Embed Size (px)
Citation preview

Psychopharmacology (1991 ) 103:17~176 003331589100006S Psychopharmacology
© Springer-Verlag 1991
9 •
Facilitory effect of A -tetrahydrocannablnol induced feeding
on hypothalamically
Weronika Trojniar* and Roy A. Wise
Center for Studies in Behavioral Neurobiology, Department of Psychology, Concordia University, Montreal, Quebec, Canada
Received May 23, 1990 / Final version August 20, 1990
Abstract. Six male Lewis rats were tested for the effect of Ag-tetrahydrocannabinol (A 9 THC) on feeding evoked by electrical stimulation of the lateral hypothalamus. Treatment with A9-THC (0.4 mg/kg IP) decreased fre- quency threshold for feeding by 20.5% (_+4.3), causing a leftward shift in the function relating stimulation fre- quency to the latency to begin eating 45-mg food pellets upon stimulation onset; there was no change in the asymptotic performance that was approached with suf- ficiently high stimulation frequencies. Naloxone (1 and 2 mg/kg) reduced the facilitory effect of A9-THC, but did so at doses that can inhibit feeding in the no-drug con- dition. These data are consistent with evidence implicat- ing endogenous opioids in feeding, and suggest (but do not confirm) that the facilitation of feeding by A9-THC may be mediated by endogenous opioids. The facilitation of stimulation-induced feeding by doses of A9-THC that have been found to facilitate brain stimulation reward is consistent with evidence suggesting common elements in the brain mechanisms of these two behavioral effects of medial forebrain bundle stimulation.
Key words: A9-tetrahydrocannabinol- A9-THC Feed- ing -Naloxone - Opioids
The main psychoactive component of marijuana and hashish is A9-tetrahydrocannabinol (Ag-THC). While this substance is of interest because of its potential for abuse, the mechanism of action of A9-THC is still un- clear (Martin 1986). It is only recently that specific bind- ing sites for A 9 THC have been identified in the central
* Present address." Department of Animal Physiology, University of Gdansk, Gdansk, Poland Offprint requests to: R.A. Wise, Department of Psychology, 1455 de Maisonneuve Boulevard West, Montreal, Quebec, Canada H3G 1 M8
nervous system (Devane et al. 1988; Herkenham et al. 1988), and little is known about the functional circuitry within which these binding sites are localized. It is of theoretical interest to know whether Ag-THC shares some properties and perhaps has an overlapping mech- anism of action - with other, better studied habit-form- ing drugs (Wise and Bozarth 1987).
While it has been difficult to demonstrate direct re- warding actions of A9THC in lower animals, Gardner et al. (1988a, b, 1989) have recently shown that Ag-THC can facilitate medial forebrain bundle brain stimulation reward. Most drugs of abuse have similar effects (Kor- netsky et al. 1979; Wise 1980; Wise and Bozarth 1987). Amphetamine (Broekkamp et al. 1975; Colle and Wise 1988b) and morphine (Broekkamp et al. 1976; Rompr6 and Wise 1989) have such effects when centrally injected at sites and doses that are known to be rewarding in their own right (Phillips and LePiane 1980; Bozarth and Wise 1981; van der Kooy et al. 1982; Carr and White 1983; Hoebet et al. 1983). Thus it has been suggested that the reward-facilitating effects of drugs of abuse reflect summation of the rewarding actions of the drugs with the rewarding actions of the brain stimulation, and that a common mechanism mediates the rewarding effects of the drugs and the stimulation (Stein and Wise 1973; Kornetsky et al. 1979; Wise 1980, 1988; Wise and Bo- zarth 1987). In this context, the findings of Gardner et al. (1988a, b, 1989) offer an animal model of habit-form- ing actions of A9-THC. Because the effect of A9-THC in this model was reversed by naloxone at doses that did not affect self-stimulation in their own right, Gardner et al. (1988a, b, 1989) have suggested the possible involvement of an opioid mechanism in the euphorigenic action of A9-THC.
