Embed Size (px)
Citation preview

Instituto Nacional de Pesquisas da Amazônia – INPA
Programa de Pós-Graduação em Genética, Conservação e Biologia
Evolutiva
Carlos Eduardo Faresin e Silva
Manaus-AM
2016
Mapeamento físico cromossômico de elementos repetitivos em
marsupiais amazônicos

Instituto Nacional de Pesquisas da Amazônia – INPA
Programa de Pós-Graduação em Genética, Conservação e Biologia
Evolutiva
Carlos Eduardo Faresin e Silva
Orientador: Eliana Feldberg, Dra.
Co-orientador: Maria Nazareth Ferreira da Silva, Dra.
Manaus-AM
2016
Mapeamento físico cromossômico de elementos repetitivos em
marsupiais amazônicos
Tese apresentada ao Programa de Pós-
Graduação em Genética, Conservação e
Biologia Evolutiva, do Instituto Nacional de
Pesquisas da Amazônia, como requisito
para a obtenção do título de Doutor em
Genética, Conservação e Biologia
Evolutiva.

Ficha catalográfica
Sinopse
No presente trabalho foram analisadas citogeneticamente 16
espécies de marsupiais, coletadas em 13 pontos da bacia hidrográfica
que compõem o bioma Amazônia. Foram realizadas comparações em
coloração convencional, distribuição da heterocromatina, regiões
organizadoras de nucléolo, DNAr 18S, sequências teloméricas e
retroelemento LINE-1. As variações cromossômicas foram analisadas
no âmbito geográfico, evidenciando padrões segregados para as
diferentes formas do cromossomo X. Na família Didelphidae,
sequências teloméricas intersticiais foram relacionadas como
componentes da heterocromatina e restritas a algumas linhagens; a
distribuição do LINE-1 mostrou padrão disperso, diferente daquele já
observado na literatura, sugerindo que esse elemento não participe
participam da inativação do cromossomo X nas espécies analisadas
S586 Silva, Carlos Eduardo Faresin e
Mapeamento físico cromossômico de elementos repetitivos em
marsupiais Amazônicos /Carlos Eduardo Faresin e Silva. --- Manaus:
[s.n.], 2016.
75 f.: il.
Tese (Doutorado) --- INPA, Manaus, 2016.
Orientador: Eliana Feldberg
Coorientador: Maria Nazareth Ferreira da Silva
Área de concentração: Genética, Conservação e Biologia
Evolutiva
1. Marsupiais - Amazônicos. 2. Citogenética. 3. Cariótipo. I. Título.
CDD 599.29811
nio

Dedicatória
Dedico este trabalho a todas
as pessoas que possuem o
comprometimento e respeito
de ausentar-se de suas
terras, visando trazer
conhecimentos dos cantos
mais remotos do mundo.
Dedico também aos
brasileiros que residem
nesses locais e auxiliam na
ocupação de nosso território
e convivem com a natureza
de forma harmônica.

Epígrafe
“A verdade é como o Sol, podes
esconder durante um tempo,
mas não desaparece”
Elvis Presley

A realização deste projeto foi possível devido:
Ao Programa de Pós-graduação em Genética, Conservação e Biologia
Evolutiva, do INPA.
Ao laboratório de Genética Animal do INPA, Coordenação de Pesquisas
em Biodiversidade, onde a maior parte deste trabalho foi desenvolvida, com
financiamento proporcionado pelos Projetos de Pesquisas Institucionais (PPIs),
Projeto Sisbiota Rede BIOPHAM (MCT/MEC/CAPES/FNDCT n◦ 47/2010), ao
Projeto CAPES Pró-Amazônia.
À Fundação de Amparo à Pesquisa do Amazonas (FAPEAM) pela
concessão da bolsa de estudo.

Agradecimentos
Agradeço à minha orientadora Dra. Eliana Feldberg, por sempre deixar
as portas de seu laboratório abertas ao longo de 10 anos, pelos incontáveis
ensinamentos (científicos e de vida) e pela fé incondicional depositada na
minha capacidade. À minha coorientadora Dra. Maria Nazareth Ferreira da
Silva, por compartilhar sua visão, pelo auxílio ao concatenar toda problemática
com a história complexa da mastofauna amazônica. A ambas pelo apoio quase
ilimitado.
Ao colega Eduardo S. Eler, pela parceria de longa data, discussões e
pela grande quilometragem que percorremos nessa selva desde 2006. À Érica
M. S. Souza, pela amizade, auxílios e sobretudo pelo entusiasmo “didelfídico”.
Aos amigos Marco Antônio A. Schetino e Rodrigo A. Andrade, pelas
contribuições e auxílios valorosos em campo e laboratório.
Aos colegas de laboratório Leila B. Ribeiro, Ramon M. Favarato, Lucas
Barros pelas sugestões.
Aos professores Dr. Fabrício Santos e Dra. Marta Svartman, por me
receberem em seus laboratórios contribuindo para meu crescimento
profissional.
Aos amigos do projeto SISBIOTA: Fabrício Bertuol, Pedro Sena,
Alexandre Almeida, Roberto Zamora e Vinícius Carvalho cujo companheirismo
foi fundamental para tornar as expedições ainda mais agradáveis e exequíveis.
E todos os auxiliares de campo (Sebastião “Sabá”, Oscírio “Juruna”,
Flamarion “Pinduca”, Jonas Paz entre outros) que doaram sua força de
trabalho, que compartilharam seu companheirismo e narraram suas
experiências de vida. Essas ocasiões são oportunidades ímpares na inserção
real de pesquisadores dentro da realidade amazônica.

Resumo
Os marsupiais sulamericanos estão representados principalmente pela família
Didelphidae considerada conservada cariotipicamente. No presente trabalho mapeamos
elementos repetitivos em 194 indivíduos, representando 16 espécies de didelfídeos,
coletadas em 13 localidades ainda não exploradas, do ponto de vista científico, na
Amazônia, para verificar possíveis padrões de evolução cariotípica. Para isso foram
utilizados métodos citogenéticos clássicos e moleculares. Variações no cromossomo X,
na distribuição da heterocromatina e no mapeamento de sequências de DNA ribossomal
18S e sequências teloméricas foram registradas. A principal variação foi na posição do
centrômero do cromossomo X de Caluromys lanatus e de Marmosa murina. Os diferentes
tipos de cromossomos X, de ambas espécies, mostraram uma separação em termos
geográficos entre leste e oeste do Brasil, com contato na Amazônia central. Para as
espécies de Marmosops spp., dois padrões de X foram evidenciados pelo método de
banda C, porém não foram separados geográficamente. As regiões organizadoras de
nucléolo foram confirmadas pelas sondas de DNAr 18S em todas as espécies, exceto a
marcação do cromossomo Y de Monodelphis brevicaudata. A distribuição desse marcador
se mostrou variável apenas no gênero Marmosa. Considerando a RON simples como um
caráter plesiomórfico sugerimos que as espécies do gênero Marmosa e Didelphis
marsupialis evoluíram independentemente para o sistema múltiplo. Em relação às
sequências teloméricas, 11 espécies foram analisadas e cinco apresentaram ITSs, todas
dentro da região de heterocromatina centromérica. A presença de ITS foi constante no
cromossomo X de indivíduos de Marmosa murina e variável nas demais espécies.
Acreditamos que a presença variável das ITSs seja uma característica particular de cada
linhagem. O retroelemento LINE-1 foi mapeado em exemplares de cinco espécies:
Caluromys philander, Gracilinanus emiliae, Marmosa murina, Marmosa demerarae e
Didelphis marsupialis. As marcações se mostraram dispersas por todos os cromossomos,
não havendo preferência por regiões específicas do cromossomo, nem mesmo pelo
cromossomo X. A variação do cromossomo X e os padrões de distribuição dos elementos
repetitivos podem ser reflexo dos eventos de especiação ocorridos nos didelfídeos.

i
Abstract
Currently, marsupials are the second largest mammals living family. The groups evolutionary history
is linked to the geological history of the continent and in South America, they are represented mainly
by Didelphidae family. This family is the only present in the Amazon and in the rest of Brazil.
Marsupial cytogenetic characterization shows a conservation of diploid numbers ranging from 10 to
32 chromosomes, and in didelphids are known karyotypes 14, 18 and 22 chromosomes. In this
context, molecular cytogenetic allows for new inferences about the chromosomal evolution, supplying
additional characters to analyze, for example, mapping repetitive regions of the genome. In this paper
we map repetitive elements in species didelphids collected in 13 new locations in the Amazon in
unexplored regions, seeking to understand evolutionary patterns of karyotypes. In all were analyzed
194 individuals of 16 species featured in conventional staining, band C, Ag-RON and FISH with
probes DNAr18S, telomeric and transposable element LINE-1. We observed variation on the X
chromosome, the distribution of heterochromatin and mapping sequences of 18S ribosomal DNA and
telomeric sequences. The main variation found in the position of the centromere was the X
chromosome of C. lanatus and Marmosa murina. Geographically, the different types of X, both
species showed a segregation between eastern and western Brazil, with contact in the central
Amazon. For species Marmosops spp. two patterns of X were evidenced by the C-band technique,
but less obvious geographical distribution. The nucleolar organizer regions were confirmed by 18S
rRNA gene probe in all species except the marking of the Y chromosome Monodelphis brevicaudata.
The distribution of this marker showed only variable in Marmosa genre. Considering RON simple as a
character plesiomorphic conclude that the species of the genus Didelphis and Marmosa evolved
independently for multiple system. Marsupials are considered chromosomally conservative, however,
given the current state of knowledge to this group, inter variations and intraspecific are observed and
this is due to the expansion of the sample and the application of more accurate techniques,
suggesting the presence of chromosomal rearrangements in evolution this animal group. Regarding
the telomeric sequences, 11 species were analyzed and compared the presence of interstitial
telomeric sequences (ITSs) with the distribution of the C band and found that STIs are coincident with
heterochromatin blocks. Of the 11 species examined, only 5 had STIs, all within the centromeric
heterochromatin region. The presence of STIs was constant on the X chromosome of Marmosa
individuals and murine variable in the other species. We believe that the variable presence of STIs is
a particular characteristic of each lineage. Our results support the inference that such sequences can
not always be interpreted as a consequence of chromosomal rearrangement. Finally, we map the
LINE-1 retroelement to five species of copies: Caluromys philander, Gracilinanus emiliae, Marmosa

ii
murina, Marmosa demerarae and Didelphis marsupialis. The markings are shown scattered all
chromosomes, with no preference for specific chromosome regions, or even the X chromosome, as
reported in the literature for other marsupials. The most informative variations were in the morphology
of the X chromosome, which verified the existence of geographic patterns and are possibly related to
different species, showing that chromosomal variation has relation to the diversification of species.
Markers of repetitive sequences (18S, telomere and L1) showed variations possibly be related to
more recent events of speciation.

iii
Sumário 1. Introdução geral ........................................................................................................................... 1
1.1. Ordem Didelphimorphia, Família Didelphidae ....................................................................... 1
1.2. Citogenética clássica em marsupiais ................................................................................... 5
1.2.1 Citogenética molecular em marsupiais ..................................................................................... 7
1.3 . Objetivo geral ........................................................................................................................... 11
2. Material e Métodos .................................................................................................................... 12
2.1. Material e locais de coleta ................................................................................................... 12
2.2 Métodos Citogenéticos ............................................................................................................ 15
2.2.1 Obtenção de cromossomos mitóticos ........................................................................................... 15
2.2.2. Bandeamentos Cromossômicos ................................................................................................... 16
2.3.1 Análise cariotípica ............................................................................................................................ 17
2.3.2. Citogenética molecular ................................................................................................................... 17
2.3.2.2. Detecção de sequências de DNA repetitivo em tandem ....................................................... 18
3. Resultados e Discussão ............................................................................................................. 21
Capítulo 1: Caracterização citogenética de marsupiais didelfídeos na Amazônia brasileira: uma
abordagem clássica sobre o cromossomo X e heterocromatina, num contexto geográfico. .............. 22
Capítulo 2: Comparação cromossômica entre heterocromatina e sequências teloméricas em 11
espécies de marsupiais amazônicos (Didelphimorphia; DIdelphidae) ............................................... 41
Capítulo 3: Mapeamento do elemento LINE-1 em cinco espécies de marsupiais amazônicos
(Didelphidae; Didelphimorphia). ........................................................................................................ 50
4. Conclusões ................................................................................................................................... 57
5. Referências ................................................................................................................................... 58
Apêndice I: ........................................................................................................................................ 72
Apêndice II: ....................................................................................................................................... 72

iv
Lista de figuras:
Introdução geral
Figura 1. Relações intergenéricas simplificadas da família Didelphidae, baseada na árvore com
dados morfológicos, cariotípicos e moleculares pelos métodos de análise Bayesiana de Voss e
Jansa (2009). Dados de número diploide conhecidos para espécies de cada gênero à frente de cada
unidade terminal. Em pontilhado a relação alternativa do gênero Glironia com o restante dos
didelfídeos, encontrada pela máxima parcimônia. ............................................................................... 5
Figura 2. Mapa da bacia amazônica, indicando os locais de coleta das espécies de Didelphidae
analisadas neste estudo. Estado do Amazonas: 1- rio Japurá, Município de Japurá; 2- rio Juruá,
Município de Juruá; 3- rio Negro, Município de Santa Isabel do Rio Negro; 4- rio Purus, Município de
Tapauá; 5- rio Cuieiras, Município de Manaus; 6- Município de Manaus (Campus da Universidade
Federal do Amazonas e Refinaria Isaac Sabá); 7- rio Aripuanã, Município de Novo Aripuanã; 8- rio
Uatumã, Município de Presidente Figueiredo; 9- rio Jatapú, Município de São Sebastião do Uatumã ;
Estado do Pará 10- rio Trombetas, Município de Oriximiná; 11- rio Tapajós, Municípios de Aveiro e
Santarém; 12- rio Jari, Município de Almeirim; Estado de Rondônia: 13- Município de Porto Velho.
Referências geográficas no apêndice I. ............................................................................................ 13
Capítulo 1
Figura 1. Cariótipos em Giemsa (I), Banda C (II), RON e DNAr 18S (III) e cromossomos sexuais
(caixas) de: a) Caluromys philander (SISTAP-M-244, caixas: CAN 34, SISTAP-M-305); b) Caluromys
lanatus (CTGA-M-701); c) Marmosa demerarae (RNL 46, caixas: MCA 27); d) Marmosa murina (RNL
69, caixa: CEF 18); e) Marmosops pinheiroi (INPA 5377, caixa: EE 192) (SISTAP-M-278, caixa:
EE107, INPA 5408); f) Metachirus nudicaudatus (SISTAP-M-302; caixa: SISSIS-M-64); g)
Gracilinanaus emiliae (SISTAP-M-243). Os espécimes testemunho encontram-se na Coleção de
Mamíferos do INPA; a sigla INPA se refere ao registro de tombamento na coleção e as demais ao
registro de campo dos indivíduos analisados. ................................................................................... 29
Figura 2 Cariótipos em Giemsa (I), Banda C (II), RON e DNAr 18S (III) e cromossomos sexuais
(caixas) de: a) Glironia venusta (BAC 80); b) Monodelphis cf. emiliae (INPA 5388); c) Monodelphis
brevicaudata (INPA 5404); d) Monodelphis sp. (CAN 44); e) Didelphis marsupialis (EE 249, caixas:
EE174). Os espécimes testemunho encontram-se na Coleção de Mamíferos do INPA; a sigla INPA
se refere ao registro de tombamento na coleção e as demais ao registro de campo dos indivíduos
analisados. ........................................................................................................................................ 30
Figura 3 Localização das formas do cromossomo X de Caluromys philander na América do Sul. Os
pontos em asterisco são da literatura e os pontos cículo cheio são do presente trabalho: (*1)
Venezuela, Reig et al. 1977; (●2) rio Purus; (●3) REMAN Manaus, presente trabalho e Souza et al.
2013; (●4) rio Trombetas; (●5) rio Tapajós; (*6) rio Jari, Souza et al. 2013; (*7) Pernambudo, Souza
et al. 1990; (*8) São Paulo, Svartman e Vianna-Morgante 1999 e Pereira et al. 2008. m =
metacêntrico; sm = submetacêntrico; a = acrocêntrico; p = puntiforme. ............................................ 34

v
Figura 4 Localização das formas do cromossomo X de Marmosa murina na América do Sul. Os
pontos em asterisco são dados da literatura (*1) Villa Vivêncio, Colômbia, Hayman e Martin 1974;
(*2) Loreto-Peru, Reig et al. 1977; (*3) Bolívar, Venezuela, Reig et al. 1977. Os pontos em ● são
dados do presente trabalho: (●4) rio Negro; (●5) rio Juruá; (●6) rio Purus; (●7) rio Uatumã; (●8) rio
Trombetas; (●9) rio Jari; (*10) Tartarugalzinho, Pará, Carvalho et al. 2002; (*11) Pacoti, Ceará,
Pagnozzi et al. 2002; (*12) Pernambuco, Souza et al. 1990; (*13) Vila Rica, Pagnozzi et al. 2002;
(*14) Tocantins, Lima 2004; (*15) UHE Corumba IV, Luziania, GO, Pereira et al. 2008; (*16) UHE
Peixe Angical, Goiás, Pereira et al. 2008; (*17) Mambai, Goiás, Carvalho et al. 2002; (*18) Espírito
Santo, Paresque 2004. ..................................................................................................................... 35
Capítulo 2
Figura 1 Locais de coleta: 1- rio Japurá; 2- rio Negro; 3- rio Purus; 4- Manaus; 5- rio Aripuanã; 6- rio
Uatumã; 7- rio Jatapu; 8- rio Jari; 9- rio Trombetas; 10- Rio Tapajós; 11- Porto Velho. ..................... 43
Figura 2 Cromossomos após hibridização de sequências teloméricas (à esquerda) e após tratamento
de banda C (à direita) de: a) Caluromys philander (SISTAP-M-244, caixa: SISTAP-M-297,
cromossomo Y SISTAP-M-305); b) Gracilinanus emiliae (SISTAP-M-344); c) Marmosops pinheiroi
(SISTAP-M-237); d) Marmosa demerarae (MCA 65), e) Marmosa murina (CEF 18); f) Metachirus
nudicaudatus (SISTAP-M-302); g) Glironia venusta (BAC 80); h) Monodelphis sp. nov. (CAN 44); i)
Didelphis marsupialis (EE 206). Siglas se referem ao registro de campo dos indivíduos analisados. 46
Capítulo 3
Figura 1. Hibridização in situ fluorescente do LINE-1 nos cromossomos de espécies de didelfídeos:
a) Caluromys philander; b) Marmosa demerarae; c) Gracilinanus emiliae; d) Marmosa murina; e)
Didelphis marsupialis. ....................................................................................................................... 52

vi
Lista de tabelas
Introdução geral
Tabela 1: Classificações da família Didelphidae por diferentes autores, de 1888 a 2009 (Voss e
Jansa 2009). ....................................................................................................................................... 3
Tabela 2. Número de espécimes analisados por espécie e por localidade. Entre parênteses o número
da localidade representada no mapa (Figura 2). ............................................................................... 14
Tabela 1: Dados cariotípicos dos espécimes aqui analisados. Em localidade, os números entre
parênteses indicam as localidades dos Apêndices I e II. A morfologia dos cromossomos é indicada
pelas letras: m-metacêntrico; sm-submetacêntrico; st-subtelocêntrico; a- acrocêntrico; 2n=número
diploide; NF=número fundamental (de braços). ................................................................................ 26
Tabela 2: Padrões de distribuição da heterocromatina do cromossomo X nas espécies do gênero
Marmosops. ...................................................................................................................................... 36
Tabela 1 Espécies analisadas neste trabalhoe enfatizando aquelas que apresentaram variação
intraespecífica no número de ITS...................................................................................................... 45

1
1. Introdução geral Os marsupiais constituem uma das três principais linhagens de mamíferos viventes e
dentre estas, representam o segundo maior grupo em número de espécies.
Taxonomicamente, as 280 espécies atuais pertencem a sete ordens: Didelphimorphia,
Paucituberculata e Microbiotheria presentes principalmente na América do Sul, sendo que
apenas Didelphimorphia pode ser encontrada nas Américas Central e do Norte. As demais
ordens: Dasyuromorphia, Diprotodontia, Notoryctemorphia e Peramelemorphia encontradas
predominantemente na Austrália, Nova Guiné e ilhas adjacentes (Aplin e Archer 1987;
Wilson e Reeder 2005).
Szalay (1982), com base na morfologia da articulação do tornozelo, propôs classificar
as ordens de marsupiais em duas coortes, Australidelphia e Ameridelphia; porém, a
condição monofilética desta última permanece controversa. Ameridelphia é representada
pelas ordens: Didelphimorphia, Paucituberculata e Microbiotheria. Estudos filogenéticos e
anatômicos indicam que esta última possui mais afinidades com marsupiais australianos que
didelfídeos (Jansa e Voss 2000; Horovitz e Sánches-Villagra 2003; Nilsson et al. 2004;
Nilsson et al. 2010).
1.1. Ordem Didelphimorphia, Família Didelphidae
O fóssil metatério mais antigo conhecido é do início do Cretáceo na China. Metatérios
do Cretáceo foram encontrados na Ásia, Europa e América do Norte (Bennett 2012). Das
três ordens de marsupiais americanos, os didelfimorfos são considerados os mais basais
dentro dos metatérios (Nilsson et al. 2010) e as espécies remanescentes descendem de
linhagem antiga do fim do Cretáceo, a qual teria colonizado quase todos os continentes
(Marshall et al. 1990; McKenna e Bell 1997). Atualmente, a distribuição geográfica dos
didelfimorfos restringe-se às Américas, desde o Canadá até o sul da Argentina.
Os didelfimorfos atuais representam formas derivadas dos mais antigos metatérios
fósseis do Paleoceno, cuja linhagem divergiu dos demais metatérios cerca de 69 milhões de
anos atrás (Simpson 1945; Szalay 1982; Reig et al. 1987; Horovitz e Sánchez-Villagra 2003;
Nilsson et al. 2004). Esta ordem possui o maior número de espécies entre os marsupiais
americanos, com 18 gêneros e 91 espécies, todas classificadas na família Didelphidae
(Nowak 1991; Gardner 1993; Groves 1993; Costa 2005; Gardner 2007). Esses gêneros

