Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Journal of South American Earth Sciences 33 (2012) 68e79
Contents lists available
Journal of South American Earth Sciences
journal homepage: www.elsevier .com/locate/ jsames
First sauropod (Dinosauria: Saurischia) remains from the Guichón Formation,Late Cretaceous of Uruguay
Matías Sotoa,*, Daniel Pereaa, Andrea Cambiasob
aDepartamento de Evolución de Cuencas, Facultad de Ciencias, 4225 Iguá, 11400 Montevideo, UruguaybAv. Corrientes 4553 12D (1195), Ciudad Autónoma de Buenos Aires, Argentina
a r t i c l e i n f o
Article history:Received 19 April 2011Accepted 18 August 2011
Keywords:SauropodaTheropodaOrnithopodaCretaceousUruguay
* Corresponding author.E-mail addresses: [email protected], sotor
[email protected] (D. Perea), [email protected] (
0895-9811/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.jsames.2011.08.003
a b s t r a c t
We report the first sauropod remains recorded in the Guichón Formation, western Uruguay. Thematerials belong to a middle-sized sauropod represented by more than one individual, and among otherremains include more than fifty caudal centra. Close to the bones, several eggshell fragments resemblingSphaerovum erbeni Mones, 1980 were found. We discuss the biostratigraphic implications of thesefindings, which for the first time allow us to confidently refer the Guichón Formation to the LateCretaceous. The combination of several synapomorphies such as a biconvex first caudal centrum,strongly procoelous middle and distal caudal centra, and a pyramidal astragalus suggests that thesauropod remains belong to a derived lithostrotian, probably related to Pellegrinisaurus powelli, Baur-utitan britoi and Alamosaurus sanjuanensis. A few isolated teeth (now lost) referred by Frederich vonHuene in 1934 to ornithomimid theropods and ornithischians are herein reinterpreted as belonging toindeterminate theropods and basal iguanodontians.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Titanosaurian sauropods, the remains of which are found inCretaceous deposits from all continents except Antarctica, includedsmall to giant-sized long-necked quadrupeds, which were taxo-nomically diverse and morphologically distinct (Curry Rogers,2005; Wilson, 2006). In South America, they were the mostabundant and diverse herbivorous vertebrates during the LateCretaceous. Dozens of genera and species from Argentina (e.g.Apesteguía, 2004; Bonaparte and Coria, 1993; Bonaparte et al.,2006; Calvo and González Riga, 2003; Calvo et al., 2007a,b;González Riga, 2003; González Riga et al., 2008; Huene, 1929b;Lydekker, 1893; Martínez et al., 2004; Powell, 1987a, 1992, 2003;Salgado, 1996; Salgado and Azpilicueta, 2000; Salgado and Coria,1993; Salgado et al., 1997a, 1997b, 2005), Brazil (e.g. Campos andKellner, 1999; Kellner and de Azevedo, 1999; Kellner et al., 2005;Powell, 1987b), Chile (Kellner et al., 2011) and Uruguay (Huene,1929a; but see Powell, 2003 concerning the impossibility to reacha specific level of determination) have been described.
Purported sauropod eggs and eggshells known so far in Uruguaycame from the Yapeyú Member of the Mercedes Formation (Goso
[email protected] (M. Soto),A. Cambiaso).
All rights reserved.
and Perea, 2004; Fig. 1A), while the bones are tentatively attrib-uted to this member (formerly part of the Asencio Formation;Huene,1929a; Perea and Ubilla, 1994; Powell, 2003) given that theyare almost always found in meteorized levels (with few exceptions;Soto, 2010a).
The remains described herein come from the Guichón Forma-tion (Fig. 1A), being the first record of the clade from this unit. Thefossil site is located about 20 km southeast of Quebracho town,Paysandú province (Fig. 1C), inside a 23-ha erosion gully (Fig. 2).Bones are disarticulated but spatially associated, slightly abradedand sometimes fractured. These features are attributable to bothancient (pre-burial weathering and transport) and modern (tram-pling) processes. Paleocurrent analysis cannot be undertaken giventhat the fossil remains do not preserve its original position. Theprovenance of the remains is from a silicified conglomeratichorizon, the weathering of which causes the bones to roll towardthe bottom of the gully.
2. Geological setting
The Guichón Formation is included in the Paysandú Group(Bossi and Navarro, 1991; Fig. 1B), along with the MercedesFormation (Bossi, 1966). Although Bossi (1966) recognized a thirdunit, the Asencio Formation, most authors include it partially ortotally in theMercedes Formation (Ford and Gancio,1988; Goso andPerea, 2004; Martínez and Veroslavsky, 2004).
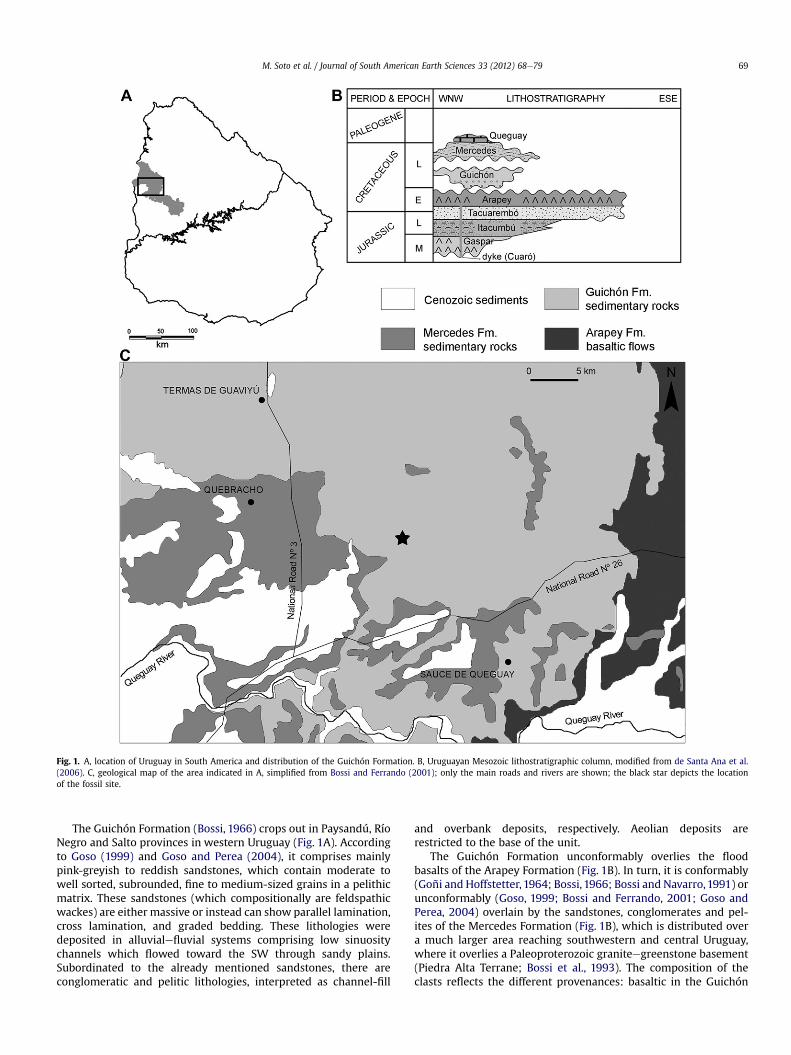
Fig. 1. A, location of Uruguay in South America and distribution of the Guichón Formation. B, Uruguayan Mesozoic lithostratigraphic column, modified from de Santa Ana et al.(2006). C, geological map of the area indicated in A, simplified from Bossi and Ferrando (2001); only the main roads and rivers are shown; the black star depicts the locationof the fossil site.
M. Soto et al. / Journal of South American Earth Sciences 33 (2012) 68e79 69
The Guichón Formation (Bossi, 1966) crops out in Paysandú, RíoNegro and Salto provinces in western Uruguay (Fig. 1A). Accordingto Goso (1999) and Goso and Perea (2004), it comprises mainlypink-greyish to reddish sandstones, which contain moderate towell sorted, subrounded, fine to medium-sized grains in a pelithicmatrix. These sandstones (which compositionally are feldspathicwackes) are either massive or instead can show parallel lamination,cross lamination, and graded bedding. These lithologies weredeposited in alluvialefluvial systems comprising low sinuositychannels which flowed toward the SW through sandy plains.Subordinated to the already mentioned sandstones, there areconglomeratic and pelitic lithologies, interpreted as channel-fill
and overbank deposits, respectively. Aeolian deposits arerestricted to the base of the unit.
The Guichón Formation unconformably overlies the floodbasalts of the Arapey Formation (Fig. 1B). In turn, it is conformably(Goñi and Hoffstetter, 1964; Bossi, 1966; Bossi and Navarro,1991) orunconformably (Goso, 1999; Bossi and Ferrando, 2001; Goso andPerea, 2004) overlain by the sandstones, conglomerates and pel-ites of the Mercedes Formation (Fig. 1B), which is distributed overa much larger area reaching southwestern and central Uruguay,where it overlies a Paleoproterozoic graniteegreenstone basement(Piedra Alta Terrane; Bossi et al., 1993). The composition of theclasts reflects the different provenances: basaltic in the Guichón

Fig. 2. A, shape of the gully (see location in Fig. 1C) inside of which the fossil remains were found. B, general view of the gully; the arrow indicates a person for scale.
Fig. 3. FC-DPV 2310, eggshell fragments. A, external view. B, radial view. Scale barequals 3 cm (A) and 1 cm (B).
M. Soto et al. / Journal of South American Earth Sciences 33 (2012) 68e7970
Formation and from the basement in theMercedes Formation. Bothunits were deposited in fluvial environments, but the latterrepresents more torrential conditions. Warm, semiarid and humidpaleoclimatic conditions, respectively, are inferred.
The deposition of the Paysandú Group began in endorrheicconditions after the extrusion of the Neocomian flood basalts (anevent regionally known as Serra Geral magmatism), and took placein the Litoral del Río Uruguay Basin (Goso, 1999), which in Uruguayoccupies about 25,000 km2, with a maximum thickness of 220 m.This sedimentary scenario continued in the Argentinean Meso-potamia, where the unit depocenter is located. The ArgentineanPuerto Yeruá Formation is correlated with both the Guichón (Gosoand Perea, 2004) and Mercedes formations.
The age of the Guichón Formation is not clear. Proposed agesrange from Aptian (Goso and Perea, 2004) to Santoniane?Campa-nian (Mones, 1997). On one hand, the geological evidence such asthe unconformity with the overlying Mercedes Formation (Cam-panianeMaastrichtian; Goso and Perea, 2004; Daners andGuerstein, 2004) and the facies similarity with the MiguesFormation (Albian; Campos et al., 1997) from the southern SantaLucía Basin, led some authors (Goso,1999; Goso and Perea, 2004) tosuggest an Early Cretaceous age for this unit. It may be noted thatthe age of the correlated Puerto Yeruá Formation (Argentina) is alsodifficult to determine beyond a Cretaceous age (De Valais et al.,2003).
