Embed Size (px)
Citation preview

FISIOLOGIA
DEFINIÇÃO ......................................... 268
HOMEOSTASE .................................... 268
Homeostase no corpo .................... 268 Líquidos corporais ............................. 268
TRANSPORTE PELA MEMBRANA ........ 269
DIFUSÃO SIMPLES ............................. 269
Difusão Não-eletrólito ................... 269 Coeficiente de partição (CA-CB) .......... 269 Coeficiente de partição (K) ................ 269 Área de superfície (A) ........................ 269
Difusão de eletrólitos ..................... 269
DIFUSÃO FACILITADA ....................... 270
TRANSPORTE ATIVO ......................... 270
Transporte ativo primário .............. 270 Bomba de sódio e potássio ................ 270 Bomba de Ca
2+ ................................... 270
Bomba de H- -K
+ ................................. 270
Transporte ativo secundário .......... 271 Co-transporte .................................... 271 Contratransporte ............................... 271
OSMOSE ........................................... 271
Osmolaridade ................................ 271 Pressão osmótica ............................... 272
CANAIS IÔNICOS ............................... 272
Dependentes de voltagem ............. 272
Dependente de ligantes ................. 272
FISIOLOGIA DO SN .............................273
BASES NEURONAIS ........................... 273
Neurônios e células da Glia ........... 273
BIOELETROGENESE ........................... 274
Potencial de repouso ..................... 274
Potencial de ação .......................... 275
SISTEMA SENSORIAL ......................... 276
Sentido mecânico .......................... 276
Controle motor .............................. 277
FISIOLOGIA CÁRDIÁCA .......................278
SISTEMA CIRCULATÓRIO .................. 278
Coração ......................................... 278 Condução elétrica no coração .......... 279 Contração ......................................... 280 Ciclo cardíaco .................................... 280 Vasos ................................................. 281
FISIOLOGIA RESPIRATÓRIA ................282
Troca gasosa.................................. 282
VOLUMES PULMONARES ................. 283
Capacidade pulmonar ................... 283
Espaço morto ................................. 283
CICLO RESPIRATÓRIO ....................... 283
Repouso ......................................... 283
Inspiração ...................................... 283
Expiração ....................................... 283
FISIOLOGIA GASTROINTESTINAL ....... 284
TUBO DIGESTÓRIO ........................... 284
Boca ............................................... 284
Faríngea ........................................ 285
Esôfago .......................................... 285
Estômago ...................................... 286
Intestino delgado .......................... 286 Duodeno ........................................... 286 Jejuno ................................................ 286 Íleo 286
Intestino grosso ............................. 287 Ceco 287 Cólon ................................................. 287 Reto 287 Anus 287
ORGÃO ACESSÓRIOS ..................... 287
Fígado ............................................ 287 Bílis 287
Pâncreas ........................................ 287 Suco pancreático ............................... 287

268
DEFINIÇÃO
Fisiologia do grego physis = natureza ou função, logia = estudo. É o estudo do funcionamento do corpo, e é o ramo da biologia que estuda as múltiplas funções mecânicas, físicas e bioquímicas nos seres vivos.
HOMEOSTASE
É a propriedade de um sistema, de regular o seu ambiente interno para manter uma condição estável, mediante ajuste de equilíbrio dinâmico controlado por mecanismo de regulação inter-relacionado. Termo criado em 1932 por Walter Brad Ford Cannon, a partir do grego Homeo = similar ou igual, stasis = estático.
Homeostase no corpo
A capacidade de sustentar a vida, no corpo humano é afetada por um leque de fatores, como a temperatura, a salinidade, o ph, ou as concentrações de nutrientes, como a glicose, íons, oxigênio, e resíduos como o dióxido de carbono e a uréia.
Líquidos corporais
No ser humano a água corresponde a 70% do peso corporal, a água corporal é dividida em dois compartimentos:
Líquido intracelular (LIC): contido dentro das
células. Seu principal cátion é o N𝐚+, e os ânios que contrabalançarão são o cloreto, Cl- e o bicarbonato HCO3.
Líquido extracelular (LEC): contido fora das células: os principais cátions do LEC são o potássio e o magnésio e os ânios que os contrabalançam, são as proteínas e os fosfatos orgânicos. O LEC é constituido pelo plasma e o líquido intersticial; o plasma é o liquido circulante nos vasos sanguíneos. O líquido intersticial banha as células e é o maior dos dois subcompartimento. Os dois estão separados pelas paredes capilares.
As diferenças de concentrações dos solutos, através da membrana, são geradas e mediadas por mecanismos de transportes consumidores de energia. A bomba de sódio e potássio transporta sódio do LIC para o LEC e, de modo simultâneo, transporta potássio do LEC para o LIC, como os dois íons são transportados contra seus gradientes, é necessário uma fonte de energia, o ATP.

269
TRANSPORTE PELA MEMBRANA
As membranas celulares são compostas por lipídios e proteínas. O componente lipídico é composto por fosfolipídios, colesterol e glicolipidios, substâncias lipossolúveis, como dióxido de carbono, oxigênio, ácidos graxos e hormônios esteroides. Também, é responsável pela baixa permeabilidade de membranas a substâncias hidrossolúveis, como íons, glicose e aminoácidos. Os componentes proteicos da membrana consistem em transportadores, canais para íons e para água. As substâncias podem ser transportadas a favor, do gradiente de concentração, ou contra este gradiente de concentração. O transporte a favor ocorre por difusão simples ou facilitada, não necessitando de energia. O transporte contra tal gradiente ocorre por transporte ativo, podendo ser primário ou secundário. No primário há necessidade de energia, no secundário, ocorre utilização indireta de energia. Outra diferença entre os mecanismos de transporte é a presença ou não de proteínas carreadoras. A difusão simples não é mediada por carreador. A difusão facilitada, o transporte ativo primário e o secundário envolvem proteínas integrais, chamadas de transporte mediado por carreador.
DIFUSÃO SIMPLES
Difusão Não-eletrólito
Ocorre como resultado de movimento térmico aleatório das moléculas, como a figura abaixo.
Figura 1: Difusão simples
Duas soluções A e B, separadas por membrana permeável ao soluto, a concentração do soluto A é, duas vezes maior do que na solução B. As moléculas estão em movimento constante, com igual probabilidade de que dada molécula atravessará a membrana para a outra solução. Contudo, como há duas vezes mais moléculas na solução A do que em B, haverá maior movimentação de moléculas de A para B do que de B para A. A difusão efetiva do soluto é chamada de fluxo e depende das seguintes variáveis: tamanho (intensidade) do gradiente de concentração, coeficiente de partição, coeficiente de difusão, espessura da membrana e área da superfície disponível para a difusão.
Coeficiente de partição (CA-CB)
É a força impulsora para a difusão efetiva. Quanto maior for à diferença de concentração do soluto entre as soluções A e B, maior a força impulsora e maior a difusão efetiva. Se as concentrações nas duas soluções forem iguais, não haverá força impulsora nem difusão efetiva.
Coeficiente de partição (K)
Descreve a solubilidade em água. Quanto maior a solubilidade em relação ao óleo, maior o coeficiente de partição e mais facilmente o soluto pode dissolver na bicamada lipídica das membranas celulares. Solutos apolares tendem a ser insolúveis em óleo e ter baixos valores de coeficiente de partição. Esse coeficiente pode ser determinado pela adição de soluto a uma mistura de azeite e água e, a seguir, dosando-se sua concentração na fase oleosa em relação á sua concentração na fase aquosa.
K=concentra ção no azeite
concentra ção na água
Área de superfície (A)
Quanto maior a área disponível da superfície da membrana, maior será a velocidade de difusão. Por exemplo, gases lipossolúveis, como o oxigênio e o dióxido de carbono, têm velocidades de difusão particularmente altas através das membranas celulares. Essas altas velocidades podem ser atribuídas a grande área de superfície para a difusão oferecida pelo componente lipídico da membrana.
Difusão de eletrólitos
Se o soluto for um íon ou um eletrólito, há duas consequências adicionais da presença de carga no soluto. 1ª se houver diferença de potencial através da membrana, essa diferença altera a velocidade de difusão efetiva de soluto com carga. Por exemplo, a difusão de íons K+ diminuirá se o K+ estiver se difundindo para área de carga positiva, e acelerará se o K+ estiver difundindo para área de carga positiva, e acelerará se o K+ estiver difundido para área de carga negativa. Esse efeito da diferença de potencial pode tanto adicionar quanto anular os efeitos diferença de potencial pode tanto adicionar quanto anular os efeitos das diferenças das concentrações, dependendo da orientação da diferença de potencial e da carga do íon difusível. Se o gradiente de concentração e o efeito de carga forem orientados no mesmo sentido através da membrana, eles se combinarão; se forem orientados em sentidos opostos, eles poderão se cancelar. 2ª quando um soluto com carga se difunde a favor do gradiente de concentração, a difusão pode, gerar diferença de potencial através da membrana, chamado potencial de difusão.

