Embed Size (px)
Citation preview

マウス精子にはウイルスを感知するToll like receptorが発現し、それは受精能を低下させる
誌名誌名 Journal of mammalian ova research = 日本哺乳動物卵子学会誌
ISSNISSN 13417738
巻/号巻/号 273
掲載ページ掲載ページ p. 136-143
発行年月発行年月 2010年10月
農林水産省 農林水産技術会議事務局筑波産学連携支援センターTsukuba Business-Academia Cooperation Support Center, Agriculture, Forestry and Fisheries Research CouncilSecretariat

J. Mamm. Ova Res. Vol. 27, 136-143,2010 136
Murine Sperm Expresses TolI胴 likeReceptor (TLR) Family that Responds to the Pathogens Released from Virus, and Decreases Fertilization Ability by the Stimuli
Toshihiro Mihara1, Keita Fukumoto1,
Tetsuji Okazaki1,2 and Masayuki Shimada1*
'Graduαte School of Biosphere Science, Hiroshima UniversifY, Hiroshima 739-8528, Japan 2Livesfock Research lnsfifufe, Oifa Prefectura/ Agriculture, Forestryαηd Fisheries Research
Center, Oita 879-7111, J.勾Dan
Abstract: Sperm express the TolI-like receptor (TLR)
family and have an innate immune function defending
against bacterial infection in semen. It has been known
that virus infection is also observed in semen with low
motility of sperm, suggesting that sperm are also able to
respond to virus infection. However, there is little
information about the expression and roles of TLRs that
recognize virus-released pathogens in semen. The
present study clθarly showed that sperm express
functional TLR9 that recognizes the unmethylated CpG
DNA sθquence. Using a specific antibody, we detected
the expression of TLR9 in sperm. However, the positivθ
band was disappeared by progesterone treatment that
induced the acrosome reaction. When sper斤1were
cultured with the TLR9 ligand, Spθrm motility significantly
decreased in a time-dθpendent manner. The treatment
of sperm with the TLR9 ligand affected the acrosomal
status and suppressed the BSA“induced capacitation of
sperm. Additionally, sperm were used for in vitro
fertilization and injection into the uterus of super-桐 ovulated
fema/e mice following pre-culture with the TLR9 ligand.
The fertilization rates were significantly suppressed both
in vitro and in vivo. From thesθresults, we conclude that
sperm express and have functional TLR9 that potentially
recognizθs virus infection in semen.
κey words: TolI綱 likereceptoにSperm,Virus infection,
Fertilization, Innate immune function
Received: March 17,2010 Accepted: April 16,2010 *To whom correspondence shou1d be addressed e-mail: [email protected]
Introduction
In the innate immune system, pathogens (known as
pathogenωassociated molecular patterns: PAMPs) are
recognized by the TolI-like receptor (TLR) family, which
is one of the pattern-recognition receptors (PPRs) [1]
Although the innate immune function has been known to
be dependent on immune cells, we have recently shown
that TLR4 is expressed on sperm, and recognizes
lipopolysaccharide (LPS), a Gram negative bacterial
factor, which results in the loss of sperm ability (motility,
apoptosis, acrosome reaction, fertilization rate) [2]
Additionally, sperm also express TLR2 that recognizes
Gram positive bacterial factor, a lipopeptide [2]. Similar
results were shown when mice sperm were cultured
with a TLR2 ligand, Pam3Cys-Seト(lys)4(Pam3Cys) as
well as LPS [2]. A significant negative correlation was
observed between the number of bacteria and sperm
motility, suggesting that TLR2 and TLR4 expressed on
sperm might play important roles in responses to
bacterial infection in semen.
The impact on male infertility of virus infection as well
as of bacterial infection has been reported. Virus DNA or
RNA are detected in semen with low sperm quality [3, 4].
The unmethylated CpG DNA sequence, single strand
RNA (ssRNA) and double stranded RNA (ds RNA) are
components of viruses that are recognized as
pathogens by immune cells [5]. In immune cells such
as macrophages, dendritic cells and natural killer cells,
virus infection is recognized by TしR3,TLR7, TLR8 and
TLR9 [6-8]. TLR3 recognizes dsRNA to induce the

secretion of proωinflammatory cytokines [6]. When
TLR7, TLR8 and TLR9 are stimulated by ssRNA or the
unmethylated CpG DNA sequence, the myeloid
differentiation factor 88 (MyD88) pathway is activated
and MyD88 interacts with 1し小receptor“associated
kinase (IRAK)-1, IRAK幽 4and TNF receptor-associated
factor (TRAド)6 [9]. This complex induces
phosphorylation and degradation of IK8, then NF叶(8is
transferred to nuclei, or activates interferon-regulatory
factor (IRF) 7 to induce target gene expressions such as
Infa or Infb [9-11]. The MyD88 dependent pathway is
also involved in the progression of apoptosis in
endothelial cells [12]. 8ased on these reports, we
hypothesized that sperm directly recognizes virus as
well as bacerial infections in semen via TしR3,TLR7,
TLR8 and TLR9, resulting in the reduction of sperm
functions, including fertilization ability. Although
Palladino et al. [13] reported the expression of the TLR
family that recognize virus pathogens in rat testes,
epididymidi and sperm, the relationship between TLRs
in sperm and the decrease of sperm fertilization ability
due to virus infection has remained unclear.
In this report, we examined whether TLR3, TLR7,
TLR8 and TLR9 were expressed in mouse sperm and
played functional roles in sperm motility and fertilization
ability. Tlr3,ηr7, Tlr8 and Tlr9 expression at the mRNA
level were detected by RT・.PCR,and the protein level
was analyzed using a specific antibody. Additionally, to
clarify the functional role of these receptors, we cultured
sperm with specific ligands and then analyzed the
motility, acrosomal status and induction of capacitation.
Finally, we also investigated whether the virus
pathogens inhibited sperm penetration to the oocyte
both in vivo and in vitro.
Materials and Methods
Materials
The TLR7 ligand, single strand RNA (ssRNA40) and
TLR9ligand, unmethylated CpG DNA (ODN1826), were
obtained from InvivoGen (San Diego, CA, USA). For
western blot analyses, antトTLR9antibody was
purchased from IMGENEX (IMG-305A, San Diego, CA,
USA), antトphospho-Tyr (P-Tyr-1 00, 9411), from Cell
Signaling Technology Inc. (Danvers, MA, USA), and
anti-かactinantibody from Sigma Chemical Co (Sigma,
St. Louis, MO). Equine chorionic gonadotropin, eCG
was purchased from Asuka Seiyaku (Tokyo, Japan).