2
foram organizados em diferentes subfamílias e tribos por diferentes autores (Tabela 1) e a
proposta filogenética mais recente é a de Voss e Jansa (2009). Esses autores revisaram a
história taxonômica da família mostrando avanços sobre a filogenia do grupo e propondo
uma nova classificação (Tabela 1), tendo por base caracteres morfológicos, cariotípicos e
moleculares.
Voss e Jansa (2009) reconhecem quatro subfamílias em Didelphidae: Glironiinae e
Hyladelphinae com apenas um gênero monotípico cada, Caluromyinae com dois gêneros e
Didelphinae com os demais gêneros. A última subfamília ainda é subdivida em quatro tribos:
Marmosini, Metachirini, Didelphini e Thylamyini (Tabela 1). As relações intergenéricas
simplificadas nesse grupo são mostradas na Figura 1.
3
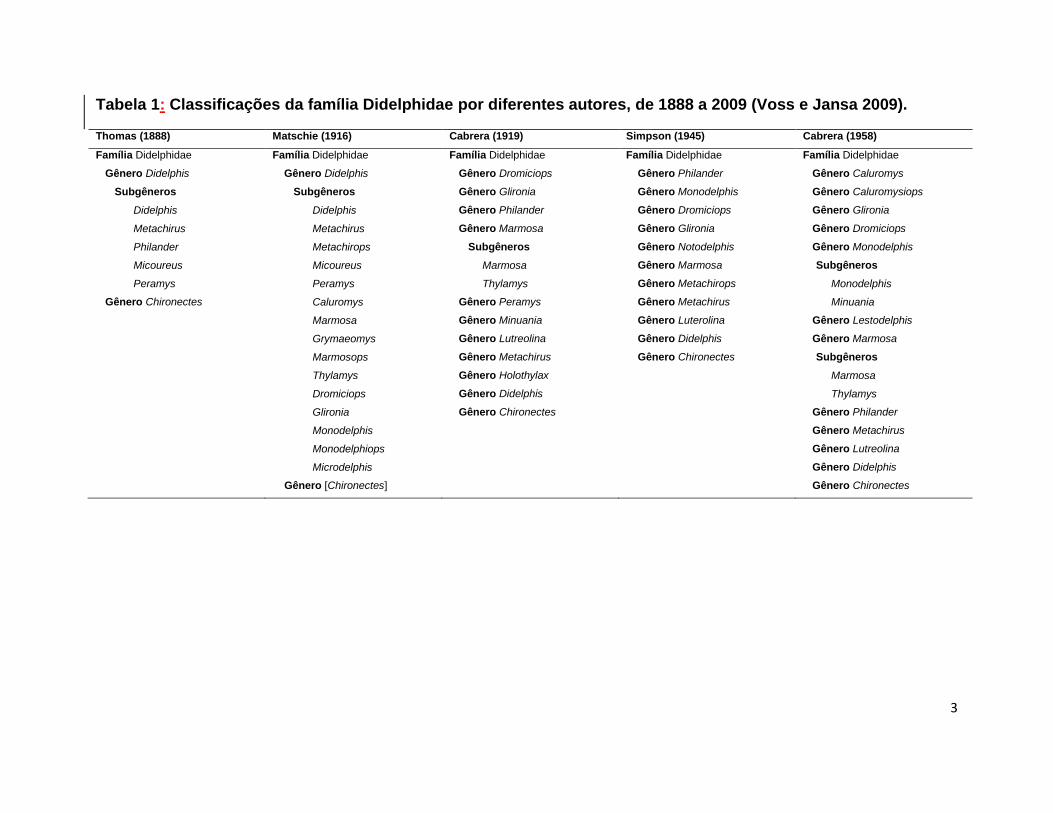
Tabela 1: Classificações da família Didelphidae por diferentes autores, de 1888 a 2009 (Voss e Jansa 2009).
Thomas (1888) Matschie (1916) Cabrera (1919) Simpson (1945) Cabrera (1958)
Família Didelphidae Família Didelphidae Família Didelphidae Família Didelphidae Família Didelphidae
Gênero Didelphis Gênero Didelphis Gênero Dromiciops Gênero Philander Gênero Caluromys
Subgêneros Subgêneros Gênero Glironia Gênero Monodelphis Gênero Caluromysiops
Didelphis Didelphis Gênero Philander Gênero Dromiciops Gênero Glironia
Metachirus Metachirus Gênero Marmosa Gênero Glironia Gênero Dromiciops
Philander Metachirops Subgêneros Gênero Notodelphis Gênero Monodelphis
Micoureus Micoureus Marmosa Gênero Marmosa Subgêneros
Peramys Peramys Thylamys Gênero Metachirops Monodelphis
Gênero Chironectes Caluromys Gênero Peramys Gênero Metachirus Minuania
Marmosa Gênero Minuania Gênero Luterolina Gênero Lestodelphis
Grymaeomys Gênero Lutreolina Gênero Didelphis Gênero Marmosa
Marmosops Gênero Metachirus Gênero Chironectes Subgêneros
Thylamys Gênero Holothylax Marmosa
Dromiciops Gênero Didelphis Thylamys
Glironia Gênero Chironectes Gênero Philander
Monodelphis Gênero Metachirus
Monodelphiops Gênero Lutreolina
Microdelphis Gênero Didelphis
Gênero [Chironectes] Gênero Chironectes
4
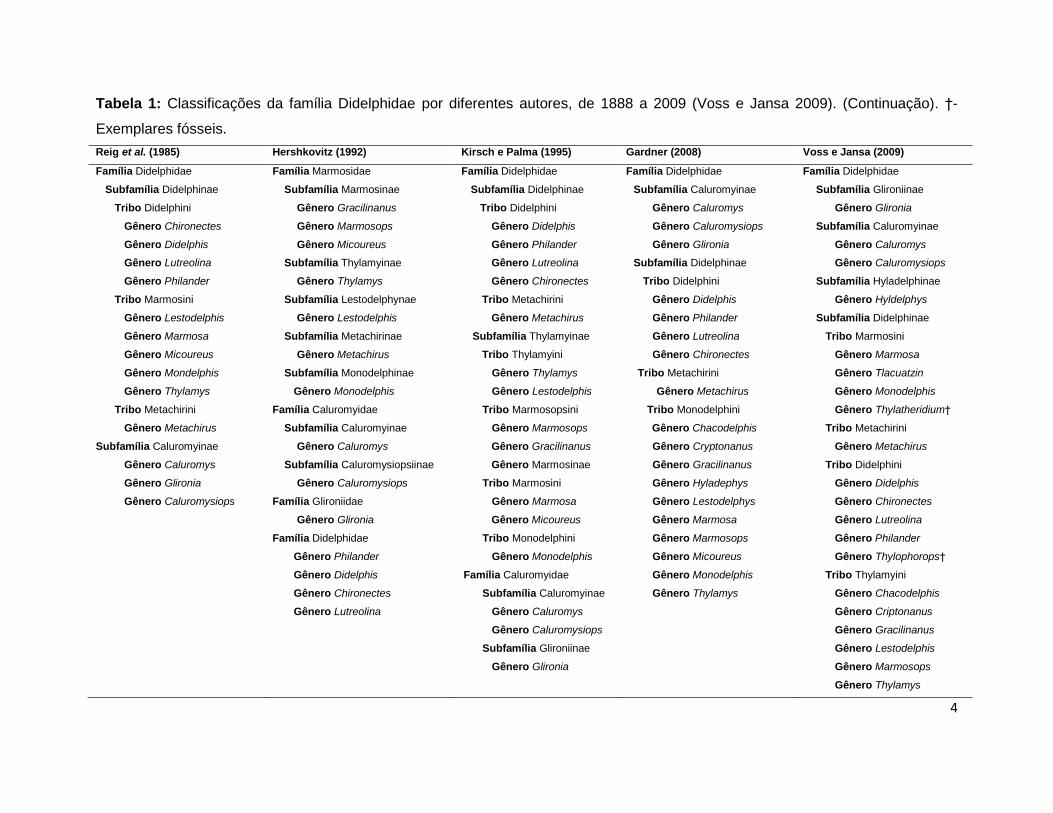
Tabela 1: Classificações da família Didelphidae por diferentes autores, de 1888 a 2009 (Voss e Jansa 2009). (Continuação). †-
Exemplares fósseis.
Reig et al. (1985) Hershkovitz (1992) Kirsch e Palma (1995) Gardner (2008) Voss e Jansa (2009)
Família Didelphidae Família Marmosidae Família Didelphidae Família Didelphidae Família Didelphidae
Subfamília Didelphinae Subfamília Marmosinae Subfamília Didelphinae Subfamília Caluromyinae Subfamília Glironiinae
Tribo Didelphini Gênero Gracilinanus Tribo Didelphini Gênero Caluromys Gênero Glironia
Gênero Chironectes Gênero Marmosops Gênero Didelphis Gênero Caluromysiops Subfamília Caluromyinae
Gênero Didelphis Gênero Micoureus Gênero Philander Gênero Glironia Gênero Caluromys
Gênero Lutreolina Subfamília Thylamyinae Gênero Lutreolina Subfamília Didelphinae Gênero Caluromysiops
Gênero Philander Gênero Thylamys Gênero Chironectes Tribo Didelphini Subfamília Hyladelphinae
Tribo Marmosini Subfamília Lestodelphynae Tribo Metachirini Gênero Didelphis Gênero Hyldelphys
Gênero Lestodelphis Gênero Lestodelphis Gênero Metachirus Gênero Philander Subfamília Didelphinae
Gênero Marmosa Subfamília Metachirinae Subfamília Thylamyinae Gênero Lutreolina Tribo Marmosini
Gênero Micoureus Gênero Metachirus Tribo Thylamyini Gênero Chironectes Gênero Marmosa
Gênero Mondelphis Subfamília Monodelphinae Gênero Thylamys Tribo Metachirini Gênero Tlacuatzin
Gênero Thylamys Gênero Monodelphis Gênero Lestodelphis Gênero Metachirus Gênero Monodelphis
Tribo Metachirini Família Caluromyidae Tribo Marmosopsini Tribo Monodelphini Gênero Thylatheridium†
Gênero Metachirus Subfamília Caluromyinae Gênero Marmosops Gênero Chacodelphis Tribo Metachirini
Subfamília Caluromyinae Gênero Caluromys Gênero Gracilinanus Gênero Cryptonanus Gênero Metachirus
Gênero Caluromys Subfamília Caluromysiopsiinae Gênero Marmosinae Gênero Gracilinanus Tribo Didelphini
Gênero Glironia Gênero Caluromysiops Tribo Marmosini Gênero Hyladephys Gênero Didelphis
Gênero Caluromysiops Família Glironiidae Gênero Marmosa Gênero Lestodelphys Gênero Chironectes
Gênero Glironia Gênero Micoureus Gênero Marmosa Gênero Lutreolina
Família Didelphidae Tribo Monodelphini Gênero Marmosops Gênero Philander
Gênero Philander Gênero Monodelphis Gênero Micoureus Gênero Thylophorops†
Gênero Didelphis Família Caluromyidae Gênero Monodelphis Tribo Thylamyini
Gênero Chironectes Subfamília Caluromyinae Gênero Thylamys Gênero Chacodelphis
Gênero Lutreolina Gênero Caluromys Gênero Criptonanus
Gênero Caluromysiops Gênero Gracilinanus
Subfamília Glironiinae Gênero Lestodelphis
Gênero Glironia Gênero Marmosops
Gênero Thylamys

5
Figura 1. Relações intergenéricas simplificadas da família Didelphidae, baseada na árvore com
dados morfológicos, cariotípicos e moleculares pelos métodos de análise Bayesiana de Voss e
Jansa (2009). Dados de número diploide conhecidos para espécies de cada gênero à frente de
cada unidade terminal. Em pontilhado a relação alternativa do gênero Glironia com o restante dos
didelfídeos, encontrada pela máxima parcimônia.
1.2. Citogenética clássica em marsupiais
Apesar de uma grande diversidade morfológica e da grande divergência entre
grupos, os metatérios exibem características citogenéticas compartilhadas por todas as
ordens: (1) baixos números diploides, com variação de 10 a 32 cromossomos; (2)
grandes cromossomos (Hayman 1990); (3) alto grau de conservação, principalmente
na eucromatina (Svartman e Vianna-Morgante 1999) e (4) ausência de região de
homologia entre cromossomos sexuais, e consequente modificação do comportamento

6
destes cromossomos durante a meiose (Solari e Bianchi 1975; Sharp 1982; Seluja et
al. 1987; Graves e Watson 1991).
A família Didelphidae se enquadra nesses padrões, exibindo apenas três
números diploides (14, 18 e 22). Desses, o número diploide mais frequente é o 2n=14,
encontrado em 15 espécies, seguido pelo 2n=18, encontrado em Monodelphis e
Glironia e o 2n=22, encontrado em oito espécies da tribo Didelphini e no gênero
Tlacuatzin. A origem desses cariótipos tem sido discutida na literatura e técnicas cada
vez mais sofisticadas têm proporcionado novos caminhos para essa discussão. Reig et
al. (1977) observaram cromossomos em coloração convencional de 14 espécies e
propuseram três hipóteses alternativas para a evolução cariotípica de didelfídeos: 1)
aumento do número diploide por fissões cêntricas, tendo como partida 2n=14
cromossomos; 2) redução e aumento dos números diploides, com fusões e fissões
cêntricas, partindo do 2n=18, e 3) redução do número diploide por fusões cêntricas a
partir do 2n=22. Dentro dessas hipóteses, ainda há discordância sobre quais eventos
foram responsáveis pela evolução desses cariótipos.
Inicialmente, Reig et al. (1977) descartaram a hipótese da ancestralidade em
2n=18 e 22 cromossomos, com base na hipótese da ortosseleção de White (1973), na
prevalência do 2n=14 em espécies australianas e americanas e também no alto grau
de derivação das espécies com 2n=22. Rofe e Hayman (1985) corroboraram a
ancestralidade do 2n=14 ao observar homeologias apresentadas pelos cromossomos
na banda G. De forma contrária, os segmentos de heterocromatina aparentam ser
espécie-específicos (Svartman e Vianna-Morgante 1999). Na impregnação com nitrato
de prata, as regiões organizadoras de nucléolo (RON) mostraram distribuição variável
nos três números diploides: em espécies com 2n=14 as RON foram observadas de
uma a quatro marcações em autossomos; nas espécies com 2n=18, foram observadas
marcações em um par autossômico e, eventualmente, no X e em espécies com 2n=22,
a quantidade de RONs ativas foi variável intraindividualmente, porém essa variação foi
de no mínimo quatro marcações e no máximo oito (Svartman e Vianna-Morgante 2003;
Svartman 2009 e referências inclusas).

7
A sobreposição dos números diploides na filogenia de Voss e Jansa (2009) não
permitiu detectar um padrão de evolução cromossômica linear (2n=14 para 2n=18 e
para 2n=22 ou o inverso), conforme a figura 1. Porém, Fantin e da Silva (2011)
mostraram que número diploide igual a 18 é compartilhado por dois gêneros
didelfídeos: Monodelphis e Glironia. Glironia localiza-se na base da filogenia da família
Didelphidae e isso pode ser uma evidência adicional à ancestralidade de um número
diploide maior que 14, porém não podemos descartar origens independentes.
1.2.1 Citogenética molecular em marsupiais
A combinação de métodos de citogenética e biologia molecular possibilitou
localizar fisicamente sequências de DNA nos cromossomos, realizando inferências
sobre a organização do genoma (Guerra 1988; 2004). Variações desses métodos
permitem identificar sequências de cópia única, sequências moderada ou altamente
repetitivas, verificar similaridade entre genomas e mapeamento (Singer 1982).
Exemplifica-se a pintura cromossômica em marsupiais australianos, que
identificou de forma precisa rearranjos cromossômicos, possibilitando inferir sobre um
modelo de cariótipo ancestral (Rens et al. 1999; O’Neill et al. 1999; De Leo et al. 1999;
Rens et al. 2003). Além disso, a hibridização in situ fluorescente (FISH) corroborou os
resultados obtidos com banda G e permitiu realizar novas discussões sobre rearranjos
cromossômicos.
A estabilidade cariotípica de marsupiais tem sido amplamente discutida e pela
citogenética clássica é possível observar uniformidade dos números diploides e
extensiva homeologia das bandas G. A pintura cromossômica e hibridização de
genomas (GISH) também mostram grande similaridade desses genomas. Nesse
âmbito a principal fonte de variabilidade cariotípica tem sido atribuída aos rearranjos
Robertsonianos e diferenças na heterocromatina, a qual mostra padrão espécie-
específico.
Todder et al. (1997) analisaram a relação entre os cariótipos de Macropus
eugenii e Wallabia bicolor e encontraram sequências do cromossomo Y de uma
espécie com sistema de determinação sexual simples em cromossomos XY1Y2 da
outra espécie com cromossomos sexuais múltiplos. Concluíram que os eventos

8
envolvidos no surgimento do sistema sexual múltiplo em uma das espécies eram um
pouco mais complexos do que o revelado pela banda G.
Pintura cromossômica no complexo Petrogale (Marsupialia; Macropodidae)
mostrou que sondas podem revelar padrões heterogêneos de hibridização ao longo de
um cromossomo, sendo que em porções distais exibiram sinais mais intensos,
enquanto nas porções proximais essa intensidade foi menor. Essa heterogeneidade foi
atribuída à divergência da heterocromatina centromérica e sequências repetitivas
dispersas na eucromatina, por amplificação diferencial de sequências contidas nessas
regiões (O’Neill et al. 1999). Esse padrão de conservação da eucromatina e
especificidade de sequências relacionadas à heterocromatina pericentromérica também
foi demonstrado em marsupiais da família Didelphidae, com a técnica de hibridização
genômica cruzada (Svartman e Vianna-Morgante 1999).
Outros elementos importantes do genoma são as sequências de DNA repetitivo,
que podem pertencer a diferentes classes e algumas têm suas funções e origens já
conhecidas, por exemplo: sequências teloméricas, unidades de DNA ribossomal e
elementos transponíveis (Brasileiro-Vidal e Guerra 2002). Essas sequências
representam um componente interessante do DNA, uma vez que há escassez de
dados relacionados a eles e também pelo papel incerto que desempenham na
organização genômica (Elder e Turner 1995).
Em didelfídeos a AgRON e o mapeamento dos sítios ribossomais 18S
mostraram que a quantidade de sítios ribossomais pode variar entre as espécies, mas
pelo menos um desses pares cromossômicos é homeólogo em todas as espécies. Isso
indica que a conservação dos cariótipos de marsupiais também se estende a algumas
sequências de DNA repetitivo (Svartman e Vianna-Morgante 2003).
As sequências teloméricas desempenham função de proteção das extremidades
coesivas dos cromossomos, impedindo que elas interajam umas com as outras. Além
disso, elas são consideradas sequências extremamente conservadas em todos os
vertebrados (McClintock 1941; Blackburn 1991). Em marsupiais, sondas dessas
sequências foram utilizadas para mostrar eventos de fusões cromossômicas,
fornecendo novos dados para a discussão da evolução cariotípica do grupo. Em

9
espécies da família Didelphidae foram encontradas sequências teloméricas intersticiais
(ITS) nos cariótipos com 2n=14 e 18, significando que estes cariótipos teriam originado
por fusão a partir de um 2n=22 (Svartman e Vianna-Morgante 1998; Carvalho e Mattevi
2000).
Pagnozzi et al. (2000; 2002) sugeriram que as ITS encontradas em didelfídeos
brasileiros podiam ser um componente da heterocromatina, ao invés de resultado de
fusões recentes. Metcalfe et al. (2004) observaram que nem todas as ITS de
Macropodidae podem ser aceitas como evidências de fusões, salvo quando não forem
os maiores constituintes da heterocromatina. Porém, Svartman (2009) argumenta que o
estado de condensação dos cromossomos pode tornar a localização das sequências
teloméricas pouco precisa por aproximar essas marcações à região de heterocromatina
pericentromérica.
Elementos transponíveis são reconhecidos como parte de repetições dispersas
pelo genoma. Eles podem pertencer a duas classes que se diferenciam pela forma de
transposição. Na classe I, temos os retrotransposons que são elementos que utilizam
um intermediário de RNA para se movimentar para novos pontos, onde serão inseridos.
Ainda dentro dessa classe, temos elementos com repetições terminais curtas (SINE -
short interspersed elements) e elementos com longas repetições terminais (LINE – long
interspersed elements). Os elementos da classe II são caracterizados por se
movimentarem com auxílio de uma enzima denominada transposase, o que elimina a
necessidade de um intermediário de RNA (Capy et al. 1998; Almeida e Carareto 2005).
O elemento LINE-1, da superfamília dos LINEs, tem sido considerado como
grande parte do genoma de mamíferos e frequentemente está relacionado com
desordens genéticas (duplicação, deleção e instabilidade cromossômica por
recombinação heteróloga), regulação da expressão gênica, inativação do X nas fêmeas
(Ostertag et al. 2001; Kazazian e Goodier 2002) e tem sido utilizado em estudos
evolutivos. Waters et al. (2007) observaram que, em mamíferos, os padrões de
divergência entre os elementos LINE-1 são específicos de cada linhagem. Porém, o
envolvimento desses elementos em alterações cromossômicas pode ser indireto, mas
ainda assim o acúmulo desses elementos pode ser associado com os padrões de

10
especiação de algumas linhagens (Dogbiny et al. 2004; Waters et al. 2004). Em
marsupiais, a distribuição desses elementos ainda foi pouco explorada, com exceção
do sequenciamento de LINE-1 em Didelphis virginiana (Dönner e Paböo 1995).
Em relação aos elementos repetitivos foi demonstrado que eles podem
corroborar padrões de especiação e estarem relacionados à plasticidade cariotípica.
Nesse campo, existem poucos estudos em marsupiais, contrastando com a realidade
das linhagens eutérias. Considerando que marsupiais e mamíferos placentários são
grupos irmãos, aqueles podem fornecer uma base de comparação como grupo externo
para o estudo dos padrões de distribuição e divergência das sequências repetitivas em
mamíferos placentários. Dessa forma o presente trabalho buscou obter padrões de
distribuição de sequências repetitivas dispersas e in tandem para espécies de
marsupiais amazônicos.

11
1.3 . Objetivo geral
Mapear fisicamente, em cromossomos metafásicos, regiões de sequências
repetitivas in tandem e dispersas em espécies de marsupiais amazônicos,
representantes dos gêneros: Caluromys, Didelphis, Glironia, Gracilinanus, Marmosa,
Marmosops, Metachirus e Monodelphis a fim de ampliar o entendimento da
organização genômica nestas espécies e buscar padrões evolutivos em marsupiais
didelfídeos.
1.4.1, Específicos
1- Comparar padrões de bandeamento cromossômico com sequências
mapeadas de espécies representantes dos gêneros de marsupiais dos
gêneros prospostos.
2- Identificar padrões intraespecíficos em espécies coletadas em diferentes
localidades em busca de variabilidade em espécies representantes dos
gêneros de marsupiais dos gêneros propostos.
3- Comparar os padrões obtidos, na tentativa de relacioná-los aos eventos de
diversificação cromossômica nas espécies representantes dos gêneros
propostos
4- Determinar a localização física de elementos do tipo LINE nos cariótipos e
verificar sua divergência entre as espécies escolhidas no presente estudo.

