Embed Size (px)
Citation preview

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis Doctoral
Fasciola hepática (Trematoda:Fasciola hepática (Trematoda:Digenea) en ganado bovino de losDigenea) en ganado bovino de losvalles cordilleranos patagónicos:valles cordilleranos patagónicos:
factores involucrados en sufactores involucrados en sutransmisión.transmisión.
Kleiman, Florencia
2004
Tesis presentada para obtener el grado de Doctor en CienciasBiológicas de la Universidad de Buenos Aires
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the Master's and Doctoral Theses Collection of the Central LibraryDr. Luis Federico Leloir, available in digital.bl.fcen.uba.ar. It should be used accompanied bythe corresponding citation acknowledging the source.
Cita tipo APA:
Kleiman, Florencia. (2004). Fasciola hepática (Trematoda: Digenea) en ganado bovino de losvalles cordilleranos patagónicos: factores involucrados en su transmisión.. Facultad deCiencias Exactas y Naturales. Universidad de Buenos Aires.http://hdl.handle.net/20.500.12110/tesis_n3746_KleimanCita tipo Chicago:
Kleiman, Florencia. "Fasciola hepática (Trematoda: Digenea) en ganado bovino de los vallescordilleranos patagónicos: factores involucrados en su transmisión.". Tesis de Doctor. Facultadde Ciencias Exactas y Naturales. Universidad de Buenos Aires. 2004.http://hdl.handle.net/20.500.12110/tesis_n3746_Kleiman

de los valles cordilleranos patagónicos:Factores involucrados en su transmisión
Florencia KLEIMAN
Tesis para optar ai títqu de Doctor de la Universidad de Buenos Aires
Unidad de Ecologia de Reservorios y Vectores de ParásitosDepartamento de Ecología, Genética y Evolución,
Facultad de Ciencias Exactas y Naturales,Universidad de Buenos Aires.
2004 V‘ Directora: María Cristina Wisnivesky

Toda nuestra ciencia,comparada con Ia realidad,
es primitiva e infantil...y sin embargo es lo más preciado
que tenemos.
ALBERT EINSTEIN
(1379-1955)

AGRADECIMIENTOS
A mi directora, la Dra. Cristina Wisnivesky, por contagiarme su pasión por la
parasitología y por permitirme trabajar con tanta libertad durante estosaños.
A la Dra. Margarita Ostrowski de Núñez por enseñarme el universo de losTrematodes.
AI Consejo Nacional de Investigaciones Científicas y Tecnológicas
(CONICET),a Ia Facultad de Ciencias Exactas y Naturales, a Ia Secretaría de
Ciencia y Técnica de la Universidad de Buenos Aires, a la Agencia Nacional
de Promoción Científica y Tecnológica y a mi directora, por otorgarme los
recursos económicos para desarrollar esta tesis.
AI Ing. Agr. Tati Iturburu y al Med. Vet. Hernán Alonso por facilitar mi
trabajo en el campo, por sus aportes y por el interés demostrado en el
tema. A ellos y a Sandra y a Paula, por las cenas, las charlas y Ia calidez con
la que me recibieron durante los muestreos.
AIMed. Vet. Hernán Alonso por realizar la toma de muestras en ganado.
A Silvia, Elsa, Pepino y Dolfi por su colaboración incondicional durante los
muestreos y por brindarme un ambiente de trabajo tan cálido.
AI Dr. Hugo Sigman, a la Dra. Silvia Gold y al Establecimiento Los Murmullos
por permitirme realizar el estudio y por el apoyo logístico.
A la Dra. Stella Maris González Cappa por facilitarme las instalaciones del
Dpto. de Microbiología, Parasitología e Inmunología de la Facultad de
Medicina, UBA, para desarrollar el trabajo de laboratorio que involucróinfección.
A Lucas Bukata y a Rafael Lim por ayudarme en las tareas técnicas de
laboratorio, por su incondicionalidad y por haberme hecho sentir cómoda
desde el primer día en un ambiente de trabajo nuevo.

A Silvia por demostrarse siempre interesada en mi trabajo, por su
dedicación, sus aportes, sus críticas, su contención y por encarar todo con
tanta energía.
A Diana por su inmensa colaboración durante la última temporada demuestreos.
A Lucila por las horas, días y casi siglos que dedicó a ayudarme a procesarlas muestras de caracoles.
A Silvia, Gerónimo y Lucila por ayudarme a procesar las muestras de
ganado.
A Aníbal, por su colaboración con el procesamiento de imágenes y el análisisde datos.
A Tato, Aníbal, Silvia, Lucila y Diana por el tiempo que dedicaron a la lectura
de esta tesis y por sus críticas y sugerencias.
A Juli Filloy, por ayudarme con mis baches estadísticos y con mis baches
espirituales.
A Darío Vezzani (Tato) por el diseño de tapa.
A Tato y a Aníbal (una vez más), porque ellos son los principales
responsables de que escribir esta tesis haya sido placentero. Por las horas
de debate científico y por las de discusiones infantiles, ambas igualmente
enriquecedoras.
A Linus, por haberme formado, acompañado y aconsejado durante el
desarrollo de mi tesis. Por haberme enseñado todo y por tratar siempre de
facilitarme el camino. Por ser un excelente maestro y compañero. Porbrindarse en forma incondicional.
A mi familia, por soportar y sobrevivir a mi intenso ritmo de muestreos y
mis ausencias. Por compartir mis logros y mis desconsuelos. Por haber sido
fundamentales durante la etapa de escritura de la tesis.
A Sofi, por haberlo resistido. Gracias.

CONTENIDO
Resumen
Abstract
1 Introducción
1.1 Conceptos generales
1.2 Ciclo de vida de Fasciola hepatica
1.3 Presentación del tema de estudio
1.4 Historia del descubrimiento del ciclo de vida de Fasciola hepatica
1.5 Rompecabezas epidemiológico. Objetivos de Ia tesis
2 ElAmbiente
2.1 Selección del área de estudio
2.1.1 Metodología
2.1.2 Resultados y Discusión2.2 Caracterización del área de estudio
2.2.1 Escala Regional. Descripción general2.2.2 Escala Local
2.3 Conclusiones
3 El Hospedador Definitivo3.1 Introducción
3.2 Objetivos
3.3 Hipótesis
3.4 Materiales y Métodos
3.4.1 El ganado: Población en estudio
3.4.2 Manejo ganadero
3.4.3 Infección en ganado
3.4.4 Infección en hospedadores silvestres3.4.5 Análisis estadístico
3.5 Resultados y Discusión
3.5.1 Método coprológico de diagnóstico
3.5.2 Infección en ganado
3.5.3 Infección en hospedadores silvestres3.6 Conclusiones
4 ElHospedador Intermediario4.1 Introducción
12
14
16
18
18
18
19
23
23
26
37
38
38
43
43
44
44
44
49
55
55
57
57
60
72
75
76
76

4.2 Objetivos
4.3 Hipótesis
4.4 Materiales y Métodos4.4.1 Períodos de estudio
4.4.2 Variables ambientales
4.4.3 El hospedador intermediario4.4.4 Análisis estadístico
4.5 Resultados y Discusión4.5.1 Variables ambientales
4.5.2 EIhospedador intermediario: Generalidades
4.5.3 El hospedador intermediario: Identificación
4.5.4 Estructura y dinámica poblacional de Lymnaea víatrix
4.5.5 Infección en Lymnaea viatrix: Fascíola hepatica
4.5.6 Infección en Lymnaea viatrix: Otros trematodes4.5.7 Consideraciones finales
4.6 Conclusiones
5 El Parásito
5.1 Metodología5.2 Resultados
5.3 Discusión
6 Discusión General
Apéndice
Bibliografía
83
83
84
84
85
88
92
94
94
109
113
113
133
137
144
147
149
149
151
152
155
170
173

RESUMEN
La fasciolosis, causada por Fasciola hepatica, es una de las enfermedades
parasitarias de mayor importancia para la actividad ganadera porque produce considerables
pérdidas económicas a escala mundial. Con el objetivo de caracterizar la transmisión en un
área de alta endemicidad como son los valles cordilleranos patagónicos, se realizaron
estudios en un establecimiento ganadero de la localidad de Cholila, Chubut, entre diciembre
de 1998 y febrero de 2002.
Se identificó al caracol que actúa como hospedador intermediario de F. hepatica, se
caracterizaron los ambientes donde se desarrolla y se estudió su dinámica temporal en
relación con las condiciones climáticas de Ia zona. Se estimaron las prevalencias en el
hospedador intermediario y en el hospedador definitivo por observación directa del parásito y
por métodos coprológicos, respectivamente. Se recopiló información sobre el tiempo de
permanencia del ganado en cada ambiente.
Lymnaea viatrix resultó ser la única especie de la Familia Lymnaeidae presente en el
área de estudio. El hallazgo de esta especie a latitudes superiores a los 41° S amplía su
rango de distribución austral mundial. Se comprobó que L. viatrix puede desarrollarse en
cualquier cuerpo de agua (temporal o permanente) que presente parches poco disturbados,
con suelo de barro cubierto por una delgada película de agua y escasa vegetación. Los
caracoles hibernaron durante el invierno, entraron en actividad durante la primavera,
ovipusieron a comienzos del verano y luego los individuos de mayor tamaño murieron a
comienzos del otoño. Esta dinámica se vio fuertemente afectada por la disponibilidad de
agua en el suelo. Se registró infección por F. hepatíca durante todo el estudio únicamente en
caracoles adultos y las prevalencias fluctuaron entre 1 y 14%. Los caracoles parasitados sólo
se detectaron en verano y otoño. La infección en caracoles no estuvo relacionado con su
abundancia en cada ambiente ni con su estructura de tamaños, pero se relacionó con el
tiempo que el ganado estuvo pastando en cada ambiente. En el ganado, la prevalencia al
comienzo del estudio resultó mayor en los animales jóvenes que en los adultos, y a pesar de
que esta tendencia desapareció luego de tres años de tratamientos antihelmínticos, los
niveles de infección general fueron cercanos al 50% en todo momento. La utilización de
drogas no tuvo efecto sobre la prevalencia en el ganado ni redujo la cantidad de huevos
eliminados al ambiente. Los resultados sugieren que el manejo sanitario basado en la
aplicación de antihelmínticos no contribuyó a disminuir la proliferación del parásito en el
ambiente, ya sea por una alta oferta de metacercarias todo el año o por una baja eficacia de
las drogas. En el área de estudio, niveles bajos de infección en caracoles mantienen altas
prevalencias en el ganado, aún con la continua aplicación de antiparasitarios. Debido a que la
mejor estrategia de control es la que combina manejo ambiental con quimioterapia
estratégicamente adecuada a las características de la transmisión en cada región, la
información microepidemiológica generada en este estudio puede ser útil para implementaruna estrategia adecuada.
Palabras claves: Fasciola hepatica, Lymnaea viatrix, fasciolosis, ganado, transmisión.

ABSTRACT
Fasciolosis in cattle, caused by Fasciola hepatica, is a worldwide disease of maj0r
economical importance. The object of this thesis was to investigate the factors involved in the
parasite transmission in a highly endemic area such as the Andean Valleys in Argentine
Patagonia. Research was conducted in a Iivestock farm located in the Locality of Cholila, Chubut,
between December 1998 and February 2002. Studies comprised the identification of the
intermediate snail host, habitat characterization, and the relationship between temporal
dynamics of the snail population and local climatic conditions. Parasite prevalences were
estimated in snails and cattle by direct parasite observation and coprological methods,
respectively. Duration of cattle grazing in each environment was registered.
Lymnaea viatrix was the only Iymnaeid species in the study area and its finding south of
parallel 41° S extends its distribution range. This snail occurred in temporary and permanent
waterbodies with scarcely disturbed patches of mud covered by a thin water layer and bare
vegetation. Snails hibernated during winter, resumed activity in spring, oviposited at the
beginning of summer and Iarge-sized individuals died at the beginning of autumn. Population
dynamics was stroneg influenced by water availability in the soil. F. hepatica was only detected
in adult snails during summer and autumn, and prevalences ranged from 1 to 14%. Snail
infection was not related with snail abundance or size structure, and was related to the duration
of cattle grazing in each environment. At the beginning of the study prevalence in cattle was
higher in young animals than in older ones, and although this tendency disappeared after three
years of anthelmintic treatment, values remained high in both groups of animals. Overall
infection levels were about 50% throughout the study.
Anthelmintic drugs had no effect on cattle prevalence or on the reduction of egg
shedding. Results suggest that sanitary management based on anthelmintics did not diminish
parasite build-up in the environment, either by a high offer of metacercariae all year round or
lack of chemotherapy efflcacy. Low infection prevalences in snails were responsible for high
levels of infection in cattle, despite the continuous administration of anthelmintics. On the basis
that the best control strategy should combine environmental management with chemotherapy
adapted to the particular regional features, miCro-epidemiological information provided hereinwould contribute to limit Iooses in live-stock.
Key words: Fasciola hepatíca, Lymnaea viatrix, fasciolosis, Iivestock, transmission.

INTRODUCCIÓN
“Fasciola hepatica is easily accessible, its distribution
¡s cosmopolitan, and its fascinating life cycle is a
rewarding challenge for zoologists, ecologists,
pathologists and biochemists interested ¡n trematoderesearch”
J. C. Boray 1969
1.1 CONCEPTOS GENERALES
La epidemiología estudia Ia distribución de las infecciones en las poblaciones
humanas y las variables que determinan su ocurrencia (Gordis 2000) y uno de sus
objetivos principales es identificar los factores de riesgo para las poblaciones afectadas.
De manera similar, la epizootiología se ocupa del estudio de Ia distribución de
infecciones parasitarias en otras especies animales (Esch & Fernández 1993). Sin
embargo, algunos autores han utilizado el término epidemiología para referirse a
cualquier tipo de población animal (Nelson 1990) y en Ia actualidad el término es
comúnmente usado para referirse al estudio de infecciones parasitarias en animales y
en el hombre. Adoptando este criterio, las variables que determinan la abundancia de
las distintas poblaciones componentes‘ del parásito son el principal objeto de estudio de
la ecoepidemiología. En el ganado, en particular, esta rama de la epidemiología trata
de dilucidar cuáles son las variables que afectan la prevalencia e intensidad de Ia
infección y eI impacto que el parásito produce en los animales, tanto en términos de
manifestaciones clínicas de la enfermedad como en pérdidas en productividad
(Torgerson & Claxton 1999).
Algunos autores proponen que Ia epidemiología puede diferenciarse en
macroepidemiología y microepidemiología. La primera abarca el estudio de los
conceptos recién desarrollados de una manera muy amplia. Estudia patrones de
distribución, prevalencia e intensidad de infección junto con estimaciones de morbilidad,
l Poblacióncomponente: conjunto de individuos de un estadio de desarrollo en un momento y lugar hospedador o ambiente- determinados (Bush et al. 1997)

mortalidad y pérdidas económicas, temas que suelen ser sintetizados con el desarrollo
de modelos matemáticos. La microepidemiología contempla la relación-hospedador
parásito desde un enfoque diferente. Dentro de los hospedadores estudia los factores
genéticos relacionados con la susceptibilidad y la respuesta inmune, y en el ambiente
estudia la forma en que el parásito se transmite y su densidad a una escala local,
contemplando las condiciones particulares de cada zona (Nelson 1990).
La comprensión de la biología del parásito o del hospedador no resultan
suficientes para dilucidar las interacciones hospedador-parásito. Es necesario conocer la
dinámica conjunta, ya que los cambios en una de las poblaciones pueden afectar
directamente la dinámica de la otra población. Dentro del marco de interés de la
ecoepidemiología, se encuentra el estudio de Ia dinámica poblacional del complejo
hospedador-parásito, que abarca los procesos que afectan conjuntamente la
abundancia de ambos en un área determinada. Es un fenómeno común que los
parásitos existan en la naturaleza constituyendo varias poblaciones diferenciadas, unas
dentro de hospedadores y otras en el ambiente. Estas poblaciones pueden aumentar su
tamaño como resultado del reclutamiento de nuevos individuos y pueden disminuirlo
como consecuencia de la muerte del parásito. Dentro del hospedador, la muerte puede
ser natural, inducida por respuesta inmune, o bien como consecuencia de la muerte del
propio hospedador. En el caso de los estadios de vida libre, los hospedadores no juegan
ningún rol en su supervivencia, sin embargo, el éxito en la transmisión se ve
enormemente influenciado por la abundancia y distribución espacial de los
hospedadores (Scott &Smith 1994).
El ambiente es un factor determinante de la dinámica poblacional del complejo
hospedador-parásito. Según la escala de estudio, el concepto de ambiente puede variar,
siendo desde un hospedador para una infrapoblación de parásitos hasta una región
geográfica para la especie. La distribución de un parásito está restringida a aquellas
zonas geográficas en las cuales la especie se encuentra permanentemente establecida,
debido a Ia presencia de sus hospedadores y de las condiciones que permiten la
transmisión. Dentro de ese rango de distribución, pueden encontrarse zonas más
favorables en las que se observan altas abundancias, y áreas de distribución en parches
con abundancias más bajas, que generalmente coinciden con los límites de las zonas
aptas para el desarrollo de la especie (Malone 1994, Parmesan et al. 2000).
Los parásitos pertenecientes a la subclase Digenea (Platyhelminthes,
Trematoda), entre los que se encuentra Fasciola hepatica (Linnaeus, 1758), se
caracterizan por presentar ciclos de vida complejos, que incluyen estadios dentro de
hospedadores intermediarios y definitivos (moluscos y vertebrados respectivamente) y
larvas de vida libre en el ambiente (Gibson 2002). Las poblaciones componentes, en
este caso, pueden diferenciarse en los huevos, los miracidios, las cercarias y las
metacercarias -en el ambiente-, las larvas dentro de los hospedadores intermediarios y
los adultos en los hospedadores definitivos (Bush et al. 1997). Exceptuando la última

población mencionada, las restantes se ven directamente afectadas por variables
ambientales -denso independientes-, como la temperatura y la humedad, ya sea
porque actúan directamente sobre los parásitos, o bien porque condicionan el desarrollo
del hospedador intermediario dentro del cual habita la población parasitaria (Torgerson
& Claxton 1999). La temperatura es considerada uno de los factores críticos más
importantes en el desarrollo de la mayoría de los organismos (Precht et al. 1973,
Ahmed & Raut 1991, Raut et al. 1992). Puede actuar como un detonador para el inicio
de un proceso biológico o bien restringir y hasta interrumpir su desarrollo (Aziz & Raut
1996). La humedad, en cambio, en el caso particular de F. hepática, es esencial para la
supervivencia de los estadios de vida libre (huevos y metacercarias) y para la movilidad
de las larvas nadadoras que emergen de los huevos y los caracoles (miracidios y
cercarias, respectivamente) (Ollerenshaw 1971a).
El desarrollo de la infección tanto dentro de los hospedadores intermediarios
como de los definitivos, depende también de variables estrechamente ligadas a
características endógenas o conductas de los hospedadores, entre las que se pueden
mencionar la resistencia natural o adquirida al parásito, el estado nutricional, el
comportamiento y los hábitos alimenticios del hospedador y la competencia
intraespecífica del parásito por los recursos del hospedador (Wisnivesky 2003).
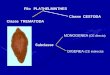
1.2 CICLODE VIDA DE Fasciola hepatica
El ciclo de vida de F. hepatica comprende estadios de desarrollo dentro de
hospedadores animales y en el ambiente (Figura 1-1). El parásito adulto alcanza la
madurez sexual en el hígado y vesícula biliar de mamíferos herbívoros ovinos, bovinos,
caprinos, camélidos, equinos, suinos y en el hombre. Los huevos que el parásito
produce son trasladados junto con la bilis hacia el intestino y eliminados con la materia
fecal. El tiempo de desarrollo embrionario hasta la eclosión, fluctúa entre 12 y 15 días
en condiciones adecuadas de temperatura, humedad, luz y oxígeno (Rowan 1956,
Ollerenshaw 1958, Luzón-Peña et al. 1994, Andrews 1999) (Figura 1-2a y b). Luego
emerge una larva nadadora llamada miracidio2 (Figura 1-2c), que busca activamente
al hospedador intermediario, un caracol perteneciente a la Familia Lymnaeidae
(Gasteropoda, Pulmonata). Dado que los estadios de vida libre no se alimentan (viven
con las reservas que generaron en el estadio anterior), los miracidios tienen
aproximadamente 24 horas para encontrar un caracol antes de morir (Hope Cawdery et
al. 1978). La penetración ocurre por la cavidad pulmonar u otras zonas del cuerpo y
dentro del hospedador intermediario se produce la reproducción asexual. Durante las
primeras 12 horas el miracidio se desarrolla en un esporoquiste; éste migra hacia el
2 Miracidio: del griego mirakídion= jovencito

hepatopáncreas, produce una primera generación de redias3 madre y muere. Las
células germinales de estas redias (Figura 1-3a) producirán nuevas generaciones de
redias hijas, que luego darán origen a las cercarias“ (Figura 1-3b). La emisión de estas
larvas al ambiente ocurre, en promedio, a partir de los 45 días postinfección (p.i)
aunque se ha registrado el desarrollo completo en menos de 30 días dependiendo de Ia
temperatura (Lee et al. 1995)-. Las cercarias abandonan las redias a través del poro de
nacimiento y llegan al agua mediante la ruptura de tejido del caracol. Estas larvas
nadadoras presentan geotropismo negativo y fototropismo positivo y una vez en el agua
buscan un sustrato donde enquistarse. Los quistes, llamados también metacercariass
(Figura 1-3c), pueden adherirse a una amplia gama de sustratos; en la naturaleza se
los encuentra en plantas, rocas y agua. En el laboratorio se adhieren a vidrio, plástico,
etc. Algunos trabajos indican que en presencia de luz intensa las cercarias tienden a
enquistarse con mayor rapidez (Ginetsinkaya 1988). En oscuridad total, las cercarias no
se enquistan y continúan nadando hasta que se les agotan las reservas de glucosa y
lípidos y mueren.
Un esporoquiste puede generar entre 5 y 6 redias madre que pueden dar nuevas
redias. Cada redia hija genera entre 15 y 20 cercarias. Con lo cual, por cada miracidio
que penetra en un caracol pueden desarrollarse aproximadamente 4000 cercarias
durante la infección (Hansen & Perry 1994).
Para poder completar el ciclo de vida, las metacercarias deben ser ingeridas por
el hospedador definitivo. Esto ocurre cuando el animal se alimenta de pastos infestados
o bebe agua con metacercarias. Dentro del hospedador definitivo éstas se desenquistan
en el tubo digestivo y los juveniles liberados atraviesan la pared del intestino delgado
(24hs. p.i.) y por vía intraperitoneal se dirigen al hígado (Andrews 1999). Allíatraviesan
la cápsula de Glisson y penetran en el tejido hepático (4 a 6 días p.i.) para llegar a los
conductos biliares (S a 6 semanas p.i.) donde alcanzan la madurez sexual (2 a 3 meses
p.i.). El adulto es hermafrodita, con lo cual un único adulto que logre instalarse en la
vesícula puede producir huevos que se liberan al ambiente.
Considerando que el juvenil que sale del quiste mide aproximadamente unos
300 pm y el adulto que se instala en la vesícula unos 3 cm de largo, el parásito
incrementa su tamaño unas 100 veces durante su recorrido dentro del hospedador
definitivo. Esto lo logra alimentándose del tejido que va atravesando.
A diferencia de los miracidios que necesitan encontrar al siguiente hospedador
en pocas horas, los huevos y metacercarias pueden vivir mucho tiempo en forma
latente y resistir a períodos de bajas temperaturas, pero no sobreviven a períodos
prolongados de temperaturas elevadas y sequías.
3 Redia: en honor al naturalista italiano F. Redi (1626-1698)4 Cercaria: del griego kerkos= cola + arius= perteneciente, parecido a5 Meta: del griego meta: después

Figura 1-1: Esquema del ciclo de vida de Fascio/a hepatica: 1) Liberación de huevos al medio,2) eclosión del miracidio y búsqueda activa del hospedador intermediario (H.I.),3) liberación de las cercarias que se enquistan en la vegetación y 4) ingestión delas metacercarias por el hospedador definitivo (H.D.), dentro del cual se desarrollael adulto que produce ios huevos.
Figura 1-2: Huevos de Fascíola hepatica: a) Sin embrión desarrollado, b) con el miracidioen desarrollo, c) miracidio emergiendo. Extraído de Bacigalupo (1934)
Figura 1-3: Estadios Iarvarios de Fasciola hepatíca dentro del hospedador intermediarioy en el ambiente: a) redia, b) cercaria y c) metacercaria
11

1.3 PRESENTACIÓN DEL TEMA DE ESTUDIO
La infección producida por F. hepatíca en los hospedadores definitivos es
conocida como fasciolosis o distomatosis. Es una parasitosis cosmopolita cuya
distribución abarca principalmente a'reas templadas de Europa y áreas templadas y
subtropicales de América. En el hemisferio norte ha sido descripta en los países nórdicos
hasta los 65° N y en el hemisferio sur hasta la provincia de Santa Cruz en nuestro país
(50° S) y en Australia e islas de Nueva Zelanda (Boray 1985, Torgerson & Claxton
1999). A una escala de mayor detalle, la distribución del parásito en el ambiente es en
parches y extremadamente variable, y depende de la presencia del hospedadorintermediario.
La fasciolosis es considerada una zoonosis" emergente por la Organización
Mundial de la Salud. Los informes más recientes estiman que 2,4 millones de personas
están infectadas en el mundo (Rim et al. 1994). En los últimos 25 años se han
registrado más de 7000 casos en 51 países de todos los continentes: 12 en Oceanía,
354 en Asia, 487 en África, 2951 en Europa y 3267 en América (Mas-Coma et al. 1999).
Cuba, por ejemplo, sufrió tres epidemias de fasciolosis humana desde la década del '40
hasta el presente (Diaz et al. 1990). Actualmente, Bolivia y Perú en Sudamérica e Irán
en Asia son los países en los cuales se han registrado las regiones con mayor número
de casos humanos (Angles et al. 1997, Esteban et al. 1997). También se han notificado
algunos casos importados por viajantes (Price et al. 1993, Graham et al. 2001). En
Argentina se desconoce la importancia de esta parasitosis, ya que no existen registros
regulares. El primer caso de infección notificado fue en Ia Provincia del Chaco, a
mediados del siglo pasado (Petraglia 1954). Durante los últimos años se registró un
caso en la Provincia de San Luis (Ale et al. 2000) y otro en Neuquén (Rubel et al. 2003).
No obstante, como esta parasitosis no es de denuncia obligatoria, su sintomatología no
es específica y el diagnóstico de certeza presenta ciertas dificultades, la cifra de
humanos afectados podría ser mucho mayor.
A nivel veterinario, es una de las enfermedades parasitarias de mayor
importancia, con considerables pérdidas económicas anuales a escala mundial. Estas
pérdidas son el reflejo del daño que produce el parásito en los animales y que incluyen
decomiso de los hígados afectados, reducción en la producción de carne y leche,
disminución en Ia calidad de la lana en ovejas y camélidos y abortos y reducción en la
fertilidad en animales con altas cargas parasitarias (Dargie 1987, Hillyer& Apt 1997).
Diversos investigadores han encarado estudios epidemiológicos de esta
parasitosis de una manera integral, intentando identificar los principales factores
responsables de que la transmisión sea tan exitosa en condiciones tan diversas
(Ollerenshaw 1971a, Malone et al. 1984, Alcaino et al. 1993, Zukowski et al. 1993,
6 Zoonosis: infección parasitaria naturalmente transmitida entre el hombre y otras especies devertebrados

Buchon et al. 1997, Yilma & Malone 1998, Fuentes et al. 2001). Sin embargo, dadas las
características particulares de cada región, dichos resultados no son siempre
extrapolables a otras zonas. En la Argentina, la infección es endémica y tiene
distribución regional, destacándose las llanuras y esteros de Buenos Aires, Entre Ríos,
Corrientes, Santa Fe y Córdoba y los valles montañosos de Mendoza, San Luis, Río
Negro, Neuquén, Chubut, Salta, Catamarca y Jujuy. A pesar de existir estudios sobre la
biología del hospedador intermediario y las prevalencias en ganado en algunas zonas
del país (Dwinger et al. 1982, Kaczorkiewicz 1983, Rossanigo et al. 1983, Venturini &
Fonrouge 1985, Moriena et al. 2001) aún no se han realizado estudios integrales en
cada región que permitan conocer qué especies actúan como hospedadores
intermediarios, cuáles son las prevalencias en hospedadores domésticos y si existen
hospedadores silvestres que participen en el ciclo de vida. Tampoco se conocen cuáles
son las variables ambientales que modulan la dinámica de transmisión del parásito en
cada región en particular.
Fascio/a hepática presenta gran especificidad’ por su hospedador intermediario,
pero una amplia gama de especies de mamíferos herbívoros pueden actuar como
hospedadores definitivos (Boray 1967). Las especies de caracoles descriptas como
hospedadores del parásito en diversas regiones pertenecen a la Familia Lymnaeidae
(Gasteropoda, Pulmonata) y nunca se encontraron especies de otras Familias
naturalmente infectadas (Malek 1985, Malone 1994). Entre los hospedadores definitivos
pueden incluirse al hombre y al ganado bovino, ovino, caprino, equino, suino y diversas
especies de camélidos, así como también una amplia variedad de especies silvestres en
las cuales el parásito puede desarrollarse exitosamente (Presidente et al. 1974, Spratt &
Presidente 1981, Boray 1985, Rickard 1992, Apt et al. 1993, Buchon et al. 1997, Mas
Coma et al. 1997, Ménard et al. 2000, Shimalov & Shimalov 2000, Ménard et al. 2001).
Por estas razones los factores limitantes más importantes para la persistencia de la
fasciolosis son los que determinan la presencia del hospedador intermediario, que en
general coinciden con los necesarios para el desarrollo del parásito en el mismo.
Existen además, muchas otras variables que pueden afectar o condicionar Ia
transmisión del parásito. La edad del hospedador puede ser un factor determinante en
la susceptibilidad a Ia infección. A medida que Ia población de hospedadores envejece,
la disponibilidad de individuos adecuados para albergar aI parásito puede modificarse.
Aún cuando la estructura etaria de la población se mantenga estable, dado que las
poblaciones son abiertas (emigración-inmigración), los individuos que se van pueden
llevarse los parásitos consigo y los que llegan pueden no estar infectados y ser más
susceptibles por la ausencia de protección inmunológica desarrollada a partir de
7 Especificidad por un hospedador se define como el grado al cual un parásito está restringido en elnúmero de especies hospederas en un determinado estadio de su ciclo de vida (Poulin 1998)

contactos previos con el parásito. Incluso si los individuos permanecen en la población,
pueden ocurrir cambios importantes en la dinámica poblacional hospedador-parásito en
el tiempo. Por ejemplo, cambios nutricionales en la población de hospedadores debido a
variaciones en la disponibilidad de alimentos durante el año con subsecuentes cambios
en la susceptibilidad a la infección. En este caso, aunque los individuos son los mismos,
su calidad como potenciales hospedadores puede cambiar. De forma similar, aunque el
número de parásitos en una población permanezca estable, los cambios en la
infectividad pueden afectar la dinámica de transmisión de la parasitosis. Dichos cambios
pueden producirse por características intrínsecas del hospedador o el parásito, como se
mencionó antes, o bien por factores externos al complejo hospedador-parásito. Por
ejemplo, el uso continuo de drogas antiparasitarias a las cuales el parásito empieza a
generar resistencia puede dar lugar a una situación en la cual una proporción de las
larvas presentes en el ambiente pierda susceptibilidad a la droga. Esto generaría que un
grupo de la población parasitaria fuera capaz de infectar a la población de hospedadores
tratados (Scott &Smith 1994).
La complejidad de los ciclos de vida de los parásitos digeneos, la gran cantidad
de factores involucrados en la transmisión y la interconexión entre los mismos
evidencian la necesidad de realizar estudios integrales para poder entender la dinámica
del complejo hospedador-parásito.
1.4 HISTORIA DEL DESCUBRIMIENTO DEL CICLO DE VIDA DE Fasciola
hepatica'
Los parásitos animales son conocidos desde hace miles de años. Los egipcios,
mil quinientos años antes de Jesucristo ya citan a las pulgas, los piojos, las tenias, los
oxiurus y los ascaris. Sin embargo, F. hepatica aparece recién mencionada en la
bibliografía en el año 1379 por Jean de Brie, quien la descubrió mientras realizaba un
tratado sobre la producción de lana y el manejo de ovejas para Carlos V de Francia.
Este autor no asoció al parásito con la enfermedad que provocaba en los animales,
conocida en ese entonces como “Iiver rot” (úlcera/podredumbre del hígado) y cuya
causa se asociaba a la ingestión de plantas tóxicas por parte de las ovejas. Desde esta
primera mención y hasta que se conoció el ciclo completo del parásito pasaron más de400 años.
Durante los siglos XVI y XVII fueron varios los investigadores que estudiaron al
parásito dentro del hospedador definitivo, pero nada se sabía aún sobre el resto del
ciclo de vida de F. hepatica. A partir de la observación de huevos dentro del útero de
individuos adultos encontrados en las vesículas biliares de ovejas, Govert Bidloo (1698)
propuso que estos animales se infectaban al ingerir el parásito adulto o sus huevos.
I Fuente: Andrews 1999

Estas observaciones fueron comentadas por Bidloo a Antony van Leeuwenhoek, quien
se puso a estudiar el tema. Leeuwenhoek supuso que el parásito vivía en el agua y que
las ovejas se enfermaban al beber el agua contaminada, pero no logró probar la
presencia de formas parasitarias en la misma. Recién hacia fines del siglo XVIII
aparecen referencias bibliográficas sobre los estadios restantes del ciclo de vida de F.
hepatíca. Johann Swammerdam, mientras disecaba caracoles para estudiar su anatomía
interna, encontró y describió organimos vivos que posteriormente se supo que
correspondían a cercarias. En 1773, Otto Müller encontró “criaturas semejantes a
renacuajos” nadando en el agua de lagunas y las nombró cercarias, nombre genérico
que les dio por su cola característica.
En 1803, Johann Zeder reportó haber observado la eclosión de huevos de
trematodes y Ia liberación de embriones nadadores ciliados, los miracidios. Algunos
años después Christian Nitzsch presenció el enquistamiento de cercarias. Mientras las
estudiaba nadando en agua, observó que luego de cierto tiempo se adherían al sustrato,
perdían la cola y se cubrían con una sustancia gelatinosa.
A pesar de los grandes avances en el conocimiento del ciclo de vida de este
parásito, lo único que se sabía con certeza era que los huevos eran liberados por los
adultos y que de ellos emergían miracidios. Aún cuando se conocían partes del ciclo y se
habían estudiado las cercarias y redias, se desconocía la conexión entre los distintos
estadios. Fue recién en 1818 que Ludwig Bojanus notó el parecido entre redias,
cercarias y gusanos adultos, y la observación de Ia emergencia de cercarias a partir de
redias fundamentó la idea de que existía una conexión entre cercarias y trematodes
adultos. Sin embargo, en el ambiente científico no prevaleció esta idea y aún se creía
que las cercarias eran formas de vida independientes. También se suponía que el
hospedador definitivo se infectaba al ingerir huevos del parásito, y a pesar de varias
teorías alternativas propuestas por diversos investigadores, recién en 1852 James
Simonds demostró que dicha teoría era incorrecta al presentar un trabajo en el cual
infectó una oveja con cientos de huevos de F. hepatica y en la necropsia realizada
varios meses después, no encontró lesiones ni rastros de trematodes adultos.
Posteriores estudios realizados por Guido Wagener (1857) permitieron observar
la penetración de miracidios en caracoles y el desarrollo de redias. EI heImintólogo
David Weinland en 1875 observó que las cercarias tendían a enquistarse en objetos por
encima del nivel del agua. Fue a partir de esta observación que conjeturó que las
cercarias se enquistaban en los pastos para luego ser comidas por las ovejas y que era
a partir de este estadio larval, que él consideró como gusanos juveniles, que se
desarrollaba la infección en el ganado. Veinte años después, Algeron Thomas y Leuckart
en forma independiente confirmaron que el hospedador intermediario en el ciclo de F.
hepatíca era un caracol del género Lymnaea.
Aún quedaba por probar que los herbívoros adquirían la infección al tragar
metacercarias y cuál era la ruta de migración del juvenil desenquistado hasta alcanzar

el hígado de su hospedador. Ambos interrogantes fueron dilucidados mediante
experimentos de laboratorio. Del primer tema se ocupó Adolpho Lutz (1892) al infectar
con éxito cobayos, un conejo, un chivo y una rata, alimentándolos con comida que
contenía metacercarias. El recorrido del parásito dentro del hospedador definitivo Io
describió Dimitry Sinitsin en 1914 y fue posteriormente confirmado por diversos
investigadores.
Finalmente, según Bacigalupo (1934), el parásito fue introducido a nuestro país
por los españoles junto con el ganado doméstico, en el siglo XV. Es notable como, a
pesar de su gran especificidad por el hospedero intermediario, F. hepatica fue capaz de
adaptarse exitosamente a las especies de Lymnaeas locales.
1.5 ROMPECABEZASEPIDEMIOLÓGICO. OBJETIVOS DE LA TESIS
Esta tesis tuvo por objetivo armar el rompecabezas del ciclo de vida de F.
hepatica en los valles cordillerano-patagónicos de Argentina. De este ciclo, algunas
piezas eran conocidas y era cuestión de mirarlas, girarlas, volver a mirarlas y hacerlas
encajar en el contexto del estudio, para luego buscar las piezas faltantes y tratar de
completar el rompecabezas. A medida que avanzó Ia investigación fui descubriendo que
el rompecabezas tenía una particularidad: cada nueva pieza que encontraba, parecía
por sí sola ser un rompecabezas complejo. Fue así que para poder completar el
rompecabezas del ciclo de vida de F. hepatica tuve que encontrar antes “las piezas que
conformaban las piezas”. Lo cual representó el mayor desafío de esta tesis.
Los interrogantes que originaron este estudio fueron:¿CuáIes son las estrategias
del parásito para mantener su ciclo en una región con un clima adverso durante gran
parte del año?, ¿Cuál o cuáles son los hospedadores intermediarios a esas Iatitudes?,
¿Con qué niveles de infección en hospedadores definitivos e intermediarios se mantiene
el ciclo?, ¿Entre que meses del año se da Ia transmisión?.
Luego surgieron preguntas más específicas vinculadas con el desarrollo del
parásito en los hospedadores definitivos: ¿Hay diferencias en los niveles de infección en
el ganado en animales con distintos tiempos de exposición a las pasturas?, y otras
relacionadas con el hospedador intermediario y su hábitat: ¿Conforman los caracoles
presentes en el área de estudio una única población, o existen poblaciones aisladas en
los distintos cuerpos de agua? Algunas de estas preguntas pudieron ser respondidas a
medida que avanzó el estudio. Otras sólo generaron nuevas preguntas. En ambos
casos, incursionar en el universo de este parásito fue una experiencia apasionante y
enriquecedora.

Losobjetivos generales de esta tesis son:
Describir la dinámica de transmisión de F. hepatica en los valles cordilleranos
patagónicos, e identificar Ios factores ambientales que Ia determinan a lo largodel año.
Desarrollar una propuesta de control integrado de la fasciolosis bovina que
incluya el manejo ambiental y la administración estratégica de antihelmínticos.
Los objetivos específicos y las hipótesis que generaron este estudio se presentan en
forma detallada en cada capítulo.
Esta tesis está estructurada en dos grandes partes. La primera (capítulos 2 a 5)
explora los distintos elementos involucrados en el rompecabezas epidemiológico, y en la
segunda (capítulo 6) se analizan conjuntamente dichos elementos y se desarrolla una
propuesta de manejo integral. En el capítqu 2 se presentan los antecedentes
epidemiológicos que llevaron a la elección del área de estudio. En los capítulos 3 y 4 se
desarrollan los estudios relacionados con los hospedadores definitivos e intermediarios,
incluido el ambiente donde ambos habitan. En el capítqu 5 se presentan los resultados
de los ensayos que permitieron confirmar la identidad taxonómica del parásito a través
de los adultos y evaluar Ia viabilidad de metacercarias y la capacidad de infectarse del
hospedador intermediario. En la segunda parte (capítulo 6) se propone un manejo del
ganado adecuado a las situaciones de riesgo de transmisión de F. hepatica en el área de
estudio, a partir de la información preexistente y la generada en la presente tesis. Se
completa el trabajo con una discusión general y las conclusiones de la tesis.

EL AMBIENTE
Las características del ambiente pueden tener un efecto directo o indirecto sobre
el hospedador intermediario, el parásito y el hospedador definitivo. También pueden
condicionar su distribución espacial y temporal. Por estos motivos, resulta un
componente fundamental en el ciclo de transmisión de Fascío/a hepatica, y es la primera
pieza del rompecabezas que se presenta.
La selección del área de estudio fue parte de un extenso trabajo preliminar que
consistió en determinar primero una región del país adecuada para el estudio (selección
a escala regional) y luego un establecimiento ganadero que cumpliera con ciertas
condiciones para llevar adelante el estudio (selección a escala local). En la primer parte
de este capítulo se presenta la metodología utilizada para llevar a cabo estas elecciones,
y los resultados obtenidos. En la segunda parte, se caracterizan - a partir de la
bibliografía disponible y de trabajo a campo - la región del país seleccionada, el
establecimiento ganadero y los distintos ambientes dentro del mismo.
2.1 SELECCIÓN DEL ÁREA DE ESTUDIO
2.1.1 Metodología
La información disponible sobre decomisos de hígados bovinos en todos los
frigoríficos del país fue facilitada por la Oficina de Estadística del Servicio Nacional de
Sanidad y Calidad Agroalimentaria (SENASA). A partir de esos datos se evaluó Ia
situación de la fasciolosis a escala nacional en los últimos 10 años y se estimó el
porcentaje de decomisos a escala regional y por provincia para los 3 años previos al
comienzo del estudio. Para el análisis de la parasitosis a escala regional se dividió al
18

país en Patagonia (Río Negro, Neuquén, Chubut, Santa Cruz y Tierra del Fuego),
Mesopotamia (Corrientes, Entre Ríos, Misiones), Buenos Aires y Santa Fe por ser las
provincias de mayor importancia ganadera, y se agruparon el resto de las provincias en
la categoría “Otras” (Fig. 2-1).
Luego del análisis de estos datos, se estableció comunicación con los Jefes de
Servicio y/o veterinarios de los frigoríficos de las provincias que en el período 1996
1998 mostraron un aumento en el porcentaje de casos de distomatosis, o bien tuvieron
los mayores valores porcentuales de decomisos anuales. Además se incluyeron
provincias de importancia ganadera como Buenos Aires y Santa Fe. En cada caso se
realizó una encuesta preliminar para conocer el nivel de preocupación que la parasitosis
generaba en los productores, los niveles de infección registrados durante las faenas y la
predisposición por parte de productores y veterinarios para colaborar con la
investigación.
El establecimiento ganadero. Una vez seleccionada Ia región de estudio se
identificaron campos con actividad ganadera y se realizó un cuestionario epidemiológico
en distintos establecimientos de las zonas más afectadas para conocer sus
características. Se buscó un campo con antecedentes de fasciolosis, donde se
produjeran los nacimientos y se realizaran la cría y el engorde del ganado —para
garantizar que la infección se produjera en el lugar -. Además, se contempló que el
tamaño de la hacienda permitiera la realización de muestreos representativos y que el
campo fuera de superficie extensa para garantizar la diversidad de ambientes y una
amplia variedad de cuerpos de agua.
2.1.2 Resultados y Discusión
Los niveles de infección en bovinos, registrados durante el periodo 1988-1998
para todo el país rondaron el 1% (Tabla 2-1). A escala regional, durante los tres años
previos al estudio se observó que las mayores prevalencias siempre se registraron en la
Patagonia (8-9,6%), y que fueron superiores a las informadas para la Región
Mesopotámica (3-5,2%), considerada zona de distomatosis por excelencia (Tabla 2-2).
Las provincias de Buenos Aires y Santa Fe presentaron los valores más bajos de
prevalencias (0,6-0,9°/o); sin embargo, aportaron la mayor proporción de animales
decomisados y faenados por región. Las provincias agrupadas en la categoría “Otras” no
tuvieron una contribución importante en los niveles de infección durante los años 1997
y 1998 (1,3% y 1,5% respectivamente), a pesar de ser grande la proporción de
animales faenados sobre el total del país. En cambio, las prevalencias fueron similares a
las de la región mesopotámica para el año 1996 (6,5%). El aporte de hígados
decomisados en la Patagonia no fue el mayor, por no ser tantos los animales que llegan
a frigorífico con relación al total de faenas del país; sin embargo, la prevalencia

registrada en la zona es de consideración. Además, los principales aportes estuvieron
dados sólo por las provincias de Neuquén, Río Negro y Chubut (Tabla 2-3).
La Tabla 2-3 presenta las prevalencias provinciales en bovinos, estimadas a
partir de decomisos en frigoríficos. La cantidad de hígados decomisados fue muy
variable entre provincias. Las mayores proporciones siempre se registraron en
provincias de la Patagonia (Neuquén, Río Negro, Chubut y Tierra del Fuego) o la
Mesopotamia (Entre Ríos, Misiones y Corrientes). Además se observó una tendencia de
aumento en los niveles de infección registrados en Córdoba, Río Negro, Mendoza y
Santiago del Estero. Las provincias que no aparecen con datos no presentaron sus
registros al SENASAese año. La actividad en frigoríficos varió a Io largo de los años en
cada provincia, como se ve reflejado en el número de frigoríficos que presentaron
registro.
A partir de los resultados y observaciones hasta aquí presentados, se estableció
contacto con frigoríficos de las siguientes provincias: Buenos Aires, Santa Fe, Entre
Ríos, Corrientes, Mendoza, Neuquén, Río Negro y Chubut.
Las diferencias observadas en el número de decomisos y de faenas entre
provincias, pone de manifiesto que la estimación de una prevalencia nacional no brinda
información real sobre la situación de esta zoonosis en el ámbito ganadero. Existen
provincias con registros de prevalencias elevadas aunque no aportan un porcentaje
importante de cabezas faenadas al total del país y otras que, aunque la proporción de
hígados decomisados sea baja, contribuyen mayoritariamente al total de animales
faenados anualmente a escala nacional. Esto genera un efecto de “dilución” de casos,
que redunda en prevalencias menores al 1% aunque haya aportes de algunas provincias
que fluctúan entre 10-80%.
El área de estudio. A partir de la información suministrada por los Jefes de
Servicio y/o veterinarios de los distintos frigoríficos contactados, resultó evidente que
los establecimientos ganaderos de la región cordillerana de la provincia del Chubut
manifestaban una preocupación especial por el problema. Se nos informó de Ia
existencia de distomatosis en varios campos ganaderos a pesar de la aplicación
frecuente de antiparasitarios y en algunos casos se registró Ia muerte de ovejas y de
caballos que al ser necropsiados presentaban adultos de F. hepatica en la vesícula y
canalículos biliares (com. pers. Med. Vet. Jorge Gomez Castañón, Frigorífico de
Trevelin). Considerando estos antecedentes, se estableció como área de estudio Ia zona
cordillerana del noroeste del Chubut (Fig. 2-2). Hasta el momento de comenzar este
estudio no existían trabajos de investigación que estimaran las prevalencias reales en
ganado, ni caracterizaran al hospedador intermediario que permite el mantenimiento delciclo a esas latitudes.
20

Figura 2-1: Mapa de la República Argentina y las provincias que conforman las distintas regiones analizadas
Referencias
i
PatagoniaI
Mesopotamia
Bs. As. - Sta. Fe
I
21

Tabla 2-1: Prevalencias en ganado bovino y ovino, estimadas a partir del número de hígados decomisadosen el total de los frigoríficos registrados del país, durante el preíodo 1988-1998. Fuente: SENASA.
OVINOS
ANO ANIMALES FAENADOS HIGADOS DECOMISADOS Pla! ANIMALES FAENADOS HIGADOS DECOMISADOS 5%!1988 9652625 117734 (1.22) 1421595 8323 (0.58)1989 2312508 113975 (4.93) 1618975 13770 (0.85)1990 10305000 119000 (1.15) 1649000 11467 (0.7)1991 10621000 111000 (1.04) 1341000 10000 (0.75)1992 10054000 110000 (1.1) 1060000 5000 (0.47)1993 10283000 121000 (1.18) 1002000 6000 (0.6)1994 10266000 129000 (1.25) 1059000 3000 (0.28)1995 10100000 131000 (1.3) 548000 800 (0.15)1996 10550949 205119 (1.94) 571595 571 (0.1)1997 10777755 114559 (1.06) 629631 1889 (0.3)1998 9470117 89328 (0.94) 517374 2070 (0.4)
Tabla 2-2: Prevalencias en bovinos estimadas a partir del número de hígados decomisados en el total de iosfrigoríficos registrados por región, durante el período 1996-1998. Fuente: SENASA.
Ramón # de hígados # de animales prevalencia # de decomisos por región/ Aporte por regióndecomisados faenados (W!) # de decomisos totales a la faena total (°/o)Patagonia 13726 155441 8.83 6.66 1.47Mesopotamia 22114 422523 5.23 10.78 4.00
1996 8s.As.-Sta.Fe 75588 8531947 0.89 36.85 80.86Resto 93691 1441038 6.50 45.68 13.66Total 205119 10550949
Patagonia 15368 160125 9.60 13.41 1.49Mesopotamia 15611 486145 3.21 13.63 4.51
1997 83.As.-Sta.Fe 62750 8558522 0.73 54.78 79.41Resto 20830 1572963 1.32 18.18 14.59Total 114559 10777755
Patagonia 11383 142186 8.01 12.74 1.50Mesopotamia 11523 367975 3.13 12.90 3.89
1998 8s.As.-Sta.Fe 45284 7556451 0.60 50.69 79.79Resto 21138 1403505 1.51 23.66 14.82Total 89328 9470117
Tabla 2-3: Prevalencias en bovinos estimadas a partir del número de hígados decomisados en el total de losfrigoríficos registrados en cada provincia, durante el período 1996-1998. Fuente: SENASA.
1991Ammus PREVALENCIA CANTIDADDE mmm msvmsncu cmnvo os ANIMALESPREVALENCIA
PROVINCIA maoaincos FAENADOS ps) meonincos FAENADOS (se) FRIGORÍFICOS FAENADOS (es)Buenos Aires 77 6241245 0.687 79 6244500 0.565 82 5529883 0.525
3 41416 0.027 3 52617 0 3 23518 1Chubut 5 14347 10.218 5 14271 7.96 5 14787 9.79Córdoba 22 706469 1.773 24 786160 1.84 25 721785 1.98Corrientes 3 18642 14.677 3 14408 3.7 3 6206 3.5Entre Rios 15 375730 4.791 20 432986 3.03 21 335396Formosa 2 32896 3.414 2 36485 2.67 2 38833 2 23La Pampa 7 209798 0.001 7 248628 0 02 7 284129 0 09Mendoza 5 218905 1.025 S 213241 1 48 5 191520 1 99Misiones 2 28151 4.891 2 38751 5 2 26373 5Neuquén 1 5800 81.638 1 7897 S7 9 1 21705 7 0Rio Negro 5 126722 5.354 5 130104 7 20 5 103994 8 09Saita 2 26989 1 263 2 37126 1 22 2 39485 1 79San Luns 2 145161 0 995 2 132861 1 06 2 58694 0 87Santa Cruz 1 2399 0 1 3648 1 1700 OSanta Fe 32 2290702 1.428 32 2314022 1 20 32 2026568 0.80Sgo. del Estero 1 27950 0.565 1 17337 0 82 1 9013 1.22T. del Fuego 1 6173 11.988 1 4205 7 34 OTucumán 3 14 0.808 3 48508 0.34 3 36528 0.76Total 189 10550949 1.238 198 10777755 1.063 202 9470117 0.9
Las provincias ausentes no presentaron datos. Las provincias sin actividad se encuentran indicadas con un cero en Iacantidad de frigoríficos.
22

