Embed Size (px)
Citation preview

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 1/150
TitleThermal tolerance of Echinolittorina species in Hong Kong:implications for their vertical distributions
Advisor(s) Williams, GA
Author(s) Li, Hoi-ting, Kathy.; “§sý.
Citation
Issued Date 2012
URL http://hdl.handle.net/10722/173874
RightsThe author retains all proprietary rights, (such as patent rights)and the right to use in future works.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 2/150
THERMAL TOLERANCE OF ECHINOLITTORINA SPECIES
IN HONG KONG:
IMPLICATIONS FOR THEIR VERTICAL DISTRIBUTIONS
by
Hoi Ting Kathy, LI
B. Sc. in Biology, The Hong Kong University of Science and Technology, Hong Kong
A thesis submitted in partial fulfillment of the requirement for
the Degree of Master of Philosophy
at The University of Hong Kong
August, 2012

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 3/150
Abstract of thesis entitled
THERMAL TOLERANCE OF ECHINOLITTORINA SPECIES
IN HONG KONG:
IMPLICATIONS FOR THEIR VERTICAL DISTRIBUTIONS
Submitted by Hoi Ting Kathy, LI
for the Degree of Master of Philosophy
at The University of Hong Kong
in August, 2012
Intertidal rocky shores represent an extremely stressful physical environment
dictated by the rise and fall of the tides. One of the major environmental stresses
over this gradient is temperature, especially towards the upper reaches of the
shore where species spend long periods out of water exposed to hot, desiccating
conditions. As a result, the thermal tolerance of intertidal species is often
positively correlated with their vertical distributions, and the physiological and
molecular limits that drive such patterns have been the subject of recent research.
Understanding these tolerance limits, from small (e.g. vertical distribution) to
large (e.g. latitudinal) spatial scales, may provide information to predict species’
success under future climate change scenarios, and thus possible changes in
community structure.
Given their abundance in the high shore, and well resolved taxonomy and
phylogeography, the littorinids Echinolittorina malaccana, E . radiata and E .

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 4/150
vidua are excellent models to investigate the relationship between thermal
tolerance and spatial distribution patterns. These littorinids are widely
distributed on Hong Kong shores and exhibit a distinct and consistent vertical
distribution that ranges from temperate to tropical regions along the western
Pacific coast. Field surveys in summer and winter at two moderately exposed
shores (Stanley and South Bay, Hong Kong) showed that E. malaccana was
distributed highest on the shore, followed by E. radiata and E. vidua respectively,
and all the three species were found ~ 0.25m lower on the shore in summer than
winter. Laboratory experiments, including determination of survival limits
(LT50), Arrhenius breakpoint temperature of heart rate (ABT of HR) and activities
of metabolic enzymes (MDH and LDH), were used to establish if the
physiological attributes of the three species were related to their distribution
patterns. The LT50 of E. malaccana were the highest of the three species
(56.47oC), followed by E . radiata (55.5oC), and finally the lower shore species E .
vidua (53.7oC); while ABT of HR in E . malaccana (48.2oC) was also higher than
E . radiata (46.5oC) and E . vidua (46.6oC). The enzyme activities did not show
any clear patterns. In terms of seasonal variation, LT50 and ABT of HR of all
three Echinolittorina species were higher in summer than winter, which showed
the potential for the littorinids to acclimate when environmental conditions
become more severe.
The present study provided a fundamental understanding of how physiological,
temperature tolerance may determine the spatial and temporal distribution patterns
of Echinolittorina species at a local scale where strong environmental gradients
vary vertically and also between seasons. Information on the tolerance limits of
physiological traits such as LT50, heart rates and enzyme functioning may direct

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 5/150
further investigations to identify the underlying causes of the survival limits of
these species to temperature variation, and whether this tolerance is genetically or
environmentally determined, for example through acclimation. Such studies will
provide insights into how a species' physiology may limit their present-day
distributions at multiple scales from local to biogeographical, but also enable
predictions of how species may respond to changing temperature regimes.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 6/150

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 7/150
ii
ACKNOWLEDGEMENTS
I would like to express my deepest gratitude to
Prof. Gray Williams, my supervisor, for his generous guidance and support,
especially during my endless thesis writing progress, as well as giving me the chance
to have lots of fun at Tsitsikamma and St. Petersburg, and BEER of course.
Dr. Stephen Cartwright, for his patience and guidance on detailed revisions on
this thesis.
Dr. Yunwei Dong, Dr. Wai-chuen Ng and Dr. Priscilla Leung, for teaching me
the techniques on enzyme works.
My field/lab helpers - June Leung (the all-the-time driver :]), Karen Villarta (the
prison breaker at Stanley), Terence Ng (the littorinids catcher), Stella Wong, Samuel
Wang, Sally Lau, Charmaine Yung, Adela Li, etc, for spending their time and
assisting me to find the little snails, either under hot & strong sun in summer or cold,
wet (by the strong wave action) & mysterious nights in winter.
All the students and staff at the Swire Institute of Marine Science (SWIMS), and
the School of Biological Sciences, The University of Hong Kong (SBS, HKU),
especially Cecily Law and Patrick Chan for their technical support.
My softball teammates at HKUST, PTIs from Mai Po, previous labmates at
CML, classmates from UST Biology, great friends from GHS, for their unlimited
supports and cheers, esp when I was desperate, and I want to say ‘I love you all!’.
My family, both real and host family from US, for letting me do whatever I want,
despite my willfulness.
Last, but most importantly, Miss Cecilia Chow, my Biology teacher at GHS, for
her generous sharing during classes that inspire my interest in the environmental field,
and lead to completion of this M.Phil. degree.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 8/150
iii
CONTENTS
Declaration ………………………………………………….…… i
Acknowledgements …………………………………………………….… ii
Contents ………………………………………………………. iii
CHAPTER 1 GENERAL INTRODUCTION ………………………..… 1
INTRODUCTION
Intertidal rocky shores as model systems to investigate
species’ distribution patterns
Temperature - a prominent factor controlling species’
distribution patterns
Littorinids as a model for studies on thermal stress
Aims of study and project outline
The Hong Kong climate
Site descriptions
CHAPTER 2 SEASONAL VARIATION IN VERTICAL DISTRIBUTION OF
ECHINOLITTORINA SPECIES ………………………..… 20
INTRODUCTION
MATERIALS AND METHODS
RESULTS
DISCUSSION

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 9/150
iv
CHAPTER 3 THERMAL TOLERANCE OF Echinolittorina SPECIES IN
HONG KONG ………………………..… 46
INTRODUCTION
Thermal tolerance and species distribution on intertidal rocky
shores
How to determine thermal tolerance of species?
Thermal tolerance and vertical distribution of Echinolittorina
species on Hong Kong rocky shores
MATERIALS AND METHODS
Preliminary studies as quality control of experimental protocol
Seasonal variation in lethal temperature
Activities of malate dehydrogenase and lactate dehydrogenase
Data analysis
RESULTS
Seasonal variation in lethal temperature
Seasonal variation in Arrhenius breakpoint temperature of
heart rate
Activities of malate dehydrogenase and lactate dehydrogenase
DISCUSSION
Lethal temperature in relation to vertical distribution
Arrhenius breakpoint temperature of heart rate in relation to
vertical distribution
Activities of malate dehydrogenase and lactate
dehydrogenase in relation to vertical distribution

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 10/150
v
Seasonal variation of LT50 and ABT of HR
CHAPTER 4 GENERAL DISCUSSION ………………………..… 104
DISCUSSION
Vertical and seasonal distribution of Echinolittorina
species in Hong Kong
Thermal tolerance of Echinolittorina species and their
seasonal variations in Hong Kong
Present study: implications for future investegations
Conclusions
LIST OF REFERENCES ………………………..… 115
APPENDIX 1 ………………………..… 138
Name used for Echinoli ttorina malaccana , E .
radiata and E . vidua in previous literature
APPENDIX 2 .1 ………………………..… 139
Heart rate of Echinolittorina species vs
temperature (Winter)
APPENDIX 2 .2 ………………………..… 140
Heart rate of Echinolittorina species vs
temperature (Summer)

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 11/150
CHAPTER 1:
GENERAL INTRODUCTION
1.1
INTRODUCTION 2
I ntertidal r ocky shores as model systems to investigate species ’
distri bution patterns
Temperature - a prominent factor controlling species’ distri bution
patterns
L ittor inids as a model for studies on thermal stress
Aims of study and project outli ne
The Hong Kong cl imate
Site descriptions

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 12/150
Chapter 1: General introduction
1.1 INTRODUCTION
Determination of the causes of species distribution patterns is a complex matter in
all types of ecosystems, as it involves the interplay of physical factors (e.g.
temperature, salinity, water availability) and biological interactions (e.g. predation,
competition, facilitation) between species (see Brown, 1984; Krebs, 2009 for
reviews). Given the complex nature of these interactions, ecologists are keen to
investigate whether individual factors can play a dominant role in determining
these distribution patterns; from large-scale latitudinal species' ranges to
small-scale microhabitat selection (see Begon et al., 1996 for review).
Identifying the factors which drive these patterns would facilitate greater
understanding of community dynamics within an ecosystem (e.g. in freshwater
systems: Welborn et al., 1996; intertidal systems: Underwood and Chapman, 1996;
also see Tilman, 1982 and Menge and Olson, 1990 for reviews).
I nter tidal r ocky shor es as model systems to investigate species ’ distri bution
patterns
Intertidal rocky shores are a model system to assess the impacts of physical
factors on species' distribution patterns, as shores experience a very extreme and
dynamic physical environment gradient determined by tidal events (see Lewis,
1964; Stephenson and Stephenson, 1972; Menge and Branch, 2001; Little et al.,
2009 for reviews). The changes brought by the rise and fall of the tides creates
alternating aquatic and terrestrial conditions which intertidal organisms have to
tolerate, and thus establishes a regularly fluctuating, severe, environmental

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 13/150
Chapter 1: General introduction
gradient along the vertical range of the shore (see Rafaelli and Hawkins, 1996;
Menge and Branch, 2001; Little et al., 2009 for reviews). The unique situation
of severe changes in environmental conditions (e.g. temperature, water
availability) and biological interactions (e.g. predation, competition) over a short
vertical gradient (often less than 10m), therefore, makes intertidal rocky shores a
model system to investigate how these factors affect the distribution of species.
With variation in the degrees of physical stress and biological interactions along
the intertidal zone, organisms exhibit different ranges of tolerance to the
conditions they can experience along this vertical environmental gradient (see
Underwood, 1979 for review). Species are unable to tolerate conditions along
the entire vertical gradient, and are assumed to have become adapted to perform
optimally to the conditions experienced at certain heights along the shore (see
Nybakken and Bertness, 2005 for review). As a result, species are found to have
their own, specific, vertical distribution patterns. The upper limit of species'
distribution along the shore is generally regarded to be set by physical factors,
including temperature and desiccation (e.g. Lewis, 1964; Stephenson and
Stephenson, 1972; Wolcott, 1973; Schonbeck and Norton, 1978; Stillman and
Somero, 2001) whilst lower limits are considered to be driven by biological
interactions, such as competition and predation (e.g. Connell, 1961a; Paine, 1974;
Wethey, 1984). These interactions are also mediated by changing environmental
conditions, for example, seasonal changes (e.g. Evans, 1948; Fraenkel, 1968);
physical disturbance (e.g. Boulding and Van Alstyne, 1993; Hutchinson and
Williams, 2003b; Sanpanich et al., 2006) and also the supply of juveniles to the
shore (Underwood and Denley, 1984; Underwood and Fairweather, 1989; Menge

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 14/150
Chapter 1: General introduction
and Branch, 2001). Ultimately, knowing how species respond to these
interactions in relation to their vertical distributions will allow a better
understanding of the processes driving intertidal community structure (Menge and
Olson, 1990; Underwood and Chapman, 1996).
Temperature - a prominent factor controlling species’ distribution patterns
Environmental temperature is widely accepted as one of the most important
physical factors associated with both latitudinal and local distribution patterns of
species in terrestrial and marine ecosystems (see Huey and Kingsolver, 1989;
Pörtner, 2002; Parmesan, 2006; Angilletta, 2009 for reviews). The temperatures
species experience can affect their distributions by altering the fitness of
organisms through influencing their crucial life functions, from biochemical to
physiological levels, hence affecting species performance such as survival,
locomotion, growth and reproduction (see Hochachka and Somero, 2002; Pörtner,
2002; Pörtner et al., 2007; Angilletta, 2009 for reviews). Since environmental
temperatures vary at a variety of spatial and temporal scales, from local to
geographic scales, studies have investigated the relationship between variation in
temperature tolerance and both small-scale, local species distributions (for
example between lizards along a 1000-m altitudinal gradient, Spellerberg, 1972;
and gastropods that occupy a narrow vertical range, often < 10m, Garrity, 1984)
as well as large-scale, latitudinal species distributions (e.g. in insects:
Addo-Bediako et al., 2000; ectotherms in both marine and terrestrial
environments, Sunday et al., 2010). Understanding how temperature tolerance
plays a role in determining the different scales of species' spatial patterns, may

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 15/150
Chapter 1: General introduction
provide information to predict species’ success, for example, under future climate
change scenarios and hence potential changes in community structure (Somero,
2010).
On intertidal rocky shores, environmental temperatures fluctuate greatly due to
the alternation of tidal events (Lewis, 1964; Stephenson and Stephenson, 1972).
Due to the variation in species tolerance limits, previous studies have often found
clear positive correlations between thermal tolerance of intertidal species and their
vertical distributions (e.g. Stirling, 1982; McMahon, 2001; Stillman and Somero,
2000; also see Underwood, 1979 for review). As well as showing changes
within the tidal cycle, environmental temperatures are often most extreme when
low tides occur during day time in hot seasons, or at low latitudes (Moore, 1972).
In Hong Kong, for example, during summer low tides in the afternoon, rock
surface temperatures can exceed 50oC (Williams, 1994b; Williams and Morritt,
1995; Williams et al, unpubl data). Such variations in environmental
temperatures, therefore, require further investigation to identify if thermal
tolerance of intertidal species can change with time (e.g. seasonal or
laboratory-based acclimation, e.g. Stirling, 1982; Obermülle et al., 2011, also see
Table 3.13 in Chapter 3). Determining a species’ thermal tolerance will allow an
understanding of the factors affecting a species’ vertical and seasonal distribution,
and also provide a scientific basis which may help predict species’ success under
long-term, global, environmental changes (Somero, 2010).

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 16/150
Chapter 1: General introduction
Littor ini ds as a model f or studies on thermal stress
Littorinids (Gastropoda: Littorinidae), also known as periwinkles, are abundant
worldwide in different types of intertidal habitats, such as mangroves and rocky
shores, where many of these species dominate the upper intertidal zone (see
Lewis, 1964; Stephenson and Stephenson, 1972; McQuaid, 1996a,b; Reid, 2007
for reviews). Due to their abundance and ubiquitous distribution, the taxonomy
of littorinids has well been documented (e.g., the genus Littoraria, Reid, 1986;
Littorina, Reid, 1996 and Echinolittorina, Reid, 2007) and, therefore, this well
resolved taxonomy provides researchers with accurate and confident
identifications of species and their phylogenetic relationships to aid comparative
studies.
From the ecological aspect, littorinids are dominant grazers on the high shore of
intertidal rocky shores (see Norton et al., 1990; McQuaid, 1996b for reviews).
The grazing activity of littorinids has been shown to reduce the biomass (e.g.
Castenholtz, 1961, 1964; Nicotri, 1977, Underwood, 1984, Mak and Williams,
1999) and alter the composition of the biofilm (e.g Lubchenco, 1978; Hunter and
Russell-Hunter, 1983, Vadas and Elner, 1992; Williams, 1994a; Stafford and
Davies, 2005). Littorinids are, therefore, key players in shaping community
structure in the high shore (see Norton et al., 1990; McQuaid, 1996b).
Living on the high shore, littorinids can experience prolonged emersion periods (~
8 days) during neap tides (McMahon, 1990; Uglow and Williams, 2001), as well
as extreme environmental temperatures (> 50oC, Williams, 1994b; Williams and

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 17/150
Chapter 1: General introduction
Morritt, 1995; Marshall et al, 2010) and are, therefore, model organisms to study
the relationship between their thermal tolerance and vertical distributions.
Species living in such stressful environmental conditions are assumed to be living
close to their physiological limits (Somero, 2010) and this is thought to be
especially true for tropical species (Tewkesbury et al, 2011), as their potential to
acclimate to higher environmental temperatures is thought to be less as compared
to temperate species (see Somero, 2005; 2010 for reviews). The majority of
previous studies have, however, been restricted to the eastern Pacific and Atlantic
coasts (e.g. Sandison, 1967; Hamby, 1975; Cannon and Hughes, 1992; Clarkes et
al., 2000b; Davenport and Davenport, 2005), and there have been few studies
along the western Pacific or on tropical shores (but see McMahon, 1990; Stirling,
1982). It is therefore important to address this knowledge gap and to investigate
responses of species found in these understudied regions, so as to determine if
there are any general patterns related to species' distribution patterns and their
acclimation potentials worldwide (see Foster, 1990).
The genus Echinolittorina (formerly Nodilittorina, see Appendix 1 for former and
present scientific names), is the largest genus in the family Littorinidae (Reid,
2007). Containing 59 recognized species, close to half of the genus can be found
in the Indo-West Pacific region (Reid, 2007). Like many other littorinids,
Echinolittorina malaccana, E . radiata and E . vidua (Figure 1.1) play an important
ecological role by controlling biofilm abundance on the high shore (Mak, 1996),
and are common along the Indo-West Pacific coast, with representatives of the
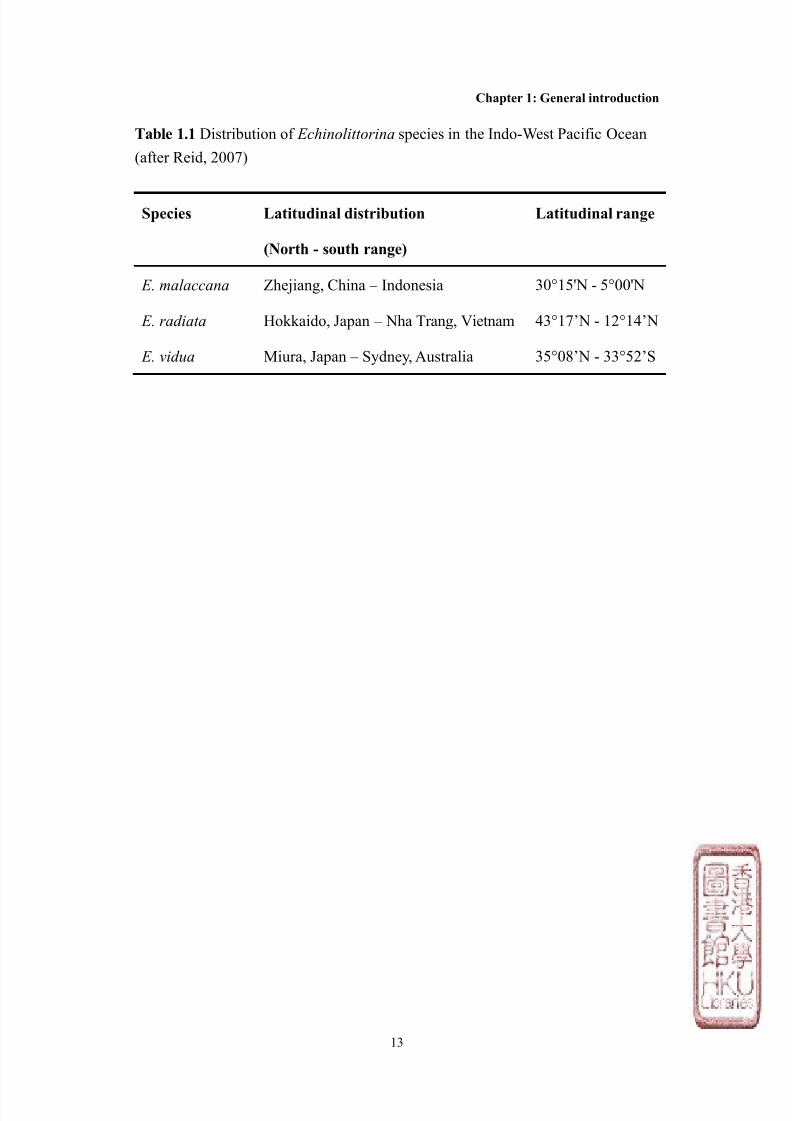
three species covering over 70o of latitude (Table 1.1). These three species are,
therefore, excellent representative candidates for determining the relationship

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 18/150
Chapter 1: General introduction
between thermal tolerance and their vertical distributions. The three species are
abundant on rocky shores in Hong Kong and their ecology has been studied in
detail by Mak (1996) and Walters (2002). All three species inhabit the mid-high
shore, between 1.3-2.7m above C.D. (Mak, 1996), where they frequently
encounter extreme fluctuations in environmental conditions driven by tidal cycles
and weather conditions. Given the fact that competition and predation pressure
on Echinolittorina species are assumed to be low in Hong Kong (see Stafford,
2002), hence the vertical distributions of these species are assumed to be related
to their tolerance to physical stresses (Stirling, 1982; Yipp et al., 1990).
Aims of study and project outli ne
The aims of this study were to measure: (i) variation in thermal tolerance between
the Echinolittorina species in Hong Kong, in relation to their local spatial and
temporal distributions; which would provide the scientific basis to direct further
investigations on larger-scale patterns (e.g. large scale latitudinal patterns and any
potential changes over temporal scales of years); and, (ii) seasonal variability of
the littorinids’ thermal tolerance, in order to determine the acclimation potential of
each species to higher environmental temperatures; ultimately to help predict and
identify relative species’ success as temperatures are proposed to increase in a
warming world.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 19/150
Chapter 1: General introduction
To achieve these goals, this study is divided into two major parts as follows:
Chapter 2 Seasonal variation in vertical distribution of Echinolittorina
species
Since the vertical distributions of Echinolittorina malaccana, E . radiata and E .
vidua in Hong Kong have been previously investigated by Mak (1996) and
Walters (2002), in this chapter, therefore, the focus was to measure the seasonal
variation in the distribution and abundance of the Echinolittorina species. Three,
monthly seasonal surveys were conducted in both winter and summer on
moderately exposed rocky shores in South Bay and Stanley, Hong Kong Island.
This data helped to determine how the distribution and abundance of the
littorinids may vary between seasons, which may be related to their tolerances to
the seasonally variable physical environment in Hong Kong.
Chapter 3 Thermal tolerance of Echinolittorina species in Hong Kong
Using a variety of physiological parameters, including lethal temperature,
Arrhenius breakpoint temperature of heart rate and activities of the enzymes
malate and lactate dehydrogenases, this chapter examined variation in thermal
tolerance of Echinolittorina species. With these information, thermal tolerance
of Echinolittorina species in relation to their vertical and seasonal distribution
could be demonstrated, and their potential to acclimate to high temperature could
also be identified.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 20/150
Chapter 1: General introduction
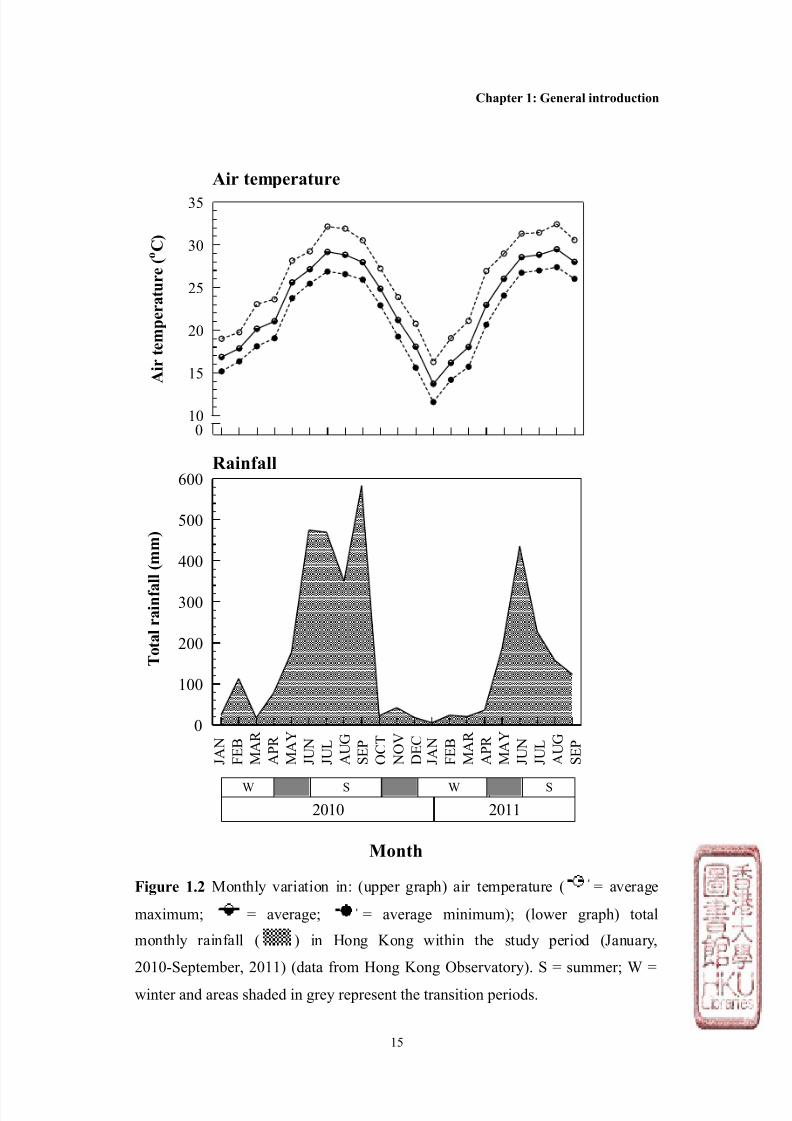
The Hong Kong cl imate
Hong Kong (22o20’ N and 114o10’E), is situated on the southern coast of China,
south of the Tropic of Cancer and has a strong seasonal, monsoon climate
(Morton and Morton, 1983). In general, between June to September, Hong
Kong experiences a hot, wet summer due to the dominant south-eastern monsoon.
In 2010 for example, the average maximum air temperature was > 30oC, average
relative humidity of ~ 82% (Hong Kong Observatory, see Figure 1.2). In
contrast, the north-eastern monsoon during the winter brings cold, dry conditions
from December to March (Kaehler and Williams, 1996), for example during 2010,
the average maximum air temperature was < 20oC, average relative humidity of ~
70% (Hong Kong Observatory, Figure 1.2). As well as differences in air
temperature and humidity, rainfall is higher in summer, with an average monthly
rainfall > 400mm in 2010, as compared with the winter average monthly rainfall
of < 50mm (Hong Kong Observatory, Figure 1.2). The periods between these
two dominant monsoon seasons, i.e. April – May and October – November, are
considered transition periods when weather conditions are variable as the
monsoon influences change (Kaehler and Williams, 1996).
Sea surface temperatures also exhibit strong seasonal variation in Hong Kong.
In summer, the Hainan current from the South China Sea brings warm water
northwards along the south China coast (Morton and Morton, 1983), and sea
surface temperature at, for example, Waglan Island (south of Hong Kong Island)
is maintained at ~ 27oC (Hong Kong Observatory). In winter, while the Taiwan
current from the East China Sea brings relatively cold water (19-23oC, Morton

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 21/150
Chapter 1: General introduction
and Morton, 1983) to the coast of Hong Kong, the Hainan current is replaced by
the warm Kuroshio current (26-29oC, Morton and Morton, 1983). The mixing of
water from the Taiwan and Kuroshio current, therefore, contributes to a relatively
warmer sea surface temperature (monthly average=18.9oC, Hong Kong
Observatory) than air temperature in winter (monthly average=15.7oC, Hong
Kong Observatory) in Hong Kong. In general wave action is more calm in
summer than winter (Apps and Chen 1973), with the exception of periods of
tropical depressions or typhoons when wave action and wind speeds are extreme
(Morton and Harper, 1995; Morton et al., 1996).
The daily tidal pattern in Hong Kong ranges between mixed, semi-diurnal, tides
during spring tides, and almost diurnal neap tides (Morton et al., 1996), with an
average tidal range of ~ 2.2m (Hutchinson and Williams, 2003a). During spring
tides, the lowest of the low tides generally occurs during late night – early
morning in winter, while during summer, the lowest of the low tides occurs in the
early afternoon and evening (Williams, 1994b). The seasonal variation in timing
of low tides, therefore, creates a strong seasonal contrast in environmental
temperatures that intertidal organisms experience (see Figure 1.2), with winters
being cool and dry and animals emersed during the night, whilst summers are hot
and wet and animals are emersed at the hottest time of the day when rock surface
temperatures can exceed 50oC (Williams, 1994b, Williams and Morritt, 1995;
Williams et al, unpubl data) .