Human chorionic gonadotropin, hCG was purchased
from Asuka Seiyaku. Oligonucleotide polyべdT)was
purchased from GE Healthcare (8uckinghamshire, UK),
Mihara, et al. 137
and AMV reverse transcriptase and Taq polymerase
were purchased from Promega (Madison, WI, USA).
80vine serum albumin (8SA, fraction V) was obtained
from Sigma (A 7888), and routine chemicals and
reagents were obtained from Nacalai Tesque, Inc.
(Kyoto, Japan) or Sigma.
Animals
Specific pathogen free (SPF) immature female
C578L16 mice, adult (8-week-old) female ICR mice and
4幽 month幽 oldmale ICR mice were obtained from Clea
Japan (Tokyo, Japan). On day 23 of age, immature
female mice were injected intraperitoneally (IP) with 4 IU
of eCG to stimulate follicular growth followed 48 h later
with 5 IU hCG to collect the ovulated cumulus oocyte
complexes (COCs) for in vitro fertilization [14]. For the
artificial insemination study, adult female ICR mice were
injected intraperitoneally (IP) with 5 IU of eCG to
stimulate follicular growth followed 48 h later with an
injection of 5 IU hCG. These mice were housed under a
16刷 hourlight!8.・hourdark schedule at the Experiment
Animal Center at Hiroshima University, and provided
food and water ad libitum.
Animals were treated in accordance with the NIH
Guide for the Care and Use of しaboratoryAnimals, as
approved by the Animal Care and Use Committee at
Hiroshima University.
Collection and culture of sperm
Spermatozoa were collected from the cauda
epididymidi of 4“month-old ICR strain mice into 500μlof
human tubal fluid (HTF) medium containing 0.4% (w/v)
8SA [14]. The sperm was cultured with 5 or 10 μg/mlof
ssRNA40 or 1 or 5μM of ODN1826 for 6 h, before
analyzing the motility. To induce the acrosome
reaction, the sperm were treated with 100 ng/ml of
progesterone (Sigma) for 60 min following pre舗
cultivation with 8SA for 60 min.
The detection of acrosome reaction
The acrosomal status of spermatozoa was measured
using FITC-Iabeled peanut agglutinin (FITC-PNA,
Sigma) and propidium iodide (PI) staining [15]. The
sperm were treated with progesterone following the
culture with 8SA.すhemixture was spread over slides
and air-dried at room temperature. Samples were then
fixed with absolute methanol for 10 min at room
temperature, and 30μ1 of FITC鵬 PNAsolution (100μg/ml)
in P8S was spread over each slide. The slides were
then incubated in a dark, moist chamber for 20 min at
370C. Following a rinse with P8S and air-drying, the
138 J. Mamm. Ova Res. Vol. 27, 2010
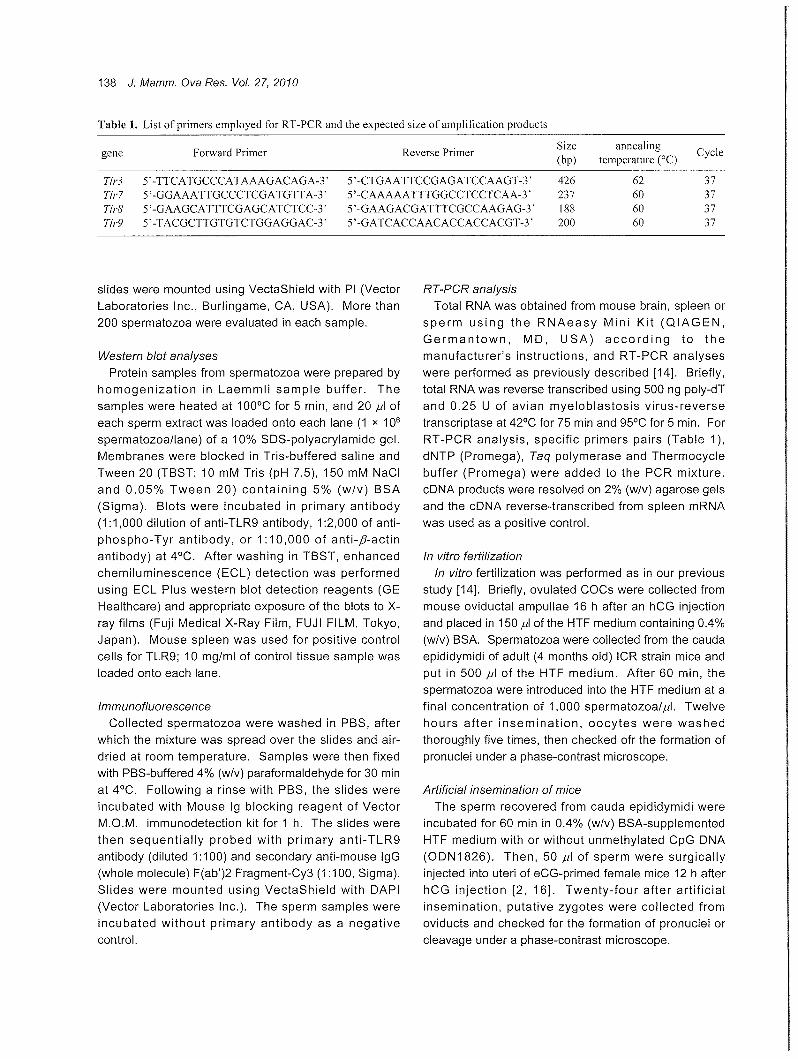
Table 1. List of primers employed for RT -PCR and the expected size of amplification products
Forward Primer Reverse Primer Size annealing
Cycle gene (bp) temperature (oC)
Tlr3 デーTTCATGCCCATAAAGACAGA時γ 5' -CTGAA TTCCGAGATCCAAGT -3' 426 62 37 Tlr7 デーGGAAATTGCCCTCGATGTτA-デ 5¥CAAAAATTTGGCCTCCTCAA-3 ' 237 60 37 Tlr8 デーGAAGCATTTCGAGCATCTCC-ゲ 5' -GAAGACGATTTCGCCAAGAG-3' 188 60 37 Tlr9 ターTACGCTτGTGTCTGGAGGAC-3' デーGATCACCAACACCACCACGT-3' 200 60 37
slides were mounted using VectaShield with PI (Vector RT剛 PCRanalysis
Laboratories Inc., Burlingame, CA, USA). More than Total RNA was obtained from mouse brain, spleen or
200 spermatozoa were evaluated in each sample. sperm using the RNAeasy Mini Kit (QIAGEN,
wθstern blot analyses
Protein samples from spermatozoa were prepared by
homogenization in Laemmli sample buffer. The
samples were heated at 1000C for 5 min, and 20μlof
each sperm extract was loaded onto each lane (1 x 106
spermatozoa/lane) of a 10% SDS-polyacrylamide gel
Membranes were blocked in Tris-buffered saline and
Tween 20 (TBST; 10 mM Tris (pH 7.5), 150 mM NaCI
and 0.05% Tween 20) containing 5% (w/v) BSA
(Sigma). Blots were incubated in primary antibody
(1: 1,000 dilution of anti-TLR9 antibody, 1 :2,000 of anti“
phospho-Tyr antibody, or 1:10,000 of antトβ'-actin
antibody) at 40C. After washing in TBST, enhanced
chemiluminescence (ECし)detection was performed
using ECL Plus western blot detection reagents (GE
Healthcare) and appropriate exposure of the blots to X-
ray films (Fuji Medical X-Ray Film, FUJI刊しM,Tokyo,
Japan). Mouse spleen was used for positive control
cells for TLR9; 10 mg/ml of control tissue sample was
loaded onto each lane.