El establecimiento ganadero. En Ia Tabla 2-4 se detallan los establecimientos
ganaderos más importantes de Ia región, los cursos de agua principales asociados, el
antihelmíntico utilizado más frecuentemente y la carga ganadera aproximada informada
por cada establecimiento. Las encuestas epidemiológicas se realizaron en 3 de los 15
establecimientos contactados, por ser los que mostraron mayor interés en que se
realizara un estudio epidemiológico sobre la parasitosis. A partir de los datos obtenidos
(Tabla 2-5), se seleccionó un establecimiento que se encuentra ubicado en Ia Localidad
de Cholila (42°32’S, 71°34’O), Departamento de Cushamen, al noroeste de Ia provincia
de Chubut (Fig. 2-2).
2.2 CARACTERIZACIÓN DEL ÁREA DE ESTUDIO
2.2.1 Escala Regional. Descripción general
EI clima que caracteriza a la Patagonia se puede explicar por factores como la
influencia oceánica, su distancia al polo, la barrera orográfica que representa Ia
Cordillera de los Andes y los centros de alta presión ubicados sobre el Océano Atlántico
y Pacifico. Así, las temperaturas máximas anuales inferiores a 0° C sólo se registran en
las zonas más elevadas de Ia Cordillera y el resto de Ia región puede considerarse como
de características templadas a templado-frías hacia el sur. Las precipitaciones están
fuertemente condicionadas por la barrera orográfica de Ia Cordillera de los Andes,
generándose una división marcada entre las zonas húmedas y las mayormente áridas
(Malvárez et al. 2004).
El área de estudio se encuentra comprendida dentro del Dominio Subantártico,
Provincia Fitogeográfica Subantártica. EI clima de este territorio es templado a frío y
húmedo, con nieve durante el invierno y heladas todo el año (Cabrera 1976). La región
presenta un régimen de precipitaciones invernales, concentrándose mayormente entre
los meses de mayo y septiembre. Durante los meses de julio y agosto, la misma suele
presentarse en forma de nieve que se deposita en los valles y principalmente en lascumbres de las montañas.
Para caracterizar los ambientes que predominan en la región es necesario
primero definir algunos términos. Se llama humedal a un conjunto de ambientes muy
diversos que integran áreas que son inundadas o saturadas por aguas superficiales o
subterráneas, con una frecuencia y duración variables. Por ende, estos ambientes
presentan suelos con características hidromórficas y vegetación adaptada al régimen
hidrológico. Un mallín o vega corresponde a un área con suelos temporaria o
permanentemente saturados con agua, con vegetación herbácea e higrofítica formada
por gramíneas y ciperáceas que aportan abundante materia orgánica a los suelos
minerales. Estos ambientes se forman en zonas con climas húmedos y baja
23

Tabla 2-4: Establecimientos ganaderos contactados en la región oeste de la provincia del Chubut
Zona Río Principal Antihelmíntico # vacasTecka* ** Tecka ‘Fasinex 2000
Esquel Esquel Fasinex 400
Río Grande* ** Grande Galgosantel 200RíoGrande Grande Galgosantel 200Gualijaina* Gualijaina ‘Fasinex 500Nantifool* Nantifool Galgosantel 200Gualijaina* Gualijaina Fasinex 400Gdor. Costa Cherque Fasinex 300
Cholila Arroyo Cordillera Galgosantel 400Cholila Arroyo Cordillera Fasinex 300
Cholila** Carrileufu Galgosantel 500Cholila Arroyo Cordillera Galgosantel 300Corinto Corinto Fasinex 300Corcovado Corcovado Ivomec F 400
Leleque* Arroyo Cordillera Fasinex 4000
* Establecimientos con animales infectados detectados mediante análisis coprológicos o necropsias. El resto de losestablecimientos detectaron Ia presencia del parásito en ganado durante la faena de animales para consumo interno.** Establecimientos en los que se realizó la encuesta epidemiológica.Galgosantel: compuesto por closantel; Fasinex: compuesto por triclabendazol; Ivomec F: ivermectina.
Tabla 2-5: Información suministrada por los responsables de los establecimientos ganaderos en los que serealizó la encuesta epidemiológica, Chubut
Y
agua encon
Enterotoxemla- (2) Enterotoxemla- Enterotoxemia-Hemosan- (2)lvermectina/Faslnex-mosca Hemogloblnurla- Galgosantel(1)/Desparasitación (2)
de vacunación/desparasitación Veces
por
24

evaporación, que generan condiciones de permanente saturación. En general, las
especies vegetales se encuentran adaptadas a condiciones extremas de bajo contenido
de oxígeno y disponibilidad de nutrientes, con aguas de pH normalmente ácido a
levemente alcalino (Roig & Roig 2004). En la región Cordillerana donde se realizó el
estudio, estas condiciones se dan en los valles, que son las zonas que reciben el agua
de escurrimiento que proviene de áreas más elevadas. Según Malvárez et al. (2004), en
la Patagonia los humedales son escasos en la mayor parte de la zona oriental
extrandina y predominantes en el extremo sudoeste de la región. Estos autores
establecieron criterios para la regionalización de los humedales, partiendo de la premisa
de que se pueden generar distintos tipos de ambientes dependiendo del régimen
climático: la estacionalidad de las precipitaciones y la relación entre éstas y la
evapotranspiración. Así, quedan definidas 6 zonas; el área de estudio se encuentra
dentro de Ia zona 4: con presencia de mallines turbosos y ocasionalmente turberas, enáreas cordilleranas.
Con respecto a la vegetación que caracteriza estos ambientes dentro del
Dominio Subantártico, en el fondo del valle (500-600 msnm) domina el bosque bajo de
ñire (Nothofagus antarctica) y Ienga (Nothofagus pumilío). Los bosques de ñire suelen
encontrarse en lugares bajos y húmedos; los de lenga son más tolerantes al frío y
ascienden por encima de los 1400 m.s.n.m. También predomina el maitén (Maytenus
boaría) en las orillas de los ríos. Asociado a los bosques, suele haber un estrato con una
bambusea de cañas macizas, el coligüe (Chusquea cu/eou). Entre los arbustos se
destacan el calafate (Berberis buxifolia) y Ia rosa mosqueta (Rosa eg/anteria). Hay
numerosas comunidades edáficas como juncales en las playas de los lagos y praderas,
generalmente inducidas por el hombre (Cabrera 1976). En relación a este último punto,
según De Pietri (1993), la presencia de manchones de pastizales en los bosques andino
patagónicos, excluyendo las praderas anegadizas (mallines), es un indicador de
actividad ganadera, esto es, de origen antrópico.
Los valles de la región son fluviales, correspondientes a antiguos valles glaciales.
El área de estudio presenta suelos desarrollados a partir de cenizas volcánicas que se
caracterizan por ser profundos, de débil desarrollo y buen drenaje y no presentan
alcalinidad ni salinidad. La textura superficial es franco arcillo arenosa (Atlas de Suelos
de la República Argentina 1985), presentando buenas condiciones de retención de agua,
factor decisivo para el desarrollo del hospedador intermediario. Estos suelos son muy
susceptibles a la erosión hídrica y/o eólica. En las laderas, los suelos son poco
profundos y pedregosos, formados a partir de los materiales originarios —pizarras de
mica, cuarzo, gneis, granito, etc. — y en muchos casos no existe desarrollo
pedogenético (De Pietri 1993).
25

2.2.2 Escala Local
2.2.2.1 Metodología
Una vez seleccionado el establecimiento ganadero, se procedió a reconocer losdistintos ambientes:
El valle. Durante una recorrida preliminar del campo ganadero, se observaron
mallines, lagunas, arroyos y charcos en Ia zona de valle y sólo unos pocos arroyos en la
zona de meseta. El acceso a las laderas fue más dificultoso por no existir sendas o
caminos, pero pudieron observarse arroyos similares a los de meseta, con mayor
corriente y más angostos. A partir de estas observaciones, se seleccionó como zona de
muestreo el terreno comprendido dentro del valle, por presentar cuerpos de agua con
condiciones teóricamente adecuadas para el desarrollo del hospedador intermediario
alta incidencia solar, corrientes suaves, poca profundidad, vegetación escasa en las
orillas- (Lombardero et al. 1979, Malone 1994) y ser accesible para la realización de los
muestreos. También se consideró que el ganado se encuentra Ia mayor parte del año en
el valle y unos pocos meses en las laderas, y que Ia zona de meseta nunca fue utilizada
para la explotación ganadera. El estudio se llevó a cabo entre diciembre de 1998 y
febrero de 2002 y los muestreos se realizaron durante la primavera, el verano y el
otoño, ya que las condiciones meteorológicas dificultan las tareas durante los mesesinvernales.
La dinámica del agua en el valle se estudió a partir de bibliografía, mapas
topográficos, fotos aéreas, imágenes satelitales, información suministrada por los
lugareños y observaciones a campo durante las fechas en que se realizaron losmuestreos.
Los cuerpos de agua. Se definieron dos tipos de cuerpos de agua: temporarios
y permanentes. Esta clasificación se consideró debido a las grandes diferencias en Ia
dinámica del agua entre esos tipos de ambientes.
Se realizaron dos muestreos preliminares en diciembre de 1998 y febrero de
1999 con el objetivo de identificar los cuerpos de agua permanentes (lagunas, ríos y
arroyos) y temporarios (mallines y charcos) presentes en el establecimiento ganadero
en una zona que abarca una superficie de aproximadamente 2000 ha. Se describió cada
cuerpo de agua según las características de su corriente, sus orillas, la profundidad de
Ia columna de agua y el tipo de vegetación. A su vez se registró el potrero dentro del
cual se encontraban y Ia accesibilidad del ganado a los mismos.
26

A partir de la información obtenida en campo y utilizando mapas topográficos,
fotos aéreas e imágenes satelitales se confeccionó un mapa del establecimiento. Los
tamaños de los cuerpos de agua se estimaron a partir de fotografías aéreas.
2.2.2.2 Resultados
EIestablecimiento ganadero en el que se realizó el estudio se halla entre los
Lagos Cholila y Rivadavia. AI este limita con el Río Carrileufú, que conecta el Lago
Cholila con el Lago Rivadavia, límite sur del campo. AI oeste se encuentra la
Cordillera y al norte linda con otro campo privado de explotación ganadera (Fig. 2
3). Abarca unas 6000 hectáreas, de las cuales 2100 ha. se encuentran en el valle,
1800 ha. en zona de meseta y 2000 ha. corresponden a Ia Pre Cordillera y
Cordillera. El valle estuvo sometido a Io largo de los años a manejos antrópicos con
el fin de generar condiciones adecuadas para Ia explotación agrícola ganadera. De
esta forma, el raleo de árboles y Ia aplicación de arbusticidas generó un valle muy
abierto, con grandes superficies de pastizales, juncales y bosques.
Descripción geomorfológica del valle. La zona de estudio se encuentra
comprendida entre los 500 y 600 metros sobre el nivel del mar y está encajonada entre
Ia Pre Cordillera y la meseta. EI valle nace en la base de Ia ladera y su nivel de menor
altitud se alcanza en el Río Carrileufú. Entre ambos extremos el ancho máximo no
supera los 2,5 Km.
La zona recibe principalmente el aporte de agua de tres arroyos de montaña que
descienden de la Cordillera, atraviesan el valle y desembocan en el río. Las áreas de
mallines se forman debido al cambio abrupto de pendiente en el terreno y a la
consecuente pérdida de velocidad del agua (Fig. 2-4).
Las formaciones de mayor altitud son un morro de aproximadamente 600
m.s.n.m. y una lomada de 560 m.s.n.m.
Ambientes estudiados. El campo fue sectorizado con alambrados en el año
2000, y a partir de entonces, los distintos ambientes quedaron contenidos en potreros.
Se individualizaron 7 cuerpos de agua permanentes (P) y 9 temporarios (T) que se
detallan a continuación y se esquematizan en la Figura 2-4.
1P.- Laguna: (Superficie aproximada: 1200 m2) Cuerpo de agua Iéntico formado sobre
un meandro (en forma de herradura) dentro del potrero 26. Aguas cálidas sin corriente.
Orillas sin vegetación tupida, con sustrato barroso y pendiente muy suave. Recibe
aportes periódicos de agua de deshielo y esporádicamente de las inundaciones del río.
Los volúmenes de agua varían desde el deshielo hacia fines del verano, habiéndose
observado a lo largo del estudio diferencias en más de 1 metro en el nivel del agua de
27

Ia costa en un período de 1 mes. Estas diferencias resultan en variaciones de las
características de las márgenes de la laguna en diferentes momentos del año. Cuando el
volumen es máximo, el agua llega a Ia altura de la vegetación herbácea permanente;
cuando disminuye queda descubierta un área barrosa, sin vegetación o con vegetación
muy escasa (Fig. 2-5). A lo largo del estudio se observó una amplia variedad de aves en
la laguna y esporádicamente coipos.
2P.- MEANDRO: (Superficie aproximada: 970 m2) Cuerpo de agua de características
similares a la laguna. Es un meandro del río Carrileufu que se encuentra aislado durante
gran parte del año y que, cuando el caudal del río aumenta, se interconecta. Por estar
tan cerca del río, su acceso es muy escarpado y su nivel está por debajo del terreno
general. Nose encuentra dentro de potreros de uso ganadero.
3P.- LAGUNAGRANDE: (Superficie aproximada: 1500 m2) Cuerpo de agua Iéntico de
gran superficie y profundidad, intersección de los potreros 35, 36 y 29. Ecotonos
abruptos definidos con juncales altos que se extienden hasta Ia orilla del lago y se
desarrollan sobre un mallín. La laguna se conecta con el río Carrileufú a través de un
arroyo de aguas claras y de corriente suave, con fondo pedregoso y bastante
vegetación. Debido a que la laguna resultó de difícil acceso sólo se muestreó el arroyo
en su nacimiento (Fig. 2-5).
4P.- RÍO CARRILEUFÚ: Río que nace en el Lago Cholila -|ago oligotrófico, de aguas
poco mineralizadas y turbias por su origen glacial- y desemboca en el Lago Rivadavia.
En la época de deshielo aumenta su caudal que disminuye hacia fines del verano. Limita
con el campo a lo largo de aproximadamente 10 km, cinco de los cuales se encuentran
alambrados y no son accesibles para el ganado. Presenta bordes escarpados, en
muchas zonas de difícil acceso. Las zonas accesibles para el ganado forman
generalmente playas de arena protegidas de la corriente (Fig. 2-5).
5P.- ARROYO 1: Arroyo que desciende de la montaña, encauzado, pedregoso y de
aguas frías. Márgenes muy arboladas. Atraviesa, de oeste a este, los potreros 21, 17,
15, 13 y 12 y desemboca en el río Carrileufu. Dentro del potrero 15 se habilita
ocasionalmente un canal de irrigación (Arroyo 1T) hacia un pequeño potrero (potrero
14) que no tiene cuerpos de agua naturales y donde en ocasiones se reúne un número
reducido de bovinos o equinos (Fig. 2-5).
6P.- ARROYO 2: Arroyo que desciende de Ia montaña en sentido oeste-este, de
características similares al arroyo 1. AI llegar al valle se ramifica formando una zona de
mallines. En el valle atraviesa los potreros 25, 29 y 27.
28

Figura 2-2:Mapa de la Provincia dei Chubut indicando el área de estudio en la Región Cordillerana
Figura 2-3: Fotografía aérea de la región noroeste de la provincia del Chubut indicando el vallede estudio, ubicado entre los lagos Rivadavia y Cholila
Lago Cholila
0.7 O 0.7 1.4 Kilometros 295:53

Figura 2-4: Mapas del valle estudiado: a) Potreros de uso ganadero y b) detalle de los cuerposde agua principales
Referencias
" PotrerosAmbientes- temporarios
permanentes
Lago RivadaviaCursos de agua
Iagmn grande
rio carrilequ
0.7 0 0.7 1.4 Kilometers5::30

7P.- ARROYO 3: Arroyo que desciende de Ia montaña en sentido oeste-este, de
características similares al arroyo 1. Desciende por el potrero 39, y al llegar al valle se
ramifica formando una zona de mallines que se extienden dentro de los potreros 36, 38
y 35, principalmente.
1T.- MALLÍN 1: (Superficie aproximada: 4500 m2) Formado a partir del arroyo 2,
abarca tres zonas principales. Dentro de los potreros 25 y 29 el arroyo forma diversos
brazos con corriente suave y zonas inundadas a su alrededor. El ambiente presenta
árboles y arbustos. Dentro del potrero 27, aguas abajo de los anteriores y habiendo
pasado el morro, se encuentra principalmente en zona de pastizal, con algunos árboles.
A pesar de ser un único ambiente, debido a que se encuentra contenido en tres potreros
distintos, para el estudio de infección se mantuvieron discriminados los distintos
sectores: dentro del potrero 29 se lo denomina mallín 1, dentro del potrero 25, mallín
1* y dentro del potrero 27, mallín 1** (Fig. 2-5).
2T.- MALLÍN 2: (Superficie aproximada: 3700 m2) Formado a partir del arroyo 1
cuando penetra en el valle y pierde velocidad. Parcialmente cubierto por un arbustal y
pastos, se encuentra dentro del potrero 17.
3T.- MALLÍN 3: (Superficie aproximada: 3500 m2) Formado a partir del arroyo 1, a
aproximadamente 800 metros aguas abajo del mallín 2. Sus características son muy
distintas a los mallines 1 y 2 ya que prácticamente sólo se observa vegetación herbácea
y el arroyo circula encauzado y ramificado y las zonas anegadizas se restringen a las
márgenes del arroyo cuando el caudal aumenta. Se encuentra ubicado dentro del
potrero 12 (Fig. 2-5).
4T.- MALLÍN 4: (Superficie aproximada: 3400 m2) Formado a partir del arroyo 3, se
encuentra alrededor de la laguna grande, en partes de los potreros 36, 38 y 35. En
apariencia más seco y heterogéneo que los mallines ya descriptos, los muestreos se
realizaron en una zona levemente más baja que sus alrededores, donde se formaba
periódicamente un piletón, con características más parecidas a una laguna que a un
mallín (ambiente con bordes definidos, sin vegetación y con sustrato barroso, de aguas
límpidas y más cálidas que las del resto del ambiente).
5T.- MALLÍN5: (Superficie aproximada: 1800 m2) Mallínformado por juncales de gran
altura, de difícilacceso. Bordea la laguna grande.
6T.- CHARCO1: (Superficie aproximada: 30 m2) Ojo de agua que se encuentra ubicado
dentro del potrero 27, a pocos metros del río. Es la parte final de un brazo del arroyo 2
que atraviesa el valle y desemboca en el río. Durante todos los muestreos realizados el
31

caudal de agua del arroyo fue bajo y el charco se encontraba desconectado o
prácticamente aislado. Presentaba en las orillas vegetación herbácea, con sustrato
barroso visible y alta incidencia solar.
7T.- CHARCO 2: (No se calculó Ia superficie) Se observó durante un único muestreo y
su formación se debió, probablemente, a intensas lluvias y la crecida del río. Se ubicó
entre el juncal y el río y presentaba aguas claras y limpias sobre las pasturas
características de gran parte del valle. Formado sobre pastos, no se observó vegetaciónacuática.
8T.- JUNCAL: (No se calculó la superficie) Mallín que se forma a partir del aporte de
agua del arroyo 2 y las inundaciones esporádicas del río. Suele mantener su humedad
durante gran parte del año. Está parcialmente cubierto por juncos de gran altura y
pastos. Ecotonos abruptos definidos.
9T.- CANAL: (Extensión aproximada: 2000 metros) Canal de irrigación artificial
construido paralelo al morro, que se alimenta a partir del arroyo 2. Tiene un ancho
aproximado de 1 metro y presenta sustrato barroso con poca vegetación y corriente
muy suave o nula, dependiendo de la época del año y del manejo agrícola (Fig. 2-5).
Las características de este valle (extenso en dirección paralela a la Cordillera y
angosto entre las montañas y el río) generan una disposición de los cuerpos de agua en
un gradiente de niveles altitudinales: desde aquellos emplazados a mayor altitud cerca
de la ladera, hasta los que están en la zona más baja junto al río (Fig. 2-6). Esta
distribución espacial se da en una superficie pequeña y en un área de pendiente muysuave.
Todos los cuerpos de agua se alimentan con el agua de deshielo que transportan
los arroyos desde las montañas y esporádicamente con las inundaciones del río. A partir
del inicio de Ia primavera, los cursos principales comienzan a aumentar su caudal,
inundando prácticamente todo el valle. Así, la mayoría de los cuerpos de agua quedan
interconectados y comienzan a individualizarse nuevamente a medida que aumentan las
temperaturas ambientales y disminuye el caudal de los arroyos. Esta dinámica se repite
año tras año pero depende fuertemente del volumen de precipitación caída durante el
invierno y de las temperaturas que se alcancen en la primavera. Si Ia cantidad de nieve
acumulada es importante y las lluvias frecuentes e intensas, es de esperarse que no
sólo queden conectados los cuerpos de agua que se encuentran alineados a distintas
altitudes, sino también aquellos ubicados a lo largo del valle a una misma altitud (Fig.
2-7). Estos últimos serán los primeros en aislarse a medida que comience el calor.
Los ambientes que se individualizaron durante este estudio presentaron
superficies variables en el tiempo. Zonas que a comienzos de la primavera estuvieron

inundadas, no pudieron ser detectadas durante las fechas de muestreos. Así, los
ambientes antes caracterizados no representan la totalidad de los existentes en el
establecimiento ganadero, pero sí los más importantes. Los cuerpos de agua
temporarios (mallines principalmente) representaron un área total mayor a la de los
cuerpos de agua permanentes (laguna, meandro y arroyos).
La accesibilidad del ganado a los distintos ambientes depende del manejo y la
rotación entre potreros. En forma general, los animales tienen posibilidad de acceso a
todos los cuerpos de agua, excepto al Río Carrileufú y a un meandro de su cauce, que
quedan excluidos de la zona de uso ganadero.

Figura 2-5: Ambientes estudiados dentro del establecimiento ganadero: a) laguna, b) río Carriieufu,c) arroyo de montaña (permanentes) y d) arroyo que nace de Ia laguna grande, e) mallín1, f) mallín 3, g) canal de irrigación (temporarios)
34

35
Figura2-6:Representacióntridimensionaldeláreadeestudio,Cholila,Chubut.Seobservaelvalleconlosprincipalescuerposde
agua,alpiedelaCordillera

Figura 2-7: Esquema de Ia dinámica del agua en el área de estudio, Cholila, Chubut
3 1 )a)
b)
c)
d)
En el esquema se representa la zona de estudio (rectángulo) en distintas situaciones con respecto a la disponibilidad de aguaen el ambiente. Los cursos de agua (arroyos, flechas azules) descienden desde las montañas y al llegar al valle alimentana los distintos cuerpos de agua (simbolizados de manera general por cuadrados azules). A favor de la pendiente, el aguaescurre a través de los distintos cuerpos de agua hacia el río (franja rectangular azul). Durante los períodos más secos del año(a) la conexión entre cuerpos de agua a un mismo nivel y a distintos niveles altitudinales es mínima. Luego del deshielo (b)el agua fluye intensamente conectando ambientes que se encuentian en el gradiente altitudinal e incluso a una misma altitud.A medida que avanza Ia primavera el caudal de los arroyos que ingresan al valle disminuye y los distintos ambientes comienzana aislarse parcialmente nuevamente (c). Finalmente se llega a la situación inicial (d) y la conexión entre cuerpos de aguavariará según las condiciones climáticas (temperaturas y precipitación) que hayan caracterizado cada ciclo.
36

2.3 CONCLUSIONES
1. Con respecto a Ia situación de Ia fasciolosis en el país durante los últimos años y
según los registros oficiales:
Las mayores prevalencias se registran en la Patagonia.
Las dos principales provincias ganaderas (Buenos Aires y Santa Fe) presentan
valores bajos de prevalencia, pero altos niveles de decomisos de hígados con
respecto al total del país.
2. La estimación de una prevalencia nacional no brinda información real sobre Ia
situación de esta zoonosis en el ámbito ganadero. A escala local, existen zonas en las
que Ia fasciolosis no resulta un problema y otras en las que las prevalencias toman
valores muy elevados. De esta manera, los valores promedio enmascaran las altas
prevalencias locales.
3. Los distintos cuerpos de agua presentes en el área de estudio no están aislados
temporal ni espacialmente. En las épocas de mayor disponibilidad de agua en el suelo,
los ambientes se interconectan. Algunos permanecen en esa situación todo el año y
otros se aíslan a medida que Ia disponibilidad hídrica disminuye.

EL HOSPEDADOR DEFINITIVO
En este capítqu se presentan los estudios que permitieron estimar los niveles de
infección en ganado bovino, en distintos grupos de interés epidemiológico y en distintos
momentos. Por otro lado, se realizó un estudio exploratorio para conocer los posibles
hospedadores silvestres que hay en la región. Para la realización de esta serie de
estudios fue necesario en primera instancia poner a punto un método de diagnóstico
coprológico confiable, cuya sensibilidad y utilidad se evalúan y discuten también en este
capítulo.
3.1 INTRODUCCIÓN
Fascio/a hepática es uno de los parásitos digeneos de ganado más ampliamente
distribuido e intensamente estudiado a escala mundial. Los hospedadores domésticos
más comunes son los bovinos y ovinos, pero el parásito se desarrolla también en
búfalos, caprinos, camélidos, suinos y equinos (Boray 1985, Rickard 1992, Apt et al.
1993, Buchon et al. 1997, Mas-Coma et al. 1997). Existen además diversas especies
silvestres en las cuales se ha documentado la presencia del parásito y en algunos casos
se ha estudiado su rol en el ciclo de transmisión: coipos, ciervos, marsupiales
australianos, Iagomorfos y pequeños roedores (Presidente et al. 1974, Spratt &
Presidente 1981, Apt et al. 1993, Ménard et al. 2000, Shimalov & Shimalov 2000,
Ménard et al. 2001).
La fasciolosis en el ganado bovino y ovino es considerada una enfermedad de
importancia en el ámbito veterinario porque produce grandes pérdidas económicas y
productivas. Esto se debe a que la enfermedad en los animales suele presentarse en
38

forma subclínica y se manifiesta como una disminución en el rendimiento (Malone
1994). El tejido hepático del hospedador es dañado durante la migración de los
juveniles de F. hepatíca y reemplazado por tejido fibroso. En los ductos o canalícuios
biliares la irritación mecánica ocasionada por los adultos produce primero inflamación y
luego el engrosamiento y en algunos casos severos la calcificación de la pared y bloqueo
del ducto biliar (Behm & Sangster 1999). Los hígados que presentan cicatrices y ductos
bloqueados son decomisados durante la faena en frigoríficos y mataderos. Según la
Asociación Americana de Parasitólogos Veterinarios, en Estados Unidos se decomisan
aproximadamente 1,5 millones de hígados anualmente (Dargie 1987). En Chile, durante
el año 1987 se decomisaron 213.921 hígados de bovinos, lo que a un peso promedio de
5 kg por hígado significó una pérdida de 1070 toneladas. (Alcaino et al. 1993). Sin
embargo, aunque el decomiso de hígados infectados es la manifestación más visible de
Ia enfermedad, las pérdidas económicas en la producción de carne exceden en alto
grado a las pérdidas producidas por decomisos. En el estado de Florida, EEUU, se
estimó que la industria de la carne pierde USIO millones cada año a causa de Ia
fasciolosis. La infección por F. hepatíca resulta en un menor incremento de biomasa que
se manifiesta en terneros más livianos al destete, crecimiento y engorde más lento de
las vaquillonas de reposición y mayores tasas de descarte de la tropa. Una infección
moderada produce una reducción de aproximadamente el 9% en la ganancia de peso y
una infección mayor puede llegar a producir pérdidas del 28%. Estas pérdidas no se
recuperan aunque los animales se traten con antiparasitarios y se elimine la infección
(Hope Cawdery et al. 1977). Se ha comprobado también que los bovinos infectados
presentan menor fertilidad, requiriendo un mayor número de inseminaciones por
concepción (2,3 vs. 1,6 para los no infectados) y que pueden producirse abortos por
liberación de toxinas o migración errática del trematode juvenil (Eddi 1991).
En las tropas de bovinos infectados se ha registrado una disminución de 10-15%
en la producción de leche, recuperándose sólo un 8% después del tratamiento
antihelmíntico (Eddi 1991, Torgerson & Claxton 1999). Incluso algunos autores sugieren
que la infección puede afectar negativamente la calidad de la leche (Black 1972).
Finalmente, a las pérdidas directas recién mencionadas deben sumársele los
gastos indirectos derivados de la necesidad de implementar el uso frecuente de
antihelmínticos, para mantener a la población parasitaria controlada y obtener los
resultados productivos esperados.
Los porcentajes de infección en ganado que permiten que el parásito se
transmita al caracol y que su ciclo se mantenga en forma exitosa, son muy variables
según las características de cada región. Restringiendo la información a los países
limítrofes con Argentina, en un amplio estudio coprológico realizado entre 1977 y 1988,
que abarcó 129 municipios del Estado de Santa Catarina en Brasil, se hallaron
prevalencias de 27,9% en vacas, 24,7% en búfalos, 16,9% en ovejas y 15,7% en
39

cabras (Maués da Serra-Freire & Nuernberg 1992). En el Altiplano Boliviano se
registraron prevalencias en el ganado que varían desde el 16,5 hasta el 60% (Buchón et
al. 1997). La información para el Uruguay deriva de las cifras oficiales de decomisos de
hígados bovinos que pasaron de 54,7% en 1981 a 59,7% en 1990 (Eddi 1991) y en
Chile, los mataderos han llegado a informar prevalencias del 90% en bovinos (Apt et al.
1993). En nuestro país, la información epidemiológica proviene de los registros de
hígados decomisados en mataderos de distintas provincias. Como se mencionó en el
capítqu 2, dichos estudios son escasos; sin embargo evidencian fluctuaciones muy
grandes en las prevalencias en distintas regiones, hallándose registros cercanos al
100% y zonas prácticamente sin infección.
Naturalmente no todas las especies descriptas como hospedadores definitivos de
F. hepatica son igualmente susceptibles a infectarse. Además, la presencia del parásito
puede producir cierto grado de resistencia en los animales, el cual varía según el
hospedador. Algunas especies muestran una resistencia innata, como es el caso de los
caballos, que son menos susceptibles que los rumiantes (Torgerson & Claxton 1999).
Con respecto a este último grupo, en general existe acuerdo en la bibliografía en que los
bovinos también presentan cierto grado de resistencia, y que los animales jóvenes,
desnutridos o preñados, junto con los ovinos, conforman el grupo de mayor
susceptibilidad al parásito (Castro 1973, Boray 1981, Dargie 1987). En ovinos no hay
evidencias de resistencia a infecciones experimentales secundarias o subsecuentes, en
cambio en bovinos se han observado niveles importantes de resistencia adquirida luego
de sucesivas infecciones. Por otro lado, se ha comprobado que en el caso del ganado
vacuno puede haber una recuperación o auto-cura luego de una infección primaria y
que no ocurre lo mismo en ovejas, animales en los cuales una infrapoblación de baja
densidad puede sobrevivir más de 11 años (Boray 1985, Keegan & Trudgett 1992). La
susceptibilidad al parásito puede variar también con la edad del hospedador. Un estudio
realizado por Gonzalez-Lanza et al. (1989) indica que la prevalencia en poblaciones
vacunas aumenta con la edad hasta los 4 años de vida y luego se mantiene constante
hasta las edades máximas (11-18 años). Con respecto a la cantidad de huevos
liberados por gramo de materia fecal (hpg), variable que algunos investigadores
consideran adecuada para estimar la carga parasitaria de helmintos (Bardie et al. 1978,
Malone 1994), se registró el máximo valor promedio en animales de entre 3 y 4 años, y
el mínimo en animales de 1 año de edad. De acuerdo a estos autores, las diferencias en
los niveles de infección entre edades serían explicadas por los distintos tiempos de
exposición que presenta cada grupo a las pasturas infestadas: el hecho de que animales
jóvenes (hasta 1 año) presenten los valores más bajos de prevalencia y de hpg serían el
reflejo de que los mismos no tuvieron oportunidad de infectarse. Por otro lado, según
Torgerson & Claxton (1999), el hecho de que a partir de cierta edad la proporción de
4o

animales infectados se mantenga constante sugiere que se alcanzó un equilibrio entre la
tasa de adquisición y muerte del parásito.
En general en todo el mundo, la fasciolosis en ganado doméstico se controla
mediante la aplicación masiva de antiparasitarios, sin discriminar entre animales
positivos y negativos. En la Argentina, el tratamiento contra esta parasitosis obedece al
mismo esquema en todo el país. Sin embargo, las características de la transmisión no
son iguales en la zona subtropical y en los valles templados. Los antiparasitarios
específicos contra F. hepatica disponibles en el mercado local están compuestos
principalmente por closantel, triclabendazol, albendazol o ivermectina y su
administración puede ser oral o inyectable, dependiendo del producto. La mayoría
matan al parásito en estado adulto, pero existen productos que lo eliminan a partir de la
primera o quinta semana postinfección. Otra de las variables entre los diversos
productos es la vida media de la droga en sangre, que puede fluctuar entre 48 horas y
aproximadamente 1 mes desde el momento de la aplicación.
En el ámbito veterinario, el principal objetivo de la aplicación de drogas es
minimizar los efectos de la parasitosis en los animales y maximizar la producción; sin
embargo, en el contexto de un estudio epidemiológico, resulta de suma importancia
poder interrumpir la oferta de huevos de F. hepatica al medio, ya que de ella depende
en gran medida la continuidad de la transmisión. Los tratamientos antihelmínticos
efectivos provocan Ia muerte de la población parasitaria dentro de los hospedadores
infectados, cortando o minimizando la liberación de huevos al medio. Los
antiparasitarios que también matan estadios juveniles del parásito ayudan a controlar la
cantidad de hospedadores potencialmente infectivos.
Existen diversas maneras de cuantificar la presencia del parásito en el
hospedador definitivo. Entre los métodos diagnósticos existentes, las técnicas
coprológicas permiten detectar a los animales con infecciones maduras que presentan
huevos en la materia fecal. Este dato es de suma importancia en un estudio
epidemiológico ya que es un buen estimador de la oferta de huevos al medio.
Posibles hospedadores silvestres en la región de estudio. Como semencionó al comienzo de esta introducción, se han descripto numerosas especies de
mamíferos que pueden actuar como hospedadores silvestres. En particular, existen
registros del hallazgo de infección por F. hepatíca en 2 especies de la Familia Leporidae
(Lagomorpha) en condiciones naturales: el conejo europeo, Orycto/agus cun/culus
(Linnaeus, 1758) con prevalencias de aproximadamente 6% en Sudamérica y 33% en
Europa (Apt et al. 1993, Ménard et al. 2000) y la liebre europea, (Lepus europaeus
Pallas 1778) con infecciones del 9% en Europa del este (Shimalov & Shimalov 2000).
También se ha estudiado el rol del coipo Myocastor coypus (Molina, 1782) como
hospedador en la naturaleza. En Brasil se detectó un 6% de infección en zona ganadera
41

endémica para el parásito (Silva Santos et al. 1992). Ménard et al. (2001) encontraron
prevalencias en Francia que fluctuaron entre 8% y 40% y comprobaron que los huevos
eliminados por los coipos continuaban su desarrollo como larvas de vida libre y luego
dentro del hospedador intermediario hasta convertirse en metacercarias tan infectivas
para los ovinos como las que provenían de los huevos liberados por parásitos
mantenidos naturalmente en las ovejas. EIcoipo es una especie nativa de Sudamérica.
En la Argentina se han descripto dos subespecies, una de las cuales se encuentra en Ia
Patagonia y Ia otra se distribuye en el norte del país (Woods et al. 1992). La liebre
europea, en cambio, fue introducida a la Argentina desde Alemania para la caza
deportiva en 1888 y pocos años después fue declarada plaga agrícola (Bonino 1986a
1986b, Silva et al. 1986). En la Patagonia es muy abundante y ocasiona daños en
explotaciones agrícolas y forestales. Por este motivo y con el objetivo de controlar Ia
densidad poblacional de la especie, en el año 1998 se sancionó en la provincia del
Chubut la Ley N° 4375‘ que autoriza a los frigoríficos la comercialización de productos
de liebre, promoviendo un manejo sustentable del recurso.
A pesar de que en la Argentina Ia fasciolosis es endémica y que tanto L.
europaeus como M. coypus son especies muy abundantes en todo el territorio, no
existen estudios para determinar su importancia en la transmisión de F. hepatica.
‘ Boletín Provincial N° 8035, 29 de junio de 1998, Chubut

3.2 OBJETIVOS
Estimar los niveles de infección por F. hepatica en el ganado al comienzo del estudio.
Luego de comenzada la investigación, se implementó un manejo sanitario regular y
mucho más riguroso en el establecimiento. Con el objetivo de evaluar cómo ese manejo
repercutió en los niveles de infección, se estimó Ia prevalencia en ganado al finalizar elestudio.
Estimar la prevalencia en los distintos grupos de interés epidemiológico.
Estudiar la incidencia del parásito en una cohorte juvenil y relacionarla con el manejo
sanitario implementado en el lugar de estudio.
Describir la ocurrencia de mamíferos herbívoros silvestres que podrían estar actuando
como hospedadores definitivos.
3.3 HIPÓTESIS
Los niveles de infección en ganado se ven afectados por el manejo al que los
animales están sometidos. El manejo antrópico es una variable de confusión inherente
al sistema epidemiológico que altera la exposición del ganado al parásito (ya sea a
través del movimiento de Ia tropa o la remoción de animales del campo) y Ia
probabilidad de que Ia infección sea exitosa (por el tratamiento antihelmíntico).
Específicamente se espera que el tratamiento antiparasitario sostenido en forma regular
en eI tiempo controle a la población de parásitos y que dicho efecto se visualice como
menores prevalencias y/o cargas parasitarias.
Las prevalencias difieren entre grupos etarios, presentando mayores niveles de
infección los anima/es jóvenes. Los animales de distintas edades presentarán distintos
niveles de infección en función del tiempo de exposición a las pasturas, la cantidad de
aplicaciones de antiparasitarios recibidas durante sus vidas y/o el manejo ganadero al
cual hayan sido sometidos. Suponiendo que los animales jóvenes ya destetados con poca
experiencia de contacto con el parásito presentarán menor resistencia a las infecciones y
reinfecciones, se espera encontrar mayores prevalencias en animales jóvenes que enadultos.
43

3.4 MATERIALES Y MÉTODOS
3.4.1 Elganado: Población en estudio
El estudio se realizó en la población de ganado bovino del establecimiento
ganadero, perteneciente a la raza Hereford (Figura 3-1a). La hacienda se clasificó en
grupos etarios según el interés epidemiológico: terneros, vaquillonas, vacas y toros. Se
definen como terneros a aquellos animales de hasta 6 meses de edad que aún no han
sido destetados; vaquillonas a las hembras de entre 6 meses y 2-3 años que aún no
tuvieron cría y vacas a las hembras luego de la primera parición (esta última categoría
puede incluir animales de entre 3-4 y 9 años de edad). Los toros son los machos
adultos. Por último, los novillos (machos de edad equivalente a las vaquillonas) no
fueron estudiados ya que durante el destete se separan los terneros hembras de los
machos y estos últimos son vendidos y salen del campo en el corto plazo.
3.4.2 Manejo ganadero
Para conocer el manejo al cual estuvo sometido el ganado, se recopiló
información a través del veterinario a cargo y del personal del establecimiento. Entre
1999 y febrero de 2002 se tuvo acceso a los registros generados por el veterinario, para
conocer el movimiento de la hacienda dentro del campo.
En forma general, el ganado permanece en la zona de estudio -valle- durante la
primavera y el verano. A comienzos del período invernal los animales son llevados a las
laderas de la Cordillera, donde permanecen hasta mediados de la siguiente primavera,
momento en el que vuelven al valle. Este manejo, común en la zona, se implementa
porque el valle se encuentra cubierto por la nieve durante los meses de invierno e
inundado luego del deshielo (hasta el mes de septiembre aproximadamente) y las
pasturas no están disponibles para los animales durante el período invernal (Fig. 3-2).
Las laderas de las montañas, en cambio, presentan gran abundancia de caña coligüe
(Chusquea cu/eou), especie vegetal que puede alcanzar los 2 metros de altura y que
protege a los bovinos de las nevadas y les permite ramonear sus hojas mientras el
suelo está nevado o congelado.
Debido al manejo ganadero de compra y venta de animales, el tamaño
poblacional fue variando durante el estudio. AI comienzo (diciembre de 1998), la
hacienda estaba compuesta por ganado bovino y ovino y no se encontraba sometida a
un manejo regular. Los animales, algunos nacidos en el campo y otros procedentes de
diversas zonas, pastaban libremente ya que el terreno no estaba sectorizado. Esta
situación se mantuvo hasta comienzos de 1999. A partir de ese momento y durante
todo el estudio, como resultado de la implementación de un nuevo manejo ganadero,
44

Figura 3-1: Ganado bovino estudiado y manejo sanitario: a) vacas de la raza Hereford, b) tratamientosanitario, c) vista de la entrada a la manga de trabajo, d) vista de ia salida de la mangade trabajo
Figura 3-2: Vista del valle donde se realizó el estudio, Choiila, Chubut: a) durante el períodoestival, b) durante el período invernal
45

sólo hubo ganado bovino en el campo. Las ovejas fueron usadas para consumo interno
durante el transcurso del primer año de estudio y los animales no se repusieron. Debido
a que el valle no se encontraba sectorizado, los animales tuvieron libre acceso a todos
los cuerpos de agua hasta mediados del año 2000, momento en que se delimitaron
potreros. A partir de entonces se pudo conocer el paradero del ganado en forma parcial
a lo largo del tiempo.
Rotación de animales. El movimiento de Ia hacienda en el valle respondió a las
necesidades ganaderas y no hubo un manejo regular y constante ni rotaciones
programadas entre potreros. A continuación se detalla Ia información recabada sobre los
potreros que estuvieron ocupados por animales a partir de mayo de 2000. Los animales
fueron divididos en: rodeo base y rodeo general, ambos rodeos de animales adultos
(RA), vaquillonas nacidas en octubre de 1998 (VQ 1998) y vaquillonas nacidas en
octubre de 1999 (VQ 1999). Los terneros siempre estuvieron junto con sus madres,
pertenecientes al RA. No se pudo conocer Ia carga animal que frecuentó cada potrero en
cada ocasión. La ausencia de información en ciertos meses o para ciertos rodeossignifica que los animales no estuvieron en el valle en dichas ocasiones.
Mayo de 2000: RAen potrero 17, VQ 1999 en potreros 20 y 21
Junio de 2000: VQ 1999 en potrero 20
Julio de 2000: VQ 1999 en potrero 20
Agosto de 2000: RAen potreros 3, 4 y 38; VQ 1998 en potrero 17; VQ 1999 en potrero
20
Septiembre de 2000: RAen potreros 2, 3, 4 y 38; VQ 1998 en potrero 17; VQ 1999 en
potrero 20
Octubre de 2000: RAen potreros 36 y 38; VQ 1998 en potrero 17; VQ 1999 en potrero
20
Noviembre de 2000: RA en potreros 3, 28, 29 y 36; VQ 1998 en potrero 27; VQ 1999
en potreros 21, 24 y 25
Diciembre de 2000: RA en potreros 3, 5, 6, 26 y 35; VQ 1998 en potrero 29; VQ 1999
en potreros 24 y 25
Enero de 2001: RA en potreros 28 y 35; VQ 1998 en potrero 29; VQ 1999 en potreros
15 y 24
Febrero de 2001: RA en potrero 35; VQ 1998 en potrero 29; VQ 1999 en potreros 12,
15 y 24
Marzo de 2001: RAen potrero 38; VQ 1999 en potreros 12, 20 y 38
Abril de 2001: VQ 1999 en potreros 13 y 15
Mayo de 2001: VQ 1999 en potrero 12
Octubre de 2001: RAen potreros 20 y 26
46

Noviembre de 2001: RA en potreros 5, 6, 14, 20 y 26, toros adultos en potrero 15,
toros de 1 año en potrero 21, VQen potrero 28
Diciembre de 2001: RAen potreros 2, 24 y 27, VQ 1999 potrero 17
Enero de 2002: RAen potreros 15, 26 y 27
La figura 3-3 presenta un esquema del movimiento recién detallado.
Manejo sanitario. Los animales recibieron tratamientos antihelmínticos
esporádicos e irregulares hasta el año 1998. A partir de ese momento, se implementó
como estrategia sanitaria la desparasitación y vacunación de toda la tropa dos veces al
año: hacia fines de Ia primavera y hacia fines del otoño. Los terneros, cuyo nacimiento
ocurre entre septiembre y octubre, no reciben tratamientos hasta el momento del
destete, aproximadamente a los 6 meses de edad y en coincidencia con el manejootoñal.
Hasta el momento en que se comenzó el estudio se utilizaba principalmente un
producto compuesto por Triclabendazol como antiparasitario interno. Esta droga elimina
parásitos de todos los estadios Iarvales, pero tiene un efecto residual de
aproximadamente una semana (Richards et al. 1990). Desde fines de 1998 y hasta la
fecha de finalización del estudio se utilizó Closantel, dosificado en función del peso de
los animales tratados. Esta droga fasciolicida actúa únicamente sobre los adultos o bien
sobre estadios juveniles avanzados (a partir de la 6a semana postinfección) y tiene un
efecto residual en sangre de aproximadamente 7 semanas (Com. pers. Depto. Técnico
Biogénesis S.A.). En general, los tratamientos sanitarios se planificaron para toda la
hacienda, pero existieron casos particulares en los que un grupo de animales fue
tratado en fechas diferentes a las de los grupos restantes. Los cronogramas generales y
particulares de desparasitación se presentan en la tabla 3-1.

Figura 3-3: Esquematización del movimiento del ganado dentro del establecimiento de estudioa partir de la delimitación de potreros
POU’GI’OS
20 21
May-00JunvOOJul-00Ago-00Sep-00Oct-00
' Nov-00Dic-00Ene-01
g Feb-01i Mar-01
Abr-01May-01Jun-01
i Jul-01Ago-01Sep-01Oct-01
E Nov-01Dic-01
Y Ene-02
Los n
* Sin** 70 RA, 28V*** VQY RA
potrero.
Fúmeros dentro de las casillas indican la carga de animales que frecuentó el Referencias:
Sin animales
acceso al cuerpo de agua principal (la Laguna) QQ
Tabla 3-1: Cronograma de desparasitación en el ganado bovino estudiado
Fecha RodeodesgarasitadoMayo 1998 todosDiciembre 1998 todos
Mayo 1999 todosSeptiembre 1999 RAy VQ 1998Diciembre 1999 todos
Mayo 2000 todosEnero 2001 todos
Mayo 2001 toros jóvenesJunio 2001 VQ 1999Noviembre 2001 toros jóvenesDiciembre 2001 todos
RA: Rodeo adulto, VQ 1998: vaquillonas nacidas en octubre de 1998, VQ1999: vaquillonas nacidas en octubrede 1999. Los toros jóvenes eran animales de 1 año de edad.
48

3.4.3 Infección en ganado
Extracción de materia fecal
Para estudiar la infección por F. hepatica en el ganado, se tomaron muestras de
materia fecal en distintos momentos durante el estudio, en función de la posibilidad de
acceder a los animales. Para Ia obtención de las muestras se aprovecharon los
momentos en que se realizaban las actividades regulares de vacunación y
desparasitación. Las fechas en que se realizaron los muestreos y el grupo de animales
que se estudió en cada ocasión se detallan a continuación:
- Diciembre de 1998: vaquillonas, vacas y toros
- Febrero de 1999: terneros (nacidos en octubre de 1998)
- Enero de 2001: vaquillonas (VQ 1999)
- Noviembre de 2001: toros jóvenes
- Diciembre de 2001: vaquillonas y vacas
Los animales fueron reunidos en un corral y se los hizo pasar por una manga de
trabajo donde se los pudo manipular en forma individual y ordenada (Figura 3-1b, c y
d). Todas las muestras fueron extraídas directamente del recto de cada individuo con
guantes descartables largos hasta el hombro, excepto en el caso de los terneros, que seobtuvieron mediante la estimulación anal —únicométodo viable en animales de esta
categoría-. En todos los casos, los animales presentaban caravanas y las muestras
fueron identificadas con el número de cada individuo. Aproximadamente 25 grs de
materia fecal se conservaron en recipientes plásticos fijados con 80 ml de formol 5%
(aproximadamente 1 volumen de materia fecal en 3 volúmenes de conservante, según
recomienda el CDC2), o frescos, congelados a —17°C. La utilización de formol como
conservante fue necesaria inicialmente debido a la imposibilidad de mantener la cadena
de frío en el campo durante los días correspondientes al primer muestreo. Para todos
los muestreos subsiguientes se congelaron las muestras hasta su procesamiento y se
prescindió del formol. La cantidad de material fresco utilizado en cada caso fue pesada
en una balanza de precisión con un error de 10'2 gramos.
Método diagnóstico
Para la detección de huevos de F. hepatica en la materia fecal se utilizó
inicialmente la técnica de sedimentación-concentración de formol-éter (Ritchie 1948,
Shore García 1973, OMS 1994). Esta técnica, a pesar de ser la más comúnmente
2 CDC.Identification and diagnosis of parasites of public health concern: diagnostic procedures for stoolspecimens. 7/10/2002. http://www.dpd.cdc.gov/DPDx/HTML/DiagnosticProcedures.html
49

utilizada en laboratorio, presentó la desventaja de requerir muchos pasos e insumir
mucho tiempo para el procesamiento y la observación de cada muestra. Además,
durante el procesamiento de las primeras muestras surgieron dudas sobre la
sensibilidad y confiabilidad del método, debido a la baja cantidad de huevos
encontrados en las muestras positivas. Por estos motivos se buscó un procedimiento
alternativo que permitiera procesar la materia fecal en menor tiempo y cuya
sensibilidad fuese mayor que Ia del método de sedimentación-concentración. Para ello
se puso a punto una técnica de tamización, basada en métodos previamente descriptos
(Giráo & Ueno 1994, Zukowski et al. 1993, Dresden 1981, Boray & Pearson 1960) y se
evaluó su utilidad relativa al método de sedimentación-concentración, comparando
sensibilidad y tiempo de desarrollo.Se utilizaron muestras de materia fecal extraídas en diciembre de 1998 a 51
bovinos (vaquillonas y vacas) nacidos y criados en el establecimiento de estudio.
Veinticinco gramos de heces de cada individuo se homogeneizaron en recipientes
plásticos que contenían 80 ml de formol al 5%. De cada muestra en suspensión, se
realizaron 2 extracciones de 8 ml —equivalentes a 2,5 gr de heces - y se asignaron al
azar a cada uno de los dos métodos.
El método de 4' ‘ " ‘ración (Shore García 1973) (Fig. 3-4a).
Cada muestra fue filtrada a través de gasas colocadas en un colador y luego 8 ml de la
suspensión se volcaron en un tubo de centrífuga de 10 ml de capacidad y se agregaron
2 ml de solución salina. Luego de homogeneizar, la suspensión fue centrifugada a 1500
RPM(174g) durante 2 minutos. El sobrenadante fue descartado y se incorporó formol al
5% hasta alcanzar nuevamente un volumen de 10 ml. Luego se homogeneizó por
agitación y se centrifugó nuevamente en las mismas condiciones. Una vez descartado el
sobrenadante, se resuspendió el sedimento en 7 ml de formol 5% y 3 ml de éter. Se
centrifugó por última vez, se descartó nuevamente el sobrenadante y el sedimento fue
observado a microscopio óptico (100x) entre cubre (20 x 20 mm) y portaobjetos.
Debido a la gran cantidad de material recuperado, cada muestra rindió entre 25 y 30
preparados. Se consideró positiva a una muestra cuando se encontró al menos un
huevo en el total de los preparados. El número de huevos recuperados fue registrado entodos los casos.
El método de tamización (Fig. 3-4b) se basó en un trabajo previo de Giráo &
Ueno (1994), modificado para simplificarlo. Ocho ml de suspensión fecal de cada
muestra se filtraron a través de un set de tres tamices encajados. Los tamaños de las
mallas son de 250, 125 y 53 pm cada uno (40, 120 y 270 U.S. Standard Size,
respectivamente), desde el primero hasta el tercero y el diámetro de los tamices es de
20cm. Los dos primeros retienen partículas grandes, mientras que el último permite
recolectar huevos de F. hepatica.
50

Luego de volcar el material sobre el primer tamiz se lavó con abundante agua
corriente a presión para permitir que el material y los huevos pasaran a través de los
tamices y que el material grande quedara retenido en cada uno, dependiendo de su
tamaño. Luego se quitó el primer tamiz y se repitió el procedimiento sobre el segundo.
Finalmente, el tamiz de 53 um se inclinó levemente y se Iavó con agua para permitir
que todo el material retenido en el mismo escurriera hacia un costado. Dicho material
fue transferido a una caja de Petri y teñido con unas gotas de azul de metileno 1% para
diferenciar los restos vegetales (teñidos de azul) de los huevos amarillos de F. hepatíca
(Boray & Pearson 1960). Se buscaron huevos del parásito bajo microscopio
estereoscópico con transiluminación a un aumento de 20x. Se contaron todos loshuevos encontrados en cada muestra.
Para evitar la contaminación entre muestras, todos los tamices fueron
vigorosamente lavados en ambos sentidos con agua corriente a presión antes de volvera utilizarlos.
Para probar la precisión del método, se agregaron aproximadamente 100 huevos
de F. hepatíca obtenidos del útero de un espécimen adulto a una suspensión de 8 ml de
heces en formol 5% que fue posteriormente procesada como se acaba de explicar. La
materia fecal utilizada para este ensayo fue obtenida de una vaca sana criada en el
predio de la Facultad de Ciencias Exactas y Naturales de Ia Universidad de Buenos Aires.
Todo el material retenido en cada tamiz y el que atravesó los mismos se examinó bajo
microscopio estereoscópico. Los huevos sólo se encontraron en el material retenido por
el tamiz de 53um. Este tamiz fue entonces lavado y se juntó el posible material que
pudiera haber quedado retenido en la malla y se examinó bajo microscopio
estereoscópico para descartar la presencia de huevos.
Dado que no se encontraron huevos que hubiesen quedado retenidos en la malla
con posterioridad al lavado, se confirmó la eficiencia del procedimiento de limpieza y lacorrecta elección de los tamices usados.
En ambos métodos dos observadores revisaron cada una de las muestras.
La determinación de los huevos de F. hepatica en materia fecal se realizó
utilizando los Medios Auxiliares para el Diagnóstico de Parasitosis Intestinales publicado
por Ia Organización Mundial de la Salud (1994).

