Embed Size (px)
Citation preview

Aus dem Zentrum für Allgemeine Molekularbiologie und Proteindesign der
Tierärztlichen Hochschule Hannover
und dem Zentrum für Pharmaforschung und medizinische Biotechnologie des
Fraunhofer Institutes für Toxikologie und Experimentelle Medizin
Genexpressions- und Metabolismusuntersuchungen der menschlichen
respiratorischen Nasenschleimhaut
INAUGURAL-DISSERTATION
zur Erlangung des Grades einer Doktorin der Veterinärmedizin
( Dr. med. vet. )
durch die Tierärztliche Hochschule Hannover
Vorgelegt von
Daniela Rahmel
aus Berlin
Hannover 2004

Wissenschaftliche Betreuung: Univ.-Prof. Dr. rer. nat. Bernd Otto
Univ.-Prof. Dr. rer. nat. Jürgen Borlak
1. Gutachter: Univ.-Prof. Dr. rer. nat. Jürgen Borlak
Univ.-Prof. Dr. rer. nat. Bernd Otto
2. Gutachter: Apl.-Prof. Dr. med. vet. Heiner Niemann
Tag der mündlichen Prüfung: 03. Juni 2004

Abkürzungsverzeichnis I
Abkürzungsverzeichnis
Acetyl-CoA Acetyl-CoenzymA
AFB1 Aflatoxin B1
AhR Aryl hydrocarbon-Rezeptor
AK Antikörper
AnH Anilin-Hydroxylase
APD Aminopyrin-N-Demethylase
AR Allergische Rhinitis
BeD Benzaldehyd-Dehydrogenase
bp Basenpaare
BROD Benzyloxyresorufin-O-Deethylase
BSA Bovines Serum Albumin
BzD Benzphetamin-N-Demethylase
bzw. beziehungsweise
CAR Constitutive Androstane Rezeptor
cDNA copy-DNA
CMT Cyclopentadienylmangantricarbonyl
CO Kohlenstoffmonoxid
CPM Counts per Minute
CYP Cytochrom P450-abhängige Monooxygenasen
DCBN 2,6-Dichlorobenzonitril
DCPIP Dichlorophenolindophenol
DEN Diethylnitrosamin
DENd Diethylnitrosamin-N-Deethylase

Abkürzungsverzeichnis II
DEPEC Diethylpyrocarbonat
DMNd Dimethylnitrosamin-N-Demethylase
DMSO Dimethylsulfoxid
DNA Desoxyribonukleinsäure
dNTP Desoxynukleosidtriphosphat
DPM Desintegrations per Minute
ECOD Ethoxycoumarin-O-Deethylase
EDTA Ethylendiamintetraessigsäure
EH Epoxid-Hydrolase
ELISA Enzyme-linked immunosorbant assay
EmD Ethylmorphin-N-Demethylase
ErD Erythromycin-N-Demethylase
EROD Ethoxyresorufin-O-Deethylase
FMO Flavinhaltige Monooxygenasen
FS Friederiken-Stift
GAPDH Glycerinaldehyd-3-phosphat-Dehydrogenase
GST Glutathion-S-Transferase
h Stunde
HMPA Hexamethylphosphoramid
HMPAd Hexamethylphosphoramid-N-Demethylase
HNEC human nasal epithelial cells
HPLC High Performance Liquid Chromatography
i.d.R. in der Regel
IgA ImmunglobulinA
IL Interleukin

Abkürzungsverzeichnis III
IR Immunoreaktivität
LE Loteprednol-Etabonat
LK Leberkontrolle
LXR Liver X Rezeptor
M Metabolit
MFO Mischfunktionelle Oxygenase
MHH Medizinische Hochschule Hannover
min Minuten
mRNA Messenger-Ribonukleinsäure
MROD Methoxyresorufin-O-Demethylase
MTBE Methyltertbutylether
NADH Nicotinamidadenindinucletid
NADPH Nicotinamidadenindinucleotidphosphat
NDEA N-Nitrosodiethylamin
NM nasal mucosa
NNK 4-(Methylnitrosamino)-1-(3-pyridyl)-1-butanon
NO Stickstoffmonoxid
NOS NO-Synthase
OM olfactory mucosa
OMP Odorant marker protein
PAGE Polyacrylamidgelelektrophorese
PAHs polyzyklische aromatische Hydrocarbone
p. appl. post applicationem
PAS periodic acid Schiff
PCR Polymerase-Kettenreaktion

Abkürzungsverzeichnis IV
pNPH p-Nitrophenol-Hydroxylase
PPAR Peroxisome proliferative activated Rezeptor
PrD Propionaldehyd-Dehydrogenase
PROD Pentoxyresorufin-O-Dealkylase
PVDF Polyvinylidendifluorid
PXR Pregnenolone X Rezeptor
RMDD retrometabolic drug design
RNA Ribonukleinsäure
RNase Ribonuklease
RT Reverse Transkriptase
s Sekunden
SDS Natriumdodecylsulfat
SDS-PAGE Natriumdodecylsulfat-Polyacrylamidgelelektrophorese
sog. Sogenannt
SOP Standard Operating Procedure
SPE solid phase extraction
TAE Tris-Essigsäure-EDTA
TBS Tris buffered saline
TCDD 2,3,7,8-Tetrachlorodibenzo-p-Dioxin
TEMED N,N,N`,N`-Tetramethylendiamin
tgl. täglich
TNF α Tumor-Nekrose-Faktor α
Tris Tris(hydroxymethyl)-aminomethan
Tween20 Polyoxyethylensorbitanmonolaurat
u.a. unter anderem

Abkürzungsverzeichnis V
UDP Uridindiphosphat
UDP-GT Uridindiphosphat-Glucuronyl-Transferase
UGT Uridindiphosphat-Glucuronyl-Transferase
Upm Umdrehungen pro Minute
usw. und so weiter
UV Ultraviolett
z.B. zum Beispiel
Zig. Zigaretten

Inhaltsverzeichnis VI
Inhaltsverzeichnis
1. Einleitung ................................................................................................ 1 1.1 Anatomie der oberen Atemwege und der Nasenmuschel.............................................2 1.2 Zellulärer Aufbau der Nasenschleimhaut ......................................................................7 1.3 Physiologie und Funktionen der oberen Atemwege....................................................11 1.4 Funktion der Nasenschleimhaut in der Biotransformation.........................................13 1.5 Nasale Applikation von Arzneimitteln - Glucocorticoide – Soft Steroide..................18 1.6 Ziel der Arbeit.............................................................................................................. 21
2. Material und Methoden........................................................................ 22
2.1 Material ........................................................................................................................22 2.1.1 Probenmaterial .........................................................................................................22 2.1.2 Chemiaklien..............................................................................................................22 2.1.3 Geräte und Verbrauchsmaterialien.............................................................................27
2.2 Methoden......................................................................................................................31 2.2.1 Gewinnung und Transport des Probenmaterials..........................................................31 2.2.2 Genexpressionsuntersuchungen.................................................................................31 2.2.3 Proteinexpressionsuntersuchungen.............................................................................35 2.2.4 Metabolismusuntersuchungen....................................................................................40
3. Ergebnisse............................................................................................. 45
3.1 Genexpression..............................................................................................................45 3.1.1 Qualität und Quantität der isolierten RNA..................................................................45 3.1.2 RT-PCR ..................................................................................................................47
3.2 Proteinexpression.........................................................................................................56 3.2.1 Poolgewichte und Bestimmung des mikrosomalen Proteingehaltes..............................56 3.2.2 Western blot.............................................................................................................56
3.3 Metabolismusuntersuchungen.....................................................................................58 3.3.1 Metabolismus von Loteprednol-Etabonat (LE)..........................................................58 3.3.2 Testosteron-Metabolismus........................................................................................69 3.3.3 EROD-Aktivitäten....................................................................................................72
4. Diskussion............................................................................................. 74
4.1 Untersuchtes Gewebe ...................................................................................................74 4.2 Exprimierte Gene .........................................................................................................75 4.3 Nachweisbares Protein.................................................................................................84 4.4 Metabolische Aktivität.................................................................................................86 4.5 Schlussfolgerungen...................................................................................................... 89
5. Zusammenfassung ................................................................................90 6. Summary...............................................................................................92

Inhaltsverzeichnis VII
7. Danksagung ..........................................................................................93 8. Literaturverzeichnis ..............................................................................94 9. Anhang .....................................................................................................I
9.1 Daten der Patienten für die Genexpression.................................................................. I 9.2 Daten der Patienten für die mikrosomalen Pools ........................................................II 9.3 Liste der verwendeten Primer....................................................................................VII 9.4 Liste der verwendeten Primär- und Sekundärantikörper.......................................... IX 9.5 Agarosegele und graphische Darstellung der Genexpression....................................X 9.6 Darstellung der Western blots ..............................................................................XLVII 9.7 HPLC-Chromatogramme von Loteprednol-Etabonat (LE) und seinen Metaboliten nach Inkubation menschlicher Nasenmuschelmikrosomen.........................................L
9.7.1 1. Versuchsabschnitt (Inkubation bei verschiedenen Inkubationszeiten): Serie 1 ...................................................................................................................... L 9.7.2 1. Versuchsabschnitt (Inkubation bei verschiedenen Inkubationszeiten): Serie 2 ....................................................................................................................LII 9.7.3 2. Versuchsabschnitt (Inkubation mit und ohne Glucuronidase-Zusatz) ...................LVII
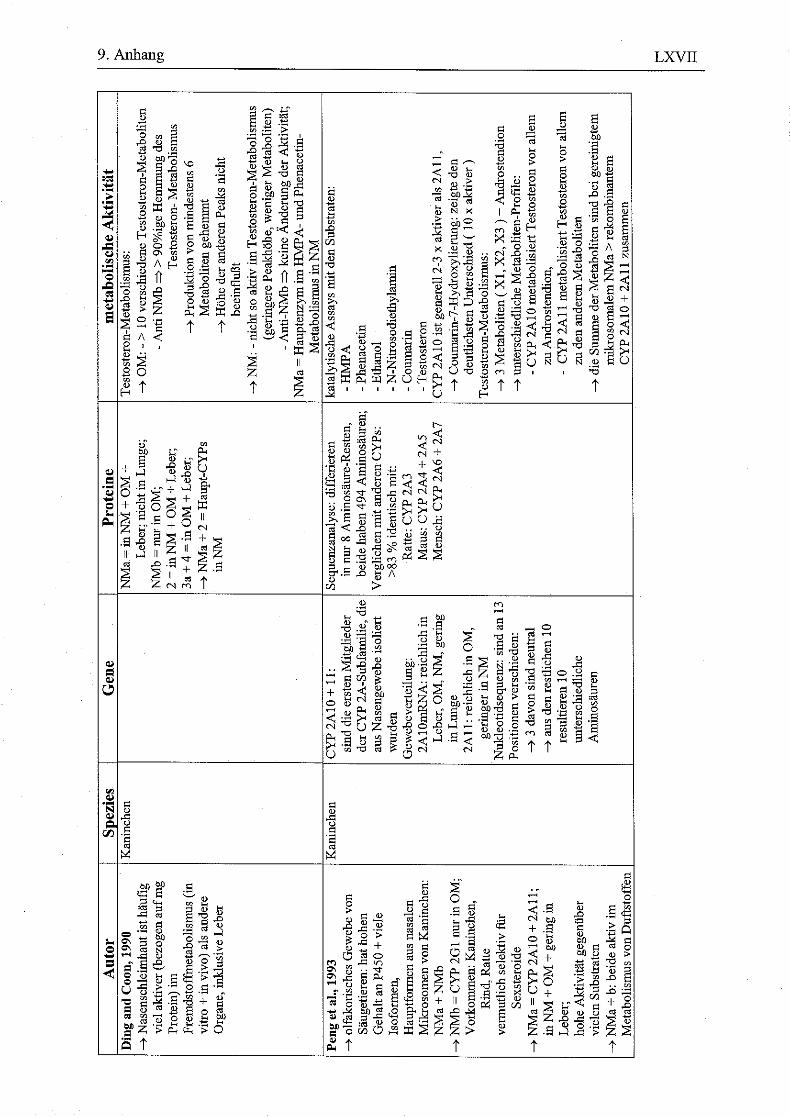
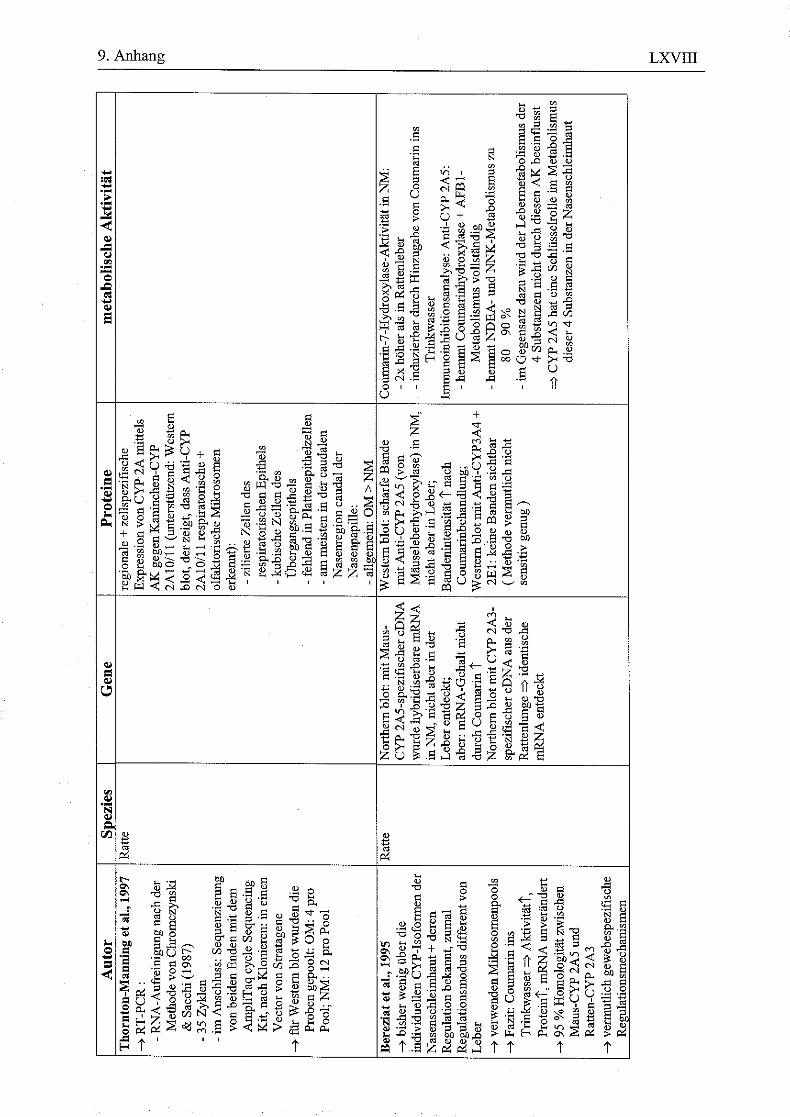
9.8 HPLC-Chromatogramme von Testosteron und seinen Metaboliten nach Inkubation menschlicher Nasenmuschelmikrosomen..............................................LXI 9.9 Vorkommen von Enzymen in der respiratorischen Nasenschleimhaut (NM) bei verschiedenen Spezies......................................................................................LXIV


1. Einleitung 1
1. Einleitung
Die nasale Arzneistoffapplikation ist aufgrund der leichten Zugänglichkeit und der guten
Durchblutung der Nasenschleimhaut nicht nur für lokal wirksame, sondern auch für
systemisch wirksame Arzneimittel attraktiv. Trotz des wachsenden Interesses der
pharmazeutischen Industrie an der nasalen Applikationsform ist die menschliche
Nasenschleimhaut bislang kaum hinsichtlich ihrer Ausstattung an Arzneistoff-
metabolisierenden Enzymen untersucht worden, was u.a. in der schwierigen Verfügbarkeit
menschlichen Nasenschleimhautgewebes für wissenschaftliche Untersuchungen begründet
sein dürfte. Ziel dieser Arbeit war es deshalb, die menschliche respiratorische
Nasenschleimhaut bezüglich vorhandener Enzymsysteme sowie deren metabolischen
Kompetenz näher zu charakterisieren. Im Fokus stand dabei der Nachweis der verschiedenen
Isoformen des Cytochrom P450 Monooxygenasesystemes (CYPs), der wichtigsten
Arzneistoff-metabolisierenden Enzymfamilie. Da das Cytochrom P450-System durch
epigenetische Faktoren beeinflusst wird, wurde zusätzlich der Einfluss von Geschlecht und
Raucher-Status auf das Genexpressionsmuster mitberücksichtigt. Darüber hinaus wurde die
Verstoffwechselung des Glucocorticoids Loteprednol-Etabonat (LE) untersucht, welches sich
derzeit in der klinischen Phase der Zulassung für die Behandlung allergischer
Atemwegserkrankungen befindet (Szelenyi and Pahl, 2002). Damit sollte geprüft werden, ob
und in welchem Ausmaß eine metabolische Inaktivierung durch die in der menschlichen
Nasenschleimhaut vorhandenen Enzymsysteme erfolgt und ob dieser Arzneistoff somit
potentiell für die nasale Applikation zur lokalen Behandlung in Frage käme.
Im Folgenden werden zunächst die anatomischen Grundlagen der oberen Atemwege und der
Nasenmuscheln, von denen die untersuchte Nasenschleimhaut stammt, beschrieben.
Anschließend wird der zelluläre Aufbau der respiratorischen Nasenschleimhaut dargestellt. Es
folgen Physiologie und Funktionen der oberen Atemwege sowie die Funktion der
Nasenschleimhaut in der Biotransformation. Schließlich wird die nasale Applikation von
Arzneimitteln erläutert, so dass über die medikamentöse Therapie allergischer
Atemwegserkrankungen der Bogen zu den Glucocorticoiden und dem Arzneistoff
Loteprednol-Etabonat geschlossen wird.

1. Einleitung 2
1.1 Anatomie der oberen Atemwege und der Nasenmuschel
Die Nasenschleimhaut ist Teil des Respirationsapparates, der sich in die folgenden drei
funktionalen Abschnitte gliedert (Blue Histology, 2002; Kristic, 1991):
1. luftleitender Abschnitt, der die charakteristische respiratorische Schleimhaut besitzt.
Hierzu zählen die Nasenhöhle mit den Nasenmuscheln und den Nasennebenhöhlen,
Nasopharynx, Larynx, Trachea und der Bronchialbaum mit seinen Bronchi, Bronchioli
und Bronchioli terminales.
2. respiratorischer Abschnitt, der für den Gasaustausch verantwortlich ist und zu dem
die Bronchioli respiratorii und das Alveolarsystem gehören.
3. Ventilationsabschnitt: Dabei handelt es sich um jene Teile des
Muskuloskelettalapparates, welche die Atembewegungen ausführen.
Die knöcherne Grundlage für die Nasenschleimhaut liefert die Nasenhöhle (Cavum nasi oder
Cavitas nasi), die gleichzeitig den Beginn des luftleitenden Abschnitts des
Respirationsapparates darstellt. Die Nasenhöhle beginnt vorne mit den beiden Nasenlöchern,
Nares, und kommuniziert hinten durch die Choanen, Choanae, mit dem nasalen Teil des
Pharynx, der auch als Nasopharynx oder Epipharynx bezeichnet wird.
Durch die Nasenscheidewand, Septum nasi, entstehen zwei getrennte Nasenhöhlen, eine
rechte und eine linke. Bei der Nasenscheidewand handelt es sich um eine mediane
schleimhautüberzogene Trennwand, die in ihrem hinteren Teil aus Knochen, Septum nasi
osseum, und in ihrem vorderen Teil aus Knorpel, Cartilago septi nasi, besteht. Das
Nasenseptum steht meist jedoch nicht genau in der Medianen, sondern ist m.o.w. stark nach
der einen oder anderen Seite verbogen (Septumdeviation). Nach Schiebler et al. (1997) tritt
die Septumdeviation sogar bei über 70% der Menschen auf.
Die Seitenwand jeder Nasenhöhle ist medial zu drei Knochenlamellen aufgeworfen, der
unteren, mittleren und oberen Nasenmuschel (Concha nasalis inferior, media et superior),
wobei die oberen beiden Teile des Ethmoids sind, während es sich bei der unteren um einen
eigenständigen Knochen handelt. Gelegentlich ist oberhalb der Concha nasalis superior noch
eine Concha nasalis suprema ausgebildet, die ebenfalls einen Ethmoidalanteil darstellt. Durch
die drei - gelegentlich vier - Nasenmuscheln ist die laterale Nasenhöhlenwand kompliziert
gestaltet (siehe Abbildung 1). Da die Nasenmuscheln von ihren Anheftungszonen an der

1. Einleitung 3
Nasenhöhlenseitenwand nach medial und unten vorragen, entstehen zwischen den lateralen
Flächen der Nasenmuscheln und der Seitenwand drei oder - bei Vorhandensein einer Concha
nasalis suprema – vier Nasengänge: Meatus nasi inferior, medius, superior und evtl.
supremus. Der Raum oberhalb der oberen Nasenmuschel wird nicht als Nasengang
bezeichnet, da er nicht durchgängig ist, sondern nach hinten durch das Keilbein, Os
sphenoidale, verschlossen wird. Er wird aus diesem Grund als Recessus spheno-ethmoidalis
bezeichnet. Die schleimhautüberzogenen Conchen erreichen i.d.R. das Nasenseptum nicht, so
dass medial von ihnen der gemeinsame Nasengang, Meatus nasi communis, entsteht, der mit
den anderen Nasengängen in Verbindung steht. Eine genauere Beschreibung der
Nasenmuscheln selbst soll am Ende dieses Kapitels erfolgen.
Concha nasalis superior
Concha nasalis media
Concha nasalis inferior
Abbildung 1: Laterale Nasenhöhlenwand mit Concha nasalis superior, media und inferior
[aus: Netter FH, Kopf und Hals. In: Atlas der Anatomie des Menschen, 2. Auflage, Novartis AG, Basel: 38,
2000]
Der Boden der Nasenhöhle, der gleichzeitig auch das Dach der Mundhöhle bildet, wird
jeweils vorne aus dem Processus palatinus maxillae und hinten aus der Lamina horizontalis
ossis palatini gebildet.
Das Dach jeder Nasenhöhle setzt sich zusammen aus Teilen des Os ethmoidale, Os frontale,
Os nasale und Os sphenoidale.

1. Einleitung 4
Grundsätzlich wird die Nasenhöhle in 3 strukturell und funktionell unterschiedliche
Abschnitte unterteilt:
1. Nasenvorhöfe, Vestibula nasi: Die unmittelbar an die Nasenlöcher, Nares,
angrenzenden Teile der Nasenhöhle stellen die ersten Zentimeter des luftleitenden
Abschnitts des Respirationsapparates dar. Sie sind ausgekleidet mit verhorntem
mehrschichtigem Plattenepithel, welches sich von der Gesichtshaut über die
Nasenöffnung bis in die Vorhöfe fortsetzt. Dieser, auch als Regio cutanea bezeichnete
Teil der Nasenhöhle, enthält außerdem besonders dicke Haare, sog. Vibrissen
(Vibrissae), sowie apokrine Knäuelsdrüsen (Glandulae vestibulares nasi) und freie,
d.h. nicht an Haare gebundene Talgdrüsen. Die Vibrissen, die im Gegensatz zu den
Haaren der Haut keinen M. arrector besitzen, verhindern das Eindringen großer
Schutzpartikel und Insekten. Seitlich befindet sich eine Epithelleiste, Limen nasi, die
in etwa dem Übergang in die eigentliche Nasenhöhle mit ihrer Regio respiratoria und
olfactoria entspricht und deshalb auch als „inneres Nasenloch“ (Ostium internum)
bezeichnet und den Nasenlöchern als Ostium externum gegenüber gestellt wird. Im
hinteren Bereich des Vestibulums werden Haare und Drüsen spärlicher. Das Epithel
verliert seine Hornschicht und geht kontinuierlich in das respiratorische Epithel über.
2. Respiratorischer Abschnitt, Regio respiratoria: Der respiratorische Abschnitt
schließt direkt an die Vorhöfe an und macht insgesamt den größten Anteil an der
Nasenhöhle aus. Das verhornte mehrschichtige Plattenepithel der Nasenvorhöfe geht
über ein unverhorntes, zunehmend dünner werdendes Plattenepithel in das
mehrreihige Flimmerepithel der Regio respiratoria über. Dieses ist charakteristisch für
alle Anteile des luftleitenden Abschnitts des Respirationsapparates und wird deshalb
respiratorisches Epithel genannt. Der genaue histologische Aufbau wird in Kapitel
1.2.1 ausführlich erläutert.
3. Olfaktorischer Abschnitt, Regio olfactoria: Dieser Abschnitt ist gekennzeichnet
durch das olfaktorische Epithel (Riechepithel, Riechschleimhaut), welches sich am
Dach der Nasenhöhle, an der oberen Nasenmuschel und am gegenüberliegenden Teil
des Nasenseptums befindet (Boenninghaus, 1996; Kristic, 1991; Speckmann and
Wittkowski, 1998). Das olfaktorische Epithel ist mit 60 µm (Kristic, 1991; Lippert,
1996) deutlich dicker als das respiratorische Epithel und durch die Einlagerung von
Lipofuszin gelbbraun gefärbt. Die Ausdehnung der Regio olfactoria wird beim
Menschen in der Literatur relativ einheitlich mit 5 cm2 angegeben (Kristic, 1991;

1. Einleitung 5
Schiebler et al., 1997; Speckmann and Wittkowski, 1998; Weiss, 1988). Im Gegensatz
zum Mensch erstreckt sich bei den Makrosmatikern die Riechschleimhaut auf eine
wesentlich größere Oberfläche. So wird sie beispielsweise beim Hund mit ca. 100 cm2
(Benninghoff, 1994) angegeben.
Zum Schluss diese Kapitels möchte ich die Anatomie der Nasenmuscheln genauer betrachten,
da von ihnen die Nasenschleimhaut stammt, die im Rahmen meiner Dissertation untersucht
werden soll. Es handelt sich dabei um muschelartige Gebilde, die von der lateralen Wand der
Nasenhöhle in das Lumen hineinragen und auf diese Weise die zuvor beschriebenen
Nasengänge, Meatus nasi, bilden (siehe Abbildung 2). Ihre Form erhält die Nasenmuschel
durch eine Knochenlamelle, die von einer m.o.w. dicken Schleimhaut überzogen ist.
Die untere Nasenmuschel, Concha nasalis inferior, ist die größte und längste. Von Lanz and
Wachsmuth (1985) geben ihre Länge mit 35 – 58 mm, im Durchschnitt 47,7 mm, an. Sie
beginnt direkt hinter dem Vestibulum nasi und hat die Form einer nach medial konvexen,
vorne stumpfen und hinten zugeschärften Platte. Die konvexe, dem Meatus nasi communis
zugewandte Fläche ist durch Anlagerung von Gefäßen und Drüsen fein modelliert, während
die konkave Fläche glatt ist. Ihr Unterrand ist leicht nach lateral eingerollt und nimmt von
hinten nach vorne an Mächtigkeit zu, was als Verstrebung aufgefasst wird.
Concha nasalis superior
Concha nasalis media
Concha nasalis inferior
Abbildung 2: computertomographische Darstellung der Nasenhöhle mit Nasenmuscheln und
Nasenseptum
[aus: Westhofen Prof Dr M, Nase, Nasennebenhöhlen, Mittelgesicht und vordere Schädelbasis. In: Hals-Nasen-
Ohrenheilkunde systematisch, UNI-MED Verlag AG, Bremen: 192, 2001]

1. Einleitung 6
Die mittlere Nasenmuschel, Concha nasalis media, ist die zweitgrößte und beginnt etwa 1,5
cm hinter dem Vorderende der unteren Nasenmuschel. Sie ist wie die untere Muschel von
einer dicken Schleimhaut überzogen und ihre mediale konvexe und ihre konkave laterale
Fläche ähneln in ihrer Oberflächenbeschaffenheit der unteren Nasenmuschel.
Die obere Nasenmuschel, Concha nasalis superior, entspringt ca. 1,5 cm hinter dem
Vorderende der Concha nasalis media und stellt die kleinste der drei Muscheln dar. Die Form
der Flächen ist mit jener der anderen beiden Conchen vergleichbar, während ihre Oberfläche
bedingt durch die marklosen Fila olfactoria durch vertikalen Rinnen gezeichnet ist. Sie besitzt
nur eine vergleichsweise dünne Schleimhaut und weist – im Gegensatz zu den unteren
Nasenmuscheln – olfaktorisches Epithel auf.
Bisweilen existiert oberhalb der Concha nasalis superior noch eine Concha nasalis suprema,
die ebenfalls vom Ethmoid gebildet wird. Von Lanz and Wachsmuth (1985) geben die
Häufigkeit dieser Nebenmuschel mit 17,7% an. In solchen Fällen entsteht noch ein weiterer
Nasengang, der Meatus nasi supremus, der – wie die anderen auch – mit dem Meatus nasi
communis und dem Meatus nasopharyngeus in Verbindung steht.

1. Einleitung 7
1.2 Zellulärer Aufbau der Nasenschleimhaut
Im Anschluss an die makroskopische Betrachtung der Nasenhöhle soll nun der
mikroskopische Aufbau der Nasenschleimhaut dargestellt werden. Die Schleimhaut, Mucosa,
des Respirationstrakts wird - wie in anderen Geweben auch - in die Lamina epithelialis
mucosae und die Lamina propria mucosae unterteilt, die durch eine Basalmembran
voneinander getrennt werden. Bei der Lamina propria mucosae, die der Einfachheit halber oft
als Propria bezeichnet wird, handelt es sich um Bindegewebe, welches als
Verbindungsgewebe zu dem darunter befindlichen Stützgewebe, Knochen bzw. Knorpel,
dient. Die Lamina epithelialis mucosae, auch Epithel genannt, vertritt die oberflächliche
Schicht der Schleimhaut und ist von einem dünnen sero-mucösen Film überzogen. Je nach
den am Aufbau des Epithels beteiligten Zellen unterscheidet man ein respiratorisches und ein
olfaktorisches Epithel und damit eine Regio respiratoria und eine Regio olfactoria, wobei
auch die Propria in beiden Regionen differiert.
Die respiratorische Schleimhaut hat ihren Namen aufgrund der Tatsache, dass sie im
gesamten Bereich des luftleitenden Abschnitts des Respirationsapparates vorkommt.
Innerhalb der Nasenhöhle befindet sie sich am Nasenhöhlenboden, an der Seitenwand im
Gebiet der unteren beiden Nasenmuscheln sowie am gegenüberliegenden Teil des
Nasenseptums. Sie kleidet außerdem die Nasennebenhöhlen aus, ist dort aber nur
vergleichsweise dünn. Am dicksten ist sie über den Conchen.
Es handelt sich hierbei um ein mehrreihiges Flimmerepithel, in dem zahlreich Becherzellen
eingelagert sind. Es werden dabei folgende Zellarten unterschieden: Flimmerepithelzellen,
Becherzellen sowie Intermediär- und Basalzellen (siehe Abbildung 3 und 4). An mechanisch
stärker beanspruchten Stellen jedoch können Inseln von Plattenepithel vorkommen (Weiss,
1988). Neuere Untersuchungen deuten darauf hin, dass auch die Vitamin A-Versorgung einen
Einfluss auf Vorkommen und Ausdehnung des Plattenepithels hat (Million et al., 2001).

1. Einleitung 8
Abbildung 3: Schematische Darstellung des respiratorischen Epithels und der Kinozilien
[aus: Rohen JW and Lütjen-Drecoll E, Epithelgewebe. In: Funktionelle Histologie, 4. Auflage, Schattauer
Verlagsgesellschaft mbH, Stuttgart: 72, 1999]
Die hochprismatischen Flimmerepithelzellen besitzen apikal feine, 5 – 10 µm lange (Linß and
Halbhuber, 1991; Rohen and Lütjen-Drecoll, 1995) und 0,2 µm dicke (Linß and Halbhuber,
1991), bewegliche Zilien (Kinozilien, siehe Abbildung 3). Die Kinozilien gehen aus den
Kinetosomen (Basalknötchen) hervor, die sich lichtmikroskopisch als feine, dunkle Linie,
sog. Basalknötchenlinie, darstellen. Diese dient als wichtiges Unterscheidungsmerkmal für
andere epitheliale Oberflächenstrukturen (z.B. Bürstensaum).
Die Kinozilienbewegung setzt sich zusammen aus einem raschen Schlag und einer langsamen
Rückschwingphase. Dabei kann eine Frequenz von bis zu 30 Hz und eine Geschwindigkeit an
der Zilienspitze von bis zu 2,5 m/s erreicht werden (Linß and Halbhuber, 1991). Beim
Vorwärtsschlag ist die Zilie aufgerichtet und schlägt mit ganzer Kraft, während sie bei der
Rückschwingphase stark gekrümmt ist, so dass sie sich praktisch unter dem Sekretfilm
hindurchzieht. Auf diese Weise entsteht ein Flüssigkeitsstrom in Schlagrichtung, wobei die
Kinozilien stets in Richtung Pharynx schlagen. Die peitschenartigen Zilienbewegungen
kommen durch ein Vorbeigleiten der Mikrotubuli innerhalb des Kinoziliums zustande. Für
den Energienachschub sorgen dabei Mitochondrien, die im apikalen Drittel der
Flimmerepithelzellen zu finden sind. Durch inter- und intrazelluläre Koordination erfolgt eine
Vereinheitlichung von Schlagrichtung und –frequenz, so dass ein wellenartiger
Bewegungsablauf (Metachronie) entsteht, der dem Transport von Flüssigkeit, Schleim, Zellen
und Fremdkörpern auf der Epitheloberfläche dient.
Flimmerepithelzelle Becherzelle Basalzelle Basalmembran seromucöse Drüsen

1. Einleitung 9
Die Becherzellen sind vor allem im hinteren Bereich der Nasenhöhle lokalisiert. Es handelt
sich dabei um hochprismatischen Zellen, die stark mit PAS-positiven Schleimgranula gefüllt
sind, so dass ihr abgeplatteter Kern und das Ergastoplasma an der Zellbasis
zusammengedrängt werden. Becherzellen können auch als endoepitheliale mucöse Drüsen
betrachtet werden, die zusammen mit den sero-mucösen Drüsen der Propria für den
Schleimfilm auf der Epitheloberfläche verantwortlich sind. Dieser Schleimfilm ist insgesamt
etwa 5 µm dick (Rohen and Lütjen-Drecoll, 1995) und setzt sich aus einer oberflächlichen
Gel-Phase und einer tieferen Sol-Phase zusammen. Die Gel-Phase ist reich an
Makromolekülen und bildet eine viskoelastische Barriere, welche einen Widerstand für den
aktiven, schnellen Schlag der Zilien gibt, die den Film dadurch verschieben können. Die Sol-
Phase, auch Hypophase genannt, ist für die Zilienbewegung wichtig. In dieser Phase ziehen
die erschlafften Zilien wieder zurück. Der Schleimfilm enthält neben wasserbindenden
Glykoproteinen noch antibakterielle Proteine (Lysozyme, Laktoferrin), Immunglobuline (vor
allem IgA) und Proteinaseinhibitoren, die für die lokale Infektionsabwehr von Bedeutung
sind.
Die undifferenzierten Basalzellen reichen nicht bis zur Epitheloberfläche und haben keine
Zilien. Wegen ihrer Teilungsfähigkeit sind sie als Stratum germinativum zu betrachten. Sie
liefern die Intermediärzellen, die sich dann entweder zu Flimmerepithelzellen oder zu
Becherzellen differenzieren.
Lamina epithelialis mucosae und Lamina propria mucosae werden durch eine auffallend
dicke und glatte Basalmembran getrennt. So können sich die Epithelzellen bei den
regenerativen Zellverschiebungen auf der Membran leicht verschieben (Rohen and Lütjen-
Drecoll, 1995).
Die Lamina propria mucosae besteht aus lockerem Bindegewebe, das mit dem Periost der
knöchernen Nasenwand bzw. dem Perichondrium der knorperligen Wandanteile fest
verbunden ist. Subepithelial befinden sich Lymphozyten und Plasmazellen, einzeln und in
Gruppen, und bilden so die lymphoide Zone. Teilweise sind diese Zellen auch intraepithelial
zu finden. Unterhalb dieser Zone ist die Drüsenzone gelegen, in der zahlreich verzweigte,
tubulo-alveoläre, sero-mucöse Drüsen, die Glandulae nasales, lokalisiert sind. Sie haben
kurze Ausführungsgänge, die auf der Epitheloberfläche münden, sowie mucöse und seröse
Drüsenanteile. In den mittleren und tieferen Schichten der Propria sind ausgedehnte venöse
Schwellkörper gelegen, die besonders an der unteren und mittleren Nasenmuschel ausgeprägt

1. Einleitung 10
sind (Plexus cavernosi concharum). Die Füllung dieser Schwellkörper hat eine erhöhte
Wärmeabstrahlung zur Folge und wird durch Dilatation der zuführenden Arteriolen sowie
durch Konstriktion der abführenden tieferen Venen, die somit als Drosselvenen oder
Sperrvenen dienen, hervorgerufen. Die Regulation erfolgt dabei autonom.
Insgesamt ist die Nasenschleimhaut gut mit Blutgefäßen versorgt. Die dickwandigen Arterien
liegen nahe am Periost in Gestalt eines Gitterwerkes, von dem arkadenartige Gefäße
abzweigen, die auf diese Weise die gesamte Propria senkrecht durchziehen. Kurz vor der
Epitheloberfläche teilen sie sich in Arteriolen, aus denen ein dichtes Netzwerk fenestrierter
Kapillaren entspringt, die teilweise die Oberfläche und teilweise die Drüsen versorgen.
Zwischen den Arterien und Venen befinden sich außerdem zahlreiche gewundene
arteriovenöse Anastomosen.
Abbildung 4: Histologischer Schnitt durch die Schleimhaut der Nase im Bereich der Regio respiratoria
(396x). Man beachte den deutlichen Flimmerbesatz (Kinozilien), die relative dicke, gerade verlaufende
Basalmembran und das lockermaschige, gefäßreiche, subepitheliale Bindegewebe, in dem sich zahlreiche
Lymphozyten und Plasmazellen befinden.
[aus: Rohen JW and Lütjen-Drecoll E, Respirationssystem. In: Funktionelle Histologie, 4. Auflage, Schattauer
Verlagsgesellschaft mbH, Stuttgart: 217, 1999]
Die Nasennebenhöhlen werden ebenfalls von Flimmerepithel ausgekleidet, wobei die
Kinozilien hier in Richtung der Ostien schlagen. Im Gegensatz zur Nasenhaupthöhle sind in
den Nebenhöhlen nur wenige Becherzellen vorhanden. Die Lamina propria mucosae ist zu
einem dünnen Bindegewebsblatt umgebildet und auffallend gefäßarm. Sero-mucösen Drüsen
sind nur vereinzelt vorhanden, venöse Schwellkörper fehlen hier vollkommen.

1. Einleitung 11
1.3 Physiologie und Funktionen der oberen Atemwege
Die Nasenhöhlen repräsentieren den ersten Teil des luftleitenden Abschnitts des
Respirationsapparates und dienen somit in erster Linie der Luftleitung. Dabei gelangt der
Hauptluftstrom von den Nasenlöchern über die Nasengänge zu den Choanen. Nur 5 – 10 %
des Inspirationsvolumens erreichen das Riechepithel, die Nasennebenhöhlen sogar nur 1 %
(Westhofen, 2001).
Neben der Luftleitung kommt den Nasenhöhlen noch eine weitere Aufgabe zu, die
Vorbehandlung der Atemluft. Durch die Nasenmuscheln ist die Oberfläche der lateralen
Nasenhöhlenwand in Falten gelegt. Die daraus resultierende Oberflächenvergrößerung
erleichtert – zusammen mit der erzeugten turbulenten Strömung – die Konditionierung der
Atemluft in Form von Erwärmung bzw. Abkühlung, Filterung bzw. Reinigung sowie in Form
des Anfeuchtens der eingeatmeten Luft. Alles zusammen dient dem Schutz der tiefen
Atemwege.
Durch unterschiedliche Blutfüllung der Schleimhaut bzw. der venösen Schwellkörper der
unteren beiden Nasenmuscheln wird die inspirierte Luft auf eine Temperatur von etwa 36°C
gebracht. Um einen effektiveren Wärmeaustausch zu erzielen, strömen Blut und Atemluft
nach dem Gegenstromprinzip.
Eine erste Filterung der eingeatmeten Luft erfolgt durch die Vibrissen der Nasenvorhöfe. Sie
fungieren als Art Reuse, die das Eindringen grober Partikel und Insekten in den
Respirationstrakt verhindert. Kleinere Partikel werden dagegen durch die mucociliäre
Clearance beseitigt. Hierbei transportieren die pharynxwärts schlagenden Kinozilien des
Flimmerepithels den oberflächlichen Schleimfilm mit den darin gelösten Fremdstoffen zum
Pharynx. Der Rachen wiederum ist reich an lymphatischen Einrichtungen, welche der Abwehr
eingedrungener Mikroorganismen dienen. Zusätzlich enthält die oberflächliche, visköse Phase
des Schleimfilmes antibakterielle Proteine, Immunglobuline sowie Proteinaseinhibitoren, die
zusammen mit den Leukozyten des Epithels und der subepithelialen Gewebeschichten für die
lokale Infektionsabwehr verantwortlich sind.
Schließlich erfolgt in den oberen Atemwegen noch das Anfeuchten der Atemluft auf 70 – 80
%. Erzielt wird dies durch Wasserverdunstung und durch Abgabe von Nasensekret, das
gleichzeitig auch die Nasenschleimhaut vor dem Austrocknen schützt.

1. Einleitung 12
Anschwellen und vermehrte Sekretion der Nasenschleimhaut können verschiedene Ursachen
haben. So kann dies reflektorisch über das vegetative Nervensystem, hormonell, durch
Entzündungen, Allergien, Arzneimittel oder aber durch mechanische, thermische oder
chemische Reize ausgelöst werden.
Im oberen hinteren Bereich der Nasenhöhle sind außerdem die Rezeptoren des
Geruchssinnes lokalisiert. Wie bereits erwähnt, erreicht normalerweise nur ein kleiner Teil
der eingeatmeten Luft die Regio olfactoria. Bei bewusster, tiefer Inspiration jedoch erhöht
sich die Geschwindigkeit des eingeatmeten Luftstromes, die turbulente Strömung verwandelt
sich in eine zunehmend laminare Strömung, und ein größerer Teil der Atemluft gelangt bis
zum Riechepithel. Durch die geruchliche Prüfung der Inspirationsluft kommt dem
olfaktorischen System somit auch noch eine Schutzfunktion zu, da viele schädliche Gase als
übelriechend empfunden werden und ihre Einatmung deswegen vermieden wird.
Interessanterweise beruht die vorübergehende Minderung des Riechvermögens (Hyposmie)
bei Schnupfen nicht auf einer Schädigung des olfaktorischen Systems, sondern auf der
Tatsache, dass die Geruchsstoffe infolge der Schleimhautschwellung nicht mehr bis zur
Riechschleimhaut gelangen.
Schließlich dient die Nasenhöhle mit ihren Nebenhöhlen beim Sprechen als Resonanzraum.
Die Konsonanten m, n und ng werden gesprochen, ohne dass der Nasenrachenraum durch das
Gaumensegel abgeschlossen ist. Somit strömt die Luft via Nasenhöhle durch die Nase aus,
weshalb genannte Konsonaten auch als Rhinophone bezeichnet werden.

1. Einleitung 13
1.4 Funktion der Nasenschleimhaut in der Biotransformation
Zusätzlich zu den in Kapitel 1.3 genannten Funktionen nimmt die Schleimhaut der oberen
Atemwege auch in Bezug auf die Xenobiotransformation eine wichtige Rolle ein. Der
Metabolismus von Xenobiotika wird in der Regel in zwei Phasen unterteilt. Die Phase I-
Reaktion, auch Funktionalisierungsreaktion genannt, beinhaltet die Einführung oder
Freisetzung funktioneller Gruppen, wie Hydroxyl-, Amino-, Carboxyl- oder Thiolgruppen.
Die Endprodukte der Phase I-Reaktion enthalten somit chemisch reaktive funktionelle
Gruppen, die von den Phase II-Enzymen benötigt werden.
Während der Phase II-Reaktion erfolgt die Konjugation der Phase I-Metaboliten an
körpereigene Stoffe, weshalb dieser Vorgang auch als Konjugationsreaktion bezeichnet wird.
Hierzu zählen Glucuronidierung, Glycosidierung, Glutathion-Konjugation, Sulfatierung,
Methylierung, Acetylierung, Aminosäuren-Konjugation sowie die Fettsäuren-Konjugation,
deren einzige gemeinsame Eigenschaft ein energiereicher oder aktivierter Zwischenmetabolit
darstellt, sei es ein aktivierter Cofaktor, wie beispielsweise UDP-Glucuronsäure bei der
Glucuronidierung oder Acetyl-CoA bei der Acetylierung, oder sei es ein aktiviertes oder
reaktives Xenobiotikum. Am Ende entstehen generell wasserlösliche und somit leicht
eleminierbare Produkte.
Die Funktionalisierung der Verbindungen im Rahmen des Phase I-Metabolismus kann durch
Oxidation, Reduktion (z.B. Reduktion von Nitrogruppen zu Aminogruppen) oder Hydrolyse
(z.B. Hydrolyse von Estern durch Esterasen) erzielt werden. Phase I-Reaktionen werden von
Monooxygenasen und anderen Oxidasen, Alkoho l- und Aldehyddehydrogenasen sowie
Esterasen und Amidasen vermittelt. Die wichtigste Komponente des oxidativen Phase-I-
Biotransformationssystems besteht aus einer Gruppe von Hämoproteinenzymen, welche
kollektiv als Cytochrom P450-abhängige Monooxygenasen (CYPs) bezeichnet werden.
Bei dem Cytochrom P450-System handelt es sich um eine Enzymsuperfamilie, die sich in
diverse Familien und Subfamilien gliedert. Derzeit sind 18 Familien der humanen Cytochrom
P450-Enzyme bekannt (Chen et al, 2002; Nelson, 2003), wobei die Anzahl sequenzierter
Gene und Pseudogene ständig weiter ansteigt.

1. Einleitung 14
Als Hämoprotein besitzt das Cytochrom P450 ein Häm in Form eines Fe-Protoporphyrins,
welches nicht-kovalent an das Apoprotein gebunden ist. Die Bezeichnung P450 stammt dabei
von der Tatsache, dass das Cytochrom (Cytochrom = Pigment = P) in reduzierter Form in
Anwesenheit von CO ein mit 450 nm spezifisches Absorptionsmaximum aufweist. Im
Gegensatz zu den Bakterien sind genannte Cytochrome bei den Säugetieren stets
membrangebunden und entweder in der inneren Membran der Mitochondrien oder aber – die
Mehrheit der Cytochrome - in der Membran des Endoplasmatischen Retikulums lokalisiert.
Das Cytochrom P450-System benötigt zusätzlich reduzierende Äquivalente, die vom NADPH
oder manchmal auch vom NADH stammen und via eines zweiten Enzyms auf das Cytochrom
übertragen werden, sowie ein Atom Sauerstoff, welches von molekularem Sauerstoff oder von
oxygenierten Substraten stammt. Da nur eines der beiden Sauerstoffatome von molekularem
Sauerstoff in das Substrat inkorporiert wird, während das andere zu Wasser reduziert wird,
werden diese Enzyme auch als Monooxygenasen bezeichnet und den Dioxygenasen, welche
beide Atome einfügen, gegenüber gestellt. Aufgrund der beschriebenen doppelten Funktion –
Oxidation des Substrates einerseits und Reduktion des Wassers andererseits – werden sie auch
als Mischfunktionelle Oxygenasen (MFO) bezeichnet.
Um die metabolische Aktivität dieser Enzymsuperfamilie untersuchen zu können, bedient
man sich bestimmter Markersubstrate, die entweder nur von einer einzelnen CYP-Isoform
bzw. von einer Subfamilie oder aber von verschiedenen CYPs umgesetzt werden. Eine sehr
häufig verwendete Gruppe sind dabei die 7-Alkoxyresorufine, zu denen 7-Ethoxyresorufin, 7-
Methoxyresorufin, 7-Benzyloxyresorufin und 7-Pentoxyresorufin gehören. Die O-
Dealkylierung des Ethoxyresorufins (EROD) ist mit den Cytochromen P450 1A1 (CYP 1A1)
und 1A2 (CYP 1A2) assoziiert, von denen CYP 1A1 das Ethoxyresorufin in einem bedeutend
größeren Ausmaß umsetzt, wenn beide Cytochrome vorhanden sind (Wardlaw et al., 1998).
Die Verstoffwechselung des Methoxyresorufins (MROD) wird dagegen einzig von CYP 1A2
ausgeführt (Wardlaw et al., 1998), während die O-Dealkylierung des Pentoxyresorufins
(PROD) wiederum den Mitgliedern der CYP 2B-Subfamilie zugeschrieben wird (Gelardi et
al., 2001; Hukkanen, 2000; Seubert et al., 2002; Wardlaw et al., 1998).
Ein weiteres Markersubstrat ist Testosteron, welches stereo- und regioselektiv von
unterschiedlichen CYP-Isoformen hydroxyliert wird und somit die funktionelle Basis für das
gleichzeitige Studium verschiedener Isozyme liefert.

