Embed Size (px)
Citation preview

INSTITUTO OSWALDO CRUZ
Mestrado em Biologia Parasitária
EFEITO DOS INIBIDORES DE ASPÁRTICO PROTEASE DO HIV SOBRE Leishmania amazonensis
LIVIA DE OLIVEIRA SANTOS
Rio de Janeiro 2009
TESE MBP -IOC* L.O. SANTOS 2009

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
INSTITUTO OSWALDO CRUZ
Pós-Graduação em Biologia Parasitária
LIVIA DE OLIVEIRA SANTOS
Efeito dos Inibidores de Aspártico Protease do HIV sobre Leishmania amazonensis
Dissertação apresentada ao Instituto Oswaldo Cruz
como parte dos requisitos para obtenção do título
de Mestre em Ciências.
Orientador: Prof. Dr. Carlos Roberto Alves
RIO DE JANEIRO
2009

iii
INSTITUTO OSWALDO CRUZ Pós-Graduação em Biologia Parasitária
AUTOR: Lívia de Oliveira Santos
EEITO DOS INIBIDORES DE ASPÁRTICO PROTEASE DO HIV
SOBRE LEISHMANIA AMAZONENSIS
ORIENTADOR (ES): Prof. Dr. Carlos Roberto Alves
Aprovada em: 16/04/2009
EXAMINADORES:
Prof. Drª. Alda Maria da Cruz - Presidente Prof. Drª. Suzana Côrte-Real Faria Prof. Drª. Helena Carla Castro
Rio de Janeiro, 13 de Julho de 2009

iv
Dedicatória
Em memória de meu pai,
que amei cada minuto da minha vida
e a quem dedico esta conquista.

v
Agradecimentos
A Jesus, pelos caminhos que percorri sob sua orientação e proteção e a Deus por tudo que tenho conquistado e por estar ao meu lado em todos os momentos da minha vida. Ao meu orientador, Dr. Carlos Roberto Alves, cujo apoio e compreensão foram essenciais para o desenvolvimento deste trabalho. A Drª Constança Britto por ter me recebido de braços abertos em seu laboratório. A todos os amigos do laboratório, sem exceções, e em especial a Dani, Karina, Bernardo, Chico, Leo, Camilinha, Lívia U., Thaisinha, Larissa, Louise e Clarinha pelo carinho especial. A Ellen e Bianca, que tanto me ajudaram neste trabalho e que me elegeram como “chefinha”, mas que na verdade eu as elegi como minhas amigas do coração. A Drª Claudia Masini d`Avila Levy e Marta Helena Branquinha de Sá por terem dado a mim a oportunidade de iniciar na vida científica. Tudo que hoje sei devo a vocês. Minhas orientadoras do coração, acima de tudo. A minha amiga querida, Fernanda de Aquino Marinho, por sua amizade verdadeira, pelos momentos de risadas, pelos momentos de desabafo, pelas horas dispensadas no auxílio à elaboração deste trabalho. Sua amizade é uma das coisas mais valiosas na minha vida. Ao Dr André Luís e a Drª Cristina Motta, da Universidade Federal do Rio de Janeiro, pelo apoio no desenvolvimento da pesquisa, bem como pelo auxílio na análise e interpretação dos experimentos. Ao corpo docente e discente e a secretaria do Programa de Pós-Graduação em Biologia Parasitária, pelos conhecimentos compartilhados e pelo zelo e auxílio constante ao aluno. A Fiocruz pelo apoio financeiro e pela oportunidade de estar em uma instituição tão séria e competente. A todos do Laboratório de Bioquímica de Insetos por serem sempre tão solícitos e permitirem o uso de seus aparelhos. A Jaime Augusto Neto, meu companheiro e meu amigo, sobretudo, sempre presente ao meu lado, por me incentivar, apoiar e acreditar nos meus sonhos e, ainda mais, por ter me ajudado a transformá-los em realidade. Te amo! Aos meus pais, irmã e avó, com quem sei que sempre poderei contar e a quem amo incondicionalmente. Obrigado a todos de coração!

vi
Epígrafe
"A dúvida é o princípio da sabedoria”
(Aristóteles)

vii
Sigas e abreviaturas:
AIDS/SIDA: Acquired Immunodeficiency Syndrome (síndrome da imunodeficiência humana)
DAN: Diazo-acetyl-norleucinemethylester (Diazoacetil-DL-norleucina metil ester)
DMEM: Dulbecco’s modified Eagle’s medium (Meio eagle modificado por Dulbecco)
DMSO: Dimetilsulfóxido
DNA/ ADN: Deoxyribonucleic acid (ácido desoxirribonucléico)
gp63: Glicoproteína 63
GPI: Glicosilfosfatidilinositol
HIV: Human Immunodeficiency Virus (Vírus da Imunodeficiência Humana)
IC50: Concentração inibitória
IL-10: interleucina-10
IP/r: Inibidor/ritonavir
IPs: Inibidores de proteases
IPs-HIV: Inibidores de protease do HIV
kDa: kilodalton
kDNA: DNA do cinetoplasto
LCD: Leishmaniose cutânea difusa
LM: Leishmaniose mucocutânea
LV: Leishmaniose visceral
mA: Miliampere
MSP: Major surface peptidase (principal peptidase de superfície)
MTT: (bromidrato de 3-(4,5-dimetiltiazol-2-il)-2,5-difeniltetrazolio)
NK: Natural Killer
OMS: Organização Mundial da Saúde
PAHO/WHO: Pan American Health Organization/World Health Organization
PBS: Solução salina tamponada com fosfato
PSP: Promastigote surface peptidase (peptidase de superfície de promastigota)
RFLP: Restriction fragment length polymorphism
rpm: Rotação por minuto
SAG: Gluconato de antimônio e sódio
SAP: Secreted aspartic protease (aspártico protease secretada)
SDS: Sodium dodecyl sulfate (dodecil sulfato de sódio)
SDS-PAGE: SDS - polyacrilamide gel electrophoresis (Eletroforese em gel de poliacrilamida
contendo SDS)

viii
SFB: Soro fetal bovino
TARV: terapia anti-retroviral de altamente efetiva
TGF-β: Transforming growth factor-β (fator transformador do crescimento-β)
TH2: T helper 2
TNF-α: Tumor necrosis factor-α (fator de necrose tumoral- α)
V: Volts

ix
INSTITUTO OSWALDO CRUZ
EFEITO DOS INIBIDORES DE ASPÁRTICO PROTEASE DO HIV
SOBRE LEISHMANIA AMAZONENSIS
RESUMO/ABSTRACT
DISSERTAÇÃO DE MESTRADO
Livia de Oliveira Santos
O protozoário Leishmania é o agente etiológico da leishmaniose, doença cuja
patogenicidade ainda não é bem compreendida. A terapêutica atual para a leishmaniose é pouco eficaz devido à toxicidade dos agentes terapêuticos disponíveis e à emergência de resistência aos medicamentos. Além disso, os países e regiões endêmicas são economicamente pobres e têm sua situação agravada devido ao aumento do número de casos de co-infecção Leishmania-HIV, devido à sobreposição das epidemias da Aids e da leishmaniose. No presente trabalho, foi investigado o efeito dos inibidores de aspártico protease do HIV (IPs-HIV) sobre a proliferação, ultraestrutura e diferenciação de Leishmania amazonensis. Além disso, foi avaliado o efeito dessas drogas sobre a interação com macrófagos, a atividade de aspártico protease e a expressão de proteases clássicas, que estão diretamente envolvidas na patogênese do parasito. Todos os IPs-HIV foram capazes de diminuir o crescimento do parasito de uma forma dose-dependente, especialmente nelfinavir e lopinavir, com o IC50 correspondendo a 15,12 µM e 16,47, respectivamente. Análises por microscopia eletrônica revelaram que o tratamento com os inibidores causou mudanças profundas na ultraestrutura do flagelado, o encolhimento do citoplasma, aumento do número de inclusões lipídicas e núcleo intimamente envolto pelo retículo endoplasmático, bem como a condensação da cromatina, sendo esta última observação um indicativo de morte por apoptose. Os IPs-HIV não foram capazes de inibir a diferenciação do parasito da forma promastigota para amastigota nas condições utilizadas neste estudo, porém foram capazes de inibir diretamente a atividade de aspártico proteases contra substratos específicos em extratos brutos do parasito. O tratamento de formas promastigotas com os IPs-HIV antes da interação induziu a uma redução drástica dos índices de associação com macrófagos murinos. O pós-tratamento também interferiu no desenvolvimento intracelular de L. amazonensis nos macrófagos, como avaliado, nos ensaios onde os IPs-HIV foram adicionados a culturas de células previamente infectadas. Apesar de todos estes efeitos benéficos, os inibidores induziram a um aumento na expressão de cisteíno proteases b (cpb) e da metaloprotease (gp63), dois fatores de virulência bem caracterizados de Leishmania spp. Diante da sobreposição leishmaniose/HIV, é fundamental compreender as sofisticadas interelações entre Leishmania, HIV e macrófagos. Além disso, há muitas questões não resolvidas, no que diz respeito ao tratamento de pacientes co-infectados com Leishmania-HIV, por exemplo, a eficácia do tratamento destinado a controlar cada patógeno nesses indivíduos continua indefinido. Os resultados in vitro apresentados adicionam novos conhecimentos sobre o vasto espectro de eficácia do IPs-HIV sobre patógenos oportunistas e sugerem que estudos adicionais devem ser realizados sobre os efeitos sinérgicos dos compostos classicamente utilizados no tratamento da leishmaniose e os inibidores da protease do HIV em macrófagos co-infectados.

x
ABSTRACT Leishmania is the etiologic agent of leishmanisais, a protozoan disease whose
pathogenic events are not well understood. Current therapy for leishmaniasis is suboptimal due to toxicity of the available therapeutic agents and the emergence of drug resistance. Furthermore, endemic countries and regions are economically poor. Compounding these problems is the increase in the number of cases of Leishmania-HIV coinfection, due to the overlap between the AIDS epidemic and leishmaniasis. In the present work, we have investigated the effect of HIV aspartyl peptidase inhibitors (PIs) on the Leishmania
amazonensis proliferation, ultrastructure, differentiation, interaction with macrophage cells, aspartyl protease activity and expression of classical proteases, which are directly involved in the Leishmania pathogenesis. All the HIV PIs impaired parasite growth in a dose-dependent fashion, especially nelfinavir and lopinavir, with an IC50 corresponding to 15,12 and 16,47, respectively. PIs treatment caused profound changes in the leishmania ultrastructure as shown by transmission electron microscopy, including cytoplasm shrinking, increase in the number of lipid inclusions and some cells presenting the nucleus closely wrapped by endoplasmic reticulum, as well as chromatin condensation, which is suggestive of apoptotic death. The PIs were not able to inhibit the promastigote-amastigote differentiation of Leishmania
amazonensis under the conditions used in this study, b ut were able to directly inhibit the activity of aspartyl proteases over specific
substrates. The pre-treatment of promastigote forms with PIs drastically reduced the association indexes during the interaction with murine macrophage cells. As well, PIs interfere with the intracellular development of Leishmania in macrophages, assessed by adding PIs in previously infected cells. Despite all these beneficial effects, the PIs induced an increase in the expression of cysteine peptidase b and the metallopeptidase gp63, two well-known virulence factors expressed by Leishmania spp. In the face of leishmaniasis/HIV overlap, it is critical to further comprehend the sophisticated interplays among Leishmania, HIV and macrophages. In addition, there are many unresolved questions related to the management of Leishmania-HIV-coinfected patients. For instance, the efficacy of therapy aimed at controlling each pathogen in coinfected individuals remains largely undefined. The results presented herein add new in vitro insight into the wide spectrum efficacy of HIV PIs and suggest that additional studies about the synergistic effects of classical antileishmanial compounds and HIV PIs in macrophages coinfected with Leishmania and HIV-1 should be performed.

xi
Lista de figuras Introdução Fig 1...........................................................................................................................................2 Fig 2...........................................................................................................................................3 Fig 3...........................................................................................................................................5 Fig 4...........................................................................................................................................6 Fig 5...........................................................................................................................................8 Fig 6.........................................................................................................................................10 Fig 7.........................................................................................................................................14 Fig 8.........................................................................................................................................21 Fig 9.........................................................................................................................................22 Fig 10.......................................................................................................................................23 Resultados Fig 11.......................................................................................................................................35 Fig 12.......................................................................................................................................36 Fig 13.......................................................................................................................................37 Fig 14.......................................................................................................................................38 Fig 15.......................................................................................................................................39 Fig 16.......................................................................................................................................40 Fig 17.......................................................................................................................................41 Fig 18.......................................................................................................................................44 Fig 19.......................................................................................................................................45 Fig 20.......................................................................................................................................46 Fig 21.......................................................................................................................................47 Fig 22.......................................................................................................................................49 Fig 23.......................................................................................................................................50 Fig 24.......................................................................................................................................52 Fig 25.......................................................................................................................................54 Fig 26.......................................................................................................................................56

xii
Índice:
I. Introdução.................................................................................................................. 1
1. Família Trypanosomatidae.......................................................................................... 1
2. Leishmania e Leishmanioses....................................................................................... 4
3. Co-infecção Leishmania-HIV..................................................................................... 9
4. Tratamento das leishmanioses..................................................................................... 12
5. Proteases...................................................................................................................... 13
6. Proteases em Leishmania............................................................................................ 15
7. Inibidores proteolíticos como quimioterápicos........................................................... 18
8. Inibidores de Protease do HIV.................................................................................... 19
9. Ação dos Inibidores de Protease do HIV sobre patógenos oportunistas..................... 23
II. Justificativa............................................................................................................... 25
III. Objetivo geral.......................................................................................................... 26
1. Objetivo específico...................................................................................................... 26
IV. Metodologia............................................................................................................. 27
1. Materiais...................................................................................................................... 27
2. Cepa utilizada e condições de cultivo......................................................................... 27
3. Obtenção de isolados infectivos.................................................................................. 27
4. Ensaio de inibição da multiplicação............................................................................ 28
5. Ensaio de inibição da diferenciação dos parasitos de promastigota para amastigota in vitro.............................................................................................................................
28
6. Microscopia eletrônica de transmissão....................................................................... 29
7. Avaliação da atividade enzimática através de dosagem.............................................. 29
8. Ensaio de viabilidade do parasito................................................................................ 30
9. Avaliação do perfil de proteases por SDS-PAGE com gelatina e por immunoblotting...............................................................................................................
31
10. Ensaio de viabilidade de macrófagos........................................................................ 32
11. Ensaio de interação Leishmania-macrófago (Pré-tratamento).................................. 32
12. Ensaio de interação Leishmania-macrófago (Pós-tratamento).................................. 33
13. Análises estatísticas................................................................................................... 33
V. Resultados................................................................................................................. 34
VI. Discussão.................................................................................................................. 57
VII. Conclusões.............................................................................................................. 67
VIII. Anexo.................................................................................................................... 68
IX. Bibliografia.............................................................................................................. 80

1
I. Introdução
1. Família Trypanosomatidae
O sub-reino Protozoa agrupa alguns protozoários de reconhecida importância médica,
veterinária e agropecuária. A família Trypanosomatidae, classificada neste grupo, possui
membros que parasitam um grande espectro de organismos, atingindo desde protozoários,
rotíferos, nematóides, anelídeos, moluscos, artrópodes e plantas, até vertebrados de quase
todas as classes (peixes, anfíbios, répteis, aves e mamíferos), incluindo o homem (Rey, 2008).
Os microrganismos desta família possuem características ultraestruturais típicas, como
flagelo único contendo uma complexa estrutura paraflagelar; mitocôndria única e ramificada
que percorre todo o corpo celular, que contém uma região com acúmulo de DNA,
denominada cinetoplasto; os glicossomos, que compartilham algumas funções metabólicas
com peroxissomos de outros organismos e que contém as enzimas da via glicolítica, e
microtúbulos subpeliculares associados à membrana plasmática (De Souza, 2002).
Os gêneros pertencentes a esta família são tradicionalmente classificados baseados em
dois critérios: formas evolutivas e ciclo de vida. Quanto a este último critério, são divididos
em tripanossomatídeos heteroxênicos e monoxênicos. Os monoxênicos são aqueles que
possuem apenas um hospedeiro durante o seu ciclo de vida, como Leptomonas, Crithidia,
Blastocrithidia, Herpetomonas e Rhynchoidomonas, e três gêneros recentemente propostos,
Wallaceina, Angomonas (Brandão et al. 2001) e Strigomonas (revisto por Santos et al, 2007).
Já os tripanossomatídeos heteroxênicos, representados pelos gêneros Trypanosoma,
Leishmania, Sauroleishmania, Endotrypanum e Phytomonas, são aqueles que possuem um
ciclo de vida complexo envolvendo mais de um hospedeiro (revisto por Santos et al, 2007)
(Figura 1). Quanto à forma evolutiva, os tripanossomatídeos podem apresentar mais de uma
morfologia celular durante o ciclo de vida, que se distinguem pelo aspecto geral da célula,
presença ou ausência de flagelo extracelular e posição do complexo flagelo - bolsa flagelar -
cinetoplasto em relação ao núcleo (Vickerman & Preston, 1976) (Figura 2).
Dentro da família Trypanosomatidae existem dois gêneros que assumem grande
importância médica, são estes: Trypanosoma, responsável pelas tripanossomíases americana
(Doença de Chagas) e africana (Doença do sono) e Leishmania, responsável pelas
leishmanioses tegumentares e viscerais (Calazar) (Rey, 2008).

2
Figura 1. Ciclos de vida dos gêneros de tripanossomatídeos. No grupo dos heteroxênicos, os
tripanossomatídeos alternam entre hospedeiros invertebrados e vertebrados (setas vermelhas)
ou plantas (setas verdes). O desenvolvimento dos monoxênicos ocorre em um único
hospedeiro invertebrado (seta azul-escuro), embora tripanossomatídeos de insetos possam ser
encontrados em plantas, e relatos recentes descrevem a presença destes tripanosomatídeos em
vertebrados (setas azul-claras). Os gêneros de tripanossomatídeos de insetos que foram
recentemente propostos estão realçados em cinza (adaptado de Santos et al, 2007).

3
Figura 2. Principais formas evolutivas encontradas nos gêneros da família Trypanosomatidae
(adaptado de Sousa, 2000).
promastigota opistomastigota paramastigota epimastigota
tripomastigota coanomastigota amastigota esferomastigota

4
2. Leishmania e Leishmanioses
As leishmanioses humanas compreendem doenças com um largo espectro de
manifestações clínicas, as quais variam desde lesões cutâneas autolimitantes até o
envolvimento visceral severo, podendo levar ao óbito (revisto por Handman and Bullen,
2002). Estima-se que haja uma prevalência de 12 milhões de indivíduos infectados no mundo
e que 2 milhões de novos casos ocorram a cada ano, dos quais 1,5 milhão sejam de
leishmaniose cutânea e 500 mil de leishmaniose visceral. Além desses números alarmantes,
estima-se que cerca de 350 milhões de pessoas encontrem-se em risco de adquirir a infecção
(PAHO/WHO, 2007).
Os agentes responsáveis pelas leishmanioses humanas são protozoários flagelados do
gênero Leishmania, que se caracterizam por apresentarem duas formas evolutivas principais
durante seu ciclo de vida. As formas amastigotas são intracelulares, e encontradas no
hospedeiro mamífero, e as promastigotas, são encontradas no insetor vetor da doença, o
flebotomíneo (Figura 3). Nas infecções do inseto vetor, gênero Phlebotomus no Velho Mundo
(África, Ásia e Europa) e Lutzomyia no Novo Mundo (Américas) (Shaw et al., 1987), podem
ser reconhecidos diferentes estágios da forma promastigota. Cada um destes estágios se
caracteriza por alterações morfológicas e funcionais, destinadas a garantir a sobrevivência do
parasito no flebotomíneo (Kamhawi, 2006).
No hospedeiro mamífero, os promastigotas metacíclicos fazem o primeiro contato com
componentes do sangue e células do sistema fagocítico mononuclear (Dominguez & Torano,
2001). Os promastigotas são rapidamente fagocitados por macrófagos do tecido, sendo
inicialmente mantidos dentro de um compartimento denominado fagossoma, que mais tarde se
funde com o lisossoma para formar o fagolisossoma, onde ocorre a diferenciação do parasito
para as formas amastigotas que residem e se multiplicam dentro do compartimento ácidíco do
fagolisossoma (Cohen-Freuea et al., 2007). Subsequentemente, ocorre a lise da célula
hospedeira com a liberação dos parasitos que podem infectar outros fagócitos adjacentes e por
via hematogênica alcançar tecidos além do ponto inicial da infecção (baço, fígado, medula
óssea ou linfonodos). O ciclo se completa quando um inseto vetor pica o hospedeiro
vertebrado infectado e ingere as formas amastigotas, que então atingem o tubo digestório,
onde novamente se diferenciam em promastigotas (revisto por Handman and Bullen, 2002)
(Figura 3).
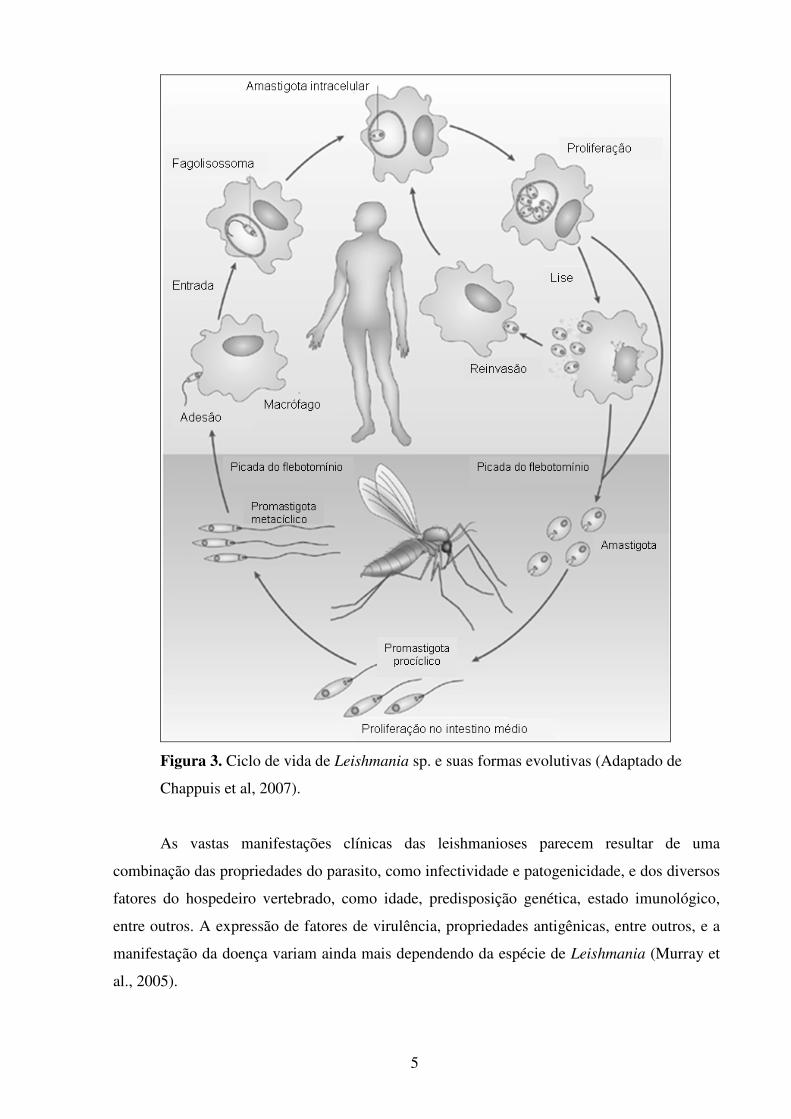
5
Figura 3. Ciclo de vida de Leishmania sp. e suas formas evolutivas (Adaptado de
Chappuis et al, 2007).
As vastas manifestações clínicas das leishmanioses parecem resultar de uma
combinação das propriedades do parasito, como infectividade e patogenicidade, e dos diversos
fatores do hospedeiro vertebrado, como idade, predisposição genética, estado imunológico,
entre outros. A expressão de fatores de virulência, propriedades antigênicas, entre outros, e a
manifestação da doença variam ainda mais dependendo da espécie de Leishmania (Murray et
al., 2005).
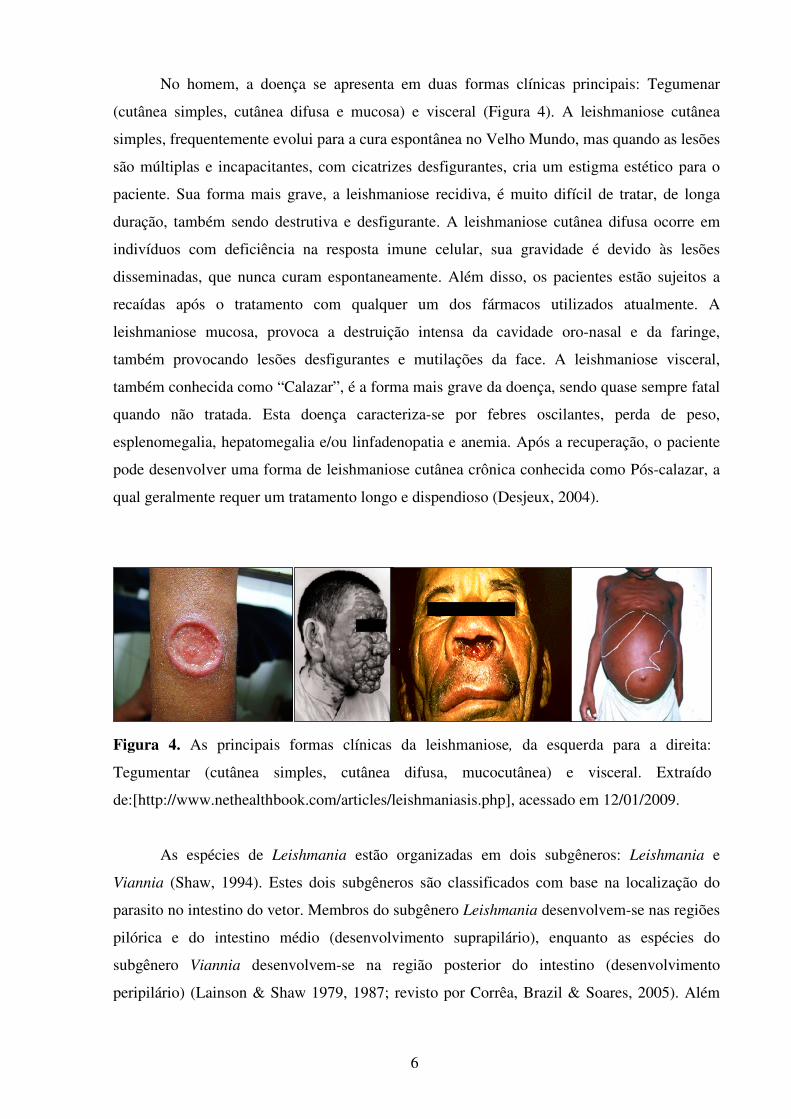
6
No homem, a doença se apresenta em duas formas clínicas principais: Tegumenar
(cutânea simples, cutânea difusa e mucosa) e visceral (Figura 4). A leishmaniose cutânea
simples, frequentemente evolui para a cura espontânea no Velho Mundo, mas quando as lesões
são múltiplas e incapacitantes, com cicatrizes desfigurantes, cria um estigma estético para o
paciente. Sua forma mais grave, a leishmaniose recidiva, é muito difícil de tratar, de longa
duração, também sendo destrutiva e desfigurante. A leishmaniose cutânea difusa ocorre em
indivíduos com deficiência na resposta imune celular, sua gravidade é devido às lesões
disseminadas, que nunca curam espontaneamente. Além disso, os pacientes estão sujeitos a
recaídas após o tratamento com qualquer um dos fármacos utilizados atualmente. A
leishmaniose mucosa, provoca a destruição intensa da cavidade oro-nasal e da faringe,
também provocando lesões desfigurantes e mutilações da face. A leishmaniose visceral,
também conhecida como “Calazar”, é a forma mais grave da doença, sendo quase sempre fatal
quando não tratada. Esta doença caracteriza-se por febres oscilantes, perda de peso,
esplenomegalia, hepatomegalia e/ou linfadenopatia e anemia. Após a recuperação, o paciente
pode desenvolver uma forma de leishmaniose cutânea crônica conhecida como Pós-calazar, a
qual geralmente requer um tratamento longo e dispendioso (Desjeux, 2004).
Figura 4. As principais formas clínicas da leishmaniose, da esquerda para a direita:
Tegumentar (cutânea simples, cutânea difusa, mucocutânea) e visceral. Extraído
de:[http://www.nethealthbook.com/articles/leishmaniasis.php], acessado em 12/01/2009.
As espécies de Leishmania estão organizadas em dois subgêneros: Leishmania e
Viannia (Shaw, 1994). Estes dois subgêneros são classificados com base na localização do
parasito no intestino do vetor. Membros do subgênero Leishmania desenvolvem-se nas regiões
pilórica e do intestino médio (desenvolvimento suprapilário), enquanto as espécies do
subgênero Viannia desenvolvem-se na região posterior do intestino (desenvolvimento
peripilário) (Lainson & Shaw 1979, 1987; revisto por Corrêa, Brazil & Soares, 2005). Além