12
2. Material e Métodos 2.1. Material e locais de coleta
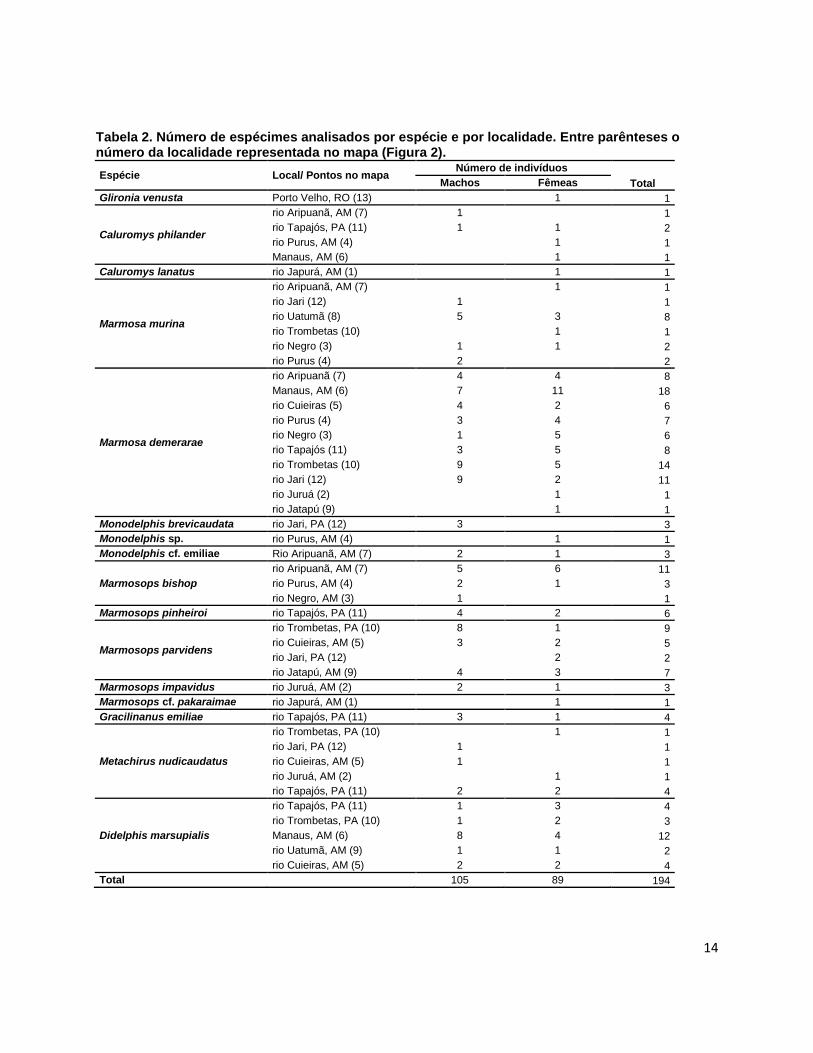
Neste estudo foram analisados 194 indivíduos de 16 espécies, representantes
de oito gêneros de marsupiais, coletadas em 13 localidades da Amazônia (Figura 2,
Tabela 2, Apêndice I). Os espécimes foram coletados sob autorizações do Instituto
Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis - IBAMA, de acordo
com as licenças de número: 02005.000642/03-11 (IBAMA); 02000.002336/2003-93
(IBAMA); 02005.002672/04 (IBAMA); 37585-5 (SISBIO); 37592-4 (SISBIO). Os
indivíduos analisados estão depositados na Coleção de Mamíferos do Instituto
Nacional de Pesquisas da Amazônia - INPA (apêndice II).
No presente trabalho foram utilizadas armadilhas “live-traps” dos tipos
“tomahawk” (14x14x40cm) e “sherman” (8x8x23cm). As armadilhas foram dispostas
aos pares (uma “tomahawk” e uma “sherman”) em 40 estações espaçadas por uma
distância de cerca de 30 metros. A disposição das armadilhas, em cada estação, foi
feita obedecendo ao seguinte critério: em estações ímpares, as “shermans” foram
instaladas no sub-bosque em árvores ou cipós que faziam alguma ligação com o
dossel, enquanto as “tomahawks” foram instaladas no chão em frente a tocas ou
troncos caídos. Em estações pares foi feito o inverso.
Em cada localidade, as armadilhas permaneceram ativadas por um período de
10 noites e foram vistoriadas a cada dia, no período matutino. As iscas foram
compostas por fatias de banana com pasta de amendoim torrado e moído, que foram
substituídas a cada dois dias ou de acordo com a necessidade.
Cada espécime coletado teve seus dados de idade, sexo, estágio reprodutivo,
peso, medidas, localidade, ambiente e condições climáticas anotados em caderno de
campo padronizado pela curadoria da Coleção de Mamíferos do INPA e foram
depositados nesta coleção, onde foram identificados pela Dra. Maria Nazareth Ferreira
da Silva, curadora da Coleção de Mamíferos do INPA. Os espécimes tiveram a pele,
crânio, esqueleto, suspensão celular preparados e seus tecidos coletados. Os
indivíduos cujas peles não foram taxidermizadas foram fixados em formol 10% e depois
preservados em álcool etílico 70%.

13
Figura 2. Mapa da bacia amazônica, indicando os locais de coleta das espécies de Didelphidae
analisadas neste estudo. Estado do Amazonas: 1- rio Japurá, Município de Japurá; 2- rio Juruá,
Município de Juruá; 3- rio Negro, Município de Santa Isabel do Rio Negro; 4- rio Purus, Município
de Tapauá; 5- rio Cuieiras, Município de Manaus; 6- Município de Manaus (Campus da
Universidade Federal do Amazonas e Refinaria Isaac Sabá); 7- rio Aripuanã, Município de Novo
Aripuanã; 8- rio Uatumã, Município de Presidente Figueiredo; 9- rio Jatapú, Município de São
Sebastião do Uatumã ; Estado do Pará 10- rio Trombetas, Município de Oriximiná; 11- rio Tapajós,
Municípios de Aveiro e Santarém; 12- rio Jari, Município de Almeirim; Estado de Rondônia: 13-
Município de Porto Velho. Referências geográficas no apêndice I.
14
Tabela 2. Número de espécimes analisados por espécie e por localidade. Entre parênteses o número da localidade representada no mapa (Figura 2).
Espécie Local/ Pontos no mapa Número de indivíduos
Total Machos Fêmeas
Glironia venusta Porto Velho, RO (13) 1 1
Caluromys philander
rio Aripuanã, AM (7) 1 1
rio Tapajós, PA (11) 1 1 2
rio Purus, AM (4) 1 1
Manaus, AM (6) 1 1
Caluromys lanatus rio Japurá, AM (1) 1 1
Marmosa murina
rio Aripuanã, AM (7) 1 1
rio Jari (12) 1 1
rio Uatumã (8) 5 3 8
rio Trombetas (10) 1 1
rio Negro (3) 1 1 2
rio Purus (4) 2 2
Marmosa demerarae
rio Aripuanã (7) 4 4 8
Manaus, AM (6) 7 11 18
rio Cuieiras (5) 4 2 6
rio Purus (4) 3 4 7
rio Negro (3) 1 5 6
rio Tapajós (11) 3 5 8
rio Trombetas (10) 9 5 14
rio Jari (12) 9 2 11
rio Juruá (2) 1 1
rio Jatapú (9) 1 1
Monodelphis brevicaudata rio Jari, PA (12) 3 3
Monodelphis sp. rio Purus, AM (4) 1 1
Monodelphis cf. emiliae Rio Aripuanã, AM (7) 2 1 3
Marmosops bishop
rio Aripuanã, AM (7) 5 6 11
rio Purus, AM (4) 2 1 3
rio Negro, AM (3) 1 1
Marmosops pinheiroi rio Tapajós, PA (11) 4 2 6
Marmosops parvidens
rio Trombetas, PA (10) 8 1 9
rio Cuieiras, AM (5) 3 2 5
rio Jari, PA (12) 2 2
rio Jatapú, AM (9) 4 3 7
Marmosops impavidus rio Juruá, AM (2) 2 1 3
Marmosops cf. pakaraimae rio Japurá, AM (1) 1 1
Gracilinanus emiliae rio Tapajós, PA (11) 3 1 4
Metachirus nudicaudatus
rio Trombetas, PA (10) 1 1
rio Jari, PA (12) 1 1
rio Cuieiras, AM (5) 1 1
rio Juruá, AM (2) 1 1
rio Tapajós, PA (11) 2 2 4
Didelphis marsupialis
rio Tapajós, PA (11) 1 3 4
rio Trombetas, PA (10) 1 2 3
Manaus, AM (6) 8 4 12
rio Uatumã, AM (9) 1 1 2
rio Cuieiras, AM (5) 2 2 4
Total 105 89 194

15
2.2 Métodos Citogenéticos
2.2.1 Obtenção de cromossomos mitóticos
Os cromossomos foram obtidos pelo método in vivo segundo Ford e Harmerton
(1956), utilizando-se colchicina diluída a 0,0125% em uma proporção de 1 mL para
cada 100 gramas de peso animal. Esta solução foi injetada intraperitonealmente no
animal vivo, que foi colocado em descanso, por um período de 30 minutos. Após esse
tempo, o animal foi eutanaziado utilizando-se inalação de dosagem elevada de
anestésico inalante (halotano). Em seguida, os fêmures foram removidos e suas
epífises foram cortadas para a obtenção da medula óssea. A medula foi lavada do
interior da diáfise femoral, com o auxílio de uma seringa de 10mL contendo solução
hipotônica de cloreto de potássio 0,075M, para uma placa de Petri até a remoção total
desse material. O material foi homogeneizado e colocado em banho-maria por 30
minutos para efetuar a hipotonização das células. A solução foi transferida para um
tubo Falcon de 15mL e foi adicionado oito a 10 gotas de fixador Carnoy (metanol-ácido
acético: 3:1) e novamente foi homogeneizado. Essa solução foi centrifugada, o
sobrenadante desprezado e novamente foi ressuspendida em fixador. Esses passos
foram repetidos duas vezes. Após a última centrifugação, o sobrenadante foi
descartado e adicionou-se fixador na proporção de 2:1 em relação à quantidade de
sedimento e homogeneizado. Por fim o material foi transferido para um tubo tipo
Eppendorf devidamente identificado com o número do espécime e foi guardado em
freezer (-10 °C) até a preparação das lâminas.
No laboratório, as lâminas foram preparadas, pingando-se duas a três gotas da
suspensão celular em uma lâmina de vidro limpa (aquecida em água destilada até 60
°C), a uma altura de 30-40cm e seca à temperatura ambiente; foram coradas com
solução de Giemsa, diluída a 5% em tampão fosfato pH 6,8, por 10 minutos.
Posteriormente foram lavadas e secas novamente ao ar.

16
2.2.2. Bandeamentos Cromossômicos
2.2.2.1. Bandeamento C segundo Sumner (1972)
A localização das regiões de heterocromatina foi feita utilizando a técnica de
banda C. Inicialmente a lâmina foi tratada com HCl 0,2N em temperatura ambiente, por
30 minutos; em seguida as lâminas foram lavadas com água destilada e secas ao ar.
As lâminas foram incubadas em solução de hidróxido de bário octahidratado
(Ba(OH)2.8H2O) a 5%, recém preparada e filtrada, por um período de cinco segundos
a um minuto em uma temperatura de 42 °C. O tempo de tratamento com hidróxido de
bário foi determinado para cada espécie. Após esse tratamento, as lâminas foram
lavadas rapidamente em solução de HCl 0,2 N a 42 °C e posteriormente foram lavadas,
várias vezes, em água destilada e novamente secas ao ar. Em seguida foram
incubadas em solução 2XSSC (cloreto de sódio 0,3M e citrato trissódico 0,06M em pH
6,8) a 60 °C, por 15 minutos. As lâminas foram lavadas e secas ao ar. Por fim foram
coradas em Giemsa diluído a 2% em tampão fosfato pH 6,8, durante 20-30 minutos ou
com Giemsa 5% por 5 a 10 minutos.
2.2.2.2. Detecção das Regiões Organizadoras de Nucléolo (RON)
segundo Howell e Black (1980)
Para a localização das regiões organizadoras de nucléolos ativas foi utilizada a
técnica de precipitação de cristais de prata (AgRON), com pequenas modificações que
consiste em: pingar sobre a lâmina com a suspensão celular, uma gota de solução
aquosa de gelatina a 2% acrescida de ácido fórmico na proporção de 1mL/100mL de
solução. Adicionar sobre a gota de gelatina, duas gotas de solução aquosa de nitrato
de prata (AgNO3) a 50%, agitando levemente a lâmina e cobrindo com lamínula;
colocar a lâmina em câmara úmida e levar ao banho-maria ou estufa à 60 °C, por um
período de dois a quatro minutos, ou até a lâmina apresentar uma coloração marrom-
dourada; lavar as lâminas em água destilada, permitindo que a lamínula e o excesso de
soluções sejam removidos e secar ao ar. As lâminas foram coradas com solução de
Giemsa diluído a 2% em tampão fosfato pH 6,8, por 20-30 segundos, lavadas e secas
ao ar.

17
2.3.1 Análise cariotípica
Os cromossomos foram analisados em microscópio de luz com objetiva de
imersão. Foram contadas pelo menos 30 metáfases de cada indivíduo, com o intuito de
estabelecer o valor modal (2n) e o número fundamental (número de braços) de cada
espécime.
As melhores metáfases foram editadas no programa Adobe Photoshop CS4. As
médias do comprimento total dos cromossomos e da razão entre os braços
cromossômicos (q/p) foram calculadas com auxílio do programa livre Image J. O
cariótipo foi montado emparelhando-se os cromossomos com base na razão entre os
braços cromossômicos e na posição do centrômero, segundo Levan et al. (1964). Os
cromossomos mitóticos foram classificados em metacêntricos (RB=1,0-1,69),
submetacêntricos (RB=1,7-2,99), subtelocêntricos (RB=3,00-7,0) e acrocêntricos
(RB>7,00).
Na determinação do número de braços (NF) foram considerados os autossomos
metacêntricos, submetacêntricos e subtelocêntricos como tendo dois braços e os
acrocêntricos como tendo apenas um braço. Os cromossomos sexuais não foram
considerados.
2.3.2. Citogenética molecular
2.3.2.1. Extração de DNA
Para a extração do DNA foi utilizado tecido muscular de espécimes de cada
espécie, seguindo o protocolo de extração descrito por Sambrook e Russell (2001),
com algumas modificações. O tampão de lise utilizado foi (Tris-HCl pH 8,0 em 10 mM,
NaCl 0,3 M, EDTA 10 mM, Urea 4 M, SDS 1%) de Estoup et al. (1993) e Asahida et al.
(1996). Posteriormente acrescentou-se: 15 μL de proteinase K (10 mg/mL) e 6 μL de
RNAse (10 mg/mL). As amostras foram incubadas para a digestão do tecido. Em
seguida lavagens sucessivas foram feitas com fenol, fenol-clorofórmio, álcool isoamílico
e clorofórmio hidratado (500μL de cada um destes reagentes). Após a lise, o DNA foi

18
separado das proteínas por precipitação salina juntamente com centrifugação a 14.000
rpm.
O sobrenadante foi precipitado com auxílio de 600 μL de isopropanol 100%, e
por centrifugação. Ao final, o DNA foi hidratado com aproximadamente 100 μL de água
milli-Q, dependendo do tamanho do pellet (DNA precipitado) formado.
Para possibilitar a análise da quantidade e integridade do material, o DNA
extraído foi quantificado por comparação com marcador de concentração conhecida,
em eletroforese padrão (com tampão Tris-Borato- EDTA 0,5X e corrida a 70 V por 40
minutos) em gel de agarose 0,8% e corado com GelRed. A visualização e análise do
DNA no gel foram feitas no fotodocumentador Easy Doc 100 (BioAgency).
2.3.2.2. Detecção de sequências de DNA repetitivo em tandem
Obtenção das sondas de DNAr 18S
Para a localização das sequências de DNAr foi utilizada uma sonda para DNAr
18S obtida via PCR a partir do DNA nuclear do marsupial Caluromys philander, usando
os iniciadores 18SF (5’-CCG CTT TGG TGA CTC TTG AT-3’) e 18SR (5’-CCG AGG
ACC TCA CTA AAC CA-3’) (Gross et al. 2010).
Obtenção das sondas teloméricas (TTAGGG)n
A sonda para detecção de sítios teloméricos foi obtida pela amplificação em até
1400 pares de base dos iniciadores (TTAGGG)n e (CCCTAA)n pela reação em cadeia
da polimerase (Polimerase Chain Reaction PCR) segundo Saiki et al. (1988) como
descrito por Ijdo et al.(1991).
Obtenção das sondas de DNAs repetitivos dispersos
Foi utilizado o iniciador (L1R, 5′-ATTCTRTTCCATTGGTCTA-3′; L1F, 5′-
CCATGCTCATSGATTGG-3′) para amplificar o retroelemento LINE-1, conforme
descrito por Waters et al. (2004).
Reações de PCR (Polimerase Chain Reaction)
As reações de PCR foram realizadas para um volume final de 25 μL consistindo

19
de 1μl de DNA genômico (100 ng), 2,5μL de tampão 10X com cloreto de magnésio
(2mM), 0,25μl de DNA Taq Polimerase (5 U/μL), 1,5μL de dNTP (1mM), 1,5μL de cada
iniciador (5mM) e água milli-Q para completar o volume. Os parâmetros das reações de
PCR seguiram: (a) para o DNAr 18S: 2 minutos a 95 °C (desnaturação); 35 ciclos por 1
minuto a 95 °C, 30 segundos a 55 °C (anelamento) e 1 minuto e 40 segundos a 72 °C
(extensão); 7 minutos a 72 °C (extensão final); (b) Sonda Telomérica: primeiros 10
ciclos de 1 minuto a 94 °C, 30 segundos a 55 °C e 1 minuto a 72 °C, seguidos por 30
ciclos de 1 minuto a 94 °C, 30 segundos a 60 °C e 1,5 minuto a 72 °C e um passo final
de 5 minutos a 72 °C; (c) para o retroelemento L1: 2 minutos a 95 °C (desnaturação);
35 ciclos por 1 minuto a 95 °C, 1 minuto a 52 °C (anelamento) e 1 minuto e 40
segundos a 72 °C (extensão); 7 minutos a 72 °C (extensão final).
Hibridização in situ fluorescente
Os produtos de PCR foram marcados com biotina-14-dATP (Biotin Nick
Translation mix; Invitrogen) e digoxigenina-11-dUTP (DigNick Translation mix; Roche)
por meio da reação de nick translation. O procedimento de hibridização foi conduzido
com base nos protocolos descritos por Pinkel et al. (1986) com 77% de estringência. O
DNA dos cromossomos mitóticos foi desnaturado em formamida 70% diluída em
2XSSC (Cloreto de Sódio 17,53g (0,29 M), 8,82g de Citrato de Sódio e água destilada
para um volume final de 1000mL, pH 7,0) a 67 °C por 10 segundos. A solução de
hibridização (100ng da sonda desnaturada, 10mg/mL de dextran-sulfato (2XSSC) e
50% de formamida em um volume final de 30 μL foi adicionada em uma lâmina e
hibridizada a 37 °C por uma ou duas noites, dependendo da sonda utilizada (uma noite
para telômeros e 2 para 18S e L1) em câmara úmida (2XSSC). As lâminas foram
lavadas após a hibridização em 2X SSC 72 °C, pH 7,0 por 5 minutos. Após essa
lavagem, as lâminas foram imersas em tampão PBD para o bloqueio (20mL de 20X
SSC, 1mL de Triton 100, 1g de leite em pó desnatado e água destilada para um volume
de 100mL, pH7,0). Para detecção da sonda foram utilizados os anticorpos FITC-
Avidina conjugada (Sigma; www.sigmaaldrich.com) em tampão C (0,1 M Bicarbonato
de Sódio) e anti-digoxigenina rodamina (Roche). As lâminas foram lavadas três vezes

20
em detergente-tampão fosfato (PBD) a 45 °C por 2 minutos. Duas séries de
amplificação foram realizadas, utilizando biotina-anti-avidina em tampão PBD (2 μL de
anti-avidina em 38 μL de tampão PBD), nas quais as lâminas foram incubadas por 5
minutos em câmara úmida a 37 °C. Cada tratamento com biotina-anti-avidina
conjugada em tampão PBD (2 μL de anti-avidina em 38 μL de PBD), na qual as
lâminas foram incubadas por 5 minutos em câmara úmida a 37 °C. Cada tratamento
com biotina-anti-avidina foi seguido pela incubação com isotiocianeto fluorescente a
0,07% (FITC)- avidina em tampão C por 8 minutos em câmara úmida a 37 °C. Seguida
de cada amplificação, as lâminas foram lavadas três vezes em PBD a 45 °C por 2
minutos. Os cromossomos foram contra-corados com DAPI (2ug/mL) diluído em
antifade (Vector; www.vectorlabs.com).
As metáfases foram analisadas em microscópio de epifluorescência Olympus
BX51, capturadas com câmera digital (Olympus DP71) através do software Image-Pro
MC 6.3 e processadas no programa Adobe Photoshop CS3.

21
3. Resultados e Discussão
Os resultados obtidos com a análise cromossômica comparativa de marsupiais
amazônicos e a discussão desses dados são apresentados na forma de três capítulos,
conforme mencionado a seguir:
3.1. Capítulo 1 – Caracterização citogenética de marsupiais didelfídeos na Amazônia
brasileira: uma abordagem clássica sobre o cromossomo X e heterocromatina, num
contexto geográfico.
3.2. Capítulo 2 – Comparação cromossômica entre heterocromatina e sequências
teloméricas em espécies de marsupiais didelfídeos (Didelphimorphia)
3.3. Capítulo 3 – Mapeamento do elemento LINE-1 em espécies de marsupiais
amazônicos (Didelphidae; Didelphimorphia).

22
Capítulo 1: Caracterização citogenética de marsupiais
amazônicos (Didelphidae; Didelphimorphia): uma abordagem
clássica sobre o cromossomo X e heterocromatina, num
contexto geográfico.
Resumo:
Foram caracterizadas citogeneticamente 16 espécies de marsupiais
amazônicos, variações no cromossomo X, na distribuição da heterocromatina e
no mapeamento de sequências de DNA ribossomal 18S. Os cariótipos de 194
indivíduos de 13 localidades da Amazônia brasileira, foram analisados
empregando-se métodos de coloração convencional, banda C, Ag-RON e FISH
com sonda de DNAr18S. A principal variação foi registrada na posição do
centrômero do cromossomo X de Caluromys philander e de Marmosa murina.
Geograficamente, os diferentes tipos de cromossomo X, de ambas espécies,
mostraram uma segregação entre leste e oeste do Brasil, com contato na
Amazônia central. Para as espécies de Marmosops spp. dois padrões de X
foram evidenciados pela técnica da banda C, porém com distribuição
geográfica mais uniforme. As regiões organizadoras de nucléolo mostraram
especificidade com a sonda de DNAr 18S em todas as espécies, exceto a
marcação do cromossomo Y de Monodelphis brevicaudata. A distribuição
desse marcador se mostrou variável apenas no gênero Marmosa (M. murina e
M. demerarae). Considerando RON simples como um caráter plesiomórfico,
concluímos que as espécies do gênero Marmosa e Didelphis marsupialis
evoluíram independentemente para o sistema múltiplo. Marsupiais são
considerados cromossomicamente conservativos; porém, dado o atual estado
de conhecimento citogenético para esse grupo, variações cromossômicas intra
e interespecíficas foram observadas. Isto se deve à ampliação da amostra junto
a aplicação dos métodos cromossômicos de bandeamento e molecular, o que
sugere a presença de rearranjos cromossômicos na evolução de marsupiais.

