Embed Size (px)
Citation preview

High Expression of PRL-3 Promotes Cancer Cell Motility and LiverMetastasis in Human Colorectal Cancer: A Predictive MolecularMarker of Metachronous Liver and Lung Metastases
Hirotaka Kato,1,2 Shuho Semba,1
Upik A. Miskad,1 Yasushi Seo,2
Masato Kasuga,2 and Hiroshi Yokozaki11Division of Surgical Pathology, Department of BiomedicalInformatics, and 2Division of Diabetes, Digestive and KidneyDiseases, Department of Clinical Molecular Medicine, KobeUniversity Graduate School of Medicine, Kobe, Japan
ABSTRACTPurpose: Overexpression of PRL-3 has been implicated
in colorectal cancer metastases. We investigated the signif-icance of PRL-3 expression in the progression and develop-ment of colorectal cancer.
Experimental Design: We transfected PRL-3-specificsmall interfering RNA into human colon cancer DLD-1 cellsand analyzed its effect on proliferation, motility, and hepaticcolonization. Using an in situ hybridization method, we ex-amined the levels of PRL-3 expression in both primary (177cases) and metastatic (92 cases) human colorectal cancersand elucidated the relationships with clinicopathological pa-rameters including the incidence of metachronous liverand/or lung metastasis after curative surgery for primarytumor.
Results: Transient down-regulation of PRL-3 expres-sion in DLD-1 cells abrogated motility (in vitro) and hepaticcolonization (in vivo), but no effect on the proliferation ofthese cells was observed. In human primary colorectal can-cers, the frequency of up-regulated PRL-3 expression incases with liver (84.4%) or lung (88.9%) metastasis wasstatistically higher than that in cases without either type ofmetastasis (liver, 35.9%; lung, 42.3%). In metastatic colo-rectal cancer lesions, high expression of PRL-3 was fre-quently detected (liver, 91.3%; lung, 100%). Interestingly,metachronous metastasis was observed more frequently inthe cases with high PRL-3 expression (P < 0.0001).
Conclusions: These results indicate that PRL-3 expres-sion in colorectal cancers may contribute to the establish-ment of liver metastasis, particularly at the step in whichcancer cells leave the circulation to extravasate into the livertissue. In addition, PRL-3 is expected to be a promisingbiomarker for identifying colorectal cancer patients at highrisk for distant metastases.
INTRODUCTIONColorectal cancer is the third most common malignant
neoplasm worldwide (1) and the second leading cause of deathdue to cancer in the United States (2). Despite recent advancesin diagnostic and therapeutic measures, the prognosis of colo-rectal cancer patients with distant metastasis still remains poor.In addition, not a few colorectal cancer patients suffer from theunexpected development of occult metastases, especially in theliver and lung, after the curative resection of their primarytumors. Therefore, it is necessary to clarify the molecular mech-anism(s) involved in metastasis and to identify the specificbiomarkers of colorectal cancer metastasis. To identify the con-sistent genetic alterations associated with the transition fromprimary colorectal cancers to liver metastases, Saha et al. (3)performed global gene expression profiles using a serial analysisof gene expression approach and found that PRL-3 (phosphataseof regenerating liver-3/PTP4A3) was frequently overexpressedin the liver metastases studied, but expressed at lower levels inprimary tumors and normal colorectal epithelium.
Protein tyrosine phosphatases play a fundamental role inregulating diverse proteins that essentially participate in everyaspect of cellular physiologic and pathogenic processes (4).PRL-1, -2, and -3 represent a novel class of protein tyrosinephosphatase superfamily members in that they possess a uniqueCOOH-terminal prenylation motif with a protein tyrosine phos-phatase-active site signature sequence CX5R (5, 6). PRLs werefound to be associated with the early endosome and plasmamembrane in their prenylated state, whereas nuclear localizationof these phosphatases may occur in the absence of prenylation(7). Although the PRLs share 75% amino acid sequence simi-larity, the ScanProsite analysis revealed that the potential sitesof phosphorylation by several kinases are quite different (6, 8).Moreover, Northern blot analysis has demonstrated that thepreferential mRNA expression pattern of these PRLs also dif-fered among organs, indicating that PRLs are quite divergent intheir functions (6, 9). PRL-1, the founding member of PRLphosphatases, was originally identified as an immediate earlygene, the expression of which was induced in mitogen-stimu-lated cells and in the regenerating liver (10, 11). Overexpressionof PRL-1 and PRL-2 has been found to transform mouse fibro-blasts and hamster pancreatic epithelial cells in culture and topromote tumor growth in nude mice, suggesting that both ofthese PRLs may participate in tumorigenesis (5, 8). Similarly,
Received 3/16/04; revised 6/22/04; accepted 7/27/04.Grant support: Japan Society for the Promotion of Science (no.15790180, S. Semba), and a Grant-in-aid for Cancer Research from theMinistry of Health, Welfare and Labor, Japan (no. 14–10, H. Yokozaki).The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.Requests for reprints: Shuho Semba, Division of Surgical Pathology,Department of Biomedical Informatics, Kobe University GraduateSchool of Medicine, 7-5-1 Kusunoki-cho, Chuo-ku, Kobe 650-0017,Japan. Phone: 81-78-382-5462; Fax: 81-78-382-5479; E-mail: [email protected].
©2004 American Association for Cancer Research.
7318 Vol. 10, 7318–7328, November 1, 2004 Clinical Cancer Research
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

PRL-3 has been found to enhance the growth of human embry-onic kidney fibroblasts (9). Although the expression of PRL-1and PRL-2 has been detected widely in various organs, humanPRL-3 is expressed predominantly in the heart, striated musclecells, and smooth muscle cells, with lower level of expression inthe pancreas (9). Zeng et al. (12) demonstrated recently thatoverexpression of PRL-3 in Chinese hamster ovary cells en-hanced the motility and invasive ability of these cells, suggest-ing that high expression of PRL-3 phosphatase may be one ofthe key alterations contributing to the metastasis of the trans-formed cells.
In the current study, we evaluated the role of PRL-3 inhuman colon cancer DLD-1 cells, especially targeting theirproliferation, motility, and hepatic colonization by down-regu-lating PRL-3 expression with small interfering RNA. We alsoexamined the levels of PRL-3 expression in both primary andmetastatic human colorectal cancers and investigated the rela-tionships with clinicopathological features, including patientoutcome.
MATERIALS AND METHODSCell Lines and Tissue Samples. Four human colon can-
cer cell lines (DLD-1, HCT-15, LoVo, and SW480) were rou-tinely maintained in RPMI 1640 (Invitrogen Co., Carlsbad, CA)supplemented with heat-inactivated 10% (v/v) fetal bovine se-rum (Invitrogen Co.) at 37°C in a humidified atmosphere of 95%air and 5% CO2.