Immunofluorescence
Collected spermatozoa were washed in PBS, after
which the mixture was spread over the slides and air-
dried at room temperature. Samples were then fixed
with PBS-buffered 4% (w/v) paraformaldehyde for 30 min
at 40C. Following a rinse with PBS, the slides were
incubated with Mouse Ig blocking reagent of Vector
M.O.M. immunodetection kit for 1 h. The slides were
then sequentially probed with primary anti-TLR9
antibody (diluted 1:100) and secondary anti-mouse IgG
(whole molecule) F(ab')2 Fragment-Cy3 (1:100, Sigma).
Slides were mounted using VectaShield with DAPI
(Vector Laboratories Inc.). The sperm samples were
incubated without primary antibody as a negative
control
Germantown, MD, USA) according to the
manufacturer's instructions, and Rτ欄 PCRanalyses
were performed as previously described [14]. Briefly,
total RNA was reverse transcribed using 500 ng poly-dT
and 0.25 U of avian myeloblastosis virus酬 reverse
transcriptase at 420C for 75 min and 950C for 5 min. For
RT-PCR analysis, specific primers pairs (Table 1),
dNTP (Promega), Taq polymerase and Thermocycle
buffer (Promega) were added to the PCR mixture
cDNA products were resolved on 2% (w/v) agarose gels
and the cDNA reverse輔 transcribedfrom spleen mRNA
was used as a positive control.
In vitro fertilization
In vitro fertilization was performed as in our previous
study [14]. Briefly, ovulated COCs were collected from
mouse oviductal ampullae 16 h after an hCG injection
and placed in 150μ1 of the HTF medium containing 0.4%
(w/v) BSA. Spermatozoa were collected from the cauda
epididymidi of adult (4 months old) ICR strain mice and
put in 500μ1 of the HTF medium. After 60 min, the
spermatozoa were introduced into the HTF medium at a
final concentration of 1,000 spermatozoalμ1. Twelve
hours after insemination, oocytes were washed
thoroughly five times, then checked ofr the formation of
pronuclei under a phase-contrast microscope.
Artificial insemination of mice
The sperm recovered from cauda epididymidi were
incubated for 60 min in 0.4% (w/v) BSA-supplemented
HTF medium with or without unmethylated CpG DNA
(ODN 1826). Then, 50μ1 of sperm were surgically
injected into uteri of eCG-primed female mice 12 h after
hCG injection [2, 16]. Twenty-four after artificial
insemination, putative zygotes were collected from
oviducts and checked for the formation of pronuclei or
cleavage under a phase-contrast microscope.
Statistical analysis
Data from three experimental trials are presented as
mean :t S.E.M. Data were analyzed using the Statistical
Analysis System Package (SAS Institute Inc., Cary, NC,
USA). AII percentage data were subjected to arcsine
transformation before ANOVA. When ANOVA revealed
a significant effect, means were compared using Fisher's
protected least significant difference post-hoc test, and
were considered to be significant when P < 0.05
Results
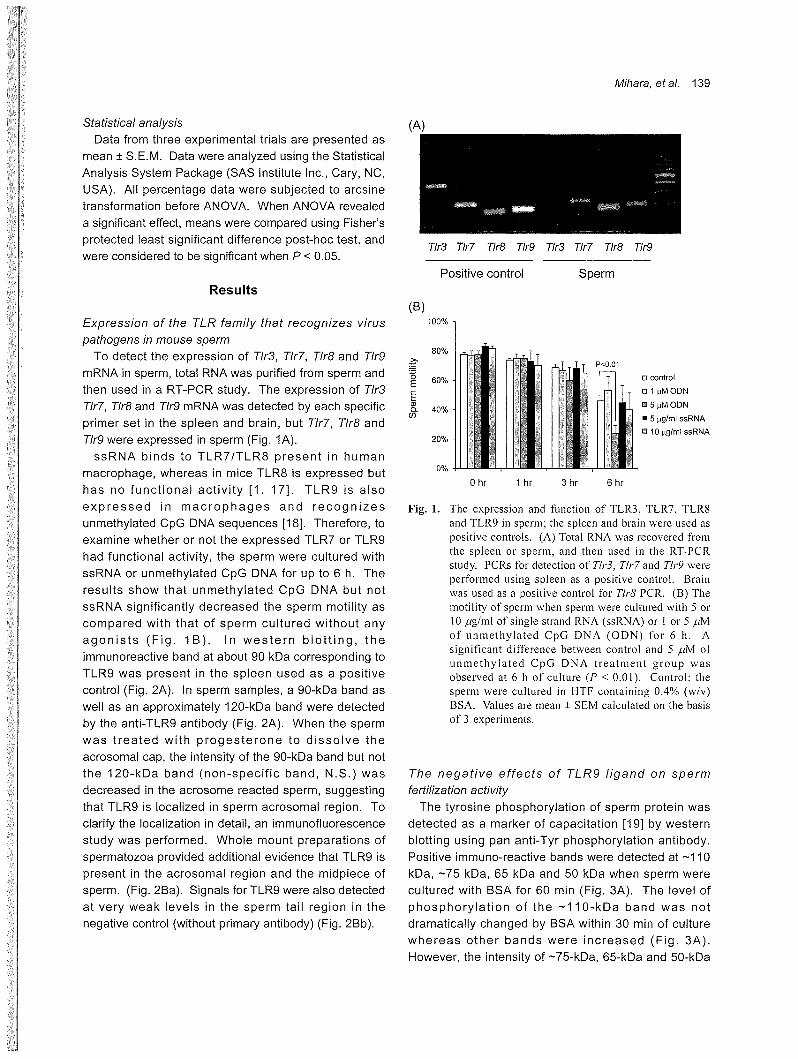
Expression of the TLR family that recognizes virus
pathogens in mouse sper斤7
To detect the expression of T/r3, T/r7, T/r8 and T/r9
mRNA in sperm, total 沢NAwas purified from sperm and
then used in a RTωPCR study. The expression of T/r3
T/r7,η'r8 and Tlr9 mRNA was detected by each specific
primer set in the spleen and brain, but T/r7, T/r8 and
ηr9 were expressed in sperm (Fig. 1A).
