Embed Size (px)
Citation preview

UNIVERSIDAD VERACRUZANA FACULTAD DE CIENCIAS BIOLÓGICAS Y AGROPECUARIAS
CAMPUS TUXPAN, VER.
TRABAJO DE INVESTIGACIÓN
“EL CICLO DE KREBS Y LA FOTOSÍNTESIS”
Tuxpan de Rodríguez Cano a 29 de Julio de 2009.
ÍNDICE
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

I Introducción………………………………………………………………………..….…...3II Objetivos…………………………………………………………………………………...4 2.1 Objetivo General…………………………………………………………………..…4 2.2 Objetivos Particulares…………………………………………………………….....4III Desarrollo de Tema ………………………………………………..……………..…..…5
3.1 Ciclo de Krebs…………………………………...…...……………….……………..83.2 Etapas del Ciclo de Krebs………………………………………….……………….63.2.1 Reacción 1: Citrato sintasa (De oxalacetato a citrato)……….………………..83.2.2 Reacción 2: Aconitasa (De citrato a isocitrato)………………………………...83.2.3 Reacción 3: Isocitrato deshidrogenasa (De isocitrato a oxoglutarato)………9 3.2.4 Reacción 4: α-cetoglutarato deshidrogenasa (De oxoglutarato a Succinil CoA)……………………………………………………………………………….10 3.2.5 Reacción 5: Succinil-CoA sintetasa (De Succinil-CoA a succinato)……......113.2.6 Reacción 6: Succinato deshidrogenasa (De succinato a fumarato).….…....123.2.7 Reacción 7: Fumarasa (De fumarato a L-malato)……….……………………133.2.8 Reacción 8: Malato deshidrogenasa (De L-malato a oxalacetato)…………133.3 Regulación…….…………………………………………………………………….153.4 Principales vías que convergen en el ciclo de Krebs……….…………………163.5 La Fotosíntesis…….………………………………………………………………..183.5.1 Fase Lumínica……………………………………………………………………193.5.2 Fase Obscura…………………………………………………………………….203.6 Cloroplasto……………………………………………………………….………….213.7 Clorofila………………………………………………………………….…………..223.8 Ciclo de Calvin…………………………………………………………….………..233.9 Fases del Ciclo de Calvin………………………………………………….………243.9.1 1ª Fase: Fijación del CO2………………………………………………….……24 3.9.2 2ª Fase: Reducción………………………………………………………………243.9.3 3ª Fase: Regeneración…………………………………………………………..253.9.4 Plantas C3 y C4…………………………………………………………………..273.9.5 Rubisco………………………………...………………………………………….29
IV Conclusiones………………….…………………………………………………….....30V Bibliografía……………………………………………………………………….……..31
I INTRODUCCIÓN
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

El ciclo de Krebs (también llamado ciclo del ácido cítrico o ciclo de los ácidos
tricarboxílicos) es una ruta metabólica, es decir, una sucesión de reacciones químicas,
que forman parte de la respiración celular en todas las células aerobicas. En organismos
aeróbicos, el ciclo de Krebs es parte de la vía catabólica que realiza la oxidación de
glúcidos, ácidos grasos y aminoácidos hasta producir CO2, liberando energía en forma
utilizable (poder reductor y GTP).
El ciclo de Krebs también proporciona precursores para muchas biomoléculas, como
ciertos aminoácidos. Por ello se considera una vía anfibólica, es decir, catabólica y
anabólica al mismo tiempo.
La fotosíntesis es la reacción impulsada por el sol que hace que las hojas de las plantas
(u otras partes que contengan clorofila) —combinando agua, sales minerales y dióxido
de carbono— fabriquen materia orgánica (azúcares) liberando oxígeno en el proceso.
La fotosíntesis es la base de nuestra cadena alimenticia, porque las plantas producen el
alimento necesario para el crecimiento de sus células y, al mismo tiempo, proporcionan
energía nutricional a los animales cuando se alimentan de ellas.
El ciclo de Calvin (también conocido como ciclo de Calvin-Benson o fase de fijación del
CO2 de la fotosíntesis) consiste en una serie de procesos bioquímicos que se realizan en
el estroma de los cloroplastos de los organismos fotosintéticos. Fue descubierto por
Melvin Calvin y Andy Benson de la Universidad de California Berkeley mediante el
empleo de isótopos radiactivos de carbono.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

II OBJETIVOS
Objetivo General
Conocer todos los procesos celulares del ciclo de Krebs y sus etapas, la Fotosíntesis
y sus dos fases asi como el ciclo de calvin en las plantas.
Objetivos Particulares
Conocer particularme el ciclo de Krebs y sus etapas en el proceso metabolico en las
plantas.
Conocer las etapas del proceso de la fotosíntesis, asi como las etapa luminosa y
obscura dentro de los cloroplastas donde se realiza esta acción.
Conocer el ciclo de Calvin y sus etapas en los procesos bioquímicos en las estomas
de las plantas
III DESARROLLO DEL TEMA
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.1 Ciclo de Krebs
El ciclo de Krebs (también llamado ciclo del ácido cítrico o ciclo de los ácidos tricarboxílicos)
es una ruta metabólica, es decir, una sucesión de reacciones químicas, que forman parte de
la respiración celular en todas las células aeróbicas. En organismos aeróbicos, el ciclo de
Krebs es parte de la vía catabólica que realiza la oxidación de glúcidos, ácidos grasos y
aminoácidos hasta producir CO2, liberando energía en forma utilizable (poder reductor y
GTP).
El metabolismo oxidativo de glúcidos, grasas y proteínas frecuentemente se divide en tres
etapas, de las cuales, el ciclo de Krebs supone la segunda. En la primera etapa, los
carbonos de estas macromoléculas dan lugar a moléculas de acetil-CoA de dos carbonos, e
incluye las vías catabólicas de aminoácidos (p. ej. desaminación oxidativa), la beta oxidación
de ácidos grasos y la glucólisis. La tercera etapa es la fosforilación oxidativa, en la cual el
poder reductor (NADH y FADH2) generado se emplea para la síntesis de ATP según la
teoría del acomplamiento quimiosmótico.