Figura 3-4: Protocolo de los métodos coprológicos de diagnóstico comparados para la detecciónde F. hepatica: a) Método de sedimentación-concentración, b) Método de tamizaciónCada muestra se subdividió y se procesó por ambos métodos
n8m.M.Eüa ®Hinata/s 7 ‘ /Y de n mFTYdsno o.| 3MB“ «by. sm Muestra
¿mg-:9 . _ m' / Hamgmaza' /Hamgmazar H
Ga» TN'IÓÓ‘ICGIw AzuldeMeüleno
I‘m ® vé V “¿a W“amd/mw ÜqÉ vw
ÜmmQI V
0'v
52

Estimación de la prevalencia al comienzo del estudio
Se realizó un primer muestreo sobre la hacienda adulta (vaquillonas, vacas y
toros) en diciembre de 1998 y sobre los terneros en febrero de 1999, al momento del
destete. La hacienda, en ese momento, estaba compuesta por 957 bovinos. Los
animales fueron agrupados en categorías de interés epidemiológico -por edades (como
ya se explicó) y según su origen-.
Los terneros (6 meses de edad) y vaquillonas (1 y 2 años) nacieron en el campo.
Las vacas (más de 5 años de edad) fueron divididas para su estudio en dos grupos: por
un lado los animales nacidos y criados en el campo (vacas NyC) y por otro las vacas que
se compraron en otros establecimientos e ingresaron al campo en mayo de 1998, 7
meses antes del estudio (vacas A). La procedencia de este grupo de animales no se
pudo identificar.
Finalmente, el grupo de los toros también estuvo formado por animales de
distinta procedencia y tiempo de estadía en el campo, pero esta información tampoco
pudo ser recopilada.
Por tratarse de un muestreo preliminar, no se estableció a priori el tamaño
muestral necesario para la estimación de Ia prevalencia, sino que la cantidad de
individuos quedó supeditada a las posibilidades en el momento de la toma de muestras.
Las vacas nacidas en el campo recibieron tratamientos antihelmínticos
esporádicos hasta el primer muestreo; las vaquillonas recibieron una única dosis.
Dentro del grupo formado por los animales que pastan (vaquillonas, vacas A, vacas NyC
y toros), las tres primeras categorías fueron desparasitadas por única o última vez en
mayo de 1998 (7 meses antes del muestreo) y los toros en noviembre del mismo año
(1 mes antes del muestreo). Los terneros nunca recibieron tratamientos
antiparasitarios.
Las muestras obtenidas fueron trasladadas al laboratorio en recipientes plásticos
con 80 mI de formol 5%. Para buscar huevos de F. hepatica se procesaron mediante la
técnica de tamización, utilizando el equivalente a 3 gramos de materia fecal. Secontaron todos los huevos encontrados en cada muestra.
Con el objetivo de comparar la prevalencia estimada por coprología con la
infección hallada durante las faenas en la zona de estudio, se recopilaron, a través del
SENASA: 1) los datos de infección en Frigoríficos de la Provincia del Chubut, para los 3
años previos al estudio (ver Tabla 2-3) y 2) los registros de decomisos de hígados
bovinos de los frigoríficos con actividad en la región Cordillerana de Ia Provincia del
Chubut, durante el segundo cuatrimestre de 1998 y el primero de 1999.
53

Estimación de la prevalencia al final del estudio
Durante los meses de noviembre y diciembre de 2001 se realizó otro muestreo
de toda la hacienda para estudiar la prevalencia general y por grupos, luego de 3 años
de tratamientos antiparasitarios regulares. La población bovina estaba compuesta, en
ese momento, por 520 animales. El grupo de las vaquillonas estaba formado por
animales de 2 y 3 años de edad, y entre las vacas había individuos de 4 y más de 5
años. Dentro del grupo de los toros había animales de 1 año y mayores de 1 año.Finalmente los terneros tenían 2 meses de edad.
Para calcular el tamaño de la muestra en esta segunda ocasión se procedió como
se describe a continuación, en función del objetivo:
1.- Se calculó el tamaño muestral necesario para estimar la prevalencia general
de la población (Zar 1999) considerando como valor teórico de prevalencia (p) la
hallada durante el primer muestreo para la hacienda general, un nivel de confianza del
95% (estadístico de prueba correspondiente: Z(._a,2,=1.96) y una mínima diferencia aser detectada del 10%.
2.- Para evaluar la existencia de diferencias (considerando las prevalencias
halladas durante el primer muestreo) en los niveles de infección entre vaquillonas y
vacas, se calculó el tamaño de muestra mínimo necesario para cada uno de los dos
grupos (Zar 1999) para tener una potencia no inferior al 80%. Se estableció un nivel de
significación del 5% y se consideraron los valores de prevalencia hallados para cada
grupo durante el primer estudio.
Los toros de 1 año de edad fueron muestreados en el mes de noviembre de
2001; habían sido destetados y desparasitados por primera y única vez en mayo delmismo año.
Las vacas y vaquillonas fueron muestreadas en el mes de diciembre. Las mismas
fueron desparasitados por última vez a fines de mayo de 2001. Las vaquillonas de 2
años (VQ1999) recibieron durante sus vidas un total de 3 desparasitaciones (Ia primera
en mayo de 2000), las vaquillonas de 3 años (VQ1998) recibieron 4 tratamientos (el
primero en mayo de 1999) y las vacas que estuvieron en el establecimiento desde el
comienzo del estudio recibieron 6 tratamientos (Tabla 3-1). Los toros mayores de 1 año
deberían haber sido muestreados también, pero por problemas operativos no se pudo
acceder a los mismos. Los terneros no pudieron ser muestreados por razones de
seguridad, ya que eran muy pequeños y podían resentirse por Ia manipulación.
Se procesaron 3 gramos de materia fecal por muestra para que los resultados
pudieran ser comparados con los del primer muestreo. Se registró la cantidad de
huevos hallados en todas las muestras positivas.
54

Estudio de una cohorte
Para conocer Ia evolución de Ia infección durante un año en un grupo de
animales jóvenes, a mediados de enero y principio de diciembre de 2001 se tomaron
muestras de materia fecal a una población de 70 vaquillonas nacidas en octubre de
1999 (VQ 1999). Se recopiló información sobre los potreros que frecuentaron los
animales y los tratamientos antiparasitarios que recibieron durante sus vidas.
Por otra parte, con el objetivo de estimar la eficacia del tratamiento
antiparasitario en estos animales, se tomaron muestras de materia fecal 24 días post
aplicación del antihelmíntico suministrado el 16 de enero de 2001. Para ello, los
animales fueron mantenidos desde esa fecha y hasta el momento de Ia toma de
muestras, en un corral cercano a la manga.
3.4.4 Infección en Hospedadores Silvestres
Durante el mes de enero de 2001 se entrevistó al personal del establecimiento
ganadero con el fin de obtener información sobre las especies de mamíferos silvestres
que habitan la zona y se recorrió el campo para verificar la presencia de liebres y
coipos, conocer el tipo de ambientes que frecuentan y constatar Ia presencia de materia
fecal de liebres -boñigas- en los distintos ambientes (inundables -temporarios y
permanentes- y no inundables).
Un estudio realizado en liebres europeas durante la zafra de 1999 demostró que
un 0,08% (127/162.328) de los animales capturados en la región cordillerana noroeste
de la provincia del Chubut presentaron fasciolas en sus hígados o canalículos biliares
(Com. pers. Med. Vet. Nirma González, SENASA Esquel). A partir de este hallazgo
surgió como objetivo a posteriori, evaluar el roI de Lepus europaeus en la transmisión
de F. hepatica en la región de estudio. Para ello, durante la siguiente visita realizada en
marzo de 2001, se buscó materia fecal en los cuerpos de agua (mallines, lagunas y
charcos), en 100 parches considerados aptos para el desarrollo del hospedador
intermediario, cuyos tamaños variaron entre 0,25 y 1 m2. Estos parches
correspondieron a una proporción de los que se utilizaron para la búsqueda de caracoles
(ver capítqu 4). Las boñigas se conservaron intactas en recipientes con formol 5 % y en
el laboratorio se pesaron y se procesaron individualmente para buscar huevos de F.
hepatica. Se contó la cantidad de huevos recuperados de cada boñiga.
3.4.5 Análisis estadístico
Para comparar la sensibilidad entre métodos diagnósticos se utilizó la prueba de
Mc. Nemar (Zar 1999) para muestras nominales pareadas.
55

Se calculó el porcentaje de concordancia (PC) entre métodos diagnósticos según
Gordis (2000) de Ia siguiente manera:
PC= (# de muestras positivas por ambos métodos/# de muestras positivas por lo
menos por un método)x100.
Se calculó un intervan de confianza para el valor de prevalencia estimado (p) en
cada muestreo, prefijando un nivel de confianza del 95% y suponiendo una distribución
aproximadamente normal (n>30, np>5 y np(1-p)>5) (Zar 1999).
La proporción de animales infectados y negativos por coprología entre los grupos
de interés epidemiológico (animales lactantes vs. animales de pastoreo y animales
jóvenes (vaquillonas) vs. adultos (vacas)), se comparó utilizando una prueba de
Homogeneidad (estadístico de prueba x2, a: 0,05) (Zar 1999). Se utilizó este mismo
método para comparar los dos grupos de vacas (vacas compradas unos meses antes del
estudio y vacas nacidas y criadas en la zona de estudio). A posteriori y en función de los
resultados obtenidos, se decidió excluir de ambos análisis al grupo de los toros.
Cuando resultó necesario, se calculó la potencia de cada prueba (hipótesis 1:
prevalencias diferentes entre lactantes y animales de pastoreo; hipótesis 2:
prevalencias diferentes entre animales jóvenes y adultos) según Zar (1999). Esta
estimación permite conocer la probabilidad de rechazar correctamente una hipótesis y
por complementariedad la probabilidad de cometer un error al no rechazarla.
La cantidad de huevos liberados por los animales infectados se comparó entre
grupos etarios y entre fechas mediante la prueba de Kruskal-Wallis (Siegel & Castellan
1995).

3.5 RESULTADOS Y DISCUSIÓN
3.5.1 Método coprológico de diagnóstico
La Tabla 3-2 presenta los resultados obtenidos a partir de la comparación de las
técnicas de sedimentación y tamización. Por sedimentación, el 21,6% (11/51) de los
animales mostraron huevos en sus heces, en tanto que por tamización, el porcentaje
fue significativamente mayor (53%, 27/51, p<0,001). Mientras que 16 animales
negativos por sedimentación fueron positivos por tamización, no se registró ningún casodonde ocurriera lo inverso.
Elporcentaje de concordancia entre métodos fue del 41% (11/(11+16)).
La técnica de tamización resultó mucho más sensible (incrementó la detección
2,5 veces —un total de 27 muestras positivas por tamización vs. 11 por sedimentación
) y requirió menor inversión de tiempo para el análisis de las muestras. La prevalencia a
lo largo del trabajo de investigación fue estimada mediante este método.
El número de huevos encontrados en las 11 muestras positivas por ambas
técnicas fue considerablemente diferente. Por sedimentación, todas las muestras
presentaron entre 1 y 3 huevos, mientras que sólo el 36% de las procesadas por
tamización arrojaron este resultado. EI64% restante presentó entre 6 y 35 huevos. Por
otra parte, de las 16 muestras discordantes entre métodos, en un 50% se encontraron
hasta 2 huevos, y en las restantes entre 3 y 16.
Tabla 3-2: Frecuencia de muestras positivas y negativas obtenidas por el método de
sedimentación-concentración y el de tamización.
TamizaciónNro.de muestras
+ - total
8C-o b 11 0 11a 3 +B :l5 E _ 16 24 4oE 8 _
'Ü .
(g g g 27 24 51
Si bien durante los últimos años se han desarrollado nuevos métodos
diagnósticos para la fasciolosis, Ia presencia del parásito es usualmente determinada
mediante la detección de huevos en heces (Ward et al. 1997, Anderson et al. 1999).

A partir del método tradicional de sedimentación, que incluye formol y éter como
solventes y centrifugación para concentrar los huevos (Ritchie 1948, Shore García 1973,
OMS 1994), se han incorporado diversas modificaciones. El método fue adaptado para
su uso en campo y en laboratorios poco sofisticados reemplazando la centrifugación por
sedimentación en conos de decantación (Dennis et al. 1954, Grock et al. 1998), lo cual
permite prescindir de electricidad, aunque insume más tiempo. También se incorporó el
uso de coladores o tamices de malla grande como primer paso del protocolo, para
eliminar las partículas de mayor tamaño de la materia fecal (Zukowski et al. 1993,
Grock et al. 1998, Anderson et al. 1999, Parr and Gray 2000).
Un trabajo realizado por Giráo & Ueno (1994) describe un método simple para
cuantificar huevos de F. hepatíca. EI mismo requiere de detergente para emulsionar la
materia fecal y una serie de tamices de mallas de distinto tamaño a través de los cuales
se filtra el material según el tamaño de las partículas, permitiendo que los huevos del
parásito queden retenidos en el último tamiz. Este material recolectado es teñido con
azul de metileno para resaltar la presencia de los huevos (Boray & Pearson 1960) y
observado bajo microscopio estereoscópico. El método de tamización puesto a punto en
esta tesis se basó en este último método, descripto por Giráo & Ueno (1994).
En función de los resultados obtenidos y considerando su mayor sensibilidad, el
método de tamización resulta más eficiente y confiable que el método tradicional de
sedimentación. Se considera que los factores principales que influyen en la detección de
huevos en los métodos coprológicos son la duración e intensidad de Ia infección, la
dificultad para obtener muestras seriadas bajo condiciones de campo y las variables
dependientes de laboratorio (Ward et al. 1997). Dado que los primeros tres items se
aplican de igual manera para ambos métodos, las diferencias entre ellos deberían estar
dadas por el grado de complejidad de cada técnica. El método de sedimentación incluye
numerosos pasos que aumentan la posibilidad de perder huevos, hecho que quedó
demostrado por el bajo número de huevos recuperados. Los huevos pueden quedar
retenidos entre los restos vegetales de materia fecal mientras el material es filtrado a
través de las gasas, o bien puede quedar en el fondo o las paredes de los recipientes y
dentro de las pipetas cuando se recupera el sedimento para la observación en
microscopio. Por el contrario, el método de tamización incluye un protocolo de un solo
paso en el cual los huevos, si están presentes, son recolectados del último tamiz.
Por otro lado, los huevos perdidos por la técnica de sedimentación pueden
atribuirse a la fatiga del observador debido al gran número de preparados requeridos
para el examen de toda una muestra, proceso que puede durar aproximadamente 2
horas. Además, los huevos pueden quedar enmascarados por partículas grandes de
restos vegetales de la materia fecal. Contrariamente, el método de tamización requiere
como máximo la observación de 2 cajas de Petri por muestra y el tiempo promedio
invertido en examinar una muestra entera se reduce a aproximadamente 15-20
58

minutos. El uso de microscopio estereoscópico junto con el hecho de que los huevos
resaltan sobre eI fondo azul, permite una búsqueda más rápida y sencilla.
El método de tamización tiene otras ventajas adicionales. Debido a su
simplicidad, reduce el tiempo necesitado para procesar numerosas muestras y facilita el
procesamiento de volúmenes grandes de suspensión si las heces están fijadas, o
diferentes cantidades, si están mantenidas en fresco.
Existen algunas diferencias operativas entre la técnica de tamización y otros
métodos coprológicos desarrollados previamente que incluyen el uso de tamices. Con
respecto al número de tamices, el tiempo de procesamiento se reduce a medida que
disminuye la cantidad de mallas utilizadas. Sin embargo, no parecería conveniente
reducir el número de tamices a 2 porque podría producirse el taponamiento de los
poros, especialmente cuando las heces son muy densas. Basándonos en las
experiencias obtenidas durante la puesta a punto de esta técnica, 3 tamices es la
cantidad adecuada para obtener resultados confiables en un tiempo de procesamientorazonable.
Los trabajos realizados por Giráo & Ueno (1994) y Zukowski et al. (1993)
proponen el uso de detergente para facilitar la separación de los huevos de las
partículas antes de la filtración. En este trabajo se ha omitido el uso de detergente ya
que se verificó que no quedaban huevos retenidos en la materia fecal durante el
procesamiento. Además, a diferencia de Zukowski et al. (1993), no se realizaron
sedimentaciones diferenciales luego del filtrado, ya que los huevos recolectados delúltimos tamiz fueron fácilmente diferenciados del resto del material.
El método de tamización incrementa las posibilidades de encontrar huevos. Por
ese motivo, es particularmente ventajoso en áreas donde la intensidad de infección y la
liberación de huevos son reducidas, ya sea por un bajo nivel de transmisión o por
tratamientos químicos regulares en los animales. Además, este método permite estimar
en forma más precisa las pérdidas económicas producidas por el parásito en los casos
en que dicha estimación se hace a partir de Ia prevalencia o de los huevos por gramo
(hpg) (Malone 1994).
Finalmente, a pesar de que el método de sedimentación es el más popular para
el diagnóstico coprológico de Ia fasciolosis, insume mucho tiempo y lleva a
subestimaciones de la prevalencia en el ganado y el nivel de huevos liberados al medio.
Actualmente existen métodos alternativos, en particular los ensayos
inmunoenzimáticos (ELISA), para el diagnóstico de F. hepatica (Abdel- Rahman et al.
1998, Ibarra et al. 1998, Castro et al. 2000, Duménigo et al. 1996). Estos métodos se
basan en la detección de antígenos o anticuerpos del parásito y son más sensibles que
el método coprológico de sedimentación, ya que pueden detectar infecciones tempranas
causadas por juveniles que aún no alcanzaron la madurez sexual y que no liberan
huevos (Duménigo et al. 2000), así como infecciones tardías con parásitos seniles. Sin
embargo, los métodos inmunológicos no proveen información sobre la cantidad de
59

huevos liberados ni sobre la proporción de hospedadores definitivos que aportan huevos
al ambiente. Estas variables son importantes para la realización de estudios
epidemiológicos, ya que la oferta de potenciales miracidios a las poblaciones locales de
caracoles modelan la dinámica de la transmisión de F. hepatica.
3.5.2 Infección en ganado
Prevalencia al comienzo del estudio
Un primer muestreo en animales de toda la hacienda permitió conocer la
situación de la parasitosis en la población del ganado del establecimiento, al momento
de iniciar el estudio. Los resultados de esta primera estimación de la prevalencia
reflejan las características de la transmisión de la parasitosis cuando los hospedadores
definitivos están sometidos a manejos sanitarios irregulares y poco frecuentes, y
cuando cohabitan con ganado ovino.
Se recolectó materia fecal de 134 animales de un total poblacional de 957
bovinos (14%). De las muestras obtenidas, 21 correspondieron a vaquillonas (7 de 1
año de edad y 14 de 2 años), 14 a vacas A, 61 a vacas NyC, 5 a toros y 33 a terneros.
La tabla 3-3 presenta el resumen de la información obtenida a partir de este
estudio: la cantidad de animales presentes en el establecimiento (frecuencia absoluta y
porcentual), divididos en categorías de interés epidemiológico; el número de animales
de cada categoría que fue muestreado y la prevalencia total y para cada categoría.
EI nivel de infección general para la hacienda fue del 50% considerando a los
terneros (Intervalo de confianza: 41°/o-58%) y del 61,4% excluyéndolos, valores mucho
mayores que el estimado a partir de la información suministrada por el SENASApara la
provincia del Chubut (prevalencia en ganado bovino para 1996: 10,2%; 1997: 8%;
1998: 15,6%; prevalencia promedio: 11%, ver Tabla 2-3). Esto permite suponer que la
prevalencia en la zona de muestreo no es la misma que la prevalencia registrada para la
provincia, y que la transmisión de la parasitosis debe ser muy heterogénea en distintas
regiones del Chubut, arrojando un valor promedio que no representa lo que pasa aescala local.
Se hallaron diferencias significativas entre las prevalencias registradas en los
terneros (5/33, 15%) y los animales de pastoreo (61/96, 61%) (p<0,001).
Como no se detectaron diferencias significativas entre las vacas compradas y las
nacidas y criadas en el campo (p>0,05), se consideró un único grupo de animales
adultos para los siguientes análisis.
Dentro del grupo de los animales que pastan -vacas y vaquillonas -, no se
hallaron diferencias significativas en cuanto a la frecuencia de animales positivos entre
60

los animales jóvenes (17/21, 81%) y los adultos (45/75, 60%) (p=0,085). La potencia
de esta prueba resultó en un 27 %, extremadamente baja.
Considerando que los rumiantes adultos pasan la mayor parte del día pastando y
que los terneros se alimentan principalmente de leche y sólo están en contacto con las
pasturas como parte del aprendizaje de alimentación, estos resultados confirman que
los terneros presentan un riesgo de infección menor que los animales adultos. Sin
embargo, resulta Ilamativamente elevado el porcentaje de lactantes infectados. Los
niveles teóricos de infección en terneros no deberían ser altos, ya que esta categoría
tiene muy corto tiempo de vida y aún pasa la mayor parte del día Iactando. Justamente
debido a este comportamiento, podría suponerse que esta categoría funcionará como un
sensor del estado de infestación de las pasturas. Si el nivel de transmisión es bajo,
debido a una baja densidad de metacercarias en el ambiente, los niveles de infección en
este grupo deberían ser cercanos a cero. Por el contrario, sólo se esperaría hallar
terneros parasitados si la infestación de las pasturas fuese muy importante. El alto
porcentaje de infección detectado durante este primer muestreo sugiere que las
pasturas se hallaban sobreinfestadas cuando se comenzó este estudio. Situaciones
similares se registraron en otras zonas de Ia Patagonia, y gracias a un seguimiento
constante de esos terneros desde su nacimiento se constató que el parásito se detecta
en materia fecal a partir de los tres meses de vida (Olaechea 1995). Un trabajo
realizado en el Uruguay en una zona endémica registró una prevalencia del 37% en
terneros mediante observación directa de los hígados (Nari & Cardozo 1976). Este valor
duplicó el hallado por coprología durante los estudios realizados en esta tesis. Estas
diferencias podrían deberse a que la detección por coprología impide detectar individuos
con infecciones tempranas, o simplemente a que los niveles de infestación en pasturas
fueron más elevados en el Uruguay. En el Altiplano Boliviano, región que presenta los
mayores valores de prevalencia en humanos, se detectó un 8% (3/37) de infección en
bovinos menores de 1 año mediante técnicas coprológicas. En este mismo estudio, se
observó que la proporción de individuos infectados ascendió a 35% en animales de
entre 1 y 2 años y luego descendió a 29% en animales de mayor edad (Buchon et al.
1997).
En coincidencia con los resultados obtenidos en el Altiplano Boliviano, dentro del
grupo de los animales que pastan se observó un mayor nivel de infección en animales
jóvenes que en adultos. Estas diferencias en las prevalencias encontradas entre
vaquillonas y vacas, aunque no resultaron significativas, fueron mucho más marcadas
que las halladas por Buchon y colaboradores (1997) y podrían estar reflejando una
susceptibilidad diferencial asociada a distintos tiempos de exposición a las pasturas. La
baja potencia de la prueba permite suponer que la probabilidad de que se haya
cometido un error al no rechazar la hipótesis planteada (igualdad en las prevalencias
entre vaquillonas y vacas) es muy grande. Esto podría deberse a que el tamaño
muestral resultó bajo, y podría estar enmascarando la verdadera diferencia que existe
61

entre las proporciones de animales infectados en cada grupo. Según Buchon et al.
(1997), la resistencia a reinfecciones o la capacidad de autocura podrían explicar porqué
la infección por F. hepatíca en ganado bovino no aumenta con la edad más allá de los 2
años. Por otro lado, esta misma tendencia fue observada en vacunos en un estudio
realizado en Cuba, pero los autores atribuyeron dicho resultado al aumento en los
controles y tratamientos a medida que los animales crecen (Percedo et al. 1987).
Tabla 3-3: Prevalencia en ganado bovino al comienzo del estudio (diciembre 1998
febrero 1999), Cholila, Chubut
Ultimotratamiento tamaño de # muestreado Prevalencia
GI’UPO Categoría antihelmíntico Ia población (No.posltlvos/No.
(°/°) ¡oo
Vaquillonas Mayo1998 252 (26,3) 21 81%Vacas (A) 80 (8,4) 14 43%
De pastoreo Vacas (NyC) 310 (32,4) 61 64%Vacas Mayo1998 390 (40,8) 75 61%Toros Noviembre 1998 15 (1,6) 5 0%
Total de PastOreo — 657 (68,7) 101 61.4%Lactantes Terneros ninguno 300 (31,3) 33 15%
TOTAL 957 (100) 134 50%
Vacas (A): vacas adquiridas en mayo de 1998, vacas (NyC): vacas nacidas y criadas en el lugar de
estudio. Se encontraron diferencias significativas entre los animales de pastoreo (excluyendo a los t0ros)
y los lactantes pero las vaquillonas y las vacas no difirieron en cuanto a la proporción de infectados en
cada grupo.
AI comparar el número de huevos liberados por gramo de materia fecal (hpg)
por las distintas clases etarias (terneros, vaquillonas y vacas) se observaron diferencias
significativas entre los grupos (p< 0,05). Las vaquillonas presentaron el mayor número
mediano de huevos en heces (3,3 hpg, 10-3o cuartil: O,85-7,7) y los terneros el menor
(0,7 hpg, 1°-3° cuartil: 0,3-1,45). Las vacas presentaron valores intermedios (1,3 hpg,
10-3o cuartil: 0,7-2,3) (Tabla 3-4). Se encontraron diferencias significativas entre el
primer grupo y los terneros (p=0,02) y las vacas (p=0,03), pero estos dos últimos no
difirieron entre sí (p=0,14). Estos resultados coinciden con los obtenidos por Claxton y
colaboradores (1997), quienes encontraron en un estudio realizado en Perú, que las
vaquillonas Iiberaban mayor cantidad de huevos por gramo de materia fecal que las
vacas y los terneros. Un estudio realizado a gran escala en España demostró que los
bovinos de entre 3 y 4 años Iiberaban en promedio mayor cantidad de huevos que los
animales más jóvenes y más viejos. Las prevalencias en ese estudio aumentaron con Ia
62

edad hasta los 4 años y luego se mantuvieron constantes hasta los 10 años
aproximadamente (González-Lanza et al. 1989).
La ausencia de huevos en la materia fecal de los toros no puede explicarse de
igual manera que en el resto de los animales, ya que es posible que el tratamiento
antiparasitario aplicado un mes antes del muestreo sea el motivo por el cual no seencontró infección en los machos adultos.
Tabla 3-4: Mediana del número de huevos por gramo de materia fecal (HPG) en
terneros, vaquillonas y vacas muestreadas al comienzo del estudio
(diciembre de 1998-febrero de 1999), Cholila, Chubut
HPG
Terneros Vacas Vaquillonasanimales positivos 5 45 17
mínimo hpg 0.3 0.3 0.3cuarti/ 1 0.3 0.8 0.85
mediana de hpg 0,7 1.3 3.3cuartil 3 1.35 2.3 7.7
máximo hpg 1.7 10 30
Un huevo en 3 gramos de materia fecal procesados equivale a 0,3 hpg.
Una de las hipótesis planteadas en este capítulo de la tesis hace referencia a las
diferencias que se esperan hallar en los niveles de infección entre animales de distintas
edades, basándose en que la susceptibilidad al parásito es mayor para animales más
jóvenes que estuvieron menos tiempo en contacto con las pasturas y no desarrollaron
resistencia a las reinfecciones (Torgerson & Claxton 1999). Durante este primer estudio
se observó una tendencia de aumento en los niveles de infección en vaquillonas
(animales más jóvenes) y disminución en vacas (animales más viejos), aunque no se
detectaron diferencias significativas entre la proporción de vaquillonas y vacas positivas.
Si esta ausencia de diferencias fuese real, estos resultados estarían indicando que la
desparasitación previa al muestreo fue efectiva y que ambos grupos de animales
tuvieron el mismo grado de exposición al parásito entre el momento de la aplicación del
antihelmíntico y el muestreo. Así, no se estaría evidenciando un efecto de resistencia a
la reinfección en animales con tiempos de exposición prolongados al parásito (vacas
adultas). De todas formas, una diferencia de un 20% en el número de animales
liberando huevos al ambiente resulta sumamente importante a nivel biológico. Si
además se considera que el grupo con mayor prevalencia es el que libera mayor
cantidad de huevos del parásito al ambiente, su aporte en el mantenimiento del ciclo
63

podría ser considerablemente mayor que el de los animales viejos. AI realizar una
estimación de dicho aporte considerando el valor mediano de hpg y que cada individuo
defeca diariamente entre 15 y 35 kg de bosta (Gürtler et al. 1979), se obtiene una
liberación diaria de 49500-115500 huevos por vaquillona, y de 19.500-45.500 huevos
por vaca. En Ia situación planteada (al comienzo del estudio) las vaquillonas
representaban un 26% de la población total y las vacas casi un 40% (Tabla 3-3).
Considerando los niveles de infección de cada grupo se podría suponer que entre las
vaquillonas había unos 202 animales liberando huevos al ambiente y entre las vacas
unos 234. Así, el primer grupo aportaba al comienzo de este estudio un mínimo de
10.000.000 de huevos diarios y el segundo grupo un mínimo de 4.000.000 de huevos
por día. La diferencia resulta importante; no obstante, ambos aportes son muy elevados
y podrían estar muy por encima del punto de saturación del ambiente. En consecuencia
las diferencias entre grupos podrían no tener implicancias epidemiológicas.
Finalmente, la prevalencia hallada en terneros junto con el alto porcentaje de
infección en los animales que pastorean, sugiere que la infestación de las pasturas es
muy alta en condiciones de manejos sanitarios irregulares y ambientes compartidos por
ganado bovino y ovino. Además, la alta infección registrada en vaquillonas evidencia
una elevada tasa de reinfección luego del tratamiento, si se considera el corto período
con condiciones climáticas favorables para el desarrollo del parásito (ver capítqu 4)
entre la fecha de la desparasitación y el momento del muestreo.
Los resultados de las faenas realizadas durante el segundo semestre de 1998 y
el primero de 1999 en el noroeste de Chubut se presentan en la tabla 3-5. En dicha
tabla figuran Ia cantidad de animales faenados por categoría y el número de hígados
decomisados totales, sin discriminar entre categorías. El principal movimiento fue de
novillos, único grupo de animales cuya prevalencia no fue estimada en el
establecimiento ganadero en diciembre de 1998. Sin embargo, sus niveles de infección
podrían ser comparados con los de las vaquillonas, animales hembras de igual edad.
Tabla 3-5: Casos de distomatosis detectados en frigoríficos de la región noroeste de Ia
Provinciadel Chubut (2° semestre de 1998-1o semestre de 1999)
Faena en Frigoríficos de Ia Provincia de Chubut. Segundo semestre de 1998Faena por categoría Faena total Casos de distomatosis
Departamento Novillos Novillitos Vacas Toros Terneros (%)Trevelin 3527 0 0 0 0 0 3527 0Futaleufu (Esquel) 2526 401 86 O 0 0 3013 311 (10.3)
Faena en Frigoríficos de la Provincia de Chubut. Primer semestre de 1999Trevelin 2434 0 0 0 0 0 2434 ¡90 (7.8)Futaleufu (Esquel) 2091 0 0 0 0 o 2091 183 (26.1)
Datos suministrados por el SENASA,D.N.F.A., Oficina de Estadística
64

Se registraron hígados con rastros de infección por F. hepatica en un 10-26% de
los novillos faenados. Estos niveles son muy inferiores al registrado en vaquillonas del
establecimiento ganadero en estudio, y sabiendo que es habitual que los frigoríficos
reciban animales de campos ganaderos de la zona de Cholila -incluso del
establecimiento en estudio- podría suponerse que los niveles de infección de esta
parasitosis, evaluados mediante los registros generados en frigoríficos, estarían siendosubestimados.
Prevalencia al final de estudio
EI estudio realizado en diciembre de 2001 aportó información sobre la
prevalencia en ganado luego de 3 años de tratamientos antiparasitarios aplicados con
regularidad dos veces por año. La hacienda en ese momento estaba formada por 520
animales, de los cuales el 52% eran vacas, el 13,5% eran vaquillonas de 2 años, el
21,5% vaquillonas de 3 años, el 9% toros de 1 año de edad y el 4% restante
correspondía a toros mayores de 3 años.
El tamaño muestral necesario para estimar la prevalencia general de Ia
población con un 95% de confianza fue de 221 animales. EI tamaño muestral mínimo
necesario por grupo para comparar las prevalencias entre vaquillonas y vacas fue de
116 y 122 animales respectivamente (establecido para detectar una diferencia del 20%
con una potencia no menor al 80%). Se tomaron muestras de materia fecal a 262
animales: 112 vacas, 46 vaquillonas de 2 años, 72 vaquillonas de 3 años (en total 118
vaquillonas) y 32 toros de 1 año.
La prevalencia general de Ia hacienda fue del 45,4% (Intervalo de confianza:
45%-45,8%). El 50,8% (60/118) de las vaquillonas, el 50,9 (57/112) de las vacas y el
6% (2/32) de los toros presentaron huevos de F. hepatica en la materia fecal (Tabla 3
6).
Tabla 3-6: Prevalencia en ganado bovino al final del estudio (noviembre-diciembre de
2001)
Ultimotratamiento Tamaño de # muestreado PrevalenciaGrupo Categoría antihelmíntico la población (No.posltlvos/No.
(°/o)
Vaquíllonas Mayo2001 182 (35) 118 50,8%De pastoreo Vacas Mayo2001 270 (52) 112 50,9%
Toros Mayo2001 50 (9,5) 32 6%TOTAL 520 262 45,4%
65

No se detectaron diferencias significativas entre los animales jóvenes
(vaquillonas) y los adultos (vacas) (p=0,1).
Al analizar las prevalencias por edad de los animales hembra (vaquillonas y
vacas) se observó que todos los grupos presentaron valores de infección similares
(Tabla 3-7). Con respecto al hpg, tanto vacas como vaquillonas presentaron un valor
mediano igual a 1 huevo por gramo de materia fecal. Resultados similares se obtuvieron
al analizar el hpg por edad (Tabla 3-8).
La aplicación de antiparasitarios dos veces por año en forma regular no tuvo un
efecto importante en la disminución de la infección luego de tres años de tratamientos.
Las vaquillonas estudiadas durante el primer muestreo habían recibido una única dosis
de antihelmíntico durante sus vidas mientras que las estudiadas en el segundo
muestreo fueron tratadas entre 3 y 4 veces. Las vacas, en forma similar, habían
recibido tratamientos poco frecuentes hasta el primer muestreo y 6 dosificaciones
regulares durante el estudio. Los resultados de este segundo muestreo indican que Ia
utilización de químicos homogeniza las prevalencias de vaquillonas y vacas, pero no
logra reducirlas de manera importante. Las prevalencias generales al comienzo y al final
del estudio fueron prácticamente iguales (p=0,5). La prevalencia de las vaquillonas se
redujoen un 30%, pero aún así fue superior al 50%; la prevalencia en vacas no varió.
Tabla 3-7: Prevalencia registrada por edades en vaquillonas y vacas en diciembre de2001
Categoría Edad (años) N Prevalencia °/o
Va uillonas 2 46 47'sq 3 72 52,8
4 42 50Vacas ,
5 o mas 70 51,4
La prevalencia en toros fue muy baja y mucho menor a los niveles de infección
registrados en los grupos restantes. El movimiento de estos animales entre potreros no
pudo establecerse, pero se sabe que por ser pocos animales y debido a que no pueden
estar en contacto permanente con las hembras, se los suele mantener en un único
potrero, o se los rota muy poco. Las diferencias registradas podrían deberse justamente
a que estos animales no frecuentan todos los ambientes que hay en el área de estudio
ni los comparten con vaquillonas y vacas. De esta forma la probabilidad de contacto con
el parásito se vería reducida. Un estudio realizado por Hope Cawdery (1984) reportó
que tanto vacas como toros presentaron los mayores niveles de decomiso de hígados,
66

descartándose de esta forma la posibilidad de que exista una tasa de infeccióndiferencial entre sexos.
Tabla 3-8: Número de huevos liberados por gramo de materia fecal (HPG) en las
vaquillonas y vacas estudiadas durante diciembre de 2001
HPG
Vaquillonas Vacas2 años 3 años Total 4 años > 5 años Total
anima/es positivos 22 38 60 21 36 57mínimo hpg 0.3 0.3 0.3 0.3 0.3 0.3
cuarti/ 1 0.3 0.3 0.3 0.7 0.7 0.7
mediana de hpg 1.7 1 1 1 1 1cuartí/ 3 6.8 2.7 3.5 1.7 2.9 2.5
máximo hgg 13.3 11 13.3 16 15.3 16
Un huevo en 3 gramos de materia fecal procesados equivale a 0,3 hpg.
El número de huevos liberados por la hacienda en general fue similar en ambos
muestreos (Tabla 3-9). No hubo modificación en el hpg en vacas aunque se notó una
leve disminución en vaquillonas que no resultó significativa (p=0,1).
El efecto de homogenización en cuanto a la cantidad de animales positivos y la
liberación de huevos que se observó en este estudio no coinciden con los resultados
obtenidos en Perú. Allí, los animales que fueron sometidos a desparasitaciones tres
veces por año mantuvieron diferencias en los hpg entre grupos etarios (Claxton et al.
1997).
Aún cuando no parece existir una relación lineal entre la cantidad de huevos
liberados con la materia fecal (hpg) y la carga parasitaria de F. hepatíca en bovinos
(Duménigo et al. 2000), una posible explicación a los resultados presentados en este
capítqu es que los grupos que liberaron mayor número de huevos pudieron albergar
mayor cantidad de parásitos adultos. Este criterio es utilizado en Estados Unidos para
estimar las pérdidas económicas producidas por el parásito en ganado bovino (Malone &
Craig 1990). A partir de la cantidad de huevos en 2 gramos de materia fecal se calcula
Ia carga parasitaria (baja, media, alta) y se evalúa el posible daño en las tropas.
Finalmente, resulta sorprendente la escasa disminución que se produjo en la
prevalencia en vaquillonas, pero más llamativo es que los tratamientos antiparasitarios
no hayan producido ningún efecto en las vacas. Este resultado parecería indicar que la
prevalencia que se puede alcanzar en este grupo de animales es independiente del uso
de antiparasitarios, y está directamente relacionada al desarrollo de resistencia con Ia
edad. Kendall y colaboradores (1978) demostraron que una infección sensibilizante
67

genera protección contra nuevas infecciones entre las 35 y las 54 semanas siguientes,
independientemente de si se aplicaron tratamientos con fasciolicidas entre las
infecciones, Io que probaría que existe resistencia adquirida.
Tabla 3-9: Número de huevos por gramo de materia fecal (HPG) liberados por toda la
hacienda al comienzo y al final del estudio (diciembre de 1998-diciembre de
2001)
HPG
1998 2001anima/es positivos 67 119
mínimo hpg 0,3 0,3cuarti/ 1 0,7 0,7
mediana de hpg 1,3 1cuartí/ 3 3,3 2,7
MUn huevo en 3 gramos de materia fecal procesados equivale a 0,3 hpg.
Estudio de una cohorte
Manejo ganadero. Las vaquillonas seleccionadas para el estudio nacieron en
octubre de 1999 y estuvieron en el sector correspondiente al potrero 17 hasta marzo de
2000 (5 meses). Luego fueron trasladadas a la ladera de las montañas hasta comienzos
de mayo de 2000 (2 meses), día en que se desparasitaron por primera vez y se
destetaron. Desde ese día se alimentaron durante un mes en el potrero 21, hasta
comienzos de junio de 2000, momento en que recibieron otro tratamiento y pasaron a
pasturas con bebederos hasta fines de octubre de 2000 (4 meses durante el invierno).
Luego fueron trasladadas al potrero 12 hasta principio de noviembre (menos de 1 mes),
permanecieron 15 días en el potrero 17 y desde mediados de noviembre hasta el día del
muestreo (2 meses) estuvieron pastando en los potreros 17 y 24 (Fig. 3-3).Prevalencia. El muestreo realizado a comienzos de enero de 2001 dio como
resultado un 40% de animales infectados (24/60). Luego de la desparasitación, ninguno
de los 24 animales previamente infectados liberaron huevos con las heces.
En octubre de 2001 ingresaron a frigorífico 40 novillos hermanos de estas
vaquillonas, con igual historia hasta enero de 2001. Los resultados de la faena,
facilitados por el Ing. Agr. Tati Iturburu y el Med. Vet. Hernán Alonso (Establecimiento
Los Murmullos), mostraron que 32 de los 40 animales faenados (80%) presentaron
adultos de F. hepatica en sus hígados.
68

La prevalencia registrada por visualización directa del parásito adulto duplicó los
valores obtenidos por la técnica coprológica. Esto podría estar indicando que en enero
parte de la infección en vaquillonas era inmadura y que por dicho motivo no se
detectaron huevos en las heces, o bien que el período de mayor riesgo de infección fue
posterior al momento del muestreo. Losanimales fueron tratados a mediados de enero,
con Io cual estuvieron teóricamente protegidos contra F. hepatica hasta principios de
marzo. Esto significa que las infecciones inmaduras existentes al momento de ser
tratados o bien aquellas que adquirieron hasta una semana postratamiento deberían
haber sido erradicadas. Por Io tanto, el alto porcentaje de novillos infectados en octubre
tiene que haber sido el producto del contacto con metacercarias entre marzo y julio,
aproximadamente.
En diciembre de 2001 se encontró infección en un 47,8% (22/46) de las
vaquillonas. Un esquema de este estudio se grafica en la Figura 3-5.
De las 46 vaquillonas que se estudiaron en diciembre, 45 pudieron ser
correctamente identificadas mediante el registro de su caravana. Al comparar la
situación de estos 45 animales en enero y diciembre de 2001 se observó que de los 18
animales positivos en enero, 14 fueron negativos en diciembre y 4 mantuvieron la
infección a pesar de haber recibido dos dosis de antihelmíntico durante el año. De los 27
animales negativos en enero, sólo 10 siguieron siendo negativos y 17 fueron positivos
en diciembre (Tabla 3-10).
Figura 3-4: Cronograma de manejo y muestreos sobre la cohorte estudiada en ganado
de Ia raza Hereford, Cholila, Chubut
Febrero
Diciembre
Ï Referencias:,, (47,8%)
Octubiie“ H Muestreoen ganado [%de infección)' Enero
- ¡d l Fecha de desporosiïocióndel ganado, (80%)
(40%) W 2(0%)fill‘ l
KNaclmlento /\ Oct.99 /\\_,/y . 2000
1 ¿Mayo
2091 destete
Junio
69

Tabla 3-10: Comparación del número de animales positivos y negativos en enero y
diciembre de 2001. Estudio realizado sobre una cohorte de vaquillonas
Ene.2001+ - Total
H
g + 4 17 21ú 14 10 240 Total 18 27 45
El estudio realizado sobre este grupo de vaquillonas demuestra que Ia
prevalencia se mantuvo constante de un año a otro a pesar de la aplicación de dos
tratamientos antiparasitarios, uno a comienzo del año y otro en junio. Aún cuando las
pasturas hubiesen estado sobre infestadas con metacercarias, la aplicación de drogas
en enero y junio pone de manifiesto la existencia de alguna falla en el manejo sanitario.
Luego de la primera aplicación, las vaquillonas deberían haber estado libres de adultos.
Efectivamente, la ausencia de huevos en la materia fecal en febrero sugiere que los
adultos fueron eliminados, pero las infrapoblaciones de parásitos jóvenes podrían haber
sobrevivido y aún no haber alcanzado la madurez sexual. Con lo cual, el hecho de que
todos los animales hayan sido negativos luego del primer tratamiento no brinda
información sobre el estado de los parásitos inmaduros. Sin embargo, suponiendo que
la droga utilizada tiene una vida media en sangre (efecto residual) de aproximadamente
50 días, podría suponerse que los juveniles que comenzaron su desarrollo antes de Ia
aplicación de la droga fueron muriendo a medida que maduraron, y que los parásitos
que alcanzaron la madurez sexual antes de comienzos de marzo fueron eliminados. Los
animales quedaron expuestos a nuevas infecciones a partir de fines de enero, ya que de
esa forma, para el momento en que los parásitos alcanzaron las 6 semanas de vida, el
antiparasitario había perdido efectividad. Nuevamente el tratamiento realizado a
principio de junio brindó protección contra el parásito hasta fines de julio, eliminando
adultos y juveniles a medida que se desarrollaban. Suponiendo que agosto y parte de
septiembre no son meses con alto riesgo de contraer Ia infección porque el suelo está
cubierto por nieve o inundado, o porque los animales no están en el valle, las
prevalencias registradas en diciembre serían el producto de infecciones adquiridas a
partir de octubre. Sin embargo, considerando que en condiciones adecuadas el parásito
puede tardar unos 2 meses en alcanzar la madurez sexual dentro del hospedador
definitivo, los tiempos entre octubre y el muestreo de diciembre son muy justos como
para haber detectado un 40% de animales con huevos en la materia fecal.
Por otro lado, este desarrollo resulta válido partiendo de la hipótesis de que las
metacercarias producidas durante el verano y el otoño sobrevivieron hasta la siguiente
70

primavera. Algunos estudios sugieren que metacercarias mantenidas a -2° C durante 8
semanas permanecen infectivas y que aproximadamente el 50% de las metacercarias
sobreviven el invierno en el Reino Unido (Andrews 1999). Un estudio realizado en
España demostró que luego del invierno, la supervivencia de metacercarias enquistadas
a comienzos del otoño fue del 15%, mientras que casi un 50% de las metacercarias
jóvenes (enquistadas avanzado el otoño) resultaron viables al comenzar Ia primavera.
Sin embargo, luego de un invierno frío (temperatura máxima del mes más frío durante
el invierno: 10° C; temperatura mínima: -2,S° C) la supervivencia y la viabilidad de las
metacercarias fueron muy bajas. El riesgo de infección para el ganado se consideró nulo
en primaveras de años cuyos inviernos fueron fríos (Luzón-Peña et al. 1994). A partir
de estos resultados y sabiendo que los inviernos en el área de estudio son aún más
desfavorables que los presentados por estos autores (ver 4.5.1), podría suponerse que
las metacercarias que se enquistan en las pasturas antes del invierno representarán un
riesgo menor para el ganado durante la primavera. Por estos motivos, una posibilidad
sería que el período de protección de Ia droga haya sido más corto que el teórico,
permitiendo el desarrollo de parte de la población de juveniles existente al momento deltratamiento.
La alta prevalencia detectada en los novillos en el mes de octubre de 2001
(registrada por observación directa del parásito en hígado) sugiere también que el
tratamiento antiparasitario no fue muy efectivo o que la probabilidad de contacto con
metacercarias fue muy elevada durante el otoño. Dichas infecciones no pudieron ser
producto del contacto con metacercarias durante la primavera temprana ya que no se
hubiese detectado un porcentaje tan alto de novillos con parásitos en los canalículos yvesícula biliar a menos de un mes de haber contraído Ia infección.
El método diagnóstico usado en estos estudios es una herramienta de gran
utilidad porque permite evaluar la oferta de huevos del parásito al ambiente. Sin
embargo, este método presenta como restricción que no permite cuantificar infecciones
inmaduras. Así, los niveles de infección aquí registrados son un buen estimador de la
proporción de animales que liberan huevos y contribuyen a la transmisión de Ia
parasitosis, pero la verdadera prevalencia podría estar siendo subestimada porque no se
contemplan las infecciones tempranas. El seguimiento realizado a una cohorte permite
suponer que dicha subestimación podría llegar a ser del 50% (40% por coprología vs.
80% por observación del adulto), aunque esto es sólo un supuesto, ya que en ningún
momento se pudieron revisar los hígados de los mismos animales que se diagnosticaron
por coprología. Además, hubo un lapso de varios meses entre las fechas en que se
muestrearon ambos grupos (vaquillonas y novillos). Sin embargo, las diferencias
registradas en este estudio entre los niveles de infección detectados por coprología y
por autopsia coinciden con las halladas en un estudio realizado en Bolivia. Hillyer y
colaboradores (1996) estudiaron Ia prevalencia en ganado bovino del Altiplano Boliviano
71

utilizando dos métodos: hallazgo de huevos en la materia fecal y detección de
anticuerpos del parásito en sangre (detecta infecciones tempranas). Hallaron que del
total de muestras que resultaron negativas por coprología, Ia mitad fue positiva por
serología.
3.5.3 Infección en Hospedadores Silvestres
Como resultado de los avistajes realizados en el establecimiento ganadero, se
comprobó la presencia y uso del ambiente por parte de coipos (Myocastor coypus) y
liebres europeas (Lepus europaeus) entre los mamíferos herbívoros potenciales
hospedadores de F. hepatica. Los coipos se observaron únicamente en Ia laguna, perono se realizaron estudios más exhaustivos.
En el establecimiento ganadero se comprobó la presencia de L. europaeus y no
se registraron otras especies del Orden Lagomorpha. Durante el estudio se observaron
ejemplares de liebres europeas en todos los ambientes en pleno día y boñigas secas
ampliamente distribuidas en pasturas no inundadas y en los alrededores de los mallines.
La figura 3-6 muestra los ambientes en los que se buscó materia fecal. De los 100
parches estudiados, se encontraron boñigas en uno, ubicado dentro de un charco
temporal en el cual había caracoles. Sobre un total de 10 boñigas recolectadas cuyo
peso varió entre 1 y 1,4 gramos, seis resultaron positivas, presentando cada una entre
uno y cinco huevos (mediana = 2,5). Se estimó una mediana de dos huevos por gramo
de materia fecal (hpg).
El rol de los lepóridos en la transmisión de F. hepatica aún no ha sido bien
establecido. Los estudios realizados en O. cunicu/us y en L. europaeus demostraron que
estas especies albergan cargas parasitarias bajas (Apt et al. 1993, Shimalov & Shimalov
2000). Además, Ménard y colaboradores (2000) comprobaron que a pesar de haber
encontrado altos porcentajes de infección en O. cuniculus, la proporción de animales
parasitados que liberan huevos al ambiente no alcanza el 20%. Finalmente, algunos
autores consideran que L. capensis no participa significativamente en la transmisión de
Ia parasitosis en el Altiplano Boliviano, basándose en que sus abundancias son muy
bajas en dicha región (Fuentes et al. 1997, Valero & Mas-Coma 2000). En el presente
estudio, la mediana de huevos liberados por gramo de materia fecal resultó similar a Ia
de los bovinos infectados en el área estudiada (incluso superior a Ia de algunos grupos,
Tablas 3-4 y 3-8) y en otras zonas de la Patagonia (Rubel et al. 2003), sugiriendo que
el número de huevos no sería por sí solo una variable limitante en la transmisión de la
parasitosis.
Con respecto a Ia proporción de animales infectados en la zona, un trabajo
realizado en un frigorífico de la región noroeste de Ia provincia del Chubut, permitió
estimar una prevalencia en liebres (registrada mediante observación directa de adultos
72

durante faenas) menor a 0,1% (Kleiman et al. 2004). A partir de estos resultados
preliminares y considerando que una liebre defeca entre 400 y 500 boñigas por día
(Novaro et al. 1992, Krebs et al. 2001, Parkes 2001), podría suponerse que cada animal
infectado liberará diariamente 600-750 huevos al ambiente. Considerando que la
densidad de esta especie en la zona noroeste de Ia Patagonia ha sido estimada en 27,9
liebres/ha para los mallines de valle durante los meses de verano (Novaro et al. 1992),
la oferta al ambiente rondaría los 1700-2000 huevos diarios por hectárea. AIrealizar un
cálcqu similar para el ganado (carga ganadera promedio: 0,3; prevalencia: 50%; hpg
general promedio: 1) se obtiene una oferta diaria y por hectárea de entre 2200 y 5200huevos.
Finalmente, aún cuando la oferta de huevos al ambiente es más importante en
ganado que en Iiebres, los Iepóridos podrían ser buenos diseminadores de los huevos de
F. hepatica, si se consideran sus altas abundancias, su gran capacidad de dispersión y
que habitan y se alimentan en los mismos ambientes donde se encuentran los caracoles
que actúan como hospedadores intermediarios del parásito (Flux & Angermann 1990).
Estos hechos, sumados a que los hábitats de L. europaeus se superponen con los que
utiliza el ganado para pastar (Bonino 1986b), indicarían que Ia liebre europea podría
estar desempeñando un rol importante en el ciclo de transmisión del parásito en la
cordillera patagónica.
Las boñigas halladas en las pasturas durante el primer recorrido se encontraban
secas y esto podría ser un factor condicionante de la supervivencia de los huevos del
parásito, ya que los mismos necesitan humedad para sobrevivir y desarrollarse en el
ambiente (Torgerson & Claxton 1999). Podría suponerse entonces, que sólo la materia
fecal depositada en un cuerpo de agua alcanzaría las condiciones necesarias para
aportar miracidios al medio. Por dicho motivo, en este primer estudio no se
consideraron las boñigas diseminadas en zonas secas y sólo se muestrearon los
ambientes aptos para caracoles. Sin embargo, las áreas inundadas varían según Ia
época del año y consecuentemente también Io hacen los ambientes adecuados para el
desarrollo de los huevos del parásito.
Este trabajo preliminar indica que es necesario investigar en forma más precisa
el aporte de las liebres al ciclo de transmisión doméstico de F. hepatica, para poder
planificarestrategias de control adecuadas.

