Embed Size (px)
Citation preview

Improved differentiation of umbilical cord blood-derivedmesenchymal stem cells into insulin-producing cellsby PDX-1 mRNA transfection
Phuc Van Pham n, Phuoc Thi-My Nguyen, Anh Thai-Quynh Nguyen, Vuong Minh Pham,Anh Nguyen-Tu Bui, Loan Thi-Tung Dang, Khue Gia Nguyen, Ngoc Kim PhanLaboratory of Stem Cell Research and Application, University of Science, Vietnam National University, Ho Chi Minh City, Viet Nam
a r t i c l e i n f o
Article history:Received 29 April 2014Received in revised form4 August 2014Accepted 18 August 2014
Keywords:Mesenchymal stem cellsUCB-MSCsInsulin producing cellsPDX-1mRNA transfection
a b s t r a c t
Numerous studies have sought to identify diabetes mellitus treatment strategies with fewer side effects.Mesenchymal stem cell (MSC) therapy was previously considered as a promising therapy; however, itrequires the cells to be trans-differentiated into cells of the pancreatic-endocrine lineage beforetransplantation. Previous studies have shown that PDX-1 expression can facilitate MSC differentiationinto insulin-producing cells (IPCs), but the methods employed to date use viral or DNA-based tools toexpress PDX-1, with the associated risks of insertional mutation and immunogenicity. Thus, this studyaimed to establish a new method to induce PDX-1 expression in MSCs by mRNA transfection. MSCs wereisolated from human umbilical cord blood and expanded in vitro, with stemness confirmed by surfacemarkers and multipotentiality. MSCs were transfected with PDX-1 mRNA by nucleofection andchemically induced to differentiate into IPCs (combinatorial group). This IPC differentiation was thencompared with that of untransfected chemically induced cells (inducer group) and uninduced cells(control group). We found that PDX-1 mRNA transfection significantly improved the differentiation ofMSCs into IPCs, with 8.372.5% IPCs in the combinatorial group, 3.2172.11% in the inducer group and 0%in the control. Cells in the combinatorial group also strongly expressed several genes related to beta cells(Pdx-1, Ngn3, Nkx6.1 and insulin) and could produce C-peptide in the cytoplasm and insulin in thesupernatant, which was dependent on the extracellular glucose concentration. These results indicatethat PDX-1 mRNA may offer a promising approach to produce safe IPCs for clinical diabetes mellitustreatment.
& 2014 International Society of Differentiation. Published by Elsevier B.V. All rights reserved.
1. Introduction
Diabetes mellitus is a highly prevalent disease estimated by theWorld Health Organization to affect approximately 500 millionpeople worldwide. However, to date, there is still no cure. All ofthe current methods used to treat diabetes mellitus aim to restore
glucose homeostasis. Cellular therapy has long been considered asa potential approach to cure this disease. However, beta cellnumbers are limited, and thus not ideal for replacement therapy.Insulin-producing cells (IPCs), on the other hand, can be differ-entiated from stem cells and offer a potential source of cells in lieuof beta cells. For this reason, numerous studies have beenconducted to establish protocols to differentiate stem cellsinto IPCs.
Various sources of stem cells have been successfully differen-tiated into IPCs, including embryonic stem cells (Hua et al., 2014;Jiang et al., 2007), induced-pluripotent stem cells (Alipio et al.,2010; Jeon et al., 2012; Zhu et al., 2011), pancreatic stem cells(Noguchi et al., 2010), mesenchymal stem cells from humanumbilical cord blood (UCB) (Parekh et al., 2009; Phuc et al.,2011), placenta (Kadam et al., 2010), bone marrow (Phadniset al., 2011), and adipose tissue (Chandra et al., 2009). Of these,UCB-derived MSCs offer several advantages, particularly becauseof the increased banking of UCB samples in recent years.
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/diff
Differentiation
http://dx.doi.org/10.1016/j.diff.2014.08.001Join the International Society for Differentiation (www.isdifferentiation.org)0301-4681/& 2014 International Society of Differentiation. Published by Elsevier B.V. All rights reserved.
Abbreviations: DMEM, Dulbecco’s modified eagle medium; GFP, green fluorescentprotein; MNC, mononuclear cell; mRNA, messenger RNA; MSC, mesenchymal stemcell; IMDM, Iscove’s modified Dulbecco’s media; IPC, insulin producing cell; PBS,phosphate buffered saline
n Corresponding author.E-mail addresses: [email protected] (P. Van Pham),
[email protected] (P. Thi-My Nguyen),[email protected] (A. Thai-Quynh Nguyen),[email protected] (V. Minh Pham),[email protected] (A. Nguyen-Tu Bui),[email protected] (L. Thi-Tung Dang), [email protected] (K. Gia Nguyen),[email protected] (N. Kim Phan).
Please cite this article as: Van Pham, P., et al., Improved differentiation of umbilical cord blood-derived mesenchymal stem cells intoinsulin-producing cells by PDX-1 mRNA transfection. Differentiation (2014), http://dx.doi.org/10.1016/j.diff.2014.08.001i
Differentiation ∎ (∎∎∎∎) ∎∎∎–∎∎∎

Consequently, UCB-MSCs have been extensively studied for IPCdifferentiation. Until now, the most successful methods to induceUCB-MSC differentiation into IPCs used nicotinamide and/orexendin-4 inducers (Phuc et al., 2011; Prabakar et al., 2012; Tsaiet al., 2012). Other studies have also successfully differentiatedUCB-MSCs into IPCs by up-regulating some of the master genesthat cause IPC differentiation (mainly PDX-1) (He et al., 2011;Wang et al., 2011). These studies have demonstrated that PDX-1 isan important factor regulating pancreatic-endocrine differentia-tion, particularly for beta cell formation and function. Further-more, PDX-1-differentiated IPCs can regulate the glucoseconcentration of diabetic mice.
The chemical induction of IPCs from MSCs, however, is gen-erally poor and, although PDX-1 up-regulation can significantlyincrease IPC production, the use of vector viruses, such as anadenovirus or a lentivirus, harbors the risk of insertional muta-genesis and immunogenicity (Dave et al., 2009; Hacein-Bey-Abinaet al., 2008; Howe et al., 2008). As such, the differentiated IPCsfrom these protocols cannot be used to treat humans in clinicalapplications. Therefore, this study aimed to develop a novel andsafe method to improve the differentiation efficiency of UCB-MSCsinto IPCs. We show improved chemical differentiation of MSCsfollowing transfection of PDX-1 mRNA.
2. Materials and methods
2.1. Isolation of UCB-MSCs
Human UCB was obtained from hospital samples with informedconsent obtained from the mother after delivery of her child. Allprocedures and manipulations were approved by our InstitutionalEthical Committee (Laboratory of Stem Cell Research and Application,University of Science, Vietnam National University, Ho Chi Minh City,Vietnam) and the Hospital Ethical Committee (Nhan Dan 115Hospital, Ho Chi Minh City, Vietnam). A bag system containing17 mL of anticoagulant (citrate, phosphate, and dextrose) was used.All UCB units were processed within 3 h after delivery. To isolatemononuclear cells (MNCs), each UCB unit was diluted 1:1 withphosphate-buffered saline (PBS) and carefully loaded onto Ficoll-Hypaque (1.077 g/mL, Sigma-Aldrich, St. Louis, MO). After densitygradient centrifugation at 3000 rpm for 20 min at room temperature,MNCs were removed from the interphase, washed twice with PBS,and resuspended in Iscove’s modified Dulbecco’s media (IMDM)with 15% fetal bovine serum (FBS) and 1% antibiotic-antimycotic(Sigma-Aldrich). MNCs were seeded in T-75 cm2
flasks at 1�105
cells/cm2 and incubated at 37 1C, 5% CO2. The medium was replacedevery 3 days. When cells reached 70–80% confluence, they were sub-cultured at a ratio of 1:3 using the same medium as primary culture.
2.2. UCB-MSC characterization
UCB-MSCs were characterized according the MSC standard setby Dominici et al. (2006). UCB-MSCs were confirmed by flowcytometry using surface marker expressions of CD14, CD34, CD45,HLA-DR, CD73, CD90 and CD105. Flow cytometry was performedon a FACSCalibur flow cytometer (BD Bioscience, San Jose, CA).UCB-MSCs were stained with anti-CD14-FITC, anti-CD34-FITC,anti-CD45-FITC, anti-HLA-DR-FTIC, anti-CD73-PE, anti-CD90-FITCand anti-CD105-FITC monoclonal antibodies. A total of 10,000 cellswere analyzed by CellQuest Pro software. Isotype controls wereused in all analyses.