ssRNA binds to TLR7/TLR8 present in human
macrophage, whereas in mice TLR8 is expressed but
has no functional activity [1, 17]. TLR9 is also
expressed in macrophages and recognizes
unmethylated CpG DNA sequences [18]. Therefore, to
examine whether or not the expressed TLR7 or TしR9
had functional activity, the sperm were cultured with
ssRNA or unmethylated CpG DNA for up to 6 h. The
results show that unmethylated CpG DNA but not
ssRNA significantly decreased the sperm motility as
compared with that of sperm cultured without any
agonists (Fig. 1 B). In western blotting, the
immunoreactive band at about 90 kDa corresponding to
TしR9was present in the spleen used as a positive
control (Fig. 2A). In sperm samples, a 90次Daband as
well as an approximately 120-1くDaband were detected
by the anti-TLR9 antibody (Fig. 2A). When the sperm
was treated with progesterone to dissolve the
acrosomal cap, the intensity of the 90次Daband but not
Mihara, et al. 139
ηr3 Tlr7 η'r8 η'r9 T/r3 ηr7 η'r8 T/r9
Positive control Sperm
(B) 100%
80% キ〉ー・'、
関E 富市ロcon!rol
EEn L40% 1il5μMODN .5μgfml ssRNA
20% i I I蹴 E 淵
, , ~YI , , 船四圃司 隠 10~gfml ssRNA
。%
o hr 1 hr 3 hr 6 hr
Fig. 1. The expression and function of TLR3, TLR7, TLR8 and TLR9 in sperm; the spleen and brain were used as positive controls. (A) Total RNA was recovered会omthe spl巴巴nor sperm, and then used in the RTωPCR study. PCRs for detection of Th・3,Tlr7 and Tlr9 were perfor・m巴dusing spleen as a positive control. Brain was used as a positive control for Tlr8 PCR. (B) The motility of sperm when sperm were cultured with 5 or 10μg/ml of single strand RNA (ssRNA) or 1 or 5μM of unmethylated CpG DNA (ODN) for 6 h. A significant difference between control and 5μM of unmethylated CpG DNA treatment group was observed at 6 h of culture (Pく 0.01). Control: the sperm were cultured in HTF containing 0.4% (w/v) BSA. Values are mean土 SEMca1culated on the basis of 3 experiments.
the 120-kDa band (non-specific band, N.S.) was The negative effects of TLR9 ligand on sperm
decreased in the acrosome reacted sperm, suggesting flθrtilization activity
that TしR9is localized in sperm acrosomal region τo The tyrosine phosphorylation of sperm protein was
clarify the localization in detail, an immunofluorescence detected as a marker of capacitation [19] by western
study was performed. Whole mount preparations of blotting using pan anti-Tyr phosphorylation antibody.
spermatozoa provided additional evidence that TLR9 is Positive immuno-reactive bands were detected at -110
present in the acrosomal region and the midpiece of kDa, -75 kDa, 65 kDa and 50 kDa when sperm were
sperm. (Fig. 2Ba). Signals for TしR9were also detected cultured with BSA for 60 min (Fig. 3A). The level of
at very weak levels in the sperm tail region in the phosphorylation of the -110・主Daband was not
negative control (without primary antibody) (Fig. 2Bb). dramatically changed by BSA within 30 min of culture
whereas other bands were increased (Fig. 3A).
However, the intensity of -75-kDa, 65-kDa and 50欄 kDa

140 J. Mamm. Ova Res. Vol. 27, 2010
(A) ぐ-150 kDa N.S.今
TLR9 .. そ-100 kDa
-(:-号、品 p:::,.0'<> ~</;-' ",t$'-c-.~'<J
C;)~Y C;)~- .C¥",O'_O cj
下,,-マ"""_q,v
,~
(8)
e
w
d
&E--VE
a吐
gf
e
。c N
nu R
I」T
Fig. 2. The localization of TLR9 in sperm. (A) The expression of TLR9 at the protein levels in sperm Sperm wer巴 culturedfor 60 min in HTF containing 0.4% (w/v) BSA and then treated with or without progesterone for 1 h. Mouse spleen was used as a positive control for TLR9. N.S.: a non-specific band of about 120 kDa was detected only in sperm, not in spleen, th巴 positivecontrol. (B) TLR9 was localized on mice sperm using anti-mous巴 monoclonalTLR9 antibody. a) TLR9 in sperm, b) the negative control for TLR9 of sperm without primary antibody. Red is the Cys3 signal conjugating anti-mouse IgG antibody and blue is DAPI staining. The sperm was fixed just aft巴rcollection and then stained by the antibody.