1. Einleitung 15
Das wohl wichtigste Enzym innerhalb des Phase II-Metabolismus ist die UDP-Glucuronyl-
Transferase (UDP-GT oder UGT), was mit der leichten Verfügbarkeit des benötigten Co-
Faktors, der UDP-Glucuronsäure, erklärbar sein dürfte, da die UDP-Glucuronsäure Teil des
intermediären Stoffwechsels ist und eng mit der Glycogensynthese verbunden ist. Auch hier
handelt es sich um eine Enzymsuperfamilie, deren Mitglieder in zwei distinkte Familien
unterteilt werden, UGT 1 und UGT 2 (King et al., 2000, Mackenzie et al., 1997).
Interessanterweise sind diese Enzyme ebenfalls in der Membran des Endoplasmatischen
Retikulums lokalisiert – was eine Ausnahme unter den Phase II-Enzymen darstellt – und
damit in enger Nachbarschaft zum Cytochrom P450-System gelegen, so dass die entstehenden
Phase I-Metaboliten direkt glucuronidiert werden können.
Ein weiteres Phase II-Enzym stellt die zytosolische Glutathion-S-Transferase (GST) dar,
welche die Konjugation mit dem endogenen Tripeptid Glutathion katalysiert und aus zwei
ähnlichen, aber nicht identischen Subunits zusammengesetzt ist. Die Glutathion-Konjugation
unterscheidet sich signifikant von der Gluconat- und der Sulfat-Konjugation, da bei
letztgenannten eine vorherige Aktivierung der Gluconat- bzw. Sulfat-Anteile notwendig ist. In
diesem Fall aber ist die einzige chemische Voraussetzung ein elektrophiles Zentrum innerhalb
des Substrates, das die Reaktion mit dem nukleophilen Glutathion ermöglicht. Somit kann die
Glutathion-Konjugation als Schutzmechanismus aufgefasst werden, bei dem potentiell
toxische, nukleophile Metaboliten einfach „aufgewischt“ werden (Gibson and Skett, 1986).
Die menschliche Nasenschleimhaut enthält alle Komponenten des Cytochrom P450-Systemes
(Gervasi et al, 1991). Der Cytochrom P450-Gehalt der respiratorischen Schleimhaut des
Menschen beträgt 25 pmol/mg Protein (Gervasi et al., 1991; Longo et al., 1989) und ist somit
vergleichbar mit jenem von Hund und Ratte, jedoch geringer als jener beim Kaninchen
(Gervasi et al., 1991). Verglichen mit anderen menschlichen Geweben entspricht diese CYP-
Menge nur etwa 5% des Cytochromgehaltes der Leber, ist dagegen aber etwa doppelt so hoch
wie in der Lunge (Gervasi et al., 1991).
Obwohl im Vergleich zur Leber die Nasenschleimhaut bezüglich ihrer Stoffwechselkapazität
ein relativ unerforschtes Gebiet ist, sind trotzdem eine Anzahl konstitutiv exprimierter CYP-
Isoformen in nasalen Geweben verschiedener Spezies gefunden worden, darunter Mitglieder
der 1A-, 2A-, 2B-, 2C-, 2E-, 3A- und 4B-Subfamilie (Longo et al., 2000; Thornton-Manning
et al., 1997). Das für die Regio olfactoria spezifische CYP 2G1 konnte bei Rind (Longo et al.,

1. Einleitung 16
1997), Schwein (Martini et al., 1998), Kaninchen (Ding and Coon, 1994) sowie bei Ratte und
Maus (Gu et al., 1997; Gu et al., 1998) nachgewiesen werden.
Besonders gut untersucht ist die CYP 2A-Subfamilie, die beim Menschen drei Mitglieder
aufweist: CYP 2A6, CYP 2A7 und CYP 2A13. CYP 2A6 und 2A7 sind reichlich in der Leber
vorhanden in einem Verhältnis von etwa 1:1 (Koskela et al., 1999), während CYP 2A13 am
höchsten in der Nasenschleimhaut exprimiert ist, gefolgt von Lunge und Trachea. Der CYP
2A13-Gehalt der Nasenschleimhaut ist etwa fünf mal höher als der CYP 2A6-Gehalt (Kosekla
et al., 1999). Auch bei anderen Spezies sind in der Nasenschleimhaut CYP 2A-Isoformen
vorhanden, beispielsweise CYP 2A10/11 beim Kaninchen, CYP 2A3 bei der Ratte und CYP
2A4/5 bei der Maus (Ding et al., 1994; Peng et al., 1993).
In der menschlichen Nasenschleimhaut ist aber nicht nur das Monooxygenase-System
vertreten, sondern auch zahlreiche nicht-oxidative Enzyme, darunter die DT-Diaphorase
(Gervasi et al., 1991), die Epoxid-Hydrolase (Gervasi et al., 1991; Green et al., 2001), die
Glutathion-S-Transferase (Aceto et al., 1989; Gervasi et al., 1991; Green et al., 2001; Krishna
et al., 1995) und das Cyanid-metabolisierende Enzyme Rhodanese (Lewis et al., 1991). Eine
Übersicht über die von Gervasi et al. (1991) untersuchten Enzymaktivitäten ve rschiedener
Phase I- und Phase II-Enzyme beim Menschen zeigen die Abbildungen 5 und 6.
Interessanterweise sind die Phase II-Enzyme zwei bis drei mal stoffwechselaktiver als das
Cytochrom P450-System, so dass reaktive Phase I-Metaboliten gar nicht erst akkumulieren
können (Gervasi et al., 1991) – eine Beobachtung, die auch Longo et al. (1991) beim Rind
gemacht haben.

1. Einleitung 17
untersuchte Parameter nasale gepoolte Mikrosomen hepatische Mikrosomen
Protein (mg/g Gewebe) 3,5 11,2
ECOD (pmol/min/mg Protein) 2,9 480
EROD (pmol/min/mg Protein) 0,5 102
AnH (pmol/min/mg Protein) 65 687
HMPAd (pmol/min/mg Protein) 92 240
DMNd (pmol/min/mg Protein) 220 2045
APD (pmol/min/mg Protein) 65 826
Abbildung 5: Monooxygenase-Aktivitäten in menschlichen nasalen und hepatischen Mikrosomen (Gervasi
et al., 1991).
untersuchte Parameter nasale gepoolte Mikrosomen hepatische Mikrosomen
cytosolisches Protein
(mg/g Gewebe)
14,5 24,8
EH (nmol/min/mg Protein) 17,4 35,6
UDP-GT1 (nmol/min/mg Protein) <0,001 2,4
GST (nmol/min/mg Protein) 56,6 476
PrD (nmol/min/mg Protein) 2,08 3,07
BeD (nmol/min/mg Protein) 12,8 1,2
DT-Diaphorase
(nmol/min/mg Protein)
6,25 0,49
Carbonylreductase
(nmol/min/mg Protein)
1,92 1,85
Abbildung 6: Epoxid Hydrolase (EH) und Phase II-Enzymaktivitäten in menschlichen nasalen und
hepatischen Mikrosomen (Gervasi et al., 1991).

1. Einleitung 18
1.5 Nasale Applikation von Arzneimitteln – Glucocorticoide – Soft Steroide
Die nasale Arzneistoffapplikation ist aufgrund der leichten Zugänglichkeit und der guten
Durchblutung der Nasenschleimhaut sowohl für lokal, als auch für systemisch wirksame
Arzneimittel eine attraktive Applikationsform. Bei den lokal wirksamen Arzneistoffen ist die
nasale Applikationstechnik bereits für verschiedene Arzneistoffgruppen etabliert, darunter
Vasokonstriktoren zur Schleimhautabschwellung ( z.B. Xylometazolin [Olynth],
Oxymetazolin [Nasivin]), Mastzellstabilisatoren zur Verhinderung der
Histaminausschüttung bei allergisch bedingten Erkrankungen (z.B. Cromoglicinsäure
[Vividrin]) sowie verschiedene Glucocorticoide. Bei Letztgenannten handelt es sich um die
wichtigste Klasse von Medikamenten zur Behandlung von Entzündungen und Allergien,
gleichzeitig aber auch um Arzneimittel mit einem sehr breiten Nebenwirkungsspektrum.
Obwohl bei nasaler Administration nur ein kleiner Teil in den Blutkreislauf gelangt, sind
systemische Wirkungen nicht ganz auszuschließen. Beispielsweise wird bei nasaler
Verabreichung von Triamcinolonacetat eine Cortisolsuppression von 8 % (Einzeldosis) bzw.
16 % (Steady state-Dosis) beobachtet (Hochhaus et al., 2002).
Glucocorticoide stellen jedoch meistens einen unverzichtbaren Therapiebestandteil bei der
Behandlung von Allergien dar, so auch bei der Behandlung der allergischen Rhinitis
(Histologischer Schnitt durch die Nasenschleimhaut eines Patienten mit allergischer Rhinitis
siehe Abbildung 7). Die allergische Rhinitis – als Allergiekorrelat der Nasenschleimhaut – ist
eine weltweit verbreitete Erkrankung und betrifft etwa 10 – 50 % der Bevölkerung (Pawankar
and Fokkens, 2001). Die steigende Prävalenz in den letzten Jahren, die häufige Kombination
mit Asthma bronchiale (Kumar and Singh, 2002; Pawankar and Fokkens, 2001) und die meist
unerlässliche Glucocorticoidtherapie haben die Suche nach neueren und sicheren
Therapiekonzepten forciert. Durch Arzneistoffkonstruktion nach dem Prinzip des RMDD
(retrometabolic drug design) ist eine neue Klasse von Steroiden entstanden, die sich durch
einen erhöhten therapeutischen Index auszeichnet, da diese Substanzen so konstruiert sind,
dass sie nach ihrer systemischen Verfügbarkeit sofort zu inaktiven Metaboliten
verstoffwechselt werden (Bodor, 1999). Erster Vertreter dieser sog. Soft Steroide ist das
Loteprednol-Etabonat (LE), das für allergische ophthalmologische Indikationen bereits
zugelassen ist und sich derzeit in der klinischen Phase der Zulassung für die Behandlung

1. Einleitung 19
allergischer Atemwegserkrankungen befindet (Szelenyi and Pahl, 2002). In Abbildung 8 sind
Design, Metabolismus und allgemeine Struktur von Soft Steroiden dargestellt.
Abbildung 7: Histologischer Schnitt durch die untere Nasenmuschel eines Patienten mit allergischer
Rhinitis. Hämatoxylin-Eosin-Färbung. Links: 100-fache Vergrößerung. Man beachte das mehrreihige
Flimmerepithel, die deutliche Basalmembran und das lockere Bindegewebe der Lamina propria, in welches in
den tieferen Schichten zahlreiche Immunzellen eingelagert sind. Rechts: 200-fache Vergrößerung.
Ausschnittsvergrößerung aus den tiefen Schichten der Nasenschleimhaut desselben Patienten. Man beachte
die deutliche Infiltration von eosinophilen Granulozyten, Lymphozyten und Plasmazellen.
[Die Aufnahmen wurden freundlicherweise von Prof. Dr. med. habil. Klaus Richter, Gemeinschaftspraxis für
Pathologie Prof. Dr. K. Richter und Dr. W. Beschow, Hannover zur Verfügung gestellt.]
200 µm 100 µm
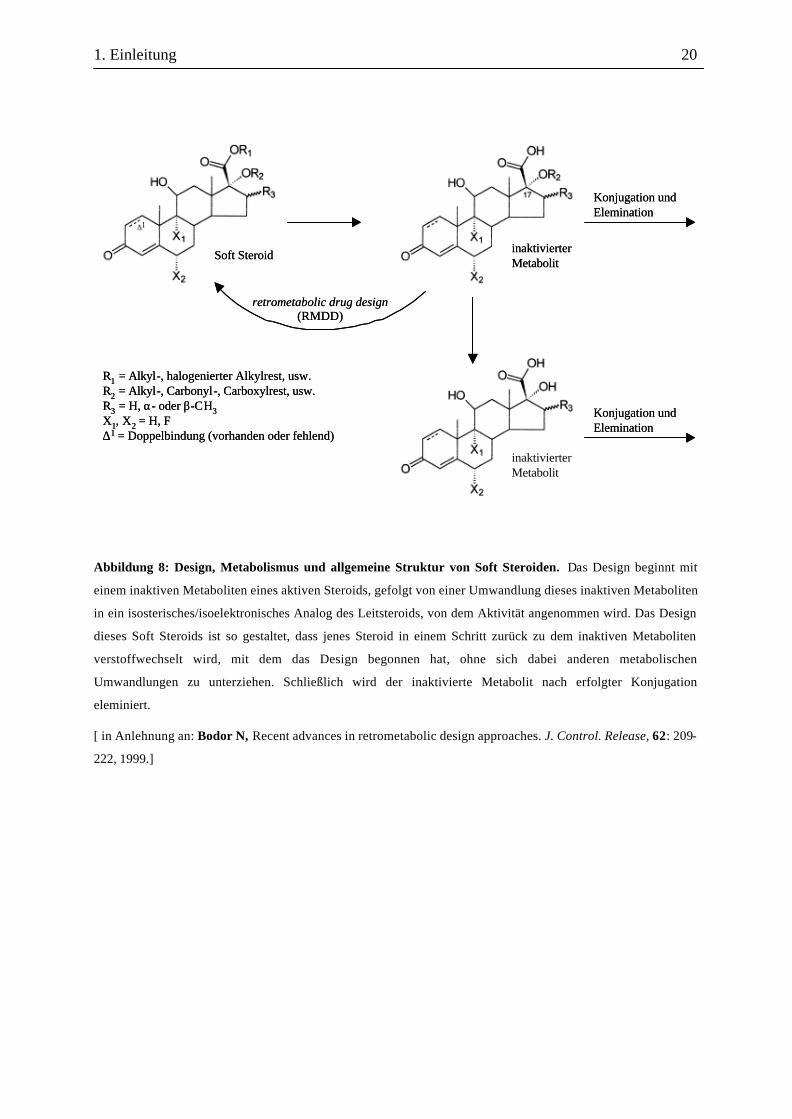
1. Einleitung 20
Abbildung 8: Design, Metabolismus und allgemeine Struktur von Soft Steroiden. Das Design beginnt mit
einem inaktiven Metaboliten eines aktiven Steroids, gefolgt von einer Umwandlung dieses inaktiven Metaboliten
in ein isosterisches/isoelektronisches Analog des Leitsteroids, von dem Aktivität angenommen wird. Das Design
dieses Soft Steroids ist so gestaltet, dass jenes Steroid in einem Schritt zurück zu dem inaktiven Metaboliten
verstoffwechselt wird, mit dem das Design begonnen hat, ohne sich dabei anderen metabolischen
Umwandlungen zu unterziehen. Schließlich wird der inaktivierte Metabolit nach erfolgter Konjugation
eleminiert.
[ in Anlehnung an: Bodor N, Recent advances in retrometabolic design approaches. J. Control. Release, 62: 209-
222, 1999.]
Soft Steroid inaktivierter Metabolit
Konjugation und Elemination
Konjugation und Elemination
retrometabolic drug design(RMDD)
R1 = Alkyl-, halogenierter Alkylrest, usw.R2 = Alkyl-, Carbonyl-, Carboxylrest, usw.R3 = H, α- oder β-CH3X1, X2 = H, F∆1 = Doppelbindung (vorhanden oder fehlend)
inaktivierter Metabolit
Soft Steroid inaktivierter Metabolit
Konjugation und Elemination
Konjugation und Elemination
retrometabolic drug design(RMDD)
R1 = Alkyl-, halogenierter Alkylrest, usw.R2 = Alkyl-, Carbonyl-, Carboxylrest, usw.R3 = H, α- oder β-CH3X1, X2 = H, F∆1 = Doppelbindung (vorhanden oder fehlend)
inaktivierter Metabolit

1. Einleitung 21
1.6 Ziel der Arbeit
Trotz des wachsenden Interesses der pharmazeutischen Industrie an der nasalen
Applikationsform ist die menschliche Nasenschleimhaut bislang kaum hinsichtlich ihrer
Arzneistoff-metabolisierenden Kompetenz untersucht. Ziel dieser Arbeit war es deshalb, die
menschliche respiratorische Nasenschleimhaut bezüglich vorhandener Enzymsysteme sowie
metabolischer Kompetenz näher zu charakterisieren. Schwerpunktmäßig wurden dabei die
verschiedenen Isoformen des Cytochrom P450-Systemes (CYPs) untersucht. Da Rauchen
nachweislich einen Einfluss auf die pulmonale CYP-Expression bei Mensch und Ratte besitzt
(Eke and Iscan, 2002; Haussmann et al., 1998; Hukkanen, 2000; Willey et al., 1997) und bei
der Ratte eine CYP-Enzyminduktion durch Zigarettenrauch auch in der nasalen Schleimhaut
beobachtet wurde (Haussmann et al., 1998; Wardlaw et al., 1998), ist davon auszugehen, dass
auch in der menschlichen Nasenschleimhaut Unterschiede im Genexpressionsmuster
abhängig vom Raucherstatus auftreten. Um dieses zu untersuchen, wurden die Spender in eine
Raucher- und eine Nicht-Raucher-Gruppe unterteilt. Die Nasenschleimhaut stammte dabei
von Patienten, die sich aufgrund einer Muschelhyperplasie einer partiellen Turbinektomie der
Concha nasalis inferior unterzogen haben.
Im ersten Versuchsabschnitt wurde die totale RNA der Schleimhaut isoliert, qualitativ und
quantitativ bestimmt und mittels RT-PCR auf die Expression verschiedener Phase I- und
Phase II-Enzyme, regulatorischer Rezeptoren sowie Entzündungsmediatoren untersucht.
Durch Ultrazentrifugation wurden im zweiten Versuchsabschnitt Mikrosomen gewonnen,
wobei aufgrund der geringen Gewebemengen die Nasenmuscheln mehrerer Patienten gepoolt
werden mussten. Auch hierbei wurde die Unterteilung in Raucher und Nicht-Raucher
beibehalten. Die Mikrosomen wurden für die anschließende Proteinbestimmung verwendet, in
deren Rahmen bestimmte Cytochrom P450 Monooxygenasen durch Western blotting weiter
charakterisiert wurden, so dass Gen- und Proteinexpression dieser Isozyme verglichen werden
konnten. Zur Untersuchung der metabolischen Kompetenz wurden die mikrosomalen
Membranen für verschiedene katalytische Assays verwendet. Der Schwerpunkt ruhte hierbei
auf der Verstoffwechselung des Soft Steroids Loteprednol-Etabonat (LE), dessen
Metabolitenprofil mittels Radio-HPLC bestimmt wurde. Abschließend wurden die
Enzymaktivitäten gegenüber den CYP-Markersubstraten Testosteron und Ethoxyresorufin
ermittelt.

2. Material und Methoden 22
2. Material und Methoden
2.1 Material
2.1.1 Probenmaterial
humane Nasenmuscheln Friederiken-Stift, Hannover;
Medizinische Hochschule Hannover
2.1.2 Chemikalien
2.1.2.1 Reagentien für die Untersuchung der Genexpression
Agarose NEEO Ultra-Qualität, Rotigarose für Carl Roth GmbH + Co, Karlsruhe
die Elektrophorese (Code 2267-3; Charge 43151566)
Bromphenolblau Sigma Chemical Company, St.Louis, USA
(Code B-8026; Charge 18H3630)
Custom Primers, 50nM, entsalzt Invitrogen GmbH, Karlsruhe
(Primerliste einschließlich PCR-Bedingungen siehe Anhang Seite VII bis VIII)
DNA ladder, 100bp Invitrogen GmbH, Karlsruhe
(Code 15628-050; Charge 1131233)
DNA Typing Grade 50X TAE Buffer Life Technologies, Paisley, Schottland
(Code 24710-030; Charge 1101351)
dNTP Mix (5mM für RT) Qiagen, Hilden
(Code 1010355; Charge 10921673)
dNTP Solution 100mM, PCR grade MBI Fermentas GmbH, St. Leon-Rot
(Code R0181; Charge 0273)
Ethanol 96% Riedel-de Haën Sigma-Aldrich, Seelze
(Code 24105; Charge 13330)

2. Material und Methoden 23
Ethidiumbromidlösung 10 mg/ml Sigma Chemical Company, St.Louis, USA
(Code E-1510; Charge 117H8509)
Glycerin Carl Roth GmbH + Co, Karlsruhe
(Code 3783.2; Charge 04252672)
HotStar Taq, 5 units/µl Qiagen, Hilden
(Code 1007837; Charge 11239460)
2-Mercaptoethanol Sigma Chemical Company, St.Louis, USA
(Code M-7522; Charge 105F-02785)
Omniscript Reverse Transcriptase 4 units/µl Qiagen, Hilden
(Code 1010890; Charge 11229931)
PCR Buffer, 10x Qiagen, Hilden
(Code 1005479; Charge 11239890)
Random Hexamers Promega, Mannheim
(Code C118A; Charge 13232211)
RNA 6000 Nano LabChip Agilent Technologies Deutschland GmbH,
(Code 5065-4476; Charge EC18BK02) Waldbronn
RNA 6000 Nano Reagents & Supplies Agilent Technologies Deutschland GmbH,
(Code 5065-4476; Charge EC21RK02) Waldbronn
RNasin Ribonuclease Inhibitor 10 000 units Promega, Mannheim
(Code N211B; Charge 13000713)
RNeasy Mini Kit Qiagen, Hilden
(Code 74104; Charge 11234541)
10 x RT Buffer Qiagen, Hilden
(Code 1010883; Charge 10922292)
Xylencyanol Sigma Chemical Company, St.Louis, USA
(Code X-4126; Charge 117H3626)

2. Material und Methoden 24
2.1.2.2 Reagentien für die mikrosomalen Untersuchungen
Acetonitril Carl Roth GmbH + Co, Karlsruhe
(Code 8825.2; Charge 20281)
30% Acrylamid / 0,8% Bisacrylamid Carl Roth GmbH + Co, Karlsruhe
(Code 3029.1; Charge 18254163)
11-α-Hydroxyprogesteron Sigma Chemical Company, St.Louis, USA
(Code H-5502; Charge 17H0285)
Ammoniumacetat Merck KG, Darmstadt
(Code 1.01116.0500; Charge A180816 932)
Ammoniumperoxodisulfat Merck KG, Darmstadt
(Code 1.01201.0100; Charge K27070601)
β-Glucuronidase (EC 3.2.1.31) Sigma Chemical Company, St.Louis, USA
(Code G-0751; Charge 120K1321)
β-NADPH, reduced form Sigma Chemical Company, St.Louis, USA
(Code N-1630; Charge 81K7067)
Bicinchoninic Acid Solution Sigma Chemical Company, St.Louis, USA
(Code B-9643; Charge 60K5300)
Bovine Serum Albumin Fraction V PAA Laboratories GmbH, Linz,
(Code K41-001-100; Charge G16112-207) Österreich
Chemicon ECL Chemicon, Temecula, USA
(Code 2230; Charge 20020402)
Dimethylsulfoxid Merck KG, Darmstadt
(Code 1.02952.1000; Charge K30558552 220)
EDTA Sigma Chemical Company, St.Louis, USA
(Code E-5134; Charge 100K0284)
Essigsäure Carl Roth GmbH + Co, Karlsruhe
(Code 6755.1; Charge 31150335)

2. Material und Methoden 25
Ethanol 96% Riedel-de Haën Sigma-Aldrich, Seelze
(Code 24105; Charge 13330)
Ethoxyresorufin Sigma Chemical Company, St.Louis, USA
(Code E-3763; Charge 22K4012)
Ethyl Acetat Mallinckrodt Baker B.V., Holland
(Code 8037; Charge 9930110002)
Glycin Carl Roth GmbH + Co, Karlsruhe
(Code 3908.2; Charge 49151490)
Heptan Fluka Chemie GmbH, Schweiz
(Code 51745; Charge 423800/1)
Kupfer-(II)-sulfat Carl Roth GmbH + Co, Karlsruhe
(Code P023.1; Charge 44046989)
[4-14C] Loteprednol Etabonate Amersham pharmacia biotech,
(Code CFQ13089) Buckinghamshire, England
Loteprednol Etabonate VIATRIS GmbH & Co KG, Frankfurt
(Code X-387; Charge 25)
Methanol Carl Roth GmbH + Co, Karlsruhe
(Code 4627.1; Charge 15253746)
Natriumchlorid Carl Roth GmbH + Co, Karlsruhe
(Code 3957.1; Charge 02146852)
Potassium Chloride Sigma Chemical Company, St.Louis, USA
(Code P-5404; Charge 87H06665)
Precision blue protein standard prestained Bio-Rad Laboratories GmbH, München
(Code 161-0372; Charge 91731)
Primärantikörper für Western blot
(Liste der verwendeten Primärantikörper siehe Anhang Seite IX)
2-Propanol Merck KG, Darmstadt
(Code 1.09634.2500; Charge K29036034 112)

2. Material und Methoden 26
Resorufin Sigma Chemical Company, St.Louis, USA
(Code R-3257; Charge 18H3639)
Roti-Block 10x Konzentrat Carl Roth GmbH + Co, Karlsruhe
(Code A151.1; Charge 06148259)
Roti-Load1 4x Konzentrat Carl Roth GmbH + Co, Karlsruhe
(Code K929.1; Charge 27045406)
Rotiszinteco plus Carl Roth GmbH + Co, Karlsruhe
(Code 0016.2; Charge 40251611)
SDS ultra pure Carl Roth GmbH + Co, Karlsruhe
(Code 2326.2; Charge 48046682)
Sekundärantikörper für Western blot Chemicon, Temecula, USA
(Liste der verwendeten Sekundärantikörper siehe Anhang Seite IX)
Sodium hydroxide Sigma Chemical Company, St.Louis, USA
(Code S-5881; Charge 127H01821)
Sucrose Sigma Chemical Company, St.Louis, USA
(Code S-9378; Charge 100K0121)
TEMED p.a. Carl Roth GmbH + Co, Karlsruhe
(Code 2367.1; Charge 06141122)
Testosteron Sigma Chemical Company, St.Louis, USA
(Code T1500; Charge 39H0638)
Tris Carl Roth GmbH + Co, Karlsruhe
(Code 4855.2; Charge 50152286)
Trizma Base Sigma Chemical Company, St.Louis, USA
(Code T-1503; Charge 11K5424)
Trizma Hydrochloride Sigma Chemical Company, St.Louis, USA
(Code T-3253; Charge 69H5435)
Tween20 Carl Roth GmbH + Co, Karlsruhe
(Code 9127.1; Charge 47046531)

2. Material und Methoden 27
Western Lightning Chemiluminescence PerkinElmer Life Sciences, Boston, USA
Reagent Plus (Code NEL105; Charge 254753)
2.1.3 Geräte und Verbrauchsmaterialien
Agilent 2100 Bioanalyzer Agilent Technologies Deutschland GmbH,
(Serien-Nr. DE01727542) Waldbronn
Analysenwaage Sartorius MC210S Sartorius AG, Göttingen
(Serien-Nr. 11410980)
Biometra Standard Power Pack P25 Biometra, Göttingen
(Serien-Nr. 4111139)
BioRad PowerPac 200 Bio-Rad Laboratories GmbH, München
(Serien-Nr. 285BR04868)
BioRad PowerPac 300 Bio-Rad Laboratories GmbH, München
(Serien-Nr. 283BR09143)
BioRad Sub-CellGT Elektrophoresekammer Bio-Rad Laboratories GmbH, München
(Serien-Nr. 61S01069)
C18-Nucleosil-Säule (250 x 4 mm, Macherey-Nagel, Düren
Partikelgröße 5 µm, Serien-Nr. 1106105)
Chirurgische Pinzette, 14,5 cm Medicalis, Garbsen
Chirurgische Schere, gerade, spitz/spitz, 14,5 cm Medicalis, Garbsen
ELISA Reader Dynatech MR5000 Dynatech, Ohio, USA
(Serien-Nr. G3112)
Emission Filter 620/10 (615-625 nm) Bio-Rad Laboratories GmbH, München
(Code 1702426)
Excitation Filter 510/10 (505-515 nm) Bio-Rad Laboratories GmbH, München
(Code 1702423)

2. Material und Methoden 28
Filterpapier Merck KG, Darmstadt
(Code 1001-931)
Flow Scintillation Analyzer 500TR Series Packard BioScience Company, USA
(Serien-Nr. 421035)
Fluorometer VersaFluor Bio-Rad Laboratories GmbH, München
(Serien-Nr. 435 BR 0769)
Gelkammer Biometra Minigel-Twin Typ G42 Biometra, Göttingen
(Serien-Nr. 1409129)
Gewebehomogenisator Dounce 7 ml und 1 ml Wheaton, USA
Glasplatte, ausgeschnitten Whatman Biometra, Göttingen
(Code 010-003)
Glasplatte mit fixierten Spacern 0,6 mm Whatman Biometra, Göttingen
(Code 010-002)
HPLC Series 1100 für Testosteronmetabolismus Hewlett Packard GmbH, Waldbronn
[ Degasser G1322A (Serien-Nr. JP 73014117)
Quat Pump G1311A (Serien-Nr. DE 83104972)
ALS G1313A (Serien-Nr. DE 82206896)
Col Comp G1316A (Serien-Nr. DE 82207725)
DAD G1315A (Serien-Nr. DE 90604736) ]
HPLC Series 1100 für Radio-HPLC Hewlett Packard GmbH, Waldbronn
[ Degasser G1322A (Serien-Nr. JP 73014143)
Quat Pump G1311A (Serien-Nr. DE 83104953)
ALS G1329A (Serien-Nr. DE 91603535)
ALS Therm G1330A (Serien-Nr. DE 82203511)
Col Comp G1316A (Serien-Nr. DE 82207724)
VWD G1314A (Serien-Nr. JP 73797282) ]
Kamm 10-zähnig 0,6 mm für Minigel und Whatman Biometra, Göttingen
Minigel-Twin (Code 010-016)
Klammern für Gelelektrophorese Whatman Biometra, Göttingen
(Code 010-007)

2. Material und Methoden 29
Kodak Digital Science Image Station 440 CF Kodak, USA
(Serien-Nr. 207102)
LS 6500 Multi-Purpos Scintillation Counter Beckmann Coulter, Palo Alto, USA
(Serien-Nr. 7070019)
Magnetrührer Heidolph MR 3000 Heidolph Instruments GmbH & Co.KG,
(Serien-Nr. 030008262) Schwabach
Mahlkugeln Wolframcarbid 5 mm F. Kurt Retsch GmbH & Co KG, Haan
(Code 05.368.0038)
Megafuge 2.0R Heraeus Kendro Laboratory Products, Osterode
(Serien-Nr. 268322)
Mini Trans-Blot Elektrophoretic Transfer Cell Bio-Rad Laboratories GmbH, München
(Serien-Nr. 37S/4382)
MS2 Minishaker IKA IKA Works, INC., Wilmington, USA
(Serien-Nr. 03.059839)
Nunc-Immuno Plates NUNC GmbH & Co KG, Wiesbaden
(Code 442 404)
Polycarbonatröhrchen Beckmann Coulter, Palo Alto, USA
(Code 355618; Charge 010115)
PVDF-Transfer-Membran NEN Life Science Products, Boston,
(Code NEF102; Charge 183321) USA
Rotilabo-Einsätze 100 µl Carl Roth GmbH + Co, Karlsruhe
(Code C516.1; Charge 69752226AS7)
Rotor 70Ti Beckmann Coulter, Palo Alto, USA
(Serien-Nr. 01E 933)
Roto-Shake Genie Scientific Industries, Inc.,
(Serien-Nr. 1580) New York, USA
Schwingmühle Retsch MM 200 F. Kurt Retsch GmbH & Co KG, Haan
(Serien-Nr. 200609018G)

2. Material und Methoden 30
Sigma Tischzentrifuge 1-15 Sigma Laborzentrifugen, Osterode
(Serien-Nr. 83054)
Silikonabdichtung 1,0 mm für Minigel und Whatman Biometra, Göttingen
Minigel-Twin (Code 010-005)
Spherisorb ODS-2-Säule (250 x 4 mm, Latek, Eppelheim
Partikelgröße 3 µm, Serien-Nr. 9802010)
Test tube heater SHT 20 Stuart Scientific Co. Ltd., Surrey, England
(Serien-Nr. 5051)
Thermocycler T3 Biometra, Göttingen
(Serien-Nr. 0904125, 0904126, 0904134)
Thermomixer compact Eppendorf-Netheler-Hinz GmbH,
(Serien-Nr. 5350 02163) Hamburg
Thermomixer comfort Eppendorf-Netheler-Hinz GmbH,
(Serien-Nr. 5355 03052) Hamburg
Trockeneis Kohlensäurewerk, Laatzen
Ultra Turrax® T8 IKA Labortechnik, Staufen
(Serien-Nr. 00.080400)
Ultrazentrifuge Optima LE-80K Beckmann Coulter, Palo Alto, USA
(Serien-Nr. COL Ø 1C27)
Verschlüsse für Polycarbonatröhrchen Beckmann Coulter, Palo Alto, USA
(Code 338824)
Waage BP 3100P Sartorius AG, Göttingen
(Serien-Nr. 11903531)
Wärmeschrank Typ UE 400 Memmert GmbH + CoKG, Schwabach
(Serien-Nr. c498-0396)
Wasserbad Julabo SW21 Julabo Labortechnik GmbH, Seelbach
(Serien-Nr. 03904420519)
Waters Oasis HLB 3cc (60mg) Extraction Waters Corporation, USA
Cartridges (Code WAT094226; Charge W2147J4)

2. Material und Methoden 31
White virgin PTFE septa, 8mm Agilent Technologies Deutschland GmbH,
(Code 5183-4434) Waldbronn
Zentrifuge Mikro 22R Hettich Zentrifugen, Tuttlingen
(Serien-Nr. 0001096-03-00)
Zentrifugenröhrchen 15 ml Sarstedt, Nümbrecht
(Code 62.554.502; Charge 2182 1603)
Zentrifugenröhrchen 50 ml Sarstedt, Nümbrecht
(Code 62.547.254; Charge 2196 7023)
2.2 Methoden
2.2.1 Gewinnung und Transport des Probenmateriales
Respiratorische Nasenschleimhaut wurde von Patienten gewonnen, die sich aufgrund einer
Muschelhyperplasie einer partiellen Turbinektomie der Concha nasalis inferior unterzogen hatten
(Auflistung der Patienten einschließlich der Patientendaten siehe Anhang Seiten I bis VI). Die
Personen wurden vor dem operativen Eingriff über das Projekt aufgeklärt und haben ihre
Einwilligung für die Untersuchung erteilt.
Innerhalb von 10 – 15 Minuten nach ihrer operativen Entfernung wurden die Nasenmuscheln mittels
Trockeneis schockgefroren und bei –80°C gelagert.
2.2.2 Genexpressionsuntersuchungen
Sämtliche Untersuchungen zur Genexpression erfolgten in den Laboren T2.09 und T2.023 des
Fraunhofer Institutes für Toxikologie und Experimentelle Medizin.

2. Material und Methoden 32
2.2.2.1 RNA-Isolation
Die RNA-Isolation aus der humanen respiratorischen Nasenschleimhaut wurde nach dem Protokoll
des Herstellers des RNeasy Mini Kits von Qiagen durchgeführt. Abweichend vom Protokoll wurde
die gewonnene RNA jedoch nicht im mitgelieferten RNase-freien Wasser, sondern in 50 µl DEPC-
(Diethylpyrocarbonat) Wasser eluiert. Um hauptsächlich epitheliales Gewebes zu erhalten, wurden
die oberflächlichen Schichten der conchalen Schleimhaut vorsichtig von den darunter liegenden
knorpeligen und knöchernen Trägerstrukturen abgekratzt. Anschließend wurde das im Buffer RLT
(lysis buffer) des RNeasy Mini Kits befindliche Gewebe mit jeweils einer Bleikugel pro Probe in
der Schwingmühle zwei Mal 60 Sekunden bei einer Frequenz von 15 Hz homogenisiert.
Die qualitative und quantitative RNA-Messung erfolgte mit dem Agilent Bioanalyzer der Agilent
Technologies Deutschland GmbH, Waldbronn nach Herstellerprotokoll, wozu 1 µl RNA je Probe
eingesetzt wurde. Zur Beurteilung der RNA-Qualität wurden die beiden ribosomalen Banden
herangezogen, deren Verhältnis (28S rRNA : 18S rRNA) im Idealfall einer reinen und intakten RNA
2:1 ist. Die Schärfe der ribosomalen Banden wurde nicht für Qualitätsbeurteilung herangezogen.
Proben mit einer RNA-Konzentration > 60 ng/µl wurden für die nachfolgende RT-(Reverse
Transkriptase) Reaktion verwendet, bei geringerer RNA-Ausbeute oder bei mangelnder Qualität
wurde die RNA-Isolation wiederholt und die RNA erneut gemessen.
2.2.2.2 RT-Reaktion und RT-PCR
Die Umschreibung der RNA in cDNA wurde mit der Omniscript Reverse Transcriptase von Qiagen
durchgeführt. Für die reverse Transkription wurde jeweils 1 µg RNA eingesetzt und mit der
entsprechenden Menge DEPC-Wasser auf ein Volumen von 12,75 µl eingestellt. Anschließend
wurde die RNA für 5 min bei 65°C denaturiert. Nach Herunterkühlen der Probe auf
Raumtemperatur wurde der Reaktionsmix bestehend aus 2 µl 10 x RT-Puffer, 2 µl dNTP Mix (5
mM), 2 µl Random Hexamers (5 µM), 0,25 µl RNasin (5 µM) und 1 µl Omniscript Reverse
Transcriptase hinzugegeben und die RT-Reaktion bei 37°C gestartet. Da bei Probe M-R 4 aufgrund
der geringen RNA-Konzentration von nur 67 ng/µl bei Einsatz von 1 µg RNA das Volumen von
12,75 µl überschritten wurde, musste bei dieser Probe der 1,5-fache RT-Ansatz gewählt werden,
d.h. die RNA wurde mittels DEPC-Wasser auf ein Volumen von 19,12 µl gebracht, die 1,5-fache
Menge des genannten Reaktionsgemisches hinzugefügt und die Reaktion gestartet. Im Anschluss an
die einstündige Inkubation bei 37°C, während der die RNA in cDNA umgeschrieben wurde, erfolgte

2. Material und Methoden 33
eine erneute Denaturierung bei 95°C für 5 min. Die entstandenen cDNA wurde bei –20°C
aufbewahrt.
Unter Zugabe von 20 µl A. bidest. bzw. von 10 µl A. bidest. im Falle des 1,5-fachen RT-Ansatzes
wurde die in der reversen Transkription gewonnene cDNA auf eine Konzentration von 25 ng/µl
verdünnt und pro PCR-Ansatz jeweils 1 µl cDNA (25 ng) eingesetzt. Die PCR wurde mit der
HotStar Taq DNA Polymerase von Qiagen durchgeführt. Zum PCR-Cocktail, bestehend aus jeweils
14,375 µl A. bidest., 2 µl 10x PCR Buffer, 0,5 µl dNTP Solution (10mM), 0,125 µl HotStar Taq
Polymerase sowie 2 µl der jeweiligen 10 µM Primergebrauchslösung ( je 10 µl 5´ bzw. 3´ Primer auf
80 µl A. bidest.) wurden pro Ansatz 1 µl cDNA hinzugegeben und die Reaktion im Thermocycler für
15 min bei 95°C gestartet. Für die zyklische Amplifizierung wurden folgende Bedingungen gewählt:
Aktivierung der Polymerase 15 min bei 95°C, Denaturierung 45 s bei 95°C, Annealing 60 s mit
primerspezifischen Temperaturen (siehe Anhang Seite VII bis VIII), Elongation 60 s bei 72°C. Je
nach Oligomer wurden unterschiedliche Zyklenzahlen verwendet. Im Anschluss an den letzten Zyklus
erfolgte stets eine Reaktion für 7 min bei 72°C sowie ein Herunterkühlen des Gerätes auf 4°C zur
Konservierung der Proben.
Parallel zu den Proben wurden bei der PCR jeweils eine Leber- und eine Negativkontrolle
mitgeführt. Bei der Leberkontrolle handelte es sich um cDNA, die aus RNA humaner
Leberzellkulturen durch reverse Transkription gewonnen wurde. Leber wurde als Kontrolle
verwendet, da es sich bei der Leber um dasjenige Organ handelt, in dem die meisten Cytochrom
P450 Monooxygenasen exprimiert sind. Nicht in der Leber exprimiert sind dagegen die Cytochrome
2A13 (Koskela et al., 1999; Su et al., 2000), 2F1 (Carr et al., 2003; Chen et al., 2002; Ding and
Kaminsky, 2002, Nishimura et al., 2003), 2S1 (Rylander et al., 2001), 3A5 (Hukkanen et al.,
2003), 4B1 (Nhamburo et al., 1989; Nishimura et al., 2003) sowie die Enzyme FMO2 (Dolphin et
al., 1998) und UGT 2A1 Jedlitschky et al., 1999). Aus methodischer Sicht wurde damit in Kauf
genommen, dass nicht bei allen untersuchten Genen in der Leberkontrolle RT-PCR Amplifikate
nachgewiesen werden konnten. Bei der Negativkontrolle wurde anstelle der cDNA 1 µl A. bidest.
eingesetzt.
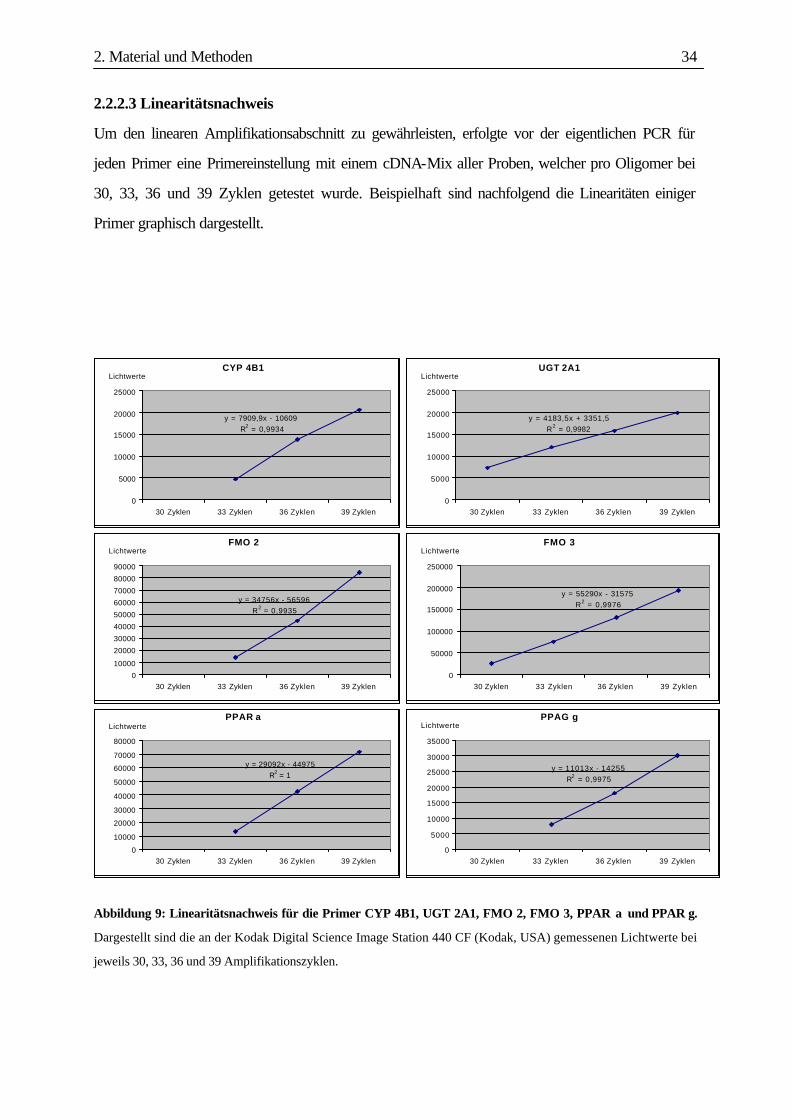
2. Material und Methoden 34
2.2.2.3 Linearitätsnachweis
Um den linearen Amplifikationsabschnitt zu gewährleisten, erfolgte vor der eigentlichen PCR für
jeden Primer eine Primereinstellung mit einem cDNA-Mix aller Proben, welcher pro Oligomer bei
30, 33, 36 und 39 Zyklen getestet wurde. Beispielhaft sind nachfolgend die Linearitäten einiger
Primer graphisch dargestellt.
Abbildung 9: Linearitätsnachweis für die Primer CYP 4B1, UGT 2A1, FMO 2, FMO 3, PPAR α und PPAR γ.
Dargestellt sind die an der Kodak Digital Science Image Station 440 CF (Kodak, USA) gemessenen Lichtwerte bei
jeweils 30, 33, 36 und 39 Amplifikationszyklen.
CYP 4B1
0
5000
10000
15000
20000
25000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 7909,9x - 10609R2 = 0,9934
UGT 2A1
0
5000
10000
15000
20000
25000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 4183,5x + 3351,5R2 = 0,9982
FMO 2
0
10000
20000
30000
40000
50000
60000
70000
80000
90000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 34756x - 56596R2 = 0,9935
FMO 3
0
50000
100000
150000
200000
250000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 55290x - 31575R2 = 0,9976
PPAR a
0
10000
20000
30000
40000
50000
60000
70000
80000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 29092x - 44975R2 = 1
PPAG g
0
5000
10000
15000
20000
25000
30000
35000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 11013x - 14255R2 = 0,9975
CYP 4B1
0
5000
10000
15000
20000
25000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 7909,9x - 10609R2 = 0,9934
UGT 2A1
0
5000
10000
15000
20000
25000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 4183,5x + 3351,5R2 = 0,9982
FMO 2
0
10000
20000
30000
40000
50000
60000
70000
80000
90000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 34756x - 56596R2 = 0,9935
FMO 3
0
50000
100000
150000
200000
250000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 55290x - 31575R2 = 0,9976
PPAR a
0
10000
20000
30000
40000
50000
60000
70000
80000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 29092x - 44975R2 = 1
PPAG g
0
5000
10000
15000
20000
25000
30000
35000
30 Zyklen 33 Zyklen 36 Zyklen 39 Zyklen
Lichtwerte
y = 11013x - 14255R2 = 0,9975

2. Material und Methoden 35
2.2.2.4 Auswertung der Amplifikate
Zur amplifizierten cDNA wurden 6 µl 6x DNA-loading buffer – bestehend aus 40 mg
Bromphenolblau, 20 mg Xylencyanol, 100,64 g Glycerin ad 200 ml A. bidest. - hinzugegeben und je
Probe 10 µl in eine Kammer auf einem 1,5%igen Agarosegel (in 100 ml 1x TAE-Puffer, versetzt mit
1 µl Ethidiumbromid) aufgetragen. Der Größenkontrolle diente dabei der in die jeweils erste Kammer
gegebene DNA-Molekulargewichtsmarker (nachfolgend Ladder genannt). Nach 45-minütiger
Auftrennung der cDNA bei 120 V in der Elektrophoresekammer erfolgte die Darstellung mittels
UV-Licht an der Kodak Digital Science Image Station 440 CF (Kodak, USA). Durch die
Verwendung spezifischer Primer ausreichender Länge und die Überprüfung der erwarteten bp-Länge
konnten die Amplifikate ausreichend verifiziert werden. Eine eigenständige Verifizierung in Form
einer Sequenzierung oder eines Restriktionsenzymverdaus wurde nicht durchgeführt.
Für die Auswertung wurden die Bandenstärken der PCR-Produkte der zu untersuchenden Gene mit
der 1D-Image Analysis Software für Windows Version 3.5.3B geldensitrometrisch ermittelt und die
Mittelwerte sowie Standardabweichungen aller Kandidatengene für die Raucher und die Nicht-
Raucher, sowie die männlichen und weiblichen Patienten berechnet. Die statistische
Signifikanzanalyse wurde mittels Mathematica4.2 der Wolfram Research, Inc. (www.wolfram.com)
durchgeführt, wobei das Programmpaket „Statistics´HypothesisTests´“ mit dem Programm
„MeanDifferenceTest“ und der Option TwoSided->True zur Anwendung kam. Voraussetzung für
die Anwendbarkeit dieses Tests ist das Vorliegen einer Normalverteilung, was vorab mit dem
Shapiro-Wilk-Test gesondert überprüft wurde.
Gemäß SOP wurde pro Patient und Primer eine RT-PCR durchgeführt, weshalb weder ein Intra-,
noch ein Inter-Assay Variationskoeffizient angegeben werden kann.
2.2.3 Proteinexpressionsuntersuchungen
Die Experimente zur Untersuchung der Proteinexpression wurden innerhalb des Fraunhofer Institutes
für Toxikologie und Experimentelle Medizin in den Räumen T2.03, T2.05, T2.09 und T2.033
durchgeführt.