A total of 197 patients who underwent surgical resection ofprimary and/or metastatic colorectal cancer between January1998 and December 2002 at Kobe University Hospital wereinvestigated. Formalin-fixed and paraffin-embedded specimensfrom 177 colorectal cancer patients who underwent surgicalresection for primary tumors and from 30 colorectal cancerpatients who underwent surgery for the resection of metastatictumors were collected (Tables 1 and 2). Both primary and
metastatic tumor specimens were collected from 10 of thesepatients. In brief, the lesions consisted of 177 primary and 92metastatic colorectal cancers (lymph node metastases, 59 cases;liver metastases, 23 cases; lung metastases, 6 cases; and perito-neal dissemination, 4 cases). Informed consent was obtainedfrom all of the patients, and no patient received any type oftherapy pre- or postsurgery. Histologic classification and clini-copathological staging were performed according to the GeneralRules for Clinical and Pathological Studies on Cancer of theColon, Rectum, and Anus (13) along with the classification ofthe International-Union Against Cancer (14).
Small Interfering RNA Transfection. For the RNA in-terference analyses, human PRL-3–specific small interfering RNA(5�-GUGACCUAUGACAAAACGCTT-3� and 5�-GCGUUUU-GUCAUAGGUCACTT-3�) and human PRL-1–specific smallinterfering RNA (5�-GAUGCAGUUCAGUUUAUAATT-3� and5�-UUAUAAACUGAACUGCAUCTT-3�) were designed andsynthesized based on the coding sequence of human PRL-3and PRL-1. Control small interfering RNA targeted Luciferase(Luc-small interfering RNA: 5�-CGUACGCGGAAUACUUC-GATT-3� and 5�-UCGAAGUAUUCCGCGUACGTT-3�), con-trol scrambled small interfering RNA for PRL-3 (scramble smallinterfering RNA 1: 5�-ACGCUAUAGCUAGAGCAACTT-3�and 5�-GUUGCUCUAGCUAUAGCGUTT-3�), and controlscrambled small interfering RNA for PRL-1 (scramble smallinterfering RNA 2: 5�-GUCAUUAAGUGUACUAGAUTT-3�and 5�-AUCUAGUACACUUAAUGACTT-3�) were also syn-thesized. All of the small interfering RNA sequences weresubjected to basic local alignment search tool search to confirmthe absence of homology to any additional known coding se-quences in the human genome. DLD-1 cells were seeded in24-well plates (1 � 105 cells/well) in RPMI 1640 plus 10% fetalbovine serum. The following day, each small interfering RNAwas added to the culture at a final concentration (0, 0.5, and 5nmol/L) in 500 �L of RPMI 1640 without antibiotics and serumin the presence of 0.8% Oligofectamine (Invitrogen Co.). Aftersmall interfering RNA transfection, cells were incubated inRPMI 1640 plus 1% fetal bovine serum. The medium wasrenewed 72 and 144 hours afterward. DLD-1 cells treated onlywith Oligofectamine were also used as a control (mock trans-fection). We confirmed that the transfection of PRL-small in-terfering RNAs and these control small interfering RNAs didnot affect the levels of �-actin using reverse transcription-PCR(RT-PCR) analyses.
Table 2 Variation of metastatic lesions in colorectal cancer patientsanalyzed in the metastatic tumor study
Metastatic tumors Number of patients
Total 30Liver 18Lung 2Dissemination 4Liver � lung 3Liver � lymph node * 2Lung � lymph node * 1
* Intra-abdominal and intrathoracic lymph node metastases wereconfirmed by histological examination. The levels of PRL-3 expressionin these lymph node metastases were also investigated.
Table 1 Variation of metastatic lesions in colorectal cancer patientsanalyzed in the primary tumor study
Primary tumors Number of patients
Total 177Metastasis (�) 100Metastasis (�) 77
Lymph node metastasis * only 36Distant metastasis† only 21
Liver 15Lung 3Dissemination 1Liver � lung 1Liver � dissemination 1
Lymph node * � distant metastasis † 20Lymph node � liver 13Lymph node � lung 3Lymph node � dissemination 2Lymph node � liver � lung 2
* Matched lymph node metastases were confirmed by histologicalexamination. The levels of PRL-3 expression in the matched lymphnode metastases were also investigated.
† Diagnosis was made pathologically in the cases with surgicalresection of metastatic tumor and made on imaging in the cases withoutsurgical treatment.
7319Clinical Cancer Research
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

Quantitative Real-Time RT-PCR Analyses. Using theRNeasy Mini kit (Qiagen, Hilden, Germany), each total RNA wasisolated from nontreated human colon cancer cell lines (DLD-1,HCT-15, LoVo, and SW480; 1 � 105) and small interferingRNA-transfected DLD-1 cells (1 � 105). Then, quantitative real-time RT-PCR analyses were performed using the ABI PRISM7700 Sequence Detection System and the QuantiTect SYBR GreenRT-PCR kit (Qiagen). Primer sets used for RT-PCR amplificationof PRL-3 and PRL-1 were as follows: PRL-3/forward, 5�-GG-GACTTCTCAGGTCGTGTC-3�; PRL-3/reverse, 5�-AGCCCCG-TACTTCTTCAGGT-3�; PRL-1/forward, 5�-ATGGCTCGAAT-GAACCGCCCAG-3�; and PRL-1/reverse, 5�-TTATTGAA-TGCAACAGTTGTTT-3�. As a control, the levels of �-actin ex-pression were also analyzed (�-actin/forward, 5�-CCACGAAAC-TACCTTCAACTCC-3�; �-actin/reverse, 5�-TCATACTCCT-GCTGCTTGCTGATCC-3�). According to the manufacturer’sinstructions, a master-mix (50 �L) of the following reaction com-ponents was prepared to the indicated end concentration: 25 �L of2 � QuantiTect SYBR Green RT-PCR Master Mix, 0.5 �mol/L ofeach forward and reverse primer, 0.5 �L of QuantiTect RT Mix, 10�L (10 ng) of total RNA, and the proper amount of RNase-freewater. After an initial incubation at 50°C for 30 minutes anddenaturation at 95°C for 15 minutes, the following cycling condi-tions (40 cycles) were used: denaturation at 94°C for 15 seconds,annealing at 60°C for 30 seconds, and elongation at 72°C for 1minute. All of the experiments were performed in triplicate.
Western Blot Analysis. To exclude the possibility thatthe protein kinase R-dependent interferon pathway activated bysmall interfering RNA transfection induced broad and compli-cating effects (15), we investigated the expression of the phos-phorylated forms of protein kinase R and the protein kinase Rsubstrate eukaryotic inhibition factor 2� in DLD-1 cells treatedwith or without small interfering RNA. For Western blotting,the cells (1 � 105) were lysed in a buffer containing 50 mmol/LTris-HCL (pH 7.4), 125 mmol/L NaCl, 0.1% Triton X-100, and5 mmol/L EDTA containing both 1% protease inhibitor (Sigma,St. Lois, MO) and 1% phosphatase inhibitor mixture II (Sigma).Protein was separated by SDS-PAGE followed by electrotrans-fer. Anti-protein kinase R, phospho-protein kinase RThr446,phospho-protein kinase RRThr451, eukaryotic inhibition factor2�, and phospho-eukaryotic inhibition factor 2�Ser51 polyclonalantibodies (1:1000 dilution; Cell Signaling, Beverly, MA) wereused in the primary reaction. Horseradish peroxidase-conjugatedgoat antirabbit IgG (Jackson ImmunoResearch LaboratoriesInc., West Grove, PA) was used as a secondary antibody.