On the other hand, the paleontological evidencewas insufficientor wrongly interpreted for decades. Dinosaur remains recovered todate (now lost) comprise a few theropod and ornithischian teethreferred to the Ornithomimidae (Huene, 1934a,b) and Iguano-dontidae (Bonaparte, 1978), respectively. These referrals, whichconstituted the basis to allocate the Guichón Formation to the LateCretaceous, are still unchallenged (Weishampel et al., 2004), whichjustify the critical review undertaken herein employing updatedsystematics. Abundant remains of the basal notosuchian Uruguay-suchus were also described for the unit (Rusconi, 1933), and newmaterials will be reported elsewhere (Soto et al., in press). From thePuerto Yeruá Formation, in turn, a ‘titanosaurid’ humerus (Powell,2003; early referred to cf. Argyrosaurus superbus by Huene,1929b), an ankylosaur osteoderm, a fragmentary theropod teeth,and an eggshell fragment referred to cf. Sphaerovum erbeni havebeen reported (De Valais et al., 2003). Hence, the sauropod remainsfrom the Guichón Formation described herein are relevant.
Institutional AbbreviationsdFC-DPV, Colección de VertebradosFósiles, Facultad de Ciencias (Universidad de la República, Uruguay).
Anatomical AbbreviationsdMeasurements of sauropod verte-brae are as follows: CL, centrum length. CtH, cotyle height. CtW,
cotyle width. CdH, condyle height. CdW, condyle width. CtR, cotyledorsoventral depression ratio, defined herein as CtH/CtW. CdR,condyle dorsoventral depression ratio, defined herein as CdH/CdW.Measurements of theropod teeth follow Smith et al. (2005): AL,apical length. CBL, crown base length. CBW, crown base width. CH,crown height. CBR, crown base ratio. DAVG, average distal serrationdensity.
3. Systematic paleontology
FAVEOLOOLITHIDAE Zhao and Ding, 1976 sensu Casadío et al.,2002SPHAEROVUM Mones, 1980SPHAEROVUM sp.(Fig. 3)
MaterialdFC-DPV 2310, several eggshell fragments.LocalitydThe material was collected 20 km SE of Quebracho
town, Paysandú province, Uruguay (31�58021.900 S, 57�44000,900 W).Stratigraphic provenance: Guichón Formation (Late Cretaceous;this contribution), base of Paysandú Group.
DescriptiondThe eggshell fragments, probably coming fromspherical eggs (Fig. 3A), measure up to 5 mm thick (Fig. 3B). Theornamentation is typically compactituberculate (i.e., externalsurface covered with dome-like tubercles; Mikhailov, 1991).Although the materials are silicified, it appears that there isa multicanaliculate pore system.
CommentsdThe above mentioned characters agree with therevised diagnosed of the oospecies S. erbeni provided by Casadíoet al. (2002): spherical egg with diameter ranging between 150and 206 mm, eggshell thickness ranging between 4.2 and 5.5. mm,presence of compactituberculate ornamentation (translated fromSpanish).
M. Soto et al. / Journal of South American Earth Sciences 33 (2012) 68e79 71
However, as mentioned by Simon (2006), given that the holo-type of S. erbeni is silicified, comparisons have to be reduced toexternal characters only (i.e. actual pore system cannot beconfirmed, which is diagnostic at the oofamily level; Mikhailov,1991). Pending detailed studies, we tentatively refer the materialsto the oogenus Sphaerovum, already recorded in several localitiesfrom Uruguay (Mercedes Formation; Mones, 1980; Verde and Soto,2005). Sphaerovum was included within the oofamily Faveolooli-thidae by Faccio and Montaña (1994), Faccio (1994) and Casadíoet al. (2002), although other authors proposed a relationship withthe Megaloolithidae (see references in Faccio and Montaña (1994)and Casadío et al. (2002)).
The relationship between big, spherical eggs and sauropods hasbeen confirmed in the Patagonian Auca Mahuevo site (e.g. Chiappeet al., 1998), although certain megaloolithids have been foundassociated with hadrosaurid ornithischians (Grigorescu et al.,2010). Sphaerovum has been long interpreted as laid by
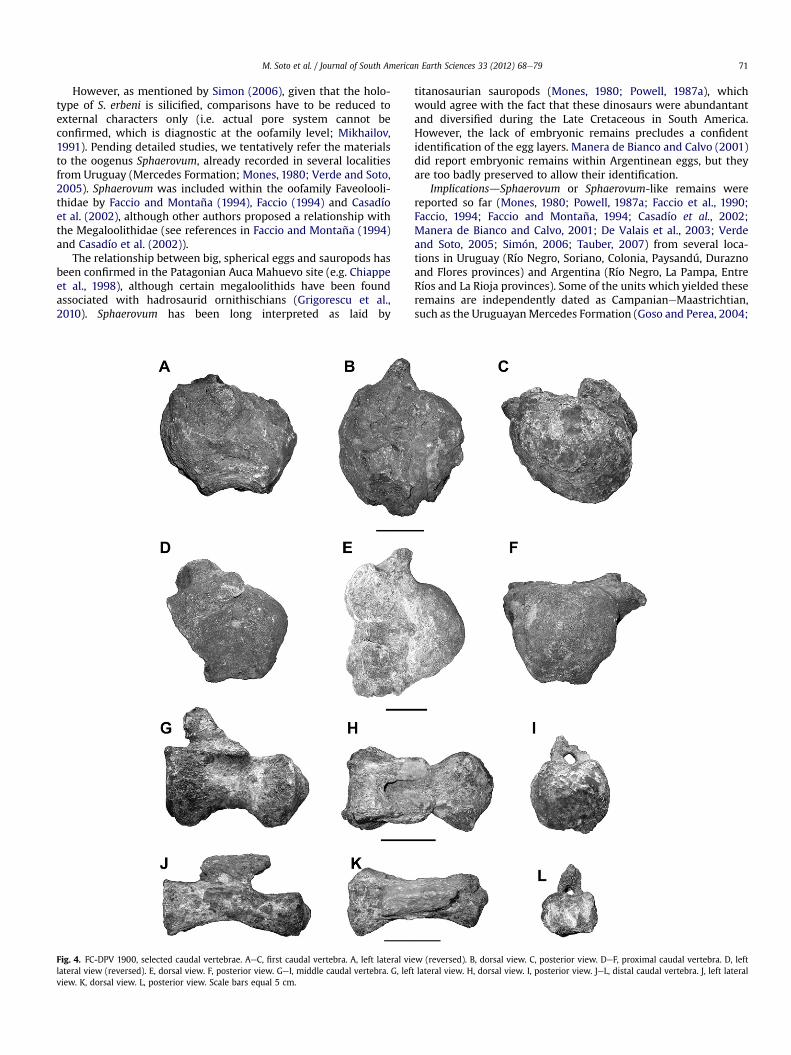
Fig. 4. FC-DPV 1900, selected caudal vertebrae. AeC, first caudal vertebra. A, left lateral vielateral view (reversed). E, dorsal view. F, posterior view. GeI, middle caudal vertebra. G, lefview. K, dorsal view. L, posterior view. Scale bars equal 5 cm.
titanosaurian sauropods (Mones, 1980; Powell, 1987a), whichwould agree with the fact that these dinosaurs were abundantantand diversified during the Late Cretaceous in South America.However, the lack of embryonic remains precludes a confidentidentification of the egg layers. Manera de Bianco and Calvo (2001)did report embryonic remains within Argentinean eggs, but theyare too badly preserved to allow their identification.
ImplicationsdSphaerovum or Sphaerovum-like remains werereported so far (Mones, 1980; Powell, 1987a; Faccio et al., 1990;Faccio, 1994; Faccio and Montaña, 1994; Casadío et al., 2002;Manera de Bianco and Calvo, 2001; De Valais et al., 2003; Verdeand Soto, 2005; Simón, 2006; Tauber, 2007) from several loca-tions in Uruguay (Río Negro, Soriano, Colonia, Paysandú, Duraznoand Flores provinces) and Argentina (Río Negro, La Pampa, EntreRíos and La Rioja provinces). Some of the units which yielded theseremains are independently dated as CampanianeMaastrichtian,such as the UruguayanMercedes Formation (Goso and Perea, 2004;
w (reversed). B, dorsal view. C, posterior view. DeF, proximal caudal vertebra. D, leftt lateral view. H, dorsal view. I, posterior view. JeL, distal caudal vertebra. J, left lateral
M. Soto et al. / Journal of South American Earth Sciences 33 (2012) 68e7972
Daners and Guerstein, 2004) and the Argentinean Allen and LosAlamitos Formations (Leanza et al., 2004 and references therein).However, no other relevant biostratigraphic data exist for theArgentinean Puerto Yeruá (De Valais et al., 2003), Colorado (Casadíoet al., 2002) and Los Llanos (Tauber, 2007) Formations.
SAUROPODOMORPHA Huene, 1932SAUROPODA Marsh, 1878TITANOSAURIA Bonaparte and Coria, 1993LITHOSTROTIA Upchurch et al., 2004SALTASAURIDAE Bonaparte and Powell, 1980SALTASAURIDAE gen. et sp. indet.(Figs. 4 and 5)
MaterialdFC-DVP 1900: forty eight caudal centra (plus frag-ments of at least another nine), two caudal neural spines, a partialcoracoid, a fragmentary ulna, metacarpal fragments, a right prox-imal fibula, a left distal tibia, two right astragali, several meta-tarsals, two non-ungual pedal phalanxes and five probableosteoderms.
LocalitydSame as for FC-DPV 2310.DescriptiondCaudal vertebrae. Among the caudal vertebrae,
most regions of the tail but the terminal one are represented. Thecentra are cylindrical and spool-shaped, given their concave ventraland lateral faces (Fig. 4). For centrum measurements see Table 1.Cotyle width is the most variable measurement (Table 1). Theshapes of most centrum articular faces can be described as sub-circular, although a few ones show higher than wide articular facesand others are slightly dorsoventrally depressed. The distal caudalcentra are moderately depressed (Fig. 4JeL).