El ciclo de Krebs también proporciona precursores para muchas biomoléculas, como ciertos
aminoácidos. Por ello se considera una vía anfibólica, es decir, catabólica y anabólica al
mismo tiempo.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

El ciclo de Krebs tiene lugar en la matriz mitocondrial en eucariotas.
El acetil-CoA (Acetil Coenzima A) es el principal precursor del ciclo. El ácido cítrico (6
carbonos) o citrato se regenera en cada ciclo por condensación de un acetil-CoA (2
carbonos) con una molécula de oxaloacetato (4 carbonos). El citrato produce en cada ciclo
una molécula de oxaloacetato y dos CO2, por lo que el balance neto del ciclo es:
Acetil-CoA + 3 NAD+ + FAD + GDP + Pi + 3 H2O → CoA-SH + 3 (NADH + H+) + FADH2 +
GTP + 2 CO2 + 3 H+
Los dos carbonos del Acetil-CoA son oxidados a CO2, y la energía que estaba acumulada
es liberada en forma de energía química: GTP y poder reductor (electrones de alto
potencial): NADH y FADH2. NADH y FADH2 son coenzimas (moléculas que se unen a
enzimas) capaces de acumular la energía en forma de poder reductor para su conversión en
energía química en la fosforilación oxidativa.
El FADH2 de la succinato deshidrogenasa, al no poder desprenderse de la enzima, debe
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

oxidarse nuevamente in situ. El FADH2 cede sus dos hidrógenos a la ubiquinona (coenzima
Q), que se reduce a ubiquinol (QH2) y abandona la enzima.
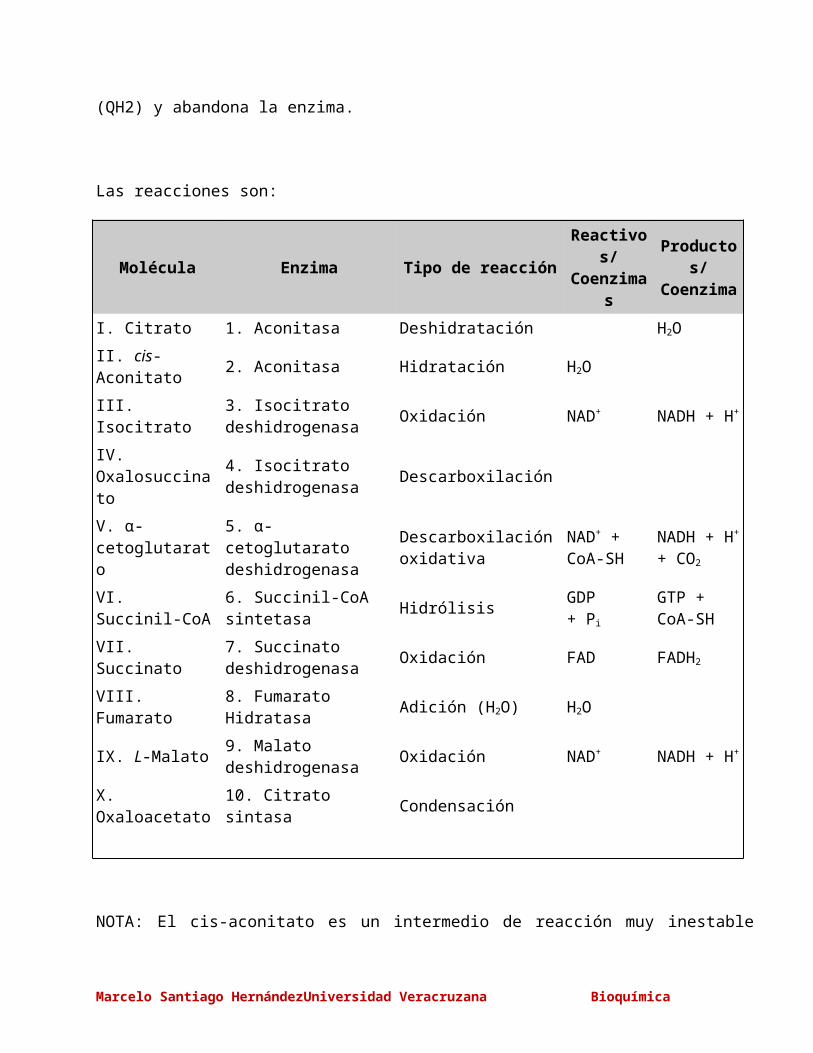
Las reacciones son:
Molécula Enzima Tipo de reacciónReactivos/Coenzimas
Productos/Coenzima
I. Citrato 1. Aconitasa Deshidratación H2O
II. cis-Aconitato 2. Aconitasa Hidratación H2O
III. Isocitrato3. Isocitrato deshidrogenasa
Oxidación NAD+ NADH + H+
IV. Oxalosuccinato
4. Isocitrato deshidrogenasa
Descarboxilación
V. α-cetoglutarato
5. α-cetoglutaratodeshidrogenasa
Descarboxilación oxidativa
NAD+ +CoA-SH
NADH + H+
+ CO2
VI. Succinil-CoA6. Succinil-CoA sintetasa
HidrólisisGDP+ Pi
GTP +CoA-SH
VII. Succinato7. Succinato deshidrogenasa
Oxidación FAD FADH2
VIII. Fumarato 8. Fumarato Hidratasa Adición (H2O) H2O
IX. L-Malato9. Malato deshidrogenasa
Oxidación NAD+ NADH + H+
X. Oxaloacetato 10. Citrato sintasa Condensación
NOTA: El cis-aconitato es un intermedio de reacción muy inestable que rápidamente se
transforma en citrato, antes de comenzar la tercera reacción.
3.2 Etapas del Ciclo de Krebs
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.2.1 Reacción 1: Citrato sintasa (De oxalacetato a citrato)
El sitio activo de la enzima, activa el acetil-CoA para hacerlo afín a un centro carbonoso del
oxalacetato. Como consecuencia de la unión entre las dos moléculas, el grupo tioéster
(CoA) se hidroliza, formando así la molécula de citrato.