UCB-MSCs were also confirmed by their potential to differenti-ate along multiple lineages. Adipogenic differentiation of MSCswas performed as described previously (Lee et al., 2004b). Briefly,UCB-MSCs at passage 5 were plated at a density of 1�104
cells/well in 24-well plates. At 70% confluence, the cells wereswitched to IMDM supplemented with 0.5 mM 3-isobutyl-1-methyl-xanthine, 1 nM dexamethasone, 0.1 mM indomethacinand 10% FBS (all from Sigma-Aldrich) and cultured for 21 days.Adipogenic differentiation was evaluated by observing the produc-tion of lipid vesicles within cells via microscopy.
For osteogenic differentiation, UCB-MSCs were plated at 1�104
cells/well in 24-well plates. At 70% confluence, the cells wereswitched to IMDM supplemented with 10% FBS, 10�7 M dexametha-sone, 50 μM ascorbic acid-2 phosphate and 10 mM β-glycerolphosphate (all from Sigma-Aldrich), and cultured for 21 days, asdescribed elsewhere (Lee et al., 2004b). Osteogenic differentiation(calcium accumulation) was confirmed by Alizarin red staining.
For chondrogenic differentiation, UCB-MSCs were inducedusing a commercial medium for chondrogenesis (StemPro Chon-drogenesis Differentiation Kit, A10071-01, Life Technologies). UCB-MSCs were differentiated in pellet form, according to manufac-turer’s guidelines. After 21 days growth, cell pellets were stainedwith an anti-aggrecan monoclonal antibody (BD Bioscience).
2.3. In vitro mRNA PDX-1 production
pcDNA3.1-hPDX-1 was amplified by PCR with 50-T7 primer(50-TAATACGACTCACTATAGGG-30) and 30-specific primer for PDX-1(50-GTCCTCCTCCTTTTTCCAC-30). pcDNA3.1-hPDX-1 was prepared inthe previous study by cutting hPDX-1 from vector pWPT-PDX1 withNotI and BamHI (Plasmid 12256, Addgene, Cambridge, MA) andinserting to vector pcDNATM 3.1 (Invitrogen, Carlsbad, CA) (Nguyenet al., 2014).
The PCR products for hPDX-1 were purified using the GenElutePCR Clean-up Kit, Sigma-Aldrich, St Louis, MO). The purified PCRproduct was employed for an in vitro transcription reaction usingthe T7 mScript Standard mRNA Production System (EpicentreBiotechnologies, Madison, WI). The mRNA concentration wasmeasured using a Nanophotometer (Eppendorf, Germany).
2.4. mRNA PDX-1 transfection
UCB-MSCs were transfected according to a previously pub-lished protocol (Arnold et al., 2012). UCB-MSCs were transfectedwith 3 μg of mRNA by nucleofection (NHDF-VPD-1001, Lonza).After transfection, these cells were plated into T-25 flasks andcultured in the medium. At 72 h, 144 h, and 216 h after nucleofec-tion, the adherent cells were transfected with “FuGENE HD”(Roche, Basel, Switzerland) according to the manufacturer’sinstructions, which was replaced with culture medium 4 h later.The ratio of “FuGENE HD” reagent and mRNA was 8 μL per 3 μg ofmRNA. Transfected samples of UCB-MSCs were evaluated forchanges in Pdx-1 expression at both transcriptional and transla-tional levels.
2.5. RNA extraction and reverse transcript real-time RT PCR
RNA was extracted from cell cultures using a Trizol extractionkit (Intron Biotechnology, Korea). mRNA was reversed trans-cribed into cDNA using an AMV reverse transcription kit (AgilentTechnologies, Santa Clara, CA). The real-time RT-PCR reactionswere carried out using Brilliant II SYBRs Green QPCR Master Mix(Agilent Technologies). The primer sequences were as follows:GAPDH, forward, 50-AGAAGGCTGGGGCTCATTTG-30, and reverse,50-AGGGGCCATCCACAGTCTTC-30; PDX-1, forward, 50-GGATGAAGTCTACCAAAGCTCACGC-30, and reverse, 50-CCAGATCTTGATGTGTCTCTCGGTC-30; INSULIN, forward, 50-AACCAACACCTGTGCGGCTCA-30; reverse, 50-TGCCTGCGGGCTGCGTCTA-30; NGN3, forward,50-CGCCGGTAGAAAGGATGAC-30, reverse: 50-GAGTTGAGGTTGTG-CATTCG-30; NKX6.1, forward: 50-CTGGAGAAGACTTTCGAACAA-30,
P. Van Pham et al. / Differentiation ∎ (∎∎∎∎) ∎∎∎–∎∎∎2
Please cite this article as: Van Pham, P., et al., Improved differentiation of umbilical cord blood-derived mesenchymal stem cells intoinsulin-producing cells by PDX-1 mRNA transfection. Differentiation (2014), http://dx.doi.org/10.1016/j.diff.2014.08.001i

reverse, 50-AGAGGCTTATTGTAGTCGTCG-30. GAPDH was used as aninternal control for the normalization of gene expression. Real-time RT-PCR was performed as per the following cycling condi-tions: 95 1C for 30 s; 60 1C for 30 s; and 72 1C for 60 s for 40 cycles.The Ct values were used to calculate gene expression according tothe 2�ΔCCt method.
2.6. IPC differentiation
In this study, UCB-MSCs were treated under three conditions:combinatorial group (PDX-1-transfected and chemically induced);inducer group (chemically induced only); and control group(no induction or transfection). At the start of the assay (hereinreferred to as day-9 of the time line), undifferentiated UCB-MSCs inthe combinatorial group were transfected with PDX-1 mRNA. Ninedays later at ‘day 0’, UCB-MSCs in the combinatorial and inducergroups were induced to differentiate into IPCs in IMDM supple-mented with 2% FBS, 100 ng/mL epidermal growth factor and 2%B27 for 3 days. From days 4 to 21, cells were incubated in IMDMsupplemented with 10 nM nicotinamide, 2% B27, 10 ng/mL betacel-lulin and 0.1 mM beta-mercaptoethanol, with fresh media replacedevery 3 days. UCB-MSCs in the control group only were culturedin basic medium (IMDM plus 15% FBS, 1% antibiotic-antimycotic).At days 7, 14, and 21, the gene expression levels of PDX-1, NGN3,NKX6.1 and INSULIN were evaluated. At day 21, the supernatantfrom all three groups was collected to analyze the amount ofinsulin production and cell extracts were used to measure C-peptide production.
2.7. Flow cytometry analysis
PDX-1-transfected cells were analyzed by flow cytometry. About5�106 cells were fixed in PFA 4% and permeabilized with 0.03%Triton X-100 (Sigma-Aldrich, St Louis, MO) diluted in PBS with 0.1%bovine serum albumin (BSA) (Sigma-Aldrich, St Louis, MO) for 1 h.Non-specific sites were blocked using 10% BSA for an additional 1 h.Cells were then incubated with anti-Pdx1 monoclonal antibody (BDBioscience) for 4 h. Stained cells were washed three times with PBSplus 0.1% BSA and then incubated with an anti-mouse secondaryIgG1-FITC for 1 h. Finally, the cells were washed three times with PBSplus 0.1% BSA and re-suspended in FACS fluid sheath for analysis andsorting on a FACSJazz flow cytometer (BD Bioscience). Isotypes wereused for this analysis.
2.8. Immunohistochemistry
For immunohistochemistry, cells were fixed with 4% paraformal-dehyde and then permeabilized by 0.03% Triton X-100 diluted in PBSwith 0.1% BSA for 1 h. Non-specific sites were blocked using 10% BSAfor an additional 1 h. Cells were then stained with primary mono-clonal antibodies against Pdx-1 (sc-390792, Santa Cruz Biotechnol-ogy, Canada) insulin (sc-52035, Santa Cruz Biotechnology, Canada).Following washing with PBS, cells were then stained with anti-mouse secondary IgG1-FITC (sc-2010, Santa Cruz Biotechnology,Canada) for 30 min, washed again and then counterstained withHoechst 33342 for 10 min (Sigma-Aldrich, St Louis, MO). Staining wasobserved under a fluorescent microscope (Cell Observer, Carl-Zeiss,Germany).
2.9. Insulin and C-peptide measurement
After 21 days of differentiation, cells were washed with PBS andthen incubated for 3 h in DMEM-LG (low glucose; 5 mM glucose).The supernatant was collected and stored at �20 1C for insulinmeasurement and the differentiated cells were harvested and thecellular extract used for C-peptide measurement. The Insulin ELISA
kit (ab100578, Abcam, Cambridge, MA) and C-peptide ELISA kit(ab178641, Abcam, Cambridge, MA) were used according to themanufacturer’s instructions. The assays were read at 450 nm in aDTX Multimode plate reader (Beckman Coulter, Fullerton, CA).