Lateral hypothalamic brain stimulation not only in- duces rewarding effects; it also induces feeding in sated animals (Wise 1974). A common medial forebrain bundle substrate is thought to be involved in self-stimulation and stimulation-induced feeding (Glickman and Schiff 1967; Hoebel 1969; Wise and Bozarth 1987; Wise 1988). Several habit-forming and reward-facilitating drugs

173
s t imulate feeding, inc lud ing morph ine , barbi tura tes , ben- zodiazepines (Wise and Bozarth 1987; Wise 1988), and even, unde r some condi t ions , amphe t amine (Ho l t zman 1974; Dobrzansk i 1976; Blundel l and L a t h a m 1980). Mos t of these drugs have been shown to facilitate s t imu- l a t ion- induced as well as depr iva t ion- induced or spon- taneous feeding (Soper and Wise 1971; Jenck et al. 1986b, 1987b; Colle and Wise 1988a).
The effects of A 9 - T H C on s t imula t ion- induced feed- ing are par t icular ly interesting, because if A 9 - T H C facili- tates the b ra in mechan i sms of na tu ra l re inforcement processes the mechan i sms of food re inforcement are am- ong the mos t likely to be facilitated. S t imula t ion of appe- tite is a wel l -known effect of m a r i j u a n a and hashish; appeti te s t imula t ing effects of m a r i j u a n a smoke or A 9 - T H C inject ion in h u m a n s have been described in n u m e r o u s anecdota l reports (Siler et al. 1933; Al len tuck and B o w m a n 1942; Tar t 1970; Haines and Green 1970; Hal ikas et al. 1971), l abora to ry studies (Hollister 1971; Greenberg et al. 1976; Fo l t in et al. 1986, 1988), and clinical invest igat ions (Noyes et al. 1976; Regulson et al. 1976; Gross et al. 1983). The present s tudy was designed to determine if A g - T H C would facilitate s t imula t ion- induced feeding, and, if so, whether the effect would be a t t enua ted by doses of na loxone that a t tenua te this agent ' s reward-faci l i ta t ing effects (Ga rdne r et al. 1989).
Materials and methods
Animals and surgery. Male Lewis rats weighing approximately 400 g at the time of surgery were used. They were housed in individual cages with free access to food and water under a normal 12 h light-12 h dark illumination cycle. Ten rats were implanted, under pentobarbital anesthesia (60 mg/kg, IP), with bilateral, monopolar, stainless steel electrodes (254 gm diameter) insulated with Formvar, except for the square-cut tip. Pellegrino et al. (1979) coordinates were: 0.8 mm posterior to bregma, 1.7 mm lateral to the midline, and 8.3-8.8 mm ventral to the dural surface. The exact depth of each placement was determined on the basis of the effects of brain stimulation during surgery. Trains of cathodal square-wave con- stant current pulses (0.1 ms pulses at 100-300 gA and 60-100 Hz) were delivered as the electrodes were lowered through the brain by 0.2-0.5 mm steps, starting from 7.0 mm below dura. Animals were observed for sniffing responses (nose and vibrissae movements ac- companied by an increase in respiration rate). Each electrode was fixed at the locus of the most vigorous stimulation-induced sniffing. The electrodes were anchored to four stainless steel skull screws with dental acrylic; a stainless steel wire wrapped around two of the screws served as the anode for electrical stimulation.
Behavioral tests. After a 1-week recovery period, the rats were screened for stimulation-induced feeding. The testing was carried out in a 250 x 350 mm box with 45-rag food pellets covering the floor. The rats were taken from their home cages, where they had free access to food, and were allowed to explore the test box for 30 min before testing to allow for habituation and complete satiation. Trains of square-wave, constant current, 0.1 ms duration cathodal pulses were conducted from the stimulator to the electrode by way of a mercury commutator and flexible wire leads. Pulse duration, pulse frequency and stimulation intensity were monitored by oscil- loscope. Screening was carried out using a fixed stimulation fre- quency of 50 Hz; current intensity was raised incrementally in 20-s trials until forward search, sniffing, and eventually eating were observed. Stimulation through each electrode was tested in a separate block of trials; the one through which stimulation induced
more reliable eating was chosen for further experiment. For each rat a stimulation intensity was determined which would, at a stimu- lation frequency of 50 Hz, induce feeding with a mean latency of 10 s; the range of such frequencies was 80-400 gA. Once deter- mined, this stimulation intensity was used for all subsequent tests. Animals that did not show signs of stimulation-induced eating within 10 days of daily screening were discarded.