La reacción es sumamente exoergónica (ΔG'°=-31.4 kJ/mol), motivo por el cual este paso es
irreversible. El citrato producido por la enzima, además, es capaz de inhibir
competitivamente la actividad de la enzima. Incluso estando la reacción muy favorecida,
porque es exoergónica, la citrato sintasa puede ser perfectamente regulada. Este aspecto
tiene una notable importancia biológica, puesto que permite una completa regulación del
ciclo de Krebs completo, convirtiendo a la enzima en una especie de marcapasos del ciclo.
3.2.2 Reacción 2: Aconitasa (De citrato a isocitrato)
La aconitasa cataliza la isomerización del citrato a isocitrato, por la formación de cis-
aconitato. La enzima cataliza también la reacción inversa, pero en el ciclo de Krebs tal
reacción es unidireccional a causa de la ley de acción de masa: las concentraciones (en
condiciones estándar) de citrato (91%), del intermediario cis-aconitato (3%) y de isocitrato
(6%), empujan decididamente la reacción hacia la producción de isocitrato.
En el sitio activo de la enzima está presente un clúster hierro-azufre que, junto a algunos
residuos de aminoácidos polares, liga el sustrato. En concreto, la unión al sustrato se
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica
asegura por la presencia de un resto de serina, de arginina, de histidina y de aspartato, que
permiten sólo la unión estereospecifica del citrato 1R,2S, rechazando la forma opuesta.
3.2.3 Reacción 3: Isocitrato deshidrogenasa (De isocitrato a oxoglutarato)
La isocitrato deshidrogenasa mitocondrial es una enzima dependiente de la presencia de
NAD+ y de Mn2+ o Mg2+. Inicialmente, la enzima cataliza la oxidación del isocitrato a
oxalsuccinato, lo que genera una molécula de NADH a partir de NAD+. Sucesivamente, la
presencia de un ión bivalente, que forma un complejo con los oxígenos del grupo carboxilo
en posición alfa, aumenta la electronegatividad de esa región molecular. Esto genera una
reorganización de los electrones en la molécula, con la consiguiente rotura de la unión entre
el carbono en posición gamma y el grupo carboxilo adyacente. De este modo se tiene una
descarboxilación, es decir, la salida de una molécula de CO2, que conduce a la formación
de α-cetoglutarato, caracterizado por dos carboxilos en las extremidades y una cetona en
posición alfa con respecto de uno de los dos grupos carboxilo.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.2.4 Reacción 4: α-cetoglutarato deshidrogenasa (De oxoglutarato a Succinil-CoA)
Después de la conversión del isocitrato en α-cetoglutarato se produce una segunda reacción de
descarboxilación oxidativa, que lleva a la formación de succinil CoA. La descarboxilación oxidativa
del α-chetoglutarato es muy parecida a la del piruvato, otro α-cetoácido.
Ambas reacciones incluyen la descarboxilación de un α-cetoácido y la consiguiente producción de
una unión tioéster a alta energía con la coenzima A. Los complejos que catalizan tales reacciones
son parecidos entre ellos.
La α-cetoglutarato deshidrogenasa (o, más correctamente, oxoglutarato deshidrogenasa),
está compuesta de tres enzimas diferentes:
* Subunidad E1: las dos cetoglutarato deshidrogenasas.
* Subunidad E2: la transuccinilasa.
(La subunidad E1 y E2 presentan una gran homología con las de la piruvato
deshidrogenasa.)
* Subunidad E3: la dihidrolipoamida deshidrogenasa, que es el mismo polipéptido presente
en el otro complejo enzimático.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.2.5 Reacción 5: Succinil-CoA sintetasa (De Succinil-CoA a succinato)
El succinil-CoA es un tioéster a alta energía (su ΔG°′ de hidrólisis está en unos -33.5 kJ mol-
1, parecido al del ATP que es de -30.5 kJ mol-1). La citrato sintasa se sirve de un
intermediario con tal unión a alta energía para llevar a cabo la fusión entre una molécula con
dos átomos de carbono (acetil-CoA) y una con cuatro (oxalacetato). La enzima succinil-CoA
sintetasa se sirve de tal energía para fosforilar un nucleósido
difosfato purinico como el GDP.
La energía procedente del tioéster viene convertida en energía ligada a una unión fosfato. El
primer paso de la reacción genera un nuevo intermediario a alta energía, conocido como
succinil fosfato. Sucesivamente, una histidina presente en el sitio catalítico remueve el
fosfato de la molécula glucídica, generando el producto succinato y una molécula de
fosfohistidina, que dona velozmente el fosfato a un nucleósido difosfato, recargándolo a
trifosfato. Se trata del único paso del ciclo de Krebs en el que se produce una fosforilación a
nivel de sustrato.
El GTP está implicado principalmente en las rutas de transducción de señales, pero su papel
en un proceso energético como el ciclo de Krebs es, en cambio, esencialmente trasladar
grupos fosfato hacia el ATP, en una reacción catalizada por la enzima nucleósid
difosfoquinasa.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.2.6 Reacción 6: Succinato deshidrogenasa (De succinato a fumarato)
La parte final del ciclo consiste en la reorganización de moléculas a cuatro átomos de
carbono hasta la regeneración del oxalacetato. Para que eso sea posible, el grupo metilo
presente en el succinato tiene que convertirse en un carbonilo. Como ocurre en otras rutas,
por ejemplo en la beta oxidación de los ácidos grasos, tal
conversión ocurre mediante tres pasos: una primera oxidación,
una hidratación y una segunda oxidación. Estos tres pasos,
además de regenerar oxalacetato, permiten la extracción ulterior
de energía mediante la formación de FADH2 y NADH.
La primera reacción de oxidación es catalizada por el complejo enzimático de la succinato
deshidrogenasa, la única enzima del ciclo que tiene como aceptor de hidrógeno al FAD en
vez de al NAD+. El FAD es enlazado de modo covalente a la enzima por un residuo de
histidina. La enzima se vale del FAD ya que la energía asociada a la reacción no es
suficiente para reducir el NAD+.