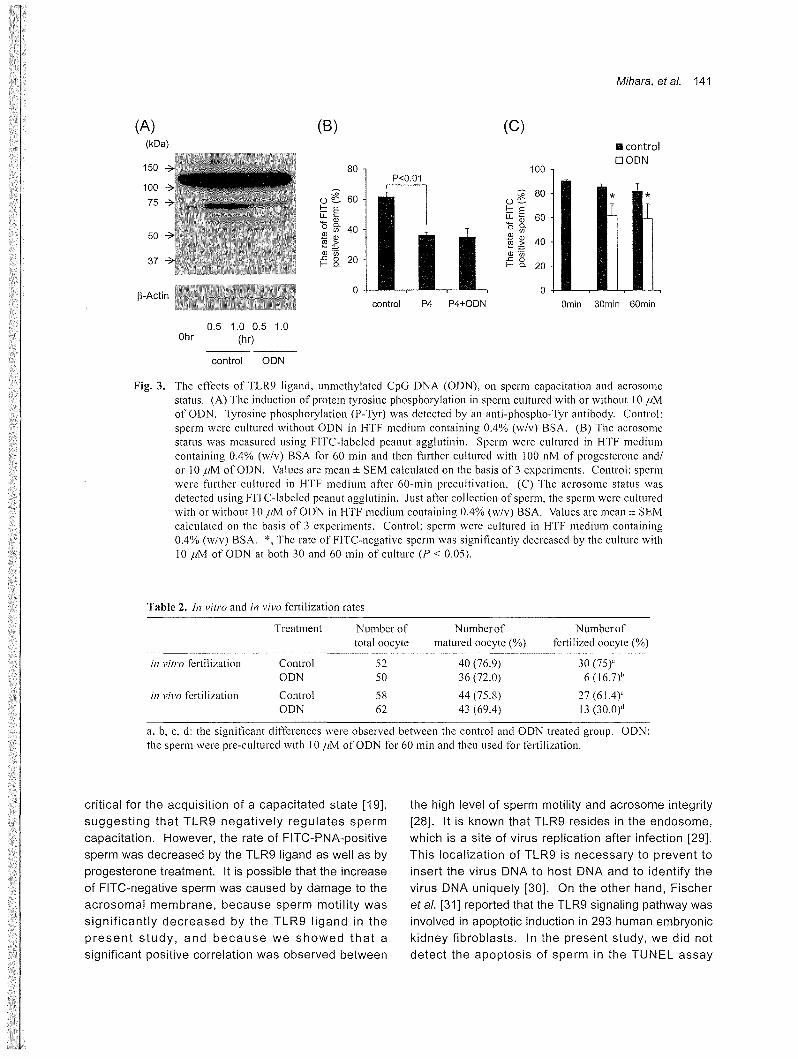
bands was dramatically decreased in sperm cultured
with unmethylated CpG DNA (Fig. 3A)
Loss of the acrosomal cap (acrosome reaction) was
detected by FITC-PNA. The rate of FITC-PNA-positive
sperm after the treatment with progesterone was
significantly decreased as compared with that for
control sperm (without progesterone treatment) (Fig
3B). When the TLR9 ligand, unmethylated CpG DNA,
was added to progesterone-containing medium, the rate
of FITC-positive sperm was not significantly changed as
compared with that for progesterone-treated sperm (Fig.
3B). However, when the sperm were cultured with
unmethylated CpG DNA alone, the rate of FITC-positive
sperm was significantly decreased as compared with
that in sperm without any agonists, and was comparable
with that for progesterone-treated sperm (Fig. 3C).
The effects of unmethylated CpG DNA on sperm
penetration to oocytes both in vitro and in vivo
Sperm recovered from the epididymidi of SPF mice
were cultured with or without unmethylated CpG DNA目
The sperm samples were then used in an in vitro
fertilization protocol or injected into the uteri of female
mice as reported previously [2]. The treatment of sperm
with unmethylated CpG DNA prior to in vitro fertilization
significantly decreased the subsequent rate of
fertilization as compared with that for sperm pre司
cultured with BSA alone (control) (Table 2).
Following this artificial insemination protocol, the
fertilization rate in control mice (without ligand
treatment) was about 60% whereas the fertilization rate
in mice injected with sperm exposed to unmethylated
CpG DNA was about 30% (Table 2).
Discussion
Sexually transmitted virus infections interfere with
male fertility. In humans, the hepatitis B and C viruses,
human immunodeficiency virus (HIV), papillomavirus
and herpes simplex virus have been detected in semen,
and infection increases the risk of infertility due to a
decrease in sperm motility [20-22]. In pig, porcine
reproductive and respiratory syndrome virus (PRRSV),
Aujeszky's disease virus and porcine parvovirus (PPV)
decrease sperm fertilization activity, which results in
economic losses in domestic animals [23-25]. The
reduction of sperm functions due to virus infections has
been thought to be caused by the immune function
accumulating leukocytes in semen [26]. Leukocytes
express TLR3, TLR7, TLR8 and TLR9 that recognize
virus-released pathogens, ssRNA, double strand RNA
or unmethylated CpG DNA, then secrete cytokines or
chemokine families [27]. However, in this study we
clearly showed that sperm also had immune functions
that recognize virus infection which decreases their own
fertilization activity.
The present study revealed that TLR9 was expressed
in sperm. Although details of the mechanism by which
sperm fertilization activity is reduced are unknown, a
TLR9 specific ligand strongly suppressed BSA-induced
tyrosine phosphorylation of sperm protein. The tyrosine
phosphorylation of proteins in sperm is thought to be
141 Mihara, et a/.
(B)
• control 口ODN
*
100
ハUn
u
n
u
n
u
n
o
c
u
d
骨
内
4
(ポ
)ε」
ω且目。〉一岩田
oa
。ト一比』口出担問」由工ト
(C)
80
n
u
n
u
n
U
6
4
2
(ポ
)ε」由且的。〉沼田口且
。ト一hL
』
Dω古川』
ωこト
(A) (kDa)
100
75
50
37
150
60min 30min Omin
。P4+0DN P4 conlrol
。
0.5 1.0 0.5 1.0 (hr)
conlrol ODN
Ohr
十Actin
Fig. 3. The effects of TLR9 ligand, unmethylated CpG DNA (ODN), on sperm capacitation and acrosome status. (A) The induction of protein tyrosine phosphorylation in sperm cultured with or without 10μM
ofODN.τyrosine phosphory1ation (P-Tyr) was detected by an anti-phospho叩Tyrantibody. Control:
sperm were cultured without ODN in HTF medium containing 0.4% (w/v) BSA. (B) The acrosome status was measured using FITC-labeled peanut agg1utinin. Sperm were CUltUl‘ed in HTF medium
containing 0.4% (w/v) BSA for 60 min and then further cultured with 100 nM of progesterone and/
or 10μM ofODN. Values are mean士SEMcalculated on the basis of 3 experiments. Control: sperm
were further cultured in HTF medium after 60-min precultivation. (C) Th巴 acrosomestatus was
detected using FITC司 labeledpeanut agglutinin. Just after collection of sperm, the sperm were cultured with or without 10μM ofODN in HTF medium containing 0.4% (w/v) BSA. Values are mean土 SEM
calculated on the basis of 3 experiments. Control: sperm were cultured in HTF medium containing
0.4% (w/v) BSA.へTherate of FITC-negative sperm was significantly decreased by the culture with
10μM of ODN at both 30 and 60 min of culture (Pく 0.05)
Table 2. 111 vitro and in vivo fertilization rates
Numberof
fertilized oocyte (%)
Numberof
matured oocyte (%) Number of total oocyte
Treatment
30 (75)"
6 (16.7)1'
27(61.4)' 13 (30.0)d
40 (76.9)
36 (72.0)
44 (75.8)
43 (69.4)
52
50
Control
ODN
Control
ODN
in vifro fertilization
58
62
in vivo ferti1ization
a, b, c, d: the significant differences were observed between the control and ODN treated group. ODN:
the sperm were pre-cultured with 10 μM of ODN for 60 min and then used for fertilization.
the high level of sperm motility and acrosome integrity
[28]. It is known that TLR9 resides in the endosome,
which is a site of virus replication after infection [29].
