Embed Size (px)
Citation preview
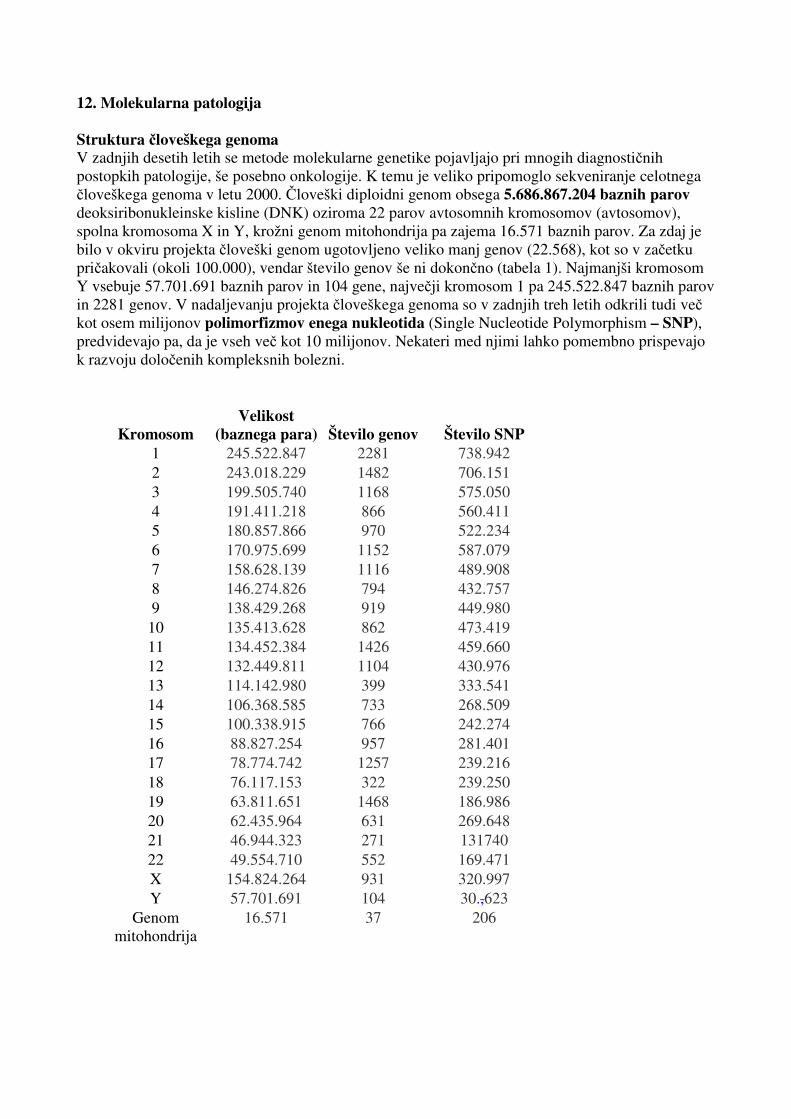
12. Molekularna patologija Struktura človeškega genoma V zadnjih desetih letih se metode molekularne genetike pojavljajo pri mnogih diagnostičnih postopkih patologije, še posebno onkologije. K temu je veliko pripomoglo sekveniranje celotnega človeškega genoma v letu 2000. Človeški diploidni genom obsega 5.686.867.204 baznih parov deoksiribonukleinske kisline (DNK) oziroma 22 parov avtosomnih kromosomov (avtosomov), spolna kromosoma X in Y, krožni genom mitohondrija pa zajema 16.571 baznih parov. Za zdaj je bilo v okviru projekta človeški genom ugotovljeno veliko manj genov (22.568), kot so v začetku pričakovali (okoli 100.000), vendar število genov še ni dokončno (tabela 1). Najmanjši kromosom Y vsebuje 57.701.691 baznih parov in 104 gene, največji kromosom 1 pa 245.522.847 baznih parov in 2281 genov. V nadaljevanju projekta človeškega genoma so v zadnjih treh letih odkrili tudi več kot osem milijonov polimorfizmov enega nukleotida (Single Nucleotide Polymorphism – SNP), predvidevajo pa, da je vseh več kot 10 milijonov. Nekateri med njimi lahko pomembno prispevajo k razvoju določenih kompleksnih bolezni.
Kromosom Velikost
(baznega para) Število genov Število SNP 1 245.522.847 2281 738.942 2 243.018.229 1482 706.151 3 199.505.740 1168 575.050 4 191.411.218 866 560.411 5 180.857.866 970 522.234 6 170.975.699 1152 587.079 7 158.628.139 1116 489.908 8 146.274.826 794 432.757 9 138.429.268 919 449.980
10 135.413.628 862 473.419 11 134.452.384 1426 459.660 12 132.449.811 1104 430.976 13 114.142.980 399 333.541 14 106.368.585 733 268.509 15 100.338.915 766 242.274 16 88.827.254 957 281.401 17 78.774.742 1257 239.216 18 76.117.153 322 239.250 19 63.811.651 1468 186.986 20 62.435.964 631 269.648 21 46.944.323 271 131740 22 49.554.710 552 169.471 X 154.824.264 931 320.997 Y 57.701.691 104 30.,623
Genom mitohondrija
16.571 37 206

Tabela 1. Sestava človeškega genoma po posameznih kromosomih z velikostmi kromosomov v baznih parih (bp), številom genov in številom polimorfizmov enega nukleotida ter sestava genoma mitohodrija. Genetske bolezni Cilj molekularnogenetskih testov v patologiji je čim bolj natančna ocena bolezni ali nagnjenosti k njej. Pri genetskih boleznih, kjer je bolezenski gen znan, je odkritje mutacije najbolj zanesljiva diagnostična metoda. To velja predvsem za tiste bolezni, ki se dedujejo po Mendlovih zakonih, in tudi za nekatere dedne oblike raka. Poznamo približno 6000 genetskih bolezni in za večino so že odkrili gene in genske okvare – mutacije, kar je prvi pogoj za uspešno in učinkovito molekularno diagnostiko. Posebna skupina genov je povezana z genetiko raka. To so predvsem onkogeni in za tumorje zaviralni geni. Onkogene so odkrili na podlagi izsledkov, da lahko nekateri virusi prek svojega genetskega gradiva povzročajo različne oblike raka pri živalih. Normalne oblike teh genov so protoonkogeni, ki so običajno nujno potrebni za pravilno delovanje celic. Mnogi med njimi uravnavajo celični cikel. V rakavih celicah so protoonkogeni mutirani, zato ne delujejo pravilno. Posledica mutacije (sprememba protoonkogena v onkogen) je običajno pridobitev nove funkcije – povečane aktivnosti – deregulacije celičnega cikla. Za spremembo aktivnosti je dovolj ena sama mutacija, zato onkogeni delujejo dominantno. Druga skupina genov so za tumorje zaviralni geni, katerih vloga v karcinogenezi je opredeljena s hipotezo dveh zadetkov. Na celični ravni sta potrebni dve mutaciji, da se pojavi inaktivacija teh genov. Ker pa gre velikokrat za redke dedne oblike raka, pri katerih je ena mutacija že podedovana, so mutacije v za tumorje zaviralnih genih največkrat povezane z avtosomno dominantno dovzetnostjo za raka. Poznamo približno 50 onkogenov in za tumorje zaviralnih genov, ki so pomembni za molekularno diagnostiko raka. Za zdaj lahko z metodami molekularne genetike uspešno diagnosticiramo približno 1600 najpogostejših genetskih bolezni. Oblike genetske nagnjenosti k bolezni Genetske bolezni lahko razdelimo v več skupin, odvisno od načina dedovanja. Ločimo avtosomno dominantno in avtosomno recesivno dedovanje, na spolna kromosoma X in Y vezano dedovanje ter mitohondrijsko dedovanje. Za avtosomne recesivne bolezni je značilno, da je za nastanek bolezni potrebna okvara obeh kopij gena (alelov). Bolniki so potomci heterozigotnih prenašalcev, ki običajno niso prizadeti. Značilni primeri teh bolezni so fenilketonurija, cistična fibroza in srpasta anemija. Pri avtosomnem dominantnem dedovanju za nastanek bolezni zadostuje že ena sama kopija okvarjenega gena. Primer tega sta Huntingtonova bolezen in ahondroplazija. Na spolni kromosom X vezane genetske bolezni so posebnost. Ker imajo moški en sam kromosom X, se pri njih bolezni izražajo dominantno. Ženske so prenašalke in zbolijo redko, na primer, če je oče bolan in mati prenašalka. Hemofilija in Alportov sindrom sta genetski bolezni, povezani z dedovanjem kromosoma X. Bolezni, povezane s kromosomom Y, podedujejo samo moški potomci. Posebna oblika dedovanja je mitohondrijsko, saj se mitohondriji na potomce prenesejo le prek citoplazme jajčne celice. Okvare genov v mitohondrijih DNK mati prenese na vse svoje potomce. Ker so mitohondriji glavni izvor celične energije prek mehanizma oksidativne fosforilacije, mitohondrijske bolezni najpogosteje prizadenejo tkiva, odvisna od zadostne preskrbe z energijo, kot so mišice in možgani.