2. Material und Methoden 36
2.2.3.1 Herstellung der Pufferlösungen
KCl-Puffer 0,15 M: 11,18 g KCl
ad 1 l A. bidest.
10x SDS-Laufpuffer 150g Tris
720g Glycin
50g SDS
ad 5 l A. bidest.
1x SDS-Laufpuffer 100 ml 10x SDS-Laufpuffer
900 ml A. bidest.
10x TBS-Puffer 121g Tris
400g NaCl
ad 5 l A. bidest.
mit 1 M HCl auf pH = 7,6 eingestellt
1x TBS-Puffer ohne Tween 100 ml 10x TBS-Puffer
900 ml A. bidest.
1x TBS-T 0,1% 1000 ml 1x TBS-Puffer ohne Tween
1 ml Tween20
10x Transfer-Puffer 151,5g Tris
720g Glycin
ad 5 l A. bidest.
1x Transfer-Puffer 100 ml 10x Transfer-Puffer
900 ml A. bidest.
pH = 8,3
TRIS-Stock 0,1 M: 6,35 g Trizma HCl
1,18 g Trizma Base
ad 500 ml A. bidest.
mit 1 M HCl auf pH = 7,4 eingestellt

2. Material und Methoden 37
TRIS-Sucrose-Puffer: 42,79 g Sucrose
100 ml TRIS-Stock
0,93 g EDTA
ad 500 ml A. bidest.
2.3.3.2 Isolation mikrosomaler Proteine durch Ultrazentrifugation
Aufgrund der geringen Gewebemengen mussten die Nasenmuscheln mehrerer Patienten gepoolt
werden, um für die nachfolgenden proteinanalytischen und metabolischen Untersuchungen
ausreichende Mengen an mikrosomalem Protein zu erhalten. So wurden insgesamt 15 Pools aus den
Nasenmuscheln von drei bis vierzehn Patienten gebildet (Patientendaten zu den einzelnen Pools siehe
Anhang Seite II bis VI). Eine Übersicht über Art und nachfolgende Verwendung der mikrosomalen
Pools gibt Abbildung 10.
Pool Art des Pools
Anzahl der
Patienten
WB LE I LE II Testost. EROD
60 min
120 min
mit Gluc.
ohne Gluc.
1 M-NR 5 X X 1 - - X - 2 M-NR 5 - X 1 X 2 - - - - 4 M-NR 6 X X 2 X 1 - - X - 5 M-NR 6 - X 2 X 1 - - - X 6 W-NR 6 X X 2 - - - X X 7 M-R 6 X X 2 X 1 - - - X 8 W-R 3 X X 2 - - - - - 9 M-NR 14 X X 2 X 2 - - X X 10 M-R 6 X X 2 X 2 - - X X 11 W-NR 5 X X 2 X 2 - - - - 12 M-NR 10 - - - X X - - 13 M-R 7 - - - X X - - 14 W-NR 10 - - - X X - - 15 W-R 5 - - - X X - - 16 W-? 4 - - - X X - -
Abbildung 10: Art und Verwendung der mikrosomalen Pools
Abkürzungen: EROD = Ethoxyresorufin-O-deethylase; LE-I = Loteprednol-Etabonat-Metabolismus Teil I
(verschiedene Inkubationszeiten) mit 1 = 1. Versuchsserie und 2 = 2. Versuchsserie; LE-II = Loteprednol-Etabonat-
Metabolismus Teil II (mit und ohne Glucuronidase-Zusatz); M = männlich; NR = Nicht-Raucher; R = Raucher;
Testost. = Testosteron-Metabolismus; W = weiblich; WB = Western blot.

2. Material und Methoden 38
Nach Bestimmung der Poolgewichte wurden die jeweiligen Organpools in der dreifachen Menge
0,15 M KCl-Puffer mit einer Schere zerkleinert und anschließend mit dem Ultra Turrax
homogenisiert. Das gewonnene Homogenat wurde in ein Polycarbonatröhrchen gefüllt, mit 0,15 M
KCl-Puffer aufgefüllt und für 30 min bei 15.000 Upm und 4°C in der Ultrazentrifuge Optima LE-
80K (Beckmann Coulter, Palo Alto, USA) zentrifugiert. Der Überstand wurde entnommen und für
60 min bei 55.000 Upm und 4°C erneut zentrifugiert. Das entstandene Pellet wurde in einigen
Millilitern 0,15 M KCl-Puffer aufgenommen und im Gewebehomogenisator Dounce (7ml)
homogenisiert. Die erhaltene Suspension wurde nach Auffüllen mit 0,15 M KCl-Puffer nochmals für
60 min bei 55.000 Upm und 4°C zentrifugiert und das gewonnene Pellet in 500 µl TRIS-Sucrose-
Puffer aufgenommen und im Gewebehomogenisator Dounce (1ml) homogenisiert. 10 µl wurden für
die anschließende Proteinbestimmung eingesetzt, der Rest bei –80°C eingefroren.
2.2.3.3 Proteinbestimmung nach Smith et al. (1985)
Die Bestimmung des Gesamtproteingehaltes wurde nach der Methode von Smith et al. (1985)
durchgeführt. Hierzu wurde von jedem Pool eine Verdünnungsreihe (1:100 bis 1:1000) hergestellt.
Zur Quantifizierung des Proteingehaltes wurde vor jeder Probenmessung eine Kalibriergerade aus
bovinem Serumalbumin (BSA) erstellt. Alle Messungen erfolgten als Doppelbestimmungen und
wurden in einer 96-Well-Mikrotiterplatte durchgeführt, wobei pro Kavität 100 µl Probe und 100 µl
frisch angesetzte Färbelösung - bestehend aus 1 Teil 4% CuSO4 und 49 Teilen Bicinchoninic Acid -
eingesetzt wurde. Zur Beschleunigung der Reaktion wurde die Mikrotiterplatte 30 min bei 60°C im
Wärmeschrank inkubiert. Die Messung des Proteingehaltes erfolgte bei 550 nm mittels eines ELISA
Reader Dynatech MR5000 (Dynatech, Ohio, USA).
2.3.3.4 Proteinexpression ausgewählter CYPs
2.3.3.4.1 Auftrennung der Proteine mittels 1D-SDS-Polyacrylamidgelelektrophorese
Die Natriumdodecylsulfat-Polyacrylamidgelelektrophorese (SDS-PAGE) wurde nach der Methode
von Laemmli (1970) durchgeführt. Die mikrosomalen Proteine von acht Pools wurden entsprechend
ihres Molekulargewichtes mittels 1D-SDS-PAGE aufgetrennt, auf eine PVDF-Membran transferiert
und durch Inkubation mit einem Primär- und Sekundärantikörper die Cytochrome 1A1, 2A, 2B6,
2E1, 3A4 und 3A5 nachgewiesen.

2. Material und Methoden 39
Pro Pool wurden 45 µg mikrosomales Protein auf ein Volumen von 12 µl eingestellt, mit je 4 µl Roti
-Load1 4x-Konz. versetzt und 5 min bei 95°C inkubiert. Auf die erste Spur des 12%igen
Polyacrylamidgeles wurde ein Marker – bestehend aus 5 µl Precision blue protein standard
prestained und 0,5 µl Chemicon ECL – aufgetragen, auf die zweite Spur eine Positivkontrolle
(humane Lebermikrosomen). Zur besseren Auflösung der Proteinbanden wurden die Proben auf ein
Sammelgel aufgetragen, welches sich über dem eigentlichen Trenngel befand. Das Trenngel bestand
aus 3 ml 30% Acrylamid / 0,8% Bisacrylamid, 1,875 ml 1,5 M Tris pH 8,8, 2,55 ml A. bidest. und
37,5 µl 20% SDS, das Sammelgel aus 850 µl 30% Acrylamid / 0,8% Bisacrylamid, 625 µl 1 M Tris
pH 6,8, 3,47 ml A. bidest. und 25 µl 20% SDS. Zum Start der Polymerisierungsreaktion wurde dem
Trenngel 75 µl 10% Ammoniumperoxodisulfat und 7,5 µl TEMED, dem Sammelgel 50 µl 10%
Ammoniumperoxodisulfat und 5 µl TEMED hinzugegeben. Die Elektrophorese erfolgte bei
Raumtemperatur in einer Minigel-Elektrophoresekammer der Firma Biometra unter Zugabe von 1x
SDS-Laufpuffer. Anfänglich betrug die Stromstärke 20 mA pro Gel, nach Erreichen des Trenngels
wurde sie für etwa 60 bis 90 min auf 25 mA pro Gel erhöht.
2.3.3.4.2 Darstellung der Proteine mittels Western blot
Direkt im Anschluss an die Elektrophorese wurde das Gel aus der Elektrophoresekammer
entnommen und 10 min in 1x Transfer-Puffer getränkt. Parallel dazu wurde die PVDF-Membran 15
s in Methanol aktiviert, 2 min in A. bidest. gewässert und ebenfalls 10 min in 1x Transfer-Puffer
getränkt. Nach Beschicken der Kassette und Einsetzen der Kassette in die Blotkammer erfolgte das
Blotten über Nacht bei 90 mA und 4°C auf einem Magnetrührer Heidolph MR3000 der Firma
Heidolph Instruments GmbH & Co.KG.
Zur Sättigung freier, unspezifischer Bindungsstellen wurde die PVDF-Membran für 1 h bei
Raumtemperatur auf einem Schüttler (Roto-Shake Genie der Firma Scientific Industries, Inc.) mit
Blocking-Reagenz – bestehend aus neun Teilen 1x TBS-Puffer ohne Tween und einem Teil
Rotiblock, 10x Konzentrat – versetzt. Nach drei fünfminütigen Waschschritten mit Waschpuffer (1x
TBS-T 0,1%) wurde die Inkubation mit dem Primärantikörper, welcher in 10 ml Diluent – bestehend
aus 99 Teilen 1x TBS-Puffer ohne Tween und einem Teil Rotiblock, 10x Konzentrat – verdünnt
wurde (Angaben zu den verwendeten Primär- und Sekundärangaben und deren Verdünnungen siehe
Anhang Seite IX), für 1 h bei Raumtemperatur auf dem Schüttler durchgeführt. Nach erneutem
Waschen folgte die Inkubation mit dem Sekundärantikörper in analoger Weise. Anschließend wurde
die PVDF-Membran wieder gewaschen und in frisch angesetztem Entwicklungsreagenz 1 min bei
Raumtemperatur geschwenkt. Nach etwa 5 min erfolgte die Exposition an der Kodak Digital

2. Material und Methoden 40
Science Image Station 440 CF (Kodak, USA). Die Lumineszenz wurde nach 1 und nach 10 min
Expositionsdauer gemessen.
2.2.4 Metabolismusuntersuchungen
Die Inkubationsexperimente wurden in den Laboren T2.06, T2.07, T2.08 sowie T2.09 des Fraunhofer
Institutes für Toxikologie und Experimentelle Medizin durchgeführt.
2.2.4.1 Metabolismus von Loteprednol-Etabonat (LE)
Um die Verstoffwechselung von Loteprednol-Etabonat, dem ersten Vertreter der sog. Soft Steroide
(Szelenyi and Pahl, 2002), zu untersuchen, wurde 14C-Loteprednol-Etabonat als Substrat verwendet
und das entstehende Metabolitenprofil mittels Radio-HPLC bestimmt. Die Messung des
Loteprednol-Etabonats und seiner Hauptmetaboliten wurde dabei durch Frau Diplom-Chemikerin
Qiong Luo am Fraunhofer Institut für Toxikologie und Experimentelle Medizin durchgeführt.
Abbildung 11: Strukturformel von Loteprednol-Etabonat (LE), dem ersten Vertreter der sog. Soft Steroide.
[aus: Bodor N, Recent advances in retrometabolic design approaches. J. Control. Release, 62: 209-222, 1999.]

2. Material und Methoden 41
2.2.4.1.1 1.Versuchsabschnitt: Inkubation bei verschiedenen Inkubationszeiten
Im ersten Versuchsabschnitt erfolgten die Inkubationen der einzelnen mikrosomalen
Nasenmuschelpools sowie der mitgeführten Negativ- (nur Puffer und Substrat) und Positivkontrollen
bei 60 und/oder 120 min (siehe Abbildung 10). Die Versuche wurden dabei in zwei Versuchsserien
aufgeteilt, wobei in der ersten Serie Rattenlebermikrosomen, in der zweiten humane
Lebermikrosomen als Positivkontrolle verwendet wurden. Die konzentrierte Mikrosomensuspension
wurde mit 0,1 M TRIS-Puffer auf 1 mg mikrosomales Protein / ml eingestellt und mit 1 mM β-
NADPH versetzt. Zum Start der Reaktion wurde je Ansatz Loteprednol-Etabonat (25 µM) mit einer
Aktivität von 1 µCi/ml hinzugegeben und die Reaktionsansätze im Wasserbad bei 37°C 60 bzw. 120
min inkubiert. Im Anschluss wurden die Proben für 5 min bei 8.000 Upm und 4°C zentrifugiert und
bis zur weiteren Aufarbeitung bei –80°C schockgefroren.
Die Metaboliten wurden durch Festphasenextraktion nach folgender Methode gewonnen: Zunächst
wurden die Kartuschen (Waters Oasis HLB 3cc (60 mg) Extraction Cartridges der Firma Waters
Corporation, USA) mit 3 ml Ethanol und 2 ml A. bidest. konditioniert, dann die Proben aufgetragen.
Nach aufeinanderfolgenden Waschschritten mit 1 ml Ethanol (15%), 1 ml A. bidest. und 1 ml
Ethylacetat/Heptan (2:98) sowie Trocknen der Säule erfolgte die Elution mit 3 ml Ethylacetat/Heptan
(35:65) und 1 ml Ethylacetat. Anschließend wurde das Eluat unter Stickstoffstrom bei
Raumtemperatur bis zur vollständigen Trocknung eingedampft und das erhaltene Extrakt in 40 µl
mobiler Phase – bestehend aus Methanol und Essigsäure (0,5%, pH 3) im Verhältnis 62:38 –
aufgenommen.
Bei einem Injektionsvolumen von 10 µl und einer Flussrate von 0,3 ml / min wurden die Metaboliten
auf einer Spherisorb ODS-2-Säule (250 x 4 mm, Partikelgröße 3 µm, Latek, Eppelheim) bei 25°C
chromatographisch aufgetrennt. Die Trennung der Metaboliten erfolgte mit folgendem Gradienten der
mobilen Phase:
I. 0-9 min mit 62% Methanol, 38 % Essigsäure (0,5%, pH 3)
II. 9-16 min mit 74% Methanol, 26 % Essigsäure (0,5%, pH 3)
III. 16-39 min mit 74% Methanol, 26 % Essigsäure (0,5%, pH 3)
IV. 39-43 min mit 62% Methanol, 38 % Essigsäure (0,5%, pH 3)
V. 43-55 min mit 62% Methanol, 38 % Essigsäure (0,5%, pH 3)
Der Detektion diente ein Flow Scintillation Analyzer 500TR Series der Firma Packard BioScience
Company, USA.

2. Material und Methoden 42
2.2.4.1.2 2. Versuchsabschnitt: Inkubation mit und ohne Glucuronidase-Zusatz
Durch zusätzlichen Glucuronidase-Verdau können Glucuronide nachgewiesen werden. Aus diesem
Grund wurden die Proben in doppelter Menge angesetzt, im Anschluss an die LE-Inkubation
gesplittet und die eine Hälfte mit, die andere Hälfte ohne Glucuronidase behandelt. Parallel zu den
Organpools wurden zwei Positiv- (humane Lebermikrosomen) und zwei Negativkontrollen (nur
Puffer und Substrat) mitgeführt (jeweils mit und ohne Glucuronidase-Zusatz).
Auch hier wurde – mit Ausnahme von Pool 15 - der mikrosomale Proteingehalt mit 0,1 M TRIS-
Stock auf 1 mg / ml eingestellt und 1 mM β-NADPH zugesetzt. Bei Pool 15 musste die
Proteinkonzentration aufgrund der geringen Proteinausbeute auf 0,5 mg / ml halbiert werden. Nach
Zugabe von 25 µM Loteprednol-Etabonat mit einer Aktivität von 1 µCi/ml wurden die Proben im
Wasserbad bei 37°C 120 min inkubiert und anschließend 5 min bei 4°C und 8.000 Upm
zentrifugiert. Die Überstände wurden abgenommen, halbiert und eine Hälfte mit, die andere ohne β-
Glucuronidase inkubiert. Die Aliquots erhielten hierzu das gleiche Volumen an
Ammoniumacetatpuffer (200 mM, pH 4,5) unter Zugabe von 5 mg / ml β-Glucuronidase bzw. ohne
β-Glucuronidase-Zusatz (Kontrollen) und wurden über Nacht bei 37°C inkubiert.
Die Festphasenextraktion der Inkubate sowie die anschließende HPLC-Messung erfolgten wie unter
2.2.4.1.1 beschrieben. Vor dem HPLC-Auftrag wurden jeweils 5% des Probenvolumens
abgenommen, mit je 4 ml Rotiszint versetzt, gemischt und die Radioaktivität des 14C-Isotops jeweils
eine Minute als DPM (Desintegrations per Minute) im LS 6500 Multi-Purpos Scintillation Counter
der Firma Beckmann Coulter, Palo Alto, USA gemessen und für die Berechnung der Wiederfindung
eingesetzt.
2.2.4.2 Testosteron-Metabolismus
Testosteron wird von unterschiedlichen CYP-Isoformen stereo- und regioselektiv hydroxyliert und
liefert auf diese Weise Informationen über die Aktivität einzelner Isozyme. Pro Ansatz wurden 50
µM Testosteron eingesetzt. Zusätzlich wurde eine Negativ- (nur Puffer und Substrat) und eine
Positivkontrolle (humane Lebermikrosomen) mitgeführt. Der mikrosomale Proteingehalt wurde mit
0,1 M TRIS-Stock auf 0,5 mg / ml eingestellt und die Inkubation unter Zusatz von 1 mM β-
NADPH bei 37°C im Wasserbad durchgeführt. Nach 120 min wurden die Proben für 5 min bei
8.000 Upm und 4°C zentrifugiert und die Überstände und Pellets bei –80°C schockgefroren.

2. Material und Methoden 43
Von den Überständen wurden 900 µl abgenommen, mit jeweils 100 µl 2-Propanol, 5 ml Ethylacetat
sowie 1 µg 11-α-Hydroxyprogesteron als internem Standard versetzt und auf dem Rotoshake 20
min bei Raumtemperatur extrahiert. Zur Trennung der Phasen wurden die Proben 20 min bei
Raumtemperatur und 2.200 Upm zentrifugiert und die obere Ethylacetat-Phase mit den darin
gelösten Metaboliten unter Stickstoffstrom eingedampft. Zur HPLC-Messung wurden die Extrakte in
je 100 µl mobiler Phase (Wasser / Methanol / Acetonitril, 60/25/15, v/v/v) aufgenommen, von denen
jeweils 80 µl in das HPLC-System (Hewlett Packard HP Series 1100) injiziert wurden. Die mobile
Phase wurde durch eine HP 1100 Quaternary Pump mit einer Flussrate von 1 ml / min durch das
System gepumpt. Die chromatographische Auftrennung der Testosteronmetaboliten 6α-
Hydroxytestosteron, 7α-Hydroxytestosteron, 6β-Hydroxytestosteron, 16α-Hydroxytestosteron,
2α-Hydroxytestosteron sowie Androstendion erfolgte auf einer C18-Nucleosil-Säule (250 x 4 mm,
Partikelgröße 5 µm, Macherey-Nagel, Düren) bei einer Ofentemperatur von 30°C.
Der Trennung der Metaboliten diente folgender Gradient der mobilen Phase:
I. 0-12 min mit 60% Wasser, 25% Methanol, 15% Acetonitril;
II. 12-15 min mit 45% Wasser, 40% Methanol, 15% Acetonitril;
III. 15-27 min mit 45% Wasser, 45% Methanol, 10% Acetonitril;
IV. 27-32 min mit 40% Wasser, 30% Methanol, 30% Acetonitril;
V. 32-35 min mit 40% Wasser, 25% Methanol, 35% Acetonitril;
VI. 35-40 min mit 30% Wasser, 20% Methanol, 50% Acetonitril;
VII. 40-45 min mit 60% Wasser, 25% Methanol, 15% Acetonitril.
Die Detektion erfolgte bei 238 nm.
2.2.4.3 EROD-Aktivitäten
Ethoxyresorufin ist ein profluoreszierendes Substrat für CYP 1A1, dessen metabolische Umsetzung
zu dem fluoreszierenden Resorufin gemessen werden kann. Hierzu wurde eine Ethoxyresorufin-
Stammlösung in DMSO mit einer Konzentration von 1 mg / ml hergestellt und durch Zugabe zu den
einzelnen Reaktionsansätzen auf die endgültige Konzentration von 10 µM verdünnt. Die konzentrierte
Mikrosomensuspension der Proben und der Positivkontrolle wurde mit 0,1 M TRIS-Stock auf 0,5
mg mikrosomales Protein / ml eingestellt und mit 1 mM β-NADPH versetzt. Mikrosomen humaner
Hepatozyten dienten dabei als Positivkontrolle. Die Inkubation erfolgte im Wasserbad bei 37°C.
Nach 120 min wurde die Reaktion mit 200 µl Acetonitril abgestoppt und die Proben 5 min bei 5.000
Upm zentrifugiert. Zur Bestimmung der Glucuronide wurden die Überstände halbiert und ein Aliquot

2. Material und Methoden 44
mit, das andere ohne Glucuronidase behandelt. Die Proben erhielten hierzu das gleiche Volumen an
Ammoniumacetatpuffer (200 mM, pH 4,5) unter Zusatz von 5 mg / ml β-Glucuronidase bzw. ohne
β-Glucuronidase-Zusatz (Kontrollen) und wurden über Nacht bei 37°C inkubiert.
Das Dealkylierungsprodukt Resorufin wurde in dem Fluorometer VersaFluor (Bio-Rad
Laboratories GmbH, München) bei einer Excitationswellenlänge von 530 nm und einer
Emissionswellenlänge von 585 nm unter Zusatz von je 500 µl Glycinpuffer und 1000 µl TRIS-Puffer
pro Probe bzw. Kalibrierlösung gemessen. Zur Erstellung der Kalibriergerade wurde Resorufin in
unterschiedlichen Konzentrationen (0, 5, 10, 20, 40, 60, 80, 100 nM) in Ethanol gelöst (siehe
Abbildung 12). Lagen die gemessenen Fluoreszenzwerte der Proben oberhalb des höchsten Wertes
der Kalibriergerade, so wurden die Proben in einem Gemisch aus Acetatpuffer, Glycinpuffer und
TRIS-Puffer (im Verhältnis 1:1:2) verdünnt und die Messung wiederholt. Bei der Berechnung der
Konzentration wurde das Ergebnis mit dem jeweiligen Verdünnungsfaktor multipliziert.
Abbildung 12: Resorufin-Kalibriergerade zur Bestimmung des Resorufin-Gehaltes in nmol.
Resorufin-Kalibriergerade
340
95
220
346
473
599y = 6,2509x - 27,578
R2 = 0,9999
0
100
200
300
400
500
600
700
0 20 40 60 80 100 120
Resorufin-Gehalt [nmol]
Fluoreszenz

3. Ergebnisse 45
3. Ergebnisse
Sämtliche Rohdaten meiner Doktorarbeit werden im Archiv des Fraunhofer Institutes für
Toxikologie und Experimentelle Medizin in Hannover gelagert.
3.1 Genexpression
3.1.1 Qualität und Quantität der isolierten RNA
Die qualitative und quantitative RNA-Messung erfolgte durch Kapillarelektrophorese mit dem
Agilent Bioanalyzer der Agilent Technologies Deutschland GmbH, Waldbronn. In Abbildung
13 ist die totale RNA des Molekulargewichtsmarkers (nachfolgend Ladder bezeichnet) sowie
die der einzelnen Proben graphisch dargestellt. Abbildung 14 zeigt die Fluoreszenzwerte in
Abhängigkeit von der Retentionszeit am Beispiel von M-NR 1. In beiden Darstellungsformen
sind die beiden Banden der 18S rRNA sowie der 28S rRNA als Indikator reiner und intakter
RNA deutlich sichtbar.
Abbildung 13: Graphische Darstellung der totalen RNA des Ladders sowie die der einzelnen Proben
gemessen durch Kapillarelektrophorese am Agilent Bioanalyzer der Agilent Technologies GmbH,
Waldbronn. Deutlich sichtbar sind die beiden ribosomalen Banden 18S rRNA und 28S rRNA als Indikator
reiner und intakter RNA.
W-NRW-R M-R M-NR
Ladder 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 7
18S rRNA
28S rRNA
6
W-NRW-R M-R M-NR
Ladder 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 7
18S rRNA
28S rRNA
6

3. Ergebnisse 46
Abbildung 14: Graphische Darstellung der 18S rRNA- und 28S rRNA-assoziierten Fluoreszenz in
Abhängigkeit von der Retentionszeit am Beispiel von M-NR 1. Die Qualität der is olierten RNA lässt sich
anhand des Verhältnisses der beiden ribosomalen Banden ( 28 S rRNA : 18S rRNA ) abschätzen. Die RNA-
Konzentration errechnet sich aus der Gesamt -RNA-Fläche in Relation zur Fläche des Ladders mit bekanntem
RNA-Gehalt.
Die RNA-Konzentration, berechnet aus der jeweiligen RNA-Fläche in Bezug zur RNA-
Fläche des Ladders mit bekanntem RNA-Gehalt, sowie die RNA-Qualität – als Verhältnis der
beiden ribosomalen Banden ( 28S rRNA : 18S rRNA ) – ist für die insgesamt 22 Patienten in
den Abbildungen 15 (Raucher) und 16 (Nicht-Raucher) tabellarisch wiedergegeben.
Probe RNA-Fläche RNA-Konz. ( ng / µl )
rRNA-Verhältnis [ 28S / 18S ]
W-R 1 616,1 357,3 2,9 W-R 2 129,7 102,0 2,1 W-R 3 181,3 105,1 3,2 W-R 4 130,7 102,8 3,0 W-R 5 142,9 112,4 2,6 M-R 1 326,6 257,0 2,5 M-R 2 575,2 452,5 2,6 M-R 3 221,9 174,6 2,4 M-R 4 116,1 67,3 1,2 M-R 5 229,8 180,8 2,3
Abbildung 15: RNA-Fläche und daraus resultierende RNA-Konzentration (in ng / µl) sowie Verhältnis
der beiden ribosomalen Banden (28S rRNA : 18S rRNA) für die Gruppe der Raucher.
19 24 29 34 39 44 49 54 59 64 69Zeit ( Sekunden )
80
70
60
50
40
30
20
10
0
Fluoreszenz
19 24 29 34 39 44 49 54 59 64 69Zeit ( Sekunden )
80
70
60
50
40
30
20
10
0
Fluoreszenz

3. Ergebnisse 47
W- NR 1 208,6 96,8 2,6 W-NR 2 674,0 390,8 2,3 W-NR 3 490,8 227,7 2,8 W-NR 4 605,0 280,6 2,0 W-NR 5 637,3 369,5 2,2 M-NR 1 452,9 210,1 2,5 M-NR 2 547,2 253,8 2,5 M-NR 3 1.091,8 633,1 2,4 M-NR 4 687,9 398,9 2,1 M-NR 5 1.165,9 676,1 2,6 M-NR 6 435,2 252,4 1,8 M-NR7 482,0 279,5 2,1
Abbildung 16: RNA-Fläche und daraus resultierende RNA-Konzentration (in ng / µl) sowie Verhältnis
der beiden ribosomalen Banden (28S rRNA : 18S rRNA) für die Gruppe der Nicht-Raucher.
3.1.2 RT-PCR
Sämtliche Agarosegele der exprimierten Gene mit dazugehöriger graphischer Auswertung
sind im Anhang auf den Seiten X bis XLVI dargestellt.
3.1.2.1 Genexpression der Housekeeping-Gene CyclophillinA und GAPDH
Genexpressionsergebnisse werden häufig in Relation zu stabil exprimierten Genen, sog.
Housekeeping-Genen, dargestellt. GAPDH, welches in der Literatur oft als Housekeeping-
Gen ausgewiesene wird (Jung et al., 1997; Mollerup et al., 1999; Jung et al., 2003), zeigte
einen signifikanten Unterschied (p<0,05) zwischen Rauchern und Nicht-Rauchern. So lag der
durchschnittliche geldensitrometrisch ermittelte RNA-Gehalt von GAPDH in der Raucher-
Gruppe signifikant höher als in der Nicht-Raucher-Gruppe (siehe Abbildung 17).
Bei CyclophillinA, einem weiteren Housekeeping-Gen (Elder et al., 2000; Yang et al., 2000),
war der beobachtete Unterschied mit p = 0,076 statistisch zwar nicht signifikant, jedoch
wurde aufgrund der Nähe zum Signifikanzniveau (p<0,05) auf eine Darstellung der Werte
relativ zu CyclophillinA ebenfalls verzichtet.
Die Genexpressionsergebnisse werden daher absolut dargestellt.

3. Ergebnisse 48
Abbildung 17: Genexpression von CyclophillinA (links) und GAPDH (rechts). Mittelwerte und
Standardabweichungen der an der Kodak Digital Science Image Station 440 CF (Kodak, USA)
gemessenen Lichtwerte für die Nicht-Raucher- (n = 12) und die Raucher-Gruppe (n = 10).
*: p = 0,076; **: p<0,05.
3.1.2.2 CYP-Genexpression
Von den insgesamt 20 untersuchten Cytochrom P450 Monooxygenasen (siehe Liste der
verwendeten Primer im Anhang auf den Seiten VII und VIII) konnten die Cytochrome 1A2,
3A7 und 4A11 auch nach zahlreichen PCR-Amplifikationen von bis zu 39 Zyklen nicht
nachgewiesen werden.
Statistisch signifikante Unterschiede (p<0,05) zwischen der Raucher- und der Nicht-Raucher-
Gruppe konnten für CYP 1A1, 1B1 und 2S1 ermittelt werden. Aus statistischer Sicht war
dieser Unterschied bei CYP 1B1 mit einer Signifikanz von p<0,00006 am stärksten
ausgeprägt, gefolgt von CYP 1A1 mit p<0,002. Alle drei Monooxygenasen waren bei den
Rauchern stärker exprimiert als bei den Nicht-Rauchern. Betrachtet man bei genannten drei
Genen zusätzlich das Geschlecht, so fällt auf, dass CYP 1A1 und 2S1 nur bei den
Raucherinnen signifikant erhöht waren. Hingegen war CYP 1B1 geschlechtsunabhängig bei
männlichen wie weiblichen Rauchern erhöht.
GAPDH
50000
60000
70000
80000
90000
100000
110000
Nicht-Raucher Raucher
LichtwerteCyclophillinA
50000550006000065000700007500080000850009000095000
100000
Nicht-Raucher Raucher
Lichtwerte
* **
GAPDH
50000
60000
70000
80000
90000
100000
110000
Nicht-Raucher Raucher
LichtwerteCyclophillinA
50000550006000065000700007500080000850009000095000
100000
Nicht-Raucher Raucher
LichtwerteGAPDH
50000
60000
70000
80000
90000
100000
110000
Nicht-Raucher Raucher
LichtwerteCyclophillinA
50000550006000065000700007500080000850009000095000
100000
Nicht-Raucher Raucher
Lichtwerte
* **

3. Ergebnisse 49
Abbildung 18: Genexpression von CYP 1B1 (links) und CYP 2S1 (rechts). Mittelwerte und
Standardabweichungen der an der Kodak Digital Science Image Station 440 CF (Kodak, USA)
gemessenen Lichtwerte für die Nicht-Raucher- (n = 12) und die Raucher-Gruppe (n = 10).
*: p<0,00006; **: p<0,03.
Ein statistisch signifikanter Geschlechtsunterschied unabhängig vom Raucherstatus konnte
nur für CYP 2J2 mit p<0,0004 ermittelt werden (siehe Abbildung 19). So exprimieren
Männer CYP 2J2 weitaus stärker als Frauen, wobei bei einer Nichtraucherin (W-NR 5) keine
CYP 2J2-Transkripte nachweisbar waren. Auch CYP 2C9 war bei Männern stärker
exprimiert, wenngleich der Unterschied mit p = 0,063 statistisch nicht signifikant war. Hinzu
kommt, dass CYP 2C9 bei nur 12 (neun Männern und drei Frauen) der 22 Probanden
erfolgreich detektiert wurde, weshalb vor einer Bewertung weitere Untersuchungen mit
größeren Patientenzahlen notwendig erscheinen.
CYP 1B1
0
20000
40000
60000
80000
100000
120000
Nicht-Raucher Raucher
LichtwerteCYP 2S1
0
20000
40000
60000
80000
100000
120000
Nicht-Raucher Raucher
Lichtwerte
* **
CYP 1B1
0
20000
40000
60000
80000
100000
120000
Nicht-Raucher Raucher
LichtwerteCYP 2S1
0
20000
40000
60000
80000
100000
120000
Nicht-Raucher Raucher
LichtwerteCYP 1B1
0
20000
40000
60000
80000
100000
120000
Nicht-Raucher Raucher
LichtwerteCYP 2S1
0
20000
40000
60000
80000
100000
120000
Nicht-Raucher Raucher
Lichtwerte
* **
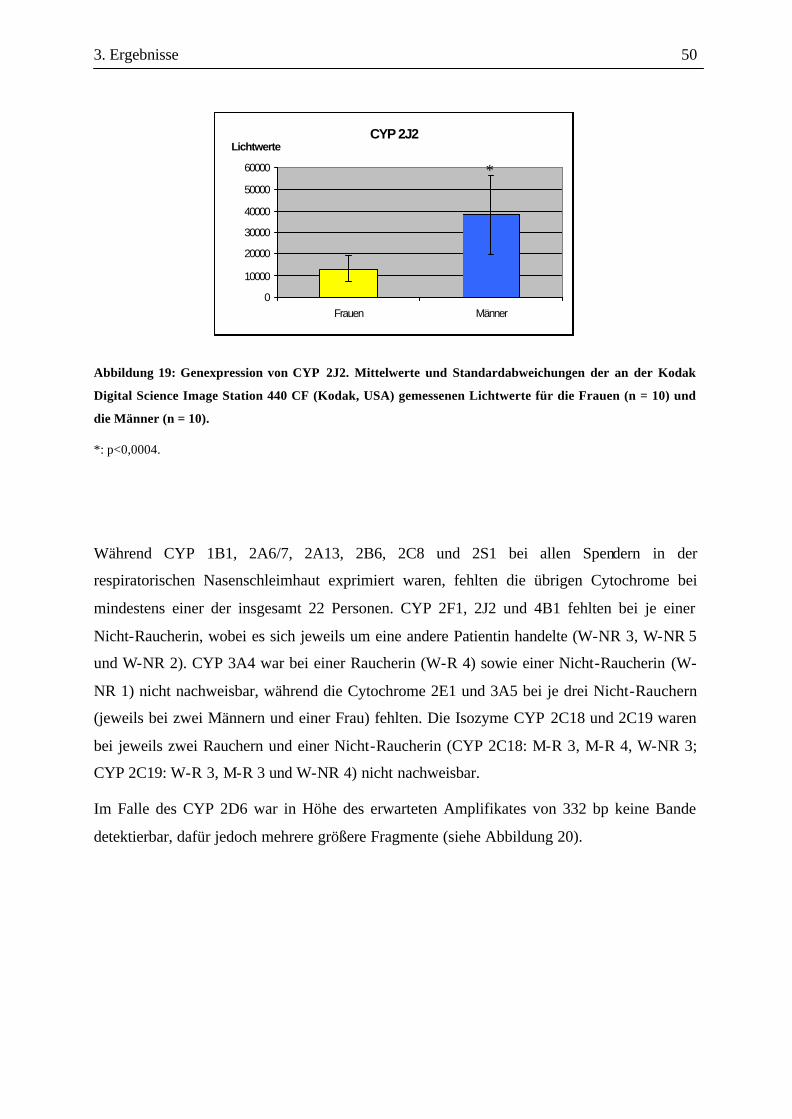
3. Ergebnisse 50
Abbildung 19: Genexpression von CYP 2J2. Mittelwerte und Standardabweichungen der an der Kodak
Digital Science Image Station 440 CF (Kodak, USA) gemessenen Lichtwerte für die Frauen (n = 10) und
die Männer (n = 10).
*: p<0,0004.
Während CYP 1B1, 2A6/7, 2A13, 2B6, 2C8 und 2S1 bei allen Spendern in der
respiratorischen Nasenschleimhaut exprimiert waren, fehlten die übrigen Cytochrome bei
mindestens einer der insgesamt 22 Personen. CYP 2F1, 2J2 und 4B1 fehlten bei je einer
Nicht-Raucherin, wobei es sich jeweils um eine andere Patientin handelte (W-NR 3, W-NR 5
und W-NR 2). CYP 3A4 war bei einer Raucherin (W-R 4) sowie einer Nicht-Raucherin (W-
NR 1) nicht nachweisbar, während die Cytochrome 2E1 und 3A5 bei je drei Nicht-Rauchern
(jeweils bei zwei Männern und einer Frau) fehlten. Die Isozyme CYP 2C18 und 2C19 waren
bei jeweils zwei Rauchern und einer Nicht-Raucherin (CYP 2C18: M-R 3, M-R 4, W-NR 3;
CYP 2C19: W-R 3, M-R 3 und W-NR 4) nicht nachweisbar.
Im Falle des CYP 2D6 war in Höhe des erwarteten Amplifikates von 332 bp keine Bande
detektierbar, dafür jedoch mehrere größere Fragmente (siehe Abbildung 20).
CYP 2J2
0
10000
20000
30000
40000
50000
60000
Frauen Männer
Lichtwerte
*
CYP 2J2
0
10000
20000
30000
40000
50000
60000
Frauen Männer
LichtwerteCYP 2J2
0
10000
20000
30000
40000
50000
60000
Frauen Männer
Lichtwerte
*

3. Ergebnisse 51
Abbildung 20: CYP 2D6-Genexpression bei Rauchern (A) und Nicht-Rauchern (B). Darstellung des 1,5
%igen Agarosegeles
Die an der Kodak Digital Science Image Station 440 CF (Kodak, USA) gemessenen mittleren
Lichtintensitäten der Cytochrome 2C8, 2C9, 2C18, 2C19, 2E1 und 3A4 waren deutlich
schwächer als jene der zugehörigen Positivkontrolle, während die mittlere CYP 2J2-Intensität
das 2,3-fache der zugehörigen Positivkontrolle betrug. Nicht in der Positivkontrolle
detektierbar waren Transkripte folgender Isoformen: CYP 2A13, 2F1, 2S1, 3A5 und 4B1.
332 bp
Ladder PK NK 1 12 4 53 2 3 4 5
Ladder
332 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
A: Raucher :
B: Nicht-Raucher:
332 bp
Ladder PK NK 1 12 4 53 2 3 4 5
Ladder
332 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
A: Raucher :
B: Nicht-Raucher:

3. Ergebnisse 52
3.1.2.3 Genexpression Flavinhaltiger Monooxygenasen (FMOs), der Epoxid-Hydrolase
(EH) sowie verschiedener Phase II-Enzyme
Von den sechs bekannten FMOs (Cashman and Zhang, 2002; Furnes et al., 2003) wurden die
Isozyme FMO 2, FMO 3 und FMO 5 auf ihre Expression in der menschlichen
respiratorischen Nasenschleimhaut untersucht. Jedes der drei Gene konnte bei allen 22
Patienten nachgewiesen werden. Die bei den Rauchern beobachtete reprimierte FMO 2-
Genexpression lag mit p=0,055 knapp unterhalb des statistischen Signifikanzniveaus von
p<0,05. Interessanterweise verursachte dabei das Rauchen bei Männer und Frauen ganz
unterschiedliche Effekte. Während die Expression von FMO 2 bei männlichen Spendern mit
p<0,05 statistisch signifikant reduziert war, konnte bei weiblichen Spendern (p<0,98) kein
Einfluss durch das Rauchen beobachtet werden (siehe Abbildung 21).
Vergleicht man die mittleren Lichtwerte der PCR-Amplifikate mit dem entsprechenden
Lichtwert der mitgeführten Positivkontrolle (humane Leber), so war der Wert für FMO 3 mit
dem der Positivkontrolle etwa vergleichbar, während der mittlere Wert für FMO 5
geringfügig darunter lag. FMO 2 dagegen konnte in der Positivkontrolle nicht nachgewiesen
werden.
Abbildung 21: Genexpression von FMO 2 bei Frauen (links) und Männern (rechts). Mittelwerte und
Standardabweichungen der an der Kodak Digital Science Image Station 440 CF (Kodak, USA)
gemessenen Lichtwerte für die Nicht-Raucherinnen (n = 5) und die Raucherinnen (n = 5) sowie für die
Nicht-Raucher (n = 7) und die Raucher (n = 5). Man beachte die unterschiedlichen Effekte durch das Rauchen
bei Männer und Frauen.
*: p<0,98; **: p<0,05.
0100002000030000400005000060000700008000090000
Nicht-Raucherinnen Raucherinnen
Lichtwerte
0100002000030000400005000060000700008000090000
Nicht-Raucher Raucher
LichtwerteFMO 2 bei Frauen FMO 2 bei Männern
***
0100002000030000400005000060000700008000090000
Nicht-Raucherinnen Raucherinnen
Lichtwerte
0100002000030000400005000060000700008000090000
Nicht-Raucher Raucher
LichtwerteFMO 2 bei Frauen FMO 2 bei Männern
0100002000030000400005000060000700008000090000
Nicht-Raucherinnen Raucherinnen
Lichtwerte
0100002000030000400005000060000700008000090000
Nicht-Raucher Raucher
Lichtwerte
0100002000030000400005000060000700008000090000
Nicht-Raucherinnen Raucherinnen
Lichtwerte
0100002000030000400005000060000700008000090000
Nicht-Raucher Raucher
LichtwerteFMO 2 bei Frauen FMO 2 bei Männern
***

3. Ergebnisse 53
Die Epoxid-Hydrolase (EH) katalysiert die Addition von Wasser an Epoxide, um das
korrespondierende Dihydrodiol zu bilden. Diese Reaktion produziert hauptsächlich
Metaboliten geringerer Reaktivität, die leicht zu konjugieren und auszuscheiden sind
(Seidegard and Ekstrom, 1997). In der untersuchten Nasenschleimhaut konnten EH-
Transkripte bei allen Personen nachgewiesen werden, wobei keine signifikanten Unterschiede
hinsichtlich Raucher-Status oder Geschlecht erkennbar waren.
Von den Phase II-Enzymen Glutathion-S-Transferase (GST) und UDP-Glucuronyl-
Transferase (UDP-GT oder UGT) wurden die Subformen GST α2, GST µ1, GST ρ1 sowie
UGT 1A1 und UGT 2A1 untersucht.
Die hier untersuchten Glutathion-S-Transferasen waren jeweils bei allen Patienten
nachweisbar. Die gemessenen Lichtwerte der PCR-Amplifikate der Spender lagen im Falle
der GST µ1 leicht oberhalb des Lichtwertes der Leberkontrolle, im Falle der GST α2
eindeutig darüber. Mit Ausnahme zweier Raucher (W-R 2 und M-R 1), die nur ein schwaches
Signal lieferten, befanden sich auch die Werte für GST ρ1 deutlich oberhalb des Wertes der
Positivkontrolle. Raucher wiesen insgesamt einen geringeren mRNA-Gehalt an GST α2 auf
(p<0,002), wie in Abbildung 22 zu sehen ist. Bei Betrachtung der Geschlechter zeigte sich
dabei, dass dieser Unterschied nur bei männlichen Spendern (p<0,001) statistisch signifikant
war, nicht jedoch bei weiblichen Spendern (p<0,33).
Abbildung 22: Genexpression von GST α2. Mittelwerte und Standardabweichungen der an der Kodak
Digital Science Image Station 440 CF (Kodak, USA) gemessenen Lichtwerte für die Nicht-Raucher- (n =
12) und die Raucher-Gruppe (n = 10) sowie Lichtwert für die Leberkontrolle.
*: p<0,002.
0
50000
100000
150000
200000
250000
Leberkontrolle Nicht-Raucher Raucher
Lichtwerte
*
GST α2 GST α2 GST α2
0
50000
100000
150000
200000
250000
Leberkontrolle Nicht-Raucher Raucher
Lichtwerte
*
GST α2 GST α2 GST α2
*
GST α2 GST α2 GST α2

3. Ergebnisse 54
Die in der Leber stark exprimierte UGT 1A1 ließ sich in der menschlichen respiratorischen
Nasenschleimhaut bei vier Rauchern und zwei Nicht-Rauchern nachweisen, allerdings erst
nach 39 PCR-Amplifikationszyklen (siehe Abbildung XXVI im Anhang Seite XXXV).
Dagegen war die auch als olfaktorische UGT bezeichnete UGT 2A1 (Jedlitschky et al., 1999)
in der Nasenschleimhaut sämtlicher 22 Spender vorhanden (siehe Abbildung XXVII im
Anhang Seite XXXVI), nicht jedoch in der Positivkontrolle (humane Leber). Signifikante
Unterschiede hinsichtlich Raucher-Status oder Geschlecht waren weder bei den drei
Glutathion-S-Transferasen, noch bei den beiden UDP-Glucuronyl-Transferasen erkennbar.
3.1.2.4 Genexpression verschiedener regulatorischer Rezeptoren
Da die transkriptorische Regulation der hier untersuchten Enzyme unter anderem über
verschiedene nukleäre Transkriptionsfaktoren bzw. Rezeptoren erfolgt, wurden ergänzend
folgende Rezeptoren in die Untersuchungen miteinbezogen: Aryl hydrocarbon-Rezeptor
(AhR), Constitutive Androstane Rezeptor β (CAR β), Liver X Rezeptor α (LXR α),
Pregnenolone X Rezeptor (PXR), Peroxisome proliferative activated Rezeptor α (PPAR α)
und Peroxisome proliferative activated Rezeptor γ (PPAR γ).
Die Rezeptoren AhR, LXR α, PXR, PPAR α und PPAR γ waren in der respiratorischen
Nasenschleimhaut aller 22 Personen exprimiert, wobei AhR, PPAR α und PPAR γ
vergleichbare Lichtintensitäten wie die Positivkontrolle (humane Leber) lieferten, während
LXR α und PXR deutlich schwächere Lichtintensitäten aufwiesen. CAR β war
interessanterweise nur bei vier weiblichen Rauchern und sechs männlichen Nicht-Rauchern
nachweisbar (siehe Abbildung XXIX im Anhang Seite XXXVIII), und zwar mit Lichtwerten
deutlich unterhalb der Positivkontrolle.
Ein statistisch signifikanter Unterschied (p<0,015) zwischen der Raucher- und der Nicht-
Raucher-Gruppe war nur bei PPAR γ (siehe Abbildung 23) vorhanden. Interessanterweise war
die beobachtete PPAR γ-Induktion nur bei den Raucherinnen aufzeigbar. PXR wiederum war
mit einer statistischen Signifikanz von p<0,005 bei weiblichen Spendern stärker exprimiert als
bei männlichen (siehe Abbildung 23).