Unfortunately, only the bases of most neural arches arepreserved. An important amount of information with potentialphylogenetic relevance (e.g. Wilson, 1999) is thus lacking. Con-cerning the transverse processes, rarely more than their bases are
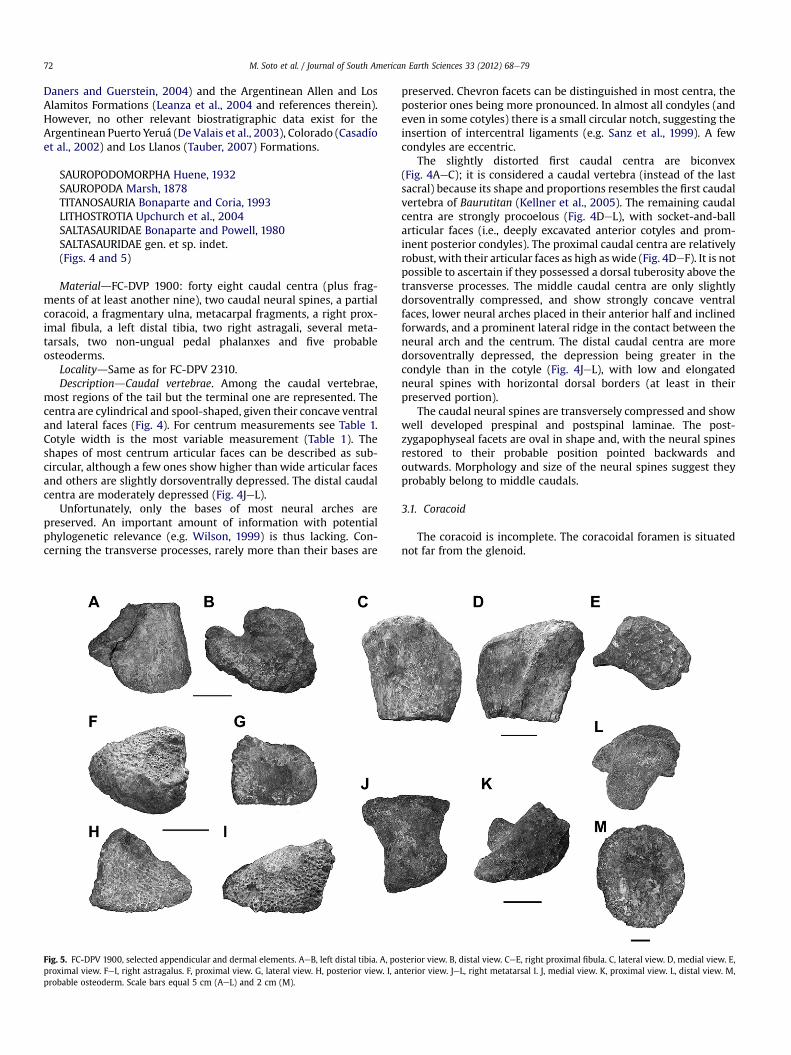
Fig. 5. FC-DPV 1900, selected appendicular and dermal elements. AeB, left distal tibia. A, poproximal view. FeI, right astragalus. F, proximal view. G, lateral view. H, posterior view. I, aprobable osteoderm. Scale bars equal 5 cm (AeL) and 2 cm (M).
preserved. Chevron facets can be distinguished in most centra, theposterior ones being more pronounced. In almost all condyles (andeven in some cotyles) there is a small circular notch, suggesting theinsertion of intercentral ligaments (e.g. Sanz et al., 1999). A fewcondyles are eccentric.
The slightly distorted first caudal centra are biconvex(Fig. 4AeC); it is considered a caudal vertebra (instead of the lastsacral) because its shape and proportions resembles the first caudalvertebra of Baurutitan (Kellner et al., 2005). The remaining caudalcentra are strongly procoelous (Fig. 4DeL), with socket-and-ballarticular faces (i.e., deeply excavated anterior cotyles and prom-inent posterior condyles). The proximal caudal centra are relativelyrobust, with their articular faces as high as wide (Fig. 4DeF). It is notpossible to ascertain if they possessed a dorsal tuberosity above thetransverse processes. The middle caudal centra are only slightlydorsoventrally compressed, and show strongly concave ventralfaces, lower neural arches placed in their anterior half and inclinedforwards, and a prominent lateral ridge in the contact between theneural arch and the centrum. The distal caudal centra are moredorsoventrally depressed, the depression being greater in thecondyle than in the cotyle (Fig. 4JeL), with low and elongatedneural spines with horizontal dorsal borders (at least in theirpreserved portion).
The caudal neural spines are transversely compressed and showwell developed prespinal and postspinal laminae. The post-zygapophyseal facets are oval in shape and, with the neural spinesrestored to their probable position pointed backwards andoutwards. Morphology and size of the neural spines suggest theyprobably belong to middle caudals.
3.1. Coracoid
The coracoid is incomplete. The coracoidal foramen is situatednot far from the glenoid.
sterior view. B, distal view. CeE, right proximal fibula. C, lateral view. D, medial view. E,nterior view. JeL, right metatarsal I. J, medial view. K, proximal view. L, distal view. M,
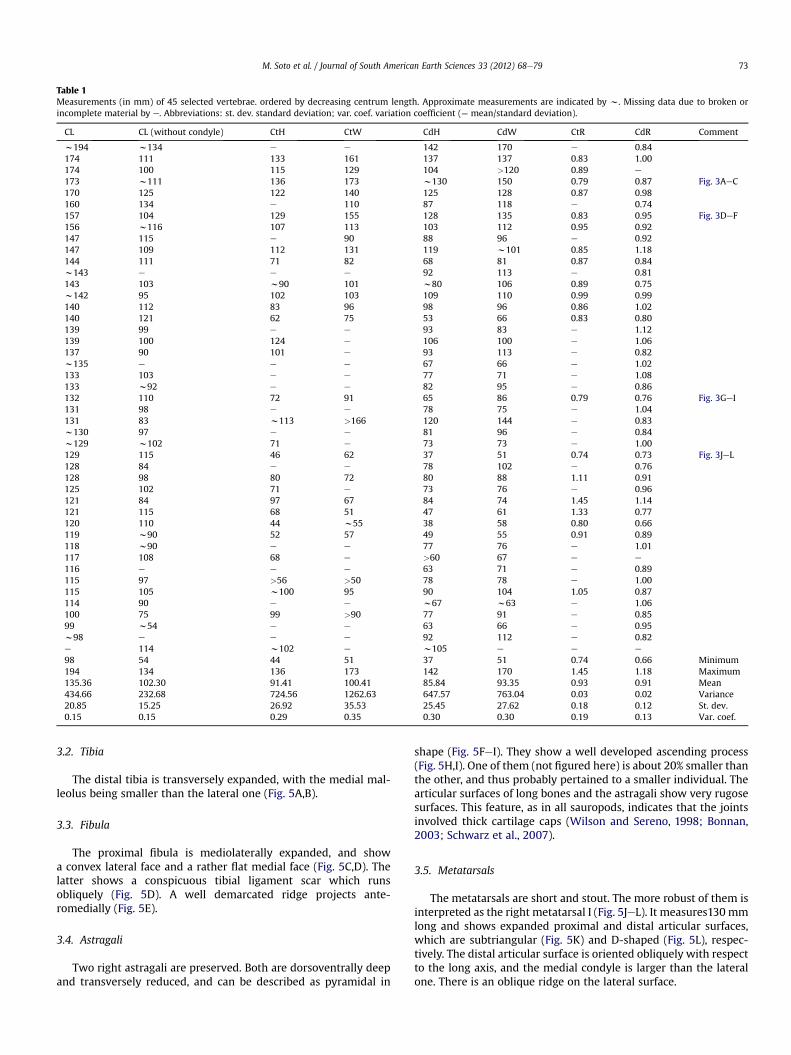
Table 1Measurements (in mm) of 45 selected vertebrae. ordered by decreasing centrum length. Approximate measurements are indicated by w. Missing data due to broken orincomplete material by e. Abbreviations: st. dev. standard deviation; var. coef. variation coefficient (¼ mean/standard deviation).
CL CL (without condyle) CtH CtW CdH CdW CtR CdR Comment
w194 w134 e e 142 170 e 0.84174 111 133 161 137 137 0.83 1.00174 100 115 129 104 >120 0.89 e
173 w111 136 173 w130 150 0.79 0.87 Fig. 3AeC170 125 122 140 125 128 0.87 0.98160 134 e 110 87 118 e 0.74157 104 129 155 128 135 0.83 0.95 Fig. 3DeF156 w116 107 113 103 112 0.95 0.92147 115 e 90 88 96 e 0.92147 109 112 131 119 w101 0.85 1.18144 111 71 82 68 81 0.87 0.84w143 e e e 92 113 e 0.81143 103 w90 101 w80 106 0.89 0.75w142 95 102 103 109 110 0.99 0.99140 112 83 96 98 96 0.86 1.02140 121 62 75 53 66 0.83 0.80139 99 e e 93 83 e 1.12139 100 124 e 106 100 e 1.06137 90 101 e 93 113 e 0.82w135 e e e 67 66 e 1.02133 103 e e 77 71 e 1.08133 w92 e e 82 95 e 0.86132 110 72 91 65 86 0.79 0.76 Fig. 3GeI131 98 e e 78 75 e 1.04131 83 w113 >166 120 144 e 0.83w130 97 e e 81 96 e 0.84w129 w102 71 e 73 73 e 1.00129 115 46 62 37 51 0.74 0.73 Fig. 3JeL128 84 e e 78 102 e 0.76128 98 80 72 80 88 1.11 0.91125 102 71 e 73 76 e 0.96121 84 97 67 84 74 1.45 1.14121 115 68 51 47 61 1.33 0.77120 110 44 w55 38 58 0.80 0.66119 w90 52 57 49 55 0.91 0.89118 w90 e e 77 76 e 1.01117 108 68 e >60 67 e e
116 e e e 63 71 e 0.89115 97 >56 >50 78 78 e 1.00115 105 w100 95 90 104 1.05 0.87114 90 e e w67 w63 e 1.06100 75 99 >90 77 91 e 0.8599 w54 e e 63 66 e 0.95w98 e e e 92 112 e 0.82e 114 w102 e w105 e e e
98 54 44 51 37 51 0.74 0.66 Minimum194 134 136 173 142 170 1.45 1.18 Maximum135.36 102.30 91.41 100.41 85.84 93.35 0.93 0.91 Mean434.66 232.68 724.56 1262.63 647.57 763.04 0.03 0.02 Variance20.85 15.25 26.92 35.53 25.45 27.62 0.18 0.12 St. dev.0.15 0.15 0.29 0.35 0.30 0.30 0.19 0.13 Var. coef.
M. Soto et al. / Journal of South American Earth Sciences 33 (2012) 68e79 73
3.2. Tibia
The distal tibia is transversely expanded, with the medial mal-leolus being smaller than the lateral one (Fig. 5A,B).
3.3. Fibula
The proximal fibula is mediolaterally expanded, and showa convex lateral face and a rather flat medial face (Fig. 5C,D). Thelatter shows a conspicuous tibial ligament scar which runsobliquely (Fig. 5D). A well demarcated ridge projects ante-romedially (Fig. 5E).
3.4. Astragali
Two right astragali are preserved. Both are dorsoventrally deepand transversely reduced, and can be described as pyramidal in
shape (Fig. 5FeI). They show a well developed ascending process(Fig. 5H,I). One of them (not figured here) is about 20% smaller thanthe other, and thus probably pertained to a smaller individual. Thearticular surfaces of long bones and the astragali show very rugosesurfaces. This feature, as in all sauropods, indicates that the jointsinvolved thick cartilage caps (Wilson and Sereno, 1998; Bonnan,2003; Schwarz et al., 2007).
3.5. Metatarsals
The metatarsals are short and stout. The more robust of them isinterpreted as the right metatarsal I (Fig. 5JeL). It measures130 mmlong and shows expanded proximal and distal articular surfaces,which are subtriangular (Fig. 5K) and D-shaped (Fig. 5L), respec-tively. The distal articular surface is oriented obliquely with respectto the long axis, and the medial condyle is larger than the lateralone. There is an oblique ridge on the lateral surface.

M. Soto et al. / Journal of South American Earth Sciences 33 (2012) 68e7974
3.6. Pedal phalanges
Among the non-ungual pedal phalanges, one of them is smalland hemispherical in shape, thus it is identified as the Ie1 phalanx.The remaining phalanxes are similar to the IIe2, IIIe2 and IVe2phalanges figured by González Riga et al. (2008).