WST-1 Cell Proliferation Assay. Cell growth and sur-vival in the presence or absence of each PRL-small interferingRNA transfection were determined using the Premix WST-1Cell Proliferation Assay System (Takara Biochemicals, Tokyo,Japan) as described elsewhere (16). Forty-eight hours after smallinterfering RNA transfection, an aliquot of 1 � 105 cells (100�L volume/well) were inoculated to triplicate wells and main-tained in phenol red-free medium for 48 hours. After incubation,10 �L of Premix WST-1 was added to each microculture well,and the plates were incubated for 30 minutes at 37°C, afterwhich absorbance at 450 nm was measured using a microplatereader. The absorbance in the cells without small interferingRNA transfection (1.272 �OD) was considered to be 100%.
Cell Motility/Invasion Assay. Cell motility and invasiveactivity were estimated using Transwells (6.5 mm in diameter;polycarbonate membrane, 8 �m pore size) coated with extra-cellular matrix gel obtained from Chemicon (Temecula, CA).Forty-eight hours after small interfering RNA transfection, analiquot of 1 � 105 cells was placed in the upper chamber with0.5 mL serum-free medium, whereas the lower chamber (24-well plate) was loaded with 1 mL of medium containing 10%fetal bovine serum. After 48 hours of incubation at 37°C with5% CO2, the cells were fixed with 4% paraformaldehyde andthen counterstained with hematoxylin. The cells that had mi-grated into the lower chamber were observed and counted undera light microscope.
Hepatic Metastasis Model. The protocol was approvedby the Kobe University Health Sciences Animal Care Commit-tee and Japanese Governmental Law 105. Eight-week–oldBALB/cA Jcl-nu female mice (housed 5 per cage) were used inthis study. Mice were locally injected to the spleen with 3 � 105
viable DLD-1 cells treated under each condition (nontreatedgroup: n 5; PRL-3–small interfering RNA group: n 5; andPRL-1–small interfering RNA group: n 5), and the mice weresacrificed under anesthesia on day 30. Liver and spleen tissueswere fixed in 10% buffered formalin (pH 7.4) and processed forroutine histology. The number and diameter of metastatic foci in5 sections per liver was determined, and the volume (V) of thesefoci was calculated using the equation V 1/2 � A � B2, whereA and B indicate long and short diameters, respectively.
In situ Hybridization Study. The specific antisense ol-igonucleotide DNA probe for PRL-3 (5�-GTTGATGGCTCCGCGGCG-3�) was designed complementary to the mRNA tran-scripts of the PRL-3 gene according to the GenBank database.The specificity of the oligonucleotide sequences was initiallydetermined by a Gen-EMBL database search using the FastAalgorithm (17), which showed minimal homology with thePRL-1 and PRL-2 genes and other nonspecific mammalian genesequences. All of the probes were synthesized with six biotinmolecules (hyperbiotinylated) at the 3� end via direct couplingusing standard phosphormidine chemistry (18, 19). We thenused multibiotinylated poly(dT)20 oligonucleotides (InvitrogenCo.) to verify the integrity of mRNA in each sample. Thelyophilized probes were reconstituted to a 1 �g/�l stock solu-tion in 10 mmol/L Tris-HCl (pH 7.6) and 1 mmol/L EDTA. Thestock solution was diluted with Brigati Probe Diluent (Invitro-gen Co.) immediately before use.
In situ hybridization was performed using manual capillaryaction technology (20) on the Microprobe Staining System(Fisher Scientific, Pittsburgh, PA). ProbeOn Plus slides (FisherScientific) were placed in the MicroProbe slide holder so as tomake a 150 �m gap, and all of the subsequent reagents wereplaced onto and drained from the slides by capillary action. Thetissue sections were dewaxed with xylene and rehydrated with1 � Tris-buffered saline-Tween 20. The tissue sections weredigested with a stable pepsin solution (DAKO, Carpinteria, CA),which was used at full strength for 3 minutes at 100°C. Hybrid-ization of the probes was carried out for 80 minutes at 60°C, andthe samples were then washed three times with 2 � SSC for 2minutes at 45°C. The samples were incubated with alkalinephosphatase-labeled avidin (Biomeda, Foster City, CA) for 30
7320 High Expression of PRL-3 in Human Colorectal Cancer
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

minutes at 45°C, briefly rinsed in 20 � Tris-buffered saline-Tween 20, rinsed with alkaline phosphatase enhancer (Invitro-gen Co.) for 1 minute, and finally incubated with the chromogensubstrate Fast Red (Biomeda) for 20 minutes at 37°C. Hybrid-ization of the samples with biotinylated poly(dT)20 probes wasalways performed to verify the integrity of mRNA. To analyzethe specificity of the hybridization signal, we performed RNasepretreatment of the tissue sections and competition assays withan unlabeled antisense probe as control procedures. Then, ascontrols for endogenous alkaline phosphatase activity, we per-formed treatment of the samples in the absence of the biotiny-lated probes and the use of chromogen in the absence of anyprobes. We confirmed that no signal was detected under any ofthese conditions.
For the evaluation of in situ hybridization reactivity, nor-mal colorectal epithelium tissue, smooth muscle cells of thevessel, and lymphocytes were used as internal controls. Thereactivity of in situ hybridization was graded as follows: highPRL-3 expression, 10% cancer cells showed PRL-3 expres-sion exceeding that of the internal controls; and low PRL-3expression, 90% cancer cells showed no increase in theexpression of PRL-3 compared with the internal controls. Thelevels of in situ hybridization reactivity were evaluated inde-pendently by three pathologists (H. K., S. S., and U. A. M.).
Prognosis Study. Among our cohort of 150 colorectalcancer patients clinically diagnosed as free of distant metastasesat the time of curative resection of their primary tumors, 104 hadcomplete follow-up information. For these 104 patients, weevaluated the associations between clinicopathological features,including the levels of PRL-3 expression in the primary tumorsand the incidence of metachronous liver and/or lung metastasisafter curative surgery for primary tumors. All of the patientsunderwent imaging examinations (computed tomography andultrasonography) at regular intervals. The median follow-upperiod was 2.8 years (range, 5 months to 6.1 years).
Statistical Analyses. The results of the in vitro assays, invivo metastasis assay, and the in situ hybridization studies wereinvestigated by �2 test. The time to the appearance of metachro-nous metastasis in cases with high and low expression of PRL-3
after surgical resection for primary tumor was compared byperforming a Kaplan-Meier analysis and testing the results withthe log-rank statistic. We also evaluated the association betweenthe incidence of metachronous metastasis and the conventionalindicators, such as primary tumor size, angiolymphatic invasion,and the presence of lymph node metastasis by Kaplan-Meieranalyses. A P � 0.05 was regarded as statistically significant.