7
disso, análises de isoenzimas são utilizadas para definir os complexos de espécies dentro de
cada subgênero (Rioux et al., 1990; Zhang et al., 2006).
A classificação das espécies no gênero Leishmania baseou-se inicialmente em critérios
ecobiológicos, tais como vetores, distribuição geográfica, tropismo, propriedades antigênicas e
manifestação clínica. No entanto, análises bioquímicas e moleculares mostraram que a
classificação de espécies baseada apenas nesses critérios nem sempre são satisfatórios para
agrupar os flagelados de acordo com suas similaridades genéticas e, portanto, outros critérios
passaram a ser utilizados para auxiliar a classificação das espécies de Leishmania, tais como
perfil de isoenzimas, sondas de DNA e anticorpos monoclonais espécie-específicos, análises
de RFLP (restriction fragment lenght polymorfism) e perfil de polimorfismo exibido pelo
kDNA (Cupolillo et al., 1998; revisto por Bañuls, Hide & Prugnolle, 2007).
No Brasil, a leishmaniose visceral é causada por Leishmania (L.) chagasi (sinonímia
L. infantum), já a leishmaniose tegumentar americana é causada por diferentes subgêneros e
espécies de Leishmania, sendo as mais importantes no Brasil: Leishmania (L.) amazonensis,
Leishmania (V.) guyanensis e Leishmania (V.) braziliensis (Ministério da Saúde, 2008). O
parasito L. amazonensis, foco deste estudo, é um dos principais causadores da leishmaniose
tegumentar americana em áreas endêmicas (Rangel & Lainson, 2003). A infecção causada por
L. amazonensis pode ser caracterizada por lesões cutâneas múltiplas e progressivas, resistência
à quimioterapia e anergia de células T específicas.
Em 1945, foi relatado pela primeira vez um caso de leishmaniose cutânea difusa
anérgica (LCD) no Brasil (revisto por Silveira, Lainson & Corbett, 2005). A partir de 1948,
autores venezuelanos descreveram alguns casos desse tipo de leishmaniose tegumentar
(revisto por Silva, 1996). Esta forma distinta de leishmaniose tegumentar americana foi
considerada, por muito tempo, uma variação clínica da infecção por L. braziliensis (revisto por
Silveira, Lainson & Corbett, 2005), porém através de estudos posteriores sobre a morfologia,
bioquímica e comportamento do parasito em culturas in vitro e em animais de laboratório, foi
demonstrado que o parasito seria um membro do complexo mexicana, recebendo a
denominação Leishmania mexicana amazonensis, salientando a sua importância médica como
a causa da LCD, uma forma potencialmente incurável da leishmaniose tegumentar americana
(Lainson & Shaw, 1972). Em uma revisão subsequente da classificação das espécies de
Leishmania neotropicais, Lainson & Shaw, em 1987, nomearam o organismo como L.
amazonensis, dentro do subgênero Leishmania (revisto por Silveira, Lainson & Corbett, 2005).
No continente americano, a leishmaniose é causada por diferentes espécies de
Leishmania e a infecção por L. amazonensis, em particular, produz um amplo espectro de
formas clínicas. As manifestações clínicas compreendem desde o envolvimento cutâneo

8
simples, forma mais comum da doença, até a forma cutânea difusa. A Leishmaniose cutânea
difusa (LCD) é uma manifestação rara da doença caracterizada pela presença de nódulos, ao
invés de lesões ulceradas, com abundância de macrófagos parasitados e ausência de resposta
imunológica mediada por células T. No Brasil, a LCD está concentrada nas regiões Norte e
Nordeste, com maior número de casos notificados no Estado do Maranhão. O parasito L.
amazonensis também já foi isolado de pacientes com leishmaniose visceral (LV) e de
pacientes com leishmaniose mucocutânea (LM), entretanto, no Brasil, essas duas
manifestações clínicas são tipicamente causadas por L. chagasi e L. braziliensis,
respectivamente (Oliveira et al., 2007).
A área geográfica onde o flagelado L. amazonensis se encontra estende-se por toda
Bacia Amazônica (compreendendo a parte brasileira, e possivelmente os países vizinhos) bem
como outros territórios, inclusive Maranhão, Ceará, Bahia, Minas Gerais e Espírito Santo,
também existem relatos nas ilhas Trinidad (Antilhas) (Rey, 2008). Descobertas recentes
indicam que este parasito está ampliando sua distribuição geográfica no Brasil, contribuindo
para quadros clínicos incomuns da doença em novas áreas de transmissão (Azeredo-Coutinho
et al., 2007) (Figura 5).
Figura 5. Mapa do Brasil mostrando a distribuição das principais espécies de Leishmania na
transmissão da leishmaniose tegumentar americana em 2005, com a adição de um caso por L.
amazonensis, recentemente descrito no Rio de Janeiro (Ministério da Saúde/Serviço de
Vigilância em Saúde/2007).

9
3. Co-infecção Leishmania-HIV
O surgimento da síndrome da imunodeficiência adquirida (AIDS) levou à
reemergência e, principalmente, ao aumento da incidência de doenças ligadas à
imunossupressão celular, causadas por micobactérias, fungos e protozoários. Inicialmente, as
principais doenças infecciosas relacionadas à aids foram a micobacteriose, histoplasmose,
pneumocistose, toxoplasmose, a criptococose e a candidíase. Em regiões onde a tuberculose
estava controlada, começaram registros de novos casos da doença, bem como relatos de
apresentações atípicas da doença de Chagas e da leishmaniose visceral (Lindoso, 2006).
O primeiro caso de leishmaniose associada à infecção com o vírus da imunodeficiência
humana (HIV) foi relatado em 1985, na Espanha, a partir de então, o número de casos
aumentou rapidamente no Sul da Europa. Atualmente, casos de co-infecção já foram relatados
em trinta e cinco países. No entanto, o impacto real da co-infecção Leishmania-HIV é
provavelmente subestimado em âmbito mundial, devido a deficiências nos sistemas de
vigilância (revisto por Alvar et al., 2008). O fato da leishmaniose não estar incluída entre as
doenças associadas a aids contribui para a escassez de informações. Por esta razão, a partir de
1998, a OMS, em colaboração com o programa para aids das Nações Unidas, criou uma rede
de vigilância ativa para determinar a real dimensão deste problema (revisto por Cruz et al,
2006). Quatro países são os principais envolvidos em casos de co-infecção no Sul da Europa,
são eles: França, Itália, Portugal e Espanha. O maior número de casos foi detectado na
Espanha, o que pode estar relacionado com a reativação de infecções assintomáticas, devido a
uma maior sobreposição geográfica entre a leishmaniose e a infecção pelo HIV, quando
comparada com a de outros países da região. No entanto, infecções recém-adquiridas têm sido
relatadas entre usuários de drogas injetáveis que partilham seringas infectadas. Em função da
sobreposição das duas doenças, casos de co-infecção têm se expandido para outros regiões do
mundo (Figura 6) (Alvar et al., 2008).

10
Figura 6. Mapa com a distribuição da Leishmaniose no mundo ( ) e países que
reportaram a co-infecção Leishmania-HIV ( ) em 2001 (Cruz et al, 2006).
A maioria dos casos de co-infecção Leishmania-HIV é descrita em adultos infectados
por HIV-1. Contudo, também têm sido reportados casos de co-infecção em pacientes HIV-2.
Nestes pacientes, a leishmaniose visceral é causada principalmente por L. infantum e L.
donovani (Alvar et al., 1997) e mais raramente por L. aethiopica (Berhe, Hailu & Gemetchu,
1995). Espécies que causam leishmaniose cutânea como L. braziliensis (Da-Cruz et al., 1992;
Romero, Taranto & Malchiodi, 2004), L. tropica (CDC, 1992; Magill et al., 1993) e L. major
(Gillis et al., 1995) também têm sido descritas como responsáveis por casos de co-infecção, de
acordo com a área geográfica afetada (revisto por Cruz et al., 2006). Várias espécies de
Leishmania dermotrópicas como L. braziliensis, L. mexicana e L. amazonensis têm sido
identificadas como o agente etiológico da leishmaniose visceral, enquanto variantes
viscerotrópicas de L. chagasi têm sido encontradas em lesões cutâneas ou pele saudável de
pacientes HIV positivos (revisto por Alvar et al, 2008).
A maioria dos casos de co-infecção registrados na América do Sul acontece no Brasil,
onde a incidência de aids aumentou de 0,8 casos por 100 mil habitantes em 1986 para 12,3
casos em 2001. Porém, os esforços aplicados na prevenção e tratamento da doença têm
ajudado a manter uma prevalência adulta nacional estável em torno de 0,5% desde 2000. Dos
150.000 casos de leishmaniose cutânea relatados de 2001 a 2005, 150 (0.1%) pacientes tinham
co-infecção com HIV. Neste mesmo período, 16.210 casos de leishmaniose visceral foram
relatados no Brasil, sendo que 315 (2%) eram pacientes co-infectados com HIV (revisto por

11
Alvar et al, 2008). Dos pacientes co-infectados, 53% e 29% dos casos foram relatados nas
regiões nordeste e sudeste, respectivamente. As formas clínicas da leishmaniose encontradas
nos casos de co-infecção são 43% mucocutânea, 37% visceral e 20% cutânea (Rabello, 2003),
padrão clínico que difere daquele encontrado no sul da Europa, onde a leishmaniose visceral
típica representa 87,3% dos casos de co-infecção, os casos atípicos devido a colonizações
intestinais, pulmonares ou outras, respondem por 7,1% dos casos, além da leishmaniose
cutânea, 4,8%, mucocutânea, 0,3% e casos combinados de leishmaniose visceral e cutânea,
0,2% (Desjeux & Alvar, 2003).
Uma vez que o parasito e o vírus se desenvolvem na mesma célula alvo, os
macrófagos, é plausível conceber que uma série de interações aconteça na célula hospedeira.
Nesse contexto, estudos recentes demonstram que a proteína Tat do HIV-1 regula a replicação
em macrófagos humanos tanto de Leishmania como de Blastocrithidia culicis, um
tripanossomatídeo monoxênico, de forma que os protozoários podem multiplicar-se de duas a
três vezes mais rápido, respectivamente, nos macrófagos infectados com HIV-1. A proteína
Tat recombinante é capaz de anular o efeito leishmanicida induzido pelo interferon-gama
(IFN-γ), o que permite a replicação de Leishmania mesmo na presença desta citocina (Barreto-
de-Souza et al., 2006; 2008). Se por um lado o HIV parece facilitar ou promover o
desenvolvimento de leishmaniose, por outro, a co-infecção pode potencializar a progressão
para a aids. Por exemplo, em pacientes co-infectados há uma taxa mais elevada de células T
CD3+ expressando CCR5, uma proteína ligada à membrana que está envolvida na entrada de
HIV-1 em células susceptíveis (Nigro et al., 2007). Além disso, o principal antígeno de
superfície de Leishmania, o LPG, pode induzir a ativação do HIV em monócitos com infecção
latente (revisto por Wolday, 1999). Além disso, foi recentemente relatado que o
estibogluconato de sódio, droga utilizada atualmente no tratamento da leishmaniose, induz o
aumento nos transcritos de HIV-1 e a replicação do vírus em células T CD4 positivas e em
histoculturas de Timo, o que pode trazer implicações clínicas para o tratamento da
leishmaniose em indivíduos infectados pelo HIV-1 (Barat et al., 2006). O parasito também é
capaz de promover o aumento de transcritos de HIV-1, e consequentemente a produção de
vírus, tanto na co-infecção de culturas ex vivo de tecido tonsilar humano, como em culturas de
macrófagos, podendo o aumento da produção de HIV-1 estar ligado à alta produção de
citocinas pro-inflamatórias induzidas pelo protozoário, como TNF-α e IL-1α (Zhao,
Papadopoulou & Tremblay, 2004a; 2004b).

12
4. Tratamento das leishmanioses
As drogas empregadas atualmente no combate das doenças parasitárias surgiram a
partir de programas de seleção empíricos. No entanto, a identificação das diferenças
bioquímicas e metabólicas entre os parasitos e seus hospedeiros mamíferos fornece
indubitavelmente uma alternativa razoável para o desenvolvimento de novos agentes
quimioterapêuticos (Taylor et al., 1994). A primeira linha de medicamentos utilizada no
tratamento da leishmaniose visceral é composta de antimoniais pentavalentes (antimoniato de
N-metilglucamina e gluconato de antimônio e sódio), com os quais é feito o tratamento
padrão. Os antimoniais, que têm sido utilizados no tratamento das leishmanioses desde o início
do século passado, produzem no hospedeiro uma ampla diversidade de efeitos colaterais, que
variam desde fraqueza, náuseas, vômitos, anorexia, tontura, palpitação, insônia, nervosismo,
cefaléia e febre, até artralgias, mialgias, dores abdominais, choque pirogênico, manifestações
clínicas de Herpes Zoster latente, insuficiência renal aguda e arritmias. Além disso, a
administração é por via parenteral, e por longos períodos, e em todas as áreas endêmicas do
mundo tem sido observada a resistência do parasito aos medicamentos (Rey, 2008). Por
exemplo, foi observado que L. donovani evade os efeitos citotóxicos da terapia com
antimoniais pentavalentes, pelo aumento do efluxo da droga através da superexpressão de
proteínas da membrana pertencentes à superfamília de transportadores ABC (“ATP-binding
cassette”) (Singh, 2006). A ausência de resposta ao primeiro tratamento tem sido constatada
em 8% dos casos na Índia, 5% na China e 2% no Quênia (Rey, 2008). A segunda linha de
medicamentos ou terapia alternativa utilizada no tratamento da leishmaniose é composta de
pentamidinas (diamidinas aromáticas) ou anfotericina B, que também apresentam sérios
efeitos colaterais e são de aplicação intravenosa. As pentamidinas destinam-se a tratar os casos
resistentes aos antimoniais e à anfotericina B ou nos casos de recidiva (Rey, 2008).
Outras modalidades de tratamento que não são normalmente empregadas no Brasil, ou
por serem menos eficazes, ou por não estarem disponíveis, ou pelo preço elevado são: a
miltefosina, as formulações lipídicas da anfotericina B e o alopurinol. A miltefosina é uma
alquilfosfocolina desenvolvida originalmente como um quimioterápico antineoplásico, e que é
reconhecida como a primeira droga oral para o tratamento da leishmaniose, com uso
recomendado para leishmaniose visceral na Índia e Etiópia, e para a leishmaniose cutânea na
Colômbia e Bolívia (revisto por Berman, 2008). Esta droga possui poucos efeitos colaterais,
porém seu custo é extremamente elevado. As formulações lipídicas da anfotericina B
apresentam efeitos colaterais mais brandos que a formulação original deste quimioterápico
(Pal, Manadal & Duttagupta, 2001; Sundar, 2001; Singh & Sivakumar, 2004), e o alopurinol

13
apresenta raros efeitos tóxicos, porém é menos ativo quando metabolizado no organismo (Rey,
2008). Recentemente, a resistência a pentamidinas tem sido reportada, bem como as
dificuldades no tratamento de pacientes imunodeprimidos, pela aids por exemplo, nos quais as
drogas convencionais são menos eficazes e doses mais altas e períodos prolongados de
tratamento são necessários (revisto por Santos et al., 2008).
O custo do medicamento é um fator extremamente limitante uma vez que a
leishmaniose é considerada uma doença dos menos favorecidos economicamente: a doença foi
descrita em 88 países, 72 dos quais são países em desenvolvimento e 13 destes são os menos
desenvolvidos do mundo. Apesar dos enormes esforços, é difícil prever os efeitos exatos da
leishmaniose na saúde pública, uma vez que diversos casos não são reportados ou são
diagnosticados erroneamente (WHO, 2006). No Brasil, em casos de co-infecção Leishmania-
HIV o tratamento recomendado é o mesmo utilizado em pacientes imunocompetentes. Os
antimoniais pentavalentes são as drogas de escolha. As alternativas disponíveis na rede pública
para o tratamento das leishmanioses são a anfotericina B e o isotionato de pentamidina. Nos
indivíduos co-infectados a terapia antiretroviral é indicada, mas o início do tratamento anti-
retroviral poderá ser instituído após o início do tratamento para leishmaniose, quando já
houver melhora dos parâmetros clínicos e laboratoriais (Ministério da Saúde, 2007). As co-
infecções com HIV estão aumentando mundialmente, porém não há tratamento e iniciativas
específicas na busca de novas terapias, e o tratamento de formas complexas de co-infecção
Leishmania-HIV ainda é inadequado (Alvar, Croft & Olliaro, 2006).
5. Proteases
As proteases, peptidases ou peptídeo-hidrolases são um grupo amplo e diversificado de
enzimas, de ocorrência geral, que podem ser encontradas nos mais diversos sistemas
biológicos, tais como plantas, animais e microrganismos. Essas enzimas desempenham papéis
fundamentais em quase todos os fenômenos biológicos e compreendem cerca de 2% do total
de proteínas presentes em todos os tipos de organismos. Várias dessas enzimas são de
importância médica, com algumas sendo utilizadas como alvo quimioterápico, tendo a seu
favor anos de análises bioquímicas, o que permitiu a descrição detalhada do seu mecanismo de
catálise, especificidade por substratos e correlação entre estrutura e função. Provavelmente,
sabe-se mais sobre a bioquímica e estrutura das proteases do que de qualquer outra família de
enzimas (McKerrow et al, 2008).
Classicamente, as peptidases são classificadas baseadas em três critérios principais, são
eles: tipo de reação catalisada; natureza química do sítio catalítico e as relações referentes a

14
sequência de aminoácidos e a estrutura da proteína (Barrett, Rawling & O`Brien, 2001). Em
relação a este último critério, o sequenciamento completo do genoma de diversos
microrganismos, levou a uma modernização do sistema de classificação das proteases, pela
necessidade de abranger o diversificado repertório catalítico encontrado na natureza, e de
catalogar e sistematizar a grande quantidade de informação produzida ao longo dos anos,
particularmente na última década (Page & Di Cera, 2008). Dessa forma, foi criado o sistema
MEROPS (http://merops.sanger.ac.uk), que agrupa as enzimas em famílias de acordo com a
homologia na sequência de aminoácidos. Por sua vez, as famílias de mesma origem ancestral
são agrupadas em clãs. Este tipo de informação é determinado pela estrutura terciária das
enzimas (Barrett, Tolle & Rawlings, 2003). Quanto ao tipo de reação catalisada, as proteases
são classificadas de acordo com a posição de clivagem do substrato (Schechter e Berger, 1967;
1968). Por este critério, as proteases são divididas em endoproteases, que degradam ligações
peptídicas internas do substrato, e exoproteases, que degradam ligações peptídicas nas
extremidades amino- ou carboxi- terminal, sendo subdivididas em aminoproteases e
carboxiproteases (revisto por Hooper, 2002). As aminoproteases podem liberar um único
resíduo de aminoácido (aminopeptidase), um dipeptídeo (dipeptidases) ou um tripeptídeo
(tripeptídeo-peptidases). As carboxiproteases atuam no terminal C da cadeia peptídica e
liberam um único aminoácido ou um dipeptídeo (Rao et al., 1998). De acordo com a natureza
química do sítio catalítico, as proteases podem ser também classificadas em seis grandes
classes: aspártico proteases, cisteíno proteases, glutâmico proteases, metalo proteases, serino
proteases e treonino proteases (Figura 7) (Xu et al., 2008).
Figura 7. Esquema de classificação das proteases quanto ao tipo de reação catalisada e a
natureza química do sítio ativo (adaptado de Bond & Butler, 1987).
aminopeptidasesHH22NN
COOHCOOH
dipeptidasesTripeptídeo-peptidases
peptidilpeptidasescarboxiproteases
endoproteases COOHCOOH
dipeptidases
...
HH22NN
COOHCOOH
COOHCOOH...
carboxiproteases
NH2
COOH
NH2
COOH
exoproteasesendoproteases
aminoproteases
serino proteasescisteíno proteasesaspártico proteasesglutâmico proteasestreonino proteasesmetalo proteases
NH2
COOH
NH2
COOH
NH2
COOH
(proteinases)
í
NH2
COOH
Proteases
HH22NN
COOHCOOH
í -
COOHCOOH...
HH22NN
COOHCOOH
COOHCOOH...
NH2
COOH
NH2
COOH
í
NH2
COOH
NH2
COOH
NH2
COOH
í
NH2
COOH
Proteases
aminopeptidasesHH22NN
COOHCOOH
dipeptidasesTripeptídeo-peptidases
peptidilpeptidasescarboxiproteases
endoproteases COOHCOOH
dipeptidases
...
HH22NN
COOHCOOH
COOHCOOH...
carboxiproteases
NH2
COOH
NH2
COOH
exoproteasesendoproteases
aminoproteases
serino proteasescisteíno proteasesaspártico proteasesglutâmico proteasestreonino proteasesmetalo proteases
NH2
COOH
NH2
COOH
NH2
COOH
(proteinases)
í
NH2
COOH
Proteases
HH22NN
COOHCOOH
í -
COOHCOOH...
HH22NN
COOHCOOH
COOHCOOH...
NH2
COOH
NH2
COOH
í
NH2
COOH
NH2
COOH
NH2
COOH
í
NH2
COOH
Proteases

15
Nos tripanossomatídeos, as proteases desempenham um papel fundamental na
interação parasito-hospedeiro, uma vez que, estão envolvidas com a invasão pela degradação
de tecido conjuntivo; com a degradação de proteínas do citoesqueleto durante invasão ou
ruptura das células do hospedeiro; com o escape ou modulação do sistema imunológico por
degradação ou ativação de moléculas do sistema imune; com a invasão de células hospedeiras;
com a intervenção na coagulação sanguínea e com o sistema fibrinolítico do hospedeiro, que
dependem de reações em cascata catalisadas por proteases; e com a variabilidade antigênica do
parasito (revisto por McKerrow et al., 1993; Sajid & McKerrow, 2002; McKerrow, 2006).
Além de serem importantes fatores de virulência nos tripanossomatídeos, as proteases
também estão envolvidas em eventos cruciais do ciclo de vida destes microrganismos, tais
como remodelação do parasito durante a transição de um estágio de vida para outro e ativação
ou degradação de enzimas ou proteínas regulatórias do parasito, além de participarem no
processamento de proteínas para fins nutricionais (revisto por McKerrow et al., 1993; Sajid &
McKerrow, 2002).
6. Proteases em Leishmania
Das seis classes de proteases, as metalo e as cisteína proteases são as mais comumente
detectadas nos tripanossomatídeos (Branquinha et al., 1996; Santos et al., 2005). No entanto,
proteases pertencentes às classes serino, treonina e aspártico também têm sido identificadas
(Burleigh et al., 1997; Paugam et al., 2003; Silva-Lopez & Giovanni-De-Simone, 2004; Alves
et al., 2005; Valdivieso et al., 2007). Em Leishmania spp., dentre os fatores moleculares que
contribuem para a virulência e patogênese dos parasitos, destaca-se uma metalo protease de
60-65 kDa, denominada MSP (major surface peptidade), gp63, leishmanolisina ou PSP
(promastigote surface peptidase), que é a glicoproteína mais abundantemente expressa na
superfície dos promastigotas (Bordier, 1987; revisto por Yao, Donelson & Wilson, 2003).
Esta glicoproteína, ligada à membrana por uma âncora de glicosilfosfatidilinositol
(GPI) (Bordier et al., 1986), é expressa em grande quantidade na superfície da forma
promastigota, infectiva ao inseto (fase logarítmica de crescimento), tem sua expressão
aumentada nas formas promastigotas metacíclicas infectivas ao homem (fase estacionária de
crescimento), e tem um baixo, porém detectável, nível de expressão na forma amastigota
intracelular (Medina-Acosta et al., 1989; Frommel et al., 1990; McGwire & Chang, 1994;
Hsiao et al, 2008). A gp63 tem sido implicada na resistência à lise mediada pelo sistema
complemento, adesão e invasão dos macrófagos, proteção da degradação dentro do
fagolisossomo de macrófagos, e inibição da quimiotaxia de monócitos e neutrófilos (revisto

16
por Yao, Donelson & Wilson, 2003). Mais recentemente, foi descrito que a gp63 promove a
evasão da morte apoptótica mediada por peptídeos antimicrobianos (Kulkarni et al, 2006) e
inibe a proliferação de células NK (Natural Killer) (Lieke at al., 2008).
As espécies de Leishmania, especialmente aquelas que fazem parte do complexo
mexicana, também expressam altos níveis de cisteíno proteases. As cisteíno proteases mais
bem caracterizadas são as similares a catepsina L (CPA e CPB) e as similares a catepsina B
(CPC), que são enzimas encontradas nos megassomas, organelas descritas como grandes
lisossomos, compostos basicamentede de cisteíno proteases, específicas do estágio amastigota
(revisto por McKerrow et al, 2006). Os genes responsáveis pela codificação destas cisteíno
proteases de baixa massa molecular (cpa, cpb e cpc) já foram identificados em diversas
espécies de Leishmania. Através da geração de mutantes nulos para cada um destes genes, foi
demonstrado que o papel da CPB talvez seja facilitar a diferenciação de promastigota para
amastigota e/ou facilitar a evasão do ataque microbicida do macrófago (revisto por Frame,
Mottram & Coombs, 2000; Mottram, Coombs & Alexander, 2004). Além disso, verificou-se
que a CPA não é essencial para a replicação de L. infantum, mas é importante para a interação
parasito-hospedeiro (Denise et al., 2006) e que a remoção dos genes ou inibição das cisteino
proteases CPA e CPB em L. mexicana não somente interfere na via de autofagia que ocorre
durante a diferenciação para promastigotas metacíclicos e amastigotas, como também impede
a metaciclogênese e transformação para amastigotas, fortalecendo a hipótese de que a
autofagia é requerida para a diferenciação celular (Williams et al., 2006).
O nível de expressão ou diversidade de isoenzimas CPB é importante para a virulência
do parasito, o que explica a presença de múltiplas cópias em tandem de genes cpb (Denise et
al., 2003). Outros estudos demonstraram que o início da doença é mais demorado em
linhagens de L. mexicana deficientes para os genes cpa e cpb. Além disso, a comparação
destas linhagens com o parasito selvagem demonstrou a formação de lesões menores, menor
carga parasitária em hamsters, a multiplicação mais lenta das formas promastigotas em
cultura, e uma infectividade menor em células fagocíticas mononucleares humanas, além de
induzirem uma produção de níveis menores das citocinas IL-10 e TGF-ß, que estão associadas
com a resposta do tipo Th2. Estes dados sugerem que o uso de Leishmania viva,
geneticamente atenuada, pode conferir imunidade protetora (Saravia et al., 2006). Além disso,
é valido ressaltar que sequências peptídicas da região COOH-terminal das CPs têm sido
avaliadas como potencial imunorregulador na leishmaniose cutânea (Alves et al., 2001; 2004).
A presença de uma serino protease de 60 kDa foi identificada na superfície celular de
L. (L.) amazonensis, assim como na membrana de compartimentos intracelulares do parasito
(Morgado-Diaz et al., 2005); e ainda, estudos de inibição sugerem a presença de uma outra