El complejo enzimático también es el único del ciclo que pasa dentro de la membrana
mitocondrial. Tal posición se debe a la implicación de la enzima en la cadena de transporte
de los electrones. Los electrones pasados sobre el FAD se introducen directamente en la
cadena gracias a la unión estable entre la enzima y el cofactor mismo
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.2.7 Reacción 7: Fumarasa (De fumarato a L-malato)
La fumarasa cataliza la adición en trans de un protón y un grupo OH- procedentes de una
molécula de agua. La hidratación del fumarato produce L-malato.
3.2.8 Reacción 8: Malato deshidrogenasa (De L-malato a oxalacetato)
La última reacción del ciclo de Krebs consiste en la oxidación del malato a oxalacetato. La
reacción, catalizada por la malato deshidrogenasa, utiliza otra molécula de NAD+ como
aceptor de hidrógeno, produciendo NADH.
La energía libre de Gibbs asociada con esta última reacción es decididamente positiva, a
diferencia de las otras del ciclo. La actividad de la enzima es remolcada por el consumo de
oxalacetato por parte de la citrato sintasa, y de NADH por parte de
la cadena de transporte de electrones.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

Visión simplificada y rendimiento del proceso.
El paso final es la oxidación del ciclo de kreps, produciendo un acetil-CoA y un CO2.
El acetil-CoA reacciona con una molécula de oxaloacetato (4 carbonos) para formar
citrato (6 carbonos), mediante una reacción de condensación.
A través de una serie de reacciones, el citrato se convierte de nuevo en oxaloacetato.
Durante estas reacciones, se substraen 2 átomos de carbono del citrato (6C) para dar
oxalacetato (4C); dichos átomos de carbono se liberan en forma de CO2
El ciclo consume netamente 1 acetil-CoA y produce 2 CO2. También consume 3 NAD+
y 1 FAD, produciendo 3 NADH + 3 H+ y 1 FADH2.
El rendimiento de un ciclo es (por cada molécula de piruvato): 1 GTP, 3 NADH, 1
FADH2, 2CO2.
Cada NADH, cuando se oxide en la cadena respiratoria, originará 2,5 moléculas de
ATP (3 x 2,5 = 7,5), mientras que el FADH2 dará lugar a 1,5 ATP. Por tanto, 7,5 + 1,5 +
1 GTP = 10 ATP por cada acetil-CoA que ingresa en el ciclo de Krebs.
Cada molécula de glucosa produce (vía glucólisis) dos moléculas de piruvato, que a
su vez producen dos acetil-CoA, por lo que por cada molécula de glucosa en el ciclo
de Krebs se produce: 4CO2, 2 GTP, 6 NADH, 2 FADH2; total 36 ATP.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.3 Regulación
Muchas de las enzimas del ciclo de Krebs son reguladas por retroalimentación negativa, por
unión alostérica del ATP, que es un producto de la vía y un indicador del nivel energético de
la célula. Entre estas enzimas, se incluye el complejo de la piruvato deshidrogenasa que
sintetiza el acetil-CoA necesario para la primera reacción del ciclo a partir de piruvato,
procedente de la glucólisis o del catabolismo de aminoácidos. También las enzimas citrato
sintasa, isocitrato deshidrogenasa y α-cetoglutarato deshidrogenasa, que catalizan las tres
primeras reacciones del ciclo de Krebs, son inhibidas por altas concentraciones de ATP.
Esta regulación frena este ciclo degradativo cuando el nivel energético de la célula es
bueno.
Algunas enzimas son también reguladas negativamente cuando el nivel de poder reductor
de la célula es elevado. El mecanismo que se realiza es una inhibición competitiva por
producto (por NADH) de las enzimas que emplean NAD+ como sustrato. Así se regulan,
entre otros, los complejos piruvato deshidrogenasa y citrato sintasa.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.4 Principales vías que convergen en el ciclo de Krebs
La mayoría de las vías catabólicas convergen en el ciclo de Krebs, como muestra el
diagrama. Las reacciones que forman intermediarios del ciclo se conocen como reacciones
anapleróticas.
El ciclo de Krebs constituye la segunda etapa del catabolismo de carbohidratos. La glucólisis
rompe la glucosa (6 carbonos) generando dos moléculas de piruvato (3 carbonos). En
eucariotas, el piruvato se desplaza al interior de la mitocondria (gracias a un transportador
específico de membrana interna). En la matriz mitocondrial, produce acetil-CoA que entra en
el ciclo de Krebs.
En el catabolismo de proteínas, los enlaces peptídicos de las proteínas son degradados por
acción de enzimas proteasas en el tubo digestivo liberando sus constituyentes
aminoacídicos. Estos aminoácidos penetran en las células, donde pueden ser empleados
para la síntesis de proteínas o ser degradados para producir energía en el ciclo de Krebs.
Para su entrada al ciclo deben eliminarse sus grupos amino (terminales y laterales) por
acción de enzimas aminotransferasas y desaminasas, principalmente.
En el catabolismo de lípidos, los triglicéridos son hidrolizados liberando ácidos grasos y
glicerol. En el hígado, el glicerol puede ser convertido en glucosa vía dihidroxiacetona
fosfato y gliceraldehído-3-fosfato, por la gluconeogénesis (ruta anabólica). En muy diversos
tejidos, especialmente en músculo cardíaco, los ácidos grasos son degradados en la matriz
mitocondrial mediante sucesivos ciclos de beta oxidación que liberan unidades de acetil-
CoA, que pueden incorporarse al ciclo de Krebs. En ocasiones, el ciclo de Krebs puede
rendir propionil-CoA (3 carbonos), que puede emplearse para la síntesis de glucosa en la
gluconeogénesis hepática.
El ciclo de Krebs siempre es seguido por la fosforilación oxidativa. Este proceso extrae la
energía en forma de electrones de alto potencial de las moléculas (Cofactores reducidos)
que son el NADH y FADH2, regenerando NAD+ y FAD, gracias a lo cual el ciclo de Krebs
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

puede continuar. Los electrones son transferidos a moléculas de O2, rindiendo H2O. Pero
esta transferencia se realiza a través de una cadena transportadora de electrones capaz de
aprovechar la energía potencial de los electrones para bombear protones al espacio
intermembrana de la mitocondria. Esto genera un gradiente electroquímico de H+, que es
utilizado para la síntesis de ATP mediante la enzima ATP sintetasa. De este modo, el ciclo
de Krebs no utiliza directamente O2, pero lo requiere al estar acoplado a la fosforilación
oxidativa.