Figura 3-6: Ambientes inundables en los que se buscaron boñigas de liebres (Lepus europaeus)A) Detalle de un mallín con vegetación baja; B) laguna y charcos formados porinundación del río; C) detalle de un mallín con pastizal; D), E) y F) primer plano deparches adecuados para el desarrollo de L. viatrix
74

3.6 CONCLUSIONES
1. La utilización del número de huevos por gramo de materia fecal como indicador de la
carga parasitaria en un animal o de las pérdidas en la producción, puede resultar
inadecuada si el método mediante el cual se estima dicha variable es poco sensible.
2. Las prevalencias estimadas al comienzo y al final del estudio superaron ampliamente
las registradas mediante faenas (SENASA)en la región Cordillerana de la provincia del
Chubut, confirmando que las prevalencias oficiales a nivel provincial no reflejan la
situación epidemiológica local.
3. La prevalencia general en el área de estudio fue del 50% independientemente del
manejo químico realizado.
4. Los niveles de infección y la cantidad de huevos por gramo de materia fecal
parecerían ser mayores en vaquillonas que en vacas cuando no existe un régimenestricto de control.
5. Los tratamientos antiparasitarios produjeron un efecto de homogenización de las
prevalencias y la liberación de huevos en animales de distintas edades.
6. Bajo las condiciones de campo los antiparasitarios podrían tener un efecto residualinferior al teórico.
7. La infección registrada en febrero en terneros nacidos en el mes de octubre indicaría
una alta infestación de las pasturas con metacercarias viables luego del invierno.
8. En la zona de estudio existen por lo menos dos potenciales hospedadores definitivos
silvestres: el coipo (Myocastor coypus) y la liebre europea (Lepus europaeus). Con
respecto a esta última especie puede decirse que podría estar jugando un rol en la
transmisión de la parasitosis.

EL HOSPEDADOR INTERMEDIARIO
En este capítulo se presentan los resultados de los estudios que permitieron en
primera instancia identificar al caracol que actúa como hospedador intermediario en la
zona de estudio y los ambientes aptos para su desarrollo, y luego estimar las
prevalencias de F. hepatica con las que se mantiene la infección en ganado. Debido a
que Ias variables ambientales tienen un efecto muy importante sobre el desarrollo del
hospedador intermediario y de los estadios Iarvales del parásito que se desarrollan en
él, en este capítulo se analizan también algunas variables del ambiente y sus
implicancias sobre la estructura y dinámica poblacional del caracol. Se presenta también
un estudio sobre digeneos parásitos de aves acuáticas, que fueron encontrados en un
ambiente infectando a los mismos caracoles que utiliza F. hepatica.
4.1 INTRODUCCIÓN
Los hospedadores intermediarios de F. hepatica son caracoles de la Familia
Lymnaeidae (Gasteropoda, Pulmonata). Las especies de más amplia distribución y las
más estudiadas en el mundo son Lymnaea tomentosa (Pfeiffer) 1855 en Australia; en la
Región Holártica L. stagna/¡s (Linnaeus, 1758), L. pa/ustris (Müller, 1774) y L.
truncatula (Müller, 1774), esta última principalmente en Europa; L. cubensis Pfeiffer,
1839 y L. bu/ímoides (Lea, 1841) en América del Norte y Central y L. columella Say,
1817 y L. víatrix Orbigny, 1835 en América del Sur (Hubendick 1951, Boray 1964,
Paraense 1982, Malone et al. 1984, Oviedo et al. 1996, Bargues & Mas-Coma 1997,
Torgerson & Claxton 1999, Durand et al. 2002, Naranjo García 2003, Cañete et al.
2004). Algunas otras especies han sido utilizadas como modelos experimentales en
varios países: L. peregra (Müller, 1774) y L. ovata (Draparnaud) en Suiza (WuIlschIeger
76

& Jokela 1999), L. luteo/a (Lamarck) en India (Misra & Raut 1993, Aziz Si Raut 1996), L.
o/lu/a Gould en Japón (Itagaki 1987), L. nata/ensis Krauss, 1848 en Tanzania (Utzinger
&Tañer 2000) y L. virídís Quoy & Gaimard, 1832 en Corea (Lee et al. 1994).
Sin embargo, Ia relevancia epidemiológica y la distribución geográfica de estos
caracoles no se conocen con precisión, y existe incertidumbre taxonómica entre las
especies del género que han sido descriptas, debido a la uniformidad morfoanatómica y
a que muchas veces dichas descripciones se han basado únicamente en las
características de la conchilla (Paraense 1976, 1982, Castellanos & Landoni 1981,
Oviedo et al. 1995, Bargues & Mas-Coma 1997, Mas-Coma 1998, Samadi et al. 2000,
Durand et al. 2002). Paraense (1976) discute cómo los diferentes criterios de
clasificación establecidos por diversos autores pueden llevar a una especie a ser
asignada a géneros o subgéneros diferentes.
En la Argentina, Lymnaea díaphana King, 1830 y Lymnaea p/¡cata Hylton Scott,
1953 fueron descriptas en Ia década del '50 (Hubendick 1951, Hylton Scott 1953), pero
rara vez citadas posteriormente (Santa Cruz 1979). La información sobre Lymnaea
colume/la y Lymnaea viatríx es mucho más abundante y actualizada. De acuerdo a la
bibliografía, la distribución de L. colume/la se restringe al noreste del país (Castellanos &
Landoni 1981, Paraense 1982, Prepelitchi et al. 2003), mientras que L. viatríx ha sido
documentada en las siguientes provincias: Salta, Jujuy (Castellanos & Landoni 1981),
Buenos Aires (Castellanos & Landoni 1981, Venturini & Fonrouge 1985), Corrientes
(Lombardero et al. 1979), San Luis (Castellanos & Landoni 1981, Rossanigo et al.
1983), Córdoba, Entre Ríos, Neuquén, Mendoza (Castellanos & Landoni 1981, Paraense
1982), y su registro más austral se remite a la localidad de Viedma (40°50’ S, 63° 0),
al este de la provincia de Río Negro (Paraense 1976). Según estos datos, L. viatríx es,
hasta ahora, el hospedador intermediario de F. hepatíca más ampliamente distribuido
en nuestro país.
Los caracoles del género Lymnaea son anfibios y suele encontrárselos en el
fango, aunque algunas especies son más acuáticas y viven a varios centímetros de
profundidad (Ollerenshaw 1971a, Castellanos & Landoni 1981). Las condiciones que
debe tener un ambiente para que el caracol pueda habitarlo y desarrollarse en él son
muy diversas y difieren para cada especie. EI conjunto de dichas condiciones es Io que
se conoce como hábitat y se define como el área con los recursos necesarios para
permitir la ocupación, supervivencia y reproducción de una especie (Krausman 1999).
Las variaciones en el tiempo y en el espacio llevaron a clasificar a los hábitats de la
siguiente forma: a) constantes en el tiempo, b) predeciblemente estacionales, con
alternancia regular de períodos favorables y desfavorables, c) impredecibles en el
tiempo, cuando los períodos favorables y desfavorables no tienen regularidad, d)
efímeros si los períodos favorables son esporádicos y están seguidos de períodos
desfavorables de duración indefinida, e) continuos en el espacio, cuando el área

favorable es más extensa que Ia que pueden cubrir los organismos, f) en mosaico,
cuando las áreas favorables y desfavorables están intercaladas, pero no resultan un
impedimento para que el organismo se traslade de un área favorable a otra y
finalmente, g) aislados, cuando las áreas favorables se encuentran demasiado alejadas
unas de otras como para que el organismo pueda dispersarse entre ellas, excepto en
determinadas y raras ocasiones (Southwood 1977). Probablemente el concepto de
hábitat sea uno de los más importantes en ecología. Sin embargo, existe gran
ambigüedad en el uso del término (Hall et al. 1997). Las definiciones aquí presentadas
son simplemente algunas de las que pueden encontrarse en la bibliografía; existen otras
más complejas e incluso definiciones derivadas del concepto de hábitat que aquí no sedesarrollan.
La heterogeneidad que puede presentar un hábitat llevó a Ia definición de
parche. Los modelos de disponibilidad de alimento proponen que un parche es un
bloque discreto del hábitat; el alimento se encuentra dentro de dichos parches y pueden
haber “espacios vacíos" entre los mismos. Una perspectiva alternativa sugiere que los
parches son subdivisiones del hábitat estadísticamente independientes, y que aunque
no sean fácilmente distinguibles se reconocen por las diferencias que se observan en
cada sitio con respecto a las variables en estudio. EI concepto de parche depende del
nivel de detalle al que se analice el ambiente; un hábitat puede ser muy parcheado a
una escala pero esencialmente uniforme a otra. Por otro lado, un hábitat heterogéneo
puede no presentar parches claramente definidos a distintas escalas. El hecho de que
no se visualicen parches o que no puedan ser claramente mapeados en un ambiente, no
quita que los mismos puedan estar bien definidos en cuanto a la disponibilidad de
alimento en un momento determinado. Finalmente, un área puede no diferir de los
espacios que Ia rodean, y sin embargo la existencia de microclimas en un momento
dado puede determinar que ciertas zonas del hábitat sean más favorables que otras
para la disponibilidad de alimento de una especie (Blackwell 2001). Más allá de las
definiciones teóricas, los conceptos de hábitat y parche deben ser definidos
cuidadosamente para cada especie, ya que los mismos pueden establecerse para un
organismo en función de los recursos y condiciones que parecen relevantes para el ojo
del investigador, y sin embargo no ser dichas variables las que realmente estén
afectando la presencia del animal en estudio.
En general, la bibliografía menciona como hábitats adecuados para Ia mayoría de
las especies del género Lymnaea, aquellos cuerpos de agua que presentan cierta
corriente que permita una aireación adecuada y sustrato con barro y poca o nula
vegetación, Io que facilita el desarrollo de las algas unicelulares que utilizan los
caracoles como alimento (Boray 1964). Se ha registrado como condición desfavorable Ia
acidez del agua y Boray (1964) encontró que ambientes en los que predominan
especies de Ia Familia Juncaceae no son adecuados para el desarrollo de L. tomentosa
debido a que las plantas impiden la llegada de luz al sustrato y acidifican el medio.
78

Las condiciones climáticas, principalmente la temperatura y la humedad, son
también cruciales para determinar si un hábitat es favorable o desfavorable. Estos
moluscos sólo pueden alimentarse, desarrollarse y reproducirse cuando el suelo se
encuentra saturado en agua, preferentemente con una delgada capa superficial. Cuando
el suelo se seca, los caracoles se retraen en su conchilla y estivan, pudiendo
permanecer vivos durante un tiempo, aunque períodos prolongados de sequía pueden
reducir las poblaciones a niveles muy bajos (Ollerenshaw 1971a). Según un trabajo
realizado por Malone et al. (1984), la humedad del suelo puede considerarse un factor
clave en la determinación de las variaciones interanuales en el riesgo de transmisión de
F. hepatica. Como se mencionó en el Capítqu 1, la temperatura es considerada un
factor ambiental crítico en la ecología de la mayoría de los organismos (Precht et al.
1973, Ahmed & Raut 1991, Raut et al. 1992). Puede detonar el comienzo de procesos
biológicos o ser un umbral por debajo del cual no hay actividad (Aziz & Raut 1996). En
particular, con respecto a los caracoles del género Lymnaea, diversos autores han
comprobado experimentalmente que existen temperaturas críticas por debajo o por
encima de las cuales los individuos no pueden completar su desarrollo, más allá de que
el resto de las condiciones sean las adecuadas, y que hay rangos óptimos de
temperatura para su crecimiento (Aziz & Raut 1996, Claxton et al. 1999). Sin embargo,
las condiciones constantes de los ensayos de laboratorio no siempre son fácilmente
extrapolables a campo, donde las fluctuaciones de temperatura son muy grandes. Se ha
reportado que en zonas con inviernos crudos, los caracoles del género Lymnaea pueden
sobrevivir enterrados en el suelo bajo la capa de nieve, e incluso en el hielo (Rognlie et
al. 1996, Andrews 1999). Esta estrategia de supervivencia, denominada hibernación,
suele ser común en caracoles de este género (Boray 1969).
La importancia del género Lymnaea en la transmisión de F. hepatica y de otros
parásitos es ampliamente reconocida (Brown 1978) y diversos autores han estudiado la
biología de distintas especies con el objetivo final de aportar información que permita
implementar medidas de control (Hunter 1975, Malone et al. 1984, Misra & Kaut 1993,
Aziz & Raut 1996). Aunque estos estudios resultan de suma utilidad, las características
del ciclo de vida del hospedador y su dinámica poblacional son particulares de cada
región, dependiendo de las condiciones meteorológicas que predominen en Ia zona, y
esto a veces impide extrapolar los resultados hallados para una especie en determinada
región, a otras especies y/o regiones. Con respecto a L. viatríx, son pocos los estudios
realizados sobre su biología y la mayoría son en condiciones de laboratorio (Venturini et
al. 1981, Mattos & Ueno 1985, Venturini & Fonrouge, 1985, Müller et al. 1998). En su
estado adulto puede desarrollarse hasta alrededor de 1 cm de largo (Bendezú & Landa
1973, Paraense 1976), pero el crecimiento de los caracoles de esta especie y de otras
está fuertemente influenciado por factores denso-dependientes y denso-independientes
como las condiciones de temperatura, humedad, pH y salinidad del medio y la oferta de

alimento, encontrándose en bibliografía información muy diversa sobre los tamaños
máximos que pueden alcanzar los individuos y los tiempos que demoran en lograrlo
(Itagaki 1987, Misra & Kaut 1993, Lee et al. 1994, Aziz & Raut 1996, Müller et al. 1998,
Tanaka et al. 1999, Cañete et al. 2004).
Los moluscos tienen una alta capacidad reproductiva y un rápido desarrollo en
condiciones favorables. Ollerenshaw (1971a) reportó que en laboratorio, en 12 semanas
y a partir de un único individuo de la especie L. truncatu/a, se obtuvieron 25000
caracoles y que en campo, 3 caracoles dieron lugar a una población de 1500 individuos
en los 4 meses de mayor temperatura. En condiciones naturales, la constante
fluctuación en las variables bióticas y abióticas recién mencionadas, la heterogeneidad
del ambiente a distintas escalas y las características biológicas de los moluscos
(capacidad reproductiva y de desarrollo) pueden generar dinámicas con patrones de
crecimiento explosivo “crash-boom" (Malone et al. 1984).
Lymnaea sp. y su rol como hospedador intermediario. Dado que en líneasgenerales el hospedador es el hábitat del parásito, comúnmente se considera que los
parásitos tienen pocos requisitos ambientales, además de la presencia de hospedadores
adecuados. Así, el supuesto de que algunos hábitats no son adecuados para el parásito
aún cuando el hospedador esté presente parece rara vez cumplirse. Sin embargo, los
parásitos con ciclos de vida complejos (varios hospedadores, estadios de vida libre)
como el de F. hepatíca, se ven restringidos a hábitats donde se encuentran todos los
hospedadores y donde están dadas las condiciones ambientales para el desarrollo de los
estadios de vida libre (Wullschleger & Jokela 1999). Las variables ambientales como la
temperatura y la humedad, entonces, no son sólo determinantes para el desarrollo del
hospedador intermediario sino también para el parásito dentro y fuera del hospedador.
Según Nari et al. (1983) la temperatura influye en dos aspectos fundamentales en el
desarrollo de F. hepatica dentro de su hospedador intermediario: determina si el ciclo
puede completarse y regula la velocidad a la que se produce. Estos autores observaron
que a medida que las temperaturas aumentan (de 15° C a 27° C) el desarrollo del
parásito llega a término en menor tiempo dentro del caracol. Además, Ollerenshaw
(1958) observó que por debajo de los 10° C el desarrollo de las larvas dentro de los
caracoles no se completa y que a temperaturas levemente superiores, el parásito tarda
tanto tiempo en desarrollarse que se encuentra expuesto a los efectos adversos de
otros factores ambientales. La temperatura también puede influir en la capacidad de los
miracidios de F. hepatica para encontrar al caracol (Christensen & Nansen 1976).
La humedad afecta directamente a los estadios de vida libre, pero, a diferencia
de Ia temperatura, sólo tiene efectos indirectos sobre el parásito dentro del hospedadorintermediario. Los huevos dentro de la materia fecal en el ambiente necesitan humedad
para mantenerse viables y mueren rápidamente si son sometidos a desecación
(Ollerenshaw 1971b). Considerando que la humedad es un factor decisivo también para
80

el caracol, estos resultados demuestran la existencia de sincronización entre la
liberación de miracidios y la actividad del hospedador intermediario.
Las condiciones en las que se encuentra el caracol en cada momento juegan
también un rol crucial en la transmisión del parásito. Por ejemplo, el estado nutricional
del caracol afecta tanto a la instalación y desarrollo de las redias como a la producción
de cercarias. Cuanto mejor es el estado nutricional del animal, mayor es Ia carga
parasitaria que puede albergar (Kendall 1950). Así, un cuerpo de agua que ofrezca
condiciones favorables para el desarrollo de los caracoles podrá sostener poblaciones de
mayor densidad, pero cuyos individuos serán de menor tamaño. Un cuerpo de agua con
menores densidades permitirá que los caracoles alcancen tamaños más grandes, y
potencialmente mayores cargas parasitarias que se desarrollen exitosamente
(Ollerenshaw 1971b). El tamaño del hospedador también puede influir en la
susceptibilidad para infectarse. Kendall (1950) observó que tanto el número de redias
establecidas como el de redias maduras aumentaba con el tamaño del hospedador en
infecciones naturales. Por otro lado, Mattos & Ueno (1986) encontraron que tanto
adultos como juveniles se infectaban y producían cercarias, pero que el primer grupo
presentaba niveles más bajos de infección y mayores tasas de mortalidad. Finalmente,
Castro et al. (2001) encontraron en campo que el porcentaje de infección aumentaba
exponencialmente con el tamaño de los caracoles, y que los niveles de infección en
caracoles pequeños eran muy bajos.
La relación entre hospedador y parásito no influye de manera positiva o negativa
únicamente sobre el parásito. En general, el efecto de esterilización del hospedador está
ampliamente reconocido, pero no es igualmente eficiente en todos los sistemas
hopedador-parásito. Las especies de parásitos que desarrollan redias son consideradas
agentes más efectivos de esterilización que las que producen esporoquistes. Esto se
debe a que los esporoquistes, al no tener canal alimentario, sólo pueden absorber
nutrientes mientras que las redias pueden ingerir pedazos de órganos. Además, los
esporoquistes producen obstrucción de Ia glándula digestiva mientras que las redias
directamente destruyen las gónadas, lo que produce una verdadera castración (Combes
1982). Se ha comprobado que F. hepatica produce efectos perjudiciales sobre el
hospedador intermediario, como castración o disminución en Ia fecundidad, aumento en
la mortalidad, destrucción de las glándulas digestivas, cambios metabólicos (mayor
energía en crecimiento y menor en reproducción, incluyendo casos de gigantismo) y
mayor sensibilidad al stress ambiental, así como alteraciones en el comportamiento
(Fernandez & Esch 1991, Graczyk & Fried 1999, Gutiérrez et al. 2000).
La instalación y el desarrollo de un esporoquiste de F. hepatica en un caracol
pueden estar condicionados por la previa existencia de otra especie parasitaria y de
complejas interacciones entre ambas (Lim & Heyneman 1972). Se han reportado casos
81

en los que la ocupación del caracol por un parásito impide el posterior desarrollo de otro
parásito (antagonismo) y situaciones en las que sólo si el caracol se encuentra infectado
por un digeneo, otro puede instalarse posteriormente (facilitación). Diversos estudios
han permitido comprobar que en infecciones dobles con especies parasitarias que
desarrollan redias y esporoquistes, el primer grupo siempre resulta dominante sobre el
segundo (Lim & Heyneman 1972). A su vez, Boray (1967) reportó que caracoles del
género Lymnaea que presentaron infecciones maduras por redias y cercarias de la
Familia Echinostomatidae, mostraron una alta resistencia a infectarse con F. hepatica.
En contraposición, se ha comprobado que existen especies de digeneos que al infectar
caracoles resistentes naturalmente a F. hepatica facilitan la posterior instalación del
parásito (Abrous et al. 1996).

4.2 OBJETIVOS
Identificar las especies de caracoles del género Lymnaea presentes en el área deestudio.
Estudiar Ia estructura y dinámica poblacional del hospedador intermediario.
Estudiar posibles asociaciones entre las fluctuaciones de temperatura y precipitación
(variable indicadora de humedad en el suelo) y el desarrollo del hospedadorintermediario.
Identificar los ambientes aptos para el desarrollo del hospedador intermediario.
Caracterizar los hábitats con presencia de caracoles.
Estudiar la infección por F. hepatica en caracoles en el tiempo.
Estudiar la presencia de otros digeneos que utilicen el mismo hospedador intermediario
que F. hepatica.
4.3 HIPÓTESIS
Debido a que la información existente indica que la zona es endémica para F. hepatica y
que se registran periódicamente casos en el ganado, deben existir una o más especies
del género Lymnaea que se desarrollan en el área de estudio.
Existen ambientes que permiten el desarrollo sostenido del hospedador intermediario.
La dinámica poblacional del caracol está modulada por la temperatura y la precipitación.
Si los distintos ambientes en los que se encuentre al hospedador intermediario
presentan diferentes condiciones para su desarrollo, esto se manifestará en la
estructura poblacional del caracol. Así, ambientes homogéneos o con características
comunes deben presentar dinámicas de desarrollo similares y ambientes heterogéneos
presentan dinámicas asincrónicas.
83

4.4 MATERIALES Y MÉTODOS
“De vuelta en Buenos Aires con una gran cantidad de
Limnaeas, más o menos 500 ejemplares de distintostamaños fueron colocadas más o menos en las mismas
condiciones en que fueron halladas”
Dr. Juan Bacigalupo 1934.
4.4.1 Períodos de estudio
Las actividades de campo se realizaron entre diciembre de 1998 y febrero de
2002 y consistieron en 11 muestreos (visitas al establecimiento ganadero) divididos en
cuatro períodos de estudio. Cada periodo de estudio se definió como el lapso entre el
momento posterior al deshielo -en primavera- y comienzos del otoño, antes de que las
condiciones adversas del invierno impidieran realizar el trabajo de campo. Las fechas en
las que se realizaron muestreos en cada período se detallan a continuación:
- Periodo 1: primavera 1998-verano 1999, muestreos en diciembre y febrero
- Periodo 2: primavera 1999-otoño 2000, muestreos en noviembre, enero, y marzo
- Periodo 3: primavera 2000-otoño 2001, muestreos en noviembre, enero, y marzo
- Periodo 4: primavera 2001-verano 2002, muestreos en octubre, noviembre, y febrero
Los muestreos realizados durante el 1° periodo fueron preliminares. En dichas
ocasiones, con la finalidad de determinar los posibles ambientes aptos para el desarrollo
del hospedador intermediario, se buscaron caracoles del género Lymnaea en todos los
cuerpos de agua del establecimiento. En los ambientes temporarios se recorrió parte de
la superficie total y en los cuerpos de agua permanentes se buscaron caracoles en las
orillas (observando aproximadamente 50 cm hacia el interior del cuerpo de agua y hacia
afuera), también en parte de su perímetro total. Además, con el objetivo de poner a
punto un diseño de recolección de caracoles para los muestreos posteriores, se
realizaron búsquedas en Ia superficie de los cuerpos de agua y bajo el suelo, de las
siguientes formas: 1) se pasó una red sobre el sustrato; 2) se recolectaron caracoles a
mano (Ollerenshaw 1971b, Utzinger &Tañer 2000, Castro et al. 2001); 3) se extrajeron
bloques de barro de 20 cm de profundidad que se tamizaron en el campo para buscar
caracoles enterrados (Malone 1984).
84

Para la realización del estudio se consideraron 3 niveles de análisis definidos de
la siguiente manera: macro escala - el establecimiento ganadero; meso escala —los
distintos cuerpos de agua dentro del establecimiento; micro escala —los hábitats en
cada cuerpo de agua. Durante todos los períodos de estudio se respetaron estos niveles
al estudiar tanto al hospedador intermediario como las variables ambientales.
En cada ocasión se estudiaron los cuerpos de agua que presentaban agua y/o a
los cuales se pudo acceder. Luego, para el análisis de los datos y en función del
objetivo, se seleccionaron distintos ambientes. Los criterios para dicha elección se
especifican en cada sección.
4.4.2 Variables ambientales
Macroescala
Los registros de temperaturas máximas, mínimas y medias diarias y los valores
de precipitación diaria fueron suministrados por el National Climatic Data Center, USA
(http://www.ncdc.noaa.gov/oa/ncdc.html) y por el Servicio Meteorológico Nacional. Los
datos correspondieron a la central meteorológica instalada en el Aeropuerto de Esquel,
Chubut, por ser la más cercana a la zona de estudio y la que presentaba información
más completa.
A partir de la información de temperaturas diarias se calcularon los promedios
estacionales y anuales para el período 1998-2001 y se estimaron las temperaturas
horarias mediante un modelo sinusoidal (Liu et al. 1995). Considerando que la
bibliografía sugiere que 10° C es una temperatura crítica para el desarrollo del caracol y
el parásito, se utilizó dicha temperatura como valor de interés biológico (Boray 1969,
Aziz & Raut 1996, Torgerson & Claxton 1999). Se calculó la cantidad de días que
durante las 24 horas tuvieron temperaturas superiores a 10° C, aquellos cuyas
temperaturas no superaron en ningún momento del día los 10o C y los días con
temperaturas medias superiores a los 10° C. Se utilizaron las temperaturas máximas
semanales para estimar los períodos de inactividad biológica para el caracol y el
parásito. Además, se calcularon Ia cantidad de horas acumuladas con temperaturas
superiores a los 10° C y a los 15° C, suponiendo que temperaturas por debajo de los
10° C inhabilitan completamente el desarrollo del caracol y del parásito y que entre 10°
y 15° podría haber crecimiento pero los caracoles difícilmente puedan completar su ciclo
biológico (Aziz & Raut 1996).
Los datos de precipitación diaria para los años 2000 y 2001 se registraron
también con un pluviómetro dentro del establecimiento ganadero y se compararon con
los registros de Esquel. Se analizó la existencia de una relación funcional lineal entre los
85

valores de precipitación mensual acumulada para Esquel y Cholila para el año 2000
(Sokal & Rohlf 1969).
Con el objetivo de analizar Ia variación conjunta de las variables temperatura y
precipitación, se construyó un balance hídrico de Thornthwaite para cada año de estudio
(1998-2001), utilizando los datos de precipitación y temperaturas medias mensuales.
Se procedió según Thornthwaite & Mather (1955) suponiendo un valor de capacidad de
campo (capacidad de retención de agua del suelo en contra de la gravedad) de 200
mm. Esta metodología permite realizar un balance entre el agua precipitada y el agua
evaporada del suelo, considerando el posible almacenamiento de agua. Así se puede
conocer la disponibilidad de agua en el suelo en cada momento del año, que se expresa
de Ia siguiente manera: Exceso, cuando la precipitación excede Ia evapotranspiración, y
el suelo se encuentra en capacidad de campo permanente y Déficit, cuando Ia relación
es inversa y falta humedad para satisfacer Ia necesidad potencial de agua de Ia
vegetación que cubre la superficie en estudio. Entre estos dos extremos pueden
presentarse situaciones de Uso o Reposición del agua del suelo (Sarmiento 1984).
Mesoescala
Durante todo el estudio, dentro de cada cuerpo de agua y en los sitios de
búsqueda y colecta de caracoles, se registraron Ia temperatura y el pH instantáneos del
agua. Las mediciones se realizaron utilizando instrumentos digitales en presencia oausencia de caracoles.
Con el objetivo de estudiar la dinámica diaria de las temperaturas del agua, se
seleccionaron cuerpos de agua representativos de los distintos tipos de ambientes. Para
los ambientes temporarios se eligieron 2 mallines de características generales muy
diferentes, y para los permanentes la laguna y un arroyo. Se registró la temperatura del
agua cada media hora en el mallín 1, el mallín 3, el arroyo 1 y Ia laguna utilizando
sensores digitales. La información se recogió durante los muestreos -Iapsos de 3 a 5
días-, en los meses de noviembre de 2000, noviembre de 2001, marzo de 2001, enero
de 2001 y octubre de 2001. Simultamente se registraron datos de temperatura del aire
dentro del establecimiento ganadero con el mismo tipo de sensores.
Microesca/a
Durante los muestreos de octubre y noviembre de 2001 y febrero de 2002 se
realizaron estudios en ambientes temporarios y permanentes, con el objetivo de
caracterizar los hábitats utilizados por los caracoles. Para ello se seleccionaron los
mallines 1, 2, 3 y el charco 1 (temporarios), y el arroyo 1 y la laguna (permanentes).
86

Se diseñó un muestreo que se adaptó para cada tipo de ambiente, según las
condiciones del mismo (Matteucci & Colma 1982). En forma general, desde un punto
elegido al azar se tomaron registros sistemáticamente de la temperatura y el pH del
agua, la cobertura vegetal, la altura mínima y máxima de Ia columna de agua y la
cantidad de caracoles presentes, ya sea en un parche natural o en un cuadradomuestral.
Los mal/ines y e/ arroyo 1. En cada ambiente se identificaron puntos en forma
sistemática (equidistantes 5 metros uno de otro) y en cada uno se determinó si el suelo
se hallaba mojado o seco. Esta característica se utilizó como un indicador de cómo varió
la superficie de suelo con agua entre fechas. Luego, desde cada punto se midió la
distancia al parche más cercano, y desde este último la distancia al parche vecino más
cercano. Se consideraron parches dentro de un ambiente aquellas zonas con poca
corriente, sustrato con barro visible y escasa vegetación, según Boray (1964). EI área
de cada parche se calculó aproximando su forma a una figura geométrica (círculo,
cuadrado, triángulo). En ambos parches se registraron la temperatura y el pH del agua,
Ia altura mínima y máxima de Ia columna de agua, el porcentaje de cobertura vegetal y
Ia cantidad de caracoles presentes.
El charco 1. Por ser un cuerpo de agua pequeño, se muestrearon todos los
parches presentes en una de sus orillas (la más accesible). La superficie de cada parche
se calculó como se describió en el punto anterior y en cada uno se registraron la
temperatura y el pH del agua, la altura mínima y máxima de Ia columna de agua y la
cantidad de caracoles presentes.
La laguna. En este ambiente no se distinguen parches sino que toda Ia orilla es
un microambiente continuo adecuado para el desarrollo del caracol. Por ese motivo, se
realizó un muestreo sistemático con cuadrados de 40 x 40 cm colocados a Io largo de Ia
orilla norte cada 5 metros y paralelos a la línea de costa (Foto 1, pág. 143). Dentro de
cada cuadrado se midieron la altura mínima y máxima de Ia columna de agua, la
temperatura y el pH del agua, el porcentaje de cobertura vegetal del suelo y la cantidad
de caracoles. Luego, por fuera de cada cuadrado, se buscaron caracoles hacia el pasto y
hacia el interior del cuerpo de agua, respetando un ancho de 40 cm (correspondiente al
ancho del cuadrado muestral). En el primer caso se muestreó hasta alcanzar la
vegetación y en el segundo hasta ausencia de visión en profundidad. En el sitio donde
se registró el caracol más alejado del cuadrado se midieron el porcentaje de cobertura
vegetal, la distancia al extremo del cuadrado muestral y Ia altura de la columna de agua
bajo la cual se encontraba el mismo (sólo en el caso de los registros tomados hacia el
interior de la laguna).
87

4.4.3 Elhospedador intermediario
La recolección de los caracoles fue realizada siempre por el mismo operadormediante tres métodos:
Búsqueda por tiempo: la abundancia relativa de Iymnaeas se estimó como el
número de caracoles colectados durante 30 minutos (extrapolado a 10 minutos) en los
parches aptos para el desarrollo de los caracoles (Rabinovich 1978). En los ambientes
negativos se extendió la búsqueda a 45 minutos (15 minutos más).
Método areal: se recolectó la totalidad de los caracoles presentes por unidad
de superficie conocida, únicamente para el estudio a microescala presentado en la
sección anterior. En los mallines, en el arroyo 1 y en el charco 1, la densidad de
caracoles se estimó como el número de individuos hallados en la superficie
correspondiente a cada parche, extrapolada a 1m2. La densidad de caracoles en la
laguna se estimó como la cantidad de caracoles dentro de la superficie del cuadrado
muestral (40 x 40 cm). Hacia el interior de la laguna se estimó como el número de
individuos hallados en la superficie comprendida por el ancho del cuadrado muestral y la
distancia máxima registrada hasta el caracol que se halló más alejado en Ia totalidad de
las mediciones realizadas. En todos los casos la densidad se extrapoló a número de
caracoles por m2.
Recolección en cantidad: para el estudio de infección, estructura y dinámica
poblacional, se juntaron caracoles de todos los cuerpos de agua en grandes cantidades
extendiendo la búsqueda arbitrariamente, más allá de los 30 minutos estipulados parael cálcqu de abundancia.
Los caracoles recolectados fueron guardados entre capas de algodones húmedos
en recipientes plásticos herméticos perfectamente rotulados y trasladados con vida allaboratorio.
De todos los caracoles que se recolectaron durante el estudio, se disecaron los
animales que llegaron con vida al laboratorio y los que murieron pero conservaban el
cuerpo en buen estado. Todos los caracoles fueron medidos, incluyendo aquellos que
sólo conservaban la conchilla. Se seleccionó un subgrupo de caracoles con vida para la
identificación taxonómica según se describe más adelante. Los tamaños se utilizaron
para el estudio de estructura y dinámica poblacional. Se seleccionaron para buscarinfección aquellos caracoles que conservaban el animal en buen estado, estuvieran
vivos o muertos. Para este último estudio, los caracoles se mantuvieron discriminados
por cuerpo de agua y según el potrero, para poder relacionar la ocurrencia del parásito
con el movimiento del ganado.
88

Identificación
Para la identificación taxonómica de los caracoles, se seleccionó de cada muestra
recolectada en campo en los distintos ambientes, un grupo de 5 ó 6 individuos de gran
tamaño (>6 mm) que no emitieron cercarias durante los primeros 10 días. Estos fueron
anestesiados en una solución de Nembutal 0,05%, matados y fijados según Paraense
(1984) (ver descripción detallada en Apéndice). Los especimenes fijados fueron
identificados por el Dr. W. Lobato Paraense (Departamento de Malacología, Instituto
Oswaldo Cruz, Brasil), según la anatomía interna de los órganos reproductivos y la
morfología de la conchilla (Paraense 1976, 1982, 1984).
Los pies de todos los individuos determinados fueron seccionados y guardados
en alcohol 70% o a —17° C y enviados al Dr. Alejandro Schijman, del Laboratorio de
Biología Molecular de la Enfermedad de Chagas, Instituto de Investigaciones en
Ingeniería Genética y Biología Molecular (INGEBI) para un estudio molecular mediante
la secuenciación del gen de la subunidad 185 del ARNribosomal.
Estructura y dinámica poblacional de los caracoles
La relación que puede existir entre tamaño y edad de los individuos es discutida
por distintos autores (Venturini et al. 1981, Mattos & Ueno 1986, Rumi & Hamman
1992, Lee et al. 1994) ya que la longitud de un caracol depende, además del tiempo de
vida, de las condiciones del ambiente a las que estuvo sometido durante su desarrollo.
Considerar que dicha relación es lineal, implica asumir muchos supuestos con respecto a
la homogeneidad en las variables ambientales que difícilmente se cumplen en
condiciones de campo. Por dicho motivo, en este estudio se utilizó la longitud de cada
caracol únicamente como estimador del tamaño. Para ello, todos los caracoles fueron
medidos, antes de ser disecados, desde el ápice hasta el margen anterior de la conchilla
(Hubendick 1951, Oviedo et al. 1995) utilizando microscopio estereoscópico con ocular
graduado. Se consideró como error de medición por parte del observador, la mitad de la
mínima unidad del ocular graduado, que correspondió a 0,08 mm. En todos los casos se
midieron la mayoría de los caracoles recolectados, excepto en la laguna, donde se midió
aproximadamente un 50% de los caracoles juntados en cada muestreo.
La distribución y Ia dinámica temporal de los cuerpos de agua (ver capítulo 2)
mostraron que existe conexión entre los mismos en algún momento del año. Este
resultado permitiría suponer que los grupos de caracoles que habitan cada ambiente no
se encuentran aislados unos de otros, y que probablemente haya transporte de
individuos desde los cuerpos de agua a mayor altitud hacia los que se encuentran más
cercanos al río en el valle. Suponiendo esto, podria considerarse que hay flujo génico
89

entre los caracoles de la especie L. viatrix en los distintos cuerpos de agua de la zona de
estudio y por ende que componen una única población.
En esta tesis se ana/izan la estructura y la dinámica de L. viatrix a macro escala,
considerando que existe una única población en el campo. Así, la distribución de
tamaños de los caracoles en cada fecha fue utilizada para determinar cambios en la
estructura poblacional y para estimar la época reproductiva. Para estudiar la proporción
de juveniles y adultos en el tiempo, se estableció como criterio que los caracoles
mayores o iguales a 4 mm han alcanzado la madurez sexual y los menores a 4 mm son
inmaduros, según lo observado en forma preliminar en el campo y sugerido por
Venturini & Fonrouge (1985) para L. viatrix en la provincia de Buenos Aires. A partir de
los datos obtenidos se realizó un análisis exploratorio entre y dentro de los distintos
años y estaciones climáticas y luego se analizaron los distintos períodos de estudio
(exceptuando el preliminar) para compararlos.
Aún cuando se considere la existencia de una única población en el área de
estudio, el aislamiento parcial entre cuerpos de agua durante ciertas épocas del año
podría estar generando una estructura y dinámica heterogéneas a meso escala. Según
el modelo de metapoblaciones, las poblaciones están espacialmente estructuradas en
subpoblaciones, y la migración entre ellas tiene efecto sobre la dinámica local,
incluyendo la posibilidad del restablecimiento de la población luego de una extinción
(Hanski & Ginin 1997). Si esto estuviese ocurriendo en la población en estudio, podría
observarse heterogeneidad entre los caracoles de distintos cuerpos de agua, generada
por el aislamiento parcial y por características particulares de cada ambiente. Con el
objetivo de estudiar la heterogeneidad en la estructura de la población a meso escala
(“subpoblaciones”) se exp/oraron los tamaños de los caracoles en cada cuerpo de agua
en las distintas fechas para buscar posibles patrones de agrupamiento.
Por otro lado, las diferencias en la dinámica del agua en ambientes temporarios
y permanentes podrían tener un efecto sobre la estructura de las subpoblaciones de
caracoles que se desarrollan en ambos tipos de ambientes. Para probar esta hipótesis
se analizó la proporción de caracoles inmaduros y maduros sexualmente en ambientes
temporarios y permanentes.
Finalmente, se exploró la relación entre los tamaños y las abundancias de los
caracoles y las siguientes variables ambientales: temperaturas máximas, medias y
mínimas, horas con temperaturas superiores a 10° C, 15° C y 22° C, precipitación y
algunos componentes del balance hídrico. Este análisis se realizó para cada tipo de
ambiente (permanente y temporario).
90

Para los últimos dos análisis se seleccionaron como ambientes representativos
de los cuerpos de agua permanentes a la laguna, y de los temporarios a los mallines 1 y
2, por ser los que tuvieron registros más frecuentes y continuos y presencia constantede caracoles.
Infección
En el laboratorio los caracoles fueron colocados, en grupos de a 10 individuos,
en recipientes plásticos de aproximadamente 100 ml de capacidad con agua declorada,
bajo una fuente de luz artificial para favorecer Ia emisión de cercarias. En los casos en
que se observaron cercarias nadando o metacercarias enquistadas dentro de algún
recipiente, los caracoles fueron separados individualmente en recipientes de 10 ml para
detectar los individuos infectados. Los moluscos que presentaron emisión espontánea
de cercarias durante los primeros días de estudio (infecciones patentes) fueron
separados y las cercarias se montaron vivas entre cubre y portaobjetos y se
identificaron bajo microscopio óptico (40x). Luego los caracoles se disecaron para
descartar infecciones dobles. Aquellos caracoles que no emitieron cercarias durante los
primeros días fueron disecados y examinados bajo microscopio estereoscópico buscando
la presencia de infecciones tempranas (infecciones prepatentes). Las cercarias de
digeneos en general y de F. hepatica en particular se fijaron en formol 5% caliente (ver
descripción detallada en Apéndice) y se identificaron mediante características
morfológicas (Dawes 1968, Odening 1971, Yamaguti 1975). Las redias inmaduras que
sólo presentaban en su interior células germinales indiferenciadas no pudieron
identificarse a nivel específico.
Para estudiar la existencia de asociación entre la presencia de infección en un
cuerpo de agua y los tamaños y abundancias de los caracoles, se compararon estas
variables por fecha considerando todos los cuerpos de agua que alguna vez presentaron
infección. Para este estudio se utilizó un subconjunto de los datos analizados en la
sección de estructura poblacional a meso escala, incorporando la información sobreinfección.
Durante el estudio de emisión de cercarias y la disección de los caracoles
recolectados en la laguna en enero y marzo del año 2000, se registró la presencia de
esporoquistes, redias, cercarias y metacercarias de digeneos diferentes de F. hepatica.
Con el objetivo de determinar los estadios larvales hallados y estudiar la abundancia de
estos parásitos en lymnaeas, se identificaron todos los estadios larvales presentes en
los caracoles de dicho cuerpo de agua, utilizando la misma metodología descripta para
F. hepatíca.
91

Además, en el campo se realizó un relevamiento de las aves acuáticas que
frecuentaban la laguna con el objetivo de explorar qué especies podrían estar actuando
como hospedadores definitivos.
4.4.4 Análisis estadístico
Se utilizaron métodos no paramétricos debido a la falta de normalidad en lasvariables consideradas.
Las distribuciones de las frecuencias acumuladas de tamaños dentro y entre
estaciones se compararon mediante una prueba de Kolmogorov Smirnov (a= 0,05)
(Siegel & Castellan 1995).
Las proporciones de caracoles inmaduros (< 4 mm) y maduros sexualmente (>4
mm) en los distintos muestreos o tipos de ambientes, así como las proporciones de
caracoles infectados y negativos en los distintos cuerpos de agua, se contrastaron
mediante pruebas de Homogeneidad con comparaciones a posteriori (estadístico de
prueba G, a: 0,05) (Fleiss 1981).
Las medidas de tendencia central para las variables superficie de los parches y la
distancia entre los mismos, así como el pH y la temperatura en cada ambiente se
compararon mediante una prueba de Mann Whitney cuando los contrastes fueron de a
pares, o Kruskall Wallis cuando se contrastaron más de 2 grupos (Siegel & Castellan
1995, Zar 1999). Estas variables se compararon entre fechas y en relación a la
presencia de caracoles.
También se utilizaron estas pruebas para realizar la comparación de medianas
de los tamaños de caracoles pertenecientes a distintos grupos (sanos vs. parasitados;
distintos cuerpos de agua; distintas estaciones climáticas). En todos los casos se fijó un
nivel de significación del 5%.
Para el estudio de la estructura y dinámica poblacional de L. víatrix se
excluyeron de cada análisis los cuerpos de agua con menos de 6 caracoles recolectados.
Se estimó el coeficiente de variación (CV= desvío estándar/media) para
comparar las densidades de caracoles entre fechas y ambientes (Sokal & Rohlf 1969).
En las comparaciones estacionales se consideró, para cada estación, el mes más
representado en los muestreos. La primavera quedó representada por el mes de
noviembre de los distintos años estudiados, el verano por el mes de enero y el otoño
por el mes de marzo.
92

Para el estudio de infección por digeneos, se compararon las proporciones de
caracoles infectados en enero y marzo con distintos estadios larvales mediante una
prueba de Z para proporciones (Fleiss 1981).
Para buscar una relación funcional lineal entre tamaño y abundancia de
caracoles y variables abióticas se utilizaron Modelos Lineales Generalizables (GLM;
procedimiento según Carbajo 2003).
Se utilizaron gráficos de cajas (Box plots) para mejor visualización e
interpretación de los datos. El centro de cada caja indica la posición de la mediana,
alrededor se encuentran los intervalos de confianza, los extremos de la caja son los
cuartiles 1 y 3 (que incluyen el 50% de los datos), luego se observan los bigotes y la
posición de los valores extremos (Norman &Streiner 1996).