17
serino protease extracelular de 56 kDa, inibida por cálcio e manganês e ativada por zinco, que
difere de todas as proteases já conhecidas e caracterizadas deste flagelado (Silva-Lopez,
Coelho & De Simone, 2005). Recentemente, identificou-se a presença de uma serino protease
em uma cepa de L. donovani (Choudhury et al., 2008) com as mesmas características
bioquímicas daquelas identificadas em L. amazonensis (Silva-Lopez, Coelho & De Simone,
2005).
Poucas proteases são reconhecidas como oligopeptidases. Essas enzimas
aparentemente desempenham funções críticas especializadas, como a modificação e destruição
de peptídeos mensageiros (Andrade et al., 1998). As serino oligopeptidases do grupo das prolil
peptidases clivam, na sua grande maioria, peptídeos após resíduos de prolina. Um subgrupo
menor dessa família cliva substratos no lado carboxila de resíduos de aminoácidos básicos, e
esta subfamília é denominada serino oligopeptidase B. Proteases pertencentes a este subgrupo
foram identificadas em L. major e L. amazonensis, e nos tripanossomas: Trypanosoma brucei,
Trypanosoma cruzi, Trypanosoma evansi e Trypanosoma congolense (Andrade et al., 1998;
Morty et al., 1999, 2005; revisto por Coetzer, Goldringa & Husona, 2008).
As aspártico proteases, foco deste estudo, estão divididas em várias famílias e
geralmente funcionam em condições ácidas. Este último aspecto limita a função das aspártico
proteases a alguns locais específicos nos diferentes organismos, o que faz com que a
ocorrência desta classe proteolítica seja menos abundante do que a de outros grupos de
proteases (Dash et al., 2003). Apesar disso, as aspártico proteases têm sido isoladas e
estudadas em uma ampla gama de organismos, desde vertebrados até plantas, fungos,
parasitos, retrovírus, e mais recentemente em bactérias (Hill & Phylip, 1997; James, 1998;
Dash et al., 2003; Takahashi et al., 2006; Alves et al., 2005; Valdivieso et al., 2007; Pinti et
al., 2007). Nos ultimos anos, as aspártico proteases têm recebido atenção como alvos para
intervenção farmacêutica visto a sua participação em vários processos fisiológicos e
patológicos importantes (Eder et al., 2007). Os principais exemplos são: o envolvimento da
renina na hipertensão, da catepsina D na metástase do câncer de mama e da protease do HIV
na AIDS (Dash et al., 2003).
As aspártico proteases ainda não estão bem caracterizadas na família
Trypanosomatidae. Em Leishmania major, essas enzimas parecem processar peptídeos de 28
kDa em lisossomas e contribuir para a degradação de cadeias invariáveis em células
apresentadoras de antígenos de ratos infectados (Zhang et al., 2000). Além disso, a atividade
de aspártico protease diminui durante a transformação de promastigotas para amastigotas em
L. amazonensis (Alves et al., 2005). Esta atividade enzimática também já foi descrita em
frações solúveis de promastigotas de Leishmania mexicana, sendo capaz de degradar um

18
substrato sintético específico para aspártico proteases do tipo catepsina D (Valdivieso et al.,
2007).
As proteases dos parasitos são de considerável interesse para pesquisa porque podem
proporcionar informações sobre a bioquímica e a biologia do parasito, assim como esclarecer
algumas facetas da interação parasito-hospedeiro. Uma vez que essas enzimas desempenham
uma série de funções críticas e essenciais para os parasitos, as peptidases são alvo para a
produção de agentes quimioterápicos (revisto por McKerrow, 1999; Vendeville et al., 2002;
Bal et al., 2003).
7. Inibidores proteolíticos como quimioterápicos
O desenvolvimento de inibidores efetivos e a descoberta de seu mecanismo de ação
podem ter influência significativa sobre estratégias terapêuticas. Desta forma, inibidores
proteolíticos têm surgido como drogas alternativas para o uso na quimioterapia de diversos
processos patológicos, como: câncer (Shaw et al., 1990; Craik et al., 1995; Zhou et al., 2002),
desordens inflamatórias (como artrite reumatóide), respiratórias (como asma) (Tanaka et al.,
1995; Fath et al., 1998), cardiovasculares (como hipertensão) (Kleinert et al., 1992; Hoover et
al., 1995; Simoneau et al., 1998), e neurodegenerativas (como mal de Alzheimer) (Vassar et
al., 1999; Hong et al., 2002). Os inibidores da aspártico protease do HIV (IPs-HIV) na
quimioterapia da AIDS constituem, indubtavelmente, o exemplo de maior impacto e sucesso
da abordagem de inibição da atividade proteolítica como forma de controlar um patógeno.
Além dos IPs-HIV empregados no tratamento da aids, outros inibidores de aspártico proteases
têm sido explorados como possíveis agentes terapêuticos, como os inibidores da plasmepsina,
para o tratamento da malária, e inibidores das SAPs (secreted aspartic peptidases), para o
tratamento da candidíase (revisto por Dash et al., 2003). Inibidores peptideomiméticos vêm
sendo desenvolvidos e testados quanto a inibição de SAPs purificadas de diferentes espécies
de Candida (Pichová et al., 2001; Majer et al., 2006). Já a plasmepsina IV, tem sido estudada
como uma enzima crítica para o desenvolvimento de inibidores para o tratamento da malária,
pelo fato de ter homologia na sequência de aminoácidos entre as quatro plasmepsinas
responsáveis por degradar a hemoglobina (Nguyen et al., 2008). Estudos de triagem, síntese e
avaliação têm identificado derivados não peptídicos que são capazes de matar os parasitos in
vitro através da inibição de proteases (Sharma, 2007).
Além dos inibidores de aspártico proteases, outras classes de inibidores, como por
exemplo, os inibidores de cisteíno proteases têm mostrado sua eficácia na terapêutica
experimental de algumas doenças causadas por tripanossomatídeos. Recentemente, foi

19
reportado que um inibidor da cruzipaína, denominado K777, é capaz de proteger cachorros de
danos no coração durante a infecção por T. cruzi (Barr et al., 2005) e que inibidores de cisteíno
proteases induzem a regressão no tamanho da lesão quando administrados em camundongos
previamente desafiados com L. major (Selzer et al., 1999). Informações estruturais sobre o
sítio ativo do receptor (ou protease) e seleção de moléculas concebidas com ferramentas de
bioinformática contribuem para a concepção de inibidores potentes e seletivos. A química
combinatória possibilita tanto a descoberta de novas moléculas para serem testadas (Dash et
al., 2003).
8. Inibidores de Protease do HIV
No HIV-1, como em todos os retrovírus, a produção da doença inevitavelmente requer
uma partícula viral com uma protease ativa. A protease do HIV é uma enzima pertencente à
classe das aspártico proteases, que consiste em uma proteína de 99 aminoácidos. No ciclo de
replicação do HIV-1, uma proteína precursora ou poliproteína é sintetizada. Esta poliproteína é
composta de proteínas estruturais (proteínas Gag) e enzimas, incluindo a transcriptase reversa,
a integrase, e a própria protease. A clivagem da poliproteína é necessária para a formação de
partículas infecciosas, na fase final do ciclo. As células contendo o DNA proviral do HIV-1
que não produzem uma protease funcional produzem partículas virais, porém estas são
imaturas e não infecciosas (Figura 8) (revisto por Eron, 2000).
A família das aspártico proteases possui dois grupos β-carboxi aspartil no sítio ativo.
Estes grupos são responsáveis por catalisar a hidrólise das ligações peptídicas. Um resíduo
aspartil auxilia na adição de uma molécula de água à carbonila da amida do substrato,
formando um intermediário tetraédrico que possui grande afinidade pelo sítio catalítico. Após
a formação deste intermediário, ocorre a quebra da ligação C-N com a formação de um ácido
carboxílico e de uma amina primária. Substâncias químicas que mimetizam este intermediário
tetraédrico impedem a hidrólise do substrato e, consequentemente, o ciclo do vírus HIV é
interrompido (Cunico, Gomes & Vellasco Junior, 2008).
Atualmente existem dez drogas disponíveis comercialmente que tem como alvo a
protease do HIV, nove delas representam estórias de sucesso no desenvolvimento racional de
moléculas peptídeo miméticas, ou seja, inibidores que se baseiam na homologia de sequências
de sítios de clivagem do substrato na poliproteína viral. O único inibidor não peptídeo
mimético da protease do HIV é o Tipranavir, o qual não possui qualquer semelhança com
peptídeos de substratos da enzima e que foi descoberto com a otimização da ligação de 4-
hidroxicomarina e 4-hidroxi-2-pirona (Tsantrizos, 2008).

20
A introdução da terapia antiretroviral de alta potência (TARV), no final de 1995 e
início de 1996, nos Estados Unidos e Oeste da Europa, mudou drasticamente o curso da
infecção pelo HIV. Esta terapia caracteriza-se pela introdução de inibidores proteolíticos ao
coquetel de tratamento pré-existente, composto por inibidores da transcriptase reversa
(análogos e não-análogos de nucleotídeos), inibidores de fusão e inibidores de entrada
(AIDSinfo, 2007). Este coquetel envolve o uso de pelo menos três agentes de duas classes
distintas de antiretrovirais: um inibidor de protease em combinação com dois inibidores de
transcriptase reversa nucleotídicos ou um inibidor de transcriptase reversa não nucleotídico
(Mastrolorenzo et al., 2007). A primeira geração de inibidores de protease do HIV (IPs-HIV)
desenvolvida compreende as drogas: saquinavir, ritonavir, indinavir, nelfinavir e amprenavir, e
a segunda geração: fosamprenavir (uma prodroga do amprenavir), lopinavir, atazanavir,
tipranavir e darunavir (Figura 9) (Mastrolorenzo et al., 2007; Hughes, Barbera & Nelsona,
2008; FDA, 2008).
O Saquinavir foi o primeiro IP-HIV a ser licenciado, em dezembro de 1995, para
terapêutica do HIV. A droga funciona simulando o estado de transição da protease do HIV e
foi desenvolvido antes dos dados de cristalografia da protease do vírus estarem disponíveis.
Apesar de representar um avanço importante até então, sua baixa biodisponibilidade oral
mostrou-se desvantajosa e a resistência ao composto foi logo identificada. A partir deste fato,
as pesquisas se concentraram na descoberta de agentes com maior potência e melhor
biodisponibilidade oral. O desenvolvimento dos IPs-HIV passou então a ser baseado nos dados
fornecidos pela estrutura terciária da peptidase, obtida pela cristalografia de raios-X (Figura
10). Dentre eles incluí-se o indinavir, ritonavir, nelfinavir, lopinavir e amprenavir.
Posteriormente, os esforços foram empenhados no desenvolvimento de drogas com menor
impacto sobre o metabolismo lipídico e maior atividade contra variantes resistentes às drogas.
Estes esforços conduziram à descoberta do atazanavir, tipranavir e darunavir. O amprenavir
foi modificado para dar origem ao fosamprenavir, que possui uma melhor absorção após a
administração oral (revisto por Martinez-Cajasa & Wainberg, 2007).
O composto ritonavir é usado, principalmente como um impulsionador em associação
com outros IPs. Os inibidores de protease potencializados com ritonavir (IP/r) oferecem maior
barreira genética à resistência do que os inibidores de transcriptase reversa análogos de
nucleosídeo. Fosamprenavir, atazanavir e lopinavir em associação com ritonavir têm sido
recomendados como esquema inicial da terapia antiretroviral para pacientes não submetidos a
tratamentos anteriores (AIDSinfo, 2008).

21
Figura 8. Esquema resumido do ciclo da replicação viral com as etapas em que atuam os
medicamentos disponíveis no mercado (Souza & Almeida, 2003).
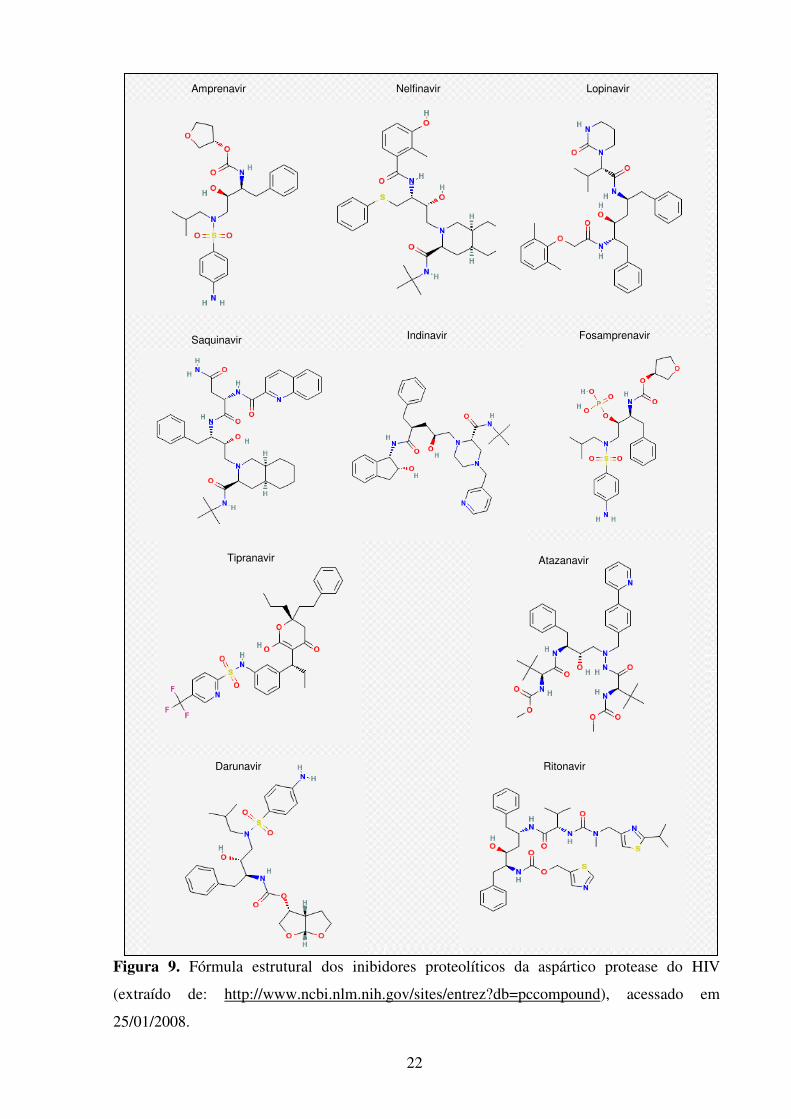
22
Figura 9. Fórmula estrutural dos inibidores proteolíticos da aspártico protease do HIV
(extraído de: http://www.ncbi.nlm.nih.gov/sites/entrez?db=pccompound), acessado em
25/01/2008.
Nelfinavir
Saquinavir Indinavir Fosamprenavir
Darunavir
Atazanavir
Ritonavir
Tipranavir
Amprenavir Lopinavir

23
Figura 10. Esquema mostrando a estrutura tridimensional da protease do HIV-1. Em A: as
duas subunidades iguais da protease (em azul e verde), os domínios da enzima (setas brancas)
e seu sítio ativo; Em B, os dois resíduos aspartil (em vermelho) (extraído de:
http://www.callutheran.edu/Academic_Programs/Departments/BioDev/omm/hiv_protease/mol
mast.htm, acessado em 02/02/2009.
9. Ação dos Inibidores de Protease do HIV sobre patógenos oportunistas
Nos pacientes com aids, a incidência, morbidade e mortalidade das co-infecções virais,
bacterianas, fúngicas e parasitárias diminuiu drasticamente após o início da TARV (Terapia
Antiretroviral), por volta de 1996 (Palella et al., 1998; Pozio & Morales, 2005). Embora na
maioria dos casos, estas melhorias tenham sido atribuídas à recuperação da imunidade do
hospedeiro, melhorias nas infecções oportunistas foram demonstradas, mesmo na ausência de
recuperação imunológica (Mele et al., 2003).
Hoegl e coloboradores (1998) publicaram um caso de um paciente que recebia a terapia
TARV e se recuperou da infecção por Candida albicans, mesmo mantendo uma baixa
contagem de celulas CD4+, sugerindo o efeito direto da terapia sobre este patógeno. Em outro
estudo, um paciente HIV positivo com criptosporidiose, e contagem de células CD4+ em torno
de 33 células/µl (paciente saudável: 800 a 1500 células/µl), rapidamente recuperou-se da
infecção após o início da terapia antiretroviral, na qual indinavir foi incluído, apesar da
contagem de células CD4+ ter aumentado somente para 84 células/µl (revisto por Pozio &
Morales, 2005). Estas observações levaram à hipótese de que os IPs-HIV teriam um efeito
direto sobre as infecções oportunistas, o que já foi demonstrado para Pneumocystis carinii,
Candida albicans, Cryptosporidium parvum, Toxoplasma gondii, Plasmodium falciparum
(revisto por Pozio & Morales, 2005), Fonsecaeae pedrosoi (Palmeira et al., 2008), Leishmania
major e Leishmania infantum (Savoia et al, 2005; Trudel et al., 2008).
Sítio ativo
Bordas (flaps)
interdigitações entre folhas beta
N e C terminaisResíduos aspartil
Sítio ativo
Bordas (flaps)
interdigitações entre folhas beta
N e C terminaisResíduos aspartilA B

24
A possível eficácia dos IPs-HIV contra infecções oportunistas pode ser explicada por
sua ação antinflamatória (Tovo, 2000), sua capacidade de bloquear o proteassoma celular
(Andre et al., 1998) e promover apoptose (Gaedicke et al., 2002). Além disso, tem sido
demonstrado um efeito sinérgico entre as drogas rotineiramente utilizadas contra as infecções
oportunistas e os IPs-HIV (Hommer, Eichholz & Petry, 2003; Casolari et al., 2004). Um
exemplo de aplicação prática da sinergia entre as diferentes drogas é a diminuição das doses
dos antimicóticos, que possuem muitos efeitos colaterais, no tratamento de infecções fúngicas
em indivíduos HIV positivos (Casolari et al., 2004).
Estudos indicam que o sucesso da resposta terapêutica na co-infecção Leishmania-HIV
está inversamente relacionada à carga viral anterior ao tratamento (Berhe et al., 1999). Outro
fato que também parece claro é que os pacientes que não recebem o TARV têm mais
possibilidades de desenvolver a patologia da leishmaniose e mostram um maior risco de falha
terapêutica no tratamento da AIDS, assim como de recaídas clínicas e parasitológicas (Alvar et
al., 1997, 2008; Pintado & López-Vélez, 2001; Pintado et al., 2001; Cruz, 2006). Além disso,
na ausência da TARV, a taxa de letalidade durante o tratamento da leishmaniose visceral era
de 24% (Alvar et al., 2008).
Embora a prevalência da leishmaniose visceral tenha diminuído desde a introdução do
TARV, tem havido um aumento na taxa de reativação da doença, o que pode ser explicado
pelo aumento da sobrevida induzida pelo tratamento. Desta forma, casos em que o TARV não
conseguiu impedir a LV também têm sido descritos. Na Espanha, 17 indivíduos HIV-positivos
em TARV com dois inibidores nucleosídicos da transcriptase reversa e um inibidor de
protease, experimentaram um primeiro episódio de LV, e cinco deles tiveram uma ou mais
recidivas. Uma pessoa HIV-positiva em TARV com o inibidor de protease indinavir, e com
uma contagem de células T CD4 + de 422 células µl-1, sofreu dois surtos de leishmaniose
cutânea causados por L. infantum cinco anos após um episódio de LV, que havia sido tratado
com anfotericina B por um ano e esplenectomia. Em uma mulher HIV-positiva com histórico
de LV, um episódio de leishmaniose cutânea pós-calazar foi documentado um ano após o
início do TARV, na qual foi incluído o indinavir. Um estudo realizado na Espanha entre os
indivíduos com infecção por L. infantum mostrou que, embora a contagem de células T CD4 +
durante episódios clínicos da LV fossem significativamente menores em indivíduos que não
recebem TARV, em comparação com aqueles que receberam o tratamento, ambos os grupos
tinham contagens de células T CD4 + inferiores a 200 células µl-1 (Revisto por Pozio e
Morales, 2005).

25
II. Justificativa
A quimioterapia utilizada no tratamento da leishmaniose é de baixa eficácia, pois os
fármacos geram severos efeitos colaterais, desde fraqueza até insuficiência renal aguda e
arritmias, principalmente devido à similaridade estrutural entre os microrganismos causadores
da doença e células humanas. As enzimas proteolíticas participam em processos biológicos
normais, e também são importantes fatores no desenvolvimento de inúmeras doenças humanas
causadas por parasitas, inclusive a leishmaniose. Inibidores de protease do HIV têm se
mostrado eficazes na diminuição da incidência, morbidade e mortalidade em pacientes co-
infectados com Leishmania e HIV. Neste contexto, os IPs-HIV, disponíveis comercialmente,
podem ser avaliados com o objetivo de se obter uma quimioterapia com curta duração, de
administração oral, mais potente e com baixo custo para o tratamento da leishmaniose.

26
III. Objetivo geral
Os objetivos deste trabalho são determinar os efeitos dos inibidores de aspártico
protease do HIV sobre a taxa de multiplicação, a diferenciação, a ultraestrutura celular e o
processo infeccioso em Leishmania amazonensis, associado com a inibição pelos IPs-HIV da
atividade das aspártico peptidases.
1. Objetivos específicos
1.1. Avaliar o efeito dos IPs-HIV sobre a taxa de multiplicação das formas promastigotas de
Leishmania amazonensis;
1.2. Determinar o IC50 das drogas que influenciam a taxa de multiplicação do parasito;
1.3. Determinar a influência dos IPs-HIV sobre a diferenciação do parasito;
1.4. Verificar o efeito dos IPs-HIV sobre a atividade enzimática de aspártico proteases no
extrato bruto das formas promastigotas de L. amazonensis;
1.5. Verificar possíveis alterações ultraestruturais nas formas promastigotas tratadas com os
IPs-HIV;
1.6. Determinar o efeito do pré- e pós-tratamento com os IPs-HIV sobre a interação com
macrófagos;
1.7. Determinar o efeito dos IPs-HIV sobre o perfil de proteases através de zimografia e
immunoblotting.

27
IV. Metodologia
1. Materiais
Os inibidores de protease do HIV (IPs-HIV) foram adquiridos das seguintes empresas:
Hoffmann-La Roche AG (Grenzach-Wyhlen, Germany) (Saquinavir e Nelfinavir), Merck
Sharp & Dohme GmbH (Haar, Germany) (Indinavir), Abbott Laboratories (Abbott Park, IL,
USA) (Lopinavir) e GlaxoSmithKline (NC, USA) (Amprenavir). O anticorpo monoclonal
anti-α-tubulina, o anticorpo secundário anti-IgG de coelho conjugado a peroxidase, soro
albumina bovina (BSA), dimetilsulfóxido (DMSO), soro fetal bovino inativado (SFB),
brometo de 3[4,5-dimetiltiazol-2-il]-2,5-difeniltetrazolio (MTT), ditiotreitol (DTT), iodeto de
propídeo e meio Schneider foram obtidos da Sigma Chemical Co. (St Louis, USA).
Constituintes de meio, componentes de tampão, reagentes usados em eletroforese e
imunoblotting foram obtidos da Bio-RAD (Califórnia, USA). O anticorpo policlonal anti-cpb,
produzido contra cisteíno protease de Leishmania mexicana, foi cedido pelo Dr. Mary Wilson
(Department of Internal Medicine, Biochemistry, Microbiology and Epidemiology, Program in
Molecular Biology, University of Iowa, USA), e o anti-gp63, produzido contra gp63 de L.
mexicana, foi cedido pelo Dr. Peter Overath (Max-Planck-Institut für Biologie, Abteilung
Membranbiochemie, Germany).
2. Cepa utilizada e condições de cultivo
Os promastigotas de Leishmania (L.) amazonensis, cepa MHOM/BR/77/LTB0016,
foram obtidos da Coleção de Leishmaniose (Fundação Oswaldo Cruz, Rio de Janeiro, RJ,
Brasil), e mantidos através de repiques semanais em garrafas de cultura de 25cm2 contendo
meio Schneider pH 7,2, suplementado com 10% de SFB a 26 ºC até o crescimento do parasito
(CEUA L-0006/07).
3. Obtenção de isolados infectivos
Para a manutenção da infectividade, a cepa de Leishmania amazonensis foi inoculada
em fêmeas de camundongos da linhagem BALB/c. Aproximadamente 1 × 107 parasitos,
obtidos da fase estacionária de crescimento, foram lavados, resssuspensos em tampão PBS
(tampão fosfato 20 mM, 150 mM NaCl, pH 7.2) estéril, e inoculados nas patas traseiras dos
camundongos. A cada 15 dias, o tamanho da pata foi aferido, com o auxílio de um paquímetro

28
de precisão de décimo de milímetro, para o controle da infecção. Cerca de 3 meses após o
inóculo, os animais foram sacrificados e os tecidos das patas foram macerados em meio
DMEM acrescido de 200 UI de penicilina e 200 µg/ml de estreptomicina). O produto deste
macerado foi inoculado em garrafas de cultura contendo 10 ml de meio Schneider,
suplementado com 20% de SFB e mantidos a 26°C até o crescimento dos parasitos.
4. Ensaio de inibição da multiplicação
Foi analisado o efeito sobre a taxa de multiplicação das formas promastigotas de L.
amazonensis de cinco inibidores distintos de aspártico protease do HIV (amprenavir, indinavir,
lopinavir, nelfinavir e saquinavir) e de um inibidor clássico de aspártico proteases (pepstatina
A). Os promastigotas foram contados em câmara de Neubauer e ressuspensos em meio
Schneider a uma concentração final de 5 × 107 promastigotas viáveis/ml. A viabilidade foi
verificada pela motilidade e exclusão do corante vital azul de tripan. O inibidor foi adicionado
à cultura em concentrações finais que variaram de 15 a 500 µM (partindo de uma solução
estoque de 20 mM em DMSO diluída seriadamente no meio de cultura). Diluições do DMSO
correspondentes às utilizadas na preparação das drogas serviram como controle. Após 24, 48,
72 e 96 horas de incubação a 26 ºC, o número de promastigotas viáveis foi quantificado por
contagem dos parasitos em câmara de Neubauer. A viabilidade foi verificada pela motilidade e
exclusão do corante vital azul de tripan. O IC50 (concentração mínima da droga capaz de
causar uma redução de 50% na sobrevivência/viabilidade do parasito em comparação com as
culturas sem o inibidor) foi determinado após 48 horas para os inibidores mais efetivos. Esses
valores foram determinados por análises de regressão linear do número de promastigotas
viáveis versus o log da concentração da droga, empregando o programa Sigma Plot para
windows versão 8.0 SPSS Inc, copyright 1986-2001.
5. Ensaio de inibição de diferenciação dos parasitos de promastigota para
amastigota in vitro
Promastigotas recém isoladas de infecção experimental em camundongos BALB/c e
criopreservadas foram recuperadas e cultivadas em meio Schneider pH 7,2 (suplementado com
10% de SFB, 200 UI de penicilina, 200 µg /ml de estreptomicina) e mantidos a temperatura de
26 ºC por quatro dias, para padronização da curva de crescimento. A diferenciação de
promastigota para amastigota foi induzida pela mimetização in vitro do ambiente do
hospedeiro vertebrado, através da acidificação do meio e aumento da temperatura. Para tal,

29
1,0x107 promastigotas/ml, obtidas de fase estacionária de crescimento, foram incubadas em
meio Schneider pH 5,5, suplementado com 20% de SFB, a 32 ºC por 24-96 h (Alves et al.,
2005). Os parasitos foram incubados na ausência ou presença de nelfinavir, amprenavir e
lopinavir nas concentrações equivalentes ao IC50 e a porcentagem de formas amastigotas-
promastigotas foi determinada por observação microscópica.
6. Microscopia eletrônica de transmissão
Os promastigotas (1 × 108) foram incubados na presença ou ausência de nelfinavir e
lopinavir na concentração do IC50 por 2, 4, 6, 8, 12 e 24 horas e então fixadas por 12 horas à 4
ºC em tampão cacodilato 0,1 M (Sigma) com 2,5% de glutaraldeído (Sigma), pH 7,2. Após a
fixação, as células foram lavadas em tampão cacodilato e pós-fixadas por 1 hora com tampão
cacodilato contendo tetróxido de ósmio 1%, ferrocianeto de potássio 0,8% e CaCl2 5 mM
(Sigma). As células foram então lavadas no mesmo tampão e desidratadas em concentrações
crescentes de acetona (50%, 70%, 90% e duas vezes 100%). Em seguida, o material foi
infiltrado por aproximadamente 12 horas em resina epóxi (Epon) diluída em acetona 1:1. Após
este período, o material foi infiltrado em resina Epon pura, onde permaneceu por cerca de 6
horas e depois foi incluído em resina Epon pura em poços de placa específica para inclusão,
onde ocorreu a polimerização a 60 ºC em forno (Biomatic) por 48 horas. Cortes ultrafinos
foram montados sobre grades de cobre de 300 mesh, coradas com acetato de uranila 5% e
citrato de chumbo e então observadas em microscópio eletrônico de transmissão Zeiss 900
(Zeiss, Oberkochen, Germany).
7. Avaliação da atividade enzimática através de dosagem proteolítica
O extrato do parasito foi obtido de acordo com metodologia descrita previamente (Alves
et al., 2005). Os promastigotas (1 × 109) foram centrifugados e lavados 3 vezes com PBS pH
7,2. As células foram ressuspensas em 1 ml de Tris–HCl 10 mM, pH 6,8, contendo 1% de
CHAPS (3-[(3-colamidopropil)-dimetilamonio]-1-propanosulfonato) e submetidas a ciclos
repetidos de congelamento e descongelamento. Os extratos foram mantidos por 40 minutos
em freezer e então centrifugados por 30 minutos, 30.000× g a 4 °C. O sobrenadante obtido foi
aliquotado, mantido em freezer a -70 ºC por até 4 dias. A concentração de proteínas foi
determinada de acordo com o método de Lowry et al., (1951), usando BSA como padrão. As
leituras foram feitas em cubeta de quartzo no espectrofotômetro (Ultrospec 1100 pro;
Amersham Biosciences, UK) com o comprimento de onda de 617 nm.