2.10. Glucose response assay
At day 21 of differentiation, cells were washed with PBS andthen incubated for 1 h in DMEM-LG. The supernatant was col-lected and stored at �20 1C. The cells were washed with PBS andincubated for an additional 1 h in DMEM-HG (high glucose;25 mM glucose) and this supernatant was also collected andstored at �20 1C. The insulin concentration was determined usingthe Insulin ELISA kit, as described above.
2.11. Statistical analysis
Significance of differences between mean values was assessedby t test and ANOVA. A P value o0.05 was considered to besignificant. All data were analyzed by Prism 6 software (GraphPadSoftware, La Jolla, CA).
3. Results
3.1. UCB-MSCs fully exhibited the MSC characteristics
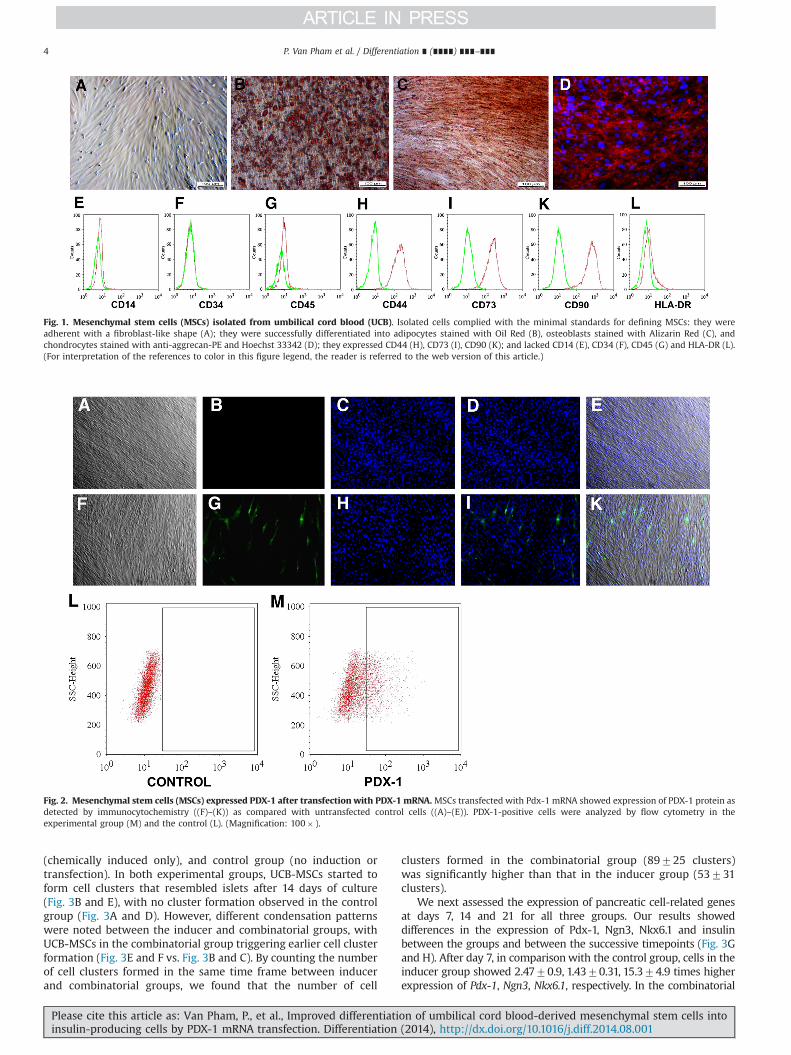
We successfully isolated MSCs from five samples of humanUCBs. All potential isolates of MSCs were confirmed according tothe guidelines set out by Dominici et al. (2006). The resultsshowed that isolated MSCs exhibited a fibroblast-like shape whencultured under adherent conditions (Fig. 1A). These cells werepositive for CD44, CD73, and CD90 and negative for CD14(a marker of monocytes), CD34 (a marker of hematopoietic stemcells), CD45 (a marker of leukocytes) and HLA-DR (Fig. 1E–L). TheMSCs could also be successfully differentiated along three differentmesenchymal lineages, including adipocyte, osteocyte and chon-drocyte lineages. For adipogenic differentiation, MSCs demon-strated a change in shape and the presence of lipid dropletswithin the cytoplasm of cells following growth in adipocyte-inducing medium. These lipid droplets were stained red by OilRed dye (Fig. 1B). For osteogenic differentiation, MSCs also demon-strated a change in shape as well as an accumulation in cytoplas-mic calcium, as determined by Alizarin red staining (Fig. 1C).Finally, MSCs were also successfully differentiated into chondro-cytes, with pellets staining positively for aggrecan, a specificmarker of chondrocytes (Fig. 1D).
3.2. PDX-1 mRNA transfection and PDX-1 expression in MSCs
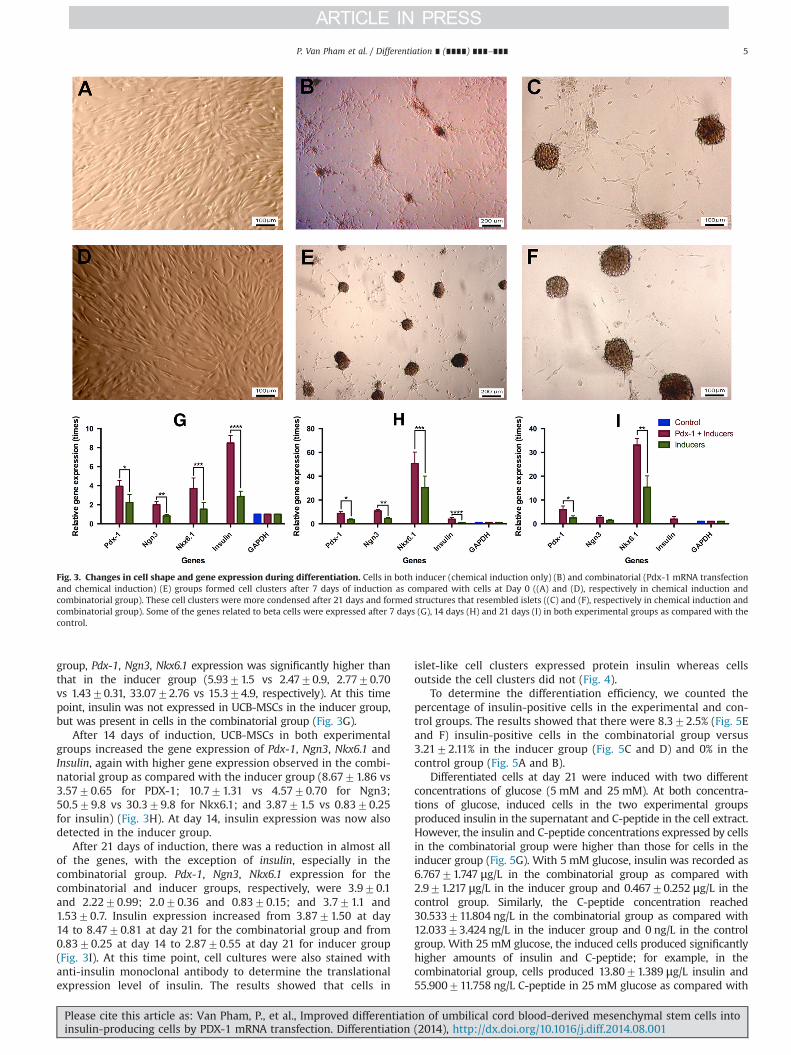
In this experiment, we transfected UCB-MSCs with PDX-1mRNA according to a published protocol (Arnold et al., 2012).We found that, compared with un-transfected control cultures(Fig. 2A–E), some of the transfected cells showed positive PDX-1expression, as determined using immunochemistry staining(Fig. 2F–K). Using flow cytometry, we determined the PDX-1-positive cell population as 12.5575.32% of the total number ofcells (Fig. 2L and M).