3. Ergebnisse 55
Abbildung 23: Genexpression von PPAR γ (links) und PXR (rechts). Mittelwerte und
Standardabweichungen der an der Kodak Digital Science Image Station 440 CF (Kodak, USA)
gemessenen Lichtwerte für die Nicht-Raucher- (n = 12) und die Raucher-Gruppe (n = 10) im Fall von
PPAR γ bzw. für Frauen (n = 10) und Männer (n = 12) im Fall von PXR.
*: p<0,015; **: p<0,005.
3.1.2.5 Genexpression verschiedener Cytokine
Da Rauchen nachweislich zu entzündlichen Atemwegsveränderungen führt (Dwyer, 2003;
Kasuga et al., 2003; Oudijk et al., 2003), wurden folgende Cytokine als Marker für mögliche
Entzündungsreaktionen (Jones et al., 2003) untersucht: IL-1β , IL-6, IL-8 und TNF α. Auf
diese Weise sollte geprüft werden, ob die zwischen Rauchern und Nicht-Rauchern
beobachteten Unterschiede in der Genexpression in Zusammenhang mit einer möglichen
Nasenschleimhautentzündung gebracht werden könnten. Unterschiede in der Expression
genannter Entzündungsmediatoren zwischen Rauchern und Nicht-Rauchern konnten jedoch
nicht festgestellt werden.
Transkripte von TNF α und IL-8 waren bei allen Patienten nachweisbar, IL-1β-Amplifikate
fehlten bei einer Nicht-Raucherin (W-NR 2). Dagegen war IL-6 nur bei sechs Rauchern (drei
weibliche und drei männliche) und vier Nicht-Rauchern (zwei männliche und zwei weibliche)
exprimiert.
0
5000
10000
15000
20000
25000
Nicht-Raucher Raucher
Lichtwerte
0
20000
40000
60000
80000
100000
120000
Frauen Männer
LichtwertePPAR γ PXR
*
**
0
5000
10000
15000
20000
25000
Nicht-Raucher Raucher
Lichtwerte
0
20000
40000
60000
80000
100000
120000
Frauen Männer
LichtwertePPAR γ PXR
0
5000
10000
15000
20000
25000
Nicht-Raucher Raucher
Lichtwerte
0
20000
40000
60000
80000
100000
120000
Frauen Männer
Lichtwerte
0
5000
10000
15000
20000
25000
Nicht-Raucher Raucher
Lichtwerte
0
20000
40000
60000
80000
100000
120000
Frauen Männer
LichtwertePPAR γ PXR
*
**

3. Ergebnisse 56
3.2 Proteinexpression
3.2.1 Poolgewichte und Bestimmung des mikrosomalen Proteingehaltes
In Abbildung 24 sind die zur Mikrosomengewinnung eingesetzten Poolgewichte der
Organpools, der jeweilige mikrosomale Proteingehalt - ermittelt durch die Proteinbestimmung
nach Smith et al. (1985) - sowie die Proteinausbeute (als mikrosomales Protein / g Gewebe)
dargestellt.
Pool-Nr. Art des Pools Poolgewicht ( g )
mikrosomales Protein
( µg / µl )
mikrosomales Protein / g Gewebe ( mg / g )
1 M-NR 1,53 5,77 1,89 2 M-NR 1,49 5,62 1,89 4 M-NR 2,7 7,74 1,43 5 M-NR 2,32 5,11 1,1 6 W-NR 1,58 5,74 1,82 7 M-R 2,6 5,94 1,14 8 W-R 1,16 3,75 1,62 9 M-NR 4,67 10,15 1,09 10 M-R 3,73 9,76 1,31 11 W-NR 2,52 5,01 0,99 12 M-NR 3,74 7,67 1,03 13 M-R 4,39 7,75 0,88 14 W-NR 4,43 6,61 0,75 15 W-R 1,16 3,04 1,31 16 W-? 1,21 4,67 1,93
Abbildung 24: Darstellung der Poolgewichte, des mikrosomalen Proteingehaltes und des mikrosomalen
Proteins / g Gewebe der insgesamt 15 Organpools.
3.2.2 Western blot
Von den mittels Western blot untersuchten Cytochromen 1A1, 2A, 2B6, 2E1, 3A4 und 3A5
konnten bei einem Proteinloading von 45 µg mikrosomalem Protein nur bei CYP 2A eine
schwache Proteinexpression beobachtet werden (siehe Abbildung 25). Die an der Kodak
Digital Science Image Station 440 CF (Kodak, USA) gemessenen durchschnittlichen
Lichtwerte der acht Organspools betrugen mit 69.424,51 und einer Standardabweichung von

3. Ergebnisse 57
20.828,68 insgesamt 21,6 % der Positivkontrolle (humane Leber), die einen Lichtwert von
321.244,48 aufwies (siehe Abbildung 26). Die Western blots aller untersuchten Cytochrome
sind im Anhang auf den Seiten XLVII bis XLIX abgebildet.
Abbildung 25: CYP 2A-Proteinexpression mittels Western blot: Messung der Lumineszenz nach 10 min
Expositionsdauer an der Kodak Digital Science Image Station 440 CF (Kodak, USA).
Man beachte die deutliche Bande der Positivkontrolle (PK) und die vergleichsweise schwachen Banden der
einzelnen Proben.
Abbildung 26: CYP 2A-Proteinexpression: an der Kodak Digital Science Image Station 440 CF (Kodak,
USA) gemessene Lichtwerte: Lichtwert der Positivkontrolle (PK) und durchschnittlicher Lichtwert und
Standardabweichung der acht Organpools
CYP 2A-Proteinexpression
0
50000
100000
150000
200000
250000
300000
350000
PK 8 Organpools
Proben
Lichtwerte
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
25 kDa
unspezifische Bande
CYP 2A
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
25 kDa
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
25 kDa
unspezifische Bande
CYP 2A
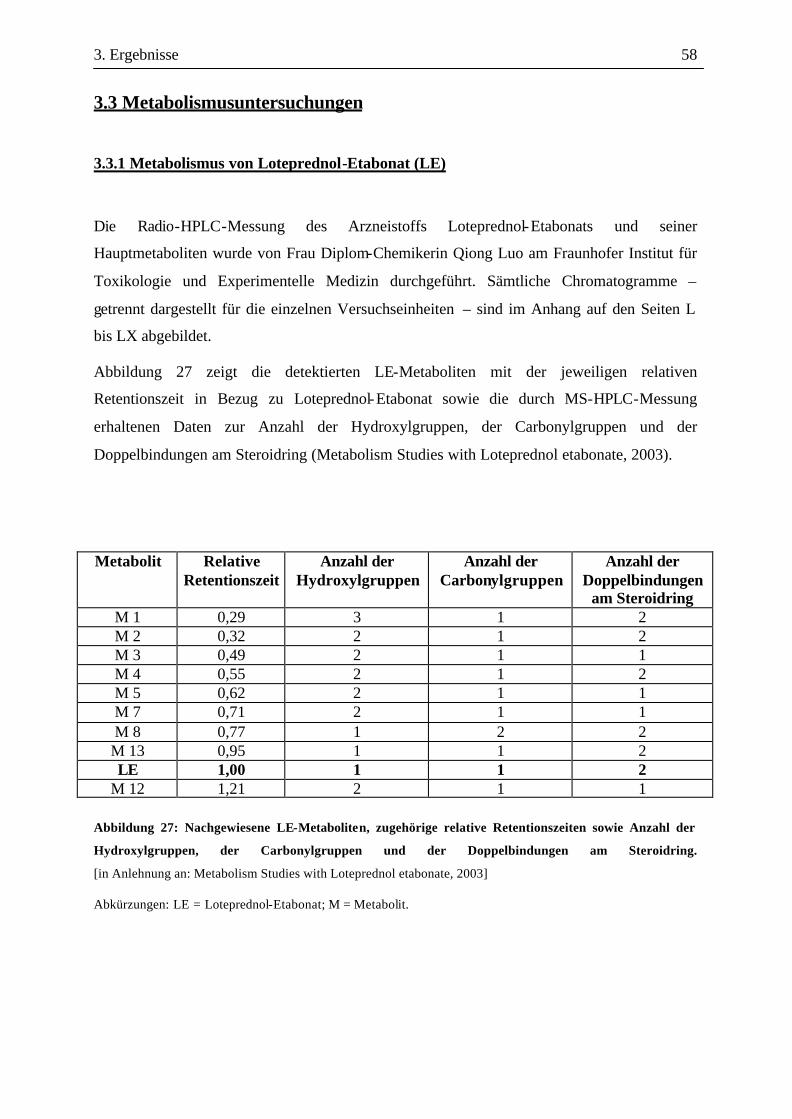
3. Ergebnisse 58
3.3 Metabolismusuntersuchungen
3.3.1 Metabolismus von Loteprednol-Etabonat (LE)
Die Radio-HPLC-Messung des Arzneistoffs Loteprednol-Etabonats und seiner
Hauptmetaboliten wurde von Frau Diplom-Chemikerin Qiong Luo am Fraunhofer Institut für
Toxikologie und Experimentelle Medizin durchgeführt. Sämtliche Chromatogramme –
getrennt dargestellt für die einzelnen Versuchseinheiten – sind im Anhang auf den Seiten L
bis LX abgebildet.
Abbildung 27 zeigt die detektierten LE-Metaboliten mit der jeweiligen relativen
Retentionszeit in Bezug zu Loteprednol-Etabonat sowie die durch MS-HPLC-Messung
erhaltenen Daten zur Anzahl der Hydroxylgruppen, der Carbonylgruppen und der
Doppelbindungen am Steroidring (Metabolism Studies with Loteprednol etabonate, 2003).
Metabolit Relative Retentionszeit
Anzahl der Hydroxylgruppen
Anzahl der Carbonylgruppen
Anzahl der Doppelbindungen
am Steroidring M 1 0,29 3 1 2 M 2 0,32 2 1 2 M 3 0,49 2 1 1 M 4 0,55 2 1 2 M 5 0,62 2 1 1 M 7 0,71 2 1 1 M 8 0,77 1 2 2 M 13 0,95 1 1 2 LE 1,00 1 1 2
M 12 1,21 2 1 1
Abbildung 27: Nachgewiesene LE-Metaboliten, zugehörige relative Retentionszeiten sowie Anzahl der
Hydroxylgruppen, der Carbonylgruppen und der Doppelbindungen am Steroidring.
[in Anlehnung an: Metabolism Studies with Loteprednol etabonate, 2003]
Abkürzungen: LE = Loteprednol-Etabonat; M = Metabolit.
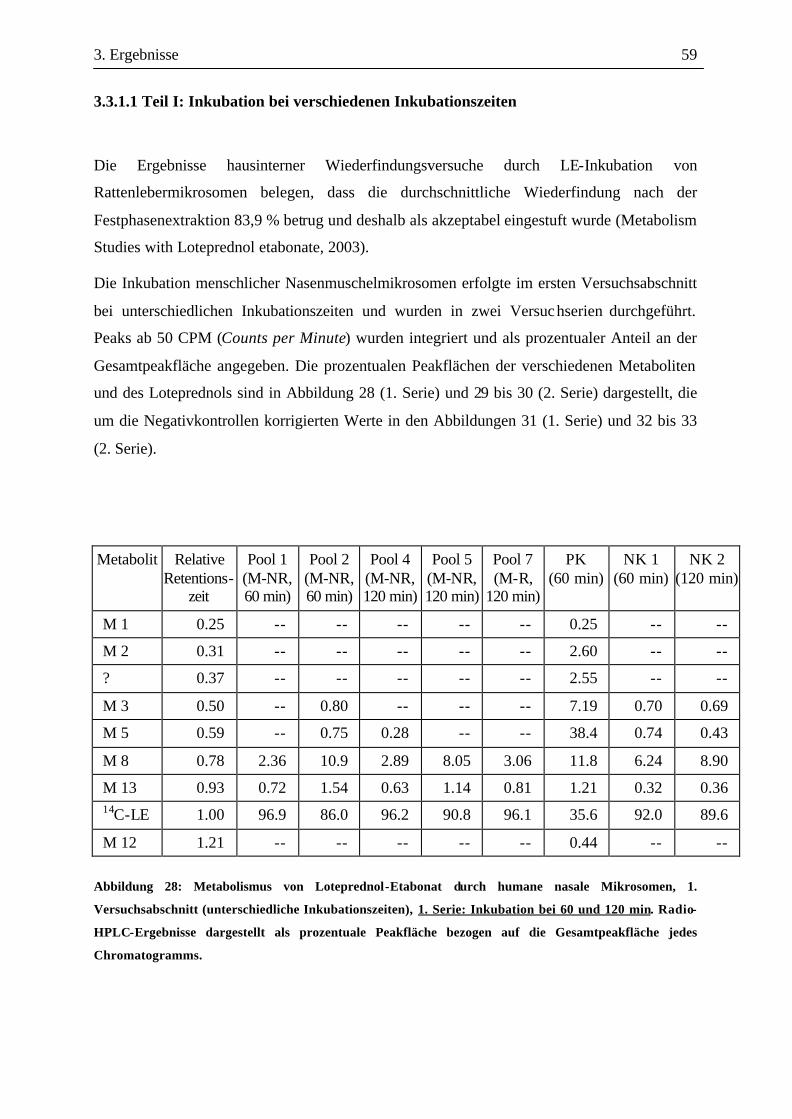
3. Ergebnisse 59
3.3.1.1 Teil I: Inkubation bei verschiedenen Inkubationszeiten
Die Ergebnisse hausinterner Wiederfindungsversuche durch LE-Inkubation von
Rattenlebermikrosomen belegen, dass die durchschnittliche Wiederfindung nach der
Festphasenextraktion 83,9 % betrug und deshalb als akzeptabel eingestuft wurde (Metabolism
Studies with Loteprednol etabonate, 2003).
Die Inkubation menschlicher Nasenmuschelmikrosomen erfolgte im ersten Versuchsabschnitt
bei unterschiedlichen Inkubationszeiten und wurden in zwei Versuchserien durchgeführt.
Peaks ab 50 CPM (Counts per Minute) wurden integriert und als prozentualer Anteil an der
Gesamtpeakfläche angegeben. Die prozentualen Peakflächen der verschiedenen Metaboliten
und des Loteprednols sind in Abbildung 28 (1. Serie) und 29 bis 30 (2. Serie) dargestellt, die
um die Negativkontrollen korrigierten Werte in den Abbildungen 31 (1. Serie) und 32 bis 33
(2. Serie).
Metabolit Relative Retentions-
zeit
Pool 1 (M-NR, 60 min)
Pool 2 (M-NR, 60 min)
Pool 4 (M-NR, 120 min)
Pool 5 (M-NR, 120 min)
Pool 7 (M-R,
120 min)
PK (60 min)
NK 1 (60 min)
NK 2 (120 min)
M 1 0.25 -- -- -- -- -- 0.25 -- --
M 2 0.31 -- -- -- -- -- 2.60 -- --
? 0.37 -- -- -- -- -- 2.55 -- --
M 3 0.50 -- 0.80 -- -- -- 7.19 0.70 0.69
M 5 0.59 -- 0.75 0.28 -- -- 38.4 0.74 0.43
M 8 0.78 2.36 10.9 2.89 8.05 3.06 11.8 6.24 8.90
M 13 0.93 0.72 1.54 0.63 1.14 0.81 1.21 0.32 0.36 14C-LE 1.00 96.9 86.0 96.2 90.8 96.1 35.6 92.0 89.6
M 12 1.21 -- -- -- -- -- 0.44 -- --
Abbildung 28: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 1.
Versuchsabschnitt (unterschiedliche Inkubationszeiten), 1. Serie: Inkubation bei 60 und 120 min. Radio-
HPLC-Ergebnisse dargestellt als prozentuale Peakfläche bezogen auf die Gesamtpeakfläche jedes
Chromatogramms.

3. Ergebnisse 60
Metabolit Relative Retentions-
zeit
Pool 4 (M-NR)
Pool 5 (M-NR)
Pool 6 (W-NR)
Pool 7 (M-R)
Pool 8 (W-R)
Pool 9 (M-NR)
Pool 10 (M-R)
Pool 11 (W-NR)
PK 1 NK 1
? 0.37 -- -- -- -- -- -- -- -- 0.29 --
M 3 0.50 -- -- -- -- -- -- -- -- 4.28 --
M 5 0.58 -- -- -- -- -- -- -- -- 27.6 --
M 8 0.77 0.98 1.36 2.62 1.47 1.33 1.82 1.64 1.91 1.67 2.52
M 13 0.95 -- -- -- -- -- -- -- -- 21.2 -- 14C-LE 1.00 99.0 98.6 97.4 98.5 98.7 98.2 98.4 98.1 45.0 97.5
Abbildung 29: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 1.
Versuchsabschnitt (unterschiedliche Inkubationszeiten), 2. Serie: Inkubation bei 60 min. Radio-HPLC-
Ergebnisse dargestellt als prozentuale Peakfläche bezogen auf die Gesamtpeakfläche jedes
Chromatogramms.
Metabolit Relative Retentions-
zeit
Pool 2 (M-NR)
Pool 9 (M-NR)
Pool 10 (M-R)
Pool 11 (W-NR)
PK 2 NK 2
? 0.37 -- -- -- -- 0.26 --
M 3 0.50 -- -- -- -- 1.54 --
M 5 0.59 -- -- -- -- 20.7 --
M 8 0.78 2.74 2.06 2.14 6.77 2.44 6.77
M 13 0.95 -- -- -- -- 10.2 -- 14C-LE 1.00 97.3 97.9 97.9 93.2 64.8 93.2
Abbildung 30: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 1.
Versuchsabschnitt (unterschiedliche Inkubationszeiten), 2. Serie: Inkubation bei 120 min. Radio-HPLC-
Ergebnisse dargestellt als prozentuale Peakfläche bezogen auf die Gesamtpeakfläche jedes
Chromatogramms.
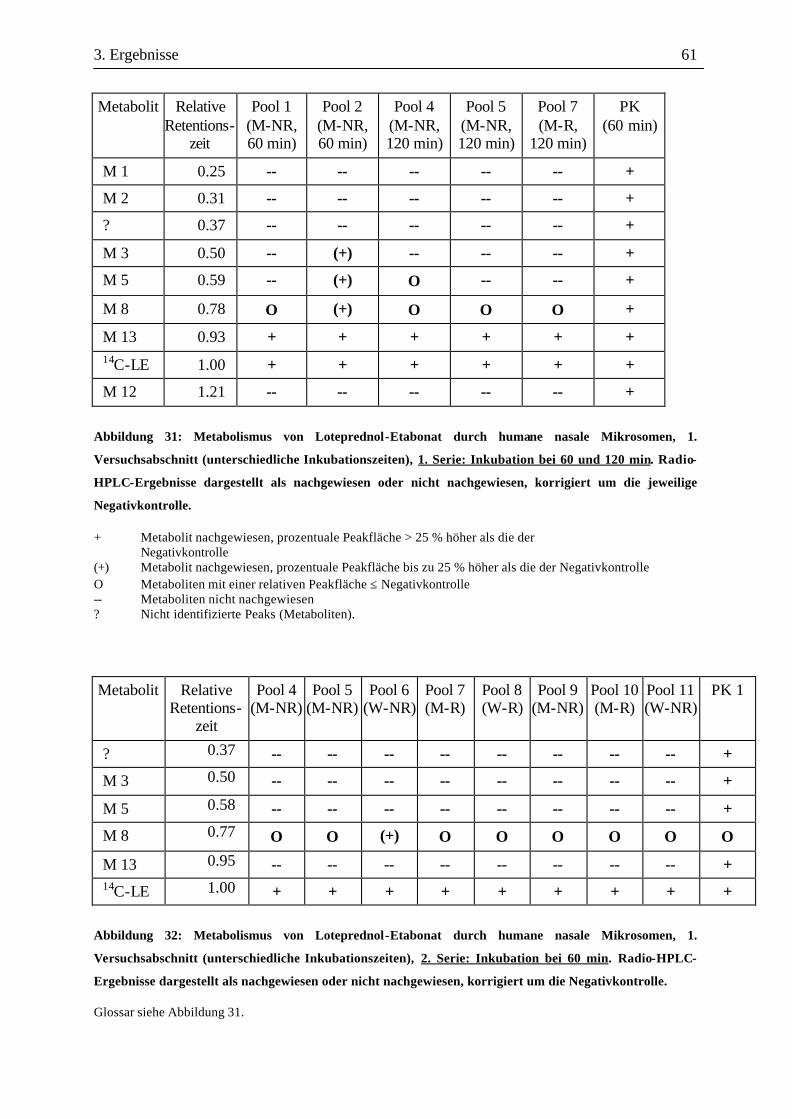
3. Ergebnisse 61
Metabolit Relative Retentions-
zeit
Pool 1 (M-NR, 60 min)
Pool 2 (M-NR, 60 min)
Pool 4 (M-NR, 120 min)
Pool 5 (M-NR, 120 min)
Pool 7 (M-R,
120 min)
PK (60 min)
M 1 0.25 -- -- -- -- -- +
M 2 0.31 -- -- -- -- -- +
? 0.37 -- -- -- -- -- +
M 3 0.50 -- (+) -- -- -- +
M 5 0.59 -- (+) Ο -- -- +
M 8 0.78 Ο (+) Ο Ο Ο +
M 13 0.93 + + + + + + 14C-LE 1.00 + + + + + +
M 12 1.21 -- -- -- -- -- +
Abbildung 31: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 1.
Versuchsabschnitt (unterschiedliche Inkubationszeiten), 1. Serie: Inkubation bei 60 und 120 min. Radio-
HPLC-Ergebnisse dargestellt als nachgewiesen oder nicht nachgewiesen, korrigiert um die jeweilige
Negativkontrolle.
+ Metabolit nachgewiesen, prozentuale Peakfläche > 25 % höher als die der Negativkontrolle (+) Metabolit nachgewiesen, prozentuale Peakfläche bis zu 25 % höher als die der Negativkontrolle Ο Metaboliten mit einer relativen Peakfläche ≤ Negativkontrolle -- Metaboliten nicht nachgewiesen ? Nicht identifizierte Peaks (Metaboliten).
Metabolit Relative
Retentions-zeit
Pool 4 (M-NR)
Pool 5 (M-NR)
Pool 6 (W-NR)
Pool 7 (M-R)
Pool 8 (W-R)
Pool 9 (M-NR)
Pool 10 (M-R)
Pool 11 (W-NR)
PK 1
? 0.37 -- -- -- -- -- -- -- -- +
M 3 0.50 -- -- -- -- -- -- -- -- +
M 5 0.58 -- -- -- -- -- -- -- -- +
M 8 0.77 Ο Ο (+) Ο Ο Ο Ο Ο Ο
M 13 0.95 -- -- -- -- -- -- -- -- + 14C-LE 1.00 + + + + + + + + +
Abbildung 32: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 1.
Versuchsabschnitt (unterschiedliche Inkubationszeiten), 2. Serie: Inkubation bei 60 min. Radio-HPLC-
Ergebnisse dargestellt als nachgewiesen oder nicht nachgewiesen, korrigiert um die Negativkontrolle.
Glossar siehe Abbildung 31.
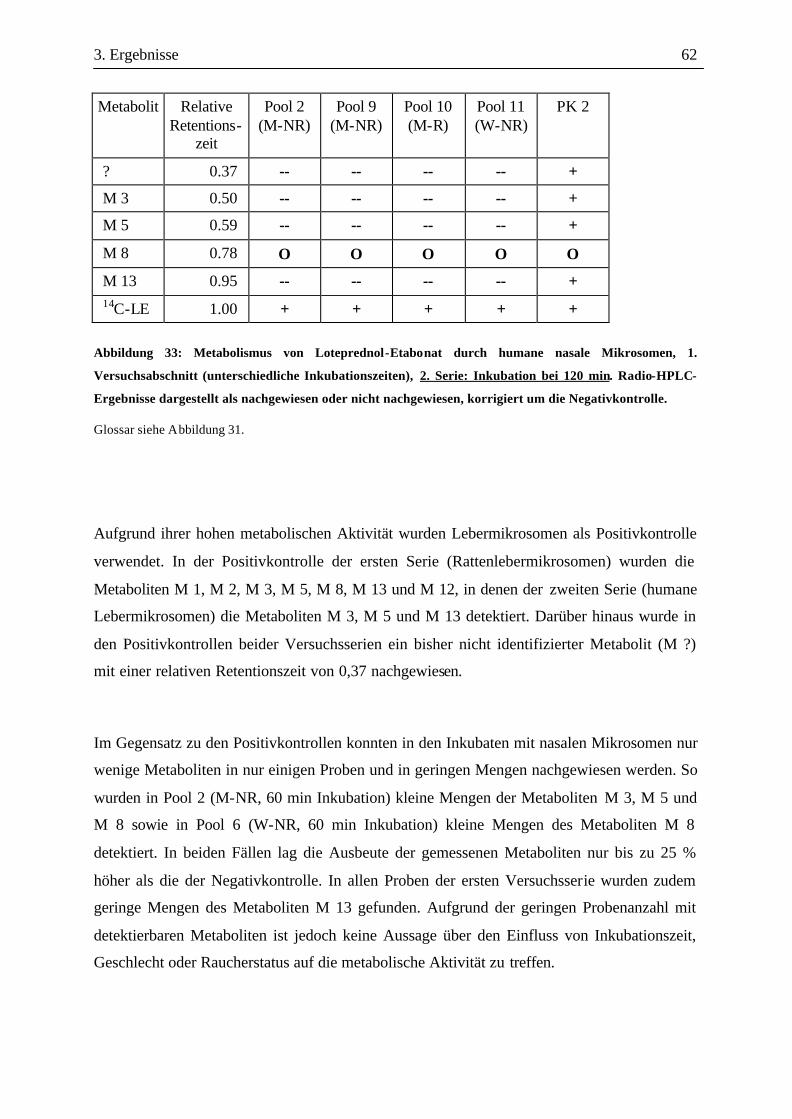
3. Ergebnisse 62
Metabolit Relative Retentions-
zeit
Pool 2 (M-NR)
Pool 9 (M-NR)
Pool 10 (M-R)
Pool 11 (W-NR)
PK 2
? 0.37 -- -- -- -- +
M 3 0.50 -- -- -- -- +
M 5 0.59 -- -- -- -- +
M 8 0.78 Ο Ο Ο Ο Ο
M 13 0.95 -- -- -- -- + 14C-LE 1.00 + + + + +
Abbildung 33: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 1.
Versuchsabschnitt (unterschiedliche Inkubationszeiten), 2. Serie: Inkubation bei 120 min. Radio-HPLC-
Ergebnisse dargestellt als nachgewiesen oder nicht nachgewiesen, korrigiert um die Negativkontrolle.
Glossar siehe Abbildung 31.
Aufgrund ihrer hohen metabolischen Aktivität wurden Lebermikrosomen als Positivkontrolle
verwendet. In der Positivkontrolle der ersten Serie (Rattenlebermikrosomen) wurden die
Metaboliten M 1, M 2, M 3, M 5, M 8, M 13 und M 12, in denen der zweiten Serie (humane
Lebermikrosomen) die Metaboliten M 3, M 5 und M 13 detektiert. Darüber hinaus wurde in
den Positivkontrollen beider Versuchsserien ein bisher nicht identifizierter Metabolit (M ?)
mit einer relativen Retentionszeit von 0,37 nachgewiesen.
Im Gegensatz zu den Positivkontrollen konnten in den Inkubaten mit nasalen Mikrosomen nur
wenige Metaboliten in nur einigen Proben und in geringen Mengen nachgewiesen werden. So
wurden in Pool 2 (M-NR, 60 min Inkubation) kleine Mengen der Metaboliten M 3, M 5 und
M 8 sowie in Pool 6 (W-NR, 60 min Inkubation) kleine Mengen des Metaboliten M 8
detektiert. In beiden Fällen lag die Ausbeute der gemessenen Metaboliten nur bis zu 25 %
höher als die der Negativkontrolle. In allen Proben der ersten Versuchsserie wurden zudem
geringe Mengen des Metaboliten M 13 gefunden. Aufgrund der geringen Probenanzahl mit
detektierbaren Metaboliten ist jedoch keine Aussage über den Einfluss von Inkubationszeit,
Geschlecht oder Raucherstatus auf die metabolische Aktivität zu treffen.
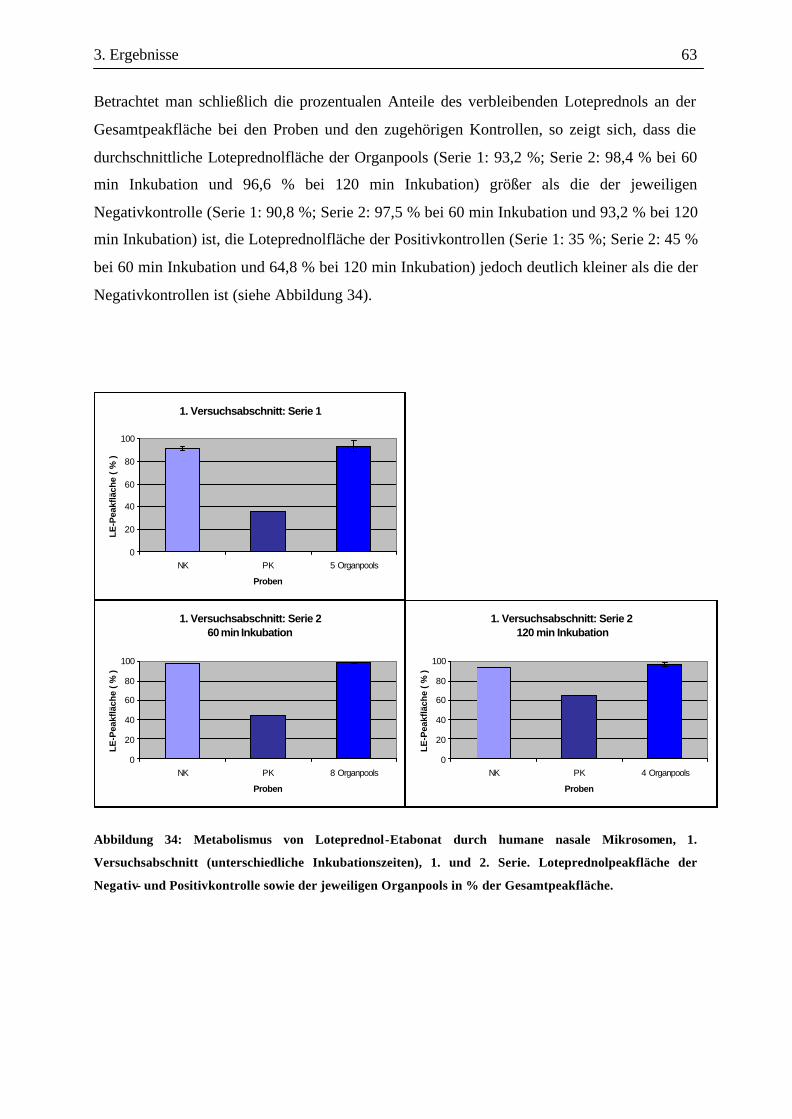
3. Ergebnisse 63
Betrachtet man schließlich die prozentualen Anteile des verbleibenden Loteprednols an der
Gesamtpeakfläche bei den Proben und den zugehörigen Kontrollen, so zeigt sich, dass die
durchschnittliche Loteprednolfläche der Organpools (Serie 1: 93,2 %; Serie 2: 98,4 % bei 60
min Inkubation und 96,6 % bei 120 min Inkubation) größer als die der jeweiligen
Negativkontrolle (Serie 1: 90,8 %; Serie 2: 97,5 % bei 60 min Inkubation und 93,2 % bei 120
min Inkubation) ist, die Loteprednolfläche der Positivkontrollen (Serie 1: 35 %; Serie 2: 45 %
bei 60 min Inkubation und 64,8 % bei 120 min Inkubation) jedoch deutlich kleiner als die der
Negativkontrollen ist (siehe Abbildung 34).
Abbildung 34: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 1.
Versuchsabschnitt (unterschiedliche Inkubationszeiten), 1. und 2. Serie. Loteprednolpeakfläche der
Negativ- und Positivkontrolle sowie der jeweiligen Organpools in % der Gesamtpeakfläche.
1. Versuchsabschnitt: Serie 1
0
20
40
60
80
100
NK PK 5 Organpools
Proben
LE-P
eakf
läch
e (
% )
1. Versuchsabschnitt: Serie 260 min Inkubation
0
20
40
60
80
100
NK PK 8 Organpools
Proben
LE
-Pea
kflä
che
( % )
1. Versuchsabschnitt: Serie 2120 min Inkubation
0
20
40
60
80
100
NK PK 4 Organpools
Proben
LE
-Pea
kflä
che
( % )
1. Versuchsabschnitt: Serie 1
0
20
40
60
80
100
NK PK 5 Organpools
Proben
LE-P
eakf
läch
e (
% )
1. Versuchsabschnitt: Serie 260 min Inkubation
0
20
40
60
80
100
NK PK 8 Organpools
Proben
LE
-Pea
kflä
che
( % )
1. Versuchsabschnitt: Serie 2120 min Inkubation
0
20
40
60
80
100
NK PK 4 Organpools
Proben
LE
-Pea
kflä
che
( % )

3. Ergebnisse 64
3.3.1.2 Teil II: Inkubation mit und ohne Glucuronidase-Zusatz
Die in diesem Versuchsabschnitt durchgeführten Wiederfindungsversuche durch Messung der
Gesamtradioaktivität vor und nach der Festphasenextraktion (siehe Abbildung 35) ergaben
eine durchschnittliche Wiederfindung nach der Festphasenextraktion von 84,5 % für die
Proben ohne Glucuronidase-Zusatz und von 84,1 % für die Proben mit Glucuronidase-Zusatz.
Damit ist die Wiederfindung mit jener der hausinternen Studie (83,9 % für Proben ohne
Glucuronidase-Zusatz, siehe Abschnitt 3.3.1.1) vergleichbar.
Abbildung 35: Gesamtradioaktivität der Kontrollen und der Organpools, dargestellt als DPM
(Desintegrations per Minute), gemessen am LS 6500 Multi-Purpos Scintillation Counter der Firma
Beckmann Coulter, Palo Alto, USA. Die oberen beiden Grafiken (jeweils ohne und mit Glucuronidase-Zusatz)
zeigen die Gesamtradioaktivität vor der SPE (solid phase extraction), die unteren beiden nach der SPE.
Gesamtradioaktivität vor SPE
0100002000030000400005000060000700008000090000
100000
NK PK Pool 12 Pool 13 Pool 14 Pool 15 Pool 16
Proben ohne Glucuronidase-Zusatz
DPMGesamtradioaktivität vor SPE
0100002000030000400005000060000700008000090000
NK PK Pool 12 Pool 13 Pool 14 Pool 15 Pool 16
Proben mit Glucuronidase-Zusatz
DPM
Gesamtradioaktivität nach SPE
0
10000
2000030000
4000050000
60000
70000
80000
NK PK Pool 12 Pool 13 Pool 14 Pool 15 Pool 16
Proben ohne Glucuronidase-Zusatz
DPMGesamtradioaktivität nach SPE
0
10000
2000030000
4000050000
60000
70000
80000
NK PK Pool 12 Pool 13 Pool 14 Pool 15 Pool 16
Proben mit Glucuronidase-Zusatz
DPM
Gesamtradioaktivität vor SPE
0100002000030000400005000060000700008000090000
100000
NK PK Pool 12 Pool 13 Pool 14 Pool 15 Pool 16
Proben ohne Glucuronidase-Zusatz
DPMGesamtradioaktivität vor SPE
0100002000030000400005000060000700008000090000
NK PK Pool 12 Pool 13 Pool 14 Pool 15 Pool 16
Proben mit Glucuronidase-Zusatz
DPM
Gesamtradioaktivität nach SPE
0
10000
2000030000
4000050000
60000
70000
80000
NK PK Pool 12 Pool 13 Pool 14 Pool 15 Pool 16
Proben ohne Glucuronidase-Zusatz
DPMGesamtradioaktivität nach SPE
0
10000
2000030000
4000050000
60000
70000
80000
NK PK Pool 12 Pool 13 Pool 14 Pool 15 Pool 16
Proben mit Glucuronidase-Zusatz
DPM

3. Ergebnisse 65
Im zweiten Versuchsabschnitt erfolgte die LE-Inkubation menschlicher nasaler Mikrosomen
bei 120 min mit und ohne Glucuronidase-Zusatz. Auch hier wurden Peaks ab 50 CPM
(Counts per Minute) integriert und als prozentualer Anteil an der Gesamtpeakfläche
dargestellt (siehe Abbildungen 36 bis 39).
Als Positivkontrolle dienten - wie in der 2. Serie des ersten Versuchsabschnittes - humane
Lebermikrosomen, bei denen in diesem Versuchsabschnitt jedoch zusätzlich zu den
Metaboliten M 3, M 5, M 13 und dem bisher nicht identifizierten Metaboliten (M ?) mit einer
relativen Retentionszeit von 0,37 des ersten Versuchsabschnittes der Metabolit M 7 sowie in
Spuren der Metabolit M 8 nachgewiesen werden konnten.
Bei allen fünf untersuchten Organpools war der Metabolit M 8 unter Zusatz des Enzyms β-
Glucuronidase deutlich gegenüber der Negativkontrolle erhöht. Ohne Glucuronidase-Zusatz
hingegen lag M 8 unterhalb der Negativkontrolle, mit Ausnahme von Pool 15 (W-R), bei dem
M 8 in dem Bereich bis zu 25 % oberhalb der Negativkontrolle zu finden war. M 8 der
Positivkontrolle befand sich ohne Glucuronidase-Zusatz ebenfalls unterhalb der
Negativkontrolle, mit Glucuronidase-Zusatz im Bereich bis zu 25 % oberhalb der
Negativkontrolle. Unter Glucuronidase-Zusatz wurde bei Pool 15 (W-R) ein weiterer
Metabolit detektiert, der mit einer relativen Retentionszeit von 0,19 bislang noch nicht
identifiziert wurde und der bei keiner der anderen Proben zu finden war.
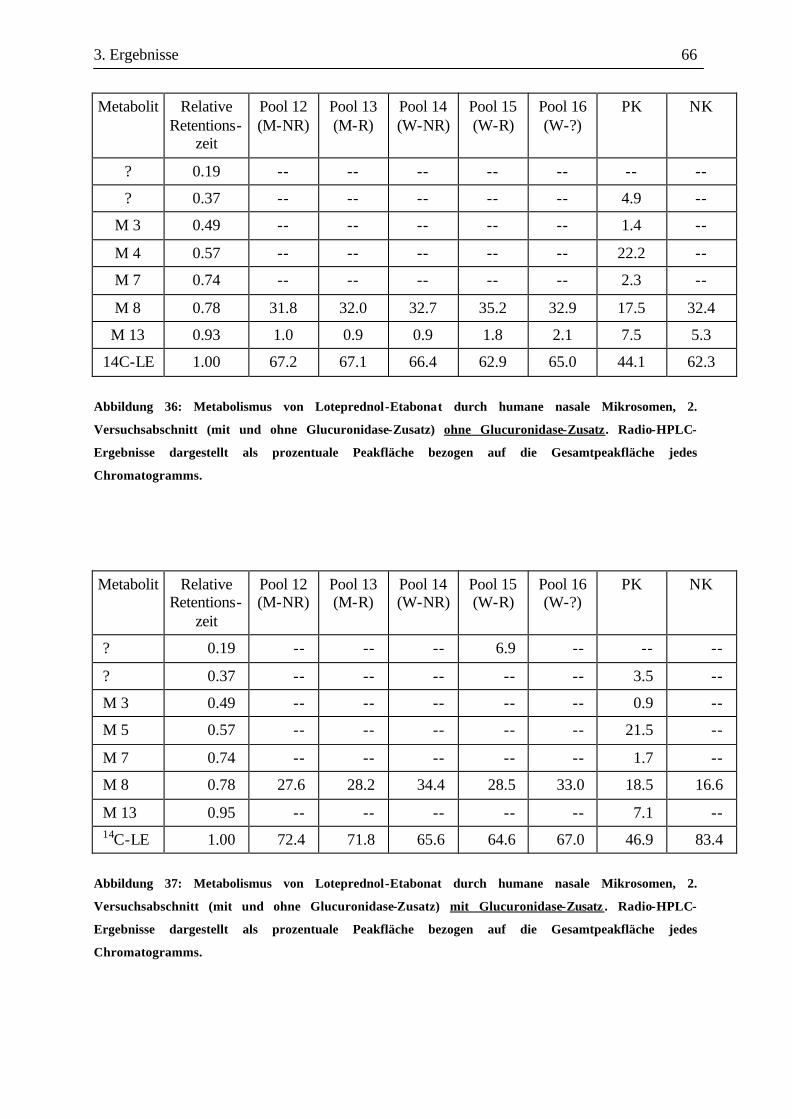
3. Ergebnisse 66
Metabolit Relative Retentions-
zeit
Pool 12 (M-NR)
Pool 13 (M-R)
Pool 14 (W-NR)
Pool 15 (W-R)
Pool 16 (W-?)
PK NK
? 0.19 -- -- -- -- -- -- --
? 0.37 -- -- -- -- -- 4.9 --
M 3 0.49 -- -- -- -- -- 1.4 --
M 4 0.57 -- -- -- -- -- 22.2 --
M 7 0.74 -- -- -- -- -- 2.3 --
M 8 0.78 31.8 32.0 32.7 35.2 32.9 17.5 32.4
M 13 0.93 1.0 0.9 0.9 1.8 2.1 7.5 5.3
14C-LE 1.00 67.2 67.1 66.4 62.9 65.0 44.1 62.3
Abbildung 36: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 2.
Versuchsabschnitt (mit und ohne Glucuronidase-Zusatz) ohne Glucuronidase-Zusatz. Radio-HPLC-
Ergebnisse dargestellt als prozentuale Peakfläche bezogen auf die Gesamtpeakfläche jedes
Chromatogramms.
Metabolit Relative Retentions-
zeit
Pool 12 (M-NR)
Pool 13 (M-R)
Pool 14 (W-NR)
Pool 15 (W-R)
Pool 16 (W-?)
PK NK
? 0.19 -- -- -- 6.9 -- -- --
? 0.37 -- -- -- -- -- 3.5 --
M 3 0.49 -- -- -- -- -- 0.9 --
M 5 0.57 -- -- -- -- -- 21.5 --
M 7 0.74 -- -- -- -- -- 1.7 --
M 8 0.78 27.6 28.2 34.4 28.5 33.0 18.5 16.6
M 13 0.95 -- -- -- -- -- 7.1 -- 14C-LE 1.00 72.4 71.8 65.6 64.6 67.0 46.9 83.4
Abbildung 37: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 2.
Versuchsabschnitt (mit und ohne Glucuronidase-Zusatz) mit Glucuronidase-Zusatz . Radio-HPLC-
Ergebnisse dargestellt als prozentuale Peakfläche bezogen auf die Gesamtpeakfläche jedes
Chromatogramms.

3. Ergebnisse 67
Metabolit Relative Retentions-
zeit
Pool 12 (M-NR)
Pool 13 (M-R)
Pool 14 (W-NR)
Pool 15 (W-R)
Pool 16 (W-?)
PK
? 0.19 -- -- -- -- -- --
? 0.37 -- -- -- -- -- +
M 3 0.49 -- -- -- -- -- +
M 5 0.57 -- -- -- -- -- +
M 7 0.74 -- -- -- -- -- +
M 8 0.78 Ο Ο Ο (+) Ο Ο
M 13 0.93 Ο Ο Ο Ο Ο + 14C-LE 1.00 + + + + + +
Abbildung 38: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 2.
Versuchsabschnitt (mit und ohne Glucuronidase-Zusatz) ohne Glucuronidase-Zusatz. Radio-HPLC-
Ergebnisse dargestellt als nachgewiesen oder nicht nachgewiesen, korrigiert um die Negativkontrolle.
+ Metabolit nachgewiesen, prozentuale Peakfläche > 25 % höher als die der Negativkontrolle (+) Metabolit nachgewiesen, prozentuale Peakfläche bis zu 25 % höher als die der Negativkontrolle Ο Metaboliten mit einer relativen Peakfläche ≤ Negativkontrolle -- Metaboliten nicht nachgewiesen ? Nicht identifizierte Peaks (Metaboliten).
Metabolit Relative Retentions-
zeit
Pool 12 (M-NR)
Pool 13 (M-R)
Pool 14 (W-NR)
Pool 15 (W-R)
Pool 16 (W-?)
PK
? 0.19 -- -- -- + -- --
? 0.37 -- -- -- -- -- +
M 3 0.49 -- -- -- -- -- +
M 5 0.57 -- -- -- -- -- +
M 7 0.74 -- -- -- -- -- +
M 8 0.78 + + + + + (+)
M 13 0.95 -- -- -- -- -- + 14C-LE 1.00 + + + + + +
Abbildung 39: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 2.
Versuchsabschnitt (mit und ohne Glucuronidase-Zusatz) mit Glucuronidase-Zusatz . Radio-HPLC-
Ergebnisse dargestellt als nachgewiesen oder nicht nachgewiesen, korrigiert um die Negativkontrolle.
Glossar siehe Abbildung 38.
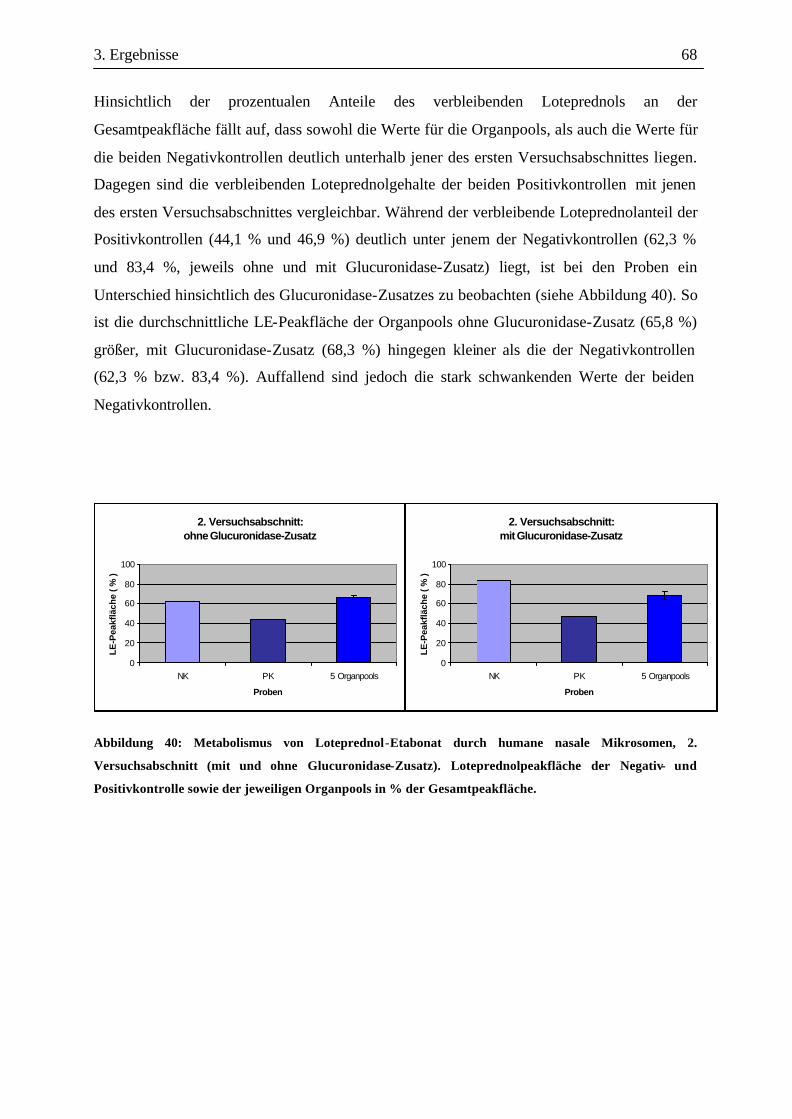
3. Ergebnisse 68
Hinsichtlich der prozentualen Anteile des verbleibenden Loteprednols an der
Gesamtpeakfläche fällt auf, dass sowohl die Werte für die Organpools, als auch die Werte für
die beiden Negativkontrollen deutlich unterhalb jener des ersten Versuchsabschnittes liegen.
Dagegen sind die verbleibenden Loteprednolgehalte der beiden Positivkontrollen mit jenen
des ersten Versuchsabschnittes vergleichbar. Während der verbleibende Loteprednolanteil der
Positivkontrollen (44,1 % und 46,9 %) deutlich unter jenem der Negativkontrollen (62,3 %
und 83,4 %, jeweils ohne und mit Glucuronidase-Zusatz) liegt, ist bei den Proben ein
Unterschied hinsichtlich des Glucuronidase-Zusatzes zu beobachten (siehe Abbildung 40). So
ist die durchschnittliche LE-Peakfläche der Organpools ohne Glucuronidase-Zusatz (65,8 %)
größer, mit Glucuronidase-Zusatz (68,3 %) hingegen kleiner als die der Negativkontrollen
(62,3 % bzw. 83,4 %). Auffallend sind jedoch die stark schwankenden Werte der beiden
Negativkontrollen.
Abbildung 40: Metabolismus von Loteprednol-Etabonat durch humane nasale Mikrosomen, 2.
Versuchsabschnitt (mit und ohne Glucuronidase-Zusatz). Loteprednolpeakfläche der Negativ- und
Positivkontrolle sowie der jeweiligen Organpools in % der Gesamtpeakfläche.
2. Versuchsabschnitt: ohne Glucuronidase-Zusatz
0
20
40
60
80
100
NK PK 5 Organpools
Proben
LE
-Pea
kflä
che
( % )
2. Versuchsabschnitt: mit Glucuronidase-Zusatz
0
20
40
60
80
100
NK PK 5 Organpools
Proben
LE
-Pea
kflä
che
( % )
2. Versuchsabschnitt: ohne Glucuronidase-Zusatz
0
20
40
60
80
100
NK PK 5 Organpools
Proben
LE
-Pea
kflä
che
( % )
2. Versuchsabschnitt: mit Glucuronidase-Zusatz
0
20
40
60
80
100
NK PK 5 Organpools
Proben
LE
-Pea
kflä
che
( % )
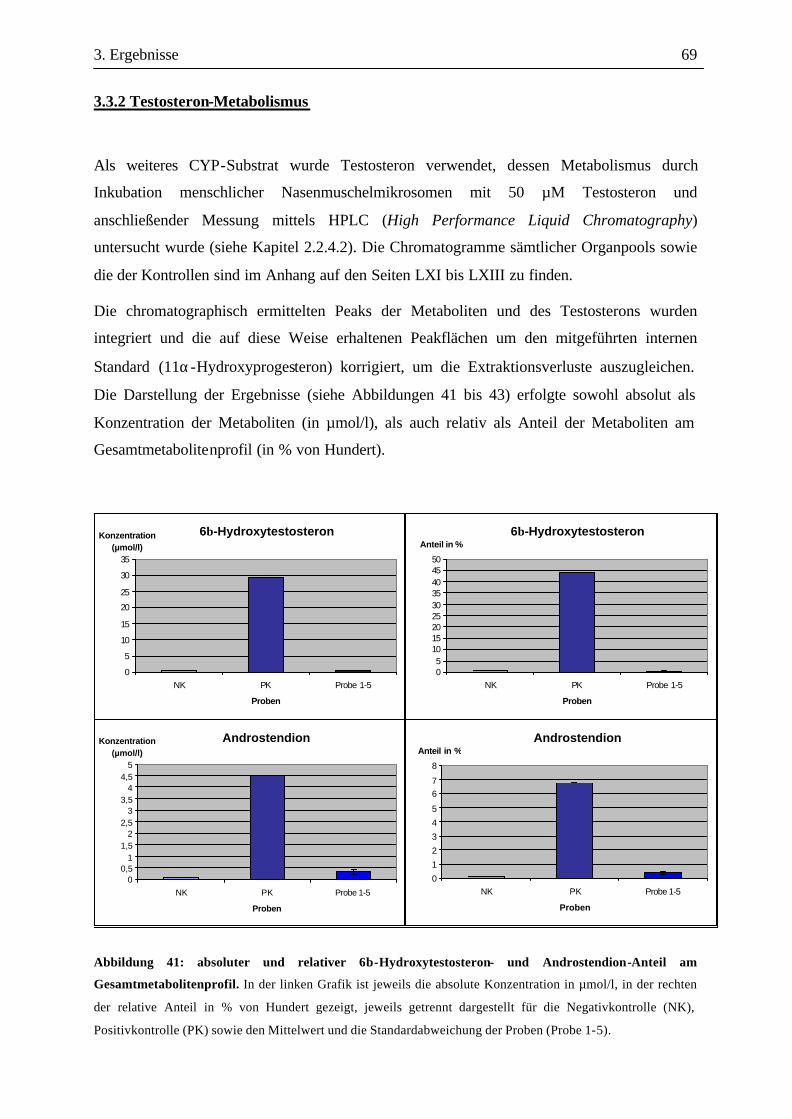
3. Ergebnisse 69
3.3.2 Testosteron-Metabolismus
Als weiteres CYP-Substrat wurde Testosteron verwendet, dessen Metabolismus durch
Inkubation menschlicher Nasenmuschelmikrosomen mit 50 µM Testosteron und
anschließender Messung mittels HPLC (High Performance Liquid Chromatography)
untersucht wurde (siehe Kapitel 2.2.4.2). Die Chromatogramme sämtlicher Organpools sowie
die der Kontrollen sind im Anhang auf den Seiten LXI bis LXIII zu finden.
Die chromatographisch ermittelten Peaks der Metaboliten und des Testosterons wurden
integriert und die auf diese Weise erhaltenen Peakflächen um den mitgeführten internen
Standard (11α-Hydroxyprogesteron) korrigiert, um die Extraktionsverluste auszugleichen.
Die Darstellung der Ergebnisse (siehe Abbildungen 41 bis 43) erfolgte sowohl absolut als
Konzentration der Metaboliten (in µmol/l), als auch relativ als Anteil der Metaboliten am
Gesamtmetabolitenprofil (in % von Hundert).
Abbildung 41: absoluter und relativer 6β-Hydroxytestosteron- und Androstendion-Anteil am
Gesamtmetabolitenprofil. In der linken Grafik ist jeweils die absolute Konzentration in µmol/l, in der rechten
der relative Anteil in % von Hundert gezeigt, jeweils getrennt dargestellt für die Negativkontrolle (NK),
Positivkontrolle (PK) sowie den Mittelwert und die Standardabweichung der Proben (Probe 1-5).
05
101520253035404550
NK PK Probe 1-5
Proben
Anteil in %6β-Hydroxytestosteron
0
5
10
15
20
25
30
35
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
6β-Hydroxytestosteron
0
1
2
3
4
5
67
8
NK PK Probe 1-5
Proben
Anteil in %Androstendion
00,5
11,5
22,5
33,5
44,5
5
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
Androstendion
05
101520253035404550
NK PK Probe 1-5
Proben
Anteil in %6β-Hydroxytestosteron
0
5
10
15
20
25
30
35
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
6β-Hydroxytestosteron
05
101520253035404550
NK PK Probe 1-5
Proben
Anteil in %6β-Hydroxytestosteron
0
5
10
15
20
25
30
35
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
6β-Hydroxytestosteron
0
1
2
3
4
5
67
8
NK PK Probe 1-5
Proben
Anteil in %Androstendion
00,5
11,5
22,5
33,5
44,5
5
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
Androstendion
0
1
2
3
4
5
67
8
NK PK Probe 1-5
Proben
Anteil in %Androstendion
00,5
11,5
22,5
33,5
44,5
5
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
Androstendion