Por cada molécula de glucosa, la energía obtenida mediante el metabolismo oxidativo, es
decir, glucólisis seguida del ciclo de Krebs, equivale a 30/32 moléculas de ATP dependiendo
del tipo de lanzadera para introducir el poder reductor dentro de la mitocondria, si es la
lanzadera de malato-aspartato son 32 y si es la de glicerol 3 fosfato, son 30.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.5 La Fotosíntesis
Fotosíntesis, proceso en virtud del cual los organismos con clorofila, como las plantas
verdes, las algas y algunas bacterias, capturan energía en forma de luz y la transforman en
energía química. Prácticamente toda la energía que consume la vida de la biosfera terrestre
la zona del planeta en la cual hay vida— procede de la fotosíntesis.
Una ecuación generalizada y no equilibrada de la fotosíntesis en presencia de luz sería:
CO2 + 2H2A → (CH2) + H2O + H2A
El elemento H2A de la fórmula representa un compuesto oxidable, es decir, un compuesto
del cual se pueden extraer electrones; CO2 es el dióxido de carbono; CH2 una generalización
de los hidratos de carbono que incorpora el organismo vivo. En la gran mayoría de los
organismos fotosintéticos, es decir, en las algas y las plantas verdes, H2A es agua (H2O);
pero en algunas bacterias fotosintéticas, H2A es anhídrido sulfúrico (H2S). La fotosíntesis con
agua es la más importante y conocida y, por tanto, será la que tratemos con detalle.
La fotosíntesis se realiza en dos etapas: una serie de reacciones que dependen de la luz y
son independientes de la temperatura, y otra serie que dependen de la temperatura y son
independientes de la luz. La velocidad de la primera etapa, llamada reacción lumínica,
aumenta con la intensidad luminosa (dentro de ciertos límites), pero no con la temperatura.
En la segunda etapa, llamada reacción en la oscuridad, la velocidad aumenta con la
temperatura (dentro de ciertos límites), pero no con la intensidad luminosa.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.5.1 Fase Lumínica
La primera etapa de la fotosíntesis es la absorción de luz por los pigmentos. La clorofila es
el más importante de éstos, y es esencial para el proceso. Captura la luz de las regiones
violeta y roja del espectro y la transforma en energía química mediante una serie de
reacciones. Los distintos tipos de clorofila y otros pigmentos, llamados carotenoides y
ficobilinas, absorben longitudes de onda luminosas algo distintas y transfieren la energía a la
clorofila A, que termina el proceso de transformación. Estos pigmentos accesorios amplían el
espectro de energía luminosa que aprovecha la fotosíntesis.
La fotosíntesis tiene lugar dentro de las células, en orgánulos llamados cloroplastos que
contienen las clorofilas y otros compuestos, en especial enzimas, necesarios para realizar
las distintas reacciones. Estos compuestos están organizados en unidades de cloroplastos
llamadas tilacoides; en el interior de éstos, los pigmentos se disponen en subunidades
llamadas fotosistemas. Cuando los pigmentos absorben luz, sus electrones ocupan niveles
energéticos más altos, y transfieren la energía a un tipo especial de clorofila llamado centro
de reacción.
En la actualidad se conocen dos fotosistemas, llamados I y II. La energía luminosa es
atrapada primero en el fotosistema II, y los electrones cargados de energía saltan a un
receptor de electrones; el hueco que dejan es reemplazado en el fotosistema II por
electrones procedentes de moléculas de agua, reacción que va acompañada de liberación
de oxígeno. Los electrones energéticos recorren una cadena de transporte de electrones
que los conduce al fotosistema I, y en el curso de este fenómeno se genera un trifosfato de
adenosina o ATP, rico en energía. La luz absorbida por el fotosistema I pasa a continuación
a su centro de reacción, y los electrones energéticos saltan a su aceptor de electrones. Otra
cadena de transporte los conduce para que transfieran la energía a la coenzima dinucleotido
fosfato de nicotinamida y adenina o NADP que, como consecuencia, se reduce a NADPH2.
Los electrones perdidos por el fotosistema I son sustituidos por los enviados por la cadena
de transporte de electrones del fotosistema II. La reacción en presencia de luz termina con
el almacenamiento de la energía producida en forma de ATP y NADPH2.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.5.2 Fase Obscura
La reacción en la oscuridad tiene lugar en el estroma o matriz de los cloroplastos, donde la
energía almacenada en forma de ATP y NADPH2 se usa para reducir el dióxido de carbono
a carbono orgánico. Esta función se lleva a cabo mediante una serie de reacciones llamada
ciclo de Calvin, activadas por la energía de ATP y NADPH2. Cada vez que se recorre el ciclo
entra una molécula de dióxido de carbono, que inicialmente se combina con un azúcar de
cinco carbonos llamado ribulosa 1,5-difosfato para formar dos moléculas de un compuesto
de tres carbonos llamado 3-fosfoglicerato. Tres recorridos del ciclo, en cada uno de los
cuales se consume una molécula de dióxido de carbono, dos de NADPH2 y tres de ATP,
rinden una molécula con tres carbonos llamada gliceraldehído 3-fosfato; dos de estas
moléculas se combinan para formar el azúcar de seis carbonos glucosa. En cada recorrido
del ciclo, se regenera la ribulosa 1,5-difosfato.
Por tanto, el efecto neto de la fotosíntesis es la captura temporal de energía luminosa en los
enlaces químicos de ATP y NADPH2 por medio de la reacción en presencia de luz, y la
captura permanente de esa energía en forma de glucosa mediante la reacción en la
oscuridad. En el curso de la reacción en presencia de luz se escinde la molécula de agua
para obtener los electrones que transfieren la energía luminosa con la que se forman ATP y
NADPH2. El dióxido de carbono se reduce en el curso de la reacción en la oscuridad para
convertirse en base de la molécula de azúcar. La ecuación completa y equilibrada de la
fotosíntesis en la que el agua actúa como donante de electrones y en presencia de luz es:
6 CO2 + 12H2O → C6H12O6 + 6O2 + 6H2O
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.6 Cloroplasto
Cloroplasto, orgánulo citoplasmático, que se encuentra en las células vegetales y en las de
las algas, donde se lleva a cabo la fotosíntesis (proceso que permite la transformación de
energía luminosa en energía química).