Six rats that ate in response to stimulation were next tested in a latency paradigm, where frequency of stimulation was varied from trial to trial. Latencies to eat were measured for each 30-s trial; stimulation was maintained for 30 s or until 5 s after the animal began to eat. Rest time of 20 s was given between trials. Four blocks of trials were given each day; stimulation frequency was progres- sively increased in the first and third blocks and decreased in the second and fourth. The between-trial increments in stimulation frequency were 5% of each previous value. The range of tested frequencies was from 18 to 50 Hz in control conditions and was adjusted as required under drug conditions. A total of 18 stimula- tion frequencies was tested per block; each block of trials took about 10 rain to complete. The four tests were averaged to obtain a daily mean latency at each stimulation frequency. Once stimula- tion-induced feeding stabilized, the animals were tested, on separate days, under drugs or drug vehicles.
Feeding threshold was defined as the stimulation frequency at which an animal began to eat with a latency of 20 s. Threshoid was derived from each rat's latency-frequency function by a method of linear interpolation.
Drugs. All drugs were administered by IP injection. The A 9 THC (DHHS, N1DA, Research Triangle Institute) was dissolved by the method of Fenimore and Loy (1971). First, A9THC was dissoived in a 20% solution of polyvinylpyrrolidone (PVP) (Sigma) prepared with ethanol. This solution was evaporated under nitrogen and the concentrated A9-THC-PVP complex was then suspended in 0.9% NaCI at a A 9 THC concentration of 1 mg/ml. A vehicle solution of 20% PVP in 0.9% NaC1 was tested on separate days (injection volumes and other parameters identical to the A9-THC injections). Vehicle and Ag-THC were each administered 15 rain before behav- ioral testing.
The animals were first tested with A9-THC at a dose of 1.5 mg/kg, the dose that Gardner et al. (1988a, b) found to signifi- cantly increase the sensitivity of their animals to rewarding brain stimulation. This dose caused a syndrome of hyper-reflexia (hyper- reactivity to environmental sounds) and catalepsy in our animals, and we subsequently tested lower doses. A dose of 0.4 mg/kg was found to be effective, both in the animals tested for stimulation- induced feeding and in other rats (of both Lewis and Long-Evans strains) tested for self-stimulation; this dose was used for subse- quent testing.
Each rat was tested once or twice under A9-THC and 1-4 times under saline and PVP. Injection days were separated by 2 days of control testing (no injections). The effect of A9-THC on stimulation induced feeding was challenged with 1 and 2 mg/kg naloxone (NAL) administered 10 min before A9-THC injection. On separate test days the naloxone effect on stimulation-induced feeding was also evaluated under vehicle injections.
Histology. After completion of behavioral testing, the animals were intracardially perfused with saline followed by 10% formalin solu- tion. The brains were removed from the skulls and stored in 10% formalin solution. The brains were frozen and sectioned at 40 lam to determine the electrode placements.