3.7. Osteoderms
Some remains of similar size (diameter ranging from 5 to 7 cm)and simple, discoidal shape (with flat dorsal and ventral surfaces)are herein interpreted as probable non-keeled osteoderms(Fig. 5M), although histological sections need to bemade in order toconfirm their nature.
CommentsdGiven the presence of two astragali of dissimilarsize and the high number of caudal centra (some lithostrotiansshow less than 35 caudal vertebrae; Wilson, 2002) it is probablethat more than one individual is represented. However, most of thebones are likely to belong to a single, medium-sized individual,probably achieving a total length of 12e14 m, which agree with thesize reported for Baurutitan (Kellner et al., 2005). Indeed, caudalcentrum in FC-DPV 1900 and Baurutitan broadly overlaps in size.
The anterior shift of neural arches in middle and distal caudalvertebrae is diagnostic of Titanosauriformes (Salgado et al., 1997a;Wilson and Sereno, 1998; Upchurch et al., 2004). The presence ofprespinal and postspinal laminae in proximal andmiddle caudals isconsidered synapomorphic of Titanosauria (Wilson and Sereno,1998; Wilson, 2002, 2005).
The presence of strongly procoelous proximal caudals(Fig. 5DeF) is a characteristic feature of lithostrotian titanosaurs(e.g. Sanz et al., 1999; Wilson, 2002), while strongly procoelousmiddle and distal caudals (Fig. 5GeL) are only present in derivedlithostrotians (e.g. Wilson, 2002; González Riga, 2003; Salgado,2003a; Upchurch et al., 2004), the single exception being theChinese eusauropod Mamenchisaurus (e.g. Wilson et al., 1999;Upchurch et al., 2004). The presence of a biconvex first caudalcentra has been proposed as a synapomorphy of the family Sal-tasauridae (Wilson, 2002, 2005, 2006).
The transverse expansion of the distal tibia is also considereda synapomorphy of Titanosauria (Wilson and Sereno, 1998; Wilson,2002, 2005).
The pyramidal astragali (Fig. 5FeH) recall the condition seen inother titanosaurs (e.g. Borsuk-Bialynicka, 1977; Salgado et al.,1997b, 2005; Powell, 2003; González Riga et al., 2008; Otero,2010). Indeed, the pyramidal shape has been proposed as a syna-pomorphy of the family Saltasauridae (Wilson, 2002, 2005, 2006).
Metatarsal shape and proportions closely resemble those ofseveral titanosaurian taxa described by other authors (You et al.,2003; Martínez et al., 2004; Bonaparte et al., 2006; González Rigaet al., 2008; Curry Rogers, 2009; Otero, 2010).
The probable osteoderms recovered in the Guichón Formation(Fig. 5M) are more similar morphologically to those of the discoidaltype (sensu D’Emic et al., 2009). Many titanosaur remains fromAfrica, Madagascar, Europe and particularly South America includeosteoderms (e.g. Huene,1929b; Jacobs et al., 1993; Curry Rogers andForster, 2001; Powell, 2003; Salgado, 2003b; for a review seeD’Emic et al. (2009)), a feature which at least in some cases can becorrelated to reduced body size.
Despite the incompleteness of the materials, comparisons withseveral lithostrotian taxa can be made. FC-DPV 1900 can be readilydifferentiated from taxawith procoelous first caudal centra, such asEpachthosaurus sciuttoi (Martínez et al., 2004), Paralititan stromeri(Smith et al., 2001), Saltasaurus loricatus (Powell, 1992), Bonitasaurasalgadoi (Apesteguía, 2004), Lirainosaurus astibiae (Sanz et al.,1999), Futalognkosaurus dukei (Calvo et al., 2007b) and
Gondwanatitan faustoi (Kellner and de Azevedo, 1999). The absenceof wide ventral fossae and/or midline ridges, and the fact thatcaudal centrum width never doubles centrum height allows toexclude the Saltasaurinae (Salgado et al., 1997a, 2005; Powell,1992;Salgado and Azpilicueta, 2000; Wilson, 2002). The absence ofanterior faces strongly inclined forward and neural archs posi-tioned very anteriorly on the centrum also allows to exclude the“Aeolosaurini” (Salgado and Coria, 1993; Salgado et al., 1997b;Kellner and de Azevedo, 1999; Franco-Rosas et al., 2004; see alsoCalvo et al., 2007a). There are no condyles that can be described asdorsally displaced as in Mendozasaurus (González Riga, 2003) norconstricted as in Lirainosaurus (Sanz et al., 1999), although in distalcaudals the high dorsoventral depression of the condyle in relationto the cotyle resemble the condition in Muyelensaurus pecheni(Calvo et al., 2007a). Finally, FC-DPV 1900 with its strongly pro-coelous tail clearly deviates from the tails of taxa such as Opistho-coelicaudia (Borsuk-Bialynicka, 1977), Rinconsaurus (Calvo andGonzález Riga, 2003), Malarguesaurus (González Riga et al., 2009)and Malawisaurus (Jacobs et al., 1993; Gomani, 2005).
Considering all the available evidence, FC-DPV 1900 is a derivedlithostrotian, probably a non-saltasaurine saltasaurid related toAlamosaurus sanjuanensis, Pellegrinisaurus powelli and Baurutitanbritoi. We are cautious with the use of Saltasauridae given that itstaxonomic content varies considerably among different phyloge-nies, ranging from very inclusive (e.g. Curry Rogers, 2005) to theopposite extreme (e.g. Upchurch et al., 2004).
ImplicationsdAs far as we know no lithostrotians with stronglyprocoelous middle and posterior caudals have been reported fromdeposits older than Cenomanian. Indeed, E. sciuttoi from the LateCenomanianeEarly Turonian of Argentina is regarded as the mostbasal lithostrotian with procoelous caudals throughout the tail(Salgado, 2003a; Martínez et al., 2004). Moreover, basal titanosaurswith amphyplatian/platycoelous middle and posterior caudalscome from ?Aptian (Jacobs et al., 1993; Gomani, 2005) to earlyCenomanian (Leanza et al., 2004) units (although rare forms arereported from the late Turonian/early Coniacian; Salgado andCalvo, 1993; Leanza et al., 2004), while taxa with slightly procoe-lous caudals come from TuronianeConiacian strata (González Riga,2003). In other regions of the world, strongly procoelous caudalsare only rarely found in units predating the Late Cretaceous(Averianov et al., 2003).
Within Lithostrotia, a biconvex first caudal centrum has onlybeen reported in taxa from late Late Cretaceous units of Argentina(P. powelli and several Titanosauria indet.; Lydekker, 1893; Huene,1929b; Salgado, 1996; Powell, 2003; Salgado et al., 2005; D’Emicand Wilson, 2011), Brazil (B. britoi; Powell, 1987b; Kellner et al.,2005), Pakistan (Titanosauria indet.; Malkani et al., 2001), and theUSA (A. sanjuanensis; Gilmore, 1946). However, as D’Emic andWilson (2011) state, in some of this forms the purported firstcaudal may be instead a last sacral, as was recently interpreted forNeuquensaurus australis (Salgado et al., 2005; D’Emic and Wilson,2011).
A procoelous first caudal is the character-state present in mosttitanosaurians for which it can be scored (e.g. Powell, 1992; Sanzet al., 1999; Smith et al., 2001; Martínez et al., 2004; Apesteguía,2004; Calvo et al., 2007a). A clade of derived, non-saltasaurinelithostrotians sharing the biconvex first caudal, including Alamo-saurus and Pellegrinisaurus (as recovered by Upchurch et al., 2004)and may be other forms, is plausible, although this hypothesis hasyet to be phylogenetically tested. In support of this, Baurutitan,Alamosaurus and FC-DPV 1900 share the presence of a dorsaltuberosity (in the contact between the neural arch and the trans-verse processes) that continues in posterior caudals as a prominentlateral ridge (Kellner et al., 2005). Pellegrinisaurus, Alamosaurus andFC-DPV 1900 shares the presence of depressed distal caudal
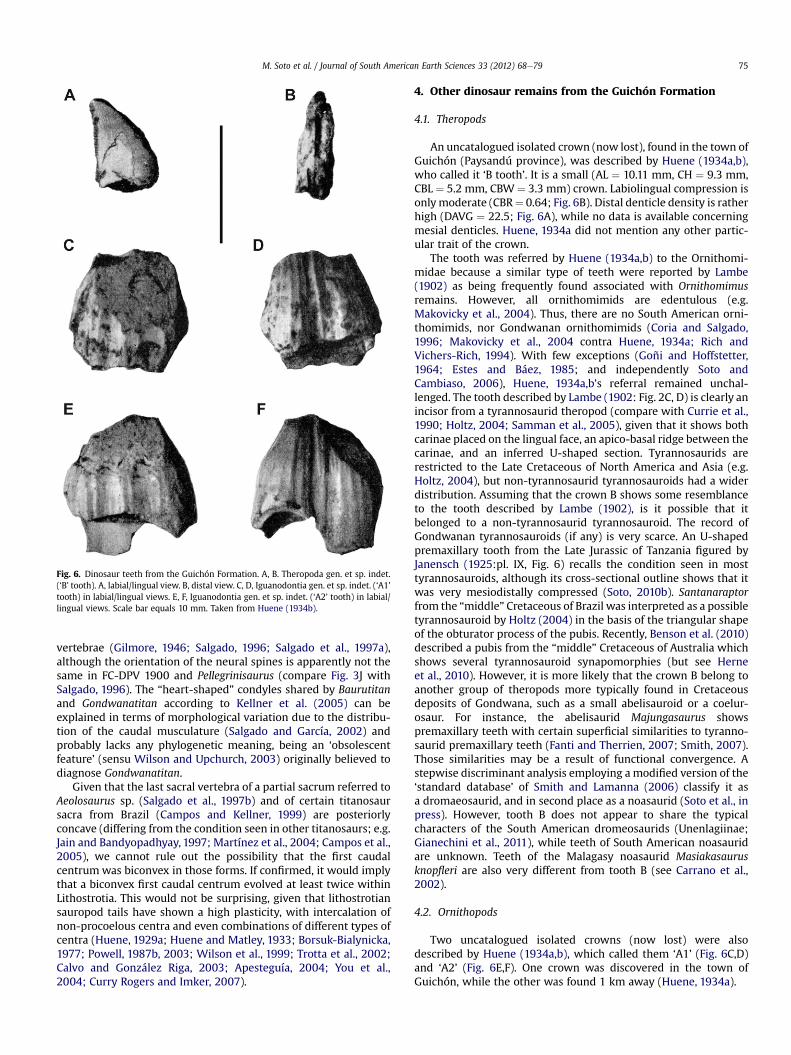
Fig. 6. Dinosaur teeth from the Guichón Formation. A, B. Theropoda gen. et sp. indet.(‘B’ tooth). A, labial/lingual view. B, distal view. C, D, Iguanodontia gen. et sp. indet. (‘A1’tooth) in labial/lingual views. E, F, Iguanodontia gen. et sp. indet. (‘A2’ tooth) in labial/lingual views. Scale bar equals 10 mm. Taken from Huene (1934b).