This localization of TLR9 is necessary to prevent to
insert the virus DNA to host DNA and to identify the
virus DNA uniquely [30]. On the other hand, Fischer
et 81. [31] reported that the TLR9 signaling pathway was
involved in apoptotic induction in 293 human embryonic
kidney fibroblasts. In the present study, we did not
detect the apoptosis of sperm in the TUNEL assay
critical for the acquisition of a capacitated state [19],
suggesting that TLR9 negatively regulates sperm
capacitation. However, the rate of FITC欄 PNA明 positive
sperm was decreased by the TLR9 ligand as well as by
progesterone treatment. It is possible that the increase
of FITC-negative sperm was caused by damage to the
acrosomal membrane, because sperm motility was
significantly decreased by the TLR9 ligand in the
present study, and because we showed that a
significant positive correlation was observed between

142 J. Mamm. Ova Res. Vol. 27, 2010
because the ligand, unmethylated CpG DNA, has involved in virus uptake are expressed in sperm, sperm
numerous terminal regions that bind to the TUN巨L is infected by viruses. To reduce the infection risk, the
probe. However, we hypothesize that the TしR9 sperm-washing technique and analysis of the
pathway induces loss of acrosomal integrity and the contamination by a high sensitivity method is required
apoptotic process, suppressing the capacitation of for not only the human HIV problem, but also large
sperm. Our previous [2] study showing the increase of animal reproductive performance.
apoptosis by the TLR2 and TLR4 pathways in sperm
supports the above hypothesis. Further study is Acknowledgements
required to clarify the mechanisms by which the TLR9
pathway downregulates sperm fertilization ability.
The expression and function of TLR9 in the
acrosomal region of sperm were revealed in this study
The innate immune function reducing sperm fertilization
activity, reduces the risk of infecting the fetus with a
virus. If semen were infected by a virus, the risk of virus
infection would increase in the female reproductive
tract. Wira and Fahey [32] in their review presented that
in a normal menstrual cycle, there is a window of
vulnerability (7-10 days following ovulation) in which the
potential for viral infection in the female reproductive
tract is enhanced. To reduce the infection risk,
especially the HIV risk, a sperm-washing technique to
remove seminal plasma that contains the virus was
reported in 1992 [33]. Sperm-washing techniques have
substantially changed the paradigm of fathering children
in serodiscordant couples with male human
immunodeficiency virus (HIV) infection, because
removing virus from the semen is beneficial for both the
fetus and mother [34]. However, the results of the
present study appear to support the possibility that not
only semen but also sperm is infected, because TLR9 is
known to localize within the cytoplasm. Thus, before
using sperm from virus-infected semen for artificial
insemination or in vitro fertilization, a highly sensitive
analysis of virus DNA purified from the washed sperm is
essential to reduce the fetal infection risk. Savasi et al.
[35] reported that the sperm from an HIV-positive men
was washed and then checked for virus contamination
by real-time PCR following artificial insemination (AI), in
vitro fertilization (IVF) or intracytoplasmic sperm
injection (ICSI). They showed that the overall
pregnancy rate (70.3%), independent of the procedure
used (AI or IVF/ICSI), justifies the efforts of the
individual patient to have a safe pregnancy [35].
In conclusion, sperm expresses and has a functional
TLR9 that potentially recognizes virus infection in
semen. The ligand for TLR9 reduces the fertilization
ability both in vitro and in vivo due to the suppression of
capacitation and a decrease in acrosome integrity.
Because TLR9 is known to localize within the
cytoplasm, and because it is known that the receptors
Thanks to Mr. N. Noma for technical assistance in
analyzing the sperm fertilization ability.
This work was supported in part by Japan Science
and Technology Agency (JST)ー.Grant(No. 12-068 and
NO.12ぺ04)(to M.S).
References
1) Akira、S.、じ巴mat側、 S.and Takeuchi, O. (2006): Pathogen
recognilion and innate immunily. CeIl, 124,783-801.
2) Okazaki, T., Fujita, Y., Mihara,丁目, Shitanaka, M., Neg凶11‘
ト1., Richards‘J.S. and Shimada, M. (2009): Expression 01'
the TolI-like receptor syslem that recognize bacterial
infection, controls fertilizalion ability in mammalian
spermalozoa. Reprod. Immllnol. Biol., 24, 174 (abslracl
13).
3) OchsendOl・f,F.R. (2008): SexlIaIly transmilled infeclions:
impact on male ferlility. Andrologia, 40, 72-75.
4) GlI己rin,B. and Pozzi, N. (2005): Virllses in boar semen:
delection and clinical as well as epidemiological
consequences regarding disease lransmission by artificial
insemination. Theriogenology, 63, 556-572
5) Koyama, S.、Ishii,K.J., Coban, C. and Akira, S. (2008):
lnnate immune response 10 viral infection. Cylokine, 43,
336-341.
6) Alexopollloll、L., Holt, A.C.今 Medzhitov,R. and FlaveIl,
R.A. (2001)・Recognitionof dOllble-stranded RNA and
aClivalion 01' NF-kappaB by ToIl-like receptor 3. Nature‘
413, 732-738.
7) Diebold, S.S.今 Kaisho句 T.,Hemmi, H., Akira今 S.and Reis, e,
SOllsa, C. (2004)・lnnateantiviral responses by means 01'
TLR7司 mediatedrecognition of single也 strandedRNA.
Science, 303, 1529-1531
8) Heil, F., Hemmi,行、 Hochrein,H., Ampenberger, F.、
Kirschning, C., Akira, S., Lipford, G., Wagner, H. and
Baller句 S.(2004): Species-specific recognition of single-
stranded RNA via toIl-like receptor 7 and 8. Science, 303,
1526-1529
9) Hacker, H., Redecke, V., Blagoev, B., Kratchmarova, 1.,
HSll, L.c., Wang, G.G., Kamps, M.P., Raz, E.、Wagner,H今
Hacker, G., Mann, M. and Karin, M. (2006): Specificity in
TolI四likereceptor signalling throllgh distinct effeclol
fllnctions ofTRAF3 and TRAF6. Nalure、439,204-207.10) Kawai、T.、Salo弓 S、lshii、K.J., Coban, c., Hemmi, H門
Yamamolo, M., Terai, K., Malsllda, M., lnoue、.1., Uemalsu,
S.. Takeuchi, O. and Akira, S. (2004): lnlerferon-alpha

induction through Toll悌 likereceptors involves a direct
interaction of IRF7 with MyD88 and TRAF6. Nat
Immunol., 5,1061-1068.