Results
St imula t ion- induced feeding was facili tated by A g - T H C ; the drug caused a leftward shift of the func t ion relat ing feeding latency to s t imula t ion frequency, decreasing the a m o u n t of s t imula t ion required to produce responding
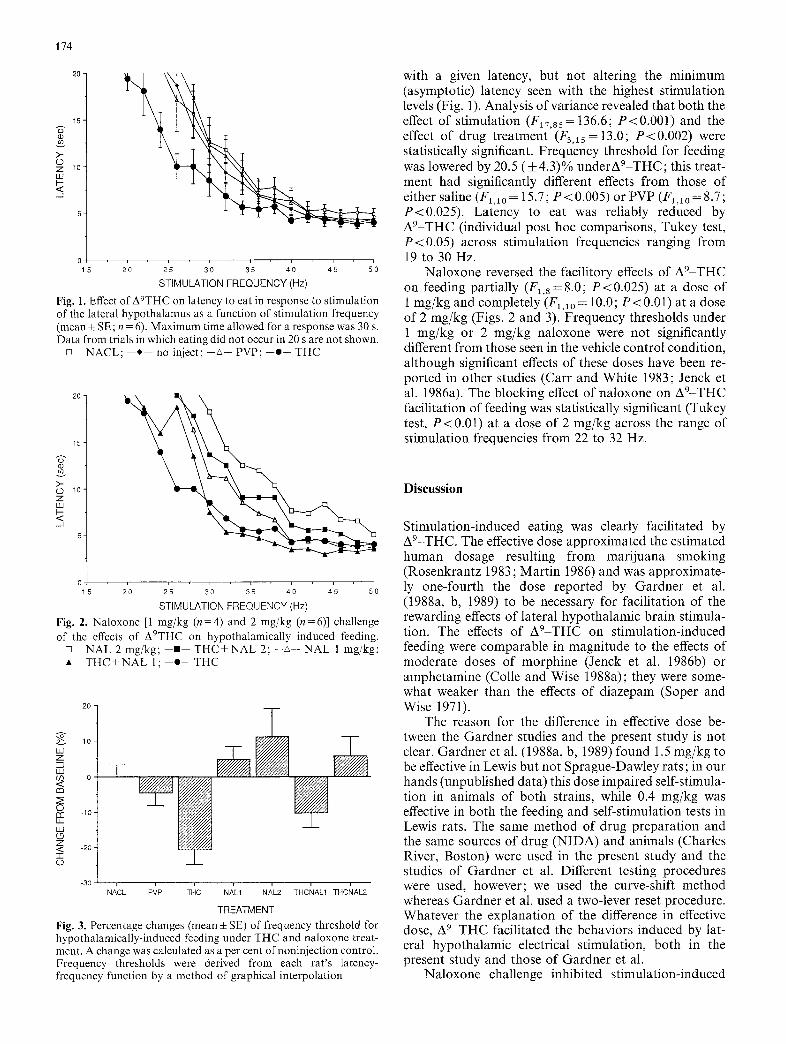
174
2 0 "
15"
>- o Z 1 0 UJ I'-" ,< _J
5
0 5 ' 2'0 ' 2'5 ' 3'0 3'5 4'0 4'5 5'0
STIMULATION FREQUENCY (Hz)
Fig. 1. Effect of A9THC on latency to eat in response to stimulation of the lateral hypothalamus as a function of stimulation frequency (mean-~ SE; n = 6). Maximum time allowed for a response was 30 s. Data from trials in which eating did not occur in 20 s are not shown.
o NACL; * no inject;--~-- PVP;--•-- THC
2 0
15'
>.- o lO.
W
< J
5
0 5 ' 2'0 ' 2'5 ' 3~0 3'5 4'0 4'5 5'0
STIMULATION FREQUENCY (Hz)
Fig. 2. Naloxone [1 mg/kg (n= 4) and 2 mg/kg (n = 6)] challenge of the effects of AgTHC on hypothalamically induced feeding.
n NAL 2 mg/kg; • THC+NAL 2; - zx-- NAL 1 mg/kg; • THC+NAL 1; • THC
IJJ Z
LU
CO :E O r r I.J._ LU
Z <
O
2 0 -
1 0
0 -V
-10 ¸
- 2 0 -
-30 j
NACL PdP ~o N2o NL2 ,H&AL1 TH&A~ TREATMENT
Fig. 3. Percentage changes (mean=l: SE) of frequency threshold for hypothalamically-induced feeding under THC and naloxone treat- ment. A change was calculated as a per cent of noninjection control. Frequency thresholds were derived from each rat's latency- frequency function by a method of graphical interpolation
with a given latency, but not altering the minimum (asymptotic) latency seen with the highest stimulation levels (Fig. 1). Analysis of variance revealed that both the effect of stimulation (F17,ss= 136.6; P<0.001) and the effect of drug treatment (F3,15 =13.0; P<0.002) were statistically significant. Frequency threshold for feeding was lowered by 20.5 (_+ 4.3)% underA9-THC; this treat- ment had significantly different effects from those of either saline (Fl,ao = 15.7; P < 0.005) or PVP (F~,lO = 8.7; P<0.025) . Latency to eat was reliably reduced by A9-THC (individual post hoc comparisons, Tukey test, P<0 .05) across stimulation frequencies ranging from 19 to 30 Hz.