M. Soto et al. / Journal of South American Earth Sciences 33 (2012) 68e79 75
vertebrae (Gilmore, 1946; Salgado, 1996; Salgado et al., 1997a),although the orientation of the neural spines is apparently not thesame in FC-DPV 1900 and Pellegrinisaurus (compare Fig. 3J withSalgado, 1996). The “heart-shaped” condyles shared by Baurutitanand Gondwanatitan according to Kellner et al. (2005) can beexplained in terms of morphological variation due to the distribu-tion of the caudal musculature (Salgado and García, 2002) andprobably lacks any phylogenetic meaning, being an ‘obsolescentfeature’ (sensu Wilson and Upchurch, 2003) originally believed todiagnose Gondwanatitan.
Given that the last sacral vertebra of a partial sacrum referred toAeolosaurus sp. (Salgado et al., 1997b) and of certain titanosaursacra from Brazil (Campos and Kellner, 1999) are posteriorlyconcave (differing from the condition seen in other titanosaurs; e.g.Jain and Bandyopadhyay, 1997; Martínez et al., 2004; Campos et al.,2005), we cannot rule out the possibility that the first caudalcentrumwas biconvex in those forms. If confirmed, it would implythat a biconvex first caudal centrum evolved at least twice withinLithostrotia. This would not be surprising, given that lithostrotiansauropod tails have shown a high plasticity, with intercalation ofnon-procoelous centra and even combinations of different types ofcentra (Huene, 1929a; Huene and Matley, 1933; Borsuk-Bialynicka,1977; Powell, 1987b, 2003; Wilson et al., 1999; Trotta et al., 2002;Calvo and González Riga, 2003; Apesteguía, 2004; You et al.,2004; Curry Rogers and Imker, 2007).
4. Other dinosaur remains from the Guichón Formation
4.1. Theropods
An uncatalogued isolated crown (now lost), found in the town ofGuichón (Paysandú province), was described by Huene (1934a,b),who called it ‘B tooth’. It is a small (AL ¼ 10.11 mm, CH ¼ 9.3 mm,CBL ¼ 5.2 mm, CBW ¼ 3.3 mm) crown. Labiolingual compression isonlymoderate (CBR¼ 0.64; Fig. 6B). Distal denticle density is ratherhigh (DAVG ¼ 22.5; Fig. 6A), while no data is available concerningmesial denticles. Huene, 1934a did not mention any other partic-ular trait of the crown.
The tooth was referred by Huene (1934a,b) to the Ornithomi-midae because a similar type of teeth were reported by Lambe(1902) as being frequently found associated with Ornithomimusremains. However, all ornithomimids are edentulous (e.g.Makovicky et al., 2004). Thus, there are no South American orni-thomimids, nor Gondwanan ornithomimids (Coria and Salgado,1996; Makovicky et al., 2004 contra Huene, 1934a; Rich andVichers-Rich, 1994). With few exceptions (Goñi and Hoffstetter,1964; Estes and Báez, 1985; and independently Soto andCambiaso, 2006), Huene, 1934a,b’s referral remained unchal-lenged. The tooth described by Lambe (1902: Fig. 2C, D) is clearly anincisor from a tyrannosaurid theropod (compare with Currie et al.,1990; Holtz, 2004; Samman et al., 2005), given that it shows bothcarinae placed on the lingual face, an apico-basal ridge between thecarinae, and an inferred U-shaped section. Tyrannosaurids arerestricted to the Late Cretaceous of North America and Asia (e.g.Holtz, 2004), but non-tyrannosaurid tyrannosauroids had a widerdistribution. Assuming that the crown B shows some resemblanceto the tooth described by Lambe (1902), is it possible that itbelonged to a non-tyrannosaurid tyrannosauroid. The record ofGondwanan tyrannosauroids (if any) is very scarce. An U-shapedpremaxillary tooth from the Late Jurassic of Tanzania figured byJanensch (1925:pl. IX, Fig. 6) recalls the condition seen in mosttyrannosauroids, although its cross-sectional outline shows that itwas very mesiodistally compressed (Soto, 2010b). Santanaraptorfrom the “middle” Cretaceous of Brazil was interpreted as a possibletyrannosauroid by Holtz (2004) in the basis of the triangular shapeof the obturator process of the pubis. Recently, Benson et al. (2010)described a pubis from the “middle” Cretaceous of Australia whichshows several tyrannosauroid synapomorphies (but see Herneet al., 2010). However, it is more likely that the crown B belong toanother group of theropods more typically found in Cretaceousdeposits of Gondwana, such as a small abelisauroid or a coelur-osaur. For instance, the abelisaurid Majungasaurus showspremaxillary teeth with certain superficial similarities to tyranno-saurid premaxillary teeth (Fanti and Therrien, 2007; Smith, 2007).Those similarities may be a result of functional convergence. Astepwise discriminant analysis employing a modified version of the‘standard database’ of Smith and Lamanna (2006) classify it asa dromaeosaurid, and in second place as a noasaurid (Soto et al., inpress). However, tooth B does not appear to share the typicalcharacters of the South American dromeosaurids (Unenlagiinae;Gianechini et al., 2011), while teeth of South American noasauridare unknown. Teeth of the Malagasy noasaurid Masiakasaurusknopfleri are also very different from tooth B (see Carrano et al.,2002).
4.2. Ornithopods
Two uncatalogued isolated crowns (now lost) were alsodescribed by Huene (1934a,b), which called them ‘A1’ (Fig. 6C,D)and ‘A2’ (Fig. 6E,F). One crown was discovered in the town ofGuichón, while the other was found 1 km away (Huene, 1934a).

M. Soto et al. / Journal of South American Earth Sciences 33 (2012) 68e7976
Although incomplete, A1 and A2 seem to have been diamond-shaped crowns, given that the mesial and distal vertices werecloser to the root than to the apex (Fig. 6CeF). Marginal denticlesare absent. The teeth show on the unworn convex face a narrowprimary ridge (Fig. 6D,F). There are four other narrow ridges,separated from the primary one by a smooth region. Strikinggrooved ridges are present on the opposite face (Fig. 6E), whichcould represent an autapomorphic trait. Maximum preservedmeasurements given by Huene for A1 and A2: maximumheight ¼ 11.5 and 12 mm, maximum length ¼ 11.7 and 12.1 mm,and maximum width ¼ 6.9 and 5.3 mm, respectively.
Huene, 1934a,b stated there were no doubt A1 and A2 belongedto an ornithischian dinosaur, finding the greatest similarities withthe ‘Camptosaurinae’, ‘Psittacosaurinae’, and ‘Leptoceratopsinae’(sensu Nopcsa, 1928). Later, Bonaparte (1978) referred them to theIguanodontidae.
Diamond-shaped crowns in which mesial and distal vertices areclose to the root recall the condition seen in Dryomorpha (Sereno,1986), while basal ornithopods show subtriangular crowns, withmesial and distal vertices closer to the apex. Marginal denticlesreduced or absent is a trait present in certain hadrosaurids (Sau-rolophinae and Bactrosaurus; Prieto-Márquez, 2010), and preciselysome Argentinean forms have been identified as saurolophines(Prieto-Márquez and Salinas, 2010). However, we consider that theabsence of marginal denticles in the Uruguayan specimens isprobably due to heavy wear. The grooved ridges, unknown in otherornithischians, could be an autapomorphic trait. The narrowprimary ridge present in the convex face of A1 recall maxillarycrowns of non-dryomorph iguanodontians and dentary crowns ofiguanodontians. Although the presence of a subtle primary ridge isa relevant character, we do not know if A1 and A2 are maxillary ordentary crowns. Both possibilities will be considered.
Maxillary crowns of non-dryomorph iguanodontians either donot show a developed primary ridge (e.g. Tenontosaurus), or theprimary ridge is not prominent and less pronounced than that ofthe dentary crowns (e.g., Muttaburrasaurus). In Dryomorpha (i.e.,Dryosaurus and more derived ornithopods) both maxillary anddentary crowns bear a primary ridge; the labial face of maxillarycrowns show a primary ridge which is more developed than that ofthe lingual face of dentary crowns (Sereno, 1986; Norman, 2004).Moreover, a diamond-shaped crown is another synapomorphy ofthe clade. In Ankylopollexia (i.e., Camptosaurus and more derivedornithopods) there is a prominent primary ridge in the labial face ofthe maxillary crowns (Sereno, 1986). This is not the case of theUruguayan specimens, the primary ridge of which is poorlydeveloped. In turn, the presence of a primary ridge in the lingualface of dentary crowns is a synapomorphy of Iguanodontia (Sereno,1986, 1999).
The presence of few narrow, longitudinal secondary ridges (asopposed to the wide secondary ridges typical of basal ornithopods)recalls the condition present in most iguanodontian. Dryosaurusand Camptosaurus show few secondary ridges, while iguano-dontians more primitive than Tenontosaurus show a higher numberof secondary ridges, like in the Uruguayan specimens.
On the basis of the presence of both a diamond-shaped crownand a primary ridge, and taking into account that it is not possibleto distinguish if A1 and A2 are maxillary or dentary crowns, themost cautious assignment would be to Iguanodontia indet.Although diamond-shaped crowns were considered a synapomor-phy of Dryomorpha (Sereno, 1986), the non-dryomorph basaliguanodontian Tenontosaurus possesses such type of crowns. Inconclusion, A1 and A2 belonged to an indeterminate iguano-dontian, (probably to a non-dryomorphan iguanodontian). Thebiochron of Iguanodontia reaches back to the Callovian (Norman,2004), although basal iguanodontians are known in South
America in deposits ranging in age from Cenomanian to Maas-trichtian (e.g., Coria and Salgado, 1996; Coria and Calvo, 2002;Novas et al., 2004; Calvo et al., 2007c).
5. Discussion
Herein it has been shown that the purported ornithomimid andiguanodontid material should be better regarded as belonging toindeterminate theropods and basal iguanodontians, respectively.Thus, they do not support a Late Cretaceous age for the GuichónFormation (contra Huene, 1934a and Bonaparte, 1978), hence theimportance of the sauropod remains herein described for sheddingsome light on the age of the Guichón Formation (and thus of thepartly correlated Puerto Yeruá Formation).
As stated in Section 3. Systematic Paleontology, within Lithos-trotia, a biconvex first caudal centrum has only been reported intaxa from late Late Cretaceous units of Argentina. Moreover,Sphaerovum or Sphaerovum-like remains have been reported so farfrom several CampanianeMaastrichtian units, leading Casadío et al.(2002) to consider it as an index fossil.
Despite the late Late Cretaceous age apparently suggested by thebones and eggshells described above, the basal position of Uru-guaysuchus within Notosuchia recovered in several recent phylo-genetic analyses (e.g. Pol and Apesteguía, 2005; Turner, 2006; Sotoet al., in press) and comparisons with the composition of LateCretaceous assemblages from the Neuquén and Baurú basinssuggest that an early Late Cretaceous age for the Guichón Forma-tion is more likely than a late Late Cretaceous one. If we admit thatthe ghost lineage of lithostrotians with biconvex first caudal mayhave reached back to the Turonian (according to recent phyloge-nies; e.g. Calvo et al., 2007a,b; González Riga et al., 2009), there isno contradiction with what have been stated before. Overall, a LateCretaceous age for the Guichón Formation is suggested by thefossils known so far, and thus the hiatus between this unit and theoverlying Mercedes Formation (CampanianeMaastrichtian) wouldbe of less relevance than proposed by Goso (1999) and Goso andPerea (2004).