3. Ergebnisse 70
Alle Testosteronmetaboliten lagen jeweils unter 0,5 % des metabolischen Gesamtprofils. Den
höchsten absoluten Wert erreichte mit 0,45 ± 0,04 % (0,38 ± 0,04 µmol/l) 6β-
Hydroxytestosteron, welches sich damit jedoch trotzdem unterhalb der Negativkontrolle mit
0,50 % (0,37 µmol/l) befand. Deutlich über der Negativkontrolle von 0,10 % (0,07 µmol/l)
lag das Androstendion mit 0,37 ± 0,12 % (0,31 ± 0,11 µmol/l).
Unterhalb der Negativkontrollen lagen die beiden Metaboliten 2α-Hydroxytestosteron und
16α-Hydroxytestosteron mit jeweils 0,16 ± 0,03 % (0,14 ± 0,02 µmol/l) und 0,18 ± 0,02 (0,15
± 0,02 µmol/l) gegenüber 0,26 % (0,20 µmol/l) und 0,32 % (0,24 µmol/l).
Abbildung 42: absoluter und relativer 2α- und 16α-Hydroxytestosteron -Anteil am
Gesamtmetabolitenprofil. In der linken Grafik ist jeweils die absolute Konzentration in µmol/l, in der rechten
der relative Anteil in % von Hundert gezeigt, jeweils getrennt dargestellt für die Negativkontrolle (NK),
Positivkontrolle (PK) sowie den Mittelwert und die Standardabweichung der Proben (Probe 1-5).
0
1
23
4
5
6
7
8
NK PK Probe 1-5
Proben
Anteil in %2α-Hydroxytestosteron
00,5
11,5
22,5
33,5
44,5
5
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
2α-Hydroxytestosteron
0
0,5
11,5
2
2,5
3
3,5
4
NK PK Probe 1-5
Proben
Konzentration (µmol/)
16α-Hydroxytestosteron
0
1
2
3
4
5
6
NK PK Probe 1-5
Proben
Anteil in %16α-Hydroxytestosteron
0
1
23
4
5
6
7
8
NK PK Probe 1-5
Proben
Anteil in %2α-Hydroxytestosteron
00,5
11,5
22,5
33,5
44,5
5
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
2α-Hydroxytestosteron
0
1
23
4
5
6
7
8
NK PK Probe 1-5
Proben
Anteil in %2α-Hydroxytestosteron
00,5
11,5
22,5
33,5
44,5
5
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
2α-Hydroxytestosteron
0
0,5
11,5
2
2,5
3
3,5
4
NK PK Probe 1-5
Proben
Konzentration (µmol/)
16α-Hydroxytestosteron
0
1
2
3
4
5
6
NK PK Probe 1-5
Proben
Anteil in %16α-Hydroxytestosteron
0
0,5
11,5
2
2,5
3
3,5
4
NK PK Probe 1-5
Proben
Konzentration (µmol/)
16α-Hydroxytestosteron
0
1
2
3
4
5
6
NK PK Probe 1-5
Proben
Anteil in %16α-Hydroxytestosteron

3. Ergebnisse 71
Nur in Spuren detektierbar waren die Metaboliten 6α- und 7α-Hydroxytestosteron mit 0,022
± 0,003 % (0,019 ± 0,003 µmol/l) und 0,0006 ± 0,0009 % (0,0005 ± 0,0007 µmol/l). Im Falle
des 6α-Hydroxytestosterons betrug die Negativkontrolle 0,018 % (0,013 µmol/l), 7α-
Hydroxytestosteron war dagegen weder in der Negativ-, noch in der Positivkontrolle
nachweisbar (siehe Abbildung 43).
Abbildung 43: absoluter und relativer 6α- und 7α-Hydroxytestosteron -Anteil am
Gesamtmetabolitenprofil. In der linken Grafik ist jeweils die absolute Konzentration in µmol/l, in der rechten
der relative Anteil in % von Hundert gezeigt, jeweils getrennt dargestellt für die Negativkontrolle (NK),
Positivkontrolle (PK) sowie den Mittelwert und die Standardabweichung der Proben (Probe 1-5).
0
0,02
0,04
0,06
0,08
0,1
0,12
0,14
NK PK Probe 1-5
Proben
Anteil in %6α-Hydroxytestosteron
00,010,020,030,040,050,060,070,080,09
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
6α-Hydroxytestosteron
00,00020,00040,00060,00080,001
0,00120,00140,00160,0018
NK PK Probe 1-5
Proben
Anteil in %7α-Hydroxytestosteron
0
0,0002
0,0004
0,0006
0,0008
0,001
0,0012
0,0014
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
7α-Hydroxytestosteron
0
0,02
0,04
0,06
0,08
0,1
0,12
0,14
NK PK Probe 1-5
Proben
Anteil in %6α-Hydroxytestosteron
00,010,020,030,040,050,060,070,080,09
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
6α-Hydroxytestosteron
0
0,02
0,04
0,06
0,08
0,1
0,12
0,14
NK PK Probe 1-5
Proben
Anteil in %6α-Hydroxytestosteron
00,010,020,030,040,050,060,070,080,09
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
6α-Hydroxytestosteron
00,00020,00040,00060,00080,001
0,00120,00140,00160,0018
NK PK Probe 1-5
Proben
Anteil in %7α-Hydroxytestosteron
0
0,0002
0,0004
0,0006
0,0008
0,001
0,0012
0,0014
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
7α-Hydroxytestosteron
00,00020,00040,00060,00080,001
0,00120,00140,00160,0018
NK PK Probe 1-5
Proben
Anteil in %7α-Hydroxytestosteron
0
0,0002
0,0004
0,0006
0,0008
0,001
0,0012
0,0014
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
7α-Hydroxytestosteron

3. Ergebnisse 72
Somit ist das Substrat Testosteron in der Positivkontrolle zu 36,76 % (24,43 µmol/l)
unverstoffwechselt geblieben, während es in der Negativkontrolle (98,80 % bzw. 74,18
µmol/l) und in den Proben (98,82 ± 0,16 % bzw. 83,37 ± 4,59 µmol/l) zu nahezu identischen
Anteilen verblieben ist (siehe Abbildung 44).
Abbildung 44: absoluter und relativer Testosteron-Anteil am Gesamtmetabolitenprofil. In der linken
Grafik ist die absolute Testosteron-Konzentration in µmol/l, in der rechten der relative Testosteron-Anteil in %
von Hundert gezeigt, jeweils getrennt dargestellt für die Negativkontrolle (NK), Positivkontrolle (PK) sowie den
Mittelwert und die Standardabweichung der Proben (Probe 1-5).
3.3.3 EROD-Aktivitäten
Aufgrund der nur geringen Mengen an mikrosomalem Protein war es nicht möglich,
klassische enzymkinetische Untersuchungen mit variablen Zeiten und variablen
Substratmengen durchzuführen (vergleiche hierzu Michaelis-Menten-Kinetik). Jedoch ließen
sich bei den Organpools auch mit den eingesetzten 500 µg mikrosomalen Proteins keine
EROD-Aktivitäten nachweisen, weshalb nur die Ergebnisse der Positivkontrolle dargestellt
sind. So konnte für die Positivkontrolle (menschliche Lebermikrosomen) eine Aktivität von
199,3 nmol/mg mikrosomales Protein/h ermittelt werden (siehe Abbildung 45). Nach
Inkubation mit β-Glucuronidase war eine Zunahme des Resorufin-Signales um 21,7 % zu
beobachten, was auf eine deutliche Glucuronidierung nach erfolgter Oxidation schließen lässt.
0102030405060708090
100
NK PK Probe 1-5
Proben
Anteil in %Testosteron
0102030405060708090
100
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
Testosteron
0102030405060708090
100
NK PK Probe 1-5
Proben
Anteil in %Testosteron
0102030405060708090
100
NK PK Probe 1-5
Proben
Konzentration (µmol/l)
Testosteron

3. Ergebnisse 73
Abbildung 45: EROD-Aktivität menschlicher Lebermikrosomen, die im Rahmen des EROD-Assays als
Positivkontrolle verwendet wurden. Dargestellt ist die Bildung des fluorimetrisch gemessenen Resorufins nach
120-minütiger Inkubation mit 10µM Ethoxyresorufin.
0
50
100
150
200
250
300
Positivkontrolle
Bild
ung
von
Res
rufin
[nm
ol/m
g m
ikro
som
ales
Pro
tein
/h]
glucuronidiertesResorufin
nicht-glucuronidiertesResorufin

4. Diskussion 74
4. Diskussion
4.1 Untersuchtes Gewebe
Ausgangsmaterial für die Untersuchungen war hypertrophe Schleimhaut der unteren Nasenmuschel,
welche bei den einzelnen Patienten aufgrund verschiedener Erkrankungen – meist chronischer
Entzündung bzw. allergischer Rhinitis – operativ entfernt werden musste. Es handelt sich dabei zwar
nicht um gesundes und damit unverändertes Gewebe, stellt aber letztendlich genau jenes Gewebe
dar, welches im Rahmen der betreffenden Erkrankung medikamentös behandelt werden soll. Bedingt
durch die Art der Gewinnung des Probenmaterials (chirurgische Entfernung der unteren
Nasenmuschel, Abkratzen der Schleimhaut von den knorpeligen und knöchernen Trägerstrukturen)
ist nicht ganz auszuschließen, dass auch einzelne Zellen adherierenden Gewebes mituntersucht
wurden.
Aufgrund ihrer anatomischen Lage gehört die untersuchte Schleimhaut zur Regio respiratoria der
Nasenschleimhaut. Nicht untersucht wurde dagegen Schleimhaut der Regio olfactoria, die für eine
topische Applikation von Arzneistoffen nicht in Frage kommt. Darüber hinaus unterscheidet sich die
olfaktorische Schleimhaut bei Kaninchen, Ratte, Hund, Schwein, Affe und vermutlich auch beim
Menschen von der respiratorischen Nasenschleimhaut durch eine deutlich höhere metabolische
Aktivität (Gervasi et al., 1991; Hadley and Dahl, 1983; Longo et al., 1989; Marini et al., 1998;
Reed, 1993) sowie durch weitere Funktionen. Hierzu gehört die Beteiligung bei der
Geruchswahrnehmung sowie der Schutz der olfaktorischen Neurone und damit des empfindlichen
Nervengewebes (Bereziat et al., 1995; Dahl and Hadley, 1991; Getchell et al., 1993; Su et al.,
1996), weshalb die olfaktorische Schleimhaut für Vergleiche insgesamt ungeeignet erscheint.
Hingegen ist die Bronchialschleimhaut der menschlichen Lunge nahezu ideal für Vergleichszwecke,
da sich die respiratorische Schleimhaut der oberen Atemwege über die Trachealschleimhaut
kontinuierlich bis in die unteren Atemwege fortsetzt und bereits mehrere Daten zur inhalativen
Arzneistoffapplikation an der menschlichen Lunge vorliegen. Ebenso für Vergleichszwecke geeignet
erscheint die respiratorische Nasenschleimhaut anderer Spezies. Im Vordergrund stehen dabei
Kaninchen, Maus und Ratte, da diese häufig für toxikologische Studien herangezogen werden und
ihre Nasenschleimhäute deshalb vergleichsweise gut charakterisiert sind. Aus den genannten Gründen
werden die Versuchsergebnisse der vorgelegten Doktorarbeit im Folgenden mit der respiratorischen

4. Diskussion 75
Nasenschleimhaut anderer Spezies sowie mit respiratorischem Gewebe der menschlichen Lunge
verglichen.
4.2 Exprimierte Gene
Mit der RT-PCR wird die mRNA verschiedener Targetgene innerhalb eines Gewebes
nachgewiesen, wobei es sich um eine Momentaufnahme des jeweiligen Gewebes handelt. Die
unterschiedlich starke Stabilisierung oder Degradierung der mRNA führt dabei zu spezifischen
Halbwertszeiten der einzelnen mRNAs. Zusätzlich kann eine mögliche Proteinbindung die Translation
behindern, weshalb vom mRNA-Gehalt nicht unmittelbar auf den Proteingehalt geschlossen werden
darf. Aus diesen Gründen wurde die Nasenschleimhaut nicht nur auf der Genexpressionsebene,
sondern auch auf der Protein- und Metabolismusebene untersucht. Die Beobachtung, dass teilweise
mRNA nachgewiesen wurde, aber kein Protein detektierbar war (wie beispielsweise im Fall der
Cytochrome 3A4 und 3A5), zeigt, dass geringe mRNA-Mengen physiologisch möglicherweise ohne
große Bedeutung sind (Koskela et al., 1999). Da die untersuchte Nasenschleimhaut von
unterschiedlichen Spendern stammte, die verschiedene Genotypen besitzen und unterschiedlichen
Umwelteinflüssen unterliegen, war es zu erwarten, dass die einzelnen Gene bei den verschiedenen
Personen unterschiedlich stark exprimiert bzw. teilweise auch gar nicht exprimiert waren. So waren
unter den untersuchten Cytochromen nur CYP 1B1, 2A6/7, 2A13, 2B6, 2C8 und 2S1 bei allen
Spendern exprimiert, während die übrigen Cytochrome bei mindestens einer der insgesamt 22
Patienten nicht exprimiert waren.
Im Anschluss an diese allgemeinen Erläuterungen wird die Expression der einzelnen Gene diskutiert.
Genexpressionsergebnisse werden häufig als Verhältnis zu stabil exprimierten Housekeeping-Genen
dargestellt. Oft werden hierfür GAPDH oder CyclophillinA eingesetzt. Da GAPDH mit einer
statistischen Signifikanz von p<0,05 in der menschlichen respiratorischen Nasenschleimhaut bei
Rauchern und Nicht-Rauchern unterschiedlich stark exprimiert war, konnte es als interne
Bezugsgröße nicht verwendet werden. Auch Deindl et al. (2002), Prieto-Alamo et al. (2003) und

4. Diskussion 76
Tricario et al. (2002) haben bereits in vorherigen Studien gezeigt, dass die Expression des häufig
verwendeten Housekeeping-Genes GAPDH innerhalb verschiedener Gewebe des Menschen
teilweise erheblich variiert und somit nicht immer als interner Standard geeignet ist. Auch
CyclophillinA wurde auf die Eignung als Housekeeping-Gen untersucht. Zwar war der beobachtete
Unterschied mit p = 0,076 statistisch nicht signifikant, jedoch wurde aufgrund der Nähe zum
Signifikanzniveau (p<0,05) auf eine Korrektur der Werte durch CyclophillinA ebenfalls verzichtet, so
dass die Genenxpressionsergebnisse absolut dargestellt wurden.
Aus der Enzymfamilie des Cytochrom P450-Systems konnten hier neben den bereits bei anderen
Spezies in der Nasenschleimhaut nachgewiesenen CYP-Isoformen der 1A-, 2A-, 2B-, 2C-, 2E-,
3A- und 4B-Subfamilie (Longo et al., 2000; Thornton-Manning et al., 1997) zusätzlich die
Expression von CYP 1B1, 2F1, 2J2 sowie des erst kürzlich beschriebenen CYP 2S1 mittels RT-
PCR gezeigt werden. Dagegen war das in Leber und Niere vorkommende CYP 4A11 (Baker et al.,
2003; Cummings et al., 2000; Savas et al., 2003) nicht in der menschlichen Nasenschleimhaut
exprimiert. Über das Vorhandensein von CYP 4A11 in anderen respiratorischen Geweben des
Menschen ist bislang auch nichts bekannt. Im Folgenden werden die Ergebnisse im einzelnen
diskutiert.
Das durch AhR-Liganden in nahezu jedem untersuchten Gewebe induzierbare CYP 1A1
(Hukkanen, 2000) ist in der menschlichen respiratorischen Nasenschleimhaut offensichtlich nicht
konstitutiv exprimiert, wohl aber durch Zigarettenrauch induzierbar. So war es bei 60 % der Raucher
exprimiert, während es bei keinem der Nicht-Raucher nachweisbar war. Eine CYP 1A1-
Enzyminduktion nach Zigarettenrauchexposition konnten Wardlaw et al. (1998) auch in der
Nasenschleimhaut von Ratten beobachten. Auch in Untersuchungen an respiratorischem Gewebe der
menschlichen Lunge erwies sich Zigarettenrauch als potenter Induktor (Anttila et al., 2000; Mollerup
et al., 1999; Willey et al., 1997), wobei quantitative RT-PCR-Messungen eine höhere Induktion bei
weiblichen als bei männlichen Rauchern belegen (Mollerup et al., 1999). Bei den hier durchgeführten
Versuchen war CYP 1A1 bei insgesamt vier weiblichen im Gegensatz zu nur zwei männlichen
Rauchern induziert, was in Übereinstimmung mit den Beobachtungen von Mollerup et al. (1999) an
der menschlichen Lunge steht. Interessanterweise war der Unterschied zwischen der Raucher- und
der Nicht-Raucher-Gruppe auch nur bei den weiblichen Personen statistisch signifikant.
Im Vergleich zu CYP 1A1 ließ sich das zweite Mitglied der CYP 1A-Subfamilie - CYP 1A2 - bei
keinem der Spender (n=22) nachweisen. Auch in der menschlichen Lunge konnte CYP 1A2 bislang

4. Diskussion 77
noch nicht detektiert werden (Shimada et al., 1996; Thum and Borlak, 2000; Wheeler et al., 1990).
Nach Hukkanen (2000) wird CYP 1A2 strikt leberspezifisch exprimiert, da bisher noch kein CYP
1A2-Protein in extrahepatischen Geweben nachgewiesen wurde. Im Gegensatz zur
Nasenschleimhaut des Menschen ist CYP 1A2 in der Nasenschleimhaut der Ratte konstitutiv
exprimiert (Longo et al., 2000). Daher liegt der Schluss nahe, dass die Ergebnisse von Tierversuchen
nicht unkritisch auf den Menschen übertragen werden können.
Mit einer Signifikanz von p<0,00006 konnte für CYP 1B1 eine statistisch hochsignifikante Induktion
für Raucher ermittelt werden. Auch diese Beobachtung steht in Einklang mit Untersuchungen an der
menschlichen Lunge (Willey et al., 1997).
Die CYP 2A-Subfamilie beinhaltet beim Menschen die Mitglieder CYP 2A6, 2A7 sowie 2A13, von
denen sich bislang nur CYP 2A6 und 2A13 als katalytisch aktiv erwiesen haben (Su et al., 2000).
CYP 2A13, das unter allen untersuchten Geweben am stärksten in der Nasenschleimhaut exprimiert
ist (Su et al., 2000), ließ sich bei allen Patienten, nicht jedoch in der menschlichen Leber nachweisen.
Dies bestätigt die früheren Ergebnisse von Koskela et al. (1999) und Su et al. (2000), die ebenfalls
CYP 2A13 nur in der Nasenschleimhaut nachweisen konnten. Im Gegensatz zu CYP 2A13 war
CYP 2A6/7 zusätzlich auch in der Leber nachweisbar. Auch dies steht in Einklang mit den
Beobachtungen von Koskela et al. (1999) und Su et al. (2000). Die CYP 2A-Isoformen des
Menschen sind hinsichtlich ihrer Nukleotidsequenz zu > 83% mit CYP 2A3 der Ratte, CYP 2A4
und 2A5 der Maus sowie CYP 2A10 und 2A11 des Kaninchens identisch (Ding et al., 1994; Peng
et al., 1993). Sie sind somit homolog zum Cytochrom P450 NMa des Kaninchens, welches zuerst in
nasalen Mikrosomen entdeckt und erst später als CYP 2A10 und 2A11 definiert wurde. Das NMa
ist eines der Hauptcytochrome in der Nasenschleimhaut des Kaninchens. Es metabolisiert mit hoher
Aktivität zahlreiche Substrate (Ding and Coon, 1990; Peng et al., 1993), weshalb sich
Untersuchungen an der Nasenschleimhaut bei verschiedenen Spezies auf die Homologe der CYP
2A-Subfamilie konzentrieren.
Übereinstimmend mit Beobachtungen an respiratorischem Gewebe der menschlichen Lunge (Willey
et al., 1997) konnten bei dem Cytochrom 2B6 auch in der Nasenschleimhaut keine Unterschiede
zwischen Rauchern und Nicht-Rauchern beobachtet werden. Auch die Mitglieder der CYP 2C-
Subfamilie zeigten weder Raucher-, noch Geschlechtsunterschiede, jedoch mussten bei der RT-PCR
von CYP 2C8, 2C9 und 2C19 sehr hohe Zyklenzahlen verwendet werden (jeweils 39 Zyklen), um

4. Diskussion 78
überhaupt Transkripte amplifizieren zu können. Die Frage, ob derart geringe mRNA-Mengen zu
einer nennenswerten Proteintranslation führen und damit physiologisch von Bedeutung sind, haben
bereits Koskela et al. (1999) diskutiert. Im Gegensatz zu den zuvor genannten Isoformen ließ sich
CYP 2C18 bereits mit 33 Zyklen amplifizieren, war aber nur bei 19 der insgesamt 22 Personen
nachweisbar. Bezogen auf die mRNA-Expression scheint CYP 2C18 in der menschlichen Lunge
eine abundante Isoform der CYP 2C-Subfamilie zu sein (Zhu-Ge et al., 2002), was wiederum in
Einklang mit meinen Untersuchungen an der respiratorischen Nasenschleimhaut steht. Obwohl
bislang wenig über spezifische CYP 2C18-Substrate bekannt ist (Zhu-Ge et al., 2002) und CYP
2C18-Protein im Gegensatz zu CYP 2C8, 2C9 und 2C19 in der Leber bisher nicht nachgewiesen
wurde (Gerbal-Chaloin et al., 2001; Hukkanen, 2000; Zhu-Ge et al., 2002), sind mit heterolog
exprimiertem CYP 2C18-Protein einige Substrate identifiziert worden, die CYP 2C18 als Mitglied
der CYP 2C-Subfamilie metabolisiert und die meist auch von den anderen Isoformen
verstoffwechselt werden. Hierzu zählen Tolbutamid (Hukkanen, 2000; Zhu-Ge et al., 2002),
Sulfaphenazolderivate (Ha-Duong et al., 2001), Bisphenol A (Niwa et al., 2001), all-trans-
Retinolsäure (Marill et al., 1999), Diclofenac (Mancy et al., 1996; Bort et al., 1999) und Omeprazol
(Hukkanen, 2000; Karam et al., 1996).
Das ebenfalls zahlreiche Arzneistoffe metabolisierende CYP 2D6-Protein ist durch eine große
interindividuelle Variabilität in der metabolischen Aktivität gekennzeichnet (Charlier et al., 2003;
Endrizzi et al., 2002; Muller et al., 2003). Begründet wird dies durch genetische Polymorphismen,
die zu unterschiedlichen Metabolisierungs-Typen führen. Der Nachweis von CYP 2D6-Transkripten
kann dabei durch Amplifikate der Pseudogene CYP 2D7P und CYP 2D8P behindert werden
(Endrizzi et al., 2002). Das Fehlen einer spezifischen Bande in Höhe der zu erwartenden 332 bp und
das Auftreten mehrerer größerer Fragmente in meinen Untersuchungen könnte auf diese Weise
erklärt werden (siehe Abbildung 20 auf Seite 51).
Mit Ausnahme dreier Nicht-Raucher war das in der Leber durch Alkohole induzierbare CYP 2E1
bei allen Spendern nachweisbar. Da über den Alkoholkonsum der getesteten Personen nichts
bekannt ist, kann somit auch die Frage nach der Induzierbarkeit von CYP 2E1 in der
respiratorischen Nasenschleimhaut nicht beantwortet werden. Während Mace et al. (1998)
berichten, dass CYP 2E1 in nur 50 % der untersuchten Lungenproben exprimiert ist, konnte in der
menschlichen Nasenschleimhaut CYP 2E1 bei 86% der Spender nachgewiesen werden (19 von 22

4. Diskussion 79
Patienten). Unterschiede in der CYP 2E1-Expression zwischen Rauchern und Nicht-Rauchern bzw.
Männern und Frauen konnten nicht festgestellt werden.
Die CYP 2F-Subfamilie ist durch wenige Isoformen charakterisiert und wird selektiv in der Lunge
und nur gering bis gar nicht in der Leber exprimiert (Carr et al., 2003; Chen et al., 2002; Ding and
Kaminsky, 2002, Nishimura et al., 2003). Mit Ausnahme einer Nicht-Raucherin konnte CYP 2F1 in
der respiratorischen Nasenschleimhaut bei allen Probanden nachgewiesen werden, während nach 33
Amplifikationszyklen CYP 2F1-Transkripte in der Leber nicht detektierbar waren. Meine
Untersuchungen belegen somit, dass es sich bei CYP 2F1 tatsächlich um ein überwiegend in
respiratorischen Geweben exprimiertes Cytochrom P450 handelt. Da CYP 2F1 eine ganze Reihe
luftgetragener Verbindungen, besonders jene aus dem Zigarettenrauch (Chen et al., 2002), zu
reaktiven und potentiell pneumotoxischen Metaboliten aktiviert (Chen et al., 2002; Carr et al.,
2003), ist zu vermuten, dass diese reaktiven Metaboliten auch in der menschlichen respiratorischen
Nasenschleimhaut gebildet werden, da die Nasenschleimhaut im Rahmen der In- und Exspiration
ebenfalls mit luftgetragenen Substanzen in Kontakt kommt.
Das abundant in cardiovasculären Geweben exprimierte und am Arachidonsäure- sowie
Linolsäuremetabolismus beteiligte CYP 2J2 (King et al., 2002; Yang et al., 2001) konnte auch in der
untersuchten menschlichen Nasenschleimhaut nachgewiesen werden. Männer waren dabei durch eine
stärkere Expression als Frauen charakterisiert (p<0,004). Auch in der menschlichen Lunge wird
CYP 2J2 exprimiert (Zeldin et al., 1996). Der Einfluss des Geschlechts auf die CYP 2J2-Expression
in der Lunge ist jedoch unbekannt.
Das erst im Jahr 2002 entdeckte CYP 2S1 ist innerhalb der CYP 2-Familie eng mit CYP 2A6,
2A13 und 2B6 verwandt (Rivera et al., 2002) und zeigt hohe Expressionsraten in Trachea, Lunge,
Magen, Dünndarm und Milz (Rylander et al., 2001). In der menschlichen alveolarepithelialen
Lungenzelllinie A549 konnte CYP 2S1 durch 2,3,7,8-Tetrachlorodibenzo-p-Dioxin (TCDD)
induziert werden. Daher vermuten Rivera et al. (2002), dass dieses Enzym eine bedeutende Rolle im
Karzinogenmetabolismus der Lunge spielt. Meine Untersuchungen belegen, dass CYP 2S1 auch in
der menschlichen respiratorischen Nasenschleimhaut exprimiert wird. Dabei könnte die beobachtete
Enzyminduktion bei Rauchern (p<0,05) als erwartete Antwort auf die Zigarettenrauchexposition
interpretiert werden und steht damit in Einklang mit den Beobachtungen von Rivera et al. (2002).
Interessanterweise war die CYP 2S1-Induktion nur bei weiblichen Rauchern statistisch signifikant
(p<0,03).

4. Diskussion 80
Aus der CYP 3A-Subfamilie, deren Mitglieder etwa die Hälfte aller gebräuchlichen Arzneistoffe
metabolisieren (De Luca et al., 2001; Hukkanen, 2000), wurden die Isoformen CYP 3A4, 3A5 und
3A7 untersucht. Erwartungsgemäß konnte Amplifikate der fetalen Form, CYP 3A7, weder in der
Leber, noch in der Nasenschleimhaut nachgewiesen werden, was den Untersuchungen von Anttila et
al. (1997) an der menschlichen Lunge entspricht. Dagegen war CYP 3A4, welches zwischen 30 –
40 % (Hukkanen, 2000) und 60 % (De Luca et al., 2001) des Gesamtcytochromgehaltes der Leber
ausmacht, nicht nur in der Leber, sondern auch in 20 der 22 Nasenschleimhautproben exprimiert.
CYP 3A5, die Haupt-CYP 3A-Form der Lunge (Anttila et al., 1997; Hukkanen et al., 2003), war
bei insgesamt 19 Patienten nachweisbar. Allerdings waren für den mRNA-Nachweis 39 (CYP 3A4)
bzw. 38 (CYP 3A5) Amplifikationszyklen notwenig, so dass erneut die Frage nach der
physiologischen Bedeutung derartig geringer mRNA-Mengen aufkommt (Koskela et al., 1997),
zumal auch im ergänzenden Western blot weder CYP 3A4-, noch 3A5-Protein nachgewiesen
wurde. Vergleicht man die benötigte Anzahl an Zyklen, so scheint in nasalem respiratorischen
Gewebe CYP 3A5 die stärker exprimierte Form zu sein. Dies entspricht den Untersuchungen an
pulmonalem respiratorischen Gewebe. Dort konnten Anttila et al. (1997) CYP 3A4- in nur 20 %
der Proben und CYP 3A5-mRNA in allen Proben nachweisen. Die Beobachtung von Hukkanen et
al. (2003), dass Rauchen zu einer verminderten CYP 3A5-Expression in der menschlichen Lunge
führt, konnte für die menschliche Nasenschleimhaut nicht bestätigt werden, steht aber in Einklang mit
Untersuchungen bronchialer Biopsien von Rauchern und Nicht-Rauchern (Borlak et al., 2004).
CYP 4B1, welches u.a. die ω-Hydroxylierung von Laurinsäure katalysiert (Imaoka et al., 2001),
war mit Ausnahme einer Nicht-Raucherin bei allen Patienten nachweisbar, nicht jedoch in der Leber.
Dies steht in Einklang mit den Untersuchungen von Nhamburo et al. (1989) und Nishimura et al.
(2003), die CYP 4B1-Transkripte nur in der menschlichen Lunge amplifizieren konnten.
Von den Flavinhaltigen Monooxygenasen (FMOs), welche bereits in der respiratorischen und
olfaktorischen Nasenschleimhaut von Ratten nachgewiesen wurden (Reed, 1993), ließen sich die
untersuchten Isoformen FMO 2, FMO 3 und FMO 5 bei allen Spendern detektieren. Dabei zeigte
nur FMO 2, bei der es sich um die Hauptform Flavinhaltiger Monooxygenasen in der Säugetierlunge
handelt (Dolphin et al., 1998; Krueger et al., 2002), einen statistisch signifikanten Unterschied
zwischen Rauchern und Nicht-Rauchern in Form einer verminderten Genexpression selektiv bei den
männlichen Rauchern. Dass der beobachtete Unterschied auch tatsächlich einen Einfluss auf den

4. Diskussion 81
Metabolismus von Arzneistoffen und anderen Xenobiotika ausübt, ist dabei aber unwahrscheinlich,
da FMO 2 für ein verkürztes und damit katalytisch inaktives Protein codiert (Dolphin et al., 1998;
Furnes et al, 2003; Whetstine et al., 2000). Das Fehlen von FMO 2-mRNA in der Leber ist zuvor
auch schon von Dolphin et al. (1998) berichtet worden.
Neben den Flavinhaltigen Monooxygenasen war in der menschlichen Nasenschleimhaut auch die
Epoxid-Hydrolase (EH) bei allen 22 Patienten nachweisbar, was die zuvor von Gervasi et al. (1991)
und Green et al. (2001) berichteten EH-Enzymaktivitäten in der menschlichen Nasenschleimhaut
bestätigt. Auch bei der Ratte (Bond, 1983; Faller et al., 2001), bei der Maus (Reed, 1993) und
beim Hund (Bond et al., 1988) konnten EH-Aktivitäten in der Nasenschleimhaut gezeigt werden,
ebenso wie in der menschlichen Lunge (Faller et al., 2001). Im pulmonalen Gewebe wurde dabei
kein Unterschied in der EH-Expression zwischen Rauchern und Nicht-Rauchern festgestellt (Willey
et al., 1997). Dies entspricht wiederum meinen Beobachtungen an nasalem Gewebe des Menschen.
Reaktive Epoxide, die auch aus einigen polyzyklischen aromatischen Hydrocarbonen (PAHs) des
Zigarettenrauchs gebildet werden können und die sich als potente Karzinogene erwiesen haben
(Hecht, 2002; Hukkanen et al., 2000; Rubin, 2001), können durch die Epoxid-Hydrolase in weniger
reaktive und leichter konjugierbare Metaboliten hydrolysiert werden, die somit leichter eleminierbar
sind (Seidegard and Ekstrom, 1997). Über diese detoxifizierende Wirkung der Epoxid-Hydrolase
besitzt die respiratorische Nasenschleimhaut des Menschen zumindest gegenüber einigen im
Zigarettenrauch enthaltenen Prokarzinogenen einen Schutzmechanismus.
Darüber hinaus konnten in der respiratorischen Nasenschleimhaut verschiedene Isoformen der
Glutathion S-Transferase nachgewiesen werden. Bereits zuvor hatten Aceto et al. (1989), Gervasi et
al. (1991), Green et al. (2001) und Krishna et al. (1995) das Vorhandensein dieser Enzymfamilie in
der menschlichen Nasenschleimhaut metabolisch bzw. immunhistochemisch nachgewiesen. Die von
Krishna et al. (1995) gemachte Beobachtung einer geschlechtsabhängigen GST α-Expression in der
Regio olfactoria, nicht jedoch in der Regio respiratoria konnte in meinen Untersuchungen für die
respiratorische Nasenschleimhaut bestätigt werden. Interessanterweise war dabei aber eine
geschlechtsabhängige Reaktion auf das Rauchen zu beobachten. So wies die Gruppe der Raucher
insgesamt geringere GST α2-Transkriptmengen auf. Bei Betrachtung der Geschlechter zeigte sich
jedoch, dass dieser Unterschied nur bei den männlichen Personen statistische Signifikanz erlangte
(p<0,002). Da die GST α2 über die Konjugation reaktiver Phase I-Metaboliten detoxifizierend und
somit zytoprotektiv wirkt (Dreij et al., 2002; Pietsch et al., 2003; Turesky et al., 2003), kann

4. Diskussion 82
vermutet werden, dass – verminderter Proteingehalt und verminderte metabolische Aktivität
vorausgesetzt - Raucher schlechter vor derartigen Substanzen geschützt sind. Da sich die GST α2
darüber hinaus als aktiv gegenüber einigen Kanzerogenen, die aus den Inhaltstoffen des
Zigarettenrauchs gebildet werden, erwiesen hat (Dreij et al., 2002), kann zusätzlich angenommen
werden, dass Raucher ein erhöhtes Risiko nasaler Tumoren besitzen. Hinweise über den
Zusammenhang zwischen Zigarettenkonsum und Tumoren der Nasenschleimhaut sind zumindest
zahlreich gegeben (Benninger, 1999; Ding and Kaminsky, 2002; Feron et al., 2001; Hildesheim et
al., 2001; Talmi et al., 2002).
Die Uridindiphosphat-Glucuronyl-Transferase 1A1 (UGT 1A1) konnte auch bei einer maximalen
Zahl von 39 Amplifikationszyklen nur bei sechs Patienten nachgewiesen werden, wobei die
ermittelten durchschnittlichen Lichtwerte dieser sechs Amplifikate nur 7,3 % vom Lichtwert der
Positivkontrolle (humane Leber) betrugen. Dies erklärt die fehlende UGT-Aktivität in der
menschlichen Nasenschleimhaut bei Gervasi et al. (1991) unter Verwendung von 1-Naphthol, einem
spezifischen UGT 1A1-Substrat (Dean et al., 2002). Die Beobachtung, dass mit 39
Amplifikationszyklen teilweise Transkripte nachgewiesen werden konnten, dies aber zu keiner
messbaren metabolischen Aktivität führt, stützt wiederum die These von Koskela et al. (1999), dass
gering exprimierte Gene physiologisch weniger von Bedeutung sind.
Im Gegensatz zu UGT 1A1 war die auch als olfaktorische UGT bezeichnete UGT 2A1 (Jedlitschky
et al., 1999) bei sämtlichen Spendern in der respiratorischen Nasenschleimhaut exprimiert, nicht
jedoch in der Positivkontrolle (humane Leber). Das Fehlen von UGT 2A1-Transkripten in der
menschlichen Leber hatten zuvor auch schon Jedlitschky et al. (1999) beobachtet. Mit diesen
Untersuchungen konnte zusätzlich zur bekannten UGT 2A1-Expression in der Regio olfactoria
(Jedlitschky et al., 1999) diese Expression nun erstmalig auch in der Regio respiratoria der
menschlichen Nasenschleimhaut gezeigt werden, weshalb die Bezeichnung olfaktorische UGT nicht
mehr korrekt erscheint.
Da die transkriptionelle Regulation der oben genannten Enzyme u.a. über Transkriptionsfaktoren und
nukleäre Rezeptoren erfolgt, wurde die Genexpression von AhR, CAR β , LXR α, PXR, PPAR α
und PPAR γ mituntersucht. Wenngleich zum Verständnis der Regulationsmechanismen
weitergehende Untersuchungen notwendig sind, können die ermittelten Expressionen regulatorischer
Transkriptionsfaktoren einen Beitrag zum verbesserten Verständnis der hier beobachteten

4. Diskussion 83
Genexpressionsprofile liefern. Da für die nasalen Schleimhäute der Ratte gewebsspezifische
Regulationsmechanismen postuliert werden (Bereziat et al., 1995; Longo and Ingelman-Sundberg,
1993; Longo et al., 2000; Wardlaw et al., 1998) und sich dies auch beim Menschen abzeichnet
(Carayol et al., 2002), sollten die für andere Gewebe bekannten Regulationsmechanismen nicht
unkritisch auf die menschliche Nasenschleimhaut übertragen werden.
Bei weiblichen Spendern war der Pregnenolone X Rezeptor (PXR) signifikant stärker exprimiert als
bei männlichen (p<0,005). Geschlechtsabhängige Unterschiede in der Genexpression wurden mit
Ausnahme von CYP 2J2 jedoch bei keinem der untersuchten Enzyme beobachtet.
Interessanterweise war bei männlichen Patienten CYP 2J2-mRNA signifikant erhöht. Der
geschlechtsspezifische Unterschied zwischen PXR und CYP 2J2 konnte jedoch durch Korrelation
der Genexpressionsbefunde nicht erklärt werden. PXR ist ein wichtiger Faktor für die Regulation der
Cytochrome der 3A-Subfamilie (Hukkanen, 2000; Kliewer et al., 1999; Waxman, 1999) sowie der
Cytochrome 2C8 und 2C9 (Hukkanen, 2000). Auch hier war keine Übereinstimmung im
Expressionsprofil mit genannten Targetgenen feststellbar.
Die PPARs spielen eine wichtige Rolle in der Karzinogenese (Cajaraville et al., 2003; Valles et al.,
2003; Vanden Heuvel et al., 2003). Es ist daher nicht verwunderlich, dass PPAR γ bei Rauchern
induziert war.
Bestandteil der transkriptorischen Regulation der CYP 1-Familienmitglieder ist der Aryl
hydrocarbon-Rezeptor (Hukkanen, 2000; Seubert et al., 2002; Waxman, 1999), dessen
Expression in der menschlichen respiratorischen Nasenschleimhaut erstaunlicherweise erheblich von
den Expressionen der Mitglieder der CYP 1-Familie abwich. Während die Genexpression des Ah-
Rezeptors unabhängig von der Zigarettenrauchexposition war, wurde CYP 1A1 bei 60 % der
Raucher exprimiert, CYP 1A2 dagegen überhaupt nicht. CYP 1B1 wiederum war bei Rauchern
hoch signifikant (p<0,00006) induziert. Gewebsspezifische Regulationsmechanismen, wie sie bereits
bei der Ratte beobachtet wurden (Bereziat et al., 1995; Longo and Ingelman-Sundberg, 1993;
Longo et al., 2000; Wardlaw et al., 1998), scheinen somit auch auf die menschliche
Nasenschleimhaut übertragbar zu sein.
Um herauszufinden, ob die beobachteten Unterschiede in der Genexpression zwischen Rauchern und
Nicht-Rauchern bei CYP 1A1, CYP 1B1, CYP 2S1, FMO 2, GST α2 und PPAR γ mit

4. Diskussion 84
entzündlichen Schleimhautveränderungen zusammenhängen, wurden die als Entzündungsmediatoren
bekannten Cytokine IL-1β (Grellner, 2002; Haspolat et al., 2002; Jones et al., 2003; Ruan et al.,
2003), IL-6 (Grellner, 2002; Jones et al., 2003), IL-8 (Beeh et al., 2003; Jones et al., 2003) und
TNF α (Grellner, 2002; Haspolat et al., 2002; Jones et al., 2003) in die Untersuchungen
miteinbezogen. Bei allen Patienten waren die Entzündungsmediatoren in unterschiedlicher
Ausprägung exprimiert, was aufgrund des medizinischen Vorberichts (chronische Entzündung bzw.
allergische Rhinitis) auch zu erwarten war. Signifikante Unterschiede zwischen Rauchern und Nicht-
Rauchern konnten jedoch bei keinem der Cytokine in der menschlichen Nasenschleimhaut
nachgewiesen werden, weshalb auch nicht von entzündungsbedingten Unterschieden in der
Expression genannter Gene auszugehen ist.
4.3 Nachweisbares Protein
In der menschlichen respiratorischen Nasenschleimhaut ließ sich mittels Western blot nur Protein der
CYP 2A-Subfamilie nachweisen, jedoch kein CYP 1A1-, 2B6-, 2E1-, 3A4- oder 3A5-Protein.
Dies bestätigt frühere Beobachtungen, wonach die CYP 2A-Enzyme in den Nasenschleimhäuten
verschiedener Spezies stark exprimiert sind, weshalb ihnen eine Schlüsselrolle im Metabolismus
zahlreicher Xenobiotika innerhalb der Nasenschleimhaut zugesprochen wird (Bereziat et al., 1995;
Ding and Coon, 1989; Ding and Coon, 1990; Putt et al., 1995; Su et al., 1996). Bereits 1993
konnten Getchell et al. mittels immunhistochemischer Untersuchungen große Mengen eines CYP 2A-
verwandten Proteins in der menschlichen Nasenschleimhaut der Regio respiratoria sowie der Regio
olfactoria nachweisen. Eine eindeutige Differenzierung der Proteine CYP 2A6 und 2A13 war und
ist jedoch nicht möglich, da aufgrund der hohen Aminosäurehomologie von 95,4 % spezifische
Antikörper nicht verfügbar sind (Ding and Kaminsky, 2002; Su et al., 2000).
Da es keine weiteren publizierten Daten zur CYP-Proteinexpression in der menschlichen
Nasenschleimhaut gibt, werden nachfolgend meine Ergebnisse mit tierexperimentell gewonnenen
Daten verglichen. So konnten Bereziat et al. (1995) ein CYP 2A-immunreaktives Protein in der
Nasenschleimhaut der Ratte detektieren, jedoch kein CYP 2E1- oder CYP 3A4-Protein. Wardlaw

4. Diskussion 85
et al. (1998) konnten nach Zigarettenrauchexposition eine CYP 1A1-Induktion bei exponierten
Ratten nachweisen, jedoch war kein CYP 1A1-Protein in den Kontrollen (nicht exponierte Tiere)
detektierbar. CYP 2B1/2-Protein war bei exponierten wie nicht exponierten Ratten vorhanden.
Auch in ergänzenden immunhistochemischen Untersuchungen konnte kein CYP 3A-Protein
nachgewiesen werden. Im Gegensatz dazu berichteten Longo et al. (2000) neben der Expression
von CYP 2A-, auch die Expression von CYP 2B-, 2C-, 2G1- sowie 3A-Protein in der
Nasenschleimhaut der Ratte, wobei aus dem Versuchsaufbau nicht genau hervorgeht, ob es sich um
respiratorisches oder olfaktorisches Nasenschleimhautgewebe handelte. Die Tatsache, dass auch
CYP 2G1-Protein nachgewiesen wurde, lässt aber darauf schließen, dass Nasenschleimhaut der
Regio olfactoria untersucht wurde, da CYP 2G1 selektiv in olfaktorischem Gewebe exprimiert wird
(Longo et al., 2000).
Die in der vorliegenden Doktorarbeit für die menschliche respiratorische Nasenschleimhaut
gewonnenen Daten sind somit mit den Ergebnissen von Bereziat et al. (1995) für die Ratte
vergleichbar. Übereinstimmend mit den Ergebnissen von Wardlaw et al. (1998) konnte kein CYP
3A-immunreaktives Protein nachgewiesen werden, ebenso kein CYP 1A1-Protein bei den Nicht-
Rauchern. Die Expression von CYP 2B-Protein ebenso wie die Expression von CYP 1A1-Protein
bei den Zigarettenrauch exponierten Tieren in den Versuchen von Wardlaw et al. (1998)
unterscheidet sich von meinen Beobachtungen an der menschlichen Nasenschleimhaut. Bezüglich des
CYP 1A1 bleibt zu bemerken, dass mittels der hoch sensitiven RT-PCR bei keinem der Nicht-
Raucher CYP 1A1-Transkripte nachgewiesen wurden, weshalb CYP 1A1-Protein in den
Nasenmuschelpools der Nicht-Raucher auch nicht zu erwarten war. Dagegen war CYP 1A1 bei
insgesamt 60 % der Raucher exprimiert, wobei die gemessenen Lichtintensitäten deutlich unterhalb
der Positivkontrolle (humane Leber) lagen. Da aufgrund der geringen Gewebemengen und des damit
verbundenen geringen Proteingehaltes der einzelnen Resektate die Nasenmuscheln mehrerer
Patienten gepoolt werden mussten, wäre es durchaus denkbar, dass die auf den Gesamtpool
bezogene nur partielle CYP 1A1-Expression und die daraus resultierende partielle CYP 1A1-
Translation zu gering ist, um im Western blot detektiert werden zu können und somit der verwendete
Western blot für den CYP 1A1-Nachweis in der menschlichen respiratorischen Nasenschleimhaut
nicht sensitiv genug ist.