Los cloroplastos son orgánulos con forma de disco, de entre 4 y 6 micrómetros de diámetro.
Aparecen en mayor cantidad en las células de las hojas, lugar en el cual parece que pueden
orientarse hacia la luz. En una célula puede haber entre 40 y 50 cloroplastos, y en cada
milímetro cuadrado de la superficie de la hoja hay 500.000 cloroplastos.
Cada cloroplasto está recubierto por una membrana doble: la membrana externa y la
membrana interna. En su interior, el cloroplasto contiene una sustancia básica denominada
estroma, la cual está atravesada por una red compleja de discos conectados entre sí,
llamados tilacoides. Muchos de los tilacoides se encuentran apilados como si fueran platillos;
a estas pilas se les llama grana. Las moléculas de clorofila, que absorben luz para llevar a
cabo la fotosíntesis, están unidas a los tilacoides. La energía luminosa capturada por la
clorofila es convertida en trifosfato de adenosina (ATP) mediante una serie de reacciones
químicas que tienen lugar en los grana. Los cloroplastos también contienen gránulos
pequeños de almidón donde se almacenan los productos de la fotosíntesis de forma
temporal.
En las plantas, los cloroplastos se desarrollan en presencia de luz, a partir de unos
orgánulos pequeños e incoloros que se llaman proplastos. A medida que las células se
dividen en las zonas en que la planta está creciendo, los proplastos que están en su interior
también se dividen por fisión. De este modo, las células hijas tienen la capacidad de producir
cloroplastos.
En las algas, los cloroplastos se dividen directamente, sin necesidad de desarrollarse a partir
de proplastos. La capacidad que tienen los cloroplastos para reproducirse a sí mismos, y su
estrecha similitud, con independencia del tipo de célula en que se encuentren, sugieren que
estos orgánulos fueron alguna vez organismos autónomos que establecieron una simbiosis
en la que la célula vegetal era el huésped.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.7 Clorofila
Clorofila, pigmento que da el color verde a los vegetales y que se encarga de absorber la luz
necesaria para realizar la fotosíntesis, proceso que transforma la energía luminosa en
energía química. La clorofila absorbe sobre todo la luz roja, violeta y azul, y refleja la verde.
La gran concentración de clorofila en las hojas y su presencia ocasional en otros tejidos
vegetales, como los tallos, tiñen de verde estas partes de las plantas. En algunas hojas, la
clorofila está enmascarada por otros pigmentos. En otoño, la clorofila de las hojas de los
árboles se descompone, y ocupan su lugar otros pigmentos.
La molécula de clorofila es grande y está formada en su mayor parte por carbono e
hidrógeno; ocupa el centro de la molécula un único átomo de magnesio rodeado por un
grupo de átomos que contienen nitrógeno y se llama anillo de porfirinas. La estructura
recuerda a la del componente activo de la hemoglobina de la sangre.
De este núcleo central parte una larga cadena de átomos de carbono e hidrógeno que une
la molécula de clorofila a la membrana interna del cloroplasto, el orgánulo celular donde
tiene lugar la fotosíntesis. Cuando la molécula de clorofila absorbe un fotón, sus electrones
se excitan y saltan a un nivel de energía superior (ver fotoquímica) esto inicia en el
cloroplasto una compleja serie de reacciones que dan lugar al almacenamiento de energía
en forma de enlaces químicos.
Hay varios tipos de clorofilas que se diferencian en detalles de su estructura molecular y que
absorben longitudes de onda luminosas algo distintas. El tipo más común es la clorofila A,
que constituye aproximadamente el 75% de toda la clorofila de las plantas verdes. Se
encuentra también en las algas verdeazuladas y en células fotosintéticas más complejas. La
clorofila B es un pigmento accesorio presente en vegetales y otras células fotosintéticas
complejas; absorbe luz de una longitud de onda diferente y transfiere la energía a la clorofila
A, que se encarga de transformarla en energía química. Algunas bacterias presentan otras
clorofilas de menor importancia.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.8 Ciclo de Calvin
El ciclo de Calvin (también conocido como ciclo de Calvin-Benson o fase de fijación del
CO2 de la fotosíntesis) consiste en una serie de procesos bioquímicos que se realizan en el
estroma de los cloroplastos de los organismos fotosintéticos. Fue descubierto por Melvin
Calvin y Andy Benson de la Universidad de California Berkeley mediante el empleo de
isótopos radiactivos de carbono.
Durante la fase luminosa o fotoquímica de la fotosíntesis, la energía lumínica ha sido
almacenada en moléculas orgánicas sencillas e inestables, que van a aportar energía para
realizar el proceso (ATP) y poder reductor, es decir, la capacidad de donar electrones
(reducir) a otra molécula (nicotín-amida dinucleótido fosfato o NADPH+H+). En general, los
compuestos bioquímicos más reducidos (simplificando la cuestión: los que tienen más
electrones) almacenan más energía que los oxidados (con menos electrones, también
simplificando) y son, por tanto, capaces de generar más trabajo (por ejemplo, aportar la
energía necesaria para el movimiento muscular). En el ciclo de Calvin se integran y
convierten moléculas inorgánicas de dióxido de carbono en moléculas orgánicas sencillas a
partir de las cuales se formará el resto de los compuestos bioquímicos que constituyen los
seres vivos. Este proceso también se puede, por tanto, denominar como de asimilación del
carbono.
La primera enzima que interviene en el ciclo y que fija el CO2 atmosférico uniéndolo a una
molécula orgánica (ribulosa-1-5-bisfosfato) se denomina RuBisCO (por las siglas de
Ribulosa bisfosfato carboxilasa-oxigenasa).