6. Conclusions
The skeletal remains described herein are clearly referrable toa derived lithostrotian sauropod, probably related to Pelle-grinisaurus, Alamosaurus and Baurutitan. The eggshell fragmentsare tentatively referred to Sphaerovum, due to their thickness andcompactituberculate ornamentation, pending a detailed compar-ison with other eggshell materials from Uruguay.
These findings are important because: (i) they constitute thefirst record of dinosaur bones and eggshells from the GuichónFormation; (ii) they represent the most complete and diagnosticdinosaur materials ever found in Uruguay; (iii) this is the first timea clear spatial association among dinosaurian eggshells and bonesis reported from Uruguay; and (iv) the simultaneous occurrence ofa derived lithostrotian and Sphaerovum provides an importantbiostratigraphic criterion to allocate the Guichón Formation (andthe correlated Puerto Yeruá Formation) to the Late Cretaceous.
Acknowledgments
Thanks are due to R. Beasley and personnel of Forestal OrientalS.A., who communicated the findings and provided fieldworksupport. S. Apesteguía, J. F. Bonaparte, P. Galton, A. Rinderknetch, L.Salgado, A. Schipilov, M. Ubilla, G. Veroslavsky, and J. A. Wilson areacknowledged for useful discussions. J. De León, R. Guzmán and R.Juárez-Valieri provided bibliographic references. J. A. Whitlock andan anonymous reviewer greatly contribute to improve an early

M. Soto et al. / Journal of South American Earth Sciences 33 (2012) 68e79 77
draft of this manuscript. This is a contribution to CSIC Project “ElMesozoico de Uruguay (Jurásico Tardío-Cretácico Tardío): biotas,estratigrafía, geocronología y ambientes”.
References
Apesteguía, S., 2004. Bonitasaura salgadoi gen. et sp. nov.: a beaked sauropod fromthe Late Cretaceous of Patagonia. Naturwissenschaften 91, 493e497.
Averianov, A., Starkov, A., Skutchas, P., 2003. Dinosaurs from the early CretaceousMurtoi Formation in Buryatia, Eastern Russia. Journal of Vertebrate Paleon-tology 23, 586e594.
Benson, R.B.J., Barrett, P.M., Rich, T.H., Vickers-Rich, P., 2010. A southern tyrantreptile. Science 327, 1613.
Bonaparte, J.F., 1978. El mesozoico de América del Sur y sus tetrápodos. Opera Lil-loana 26, 1e596.
Bonaparte, J.F., Coria, R.A., 1993. Un nuevo y gigantesco saurópodo Titanosaurio dela Formación Río Limay (AlbianoeCenomaniano) de la provincia del Neuquén,Argentina. Ameghiniana 30, 271e282.
Bonaparte, J.F., González Riga, B.J., Apesteguía, S., 2006. Ligabuesaurus leanzai nov.gen. et sp., a new titanosaur from the Aptian of Patagonia, Argentina. Creta-ceous Research 27, 364e376.
Bonaparte, J.F., Powell, J.E., 1980. A continental assemblage of tetrapods from theUpper Cretaceous beds of El Brete, northwestern Argentina (Sau-ropodaeCoelurosauriaeCarnosauriaeAves). Memoire de la Societé Geologiquede France, N. S. 139, 19e28.
Bonnan, M.F., 2003. The evolution of manus shape in sauropod dinosaurs: impli-cations for functional morphology, forelimb orientation, and phylogeny. Journalof Vertebrate Paleontology 23, 595e613.
Borsuk-Bialynicka, M., 1977. A new camarasaurid sauropod Opisthocoelicaudiaskarzynskii, gen. n., sp. n. from the Upper Cretaceous of Mongolia. Acta Palae-ontologica Polonica 37, 1e64.
Bossi, J., 1966. Geología del Uruguay. Departamento de Publicaciones de la Uni-versidad de la República, Montevideo, p. 469.
Bossi, J., Ferrando, L., 2001. Carta geológica del Uruguay. Escala 1:500.000 EdiciónGeoeditores SRL (CDeROM). Montevideo.
Bossi, J., Navarro, R., 1991. Geología del Uruguay. Departamento de Publicaciones dela Universidad de la República, Montevideo, p. 967.
Bossi, J., Preciozzi, F., Campal, N., 1993. Predevoniano del Uruguay e Parte I: TerrenoPiedra Alta. DINAMIGE, Montevideo, p. 50.
Calvo, J.O., González Riga, B.J., 2003. Rinconsaurus caudamirus gen. et sp. nov., a newtitanosaurid (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia,Argentina. Revista Geológica de Chile 30, 333e353.
Calvo, J.O., González Riga, B.J., Porfiri, J.D., 2007a. A New Titanosaur Sauropod fromthe Late Cretaceous of Neuquén, Patagonia, Argentina, vol. 65. Arquivos doMuseu Nacional, Rio do Janeiro. 485e504.
Calvo, J.O., Porfiri, J.D., González Riga, B.J., Kellner, A.W.A., 2007b. Anatomy ofFutalognkosaurus dukei Calvo, Porfiri, González Riga and Kellner, 2007 (Dino-sauria, Titanosauridae) from the Neuquén Group (Late Cretaceous), Patagonia,Argentina, vol. 65. Arquivos do Museu Nacional, Rio do Janeiro. 511e526.
Calvo, J.O., Porfiri, J.D., Novas, F.E., 2007c. Discovery of a New Ornithopod Dinosaurfrom the Portezuelo Formation (Upper Cretaceous), Neuquén, Patagonia,Argentina, vol. 65. Arquivos do Museu Nacional, Rio do Janeiro. 471e483.
Campos, C., García, J., Dino, R., Veroslavsky, G., 1997. Registro de palinomorfos naFormação Migues (Poço SL11eSP1), Bacia de Santa LucíaeUruguai, vol. II.Revista Universidade Guarulhos, Geociências. 85e97.
Campos, A.D., Kellner, A.W.A., 1999. On some (Titanosauridae) pelves from thecontinental Cretaceous of Brazil. In: Tomida, Y., Rich, T.H., Vickers-Rich, P. (Eds.),Proceedings of the Second Gondwanan Dinosaur Symposium, National ScienceMuseum Monographs, vol. 15, pp. 143e166.
Campos, D.A., Kellner, A.W.A., Bertini, R.J., Santucci, R.M., 2005. On a titanosaurid(Dinosauria, Sauropoda) vertebral column from the Bauru Group, Late Creta-ceous of Brazil. Arquivos do Museu Nacional 63, 565e593.
Carrano, M.T., Sampson, S.D., Forster, C.A., 2002. The osteology of Masiakasaurusknopfleri, a small abelisauroid (Dinosauria: Theropoda) from the Late Creta-ceous of Madagascar. Journal of Vertebrate Paleontology 22, 510e534.
Casadío, S., Manera, T., Parras, A., Montalvo, C.I., 2002. Huevos de dinosaurios(Faveoloolithidae) del Cretácico Superior de la cuenca del Colorado, provinciade La Pampa, Argentina. Ameghiniana 39, 285e293.
Chiappe, L.M., Coria, R.A., Dingus, L., Jackson, F., Chinsamy, A., Fox, M., 1998.Sauropod dinosaur embryos from the Late Cretaceous of Patagonia. Nature 396,258e261.
Coria, R.A., Calvo, J.O., 2002. A new iguanodontian ornithopod from Neuquén Basin,Patagonia, Argentina. Journal of Vertebrate Paleontology 22, 503e509.
Coria, R.A., Salgado, L., 1996. “Loncosaurus argentinus” Ameghino, 1899 (Ornithi-schia, Ornithopoda): a revised description with comments on its phylogeneticrelationships. Ameghiniana 33, 373e376.
Currie, P.J., Rigby Jr., J.K., Sloan, R.E., 1990. Theropod teeth from the Judith RiverFormation of southern Alberta, Canada. In: Carpenter, K., Currie, P.J. (Eds.),Dinosaur Systematics: Perspectives and Approaches. Cambridge UniversityPress, Cambridge, pp. 107e125.
Curry Rogers, K.A., 2005. Titanosauria: a phylogenetic overview. In: Curry Roger-s, K.A., Wilson, J.A. (Eds.), The Sauropods: Evolution and Paleobiology. IndianaUniversity Press, pp. 50e103.
Curry Rogers, K.A., 2009. The postcranial osteology of Rapetosaurus krausei (Sau-ropoda: Titanosauria) from the Late Cretaceous of Madagascar. Journal ofVertebrate Paleontology 29, 1046e1086.
Curry Rogers, K.A., Forster, C., 2001. The last of the dinosaur titans: a new sauropodfrom Madagascar. Nature 412, 530e534.
Curry Rogers, K.A., Imker,M., 2007. Newdata on ‘Malagasy taxon B’, a titanosaur fromthe Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 27, 64A.
Daners, G., Guerstein, R., 2004. Dinoflagelados del Maastrichtiense-Paleógeno en laFormación Gaviotín, cuenca Punta del Este. In: Veroslavsky, G., Ubilla, M.,Martínez, S. (Eds.), Cuencas Sedimentarias de Uruguay: Geología, Paleontologíay recursos naturales e Cenozoico. DIRACeFacultad de Ciencias, Montevideo,pp. 37e62.
D’Emic, M.D., Wilson, J.A., Chatterjee, S., 2009. The titanosaur (Dinosauria: Sau-ropoda) osteoderm record: review and first definitive specimen from India.Journal of Vertebrate Paleontology 29, 165e177.
D’Emic, M.D., Wilson, J.A., 2011. New remains attributable to the holotype of thesauropod dinosaur Neuquensaurus australis, with implications for saltasaurinesystematics. Acta Paleontologica Polonica 56, 61e73.
de Santa Ana, H., Veroslavsky, G., Fúlfaro, V., Rossello, E., 2006. Cuenca Norte:evolución tectónica y sedimentaria del Carbonífero-Pérmico. In: Veroslavsky, G.,Ubilla, M., Martínez, S. (Eds.), Cuencas sedimentarias de Uruguay: geología,paleontología y recursos naturales. Paleozoico. DIRAC, Montevideo,pp. 209e256.
De Valais, S., Apesteguía, S., Udrizar Sauthier, D., 2003. Nuevas evidencias dedinosaurios de la Formación Puerto Yeruá (Cretácico), Provincia de Entre Ríos,Argentina. Ameghiniana 40, 507e511.
Estes, R., Báez, A.M., 1985. Herpetofaunas of North and South America during theLate Cretaceous and Cenozoic: evidence for interchange? In: Stehli, F.G.,Webb, S.D. (Eds.), The Great American Biotic Interchange. Plenum Press, NewYork, pp. 139e197.
Faccio, G., 1994. Dinosaurian eggs from the Upper Cretaceous of Uruguay. In:Carpenter, K., Horner, J., Hirsch, K.F. (Eds.), Dinosaur Eggs and Babies. CambridgeUniversity Press, pp. 7e55.