4.5 RESULTADOSY DISCUSIÓN
“Sabiendo que las larvas del Saguaypé viven en
caracoles, hicimos buscar en terrenos infectados por el
mal, caracoles de todas las clases existentes, y
pudimos convencernos que en todos los terrenos
invadidos, son sobre todo dos clases de caracoles
pequeños los que encierran las formas ¡ntermediarias;
en tan gran número se hallan los caracoles infectados,
que es más fácil hallar caracoles con hígados enfermos
que con hígados sanos”
Wernicke 1888.
4.5.1 Variables ambientales
Macroescala
Temperatura. Las temperaturas mínimas, medias y máximas estacionales y
anuales registradas durante los años de estudio se presentan en Ia Tabla 4-1. Las
temperaturas mínimas anuales fluctuaron entre aproximadamente 2-30 C, las medias
entre 8-90 C y las máximas entre 14-16o C.
Tabla 4-1: Temperaturas mínimas, medias y máximas estacionales y anuales. Estación
meteorológica de Esquel, Chubut
Año/ Verano Otoño Invierno Primavera Anual
Temp. (°C) Minima Media Máxima Mínima Media Máxima Mínima Media Máxima Mínima Media Máxima Mínima Media Máxima1998 6.7 14.0 21.5 1.8 6.8 12.4 -1.3 4.8 10.3 4.6 11.5 18.8 2.9 9.2 15.71999 7.1 14.8 22.9 -1.6 4.7 10.8 -2.2 3.8 8.8 5.0 13.1 19.3 2.0 9.0 15.42000 6.2 13.9 19.7 -0.4 6.2 11.6 -2.6 2.9 7.1 4.1 9.7 16.6 1.3 3.1 13.7
2001 6.6 12.8 19.9 -0.S 5.3 11.7 -2.4 2.7 8.5 4.7 11.8 18.9 2.1 8.2 14.8
La Figura 4-1 presenta las temperaturas medias semanales registradas durante
el período 1998-2001. EIaño 1998 tuvo un invierno más cálido que los años 1999, 2000
y 2001. En ningún momento se registraron temperaturas medias inferiores a los 2° C.
Durante los inviernos de 1999, 2000 y 2001, en cambio, se registraron temperaturas

medias inferiores a los —3° C. La primavera más cálida correspondió al año 1999,
registrándose las menores temperaturas medias durante el período estival del año
2000. Durante los meses de verano y hasta mitad del otoño, las temperaturas medias
fueron similares para 1998, 1999, 2000 y 2001.
La Figura 4-2 muestra el curso de temperaturas semanales máximas, medias y
mínimas para el período 1998-2001. Puede observarse, para cada año, el lapso en el
que las temperaturas máximas no superaron los 10° C, es decir los días que tuvieron 24
horas con condiciones adversas para el desarrollo del caracol y el parásito. Para 1998
dicho período comenzó en la semana 22 (fines de mayo) y se extendió durante 12
semanas (principio de agosto). El año 1999 presentó, desde la semana 20 (mediados
de mayo), 18 semanas con temperaturas por debajo de los 10° C (mediados de
septiembre), el año 2000 tuvo 14 semanas con las condiciones mencionadas, a partir
de la semana 21 (entre mediados de mayo y fines de agosto) y el año 2001 tuvo 12
semanas en las que la temperatura máxima no superó los 10° C (desde la semana 20,
hasta la semana 32) (Tabla 4-2). Estos resultados muestran que los años que
presentaron menos semanas con temperaturas adversas para el desarrollo del
hospedador intermediario y el parásito fueron 1998 y 2001. Durante el 2000, el período
con temperaturas inferiores a los 10° C comenzó a principios del invierno, mientras que
los años restantes empezó durante el otoño. Con respecto a las temperaturas mínimas,
el año 1998 tuvo 18 días en los cuales la temperatura excedió los 10° C durante las 24
horas. Los mismos se registraron en los meses de enero, febrero y diciembre. Durante
1999 se registraron 26 días con estas características; Ia diferencia con el año anterior
radicó en que hubo días durante el mes de noviembre en los que las temperaturas
mínimas excedieron los 10° C. El año 2000 sólo presentó 12 días con condiciones
favorables todo el día. Los mismos se registraron durante enero, febrero, marzo y
diciembre. Para el año 2001 se encontraron 16 días con temperaturas adecuadas para
el desarrollo de hospedador intermediario y el parásito entre enero y marzo, un día
durante el mes de noviembre y 12 durante diciembre. AI analizar las temperaturas
medias, se observó que para el año 1998, las mismas fueron inferiores a los 10° C
entre Ia semana 15 (comienzos de abril) y la 45 (comienzos de noviembre). Durante
1999 las temperaturas medias no superaron los 10° C entre Ia semana 13 (fines de
marzo) y la 41 (principios de octubre), en el 2000 entre las semanas 18 (fines de abril)
y 43 (mediados de octubre) y en el 2001 entre la 16 (mediados de abril) y Ia 42
(mediados de octubre).
Con respecto a los períodos favorables y desfavorables para el desarrollo del
caracol, si se consideran las temperaturas medias inferiores a los 10° C (OIIerenshaw
1971a), podría suponerse que el reposo invernal (hibernación) ocurre entre marzo-abril
y octubre y que los meses restantes son los de actividad biológica. Si además se toman
las fechas en las que las temperaturas son inferiores a los 10o C durante todo el día
(OIIerenshaw 1971a, Aziz & Raut 1996, Claxton et al. 1999), el período de reposo se

Figura 4-1: Temperaturas medias semanales registradas en Esquel, Chubut (1998-2001)
E3
TempMediasSemanales(oC)
nE5
<
3
No h ->
13 57911131517192!!!
primavera
Eleje de las abscisas indica las semanas del año (entre 1 y 52). Las flechas indican los momentos aproximadosen los que las temperaturas pasan a ser inferiores o superiores a los 10°C.
Figura 4-2: Temperaturas máximas, medias y mínimas semanales registradas en Esquel, Chubutdurante el período 1998-2001
Temp.Semanalespromedio(oC)
Los registros se presentan desde enero de 1998 hasta diciembre de 2001.
Temperaturas máximas (azul), medias (rojo) y mínimas (amarillo) expresadas semanalmente. 96

restringe a mayo-septiembre. De esta forma, a partir de los datos de temperatura
ambiental se puede concluir que el período de hibernación para los caracoles
comenzaría en el rango que va entre fines de marzo y mediados de mayo y finalizaría
entre comienzos de septiembre y octubre. Por supuesto, esta dinámica a macro escala
puede presentar diferencias entre ambientes, pero siempre serán dentro del rango
presentado.
Tabla 4-2: Cantidad de semanas por año cuyas temperaturas máximas no superaron los
10° C. Estación meteorológica de Esquel, Chubut
# de semanas con Lapso (en semanas delAño ,,
Temp. Max. < 100 C ano -entre 1 y 52-)1998 12 22-341999 18 20-382000 14 21-352001 12 20-32
La estimación de las temperaturas horarias permitió calcular la cantidad de
horas acumuladas por encima de ciertas temperaturas de interés biológico. Excluyendo
los meses en los cuales las temperaturas son inadecuadas para el desarrollo del caracol
y el parásito (entre mayo y septiembre según se desprende de los resultados
presentados en la Tabla 4-2), durante el primer período de estudio, entre septiembre y
mayo se acumularon 3221 horas con temperaturas superiores a los 10° C, en el
segundo período de estudio 3280 horas, durante el tercer período de estudio 2922
horas y durante el cuarto período 2654 horas entre septiembre y febrero, momento en
que finalizó esta investigación. Contemplando la ausencia de datos en marzo y abril del
último período de estudio, en la Tabla 4-3 se presenta la cantidad de horas con
temperaturas superiores a los 10° C para cada período, pero subdivididos de la
siguiente forma: entre septiembre y febrero, y marzo-abril. Así, haciendo posible la
comparación entre períodos, se observa que entre septiembre y febrero, la mayor
cantidad de horas acumuladas con temperaturas superiores a los 10° C se registró en el
4° período de estudio (primavera 2001-verano 2002) y la menor durante el tercero
(primavera 2000-verano 2001) (Tabla 4-3).
Finalmente, al realizar un análisis semejante pero por año, se observa que la
cantidad de horas acumuladas por encima de los 10° C fue similar entre los meses de
enero y abril para todos los años, pero como consecuencia de que el invierno de 1998 y
la primavera de 1999 fueron más cálidos que los de 2000 y 2001, estos primeros dos
años de estudio acumularon un total de horas con temperaturas favorables mayor (Fig.
97

4-3). AI realizar el mismo análisis con temperaturas acumuladas por encima de los 15°
C, la diferencia entre años se acentúa desde el comienzo del año (Fig. 4-4).
Tabla 4-3: Cantidad de horas acumuladas por encima de los 10° C entre septiembre y
mayo de cada período de estudio
Período Sep-Feb Mar-Abril Total10 2491 73o 32212° 2543 737 32803° 2209 713 29224° 2654 s/d 2654
S/d= sin dato.
Precipitación. Los registros de lluvias tomados dentro del establecimiento de
estudio durante los años 2000 y 2001 mostraron niveles de precipitación superiores a
los registrados para Esquel en esos mismos años. Este resultado es reflejo del marcado
gradiente de precipitaciones oeste-este, producto de la existencia de la Cordillera. Por
esta razón, se consideró que el sesgo en la información sería mayor utilizando los
registros de Esquel que estimando la verdadera precipitación de Cholila mediante un
análisis de regresión lineal. Al estudiar la relación funcional entre la precipitación en
ambas zonas (Cholila en función de Esquel) durante el año 2000, se encontró una
regresión significativa (R2=0,98, p<0,01), explicada por la siguiente ecuación:
Y= 33,837+ 1,7107 X
A partir de esta recta estimada se transformaron los datos de precipitación
obtenidos en Esquel y se utilizaron los valores transformados como estimación de la
precipitación en la zona de estudio, para los dos años con información faltante (1998
1999).
La precipitación acumulada mensualmente durante el período de estudio se
presenta en la Figura 4-5. Esta variable resultó muy heterogénea para un mismo mes
en los distintos años. El año 2001 fue el que registró mayores precipitaciones en Cholila
y el año 1999 fue el más seco. Durante el año 2001, los mayores aportes de agua se
registraron durante los meses de mayo, junio y julio. Sin embargo, prácticamente
durante todo el año las precipitaciones acumuladas superaron a las de los años
restantes. Solamente en enero de 2001 cayeron 190 mm de agua, mientras que en
enero de 1998 llovieron unos 67 mm, 34 mm en 1999 y durante los meses de enero de
2000 y 2002 aproximadamente 20 mm. A la vez, durante mayo y julio de 2001
cayeron, cada mes, aproximadamente 130 mm más que durante los mismos meses del

Figura 4-3:Cantidad de horas acumuladas con temperaturas ambientales superiores a los 10°Cen Esquel, Chubut (1998-2001)
3500
3000
2500
2000
1500
1000
Figura4-4:Cantidad de horas acumuladas con temperaturas ambientales superiores a los 15°Cen Esquel, Chubut (1998-2001)
2500
1500
1000
99

Figura 4-5: Precipitación acumulada mensualmente en Cholila, Chubut durante el período 1998-2001
Pp.Acumulada(mm)
Figura 4-6: Interpretación de los balances hídricos para el período 1998-2001, Cholila, Chubut
MESE F M A M J J A S 0 N D
1998 u d d r r u e e e u u u1999 d d d d d r r r r u u u
zz 2000 d r d d r e e e e e u u2001 e u e u e e e e u u u d2002 r r
Para cada mes de cada año se indica la situación de disponibilidad de agua en el suelo: u- uso, d- déficit,r- reposición, e- exceso.
100

año anterior. Gracias a estos aportes extraordinarios, la precipitación acumulada
durante el año 2001 fue de 1178 mm contra 1050 mm para 1998, 739 mm para 1999 y
897 mm caídos durante 2000. A pesar de esto, los meses de septiembre a diciembre de
2001 fueron más secos (43 mm acumulados) que los del año previo (196 mm
acumulados).
Balance Hídrico. El resultado del análisis conjunto del efecto de las
precipitaciones y las temperaturas sobre Ia disponibilidad de agua en el suelo se
presenta en la Figura 4-6. Dicha figura representa una esquematización del balance
hídrico anual correspondiente a Ia región de estudio. Puede apreciarse que la situación
de disponibilidad de agua fue muy heterogénea dentro de una misma estación climática
durante los distintos años estudiados. Todos los veranos comenzaron (diciembre) con
humedad en el suelo, excepto el verano de 2001. Sin embargo, dichas condiciones sólo
se mantuvieron entrado el verano (enero, febrero) de 2001. Los restantes presentaron
situaciones de estrés hídrico (déficit o reposición). El otoño fue homogéneo (déficit de
agua en el suelo) en 1998, 1999 y 2000, pero el 2001 fue marcadamente más húmedo,
con disponibilidad de agua en el suelo durante toda la temporada. La primavera
comenzó con abundante agua en el suelo (producto de deshielos importantes) exceptoen 1999.
El análisis de las variables temperatura ambiente, precipitación y disponibilidad
de agua en el suelo muestran que las estaciones climáticas no son uniformes entre años
para las tres variables de interés. La temperatura presenta cierta homogeneidad
durante los meses cálidos, pero los inviernos son muy distintos. Además, la
precipitación fluctúa mucho entre meses y entre años, generando así un balance hídrico
con características particulares para cada estación de cada año estudiado.
Mesoescala
Los pH y las temperaturas del agua, medidos en los ambientes en los que se
encontraron caracoles activos, resultaron muy variables en cada cuerpo de agua y entre
los mismos durante el estudio. EI pH fluctuó entre 5,3 y 9,5 y las temperaturas entre
7°Cy 29°C y no se observó ninguna relación dentro o entre cuerpos de agua.
Los registros de temperaturas del agua para el mallín 1, el mallín 2, la laguna y
el arroyo 1 (en algunas ocasiones comparadas con las temperaturas ambientales) se
presentan en Ia Figura 4-7. En noviembre de 2000 las temperaturas del agua del mallín
1 y el mallín 3 fueron similares durante el período analizado, y generalmente inferiores
a las temperaturas registradas en Ia laguna. Sus temperaturas mínimas fueron siempre
superiores a las del aire y las máximas menores. En el caso de la laguna, las
101

temperaturas mínimas fueron siempre superiores a las del aire pero se alcanzaron
temperaturas máximas similares y sin ningún desfasaje (Fig.4-7a). En noviembre de
2001 se observaron temperaturas mínimas similares entre el mallín 1 y la laguna, pero
esta última alcanzó nuevamente temperaturas máximas superiores (Fig.4-7b). En enero
de 2001 las temperaturas en el mallín 1 fueron superiores a las registradas en el mallín
3 (Fig.4-7c). Finalmente, en marzo de 2001 las temperaturas de los mallines 1 y 3
fueron similares y las de Ia laguna generalmente superiores, registrándose una situación
parecida a la descripta para noviembre de 2000 (Fig. 4-7d).
Esta información, aunque fragmentaria, muestra las siguientes tendencias:
Durante los meses en los que el clima no es tan cálido (noviembre y marzo) los mallines
se comportan de manera similar entre sí. Sus temperaturas mínimas son superiores a
las ambientales, pero sus temperaturas máximas difícilmente alcanzan las del aire.
Durante el verano (enero), momento de más calor, el mallín 1 tiene mayor capacidad
de calentarse, alcanzando siempre temperaturas más altas que las del mallín 3, con
mínimas y máximas superiores también a las del aire. La laguna presenta aguas más
cálidas en todo momento, alcanzando temperaturas máximas mucho más elevadas que
las de los mallines y temperaturas mínimas generalmente superiores o similares. El
único registro del arroyo 1 muestra un patrón muy homogéneo en el tiempo, sugiriendo
la ausencia de una influencia importante por parte de las temperaturas ambientales.
Microescala
Durante el estudio de variables ambientales realizado a micro escala se
obtuvieron registros en: 10 parches en el arroyo 1, 42 en el mallín 1, 20 en el mallín 2,
16 en el mallín 3 y 12 en el charco 1 durante los meses de octubre y noviembre de
2001. Los resultados y el análisis de los datos obtenidos en la laguna se presentan por
separado por no haberse realizado sobre parches.
La superficie con suelo mojado en los mallines disminuyó entre octubre y
noviembre (entre el deshielo y a medida que aumentaron las temperaturas
ambientales). En octubre el 73,7% (28/38) de los puntos medidos presentaban
humedad, versus un 56,5% (13/23) en noviembre.
AI analizar conjuntamente la superficie de los parches de los mallines, el arroyo
1 y el charco 1, se encontraron diferencias significativas entre fechas (p=0,002),
hallándose parches más grandes durante el mes de noviembre (Fig. 4-8). La distancia
entre parches también difirió significativamente entre fechas (p<0,001), siendo
mayores en noviembre (Fig. 4-9). AIanalizar la superficie de los parches en cada fecha,
no se encontraron diferencias significativas entre el arroyo 1, el mallín 1, el mallín 2, el
mallín 3 y el charco 1 en octubre de 2001 (p=0,2), ni entre el arroyo 1 y el mallín 1 en
noviembre de 2001 (p=0,83, Fig. 4-10). Sin embargo, las distancias entre parches

Figura 4-7: Temperaturas horarias del agua en los distintos ambientes estudiados, Cholila, Chubut
a .Referenc1as+Laguna
6 1+Mallín 1É —0-—Maliin 3.5, Mar-v Arroyo 1E +Aire8.Ep
b30
25G3 208
«E 15aE 10
¡2 5
0
i I 13-11-2001 I 14-11-2001 l 1511-21101 i |
C
Ü.9,II
L5EaE¡2
116-171-2001} 17-0141101 [ 10-01-2001 i 1901-20111 120-111-2001]
d
Temperatura(oC)
l l 20-03-2001 ¡Hum | 22-0321101 | ¡Ha-¡nm I 24-03-2001 |
Las temperaturas se registraron cada 30 minutos. a) Temperaturas del agua (Laguna, mallín 1, mallín 3) ydei aire entre ei 13 y el 16 de noviembre de 2000; b) Temperaturas del agua registradas en la Laguna, elmallín 1 y el arroyo 1 entre ei 12 y ei 16 de noviembre de 2001; c) Temperaturas del agua (mailines 1 y 3)y del aire entre 16 y el 19 de enero de 2001; d) Temperaturas del agua (Laguna, mallín 1, mallín 3) y delaire entre el 19 y el 24 de marzo de 2001.
103

Figura 4-8: Superficie de los parches en los hábitats para caracoles (maliines, arroyo 1 y charco 1)en el área de estudio, Cholila, Chubut (2001), (p < 0,01)
Superficie(cm2)
.
Octubre Noviembre
Figura 4-9: Distancia entre parches en los hábitats para caracoles (maliines, arroyo 1 y charco 1)en el área de estudio, Cholila, Chubut (2001), (p < 0,001)
rs," Mi
Distancia(cm)
g
Octubre Noviembre
Figura 4-10: Superficie de los parches en los hábitats para caracoles, para cada uno de los mesesestudiados (2001) (p= 0,2 para octubre; p= 0,83 para noviembre)
dcïuBre
8000 - . .t
ST 3000 E3
.9 - ‘QÉ, Ñ VIemBregw - nooo
' 3000
Arroyo 1 Mallín 3 Charco 1 Mallín 2 Mallín 1
Loscírculos negros representan Ia mediana, ubicada en el centro del Intervalo de confianza (gris oscuro). Los rectángulos representanlos cuartiles 1 y 3 y en su Interior están contenidos el 50% de los datos. Los CIrculosblancos son valores extremos (ver Mat. y Met., 4.4.4).
104

cercanos fueron significativamente distintas entre los ambientes en octubre (p<0,001) y
en noviembre (p<0,001). Las mayores distancias entre parches siempre se encontraron
en el arroyo 1. En el mes de noviembre el mallín 3 presentó distancias entre parches
similares a las registradas en el arroyo 1. El charco 1 tuvo distancias entre parches
intermedias, seguido por el mallín 2 (Fig. 4-11). El ambiente con parches más cercanos
fue el mallín 1. Estos resultados indicarían que la superficie de los parches no es una
variable que condicione la presencia de caracoles en los distintos ambientes en un
momento dado. Sin embargo, las variaciones temporales podrían jugar un rol
importante. La dinámica del agua a medida que se secan los ambientes (entre la época
de deshielo y Ia llegada del verano) parecería estructurar los parches de Ia siguiente
forma: parches más pequeños y cercanos (mayor conectividad) cuando hay más agua y
parches más grandes y alejados a medida que los ambientes se van secando.
Se encontraron caracoles en 20 de los 100 parches estudiados (1 en el charco 1
y 3 en el mallín 2 en octubre, y los restantes en el mallín 1, en octubre y noviembre).
Por esta razón, los análisis asociados a la presencia de caracoles que se presentan a
continuación, se realizaron con los datos de los mallines exclusivamente.
Los parches con individuos fueron significativamente más grandes que los no
utilizados (p<0,05, Tabla 4-4). EI60% de los caracoles hallados estaban en parches de
superficie mayor a los 1000 cmz. La distancia entre parches más cercanos no difirió
entre los que tenían caracoles y los que estaban vacíos (p=0,06), aunque Ia diferencia
fue estadísticamente marginal (Fig. 4-12). Por este motivo se analizaron dichas
variables por ambiente y por fecha y en ninguno de los casos estudiados se encontraron
diferencias significativas (p>0,05 en todos los casos).
Con respecto a las profundidades a las que se encuentran los caracoles, las
mismas oscilaron entre cero (lo que se llamará “playa”) y 6 cm. Sólo se encontraron
caracoles en parches cuya columna de agua mínima fue de cero centímetros (Fotos 2 y
3, pág. 143). En los parches con presencia de playas, las alturas máximas de las
columnas de agua no parecieron condicionar Ia presencia de caracoles ya que no se
encontraron diferencias significativas entre dicha variable y Ia columna máxima en cada
parche (p=0,4).
Los parches con presencia de caracoles presentaron pH significativamente más
bajos que los sitios sin caracoles (p<0,01, Fig. 4-13). Los caracoles del género Lymnaea
se encontraron en microambientes cuya mediana de pH fue de 6,3 (1o -3° cuartil= 6,2
6,6). No se encontraron diferencias significativas en las temperaturas registradas en
parches con y sin caracoles (p= 0,21, Fig. 4-14).La variabilidad relativa en Ia densidad de caracoles fue similar en ambas fechas
(3,3 en octubre vs. 3,7 en noviembre). Los coeficientes de variación fueron de 1,5
(desvío: 26, media= 16,8) para el mallín 1, y 3 (desvío: 5,2, media: 1,75) para el
105

Figura 4-11: Distancia entre parches en los diversos hábitats para caracoles,Chubut (2001)
por mes. Cholila,
400- fi y.300
200
.A o0l
Distancia(cm)
o
Octubre
' 3000
‘ 1000,4 7 ., .m
Arro'yo 1 Máliin 3 Cha'rco1 Mállínz Mállin1
Tabla 4-4: Superficie de los parches utilizados y no utilizados por los caracoles en todos losambientes estudiados, Cholila,Chubut (Octubre-noviembre de 2001)
Superficie (cm?)Parches c/ca racoles Parches s/caracoles
250Mínima1er cuartil 434.97 270Mediana 1016.3a saab
3er cuartil 3488.4 1300Máxima 9225 6939.8
N 20 80
Letras distintas indican diferencias significativas.
Figura 4-12: Distancia entre parches cercanos utilizados y no utilizados por los caracoles en todoslos ambientes estudiados, Cholila,Chubut (Octubre-noviembre de 2001) (p > 0,05)
Distancia(cm)
400’ .7 P
I
Parches sincaracoles
Para interpretación de los gráficos de caja ver Figura 4-10 o Materiales y Métodos (4.4.4).
Parchescon caracoles
106

Figura 4-13: Valores de pH en los parches utilizados y no utilizados por los caracoles en todoslos ambientes estudiados, Cho|i|a, Chubut (Octubre-noviembre de 2001) (p > 0,05)
7.1 “
pH 6.6
5.1 l
Parches slncaracoles Parches con caracoles
Figura 4-14: Temperatura del agua en los parches utilizados y no utilizados por los caracoles entodos los ambientes estudiados, Cho|i|a, Chubut (Octubre-noviembre de 2001) (p > 0,05)
,,,,,,,,,,,,,,,,,,,,,,,,,,,,,, ..
25‘
20‘
Temperatura(oC)
Z‘nl
0 l I
Parches SIl'lcaracoles Parches con COYClCOIGS
Para interpretación de los gráficos de caja ver Figura 4-10 o Materiales y Métodos (4.4.4).
107

mallín 2, durante el mes de octubre. En noviembre el mallín 1 presentó un coeficiente
de variación igual a 2,8 (desvío= 11,9, media= 4,3). La distribución de estos animales
resultó agrupada; Ia mayoría de los parches estaban vacíos, y algunos pocos contenían
gran cantidad de caracoles (Tabla 4-5). Por esta razón, se compararon las densidades
excluyendo los parches vacíos. Durante el mes de octubre, el mallín 1 (n= 10) presentó
una densidad mediana de 23,1 caracoles por metro2 de hábitat (1o -3° cuartil: 11,3-67)
y el mallín 2 (n= 3) una densidad mediana de 11,5 caracoles por metro2 (1o -3° cuartil:
2,7-20,8). En noviembre la densidad mediana para el mallín 2 (n= 5) fue de 7,9 (1° -3°
cuartil: 2,5-40,6).
La cobertura vegetal de los parches con caracoles nunca superó el 50% de la
superficie, permitiendo en todos los casos que la luz alcanzara el sustrato.
La laguna. En febrero de 2002 el nivel del agua en la laguna había disminuido
con respecto a los meses de primavera, quedando al descubierto una orilla barrosa
(Foto 4, pág. 143). En dicha ocasión, se tomaron registros en 30 cuadrados muestrales
equidistantes, y se observó el ecotono de la laguna como un ambiente homogéneo a
simple vista.Se encontraron caracoles en 26 de los 30 cuadrados muestrales estudiados. En
todos los casos hubo playas y la altura máxima de la columna de agua dentro de los
cuadrados fue de 1,2 cm. No se encontraron diferencias significativas entre esta última
variable y la presencia de caracoles (p=0,15). AI alejarse del cuadrado muestral hacia
las pasturas, sólo se encontraron 4 caracoles a distancias que nunca superaron los 90
cm desde el ecotono de la laguna, y siempre que el suelo mantenía humedad y no
estaba cubierto por pastos. Hacia el interior del cuerpo de agua, en el 90% de los sitios
muestreados (27/30) se encontraron caracoles sumergidos hasta 6,7 cm de profundidada no más de 65 cm de la orilla.
El coeficiente de variación para la densidad de caracoles fue de 1,01 (desvío=
73,9, media: 72,5) en los cuadrados muestrales y de 1,43 (desvío= 62,7, media=
43,8) hacia el interior de la laguna. Este resultado pone de manifiesto que la
distribución de caracoles es al azar en la orilla de la laguna y con cierto grado de
agrupamiento hacia el interior de la misma (Tabla 4-6).
La cobertura vegetal dentro de los cuadrados muestrales nunca superó el 10%
de la superficie, y el 90% (27/30) de los cuadrados no presentó vegetación en absoluto
(Foto 5, pág. 143). Algo similar se observó hacia el interior de la laguna, donde
exceptuando un cuadrado con el 80% de su superficie cubierta, los restantes
presentaron a lo sumo el 5% de cobertura vegetal.
El pH en este ambiente no parece tener asociación con la presencia de caracoles,
ya que no se encontraron diferencias significativas entre los valores de acidez y la
aparición de individuos en la orilla, dentro de un cuadrado muestral (p=0,6, Fig. 4-15).
Sin embargo, a diferencia de lo encontrado en los ambientes temporarios, las
108

temperaturas registradas en los cuadrados muestrales con y sin caracoles difirieron
significativamente (p= 0,008), registrándose mayores valores en los cuadrados
muestrales con caracoles (Mediana=18,1° C; 1°-3° cuartil: 16,3-20,2° C versus Me=
11,70 c; 10-3° cuartil: 9,9-15,4° c; Fig. 4-16).
En ambos tipos de ambientes, la presencia de sustrato barroso con playa, baja
cobertura vegetal y alta incidencia solar caracterizaron los sitios con caracoles. Estas
condiciones probablemente sean necesarias para permitir el desarrollo de alimentos
(Boray 1964) o generar microambientes con temperaturas y pH óptimos (Aziz & Raut
1996, Claxton et al. 1999, Cañete et al. 2004).
El pH es una de las principales variables que condicionan la presencia de los
caracoles del género Lymnaea en un determinado ambiente a macro escala (Malone
1994). A ese nivel de análisis, el área de estudio presenta suelos adecuados para el
desarrollo del hospedador intermediario (ver capítqu 2). A una escala de mayor detalle,
una recopilación bibliográfica realizada por Boray (1969) indica que en Australia, L.
tomentosa puede vivir en ambientes cuyos pH fluctúan entre 5 y 8, aunque el 90% de
los ambientes adecuados tienen pH entre 6 y 7. Este autor no encontró caracoles en
ambientes con pH por debajo de 5 o encima de 8. Durante el desarrollo de esta tesis,
los pH medidos en forma instantánea en todos los cuerpos de agua estudiados (meso
escala) presentaron un rango de variación mucho mayor (5,3-9,5), indicando que en el
área de estudio, el hospedador intermediario puede adaptarse a condiciones muy
diversas. Además, se encontró asociación entre el pH y la presencia del caracol sólo en
un tipo de ambientes (temporario), sugiriendo que a micro escala, las características de
cada cuerpo de agua (temperatura, corriente, tamaño de los parches, conectividad
entre los mismos, existencia de parches o hábitas continuos) pueden modificar Ia
importancia que puede tener el pH sobre el caracol.
Con respecto a la temperatura, sólo se encontraron diferencias en la laguna. En
ese ambiente, los sectores con caracoles presentaron una mediana de temperatura
mucho más elevada que las zonas sin caracoles. Este resultado sugiere que con las
temperaturas ocurre algo similar al pH, y que en un ambiente donde hay mucha
variabilidad de temperaturas los caracoles se desplazarían pequeñas distancias haciasectores más cálidos.
4.5.2 Elhospedador intermediario: Generalidades
Durante el estudio se recolectaron aproximadamente 4000 caracoles de la
Familia Lymnaeidae. Los individuos siempre se hallaron sobre superficies húmedas o
semienterrados en el barro y se distinguieron fácilmente a simple vista (Fotos 5, 6 y 7,
pág. 143). En las ocasiones en las que se procedió a buscar caracoles en bloques de
109

barro de hasta 20 cm de profundidad o con coladores enterrados en el fango, nunca se
hallaron individuos por debajo de la capa de barro superficial. Por otro lado, las
características de los ambientes donde se encontraron las lymnaeas imposibilitaron
utilizar redes para recolectarlos. Por estos motivos, los caracoles se juntaron con
colador y a mano, en vez de utilizar red y tamices.
Debido a que el colador no se pasó en forma sistemática por todo el cuerpo de
agua, sino que se lo utilizó en los parches donde se detectaba algún caracol a simple
vista, podría suponerse que si el método tiene sesgo, el mismo será sobre los caracoles
de menor tamaño (<2 mm) que son los que a simple vista resultan más difíciles de
observar. Ollerenshaw (1971b) plantea que ante una situación semejante, el hecho de
encontrar caracoles pequeños en ciertas épocas del año, aunque sea en cantidades
menores a las que realmente hay en campo, es un indicio de que el muestreo presenta
un sesgo pequeño. El hecho de no encontrar caracoles pequeños podría estar indicando
su real ausencia, o bien que sus densidades son demasiado bajas como para detectarlos
por este método.
Se estudiaron en forma regular un cuerpo de agua permanente (laguna) y dos
temporarios (mallín 1 y 2) y de manera esporádica el arroyo 1, el canal, el charco 1, el
juncal, el arroyo que nace en Ia laguna grande (arroyito), el meandro y tres mallines (3,
4 y 5) (ver definiciones de los ambientes en el capítulo 2). En Ia Tabla 4-7 se presentanla cantidad de caracoles medidos de los recolectados en cada ambiente. Las celdas sin
dato y con guión indican las fechas en las que no se tomaron registros y los ceros
significan ausencia de caracoles o que el cuerpo de agua estaba seco. Entre noviembre
de 1999 y febrero de 2002 se encontraron caracoles del género Lymnaea aunque sea
una vez en todos los mallines, en las lagunas (en la laguna y en el arroyo que nace en
la laguna grande), en el meandro, en el charco 1 y en el arroyo 1. En este último cuerpo
de agua la presencia de individuos fue esporádica y siempre se los encontró en
pequeñas playitas con suelo de barro, protegidos de la corriente. Nunca se encontraron
caracoles vivos en el río, pero sí conchillas. Las mismas se hallaron semienterradas en
las playitas sin corriente que se forman en la orilla, en diversos sitios a Io largo de los 2
kilómetros estudiados. En muchos casos se encontraron conchillas agrupadas en una
playita, coincidiendo con la desembocadura de algún mallín o arroyo.
Los mallines presentaron características generales muy distintas (vegetación
esencialmente arbustiva, herbácea escasa o herbácea tupida —juncaI-)y sin embargo se
encontraron caracoles en los diferentes ambientes, pero siempre en zonas en las que la
luz llegaba al nivel del suelo (Foto 8, pág. 143).
110

Tabla 4-5: Densidad de caracoles en los parches analizados en los mallines, Cholila, Chubut (2001)
# de Parches
Dens'dad de “faciles Octubre 2001 Noviembre 2001(# individuos/m )
O 60 20
<10 4 3
10-20 4 0
20-30 2 0
30-40 1 1
>40 4 1
Total 75 25
Tabla 4-6: Densidad de caracoles en la laguna, Cholila, Chubut (2002)
# de cuadrados
Densidad de caracoles Febrero de 2002
(# individuos/m2) A B0 4 3
<10 3 7
10-20 1 9
20-30 1 0
30-40 3 0
40-50 1 2
>50 17 9
Total 30 30
A: en Ia orilla de la laguna, dentro de los cuadrados muestrales, B: hacia el interior de la laguna.
Figura 4-15: Valores de pH del agua en la Laguna dentro de los cuadrados muestrales con y sincaracoles. Cholila, Chubut, Febrero de 2002 (p= 0,6)
pH
I l
Cuadrados sincaracoles Cuadrados con caracoles 111Para interpretación de los gráficos de caja ver Figura 4-10 o Materiales y Métodos (4.4.4).

Figura 4-16: Temperatura del agua en los cuadrados muestrales con y sin caracoles. Cholila,Chubut, Febrero de 2002 (p < 0,01)
25
20‘
10'l
Cuadrados sin caracolesl
Cuadrados con caracoles
Para interpretación de los gráficos de caja ver Figura 4-10 o Materiales y Métodos (4.4.4).
Tabla4-7: Frecuencia absoluta de caracoles medidos de los recolectados en cada cuerpo de agua
Las celdas sin datos y con guión indican las fechas en las que no se tomaron registros para esos cuerpos deagua y los ceros significan ausencia de caracoles o que el cuerpo de agua estaba seco.
112

4.5.3 Elhospedador intermediario: Identificación
Todos los individuos examinados correspondieron a la especie Lymnaea viatrix
Orbígny, 1835, por observación de su genitalia y su conchilla. El resto de los caracoles
resultaron idénticos por su m0rfología externa a los determinados. En la Figura 4-17
puede observarse el aspecto externo de un individuo.
Los estudios de la secuenciación del gen de la subunidad 188 del ARNribosomal,
realizados por el Dr. Alejandro Schijman, del Laboratorio de Biología Molecular de Ia
Enfermedad de Chagas, Instituto de Investigaciones en Ingeniería Genética y Biología
Molecular (INGEBI), brindaron más información sobre la especie. El fragmento de ADN
secuenciado resultó idéntico entre individuos. Además, las secuencias obtenidas fueron
contrastadas con las de especimenes de L. viatrix recolectados en la provincia de
Córdoba y no se encontraron diferencias entre ambas poblaciones. Algunos autores
sugieren que existen problemas de determinación a nivel específico debido a launiformidad tanto de las conchillas como de la anatomía de la masa visceral entre
algunas especies (Oviedo et al. 1995). La coincidencia entre la identificación morfológica
y molecular de la especie brinda un resultado sumamente confiable, y demuestra la
utilidad, en este caso, del estudio morfológico como herramienta para la determinación
específica de los caracoles. Los resultados obtenidos dieron lugar a una tesis de
licenciatura y se presentan en forma detallada en la misma (Matos 2001).
Este resultado es Ia primera referencia que se tiene del hallazgo de la especie en
la provincia del Chubut. En la bibliografía no existen registros de su presencia a estas
latitudes, siendo el más austral el correspondiente a la ciudad de Viedma, provincia de
Río Negro, al norte del paralelo 41° S (Paraense 1976) (Fig. 4-18). Considerando que
mundialmente se han documentado casos endémicos de fasciolosis entre los 65° N y los
50° S (Boray 1985, Torgerson & Claxton 1999) y que Ia presencia de L. viatrix ha sido
descripta desde México (aproximadamente 20° N) hasta el sur argentino
(aproximadamente 40° S) (Paraense 1982, Naranjo García 2003), el hallazgo de esta
especie en la localidad de Cholila (42° S) indicaría que a esta latitud la especie seencuentra en el extremo austral de su distribución mundial.
4.5.4 Estructura y dinámica poblacional de Lymnaea viatrix
Marcoescala
Las distribuciones de tamaños en una misma estación en distintos años,
difirieron significativamente (p< 0,05 para todas las comparaciones, Fig. 4-19).
AI comparar las proporciones de caracoles inmaduros (< 4 mm) y maduros
sexualmente (> 4 mm) entre fechas, se hallaron diferencias significativas para todo el
113

grupo (G ag.= 213,26, p<0,05). Sin embargo, Ia proporción de caracoles inmaduros y
maduros sexualmente resultó homogénea entre noviembre de 1999, 2000, 2001
(primaveras) y febrero de 2002 (verano) (G = 8,38, p>0,05), entre marzo de 2000,
2001 (otoños) y octubre de 2001 (primavera) (G = 2,5, p>0,05) y entre ambos eneros
(2000, 2001) (veranos) (G = 5,27, p> 0,05, Tabla 4-8). La mayor proporción de
adultos siempre se encontró durante las primaveras, momentos en los que entre el 95%
y el 100% de los caracoles fueron mayores a 4 mm. En los eneros el porcentaje se
redujo a un 70% de maduros vs. 30% de inmaduros y durante los muestreos realizados
en marzo el porcentaje de caracoles adultos varió entre 85-90%.Las diferencias en las distribuciones de tamaños dentro de cada estación durante
el período 1998-2001 podrían ser el reflejo de Ia variabilidad en las condiciones
ambientales estudiadas a macro escala. El hecho de que las primaveras, veranos,
otoños e inviernos hayan presentado temperaturas y valores de precipitación tan
disímiles entre años estaría explicando Ia falta de homogeneidad en los tamaños de los
caracoles. Un resultado semejante se encontró al estudiar la dinámica poblacional de
Lymnaea bu/¡moides en Estados Unidos (Malone et al. 1984). Estos autores hallaron que
los tamaños promedio de los caracoles en cada mes y el esfuerzo reproductivo variaban
entre años dependiendo de las condiciones climáticas.
Sin embargo, a pesar de las diferencias registradas en las variables abióticas, al
analizar la estructura poblacional en función de la madurez sexual de los individuos, se
encontró homogeneidad dentro de cada estación. Estos resultados sugieren que la
variabilidad en el ambiente se refleja en variabilidad en los tamaños de los caracoles,
pero que en líneas generales, las condiciones en cada estación pueden estructurar a la
población en cuanto a su desarrollo (madurez/inmadurez sexual).
Con respecto a los tamaños, se observaron diferencias significativas entre
estaciones (p<0,001), encontrándose el mayor valor mediano de tamaño de caracoles
en primavera (N= 395, Me= 7,9, 1°-3° cuartil= 6,8-9,4) y el menor en otoño (794,
Me: 4,9, 1°-3° cuarti|= 4,4-5,6), presentando el verano valores intermedios (1049,
Me: 6,2, 1°-3° cuartil= 4-8, Fig. 4-20).
Durante el segundo período de estudio (primavera de 1999 —otoño de 2000)
se encontraron diferencias significativas en los tamaños de los caracoles entre
estaciones (p=0,0001, Tabla 4-9). En noviembre de 1999 (N = 92) el 67% de los
individuos presentó tamaños mayores a 8 mm y no se encontraron caracoles menores a
4 mm, indicando que a la salida de la hibernación, todos los individuos habían alcanzado
la madurez sexual y Ia mayoría eran muy grandes (>8 mm). En enero de 2000 (N =
653) la proporción de caracoles mayores a 8 mm se redujo (34%) y aparecieron
individuos de tamaños menores a 4 mm (22,5%), de los cuales más de la mitad fueron
muy pequeños (<3 mm, 14%), indicando que entre noviembre y enero se produjo el
reclutamiento de una nueva población. En marzo de 2000 (N = 490) ya no se
114

Flgura 4-17: Aspecto externo de un Individuo de Lymnaea viatrix de Cholila, Chubut
Figura 4-18: Mapa de parte de la Patagonia Argentina indicando la localidad donde se encontró aL. viatrix (Cholila) y el registro más austral (Viedma)
Rio Ncgm
O Chulilu
Chubul
115

Figura 4-19: Frecuencia porcentual acumulada de tamaños de caracoles recolectados en cadafecha de estudio: a) Muestreos de primavera (noviembre de 1999, noviembre de2000 y noviembre de 2001), b) Muestreos de verano (enero de 2000 y enero de2001), c) Muestreos de otoño (marzo de 2000 y marzo de 2001). Cholila, Chubut
DJ V
50ï40<
30
20
10
+ Nov-99+ Nov-00
l + Nov-010 :—v i . r
Frecuenciaporcentualacumulada
Tamaños (mm)
UV
100
90
80 e
Frecuenciaporcentualacumulada
U1O
6Tamaños (mm)
30“
Frecuenciaporcentualacumulada
I
6Tamaños (mm)

Tabla4-8: Cantidad de caracoles inmaduros sexualmente (<4 mm) y maduros sexualmente(>4 mm) por fecha. Cholila, Chubut (1999-2002)
Novlembre Octubre Enero Fabrero _—____Ma"Z°tamaños (mm) 1999a 2000a 2001a 2001c 2000” 2001':| 2902a 2000c 2001c
<4.0 0 1 7 8 148 115 8 57 46
>4.0 9-2 129 16__6 70 505 281 270 433 258
total 9-2 130 173 78 653 396 278 490 304
Las letras distintas indican diferencias significativas en las proporciones entre fechas (p< 0.05).
Figura 4-20: Tamaño de los caracoles por estación (Otoño representado por los meses de marzo,primavera por los de noviembre y verano por los meses de enero). Cholila, Chubut(1999-2002)
Tamaño(mm)
0 l l l
Otoño Primavera Verano
Para interpretación de los gráflcos de caja ver Figura 4-10 o Materiales y Métodos (4.4.4).
Tabla 4-9: Medidas de posición central y dispersión características para el tamaño de caracolespor fecha (1999-2002). Cholila, Chubut
Tamaño Mediano (mm) Cuartil 1 Cuartil 3 Tamaño mínimo Tamaño máximo NPrimer Nov. 1999 Primavera 9.2a 7.5 10.3 4.7 12.4 92
Período de Ene. 2000 Verano 7.2" 4.3 8.5 2 12 653Estudio Mar. 2000 Otoño 4.8“ 4.4 5.2 2.2 7.6 490
Segundo Nov. 2000 Primavera 7.9“ 7 8.7 3.1 11.2 130Período de Ene. 2001 Verano 5.2" 3.9 6.7 1.9 11 396
Estudio Mar. 2001 Otoño 5-4" 4.4 6.4 1.7 13.4 304Tercer Oct. 2001 Primavera 5.9' 5 7.9 2.3 10.4 78
Período de Nov.2001 Primavera 7.4" 6.1 9.4 2 13.7 173Estudio Feb. 2002 Verano 6.3' 5.2 7.2 2.4 10.9 278
Las letra distintas indican diferencias significativas entre meses, dentro de cada período de estudio.
117

encontraron caracoles mayores a 8 mm. Los caracoles menores a 4 mm representaron
el 11% del total pero dentro de ese intervalo sólo 2 fueron menores a 3 mm. El 88,4%
de los individuos fueron adultos cuyos tamaños variaron entre 4-8 mm, concentrándose
el 82% entre los 4-6 mm, sugiriendo que ese es el rango de tamaños al cual acceden la
mayoría de los caracoles antes de entrar en período de reposo (Fig. 4-21).
El tercer período de estudio (primavera de 2000 - otoño de 2001) mostró
diferencias significativas entre primavera y verano (p = 0,0001) y entre primavera y
otoño (p = 0,0001), pero verano y otoño no presentaron diferencias significativas en
cuanto a los tamaños de caracoles (p = 0,1, Tabla 4-9). En forma similar al primer
período de estudio, en noviembre de 2000 (N = 130) prácticamente el total de los
caracoles hallados fueron adultos, y entre ellos el 45% presentó tamaños mayores a 8
mm. En enero de 2001 (N = 396) el porcentaje de individuos mayores a 8 mm se
redujo al 13%. Un 11% presentó tamaños menores a 4 mm, de entre los cuales el 9%
fue menor a 3 mm, indicando nuevamente que el período reproductivo fue a comienzos
del verano. Prácticamente el 60% de la población fueron adultos de tamaños
intermedios (4-8 mm). Finalmente, en marzo de 2001 (N = 304) sólo el 9,5% de los
caracoles superó los 8 mm y por debajo de los 4 mm se halló un 15%, pero sólo un 3%
fue menor a 3 mm. En el intervalo 4-8 mm se encontró al 75% de los individuos,
sugiriendo nuevamente que los caracoles adultos que sobreviven el período invernal y
entran en actividad luego de la hibernación, mueren hacia comienzos del otoño y que
los que nacen a principio del verano alcanzan tamaños intermedios antes de entrar en
período de reposo (Fig. 4-21).
En el cuarto período de estudio (primavera de 2001 - verano de 2002) los
tamaños de los caracoles difirieron significativamente entre octubre y noviembre de
2001 (p = 0,0001) y entre noviembre de 2001 y febrero de 2002 (p=0,0001), pero
octubre y febrero no presentaron diferencias significativas (p = 0,92, Tabla 4-9). En el
muestreo de octubre de 2001 el 24% de los caracoles (N=78) resultó mayor a 8 mm y
se encontró un 10% menor a los 4 mm, pero sólo 1 individuo estuvo por debajo de los 3
mm. Los restantes (66%) fueron adultos de tamaños intermedios y el intervan más
frecuente fue el de 5-6 mm, conteniendo el 32% de los caracoles. En noviembre de
2001 (N = 173), el 42% de los individuos superaron los 8 mm y un 54% fueron adultos
de tamaños intermedios (4-8 mm). A diferencia de los primeros períodos de estudio, se
encontró un 8% de caracoles menores a 4 mm, pero sólo 2 fueron de tamaños menores
a los 3 mm. En el muestreo de verano, realizado en febrero (N = 278), el 14% de los
caracoles recolectados fue mayor a 8 mm y un 3% (8 caracoles) se encontró por debajo
de los 4 mm, de los cuales 2 individuos fueron de tamaño menor a los 3 mm. El resto
fueron adultos de tamaños intermedios (Fig. 4-21).
118

Figura 4-21: Estructura poblacional de L. viatrix: frecuencia porcentual de individuos en cadaclase de tamaños por fecha (1999-2002). Cholila, Chubut
100%
80%
60%
40°/o
20%
0%
I>8rrm¡214.0-8 nml3-4.0 n’mEl<3rrm
Nov 99 Ene 00 Mar OONov 00 Ene 01 Mar 01 Oct-01 Nov 01 Feb-02
Individuos de tamaños menores a 4 mm se consideran juveniles y mayores a 4 mm adultos.
119

A macro escala, se observan patrones de desarrollo sincronizados a lo largo de
los años. De manera general, los individuos adultos que durante la primavera salen de
la hibernación, oviponen durante esa temporada y luego van muriendo hacia el otoño,
como lo demuestra la disminución, entre noviembre y marzo, del porcentaje de
caracoles de gran tamaño. La nueva generación nacida en verano alcanza tamaños
intermedios para la época en que entran en reposo, y a Ia salida de la hibernación
prácticamente el total de la población está sexualmente madura. Esto indicaría que
estos animales tienen un ciclo de vida que no supera los 15-16 meses y que durante la
primera mitad del mismo alcanzan Ia madurez sexual, Io que les garantiza poder dejar
descendencia durante la temporada de actividad previa a morir. Esta dinámica es
similar a la reportada para L. truncatu/a en Escocia (Kendall 1950).
En ningún momento se pudieron extender los muestreos hasta entrado el otoño
(mayo) o bien recién comenzada Ia primavera (septiembre). Esta restricción impidió
conocer los momentos exactos en los que los caracoles entran en reposo y nuevamente
en actividad, para confirmar así los supuestos realizados a partir de las temperaturas
ambientales (ver 4.5.1, temperaturas a macro escala). Los tamaños de los caracoles
antes de entrar en hibernación y al volver a Ia actividad se estimaron utilizando los
datos de marzo y noviembre, respectivamente. Si se considera que dichas mediciones
pueden llegar a diferir de las correspondientes a los momentos exactos de entrada y
salida de Ia hibernación a lo sumo en un poco menos de dos meses, podría suponerse
que el error en Ia estimación será pequeño. Únicamente durante el cuarto período de
estudio se pudo realizar un muestreo en el mes de octubre y en dicha ocasión Ia
proporción de caracoles ¡nmaduros y maduros sexualmente no se asemejó a las
encontradas durante los meses de noviembre sino a las de marzo del período anterior.
Además, Ia estructura de tamaños fue similar en marzo y octubre de 2001,
encontrándose el tamaño modal en el mismo intervan en ambos casos, cosa que no
ocurrió entre marzo y noviembre de 2000 (Fig. 4-22). Así, cuanto más cercano es el
muestreo al momento en el que los caracoles entran en actividad, más parecidos son
sus tamaños a los que se registraron justo antes de entrar en hibernación. En
coincidencia con otros autores, este resultado sugeriría que durante el período invernal
y sometidos a las rigurosas condiciones climáticas de la región (temperaturas medias
anuales inferiores a los 10° C y temperaturas máximas que apenas alcanzan los 15° C),
los caracoles interrumpen su desarrollo o crecen a una tasa muy baja (Aziz & Raut
1996, Claxton et al. 1999). Como ejemplo, Hodasi (1976) estudió la influencia del frío
en L. truncatu/a y comprobó que caracoles de 2 mm de longitud promedio mantenidos a
5° C durante tres meses, no incrementaron su longitud durante ese tiempo en más de
0,4 mm promedio.
La proporción de caracoles ¡nmaduros sexualmente (3-4 mm) registrada en
octubre estaría indicando que existe cierto porcentaje de individuos que pasan el
invierno sin haber alcanzado la madurez, pero que maduran rápidamente ni bien
120

mejoran las condiciones climáticas. Los resultados obtenidos en este estudio difieren de
los hallados por Boray (1969) con L. tomentosa. Los tamaños promedio de la población
estudiada por este autor disminuyeron a Ia salida de la estivación (6 mm vs. 3,5 mm),
sugiriendo que los caracoles jóvenes (de menor tamaño) son los más resistentes a Ia
desecación. Aún cuando Ia estivación y la hibernación son períodos modulados por
variables distintas, en ambas situaciones el caracol se encuentra sometido a estrés,
interrumpiendo su actividad. En la población estudiada en Ia provincia de Chubut, los
tamaños promedio siempre fueron mayores a la salida de Ia hibernación (4,8 mm vs.
7,9 mm en marzo y noviembre de 2000 respectivamente, y 5,6 mm vs. 6,4 mm en
marzo y octubre de 2001), sugiriendo que no hay una mortalidad diferencial de
individuos de mayores tamaños durante el invierno.
La Figura 4-22 presenta Ia dinámica de L. viatrix durante el período estudiado.
Las distintas clases de tamaños fueron armadas de manera sistemática con amplitudes
de 1 mm (excepto la de caracoles más pequeños) y permiten observar los tamaños más
representados (tamaños modales) en cada fecha, a partir de los cuales y en forma
aproximada se diferenciaron las distintas cohortes (indicadas por una línea negra) y la
superposición temporal de generaciones. A pesar de que en todo momento la
variabilidad de tamaños es muy grande, en parte por Ia diferencia que puede haber en
la tasa de crecimiento entre individuos, la evidencia más contundente de superposición
de generaciones parece observarse durante el verano, donde los individuos seniles
cohabitan con Ia generación de caracoles recién nacidos. En otoño los individuos
pertenecientes a Ia generación anterior ya murieron y en primavera aún no hubo
reclutamiento de una nueva generación.
AIcomparar los tamaños de los caracoles entre los meses de cada estación (Fig.
4-22) puede observarse que: 1) Ia dispersión de tamaños es mayor en noviembre de
2001, intermedia en noviembre de 1999 y mínima en noviembre de 2000, 2) en enero
de 2001 Ia población se encuentra más concentrada alrededor de tamaños grandes, en
cambio en enero de 2000 todos los tamaños están representados, 3) la dispersión de
tamaños es mayor en marzo de 2001 que en marzo de 2000, momento en que la
mayoría de la población se encuentra representada por tamaños intermedios. El análisis
del balance hídrico de cada año (Fig. 4-6) demuestra que cuanto mayor es la
disponibilidadde agua (exceso constante) durante los meses previos a cada observación
(otoño para las observaciones de primavera y primavera para las de verano), mayor es
Ia variabilidad de tamaños en Ia población. Esto sugiere que las condiciones de
constante humedad en el suelo durante las temporadas de actividad, generan
condiciones Iaxas y más estables en el ambiente. De esta forma, el desarrollo de los
individuos depende de las características particulares del parche en el que habiten y al
haber agua disponible a macro escala hay muchos cuerpos de agua aislados o semi
aislados entre sí que resultan hábitats adecuados. En cambio una situación de estrés en
121

la disponibilidad de agua en el suelo condiciona el desarrollo de Ia población a los
momentos y a los pocos ambientes favorables, observándose pulsos de crecimiento
sincronizados. La mayor variabilidad, entonces, se debería a que las subpoblaciones de
los distintos cuerpos de agua crecen en forma asincrónica y estructuran a la población
general de manera distinta en función de las particularidades de cada ambiente.
En la Figura 4-22 se observa también que, en campo y sometidos a condiciones
ambientales heterogéneas en el tiempo, los caracoles crecen aproximadamente 1 mm
entre primavera y verano (noviembre-enero, 2 meses) y entre verano y otoño (enero
marzo, 2 meses), y 2 mm entre noviembre y febrero (3 meses), lo cual implicaría una
tasa de desarrollo de aproximadamente 0,5 mm por mes en período favorable.
Un estudio realizado utilizando a L. viridis como modelo experimental (Lee et al.
1994), demostró que el estado nutricional y la calidad del alimento influyen fuertemente
en el desarrollo de los caracoles, medido como tamaños alcanzados. En otro estudio,
individuos de la especie L. víatrix criados a temperaturas entre 240-26o C, aumentaron
su tamaño, en promedio, unos 4 mm en 36 días, mientras que caracoles criados en
condiciones de menor densidad pero a 200-22o C, crecieron en promedio 1 mm en 48
días (Venturini et al. 1981). A pesar de que las observaciones realizadas a campo
durante el curso de esta tesis permiten suponer que el alimento no es un factor
restrictivo en ninguno de los ambientes en los que se encontraron caracoles, la falta de
uniformidad en las variables del ambiente (por ejemplo temperatura del agua) podríanestar afectando la tasa de crecimiento de L. víatrix. En coincidencia con el criterio
adoptado por Malone et al. (1984), se consideró que caracoles muy pequeños (< 2 mm)
son individuos recién nacidos y que los mayores a 8 mm cumplieron un ciclo de vida
completo. Podría suponerse que la mayor distorsión se producirá en tamaños
intermedios (4-8 mm aproximadamente) donde la variabilidad podría estar modulada
tanto por el tiempo de vida como por las condiciones a las que estuvieron expuestos los
caracoles durante ese período. Cualesquiera que sean los factores que condicionan el
crecimiento, se puede suponer que la madurez sexual se alcanza aproximadamente a
partir de los 4 mm (Venturini & Fonrouge 1985), ya sea que este tamaño se alcance a
mayor tiempo en condiciones subóptimas, o a tiempos muy cortos de vida gracias a
condiciones de desarrollo muy favorables.