Naloxone reversed the facilitory effects of A9-THC on feeding partially (Fx, 8 = 8.0; P<0.025) at a dose of 1 mg/kg and completely (F1, so = 10.0; P < 0.01) at a dose of 2 mg/kg (Figs. 2 and 3). Frequency thresholds under 1 mg/kg or 2 mg/kg naloxone were not significantly different from those seen in the vehicle control condition, although significant effects of these doses have been re- ported in other studies (Carr and White 1983; Jenck et al. 1986a). The blocking effect of naloxone on Ag-THC facilitation of feeding was statistically significant (Tukey test, P<0 .01) at a dose of 2 mg/kg across the range of stimulation frequencies from 22 to 32 Hz.
Discussion
Stimulation-induced eating was clearly facilitated by A9-THC. The effective dose approximated the estimated human dosage resulting from marijuana smoking (Rosenkrantz 1983; Martin 1986) and was approximate- ly one-fourth the dose reported by Gardner et al. (1988a, b, 1989) to be necessary for facilitation of the rewarding effects of lateral hypothalamic brain stimula- tion. The effects of A 9 THC on stimulation-induced feeding were comparable in magnitude to the effects of moderate doses of morphine (Jenck et al. 1986b) or amphetamine (Colle and Wise 1988a); they were some- what weaker than the effects of diazepam (Soper and Wise 1971).
The reason for the difference in effective dose be- tween the Gardner studies and the present study is not clear. Gardner et al. (1988a, b, 1989) found 1.5 mg/kg to be effective in Lewis but not Sprague-Dawley rats; in our hands (unpublished data) this dose impaired self-stimula- tion in animals of both strains, while 0.4 mg/kg was effective in both the feeding and self-stimulation tests in Lewis rats. The same method of drug preparation and the same sources of drug (NIDA) and animals (Charles River, Boston) were used in the present study and the studies of Gardner et al. Different testing procedures were used, however; we used the curve-shift method whereas Gardner et al. used a two-lever reset procedure. Whatever the explanation of the difference in effective dose, A9-THC facilitated the behaviors induced by lat- eral hypothalamic electrical stimulation, both in the present study and those of Gardner et al.
Naloxone challenge inhibited stimulation-induced

175
feeding, but - while the effects on baseline feeding were not statistically reliable - it did so to approximately the same extent in the vehicle conditions and in the A9-THC conditions. Since statistically reliable effects on baseline stimulation-induced feeding have been reported by others with similar or lower doses of naloxone, it would not be prudent to ignore the statistically insignificant effects on baseline responding seen in the present study. It is established in the literature that naloxone exerts a small but repeatable effect on stimulation-induced eating at doses as low as 0.2 (Carr and Simon 1983) or 0.5 mg/kg (Jenck et al. 1986a). Thus it should not be quickly concluded f rom our data that the effects of A9-THC were mediated by an endogenous opioid mech- anism. Rather, the conservative interpretation would be that stimulation-induced feeding is, itself, partially de- pendent on an endogenous opioid mechanism and that naloxone eliminates the contribution of that mechanism to performance under both baseline and A9-THC con- ditions.