23
Introdução
Historicamente, os primeiros dados citogenéticos de marsupiais provém
do trabalho de Jordan (1911) apud Reig et al. (1977) sobre a espermatogênese
de Didelphis virginiana. Desde então, dados citogenéticos de espécies de
marsupiais australianos e americanos têm sido ampliados e atualmente são
conhecidos os cariótipos de aproximadamente 180 espécies (Hayman 1990).
Citogeneticamente, marsupiais mostram pouca variação quando comparados
às outras ordens de mamíferos, como por exemplo, Rodentia e Chiroptera
(Baker 1967; Baker e Bickham 1980; Moratelle e Morielle-Versute 2007;
Nagamachi et al. 2015 e referências).
Extensas comparações de marcadores citogenéticos (banda-G, -C e
região organizadora do nucléolo) entre espécies americanas e australianas
revelam que a estabilidade cromossômica é verificada desde número diploide
até padrões de bandeamento longitudinal (Yonenaga-Yassuda et al. 1982; Rofe
e Hayman 1985; Casartelli et al. 1986; Souza et al. 1990; Svartman e Vianna-
Morgante 1999). Três principais números diploides estão presentes entre
espécies de ambos os continentes: 14, 18 e 22 numa amplitude de 10 a 32
cromossomos. Dentre todas as famílias, Macropodidae (ordem Diprotodontia) é
a que possui maior diversidade de cariótipos, variando de 2n=10 a 32,
enquanto que a família Didelphidae (ordem Didelphimorphia) mostra apenas os
três números diploides predominantes (14, 18 e 22); dentre esses, o 2n=14 é o
mais frequente tanto nesta família como nos demais marsupiais (Reig et al.
1977; Hayman 1990, Palma e Yates 1996; Carvalho et al. 2002). Este número
diploide vem sendo sugerido como ancestral para os marsupiais (Reig et al.
1977; O’Neill et al. 1997), embora a ancestralidade desse grupo ainda seja
motivo de discussão (Svartman e Vianna-Morgante 1998; Carvalho e Mattevi
2000).
No bioma amazônico, a grande abrangência territorial, aliada à carência
de boas amostragens, torna o conhecimento da fauna de mamíferos incipiente
(Ab’Saber 1977; Voss e Emmons 1996; INPE 1999). Especificamente para as
27 espécies amazônicas de marsupiais (Paglia et al. 2012), dados
citogenéticos estão disponíveis apenas para 17 (Brandão et al. 2015;

24
Nagamachi et al. 2015). A representatividade desses dados está sujeita às
mudanças taxonômicas indicadas por estudos mais recentes, envolvendo
dados filogenéticos, dificultando o reconhecimento do avanço no conhecimento
citogenético, uma vez que frequentemente falta conexão entre os dados do
espécime, identificação da espécie e o cariótipo. Nesse âmbito, alguns fatores
limitam o conhecimento do grupo, como o número de espécies analisadas e a
restrição frequente das análises citogenéticas apenas aos números diploide e
fundamental (Nagamachi et al. 2015).
Ainda nesse contexto, as principais discussões têm centrado na
estrutura dos autossomos, enquanto que os cromossomos sexuais têm sido
estudados mais em relação ao comportamento em núcleos meióticos (Solari e
Bianchi 1975; Sharp 1982; Seluja et al. 1987; Graves e Watson 1991).
Neste trabalho discutimos as principais diferenças morfológicas
encontradas nos cromossomos sexuais e nos padrões de banda C de 16
espécies de didelfídeos coletadas na Amazônia e. Ainda, discutimos esses
padrões em um contexto geográfico amplo.
Material e Métodos
Neste estudo foram analisadas 194 indivíduos de 16 espécies,
pertencentes a oito gêneros de marsupiais, coletadas em 13 localidades da
Amazônia brasileira (Tabela 1, Apêndice I). Os espécimes foram coletados sob
autorizações do Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais
Renováveis - IBAMA, de acordo com os números de licença: 02005.000642/03-
11 (IBAMA); 02000.002336/2003-93 (IBAMA); 02005.002672/04 (IBAMA);
37585-5 (SISBIO); 37592-4 (SISBIO). Os indivíduos foram depositados na
Coleção de mamíferos do INPA (Instituto Nacional de Pesquisas da Amazônia)
(Apêndice II).
Os cromossomos mitóticos foram obtidos a partir da medula óssea,
segundo Ford e Harmerton (1956). Os padrões de banda-C e da região
organizadora de nucléolo foram determinados segundo as técnicas descritas
por Sumner (1972) e Howell e Black (1980), respectivamente. Os cariótipos
foram organizados, emparelhando os cromossomos com base na razão de

25
braços cromossômicos e na posição do centrômero, segundo Levan et al.
(1964).
A localização da sequência dos cístrons ribossomais foi determinada por
meio da hibridização in situ fluorescente (Pinkel et al. 1986) utilizando a sonda
DNAr 18S, obtida via PCR a partir do DNA nuclear do marsupial Caluromys
philander, usando os iniciadores 18SF (5’-CCG CTT TGG TGA CTC TTG AT-
3’) e 18SR (5’-CCG AGG ACC TCA CTA AAC CA-3’) (Gross et al. 2010) e a
conforme descrito em.
Resultados
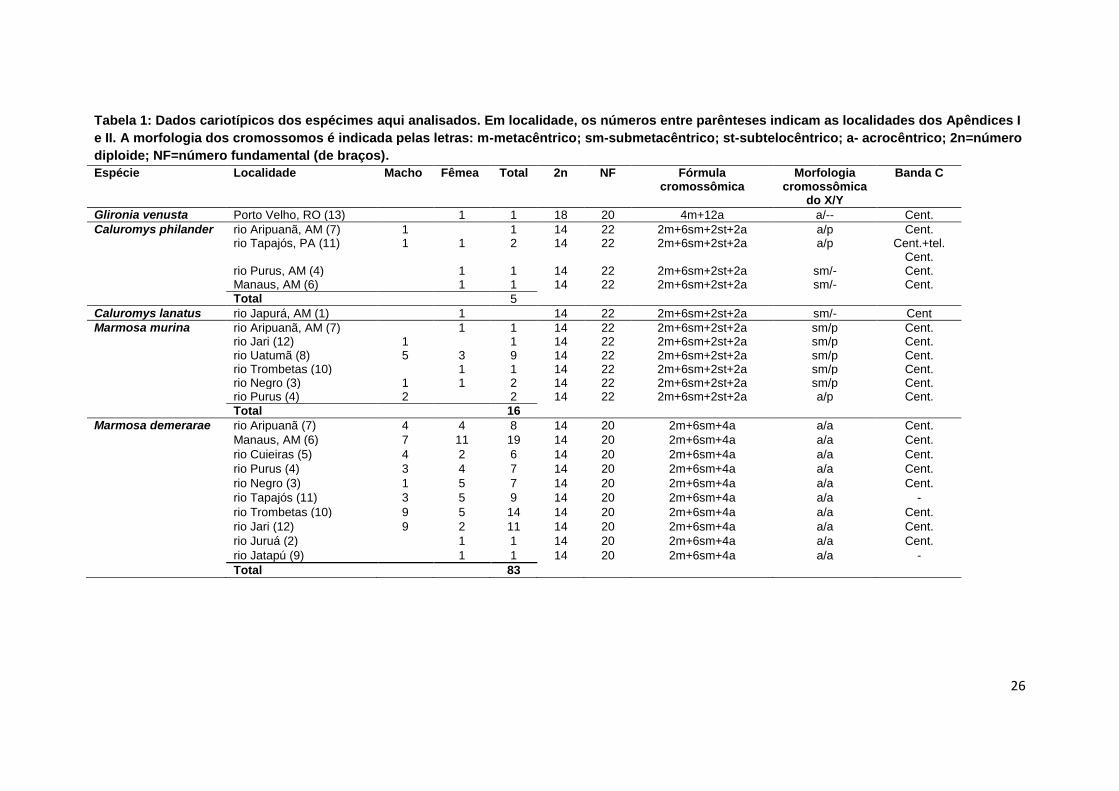
Dentre as 16 espécies analisadas, 11 apresentaram 2n=14, quatro
2n=18 e uma 2n=22 cromossomos (Tabela 1).
Entre as espécies com 2n=14, observou-se uma variação no número
fundamental de braços (NF), ocasionando mudanças nas fórmulas
cromossômicas (Tabela 1), sendo NF=20 em Marmosa demerarae (Fig. 1c-I) e
Metachirus nudicaudatus (Fig. 1f-I), NF=22 em Caluromys philander (Fig. 1a-I),
Caluromys lanatus (Fig. 1b-I), Marmosa murina (Fig. 1d-I) e Gracilinanus
emiliae (Fig.1g-I) e NF=24 nas espécies pertencentes ao gênero Marmosops
(M. bishopi, M. pinheiroi, M. parvidens, M. impavidus, M. cf. pakaraimae).
Todas as espécies de Marmosops analisadas apresentaram características
cariotípicas similares (Fig. 1e-I – mostrado apenas M. pinheiroi).
26
Tabela 1: Dados cariotípicos dos espécimes aqui analisados. Em localidade, os números entre parênteses indicam as localidades dos Apêndices I
e II. A morfologia dos cromossomos é indicada pelas letras: m-metacêntrico; sm-submetacêntrico; st-subtelocêntrico; a- acrocêntrico; 2n=número
diploide; NF=número fundamental (de braços).
Espécie Localidade Macho Fêmea Total 2n NF Fórmula cromossômica
Morfologia cromossômica
do X/Y
Banda C
Glironia venusta Porto Velho, RO (13) 1 1 18 20 4m+12a a/-- Cent.
Caluromys philander rio Aripuanã, AM (7) 1 1 14 22 2m+6sm+2st+2a a/p Cent. rio Tapajós, PA (11) 1 1 2 14 22 2m+6sm+2st+2a a/p Cent.+tel.
Cent. rio Purus, AM (4) 1 1 14 22 2m+6sm+2st+2a sm/- Cent. Manaus, AM (6) 1 1 14 22 2m+6sm+2st+2a sm/- Cent.
Total 5
Caluromys lanatus rio Japurá, AM (1) 1 14 22 2m+6sm+2st+2a sm/- Cent
Marmosa murina rio Aripuanã, AM (7) 1 1 14 22 2m+6sm+2st+2a sm/p Cent. rio Jari (12) 1 1 14 22 2m+6sm+2st+2a sm/p Cent. rio Uatumã (8) 5 3 9 14 22 2m+6sm+2st+2a sm/p Cent. rio Trombetas (10) 1 1 14 22 2m+6sm+2st+2a sm/p Cent. rio Negro (3) 1 1 2 14 22 2m+6sm+2st+2a sm/p Cent. rio Purus (4) 2 2 14 22 2m+6sm+2st+2a a/p Cent.
Total 16
Marmosa demerarae rio Aripuanã (7) 4 4 8 14 20 2m+6sm+4a a/a Cent.
Manaus, AM (6) 7 11 19 14 20 2m+6sm+4a a/a Cent.
rio Cuieiras (5) 4 2 6 14 20 2m+6sm+4a a/a Cent.
rio Purus (4) 3 4 7 14 20 2m+6sm+4a a/a Cent.
rio Negro (3) 1 5 7 14 20 2m+6sm+4a a/a Cent.
rio Tapajós (11) 3 5 9 14 20 2m+6sm+4a a/a -
rio Trombetas (10) 9 5 14 14 20 2m+6sm+4a a/a Cent.
rio Jari (12) 9 2 11 14 20 2m+6sm+4a a/a Cent.
rio Juruá (2) 1 1 14 20 2m+6sm+4a a/a Cent.
rio Jatapú (9) 1 1 14 20 2m+6sm+4a a/a -
Total 83
27
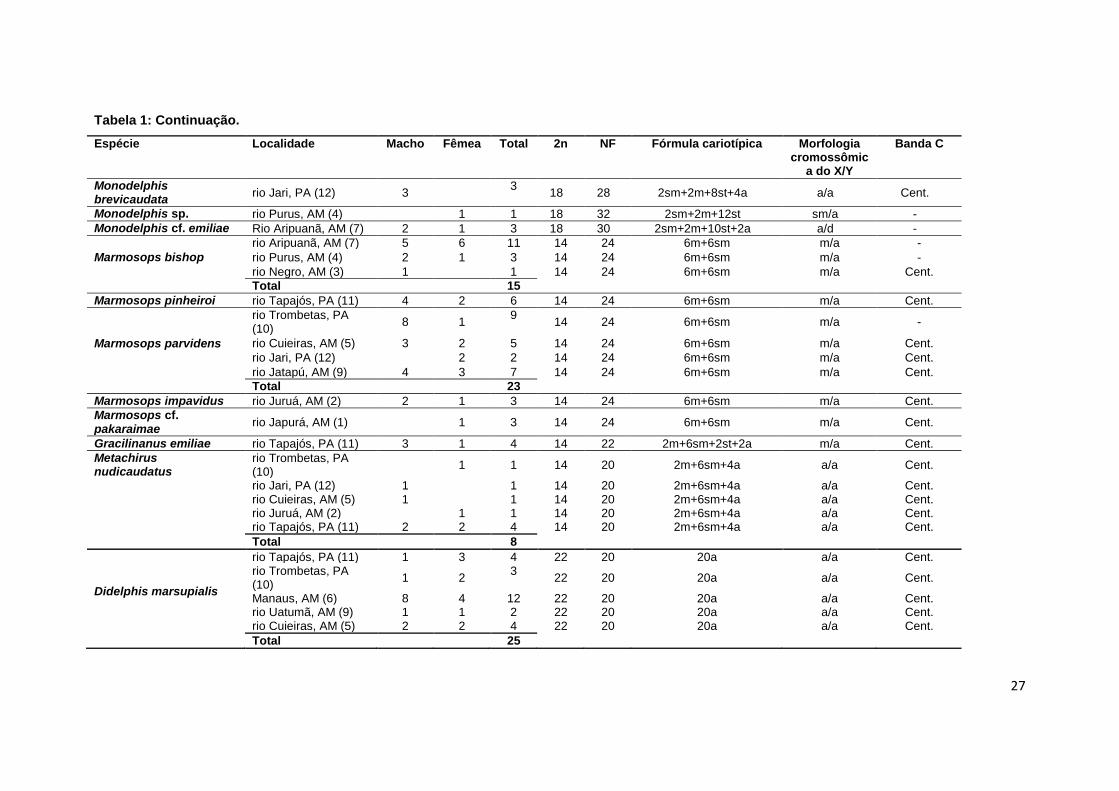
Tabela 1: Continuação.
Espécie Localidade Macho Fêmea Total 2n NF Fórmula cariotípica Morfologia cromossômic
a do X/Y
Banda C
Monodelphis brevicaudata
rio Jari, PA (12) 3
3 18 28 2sm+2m+8st+4a a/a Cent.
Monodelphis sp. rio Purus, AM (4)
1 1 18 32 2sm+2m+12st sm/a -
Monodelphis cf. emiliae Rio Aripuanã, AM (7) 2 1 3 18 30 2sm+2m+10st+2a a/d -
Marmosops bishop
rio Aripuanã, AM (7) 5 6 11 14 24 6m+6sm m/a -
rio Purus, AM (4) 2 1 3 14 24 6m+6sm m/a -
rio Negro, AM (3) 1 1 14 24 6m+6sm m/a Cent.
Total 15
Marmosops pinheiroi rio Tapajós, PA (11) 4 2 6 14 24 6m+6sm m/a Cent.
Marmosops parvidens
rio Trombetas, PA (10)
8 1 9
14 24 6m+6sm m/a -
rio Cuieiras, AM (5) 3 2 5 14 24 6m+6sm m/a Cent.
rio Jari, PA (12) 2 2 14 24 6m+6sm m/a Cent.
rio Jatapú, AM (9) 4 3 7 14 24 6m+6sm m/a Cent.
Total 23
Marmosops impavidus rio Juruá, AM (2) 2 1 3 14 24 6m+6sm m/a Cent.
Marmosops cf. pakaraimae
rio Japurá, AM (1) 1 3 14 24 6m+6sm m/a Cent.
Gracilinanus emiliae rio Tapajós, PA (11) 3 1 4 14 22 2m+6sm+2st+2a m/a Cent.
Metachirus nudicaudatus
rio Trombetas, PA (10)
1 1 14 20 2m+6sm+4a a/a Cent.
rio Jari, PA (12) 1
1 14 20 2m+6sm+4a a/a Cent. rio Cuieiras, AM (5) 1
1 14 20 2m+6sm+4a a/a Cent.
rio Juruá, AM (2)
1 1 14 20 2m+6sm+4a a/a Cent. rio Tapajós, PA (11) 2 2 4 14 20 2m+6sm+4a a/a Cent.
Total 8
Didelphis marsupialis
rio Tapajós, PA (11) 1 3 4 22 20 20a a/a Cent. rio Trombetas, PA (10)
1 2 3
22 20 20a a/a Cent.
Manaus, AM (6) 8 4 12 22 20 20a a/a Cent. rio Uatumã, AM (9) 1 1 2 22 20 20a a/a Cent. rio Cuieiras, AM (5) 2 2 4 22 20 20a a/a Cent.
Total 25

28
Em relação aos cromossomos sexuais, o X apresentou-se com três
morfologias: metacêntrico em G. emiliae e em Marmosops spp.;
submetacêntrico em uma fêmea C. lanatus), acrocêntrico em M. demerarae e
M. nudicaudatus. Em duas espécies, Caluromys philander e Marmosa murina,
a morfologia do cromossomo X variou em acrocêntrico ou submetacêntrico, em
uma mesma ou em diferentes localidades. Já, o Y apresentou-se como
acrocêntrico em G. emiliae, Marmosops spp., M. demerarae e M. nudicaudatus
e puntiforme em C. philander e M. murina (Fig. 1).

29
Figura 1. Cariótipos em Giemsa (I), Banda C (II), DNAr 18S e RON (III) e cromossomos
sexuais (caixas) de: a) Caluromys philander (SISTAP-M-244, caixas: CAN 34, SISTAP-M-
305); b) Caluromys lanatus (CTGA-M-701); c) Marmosa demerarae (RNL 46, caixas: MCA
27); d) Marmosa murina (RNL 69, caixa: CEF 18) (IV) variação no mapeamento de 18S
encontrada nos indivíduos do rio Purus, Tapauá, Amazonas; e) Marmosops pinheiroi
(INPA 5377, caixa: EE 192) (SISTAP-M-278, caixa: EE107, INPA 5408); f) Metachirus
nudicaudatus (SISTAP-M-302; caixa: SISSIS-M-64); g) Gracilinanaus emiliae (SISTAP-M-
243). Os espécimes testemunho encontram-se na Coleção de Mamíferos do INPA; a sigla
INPA se refere ao registro de tombamento na coleção e as demais ao registro de campo
dos indivíduos analisados.

30
Entre as espécies com 2n=18 observou-se uma variação no número
fundamental de braços (NF), sendo NF=20 em Glironia venusta (Fig. 2a-I),
NF=28 em Monodelphis brevicaudata (Fig. 2c-I), NF=30 em Monodelphis cf.
emiliae (Fig. 2b-I) e NF=32 em Monodelphis sp. (Fig. 2d-I). Em M. cf. emiliae e
em M. brevicaudata o X foi acrocêntrico e em Monodelphis sp. foi
submetacêntrico. O Y foi acrocêntrico em Monodelphis brevicaudata e em
Monodelphis sp. e puntiforme em M. cf. emiliae.
Didelphis marsupialis foi a única espécie que apresentou 2n=22
cromossomos e NF=20 (Fig. 2e-I), sendo o X e Y acrocêntricos.
Figura 2 Cariótipos em Giemsa (I), Banda C (II), DNAr 18S e RON (III) e cromossomos sexuais (caixas) de: a) Glironia venusta (BAC 80); b) Monodelphis cf. emiliae (INPA 5388); c) Monodelphis brevicaudata (INPA 5404); d) Monodelphis sp. (CAN 44); e) Didelphis marsupialis (EE 249, caixas: EE174). Os espécimes testemunho encontram-se na Coleção de Mamíferos do INPA; a sigla INPA se refere ao registro de tombamento na coleção e as demais ao registro de campo dos indivíduos analisados.

31
A localização da heterocromatina nas espécies com 2n=14 foi
centromérica, apresentando-se de forma conspícua em M. demerarae (Fig. 1c-
II), M. murina (Fig. 1d-II), M. pinheiroi (Fig 1e-II) e G. emiliae (Fig. 1g-II). Em C.
philander e C. lanatus, os blocos heterocromáticos foram difusos, sendo que na
primeira foi detectada heterocromatina na região telomérica (Fig. 1a-II e 1b-II).
O cromossomo X em C. philander foi totalmente heterocromático, exceto por
uma faixa distal nos braços longos (Fig. 1a-II); já em M. demerarae foi
totalmente heterocromático, exceto por uma faixa proximal nos braços longos
(Fig. 1c-II); e centromérica em M. murina (Fig. 1d-II), M. nudicaudatus (Fig. 1f-
II) e G. emiliae (Fig. 1g-II).
Para Marmosops spp. foram detectados dois padrões de banda C para o
cromossomo X. No padrão 1, o X foi totalmente heterocromático, exceto por
uma faixa proximal nos braços longos (Fig. 1e - caixa); no padrão 2, a
heterocromatina concentrou-se nos braços curtos e centrômero (Fig. 1e -
caixa). Ambos padrões estiveram presentes em M. parvidens e M. bishopi,
enquanto que apenas o padrão 1 foi observado em M. cf. pakaraimae, M.
impavidus e M. pinheiroi (Tabela 2). O Y foi totalmente heterocromático em
todas as espécies.
Nas espécies com 2n=18 cromossomos, a heterocromatina foi
centromérica em G. venusta (Fig. 2a-II), M. emiliae (Fig. 2b-II) e Monodelphis
brevicaudata (Fig. 2c-II). O Y foi completamente heterocromático nas duas
espécies de Monodelphis (Fig. 2b-II e 2c-II). Em Monodelphis sp. não foi
possível determinar o padrão de banda C.
As regiões organizadoras de nucléolo (RON), confirmadas pela FISH
com sonda DNAr 18S, estiveram presentes nos braços curtos do par 6 em
todas as espécies com 2n=14. M. demerarae e M. murina tiveram o par 5
também marcado em posição terminal nos braços longos (Fig. 1, c-III e d-III).
Nas espécies com 2n=18, a RON foi simples e confirmada pela sonda DNAr
18S. Em M. emiliae, a RON localizou-se nos braços curtos do par 7 (Fig. 2b-III)
e em M. brevicaudata localizou-se nos cromossomos X e Y, embora nenhum
sítio de 18S tenha sido detectado no Y (Fig. 2c-III).
Em D. marsupialis, a RON foi evidenciada, tanto pela prata quanto pela
sonda DNAr 18S, em três pares cromossômicos, os pares putativos 5, 7 e 8.