4. Diskussion 86
4.4 Metabolische Aktivität
Neben der Gen- und Proteinexpression wurde auch die Enzymaktivität der menschlichen
respiratorischen Nasenschleimhaut untersucht. Hierzu wurden mikrosomale Membranen mit den
Substraten Loteprednol-Etabonat, Testosteron und Ethoxyresorufin inkubiert. Nach Inkubation
gepoolter Nasenmuschelmikrosomen mit Loteprednol-Etabonat konnte nur eine geringfügige
Loteprednol-Verstoffwechselung ermittelt werden. M 8 stellte dabei den Hauptmetaboliten dar.
Da keine Untersuchungen zur Verstoffwechselung des bislang nur ophthalmologisch zugelassenen
Loteprednol-Etabonates in extraokulären Geweben beim Menschen existieren, sind lediglich
Vergleiche mit den Ergebnissen einer hausinternen Studie möglich (Metabolism Studies with
Loteprednol etabonate, 2003), in deren Rahmen auch Inkubationen mit rekonstituierten
Enzymsystemen durchgeführt wurden. CYP 3A4, eines der Haupt-Cytochrome der Leber (De Luka
et al., 2001; Hukkanen, 2000), war dabei maßgeblich für die Biotransformation des Loteprednols
verantwortlich.
In den vorausgegangen proteinanalytischen Untersuchungen war in der menschlichen
Nasenschleimhaut nur Protein der CYP 2A-Subfamilie nachweisbar, wobei aufgrund der fehlenden
Verfügbarkeit spezifischer Antikörper (Ding and Kaminsky, 2002; Su et al., 2000) eine
Unterscheidung von CYP 2A6 und 2A13 nicht möglich war. In den Experimenten mit
rekonstituiertem und funktionell aktivem CYP 2A6 war keine nennenswerte Metabolisierung des
Loteprednols zu beobachten (Metabolism Studies with Loteprednol etabonate, 2003), wobei das
entstehende Metabolitenprofil dem der Inkubationen mit Nasenmuschelmikrosomen ähnelte.
Insgesamt betrachtet korreliert die deutliche metabolische Umsetzung des Loteprednols in den
verwendeten Positivkontrollen (Lebermikrosomen der Ratte und des Menschen) mit den Daten
durch separate Inkubation mit CYP 3A4, dem Haupt-Cytochrom der Leber. Ebenso stimmt die nur
geringe Loteprednolverstoffwechselung durch menschliche nasale Mikrosomen mit den
Beobachtungen überein, dass in der menschlichen respiratorischen Nasenschleimhaut nur Protein der
CYP 2A-Subfamilie detektierbar war und dass dessen Mitglied CYP 2A6 in den Untersuchungen
mit Supersomen gegenüber Loteprednol nur geringe Aktivitäten zeigte.

4. Diskussion 87
Bei der Betrachtung der Ergebnisse des 2. Versuchsabschnittes (mit Zugabe von
Ammoniumacetatpuffer) fällt zunächst auf, dass das Substrat Loteprednol-Etabonat insgesamt sehr
viel stärker zerfällt als im 1. Versuchsabschnitt (ohne Zugabe von Ammoniumacetatpuffer). So liegt
der relative Anteil des verbleibenden Loteprednols in den Negativkontrollen des 2. Abschnittes um
durchschnittlich 21,7 % niedriger als in den Negativkontrollen des 1. Abschnittes. Wenngleich die
Zugabe von Ammoniumacetatpuffer (200 mM, pH 4,5) für den β-Glucuronidase-Verdau notwendig
ist, scheint dessen Zusatz zu einer Instabilität des Loteprednols zu führen. Darüber hinaus sind die
deutlich schwankenden Werte zwischen den beiden Negativkontrollen des 2. Abschnittes auffällig,
weshalb die Daten dieses Abschnittes insgesamt nur qualitativ betrachtet werden.
Im 2. Versuchsabschnitt wurde zum Nachweis möglicher Glucuronide ein zusätzlicher
Glucuronidase-Verdau durchgeführt. Durch Zugabe des Enzyms β-Glucuronidase sollte die
Abspaltung des Glucuronsäure-Restes erzielt werden, womit der Nachweis der Glucuronidierung
erbracht wird. Der einzige Metabolit, der in den Inkubaten ohne Glucuronidase-Zusatz auftrat, war –
wie im 1. Versuchsabschnitt – M 8, der wiederum nur in einem Pool (Pool 15) oberhalb der
Negativkontrolle lag. Somit stellt M 8 bei Pool 15 den einzigen Metaboliten dar, von dem eine
Glucuronidierung hätte nachgewiesen werden können. Interessanterweise wurde nach
Glucuronidase-Verdau auch nur bei Pool 15 (W-R) ein weiterer Metabolit detektiert. Dabei
handelte es sich um einen neuen, bislang unidentifizierten Metaboliten mit einer relativen
Retentionszeit von 0,19. Dies lässt auf eine Glucuronidierung von M 8 schließen.
Darüber hinaus war auffällig, dass M 8 in den Inkubaten mit Glucuronidase-Zusatz gegenüber der
Negativkontrolle erhöht erschien. Betrachtet man dagegen die Metabolitenprofils der einzelnen
Nasenmuschelpools mit und ohne Enzymzusatz unabhängig von der jeweiligen Negativkontrolle, so
ist kaum ein Unterschied erkennbar. Dafür weist die Negativkontrolle mit Glucuronidase-Zusatz
deutlich geringere Mengen an Zerfallsprodukten auf, die nicht erklärbar sind. Korrigiert man nun die
absoluten M 8-Anteile der Organpools um die jeweilige Negativkontrolle, erscheint M 8 in den
Inkubaten mit Enzymzusatz erhöht. Die Überprüfung der Ergebnisse des 2. Versuchsabschnittes war
jedoch nicht möglich, da keine weiteren Mikrosomen menschlicher Nasenschleimhaut zur Verfügung
standen und die Gewinnung des Materials ausgesprochen schwierig und langwierig ist.
Zusätzlich wurden Metabolismusuntersuchungen mit dem endogenen Steroid Testosteron
durchgeführt. Betrachtet man die relativen Anteile des verbleibenden Testosterons in den

4. Diskussion 88
Nasenmuschelpools und in der Negativkontrolle, so scheint so gut wie keine Verstoffwechselung des
Testosterons stattzufinden. Zieht man jedoch die geringe Verschiebung des Metabolitenprofils
gegenüber der Negativkontrolle in Betracht, so dürfte zumindest von einem geringen Testosteron-
Metabolismus auszugehen sein. So lagen nach 120-minütiger Inkubation der mikrosomalen
Organpools mit 50 µM Testosteron alle untersuchten Testosteronmetaboliten jeweils unter 0,5 % des
metabolischen Gesamtprofils. Oberhalb der Negativkontrolle befanden sich dabei Androstendion mit
0,37 % sowie in Spuren 6α-Hydroxytestosteron und 7α-Hydroxytestosteron.
7α-Hydroxytestosteron war dabei der einzige Metabolit, der weder in der Negativ-, noch in der
Positivkontrolle (humane Leber) detektierbar war. Dagegen war er in äußerst geringen Mengen in
allen Nasenmuschelinkubaten nachweisbar. Somit scheint es sich um einen Metaboliten zu handeln,
der durch menschliche nasale Enzyme gebildet wird, welche nicht parallel auch in der Leber
vorhanden sind. Inwieweit der Testosteronmetabolit durch die Cytochrome 2A13 und 2S1 gebildet
wird, muss noch weitergehend untersucht werden.
Für den Menschen ist über den Metabolismus von Testosteron in der respiratorischen
Nasenschleimhaut nichts bekannt. Dagegen wurden beim Rind in der Regio respiratoria insgesamt
acht Testosteronmetaboliten nachgewiesen, wobei die Testosteronoxidationsrate um insgesamt etwa
15 % niedriger als in der Regio olfactoria lag (Longo et al., 1991). Mit heterolog exprimiertem
CYP 2A verschiedener Spezies wurden unterschiedliche Beobachtungen hinsichtlich der
Verstoffwechselung von Testosteron gemacht. So wurden beim Kaninchen sowohl mit gereinigtem
mikrosomalem nasalem NMa (entspricht CYP 2A10/11), als auch mit rekombinantem CYP 2A10
und 2A11 neben Androstendion noch 3 weitere, nicht näher charakterisierte Testosteronmetaboliten
X1, X2 und X3 detektiert (Peng et al., 1993), während bei der Ratte mit heterolog exprimiertem
CYP 2A3 insgesamt 15 verschiedene α-Hydroxytestosteronmetaboliten nachgewiesen wurden (Liu
et al., 1996). Dagegen zeigte heterolog exprimiertes humanes CYP 2A6, welches homolog zu CYP
2A10/11 des Kaninchens und zu CYP 2A3 der Ratte ist (Ding et al., 1994; Peng et al., 1993),
keine Aktivität gegenüber Testosteron (Liu et al., 1996). Dies könnte wiederum die geringe
Testosteronoxidationsrate menschlicher respiratorischer Nasenschleimhaut im Gegensatz zur
vergleichsweise hohen Aktivität bei anderen Spezies (Ding and Coon, 1990; Longo et al., 1991)
erklären, da CYP 2A-Protein in meinen Untersuchungen das einzige Cytochrom war, welches sich
auch auf Proteinebene nachweisen ließ. Wie bereits zuvor erwähnt, war aufgrund der fehlenden

4. Diskussion 89
Verfügbarkeit spezifischer Antikörper (Ding and Kaminsky, 2002; Su et al., 2000) eine
Unterscheidung von CYP 2A6 und 2A13 nicht möglich. Der Nachweis von CYP 2A13-
Transkripten in der menschlichen respiratorischen Nasenschleimhaut (Koskela et al., 1999; Su et al.,
2000) in Kombination mit dem Nachweis der Funktionsfähigkeit von CYP 2A13 (Su et al., 2000)
könnte die Beobachtung erklären, dass trotz fehlender Aktivität von CYP 2A6 gegenüber
Testosteron eine – wenn auch nur geringe – Testosteronverstoffwechselung nachgewiesen werden
konnte.
Das CYP 1A1-Markersubstrat Ethoxyresorufin (Burke et al., 1994) wurde von keinem der
insgesamt fünf untersuchten Nasenmuschelpools (zwei Raucher- und drei Nicht-Raucher-Pools)
umgesetzt. Die fehlende EROD-Aktivität bei den Nicht-Rauchern war erwartungsgemäß, da bei den
Nicht-Rauchern in der respiratorischen Nasenschleimhaut weder CYP 1A1-Transkripte, noch CYP
1A1-Protein nachgewiesen werden konnte. Bei Rauchern war CYP 1A1 zwar in 60 % der Fälle
exprimiert, jedoch konnte kein CYP 1A1-Protein detektiert werden, was wiederum mit der
fehlenden Ethoxyresorufin-Verstoffwechselung übereinstimmt.
4.5 Schlussfolgerungen
Insgesamt betrachtet sind somit in der menschlichen respiratorischen Nasenschleimhaut verschiedene
Cytochrom P450 Monooxygenasen (CYPs) und weitere fremdstoffmetabolisierende Enzyme
exprimiert, wenngleich im Western blot nur CYP 2A-Protein nachgewiesen werden konnte und die
metabolische Aktivität gegenüber den getesteten Substraten Loteprednol-Etabonat (LE) und
Testosteron deutlich geringer als in der Leber war. Bei den durchgeführten in vitro-Versuchen konnte
gezeigt werden, dass der untersuchte Arzneistoff Loteprednol-Etabonat so gut wie keiner
Verstoffwechselung durch menschliche Nasenmuschelmikrosomen unterliegt. Folgerichtig erscheint
eine topische Applikation von Loteprednol-Etabonat mit keinem komplexen und in der menschlichen
respiratorischen Nasenschleimhaut ortsgebundenen Metabolismus verbunden zu sein, was zu
weiteren Untersuchungen ermutigt.

5. Zusammenfassung 90
5. Zusammenfassung
Daniela Rahmel
Genexpressions- und Metabolismusuntersuchungen der menschlichen respiratorischen
Nasenschleimhaut
Ziel dieser Arbeit war es, die menschliche respiratorische Nasenschleimhaut hinsichtlich ihrer
enzymatischen Ausstattung sowie metabolischen Kompetenz näher zu charakterisieren, wobei
schwerpunktmäßig die verschiedenen Isoformen des Cytochrom P450 Monooxygenasesystemes
(CYPs) untersucht wurden. Die respiratorische Nasenschleimhaut stammte dabei von Patienten, die
sich aufgrund einer Schleimhauthyperplasie einer partiellen Turbinektomie der unteren Nasenmuschel
unterzogen haben.
Mittels RT-PCR konnten zahlreiche Cytochrom P450 Monooxygenasen nachgewiesen werden,
darunter CYP 1A1, 1B1, 2A6/7, 2A13, 2B6, 2C8, 2C9, 2C18, 2C19, 2E1, 2F1, 2J2, 2S1, 3A4,
3A5 und 4B1. Nicht detektierbar waren hingegen CYP 1A2, 2D6, 3A7 und 4A11. Darüber hinaus
konnte die Expression der Epoxid-Hydrolase (EH), der Flavinhaltigen Monooxygenasen 2, 3 und 5
(FMO 2, FMO 3 und FMO 5), der Glutathion-S-Transferasen α2, µ1 und ρ1 (GST α2, GST µ1
und GST ρ1) sowie der UDP-Glucuronyl-Transferase 2A1 (UGT 2A1) in der menschlichen
respiratorischen Nasenschleimhaut belegt werden. Rauchen führte zu einer verstärkten Expression
von CYP 1A1, 1B1 und 2S1, während GST α2 vermindert exprimiert wurde. Der Einfluss des
Geschlechts konnte nur für CYP 2J2 in Form einer stärkeren Expression bei männlichen Spendern
ermittelt werden. Mittels Western blot konnte immunreaktives CYP 2A-, nicht jedoch CYP 1A1-,
2B6-, 2E1-, 3A4- oder 3A5-Protein nachgewiesen werden. In den mikrosomalen Assays mit
Nasenmuschelpools wurde das CYP 1A-Substrat Ethoxyresorufin nicht umgesetzt, wohingegen
Testosteron und Loteprednol-Etabonat (LE) in geringem Umfang metabolisiert wurden.
Somit konnte für die menschliche respiratorische Nasenschleimhaut die Expression zahlreicher CYP-
Isoformen nachgewiesen werden, wenngleich nur CYP 2A-Protein translatiert erschien und die
metabolische Aktivität gegenüber Loteprednol-Etabonat und Testosteron deutlich geringer als in der
Leber ausfiel. Bei den durchgeführten in vitro-Versuchen konnte gezeigt werden, dass der
untersuchte Arzneistoff Loteprednol-Etabonat so gut wie keiner Verstoffwechselung durch die
verwendeten menschlichen Nasenmuschelmikrosomen unterliegt, weshalb nach lokaler Applikation in

5. Zusammenfassung 91
der respiratorischen Nasenschleimhaut beim Menschen mit keinem nennenswerten metabolischen
Abbau zu rechnen ist.

6. Summary 92
6. Summary
Daniela Rahmel
Investigations of gene expression and metabolic activity of the human respiratory nasal
mucosa
The objective of this study was to characterize the expression of xenobiotic metabolizing enzymes as
well as the metabolic capacity of the human respiratory nasal mucosa. In particular, the various
isoforms of the cytochrome P450 monooxygenases (CYPs) were investigated. The respiratory nasal
mucosa samples were taken from patients who had undergone a partial turbinectomy of the lower
nasal concha after mucosal hyperplasia had been diagnosed.
Several cytochrome P450 monooxygenases could be identified using RT-PCR, including CYP 1A1,
1B1, 2A6/7, 2A13, 2B6, 2C8, 2C9, 2C18, 2C19, 2E1, 2F1, 2J2, 2S1, 3A4, 3A5 and 4B1, albeit
at different expression levels. The enzymes CYP 1A2, 2D6, 3A7 and 4A11 were below the limit of
quantification. Furthermore, the expression of epoxide hydrolase (EH), the flavin containing
monooxygenases 2, 3 and 5 (FMO 2, FMO 3 and FMO 5), the glutathione-S-transferases α2, µ1
and ρ1 (GST α2, GST µ1 and GST ρ1) as well as the UDP-glucuronyltransferase 2A1 (UGT 2A1)
could be demonstrated in human respiratory nasal mucosa. Abundant expression of CYP 1A1, 1B1
and 2S1 was found in patients that smoked, while GST α2 was expressed at lower levels in these
patients. Male patients showed a higher expression level of CYP 2J2 than female patients, otherwise
no other sex-related differences were observed. Immune reactive CYP 2A protein was confirmed
by Western blot analysis, but not CYP 1A1, 2B6, 2E1, 3A4 or 3A5 protein. In microsomal assays
with pooled nasal concha, the CYP 1A substrate ethoxyresorufin was not metabolized, while
testosterone and loteprednol-etabonate (LE) were metabolized to a limited extent.
Thus, the expression of several CYP isoforms could be demonstrated in human respiratory nasal
mucosa, although only the CYP 2A protein appeared to be translated. The metabolic activities
shown for testosterone and loteprednol-etabonate were markedly lower in nasal mucosa than in the
liver. The drug loteprednol-etabonate was found to undergo only minimal metabolism in the
experiments conducted with human nasal microsomal preparations, and for this reason may be
suitable for therapy using local administration on the nasal mucosa in man.

7. Danksagung 93
7. Danksagung
Mein Dank gilt in erster Linie dem Institut für Toxikologie und Experimentelle Medizin der
Fraunhofer Gesellschaft, das durch die Bereitstellung der Räumlichkeiten, Geräte und Materialien die
Voraussetzung für diese Dissertation geschaffen hat.
Bei Herrn Prof. Dr. Borlak möchte ich mich für die wissenschaftliche Betreuung bedanken, ebenso
bei Herrn Prof. Dr. Otto, der freundlicherweise das Koreferat von Seiten der Tierärztlichen
Hochschule Hannover übernommen hat.
Prof. Dr. Lamm und Dr. Lamm sowie dem gesamten OP-Team des Friederiken-Stifts Hannover
danke ich für die Unterstützung beim Sammeln der Nasenmuscheln, ohne deren Hilfe dieses Projekt
nicht zustande gekommen wäre.
Bei allen technischen Assistenten der Abteilung bedanke ich mich dafür, dass sie mir stets mit Rat
und Tat zur Seite gestanden haben. Dies waren insbesondere Marlies und Antje, die mir beim
Sammeln der Nasenmuscheln geholfen haben, Rong und Kerstin, die mich in die Methodik der RT-
PCR eingewiesen haben, Andrea, die mir die Western blots näher gebracht hat, sowie Gitta, die mir
im Rahmen der metabolischen Assays bei der Durchführung und der anschließenden Auswertung
behilflich war.
Den Post-docs Thomas Thum, Tanja Hansen und Volker Zelinski möchte ich für die Unterstützung
bei der jeweiligen Versuchsplanung, -durchführung und -auswertung danken.
Herrn Dr. Holländer bin ich für die statistische Auswertung und seine ausführlichen Erläuterungen zu
Dank verpflichtet.
Prof. Dr. Richter danke ich für die Bereitstellung der Aufnahmen der histologischen Schnitte eines
Patienten mit allergischer Rhinitis.
Bei Frau Ockert möchte ich mich für die Hilfe bei der englischen Übersetzung bedanken.
Bei meinen Eltern bedanke ich mich für die moralische und finanzielle Unterstützung. Mein
besonderer Dank gilt dabei meinem Vater, der durch seine Sicht als Chemiker und durch die stets
sachlichen Diskussionen zur Klärung vieler Fragen beigetragen hat. Darüber hinaus möchte ich allen
meinen Freunden danken, die stets ein offenes Ohr für mich hatten und mich immer im richtigen
Moment wieder motiviert haben.

8. Literaturverzeichnis 94
8. Literaturverzeichnis
Aceto A, Di Ilio C, Angelucci S, Longo V, Gervasi PG and Federici G, Glutathione
transferases in human nasal mucosa. Arch. Toxicol., 63: 427-431, 1989.
Anttila S, Hukkanen J, Hakkola J, Stjernvall T, Beaune P, Edwards RJ, Boobis AR,
Pelkonen O and Raunio H, Expression and localization of CYP3A4 and CYP3A5 in human lung.
Am. J. Respir. Cell Mol. Biol., 16: 242-249, 1997.
Anttila S, Lei XD, Elovaara E, Karjalainen A, Sun W, Vainio H and Hankinson O, An
uncommon phenotype of poor inducibility of CYP1A1 in human lung is not ascribable to
polymorphisms in the AHR, ARNT, or CYP1A1 genes. Pharmacogenetics, 10: 741-751, 2000.
Baker JR, Satarug S, Edwards RJ, Moore MR, Williams DJ and Reilly PE, Potential for
early involvement of CYP isoforms in aspects of human cadmium toxicity. Toxicol. Lett., 137: 85-
93, 2003.
Beeh KM, Beier J, Kornmann O, Mander A and Buhl R, Long-term repeatability of induced
sputum cells and inflammatory markers in stable, moderately severe COPD. Chest., 123: 778-783,
2003.
Benninger MS, The impact of cigarette smoking and environmental tobacco smoke on nasal and
sinus disease: a review of the literature. Am. J. Rhinol., 13: 435-438, 1999.
Benninghoff A, Nervensystem, Haut, Paraganglien. In: Anatomie Makroskopische Anatomie,
Embryologie und Histologie des Menschen Band 2, 15. Auflage, Urban & Schwarzenberg,
München: 784-790, 1994.
Bereziat JC, Raffalli F, Schmezer P, Frei E, Geneste O and Lang MA, Cytochrome P450 2A
of nasal epithelium: regulation and role in carcinogen metabolism. Mol. Carcinog., 14: 130-139,
1995.
Bianco A, Whiteman SC, Sethi SK, Allen JT, Knight RA and Spiteri MA, Expression of
intercellular adhesion molecule-1 (ICAM-1) in nasal epithelial cells of atopic subjects: a mechanism
for increased rhinovirus infection? Clin. Exp. Immunol., 121: 339-345, 2000.

8. Literaturverzeichnis 95
Blanchard KT and Morris JB, Effects of m-xylene on rat nasal cytochrome P450 mixed function
oxidase activities. Toxicol. Lett., 70: 253-259, 1994.
Blue Histology – Respiratory System aus: http://www.lab.anhb.uwa.edu.au/mb140/CorePag
es/Respiratory/Respir.htm, 2002.
Bodor N, Recent advances in retrometabolic design approaches. J. Control. Release, 62: 209-
222, 1999.
Boenninghaus HG, Nase, Nebenhöhlen und Gesicht. In: Hals-Nasen-Ohrenheilkunde für
Studierende der Medizin, 10. Auflage, Springer Verlag, Berlin Heidelberg New York: 161-173,
1996.
Bond JA, Some biotransformation enzymes responsible for polycyclic aromatic hydrocarbon
metabolism in rat nasal turbinates: effects on enzyme activities of in vitro modifiers and intraperitoneal
and inhalation exposure of rats to inducing agents. Cancer Res., 43: 4805-4811, 1983.
Bond JA, Harkema JR and Russell VI, Regional distribution of xenobiotic metabolizing enzymes
in respiratory airways of dogs. Drug Metab. Dispos., 16: 116-124, 1988.
Borlak J, Moeller J, Erpenbeck VJ, Krug N and Thum T, Expression of xenobiotic
metabolising enzymes in bronchial biopsies of BALs of smokers. Am. J. Respir. Crit. Care Med. (in
press), 2004.
Bort R, Macé K, Boobis A, Gómez-Lechón MJ, Pfeifer A and Castell J, Hepatic metabolism
of diclofenac: role of human CYP in the minor oxidative pathways. Biochem. Pharmacol., 58: 787-
796, 1999.
Burke MD, Thompson S, Weaver RJ, Wolf CR and Mayer RT, Cytochrome P450
specificities of alkoxyresorufin O-dealkylation in human and rat liver. Biochem. Pharmacol., 48:
923-936, 1994.
Cajaraville MP, Cancio I, Ibabe A and Orbea A, Peroxisome proliferation as a biomarker in
environmental pollution assessment. Microsc. Res. Tech., 61: 191-202, 2003.
Carayol N, Vachier I, Campbell A, Crampette L, Bousquet J, Godard P and Chanez P,
Regulation of E-Cadherin expression by dexamethasone and tumour necrosis factor-alpha in nasal
epithelium. Eur. Respir. J., 20: 1430-1436, 2002.

8. Literaturverzeichnis 96
Carr BA, Wan J, Hines RN and Yost GS, Characterization of the human lung CYP2F1 gene and
identification of a novel lung-specific binding motif. J. Biol. Chem., 278: 15473-15483, 2003.
Cashman JR and Zhang J, Interindividual differences of human flavin-containing monooxygenase
3: genetic polymorphisms and functional variation. Drug. Metab. Dispos., 30: 1043-1052, 2002.
Charlier C, Broly F, Lhermitte M, Pinto E, Ansseau M and Plomteux G, Polymorphisms in
the CYP 2D6 gene: association with plasma concentrations of fluoxetine and paroxetine. Ther. Drug
Monit., 25: 738-742, 2003.
Chen N, Whitehead SE, Caillat AW, Gavit K, Isphording DR, Kovacevic D, McCreary
MB and Hoffman SMG, Identification and cross-species comparisons of CYP2F subfamily genes
in mammals. Mutat. Res., 499: 155-161, 2002.
Cummings BS, Lasker JM and Lash LH, Expression of glutathione-dependent enzymes and
cytochrome P450s in freshly isolated and primary cultures of proximal tubular cells from human
kidney. J. Pharmacol. Exp. Ther., 293: 677-685, 2000.
Dahl AR and Hadley WM, Nasal cavitiy enzymes involved in xenobiotic metabolism: effects on
the toxicity of inhalants. Crit. Rev. Toxicol., 21: 345-372, 1991.
Dean B, Chang S, Stevens J, Thomas PE and King C, Isolation and characterization of a UDP-
glucuronosyltransferase (UGT1A01) cloned from female rhesus monkey. Arch. Biochem. Biophys.,
402: 289-295, 2002.
Deindl E, Boengler K, van Royen N and Schaper W, Differential expression of GAPDH and
beta3-actin in growing collateral arteries. Mol. Cell Biochem., 236: 139-146, 2002.
De Luca A, Gysling E, Bonnabry P, Masche U und Meier-Abt P, Revision 2001: Zytochrome
und ihre Bedeutung für Arzneimittelinteraktionen. In: Pharma-kritik, Infomed-Verlags AG, CH-Wil,
2001.
Ding X and Coon MJ, Immunochemical characterization of multiple forms of cytochrome P-450 in
rabbit nasal microsomes and evidence for tissue-specific expression of P-450s NMa and NMb.
Mol. Pharmacol., 37: 489-496, 1990.
Ding X and Coon MJ, Steroid metabolism by rabbit olfactory-specific P450 2G1. Arch.
Biochem. Biophys., 315: 454-459, 1994.

8. Literaturverzeichnis 97
Ding X and Kaminsky LS, HUMAN EXTRAHEPATIC CYTOCHROMES P450: Function in
Xenobiotic Metabolism and Tissue-Selective Chemical Toxicity in the Respiratory and
Gastrointestinal Tracts. Annu. Rev. Pharmacol. Toxicol., 43: 149-173, 2002.
Ding X, Peng HM and Coon MJ, Structure-function analysis of CYP2A10 and CYP2A11, P450
cytochromes that differ in only eight amino acids but have strikingly different activities toward
testosterone and coumarin. Biochem. Biophys. Res. Commun., 203: 373-378, 1994.
Dolphin CT, Beckett DJ, Janmohamed A, Cullingford TE, Smith RL, Shephard EA and
Phillips IR, The flavin-containing monooxygenase 2 gene (FMO2) of humans, but not of other
primates, encodes a truncated, nonfunctional protein. J. Biol. Chem., 273: 30599-30607, 1998.
Dreij K, Sundberg K, Johansson AS, Nordling E, Seidel A, Persson B, Mannervik B and
Jernstrom B, Catalytic activities of human alpha class glutathione transferases toward carcinogenic
dibenzo[a,l]pyrene diol epoxides. Chem. Res. Toxicol., 15: 825-831, 2002.
Dwyer TM, Cigarette smoke-induced airway inflammation as sampled by the expired breath
cendensate. Am. J. Med. Sci., 326: 174-178, 2003.
Eke BC and Iscan M, Effects of cigarette smoke with different tar contents on hepatic and
pulmonary xenobiotic metabolizing enzymes in rats. Hum. Exp. Toxicol., 21: 17-23, 2002.
Elder DA, Karayal AF, D̀ Ercole AJ and Calikoglu AS, Effects of hypothyroidism on insulin-
like growth factor-I expression during brain development in mice. Neurosci. Lett., 293: 99-102,
2000.
Endrizzi K, Fischer J, Klein K, Schwab M, Nussler A, Neuhaus P, Eichelbaum M and
Zanger UM, Discriminative quantification of cytochrome P4502D6 and 2D7/8 pseudogene
expression by TaqMan real-time reverse transcriptase polymerase chain reaction. Anal. Biochem.,
300: 121-131, 2002.
Faller TH, Csanady GA, Kreuzer PE, Baur CM and Filser JG, Kinetics of propylene oxide
metabolism in microsomes and cytosol of different organs from mouse, rat, and humans. Toxicol.
Appl. Pharmacol., 172: 62-74, 2001.
Feron VJ, Arts JH, Kuper CF, Slootweg PJ and Woutersen RA, Health risks associated with
inhaled nasal toxicants. Crit. Rev. Toxicol., 31: 313-347, 2001.

8. Literaturverzeichnis 98
Furnes B, Feng J, Sommer SS and Schlenk D, Identification of novel variants of the flavin-
containing monooxygenase gene family in african americans. Drug. Metab. Dispos., 31: 187-193,
2003.
Gelardi A, Morini F, Dusatti F, Penco S and Ferro M, Induction by xenobiotics of phase I and
phase II enzyme activities in the human keratinocyte cell line NCTC 2544. Toxicol. In Vitro, 15:
701-711, 2001.
Gerbal-Chaloin S, Pascussi JM, Pichard-Garcia L, Daujat M, Waechter F, Fabre JM,
Carrere N and Maurel P, Induction of CYP2C genes in human hepatocytes in primary culture.
Drug. Metab. Dispos., 29: 242-251, 2001.
Gervasi PG, Longo V, Naldi F, Panattoni G and Ursino F, Xenobiotic-metabolizing enzymes in
human respiratory nasal mucosa. Biochem. Pharmacol., 41: 177-184, 1991.
Getchell ML, Chen Y, Ding X, Sparks DL and Getchell TV, Immunohistochemical localization
of a cytochrome P-450 isozyme in human nasal mucosa: age-related trends. Ann. Otol. Rhinol.
Laryngol., 102: 368-374, 1993.
Gibson GG and Skett P, Enzymology and molecular mechanisms of drug metabolism. In:
Introduction to drug metabolism, Chapman and Hall Ltd., London: 66-69, 1986.
Giorgi M, Marini S, Longo V, Mazzaccaro A, Amato G and Gervasi PG, Cytochrome P450-
dependent monooxygenase activities and their inducibility by classic P450 inducers in the liver,
kidney, and nasal mucosa of male adult ring-necked pheasants. Toxicol. Appl. Pharmacol., 167:
237-245, 2000.
Green T, Lee R, Toghill A, Meadowcroft S Lund V and Foster J, The toxicity of styrene to the
nasal epithelium of mice and rats: studies on the mode of action and relevance to humans. Chem.
Biol. Interact., 137: 185-202, 2001.
Grellner W, Time-dependent immunohistochemical detection of proinflammatory cytokines (IL-
1beta, IL-6, TNF-alpha) in human skin wounds. Forensic Sci. Int., 130: 90-96, 2002.
Gu J, Walker VE, Lipinskas TW, Walker DM and Ding X, Intraperitoneal administration of
coumarin causes tissue-selective depletion of cytochromes P450 and cytotoxicity in the olfactory
mucosa. Toxicol. Appl. Pharmacol., 146: 134-143, 1997.

8. Literaturverzeichnis 99
Gu J, Zhang QY, Genter MB, Lipinskas TW, Negishi M, Nebert DW and Ding X,
Purification and characterization of heterologously expressed mouse CYP2A5 and CYP2G1: role in
metabolic activation of acetaminophen and 2,6-dichlorobenzonitrile in mouse olfactory mucosal
microsomes. J. Pharmacol. Exp. Ther., 285: 1287-1295, 1998.
Hadley WM and Dahl AR, Cytochrome P-450-dependent monooxygenase activity in nasal
membranes of six species. Drug Metab. Dispos., 11: 275-276, 1983.
Ha-Duong NT, Dijols S, Marques-Soares C, Minoletti C, Dansette PM and Mansuy D,
Synthesis of sulfaphenazole derivates and their use as inhibitors and tools for comparing the active
sites of human liver cytochromes P450 of the 2C subfamily. J. Med. Chem., 44: 3622-3631, 2001.
Haspolat S, Mihci E, Coskun M, Gumuslu S, Ozbenm T and Yegin O, Interleukin-1beta,
tumor necrosis factor-alpha, and nitrite levels in febrile seizures. J. Child. Neurol., 17: 749-751,
2002.
Haussmann HJ, Anskeit E, Becker D, Kuhl P, Stinn W, Teredesai A, Voncken P and Walk
RA, Comparison of fresh and room-aged cigarette sidestream smoke in a subchronic inhalation
study on rats. Toxicol. Sci., 41: 100-116, 1998.
Hecht SS, Tobacco smoke carcinogens and breast cancer. Environ. Mol. Mutagen., 39: 119-126,
2002.
Heydel J, Leclerc S, Bernard P, Pelczar H, Gradinaru D, Magdalou J, Minn A, Artur Y and
Goudonnet H, Rat olfactory bulb and epithelium UDP-glucuronosyltransferase 2A1 (UGT2A1)
expression: in situ mRNA localization and quantitative analysis. Brain Res. Mol. Brain Res., 90: 83-
92, 2001.
Hildesheim A, Dosemeci M, Chan CC, Chen CJ, Cheng YJ, Hsu MM, Chen IH, Mittl BF,
Sun B, Levine PH, Chen JY, Brinton LA and Yang CS, Occupational exposure to wood,
formaldehyde, and solvents and risk of nasopharyngeal carcinoma. Cancer Epidemiol. Biomarkers
Prev., 10: 1145-1153, 2001.
Hochhaus G, Gonzalez MA, Dockhorn RJ, Shilstone J and Karafilidis J, A new solution-
based intranasal triamcinolone acetonide formulation in patients with perennial allergic rhinitis: how
does the pharmacokinetic/pharmacodynamic profile for cortisol suppression compare with an
aqueous suspension-based formulation? J. Clin. Pharmacol., 42: 662-669, 2002.

8. Literaturverzeichnis 100
Hukkanen J, Xenobiotic-metabolizing cytochrome P450 enzymes in human lung. Academic
Dissertation, Department of Pharmacology and Toxicology, University of Oulu, 2000.
Hukkanen J, Lassila A, Päivärinta K, Valanne S, Sarpo S, Hakkola J, Pelkonen O and
Raunio H, Induction and regulation of xenobiotic-metabolizing cytochrome P450s in the human
A549 lung adenocarcinoma cell line. Am. J. Respir. Cell Mol. Biol., 22: 360-366, 2000.
Hukkanen J, Vaisanen T, Lassila A, Piipari R, Anttila S, Pelkonen O, Raunio H and
Hakkola J, Regulation of CYP3A5 by glucocorticoids and cigarette smoke in human lung-derived
cells. J. Pharmacol. Exp. Ther., 304: 745-752, 2003.
Imaoka S, Hayashi K, Hiroi T, Yabusaki Y, Kamataki T and Funae Y, A transgenic mouse
expressing human CYP4B1 in the liver. Biochem. Biophys. Res. Commun., 284: 757-762, 2001.
Jedlitschky G, Cassidy AJ, Sales M, Pratt N and Burchell B, Cloning and characterization of a
novel human olfactory UDP-glucuronosyltransferase. Biochem. J., 340: 837-843, 1999.
Jones MA, Totemeyer S, Maskell DJ, Bryant CE and Barrow PA, Induction of
Proinflammatory Responses in the Human Monocytic Cell Line THP-1 by Campylobacter jejuni.
Infect. Immun., 71: 2626-2633, 2003.
Jung M, Romer A, Keyszer G, Lein M, Kristiansen G, Schnorr D, Loening SA and Jung K,
mRNA expression of the five membrane-type matrix metalloproteinases MT1-MT5 in human
prostatic cell lines and their down-regulation in human malignant prostatic tissue. Prostate, 55: 89-
98, 2003.
Karam WG, Goldstein JA, Lasker JM and Ghanayem BI, Human CYP2C19 is a major
omeprazole 5-hydroxylase, as demonstrated with recombinant cytochrome P450 enzymes. Drug.
Metab. Dispos., 24: 1081-1087, 1996.
Kasuga I, Pare PD, Ruan J, Connett JE, Anthonisen NR and Sandford AJ, Lack of
association of group specific component haplotypes with lung function in smokers. Thorax, 58: 790-
793, 2003.
King CD, Rios GR, Green MD and Tephly TR, UDP-glucuronosyltransferases. Curr. Drug.
Metab. 1: 143-161, 2000.

8. Literaturverzeichnis 101
King LM, Ma J, Srettabunjong S, Graves J, Bradbury JA, Li L, Spiecker M, Liao JK,
Mohrenweiser H and Zeldin DC, Cloning of CYP2J2 genes and identification of functional
polymorphisms. Mol. Pharmacol., 61: 840-852, 2002.
Kiewer SA, Lehmann JM, Milburn MV and Willson TM, The PPARs and PXRs: nuclear
xenobiotic receptors that define novel hormone signaling pathways. Recent Prog. Horm. Res., 54:
345-367, 1999.
Kobayashi N, Terada N, Hamano N, Numata T and Konno A, Transepithelial migration of
activated eosinophils induces a decrease of E-cadherin expression in cultured human nasal epithelial
cells. Clin. Exp. Allergy, 30: 807-817, 2000.
Koskela S, Hakkola J, Hukkanen J, Pelkonen O, Sorri M, Saranen A, Anttila S,
Fernandez-Salguero P, Gonzalez F and Raunio H, Expression of CYP2A genes in human liver
and extrahepatic tissues. Biochem. Pharmacol., 57: 1407-1413, 1999.
Krishna NS, Getchell TV, Dhooper N, Awasthi YC and Getchell ML, Age- and gender-
related trends in the expression of glutathione S-transferases in human nasal mucosa. Ann. Otol.
Rhinol. Laryngol., 104: 812-822, 1995.
Kristic RV, Respiratory System. In: Human Microscopic Anatomy, Springer Verlag, Berlin
Heidelberg: 124-133, 1991.
Krueger SK, Martin SR, Yueh MF, Pereira CB and Williams DE, Identification of active
flavin-containing monooxygenase isoform 2 in human lung and characterization of expressed protein.
Drug Metab. Dispos., 30: 34-41, 2002.
Kumar L and Singh M, Respiratory allergy. Indian J. Pediatr., 69: 237-244, 2002.
Laemmli UK, Cleavage of structural proteins during the assembly of the head of bacteriophage T4.
Nature, 227: 680-685, 1970.
Lazard D, Zupko K, Poria Y, Nef P, Lazarovits J, Horn S, Khen M and Lancet D, Odorant
signal termination by olfactory UDP glucuronosyl transferase. Nature, 349: 790-793, 1991.
Lewis JL, Rhoades CE, Gervasi PG, Griffith WC and Dahl AR, The cyanide-metabolizing
enzyme rhodanese in human nasal respiratory mucosa. Toxicol. Appl. Pharmacol., 108: 114-120,
1991.

8. Literaturverzeichnis 102
Linß W und Halbhuber K, Atmungsapparat. In: Histologie und mikroskopische Anatomie, 17.
Auflage, Thieme Verlag, Stuttgart: 179-181, 1991.
Lippert H, Kopf II und Hals. In: Lehrbuch Anatomie, 4. Auflage, Urban & Schwarzenberg,
München: 567-571, 1996.
Liu C, Zhuo X, Gonzalez FJ and Ding X, Baculovirus-mediated expression and characterization
of rat CYP2A3 and human CYP2a6: role in metabolic activation of nasal toxicants. Mol.
Pharmacol., 50: 781-788, 1996.
Longo V and Ingelman-Sundberg M, Acetone-dependent regulation of cytochromes P4502E1
and P4502B1 in rat nasal mucosa. Biochem. Pharmacol., 46: 1945-1951, 1993.
Longo V, Citti L and Gervasi PG, Metabolism of diethylnitrosamine by nasal mucosa and hepatic
microsomes from hamster and rat: species specificity of nasal mucosa. Carcinogenesis, 7: 1323-
1328, 1986.
Longo V, Pacifici GM, Panattoni G, Ursino F and Gervasi PG, Metabolism of
diethylnitrosamine by microsomes of human respiratory nasal mucosa and liver. Biochem.
Pharmacol., 38: 1867-1869, 1989.
Longo V, Mazzaccaro A, Naldi F and Gervasi PG, Drug-metabolizing enzymes in liver,
olfactory, and respiratory epithelium of cattle. J. Biochem. Toxicol., 6: 123-128, 1991.
Longo V, Amato G, Santucci A ang Gervasi PG, Purification and characterization of three
constitutive cytochrome P-450 isoforms from bovine olfactory epithelium. Biochem. J., 323: 65-70,
1997.
Longo V, Ingelman-Sundberg M, Amato G, Salvetti A and Gervasi PG, Effect of starvation
and chlormethiazole on cytochrome P450s of rat nasal mucosa. Biochem. Pharmacol., 59: 1425-
1432, 2000.
Mace K, Bowman ED, Vautravers P, Shields PG, Harris CC and Pfeifer AM,
Characterization of xenobiotic-metabolising enzyme expression in human bronchial mucosa and
peripheral lung tissues. Eur. J. Cancer, 34: 914-920, 1998.

8. Literaturverzeichnis 103
Mackenzie PI, Owens IS, Burchell B, Bock KW, Bairoch A, Belanger A, Fourel-Gigleux S,
Green M, Hum DW, Iyanagi T, Lancet D, Louisot P, Magdalou J, Chowdhury JR, Ritter
JK, Schachter H, Tephly TR, Tipton KF and Nebert DW, UDP glycosyltransferase gene
superfamily: recommended nomenclature update based on evolutionary divergence.
Pharmacogenetics, 7: 255-269, 1997.
Mancy A, Antignac M, Minoletti C, Dijols S, Mouries V, Duong NT, Battioni P, Dansette
PM and Mansuy D, Diclofenac and its derivates as tools for studying human cytochromes P450
active sites: particular efficiency and regioselectivity of P450 2Cs. Biochemistry, 38: 14264-14270,
1999.
Marill J, Cresteil T, Lanotte M and Chabot GG, Identification of human cytochrome P450s
involved in the formation of all-trans-retinoic acid principal metabolites. Mol. Pharmacol., 58:
1341-1348, 2000.
Marini S, Longo V, Mazzaccaro A and Gervasi PG, Xenobiotic-metabolizing enzymes in pig
nasal and hepatic tissues. Xenobiotica, 28: 923-935, 1998.
Million K, Tournier F, Houcine O, Ancian P, Reichert U and Marano F, Effects of retinoic
acid receptor-selective agonists on human nasal epithelial cell differentiation. Am. J. Respir. Cell
Mol. Biol.,25: 744-750, 2001.
Mollerup S, Ryberg D, Hewer A, Phillips DH and Haugen A, Sex differences in lung CYP1A1
expression and DNA adduct levels among lung cancer patients. Cancer Res., 59: 3317-3320,
1999.
Muller B, Zopf K, Bachofer J and Steimer W, Optimized strategy for rapid cytochrome P450
2D6 genotyping by real-time long PCR. Clin. Chem., 49: 1624-1631, 2003.
Nelson DR, Comparison of P450s from human and fugu: 420 million years of vertebrate P450
evolution. Arch. Biochem. Biophys., 409: 18-24, 2003.
Netter FH, Kopf und Hals. In: Atlas der Anatomie des Menschen, 2. Auflage, Novartis AG,
Basel: 38, 2000.
Nhamburo PT, Gonzalez FJ, McBride OW, Gelboin HV and Kimura S, Identification of a new
P450 expressed in human lung: complete cDNA sequence, cDNA-directed expression, and
chromosome mapping. Biochemistry, 28: 8060-8066, 1989.

8. Literaturverzeichnis 104
Nishimura M, Yaguti H, Yoshitsugu H, Naito S and Satoh T, Tissue distribution of mRNA
expression of human cytochrome P450 isoforms assessed by high-sensitivity real-time reverse
transcription PCR. Yakugaku Zasshi, 123: 369-375, 2003.
Niwa T, Fujimoto M, Kishimoto K, Yabusaki Y, Ishibashi F and Katagiri M, Metabolism and
interaction of bisphenol A in human hepatic cytochrome P450 and steroidogenic CYP17. Biol.
Pharm. Bull., 24: 1064-1067, 2001.
Oudijk EJ, Lammers JW and Koenderman L, Systemic inflammation in chronic obstructive
pulmonary disease. Eur. Respir. J. Suppl., 46: 5-13, 2003.
Pawankar R and Fokkens W, Evidence-based treatment of allergic rhinitis. Curr. Allergy
Asthma Rep., 1: 218-226, 2001.
Peng HM, Ding X and Coon MJ, Isolation and heterologous expression of cloned cDNAs for
two rabbit nasal microsomal proteins, CYP2A10 and CYP2A11, that are related to nasal
microsomal cytochrome P450 form a. J. Biol. Chem., 268: 17253-17260, 1993.
Pietsch EC, Hurley AL, Scott EE, Duckworth BP, Welker ME, Leone-Kabler S, Townsend
AJ, Torti FM and Torti SV, Oxathiolene oxides: a novel family of compounds that induce ferritin,
glutathione S-transferase, and other proteins of the phase II response. Biochem. Pharmacol., 65:
1261-1269, 2003.
Prieto-Alamo MJ, Cabrera-Luque JM and Pueyo C, Absolute quantitation of normal and ROS-
induced patterns of gene expression: an in vivo real-time PCR study in mice. Gene Expr., 11: 23-
34, 2003.
Putt DA, Ding X, Coon MJ and Hollenberg PF, Metabolism of aflatoxin B1 by rabbit and rat
nasal mucosa microsomes and purified cytochrome P450, including isoforms 2A10 and 2A11.
Carcinogenesis, 16: 1411-1417, 1995.
Reed CJ, Drug metabolism in the nasal cavity: relevance to toxicology. Drug Metab. Rev., 25:
173-205, 1993.
Rivera SP, Saarikoski ST and Hankinson O, Identification of a novel dioxin-inducible
cytochrome P450. Mol. Pharmacol., 61: 255-259, 2002.
Rohen JW und Lütjen-Drecoll E, Epithelgewebe. In: Funktionelle Histologie, 4. Auflage,
Schattauer Verlagsgesellschaft mbH, Stuttgart: 71-74, 1999.

8. Literaturverzeichnis 105
Rohen JW und Lütjen-Drecoll E, Respirationssystem. In: Funktionelle Histologie, 4. Auflage,
Schattauer Verlagsgesellschaft mbH, Stuttgart: 216-221.
Ruan XZ, Moorhead JF, Fernando R, Wheeler DC, Powis SH and Varghese Z, PPAR
agonists protect mesangial cells from Interleukin 1beta-induced intracellular lipid accumulation by
activating the ABCA1 cholesterol efflux pathway. J. Am. Soc. Nephrol., 14: 593-600, 2003.
Rubin H, Synergistic mechanisms in carcinogenesis by polycyclic aromatic hydrocarbons and by
tobacco smoke: a bio-historical perspective with updates. Carcinogenesis, 22: 1903-1930, 2001.
Rylander T, Neve EP, Ingelman-Sundberg M and Oscarson M, Identification and tissue
distribution of the novel human cytochrome P450 2S1 (CYP2S1). Biochem. Biophys. Res.
Commun., 281: 529-535, 2001.
Savas U, Hsu MH and Johnson EF, Differential regulation of human CYP4A genes by
peroxisome proliferators ad dexamethasone. Arch. Biochem. Biophys., 409: 212-220, 2003.
Schiebler TH, Schmidt W und Zilles K, Kopf und Hals. In: Anatomie, 7. Auflage, Springer
Verlag, Berlin: 404-425, 1997.
Seidegard J and Ekstrom G, The role of human glutathione transferases and epoxide hydrolases in
the metabolism of xenobiotics. Environ. Health Perspect., 105: 791-799, 1997.
Seubert JM, Sinal CJ and Bend JR, Acute sodium arsenite administration induces pulmonary
CYP1A1 mRNA, protein and activity in the rat. J. Biochem. Mol. Toxicol., 16: 84-95, 2002.
ShimadaT, Yamazaki H, Mimura M, Wakamiya N, Ueng YF, Guengerich FP and Inui Y,
Characterization of microsomal cytochrome P450 enzymes involved in the oxidation of xenobiotic
chemicals in human fetal liver and adult lungs. Drug Metab. Dispos., 24: 515-522, 1996.
Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, Fujimoto
EK, Goeke NM, Olson BJ and Klenk DC, Measurement of protein using bicinchoninic acid.
Anal. Biochem., 150: 76-85, 1985.
Speckmann EJ und Wittkowski W, Sensorisches System. In: Bau und Funktionen des
menschlichen Körpers, 19. Auflage, Urban 6 Schwarzenberg, München: 137-139, 1998.
Su T, Sheng JJ, Lipinskas TW and Ding X, Expression of CYP2A genes in rodent and human
nasal mucosa. Drug Metab. Dispos., 24: 884-890, 1996.