Faccio, G., Ford, I., Gancio, F., 1990. Primer registro fósil in situ de huevos dedinosaurios del Uruguay (Fm. Mercedes). Facultad de Agronomía, Boletín deInvestigación 26, 1e20.
Faccio, G., Montaña, J., 1994. Registro fósil in situ de huevos de dinosaurios del Cre-tácico Superior deDuraznoy la presencia de Faveoloolithidae (Zhao et Ding 1976)en huevos de dinosaurios del Uruguay. Paleociencias Serie Didáctica 2, 16e18.
Fanti, F., Therrien, F., 2007. Theropod tooth assemblages from the Late CretaceousMaevarano Formation and the possible presence of dromaeosaurids in Mada-gascar. Acta Paleontologica Polonica 52, 155e166.
Ford, I., Gancio, F., 1988. Asociación caolinita-montmorillonita en un paleosuelodel Terciario Inferior del Uruguay. Facultad de Agronomía, Boletín de Inves-tigación 12.
Franco-Rosas, A.C., Salgado, L., Rosas, C.F., Carvalho, I. de S., 2004. Nuevos materialesde titanosaurios (Sauropoda) en el Cretácico Superior de Mato Grosso, Brasil.Revista Brasileira de Paleontologia 7, 329e336.
Gianechini, F.A., Makovicky, P.J., Apesteguía, S., 2011. The teeth of the unenlagiinetheropod Buitreraptor from the Cretaceous of Patagonia, Argentina, and theunusual dentition of the Gondwanan dromaeosaurids. Acta PalaeontologicaPolonica 56, 279e290.
Gilmore, C.W., 1946. Reptilian fauna of the North Horn Formation of central Utah.Geological Survey Professional Paper 210C, 29e52.
Gomani, E.M., 2005. Sauropod dinosaurs from the Early Cretaceous of Malawi,Africa. Palaeontologia Electronica 8 (1), 1e37. http://palaeoelectronica.org/paleo/2005_1/gomani27/issue1_05.htm.
González Riga, B.J., 2003. A new titanosaur (Dinosauria, Sauropoda) from the UpperCretaceous of Mendoza Province, Argentina. Ameghiniana 40, 155e172.
González Riga, B.J., Calvo, J.O., Porfiri, J., 2008. An articulated titanosaur from Pata-gonia (Argentina): new evidences of the pedal evolution. Palaeoworld 17, 33e40.
González Riga, B.J., Previtera, E., Pirrone, C., 2009. Malarguesaurus florenciae gen. etsp. nov., a new titanosauriform (Dinosaurioa, Sauropoda) from the UpperCretaceous of Mendoza, Argentina. Cretaceous Research 30, 135e148.
Goñi, J.C., Hoffstetter, R., 1964 (Amérique Latine, 9�). Uruguay. Lexique Strati-graphique International, vol. 5. CNRS, Paris. 1e200.
Goso, C., 1999. Análise estratigráfica do Grupo Paysandú (Cretáceo) na Bacia doLitoral Uruguai. IGCEeUNESP, Rio Claro, Unpublished PhD Dissertation, p. 184.
Goso, C., Perea, D., 2004. El Cretácico post-basáltico de la Cuenca litoral del ríoUruguay: geología y paleontología. In: Veroslavsky, G., Ubilla, M., Martínez, S.(Eds.), Cuencas sedimentarias de Uruguay: geología, paleontología y recursosnaturales. Mesozoico, second ed. DIRAC, Montevideo, pp. 143e171.
Grigorescu, D., Garcia, G., Csiki, Z., Codrea, V., Bojar, A.-V., 2010. Uppermost Creta-ceous megaloolithid eggs from the Hateg Basin, Romania, associated withhadrosaur hatchlings: Search for explanation. Palaeogeography, Palae-oclimatology, Palaeoecology 293, 360e374.
Herne, M.C., Nayr, J.P., Salisbury, S.W., 2010. Comment on “a southern tyrantreptile”. Science 329, 1013.
Holtz Jr., T.R., 2004. Tyrannosauroidea. In: Weishampel, D.B., Dodson, P.,Osmólska, H. (Eds.), The Dinosauria, second Edition. University of CaliforniaPress, Berkeley, pp. 111e136.
Huene, F. von, 1929a. Terrestrische Oberkreide in Uruguay. Zentralblatt für Geologie,Mineralogie und Paläontologie, Ab. B. 4, 107e112.
Huene, F. von, 1929b. Los Saurisquios y Ornitisquios del Cretáceo Argentino. Analesdel Museo de La Plata 3 (2a serie), 1e196.

M. Soto et al. / Journal of South American Earth Sciences 33 (2012) 68e7978
Huene, F. von, 1932. Die fossile ReptileOrdnung Saurischia, ihre Entwicklung undGeschichte. Monographie Geologie Paläontologie 1941B, 1e34.
Huene, F. von, 1934a. Nuevos dientes de Saurios del Cretáceo del Uruguay. Boletíndel Instituto Geológico de Uruguay 21, 13e20.
Huene, F. von, 1934b. Neue Saurier-Zähne aus der Kreide von Uruguay. Centralblattfür Mineralogie, Geologie und Paläontologie, Abteilung B: Geologie und Pal-äontologie 4, 183e189.
Huene, F. von, Matley, C.A., 1933. The Cretaceous Saurischia and Ornithischia of thecentral province of India. Palaeontologia Indica 21, 1e74.
Jacobs, L., Winkler, D.A., Downs, W.R., Gomani, E.M., 1993. New material of an EarlyCretaceous titanosaurid sauropod dinosaur from Malawi. Palaeontology 36,523e534.
Jain, S.L., Bandyopadhyay, S., 1997. New titanosaurid (Dinosauria: Sauropoda) fromthe Late Cretaceous of central India. Journal of Vertebrate Paleontology 17,114e136.
Janensch, W., 1925. Die Coelurosaurier und Theropoden der TendagurueSchichtenDeutscheOstafrikas. Palaeontographica VIII (Suppl.), 1e100.
Kellner, A.W.A., de Azevedo, S.A.K., 1999. A new sauropod dinosaur (Titanosauria)from the Late Cretaceous of Brazil. In: Tomida, Y., Rich, T.H., VickerseRich, P.(Eds.), Proceedings of the Second Gondwana Dinosaur Symposium, NationalScience Museum Monographs, vol. 15, pp. 111e142.
Kellner, A.W.A., Campos, D.A., Trotta, M.N.F., 2005. Description of a TitanosauridCaudal Series from the Bauru Group, Late Cretaceous of Brazil, vol. 63. Arquivosdo Museu Nacional, Rio de Janeiro. 529e564.
Kellner, A.W.A., Rubilar-Rogers, D., Vargas, A., Suárez, M., 2011. A new titanosaursauropod from the Atacama Desert, Chile. Anais da Academia Brasileira deCiências 83, 211e219.
Lambe, L., 1902. New genera and species from the Belly River Series (mid-Creta-ceous). Contributions to Canadian Paleontology 3, 25e81.
Leanza, H.A., Apesteguía, S., Novas, F.E., de la Fuente, M.S., 2004. Cretaceousterrestrial beds from the Neuquén Basin (Argentina) and their tetrapodassemblages. Cretaceous Research 25, 61e87.
Lydekker, R., 1893. Contributions to the study of the fossil vertebrates of Argentina.I. The dinosaurs of Patagonia. Anales del Museo de La Plata 2, 1e14.
Makovicky, P.J., Kobayashi, Y., Currie, P.J., 2004. Ornithomimosauria. In:Weishampel, D.B., Dodson, P., Osmólska, H. (Eds.), The Dinosauria, SecondEdition. University of California Press, Berkeley, pp. 137e150.
Malkani, M.S., Wilson, J.A., Gingerich, P.D., 2001. First dinosaurs from Pakistan.Journal of Vertebrate Paleontology 21 (Suppl.3), 77A.
Manera de Bianco, T., Calvo, J.O., 2001. Primer registro de restos embrionarios dedinosaurios en la provincia de Río Negro. Ameghiniana 38 (Suppl.), 12R. XVIIJornadas Argentinas de Paleontología de Vertebrados.
Martínez, R.D., Giménez, O., Rodríguez, J., Luna, M., Lamanna, M.C., 2004. Anarticulated specimen of the basal titanosaurian (Dinosauria: Sauropoda)Epachthosaurus sciuttoi from the early Late Cretaceous Bajo Barreal formation ofChubut province, Argentina. Journal of Vertebrate Paleontology 24, 107e120.
Marsh, O.C., 1878. Principal characters of American Jurassic dinosaurs. Part I.American Journal of Science (Third Series) 21, 411e416.
Martínez, S., Veroslavsky, G., 2004. Registros no depositacionales del TerciarioTemprano de Uruguay. In: Veroslavsky, G., Ubilla, M., Martínez, S. (Eds.),Cuencas sedimentarias de Uruguay: geología, paleontología y recursos natu-rales e Cenozoico. DIRACeFacultad de Ciencias, Montevideo, pp. 63e82.
Mikhailov, K.E., 1991. Classification of fósil eggshells of amniotic vertebrales. ActaPaleontologica Polonica 36, 193e238.
Mones, Á., 1980. Nuevos elementos de la paleoherpetofauna del Uruguay (Crocodiliay Dinosauria). II Congreso Argentino de Paleontología y Bioestratigrafía y ICongreso Latinoamericano de Paleontología, Buenos Aires, Actas 1, pp. 265e277.
Mones, Á., 1997. Los vertebrados mesozoicos del Uruguay y sus relaciones con los deáreas vecinas. In: Arroyo Cabrales, J., Polaco, O.J. (Eds.), Homenaje al ProfesorTicul Álvarez, vol. 357. Instituto Nacional de Antropología e Historia, México,pp. 205e222. Colección Científica.
Nopcsa, F., 1928. The genera of reptiles. Palaeobiologica 1, 163e188.Norman, D.B., 2004. Basal Iguanodontia. In: Weishampel, D.B., Dodson, P.,
Osmólska, H. (Eds.), The Dinosauria, second ed. University of California Press,Berkeley, pp. 413e437.
Novas, F.E., Cambiaso, A.V., Ambrioso, A., 2004. A newbasal iguanodontian (Dinosauria,Ornithischia) from the Upper Cretaceous of Patagonia. Ameghiniana 41, 75e82.
Otero, A., 2010. The appendicular skeleton of Neuquensaurus, a Late Cretaceoussaltasaurine sauropod from Patagonia, Argentina. Acta Palaeontologica Polonica55, 399e426.
Perea, D., Ubilla, M., 1994. Tetrápodos pre-cenozoicos del Uruguay II: comentariossobre nuevos restos de Titanosauridae. In: Paleociencias: Serie Didáctica, vol. 2.Depto. de Río Negro. 13e14.
Pol, D., Apesteguía, S., 2005. New Araripesuchus remains from the early LateCretaceous (CenomanianeTuronian) of Patagonia. American Museum Novitates3490, 1e38.
Powell, J.E., 1987a. The Late Cretaceous fauna of Los Alamitos, Patagonia, Argentina.Part VI e The titanosaurids. Revista del Museo Argentino de Ciencias NaturalesIII, 147e153.
