompartiment auftritt.
Erneut ergaben sich keine nennenswerten Unterschiede zwischen den Altersgruppen. Dieser
Befund deckt sich mit den Schätzungen der Vesikelvolumina aus den Glucose-
Ausgleichswerten, die ebenfalls keinen Alterseffekt aufwiesen (Abbildung 8).
Insgesamt ist also davon auszugehen, dass für alle Tiergruppen ausreichend angereicherte
BSMV-Suspensionen mit adäquater funktioneller Integrität zur Bestimmung der kinetischen
Parameter des Ca2+-Transportes durch duodenale Bürstensaummembranen vorlagen.
81

Diskussion Bei der Beurteilung der Ca2+-Aufnahmeraten in BSMV ist des Weiteren zu beachten, dass
mittels der Schnellfiltrationsmethode zunächst der Gesamttransport ermittelt wird, der sich
seinerseits aus einer sättigbaren und einer nicht sättigbaren Komponente zusammensetzt
(SCHEDL u. WILSON 1985, GHISHAN et al. 1989, KAUNE 1992). Einen wichtigen Anteil
an der unspezifischen Komponente des Ca2+-Transportes bildet die Proteinbindung von Ca2+,
die sowohl an der Außen- als auch Innenseite der isolierten Bürstensaummembran erfolgen
kann. Diese Proteinbindung wurde durch den Vergleich der aus Glucose- und Ca2+-
Aufnahmeversuchen berechneten Vesikelvolumina geschätzt und betrug bei MILLER u.
BRONNER (1981) 50%, bei KASSIANOFF (1989) 90 % und bei MERRIL et al. (1986)
100 %.
Die sättigbare Komponente kann im Gegensatz zur nicht sättigbaren Komponente mit dem
aktiven Ca2+-Transport korreliert werden (SCHEDL u. WILSON 1985), und es lassen sich
aus den Daten der Ca2+-Aufnahmeraten als Funktion von [Ca2+]f nach der Theorie von
Michaelis-Menten die kinetischen Parameter Vmax und Km kalkulieren. Dabei muss beachtet
werden, dass die „schnellen“ Ca2+-Aufnahmeraten innerhalb der ersten Minute für die
Berechnung berücksichtigt werden, da später gemessene Werte für die gewählte
Konzentration nicht mehr repräsentativ sind (GHIJSEN et al. 1987). Die apparenten Km-
Werte für die Absetz- und Mastferkel lagen bei 1,1 mmol.l-1 und waren damit signifikant von
den Km-Werten der Saugferkel verschieden, die bei 1,4 mmol.l-1 lagen. Ähnliche Km-Werte
wurden für BSMV aus Hühnerenterozyten beschrieben (RASMUSSEN et al. 1979). Eine
Überprüfung der kinetischen Daten in Scatchard-Plots zeigte eine lineare Beziehung und gab
damit keinen Hinweis auf die Existenz von mehreren, sich deutlich unterscheidenden
Transportsystemen. Es soll darauf hingewiesen werden, dass die hier kalkulierten Km-Werte
je nach Ausmaß der Präsenz von ECaC1 oder ECaC2 bzw. Koexistenz beider Systeme in der
Bürstensaummembran die Ca2+-Affinität entweder des einen oder anderen oder den
Mittelwert beider Systeme darstellen können. Insofern können die unterschiedlichen Km-
Werte als Hinweis auf die unterschiedliche Expression von ECaC1 und/oder ECaC2 bei
Absetz-/Mastferkeln bzw. Saugferkeln gewertet werden. Jedoch sind die Km-Werte trotz der
statistischen Unterschiede aus transporterphysiologischer Sicht so ähnlich, dass eine
Unterscheidung der Ca2+-Kanäle anhand der Km-Werte nicht möglich ist. Hierfür müssen
erfahrungsgemäß Unterschiede bei den Km-Werten von mindestens einer Zehnerpotenz
82

Diskussion bestehen. Erst dann ist eine Unterscheidung unter Verwendung eines „Two-side-binding“-
Modells (Annahme der Beteiligung von 2 unterschiedlichen Transportersystemen)
rechnerisch sinnvoll und möglich. Im vorliegenden Fall wurde nach KAUNE et al. (1992) ein
„One-side-binding“-Modell zur Berechnung von Vmax und Km verwendet, dass den Gesamt-
Vmax-Wert aller potenziellen Ca2+-Transportsysteme angibt und für den altersabhängigen
Gruppenvergleich verwendet werden konnte.
Im Gegensatz zu den hier dargestellten Werten haben KAUNE et al. (1992) eine apparente
Km für das Absetzferkel von 0,03 mmol.l-1 ermittelt. Diese Abweichung kann am Besten
dadurch erklärt werden, dass bei den Untersuchungen von Kaune die Km-Werte in
Anwesenheit eines vesikelauswärts gerichteten Kalium-Gradienten ermittelt wurden, was zu
einer transmembranalen Potentialdifferenz (Vesikelinneres negativ) führte. Inzwischen ist
gesichert, dass intestinale Ca2+-Kanäle spannungssensitiv sind (HOENDEROP et al. 1999b),
und es ist damit davon auszugehen, dass die von KAUNE et al. vorgelegte Kalium-
Spannungsklemme einen signifikanten Einfluss auf die Kinetik des Ca2+-Transportes hatte. Es
soll an dieser Stelle darauf hingewiesen werden, dass in zukünftigen Experimenten zur
Charakterisierung des Ca2+-Transportes über die duodenale Bürstensaummembran bei der
Auswahl der Versuchsbedingungen eine Vereinheitlichung der membranalen
Spannungsverhältnisse notwendig erscheint.
Für die beiden bisher bekannten Ca2+-Kanäle sind die Km-Werte mit ECaC-exprimierenden
Oozyten von Xenopus laevis bestimmt worden. Sie variierten beim ECaC1 von 0,2 mmol.l-1
(HOENDEROP et al. 1999a) bis 0,66 mmol.l-1 (PENG et al. 2000b) und beim ECaC2 von
0,25 mmol.l-1 (PENG et al. 2000a) bis 0,44 mmol.l-1 (PENG et al. 1999), was in der
Größenordnung zumindest teilweise eher den in dieser Arbeit gezeigten Km-Werten entspricht
als den von KAUNE et al. beschriebenen.
Die Vmax-Werte für die Absetz- und Mastferkel lagen bei 1,89 und 2,12 nmol.mg-1 Protein und
waren signifikant von dem Vmax-Wert der Saugferkel verschieden, der bei 3,64 nmol.mg-1
Protein lag, was als Hinweis auf eine erhöhte Anzahl von ECaC gewertet werden kann.
Unterstützt wird diese Annahme durch Befunde aus Versuchen mit duodenalen Epithelien in
Ussing-Kammern, bei denen ein tendenziell erhöhter, transepithelialer Ca2+-Nettofluss beim
Saugferkel gegenüber Absetzferkeln gezeigt werden konnte (SCHRÖDER et al. 1993,
1998a,b).
83

Diskussion 5.2 Untersuchungen zur Struktur der Ca2+-Kanäle
Zunächst wurde mit Hilfe der RT-PCR untersucht, ob auch beim Schwein wie bei den bisher
untersuchten Spezies spezifische Ca2+-Kanäle am transepithelialen Ca2+-Transport im
Duodenum beteiligt sind. Für diesen Nachweis standen Primer, die vom ECaC1 des
Kaninchens bzw. ECaC2 des Menschen abgeleitet wurden, zur Verfügung. Damit konnte
zwar erstmals die Existenz von Ca2+-Kanälen im Schweinedünndarm nachgewiesen werden,
aber eine Typisierung war nicht möglich, da die identifizierte Teilsequenz aus einem Bereich
stammte, in dem sich ECaC1 und ECaC2 nur geringfügig unterscheiden. Deshalb wurden
anschließend Teilsequenzen identifiziert, die dies nun ermöglichen. Davon hatte die eine eine
95%ige Homologie zum ECaC1 der Ratte und nur eine 33%ige Homologie zum ECaC2 des
Menschen. Die entsprechende Aminosäuresequenz zeigte eine 97%ige Homologie zum
ECaC1 der Ratte und eine 33%ige Homologie zum ECaC2 der Ratte. Daraus kann
geschlossen werden, dass im Dünndarm des Schweines die mRNA, die für den ECaC1
codiert, präsent ist. Eine andere Teilsequenz überschnitt diese in 12 Aminosäuren, von denen
allerdings 5 unterschiedlich waren, so dass davon ausgegangen werden kann, dass diese
Teilsequenz dem ECaC2 zuzuordnen ist. Es gilt demnach als wahrscheinlich, dass im
Dünndarm des Schweines ebenfalls die mRNA, die für den ECaC2 codiert, präsent ist. Dies
wird dadurch untermauert, dass bei der Basensequenz eine höhere Homologie zum ECaC2
(88 % im Vergleich zum ECaC2 des Menschen) als zum ECaC1 (81 % im Vergleich zum
ECaC1 der Ratte) vorlag und beide Primer, die zu diesem PCR-Produkt führten, von einem
ECaC2 abgeleitet wurden.
84