Genetske bolezni, ki so v glavnem posledica mutacij v enem genu, so monogenske in sledijo Mendlovim zakonom dedovanja. Večina predvsem najbolj pogostih bolezni pa je multifaktorskih – kompleksnih, kar je posledica multiplih genov in interakcije različnih zunanjih dejavnikov. Tako ločimo genetske in zunanje dejavnike tveganja. Genetski dejavniki tveganja se nanašajo predvsem na določene funkcijske polimorfizme, polimorfizme enega nukleotida (SNP-je) oziroma posebne kombinacije teh polimorfizmov, ki so povezani s povečanim tveganjem za razvoj bolezni. Vemo, da lahko nekateri pomembno prispevajo k razvoju nekaterih kompleksnih bolezni. Take t. i. funkcijske SNP-je vključujemo v molekularno diagnostiko, saj nam pomagajo pri oceni genetskih dejavnikov tveganja. Primeri nekaterih takih SNP-jev so SNP C804A v genu TNF-beta, kadar poveča tveganje za akutni miokardni infarkt, SNP N219S v genu za lipoprotein lipazo, eden od večih SNP-jev, ki tudi lahko prispeva k povečanemu tveganju za Alzheimerjevo bolezen, SNP G894T v genu za endotelijsko dušikov monoksid sintazo 3 poveča tveganje za povišan krvni tlak, SNP G2046T v genu za kolagen COL1A1 poveča tveganje za osteoporozo in SNP C13910T v genu laktaze poveča tveganje za intoleranco na laktozo. MOLEKULARNE TEHNIKE Osamitev nukleinskih kislin Vse naše celice z jedrom vsebujejo deoksiribonukleinsko kislino (DNK) in ribonukleinsko kislino (RNK). DNK je mogoče osamiti iz zelo različnih kliničnih vzorcev periferne krvi, kostnega mozga, seča, blata, likvorja, citoloških razmazov, sveže zamrznjenih tkivnih vzorcev ali iz v parafin vklopljenih tkivnih vzorcev, starih več let. Klinične vzorce za osamitev DNK iz krvi ali kostnega mozga običajno zberemo v epruvete, ki vsebujejo antikoagulant, ki je lahko etilendiaminotetraocetna kislina (EDTA) ali citrat. Če za molekularnogenetsko analizo potrebujemo RNK, moramo osamitev iz kliničnega vzorca opraviti takoj ali tkivo zamrzniti v tekočem dušiku. DNK osamimo iz celic z uporabo litičnih encimov za liziranje celičnih sten (proteinaze), nato z organskimi topili (fenol, kloroform) oborimo beljakovine in DNK izločimo z obarjanjem z amonijevim acetatom in etanolom. Vedno pogosteje uporabljajo majhne kromatografske kolone na osnovi afinitetne kromatografije nukleinskih kislin, ki jih je mogoče dobiti v kompletih komercialnih podjetij in so primerne predvsem za osamitve DNK iz manjših vzorcev. DNK je v primerjavi z RNK veliko bolj stabilna, ker je mogoče katalitične encime DNKaze, ki jo razgrajujejo, precej preprosto inaktivirati. Že prisotnost EDTA ali toplote inaktivira večino DNKaz, v nasprotju z njimi pa so RNKaze precej bolj stabilne tudi pri povišani temperaturi. Posledično je osamitev RNK bistveno bolj težka. Zato si pri tem pomagamo z litični pufri, ki vsebujejo inhibitorje RNKaz, na primer gvanidinijev klorid. Kljub temu je RNK kakovostno mogoče osamiti le iz svežih ali primerno zamrznjenih kliničnih vzorcev, fiksirani ali arhivski vzorci pa povzročajo precej težav. Z novejšimi metodami so kljub temu uspeli osamitvi RNK, ki pa je navadno degradirana in omogoča pomnoževanje le zelo kratkih predelov, običajno do 100 baznih parov. Čistost DNK lahko preverimo z merjenjem optične gostote v ultravijoličnem delu spektra. Primerjamo absorpcijo pri valovni dolžini 260 nm in 280 nm, ki je značilna za proteine. Razmerje, ki je v območju od 1,7 do 2 pomeni zelo čisto DNK, kar navadno zadostuje za običajne molekularnogenetske analize. DNK slabše kakovosti je mogoče dodatno čistiti s posebnimi kolonami na podlagi afinitetne kromatografije. V posebnih primerih lahko DNK ali dele DNK ločujemo z gelsko elektroforezo. Mikrodisekcija V novejšem času se zaradi očitnih prednosti vedno bolj uveljavlja tudi molekularna analiza na tarčnem tkivu celic, pridobljenih z mikrodisekcijo. Obstaja več vrst mikrodisekcije, in sicer od

preproste ročne do sodobne laserske mikrodisekcije, ki je hitrejša in veliko bolj natančna od ročne, možnost onesnaženja tako pridobljenega vzorca s tujo DNK, mRNa ali z RNAzami pa je precej manjša. Sistem za lasersko mikrodisekcijo sestavljajo mikroskop, laser in visokoločljivostna kamera, ki so povezani z osebnim računalnikom. Laserski žarek ima zelo majhen premer (okoli 1 µm) in ga je mogoče zelo natančno voditi po tkivni rezini. Z laserskim žarkom ločimo tarčne celice od ostalega tkiva in jih prenesemo v reakcijsko posodico. Laserska mikrodisekcija torej omogoča izolacijo nekaterih struktur (na primer glomerulov) oziroma skupin celic (kot so tumorske celice) ali celo posameznih celic (denimo Reed-Sternbergove velikanke) iz okolnega tkiva brez primesi drugih sestavin tkiva (kot so tubuli, intersticij, žile, vnetni infiltrati, tumorska stroma). Mikrodisekcijo lahko uporabimo na širokem spektru tkiv in celic in tudi na rezinah v parafin vklopljenih tkiv ter citoloških razmazih. Odvisno od izhodnega vzorca za mikrodisekcijo je mogoče osamiti DNK, dovolj dobro mRNK za študije izražanja genov in tudi proteine. V prihodnje bo laserska mikrodisekcija omogočila ugotavljanje “genetskih prstnih odtisov” specifičnih patoloških lezij predvsem v molekularni onkologiji. Glavna prednost mikrodisekcije je, da dobimo relativno čist vzorec morfološko preverjene populacije izbranih celic. To je včasih zelo pomembno za točno ugotovitev genetske spremembe ali za samo kvantificiranje genetske spremembe, posebno kadar preiskovani vzorec vsebuje obilno stromo ali limfoidni infiltrat, ki lahko vplivata na rezultat molekularne analize. Pri določevanju izgube heterozigotnosti (loss of