32
Nos pares 5 e 8 as marcações foram observadas em posição terminal dos
braços longos, enquanto que no par 7 foram biteloméricas (Fig. 2e-III). Porém,
quanto à atividade, houve variação de quatro a oito marcações de Ag-RON.
Discussão
Nos últimos anos, avanços nos estudos sistemáticos e taxonômicos da
família Didelphidae introduziram mudanças na nomenclatura para muitas de
suas espécies (Jansa e Voss 2000; Voss e Jansa 2003; 2009; Gutiérrez et al.
2010). Aqui utilizamos a nomenclatura de Voss e Jansa (2009) e relacionamos
nossos dados citogenéticos das 16 espécies analisadas, com a árvore
consenso do referido trabalho. Observando principalmente a estrutura
cromossômica dos autossomos, nossos resultados corroboram o padrão
cariotípico conservado (número diploide e fórmula cromossômica) descrito na
literatura para esta família, principalmente para as espécies: Didelphis
marsupialis, Marmosa demerarae, Metachirus nudicaudatus, Monodelphis
brevicaudata, Monodelphis cf. emiliae e para as espécies de Marmosops
(Biggers et al. 1965; Reig et al. 1977; Yonenaga-Yassuda et al. 1982; Seluja et
al. 1984; Casartelli et al. 1986; Hayman 1990; Souza et al. 1990; Palma e Yates
1996; Svartman e Vianna-Morgante 1998; 1999; Carvalho et al. 2002).
É interessante observar que, apesar dos marsupiais serem considerados
cariotipicamente conservados, ao compararmos os cariótipos de alguns
gêneros, é possível associá-los a uma dada espécie, devido a presença de
caracteres diagnósticos. Por exemplo, M. demerarae e M. murina diferem no
número fundamental, morfologia e tamanho dos cromossomos sexuais. Já as
espécies de Monodelphis, o NF é específico para cada espécie. Entretanto, o
mesmo não acontece para o gênero Marmosops, no qual as cinco espécies
apresentam macroestrutura cromossômica muito semelhante.
Especificamente no caso do gênero Marmosa, a taxonomia é
considerada complicada é considerado de taxonomia problemática e muitas
espécies, inclusive Micoureus demerarae (Tate 1933) Recentemente
Micoureus foi reincorporado ao gênero Marmosa (Voss e Jansa 2009), sendo
atualmente considerado sub-gênero. Vale ressaltar que nesse processo de
reorganização taxonômica em Didelphidae, uma defasagem pode ter ocorrido

33
quanto à nomenclatura dos espécimes já cariotipados, gerando dúvidas sobre
quantas e quais espécies foram de fato cariotipadas, uma vez que
determinados indivíduos podem ter sido reclassificados e espécies divididas
em duas ou mais táxons válidos. Dessa forma uma revisão da nomenclatura
das espécies com caiótipos já apresentados na literatura se faz necessário.
Em relação à estrutura do cromossomo X, verificamos variações
intraespecíficas em Caluromys philander e Marmosa murina, ao longo de suas
distribuições, já considerando o grau de condensação cromossômica.
Considerando primeiramente C. philander, observamos que o
cromossomo X foi acrocêntrico ou submetacêntrico, visto tanto em homozigose
como em heterozigose em fêmeas (Fig. 3). Os nossos dados complementam
os padrões de distribuição geográfica das formas apresentadas do
cromossomo X dessa espécie (Souza et al. 2013). Em uma escala geográfica
maior, a forma acrocêntrica do X está presente nas regiões sudeste e nordeste
do Brasil, assim como no leste amazônico (localidades 3 a 6). A forma
submetacêntrica é encontrada em áreas do oeste, central e leste da Amazônia
brasileira (Fig. 3) (Reig et al. 1977; Svartman e Vianna-Morgante 1999; Pereira
et al. 2008), onde se observa homozigose assim como heterozigose em
fêmeas. Os poucos registros de heterozigotos para o X podem ser explicados
pela baixa quantidade de indivíduos capturados e/ou analisados
citogeneticamente (Peres 1999).

34
Figura 3 Localização das formas do cromossomo X de Caluromys philander na América
do Sul. Os pontos em asterisco são da literatura e os pontos cículo cheio são do
presente trabalho: (*1) Venezuela, Reig et al. 1977; (●2) rio Purus; (●3) REMAN Manaus,
presente trabalho e Souza et al. 2013; (●4) rio Trombetas; (●5) rio Tapajós; (*6) rio Jari,
Souza et al. 2013; (*7) Pernambuco, Souza et al. 1990; (*8) São Paulo, Svartman e Vianna-
Morgante 1999 e Pereira et al. 2008. m = metacêntrico; sm = submetacêntrico; a =
acrocêntrico; p = puntiforme.
Em M. murina o X submetacêntrico é encontrado em indivíduos
provenientes de duas localidades do centro-oeste do Brasil, situadas
marginalmente na distribuição da espécie. É encontrado também em todas as
localidades da Amazônia brasileira analisadas, exceto no rio Purus (Fig. 4,
localidade 6). Adicionalmente, cromossomos bibraquiais de forma metacêntrica
são também encontrados na Amazônia, mais a oeste e ao norte das
localidades com submetacêntrico (Fig. 4, localidades 1 a 3) (Carvalho et al.
2002; Reig et al. 1977). Já, o X acrocêntrico é encontrado nas regiões centro-
oeste, sudeste e nordeste do Brasil (Souza et al. 1990; Palma e Yates 1996),
assim como no rio Purus, Amazonas (presente trabalho) (Fig. 4). Assim,
observa-se alta estruturação geográfica na distribuição das formas
morfológicas do cromossomo X em Marmosa murina, com o X
submetacêntrico, apresentando ocorrência restrita à bacia amazônica (Fig. 4).
Em contraste, não observamos um padrão de variação para as formas

35
morfológicas do cromossomo X de M. murina, mesmo com um maior número
de exemplares cariotipados por localidade, não tendo sido detectado até o
momento polimorfismo das populações.
Figura 4 Localização das formas do cromossomo X de Marmosa murina na América do
Sul. Os pontos em asterisco são dados da literatura e os pontos em ● são dados do
presente trabalho: (*1) Villa Vivêncio, Colômbia, Hayman e Martin 1974; (*2) Loreto-Peru,
Reig et al. 1977; (*3) Bolívar, Venezuela, Reig et al. 1977; (●4) rio Negro; (●5) rio Juruá;
(●6) rio Purus; (●7) rio Uatumã; (●8) rio Trombetas; (●9) rio Jari; (*10) Tartarugalzinho,
Pará, Carvalho et al. 2002; (*11) Pacoti, Ceará, Pagnozzi et al. 2002; (*12) Pernambuco,
Souza et al. 1990; (*13) Vila Rica, Pagnozzi et al. 2002; (*14) Tocantins, Lima 2004; (*15)
UHE Corumba IV, Luziania, GO, Pereira et al. 2008; (*16) UHE Peixe Angical, Goiás,
Pereira et al. 2008; (*17) Mambai, Goiás, Carvalho et al. 2002; (*18) Espírito Santo,
Paresque 2004.
Distribuição de Heterocromatina
O padrão da heterocromatina em 8 das 16 espécies (C. lanatus, G.
venusta, D. marsupialis, M. brevicaudata, M. emiliae, Monodelphis sp., G.
emiliae, M. nudicaudatus) analisadas no presente trabalho foi conservado e se
apresentou como pequenos blocos centroméricos, de coloração tênue em
todos os autossomos. Em C. philander foram encontrados blocos
heterocromáticos na região telomérica dos autossomos, divergindo do que já
havia sido registrada na literatura. Já em Marmosa demerarae, Marmosa

36
murina e as cinco espécies de Marmosops spp. se destacaram por apresentar
grandes blocos de heterocromatina nos centrômeros dos autossomos. Estas
diferenças nos padrões da heterocromatina geralmente são explicadas pela
diferença na composição (tipos de sequências) do DNA desta região, que varia
tanto inter como intraespecificamente (Sumner 2003). Sendo que as regiões
heterocromáticas geralmente são compostas por sequências repetitivas de
DNA, especialmente DNA satélite e transposons (John 1988; Grewal e Jia
2007). Essas diferenças de composição da heterocromatina podem ser
responsáveis pelos padrões observados em C. philander, M. murina, M.
demerarae e Marmosops spp.
Quanto a Marmosops spp., os diferentes padrões de banda C do
cromossomo X, distribuídos pela bacia amazônica, ocorrem em simpatria na
área compreendida entre os eixos Negro-Purus e Trombetas-Tapajós,
flanqueada pelo padrão 1 a oeste e pelo padrão 2 a leste (Tabela 2). A
correlação desses padrões com possíveis espécies crípticas ainda não foi
possível de ser estabelecida, dependendo ainda de análises moleculares e
morfológicas mais amplas (da Silva, comunicação pessoal). No entanto, a
diversidade de padrões heterocromáticos do cromossomo X em Marmosops
spp. pode também ser atribuída a variações presentes ao longo da bacia
amazônica.
Tabela 2: Padrões de distribuição da heterocromatina do cromossomo X nas espécies do gênero Marmosops.
Espécie Localidade Padrão
1 2
Marmosops bishopi Rio Purus 1 macho 2 machos; 2 fêmeas
Rio Negro 1 macho
Rio Aripuanã 2 machos; 4 fêmeas 1 macho; 1 fêmea
Marmosops parvidens Rio Cuieiras 2 machos 2 fêmeas
Rio Trombetas 5 machos 4 machos; 1 fêmea
Rio Jari 2 fêmeas
Marmosops pinheiroi Rio Cuieiras 1 macho
Rio Tapajós 4 machos
Marmosops impavidus Rio Juruá 1 macho
Marmosops cf. pakaraimae Rio Japurá 1 fêmea
Segundo Sumner (2003), a identificação do padrão heterocromático é
um aspecto essencial para a caracterização do cariótipo de uma espécie,
mesmo porque diferenças podem ser detectadas entre populações de uma
mesma espécie. A heterocromatinização também é um processo utilizado para

37
o silenciamento de genes, portanto, o padrão heterocromático tanto pode servir
como caráter citotaxonômico, como também ser útil para compreensão dos
mecanismos de evolução cromossômica e de regulação da expressão gênica.
No entanto, no presente trabalho observamos que, com exceção de
Marmosops spp., as espécies aqui analisadas apresentaram pouca variação
intraespecífica em seus padrões de heterocromatina, inclusive do X, o que
sugere que o padrão de distribuição de heterocromatina, isoladamente, não
permite a distinção populacional em didelfídeos. Todavia, quando comparamos
este caráter em nível específico, a distribuição da heterocromatina torna-se um
marcador eficiente para caracterizá-las, como é o caso de M. demerarae e M.
murina, uma vez que a primeira apresenta blocos heterocromáticos
centroméricos maiores do que a segunda, permitindo melhor diferenciação dos
táxons. Isto também foi observado entre as espécies congenéricas C. philander
e C. lanatus, ambas com 2n=14 e NF=24, porém com padrão heterocromático
distinto.
Quanto à região organizadora do nucléolo (RON) em Didelphidae, esta
pode ser simples ou múltipla. Segundo Hsu et al. (1975), a RON num cístron
único e longo seria um caráter ancestral em mamíferos, sendo que rearranjos
subsequentes levariam ao surgimento de RONs múltiplas em grupos derivados.
Deste modo, os gêneros Glironia, Monodelphis, Caluromys, Gracilinanus e
Marmosops possivelmente apresentam uma condição plesiomórfica, já que nas
espécies desses gêneros, a RON está presente somente no par autossômico
6, confirmada pela técnica de FISH com a sonda DNAr 18S. Por outro lado, as
espécies de Marmosa e Didelphis, nas quais as RONs são múltiplas,
representariam uma condição derivada. O mesmo foi verificado para espécies
do gênero Philander (Yonenaga-Yassuda et al. 1982; Svartman e Vianna-
Morgante 2003).
Quando analisamos este caráter em Monodelphis, encontramos sítios de
RON presentes no par 7 e no cromossomo sexual X. No presente trabalho
detectamos RON simples, presentes no par 7 em Monodelphis cf. emiliae e
Monodelphis sp., RON simples presente nos cromossomos X e Y em
Monodephis brevicaudata, sendo esta última uma novidade para o gênero.
Genes ribossomais em cromossomos sexuais de mamíferos já foram relatados

38
numa espécie de morcego, Carollia castanea (Hsu et al. 1975), mas os
mesmos autores ressaltam que RON no cromossomo X pode gerar problemas
de compensação de dose em mamíferos. A presença de RON em
cromossomos sexuais pode ser considerada um caráter derivado, uma vez que
estaria presente originalmente em autossomos e por rearranjos, tipo
translocação ou transposição, passaria a ocupar o cromossomo X.
A precipitação de Nitrato de Prata pode ocorrer também em regiões de
heterocromatina, levando a uma interpretação errônea sobre a distribuição
desse marcador (Sumner 2003; Schneider et al. 2012). Situação semelhante foi
verificada no cromossomo Y de M. brevicaudata, cuja marcação não foi
confirmada pela FISH, o que nos leva a sugerir não se tratar de um sítio
ribossomal, mas sim um bloco de heterocromatina.
Se sobrepusermos o caráter RON à filogenia de Voss e Jansa (2009)
(não mostrado) verificamos que RONs múltiplas estão distribuídas em duas
linhagens derivadas, indicando surgimentos independentes; a primeira no
gênero Marmosa e a segunda nas espécies com 22 cromossomos, como
exemplo Didelphis e Philander. Assim, os dados de RON nos marsupiais
corroboram a hipótese de Hsu et al. (1975), onde o sistema simples teria sido
retido em Caluromys philander, C. lanatus, Gracilinanus emiliae, Marmosops
spp., Monodelphis brevicaudata, Monodelphis kunsi, Monodelphis dimidiata e
Metachirus nudicaudatus (Souza et al. 1990; Palma e Yates 1996; Carvalho et
al. 2002, Svartman e Vianna-Morgante 2003; Pereira et al. 2008, Souza et al.
2011).
Interessante notar que para RONs múltiplas foram identificadas
variações intraespecíficas tanto em M. murina como em M. demerarae. As
RONs múltiplas de M. murina descritas no presente trabalho contrastam com
dados bibliográficos, uma vez que em indivíduos coletados em Goiás, a RON é
simples nos braços curtos do par 6 (Palma e Yates 1996; Carvalho et al. 2002),
enquanto que indivíduos de Pernambuco apresentam marcações adicionais
nos braços longos dos pares 3 e 5 (Souza et al. 1990). Ainda, em M. murina, os
dois indivíduos do rio Purus diferiram tanto em relação aos cromossomos
sexuais quanto às RONs dos demais indivíduos dessa espécie.

39
Já, em M. demerarae, os indivíduos aqui analisados não tiveram os
cístrons ribossomais nos braços curtos do par 5, como registrados para
espécimes da Mata Atlântica (Svartman e Vianna-Morgante 2003).
Em Didelphis marsupialis apenas a atividade das RONs variou,
conforme já registrado em espécimes da Mata Atlântica (Yonenaga-Yassuda et
al. 1982; Svartman e Vianna-Morgante 2003).
Assim, entre os marsupiais da região amazônica analisados, chamam a
atenção as variações relacionadas à posição do centrômero e padrões de
heterocromatina do cromossomo X. Souza et al. (2013) sugeriram que
inversões pericêntricas no cromossomo X de Caluromys philander ocorreram,
alterando sua morfologia. Os padrões aqui observados complementam essas
observações.
Para M. murina as diferentes morfologias do X (m, sm, a) podem ser
devidas a um reposicionamento do centrômero sem a presença de rearranjos,
o que já foi observado em mamíferos e isso pode ser relacionado a três
principais regiões do cromossomo: subtelomérica, próxima ao centrômero
nativo e no limite entre heterocromatina e eucromatina (Rochi et al. 2012;
Burrack e Berman 2012).
Dutrillaux (1979) argumenta que o conhecimento acerca dos cariótipos
das espécies é limitado devido à baixa amostragem de indivíduos e que o
incremento na amostra pode identificar variações adicionais. Acreditamos que
a ampliação da amostra e aplicação de análises integrativas e novas
metodologias poderão fundamentar melhor o significado dessas variações
cromossômicas. Entretanto, para a região amazônica, a grande limitação dos
estudos citogenéticos ainda reside nas coletas de material, dada a sua enorme
extensão e dificuldades de acesso.
A irradiação de espécies da família Didelphidae ocorreu no continente
sul americano. Jansa et al. (2014) encontraram evidências consistentes, que
corroboram a origem dessas espécies em florestas úmidas como as florestas
pluviais atuais, que existem desde o Eoceno (Morley 2000; Burnham e Johnson
2004). Se as espécies de didelfídeos tiveram origem onde atualmente se
encontra a Amazônia e tiveram uma dispersão para o restante do continente,
possivelmente os padrões do cromossomo X encontrados em C. philander e M.

40
murina refletem essa dispersão, onde o reposicionamento do centrômero
aconteceu na transição entre a faixa central-leste desse bioma.
O acúmulo de dados citogenéticos de marsupiais permite identificar
variações em marcadores cromossômicos e o uso desses marcadores permite
ainda identificar padrões de estratificação geográfica em sua distribuição,
mesmo que ainda existam áreas (principalmente entre Amazônia e Cerrado),
que necessitem de amostras para interpretar essas variações de forma
contínua. Contudo, dada a amplitude geográfica, a variação nesses caracteres
pode representar gradações intraespecíficas, mas podem também estar
relacionadas a linhagens ou espécies ainda não identificadas.

41
Capítulo 2: Comparação cromossômica entre heterocromatina e sequências teloméricas em 11 espécies de marsupiais amazônicos (Didelphimorphia; DIdelphidae)
Resumo: Sequências teloméricas intersticiais (ITSs) têm sido mapeadas em
espécies de marsupiais americanas e australianas, mostrando que nem
sempre podem ser interpretadas como resquícios de fusão cromossômica.
Neste trabalho analisamos o cariótipo de 11 espécies de didelfídeos
amazônicos, comparando a distribuição da banda C com a das marcações
teloméricas e verificamos que as ITS são coincidentes com blocos de
heterocromatina. Das 11 espécies analisadas, seis apresentaram ITSs, todas
dentro da região de heterocromatina centromérica. A presença de ITS foi
constante no cromossomo X de indivíduos de Marmosa murina e variável nas
demais espécies. Acreditamos que a presença variável das ITSs seja uma
característica particular de cada linhagem. Nossos resultados corroboram as
inferências de que essas sequências nem sempre podem ser interpretadas
como consequência de rearranjo cromossômico.
Introdução
Sequências teloméricas são altamente conservadas entre entre
eucariotos e desempenham a função de proteção das extremidades coesivas
dos cromossomos, impedindo a sua interação com outras cromátides e ação
de DNAses (McClintock 1941; Meyne et al. 1989; Blackburn 1991). Entretanto,
essas sequências podem ser encontradas também em uma posição intersticial
nos braços cromossômicos ou na região centromérica. Essa situação tem sido
interpretada tanto como respostas a danos no DNA ou à ação de repetições
dispersas quanto à reorganização cromossômica (Azzalin et al. 2001; Ruiz-
Herrera et al. 2008; Sánchez et al. 2010). Independente de sua origem, as ITSs
podem ser fixadas e serem interpretadas no âmbito evolutivo (Nergadze et al.
2004).
Na família Didelphidae o mapeamento de ITS em cariótipos com 2n=14
e 18 levou alguns autores a sugerirem que os mesmos teriam originado por
rearranjos cromossômicos do tipo fusão a partir de um cariótipo com 2n=22
(Svartman e Vianna-Morgante 1998; Carvalho e Mattevi 2000). Já, Pagnozzi et

42
al. (2000; 2002), analisando algumas espécies de didelfídeos brasileiros,
sugeriram que as ITS encontradas nas mesmas seriam um componente da
heterocromatina, e não o resultado de fusões recentes. Metcalfe et al. (2004)
também concordam que nem todas as ITS de marsupiais australianos da
família Macropodidae podem ser aceitas como evidências de fusões, salvo
quando não forem os maiores constituintes da heterocromatina. Porém,
Svartman (2009) novamente argumenta que o estado de condensação dos
cromossomos pode tornar a localização das sequências teloméricas pouco
precisa, por aproximar essas marcações à região de heterocromatina
pericentromérica.
Marsupiais americanos e australianos têm se mostrado um grupo
interessante para estudo, dada sua história evolutiva e o compartilhamento do
padrão cromossômico. Portanto, mesmo sob o modelo da estabilidade
cromossômica, no presente estudo fizemos uma análise comparativa da
distribuição da heterocromatina em relação ao padrão de hibridização de
sequências teloméricas em 11 espécies de marsupiais didelfídeos amazônicos.
Material e Métodos
Neste estudo foram analisadas 11 espécies de marsupiais amazônicos,
representantes de oito gêneros de marsupiais, coletadas em 11 localidades da
Amazônia (Figura 1, Apêndice I). Os espécimes foram coletados sob
autorizações do Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais
Renováveis - IBAMA, de acordo com os números de licença: 02005.000642/03-
11 (IBAMA); 02000.002336/2003-93 (IBAMA); 02005.002672/04 (IBAMA);
37585-5 (SISBIO); 37592-4 (SISBIO). Os indivíduos foram depositados na
Coleção de mamíferos do INPA (Instituto Nacional de Pesquisas da Amazônia)
(Apêndice II).