3.3. Differentiation of UCB-MSCs into IPCs by chemical inductionwith or without PDX-1 mRNA transfection
To evaluate the efficacy of PDX-1 mRNA transfection on IPCdifferentiation of UCB-MSCs, we set up two experimental groupsand a control group herein referred to as the combinatorialgroup (Pdx-1-transfected and chemically induced), inducer group
P. Van Pham et al. / Differentiation ∎ (∎∎∎∎) ∎∎∎–∎∎∎ 3
Please cite this article as: Van Pham, P., et al., Improved differentiation of umbilical cord blood-derived mesenchymal stem cells intoinsulin-producing cells by PDX-1 mRNA transfection. Differentiation (2014), http://dx.doi.org/10.1016/j.diff.2014.08.001i
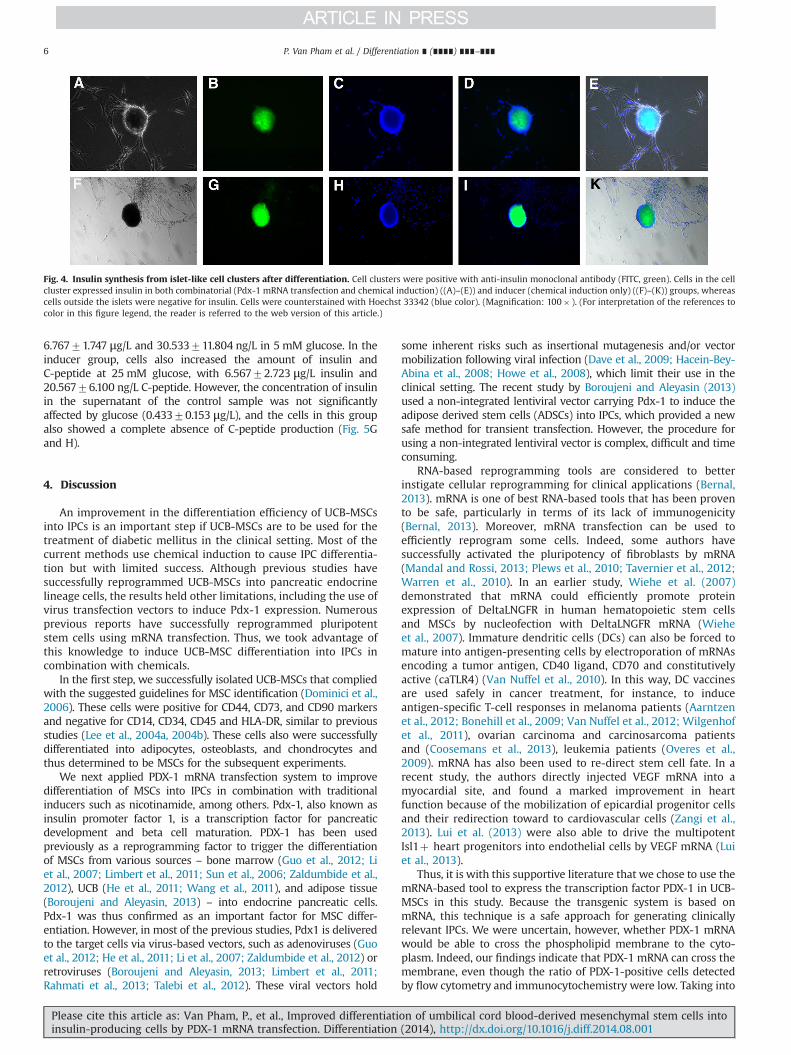
(chemically induced only), and control group (no induction ortransfection). In both experimental groups, UCB-MSCs started toform cell clusters that resembled islets after 14 days of culture(Fig. 3B and E), with no cluster formation observed in the controlgroup (Fig. 3A and D). However, different condensation patternswere noted between the inducer and combinatorial groups, withUCB-MSCs in the combinatorial group triggering earlier cell clusterformation (Fig. 3E and F vs. Fig. 3B and C). By counting the numberof cell clusters formed in the same time frame between inducerand combinatorial groups, we found that the number of cell
clusters formed in the combinatorial group (89725 clusters)was significantly higher than that in the inducer group (53731clusters).
We next assessed the expression of pancreatic cell-related genesat days 7, 14 and 21 for all three groups. Our results showeddifferences in the expression of Pdx-1, Ngn3, Nkx6.1 and insulinbetween the groups and between the successive timepoints (Fig. 3Gand H). After day 7, in comparisonwith the control group, cells in theinducer group showed 2.4770.9, 1.4370.31, 15.374.9 times higherexpression of Pdx-1, Ngn3, Nkx6.1, respectively. In the combinatorial
Fig. 2. Mesenchymal stem cells (MSCs) expressed PDX-1 after transfection with PDX-1 mRNA.MSCs transfected with Pdx-1 mRNA showed expression of PDX-1 protein asdetected by immunocytochemistry ((F)–(K)) as compared with untransfected control cells ((A)–(E)). PDX-1-positive cells were analyzed by flow cytometry in theexperimental group (M) and the control (L). (Magnification: 100� ).
Fig. 1. Mesenchymal stem cells (MSCs) isolated from umbilical cord blood (UCB). Isolated cells complied with the minimal standards for defining MSCs: they wereadherent with a fibroblast-like shape (A); they were successfully differentiated into adipocytes stained with Oil Red (B), osteoblasts stained with Alizarin Red (C), andchondrocytes stained with anti-aggrecan-PE and Hoechst 33342 (D); they expressed CD44 (H), CD73 (I), CD90 (K); and lacked CD14 (E), CD34 (F), CD45 (G) and HLA-DR (L).(For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
P. Van Pham et al. / Differentiation ∎ (∎∎∎∎) ∎∎∎–∎∎∎4
Please cite this article as: Van Pham, P., et al., Improved differentiation of umbilical cord blood-derived mesenchymal stem cells intoinsulin-producing cells by PDX-1 mRNA transfection. Differentiation (2014), http://dx.doi.org/10.1016/j.diff.2014.08.001i
group, Pdx-1, Ngn3, Nkx6.1 expression was significantly higher thanthat in the inducer group (5.9371.5 vs 2.4770.9, 2.7770.70vs 1.4370.31, 33.0772.76 vs 15.374.9, respectively). At this timepoint, insulin was not expressed in UCB-MSCs in the inducer group,but was present in cells in the combinatorial group (Fig. 3G).
After 14 days of induction, UCB-MSCs in both experimentalgroups increased the gene expression of Pdx-1, Ngn3, Nkx6.1 andInsulin, again with higher gene expression observed in the combi-natorial group as compared with the inducer group (8.6771.86 vs3.5770.65 for PDX-1; 10.771.31 vs 4.5770.70 for Ngn3;50.579.8 vs 30.379.8 for Nkx6.1; and 3.8771.5 vs 0.8370.25for insulin) (Fig. 3H). At day 14, insulin expression was now alsodetected in the inducer group.
After 21 days of induction, there was a reduction in almost allof the genes, with the exception of insulin, especially in thecombinatorial group. Pdx-1, Ngn3, Nkx6.1 expression for thecombinatorial and inducer groups, respectively, were 3.970.1and 2.2270.99; 2.070.36 and 0.8370.15; and 3.771.1 and1.5370.7. Insulin expression increased from 3.8771.50 at day14 to 8.4770.81 at day 21 for the combinatorial group and from0.8370.25 at day 14 to 2.8770.55 at day 21 for inducer group(Fig. 3I). At this time point, cell cultures were also stained withanti-insulin monoclonal antibody to determine the translationalexpression level of insulin. The results showed that cells in
islet-like cell clusters expressed protein insulin whereas cellsoutside the cell clusters did not (Fig. 4).
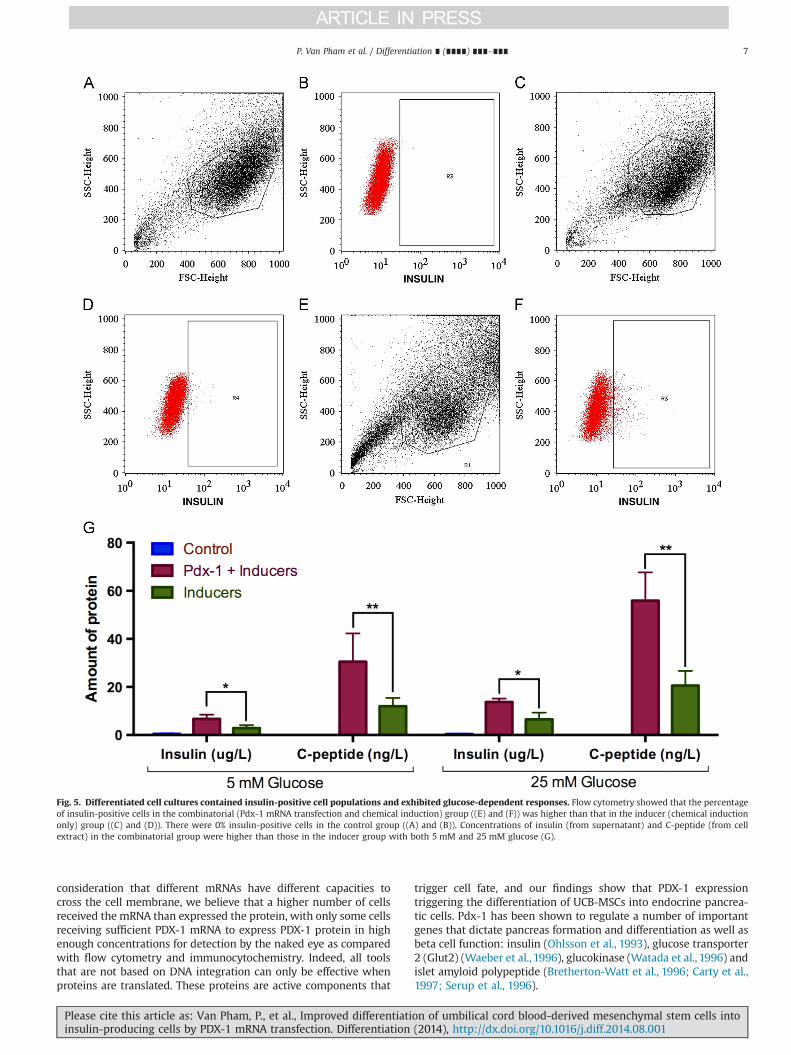
To determine the differentiation efficiency, we counted thepercentage of insulin-positive cells in the experimental and con-trol groups. The results showed that there were 8.372.5% (Fig. 5Eand F) insulin-positive cells in the combinatorial group versus3.2172.11% in the inducer group (Fig. 5C and D) and 0% in thecontrol group (Fig. 5A and B).