8. Literaturverzeichnis 106
Su T, Bao Z, Zhang QY, Smith TJ, Hong JY and Ding X, Human cytochrome P450 CYP2A13:
predominant expression in the respiratory tract and its high efficiency metabolic activation of a
tobacco-specific carcinogen, 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone. Cancer Res., 60:
5074-5079, 2000.
Szelenyi I and Pahl A, Drug therapy in asthma bronchiale in the new millennium. Pharmazie, 57:
83-86, 2002.
Talmi YP, Wolf M, Horowitz Z, Bedrin L, Kronenberg J and Pfeffer MR, Smoking-induced
squamous-cell cancer of the nose. Arch. Environ. Health, 57: 422-424, 2002.
Thornton-Manning JR, Nikula KJ, Hotchkiss JA, Avila KJ, Rohrbacher KD, Ding X and
Dahl AR, Nasal cytochrome P450 2A: identification, regional localization, and metabolic activity
toward hexamethylphosphoramide, a known nasal carcinogen. Toxicol. Appl. Pharmacol., 142:
22-30, 1997.
Thum T and Borlak J, Gene expression in distinct regions of the heart. Lancet 355: 979-983,
2000.
Tricarico C, Pinzani P, Bianchi S, Paglierani M, Distante V, Pazzagli M, Bustin SA and
Orlando C, Quantitative real-time reverse transcription polymerase chain reaction: normalization to
rRNA or single housekeeping genes is inappropriate for human tissue biopsies. Anal. Biochem.,
309: 293-300, 2002.
Turesky RJ, Richoz J, Constable A, Curtis KD, Dingley KH and Turteltaub KW, The
effects of coffee on enzymes involved in metabolism of the dietary carcinogen 2-amino-1-methyl-6-
phenylimidazo[4,5-b]pyridine in rats. Chem. Biol. Interact., 145: 251-265, 2003.
Valles EG, Laughter AR, Dunn CS, Cannelle S, Swanson CL, Cattley RC and Corton JC,
Role of the peroxisome proliferator-activated receptor alpha in responses to diisononyl phthalate.
Toxicology, 191: 211-225, 2003.
Vanden Heuvel JP, Kreder D, Belda B, Hannon DB, Nugent CA, Burns KA and Taylor
MJ, Comprehensive analysis of gene expression in rat and human hepatoma cells exposed to the
peroxisome proliferator WY14,643. Toxicol. Appl. Pharmacol., 188: 185-198, 2003.

8. Literaturverzeichnis 107
Voigt JM, Guengerich FP and Baron J, Localization and induction of cytochrome P450 1A1 and
aryl hydrocarbon hydroxylase activity in rat nasal mucosa. J. Histochem. Cytochem., 41: 877-885,
1993.
Von Lanz T und Wachsmuth W, Splanchnocranium. In: Praktische Anatomie 1. Band Teil 1A,
Springer Verlag, Berlin Heidelberg: 164-170, 1985.
Wardlaw SA, Nikula KJ, Kracko DA, Finch GL, Thornton-Manning JR and Dahl AR,
Effect of cigarette smoke on CYP1A1, CYP1A2 and CYP2B1/2 of nasal mucosa in F344 rats.
Carcinogenesis, 19: 655-662, 1998.
Waxman DJ, P450 gene induction by strucurally diverse xenochemicals: central role of nuclear
receptors CAR, PXR, and PPAR. Arch. Biochem. Biophys., 369: 11-23, 1999.
Weiss L, The Respiratory System. In: Cell and Tissue Biology A Textbook of Histology, 6.
Edition, Urban & Schwarzenberg, Baltimore Munich: 753-758, 1988.
Westhofen Prof Dr M, Nase, Nasennebenhöhlen, Mittelgesicht und vordere Scgädelbasis. In:
Hals-Nasen-Ohrenheilkunde systematisch, UNI-MED Verlag AG, Bremen: 192, 2001.
Wheeler CW, Park SS and Guenther TM, Immunochemical analysis of a cytochrome P-450IA1
homologue in human lung microsomes. Mol. Pharmacol., 38: 634-643, 1990.
Whetstine JR, Yueh MF, McCarver DG, Williams DE, Park CS, Kang JH, Cha YN,
Dolphin CT, Shephard EA, Phillips IR and Hines RN, Ethnic differences in human flavin-
containing monooxygenase 2 (FMO2) polymorphisms: detection of expressed protein in African-
Americans. Toxicol. Appl. Pharmacol., 168: 216-224, 2000.
Willey JC, Coy EL, Frampton MW, Torres A, Apostolakos MJ, Hoehn G, Schuermann
WH, Thilly WG, Olson DE, Hammersley JR, Crespi CL and Utell MJ, Quantitative RT-PCR
measurement of cytochromes p450 1A1, 1B1, and 2B7, microsomal epoxide hydrolase, and
NADPH oxidoreductase expression in lung cells of smokers and nonsmokers. Am. J. Respir. Cell
Mol. Biol., 17: 114-124, 1997.
Yang Y, Sanchez D, Figarella C and Lowe ME, Discoordinate expression of pancreatic lipase
and two related proteins in the human fetal pancreas. Pediatr. Res., 47: 184-188, 2000.

8. Literaturverzeichnis 108
Yang B, Graham L, Dikalov S, Mason RP, Falck JR, Liao JK and Zeldin DC,
Overexpression of cytochrome P450 CYP2J2 protects against hypoxia-reoxygenation injury in
cultured bovine aortic endothelial cells. Mol. Pharmacol., 60: 310-230, 2001.
Zeldin DC, Foley J, Ma J, Boyle JE, Pascual JM, Moomaw CR, Tomer KB, Steenbergen
C and Wu S, CYP2J subfamily P450s in the lung: expression, localization, and potential functional
significance. Mol. Pharmacol, 50: 1111-1117, 1996.
Zhu-Ge J, Yu YN, Qian YL and Li X, Establishment of a transgenic cell line stably expressing
human cytochrome P450 2C18 and identification of a CYP2C18 clone with exon 5 missing. World
J. Gastroenterol., 8: 888-892, 2002.

9. Anhang I
9. Anhang
9.1 Daten der Patienten für die Genexpression Patient Geschlecht
( M / W ) Raucher ( R / NR )
Alter (Jahre)
Medikamente Allergien OP-Datum (OP-Ort)
W-R 1 W R ( 15 Zig. tgl. )
34 - Katzen 10.09.2002 (FS)
W-R 2 W R ( 3 Zig. tgl. )
20 - - 25.07.2002 (FS)
W-R 3 W R ( viele ) 39 Pille Nickel 09.07.2002 (FS)
W-R 4 W R ( 15-20 Zig. tgl. )
34 Ibuprofen, Levofloxacin, Pille
Gräser, Hausstaub, Katze
27.06.2002 (FS)
W-R 5 W R ( 20-25 Zig. tgl. )
56 Clirogest N1000, Dolormin extra, Togal, Atrex duo
- 12.06.2002 (MHH)
M-R 1 M R ( 3-4 Zig. tgl. )
36 - - 27.08.2002 (FS)
M-R 2 M R ( 3-4 Zig. tgl. )
30 Eutyrox 200 Nahrungsmittel 20.08.2002 (FS)
M-R 3 M R ( 50 Zig. tgl. )
45 - - 06.08.2002 (FS)
M-R 4 M R ( <15 Zig. tgl. )
28 - Heuschnupfen, Katzenhaare
25.07.2002 (FS)
M-R 5 M R ( 15 Zig. tgl. )
51 - Heuschnupfen 15.05.2002 (MHH)
W-NR 1 W NR 30 - - 05.09.2002 (FS)
W-NR 2 W NR 26 Nasonex, Vividrin, Aerius
Penicillin, Gräser, Pollen
18.07.2002 (FS)
W-NR 3 W NR 62 Augentropfen (Arteoptic)
Gräser 07.05.2002 (MHH)
W-NR 4 W NR 23 Pille Penicillin 07.05.2002 (MHH)
W-NR 5 W NR 19 - - 02.07.2002 (FS)
M-NR 1 M NR 47 Paracetamol - 10.09.2002 (FS)
M-NR 2 M NR 61 Vitamine, Prostata
- 05.09.2002 (FS)
M-NR 3 M NR 33 - - 03.09.2002 (FS)
M-NR 4 M NR 41 für Heuschnupfen Heuschnupfen 03.09.2002 (FS)

9. Anhang II
Patient Geschlecht ( M / W )
Raucher ( R / NR )
Alter (Jahre)
Medikamente Allergien OP-Datum (OP-Ort)
M-NR 5 M NR 20 - Hausstaub 03.09.2002 (FS)
M-NR 6 M NR 27 - Penicillin 17.09.2002 (FS)
M-NR 7 M NR 50 Bluthochdruck - 17.09.2002 (FS)
9.2 Daten der Patienten für die mikrosomalen Pools Pool Geschlecht
( M / W ) Raucher ( R / NR )
Alter (Jahre)
Medikamente Allergien OP-Datum (OP-Ort)
M NR 42 Sortis, Nexium Pollen 23.07.2002 (FS)
M NR 27 - Pollen 19.09.2002 (FS)
M NR 60 Bluthochdruck - 24.09.2002 (FS)
M NR 51 - Pflaster 01.10.2002 (FS)
1
M NR 16 - - 01.10.2002 (FS)
M NR 37 Antidepressivum - 09.07.2002 (FS)
M NR 34 - - 30.07.2002 (FS)
M NR 47 Paracetamol - 10.09.2002 (FS)
M NR 62 - Latex 12.09.2002 (FS)
2
M NR 27 - Penicillin 17.09.2002 (FS)
M NR 14 - - 12.06.2002 (MHH)
M NR 46 - - 18.06.2002 (FS)
M NR 62 - - 20.08.2002 (FS)
M NR 61 - - 27.08.2002 (FS)
M NR 53 - - 29.08.2002 (FS)
4
M NR 61 Vitamine, Prostata
- 05.09.2002 (FS)

9. Anhang III
Pool Geschlecht ( M / W )
Raucher ( R / NR )
Alter (Jahre)
Medikamente Allergien OP-Datum (OP-Ort)
M NR 14 - - 20.06.2002 (FS)
M NR 62 - - 27.06.2002 (FS)
M NR 52 - Jod 09.07.2002 (FS)
M NR 62 - - 25.07.2002 (FS)
M NR 62 - - 30.07.2002 (FS)
5
M NR 57 Sempocort, Oneep
Aspirin 20.08.2002 (FS)
W NR 62 Augentropfen (Arteoptic)
Gräser 07.05.2002 (MHH)
W NR 23 Pille Penicillin 07.05.2002 (MHH)
W NR 19 - - 02.07.2002 (FS)
W NR 30 - - 05.09.2002 (FS)
W NR 16 - Heuschnupfen 08.10.2002 (FS)
6
W NR 58 - Haselnuß 10.10.2002 (FS)
M R ( 15 Zig. tgl. )
51 - Heuschnupfen 15.05.2002 (MHH)
M R ( <15 Zig. tgl. )
28 - Heuschnupfen, Katzenhaare
25.07.2002 (FS)
M R ( 50 Zig. tgl. )
45 - - 06.08.2002 (FS)
M R ( 3-4 Zig. tgl. )
30 Eutyrox 200 Nahrungsmittel 20.08.2002 (FS)
M R ( 3-4 Zig. tgl. )
36 - - 27.08.2002 (FS)
7
M R 31 - - 10.10.2002 (FS)
W R ( 20-25 Zig. tgl. )
56 Clirogest N1000, Dolormin extra (1-2 Tabl.), Togal (1-2 Tabl.), Atrex duo (1-2 Tabl.)
- 12.06.2002 (MHH)
W R ( 15-20 Zig. tgl. )
34 Ibuprofen, Levofloxacin, Pille
Gräser, Hausstaub, Katze
27.06.2002 (FS)
8
W R ( 3 Zig. tgl. )
20 - - 25.07.2002 (FS)

9. Anhang IV
Pool Geschlecht ( M / W )
Raucher ( R / NR )
Alter (Jahre)
Medikamente Allergien OP-Datum (OP-Ort)
M NR 55 Augensalbe - 27.06.2002 (FS)
M NR ( seit Dezember )
33 - - 30.07.2002 (FS)
M NR 44 - - 01.08.2002 (FS)
M NR 33 - - 03.09.2002 (FS)
M NR 53 - Pollen 15.10.2002 (FS)
M NR 63 Bluthochdruck Penicillin 17.10.2002 (FS)
M NR 36 - - 29.10.2002 (FS)
M NR 43 - - 29.10.2002 (FS)
M NR 32 - Pollen 05.11.2002 (FS)
M NR 55 Eupkylong Pflaster 05.11.2002 (FS)
M NR 55 - Penicillin 07.11.2002 (FS)
M NR 62 Bluthochdruck - 12.11.2002 (FS)
M NR 38 - - 19.11.2002 (FS)
9
M NR 59 Antiallergicum Tierhaare 19.11.2002 (FS)
M R ( 3-4 Zig. tgl. )
16 - Penicillin, Pollen
27.06.2002 (FS)
M R 31 - Hausstaub, Katzenhaare
15.10.2002 (FS)
M R ( 10 Zig. tgl. )
29 - - 29.10.2002 (FS)
M R ( 30 Zig. tgl. )
59 Asthma - 29.10.2002 (FS)
M R ( 20 Zig. tgl. )
35 - - 05.11.2002 (FS)
10
M R ( 20 Zig. tgl. )
52 Bluthochdruck - 05.11.2002 (FS)
W NR 30 Pille Penicillin 17.10.2002 (FS)
W NR 31 Antiallergicum Hausstaub 12.11.2002 (FS)
W NR 55 - - 14.11.2002 (FS)
11
W NR 20 - Penicillin 21.11.2002 (FS)
9. Anhang V
Pool Geschlecht ( M / W )
Raucher ( R / NR )
Alter (Jahre)
Medikamente Allergien OP-Datum (OP-Ort)
11 W NR 38 - - 26.11.2002 (FS)
M NR 58 - - 07.01.2003 (FS)
M NR 32 - - 14.01.2003 (FS)
M NR 45 - - 28.01.2003 (FS)
M NR 35 - - 28.01.2003 (FS)
M NR 56 Bluthochdruck Pollen 06.02.2003 (FS)
M NR 29 - Hausstaub, Milben
18.02.2003 (FS)
M NR 32 Allergie alles 20.02.2003 (FS)
M NR 42 - - 20.02.2003 (FS)
M NR 25 - Hausstaub 06.03.2003 (FS)
12
M NR 45 - Pollen, Getreide
06.03.2003 (FS)
M R ( 10 Zig. tgl. )
29 - Pollen, Asthma
21.01.2003 (FS)
M R ( 1 Schachtel tgl. )
26 - - 28.01.2003 (FS)
M R (gelegentlichHasch )
22 - Pollen, Hausstaub, Tierhaare, Penicillin, Amoxycillin
06.02.2003 (FS)
M R ( 5-10 Zig. tgl. )
22 Pulmicord Pollen 11.02.2003 (FS)
M R ( Pfeife ) 41 - - 20.02.2003 (FS)
M R ( 1 Schachtel tgl. )
20 - - 11.03.2003 (FS)
13
M R ( 10 Zig. tgl. )
35 - - 11.03.2003 (FS)
W NR 28 Pille Pflaster 14.01.2003 (FS)
W NR 66 Bluthochdruck, Diabetes
- 21.01.2003 (FS)
14
W NR 30 - Neurodermitis, Lokalanästhesie
21.01.2003 (FS)

9. Anhang VI
Pool Geschlecht ( M / W )
Raucher ( R / NR )
Alter (Jahre)
Medikamente Allergien OP-Datum (OP-Ort)
W NR 24 - - 23.01.2003 (FS)
W NR 42 - - 06.02.2003 (FS)
W NR 24 - Erdnuss 04.03.2003 (FS)
W NR 19 - - 04.03.2003 (FS)
W NR 72 Schilddrüse Pflaster, Jod
11.03.2003 (FS)
W NR 44 - Pflaster 13.03.2003 (FS)
14
W NR 27 - - 13.03.2003 (FS)
W R 24 Pille - 17.10.2002 (FS)
W R ( 10 – 15 Zig. tgl. )
41 - Wolle 14.11.2002 (FS)
W R ( 15 Zig. tgl. )
40 Schilddrüse Nickel 28.01.2003 (FS)
W R ( 15 Zig. tgl. )
23 - Hausstaub, Milben
18.02.2003 (FS)
15
W R ( 15 Zig. tgl. )
22 - - 11.03.2003 (FS)
W ? 24 Pille Penicillin 13.02.2003 (FS)
W ? 26 - - 13.02.2003 (FS)
W ? 32 L-Thyroxin - 13.02.2003 (FS)
16
W ? 62 Schilddrüse Pflaster, Wespen
18.02.2003 (FS)
Abkürzungen: M = männlich; NR = Nicht-Raucher; R = Raucher; tgl. = täglich; W = weiblich; Zig. = Zigaretten; ? = keine Angabe

9. Anhang VII
9.3 Liste der verwendeten Primer
Primer Annealing
Temperatur Zyklen Länge Sequenz Genbank
CyclophillinA 55°C 33 347 5´CTTGCCATTCCTGGACCCAA 3´TTTCGTGCTCTGAGCACTGG
NM_021130
GAPDH 55°C 33 353 5´GGCCAAGGTCATCCATGA 3´TCAGTGTAGCCCAGGATG
NM_002046
CYP 1A1 58°C 36 432 5´TCACAGACAGCCTGATTGAG 3´GATGGGTTGACCCATAGCTT
NM_000499
CYP 1A2 55°C - 308 5´TGGCTTCTACATCCCCAAGAAAT 3´TTCATGGTCAGCCCGTAGAT
NM_000761
CYP 1B1 55°C 33 301 5´GCAGAATTGGATCAGGTCGT 3´TGGTCAGGTCCTTGTTGATG
NM_000104
CYP 2A6/7 55°C 36 1151 5´GTGTGGACATGATGCCGT 3´AGGACTTGAGGCGGAAGT
NM_000762 (2A6) NM_000764 (2A7)
CYP 2A13 55°C 36 320 5´ACAAAGAGTTCCTGTCACTG 3´CCGCAAAGAAGAGGTTCAGG
NM_000766
CYP 2B6 55°C 33 357 5´CCATACACAGAGGCAGTCAT 3´GGTGTCAGATCGATGTCTTC
NM_000767
CYP 2C8 58°C 39 311 5´GATCATGTAATTGGCAGACACA 3´CCTGCTGAGAAAGGCATGAAG
NM_000770
CYP 2C9 58°C 39 437 5´AGCTTGGAAAACACTGCAGT 3´CCTGCTGAGAAAGGCATGAAG
NM_000771
CYP 2C18 55°C 33 431 5´CTGTAACTGATATGTTTGGG 3´CCTGCTGAGAAAGGCATGAAG
NM_000772
CYP 2C19 58°C 39 431 5´GTAATCACTGCAGCTGACTTAC 3´CCTGCTGAGAAAGGCATGAAG
NM_000769
CYP 2D6 58°C 38 332 5´TGATGAGAACCTGCGCATAG 3´ACCGATGACAGGTTGGTGAT
NM_000106
CYP 2E1 55°C 36 365 5´AGCACAACTCTGAGATATGG 3´ATAGTCACTGTACTTGAACT
NM_000773
CYP 2F1 55°C 33 283 5´ATGAACTTGCCGCACCGCGT 3´ACAGGCTCCACTTACGGTGC
NM_000774
CYP 2J2 55°C 33 389 5´CCCACCAAACTCTCTTCAGC 3´CATTCTCTGCACCTCATGGA
NM_000775
CYP 2S1 55°C 33 312 5´ACCCCAACATCTTCAAGCAC 3´TTCATCTGGTCTGCGTGGT
NM_030622
CYP 3A4 55°C 39 324 5´CCAAGCTATGCTCTTCACCG 3´TCAGGCTCCACTTACGGTGC
NM_017460
CYP 3A5 55°C 38 472 5´TGTCCAGCAGAAACTGCAAA 3´TTGAAGAAGTCCTTGCGTGTC
NM_000777
CYP 3A7 55°C - 475 5´CTATGATACTGTGCTACAGT 3´TCAGGCTCCACTTACGGTCT
NM_000765
CYP 4A11 55°C - 280 5´CAAGTGACCTCCCTGCTCAT 3´CTGATCTCCCCAGAATCACC
NM_000778
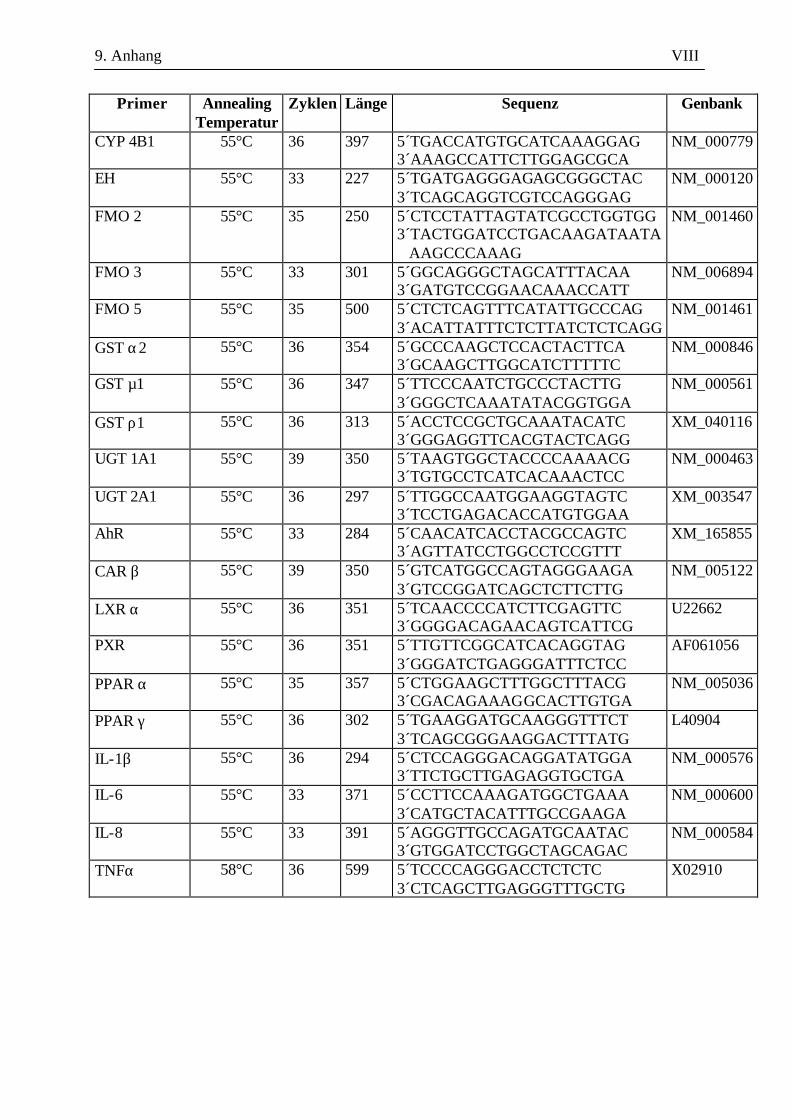
9. Anhang VIII
Primer Annealing Temperatur
Zyklen Länge Sequenz Genbank
CYP 4B1 55°C 36 397 5´TGACCATGTGCATCAAAGGAG 3´AAAGCCATTCTTGGAGCGCA
NM_000779
EH 55°C 33 227 5´TGATGAGGGAGAGCGGGCTAC 3´TCAGCAGGTCGTCCAGGGAG
NM_000120
FMO 2 55°C 35 250 5´CTCCTATTAGTATCGCCTGGTGG 3´TACTGGATCCTGACAAGATAATA AAGCCCAAAG
NM_001460
FMO 3 55°C 33 301 5´GGCAGGGCTAGCATTTACAA 3´GATGTCCGGAACAAACCATT
NM_006894
FMO 5 55°C 35 500 5´CTCTCAGTTTCATATTGCCCAG 3´ACATTATTTCTCTTATCTCTCAGG
NM_001461
GST α2 55°C 36 354 5´GCCCAAGCTCCACTACTTCA 3´GCAAGCTTGGCATCTTTTTC
NM_000846
GST µ1 55°C 36 347 5´TTCCCAATCTGCCCTACTTG 3´GGGCTCAAATATACGGTGGA
NM_000561
GST ρ1 55°C 36 313 5´ACCTCCGCTGCAAATACATC 3´GGGAGGTTCACGTACTCAGG
XM_040116
UGT 1A1 55°C 39 350 5´TAAGTGGCTACCCCAAAACG 3´TGTGCCTCATCACAAACTCC
NM_000463
UGT 2A1 55°C 36 297 5´TTGGCCAATGGAAGGTAGTC 3´TCCTGAGACACCATGTGGAA
XM_003547
AhR 55°C 33 284 5´CAACATCACCTACGCCAGTC 3´AGTTATCCTGGCCTCCGTTT
XM_165855
CAR β 55°C 39 350 5´GTCATGGCCAGTAGGGAAGA 3´GTCCGGATCAGCTCTTCTTG
NM_005122
LXR α 55°C 36 351 5´TCAACCCCATCTTCGAGTTC 3´GGGGACAGAACAGTCATTCG
U22662
PXR 55°C 36 351 5´TTGTTCGGCATCACAGGTAG 3´GGGATCTGAGGGATTTCTCC
AF061056
PPAR α 55°C 35 357 5´CTGGAAGCTTTGGCTTTACG 3´CGACAGAAAGGCACTTGTGA
NM_005036
PPAR γ 55°C 36 302 5´TGAAGGATGCAAGGGTTTCT 3´TCAGCGGGAAGGACTTTATG
L40904
IL-1β 55°C 36 294 5´CTCCAGGGACAGGATATGGA 3´TTCTGCTTGAGAGGTGCTGA
NM_000576
IL-6 55°C 33 371 5´CCTTCCAAAGATGGCTGAAA 3´CATGCTACATTTGCCGAAGA
NM_000600
IL-8 55°C 33 391 5´AGGGTTGCCAGATGCAATAC 3´GTGGATCCTGGCTAGCAGAC
NM_000584
TNFα 58°C 36 599 5´TCCCCAGGGACCTCTCTC 3´CTCAGCTTGAGGGTTTGCTG
X02910

9. Anhang IX
9.4 Liste der verwendeten Primär- und Sekundärantikörper Primärantikörper Verdünnung Sekundärantikörper Verdünnung CYP 1A1 (Code MAb 1A3-03, rubitec GmbH, Bochum)
1 : 400 α-mouse (Code AP 160 P; Charge 20040081, Chemicon, Temecula, USA)
1 : 5.000
CYP 2A (Code sc-9896; Charge C280, Santa Cruz Biotechnology, Santa Cruz, USA)
1 : 100 α-sheep (Code AP 147 P; Charge 20021225, Chemicon, Temecula, USA)
1 : 10.000
CYP 2B6 (Code AB-1268; Charge 20120789, Chemicon, Temecula, USA)
1 : 4.000 α-sheep (Code AP 147 P; Charge 20021225, Chemicon, Temecula, USA)
1 : 10.000
CYP 2E1 (Code AB-1274; Charge 2210034, Chemicon, Temecula, USA)
1 : 2.000 α-sheep (Code AP 147 P; Charge 20021225, Chemicon, Temecula, USA)
1 : 10.000
CYP 3A4 (Code AB-1278; Charge 19101552, Chemicon, Temecula, USA)
1 : 1.000 α-sheep (Code AP 147 P; Charge 20021225, Chemicon, Temecula, USA)
1 : 10.000
CYP 3A5 (Code AB-1279; Charge 2103001, Chemicon, Temecula, USA)
1 : 1.000 α-rabbit (Code AP 132 P; Charge 20070556, Chemicon, Temecula, USA)
1 : 10.000
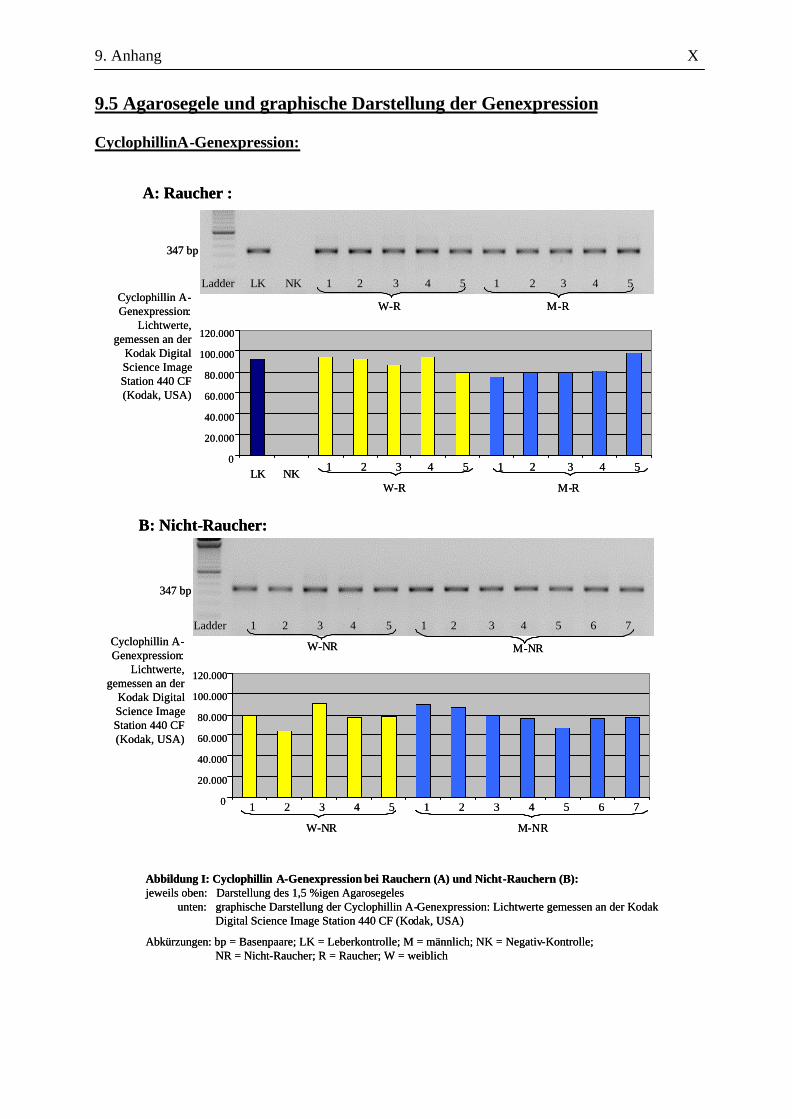
9. Anhang X
9.5 Agarosegele und graphische Darstellung der Genexpression CyclophillinA-Genexpression:
347 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung I: Cyclophillin A-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der Cyclophillin A-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
347 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
Cyclophillin A-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
Cyclophillin A-Genexpression:
Lichtwerte, gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
347 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung I: Cyclophillin A-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der Cyclophillin A-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
347 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
Cyclophillin A-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
Cyclophillin A-Genexpression:
Lichtwerte, gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
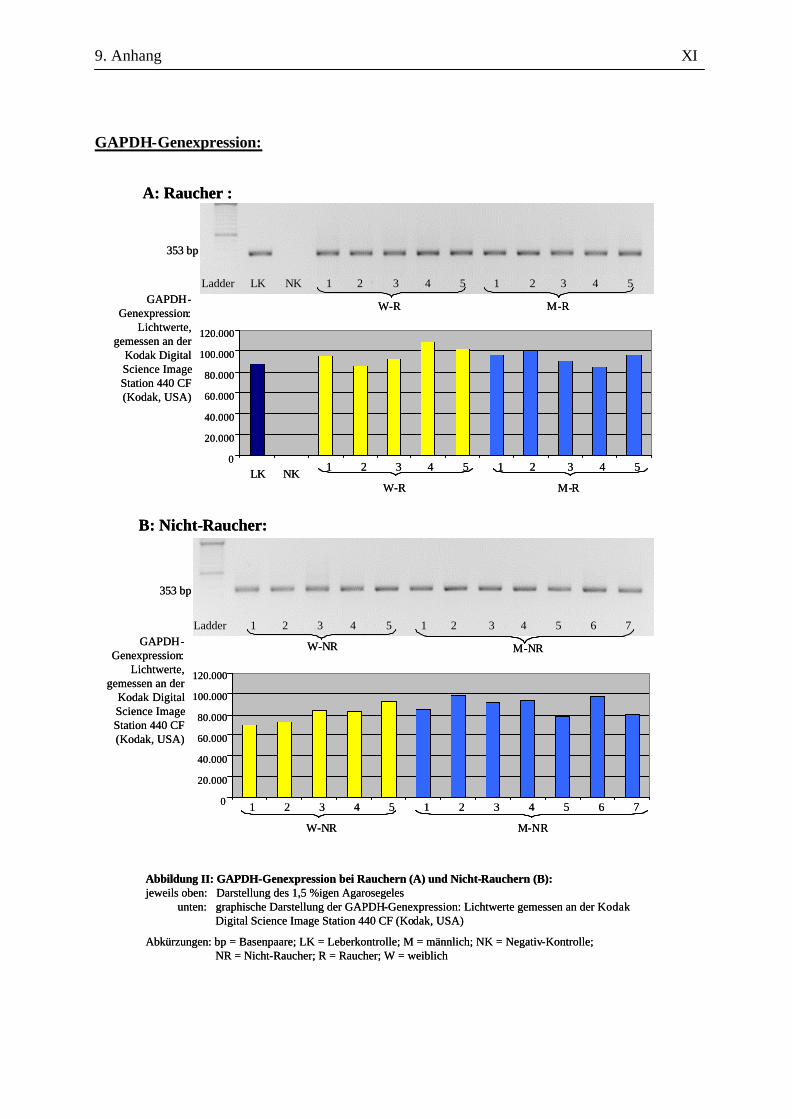
9. Anhang XI
GAPDH-Genexpression:
353 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung II: GAPDH-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der GAPDH-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
353 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
GAPDH-Genexpression:
Lichtwerte, gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
GAPDH-Genexpression:
Lichtwerte, gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
353 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung II: GAPDH-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der GAPDH-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
353 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
GAPDH-Genexpression:
Lichtwerte, gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
GAPDH-Genexpression:
Lichtwerte, gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XII
CYP 1A1-Genexpression:
432 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung III: CYP 1A1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 1A1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
432 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
A: Raucher :
B: Nicht-Raucher:
CYP 1A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 1A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
0
10.000
20.000
30.000
40.000
50.000
60.000
70.000
10.000
0
20.000
30.000
40.000
50.000
60.000
70.000
432 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung III: CYP 1A1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 1A1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
432 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
A: Raucher :
B: Nicht-Raucher:
CYP 1A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 1A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
0
10.000
20.000
30.000
40.000
50.000
60.000
70.000
10.000
0
20.000
30.000
40.000
50.000
60.000
70.000

9. Anhang XIII
CYP 1B1-Genexpression:
301 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung IV: CYP 1B1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 1B1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
301 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 1B1-Genexpression:
Lichtwerte, gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 1B1-Genexpression:
Lichtwerte, gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
301 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung IV: CYP 1B1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 1B1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
301 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 1B1-Genexpression:
Lichtwerte, gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 1B1-Genexpression:
Lichtwerte, gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
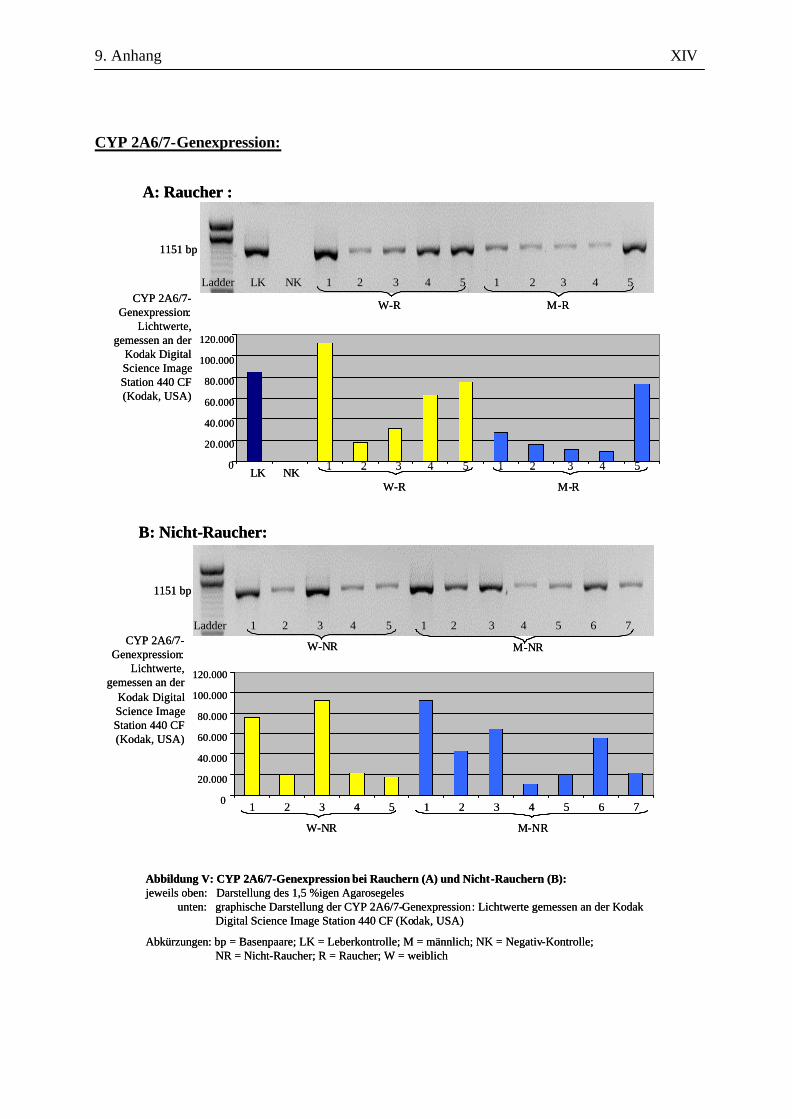
9. Anhang XIV
CYP 2A6/7-Genexpression:
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
1151 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung V: CYP 2A6/7-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2A6/7-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
1151 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
A: Raucher :
B: Nicht-Raucher:
CYP 2A6/7-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2A6/7-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
0
20.000
40.000
60.000
80.000
100.000
120.000
0
20.000
40.000
60.000
80.000
100.000
120.000
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
1151 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung V: CYP 2A6/7-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2A6/7-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
1151 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
A: Raucher :
B: Nicht-Raucher:
CYP 2A6/7-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2A6/7-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
0
20.000
40.000
60.000
80.000
100.000
120.000
0
20.000
40.000
60.000
80.000
100.000
120.000

9. Anhang XV
CYP 2A13-Genexpression:
320 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung VI: CYP 2A13-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2A13-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
320 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
0
0
A: Raucher :
B: Nicht-Raucher:
CYP 2A13-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2A13-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
20.000
40.000
60.000
80.000
100.000
120.000
140.000
20.000
40.000
60.000
80.000
100.000
120.000
140.000
320 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung VI: CYP 2A13-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2A13-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
320 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
0
0
A: Raucher :
B: Nicht-Raucher:
CYP 2A13-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2A13-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
20.000
40.000
60.000
80.000
100.000
120.000
140.000
20.000
40.000
60.000
80.000
100.000
120.000
140.000
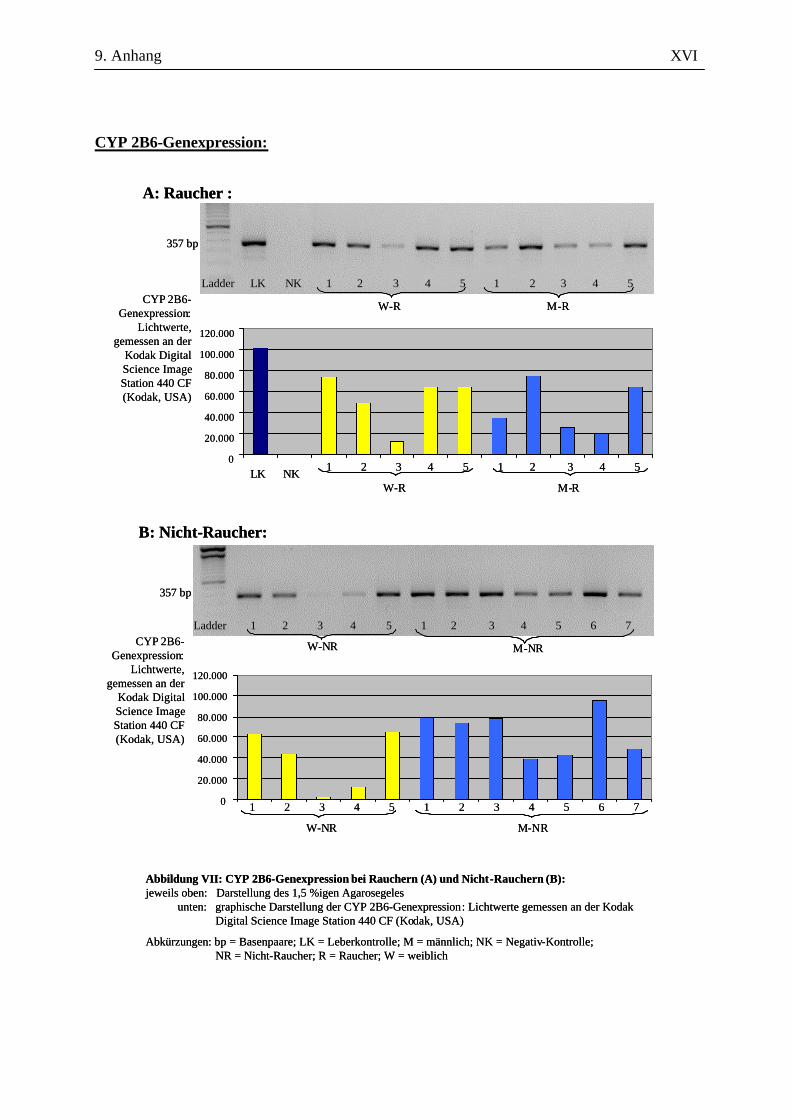
9. Anhang XVI
CYP 2B6-Genexpression:
357 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung VII: CYP 2B6-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2B6-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
357 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2B6-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2B6-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
357 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung VII: CYP 2B6-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2B6-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
357 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2B6-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2B6-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XVII
CYP 2C8-Genexpression:
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
311 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung VIII: CYP 2C8-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2C8-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
311 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2C8-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2C8-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
311 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung VIII: CYP 2C8-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2C8-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
311 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2C8-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2C8-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XVIII
CYP 2C9-Genexpression:
437 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung IX: CYP 2C9-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2C9-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
437 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2C9-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2C9-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
437 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung IX: CYP 2C9-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2C9-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
437 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2C9-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2C9-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
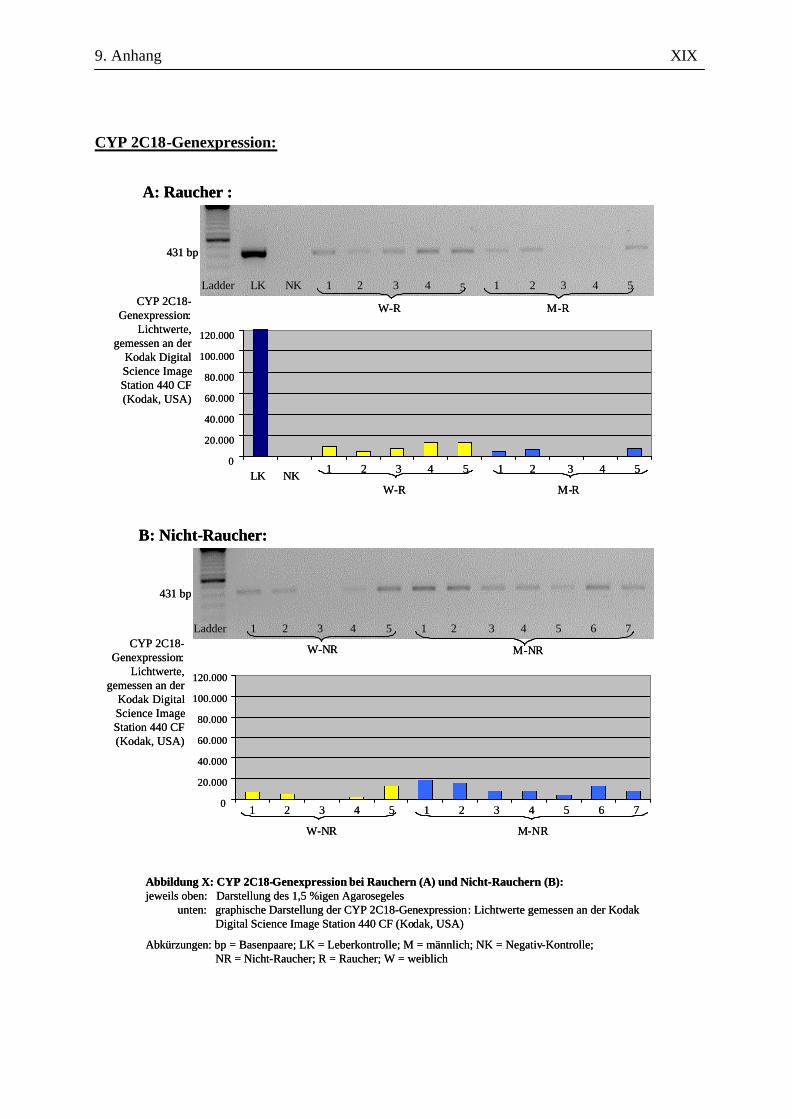
9. Anhang XIX
CYP 2C18-Genexpression:
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
431 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung X: CYP 2C18-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2C18-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
431 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2C18-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2C18-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
431 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung X: CYP 2C18-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2C18-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
431 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2C18-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2C18-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
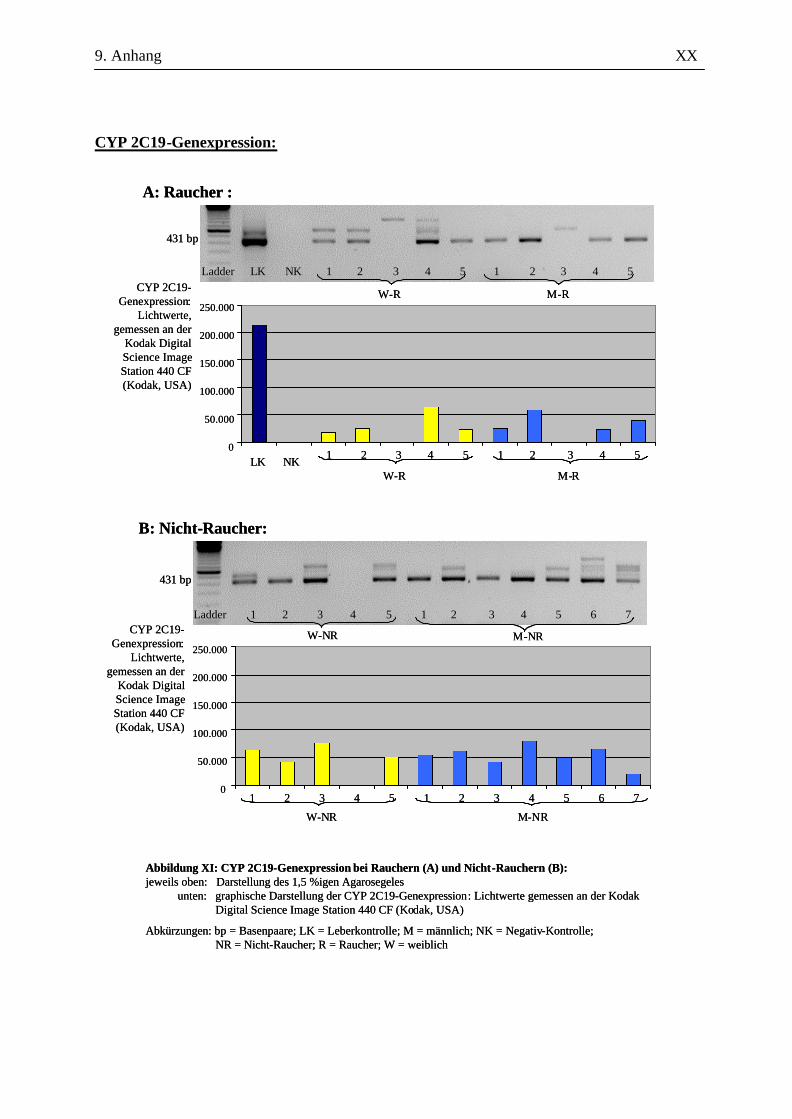
9. Anhang XX
CYP 2C19-Genexpression:
431 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XI: CYP 2C19-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2C19-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
431 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2C19-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2C19-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
431 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XI: CYP 2C19-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2C19-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
431 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2C19-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2C19-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XXI
CYP 2E1-Genexpression:
365 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XII: CYP 2E1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2E1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
365 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2E1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2E1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
365 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XII: CYP 2E1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2E1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
365 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2E1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2E1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XXII
CYP 2F1-Genexpression:
283 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XIII: CYP 2F1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2F1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
283 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
A: Raucher :
B: Nicht-Raucher:
CYP 2F1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2F1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
0
20.000
60.000
40.000
80.000
100.000
120.000
0
20.000
40.000
60.000
80.000
100.000
120.000
283 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XIII: CYP 2F1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2F1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
283 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
A: Raucher :
B: Nicht-Raucher:
CYP 2F1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2F1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
0
20.000
60.000
40.000
80.000
100.000
120.000
0
20.000
40.000
60.000
80.000
100.000
120.000