This appears to be quite different f rom the case of self-stimulation. In general, stimulation-induced eating seems considerably more sensitive to naloxone challenge than is self-stimulation; naloxone does not have clear-cut effects on lateral hypothalamic self-stimulation until doses of 5 mg/kg or higher (Wauquier et al. 1974; van der Kooy et al. 1977; Lorens and Sainati 1978; Esposito et al. 1980; Perry et al. 1981 ; Seeger et al. 1981 ; West and Wise 1988; Gardner et al. 1989), while it begins to have marked effects on stimulation-induced feeding at doses an order of magnitude lower (Carr and Simon 1983; Jenck et al. 1986a). It is possible that the difference merely reflects the greater variability of self-stimulation, which may obscure real effects at lower doses than are usually used. On the other hand, it may be that there is some degree of difference in the components of the medial forebrain bundle mechanisms of self-stimulation and stimulation-induced feeding. While the two behav- iors are similarly altered by most o f the same drug treat- ments and are thus thought to involve common or at least overlapping mechanisms (Hoebel 1969; Wise and Bozarth 1987; Wise 1989), there are reported pharma- cological differences that involve opioids. Both morphine and the kappa opioid U50,488H facilitate stimulation- induced feeding (Jenck et al. 1987b) while only morphine and not the kappa agonist facilitate self-stimulation (Jenck et al. 1987a). Thus there may be a kappa opioid contribution to stimulation-induced feeding that does not interact with self-stimulation. Still this possibility does not offer a very good explanation for the greater sensitivity of feeding to naloxone; kappa-media ted ef- fects are less sensitive to naloxone than are mu- or delta- mediated effects, and if kappa opioids are involved in feeding but not self-stimulation then naloxone would be expected to be less rather than more sensitive to nal- oxone.
While the role of opioid mechanisms remains to be elucidated, the effects of A 9 T H C on stimulation- induced feeding add yet another t reatment to the list of treatments that similarly affect self-stimulation and stim- ulation-induced eating, While neither the present finding
nor the accumulated findings can establish that a com- mon mechanism mediates the two effects of lateral hypothalamic stimulation, the accumulating evidence does suggest that the two behaviors are likely to depend on at least partially common medial forebrain bundle components.
References
Allentuck S, Bowman KM (1942) The psychiatric aspects of mari- juana intoxication. Am J Psychiatry 99:248-251
Blundell JE, Latham CJ (1980) Characterization of adjustments to the structure of feeding behavior following pharmacological treatment: effects of amphetamine and fenfluramine and the antagonism produced by pimozide and methergoline. Pharma- col Biochem Behav 12:71%722
Bozarth MA, Wise RA (1981) Intracranial self-administration of morphine into the ventral tegmental area of rats. Life Sci 28: 551-555
Broekkamp CLE, Pijnenburg AJJ, Cools AR, Van Rossum JM (1975) The effect of microinjections of amphetamine into the neostriatum and the nucleus accumbens on self-stimulation be- havior. Psychopharmacologia 42: 179-183
Broekkamp CLE, Van den Bogaard JH, Heijnen HJ, Rops RH, Cools AR, Van Rossum JM (1976) Separation of inhibiting and stimulating effects of morphine on self-stimulation behavior by intracerebral microinjections. Eur J Pbarmacol 36:443-446
Carr GD, White NM (1983) Conditioned place preference from intra-accumbens but not intra-caudate amphetamine injections. Life Sci 33:2551-2557
Carr KD, Simon EJ (1983) Effects of naloxone and its quarternary analogue on stimulation-induced feeding. Neuropharmacology 22:127-130