11 ) じ巴matsu,S., Sato, S., Yamamoto, M., Hirotani, T., Kato,
H., Takeshita, F., Matsudaラ M.,Coban, C., Ishii, K.J.,
Kawai, T., Takeuchi, O. and Akira, S. (2005): Interleukin-I
receptor-associated kinase輸 1plays an essential role fOl・
Toll-like receptor (TLR)7同 andTLR9-m巴diatedinterferon-
{alpha} induction. J. Exp. Med., 201, 915-923.
12) Bannerman, D.D., Erwert, R.D., Winn, R.K. and Harlan,
J.M. (2002): TIRAP mediates endotoxin-induced NF-
kappaB activation and apoptosis in endothelial cells
Biochel11. Biophys. Res. Coml11u日刊 295、157-162.
13) Palladino, M.A., Savarese, M.A., Chapl11an, J.し, Dughi,
M.K. and Plaska, D. (2008): Localization of Toll-like
receptors on epididYl11al epithelial cells and sperl11atozoa
Al11. J. Reprod. Immunol., 60, 541-555
14) Shimada, M., Yanai, Y円 Okazaki,T., Noma, N円
Kawashima, 1., Mori,τ. and Richards, J.S. (2008):
Hyaluronan fragments generated by sperl11-secreted
hyaluronidase stimulate cytokine/chemokine production
via the TLR2 and TLR4 pathway in cumulus cells of
ovulat巴dCOCs, which l11ay enhance fertilization
Development, 135, 2001-2011.
15) Zeng, W.X., Shimada, M., Isobe, N. and Terada, T. (2001):
Survival of boar sp巴rl11atozoa仕ozenin diluents of varying
osmolality. Theriogenology, 56, 447-458
16) Kile, J.c. Jr. (1951): An improved l11ethod for th巴artificial
insemination of mice. Anat. Rec., 109, 109-117.
17) Delgado, M.A., Elmaoued, R.A.ラ Davisラ A.S.,Kyei, G. and
Deretic, V. (2008): Toll-lik巴 receptorscontrol autophagy.
EMBO J., 27,1110-1121.
18) Latz, E., Schoenemeyer, A., Visintin, A., Fitzgerald, K.Aーラ
Monks, B.G., Knetter, C目F.,Lien, E., Nilsen, N.J., Espevik,
T. and Golenbock, D.T. (2004): TLR9 signals after
translocating from the ER to CpG DNA in the Iysosome.
Nat. Immunol., 5, 190-198.
19) Naz, R.K., Ahmad, K. and Kumar, R. (1991): Role of
me口lbranephosphotyrosine proteins in human
spermatozoal function. J. Cell Sci., 99う 157-165.
20) Moretti, E., Federico, M.G., Giannerini, V. and Collodel,
G. (2008): Sperm ultrastructure and meiotic segregation in
a group of patients with chronic hepatitis B and C.
Andrologia, 40, 286-291.
21) Foresta, c., Garolla, A., Zuccarello, D., Pizzol, D., Moretti,
A., Barzol1, L. and P
Mihara, et al. 143
susceptibility of Streptococcus suis isolated from
slaughtered pigs. Zentralbl. Veterinarm巴d.B, 41, 608-617
24) Hall, L.B. Jr, Kluge, J.P., Evans, L.E. and Hill, H.T
(1984): The effect of pseudorabies (Aujeszky's) virus
infection 011 young mature boars and boar fertility. Can. J.
Comp. Med., 48,192-197
25) Guerin, B. and Pozzi, N. (2005): Viruses in boar semen: detection and clinical昌swell as epidemiological
consequ巴nc巴sregarding disease transmissiol1 by artificial
insel11ination. Theriogenology, 63, 556-572.
26) Denny, T.N., Scolpino, A., Garc問、 A.,Polyak, A円 Weiss,
S.N., Skurnick, J.H., Passannante, M.R. and Colon, J.
(1995): Evaluation of T -IYl11phocyt巴 subsetspresent in
semen and peripheral blood of healthy donors: a report
合omthe heterosexual transmission study. Cytometry, 20,
349-355.
27) Zarember, K.A. and Godowski, P.J. (2002): Tissue
expression of human Toll-like receptors and diffel巴ntial
regulation of Toll-like receptor mRNAs in leukocytes in
response to microb巴s,their products, and cytokines. J.
Immunol., 168, 554-561.
28) Okazaki, T., Abe, S., Yoshida, S. and Shimada, M. (2009):
Seminal plasma damages sperm during cryopreservation, but its presence during thawing il11proves semen quality
and conc巴ptionrates in boars with poor post-thaw semen
quality. Theriogenology, 71, 49ト刊498.
29) Sapp, M. and Bienkowska-Haba, M. (2009): Viral entry
mechanisms: human papillomavirus and a long journey
from extracellular matrix to the nucleus. FEBS J., 276,
7206-7216.
30) Barton, G.M., Kagan, J.C. and Medzhitov, R. (2006):
Intracellular localization of Toll四 likereceptor 9 prevents
recognition of self DNA but facilitates access to viral
DNA. Nat. Immunol., 7, 49-56.
31) Fischer, S.F., Rehm, M., Bauer, A., Hofling, F., Kirschn巴k,
S., Rutz, M., Bauer, S.ヲ Wagner,H. and Hacker, G. (2005):
Tolトlikereceptor 9 signaling can sensitize fibroblasts for
apoptosis. Immunol. Lett., 97, 115-122.
32) Wira, C.R. and Fahey, J.V. (2008): A new strategy to
understand how HIV infects women: identification of a
window of vulnerability during th巴m巴nstrualcycle. AIDS,
22ヲ 1909-1917.
33) Huyser, C., Fourie, Fし, Oosthuizen, M. and Neethling, A.
(1991): Microbial flora in sel11en during in vitro
fertilization. J. In Vitro Fert. Embryo Transf., 8, 260-264.