43
Figura 1 Locais de coleta: Estado do Amazonas: 1- rio Japurá, Município de Japurá; 2-
rio Negro, Santa Isabel do Rio Negro; 3- rio Purus, Município de Tapauá; 4- rio Negro,
Município de Manaus; 5- rio Aripuanã, Município de Novo Aripuanã; 6- rio Uatumã,
Municipio de Prestidente Figueiredo; 7- rio Jatapu, Município de São Sebastião do
Uatumã; Estado do Pará; 8- rio Jari, Município de Almeirim; 9- rio Trombetas, Município
de Oriximiná; 10- rio Tapajós, Municípios de Aveiros e Santarém; Estado de Rondônia:
11- Município de Porto Velho.
Os cromossomos mitóticos foram obtidos a partir da medula óssea pelos
métodos in vivo e in vitro segundo Ford e Harmerton (1956) e Tjio e Whang
(1962), respectivamente. O padrão de banda-C foi determinado segundo a
técnica descrita por Sumner (1972). As sequências teloméricas foram
mapeadas pela técnica de hibridização in situ fluorescente (FISH) conforme
Pinkel et al. (1986), cuja sonda foi obtida por PCR (Ijdo et al.1991).
Resultados
O padrão de banda C utilizado nesta comparação foi descrito em Silva et
al. (em preparação, capítulo 1 desta tese), para 10 espécies (Tabela 1). A
sonda telomérica hibridizou nas extremidades de todos os cromossomos de
todas as espécies (Fig. 2). Destas, as espécies Marmosops pinheiroi,
Marmosops cf. pakaraimae, Marmosops parvidens, Marmosa demerarae,

44
Marmosa murina, e Monodelphis sp. tiveram esta sequência hibridizada
também nas regiões centroméricas, caracterizando assim regiões teloméricas
intersticiais (ITS). As ITSs estiveram co-localizadas com blocos de
heterocromatina em todas as espécies (Fig. 2).
Marmosops pinheiroi (Figura 2c) e Marmosops cf. pakaraimae (dados
não mostrados) foram as espécies que apresentaram o maior número de
cromossomos, tanto autossomos quanto o cromossomo X, com marcações
intersticiais centroméricas e com um bloco conspícuo no centrômero do
cromossomo X. Em Marmosops parvidens (dados não mostrados) observamos
uma variação na quantidade de ITS entre os indivíduos, com cariótipos
apresentando zero, duas, cinco e 13 marcações intersticiais (Tabela 1).
Variação interindividual também foi observada em Marmosa demerarae, que
apresentou ITSs centroméricas em todos os cromossomos, exceto nos sexuais
(Fig. 2d), apresentando indivíduos com zero, quatro, cinco, seis, sete, oito, 10 e
12 marcações (Tabela 1). Em Marmosa murina, a ITS localizou-se no
centrômero do segundo par autossômico e do cromossomo X (Fig. 2e). Apenas
um indivíduo de Caluromys philander, coletado no rio Tapajós, mostrou uma
ITS pericentromérica no par autossômico 3 (Fig. 2a, caixa). Monodelphis sp.
nov. apresentou uma ITS que foi detectada no maior par autossômico (Fig 2h).
Nas demais espécies (Gracilinanus emiliae, Metachirus nudicaudatus, Glironia
venusta e Didelphis marsupialis) não foram detectadas ITS (Fig. 2b, f, g, i). Por
outro lado, em Didelphis marsupialis, os sítios teloméricos foram menores do
que os observados nas outras espécies (Fig. 2i).
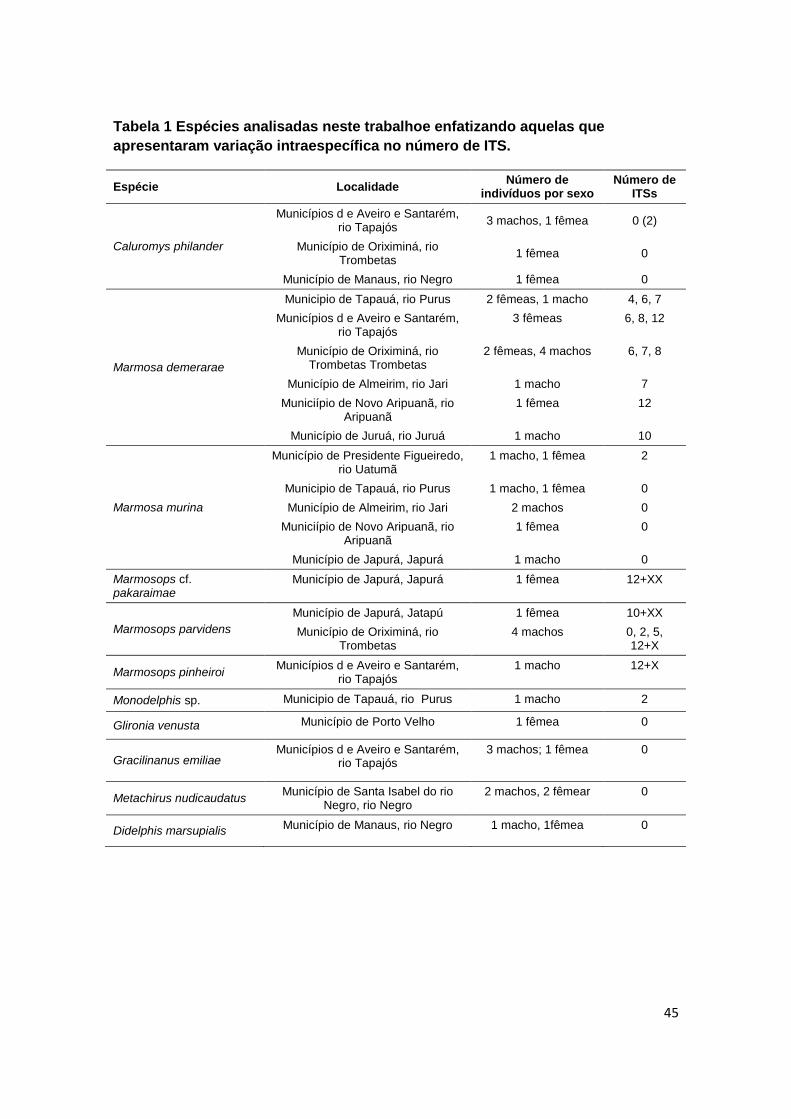
45
Tabela 1 Espécies analisadas neste trabalhoe enfatizando aquelas que
apresentaram variação intraespecífica no número de ITS.
Espécie Localidade Número de
indivíduos por sexo Número de
ITSs
Caluromys philander
Municípios d e Aveiro e Santarém, rio Tapajós
3 machos, 1 fêmea 0 (2)
Município de Oriximiná, rio Trombetas
1 fêmea 0
Município de Manaus, rio Negro 1 fêmea 0
Marmosa demerarae
Municipio de Tapauá, rio Purus 2 fêmeas, 1 macho 4, 6, 7
Municípios d e Aveiro e Santarém, rio Tapajós
3 fêmeas 6, 8, 12
Município de Oriximiná, rio Trombetas Trombetas
2 fêmeas, 4 machos 6, 7, 8
Município de Almeirim, rio Jari 1 macho 7
Municiípio de Novo Aripuanã, rio Aripuanã
1 fêmea 12
Município de Juruá, rio Juruá 1 macho 10
Marmosa murina
Município de Presidente Figueiredo, rio Uatumã
1 macho, 1 fêmea 2
Municipio de Tapauá, rio Purus 1 macho, 1 fêmea 0
Município de Almeirim, rio Jari 2 machos 0
Municiípio de Novo Aripuanã, rio Aripuanã
1 fêmea 0
Município de Japurá, Japurá 1 macho 0
Marmosops cf. pakaraimae
Município de Japurá, Japurá 1 fêmea 12+XX
Marmosops parvidens
Município de Japurá, Jatapú 1 fêmea 10+XX
Município de Oriximiná, rio Trombetas
4 machos 0, 2, 5, 12+X
Marmosops pinheiroi Municípios d e Aveiro e Santarém,
rio Tapajós 1 macho 12+X
Monodelphis sp. Municipio de Tapauá, rio Purus 1 macho 2
Glironia venusta Município de Porto Velho 1 fêmea 0
Gracilinanus emiliae Municípios d e Aveiro e Santarém,
rio Tapajós 3 machos; 1 fêmea 0
Metachirus nudicaudatus Município de Santa Isabel do rio
Negro, rio Negro 2 machos, 2 fêmear 0
Didelphis marsupialis Município de Manaus, rio Negro 1 macho, 1fêmea 0

46
Figura 2 Cromossomos após hibridização de sequências teloméricas (à esquerda) e
após tratamento de banda C (à direita) de: a) Caluromys philander (SISTAP-M-244, caixa:
SISTAP-M-297, cromossomo Y SISTAP-M-305); b) Gracilinanus emiliae (SISTAP-M-344);
c) Marmosops pinheiroi (SISTAP-M-237); d) Marmosa demerarae (MCA 65), e) Marmosa
murina (CEF 18); f) Metachirus nudicaudatus (SISTAP-M-302); g) Glironia venusta (BAC
80); h) Monodelphis sp. (CAN 44); i) Didelphis marsupialis (EE 206). Siglas se referem ao
registro de campo dos indivíduos analisados.

47
Discussão
A uniformidade dos números diploides em marsupiais gera
questionamentos acerca dos eventos das mudanças cromossômicas nesses
organismos. Cariótipos com 2n=14 estão presentes em espécies atuais de
ambos os continentes (americano e australiano). Entretanto, diferem quanto ao
número fundamental devido a rearranjos do tipo inversões pericêntricas. Ainda,
evidências também apontam para fissão cêntrica, gerando números diploides
maiores (Rofe e Hayman 1985). O papel desse rearranjo como principal evento
para o aumento no número diploide é corroborado, quando dados de pintura
cromossômica e filogenia são analisados em conjunto, concluindo que 2n=14
realmente é o candidato mais provável para o número diploide ancestral dos
marsupiais (Westerman et al. 2010).
Ainda do ponto de vista filogenético, Westerman et al. (2010, material
suplementar 2) observaram que as ITSs estão presentes em cromossomos de
poucas espécies da família Didelphidae, sendo encontradas apenas nos
gêneros Marmosa, Monodelphis, Gracilinanus e Marmosops (Svartman e
Vianna-Morgante 1998; Carvalho e Mattevi 2000; Pagnozzi et al. 2002).
Por outro lado, se sobrepusermos esses dados na filogenia de Voss e
Jansa (2009), Glironia venusta (2n=18) e Caluromys spp. (2n=14), táxons
considerados basais, não apresentam ITS, o que sugere evento de fissão, pois
para justificar evento de fusão teríamos que interpretar os resquícios de ITS
como não retidos pelos táxons basais da família ou não detectáveis devido a
limitações técnicas. Isto porque as sequências devem apresentar extensão
suficiente para serem detectadas ao microscópio (Azzalin et al. 1997; Bolzán
2012). Ao sobrepormos dos dados na filogenia de Voss e Jansa (2009)
observamos que os padrões de ITSs registrados estão restritos a dois clados
dentro da família Didelphidae: as tribos Marmosini e Thylamyini, ambos da
subfamília Didelphinae. Para cada um desses dois agrupamentos percebe-se
que a distribuição das ITSs parece aumentar nas espécies mais derivadas e
encontram-se reduzidas nas basais, como por exemplo em Marmosa murina
com ITS no cromossomo X e M. demerarae com marcações em quase todos
os centrômeros. Outro exemplo dessa situação são as espécies Gracilinanus,

48
onde G. emiliae não mostra marcações e G. microtarsus mostrou pelo menos em um
par (Carvalho e Mattevi 2000; presente trabalho).
Acerca das variações intraespecíficas nas ITSs, em Marmosa
demerarae registramos ITS em 12 cromossomos, superando as 8 ITSs já
registrado por Pagnozzi et al. (2000), chegando a. Essa variação não se
restringe a esse táxon, pois observamos situação similar em Marmosops
parvidens (Tabela 1). Tal variação parece estar correlacionada a espécies em
processo recente de especiação, mas também a rearranjos cromossômicos
não Robertsonianos, como inversões e/ou translocações.
Marmosa murina e C. philander desse estudo tiveram variações de ITS
mais discretas. Para a primeira, ITSs foram observadas apenas em nossos
exemplares, enquanto que essas marcações não foram detectadas em
exemplares de outras localidades examinadas em outros estudos (Carvalho e
Mattevi 2000; Pagnozzi et al. 2002). Já, para C. philander, os registros
restringem-se aos exemplares amazônicos analisados por Souza et al. (2013) e
neste estudo. Mesmo com acréscimo de indivíduos analisados, detectamos
ITSs em apenas um indivíduo, mostrando que existe variação menor para essa
espécie.
Quanto à sua organização, as sequências teloméricas intersticiais têm
sido classificadas em het-ITSs, aquelas correlacionadas à heterocromatina e s-
ITSs, sequências curtas em posições intersticiais (Ruiz-Herrera et al. 2008; Lin
e Yan 2008). Considerando essas classes, podemos relacionar as ITSs
observadas no presente trabalho ao tipo het-ITS, que são sequências maiores
e encontradas em associação com a heterocromatina. Situação similar foi
observada em marsupiais australianos (Metcalfe et al. 2004; 2007).
Propomos aqui que, possivelmente que a het-ITS pode ter surgido como
uma sequência curta por fatores do tipo: inserção pelo mecanismo de reparo
(Nergadze et al. 2004, 2007), fusão (Bolzán e Bianchi 2006) ou transposição
(Bouffler et al. 1993; Nergadze et al. 2007). Posteriormente, essas sequências
aumentaram em comprimento, via duplicação por eventos específicos e
independentes para cada espécie observada, integrando à heterocromatina e
tornando-se detectáveis pela FISH. Essa heterocromatinização serviria como
alternativa para “proteção” cromossômica, o que explicaria a associação das

49
ITSs, por nós observadas, com os blocos heterocromáticos maiores e
fortemente marcados na Banda C.
Neste trabalho, sugerimos que a variação no padrão de sequências
teloméricas em didelfídeos é principalmente intraespecífica. Entretanto, é
necessário a inclusão dos demais gêneros de didelphideos à essa análise para que
possamos inferir sobre um padrão evolutivo das sequências teloméricas intersticiais na
família. Por outro lado, sugerimos que nesse grupo, essas ITSs não são o resultado de
eventos de fusão cromossômica, hipótese essa também corroborada pela
sobreposição dos dados de Its na filogenia de Voss e Jansa (2009), na qual táxons
basais apresentam menor número diploide e ainda, citogeneticamente, a associação
dessas sequências com a heterocromatina indicam que as ITSs fazem parte do DNA.
Entretanto, ainda não podemos formular uma hipótese para a origem destas ITS aqui
encontradas e nem tão pouco para a sua variação.

50
Capítulo 3: Mapeamento do elemento LINE-1 em cinco espécies de marsupiais
amazônicos (Didelphidae; Didelphimorphia).
Resumo: No presente trabalho mapeamos o retroelemento LINE-1 em espécies de
didelfídeos, buscando entender a dinâmica desse elemento na evolução cariotípica de
marsupiais amazônicos. Foram analisados exemplares de cinco espécies: Caluromys
philander, Gracilinanus emiliae, Marmosa murina, Marmosa demerarae e Didelphis
marsupialis. As marcações se mostraram dispersas por todos os cromossomos, não
havendo preferência por regiões específicas do cromossomo, nem mesmo pelo
cromossomo X, como já registrado na literatura para outros marsupiais.
Introdução
Elementos transponíveis (ET) são sequências de DNA dotadas de mobilidade,
o que determina sua configuração dispersa pelo genoma. A movimentação desses
elementos pelo genoma pode ocorrer de forma direta via DNA-DNA ou por meio de um
intermediário de RNA. Essa característica, por sua vez, pode se relacionar com
funções biológicas como: transferência horizontal, regulação gênica, mudanças
cromossômicas, inativação do X em fêmeas (Lyon 1998; Ostertag et al. 2001 Kazazian
e Goodier 2002; Chow et al. 2010).
Elementos transponíveis estão presentes em todos os organismos, abrangendo
procariotos e eucariotos e correspondem a uma grande fração dos genomas dos
hospedeiros (Lander et al. 2001). Em mamíferos, os LINEs (Long Interspersed Nuclear
Elements) e SINEs (Short Interspersed Nuclear Elements) são os ETs mais bem
representados. Em Monodelphis domestica os LINEs representam 20%, em Mus
musculus 18,7% e em humanos aproximadamente 20% (Gentles et al. 2007;
Rebuzzini et al. 2009).
Waters et al. (2007) observaram que, em mamíferos, os padrões de
divergência entre os elementos LINE-1 são específicos de cada linhagem. Porém, o
envolvimento desses elementos em alterações cromossômicas pode ser indireto, mas
ainda assim o seu acúmulo pode ser associado com os padrões de especiação de
algumas linhagens (Dobigny et al. 2004; Waters et al. 2004). Em marsupiais, a
distribuição desses elementos em cromossomos ainda foi pouco explorada,
restringindo-se ao sequenciamento de LINE-1 em Didelphis virginiana e hibridização in
situ fluorescente de retrovírus endógenos de cangurus (KERV) em espécies de
wallaby do continente australiano (Dönner e Paböo 1995; O’Neill et al. 1998).

51
Mapeamento cromossômico desse elemento em mamíferos tem possibilitado
realizar inferências sobre a estrutura e reorganização da cromatina em
mamíferos Referências.
Deste modo, o presente estudo teve como objetivo mapear a distribuição
dos elementos LINE-1 nos cromossomos de cinco espécies de didelfídeos
amazônicos para conhecer sua organização.
Material e Métodos
A distribuição do elemento transponível LINE-1 foi mapeada nos
cromossomos de cinco espécies de didelfídeos amazônicos: Caluromys
philander (1 macho e 2 fêmeas), Marmosa murina (1 macho e 1 fêmea),
Marmosa demerarae (1 macho e 2 fêmeas), Gracilinanus emiliae (1 fêmea) e
Didelphis marsupialis (1 macho e 1 fêmea), cujas preparações cromossômicas
foram obtidas segundo Ford e Hammenrton (1956). Para a obtenção da sonda
utilizamos os primers desenvolvidos por Dobigny (2002): L1-F 5' CCA-TGC-
TCA-TSG-ATT-GG-3', L1-R 5' ATT-CRT-TCC-CAT-TGG-TCTA 3', seguindo os
parâmetros de Waters et al. (2004), aumentando apenas o tempo de
anelamento para um minuto o que permitiu melhores resultados. As sondas
foram marcadas por nick translation com kits de marcação DIG-Nick, da Roche,
seguindo as recomendações do fabricante. A hibridização in situ fluorescente
(FISH) seguiu o protocolo descrito por Pinkel et al. (1986), com estringência de
70%, onde foram obtidos os melhores resultados.
Resultados
Das cinco espécies analisadas, nas quais o LINE-1 foi mapeado quatro
têm 2n=14 cromossomos e uma 2n=22 (Figura 1). Em todas as espécies o
LINE-1 se mostrou disperso. Entretanto, Caluromys philander (Figura 1a) e
Marmosa demerarae (Figura 1b) apresentaram uma maior concentração de
marcações, distribuídas por todas as cromátides. Em contraste, em G. emiliae
(Figura 1c) e em M. murina (Figura 1d) a densidade pareceu ser bem menor.
Didelphis marsupialis (Figura 1e) mostrou um padrão intermediário.
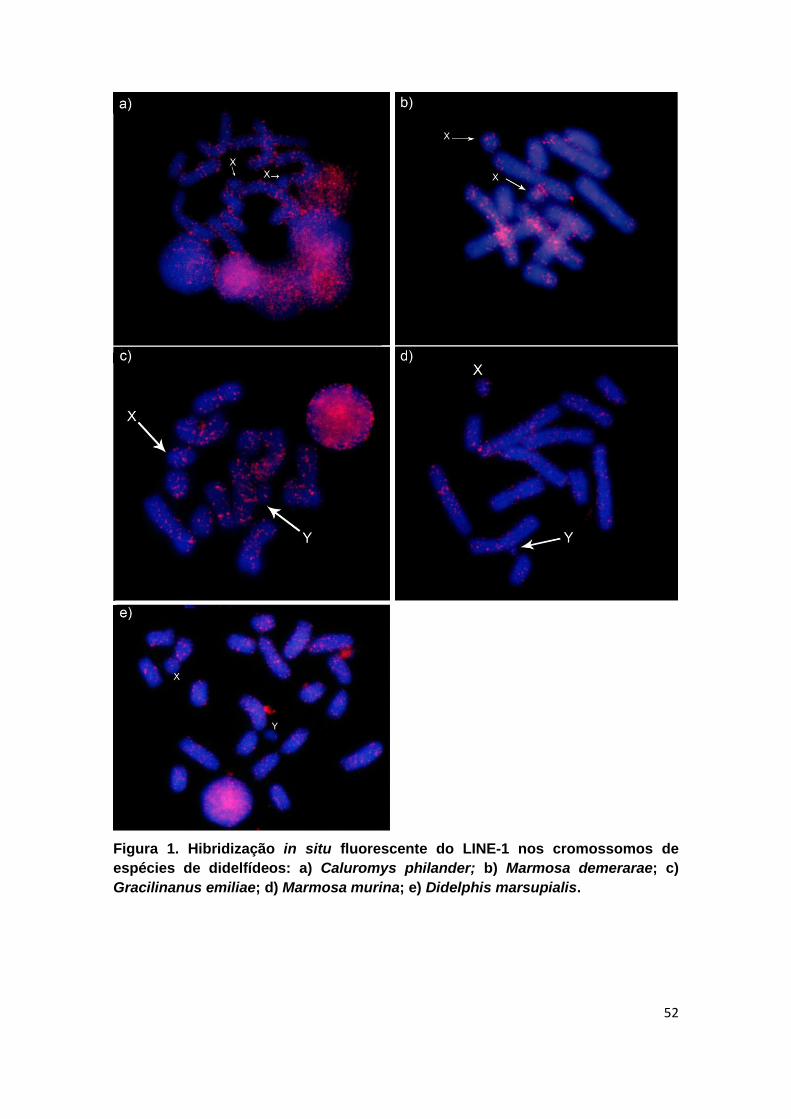
52
Figura 1. Hibridização in situ fluorescente do LINE-1 nos cromossomos de
espécies de didelfídeos: a) Caluromys philander; b) Marmosa demerarae; c)
Gracilinanus emiliae; d) Marmosa murina; e) Didelphis marsupialis.
e)

53
Discussão
Padrões de hibridização de LINE-1 (L1) nos cromossomos de mamíferos
foram observados em pelo menos 10 ordens: Tubulidentata, Proboscidea,
Macrocelidea, Xenarthra, Sirenia, Chiroptera, Cetacea, Rodentia, Primates e
Diprotodontia (O’Neill et al. 1998; Parish et al. 2002; Waters et al. 2004; Acosta
et al. 2008; Rebuzzini et al. 2009; Bonifácio et al. 2012). Dentre essas, apenas
Diprotodontia pertence à infraclasse Metatheria, onde estão classificados os
marsupiais (Wilson e Reeder 2005).
Em Macropus eugenii, marsupial australiano, a distribuição de elemento
transponível (ET) mostrou-se dispersa pelos braços cromossômicos e ausente
nos centrômeros, sendo que a dispersão do L1 foi limitada pela presença do
retroelemento KERV-1 (Kangaroo Endogenous RetroVirus) (Ferreri et al. 2011).
Apesar de não compararmos a distribuição do L1 com a de outros ET, a
dispersão desse elemento não apresentou limitações claras entre regiões das
cromátides. Portanto, espécies com clusters mais definidos podem apresentar
esse padrão devido interações entre dois retroelementos, confinando-os à
regiões específicas e separadas (como exemplo: Marmosa murina e
Gracilinanus emiliae).
Em relação aos eutérios, os padrões de hibridização de L1 estão
disponíveis para Cetacea, Tubulidentata, Proboscidea, Macrocelidea,
Xenarthra e Sirenia. Os padrões obtidos neste estudo assemelham-se mais
aos padrões do xenartro amazônico Bradypus tridacylus, cujos sinais de
hibridização foram mais dispersos (Waters et al. 2004;). Esse tipo de situação é
contrário ao observado em roedores, onde a inserção pode ser alocada em
bandas-G positiva ou dispersa por toda a cromátide, exceto no centrômero. Em
espécies do gênero Taterillus, por exemplo, a dispersão desse elemento pelos
cromossomos foi maior em espécies que apresentam divergência mais recente.
Enquanto que no gênero Mus, essa dispersão foi constante em todas as
subespécies investigadas (Dobigny et al. 2004; Rebuzzini et al. 2009).
Ao contrário de Taterillus, não podemos atribuir a dispersão do LINE-1
de nossos espécimes com estimativas de tempos de divergência entre
espécies, pois mesmo que a irradiação dos didelfídeos atuais seja mais recente

54
do que o esperado (Jansa et al. 2015), a escala temporal é muito maior do que
a verificada em Taterillus.
Os padrões de hibridização do L1 de espécies de elefantes, Ardvaark e
xenartros, levaram Waters et al. (2004) a inferirem que padrões de L1
semelhantes a padrões de bandeamento G são característicos de
Euarchotonglires (coelhos, roedores e primatas). Porém, essa organização não
parece ser exclusiva desse grupo, pois registros no cetáceo Inia geoffrensis e
no quiróptero Carollia brevicauda também exibiram o mesmo padrão (Parish et
al. 2002; Bonifácio et al. 2012). No caso de quirópteros (Tonatia saurophila e
Mimon crenulatum) foi observado um enriquecimento de L1 na região
pericentromérica dos cromossomos. Isso demonstra que não existe padrão
único para cada linhagem de mamífero, dado que um mesmo padrão pode ser
encontrado em linhagens distantes e dois padrões podem ser vistos em um
mesmo grupo.
Nossos resultados também divergem da afirmativa de que exista um
padrão preferencial para a inserção do L1 em quaisquer regiões
cromossômicas, pois não pudemos relacionar a distribuição de padrões
considerados semelhantes com algum tipo de estrutura cromossômica
evidenciada por coloração convencional ou bandeamentos.
Ainda, foi demonstrado que LINE-1 propaga-se verticalmente entre as
linhagens de mamíferos, refletindo sua história evolutiva (Waters et al. 2007).
Apesar de, essa afirmação ser um elemento-chave para compreensão dos
padrões evolutivos desse elemento em mamíferos, ainda falta estabelecer uma
relação clara de como a compartimentalização foi estabelecida em cada
linhagem de mamíferos.
Em termos de organização genômica, os bandeamentos cromossômicos
(-G, -R e –C) permitem reconhecer regiões dos cromossomos em diferentes
níveis de compactação da cromatina. Para didelfídeos, esses métodos são
bem explorados na literatura e recentemente descrevemos padrões cariotípicos
para 16 espécies, coletadas em 13 localidades da Amazônia (Rofe e Hayman
1985; Palma e Yates 1996; Svartman e Vianna-Morgante 1999; Carvalho e
Mattevi 2002; Svartman 2009; Silva et al. em preparação [capítulo 1 do
presente trabalho]).