Diskussion 5.3 Untersuchungen zur Quantifizierung der Ca2+-Kanäle
Um die jeweiligen quantitativen Anteile von ECaC1 und ECaC2 im Duodenum von Ferkeln
während der postnatalen Entwicklung näher zu charakterisieren, wurden zunächst Northern-
Blot-Analysen durchgeführt. Es gelang allerdings nicht, die mit den verschiedenen Sonden
dargestellten Banden dem ECaC1 oder ECaC2 zuzuordnen. Obwohl die „ECaC2-Sonde“ und
in besonderem Maße die „ECaC1-Sonde“ aus dem Bereich gewählt wurde, in dem sich
ECaC1 und ECaC2 deutlich unterscheiden müssten, entstanden bei beiden Sonden jeweils
zwei Banden und die „ECaC-Sonde“, die beide Kanäle hätte darstellen sollen, stellte nur eine
Bande dar. Die Ursachen für dieses paradoxe Phänomen sind nicht bekannt.
Mit den Ergebnissen entlang der cranial-caudalen Darmachse, die die höchste Expression des
ECaC im vorderen Dünndarm zeigten, lässt sich erstmals auf mRNA-Ebene bestätigen, dass
der vordere Dünndarm der Hauptort der aktiven Ca2+-Absorption ist, wie es von
Untersuchungen auf funktioneller Ebene seit langem bekannt ist (PARTRIDGE 1978, VAN
OS 1987, BRONNER 1992). Auch das Colon scheint beim Schwein zum aktiven Ca2+-
Transport befähigt, da auch in diesen Darmabschnitten die für die Ca2+-Kanäle codierende
mRNA nachgewiesen werden konnte.
Mit dem Verfahren der Real-Time-PCR war es möglich, die ECaC1- und ECaC2- Sequenzen
zu unterscheiden. Für den ECaC1 ergaben sich Ct-Werte, die größer als 37 waren, und somit
im Bereich der Nachweisgrenze lagen. Der Nachweis gelang bei 4 Saug- und 5 Absetzferkeln.
Für jeweils 5 Saug- und 5 Absetzferkel konnte der Nachweis auch für den ECaC2
durchgeführt werden. Es ergaben sich dabei Ct-Werte, die bei 21 lagen, und damit auf eine
höhere Präsenz von ECaC2 als von ECaC1 im Dünndarm von Saug- und Absetzferkeln
hinweisen. Dies deckt sich mit Untersuchungen, die PENG at al. (2001) durchführten, wobei
sie in humanem Dünndarmgewebe nur den ECaC2 nachweisen konnten und der ECaC1
unterhalb der Nachweisgrenze lag. Real-Time Untersuchungen sind jedoch nur bedingt
geeignet, Aussagen zu treffen über den direkten Vergleich der mRNA-Gehalte von ECaC1
und ECaC2, da bei den PCR-Reaktionen unterschiedliche Bedingungen vorherrschen, wie
z. B. eine unterschiedliche Primer- oder MgCl2-Konzentration und die zu amplifizierenden
Stücke eine unterschiedliche Länge und Basensequenz haben.
85

Diskussion Um die Präparationen der Tiere miteinander vergleichen zu können, wurden die ermittelten
mRNA-Gehalte auf den internen Standard β-Aktin bezogen (PEIRCE u. CHEN 2001, DA
COSTA et al. 2002). Vom β−Aktin wird vorausgesetzt, dass es in gleicher Menge in den
Geweben bei den verschiedenen Tieren vorkommt (BISHOP et al. 2000, DA COSTA et al.
2002, KLEMCKE et al. 2001, LABUHN u. BRACK 1997, OUE et al. 1999, PEIRCE u.
CHEN 2001). Dadurch konnten Proben verglichen werden, deren Menge an vorhandener
cDNA nicht exakt gleich war.
Mit Hilfe der relativen ECaC1 und ECaC2 mRNA-Gehalte wurde der Alterseffekt untersucht.
Die mRNA-Gehalte für den ECaC1 unterschieden sich dabei nicht nennenswert zwischen den
Saug- und Absetzferkeln. Demnach scheint der ECaC1 auf m-RNA-Ebene nicht
altersabhängig reguliert zu werden. Ein deutlicher Alterseffekt, der aber aufgrund der hohen
Streuung innerhalb der Gruppen statistisch nicht gesichert werden konnte, deutet sich aber in
Bezug auf die mRNA-Gehalte des ECaC2 an. Beim Saugferkel war die Präsenz des ECaC2
um 2/3 niedriger als beim Absetzferkel.
5.4 Schlussbetrachtung und Ausblick
Möglicherweise wird mit dem ECaC1 eine Basisversorgung des Organismus mit Ca2+
erreicht, die in besonderem Maße beim Saugferkel, das mit einer höheren Ca2+-Konzentration
in der Nahrung versorgt wird, ausreichend ist, um die Ca2+-Homöostase aufrecht zu erhalten.
Mit zunehmendem Alter sinkt die Transporterkapazität am Epithel und in den BSMV,
gleichzeitig wird der mRNA-Gehalt für den ECaC2 in den Enterozyten hochreguliert. Dies
deutet auf eine Regulation auf Translationsebene, die noch untersucht werden muss. Da aber
der mRNA-Gehalt mit zunehmendem Alter ansteigt, liegt die Vermutung nahe, dass der
ECaC2 im Dünndarm der Schweine, wie es ebenfalls für Caco-2-Zellen gezeigt (WOOD et al.
2001) wurde, durch Calcitriol reguliert wird, da aus früheren Untersuchungen bekannt ist,
dass die Ca2+-Versorgung beim Schwein erst altersabhängig mehr und mehr unter die
Kontrolle dieses Hormons kommt. Dabei bleibt allerdings die Konzentration des Calcitriols
altersunabhängig konstant, jedoch steigt mit zunehmendem Alter die maximale
Bindungskapazität des Vitamin D-Rezeptors (SCHRÖDER et al. 1993). Demnach würde der
ECaC2 für die Feinregulation der Ca2+-Homöostase von Bedeutung sein. Diese Hypothesen
86

Diskussion über die physiologische Bedeutung der verschiedenen Ca2+-Kanäle müssen jedoch noch auf
der Proteinebene untersucht werden und können auch dadurch verifiziert werden, dass die
Regulation auf mRNA-Ebene bei verschiedenen Behandlungsmodellen mit Calcitriol
betrachtet wird. Für zukünftige Untersuchungen zur Charakterisierung der Rolle der Ca2+-
Kanäle bei der postnatalen Entwicklung des duodenalen Ca2+-Transportes beim Schwein
stehen nun Techniken zur Verfügung.
87

Zusammenfassung 6 ZUSAMMENFASSUNG Thomas Hinterding Expression der Ca2+-Kanäle ECaC1 und ECaC2 im Dünndarm von Saug- und
Absetzferkeln
Es wurden funktionelle Daten zur postnatalen Entwicklung des intestinalen Ca2+-Transportes
bei Schweinen erhoben und anschließend mit molekularbiologischen Methoden diejenigen
Ca2+-Kanäle, die für die Aufnahme von Ca2+ durch die Bürstensaummembran der Enterozyten
verantwortlich sind, qualitativ und altersabhängig quantitativ untersucht.
Zur Erfassung der funktionellen Daten wurden Bürstensaummembranvesikel von Enterozyten
aus dem vorderen Dünndarm von Saug-, Absetz- und Mastferkeln mittels Mg2+-Präzipitation
und Differentialzentrifugation hergestellt und die Ca2+-Aufnahmeraten mittels
Schnellfiltrationstechnik unter Verwendung des Radiotracers 45Ca untersucht.
Der qualitative Nachweis der epithelialen Ca2+-Kanäle (ECaC) erfolgte mit der RT-PCR und
Primern in Anlehnung an bekannte Sequenzen anderer Spezies.
Zur relativen Quantifizierung der ECaC auf mRNA-Ebene wurden Untersuchungen bei Saug-
und Absetzferkeln mit Hilfe von Northern-Blots und Real-Time-PCRs durchgeführt.
Die Untersuchungen führten zu folgenden Ergebnissen:
(1) Die Kinetik der Ca2+-Aufnahme in duodenale BSMV des Ferkels konnte am Besten mit
einem „One-site-binding“-Modell angepasst werden. Die daraus erhaltenen kinetischen
Daten ergaben keinen unmittelbaren Hinweis für die Präsenz verschiedener ECaC im
Schweinedünndarm.
(2) Saugferkel hatten eine signifikant höhere maximale Ca2+-Aufnahmerate (Vmax) als
Absetzferkel (p < 0,01) oder Mastferkel (p < 0,05). Die Vmax-Werte der Saugferkel
betrugen 3,64 ± 0,45 nmol.mg-1 Protein.30s-1, die der Absetzferkel 1,89 ± 0,20 nmol.mg-1
Protein.30s-1 und die der Mastferkel 2,13 ± 0,20 nmol.mg-1 Protein.30s-1 ( x ± SEM,
n = 5).
88