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 22/150
Chapter 1: General introduction
Site descri ptions
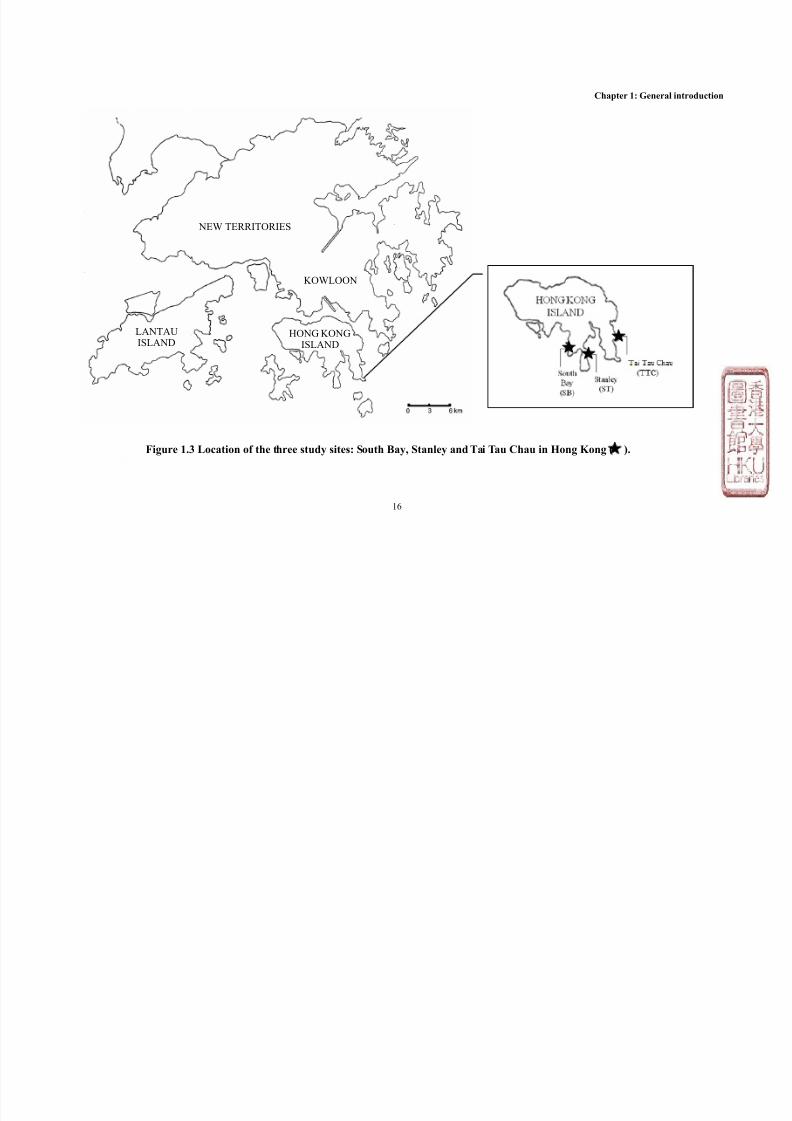
All field surveys and animal collections were conducted at South Bay (SB,
22°13’N, 114°11’E), Stanley (ST, 22°12’N, 114°13’E) and Tai Tau Chau (TTC,
22°13’N,114°15’E). All the three sites are located within the eastern, oceanic
waters of Hong Kong (Figure 1.3), to minimize the confounding effect of salinity
variation brought by the Pearl River Estuary which influences Hong Kong's
western waters. The three sites experience similar levels of wave exposure and
can be categorized as moderately exposed shores as defined by the distribution of
flora and fauna (Morton and Morton, 1983, Kaehler and Williams, 1996). While
South Bay is composed of a nearly vertical rock cliff (see Figure 1.4 a), and faces
south-west; Stanley and Tai Tau Chau are composed of more flattened rock
platforms (Figures 1.4 b & c), and face north and east respectively.
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 23/150
Chapter 1: General introduction
Table 1.1 Distribution of Echinolittorina species in the Indo-West Pacific Ocean
(after Reid, 2007)
Species Latitudinal distribution
(North - south range)
Latitudinal range
E. malaccana Zhejiang, China – Indonesia 30°15'N - 5°00'N
E. radiata Hokkaido, Japan – Nha Trang, Vietnam 43°17’N - 12°14’N
E. vidua Miura, Japan – Sydney, Australia 35°08’N - 33°52’S

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 24/150
Chapter 1: General introduction
E . malaccana
E . radiata
E . vidua
Figure 1.1 The three common rocky shore Echinolittorina species in Hong Kong.
5 mm
5 mm
5 mm
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 25/150
Chapter 1: General introduction
10
15
20
25
30
35
0
100
200
300
400
500
600
W S W
A i r t e m p e r a t u r e ( o C )
T o t a l r a i n f a
l l ( m m )
S
2010 2011
J A N
F E B
M A R
A P R
M A Y
J U N J U L A U G
S E P
O C T
N O V
D E C
J A N
F E B
M A R
A P R
M A Y
J U N J U L A U G
S E P
Month
0
Air temperature
Rainfall
Figure 1.2 Monthly variation in: (upper graph) air temperature ( = average
maximum; = average; = average minimum); (lower graph) total
monthly rainfall ( ) in Hong Kong within the study period (January,
2010-September, 2011) (data from Hong Kong Observatory). S = summer; W =
winter and areas shaded in grey represent the transition periods.
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 26/150
Chapter 1: General introduction
16
Figure 1.3 Location of the three study sites: South Bay, Stanley and Tai Tau Chau in Hong Kong ( ).
NEW TERRITORIES
KOWLOON
HONG KONGISLAND
LANTAUISLAND

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 27/150
Chapter 1: General introduction
17
Figure 1.4 a Photograph of study site at South Bay (SB) in summer.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 28/150
Chapter 1: General introduction
18
Figure 1.4 b Photograph of study site at Stanley (ST) in winter.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 29/150
Chapter 1: General introduction
19
Figure 1.4 c Photograph of study site at Tai Tau Chau (TTC) in summer.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 30/150
CHAPTER 2:
SEASONAL VARIATION IN VERTICAL
DISTRIBUTION OF ECHINOLITTORINA
SPECIES
2.1 INTRODUCTION 21
2.2 MATERIALS AND METHODS 26
2.3 RESULTS 29
2.4 DISCUSSION 40

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 31/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
2.1 INTRODUCTION
Intertidal ecologists have been trying to understand how physical factors and
biological interactions drive distribution patterns using sessile species (in
barnacles: Connell, 1961a,b; Wethey, 1984; mussels: Paine, 1974; algae:
Schonbeck and Norton 1978, 1980; Underwood, 1980; also see Underwood, 1979
for reviews). The lower limit of distribution of sessile species has been proposed
to be driven by biological interactions, such as competition and predation
(Connell, 1961a,b; Wethey, 1984; Paine, 1974; Schonbeck and Norton, 1980).
Connell (1961a), for example, showed that the rapid growth in the lower shore
Balanus balanoides was responsible for mortality of the upper shore barnacle,
Chthamalus stellatus, when they were found low on the shore, and proposed that
the lower limit of C. stellatus was determined by competition. The upper limit
of species' distributions, on the other hand, is generally driven by physical factors,
such as temperature and desiccation stresses (Connell, 1961a; Schonbeck and
Norton, 1978; Underwood, 1980). Three species of fucoid algae ( Pelvetia
canaliculata, Fucus spiralis and Ascophyllum nodosum), for example, showed
clear tissue damage at their maximum height of distribution when they were
emersed for over a 20-day period (Schonbeck and Norton, 1978), illustrating the
important roles played by temperature and desiccation in determining the upper
limit of species' distributions. Sessile species are also unable to escape from
extreme environmental conditions during emersion, especially in summer.
During hot summer months, environmental conditions can exceed the survival
limits of sessile species, often resulting in ‘high shore kills’ of these species (e.g.
Connell, 1961a,b; Hawkins and Hartnoll, 1985; Chan et al., 2006).

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 32/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
Unlike sessile species, mobile species have the advantage to avoid extreme
environmental conditions by moving to microhabitat refuges, such as crevices,
cracks and rock pools (Garrity, 1984; Williams and Morritt, 1995; Helmuth and
Hofmann, 2001), or migrating down shore to avoid hot summer conditions (Lewis,
1954; Branch, 1975; Liu, 1994; Williams and Morritt, 1995; Takada, 1996; Harper
and Williams, 2001; see Underwood, 1979 for review). Given the ability of
mobile species to use behavioural mechanisms to avoid extreme environments,
mobile species, therefore, may have less risk of mortality as compared to sessile
species. The upper limit of mobile species distribution is, however, in a similar
manner to sessile species, largely determined by physical factors (for example, in
limpets: Wolcott, 1973; gastropods: Britton and McMahon, 1990; McMahon,
1990; 2001; porcelain crabs: Stillman and Somero, 2001; see Underwood, 1979
for review). These studies generally showed that the physiological tolerance of
mobile species to physical stresses was positively correlated to their
corresponding shore heights. The behavioural and physiological strategies
exhibited by mobile intertidal species cannot be placed into context without a
basic understanding of the vertical distribution of a species (Underwood et al.,
2000). Understanding the vertical level that a species inhabits allows the
description or quantification of the physical environment that they will experience
and the ranges of environmental conditions that they will have to tolerate. To fully
appreciate the ecology of intertidal species, therefore, requires an initial
assessment of their vertical distribution patterns (Underwood et al., 2000).
Generally categorized as high shore gastropods, littorinids are abundant at the

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 33/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
upper edge of intertidal habitats (e.g. rocky shores and mangroves) throughout the
world (see Lewis, 1964; Stephenson and Stephenson, 1972; McQuaid, 1996a,b;
Reid, 2007 for reviews). Littorinids are commonly restricted to areas above the
high water mark and, therefore, experience prolonged periods of emersion, which
can last longer than 8 days during neap tides (McMahon, 2001; Uglow and
Williams, 2001). Throughout the prolonged emersion period, littorinids
frequently experience extreme environmental conditions (McMahon, 1990;
Williams, 1994b; McQuaid, 1996b; Uglow and Williams, 2001). To survive the
harsh conditions on the high shore, littorinids are well-adapted to this environment
through morphological (Vermeij, 1973; McQuaid and Scherman, 1988; Lee and
Lim, 2009), behavioural (e.g. Vermeij, 1971; McQuaid and Scherman, 1988;
Muñoz et al., 2005), and physiological (e.g. Evans, 1948; Stirling, 1982;
McMahon, 2001) strategies. Morphologically, many species have light-coloured
and nodulated shells, which help reflect heat (Vermeij, 1973; McQuaid and
Scherman, 1988; Lee and Lim, 2009). In terms of behavioural aspects, most
littorinids withdraw their foot, and attach themselves via a mucus thread when the
substratum reaches a high temperature (Vermeij, 1971; McQuaid and Scherman,
1988). This 'standing' behaviour reduces their direct contact with the substratum
and, therefore, minimizes heat uptake through conduction from the rock surface
(Marshall et al., 2010). Such strategies, therefore, allow littorinids to dominate
the physically harsh environment high on intertidal shores.
Littorinids, similar to other intertidal gastropods (limpets Liu, 1994; Williams and
Morritt, 1995; top shells: Takada, 1996; various intertidal gastropods: Harper and

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 34/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
Williams, 2000), also show seasonal variation in their vertical distribution, which
has been suggested to minimize physical stresses (Lambert and Farley, 1968;
Hannaford Ellis, 1985; Ohgaki, 1988b; Mak, 1996). These studies, in general,
observed that adult individuals are distributed lower on shores during hot periods
of the year (i.e. summer) than during cooler, winter periods. Such migrations
may, however, be related to other factors, such as wave exposure, food availability
and reproduction (Lambert and Farley, 1968; Williams and Ellis, 1975). For
example, Ohgaki (1988b; 1989; Ito et al., 2002) suggested that the downshore
migration of Echinolittorina radiata in Japan was related to their spawning
pattern. Mak (1996) observed a similar pattern in Hong Kong, which he
suggested may be related to seasonal variation in abundance of microalgae.
While explanations for the cause of migration patterns remain controversial,
determination of such patterns may be important in understanding the physical
environment species face, and the forces which drive these migration patterns.
Echinolittorina malaccana, E . radiata and E . vidua are three common littorinds
that are widely distributed on Hong Kong rocky shores (Ohgaki, 1985, Williams,
1994; Mak 1996; Walters, 2002). Given that these Echinolittorina species live
close to the upper limit of the intertidal zone, they show distinct and consistent
vertical distribution patterns over shores of varying wave exposure (Ohgaki, 1985;
Mak, 1996), as well as seasonal vertical migration (Mak, 1996). On all three
shores of varying wave exposure that Mak (1996) studied, E. malaccana inhabited
the highest level of the intertidal zone (mean height = 2.2-2.7 m C.D.), followed
by E. radiata (1.8-2.5 m C.D.) and E. vidua (1.3-2.0 m C.D.). From his surveys,

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 35/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
Mak also found that all three Echinolittorina species were distributed at lower
shore levels in summer as compared to winter (Mak, 1996). There are several
explanations or theories/hypotheses which can be applied to explain the
distribution patterns of Hong Kong Echinolittorina species. Understanding both
the spatial and temporal distribution of the littorinids, however, is an essential
starting point to help generate hypotheses about how a specific factor may play a
role in determining the observed distributions (Underwood et al., 2000).
In this study, as well as determining the species' vertical distributions (which have
been previously described in earlier studies, Mak, 1996), variation in the seasonal
distribution and abundance of E . malaccana, E . radiata and E . vidua in Hong
Kong were also quantified. Seasonal surveys were conducted in winter and
summer months on moderately exposed rocky shores at South Bay and Stanley,
Hong Kong (see Chapter 1). These surveys aimed to establish the physical
environment these snails experienced to inform further investigations on the
relative physiological tolerances of these species to physical factors, such as
temperature, and how these may be related to the spatial and temporal
distributions of Echinolittorina species in Hong Kong (see Chapter 2).

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 36/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
2.2 MATERIALS AND METHODS
Field surveys were conducted to determine the vertical distributions of
Echinolittorina malaccana, E . radiata and E . vidua in winter and summer.
Surveys were carried out on three, randomly selected, replicate days in winter
(Dec 2010-Feb, 2011) and summer (Jun-Aug, 2011, as defined by Kaehler and
Williams, 1996) at the two study sites (refer to Chapter 1). Average monthly air
temperatures were between 13.7-20.2oC during winter, and 27.1-29.5oC in
summer (Hong Kong Observatory, see Chapter 1). Surveys were conducted
during low water on spring tides (tidal height ≦ 1.20m above Chart Datum, C.D.),
when the littorinids were emersed and inactive (Mak, 1996). All surveys were
conducted during dry days, to minimize potential disturbance to the littorinids’
distributions due to dislodgement by rain (Ohgaki, 1988a).
The survey method was adopted from Mak (1996), who also conducted surveys
on Echinolittorina species in Hong Kong. At each site, a horizontal, 10m stretch
of shore was vertically divided into 25cm height intervals from 1.5-3.25m above
C.D., which is the normal vertical range for Echinolittorina species (Mak, 1996).
At each height, 10, 25x25cm quadrats were randomly placed, and the total
number of each Echinolittorina species in each quadrat was recorded. The
vertical distribution for each Echinolittorina species was presented in terms of
mean number and mean percentage of individuals at different tidal heights from
the three replicate days during each season, as individual day to day differences
were not of interest in this study. The percentage of individuals at different tidal
heights was calculated as follows:

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 37/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
The overall mean height of each species was calculated following Harper and
Williams (2001):
The sample area covered at each height was 6250cm2 [25 x 25cm (area of each
quadrat) x 10 quadrats], whereas the total area covered on each sampling day was
50000cm
2
[Total area= 6250cm
2
(area of each height) x 8 heights].
Due to inherent local variability (e.g. wave exposure, topography, inclination), the
vertical distribution of a species would be expected to show some degree of
variation between different shores (Mak, 1996). As the present study aimed to
obtain a species-level view of overall distribution patterns of the three
Echinolittorina species, between shore variation was not analyzed, and a two-way
Analysis of Variance (ANOVA) was used to test for any possible variation in
vertical distribution and total abundance of Echinolittorina species between
seasons at South Bay and Stanley separately, using WinGMAv5 (EICC, The
University of Sydney), with species (sp) [three levels: E . malaccana, E . radiata
and E . vidua] and seasons (se) [two levels: Summer and Winter] treated as fixed

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 38/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
factors. Cochran’s test was used to check for homogeneity of variances
(Cochran, 1951) and data were transformed, where possible, if they were
heterogeneous. Analyses were still conducted when data failed tests for
homogeneity of variances after transformation, as ANOVA is considered to be
robust to such heterogeneity, provided the error degrees of freedom are relatively
large (Underwood, 1997). Where variances were homogenous, to minimize the
chance of Type I errors, p-values were adjusted to a more conservative level ( p <
0.01, Underwood, 1981). Significant difference(s) for fixed factors were further
analysed by post -hoc Student-Newman-Keuls (SNK) tests to identify potential
differences in treatment means.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 39/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
2.3 RESULTS
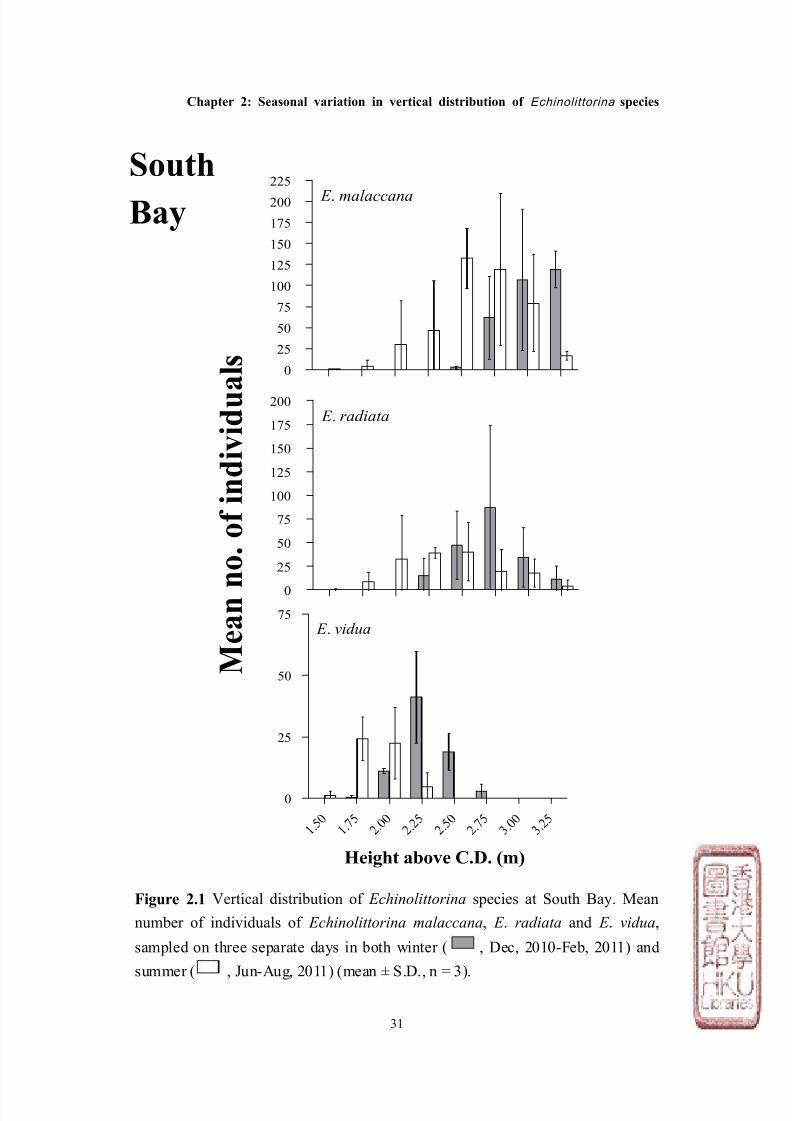
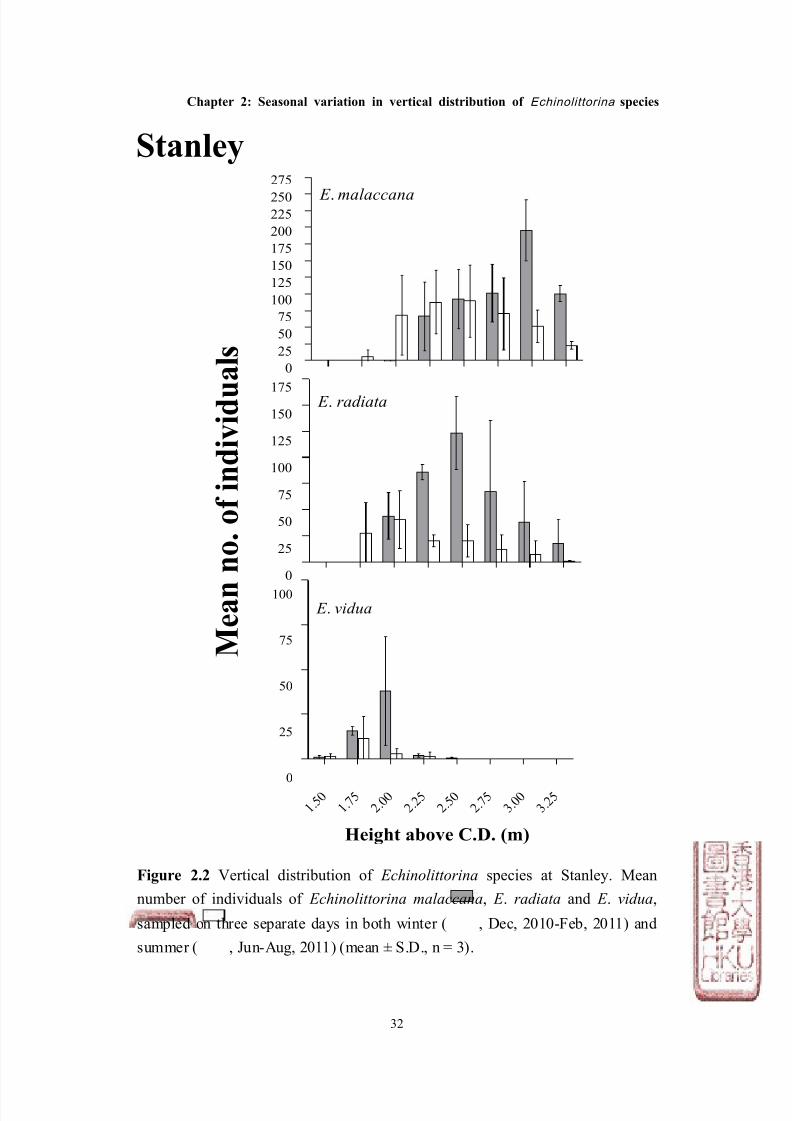
The distribution of Echinolittorina malaccana, E . radiata and E . vidua showed
consistent vertical patterns at both South Bay and Stanley. In general, E .
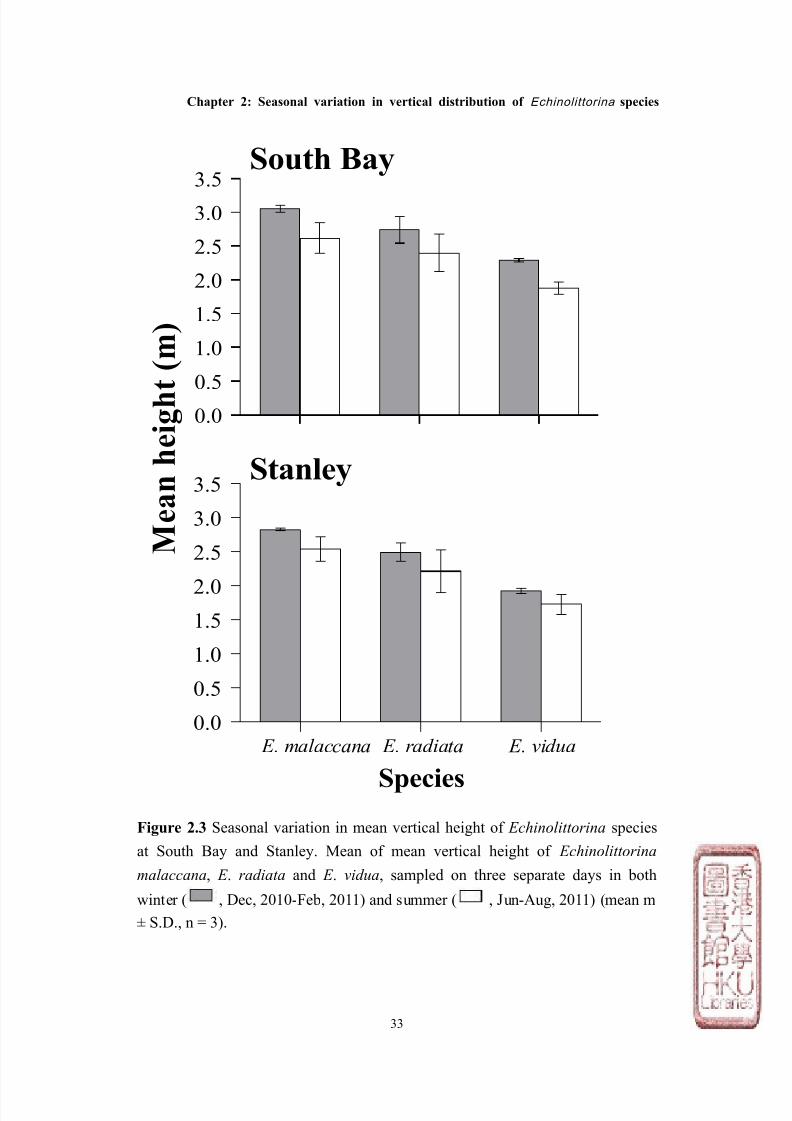
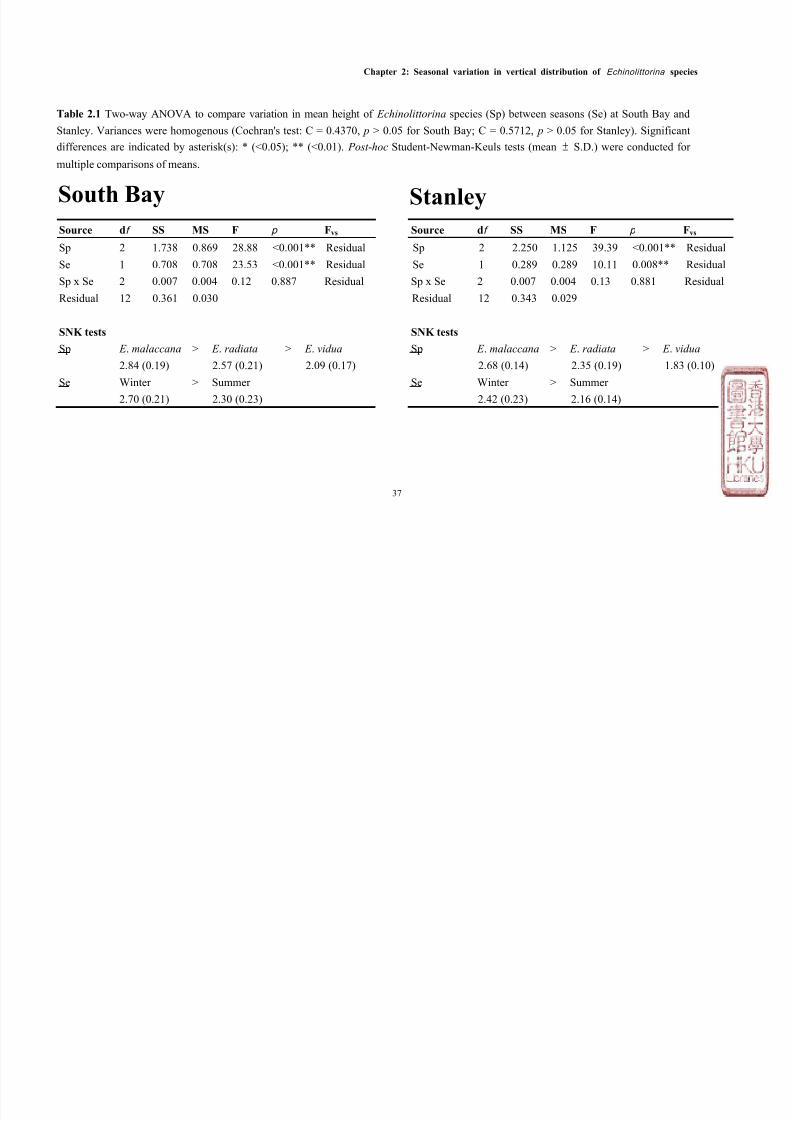
malaccana occupied the highest position on both shores (mean height ± S.D. =
2.76m above C.D. ± 0.24), which overlapped with E . radiata which was found
slightly lower on the shore (2.46m above C.D. ± 0.29), and finally E . vidua which
overlapped E. radiata, but was found lowest on the shore (1.96m above C.D. ±
0.23, see Figures 2.1 & 2.2). The vertical distribution ranges for the three
species between the two shores were similar, although the average mean height
for all species in Stanley, was ~ 0.25m lower than at South Bay (Figure 2.3).
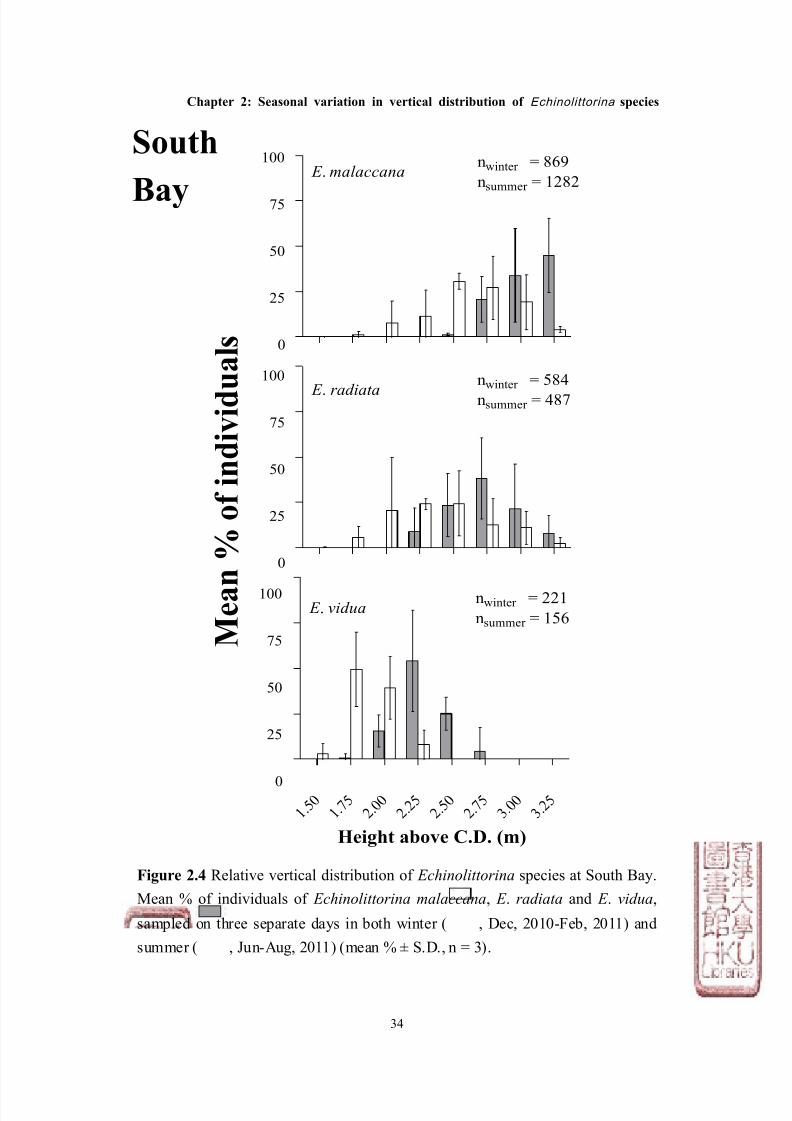
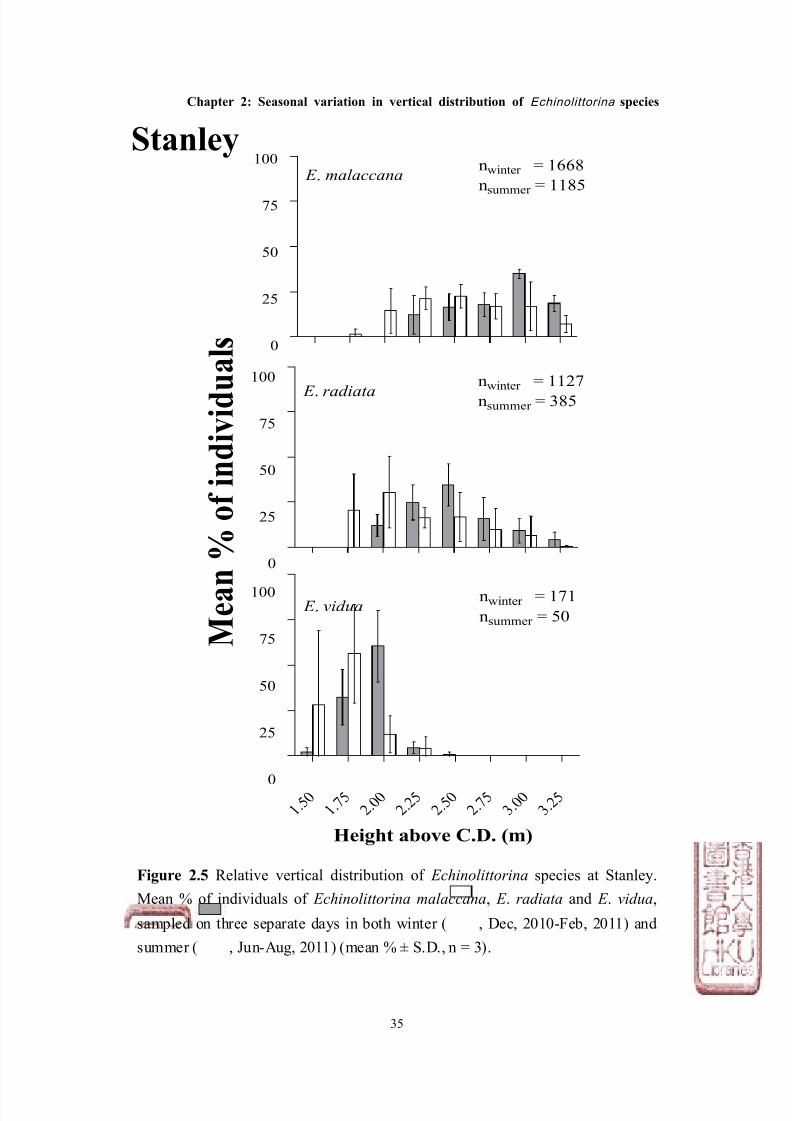
The abundance of Echinolittorina species also showed a consistent pattern at the
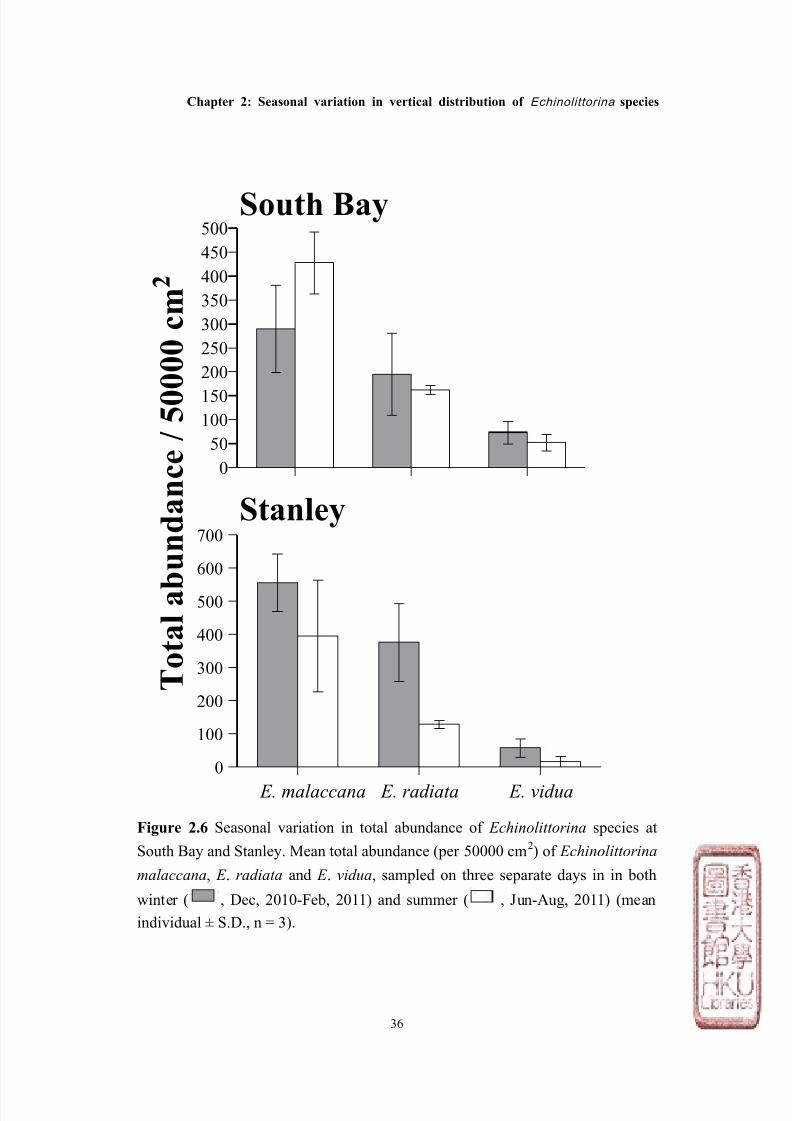
two shores (Figures 2.4 & 2.5, Tables 2.2 & 2.3). E . malaccana was the most
abundant among the three species (total abundance from all surveys: 2,151
individuals for South Bay; 2,853 for Stanley), whilst E . radiata was ~ 50% less
abundant on both shores (total abundance: 1,071 for South Bay; 1,512 for Stanley),
and E . vidua was the least abundant (total abundance: 377 for South Bay; 221 for
Stanley, see Figures 2.4 & 2.5).
Both the vertical distribution and abundance of the individual species varied
between seasons (Tables 2.1-2.3). The vertical distributions of E . malaccana, E .
radiata and E . vidua were generally lower by at least 0.25m during the summer
than in the winter months at both sites (Figures 2.1 & 2.2), with the exception of E .
vidua at Stanley which showed a similar distribution between seasons. The