RESULTSExpression of PRL-3 in Human Colon Cancer Cell
Lines. We first examined the expression of PRL-3 and PRL-1in 4 human colon cancer cell lines and normal colonic epithe-lium by quantitative real-time RT-PCR analyses. AlthoughPRL-3 and PRL-1 were expressed in all of the cell lines, theexpression levels varied significantly. The DLD-1 cells demon-strated the highest level of PRL-3 expression, which was equiv-alent to about three times that of the normal colonic epithelium,whereas the LoVo and SW480 cells showed low levels of PRL-3expression that were almost equal to that of the normal colonicepithelium (Fig. 1A). As shown in Fig. 1B, DLD-1 cells alsodemonstrated high PRL-1 expression equivalent to about twicethat of the normal colonic epithelium, and therefore we per-formed the following RNA interference studies using theDLD-1 cells.
Transfection of PRL-3-Small Interfering RNA Down-Regulates Endogenous PRL-3 without Activating the ProteinKinase R-Dependent Interferon Pathway. Transfection ofPRL-3 and PRL-1–small interfering RNAs into DLD-1 cellsdown-regulated the level of the expression of each type of PRLin a concentration-dependent manner (Fig. 2, A and B). After thetransfection of PRL-3– and PRL-1–small interfering RNAs (5nmol/L), the expression of each PRL reached a minimal level at48 to 96 hours, which gradually recovered to almost the baselinelevel at 168 hours (Fig. 2, C and D). According to these results,we performed Western blotting analysis, WST-1 cell prolifera-tion assay, cell motility/invasion assay, and hepatic metastasisassay experiments using DLD-1 cells 48 hours after smallinterfering RNA (5 nmol/L) transfection. Western blotting anal-ysis showed that the transfection of each small interfering RNA
Fig. 1 The levels of PRL-3 and PRL-1 expression in human colon cancer cell lines. A and B, various levels of PRL-3 and PRL-1 expression weredetected by quantitative real-time RT-PCR analyses. The results are shown as the ratio between each PRL and �-actin.
7321Clinical Cancer Research
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

(5 nmol/L) had no influence on the levels of expression of thephosphorylated forms of protein kinase R and eukaryotic inhi-bition factor 2� (Fig. 2E), thereby demonstrating that the exog-enously transfected small interfering RNAs (5 nmol/L) did notactivate the protein kinase R-dependent interferon pathway inthe DLD-1 cells. In contrast, the DLD-1 cells treated with 0.1�mol/L Calyculin A (serine/threonine phosphatase inhibitor;Cell Signaling) demonstrated quite high levels of expression ofthe phosphorylated forms of protein kinase R and eukaryoticinhibition factor 2� (Fig. 2E).
Transfection of PRL-3-Small Interfering RNA Sup-presses Cancer Cell Motility Accompanied with Morpho-logic Alterations. As a starting point of our attempt to deter-mine the significance of PRL-3 in the progression anddevelopment of human colorectal cancer, we evaluated the rolesof PRL-3 in cell proliferation and motility/invasion using anRNA interference technique. First, we attempted to analyze theeffect of PRL-3 and PRL-1 on cell growth in vitro. Transfectionof these PRL-small interfering RNAs did not alter the cellgrowth of DLD-1 cells, as compared with the nontreated control
Fig. 2 Specific suppression of endogenous PRLs by small interfering RNA transfection in human colon cancer DLD-1 cells. A and B, theconcentration-dependent effects of PRL-3- and PRL-1-small interfering RNAs. The levels of PRL-3 and PRL-1 expression in DLD-1 cells wereexamined 48 hours after small interfering RNA transfection at different concentrations (0, 0.5, and 5 nmol/L) using quantitative real-time RT-PCR.As controls, the levels of PRL-3 and PRL-1 expression in DLD-1 cells transfected with Luciferase-small interfering RNA (Luc-si RNA), scrambledsmall interfering RNA for PRL-3 (scramble 1), and scrambled small interfering RNA for PRL-1 (scramble 2) were also examined. C and D, the timecourse of PRL-3 and PRL-1 expression after small interfering RNA transfection. The levels of PRL-3 and PRL-1 expression in DLD-1 cells wereexamined at the indicated time after small interfering RNA (5 nmol/L) transfection using quantitative real-time RT-PCR. The expression of each PRLwas analyzed 0 to 168 hours after small interfering RNA transfection. As controls, the levels of PRL-3 and PRL-1 expression in DLD-1 cells treatedwith only Oligofectamine (mock), Luciferase-small interfering RNA (Luc-siRNA), scrambled small interfering RNA for PRL-3 (scramble 1), andscrambled small interfering RNA for PRL-1 (scramble 2) were also examined. E, phosphorylation of PKR and eIF2� in response to small interferingRNA transfection. The expression of phosphorylated forms of PKR and the PKR substrate eIF2� in DLD-1 cells with or without small interferingRNA transfection were examined by Western blotting analyses. As a control, the DLD-1 cells treated with 0.1 �mol/L of the serine/threoninephosphatase inhibitor Calyculin A were used. (PKR, protein kinase R; eIF2�, eukaryotic inhibition factor 2�)
7322 High Expression of PRL-3 in Human Colorectal Cancer
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from
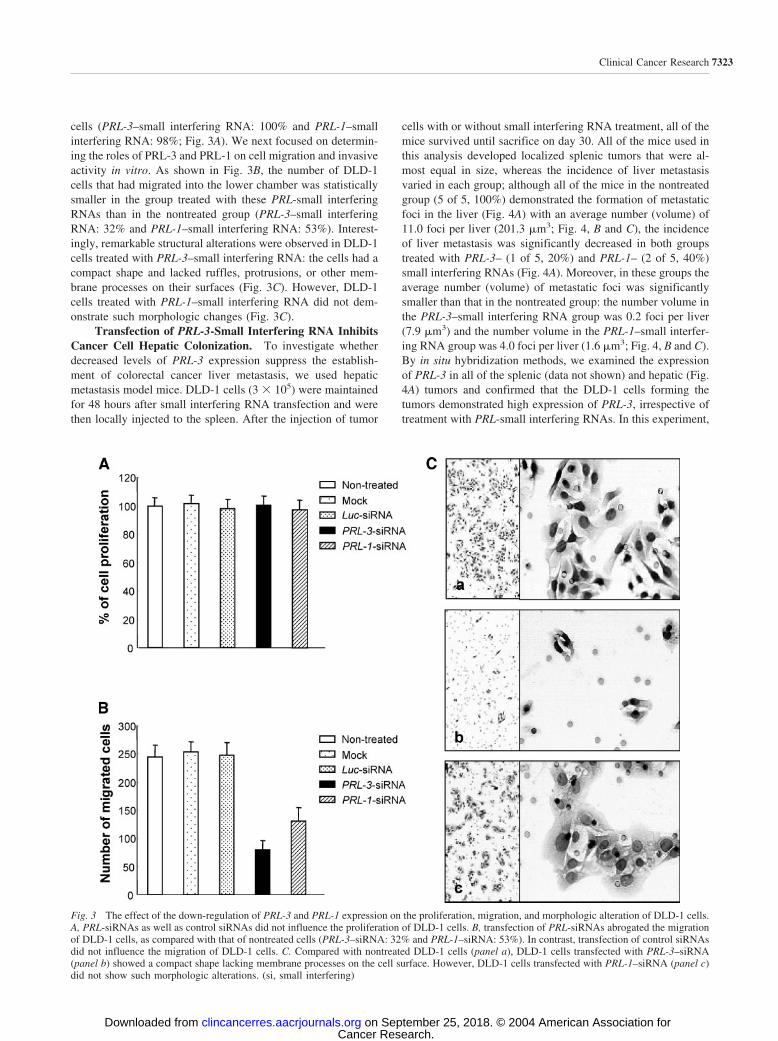
cells (PRL-3–small interfering RNA: 100% and PRL-1–smallinterfering RNA: 98%; Fig. 3A). We next focused on determin-ing the roles of PRL-3 and PRL-1 on cell migration and invasiveactivity in vitro. As shown in Fig. 3B, the number of DLD-1cells that had migrated into the lower chamber was statisticallysmaller in the group treated with these PRL-small interferingRNAs than in the nontreated group (PRL-3–small interferingRNA: 32% and PRL-1–small interfering RNA: 53%). Interest-ingly, remarkable structural alterations were observed in DLD-1cells treated with PRL-3–small interfering RNA: the cells had acompact shape and lacked ruffles, protrusions, or other mem-brane processes on their surfaces (Fig. 3C). However, DLD-1cells treated with PRL-1–small interfering RNA did not dem-onstrate such morphologic changes (Fig. 3C).