ow:
(Lum)sogewel
Oflhümv"
Figura4-22:DinámicaPoblacionaldeL.viatrixenCholila,Chubut,duranteelperíodo1999-2002
N=653
N=490
N=130N=396N=304N=78N=173N=278
V
Nov1999Ene.2000Mar.2000Nov.2000Ene.2001Mar.2001Oct.2001Nov.2001Feb.2002
Elanchodelasbarrasindicalafrecuenciadecaracolesencadacategoríadetamaños.Laslíneasindicanlaevolucióntemporaldecadacohorte
123

Mesoescala
La abundancia de caracoles a meso escala fluctuó mucho entre cuerpos de agua
y en el tiempo, lo cual dificultó el análisis de los datos. La Tabla 4-10 muestra Ia
cantidad de caracoles encontrados en 10 minutos de búsqueda por ambiente y por
fecha. Las celdas sombreadas indican los cuerpos de agua en los que se buscaron
caracoles y las celdas sombreadas vacías los momentos en los que no se realizaron
estimaciones. La abundancia de caracoles por cuerpo de agua presentó 3 registros
extremos por encima de los 25 caracoles en 10 minutos de búsqueda. Estos valores
correspondieron a la laguna en otoño (marzo de 2000) y verano (febrero de 2002) y al
mallín 4 en verano (enero de 2001). Los ambientes que presentaron abundancias entre
20 y 40 caracoles en algún momento, fueron el meandro y el charco 1. Las abundancias
fueron predominantemente menores a 10 en todos los ambientes y fechas restantes. En
el mallín 4, a diferencia de los otros mallines, los caracoles siempre se encontraron en
una zona más baja del ambiente donde se formaba una pequeña laguna que cuando
comenzaba a secarse era similar a micro escala a las orillas de la laguna y el meandro.
Los caracoles se hallaron en los bordes con barro y sin vegetación. A partir de estas
diferencias entre la zona estudiada del mallín 4 y los restantes mallines, podría
suponerse que una característica común a los cuerpos de agua que tuvieron las
mayores abundancias es que todos ellos se presentaron como hábitats continuos
(laguna, meandro, mallín 4) o muy conectados entre si (charco 1). Los cuerpos de agua
cuyas abundancias se mantuvieron siempre por debajo de 10 caracoles tuvieron un
hábitat en parches. Entre ellos, el que presentó los mayores valores de abundancia y la
mayor conectividad entre parches fue el mallín 1. El mallín 3 presentó, junto con el
arroyo 1 los parches más distantes entre sí (ver 4.5.1 micro escala).
AI comparar las abundancias de los dos cuerpos de agua con registros más
frecuentes (la laguna -cuerpo de agua permanente- y el mallín 1 -cuerpo de agua
temporario-) se observaron tendencias inversas. En el caso del cuerpo de agua
permanente, no se encontraron caracoles en primavera (noviembre de 1999, noviembre
de 2000 y octubre de 2001) o bien las abundancias fueron bajas (noviembre de 2001) y
aumentaron hacia el otoño (Fig. 4-23a). La tendencia observada en el cuerpo de agua
temporario fue de disminución de las abundancias desde primavera hacia el otoño (Fig.
4-23b). La descripción de los ambientes (capítulo 2) y los estudios realizados a micro
escala en la laguna (4.5.1), permiten ofrecer una explicación para esta tendencia
observada. Los caracoles habitan sobre superficie de barro con playas y cobertura
vegetal mínima o nula. Luego del deshielo, el nivel de agua en la laguna es máximo y
no se observan dichas condiciones en sus orillas. El volumen de agua disminuye a
medida que las temperaturas aumentan, y de esa forma queda al descubierto un suelo
con barro y sin vegetación, que favorecería el desarrollo del caracol. En los mallines se
observó que las densidades tendieron a disminuir a medida que avanzó el verano.
124

Tabla 4-10: Abundancia de caracoles estimada como el número de individuos recolectados en10 minutos de búsqueda.
Canal Juncal Meandro
2.72.3
Las celdas sombreadas corresponden a las ocasiones en que se recolectaron caracoles. Las celdas sombreadasvacías indican los momentos en que no se registró Ia abundancia y las celdas con guiones las fechas en lasque no se tomaron registros.
Figura 4-23: Abundancias de L. viatrix registradas en a) la laguna (ambiente permanente) yb) el mallín 1 (ambiente temporario), en el tiempo
.5
NUU’IOÑQÚO0°000000
.a
00Abundancla(IIcaracolesan10')
éo
a) Laguna MIJOCIICOO
á
Abundancla(¡Icaracolesan10')
N
b) Mallln1 Mmm“
No se mantuvieron las escalas para una mej0r visualización de las tendencias de cada gráfico. Las fechas demuestreos están Ordenadas cronológicamente (1: noviembre de 1999 (primavera), 2: enero de 2000 (verano),3: marzo de 2000 (otoño), 4: noviembre de 2000 (primavera), 5: enero de 2001 (verano), 6: marzo de 2001(otoño), 7: octubre de 2001 (primavera), 8: noviembre de 2001 (primavera), 9: febrero de 2002 (verano)).
125

Podría suponerse que cuando las temperaturas aumentan y la disponibilidad de
agua disminuye, Ia cantidad de parches adecuados para el desarrollo de L. viatrix
decrece. Aún cuando el estudio a micro escala demostró que los parches de mayor
tamaño son más utilizados que los más pequeños y que se generan parches más
grandes a medida que los ambientes se van secando, las tendencias de las abundancias
permiten suponer que dichas condiciones no son suficientes para el desarrollo del
caracol. La disminución de la conectividad entre parches, con el consiguiente
aislamiento en condiciones subóptimas, podría ser un factor importante.
EIdiseño de muestreo para este estudio consistió en recolectar caracoles de una
pequeña área de cada ambiente estudiado. Por tal motivo, y en coincidencia con el
criterio adoptado por otros autores, puede suponerse que el muestreo no tuvo un efecto
en la abundancia total los individuos de cada cuerpo de agua (Claxton et al. 1997). En
el único sitio en eI que se puede considerar que las extracciones realizadas afectaron a
Ia subpoblación es en eI charco 1. Este cuerpo de agua fue prácticamente censado
durante noviembre de 1999, y no se volvieron a encontrar caracoles hasta noviembre
de 2000. Esto demuestra que los individuos no habían comenzado el período
reproductivo al momento de la extracción, con Io cual no se registró una nueva
generación de caracoles en marzo de 2000, dos meses más tarde. Si se tiene en cuenta
que este charco se encuentra prácticamente aislado entre las épocas de deshielo, el
hallazgo de caracoles nuevamente en noviembre de 2001 estaría sugiriendo que hubo
recolonización del ambiente a través de la migración de individuos transportados desde
ambientes aguas arriba.
Al analizar la estructura de tamaños en cada cuerpo de agua por fecha, no se
observó un patrón definido que indique Ia existencia de grupos de ambientes con
condiciones homogéneas (Fig. 4-24). El análisis estadístico para cada muestreo mostró,
con un nivel de significación del 10% (ajustado según el criterio de Bonferroni para cada
caso), que hay evidencias para suponer que:
En noviembre de 1999 el tamaño mediano de los caracoles encontrados en el
charco 1 es estadísticamente mayor al de los individuos del mallín 1 (Fig. 4-24a).
En enero de 2000 el tamaño mediano de los caracoles hallados en Ia laguna es
estadísticamente mayor al de los individuos del mallín 1 (Fig. 4-24b).
En noviembre de 2000 el tamaño mediano de los caracoles del charco 1 y eI
mallín 1 es mayor al de los individuos del mallín 2 (Fig. 4-24c).
126

En enero de 2001 los tamaños medianos de los caracoles encontrados en Ia
laguna y el meandro difieren significativamente de los de los mallines 2 y 5. Estos
últimos presentaron tamaños medianos menores (Fig. 4-24d).
En marzo de 2001 la longitud mediana de los caracoles del arroyo 1 es
significativamente menor a las del resto de los cuerpos de agua.
No se encontraron diferencias significativas entre el juncal y el mallín 1*, ni
entre el mallín 1 y Ia laguna, ni entre el meandro y el mallín 1**. Como se definió en el
capítulo 2, el mallín 1 abarca 3 potreros. Debido a que los potreros están delimitados
por alambrado, esta separación representa una división del ambiente para el ganado,
pero no para los caracoles. De esta forma, se puede considerar que el mallín 1, el mallín
1* y el mallín 1** son un mismo hábitat -sin barreras- para el hospedador
intermediario, más allá de Ia existencia de parches que podrían hacer del ambiente un
hábitat en mosaico (Southwood 1977). Para la exploración de los datos a meso escala,
los datos del mallín 1 se mantuvieron discriminados por potrero para poder
compararlos, suponiendo a priori que no existirían diferencias significativas en los
tamaños medianos de los caracoles que allí se desarrollan (Fig. 4-24e).
En octubre de 2001 no hay diferencias entre los tamaños medianos de los
caracoles del arroyo y del mallín 1 (Fig. 4-24f).
En noviembre de 2001 no se encontraron diferencias significativas entre los
tamaños medianos de los individuos hallados en Ia laguna y el mallín 5, pero sí entre
este grupo y el mallín 1 y el meandro (Fig. 4-24g).
En febrero de 2002, el tamaño mediano de los caracoles provenientes del mallín
1 es significativamente mayor al de los individuos de la laguna (Fig. 4-24h).
Las estructuras de tamaños por fecha en las subpoblaciones de cada tipo de
ambiente (temporarios, representados por los mallines 1 y 2; permanentes,
representados por la laguna) se presentan en Ia Figura 4-25. Durante el estudio nunca
se encontraron caracoles en la laguna en el muestreo realizado a Ia salida de Ia
hibernación (noviembre para los años 1999 y 2000 y octubre para el 2001), aún cuando
en los ambientes temporarios ya había actividad. La proporción de caracoles inmaduros
sexualmente en los cuerpos de agua temporarios siempre fue nula o despreciable
durante los meses de noviembre. EI reclutamiento de una nueva subpoblación,
representado por Ia aparición de individuos de tamaños pequeños, siempre se observó
en enero en los ambientes temporarios. En los permanentes, el reclutamiento fue en
enero en el año 2000, pero hubo un corrimiento hacia marzo en el 2001.
127

Figura 4-24: Mediana del tamaño de los caracoles hallados en cada ambiente, por fecha (1999-2002).
a)
b)
d)
Yamaha(mm)
Tamaño(mm)
Tamaño(mm)
Tamaño(mm)
Cholila, Chubut
b Noviembre 1999 ‘ Marzo 2001
12 ‘
12 ‘
10 ‘
'E‘e 8‘
a — .2uE.2
e < 4 .
Quiz?4 ,
N=0‘ ' V ' ’ u I I I I I l I
mallín3 diam“ laguna mallln2 mallln1 arroyol juncal laguna mallln1 maandro mallín1' mallín 1"Ambiente Amen”
a. . . , Enero zooo Octubre 200111 a
11 n W "
b 9 .
i 'E‘5 i
7 ' a 7 .cnE¡1' .
5 - 5 .
3 _ N=29 ‘ i) y N“z 3 . N=17
N- 19 rrrrrrr rr lt - .6 N=51
1 N=0
mallln3 laguna mallin 2 mailln 1 a"W" 1' mm“ 1 laguna mm" 2 mallln 1Ambiente Ambkme
Noviembre 200111 b b.7" " b ,.+_
12 ' . . ,9
' E
7 -; É.2 BuE¡1’
5 .
N=12 ‘ '3fg _ _ O
N-4 N-31 :71NSO
1 l v v v . . . . . .
arroyo 1 mallín 3 charco 1 laguna mallin 2 mallÍn 1 charco 1 laguna mallln 2 meanaro mallln 1 mallln 5Ambiente ¡WWW
l?7 Enero 2001 Febrero 2002
10 '
9 .
É6. g o. o' í
N-6 s,_
4 . I2_ 3- ¡”777%7 y g x7777777fi N=2
N=233 Nm
u . . y . u l u r . . . .
canal nrroyito laguna mallln 2 meandro mallin 5 mnllin 6 laguna mallln 3 mallln1 mallln 2
AmbienteAmbiente
e)
9)
Las letras distintas indican diferencias significativas en los tamaños medianos entre cuerpos de agua, dentrode cada fecha (p< 0.05). Los ambientes con 6 o menos observaciones se excluyeron del análisis estadístico.
128

Un análisis del balance hídrico (Fig. 4-6) de cada año evidenció algunas
diferencias que podrían explicar los resultados recién presentados. Con respecto a los
ambientes permanentes, el desfasaje en el reclutamiento entre el año 2000 y el 2001
podría deberse a la situación de saturación de agua en el suelo (exceso) hasta mucho
más avanzada Ia primavera del 2000 que de otros años. Además, en esa primavera el
suelo perdió mucha menos agua que en 1998, 1999 y 2001, como se ve reflejado en la
disponibilidad de agua durante el verano del año 2001 (exceso o uso, vs. déficit los
otros años). La disponibilidad de agua en el suelo siempre fue homogénea en noviembre
(uso). La actividad de los caracoles se adelantó durante el último período de estudio y
en el mes de noviembre de 2001 se encontraron caracoles en el ambiente permanente.
Al analizar el balance hídrico para dicho año, puede observarse que la situación de uso
comenzó en septiembre, antes que los años restantes. Esto estaría indicando que las
temperaturas favorables se anticiparon con respecto a otros años. Efectivamente, el
período con temperaturas desfavorables para el desarrollo del caracol ese año fue el
más corto y el que finalizó antes (Tabla 4-2).
Durante el segundo y tercer período de estudio el reclutamiento fue evidente en
enero, pero en el cuarto período no se encontraron porcentajes altos de individuos
inmaduros en ningún momento. Esto podría deberse a que durante ese último período
no se realizó un muestreo en el mes de enero, y si el reclutamiento hubiese ocurrido a
principio de año podría esperarse que la proporción de caracoles pequeños ya no se
detectase en febrero. Otra posibilidad sería que la rara situación de déficit hídrico en
diciembre del año 2001 haya alterado Ia dinámica reproductiva de los caracoles eseaño.
Evidentemente, la disponibilidad de agua en el suelo condiciona el desarrollo de
L. víatríx y las particularidades de Ia dinámica del agua en ambientes temporarios y
permanentes pueden provocar diferencias en la estructura de la población a esa escala
de análisis. La evapotranspiración real (El'Pr) explicó aproximadamente un 84,4% de la
variabilidad de las abundancias de caracoles en la laguna (ambiente permanente con
hábitats continuos) (p< 0,01). EImodelo encontrado predice que las abundancias serán
mayores en los momentos en que la ErPr sea menor (relación inversa). En los
ambientes temporarios con hábitats en parches, la diferencia entre la precipitación y la
ETPr (PP-El'Pr) explicó aproximadamente un 71,1% de la variabilidad de los tamaños
medios de los caracoles hallados en los mallines (p< 0,01). Nuevamente la relación fue
inversa, sugiriendo que se hallarán tamaños medios mayores cuando la diferencia PP
El'Pr sea menor. Estos modelos apoyan las observaciones hechas a campo y los
resultados presentados hasta ahora.
Respecto a la longitud que puede alcanzar L. viatrix, en condiciones
experimentales se obtuvieron tamaños máximos de 11 mm y 14 mm (Lara et al. 1988,
Mülleret al. 1998). En condiciones de campo, en el Uruguay se encontraron ejemplares
129

de tamaños de hasta 10 mm (Castro et al. 2001). Venturini & Fonrouge (1985)
realizaron un exhaustivo estudio en la provincia de Buenos Aires, y encontraron que el
mes en el que se hallaron los mayores tamaños de caracoles, la población presentó un
tamaño promedio de 4,23 mm. Paraense (1976) encontró tamaños máximos de 12,6
mm en caracoles recolectados en Viedma, Río Negro. En esta tesis, los datos de campo
muestran que en la región estudiada es frecuente encontrar caracoles de tamaños
mayores a 8 mm y que el máximo tamaño registrado durante el estudio fue de 13,7
mm, superior a lo documentado en la bibliografia. Dicho tamaño se registró en la laguna
en Ia última primavera (noviembre de 2001). Además, todos los individuos mayores a
12 mm (n=11) se encontraron en ambientes permanentes (la laguna o el meandro), o
bien en cuerpos de agua temporarios con características particulares (el charco 1 y el
mallín 4) (Tabla 4-11).
Desafortunadamente, en bibliografía no se pudieron encontrar datos sobre los
tamaños máximos alcanzados por la especie en condiciones naturales en países
situados a latitudes menores (Brasil, Bolivia, Perú, México), que permitieran comparar
la capacidad de desarrollo de los caracoles en regiones más cálidas y con condiciones
ambientales menos adversas que las que presenta la zona de estudio (Paraense 1982,
2003, Claxton et al. 1997, Durand et al. 2002). La Figura 4-26 muestra los distintos
tamaños de L. víatríx recolectados en campo, desde los más pequeños hasta los más
grandes.
La Figura 4-27 presenta la estructura general de tamaños de L. víatríx para cada
tipo de ambiente, independientemente de la fecha. La proporción de caracoles juveniles
(<4 mm) en los ambientes permanentes duplicó el valor obtenido para los cuerpos de
agua temporarios (15% en permanentes vs. 7% en temporarios, Fig. 4-27a),
principalmente porque se hallaron individuos menores a 2 mm (Fig. 4-27b). Los
individuosadultos entre 4 y 8 mm estuvieron igualmente representados en ambos tipos
de ambientes (Fig. 4-27a) aunque las proporciones fueron heterogéneas entre las clases
de tamaños dentro del grupo (Fig. 4-27b). La proporción de individuos mayores a 8 mm
fue levemente superior en los ambientes temporarios principalmente por un aporte
mayor de caracoles de 8-10 mm. Sólo se detectaron diferencias significativas entre las
proporciones de caracoles inmaduros (p< 0,05), sugiriendo que L. viatrix puede
desarrollarse de manera similar en ambientes temporarios (con hábitats en parche) y
permanentes (con hábitats continuos). Este resultado difiere del obtenido por
Ollerenshaw (1971b), quien observó que cuerpos de agua con condiciones más
favorables para el desarrollo de los caracoles pueden sostener poblaciones de mayor
densidad, pero con individuos de menor tamaño, y que cuerpos de agua con menores
densidades permiten que los caracoles alcancen tamaños más grandes.
130

Figura 4-25: Estructura de tamaños de las subpoblaciones de L. viatrix en ambientes temporariosy permanentes durante el período 1999-2002, Cholila, Chubut
100 ' TamañosI90 >4mm
El<4 mm
Frecuenciadecaracoles(%)
1 2 1 2°i112No v-99
1 l 2
Oct-01
1|21I2 1'21I2Ene-00 Mar-00 Nov-00 Ene-01 Mar-01 Nov-01 Feb-02
1: Enambientes temporarios (representados por un mallín) y 2: en ambientes permanentes (representadospor la Laguna).
Tabla 4-11: Ambientes y fechas donde se registraron los caracoles de mayor tamaño en el período1999-2002. Cholila, Chubut
l Tamaño (mm) # de individuos Ambiente Fecha ll 13.7 1 laguna Nov-01 ‘
l 13.4 1 meandro Mar-01‘ 13.3 1 meandro Mar-01
l 12.8 1 meandro Mar-01
12.7 1 mallín 4 Nov-01
, l 12.5 1 meandro Mar-013 12.4 1 charco 1 Nov-99
l 12.2 1 meandro Mar-0112.1 1 mallln 4 Nov-01
; 12 2 laguna En-00/Nov-01
k 1 1.9 1 charco 1 Nov-9911.8 1 charco 1 Nov-99
‘ 11.7 2 laguna Nov-O1l 11.6 1 charco 1 Nov-99
i 11.5 2 charco 1/Iaguna Nov-99/Nov-O1¡ 11.4 2 laguna En-OO l
11i3 2 charco 1/mallín1 Nov-99/Nov-01’ 11.2 5 charco 1/laguna/mallín2 Nov-99/En-00lNov-00
‘ 11.1 6 laguna En-OO/Nov-01
‘ 11 10 charco 1/laguna/mallín2 Nov-991En-01/Nov-O1
1 mallín 1/mallín 4
131

Figura 4-26: Ejemplares de L. víatrix hallados en el campo en la localidad de Cholila, Chubut
Figura 4-27: Estructura de tamaños de las subpoblaciones de L. viatrix en ambientes temporariosy permanentes durante todo el estudio (1999-2002), Cholila,Chubut
a)
DAnbientesTenporarlos
AnblentesPemanentes
FrecuenciaPorcentual
U'V
0-1 1-2 2-3 3-4 4-5 5-6 6-7 7-6 5-9 9-10 10-11 11-12 >12
Tamaños (mm)
132

4.5.5 Infección en Lymnaea víatrix: Fasciola hepatica
Durante el estudio, de los 2594 caracoles medidos, se disecaron 1633 (63%) y
se confirmó la infección por F. hepatica en el 0,67% (11/1633) de los individuos. Se
encontraron caracoles infectados tanto en cuerpos de agua permanentes como en
temporarios. En Ia Tabla 4-12 se presentan el número de caracoles recolectados,
medidos, disecados e infectados en cada cuerpo de agua y por potrero, durante el
estudio. Se encontraron redias con cercarias maduras de F. hepatica en 5,9% (7/119)
de los caracoles recolectados en enero de 2000 y 0,9% (2/222) en febrero de 2002 en
Ia laguna, y en marzo de 2001 se encontró un 2% (1/53) de infección en el mallín 1
dentro del potrero 29 y un 14% (1/7) en el arroyo 1 en la zona del casco delestablecimiento.
De los 11 caracoles infectados con F. hepatíca, 10 fueron medidos. Los tamaños
de los individuos fluctuaron entre 4,5 y 9,4 mm con una mediana de 7,55 mm. Un único
caracol positivo encontrado en el mallín 1 en marzo de 2001 midió 4,5 mm y los demás
superaron los 6,7 mm (Tabla 4-13). Según estos tamaños y los criterios adoptados,
todos los caracoles infectados correspondieron a individuos sexualmente maduros que
habrían nacido en la temporada reproductiva anterior. La importancia de los caracoles
de gran tamaño en la transmisión del parásito es discutida por Ollerenshaw (1971b).
Este autor encontró que los individuos de la especie L. truncatu/a mayores a 5 mm
representaron sólo el 25% de la población estudiada, pero que el 85% de los caracoles
infectados perteneció a esta categoría, demostrando la relevancia de individuos de gran
tamaño en la producción de metacercarias.
Se encontraron también 50 caracoles infectados con redias inmaduras que no
pudieron ser determinadas mediante observación directa. Dichos estadios larvales
podrían pertenecer a F. hepatica o bien a digeneos de la Familia Echinostomatidae (ver
4.5.6). El 50% de estos caracoles (25/50), se encontraron en los mismos momentos y
cuerpos de agua en los que se encontró infección por F. hepatica: 6 aparecieron en
enero de 2000 y 19 en febrero de 2001 en la laguna.
AI estudiar los tamaños y las abundancias de los caracoles en los cuerpos de
agua y fechas donde alguna vez se registró infección (laguna, mallín 1 y arroyo 1; enero
de 2000, marzo de 2001 y febrero de 2002) se encontró que: en enero de 2000, el
tamaño mediano de los caracoles de la laguna (donde se registró infección) fue
significativamente mayor que el de los individuos hallados en el mallín 1 (p< 0,05).
Además, la abundancia fue también mayor (23 caracoles en 10 minutos de búsqueda en
la laguna vs. 3/10' en el mallín 1). En marzo de 2001 no se encontraron diferencias
significativas entre los tamaños de los caracoles de la laguna y el mallín 1 (donde se
registró infección) (p> 0,05), pero los del arroyo fueron significativamente menores al
resto. Durante este muestreo, las abundancias registradas fueron de 9/10' en la laguna
133

y 4,5/10' en el mallín. Finalmente, en febrero de 2002 el tamaño mediano de los
caracoles de la laguna (donde se registró infección) fue significativamente menor al del
mallín 1 y la abundancia fue mayor en la laguna (100 caracoles en 10' en la laguna vs.
2,3/10' en el mallín) (Fig. 4-24 y Fig. 4-25). Para mejor visualización de los resultados,
la Figura 4-28 resume los valores de tamaños medianos y abundancias de caracoles que
caracterizaron a los cuerpos de agua en los que alguna vez se registró infección por F.
hepatíca. La abundancia siempre tomó valores mayores en la laguna, Io que sugiere la
falta de asociación entre dicha variable y la aparición de infección en caracoles.
Tampoco parece haber una relación entre Ia ocurrencia de infección y la estructura de la
población por ambiente, ya que en un caso el cuerpo de agua con individuos parasitados
presentó tamaños medianos mayores (enero de 2000), en otra ocasión menores
(febrero de 2002) y en marzo de 2001 no hubo diferencias en los tamaños medianosentre ambientes.
Llamativamente, la mayor prevalencia durante el estudio se registró en el arroyo
1 (14%), en un tramo de su curso al cual no tiene acceso el ganado. La proporción de
caracoles infectados en dicha ocasión fue mucho mayor a las halladas en otros cuerpos
de agua durante el estudio. Más allá de este hallazgo, la mayor prevalencia se encontró
en la laguna. Sin embargo, no se encontraron diferencias significativas entre la
proporción de caracoles infectados en la laguna en enero de 2000 (7/119, 5,9%) y en el
mallín 1 en marzo de 2001 (1/53, 2%) (629.: 1,57, p> 0,05), ni entre este último y la
proporción de caracoles infectados en la laguna en febrero de 2002 (2/222, 0,9%)
(629.: 0,33, p> 0,05).
Nunca se encontraron caracoles infectados durante Ia primavera. Las infeccionesmaduras fueron recién evidentes durante los muestreos de verano u otoño. Si se
considera que en condiciones adecuadas de temperatura (superiores a los 10° C) el
parásito necesita entre 45 días y 2 meses para desarrollarse desde esporoquiste hasta
cercarias maduras, podría suponerse que las infecciones de verano son el resultado del
contacto entre caracol y miracidio a partir de octubre. Otra posibilidad sería que el
caracol haya hibernado con la infección que contrajo durante el otoño anterior. Si el
período de hibernación encontró al caracol con una infección reciente (inmadura), estos
resultados indicarían que las condiciones de temperatura o humedad que caracterizan a
la región no permiten que el parásito termine de desarrollarse exitosamente antes del
verano siguiente. Si, en cambio, el caracol entró en hibernación con una infección ya
madura adquirida durante el verano anterior, evidentemente la probabilidad de que
sobreviva el período invernal es muy baja (debido al efecto conjunto de la invasión
parasitaria y el estrés ambiental), ya que nunca se encontraron caracoles infectados
durante octubre o noviembre, a la salida del período de reposo. Estos resultados
concuerdan con los obtenidos por Ollerenshaw (1971a) en Gran Bretaña. Este autor
134

Tabla4-12:CantidaddecaracolesdelaespecieLymnaeavíatrixrecolectados,medidos,disecadoseinfectadosduranteelestudio(1999-2002).Cholila,Chubut
cuerpodeagua
Lasceldasconguiónindicanlosambientesymomentosenlosquenosetomaronregistros;losasteriscosindicanloscuerposdeaguaqueestabansecosodondenose encontraroncaracoles.LasletrasindicanlacantidaddecaracolesR:recolectados,M:medidos,D:disecados,I:infectados(expresadoenporcentaje).Seindicanlasfechas enlasqueseencontraroncaracolesinfectadosconF.hepatica(+)yenlasqueno(-).
135

Tabla 4-13: Tamaño de los caracoles de Ia especie L. víatrix infectados con F. hepatíca, Cholila, Chubut
FECHA CUERPO DE AGUA TAMAÑO (mm) mediana (1er-3ercuartil)6.8
6.97.4
Enero 2000 Laguna 7.7 7.7 (7.15-8.45)7.9 7.55
9 (6.82-7.97)9.1
Marzo 2001 A"°y° 1 5'5 5.05‘ Mallln 1 4.6
l Febrero 2002 La_gu_na 8 8
En febrero de 2002 se encontraron 2 caracoles infectados pero uno no pudo ser medido.
Figura 4-28: Tamaños y abundancias de los caracoles por cuerpo de agua: a) durante enero de2000, b) durante marzo de 2001 y c) durante febrero de 2002. Sólo se incluyen losambientes donde alguna vez se registró infección (laguna, mallín 1 y arroyo 1).
Elmo 2000
Tlmlflo(mm)
N=619 77l - A=23
Turran(mm)
e
r“. N=27.
mallln 1' mallln 1"
Febrero 2902
“Inma(mm)
q
UI
" 74:37#A=2.3
laguna mallln |
Ambiente
N: tamaño de la muestra, A: abundancia de caracoles.Para interpretación de los gráficos de caja ver Figura 4-10 o Materiales y Métodos (4.4.4).
136

realizó un estudio epidemiológico durante 3 años y concluyó que la infección en
caracoles que sobreviven al invierno es generalmente mucho menor y de menor
importancia epidemiológica que la de verano. Por otro lado, ensayos de laboratorio
confirmaron que Ia supervivencia de individuos de la especie Fossaría cubensis
(Gastropoda: Lymnaeidae) infectados con F. hepatíca fue mucho menor que la de los
controles sanos (Gutiérrez et al. 2000).
4.5.6 Infección en Lymnaea viatrix: Otros trematodes
Durante los meses de enero y marzo del año 2000, en los caracoles hallados en
Ia laguna, se detectaron esporoquistes, cercarias y metacercarias pertenecientes a una
especie del género Coty/urus sp. (Digenea: Strigeioidea) y redias, cercarias y
metacercarias pertenecientes a dos géneros de Ia familia Echinostomatidae (Digenea:
Echinostomatoidea), que para este estudio no fueron discriminados. Estos parásitos
utilizan aves como hospedadores definitivos y moluscos como hospedadores primarios
(H1) donde se desarrollan las cercarias y como hospedadores secundarios (H2) donde
se enquistan las metacercarias que luego son ingeridas por las aves (Basch 1969,
Prepelitchi 2002) (Figura 4-29).
Un relevamiento hecho en la laguna en enero y marzo de 2000 permitió
reconocer las siguientes especies de aves acuáticas o semiacuáticas que se alimentan
ocasionalmente de caracoles y que podrían estar actuando como hospedadores
definitivos de los digeneos hallados: Fulica sp. —gallareta-, Cery/e torquata -martín
pescador-, Anas flavirostris, Anas georgica -patos- Theristicus caudatus —bandurria-.
Para este estudio se utilizaron 382 caracoles, de los cuales 119 fueron los
disecados durante enero (Tabla 4-11); los restantes pertenecieron a un subgrupo de los
445 caracoles disecados en marzo (n= 263).
Sobre el total de caracoles procesados, 35 (9%) estuvieron parasitados por
redias y cercarias de la familia Echinostomatidae, 226 (59%) por esporoquistes y
cercarias del género Coty/urus 5p., 132 (34%) presentaron metacercarias de
Echinostomatidae y 75 (20%) metacercarias de Coty/urus sp. El total discriminado por
tipo de infección supera el total de caracoles procesados porque se encontraron
infecciones simultáneas por cercarias y metacercarias en un mismo individuo.
El porcentaje de infección general para cada muestreo fue muy elevado. En
enero, 106 caracoles sobre un total de 119 disecados (89%) estuvieron parasitados con
alguno de los digeneos hallados en cualquiera de los estadios Iarvales posibles; además
7 caracoles estaban parasitados únicamente con larvas de F. hepatica y sólo 6 (5%)
resultaron negativos. El 57% (60/106) de los caracoles positivos (sin incluir los
infectados con F. hepatíca) presentó infecciones por cercarias (H1) y el 93% (99/106)
137

por metacercarias (H2). Se encontraron 7 (6,6%) caracoles actuando como H1
únicamente, 46 (43%) actuando únicamente como H2 y 53 (50%) que presentaron
infecciones primarias y secundarias simultáneamente (Tabla 4-14). En marzo se
encontraron estadios larvales de parásitos en 424 caracoles sobre 445 individuos
disecados (95,3%). Sin embargo, la correcta disección y clasificación pudo realizarse
sobre 242 individuos de los 424 parasitados. Entre estos caracoles, un 85% (205/242)
se encontraba actuando como H1 y un 23% (56/242) como H2. Se encontraron 185
(76%) caracoles infectados únicamente con cercarias (H1) (entre los que se incluyen 11
caracoles infectados con redias inmaduras), 23 (9,5%) únicamente con metacercarias
(H2) y 34 (14%) caracoles presentaron infecciones primarias (H1) y secundarias (H2)
simultáneamente (entre los que se incluyen 4 infectados con redias inmaduras) (Tabla
4-14).
En enero el tamaño mediano de los caracoles sanos (6/119) fue de 7,3 mm con
un rango que varió entre 6,7-10,5 mm y el de los infectados con redias o esporoquistes
y cercarias de cualquiera de los digeneos hallados (incluyendo a F. hepatíca) (67/119)
fue de 7,2 mm (3,3-11,2 mm). En marzo el tamaño mediano de los caracoles sanos
(21/263) fue de 4,8 mm con un rango que varió entre 2,2-6 mm y el de los individuos
parasitados con redias o esporoquistes y cercarias (242/263) fue de 4,9 mm (2,8-7,3
mm). No se hallaron diferencias significativas entre los tamaños de los animales sanos y
parasitados para el mes de marzo (p=0,07). Para enero no se pudo aplicar el test
debido a que la cantidad de animales en el grupo de individuos sanos fue muy baja.
Infecciones simultáneas. Durante este estudio sólo se encontraron 3
caracoles con infecciones dobles sobre un total de 382 (119+263) caracoles procesados.
Todos los casos se registraron en el mes de marzo de 2000. Uno de ellos (tamaño: 5,5
mm) presentó cercarias de Cotylurus sp. y Echinostomatidae y los otros dos (2,8 y 4,9
mm) presentaron infecciones maduras con esporoquistes y cercarias de Cotylurus sp. y
redias inmaduras. En todos los demás casos sólo se observaron infecciones simples por
esporoquistes o redias con cercarias. En coincidencia con los resultados obtenidos por
Boray (1967) la interacción antagónica entre digeneos parecería ocurrir únicamente
cuando el caracol se encuentra infectado con estadios larvales correspondientes a una
infección primaria (redias o esporoquistes) y no cuando el único tipo de infección es por
metacercarias. Este autor encontró que caracoles de Ia especie L. tomentosa que
albergaban infecciones maduras de Echinostomatideos eran resistentes a infectarse con
F. hepatíca. Durante el desarrollo de esta tesis, nunca se encontraron infecciones
maduras de F. hepatíca cuando hubo otro digeneo parasitando un caracol. Pero
llamativamente, los dos caracoles que presentaron infecciones dobles con uno de los
digeneos de aves desarrollado y el otro inmaduro, albergaban esporoquistes maduros y
redias inmaduras. La bibliografía sugiere que, debido a la presencia de una faringe en
las redias, éstas pueden depredar a los esporoquistes
138

Figura 4-29: Ciclode vida de digeneos parásitos de aves hallados en L. víatrix en la laguna.Modificado de Haas et al. 1995.
Adulto en aves
Huevo en el ambiente
Metacerca rla
MIracldio Invade al primerhospedador lntermediario
Cercarla invade al Segundo Esporoqulstehospedador lntermediarlo
Generación de redias
Tabla 4-14: Infección por digeneos de aves y por F. hepatica en caracoles de Ia laguna duranteenero y marzo de 2000.
Infección por digeneos de aves Infección por Tamaño mediano (rango) Sin Tamaño mediano (rango) Total
FeCha exclusiva H1 exclusiva H2 simultánea Total Infectados F. hepatica caracoles Infectados H1 infección caracoles sanos dlsecados
Enero 2000 7 46 53 106 7 7.2 (3.3-11.2) mm 6 7.3 (6.7-10.5) mm 119
Marzo 2000 185 23 34 242 0 4.9 (2.8-7.3) mm 21 4.8 (2.2-6) mm 263
Se registraron las infecciones por esporoquistes o redias con cercarias, donde el caracol actúa como hospedadorprimario (H1) y las infecciones por metacercarias, donde el caracol actúa como hospedador secundario (H2).
139

(Combes 1982). No se ha documentado que una especie que desarrolla esporoquistes
se imponga sobre una con redias, ni que Ia ocupación del caracol por esporoquistes
impida el desarrollo de infecciones posteriores por redias, a excepción de un trabajo
realizado en 1971 por Owyang & Lie, citado por Combes (1982).
Sobre el total de caracoles estudiados, el porcentaje de individuos infectados con
redias y cercarias de Echinostomatidae (H1) fue del 11% (13/119) en enero (sin contar
las infecciones por redias inmaduras (n=5)) y del 8% (22/263) en marzo, y no se
encontraron diferencias significativas entre dichas proporciones (p= 0,78). La infección
por metacercarias (H2) alcanzó un 85% (101/119) en enero y disminuyó a un 8%
(21/263) en marzo y las diferencias entre ambas proporciones fueron significativas (p<
0,05).
La Figura 4-30 muestra la prevalencia de Echinostomatidae y Coty/urus sp. en
caracoles de distintos tamaños, en los muestreos realizados en enero y marzo del año
2000. En enero los caracoles menores a 4 mm no presentaron infección por redias y
cercarias de Echinostomatidae pero la infección por metacercarias alcanzó a todos los
tamaños. En marzo todas las clases de tamaños presentaron infección por redias y
cercarias (aunque en una proporción muy baja en caracoles de tamaños <4 mm y entre
7-8 mm) y las metacercarias se hallaron sólo en individuos de tamaños superiores a 4
mm (Fig. 4-30a). Puede observarse que en el mes de enero el 10% de los caracoles
emitiendo cercarias (que son los de mayor tamaño) es suficiente para que haya un alto
porcentaje de infección por metacercarias en todos los tamaños, pero no ocurre Io
mismo en marzo. A partir de estos resultados podría suponerse también que existieron
dos momentos de infección por miracidios; el primero fue previo al reclutamiento de
una nueva población de caracoles, motivo por el cual los individuos muy pequeños no
estuvieron infectados en enero, y el segundo fue entre enero y marzo, ya que para el
segundo muestreo todas las clases de edades presentaron infecciones por redias.
La Figura 4-30b muestra los resultados obtenidos para Coty/urus sp. La
prevalencia total por esporoquistes y cercarias fue del 35% (42/119) en enero y se
duplicó en marzo (70%, 184/263), hallándose diferencias significativas entre
proporciones (p<0,05). La prevalencia de metacercarias también difirió
significativamente entre ambos meses (p< 0,05). En enero fue del 32% (38/119) y en
marzo del 14% (37/263). La infección por cercarias alcanzó a todos los tamaños decaracoles tanto en enero como en marzo. Se encontraron metacercarias en todas las
categorías de tamaños en enero pero en marzo ya no se registraron en caracoles
grandes (>6 mm). Se sabe que las especies pertenecientes a este género se desarrollan
en aproximadamente 2 meses dentro del caracol, pero aproximadamente en una
semana dentro del hospedador definitivo, y que el tiempo de liberación de huevos es
muy corto (Basch 1969). Podría suponerse entonces, que los hospedadores definitivos
140

se ¡nfectaron durante enero y que una posterior liberación de huevos permitió la
reinfección de los caracoles, incrementando la prevalencia como H1 dos meses después.
Las metacercarias del género Cotylurus sp. aumentan mucho su tamaño dentro
del caracol entre el momento en que penetran las cercarias, que son muy pequeñas, y
eI momento en que se enquistan. Prácticamente todas las metacercarias encontradas en
ambos muestreos aún no se habían enquistado, pero eran de gran tamaño. En el mes
de marzo, que es cuando hay mayor número de caracoles emitiendo cercarias, el
porcentaje de metacercarias encontrado resultó bajo, probablemente porque las
infecciones recientes, con metacercarias pequeñas sin enquistar, no pudieronobservarse durante las disecciones.
Finalmente, la proporción de caracoles con infecciones primarias (H1) de
cualquiera de los digeneos, fue significativamente mayor en marzo que en enero (p<
0,05) y lo contrario ocurrió con las infecciones por metacercarias (H2).
La ausencia de caracoles de gran tamaño en marzo, las diferencias en las épocas
de infección y la competencia interespecífica, entre otros factores, podrían explicar
parte de la fluctuación en las infecciones por estos digeneos. Las altas cargas
parasitarias registradas en todos los caracoles infectados con redias o esporoquistes
podrían estar jugando algún rol en Ia población de caracoles, por ejemplo causándoles
Ia muerte. En principio no podría pensarse en Ia ocurrencia de un fenómeno de
gigantismo a partir de estos resultados, debido a que no se hallaron diferencias entre
los tamaños de los caracoles infectados y los caracoles sanos. Un estudio más
prolongado sobre la dinámica de estos parásitos podría permitirnos conocer mejor cómo
es la transmisión en Ia naturaleza, cómo influye el parasitismo en la supervivencia de
los caracoles y si está interfiriendo de alguna manera en la instalación y transmisión de
F. hepatica.

Figura 4-30: Infección por digeneos de aves en L. viatrix de la laguna: a) Prevalencia de Echinostomatidae(Echino) y b) Cotylurus sp. (Cot.) en enero y marzo de 2000, Cholila, Chubut
a)4o 4o so
3 3 W -- 45 mg 35 ' 3 8 35 -- 2
U <- O
3. 30- e 5 30 .c 4° g1'! 8 5 -- 35 gi" 25 ' o 25 -- Uv 'u 3 -- 30 goFZO' a‘ï °\° 20-- --2s ¡e
15 . LJ ,9 -- 20 flC 5 g 15 "' É“¡’10- a u lo" --15 gu ° a -- 1o UE El E o E"' E 5 " -- s "
o- 0 :"ma : : 0.. <4 4401-5 5.01-6 6.01-7 7,01—8 8401-9 >9
Tamanos (mm) Tamaños (mm)Enero Marzo
H 1 Echino
l:l H2 Echino
+Pob|ación decaracoles
b),,, 4o 50 u, 4o 50 m8 a 8 - 45 a5 35" 3 s 35 4o TaU ..E 3o «- g 2' 3o E'o n, 'o -- 35 33 25 " z 3 25 -- 30 oe 20 -- v \° 20 .. 25 lgo e o e}-— 15 -- Í. .E 15 " 2° mu - u .. 15 5I: u c Cg 1° " 5 g 1° -- lo 3u _P 8 u 5 .. °dl 5 g a 5 o" ¡. I. > É¡L o ' 5‘ . . , o IL u. 0 . : : : : : : o
<4 ¿01.5 5.015 5.01.7 7.014 8_01_g >g <4 4.01-5 501-6 6‘01-7 7‘01-8 8.01-9 >9
Tamaños (mm) Tamanos (mm)
Enero MarzoEl H1 Cot.
H 2 Cot.
+Población decaracoles
H1: infección por cercarias (hospedador primario), H2: infección por metacercarias (hospedador secundario).
142

Foto 1
FOÏO 3 Foto 4
Foto 7 Foto 8
Foto 1:cuadrado muestral utilizado en la laguna para el estudio de microambientes. Fotos 2 y 3: parches con playas.Foto 4: orillas de Ia laguna mostrando la disminución del nivel de agua con respecto a meses de mayor disponibilidadde agua. Fotos 5 y 6: L. viatrix en su hábitat caracterizado por sustrato de barro, baja o nula cobertura vegetal y delgadaspelículas de agua. Foto 7: L. viatrix tomando sol... Foto 8: pequeño parche en un ambiente con vegetación densa.
143

4.5.7 Consideraciones finales
Los estudios realizados y presentados en este capítulo aportaron información
sobre el hospedador intermediario de F. hepatica a distintas escalas de análisis. Las
características de la estructura y dinámica poblacional a macro escala permitieron
establecer un patrón general de desarrollo de L. víatrix, modulado principalmente por la
disponibilidad de agua en eI suelo (determinada por Ia temperatura y la precipitación) y
por las temperaturas ambientales. Esta escala de análisis es de suma importancia para
el estudio ecoepidemiológico de la fasciolosis, ya que aunque los caracoles presenten
dinámicas de desarrollo asincrónicas a escalas de mayor detalle, los factores
ambientales recién mencionados restringen Ia actividad de los moluscos a ciertas
épocas del año independientemente de lo que ocurra en cada ambiente.
La dispersión activa y pasiva de los caracoles del género Lymnaea es discutida
por diversos autores (recopilado por Boray 1969). Algunos estudios permiten suponer
que la dispersión durante ciertas épocas del año desde ambientes desfavorables hacia
zonas favorables, no es un evento común en L. truncatula y L. tomentosa. Sin embargo,
ensayos experimentales realizados a campo demostraron que los caracoles pueden ser
transportados aguas abajo con Ia corriente —dispersión pasiva- (Boray 1969). El
hallazgo de conchillas en el río, agrupadas en la desembocadura de algún mallín o
arroyo; Ia ocurrencia permanente de caracoles en el mallín 2, pero esporádica en el
arroyo 1 y el mallín 3 (ambos aguas abajo del mallín 2) y el evidente evento de
recolonización del charco 1 luego de la primera extracción de caracoles, manifiestan que
los caracoles de esta población se dispersan de manera pasiva entre ambientes,
probablemente llevados por la corriente.
Todos los ambientes presentes en el área de estudio, a excepción del río,
pueden potencialmente albergar caracoles. Sin embargo, en algunos la presencia de L.
viatrix fue permanente durante los meses estudiados y en otros sólo se hallaron unos
pocos individuos de manera irregular en el tiempo. Este resultado permite suponer que
no todos los cuerpos de agua son hábitats igualmente aptos para eI desarrollo sostenido
del caracol. Por ejemplo, las mayores abundancias siempre se encontraron en hábitats
continuos o con parches muy conectados, a pesar de que no se detectó un patrón claro
entre ambientes en el tiempo. La ausencia de subpoblaciones establecidas en el río y el
arroyo 1 podría deberse a que, según Boray (1969), ambientes en zonas con mucha
pendiente no suelen desarrollar poblaciones grandes de caracoles debido a que el flujo
de agua lava el suelo de barro y arrastra a los caracoles. Cuerpos de agua con
velocidades medias permiten el desarrollo de algas, la migración activa y la
reproducción de los caracoles.
144

El porcentaje de infección por F. hepatica en caracoles fluctuó entre un 1% y un
14% durante todo el estudio. Las implicancias de estas prevalencias, así como Ia
importancia de cada tipo de ambiente en Ia transmisión del parásito se discuten
relacionadas al manejo ganadero en el capítqu 6. Sin embargo, el hecho de que las
infecciones sólo se hayan registrado en animales grandes (mediana= 7,55 mm) y que Ia
proporción de individuos de gran tamaño haya sido homogénea en ambientes
temporarios y permanentes sugiere que ante una oferta similar de miracidios, podría
esperarse igual probabilidad de infección en ambos tipos de ambientes. Con respecto a
la elevada proporción de caracoles infectados en el arroyo 1 (1/7, 14%), este hallazgo
puede considerarse un evento poco común, ya que el único caracol infectado (5,5 mm)
de un total de 7 individuos hallados, probablemente contrajo la infección aguas arriba,
en el mallín 2, y fue transportado por el arroyo hasta detenerse en un pequeño
remanso. Un único caracol además del infectado superó los 5 mm de longitud. Los
restantes fueron pequeños (< 3,5 mm), sugiriendo que probablemente los dos
individuos adultos fueron transportados por la corriente y dejaron descendencia en elsitio donde fueron recolectados.
Un trabajo realizado por Boray (1967) demostró que caracoles del género
Lymnaea infectados con un digeneo de Ia familia Echinostomatidae no se infectaron
posteriormente con F. hepatica. En ese caso, ambos parásitos estudiados desarrollan
redias dentro del hospedador intermediario y podría suponerse que la voracidad del
primer colonizador podría inhibir Ia instalación del siguiente parásito en el mismo
caracol (Boray 1967, Combes 1982). Este tipo de competencia entre especies se
considera una interferencia directa, ya que la especie dominante se alimenta de Ia
especie subordinada, eliminándola e impidiendo el establecimiento de posteriores
infecciones. EI efecto de interferencia se ha observado también entre especies que
desarrollan solamente esporoquistes, pero en estos casos la competencia se considera
indirecta, ya que el desplazamiento no se produce por ingestión de una especie a Ia
otra. En los casos en los que se ha observado competencia entre una especie con redias
y una con esporoquistes, siempre se ha asumido que la primera es dominante sobre la
segunda, aún cuando la especie subordinada logre persistir un tiempo antes de ser
desplazada (Kuris 1990).
En los estudios a campo realizados en esta tesis se comprobó que los caracoles
que intervienen en el ciclo natural de F. hepatica actúan también como hospedadores de
otros parásitos de aves, y que sus prevalencias pueden ser altas. Se observó que las
infecciones dobles fueron muy raras, y que en los pocos casos en los que se
presentaron, en una única ocasión hubo emisión de cercarias de ambas especies y en
los dos casos restantes Ia especie con esporoquistes predominó sobre las especies con
redias. Este resultado podría estar reflejando Ia situación en Ia que Ia especie
dominante (con desarrollo de redias) que invadió al caracol en segundo lugar, aún no
145

desplazó a la especie subordinada (Cotylurus 5p.), pero que eventualmente la especie
voraz prevalecerá. Durante estos estudios resultó llamativa la ausencia de infección por
F. hepatica en la laguna en marzo de 2000, habiéndose encontrado caracoles infectados
dos meses antes. Aún cuando durante marzo hubo un marcado incremento (con
respecto a enero) en la prevalencia (como H1) de los digeneos de aves, el mayor
porcentaje de infección fue por Cotylurus sp. (especie con esporoquistes). Podría
suponerse que el desarrollo de las redias de F. hepatíca dentro de un caracol podría
haberse visto demorado por la ocurrencia previa de otra infección. Sin embargo,
deberían haberse encontrado mayor cantidad de infecciones dobles con esporoquistes
maduros y redias inmaduras, que permitieran suponer un desarrollo lento de F. hepatica
como consecuencia de una interacción indirecta débil. Alternativamente, podría
pensarse que la invasión del hospedador por parte de una especie que desarrolla
esporoquistes puede perjudicar el posterior desarrollo de otro parásito (interacción
antagónica indirecta), independientemente de la voracidad de cada especie.
Finalmente, el hecho de que L. viatrix sea la única especie del género en la zona
y que se hayan encontrado ejemplares infectados durante el estudio, indicaría que es el
único hospedador intermediario de F. hepatíca y que el éxito en la transmisión depende
exclusivamente de su presencia.