9. Anhang XXIII
CYP 2J2-Genexpression:
389 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XIV: CYP 2J2-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2J2-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
389 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
40.000
30.000
20.000
10.000
0
40.000
30.000
20.000
10.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2J2-Genexpression:L
ichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2J2-Genexpression:L
ichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
50.000
60.000
70.000
50.000
60.000
70.000
389 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XIV: CYP 2J2-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2J2-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
389 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
40.000
30.000
20.000
10.000
0
40.000
30.000
20.000
10.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2J2-Genexpression:L
ichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2J2-Genexpression:L
ichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
50.000
60.000
70.000
50.000
60.000
70.000
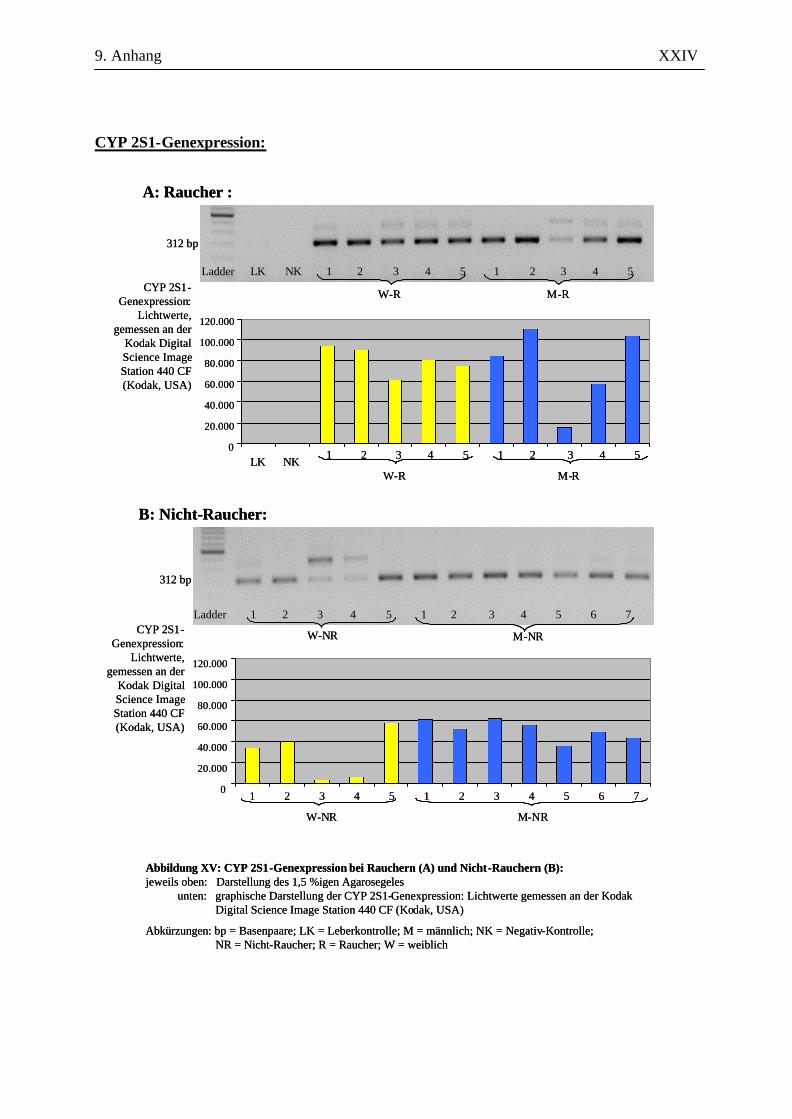
9. Anhang XXIV
CYP 2S1-Genexpression:
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
312 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XV: CYP 2S1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2S1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
312 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2S1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2S1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
312 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XV: CYP 2S1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 2S1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
312 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 2S1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 2S1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XXV
CYP 3A4-Genexpression:
324 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XVI: CYP 3A4-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 3A4-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
324 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 3A4-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 3A4-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
324 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XVI: CYP 3A4-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 3A4-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
324 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 3A4-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 3A4-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XXVI
CYP 3A5-Genexpression:
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
472 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XVII: CYP 3A5-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 3A5-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
472 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
80.000
60.000
40.000
20.000
0
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 3A5-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 3A5-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
472 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XVII: CYP 3A5-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 3A5-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
472 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
80.000
60.000
40.000
20.000
0
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 3A5-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 3A5-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XXVII
CYP 4B1-Genexpression:
397 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XVIII: CYP 4B1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 4B1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
397 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 4B1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 4B1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
397 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XVIII: CYP 4B1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CYP 4B1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
397 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
CYP 4B1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CYP 4B1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
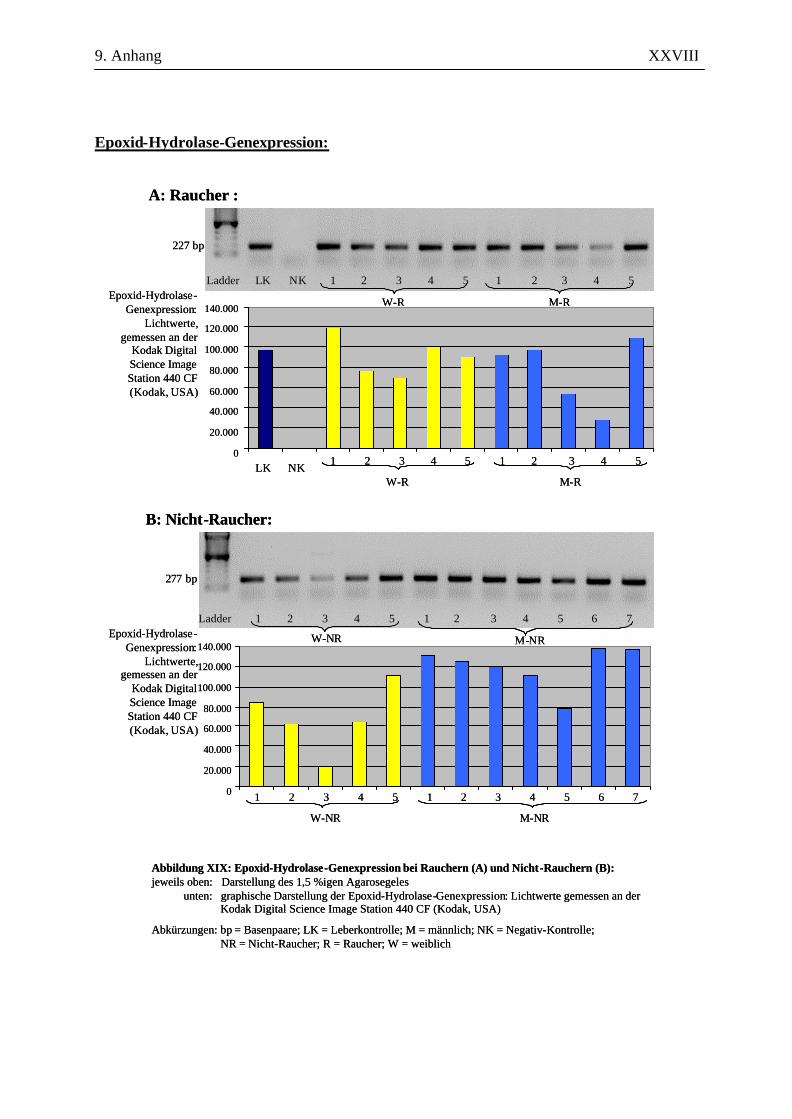
9. Anhang XXVIII
Epoxid-Hydrolase-Genexpression:
227 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XIX: Epoxid-Hydrolase-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der Epoxid-Hydrolase-Genexpression: Lichtwerte gemessen an derKodak Digital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
277 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
0
0
A: Raucher :
B: Nicht-Raucher:
Epoxid-Hydrolase-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
Epoxid-Hydrolase-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
20.000
40.000
60.000
80.000
100.000
120.000
140.000
20.000
40.000
60.000
80.000
100.000
120.000
140.000
227 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XIX: Epoxid-Hydrolase-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der Epoxid-Hydrolase-Genexpression: Lichtwerte gemessen an derKodak Digital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
277 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
0
0
A: Raucher :
B: Nicht-Raucher:
Epoxid-Hydrolase-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
Epoxid-Hydrolase-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
20.000
40.000
60.000
80.000
100.000
120.000
140.000
20.000
40.000
60.000
80.000
100.000
120.000
140.000

9. Anhang XXIX
FMO 2-Genexpression:
250 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XX: FMO2-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der FMO2-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
250 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
100.000
80.000
60.000
20.000
0
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
FMO2-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
FMO2-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA) 40.000
250 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XX: FMO2-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der FMO2-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
250 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
100.000
80.000
60.000
20.000
0
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
FMO2-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
FMO2-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA) 40.000

9. Anhang XXX
FMO 3-Genexpression:
301 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXI: FMO3-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der FMO3-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
301 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
80.000
40.000
0
120.000
80.000
40.000
0
A: Raucher :
B: Nicht-Raucher:
FMO3-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
FMO3-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
160.000
160.000
301 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXI: FMO3-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der FMO3-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
301 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
80.000
40.000
0
120.000
80.000
40.000
0
A: Raucher :
B: Nicht-Raucher:
FMO3-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
FMO3-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
160.000
160.000

9. Anhang XXXI
FMO 5-Genexpression:
500 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXII: FMO5-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der FMO5-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
500 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
A: Raucher :
B: Nicht-Raucher:
FMO5-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
FMO5-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
0
20.000
40.000
60.000
80.000
100.000
120.000
140.000
0
20.000
40.000
60.000
80.000
100.000
120.000
140.000
500 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXII: FMO5-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der FMO5-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
500 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
A: Raucher :
B: Nicht-Raucher:
FMO5-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
FMO5-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
0
20.000
40.000
60.000
80.000
100.000
120.000
140.000
0
20.000
40.000
60.000
80.000
100.000
120.000
140.000

9. Anhang XXXII
GST α2-Genexpression:
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
354 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXIII: GST αα 2-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der GST αα 2-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
354 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
GST αα 2-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
GST αα2-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
354 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXIII: GST αα 2-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der GST αα 2-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
354 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
250.000
200.000
150.000
100.000
50.000
0
250.000
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
GST αα 2-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
GST αα2-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
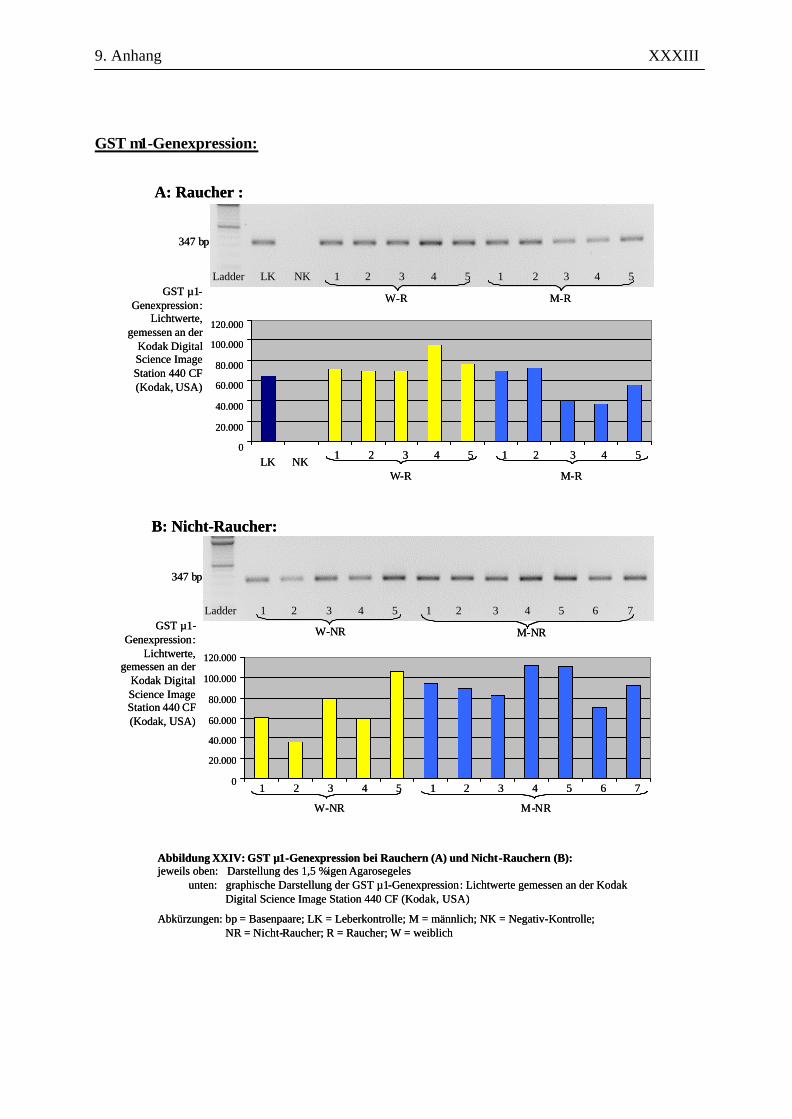
9. Anhang XXXIII
GST µ1-Genexpression:
347 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXIV: GST µ1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der GST µ1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
347 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
0
0
A: Raucher :
B: Nicht-Raucher:
GST µ1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
GST µ1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
20.000
40.000
60.000
80.000
100.000
120.000
20.000
40.000
60.000
80.000
100.000
120.000
347 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXIV: GST µ1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der GST µ1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
347 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
0
0
A: Raucher :
B: Nicht-Raucher:
GST µ1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
GST µ1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
20.000
40.000
60.000
80.000
100.000
120.000
20.000
40.000
60.000
80.000
100.000
120.000
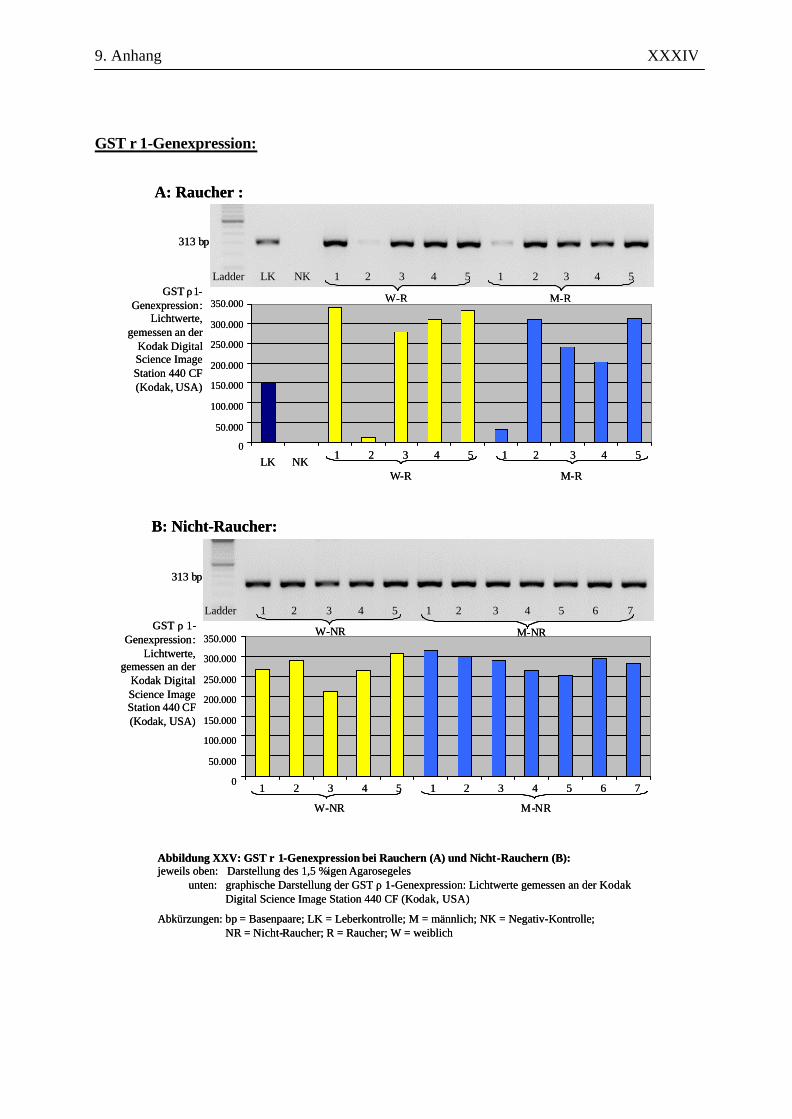
9. Anhang XXXIV
GST ρ1-Genexpression:
313 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXV: GST ρ 1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der GST ρ 1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
313 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
0
A: Raucher :
B: Nicht-Raucher:
GST ρ 1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
GST ρ1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
50.000
100.000
150.000
200.000
250.000
300.000
350.000
0
50.000
100.000
150.000
200.000
250.000
300.000
350.000
313 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXV: GST ρ 1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der GST ρ 1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
313 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
0
A: Raucher :
B: Nicht-Raucher:
GST ρ 1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
GST ρ1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
50.000
100.000
150.000
200.000
250.000
300.000
350.000
0
50.000
100.000
150.000
200.000
250.000
300.000
350.000

9. Anhang XXXV
UGT 1A1-Genexpression:
350 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXVI: UGT 1A1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der UGT 1A1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
350 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
300.000
250.000
200.000
0
350.000
300.000
250.000
200.000
0
A: Raucher :
B: Nicht-Raucher:
UGT 1A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
UGT 1A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
350.000
50.000
100.000
150.000
50.000
100.000
150.000
350 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXVI: UGT 1A1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der UGT 1A1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
350 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
300.000
250.000
200.000
0
350.000
300.000
250.000
200.000
0
A: Raucher :
B: Nicht-Raucher:
UGT 1A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
UGT 1A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
350.000
50.000
100.000
150.000
50.000
100.000
150.000

9. Anhang XXXVI
UGT 2A1-Genexpression:
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
297 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXVII: UGT 2A1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der UGT 2A1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
297 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
200.000
150.000
100.000
50.000
0
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
UGT 2A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
UGT 2A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
297 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXVII: UGT 2A1-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der UGT 2A1-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
297 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
200.000
150.000
100.000
50.000
0
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
UGT 2A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
UGT 2A1-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XXXVII
AhR-Genexpression:
284 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXVIII: AhR-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der AhR-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
284 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
AhR-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
AhR-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
284 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXVIII: AhR-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der AhR-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
284 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
AhR-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
AhR-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XXXVIII
CAR β -Genexpression:
350 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXIX: CAR ß -Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CAR ß-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
350 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
100.000
75.000
50.000
25.000
0
100.000
175.000
50.000
25.000
0
A: Raucher :
B: Nicht-Raucher:
CAR ß-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CAR ß-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
125.000
150.000
125.000
150.000
350 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXIX: CAR ß -Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der CAR ß-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
350 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
100.000
75.000
50.000
25.000
0
100.000
175.000
50.000
25.000
0
A: Raucher :
B: Nicht-Raucher:
CAR ß-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
CAR ß-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
125.000
150.000
125.000
150.000

9. Anhang XXXIX
LXR α -Genexpression:
351 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXX: LXR α -Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der LXR α -Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
351 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
70.000
60.000
50.000
40.000
30.000
20.000
10.000
0
70.000
60.000
50.000
40.000
30.000
20.000
10.000
0
A: Raucher :
B: Nicht-Raucher:
LXR α -Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
LXR α-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
351 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXX: LXR α -Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der LXR α -Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
351 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
70.000
60.000
50.000
40.000
30.000
20.000
10.000
0
70.000
60.000
50.000
40.000
30.000
20.000
10.000
0
A: Raucher :
B: Nicht-Raucher:
LXR α -Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
LXR α-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XL
PXR-Genexpression:
351 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXI: PXR-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der PXR-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
351 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
0
0
A: Raucher :
B: Nicht-Raucher:
PXR-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
PXR-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
20.000
40.000
60.000
80.000
100.000
120.000
140.000
20.000
40.000
60.000
80.000
100.000
120.000
140.000
351 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXI: PXR-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der PXR-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
351 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
0
0
A: Raucher :
B: Nicht-Raucher:
PXR-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
PXR-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
20.000
40.000
60.000
80.000
100.000
120.000
140.000
20.000
40.000
60.000
80.000
100.000
120.000
140.000
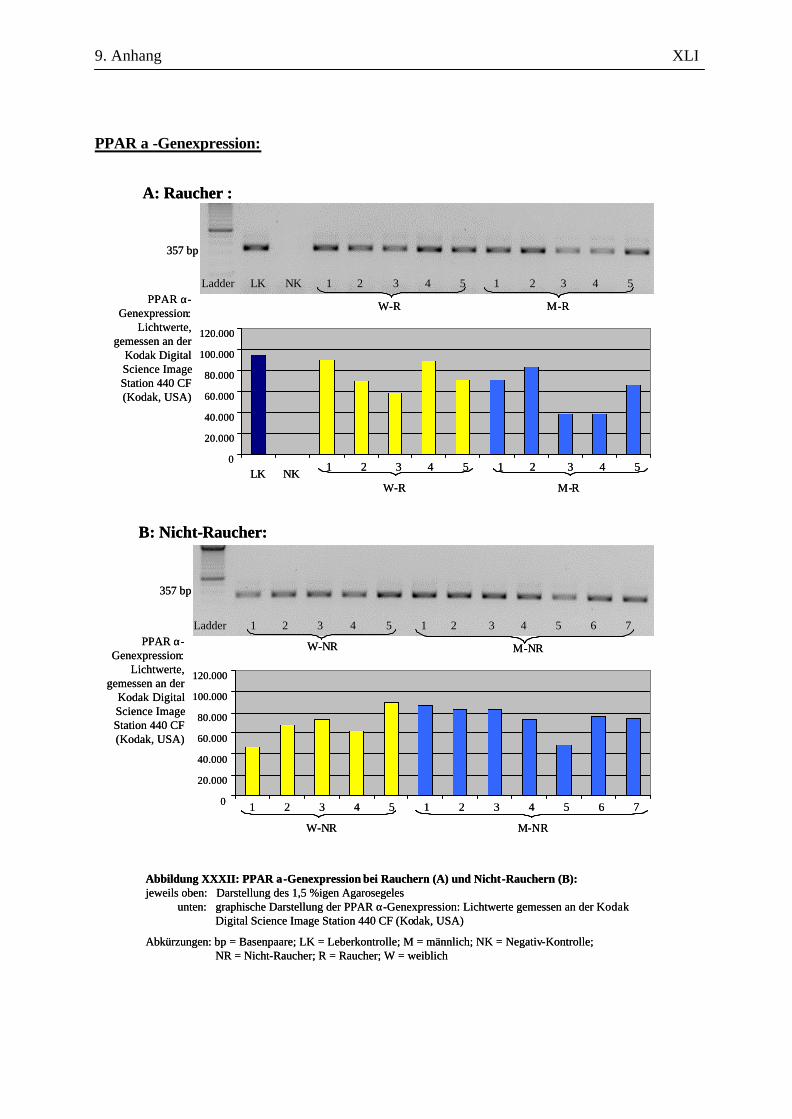
9. Anhang XLI
PPAR α -Genexpression:
357 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXII: PPAR α-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der PPAR α-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
357 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
PPAR α-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
PPAR α-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
357 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXII: PPAR α-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der PPAR α-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
357 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
PPAR α-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
PPAR α-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
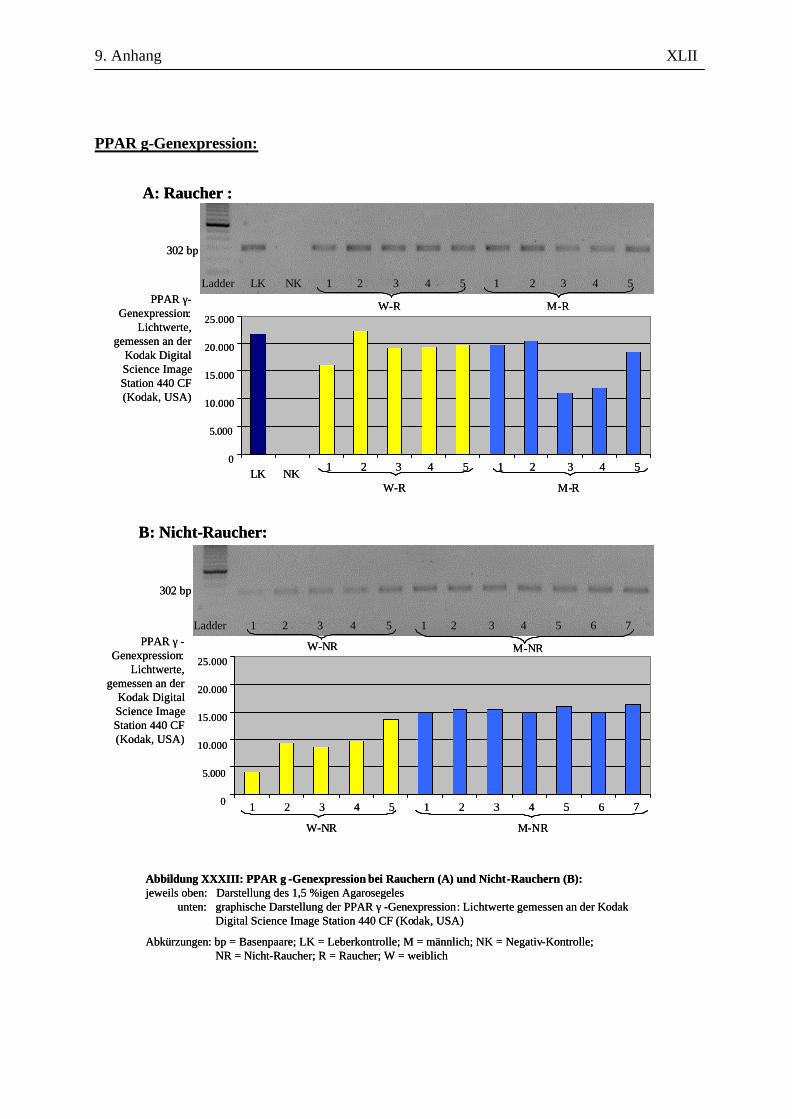
9. Anhang XLII
PPAR γ-Genexpression:
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
302 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXIII: PPAR γ -Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der PPAR γ -Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
302 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
25.000
20.000
15.000
10.000
5.000
0
25.000
20.000
15.000
10.000
5.000
0
A: Raucher :
B: Nicht-Raucher:
PPAR γ -Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
PPAR γ-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
302 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXIII: PPAR γ -Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der PPAR γ -Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
302 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
25.000
20.000
15.000
10.000
5.000
0
25.000
20.000
15.000
10.000
5.000
0
A: Raucher :
B: Nicht-Raucher:
PPAR γ -Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
PPAR γ-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XLIII
IL-1β -Genexpression:
294 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXIV: IL-1β -Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der IL-1β -Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
294 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
60.000
50.000
40.000
30.000
20.000
10.000
0
60.000
50.000
40.000
30.000
20.000
10.000
0
A: Raucher :
B: Nicht-Raucher:
IL-1β -Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
IL-1β-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
294 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXIV: IL-1β -Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der IL-1β -Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
294 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
60.000
50.000
40.000
30.000
20.000
10.000
0
60.000
50.000
40.000
30.000
20.000
10.000
0
A: Raucher :
B: Nicht-Raucher:
IL-1β -Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
IL-1β-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
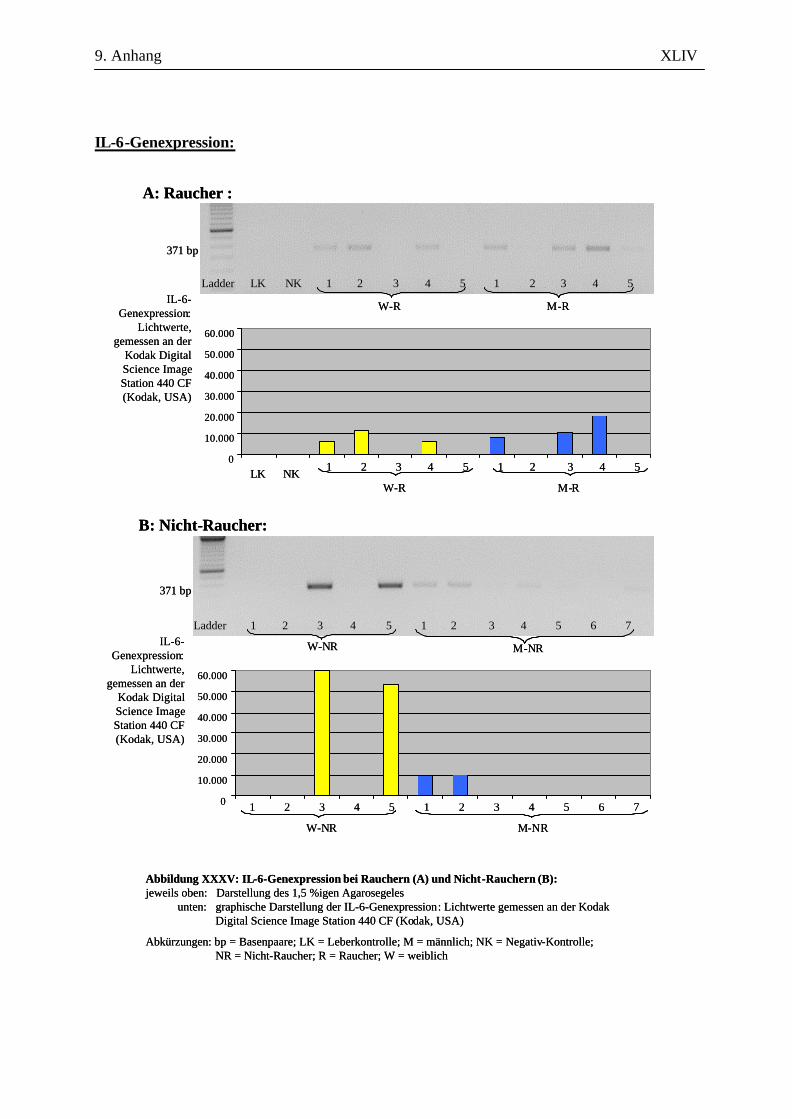
9. Anhang XLIV
IL-6-Genexpression:
371 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXV: IL-6-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der IL-6-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
371 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
60.000
50.000
40.000
30.000
20.000
10.000
0
60.000
50.000
40.000
30.000
20.000
10.000
0
A: Raucher :
B: Nicht-Raucher:
IL-6-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
IL-6-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
371 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXV: IL-6-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der IL-6-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
371 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
60.000
50.000
40.000
30.000
20.000
10.000
0
60.000
50.000
40.000
30.000
20.000
10.000
0
A: Raucher :
B: Nicht-Raucher:
IL-6-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
IL-6-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XLV
IL-8-Genexpression:
391 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXVI: IL-8-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der IL-8-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
391 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
200.000
150.000
100.000
50.000
0
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
IL-8-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
IL-8-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
391 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXVI: IL-8-Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der IL-8-Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
391 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
200.000
150.000
100.000
50.000
0
200.000
150.000
100.000
50.000
0
A: Raucher :
B: Nicht-Raucher:
IL-8-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
IL-8-Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XLVI
TNF α -Genexpression:
599 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXVII: TNFα -Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der TNFα -Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
599 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
TNFα -Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
TNFα-Genexpression:L
ichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
599 bp
Ladder LK NK 1 12 4 53 2 3 4 5
Abbildung XXXVII: TNFα -Genexpression bei Rauchern (A) und Nicht-Rauchern (B):jeweils oben: Darstellung des 1,5 %igen Agarosegeles
unten: graphische Darstellung der TNFα -Genexpression: Lichtwerte gemessen an der KodakDigital Science Image Station 440 CF (Kodak, USA)
Abkürzungen: bp = Basenpaare; LK = Leberkontrolle; M = männlich; NK = Negativ-Kontrolle; NR = Nicht-Raucher; R = Raucher; W = weiblich
LK NKW-R M-R
Ladder
599 bp
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
W-R M-R
1 2 3 4 5 1 2 3 4 5
1 2 3 4 5 1 2 3 4 5 6 7
W-NR M-NR
120.000
100.000
80.000
60.000
40.000
20.000
0
120.000
100.000
80.000
60.000
40.000
20.000
0
A: Raucher :
B: Nicht-Raucher:
TNFα -Genexpression:
Lichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)
TNFα-Genexpression:L
ichtwerte,gemessen an der
Kodak Digital Science Image Station 440 CF (Kodak, USA)

9. Anhang XLVII
9.6 Darstellung der Western blots CYP 1A1
Abbildung XL: CYP 1A1-Proteinexpression mittels Western blot: Messung der Lumineszenz nach 10 min Expositionsdauer an der Kodak Digital Science Image Station 440 CF (Kodak, USA).
Abkürzungen: M = männlich; NR = Nicht-Raucher; PK = Positiv-Kontrolle; R = Raucher; W = weiblich CYP 2A
Abbildung XLI: CYP 2A-Proteinexpression mittels Western blot: Messung der Lumineszenz nach 10 min Expositionsdauer an der Kodak Digital Science Image Station 440 CF (Kodak, USA).
Abkürzungen: M = männlich; NR = Nicht-Raucher; PK = Positiv-Kontrolle; R = Raucher; W = weiblich
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
25 kDa
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
25 kDa

9. Anhang XLVIII
CYP 2B6
Abbildung XLII: CYP 2B6-Proteinexpression mittels Western blot: Messung der Lumineszenz nach 10 min Expositionsdauer an der Kodak Digital Science Image Station 440 CF (Kodak, USA).
Abkürzungen: M = männlich; NR = Nicht-Raucher; PK = Positiv-Kontrolle; R = Raucher; W = weiblich CYP 2E
Abbildung XLIII: CYP 2E-Proteinexpression mittels Western blot: Messung der Lumineszenz nach 10 min Expositionsdauer an der Kodak Digital Science Image Station 440 CF (Kodak, USA).
Abkürzungen: M = männlich; NR = Nicht-Raucher; PK = Positiv-Kontrolle; R = Raucher; W = weiblich
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa
75 kDa
50 kDa
37 kDa
25 kDa
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa
75 kDa
50 kDa
37 kDa
25 kDa
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
25 kDa
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
25 kDa

9. Anhang XLIX
CYP 3A4
Abbildung XLIV: CYP 3A4-Proteinexpression mittels Western blot: Messung der Lumineszenz nach 10 min Expositionsdauer an der Kodak Digital Science Image Station 440 CF (Kodak, USA).
Abkürzungen: M = männlich; NR = Nicht-Raucher; PK = Positiv-Kontrolle; R = Raucher; W = weiblich CYP 3A5
Abbildung XLV: CYP 3A5-Proteinexpression mittels Western blot: Messung der Lumineszenz nach 10 min Expositionsdauer an der Kodak Digital Science Image Station 440 CF (Kodak, USA).
Abkürzungen: M = männlich; NR = Nicht-Raucher; PK = Positiv-Kontrolle; R = Raucher; W = weiblich
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
25 kDa
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
25 kDa
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
25 kDa
Ladder PKPool 8(W-R)
Pool 7(M-R)
Pool 10(M-R)
Pool 6(W-NR)
Pool 11(W-NR)
Pool 1(M-NR)
Pool 4(M-NR)
Pool 9(M-NR)
100 kDa75 kDa
50 kDa
37 kDa
25 kDa

9. Anhang L
9.7 HPLC-Chromatogramme von Loteprednol-Etabonat (LE) und seinen Metaboliten nach Inkubation menschlicher Nasenmuschelmikrosomen 9.7.1 1.Versuchsabschnitt ( Inkubation bei verschiedenen Inkubationszeiten ) Serie 1
Negativkontrolle 1 ( Puffer + Substrat ) Inkubation 60 min
Negativkontrolle 2 ( Puffer + Substrat )Inkubation 120 min
Positivkontrolle ( Rattenlebermikrosomen ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 1 ( M-NR ) Inkubation 60 min
Negativkontrolle 1 ( Puffer + Substrat ) Inkubation 60 min
Negativkontrolle 2 ( Puffer + Substrat )Inkubation 120 min
Positivkontrolle ( Rattenlebermikrosomen ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 1 ( M-NR ) Inkubation 60 min
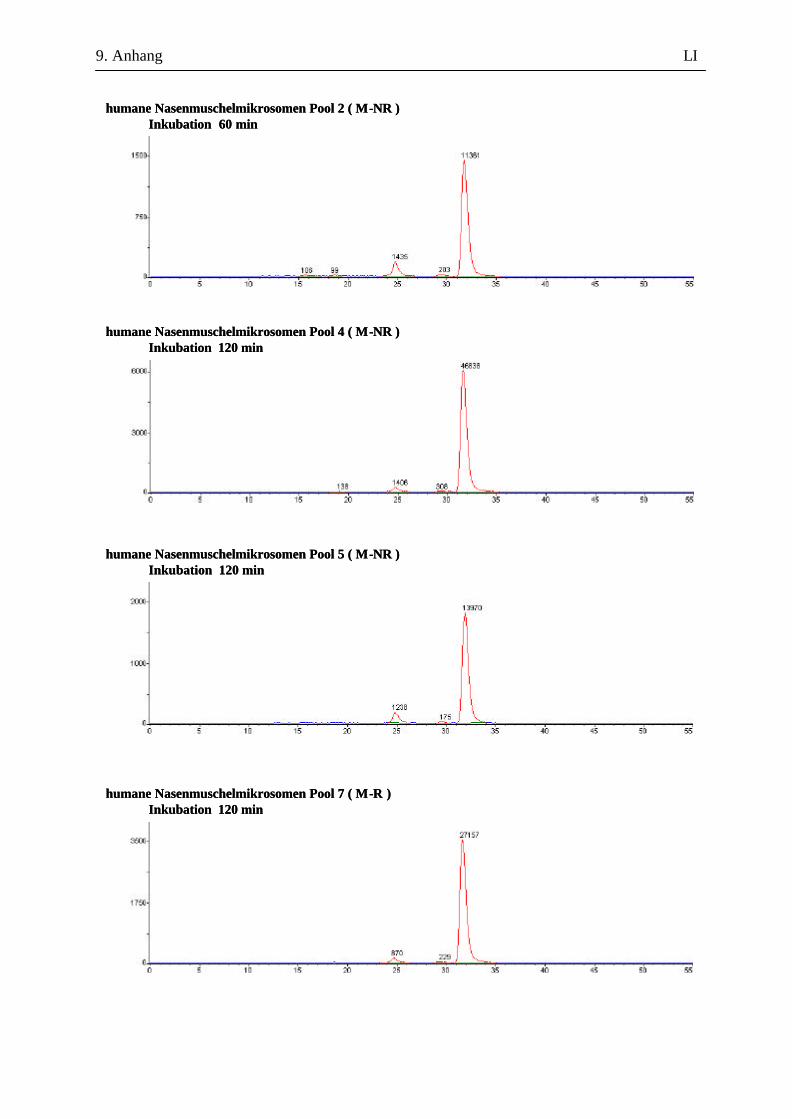
9. Anhang LI
humane Nasenmuschelmikrosomen Pool 2 ( M-NR ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 4 ( M-NR ) Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 5 ( M-NR ) Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 7 ( M-R ) Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 2 ( M-NR ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 4 ( M-NR ) Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 5 ( M-NR ) Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 7 ( M-R ) Inkubation 120 min

9. Anhang LII
9.7.2 1.Versuchsabschnitt ( Inkubation bei verschiedenen Inkubationszeiten ) Serie 2
Negativkontrolle ( Puffer + Substrat ) Inkubation 60 min
Positivkontrolle ( humane Lebermikrosomen )Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 4 ( M-NR ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 5 ( M-NR ) Inkubation 60 min
Negativkontrolle ( Puffer + Substrat ) Inkubation 60 min
Positivkontrolle ( humane Lebermikrosomen )Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 4 ( M-NR ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 5 ( M-NR ) Inkubation 60 min

9. Anhang LIII
humane Nasenmuschelmikrosomen Pool 6 ( W-NR ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 7 ( M-R )Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 8 ( W-R ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 9 ( M-NR ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 6 ( W-NR ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 7 ( M-R )Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 8 ( W-R ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 9 ( M-NR ) Inkubation 60 min

9. Anhang LIV
humane Nasenmuschelmikrosomen Pool 10 ( M-R ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 11 ( W-NR )Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 10 ( M-R ) Inkubation 60 min
humane Nasenmuschelmikrosomen Pool 11 ( W-NR )Inkubation 60 min

9. Anhang LV
Negativkontrolle ( Puffer + Substrat ) Inkubation 120 min
Positivkontrolle ( humane Lebermikrosomen )Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 2 ( M-NR ) Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 9 ( M-NR ) Inkubation 120 min
Negativkontrolle ( Puffer + Substrat ) Inkubation 120 min
Positivkontrolle ( humane Lebermikrosomen )Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 2 ( M-NR ) Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 9 ( M-NR ) Inkubation 120 min

9. Anhang LVI
humane Nasenmuschelmikrosomen Pool 10 ( M-R ) Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 11 ( W-NR )Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 10 ( M-R ) Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 11 ( W-NR )Inkubation 120 min

9. Anhang LVII
9.7.3 2.Versuchsabschnitt ( Inkubation mit und ohne Glucuronidase-Zusatz )
Negativkontrolle ( Puffer + Substrat ) mit Glucuronidase-ZusatzInkubation 120 min
Positivkontrolle ( humane Lebermikrosomen ) mit Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 12 ( M-NR ) mit Glucuronidase-Zusatz Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 13 ( M-R ) mit Glucuronidase-Zusatz Inkubation 120 min
Negativkontrolle ( Puffer + Substrat ) mit Glucuronidase-ZusatzInkubation 120 min
Positivkontrolle ( humane Lebermikrosomen ) mit Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 12 ( M-NR ) mit Glucuronidase-Zusatz Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 13 ( M-R ) mit Glucuronidase-Zusatz Inkubation 120 min
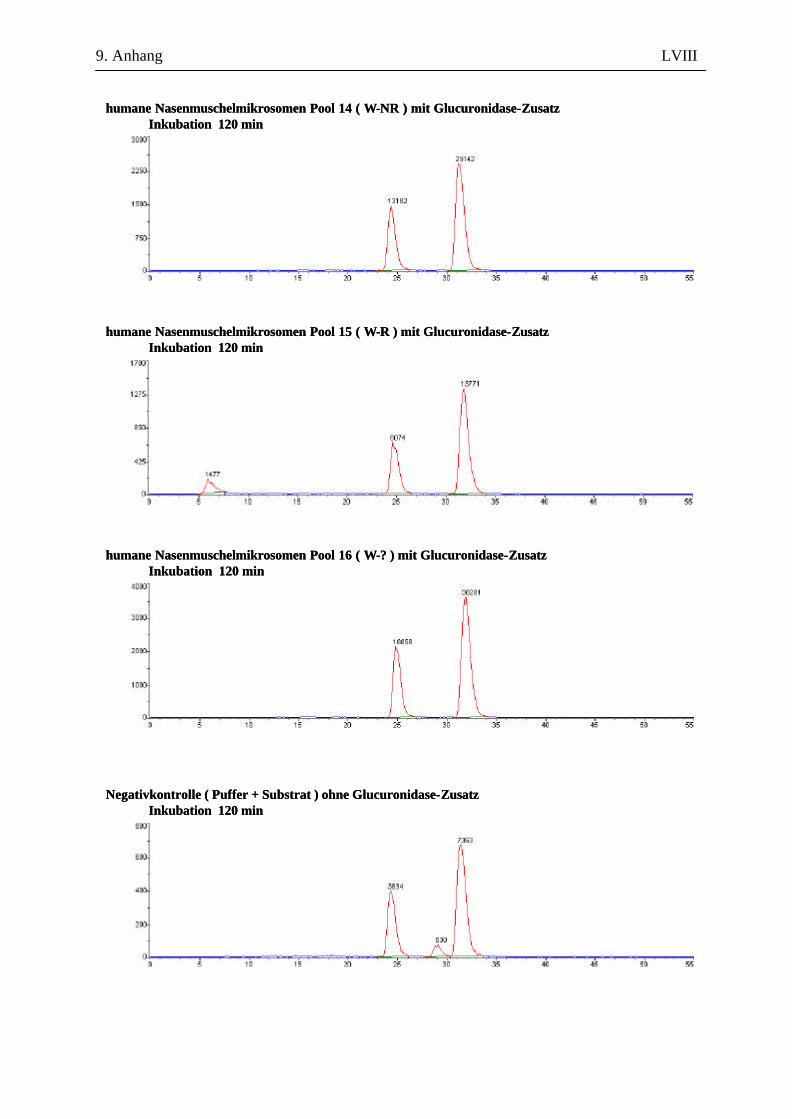
9. Anhang LVIII
humane Nasenmuschelmikrosomen Pool 14 ( W-NR ) mit Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 15 ( W-R ) mit Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 16 ( W-? ) mit Glucuronidase-Zusatz Inkubation 120 min
Negativkontrolle ( Puffer + Substrat ) ohne Glucuronidase-Zusatz Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 14 ( W-NR ) mit Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 15 ( W-R ) mit Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 16 ( W-? ) mit Glucuronidase-Zusatz Inkubation 120 min
Negativkontrolle ( Puffer + Substrat ) ohne Glucuronidase-Zusatz Inkubation 120 min
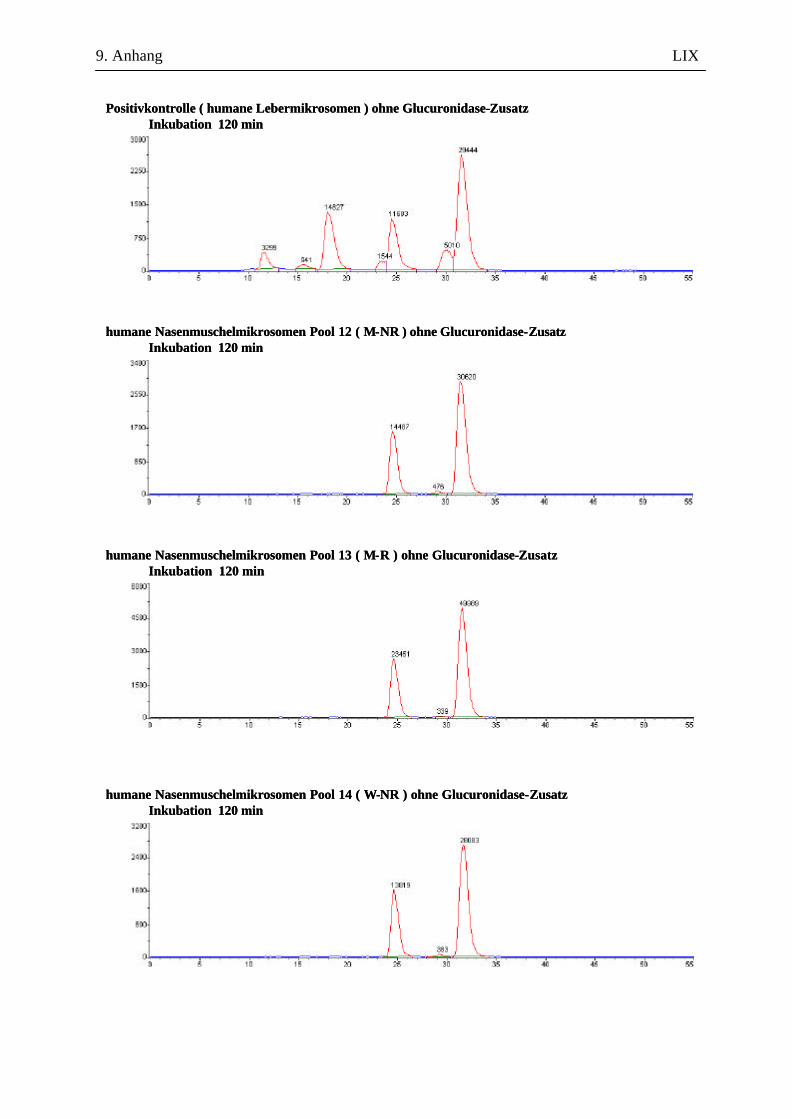
9. Anhang LIX
Positivkontrolle ( humane Lebermikrosomen ) ohne Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 12 ( M-NR ) ohne Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 13 ( M-R ) ohne Glucuronidase-Zusatz Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 14 ( W-NR ) ohne Glucuronidase-Zusatz Inkubation 120 min
Positivkontrolle ( humane Lebermikrosomen ) ohne Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 12 ( M-NR ) ohne Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 13 ( M-R ) ohne Glucuronidase-Zusatz Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 14 ( W-NR ) ohne Glucuronidase-Zusatz Inkubation 120 min

9. Anhang LX
humane Nasenmuschelmikrosomen Pool 15 ( W-R ) ohne Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 16 ( W-? ) ohne Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 15 ( W-R ) ohne Glucuronidase-ZusatzInkubation 120 min
humane Nasenmuschelmikrosomen Pool 16 ( W-? ) ohne Glucuronidase-ZusatzInkubation 120 min

9. Anhang LXI
9.8 HPLC-Chromatogramme von Testosteron und seinen Metaboliten nach Inkubation menschlicher Nasenmuschelmikrosomen
Negativkontrolle ( Puffer + Substrat )Inkubation 120 min
Positivkontrolle ( humane Lebermikrosomen )Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 1 ( M-NR )Inkubation 120 min
Negativkontrolle ( Puffer + Substrat )Inkubation 120 min
Positivkontrolle ( humane Lebermikrosomen )Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 1 ( M-NR )Inkubation 120 min

9. Anhang LXII
humane Nasenmuschelmikrosomen Pool 4 ( M -NR )Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 6 ( W-NR )Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 9 ( M -NR )Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 4 ( M -NR )Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 6 ( W-NR )Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 9 ( M -NR )Inkubation 120 min

9. Anhang LXIII
humane Nasenmuschelmikrosomen Pool 10 ( M-R )Inkubation 120 min
humane Nasenmuschelmikrosomen Pool 10 ( M-R )Inkubation 120 min