heterozygosity – LOH) za tumorje zaviralnih genov lahko primes normalnih celic v vzorcu zakrije dejansko stanje preiskovanega vzorca, zato izgube heterozigotnosti velikokrat ne moremo dovolj natančno določiti. Prav tako je mikrodisekcija zelo pomembna, ko želimo zelo natančno določiti izražanje kakega gena. Tak je primer Hodgkinovega limfoma, kjer brez mikrodisekcije neoplastičnih Reed-Sternbergovih celic, ki so potrebne za nadaljnjo molekularno analizo, zaradi obilne primesi reaktivnih limfocitov, eozinofilnih granulocitov, plazmatk in makrofagov tako rekoč ne bi mogli opraviti osamitve. Laserska mikrodisekcija je v veliko pomoč pri študiju ekspresije genov v kompleksnih organih, kot je ledvica, ki jo sestavlja več kakor 20 različnih tipov celic, bolezenski proces pa velikokrat prizadene le nekatere od njih. Analiza DNK z gelsko elektroforezo Gelska elektroforeza je analitska in preparativna metoda za analizo ali čiščenje DNK. Gele običajno pripravimo s pufri blizu nevtralnega pH, pri katerem so fosfatne skupine nukleinske kisline negativno nabite in zato DNK potuje proti pozitivni elektrodi (anodi). Hitrost, s katero se molekule pomikajo proti elektrodi, je odvisna od velikosti DNK in por gela. Manjši odseki DNK potujejo hitreje, večji pa počasneje. Agarozni geli so primerni za ločevanje fragmentov DNK velikosti od nekaj 100 do 20.000 baznih parov. Za manjše fragmente, od 100 do 500 baznih parov, pa je bolj primerna elektroforeza na akrilamidnih gelih. Vizualizacijo DNK izvedemo običajno v ultravijolični svetlobi po reakciji s kemičnimi reagenti, na primer etidijevim bromidom, ki v tej svetlobi fluorescirajo, ali z reagenti, ki se vežejo na DNK v vidni svetlobi, na primer sybr-zeleno, akridinsko oranžno, metilensko modro in podobno. Verižna reakcija s polimerazo (polymerase chain reaction – PCR) Metoda verižne reakcije s polimerazo je postala osnovna metoda molekularne diagnostike, saj omogoča pomnoževanje poljubnega odseka DNK, dolgega tudi do več 1000 baznih parov v zadostni množini (do 1 mikrograma), ki je potrebna za nadaljnje diagnostične postopke (na primer reakcijo z restrikcijsko endonukleazo, hibridizacijo, analizo mutacij s SSCP, analizo heterodupleksov ali drugo metodo, dot-blot analizo ali sekvenčno reakcijo). Predvsem pri diagnostiki pomembnih virusov in mikroorganizov poznamo diagnostične komplete in skoraj popolnoma avtomatizirane instrumente in postopke, ki omogočajo diagnostiko več 100 vzorcev v

zelo kratkem času. To velja pri diagnosticiranju virusov, kot so virus citomegalije (CMV), virus Epstein-Barr (EBV), virus hepatitisa B (HBV), virus hepatitisa C (HCV), virus herpesa simpleksa (HSV), človeški herpesvirus 6 (HHV-6), virus človeške imunske pomanjkljivosti, tip 1 (HIV-1), humani papilomski virus (HPV) in virus varicella zoster (VZV), ter nekaterih bakterij, kot so na primer Chlamydia trachomatis, Neisseria gonorrhoea in Mycobacterium tuberculosis. Poleg osnovnega PCR obstaja še več specifičnih oblik pomnoževanja DNK v verižni reakciji s polimerazo. Pri multipleksnem PCR uporabimo dva ali več parov začetnih oligonukleotidov v isti reakcijski zmesi. Tako lahko pomnožimo več odsekov DNK naenkrat in v veliko primerih tudi analiziramo v nadaljnjih stopnjah. Tako prihranimo čas predvsem pri analizah genov s številnimi eksoni. V nekaterih primerih je težko izbrati ustrezne pogoje pomnoževanja, da se določeni odsek DNK v končni zmesi ne bi pojavil v preveliki koncentraciji. Zato je v številnih primerih potrebna natančna optimizacija tako glede izbire začetnih oligonukleotidov, kot tudi glede pogojev pomnoževanja. Prav tako moramo za pomnoževanje izbrati fragmente DNK različnih dolžin, da jih lahko po koncu razlikujemo na gelski elektroforezi. Postopek vgnezdenega PCR uporabimo, ko glede na dano sekvenco DNK, ki jo želimo pomnožiti, ne uspemo izbrati dovolj učinkovitega para začetnih nukleotidov. Pri vgnezdenem postopku PCR najprej pomnožimo daljši fragment DNK v prvih 25 ciklih. V drugi stopnji pa to uporabimo kot tarčna DNK za drugi začetni par olikonukleotidov, ki bodo pomnožili krajši odsek DNK. Metoda izboljša specifičnost, ker uporabimo dva para začetnih oligonukleotidov, in tudi občutljivost, ker je pomnoževanje v vsaki stopnji vsaj 25 ciklov dvostopenjsko. Določanje mutacij in polimorfizmov Mutacija pomeni vsako spremembo v zaporedju nukleotidov DNK, ki se praviloma izraža v bolezenskem fenotipu. Posledica mutacij so različni zapisi DNK pri določenem genu ali aleli. Če se alela homolognega genskega para razlikujeta, pravimo, da gre za heterozigotno stanje, če sta enaka, pa govorimo o homozigotu. Homozigotna stanja so povezana predvsem z genetskimi boleznimi, ki se dedujejo recesivno, heterozigotna stanja pa z boleznimi, ki se dedujejo dominantno. Prve mutacije in polimorfizme so ugotavljali z uporabo restrikcijskih nukleaz. Restrikcijske nukleaze so posebni encimi, ki so jih uspeli pridobiti iz številnih mikroorganizmov, in cepijo DNK. Znanih je več kot 300 restrikcijskih endonukleaz. Encimi prepoznajo t. i. palindromska zaporedja na dvoverižni DNK in na takem mestu cepijo DNK. Tako na primer restrikcijska endonukleaza Msp I prepozna naslednje zaporedje dvoverižne DNK 5'-C C G G -3' in cepi DNK za prvim C in pred zadnjim G. Če je kateri koli C ali G mutiran v drugačen nukleotid, taka točkasta mutacija uniči to restrikcijsko mesto, zato do cepitve DNK na tem mestu ne pride. Restrikcijske endonukleaze veliko uporabljajo pri ugotavljanju polimorfizmov enega baznega para (SNP-jev) in točkastih mutacij (slika 12.1).