Transfection of PRL-3-Small Interfering RNA InhibitsCancer Cell Hepatic Colonization. To investigate whetherdecreased levels of PRL-3 expression suppress the establish-ment of colorectal cancer liver metastasis, we used hepaticmetastasis model mice. DLD-1 cells (3 � 105) were maintainedfor 48 hours after small interfering RNA transfection and werethen locally injected to the spleen. After the injection of tumor
cells with or without small interfering RNA treatment, all of themice survived until sacrifice on day 30. All of the mice used inthis analysis developed localized splenic tumors that were al-most equal in size, whereas the incidence of liver metastasisvaried in each group; although all of the mice in the nontreatedgroup (5 of 5, 100%) demonstrated the formation of metastaticfoci in the liver (Fig. 4A) with an average number (volume) of11.0 foci per liver (201.3 �m3; Fig. 4, B and C), the incidenceof liver metastasis was significantly decreased in both groupstreated with PRL-3– (1 of 5, 20%) and PRL-1– (2 of 5, 40%)small interfering RNAs (Fig. 4A). Moreover, in these groups theaverage number (volume) of metastatic foci was significantlysmaller than that in the nontreated group: the number volume inthe PRL-3–small interfering RNA group was 0.2 foci per liver(7.9 �m3) and the number volume in the PRL-1–small interfer-ing RNA group was 4.0 foci per liver (1.6 �m3; Fig. 4, B and C).By in situ hybridization methods, we examined the expressionof PRL-3 in all of the splenic (data not shown) and hepatic (Fig.4A) tumors and confirmed that the DLD-1 cells forming thetumors demonstrated high expression of PRL-3, irrespective oftreatment with PRL-small interfering RNAs. In this experiment,
Fig. 3 The effect of the down-regulation of PRL-3 and PRL-1 expression on the proliferation, migration, and morphologic alteration of DLD-1 cells.A, PRL-siRNAs as well as control siRNAs did not influence the proliferation of DLD-1 cells. B, transfection of PRL-siRNAs abrogated the migrationof DLD-1 cells, as compared with that of nontreated cells (PRL-3–siRNA: 32% and PRL-1–siRNA: 53%). In contrast, transfection of control siRNAsdid not influence the migration of DLD-1 cells. C. Compared with nontreated DLD-1 cells (panel a), DLD-1 cells transfected with PRL-3–siRNA(panel b) showed a compact shape lacking membrane processes on the cell surface. However, DLD-1 cells transfected with PRL-1–siRNA (panel c)did not show such morphologic alterations. (si, small interfering)
7323Clinical Cancer Research
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

formation of metastasis was not observed in any organs exceptthe liver.
High Expression of PRL-3 Is Closely Correlated withLiver and Lung Metastases in Human Colorectal Cancer.We next investigated the levels of PRL-3 expression in humancolorectal cancers using in situ hybridization methods. Weak
PRL-3 expression was detected in every sample of normalcolorectal epithelium analyzed, and the levels were almost thesame in each sample (Fig. 5). However, in the primary colorec-tal cancer cells, PRL-3 expression varied significantly amongcases and was distributed heterogeneously. The results are sum-marized in Table 3. In general, a high level of PRL-3 expression
Fig. 4 The effect of the down-regulation of PRL-3 and PRL-1 expression on DLD-1 cells hepatic colonization in mice. A, representative sectionsof hepatic foci formed by nontreated DLD-1 cells are shown in (panel a, �20, H&E; panel b, �200, H&E; and panel c, �200, in situ hybridization).Representative sections of hepatic foci formed by PRL-3–small interfering RNA-transfected DLD-1 cells (panel d, �20, H&E; panel e, �200, H&E;and panel f, �200, in situ hybridization) and by PRL-1–small interfering RNA-transfected DLD-1 cells (panel g, �20, H&E; panel h, �200, H&E;and panel i, �200, in situ hybridization) are also exhibited. T and H, tumor and hepatocytes, respectively. B and C, compared with the control groupinjected with nontreated DLD-1 cells, both the number and volume of hepatic metastatic foci decreased significantly in the mice injected withPRL-small interfering RNA-treated DLD-1 cells. Especially in terms of the number of metastatic foci, PRL-3–small interfering RNA demonstratedmore remarkable abrogation than PRL-1–small interfering RNA. The data represent average values from 5 mice per group SD.