4.6 CONCLUSIONES
1. La principal y aparentemente única especie que actúa como hospedador
intermediario en el ciclo de transmisión de F. hepatica es Lymnaea viatrix.
2. EI hallazgo de esta especie a latitudes superiores a los 41° S amplía su rango dedistribución austral mundial.
Con respecto a los parches que utiliza L. viatrix se encontró que:
3. Son más pequeños y presentan mayor conectividad cuando hay más agua
disponible en el suelo y aumentan de tamaño y disminuye su conectividad a medida quelos ambientes se van secando.
4. Las superficies de los parches no difieren entre ambientes, pero su conectividad sí.
5. Los parches utilizados por L. viatrix fueron de mayor tamaño que los no utilizados.
6. Todos los parches con caracoles presentaron playas (columna mínima de agua
nula) independientemente de la máxima profundidad, que nunca superó los 6 cm.
7. El valor mediano de pH en los ambientes utilizados fue de 6,3, menor que el de los
parches no utilizados. En los ambientes de hábitat continuo el pH no difirió entre
parches utilizados y vacíos.
8. La cobertura vegetal en los parches con caracoles nunca superó el 50%.
9. Tanto los ambientes temporales como los permanentes resultaron adecuados para eldesarrollo sostenido del caracol.
10. El río fue el único ambiente estudiado no apto para el desarrollo de L. viatrix,
probablemente por presentar parches muy disturbados.
11. Con respecto a la dinámica poblacional de L. viatrix se concluye que: 1) los
caracoles hibernan durante los meses fríos, entran en actividad y oviponen hacia
principio de verano y luego mueren antes del comienzo del otoño siguiente; 2) el
reclutamiento de una nueva población ocurre entre noviembre y enero, dependiendo de
las condiciones ambientales y los caracoles que nacen a fines de primavera o durante el
verano alcanzan tamaños intermedios (4-6 mm) antes de entrar en período de latencia
147

invernal; 3) en ciertas épocas del año puede observarse Ia superposición de dos
generaciones.
12. Los hábitats continuos para el hospedador intermediario presentaron mayoresabundancias de caracoles.
13. Las infecciones patentes por F. hepatíca en el hospedador intermediario se
registraron a partir del verano y en otoño, pero nunca en primavera.
14. La ocupación de un caracol por parte de un parásito parecería impedir la instalación
y desarrollo de otra especie parasitaria.
15. Existe transporte pasivo de caracoles y podría existir entonces acarreo de Iainfección.

EL PARÁSITO
Fascio/a hepatica puede considerarse una pieza más del rompecabezas
epidemiológico, pero es ciertamente parte de todas y cada una de ellas. Hasta aquí, ha
sido presentada en los capítulos 3 y 4 junto con los estudios realizados en los
hospedadores definitivos e intermediarios. En dichos capítulos se estudiaron Ia
prevalencia y parte de la dinámica parásito-hospedador tanto en ganado como en
caracoles. Además, se describieron los métodos utilizados para realizar la identificación
taxonómica de los estadios Iarvales encontrados en el hospedador definitivo (huevos) y
en el hospedador intermediario (cercarias). En este capítulo se describen una serie de
ensayos complementarios al trabajo realizado en campo, que permitieron realizar la
certificación taxonómica de la especie mediante la obtención experimental de adultos y
confirmar la infectividad de las metacercarias obtenidas a partir de infecciones
naturales. Finalmente, los huevos recuperados de los animales parasitados fueron
cultivados para realizar infecciones en Lymnaea viatrix.
5.1 METODOLOGÍA
Se realizaron infecciones experimentales en ratones y ratas para confirmar si las
cercarias de F. hepatíca emitidas por los caracoles de campo eran viables e infectivas y
para obtener parásitos adultos a partir de las larvas encontradas en el ambiente. Para
ello, en distintas oportunidades a Io largo del estudio se recolectaron metacercarias de
los recipientes que contenían a los caracoles traídos de campo y se mantuvieron en
agua declorada y limpia a 4°C hasta su posterior uso. Antes de la administración de las
metacercarias se descartaron los quistes dañados y se eligieron los que en apariencia
eran viables. Se utilizó como criterio de viabilidad de los quistes la refringencia de los
149

gránulos excretores y el estado general de cada metacercaria (Boray 1969, Valero et al.
1996, Mc. Kown et al. 2000).
Ensayos en ratones. Grupo 1.- Diez ratones hembras de la cepa CF1 de 30 días de
edad recibieron vía oral 10 metacercarias cada uno. Luego de la infección y hasta su
muerte, se mantuvieron en jaulas acondicionadas, con alimento ad libitum y en
condiciones de luz-oscuridad: 12hs-12hs. Se planificó la búsqueda de huevos del
parásito en la materia fecal a partir de los 30 días post infección (p.i.). Los individuos
que murieron antes de esa fecha fueron autopsiados. Grupo 2.- Este ensayo se diseñó a
partir de los resultados obtenidos con el grupo 1. Se modificaron las dosis de
metacercarias administradas y el tiempo p.i. en que se comenzó a buscar indicios de la
infección. Nueve ratones de Ia misma cepa y edad recibieron las siguientes dosis: 3
individuos fueron infectados con 5 metacercarias cada uno, 2 con 10, 2 con 15 y 2 con
20 metacercarias cada uno. Se planificó la búsqueda de huevos del parásito en la
materia fecal a partir de los 14 días p.i.
Ensayos en ratas. Se infectaron por vía oral 8 ratas hembras Wistar de 27 días de
edad. Un grupo de 4 animales recibió 3 metacercarias y otro grupo 5 metacercarias
cada uno. Luego de la infección los animales se mantuvieron en condiciones similares a
las de los ratones. Se planificó la búsqueda de huevos del parásito en la materia fecal a
partir de la octava semana p.i.
En todos los casos Ia materia fecal se procesó utilizando el método de tamización
descripto en 3.4.4.
Los animales positivos por coprología fueron sacrificados utilizando un exceso de
anestesia a base de éter y disecados para buscar juveniles y adultos de F. hepatíca en
el hígado y en los conductos biliares. Para ello, en cada caso se extrajo el hígado del
animal y en una caja de Petri con solución fisiológica se realizaron cortes finos del
órgano y se presionó el material para facilitar la liberación de los parásitos. Los
juveniles y adultos recuperados se fijaron en formol 5% caliente y se determinaron
morfológicamente (Dawes 1968, Yamaguti 1975).
Ensayos en caracoles. Se recolectaron huevos de F. hepatica de Ia materia fecal de
los animales infectados experimentalmente y se mantuvieron en agua fresca a 25°C y a
la oscuridad durante 13 días, momento en el que se expusieron a luz intensa hasta la
eclosión de los miracidios pocas horas después.
Para realizar las infecciones se utilizaron 21 caracoles de la especie Lymnaea
viatríx (rango de tamaños: 3-S.6mm) nacidos y criados en el laboratorio a partir de
individuos traídos de campo. Se infectaron 3 grupos de caracoles con 5 miracidios cada
caracol. Los dos primeros grupos pertenecían a la filial 2 de caracoles originalmente
recolectados en el lugar de estudio (Localidad de Cholila): el primer grupo (GI) formado
150

por 7 caracoles (tamaños entre 4,9 y 5,6mm) cuyos progenitores fueron recogidos del
mallín 2 y el segundo grupo (G2) de 5 caracoles (tamaños entre 3,5 y 4,8mm)
originalmente extraídos del charco 1. El tercer grupo (G3), de 9 caracoles (tamaños
entre 3 y 4,5mm), era la filial 2 obtenida a partir de material recolectado en una
acequia en Ia provincia de Córdoba, Argentina.
Cada caracol fue colocado individualmente en un recipiente de 5m| de capacidad
con agua declorada y se mantuvo en contacto con los miracidios durante 3 V2horas a
temperatura ambiente y bajo luz natural. Los recipientes se mantuvieron tapados para
evitar que los caracoles quedaran fuera del agua. Durante el transcurso de las 3 V2
horas se revisaron periódicamente los recipientes para confirmar que los miracidios
hubiesen penetrado en los caracoles. La mayoría de las larvas ya no se observaron
luego de 1 hora de haber sido expuestas a los caracoles. A partir de los 34 días p.i. se
comenzaron a revisar todos los caracoles periódicamente (según se describe en 4.4.3).
En cada ocasión se contó el número de animales con infecciones maduras por emisión
de cercarias y se los separó de los individuos que aún no emitían.
5.2 RESULTADOS
Ensayos en ratones
Grupo 1.- Veinticuatro días post-infección (p.i.) y antes de alcanzar el período de
patencia murieron 9 ratones que no pudieron ser autopsiados. El individuo restante fue
sacrificado y se recuperaron 6 fasciolas juveniles del hígado.
Grupo 2.- Catorce días p.i. se sacrificó el primer ratón perteneciente al grupo
que había recibido la mayor dosis de metacercarias y se recuperaron del hígado 15/20
fasciolas juveniles, pero no se detectaron adultos instalados en la vesícula. Veinticinco
días p.i. murieron 7 ratones. Cinco de estos animales pudieron ser autopsiados y en
todos los casos se observaron los lóbulos hepáticos dañados y la vesícula biliar
agrandada y con coágulos en su interior, pero no se encontraron parásitos. El ratón
sobreviviente, infectado con 5 metacercarias, fue sacrificado. No se recuperaron
parásitos ni se observaron signos de infección.
Ensayos en ratas
Todos los animales llegaron con vida y en buen estado general de salud al
momento en que se comenzó a revisar la materia fecal, ocho semanas p.i. Para esa
fecha todos los individuos resultaron positivos por coprología. Se sacrificaron 2 animales
y se recuperaron adultos de F. hepatica de los conductos biliares. El resto de los
151

animales se conservaron con vida para el mantenimiento del ciclo de vida del parásitoen el laboratorio.
Ensayos en Caracoles
A los 34 días p.i. ya se encontraron caracoles emitiendo cercarias de F. hepatica,
con Io cual no se pudo establecer exactamente el período de prepatencia. Se registraron
2/7 caracoles del grupo 1 (G1) infectados y 3/9 del grupo 3 (G3). Los caracoles del
grupo 2 (G2) no emitieron durante esa primera exposición. Luego de 40 días p.i.
emitían cercarias 2/7 (28.6%) caracoles del G1, 4/9 (44.4%) caracoles del G3 y
ninguno del G2. Hasta los 90 días p.i., momento en que se finalizó el ensayo, se
obtuvieron 13/21 (61.9%) caracoles infectados: 6/7 (85.7%) caracoles del G1, 2/5
(40%) caracoles del G2 y 5/9 (55.6%) caracoles del G3 (Tabla 5-1). EI porcentaje de
mortalidad fue del 66% (6/9) para los caracoles del G3, 40% (2/5) para los del G2 y
0% para los pertenecientes al Gl. Todos los individuos que murieron resultaron
positivos al ser disecados excepto uno de G3 que no emitió en vida y no pudo serdisecado.
Tabla 5-1: Cantidad de nuevos casos de infección por Fascio/a hepatica en Lymnaea
viatrix infectadas experimentalmente
Días p.i. 34 40 90 Total (%) Mortaudad 0/0
G1 (N=7) 2 o 4 6 (85.7) o
GZ (N=5) 0 0 2 2 (40) 40
G3 (N=9) 3 1 1 5 (55.6) 66
Total (N=21) 5 1 7 13 (61.9) 38
El total de caracoles infectados y la mortalidad se midieron a los 90 días p.i., momento en que finalizó el
ensayo. La positividad de un caracol se midió como emisión de cercarias. G1: filial 2 del mallín 2; GZ:
filial 2 del charco 1; G3: filial 2 de la Provincia de Córdoba.
5.3 DISCUSIÓN
Ninguno de los ratones infectados sobrevivió más de 25 días desde el momento
en que fueron infectados, independientemente de la dosis de metacercarias
suministrada. Aunque en algunos casos no se recuperaron parásitos de los individuos
muertos o sacrificados, las lesiones observadas en el tejido hepático fueron una
evidencia del éxito del desarrollo (aunque en forma incompleta) de las metacercarias
desenquistadas en los ratones. Evidentemente, el daño hepático producido por F.
152

hepatíca durante la migración de las fasciolas juveniles fue tan importante que provocó
la muerte de los animales antes de alcanzar el período de prepatencia, con lo cual el
ciclo no pudo establecerse utilizando ratones como hospedadores definitivos (Behm &
Sangster 1999). Estos resultados coinciden con los obtenidos por otros autores, que
sugieren que los ratones no son buenos hospedadores experimentales de F. hepatíca y
que mueren dentro de los 30 días post-infección aún con infecciones de sólo 2 parásitos
(Mulcahy et al. 1999).
Contrariamente a Io ocurrido en ratones, las ratas soportaron bien las
infecciones. Diversos autores han demostrado experimentalmente que estos roedores
son buenos hospedadores definitivos del parásito (Valero et al. 1996, Díaz et al. 1998,
Valero & Mas-Coma 2000) e incluso algunos trabajo sugieren que pueden convivir con Ia
infección toda su vida (discutido en Keegan &Trudgett 1992 y Mulcahy et al. 1999).
El período de prepatencia fue menor al informado por Díaz y colaboradores
(1998). Según estos autores, la eliminación de huevos en la materia fecal de ratas
Wistar infectadas con 25 metacercarias cada una fue evidente a partir de la 10a semana
p.i. Sin embargo, en los ensayos realizados en esta tesis, los animales ya resultaron
coprológicamente positivos a la 8a semana p.i. La elección del momento a partir del cual
iniciar Ia búsqueda de huevos en materia fecal se estableció en función de los resultados
obtenidos por estos autores, y debido a que la infección se desarrolló mucho más
rápido, no pudo establecerse con certeza la duración del período de prepatencia.
Independientemente de la capacidad de resistir la infección por F. hepática de
uno y otro tipo de hospedador, el éxito de infección de las metacercarias obtenidas a
partir de caracoles de campo estaría demostrando la viabilidad de estas larvas y su alta
capacidad infectiva.
La viabilidad de los huevos liberados por los adultos desarrollados en las ratas y
la infectividad de los miracidios eclosionados también pudieron confirmarse mediante
los ensayos preliminares realizados en L. viatrix. EI tiempo de desarrollo de los huevos
se correspondió con lo reportado en bibliografía (Rowan 1956, Ollerenshaw 1958,
Mattos & Ueno 1986, Luzón-Peña et al. 1994, Andrews 1999) y el éxito de infección fue
de aproximadamente el 62%, valor que no resulta bajo si se considera que los tamaños
de los caracoles no fueron estrictamente homogéneos y que la susceptibilidad puede
depender de esta variable (Mattos & Ueno 1986). En estos ensayos se utilizaron por un
lado caracoles con “experiencia” con el parásito (Localidad de Cholila) y otros que nunca
antes habían tenido contacto con el parásito que se desarrolla en el área de estudio
(Provincia de Córdoba). El éxito de infección no pareció diferir entre ambas poblaciones
(8/12, 66.7% en Cholila vs. 5/9, 55.6% en Córdoba), pero la mortalidad resultó mayor
en los caracoles infectados pertenecientes a la población de Córdoba (66% vs. 40%).
Sin embargo, no era el objetivo de estos ensayos preliminares responder a estos
interrogantes, con lo cual, para estudiar en detalle estas tendencias observadas, debería
153

realizarse un estudio con mayor control sobre las variables involucradas, con tamaños
muestrales mayores, réplicas y controles adecuados.
Finalmente, la obtención experimental del adulto y su estudio taxonómico
permitieron confirmar su identificación específica como F. hepatica.

DISCUSIÓN GENERAL
La fasciolosis es una de las principales infecciones parasitarias del ganado
doméstico en nuestro país. En Ia región de estudio, los productores manifestaron un
importante nivel de preocupación por los daños que causa Fasciola hepatica a escala
local. Sin embargo, a nivel nacional las prevalencias registradas oficialmente fueron
muy bajas como resultado probablemente de información recabada en forma irregular,
poco rigurosa y con distintos criterios en cada región. La fasciolosis es la cuarta
parasitosis en importancia en ganado bovino, según los registros del SENASA. La
evidente subestimación de la prevalencia general de esta parasitosis permitesuponer que la cantidad de casos anuales en el país debe superar ampliamentelos registrados oficialmente, y que sus implicancias en la producción ganaderadeben ser mayores a las estimadas.
La decisión de implementar medidas de control local no puede basarse en las
prevalencias reportadas a escala provincial o regional, ya que el promedio de los casos
detectados en una zona geográfica amplia, puede enmascarar la importancia del
problema en áreas más pequeñas. En la Argentina se han realizado pocas
investigaciones que permitan estimar las prevalencias en ganado de manera precisa,
pero en dichas ocasiones los niveles de infección registrados fueron altos. En un estudio
realizado en la provincia de Salta se encontró que sólo un 3% de los bovinos presentó
huevos en la materia fecal, pero un 14% de los animales necropsiados estuvo infectado
con F. hepatica (Dwinger et al. 1982). En la provincia de Neuquén, un estudio específico
realizado en mataderos detectó un 94% de infección durante el primer semestre de
1981 (Kaczorkiewicz 1983).
Es necesaria una correcta y rigurosa re-evaluación de la importancia de la
fasciolosis que permita cuantificar las pérdidas reales producidas por el decomiso de
hígados afectados por el parásito. Una campaña unificada que permitiera generar
155

registros en distintos frigoríficos de diversas provincias en una misma época del año,
podría ser un buen indicador de si existe realmente una subestimación de esta
parasitosis y de su relevancia.
Para identificar los factores involucrados en la transmisión de F. hepatica en los
valles cordilleranos patagónicos, primero fue necesario identificar las especies de
lymneidos presentes en la región y su posible participación en el ciclo de vida del
parásito. Luego, para el estudio del hospedador intermediario, se definieron distintas
escalas de análisis y los resultados obtenidos permitieron conocer algunas
características poblacionales y relacionarlas con el riesgo de transmisión al ganado. Así,
pudo demostrarse que la única especie de caracoles que actúa como hospedadorintermediario de F. hepatica en el área de estudio es Lymnaea víatrix y presentaun desarrollo sincronizado por la temperatura ambiental y la disponibilidad de agua en
el suelo; su actividad está restringida a unos pocos meses entre la primavera y el
otoño. La postura de huevos suele ocurrir durante el verano y esa nueva generación de
individuos necesita sobrevivir al invierno para poder dejar descendencia la temporada
siguiente a su nacimiento. El ciclo se completa en aproximadamente 15 meses. L.
víatrix demostró ser un excelente hospedador y estar bien adaptada a climas adversos,
a pesar de hallarse en el extremo austral de su distribución, posiblemente sometida a
situaciones de estrés climático. Sin embargo, la tasa de crecimiento durante los
periodos más favorables resultó baja en comparación con otras especies del género de
similar desarrollo. Por ejemplo, en Australia L. tomentosa (cuyo tamaño máximo es
apenas superior al de L. víatrix) en laboratorio y bajo condiciones favorables de
temperatura alcanzó una longitud de 3 mm en 6 días, 4 mm en 14 días y 5,8 mm en 17
días luego de eclosionar. Caracoles de 20-27 días produjeron huevos, y en campo, el
tiempo generacional puede ser de 1 mes entre primavera y otoño (Boray 1969). En
regiones de clima tropical, L. víatrix demora en promedio desde su nacimiento, 27 días
en comenzar la oviposición (Lara et al. 1988, Müller et al. 1998). A latitudes superiores
a los 41° S como es el caso del área de estudio, y sometidos a periodos cortos de
temperaturas adecuadas para su desarrollo, los caracoles requirieron entre 3 y 4 meses
para alcanzar la madurez sexual, y aún así, el período de oviposición se vio demorado
hasta la siguiente temporada.
A una escala de mayor detalle se observó que la población de caracoles está
dividida en subpoblaciones que habitan distintos cuerpos de agua. La estructura de cada
una de estas subpoblaciones depende de las características particulares de cada
ambiente. Dentro de cada cuerpo de agua los caracoles se desarrollan en pequeños
parches y, en definitiva, son estos microambientes los que condicionan la presencia o
abundancia de L. víatrix en los distintos ambientes de la región de estudio.
La gran capacidad de adaptación del hospedador intermediario a distintos
ambientes fue evidente al encontrar, durante todo el estudio, subpoblaciones
establecidas o efímeras tanto en cuerpos de agua permanentes (pequeñas lagunas,
156

arroyos) como temporarios (mallines, charcos, canales de irrigación). En general se
observóque L. viatrix habita ambientes que presentan parches con sustrato debarro, poca vegetación y alta incidencia solar. Estos parches pueden encontrarse
dentro de ambientes óptimos o subóptimos, y de ello dependerá el desarrollo efímero o
permanente de las subpoblaciones de caracoles. Aquellos ambientes que presentaron
parches grandes y muy conectados entre si albergaron mayores abundancias de
caracoles. Además se comprobó que la ocurrencia del hospedador intermediarioestuvo asociada a la presencia de “playas” (sectores con columna de aguamínima nula) dentro de los parches. Este resultado permite suponer que los
caracoles no entrarán en actividad luego del deshielo hasta que el retraimiento del agua
genere ambientes húmedos pero no completamente inundados. Estas condiciones se
dieron a partir de octubre-noviembre en el 2000 y entre agosto y septiembre en el 2001
y, durante este último año, la subpoblación de caracoles de la laguna entró en actividad
antes que en el 2000. El año 1999 presentó un invierno y una primavera mucho más
secos que los años restantes y en ningún momento se pasó de una situación de exceso
de agua en el suelo a una situación de menor humedad. En este caso no se encontró
una relación entre la actividad de los caracoles y esas particulares condicionesambientales.
La importancia de cada tipo de ambiente con respecto a la colonización y el
desarrollo de L. viatrix puede discutirse considerando las características de las
subpoblaciones que albergan y la superficie relativa de cada tipo de ambiente en el área
de estudio. Durante este trabajo se encontró que los ambientes con hábitats continuos
(máximo tamaño y conectividad) para el hospedador intermediario presentaron
generalmente las mayores abundancias. La laguna y un meandro del río fueron algunos
de los cuerpos de agua con esta característica, junto con unos pocos ambientes
temporarios (algún charco o sector de un mallín). Las altas densidades que puede
desarrollar L. viatrix en los ecotonos de estos ambientes permiten suponer que los
hábitats continuos serían los más riesgosos para el hospedador definitivo. En estos
ambientes suelen darse simultáneamente todos los factores necesarios para la
transmisión: 1) la presencia concentrada del caracol, 2) la frecuente visita del ganado
para beber, 3) la defecación de los animales infectados en un área húmeda que asegura
la supervivencia de los huevos y 4) la alta probabilidad de encuentro entre miracidios y
caracoles. Sin embargo, este tipo de ambientes representa una superficie muy pequeña
del total de los cuerpos de agua de la zona. Los mallines, en cambio, generalmente
albergaron abundancias de caracoles más bajas, y la probabilidad de ocurrencia
simultánea de todos los factores recién detallados es mucho menor que en un ambiente
permanente. Pero estos cuerpos de agua ocupan una superficie muy extensa en toda la
zona Cordillerana Patagónica.
En general se considera que los ambientes que permiten el desarrollo de altas
abundancias de caracoles, pueden actuar como fuente de dispersión hacia otros sitios
en los que las subpoblaciones se mantienen gracias a constantes eventos de

recolonización, resultando en un sistema del tipo fuente-sumidero o de metapoblaciones
(Hanski & Gi|pin 1997). Ambientes con hábitats continuos para L. viatrix podrían actuar
como “fuentes” en el área de estudio. Dentro del establecimiento ganadero, sin
embargo, Ia laguna se encuentra en la zona más baja del gradiente altitudinal, con Io
cual las subpoblaciones que allí se desarrollan difícilmente podrían dispersarse a otros
ambientes que se encuentran más elevados.
Hasta aquí entonces, se analizaron la densidad de caracoles que puede sostener
un ambiente y la superficie relativa de dicho ambiente en Ia región de interés; ambas
variables en función del riesgo para el ganado. Existe una tercera variable que está
relacionada con el desarrollo del parásito dentro del hospedador intermediario: el
tamaño que este último puede alcanzar. La infección por F. hepatica siempre se
registró en caracoles grandes. Además, tanto en la laguna como en los mallines, las
proporciones de individuos de gran tamaño y las de infectados no difirieron
significativamente entre ambientes. Sin embargo, podría suponerse que hábitats más
adecuados (como es uno continuo con respecto a uno parcheado) podrían permitir el
desarrollo de mayores proporciones de caracoles de gran tamaño y consecuentemente
de mayores niveles de infección, siempre y cuando las altas densidades no generen un
efecto negativo en el desarrollo de los caracoles. Evidentemente deben existir factores
que juegan un rol en la tasa de crecimiento de L. viatrix y en los niveles de transmisión
que se pueden alcanzar en cada tipo de ambiente. Podría suponerse que las altas
abundancias de otras especies de digeneos halladas parasitando a los caracoles de la
laguna podrían provocar una mortalidad diferencial de individuos grandes muy
parasitados (Gutiérrez et al. 2000); esto podría justificar porqué no se encontró una
mayor proporción de individuos de gran tamaño en ese cuerpo de agua. A pesar de no
haber diferencias en los niveles de infección, podría suponerse que el ambiente con
mayores densidades contendrá mayor cantidad absoluta de individuos de gran tamaño
parasitados, lo cual permitirá desarrollar niveles más altos de infestación de las
pasturas.
Finalmente quedan definidas dos situaciones muy distintas con respectoal riesgo de infección para el ganado. Por un lado la alta concentración demetacercarias en las orillas de los ambientes como la laguna y por el otro ladistribución parcheada y diluida de los quistes en las extensas superficies demallines.
Un análisis de la relación entre Ia presencia de ganado y Ia infección en L. viatrix
permitirá comparar ambos tipos de ambientes:
Se encontraron caracoles infectados en la laguna en dos ocasiones durante el
estudio. La mayor prevalencia se registró en enero de 2000 cuando el acceso de los
animales al cuerpo de agua era libre ya que el valle no se encontraba aún sectorizado.
En ese momento, no se encontró infección en los mallines 1 y 3, que también eran
visitados en forma irrestricta por los animales. Sin embargo en estos casos debe
contemplarse que la cantidad de caracoles hallados fue baja. Luego de este muestreo y
158

debido a la alta prevalencia encontrada, se decidió en el establecimiento interrumpir el
acceso de los animales a la laguna, y la misma estuvo aislada hasta fines de septiembre
de 2001. A partir de ese momento y aproximadamente durante 2 meses, un centenar
de animales adultos tuvieron acceso a las orillas del cuerpo de agua. Los huevos que
probablemente liberaron los individuos parasitados, infectaron exitosamente a los
caracoles que entraron en actividad durante la primavera, y dichas infecciones
maduraron y fueron evidentes en febrero de 2002. De esta observación se desprende
que unos 100 animales frecuentando dicho ambiente durante 2 meses permitieron que
se registrara infección en los caracoles. La prevalencia en el hospedador intermediario
fue significativamente menor que la encontrada en enero de 2000 (2/222 vs. 7/119).
Esto podría deberse a que durante prácticamente 2 temporadas la laguna estuvo libre
de infección y a que los caracoles infectados en febrero de 2002 fueron el producto del
contacto con miracidios unos meses antes ya que no pudo haber caracoles que
aportaran infecciones inmaduras de la temporada anterior. Por otro lado, los bovinos
que frecuentaron la laguna entre octubre y noviembre de 2001 habían sido
desparasitados en mayo del mismo año. Suponiendo que la probabilidad de reinfectarse
durante el invierno es baja (baja disponibilidad de metacercarias viables accesibles), no
se esperaría encontrar animales con infecciones maduras a comienzos de la primavera
(septiembre). Así, podría pensarse que el tratamiento en realidad no fue 100% efectivo,
y que un grupo de animales se mantuvo infectado aún después de la aplicación del
antiparasitario. De esta forma, una proporción baja de animales infectivos remanentes
fueron suficientes para reinfectar los caracoles de la laguna, aunque con prevalenciasmenores al 1%.
En los mallines sólo se encontró infección una vez durante los 3 años de estudio.
En dicha ocasión (marzo de 2001, prevalencia: 2%, 1/53) el ganado había estado
pastando en el mallín 1 -dentro del potrero 29- durante los cuatro meses previos al
hallazgo (noviembre y diciembre 2000 y enero y febrero 2001) (Fig. 3-3). En el mallín 2
hubo ganado pastando en mayo, agosto, septiembre, octubre y parte de noviembre de
2000 (durante el invierno estuvieron en Ia ladera de la montaña), y no se encontraron
caracoles infectados (n= 86) en el muestreo realizado en noviembre. En principio, y a
partir de los registros a los que se pudo acceder, estas fueron las únicas situaciones en
las que hubo ganado en gran cantidad y durante tanto tiempo en un mallín.
Aún cuando Ia presencia de infección en el mallín 1 se confirmó en un único
individuo y Ia cantidad de caracoles revisados en el mallín 2 no fue muy elevada, si se
supone que el resultado obtenido es el reflejo de situaciones epidemiológicas distintas y
no del azar, podría esperarse hallar una explicación al analizar lo que pasó en cadaambiente antes de marzo de 2001:
1.- En el mallín 1 (donde se encontró infección) Ia carga ganadera habría sido
mayor: en noviembre de 2000 pastaban vacas y en diciembre de 2000, enero y febrero
de 2001, vacas y vaquillonas, mientras que en el mallín 2 en mayo de 2000 pastaron
vacas y en agosto, septiembre, octubre y noviembre sólo vaquillonas.
159

2.- Las épocas del año en las que se produjo la oferta de huevos al medio fue
distinta: en el mallín 1 el ganado liberó huevos durante la primavera y el verano y el
muestreo en caracoles se realizó a comienzos del otoño, mientras que en el mallín 2 Io
mismo ocurrió principalmente durante el otoño y el invierno y a comienzos de la
primavera, y el muestreo en caracoles ocurrió a mediados de la primavera.
Evidentemente ambas diferencias podrían haber aportado al resultado final
observado, ya que a mayor carga ganadera hay mayor oferta de huevos al medio y
mayor probabilidad de encuentro entre miracidios y caracoles. Además la época del año
en que se da dicha oferta es también importante. Un estudio realizado en Chile (35° S)
permitió conocer los tiempos que demoran los huevos de F. hepatica en desarrollarse a
temperaturas ambientales. Los tiempos fueron máximos para los huevos liberados en
otoño y mínimos para los de fines de la primavera. Los huevos liberados al ambiente en
marzo, abril o mayo, eclosionaron recién en septiembre. Por otro lado, los huevos que
estuvieron a la intemperie a partir de noviembre, diciembre o enero, demoraron a lo
sumo 2 meses en eclosionar. En forma general, los huevos no eclosionaron mientras
estuvieron expuestos a temperaturas medias inferiores a los 10° C (Alcaíno et al.
1993). En otro estudio se observó que los huevos sin embrionar que se liberaron al
ambiente en otoño, no sobrevivieron el invierno, mientras que todos los huevos que
llegaron embrionados al invierno, eclosionaron exitosamente durante la siguiente
primavera (Luzón-Peña et al. 1994). A partir de estas observaciones y considerando las
condiciones climáticas durante el periodo 1998-2002 (4.5.1), podría suponerse que enel área de estudio los huevos no estarían en condiciones de eclosionar entre fines de
marzo y fines de septiembre. En el caso del mallín 1, los huevos fueron liberados en la
época del año con condiciones climáticas más favorable para su desarrollo y en
momentos en los que los caracoles estaban en actividad. En cambio, en el mallín 2, los
huevos que fueron liberados durante los meses de otoño e invierno probablemente no
encontraron las condiciones adecuadas para desarrollarse hasta la primavera, y aunque
algunos lo hubiesen logrado, los caracoles estaban en período de hibernación. Si hubo
coincidencia entre la oferta de huevos viables y el período de actividad de los caracoles,
esto pudo haber ocurrido únicamente durante el mes de octubre y comienzos de
noviembre. Considerando que el desarrollo de redias inmaduras ocurre a partir de los 9
días postinfección en condiciones de laboratorio (Mattos & Ueno 1989), podría
suponerse que si los caracoles se hubieran infectado durante octubre y noviembre,
habría sido posible observar redias inmaduras para el momento en que se realizó el
muestreo y se disecaron los caracoles (noviembre de 2000). A pesar de que todas las
infecciones podrían haber estado maduras para enero de 2001, en ese momento
tampoco se encontraron caracoles parasitados (n=32).
La ausencia de infección en los mallines 1 (n= 23), 2 (n= 6) y 3 (n=2) en
noviembre de 1999 y en los mallines 1 (n= 29) y 3 (n=5) en enero de 2000 no pudo
relacionarse con el manejo ganadero, ya que no fue posible obtener información
durante ese período. De todas formas la cantidad de caracoles hallados fue muy baja
160

excepto en el mallín 1. En este último ambiente, desde el momento en que se tienen
registros (mayo de 2000) y hasta noviembre de 2000, no hubo ganado en los potreros
que Io contienen.
Finalmente cabe resaltar que, a diferencia de lo observado en la laguna, la
presencia de ganado durante menos de 4 meses en los mallines no permitió registrarinfección en los caracoles.
En el arroyo 1 se encontraron ejemplares de L. víatríx en forma esporádica y
siempre en muy bajas cantidades (marzo (n= 0) y noviembre de 2000 (n= 5), enero
(n= 5), marzo (n= 18), octubre (n= 0) y noviembre de 2001 (n= 0) y febrero de 2002
(n=0)), generalmente agrupados todos en un mismo parche. El hallazgo de caracoles
infectados en este arroyo estaría demostrando la importancia del transporte pasivo del
hospedador intermediario y del parásito (Boray 1969, Rondelaud et al. 2004), y
confirma que la liberación de metacercarias puede ocurrir en un ambiente distinto (por
sus características y por su disposición espacial) al ambiente donde se encuentra el
ganado infectado y donde pudieron infectarse los caracoles. Con respecto a este
hallazgopodría concluirseque no debe minimizarse la importancia epidemiológicade un ambiente que no permite el desarrollo del caracol en forma sostenida,pero que podría actuar como un corredor de dispersión y conectividad entrehábitats favorables.
EI hallazgo de huevos de F. hepatíca en materia fecal de liebres confirma la
participación potencial de hospedadores silvestres en el ciclo de transmisión. Un estudio
sobre la viabilidad de los huevos liberados por los parásitos que se desarrollan dentro
de este hospedador permitiría evaluar si es factible que se infecten caracoles en un
ambiente, en ausencia del ganado. Conocer el rol de este hospedador en la transmisión
de F. hepatica sería de suma utilidad a la hora de planificar medidas de control, ya que
así podría determinarse si es posible que exista infección en caracoles y
consecuentemente metacercarias en las pasturas dentro de un potrero, aún cuando el
ganado no haya estado presente en dicho ambiente.
El estudio de la población del hospedador intermediario resultó necesario para
interpretar los mecanismos involucrados en la transmisión de F. hepatica. Los
resultados obtenidos demostraron que es muy difícil predecir pequeños cambios en la
dinámica de L. víatríx, aunque se puede conocer fácilmente la dinámica general para
cada año. Se encontró una asociación entre variables ambientales y las abundancias y
los tamaños de los caracoles en la laguna y en los mallines, respectivamente, aunque
dicha relación fue instantánea (variables ambientales y variables del hospedador
medidas en un mismo momento). Con lo cual este modelo carece de valor predictivo.
Sin embargo, puede concluirse que los años que presenten períodos contemperaturas desfavorables cortos, junto con condiciones de exceso en la
161

disponibilidad de agua durante el invierno, permitirán la actividad delhospedador intermediario de manera anticipada.
La infección en el hospedador intermediario se evidenció recién a partirdel verano. Considerando que las temperaturas son desfavorables para el desarrollo
del caracol y el parásito aproximadamente entre mayo y fines de agosto, y que las
temperaturas medias no exceden los 10° C entre abril y fines de octubre, podría
suponerse que una infección inmadura que fue adquirida la temporada anterior, podría
comenzar la maduración dentro del hospedador intermediario recién a partir de
noviembre. Por otro lado, si los huevos en el ambiente pueden eclosionar recién a partir
de octubre, el contacto entre miracidio y caracol en ese momento podría producir unainfección madura recién hacia fines de diciembre.
Los niveles de infección registrados en las subpoblaciones de L. víatrix fluctuaron
durante el estudio pero fueron bajos para las altas prevalencias que se registraron
constantemente en el ganado. Sin considerar el hallazgo de infección en el arroyo 1, los
valores de infección por F. hepatíca en L. víatrix fluctuaron entre aproximadamente un 1
y un 6%. Una recopilación bibliográfica realizada por Hurtrez-Bousses y colaboradores
(2001) presenta prevalencias en especies del género Lymnaea de 12% en Francia,
entre 0,1 y 3,1% en Estados Unidos y hasta 80% en el Altiplano Boliviano. Sin embargo
estos estudios no permiten relacionar dichos niveles de infección con los del ganado.
Otro estudio realizado durante 4 años permitió conocer la evolución de la infección en
ganado y caracoles cuando se utilizan antiparasitarios en forma estratégica y muy
frecuente. AI comienzo, el 95% de la hacienda eliminaba huevos de F. hepatica en la
materia fecal y se halló un 12% de infección en caracoles. Durante el segundo año de
estudio la prevalencia en ganado se redujo al 50% y en caracoles al 2,5%. Hacia fines
del tercer año hubo un pico en los niveles de infección en los rumiantes (71%); sin
embargo la infección no superó el 1% en el hospedador intermediario. Finalmente, no
se encontraron huevos en la materia fecal durante el último año y la infección en
caracoles se redujo al 0,4% (Parr & Gray 2000).
Durante el presente estudio se comprobó que niveles bajos de infecciónen caracoles pueden mantener altas prevalencias en ganado, aún con laaplicación frecuente de antiparasitarios (Fig.6-1). Este resultado demuestra que la
transmisión de F. hepatica es altamente eficiente en los valles cordilleranos
patagónicos. Los ensayos realizados en el laboratorio demostraron que las
metacercarias obtenidas a partir de caracoles de campo resultaron ¡nfectivas para
hospedadores definitivos experimentales y que los huevos de los adultos que sedesarrollaron infectaron exitosamente a L. víatrix.
A pesar de que se observó una disminución en la prevalencia en caracoles en
uno de los cuerpos de agua entre enero de 2000 y febrero de 2002, esta diferencia no
puede atribuirse directamente a la reducción de la infección en el ganado, ya que el
tiempo que fue utilizado dicho ambiente previo a cada hallazgo y la cantidad de
animales que lo frecuentaron no fueron homogéneos en ambos momentos.
162

Los bovinos mostraron una aparente respuesta diferencial por edades a la
infección mientras estuvieron sometidos a un manejo sanitario poco frecuente. Esta
diferencia desapareció luego de 3 años de tratamientos antiparasitarios regulares,
produciendo un efecto de homogenización de los niveles de infección pero no de
reducción.Las prevalencias en ganado fueron cercanas al 50% en todos losmomentos en los que se generaron registros (Fig. 6-1). Lo mismo ocurrió con el
número de huevos liberados por gramo de materia fecal (hpg), aunque esta variable
tomó valores muy bajos en comparación con estudios realizados en otros países. Por
ejemplo, en una investigación realizada en 81 establecimientos ganaderos italianos, se
encontraron huevos de F. hepatíca en 1,8% (N=975) de los animales y el hpg presentó
un valor promedio de 27 (rango: 10-100) (Cringoli et al. 2002). Otro trabajo llevado a
cabo en España en una extensa región arrojó una prevalencia del 29,5% (N=1301) con
un hpg promedio de 50 (rango: 10-800) (González-Lanza et al. 1989).
La ausencia de efecto aparente de los tratamientos (que debería haberse
visualizado como disminución en las prevalencias y/o el hpg) resulta llamativa. Las
posibles explicaciones para estos resultado son dos: o bien la oferta de metacercarias
viables es permanente durante todo el año, o existe algún problema con los
tratamientos suministrados. El estudio de una cohorte juvenil permitió estudiar en
mayor detalle ambas posibilidades. Estos animales nacieron en octubre de 1999 y
durante el primer año de vida recibieron dos tratamientos antiparasitarios (principio de
mayo y mediados de junio del año 2000). Podría suponerse que la quimioterapia eliminó
completamente las potenciales infrapoblaciones parasitarias adquiridas durante el
primer semestre de ese año. Luego, durante el invierno y parte de la primavera (julio
octubre), las vaquillonas fueron alimentadas con pasturas y bebederos, con Io cual la
infección detectada en enero de 2001 (40%) podría ser el resultado del contacto con
metacercarias durante noviembre (dándoles a los parásitos un mes y medio para
desarrollarse hasta alcanzar Ia madurez sexual). Ese mes los animales estuvieron 15
días en el potrero 12 y otros 15 días en el potrero 17. Sin embargo, durante los
estudios realizados en noviembre de 2000, no se detectaron caracoles en el mallín 3
dentro del potrero 12 y no se encontraron individuos infectados (n= 86) en el mallín 2
dentro del potrero 17. Tampoco se encontró infección por F. hepatíca dos meses
después. Estos resultados darían peso a la hipótesis de que las vaquillonas adquirieron
Ia infección a partir de metacercarias que sobrevivieron el invierno. Aún cuando esta
hipótesis es viable, los tiempos entre el momento de ingestión de las metacercarias
(todo noviembre) y Ia eliminación de huevos (mediados de enero) resultan muy justos
para permitir el completo desarrollo del parásito. Una infección adquirida a fines denoviembre difícilmente hubiese sido detectada a mediados de enero mediante métodos
coprológicos. Con Io cual, descartando que el ganado haya podido infectarse durante el
invierno con alimentación artificial, podría pensarse que incluso dos tratamientos
antiparasitarios no lograron eliminar completamente las infecciones. Así, la prevalencia
detectada en enero podría ser el resultado combinado de infecciones remanentes
163

mantenidas desde el verano anterior y nuevas adquiridas a partir de metacercarias quesobrevivieron el invierno.
En enero y junio de 2001 los animales fueron tratados nuevamente.
Independientemente de lo que haya ocurrido durante el primer semestre con las
infecciones, podría suponerse que el tratamiento recibido en el mes de junio debió
eliminar todos los parásitos de la población de vaquillonas. Sin embargo, en diciembre
de ese año casi un 50% de los animales Iiberaba huevos del parásito con sus heces
(Fig. 6-1). Suponiendo que un alto porcentaje de las metacercarias formadas durante el
verano y el otoño hubiesen permanecido viables después del invierno, los animales
podrían haberse infectado luego de la desparasitación de junio de 2001. Si se considera
que ese año el valle permaneció cubierto por nieve o agua hasta fines de agosto (Fig. 4
6), el ganado podría haber accedido a las metacercarias a partir de septiembre. De esta
manera, si una alta proporción de parásitos logró alcanzar la madurez sexual en 2
meses, podría justificarse la prevalencia hallada a principio de diciembre. Si las
metacercarias hubiesen estado disponibles recién a partir de octubre o si el tiempo de
desarrollo de los parásitos hubiese sido levemente mayor a 2 meses dentro del
hospedador definitivo, no se explicarían las altas tasas de infección registradas.
La viabilidad de los quistes depende del nivel de humedad y de las temperaturas
del ambiente (Andrews 1999). En el área de estudio, las metacercarias producidas
durante el verano encuentran condiciones adecuadas para su supervivencia y puede
suponerse que la mayoría permanecerán viables hasta el otoño. Durante el invierno, la
humedad no es un factor limitante, pero las temperaturas suelen ser críticas. Algunos
autores sugieren que las metacercarias que se enquistan antes de un invierno crudo no
son un riesgo para el ganado durante la primavera siguiente (Luzón-Peña et al. 1994).
Sin embargo otros estudios demostraron que las metacercarias permanecieron viables
cuando se las mantuvo a -2° C durante 8 semanas (Andrews 1999). Los resultados del
presente estudio en una cohorte juvenil sugieren que un alto porcentaje de
metacercarias podría sobrevivir bajo la nieve y producir en el ganado infecciones
importantes en primavera. La prevalencia hallada al comienzo de la investigación
(febrero de 1999) en terneros nacidos en octubre de 1998 confirma que el ganado
puede contraer infecciones luego del deshielo, antes de Ia época en la que se generan
quistes nuevos.
Por otro lado, la hipótesis de Ia existencia de alguna falla en el manejo sanitario
se encuentra apoyada por el hecho de que en el transcurso de 3 años las prevalencias
prácticamente no se redujeron y no se observó una disminución en la cantidad de
huevos liberados por los animales. El resultado de los manejos sanitarios puede verse
afectado por variables como el estado nutricional del animal (crítico luego de un período
climático adverso), la existencia de otras infecciones parasitarias que alteren el estado
inmunológico del hospedador, la frecuencia de los tratamientos antiparasitarios o Ia
incorrecta dosificación de Ia droga. Por otro lado, los bovinos pueden presentar cierto
164

grado de resistencia adquirida a F. hepatica (Castro 1973, Boray 1981, Dargie 1987,
Roberts &Suhardono 1996).
Aunque los niveles registrados actualmente son bajos, debe considerarse que Ia
resistencia a los antihelmínticos puede diseminarse rápidamente en una población sin
manifestaciones evidentes al principio, ya que la frecuencia inicial de aparición es muy
baja y la forma de crecimiento es Iogarítmica (Hastings 2001). Así, el problema no se
hace evidente hasta que ya está completamente instalado en la población. En varios
países de Europa, por ejemplo, el ganado presenta una resistencia creciente contra el
Triclabendazol, que actúa tanto sobre fasciolas juveniles como sobre maduras (Mollet
al. 2000). Se ha comprobado que Ia eficacia del tratamiento con Triclabendazol y
Closantel varía entre un 80 y 90% en animales naturalmente infectados, pero que en
ambos casos luego de unas semanas el conteo de huevos en heces aumenta. Esto
refleja que algunas fasciolas inmaduras sobrevivieron hasta alcanzar la madurez y
comenzaron a eliminar huevos (Ibarra et al. 2002). Por Io tanto, el ganado bovino
tratado alberga cargas parasitarias bajas sin manifestación clínica, pero suficientes para
mantener el ciclo biológico del parásito en el ambiente y causar daño hepático con
pérdidas económicas por decomiso de los órganos afectados (Geerts et al. 1997).
Alternativamente, podría ocurrir que el efecto residual de la droga no sea el teórico,
debido a problemas de subdosificación o a variables intrínsecas del animal que reducensu efecto residual.
Los resultados de esta tesis permiten suponer que ninguna de las dos posibles
causas planteadas podría justificar por sí sola las altas y constantes prevalencias
registradas. Los niveles de infestación de las pasturas deberían ser muy altos durante la
primavera y el contacto entre el hospedador y los quistes muy eficiente, para lograr que
en tan sólo 2 meses se detecte al 50% de la población liberando huevos. Por otro lado,
aunque hubiese un problema de resistencia a los antiparasitarios, podría suponerse que
con dos tratamientos anuales, el porcentaje de animales coprológicamente negativos
debería superar ampliamente el 50%. Un seguimiento más frecuente de la infección en
el ganado sería necesario para poder probar estas hipótesis.
Ya sea como resultado de una alta oferta de metacercarias todo el año o por
baja eficacia de las drogas, resulta evidente que el manejo sanitario basado en la
aplicación de antihelmínticos no contribuye a disminuir la proliferación delparásito en el ambiente ni reduce la infección en el ganado. Se puede concluirque, debido a la elevada proporción de metacercarias que podrían sobrevivir elinvierno y a la oferta de quistes nuevos durante el verano, el ganado seencontraría constantemente expuesto a las reinfecciones y el único momentodel año con bajo riesgo de infección sería el invierno.
165

Figura 6-1: Infección por Fasciola hepatica en ganado bovino y en Lymnaea viatrix durante el período1998-2002 en los valles cordilleranos patagónícos de la provincia del Chubut
45%
Ó ¿6/0?¿l0 DIC .Ene.
Febϑ(6,5%) 2001 (0,9%)
Referencias:
* Comienzo del Períodoprimaveral
* Comienzodel Períodoesfival
ü Comienzodel Períodootoñal
% Comienzo del PeríodoinvernalW Muefi'eoe” ganado (7°de ¡“ÍGCCÍÓnl fi Alimentaciónanificial
. Caracolessininfección
b Caracoles infectados (0/ode infección)
,L Fecha de desparasiiacióndel ganado
Elganado y las desparasitaciones en verde indican el seguimiento y desparasitación de un grupo de vaquillonas166