Slika 12.1. Primer uporabe restrikcijske endonukleaze MspI pri posredni diagnostiki avtosomne recesivne dedne bolezni cistične fibroze. Po pomnoževanju 414 bp dolgega odseka DNK v predelu gena MetH v verižni reakciji s polimerazo za vseh šest članov družine so izvedli reakcijo z restrikcijsko endonukleazo MspI. Odsek DNK ima mesto cepitve za encimom MspI pri 200 bp, tako da nastanejo v primeru heterozigotov trije fragmenti (414, 220 in 194 bp), pri homozigotu, ki nima restrikcijskega mesta, ostane začetni fragment 414 bp, medtem ko se pri homozigotu z restrikcijskim mestom oba alela polimorfizma razcepita v dva nova fragmenta DNK (220 in 194 bp). Prazen simbol pomeni zdravega posameznika, polovično polni simboli pomenijo prenašalce, polni simbol pa bolnika s cistično fibrozo. Analiza konformacij enoverižnih DNK (single stranded conformational polymorphism – SSCP) in analiza heterodupleksov (heteroduplex analysis – HA) sta po pomnoževanju določenega odseka DNK v verižni reakciji s polimerazo med najpogostejšimi tehnikami za odkrivanje napak v bolezenskih genih. Enoverižna DNK se v nedenaturacijski raztopini zvije v specifično sekundarno strukturo, odvisno od njenega zaporedja. Verige DNK, ki se ločijo celo za 1 nukleotid, lahko zavzamejo drugačno konformacijo – trodimenzionalno strukturo enoverižne DNK. Te razlike v strukturi lahko opazimo v različnem potovanju verig – konformacij po elektroforezi na poliakrilamidnem gelu v primerjavi s kontrolno DNK, brez mutacije. Njeni prednosti sta predvsem hitrost in nezapletenost, vendar pa ni stoodstotno občutljiva in jo je včasih težko interpretirati, še zlasti, kadar se pojavijo metastabilne konformacije.
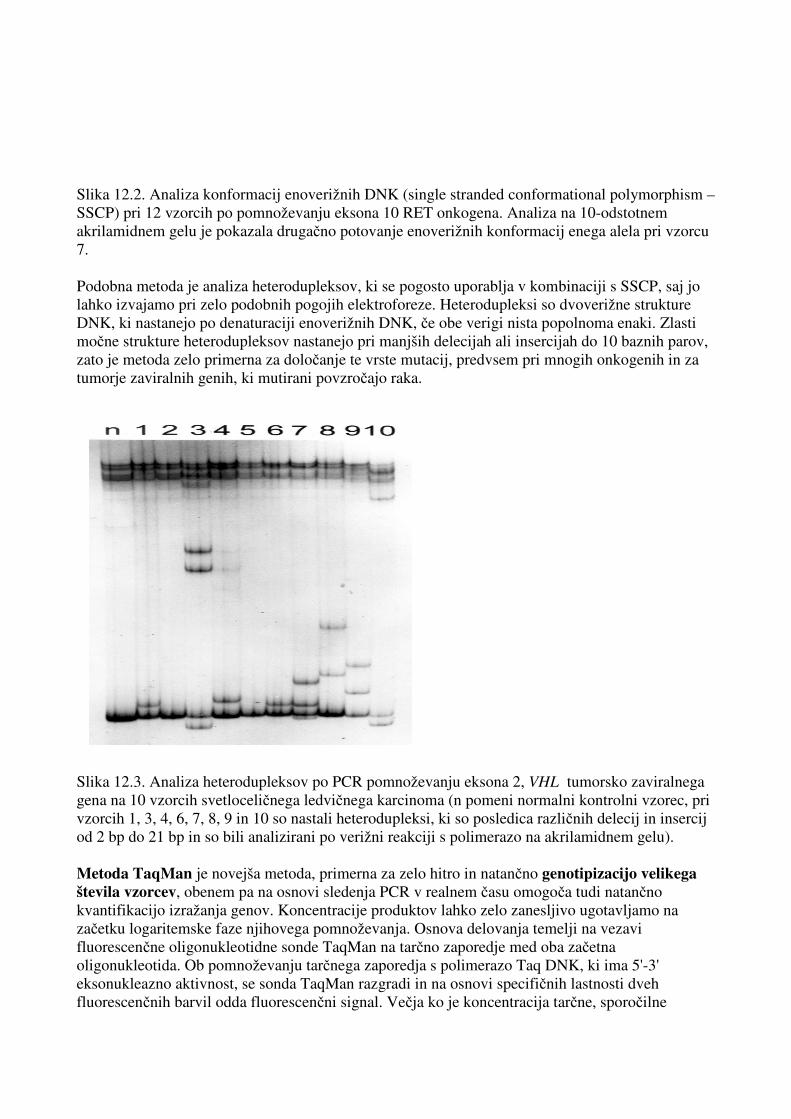
Slika 12.2. Analiza konformacij enoverižnih DNK (single stranded conformational polymorphism – SSCP) pri 12 vzorcih po pomnoževanju eksona 10 RET onkogena. Analiza na 10-odstotnem akrilamidnem gelu je pokazala drugačno potovanje enoverižnih konformacij enega alela pri vzorcu 7. Podobna metoda je analiza heterodupleksov, ki se pogosto uporablja v kombinaciji s SSCP, saj jo lahko izvajamo pri zelo podobnih pogojih elektroforeze. Heterodupleksi so dvoverižne strukture DNK, ki nastanejo po denaturaciji enoverižnih DNK, če obe verigi nista popolnoma enaki. Zlasti močne strukture heterodupleksov nastanejo pri manjših delecijah ali insercijah do 10 baznih parov, zato je metoda zelo primerna za določanje te vrste mutacij, predvsem pri mnogih onkogenih in za tumorje zaviralnih genih, ki mutirani povzročajo raka. Slika 12.3. Analiza heterodupleksov po PCR pomnoževanju eksona 2, VHL tumorsko zaviralnega gena na 10 vzorcih svetloceličnega ledvičnega karcinoma (n pomeni normalni kontrolni vzorec, pri vzorcih 1, 3, 4, 6, 7, 8, 9 in 10 so nastali heterodupleksi, ki so posledica različnih delecij in insercij od 2 bp do 21 bp in so bili analizirani po verižni reakciji s polimerazo na akrilamidnem gelu). Metoda TaqMan je novejša metoda, primerna za zelo hitro in natančno genotipizacijo velikega števila vzorcev, obenem pa na osnovi sledenja PCR v realnem času omogoča tudi natančno kvantifikacijo izražanja genov. Koncentracije produktov lahko zelo zanesljivo ugotavljamo na začetku logaritemske faze njihovega pomnoževanja. Osnova delovanja temelji na vezavi fluorescenčne oligonukleotidne sonde TaqMan na tarčno zaporedje med oba začetna oligonukleotida. Ob pomnoževanju tarčnega zaporedja s polimerazo Taq DNK, ki ima 5'-3' eksonukleazno aktivnost, se sonda TaqMan razgradi in na osnovi specifičnih lastnosti dveh fluorescenčnih barvil odda fluorescenčni signal. Večja ko je koncentracija tarčne, sporočilne
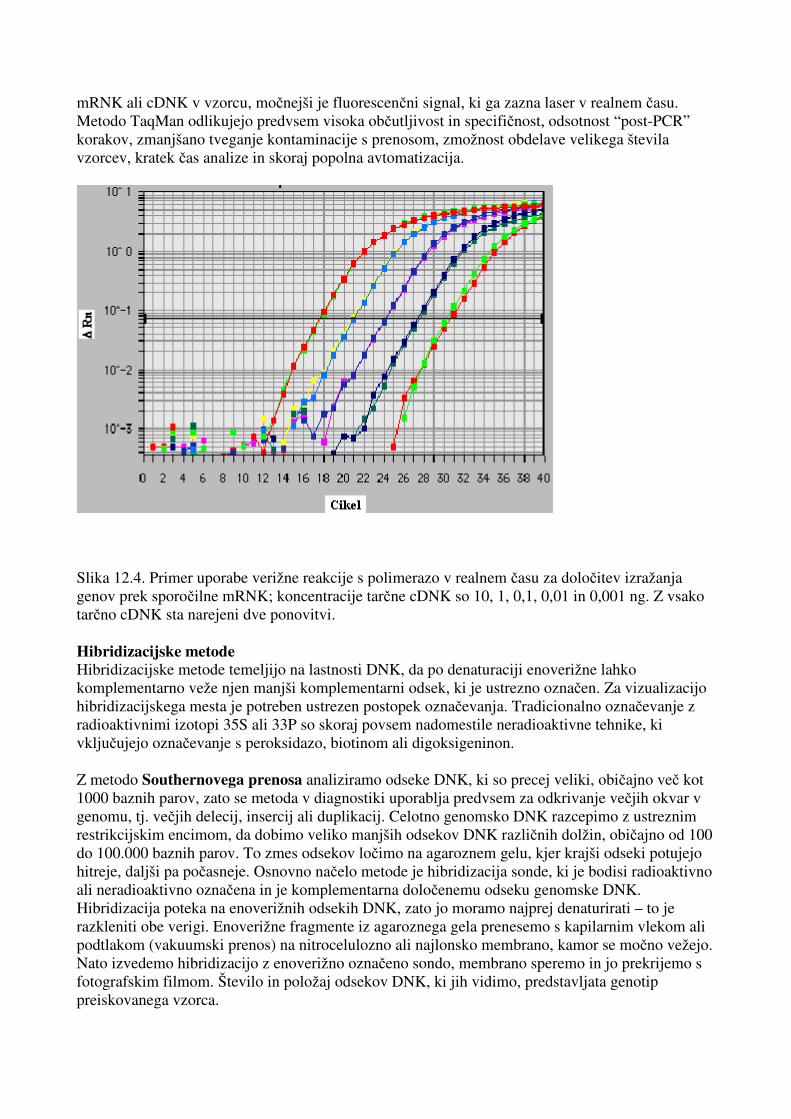
mRNK ali cDNK v vzorcu, močnejši je fluorescenčni signal, ki ga zazna laser v realnem času. Metodo TaqMan odlikujejo predvsem visoka občutljivost in specifičnost, odsotnost “post-PCR” korakov, zmanjšano tveganje kontaminacije s prenosom, zmožnost obdelave velikega števila vzorcev, kratek čas analize in skoraj popolna avtomatizacija.