7324 High Expression of PRL-3 in Human Colorectal Cancer
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

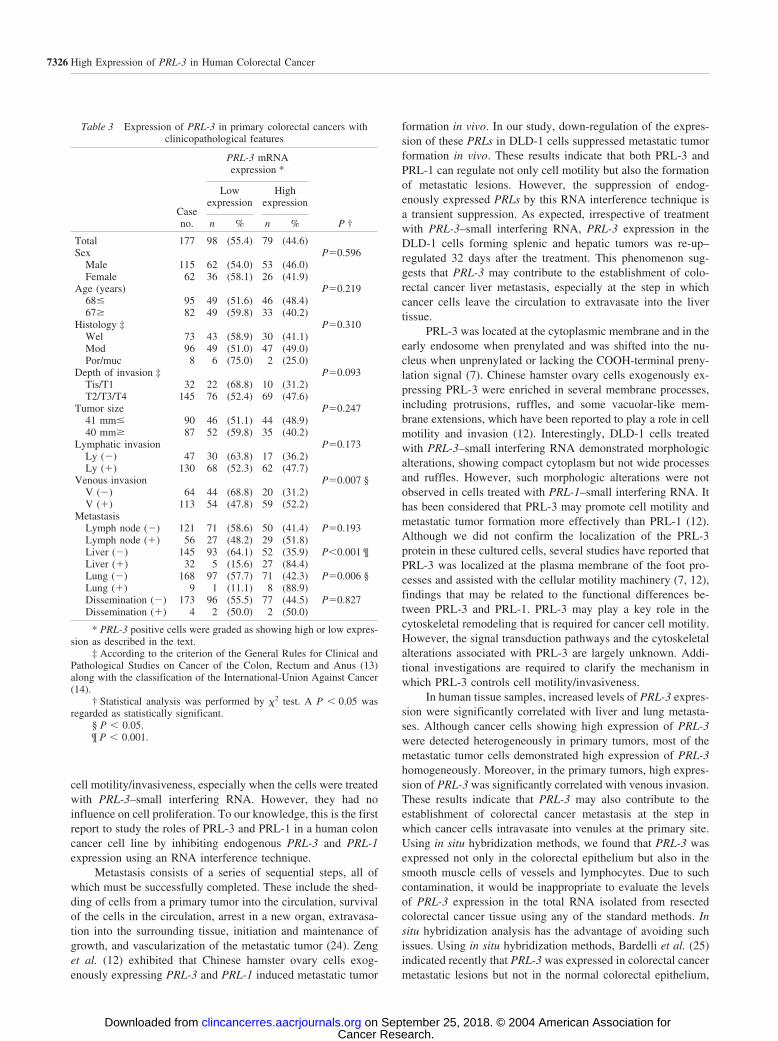
was observed in 79 (44.6%) of 177 primary colorectal cancers.The frequency of high expression of PRL-3 in cases with distantmetastasis (liver: 84.4% and lung: 88.9%) was statisticallyhigher than that in the cases without distant metastasis [liver:35.9% (P � 0.001) and lung: 42.3% (P 0.006)]. Simulta-neously, high expression of PRL-3 was detected more frequentlyin the cases with venous invasion (52.2%) than in the caseswithout venous invasion (31.2%, P 0.007). Cancer cellsforming intravenous tumor emboli showed strikingly increasedPRL-3 expression (Fig. 6A). However, the levels of PRL-3expression showed no correlation with any other clinicopatho-logical features such as depth of invasion, tumor size, lymphaticinvasion, or the presence of lymph node metastasis.
In metastatic colorectal cancer lesions, high expression ofPRL-3 was found in 21 of 23 (91.3%) cases of liver metastasisand in 6 of 6 (100%) cases of lung metastasis (Table 4).However, in lymph node metastasis and peritoneal dissemina-tion, high expression of PRL-3 was observed in only 47.5% and50.0% of the total number of cases, respectively. In liver andlung metastatic lesions, almost all of the colorectal cancer cellshomogeneously demonstrated high expression of PRL-3. Re-garding the 10 cases for which serial analyses of both primaryand metastatic tumors could be performed, all of the specimensshowed high expression of PRL-3 (Fig. 6, C–F).
PRL-3 Is a Predictive Molecular Marker of Liver andLung Metastases after Curative Surgery for Primary Colo-rectal Cancer. Finally, we performed a prognosis study todetermine whether PRL-3 could be used as a biomolecularmarker to monitor the risk of metachronous metastasis aftercurative surgery for primary colorectal cancer. Overall, postop-erative development of occult liver and/or lung metastasis ap-
peared in 14 of 104 cases (13.5%). As shown in Fig. 7, Kaplan-Meier analysis showed that cases with high PRL-3 expressionhad a greater risk for the development of metachronous metas-tasis than those with low PRL-3 expression (P � 0.0001).However, the incidence of metachronous metastasis was notstatistically related to the conventional indicators such as pri-mary tumor size, angiolymphatic invasion, and the presence oflymph node metastasis (data not shown).
DISCUSSIONUsing serial analysis of gene expression technology, which
is a powerful strategy for the detection of altered gene expres-sion, PRL-3 was recognized as the most important molecule thatwas consistently and specifically activated in liver metastases ofhuman colorectal cancers (3). As a member of the PRL familyof phosphatases, PRL-3 has a catalytic active signature motif(C104S; refs. 9, 21). Stable expression of wild-type activePRL-3 dramatically enhanced cell migration, whereas the cata-lytically inactive PRL-3 (C104S) mutant greatly reduced thepromotion of cell migration (12). These results indicated that theability of PRL-3 to promote cell migration depended on itsphosphatase activity. Protein prenylation is important in target-ing proteins to intracellular membranes and in protein-proteininteractions (22, 23). PRL-3 has been reported to be a memberof the prenylated protein phosphatase family, and the metastaticproperties of PRL-3, like those of PRL-1, are dependent on itsprenylation activity (7, 12). In the current study, we investigatedthe roles of PRL-3 and PRL-1 in human colon cancer DLD-1cells, targeting in particular their metastasis-related activities.Decreased levels of the expression of these PRLs suppressed
Fig. 5 Evaluation of PRL-3expression in primary colorec-tal cancer. The expression ofPRL-3 was assessed using insitu hybridization methods. A,low expression of PRL-3 incancer cells. No increase in thereactivity of the cancer cellswas observed when they werecompared with the adjacentnormal colorectal epithelium.B, serial section of A, stainedwith H&E. C, high expressionof PRL-3 in cancer cells. Thereactivity in the cancer cellsclearly exceeded that of the ad-jacent normal colorectal epithe-lium. D, serial section of C,stained with H&E. T and N, tu-mor and normal colorectal epi-thelium, respectively. Bars 50 �m.
7325Clinical Cancer Research
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from
cell motility/invasiveness, especially when the cells were treatedwith PRL-3–small interfering RNA. However, they had noinfluence on cell proliferation. To our knowledge, this is the firstreport to study the roles of PRL-3 and PRL-1 in a human coloncancer cell line by inhibiting endogenous PRL-3 and PRL-1expression using an RNA interference technique.
Metastasis consists of a series of sequential steps, all ofwhich must be successfully completed. These include the shed-ding of cells from a primary tumor into the circulation, survivalof the cells in the circulation, arrest in a new organ, extravasa-tion into the surrounding tissue, initiation and maintenance ofgrowth, and vascularization of the metastatic tumor (24). Zenget al. (12) exhibited that Chinese hamster ovary cells exog-enously expressing PRL-3 and PRL-1 induced metastatic tumor
formation in vivo. In our study, down-regulation of the expres-sion of these PRLs in DLD-1 cells suppressed metastatic tumorformation in vivo. These results indicate that both PRL-3 andPRL-1 can regulate not only cell motility but also the formationof metastatic lesions. However, the suppression of endog-enously expressed PRLs by this RNA interference technique isa transient suppression. As expected, irrespective of treatmentwith PRL-3–small interfering RNA, PRL-3 expression in theDLD-1 cells forming splenic and hepatic tumors was re-up–regulated 32 days after the treatment. This phenomenon sug-gests that PRL-3 may contribute to the establishment of colo-rectal cancer liver metastasis, especially at the step in whichcancer cells leave the circulation to extravasate into the livertissue.