30
Para o ensaio de atividade enzimática, cerca de 10 µg do extrato bruto dos parasitos
foram incubados (37 °C, 60 minutos) com 2 µM de substrato fluorogênico específico para
aspártico protease do HIV-1 (Arg-Glu(EDANS)-Ser-Gln-Asn-Tyr-Pro-Ile-Val-Gln-
Lys(DABCYL)-Arg e para Renina (Arg-Glu(EDANS)-Ile-His-Pro-Phe-His-Leu-Val-Ile-His-
Thr-Lys(DABCYL)-Arg) em um volume final de 60 µl, em tampão de ativação (acetato de
sódio 0.1 M, cloreto de sódio 1 M, EDTA 1mM , DTT 1 mM, DMSO 10%, BSA 1mg/ml, pH
4,7) ( Matayoshi et al., 1990). Para os ensaios de inibição, o extrato bruto foi pré-incubado (25
°C, 5 minutos) com os inibidores de aspártico protease do HIV (nelfinavir e amprenavir 1 e 10
µM, lopinavir 0,1; 1 e 10 µM) e um inibidor clássico de aspártico proteases (Pepstatina A 1
µM). A variação da fluorescência foi monitorada durante 60 minutos e a velocidade da reação
calculada usando a fórmula v = [s – so]/(t - to), onde v = velocidade, [s - so] = concentração de
substrato final subtraída da concentração de substrato inicial e (t - to) = tempo final subtraído
do tempo inicial. Os ensaios foram controlados verificando a auto-liberação do conjugado
fluorogênico nos mesmos intervalos de tempo. A atividade enzimática foi expressa em
fluorescência do produto gerado (EDANS) por minuto por miligrama de proteína (abs min -1
mg ptn -1). A fluorescência foi medida continuamente em fluorímetro (SpectraMax Gemini
xps, Molecular Devices, CA, USA) a 340 nm de excitação e 490 nm de emissão.
8. Ensaio de viabilidade do parasito
Após a contagem dos parasitos em câmara de Neubauer o número de células foi
ajustado para uma concentração final de 1 × 106 em 100 µl na presença ou ausência dos IPs-
HIV em concentrações que variaram de 50 a 250 µM. As células foram incubadas por 1 a 4
horas, centrifugadas, lavadas duas vezes com PBS e incubadas durante 15 minutos em uma
solução contendo 3 mM de iodeto de propídeo diluído em tampão de marcação (Tris 100 mM,
pH 7,4, NaCl 150 mM, 1 mM de CaCl2, 0,5 mM MgCl2). Para controle de morte celular, uma
alíquota dos parasitos foi aquecida a 100 oC por 5 minutos, e tratada com a solução de iodeto
de propídeo. A detecção da viabilidade foi determinada em citômetro de fluxo (FACS Calibur,
Becton Dickinson Immunocytometry System, San Jose, CA, USA), empregando o software
Cell Quest iPro (BD Immunocytometry. Systems, San Jose, CA). As populações celulares
analisadas foram reconhecidas por meio das propriedades de espalhamentos frontais – FSC – e
dos espalhamentos laterais – SSC das mesmas. A fluorescência vermelha do iodeto de
propídeo foi mensurada a 585±42nm (FL2).

31
9. Avaliação do perfil de proteases por SDS-PAGE com gelatina e por
immunoblotting
Os promastigotas (1 × 108 células) foram incubados com os IPs-HIV por 4 horas a 26
ºC na concentração de 100 µM. Os parasitos tratados mantiveram mais de 90% das células
viáveis, de acordo com a metodologia descrita acima. As formas promastigotas tratadas ou não
com os IPs-HIV foram lisadas pela adição de tampão de amostra para SDS-PAGE (Tris 125
mM, pH 6,8, SDS 4%, glicerol 20% e azul de bromofenol 2%). Em cada “slot” foi aplicado o
extrato do parasito correspondente a 5 x 106 células e a eletroforese (cuba de eletroforese
vertical, Bio Rad) foi realizada a 60 mA, 4 ºC por 2 horas. Após a eletroforese, o SDS foi
removido por incubação com Triton X-100 2,5 % por 1 hora. Subsequentemente, os géis
foram incubados em tampão fosfato de sódio 50 mM, pH 5,5, suplementado com DTT 2 mM a
37 ºC. Após incubação por 48 horas, os géis foram lavados 2 vezes com água destilada,
corados por 2 horas com azul de comassie R-250 em metanol-ácido acético-água (50:10:40), e
descorados “overnight” em solução contendo metanol-ácido acético-água (5:10:85). Os géis
foram secos, fotografados e a densitometria dos halos de degradação foi realizada empregando
o programa Scion Image (Scion Corporation, base don NIH for MacIntosh by Wayne
Rasbanol, NIH, USA).
Alternativamente, o extrato dos parasitos obtido como descrito acima foi adicionado de
β-mercaptoetanol 10%, seguido de desnaturação a 100 ºC por 5 minutos, e as proteínas foram
separadas eletroforeticamente (SDS-PAGE 12%), e posteriormente transferidas a 4 ºC, 100
V/300mA por 2 horas para membranas de nitrocelulose (0,22 µm). A membrana foi bloqueada
em solução de 5% de leite em pó desnatado diluído em PBS contendo Tween 20 0,5% por 1
hora em temperatura ambiente. As membranas foram então lavadas 3 vezes (10 minutos cada)
com solução de lavagem (5% de leite em pó desnatado diluído em PBS) e incubadas por 2
horas com os seguintes anticorpos policlonais: anti-cpb (1:1000), anti-gp63 (1:2000) e o
anticorpo monoclonal anti-α-tubulina (1:500). O anticorpo secundário utilizado foi o anti-IgG
de coelho conjugado a peroxidase (1:20000) seguido da imunodetecção por
quimioluminescência. A massa molecular relativa dos polipeptídeos reativos foi calculada por
comparação com padrões de peso molecular. Os filmes de raio-x foram fotografados e a
densitometria das bandas reativas foi realizada empregando o programa Scion Image (Scion
Corporation, base don NIH for MacIntosh by Wayne Rasbanol, NIH, USA).

32
10. Ensaio de viabilidade de macrófagos
O efeito dos IPs-HIV sobre a viabilidade de macrófagos murinos foi avaliada pelo
ensaio de MTT, conforme descrito previamente (Dutta et al., 2006). Macrófagos peritoniais de
fêmeas de camundongo BALB/c (6 a 8 meses de idade) foram coletados em PBS gelado e 5 ×
105 células foram colocadas para aderir em microplacas de 96 poços por 1 hora a 37 °C, em
atmosfera de 5% de CO2. As células não aderidas foram removidas através de lavagem com
PBS, e os poços foram preenchidos com meio DMEM suplementado com 10% de SFB. Os
macrófagos foram então incubados com concentrações crescentes dos IPs-HIV (1,56 a 25
µM), em triplicata. Após 24 horas, o meio foi retirado, o MTT adicionado (5mg/ml em PBS,
50 µg/poço) e as células incubadas por 3 horas ao abrigo de luz, a 37 ºC. As microplacas
foram subsequentemente centrifugadas a 190 g por 7 minutos, o sobrenadante foi removido e o
“pellet” dissolvido em 200 µl de DMSO. A absorbância foi mensurada em leitor de ELISA a
490 nm (Bio-Tek Instruments). Os macrófagos incubados sem a droga foram utilizados como
controle. As concentrações dos IPs-HIV capazes de manter 95% da viabilidade dos
macrófagos foram utilizadas nos ensaios de interação. Os experimentos foram realizados de
acordo com protocolos aprovados pelo Comitê Institucional de Uso e Cuidado com Animais
da Universidade Federal do Rio de Janeiro.
11. Ensaio de interação Leishmania-macrófago (Pré-tratamento)
Os promastigotas foram pré-tratados por 1 hora com os inibidores em concentrações
que variaram de 50 a 250 µM. Após esse período, os parasitos foram contados em câmara de
Neubauer e as concentrações capazes de manter 90% do número de células viáveis em relação
ao controle foram utilizadas no ensaio de interação. Os parasitos foram então pré-tratados
com os IPs-HIV por 1 hora com concentrações subinibitórias da droga (50 e 100 µM de
nelfinavir e lopinavir; 100 e 200 µM de amprenavir) e adicionados às placas contendo a
cultura de macrófagos. Após 1 hora de interação, as células foram fixadas em metanol, coradas
com Giemsa, desidratadas em soluções de acetona progressivamente substituídas por xilol e a
montagem das lâminas feita com Permount. A porcentagem de macrófagos infectados foi
determinada pela contagem de pelo menos 200 células em cada lamínula da triplicata. O índice
de associação foi obtido multiplicando-se a porcentagem de macrófagos infectados pelo
número de amastigotas por macrófagos infectados (Palmeira et al., 2008).

33
12. Ensaio de interação Leishmania-macrófago (Pós-tratamento)
Promastigotas obtidos da fase estacionária de crescimento foram lavados com PBS,
contados em câmara de Neubauer e adicionados as placas de 24 poços contendo lamínulas de
vidro com os macrófagos aderidos. Os parasitos foram adicionados em uma proporção
parasito/macrófago de 10:1, e incubados por 2 horas a 37 ºC em uma atmosfera de 5% de CO2.
Os parasitos livres foram removidos por lavagens sucessivas com PBS. Posteriormente, as
células infectadas com Leishmania foram tratadas por 24 horas com os IPs-HIV que tiveram
efeito sobre a multiplicação dos promastigotas, com concetrações finais de 6,25, 12,5 e 25 µM
de nelfinavir e 3,125, 6,25 e 12,5 µM de amprenavir e lopinavir (concentrações dos IPs-HIV
que mantem 95% da viabilidade dos macrófagos). Após a interação, as células foram lavadas
com PBS a 37 ºC, as lamínulas de vidro foram fixadas com metanol e coradas como descrito
acima.
13. Análises estatísticas
Todos os experimentos foram realizados no mínimo em triplicata e repetidos pelo
menos 2 vezes. Os dados foram analisados estatisticamente usando o teste t de Student usando
o programa Sigma Plot 2002 (SPSS Incorporated). Os valores de P iguais ou inferiores a 0,05
foram considerados estatisticamente significativos.

34
V. Resultados
1. Efeito dos inibidores de protease do HIV sobre a taxa de mutiplicação de
Leishmania amazonensis
Com o objetivo de avaliar se os inibidores de aspártico protease do HIV têm efeito
sobre a taxa de multiplicação das formas promastigotas de L. amazonensis, os IPs-HIV
(nelfinavir, lopinavir, amprenavir, saquinavir e indinavir), na concentração de 50 µM, foram
adicionados às formas promastigotas (5 × 107 células) e o crescimento celular in vitro foi
monitorado por 4 dias. Somente lopinavir e nelfinavir reduziram significativamente a taxa de
multiplicação do parasito, na concentração de 50 µM, a partir de 48 horas de cultivo (P < 0,05)
(Figura 11). A partir desta análise preliminar, cada inibidor foi testado na faixa de
concentração apropriada, ou seja, concentrações maiores que 50 µM para amprenavir,
indinavir e saquinavir e menores que 50 µM para nelfinavir e lopinavir. Todos os IPs-HIV
inibiram a multiplicação do parasito de forma dose-dependente (Figuras 12-16), porém a
diferença de susceptibilidade do parasito a cada inibidor foi marcante. O nelfinavir e o
lopinavir induziram a uma redução significativa na taxa de crescimento celular a partir de 48
horas nas concentrações acima de 25 µM (Figuras 12 e 13). O IC50 após 48 horas foi 15,12 ±
1,1µM e 16,47 ± 0,8 µM, respectivamente. O amprenavir a 250 µM também reduziu
significativamente (P≤ 0,05) a multiplicação do parasito após 48 horas de incubação. Com esta
droga a 500 µM, observou-se a morte predominante do parasito (Figura 14). O IC50 após 48
horas correspondeu a 62,0 µM ± 2,1 µM. O Indinavir alterou significativamente a taxa de
multiplicação do parasito somente na maior concentração e após 72 horas de cultivo (Figura
15). Já o saquinavir mesmo na maior concentração usada não diminuiu significativamnete a
proliferação do parasito (Figura 16). A pepstatina A, um inibidor clássico de aspártico
proteases, foi testado em paralelo para fins comparativos. Este inibidor diminuiu de forma
significativa o crescimento do parasito a 50 µM (Figura 17). O DMSO, solvente utilizado para
diluir as drogas, não afetou significativamente a multiplicação do parasito em relação ao
controle (Figuras 11-17).

35
Figura 11. Efeito dos inibidores de protease do HIV sobre a taxa de multiplicação de
Leishmania amazonensis. Os promastigotas (5 × 107 células) foram incubados a 26 ºC por 96
horas na ausência ou presença de amprenavir, saquinavir, nelfinavir, lopinavir e indinavir na
concentração de 50 µM. Os inibidores foram adicionados no dia 0 e as células foram contadas
diariamente em câmara de Neubauer. O dimetilsulfóxido foi utilizado na diluição
correspondente a concentração de 50 µM das drogas. Os resultados representam a média de
dois experimentos independentes, realizados em triplicata. As barras indicam o desvio padrão.
4
5
6
7
8
9
10
0 24 48 72 96
Tempo (h)
Número de promastigotasx 107
Controle
DMSO
Amprenavir
Saquinavir
Nelfinavir
Lopinavir
Indinavir
4
5
6
7
8
9
10
0 24 48 72 96
Tempo (h)
Número de promastigotasx 107
Controle
DMSO
Amprenavir
Saquinavir
Nelfinavir
Lopinavir
Indinavir
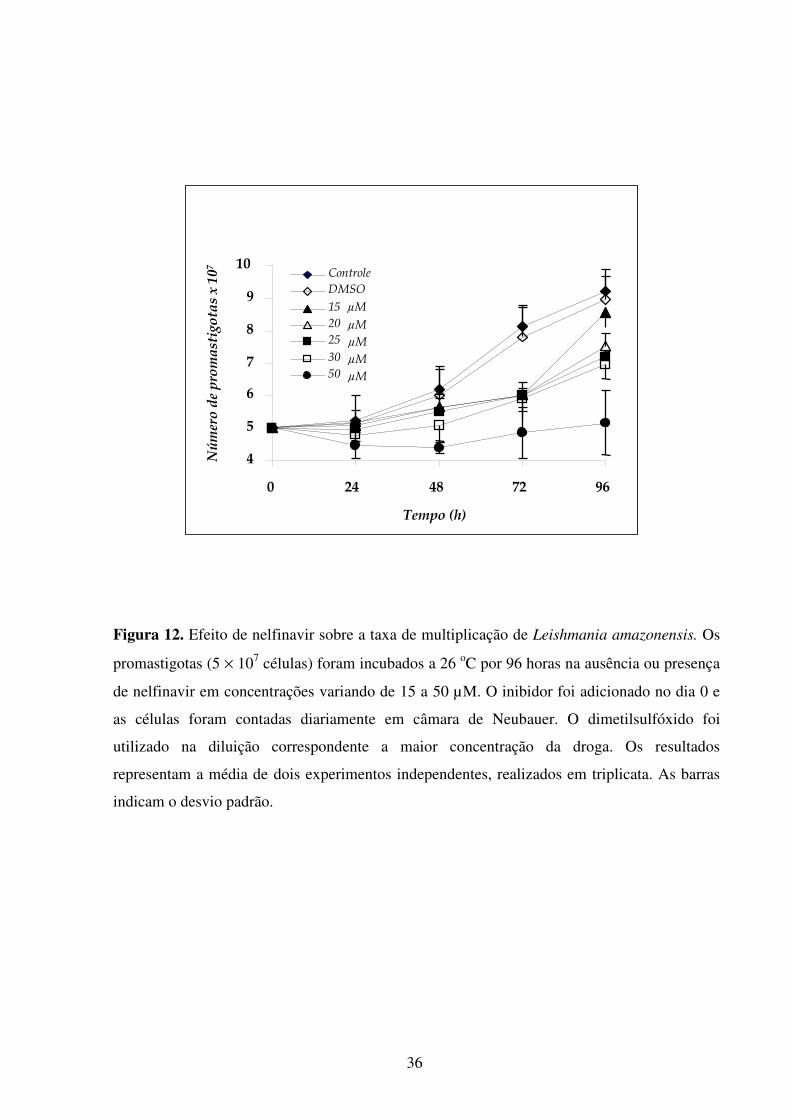
36
Figura 12. Efeito de nelfinavir sobre a taxa de multiplicação de Leishmania amazonensis. Os
promastigotas (5 × 107 células) foram incubados a 26 oC por 96 horas na ausência ou presença
de nelfinavir em concentrações variando de 15 a 50 µM. O inibidor foi adicionado no dia 0 e
as células foram contadas diariamente em câmara de Neubauer. O dimetilsulfóxido foi
utilizado na diluição correspondente a maior concentração da droga. Os resultados
representam a média de dois experimentos independentes, realizados em triplicata. As barras
indicam o desvio padrão.
4
5
6
7
8
9
10
0 24 48 72 96
Tempo (h)
Número de promastigotasx 10
7
Controle
DMSO
15 µM
20
25
30
50
µM
µM
µM
µM
4
5
6
7
8
9
10
0 24 48 72 96
Tempo (h)
Número de promastigotasx 10
7
Controle
DMSO
15 µM
20
25
30
50
µM
µM
µM
µM

37
Figura 13. Efeito de lopinavir sobre a taxa de multiplicação de Leishmania amazonensis. Os
promastigotas (5 × 107 células) foram incubados a 26 oC por 96 horas na ausência ou presença
de lopinavir em concentrações variando de 15 a 50 µM. O inibidor foi adicionado no dia 0 e as
células foram contadas diariamente em câmara de Neubauer. O dimetilsulfóxido foi utilizado
na diluição correspondente a maior concentração da droga. Os resultados representam a média
de dois experimentos independentes, realizados em triplicata. As barras indicam o desvio
padrão.
Lopinavir
4
5
6
7
8
9
10
0 24 48 72 96
Controle
DMSO
15 µM
20
25
30
50
Tempo (h)
Número de promastigotasx 10
7
µM
µM
µM
µM
Lopinavir
4
5
6
7
8
9
10
0 24 48 72 96
Controle
DMSO
15 µM
20
25
30
50
Tempo (h)
Número de promastigotasx 10
7
µM
µM
µM
µM
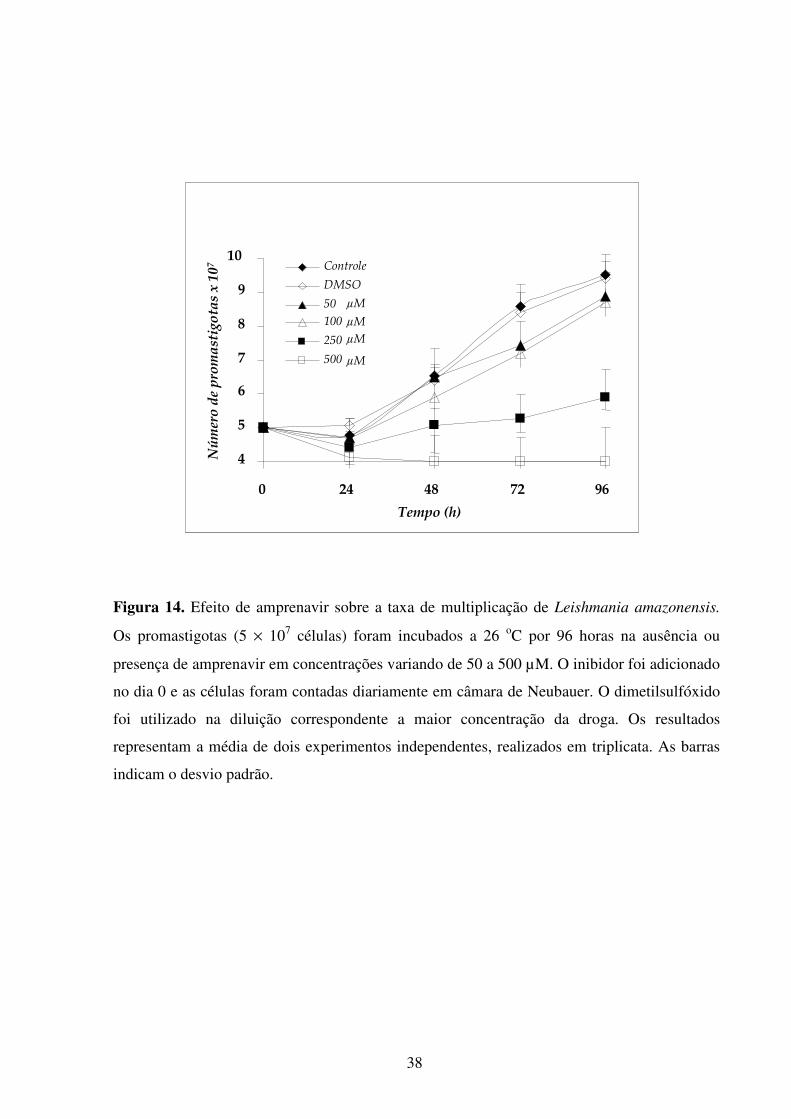
38
Figura 14. Efeito de amprenavir sobre a taxa de multiplicação de Leishmania amazonensis.
Os promastigotas (5 × 107 células) foram incubados a 26 oC por 96 horas na ausência ou
presença de amprenavir em concentrações variando de 50 a 500 µM. O inibidor foi adicionado
no dia 0 e as células foram contadas diariamente em câmara de Neubauer. O dimetilsulfóxido
foi utilizado na diluição correspondente a maior concentração da droga. Os resultados
representam a média de dois experimentos independentes, realizados em triplicata. As barras
indicam o desvio padrão.
4
5
6
7
8
9
10
0 24 48 72 96
Controle
DMSO
50
100
250
500 µM
µM
µM
µM
Tempo (h)
Número de promastigotasx 10
7
4
5
6
7
8
9
10
0 24 48 72 96
Controle
DMSO
50
100
250
500 µM
µM
µM
µM
Tempo (h)
Número de promastigotasx 10
7

39
Figura 15. Efeito de indinavir sobre a taxa de multiplicação de Leishmania amazonensis. Os
promastigotas (5 × 107 células) foram incubados a 26 oC por 96 horas na ausência ou presença
de nelfinavir em concentrações variando de 50 a 500 µM. O inibidor foi adicionado no dia 0 e
as células foram contadas diariamente em câmara de Neubauer. O dimetilsulfóxido foi
utilizado na diluição correspondente a maior concentração da droga. Os resultados
representam a média de dois experimentos independentes, realizados em triplicata. As barras
indicam o desvio padrão.
4
5
6
7
8
9
10
0 24 48 72 96
Tempo (h)
Número de promastigotasx 10
7
Controle
DMSO50
100
250
500 µM
µM
µM
µM
4
5
6
7
8
9
10
0 24 48 72 96
Tempo (h)
Número de promastigotasx 10
7
Controle
DMSO50
100
250
500 µM
µM
µM
µM

40
Figura 16. Efeito de saquinavir sobre a taxa de multiplicação de Leishmania amazonensis. Os
promastigotas (5 × 107 células) foram incubados a 26 oC por 96 horas na ausência ou presença
de saquinavir em concentrações variando de 50 a 500 µM. O inibidor foi adicionado no dia 0 e
as células foram contadas diariamente em câmara de Neubauer. O dimetilsulfóxido foi
utilizado na diluição correspondente a maior concentração da droga. Os resultados
representam a média de dois experimentos independentes, realizados em triplicata. As barras
indicam o desvio padrão.
4
5
6
7
8
9
10
0 24 48 72 96
Controle
DMSO
50
100
250
500 µM
µM
µM
µM
Tempo (h)
Número de promastigotasx 10
7
4
5
6
7
8
9
10
0 24 48 72 96
Controle
DMSO
50
100
250
500 µM
µM
µM
µM
Tempo (h)
Número de promastigotasx 10
7
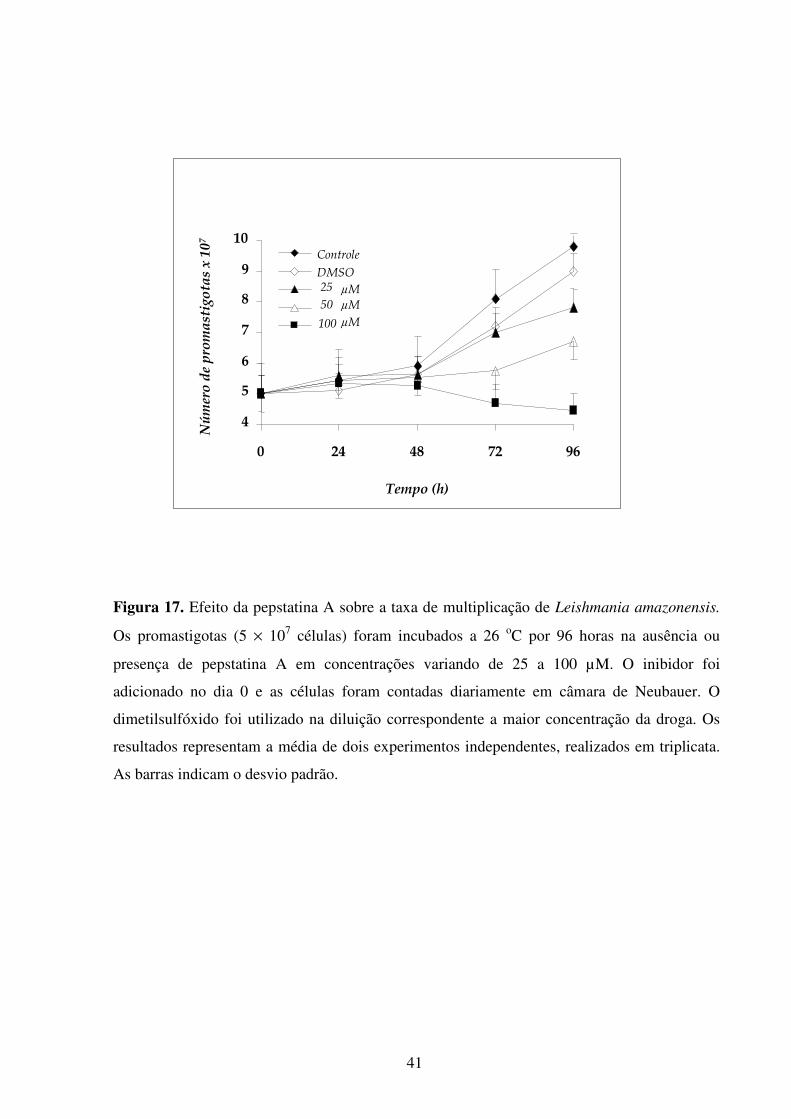
41
Figura 17. Efeito da pepstatina A sobre a taxa de multiplicação de Leishmania amazonensis.
Os promastigotas (5 × 107 células) foram incubados a 26 oC por 96 horas na ausência ou
presença de pepstatina A em concentrações variando de 25 a 100 µM. O inibidor foi
adicionado no dia 0 e as células foram contadas diariamente em câmara de Neubauer. O
dimetilsulfóxido foi utilizado na diluição correspondente a maior concentração da droga. Os
resultados representam a média de dois experimentos independentes, realizados em triplicata.
As barras indicam o desvio padrão.
4
5
6
7
8
9
10
0 24 48 72 96
Tempo (h)
Número de promastigotasx 10
7
Controle
DMSO25
50
100
µM
µM
µM
4
5
6
7
8
9
10
0 24 48 72 96
Tempo (h)
Número de promastigotasx 10
7
Controle
DMSO25
50
100
µM
µM
µM

42
2. Efeito dos inibidores sobre a diferenciação de formas promastigotas para
amastigotas de L. amazonensis:
As formas promastigotas dos parasitos (1 × 107 células/ml) foram incubadas em
condições que mimetizam o ambiente da célula hospedeira (pH 5,5 e temperatura a 32 ºC) para
induzir a diferenciação in vitro para as formas amastigotas. A fim de observar o efeito dos IPs-
HIV sobre a diferenciação, os compostos que tiveram efeito marcante sobre a proliferação
celular (nelfinavir, lopinavir e amprenavir) foram testados nas concentrações correspondentes
ao IC50 e a duas vezes o IC50: nelfinavir: 15,12 e 30,24 µM; lopinavir: 16,47 e 32,94 µM;
amprenavir: 62,0 e 124 µM. Os IPs-HIV não foram capazes de interferir na diferenciação dos
parasitos, sob estas condições experimentais.
3. Efeito dos IPs-HIV sobre a atividade enzimática de aspártico proteases de L.
amazonensis:
Através do emprego de substratos peptídicos seletivos para aspártico protease do HIV-
1 [Arg-Glu(EDANS)-Ser-Gln-Asn-Tyr-Pro-Ile-Val-Gln-Lys(DABCYL)-Arg] e para renina
[Arg-Glu(EDANS)-Ile-His-Pro-Phe-His-Leu-Val-Ile-His-Thr-Lys(DABCYL)-Arg] foi
possível demonstrar a presença de atividade de aspártico protease nos extratos de
promastigotas de L. amazonensis. A figura 18 mostra a degradação do substrato para aspártico
protease do HIV em pH 4,7 pelas proteínas solúveis de L. amazonensis e pela Pepsina, uma
aspártico protease conhecida, ao longo de 20 minutos (Figura 18). A partir deste resultado, a
degradação dos substratos para a protease do HIV e para a renina foi comparada sob as
mesmas condições experimentais. Apesar do peptídeo sintético para renina ter sido mais bem
hidrolisado do que o substrato para aspártico protease do HIV-1, a diferença não foi
estatisticamente significativa (Figura 19).
Em seguida, para caracterizar a atividade proteolítica detectada, o extrato de L.
amazonensis foi incubado com os IPs-HIV e pepstatina A. Inicialmente, todos os inibidores
na concentração de 1 µM foram testados com o substrato do HIV-1. O lopinavir e a pepstatina
A inibiram drasticamente a degradação do substrato em 91,5 % e 83,5 %, respectivamente. Já
com o substrato para renina, apenas pepstatina A foi capaz de inibir a atividade enzimática na
concentração de 1 µM. Apartir desses dados, avaliamos o efeito sobre a hidrólise do substrato
para aspártico protease do HIV-1, de concentrações dez vezes maiores de nelfinavir e
amprenavir. Os resultados indicaram que nelfinavir a 10 µM reduziu a atividade proteolítica
em 98 %, enquanto amprenavir não exerceu nenhum efeito. Tendo em vista a drástica inibição

43
por lopinavir a 1 µM, testamos concentrações menores deste composto, que não foi capaz de
inibir a degradação do substrato a 0,1 µM (Figura 20).
O substrato para renina não teve a degradação inibida por nenhum IP-HIV a 1 µM.
Apenas nelfinavir na concentração de 10 µM foi capaz de interferir na hidrólise do substrato
(64,6 % de inibição) (Figura 21).