Differentiated cells at day 21 were induced with two differentconcentrations of glucose (5 mM and 25 mM). At both concentra-tions of glucose, induced cells in the two experimental groupsproduced insulin in the supernatant and C-peptide in the cell extract.However, the insulin and C-peptide concentrations expressed by cellsin the combinatorial group were higher than those for cells in theinducer group (Fig. 5G). With 5 mM glucose, insulin was recorded as6.76771.747 μg/L in the combinatorial group as compared with2.971.217 μg/L in the inducer group and 0.46770.252 μg/L in thecontrol group. Similarly, the C-peptide concentration reached30.533711.804 ng/L in the combinatorial group as compared with12.03373.424 ng/L in the inducer group and 0 ng/L in the controlgroup. With 25 mM glucose, the induced cells produced significantlyhigher amounts of insulin and C-peptide; for example, in thecombinatorial group, cells produced 13.8071.389 μg/L insulin and55.900711.758 ng/L C-peptide in 25 mM glucose as compared with
Fig. 3. Changes in cell shape and gene expression during differentiation. Cells in both inducer (chemical induction only) (B) and combinatorial (Pdx-1 mRNA transfectionand chemical induction) (E) groups formed cell clusters after 7 days of induction as compared with cells at Day 0 ((A) and (D), respectively in chemical induction andcombinatorial group). These cell clusters were more condensed after 21 days and formed structures that resembled islets ((C) and (F), respectively in chemical induction andcombinatorial group). Some of the genes related to beta cells were expressed after 7 days (G), 14 days (H) and 21 days (I) in both experimental groups as compared with thecontrol.
P. Van Pham et al. / Differentiation ∎ (∎∎∎∎) ∎∎∎–∎∎∎ 5
Please cite this article as: Van Pham, P., et al., Improved differentiation of umbilical cord blood-derived mesenchymal stem cells intoinsulin-producing cells by PDX-1 mRNA transfection. Differentiation (2014), http://dx.doi.org/10.1016/j.diff.2014.08.001i
6.76771.747 μg/L and 30.533711.804 ng/L in 5 mM glucose. In theinducer group, cells also increased the amount of insulin andC-peptide at 25 mM glucose, with 6.56772.723 μg/L insulin and20.56776.100 ng/L C-peptide. However, the concentration of insulinin the supernatant of the control sample was not significantlyaffected by glucose (0.43370.153 μg/L), and the cells in this groupalso showed a complete absence of C-peptide production (Fig. 5Gand H).
4. Discussion
An improvement in the differentiation efficiency of UCB-MSCsinto IPCs is an important step if UCB-MSCs are to be used for thetreatment of diabetic mellitus in the clinical setting. Most of thecurrent methods use chemical induction to cause IPC differentia-tion but with limited success. Although previous studies havesuccessfully reprogrammed UCB-MSCs into pancreatic endocrinelineage cells, the results held other limitations, including the use ofvirus transfection vectors to induce Pdx-1 expression. Numerousprevious reports have successfully reprogrammed pluripotentstem cells using mRNA transfection. Thus, we took advantage ofthis knowledge to induce UCB-MSC differentiation into IPCs incombination with chemicals.
In the first step, we successfully isolated UCB-MSCs that compliedwith the suggested guidelines for MSC identification (Dominici et al.,2006). These cells were positive for CD44, CD73, and CD90 markersand negative for CD14, CD34, CD45 and HLA-DR, similar to previousstudies (Lee et al., 2004a, 2004b). These cells also were successfullydifferentiated into adipocytes, osteoblasts, and chondrocytes andthus determined to be MSCs for the subsequent experiments.
We next applied PDX-1 mRNA transfection system to improvedifferentiation of MSCs into IPCs in combination with traditionalinducers such as nicotinamide, among others. Pdx-1, also known asinsulin promoter factor 1, is a transcription factor for pancreaticdevelopment and beta cell maturation. PDX-1 has been usedpreviously as a reprogramming factor to trigger the differentiationof MSCs from various sources – bone marrow (Guo et al., 2012; Liet al., 2007; Limbert et al., 2011; Sun et al., 2006; Zaldumbide et al.,2012), UCB (He et al., 2011; Wang et al., 2011), and adipose tissue(Boroujeni and Aleyasin, 2013) – into endocrine pancreatic cells.Pdx-1 was thus confirmed as an important factor for MSC differ-entiation. However, in most of the previous studies, Pdx1 is deliveredto the target cells via virus-based vectors, such as adenoviruses (Guoet al., 2012; He et al., 2011; Li et al., 2007; Zaldumbide et al., 2012) orretroviruses (Boroujeni and Aleyasin, 2013; Limbert et al., 2011;Rahmati et al., 2013; Talebi et al., 2012). These viral vectors hold
some inherent risks such as insertional mutagenesis and/or vectormobilization following viral infection (Dave et al., 2009; Hacein-Bey-Abina et al., 2008; Howe et al., 2008), which limit their use in theclinical setting. The recent study by Boroujeni and Aleyasin (2013)used a non-integrated lentiviral vector carrying Pdx-1 to induce theadipose derived stem cells (ADSCs) into IPCs, which provided a newsafe method for transient transfection. However, the procedure forusing a non-integrated lentiviral vector is complex, difficult and timeconsuming.
RNA-based reprogramming tools are considered to betterinstigate cellular reprogramming for clinical applications (Bernal,2013). mRNA is one of best RNA-based tools that has been provento be safe, particularly in terms of its lack of immunogenicity(Bernal, 2013). Moreover, mRNA transfection can be used toefficiently reprogram some cells. Indeed, some authors havesuccessfully activated the pluripotency of fibroblasts by mRNA(Mandal and Rossi, 2013; Plews et al., 2010; Tavernier et al., 2012;Warren et al., 2010). In an earlier study, Wiehe et al. (2007)demonstrated that mRNA could efficiently promote proteinexpression of DeltaLNGFR in human hematopoietic stem cellsand MSCs by nucleofection with DeltaLNGFR mRNA (Wieheet al., 2007). Immature dendritic cells (DCs) can also be forced tomature into antigen-presenting cells by electroporation of mRNAsencoding a tumor antigen, CD40 ligand, CD70 and constitutivelyactive (caTLR4) (Van Nuffel et al., 2010). In this way, DC vaccinesare used safely in cancer treatment, for instance, to induceantigen-specific T-cell responses in melanoma patients (Aarntzenet al., 2012; Bonehill et al., 2009; Van Nuffel et al., 2012; Wilgenhofet al., 2011), ovarian carcinoma and carcinosarcoma patientsand (Coosemans et al., 2013), leukemia patients (Overes et al.,2009). mRNA has also been used to re-direct stem cell fate. In arecent study, the authors directly injected VEGF mRNA into amyocardial site, and found a marked improvement in heartfunction because of the mobilization of epicardial progenitor cellsand their redirection toward to cardiovascular cells (Zangi et al.,2013). Lui et al. (2013) were also able to drive the multipotentIsl1þ heart progenitors into endothelial cells by VEGF mRNA (Luiet al., 2013).
Thus, it is with this supportive literature that we chose to use themRNA-based tool to express the transcription factor PDX-1 in UCB-MSCs in this study. Because the transgenic system is based onmRNA, this technique is a safe approach for generating clinicallyrelevant IPCs. We were uncertain, however, whether PDX-1 mRNAwould be able to cross the phospholipid membrane to the cyto-plasm. Indeed, our findings indicate that PDX-1 mRNA can cross themembrane, even though the ratio of PDX-1-positive cells detectedby flow cytometry and immunocytochemistry were low. Taking into
Fig. 4. Insulin synthesis from islet-like cell clusters after differentiation. Cell clusters were positive with anti-insulin monoclonal antibody (FITC, green). Cells in the cellcluster expressed insulin in both combinatorial (Pdx-1 mRNA transfection and chemical induction) ((A)–(E)) and inducer (chemical induction only) ((F)–(K)) groups, whereascells outside the islets were negative for insulin. Cells were counterstained with Hoechst 33342 (blue color). (Magnification: 100� ). (For interpretation of the references tocolor in this figure legend, the reader is referred to the web version of this article.)