55
Segundo Sumner (1998; 2003), uma das características das regiões G-
positivas é serem mais ricas em genes expressos e em elementos do tipo
LINE. Porém, esse acúmulo não foi verificado nas espécies aqui estudadas.
Considerando que os LINEs podem participar da regulação da expressão
gênica (Lippman et al. 2004; Lunyak et al. 2007), sugerimos que a falta de
acúmulo de LINE nessas regiões indica que a participação desses elementos
na regulação gênica, não exije uma localização física entre genes expressos e
cópias do transponível.
Tanto metatérios como eutérios possuem sistema de determinação
sexual do tipo XX/XY, sendo que um dos cromossomos X é inativado no sexo
homogamético (Lyon 1961). Porém, em marsupiais existem algumas
peculiaridades em relação aos cromossomos sexuais, que abrangem: o
tamanho desses cromossomos em relação ao restante do genoma, diferenças
estruturais, como ausência de região de homologia entre estes, e ausência do
complexo sinaptonêmico na meiose (Solari e Bianchi 1975; Sharp 1982; Seluja
et al. 1987; Graves e Watson 1991; Graves 1996). Adicionalmente, podemos
somar a essas diferenças, a ausência do acúmulo dos elementos LINE/L1 na
inativação de um dos cromossomos X, como impulsionadores desse processo
em eutérios (Lyon 1998; 2000). Nossos dados mostram essa falta de acúmulo
de L1 no X. Com o sequenciamento completo do genoma de Monodelphis
domestica observa-se que uma quantidade significativamente menor do ET
esteve presente no cromossomo X (22%), em relação ao encontrado para
camundongos (35%) e até mesmo em humanos (O’Neill et al. 1998; Mikkelsen
et al. 2007).
Provavelmente não haja necessidade da participação desses elementos,
pois, dentre as inúmeras particularidades que diferem a inativação do X entre
os dois táxons de placentários, verifica-se a presença de um gene (RSX), que
substitui a função do gene Xist de eutério, responsável pela inativação
(Davidow et al. 2007; Hore et al. 2007). É possível que a atuação do gene RSX
não necessita da participação desse ET, durante o processo de inativação.
Portanto, ambas informações suportam as evidências obtidas pela FISH da
falta de acúmulo de LINE nos cromossomos X de marsupiais.

56
Ainda sobre o cromossomo X, cada vez mais evidências apontam para
uma dinâmica evolutiva diferenciada deste, em didelfídeos. Previamente,
registramos variações intraespecíficas na posição do centrômero do X de
Marmosa murina e Caluromys philander e também na distribuição da
heterocromatina em espécies de Marmosops (Silva et al. em preparação).
Assim, acreditamos que os cromossomos sexuais desses organismos ainda
ofereçam um vasto campo a ser explorado tanto no âmbito molecular quanto
em estudos citomoleculares.
Diferente dos eutérios, sobretudo euarchontoglires (lagomorfos, primatas
e roedores), marsupiais didelfídeos não mostram padrão de distribuição
preferencial dos retroelementos LINE-1 e os padrões observados em cada
espécie de mamífero deve seguir uma dinâmica própria dentro de cada
linhagem. Assim, a aparente falta de padrão dos didelfídeos deste estudo, tanto
pode ser fruto de uma irradiação recente de espécies quanto uma interação
particular dentro da família Didelphidae

57
4. Conclusões 1- Cariótipos dos didelfídeos mostram estabilidade, porém essa
característica marcante quanto ao número diploide é atenuada, dado
que é possível detectar variações no que se refere aos marcadores
cromossômicos (banda-C, RON, DNAr 18S). Essas variações tornam-se
mais evidentes ao incluir os cromossomos sexuais, principalmente o X
nas análises.
2- A observação das variações de marcadores cromossômicos mostra
estruturação geográfica em algumas espécies de didelfídeos. Os
agrupamentos formados podem ser indicativos da existência de
diferentes linhagens ou espécies crípticas.
3- Considerando que as sequências teloméricas intersticiais estão
presentes em poucas linhagens dentro dessa família, esse marcador
parece insuficiente como fonte de atribuição de fusões como principal
gerador dos números diploides. Isso fica ainda mais evidente quando
expandimos essas observações para as espécies basais.
4- A variação intraespecífica nas ITSs é maior do que o já registrado na
literatura. Essa variação mostra uma forte correlação com a
heterocromatina, indicando que essas sequências podem ser apenas
um componente do DNA satélite.
5- O ET- LINE-L1 não mostrou relação com regiões específicas dos
cromossomos, fortalecendo a hipótese de que sua distribuição está
relacionada à dinâmica evolutiva de cada grupo.
6- A falta de acúmulo de L1 no cromossomo X dos marsupiais pode ser um
resultado do mecanismo de inativação do cromossomo X nesses
organismos, que difere dos demais mamíferos placentários.
7- A variação do cromossomo X e os padrões de distribuição dos
elementos repetitivos podem ser reflexo dos eventos de especiação
ocorridos nos didelfídeos.

58
5. Referências
Ab’Saber, A.N. 1977. Os domínios morfoclimáticos na América do Sul: primeira
aproximação. Geomorfologia, 52: 1-23.
Acosta, M. J.; Marchal, J. A.; Fernandez-Espartero, C. H.; Bullejos, M.;
Sanchez, A. 2008. Retroelements (LINEs and SINEs) in vole genomes:
Differential distribution in the constitutive heterochromatin. Chromosome
Research, 16: 949-959.
Almeida, L. M.; Carareto, C. M. A. 2005. Origem, proliferação e extinção de
elementos transponíveis: qual seria a importância da transferência
horizontal na manutenção deste ciclo? - Série Monografias SBG. Ribeirão
Preto: SBG, 43p.
Aplin, K. P.; Archer, M. 1987. Recent advances in marsupial systematics with a
new syncretic classification. In: Archer, M. (ed.), Possums and opossums:
studies in evolution. Vol. 1, xv–lxxii. Sydney: Surrey Beatty.
Asahida, T.; Kobayashi, T.; Saitoh, K.; Nakayama, I. 1996. Tissue preservation
and total DNA extraction from fish stored at ambient temperature using
buffers containing high concentration of urea. Fisheries Science, 62(5): 727-
730.
Azzalin, C.M.; Mucciolo, E.; Bertoni, L.; Giulotto, E. 1997. Fluorescence in situ
hybridization with a synthetic (T2AG3)n polynucleotide detects several
intrachromosomal telomere-like repeats on human chromosomes.
Cytogenetics and Cell Genetics, 78: 112 – 125.
Azzalin, C.M.; Nergadze, S.G.; Giulotto, E. 2001. Human intrachromosomal
telomeric-like repeats: sequence organization and mechanisms of origin.
Chromosoma, 110: 75–82.
Baker, R. J.1967. Karyotypes of bat of the Phyllostomidae and their taxonomic
implications. South West Naturalist, 12: 407-428.
Baker, R. J.; Bickham, J. W. 1980. Karyotypic evolution: Evidence of extensive
and conservative chromosomal evolution in closely related taxa. Systematic
Zoology, 29(3): 239-253.
Biggers, J. D; Fritz, W. C.; Hare, W. C. D.; McFeely, R. A. 1965. Chromosomes
of American marsupials. Science, 148: 1602-1603.

59
Blackburn, E. H. 1991. Structure and function of telomeres. Nature, 350: 569–
573.
Bolzán, A. D.; Bianchi, M. S. 2006. Telomeres, interstitial telomeric repeat
sequences, and chromosomal aberrations. Mutation Research, 612: 189–214.
Bolzán, A. D. 2012. Chromosomal aberrations involving telomeres and
interstitial telomeric sequences. Mutagenesis, 27(1): 1-15.
Bonifácio, H. L.; da Silva, V. M. F; Martin, A. R.; Feldberg, E. 2012. Molecular
cytogenetic characterization Amazon river dolphin Inia geofrensis.
Genetica, 140(7): 307:315.
Bouffler, S.; Silver, A.; Papworth, D.; Coates, J.; Cox, R. 1993. Murine radiation
myeloid leukaemogenesis: relationship between interstitial telomere-like
sequences and chromosome 2 fragile sites. Genes, Chromosomes and Cancer, 6:
98-106.
Brandão, M. V.; Rossi, R. V.; Semedo, T. B. F.; Pavan, S. E. 2015. Diagnose e
distribuição geográfica dos marsupiais da Amazônia brasileira. In:
Pequenos mamíferos não-voadores da Amazônia brasileira (Mendes-
Oliveira, A.C. and Miranda, C.L.). Sociedade Brasileira de Mastozoologia.
ISBN: 978-85-63705-02-0, 336 p.
Brasileiro-Vidal, A. C.; Guerra, M. 2002. Citogenética molecular em cereais, p.
277-298. In: Brammer, S.P.; Iorczeski, E.J. Atualização em técnicas
celulares e moleculares aplicadas ao melhoramento genético vegetal.
Embrapa, Passo fundo, MG.
Burnham, R.; Johnson, K. 2004. South American palaeobotany and the origins
of neotropical rainforests. Philosophical Transations Of Royal Society
London. Series, 359: 1595–1610.
Burrack, L. S.; Berman, J. 2012. Flexibility of centromere and kinetochore
structures. Trends in Genetiscs, 28(5): 204-212.
Cabrera, A. 1919. Genera mammalium: Monotremata, Marsupialia. Madrid:
Museo Nacional de Ciencias Naturales.
Cabrera A. 1958 (‘‘1957’’). Catálogo de los mamíferos de América del Sur [part
1]. Revista del Museo Argentino de Ciencias Naturales (Ciencias
Zoológicas), 4(1): 1–307.

60
Capy, P.; Bazin, C.; Higuet, D.; Langrin, T. 1998. Dynamics and Evolution of
Transposible Elements. Austin, TX: Springer-Verlag.
Carvalho, B. A.; Mattevi, M. 2000. (T2AG3)n telomeric sequence hybridization
suggestive of centric fusion in karyotype marsupials evolution. Genetica,
108: 205–210.
Carvalho, B. A.; Oliveira, L. F.; Nunes, A. P.; Mattevi, M. S. 2002. Karyotypes of
nineteen marsupial species from Brazil. Journal of Mammalogy, 83(1): 58-
70.
Casartelli, C.; Rogatto, S. R.; Ferrari, I. 1986. Cytogenetic analysis of some
Brazilian marsupials (Didelphidae: Marsupialia). Canadian Journal of
Genetics and Cytology, 28: 21-29.
Chow J. C.; Ciaudo, C.;Fazzari, M. J.; Mise, N.; Servant, N.; Glass, J. L.;
Attreed, M.; Avner, P.; Wutz, A.; Barillot, E.; Greally, J. M.; Voinnet, O.;
Heard, E. 2010. LINE-1 activity in facultative heterochromatin formation
during X chromosome inactivation. Cell, 11141(6): 956-69.
Costa, L. P. 2005. Filogenia de marsupiais didelfídeos e implicações para
taxonomia, p. 699-736. In: Cáceres, N. C.; Monteiro-Filho, E.L.A. (Org.). Os
marsupiais do Brasil: biologia, ecologia e evolução. UNIDERP e
Universidade Federal do Mato Grosso do Su. Curitiba, PR.
Davidow, L. S.; Breen, M.; Duke, S. E.; Samollow, P. B.; McCarrey, J. R.; Lee,
J. T. 2007. The search for a marsupial XIC reveals a break with vertebrate
synteny. Chromosome Research, 15(2): 137-146.
De Leo, A. A.; Guedelha, N.; Toder, R.; Voullaire, L; Ferguson-Smith, M. A.;
O'Brien, P. C. M.; Graves, J. A. M. 1999. Comparative chromosome
painting between marsupial orders: relationships with a 2n=14 ancestral
marsupial karyotype. Chromosome Research, 7: 509-517.
Dobigny, G. 2002. Speciation Chromosomique chez les Especes Jumelles
Ouest-Africaines du Genre Taterillus (Rodentia, Gerbillinae): Implications
Syste´matiques et Biogeographiques, Hypothe`ses Ge´nomiques. PhD
thesis, Muse´um National d’Histoire Naturelle, Paris.
Dobigny, G.; Ozouf-Costaz, C.; Waters, P. D.; Bonillo, C.; Coutanceau, J. P.;
Volobouev, V. 2004. LINE-1 amplication accompanies explosive genome
repatterningin rodents. Chromosome Research, 12: 787–793.

61
Dönner, M.; Paböo, S. 1995. Nucleotide sequence of a marsupial LINE-1
element and evolution of placental mammals. Molecular Biology Evolution,
12 (5): 944-948.
Dutrillaux, B. 1979. Chromosomal evolution in primates: tentative phylogeny
from Microcebus murinus (Prosimian) to man. Human Genetics, 48: 251-
314.
Elder, J. F. Jr.; Turner, B. J. 1995. Concerted evolution of repetitive DNA
sequences in eukaryotes. Quarterly Review of Biology, 70(3): 297-320.
Estoup, A.; Presa, P.; Krieg, F.; Vaiman, D.; Guyomard, R. 1993. (CT)n and
(GT)n microsatellites: a new class of genetic markers for Salmo truta L.
(brown trout). Heredity, 71: 488-496.
Fantin, C.; da Silva, M. N. F. 2011. The Karyotype of a rare South American
marsupial. The bushy-tailed opossum genus Glironia (Didelphimorphia:
Didelphidae). Mastozoologia Neotropical, 18(1): 125-130.
Ferreri, G. C; Brown, J. D.; Obergfell, C.; Jue, N.; Finn, C. E.; O’Neill, M. J.;
O’Neill, R. 2011. Recent Amplification of the Kangaroo Endogenous
Retrovirus, KERV, Limited to Centromere. Journal of Virology, 85(10):
4761-4771.
Ford, C.; Hamerton, J. 1956. A colchicines hypothonic citrate squash sequence
for mammalian chromosomes. Stain Technology, 31: 247-251.
Gardner, A. L. 1993. Order Didelphimorphia, p. 15-23. In: Wilson, D.E.; Reeder,
D.M. (eds.) Mammal species of the world: a taxonomic and geographic
reference. Smithsonian Institution Press, Washington, D.C.
Gardner, A. L. 2007. Cohort Marsupialia, p. 669. In: Gardner, A.L. The
Mammals of South America. University of Chicago Press, Chicago.
Gardner, A. L. 2008. Mammals of South America. Volume I: Marsupials,
Xenarthrans, Shrews, and Bats. The University of Chicago Press. 669p.
Gentles, A. J.; Wakefield, M. J.; Kohany, O.; Gu, W.; Batzer, M. A.; Pollock, D.
D.; Jurka, J. 2007. Evolutionary dynamics of transposable elements in the
short-tailed opossum Monodelphis domestica. Genome Research, 17: 992–
1004.
Graves, J. A. M.; Watson, J. M. 1991. Mammalian sex chromosomes: evolution
of organization and function. Chromosoma, 101: 63-68.

62
Graves, J. A. M. 1996. Mammals that break the rules: Genetics of Marsupials
and monotremes. Annual reviews in Genetics, 30: 233-240.
Grewal, S. I. S.; Jia, S. 2007. Heterochromatin revisited. Nature. Reviews in.
Genetics, 8: 35-46.
Gross, M. C.; Schneider, C. H.; Valente, G. T.; Martins, C.; Feldberg, E. 2010.
Variability of 18S rDNA locus among Symphysodon fishes: chromosomal
rearrangements. Journal of Fish Biology, 76: 1117–1127.
Groves, C. P. 1993. Order Dasyuromorphia, Peramelemorphia,
Notoryctemorpha, Diprodontia, p. 29–62. In: Wilson, D.E.; Reeder, D.M.
(eds). Mammal species of the world: a taxonomic and geographic
reference. Smithsonian Institution Press, Washington, D.C.
Guerra, M.S. 1988. Introdução à citogenética geral. Guanabara, 144pp.
Guerra, M.S. 2004. FISH. Conceitos e Aplicações na citogenética. Sociedade
Brasileira de Genética, 176pp.
Gutiérrez, E. E.; Jansa, A. S.; Voss, R. S. 2010. Molecular systematics of
mouse opossums (Didelphidae: Marmosa): assessing species limits using
mitochondrial DNA sequences, with comments on phylogenetic
relationships and biogeography. American Museum Novitates, 3692: 22p.
Hayman, D. L.; Martin, P. G. 1974. Mammalia I: Monotremata and Marsupialia.
In: Animal cytogenetics. Gebru¨der Borntraeger, Berlin, Germany, 1-110p.
Hayman, D. L. 1990. Marsupial cytogenetics. Australian Journal of Zoology, 37:
331-349.
Hershkovitz, P. 1992. The South American gracile mouse opossums, genus
Gracilinanus Gardner and Creighton, 1989 (Marmosidae, Marsupialia): a
taxonomic review with notes on general morphology and relationships.
Fieldiana Zoology (New Series) 39: i–vi, 1–56.
Hore, T. A.; Koina, E.; Wakefield, M. J.; Graves, J. A. M. 2007. The region
homologous to the X-chromosome inactivation centre has been disrupted in
marsupial and monotreme mammals. Chromosome Research, 15: 147–
161.
Horovitz, I.; Sánchez-Villagra, M. R. 2003. A morphological analysis of
marsupial mammal higher-level phylogenetic relationships. Cladistics, 19:
181-212.

63
Howell, W. M.; Black, D. A. 1980. Controlled silver-staining of nucleolus
organizer region with a protective colloidal developer: a 1-step method.
Experientia, 36: 1014-1015.
Hsu, T. C.; Spirito, S. E.; Pardue, M. L. 1975. Distribution of 18+28S ribosomal
genes in mammalian genomes. Chromosoma, 53: 25-36.
Ijdo, J. W.; Wells, R. A.; Baldini, A.; Reeders, S. T. 1991. Improved telomere
detection using a telomere repeat probe (TTAGGG)n generated by PCR.
Nucleic Acid. Research, 17: 4780.
INPE, 1999. Relatório Projeto PRODES (http://www.inpe.br/
Informações_Eventos/ amz/amz.html). Acesso 10/11/2012.
Jansa, A. S.; Voss, R. S. 2000. Phylogenetic studies on didelphid marsupials I.
Introduction and preliminary results from nuclear IRBP gene sequences.
Journal of Mammalian Evolution, 7(1): 43-77.
Jansa, S. A.; Barker, F. K.; Voss, R. S. 2013. The early diversification history of
didelphid marsupials: A window into South America’s “splendid isolation”.
Evolution, 68(3): 684-695.
Jansa, S. A.; Baker, F. K.; Voss, R. S. 2014. The early diversification history of
didelphid marsupials: a window into South America’s “splendid isolation”.
Evolution, 68(3): 684-695.
John, B. 1988. The biology of heterochromatin. 1-147p. In: Verma, R. S.
Heterochromatin: Molecular and structural aspects. xv+301p. Cambridge
Press. ISBN: 0-521-33480-2
Jordan, H. E. 1911. The spermatogenesis of the opossum, with special
reference to the accessory chromosome and the chondriosomes. Archiv fur
Zellforschung, 7: 41-86.
Kazazian Jr., H. H.; Goodier, J. L. 2002. LINE drive retrotransposition and
genome instability. Cell, 110: 277-280.
Kirsch, J. A. W.; Palma. R. E. 1995. DNA/DNA hybridization studies of
carnivorous marsupials. V. A further estimate of relationships among
opossums (Marsupialia: Didelphidae). Mammalia, 59: 403–425.
Lander, E. S.; Linton, L. M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; et
al. 2001. Initial sequencing and analysis of the human genome. Nature,
409: 860–921.

64
Levan, A.; Fredga, K.; Sandberg, A. A. 1964. Nomenclature for centromeric
position on chromosomes. Hereditas, 52: 201-220.
Lima, J. F. S. 2004. Cariótipos e Regiões Organizadoras de Nucléolo (RON) de
Marmosa e Didelphis (Didelphidae) do estado do Tocantins, Brasil. Revista
Nordestina de Biologia, 18(2): 87-93.
Lin, K. W.; Yan, J. 2008. Endings in the middle: Current knowledge of interstitial
telomeric sequences. Mutation Research, 658: 95–110.
Lippman, Z; Gendrel, A. V.; Black, M.; Vaughan, M. V.; Dedhia, N.; McCombie,
R.; Lavine, K.; Mittal, V.; May, B.; Kasschau, K. D.; Carrington, J. C.;
Doerge, R. W.; Colotz, V.; Martienssen, R. 2004. Role of transposable
elements in heterochromatin and epigenetic control. Nature, 430: 471-476.
Lunyak, V. V.; Prefontaine, G. G.; Núñez, E.; Cramer, T.; Ju, B.; Ohgi, K. A.;
Hutt, K.; Roy, R.; García-Díaz, A.; Zhu, X.; Yung, Y.; Montoliu, L.; Glass, C.
K.; Rosenfeld, M. G. 2007. Developmentally regulated activation of a SINE
B2 repeat as a domain boundary in organogenesis. Science, 5835(317):
248-251.
Lyon, M. F. 1961. Gene action in the X-chromosome of the mouse (Mus
musculus L.). Nature, 190: 372–373.
Lyon, M. F. 1998. X-chromosome inactivation: a repeat hypothesis. Cytogenetic
and Cell Genetics, 80: 133–137.
Lyon, M. F. 2000. LINE-1 elements and X chromosome inactivation: A function
for “junk” DNA? PNAS, 97(12):6248-6249, doi:10.1073/pnas.97.12.6248
Marshall, L. G.; Case, J. A.; Woodburne, M. O. 1990. Phylogenetic relationships
of the families of marsupials, p. 433–506. In: Current Mammalogy.
Genoways, H.H. New York: Plenum Press.
Matschie, P. 1916. Bemerkungen uber die Gattung Didelphis L.
Sitzungsberichte der gesellschaft naturforschender freunde zu Berlin 1916:
259-272.
McClintock, B. 1941. The stability of broken ends of chromosomes of Zea mays.
Genetics, 23: 234–282.
McKenna, M. C.; Bell, S. K. 1997. Classification of mammals above the species
level. New York: Columbia University Press, xii+631pp.