Zusammenfassung (3) Saugferkel wiesen eine signifikant höhere apparente Km auf als Absetzferkel oder
Mastferkel (p < 0,05). Die Km-Werte der Saugferkel betrugen 1,44 ± 0,06 mmol.l-1 , die
der Absetzferkel 1,14 ± 0,05 mmol.l-1 und die der Mastferkel 1,13 ± 0,09 mmol.l-1
( x ± SEM, n = 5).
(4) Es wurden Teilsequenzen von Ca2+-Kanälen ermittelt, die sich teilweise überlagerten, was
auf die Präsenz zweier verschiedener Ca2+-Kanäle hinweist. Eine dieser Teilsequenzen
hatte eine deutlich höhere Homologie zum ECaC1 der Ratte (95 %) als zum ECaC2 des
Menschen (33 %). Die andere Teilsequenz wies eine höhere Homologie zum ECaC2 des
Menschen (88 %) als zum ECaC1 der Ratte auf (81 %).
(5) Mit der Real-Time-PCR-Methode konnte zwischen ECaC1 und ECaC2 differenziert
werden. Die mRNA für den ECaC1 war bei Saug- und Absetzferkeln in sehr geringem
Umfang vorhanden, der aber nicht unterschiedlich war. Ein Unterschied in Abhängigkeit
des Alters konnte nicht festgestellt werden. Der mRNA-Gehalt für den ECaC2 war beim
Saugferkel gegenüber dem Absetzferkel um 2/3 erniedrigt.
Die Ergebnisse zeigen, dass wie bei anderen bislang untersuchten Säugerspezies auch beim
Schwein zwei verschiedene Ca2+-Kanäle am intestinalen Ca2+-Transport beteiligt sind. Der
mRNA-Gehalt dieser Kanäle lässt sich über die Real-Time-PCR quantifizieren und zeigt für
den ECaC2 einen altersabhängigen Effekt.
89

Summary 7 SUMMARY Thomas Hinterding
Expression of the Ca2+-channels ECaC1 and ECaC2 in small intestines of suckling and
weaned piglets
Functional data from postnatal development of the intestinal Ca -absorption in pigs were
collected and the Ca -channels which are responsible for the Ca -uptake across the brush
border membrane of enterocytes were investigated qualitatively and age-dependent
quantitatively.
2+
2+ 2+
To collect the functional data brush border membrane vesicles were prepared from
enterocytes of the proximal small intestine of suckling and weaned piglets as well as porkers
by combined Mg2+-precipitation method and differential centrifugation and the Ca2+-uptakes
were investigated by the rapid filtration technique using the radiotracer 45Ca.
Epithelial Ca2+-channels (ECaC) were identified with RT-PCR using primers deduced from
well known sequences of other species.
For the relative quantification of ECaC on mRNA levels investigations were carried out with
preparations of suckling and weaned piglets by northern blots and real-time-PCRs.
Following results are to be emphasised:
(1) The kinetic of the Ca2+-uptake into the duodenal brush border membrane vesicles could
be fitted best with a “one-site-binding” model. The results did not support directly the
assumption of the presence of different ECaCs in the small intestine of pigs.
(2) Suckling piglets had a significantly higher maximal Ca2+-uptake (Vmax) compared with
weaned piglets (p < 0.01) or porkers (p < 0.05). Vmax values were 3.64 ± 0.45 nmol.mg-1
protein.30s-1 in suckling piglets, 1.89 ± 0.20 nmol.mg-1 protein.30s-1 in weaned piglets
and 2.13 ± 0.20 nmol.mg-1 protein.30s-1 in porkers ( x ± SEM, n = 5).
90

Summary (3) Suckling piglets had a significantly higher apparent Km compared with weaned piglets or
porkers (p < 0.05). Km values were 1.44 ± 0.06 mmol.l-1 in suckling piglets,
1.14 ± 0.05 mmol.l-1 in weaned piglets and 1.13 ± 0.09 mmol.l-1 in porkers ( x ± SEM,
n = 5).
(4) Partial sequences from the Ca2+-channels were detected which show to some extent
sequence overlay. That can be analysed as an evidence for the presence of two different
Ca2+-channels in porcine duodenum. One of the partial sequences had a higher homology
to the ECaC1 of rat (95 %) compared with ECaC2 of human (33 %). The other partial
sequence exhibited a higher homology to the hECaC2 (88 %) compared with the rECaC1
(81 %).
(5) With the real-time-PCR method differentiation of ECaC1 and ECaC2 was enabled. The
mRNA for ECaC1 was present in suckling and weaned piglets in rather small extent but
without a difference. The mRNA level for ECaC2 was lower by 66 % in suckling piglets
compared with weaned piglets.
In summary, the results show that two different Ca2+-channels are involved in Ca2+-absorption
from the small intestines, similarly to those shown recently in other mammalian species. The
mRNA levels of these channels can be quantified using the real-time-PCR method and the
results for the ECaC2 point to same kind of postnatal adaptation, which should be further
elucidated in following investigations.
91

Literaturverzeichnis 8 LITERATURVERZEICHNIS AHEARN, G.A. und H. MURER (1984): Functional roles of Na+ and H+ in SO4
2- transport by rabbit ileal brush border membrane vesicles. J. Membrane Biol. 78, 177-18. AUDRAN, M. und R. KUMAR, (1985): The physiology and pathophysiology of vitamin D. Mayo. Clin. Proc. 60, 851-866. AVIV, H. und P. LEDER (1972): Purification of biologically active globin messenger RNA by chromatography on oligothymidylic acid-cellulose. Proc. Natl. Acad. Sci. USA 69(6), 1408-1412. BARLEY, N.F., A. HOWARD, D. O´CALLAGHAN, S. LEGON und J.R.F. WALTERS (2001): Epithel calcium transporter expression in human duodenum. Am. J. Physiol. Gastrointest. Liver. Physiol. 280, G285-G290. BECKER, A., A. REITH, J. NAPIWOTZKI und B. KADENBACH (1996): A quantitative method of determining initial amounts of DNA by polymerase chain reaction cycle titration using digital imaging and novel DNA stain. Anal. Biochem. 237(2), 204-207. BEESON, W.M., E.W. CRAMPTON, T.J. CUNHA, N.R. ELLIS und R.W. LUECKE (1953): Nutrient requirements of swine. Natl. Research Council Publ. 295, 19. BELL, D.A. und D.M. DEMARINI (1991): Excessive cycling converts PCR products to random-length higher molecular weight fragments. Nucleic Acids Res. 19(18), 5079. BERNER, W., R. KINNE und H. MURER (1976): Phosphate transport into brush-border membrane vesicles isolated from rat small intestine. Biochem. J. 160, 467-474. BIKLE, D.D., S. MUNSON und D.T. ZOLOCK (1983): Calcium flux across chick duodenal brush border membrane vesicles: regulation by 1,25-dihydroxyvitamin D. Endocrinology 113(6), 2072-2080.
92

Literaturverzeichnis BINDELS, R.J.M., A. HARTOG, J. TIMMERMANS und C.H. VAN OS (1991): Active Ca2+ transport in primary cultures of rabbit kidney CCD: stimulation by 1,25-dihydroxyvitamin D3 and PTH. Am. J. Physiol. 261, F799-F807. BISHOP, C.M., C.J. MCCABE, N.J. GITTOES, P.J. BUTLER und J.A. FRANKLYN (2000): Tissue-specific regulation of thyroid hormone receptor mRNA isoforms and target gene proteins in domestic ducks. J. Endocrinol. 165(3), 607-615. BLIND, E., F. RAUE, T. MEINEL, M. PECHERSTORFER, U. RÄTH, H. SCHMIDT-GAYK, B. KOHL und R. ZIEGEL (1993): Diagnostische Bedeutung von Parathormone-related-Protein bei Tumorpatienten mit Hypercalcämie. Dtsch. Med. Wschr. 118, 330-335. BOROWITZ, S.M. und F.K. GISHAN (1985): Maturation of jejunal phosphate transport by rat brush border membrane vesicles. Pediatr. Res. 19, 1308-1312. BRADFORD, M.M. (1976): A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248-254. BRANDIS, M., J. HARMEYER, R. KAUNE, M. MOHRMANN, H. MURER und Z. ZIMOLO (1987): Phosphate transport in brush-border membranes from control and rachitic pig kidney and small intestine. J. Physiol. 384, 479-490. BREEMEN VAN, C. und K. SAIDA (1989): Cellular mechanisms regulating [Ca2+]i smooth muscle. Annu. Rev. Physiol. 51, 315-329. BRONNER, F. (1992): Current concepts of calcium absorption: an overview. J. Nutr. 122(1), 641-643. BRONNER,F., D. PANSU und W.D. STEIN (1986): An analysis of intestinal calcium transport across the rat intestine. Am. J. Physiol. 250, G561-G569.
93