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 40/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
mean height of all species was significantly lower in summer than winter at both
sites (Figure 2.3, Table 2.1). The total abundance of Echinolittorina species,
although not significantly different within species (except for E . malaccana at
South Bay, see Tables 2.2 & 2.3) was, in general, less in summer than winter at
both sites (Figure 2.6, Table 2.2). Both E . radiata and E . vidua were ~ 25% less
abundant in summer than winter at South Bay, and more than 50% less abundant
at Stanley. This temporal pattern was also observed for E . malaccana at Stanley,
which was also ~ 25% less abundant during summer as compared with winter.
In contrast, however, E . malaccana were significantly more abundant during
summer than winter at South Bay by ~ 40%.
To conclude, consistent patterns were observed for both the vertical distribution
and abundance of Echinolittorina species at South Bay and Stanley.
Echinolittorina malaccana occupied the highest tidal level and was the most
abundant species, followed by E . radiata which dominated relatively lower tidal
levels and was less abundant, and finally E . vidua, which was found the lowest on
the shore and was the least abundant among the three species. In terms of
temporal variation, all three species were less abundant and found at lower shore
levels in summer than winter, with the exception of E . malaccana at South Bay.
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 41/150
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 42/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
0
25
50
75
100
125
150
175
200
225
250
275
E . malaccana
0
25
50
75
100
125
150
175
E . radiata
1. 5 0
1. 7 5
2. 0 0
2. 2 5
2. 5 0
2. 7 5
3. 0 0
3. 2 5
0
25
50
75
100
E . vidua
Height above C.D. (m)
M e a n
n o . o f i n d i v i d u
a l s
Figure 2.2 Vertical distribution of Echinolittorina species at Stanley. Mean
number of individuals of Echinolittorina malaccana, E . radiata and E . vidua,
sampled on three separate days in both winter ( , Dec, 2010-Feb, 2011) and
summer ( , Jun-Aug, 2011) (mean ± S.D., n = 3).
Stanley
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 43/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
M e a n h e i g h t ( m )
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
E. malaccana E. radiata E. vidua
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
Species
South Bay
Stanley
Figure 2.3 Seasonal variation in mean vertical height of Echinolittorina species
at South Bay and Stanley. Mean of mean vertical height of Echinolittorina
malaccana, E . radiata and E . vidua, sampled on three separate days in both
winter ( , Dec, 2010-Feb, 2011) and summer ( , Jun-Aug, 2011) (mean m
± S.D., n = 3).
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 44/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
0
25
50
75
100
E . malaccananwinter = 869
nsummer = 1282
0
25
50
75
100
E . radiata
nwinter = 584
nsummer = 487
1. 5 0
1. 7 5
2. 0 0
2. 2 5
2. 5 0
2. 7 5
3. 0 0
3. 2 5
0
25
50
75
100
E . viduanwinter = 221
nsummer = 156
Height above C.D. (m)
M e a n % o
f i n d i v i d u a l s
Figure 2.4 Relative vertical distribution of Echinolittorina species at South Bay.
Mean % of individuals of Echinolittorina malaccana, E . radiata and E . vidua,
sampled on three separate days in both winter ( , Dec, 2010-Feb, 2011) and
summer ( , Jun-Aug, 2011) (mean % ± S.D., n = 3).
South
Bay
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 45/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
0
25
50
75
100
E . malaccananwinter = 1668
nsummer = 1185
0
25
50
75
100
E . radiata
nwinter = 1127
nsummer = 385
1. 5 0
1. 7 5
2. 0 0
2. 2 5
2. 5 0
2. 7 5
3. 0 0
3. 2 5
0
25
50
75
100 E . vidua
nwinter = 171
nsummer = 50
Height above C.D. (m)
M e a n % o
f i n d i v i d u a l s
Figure 2.5 Relative vertical distribution of Echinolittorina species at Stanley.
Mean % of individuals of Echinolittorina malaccana, E . radiata and E . vidua,
sampled on three separate days in both winter ( , Dec, 2010-Feb, 2011) and
summer ( , Jun-Aug, 2011) (mean % ± S.D., n = 3).
Stanley
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 46/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
0
50
100
150
200
250
300
350
400
450
500
E. malaccana E. radiata E. vidua
0
100
200
300
400
500
600
700
T o t a l a b
u n d a n c e / 5 0 0 0 0 c m
2
South Bay
Stanley
Figure 2.6 Seasonal variation in total abundance of Echinolittorina species at
South Bay and Stanley. Mean total abundance (per 50000 cm2) of Echinolittorina
malaccana, E . radiata and E . vidua, sampled on three separate days in in both
winter ( , Dec, 2010-Feb, 2011) and summer ( , Jun-Aug, 2011) (mean
individual ± S.D., n = 3).
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 47/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
37
Table 2.1 Two-way ANOVA to compare variation in mean height of Echinolittorina species (Sp) between seasons (Se) at South Bay and
Stanley. Variances were homogenous (Cochran's test: C = 0.4370, p > 0.05 for South Bay; C = 0.5712, p > 0.05 for Stanley). Significant
differences are indicated by asterisk(s): * (<0.05); ** (<0.01). Post -hoc Student-Newman-Keuls tests (mean ±
S.D.) were conducted for
multiple comparisons of means.
Source df SS MS F p Fvs Source df SS MS F p Fvs Sp 2 1.738 0.869 28.88 <0.001** Residual Sp 2 2.250 1.125 39.39 <0.001** Residual
Se 1 0.708 0.708 23.53 <0.001** Residual Se 1 0.289 0.289 10.11 0.008** Residual
Sp x Se 2 0.007 0.004 0.12 0.887 Residual Sp x Se 2 0.007 0.004 0.13 0.881 Residual
Residual 12 0.361 0.030 Residual 12 0.343 0.029
SNK tests SNK tests
Sp E . malaccana > E . radiata > E . vidua Sp E . malaccana > E . radiata > E . vidua
2.84 (0.19) 2.57 (0.21) 2.09 (0.17) 2.68 (0.14) 2.35 (0.19) 1.83 (0.10)
Se Winter > Summer Se Winter > Summer
2.70 (0.21) 2.30 (0.23) 2.42 (0.23) 2.16 (0.14)
StanleySouth Bay

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 48/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
Table 2.2 Two-way ANOVA to compare variation in total abundance of
Echinolittorina species (Sp) between seasons (Se) at South Bay. Variances were
homogenous (Cochran's test: C = 0.4039, p > 0.05). Significant differences are
indicated by asterisk(s): * (<0.05); ** (<0.01). Post -hoc Student-Newman-Keuls
tests (mean ± S.E.) were conducted for multiple comparisons of means.
Source df SS MS F p Fvs
Sp 2 266395.1 133197.6 38.60 <0.001** Residual
Se 1 3500.1 3500.1 1.01 0.334 Residual
Sp x Se 2 27200.4 13600.2 3.94 0.048* Residual
Residual 12 41403.3 3450.3
SNK tests
Se (Sp) E . malaccana
Winter < Summer
289.67 (91.45) 427.33 (69.25)
E . radiata
Winter = Summer
194.67 (85.34) 162.33 (9.28)
E . vidua Winter = Summer
73.67 (23.76) 52.00 (17.44)
Sp (Se) Summer
E . malaccana > E . radiata > E . vidua
427.33 (69.25) 162.33 (9.28) 52.00 (17.44)
Winter
E . malaccana = E . radiata > E . vidua
289.67 (91.45) 194.67 (85.34) 73.67 (23.76)

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 49/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
Table 2.3 Two-way ANOVA to compare variation in total abundance of
Echinolittorina species (Sp) between seasons (Se) at Stanley. Variances were
homogenous (Cochran's test: C = 0.5521, p > 0.05). Significant differences are
indicated by asterisk(s): * (<0.05); ** (<0.01). Post -hoc Student-Newman-Keuls
tests (mean ± S.D.) were conducted for multiple comparisons of means.
Source df SS MS F p Fvs
Sp 2 577354.8 288677.4 34.20 <0.001** Residual
Se 1 100650.9 100650.9 11.92 0.005** Residual
Sp x Se 2 32431.4 16215.7 1.92 0.189 Residual
Residual 12 101300.0 8441.7
SNK tests
Sp E . malaccana > E . radiata > E . vidua
475.50 (104.93) 252.00 (109.48) 36.83 (21.01)
Se Winter > Summer
329.56 (133.47) 180.00 (108.69)

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 50/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
2.4 DISCUSSION
In Hong Kong, clear vertical zonation patterns of three Echinolittorina species
were consistently observed on two moderately exposed rocky shores. E .
malaccana was distributed highest on the shore, and its range overlapped with E .
radiata, which was dominant slightly lower on the shore, while E . vidua was
found lowest on the shore. The vertical zonation pattern of the three
Echinolittorina species matches previous observations on Hong Kong shores
(Ohgaki, 1985; Cleland and McMahon, 1990; Dudgeon and Yipp, 1990; Williams,
1994; Mak, 1996; Walters, 2002). Cleland and McMahon (1990), for example,
also showed that the vertical distribution of E . malaccana and E . radiata
overlapped, and E . radiata were also found ~ 0.5m lower on the shore than E .
malaccana. A similar pattern was also observed for E . malaccana and E . radiata
in Singapore (Lee and Lim, 2009), while in Tanabe Bay (Japan) and the Gulf of
Thailand, E . vidua was found below E . radiata and E . malaccana respectively and
overlapped with the vertical distribution of barnacles, Tetraclita spp. (Habe, 1958;
Tsuchiya and Lirdwitayapasit, 1986; also see Reid, 2007 for review).
Authors working in Hong Kong have proposed that Echinolittorina species
experience reduced biological interactions, with low predation risk (Mak, 1996;
Stafford, 2002; Li, personal observation) and competition pressures (Dudgeon and
Yipp, 1990) by living high on the shore. These potential benefits may be offset
by the high physical stresses which the littorinids will experience living in the
high-shore. Species specific variation in tolerance to thermal and desiccation
stresses may positively match with the vertical distributions of the three

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 51/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
Echinolittorina species. Laboratory experiments, for example, suggest that E .
malaccana are more tolerant to desiccation stress than E . radiata in Hong Kong,
given that the LT (lethal time) 50 in air was 34 days more for E. malaccana than E .
radiata (Yipp et al., 1986), although such exposure times are highly unrealistic.
Similarly, thermal tolerance, in terms of heat coma and lethal temperature, were
higher in E . malaccana than E . radiata according to studies conducted in Hong
Kong by Stirling (1982) and McMahon (2001). These data, although not able to
fully explain the zonation patterns of Echinolittorina species, do illustrate the
importance of physical factors in relation to species' distribution, and hence
implications for community structure and ecological processes (Menge and Olson,
1990; Underwood and Chapman, 1996).
As was recorded by Mak (1996), Echinolittorina malaccana, E . radiata and E .
vidua, showed seasonal variation in their vertical distribution, with all three
species generally being distributed higher on the shore during winter than summer.
This phenomenon has been interpreted to enhance the reproductive success of
littorinids (Kojima, 1959; Hannaford Ellis, 1985; Ohgaki, 1988a; Mak, 1996), as
downward migration of the littorinids generally coincides with their spawning
periods. Moving downshore, therefore, was suggested to increase the chance for
the littorinids to release their eggs in water by increasing their duration of
immersion. The immersion period can, for example, be up to 2 hours longer at
1.5m C.D. than 1.75m during summer neap tides on Hong Kong shores (Ng,
2007). Most of these studies, however, failed to provide any evidence of the
actual benefits of an increased duration to release eggs when littorinids were

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 52/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
immersed in support of this theory. Other littorinid species, such as Littorina
unifasciata and L. cincta, for example, do not migrate downwards during their
spawning period (Pilkington, 1971), therefore, the downward movement of
littorinids in summer may not be fully explained by spawning migrations. The
downward migration of littorinids may also possibly be related to food availability
(Mak, 1996; Ito et al., 2002), or perhaps seasonal variation in tidal height. The
lowest tides are generally lower in the summer than winter (Kaehler and Williams,
1996) which may account for the lower distribution of littorinids at this time of
year (Ohgaki, 1989). The littorinids may also migrate downshore to avoid
extreme physical stress by minimizing emersion duration (Lambert and Farley,
1968; Williams and Ellis, 1975), as described for other intertidal species in Hong
Kong (Harper and Williams, 2001).
Seasonal variation in the abundance of Echinolittorina species was also detected
at Stanley. The lower abundances recorded during summer months matches with
Mak (1996)’s monthly surveys at three shores of varying exposures in Hong Kong
(Big Wave Bay, Cape d’ Aguilar and South Bay). Similar patterns of seasonal
abundance were also observed for E . radiata and E . vidua at South Bay, although
these were not statistically significant. A decrease in abundance of littorinids
during the summer months may be related to mortality associated with conditions
at this time of year; such as heat stress (Williams, 1994b; Mak, 1996).
Dislodgement of littorinids caused by heavy rainfall has also been proposed to
affect populations in both temperate Japan (Ohgaki, 1988b) and Hong Kong due
to tropical storms in summer (Mak, 1996).

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 53/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
Authors have suggested that a decrease in abundance of intertidal species often
occurs following tropical storms (Kohn, 1980; Boulding and Van Alstyne, 1993;
Mak, 1996; Hutchinson and Williams, 2003b; Sanpanich et al., 2006). During
tropical storms, intertidal gastropods may move to refuges, such as cracks and
crevices, to avoid dislodgement by strong wave action, and if they fail to find
these refuges they may be washed away resulting in mortality (Kohn, 1980). In
2011, the first tropical storm (Sarika) arrived on 9 th June after the first summer
survey was conducted, and rainfall was not heavy during the storm period
(accumulated rainfall 7th – 11th June < 20 mm, Hong Kong Observatory). This
storm did not seem to have any observable effect on littorinid abundance as the
abundance of E . malaccana and E . radiata was observed to have increased, or
was similar, in the second summer survey. E .vidua, did, however, decline in
abundance between the first and second survey, and this continued between the
second and third survey, although there were no further storms. Mortality due to
tropical storms is, therefore, unlikely to explain the reduction in littorinid
abundance in the present study, as also noted by Walters working on
Echinolittorina in Hong Kong (2002).
Despite the potential effect of tropical storms, heavy rainfall (i.e. > 46mm per day,
Ohgaki, 1988b), may also contribute towards the decrease in abundance of
Echinolittorina species in summer. Ohgaki (1988b) showed that heavy rainfall
would increase detachment of mucus threads in E . radiata, thus dislodging the
littorinids. As the first heavy rainfall (total rainfall per day: 69.8.mm, Hong

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 54/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
Kong Observatory) was reported on 22nd May, 2011, this effect may explain the
lower abundance observed in, for example, E . malaccana in the first field survey
in both South Bay (~ 30%) and Stanley (~ 80%) as compared with the second
survey. Heavy rainfall, may, therefore, have been responsible for the decline in
littorinid abundance in summer, although no direct evidence is available to link
this with snail mortality, since no tagging experiments have been conducted on
Echinolittorina species to determine their survival in relation to rainfall (see Mak,
1996).
The major link to the reduction in abundance of the three Echinolittorina species
during summer in Hong Kong may be associated with the stressful physical
environment experienced on rocky shores during this time. In summer the
lowest tides occur during the afternoon, when rock surface temperatures may
exceed 50oC (see previous references). Such extreme high temperature
conditions during summer can cause 50% mortality of mobile species on Hong
Kong shores (Williams et al., unpubl data). Large mortality events are
commonly observed, for example, in the limpet, Cellana toreuma (Firth and
Williams, 2009) and barnacle, Tetraclita japonica (Chan et al., 2006), when
physical stresses exceed species tolerance limits (Williams and Morritt, 1995;
Dong and Williams, 2011) and attempting to reduce this stress by migrating
downshore may explain the seasonal variation in the littorinids vertical
distribution (Williams and Morritt, 1995; Harper and Williams, 2001). Little is
known, however, about possible seasonal variation in littorinids’ tolerance to
physical stresses in Hong Kong. While the reduction in littorinid abundance in

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 55/150
Chapter 2: Seasonal variation in vertical distribution of Echinolittorina species
summer may be caused by mortality, the increased abundance in winter was
suggested to be a result of recruitment between October to January (Mak, 1996).
The contrary lower abundance of E . malaccana in winter than summer at South
Bay is, however, difficult to explain and was not recorded by Mak (1996),
suggesting this may be an anomaly. This anomaly may also be related to the
recruitment patterns of E . malaccana at South Bay (Menge and Branch, 1999), for
example, variation in sporadic recruitment and recruitment failure.
In conclusion, Echinolittorina malaccana, E . radiata and E . vidua, as in many
previous studies, showed a consistent vertical pattern on rocky shores in Hong
Kong. In general, Echinolittorina species were found lower down the shore, in
reduced numbers in summer than winter. While the downward migration in
summer may be a response to enhance reproductive success, the decline in
abundance may be related to mortality/ dislodgement of littorinids during heavy
rainfall, however there is little direct evidence to support these explanations. As
these Echinolittorina species will experience relatively low negative biological
interactions, such as predation and competition (Stafford, 2002), living in the high
shore on Hong Kong shores; it seems logical to assume that physical factors, e.g.
temperature and water availability, may play important roles in determining
spatial and temporal variation in their vertical distribution and abundance. As a
result, a series of laboratory based assays are required to try and establish whether
there is a correlation between their physical tolerances and spatial and temporal
patterns, and these are reported in the next chapter.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 56/150
CHAPTER 3:
THERMAL TOLERANCE OF
ECHINOLITTORINA SPECIES IN HONG KONG
3.1 INTRODUCTION 48
Thermal tolerance and species distribution on intertidal rocky
shores
How to determine thermal tolerance of species?
Thermal tolerance and vertical distribution ofEchinolittorina
species on Hong Kong rocky shor es
3.2 MATERIALS AND METHODS 58
Preliminary studies as quali ty control of experimental protocol
Seasonal var iati on in lethal temperature
Seasonal var iation in Ar rhenius breakpoin t temperatur e of heart
rate
Acti vit ies of malate dehydrogenase and l actate dehydrogenase
Data analysis
3.3 RESULTS 72
Seasonal var iati on in lethal temperature
Seasonal var iation in Ar rhenius breakpoin t temperatur e of heart
rate

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 57/150
Acti vit ies of malate dehydrogenase and l actate dehydrogenase
3.4 DISCUSSION 84
Lethal temperature in r elation to verti cal distri bution
Arrhenius breakpoint temperature of heart rate in relation to
ver tical distri bution
Acti vit ies of malate dehydrogenase and lactate dehydrogenase in
relation to vertical distri bution
Seasonal variati on of LT 50 and ABT of HR

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 58/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
3.1. INTRODUCTION
Thermal tolerance and species distributi on on intertidal rocky shores
The intertidal zone is an extreme and dynamic environmental gradient, where
species can experience severe environmental stress over a short vertical distance
(< 10m, see Chapter 1). This is especially true of intertidal rocky shores, as the
rock surface is a two-dimensional habitat which offers little protection against
physical stresses, which are generally suggested to set the upper limit of species
distribution (e.g. Connell, 1961a; Newell, 1970; Moore, 1972; Wolcott, 1973; also
see Underwood, 1979 for review). The linkage between thermal tolerance and
vertical distribution of species has been widely studied, with comparative studies
of genera distributed at different shore levels being relatively common. For
example, McMahon (2001) investigated heat coma temperature (see below for
definition) for 60 species of gastropods collected from various types of intertidal
habitats from different geographic areas, and found a strong positive relationship
between thermal tolerance and corresponding shore height. A similar trend, with
thermal tolerance being correlated with tidal height, was also observed by Stirling
(various gastropods, 1982) and Davenport and Davenport (various gastropods,
barnacles & bivalves, 2005). Although these studies compared thermal tolerance
across genera to illustrate this linkage, such comparisons may be confounded by
the role phylogenetic relationships play in determining the distribution of species
(see Somero, 2002). Somero (2010), therefore, suggested studies which focus
on comparative physiology of congeneric species instead of across different
genera to overcome this problem. Using this approach, Stillman and Somero
(2000) showed that upper shore porcelain crabs in the genus Petrolisthes, tended

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 59/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
to have greater thermal tolerance than lower shore species in the same genera.
This pattern was also supported by Tomanek and Somero (1999) and Somero
(2002) for the top shells Chlorostoma (previously Tegula) and littorinids in the
genus Littorina respectively.
Temporal variation in environmental temperatures on intertidal rocky shores, for
example between seasons, also poses the question of whether the intertidal species
have the ability to acclimate to the changes in environmental temperatures, thus
altering their thermal tolerance. Such potential impacts on thermal tolerance
have been investigated either by seasonal field-based (e.g. Evans, 1948; Fraenkel,
1968; Stirling, 1982) and/or laboratory-based acclimation experiments (e.g.
Newell, 1971; Wolcott, 1973; McMahon et al., 1995; Tomanek and Somero, 1999;
Stillman and Somero, 2000; Stenseng et al., 2005; Hopkin et al., 2006).
Fraenkel (1968), for example, tested thermal tolerance of Littorina littorea
between seasons, and found that thermal tolerance was greater in summer than
winter; whilst Stillman and Somero (2000) acclimated three porcelain crab
species in the laboratory at 8oC and 18oC, and reported that thermal tolerance of
warmer acclimated crabs was greater than those acclimated to cooler
temperatures.
Species that have less potential to acclimate for greater thermal tolerance may
exhibit other strategies to avoid or minimize this stress such as seeking refuge in
benign microhabitats (Garrity, 1984; McQuaid and Scherman, 1988; Helmuth and
Hoffman, 2002), orientating their body positions (Muñoz et al., 2005) or even
migrating downshore (Williams and Morritt, 1995; Harper and Williams, 2001) to

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 60/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
minimize exposure to stressful conditions (also see Chapter 2). However, in
terms of longer temporal scales, for example, under climate change scenarios
where environmental temperature is predicted to be elevated by 2-6o
C by 2100
(McCarthy, 2001), knowing the acclimation potential of intertidal species would
allow us to identify which species are more likely to survive or become locally
extinct, and therefore, possible impacts on community structures may also be
understood (Somero, 2002, 2010).
How to determine thermal tolerance of species?
Thermal tolerance generally refers to the survival limit of species under a given
set of thermal conditions (see Angilletta, 2009 for review). Traditional
approaches to determine species' thermal tolerances in intertidal ecology are to
measure species lethal (LT50) or heat coma temperatures (HCT) (e.g. Evans, 1948;
Newell, 1958, Southward, 1958; Fraenkel, 1960; Sandison, 1967; Stirling, 1982;
Urban, 1994; McMahon, 2001; Davenport and Davenport, 2005). LT50 is
defined as the temperature at which 50% mortality of individuals of a species is
observed within the tested population, where mortality is determined as when
individuals fail to give any response to external stimulus and fail to recover after
return to ambient conditions (Evans, 1948; Fraenkel, 1960). HCT is the
temperature at which 50% of individuals in the population start to lose their
normal neural function (i.e. enter a coma), and under such conditions, an
individual ceases locomotion and fails to attach to the substratum (Evans, 1948;
Southward, 1958; Stirling, 1982; McMahon, 2001).

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 61/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Most studies have measured LT50 and/or HCT of various intertidal species while
immersed in water (see Table 3.1), in order to minimize the confounding effect of
desiccation, and generally showed a positive correlation between thermal
tolerance of species and their vertical distribution (see Underwood 1979 for
review). Most species, however, and especially those which live on the upper
shore such as littorinids, do not experience extreme temperatures while they are
immersed but when they are emersed (Marshall et al., 2010), and may respond
differently when stressed under unnatural conditions. For example, Sandison
(1976) measured HCT and LT50 for four Littorina species and Thais lapillus in air
and water and, in general, found both HCT and LT50 to be higher when
determined in air than water. It is, therefore, important to select an
environmentally realistic heating condition when determining species’ thermal
tolerances.
When determining species’ thermal tolerance, although much of the past focus has
investigated survival limits, the tolerance of physiological systems from the
biochemical (e.g. protein synthesis, enzyme stability) to organ (e.g. heart function,
respiratory response) levels will provide important insights in relation to species'
survival limits, and thus their distributions (see Somero, 2002 for review).
Using four congeneric species of porcelain crabs ( Petrolisthes) , that live at
different shore levels, Stillman (2003) found that the Arrhenius breakpoint
temperature of heart rate (ABT of HR, see below for definition) was positively
correlated with the species LT50, and hence their vertical distributions. Similarly,
at the biochemical level, the stability and function of the metabolic enzyme,
malate dehydrogenase, in five species of Lottia was found to be correlated with

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 62/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
the respective species vertical and geographic distributions (Dong and Somero,
2009).
Heart rate, at the organ-level, has traditionally been used as a physiological
indicator of intertidal species response to thermal conditions (e.g. Pickens, 1965;
Trueman, 1967; Jones, 1970; Marshall and McQuaid, 1992; Santini et al., 1999;
Williams et al., 2005, 2010; Marshall et al., 2010; Dong and Williams, 2011; also
see Underwood, 1979; Somero, 2010 for reviews), as heart rate is known to be a
thermally sensitive measure of metabolic performance in many marine
invertebrates (see Hochachka and Somero, 2002). As well as measuring species’
metabolic performance, heart rate has shown to be positively related to oxygen
consumption of an organism (Marshall and McQuaid, 1992; see Pörtner, 2001;
2010 for reviews). This implies that once heart function of an individual starts
dropping drastically at high temperatures, individuals switch to a less-energy
producing anaerobic metabolism, which ultimately will result in mortality (see
Pörtner, 2001 for review). It can be illustrated in the case of Stillman's work on
porcelain crabs, where the temperature breakpoint of heart rate (see below for
definition) was shown to be positively correlated to an individuals' lethal
temperature (reviewed in Somero, 2010). Heart rate can, therefore, be used as a
sensitive, sub-lethal, indicator to determine thermal tolerance of intertidal species.
To determine thermal tolerance using heart rate, the Arrhenius breakpoint
temperature (ABT of HR) of heart rate (defined as the temperature at which heart
rate of an individual drops dramatically, Stillman and Somero, 1996), is
commonly determined to assess the initiation of breakdown of heart function

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 63/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
(Tomanek and Somero, 1999; Stillman, 2003; Stenseng et al., 2005; Braby and
Somero, 2006; Marshall et al., 2010, 2011; Dong and Williams, 2011). Similar
to survival limits, the ABT of HR for congeneric intertidal species has also been
found to be positively correlated to their vertical distributions (porcelain crabs:
Stillman and Somero, 1996; top shells: Stenseng et al., 2005; limpets: Dong and
Williams, 2011).
Enzyme function is also known to be thermally labile, with function breaking
down at temperatures when enzymes denature (see Hochachka and Somero, 2002).
Therefore, to study thermal tolerance limits in marine invertebrates at the
biochemical level, the activities of a suite of metabolic enzymes have been widely
determined (e.g. Sokolova and Pörtner, 2001, 2003; Dahlhoff et al., 2002; Lee and
Lim, 2009; see Pörtner, 2001, 2002; Hochachka and Somero, 2002; Somero, 2002,
2010; Dahlhoff, 2004 for reviews). Malate dehydrogenase (MDH) and lactate
dehydrogenase (LDH) are common metabolic enzymes used to investigate
thermal tolerance of species, as their kinetics and stability to temperature have
been found to vary between several congeneric species (e.g. Dahlhoff and Somero,
1993; Fields et al., 2002, 2006; Dong and Somero, 2009).
Malate dehydrogenase (MDH) in general, contains two isoforms, mitochondrial
malate dehydrogenase (mMDH) and cytosolic malate dehydrogenase (cMDH)
and serves several functions involved in aerobic metabolism (see Goward and
Nicholls, 1994; Mathews et al., 2000 for reviews). Aerobic metabolism refers to
the cellular energy production mechanism that oxidizes glucose through
glycolysis, the Krebs cycle and the electron transport chain, under the presence of

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 64/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
oxygen, which serves as the final electron acceptor in the electron transport chain
to synthesize adenosine triphosphate (ATP) as a temporary energy store (see
Mathews et al., 2000; Hochachka and Somero, 2002 for reviews).
LDH, on the other hand, is a terminal enzyme in anaerobic glycolysis, catalyzing
the conversion of pyruvate into lactate (see Mathews et al., 2000). It is often
used to monitor the anaerobic capacity of species (e.g. Childress and Somero,
1979; Wu and Lam, 1997; Rinke and Lee, 2009; Ombres et al., 2011; see
Hochachka and Somero, 2002; Dahlhoff, 2004 for reviews), which is particularly
critical at high temperatures when oxygen transportation is limited due to a
reduction in heart function (see Pörtner, 2001; Hochachka and Somero, 2002 for
review). Both enzymes have been shown to demonstrate adaptive variation with
temperature in several marine invertebrates (abalone: Dahlhoff and Somero, 1993;
mussels: Fields et al., 2006; littorinids: Sokolova and Pörtner, 2001; limpets:
Dong and Somero, 2009), which has been correlated with species biogeographic
and vertical distributions (e.g. Dahlhoff and Somero, 1993).
Most studies to determine thermal tolerance of intertidal species in relation to
their vertical distribution have, however, been conducted on the eastern Pacific
coast, therefore, more research needs to be conducted on the western Pacific and
especially the tropics to determine if there are any general patterns which emerge
with relation to species' responses to thermal stress (see Foster, 1990).