Powell, J.E., 1987b. Morfología del esqueleto axial de los dinosaurios titanosáuridos(Saurischia, Sauropoda) del estado de Minas Gerais, Brasil. Anais do X CongresoBrasileiro de Paleontologia 1, 155e171.
Powell, J.E., 1992. Osteología de Saltasaurus loricatus (Sauropoda: Titanosauridae)del Cretácico Superior del Noroeste Argentino. In: Sanz, J.L., Buscalioni, A.D.
(Eds.), Los dinosaurios y su entorno biótico. Instituto Juan de Valdéz, Cuenca,pp. 165e230.
Powell, J.E., 2003. Revision of South American titanosaurid dinosaurs: palae-obiological, palaeobiogeographical and phylogenetic aspects. Records of theQueen Victoria Museum 111, 1e173.
Prieto-Márquez, A., 2010. Global phylogeny of hadrosauridae (Dinosauria: Orni-thopoda) using parsimony and Bayesian methods. Zoological Journal of theLinnean Society 159, 435e502.
Prieto-Márquez, A., Salinas, G.C., 2010. A re-evaluation of Secernosaurus koerneri andKritosaurus australis (Dinosauria, Hadrosauridae) from the Late Cretaceous ofArgentina. Journal of Vertebrate Paleontology 30, 813e837.
Rich, T.H., Vichers-Rich, P.V., 1994. Neoceratopsians & ornithomimosaurs: dinosaursof Gondwana origin? Research & Exploration 10, 129e131.
Rusconi, C., 1933. Sobre reptiles cretáceos del Uruguay (Uruguayschus Aznarezi [sic]n. g. n. sp.) y sus relaciones con los notosúquidos de Patagonia. Boletín delInstituto Geológico del Uruguay 19, 1e64.
Salgado, L., 1996. Pellegrinisaurus powelli nov. gen. et sp. (Sauropoda, Titanosaur-idae) from the Upper Cretaceous of Lago Pellegrini, northwestern Patagonia,Argentina. Ameghiniana 33, 355e365.
Salgado, L., 2003a. Should we abandon the name Titanosauridae?: some commentson the taxonomy of titanosaurian sauropods (Dinosauria). Revista Española dePaleontología 18, 15e21.
Salgado, L., 2003b. Considerations on the bony plates assigned to titanosaurs(Dinosauria, Sauropoda). Ameghiniana 40, 441e456.
Salgado, L., Apesteguía, S., Heredia, S., 2005. A new specimen of Neuquensaurusaustralis, a Late Cretaceous saltasaurine titanosaur from North Patagonia.Journal of Vertebrate Paleontology 25, 623e634.
Salgado, L., Azpilicueta, C., 2000. Un nuevo saltasaurino (Sauropoda, Titanosaur-idae) de la Provincia de Rio Negro (Formación Allen, Cretácico Superior),Patagonia, Argentina. Ameghiniana 37, 259e264.
Salgado, L., Calvo, J.O., 1993. Report of a sauropod with amphiplatyan mid-caudalvertebrae from the Late Cretaceous of Neuquén Province (Argentina). Ame-ghiniana 30, 215e218.
Salgado, L., Coria, R.A., 1993. El género Aeolosaurus (Sauropoda, Titanosauridae) enla Formación Allen (CampanianoeMaastrichtiano) de la Provincia de Río Negro,Argentina. Ameghiniana 30, 119e128.
Salgado, L., Coria, R.A., Calvo, J.O., 1997a. Evolution of titanosaurid sauropods. I:phylogenetic analysis based on the postcranial evidence. Ameghiniana 34, 3e32.
Salgado, L., Coria, R.A., Calvo, J.O., 1997b. Presencia del género Aeolosaurus (Sau-ropoda: Titanosauridae) en la Formación Los Alamitos, Cretácico Superior de laprovincia de Río Negro, Argentina. Revista Universidade Guarulhos. GeociênciasII, 44e49.
Salgado, L., García, R., 2002. Variación morfológica en la secuencia de vértebrascaudales de algunos saurópodos titanosaurios. Revista Española de Paleon-tología 17, 211e216.
Samman, T., Powell, G.L., Currie, P.J., Hills, L.V., 2005. Morphometry of the teeth ofwestern North American tyrannosaurids and its applicability to quantitativeclassification. Acta Palaeontologica Polonica 50, 757e776.
Sanz, J.L., Powell, J.E., Loeuff, J., Le, Martínez, R., Pereda-Suberbiola, X., 1999.Sauropod remains from the Upper Cretaceous of Laño (northcentral Spain),titanosaur phylogenetic relationships. Estudios del Museo de Ciencias Naturalesde Álava 14, 235e255.
Schwarz, D., Wings, O., Meyer, C.A., 2007. Super sizing the giants: first cartilagepreservation at a sauropod dinosaur limb joint. Journal of the Geological Society164, 61e65.
Sereno, P., 1986. Phylogeny of the bird-hipped dinosaurs (Order Ornithischia).National Geographic Research 2, 234e256.
Sereno, P., 1999. The evolution of dinosaurs. Science 284, 2137e2147.Simón, M.E., 2006. Cáscaras de huevos de dinosaurios de la Formación Allen
(CampanianoeMaastrichtiano), en Salitral Moreno, provincia de Río Negro,Argentina. Ameghiniana 43, 513e528.
Smith, J.B., 2007. Dental morphology and variation in Majungasaurus crenatissimus(Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. In:Sampson, S.D., Krause, D.W., (Eds.), Majungasaurus crenatissimus (Theropoda:Abelisauridae) from the Late Cretaceous of Madagascar. Journal of VertebratePaleontology, 27(Suppl. 3), pp. 103e126.
Smith, J.B., Lamanna, M.C., 2006. An abelisaurid from the Late Cretaceous of Egypt:implications for theropod biogeography. Naturwissenschaften 93, 242e245.
Smith, J.B., Lamanna, M.C., Lacovara, K.J., Dodson, P., Smith, J.R., Poole, J.C.,Giegengack, R., Attia, Y., 2001. A giant sauropod dinosaur from an UpperCretaceous mangrove deposit in Egypt. Science 292, 1704e1706.
Smith, J.B., Vann, D.R., Dodson, P., 2005. Dental morphology and variation intheropod dinosaurs: implications for the taxonomic identification of isolatedteeth. The Anatomical Record Part A 285, 699e736.
Soto, M., 2010a. New titanosaurian (Dinosauria, Sauropoda) remains from theMercedes Formation (Late Cretaceous) of Uruguay. X Congreso Argentino dePaleontología y Bioestratigrafía y VII Congreso Latinoamericano de Paleon-tología, Resúmenes, p. 211.
Soto, M., 2010b. Estudio de los dientes de terópodos (Dinosauria, Saurischia) de laFormación Tacuarembó (Jurásico TardíoeCretácico Temprano), Uruguay. Facul-tad de CienciasePEDECIBA, Unpublished Master Thesis, p. 266.
Soto, M., Cambiaso, A., 2006. Reinterpretación de los dientes de dinosaurios de laFormación Guichón, Uruguay: iguanodontes basales y terópodos no ornitomí-midos. Ameghiniana 43 (Suppl.), 55Ae56A.

M. Soto et al. / Journal of South American Earth Sciences 33 (2012) 68e79 79
Soto, M., Pol, D., Perea, D. A new specimen of Uruguaysuchus aznarezi (Croc-odyliformes: Notosuchia) from the Cretaceous of Uruguay and its phylogeneticrelationships. Zoological Journal of the Linnean Society, in press.
Tauber Jr., A.A., 2007. Primer yacimiento de huevos de dinosaurios (CretácicoSuperior) de la provincia de La Rioja, Argentina. Ameghiniana 44, 11e28.
Trotta, M.N.F., Campos, D.A., Kellner, A.W.A., 2002. Unusual caudal vertebral centraof a titanosaurid (Dinosauria, Sauropoda) from the continental Upper Creta-ceous of Brazil. Boletim do Museo Nacional, nova série, Geologia 64, 1e11.
Turner, A.H., 2006. Osteology and phylogeny of a new species of Araripesuchus(Crocodyliformes: Mesoeucrocodylia) from the Late Cretaceous of Madagascar.Historical Biology 18, 255e369.
Upchurch, P., Barret, P.M., Dodson, P., 2004. Sauropoda. In: Weishampel, D.B.,Dodson, P., Osmólska, H. (Eds.), The Dinosauria, second ed. University of Cal-ifornia Press, Berkeley, pp. 259e324.
Verde, M., Soto, M., 2005. Sobre una asociación de huevos de dinosaurios y cop-rolitos del Cretácico Superior de Uruguay. XXI Jornadas Argentinas de Paleon-tología de Vertebrados, Plaza Huincul, Actas, p. 42.
Weishampel, D.B., Barrett, P.M., Coria, R.A., Le Loeuff, J., Xu, X., Zhao, X., Sahni, A.,Gomani, E.M.P., Noto, C.R., 2004. Dinosaur distribution. In: Weishampel, D.B.,Dodson, P., Osmólska, H. (Eds.), The Dinosauria, second ed. University of Cal-ifornia Press, Berkeley, pp. 517e606.
Wilson, J.A., 1999. A nomenclature for vertebral laminae in sauropods and othersaurischian dinosaurs. Journal of Vertebrate Paleontology 19, 639e653.
Wilson, J.A., 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis.Zoological Journal of the Linnean Society 136, 217e276.
Wilson, J.A., 2005. Overview of sauropod phylogeny and evolution. In: CurryRogers, K.A., Wilson, J.A. (Eds.), The Sauropods: Evolution and Paleobiology.University of California Press, Berkeley, pp. 15e49.
Wilson, J.A., 2006. An overview of titanosaur evolution and phylogeny. In:Torcida, F., Huerta, P. (Eds.), 3rd Internacional Symposio de Dinosaurios y suEntorno Biótico. Burgos. Museo de Salas de los Infantes, pp. 169e190.
Wilson, J.A., Martínez, R.N., Alcober, O., 1999. Distal tail segment of a titanosaur(Dinosauria: Sauropoda) from the Upper Cretaceous of Mendoza, Argentina.Journal of Vertebrate Paleontology 19, 591e594.
Wilson, J.A., Sereno, P.C., 1998. Early evolution and higher-level phylogeny ofsauropod dinosaurs. Society of Vertebrate Paleontology, Memoir 5, 1e68.
Wilson, J.A., Upchurch, P., 2003. A revision of Titanosaurus Lydekker (Dinosauria eSauropoda), the first dinosaur genus with a ‘Gondwanan’ distribution. Journalof Systematic Palaeontology 1, 125e160.
You, H., Ji, Q., Lamanna, M.C., Li, Y., 2004. A titanosaurian sauropod dinosaur withopisthocoelous caudal vertebrae from the early Late Cretaceous of LiaoningProvince, China. Acta Geologica Sinica 78, 907e911.
You, H., Tang, F., Luo, Z., 2003. A new basal titanosaur (Dinosauria: Sauropoda) fromthe Early Cretaceous of China. Acta Geologica Sinica 77, 424e429.
Zhao, Z.K., Ding, S.R., 1976. Discovery of the dinosaurs eggs from Alashanzuoqi andits stratigraphical meaning. Vertebrata PalAsiatica 14, 42e44.