Slika 12.4. Primer uporabe verižne reakcije s polimerazo v realnem času za določitev izražanja genov prek sporočilne mRNK; koncentracije tarčne cDNK so 10, 1, 0,1, 0,01 in 0,001 ng. Z vsako tarčno cDNK sta narejeni dve ponovitvi. Hibridizacijske metode Hibridizacijske metode temeljijo na lastnosti DNK, da po denaturaciji enoverižne lahko komplementarno veže njen manjši komplementarni odsek, ki je ustrezno označen. Za vizualizacijo hibridizacijskega mesta je potreben ustrezen postopek označevanja. Tradicionalno označevanje z radioaktivnimi izotopi 35S ali 33P so skoraj povsem nadomestile neradioaktivne tehnike, ki vključujejo označevanje s peroksidazo, biotinom ali digoksigeninon. Z metodo Southernovega prenosa analiziramo odseke DNK, ki so precej veliki, običajno več kot 1000 baznih parov, zato se metoda v diagnostiki uporablja predvsem za odkrivanje večjih okvar v genomu, tj. večjih delecij, insercij ali duplikacij. Celotno genomsko DNK razcepimo z ustreznim restrikcijskim encimom, da dobimo veliko manjših odsekov DNK različnih dolžin, običajno od 100 do 100.000 baznih parov. To zmes odsekov ločimo na agaroznem gelu, kjer krajši odseki potujejo hitreje, daljši pa počasneje. Osnovno načelo metode je hibridizacija sonde, ki je bodisi radioaktivno ali neradioaktivno označena in je komplementarna določenemu odseku genomske DNK. Hibridizacija poteka na enoverižnih odsekih DNK, zato jo moramo najprej denaturirati – to je razkleniti obe verigi. Enoverižne fragmente iz agaroznega gela prenesemo s kapilarnim vlekom ali podtlakom (vakuumski prenos) na nitrocelulozno ali najlonsko membrano, kamor se močno vežejo. Nato izvedemo hibridizacijo z enoverižno označeno sondo, membrano speremo in jo prekrijemo s fotografskim filmom. Število in položaj odsekov DNK, ki jih vidimo, predstavljata genotip preiskovanega vzorca.

Metoda prenosa po Northernu je precej podobna, le da pri njej uporabimo RNK namesto DNK. Imamo tri različne vrste RNK: tRNK (transportna RNK, ki je aktivna pri sestavljanju polipeptidnih verig), rRNK (ribosomska RNK, ki je del strukture ribosomov) in mRNK (sporočilna RNK, ki je rezultat prepisovanja DNK in vsebuje zapise za proteine). Sporočilno mRNK z ustreznimi postopki osamimo iz preiskovanega vzorca in analiziramo na agaroznem gelu. Podobno kakor pri DNK se tudi tu sporočilne RNK različnih dolžin ustrezno ločijo, ker so tudi geni različno dolgi. Ker je RNK enoverižna, jo lahko prenesemo neposredno na membrano brez predhodne obdelave. Hibridizacija z označeno sondo, ki se nanaša na ustrezen gen, poteka zelo podobno kakor pri Southernovem prenosu. Enako poteka vizualizacija, ki jo izvedemo bodisi z izotopom ali neradioaktivno. Slika 12.5. Shematski prikaz prenosa po Northernu. Pri sporočilni mRNK, ki smo jo osamili iz pljuč (P), možganov (M) in jeter (J), smo s sondama A in B ugotovili enako velikost prepisa, pri čemer je izražanje tega gena najmočnejše v jetrih. Pri sondi C je velikost prepisa, izraženega v možganih in jetrih, krajša, pri sondi D pa je prepis daljši, in se izraža samo v pljučih. Metoda prenosa Western se uporablja za proteine. Omogoča določitev molekulske mase proteina ali meritev relativne množine proteina v preiskovanem vzorcu. Proteine osamimo iz preiskovanega vzorca in ločimo v postopku elektroforeze na poliakrilamidnem gelu z natrijevim dodecilsulfatom. Proteini se na gelu ločijo po velikosti, nato jih prenesemo na nitrocelulozno membrano in inkubiramo z generičnimi proteini (na primer mlečnimi), da zapolnimo neproteinska mesta na membrani. Sledi inkubacija s specifičnim označenim protitelesom, ki se veže samo na določen protein. V procesu razkrivanja ugotavljamo mesto vezave protitelesa na protein. Hibridizacija in situ (in situ hybridisation – ISH) je v zadnjih letih v molekularni patologiji postala povsem avtomatizirana. V primerjavi s klasično hibridizacijo na membranah poteka tu postopek hibridizacije v celicah (in situ). Zato je ISH posebno uporabna za dokazovanje prisotnosti tujih genomov virusov in mikroorganizmov. Po vezavi specifično označene sonde, ki je komplementarna tarčni DNK ali RNK, mesto vezave razkrijemo z avidinom ali antibiotinskimi protitelesi, ki so označena z alkalno fosfatazo ali hrenovo peroksidazo.