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 65/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Thermal tolerance and vertical distribution ofEchinolittorina species on Hong
Kong rocky shor es
Littorinids are, among the intertidal gastropods, the most tolerant family to high
temperature in different intertidal habitats from temperate to tropical regions
(Evans, 1948; Newell, 1958, Sandison, 1967; Stirling, 1982; McMahon, 2001;
Davenport and Davenport, 2005). Within this family, the high-shore
Echinolittorina species are the most thermally tolerant species on Hong Kong
shores (Stirling, 1982; McMahon, 2001). Given the well-studied taxonomy and
ecology of this genus (Reid, 2007; Mak, 1996), Echinolittorina is an excellent
genus to study how their thermal tolerance, measured at different organization
levels, can be related to species' distribution patterns.
In Hong Kong, the three high-shore Echinolittorina species ( E. malaccana, E.
radiata and E. vidua), as well as showing distinct vertical distribution patterns
(see Chapter 2), also exhibit strong seasonal variation in their vertical distribution
(see Mak, 1996; Chapter 2). All three Echinolittorina species are found higher
on the shore during winter, and move down-shore in summer, with their average
tidal heights varying by ~ 0.5m between seasons (see Chapter 2; Mak, 1996).
Many studies have related this phenomenon to spawning behaviour of littorinids
(Hannaford Ellis, 1985; Ohgaki, 1988b), however, the cause and benefits of this
migration behaviour remain unclear. Migrating down-shore may be a strategy
for these littorinids to avoid thermal stress at higher shore levels during the hot,
wet summer, where rock temperature can reach close to 58°C (Williams et al.,
unpubl data) and exceed their optimal range, or even be close to their survival

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 66/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
limits (Marshall et al., 2011). Determining variation in the littorinids’ thermal
tolerance between season, may also allow an understanding of their potential to
acclimate due to the seasonal differences in environmental temperatures (see
Chapter 1).
This study, as well as investigating the thermal tolerance of the three
Echinolittorina species in relation to their vertical distribution, also aimed to test
if there was any seasonal variation in thermal tolerance of these littorinids, which
may be linked to their seasonal migration patterns, as well their potential to
acclimate to higher temperatures. Thermal tolerances in the littorinids were
assessed through lethal temperature (LT50), Arrhenius breakpoint temperature of
heart rate (ABT of HR) and activities of malate dehydrogenase and lactate
dehydrogenase. Using such an integrated approach at the organismal,
physiological and biochemical levels, was intended to provide more detailed
insights into the factors which drive survival limits in these species.
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 67/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
57
Table 3.1 Studies on thermal tolerance of intertidal species in relation to different factors, heating condition, and parameters used to determine
survival limits.
Aim of study
(Factor(s) in relation to thermal tolerance)
Species (no. of species tested) Heating
Condition
Thermal
Tolerance
Reference
Water Air HCT LT50
Vertical distribution Molluscs 11 Evans, 1948
Environmental temperatures and geographical distribution Molluscs and barnacles 15 Southward, 1958
Illustration of temperature adaptation Cod, littorinid and hermit crab 3 Fraenkel, 1960
Geographical distributions Dog whelks and littorinids 5 Sandison, 1967
Season, vertical and geographical distribution Gastropods 22 Stirling, 1982
Vertical distributions Gastropods 9 Cleland and McMahon, 1990
El Nino effect Bivalves 10 Urban, 1994
Shore height and geographical distribution Littorinids 7 Clarke et al, 2000b
Habitats types, vertical and geographical distribution Gastropods 60 McMahon, 2001
Shore height, wave exposure, geographical distance Gastropods, bivalves & barnacles 10 Davenport and Davenport, 2005
Vertical distributions Littorinids 3 Lee and Lim, 2009

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 68/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
3.2. MATERIALS AND METHODS
Preliminary studies as qual ity control of experimental protocol
To assure better quality of data and to minimize the possible confounding effects
that might contribute to significant differences between the assessments of
thermal tolerance, the effect on littorinid size, sample size, duration of short-term
acclimation, and time for recovery were tested through LT50 measurements using
E . malaccana (see Table 3.2) to aid development of standard protocols. All
experiments from the preliminary studies were conducted based on Stirling (1984)
with modifications of heating condition and rate of heating. Potential
differences in LT50 with littorinid size, sample size, short-term acclimation and
recovery time were tested using Student’s t-tests.
Preliminary studies showed that significant differences were only detected
between different sized littorinids, but there was no difference in LT50 with sample
size, duration of short-term acclimation and recovery time, thus a standard
protocol was developed according to these results. Firstly, given the size range
of 7-8mm is more commonly used for studies on Echinolittorina species than the
smaller range of 6-7mm (e.g. Lee and Lim, 2009; Marshall et al., 2010), therefore,
7-8mm littorinids were collected for the experiments. Secondly, to avoid
over-collection of littorinids that might affect populations at the collection site, a
sample size of 5 individuals per replicate (see below for details) was chosen.
Thirdly, to facilitate experimental work, littorinids were not acclimated before the
experiment. Lastly, 12 hours recovery was chosen as opposed to 24 hours,

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 69/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
which was commonly used in previous studies (e.g. Fraenkel, 1968; Stirling,
1982), since there was no significant difference in LT50 assessments between the
two recovery times.
Apart from the factors tested, spatial variation in LT50 of the three Echinolittorina
species in Hong Kong was also tested to assure generality of the data collected for
sites in Hong Kong (Figure 3.1, Table 3.3). To test for possible variation
between sites, a nested Analysis of Variance (ANOVA) was employed, with sites
(si) nested within species (Sp) [three levels: E . malaccana (Tai Miu Wan and
Stanley beach), E . radiata (Tai Miu Wan and Tai Tau Chau) and E . vidua (Stanley
prison and South Bay)]. The LT50 of the three Echinolittorina species showed
significant differences with species but no effect of sites (Figure 3.1, Table 3.3),
which leads to the assumption that there is little spatial variation in thermal
tolerance, determined using LT50, of Echinolittorina species on moderately
exposed shores of Hong Kong
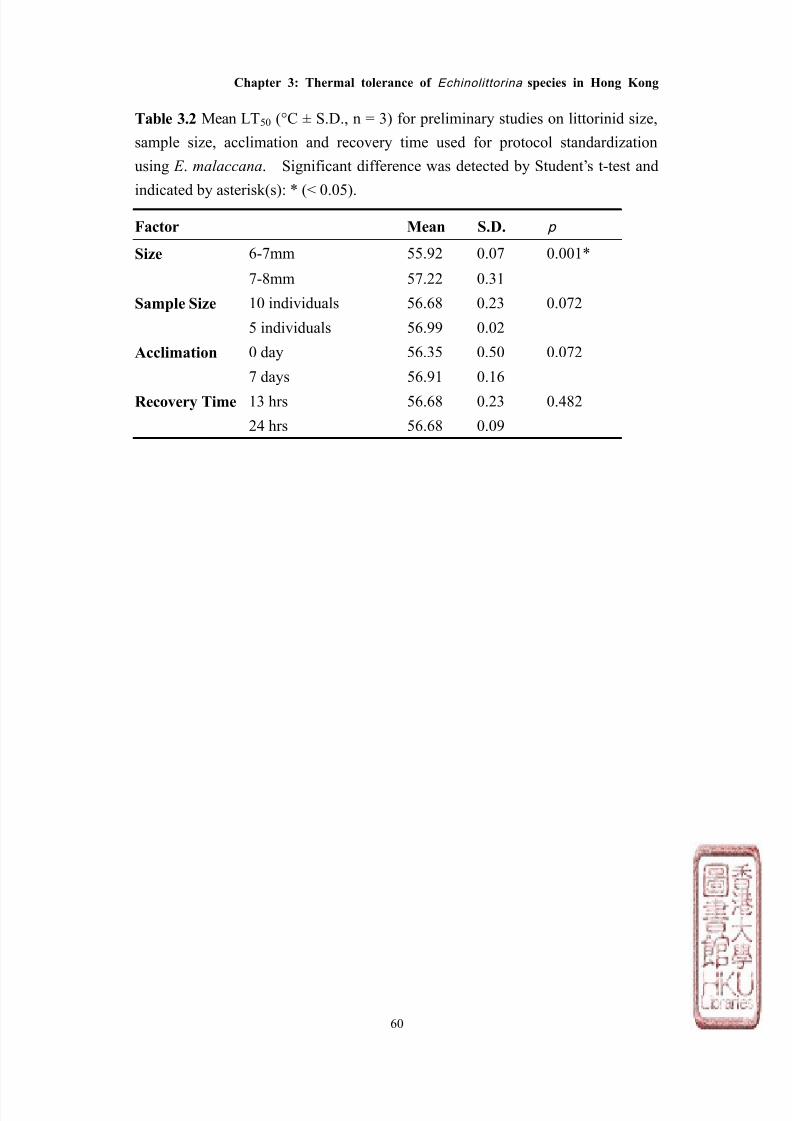
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 70/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Table 3.2 Mean LT50 (°C ± S.D., n = 3) for preliminary studies on littorinid size,
sample size, acclimation and recovery time used for protocol standardization
using E . malaccana. Significant difference was detected by Student’s t-test and
indicated by asterisk(s): * (< 0.05).
Factor Mean S.D. p
Size 6-7mm 55.92 0.07 0.001*
7-8mm 57.22 0.31
Sample Size 10 individuals 56.68 0.23 0.072
5 individuals 56.99 0.02
Acclimation 0 day 56.35 0.50 0.072
7 days 56.91 0.16
Recovery Time 13 hrs 56.68 0.23 0.482
24 hrs 56.68 0.09
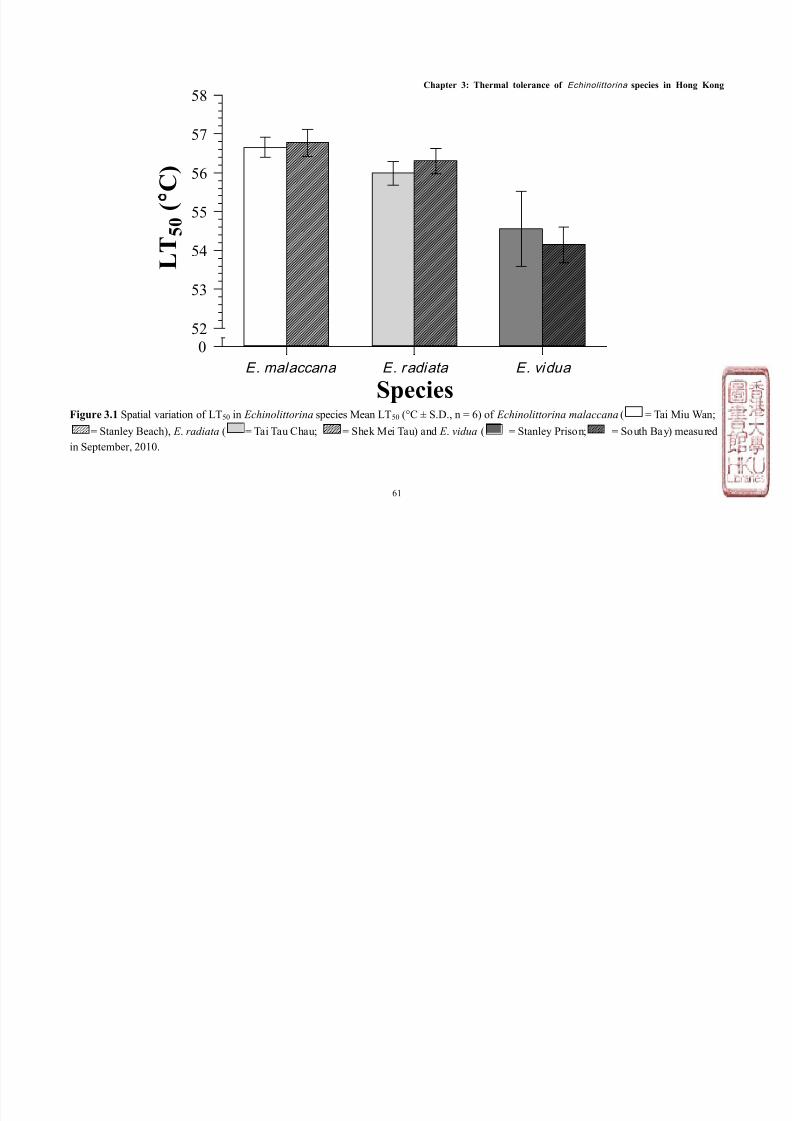
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 71/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
61
E. malaccana E. radiata E. vidua
0
52
53
54
55
56
57
58
Species
L T 5 0 (
C )
Figure 3.1 Spatial variation of LT50 in Echinolittorina species Mean LT50 (°C ± S.D., n = 6) of Echinolittorina malaccana ( = Tai Miu Wan;
= Stanley Beach), E . radiata ( = Tai Tau Chau; = Shek Mei Tau) and E . vidua ( = Stanley Prison; = South Bay) measured
in September, 2010.
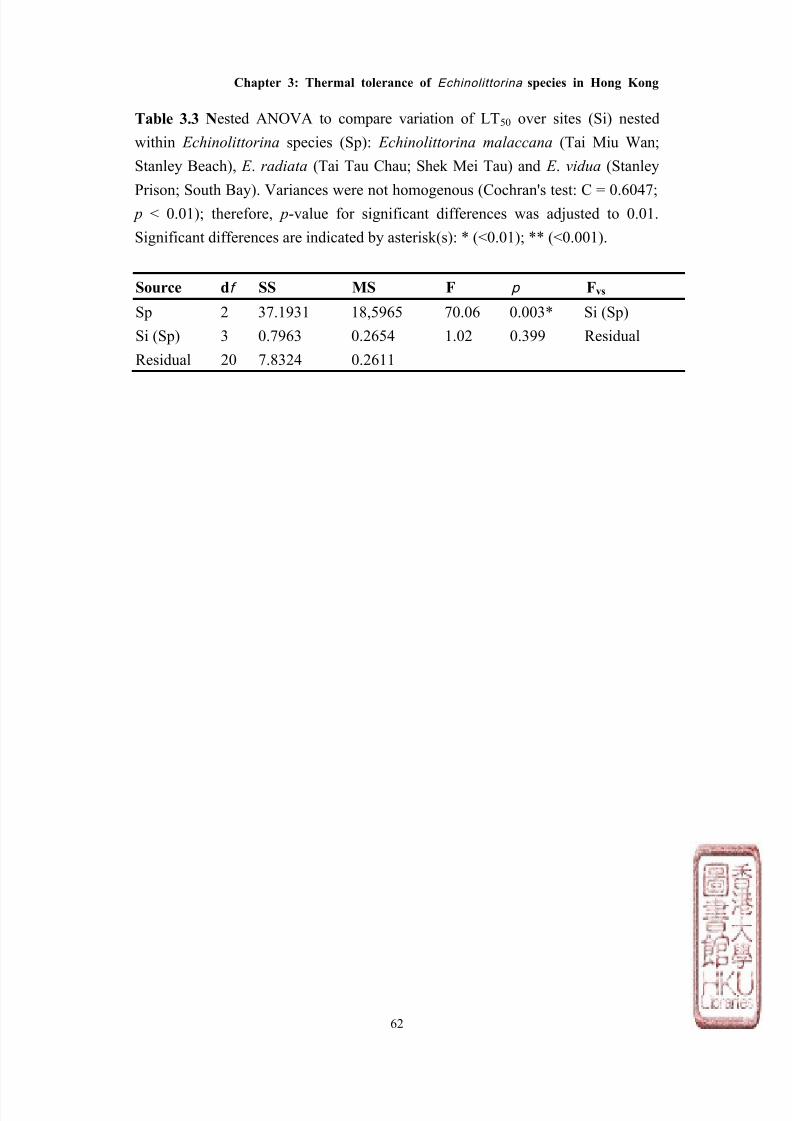
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 72/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Table 3.3 Nested ANOVA to compare variation of LT50 over sites (Si) nested
within Echinolittorina species (Sp): Echinolittorina malaccana (Tai Miu Wan;
Stanley Beach), E . radiata (Tai Tau Chau; Shek Mei Tau) and E . vidua (Stanley
Prison; South Bay). Variances were not homogenous (Cochran's test: C = 0.6047; p < 0.01); therefore, p-value for significant differences was adjusted to 0.01.
Significant differences are indicated by asterisk(s): * (<0.01); ** (<0.001).
Source df SS MS F p Fvs
Sp 2 37.1931 18,5965 70.06 0.003* Si (Sp)
Si (Sp) 3 0.7963 0.2654 1.02 0.399 Residual
Residual 20 7.8324 0.2611

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 73/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Seasonal var iati on in lethal temperature
Echinolittorina species were collected from South Bay and Stanley (see Chapter 1
for site description) on two randomly selected, replicate, days in each season,
during non-rainy, low water spring tide as applied for the field surveys.
Littorinids were collected from the height at which each species was most
abundant, i.e. their average mean height ± S.D. as determined from the field
survey (Chapter 2, E . malaccana: 2.76m ± 0.24; E . radiata: 2.46m ± 0.29; E .
vidua: 1.96m ± 0.23). To minimize variation that might be related to size (Clarke
et al., 2000a; also see Table 3.1), individuals with shell length of 7-8mm
(measured from the apex to the base of the columella, see Figure 3.2 modified
from Mak, 1996), were collected for all three species. All littorinids were
collected ~ 10m outside the population sampling sites, to ensure collection would
not impact the vertical distribution study. At each site, ~ 60-100 individuals of
each species were taken dependant on availability of individuals of the required
size range. Animals were kept moist in unsealed plastic bags that were placed
inside an insulated box, and were immediately transported to the Swire Institute of
Marine Science (Cape d’Aguilar, Hong Kong) within 70 minutes of collection.
Littorinids were rehydrated in Petri dishes for two hours in natural seawater
collected from the respective site.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 74/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Figure 3.2 Shell length of E . radiata (Mak, 1996) for measurement of littorinids.
The lethal temperature (LT50) of Echinolittorina species was determined for each
of the two days within summer and winter, using modifications of the method of
Stirling (1982). Only active littorinids, as defined by having their foot attached
to the substratum when immersed, were selected and blotted dry. Five
individuals were placed in labelled, cylindrical screw-top plastic vials (diameter
2.7cm and height 5.7cm) in air, with caps loosely fitted to allow air exchange.
Vials were randomly positioned in a grid and immersed in a Grant GP200
programmable water bath at 25C. The temperatures of the water bath (internal
thermometer) and inside a test vial, in a position where the littorinids were located
(K-type thermocouples, C ± 0.05, Omega, recorded using digital thermometers,
Lutron TM-903A, Taiwan), were continuously monitored throughout the
experiment. Temperature differences between the water bath and the vial were <
0.7C during the experiment.
Temperature was initially increased at a rate of 5C / 10 minutes until 50C, when
the heating rate was reduced to 1C / 10 minutes, and the duration of heating after
Shell length

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 75/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
50C varied between species as preliminary trials showed E. vidua to be less
tolerant than E. malaccana and E. radiata (∑ Duration of exposure = ( E .
malaccana & E. radiata) (5 x 10) + (8 x 10) = 130 minutes; ( E . vidua) (5 x 10) +
(5 x 10) = 100 minutes). When temperature inside the test vial reached 52, 53,
54 and 55C (for E . vidua) or 53, 54, 55, 56, 57 and 58C (for E . malaccana and
E . radiata), three randomly selected vials (= three replicates), were removed from
the water bath at each temperature interval, and cooled at ambient laboratory
temperature (in summer: ~ 23-26C; in winter: ~ 16-19C) for 5 minutes. For
each vial, littorinids were then removed and gently placed into a labelled Petri
dish (8.8 cm diameter) with a film of natural seawater (~ 2-3mm in depth)
collected from the site at ambient temperatures for a 12-hour recovery period (see
Table 3.2). After 12 hours, individuals that had their foot extended and attached
to the substratum, or responded to gentle pricking by a blunt forceps were
recorded as alive (Evans, 1948), while the remaining littorinids were assumed to
be dead.
LT50 of littorinids, which represents 50% mortality in the tested population, was
determined according to the following protocol. First, the number of dead snails
in each vial was converted to a proportion, i.e. percentage of mortality. Then, to
obtain three LT50 values from each set of experiments, the percentage mortality of
individual vials at each temperature point were randomly assigned to obtain three
sets of equal temperature groups, each with one value for the temperatures: 52, 53,
54 and 55C (for E . vidua) or 53, 54, 55, 56, 57 and 58C (for E . malaccana and
E . radiata). Finally, three LT50 values were calculated by fitting the proportion

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 76/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
of mortality data of each of the three temperature groups to a sigmoidal
log(dose)-response function (variable slope) using GraphPad Prism version 5.00
for Windows (GraphPad Software, San Diego California USA) for each of the two
days within each season.
Seasonal variati on in Arrhenius breakpoint temperature of heart rate
Variation of heart rate in Echinolittorina species with temperature was monitored
according to Marshall and McQuaid (2011) with modification of the rate of
heating, while determination of Arrhenius breakpoint temperature of heart rate
(ABT of HR) was based on the method used by Stillman and Somero (1996).
Echinolittorina species were collected from Tai Tau Chau (see Chapter 1 for site
description) between February – March 2011 and June – August 2011 during low
tide (tidal height ≦ 1.2m above CD). Approximately 20 individuals of each
species were collected following the criteria described above and transported to
the Swire Institute of Marine Science under the same procedures for the LT50
experiment (see above), and rehydration of snails was carried out within a period
of 24 hours.
Littorinids were placed in a 30C oven (Binder FD115) for 30 minutes to stabilize
heart rate, in order to facilitate heart rate measurements (DJ Marshall; pers
comm.). Preliminary studies showed no mortality after drying E . malaccana in
the oven for 4 days (Marshall and McQuaid, 2011). Littorinids were then fitted
with infrared sensors (Vishay Semiconductors, CNY70), secured with Blu-tack®

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 77/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
(Bostik), and placed in plastic vials (see LT50 experiment). Signals were
amplified and filtered using a custom-made pre-amplifier (a modified version of
Chelazzi et al., 1999, and used by Marshall and McQuaid, 2011 in E . malaccana),
and data were digitally logged using a data logger (PowerLab 8/30, ADI
Instruments, Figure 3.3). Littorinids were heated in a programmable water bath
(Grant GP200) at a rate of 0.5C/ minute (initial heating rate for LT50 experiment),
as a constant heating rate is commonly used when determining ABT of HR in
intertidal species (e.g. Stenseng et al., 2005; Marshall et al., 2010), between
30-55C after an initial 10 minutes at 30C (i.e. total duration = 60 minutes), and
heart rate was logged every minute and expressed as beats per minute (bpm).
Arrhenius breakpoint temperature (ABT) of heart rate, which is defined as the
temperature that shows a sharp discontinuity in the Arrhenius plot, indicates the
initial breakdown of heart function of the snail (Stillman and Somero, 1996).
Plots of heart rate versus temperature were constructed with heart rate (beat per
minute, bpm) versus 1000/temperature (Kelvin scale). By fitting the two best-fit
regression lines before and after the distinctive change in heart rate, the Arrhenius
breakpoint temperature could be determined by back-calculating the intersection
point of the two regression lines and converting this point on the x axis to the
Celsius scale (see Stillman & Somero, 1996; Stenseng et al., 2005).
Acti vit ies of malate dehydrogenase and lactate dehydrogenase
Echinolittorina species were collected from Tai Tau Chau on 15th July, 2011
during low tide (tidal height ≦ 1.2m above C.D.). Approximately 120

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 78/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
individuals of each species were collected following the criteria described above
and transported to the Swire Institute of Marine Science and prepared for heating
under the same procedures carried out in the LT50 experiment (see above).
Temperature was increased at a rate of 5C/ 10 minutes from 30C to 55C (i.e.
total duration = 60 minutes). When temperature reached 30, 35, 40, 45, 50 and
55C, three randomly selected vials, each containing five individuals, were
removed from the water bath at each temperature interval. Littorinids were then
immediately placed in 2 ml eppendorf tubes and frozen in liquid nitrogen, and
were stored in – 80C freezer (Sanyo, Japan) prior to enzyme measurements.
For sample preparation, foot muscle tissue, with the operculum removed, was
dissected from 5 individuals (approx. 0.2 g of tissue) in each vial and pooled.
The samples were pooled to minimize possible variations in enzyme activities
between individuals treated at the same temperature, which may be for example
caused by variation in food availability (Yang and Somero, 1993). The tissue
was then homogenised in 500 µl homogenization buffer (50 mmol l-1 potassium
phosphate, pH 6.8 at 20C; 1 mmol l-1 DL-Dithiothreitol) on ice with a
rotor-stator homogenizer (IKA Works, Staufen, Germany). After
homogenisation, the homogenate was centrifuged at 12,500 rpm at 4C for 10
minutes (Eppendorf centrifuge 5810 R, Eppendorf-Netheler-Hinz, Germany), and
the supernatant was stored at – 80C for determination of enzyme activities and
protein quantification.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 79/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Activities of malate dehydrogenase (MDH, E.C.1.1.1.37) and lactate
dehydrogenase (LDH, E.C.1.1.1.27) were measured according to Yang & Somero
(1993) with modifications in concentration of chemicals and expression unit for
enzyme activity, i.e. (μmol substrate converted to product min−1) per gram protein.
To monitor enzyme activities, the homogenate was thawed on ice at room
temperature, and diluted by a factor of 25 (determined from preliminary trials)
using homogenisation buffer for MDH assay, while no dilution was made for
LDH. MDH was then assayed in 200mmol l-1 imidazole-HCl (pH 7.0 at 20C),
150 µmol l-1 NADH and 0.2mmol l-1 oxaloacetate; while LDH was assayed in 200
mmol l-1 imidazole-HCl (pH 7.5 at 20C), 150µmol l-1 NADH and 4mmol l-1
pyruvate. The activities of MDH and LDH were assayed at 30C ± 0.05 for all
samples in 96-well microplates (Serological plates SLP-000-09C, Jet Biofil), and
changes in absorbance at 340nm measured using a SpectraMax M2©
spectrophotometer (Molecular Devices) equipped with a temperature-controlled
micro-plate reader. The protein content of each sample was quantified using the
procedures adopted in the Bio-Rad Bradford Protein Assay (Bio-Rad©, United
States). The homogenate was diluted by a factor of 25 times (determined from
preliminary trials) using homogenisation buffer, and was assayed in Protein Dye
Reagent Concentrate (Bio-Rad©, United States) using the 96-well microplate at
30C ±
0.05. The absorbance was measured at 595 nm using the
spectrophotometer as described, and protein content was obtained by comparing
the value to a standard curve, which was constructed using bovine serum albumin
standards at concentrations of 12.5, 25, 50, 100, 200 and 400ng/µl. Enzyme
assays and protein quantifications were done in duplicate for all samples, and the

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 80/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
mean value was taken for statistical analyses.
Data analysis
To test for any possible variation in lethal temperature of Echinolittorina species
(n = 3 from each set of experiment x 2 sampling days = 6) between seasons, a
two-way Analysis of Variance (ANOVA) was employed using WinGMAv5 (EICC,
The University of Sydney), with species (Sp) [three levels: E . malaccana, E .
radiata and E . vidua] and seasons (Se) [two levels: summer & winter] treated as
fixed factors. The two replicate days were pooled to increase statistical power,
as within season variation was not of interest in this comparison. Cochran’s test
was used to check for homogeneity of variances (Cochran, 1951). Data were
transformed, if they were heterogeneous, however, where data failed tests for
homogeneity of variances after transformation, analyses were still conducted as
ANOVA is considered to be robust to such heterogeneity provided there is a large
error degrees of freedom (Underwood, 1997). To be conservative in
interpretation of such ANOVA results p-values for significant differences
(originally p < 0.05) were adjusted to a lower value ( p < 0.01, Underwood, 1981).
If any significant difference(s) was detected, post-hoc Student-Newman-Keuls
(SNK) tests were performed to identify the difference by multiple comparisons of
treatment means.
Potential difference(s) in Arrhenius breakpoint temperature of heart rate between
seasons and activities of malate dehydrogenase and lactate dehydrogenase among