Para un total de 6 moléculas de CO2 fijado, la estequiometría final del ciclo de Calvin se
puede resumir en la ecuación:
6CO2 + 12NADPH + 18 ATP → C6H12O6P + 12NADP+ + 18ADP + 17 Pi
que representaría la formación de una molécula de azúcar-fosfato de 6 átomos de carbono
(hexosa) a partir de 6 moléculas de CO2.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.9 Fases del Ciclo de Calvin
3.9.1 1ª Fase: Fijación del CO2
La RuBisCO cataliza la reacción entre la ribulosa bisfosfato (una pentosa, es decir un
monosacárido de 5C, RuBP) con el CO2, para crear 1 molécula de 6 carbonos, la cual al ser
inestable termina por separarse en 2 moléculas que contienen 3 átomos de carbono cada
una, el fosfoglicerato (PGA). La importancia de la RuBisCo queda indicada por el hecho de
ser el enzima más abundante en la naturaleza.
3.9.2 2ª Fase: Reducción
Primero ocurre un proceso de activación en el cual una molécula de ATP, proveniente de la
fase fotoquímica, es usada para la fosforilización del PGA, transformándolo en
difosfoglicerato. Esa transferencia de un enlace fosfato permite que una molécula de
NADPH+H+ reduzca el PGA, mediante la acción de la enzima gliceraldehído-3-fosfato-
deshidrogenasa, para formar gliceraldehído-3-fosfato (PGAL). Esta última molécula es una
triosa-fosfato, un azúcar de tipo aldosa con 3C, que es una molécula estable y con mayor
energía libre (capaz de realizar mayor cantidad de trabajo) que las anteriores. Parte de
PGAL se transforma en su isómero dihidroxiacetona-fosfato (cetosa de 3C). Estas dos
triosas-fosfato serán la base a partir de la cual se formen el resto de azúcares (como la
fructosa y glucosa), oligosacáridos (como la sacarosa o azúcar de caña) y polisacáridos
(como la celulosa o el almidón). También, a partir de estos azúcares, se formarán directa o
indirectamente las cadenas de carbono que componen el resto de biomoléculas que
constituyen los seres vivos (lípidos, proteínas, ácidos nucleicos, etc.).
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.9.3 3ª Fase: Regeneración
El ciclo continua a lo largo de una serie de reacciones hasta formar ribulosa-fosfato, que
mediante el consumo de otra molécula de ATP, regenera la ribulosa bisfosfato (RuBP)
original, dejándola disponible para que el ciclo se repita nuevamente.
Por tanto, por cada vuelta del ciclo se incorpora una molécula de carbono fijado (CO2) a otra
molécula preexistente de 5 átomos de carbono (ribulosa bisfosfato), el resultado final es la
regeneración de la molécula de 5 átomos de carbono y la incorporación de un nuevo
carbono en forma orgánica C(H2O). Para comprenderlo hay que tener en cuenta que el
producto fundamental del ciclo de Calvin es el gliceraldehído-3-fosfato (de 3 átomos de
carbono), molécula que sirve como base para la síntesis del resto de carbohidratos. Tras 3
vueltas del ciclo, una nueva molécula de PGAL sale de éste y puede ser posteriormente
utilizada para la formación de otras moléculas.
Durante años se pensó que el ciclo de Calvin era independiente de la luz y se denominó
"fase oscura de la fotosíntesis". Hoy en día se conoce perfectamente que tanto la actividad
de RuBisCO como de otras enzimas clave del ciclo es regulada por la luz, desactivándose
en condiciones de oscuridad y reactivándose en condiciones de iluminación.
A bajas concentraciones de CO2 (como cuando se cierran los estomas para evitar pérdida
de agua en la planta), la Rubisco reaccionara con O2 en vez de CO2. Esta reacción provoca
una disminución del porcentaje de carbono fijado y está asociada al fenómeno denominado
fotorrespiración. Estos procesos son más graves a temperaturas relativamente altas,
disminuyendo la tasa de fotosíntesis (una medida de la capacidad de la planta para asimilar
CO2).
Por ello plantas adaptadas a climas cálidos han desarrollado estrategia para optimizar la
capacidad de asimilación de dióxido de carbono (plantas C-4 y plantas CAM). Las plantas
C4 usan inicialmente la enzima PEP carboxilasa (fosfoenolpiruvato carboxilasa), que
convierte el fosfoenolpiruvato (compuesto de 3C) en oxalacetato (compuesto de 4C) a partir
de bicarbonato que se forma por reacción del CO2 con agua (facilitado por la presencia de la
enzima anhidrasa carbónica que cataliza esta reacción). La PEP carboxilasa tiene una
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

afinidad muy alta por el bicarbonato, mayor que RubisCO por el CO2. El nombre de este tipo
de fotosíntesis proviene, precisamente, de que el primer compuesto orgánico formado
(oxalacetato) tiene 4 átomos de carbono.
A partir de oxalacetato se produce malato (un compuesto más reducido) lo que conlleva una
pérdida del poder reductor acumulado en la fotosíntesis. El malato formado desprende el
carbono fijado en las inmediaciones de RubisCO, aumentando la concentración de dióxido
de carbono respecto a oxígeno en el entorno de esta última enzima. A partir de aquí el
proceso es similar al descrito anteriormente (plantas C-3, en las que el primer producto de la
asimilación de CO2,es el PGA de 3 átomos de carbono). De esta forma se consigue evitar la
actividad oxigenasa de la Rubisco.
La formación de malato y su descomposición ocurren en células diferentes, cada una de
ellas provistas de cloroplastos especializados en llevar a cabo cada una de las dos
funciones. Otro tipo de adaptación es el de las plantas con fotosíntesis CAM (de las siglas
en inglés "Crassulacean acid metabolism", metabolismo de plantas crasuláceas) frecuente
en plantas xerófitas (plantas adaptadas a ambientes áridos) y que les permite mantener
cerrados los estomas evitando la pérdida de agua.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.9.4 Plantas C3 y C4.
Los isótopos de un elemento químico son las variedades en las que se suelen presentar sus
átomos. Existen en la naturaleza tres isótopos del carbono: el 12C, el 13C y el 14C. Son tres
variedades de un mismo elemento químico, el carbono, cuyos núcleos contienen el mismo
número de protones (seis), pero un número diferente de neutrones (seis, siete y ocho), lo
que les hace, a pesar de tener propiedades químicas semejantes, tener una masa atómica
diferente: doce, trece y catorce.