44
Pepsina Extrato total de
L. amazonensis
0
20
40
60
80
100
120
Atividade enzim
ática (abs min
-1 m
g ptn
-1)
Figura 18. Detecção da atividade de aspártico protease no extrato bruto das formas
promastigotas de L. amazonensis e de uma enzima pura (Pepsina) contra substrato específico
para aspártico protease do HIV-1 [Arg-Glu(EDANS)-Ser-Gln-Asn-Tyr-Pro-Ile-Val-Gln-
Lys(DABCYL)-Arg]. A atividade do extrato protéico solúvel de L. amazonensis (10 µg) foi
determinada a 37 oC em pH 4,7 por 20 minutos, conforme descrito na metodologia. Os
resultados representam a média de dois experimentos independentes, realizados em
quadruplicata. As barras indicam o erro padrão.
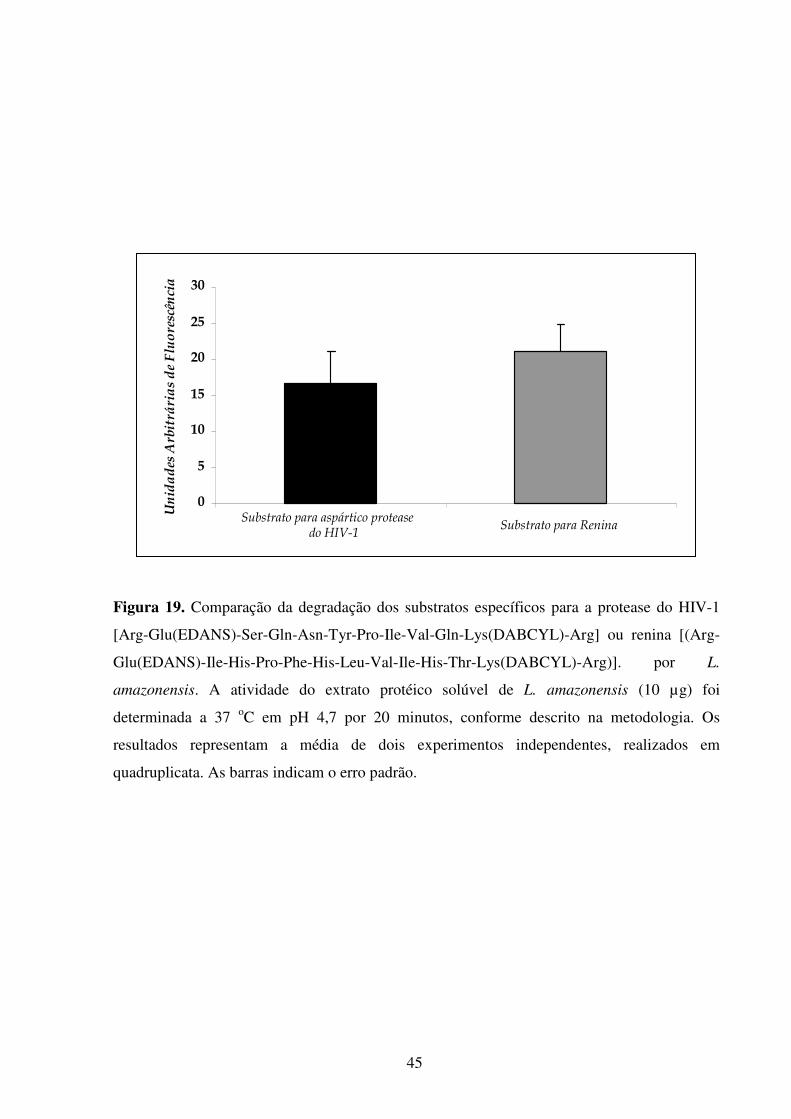
45
Figura 19. Comparação da degradação dos substratos específicos para a protease do HIV-1
[Arg-Glu(EDANS)-Ser-Gln-Asn-Tyr-Pro-Ile-Val-Gln-Lys(DABCYL)-Arg] ou renina [(Arg-
Glu(EDANS)-Ile-His-Pro-Phe-His-Leu-Val-Ile-His-Thr-Lys(DABCYL)-Arg)]. por L.
amazonensis. A atividade do extrato protéico solúvel de L. amazonensis (10 µg) foi
determinada a 37 oC em pH 4,7 por 20 minutos, conforme descrito na metodologia. Os
resultados representam a média de dois experimentos independentes, realizados em
quadruplicata. As barras indicam o erro padrão.
0
5
10
15
20
25
30
HIV-1 Peptidase Renina
Unidades Arbitrárias de Fluorescência
Substrato para aspártico protease do HIV-1
Substrato para Renina
0
5
10
15
20
25
30
HIV-1 Peptidase Renina
Unidades Arbitrárias de Fluorescência
Substrato para aspártico protease do HIV-1
Substrato para Renina
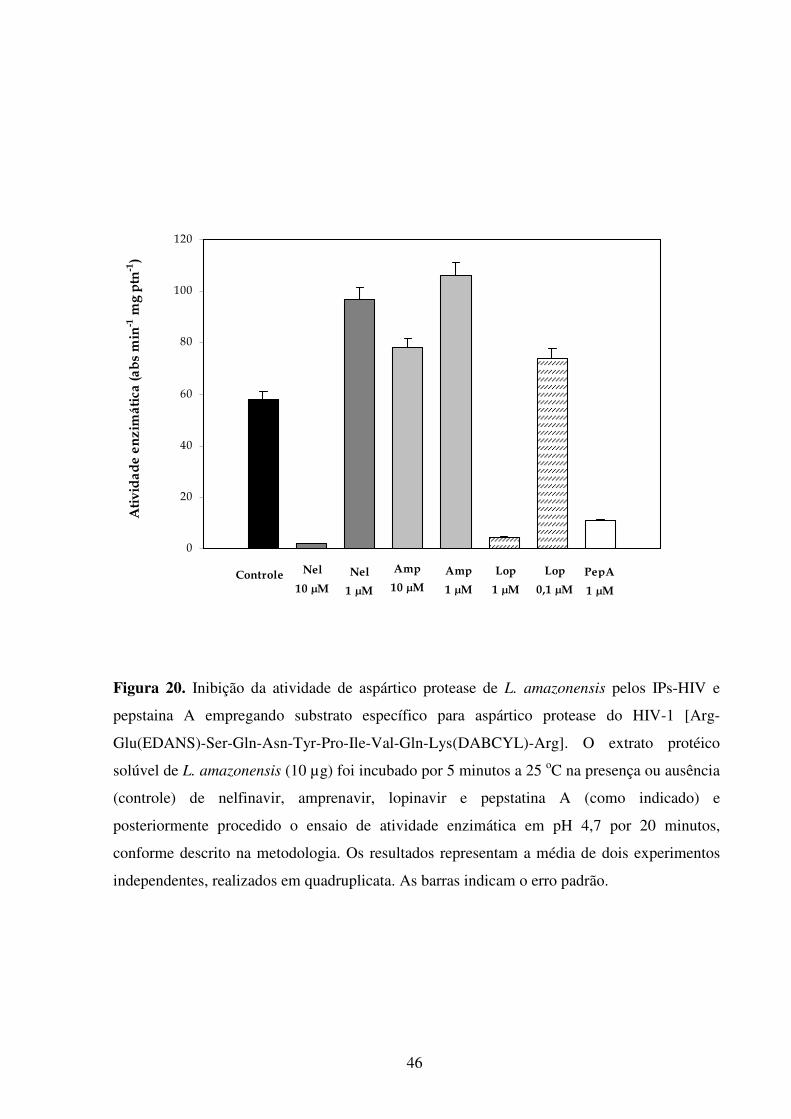
46
Controle Nel
10 µM
Nel
1 µM
Amp
10 µM
Amp
1 µM
Lop
1 µM
Lop
0,1 µM
PepA
1 µM
0
20
40
60
80
100
120
Atividade enzim
ática (abs min
-1 m
g ptn
-1)
Figura 20. Inibição da atividade de aspártico protease de L. amazonensis pelos IPs-HIV e
pepstaina A empregando substrato específico para aspártico protease do HIV-1 [Arg-
Glu(EDANS)-Ser-Gln-Asn-Tyr-Pro-Ile-Val-Gln-Lys(DABCYL)-Arg]. O extrato protéico
solúvel de L. amazonensis (10 µg) foi incubado por 5 minutos a 25 oC na presença ou ausência
(controle) de nelfinavir, amprenavir, lopinavir e pepstatina A (como indicado) e
posteriormente procedido o ensaio de atividade enzimática em pH 4,7 por 20 minutos,
conforme descrito na metodologia. Os resultados representam a média de dois experimentos
independentes, realizados em quadruplicata. As barras indicam o erro padrão.

47
Controle Nel
10 µM
Nel
1 µM
Amp
10 µM
Amp
1 µM
Lop
10 µM
Lop
1 µM
PepA
1 µM
0
20
40
60
80
100
120
140
160
180
Atividade enzim
ática (abs m
in-1 m
g ptn
-1)
Figura 21. Inibição da atividade de aspártico protease de L. amazonensis pelos IPs-HIV e
pepstaina A empregando substrato específico para renina [(Arg-Glu(EDANS)-Ile-His-Pro-
Phe-His-Leu-Val-Ile-His-Thr-Lys(DABCYL)-Arg)]. O extrato protéico solúvel de L.
amazonensis (10 µg) foi incubado por 5 minutos a 25 oC na presença ou ausência (controle) de
nelfinavir, amprenavir, lopinavir e pepstatina A (como indicado) e posteriormente procedido o
ensaio de atividade enzimática em pH 4,7 por 20 minutos, conforme descrito na metodologia.
Os resultados representam a média de dois experimentos independentes, realizados em
quadruplicata. O controle é indicado como 100%. As barras indicam o erro padrão.

48
4. Efeito dos IPs-HIV sobre a ultraestrutura de L. amazonensis:
Tendo em vista a eficácia dos IPs-HIV sobre a taxa de mutiplicação de L. amazonensis,
principalmente nelfinavir e lopinavir, avaliou-se o efeito destes inibidores na ultraestrutura de
L. amazonensis, por microscopia eletrônica de transmissão. A morfologia de células não-
tratadas (Figura 22 e 23; A) foi comparada com a ultraestrutura do protozoário tratado com os
inibidores (Figura 22, B-I; Figura 23; B-E). Os resultados mostraram pedaços de membrana
sendo liberados em toda a superfície celular (Figura 22; B-E cabeça de setas), incluindo a
membrana do flagelo (Figura 22; D cabeça de setas), após 4 horas de tratamento usando
concentrações correspondentes ao IC50 (15,12 µM para nelfinavir e 16,47 µM para lopinavir).
As células tratadas com nelfinavir mostraram também uma retração do citoplasma (Figura 22;
B, E �), um alto número de vesículas que, de acordo com sua eletrodensidade, provavelmente
correspondem a compartimentos ricos em lipídeos (Figura 22; E-F v) ou acidocalcissomas
(Figura 22; G �). As células tratadas com ambas as drogas apresentaram seus núcleos
intimamente envoltos por retículo endoplasmático (Figura 22; G-H e Figura 23; E setas
pretas), mitocôndria túrgida (Figura 22; F cabeça de seta branca) e estruturas similares à
mielina (Figura 22; H seta maior). Após 24 horas de tratamento, algumas alterações
visualizadas foram exclusivamente induzidas pelo inibidor lopinavir, como vesículas com
tamanho aumentado (Figura 23, B �) além de vários blocos de cromatina condensada
próximos ao envelope nuclear (Figura 23; B-D setas brancas). Esta alteração é sugestiva de
morte celular induzida por apoptose. Ainda com 24 horas de tratamento, foi possível verificar
uma desorganização total da ultraestrutura celular, culminando com a morte do parasito, como
observado tanto para nelfinavir quanto lopinavir (figura 22, I).

49
Figura 22. Alterações ultraestruturais em Leishmania amazonensis induzidas pelo
tratamento com nelfinavir. Os parasitos (1 × 108 celulas) obtidos de fase log de cultivo foram
inoculados em meio estéril na ausência (A) ou presença (B−I) de nelfinavir a 15,12 µM (IC50),
incubadas por 4 (B−E), 6 (F−G), 8 (H) e 24 horas (I), e processadas para microscopia
eletrônica de transmissão. Algumas alterações ultraestruturais foram evidenciadas tanto com
nelfinavir como com lopinavir, e são ilustradas com os parasitos tratados com nelfinavir, como
indicado no texto. As letras minúsculas significam: c−cinetoplasto; f−flagelo; m−mitocôndria
e n−núcleo. Pedaços de membrana (B-E-D cabeça de setas); retração do citoplasma (B, E �),
vesículas (E-F v) ou acidocalcissomas (G �), núcleos intimamente envoltos por retículo
endoplasmático (G-H), mitocôndria túrgida (F cabeça de seta branca) e estruturas similares à
mielina (H seta maior),

50
Figura 23. Alterações ultraestruturais em Leishmania amazonensis induzidas pelo
tratamento com lopinavir. Os parasitos (1 × 108 celulas) obtidos de fase log de cultivo foram
inoculados em meio estéril na ausência (A) ou presença (B−E) de lopinavir a 16,47 µM (IC50),
incubadas por 24 (B−D) e 6 horas (E) e processadas para microscopia eletrônica de
transmissão. As alterações ultraestruturais em B, C e D foram evidenciadas exclusivamente
com lopinavir. As letras minúsculas significam: c−cinetoplasto; f−flagelo; m−mitocôndria e
n−núcleo. Núcleos intimamente envoltos por retículo endoplasmático (E setas pretas)
vesículas com tamanho aumentado (B �), blocos de cromatina condensada próximos ao
envelope nuclear (B-D setas brancas).

51
5. Efeito dos IPs-HIVsobre a interação de Leishmania com macrófagos (pré-tratamento)
O efeito do tratamento das formas promastigotas com os IPs-HIV antes da interação
com macrófagos é mostrado na figura 24. Inicialmente, os inibidores nelfinavir, lopinavir e
amprenavir que mostraram maior efeito sobre a proliferação in vitro, foram incubadas por 1
hora com os inibidores nas concentrações que variaram de 50 a 250 µM, em intervalos de 50
µM. Nos experimentos de interação, foram utilizadas apenas as concentrações capazes de
manter mais de 95% dos parasitos viáveis, como observado através da sua morfologia,
motilidade, contagem em câmara de neubauer e exclusão de iodeto de propídeo. A partir deste
ensaio, as formas promastigotas foram tratadas por 1 hora com concentrações sub-inibitórias
dos IPs-HIV, que foram posteriormente removidos por lavagem. Os parasitos foram colocados
para interagir com os macrófagos por 1 hora, e após esse período, os flagelados livres foram
removidos e as células incubadas por mais 2 horas, e o índice de associação foi determinado.
O inibidor que causou uma redução mais drástica no índice de associação de L. amazonensis
com macrófagos foi nelfinavir, com um perfil de inibição dose-dependente, onde a inibição
aumentou de 86 % para 93 %, em relação ao controle, nas concentrações de 50 e 100 µM.
Quando os parasitos foram pré-tratados com amprenavir a 100 ou 200 µM, os índices de
associação foram, respectivamente, 75 % e 81% menores quando comparados ao controle. O
lopinavir não apresentou um perfil de inibição dose-dependente tão proeminente, ficando em
torno de 80% tanto para 50 µM como para 100 µM (Figura 24).

52
Figura 24. Efeito do tratamento das formas promastigotas de Leishmania amazonensis com os
IPs-HIV antes da interação com macrófagos. Os promastigotas de L. amazonensis foram
tratados ou não (controle), com nelfinavir, amprenavir ou lopinavir (como indicado) durante 1
hora antes da interação parasito-macrófago e, em seguida, as células foram lavadas com PBS.
O parasito manteve sua viabilidade nestas condições experimentais (como descrito na
Metodologia). Os macrófagos foram, então, infectados com as formas promastigotas por 1
hora a 37 ° C, e as monocamadas foram lavadas com PBS para a remoção dos parasitos livres.
O índice de associação foi determinado após 2 horas de infecção por microscopia de luz,
contando pelo menos 200 células de cada triplicata. Cada barra representa a média ± erro
padrão de pelo menos três experimentos independentes.
Association Index
0
10
20
30
40
50
60
70
controlecontrolenelfinavirnelfinavir amprenaviramprenavir lopinavirlopinavir
5050 100100 100100 200200 5050 100100 µµMM
IPsIPs
P P < 0.05< 0.05
Índ
ice
de
asso
ciaç
ão d
e L
. am
azo
nen
sis
com
mac
rófa
go
s (%
)
Controle 50 100
Nelfinavir
100 200
Amprenavir
50 100
lopinavir
µM
IPs
70
60
50
40
30
20
10
0
Association Index
0
10
20
30
40
50
60
70
controlecontrolenelfinavirnelfinavir amprenaviramprenavir lopinavirlopinavir
5050 100100 100100 200200 5050 100100 µµMM
IPsIPs
P P < 0.05< 0.05
Índ
ice
de
asso
ciaç
ão d
e L
. am
azo
nen
sis
com
mac
rófa
go
s (%
)
Controle 50 100
Nelfinavir
100 200
Amprenavir
50 100
lopinavir
µM
IPs
Association Index
0
10
20
30
40
50
60
70
controlecontrolenelfinavirnelfinavir amprenaviramprenavir lopinavirlopinavir
5050 100100 100100 200200 5050 100100 µµMM
IPsIPs
P P < 0.05< 0.05
Índ
ice
de
asso
ciaç
ão d
e L
. am
azo
nen
sis
com
mac
rófa
go
s (%
)
Controle 50 100
Nelfinavir
100 200
Amprenavir
50 100
lopinavir
µM
IPs
70
60
50
40
30
20
10
0

53
6. Efeito dos IPs-HIV sobre a interação de Leishmania amazonensis com macrófagos
(pós-tratamento)
Primeiramente, foi testado o efeito isolado dos inibidores sobre a viabilidade dos
macrófagos peritoniais de camundongo BALB/c através do ensaio com MTT. Um efeito
deletério significativo foi observado somente para lopinavir a 25 µM, enquanto amprenavir,
também nesta concentração, ficou no limite entre os efeitos não tóxicos e a toxicidade para os
macrófagos (Figura 25, A). A partir destas observações, foram utilizadas concentrações dos
IPs-HIV, que mantivessem, mais de 95% dos macrófagos viáveis durante 24 horas de
tratamento para os ensaios de interação de L. amazonensis com macrófagos na presença dos
inibidores. Os macrófagos foram pré-infectados com L. amazonensis por 1 hora, os parasitos
não aderidos ou internalizados foram removidos, e a cultura foi então tratada com nelfinavir,
lopinavir e amprenavir por 24 horas. Os resultados evidenciaram que todos os IPs-HIV
testados reduziram de forma significativa a sobrevivência intracelular de L. amazonensis nos
macrófagos, de maneira dose dependente (Figura 25). Os inibidores amprenavir e lopinavir, já
na concentração de 3.125 µM, mostraram uma redução do índice de associação de 20% e 60%,
respectivamente. Com a concentração de 6.25 µM houve uma redução do índice de associação
de 23% para nelfinavir, 35% para amprenavir e 75% para lopinavir. O lopinavir a 12.5 µM
reduziu drasticamente o índice de associação em torno de 90 %, assim como nelfinavir, que
promoveu, aproximadamente, uma inibição de 70% na concentração de 25 µM.

54
Figura 25. Toxicidade dos IPs-HIV sobre macrófagos peritoniais de camundongo BALB/c e
susceptibilidade dos parasitos intracelulares aos IPs-HIV. Inicialmente, os macrófagos (1 x 105
células) foram incubados em placas de 96 poços durante 24 horas na ausência ou na presença
de diferentes concentrações (como indicado) de nelfinavir, amprenavir e lopinavir. Após este
período, a viabilidade das células de macrófagos foi determinada espectrofotometricamente a
490 nm, através de ensaio de MTT. A linha pontilhada separa o gráfico em duas porções: < ou
≥ a 95% dos macrófagos viáveis. O controle representa os macrófagos sem inibidor (A). Os
macrófagos foram infectados com promastigotas de Leishmania amazonensis por 1 hora a 37 °
C, seguido de exaustiva lavagem com PBS e, em seguida, a interação foi incubada por mais 24
horas na ausência (controle), ou na presença de nelfinavir, amprenavir ou lopinavir. O índice
de associação foi determinado por microscopia de luz, contando pelo menos 200 células de
cada triplicata. O controle é indicado como 100%. Cada barra representa a média ± erro
padrão de pelo menos três experimentos independentes (B).
ABS 490
0,0
0,5
1,0
1,5
2,01.562 µM3.125 µM6.25 µM12.5 µM25 µM
Controle NelfinavirNelfinavir AmprenavirAmprenavir LopinavirLopinavir
A
% Association index
0
20
40
60
80
100
120
6.256.25 12.512.5 µµMM2525 3.123.12 6.256.25 12.512.5 3.123.12 6.256.25 12.512.5controlecontrole
NelfinavirNelfinavir AmprenavirAmprenavir LopinavirLopinavir
BP P < 0.05< 0.05
Controle
Índice de associação de
L. amazonensiscom macrófagos (%)
1.562 µM3.125 µM6.25 µM12.5 µM25 µM
Absorbância490
Nelfinavir Amprenavir Lopinavir
Nelfinavir Amprenavir Lopinavir
Controle
IPs
ABS 490
0,0
0,5
1,0
1,5
2,01.562 µM3.125 µM6.25 µM12.5 µM25 µM
Controle NelfinavirNelfinavir AmprenavirAmprenavir LopinavirLopinavir
A
ABS 490
0,0
0,5
1,0
1,5
2,01.562 µM3.125 µM6.25 µM12.5 µM25 µM
Controle NelfinavirNelfinavir AmprenavirAmprenavir LopinavirLopinavir
A
% Association index
0
20
40
60
80
100
120
6.256.25 12.512.5 µµMM2525 3.123.12 6.256.25 12.512.5 3.123.12 6.256.25 12.512.5controlecontrole
NelfinavirNelfinavir AmprenavirAmprenavir LopinavirLopinavir
BP P < 0.05< 0.05
Controle
Índice de associação de
L. amazonensiscom macrófagos (%)
1.562 µM3.125 µM6.25 µM12.5 µM25 µM
Absorbância490
Nelfinavir Amprenavir Lopinavir
Nelfinavir Amprenavir Lopinavir
Controle
IPs