34) Bostan, A., Vannin, A.S., Emiliani, S., Debais

163
マウス精子にはウイルスを感知する TolIlike receptorが発現し,
それは受精能を低下させる……....・H ・........・H ・........・H ・.....…....・H ・.......・H ・....…H ・H ・.......・H ・.136-143
三原敏敬福本景太 l・附崎哲司1.2・島問昌之広烏大学大学院生物園科学研究科,
東広鳥rI1 〒 739-8528,2大分県農林水産センター,壊後大野市 〒 879叩 7111
細菌感染した精液では,精子自身か、細菌が放出する内毒素を TolI
like receptor (τLR) を介して感知し,自身の運動伎を低下させる
ウイルス感染した精液においても,精子の低運動性が認められるこ
とから,精子はウイルス感染も感知している可能性がある しかし,
ウイルスが放出する病原体に対する TLRfamilyについて,精子に
おける発現とその役割に演する報告はほとんどない.本研究におい
て,精子に脱メチル化 CpG!ie列に反応する TLR9が発現している
ことが示された.ウエスタンフロッティング解析の結果, TLR9の
タンパク質レベルでの発現も検出されたが,先体反応を誘起した精
子では,そのバンドが、落くなっていたことから, TLR9が精子頭部
に局在していると推察された.τLR9のリガンドは,先体の擦傷を
促し,臼SAによる受精能獲得誘起を抑制した. さらに,体外受精
および人工授精試験の結果, TLR9 1):ガンドで前士官蒸した精子の体
内,体外における受精能が有意に低下していた.以よの結果から,
精子にはウイルスの DNAである脱メチルイヒ CpG配列を感知する
TLR9が発現し,精液中のウイルス感染を感知可能であることが示
唆された.
キーワード:TolI様受容体,精子,ウイルス感染,受精,初期免疫
スナネズミ粧のパリン代謝….....・H ・-…............................................................................司....144-149
小i憾隆一郎・辻-Ji二弘J忠 信州大学農学部動物発生工学研究室,上伊那郡 守 399-4598
スナネズミ初期怪の体外培養はらcell期に起こる cellblockで
既知培養液での培養(;1:困難であった 我々は,修正 M16士宮餐液に
パリンを添加することで 2納胞期限からの怪盤胞への発生に成功
した本研究は,スナネスミ初期医発生におけるハリンの役部を放
射性向位体を用いて調べた.過排卵誘起したスナネズミより 1,2,
8細胞期医,桑実期怪,怪盤総賂庇を採取した.修正 M16滋養液
を用いてバリン添加・非添加持の 14C_グルコース取り込みと酸化,
14C_バリンの取り込みと酸化を測定した 14C_J~ 1)ンの取り込み
は8細胞期肢で,叫C-バリンの酸化は 8細胞期および怪盤胞湖底
で有窓に増加した 14C剛グルコースの取り込みは)ff滋飽期阪では
J~ 1)ン添加IKが,酸化は B級施期阪ではバリン非添加区が有慈に高
かった. また,パリンの代言語を遺跡調主ましたところ 0.8%が脂質に
代謝されていた.これらの結果からパリンはスナネズミ初期医にお
いてエネルギー源として利用されている可能性が示唆された.
キーワート、:バリン,アミノ酸代童話, fi主,スナネズミ
ピエゾ ICSIにおいて縮胞摸が低伸展性を示すとト卵子のリスク評価…H ・H ・..............……….150-156
岩山 広 1・1泉地良一 2・山下正紀山下レディースクリニック,神戸市 千 651-0086,
2信州大学繊維学者[5,上田市 〒 386-8567
ピエゾ ICSIで周いられるインジェクションピペットは先端が平
沼であるために卵細胞膜の穿破が先端形状に影饗されることなく,
その伶展性を正確に評価することができる 本研究では,細胞膜が
低伸展性を示す卵子における膝発生能と議床能を,通常の膜性を示
す卵子と比較検討した 細胞膜が穿刺点より卵子直径の 75%未満
の伸渓でどエゾパルスなして穿破された卵子を「低伸展性」と定義
し, 75%以上伸展しピエゾパルスを適尽し穿破された卵子を「高
伸展性jと定義した.版発生能については, ICSI後の生存率およ
び)ff盤胞発生E容を比較した.査費床能については,多変緩解析を用い
て f低伸展性j卵子の栂対リスクを検討した. ["低伸展性j卵子の
ICSI後の生存伎は f高伸震性jのそれに比べ有意に低下した し
かし,目壬盤胞発生率については同様であった. また,古着床において
は卵細胞緩の「低伸震性jが危検因子として検出され,移植怪を選
択する際の基準の 1つとなり得ることか、示唆された
キーワート、:ヒト卵子,着床,卵細胞膜の伸展性, ピエゾ ICSI,
生存性
体外受精で得られた水牛初期腔における染色体異常の出現率.…...・H ・......・H ・-…H ・H ・-……….157-160
省i翠 緑 1,2・Ul10aUl1oa, C.M.1. 2・Hufana叩 Ouran,0.3.4.5・Atabay,E?' 4・Ouran,p.G.3.4・Cruz,L.C?' 4・金子t幸雄 5.
高橋芳さ幸存字吉都15宮大学}農箆学部生物生産科学科 字者都F宮市 〒 32引1嶋 8お50出5. 東京l縫立工大学大学院迷合}縫箆学研究科,府
中干市打 宇 18回3-8白509,3ブイリピンカラパオ水t牛ドセンタ一(伊PCCο),ヌエパ.エシ/ハ、サ1+札卜
5筑i波皮大学大学院生命z環策境科学研究科 つくば市 =守f30伯5-8邸57η2. 6北i海毎道大学大学l院淀釦i獣滋医学研究科,札申幌晃市 干 06ωOー0818
体外受精水牛座の低受胎主義の原因究明のために,分割初期妊に
おける染色体異常の出現率を明らかにした.フィリどン,インドの
と場入手の汚川製水牛卵巣の直径 2-8mm卵胞から卵子を吸引採
取, 23-24時間成熟培養し,河川型水牛の凍結精子を最終濃度 1x
106 sperm/mlで媒精した.媒精 12-18時局後に体外培養用培地へ
移し 36-40時間卵丘細胞と共培養し, 2-8細胞期怪を 100ng/ml
のビンプラスチンで 6-10時間処返した後,吉j察ら (1998)のウ
シ体外受精涯の方法に準じて染色体襟本を作製した. 206館の阪の
51.0% (105/206) が中期像を脊し, 87.6% (92パ05) で分析可
能であり,染色体異常の出現率は 47.8% (44/92) で,多倍数体は
23.9% (22192),昆倍数{本と半数体は各 12% (11/92) であり,多
倍数体や混倍数{本は多精子侵入が原因と考えられた
キーワード:水牛, )ff,体外受精,染色体異常,多精受精