P M J P M J P M J P M J
Sonda A Sonda B Sonda C Sonda D
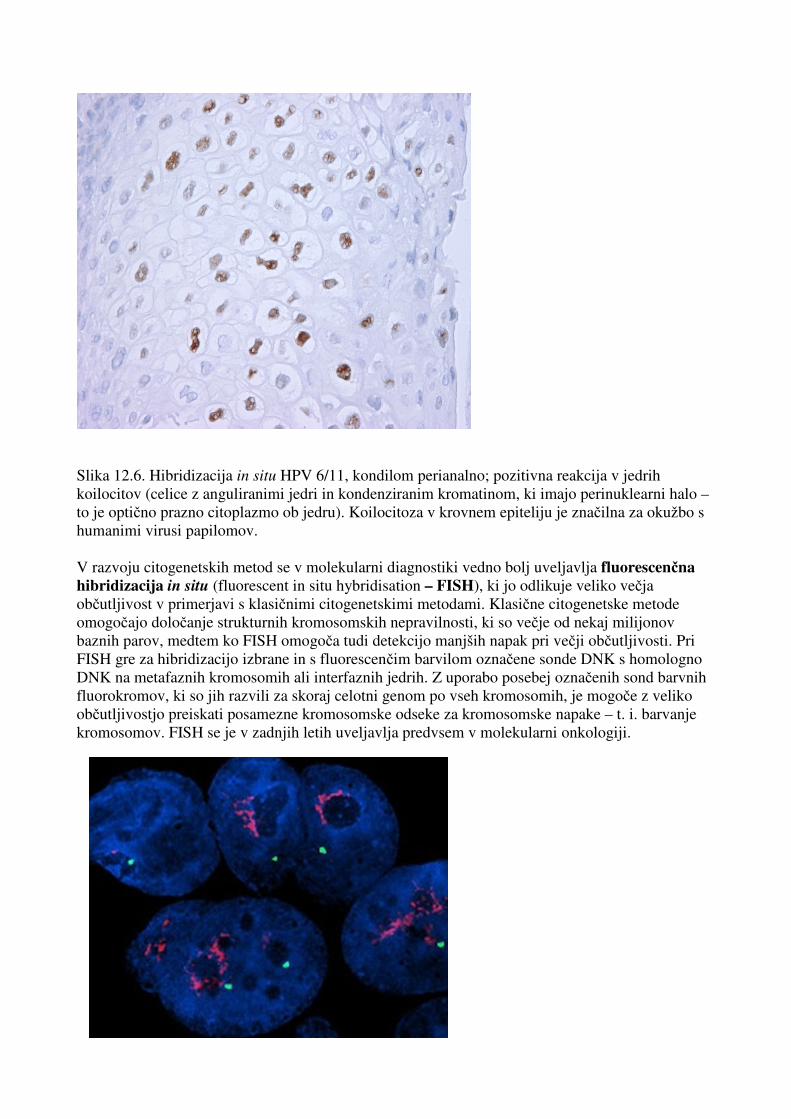
Slika 12.6. Hibridizacija in situ HPV 6/11, kondilom perianalno; pozitivna reakcija v jedrih koilocitov (celice z anguliranimi jedri in kondenziranim kromatinom, ki imajo perinuklearni halo – to je optično prazno citoplazmo ob jedru). Koilocitoza v krovnem epiteliju je značilna za okužbo s humanimi virusi papilomov. V razvoju citogenetskih metod se v molekularni diagnostiki vedno bolj uveljavlja fluorescenčna hibridizacija in situ (fluorescent in situ hybridisation – FISH), ki jo odlikuje veliko večja občutljivost v primerjavi s klasičnimi citogenetskimi metodami. Klasične citogenetske metode omogočajo določanje strukturnih kromosomskih nepravilnosti, ki so večje od nekaj milijonov baznih parov, medtem ko FISH omogoča tudi detekcijo manjših napak pri večji občutljivosti. Pri FISH gre za hibridizacijo izbrane in s fluorescenčim barvilom označene sonde DNK s homologno DNK na metafaznih kromosomih ali interfaznih jedrih. Z uporabo posebej označenih sond barvnih fluorokromov, ki so jih razvili za skoraj celotni genom po vseh kromosomih, je mogoče z veliko občutljivostjo preiskati posamezne kromosomske odseke za kromosomske napake – t. i. barvanje kromosomov. FISH se je v zadnjih letih uveljavlja predvsem v molekularni onkologiji.
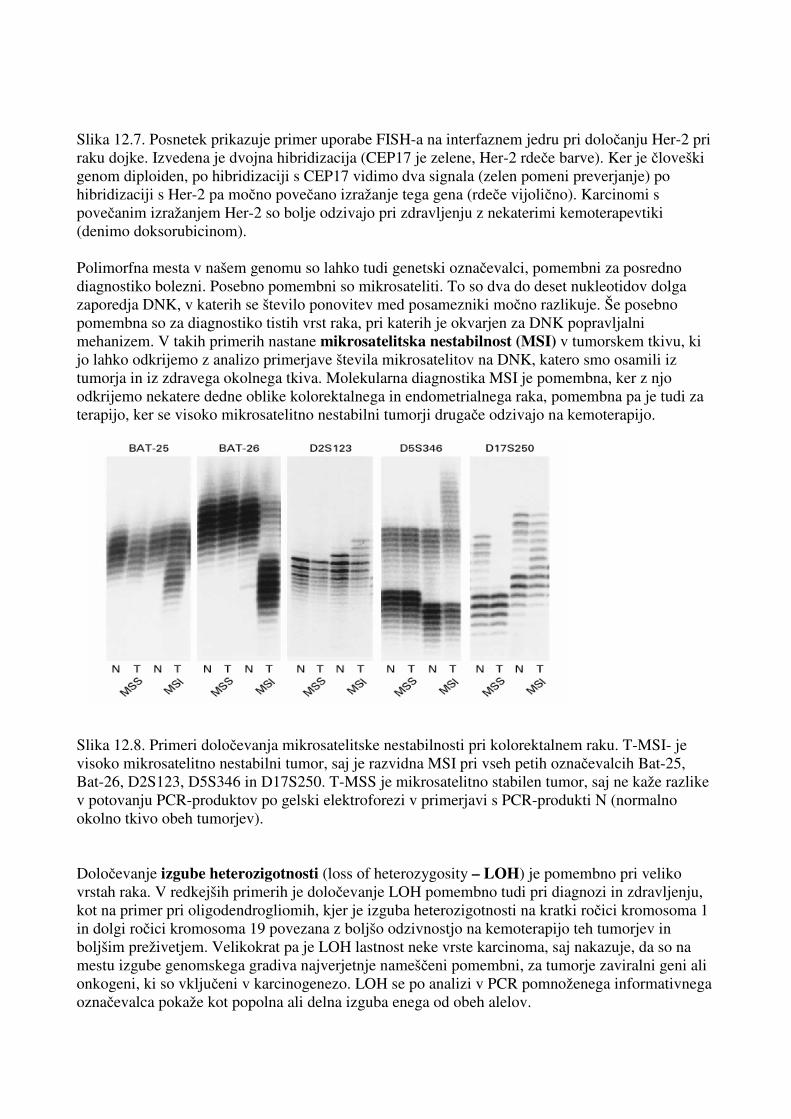
Slika 12.7. Posnetek prikazuje primer uporabe FISH-a na interfaznem jedru pri določanju Her-2 pri raku dojke. Izvedena je dvojna hibridizacija (CEP17 je zelene, Her-2 rdeče barve). Ker je človeški genom diploiden, po hibridizaciji s CEP17 vidimo dva signala (zelen pomeni preverjanje) po hibridizaciji s Her-2 pa močno povečano izražanje tega gena (rdeče vijolično). Karcinomi s povečanim izražanjem Her-2 so bolje odzivajo pri zdravljenju z nekaterimi kemoterapevtiki (denimo doksorubicinom). Polimorfna mesta v našem genomu so lahko tudi genetski označevalci, pomembni za posredno diagnostiko bolezni. Posebno pomembni so mikrosateliti. To so dva do deset nukleotidov dolga zaporedja DNK, v katerih se število ponovitev med posamezniki močno razlikuje. Še posebno pomembna so za diagnostiko tistih vrst raka, pri katerih je okvarjen za DNK popravljalni mehanizem. V takih primerih nastane mikrosatelitska nestabilnost (MSI) v tumorskem tkivu, ki jo lahko odkrijemo z analizo primerjave števila mikrosatelitov na DNK, katero smo osamili iz tumorja in iz zdravega okolnega tkiva. Molekularna diagnostika MSI je pomembna, ker z njo odkrijemo nekatere dedne oblike kolorektalnega in endometrialnega raka, pomembna pa je tudi za terapijo, ker se visoko mikrosatelitno nestabilni tumorji drugače odzivajo na kemoterapijo. Slika 12.8. Primeri določevanja mikrosatelitske nestabilnosti pri kolorektalnem raku. T-MSI- je visoko mikrosatelitno nestabilni tumor, saj je razvidna MSI pri vseh petih označevalcih Bat-25, Bat-26, D2S123, D5S346 in D17S250. T-MSS je mikrosatelitno stabilen tumor, saj ne kaže razlike v potovanju PCR-produktov po gelski elektroforezi v primerjavi s PCR-produkti N (normalno okolno tkivo obeh tumorjev). Določevanje izgube heterozigotnosti (loss of heterozygosity – LOH) je pomembno pri veliko vrstah raka. V redkejših primerih je določevanje LOH pomembno tudi pri diagnozi