P. Van Pham et al. / Differentiation ∎ (∎∎∎∎) ∎∎∎–∎∎∎6
Please cite this article as: Van Pham, P., et al., Improved differentiation of umbilical cord blood-derived mesenchymal stem cells intoinsulin-producing cells by PDX-1 mRNA transfection. Differentiation (2014), http://dx.doi.org/10.1016/j.diff.2014.08.001i
consideration that different mRNAs have different capacities tocross the cell membrane, we believe that a higher number of cellsreceived the mRNA than expressed the protein, with only some cellsreceiving sufficient PDX-1 mRNA to express PDX-1 protein in highenough concentrations for detection by the naked eye as comparedwith flow cytometry and immunocytochemistry. Indeed, all toolsthat are not based on DNA integration can only be effective whenproteins are translated. These proteins are active components that
trigger cell fate, and our findings show that PDX-1 expressiontriggering the differentiation of UCB-MSCs into endocrine pancrea-tic cells. Pdx-1 has been shown to regulate a number of importantgenes that dictate pancreas formation and differentiation as well asbeta cell function: insulin (Ohlsson et al., 1993), glucose transporter2 (Glut2) (Waeber et al., 1996), glucokinase (Watada et al., 1996) andislet amyloid polypeptide (Bretherton-Watt et al., 1996; Carty et al.,1997; Serup et al., 1996).
Fig. 5. Differentiated cell cultures contained insulin-positive cell populations and exhibited glucose-dependent responses. Flow cytometry showed that the percentageof insulin-positive cells in the combinatorial (Pdx-1 mRNA transfection and chemical induction) group ((E) and (F)) was higher than that in the inducer (chemical inductiononly) group ((C) and (D)). There were 0% insulin-positive cells in the control group ((A) and (B)). Concentrations of insulin (from supernatant) and C-peptide (from cellextract) in the combinatorial group were higher than those in the inducer group with both 5 mM and 25 mM glucose (G).
P. Van Pham et al. / Differentiation ∎ (∎∎∎∎) ∎∎∎–∎∎∎ 7
Please cite this article as: Van Pham, P., et al., Improved differentiation of umbilical cord blood-derived mesenchymal stem cells intoinsulin-producing cells by PDX-1 mRNA transfection. Differentiation (2014), http://dx.doi.org/10.1016/j.diff.2014.08.001i

We showed that the combinatorial group was able to signifi-cantly increase the expression of genes related to beta cells overcells subjected to chemical induction alone, demonstrating theutility of PDX-1 mRNA transfection. The results illustrate thatalthough the chemical inducers have a positive effect on pancrea-tic differentiation, the active expression of PDX-1 helps to facilitateand accelerate this process. The results from the gene analysisshowed that PDX-1 expression initiated this process. In the controlgroup, UCB-MSCs expressed extremely low levels of PDX-1 andthus were unable to differentiate into IPCs. In contrast, UCB-MSCsexpressed Pdx-1 mRNA at the transcriptional level in both experi-mental groups. By quantitative analysis, PDX-1 mRNA expressionin the combinatorial group was higher than the endogenousexpression of PDX-1 identified in cells in the inducer group. Thisdifference in the expression level of PDX-1 between these twogroups subsequently caused a difference in the gene expressionlevels of Ngn3, Nkx6.1, and insulin. In addition, the difference ininsulin transcription because of these levels of PDX-1 affected theC-peptide concentration. These results were similar to those ofanother previously published study (Yuan et al., 2012). Yuan et al.(2012) showed that the expression of Pdx-1 correlated with thelevel of insulin at both the transcriptional and translational levels(Yuan et al., 2012). Our results from the analysis of insulin-positivecells after 21 days of induction also indicated that the percentageof insulin-positive cells (8.372.5%) was higher than culturessubjected to chemical induction only. This finding was similar tothat reported for human embryonic stem cell differentiation intoIPCs (Jiang et al., 2007). Interestingly, our insulin production levelswere higher than those achieved by transfecting bone marrow-derived mesenchymal stem cells with plasmid vectors containingPdx-1 and Betacellulin, with only �5% insulin-positive cells in thatstudy (Li et al., 2008); this was similar to the transduction of PDX-1in UCB-MSCs with an adenovirus vector (11.6174.83% insulin-positive cells) (Wang et al., 2011). However, all of these results,including our own study, showed a lower expression of insulin ascompared with PDX-1 transduction by lentiviral vector (28.23%insulin-positive cells) (Sun et al., 2006).
Although we also detected some insulin in the control group,we did not detect C-peptide in this group. It is thus possible thatthe insulin detected in this group could be from cross-reactionbetween bovine insulin in the FBS. This study showed that PDX-1mRNA transfection could not only increase the percentage of IPCsbut also produce functional IPCs. IPCs in both experimental groupscould produce insulin in a glucose-dependent manner. Thus, PDX-1 mRNA transfection increased the differentiation efficiency with-out interfering with the normal pancreatic differentiation process.However, the level of C-peptide as well as insulin produced fromIPCs had not been compared to them in islets of Langerhans.Another limitation was insulin production of IPCs had not beenevaluated for a long time. Quality of IPCs needs to be evaluatedbefore they can be used in preclinical and clinical trials.
5. Conclusions
The establishment of an efficient and safe protocol for UCB-MSC differentiation into IPCs is a crucial step for the utility of MSCsin the treatment of clinical diabetes mellitus. This study showedthat PDX-1 mRNA transfection in combination with chemicalinducers is a safe and efficient method to improve UCB-MSCdifferentiation into IPCs. PDX-1 mRNA transfection significantlyincreased the percentage of IPCs in the differentiated cell popula-tion as compared with the use of chemical induction alone. Thesedifferentiated cells strongly expressed some of the genes related tobeta cell function and produced insulin and C-peptide in a
glucose-dependent manner. These results provide a new methodfor the potential clinical application of IPCs from UCB-MSCs.
Authors’ contributions
PVP carried out studies including primary culture of MSCs, MSCtransfection, flow cytometry analysis, gene analysis. PTMN andATQN collected umbilical cord blood; prepared and isolation ofplasmid containing Pdx-1 gene; PCR product purification. VMP,ANTB, LTTD, KGN take care MSCs before and after transfection,ELISA analysis, PCR preparation. NKP revised the manuscript,spelling and grammatical fixation.
Competing interests
The authors declare that they have no competing interests.
Acknowledgements
This work was funded by grants from Vietnam National Uni-versity, Ho Chi Minh city, Vietnam (B2010-18-02TD), and Ministryof Science and Technology, Vietnam (DTDL.2012.-G/2323).
References
Aarntzen, E.H., Schreibelt, G., Bol, K., Lesterhuis, W.J., Croockewit, A.J., de Wilt, J.H.,van Rossum, M.M., Blokx, W.A., Jacobs, J.F., Duiveman-de Boer, T., Schuurhuis, D.H., Mus, R., Thielemans, K., de Vries, I.J., Figdor, C.G., Punt, C.J., Adema, G.J., 2012.Vaccination with mRNA-electroporated dendritic cells induces robust tumorantigen-specific CD4þ and CD8þ T cells responses in stage III and IVmelanoma patients (An Official Journal of The American Association for CancerResearch). Clin. Cancer Res. 18, 5460–5470.
Alipio, Z., Liao, W., Roemer, E.J., Waner, M., Fink, L.M., Ward, D.C., Ma, Y., 2010.Reversal of hyperglycemia in diabetic mouse models using induced-pluripotentstem (iPS)-derived pancreatic beta-like cells. Proc. Nat. Acad. Sci. U.S.A. 107,13426–13431.
Arnold, A., Naaldijk, Y.M., Fabian, C., Wirth, H., Binder, H., Nikkhah, G., Armstrong, L.,Stolzing, A., 2012. Reprogramming of human huntington fibroblasts usingmRNA. ISRN Cell Biol. 2012, 12.
Bernal, J.A., 2013. RNA-based tools for nuclear reprogramming and lineage-conver-sion: towards clinical applications. J. Cardiovasc. Transl. Res. 6, 956–968.
Bonehill, A., Van Nuffel, A.M., Corthals, J., Tuyaerts, S., Heirman, C., Francois, V.,Colau, D., van der Bruggen, P., Neyns, B., Thielemans, K., 2009. Single-stepantigen loading and activation of dendritic cells by mRNA electroporation forthe purpose of therapeutic vaccination in melanoma patients (An OfficialJournal of The American Association for Cancer Research). Clin. Cancer Res.15, 3366–3375.
Boroujeni, Z.N., Aleyasin, A., 2013. Insulin producing cells established using non-integrated lentiviral vector harboring PDX1 gene. World J. Stem Cells 5,217–228.