270
DIFUSÃO FACILITADA
Parecida com a difusão simples, ocorrendo a favor do gradiente de potencial eletroquímico, não necessitando de suporte de energia metabólica, a difusão facilitada utiliza um carreador de membrana, isso é a que difere da difusão simples. A difusão facilitada ocorre mais rapidamente do que a difusão simples devido à função do carreador.
TRANSPORTE ATIVO
Transporte ativo primário
Nele um ou mais soluto se movem contra um gradiente de concentração. Devido ao fato de se movimentar contra o gradiente é necessário o uso de energia na forma de ATP. O ATP é hidrolisado em difosfato de adenosina ADP e o fosfato inorgânico P, liberando energia da ligação fosfato terminal de alta energia do ATP. Quando o fosfato terminal é liberado, ele se transfere para a proteína transportadora, iniciando ciclo de fosforilação e desfosforilação. Quando a fonte de energia do ATP é acoplada ao processo de transporte, ela é chamada de proteína de transporte primário.
Bomba de sódio e potássio
Está presente nas membranas de todas as células, bombeando sódio do LIC para o LEC e potássio do LEC para o LIC. Para cada três íons de sódio bombeando para fora, dois íons de potássio são bombeados para dentro da célula. A bomba de sódio e potássio consiste em subunidade α e β. A subunidade de α contém a atividade ATPásica, assim como os locais de ligação para os íons transportado, Na+ e K+. a bomba de sódio e potássio se alterna entre os dois principais estados conformacionais, E1 e E2 o estado E1 os sítios de ligação para Na+ e K+. A função extracelular e enzima tem alta afinidade por K+ à função transportadora de íons dessa enzima se baseia nos ciclos entre os estados E1 e E2 e recebe sua energia da hidrolise do ATP.
O ciclo de transporte se inicia com a enzima no estado E1 que se liga ao ATP. Nesse estado, os
locais de ligação do íon estão voltados para o líquido intracelular, e a enzima tem alta afinidade por Na+, três íons Na+ se ligam e o ATP é hidrolisado, e o fosfato terminal é transferido para a enzima, gerando uma forma de alta energia. Agora ocorre uma alteração conformacional, e a enzima se transfere de E1 para E2. No estado E2 os sítios de ligação iônicos estação voltados para o líquido extracelular, a afinidade para Na+ e baixa afinidade para o K+ é alta. Os três íons de Na+ sã liberados da enzima para o líquido extracelular, são ligados dois íons K+ e alta. Os três íons Na+ e fosfato inorgânico são liberados de E2. A enzima se liga ao ATP no lado intracelular da membrana, e passa por outra alteração conformacional, o que faz com que retorne à sua forma original; os dois íons K+ são liberados para o líquido intracelular e as enzimas estão prontas para começar o ciclo.
Figura 2: bomba de Na
+ K
+ das membranas celulares. ADP,
difosfato de adenosina ATP, trifosfato de adenosina; E, Na Na
+ K
+ ATPase, E~P, Na
+ K
+ ATPase fosforilada; P, fosfato
inorgânico.
Bomba de Ca2+
A maioria das membranas celulares contém uma Ca2+ ATPase ou Ca2+ da cpelula contra o gradiente eletroquímico; um íon Ca2+ é expulso para cada ATP hidrolisado. É responsável por manter muito baixa a concentração intracelular de Ca2+. O retículo sarcoplasmático das células musculares e o retículo endoplasmático das outras células contêm variantes da Ca2+ ATPase que bombeiam dois íons Ca2+ do líquido intracelular para o interior do retículo sarcoplasmático ou endoplasmático, isto é, ocorre sequestro de Ca2+. Essa variantes são chamadas de Ca2+ ATPase dos retículos sarcoplasmático e endoplasmático.
Bomba de H- -K+
Esta bomba é encontrada nas células parietais da mucosa gastrica e nas células intercaladas α do ducto coletor renal. No estômago, ela bombeia H+ do LIC das células parietais para o lúmen do estõmago, onde acidifica os conteúdos gástricos.

271
Transporte ativo secundário
São aqueles nos quais é acoplado o transporte de dois ou mais solutos geralmente sódio, se movendo de acordo com seu gradiente de concentração. O movimento sem gasto de energia do sódio fornece energia para o movimento com gasto de energia do outro soluto. Sendo assim o ATP, não é utilizado diretamente, mas é fornecida de modo indireto pelo gradiente de concentração do sódio através da membrana celular. O nome secundário se refere à utilização indireta de ATP como fonte de energia. Há dois tipos de transporte secundário, distinguido pela direção do moviemento do soluto Uphill. Se o soluto se move no mesmo sentido que o Na+, é chamado co-transporte ou simporte. Se o soluto se move em sentido oposto ao do Na+, é chamado contratransporte, antiporte ou troca.
Co-transporte
É uma forma de transporte ativo secundário na quais todos os solutos são transportados no mesmo sentido através da membrana celular. O sódio se move para dentro da célula por meio do carreador, de acordo com a gradiente e os solutos, co-transportados com o sódio, se movam para dentro das células. O co-transporte está envolvido em vários processos fisiológicos, principalmente no epitelio absortivo do intestino delgado e do túblo renal. Por exemplo, os co-transportes de Na+ glicose e de Na+ aminoácidos estão presentes nas membranas luminais das células epiteliais tanto do intestino delgado quanto no túbulo proximal do rim.
Figura 3: Co-transporte de Na
+ -glicose em uma célula
epitelial do intestino. ATP, SGLT1, proteína 1 transportadora de Na
+ -glicose.
Contratransporte
É a forma de transporte ativo secundário na qual os solutos se movem em sentido oposto através da membrana celular. O Na+ se move para dentro da célula por meio do carreador a favor de seu gradiente eletroquímico: sos solutos que são contratransportados ou trocados pelo Na+ se movem para fora da célula.
Figura 4: Contratransporte (troca) de Ca
2+-Na
+ em uma célula
muscular.
OSMOSE
É o fluxo de água através da membrana semipermeável, pela diferença de concentração de soluto. A osmose da água não é difusão da água, a osmose ocorre devido à diferença de pressão, enquanto a difusão ocorre devido a diferência de concentração da água.
Osmolaridade
É a sua concentração de partículas osmóticamente ativas. Para calcular a osmolaridade, é necessário conhecer a concentração do soluto e se o soluto se dissocia na solução. Por exemplo, a glicose não se dissocia em solução. O Nacl se dissocia em duas particulas, o CaCl2 se dissocia em três partículas. O símbolo g fornece o número de partículas em solução e também explica se há dissociação completa ou somente parcial. Assim, se o NaCl se dissocia completamente em duas partículas, g é igual a 2,0; se NaCl se dissocia somente de modo parcial, então g declina, ficando entre 1,0 e 2,0. A osmolaridade é calculada como se segue:
Osmolaridade=g∙C Onde:
Osmolaridade= concentração de partículas;
S=número de partículas por mol em solução;
C= concentração. Se a solução tem a mesma osmolaridade calculada, elas são chamadas de isosmóticas. Se duas soluções têm diferentes osmolaridade é chamada de hiperosmótica, e a solução com a menor osmolaridade, de hiposmótica.

272
Pressão osmótica
Osmose é o fluxo de água através de membrana semipermeável, a concentração de soluto gera diferença de pressão osmótica através da membrana, e essa diferença é a força impulsora para o fluxo osmótico de água. Na figura abaixo temos duas soluções aquosas, abertas à atmosfera, são mostrada. A membrana que separa as soluções é permeável à água, mas é impermeável ao soluto. Inicialmente, o soluto está presente apenas na solução 1. O soluto na solução produz pressão osmótica e causa, pela interação hidrostática da solução 1. A diferença resultante, na pressão hidrostática através da membrana, provoca, o fluxo de água da solução 2 para a solução 1. Com o tempo, o fluxo de água da solução 1. Com o tempo, o fluxo de água causa aumento do volume da solução e diminuição do volume da solução 2.
Figura 5: osmose através de membrana semipermeável: o soluto está presente em um lado de uma membrana semipermeável; com o tempo a pressão osmótica, gerada pelo soluto, causa o fluxo de água da solução 2 para a solução 1.
CANAIS IÔNICOS
São proteínas de membranas integrais que, permitem a passagem de certos íons. Eles são seletivos e permitem que íons com características especificas, se movam por eles, seletivamente baseada no tamanho do canal quanto nas cargas que os revestem. Por exemplo, um canal revestido por carga negativa, seletivo para cátions, deve permitir a passagem de sódio, mas exclui potássio, outro canal seletivo para cátions, deve ter menos seletividade e permitir a passagem de vários pequenos cátions diferente. Os canais iônicos são controlados por comportas e dependente de sua posição. Os canais podem abrir ou fechar quando um canal se abre, os íons para os quais ele é seletivo podem fluir. Os canais ionicos são controlados por comporta e, dependendo da sua posição, os canais podem abrir ou fechar. Quando um canal se abre, os íons para os quais ele é seletivo podem fluir por ele, movido pelo gradiente eletroquímico existente. Quando o canal se fecha, os íons não podem fluir por ele, não importando a grandeza do gradiente eletroquimico. Dois tipos de comportas controlam a abertura e o fechamento dos canais iônicos.
Os canais dependentes de voltagem abrem e fecham em respostas à ligação de ligantes, como
hormônios, neurotransmissores ou segundos mensageiros.
Figura 6: Canal iônico dependente de ativador.
Dependentes de voltagem
Têm comportas que são controladas por alterações do potencial de membrana. Por exemplo, a comporta de ativação do canal de Na+ no nervo é aberta pela despolarização da membrana celular do nervo; a abertura desse canal é responsável pelo curso ascendente de potencial de ação.
Figura 7: Canal iônico dependente de voltagem.
Dependente de ligantes
Têm comportas que são controlados por hormônios, por neurotransmissores e por segundos mensageiros. Por exemplo, o receptor nicotinico da placa motora é relamente um canal iônico que se abre quanto a acetilcolina se liga a ele; quando aberto, ele é permeavel aos íons Na+ e K+.
Figura 8: Canal iónico dependente de ligante.