55
7. Efeito dos IPs-HIV sobre o perfil de proteases em Leishmania através de zimografia e
immunoblotting.
Nesta parte dos experimentos, o objetivo foi avaliar se o estresse celular induzido pelos
IPs-HIV poderia levar a uma mudança no padrão de produção de proteases por L.
amazonensis. Sendo assim, as células foram incubadas por 4 horas na presença de nelfinavir,
lopinavir e amprenavir a 100 µM e seu perfil proteolítico avaliado por zimografia. Nestas
condições experimentais, no mínimo 95% dos parasitos mantiveram sua viabilidade, como
observado através de sua morfologia, motilidade, contagem em câmara de neubauer e exclusão
de iodeto de propídio. O perfil proteolítico de L. amazonensis, determinado através de
zimografia, é composto por cisteíno e metaloproteases (Yao et al., 2003; Mottram et al., 2004).
A comparação da expressão das proteases por SDS-PAGE, de parasitos incubados por 4 horas
com os IPs-HIV, revelou que uma metaloprotease de 63 kDa e as cisteíno proteases de baixo
peso molecular, tiveram suas atividades aumentadas.
Tendo em vista a importância das metalo proteases e císteino proteases,
particularmente a gp63 e cpb, como fatores de virulência em Leishmania, objetivou-se
determinar a influência dos IPs-HIV sobre a produção dessas proteínas pelo parasito, através
da análise por western blotting dessas moléculas empregando anticorpos contra gp63 e cpb.
Com este intuito, as proteínas de L. amazonensis foram separadas eletroforeticamente,
transferidas para uma membrana de nitrocelulose, e incubadas com os anticorpos anti-gp63 e
anti-cpb. Os resultados demonstraram um aumento na expressão dessas moléculas (Figura 26).
Em todas as análises empregadas, lopinavir foi o inibidor que promoveu o efeito mais
pronunciado. Como controle, o western blotting foi revelado com anticorpo anti-α-tubulina, o
qual não mostrou diferença entre as drogas e o controle (Figura 26).
Análises de densitometria mostraram uma diferença estatisticamente significativa entre
o controle e parasitos tratados com amprenavir e lopinavir para a gp63 e com todos os
inibidores para a cpb (Figura 26).
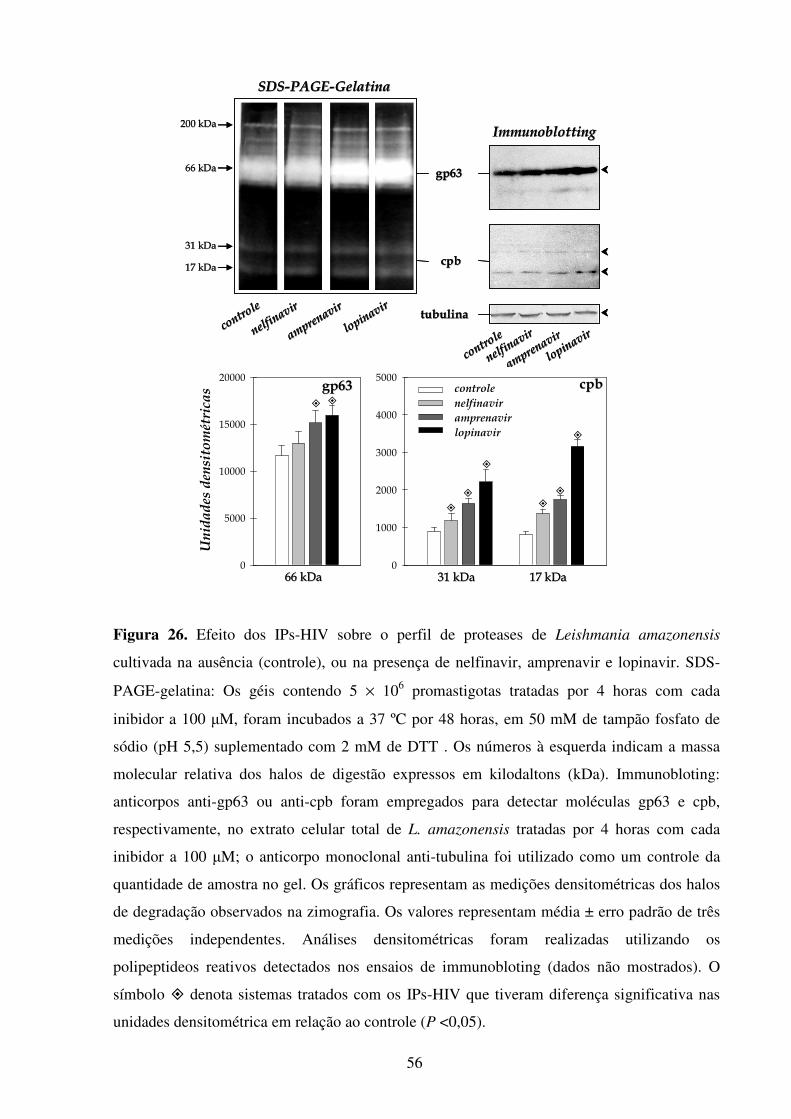
56
Figura 26. Efeito dos IPs-HIV sobre o perfil de proteases de Leishmania amazonensis
cultivada na ausência (controle), ou na presença de nelfinavir, amprenavir e lopinavir. SDS-
PAGE-gelatina: Os géis contendo 5 × 106 promastigotas tratadas por 4 horas com cada
inibidor a 100 µM, foram incubados a 37 ºC por 48 horas, em 50 mM de tampão fosfato de
sódio (pH 5,5) suplementado com 2 mM de DTT . Os números à esquerda indicam a massa
molecular relativa dos halos de digestão expressos em kilodaltons (kDa). Immunobloting:
anticorpos anti-gp63 ou anti-cpb foram empregados para detectar moléculas gp63 e cpb,
respectivamente, no extrato celular total de L. amazonensis tratadas por 4 horas com cada
inibidor a 100 µM; o anticorpo monoclonal anti-tubulina foi utilizado como um controle da
quantidade de amostra no gel. Os gráficos representam as medições densitométricas dos halos
de degradação observados na zimografia. Os valores representam média ± erro padrão de três
medições independentes. Análises densitométricas foram realizadas utilizando os
polipeptideos reativos detectados nos ensaios de immunobloting (dados não mostrados). O
símbolo � denota sistemas tratados com os IPs-HIV que tiveram diferença significativa nas
unidades densitométrica em relação ao controle (P <0,05).
66 66 kDakDa
17 17 kDakDa
31 31 kDakDa
200 200 kDakDa
gp63gp63
cpbcpb
��������
��������
��������
��������tubulinatubulinacontrole
controle
nelfinavir
nelfinavir
amprenavir
amprenavir
lopinavir
lopinavir
controle
controle
nelfinavir
nelfinavir
amprenavir
amprenavir
lopinavir
lopinavir
SDSSDS--PAGEPAGE--GelatinaGelatina
ImmunoblottingImmunoblotting
Densitometric Units
0
5000
10000
15000
20000
0
1000
2000
3000
4000
5000control
nelfinavir
amprenavir
lopinavir
gp63gp63 cpbcpb
66 66 kDakDa 31 31 kDakDa 17 17 kDakDa
� �
�
�
�
�
�
�
Unidades densitométricas controle
nelfinaviramprenavirlopinavir
66 66 kDakDa
17 17 kDakDa
31 31 kDakDa
200 200 kDakDa
gp63gp63
cpbcpb
��������
��������
��������
��������tubulinatubulinacontrole
controle
nelfinavir
nelfinavir
amprenavir
amprenavir
lopinavir
lopinavir
controle
controle
nelfinavir
nelfinavir
amprenavir
amprenavir
lopinavir
lopinavir
SDSSDS--PAGEPAGE--GelatinaGelatina
ImmunoblottingImmunoblotting
Densitometric Units
0
5000
10000
15000
20000
0
1000
2000
3000
4000
5000control
nelfinavir
amprenavir
lopinavir
gp63gp63 cpbcpb
66 66 kDakDa 31 31 kDakDa 17 17 kDakDa
� �
�
�
�
�
�
�
Unidades densitométricas controle
nelfinaviramprenavirlopinavir

57
VI. Discussão
A leishmaniose é uma doença tropical negligenciada (WHO, 2007), e a pandemia da
AIDS tem contribuído para a mudança do perfil desta doença no mundo (Alvar et al., 1997).
O número de co-infecções Leishmania-HIV tem aumentado de forma considerável,
especialmente devido à sobreposição de ambas epidemias (Alvar et al., 2008). Além disso, o
impacto real da co-infecção Leishmania-HIV é provavelmente subestimada em escala global
devido à deficiência nos sistemas de vigilância. O fato da leishmaniose não estar incluída entre
as “doenças oportunistas associadas à aids” contribui para a escassez de informações (Cruz et
al., 2006). A infecção pelo HIV pode aumentar o risco de desenvolvimento da leishmaniose
visceral na ordem de 100 a 1000 vezes em áreas endêmicas (Alvar et al., 2008). Além disso, a
visceralização de cepas de Leishmania classicamente cutâneas, como Leishmania
amazonensis, tem sido observada (Barral et al., 1986, 1987; Ramos-Santos et al., 2000; revisto
por Alvar et al., 2008). A infecção pelo HIV também reduz a probabilidade de sucesso da
quimioterapia, e aumenta acentuadamente a probabilidade de reativação da doença (Gradoni et
al., 1996; Lopez-Velez et al., 1998). Ao mesmo tempo, a leishmaniose visceral promove a
progressão clínica para AIDS e o desenvolvimento das condições que caracterizam a doença.
Ambas as enfermidades exercem um efeito sinérgico danoso sobre a resposta imune celular
pois compartilham o mesmo alvo celular do sistema imunológico, isto é, células do sistema
fagocítico mononuclear (Alvar et al., 2008).
Pacientes com infecção pelo HIV têm vivido por mais tempo e com melhor qualidade
de vida como resultado da introdução do TARV, que se caracteriza por esquemas que
consistem da combinação de inibidores de protease com pelo menos duas outras drogas
antiretrovirais (Andrews & Friedland, 2000; Pernerstorfer-Schoen et al., 2001). Os IPs-HIV
ligam-se competitivamente ao sítio de clivagem da aspártico protease do HIV-1. Eles tornam a
enzima não funcional e levam à liberação de partículas virais não infecciosas e imaturas (Saag
& Schooly, 1998). Além disso, foi demonstrado que os IPs-HIV têm efeitos vantajosos sobre
infecções oportunistas causadas por fungos patogênicos, e também por alguns protozoários,
principais causadores de morbidade e mortalidade em indivíduos com AIDS. Estes efeitos são
devidos à restauração do sistema imunológico do hospedeiro e também a uma ação direta
desses compostos sobre os patógenos oportunistas. Apesar da clara sobreposição da pandemia
da AIDS com a endemia da leishmaniose, algumas particularidades da epidemiologia,
patogênese, profilaxia, e especialmente do tratamento da co-infecção Leishmania-HIV são
pouco claras à indefinidas (WHO, 2007; Marques et al., 2007). Neste contexto, o presente
estudo foi realizado com o intuito de avaliar a influência de diferentes IPs-HIV sobre o

58
protozoário parasito L. amazonensis, com foco nos efeitos sobre a atividade enzimática de
aspártico proteases, proliferação do parasito, ultraestrutura, interação com células hospedeiras
e expressão de fatores de virulência.
As formas promastigotas de L. amazonensis foram cultivadas com pepstatina A,
nelfinavir, indinavir, saquinavir, lopinavir e amprenavir e a taxa de multiplicação do parasito
analisada ao longo de 96 horas. O inibidor pepstatina A promoveu uma forte redução da
multiplicação de L. amazonensis. Apesar deste dado indicar indiretamente, que aspártico
proteases de L. amazonensis possam ser um alvo em potencial para o emprego de inibidores
dessa classe enzimática no tratamento das leishmanioses, drogas semelhantes à pepstatina A
não são usadas clinicamente devido ao seu metabolismo no fígado e a rápida eliminação do
sangue (Rüchel, Ritter & Schajrinski, 1990). Os inibidores nelfinavir e lopinavir inibiram
acentuadamente a multiplicação do parasito a 25 µM, em contraste com os outros inibidores
testados, que só apresentaram efeitos significativos em concentrações consideravelmente
maiores. Deve-se notar que o efeito inibitório dos IPs-HIV in vitro foi observado em
concentrações substanciais (na faixa de µM), bem mais altas do que as necessárias para a
inibição da protease do HIV (na ordem de nM) (Flexner, 1998). Este dado provavelmente
reflete uma afinidade bem menor dessas drogas para alvos ainda não identificados em
Leishmania quando comparada com sua alta afinidade à protease do HIV (Flexner, 1998).
Os protozoários do gênero Leishmania são um grupo biologicamente diverso de
microrganismos. Estudos taxonômicos de isolados de Leishmania indicam uma grande
diversidade dentro do gênero (Cupolillo et al., 2001), e a diversidade genética pode ter um
impacto direto sobre propriedades relevantes dos organismos, incluindo manifestações clínicas
distintas e susceptibilidade a drogas (Chang, Akman & Nielsen, 1999). Desta forma, é
esperado que o efeito dos IPs-HIV sobre espécies de Leishmania responsáveis por
leishmaniose visceral ou mucosa seja bastante distinto. De fato, Savoia e colaboradores
(Savoia, Allice & Tovo, 2005) relataram o IC50 para promastigotas de Leishmania major na
faixa de 10 µM para indinavir e saquinavir após 24 horas de crescimento, enquanto
Leishmania infantum, que causa leishmaniose visceral, foi inibida somente parcialmente (em
torno de 30%) na concentração mais alta testada dessas drogas (50µM) e somente após 72
horas de crescimento (Savoia, Allice & Tovo, 2005). Recentemente, Trudel e colaboradores
(Trudel et al., 2008) relataram o efeito de nelfinavir, ritonavir e saquinavir sobre a taxa de
multiplicação de promastigotas de L. infantum. Nenhuma alteração significativa na taxa de
multiplicação do parasito foi observada nas concentrações até 25 µM, após 72 horas de
incubação (Trudel et al., 2008).

59
No presente trabalho, o indinavir somente alterou significativamente a taxa de
multiplicação do parasito após 72 horas de cultivo e na concentração de 500 µM, ao contrário
do que foi observado em L. major por Savoia e colaboradores, que relataram um efeito
inibitório com concentração 10 vezes menor do que a utilizada neste trabalho. O saquinavir,
corroborando com os resultados de L. infantum (Trudel et al., 2008), não diminuiu de forma
significativa a taxa de multiplicação do parasito mesmo na maior concentração utilizada. Esses
dados reforçam as diferenças de susceptibilidade aos IPs-HIV das diferentes espécies de
Leishmania. Além disso, uma cepa de Leishmania donovani, resistente ao estibugluconato de
sódio, é igualmente susceptível aos IPs-HIV quando comparada a uma cepa sensível,
sugerindo que a resistência ao quimioterápico para leishmaniose não resulta em uma
resistência cruzada aos IPs-HIV (Trudel et al., 2008).
O gênero Leishmania compreende protozoários parasitos com ciclo de vida complexo,
envolvendo estágios distintos de desenvolvimento. Esses diferentes estágios representam uma
adaptação às mudanças ambientais encontradas pelos parasitos dentro de seus hospedeiros: um
mamífero, para o qual eles são patogênicos, e um inseto vetor (Besteiro et al., 2007). Durante a
transição entre os ambientes extra e intracelular, a Leishmania é exposta a mudanças
ambientais importantes, que vão desde variações na disponibilidade de nutrientes, pH,
temperatura, bem como na disponibilidade de oxigênio. A estratégia adotada pelos parasitos
para sobreviver a essas mudanças é o desenvolvimento de formas altamente adaptadas e
especializadas (Besteiro et al., 2007). Alguns dos marcadores estágio-específicos mais bem
definidos incluem as proteases, algumas das quais são associadas com a virulência de
Leishmania em mamíferos (Mottram et al., 2004). Recentemente, foi demonstrado que a
atividade de proteases é profundamente alterada durante o processo de diferenciação (Alves et
al., 2005). Tendo em vista a possibilidade de envolvimento de aspártico proteases no processo
de diferenciação de Leishmania, o efeito dos IPs-HIV foi testado sob condições específicas
que mimetizam o ambiente celular do hospedeiro mamífero, porém sob as condições
experimentais utilizadas neste estudo, não foi possível detectar interferência dos IPs-HIV neste
processo. É possível que aspártico proteases possam ter suas funções voltadas para etapas que
precedem a diferenciação ou que sejam reguladas negativamente em formas amastigotas
(Alves et al., 2005). É importante ressaltar também que as concentrações empregadas das
drogas podem ter sido insuficientes em função de uma possível baixa afinidade com o alvo,
uma vez que os IPs-HIV não foram especificamente desenhados para um alvo em Leishmania.
A eficiência dos IPs-HIV em tratar infecções parasitárias pode estar associada à sua
capacidade de modular ou bloquear o proteassoma celular ou promover apoptose (Pozio &
Morales, 2005). Alternativamente, eles poderiam atuar diretamente sobre aspártico proteases

60
produzidas pelos protozoários. Esta classe proteolítica já foi descrita em L. amazonensis
(Alves et al., 2005) e L. mexicana (Valdivieso, Dagger & Rascón, 2007). Recentemente,
também foi demonstrado que diazoacetil-DL-norleucina metil éster (DAN), um inibidor de
aspártico protease, reduz significativamente a multiplicação de promastigotas de Leishmania
mexicana. Apesar de parecer razoável assumir que aspártico proteases e/ou o proteassoma de
Leishmania sejam o alvo dos IPs-HIV, a possibilidade de efeitos indiretos sobre L.
amazonensis devem ser considerados, como efeitos tóxicos generalizados ou não específicos
sobre os parasitos. Neste contexto, a observação por microscopia eletrônica de transmissão
dos parasitos expostos a nelfinavir ou lopinavir revelou algumas alterações peculiares nas
estruturas vitais da célula, como estruturas celulares internas e membrana citoplasmática,
sugerindo prejuízos metabólicos irreversíveis que culminaram com a morte celular do parasito.
Algumas alterações observadas em L. amazonensis, tais como aumento no número de
vesículas e envolvimento do núcleo por retículo endoplasmático, são sugestivos de autofagia.
Dados recentes indicam a existência de diferentes formas de morte celular programada. Além
da bem caracterizada morte celular por apoptose, a autofagia também tem sido descrita em
células eucariotas (Kissová et al., 2006). A autofagia é um processo de degradação normal que
existe em todas as células eucarióticas e é estimulado em resposta a uma série de estresses
ambientais, os quais necessitam do uso do mecanismo autofágico para possibilitar a
sobrevivência celular (Dunn, 1994; Seglen & Bohley, 1992). A autofagia envolve o
“sequestro” de organelas citoplasmáticas ou citosol dentro de membranas duplas, criando
autofagossomas (também denominados vacúolos autofágicos). Os autofagossomas
subsequentemente se fundem com endossomas e eventualmente com lisossomas, criando então
autofagolisossomas ou autolisossomas. No lúmem dessas estruturas tardias, enzimas
lisossomais operam a baixo pH e catabolizam o material autofágico (Levine & Klionsky,
2004; Shintani & Klionsky, 2004). Níveis basais de autofagia são responsáveis pela regulação
de eventos essenciais nas células e, portanto uma melhor sobrevivência das células, enquanto
que níveis anormais de autofagia podem ser responsáveis pela morte celular (Shintani &
Klionsky, 2004). Até o momento, os eventos responsáveis pela desregulação da atividade
autofágica não foram bem caracterizados (González-Polo et al., 2005). Além disso, em
contraste com as células de mamíferos, o processo autofágico em organismos unicelulares é
pouco estudado. Por fim, o acúmulo de vacúolos autofágicos pode preceder a morte celular
por apoptose (González-Polo et al., 2005). Curiosamente, o crescimento de Leishmania na
presença de lopinavir por 24 horas exibe uma condensação de cromatina, a qual é um dos
indicadores de apoptose. Por estas razões, seria interessante explorar mais a fundo o
mecanismo de morte celular induzido pelos IPs-HIV em Leishmania.

61
Em infecções oportunistas pelo fungo patogênico Cryptococcus neoformans, o
indinavir inibe de forma seletiva a produção de alguns fatores de virulência, tais como urease e
protease. Adicionalmente, indinavir também interfere na formação da cápsula polissacarídica,
o principal fator de virulência produzido por este microrganismo (Monari et al., 2005). Em
Candida albicans, estudos bioquímicos demonstraram que os IPs-HIV foram capazes de inibir
diretamente as aspártico proteases secretadas por este fungo (Munro & Hube, 2002). Andrews
e colaboradores, em 2006, também relataram que as enzimas plasmepsina II e IV de
Plasmodium falciparum foram diretamente inibidas pelos IPs-HIV, saquinavir, ritonavir, e
lopinavir (Andrews et al., 2006). Neste contexto, nosso objetivo foi avaliar se os IPs-HIV
induziriam a alguma mudança na expressão de proteases por L. amazonensis. Com o passar
dos anos, ficou claro que as proteases de tripanossomatídeos patogênicos desempenham um
importante papel em diversas etapas da infecção no hospedeiro, tais como: adsorção,
penetração, sobrevivência intracelular, replicação, diferenciação, infectividade, evasão da
resposta imune e nutrição. Os tripanossomatídeos apresentam um amplo e variado conjunto de
proteases extracelulares e/ou intracelulares, as quais regulam a expressão de funções
específicas requeridas nos estágios de vida do parasito (Vermelho et al., 2007).
As espécies de Leishmania contêm múltiplas atividades de cisteíno proteases
intracelulares altamente ativas e uma metaloprotease abundantemente expressa na superfície
do parasito, denominada gp63 ou leishmaniolisina (Mottram, Coombs & Alexander, 2004;
Vermelho et al., 2007; Yao et al., 2003). A maioria dos estudos até agora têm sido feitos com
apenas três tipos de cisteíno proteases designadas como CPA, CPB e CPC (Mottram, Coombs
& Alexander, 2004). A geração de mutantes nulos para os genes cpa, cpb e cpc em L.
mexicana forneceu o primeiro suporte genético para o papel de cisteíno proteases de
Leishmania na virulência do parasito, e consequentemente sua validade como alvo para drogas
(Mottram, Coombs & Alexander, 2004). De acordo, cisteíno proteases são expressas
preferencialmente em promastigotas virulentos, em oposição a promastigotas avirulentos de L.
amazonensis (Soares et al., 2003). As múltiplas enzimas CPBs têm importantes papéis
imunomodulatórios para facilitar a infecção pelo parasito. As Amastigotas de L. mexicana
deficientes na produção de CPB, embora tão infectivos para os macrófagos in vitro quanto o
tipo selvagem do parasito, têm capacidade reduzida de induzir o crescimento da lesão em
camundongos BALB/c. Além disso, mutantes para cpa mostraram-se menos infectivos que
mutantes para cpb, sugerindo que CPA também é um fator de virulência, além de ter
sobreposição de funções com CPB. Juntos cpa, cpb1 e cpb2 (isoformas de cpb) são expressos
em promastigotas metacíclicos, que são as formas infectivas de Leishmania para os mamíferos
(revisto por Mottram, Coombs, Alexander, 2004).

62
A gp63 também está envolvida na virulência de Leishmania. A expressão desta enzima
aumenta em promastigotas metacíclicos e a gp63 pode ser um dos ligantes envolvidos na
interação do parasito com sistemas de defesa do hospedeiro, incluindo componentes do
sistema complemento e da superfície de macrófagos. Esta enzima também pode ter um papel
na sobrevivência intracelular de amastigotas dentro de fagolisossomas. Porém, dados de
“knockout” para gp63 são conflitantes, visto que a restauração dos níveis de expressão da
gp63, ao mesmo nível das cepas selvagens, devolve apenas parcialmente a capacidade
infectiva do parasito em macrófagos, assim como sua sensibilidade ao complemento e a
capacidade de formar lesões. As possíveis explicações para essa observação são que a gp63
contribui parcialmente para a sensibilidade ao complemento e que outros fatores são
necessários para a restauração completa da virulência e para complementar a resistência do
parasito. Finalmente, a perda total da molécula pode contribuir para uma perturbação da
superfície de Leishmania, fazendo com que o parasito fique mais vulnerável à lise mediada
pelo sistema complemento (Yao, Donelson & Wilson, 2003).
Desta forma, em contraste à visão de que os IPs-HIV conduziriam à uma redução dos
níveis de protease, como descrito para fatores de virulência em fungos, como Cryptococcus
neoformans (Monari et al., 2005), Pneumocystis carinii, Candida albicans (revisto por Pozio
& Morales, 2005), Fonsecaeae pedrosoi (Palmeira et al., 2008), o presente estudo revelou um
aumento significativo na expressão destas moléculas quando os parasitos foram sujeitos à ação
dos IPs-HIV antes da extração das proteínas. Curiosamente, um efeito similar foi descrito
quando L. amazonensis foi tratada com partenolida, uma lactona sesquiterpeno purificada de
extratos hidroalcoolicos de partes aéreas de Tanacetum parthenium (Tiuman et al., 2005).
Tendo em vista que as cisteíno proteases, CPA e CPB de L. mexicana são essenciais para
autofagia e diferenciação do parasito (Williams, Tetley & Coombs, 2006), e levando-se em
consideração que promastigotas tratados com os IPs-HIV apresentaram alterações
ultraestruturais que assemelham-se a processos autofágicos, o aumento da produção de
proteases poderia estar correlacionado com este processo. Alternativamente, a atividade
exocítica intensa observada na região da bolsa flagelar em promastigotas tratados com
nelfinavir e lopinavir poderiam indicar um processo de exacerbação da produção de proteínas
pela célula como tentativa de sobrevivência. Contudo, a maior expressão de fatores de
virulência não foi acompanhada pelo aumento da capacidade de infecção, como evidenciado
nos experimentos de interação.
As aspártico proteases foram isoladas e estudadas de uma gama de organismos,
variando desde vertebrados, plantas, fungos, parasitos, retrovírus e bactérias. Essas enzimas
têm despertado interesse devido ao seu papel em doenças humanas como, o envolvimento da

63
renina na hipertensão, da catepsina D na metástase do câncer de mama, da β-Secretase na
doença de Alzheimer, das plasmepsinas na malária, da peptidase do HIV-1 na AIDS e das
aspártico proteases secretadas em infecções por Candida (revisto por Dash et al., 2003). Além
disso, recentemente, foi demonstrada e caracterizada atividade de aspártico proteases em L.
amazonensis e L. mexicana através do uso combinado de inibidores e substratos cromogênicos
e fluorogênicos específicos para esta classe de enzimas (Alves et al., 2005; Valdivieso, Dagger
e Rascón, 2007).
Com o objetivo de detectar atividade de aspártico protease em promastigotas de L.
amazonensis e avaliar se os IPs-HIV são capazes de inibir diretamente esta(s) enzima(s),
substratos fluorogênicos específicos para aspártico protease do HIV-1 e para a renina foram
utilizados. O substrato de ambas enzimas é uma seqüência de peptídeo sintético que contém o
sítio de clivagem (Tyr-Pro) para a protease do HIV e o sítio (Leu-Val) para a renina, bem
como dois aminoácidos covalentemente modificados para a detecção da clivagem (Matayoshi
et al., 1990). Ambos substratos foram degradados pelo extrato protéico de L. amazonensis,
porém o substrato para renina teve uma taxa de hidrólise um pouco mais acentuada. A
pepstatina A, um inibidor clássico de aspártico proteases, inibiu fortemente a atividade
enzimática a 1µM, ao contrário do que foi relatado por Valdivieso e colaboradores, que não
detectaram qualquer inibição da aspártico protease de L. mexicana por pepstatina A a 5 µM,
porém foi verificada a completa inibição por outro inibidor de aspártico proteases, o DAN
(Valdivieso, Dagger e Rascón, 2007). Porém, diferenças de sensibilidade a inibidores de
aspártico proteases já vem sendo descritas para outros organismos como bactérias e
nematódeos (Oda et al., 1987; Toogood et al., 1993, 1995; Sato et al., 1993). Esse resultado
também sugere a possibilidade de diferença dessas enzimas dentro do próprio gênero
Leishmania. Os IPs-HIV nelfinavir e lopinavir também foram capazes de inibir a atividade
enzimática de Leshmania amazonensis, mostrando que as aspártico proteases em Leishmania
são um alvo em potencial para esses inibidores e que estudos adicionais são necessários para
melhor caracterizar essas proteases, e para tentar identificar o alvo intracelular dos IPs-HIV.
Curiosamente, a inibição enzimática por lopinavir e nelfinavir, a 1 e 10 µM, respectivamente,
se mostrou muito mais proeminente nos ensaios realizados com o substrato para aspártico
protease do HIV-1 do que para o substrato da renina. Essa diferença acentuada no percentual
de inibição enzimática para os ensaios realizados com diferentes substratos, talvez esteja
relacionada à degradação por aspártico proteases diferentes, que exibem afinidade distinta
pelos IPs-HIV.
A infecção murina é um dos melhores modelos experimentais caracterizados para o
estudo da interação Leishmania-células hospedeiras de mamíferos (Pereira & Alves, 2008).