Literaturverzeichnis BROWN, E.M. (1991): Extracellular Ca2+ sensing, regulation of parathyroid cell function and role of Ca2+ and other ions as extracellular (first) messengers. Physiol. Rev. 71, 371-411. BUDDINGTON, R.K. (1994): Nutritition and ontogenetic development of the intestine. Can. J. Physiol. Pharmacol. 72(3), 251-259. BUDDINGTON, R.K. und J.M. DIAMOND (1989): Ontogenetic development of intestinal nutrient transporters. Annu. Rev. Physiol. 51, 601-619. BUDDINGTON, R.K. und C. MALO (1996): Intestinal brush-border membrane enzyme activities and transport functions during prenatal development of pigs. J. Pediatr. Gastroenterol. Nutr. 23(1), 51-64. BUDDINGTON, R.K., J. ELNIF, A.A. PUCHAL-GARDINER und P.T. SANGLID (2001): Intestinal apical amino acid absorption during development of the pig. Am. J. Physiol. Regulatory Integrative Com. Physiol. 280, R241-R247. CAMPBELL, A.K. (1988): Calcium as an intracellular regulator. In: Nordin, B.E.C. (Hrsg): Calcium in human biology. Springer Verlag, London, 261-316. CARAFOLI, E. (1987): Intracellular calcium homeostasis. Ann. Rev. Biochem. 56, 395-433. CARLBERG, C. (1995): Mechanisms of nuclear signalling by vitamin D3. Interplay with retinoid and thyroid hormone signalling. Eur. J. Biochem. 231, 517-527. CERA, K.R., D.C. MAHAN, R.F. CROSS, G.A. REINHART und R.E. WHITMOYER (1988): Effect of age, weaning and postweaning diet on small intestinal growth and jejunal morphology in young swine. J. Anim. Sci. 66, 574-584. CHIEN, A., D.B. EDGAR und J.M. TRELA (1976): Deoxyribonucleic acid polymerase from the extreme thermophile Thermus aquaticus. J. Bacteriol. 127(3), 1550-1557.
94

Literaturverzeichnis CHRISTAKOS, S., M. RAVAL-PANDYA, R.P. WERNYJ und W. YANG (1996): Genomic mechanisms involved in the pleiotropic actions of 1,25-dihydroxyvitamin D3. Biochem. J. 316, 361-371. CLINE, J., J.C. BRAMAN und H.H. HOGREFE (1996): PCR fidelity of pfu DNA polymerase and other thermostable DNA polymerases. Nucleic. Acids. Res. 24(18), 3546-3551. CORRADINO, R.A. u. R.H. WASSERMAN (1968): Actinomycin D inhibition of Vitamin D3-induced calcium-binding protein (CaBP) formation in chick duodenal mucosa. Arch. Biochem. Biophys. 126(3), 957-960. CORVEN VAN, E.J., M.D. DE JONG und C.H. VAN OS (1987a): The adenosine triphosphate-dependent calcium pump in rat small intestine: effects of vitamin D deficiency and cell isolation methods. Endocrinology 120(3), 868-873. CORVEN VAN, E.J., P.M. VERBOST, M.D. DE JONG und C.H. VAN OS (1987b): Kinetics of ATP-dependent Ca2+ uptake by permeabilized rat enterocytes. Effects of inosiol 1,4,5-trisphosphate. Cell calcium 8(3), 197-206. CROMPHAUT VAN, S.J., M. DEWERCHIN, J.G.J. HOENDEROP, I. STOCKMANS, E. VAN HERCK, S. KATO, R.J.M. BINDELS, D. COLLEN, P. CARMELIET, R. BOUILLON und G. CARMELIET (2001): Duodenal calcium absorption in vitamin D receptor-knockout mice: functional and molecular aspects. Proc. Natl. Acad. Sci. USA 98, 13324-13329. DA COSTA, N., R. BLACKLEY, H. ALZUHERRI und K.C. CHANG (2002): Quantifying the temporospatial expression of postnatal porcine skeletal myosin heavy chain genes. J. Histochem. Cytochem. 50(3), 353-364. DAHL, M.R. (1999): Die postnatale Entwicklung der duodenalen Calcium- und Phosphatabsorption beim Ferkel. Inaugural-Dissertation med. vet., Justus-Liebig-Universität Gießen DANISI, G., H. MURER und R.W. STRAUB (1984): Effect of pH on phosphate transport into intestinal brush-border membrane vesicles. Am. J. Physiol. 246(2), G180-G186. DELUCA, H.F. (1986): The metabolism and functions of vitamin D. Adv. Exp. Med. Biol. 196, 361-375.
95

Literaturverzeichnis DONOWITZ, M. und M.J. WELSH (1986): Ca2+ and cyclic AMP in regulation of intestinal Na, K and Cl transport. Ann. Rev. Physiol. 48, 135-150. DUNSFORD, B.R., D.A. KNABE und W. E. HAENSLY (1989): Effect of dietary soybean meal on the microscopic anatomy of the small intestine in the early-weaned pig. J. Anim. Sci. 67(7), 1855-1863. FAVUS, M.J. (1985): Factors that influence absorption and secretion of calcium in the small intestine and colon. Am. J. Physiol. 248, G147-G157. FEHER J.J., C.S. FULLMER und R.H. WASSERMAN (1992): Role of faciliated diffusion of calcium by calbindin in intestinal calcium absorption. Am. J. Physiol. 262, C517-C526. FEINBERG, A.P. und B. VOGELSTEIN (1984): A technique for radiolabeling DNA restriction endonuclease fragments to high specific activity. Addendum. Anal. Biochem. 137(1), 266-267. FINCKH, U., P.A. LINGENFELTER und D. MYERSON (1991): Producing single-stranded DNA probes with the Taq DNA polymerase: a high yield protocol. Biotechniques 10(1), 35-36, 38-39. FULLMER, C.S. (1992): Intestinal calcium absorption: calcium entry. J. Nutr. 122(2), 644-650. FUJITA, M., H. MATSUI, K. NAGNAO und M. NAKAO (1971): Asymmetric distribution of ouabain-sensitive ATPase activity in rat intestinal mucosa. Biochim. Biophys. Acta 233(2), 404-408. GHIJSEN, W.E., U. GANGULI, G. STANGE, P. GMAJ und H. MURER (1987): Calcium uptake into rat small intestinal brush border membrane vesicles: characterization of transmembrane calcium transport at short initial incubation times. Cell Calcium 8(2), 157-169. GHISHAN, F.K., N. ARAB und W. NYLANDER (1989): Characterisation of calcium uptake by brush border membrane vesicles of human small intestine. Gastroenterology 96, 122-129.
96

Literaturverzeichnis GLENNY, J.R. und K. WEBER (1980): Calmodulin-binding proteins of microfilaments in isolated brush borders and microvilli of intestinal epithelial cells. J. Biol. Chem. 255, 10551-10554. GRAND, R.J., S.V. PERRY und R.A. WEEKS (1979): Troponin C-like proteins (calmodulins) from mammalian smooth muscle and other tissues. Biochem. J. 177(2), 521-529. HALL, G.A. und T.F. BYRNE (1989): Effects of age and diet on small intestinal structure and function in gnotobiotic piglets. Res. Vet. Sci. 47(3), 387-392. HAMPSON, D.J. (1986): Alteration in piglet small intestinal structure at weaning. Res. Vet. Sci. 40(1), 32-40. HARMEYER, J. und R. KAUNE (1988): Wie steuert Vitamin D-Hormon die intrazelluläre Calcium-Homöostase? Biologie in unserer Zeit 18, 110-115. HATTENHAUER, O.G. (1998): In-vitro Untersuchungen zur Ontogenese des intestinalen Phosphat-Transportes beim Schwein. Inaugural-Dissertation med. vet., Justus-Liebig-Universität Gießen. HENNING, U., J. WÜNSCHE, F. KREIENBRING, E. MEHNERT und U. HERRMANN (1988): Ernährungsphysiologische Untersuchungen an Schweinen mit Ileo-Rektal-Anastomosen. Arch. Anim. Nutr. 38, 585-601. HENRY, H.L. und A.W. NORMAN (1984): Vitamin D: metabolism and biological actions. Annu. Rev. Nutr. 4, 493-520. HENRY, H.L. und A.W. NORMAN (1991): Metabolism of vitamin D. In: Coe F.L., M.J. Favus (Hrsg): Disorders of bone and mineral metabolism. Raven Verlag, New York, 149-162. HIGUCHI, R., C. FOCKLER, G. DOLLINGER und R. WATSON (1993): Kinetic PCR analysis: real-time monitoring of DNA amplification reactions. Biotechnology 11(9), 1026-1030.
97