Propuesta de un plan de manejo integral
La epidemiología de trematodes de rumiantes es la base a partir de la cual se
diseñan programas de control estratégicos. Sin Ia información generada por esta
disciplina no se pueden utilizar los antihelmínticos para proveer beneficios óptimos en el
control de los gusanos adultos y de los estadios Iarvales en el ambiente. La ausencia de
programas de control generalmente resulta en el uso de quimioterapia según el criterio
y la conveniencia de cada productor. Esto suele tener un impacto muy bajo sobre Ia
población parasitaria (Stromberg &Averbeck 1999).
La erradicación de las infecciones parasitarias es rara vez una opción práctica y
el objetivo del control debe ser Ia reducción de los efectos de las parasitosis en los
animales, para permitir una adecuada producción económica ganadera. Los objetivos
específicos de los programas de control deben ser prevenir Ia proliferación de los
parásitos en el ambiente y evitar áreas con pasturas altamente contaminadas
(Brunsdon 1980).
EItipo de programa de control o manejo que se puede recomendar depende en
gran medida de las características ganaderas locales, de las condiciones climáticas y de
las posibilidades socio-económicas de cada productor. La mejor estrategia de control es
la que combina manejo ambiental con quimioterapia estratégicamente adecuada a las
características de la transmisión en cada región. No resulta rentable utilizar
constantemente antiparasitarios ya que son muy costosos y posiblemente no son lo
suficientemente efectivos. Por otro lado, la información generada en el presente estudio
permite suponer que un manejo exclusivamente ambiental no resultará viable en una
región donde el mejor momento para la transmisión del parásito coincide con Ia época
de mayor productividad de pasturas en los valles. Así, sólo se puede proponer un plan
de dosificación de los animales en las épocas y con el tipo de antiparasitario más
adecuados y sugerir un programa de rotación de potreros que minimice el contacto
entre el ganado y las metacercarias y entre los caracoles y los miracidios. De esta forma
se intenta reducir los daños hepáticos en los animales y la disponibilidad de larvasinfectantes en el ambiente.
Movimiento del ganado. Entre los meses de octubre y marzo se sugiere
encerrar a los animales en un potrero con mallín durante el lapso de a lo sumo 2 meses
(ya que la probabilidad de que los caracoles se infecten es baja en esa situación). Luego
dejar descansar el ambiente hasta la primavera siguiente, momento en que se podría
suponer que los caracoles infectados habrán muerto durante el invierno y si el clima fue
adverso las metacercarias viables representarán un riesgo bajo.
Si fuese necesario mantener a los animales un tiempo mayor de 2 meses en un
mismo potrero, entonces sería conveniente reducir Ia carga ganadera.
Sería recomendable evitar que los animales accedan a los ambientes con
pequeñas lagunas, ya que estos son los de mayor riesgo para el ganado. De no ser
167

posible, deberia administrarse una dosis de antihelmíntico a base de Triclabendazol
justo antes de permitirles el acceso. Si este esquema se repitiera en forma sistemática
cada vez que los animales fuesen a permanecer en ambientes de alto riesgo, se
impediría la eliminación de huevos al ambiente y la consiguiente infección de loscaracoles.
Quimioterapia. Con respecto a los antiparasitarios, sería recomendable
estudiar si existe un efecto de resistencia a los mismos o algún problema en las
estrategias de dosificación. Se han documentado diversos factores que seleccionan la
resistencia, entre los que pueden mencionarse el uso frecuente de los antihelmínticos
sin discriminar entre animales parasitados y sanos, la utilización de una misma droga
durante largos períodos y la subdosificación (Geerts et al. 1997). En principio y a partir
de los resultados obtenidos en esta tesis, los antiparasitarios a base de Closantel
parecerían no ser muy efectivos contra las formas inmaduras que alcanzan la vesícula
dentro de las 7 semanas de vida media de Ia droga.
Las dos principales drogas en el mercado, específicas contra F. hepatica, son el
Triclabendazol y el Closantel. La primera elimina todos los estadios larvales y tiene un
efecto residual que no supera la semana. La segunda sólo mata gusanos que están por
alcanzar la madurez sexual pero tiene una vida media mayor. Sin embargo, un análisis
teórico de la evolución de la infección dentro del hospedador definitivo, permite suponer
que no existen diferencias importantes en cuanto a los efectos de ambos tipos de
drogas:
1.- Un grupo de animales con infecciones de distintos tiempos es tratado con
Triclabendazol. Se espera que los individuos queden libres de parásitos y estén
protegidos contra nuevas infecciones durante 4-7 días. Luego, cualquier metacercaria
ingerida podrá desarrollarse sin ser afectada por la droga.
2.- Un grupo de animales con infecciones de distintos tiempos es tratado con
Closantel. Se espera que los individuos queden libres de parásitos adultos, y que las
larvas de menos de 6 semanas de vida que ya se encontraban dentro del hospedador,
vayan muriendo a medida que superen la 6ta semana postinfección (momento en que
alcanzan los ductos biliares). De esta forma, todos los parásitos que estaban dentro del
animal al momento de tratarse con el antiparasitario serán eliminados eventualmente,
pero los daños hepáticos producidos serán mayores al permitir la evolución de los
juveniles hasta haber alcanzado los conductos biliares. Las características de esta droga
permiten pensar que las metacercarias ingeridas una semana o 10 días después de Ia
aplicación podrán desarrollarse sin ser afectadas por el antiparasitario, ya que para el
momento en que alcancen las 6 semanas, la droga ya no estará surtiendo efecto.
A partir de este razonamiento se sugiere utilizar Triclabendazol en los momentos
que se sospeche que el ganado pueda estar infectado con parásitos de distinto tiempos
168

de vida, y Triclabendazol o Closantel siempre que se sepa que las infecciones son todasmaduras.
Una desparasitación en abril antes del invierno, resultaría adecuada para
mejorar el estado sanitario de los animales antes de la época de mayor estrés. Debido a
que en ese momento los animales podrían estar parasitados con adultos (producto del
contacto con metacercarias a partir de enero) y larvas migrantes (infecciones contraídas
en marzo y abril) y suponiendo que no hay riesgo de infección en la invernada, seríarecomendable tratarlos con Triclabendazol.
Para la primavera podrían plantearse dos estrategias distintas. Si se sospecha o
se comprueba (mediante el diagnóstico coprológico de un grupo de animales) que el
tratamiento de otoño no fue 100% efectivo, sería conveniente realizar una dosificación
con Closantel o Triclabendazol en ese momento (octubre). Si se cree que los animales
están limpios, entonces podrían ser tratados a comienzos del verano con Triclabendazol,
para eliminar posibles infecciones contraídas a partir de metacercarias remanentes.
Los resultados de esta tesis indican que los manejos no deben realizarse en
forma flexible y que el estricto cumplimiento de las fechas pautadas es recomendable.
En áreas con altas prevalencias en ganado y en las que el efecto de los
tratamientos no es el esperado, podría ser conveniente aumentar el número de
tratamientos durante el primer año, para impedir períodos prolongados de eliminación
de huevos al ambiente (Parr & Gray 2000). La alternancia de drogas puede ser también
conveniente para demorar el efecto de resistencia (Geerts et al. 1997, Sangster 2001).
Finalmente, las fluctuaciones interanuales en el clima pueden anticipar o
demorar Ia entrada en actividad de los caracoles. Así, la evaluación del momento exacto
en el que se realice el tratamiento antiparasitario debería contemplar estos cambios en
Ia población del hospedador intermediario.
(jul-/ Q

APÉNDICE
FIJACIÓN DE MOLUSCOSDEAGUADULCE(adaptado al género Lymnaea sp.)
a) Cómo Anestesiarlos
Colocar a los caracoles en un recipiente con Nembutal al 0.05% en agua
corriente (1 pastilla en 1/2litro de agua o 0.1 de nembutal + 200ml de agua) durante
aproximadamente 12 horas (no dejarlos muchas más horas porque se mueren). Evitar
que el anestésico esté muy concentrado cuando se incorpora a los animales ya que se
contraen en vez de relajarse.
En ausencia de nembutal, puede utilizarse otro barbitúrico, siempre y cuando seconozcan o establezcan las condiciones adecuadas. Otra forma de anestesiarlos es
colocando a los caracoles en un recipiente con agua sin cloro (nunca destilada) y luego
agregar un par de cristales de mentol (la solución anestésica a base de nembutal es
preferida sobre este método).
Importante: si los anestésicos están muy concentrados los caracoles se contraerán en
vez de relajarse. Es importante observarlos durante los primeros minutos luego de
sumergirlos en el anestésico y en el caso de no obtenerse el efecto deseado, sacar a los
animales del recipiente con el anestésico de inmediato, ponerlos en agua nuevamente
para que se recuperen y probar con otras diluciones.
b) Cómo Matarlos
Una vez completamente relajados (con la región céfan-pedal fuera de la
conchilla y al tocarles el pie los animales no reaccionan) se sumerge a los caracoles en
agua a 70° C durante 40 segundos (o durante un tiempo proporcional al tamaño del
ejemplar -cerca de 45 segundos para planórbidos de aproximadamente 20mm de
diámetro, u otros moluscos de tamaño equivalente; cerca de 30 segundos para
planórbidos pequeños-). Este procedimiento se puede realizar individualmente
sosteniéndolos con una pinza, o en grupo dentro de un colador metálico. Luego de
transcurridos 40 segundo se los sumerge en agua fría. No sobrepasar la temperatura
adecuada ni el tiempo de inmersión, para evitar que se dañen los tejidos y se
perjudique la disección.
Sumergir el animal muerto en agua fría y luego desprenderlo de la conchilla
sosteniéndolo por la parte posterior de la cabeza (por detrás del pie) con una pinza
adecuada y empujando con suavidad para desprender la inserción del múscqu
170

columelar. A medida que se retira el animal, Ia conchilla es ocupada por el agua, y
facilita la liberación del cuerpo.
Guardar la conchilla perfectamente rotulada habiéndola dejado secar
previamente.
c) Técnicas moleculares
Si se desea conservar ADNde los caracoles para realizar técnicas moleculares
para su identificación, cortar el pie antes de fijarlos y conservarlo en alcohol 70%
(proceder en todo momento con guantes, para no contaminar la muestra).
Todos los elementos que se utilicen en este procedimiento deben estar
perfectamente lavados, autoclavados o Iimpiadoscon alcohol, según corresponda.
d) Cómo Fijarlos
Los animales ya sin su caparazón se sumergen en un fijador (puede ser Raillet
Henry). La cantidad de fijador no debe ser menor que 10 veces el volumen del material
a ser fijado. El fijador debe ser cambiado luego de 24hs y la cantidad de fijador puedeser entonces reducida.
Raillet-Henry: esta solución es fijadora y a la vez diafanizador, especialmente indicado
para especimenes de dimensiones reducidas.
Fórmula: Solución fisiológica 0.8% (CINa)........... .. 93 ml
Formolconcentrado ............................. .. 5 ml
Acido acético 2 ml
Dejar las partes blandas del animal por lo menos 24hs. en fijador antes de
proceder a disecar. El material puede conservarse por tiempo indefinido en el fijador. Si
hay urgencia por hacer Ia disección, realizar la fijación en estufa a 40-45o C durante12hs.
Comentario: Ante Ia imposibilidad de utilizar un relajante adecuado, proceder de Ia
siguiente manera: tomar con una pinza un espécimen que se encuentre moviéndose
libremente, con la cabeza y el pie expuestos, teniendo mucho cuidado de que no se
retraiga durante la manipulación. Manteniendo la abertura de la conchilla hacia arriba,
sumergir gradualmente el molusco en agua caliente, como ya se describió. Mantener la
conchilla inmersa en el agua hasta casi el nivel de apertura durante los primeros 15
segundos, sumergiendo luego el espécimen completamente durante el tiempo restante.
171

Proseguir conforme a las recomendaciones del protocolo. Los animales fijados sin previa
relajación presentan desventajas para un buen estudio anatómico.
FIJACIÓN DE TREMATODES LARVALES DE CARACOLES
Colocar al caracol infectado en un recipiente pequeño con agua corriente sin
cloro bajo luz artificial para favorecer la emisión de cercarias. Cuando se registra
emisión proceder de Ia siguiente manera: sacar el caracol y ponerlo en un nuevo
recipiente. Utilizar el agua con cercarias en el recipiente original para fijar las larvas.
Calentar formol al 10% en un tubo de ensayo a baño María hasta el primer
hervor y volcarlo en el recipiente con agua que contiene las cercarias recuperadas de los
caracoles. Guardar en esa misma solución hasta su posterior determinación.
Si se van a utilizar las cercarias para realizar PCR volcarlas en alcohol 70% frío
en vez de proceder como se acaba de detallar.

AbdeI-Rahman S.M., O'Reilly K.L. & Malone J.B. 1998. Evaluation of a diagnostic
monoclonal antibody-based capture enzyme-Iinked immunosorbent assay for
detection of a 26- to 28-kd Fascíola hepatica coproantigen in cattle. American
Journal of Veterinary Research, 59 (5): 533-537.
Abrous M., Rondelaud D. & Dreyfuss G. 1996. Paramphistomum daubneyí and Fascíola
hepatica: the effect of dual infection on prevalence and cercarial shedding in
preadult Lymnaea g/abra. Journal of Parasitology, 82(6): 1026-1029.
Ahmed M. & Raut S.K. 1991. Influence of temperature on the growth of the pestiferous
land snail Achatína fu/ica (Gastropoda: Achatinídae). Walkerana, S: 33-62.
Alcaíno H., Vega F. & Gorman T. 1993. Epidemiología de la fasciolasis hepática en la VII
Región, Chile. Parasitología al día, 17: 99-106.
Ale D., Merciai G. & Gil Echeverría M. 2000. Fascio/a hepatica. Intensificación de su
búsqueda en pacientes con eosinofilia y epidemiología positiva. En: Resumenes
del III Congreso Argentino de Parasitología. Mar del Plata: Sociedad Argentina de
Parasitología; Pp. 421.
Anderson N., Luong T.T., Vo N.G., Bui K.L., Smooker P.M. & Spithill T.W. 1999. The
sensitivity and specificity of two methods for detecting Fascio/a infections in cattle.
Veterinary Parasitology 83, 15-24.
Andrews S.J. 1999. The life cycle of Fasciola hepatica, Chapter 1, 1-29. En: Fascio/osis,
ed. J.P.Da|ton, Dublin City University, Ireland. CABIPublishing.
Angles R., Strauss W., Ramirez S., Esteban J.G. & Mas-Coma S. 1997. Human
fascioliasis in Bolivia: coprological surveys in different provinces of the
Department of La Paz. Research and Reviews in Parasitology, 57233-37.
Apt W., Aguilera X., Vega F., Alcaíno H., Zulantay I., Apt P., González V., Retamal C.,
Rodríguez J. & Sandoval J. 1993. Prevalencia de fascioliasis en humanos, caballos,
cerdos y conejos silvestres en tres provincias de Chile. Boletín de la Oficina
Sanitaria Panamericana, 115(5): 405-414.
173

Atlas de Suelos de la República Argentina, 1985. Tomo I, Provincia de Chubut.
Secretaría de Agricultura, Ganadería y Pesca / INTA.
Aziz A. & Raut S.K. 1996. Thermal effect on the life-cycle parameters of the medically
important freshwater snail species Lymnaea (Radix) luteola (Lamarck). Memorias
do Instituto Oswaldo Cruz, 91(1): 119-128.
Bacigalupo J. 1934. Distomatosis por Fasciola hepatíca. Su ciclo evolutivo en la
República Argentina. Ed. A. Lopez, Buenos Aires, Argentina.
Bardie J., Nari A. & Cardozo H. 1978. Correlación entre niveles de infección de Fasciola
hepatíca y contaje de huevos en ovinos. Veterinaria, 14(68): 143-154.
Bargues M.D. & Mas-Coma S. 1997. Phylogenetic analysis of Lymnaeid snails based on
185 rDNAsequences. Molecular Biology and Evolution, 14(5): 569-577.
Basch P.F. 1969. Coty/urus Iutzí sp. N. (Trematoda: Strigeidae) and its life cycle. The
Journal of Parasitology, 55(3): 527-539.
Behm C.A. & Sangster N.C. 1999. Pathology, pathophysiology and clinical aspects, 185
224. En: Fasciolosis, ed. J.P.Dalton, Dublin City University, Ireland. CABI
Publishing.
Bendezú P.B. & Landa H. 1973. Distomatosis hepática. Epidemiología y Control. IVITA,
Universidad Nacional Mayor de San Marcos, 14: 1-32.
Black N.M. 1972. The possible influence of liver fluke infestation on milk quality. The
Veterinary Record, January: 71-72.
Blackwell P.G. 2001. Heterogeneity, Patchiness and Correlation of Resources. Reserch
Report, Department of Probability and Statistics, University of Sheffield, U.K.,
19p.
Bonino N. 1986a. La liebre europea I. Aspectos bio-ecológicos e importancia económica.
Series Folletines Fauna Silvestre INTA,Argentina, 4pp.
Bonino N. 1986b. La liebre europea II. La liebre como problema —Métodos de control.
Series Folletines Fauna Silvestre INTA,Argentina, 4pp.
174

Boray J.C. & Pearson LG. 1960. The anthelmintic efficiency of tetrachIorodifluoroethane
in sheep infested with Fascio/a hepatica. The Australian Veterinary Journal,
august: 331-336.
Boray J.C. 1964. Studies on the ecology of Lymnaea tomentosa, the intermediate host
of Fasciola hepatica. I. History, geographical distribution and environment.
Australian Journal of Zoology, 12(2): 217-230.
Boray J.C. 1967. Host-Parasite relationship between Iymnaeid snails and Fasciola
hepatica. In: The reaction of the host to parasitism. Veterinary Medical Review,132-140.
Boray J.C. 1969. Experimental fascioliasis in Australia. Advances in Parasitology, 7: 95
210.
Boray J.C. 1981. Fasciolasis and other trematode infections. I. Recent advances in
research on fasciolasis and other trematodes of animals. In: W. Slusarski (Editor),
Review of Advances in Parasitology. PWN-Polish Scientific Publishers, Warsaw,
Poland, pp.317-339.
Boray J.C. 1985. Flukes of domestic animals. In: Parasites, pests and predators. S.M.
Gaafar, W.E. Howard & R.E. Marsh (eds.). World Animal Science, Elsevier.
Brown D.S. 1978. Pulmonate mollusks as intermediate hosts for digenetic trematodes.
En: Pulmonates, Vol. 2A. Systematics, evolution and ecology. V. Fretter & J.
Peake editores. Academic Press, London.
Brunsdon R.V. 1980. Principles of helminth control. Veterinary Parasitology, 6:185-215.
Buchon P., Cuenca H., Quiton A., Camacho A.M. Bi Mas-Coma S. 1997. Fascioliasis in
cattle in the human high endemic region of the Bolivian Altiplano. Research and
Reviews in Parasitology, 57 (2): 71-83.
Bush A.0., Lafferty K.D., Lotz J.M. & Shostak A.W. 1997. Parasitology meets ecology on
its own terms: Margolis et. al revisited. Journal of Parasitology, 83 (4): 575-583.
Cabrera A.L. 1976. Regiones fitogeográficas argentinas. Parodi L.R. (ed.), Enciclopedia
argentina de agricultura y jardinería, 2a edición, 2(1). Editorial Acmé, Buenos
Aires. Pp. 1-85
175

Cañete R., Yong M., Sánchez J., Wong L. & Gutiérrez A. 2004. Population dynamics of
intermediate snail hosts of Fascio/a hepatica and some environmental factors in
San Juan y Martinez Municipality, Cuba. Memorias do Instituto Oswaldo Cruz,
99(3): 257-262.
Carbajo A.E. 2003. Distribución espacio-temporal de Aedes aegypti (Diptera: Culicidae).
Su relación con el ambiente urbano y el riesgo de transmisión del virus dengue en
la Ciudad de Buenos Aires. Departamento de Ecología, Genética y Evolución,
Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Tesis para
optar al títqu de Doctor de Ia Universidad de Buenos Aires.
Castellanos Z.A. & Landoni N.A. 1981. Mollusca, Gasteropoda. Lymnaeidae. En Fauna de
Agua Dulce de la República Argentina, Profadu (CONICET),voI.15-fasc.5.
Castro E.R. 1973. Distomatosis de los rumiantes. “Saguaype”. Almanaque del Banco de
Seguros del Estado.
Castro E., Freyre A. & Hernández Z. 2000. Serological responses of cattle after
treatment and during natural re-¡nfection with Fascíola hepatica, as measured
with a dot-ELISAsystem. Veterinary Parasitology 90, 201-208.
Castro 0., Heinzen T. & Carballo M. 2001. Dinámica de infección de Lymnaea Viator con
Fascíola hepatica en condiciones naturales en Uruguay. Veterinaria (Montevideo),
36(142): 13-20.
Christensen N.O. & Nansen P. 1976. The influence of temperature on the infectivity of
Fascio/a hepatica miracidio to Lymnaea truncatula. The Journal of Parasitology,62: 698-701.
Claxton J.R., Sutherst J., Ortiz P. & Clarkson M.J. 1999. The effect of cyclic
temperatures on the growth of Fascio/a hepatica and Lymnaea viatríx. The
Veterinary Journal, 157, 166-171.
Claxton J.R., Zambrano H., Ortiz P., Amorós C., Delgado E., Escurra E. & Clarkson M.J.
1997. The epidemiology of fasciolosis in the Inter-Andean valley of Cajamarca,
Peru. Parasitology International, 46:281-288.
Combes C. 1982. Trematodes: antagonism between species and sterilizing effects on
snails in biological control. Parasitology, 84: 151-175.
176

Dargie J. 1987. The impact on production and mechanism of pathogenesis of trematode
infections in cattle and sheep. International Journal for Parasitology 17: 453-463.
Dawes B. 1968. The Trematoda. Cambridge University Press.
Dennis W.R., Stone W.M. & Swanson L.E. 1954. A new laboratory and field diagnostic
test for fluke ova in feces. Journal of American Veterinary Medical Association,124: 47-50.
De Pietri D.E. 1993. Dinámica de alteraciones por fuego y ganadería en un sistema
forestal del Parque Nacional Los Alerces, Chubut, Argentina. Laboratorio de
Ecología Ambiental, Facultad de Ciencias Exactas y Naturales, UBA. Tesis
Doctoral.
Díaz A., Li-Elías 0., Otero 0., García C. & Espino, A.M. 1998. Identificación, mediante
Western Blot, de inmunógenos de Fascio/a hepatica, reconocidos por los sueros de
ratas infectadas experimentalmente. Revista Cubana de Medicina Tropical,
50(1):12-17.
Diaz J., Piña B., Lastre M., Rivera L. & Perez 0. 1990. Fascioliasis humana epidémica.
Cuba 1983 VI. Estudio clínico de 40 niños del Hospital Provincial de Sagua la
Grande. Revista de Ia Sociedad Venezolana de Gastroenterología, 44 (4): 385388.
Dresden M.H. 1981. A sieving method for the collection of Schistosome eggs from
mouse intestines. Journal of Parasitology, 67(3), 450-452.
Duménigo B.E., Espino A.M. & Finlay C.M. 1996. Detection of Fascio/a hepatica antigen
¡n cattle faeces by a monoclonal antibody-based sandwich immunoassay.
Research in Veterinary Science 60, 278-279.
Duménigo B.E., Espino A.M., Finlay C.M. 8:.Mezo, M. 2000. Kinetics of antibody-based
antigen detection in serum and faeces of sheep experimentally infected with
Fascio/a hepatica. Veterinary Parasitology, 89: 153-161.
Durand P., Pointier J.P., Escoubeyrou K., Arenas J.A., Yong M., Amarista M., Bargues
M.D., Mas-Coma S. & Renaud F. 2002. Occurrence of a sibling species complex
within neotropical Iymnaeids, snail intermediate hosts of fascioliasis. Acta Tropica,83: 233-240.
177

Dwinger R.H., Le Riche P.D. & Kühne G.I. 1982. Fascioliasis in beef cattle in north-west
Argentina. Trop. Anim. Hlth. Prod., 14: 167-171.
Eddi C. 1991. Distomatosis. Aspectos epidemiológicos y económicos de esta zoonosis.
Publicación del Instituto de Patobiología, Centro de Investigaciones en Ciencias
Veterinarias, INTA, 13 p.
Esch G.W. & Fernández J.C. 1993. A Functional Biology of Parasitism. Ecological and
evolutionary implications, Chapman and Hall, 337pgs.
Esteban J.G., Flores A., Aguirre C., Strauss W., Angles R. & Mas-Coma S. 1997.
Presence of very high prevalence and intensity of infection with Fasciola hepatíca
among Aymara children from the Northern Bolivian Altiplano. Acta Tropica, 66:114.
Fernandez J. & Esch G.W. 1991. Effect of parasitism on the growth rate of the
pulmonate snail He/isoma anceps. Journal of Parasitology, 77(6): 937-944.
Fleiss J.L. 1981. Statistical Methods for Rates and Proportions. John Wiley & Sons,Estados Unidos.
Flux J.E.C. & Angermann R. 1990. The hares and Jackrabbits. In: J.A. Chapman and
J.E.C. Flux (eds) Rabits, Hares and Pikas: status survey and conservation action
plan: chapter 4, 61-94. International Union for Conservation of Nature and
Natural Resources, Gland, Switzerland.
Fuentes M.V., Coello J.R., Bargues M.D., Valero M.A., Esteban J.G., Fons R. & S. Mas
Coma. 1997. Small mammals (Lagomorpha and Rodentia) and fascioliasis
transmission in the Northern Bolivian Altiplano endemic zone. Research and
Reviews in Parasitology, 57 (2): 115-121.
Fuentes M.V., Malone J.B. & Mas-Coma S. 2001. Validation of a mapping and prediction
model for human fasciolosis transmission in Andean very high altitude endemic
areas using remote sensing data. Acta Tropica, 79: 87-95.
Geerts S., Coles G.C. & Gryseels B. 1997. Anthelmintic resistance in human helminths:
learning from the problem with worm control in Iivestock. Parasitology Today, 13
(4): 149-151.
Ginetsinkaya T.A. 1988. Trematodes, their Life Cycles, Biology and Evolution. Amerind
Publishing Co., Pvt. Ltd, New Delhi, 559 pp.
178

Giráo E.S. & Ueno H. 1994. Técnica Quatro Tamices Metálicos. En: Ueno, H.; Gongalves,
P.C. Manual para diagnóstico das he/mintoses de ruminantes. 3 ed. Japan
International Cooperation Agency and Universidad Federal do Rio Grande do Sul,
P0rto Alegre, Brazil, p.62.
Gordis L. 2000. Epidemiology; W.B. Saunders Company, 2"“ ed., 307 pp.
Gonzalez-Lanza C., Manga-Gonzalez Y., Del Pozo Carnero P. & Hidalgo Argüello R. 1989.
Dynamics of elimination of the eggs of Fasciola hepatica (Trematode, Digenea) in
the faeces of cattle in the Porma Basin, Spain. Veterinary Parasitology, 34:35-43.
Graczyk T.K. & Fried B. 1999. Development of Fasciola hepatica in the intermediate
host, pp. 31-46. En: Fascio/osís, ed. J.P. Dalton, Dublin City University, Ireland.
CABI Publishing.
Graham C.S., Brodie S.B. & Weller P.F. 2001. Imported Fasciola hepatica infection in the
United States and Treatment with Triclabendazole. Clinical Infectious Diseases,33: 1-6.
Grock R., Morales G., Vaca J.L. & Mas-Coma S. 1998. Fascioliasis in sheep in the human
high endemic region of the northern Bolivian Altiplano. Research and Reviews in
Parasitology, 58 (2), 95-101.
Gürtler H, Ketz H.A., Kolb E., Schroder L. & Seidel H. 1979. Las heces. En: Kolb E (ed)
Fisiología Veterinaria: 377. Editorial Acribia, Zaragoza, España.
Gutierrez A., Perera G., Yong M., Sánchez J. & Wong L. 2000. Life-history traits of
Fossaria cubensis (Gastropoda: Lymnaeidae) under experimental exposure to
Fasciola hepatíca (Trematoda: Digenea). Memorias do Instituto Oswaldo Cruz,
95(5): 747-752.
Haas W., Haberl B., Kalbe M. & Corner M. 1995. SnaiI-host-finding by miracidia and
cercariae: chemical host cues. Parasitology Today, 11(12): 468-472.
Hall L.S., Krausman P.R. & Morrison M.L. 1997. The habitat concept and a plea for
standard terminology. WildlifeSociety Bulletin, 25: 173-182.
Hansen J. & Perry B. 1994. The Epidemiology, Diagnosis and Control of Helminth
Parasites of Ruminants. ILRAD,Nairobi, Kenia.
179

Hanski I. & Gilpin M.E. 1997. Metapopulation biology: ecology, genetics and evolution.
Academic Press, London.
Hastings I.M. 2001. Modelling parasite drug resistance: lessons for management and
control strategies. Tropical Medicineand International Health, 6: 883-890.
Hillyer G.V. & Apt W. 1997. Food-borne trematode infections in the Americas.
Parasitology Today, 13 (3): 87-88.
Hillyer G.V., Soler de Gaianes M., Buchón P. & Bjorland J. 1996. Herd evaluation by
enzyme-Iinked immunosorbent assay for the detection of Fascio/a hepatica
infection in sheep and cattle from the Altiplano of Bolivia. Veterinary Parasitology
61, 211-220.
Hodasi J.K. 1976. The effects of low temperature on Lymnaea truncatu/a. Z.Parasitenkd. 48: 281-286.
Hope Cawdery M.J, Strickland K.L., Conway A. & Crowe P.J. 1977. Production effects of
liver fluke in cattle. I. The effects of infection on live weight gain, food intake and
food conversion efficiency in beef cattle. British Veterinary Journal, 133: 145-159.
Hope Cawdery M.J. 1984. Review of the economic importance of fascioliasis in sheep
and cattle. Irish Veterinary News (September): 14-22.
Hubendick B. 1951. Recent Lymnaeidae: their morphology, taxonomy nomenclature anddistribution. K. Svenska Vetensk. Akad. Hand 3: 1 - 223.
Hunter R.D. 1975. Growth, fecundity and bioenergetics in three population of Lymnaea
palustris in upstate New York. Ecology, 56: 50-63.
Hurtrez-Boussés S., Meunier C., Durand P. & Renaud F. 2001. Dynamic of host-parasite
interactions: the example of population biology of the liver fluke (Fascio/a
hepatica). Microbes and Infection, 3: 841-849.
Ibarra F., Montenegro N., Vera Y., Boulard Ch., Quiroz H., Flores J. & Ochoa P. 1998.
Comparison of three ELISA tests for seroepidemiology of bovine fascioliasis.
Veterinary Parasitology 77: 229-236.
180

Ibarra F., Montenegro N., Vera Y., Castillo R., Hernández A. & Ochoa P. 2002.
Comparative efficacy of an experimental fasciolicide, triclabendazole and closantel
in cattle naturally infected with Fasciola hepatíca. Veterinaria México 33(3): 237245.
Itagaki T. 1987. Influence of food on the growth and fecundity of Lymnaea ollu/a, the
intermediate host of the Iiver fluke. The Japanese Journal of Parasitology 36(1):30-35.
Kaczorkiewicz A.J. 1983. Distomatosis en la provincia del Neuquén. Revista Médica
Veterinaria, 64 (5/6): 354-356.
Keegan P.S. & Trudgett A. 1992. Fasciola hepatica in the rat: immune responses
associated with the development of resistance to infection. Parasite Immunology,14: 657-669.
Kendall S.B. 1950. Snail hosts of Fasciola hepatica in Britain. Journal of Helminthology,
24 (1-2): 63-74.
Kendall S.B., Sinclair I.J., Everett G. & Parfitt J.W. 1978. Resistance to Fasciola hepatíca
in cattle. I. Parasitological and serologicai observations. Journal of Comparative
Pathology, 88: 115-122.
Kleiman F., González N., Rubel D. & Wisnivesky C. 2004. Fasciola hepatíca (Linnaeus,
1758) (Trematoda, Digenea) en Iiebres europeas (Lepus europaeus, Pallas 1778)
(Lagomorpha, Leporidae) en la región Cordillerana Patagónica, Chubut, Argentina.
Parasitología Latinoamericana, 59: 68-71.
Krausman P.R. 1999. Some basic principles of habitat use. En: Grazing behavior of
livestock and wildlife. Launchbaugh K.L., Mosley J.C. & Sanders K.D. (eds.),
Pacific Northwest Range Short Course, Universidad de Idaho, Idaho, EstadosUnidos.
Krebs C.J., Boonstra R, Nams V., O’Donoghue M., Hogdes K.E. & Boutin S. 2001.
Estimating snowshoe hare population density from pellet plots: a further
evaluation. Canadian Journal of Zoology, 79: 1-4.
Kuris A. 1990. Guild structure of Iarval Trematodes in molluscan hosts: prevalence,
dominance and significance in competition. En: Parasite communities: patterns
181

and processes. Esch G.W., Bus A.O. & Aho J.M. (eds.), London, New York,
Chapman and Hall.
Lara S.I.M., Müller G., Fernández F.G. & Silveira Júnior P. 1988. Biología de Lymnaea
viatrix, 0rb., 1835. II. Desenvolvimento, Postura e Longevidade. Arquivo
Brasileiro de Medicina Veterinaria e Zootecnia, 40(2): 101-113.
Lee C.G., Cho S.H. & Lee C.Y. 1995. Metacercarial production of Lymnaea viridis
experimentally infected with Fascio/a hepatica. Veterinary Parasitolgoy 58: 313318.
Lee C.G., Kim S.K. & Lee C.Y. 1994. Rapid growth of Lymnaea viridis, the intermediate
host of Fasciola hepatica, under laboratory conditions. Veterinary Parasitology,51: 327-331.
Lim H. & Heyneman D. 1972. Intramolluscan inter-trematode antagonism: a review of
factors influencing the host-parasite system and its possible role in biological
control. Advances in Parasitology, 10: 191-268.
Liu S.S., Zhang G.M. & Zhu J. 1995. Influence of Temperature Variations on Rate of
Development in Insects: Analysis of Case Studies from Entomological Literature.
Annals of the Entomological Society America 88(2): 107-119.
Lombardero 0., Moriena R., Racioppi 0., Quiroz L. 1979. Bionomía de Lymnaea sp. en la
provincia de Corrientes. Veterinaria (Corrientes), 2:5-20.
Luzón-Peña M., Rojo-Vázquez F.A. & Gómez-Bautista M. 1994. The overwintering of
eggs, intramolluscal stages and metacercariae of Fascio/a hepatica under the
temperatures of a Mediterranean area (Madrid, Spain). Veterinary Parasitology,55: 143-148.
Malek E.A. 1985. Fascioliasis and its snail hosts. En: Snails host of Schistosomiasis and
other snail-transmitted diseases, PAHO,ed., Washington. Chapter IV, 108-128.
Malone J.B., Loyacano A., Armstrong D.A. & Archbald L.F. 1982. Bovine fascioliasis:
economic impact and control in Gulf Coast cattle based on seasonal transmission.
The Bovine Practitioner, 17:126-133.
182

Malone J.B., Loyacano A.F., Hugh-Jones M.E. & Corkum K.C. 1984. A three year study
on seasonal transmission of Fascíola hepatica of cattle in Louisiana. Preventive
Veterinary Medicine, 3:131-141.
Malone J.B. & Craig T.M. 1990. Cattle Iiver flukes: risk assessment and control.
Compendium on Continuing Education for the Practicing Veterinarian, 12:747-754.
Malone J.B. 1994. The Iandscape epidemiology of fasciolosis: geographic determinants
of disease risk. In: J.C. Boray (Ed.), Immunology, pathobiology and control of
fasciolosis. Round Table Conference, ICOPAVIII, Izmir, pp. 65-81.
Malvárez A.I., Kandus P. & Carbajo A. 2004. Distribución regional de los turbales en
Patagonia (Argentina y Chile). En: Los Turbales de la Patagonia. Bases para su
inventario y la conservación de su biodiversidad. Blanco D.E. & Balze V.M.
editores, pp. 23-32. Wetlands International, Buenos Aires, Argentina.
Mas-Coma S., Rodriguez A., Bargues M.D., Valero M.A., Coello J.R. & R. Angles 1997.
Secondary reservoir role of domestic animals other than sheep and cattle in
fascioliasis transmission in the Northern Bolivian Altiplano. Research and Reviews
in Parasitology, 57 (1): 39-46.
Mas-Coma S. 1998. Human fascioliasis in Europe and Latin America. En: Infectious
Diseases and Public Health. Mario Angelico and Giovanni Rocchi, Editors. Balaban
Publishers, Philadelphia.
Mas-Coma M.S., Esteban J.G. & Bargues M.D. 1999. Epidemiology of human fascioliasis:
a review and proposed new classification. Bulletin of the World Health
Organization, 77 (4): 340-346.
Matos M.L. 2001. Análisis sistemático molecular de Iymnaeidos de la República
Argentina. Tesis de licenciatura. Facultad de Ciencias Exactas y Naturales,
INGEBI, Universidad de Buenos Aires. Director: Dr. Alejandro Schijman.
Matteuci S.D. & Colma A. 1982. Metodología para el estudio de la vegetación. Secretaría
General de Ia Organización de los Estados Americanos, Programa Regional de
Desarrollo Científico y Tecnológico, Washington D.C., Estados Unidos.
Mattos M.J. & Ueno H. 1985. Manutencao de Lymnaea víatrix Orbigny, 1835 em
condicoes laboratoriais. A Hora Veterinaria, 5(26): 48-50.
183

Mattos M.J. & Ueno H. 1986. Relacao entre idade e susceptibilidade de Lymnaea
colume/la frente ás infeccoes experimentais por Fasciola hepatica. A Hora
Veterinaria, 6(33): 16-20.
Mattos M.J. & Ueno H. 1989. Susceptibilidade de Lymnaea viatríx e L. colume/Ia as
infeccoes experimentais com Fasciola hepatica. Pesq. Agropec. Bras. Brasilia,
24(5):615-622.
Maués da Serra-Freire N. & Nuernberg S. 1992. Geopolítical dispersion of the
occurrence of Fasciola hepatica in the State of Santa Catarina, Brazil. Memorias do
Instituto Oswaldo Cruz, 87: 263-269.
Mc Kown R.D., Ridley R.K. & Kennedy G.A. 2000. The hispid cotton rat (Sigmodon
hispídus) as an experimental host for the bovine fluke (Fasciola hepatica).
Veterinary Parasitology 87: 125-132.
Ménard A., Agouion A., L'Hostis M., Rondelaud D., Collard S. & Chauvin A. 2001.
Myocastor coypus as a reservoir host of Fasciola hepatíca in France. VeterinaryResearch 32: 499-508.
Ménard A. L'Hostis M., Leray G., Marchandeau S., Pascal M., Roudot N., Michel V. 81
Chauvin A. 2000. Inventory of wild rodents and Iagomorphs as natural hosts of
Fasciola hepatíca on a farm located in a humid area ¡n Loire Atlantique (France).
Parasite, 7: 77-82.
Misra T.K. & Raut S.K. 1993. Dynamic of population-dynamics in a medically important
snail species Lymnaea (Radix) luteo/a (Lamarck). Memorias do Instituto Oswaldo
Cruz, 88(3): 469-485.
Moriena R.A., Racioppi 0., Alvarez J.D. & Lombardero O.J. 2001. Fasciola hepatíca y
otros trematodes en bovinos: prevalencia según decomisos en plantas faenadoras
en la Provincia de Corrientes (Argentina). Veterinaria Argentina, XVIII(177): 493498.
Mulcahy G., Joyce P. 8: Dalton J.P. 1999. Immunology of Fasciola hepatica infection,
Chapter 10, 341-375. En: Fasciolosís, ed. J.P.Dalton, Dublin City University,
Ireland. CABI Publishing.
184

Müller G., Lara S.I.M., Silveira P. & Antunes P. 1998. Acompanhamento Iaboratorial do
ciclo biologico de Lymnaea viatrix, hospedeiro intermediario de Fasciola hepatíca.
Revista brasilera de agrociencia, 4(3): 172-176.
Naranjo García E. 2003. Moluscos continentales de México: dulceacuícolas. Revista de
BiologíaTropical 51(3): 495-505.
Nari A. & Cardozo H. 1976. Prevalencia y distribución geográfica de la fasciolosis
hepato-biliar en bovinos de carne del Uruguay. Revista Veterinaria, 13(63): 1116.
Nari A., Cardozo H., Acosta D., Solari M. & Petraccia C. 1983. Efecto de la temperatura
en el desarrollo de Fascio/a hepatíca en su huesped intermediario Lymnaea viatrix
D Orbigny. Veterinaria, 19: 36-39.
Nelson G.5. 1990. Microepidemiology, the key to the control of parasitic infections.
Transactions of the Royal Society of Tropical Medicine and Hygene, 84: 3-13.
Norman G.R. & Streiner D.L. 1996. Bioestadística. Mosby-Doyma Libros, España.
Novaro A.J., Capurro A.F., Travaini A., Funes M.C. & Rabinovich J.E., 1992. Pellet-count
sampling based on spatial distribution: a case study of the European hare in
Patagonia. Ecología Austral, 2:11-18.
Odening K. 1971. Der Grosse Leberegel und seine Verwandten. Die Neue Brehm
Bücherei. A. Ziemsen Verlag-Wittenberg Lutherstadt. Pp. 5-127.
Olaechea F.V. 1995. Fasciola hepatíca. Sistema Regional de Salud Animal, Estación
Experimental Agropecuaria Bariloche, Unidad de Salud Animal.
OIIerenshaw C.B. 1958. Climate and Liver Fluke, Agriculture, 65, 231-5
OIIerenshaw C.B. 1971a. The influence of climate on the life cycle of Fascio/a hepatíca in
Britain with some observations on the relationship between climate and the
incidence of fascioliasis in The Netherlands. Symposium of The Parasitological
Department of the Central Diesgeneeskunder Institut, Lelystad, 41-63.
OIIerenshaw C.B. 1971b. Some observations on the epidemiology of Fascioliasis in
relation to the timing of molluscicide applications in the control of the disease. The
Veterinary Record, February.
185

OMS, Organización Mundial de Ia Salud, 1994. Medios auxiliares para el diagnóstico de
las parasitosis intestinales.
Oviedo J.A., Bargues M.D. & Mas-Coma S. 1996. The intermediate snail host of Fasciola
hepatica in the Mediterranean island of Corsica. Research and Reviews in
Parasitology, 56(4): 217-220.
Paraense W.L. 1976. Lymnaea viatríx: a study of topotypic specimens (Mollusca:
Lymnaeidae). Revista Brasileira de Biologia, 36(2):419-428.
Paraense W.L. 1982. Lymnaea víatrix and Lymnaea co/umeI/a in the neotropical region:
a distributional outline. Memorias do Instituto Oswaldo Cruz, Río de Janeiro,
77(2): 181-188.
Paraense W.L. 1984. Lymnaea diaphana: a study of topotypic specimens (Pulmonata:
Lymnaeidae). Memorias do Instituto Oswaldo Cruz, 79(1): 75-81.
Paraense W.L. 2003. Planorbidae, Lymnaeidae and Physidae of Peru (Mollusca:
Basommatophora). Memorias do Instituto Oswaldo Cruz, 98(6): 767-771.
Parmesan C., Root T.L. & Willig M.R. 2000. Impacts of extreme weather and climate on
terrestrial biota. Bulletin of the American Meteorological Society, 81(3): 443-450.
Parr S.L. & Gray J.S. 2000. A strategic dosing scheme for the control of fasciolosis in
cattle and sheep in Ireland. Veterinary Parasitology, 88, 187-197.
Parkes J. 2001. Methods to monitor the density and impact of hares (Lepus europaeus)
in grassland in New Zeland. DOC Science Internal Series 8, Department of
Conservation, New Zeland.
Percedo M.I., Mesa J.J., Leyva D. & Barroso A. 1987. Infestación por Fasciola hepatica
en bovinos jóvenes en condiciones semiintensivas de producción. Revista de Salud
Animal, 9: 71-76.
Petraglia A.A. 1954. Parasitosis humana por Fasciola hepatíca. Primer caso que se
describe en el Noreste Argentino. Actas y Trabajos de la Asociación Argentina para
el Estudio de Enfermedades Transmisibles, 3 (2): 47-48.
186

Precht H., Christopherson J., Hensel H. & Larcher A. 1973. Temperature and life.
Springer-Verlag, Berlin, XI: 315pp.
Prepelitchi L. 2002. Ciclo evolutivo parcial de dos especies de Echinostomatidae
(Trematoda: Digenea) halladas en Lymnaea viatrix de Cholila, Chubut. Tesis de
Licenciatura. Facultad de Ciencias Exactas y Naturales, Universidad de Buenos
Aires, Argentina.
Prepelitchi L., Kleiman F., Pietrokovsky S.M., Moriena, R.A., Racioppi 0., Alvarez J. &
Wísnivesky-Colli C. 2003. First report of Lymnaea colume/la Say, 1817
(Pulmonata: Lymnaeidae) naturally infected with Fascio/a hepatica (Linnaeus,
1758) (Trematoda: Digenea) in Argentina. Memorias do Instituto Oswaldo Cruz,
98(7): 889-891.
Presidente P.J.A., Mc Craw B.M. & Lumsden J.H. 1974. Early pathological changes
associated with Fascio/a hepatica infection in white-tailed deer. Canadian Journal
of Comparative Medicine 38: 271-279.
Price T.A., Tuazon C.U. & Simon G.L. 1993. Fascioliasis: case reports and review.
ClinicalInfectious Diseases, 17: 426-430.
Poulin R. 1998. Evolutionary Ecology of Parasites. Chapman and Hall, New York.
Rabinovich J.E.1978. Ecología de poblaciones animales. Organización de los Estados
Americanos. Monografía N° 21, 110 p.
Raut S.K., Rahaman M.S. & Samanta S.K. 1992. Influence of temperature on survival,
growth and fecundity of the freshwater snail Indop/anorbis exustus (Deshayes).
Memorias do Instituto Oswaldo Cruz, 87: 15-19.
Richards R.J., Bowen F.L., Essenwein F., Steiger R.F. & Büscher G. 1990. The efficacy of
triclabendazole and other anthelmintics against Fasciola hepatica in controlled
studies in cattle. The Veterinary Record, 126: 213-216.
Rickard L. 1992. Llama parasites. Large Animal Veterinarian, September-October: 6-13.
Rim H-J., Farag H.F., Sornmani S. & Cross J.H. 1994. Food-borne trematodes: ignored
or emerging? Parasitology Today, 10(6): 207-209.
187

Ritchie L.S. 1948. An ether sedimentation technique for routine stool examinations.
Bulletin of the U.S. Army Medical Department, 8, 326.
Roberts J.A. & Suhardono 1996. Approaches to the control of fasciolosis in ruminants.
International Journal for Parasitology, 26: 971-981.
Rognlie M.C., Dimke K.L., Potts R.S. & Knapp S.E. 1996. Seasonal transmission of
Fascio/a hepatíca in Montana, USA, with detection of infected intermediate hosts
using a DNA-based assay. Veterinary Parasitology, 65: 297-305.
Roig C. & Roig F. 2004. Consideraciones generales. En: Los Turbales de la Patagonia.
Bases para su inventario y la conservación de su biodiversidad. Blanco D.E. &
Balze V.M.editores, pp. 5-22. Wetlands International, Buenos Aires, Argentina.
Rossanigo C.E., Avila J.D., Vasquez R. & Sager R.L. 1983. Incidencia, distribución e
identificación del huésped intermediario de la distomatosis bovina en la provincia
de San Luis. Gaceta Veterinaria, 382: 739-746.
Rowan W.B. 1956. The mode of hatching of the egg of Fascio/a hepatica. Experimental
Parasitology, 5: 118-137.
Rubel D., Prepelitchi L., Kleiman F., Carnevale S. & Wisnivesky-Colli C. 2003. Estudio de
foco a partir de un caso humano de Fasciolosis en la localidad de Loncopué,
Neuquen, Argentina. XVI Congreso Latinoamericano de Parasitología, La Paz,
Bolivia.
Rumi A. & Hamman M.I. 1992. Spatial and seasonal trends of a natural population of
Biompha/aría occidenta/¡s in Northeastern Argentina. Memorias do Instituto
Oswaldo Cruz, 87(2): 181-186.
Samadi S., Roumégoux A., Bargues M.D., Mas-Coma S., Yong M. & Pointier J.P. 2000.
Morphologicalstudies of Iymnaeid snails from the human fascioliasis endemic zone
of Bolivia. Journal of Molluscan Studies, 66: 31-44.
Santa Cruz A.C.M. 1979. Comportamiento de Límnaea díaphana King en condiciones de
laboratorio. Veterinaria, II (3): 29-40.
Sangster N.C. 2001. Managing parasiticide resistance. Veterinary Parasitology, 98: 89109.
188

Sarmiento G. 1984. Los ecosistemas y la ecosfera. Blume Ecología, Barcelona.
Scott M.E. & Smith G., 1994. Parasitic and Infectious Diseases. Epidemiology and
Ecology. Academic Press, Inc. U.S.A.
Shimalov V.V. & Shimalov V.T. 2000. Findings of Fascio/a hepatica Linnaeus 1758, in
wild animals in Belorussian Polesye. Parasitology Research, 86: 342.
Shore García A. 1973. Manual de Laboratorio Clínico, Editorial Médica Panamericana.
Siegel S. & Castellan N.J. 1995. Estadística no paramétrica: aplicada a las ciencias de la
conducta. Editorial Trillas, México.
Silva J.R. Raty P & Bifani V. 1986. Evaluación de canales de liebre (Lepus europaeus
Pallas). Revista Argentina de Producción Animal, 6: 731-737.
Silva Santos I.C., Scaini C.J. & Rodríguez I.A.F. 1992. Myocastor coypus (Rodentia
Capromyidae) como reservatorio silvestre de Fascíola hepatica (Lineu, 1758).
Revista Brasilera de Parasitología Veterinaria, 1, 1: 27-30.
Sokal R.R. & Rohlf F.J. 1969. Biometry. Freeman and Company, San Francisco, Estados
Unidos.
Southwood T.R.E. 1977. Habitat, the templet for ecological strategies? Journal of Animal
Ecology, 46: 337-365.
Spratt D.M. & Presidente P.J.A. 1981. Prevalence of Fascio/a hepatíca infection in native
mammals in Southeastern Australia. Ajebak, 59: 713-721.
Stromberg B.E. & Averbeck G.A. 1999. The role of parasite epidemiology in the
management of grazing cattle. International Journal for Parasitology, 29: 33-39.
Tanaka K., Watanabe T., Miyamoto H.H.D., Yusa Y., Kiyonaga T., Kiyota H., Suzuki Y. &
Wada T., 1999. Density-dependent growth and reproduction of the apple snail,
Pomacea cana/¡culata: a density manipulation experiment in a paddy field.
Researches on Population Ecology, 41: 253-262.
Thornthwaite C.W. & Mather J.R. 1955. The water balance. Publications ín Climatology
VIII, (1):104 p. Drexel Institute of Technology, New Jersey USA.
189

Torgerson P. & Claxton J. 1999. Epidemiology and Control, pp. 113-149. En: Fasciolosis,
ed. J.P.Da|ton, Dublin City University, Ireland. CABIPublishing.
Utzinger J. & Tanner M. 2000. Microhabitat preferences of Biomphalaria pfeifferí and
Lymnaea nata/ensis in a natural and a man-made habitat in Southeastern
Tanzania. Memorias do Instituto Oswaldo Cruz, 95(3): 287-294.
Valero M.A., Marcos M.D. & Mas-Coma S. 1996. A mathematical model for the ontogeny
of Fasciola hepatica in the definitive host. Research and Reviews in Parasitology,
56(1): 13-20.
Valero M.A. & Mas-Coma S. 2000. Comparative infectivity of Fasciola hepatica
metacercariae from isolates of the main and secondary reservoir animal host
species in the Bolivian Altiplano high human endemic region. Folia Parasitologica;47: 17-22.
Venturini L.M., Led J.E., Valenzuela M.L., Radman N.E. & Fonrouge R.F. 1981. Cría de
Lymnaea Viator en laboratorio. Revista Médica Veterinaria (Buenos Aires), 62 (4).
Venturini L. & Fonrouge R. 1985. Lymnea Viator (Dórbigny1835): aspectos de su
biología en condiciones naturales. Revista Médica Veterinaria (Buenos Aires) 66:340-345.
Ward M.P., Lyndal-Murphy M. & Baldock F.C. 1997. Evaluation of a composite method
for counting helminth eggs in cattle faeces. Veterinary Parasitology, 73: 181-187.
Wisnivesky C. 2003. Control de las poblaciones parasitarias. En: Ecología y
Epidemiología de las Infecciones Parasitarias. Libro Universitario Regional (EULAC
GTZ) ed., Cartago, Costa Rica. Capítulo S, 111-140.
Woods C.A., Contreras L., Willner-Chapman G. & Whidden P. 1992. Myocastor coypus.
Mammalian Species. The American Society of Mammalogists, 389:1-8.
Wullschleger E. & Jokela J. 1999. Does habitat-specific variation in trematode infection
risks influence habitat distribution of two closely related freshwater snails?.
Oecología, 121: 32-38.
Yamaguti S. 1975. A synoptical review of life histories of digenetic trematodes of
vertebrates. Keigaku Publishing Co., Tokyo.
190

Yiima J.M. & Malone J.B. 1998. A geographic Information system forecast model for
strategic control of fasciolosls in Ethiopia. Veterinary Parasltology, 78: 103-127.
Zar J.H. 1999. Biostatistical Analysis. Fourth edition, Prentice-Hall, USA.
Zukowskl S.H., Wayne Wilkerson G. & Malone J.B. 1993. Fasclolosis in cattle in
Louisiana. II. Development of a system to use soil maps in a geographic
information system to estimate disease risk on Louisiana coastal marsh rangeland.
Veterinary Parasltology, 47: 51-65.
, Hifi/09"

![ELMINTI [metazoi]. Classificazione Phylum Platyhelminthes Classe Trematoda (vermi piatti) Sottoclasse Digenea Classe Cestoda (vermi a nastro) Sottoclasse](https://img.pdfslide.tips/doc/110x75/5542eb75497959361e8dee74/elminti-metazoi-classificazione-phylum-platyhelminthes-classe-trematoda-vermi-piatti-sottoclasse-digenea-classe-cestoda-vermi-a-nastro-sottoclasse.jpg)