in zdravljenju, kot na primer pri oligodendrogliomih, kjer je izguba heterozigotnosti na kratki ročici kromosoma 1 in dolgi ročici kromosoma 19 povezana z boljšo odzivnostjo na kemoterapijo teh tumorjev in boljšim preživetjem. Velikokrat pa je LOH lastnost neke vrste karcinoma, saj nakazuje, da so na mestu izgube genomskega gradiva najverjetnje nameščeni pomembni, za tumorje zaviralni geni ali onkogeni, ki so vključeni v karcinogenezo. LOH se po analizi v PCR pomnoženega informativnega označevalca pokaže kot popolna ali delna izguba enega od obeh alelov.
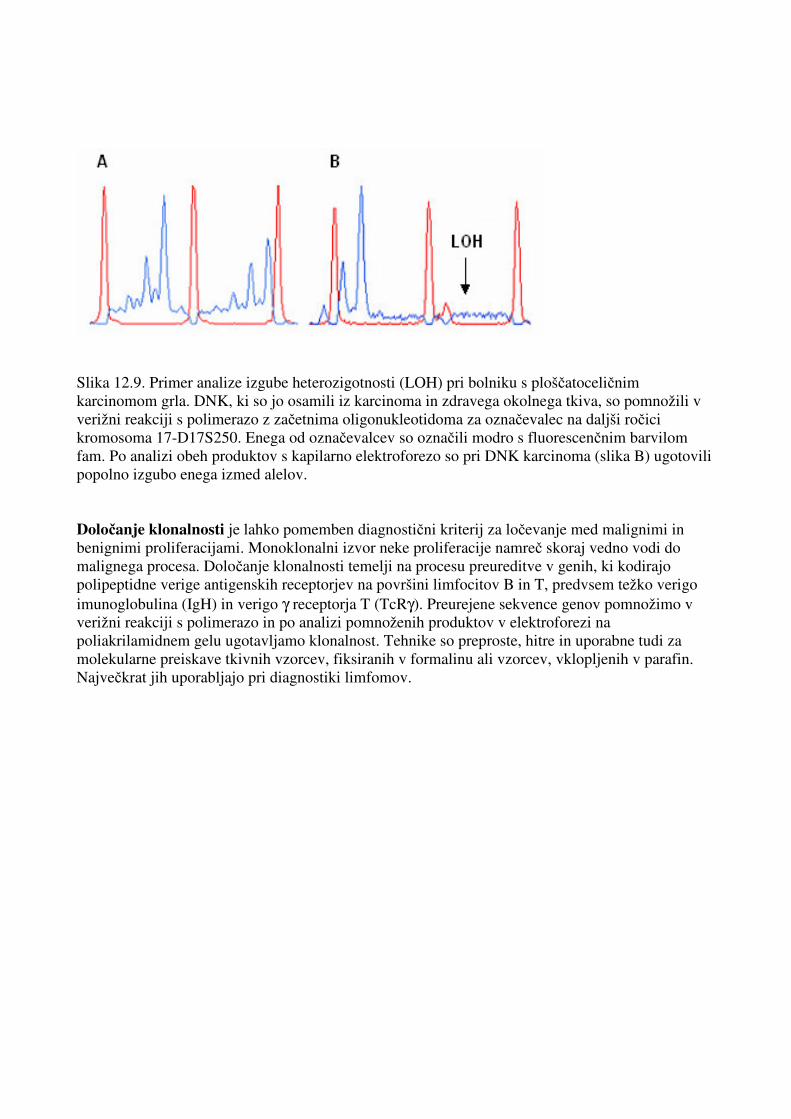
Slika 12.9. Primer analize izgube heterozigotnosti (LOH) pri bolniku s ploščatoceličnim karcinomom grla. DNK, ki so jo osamili iz karcinoma in zdravega okolnega tkiva, so pomnožili v verižni reakciji s polimerazo z začetnima oligonukleotidoma za označevalec na daljši ročici kromosoma 17-D17S250. Enega od označevalcev so označili modro s fluorescenčnim barvilom fam. Po analizi obeh produktov s kapilarno elektroforezo so pri DNK karcinoma (slika B) ugotovili popolno izgubo enega izmed alelov. Določanje klonalnosti je lahko pomemben diagnostični kriterij za ločevanje med malignimi in benignimi proliferacijami. Monoklonalni izvor neke proliferacije namreč skoraj vedno vodi do malignega procesa. Določanje klonalnosti temelji na procesu preureditve v genih, ki kodirajo polipeptidne verige antigenskih receptorjev na površini limfocitov B in T, predvsem težko verigo imunoglobulina (IgH) in verigo γ receptorja T (TcRγ). Preurejene sekvence genov pomnožimo v verižni reakciji s polimerazo in po analizi pomnoženih produktov v elektroforezi na poliakrilamidnem gelu ugotavljamo klonalnost. Tehnike so preproste, hitre in uporabne tudi za molekularne preiskave tkivnih vzorcev, fiksiranih v formalinu ali vzorcev, vklopljenih v parafin. Največkrat jih uporabljajo pri diagnostiki limfomov.

Slika 12.10. Določitev klonalnosti pri B-celičnem limfomu s pomnoževanjem IgI-lambda lahke verige v verižni reakciji s polimerazo in analizo po elektroforezi na poliakrilamidnem gelu (M – označevalec velikosti, 1 – negativna kontrola, 2, 3, 4 – monoklonalni B-celični limfomi, 5, 6 in 7 – reaktivne bezgavke). Edina metoda, s katero neposredno dokažemo mesto mutacije, je določevanje nukleotidnega zaporedja oziroma sekvenčna analiza. Kljub pomembnemu napredku v zadnjih letih zaradi avtomatizacije in hitrosti izvedbe ter neradioaktivnim postopkom je metoda še vedno precej draga in zamudna, zato se v molekularni diagnostiki običajno uporablja v zadnji stopnji, ko je treba vzorec DNK, v katerem smo odkrili mutacijo z drugimi metodami, potrditi oziroma točno določiti mesto okvare. Še vedno se uporablja metoda po Sangerju, ki je nekoliko spremenjena, in sicer namesto radioaktivnega izotopa uporabljamo s štirimi različnimi fluorescenčnimi barvili (Joe-ddA – zelena, Rox-ddT – rdeča, Fam-ddC – modra in Tamra-ddG – črna) označene dideoksinukleotidne terminatorje, ki naključno v različnih položajih končujejo verigo. Ker so vsi štirje različno označeni, jih lahko dodamo v reakcijsko zmes hkrati, kar pri izotopu ni bilo mogoče. Barva dideoksinukleotida nam po ločitvi verig v kapilarni elektroforezi in laserski detekciji pove zaporedje nukleotidov v verigi DNK oziroma sekvenco.