Literaturverzeichnis HOENDEROP, J.G.J., A.W.C.M. VAN DER KEMP, A. HARTOG, S.F.J. VAN DE GRAAF, C.H. VAN OS, P.H.G.M. WILLEMS und R.J.M. BINDELS (1999a): Molecular identification of the apical Ca2+channel in 1,25-dihydroxivitamin D3-responsive epithelia. J. Biol. Chem. 274(13), 8375-8378. HOENDEROP, J.G.J., A.W.C.M. VAN DER KEMP, A. HARTOG, C.H. VAN OS, P.H.G.M. WILLEMS und R.J.M. BINDELS (1999b): The epithelial calcium channel, ECaC, is activated by hyperpolarization and regulated by cytosolic calcium. Biochem. Biophys. Res. Commun. 261, 488-492. HOENDEROP, J.G.J., D. MÜLLER, M. SUZUKI, C.H. VAN OS und R.J.M. BINDELS (2000a): Epithelial calcium channel: gate-keeper of active calcium reabsorption. Curr. Opin. Nephrol. Hypertens. 9, 335-340. HOENDEROP, J.G.J., P.H.G.M. WILLEMS und R.J.M. BINDELS (2000b): Toward a comprehensive molecular model of active calcium reabsorption. Am. J. Physiol. Renal Physiol. 278, F352-F360. HOENDEROP, J.G.J, A. HARTOG, M. STRUIVER, A. DOUCET, P.H.G.M. WILLEMS und R.J.M. BINDELS (2000c): Localization of epithel Ca2+channel in rabbit kidney and intestine. J. Am. Soc. Nephrol. 11(7), 1171-1178. HOENDEROP, J.G.J., R. VENNEKENS, D. MÜLLER, J. PRENEN, G. DROOGMANS, R.J.M. BINDELS und B. NILIUS (2001): Function and expression of the epithelial Ca2+channel family: comparison of mammalian ECaC1 and 2. J. Physiol. 537(3), 747-761. HOPFER, U., K. NELSON, J. PEROTTO und K.J. ISSELBACHER (1973): Glucose transport in isolated brush border membrane from rat small intestine. J. Biol. Chem. 248(1), 25-32. HOPFER, U., K. SIGRIST-NELSON, E. AMMAN und H. MURER (1976): Differences in neutral amino acid and glucose transport between brush border and basolaterale plasma membrane of intestinal epithelial cells. J. Cell Physiol. 89(4), 805-810. ISHIBASHI, K., M. SUZUKI und M. IMAI (2000): Molecular cloning of a novel form (two-repeat) protein related to voltage-gated sodium and calcium channels. Biochem. Biophys. Res. Commun. 270(2), 370-376.
98

Literaturverzeichnis JOHNSON, J.A. und R. KUMAR (1994): Renal and intestinal calcium transport: roles of vitamin D and vitamin D-dependent calcium binding proteins. Sein. Nephrol. 14(2), 119-128. KASSIANOFF, I. (1989): Die Wirkung von Vitamin D auf die Calciumaufnahme im Dünndarm. Inaugural-Dissertation med. vet., Tierärztliche Hochschule Hannover. KAUNE, R. (1992): Die Regulation des intestinalen aktiven Calciumtransportes durch Calcitriol. Habilitationsschrift Tierärztliche Hochschule Hannover. KAUNE, R., B. SCHRÖDER und J. HARMEYER (1990): Binding properties of plasma vitamin D-binding protein and intestinal 1,25-dihydroxyvitamin D3 receptor in piglets with pseudo-vitamin D-deficiency rickets, type I: treatment effects with pharmacological doses of vitamin D3. Arch. Biochem. Biophys. 282(2), 326-332. KAUNE, R., I. KASSIANOFF, B. SCHRÖDER und J. HARMEYER (1992): The effects of 1,25-dihydroxyvitamin D3 deficiency on Ca2+-transport and Ca2+-uptake into brush-border membrane vesicles from pig small intestine. Biochemical et Biophysica Acta 1109, 187-194. KAWADE, M. (1964): An automatic method for determination of alkaline Phosphatase in serum using p-nitrophenylphosphat as substrate. Mie. Med. J. 14(1), 41-46. KELJO, D.J., R.J. MACLEOD, M.H. PERDUE, D.G. BUTLER und J.R. HAMILTON (1985): D-glucose transport in piglet jejunal brush-border membranes: insights from a disease model. Am. J. Physiol. 249, G751-G760. KELLY, D., J.A. SMYTH und K.J. MCCRACKEN (1990): Effect of creep feeding on structural and functional changes of the gut of early weaned pigs. Res. Vet. Sci. 48(3), 350-356. KEMP, D.J., D.B. SMITH, S.J. FOOTE, N. SAMARAS und M.G. PETERSON (1989): Colorimetric detection of specific DNA segments amplified by polymerase chain reactions. Proc. Natl. Acad. Sci. U S A 86(7), 2423-2427. KIRCHGESSNER, M. (1992): Mineral- und Wirkstoffe. Tierernährung, 10. Auflage DLG-Verlag, Frankfurt am Main.
99

Literaturverzeichnis KLEIN, C. (1999): Einfluß von PTHrP (Parathyroid Hormone related Peptid) in der Milch auf den intestinalen Calciumtransport bei Saugferkeln. Inaugural-Dissertation med. vet., Tierärztliche Hochschule Hannover. KLEMCKE, H.G., J.L. VALLET, R.K. CHRISTENSON und P.L. PEARSON (2001): Erythropoitin mRNA expression in pig embryos. Anim. Reprod. Sci. 66(1-2), 93-108. KÖMÜVES, L.G. und J.P. HEATH (1992): Uptake of maternal immunglobulins in the enterocytes of suckling piglets: improved detection with a streptavidin-biotin bridge gold technique. J. Histochem. Cytochem. 40(11), 1637-1646. KOLB, E. (1989): Lehrbuch der Physiologie der Haustiere Teil I, 5. Auflage Verlag Fischer, Stuttgart. KOLDOVSKY, O., P. SUNSHINE und N. KRETCHMER (1966): Cellular migration of intestinal epithelia in suckling and weaned rats. Nature 212(68), 1389-1390. KOVARIK, J. (1983): Zur Physiologie von Parathormon und Kalzitonin. Pädiatrie und Pädologie 18(2), 161-169. KREUTTER, D., D.C. LAFRENIERE und H. RASMUSSEN (1984): The role of fatty acids in the regulation of brush-border Ca2+ transport. Biochim. Biophys. Acta 777, 261-266. LABUHN, M. und C. BRACK (1997): Age-related changes in the mRNA expression of actin isoforms in Drosophila melanogaster. Gerontology 43(5), 261-267. LACHENMAIER-CURRLE, U. und J. HARMEYER (1988): Intestinal absorption of calcium in newborn piglets. Role of vitamin D. Biol. Neonate 53(6), 327-335. LARSEN, T. und B. SANDSTRÖM (1993): Effects of dietary calcium level on mineral and trace element utilization from a rapeseed (Brassica napus L.) diet fed to ileum-fistulated pigs. Br. J. Nutr. 69(1), 211-224.
100

Literaturverzeichnis LIANG, C.T., J.B. BARNES, R.A. BALAKIR und B. SACKOR (1986): Characterization of 1,25-dihydroxyvitamin D3-dependent calcium uptake in isolated chick duodenal cells. J. Membrane Biol. 90, 145-156. LOTTSPEICH, F. und H. ZORBAS (1998): Bioanalytic. Spektrum Akademischer Verlag, Heidelberg, Berlin. LYONS, J. (1992): The polymerase chain reaction and cancer diagnostics. Cancer 69 (6 Suppl.), 1527-1531. MADSEN, K.L., M.M. TAVERNINI, C. YACHIMEC, D.L. MENDRICK, P.J. ALFONSO, M. BUERGIN, H.S. OLSEN, M.J. ANTONACCIO, A.B. THOMSON und R.N. FEDORAK (1998): Stanniocalcin: a novel protein regulating calcium and phosphate transport across mammalian intestine. Am. J. Physiol. 274(1 Pt 1), G96-G102. MARTINSSON,K. und L. JÖNSSON (1976): The uptake of macromolecules in the ileum of piglets after intestinal “closure”. Zentralbl. Veterinärmed. A 23, 277-282. MCCANCE, R.A. (1974): The effect of age on the weights and lengths of pigs’ intestines. J. Anat. 117, 475-479. MERRILL, A.R., P. PROULX und A.G. SZABO (1986): Studies on calcium binding to brush-border membranes from rabbit small intestine. Biochim. Biophys. Acta 859, 237-245. MEYER, H. und J. KAMPHUES (1990): Grundlagen der Ernährung von Neugeborenen. In: Walser, K. und H. Bostedt: Neugeborenen- und Säuglingskunde der Tiere. 1. Auflage Enke Verlag, Stuttgart. MILLER, A. und F. BRONNER (1981): Calcium uptake in isolated brush-border vesicles from rat small intestine. Biochem. J. 196, 391-401. MILLER, B.G., T.J. NEWBY, C.R. STOKES, D.J. HAMPSON, P.J. BROWN und F.J. BOURNE (1984): The importance of dietary antigen in the cause of postweaning diarrhea in pigs. Am. J. Vet. Res. 45(9), 1730-1733.
101