273
FISIOLOGIA DO SN
Tem três funções básicas:
Processar informações que chegam; (sistema sensorial);
Organizar a função motora; (sistema somático);
Processar as tomadas de decisões (funções corticais elaboradas).
O sistema inclui neurônios sensoriais e neurônios motores, e diversos inter-neurônios no SNC que interconectam as partes e estão envolvido em processos mais avançados e complexos do SN.
BASES NEURONAIS
Neurônios e células da Glia
O neurônio é a célula funcional do SN, composto por três porções: soma, axônio e dentritos. Pequenos botões chamados terminações pré-sinápticas estão sobre a superfície dos dentritos e da soma. Essas terminações podem ser excitatórias, quando secretam substâncias que existam os neurônios pré-sinápticos, ou podem ser excitatórias, quando secretam substâncias que existem nos neurônios pós-sinápticos, ou podem ser inibitórias, quando o inibem. Os neurônios interconectados constituem os circuitos neurais, que são redes de comunicações feitas através de potenciais de ação. As informações passam por transmissões sinápticas, onde o terminal pré-sináptico libera substâncias neurotransmissoras na fenda sináptica. Os neurônios da medula e do cérebro diferem dos neurônios motores no tamanho da soma, comprimento, tamanho e número de dentritos, comprimentos e tamanho do axônio e número de terminações pré-sinápticas. Pelo axônio há o transporte de vesículas contendo proteínas, lipídeos, neurotransmissores, açucares e etc. de forma retrógrada, da periferia para o corpo celular, há o transporte de toxinas, vírus e fatores de crescimento de nervos. As terminações pré-sinápticas têm duas estruturas internas: vesículas com neurotransmissores e mitocôndrias, que fornecem ATP para síntese das substâncias transmissoras liberadas causa a alteração da permeabilidade da membrana pré-sináptica. A membrana pré-sináptica contém muitos canais de cálcio sensíveis à voltagem, quando o potencial de ação despolariza a terminação, muitos cálcios flui para dentro do botão, permitindo que as terminações e as vesículas com neurotransmissores sejam excitadas. Na sinapse, esse conteúdo se liga aos receptores de membrana no neurônio pós-sináptico para alterar o funcionamento dessa célula.
Figura 9: os neurotransmissores são a mensagem em uma
sinapse química. (A) axônio dos neurônios pré-sináptico; (B)
neurônio pós-sináptico; (C) membrana pós-sináptica; (D)
receptores; (E) meurotransmissor; (F) fenda sináptica; (G)
vesículas sinápticas; (H) terminal axônio; (I) mitocôndria.
Transporte axonal. (A) Peptídeos sintetizados e empacotados;
(B) Transporte axonal rápido ao longo da rede de microtúbulos;
(C) conteúdo das vesículas liberados por exocitose; (D)
Reciclagem das vesículas sinápticas; (E) transporte axonal
rápido retrógrado; (F) componentes velhos da membrana são
digeridos nos lisossomos. (a) RER; (b) soma; (c) lisossomo; (d)
aparelho de Golgi; (e) Vesícula sináptica.
Cálcio é o sinal de liberação dos neurotransmissores na sinapse. A liberação de neurotransmissores na fenda sináptica ocorre por exocitose. Quando a despolarização de um potencial de ação alcança o terminal axônico, mudança no potencial de membrana do terminal axônico possui canais de Ca+2 controlados por voltagem que se abrem em resposta à despolarização. Como os íons cálcio são mais concentrados no líquido extracelular do que no citosol, eles se movem para dentro da célula. O Ca2+ liga-se a proteína reguladora e inicia a exocitose. A membrana da vesícula sináptica se funde com a membrana celular, com o auxílio de várias proteínas da membrana. A área fundida se abre e os neurotransmissores se movem de dentro da vesícula sináptica para fenda sináptica. As moléculas do neurotransmissor se difundem através da fenda para se ligarem com receptores na membrana da célula pós-sináptica. Quando os neurotransmissores se ligam aos seus receptores, uma resposta é iniciada na célula pós-sináptica.
Figura 10: transferência de informações na sinapse. (A) um
potencial de ação despolariza o terminal axônico; (B) a
despolarização abre canais de Ca2+
controlados por voltagem e
o Ca2+
entra na célula; (C) a entrada do cálcio inicia a exocitose
do conteúdo das vesículas sinápticas; (D) o neurotransmissor
se difunde através da fenda sináptica e se liga aos receptores
na célula pós-sináptica; (E) a ligação do neurotransmissor inicia
uma resposta na célula pós-sináptica. (a) terminal axônico; (b)
vesículas sináptica; (c) proteína de ancoragem; (d) resposta
celular; (e) célula pós-sinaptica; (f) canal de Ca2+
controlado por
voltagem; (g) receptor; (h) fenda sináptica; (i) molécula do
neurotransmissor.

274
O restante do conteúdo da fenda é captada por células a Glia, que rodeiam os neurônios. No SNC, as células da Glia são Oligodendrócitos, Astrócitos, Microglia e células ependimárias. No SNC, os axônios são envolvidos pelo Oligodentrócitos que constituem a bainha de mielina, camadas lipídicas concêntricas ao redor dos axônios. A bainha de mielina atua como um isolante contra corrente iônica e as interrupções são chamados Nódulos de Ranvier.
Figura 11: (A) a célula de Schwann se enrola muitas vezes ao
redor do axônio; (B) o núcleo da célula de Schwann é
empurrado para fora da bainha de mielina; (C) a mielina
consiste em múltiplas camadas de membranas celulares; (D) o
nó de Ranvier é uma porção não mielinizada da membrana do
axônio entre duas células de Schwann. (1) formação da mielina
no SNP; (2) cada célula de Schwann forma mielina ao redor de
um pequeno segmento de um axônio. (a) núcleo; (b) axônio; (c)
corpo celular.
Os Astrócitos possuem prolongamento que isolam as sinapses, metabolizam os neurotransmissores jogados na fenda sináptica, fazem o tamponamento de íons potássio e hidrogênio, servem de sustentação para axônios e formam cicatrizes após agressões ao SNC. As Microglia são fagocitárias em potencial, auxiliando na remoção de resíduos que invadem o SNC. As células ependimárias formam o epitélio que separa o SNC do líquido cefalorraquidiano, revestindo o espaço vazio do SNC. O líquido cefalorraquidiano é secretado, por células, que fazem parte do plexo coroide. As células da Glia do SNP são as células de Schwann, que produzem a bainha de mielina, e as células ganglionares das raízes dorsais e dos nervos cranianos.
BIOELETROGENESE
O transporte de íons através da membrana é realizado por proteínas especializadas localizadas na membrana celular. Se a bicamada lipídica estiver ausente de proteínas, a membrana será altamente permeável, mesmo a íons muito pequenos. A diferença de cargas geradas é o potencial elétrico, que tem como medida volt (V). A corrente elétrica é o fluxo entre esses dois pontos. Essa diferença de potencial elétrico é mantida na célula pela ação da bomba Na+ e K+ e pela busca constante do equilíbrio eletroquímico pelos íons envolvidos. Por esse sistema de bombeamento, mais a difusão de potencial entre o meio interno e o externo, ou seja, cria energia potencial: Bioeletrogenese.
Potencial de repouso
É a diferença de potencial que existe através da membrana celular excitável, como as células nervosas e as musculares no período entre dois potenciais de ação. O potencial de repouso é estabelecido pelos potenciais de difusão, resultante das diferenças de concentração para vários íons, através da membrana celular. Cada íon tenta impulsionar o potencial de membrana em direção a seu próprio potencial de equilíbrio. Os íons mais permeáveis em repouso terão contribuições menores terão pouca ou nenhuma contribuição. O potencial de repouso dessas células está entre -70 a -80 mV. O potencial de repouso está próximo do potencial de equilíbrio do potássio e do cloro, pois a permeabilidade a esses íons em repouso é alta. O potencial de repouso está longe dos potenciais de equilíbrio do sódio e cálcio, pois a permeabilidade a esses íons em repouso é baixa.
Figura 12: (A) no potencial de repouso, os canais de sódio estão
fechados; (B) quando a membrana é despolarizada, mudanças
conformacionais abrem os canais de sódio.

275
Potencial de ação
As células possuem um potencial eletroquímico gerado por elas mesmas. A alteração rápida deste potencial, seguida pela restauração rápida deste potencial, seguida pela restauração do mesmo, é chamada de potencial de ação. É um fenômeno das células excitáveis, como as nervosas e as musculares, que consiste na rápida despolarização, seguida por repolarização do potencial de membrana. São os mecanismos básicos para a transmissão da informação no sistema nervoso e em todos os tipos de músculos. Despolarização: É o processo que torna o potencial de membrana menos negativo. Como observado, o potencial de repouso das células excitáveis é orientado com o interior da célula negativa. A despolarização torna o interior da célula menos negativa, até mesmo, fazer com que o interior da célula fique positivo. Hiperpolarização: É o processo que torna o potencial de membrana mais negativa. Semelhante à despolarização, os termos, aumento ou diminuindo, não deveriam ser utilizado para descrever a variação que torna o potencial de membrana mais negativa. Corrente de influxo: É o fluxo de carga positiva para fora da célula, corrente de efluxo hiperpolarizam o potencial de membrana. ex.: a corrente de influxo é o fluxo onde o sódio para dentro da célula, durante o curso ascendente do potencial de ação. Corrente de efluxo: É o fluxo de carga positiva para fora das células corrente de efluxo hiperpolarizam o potencial de membrana. Um exemplo de corrente de efluxo é o fluxo de potássio para fora da célula, durante a fase de despolarização do potencial de ação.
Figura 13: condução do potencial de ação. (A) um potencial
graduado acima d limiar alcança a zona de disparo; (B) canal de
Na+ controlados por voltagem se abrem e o Na
+ entra MP
axônio; (C) cargas positivas fluem para o segmentos adjacentes
do neurônio por fluxo de corrente local; (D) o fluxo de corrente
local da região ativa faz com que os outros segmentos da
membrana se despolarizam; (E) o período refratário impede a
condução retrógrada. A perda de K+ do citoplasma repolariza a
membrana. (a) região refratária; (b) região ativa (c) região
inativa.
Limiar É o potencial de membrana no qual é inevitável a ocorrência do potencial de ação. Quando se atinge o limiar, abrem-se canais de sódio, e muito sódio do meio extracelular flui para o meio intracelular. A célula recebe muita carga positiva do sódio e eleva seu potencial elétrico até 20mv. Essa fase ascendente é chamada despolarização. Quando houver aumento na condutância para o Na+, ele entrará na célula e alcançará o limiar de desencadeamento do potencial de ação. Esse limiar é o valor elétrico que cada célula possui para provocar a abertura de canais dependentes de voltagens. Assim, quando essas células atingem esse valor, novos canais do sódio e potássio abrem-se permitindo o fluxo de sódio e efluxo de potássio. Quando os canais de potássio abrem-se e ocorre acentuado fluxo de potássio para o meio extracelular. Assim, a célula perde carga positiva e retorna seu potencial de membrana ao valor mais negativa esta fase chama-se repolarização. Algumas vezes o canal de potássio tarda a se fechar e o fluxo de potássio ocorre em demasia, então à célula fica mais negativa do que durante o repouso, tendo essa fase o nome de hiperpolarização. Durante o período da repolarização, nenhum novo estimulo pode voltar a alterar os canais voltagem dependendo da célula. A este período damos o nome de período refratório absoluto. No final da repolarização, há o período refratário relativo, no qual um estímulo forte pode gerar novo potencial de ação.
Figura 14: (1) canais de Na
+ e k
+. (A) ambos os canais fechados;
(B) os canais de Na+
se abrem; (D) os canais de Na+ voltam à
posição original, ao passo que os canais de K+ permanecem
abertos; (E) ambos os canais estão fechados. (a) período
refratário absoluto; (b) período refratário relativo.