Bretherton-Watt, D., Gore, N., Boam, D.S., 1996. Insulin upstream factor 1 and anovel ubiquitous factor bind to the human islet amyloid polypeptide/amylingene promoter. Biochem. J. 313, 495–502.
Carty, M.D., Lillquist, J.S., Peshavaria, M., Stein, R., Soeller, W.C., 1997. Identificationof cis- and trans-active factors regulating human islet amyloid polypeptidegene expression in pancreatic beta-cells. J. Biol. Chem. 272, 11986–11993.
Chandra, V.G S., Phadnis, S., Nair, P.D., Bhonde, R.R., 2009. Generation of pancreatichormone-expressing islet-like cell aggregates from murine adipose tissue-derived stem cells. Stem Cells (Dayton, Ohio) 27, 1941–1953.
Coosemans, A., Vanderstraeten, A., Tuyaerts, S., Verschuere, T., Moerman, P.,Berneman, Z., Vergote, I., Amant, F., Van Gool, S.W., 2013. Immunologicalresponse after WT1 mRNA-loaded dendritic cell immunotherapy in ovariancarcinoma and carcinosarcoma. Anticancer Res. 33, 3855–3859.
Dave, U.P., Akagi, K., Tripathi, R., Cleveland, S.M., Thompson, M.A., Yi, M., Stephens, R.,Downing, J.R., Jenkins, N.A., Copeland, N.G., 2009. Murine leukemias with retro-viral insertions at Lmo2 are predictive of the leukemias induced in SCID-X1patients following retroviral gene therapy. PLos Genet. 5, e1000491.
Dominici, M., Le Blanc, K., Mueller, I., Slaper-Cortenbach, I., Marini, F., Krause, D.,Deans, R., Keating, A., Prockop, D., Horwitz, E., 2006. Minimal criteria fordefining multipotent mesenchymal stromal cells. The International Society forCellular Therapy position statement. Cytotherapy 8, 315–317.
Guo, Q.S., Zhu, M.Y., Wang, L., Fan, X.J., Lu, Y.H., Wang, Z.W., Zhu, S.J., Wang, Y.,Huang, Y., 2012. Combined transfection of the three transcriptional factors,PDX-1, NeuroD1, and MafA, causes differentiation of bone marrow mesenchy-mal stem cells into insulin-producing cells. Exp. Diabetes Res. 2012, 672013.
P. Van Pham et al. / Differentiation ∎ (∎∎∎∎) ∎∎∎–∎∎∎8
Please cite this article as: Van Pham, P., et al., Improved differentiation of umbilical cord blood-derived mesenchymal stem cells intoinsulin-producing cells by PDX-1 mRNA transfection. Differentiation (2014), http://dx.doi.org/10.1016/j.diff.2014.08.001i

Hacein-Bey-Abina, S., Garrigue, A., Wang, G.P., Soulier, J., Lim, A., Morillon, E., Clappier,E., Caccavelli, L., Delabesse, E., Beldjord, K., Asnafi, V., MacIntyre, E., Dal Cortivo, L.,Radford, I., Brousse, N., Sigaux, F., Moshous, D., Hauer, J., Borkhardt, A., Belohradsky,B.H., Wintergerst, U., Velez, M.C., Leiva, L., Sorensen, R., Wulffraat, N., Blanche, S.,Bushman, F.D., Fischer, A., Cavazzana-Calvo, M., 2008. Insertional oncogenesis in4 patients after retrovirus-mediated gene therapy of SCID-X1. J. Clin. Invest. 118,3132–3142.
He, D., Wang, J., Gao, Y., Zhang, Y., 2011. Differentiation of PDX1 gene-modifiedhuman umbilical cord mesenchymal stem cells into insulin-producing cellsin vitro. Int. J. Mol. Med. 28, 1019–1024.
Howe, S.J., Mansour, M.R., Schwarzwaelder, K., Bartholomae, C., Hubank, M.,Kempski, H., Brugman, M.H., Pike-Overzet, K., Chatters, S.J., de Ridder, D.,Gilmour, K.C., Adams, S., Thornhill, S.I., Parsley, K.L., Staal, F.J., Gale, R.E., Linch,D.C., Bayford, J., Brown, L., Quaye, M., Kinnon, C., Ancliff, P., Webb, D.K., Schmidt,M., von Kalle, C., Gaspar, H.B., Thrasher, A.J., 2008. Insertional mutagenesiscombined with acquired somatic mutations causes leukemogenesis followinggene therapy of SCID-X1 patients. J. Clin. Invest. 118, 3143–3150.
Hua, X.F., Wang, Y.W., Tang, Y.X., Yu, S.Q., Jin, S.H., Meng, X.M., Li, H.F., Liu, F.J., Sun,Q., Wang, H.Y., Li, J.Y., 2014. Pancreatic insulin-producing cells differentiatedfrom human embryonic stem cells correct hyperglycemia in SCID/NOD mice, ananimal model of diabetes. PLoS One 9, e102198.
Jeon, K., Lim, H., Kim, J.H., Thuan, N.V., Park, S.H., Lim, Y.M., Choi, H.Y., Lee, E.R., Kim,J.H., Lee, M.S., Cho, S.G., 2012. Differentiation and transplantation of functionalpancreatic beta cells generated from induced pluripotent stem cells derivedfrom a type 1 diabetes mouse model. Stem Cells Dev. 21, 2642–2655.
Jiang, J., Au, M., Lu, K., Eshpeter, A., Korbutt, G., Fisk, G., Majumdar, A.S., 2007.Generation of insulin-producing islet-like clusters from human embryonic stemcells. Stem Cells (Dayton, Ohio) 25, 1940–1953.
Kadam, S., Muthyala, S., Nair, P., Bhonde, R., 2010. Human placenta-derivedmesenchymal stem cells and islet-like cell clusters generated from these cellsas a novel source for stem cell therapy in diabetes. Rev. Diabetic Stud.: RDS 7,168–182.
Lee, M.W., Choi, J., Yang, M.S., Moon, Y.J., Park, J.S., Kim, H.C., Kim, Y.J., 2004a.Mesenchymal stem cells from cryopreserved human umbilical cord blood.Biochem. Biophys. Res. Commun. 320, 273–278.
Lee, O.K., Kuo, T.K., Chen, W.M., Lee, K.D., Hsieh, S.L., Chen, T.H., 2004b. Isolation ofmultipotent mesenchymal stem cells from umbilical cord blood. Blood 103,1669–1675.
Li, L., Li, F., Qi, H., Feng, G., Yuan, K., Deng, H., Zhou, H., 2008. Coexpression of Pdx1and betacellulin in mesenchymal stem cells could promote the differentiationof nestin-positive epithelium-like progenitors and pancreatic islet-like spher-oids. Stem Cells Dev. 17, 815–823.
Li, Y., Zhang, R., Qiao, H., Zhang, H., Wang, Y., Yuan, H., Liu, Q., Liu, D., Chen, L., Pei, X.,2007. Generation of insulin-producing cells from PDX-1 gene-modified humanmesenchymal stem cells. J. Cell. Physiol. 211, 36–44.
Limbert, C., Path, G., Ebert, R., Rothhammer, V., Kassem, M., Jakob, F., Seufert,J., 2011. PDX1- and NGN3-mediated in vitro reprogramming of human bonemarrow-derived mesenchymal stromal cells into pancreatic endocrine lineages.Cytotherapy 13, 802–813.
Lui, K.O., Zangi, L., Silva, E.A., Bu, L., Sahara, M., Li, R.A., Mooney, D.J., Chien, K.R.,2013. Driving vascular endothelial cell fate of human multipotent Isl1þ heartprogenitors with VEGF modified mRNA. Cell Res. 23, 1172–1186.
Mandal, P.K., Rossi, D.J., 2013. Reprogramming human fibroblasts to pluripotencyusing modified mRNA. Nat. Protoc. 8, 568–582.
Nguyen, P.T.-M., Nguyen, A.T.-Q., Nguyen, N.T., Nguyen, N.T.-M., Duong, T.T., Truong,N.H., Phan, N.K., 2014. Human umbilical cord blood derived mesenchymal stemcells were differentiated into pancreatic endocrine cell by Pdx-1 electrotransfer.Biomed. Res. Ther. 1, 50–56.
Noguchi, H., Naziruddin, B., Shimoda, M., Fujita, Y., Chujo, D., Takita, M., Peng, H.,Sugimoto, K., Itoh, T., Tamura, Y., Olsen, G.S., Kobayashi, N., Onaca, N., Hayashi, S.,Levy, M.F., Matsumoto, S., 2010. Induction of insulin-producing cells from humanpancreatic progenitor cells. Transplant. Proc. 42, 2081–2083.
Ohlsson, H., Karlsson, K., Edlund, T., 1993. IPF1, a homeodomain-containingtransactivator of the insulin gene. EMBO J. 12, 4251–4259.