Literaturverzeichnis MIRCHEFF, A.K. und E.M. WRIGHT (1976): Analytical isolation of plasma membranes of intestinal epithel cells: identification of Na, K-ATPase rich membranes and the distribution of enzyme activities. J. Membr. Biol. 28(4), 309-333. MONTELL, C., L. BIRNBAUMER, V. FLOCKERZI, R.J. BINDELS, E.A. BRUFORD, M.J. CATERINA, D.E. CLAPHAM, C. HARTENECK, S. HELLER, D. JULIUS, I. KOJIMA, Y. MORI, R. PENNER, D. PRAWITT, A.M. SCHARENBERG, G. SCHULTZ, N. SHIMIZU und M.X. ZHU (2002): A unified nomenclature for the superfamiliy of TRP cation channels. Molecular Cell 9, 229-231. MOON, H.W. (1971): Epithelial cell migration in the alimentary mucosa of the suckling pig. Proc. Soc. Exp. Biol. Med. 137(1), 151-154. MOON, H.W. (1972): Vacuolated villous epithelium of small intestine of young pigs. Vet. Pathol. 9(1), 3-21. MORIN, R.J. (1980): The role of phospholipids in platetet function. Ann. Clin. Lab. Sci. 10(6), 463-473. MORRISON, T.B., J.J. WEIS und C.T. WITTWER (1998): Quantification of low-copy-transcripts by continuous SYBR® Green I monitoring during amplification. Biotechniques 24, 954-958, 960,962. MOSSA, A.C., L.M. GOVEN und J.G. ATWOOD (1991): Thermal cycler for automatic performance of the PCR with close temperature control. Eur. Pat. Appl. No 0488768 MUALLEM, S. (1989): Calcium transport pathways of pancreatic acinar cells. Annu. Rev. Physiol. 51, 83-105. MUELHARDT, C. (2002): Der Experimentator: Molekularbiologie. 3. Auflage Verlag Fischer, Stuttgart. MÜLLER, D., J.G.J. HOENDEROP, G.F.M. MERKX, C.H. VAN OS und R.J.M. BINDELS (2000a): Gene structure and chromosomal mapping of human epithelial calcium channel. Biochem. Biophys. Res. Commun. 275, 47-52.
102

Literaturverzeichnis MÜLLER, D., J.G.J. HOENDEROP, I.C. MEIJ, L.P.J. VAN DEN HEUVEL, N.V.A.M. KNOERS, A.I. DEN HOLLANDER, P. EGGERT, V. GARCIA-NIETO, F. CLAVERIE-MARTIN und R.J.M. BINDELS (2000b): Molecular cloning, tissue distribution and chromosomal mapping of the human epithelial Ca2+channel (ECaC1). Genomics 67, 48-53. MULLIS, K.B. und F.A. FALOONA (1987): Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods in Enzymology 155, 335-350. MURER, H., P. GMAJ, B. STIEGER und B. HAGENBUCH (1989): Transport studies with renal proximal tubular and small intestinal brush border and basolateral membrane vesicles: vesicle heterogeneity, coexistence of transport systems. Methods in Enzymology 172, 346-364. MYERS, T.W. und D.H. GELFAND (1991): Reverse transcription and DNA amplification by a Thermus thermophilus DNA polymerase. Biochemistry 30(31), 7661-7666. NASSAR, C.F., E.W. KHAIRALLAH, M.G. NASSAR und Z.M. HABBAL (1988): Cytoskeletal control of calcium absorption across the rat small intestine. Comp. Biochem. Physiol. 89A, 197-201. NEDELMAN, J., P. HEAGERTY und C. LAWRENCE (1992): Quantitative PCR with internal controls. Comput. Appl. Biosci. 8(1), 65-70. NELLANS, H.N. (1990): Intestinal calcium absorption. Interplay of paracellular and cellular pathway. Miner. Electrol. Metab. 16, 101-108. NEMERE, I., Y. YOSHIMOTO und A.W. NORMAN (1984): Calcium transport in perfused duodena from normal chicks: enhancement within fourteen minutes of exposure to 1,25-dihydroxyvitamin D3. Endocrinology 115(4), 1476-1483. NEMERE, I., V. LEATHERS und A.W. NORMAN (1986): 1,25-dihydroxivitamin D3-mediated intestinal calcium transport. Biochemical identification of lysosomes containing calcium and calcium-binding protein (calbindin-D28k). J. Biol. Chem. 261, 16106-16114. NEMERE, I. und A.W. NORMAN (1988): 1,25-dihydroxyvitamin D3-mediated vesicular transport of calcium in intestine: time course-studies. Endocrinology 122(6), 2962-2969.
103

Literaturverzeichnis NEMERE, I. und A.W. NORMAN (1990): Transcaltachia, vesicular calcium transport, and microtubule-associated calbindin-D28k: emerging views of 1,25-dihydroxyvitamin D3-mediated intestinal calcium absorption. Miner. Electrolyte Metab. 16, 109-114. NEMERE, I. und A.W. NORMAN (1991): Transport of Calcium. In: S. G. Schulz: Handbook of Physiology, section 6, The Gastrointestinal System, Band IV, Intestinal Absorption and Secretion, Am. Physiological Society, Bethesda, MD, USA, 337-360. NEMERE, I. und M.C. FARACH-CARSON (1998): Membrane receptors for steroid hormones: a case for specific cell surface binding sites for vitamin D metabolites and estrogens. Biochem. Biophys. Res. Commun. 248(3), 443-449. NEMERE, I., D. LARSSON und K. SUNDELL (2000): A specific binding moiety for 1,25-dihydroxyvitamin D3 in basal lateral membranes of carp enterocytes. Am. J. Physiol. Endocrinol. Metab. 279, E614-E621. NIEMEYER, B.A., C. BERGS, U. WISSENBACH, V. FLOCKERZI und C. TROST (2001): Competitive regulation of CaT-like-mediated Ca2+ entry by protein kinase C and calmodulin. Proc. Natl. Acad. Sci. USA 98(6), 3600-3605. NILIUS, B., R. VENNEKENS, J. PRENEN, J.G.J. HOENDEROP, R.J.M. BINDELS und G. DROOGMANS (2000): Whole-cell and single channel monovalent cation currents through the novel rabbit epithelial Ca2+channel ECaC. J. Physiol. 527, 239-248. NILIUS, B., J. PRENEN, R. VENNEKENS, J.G. HOENDEROP, R.J. BINDELS und G. DROOGMANS (2001a): Modulation of the epithelial calcium channel, ECaC, by intracellular Ca2+. Cell calcium 29(6), 417-428. NILIUS, B., J. PRENEN, R. VENNEKENS, J.G. HOENDEROP, R.J. BINDELS und G. DROOGMANS (2001b): Pharmacological modulation of monovalent cation currents through the epithelial Ca2+-channel ECaC1. Br. J. Pharmacol. 134(3), 453-462. NORMAN, A.W. (1987): Studies on the vitamin D endocrine system in the avian. J. Nutr. 117, 797-807.
104

Literaturverzeichnis NORMAN, A.W. und F.P. ROSS (1979): Vitamin D seco-steroids: unique molecules with both hormone and possible membranophilic properties. Life Sci. 24(9), 759-769. NORMAN, A.W., X. SONG, L. ZANELLO, C. BULA und W.H. OKAMURA (1999): Rapid and genomic response are mediated by different shapes of agonist steroid hormone, 1,25(OH)2vitaminD3. Steroids 64, 120-128. OLTJEN, R.T., J.P. FONTENOT, C.B. AMMERMAN, C.E. COPPOCK, P.W. MOE, Q.R. ROGERS, G.L. RUMSEY, V.C. SPEER und J.D. SUMMERS (1979): Nutrient requirements of swine. Eight revised edition. Natl. Research Council Publ. No 2, 23. OS VAN, C.H. (1987): Transcellular calcium transport in intestinal and renal epithelial cells. Biochim. Biophys. Acta 906, 195-222. OUE, T., Y. TAIRA, H. SHIMA, E. MIYAZAKI und P. PURI (1999): Effect of antenatal glucocorticoid administration on insulin-like growth factor I and II levels in hypoplastic lung in nitrofen-induced congenital diaphragmatic hernia in rats. Pediatr. Surg. Int. 15, 175-179. PAHUJA, D.N. und H.F. DELUCA (1981): Stimulation of intestinal calcium transport and bone calcium mobilization by prolactin in vitamin D-deficient rats. Science 214, 1038-1039. PANSU, D., C. BELLATON und F. BRONNER (1981): Effect of Ca intake on saturable and nonsaturable components of duodenal Ca transport. Am. J. Physiol. 240(1), G32-G37. PANSU, D., C. BELLATON und F. BRONNER (1983a): Developmental changes in the mechanisms of duodenal calcium transport in the rat. Am. J. Physiol. 244(2), G20-G26. PANSU, D., C. BELLATON, C. ROCHE und F. BRONNER (1983b): Duodenal and ileal calcium absorption in the rat and effects of vitamin D. Am. J. Physiol. 244(2), G695-G700. PARTRIDGE, I.G. (1978): Studies on digestion and absorption in the intestines of growing pigs. Br. J. Nutr. 39, 527-537.
105

Literaturverzeichnis PECHET, M.M., E. BOBADILLA, E.L. CRROL und R.H. HESSE (1967): Regulation of bone resorption and formation. Influences of thyrocalcitonin, parathyroid hormone, neutral phosphate and vitamin D3. Am. J. Med. 43(5), 696-710. PEIRCE, S.K. und W.Y. CHEN (2001): Quantification of prolactin receptor mRNA in multiple human tissues and cancer cell lines by real time RT-PCR. PENG, J.B., X.Z. CHEN, U.V. BERGER, P.M. VASSILEV, H. TSUKAGUCHI, E.M. BROWN und M.A. HEDIGER (1999): Molecular cloning and characterization of a channel-like transporter mediating intestinal calcium absorption. J. Biol. Chem. 274(32), 22739-22746. PENG, J.B., X.Z. CHEN, U.V. BERGER, S. WEREMOWICZ, C.C. MORTON, P.M. VASSILEV, E.M. BROWN und M.A. HEDIGER (2000a): Human calcium transport protein CaT1. Biochem. Biophys. Res. Commun. 287(2), 326-332. PENG, J.B., X.Z. CHEN, U.V. BERGER, P.M. VASSILEV, E.M. BROWN und M.A. HEDIGER (2000b): A rat kidney-specific calcium transporter in the distal nephron. J. Biol. Chem. 275(36), 28186-28194. PENG, J.B., E.M. BROWN und M.A. HEDIGER (2001): Structural conservation of the genes encoding CaT1, CaT2 and related cation channels. Genomics 76, 99-109. PFAFFL, M.W. (2001): A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Research 29(9), 2002-2007. PUCHAL, A.A. und R.K. BUDDINGTON (1992): Postnatal development of monosaccharide transport in pig intestine. Am. J. Physiol. 262, G895-G902. RADDE, I.C.,D. DAVIS, J. SHEEPERS und H.G. MCKERCHER (1980): Bidirectional transmucosal 45Ca and 32P fluxes across the small intestine of young piglet: Relationship to intestinal Ca2+-Mg2+-ATPase activity and postnatal age. In: DeLuca, H.F. and Anast, C.S. (eds); Pediatric Diseases related to Calcium. Blachwell Cientific Publ., Oxford, S. 153-163.
106

Literaturverzeichnis RASMUSSEN, H., O. FONTAINE, E.E. MAX und D.B.P. GOODMAN (1979): The effect of 1α-hydroxyvitamin D3 administration on calcium transport in chick intestine brush border membrane vesicles. J. Biol. Chem. 254, 2993-2999. REMICK, D.G., S.L. KUNKEL, E.A. HOLBROOK und C.A. HANSON (1990): Theory and applications of the polymerase chain reaction. Am. J. Clin. Pathol. 93(4 Suppl.), S49-S54. RÜBELT, A.C. (1995): Untersuchungen zur Phosphataufnahme durch die intestinale Bürstensaummembran von Ziegen. Inaugural-Dissertation med. vet., Justus-Liebig-Universität Gießen. RYCHLIK, W., W.J. SPENCER und R.E. RHOADS (1990): Optimization of the annealing temperature for DNA amplification in vitro. Nucleic Acids Res. 18(21), 6409-6412. SAIKI, R.K., D.H. GELFAND, S. STOFFEL, S.J. SCHARF, R. HIGUCHI, G.T. HORN, K.B. MULIS und H.A. ERLICH (1988): Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 239 (4839), 487-491. SAUER, W.C., A. JUST und H. JORGENSEN (1982): The influence of daily intake on the apparent digestibility of crude protein, amino acids, calcium and phosphorus at the terminal ileum and overall in pigs. Z. Tierphysiol., Tierernährg. u. Futtermittelkde. 48, 177-182. SCHEDL, H.P. und H.D. WILSON (1985): Calcium uptake by intestinal brush border membrane vesicles. Comparison with in vivo calcium transport. J. Clin. Invest. 76, 1871-1878. SCHOENMAKERS, T.J.M., G.J. VISSER, G. FLIK und A.P.R. THEUVENET (1992): Chelator: An improved method for computing metal ion concentrations in physiological solutions. Biotechniques 12(6), 870-879. SCHRÖDER, B. (1996): Vergleichende Physiologie der gastrointestinalen Calcium- und Phosphatabsorption bei Schweinen und kleinen Wiederkäuern. Habilitationsschrift Justus-Liebig-Universität Gießen.
107

Literaturverzeichnis SCHRÖDER, B., R. KAUNE und J. HARMEYER (1990): Studies of porcine intestinal calcitriol receptor in pseudo-vitamin D deficiency rickets type I. Clin. Sci. 79, 409-414. SCHRÖDER, B., R. KAUNE, C. SCHLUMBOHM, G. BREVES und J. HARMEYER (1993): Evidence for vitamin D-independent active calcium absorption in newborn piglets. Calcif. Tissue Int. 52, 305-309. SCHRÖDER, B., C. SCHLUMBOHM, R. KAUNE und G. BREVES (1996): Role of calbindin-D9k in buffering cytosolic free Ca2+ ions in pig duodenal enterocytes. J. Physiol. London 492, 715-722. SCHRÖDER, B., M.R. DAHL und G. BREVES (1998a): Duodenal Ca2+ absorption is not stimulated by calcitriol during early postnatal development of pigs. Am. J. Physiol. 275, G305-G313. SCHRÖDER, B., M.R. DAHL und G. BREVES (1998b): Zur Rolle des Cytoskeletts bei der intestinalen Ca2+-Absorption in der frühen postnatalen Lebensphase. Z. Gastroenterol Suppl.1, 44-48. SCHRÖDER, B., O. HATTENHAUER und G. BREVES (1998c): Phosphate transport in pig proximal small intestines during postnatal development: lack of modulation by calcitriol. Endocrinology 139(4), 1500-1506. SCHWARTZ, Z., V.L. SYLVIA, D. LARSSON, I. NEMERE, D. CASASOLA, D.D. DEAN und B.D. BOYAN (2002): 1α,25(OH)2D3 regulates chondrocyte matrix vesicle protein kinase C (PKC) directly via G-protein-dependent mechanisms and indirectly via incorporation of PKC during matrix vesicle biogenesis. J. Biol. Chem. 277(14), 11828-11837. SHIRAZI-BEECHEY, S.P., R.B. BEECHEY, J. PENNY, S. VAYRO, W. BUCHAN und D. SCOTT (1991): Mechanisms of phosphate transport in sheep intestine and parotid gland: response to variation in dietary phosphate supply. Exp. Physiol. 76(2), 231-241. SHULTZ, T.D., S. BOLLMAN und R. KUMAR (1982): Decreased intestinal calcium absorption in vitro and normal brush border membrane vesicle calcium uptake in cortisol-treated chickens: evidence for dissociation of calcium absorption from brush border vesicle uptake. Proc. Natl. Acad. Sci. USA 79, 3542-3546.
108

Literaturverzeichnis SLEPCHENKO, B.M. und F. BRONNER (2001): Modeling of transcellular Ca transport in rat duodenum points to coexistence of two mechanisms of apical entry. Am. J. Physiol. Cell. Physiol. 281, C270-C281. SOUTHERN; E.M. (1975): Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98(3), 503-517. SUZUKI, M., K. ISHIBASHI, G. OOKI, S. TSURUOKA und M. IMAI (2000): Electrophysiologic characteristics of the ca-permeable channels, ECaC and CaT, in the kidney. Biochem. Biophys. Res. Commun. 274, 344-349. TAYLOR, A.N. und R.H. WASSERMAN (1967): Vitamin D3-induced calcium binding protein: partial purification, electrophoretic visualisation and tissue distribution. Arch. Biochem. Biophys. 119, 536-540. TIMMERMANS, J.A.H., R. KAUNE, R.J.M. BINDELS und C.H. VAN OS (1991): Quantification of Ca2+-ATPases in porcine duodenum. Effects of 1,25(OH)2D3 deficiency. Biochim. Biophys. Acta 1065, 177-184. TINDALL, K.R. und T.A. KUNKEL (1988): Fidelity of DNA synthesis by Thermus aquaticus DNA polymerase. Biochemistry 27(16), 6008-6013. VENNEKENS, R., J.G.J. HOENDEROP, J. PRENEN, M. STRIVER, P.H.G.M. WILLEMS, G. DROOGMANS, B. NILIUS und R.J.M. BINDELS (2000): Permeation and gatting properties of the novel epithelial Ca2+channel. J. Biol. Chem. 275(6), 3963-3969. WALTER, F.C. (1999): Untersuchungen zur funktionellen und strukturellen Charakterisierung renaler Phosphattransportsysteme bei kleinen Wiederkäuern. Inaugural-Dissertation zur Erlangung des Doktorgrades im Fachbereich Agrarwissenschaften, Ökotrophologie und Umweltmanagement der Justus-Liebig-Universität Gießen. WANG, C.H. und A.L. KOCH (1978): Constancy of growth on simple and complex media. J. Bacteriol.. 136(3), 969-975. WANG, A.M., M.V. DOYLE und D.F. MARK (1989): Quantitation of mRNA by the polymerase chain reaction. Proc. Natl. Acad. Sci. U S A 86, 9717-9721.
109

Literaturverzeichnis WAREMBOURG, M., C. PERRET und M. THOMASSET (1986): Distribution of vitamin D-dependent calcium binding protein messenger ribonucleic acid in rat placenta and duodenum. Endocrinology 119(1), 176-184. WASSERMAN, R. H., A.N. TAYLOR und F.A. KALLFELZ (1966): Vitamin D and transfer of plasma calcium to intestinal lumen in chicks and rats. Am. J. Physiol. 211(2), 419-423. WASSERMAN, R.H. und A.N. TAYLOR (1966): Vitamin D3-induced calcium-binding protein in chick intestinal mucosa. Science 152, 791-793. WASSERMAN, R.H., C.A. SMITH, M.E. BRINDAK, N. DE TALAMONI, C.S. FULLMER, J.T. PENNISTON und R. KUMAR (1992a): Vitamin D and mineral deficiencies increase the plasma membrane calcium pump of chicken intestine. Gastroenterology 102(3), 886-894. WASSERMAN, R.H., J.S. CHANDLER, S.A. MEYER, C.A. SMITH, M.E. BRINDAK, C.S. FULLMER, J.T. PENNISTON und R. KUMAR (1992b): Intestinal calcium transport and calcium extrusion processes at the basolateral membrane. J. Nutr. 122(Suppl. 3), 662-671. WEBER, K., R.G. ERBEN, A. RUMP und J. ADAMSKI (2001): Gene structure and regulation of murine epithelial calcium channels ECaC1 and 2. Biochem. Biophys. Res. Commun. 289(5), 1287-1294. WILSON, P.W. und D.E.M. LAWSON (1980): Calcium binding activity by chick intestinal brush-border membrane vesicles. Europ. J. Physiol. Pflügers Arch. 389, 69-74. WILSON, H.D., H.P. SCHEDL und K. CHRISTENSEN (1989): Calcium uptake by brush-border membrane vesicles from rat intestine. Am. J. Physiol. 257, F447-F453. WISSENBACH, U., B.A. NIEMEYER, T. FIXEMER, A. SCHNEIDEWIND, C. TROST, A. CAVALIÉ, K. REUS, E. MEESE, H. BONKHOFF und V. FLOCKERZI (2001): Expression of CaT-like, a novel calcium-selective channel, correlates with the malignancy of prostate cancer. J. Biol. Chem. 276, 19461-19468.
110

Literaturverzeichnis WOO, T.H., B.K. PATEL, M. CINCO, L.D. SMYTHE, M.L. SYMONDS, M.A. NORRIS und M.F. DOHNT (1998): Real-time homogeneous assay of rapid cycle polymerase chain reaction product for identification of Leptonema illini. Anal. Biochem. 259(1), 112-117. WOOD, R.J., L. TCHACK und S. TAPARIA (2001): 1,25-dihydroxyvitamin D3 increases the expression of the CaT1 epithelial calcium channel in the Caco-2 human intestinal cell line. BMC Physiol. 1(1), 1-11. XU, R.J., D.J. MELLOR, P. TUNGTHANATHNICH, M.J. BIRTLES, G.W. REYNOLDS und H.V. SIMPSON (1992): Growth and morphological changes in the small and large intestine in piglets during the first three days after birth. J. Dev. Physiol. 18(4), 161-172. YUE, L., J.B. PENG, M.A. HEDIGER und D.E. CLAPHAM (2001): CaT1 manifests the pore properties of the calcium-release-activated calcium channel. Nature 410, 705-509. ZHANG, H., C. MALO und R.K. BUDDINGTON (1997): Suckling induces rapid intestinal growth and changes in brush border digestive functions of newborn pigs. J. Nutr. 127(3), 418-426. ZHANG, H., C. MALO, C.R. BOYLE und R.K. BUDDINGTON (1998): Diet influences development of the pig (sus scrofa) intestine during the first 6 hours after birth. J. Nutr. 128(8), 1302-1310. ZUO, Q., D. CLAVEAU, G. HILAL, M. LECLERC und M.G. BRUNETTE (1997): Effect of calcitonin on calcium transport by the luminal and basolateral membranes of the rabbit nephron. Kidney int. 51, 1991-1999.
111

Hannover, den 25.August 2002
ERKLÄRUNG Hiermit erkläre ich, dass ich die Dissertation mit dem Titel: „Expression der Ca2+-Kanäle ECaC1 und ECaC2 im Dünndarm von Saug- und Absetzferkeln“ selbständig verfasst habe. Bei der Anfertigung wurden folgende Hilfen Dritter in Anspruch genommen: keine. Ich habe keine entgeltliche Hilfe von Vermittlungs- bzw. Beratungsdiensten (Promotionsberater oder anderen Personen) in Anspruch genommen. Niemand hat von mir unmittelbar oder mittelbar entgeltliche Leistungen für Arbeiten erhalten, die im Zusammenhang mit dem Inhalt der vorgelegten Dissertation stehen. Ich habe die Dissertation am Physiologischen Institut der Tierärztlichen Hochschule Hannover angefertigt. Die Dissertation wurde bisher nicht für eine Prüfung oder Promotion oder für einen ähnlichen Zweck zur Beurteilung eingereicht. Ich versichere, dass ich die vorstehenden Angaben nach bestem Wissen vollständig und der Wahrheit entsprechend vorgenommen habe. ________________________
Thomas Hinterding

Danksagung Ich habe die Danksagung am Ende einer Dissertation immer für die Pflichterfüllung des
jeweiligen Doktoranden gegenüber den Angehörigen des Institutes, Freunden und
Verwandten angesehen. Heute weiss ich, dass sie wirklich Herzenssache ist aus Dankbarkeit
zum Überstehen einer schwierigen Lebensphase. Durch diese Zeit haben mich sehr viele
Menschen begleitet, von denen ich an dieser Stelle einige besonders hervorheben möchte.
Zuerst möchte ich mich bei Prof. Dr. G. Breves für die freundliche Aufnahme in der
Arbeitsgruppe bedanken.
In einem ganz besonderem Maße möchte ich mich bei Prof. Dr. Bernd Schröder für die
Überlassung des Themas und die nette Betreuung bedanken. Er hat sich immer die Zeit
genommen, Fragen und Probleme zu diskutieren und hat mich mit unzähligen hilfreichen
Ratschlägen immer ein Stück weiter gebracht auf dem Weg, diese Dissertation zu erstellen.
Bedanken möchte ich mich auch bei Frau Dr. Korinna Huber, die mich in die Geheimnisse
der molekularbiologischen Techniken eingeführt hat und unter deren kritischen Blicken ich
gelernt habe, wissenschaftlich zu denken.
Frau Becker, Christina, Marion, Katrin und Kerstin danke ich für die gute Zusammenarbeit im
Labor und die Unterstützung, die ich durch sie erhalten habe.
Bedanken möchte ich mich natürlich auch bei den anderen Doktoranden (Alexandra, Eva,
Hakan, Magnus, Julia, Uta), mit denen ich in der ganzen Zeit sehr viel Spaß hatte.
Bei Yvonne, Michael und Maike möchte ich mich dafür bedanken, dass ich auch im Institut
dem Institut entkommen konnte, wenn es nötig war.
René J. M. Bindels und Joost G. J. Hoenderop danke ich ebenso wie ihrer gesamten
Arbeitsgruppe für die freundliche Aufnahme und die fachliche Unterstützung.
Mein Dank gilt auch dem Institut für Parasitologie, insbesondere Prof. Dr. G. von Samson,
dessen technische Ausrüstung ich nutzen durfte.
Der H. Wilhelm Schaumann Stiftung danke ich für die finanzielle Unterstützung.
Nicht zu letzt, aber an besonders exponierter Stelle, danke ich Corinna --- für alles! ;-)