64
Dados de infecção de macrófagos peritoneais de camundongo por L. amazonensis revelaram
que parasitos pré-tratados com inibidores de protease têm índices de associação
consideravelmente menores em comparação aos sistemas controle, com parasitos não tratados.
Estes dados sugerem a interferência nas etapas iniciais da infecção de macrófagos, assim como
na modulação de fatores de virulência ainda não identificados ou por efeitos tóxicos não
específicos diretos dos inibidores nas formas promastigotas. Deve-se destacar que estes perfis
inibitórios da interação não foram causados pela diminuição da viabilidade celular de L.
amazonensis, uma vez que controles de viabilidade foram realizados como, observação de
motilidade, contagem em câmara de neubauer e exclusão do iodeto de propídeo.
Anteriormente, foi demonstrado que as atividades de aspártico, metalo e serino proteases são
reguladas negativamente durante a transformação induzida por choque térmico de
promastigotas para amastigotas in vitro (Alves et al., 2005). Contudo, a identificação do
mecanismo molecular pelo qual os IPs-HIV interferem na interação com células de mamíferos
ainda é uma questão em aberto. É possível que aspártico proteases estejam envolvidas nas
etapas iniciais da interação com macrófagos e/ou na diferenciação dos parasitos. Contudo, o
efeito dos IPs-HIV na diferenciação não foi constatado nas concentrações utilizadas neste
trabalho.
A efetividade de inibidores contra Leishmania pode depender da fase de
desenvolvimento do parasito. Por exemplo, amastigotas de L. mexicana em desenvolvimento
dentro de macrófagos durante a infecção humana são mais sensíveis à lactacistina, um inibidor
de proteassoma, do que promastigotas que se desenvolvem em meio de cultura (Robertson,
1999), o que pode ser explicado pela ação distinta das drogas sobre promastigotas isolados e
amastigotas no ambiente intracelular, ou pela concentração das drogas pelos macrófagos. A
fim de avaliar se o mesmo acontece com os IPs-HIV, o efeito de nelfinavir, amprenavir e
lopinavir sobre a multiplicação de Leishmania no interior das células alvo foi verificado. As
concentrações da droga empregadas neste ensaio não mostraram toxicidade direta sobre
macrófagos e reduziram drasticamente o índice de associação. A maneira como atuam os
inibidores, seja diretamente sobre os amastigotas, ou indiretamente, através da modulação da
capacidade do macrófago de matar o parasito, permanece uma questão em aberto. Além disso,
os macrófagos poderiam concentrar níveis elevados dos IPs-HIV. Por exemplo, foi mostrado
para L. chagasi, que concentrações mais elevadas da droga são necessárias para interferir no
desenvolvimento de amastigotas axênicos quando comparados com amastigotas intracelulares
(Trudel et al., 2008). Em adição, mesmo as menores doses necessárias para inibir o
desenvolvimento de amastigota são mais elevadas do que aquelas necessárias para a inibição
da progressão do HIV em humanos. No entanto, deve-se considerar que o TARV consiste de

65
uma combinação de medicamentos anti-retrovirais, e a farmacodinâmica de um modelo in
vitro é muito diferente do que ocorre em seres humanos. Além disso, os IPs-HIV foram
projetados para se ajustarem a protease viral e podem, assim, ter uma baixa afinidade para
aspártico proteases de L. amazonensis. Estudos adicionais in vivo podem ajudar a elucidar essa
questão, e também mais esforços devem ser direcionados para encontrar novos inibidores que
sejam mais específicos para as enzimas do parasito.
No Brasil, as forma clínica encontrada em casos de co-infecção é majoritariamente
visceral. Em face da distribuição da leishmaniose no Brasil, sua expansão para importantes
cidades e sua sobreposição com indivíduos infectados pelo HIV, é importante entender as
sofisticadas inter-relações entre Leishmania, HIV e macrófagos. Além disso, existem várias
questões não resolvidas relativas à conduta com pacientes co-infectados. Por exemplo, a
eficácia da terapia visando o controle de cada patógeno em indivíduos co-infectados
permanece indefinida. Os resultados apresentados neste estudo indicam que L. amazonensis é
susceptível aos IPs-HIV, e acrescentam mais um microrganismos à lista de patógenos
oportunistas susceptíveis a esses compostos. Dessa forma, seria interessante avaliar o efeito
sinérgico de compostos antileishmania clássicos e os IPs-HIV em macrófagos co-infectados
com Leishmania e HIV-1. Estas buscas são relevantes especialmente, em meio a descoberta
de que o estibogluconato de sódio pode estimular a replicação do HIV-1 in vitro (Barat et al.,
2007), e que a infecção por Leishmania também aumenta a replicação do HIV-1 (Zhao,
Papadopoulou & Tremblay, 2004; Zhao et al., 2006), do mesmo modo que macrófagos co-
infectados permitem um crescimento maior de Leishmania, em comparação com macrófagos
livres do HIV-1 (Barreto-de-Souza et al., 2006). O efeito do HIV-1 em facilitar a progressão
da infecção da leishmânia nos macrófagos é bem ilustrado pelo fato de Blastocrithidia culicis,
um protozoário tipicamente não patogênico, ser capaz de infectar macrófagos infectados com
HIV-1 (Barreto-de-Souza et al., 2008).
Por fim, foi recentemente relatado que promastigotas de L. infantum só aumentam a
replicação do HIV-1 em macrófagos derivados de monócitos nas etapas finais da curva de
produção do vírus, mas também, surpreendentemente, é observada uma redução na produção
de HIV-1 durante os dias iniciais após a infecção. Este efeito primário é causado pela inibição
da entrada do vírus dentro dos macrófagos através da ação da lipofosfoglicana (LPG), o
principal glicolipídeo de superfície dos promastigotas de Leishmania, o que sugere o
estabelecimento de interações complexas em células hospedeiras que são comuns ao HIV e a
Leishmania (Garg et al., 2008). Nesse contexto, o estudo das aspártico proteases em
Leishmania spp. se faz importante para a melhor compreensão da patogenia da leishmaniose e

66
da co-infecção com HIV, além de gerar conhecimentos para a otimização da quimioterapia nos
pacientes co-infectados.

67
VII. Conclusões
• O tratamento de Leishmania amazonensis com os IPs-HIV (nelfinavir, lopinavir, e
amprenavir) promoveu a queda da proliferação celular, especialmente quando concentrações
mais elevadas das drogas foram utilizadas.
• Os IPs-HIV foram capazes de causar alterações ultraestruturais marcantes, tais como,
aumento de vesículas, encolhimento de citoplasma, condensação de cromatina, aumento de
retículo em volta do núcleo e intensa liberação de pedaços de membrana, o que culminou com
a morte celular.
• Análises bioquímicas revelaram que a expressão de proteases em Leishmania
amazonensis, comprovadamente fatores de virulência, tais como gp63 e cpb, foi aumentada
após o tratamento do parasito com os IPs-HIV.
• A atividade de aspártico protease foi detectada contra substratos específicos e inibida
pelos IPs-HIV, o que sugere que estas enzimas possam ser um alvo potencial para estes
compostos.
• Nos ensaios de interação, o índice de associação do parasito com macrófagos foi
reduzido, tanto no pré-tratamento, quanto no pós-tratamento com os IPs-HIV, mostrando que
de alguma forma, esses inibidores interferem na relação parasito-hospedeiro, quer seja,
diretamente sobre o parasito, quer seja pela modulação da ação dos macrófagos.
• Considerados em conjunto, os resultados indicam que os IPs-HIV podem ser
quimioterápicos promissores no tratamento da leishmaniose. Isto porque, estas drogas parecem
interferir no metabolismo de Leishmania amazonensis e nas relações de interação entre o
parasito e sua célula hospedeira.

68
VIII. Anexo:
1. Produção bibliográfica relacionada à dissertação de mestrado 1.1. Artigo publicado
Santos, L.O., Marinho, F.A., Altoé, E.F., Vitório, B.S., Alves, C.R., Britto, C., Motta, M.C., Branquinha, M.H., Santos, A.L.S., d`Ávila-levy, C.M. 2009. HIV Aspartyl peptidase inhibitors interfere with cellular proliferation, ultraestructure and macrophage infection of Leishmania amazonensis. 1.2. Trabalhos selecionados para apresentação oral em congressos, simpósios ou
seminários Simpósio Jovem Cientista do Cone Sul 2009. trabalho aceito para apresentação oral no congresso brasileiro da sociedade de bioquímica e biologia molecular (SBBq). Simpósio Jovem Cientista do Cone Sul 2009. Santos, L.O., Marinho, F.A., Altoé, E.F., Vitório, B.S., Alves, C.R., Britto, C., Motta, M.C., Branquinha, M.H., Santos, A.L.S., d`Ávila-levy, C.M. 2009. HIV Aspartyl peptidase inhibitors interfere with cellular proliferation, ultraestructure and macrophage infection of Leishmania amazonensis.
1.3. Trabalhos publicados em anais de congressos
Santos, L.O., Branquinha, M.H., Santos, A.L.S., Britto, C., Alves, C.R., Martins, L.Z., Vitório, B.S., d`Ávila-levy, C.M. 2008. Effect of HIV protease inhibitors on promastigote forms os Leishmania amazonensis. In: XXXVII Reunião anual da sociedade brasileira de bioquímica e biologia molecular (SBBq). Águas de Lindóia.
Santos, L.O., Martins, L.Z., Vitório, B.S., Motta, M.C., Britto, C., Branquinha, M.H., Santos, A.L.S., Alves, C.R., d`Ávila-levy, C.M. 2008. Antileishmanial activity of HIV protease inhibitors. In: XXIV Reunião da sociedade de protozoologia e XXXV Reunião para pesquisa básica em doença de chagas (SBPz). Águas de Lindóia. Santos L.O. 2008. Atividade leishmanicida de inibidores de protease do HIV. In: Bienal de pesquisa, XVI Raic. Rio de Janeiro.
Santos, L.O., Branquinha, M.H., Santos, A.L.S., Alves, C.R., d`Ávila-levy, C.M. 2007. Effect of nelfinavir and indinavir on promastigote forms of Leishmania amazonensis. In: XXII Reunião anual da sociedade brasileira de protozoologia (SBPz), Caxambu.





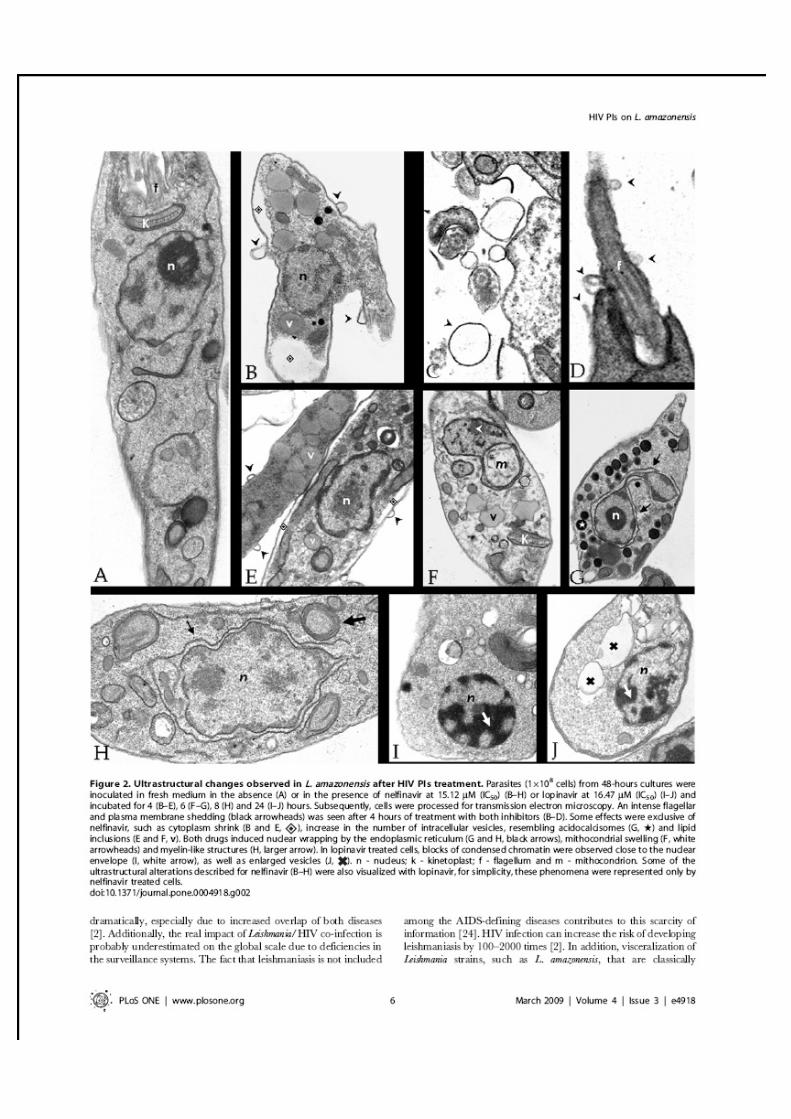

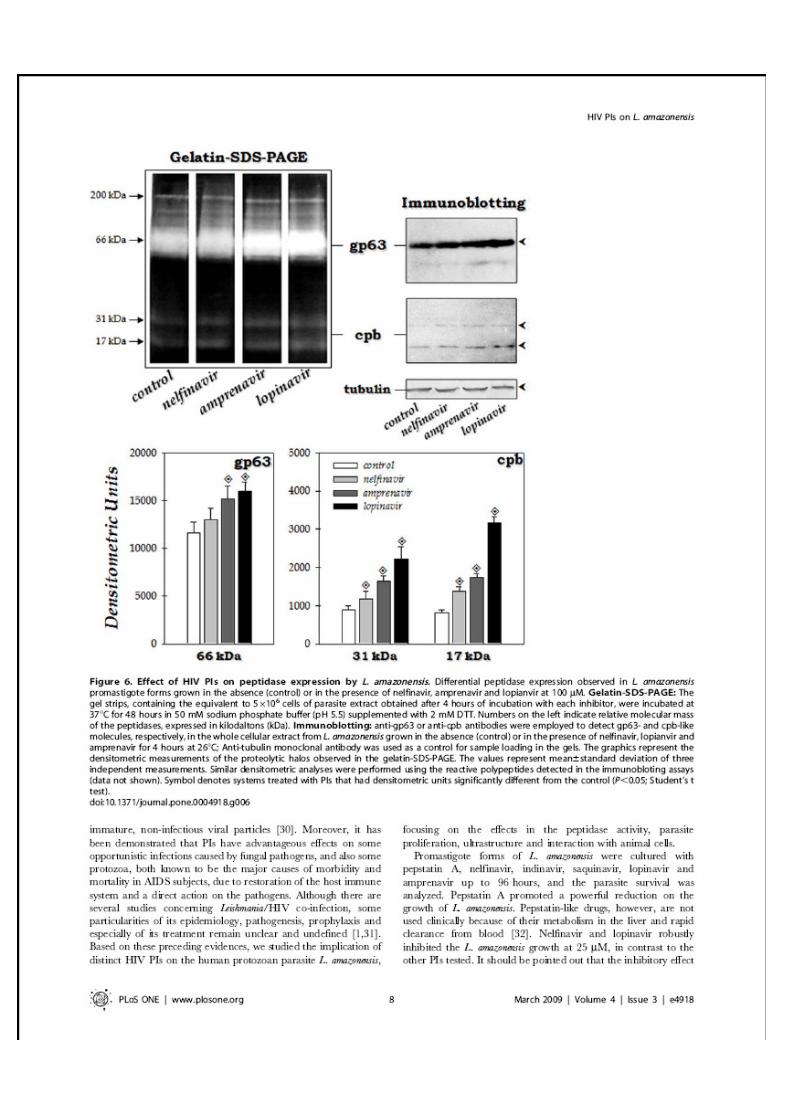




80
IX. Bibliografia:
Aidsinfo, Offering information on HIV/AIDS Treatment, Prevention and Research [homepage
na Internet]. Rockville: US Department of health and human services; c2007. [acesso em 2008 Dez 15]. Treatment of HIV Infection; [aproximadamente 2 telas]. Disponível em: http://www.niaid.nih.gov/factsheets/treat-hiv.htm
Aidsinfo, Offering information on HIV/AIDS Treatment, Prevention and Research [homepage
na Internet]. Rockville: US Department of health and human services; c2008. [acesso em 2008 Dez 15]. Panel on Antiretroviral Guidelines for Adults and Adolescents. Guidelines for the use of antiretroviral agents in HIV-1-infected adults and adolescents; [aproximadamente 2 telas]. Disponível em: http://www.aidsinfo.nih.gov/contentfiles/adultandadolescentgl.pdf.
Alvar J, Aparicio P, Aseffa A, Den Boer M, Cañavate C, Dedet JP et al. The Relationship
between Leishmaniasis and AIDS: the Second 10 Years. Clin Microbiol Rev. 2007; 21(2): 334-59.
Alvar J, Cañavate C, Guitérrez-Solar B, Jiménez M, Laguna F, López-Vélez R et al.
Leishmania and human immunodeficiency virus coinfection: the first 10 years. Clin Microbiol Rev. 1997; 10(2): 298-318.
Alvar J, Croft S, Olliaro P. Chemotherapy in the treatment and control of leishmaniasis. Adv
Parasitol. 2006; 6: 223-274. Alves CR, Benevolo-De-Andrade TC, Alves JL, Pirmez C. Th1 and Th2 immunological
profile induced by cysteine proteinase in murine leishmaniasis. Parasite Immunol. 2004; 26(3): 127-35.
Alves CR, Corte-Real S, Bourguignon SC, Chaves CS, Saraiva EM. Leishmania
amazonensis: early proteinase activities during promastigote-amastigote differentiation in vitro. Exp. Parasitol. 2005; 109(1): 38-48.
Alves CR, Pontes de Carvalho LC, Souza AL, De Simone SG. A strategy for the
identification of T-cell epitopes on Leishmania cysteine proteinases. Cytobios. 2001; 104(405): 33-41.
Andrade ASR, Santoro MM, Melo MN, Mares-Guia M. Leishmania (Leishmania)
amazonensis: Purification and enzymatic characterization of a soluble serine oligopeptidase from promastigotes. Exp Parasitol. 1998; 89(2): 153-160.
André P, Groettrup M, Klenerman P, de Giuli R, Booth BL Jr, Cerundolo V, Bonneville M,
Jotereau F, Zinkernagel RM, Lotteau V. An inhibitor of HIV-1 protease modulates proteasome activity, antigen presentation, and T cell responses. Proc Natl Acad Sci U S A. 1998; 95(22): 13120-4.
Andrews KT, Fairlie DP, Madala PK, Ray J, Wyatt DM, Hilton PM et al. Potencies of human
immunodeficiency virus protease inhibitors in vitro against Plasmodium falciparum and in vivo against murine malaria. Antimicrob Agents Chemother. 2006; 50(2): 639-48.

81
Andrews L, Friedland G. Progress in HIV therapeutics and the challenges of adherence to antiretroviral therapy. Infect Dis Clin North Am. 2000; 14(4): 901-28.
Azeredo-Coutinho RB, Conceicao-Silva F, Schubach A, Cupolillo E, Quintella LP, Madeira
MF et al. First report of diffuse cutaneous leishmaniasis and Leishmania amazonensis infection in Rio de Janeiro State, Brazil. Trans R Soc Trop Med Hyg. 2007; 101(7): 735-7.
Bal G, Van der Veken P, Antonov D, Lambeir AM, Grellier P, Croft SL et al. A
Prolylisoxazoles: potent inhibitors of prolyloligopeptidase with antitrypanosomal activity. Bioorg Med Chem Lett. 2003; 13(17): 2875-8.
Bañuls AL, Hide M, Prugnolle F. Leishmania and the leishmaniases: a parasite genetic update
and advances in taxonomy, epidemiology and pathogenicity in humans. Adv Parasitol. 2007; 64:1-109. Review.
Barat C, Zhao C, Ouellette M, Tremblay MJ. HIV-1 replication is stimulated by sodium
stibogluconate, the therapeutic mainstay in the treatment of leishmaniasis. J Infect Dis. 2007; 195(2): 236-45.
Barr SC, Warner KL, Kornreic BG, Piscitelli J, Wolfe A, Benet L, McKerrow JH. A cysteine
protease inhibitor protects dogs from cardiac damage during infection by Trypanosoma
cruzi. Antimicrob Agents Chemother. 2005; 49(12): 5160-1. Barral A, Badaró R, Barral-Netto M, Grimaldi G Jr, Momem H, Carvalho EM. Isolation of
Leishmania mexicana amazonensis from the bone marrow in a case of American visceral leishmaniasis. Am J Trop Med Hyg 1986; 35: 732-4.
Barral A, Pedral-Sampaio D, Grimaldi G Jr, Momen H, McMahon-Pratt D, Ribeiro de Jesús
A, et al. Leishmaniasis in Bahia, Brazil: evidence that Leishmania amazonensis produces a wide spectrum of clinical disease. Am J Trop Med Hyg 1991; 44: 536-46.
Barreto-de-Souza V, Pacheco GJ, Silva AR, Castro-Faria-Neto HC, Bozza PT, Saraiva EM et
al. Increased Leishmania replication in HIV-1-infected macrophages is mediated by tat protein through cyclooxygenase-2 expression and prostaglandin E2 synthesis. J. Infect. Dis. 2006; 194(6): 846–54.
Barreto-de-Souza V, Xavier Medeiros T, Machado Motta MC, Bou-Habib DC, Saraiva EM.
HIV-1 infection and HIV-1 Tat protein permit the survival and replication of a non-pathogenic trypanosomatid in macrophages through TGF-beta1 production. Microbes Infect. 2008; 10(6): 642-9.
Barrett AJ, Rawlings ND, O'Brien EA. The MEROPS database as a protease information
system. J Struct Biol. 2001; 134(2-3):95-102. Barrett AJ, Tolle DP, Rawlings ND. Managing peptidases in the genomic era. Biol Chem.
2003; 384(6): 873-82. Berhe N, Hailu A, Gemetchu T. Human immunodeficiency virus and recurrence of cutaneous
leishmaniasis long after healed localized cutaneous leishmaniasis due to Leishmania
aethiopica. Trans R Soc Trop Med Hyg. 1995; 89(4): 400-1.

82
Berhe N, Wolday D, Hailu A, Abraham Y, Ali A, Gebre-Michael T et al. HIV viral load and response to antileishmanial chemotherapy in co-infected patients. AIDS. 1999; 13(14): 1921-1925.
Berman JJ. Treatment of leishmaniasis with miltefosine: 2008 status. Expert Opin Drug Metab Toxicol. 2008; 4(9): 1209-16.
Besteiro S, Williams RA, Coombs GH, Mottram JC. Protein turnover and differentiation in
Leishmania. Int J Parasitol. 2007; 37(10): 1063-75. Review. Bond JS, Butler PE. Intracellular proteases. Annu Rev Biochem. 1987; 56: 333-64. Bordier C, Etges RJ, Ward J, Turner MJ, Cardoso de Almeida ML. Leishmania and
Trypanosoma surface glycoproteins have a common glycophospholipid membrane anchor. Proc Natl Acad Sci U S A. 1986; 83(16): 5988-91.
Bordier C. The promastigote surface protease of Leishmania. Parasitol Today. 1987; 3(5):
151-3. Brandão AA, Miranda A, Degrave WM, Sousa MA. The heterogeneity of choanomastigote-
shaped trypanosomatids as analyzed by their kDNA minicircle size: taxonomic implicaitons. Parasitol Res. 2000; 86 (10):809-12.
Branquinha MH, Vermelho AB, Goldenberg S, Bonaldo MC. Ubiquity of cysteine- and
metalloproteinase activities in a wide range of trypanosomatids. J Eukaryot Microbiol. 1996; 43(2): 131-5.
Burleigh BA, Caler EV, Webster P, Andrews N. A cytosolic serine endopeptidase from
Trypanosoma cruzi is required for the generation of Ca+2 signaling in mammalian cells. J Cell Sci. 1997; 136(3): 609-20.
Califórnia Lutheran University [homepage na Internet]. Califórnia: c2009 [acesso em 2009
fev 02]. HIV-1 protease; [aproximadamente 2 telas]. Disponível em: http://www.callutheran.edu/Academic_Programs/Departments/BioDev/omm/hiv_protease/molmast.htm
Casolari C, Rossib T, Baggio B, Coppic A, Zandomeneghic G, Rubertoc AI et al. Interaction
between saquinavir and antimycotic drugs on C. Albicans and C. Neoformans strains. Pharmacol Res. 2004; 50(6): 605–10.
Cazzulo JJ, Stoka V, Turk V. The major cysteine proteinase of Trypanosoma cruzi: a valid
target for chemotherapy of Chagas disease. Curr Pharm Des. 2001; 7(12): 1143-56. CDC (Centers for Disease Control). Viscerotropic leishmaniasis in persons returning from
operation Desert Storm 1990-1991. Morbid Mortal Wkly Rep. 1992; 41(8): 131-4. Chang KP, Akman L, Nielsen JS. Leishmania virulence and genetic heterogeneity. Clin
Dermatol. 1999; 17(3): 269-73. Review. Chappuis F, Sundar S, Hailu A, Ghalib H, Rijal S, Peeling RW et al. Visceral leishmaniasis:
what are the needs for diagnosis, treatment and control? Nat Rev Microbiol. 2007; 5(11):873-82. Review.

83
Choudhury R, Bhaumik SK, De T, Chakraborti T. Identification, purification, and characterization of a secretory serine protease in an Indian strain of Leishmania donovani. Mol Cell Biochem. 2009; 320(1-2): 1-14.
Coetzer THT, Goldringa JPD, Husona LEJ. Cellular proteolysis Oligopeptidase B: A
processing peptidase involved in pathogenesis. Biochimie. 2008; 90(2): 336-344. Cohen-Freuea G, Holzerb TR, Forneyb JD, McMaster WR. Global gene expression in
Leishmania. Int J Parasitol. 2007; 37(10): 1077-86. Corrêa JR, Brazil RP, Soares MJ. Leishmania (Viannia) lainsoni (Kinetoplastida:
Trypanosomatidae), a divergent Leishmania of the Viannia subgenus--a mini review. Mem Inst Oswaldo Cruz. 2005; 100(6): 587-92.
Craik M, Debouck C. In: Perspectives in drug discovery and design. McKerrow JH, James
MNG, editors. Leiden: ESCOM. 1995; 2: 1-125. Cruz I, Nieto J, Moreno J, Canavate C, Desjeux P, Alvar J. Leishmania/HIV co-infections in
the second decade. Indian J Med Res. 2006; 123(3): 357-88. Cunico W, Gomes CRB, Vellasco WT Jr. HIV – recentes avanços na pesquisa de fármacos.
Quim Nova. 2008; 31(8): 2111-7. Review. Cupolillo E, Aguiar Alves F, Brahim LRN, Naiff MF, Pereira LO, Oliveira-Neto MP et al.
Recent advances in the taxonomy of the new world leishmanial parasites. Med Microbiol Immunol. 2001; 190(1-2): 57-60.
Cupolillo E, Momen H, Grimaldi G Jr. Genetic diversity in natural populations of New World
Leishmania. Mem Inst Oswaldo Cruz. 1998; 93(5): 663-8. Da-Cruz AM, Machado ES, Menezes JA, Rutowitsch MS, Coutinho SG. Cellular and
humoral immune responses of a patient with American cutaneous leishmaniasis and AIDS. Trans R Soc Trop Med Hyg. 1992; 86: 511-512.
Dash C, Kulkarni A, Dunn B, Rao M. Aspartic peptidase inhibitors: implications in drug
development. Crit Rev Biochem Mol Biol. 2003; 38(2): 89-119. Review. De Araújo Soares RM, Santos ALS, Bonaldo MC, Braga AF, Alviano CS, Angluster J et al.
Leishmania (Leishmania) amazonensis: differential expression of proteinases and cell-surface polypeptides in virulent and avirulent promastigotes. Exp Parasitol. 2003; 104(3-4): 104-12.
De Souza W. Special organelles of some pathogenic protozoa. Parasitol Res. 2002; 88(12):
1013-1025. Review. Denise H, McNeil K, Brooks DR, Alexander J, Coombs GH, Mottram JC. Expression of
multiple CPB genes encoding cysteine proteases is required for Leishmania mexicana virulence in vivo. Infect Immun. 2003; 71(6): 3190-5.
Denise H, Poot J, Jimenez M, Ambit A, Herrmann DC, Vermeulen AN et al. Studies on the
CPA cysteine peptidase in the Leishmania infantum genome strain JPCM5. BMC Mol Biol. 2006; 7: 42-55.

84
Desjeux P, Alvar J. Leishmania/HIV co-infections: epidemiology in Europe. Ann. Trop. Med.
Parasitol. 2003; 97: 3–15. Desjeux P. Leishmaniasis: current situation and new perspectives. Comp Immunol Microbiol
Infect Dis. 2004; 27(5): 305-18. Dominguez M, Torano A. Leishmania immune adherence reaction in vertebrates. Parasite
Immunol. 2001; 23: 259-65. Dunn WA Jr. Autophagy and related mechanisms of lysosome-mediated protein degradation.
Trends Cell Biol. 1994; 4(4): 139–43. Dutta A, Bandyopadhyay S, Mandal C, Chatterjee M. Development of a modified MTT assay
for screening antimonial resistant field isolates of Indian visceral leishmaniasis. Parasitol Int. 2005; 54(2): 119-22.
Eder J, Hommel U, Cumin F, Martoglio B, Gerhartz B. Aspartic proteases in drug discovery.
Curr Pharm Des. 2007; 13(3): 271-85. Review. Eron JJ Jr. HIV-1 protease inhibitors. Clin Infect Dis. 2000; 30(2): 160-70. Review. Fath M, Wu X, Hileman R, Linhardt R, Kashem M, Nelson R et al. Interaction of secretory
leukocyte protease inhibitor with heparin inhibits proteases envolved in asthma. J Biol Chem. 1998; 273: 13563-9.
FDA, U.S. Food and Drug Administration [homepage na Internet]. Silver Spring: US
Department of health and human services; c2008. [acesso em 2008 Dez 18]. Drugs used in the treatment of HIV infection; [aproximadamente 2 telas]. Disponível em: http://www.fda.gov/oashi/aids/virals.html
Flexner C. HIV-protease inhibitors. N Engl J Med. 1998; 338(18): 1281-1292. Frame MJ, Mottram JC, Coombs GH. Analysis of the roles of cysteine proteinases of
Leishmania mexicana in the host-parasite interaction. Parasitology. 2000; 121(Pt 4): 367-377.
Frommel TO, Button LL, Fujikura Y, McMaster WR. The major surface glycoprotein (GP63)
is present in both life stages of Leishmania. Mol Biochem Parasitol. 1990; 38(1): 25-32. Gaedicke S, Firat-Geier E, Constantiniu O, Lucchiari-Hartz M, Freudenberg M, Galanos C. et
al. Antitumor effect of the human immunodeficiency virus protease inhibitor ritonavir: induction of tumor-cell apoptosis associated with perturbation of proteasomal proteolysis. Cancer Res. 2002; 62: 6901-8.
Garg R, Lodge R, Descoteaux A, Tremblay MJ. Leishmania infantum promastigotes reduce
entry of HIV-1 into macrophages through a lipophosphoglycan-mediated disruption of lipid rafts. J Infect Dis. 2008; 197(12): 1701-8.
Gillis D, Klaus S, Schnur LF, Piscopos P, Maayan S, Okon E, Engelhard D. Diffusely
disseminated cutaneous Leishmania major infection in a child with acquired immunodeficiency syndrome. Pediatr Infect Dis. 1995; 14(3): 247-9.