Overes, I.M., Fredrix, H., Kester, M.G., Falkenburg, J.H., van der Voort, R., de Witte, T.M.,Dolstra, H., 2009. Efficient activation of LRH-1-specific CD8þ T-cell responses fromtransplanted leukemia patients by stimulation with P2�5 mRNA-electroporateddendritic cells. J. Immunother. (Hagerstown, Md.: 1997) 32, 539–551.
Parekh, V.S., Joglekar, M.V., Hardikar, A.A., 2009. Differentiation of human umbilicalcord blood-derived mononuclear cells to endocrine pancreatic lineage. Differ.Res. Biol. Divers. 78, 232–240.
Phadnis, S.M., Joglekar, M.V., Dalvi, M.P., Muthyala, S., Nair, P.D., Ghaskadbi, S.M.,Bhonde, R.R., Hardikar, A.A., 2011. Human bone marrow-derived mesenchymalcells differentiate and mature into endocrine pancreatic lineage in vivo.Cytotherapy 13, 279–293.
Phuc, P.V., Nhung, T.H., Loan, D.T., Chung, D.C., Ngoc, P.K., 2011. Differentiating ofbanked human umbilical cord blood-derived mesenchymal stem cells intoinsulin-secreting cells. In vitro cellular and developmental biology. Animal 47,54–63.
Plews, J.R., Li, J., Jones, M., Moore, H.D., Mason, C., Andrews, P.W., Na, J., 2010.Activation of pluripotency genes in human fibroblast cells by a novel mRNAbased approach. PLoS One 5, e14397.
Prabakar, K.R., Dominguez-Bendala, J., Molano, R.D., Pileggi, A., Villate, S., Ricordi, C.,Inverardi, L., 2012. Generation of glucose-responsive, insulin-producing cellsfrom human umbilical cord blood-derived mesenchymal stem cells. CellTransplant. 21, 1321–1339.
Rahmati, S., Alijani, N., Kadivar, M., 2013. In vitro generation of glucose-responsiveinsulin producing cells using lentiviral based pdx-1 gene transduction of mouse(C57BL/6) mesenchymal stem cells. Biochem. Biophys. Res. Commun. 437,413–419.
Serup, P., Jensen, J., Andersen, F.G., Jorgensen, M.C., Blume, N., Holst, J.J., Madsen, O.D.,1996. Induction of insulin and islet amyloid polypeptide production in pancreaticislet glucagonoma cells by insulin promoter factor 1. Proc. Nat. Acad. Sci. U.S.A.93, 9015–9020.
Sun, J., Yang, Y., Wang, X., Song, J., Jia, Y., 2006. Expression of Pdx-1 in bone marrowmesenchymal stem cells promotes differentiation of islet-like cells in vitro. Sci.China, Ser. C Life Sci./Chin. Acad. Sci. 49, 480–489.
Talebi, S., Aleyasin, A., Soleimani, M., Massumi, M., 2012. Derivation of islet-likecells from mesenchymal stem cells using PDX1-transducing lentiviruses.Biotechnol. Appl. Biochem. 59, 205–212.
Tavernier, G., Wolfrum, K., Demeester, J., De Smedt, S.C., Adjaye, J., Rejman, J., 2012.Activation of pluripotency-associated genes in mouse embryonic fibroblasts bynon-viral transfection with in vitro-derived mRNAs encoding Oct4, Sox2, Klf4and cMyc. Biomaterials 33, 412–417.
Tsai, P.J., Wang, H.S., Shyr, Y.M., Weng, Z.C., Tai, L.C., Shyu, J.F., Chen, T.H., 2012.Transplantation of insulin-producing cells from umbilical cord mesenchymalstem cells for the treatment of streptozotocin-induced diabetic rats. J. Biomed.Sci. 19, 47.
Van Nuffel, A.M., Benteyn, D., Wilgenhof, S., Corthals, J., Heirman, C., Neyns, B.,Thielemans, K., Bonehill, A., 2012. Intravenous and intradermal TriMix-dendritic cell therapy results in a broad T-cell response and durable tumorresponse in a chemorefractory stage IV-M1c melanoma patient. Cancer Immu-nol., Immunother.: CII 61, 1033–1043.
Van Nuffel, A.M., Corthals, J., Neyns, B., Heirman, C., Thielemans, K., Bonehill, A.,2010. Immunotherapy of cancer with dendritic cells loaded with tumorantigens and activated through mRNA electroporation. Methods Mol. Biol.(Clifton, N.J.) 629, 405–452.
Waeber, G., Thompson, N., Nicod, P., Bonny, C., 1996. Transcriptional activation ofthe GLUT2 gene by the IPF-1/STF-1/IDX-1 homeobox factor. Mol. Endocrinol.(Baltimore, Md.) 10, 1327–1334.
Wang, J., Gao, Y., Lu, Y., Tang, X., He, D., Zhang, Y., 2011. Induced differentiation ofhuman umbilical cord mesenchymal stem modified by cells Pdx1gene into isletbeta-like cells in vitro. Sheng wu yi xue gong cheng xue za zhi¼ J. Biomed.Eng.¼Shengwu yixue gongchengxue zazhi 28 (1175–1180), 1206.
Warren, L., Manos, P.D., Ahfeldt, T., Loh, Y.H., Li, H., Lau, F., Ebina, W., Mandal, P.K.,Smith, Z.D., Meissner, A., Daley, G.Q., Brack, A.S., Collins, J.J., Cowan, C.,Schlaeger, T.M., Rossi, D.J., 2010. Highly efficient reprogramming to pluripo-tency and directed differentiation of human cells with synthetic modifiedmRNA. Cell Stem Cell 7, 618–630.
Watada, H., Kajimoto, Y., Umayahara, Y., Matsuoka, T., Kaneto, H., Fujitani, Y.,Kamada, T., Kawamori, R., Yamasaki, Y., 1996. The human glucokinase genebeta-cell-type promoter: an essential role of insulin promoter factor 1/PDX-1 inits activation in HIT-T15 cells. Diabetes 45, 1478–1488.
Wiehe, J.M., Ponsaerts, P., Rojewski, M.T., Homann, J.M., Greiner, J., Kronawitter, D.,Schrezenmeier, H., Hombach, V., Wiesneth, M., Zimmermann, O., Torzewski, J.,2007. mRNA-mediated gene delivery into human progenitor cells promoteshighly efficient protein expression. J. Cell. Mol. Med. 11, 521–530.
Wilgenhof, S., Van Nuffel, A.M., Corthals, J., Heirman, C., Tuyaerts, S., Benteyn, D.,De Coninck, A., Van Riet, I., Verfaillie, G., Vandeloo, J., Bonehill, A., Thielemans,K., Neyns, B., 2011. Therapeutic vaccination with an autologous mRNA electro-porated dendritic cell vaccine in patients with advanced melanoma. J. Immun-other. (Hagerstown, Md.: 1997) 34, 448–456.
Yuan, H., Liu, H., Tian, R., Li, J., Zhao, Z., 2012. Regulation of mesenchymal stem celldifferentiation and insulin secretion by differential expression of Pdx-1. Mol.Biol. Rep. 39, 7777–7783.
Zaldumbide, A., Carlotti, F., Goncalves, M.A., Knaan-Shanzer, S., Cramer, S.J., Roep, B.O.,Wiertz, E.J., Hoeben, R.C., 2012. Adenoviral vectors stimulate glucagon transcrip-tion in humanmesenchymal stem cells expressing pancreatic transcription factors.PLoS One 7, e48093.
Zangi, L., Lui, K.O., von Gise, A., Ma, Q., Ebina, W., Ptaszek, L.M., Spater, D., Xu, H.,Tabebordbar, M., Gorbatov, R., Sena, B., Nahrendorf, M., Briscoe, D.M., Li, R.A.,Wagers, A.J., Rossi, D.J., Pu, W.T., Chien, K.R., 2013. Modified mRNA directs thefate of heart progenitor cells and induces vascular regeneration after myocar-dial infarction. Nat. Biotechnol. 31, 898–907.
Zhu, F.F., Zhang, P.B., Zhang, D.H., Sui, X., Yin, M., Xiang, T.T., Shi, Y., Ding, M.X., Deng,H., 2011. Generation of pancreatic insulin-producing cells from rhesus monkeyinduced pluripotent stem cells. Diabetologia 54, 2325–2336.
P. Van Pham et al. / Differentiation ∎ (∎∎∎∎) ∎∎∎–∎∎∎ 9
Please cite this article as: Van Pham, P., et al., Improved differentiation of umbilical cord blood-derived mesenchymal stem cells intoinsulin-producing cells by PDX-1 mRNA transfection. Differentiation (2014), http://dx.doi.org/10.1016/j.diff.2014.08.001i





