Casi el 99 % del CO2 atmosférico es del tipo que contiene el carbono ligero 12C. Una
pequeña parte, el 1,1 % del CO2 , es algo más pesado, ya que contiene 13C. Y finalmente
existe también en la atmósfera, en muy pequeña proporción, un tipo de CO2 que contiene
14C , que es radiactivo e inestable, y cuyas aplicaciones han solido ser fundamentalmente
paleocronológicas.
Veamos a continuación dónde radica el interés del 13C en la investigación paleoclimática.
En el proceso de absorción fotosintética de CO2 , la vegetación terrestre y el fitoplancton
marino practican una discriminación en contra de las moléculas pesadas, prefiriendo el 12C
al 13C. De esta forma el carbono atrapado en los vegetales continentales contiene una
menor proporción de 13C que el carbono del CO2 atmosférico.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

De igual manera, en el océano, el carbono orgánico del plancton también tiene una
proporción menor de 13C que la que tiene el carbono inorgánico disuelto en el océano (DIC,
dissolved inorganic carbon).
Esta discriminación practicada en la fotosíntesis es, sin embargo, variable, y depende de los
niveles existentes de CO2 , tanto en el aire como en el mar. La discriminación es mayor
cuanto mayor sea la concentración de CO2 en la atmósfera o en el mar.
El signo d13C simboliza la desviación de la concentración isotópica de 13C en cualquier
muestra, viva o fósil, con respecto a una medida standard, que suele ser el carbono
contenido en el carbonato cálcico de la concha de un determinado fósil marino denominado
PDB (Pee Dee Belemnite), o VPDB, perteneciente a una formacíón geológica del Cretácico
en Carolina del Norte, y cuyo valor ha sido establecido por la Agencia Internacional de la
Energía Atómica, con sede en Viena.
La fórmula de d13C (en ‰) es la siguiente:
(13C/12C)muestra – (13C/12C)standard
——————————————––––––––––––––– x 1.000
(13C/12C)standard
siendo (13C/12C)standard la ratio isotópica de PDB . La ratio absoluta13C/12C de la
muestra standard VPDB es 0.0112372. Los materiales con ratios 13C/12C > 0.00112372
tienen valores delta positivos, y aquellos con ratios 13C/12 C < 0.00112372 tienen valores
delta negativos.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

3.9.5 Rubisco
RuBisCO es la forma abreviada con que normalmente se designa a la enzima cuyo nombre
completo es ribulosa-1,5-bisfosfato carboxilasa oxigenasa. Esta enzima tiene un doble
comportamiento que justifica su nombre, catalizando dos procesos opuestos. Primero la
fijación del CO2 a una forma orgánica, lo que justifica su clasificación como carboxilasa.
Segundo, la fotorrespiración, en la que actúa como oxigenasa del mismo sustrato. La
RuBisCO es la proteína más abundante en la biosfera.
La RuBisCO que se observa en los plastos es una proteína oligómera formada por 16
polipéptidos, que son de dos tipos, uno grande (subunidad L) y otro pequeño (subunidad S).
Esta estructura se observa también en cianobacterias, lo que no sorprende, ya que al fin y al
cabo los plastos eucarióticos derivan evolutivamente de ellas por endosimbiosis. En
procariontes distintos de las cianobacterias la RuBisCO forma agregados más sencillos,
típicamente de dos subunidades grandes y dos pequeñas.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

IV CONCLUSION
Las conclusiones del trabajo de investigación son las siguientes, que ell ciclo de Krebs es la vía
catabólica que realiza la oxidación de glúcidos, ácidos grasos y aminoácidos hasta producir CO2,
liberando energía en forma utilizable, ademas también proporciona precursores para muchas
biomoléculas, como ciertos aminoácidos.
Que la fotosíntesis es la reacción impulsada por el sol que hace que las hojas de las plantas que
contengan clorofila, que combinando agua, sales minerales y dióxido de carbono fabriquen
materia orgánica (azúcares) liberando oxígeno en el proceso.
Ademas la fotosíntesis es un proceso que se desarrolla en dos etapas. La primera es un
proceso dependiente de la luz (etapa clara), requiere de energía de la luz para fabricar
moléculas portadoras de energía a usarse en la segunda etapa. En la etapa independiente
de la luz (etapa oscura) los productos de la primera etapa son utilizados para formar los
enlaces C-C de los carbohidratos. Las reacciones de la etapa oscura usualmente ocurren en
la oscuridad si los transportadores de energía provenientes de la etapa clara están
presentes. Evidencias recientes sugieren que la enzima más importante de la etapa oscura
esta estimulada indirectamente por la luz, de ser así el termino no sería correcto
denominarla "etapa oscura". La etapa clara ocurre en la grana y la oscura en el estroma de
los cloroplastos para reactiva al siguiente proceso celular del ciclo de Calvin entre sus 8
fases de este para la obtención de energia para las plantas vasculares.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica

V BIBLIOGRAFIA
1. J. Azcón-Bieto, M. Talón (eds.). Fundamentos de Fisiología Vegetal. Madrid:
McGraw-Hill/Interamericana, Edicions Universitat de Barcelona, 2000.
2. B.B. Buchanan, W. Gruissem, R. Jones. Biochemistry and Molecular Biology of
plants. Rockville (USA): American Society of Plant Physiologists, 2000.
3. D. T. Dennis and D.H. Turpin (eds). Plant metabolism. Plant physiology,
Biochemistry, and Molecular Biology. Orlando, USA: Academic Press, 1998.
4. H.W. Heldt. Plant Biochemistry and Molecular Biology. Oxford (U.K.): Oxford
University Press, 2004.
5. Frank B. Salisbury, Cleon W. Ross. Fisiología Vegetal. México: Grupo Editorial
Iberoamericana, 1994. (traducción de la 4ª edición original en inglés: Plant
Physiology.
6. Wikipedia Enciclopedia Libre, la fotosíntesis.
7. Lehninger. Fundamentos de Bioquimica, 2002.
Marcelo Santiago Hernández Universidad Veracruzana Bioquímica





