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 81/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
the Echinolittorina species were analysed similarly using WinGMAv5 (EICC, The
University of Sydney). For Arrhenius breakpoint temperature of heart rate (n =
14), a two-way orthogonal ANOVA was performed, with species (Sp) [three levels:
E . malaccana, E . radiata and E . vidua] and seasons (Se) [two levels: summer &
winter] as fixed factors; while for activities of malate dehydrogenase and lactate
dehydrogenase (n = 3), a two-way orthogonal ANOVA was performed, with
species (Sp) [three levels: E . malaccana, E . radiata and E . vidua] and
temperature (Te) [temperature: 30, 35, 40, 45, 50 and 55C] as fixed factors.
Cochran’s test was used to check for homogeneity of variances (Cochran, 1951)
and data were treated as above where variances were heterogeneous. If any
significant difference(s) for fixed factors was detected, post-hoc
Student-Newman-Keuls (SNK) tests were performed to identify differences by
multiple comparisons of treatment means.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 82/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
3.3 RESULTS
Seasonal var iati on in lethal temperature
In the analysis of lethal temperature (LT50) of Echinolittorina species, an
interaction was found between species and season (Figure 3.3, Tables 3.4 & 3.5).
E . malaccana, which occupies the highest location on the shore (Chapter 2), in
general, had the highest LT50, followed by E . radiata and E . vidua in both seasons
at South Bay (Table 3.4) and Stanley (Table 3.5). Similar patterns of LT50 were
observed between summer and winter, however, the LT50 for each Echinolittorina
species was higher in summer than winter, and the response was greatest for E .
radiata, (1.15°C, South Bay and 1.53°C Stanley), followed by E . malaccana
(0.74°C, South Bay and 1.14°C Stanley), with E . vidua having the smallest
difference (0.57°C South Bay and 0.5°C Stanley).
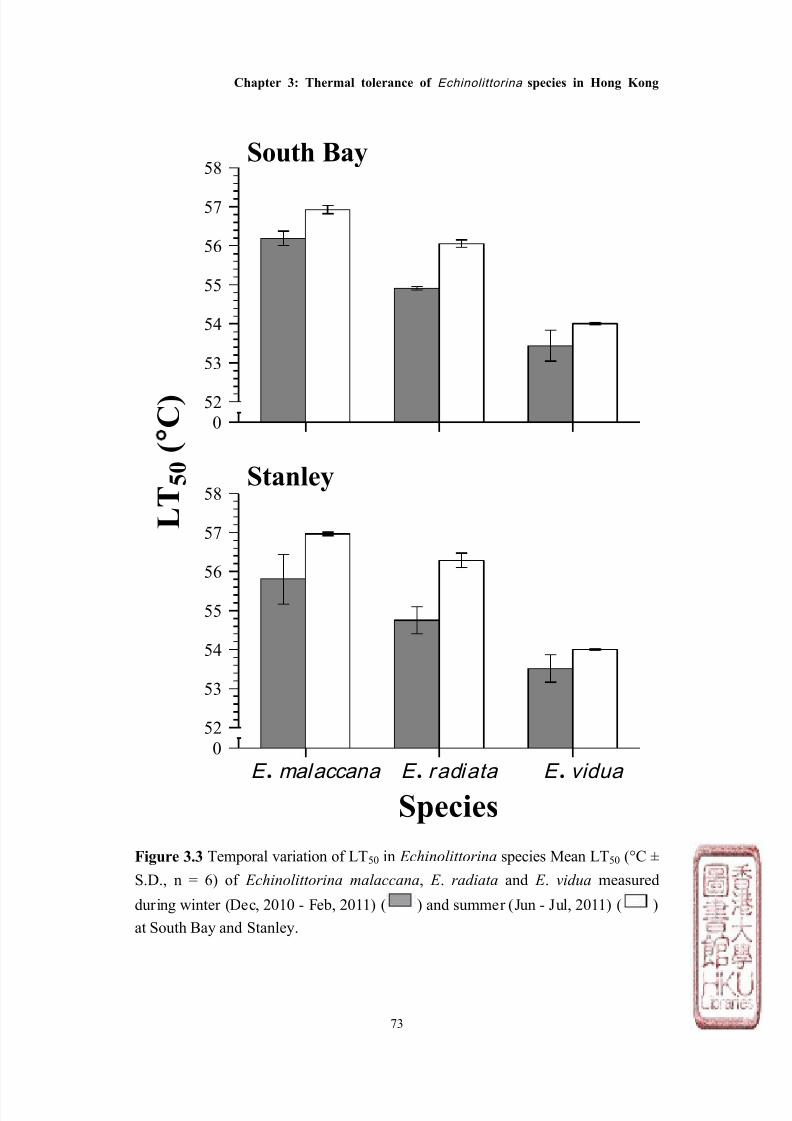
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 83/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
0
52
53
54
55
56
57
58South Bay
L T 5 0 ( C )
0
52
53
54
55
56
57
58Stanley
E . malaccana E . vidua E . radiata
Species
Figure 3.3 Temporal variation of LT50 in Echinolittorina species Mean LT50 (°C ±
S.D., n = 6) of Echinolittorina malaccana, E . radiata and E . vidua measured
during winter (Dec, 2010 - Feb, 2011) ( ) and summer (Jun - Jul, 2011) ( )
at South Bay and Stanley.
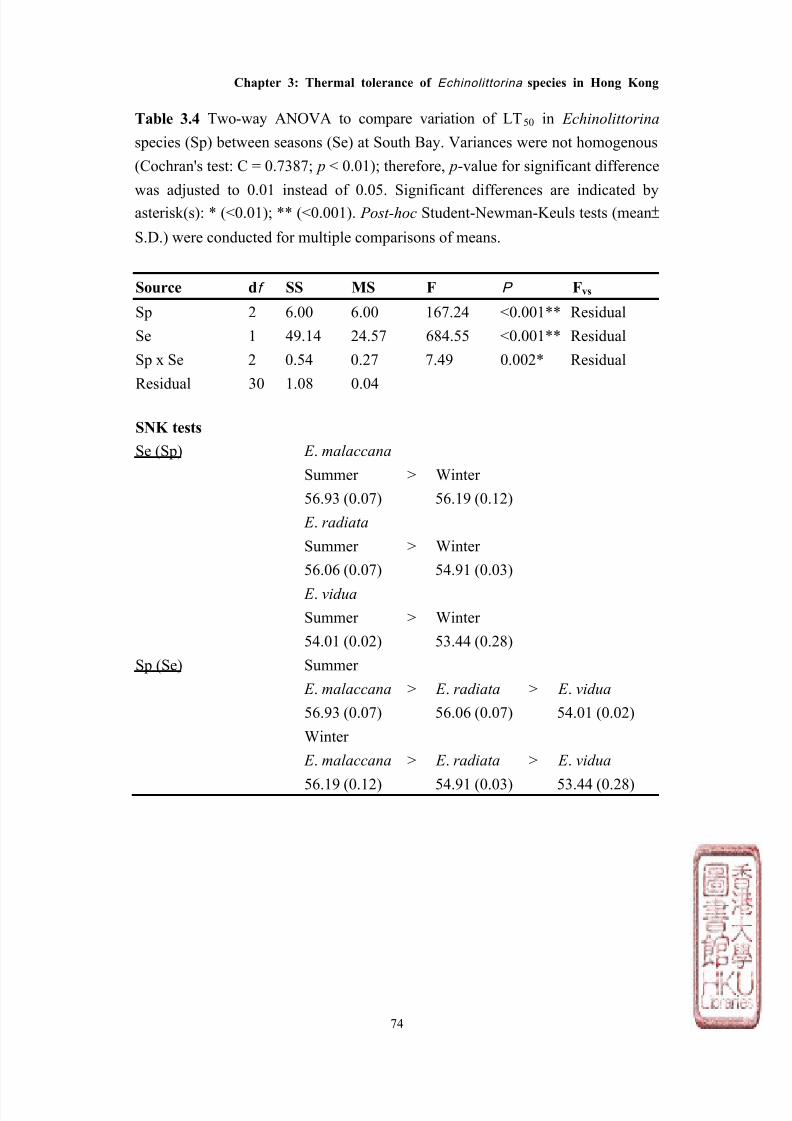
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 84/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Table 3.4 Two-way ANOVA to compare variation of LT50 in Echinolittorina
species (Sp) between seasons (Se) at South Bay. Variances were not homogenous
(Cochran's test: C = 0.7387; p < 0.01); therefore, p-value for significant difference
was adjusted to 0.01 instead of 0.05. Significant differences are indicated byasterisk(s): * (<0.01); ** (<0.001). Post-hoc Student-Newman-Keuls tests (mean±
S.D.) were conducted for multiple comparisons of means.
Source df SS MS F P Fvs
Sp 2 6.00 6.00 167.24 <0.001** Residual
Se 1 49.14 24.57 684.55 <0.001** Residual
Sp x Se 2 0.54 0.27 7.49 0.002* Residual
Residual 30 1.08 0.04
SNK tests
Se (Sp) E . malaccana
Summer > Winter
56.93 (0.07) 56.19 (0.12)
E . radiata
Summer > Winter
56.06 (0.07) 54.91 (0.03)
E . vidua
Summer > Winter
54.01 (0.02) 53.44 (0.28)
Sp (Se) Summer
E . malaccana > E . radiata > E . vidua
56.93 (0.07) 56.06 (0.07) 54.01 (0.02)
Winter
E . malaccana > E . radiata > E . vidua
56.19 (0.12) 54.91 (0.03) 53.44 (0.28)
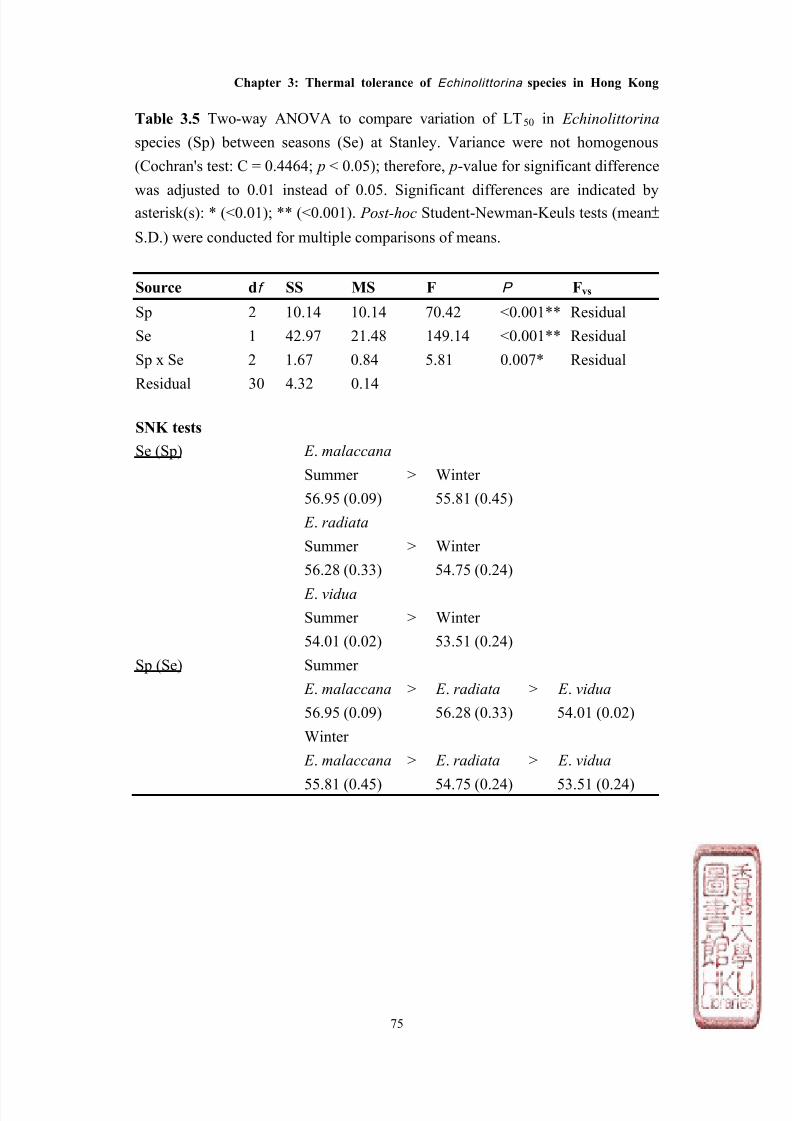
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 85/150

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 86/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Seasonal variati on in Arrhenius breakpoint temperature of heart rate
The Arrhenius breakpoint temperature (ABT) of heart rate (HR) showed an
interaction between Echinolittorina species and seasons (Figure 3.4, Table 3.6).
During summer, the mean ABT of HR for E . malaccana was the highest, followed
by E . radiata, and E . vidua. In winter, however, the pattern of ABT differed
from summer, with E . malaccana having the highest mean ABT, followed by E .
vidua, and E . radiata. The ABT for E . radiata and E . vidua were generally
greater in summer than winter (Table 3.6), whereas the ABT for E . malaccana
was similar between summer and winter.
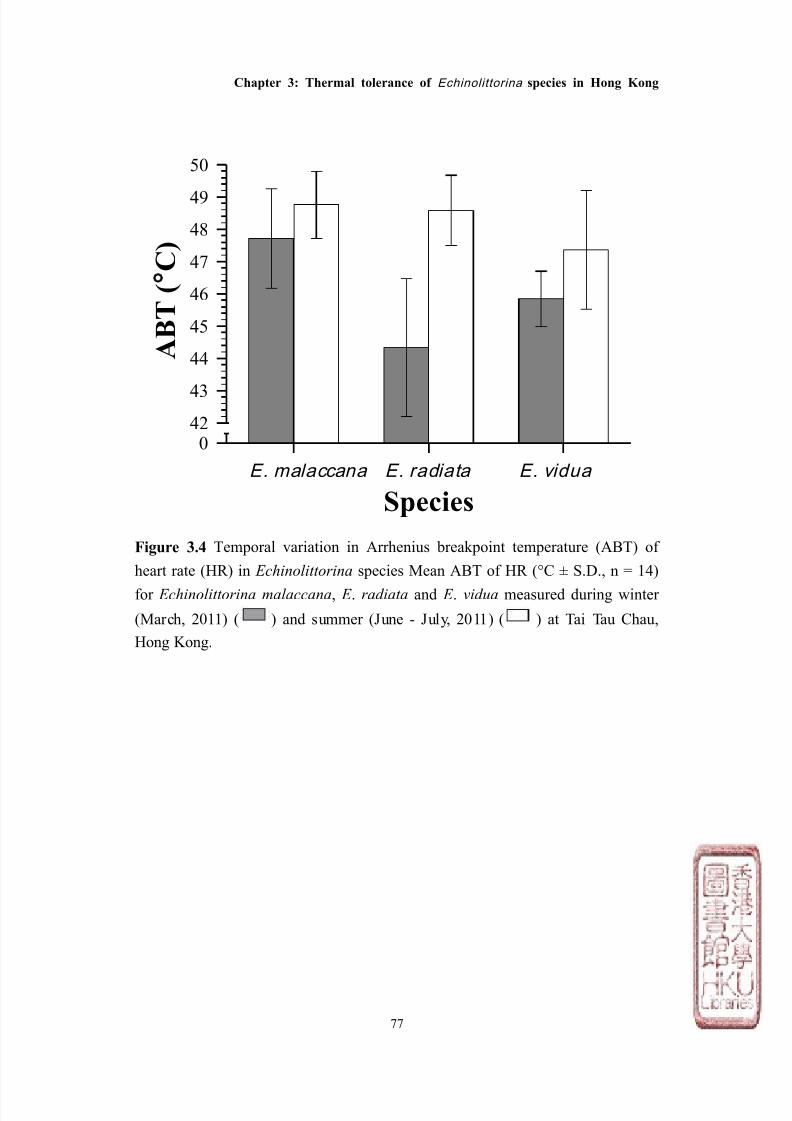
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 87/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
E. malaccana E. radiata E. vidua
042
43
44
45
46
47
48
49
50
Species
A B T ( C )
Figure 3.4 Temporal variation in Arrhenius breakpoint temperature (ABT) of
heart rate (HR) in Echinolittorina species Mean ABT of HR (°C ± S.D., n = 14)
for Echinolittorina malaccana, E . radiata and E . vidua measured during winter
(March, 2011) ( ) and summer (June - July, 2011) ( ) at Tai Tau Chau,
Hong Kong.
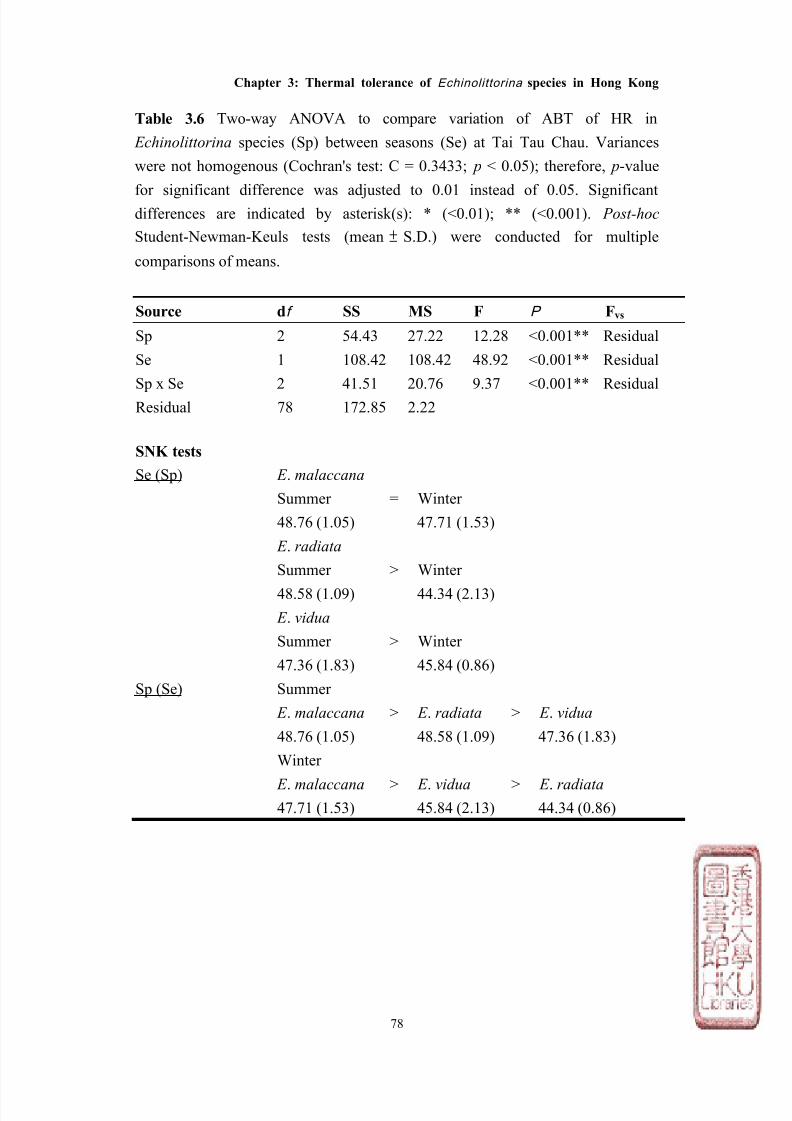
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 88/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Table 3.6 Two-way ANOVA to compare variation of ABT of HR in
Echinolittorina species (Sp) between seasons (Se) at Tai Tau Chau. Variances
were not homogenous (Cochran's test: C = 0.3433; p < 0.05); therefore, p-value
for significant difference was adjusted to 0.01 instead of 0.05. Significantdifferences are indicated by asterisk(s): * (<0.01); ** (<0.001). Post-hoc
Student-Newman-Keuls tests (mean ± S.D.) were conducted for multiple
comparisons of means.
Source df SS MS F P Fvs
Sp 2 54.43 27.22 12.28 <0.001** Residual
Se 1 108.42 108.42 48.92 <0.001** Residual
Sp x Se 2 41.51 20.76 9.37 <0.001** Residual
Residual 78 172.85 2.22
SNK tests
Se (Sp) E . malaccana
Summer = Winter
48.76 (1.05) 47.71 (1.53)
E . radiata
Summer > Winter
48.58 (1.09) 44.34 (2.13)
E . vidua
Summer > Winter
47.36 (1.83) 45.84 (0.86)
Sp (Se) Summer
E . malaccana > E . radiata > E . vidua
48.76 (1.05) 48.58 (1.09) 47.36 (1.83)
Winter
E . malaccana > E . vidua > E . radiata 47.71 (1.53) 45.84 (2.13) 44.34 (0.86)

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 89/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Acti vit ies of malate dehydrogenase and lactate dehydrogenase
Activities of both malate dehydrogenase (MDH) and lactate dehydrogenase (LDH)
varied with temperature, however, only MDH was shown to vary between species
(Figures 3.5 & 3.6, Tables 3.7 & 3.8). MDH activity was highest for E . vidua,
followed by E . malaccana, and then E . radiata (Figure 3.5, Table 3.7). While
LDH activity showed no difference among the species, activity of this enzyme
was generally higher for E . malaccana than E . vidua and E . radiata (Figure 3.6,
Table 3.8). Both activities of MDH and LDH were highest in the three littorinid
species when temperatures reached 55oC (Figures 3.5 & 3.6, Tables 3.7 & 3.8).
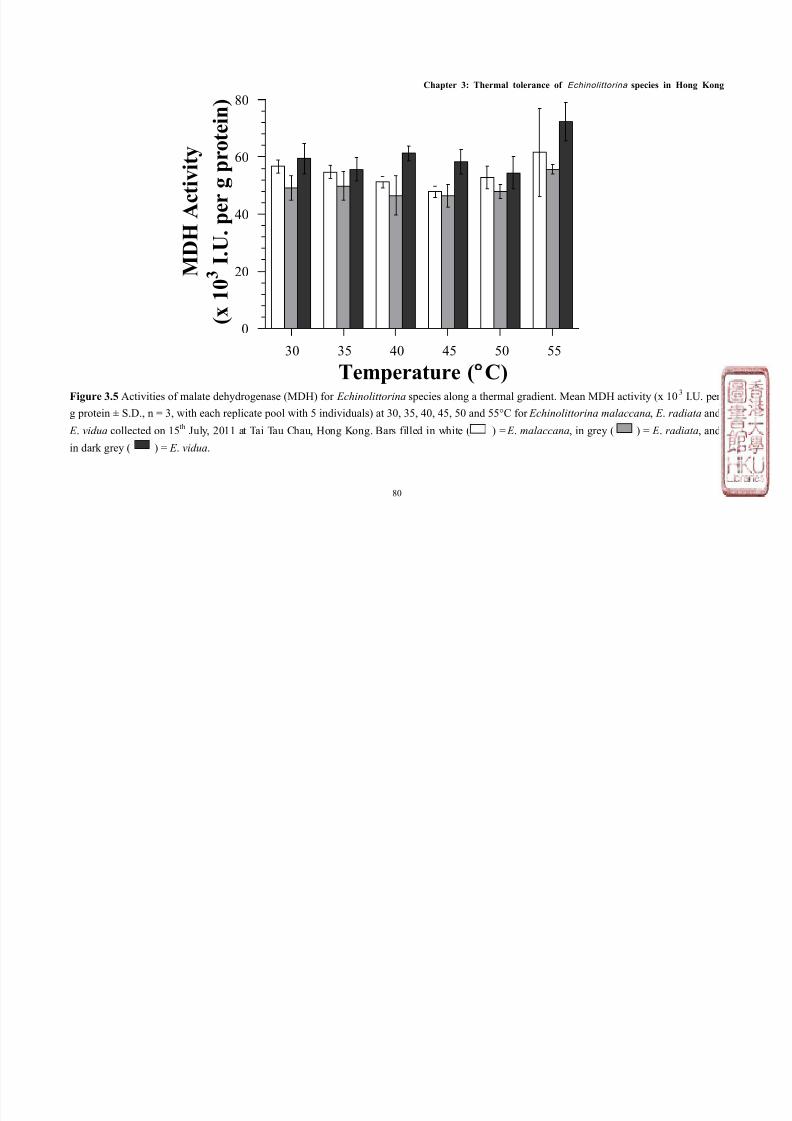
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 90/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
80
Figure 3.5 Activities of malate dehydrogenase (MDH) for Echinolittorina species along a thermal gradient. Mean MDH activity (x 10 3 I.U. per
g protein ± S.D., n = 3, with each replicate pool with 5 individuals) at 30, 35, 40, 45, 50 and 55°C for Echinolittorina malaccana, E . radiata and
E . vidua collected on 15th July, 2011 at Tai Tau Chau, Hong Kong. Bars filled in white ( ) = E . malaccana, in grey ( ) = E . radiata, and
in dark grey ( ) = E . vidua.
30 35 40 45 50 55
0
20
40
60
80
Temperature ( C)
M D H A c t i v i t y
( x 1 0 3
I . U . p e r
g p r o t e i n )
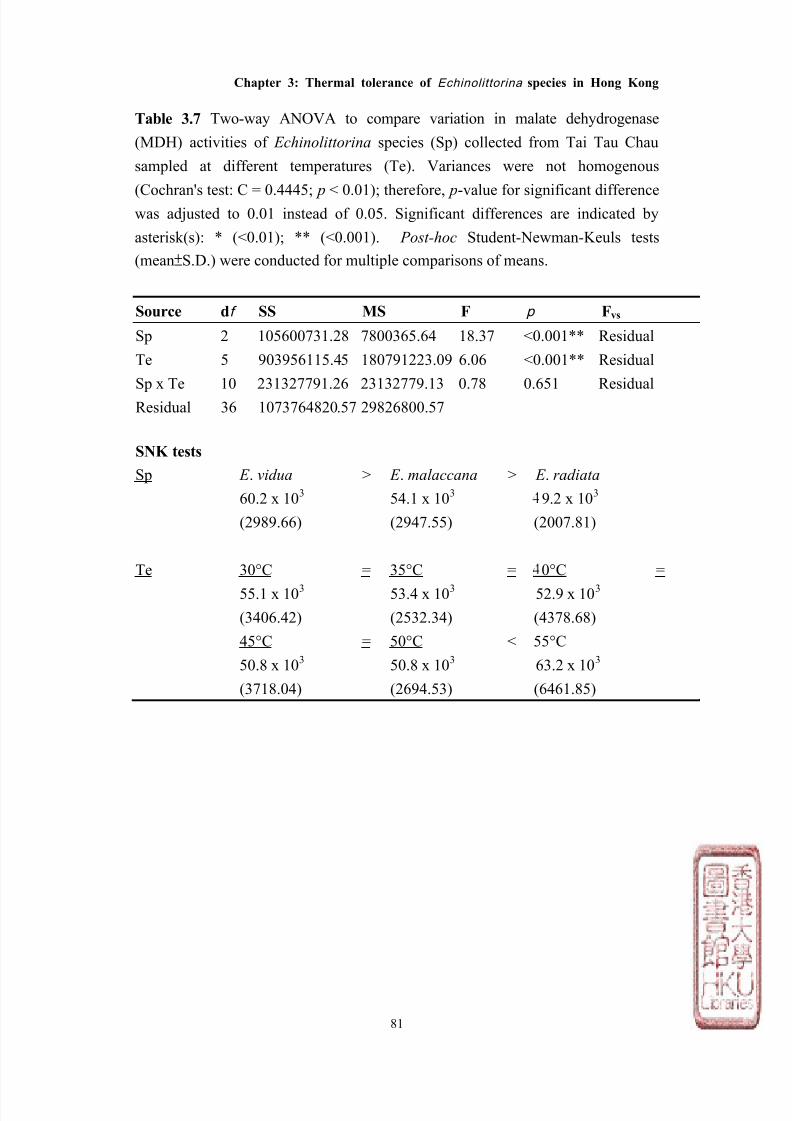
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 91/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Table 3.7 Two-way ANOVA to compare variation in malate dehydrogenase
(MDH) activities of Echinolittorina species (Sp) collected from Tai Tau Chau
sampled at different temperatures (Te). Variances were not homogenous
(Cochran's test: C = 0.4445; p < 0.01); therefore, p-value for significant differencewas adjusted to 0.01 instead of 0.05. Significant differences are indicated by
asterisk(s): * (<0.01); ** (<0.001). Post-hoc Student-Newman-Keuls tests
(mean±S.D.) were conducted for multiple comparisons of means.
Source df SS MS F p Fvs
Sp 2 105600731.28 7800365.64 18.37 <0.001** Residual
Te 5 903956115.45 180791223.09 6.06 <0.001** Residual
Sp x Te 10 231327791.26 23132779.13 0.78 0.651 Residual
Residual 36 1073764820.57 29826800.57
SNK tests
Sp E . vidua > E . malaccana > E . radiata
60.2 x 103
(2989.66)
54.1 x 103
(2947.55)
9.2 x 103
(2007.81)
Te 30°C = 35°C = 0°C =
55.1 x 103
(3406.42)
53.4 x 103
(2532.34)
52.9 x 103
(4378.68)
45°C = 50°C < 55°C
50.8 x 103
(3718.04)
50.8 x 103
(2694.53)
63.2 x 103
(6461.85)
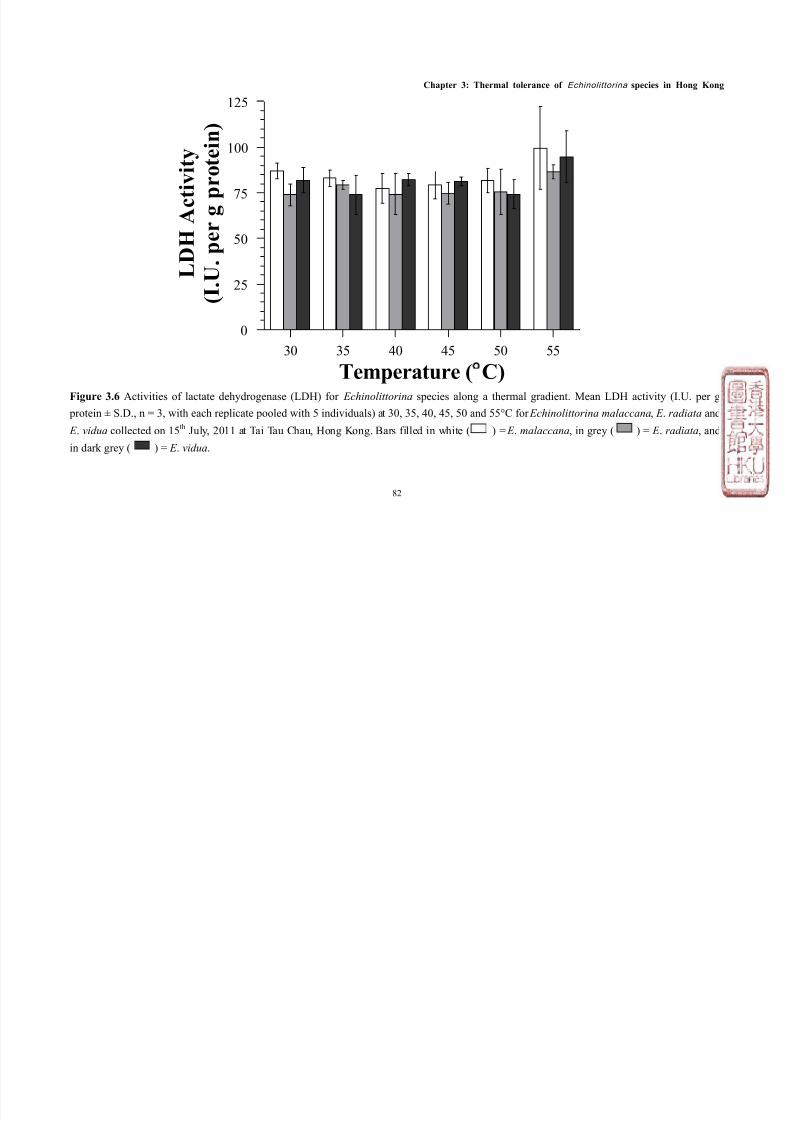
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 92/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
82
Figure 3.6 Activities of lactate dehydrogenase (LDH) for Echinolittorina species along a thermal gradient. Mean LDH activity (I.U. per g
protein ± S.D., n = 3, with each replicate pooled with 5 individuals) at 30, 35, 40, 45, 50 and 55°C for Echinolittorina malaccana, E . radiata and
E . vidua collected on 15th July, 2011 at Tai Tau Chau, Hong Kong. Bars filled in white ( ) = E . malaccana, in grey ( ) = E . radiata, and
in dark grey ( ) = E . vidua.
30 35 40 45 50 55
0
25
50
75
100
125
Temperature (
C)
L D H A
c t
i v i t y
( I . U . p e r g p
r o t e i n )
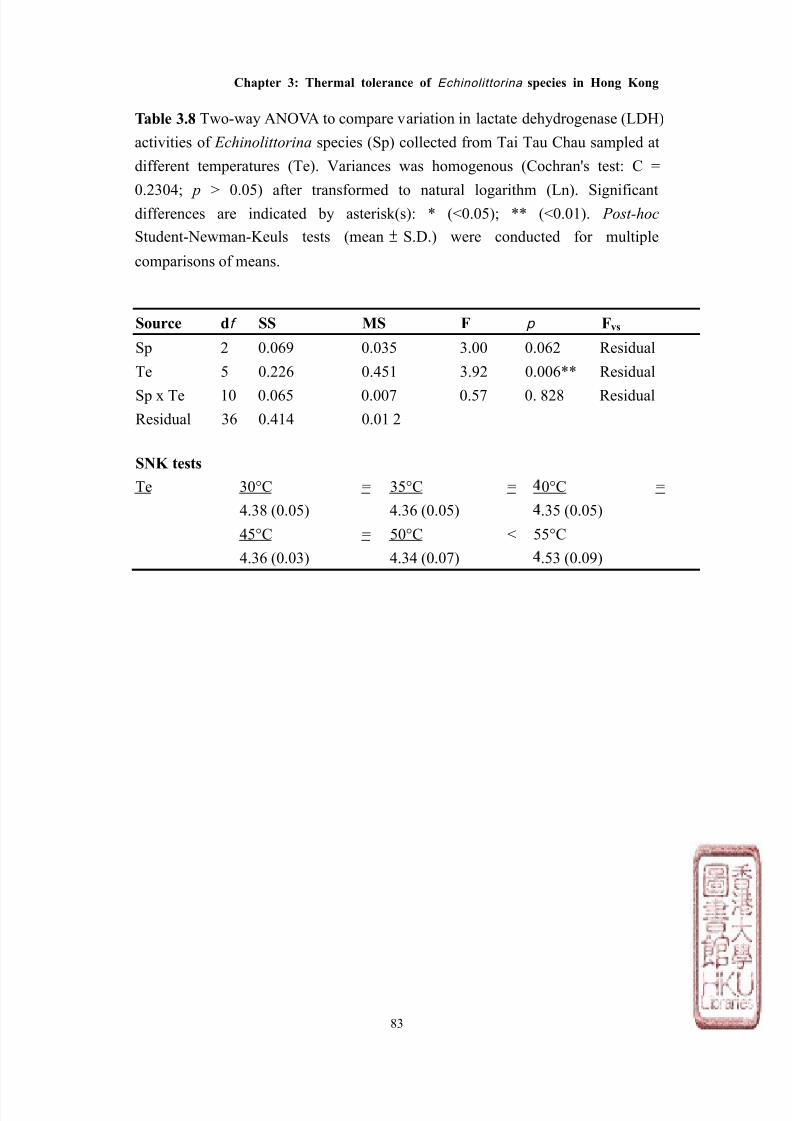
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 93/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Table 3.8 Two-way ANOVA to compare variation in lactate dehydrogenase (LDH)
activities of Echinolittorina species (Sp) collected from Tai Tau Chau sampled at
different temperatures (Te). Variances was homogenous (Cochran's test: C =
0.2304; p > 0.05) after transformed to natural logarithm (Ln). Significantdifferences are indicated by asterisk(s): * (<0.05); ** (<0.01). Post-hoc
Student-Newman-Keuls tests (mean ± S.D.) were conducted for multiple
comparisons of means.
Source df SS MS F p Fvs
Sp 2 0.069 0.035 3.00 0.062 Residual
Te 5 0.226 0.451 3.92 0.006** Residual
Sp x Te 10 0.065 0.007 0.57 0. 828 Residual
Residual 36 0.414 0.01 2
SNK tests
Te 30°C = 35°C = 0°C =
4.38 (0.05) 4.36 (0.05) .35 (0.05)
45°C = 50°C < 55°C
4.36 (0.03) 4.34 (0.07) .53 (0.09)