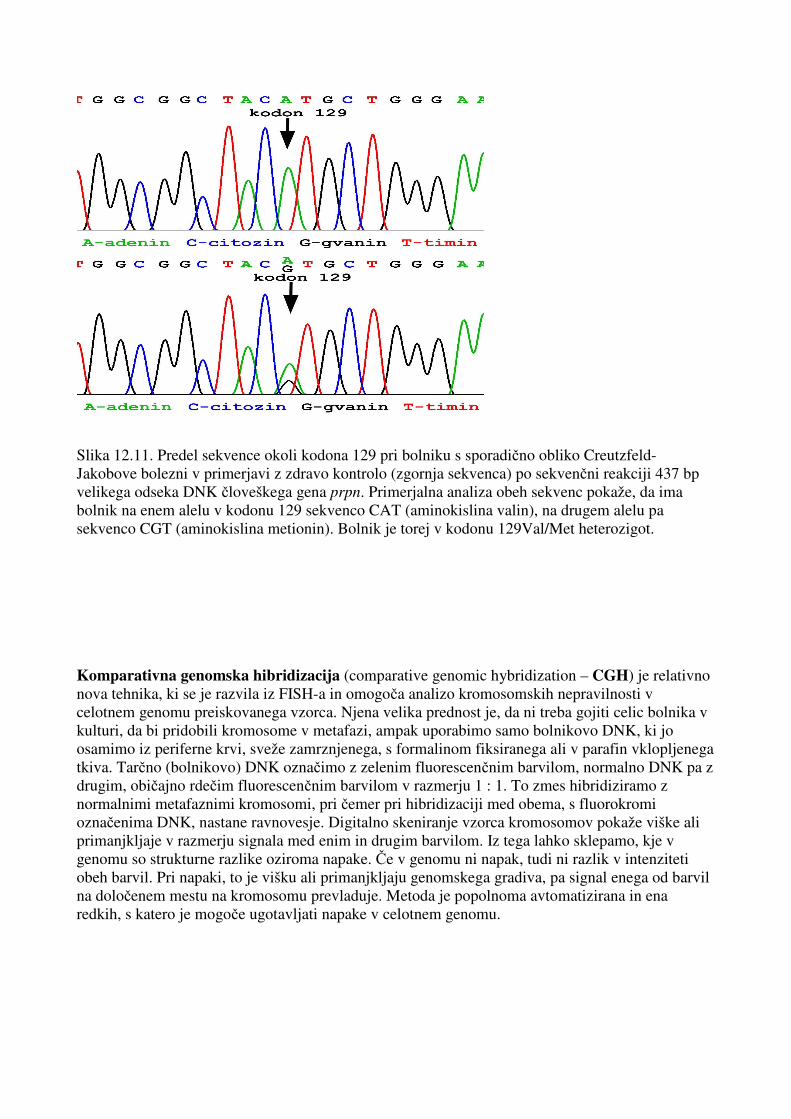
Slika 12.11. Predel sekvence okoli kodona 129 pri bolniku s sporadično obliko Creutzfeld-Jakobove bolezni v primerjavi z zdravo kontrolo (zgornja sekvenca) po sekvenčni reakciji 437 bp velikega odseka DNK človeškega gena prpn. Primerjalna analiza obeh sekvenc pokaže, da ima bolnik na enem alelu v kodonu 129 sekvenco CAT (aminokislina valin), na drugem alelu pa sekvenco CGT (aminokislina metionin). Bolnik je torej v kodonu 129Val/Met heterozigot. Komparativna genomska hibridizacija (comparative genomic hybridization – CGH) je relativno nova tehnika, ki se je razvila iz FISH-a in omogoča analizo kromosomskih nepravilnosti v celotnem genomu preiskovanega vzorca. Njena velika prednost je, da ni treba gojiti celic bolnika v kulturi, da bi pridobili kromosome v metafazi, ampak uporabimo samo bolnikovo DNK, ki jo osamimo iz periferne krvi, sveže zamrznjenega, s formalinom fiksiranega ali v parafin vklopljenega tkiva. Tarčno (bolnikovo) DNK označimo z zelenim fluorescenčnim barvilom, normalno DNK pa z drugim, običajno rdečim fluorescenčnim barvilom v razmerju 1 : 1. To zmes hibridiziramo z normalnimi metafaznimi kromosomi, pri čemer pri hibridizaciji med obema, s fluorokromi označenima DNK, nastane ravnovesje. Digitalno skeniranje vzorca kromosomov pokaže viške ali primanjkljaje v razmerju signala med enim in drugim barvilom. Iz tega lahko sklepamo, kje v genomu so strukturne razlike oziroma napake. Če v genomu ni napak, tudi ni razlik v intenziteti obeh barvil. Pri napaki, to je višku ali primanjkljaju genomskega gradiva, pa signal enega od barvil na določenem mestu na kromosomu prevladuje. Metoda je popolnoma avtomatizirana in ena redkih, s katero je mogoče ugotavljati napake v celotnem genomu.

Slika 12.12. Primer določevanja pentasomije kromosoma 21 pri bolniku z levkemijo dendritičnih celic. Po hibridizaciji na metafazne kromosome sta zaradi amplifikacije vidna zeleno obarvana kromosoma 21. Ker je kariotip moški, je kromosom X rdeč (primanjkljaj genoma v primerjavi z ženskim, referenčnim genomom), kromosom Y pa je modre barve (DAPI), ker v heterokromatski regiji ni hibridizacije. DNK-mikromreže (DNA microarrays) omogočajo analizo celotnega genoma. Hibridizacije potekajo na t. i. DNK-čipih z visoko ločljivostjo. Ločimo DNK-mikromreže, s katerimi ugotavljamo razlike v izražanju posameznih genov, in DNK-mikromreže, s katerimi določamo polimorfizme in mutacije v posameznih genih ali skupini genov.

Slika 12.12. DNK-mikromreža za določanje mutacij v genu ABCR, ki povzročajo dedno obliko Stargardtovega sindroma. Klasična genetska analiza gena ABCR, ki vsebuje 50 eksonov in je genetsko zelo heterogen, je izredno zamudna, z DNK-mikromrežo pa je končana v nekaj urah. Hkrati lahko analiziramo vseh 438 znanih mutacij, ki povzročajo to bolezen. Barvni signal posameznega nukleotida na mreži je: adenozin je rumen, citozin rdeč, gvanozin zelen, timin pa moder.







![DUCHENNOVA SVALOVÁ DYSTROFIA: Diagnostika a lieč...Duchennova svalová dystrofia: Diagnostika a liečba, Príručka pre rodiny; marec 2010 [2] 1. ÚVOD Táto príručka je určená](https://img.pdfslide.tips/doc/110x75/60a0bab76939665ac0035e1a/duchennova-svalov-dystrofia-diagnostika-a-lie-duchennova-svalov-dystrofia.jpg)





