276
Figura 15: Modelo do canal de Na
+ controlado por voltagem. (A)
durante o potencial de membrana em repouso o portão de
ativação fecha o canal; (B) o estimulo despolarizante chega ao
canal. Portão de ativação abre; (C) com o portão de ativação
aberto, o Na+ entra na célula; (D) o portão de inativação se fecha
e a entrada de Na+ para; (E) durante a repolarização causada
pela saída do K+ da célula, os dois portões voltam ás suas
posições iniciais.
Figura 16: (1) potencial de membrana em repouso; (2) estímulo
despolarizante; (3) membrana despolarizada até o limiar. Os
canais do Na+ controlados por voltagem se abrem rapidamente
e o Na+ entra na célula de K
+ controlado por voltagem começam
a se abrir lentamente; (4) a entrada rápida de Na+ despolariza a
célula; (5) os canais de Na+ se fecham e os canais de K
+ mais
lentos se abrem; (6) o K+ se move da célula para o líquido
extracelular; (7) os canais de K+ continuam abertos e mais K
+
deixa a célula, hiperpolarizando-a; (8) os canais de K+
controlados por voltagem se fecham e menos K+ sai da célula;
(9) a célula em repouso.
SISTEMA SENSORIAL
Nos animais, precisamos receber informações do ambiente onde vivemos, para podermos detectar alterações ambientais e responder a elas. Os órgãos do sentido constituem a porta de entrada dos sinais ambientais. O órgão sensorial serve como um transdutor que converte o sinal externo em uma alteração do potencial de membrana da célula receptora. Assim, sinais mecânicos químicos ou luminosos são transformados em potenciais de ação e podem ser interpretados pelo SNC. Alterações na frequência dos potenciais de ação no axônio podem ser usadas como indicador de intensidade do estimulo, quanto mais forte os estímulos, maior será o número de potenciais de ação enviados ao SNC. Em todos os sentidos, o principio básico é o mesmo:
Cada célula sensorial capta apenas um tipo de sinal, sem especificar para ele;
O órgão sensorial serve como transdutor em que o sinal externo gera potenciais receptores;
A informação é transmitida pelos nervos sensoriais na forma de potenciais de ação;
O potencial de ação em todos os nervos sensoriais é da mesma intensidade e magnitude;
A intensidade do estimulo é codificada pela modulação da frequência dos potenciais de ação.
Sentido mecânico
Os mecanaceptores respondem a estímulos mecânicos como vibração, toque, gravidade, estriamento. O sentido tátil corresponde à sensação sobre a pele. Há dois tipos de receptores para o tato: receptores de adaptação rápida e lenta. Os receptores de adaptação rápida: levam sinais no inicio e no final da estimulação, mas param de responder o estimulo e continua. Quando trocamos de roupa pela manhã,quando colocamos anel ou óculos, inicialmente percebemos a presença do objeto, mas ao longo do tempo esquecemos que o objeto está lá, pois os receptores sensoriais param de mandar sinais quando são continuamente estimulados. Os receptores de adaptação lenta: mandam sinais sensoriais ininterruptos, mantendo o SNC informado do estimulo se a presença deste for prolongada. As regiões mais sensíveis ao toque possuem mais receptores sensoriais, como a boca e as patas/mão. Nos animais também temos receptores sensíveis à temperatura (termoceptores) que respondem a frio (entre 10 e 25 ºC) e ao calor (entre 30 e 45 ºC).

277
O sentido da dor é percebido por nociceptores. Há dois tipos de dor levadas ao SNC por dois tipos de nociceptores: A dor rápida, sinalizada como uma pontada e percebida imediatamente ao dano tecidual é carregado ao SNC por neurônios mielinizados e a rota até o córtex conta com poucas sinapses. A dor lenta, em queimação, que é levada por uma rede de neurônios amielinicos, chegando lentamente ao SNC. Durante a passagem até o córtex sensorial, os sinais da dor passam pelo tálamo.
Controle motor
A coordenação de contração muscular para execução de tarefas é gerada dentro do SN, pela chegada de impulsos na junção neuromuscular e geração de potencial de ação na unidade motora. Quando um potencial de ação se inicia no neurônio motor, a excitação da membrana da unidade gera resposta de todas as fibras musculares envolvidas, gerando contração da unidade motora. Os músculos de vertebrados possuem diferentes tipos de fibras de modo que o SN pode modular quais fibras e quantas fibras estarão ativas em determinadas atividades. Por causa disso, podemos segurar um copo de plástico cheio de água numa mão enquanto na outra seguramos um tijolo. Todos os comportamentos são controlados por sinais motores vindos do SNC. A atividade motora voluntária ou em resposta a determinado estimulo ambiental depende da organização dos circuitos neuronais e do modo como os neurônios processam e integram as informações. As redes neuronais mais simples são os arcos-reflexos neste caso, uma informação aferente chega ao SNC e imediatamente uma informação eferente sai, levando à contração coordenada do músculo e resposta motora um exemplo, é a retirada da mão quando tocamos um objeto quente.
.

278
FISIOLOGIA CÁRDIÁCA
Sua principal função é entregar sangue aos tecidos, fornecendo nutrientes às células para seu metabolismo e removendo os dejetos metabólicos das células. O coração funciona como uma bomba. Esse sistema também está envolvido em várias funções homeostáticas, participa da regulação da pressão sanguínea arterial, leva hormônios regulares das glândulas endócrinas para seus locais de ação. Participam da temperatura corporal.
SISTEMA CIRCULATÓRIO
O sistema circulatório (SC) transporta materiais para, e de todas as partes do corpo. As substâncias transportadas pelo SC podem ser divididas em: nutrientes, água e gases que entram no corpo a partir do meio externo; materiais que movem-se de células para células e resíduos que as células eliminam. Em animais de porte relativamente grande possuem vasos pelos quais o fluido sanguíneo flui e leva os gases e nutriente às partes do corpo, enquanto animais menores ainda usam difusão simples porque a distância a ser percorrida é pequena. O SC aberto é característico de invertebrados, no qual não há vasos conectando artérias e veias. No SC fechado, o sangue fica confinado nos vasos ao longo de todo o percurso pelo sistema vascular. Neste sistema, o coração bombeia sangue pelas artérias que ramificam até arteríolas e uma rede de capilares. O sangue deixa os capilares, entra em vênulas, veias e retorna ao coração. O SC também recolhe os resíduos metabólitos e o dióxido de carbono liberado pelas células e os transporta para os pulmões e rins, de onde serão excretados. Alguns produtos são transportados até o fígado para serem processados antes de serem excretados na urina e nas fezes.
Figura 17: (A) cabeça e encéfalo; (B) braços; (C) pulmões; (D)
tronco; (E) fígado; (F) TG; (G) rins; (H) pelve e pernas. (a)
capilares; (b) artérias; (c) artérias ascendentes; (d) parte
abdominal da aorta; (e) artérias descendentes; (f) valvas
venosas; (g) veias ascendentes; (h) veia cava inferior; (i) veia
cava superior; (j) veias; (l) átrio direito; (m) artérias pulmonares;
(n) veias pulmonares; (o) átrio esquerdo; (p) artérias coronárias;
(q) ventrículo esquerdo; (r) coração; (s) artéria hepática; (t) veia
porta do fígado; (u) veia hepática; (v) artérias renais; (x) veias
renais; (z) ventrículo direito.
Coração
O coração é um músculo que contrai continuamente, descansando somente nas pausas entre os batimentos. Estima-se que o trabalho do coração, em um minuto, seja equivalente a levantar 3kg a uma altura de 30cm. A energia necessária para este trabalho requer um suprimento contínuo de nutriente e oxigênio. O trabalho cardíaco é dividido em sístole (contração) e diástole (relaxamento). As fibras musculares cardíacas são interconectadas entre si, formando um sincício. O sincício atrial é separado do sincício ventricular por tecido fibroso. Assim todo o átrio contrai num só tempo e todo ventrículo também. Coração é constituído por duas bombas distintas:
O coração direito; bombeia sangue para os pulmões.
O coração esquerdo; bombeia sangue para órgãos periféricos.
Cada um desses corações é uma bomba pulsátil de duas câmaras compostas de um átrio e um ventrículo.
Figura 18: (a) aorta; (b) valva semilunar pulmonar; (c) artéria
pulmonar esquerda; (d) veias pulmonares esquerda; (e) átrio
esquerdo; (f) folheto da valva AV esquerda; (g) cordas
tendíneas; (h) músculos papilares; (i) ventrículo esquerdo
(biscúpide); (j) parte descendente da aorta, (l) veia cava inferior;
(m) ventrículo direito; (n) folheto da valva AV direita (tricúspide);
(o) átrio direito; (p) veia cava superior; (q) artéria pulmonar
direita.