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 94/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
3.4 DISCUSSION
Lethal temperature in relation to verti cal distri bution
Littorinids inhabiting the high shore (reviewed by Lewis, 1972; see Chapter 1 for
definition) of intertidal habitats have been well documented to have a stronger
ability to tolerate extreme high temperatures than other gastropod taxa (Evans,
1948; Southward, 1958; Fraenkel, 1966; Stirling, 1982; McMahon and Britton,
1985; Cleland and McMahon, 1990; McMahon, 1990, 2001; Britton, 1992).
McMahon (2001), for example, categorized 60 species of gastropods from various
intertidal habitats, including rocky shores, salt marshes and mangroves, into two
groups, the littorinids and nonlittorinids, and showed that the mean HCT for
littorinids was significantly higher than the nonlittorinids by ~ 2.5°C, and that this
group generally went into coma only when temperatures reached > 40°C. This is
also true for littorinids in Hong Kong (see Table 3.9), and in this study, lethal
temperatures determined for E . malaccana, E . radiata and E .vidua, in general,
exceeded 53°C. This lethal temperature is slightly below the maximum rock
surface temperatures recorded at +2 m above CD (~ 57.6°C) in summer at Cape
d’Aguilar, Hong Kong (Williams et al., unpubl i-button data).
Echinolittorina malaccana, E . radiata and E . vidua in Hong Kong have a strong
tolerance to high temperatures and showed a consistent pattern in their tolerance
which matched their vertical distributions at South Bay and Stanley (see Chapter 2;
Williams, 1994a; Mak, 1996). E . malaccana, which occupies the highest level

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 95/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
of the high shore, has the highest lethal temperatures in both summer and winter,
followed by the lower-level E . radiata and E . vidua, respectively. This pattern
matches with the HCT determined by Stirling (1982) and McMahon (2001, see
Table 3.9), who conducted similar thermal stress experiments, but in water, on E .
malaccana and E . radiata during summer in Hong Kong. In terms of LT50,
Stirling (1984) showed no difference between E . malaccana and E . radiata during
summer in Hong Kong. Stirling's results are similar to those obtained in the
present study, with the LT50 of E . malaccana being only ~ 0.5oC higher than that
of E . radiata (see Table 3.9). The LT50 for E . malaccana and E . vidua, however,
demonstrated a greater difference, with the relatively higher shore E . malaccana
LT50s being ~ 2oC greater than E . vidua in the present study, as was also recorded
by Lee and Lim in Singapore (2009, see Table 3.10). Based on the survival
limits determined, therefore, thermal tolerance of Echinolittorina species in Hong
Kong generally increases with increasing shore levels.
Despite the close concordance of patterns of thermal tolerance of Echinolittorina
species with previous results, there is likely to be some degree of variation
introduced by the methodologies used, such as heating conditions and rates
(Terblanche et al., 2011; also see Table 3.10), which makes absolute comparison
with other established work difficult (Clarke et al., 2000a). Many studies (see
Table 3.1), to minimize the effect of desiccation, have determined survival limits
while organisms were immersed in seawater. There are only a few studies
(Sandison, 1976; Davenport and Davenport, 2005, 2007; present study) that have
measured limits whilst animals are emersed, which is the normal condition that

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 96/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
animals will experience thermal stress. During summer low tide periods, for
example, rock surface temperatures in Hong Kong rocky shores can exceed 50oC
(Williams and Morritt, 1995; Williams, unpubl data). Intertidal animals living at
the high shore are more likely to experience such extreme thermal conditions, and
can spend more than 85% of their time emersed between successive spring tides
(McMahon, 2001; Uglow and Williams, 2001; Lee and Boulding, 2010). Due to
variations in the environmental medium and thermal conditions, animals may,
therefore, exhibit different respiratory rates in air and water. Sandison (1967),
for example, showed a greater oxygen uptake in air than water in Littorina species
and Thais lapillus.
Based on this finding, Lee and Boulding (2010) pointed out that HCT determined
in water is positively correlated to that in air, therefore, implying that heating
condition is not critically important in determining species’ survival limits, and
allowing direct comparison with previous studies. Lee and Boulding, however,
did not supply any evidence to support this view (2010). Despite being
positively correlated, realistically, different heating conditions may still contribute
to variation in recorded LT50 values (Sandison, 1967; Davenport and Davenport,
2007). The lethal temperature of Littorina littoralis, L. littorea and Thais
lapillus were, for example, found to be 1-3˚C higher in air than water, while no
difference was found in L. neritoides and L. saxatilis (Sandison, 1967). Similar
to Littorina littoralis, L. littorea and Thais lapillus, in the present study, the LT50
of E . malaccana was significantly lower in water than air and a similar, but not
significant, trend was also observed in E . radiata (Table 3.11). This implies that

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 97/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
when comparing the LT50 as a measure of thermal stress in Echinolittorina species,
heating conditions have to be noted, as these may confound potential differences
or similarities between species.
Apart from heating conditions, differences in heating rates may also result in
variation in measured species’ survival limits. Stirling (1982), for example, used
two different approaches (i.e. 6˚C/hr and constant ˚C/hr) to determine the LT50 of
intertidal gastropods in Dar es Salaam, Tanzania, and showed that constant
heating at a fixed temperature led to lower LT50 values than an increasing rate.
Similarly the above results also apply to the limpets Patella vulgata, P . depressa
and P . athletica (Evans, 1948), where survival limits were found to be lower at a
slower heating rate. Conclusions made from the above studies also hold true for
E . radiata, where the LT50 was positively related to heating rate, although this was
not true in E . malaccana (see Table 3.11). When comparing survival limits in
Echinolittorina species, therefore, heating rate should also be considered as it may
potentially influence the results obtained (Terblanche et al., 2011).
In general, regardless of differences in methodology, such as heating rate, sizes of
littorinids, heating conditions (see Table 3.10) that make absolute comparison
with other research difficult (Clarke et al., 2000a), the pattern of survival limits to
high temperatures in the three Echinolittorina species remains consistent, with E .
malaccana having greater survival limits than E . radiata, and followed by E .
vidua, i.e., the LT50 of Echinolittorina species increases with higher shore heights.
The positive relationship between survival limits and vertical distribution of the

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 98/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Echinolittorina species also agrees with most previous work on intertidal species,
either on non-congeneric (Broekhuysen,1940; Evans, 1948; Stirling, 1982;
Cleland and McMahon, 1990; Clarke et al., 2000b; McMahon, 2001; Davenport
and Davenport, 2005; Lee and Lim, 2009), or on congeneric species (in littorinids:
Markel, 1971; top shells: Tomanek and Somero, 1999; porcelain crabs: Stillman
and Somero, 2000; littorinids: Somero, 2002).
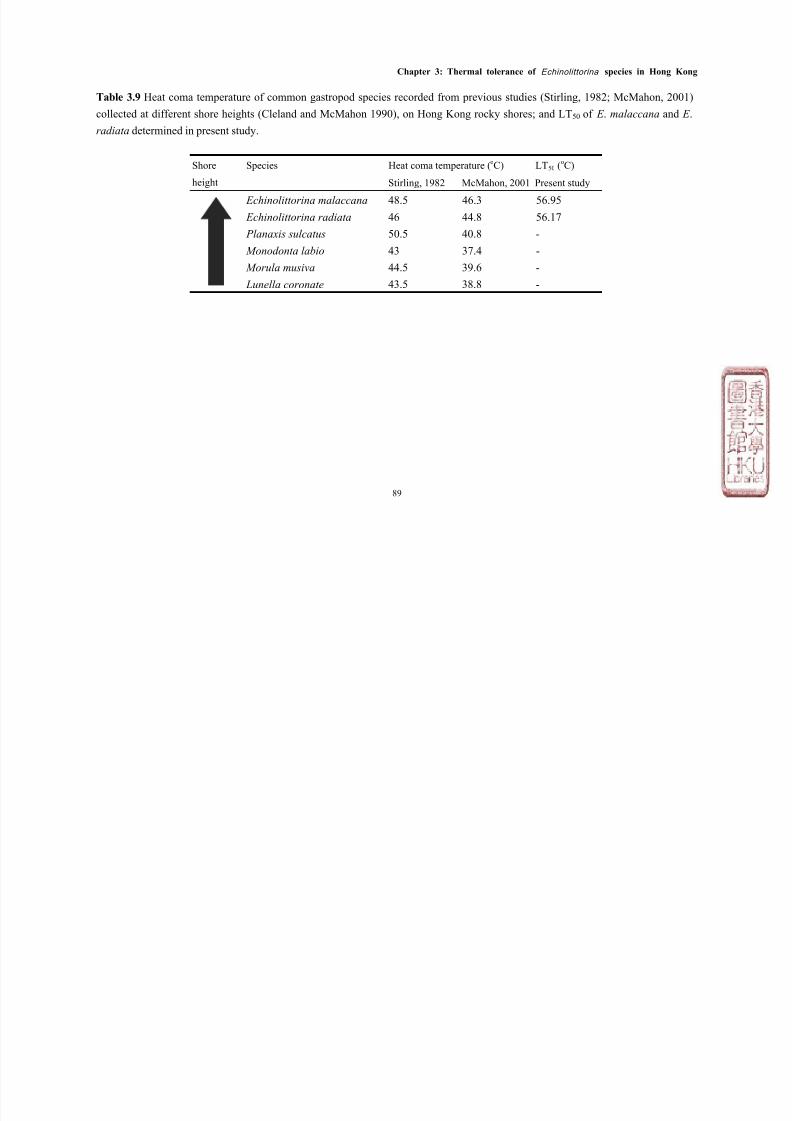
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 99/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
89
Table 3.9 Heat coma temperature of common gastropod species recorded from previous studies (Stirling, 1982; McMahon, 2001)
collected at different shore heights (Cleland and McMahon 1990), on Hong Kong rocky shores; and LT 50 of E . malaccana and E .
radiata determined in present study.
Shore
height
Species Heat coma temperature (oC) LT50 (
oC)
Stirling, 1982 McMahon, 2001 Present study
Echinolittorina malaccana 48.5 46.3 56.95
Echinolittorina radiata 46 44.8 56.17
Planaxis sulcatus 50.5 40.8 -
Monodonta labio 43 37.4 -
Morula musiva 44.5 39.6 -
Lunella coronate 43.5 38.8 -
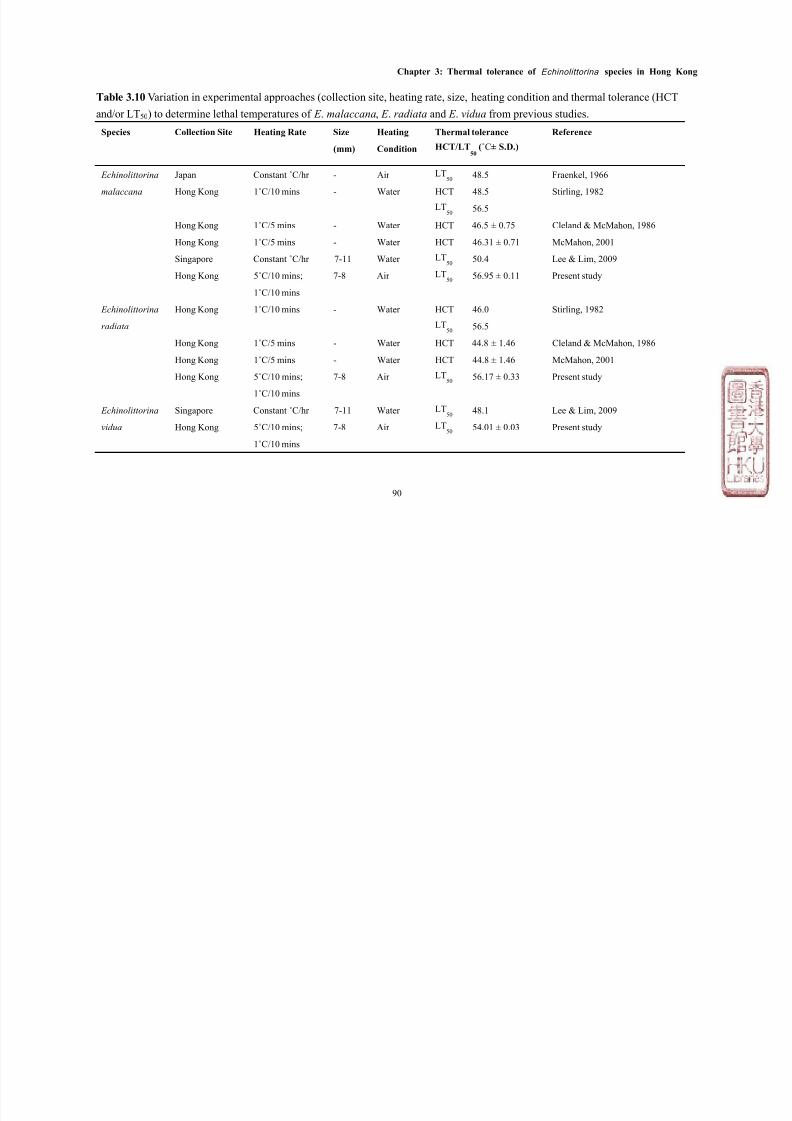
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 100/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
90
Table 3.10 Variation in experimental approaches (collection site, heating rate, size, heating condition and thermal tolerance (HCT
and/or LT50) to determine lethal temperatures of E . malaccana, E . radiata and E . vidua from previous studies.
Species Collection Site Heating Rate Size
(mm)
Heating
Condition
Thermal tolerance
HCT/LT50
(˚C± S.D.)
Reference
Echinolittorina
malaccana
Japan Constant ˚C/hr - Air LT50
48.5 Fraenkel, 1966
Hong Kong 1˚C/10 mins - Water HCT 48.5 Stirling, 1982LT
50 56.5
Hong Kong 1˚C/5 mins - Water HCT 46.5 ± 0.75 Cleland & McMahon, 1986
Hong Kong 1˚C/5 mins - Water HCT 46.31 ± 0.71 McMahon, 2001
Singapore Constant ˚C/hr 7-11 Water LT50
50.4 Lee & Lim, 2009
Hong Kong 5˚C/10 mins;
1˚C/10 mins
7-8 Air LT50
56.95 ± 0.11 Present study
Echinolittorina
radiata
Hong Kong 1˚C/10 mins - Water HCT 46.0 Stirling, 1982
LT50
56.5
Hong Kong 1˚C/5 mins - Water HCT 44.8 ± 1.46 Cleland & McMahon, 1986
Hong Kong 1˚C/5 mins - Water HCT 44.8 ± 1.46 McMahon, 2001
Hong Kong 5˚C/10 mins;
1˚C/10 mins
7-8 Air LT50
56.17 ± 0.33 Present study
Echinolittorina
vidua
Singapore Constant ˚C/hr 7-11 Water LT50
48.1 Lee & Lim, 2009
Hong Kong 5˚C/10 mins;
1˚C/10 mins
7-8 Air LT50
54.01 ± 0.03 Present study

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 101/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Table 3.11 Lethal temperature (LT50) of (a) E . malaccana and (b) E . radiata
under different heating conditions and heating rates. Student’s t-tests were
used to compare variation in LT50 between heating conditions; while one-way
ANOVA was used to compare LT50 difference(s) in heating rates.Significant differences are indicated by asterisk(s): * (< 0.05).
(a) E . malaccana
Factor Mean S.D. n p
Condition Air 56.99 0.02 3<0.001*
Water 55.99 0.06 3
Rate 5°C /10 min 56.89 0.13 3
0.2991°C /5 min 56.95 0.02 35°C /10 min (25-50°C);
1°C / 10 min (50-58°C)56.99 0.02 3
(b) E . radiata
Factor Mean S.D. n p
Condition Air 56.59 0.49 3 0.077
Water 55.95 0.06 3
Rate 5°C /10 min 56.98 0.04 3 0.02*
1°C /5 min 56.05 0.02 3
5°C /10 min (25-50°C);
1°C / 10 min (50-58°C)56.59 0.49 3

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 102/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Arrhenius breakpoint temperature of heart rate in relation to vertical
distribution
In Echinolittorina species in Hong Kong, the pattern of ABT of HR is similar to
their LT50, and was positively related to species' vertical distributions. The
relatively higher species, E . malaccana, in general, has the highest ABT of HR,
than the two relatively lower species, E . radiata and E . vidua. This result
matched with studies on congeneric intertidal species (e.g. porcelain crabs, top
shells, limpets) that also showed that the ABT of HR for species was positively
correlated to their corresponding shore heights (Stillman and Somero, 1996;
Stenseng et al., 2005; Dong and Williams, 2011). When comparing the ABT of
HR in congeneric top shells (Tegula = genus Chlorostoma), Stenseng and
colleagues (2005), for example, found that the ABT of HR in the relatively higher
shore Tegula funebralis was > 6o
C higher than the lower shore Tegula brunnea
and Tegula monteryi. In general, therefore, congeneric species inhabiting higher
shore heights tend to have higher ABT of HRs than lower shore species.
The ABT of HR refers to the temperature that heart rate of an individual drops
dramatically, and is used to indicate the initiation of loss of the ability for efficient
oxygen transportation of an individual (Stillman and Somero, 1996). With a
higher ABT of HR, therefore, E . malaccana can maintain efficient oxygen
transportation at a higher temperature than E . radiata and E . vidua. This
matches the higher LT50 determined for E . malaccana than the other two species.
A weak positive correlation between ABT of HR and LT50 for Echinolittorina

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 103/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
species (see Figure 3.7), however, matches with the trend shown in four
congeneric porcelain crabs Petrolisthes (summarized by Somero, 2010), which
suggests a general positive relationship between the two parameters. Both ABT
of HR and LT50 of Echinolittorina species, therefore, play an important role in
determining the vertical distribution of the species (see Somero, 2010 for review).
Although there have been no comparative studies on ABT of HR of congeneric
littorinid species, the heart rate of E . malaccana has recently been studied
(Marshall and McQuaid, 2011; Marshall et al., 2010, 2011). After a slower
heating rate (i.e. 1oC per 5 minutes) the ABT of HR for E . malaccana in Brunei
Darussalam (see Marshall and McQuaid, 2011; Marshall et al., 2011) was 1-3oC
higher than recorded in the present study. Such variation in ABT of HR may be
due to latitudinal differences, as shown in previous studies on intertidal
gastropods (e.g. McMahon, 2001; Stillman, 2002; Kuo and Sanford, 2009) where
thermal tolerance of species increases with decreasing latitude. However, such a
conclusion can only be drawn when the experimental protocols used are similar,
in order to reduce the confounding effects of experimental factors, such as animal
sizes, heating conditions and rates (se LT50 section).
To conclude, ABT of HR in Echinolittorina species in Hong Kong, like many
other intertidal species from other parts of the world, was found to be positively
correlated to their vertical heights. The positive correlation between ABT of HR
for Echinolittorina species and LT50 may, therefore, help explain that thermal
tolerance of the littorinid’s heart is linked to their survival limits under thermal

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 104/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
stresses.
Acti vit ies of malate dehydrogenase and lactate dehydrogenase in relation to
verti cal distri bution
In the present study, the two isoforms of MDH were not isolated for the enzyme
assays, thus, the activity measured refers to the overall activities of the two
isoforms. It has been reported that the overall activities of the isoforms are
positively related to oxygen consumption (in mussels, Mytilus californianus and
dogwhelks, Nucella ostrina Dahlhoff et al., 2002), and so this enzyme has been
used as an indicator for aerobic metabolism (e.g. Childress and Somero, 1979;
Yang and Somero, 1993; Thuesen and Childress, 1994; Dahlhoff et al., 2002;
Ombres et al., 2011).
The in vivo MDH and LDH activities of the Echinolittorina species, in general,
did not show any clear trend with increasing temperature, except for the relatively
higher activities detected when temperatures reached 55°C. This lack of
variation with temperature implies that the amount of active MDH and LDH in
the littorinids remained relatively constant as temperatures increased, even beyond
the ABT of HR, which is assumed to mark the start of the breakdown in aerobic
metabolism (Stillman and Somero, 1996; Stenseng et al., 2005; Marshall et al.,
2010, 2011; Dong and Williams, 2011).
Enzyme activity is generally expected to decrease with increasing temperature
once temperatures exceed the enzymes' optimal temperature, as enzymes are

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 105/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
thermally liable and denature at extreme temperatures (see Mathews et al., 2000;
Hochachka and Somero, 2002; Angilletta, 2009). Sokolova and Pörtner (2001),
determined the in vivo activities of two metabolic enzymes, citrate synthase (CS)
and NADP-dependent isocitrate dehydrogenase (NADP-IDH), in Littorina
saxatillis and L. obtusata, and found that activity levels decreased after heating
the littorinids from 20-25°C to 45°C at a constant rate of 1°C per 5 minutes. The
contrasting results obtained in this study, where there was little change in enzyme
activity with increasing temperatures, could possibly be explained by MDH and
LDH being thermally stable over the high temperatures experienced by
Echinolittorina species. This hypothesis, however, would need to be confirmed
using in vitro methods (e.g. Sokolova and Pörtner, 2003; Lee and Lim, 2009) by
measuring the activities of MDH and LDH along the temperature gradient using
cell extracts. When heating the littorinids in vivo, individuals may respond by
continuously inducing production of MDH and LDH to maintain the activity
levels of these enzymes. The in vitro method would, therefore, help to establish
whether the stable activity levels of the enzymes along the temperature gradient
was due to the stability of enzyme structure, and not a result of increased enzyme
production rates.
When comparing activity among species, it was found that E . vidua, which
inhabits the lowest shore level, had the highest MDH activity, and this implies this
species has the greatest aerobic capacity, followed by the two upper shore species,
E . malaccana and E . radiata respectively. The higher enzyme activity of MDH
in E . vidua than E . malaccana matches the pattern seen in a study on the activity

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 106/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
of glutamate oxaloacetate transaminase (GOT, which is responsible for ammonia
detoxification and nitrogen excretion), between the two species (Lee and Lim,
2009). This finding does not, however, support the hypothesis that E . vidua has
a greater aerobic capacity than E . malaccana as GOT is not involved in energy
metabolism, but does indicate that E . vidua may have a higher metabolic rate than
E . malaccana. This appears to be true, as the basal heart rate for E . vidua, in
general, is slightly higher than that of E . malaccana (see Appendix 2.1 and 2.2).
Enzyme activity is commonly used as an indicator in determination of metabolic
rate (see Dahlhoff, 2004 for review), as it has been reported to be positively
correlated with metabolic rate in pelagic fishes (Childress and Somero, 1979;
Sullivan and Somero, 1983; Torres and Somero, 1988a,b; Yang and Somero,
1993). If this is the case, the lower metabolic rate of E . malaccana may result
from their enhanced capability for metabolic depression (Marshall et al., 2011).
In some cases, E . malaccana is known to depress metabolism, and their heart rate
at 40°C has been shown to be lower than at 30°C during constant heating
(Marshall and McQuaid, 2011; Marshall et al., 2011). Possibly due to different
heating approaches, metabolic depression was not observed in the Echinolittorina
species in the present study (see Appendix 2) as well as the study by Matumba
and colleagues (unpubl data), who also worked on E . malaccana and E . vidua.
Yet, both studies showed that the heart rate of E . malaccana was relatively
independent of temperature, i.e. the HR did not show much increase as
temperature increased as compared with E . vidua. This phenomenon seen in the
heart rate for E . malaccana may due to metabolic depression, which may also

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 107/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
apply to E . radiata which showed a lower level of MDH activity than E . vidua.
Such physiological mechanisms may, therefore, help E . malaccana to conserve
energy to protect themselves from thermal stress (e.g. to be able to produce heat
shock proteins, Marshall and McQuaid, 2011; Marshall et al., 2011).
The activity of LDH between the species was slightly different from MDH,
although these differences were not significant. Activity was highest in E .
malaccana, while E . radiata and E . vidua showed similar LDH activities. High
LDH activity means that, despite having the ability to undergo metabolic
depression, the high shore E . malaccana also has the greatest anaerobic capacity
when compared with E . radiata and E . vidua. High shore species have, in
general, been proposed to have a greater capacity for anaerobic metabolism than
lower shore species (Simpfendörfer et al., 1995; Sokolova and Pörtner, 2001;
Rinke and Lee, 2009). Sokolova and Pörtner (2001), for example, compared the
anaerobic capacity of two Littorina species, through the ratio of pyruvate kinase
(PK) and phosphoenolpyruvate carboxykinase (PEPCK). The higher shore L.
saxatilis had a lower PK/PEPCK activity ratio than the lower shore L. obtusata,
and Sokolova and Pörtner suggested that L. saxatilis had a greater anaerobic
capacity than L. obtusata. Similarly, in the present study, although using a
different anaerobic enzyme, the higher shore species tend to have greater
anaerobic capacity than the lower species. Therefore, even when rock surface
temperature reaches close to their ABT of HR, when oxygen transportation is
limited, the higher shore species are still able to produce energy through anaerobic
metabolism, in order to maintain their basal needs for survival.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 108/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Seasonal variati on of LT 50 and ABT of HR
Seasonal impacts on thermal tolerance, including survival limits and heart rate, in
intertidal species, especially gastropods, have been previously studied (for heart
rate: Segal, 1956; survival limit: see Table 3.13). These studies have shown that
thermal tolerance was higher when animals were collected at higher air
temperature environments during summer than winter. Evans (1948), for
example, determined the seasonal HCT in the limpets Patella vulgata, P . depressa
and P . athletica, and found that their HCTs in summer were ~ 0.7-1.6 oC higher
than those recorded in winter. Such observations, in general, are also true for the
LT50 and ABT of HR in the three Echinolittorina species in Hong Kong, where the
LT50 was ~ 0.5-1.5oC higher in summer, and ABR of HR was ~ 0.1-4.2oC higher
than in winter.
Seasonal migration patterns are a common phenomena observed in littorinids
which, apart from being linked with reproductive success (e.g Kojima, 1959;
Fretter and Graham, 1962; Hannaford Ellis, 1985; Ohgaki, 1988b; Mak, 1996),
have also been suggested to be related to environmental factors (e.g. Lambert and
Farley, 1968; Williams and Ellis, 1975; Harper and Williams, 2001), such as
temperature. Lambert and Farley (1968) and Williams and Ellis (1975), for
example, suggested that the downward migration observed in Littorina littorea
during winter allowed it to escape from cold conditions high on the shore. The
three Echinolittorina species in Hong Kong exhibit seasonal migration, moving
down shore in summer (Mak, 1996; see Chapter 2). Data on thermal tolerance
suggest that this behaviour may not be driven by exposure to high temperatures in

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 109/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
summer, as the littorinids are more tolerant to high temperature in summer than
winter. This behaviour may, however, be to enhance their reproductive success
and increase duration of feeding (Ohgaki, 1988a; Mak, 1996; Ito et al., 1998).
To enhance littorinids’ reproductive success, for example, seasonal migration of
Echinolittorina species in Hong Kong was found to coincide with their spawning
period (Mak, 1996). As the low tides are lower in Hong Kong in summer than
winter (Kaehler and Williams, 1996), the seasonal migration may, therefore, help
facilitate the littorinids’ chance to release their eggs whilst immersed, as has been
observed for E . radiata in Shirahama, Japan, (Ohgaki, 1988a).
The cause of the seasonal variation in thermal tolerance of the Echinolittorina
species in Hong Kong was not investigated in the present study. It may, however,
be linked to the littorinids’ ability to acquire acclimation-induced heat shock
responses, as shown in other intertidal species (see Feder and Hofmann, 1999;
Dahlhoff, 2004; Tomanek, 2010; Somero, 2010 for reviews), which help the
species to survive in extreme environmental conditions during the hot summer.
Heat shock response, as defined by Tomanek (2010), is the production of a group
of heat shock proteins (hsp) that help stabilize denatured proteins as well as to
refold denatured proteins to correct suboptimal conformations produced under
stressful environmental conditions, such as high temperatures. Seasonal or
laboratory-based acclimation to higher temperature can coincide with an increase
in levels of hsps in many intertidal species (e.g., topshells, Chlorostoma: Tomanek
and Somero, 1999; Tomanek, 2002; 2008; mussels, Mytilus: Ioannou et al., 2009;
multiple species: Barua and Heckathron, 2004). Both hsp 38 and 70, for

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 110/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
example, showed increases when top shells Tegula brunnea (=genus Chlorostoma)
and T . funebralis were acclimated separately at 13 and 18oC (Tomanek and
Somero, 1999), which implied that the top shells are more readily protected from
extreme high thermal conditions when acclimated at higher temperatures.
Physiological systems may, therefore, be protected by increased levels of hsps,
which convey a greater ability to survive thermal stresses. This can also be seen
by the fact that the ABT of HR in T . brunnea and T . funebralis were higher when
the snails were acclimated at higher temperatures (Stenseng et al., 2005), thus,
enhancing an individual’s survival limits. The higher ABT of HR recorded when
organisms were acclimated to higher temperatures was also demonstrated by
Stillman (2000) where the LT50 of porcelain crabs Petrolisthes cinctipes, P .
eriomerus, and P . manimaculis increased after they had been acclimated at a
higher temperature. Acclimation-induced heat shock responses of
Echinolittorina species were, although, not investigated in the present study,
investigated by Marshall and colleagues (2011) who found that concentration of
hsp 70 was doubled in E . malaccana on reaching high temperatures, i.e. 50oC
after an initial stage of metabolic depression. This result implies E . malaccana’s
ability to induce heat-shock response even under a short heating period, and may
reflect acclimation-induced heat shock responses which may explain their higher
thermal tolerance in summer than winter.
In summary, the LT50 and ABT of HR of Echinolittorina malaccana, E . radiata
and E . vidua in Hong Kong displayed a distinct pattern in relation to their vertical
distribution. Variations in the activities of MDH and LDH for the littorinids,