PRL-3 was located at the cytoplasmic membrane and in theearly endosome when prenylated and was shifted into the nu-cleus when unprenylated or lacking the COOH-terminal preny-lation signal (7). Chinese hamster ovary cells exogenously ex-pressing PRL-3 were enriched in several membrane processes,including protrusions, ruffles, and some vacuolar-like mem-brane extensions, which have been reported to play a role in cellmotility and invasion (12). Interestingly, DLD-1 cells treatedwith PRL-3–small interfering RNA demonstrated morphologicalterations, showing compact cytoplasm but not wide processesand ruffles. However, such morphologic alterations were notobserved in cells treated with PRL-1–small interfering RNA. Ithas been considered that PRL-3 may promote cell motility andmetastatic tumor formation more effectively than PRL-1 (12).Although we did not confirm the localization of the PRL-3protein in these cultured cells, several studies have reported thatPRL-3 was localized at the plasma membrane of the foot pro-cesses and assisted with the cellular motility machinery (7, 12),findings that may be related to the functional differences be-tween PRL-3 and PRL-1. PRL-3 may play a key role in thecytoskeletal remodeling that is required for cancer cell motility.However, the signal transduction pathways and the cytoskeletalalterations associated with PRL-3 are largely unknown. Addi-tional investigations are required to clarify the mechanism inwhich PRL-3 controls cell motility/invasiveness.
In human tissue samples, increased levels of PRL-3 expres-sion were significantly correlated with liver and lung metasta-ses. Although cancer cells showing high expression of PRL-3were detected heterogeneously in primary tumors, most of themetastatic tumor cells demonstrated high expression of PRL-3homogeneously. Moreover, in the primary tumors, high expres-sion of PRL-3 was significantly correlated with venous invasion.These results indicate that PRL-3 may also contribute to theestablishment of colorectal cancer metastasis at the step inwhich cancer cells intravasate into venules at the primary site.Using in situ hybridization methods, we found that PRL-3 wasexpressed not only in the colorectal epithelium but also in thesmooth muscle cells of vessels and lymphocytes. Due to suchcontamination, it would be inappropriate to evaluate the levelsof PRL-3 expression in the total RNA isolated from resectedcolorectal cancer tissue using any of the standard methods. Insitu hybridization analysis has the advantage of avoiding suchissues. Using in situ hybridization methods, Bardelli et al. (25)indicated recently that PRL-3 was expressed in colorectal cancermetastatic lesions but not in the normal colorectal epithelium,
Table 3 Expression of PRL-3 in primary colorectal cancers withclinicopathological features
Caseno.
PRL-3 mRNAexpression *
P †
Lowexpression
Highexpression
n % n %
Total 177 98 (55.4) 79 (44.6)Sex P0.596
Male 115 62 (54.0) 53 (46.0)Female 62 36 (58.1) 26 (41.9)
Age (years) P0.21968� 95 49 (51.6) 46 (48.4)67� 82 49 (59.8) 33 (40.2)
Histology ‡ P0.310Wel 73 43 (58.9) 30 (41.1)Mod 96 49 (51.0) 47 (49.0)Por/muc 8 6 (75.0) 2 (25.0)
Depth of invasion ‡ P0.093Tis/T1 32 22 (68.8) 10 (31.2)T2/T3/T4 145 76 (52.4) 69 (47.6)
Tumor size P0.24741 mm� 90 46 (51.1) 44 (48.9)40 mm� 87 52 (59.8) 35 (40.2)
Lymphatic invasion P0.173Ly (�) 47 30 (63.8) 17 (36.2)Ly (�) 130 68 (52.3) 62 (47.7)
Venous invasion P0.007 §V (�) 64 44 (68.8) 20 (31.2)V (�) 113 54 (47.8) 59 (52.2)
MetastasisLymph node (�) 121 71 (58.6) 50 (41.4) P0.193Lymph node (�) 56 27 (48.2) 29 (51.8)Liver (�) 145 93 (64.1) 52 (35.9) P�0.001 ¶Liver (�) 32 5 (15.6) 27 (84.4)Lung (�) 168 97 (57.7) 71 (42.3) P0.006 §Lung (�) 9 1 (11.1) 8 (88.9)Dissemination (�) 173 96 (55.5) 77 (44.5) P0.827Dissemination (�) 4 2 (50.0) 2 (50.0)
* PRL-3 positive cells were graded as showing high or low expres-sion as described in the text.
‡ According to the criterion of the General Rules for Clinical andPathological Studies on Cancer of the Colon, Rectum and Anus (13)along with the classification of the International-Union Against Cancer(14).
† Statistical analysis was performed by �2 test. A P � 0.05 wasregarded as statistically significant.
§ P � 0.05.¶ P � 0.001.
7326 High Expression of PRL-3 in Human Colorectal Cancer
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

nonmetastatic colorectal cancer, or gastric cancer. In the presentstudy, on the other hand, we observed PRL-3 expression in thenormal colorectal epithelium and in nonmetastatic colorectalcancer tissue. This inconsistency may have been due to differ-ences in the number of cases studied, the criteria for the eval-uation of in situ hybridization signals, or the specificity of theantisense DNA probe for PRL-3. However, our in situ hybrid-ization analyses enabled the detection of slight differences inPRL-3 expression among primary colorectal cancer cases, andwe also found PRL-3 expression in cases of gastric cancersusing RT-PCR and in situ hybridization analyses (data notshown).
Unexpectedly, colorectal cancer patients develop meta-chronous liver (8.9%) and/or lung (6.0%) metastasis after cura-tive surgery for the primary tumors (26). Thus, it would be ofgreat value to identify a promising biomarker for the metachro-nous metastasis of colorectal cancer. In addition to our in situhybridization analyses of colorectal cancer tumor samples, ourprognosis study suggested that the level of PRL-3 expression in
the primary colorectal cancer lesion is a more promising pre-dictor of the postoperative development of metachronous liverand/or lung metastasis than such conventional predictors astumor size, angiolymphatic invasion, or the presence of lymphnode metastasis. High expression of PRL-3 in surgical and
Table 4 Expression of PRL-3 in metastatic lesions
Case no.
PRL-3 mRNA expression *
Lowexpression
Highexpression
N % N %
Lymph node 59 31 (52.5) 28 (47.5)Liver 23 2 (8.7) 21 (91.3)Lung 6 0 (0) 6 (100)Dissemination 4 2 (50.0) 2 (50.0)
* PRL-3 positive cells were graded as showing high or low expres-sion as described in the text.
Fig. 6 In situ hybridizationanalyses of the expression ofPRL-3 in cases of colorectalcancer with intensive venousinfiltration and distant metasta-sis. A, representative case ofrectal cancer with intensive ve-nous invasion. Cancer cellsforming intravenous tumor em-boli (T) showed significantlyincreased PRL-3 expression. Vand A, vein and artery, respec-tively. B, serial section of A,stained with H&E. C and D,representative case of sigmoidcolon cancer with liver metas-tasis. Cancer cells with highPRL-3 expression were de-tected heterogeneously in theprimary tumor (C), whereas inthe liver metastasis (D) almostall of the cancer cells homoge-neously demonstrated high ex-pression of PRL-3. E and F,representative case of rectalcancer with lung metastasis.Cancer cells with high PRL-3expression were detected heter-ogeneously in the primary tu-mor (E), whereas in the lungmetastasis (F) almost all of thecancer cells homogeneouslydemonstrated high expressionof PRL-3. Bars 50 �m.