Colle LM, Wise RA (1988a) Concurrent facilitory and inhibitory effects of amphetamine on stimulation-induced eating. Brain Res 459: 356360
Colle LM, Wise RA (t988b) Effects of nucleus accumbens am- phetamine on lateral hypothalamic brain stimulation reward. Brain Res 459: 361-368
Devane AF, Dysarz A, Johnson MR, Melvin LS, Howlett C (1988) Determination and characterization of a cannabinoid receptor in the rat brain. Mol Pharmacol 34:605
Dobrzanski S (1976) The effects of (+)-amphetamine and fen- fluramine on feeding in starved and satiated mice. Psycho- pharmacology 48: 283-286
Esposito RU, Perry W, Kornetsky C (1980) Effects of d-am- phetamine and naloxone on brain stimulation reward. Psycho- pharmacology 69:187-191
Fenimore DC, Loy PR (1971) Injectible dispersion of A 9- tetrahydrocannabinol in saline using polyvinylpirrolidone. J Pharmacol Exp Ther 23:310
Foltin RW, Brady JV, Fischman MW (1986) Behavioral analysis of marijuana effects on food intake in humans. Pharmacol Biochem Behav 25: 577-582
Foltin RW, Fischman MW, Byrne MF (1988) Effects of smoked marijuana on food intake and body weight in humans living in a resident laboratory. Appetite 11 : 1-14
Gardner EL, Paredes W, Smith D, Donner A, Milling C, Cohen D, Morrison D (1988a) Facilitation of brain stimulation reward by Ag-tetrahydrocannabinol. Psychopharmacology 96:142-144
Gardner EL, Paredes W, Smith D, Seeger TF, Donner A, Milling C, Cohen D, Morrison D (1988b) Strain-specific sensitization of brain stimulation reward by Ag-tetrahydrocannabinol in lab- oratory rats. Psychopharmacology [Suppl] 96:365
Gardner EL, Paredes W, Smith D, Zukin RS (1989) Facilitation of brain stimulation reward by A9-tetrahydrocannabinol is mediated by an endogenous opioid mechanism. Adv Biosci 175: 671-674

176
Glickman SE, Schiff BB (1967) A biological theory of reinforce- ment. Psychol Rev 74:81-109
Greenberg I, Kuehnle J, Mendelson JH, Bernstein JG (1976) Effects of marihuana use on body weight and caloric intake in humans. Psychopharmacology 49: 79-84
Gross H, Ebert MH, Faden VB, Goldbert SC, Kaye WH, Caine ED, Hawks R, Zinberg N (1983) A double-blind trial of delta-9- tetrahydrocannabinol in primary anorexia nervosa. J Clin Psychopharmacol 3:165-171
Haines L, Green W (1970) Marijuana use patterns. Br J Addict 65 : 347-362
Halikas JA, Goodwin DW, Gruze SB (1971) Marijuana effects - a survey of regular users. JAMA 217:692-694
Herkenham M, Little MD, Johnson MR, Melvin LS, Howlett AC, Rothman RB, deCosta B, Rice KC (1988) Localization of can- nabinoid receptors in brain. Soc Neurosci Abstr 14:104
Hoebel BG (1969) Feeding and self-stimulation. Ann NY Acad Sci 157 : 758-778
Hoebel BG, Monaco AP, Hernandez L, Aulisi EF, Stanley BG, Lenard L (1983) Self-injection of amphetamine directly into the brain. Psychopharmacology 81:158-163
Hollister LE (1971) Hunger and appetite after single doses of mari- juana, alcohol, and dextroamphetamine. Clin Pharmacol Ther 12 : 44-49
Holtzman SG (1974) Behavioral effects of separate and combined administrations of naloxone and d-amphetamine. J Pharmacol Exp Ther 189:51-60
Jenck F, Gratton A, Wise RA (1986a) Effects of pimozide and naloxone on latency for hypothalamically induced eating. Brain Res 375:329-337
Jenck F, Gratton A, Wise RA (1986b) Opposite effects of ventral tegmental and periaqueductal gray morphine injections on lat- eral hypothalamic stimulation-induced feeding. Brain Res 399 : 24-32
Jenck F, Gratton A, Wise RA (1987a) Opioid receptor subtypes associated with ventral tegmental facilitation of lateral hypo- thalamic brain stimulation reward. Brain Res 423 : 34.38
Jenck F, Quirion R, Wise RA (1987b) Opioid receptor subtypes associated with ventral tegmental facilitation and periaqueduct- ai gray inhibition of feeding. Brain Res 423 : 39-44