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 111/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
however, were less clear and require further in vitro experiments to confirm the
present results. In addition, activities of more enzymes are suggested to be
examined to acquire a better understanding of the aerobic and anaerobic scope of
energy metabolism of the Echinolittorina species in relation to their distribution
patterns. Seasonal variation of LT50 and ABT of HR for Echinolittorina species
were also detected in the present study, which, in general, were higher in summer
than winter. These variations are unlikely to contribute to the seasonal
down-shore migration pattern commonly observed in these littorinids, which is
probably related to other factors, such as spawning and food availability. The
cause of the variation may possibly be brought by the ability of the littorinids to
acquire acclimation-induced heat shock responses, which enhance the thermal
tolerance of physiological systems of individuals, and thus their survival, although
there is currently little evidence for this, and this is an area for further
investigation.
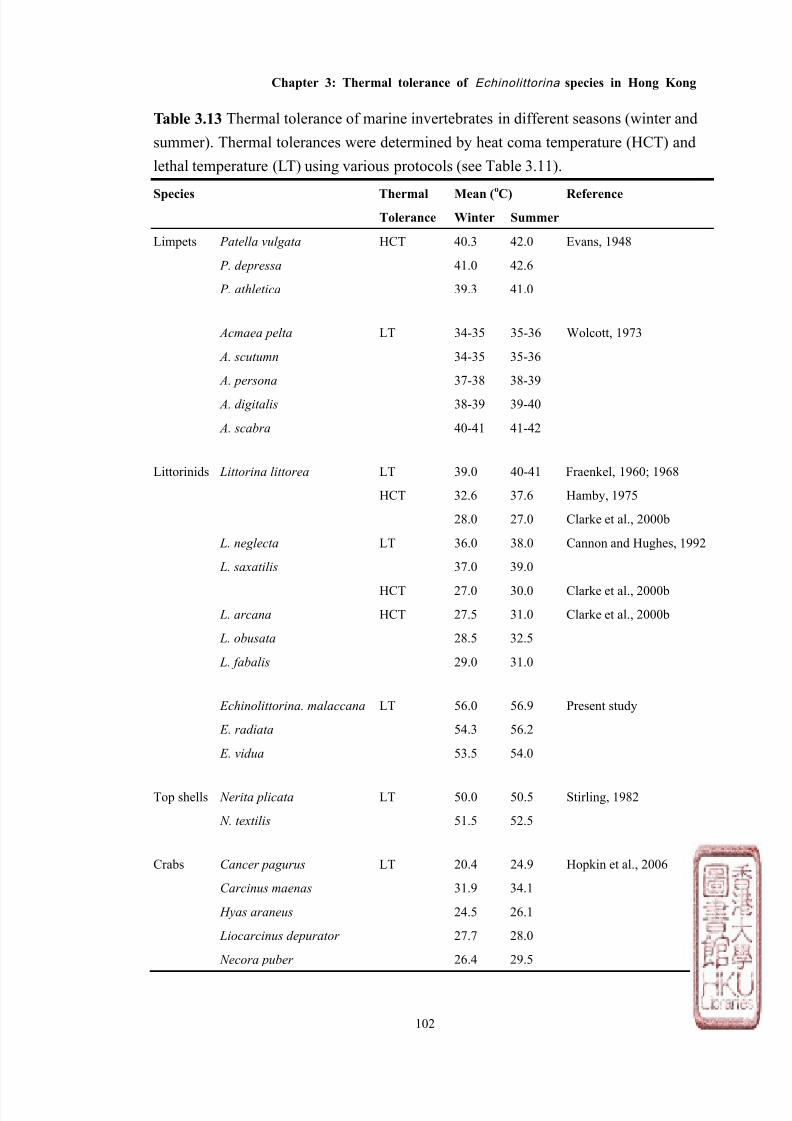
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 112/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
Table 3.13 Thermal tolerance of marine invertebrates in different seasons (winter and
summer). Thermal tolerances were determined by heat coma temperature (HCT) and
lethal temperature (LT) using various protocols (see Table 3.11).
Species Thermal
Tolerance
Mean (o
C) Reference
Winter Summer
Limpets Patella vulgata HCT 40.3 42.0 Evans, 1948
P. depressa 41.0 42.6
P. athletica 39.3 41.0
Acmaea pelta LT 34-35 35-36 Wolcott, 1973
A. scutumn 34-35 35-36
A. persona 37-38 38-39
A. digitalis 38-39 39-40
A. scabra 40-41 41-42
Littorinids Littorina littorea LT 39.0 40-41 Fraenkel, 1960; 1968
HCT 32.6 37.6 Hamby, 1975
28.0 27.0 Clarke et al., 2000b
L. neglecta LT 36.0 38.0 Cannon and Hughes, 1992
L. saxatilis 37.0 39.0
HCT 27.0 30.0 Clarke et al., 2000b
L. arcana HCT 27.5 31.0 Clarke et al., 2000b
L. obusata 28.5 32.5
L. fabalis 29.0 31.0
Echinolittorina. malaccana LT 56.0 56.9 Present study
E. radiata 54.3 56.2
E. vidua 53.5 54.0
Top shells Nerita plicata LT 50.0 50.5 Stirling, 1982
N. textilis 51.5 52.5
Crabs Cancer pagurus LT 20.4 24.9 Hopkin et al., 2006
Carcinus maenas 31.9 34.1
Hyas araneus 24.5 26.1
Liocarcinus depurator 27.7 28.0
Necora puber 26.4 29.5
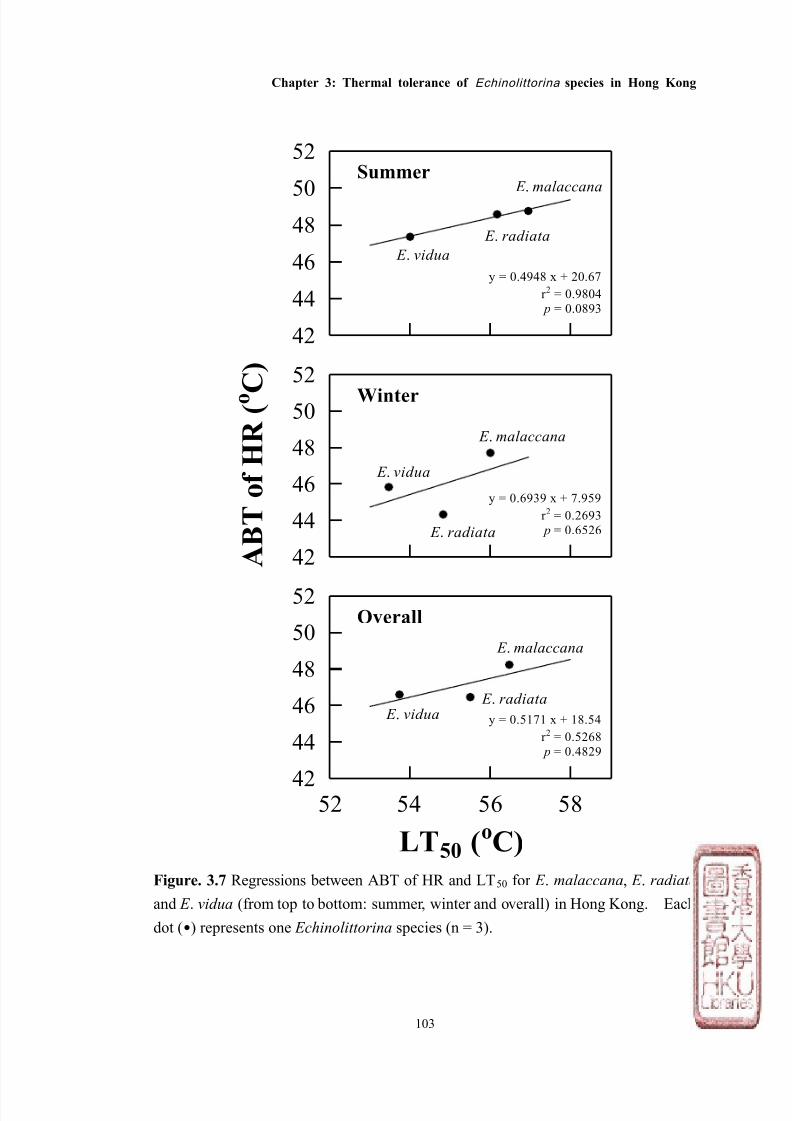
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 113/150
Chapter 3: Thermal tolerance of Echinolittorina species in Hong Kong
42
44
46
4850
52
E . vidua
E . radiata
E . malaccana
y = 0.4948 x + 20.67
r 2 = 0.9804
p = 0.0893
42
44
46
48
50
52
E . vidua
E . radiata
E . malaccana
y = 0.6939 x + 7.959
r 2 = 0.2693
p = 0.6526
42
44
46
48
50
52
E . vidua E . radiata
E . malaccana
y = 0.5171 x + 18.54
r 2 = 0.5268
p = 0.4829
54 56 5852
A B T o f H R ( o C
)
Summer
Winter
Overall
LT50 (oC)
Figure. 3.7 Regressions between ABT of HR and LT50 for E . malaccana, E . radiata
and E . vidua (from top to bottom: summer, winter and overall) in Hong Kong. Each
dot (•) represents one Echinolittorina species (n = 3).

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 114/150
CHAPTER 4:
GENERAL DISCUSSION
4.1 DISCUSSION 105
Vertical and seasonal distribution of Echinolittorina species
in Hong Kong
Thermal tolerance of Echinolittorina species and their
seasonal variation in Hong Kong
Present study: impli cations for fu ture investigations
Conclusions

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 115/150
Chapter 4: General discussion
4.1 DISCUSSION
Vertical and seasonal distribution of Echinolittorina species in Hong Kong
On two moderately exposed shores, i.e. Stanley and South Bay, in Hong Kong,
Echinolittorina malaccana, E . radiata and E . vidua, showed a consistent vertical
pattern and were located between 1.75 to 3.25m above C.D. (Chapter 2). E .
malaccana was found highest on the high shore (mean height ~2.76m above C.D.),
and overlapped with E . radiata which dominated ~ 0.3m lower down the shore
(~2.46m above C.D.) than E . malaccana, and E . vidua which was found the
lowest (~1.96m above C.D.) among the three species, living at the edge between
high and mid shore. Through field-manipulation experiments, Yipp and
Dudgeon (1990) showed that vertical distributions of E.malaccana and E . radiata
did not vary with the existence of the other species, therefore suggesting that
competition between the species may not be the major factor determining the
differences in their vertical distributions. Predation is also unlikely to drive the
vertical distribution of Echinolittorina species on Hong Kong shores (Mak, 1996;
Stafford, 2002; Walters, 2002; Li, personal observation) as the density of their
potential predators, for example, the Pacific reef egret Egretta sacra, is not high
enough to affect their distribution. With limited competition and predation
pressure observed in Hong Kong (Stafford, 2002), it can be assumed that their
vertical distribution is, therefore, related to their tolerance to physical stresses
(Stirling, 1982; Yipp et al., 1990).
Each of the three Echinolittorina species was also found lower on the shore by ~

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 116/150
Chapter 4: General discussion
0.25-0.5m in summer than winter. The seasonal variation in the littorinids’
distribution have been commonly proposed to enhance their reproductive success
by increasing their chance to spawn while immersed (e.g. Kojima, 1959;
Hannaford Ellis, 1985; Ohgaki, 1988a). However, this phenomena, as observed
in other Echinolittorina species, has also been suggested to be related to a
seasonal shift in food availability (e.g. Ito et al., 2002), seasonal variation in wave
action (e.g. Ohgaki, 1989), as well as physical stresses (e.g. Lambert and Farley,
1968).
In terms of abundance, Echinolittorina species were, in general, less abundant in
summer than winter on both shores. The lower abundance in summer was
possibly related to mortality due to thermal stress as suggested for other intertidal
species in Hong Kong (e.g. limpets: Firth and Williams, 2009; barnacles: Chan et
al., 2006) or dislodgement of littorinids during heavy rainfall (Ohgaki, 1988b;
Mak, 1996). Loss of snails due to tropical storms, as described in previous
studies (e.g. Kohn, 1980; Boulding and Van Alstyne, 1993), with the littorinids
being washed away by strong wave action is, however, thought to be unlikely in
the present study as there were few storms in 2010. Mortality may also be
increased in summer due to a decrease in food availability, as the biofilm is known
to be less abundant in summer months (Nagarkar and Williams, 1999). A
decrease in food supply, in tandem with increased physical stress, may lead to
increased mortality rates and hence reduced numbers of littorinids. In winter,
however, the increase in abundance of Echinolittorina species was associated with
the recruitment period of the littorinids (Mak, 1996), with the exception of E .
malaccana, which was less abundant in winter than summer in South Bay, a factor

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 117/150
Chapter 4: General discussion
which is difficult to explain.
Since these Echinolittorina species experience relatively low negative biological
interactions, such as predation and competition, on Hong Kong shores (Mak, 1996;
Walters, 2002; Stafford, 2002; Li, personal observation), it seems logical to
assume that physical factors, e.g. temperature and water availability, may play
important roles in determining spatial and temporal variation in their vertical
distribution and abundance. The physiological tolerances of the littorinids to
temperature were, therefore, determined through laboratory based experiments
(Chapter 3) to try and establish possible correlations with their spatial and
temporal distribution in directing further investigations to understand larger-scale
spatial patterns as well as to predict species’ success under long-term, climate
change.
Thermal tolerance of Echinolittorina species and their seasonal variation in
Hong Kong
Thermal tolerance of Echinolittorina species was assessed using an integrated
approach at the organismic, physiological and biochemical level by lethal
temperature (LT50), Arrhenius breakpoint temperature of heart rate (ABT of HR)
and activities of malate dehydrogenase (MDH) and lactate dehydrogenase (LDH).
The LT50 and ABT of HR of E . malaccana, E . radiata and E . vidua, as in other
congeneric intertidal species (e.g. in top shells: Tomanek and Somero, 1999;
porcelain crabs: Stillman and Somero, 2000), displayed a distinct pattern in

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 118/150
Chapter 4: General discussion
relation to their vertical distribution (Chapter 3). In summer, for example, E .
malaccana which lived highest on the shore, had the greatest LT50 value (56.95oC,
see Table 3.10, Chapter 3) close to 0.8o
C higher than the slightly lower E . radiata
(56.17oC), while the lowest shore E . vidua had the lowest LT50 (54.01oC). The
LT50 pattern of Echinolittorina species in Hong Kong also matched with previous
studies on similar species in Hong Kong (e.g. Stirling, 1982) and Singapore (Lee
and Lim, 2009), regardless of differences in methodology used to calculate the
value (see Table 3.11) that make absolute comparison difficult (Clarke et al.,
2000a). Similar to LT50 that showed a positive correlation with their vertical
distribution, the ABT of HR was also higher in E . malaccana (48.2oC) than the
two lower shore E . radiata and E . vidua (46.5 and 46.6oC respectively) (see Table
3.6, Chapter 3), which implied that E . malaccana can maintain efficient oxygen
transportation for aerobic metabolism under higher environmental temperatures
than E . radiata and E . vidua. The weak, but positive relationship between ABT
of HR and LT50, (Figure 3.7, Chapter 3) suggest that thermal tolerance at the
physiological level may be linked to their survival limits under thermal stress, as
reviewed by Somero (2010) in other intertidal species (e.g. top shells Tegula (=
genus Chlorostoma) and porcelain crabs, Petrolisthes).
The activities of malate dehydrogenase (MDH) and lactate dehydrogenase (LDH)
did not show a clear pattern in relation to the vertical distribution patterns of the
Echinolittorina species in Hong Kong, which was in contrast to the clear patterns
seen for LT50 and ABT of HR. Activity of MDH in E . vidua, on the contrary,
was higher than the two upper shore species E . malaccana and E . radiata at all
sampling temperatures (Chapter 3). This result cannot confidently confirm that,

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 119/150
Chapter 4: General discussion
in general, E . vidua has the greatest aerobic capacity than the other two species,
however, the metabolic rate of E . vidua has been suggested to be higher than E .
malaccana as illustrated by the higher activity of glutamate oxaloacetate
transaminase (GOT, Lee and Lim, 2009) and differences in basal heart rates (see
Appendices 2.1 & 2.2) in E . vidua and E . malaccana. The higher metabolic rate
in E . vidua, however, does not give a direct explanation to explain their thermal
tolerance limits. In contrast to MDH, LDH activity was similar between the
three species littorinids, although E . malaccana had a slightly greater activity than
E . radiata and E . vidua (Chapter 3). This implies the possibility that E .
malaccana has a higher anaerobic capacity than the other two species, therefore,
when environmental temperatures reach close to their ABT of HR, it would still
be able to produce energy through anaerobic metabolism to maintain basal needs,
which may attribute to their greater thermal tolerance limits.
However, as suggested (Chapter 3), in vitro methods should be conducted to
confirm the current results on activity of MDH and LDH, for example, activities
of CS and NADP-IDH in L. saxatilis and L. obtusata were measured by both in
vivo and in vitro methods by Sokolova and Pörtner (2001), and both methods
showed decreases in activities of the enzymes. Further, detailed examination on
enzyme function, for example, by determining enzyme kinetics and stability and
by measuring resistance to denaturing at high temperatures (e.g. Dahlhoff and
Somero, 1993; Fields et al., 2006; Dong and Somero, 2009) is recommended, as
such approaches may also help in understanding the causes of the vertical
distribution pattern of these species at the biochemical level. As in these studies,
Dong and Somero (2009) for example, used congeneric limpets, Lottia species, to

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 120/150
Chapter 4: General discussion
show that enzyme resistance to denaturing of cMDH between species increased
with increasing shore levels. Such differences could be attributed to the structure
of cMDH, as for L. digitalis (lower shore) and L. austrodigitalis (higher shore), as
an extra hydrogen bond exists in the cMDH of L. austrodigitalis, and therefore,
contributes to a more stable conformation for enzymatic reactions. Activities of
more metabolic enzymes that are involved in aerobic and anaerobic metabolism,
such as citrate synthase, pyruvate kinase, phosphoenolpyruvate carboxykinase,
NADP-dependent isocitrate dehydrogenase, should also be tested (e.g. Childress
and Somero, 1979; Sokolova and Pörtner, 2001; Rinke and Lee, 2009; Ombres et
al., 2011), in order to acquire a more general picture of how their activities
contribute to an individual’s overall response to thermal stress.
Thermal tolerance, in terms of LT50 and ABT of HR, was also found to vary
seasonally for the Echinolittorina species (Chapter 3), in which both LT50 and
ABT of HR for individual species were higher in summer than winter by ~
0.5-1.5oC and ~ 1-4oC respectively (see Chapter 3). A higher thermal tolerance
in summer than winter has also been recorded in other intertidal species, such as
limpets, top shells and crabs (see Chapter 3, Table 3.13). This seasonal variation
may not well be linked to the seasonal distribution patterns of the Echinolittorina
species (Chapter 2), however, the variation in thermal tolerance between seasons
showed that Echinolittorina species have the potential to acclimate to higher
environmental temperatures, which may lead to a higher survival limit to
increasing temperatures (as has been noted for other species, Stillman and Somero,
2002; Stenseng et al., 2005). The cause of this seasonal variation, although not
investigated in the present study, like many other intertidal species (e.g. Tomanek

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 121/150
Chapter 4: General discussion
and Somero, 1999; Ioannou et al., 2009), may be due to the littorinids acquiring
acclimation-induced heat shock responses to increase the production of heat shock
proteins, which may enhance the thermal tolerance of the physiological systems of
individuals, and thus their survival. There is currently little evidence for this,
and this, therefore, requires further investigation. One exception is the work by
Marshall and colleagues (2011) who showed that at higher temperatures, after a
period of metabolic depression, E . malaccana metabolism increased to induce the
production of heat shock proteins, so these Echinolittorina species do possess the
ability to produce these heat shock responses.
Present study: implications for future investigations
The present study has demonstrated a positive relationship between the vertical
distributions of Echinolittorina species with their thermal tolerance on two
moderately exposed shores in Hong Kong. Such results may, therefore, be able
to direct future investigations on larger-scale spatial patterns (e.g. between local
shores, latitudes). Latitudinal species distributions, for example, have been
hypothesized to give similar trends, in which, thermal tolerance of intertidal
species tends to increase with decreasing latitude. Although such trends between
thermal tolerance and latitude have been shown in previous studies (e.g.
McMahon, 2001; Stillman, 2002), they are often, however, not linear with latitude
in some intertidal species (in littorinids: Clarke et al., 2001; Lee and Boulding,
2010; dog whelks: Sorte et al., 2011), with thermal tolerance of species sometimes
being found to be greater at a higher latitude. Such unique places along the
latitudinal gradient were termed ‘hot spot’s by Helmuth and colleagues (2002), as

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 122/150
Chapter 4: General discussion
they found that environmental temperatures experienced by the mussel, Mytilus
californianus, were unexpectedly higher in some higher latitude sites than at
places at lower latitudes due to local variation in the timing of low tides at these
‘hot spot’s. Such observations, therefore, lead to the question of whether the
patterns of thermal responses seen in the Echinolittorina species from Hong Kong
shores can be applied over wider latitudinal scales. Any abnormality observed in
the patterns may imply that ‘hot spot’s exist, not only at a latitudinal scale, but
also at a local (within Hong Kong) spatial scale. If such phenomenon could be
observed locally, it might help to understand the possible environmental
conditions which may cause these ‘hot spot’s other than being due to variation in
the timing of low tides. Based on the preliminary studies, for example, on
spatial variation of LT50, the littorinids showed little spatial variation between
different shores in Hong Kong (see Chapter 3, Figure 3.1). This may not hold
true over larger spatial gradients, if thermal tolerance is solely determined by
genetics, as E. malaccana is thought to have a number of clades distributed over
various latitudes, and there is some question as to whether E . vidua also varies
genetically along its latitudinal distribution range (see Reid, 2007 for review).
To give possible clues to answer this question, potential factors, for example,
genetic variation (Kuo and Sanford, 2009), response to wave exposure (Davenport
and Davenport, 2005), and local environmental factors such as the timing of low
tides (Helmuth et al., 2002), all of which may drive variations in thermal tolerance
of Echinolittorina species, should be considered and studied.
The fundamental goal of this study was to understand thermal tolerance of
Echinolittorina species in relation to their vertical and seasonal distribution. A

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 123/150
Chapter 4: General discussion
secondary purpose was to provide basic information on species distribution limits
and acclimation potential of these species. Such information will allow
predictions of the littorinid’s response to changing environmental temperatures
such as under future climate change scenarios (see Somero, 2002, 2010 for
reviews). In his reviews, Somero suggested that warm-adapted species with
narrow distribution ranges, with little genetic variation or capacity to acclimate
would be most vulnerable towards climate warming; while species with wide
distribution range and with both genetic variability and phenotypical plasticity
would be less vulnerable. Understanding which species will be ‘winners’ or
‘losers’, he suggested, would enable prediction of future changes in species
distribution patterns and hence community structure.
In terms of acclimation ability, from the present study, E . radiata had the greatest
ability (Chapter 3) and are, therefore, more likely to be able to cope with
environmental changes than E . malaccana and E . vidua. With the little
information available, however, the present study alone cannot help conclude
which of the three Echinolittorina species may be ‘winner s’ or ‘loser s’ ( sensu
Somero, 2010). Further investigations and detailed examination of the littorinids’
phylogeny related to their thermal tolerance, as well as their phenotypic potentials
over a longer time scale are therefore required to predict species’ success under
climate change scenarios.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 124/150
Chapter 4: General discussion
Conclusions
The positive correlation between thermal tolerance of Echinolittorina species and
their vertical distribution on two moderately exposed shores in Hong Kong has
illustrated that temperature, as a physical factor, may play an important role in
determining the vertical distribution of these species. Thermal tolerance of
Echinolittorina species was also found to vary seasonally in the present study,
suggesting their potential to acclimate to higher environmental temperatures.
These results provide a basic understanding on small-scale vertical and seasonal
patterns in thermal tolerance of Echinolittorina species which, linked to their
larger-scale spatial distribution patterns, may help predict which species will be
successful under long-term changes in the environment.

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 125/150
115
LIST OF REFERENCES
Addo-Bediako A, Chown SL, Gaston KJ (2000) Thermal tolerance, climatic
variability and latitude. Proc R Soc B 267:739-745
Angilletta MJ (2009) Thermal adaptation: a theoretical and empirical synthesis.
Oxford University Press, New York, USA
Apps RF, Chen TY (1973) Sea waves at Waglan Island, Hong Kong. Royal
Observatory, Hong Kong Technical Note 36
Barua D, Heckathorn SA (2004) Acclimation of the temperature set-points of the
heat-shock response. J Therm Biol 29:185-193
Begon M, Harper JL, Townsend CR (1996) Ecology: individuals, populations, and
communities. Blackwell Science
Boulding EG, Van Alstyne KL (1993) Mechanisms of differential survival and growth
of two species of Littorina on wave-exposed and on protected shores. J Exp
Mar Biol Ecol 169:139-166
Brown JH (1984) On the relationship between abundance and distribution of species.
Am Nat 124: 255-279
Braby CE, Somero GN (2006) Following the heart: temperature and salinity effects
on heart rate in native and invasive species of blue mussels (genus Mytilus). J
Exp Biol 209:2554-2566
Branch GM (1975) Mechanisms reducing intraspecific competition in Patella spp.:
migration, differentiation and territorial behaviour. J Anim Ecol 44:575-600

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 126/150
List of references
116
Britton JC (1992) Evaporative water loss, behaviour during emersion, and upper
thermal tolerance limits in seven species of eulittoral-fringe Littorinidae
(Mollusca: Gastropoda) from Jamaica. In: Grahame J., Mill PJ, Reid DG
(eds.), Proceedings of the Third International Symposium on Littorinid
Biology. The Malacological Society of London, London, UK, pp. 69 – 83
Britton JC, McMahon RF (1990) The relationship between vertical distribution,
evaporative water loss rate, behaviour, and some morphometric parameters in
four species of rocky intertidal gastropods from Hong Kong. In: Morton B (ed)
Proceedings of the Second International Marine Biological Workshop:
The Marine Flora and Fauna of Hong Kong and Southern China II. Hong
Kong University Press, Hong Kong, pp 1153-1171
Broekhuysen GJ (1940) A preliminary investigation of the importance of desiccation,
temperature, and salinity as factors controlling the vertical distribution of
certain intertidal marine gastropods in False Bay, South Africa. Trans R Soc S
Afr 28:255-292
Cannon JP, Hughes RN (1992) Resistance to environmental stressors in Littorina
saxatilis (Olivi) and L. neglecta (Bean). In: Grahame J, Mill PJ, and Reid DG
(eds) Proceedings of the Third International Symposium on Littorinid
Biology: The Malacological Society of London, London, UK, pp 61-68
Castenholz RW (1961) An experimental study of the vertical distribution of littoral
marine diatoms. Limnol Oceanog 8:450-462
Castenholz RW (1964) The effect of grazing on marine littoral diatom populations.
Ecology 42: 783-794

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 127/150
List of references
117
Catesby SM, McKillup SC (1998) The importance of crevices to the intertidal snail
Littoraria articulata (Philippi) in a tropical mangrove forest. Hydrobiologia
367:131-138
Chan BKK, Morritt D, De Pirro M, Leung KMY, Williams GA (2006) Summer
mortality: effects on the distribution and abundance of the acorn barnacle
Tetraclita japonica on tropical shores. Mar Ecol Prog Ser 328:195-204
Chelazzi G, Williams GA, Gray DG (1999) Field and laboratory measurement of
heart rate in a tropical limpet, Cellana grata. J Mar Biol Assoc UK 79:749-
751
Childress JJ, Somero GN (1979) Depth-related enzymic activities in muscle, brain
and heart of deep-living pelagic marine teleosts. Mar Biol 52:273-283
Clarke AP, Mill PJ, Grahame J (2000a) The nature of heat coma in Littorina littorea
(Mollusca: Gastropoda). Mar Biol 137:447-451
Clarke AP, Mill PJ, Grahame J, McMahon RF (2000b) Geographical variation in heat
coma temperatures in Littorina species (Mollusca: Gastropoda). J Mar Biol
Ass UK 80:855-863
Cleland JD, McMahon RF (1990) Upper thermal limit of nine intertidal gastropod
species from a Hong Kong rocky shore in relation to vertical distribution and
desiccation associated with evaporative cooling. In: Morton B (ed)
Proceedings of the Second International Marine Biological Workshop: The
Marine Flora and Fauna of Hong Kong and Southern China II. Hong Kong
University Press, Hong Kong, pp 1141-1152
Cochran WG (1951) Testing a linear relation among variances. Biometrics 7: 17-32

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 128/150
List of references
118
Connell JH (1961a) Effects of competition, predation by Thais lapillus, and other
factors on natural populations of the barnacle Balanus balanoides. Ecol
Monogr 31:61-104
Connell JH (1961b) The influence of interspecific competition and other factors on
the distribution of the barnacle Chthamalus stellatus. Ecology 42:710-723
Dahlhoff EP, Somero GN (1993) Kinetic and structural adaptations of cytoplasmic
malate dehydrogenases of eastern Pacific abalone (genus Haliotis) from
different thermal habitats: biochemical correlates of biogeographical
patterning. J Exp Biol 185:137-150
Dahlhoff EP (2004) Biochemical indicators of stress and metabolism: applications for
marine ecological studies. Annu Rev Physiol 66:183-207
Dahlhoff EP, Stillman JH, Menge BA (2002) Physiological community ecology:
variation in metabolic activity of ecologically important rocky intertidal
invertebrates along environmental gradients. Integr Comp Biol 42:862-871
Davenport J, Davenport JL (2005) Effects of shore height, wave exposure and
geographical distance on thermal niche width of intertidal fauna. Mar Ecol
Prog Ser 292:41-50
Davenport J, Davenport JL (2007) Interaction of thermal tolerance and oxygen
availability in the eurythermal gastropods Littorina littorea and Nucella
lapillus. Mar Ecol Prog Ser 332:167-170
Dong YW, Williams GA (2011) Variations in cardiac performance and heat shock
protein expression to thermal stress in two differently zoned limpets on a
tropical rocky shore. Mar Biol 158:1223-1231

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 129/150
List of references
119
Dong YW, Somero GN (2009) Temperature adaptation of cytosolic malate
dehydrogenases of limpets (genus Lottia): differences in stability and function
due to minor changes in sequence correlate with biogeographic and vertical
distributions. J Exp Biol 212:169-177
Evans RG (1948) The lethal temperatures of some common british littoral molluscs. J
Anim Ecol 17:165-173
Feder ME, Hofmann GE (1999) Heat-shock proteins, molecular chaperones, and the
stress response: evolutionary and ecological physiology. Annu Rev Physiol
61:243-282
Fretter V, Graham A (1962) The Bristish prosobranch molluscs. Ray Society, London,
UK
Fields PA, Kim YS, Carpenter JF, Somero GN. (2002) Temperature adaptation in
Gillichthys (Teleost: Gobiidae) A4-lactate dehydrogenases: identical primary
structures produce subtly different conformations. J Exp Biol 205:1293-1303
Fields PA, Rudomin EL, Somero GN (2006) Temperature sensitivities of cytosolic
malate dehydrogenases from native and invasive species of marine mussels
(genus Mytilus): sequence-function linkages and correlations with
biogeographic distribution. J Exp Biol 209:656-667
Firth LB, Williams GA (2009) The influence of multiple environmental stressors on
the limpet Cellana toreuma during the summer monsoon season in Hong
Kong. J Exp Mar Biol Ecol 375:70-75
Foster MS (1990) Organization of macroalgal assemblages in the Northeast Pacific:
the assumption of homogeneity and the illusion of generality. Hydrobiologia
192: 21-33

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 130/150
List of references
120
Fraenkel G (1968) The heat resistance of intertidal snails at Bimini, Bahamas; Ocean
Springs, Mississippi; and Woods Hole, Massachusetts. Physiol Zool 41:1-13
Garrity SD (1984) Some adaptations of gastropods to physical stress on a tropical
rocky shore. Ecology 65:559-574
Goward CR, Nicholls DJ (1994) Malate dehydrogenase: A model for structure,
evolution, and catalysis. Protein Science 3:1883-1888
Habe T (1958) A study on the productivity of the Tanabe Bay (Part 1) VI. Zonal
arrangement of intertidal benthic animals in the Tanabe Bay. Records of
Oceanographic Works in Japan Special Vol. 2:43-49
Hannaford Ellis CJ (1985) The breeding migration of Littorina arcana Hannaford
Ellis, 1978 (Prosobranchia: Littorinidae). Zool Linn Soc 84:91-96
Harper KD, Williams GA (2001) Variation in abundance and distribution of the chiton
Acanthopleura japonica and associated molluscs on a seasonal, tropical,
rocky shore. J Zool 253:293-300
Hawkins SJ, Hartnoll RG (1983) Changes in a rocky shore community: An evaluation
of monitoring. Mar Env Res 9:131-181
Hawkins SJ, Hartnoll RG (1985) Factors determining the upper limits of intertidal
canopy-forming algae. Mar Ecol Prog Ser 20:265-271
Helmuth BST, Harley CDG, Halpin PM, O'Donnell M, Hofmann GE, Blanchette CA
(2002) Climate change and latitudinal patterns of intertidal thermal stress.
Science 298:1015-1017

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 131/150
List of references
121
Helmuth BST, Hofmann GE (2001) Microhabitats, thermal heterogeneity, and
patterns of physiological stress in the rocky intertidal zone. Biol Bull 201:374-
384
Hochachka PW, Somero GN (2002) Biochemical adaptation: mechanism and process
in physiological evolution. Oxford University Press, New York, USA
Hopkin RS, Qari S, Bowler K, Hyde D, Cuculescu M. (2006) Seasonal thermal
tolerance in marine Crustacea. J Exp Mar Biol Ecol 331:74-81
Huey RB, Kingsolver JG (1989) Evolution of thermal sensitivity of ectotherm
performance. Trends Ecol Evol 4:131-135
Hunter RD and Russel-Hunter WD (1983) Bioenergetic and community changes in
intertidal aufwuchs grazed by Littorina littorea. Ecology 64: 761-769
Hutchins LW (1947) The bases for temperature zonation in geographical distribution.
Ecol Monogr 17:325-335
Hutchinson N, Williams GA (2003a) An assessment of variation in molluscan
grazing pressure on Hong Kong rocky shores. Mar Biol 142:495-507
Hutchinson N, Williams GA (2003b) Disturbance and subsequent recovery of mid-
shore assemblages on seasonal, tropical, rocky shores. Mar Ecol Prog Ser
249:25-38
Ioannou S, Anestis A, Pörtner HO, Michaelidis B (2009) Seasonal patterns of
metabolism and the heat shock response (HSR) in farmed mussels Mytilus
galloprovincialis. J Exp Mar Biol Ecol 381:136-144