7327Clinical Cancer Research
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

biopsied colorectal cancer specimens may provide cliniciansuseful information not only for identifying occult metastases butalso for initiating adjuvant chemotherapy after the surgicaltreatment of the primary tumor. Moreover, PRL-3 may providea novel therapeutic target for intractable colorectal cancer me-tastasis. Although the specific protein substrate for PRL-3 hasnot yet been identified, the function of PRL-3 in metastasiscould be blocked or reduced by inhibiting prenylation and/orinactivating the catalytic function of the PRL-3 phosphataseactive site (12). Additional investigations will be necessary toclarify the role(s) of PRL-3 in the process of colorectal cancermetastasis and to develop inhibitors against PRL-3 itself.
ACKNOWLEDGMENTSThe authors would like to thank Dr. Hiroki Kuniyasu (Department
of Oncological Pathology, Nara Medical University) for his valuableinput, and Ms. Akiko Obata for her technical assistance.
REFERENCES1. Shike M, Winawer SJ, Greenwald PH, Bloch A, Hill MJ, SwaroopSV. Primary prevention of colorectal cancer. The WHO CollaboratingCenter for the Prevention of Colorectal Cancer. Bull World HealthOrgan 1990;68:377–85.2. Winawer SJ, Fletcher RH, Miller L, et al. Colorectal cancer screen-ing: clinical guidelines and rationale. Gastroenterology 1997;112:594–642.3. Saha S, Bardelli A, Buckhaults P, et al. A phosphatase associatedwith metastasis of colorectal cancer. Science (Wash DC) 2001;294:1343–6.4. Zhang ZY, Zhou B, Xie L. Modulation of protein kinase signaling byprotein phosphatases and inhibitor. Pharmacol Ther 2002;93:307–17.5. Cates CA, Michael RL, Stayrook KR, et al. Prenylation of oncogenichuman PTP (CAAX) protein tyrosine phosphatases. Cancer Lett 1996;110:49–55.
6. Zeng Q, Hong W, Tan YH. Mouse PRL-2 and PRL-3, two poten-tially prenylated protein tyrosine phosphatases homologous to PRL-1.Biochem Biophys Res Commun 1998;244:421–7.7. Zeng Q, Si X, Horstmann H, Xu Y, Hong W, Pallen CJ. Prenylation-dependent association of protein-tyrosine phosphatases PRL-1, -2, and-3 with the plasma membrane and the early endosome. J Biol Chem2000;275:21444–52.8. Diamond RH, Cressman DE, Laz TM, Abrams CS, Taub R. PRL-1,a unique nuclear protein tyrosine phosphatase, affects cell growth. MolCell Biol 1994;14:3752–62.9. Matter WF, Estridge T, Zhang C, et al. Role of PRL-3, a humanmuscle-specific tyrosine phosphatase, in angiotensin-II signaling. Bio-chem Biophys Res Commun 2001;83:1061–8.10. Mohn KL, Laz TM, Hsu JC, Melby AE, Bravo R, Taub R. Theimmediate-early growth response in regenerating liver and insulin-stimulated H35 cells comparison with serum stimulated 3T3 cells andidentification of 41 novel immediate-early genes. Mol Cell Biol 1991;11:381–90.11. Montagna M, Serova O, Sylla BS, Feunteun J, Lenior GM. A100-kb physical and transcriptional map around the EDH17B2 gene:identification of three novel genes and a pseudogene of a humanhomologue of the rat PRL-1 tyrosine phosphatase. Hum Genet 1995;96:532–8.12. Zeng Q, Dong J, Guo K, et al. PRL-3 and PRL-1 promote cellmigration, invasion and metastasis. Cancer Res 2003;63:2716–22.13. Japanese Society for Cancer of Colon and Rectum. General rules forclinical and pathological studies on cancer of the colon, rectum andanus. 6th ed. Tokyo: Kanehara Public Co; 1998.14. Sobin LH, Wittekind CH. UICC TNM Classification of malignanttumors. 5th ed. New York: John Wiley & Sons, Inc; 1997.15. Carol AS, Michelle H, Michael JdV, Robert HS, Bryan RGW.Activation of the interferon system by short-interfering RNAs. Nat CellBiol 2003;5:834–9.16. Semba S, Itoh N, Ito M, Harada M, Yamakawa M. The in vitro andin vivo effects of 2-(4-morpholinyl)-8-phenylchrmone (LY294002), aspecific inhibitor of phosphatidyl inositol 3�-kinase in human coloncancer cells. Clin Cancer Res 2002;8:1957–63.17. Pearson WR, Lipman DJ. Improved tools for biological sequencecomparison. Proc Natl Acad Sci USA 1988;85:2444–8.18. Park CS, Manahan LJ, Brigati DJ. Automated molecular pathology:one hour in situ DNA hybridization. J Histotechnol 1991;14:219–29.19. Caruthers MH, Becaucage SL, Efcavitch JW, et al. Chemical syn-thesis and biological studies on mutated gene-control region. ColdSpring Harbor Symp Quant Biol 1982;47:411–8.20. Reed JA, Manahan LJ, Park CS, Brigati DJ. Complete one-hourimmunocytochemistry based on capillary action. Biotechniques 1992;13:434–43.21. Zhao Z, Lee CC, Monckton DG, et al. Characterization andgenomic mapping of genes and pseudogenes of a new human proteintyrosine phosphatase. Genomics 1996;35:172–81.22. Seabra MC. Review: Membrane association and targeting of pre-nylated Ras-like GTPases. Cell Signalling 1998;10:167–72.23. Si X, Zeng Q, Ng CH, Hong W, Pallen CJ. Interaction of farnesy-lated PRL-2, a protein-tyrosine phosphatase, with the beta-submit ofgeranylgeranyltransferase II. J Biol Chem 2001;276:32875–82.24. Chambers AF, Groom AC, MacDonald IC. Dissemination andgrowth of cancer cells in metastatic sites. Nat Rev Cancer 2002;2:563–72.25. Bardelli A, Saha S, Sager JA, et al. PRL-3 expression in metastaticcancers. Clin Cancer Res 2003;9:5607–15.26. Sadahiro S, Suzuki T, Ishikawa K, et al. Recurrence patterns aftercurative resection of colorectal cancer in patients followed for a mini-mum of ten years. Hepato-Gastroenterology 2003;50:1362–6.
Fig. 7 Kaplan-Meier analysis of metastasis-free survival according toPRL-3 levels in surgically resected primary colorectal cancers. Highexpression of PRL-3 was associated with metachronous liver and/orlung metastasis after curative surgery for the primary tumor.
7328 High Expression of PRL-3 in Human Colorectal Cancer
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

2004;10:7318-7328. Clin Cancer Res Hirotaka Kato, Shuho Semba, Upik A. Miskad, et al. MetastasesMolecular Marker of Metachronous Liver and LungLiver Metastasis in Human Colorectal Cancer: A Predictive
Promotes Cancer Cell Motility andPRL-3High Expression of
Updated version
http://clincancerres.aacrjournals.org/content/10/21/7318
Access the most recent version of this article at:
Cited articles
http://clincancerres.aacrjournals.org/content/10/21/7318.full#ref-list-1
This article cites 24 articles, 9 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/10/21/7318.full#related-urls
This article has been cited by 16 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://clincancerres.aacrjournals.org/content/10/21/7318To request permission to re-use all or part of this article, use this link
Cancer Research. on September 25, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from