279
Condução elétrica no coração
O potencial de ação do músculo cardíaco dura cerca de três décimos de segundos. Portanto, a duração de contração cardíaca é maior do que a do músculo esquelético. Um pequeno grupo de fibras cardíacas na parede superior do átrio direito que formam o nó sinoatrial (SA). Esse nodo é capaz de contração rítmica. As membranas das fibras do nó SA são muito permeáveis ao sódio, que faz com que o potencial de repouso se desvie sempre para valores mais positivos. Ao seu término, a membrana fica temporariamente menos permeável ao sódio e mais permeável ao potássio, o que caracteriza uma hiperpolarização. Daí, a permeabilidade ao sódio retorna ao normal e um novo potencial de ação pode ser gerado. Isso dura sem interrupção por toda a vida, o que dá à excitação rítmica das fibras do nó SA. O fato da duração da contração cardíaca ser maior do que o músculo esquelético se deve à lentidão da membrana das células em se repolarizar após a despolarização. Isso leva um platô no pico do potencial de ação de cerca de 0,3s. Durante a despolarização, entram íons de sódio e de cálcio em grande número. Os canais de cálcio são lentos e então, este íon continua entrando após cessar a entrada de sódio. Após alguns décimos de segundo, os canais de cálcio fecham-se e tem inicio a repolarização. A duração da contração é semelhante à duração do potencial de ação. Há dois tipos de potencial de ação: os de resposta rápida e o de resposta lenta. A resposta rápida ocorre no átrio, nos miócitos ventriculares e nas fibras especializadas de Purkinje. A resposta lenta ocorre nos nodos AS e atrioventricular (AV). Nessas células o potencial de ação se propaga mais lentamente e a probabilidade de bloqueio da condução é maior. O nodo AS é considerado marca-passo do coração. A atividade elétrica do marca-passo atinge primeiros os átrios e depois os ventrículos. Devido à separação dos átrios e ventrículos por tecidos fibrosos que não deixam o impulso elétrico passar, há a necessidade de um feixe de condução entre eles: o feixe atrioventricular. O feixe leva o impulso do nó SA até o nó AV. o ventrículo, por ser maior, depende de uma rede de fibras para poder contrair em uníssono para o eficaz bombeamento do sangue. Esse arranjo permite que a contração inicie no ápice e espalhe pelo tecido empurrando o sangue para a artéria de modo eficiente.
Figura 19: condução elétrica no coração. (A) o nó AS
despolariza; (B) a atividade elétrica vai para o nó AV pelas vias
internodais; (C) a despolarização se espalha mais lentamente
através do átrio. A condução atrasa através do nó AV; (D) a
despolarização move-se rapidamente pelo sistema de condução
ventricular para o ápice do coração; (E) a onda de
despolarização se espalha para cima a partir do ápice. (a) nó
AS; (b) nó AV; (f) vias internodais; (g) fascioulo AV; (h) ramos
do fascículo; (i) ramos subendocárdicos (fibras de Purkinje).
O potencial no interior da célula cardíaca é -90mV em relação ao meio externo. Durante a despolarização, o potencial se inverte e excede o externo em 20mV. Depois da despolarização rápida da fibra, segue-se uma breve repolarização precoce parcial, e então há um platô por 0,1 a 0,2s, período em que há influxo de cálcio e efluxo de potássio. A despolarização rápida ocorre com o desenvolvimento da força-cardiáca. A repolarização completa coincide com o pico da força. O relaxamento ocorre quando o potencial volta ao repouso.
Figura 20: potencial de ação de uma célula cardíaca contrátil. (0)
canais de Na+ abertos; (1) canais de Na
+ fechados; (2) canais de
Ca2+
abertos; canais de K+ rápidos fechados; (3) canais de Ca
2+
abertos; (4) potencial de repouso.

280
Contração
A maior parte das células musculares cardíacas é contrátil, cerca de 1% é especializada em gerar potenciais de ação espontaneamente. O coração pode se contrair sem uma conexão com outras partes do corpo, pois o sinal da contração é originado dentro do próprio músculo cardíaco. O sinal para a contração vem de células conhecidas como células autoexcitáveis. Essas células são conhecidas como marca-passo porque determinam a frequência dos batimentos cardíacos. A entrada do cálcio é uma do acoplamento excitação-contração (EC) cardíaca. No músculo cardíaco, um potencial de ação se origina espontaneamente nas células de marca-passo do coração e se propaga para as células contrateis pelas junções comunicantes. Um potencial de ação entra numa célula contrátil se move pelo sarcolema e entro nos túbulos T, onde abre os canais de Ca2+ controlados por voltagem, na membrana das células. O Ca2+ entra nas células e abre os canais liberadores de Ca2+ receptores de rianodina (RYR) no retículo sarcoplasmático. Quando os canais RYR se abrem, o Ca2+ entra nas células para fora do retículo sarcoplasmático e para dentro do citosol, criando um pulso de Ca2+, varias faíscas de diferentes canais de RYR se somam para criar um sinal de Ca2+. Este processo de acoplamento E-C no músculo cardíaco é também chamado de liberação de Ca2+ induzido por Ca2+. A liberação de cálcio do reticulo sarcoplasmático fornece, 90% do Ca2+ necessário para a contração muscular, sendo que os 10% restantes entram na célula aparti do líquido extracelular. O cálcio se difunde pelo citosol para os elementos contráteis, onde se liga à troponina e inicia o ciclo de formação de pontes cruzadas e o movimento. Quando as concentrações citoplasmáticas de Ca2+ diminuem o Ca2+ desliga-se da troponina, a miosina libera a actina e os filamentos contráteis deslizam de volta para sua posição de relaxamento. No músculo cardíaco o Ca2+ também é removido da célula na troca por Na+ via trocador de antiporte Na+, -Ca2+ (NCX). Cada Ca2+ sai da célula contra seu gradiente eletroquímico em troca por 3Na+ que entram na célula a favor do seu gradiente eletroquímico. O sódio que entra na célula durante essa transferência é removida pela Na+ -K+ -ATPase.
Figura 21: acoplamento excitação-contração (E-C) e
relaxamento no músculo cardíaco. (1) contração; (2)
relaxamento. (a) Actina; (b) miosina. (A) o potencial de ação
chega proveniente de células adjacentes; (B) canais de Ca2+
controlados por voltagens se abrem. O Ca2+
entra na célula; (C)
o Ca2+
induz a liberação de Ca2+
pelos canais receptores de
rinodina (RYR); (D) liberação local causa faísca de Ca2+
; (E) os
íons de Ca2+
se ligam na troponina para iniciar a contração; (F)
os íons de ca2+
se desliga da troponina; (G) Ca2+
é bombeado de
volta para dentro do retículo sarcoplasmático para se
armazenado; (H) o Ca2+
é trocado por Na2+
pelo trocador de
antiporte NCX; (I) o gradiente de Na2+
é mantido pelo Na+ -K
+ -
ATPase.
Ciclo cardíaco
Cada ciclo possui duas fases: diástole, período durante o qual o músculo cardíaco relaxa, e sístole, período durante o qual o músculo está contraindo. O sangue flui de uma área de pressão mais alta para uma área de pressão mais baixa. A contração aumenta a pressão, ao passo que o relaxamento diminui a pressão. Coração em repouso: diástole atrial e ventricular: os átrios estão se enchendo com o sangue vindo das veias, os ventrículos acabaram de completar uma contração. À medida que os ventrículos relaxam, as valvas AV entre os átrios e os ventrículos relaxam, as valvas AV entre os átrios e os ventrículos se abrem e o sangue flui por gravidade dos átrios para os ventrículos. Término do enchimento ventricular: a sístole atrial. A maior parte do sangue entra nos ventrículos enquanto os átrios estão relaxados, os restantes 20% de enchimento dos ventrículos ocorrem quando os átrios constroem e empurram o sangue para os ventrículos. A sístole inicia seguindo a onda de despolarização que percorre rapidamente os átrios, a pressão aumentada que acompanha a contração empurra o sangue para dentro dos ventrículos. Contração ventricular inicial e o primeiro som cardíaco: enquanto os átrios se contraem, a onda de despolarização se move lentamente pelas células condutoras do nó AV e rapidamente segue pelos ramos das fascículo AV- fibras de Purkinje até a ápice do coração. A sístole ventricular inicia no ápice do coração quando as bandas musculares em espiral empurram o sangue para cima em direção à base. O sangue empurrado contra a porção inferior das valvas AV faz com que elas se fechem de modo que não haja refluxo para os átrios.