Kornetsky C, Esposito RU, McLean S, Jacobson JO (1979) In- tracranial self-stimulation thresholds: a model for the hedonic effects of drugs of abuse. Arch Gen Psychiatry 36:289- 292
Lorens SA, Sainati SM (1978) Naloxone blocks the excitatory effect of ethanol and chlordiazepoxide on lateral hypothalamic self- stimulation in the rat. Life Sci 23:1359-1364
Martin BR (1986) Cellular effects of cannabinoids. Pharmacol Rev 38: 45-74
Noyes R, Brunk SF, Avery DH, Canter A (1976) Psychologic effects of oral delta-9-tetrahydrocannabinol in advanced cancer pa- tients. Comp Psychiatry 17:641-646
Pellegrino LJ, Pellegrino AS, Cushman AJ (1979) A stereotaxic atlas of the rat brain. Plenum Press, New York
Perry W, Esposito RU, Kornetsky C (198t) Effects of chronic
naloxone treatment on brain-stimulation reward. Pharmacol Biochem Behav 4 : 247-249
Phillips AG, LePiane FG (1980) Reinforcing effects of morphine microinjection into the ventral tegmental area. Pharmacol Biochem Behav 12:965-968
Regulson W, Buther JR, Schultz J, Kirk T, Peek KL, Green ML, Zalis MO (1976) Ag-tetrahydrocannabinol as an effective anti- depressant and appetite-stimulating agent in advanced cancer patients. In: Szara S (ed) The pharmacology of marihuana. Raven Press, New York, pp 763-776
Rompr6 P-P, Wise RA (1989) Opioid-neuroleptic interaction in brainstem self-stimulation. Brain Res 477: 144-151
Rosenkrantz H (1983) Cannabis, marijuana and cannabinoid toxi- cological manifestations in man and animals. In: Fehr KO, Katant H (eds) Adverse health and behavioral consequences of cannabis use. Addiction Research Foundation, Toronto, pp 91-175
Seeger TF, Carlson KR, Nazarro JM (1981) Pentobarbitol induces a naloxone-reversible decrease in mesolimbic self-stimulation threshold. Pharmacol Biochem Behav 15:583-586
Siler JF, Sheep WL, Bastes LB, Clark GW, Cook GW, Smith WA (1933) Marihuana smoking in Panama. Milit Surg 73:269-280
Soper WY, Wise RA (1971) Hypothalamically induced eating: eating from "non-eaters" with diazepam. Tower-Int-Tech- nomed J Life Sci 1 : 79-84
Stein L, Wise CD (1973) Amphetamine and noradrenergic reward pathways. In: Usdin E, Snyder SH (eds), Frontiers in catechol- amine research. Pergamon Press, New York, pp 963-968
Tart CT (1970) Marijuana intoxication: common experiences. Nature 226:701-704
van der Kooy D, LePiane FG, Phillips AG (1977) Apparent in- dependence of opiate reinforcement and electrical self-stim- ulation systems in rat brain. Life Sci 20:981-986
van der Kooy D, Mucha RF, O'Shaughnessy M, Bucenieks P (1982) Reinforcing effects of brain microinjections of morphine re- vealed by conditioned place preference. Brain Res 243 : 107-117
Wauquier A, Niemegeers CJE, Lal H (1974) Differential antagon- ism by naloxone of inhibitory effects of haloperidol and mor- phine on brain self-stimulation. Psychopharmacologia 37 : 303-310
West TEG, Wise RA (1988) Effects ofnaltrexone on nucleus accum- bens lateral hypothalamic, and ventral tegmental self-stimu- lation rate-frequency functions. Brain Res 426:126-133
Wise RA (1974) Lateral hypothalamic electrical stimulation: does it make animals hungry? Brain Res 67:187-209
Wise RA (1980) Action of drugs of abuse on brain reward systems. Pharmacol Biochem Behav 13 : 213-223
Wise RA (1988) The neurobiology of craving: implications for the understanding and treatment of addiction. J Abnorm Psychol 97:118-132
Wise RA (1989) The brain and reward. In: Liebman JM, Cooper SJ (eds) The neuropharmacological basis of reward. Oxford University Press, Oxford, pp 377-424
Wise RA Bozarth MA (1987) A psychomotor stimulant theory of addiction. Psychol Rev 94:469-492





