65
Metcalfe, C. J.; Eldridge, M. D. B.; Johnston, P. G. 2004. Mapping the
distribution of the telomeric sequence (T2AG3)n in the 2n=14 ancestral
marsupial complement and in the macropodines (Marsupialia:
Macropodidae) by fluorescence in situ hybridization. Chromosome
Research, 12: 405-414.
Metcalfe, C. J.; Eldridge, M. D. B.; Johnston, P. G. 2007. Mapping the
distribution of the telomeric sequence (T2AG3)n in the Macropodoidea
(Marsupialia) by fluorescence in situ hybridization.II. The ancestral 2n = 22
macropodid karyotype. Cytogenetics and Genome Research, 116: 212–
217.
Meyne, J.; Ratliff, R. L.; Moyzis, R. K. 1989. Conservation of the human
telomere sequence (TTAGGG)n among vertebrates. Proceedings of the
National Academy of Sciences USA, 86: 7049–7053.
Mikkelsen, T. S.; Wakefield, M. J.; Aken, B.; Amemiya, C. T.; et al. 2007.
Genome of the marsupial Monodelphis domestica reveals innovation in
non-coding sequences. Nature, 447(7141): 167-177.
Morley, R. J. 2000. Origin and evolution of tropical rain forests. John Wiley &
Sons Inc, Chichester, England.
Moratelli, R.; Morielle-Versute, E. 2007. Métodos e aplicações da citogenética
na taxonomia de morcegos brasileiros. Pp. 197-218. In: Morcegos do Brasil
(Reis, N. R.; Peracchi, A. L.; Pedro, W. A.; Lima, I. P.). 253p. ISBN:978-85-
906395-1-0.
Nagamachi, C. Y.; Feldberg, E.; Pieczarka, J.; Pereira, A. L.; Silva, C. E. F;
Rosa, C. C.; Souza, E. M. S.; Pinto, J. A.; da Costa, M. J. R.; Malcher, S.
M.; Paixão, V. S.; Silva, W. O. 2015. Citogenética de pequenos mamíferos
não-voadores da Amazônia brasileira. In: Pequenos mamíferos não-
voadores da Amazônia brasileira (Mendes-Oliveira, A. C., Miranda, C. L.).
Sociedade Brasileira de Mastozoologia. ISBN: 978-85-63705-02-0, 336 p.
Nergadze, S. G.; Rocchi, M.; Azzalin, C. M.; Mondello, C.; Giulotto, E. 2004.
Insertion of telomeric repeats at intrachromosomal break sites during
primate evolution. Genome Research, 14: 1704–1710.
Nergadze, S. G.; Santagostino, M. A.; Salzano, A.; Mondello, C.; Giulotto, E.
2007. Contribution of telomerase RNA retrotranscription to DNA double-

66
strand break repair during mammalian genome evolution. Genome Biology,
8: 260.
Nilsson, M. A.; Arnasona, U.; Spencerb, P. B. S.; Jankea, A. 2004. Marsupial
relationships and a timeline for marsupial radiation in South Gondwana.
Gene, 340: 189–196.
Nilsson, M.A.; Churakov, G.; Sommer, M.; Tran, N.V.; Zemann, A.; Brosius, J.;
Schmitz, J. 2010. Tracking marsupial evolution using archaic genomic
retroposon insertions. PLoS Biol 8(7): e1000436. doi:
10.1371/journal.pbio.1000436
Nowak, R. M. 1991. Walker’s Mammals of the World. The John Hopkins
University Press 5ª ed., 1629pp.
O’Neill, R. J. W.; O’Neill, M. J.; Marshall, G. J. A. 1998. Undermethylation
associated with retroelement activation and chromosome remodelling in an
interspecific mammalian hybrid. Nature, 393: 68-72.
O’Neill, R. J. W.; Eldridge, M. D. B.; Toder, R.; Ferguson-Smith, M. A.; O’Brien,
P. C.; Graves, J. A. M. 1999. Chromosome evolution in kangaroos
(Marsupialia: Macropodidae): Cross species chromosome painting between
the tammar wallaby and rock wallaby spp. with the 2n=22 ancestral
macropodid karyotype. Genome, 42: 525–530.
Ostertag, E. M.; Kazazian Jr., H. H. 2001. Biology of mammalian L1
retrotransposons. Annual Reviews of Genetics, 35: 501-538.
Paglia, A.P., G.A.B. Fonseca, A.B. Rylands, G. Herrmann, L.M.S. Aguiar, A.G.
Chiarello, Y.L.R. Leite, L.P. Costa, S. Siciliano, M.C.M. Kierulff, S.L.
Mendes, V.C. Tavares, R.A. Mittermeier and J.L. Patton. 2012. Lista
Anotada dos Mamíferos do Brasil / Annotated Checklist of Brazilian
Mammals. 2nd Edition. Occasional Papers in Conservation Biology, 6:1–76
Pagnozzi, J. M.; Silva, M. J. J.; Yonenaga-Yassuda, Y. 2000. Intraspecific
variation in the distribution of the interstitial telomeric (TTAGGG)n
sequences in Micoureus demerarae (Marsupialia: Didelphidae).
Chromosome Research, 8: 585–591.
Pagnozzi, J. M.; Ditchfield, A. D.; Yonenaga-Yassuda, Y. 2002. Mapping the
distribution of the interstitial telomérica (TTAGGG)n sequences in eight
species of Brazilian marsupials (Didelphidae) by FISH and the correlation

67
with constitutive heterochromatin. Do ITS represent evidence for fusion
events in American marsupials? Cytogenetics and Genome Research, 98:
278–284.
Palma, R. E.; Yates, T. L. 1996. The chromosomes of Bolivian didelphid
marsupials. Occasional Papers of Museum of Texas University, 162: 1-20.
Parish, D. A.; Vise, P.; Wichman, H. A.; Bull, J. J.; Baker, R. J. 2002.
Distribution of LINEs and other repetitive elements in the karyotype of the
bat Carollia: implications for X chromosome inactivation. Cytogenetic and
Genome Research, 96: 191-197.
Paresque, R.; Souza, W. P.; Mendes, S. L.; Fagundes V. 2004. Composição
cariotípica da fauna de roedores e marsupiais de duas áreas de Mata
Atlântica do Espírito Santo, Brasil. Boletim do Museu de Biologia Mello
Leitão, 17: 5-33.
Pereira, N. P.; Ventura, V.; Silva, M. C. J.; Silva, D. M.; Yonenaga-Yassuda, Y.;
Pellegrino, C. M. 2008. Karyotype characterization and nucleolar organizer
regions of marsupial species (Didelphidae) from areas of Cerrado and
Atlantic Forest in Brazil. Genetics and Molecular Biology, 31(4): 887–892.
Peres, C. 1999 The structure of non volant mammal communities in different
Amazonian forest types. Pp: 564-581. In: Eisenberg, J.; Redford, K.
Mammals of the Neotropics, the central Neotropics: Ecuador, Peru, Bolivia
and Brazil. Illinois: University of Chicago Press.
Pinkel, D.; Straume, T.; Gray, J. W. 1986. Cytogenetic analysis using
quantitative, highsensitivity, fluorescence hybridization. Proceedings of the
National Academy of Sciences, 83: 2934-2938.
Rebuzzini, P.; Castiglia, R.; Nergadze, S. G.; Mitsainas, G.; Munclinger, P.;
Zuccotti, M.; Capanna, E.; Redi, C. A.; Garagna, S. 2009. Quantitative
variation of LINE-1 sequences in five species and three subspecies of the
subgenus Mus and in five Robertsonian races of Mus musculus
domesticus. Chromosome Research, 17: 65-76.
Reig, O. A.; Gardner, A. L.: Bianchi, N. O.; Patton, J. L. 1977. The
chromosomes of Didelphidae (Marsupialia) and their evolutionary
significance. Biology Journal of Linnean Society, 9: 191-216.

68
Reig, O. A; Kirsch, J. A. W.; Marshall, L. G. 1985. New conclusions on the
relationship of opossum-like marsupials, with an annotated classification of
the Didelphimorphia. Amerighiniana, 21: 335-343.
Reig, O. A.; Kirsch, J. A. W.; Marshall, L. G. 1987. Systematic relationships of
the living and Neocenic American “opossum-like” marsupials (suborder
Didelphimorphia), with comments on the classification of these and of the
Cretaceous and Paleogene New World and European metatherian. In: M.
Archer (editor), Possums and opossums; studies in evolution. 1: 1-89.
Rens, W.; O’Brien, P. C. M.; Yang, F.; Graves, J. A. M.; Ferguson-Smith, M. A.
1999. Karyotype relationships between four distantly related marsupials
revealed by reciprocal chromosome painting. Chromosome Research, 7:
461-474.
Rens, W.; O’Brien, P. C. M.; Fairclough, H.; Harman, L.; Graves, J. A. M.;
Ferguson-Smith, M. A. 2003. Reversal and convergence in marsupial
chromosome evolution. Cytogenetics and Genome Research, 102: 282–
290.
Rocchi, M.; Archidiacono, N.; Schemp, W.; Capozzi, O.; Stanyon, R. 2012.
Centromere repositioning in mammals. Heredity, 108: 59–67.
Rofe, R.; Hayman, D. 1985. G-banding evidence for a conserved complement
in the Marsupialia. Cytogenetics and Cell Genetics, 39: 40-50.
Saiki, R. K.; Gelfand, D. H.; Stoffel, S.; Scharf, S. J.; Higuchi, R.; Horn, G.
T.;Mullis, K. B.; Erlich, H. A. 1988. Primer-directed enzymatic amplification
of DNA with a thermostable DNA polymerase. Science, 239: 487-491.
Sambrook, J.; Russell, D. W. 2001. Molecular cloning: a laboratory manual.
Cold Spring Harbor Laboratory Press, New York, EUA. p. 633–664.
Ruiz-Herrera, A.; Nergadze, S. G.; Santagostino, M.; Giulotto, E. 2008.
Telomeric repeats far from the ends: mechanisms of origin and role in
evolution. Cytogenetics and Genome Research, 122: 219–228.
Sánchez, J.; Bianchi, M. S.; Bolzán, A. D. 2010. Relationship between
heterochromatic interstitial telomeric sequences and chromosome damage
induced by the radiomimetic compound streptonigrin in Chinese hamster
ovary cells. Mutation research/Fundamental and Molecular Mecanisms of
Mutagenesis, 684: 90-97.

69
Scheneider, C. H.; Gross, M. C.; Terencio, M. L.; Artoni, R. F.; Vicari, M. R.;
Martins, C.; Feldberg, E. 2013. Chromosomal evolution of neotropical
cichlids: the role of repetitive DNA sequences in the organization and
structure of karyotype. Revision in Fish Biology and Fisheries, 23(2): 201-
214.
Seluja, G.; Roche, L.; Solari, A. J. 1987. Male meiotic prophase in Didelphis
albiventris. Journal of Heredity, 78: 218-222.
Sharp, P. 1982. Sex chromosome pairing during male meiosis in. marsupials.
Chromosoma, 86: 27–47.
Singer, M. F. 1982. Highly repeated sequences in mammalian genome.
International Review of Cytology, 76: 667-1112.
Solari, A. J.; Bianchi, N. O. 1975. The synaptic behavior of the X and Y
chromosomes in the marsupial Monodelphis dimidiata. Chromosoma, 52:
11-25.
Sotero-Caio, C.G.; Volleth, M.; Hoffmann, F.G.; Scott, L.; Wichman, H.A.; Yang,
F.; Baker, R.J. 2015. Integration of molecular cytogenetics, dated molecular
phylogeny, and model-based predictions to understand the extreme
chromosome reorganization in the Neotropical genus Tonatia (Chiroptera:
Phyllostomidae). Evolutionary Biology, 15:220-235. DOI 10.1186/s12862-
015-0494-y.
Souza, M. J.; Maia, V.; Santos, J. F. 1990. Nucleolar organizer regions, G- and
C-band in some Brazilian species of Didelphidae. Revista Brasileira de
Genética, 13: 767-775.
Souza, E. M. S.; Silva, C. E. F.; Eler, E. S.; da Silva, M. N. F.; Feldberg, E.
2013. Variations of chromosomal structures in Caluromys philander
(Didelphimorphia: Didelphidae) from the Amazon region. Genetica, 141(1-
3): 89-93.
Sumner, A. T. 1972. A simple technique for demonstrating centromeric
heterochromatin. Experimental Cell Research, 75: 304-306.
Sumner, A. T. 1998. The mitotic chromosome. Advances in Genome Biology,
5A, 211-261.
Sumner, A. T. 2003. Chromosomes organization and function. Blackwell
Publishing, Malden. 287 p.

70
Svartman, M.; Vianna-Morgante, A. M. 1998. Karyotype evolution of marsupials:
from higher to lower diploid numbers. Cytogenetics and Cell Genetics, 82:
263–266.
Svartman, M.; Vianna-Morgante, A. M. 1999. Comparative genome analysis in
American marsupials: chromosomes banding and in-situ hybridization.
Chromosome Research, 7: 267-275.
Svartman, M.; Vianna-Morgante, A. M. 2003. Conservation of nucleolus
organizer in American marsupials (Didelphidae). Genetica, 118: 11-16.
Svartman, M. 2009. American marsupial chromosomes: Why study them?
Genetics and Molecular Biology, 32(4): 675-687.
Thomas, O. 1888. Catalogue of the Marsupialia and Monotremata in the
collection of the British Museum (Natural History). London: Trustees of the
British Museum (Natural History).
Tjio, J. H.; Whang, J. 1962. Chromosome preparations of bone marrow cells
without prior in vitro culture ou in vivo colchicine administration. Stain
technology, 37(1): 17-20.
Toder, R.; O'Neill, R. J.W. ; Wienberg, J.; O'Brien, P. C.M.; Voullaire, L.;
Graves, J. A. M. 1997. Comparative chromosome painting between two
marsupials: origins of an XX/XY1Y 2 sex chromosome system. Mammalian
Genome 8, 418-422
Voss, R.L; Emmons, L.H. 1996. Mammalian diversity in Neotropical lowland
rainforests: a preliminary assessment. Bulletin of the American Museum of
Natural History, 230: 1-115.
Voss, R. S.; Jansa, S. A. 2003. Phylogenetic studies on didelphid marsupials II.
Non-molecular data and new IRBP sequences: separate and combined
analyses of didelphine relationships with denser taxon sampling. Bulletin of
the American Museum of Natural History, 276: 82p. ISSN 0003-0090
Voss, R.; Jansa, S. 2009. Phylogenetic relationships and classification of
didelphid marsupials, an extant radiation of new world metatherian
mammals. Bulletin of the American Museum of Natural History, 322: 1-177.
ISSN 0003-0090.

71
Yonenaga-Yassuda, Y.; Kasahara, S.; Souza, M. J.; L’abbate, M. 1982
Constitutive heterochromatin, G-bands and nucleolus-organizer regions in
four species of Didelphidae (Marsupialia). Genetica, 58: 71–77.
Waters, P. D.; Dobigny, G.; Pardini, A. T.; Robinson, T. J. 2004. LINE-1
distribution in Afrotheria and Xenarthra: implications for understanding the
evolution of LINE-1 in eutherian genomes. Chromosoma, 113: 137-144.
Waters, P. D.; Dogbiny, G.; Wadell, P. J.; Robinson, T. J. 2007. Evolutionary
history of LINE-1 in the major clades of placental mammals. Plos One, 2(1):
158.
Westerman, M.; Meredith, R. W.; Springer, M. S. 2010. Cytogenetics Meets
Phylogenetics: A Review of Karyotype Evolution in Diprotodontian
Marsupials. Journal of Heredity,101(6): 690–702.
White, M. J. D. 1973. Animal cytology and evolution. 3ª Edição, Cambridge
University Press. 961pp.
Wilson, D. E.; Reeder, D. M. 2005. Mammal Species of the World. Johns
Hopkins University Press. 2142 pp.

72
Apêndice I: Coordenadas geográficas para cada local de coleta no formato de grau decimal,
coletadas no sistema de referência WGS 84:
1- rio Japurá (1.84341666667°S, -69.0264722222°W); 2- rio Juruá (3.64151111111°S,
66.1006916667°W); 3- rio Negro (0.57725000000° S, 64.8976944444°W); 4- rio Purus
(4.98066666667° S; 62.9770000000° W); 5- rio Cuieiras (2.70708611111°S,
60.3738388889°W); 6- Manaus (UFAM campus: 3.100548° S, 59.974595° W; Refinaria Isaac
Sabá: 3.15264166667°S, 59.9843444444°W); 7- rio Aripuanã (6.00000000000° S,
60.1666666667° W); 8- rio Uatumã (1.84998888889°S, 59.4402000000°W); 9- rio Jatapú
(2.017940° S, 58.203228° W); 10- rio Trombetas (1.48163888889° S, 56.4573333333° W); 11-
rio Tapajós (3.35486111111° S; 55.2031666667° W); 12- rio Jari (0.7000000000° S,
52.6666666667° W); 13- Porto Velho (8.87416666667° S, 64.0077777778° W).
Apêndice II: Material testemunho: Todos os espécimes analisados foram depositados na
Coleção de Mamíferos do Instituto Nacional de Pesquisas da Amazônia; os
espécimes estão indicados pela espécie, sítios de coleta, gênero, e número de
coletor, seguidos pelo registro do INPA (em parênteses) quando disponível.
Voucher specimens: Glironia venusta: (BAC 80) - Caluromys philander:
rioTapajós (male: SISTAP-M-297; SISTAP-M-305; SISTAP-M-318; SISTAP-M-382;
female: SISTAP-M-244); rio Trombetas (female: CTGA-M-652); rio Purus (female: CAN
34); Manaus (female: MSN 01); (female: BAC 102) - Caluromys lanatus: rio Japurá
(female: CTGA-M-701) - Marmosops sp.: rio Aripuanã ( female: MCA 3; MCA 7; MCA
8; MCA 26; MCA 31; MCA 35; male: MCA 4; MCA 16; MCA 38; MCA 39); rio Jari
(female: TAG 3459; RNL 70); rio Juruá (male: EE 107; EE 139; female: EE135); rio
Cuieiras (female: EE 198; EE 211; male: EE 192; EE 201; EE216) - Marmosops
bishopi: rio Negro (male: SISIS-M-127); rio Purus (male: SISPUR-M-135; SISPUR-M-
157; SISPUR-M-160; SISPUR-M-164; SISPUR-M-135; CAN 30; CAN 51; female: CAN
48) - Marmosops pinheiroi: rio Tapajós (male: SISTAP-M-237; SISTAP-M-278;
female: SISTAP-M-268; SISTAP-M-277) - Marmosops parvidens: rio Trombetas
(male: CTGA-M-501; CTGA-M-516; CTGA-M-531; CTGA-M-532; CTGA-M-551;
CTGA-M-555; CTGA-M-581; CTGA-M-600; female: CTGA-M-533) - Marmosops
impavidus: rio Purus (male: SISPUR-M-149) - Marmosops cf. pakaraime: rio Japurá
(male: SISJAP-M-705) - Marmosa murina: rio Jari (male: RNL 45); rio Uatumã (male:
CEF 4; CEF 8; CEF 18; CEF 27; CEF 28; CEF 32; female: CEF 16; CEF 34; CTGA-M-
8; CTGA-M-22; CTGA-M-41;), rio Negro (male: SISIS-M-57; SISIS-M-63); rio
Trombetas ( female: CTGA-M—519); rio Purus (male: CAN 43); rio Japurá (male:
CTGA-M-708) - Marmosa cf. lepida: rio Aripuanã (female: MCA12, rio Japurá (male:
SISJAP-M-764)- Gracilinanus emiliae: rio Tapajós: (male: SISTAP- M-245; SISTAP-
M-343; SISTAP- M-344; SISTAP- M-345) - Micoureus demerarae: rio Aripuanã
(female: MCA 27; MCA 36; MCA 58; MCA 65; male: MCA 21; MCA59); rio Jari (female:
RNL 31; RNL 48; male: RNL 30; MCA 32; MCA 46; MCA 49; MCA 58; MCA 61; MCA
64; MCA 66; MCA 67) rio Juruá (female: EE136; male: EE 143); Manaus (female: EE
149: EE 150; EE 151; EE 154; EE 158; EE 159; EE 169; EE 222; EE 228; 229; EE
234; male: EE 157; EE 167; EE 170; EE 176; EE 189; EE 194; EE 196; EE 202; EE
215; EE 220; EE 235); rio Cuieiras (female: EE 193; EE 219); rio Tapajós ( female:
SISTAP-M-229; SISTAP-M-241; SISTAP-M-321; SISTAP-M-333; SISTAP-M-369;

73
male: SISTAP-M-267; SISTAP-M-279; SISTAP-M-322); rio Trombetas (female: CTGA-
M-579; CTGA-M-590; CTGA-M-622; CTGA-M-667; CTGA-M-672; male: CTGA-M-535;
CTGA-M-539; CTGA-M-557; CTGA-M-558; CTGA-M-572; CTGA-M-573; CTGA-M-
578; CTGA-M-580; CTGA-M-613); rio Negro (female: SISIS-M-85; SISIS-M-110;
SISIS-M-117; SISIS-M-128; male SISIS-M- 86); rio Purus (female: SISPUR-M-145;
CAN 25; CAN 31; CAN 50: male: SISPUR-M-144; SISPUR-M-147; SISPUR-M-148) -
Monodelphis brevicaudata: rio Jari (male: TAG 2731; RNL 68) - Monodelphis sp.:
rio Purus: (male: CAN 44) - Monodelphis cf. emiliae: rio Aripuanã (female: MCA 31) -
Metachirus nudicaudatus: rio Jari: (RNL 47); rio Cuieiras: (fêmas: EE 200); rio
Tapajós (ind: SISTAP-M-230; fêmas: SISTAP-M-230; male: SISTAP-M-251; SISTAP-
M-269); rio Trombetas: (female: CTGA-M-655); rio Jatapú: (female: CTGA-M-52;
CTGA-M-58); rio Negro: (female: SISIS-M-64; SISIS-M-78; male: SISIS-M-84; SISIS-
M-116); rio Purus: (male: CAN 33) - Didelphis marsupialis: rio Jari: (female: RNL 44;
RNL 53; RNL 59; male: RNL 52; RNL 55; RNL 62; RNL 63); Manaus: female (EE 174;
EE 197; EE 204; EE 224; EE 227; EE 246; EE 250; EE 155; EE 155; EE 173; EE 183;
EE 190; EE 203; EE 205; EE 206; EE 223; EE 232; EE 233; EE 237; EE 247;EE 248;
EE249; EE 190); rio Uatumã (female: CEF 5; male: CEF 13); rio Trombetas (female:
CTGA-M-594; CTGA-M-606; male: CTGA-M-607); rio Purus (male: SISPUR-M-185);
rio Negro (male: SISIS-M-73): rio Tapajós (female: SISTAP-M-324; SISTAP-M-346;
SISTAP–M-347;male: SISTAP-M-243); rio Japurá: (male: CTGA-M-732).





