281
As vibrações geradas pelo fechamento das valvas AV criam o primeiro som cardíaco, S1 o Tum do Tum-tá. Com ambos os conjuntos de valvas AV e semilunares fechados, o sangue é pressionado da mesma forma que apertaria um balão cheio de água com as aos. O coração bombeia ejeção ventricular: quando os ventrículos contraem, eles geram pressão suficiente para abrir as valvas semilunares e empurrar o sangue para as artérias. A pressão criada pela contração ventricular se torna a força propulsora do fluxo sanguíneo. O sangue sobre alta pressão é forçado para dentro das artérias, deslocando o sangue sobre baixa pressão que o preenche e empurrando-o para adiante na vasculatura. Durante essa fase, as valvas AV permanecem fechadas os átrios continuam se enchendo. Relaxamento ventricular e o segundo som cardíaco: no final da ejeção ventricular, os ventrículos começam a repolarizar e a relaxar, diminuindo a pressão ventricular. Uma vez que a pressão ventricular cai abaixo da pressão nas artérias, o sangue começa a fluir de volta para o coração. Este fluxo retrógrado enche os folhetos (cúspide) em forma de taça das valvas semilunares, forçando-os para a posição fechada, fechando as valvas. As vibrações geradas pelo fechamento das valvas semilunares ao o segundo som cardíaco S2, tã do Tum-tã. Uma vez que as valvas semilunares se fecham, os ventrículos novamente se tornam câmaras isoladas. As valvas AV permanecem fechadas devido à pressão ventricular que, ainda é maior que a pressão nos átrios. Esse período é chamado de relaxamento ventricular isovolumétrico porque o volume de sangue nos ventrículos não está mudando. Quando o relacionamento do ventrículo faz com que a pressão nos átrios durante a contração ventricular. O ciclo cardíaco começou novamente.
Figura 22: ciclo cardíaco. (A) final da diástole: ambos os
conjuntos de câmaras estão relaxados e os ventrículos enchem-
se passivamente; (B) sístole atrial: a contração atrial força uma
pequena quantidade de sangue adicional para dentro dos
ventrículos; (C) contração ventricular isovolumétrica: a primeira
fase da contração ventricular empurra as valva AV e elas se
fecham, mas não cria pressão suficiente para abrir as valvas
semilunares; (D) ejeção ventricular: como a pressão ventricular
aumenta e excede a pressão nas artérias, as valvas semilunares
se abrem e o sangue é ejetado; (E) relaxamento ventricular
isovolumétrico: quando os ventrículos relaxam, a pressão nos
ventrículos cai, o sangue flui de volta para as cúspides das
valvas semilunares e elas se fecham.
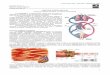
Vasos
São tubos de diferentes tamanhos com paredes elásticas e muscular para facilitar a alteração do diâmetro quando recebe o sangue. No sistema fechado, os vasos se ramificam muito, até atingirem calibres, de micrômetros e depois coalescem e voltam a ganhar diâmetro no retorno ao coração. Os vasos são classificados de acordo com suas estruturas: Artérias: são tubos cilindróides, elásticos, sem válvulas, que conduzem o sangue que sai do coração. Arteríolas: são vasos menores, mais estreitos e com grande resistência a passagem de sangue; Capilares: são vasos ainda menores, compostos por uma camada de endotélio, que permite a troca de gases e nutrientes; Vênulas e veias: são vasos que aparecem no retorno do sangue ao coração, depois das trocas teciduais.
Figura 23: estrutura do vaso sanguíneo. (A) artéria; (B) arteríola;
(C) capilar; (D) vênula; (E) veia.
Os vasos de trocas são os capilares, tubos com calibre extremamente pequeno isso facilita a troca de gases, nutrientes, metabólitos, calor e etc. O sistema venoso tem grande influencia da pressão hidrostática. As veias são tubos cilindróides, pouco elástico e repletos de válvulas que auxiliam na impulsão do sangue em direção ao coração, o calibre aumenta, a pressão sanguínea decresce progressivamente e a velocidade do fluxo diminui. O sistema vascular tem a função de transportar e distribuir substâncias essenciais e remover produtos do metabolismo das células, manter a homeostase, regular a temperatura, fazer a manutenção dos fluidos e o suprimento de oxigênio e nutriente. A pressão de pulso (PP) é a diferença entre a pressão sistólica e a diastólica o débito sistólico que é o volume de sangue bombeando a cada batimento cardíaco, pode alterar a PP.
Figura 24: Pressão arterial média x débido cardíaco x
resistência. (A) débito cardíaco; (B) resistência variável. (a)
vetrículo esquerdo; (b) artérias elásticas; (c) arteríolas.
O coração e a rede vascular são regulados por fatores neurais e humorais. Mecanismos neuronais envolvem os ramos simpáticos adrenérgicos e parassimpáticos colinérgicos do SNA. Em geral o sistema simpático estimula o coração e contrai os vasos, resultando em aumento de pressão arterial. O parassimpático deprime a função cardíaca, provoca a queda da pressão.

282
FISIOLOGIA RESPIRATÓRIA
A respiração é o processo de utilização de O2 pelos organismos. Os pulmões auxiliam nesse caso, já que são áreas invaginadas altamente vascularizadas. Nos mamíferos, milhões de sacos chamados alvéolos revestem a parte de troca gasosa pulmonar. Cada alvéolo é muito vascularizado, levando grande quantidade de O2
para o sangue o ar pulmonar. De todo o ar inspirado, apenas uma parte menor chega aos alvéolos e o O2 é retirado. Grande parte fica presa nas áreas onde não há troca essa regiões são chamadas de espaço morto. A relação entre ventilação e perfusão representa o maior determinante da troca gasosa normal. A ventilação é o volume de ar que entra ou sai do órgão respiratório em um determinado tempo, e é o processo pelo qual os novos gases se movem para dentro e para fora do organismo. A perfusão é o processo pelo qual o sangue desoxigenado torna-se oxigenado durante o contato entre o órgão respiratório e o sistema circulatório. A relação entre ventilação e perfusão é definida como relação entre o fluxo de ar que entra e sai do organismo respiratório e o sistema circulatório. A relação entre ventilação e perfusão é definida como relação entre o fluxo de ar que entra e sai do organismo e o fluxo o sanguíneo que passa pelo organismo respiratório afim de capturar O2 e eliminar CO2.
Figura 25: visão geral da respiração celular e respiração interna.
(a) vias aéreas; (b) alvéolos dos pulmões; (c) circulação
pulmonar; (d) circulação sistêmica. (A) troca I: entre a atmosfera
e o pulmão (ventilação); (B) troca II: entre o pulmão e o sangue;
(D) troca III: entre o sangue e a células. (1) célula; (2) nutrientes.
Troca gasosa
A respiração é normalmente involuntário, controlado pelo centro da respiração no TE. Neurônios localizados ao redor das vias aéreas, e no SNC podem perceber a quantidade de CO2 circulante e estimular ou inibir o centro de controle. O CO2 é o responsável porque seu excesso causa a diminuição do pH. Em todos os vertebrados há proteínas chamadas pigmentos respiratórios que auxiliam no transporte de O2 e CO2 no fluido corpóreo. A hemoglobina é o pigmento respiratório mais comum, uma proteína que contém ferro e dá a cor vermelha ao sangue. Essa proteína tem alta afinidade com O2 e carrega o gás pelo sangue para os tecidos. O transporte de O2 e CO2 definido pela difusão de gases e sua solubilidade. Enquanto o mecanismo de transporte de O2 e CO2 é ligado à hemoglobina, o CO2 é transportado dentro das hemácias na forma de bicarbonato (HCO3-). O O2 liga-se a grupamentos heme de hemoglobinas e sua afinidade pelo O2 pode ser alterado na presença de CO2-.
Figura 26: o O2 e o CO2 se movem para dentro e para fora do
sangue nos capilares pulmonares e nos capilares. (1) circulação
pulmonar; (2) circulação sistêmica; (A) troca de O2 na interfase
capilar-alveolar; (B) transporte de oxigênio; (C) troca de O2 nas
células; (D) troca de CO2 nas células; (E) transporte de CO2; (F)
troca de CO2 na interfase capilar alveolar.

283
VOLUMES PULMONARES
Os volumes dos pulmões são medidos por um Espirômentro. A respiração normal é suave corresponde à inspiração e à expiração de volume corrente Vr. O volume normal é de cerca de 500mL. Com o volume adicional que pode ser inspirado acima do volume corrente é chamado de volume inspiratório de reserva e é de quase 3.000ml. o volume adicional que pode ser expirado abaixo do volume corrente é chamado de volume expiratório de reserva, e é de cerca de 1,200ml. O volume de gás que fica nos pulmões após expiração máxima forçada e o volume residual (VR) que é de aproximadamente 1,200 ml.
Capacidade pulmonar
A capacidade inspiratória (CI) é composta pelo volume corrente mais o volume inspiratório de reserva, e é de cerca de 3.500ml. A capacidade funcional residual (CFR) é composta pelo volume expiratório de reserva (VER) mais o volume residual. A CFR é o volume que permanece nos pulmões após ser expirado o volume corrente normal, e pode ser considerado o volume que pode ser expirado após a inspiração máxima. A capacidade pulmonar total (CPT) inclui todos os volumes pulmonares, é a capacidade vital mais o volume residual.
Espaço morto
É o volume das vias aéreas e dos pulmões que não participam da troca de gases. O espaço morto pode ser considerado como os alvéolos que não participam das trocas gasosas. Eles não participam por causa de um desencontro entre a ventilação e a perfusão.
CICLO RESPIRATÓRIO
É dividido em três fases: repouso, Inspirações e expiração.
Repouso
Período entre os ciclos respiratório, quando o diafragma está em sua posição de equilíbrio, no repouso, nenhum ar está se movendo para dentro ou para fora dos pulmões. A pressão alveolar é igual à atmosférica, diz-se que a pressão alveolar é zero.
Inspiração
Durante a inspiração, o diafragma se contrai, fazendo com que aumente o volume do tórax. Quando o volume do pulmão aumenta, a pressão pulmonar tem de diminuir. No meio do caminho na inspiração a pressão alveolar cai abaixo da pressão atmosférica. O ar se dirige aos pulmões até que, ao final da inspiração, a pressão alveolar, mais uma vez, é igual a pressão atmosférica.
Expiração
A pressão alveolar torna-se positiva porque as forças elásticas dos pulmões e os volumes pulmonares voltam à CFR.
Figura 27: (A) em repouso, o diafragma está relaxado; (B)
quando o diafragma contrai, o volume torácico aumenta; (C)
quando o diafragma relaxa o volume torácico diminui.

284
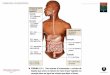
FISIOLOGIA GASTROINTESTINAL
O trato gastrointestinal (TGI) é um tubo longo e oco que passa através do corpo. Os animais usam o alimento para conseguir matéria prima para a formação e manutenção celular. Os alimentos vindos de animais ou vegetais precisam ser degradados, para fornecer os compostos ideais para atividade celular. Todos os mamíferos são sugadores no inicio da vida, quando se alimentam apenas do leite materno. Alguns animais usam a raspagem e mastigação antes de engolir grandes bolos alimentares. Os dentes, que auxiliam na mastigação podem estar presentes em diferentes formatos e quantidades. A mastigação favorece a ação enzimática no TGI. O estômago também serve como armazenador, porem tem alguma função na digestão de proteínas devido à presença da enzima pepsina. Depois do estômago, o intestino faz o papel final da digestão e absorção de nutrientes.
TUBO DIGESTÓRIO
O TGI é formado por uma estrutura básica. É um tubo oco com camadas de tecido que o rodeia, onde temos glândulas, vasos, músculo e tecidos nervosos e conectivos em vertebrado, a estrutura do TGI é formada por quatro camadas: Túnica serosa: Camada mais externa e consiste no peritônio visceral e em tecido conjuntivo limitado subjacente a ela. Túnica mucosa: Essa camada possui invaginações que forma uma infinidade de vilosidades que aumentam a superfície de contato com o alimento. Túnica submucosa: Camada de tecido conjuntivo frouxo no qual são encontrados vasos sanguíneos e nervos. Contém pequenas glândulas que lançam suas secreções em direção à luz do esôfago. Substancias que combatem os agentes infecciosos do meio externo. Túnica muscular: Bem desenvolvida, importante para a mobilidade do sistema digestório.
Figura 28: (a) vilosidade; (b) criptas; (c) mucosa; (d) muscular
da mucosa; (e) submucosa; (f) muscular externa; (1) camada
circular; (2) camada longitudinal; (g) serosa; (h) artéria e veia
submucosa; (i) placa de Peyer; (j) plexo mioentérico; (l) plexo
submucoso; (m) vaso linfático.
Boca
A mastigação é o primeiro movimento voluntário e parte reflexa. Este movimento tem a intenção de triturar o alimento e misturá-lo às secreções salivares. Processo de mastigação. É causada pelo reflexo da mastigação; a presença de um bolo alimentar na boca dá início à inibição reflexa dos músculos da mastigação, o que permite que a mandíbula caia. Essa queda inicia um reflexo de estiramento dos músculos da mandíbula, o que leva uma contração de rebote. Isso eleva a mandíbula, provocando o fechamento dos dentes, mas também comprime o bolo alimentar novamente contra as paredes da boca, o que inibe de novo os músculos da mandíbula, propiciando a queda da mandíbula, e outra vez, o rebote. A mastigação auxilia na digestão do alimento por uma simples razão, as enzimas digestivas atuam apenas na superfície das partículas alimentares, a velocidade da digestão depende muito da área total da superfície exposta às secreções intestinais. O próximo movimento é a deglutição. A primeira fase da deglutição é voluntária (fase oral): a língua move o bolo de alimento e o força pela faringe. A segunda fase é a faringe, começa quando o palato mole é empurrado para cima e a faringe é movida contra a epiglote.
Figura 29: reflexo da deglutição. (A) a língua empurra o bolo
contra o palato mole e a parte posterior da cavidade oral,
disparando o reflexo da deglutição. (a) palato mole; (b) palato
duro; (c) língua; (d) bolo alimentar; (e) epiglote; (f) glote; (g)
laringe; (h) esfíncter esofágico superior contraído. (B) o
esfíncter esofágico superior relaxa enquanto a epiglote fecha
para manter o material deglutido fora as vias aéreas. (C) o
alimento move-se para baixo no interior do esôfago, propelido
por ondas peristálticas e auxiliados pela gravidade.

285
Faríngea
À medida que o bolo alimentar entra na parte superior da boca e na faringe, ele estimula as áreas receptoras da deglutição situadas ao redor da abertura da faringe. Impulsos dessas áreas vão para o tronco cerebral para dar início a uma série de contrações automáticas da musculatura da faringe da seguinte forma:
O palato mole é empurrado para cima, fechando a parte superior das narinas.
As pregas palatofaríngeas são puxadas medialmente e se aproximam.
As cordas vogais são puxadas, e a faringe é puxada para cima e para frente pelos músculos do pescoço. Essas ações fazem com que a presença desses ligamentos impeça o movimento da epiglote para cima, fazendo com que essa se dobre para trás, recobrindo a abertura da faringe. O movimento da laringe para cima também traciona para cima a abertura do esôfago. Ao mesmo tempo, as paredes musculares superior do esôfago denominadas de esfíncter esofágico superior relaxam-se, permitindo que o alimento se desloque fácil e livremente da porção posterior da faringe para a parte superior do esôfago. Ao mesmo tempo em que a laringe é elevada e os esfíncter faringo-esofágico se relaxa, começando em sua parte superior muscular da faringe, começando em sua parte superior e propagando-se para baixo como uma rápida onda peristáltica que passa pelas porções médias e inferior da faringe e alcança o esôfago, impulsionando o alimento para este órgão. A mecânica da etapa faríngea da deglutição inclui o fechamento da traqueia, abertura do esôfago e uma rápida onda peristáltica originada na faringe que força o bolo alimentar para o esôfago.
Esôfago
O esôfago conduz o alimento da faringe para o estômago. O esôfago apresenta dois tipos de movimentos peristálticos. Peristáltico primário: É a continuação da onda peristáltica iniciada na faringe e se propaga para o esôfago durante a etapa faríngea da deglutição. Peristáltico secundário: Caso a onda primária não consiga empurrar todo o alimento para o estômago, surgem as ondas peristálticas secundárias resultantes da distensão do esôfago pelo alimento retido. Esfíncter cárdia: Situado na parte inferior do esôfago, um pouco acima de sua junção com estomago, o músculo esofágico, funciona como esfíncter. Ele permanece contraído, encontraste com a porção média do esôfago, que em geral permanece relaxada. Quando uma onda peristáltica de deglutição se propaga pelo esôfago o relaxamento receptivo, relaxa o esfíncter à frente da onda peristáltica, permitindo a propulsão do alimento para o estômago. Ele também evita o refluxo do bolo alimentar.
Figura 30: contração peristáltica e segmentares no TGI. (A) as
contrações peristálticas são responsáveis pelo movimento para
adiante; (a) tempo zero; (b) contração; (c) direção do
movimento; (d) segmento receptor; (e) segundos depois. (B) as
contrações resultantes para adiante responsáveis pela mistura;
(a) nenhum movimento resultante para adiante.

286
Estômago
O estômago tem como função motora armazenar o alimento, misturá-lo com secreções gástricas até formar o quimo e também é a passagem do alimento para o intestino delgado. O suco digestivo é secretado pelas glândulas gástricas, que recobrem quase toda a a parede do corpo do estômago. Essas secreções entram em contato com o alimento armazenado. No mesmo tempo ondas peristálticas, deslocam-se em direção ao piloro, ao longo da parede do estômago. A medida que essas ondas percorrem a parede do estômago, elas misturam e propulsionam o alimento em direção ao fundo. aparti daí as ondas impulsionam o alimento em direção ao piloro. Após a mistura do alimento com as secreções gástricas, a mistura resultante que se desloca pelo intestino é o quimo. O suco gástrico é um líquido incolor e ácido. Possui componentes como enzimas digestórias e ácido clorídrico (HCl) o papel do HCl é: Tornar o suco gástrico ácido com ph 2,0 fato importante para a ativação da pepsina. Tem ação antisséptica, promovendo a morte ou inibição de vários microrganismos que penetram no tubo digestivo junto com o alimento. Atua sobre a mucosa gástrica, permitindo a abertura da válvula pilórica. Fato importante por permitir a passagem gradativa do alimento do estômago para o duodeno. A principal enzima gástrica é a pepsina. Ela é lançada na cavidade gástrica como pepsinogênio. Em presença de HCl. Ela se transforma em pepsina ativa. Ela digere as proteínas.
Figura 31: (1) estômago. (a) antro; (b) corpo; (c) diafragma; (d)
esôfago; (e) fundo; (f) piloro; (g) rugas: superfície pregueada
que aumenta a área. (2); (a) abertura da glândula gástrica; (b)
epitélio; (c) vaso linfático; (d) lâmina própria; (e) muscular da
mucosa; (f) submucosa; (g) muscular externo: (1) camada
obliqua; (2) camada circular; (3) camada longitudinal; (h)
serosa; (i) plexo mientérico; (l) artéria; (m) mucosa.
.
Intestino delgado
O quimo é impulsionado no intestino delgado por ondas peristálticas. O intestino é dividido em três porções:
Duodeno
É um tubo que liga o estômago ao intestino delgado, é nele que o suco biliar vindo da vesícula, junta-se ao alimento. Pela mudança de ph, diversas enzimas digestivas são ativadas. É a porção primária do intestino delgado e possui diversas vilosidades. É onde se tem espaço para maior parte do processo digestório.
Jejuno
É a porção do intestino delgado que está entre o duodeno e o íleo. É um órgão responsável pela absorção de carboidratos e aminoácidos digeridos parcialmente pelo estômago e o duodeno.
Íleo
É onde continua a digestão química após passar pelo duodeno e jejuno, recebe esse nome porque está apoiado sobre o osso íleo. Absorve bastante vitamina B12.
Figura 32: vilosidade e uma cripta no intestino delgado. (a)
borda em escova; (b) microvilocidade; (c) os enterócitos
transportam nutrientes e íons; (d) os capilares transportam a
maioria dos nutrientes absorvidos; (e) as células caliciformes
secretam muco; (f) o ducto lactifero transporta a maior parte
das gorduras para a linha; (g) lâmina própria; (h) células
endócrinas; (i) muscular de mucosa; (j) lúmen da cripta.

287
Intestino grosso
Tubo muscular com cerca de 1mm de comprimento e 7 cm de diâmetro, ele inicia-se na válvula íleo cecal e termina na válvula anal divide-se em três segmentos:
Ceco
É a primeira parte do intestino grosso que recebe o conteúdo do intestino delgado. É onde se localiza o apêndice. Sua única função é a reabsorção de água e nutrientes.
Cólon
É a parte mais longa do intestino grosso e subdivide-se em cólon ascendente, cólon transverso, cólon descendente e cólon sigmoide. As principais funções do intestino grosso são: Reabsorção da água. Formação e acúmulo de fezes.
Reto
É a porção final do intestino grosso, a qual se inicia no ângulo obtuso da alça sigmoide e termina no canal anal. Sua função é acumular as fezes, para absorção final de nutrientes, antes de serem eliminadas pelo organismo, através do anus.
Anus
É o orifício final do intestino grosso por onde são eliminadas as fezes e gazes intestinais. A musculatura de suporte do ânus é a do períneo, juntamente com o esfíncter interno e externo da região.
Figura 33: (1) intestino delgado. (a)veia porta do fígado; (b) veia
cava inferior; (c) colo transverso; (d) colo ascendente; (e) papila
ilial; (f) ceco; (g) apêndice vermiforme; (h) íleo; (i) reto; (j) colo
sismóide; (l) saculação do colo; (m) colo: tênia do colo; (n)
aorta. (2) corte do intestino grosso. (a) lifonodo; (b) glândula
intestinal; (c) muscular da mucosa; (d) submucosa; (e) muscular
externa: (1) camada longitudinal; (2) camada circular. (3) reto.
(a) esfíncter interno do ânus; (b) esfíncter externo do ânus; (c)
ânus.
ORGÃO ACESSÓRIOS
Fígado
Bílis
É produzido pelo fígado e armazenado na vesícula biliar, de onde é liberada para o duodeno, através do canal colédoco. A bílis apresenta os sais biliares que não só emulsionam lipídios, também tornam solúveis os ácidos graxos e os gliceróis
Pâncreas
É uma glândula, anexa ao duodeno, ela produz o suco pancreático, um líquido incolor, rico em enzimas e básicos, com ph ao redor de 9. As principais enzimas pancreáticas são: Tripsina: quebra proteínas. Amilopsina: quebra amido. Lípase: quebra lipídios. Nuclease: quebra ácidos nucleicos. A digestão no intestino delgado ocorre graças à ação conjunta de três sulcos digestório:
Suco pancreático
O suco pancreático é rico em bicarbonato, que neutralizam no intestino a acidez do quimo gástrico. A atividade do pâncreas como produtor de suco pancreático sofre poderosa influência hormonal. Isso porque o HCl gástrico estimula no duodeno, a produção de dois hormônios que ativam a secreção do suco pancreático. . SECRETINA Hormônio que provoca a produção de um grande volume de suco pancreático alcalina, porém pobre em enzimas digestivas. PANCREOZIMINA Hormônio que provoca a produção de um pequeno volume de suco pancreático rico em enzima digestória.

288
SISTEMA URINÁRIO O sistema urinário humano é constituído por: Rins (a). Ureteres (b). Bexiga urinária (c). Uretra (d). Pela parte côncava do rim (hilo) penetra a artéria renal, transportando o sangue que deve ser purificado.Osangue drenado sai do rim também por essa região, através da veia renal.
ELEMENTOS VASCULARES DO RIM O sangue entra no rim pela artéria renal, o sangue flui apartir da artéria pela eferente para
uma rede de capilares semelhante a uma bola, chamada de glomérulo. O sangue deixa o glomérulo flui para uma arteríola eferente e depois para um segundo conjunto de capilares. CONSTITUIÇÃO DOS RINS Os rins em especial são responsáveis pela filtração e secreção de hormônios, como renina, que participa da regulação da pressão sanguínea, e a eritropoietina, que estimula a produção da ativação da vitamina D3 e um pró-hormônioesteroide, no hormônio ativo. Órgão com forma de feijão, na sua região côncava encontra-se o hilo, onde entram e saem vasos sanguíneos, nervos e saem osureteres. O hilo contém dois ou três cálices, que se unem para formar a pélvis renal. O rim é constituído poruma zonacortical e outra zona medular.
ZONA CORTICAL Formado pelos corpúsculos renais, as porções retas e contorcidas dos nefrons, vasos e os raios medulares.
FILTRAÇÃO RENAL É o movimento de líquido do sangue para o lúmen do néfron, ela ocorre apenas no corpúsculo renal, onde as paredes são modificadas para permitir o fluxo de massa do líquido. ZONA MEDULAR Sãoformados de 10 a 18 pirâmides medulares (de Malpighi), seus vértices fazem saliência nos cálices renais. Da base de cada pirâmide partem os raios medulares, que penetram no córtex. Região onde é encontrado a alça de Henle e os túbulos coletores

289
ELEMENTOS TUBULARES DO RIM O néfron se inicia com a cápsula de Bowman que circunda o glomérulo. O endotélio do glomérulo é fundido ao epitélio da cápsula de Bowman de modo que o líquido da capsula passa diretamente para dentro do lúmen do túbulo da capsula de Bowman, o líquido filtrado segue para dentro do túbulo proximal e depois para a alça de Henle, um segmento em forma de U que vai em direção à medula e depois volta em direção ao córtex. O líquido passa então para o túbulo distal. Os túbulos distais de até oito Néfrons drenam para um único tubo maior chamado ducto colector. Eles passam do córtex para a medula, e drenam na pelve renal, o líquido filtrado, agora chamado de urina, flui para o ureter. NÉFRONS São as unidades morfofuncionais dos rins, são os responsáveis pela eliminação e absorção de nutrientes. São constituídos por: Corpúsculo renal (glomérulo e cápsula de Bowman) Túbulo contorcido proximal. Alça de Henle. Túbulo contorcido distal.
FILTRAÇÃO GLOMERULAR É determinado pelo equilíbrio entre pressões hidrostáticas e coloidosmótica que atuam através da membrana capilar e pelo coeficiente de filtração capilar. No ser humano adulto filtração glomerular é em média, 180 litros por dia. As pressões efetivas da filtração incluem: A pressão hidrostática nointerior dos capilares glomerulares que promovem a filtração. A pressão hidrostática na cápsula de Bowman para fora dos capilares, que se apõe à filtração.
REABSORÇÃO RENAL É necessário para que o corpo recupere os componentes eferentes envolve os túbulos renais, capturando os nutrientes uteis ao organismo que se encontram no filtrado. TÚBULO PROXIMAL Absorve cerca de 85% da água filtrada retornando-a para o sangue. Moléculas de glicose, aminoácidos e vitaminas, entre outras substâncias, retornam aos capilares sanguíneos em grandes partes através de um mecanismo de transporte ativo.
ALÇA DE HENLE Em sua porção ascendente, tem absorção de N𝑎+, por transporte ativoseguindo o gradiente elétrico, ocorre com um considerável fluxo de C𝑙− do filtrado para o sangue. TÚBULO DISTAL Com sangue mais concentrado,deve ocorre um fluxo osmótico de água para o interior dotúbulo distal depende da presença de ADH, Hormônio Antidiurético. Hormônio produzido no hipotálamo é armazenado e liberado pela neuroipófise no sangue, tem a propriedade de aumentar a permeabilidade dotúbulo distal e, determinar a maior reabsorção da água. Em situações em que aingestão da água é muito baixa, a neuroipófise promove grande liberação de ADH no sangue resultado em uma alta reabsorção de água no túbulo distal. Consequentemente, a urina se tornaescassas e concentrada, determinando, uma acentuada economia hídrica para o individuo em situações inversas, a liberação de ADH no sangue resultando em uma alta reabsorção de água notúbulo distal consequentemente, a urina se torna escassa e concentrada, determinando uma economia hídrico para o individuo em situações inversas,a liberação de ADH no sangue diminui, acarretando uma baixa reabsorção de água no túbulo distal e a formação de urina volumosa diluída.

290
SECREÇÃO Remove moléculas específicas do sangue e as adiciona ao filtrado glomerular movem substâncias do sangue para dentro do túbulo a secreção é um processo mais seletivo que geralmente usa proteína de membrana para transportar as moléculas através do epitélio tubular. MICÇÃO Uma vez que o filtrado deixa os ductos coletores, sua composição não muda. O filtrado agora é chamado de urina, flui para a pelve renal e então desce pelo ureter em direção a bexiga com ajuda de contrações rítmicas de músculo liso. A bexiga urinária é um órgão oco cujas paredes contêmcamadas bem desenvolvidas de músculos lisos. A bexiga urinária pode se expandir até conter um volume aproximado de 500mL. O colo da bexiga é contínuo com a uretra, um tubo púnico pelo qual a urina passa para alcançar o meio externo. A abertura entre a bexiga urinária e a uretra é fechada por dois anéis de músculos denominados esfíncteres: O esfíncter interno da uretra é uma continuação da parede da bexiga e é constituído de músculo liso. O esfíncter externo da uretra é um anel de músculo esquelético controlado por neurônios motores somáticos. A micção é um reflexo espinal simples que está sujeito aos controles consciente e inconsciente pelos centros superiores do encéfalo. A medida que a bexiga se enche com urina e suas paredes se expandem, receptores de estiramento enviam sinais via neurônios sensoriais para a medula espinal.