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 132/150
List of references
122
Ito A, Ilano AS, Goshima S, Nakao S (2002) Seasonal and tidal-height variations in
body weight and radular length in Nodilittorina radiata (Eydoux & Souleyet,
1852). J Moll Stud 68:197-203
Jones H (1970) Hydrostatic pressures within the heart and pericardium of Patella
vulgata L. Comp Biochem Physiol 34:263-272
Kaehler S, Williams GA (1996) Distribution of algae on tropical rocky shores: spatial
and temporal patterns of non-coralline encrusting algae in Hong Kong. Mar
Biol 125:177-187
Krebs CJ (2009) Ecology: the experimental analysis of distribution and abundance (6
ed.) Pearson Benjamin Cummings, San Francisco, USA
Kohn A (1980) Populations of tropical intertidal gastropods before and after a
typhoon. Micronesica 16:215-228
Kojima Y (1959) The relation between seasonal migration and spawning of a
periwinkle, Littorina brevicula (Philippi). Bull Mar. Biol. St. Asamushi 9:
183-186
Kuo E, Sanford E (2009) Geographic variation in the upper thermal limits of an
intertidal snail: implications for climate envelope models. Mar Ecol Prog Ser
388:137-146
Lambert TC, Farley J (1968) The effect of parasitism by the trematode Cryptocotyle
lingua (Creplin) on zonation and winter migration of the common periwinkle,
Littorina littorea (L.). Can J Zool 46:1139-1147

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 133/150
List of references
123
Lee HJ, Boulding EG. (2010) Latitudinal clines in body size, but not in thermal
tolerance or heat-shock cognate 70 ( HSC70), in the highly-dispersing
intertidal gastropod Littorina keenae (Gastropoda: Littorinidae). Biol J Linn
Soc 100:494-505
Lee SL, Lim SSL (2009) Vertical zonation and heat tolerance of three littorinid
gastropods on a rocky shore at Tanjung Chek Jawa, Singapore. Raff Bull Zool
57:551-560
Lewis JR (1954) Observations on a high-level population of limpets. J Anim Ecol
23:85-100
Lewis JR (1964) The ecology of rocky shores. English Universities Press.
Little C, Williams GA, Trowbridge C (2009) The biology of rocky shores. 2nd
edition.
Oxford University Press, New York, USA
Liu JH (1994) The ecology of the Hong Kong limpets Cellana grata (Gould, 1859)
and Patelloida pygmaea (Dunker, 1860)-distribution and population-dynamics.
J Moll Stud 60:55-67
Louw G (1993) Physiological animal ecology. 1st edition. Longman Scientific &
Technical, Hong Kong
Lubchenco J (1978) Plant species diversity in a marine intertidal community:
importance of herbivore food preference and algal competitive abilities.
Am Nat 112:23-39
Mak YM (1996) The ecology of the high-zoned littorinids, Nodilittorina trochoides,
N. radiata and N. vidua, on rocky shores in Hong Kong. PhD Thesis, The
University of Hong Kong, Hong Kong, China

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 134/150
List of references
124
Mak YM, Williams GA (1999) Littorinids control high intertidal biofilm abundance
on tropical, Hong Kong rocky shores. J Exp Mar Biol Ecol 233:81-94
Marshall DJ, Dong YW, McQuaid CD, Williams GA (2011) Thermal adaptation in
the intertidal snail Echinolittorina malaccana contradicts current theory by
revealing the crucial roles of resting metabolism. J Exp Biol 214:3649-3657
Marshall DJ, McQuaid CD (1992) Relationship between heart-rate and oxygen-
consumption in the intertidal limpets Patella granularis and Siphonaria
oculus. Comp Biochem Physiol A103:297-300
Marshall DJ, McQuaid CD (2011) Warming reduces metabolic rate in marine snails:
adaptation to fluctuating high temperatures challenges the metabolic theory of
ecology. Proc R Soc B 278:281-288
Marshall DJ, McQuaid CD, Williams GA (2010) Non-climatic thermal adaptation:
implications for species' responses to climate warming. Biol Lett 6:669-673
Mathews CK., van Holde KE, Ahern KG (2000) Biochemistry. 3rd
edition. Prentice
Hall, San Francisco, USA
McCarthy JJ (2001) Climate Change 2001: Impacts, Adaptation, and Vulnerability :
Contribution of Working Group II to the Third Assessment Report of the
Intergovernmental Panel on Climate Change, Intergovernmental Panel on
Climate Change. Working Group II. Cambridge University Press, Cambridge,
UK
McMahon R, Russell-Hunter W, Aldridge D (1995) Lack of metabolic temperature
compensation in the intertidal gastropods, Littorina saxatilis (Olivi) and L.
obtusata (L.). Hydrobiologia 309:89-100

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 135/150
List of references
125
McMahon RF (1990) Thermal tolerance, evaporative water loss, air-water oxygen
consumption and zonation of intertidal prosobranchs: a new synthesis.
Hydrobiologia 193:241-260
McMahon RF (2001) Acute thermal tolerance in intertidal gastropods relative to
latitude, superfamily, zonation and habitat with special emphasis on the
littorinoidea. J Shell Res 20: 459-467
McMahon RF, Britton JC (1985) The relationship between vertical distribution,
thermal tolerance, evaporative water loss rate, and behaviour on emergence in
six species of mangrove gastropods from Hong Kong. In: Morton B,
Dudgeon D (eds) Proceedings of the Second International Workshop on the
Malacofauna of Hong Kong and Southern China, Hong Kong, 1983.
Hong Kong University Press, Hong Kong, pp 565-582
McQuaid CD (1996a) Biology of the gastropod family Littorinidae. I. Evolutionary
Aspects. Oceanogr Mar Biol Annu Rev 34: 233-262
McQuaid CD (1996b) Biology of the gastropod family Littorinidae. II. Role in the
ecology of intertidal and shallow marine ecosystems. Oceanogr Mar Biol
Annu Rev 34: 263-301
McQuaid CD, Scherman PA (1988) Thermal stress in a high shore intertidal
environment: morphological and behavioural adaptations of the gastropod
Littorina africana. In: Chelazzi G, Vannini M (eds) Behavioural Adaptation to
Intertidal Life. Plenum Press, New York, USA, pp 213-224
Menge BA, Branch GM (2001) Rocky intertidal communities. In Bertness MD,
Gaines SD, Hay M (eds) Marine community ecology. Sinauer Associates, pp
221-251

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 136/150
List of references
126
Menge BA, Olson AM (1990) Role of scale and environmental factors in regulation
of community structure. Trends Ecol Evol 5:52-57
Moore HB (1972) Aspects of Stress in the tropical marine environment. Adv Mar
Biol 10: 217-269
Morton B, Harper E (1995) An introduction to the Cape d’Aguilar Marine Reserve,
Hong Kong. Hong Kong University Press, Hong Kong
Morton B, Morton J (1983) The seashore ecology of Hong Kong. Hong Kong
University Press, Hong Kong
Morton B, Williams GA, Lee SY (1996) The benthic marine ecology of Hong Kong:
a dwindling heritage? In: Civil Engineering Department (ed) Coastal
Infrastructure Development in Hong Kong: a Review. Hong Kong
Government, Hong Kong, pp 233-267
Muñoz JLP, G Randall Finke, PA Camus, F Bozinovic (2005) Thermoregulatory
behavior, heat gain and thermal tolerance in the periwinkle Echinolittorina
peruviana in central Chile. Comp Biochem Physiol A 142:92-98
Newell RC (1979) Biology of intertidal animals. Marine Ecological Surveys
Newell RC, Pye VI (1970) Seasonal changes in the effect of temperature on the
oxygen consumption of the winkle Littorina littorea (L.) and the mussel
Mytilus edulis (L.) Comp Biochem Physiol 34:367-383
Newell RC, Pye VI, Ahsanull M (1971) Effect of thermal acclimation on heat
tolerance of intertidal prosobranchs Littorina littorea (L) and Monodonta
lineata (Da Costa). J Exp Biol 54:525-533

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 137/150
List of references
127
Ng SSJ (2007) Resource partitioning and coexistence of molluscan grazers on Hong
Kong rocky shores. PhD Thesis, The University of Hong Kong, Hong Kong,
China
Nicotri ME (1977) Grazing effects of four marine intertidal herbivores on the
microflora. Ecology 58: 1020-1032
Norton TA, Hawkins SJ, Manley NL, Williams GA, Watson DC (1990) Scraping a
living: a review of littorinid grazing. Hydrobiologia 193: 117-138
Nybakken JW, Bertness MD (2005) Marine biology: an ecological approach.
Pearson/Benjamin Cummings, San Francisco, USA
Ohgaki S (1985) Distribution of the family Littorinidae (Gastropoda) on Hong Kong
rocky shores. In: Morton B, Dudgeon D (eds) The Malacofauna of Hong
Kong and Southern China II. Hong Kong University Press, Hong Kong, pp
457-464
Ohgaki S (1988a) Rain and the distribution of Nodilittorina exigua (Dunker)
(Gastropoda: Littorinidae). J Exp Mar Biol Ecol 122:213-223
Ohgaki S (1988b) Vertical migration and spawning in Nodilittorina exigua
(Gastropoda: Littorinidae). J Ethol 6:33-38
Ohgaki S (1989) Vertical movement of the littoral fringe periwinkle Nodilittorina
exigua in relation to wave height. Mar Biol 100:443-448
Ombres EH, Donnelly J, Clarke ME, Harms JH, Torres JJ (2011) Aerobic and
anaerobic enzyme assays in Southern California rockfish: proxies for
physiological and ecological data. J Exp Mar Biol Ecol 399:201-207

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 138/150
List of references
128
Paine RT (1974) Intertidal community structure - experimental studies on relationship
between a dominant competitor and its principal predator. Oecologia 15:93-
120
Parmesan C (2006) Ecological and evolutionary responses to recent climate change.
Annu Rev Ecol Evol Syst 37:637-669
Pickens PE (1965) Heart rate of mussels as a function of latitude, intertidal height,
and acclimation temperature. Physiol Zool 38:390-405
Pilkington MC (1971) Eggs, larvae, and spawning in Melarapha cincta (Quoy &
Gaimard) and M . oliveri finlay (Littorinidae, Gastropoda). Aust J Mar Freshw
Res 22: 79-90
Pörtner HO (2001) Climate change and temperature-dependent biogeography: oxygen
limitation and thermal tolerance limits in animals. Naturwissenschaften
88:137-146
Pörtner, HO (2002) Climate variations and the physiological basis of temperature
dependent biogeography: systematic to molecular hierarchy of thermal
tolerance in animals. Comp Biochem Physiol A 132:739 – 761
Pörtner, HO (2010) Oxygen- and capacity-limitation of thermal tolerance: a matrix
for integrating climate-related stressor effects in marine ecosystems. J Exp
Biol 213:881-893
Pörtner HO, Peck L, Somero G (2007) Thermal limits and adaptation in marine
Antarctic ectotherms: an integrative view. Phil Trans R Soc B 362: 2233-2258

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 139/150
List of references
129
Raffaelli DG, Hawkins SJ (1996) Intertidal ecology. Chapman & Hall, London, UK
Reid DG (1986) The littorinid molluscs of mangrove forests in the Indo-Pacific
region: the genus Littoraria British Museum (Natural History), London, UK
Reid DG (1996) Systematics and evolution of Littorina. Ray Society, London, UK
Reid DG (2007) The genus Echinolittorina Habe, 1956 (Gastropoda: Littorinidae) in
the Indo-West Pacific Ocean. Zootaxa 1420:1-161
Rinke C, Lee R (2009) Pathways, activities and thermal stability of anaerobic and
aerobic enzymes in thermophilic vent paralvinellid worms. Mar Ecol Prog Ser
382:99-112
Sandison EE (1967) Respiratory response to temperature and temperature tolerance
of some intertidal gastropods. J Exp Mar Biol Ecol 1:271-281
Sanpanich, K., Wells FE, Chitramvong Y (2006) Effects of the 26 December 2004
tsunami on littorinid molluscs near Phuket, Thailand. J Moll Stud 72: 311-313
Santini G, De Pirro M, Chelazzi G (1999) In situ and laboratory assessment of heart
rate in a Mediterranean limpet using a noninvasive technique. Physiol
Biochem Zool 72:198-204
Schonbeck MW, Norton TA (1978) Factors controlling the upper limits of fucoid
algae on the shore. J Exp Mar Biol Ecol 31:303-313
Schonbeck MW, Norton TA (1980) Factors controlling the lower limits of fucoid
algae on the shore. J Exp Mar Biol Ecol 43:131-150

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 140/150
List of references
130
Simpfendörfer RW, Vial MV, López DA, Verdala M, González ML (1995)
Relationship between the aerobic and anaerobic metabolic capacities and the
vertical distribution of three intertidal sessile invertebrates: Jehlius cirratus
(Darwin) (Cirripedia), Perumytilus purpuratus (Lamarck) (Bivalvia) and
Mytilus chilensis (Hupé) (Bivalvia). Comp Biochem Physiol B 111:615-623
Sokolova IM, Pörtner HO (2001) Temperature effects on key metabolic enzymes in
Littorina saxatilis and L. obtusata from different latitudes and shore levels.
Mar Biol 139:113-126
Sokolova IM, Pörtner HO (2003) Metabolic plasticity and critical temperatures for
aerobic scope in a eurythermal marine invertebrate ( Littorina saxatilis,
Gastropoda: Littorinidae) from different latitudes. J Exp Biol 206:195-207
Somero GN (2002) Thermal Physiology and Vertical Zonation of Intertidal Animals:
Optima, Limits, and Costs of Living. Integr Comp Biol 42:780-789
Somero GN (2005) Linking biogeography to physiology: evolutionary and
acclimatory adjustments of thermal limits. Front Zool 2:1
Somero GN (2010) The physiology of climate change: how potentials for
acclimatization and genetic adaptation will determine 'winners' and 'losers'. J
Exp Biol 213:912-920
Sorte CJB, Jones SJ, Miller LP (2011) Geographic variation in temperature tolerance
as an indicator of potential population responses to climate change. J Exp Mar
Biol Ecol 400: 209-217
Southward A J (1958) Note on the temperature tolerances of some intertidal animals
in relation to environmental temperatures and geographical distribution. J Mar
Biol Ass UK 37:49-66

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 141/150
List of references
131
Spellerberg IF (1972) Temperature tolerances of Southeast Australian reptiles
examined in relation to reptile thermoregulatory behaviour and distribution.
Oecologia 9:23-46
Stafford R (2002) The role of environmental stress and physical and biological
interavtions on the ecology of high shore littorinids in a temperate and a
tropical region. PhD Thesis, University of Sunderland, UK
Stafford R, Davies MS (2005) Spatial patchiness of epilithic biofilm caused by
refuge-inhabiting high shore gastropods. Hydrobiologia 545: 279-287
Stenseng E, Braby CE, Somero GN (2005) Evolutionary and acclimation-induced
variation in the thermal limits of heart function in congeneric marine snails
(genus Tegula): implications for vertical zonation. Biol Bull 208:138-144
Stephenson TA, Stephenson A (1972) Life between tidemarks on rocky shores. WH
Freeman, San Francisco, USA
Stillman JH (2003) Acclimation capacity underlies susceptibility to climate change.
Science 301:65
Stillman JH, Reeb C (2001) Molecular phylogeny of eastern Pacific porcelain crabs,
genera Petrolisthes and Pachycheles, based on mtDNA sequence:
phylogeographic and systematic implications. Mol Phylogenet Evol 19:236-
245
Stillman JH, Somero GN (1996) Adaptation to temperature stress and aerial exposure
in congeneric species of intertidal porcelain crabs (genus Petrolisthes):
correlation of physiology, biochemistry and morphology with vertical
distribution. J Exp Biol 199:1845-1855

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 142/150
List of references
132
Stillman JH, Somero GN (2000) A comparative analysis of the upper thermal
tolerance limits of eastern Pacific porcelain crabs, genus Petrolisthes :
influences of latitude, vertical zonation, acclimation, and phylogeny. Physiol
Biochem Zool 73:200 - 208
Stirling HP (1982) The upper temperature tolerance of prosobranch gastropods of
rocky shores at Hong Kong and Dar Es Salaam, Tanzania. J Exp Mar Biol
Ecol 63:133-144
Sullivan KM, Somero GN (1983) Size- and diet-related variations in enzymic activity
and tissue composition in the sablefish, Anoplopoma fimbria. Biol Bull
164:315-326
Sunday JM, Bates AE, Dulvy NK (2010) Global analysis of thermal tolerance and
latitude in ectotherms. Proc R Soc B 278: 1823-1830
Takada Y (1996) Vertical migration during the life history of the intertidal gastropod
Monodonta labio on a boulder shore. Mar Ecol Prog Ser 130:117-123
Terblanche JS, Hoffmann AA, Mitchell KA, Rako L, le Roux PC, Chown SL (2011)
Ecologically relevant measures of tolerance to potentially lethal temperatures.
J Exp Biol 214:3713-3725
Thuesen EV, Childress JJ (1994) Oxygen consumption rates and metabolic enzyme
activities of oceanic California Medusae in relation to body size and habitat
depth. Biol Bull 187:84-98
Tilman D (1982) Resource Competition and Community Structure. Princeton
University Press, Princeton, USA

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 143/150
List of references
133
Tomanek L (2002) The heat-shock response: its variation, regulation and ecological
importance in intertidal gastropods (genus Tegula). Integr Comp Biol 42:797-
807
Tomanek L (2008) The importance of physiological limits in determining
biogeographical range shifts due to global climate change: the heat-shock
response. Physiol Biochem Zool 81:709-717
Tomanek L (2010) Variation in the heat shock response and its implication for
predicting the effect of global climate change on species' biogeographical
distribution ranges and metabolic costs. J Exp Biol 213:971-979
Tomanek L, Somero GN (1999) Evolutionary and acclimation-induced variation in
the heat-shock responses of congeneric marine snails (genus Tegula) from
different thermal habitats: implications for limits of thermotolerance and
biogeography. J Exp Biol 202:2925- 2936
Torres JJ, Somero GN (1988a) Metabolism, enzymic activities and cold adaptation in
Antarctic mesopelagic fishes. Mar Biol 98:169-180
Torres JJ, Somero GN (1988b) Vertical distribution and metabolism in Antarctic
mesopelagic fishes. Comp Biochem Physiol B 90:521-528
Trueman ER (1967) Activity and heart rate of bivalve molluscs in their natural habitat.
Nature 214:832-833
Tsuchiya M, Lirdwitayapasit T (1986) Distribution of intertidal animals on rocky
shores of the Sichang Islands, the Gulf of Thailand. Galaxea 5:15-25
Uglow RF, Williams GA (2001) The effect of emersion on ammonia efflux of three
Hong Kong Nodilittorina species. J Shell Res 20:489-493

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 144/150
List of references
134
Underwood AJ (1979) The ecology of intertidal gastropods. Adv Mar Biol 16:111-
210
Underwood AJ (1980) The effects of grazing by gastropods and physical factors on
the upper limits of distribution of intertidal macroalgae. Oecologia 46:201-
213
Underwood AJ (1981) Structure of a rocky intertidal community in New South Wales:
patterns of vertical distribution and seasonal changes. J Exp Mar Biol Ecol
51:57-85
Underwood AJ (1984) Vertical and seasonal patterns in competition for microalgae
between intertidal gastropods. Oecologia 64: 211-222
Underwood AJ (1997) Experiments in ecology: their logical design and interpretation
using analysis of variance. Cambridge University Press, Cambridge, UK
Underwood AJ, Chapman MG (1996) Scales of spatial patterns of distribution of
intertidal invertebrates. Oecologia 107:212-224
Underwood AJ, Chapman MG, Connell SD (2000) Observations in ecology: you
can’t make progress on processes without understanding the patterns. J Exp
Mar Biol Ecol 250:97-115
Underwood AJ, Denley EJ (1984) Paradigms, explanations, and generalizations
in models for the structure of intertidal communities on rocky shores. In:
Simberloff D, Strong DR, Abele L, Thistle AR (eds) Ecological communities:
Conceptual issues and the evidence. Princeton University Press, Princeton,
USA

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 145/150
List of references
135
Underwood AJ, Fairweather PG (1989) Supply-side ecology and benthic marine
assemblages. Trends Ecol Evol 4: 16-20
Urban HJ (1994) Upper temperature tolerance of ten bivalve species off Peru and
Chile related to El Nino. Mar Ecol Prog Ser 107:139-145
Vermeij GJ (1971) Temperature relationships of some tropical Pacific intertidal
gastropods. Mar Biol 10:308-314
Vermeij GJ (1973) Morphological patterns in high-intertidal gastropods: adaptive
strategies and their limitations. Mar Biol 20:319-346
Walters A (2002) The zonation of three Hong Kong high shore littorinids:
Nodilittorina trochoides , Nodilittorina radiata, and Nodilittorina vidua.
Senior thesis, Princeton University, USA
Wellborn GA, Skelly DK, Werner EE (1996) Mechanisms creating community
structure across a freshwater habitat gradient. Annu Rev Ecol Syst 27: 337-
363
Wethey DS (1984) Sun and shade mediate competition in the barnacles
Chthamalusand semibalanus: a field experiment. Biol Bull 167:176-185
Williams GA (1994a) Grazing by high-shore littorinids on a moderately exposed
tropical rocky shore. In: Morton B (ed) The Malacofauna of Hong Kong and
Southern China III. Hong Kong University Press, Hong Kong, pp 379-390
Williams GA (1994b) The relationship between shade and molluscan grazing in
structuring communities on a moderately-exposed tropical rocky shore. J Exp
Mar Biol Ecol 178:79-95

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 146/150
List of references
136
Williams GA, De Pirro M, Cartwright S, Khangura K, Ng WC, Leung PTY, Morritt D
(2010) Come rain or shine: the combined effects of physical stresses on
physiological and protein-level responses of an intertidal limpet in the
monsoonal tropics. Funct Ecol 25:101-110
Williams GA, Morritt D (1995) Habitat partitioning and thermal tolerance in a
tropical limpet, Cellana grata. Mar Ecol Prog Ser 124:89-103
Williams GA, De Pirro M, Leung KMY,, Morritt D (2005) Physiological responses to
heat stress on a tropical shore: the benefits of mushrooming behaviour in the
limpet Cellana grata. Mar Ecol Prog Ser 292:213-224
Williams IC, Ellis C (1975) Movements of the common periwinkle, Littorina littorea
(L.), on the Yorkshire coast in winter and the influence of infection with larval
Digenea. J Exp Mar Biol Ecol 17:47-58
Wolcott TG (1973) Physiological ecology and intertidal zonation in limpets ( Acmaea):
a critical look at 'limiting factors'. Biol Bull 145:389-422
Wu RSS, Lam PKS (1997) Glucose-6-phosphate dehydrogenase and lactate
dehydrogenase in the green-lipped mussel ( Perna viridis): Possible
biomarkers for hypoxia in the marine environment. Water Research 31:2797-
2801
Yang TH, Somero GN (1993) Effects of feeding and food deprivation on oxygen
consumption, muscle protein concentration and activities of energy
metabolism enzymes in muscle and brain of shallow-living (Scorpaena
guttata) and deep-living (Sebastolobus alascanus) scorpaenid fishes. J Exp
Biol 181:213-232

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 147/150
List of references
137
Yipp MW, Dudgeon D (1990) Does competition determine the vertical zonation of
high-intertidal Littorinidae (Gastropoda) in Hong Kong? In: Morton B (ed)
Proceedings of the Second International Marine Biological Workshop:
The Marine Flora and Fauna of Hong Kong and Southern China II. Hong
Kong University Press, Hong Kong, pp 795-802
Yipp MW, Dudgeon D, Cha MW (1990) Respiratory adaptations and survival of three
high-intertidal littorinids (Gastropoda) from Hong Kong rocky shores. In:
Morton B (ed) Proceedings of the Second International Marine Biological
Workshop: The Marine Flora and Fauna of Hong Kong and Southern China II.
Hong Kong University Press, Hong Kong, pp 1041-1054

8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 148/150
138
APPENDIX 1
NAME USED FOR ECHI NOLITTORINA MALACCANA, E . RADIATA AND E . VIDUA
IN PREVIOUS LITERATURE
Scientific name Reference
Current Past
Echinolittorina malaccana Nodilittorina pyramidalis Fraenkel, 1966
Stirling, 1982
Ohgaki, 1985
Yipp and Dudgeon, 1990
Yipp et al., 1990
Cleland and McMahon, 1990
McMahon, 2001
Nodilittorina trochoides Williams, 1994a
Mak, 1996
Walters, 2002
Echinolittorina radiata Nodilittorina exigua Ohgaki, 1985, 1988a,b, 1989
Yipp and Dudgeon, 1990
Yipp et al., 1990
Ohgaki, 1988, 1989
McMahon, 2001
Nodilittorina millegrana Stirling, 1982
Nodilittorina radiata Williams, 1994a
Mak, 1996
Walters, 2002
Echinolittorina vidua Nodilittorina millegrana Ohgaki, 1985
Nodilittorina vidua Williams, 1994a
Mak, 1996
Walters, 2002
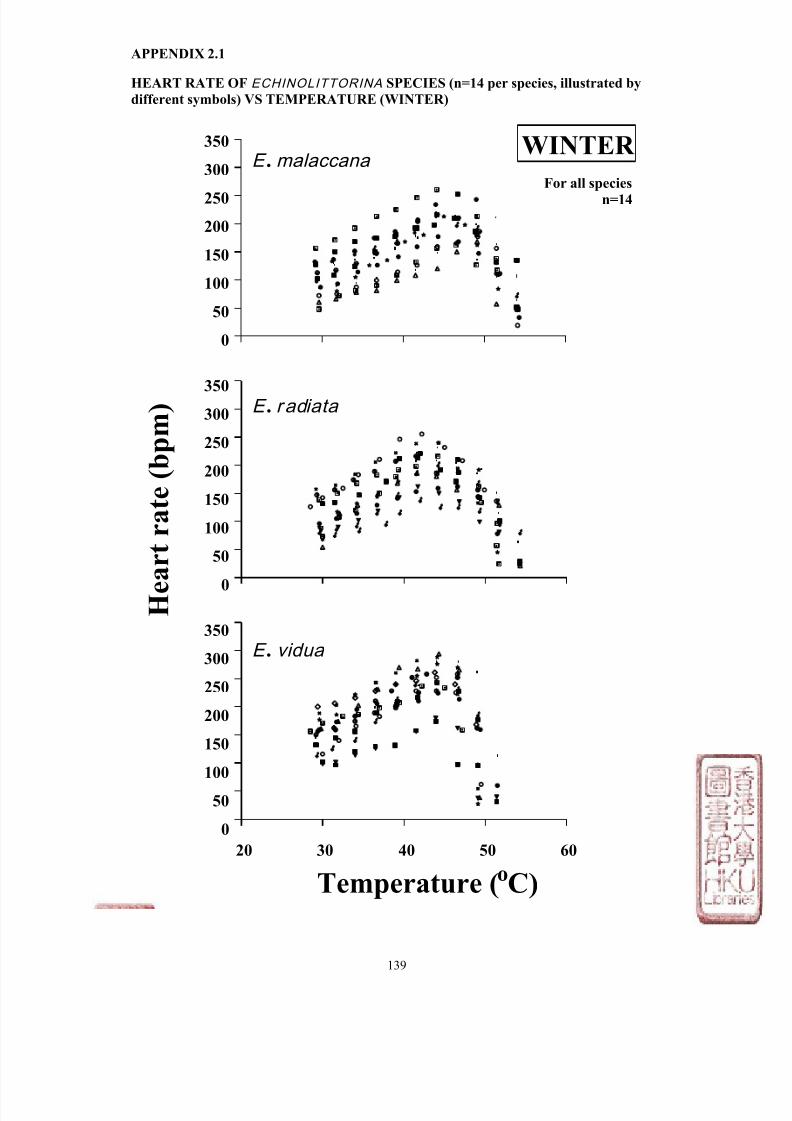
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 149/150
139
APPENDIX 2.1
HEART RATE OF ECHINOLITTORINA SPECIES (n=14 per species, illustrated by
different symbols) VS TEMPERATURE (WINTER)
0
50
100
150
200
250
300
350
0
50
100
150
200
250
300350
0
50
100
150
200
250
300
350
H e
a r t r a t e ( b p m )
20 30 40 50 60
Temperature (oC)
E . malaccana
E . r adiata
E . vidua
WINTER
For all species
n=14
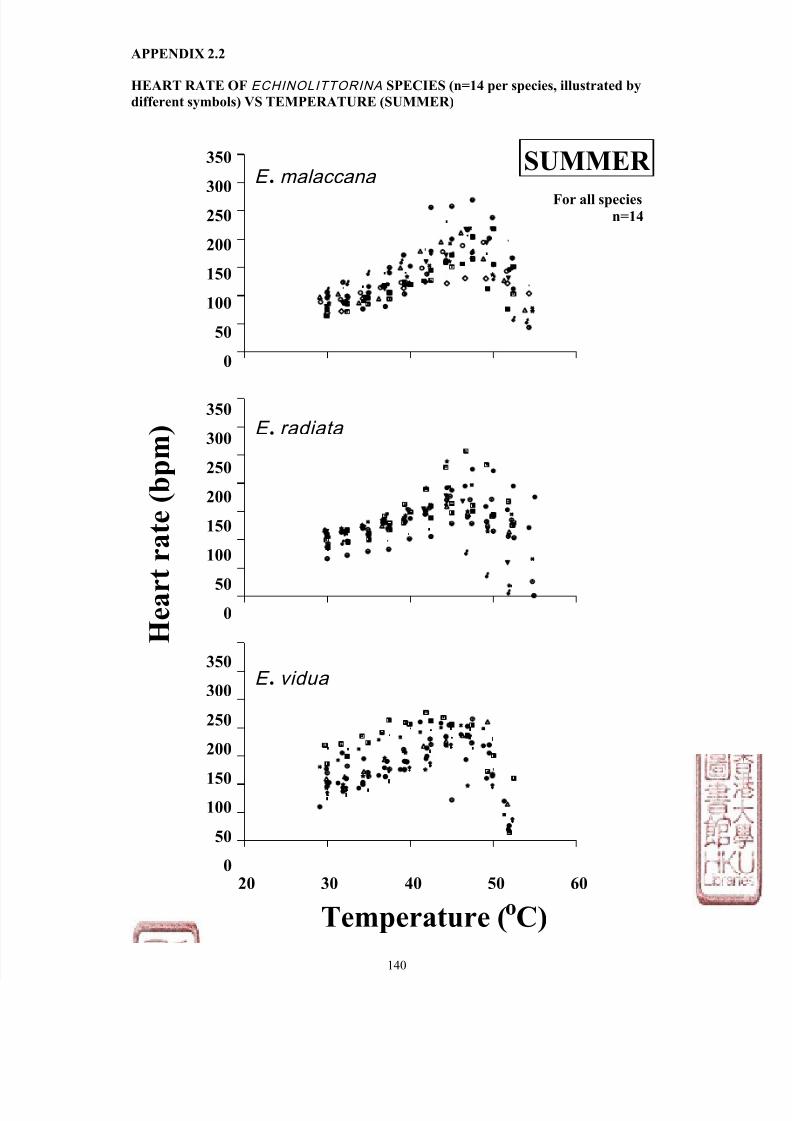
8/17/2019 FullText (15)
http://slidepdf.com/reader/full/fulltext-15 150/150
APPENDIX 2.2
HEART RATE OF ECHINOLITTORINA SPECIES (n=14 per species, illustrated by
different symbols) VS TEMPERATURE (SUMMER)
0
50
100
150
200
250
300
350
150
200
250
300
350
a t e ( b p m )
E . malaccana
E . radiata
SUMMER
For all speciesn=14





