85
González-Polo RA, Boya P, Pauleau AL, Jalil A, Larochette N, Souquère S et al. The
apoptosis/autophagy paradox: autophagic vacuolization before apoptotic death. J Cell Sci. 2005; 118(14): 3091-102.
Gradoni L, Scalone A, Gramiccia M, Troiani M. Epidemiological surveillance of
leishmaniasis in HIV-1-infected individuals in Italy. AIDS. 1996; 10(7): 785-91. Handman E, Bullen DV. Interaction of Leishmania with the host macrophage. Trends
Parasitol. 2002; 18(8): 332-4. Hill J, Phylip L. Bacterial aspartic proteinases. FEBS Lett. 1997; 409(3): 357–360. Hoegl L, Thoma-Greber E, Rocken M, Korting HC. Shift from persistent oral
pseudomembranous to erythematous candidosis in a human immunodeficiency virus (HIV)-infected patient upon combination treatment with an HIV protease inhibitor. Mycoses. 1998; 41(5-6): 213-7.
Hommer V, Eichholz J, Petry F. Effect of antiretroviral protease inhibitors alone, and in
combination with paromomycin, on the excystation, invasion and in vitro development of Cryptosporidium parvum. J Antimicrob Chem. 2003; 52(3): 359–64. Corrigido e republicado: J Antimicrob Chemother. 2003; 52(3): 535.
Hong L, Turner RT, Koelsch G, Shin D, Ghosh AK, Tang J. Crystal structure of memapsin 2
(β-secretase) in complex with an inhibitor OM00-3. Biochem. 2002; 41: 10963-7. Hooper NM. Proteases: a primer. Essays Biochem. 2002; 38: 1-8. Review. Hoover D, Lefker B, Rosati R, Wester R, kleinman E, Bindra J et al. Discovery of inhibitors
of human rennin with high oral biovailability. Adv Exp Med Biol. 1995; 362: 167-80. Hsiao CH, Yao C, Storlie P, Donelson JE, Wilson ME. The major surface protease (MSP or
GP63) in the intracellular amastigote stage of Leishmania chagasi. Mol Biochem Parasitol. 2008; 157(2): 148-59.
Hughes A, Barber T, Nelson M. New treatment options for HIV salvage patients: an overview
of second generation PIs, NNRTIs, integrase inhibitors and CCR5 antagonists. J Infect. 2008; 57(1): 1-10. Review.
James M (editor). Structure and function of aspartic protease: retroviral and cellular enzymes.
Plenum Press. New York; 1998. Kamhawi S. Phlebotomine sand flies and Leishmania parasites: friends or foes? Trends
Parasitol. 2006; 22(9): 439-45. Kissová I, Plamondon LT, Brisson L, Priault M, Renouf V, Schaeffer et al. Evaluation of the
roles of apoptosis, autophagy, and mitophagy in the loss of plating efficiency induced by Bax expression in yeast. J Biol Chem. 2006; 281(47): 36187-97.
Kleinert H, Stein H, Boyd S, Fung A, Baker W, Verburg K et al. Discovery of a well-
absorbed, efficatious rennin inhibitor, a-74273. Hypertension. 1992; 20: 768-75.

86
Kulkarni MM, McMaster WR, Kamysz E, Kamysz W, Engman DM, McGwire BS. The major surface-metalloprotease of the parasitic protozoan, Leishmania, protects against antimicrobial peptide-induced apoptotic killing. Mol Microbiol. 2006; 62(5): 1484-97.
Lainson R, Shaw JJ. Evolution, classification and geographical distribution. In: Killick-
Kendric R, Peters W, editores. The Leishmaniases in Biology and Medicine. London: Academic Press; 1987. p. 1-120
Lainson R, Shaw JJ. Leishmaniasis of the New World: taxonomic problems. Brit Med Bull.
1972; 28: 44-48. Lainson R, Shaw JJ. The role of animals in the epidemiology of South American
leishmaniasis. In: Lumsden WHR, Evans DA, editores. Biology of the Kinetoplastida. Vol. 2, London and New York: Academic Press; 1979. p. 1-116.
Levine B, Klionsky DJ. Development by self-digestion: molecular mechanisms and biological
functions of autophagy. Dev Cell. 2004; 6(4): 463–77. Lieke T, Nylén S, Eidsmo L, McMaster WR, Mohammadi AM, Khamesipour A, Berg L,
Akuffo H. Leishmania surface protein gp63 binds directly to human natural killer cells and inhibits proliferation. Clin Exp Immunol. 2008; 153(2): 221-30.
Lindoso, JA. Comportamento Oportunista das Leishmanioses/Opportunistic Behavior of
Leishmaniosis. BEPA. 2006; 3(32): 8-12. Lopez-Velez R, Perez-Molina JA, Guerrero A, Baquero F, Villarrubia J, Escribano L et al.
Clinicoepidemiologic characteristics, prognostic factors, and survival analysis of patients coinfected with human immunodeficiency virus and Leishmania in an area of Madrid, Spain. Am J Trop Med Hyg. 1998; 58(4): 436-43.
Magill AJ, Grögl M, Gasser RA Jr, Sun W, Oster CN. Visceral infection caused by
Leishmania tropica in veterans of Operation Desert Storm. N Engl J Med. 1993; 328(19): 1383-7.
Majer F, Pavlícková L, Majer P, Hradilek M, Dolejsí E, Hrusková-Heidingsfeldová O,
Pichová I. Structure-based specificity mapping of secreted aspartic proteases of Candida parapsilosis, Candida albicans, and Candida tropicalis using peptidomimetic inhibitors and homology modeling. Biol Chem. 2006; 387(9): 1247-54.
Marques N, Cabral S, Sá R, Coelho F, Oliveira J, Saraiva da Cunha JG et al. Visceral
leishmaniasis and HIV infection in the HAART era. Acta Med Port. 2007; 20(4): 291-8. Martinez-Cajas JL, Wainberg MA. Protease inhibitor resistance in HIV-infected patients:
molecular and clinical perspectives. Antiviral Res. 2007; 76(3): 203-21. Review. Mastrolorenzo A, Rusconi S, Scozzafava A, Barbaro G, Supuran CT. Inhibitors of HIV-1
protease: current state of the art 10 years after their introduction. From antiretroviral drugs to antifungal, antibacterial and antitumor agents based on aspartic protease inhibitors. Curr Med Chem. 2007; 14(26): 2734-48. Review.
Matayoshi ED, Wang GT, Krafft GA, Erickson J. Novel fluorogenic substrates for assaying
retroviral proteases by resonance energy transfer. Science. 1990; 247(4945): 954-8.

87
McGwire B, Chang KP. Genetic rescue of surface metalloproteinase (gp63)-deficiency in Leishmania amazonensis variants increases their infection of macrophages at the early phase. Mol Biochem Parasitol. 1994; 66(2): 345-7.
McKerrow JH, Caffrey C, Kelly B, Loke P, Sajid M. Proteases in parasitic diseases. Annu
Rev Pathol. 2006; 1: 497-536. Review. McKerrow JH, Rosenthal PJ, Swenerton R, Doyle P. Development of protease inhibitors for
protozoan infections. Curr Opin Infect Dis. 2008; 21(6): 668-72. Review. Mckerrow JH, Sun E, Rosenthal PJ, Bouvier J. The proteases and pathogenicity of parasitic
protozoa. Annu. Rev. Microbiol. 1993; 47: 821-853. Mckerrow JH. Development of cysteine protease inhibitors as chemoterapy for parasitic
diseases: insights on safety, target validation, and mechanisms of action. Int J Parasitol. 1999; 29(6): 833-7.
Medina-Acosta E, Karess RE, Schwarz H, Russell DG. The promastigote surface protease
(gp63) of Leishmania is expressed but differentially processed and localized in the amastigote stage. Mol Biochem Parasitol. 1989; 37(2): 263-73.
Mele R, Morales MA, Tosini F, Pozio E. Indinavir reduces Cryptosporidium parvum infection
in both in vitro and in vivo models. Int J Parasitol. 2003; 33(7): 757-64. Ministério da Saúde. [homepage na Internet]. Brasília D.F.: Serviço de Vigilância em Saúde;
c2007 [acesso em 2008 Dez 16]. Manual de controle da leishmaniose tegumentar americana; [aproximadamente 3 telas]. Disponível em: http://portal.saude.gov.br/portal/arquivos/pdf/manual_lta_2ed.pdf.
Ministério da Saúde. [homepage na Internet]. Brasília: Serviço de Vigilância em Saúde;
[acesso em 2008 Dez 16]. Vigilância em saúde; [aproximadamente 3 telas]. Disponível em: http://portal.saude.gov.br/portal/saude/visualizar_texto.cfm?Idtxt=27239.
Monari C, Pericolini E, Bistoni G, Cenci E, Bistoni F, Vecchiarelli A. Influence of indinavir
on virulence and growth of Cryptococcus neoformans. J Infect Dis. 2005; 191(2): 307–11. Morgado-Diaz JA, Silva-Lopez RE, Alves CR, Soares MJ, Corte-Real S, De Simone SG.
Subcellular localization of an intracellular serine protease of 68 kda in Leishmania (Leishmania) amazonensis promastigotes. Mem Inst Oswaldo Cruz. 2005; 100(4): 377-83.
Morty RE, Lonsdale-Eccles JD, Morehead J, Caler EV, Mentele R, Auerswald EA et al.
Oligopeptidase B from Trypanosoma brucei, a new member of an emerging subgroup of serine oligopeptidases. J Biol Chem. 1999; 274(37): 26149-26156.
Morty RE, Pelle R, Vadasz I, Uzcanga GL, Seeger W, Bubis J. Oligopeptidase B from
Trypanosoma evansi. A parasite peptidase that inactivates atrial natriuretic factor in the bloodstream of infected hosts. J Biol Chem. 2005; 280(12): 10925-10937.
Mottram JC, Coombs GH, Alexander J. Cysteine peptidases as virulence factors of
Leishmania. Curr Opin Microbiol. 2004; 7(4): 375-381.

88
Munro CA, Hube B. Anti-fungal therapy at the HAART of viral therapy. Trends Microbiol. 2002; 10(4): 173-7.
Murray HW, Berman JD, Davies CR, Saravia NG. Advances in leishmaniasis. Lancet. 2005;
366: 1561–77. NCBI [homepage na Internet]. Rockville Pike (Bethesda): National Center for Biotechnology
Information; [atualizada em 2009 jan 8; acesso em 2009 jan 25]. Pubchem compoud; [aproximadamente 2 telas]. Disponível em: http://www.ncbi.nlm.nih.gov/sites/entrez?db=pccompound.
Nguyen JT, Hamada Y, Kimura T, Kiso Y. Design of potent aspartic protease inhibitors to
treat various diseases. Arch Pharm (Weinheim). 2008; 341(9): 523-35. Review. Nigro L, Rizzo ML, Vancheri C, La Rosa R, Mastruzzo C, Tomaselli V et al. Med Microbiol
Immunol. 2007; 196(4): 253-5. Oda K, Sugitani M, Fukuhara K, Murao S. Purification and properties of a pepstatin-
insensitive carboxyl proteinase from a gram-negative bacterium. Biochim Biophys Acta. 1987; 923(3): 463-9.
Oliveira JPC, Fernandes F, Cruz AK, Trombela V, Monteiro E, Camargo AA et al. Genetic
diversity of Leishmania amazonensis strains isolated in northeastern Brazil as revealed by DNA sequencing, PCR-based analyses and molecular karyotyping. Kinetoplastid Biol Dis. 2007; 6: 5.
Page MJ, Di Cera E. Evolution of peptidase diversity. J Biol Chem. 2008; 283(44): 30010-4. Pal S, Mandal A, Duttagupta S. Studies on stibanate resistant Leishmania donovani isolate of
Indian origin. Indian J Exp Biol. 2001; 39(3): 249-54. Palella FJ Jr, Delaney KM, Moorman AC, Loveless MO, Fuhrer J, Satten GA et al. Declining
morbidity and mortality among patients with advanced human immunodeficiency virus infection. HIV Outpatient Study Investigation. N Engl J Med. 1998; 338(13): 853-60.
Palmeira VF, Kneipp LF, Rozental S, Alviano CS, Santos AL. Beneficial effects of HIV
peptidase inhibitors on Fonsecaea pedrosoi: promising compounds to arrest key fungal biological processes and virulence. PLoS ONE. 2008; 3(10): 3382.
Pan American Health Oganization (PAHO) [homepage na Internet]. Washington D.C.:
Regional Office of the World Health Organization; c2007-2009 [acesso em 2009 jan 28]. Leishmaniasis: 2007 Update; [aproximadamente 3 telas]. Disponível em: http://www.paho.org/English/AD/DPC/CD/leish-2007.htm.
Paugam A, Bulteau AL, Dupouy-Camet J, Creuzet C, Friguet B. Characterization and role of
protozoan parasite proteasomes. Trends Parasitol. 2003; 19(2): 55-9. Pereira BA, Alves CR. Immunological characteristics of experimental murine infection with
Leishmania (Leishmania) amazonensis. Vet Parasitol. 2008; 158(4): 239–55. Pernerstorfer-Schoen H, Jilma B, Perschler A, Wichlas S, Schindler K, Schindl A et al. Sex
differences in HAART-associated dyslipidaemia. AIDS. 2001; 15(6): 725-34.

89
Pichová I, Pavlícková L, Dostál J, Dolejsí E, Hrusková-Heidingsfeldová O, Weber J et al. Secreted aspartic proteases of Candida albicans, Candida tropicalis, Candida parapsilosis and Candida lusitaniae. Inhibition with peptidomimetic inhibitors. Eur J Biochem. 2001; 268(9): 2669-77.
Pintado V, López-Vélez R. Visceral leishmaniasis associated with human immunodeficiency
virus infection. Enferm Infecc Microbiol Clin. 2001; 19(7): 353-7. Pintado V, Martín-Rabadán P, Rivera ML, Moreno S, Bouza E. Visceral leishmaniasis in
humanimmunodeficiency virus (HIV)-infected and non hivinfected patients. Medicine. 2001; 80(1): 54-73.
Pinti M, Orsi CF, Gibellini L, Esposito R, Cossarizza A, Blasi E et al. Identification and
characterization of an aspartyl protease from Cryptococcus neoformans. FEBS Lett. 2007; 581(20): 3882-6.
Pozio E, Morales MA. The impact of HIV-protease inhibitors on opportunistic parasites.
Trends Parasitol. 2005; 21(2): 58-63. Review. Rabello A, Orsini M, Disch J. Leishmania/HIV co-infection in Brazil: an appraisal. Ann.
Trop. Med. Parasitol. 2003; 97(1): 17–28. Ramos-Santos C, Hernández-Montes O, Sánchez-Tejeda G, Monroy-Ostria A. Visceral
leishmaniosis caused by Leishmania (L.) mexicana in a Mexican patient with human immunodeficiency virus infection. Mem Inst Oswaldo Cruz 2000; 95: 733-7.
Rangel EF, Lainson R. Ecologia das Leishmanioses. In: Rangel EF, Lainson R.
Flebotomíneos do Brasil. Rio de Janeiro: Fiocruz; 2003. p. 291-305. Rao MB, Tanksale AM, Ghatge MS, Deshpande VV. Molecular and biotechnological aspects
of microbial proteases. Microbiol Mol Biol Rev. 1998; 62(3): 597-635. Review. Rey L. Parasitologia: parasitos e doenças parasitárias do homem nos trópicos ocidentais. Rio
de Janeiro: Guanabara koogan; 2008. Rioux JA, Lanotte G, Serres E, Pratlong F, Bastien P, Perieres J. Taxonomy of Leishmania.
Use of isoenzymes. Suggestions for a new classification. Ann Parasitol Hum Comp. 1990; 65(3): 111-25. Review.
Robertson CD. The Leishmania mexicana proteasome. Mol Biochem Parasitol. 1999; 103(1):
49–60. Romero HD, Taranto NJ, Malchiodi EL. American tegumentary leishmaniasis associated with
AIDS in Argentina. Medicina (B Aires). 2004; 64(1): 47-50. Rüchel R, Ritter B, Scha¡rinski M. Modulation of experimental systemic murine candidosis
by intravenous pepstatin A. Zentralbl Bakteriol. 1990; 273(3): 391-403. Saag MS, Schooly RT. Antiretroviral chemotherapy. In: Remington J, Schwartz MN,
editores. Current Clinical Topics in Infectious Disease. Boston: McGraw Hill; 1998. p. 154–79.

90
Sajid M, McKerrow JH. Cysteine proteases of parasitic organisms. Mol Biochem Parasitol. 2002; 120(1):1-21. Review. Corrigido e republicado: Mol Biochem Parasitol. 2002; 121(1): 159.
Santos AL, d'Avila-Levy CM, Elias CG, Vermelho AB, Branquinha MH. Phytomonas
serpens: immunological similarities with the human trypanosomatid pathogens. Microbes Infect. 2007; 9(8): 915-921. Review.
Santos ALS, Abreu CM, Alviano CS, Soares RMA. Use of proteolytic enzymes as an
additional tool for trypanosomatid identification. Parasitology. 2005; 130 (1): 79-88. Santos DO, Coutinho CE, Madeira MF, Bottino CG, Vieira RT, Nascimento SB et al.
Leishmaniasis treatment--a challenge that remains: a review. Parasitol Res. 2008; 103(1): 1-10.
Saravia NG, Escorcia B, Osorio Y, Valderrama L, Brooks D, Arteaga L et al. Pathogenicity
and protective immunogenicity of cysteine proteinase-deficient mutants of Leishmania
mexicana in non-murine models. Vaccine. 2006; 24(19): 4247-4259. Sato K, Nagaib Y, Suzukic M. Purification and partial characterization of an acid proteinase
from Dirofilaria immitis. Mol Biochem Parasitol. 1993; 58(2): 293-9. Savoia D, Allice T, Tovo PA. Antileishmanial activity of HIV protease inhibitors. Int J
Antimicrob Agents. 2005; 26(1): 92– 4. Schechter I, Berger A. On the active site of proteases. 3. Mapping the active site of papain;
specific peptide inhibitors of papain. Biochem Biophys Res Commun. 1968; 32(5): 898-902.
Schechter I, Berger A. On the size of the active site in proteases. I. Papain. Biochem Biophys
Res Commun. 1967; 27(2): 157-62. Schilling RMD. NetHealthBook.com [homepage na Internet]. Kelowna (Canadá): NetHealth
Holdings Inc.; c2008 [atualizada em 2009 jan 10; acesso em 2009 jan 12]. Leishmaniasis; [aproximadamente 2 telas]. Disponível em: http://www.nethealthbook.com/articles/leishmaniasis.php.
Seglen PO, Bohley P. Autophagy and other vacuolar protein degradation mechanisms.
Experientia. 1992; 48(2): 158–72. Selzer PM, Pingel S, Hsieh I, Ugele B, Chan VJ, Engel JC et al. Cysteine inhibitors as
chemotherapy: lessons from a parasite target. Proc Natl Acad Sci U.S.A. 1999; 96: 11015-22.
Sharma A. Malarial protease inhibitors: potential new chemotherapeutic agents. Curr Opin
Investig Drugs. 2007; 8(8): 642-52. Review. Shaw E. Cystenyl proteinases and their selective inactivation. Adv Enzymol Relat Areas Mol
Biol. 1990; 63: 271-347.

91
Shaw JJ, Lainson R, Ryan L, Braga RR, McMahon-Pratt D, David JR. Leishmaniasis in Brazil: XXIII. The identification of Leishmania braziliensis braziliensis in wild-caught neotropical sandflies using monoclonal antibodies. Trans R Soc Trop Med Hyg. 1987; 81(1): 69-72.
Shaw JJ. Taxonomy of the genus Leishmania: present and future trends and their
implications. Mem Inst Oswaldo Cruz. 1994; 89(3):471-478. Shintani T, Klionsky DJ. Autophagy in health and disease: a double-edged sword. Science.
2004; 306(5698): 990–5. Silva D. Leishmaniose anérgica hansenóide no Estado do Pará, Brasil. Na Bras Dermatol.
1996; 71(3): 253-6. Silva-Lopez RE, Coelho MG, De Simone SG. Characterization of an extracellular serine
protease of Leishmania (Leishmania) amazonensis. Parasitology. 2005; 131(1): 85-96. Silva-Lopez RE, Giovanni-De-Simone S. Leishmania (Leishmania) amazonensis: purification
and characterization of a promastigote serine protease. Exp Parasitol. 2004; 107(3-4): 173-82.
Silveira FT, Lainson R, Corbett CE. Further observations on clinical, histopathological, and
immunological features of borderline disseminated cutaneous leishmaniasis caused by Leishmania (Leishmania) amazonensis. Mem Inst Oswaldo Cruz. 2005; 100(5): 525-34.
Simoneau B, Lavallee P, Anderson PC, Bailey M, Bantle G, Berthiaume S et al. Discovery of
non-peptidic P2-P3 butanediamide rennin inhibitors with oral efficacy. Bioorg Med Chem. 1999; 7: 489-508.
Singh N. Drug resistance mechanisms in clinical isolates of Leishmania donovani. Indian J
Med Res. 2006; 123(3): 411-22. Singh S, Sivakumar R. Challenges and new discoveries in the treatment of leishmaniasis. J
Infect Chemother. 2004; 10(6): 307-15. Souza MV, Almeida MV. Drogas anti-HIV: passado, presente e perspectivas futuras. Quim
Nova. 2003; 26(3): 366-72. Sundar S. Drug resistence in Indian visceral leishmaniasis. Trop Med Int Health. 2001; 6(11):
849-54. Takahashi S, Ogasawara H, Hiwatashi K, Hori K, Hata K, Tachibana T et al. A novel D-
aspartyl endopeptidase from Paenibacillus sp. B38: purification and substrate specificity. J Biochem. 2006;139(2): 197-202.
Tanaka RD, Clark JN, Warne RL, Abraham WM, Katsuoka K. Mast cell tryptase: a new
target for therapeutic intervention in asthma. Int Arch Allergy Immunol. 1995; 107: 408-9.
Taylor MC, Kelly JM, Chapman CJ, Fairlamb AH, Miles MA. The structure, organization,
and expression of the Leishmania donovani gene encoding trypanothione reductase. Mol Biochem Parasitol. 1994; 64(2): 293-301.

92
Tiuman TS, Ueda-Nakamura T, Garcia Cortez DA, Dias Filho BP, Morgado-Díaz JA, de Souza W et al. Antileishmanial activity of parthenolide, a sesquiterpene lactone isolated from Tanacetum parthenium. Antimicrob Agents Chemother. 2005; 49(1): 176–82.
Toogood HS, Prescott M, Daniel RM. A pepstatin-insensitive aspartic proteinase from a
thermophilic Bacillus sp. Biochem J. 1995; 307 ( Pt 3): 783-9. Tovo PA. Highly active antiretroviral therapy inhibits cytokine production in HIV-uninfected
subjects. AIDS. 2000; 14(6): 743-4. Trudel N, Garg R, Messier N, Sundar S, Ouellette M, Tremblay MJ. Intracellular survival of
Leishmania species that cause visceral leishmaniasis is significantly reduced by HIV-1 protease inhibitors. J Infect Dis. 2008; 198(9): 1292-9.
Tsantrizos YS. Peptidomimetic therapeutic agents targeting the protease enzyme of the human
immunodeficiency virus and hepatitis C virus. Acc Chem Res. 2008; 41(10): 1252-63. Valdivieso E, Dagger F, Rascón A. Leishmania mexicana: identification and characterization
of an aspartyl proteinase activity. Exp Parasitol. 2007; 116(1): 77-82. Vassar R, Bennett BD, Babu-Khan S, Mendiaz EA, Denis P, Teplow DB et al. β-secretase
cleavage of Alzheimer’s amyloid precursor protein by the transmembrane aspartic protease BACE. Science. 1999; 286: 735-41.
Vendeville S, Goossens F, Debreu-Fontaine MA, Landry V, Davioud-Charvet E, Grellier P et
al. Comparison of the inhibition of human and Trypanosoma cruzi prolyl endopeptidases. Bioorg Med Chem. 2002; 10(6): 1719-29.
Vermelho AB, Giovanni-de-Simone S, d' Avila-Levy CM, Santos ALS, Melo ACN, Silva FP
Jr et al. Trypanosomatidae peptidases: a target for drug development. Curr Enz Inhib. 2007; 3(1): 19–48.
Vickerman K, Preston TM. Comparative cell biology of the kinetoplastids flagellates. In:
Lumsdem WHR, Evans DA, editores: Biology of the Kinetoplastida. New York: Academic Press; 1976. p. 35-130.
Williams RA, Tetley L, Mottram JC, Coombs GH. Cysteine peptidases CPA and CPB are
vital for autophagy and differentiation in Leishmania mexicana. Mol Microbiol. 2006; 61(3): 655-674.
Williams RA, Tetley L, Mottram JC, Coombs GH. Cysteine peptidases CPA and CPB are
vital for autophagy and differentiation in Leishmania mexicana. Mol Microbiol. 2006; 61(3): 655–674.
Wolday D, Berhe N, Akuffo H, Britton S. Leishmania-HIV interaction: immunopathogenic
mechanisms. Parasitol Today. 1999; 15(5): 182-7. World Health Organization (WHO). [homepage na Internet]. Geneva: Special Programme for
Research and Training in Tropical Diseases (TDR); c2009. [acesso em 2008 Dez 15]. Leishmaniasis; [aproximadamente 4 telas]. Disponível em: http://www.who.int/tdr/svc/diseases/leishmaniasis.

93
Xu X, Yu D, Fang W, Cheng Y, Qian Z, Lu W, Cai Y, Feng K. Prediction of peptidase category based on functional domain composition. J Proteome Res. 2008; 7(10): 4521-4.
Yao C, Donelson JE, Wilson ME. The major surface protease (MSP or GP63) of Leishmania
sp. Biosynthesis, regulation of expression, and function. Mol Biochem Parasitol. 2003; 132(1): 1–16.
Zhang T, Maekawa Y, Yasutomo K, Ishikawa H, Fawzy Nashed B, Dainichi T et al.
Pepstatin A-sensitive aspartic proteases in lysosome are involved in degradation of the invariant chain and antigen-processing in antigen presenting cells of mice infected with Leishmania major. Biochem Biophys Res Commun. 2000; 276(2): 693-701.
Zhang WW, Miranda-Verastegui C, Arevalo J, Ndao M, Ward B, Llanos-Cuentas A,
Matlashewski G. Development of a genetic assay to distinguish between Leishmania
viannia species on the basis of isoenzyme differences. Clin Infect Dis. 2006; 42(6): 801-9. Zhao C, Papadopoulou B, Tremblay MJ. Leishmania infantum enhances human
immunodeficiency virus type-1 replication in primary human macrophages through a complex cytokine network. Clin Immunol. 2004b; 113(1): 81-8.
Zhao C, Papadopoulou B, Tremblay MJ. Leishmania infantum promotes replication of HIV
type 1 in human lymphoid tissue cultured ex vivo by inducing secretion of the proinflammatory cytokines TNF-alpha and IL-1 alpha. J Immunol. 2004a; 172(5): 3086-93.
Zhao C, Thibault S, Messier N, Ouellette M, Papadopoulou B, Tremblay MJ. In primary
human monocyte-derived macrophages exposed to Human immunodeficiency virus type 1, does the increased intracellular growth of Leishmania infantum rely on its enhanced uptake? J Gen Virol. 2006; 87(Pt 5): 1295–302.
Zhou H, Zheng C, Zou G, Tao D, Gong J. G1-phase specific apopitosis in liver carcinoma cell
line induced by copper-1, 10-phenanthroline. Intern J Biochem Cell Biol. 2002; 34: 678-84.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo