Embed Size (px)
Citation preview

1
TẠP CHÍKHOA HỌC CÔNG NGHỆNÔNG NGHIỆP VIỆT NAM
Journal of Vietnam Agricultural Science and Technology
NĂM THỨ MƯỜI HAI
SỐ 7 NĂM 2017
TỔNG BIÊN TẬPEditor in chief
GS.TS. NGUYỄN VĂN TUẤT
PHÓ TỔNG BIÊN TẬPDeputy Editor
GS.TS. BÙI CHÍ BỬUTS. TRẦN DANH SỬU
TS. NGUYỄN THẾ YÊN
THƯỜNG TRỰCThS. PHẠM THỊ XUÂN - THƯ KÝ
TÒA SOẠN - TRỊ SỰBan Thông tin
Viện Khoa học Nông nghiệp Việt Nam Vĩnh Quỳnh, Thanh Trì, Hà Nội
Điện thoại: (024) 36490503; (024) 36490504; 0949940399
Fax: (024) 38613937;Website: http//www.vaas.org.vn
Email: [email protected];[email protected]
ISSN: 1859 - 1558Giấy phép xuất bản số:
1250/GP - BTTTTBộ Thông tin và Truyền thôngcấp ngày 08 tháng 8 năm 2011
MỤC LỤC1. Hô Văn Đươc, Nguyên Thi Lang, Bùi Phước Tâm,
Phạm Thi Bé Tư. Ứng dụng chỉ thị phân tử chọn giống lúa có hàm lượng amylose thấp trên quần thể lai hồi giao OM6976/Jasmine 85//OM6976
2. Hô Công Trực, Nguyên Thi Thảo Nhung, Trương Văn Bình, Đoàn Văn Thanh. Kết quả khảo nghiệm một số giống lúa chịu hạn có triển vọng tại huyện Lắk, Đắk Lắk
3. Lưu Quang Huy, Nguyên Thi Ngọc Huệ, Vũ Linh Chi, Dương Hông Mai, Vũ Đăng Toàn, Bùi Thi Thu Huyền, Hà Minh Loan, Trần Danh Sửu. Khả năng chịu hạn của một số nguồn gen lúa địa phương đang được lưu giữ tại Ngân hàng gen cây trồng Quốc gia
4. Dương Trung Dũng. Nghiên cứu khả năng sinh trưởng, phát triển và năng suất của một số giống đậu tương trên đất nương rẫy tại huyện Hoàng Su Phì, tỉnh Hà Giang
5. Lâm Văn Hà. Ảnh hưởng của phân khoáng N, P và phân hữu cơ đến năng suất cà phê vối (Coffea Canephora Pierre) trên đất đỏ bazan vùng cao nguyên Di Linh, tỉnh Lâm Đồng
6. Hô Hữu Huân, Trình Công Tư. Đánh giá hiệu quả của phân Polysulphate đối với cà phê vối trên đất nâu đỏ bazan tỉnh Lâm Đồng
7. Dương Trung Dũng, Trần Xuân Hoàng. Nghiên cứu ảnh hưởng của một số loại phân hữu cơ vi sinh đến sinh trưởng, phát triển giống chè Shan tại Thuận Châu, Sơn La
8. Nguyên Xuân Cảnh, Nguyên Thi Khánh, Phạm Hông Hiển. Nghiên cứu chủng xạ khuẩn có khả năng đối kháng với vi khuẩn Erwinia carotovora gây bệnh thối nhũn trên một số loại cây trồng
9. Nguyên Văn Giang, Đinh Văn Lơi, Phạm Hông Hiển. Ảnh hưởng của điều kiện nuôi cấy, môi trường dinh dưỡng đến khả năng sinh trưởng của chủng xạ khuẩn GL30
10. Nguyên Thi Thanh Thuỷ, Phí Quyết Tiến. Ảnh hưởng của nhiệt độ sấy và chế độ ngâm chiết tới sự biến đổi thành phần hoá học và cảm quan của giá thể sau thu hoạch đông trùng hạ thảo
11. Nguyên Thi Mai Hương, Hô Tuấn Anh. Nghiên cứu hoạt tính kháng khuẩn của một số loại tinh dầu
3
9
15
20
26
33
36
41
47
52
57

2
TẠP CHÍKHOA HỌC CÔNG NGHỆNÔNG NGHIỆP VIỆT NAM
Journal of Vietnam Agricultural Science and Technology
NĂM THỨ MƯỜI HAI
SỐ 7 NĂM 2017
TỔNG BIÊN TẬPEditor in chief
GS.TS. NGUYỄN VĂN TUẤT
PHÓ TỔNG BIÊN TẬPDeputy Editor
GS.TS. BÙI CHÍ BỬUTS. TRẦN DANH SỬU
TS. NGUYỄN THẾ YÊN
THƯỜNG TRỰCThS. PHẠM THỊ XUÂN - THƯ KÝ
TÒA SOẠN - TRỊ SỰBan Thông tin
Viện Khoa học Nông nghiệp Việt Nam Vĩnh Quỳnh, Thanh Trì, Hà Nội
Điện thoại: (024) 36490503; (024) 36490504; 0949940399
Fax: (024) 38613937;Website: http//www.vaas.org.vn
Email: [email protected];[email protected]
ISSN: 1859 - 1558Giấy phép xuất bản số:
1250/GP - BTTTTBộ Thông tin và Truyền thôngcấp ngày 08 tháng 8 năm 2011
12. Vũ Thi Hạnh Nguyên, Nguyên Thi Thanh Thủy, Phí Quyết Tiến. Nâng cao khả năng sinh tổng hợp validamycin-A từ chủng Streptomyces hygroscopicus 11405 bằng đột biến tế bào trần
13. Nguyên Văn Giang, Chu Đức Hà, Nguyên Thi Thu. Khảo sát một số đặc điểm của chủng xạ khuẩn HT1 có khả năng kháng vi khuẩn Streptococcus agalactiae gây bệnh trên cá rô phi
14. Nguyên Thi Thanh Thủy, Hô Tuấn Anh. Tối ưu hóa điều kiện tự phân tế bào nấm men bia thải
15. Nguyên Hoàng Anh, Hô Tuấn Anh. Tuyển chọn chủng vi khuẩn Lactic có khả năng sinh enzyme β - galactosidase chịu axit (pH 2 - 3)
16. Trần Văn Huy, Phạm Văn Nhạ, Nguyên Thi Nga, Nguyên Mạnh Cường, Vũ Xuân Trung, Phạm Việt Hông, Lê Thi Thu Hiền, Nguyên Trường Phi. Cải tiến công nghệ sản xuất chế phẩm nấm xanh Metarhizium anisopliae để phòng trừ rầy nâu hại lúa
17. Mai Văn Quân, Dương Thi Nguyên. Kết quả giám định và một số đặc điểm của nấm Botrytis cinerea Pers. gây bệnh thối xám trên hoa thược dược (Dahia pinnata Cav.) tại Việt Nam
18. Nguyên Văn Toản, Nguyên Văn Huế. Nghiên cứu ảnh hưởng của chế độ sấy (nhiệt độ, thời gian) bằng phương pháp sấy bơm nhiệt đến sản phẩm tinh bột nghệ
19. Nguyên Văn Toản, Nguyên Hông Phúc. Nghiên cứu ảnh hưởng của Aminoethoxyvinylglycine (AVG) kết hợp xử lý nước nóng đến quá trình chín quả bơ (Booth7) sau thu hoạch
20. Nguyên Quang Vĩnh, Nguyên Thi Phương, Bùi Tuấn Anh, Trần Văn Tú, Vũ Thi Hải, Lê Huy Công. Nghiên cứu xây dựng tiêu chuẩn cơ sở dược liệu Tam thất hoang
21. Nguyên Quang Vĩnh, Nguyên Thi Phương, Bùi Tuấn Anh, Trần Văn Tú, Vũ Thi Hải, Lê Huy Công. Nghiên cứu xây dựng tiêu chuẩn cơ sở dược liệu Hoàng liên ô rô
22. Đông Huy Giới. Bước đầu sử dụng nano bạc trong phòng trừ bệnh thối nâu do vi khuẩn Gluconobacter frateurii gây ra trên quả nhãn
23. Nguyên Thanh Hiệu, Dương Nhựt Long, Lam Mỹ Lan. Ảnh hưởng của thức ăn đến sự thành thục của cá heo (Botia modesta Bleeker, 1865)
63
69
73
79
83
87
92
96
102
105
110
114

3
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀTrong những năm gần đây, nhu cầu thị trường
gạo có nhiều thay đổi về số lượng lẫn chất lượng. Trong đó, các giống lúa phẩm chất cao, mềm dẻo, có mùi thơm đang chiếm thị phần ngày càng tăng trong canh tác và xuất khẩu của vùng. Tuy nhiên, thực tế sản xuất của ĐBSCL đang gặp nhiều khó khăn, biến đổi khí hậu làm giảm năng suất lúa, các giống chống chịu tốt hơn nhưng phẩm chất thấp và giá trị thương phẩm không cao. Chính vì vậy, việc nghiên cứu, lai tạo và phát triển các giống lúa mới ngon, dẻo lại thích nghi đa dạng và cho năng suất cao đang là nhu cầu cấp bách của sản xuất lúa gạo trong vùng.
Nhiều nghiên cứu cho rằng tính chất mềm xốp của cơm phụ thuộc chủ yếu vào hàm lượng amylose (AC) trong thành phần tinh bột của hạt gạo (Hu et al., 2004; Martin and Smith, 1995). Gen waxy, nằm trên nhiễm sắc thể số 6, là gen điều khiển chỉ tiêu AC trong hạt (Denyer et al., 2001; Nakamura and Yuki, 1992; Nakamura et al., 1989). Các giống lúa được phân ra thành hai nhóm phẩm chất: waxy (hàm lượng amylose 1-2%) và nonwaxy (hàm lượng amylose > 2%). Đối với nhóm nonwaxy, hàm lượng amylose thấp khi AC = < 20%, hàm lượng amylose trung bình khi AC = 20 - 25% và hàm lượng amylose cao khi AC > 25% (Frei et al., 2003; Coffman and Juliano, 1987).
Ứng dụng chỉ thị phân tử trong chọn giống lai hồi giao (MABC) là phương pháp chuyển một gen mục tiêu từ giống cho gen sang giống nhận gen trong khi
vẫn giữ lại các đặc tính quan trọng của giống nhận thông qua lai hồi giao. Việc sử dụng các chỉ thị phân tử cho phép giải mã di truyền của con lai ở mỗi thế hệ, làm tăng tốc độ của quá trình chọn tạo, do đó tăng hiệu quả chọn lọc gen trên một đơn vị thời gian (Hospital, 2003). MABC được sử dụng trong nhiều nghiên cứu tạo chọn giống lúa chất lượng cao trước đây (Hasan et al., 2015; Hồ Văn Được và ctv., 2015; Nguyễn Thị Lang, 2004; Zhou et al., 2003).
Xuất phát từ những vấn đề đặt ra, nghiên cứu được thực hiện nhằm khai thác phương pháp MABC trong lai tạo giống lúa có hàm lượng amylose thấp (< 20%), năng suất cao phù hợp với nhu cầu về giống lúa cũng như điều kiện canh tác của ĐBSCL hiện nay.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu - Giống lúa OM6976: có năng suất cao (7-8 tấn/
ha), thích nghi rộng nhưng hàm lượng amylose cao (24 - 25%).
- Giống lúa Jasmine 85: năng suất khá, mặt gạo đẹp, cơm dẻo (16 - 18%).
- Các chỉ thị phân tử: Wx là chỉ thị phân tử đặc hiệu sử dụng cho đánh dấu gen waxy quy định chỉ tiêu hàm lượng amylose trên lúa. RM240, RM162, RM256 và RM257 sử dụng đánh dấu trên giống mẹ OM6976 liên quan đến gen quy định các thành phần năng suất và năng suất (Bảng 1).
1 Trường Đại học Cần Thơ; 2 Viện Lúa Đồng bằng sông Cửu Long
ỨNG DỤNG CHỈ THỊ PHÂN TỬ CHỌN GIỐNG LÚA CÓ HÀM LƯỢNG AMYLOSE THẤP TRÊN QUẦN THỂ LAI HỒI GIAO OM6976/JASMINE 85//OM6976
Hồ Văn Được1, Nguyễn Thị Lang1, Bùi Phước Tâm2, Phạm Thị Bé Tư2
TÓM TẮTChiến lược chọn tạo giống lúa phẩm chất liên quan mật thiết đến hàm lượng amylose thấp. Phương pháp chọn
tạo giống nhờ vào chỉ thị phân tử là phương pháp hiện đại đạt được nhiều thành công qua nhiều nghiên cứu trên cây lúa. Trong nghiên cứu này, 71 giống lúa cao sản được đánh giá về hàm lượng amylose, năng suất và thành phần năng suất và chọn ra cá thể tốt nhất để thực hiện lai hồi giao. OM6976, giống lúa có năng suất cao, được chọn làm mẹ và Jasmine 85, giống lúa có hàm lượng amylose thấp, được chọn làm bố. Các cá thể con lai được lai hồi giao và chọn lọc liên tục qua các thế hệ kết hợp với phương pháp sử dụng chỉ thị phân tử đến thế hệ BC4. 41 chỉ thị phân tử được sử dụng để đánh giá đa dạng di truyền của giống bố mẹ, trong đó, 1 chỉ thị phân tử (Wx) đánh dấu gen quy định hàm lượng amylose và 4 chỉ thị (RM420, RM162, RM256 và RM257) liên quan đến tính trạng năng suất và thành phần năng suất cho kết quả đa hình. Ở thế hệ BC4F3, 10 dòng cho hàm lượng amylose thấp từ 17,5 - 20,6%. Trong đó, 4 dòng ưu tú là dòng D75, D131, D142 và D150 vừa có hàm lượng amylose thấp và vừa cho năng suất cao.
Từ khóa: Lúa, amylose, lai hồi giao, chỉ thị phân tử, năng suất, con lai

4
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
2.2. Phương pháp nghiên cứu
2.2.1. Phát triển quần thể lai hồi giao OM6976/Jasmine 85//OM6976
Hình 1. Sơ đồ phát triển quần thể lai hồi giao BC4F3 bằng chỉ thị phân tử
2.2.2. Phương pháp phân tích hàm lượng amylose, độ trở hồ và độ bền thể gel
Phương pháp đánh giá các chỉ tiêu phẩm chất (hàm lượng amylose (%), độ bền gel (mm), độ trở hồ (cấp) được thực hiện theo phương pháp của IRRI (1996).
2.2.3. Các chỉ tiêu về nông họcThời gian sinh trưởng (ngày) được tính từ khi
gieo hạt đến khi bông lúa chín (85% số hạt trên bông chín). Chiều cao cây (cm) được đo từ mặt đất đến đỉnh bông cái.
Năng suất và thành phần năng suất: số bông/bụi [P/số bụi thu thập]; số hạt chắc/bông [(f/v) ˟ (W+w)/P]; khối lượng 1000 hạt (g) [(W/f) ˟ 1000]; năng suất (tấn/ha) được qui về 14% ẩm độ. Trong đó: P là tổng số bông đếm được trên các bụi lúa đã chọn làm mẫu, f: Tổng số hạt chắc/bông cái, W: Khối lượng hạt chắc trên tất cả bông lúa, v: khối lượng tổng số hạt chắc, w: khối lượng 1000 hạt lấy ra từ bông cái.
2.2.4. Phương pháp ly trích DNA và phản ứng PCR-SSR
Theo phương pháp cải tiến của Nguyễn Thị Lang (2002).
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Đánh giá vật liệu bố mẹBảy mươi mốt giống lúa cao sản được thu thập từ
Ngân hàng gen của Viện Lúa Đồng bằng sông Cửu Long. Các giống lúa này lần lượt được đánh giá hàm lượng amylose, các tính trạng nông học, các thành phần năng suất và năng suất. Jasmine 85 có thời gian sinh trưởng ngắn (103,3 ngày) trong khi OM6976 là 110,0 ngày. Về năng suất, Jasmine 85 đạt 7,43 tấn/ha và OM6976 đạt 7,67 tấn/ha, cao hơn đối chứng AS996. Về hàm lượng amylose, Jasmine 85 là 18,30% trong khi OM6976 có hàm lượng amylose cao (26,38%) (Bảng 2). Do đó, OM6976 được chọn làm mẹ vì có nhiều đặc tính phù hợp như năng suất cao, ổn định, thích nghi rộng, khá ngắn ngày. Jasmine 85 được chọn làm bố vì có hàm lượng amylose thấp, ngoài ra giống này còn đặc tính ngắn ngày và năng suất khá cao. Hai giống này được chọn làm bố mẹ nhằm tạo ra thế hệ con lai có hàm lượng amylose thấp, vừa ngắn ngày và đạt năng suất cao.
Bảng 1. Các chỉ thị phân tử đa hình dùng trong thí nghiệm
Ghi chú: NS: năng suất; TPNS: thành phần năng suất
Chỉ thi NST Trình tự (5’-3’) Kích thước (bp) Tính trạng liên kết
Wx 6 ctttgtctatctcaagacacttgcagatgttcttcctgatg 210-220 Hàm lượng amylose
RM240 2 ccttaatgggtagtgtgcactgtaaccattccttccatcc 150-200 NS và TPNS
RM162 6 gccagcaaaaccagggatccggcaaggtcttgtgcggcttgcgg 250-300 NS và TPNS
RM256 8 gacagggagtgattgaaggcgttgatttcgccaagggc 100-200 NS và TPNS
RM257 9 cagttccgagcaagagtactcggatcggacgtggcatatg 150-250 NS và TPNS
OM6976 Jasmine 85
OM6976
OM6976
Wx
RM240, RM162,RM256, RM257
x
x
x
F1
BC1F1 (100-200 cây)
BC1F1 (50-100 cây Dị hợp tử)
BC1F1(1-20 cây)
BC4F3
BC4F1BC2F1Tương tự BC1
Đánh giá và chọn lọc
..........
......................................................
...
......................................................

5
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bốn mươi mốt chỉ thị phân tử được sử dụng để đánh giá đa dạng di truyền giữa các giống lúa bố mẹ. Trong đó, 1 chỉ thị phân tử (Wx) đánh dấu gen quy định hàm lượng amylose và 4 chỉ thị (RM240, RM162, RM256 và RM257) liên quan đến các thành phần năng suất và năng suất cho kết quả đa hình giữa giống bố và giống mẹ.
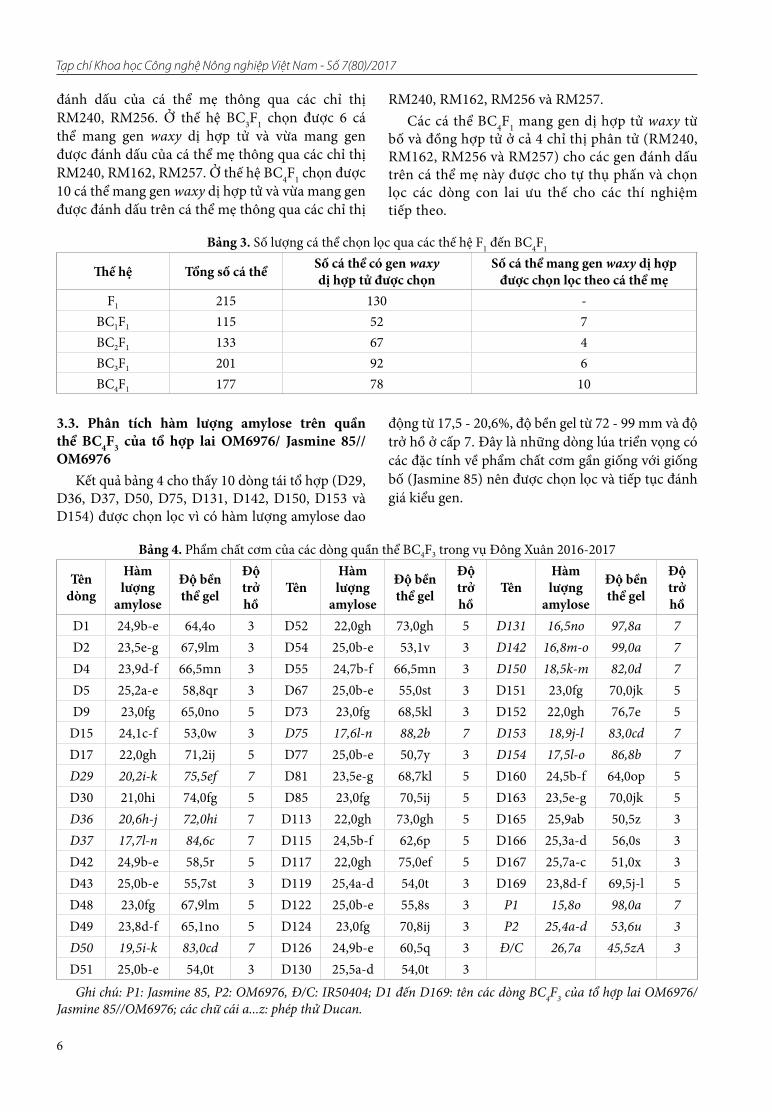
Với chỉ thị Wx, kết quả khuếch đại PCR cho băng hình ở hai kích thước khác nhau 210bp và 220bp (Hình 2). Ở kích thước 220bp, các giống KDML105, Jasmine85 và OM7347 thể hiện băng hình ở vị trí này, đây cũng là kích thước của gen waxy. Các giống như IR64, OM5930, OM6073 và OM6976 cho băng hình ở kích thước 210bp. Các giống này biểu hiện không mang gen waxy.
Hình 2. Sản phẩm PCR của các giống lúa bố mẹ với chỉ thị Wx trên gel agarose 3%
Ghi chú: M: Thang chuẩn DNA (1Kb)
Bảng 2. Các đặc tính của các giống OM6976 và Jasmine 85 trong vụ Đông Xuân 2013-2014
Hình 3. Kết quả đa hình của các giống bố mẹ với các chỉ thị cho gen liên quan đến các thành phần năng suất và năng suất trên gel agarose 3%.
Ghi chú: 1: OM6976, 2: OM6073, 3: OM5930, 4: KDML 105, 5: Jasmine 85, 6: OM7347, M: Thang chuẩn DNA (1Kb)
TT Tên giốngTGST Chiều
cao cây
Số bông/
bụi
Chiều dài
bông
Số hạt chắc/ bông
Tỷ lệ hạt lép/
bông
Khối lương
1000 hạt
Năng suất
Hàm lương
amylosengày cm bông cm hạt % g tấn/ha %
1 AS996 (đc) 90,0 96,5 9,4 21,20 72,0 20,29 29,34 2,33 24,912 Jasmine 85 103,3 101,7 11,9 28,10 100,3 24,37 27,00 7,43 18,303 OM6976 110,0 104,7 11,3 29,60 226,0 24,90 27,00 7,67 26,38
Qua đánh giá các chỉ thị phân tử liên quan đến năng suất và thành phần năng suất, với chỉ thị RM240 thể hiện băng ở kích thước 150bp (OM6976) và 200bp (Jasmine 85). Tương tự, RM162 ở 250bp (Jasmine 85) và 300bp (OM6976), RM256 ở 100bp (OM6976) và 200 (Jasmine 85), RM257 ở 150bp (OM6976) và 250bp (Jasmine 85). Các chỉ thị này dùng để đánh dấu trên cá thể mẹ (OM6976) trong nghiên cứu này.
3.2. Kết quả lai tạo quần thể hôi giao OM6976/Jasmine 85//OM6976
Quần thể OM6976/Jasmine85//OM6976 bắt đầu được lai tạo từ năm 2014 và các cá thể con lai ở liên tục được chọn lọc dựa trên chỉ thị phân tử cho đến thế hệ BC4 (Bảng 3). Các cá thể ở mỗi thế hệ được
lựa chọn dựa trên gen dị hợp tử waxy từ bố và đồng hợp tử ở 4 chỉ thị phân tử (RM240, RM162, RM256 và RM257) cho các gen đánh dấu trên cá thể mẹ.
Ở thế hệ F1, 215 cá thể được thu hoạch. Tuy nhiên, qua kiểm tra kiểu gen các cá thể này với chỉ thị Wx chỉ có 130 cá thể thể hiện mang gen waxy dị hợp tử. Các cá thể có kiểu hình tốt nhất được chọn để lai tạo cho các thế hệ kế tiếp.
Ở thế hệ BC1F1, số cá thể được thu hoạch là 115. Trong đó, 52 cá thể thể hiện gen waxy dị hợp tử, tuy nhiên, chỉ có 7 cá thể tốt nhất được chọn vì vừa mang gen waxy dị hợp tử và vừa mang gen được đánh dấu của cá thể mẹ thông qua các chỉ thị RM240, RM162.
Tương tự, ở thế hệ BC2F1 chọn được 4 cá thể mang gen waxy dị hợp tử và vừa mang gen được
210 bp
M
OM
7347
OM
6073
OM
6976
Jasm
ine8
5
OM
5930
IR64
KD
ML1
05
220 bp
6
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
đánh dấu của cá thể mẹ thông qua các chỉ thị RM240, RM256. Ở thế hệ BC3F1 chọn được 6 cá thể mang gen waxy dị hợp tử và vừa mang gen được đánh dấu của cá thể mẹ thông qua các chỉ thị RM240, RM162, RM257. Ở thế hệ BC4F1 chọn được 10 cá thể mang gen waxy dị hợp tử và vừa mang gen được đánh dấu trên cá thể mẹ thông qua các chỉ thị
RM240, RM162, RM256 và RM257.Các cá thể BC4F1 mang gen dị hợp tử waxy từ
bố và đồng hợp tử ở cả 4 chỉ thị phân tử (RM240, RM162, RM256 và RM257) cho các gen đánh dấu trên cá thể mẹ này được cho tự thụ phấn và chọn lọc các dòng con lai ưu thế cho các thí nghiệm tiếp theo.
Bảng 3. Số lượng cá thể chọn lọc qua các thế hệ F1 đến BC4F1
Bảng 4. Phẩm chất cơm của các dòng quần thể BC4F3 trong vụ Đông Xuân 2016-2017
Ghi chú: P1: Jasmine 85, P2: OM6976, Đ/C: IR50404; D1 đến D169: tên các dòng BC4F3 của tổ hợp lai OM6976/Jasmine 85//OM6976; các chữ cái a...z: phép thử Ducan.
Thế hệ Tổng số cá thể Số cá thể có gen waxy di hơp tử đươc chọn
Số cá thể mang gen waxy di hơp đươc chọn lọc theo cá thể mẹ
F1 215 130 -BC1F1 115 52 7BC2F1 133 67 4BC3F1 201 92 6BC4F1 177 78 10
3.3. Phân tích hàm lương amylose trên quần thể BC4F3 của tổ hơp lai OM6976/ Jasmine 85// OM6976
Kết quả bảng 4 cho thấy 10 dòng tái tổ hợp (D29, D36, D37, D50, D75, D131, D142, D150, D153 và D154) được chọn lọc vì có hàm lượng amylose dao
động từ 17,5 - 20,6%, độ bền gel từ 72 - 99 mm và độ trở hồ ở cấp 7. Đây là những dòng lúa triển vọng có các đặc tính về phẩm chất cơm gần giống với giống bố (Jasmine 85) nên được chọn lọc và tiếp tục đánh giá kiểu gen.
Têndòng
Hàm lương
amylose
Độ bền thể gel
Độ trở hô
TênHàm lương
amylose
Độ bền thể gel
Độ trở hô
TênHàm lương
amylose
Độ bền thể gel
Độ trở hô
D1 24,9b-e 64,4o 3 D52 22,0gh 73,0gh 5 D131 16,5no 97,8a 7D2 23,5e-g 67,9lm 3 D54 25,0b-e 53,1v 3 D142 16,8m-o 99,0a 7D4 23,9d-f 66,5mn 3 D55 24,7b-f 66,5mn 3 D150 18,5k-m 82,0d 7D5 25,2a-e 58,8qr 3 D67 25,0b-e 55,0st 3 D151 23,0fg 70,0jk 5D9 23,0fg 65,0no 5 D73 23,0fg 68,5kl 3 D152 22,0gh 76,7e 5
D15 24,1c-f 53,0w 3 D75 17,6l-n 88,2b 7 D153 18,9j-l 83,0cd 7D17 22,0gh 71,2ij 5 D77 25,0b-e 50,7y 3 D154 17,5l-o 86,8b 7D29 20,2i-k 75,5ef 7 D81 23,5e-g 68,7kl 5 D160 24,5b-f 64,0op 5D30 21,0hi 74,0fg 5 D85 23,0fg 70,5ij 5 D163 23,5e-g 70,0jk 5D36 20,6h-j 72,0hi 7 D113 22,0gh 73,0gh 5 D165 25,9ab 50,5z 3D37 17,7l-n 84,6c 7 D115 24,5b-f 62,6p 5 D166 25,3a-d 56,0s 3D42 24,9b-e 58,5r 5 D117 22,0gh 75,0ef 5 D167 25,7a-c 51,0x 3D43 25,0b-e 55,7st 3 D119 25,4a-d 54,0t 3 D169 23,8d-f 69,5j-l 5D48 23,0fg 67,9lm 5 D122 25,0b-e 55,8s 3 P1 15,8o 98,0a 7D49 23,8d-f 65,1no 5 D124 23,0fg 70,8ij 3 P2 25,4a-d 53,6u 3D50 19,5i-k 83,0cd 7 D126 24,9b-e 60,5q 3 Đ/C 26,7a 45,5zA 3D51 25,0b-e 54,0t 3 D130 25,5a-d 54,0t 3

7
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Qua kết quả đánh giá phẩm chất các dòng BC4F3 (Bảng 4) cho thấy các dòng có hàm lượng amylose thấp (<20%) và có độ bền thể gel mềm (>60 mm) như: Dòng 29, 36, 37, 50, 75, 131, 142, 150, 153, 154 tương đương với giống đối chứng Jasmine 85 thể hiện tính trạng cơm ướt, ngon dẻo, bóng láng khi nấu chín và có tính gel mềm khi để nguội. Kết quả trên cho giá trị khác biệt có ý nghĩa thống kê so với giống OM6976 và giống đối chứng IR50404. Thêm vào đó, độ trở hồ của các dòng chọn lọc trên ở cấp 3 tương đương với đối chứng Jasmine 85. Điều này cho thấy các dòng được chọn từ quần thể BC4F3 có độ trở hồ trung bình cho phẩm chất gạo tốt.
3.4. Phân tích kiểu gen trên quần thể con lai BC4F3 của tổ hơp OM6976/ Jasmine 85// OM6976
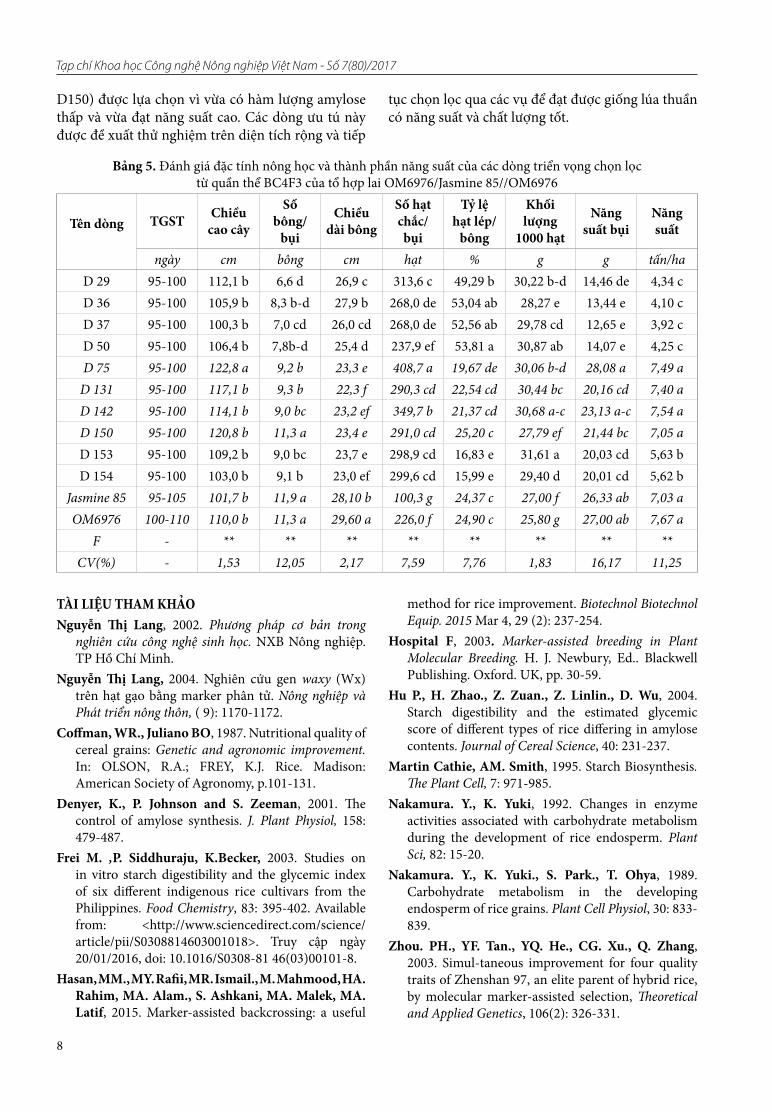
Kết quả thể hiện ở hình 4 cho thấy các băng hình của cá thể số 1 đến 10 (BC4F3) ở vị trí 220bp (giống với Jasmine 85), điều này chứng tỏ các dòng này đều mang gen waxy, qui định hàm lượng amylose thấp. Sau đó, 10 cá thể tái tổ hợp mang gen waxy này sẽ được sử dụng trong thí nghiệm xác định cá thể có
nền di truyền giống với giống nhận gen (OM6976). Kết quả kiểu gen với 4 chỉ thị RM240, RM257, RM162 và RM256 đều thể hiện đa hình và 10 dòng trên đều biểu hiện giống với giống mẹ OM6976 ở các vị trí đánh dấu trên bộ nhiễm sắc thể (Hình 5).
Qua đánh giá kiểu hình và kiểu gen liên quan đến hàm lượng amylose thấp, các dòng triển vọng được chọn lọc là D29, D36, D37, D50, D75, D131, D142, D150, D153 và D154.
Hình 4. Kết quả điện di sản phẩm PCR của các dòng BC4F3 ở tổ hợp OM6976/Jasmine 85//OM6976
với chỉ thị Wx trên gel agarose 3%Ghi chú: M: thang chuẩn; P1: OM6976; P2: Jasmine 85;
1-10: các cá thể BC4F3 lần lượt là: D29, D36, D37, D50, D75, D131, D142, D150, D153 và D154.
3.5. Chọn dòng triển vọng hàm lương amylose thấp trên quần thể OM6976/ Jasmine 85// OM6976
Mười dòng triển vọng được xác định có hàm lượng amylose thấp thông qua đánh giá kiểu hình và kiểu gen tiếp tục được đánh giá năng suất và các thành phần năng suất trong vụ Đông Xuân 2016 - 2017 (Bảng 5).
Kết quả đánh giá từ bảng 5 cho thấy, hầu hết các dòng chọn lọc đều có thời gian sinh trưởng ngắn từ 95 - 100 ngày. Chiều cao cây không khác biệt có ý nghĩa so với dạng hình bố mẹ trừ Dòng 75 cao trội hơn so với bố mẹ. Chiều dài bông của các dòng chọn lọc tương đối ngắn hơn so với giống bố mẹ tuy nhiên có số hạt chắc/bụi cao, tỉ lệ hạt lép/bông thấp và khối lượng 1000 hạt rất cao (27,79 - 30,87 g) cao hơn giống bố mẹ khác biệt ở mức ý nghĩa 1%. Do đó,
năng suất/bụi cao và cho năng suất cao tương đương giống bố mẹ như các dòng 75, 131, 142 và 150.
Xét về năng suất, 4 dòng tốt nhất được chọn là dòng D75, D131, D142 và D150. Các dòng này cho năng suất cao so với đối chứng (Jasmine 85) và hàm lượng amylose thấp (<20%).
IV. KẾT LUẬNPhương pháp ứng dụng chỉ thị phân tử trong
chọn tạo giống lúa có hàm lượng amylose thấp trên tổ hợp hồi giao OM6976/Jasmine 85//OM6976 bước đầu đạt được các dòng triển vọng. Qua phân tích các cá thể BC4F3 của tổ hợp lai hồi giao OM6976/Jasmine 85//OM6976 ghi nhận 10 dòng có hàm lượng amylose thấp dao động từ 17,50 - 20,60%. Trong đó, 4 dòng tốt nhất (D75, D131, D142 và
Hình 5. Kết quả kiểm tra di truyền các cá thể BC4F3 ở tổ hợp OM6976/Jasmine 85//OM6976 trên gel agarose 3% bằng các chỉ thị phân tử RM240, RM257, RM162, RM256
Ghi chú: M: thang chuẩn; P1: OM6976; P2: Jasmine 85; 1-10: các cá thể BC4F3 lần lượt là: D29, D36, D37, D50, D75, D131, D142, D150, D153 và D154.
8
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 5. Đánh giá đặc tính nông học và thành phần năng suất của các dòng triển vọng chọn lọc từ quần thể BC4F3 của tổ hợp lai OM6976/Jasmine 85//OM6976
Tên dòng
TGST Chiều cao cây
Số bông/
bụi
Chiều dài bông
Số hạt chắc/ bụi
Tỷ lệ hạt lép/
bông
Khối lương
1000 hạt
Năng suất bụi
Năng suất
ngày cm bông cm hạt % g g tấn/haD 29 95-100 112,1 b 6,6 d 26,9 c 313,6 c 49,29 b 30,22 b-d 14,46 de 4,34 cD 36 95-100 105,9 b 8,3 b-d 27,9 b 268,0 de 53,04 ab 28,27 e 13,44 e 4,10 cD 37 95-100 100,3 b 7,0 cd 26,0 cd 268,0 de 52,56 ab 29,78 cd 12,65 e 3,92 cD 50 95-100 106,4 b 7,8b-d 25,4 d 237,9 ef 53,81 a 30,87 ab 14,07 e 4,25 cD 75 95-100 122,8 a 9,2 b 23,3 e 408,7 a 19,67 de 30,06 b-d 28,08 a 7,49 a
D 131 95-100 117,1 b 9,3 b 22,3 f 290,3 cd 22,54 cd 30,44 bc 20,16 cd 7,40 aD 142 95-100 114,1 b 9,0 bc 23,2 ef 349,7 b 21,37 cd 30,68 a-c 23,13 a-c 7,54 aD 150 95-100 120,8 b 11,3 a 23,4 e 291,0 cd 25,20 c 27,79 ef 21,44 bc 7,05 aD 153 95-100 109,2 b 9,0 bc 23,7 e 298,9 cd 16,83 e 31,61 a 20,03 cd 5,63 bD 154 95-100 103,0 b 9,1 b 23,0 ef 299,6 cd 15,99 e 29,40 d 20,01 cd 5,62 b
Jasmine 85 95-105 101,7 b 11,9 a 28,10 b 100,3 g 24,37 c 27,00 f 26,33 ab 7,03 aOM6976 100-110 110,0 b 11,3 a 29,60 a 226,0 f 24,90 c 25,80 g 27,00 ab 7,67 a
F - ** ** ** ** ** ** ** **CV(%) - 1,53 12,05 2,17 7,59 7,76 1,83 16,17 11,25
D150) được lựa chọn vì vừa có hàm lượng amylose thấp và vừa đạt năng suất cao. Các dòng ưu tú này được đề xuất thử nghiệm trên diện tích rộng và tiếp
tục chọn lọc qua các vụ để đạt được giống lúa thuần có năng suất và chất lượng tốt.
TÀI LIỆU THAM KHẢO Nguyên Thi Lang, 2002. Phương pháp cơ bản trong
nghiên cứu công nghệ sinh học. NXB Nông nghiệp. TP Hồ Chí Minh.
Nguyên Thi Lang, 2004. Nghiên cứu gen waxy (Wx) trên hạt gạo bằng marker phân tử. Nông nghiệp và Phát triển nông thôn, ( 9): 1170-1172.
Coffman, WR., Juliano BO, 1987. Nutritional quality of cereal grains: Genetic and agronomic improvement. In: OLSON, R.A.; FREY, K.J. Rice. Madison: American Society of Agronomy, p.101-131.
Denyer, K., P. Johnson and S. Zeeman, 2001. The control of amylose synthesis. J. Plant Physiol, 158: 479-487.
Frei M. ,P. Siddhuraju, K.Becker, 2003. Studies on in vitro starch digestibility and the glycemic index of six different indigenous rice cultivars from the Philippines. Food Chemistry, 83: 395-402. Available from: <http://www.sciencedirect.com/science/article/pii/S0308814603001018>. Truy cập ngày 20/01/2016, doi: 10.1016/S0308-81 46(03)00101-8.
Hasan, MM., MY. Rafii, MR. Ismail., M. Mahmood, HA. Rahim, MA. Alam., S. Ashkani, MA. Malek, MA. Latif, 2015. Marker-assisted backcrossing: a useful
method for rice improvement. Biotechnol Biotechnol Equip. 2015 Mar 4, 29 (2): 237-254.
Hospital F, 2003. Marker-assisted breeding in Plant Molecular Breeding. H. J. Newbury, Ed.. Blackwell Publishing. Oxford. UK, pp. 30-59.
Hu P., H. Zhao., Z. Zuan., Z. Linlin., D. Wu, 2004. Starch digestibility and the estimated glycemic score of different types of rice differing in amylose contents. Journal of Cereal Science, 40: 231-237.
Martin Cathie, AM. Smith, 1995. Starch Biosynthesis. The Plant Cell, 7: 971-985.
Nakamura. Y., K. Yuki, 1992. Changes in enzyme activities associated with carbohydrate metabolism during the development of rice endosperm. Plant Sci, 82: 15-20.
Nakamura. Y., K. Yuki., S. Park., T. Ohya, 1989. Carbohydrate metabolism in the developing endosperm of rice grains. Plant Cell Physiol, 30: 833-839.
Zhou. PH., YF. Tan., YQ. He., CG. Xu., Q. Zhang, 2003. Simul-taneous improvement for four quality traits of Zhenshan 97, an elite parent of hybrid rice, by molecular marker-assisted selection, Theoretical and Applied Genetics, 106(2): 326-331.

9
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Application of molecular marker for rice breeding with low amylose content in the backcross population of OM6976/Jasmine 85//OM6976
Ho Van Duoc, Nguyen Thi Lang, Bui Phuoc Tam, Pham Thi Be TuAbstractThe strategy of selecting quality rice cultivars is closely related to low amylose content. The breeding method using the molecular markers is a modern method that has been successful in many previous researches on rice. In this study, 71 high yielding rice varieties were evaluated for amylose content, yield and yield components, and the best individuals were selected for the backcrossing. OM6976, the high yielding variety was selected as a recipient (mother) and Jasmine 85, the low amylose variety was chosen as donor (father). Progeny plants have been continuously backcrossed and selected through generations combined with the use of molecular markers to the BC4 generation. 41 molecular markers were used to evaluate the genetic diversity of parental varieties in which a molecular marker (Wx) marked the gene for amylose content and four markers (RM420, RM162, RM256 and RM257) related to yield and yield components for results of polymorphism. In the BC4F3 generation, 10 lines had low amylose content of 17.5 - 20.6%. Of these, the four best lines including D75, D131, D142 and D150 had low amylose content and high yield.Key words: Rice, amylose, backcrossing, marker, yield, progeny
Ngày nhận bài: 16/6/2017Ngày phản biện: 20/6/2017
Người phản biện: TS. Trần Danh SửuNgày duyệt đăng: 25/6/2017
1 Trung tâm Nghiên cứu Đất, Phân bón và Môi trường Tây Nguyên, Viện Thổ nhưỡng Nông hóa2 Liên hiệp các Hội Khoa học và Kỹ thuật tỉnh Đăk Lăk
KẾT QUẢ KHẢO NGHIỆM MỘT SỐ GIỐNG LÚA CHỊU HẠN CÓ TRIỂN VỌNG TẠI HUYỆN LẮK, ĐẮK LẮK
Hồ Công Trực1, Nguyễn Thị Thảo Nhung1, Trương Văn Bình1, Đoàn Văn Thanh2
TÓM TẮTThí nghiệm so sánh các giống lúa chịu hạn có triển vọng đã được thực hiện tại ba điểm trên vùng khó khăn về
nước tưới của huyện Lắk, tỉnh Đắc Lắk trong hai vụ Đông Xuân và Hè Thu năm 2015, 2016. Các giống tham gia thí nghiệm bao gồm: CH207, CH208, CH19, LCH37, DH39, P6ĐB, OM4900, CXT30, trong đó giống IR64 trồng phổ biến tại địa phương làm đối chứng. Kết quả thí nghiệm cho thấy các giống đưa vào khảo nghiệm đều thích nghi và cho năng suất cao hơn so với giống đối chứng IR64 (là giống phổ biến tại địa phương). Giống lúa chịu hạn LCH37 và giống né vụ P6ĐB cho năng suất cao nhất (P6ĐB đạt 59,0 - 72,5 tạ/ha vụ Hè Thu, 52,3 - 58,7 tạ/ha vụ Đông Xuân tăng 15,2 - 24,4% so đối chứng; LCH37 đạt 54,7 - 68,3 tạ/ha vụ Hè Thu, 51,8 - 57,5 tạ/ha vụ Đông Xuân tăng 11,7 - 17,8% so đối chứng).
Từ khóa: Giống lúa, khảo nghiệm, chịu hạn, thích nghi, huyện Lắk, tỉnh Đắc Lắk
I. ĐẶT VẤN ĐỀHuyện Lắk là vựa lúa của tỉnh Đắk Lắk với diện
tích đất trồng lúa năm 2013 là 10.467 ha, là một trong bốn huyện có diện tích lúa lớn nhất tỉnh trong số này diện tích chủ động nước chỉ chiếm khoảng 60% phân bố ở lưu vực sông Krông Ana, Krông Nô và hồ Lắk còn lại là đất không chủ động nước. Đặc điểm khí hậu của huyện mang đậm nét khí hậu Tây Trường Sơn, phân biệt mùa mưa và mùa khô rõ rệt, lượng mưa trung bình hàng năm ở đây vào khoảng 1.800 - 2.000 mm nhưng tập trung chủ yếu vào các tháng 5 - 10 (vụ mùa), thời gian còn lại hầu như không có mưa.
Theo báo cáo tổng kết của Sở Nông nghiệp và PTNT tỉnh Đắk Lắk (2013), sản lượng lúa toàn huyện năm 2013 đạt khoảng 53,7 nghìn tấn. Năng suất lúa trung bình của huyện chỉ đạt 51,3 tạ/hạ. Cơ cấu giống lúa hiện nay của huyện còn nghèo nàn, chủ yếu là các giống lúa thuần như Khang dân 18, Xi23, IR64. Diện tích lúa không chủ động nước rất lớn, các giống lúa chịu hạn hiện nay chủ yếu là LC93-1, LC93-4 chất lượng gạo khá nhưng năng suất thấp. Vụ Đông Xuân 2012 - 2013 với diện tích lúa nước bị hạn 1.740 ha tập trung chủ yếu tại các xã Bông Krang, Đăk Phơi, Yang Tao. Đây là các xã nghèo, kinh tế chủ yếu là sản xuất nông nghiệp, mức

10
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
sống của người dân còn thấp, đồng bào dân tộc thiểu số tại chỗ là chủ yếu (>80%). Vụ Đông Xuân thường bị thiếu nước đặc biệt là những diện tích lúa thường xuyên bị hạn, dẫn đến năng suất lúa trung bình của các xã này còn thấp so với các xã khác trong địa bàn huyện. Ngoài ra việc áp dụng các biện pháp kỹ thuật như bón phân, mật độ gieo sạ và các biện pháp canh tác khác còn nhiều hạn chế.
Xuất phát từ những vấn đề thực tế nêu trên, nhằm khai thác tiềm năng cho những vùng đất khó khăn phục vụ phát triển kinh tế xã hội của địa phương, giúp bà con nông dân xóa đói giảm nghèo, đồng thời giúp giảm bớt những khó khăn trong sản xuất lúa, cung cấp nguồn giống lúa tại chỗ và các hỗ trợ về kỹ thuật, xây dựng quy trình canh tác thích hợp với điều kiện canh tác tại địa phương đề tài “Nghiên cứu đánh giá một số giống lúa có khả năng chịu hạn cho vùng khó khăn về nước tưới tại huyện Lắk, tỉnh Đắk Lắk” đã được triển khai thực hiện.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứuSử dụng các giống lúa: CH207; CH208; CH19;
P6ĐB; LCH37 (Viện Cây lương thực và Cây thực phẩm chọn tạo); DH39 (Viện Khoa học Kỹ thuật Nông nghiệp Duyên hải Nam Trung bộ chọn tạo); OM4900 (Viện Lúa Đồng bằng sông Cửu Long chọn tạo); CXT30 (Viện Khoa học Kỹ thuật Nông nghiệp Việt Nam chọn tạo) và IR64 giống phổ biến tại địa phương làm đối chứng.
2.2. Phương pháp nghiên cứuThí nghiệm được bố trí theo khối ngẫu nhiên đầy
đủ (RCBD) 3 lần nhắc lại với 9 nghiệm thức. Thí nghiệm thực hiện ở 3 vùng khó khăn về nước là xã Đắk Phơi, xã Yang Tao, xã Bông Krang. Bố trí 2 vụ: Hè Thu và Đông Xuân năm 2015 và 2016. Các chỉ tiêu theo dõi và phương pháp đánh giá áp dụng theo Quy chuẩn kỹ thuật Quốc gia về khảo nghiệm giá trị canh tác và giá trị sử dụng của giống lúa (ký hiệu: QCVN 01-55:2011/BNNPTNT).
Phân bón cho 1 ha: 10 tấn phân chuồng + 80 kg N + 80 kg P2O5 + 80 kg K2O + 400 kg vôi bột, lượng giống gieo 160 kg/ha (Hồ Công Trực và ctv., 2014). Chế độ nước sử dụng nước mưa tự nhiên.
2.3. Thời gian và đia điểm nghiên cứuNghiên cứu được thực hiện năm 2015, 2016 tại
xã Yang Tao, Đắk Phơi, Bông Krang, huyện Lắk, tỉnh Đắk Lắk.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Đặc điểm nông học và sinh trưởng phát triển của các giống lúa khảo nghiệm tại huyện Lắk
Kết quả nghiên cứu về đặc điểm nông học của các giống lúa khảo nghiệm vụ Hè Thu và Đông Xuân năm 2015 tại các xã Yang Tao, Bông Krang, Đắk Phơi, huyện Lắk (Bảng 1, 2, 3) cho thấy các giống lúa đều thuộc nhóm giống ngắn ngày, vụ Hè Thu từ 82 - 113 ngày, vụ Đông Xuân từ 87 - 125 ngày; trong đó giống P6ĐB là giống ngắn ngày nhất, vụ Hè Thu 82 - 83 ngày, vụ Đông Xuân 87 - 88 ngày.
Các giống có thời gian trổ có độ dài ở mức trung bình (5 ngày), đa số các giống đều có độ cứng cây ở mức 1 (không đổ ngã) và độ tàn lá ở mức trung bình.
Bảng 1. Đặc điểm nông học của các giống lúa vụ Hè Thu và Đông Xuân năm 2015 tại xã Yang Tao, huyện Lắk
Giống lúaTGST (ngày)
Độ dài giai đoạn trổ (ngày)
Độ cứng cây (điểm)
Độ tàn lá (điểm)
Chiều cao cây (cm)
HT ĐX HT ĐX HT ĐX HT ĐX HT ĐXCH207 110 122 5 5 3 3 5 5 102,9 98,5CH208 112 125 5 5 3 3 5 5 116,8 113,5CH19 100 120 5 5 1 3 5 5 109,2 102,0P6ĐB 82 87 5 5 1 1 5 5 101,2 102,2
LCH37 98 110 5 5 1 1 5 5 106,0 100,5DH39 100 112 5 5 1 3 5 5 103,9 103,8
OM4900 96 100 5 5 1 1 5 5 111,1 110,7CXT30 86 90 5 5 1 3 5 5 97,1 100,4
IR64 95 100 5 5 1 3 5 5 99,4 97,9LSD0,05 1,66 3,44CV(%) 0,92 1,93

11
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 2. Đặc điểm nông học của các giống lúa vụ Hè Thu và Đông Xuân năm 2015 tại xã Bông Krang, huyện Lắk
Bảng 3. Đặc điểm nông học của các giống lúa vụ Hè Thu và Đông Xuân năm 2015 tại xã Đắk Phơi, huyện Lắk
Hình 1. Biểu đồ số liệu KT 2015 tại huyện Lắk Hình 2. Biểu đồ số liệu KT 2016 tại huyện Lắk
0
20
40
60
80
100
0
100
200
300
400
1 2 3 4 5 6 7 8 9 10 11 12
Mưa (mm) Bay hơi (mm)Nhiệt độ KK TB (oC) Ẩm độ (%)
0
20
40
60
80
100
0
100
200
300
400
500
1 2 3 4 5 6 7 8 9 10 11 12
Mưa (mm) Bay hơi (mm)Nhiệt độ KK TB (oC) Ẩm độ (%)
Giống lúaTGST (ngày)
Độ dài giai đoạn trổ (ngày)
Độ cứng cây (điểm)
Độ tàn lá (điểm)
Chiều cao cây (cm)
HT ĐX HT ĐX HT ĐX HT ĐX HT ĐXCH207 110 125 5 5 3 3 5 5 102,9 100,2CH208 112 125 5 5 3 3 5 5 118,3 111,7CH19 100 120 5 5 1 1 5 5 122,5 122,5P6ĐB 83 87 5 5 1 1 5 5 97,4 92,2
LCH37 98 110 5 5 1 1 5 5 104,6 102,8DH39 100 112 5 5 1 1 5 5 104,7 104,7
OM4900 95 100 5 5 1 1 5 5 114,5 112,0CXT30 86 92 5 5 1 1 5 5 98,6 98,6
IR64 98 100 5 5 1 3 5 5 100,7 99,5LSD0,05 3,49 3,49CV(%) 1,88 1,90
Giống lúaTGST (ngày) Độ dài giai
đoạn trổ (ngày)Độ cứng cây
(điểm)Độ tàn lá
(điểm)Chiều cao cây
(cm)HT ĐX HT ĐX HT ĐX HT ĐX HT ĐX
CH207 110 122 5 5 1 3 5 5 108,4 99,2CH208 113 123 5 5 1 3 5 5 115,2 113,6CH19 100 120 5 5 1 1 5 5 101,1 98,2P6ĐB 82 88 5 5 1 1 5 5 101,2 92,2
LCH37 100 110 5 5 1 1 5 5 100,7 98,3DH39 105 112 5 5 1 3 5 5 100,4 97,9
OM4900 96 100 5 5 1 1 5 5 113,7 111,3CXT30 86 90 5 5 1 1 5 5 97,5 98,4IR64 95 105 5 5 1 3 5 5 99,1 96,4
LSD0,05 4,76 1,91CV(%) 2,64 1,09

12
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Chiều cao cây của các giống ở mức trung bình, vụ Hè Thu có chiều cao từ 97 - 122 cm, vụ Đông Xuân từ 96 - 122 cm. Các giống lúa có chiều cao cây cao hơn là CH19, CH208, OM4900; Các giống có chiều cao thấp hơn là CXT30 và IR64.
Các đặc điểm nông học của các giống lúa đưa vào nghiên cứu thử nghiệm tại các xã Yang Tao, Bông Krang và Đắk Phơi tại huyện Lắk ở các vụ Hè Thu và Đông Xuân năm 2015 cho thấy là khá phù hợp với điều kiện đất đai và khí hậu của vùng. Kết quả này phù hợp với kết quả nghiên cứu một số giống lúa chịu hạn tại Tây Nguyên của Lại Đình Hòe và cộng tác viên (2013).
3.2. Các yếu tố cấu thành năng suất và năng suất các giống lúa
Các yếu tố cấu thành năng suất và năng suất thực thu của các giống lúa (bảng 4, 5, 6) qua theo dõi ở các vụ Hè Thu và Đông Xuân 2015 cho thấy:
- Số bông/m2: Các giống đều có số lượng bông/m2 khá cao (tại xã Yang Tao vụ Hè Thu từ 338 - 352 bông/m2, vụ Đông Xuân 317 - 335 bông/m2; tại xã Bông Krang vụ Hè Thu từ 328 - 341 bông/m2, vụ Đông Xuân 320 - 338 bông/m2; tại xã Đắk Phơi vụ Hè Thu từ 316 - 324 bông/m2, vụ Đông Xuân 321 - 324 bông/m2).
Bảng 4. Các yếu tố cấu thành năng suất và năng suất thực thu các giống lúa vụ Hè Thu và Đông Xuân năm 2015 tại xã Yang Tao, huyện Lắk
- Hạt chắc/bông: Số hạt chắc/bông ở các vụ Hè Thu và Đông Xuân của các giống không có khác biệt nhau đáng kể, vụ Hè Thu cao hơn vụ Đông Xuân (tại xã Yang Tao vụ Hè Thu từ 89 - 94 hạt/bông, vụ Đông Xuân 79 - 84 hạt/bông; tại xã Bông Krang vụ Hè Thu từ 89 - 92 hạt/bông, vụ Đông Xuân 80 - 84 hạt/bông; tại xã Đắk Phơi vụ Hè Thu từ 87 - 90, vụ Đông Xuân 85 - 87 hạt chắc/bông).
- Tỷ lệ hạt lép: Các giống khảo nghiệm có tỷ lệ hạt lép ở xã Yang Tao vụ Hè Thu từ 15,3 - 16,9%, vụ Đông Xuân từ 14,4 - 16,0%; xã Bông Krang vụ Hè Thu 14,7 - 16,5%, vụ Đông Xuân 12,2 - 14,5%; xã Đắk Phơi vụ Hè Thu 15,3 - 18,2%, vụ Đông Xuân 14,7 - 16,5%. Giữa các giống có tỷ lệ khác biệt không có ý nghĩa thống kê.
- Khối lượng 1.000 hạt: Khối lượng 1.000 hạt ở các giống có sự khác biệt ý nghĩa thống kê giống P6ĐB và giống LCH37 có khối lượng hạt lớn hơn các giống khác. Khối lượng 1.000 hạt của các giống
ở xã Yang Tao vụ Hè Thu từ 24,3 - 25,7 g, vụ Đông Xuân từ 23,8 - 25,5 g; xã Bông Krang vụ Hè Thu 23,0 - 24,9 g, vụ Đông Xuân 23,5 - 25,3 g; xã Đắk Phơi vụ Hè Thu 23,7 - 25,3 g, vụ Đông Xuân 23,3 - 24,7 g.
- Năng suất thực thu của các giống ở vụ Hè Thu và Đông Xuân 2015 cho thấy các giống lúa có năng suất đạt trung bình từ 51,5 - 72,5 tạ/ha (vụ Hè Thu), các giống khảo nghiệm cho năng suất cao hơn đối chứng từ 2,0 - 13,2 tạ/ha, tăng từ 3,7 - 24,4%; Vụ Đông Xuân năng suất trung bình đạt 50,5 - 58,7 tạ/ha, cao hơn so đối chứng 0,9 - 7,8 tạ/ha, tăng từ 1,8 - 15,2%. Trong đó giống lúa chịu hạn LCH37 và giống né vụ P6ĐB cho năng suất cao nhất.
Kết quả khảo nghiệm vụ Hè Thu và Đông Xuân 2016 trên 2 giống lúa P6ĐB và LCH37 cho kết quả năng suất thu hoạch (bảng 8) đạt năng suất trung bình từ 52,7 - 60,2 tạ/ha. Trong đó giống lúa P6ĐB cho năng suất vụ Hè Thu 60,2 tạ/ha (tăng 19,7% so đối chứng), vụ Đông Xuân 53,8 tạ/ha (tăng 19,6%
Giống lúaSố bông HH
(bông/m2) Hạt chắc/bông Tỷ lệ hạt lép (%)
Khối lương 1000 hạt (g)
NSTT (tạ/ha)
HT ĐX HT ĐX HT ĐX HT ĐX HT ĐXCH207 339 323 91 80 16,9 15,3 24,3 24,5 59,8 52,0CH208 338 321 90 81 15,8 14,9 24,8 25,1 60,4 54,2CH19 338 320 89 81 15,3 16,0 24,3 24,6 58,3 52,0P6ĐB 349 332 91 83 15,5 14,4 25,7 25,5 72,5 58,5
LCH37 352 335 94 84 15,4 14,8 24,5 23,9 68,3 57,1DH39 339 322 89 81 15,7 14,8 24,7 24,2 61,4 51,6
OM4900 343 322 90 81 15,7 15,8 24,6 24,7 60,7 53,4CXT30 341 317 91 79 15,3 14,6 24,8 24,7 64,7 52,9
IR64 341 318 89 81 16,3 14,6 24,0 23,8 57,6 50,4LSD0,05 14,2 22,9 2,3 2,7 2,21 1,38 0,40 0,34 2,43 3,08CV(%) 2,4 4,1 1,5 1,9 8,12 5,31 0,95 0,81 2,25 3,32

13
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 5. Các yếu tố cấu thành năng suất và năng suất thực thu các giống lúa vụ Hè Thu và Đông Xuân năm 2015 tại xã Bông Krang, huyện Lắk
Giống lúaSố bông HH
(bông/m2) Hạt chắc/bông Tỷ lệ hạt lép (%)
Khối lương1000 hạt (g)
NSTT (tạ/ha)
HT ĐX HT ĐX HT ĐX HT ĐX HT ĐXCH207 333 321 90 82 16,0 12,5 24,4 24,1 53,9 52,7CH208 328 320 91 84 15,9 12,1 24,9 24,3 56,5 54,5CH19 330 328 92 80 16,1 14,2 23,8 23,9 54,8 52,4P6ĐB 341 338 91 81 14,7 12,2 24,7 25,3 66,5 58,7
LCH37 340 335 92 83 16,5 12,4 24,2 24,6 62,9 57,5DH39 332 328 90 83 15,1 12,9 24,0 23,8 57,3 52,7
OM4900 333 323 89 81 15,9 14,5 23,4 24,2 57,4 54,3CXT30 336 326 90 82 15,8 13,1 23,4 24,1 57,3 54,6
IR64 337 331 89 81 14,8 13,2 23,0 23,5 53,5 51,8LSD0,05 16,3 12,1 4,4 2,3 0,38 0,42 0,33 0,46 2,97 2,85CV(%) 2,8 2,1 2,8 1,6 5,55 6,72 0,80 1,01 2,98 3,02
Bảng 6. Các yếu tố cấu thành năng suất và năng suất thực thu các giống lúa vụ Hè Thu và Đông Xuân năm 2015 tại xã Đắk Phơi, huyện Lắk
Bảng 7. Năng suất thực thu của giống lúa P6ĐB và LCH37 vụ Hè Thu và Đông Xuân 2016 (tạ/ha)
Giống lúaSố bông HH
(bông/m2) Hạt chắc/bông Tỷ lệ hạt lép (%)
Khối lương 1000 hạt (g)
NSTT (tạ/ha)
HT ĐX HT ĐX HT ĐX HT ĐX HT ĐXCH207 316 321 88 85 17,9 15,2 24,7 23,9 54,9 52,6CH208 319 323 89 85 16,8 15,8 24,5 24,3 56,1 53,4CH19 319 322 88 86 18,2 16,1 24,4 23,8 53,3 52,5P6ĐB 322 324 90 87 15,3 15,7 25,3 24,7 63,3 57,5
LCH37 322 324 90 87 17,6 15,5 24,7 24,6 60,4 55,1DH39 322 323 87 86 17,0 15,0 24,2 23,9 52,9 52,5
OM4900 322 323 88 85 17,3 16,5 24,5 24,1 55,3 53,9CXT30 323 324 87 86 17,0 14,7 24,2 24,1 54,9 54,0
IR64 324 324 89 86 15,9 14,9 23,7 23,3 51,5 52,5LSD0,05 11,4 15,9 2,8 2,4 0,35 0,22 0,38 0,27 3,43 3,09CV(%) 2,0 2,8 1,8 1,6 4,90 3,25 0,91 0,66 3,55 3,32
so đối chứng); Giống LCH37 cho năng suất vụ Hè Thu 55,7 tạ/ha (tăng 10,7% so đối chứng), vụ Đông Xuân 52,7 tạ/ha (tăng 17,1% so đối chứng), phù
hợp với kết quả nghiên cứu của Đỗ Việt Anh và cộng tác viên (2013, 2016).
Giống lúa Thời vụ Yang Tao Bông Krang Đắk Phơi Trung bình % so với ĐC
P6ĐBHè Thu 59,6 59,0 62,1 60,2 119,7
Đông Xuân 53,4 52,3 55,7 53,8 119,6
LCH37Hè Thu 55,9 54,7 56,5 55,7 110,7
Đông Xuân 53,1 51,8 53,3 52,7 117,1
IR64 (ĐC)Hè Thu 50,2 50,5 50,1 50,3 100,0
Đông Xuân 45,8 44,9 44,3 45,0 100,0

14
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luận- Các giống lúa khảo nghiệm thuộc nhóm ngắn
ngày, có thời gian sinh trưởng vụ Hè Thu từ 80 - 112 ngày, vụ Đông Xuân từ 87 - 127 ngày, có khả năng chịu và tránh hạn phù hợp với cơ cấu thời vụ và điều kiện đất đai, khí hậu tại huyện Lắk, tỉnh Đắk Lắk.
- Năng suất thực thu các giống lúa đưa vào khảo nghiệm đạt 51,5 - 72,5 tạ/ha (vụ Hè Thu), các giống khảo nghiệm cho năng suất cao hơn đối chứng (IR64) từ 2,0 - 13,2 tạ/ha, tăng từ 3,7 - 24,4%; Vụ Đông Xuân năng suất trung bình đạt 50,5 - 58,2 tạ/ha, cao hơn so đối chứng 0,9 - 7,8 tạ/ha, tăng từ 1,8 - 15,2%.
- Giống lúa chịu hạn LCH37 và giống né vụ P6ĐB cho năng suất cao nhất. Giống lúa LCH37 có TGST vụ Hè Thu là 82 - 83 ngày, cho năng suất đạt 54,7 - 68,3 tạ/ha; TGST vụ Đông Xuân là 87 - 88 ngày, cho năng suất đạt 51,8 - 57,5 tạ/ha. Giống lúa P6ĐB có TGST vụ Hè Thu là 98 - 100 ngày, cho năng suất đạt 59,0 - 72,5 tạ/ha; vụ Đông Xuân có TGST là 110 - 112 ngày, năng suất đạt 52,3 - 58,7 tạ/ha.
4.2. Đề nghiĐưa giống lúa LCH37 và P6ĐB vào trồng diện
rộng ở các vùng đất khó khăn về nước tại khu vực huyện Lắk, tỉnh Đắk Lắk trong những năm tới.
TÀI LIỆU THAM KHẢO Đỗ Việt Anh, Nguyên Xuân Dũng, 2013. Kết quả bước
đầu về nghiên cứu chọn tạo giống lúa chịu hạn cho vùng đất cạn và vùng sinh thái có điều kiện khó khăn. Hội thảo quốc gia về Khoa học cây trồng lần thứ nhất. Viện Khoa học Nông nghiệp Việt Nam.
Đỗ Việt Anh, Nguyên Xuân Dũng, Trần Văn Tứ, Nguyên Anh Dũng, Nguyên Văn Chinh, 2016. Kết quả nghiên cứu, chọn tạo giống lúa chịu hạn cho vùng đất cạn nhờ nước trời và vùng khó khăn về nước. Hội thảo quốc gia về Khoa học cây trồng lần thứ hai. Viện Khoa học Nông nghiệp Việt Nam.
Lại Đình Hòe, Đặng Bá Đàn, Hô Công Trực, 2013. Nghiên cứu tuyển chọn giống lúa chịu hạn và kỹ thuật canh tác cho vùng duyên hải Nam Trung bộ, Tây Nguyên. Hội thảo quốc gia về Khoa học cây trồng lần thứ nhất. Viện Khoa học Nông nghiệp Việt Nam.
Sở Nông nghiệp và PTNT tỉnh Đắk Lắk, 2013. Báo cáo tổng kết sản xuất nông nghiệp của tỉnh Đắk Lắk năm 2013.
Hô Công Trực, Nguyên Thi Thúy, Nguyên Thi Kim Thu, Lại Đình Hòe, 2014. Nghiên cứu lượng giống gieo sạ và liều lượng phân bón thích hợp cho giống lúa lai BTE1 và giống CH208 ở vùng Tây Nguyên. Kết quả 45 năm nghiên cứu khoa học và chuyển giao công nghệ. Viện Thổ nhưỡng Nông hóa.
Testing of drought-tolerant rice varieties in Lak district, Dak Lak provinceHo Cong Truc, Nguyen Thi Thao Nhung,
Truong Van Binh, Doan Van ThanhAbstractExperiments of testing were carried out in three different locations with difficult watering in Lak district, Dak Lak province during 2 seasons (summer-autumn and winter-spring seasons) of 2015 and 2016. The tested rice varieties included drought tolerant varieties (CH207, CH208, CH19, LCH37, DH39), short duration P6DB and OM4900, CXT30 varieties and popular rice variety IR64 as control one. The result showed that all tested rice varieties were adaptable to local condition and had the yield higher than that of the control. Drought-tolerant rice variety LCH37 and short duration variety P6DB showed the highest yield (P6DB reached 59.0 - 72.5 quintal ha-1 in summer-autumn season, 52.3 - 58.7 quintal ha-1 in winter-spring season, increasing by 15.2 - 24.4% compared with control while LCH37 reached 54.7 - 68.3 quintal ha-1 in summer-autumn season, 51.8 - 57.5 quintal ha-1 in winter-spring season, 11.7 - 17.8% in comparison with control).Key words: Rice, testing, drought tolerant rice variety, adaptable, Lak district, Dak Lak province
Ngày nhận bài: 9/6/2017Ngày phản biện: 19/6/2017
Người phản biện: TS. Vũ Tiến KhangNgày duyệt đăng: 25/6/2017

15
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀLúa (Oryza sativa L.) là cây lương thực quan
trọng nhất của Việt Nam. Năm 2014, sản lượng lúa của Việt Nam đạt trên 44 triệu tấn, đứng thứ 5 thế giới sau Trung Quốc, Ấn Độ, Banglades và Indonesia (FAO, 2015). Tuy nhiên, những năm gần đây, sự biến đổi khí hậu toàn cầu và suy thoái môi trường đang làm ảnh hưởng đến sản xuất nông nghiệp trên toàn thế giới. Khô hạn sẽ là yếu tố quan trọng bậc nhất ảnh hưởng đến an ninh lương thực của thế giới, gây thiệt hại lớn nhất đối với năng suất lúa (Dorner et al.,1989) vì lúa là cây trồng rất mẫn cảm với hạn do hệ thống rễ nhỏ, khí khổng rất nhạy cảm và lá nhanh bị già hóa khi gặp hạn.
Ở Việt Nam, nghiên cứu về lúa chịu hạn thời gian qua đã tập trung vào đánh giá các đặc điểm hình thái, sinh lý sinh hóa và di truyền liên quan đến tính chịu hạn của cây lúa đồng thời chọn tạo, đưa ra các giống lúa chịu hạn phục vụ sản xuất (Vũ Tuyên Hoàng và ctv., 1992; Nguyễn Hữu Cường và ctv., 2003; Nguyễn Thị Thu Hoài, 2005; Phạm Anh Tuấn và ctv., 2008). Đến nay, kết quả nhiều công trình nghiên cứu khoa học đã được công bố và hàng chục giống lúa chịu hạn đã được Viện Khoa học Nông nghiệp Việt Nam chọn tạo ra như LC93-1, BC12, CH2, CH3, CH 133, CH5… đang được trồng rộng rãi ở vùng Trung du miền núi phía Bắc, Trung bộ, Đông Nam bộ và Tây Nguyên. Tuy nhiên, đến nay chương trình chọn tạo, nhân giống lúa tập trung vào khả năng chịu hạn nhưng chưa tạo ra được nhiều giống lúa chịu hạn, năng suất cao phục vụ sản xuất. Vì vậy, việc tiếp tục nghiên cứu khả năng chịu hạn của tập đoàn lúa địa
phương đang được lưu giữ tại Ngân hàng gen cây trồng Quốc gia, từ đó phân nhóm và xác định các nguồn gen chịu hạn tốt đề xuất như nguồn vật liệu khởi đầu phục vụ chọn tạo giống lúa chịu hạn hoặc giới thiệu trực tiếp cho sản xuất ở những vùng khó khăn về nước tưới là yêu cầu cấp thiết.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu - 100 mẫu giống lúa đang được lưu giữ tại Ngân
hàng gen cây trồng Quốc gia có nguồn gốc từ các tỉnh Bắc Trung Bộ (Thanh Hóa - 54 mẫu, Nghệ An - 22 mẫu, Hà Tĩnh - 17 mẫu, Quảng Bình - 7 mẫu) và giống lúa cạn CH5 làm đối chứng.
- Dung dịch Polyethelen Glycol (PEG) 6000 (40%).- Dung dịch Kimura B (Yoshida and Forno,
1971): Bao gồm các nguyên tố đa lượng (NH4No3; NaH2PO4.2H2O; K2SO4; CaCl2.2H2O; MgSO4.7H2O) và vi lượng (MnCl2.4H2O; (NH4)6.Mo7O24.4H2O; ZnSO4.H2O; H3PO3; CuSo4.5H2O; FsCl3.6H2O; C6H8O4.H2O; C6H8O4.H2O).
2.2. Phương pháp nghiên cứu
2.2.1. Bố trí thí nghiệm- Thí nghiệm 1: Đánh giá gián tiếp khả năng
chịu hạn của các mẫu giống lúa thông qua tỷ lệ nảy mầm của hạt, sau khi được xử lý bằng dung dịch Polyethelen Glycol (PEG) 6000 (40%).
Thí nghiệm được bố trí theo kiểu ngẫu nhiên hoàn toàn, 3 lần nhắc lại, mỗi lần nhắc 20 hạt. Hạt mẫu giống được ngâm trong dung dịch PEG 6000
1 Trung tâm Tài nguyên thực vật; 2 Viện Khoa học Nông nghiệp Việt Nam3 Hội Giống cây trồng Việt Nam
KHẢ NĂNG CHỊU HẠN CỦA MỘT SỐ NGUỒN GEN LÚA ĐỊA PHƯƠNG ĐANG ĐƯỢC LƯU GIỮ TẠI NGÂN HÀNG GEN CÂY TRỒNG QUỐC GIA
Lưu Quang Huy1, Nguyễn Thị Ngọc Huệ3, Vũ Linh Chi1, Dương Hồng Mai1, Vũ Đăng Toàn1, Bùi Thị Thu Huyền1,
Hà Minh Loan1,Trần Danh Sửu2
TÓM TẮTNghiên cứu nhằm xác định khả năng chịu hạn và khả năng phục hồi của 100 mẫu giống lúa địa phương được thu
thập tại các tỉnh vùng Bắc Trung bộ theo phương pháp gây hạn nhân tạo ở các giai đoạn sinh trưởng khác nhau. Kết quả cho thấy các mẫu giống lúa có phản ứng khác nhau với điều kiện hạn ở mỗi giai đoạn sinh trưởng phát triển. 100 mẫu giống lúa đã được phân nhóm theo mức độ chịu hạn, khả năng phục hồi ở các giai đoạn sinh trưởng phát triển. Xác định được bốn mẫu giống lúa chịu hạn tốt (điểm 1) qua tất cả các giai đoạn là Tứ thời Thanh Hóa (SĐK 12), Tám đỏ Thanh Hóa (SĐK 299), Nếp Lốc Thanh Hóa (SĐK 325) và Mùa trắng Thanh Hóa (SĐK 585). Ngoại trừ mẫu giống Tám đỏ Thanh Hóa (SĐK 299), 03 giống còn lại có năng suất cao hơn hoặc tương đương đối chứng CH5 (4,18 tấn/ha) trong điều kiện hạn nhân tạo.
Từ khóa: Lúa địa phương, gây hạn nhân tạo, chịu hạn, đánh giá, khả năng phục hồi

16
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
(40%) trong 48 h, sau đó, rửa sạch bằng nước trung tính rồi chuyển sang đĩa petri có lót giấy lọc ẩm để hạt nảy mầm. Khả năng chịu hạn được đánh giá dựa vào tỷ lệ (%) hạt nảy mầm.
- Thí nghiệm 2: Nghiên cứu ảnh hưởng của điều kiện hạn nhân tạo đến 100 mẫu giống lúa và giống đối chứng CH5 ở giai đoạn mạ 3 lá trong nhà lưới.
Bố trí thí nghiệm 1 nhân tố với 3 lần nhắc lại. Hạt mẫu giống được khử trùng bằng dung dịch ethanol 10% trong 3 phút và NaOCl 5% trong 30 phút, sau đó ngâm vào nước 30oC trong 2 ngày cho tới khi nứt nanh. Tiếp theo, các hạt nảy mầm được gieo vào các khay mạ chứa bùn ruộng dày 5cm. Khi mạ được 3 lá thật, trồng cây vào các chậu (55 cm ˟ 38 cm ˟ 18 cm) chứa 16 lít dung dịch Kimura B và được cố định bằng xốp mút, với phần rễ ngập trong dung dịch. Độ pH = 5 được duy trì trong suốt thời gian sinh trưởng (sử dụng NaOH 10% hoặc HCl 10% để hiệu chỉnh). Thay mới dung dịch dinh dưỡng 7 ngày/lần. ất cả các cây của 100 nguồn gen (mẫu giống) và đối chứng trồng trong chậu với 10 cây/giống/1 lần nhắc được gây hạn nhân tạo bằng cách bổ sung PEG 6000 (20%) (Money, 1989). Sau 7 ngày tiến hành đánh giá khả năng chịu hạn. Sau đó tiếp tục cung cấp đầy đủ nước, theo dõi và đánh giá khả năng phục hồi của các mẫu giống; Đánh giá khả năng chịu hạn qua theo dõi độ cuốn lá theo thang điểm 0 - 9; Đánh giá khả năng phục hồi mạ sau các ngày gây hạn theo thang điểm 1 - 9 ( IRRI, 2002).
- Thí nghiệm 3: Đánh giá chịu hạn giai đoạn sinh trưởng sinh dưỡng và sinh trưởng sinh thực trong điều kiện nhà lưới theo phương pháp của Fischer và cộng tác viên (IRRI, 2003).
Bước 1: Chuẩn bị mạ cho tới khi cây có 3 lá thật.Bước 2: Cấy mạ mỗi mẫu giống vào 3 xô nhựa
(mỗi xô 3 cây cấy hình tam giác). Theo dõi và chăm sóc cùng điều kiện dinh dưỡng, nếu cạn nước thì phải bổ sung nước.
Bước 3: Gây hạn nhân tạo: Sau cấy 3 tuần, tiến hành rút nước gây hạn nhân tạo.
Bước 4: Đánh giá khả năng chịu hạn ở giai đoạn đẻ nhánh: Sau gây hạn 14 - 21 ngày đánh giá khả năng chịu hạn theo thang điểm 0 - 9 (lúc này triệu chứng lá cuốn thể hiện rõ nhất). Đánh giá xong tiếp tục cung cấp đầy đủ nước, theo dõi và đánh giá khả năng phục hồi của các mẫu giống theo thang điểm 1 - 9.
Bước 5: Đánh giá khả năng chịu hạn ở giai đoạn phân hóa đòng đến giai đoạn trỗ bông, vào chắc.
Tiến hành gây hạn nhân tạo và đánh giá khả năng chịu hạn sau 10 ngày (lúc triệu chứng lá cuốn thể hiện rõ nhất) theo thang điểm 0 - 9. Sau khi đánh giá xong tiếp tục cung cấp đầy đủ nước và theo dõi khả năng phục hồi (điểm 1 - 9) và sinh trưởng, phát triển của cây lúa đến khi thu hoạch.
2.2.2. Các chỉ tiêu theo dõiTỷ lệ hạt nảy mầm (GR%) = Số hạt nảy mầm (n)/
Số hạt được gieo (N) ˟ 100.Ngày gieo, ngày cấy, ngày bắt đầu đẻ nhánh, ngày
kết thúc đẻ nhánh, ngày trỗ 10%, ngày trỗ 80%, ngày trỗ hoàn toàn. Số nhánh tối đa, số nhánh hữu hiệu và chiều cao cây. Một số đặc điểm hình thái chính của lá, bông và hạt). Năng suất lý thuyết và các yếu tố cấu thành đến năng suất. Đánh giá khả năng chống chịu hạn, khả năng phục hồi, khi có hạn xảy ra theo thang điểm SES của IRRI (2002).
2.2.3. Xử lý số liệuSố liệu thí nghiệm được xử lý theo phương pháp
thống kê trong phần mềm Excel.
2.3. Thời gian và đia điểm nghiên cứuNghiên cứu được thực hiện tại Trung tâm Tài
nguyên thực vật, Hoài Đức, Hà Nội, năm 2016.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Khả năng nảy mầm của các mẫu giống sau khi ngâm ủ
Những giống có khả năng chịu hạn là những giống có khả năng nảy mầm tốt trong điều kiện thiếu nước (Nguyễn Thị Thu Hoài, 2005). Kết quả đánh giá gián tiếp khả năng chịu hạn trong điều kiện hạn nhân tạo, cho thấy 100% mẫu giống nghiên cứu đều nảy mầm trong môi trường nước cất với tỉ lệ từ 75 - 90%, trong khi đó, tỷ lệ nảy mầm của 100 mẫu giống khi được xử lý bằng dung dịch PEG 6000 40% thấp hơn rất nhiều, chỉ từ 1,7 - 35%, và có 10 mẫu không nảy mầm. Như vậy, dung dịch PEG6000 ở nồng độ 40% đã có khả năng gây hạn nhân tạo cho cây lúa. Qua đánh giá cho thấy, sau khi được xử lý hạt bằng dung dịch PEG 6000 (40%), 21 mẫu giống (21%) có tỷ lệ nảy mầm cao hơn (23,3 - 35,0%) và 5 mẫu giống (5%) có tỷ lệ nảy mầm tương đương giống đối chứng chịu hạn CH5 là 21,7 % (Hình 1). Điển hình ba mẫu giống Dự Thanh Hóa (SĐK 134), Tám thơm Thanh Hóa (SĐK 89) và Mùa trắng Thanh Hóa (SĐK585) đều có tỷ lệ nảy mầm cao nhất 35,0% sau khi được xử lý dung dịch PEG 6000 (40%).

17
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Hình 1. Tỷ lệ nảy mầm của 100 mẫu giống sau khi xử lý PEG6000 40%
3.2. Khả năng chiu hạn, phục hôi của các mẫu giống lúa ở giai đoạn mạ 3 lá
3.2.1. Phân nhóm các mẫu giống lúa theo mức độ chịu hạn
Dựa vào nguyên lý về tính chống chịu hạn và cơ chế điều chỉnh thẩm thấu, đã nghiên cứu ảnh hưởng của PEG 6000 (20%) đến mức độ cuốn lá ở giai đoạn mạ 3 lá của 100 mẫu giống.
Bảng 1. Phân nhóm 100 mẫu giống lúa theo mức độ chịu hạn ở giai đoạn 3 lá
Từ kết quả đánh giá đã phân lập 100 mẫu giống lúa thành 3 nhóm giống: Chịu hạn tốt với 32 mẫu giống (chiếm 32% tổng số mẫu nghiên cứu) khi hình thái và màu sắc lá vẫn biểu hiện bình thường hay bị cuốn nhẹ theo hình chữ V (điểm 1 - 3); Chịu hạn trung bình với 31 giống (chiếm 31%), khi bộ lá bị biến dạng theo hình chữ V sâu hoặc nặng hơn như hình chữ U (điểm 5); và chịu hạn kém và rất kém với 37 giống (chiếm 37%) (điểm 7 và 9), là những mẫu giống có bộ lá bị biến dạng một phần hay toàn bộ mà đặc trưng chính là cuốn theo hình chữ O hoặc bó chặt lại (Bảng 1).
3.2.2. Kết quả đánh giá khả năng phục hồi sau khi gây hạn ở giai đoạn mạ 3 lá
Khả năng phục hồi được đánh giá sau 10 ngày cho nước vào, thang điểm được ghi nhận từ 1 - 9 (tương đương với tỷ lệ cây phục hồi từ 100% - 0%).
Kết quả đánh giá cho thấy có 36/100 mẫu giống có khả năng phục hồi tốt (hình 2), trong đó có 31/100 mẫu giống vừa có khả năng chịu hạn tốt (điểm 0 - điểm 1) vừa có khả năng phục hồi tốt sau chịu hạn (điểm 1 - 3), đạt 31% tổng số mẫu nghiên cứu.
Hình 2. Khả năng chịu hạn, phục hồi của 100 mẫu giống lúa ở giai đoạn 3 lá
Phát hiện thú vị là 31 mẫu giống này đều nằm trong số 32 mẫu giống được phân nhóm chịu hạn tốt, đó là các mẫu giống có số đăng ký: SĐK12, SĐK21, SĐK46, SĐK106, SĐK134, SĐK136, SĐK289, SĐK299, SĐK325, SĐK473, SĐK584, SĐK585, SĐK593, SĐK596, SĐK601, SĐK603, SĐK607, SĐK613, SĐK633, SĐK643, SĐK648, SĐK651, SĐK657, SĐK658, SĐK671, SĐK686, SĐK688, SĐK730, SĐK731, SĐK732, SĐK737. Trong đó, có một số giống được đánh giá khá tốt ( điểm 1) như Tứ thời Thanh Hóa (SĐK 12), Ba tháng nước Nghệ An (SĐK 21), Dự Thanh Hóa (SĐK 134), Mai hương Hà Tĩnh (SĐK 136), Tám thơm Thanh Hóa (SĐK 289), Tám đỏ Thanh Hóa (SĐK 299), Mùa Trắng Thanh Hóa (SĐK 585).
3.3. Khả năng chiu hạn, khả năng phục hôi của các mẫu giống lúa ở giai đoạn sinh trưởng sinh dưỡng, sinh trưởng sinh thực trong điều kiện nhà lưới
3.3.1. Khả năng chịu hạn, khả năng phục hồi của các mẫu giống lúa ở giai đoạn sinh trưởng sinh dưỡng (giai đoạn đẻ nhánh)
Với thời gian xử lý dung dịch PEG6000 40% gây hạn kéo dài 15 ngày ở giai đoạn đẻ nhánh, kết quả đánh giá tính chịu hạn theo mức độ biến dạng bộ lá của 100 mẫu giống đã phân lập được 3 nhóm giống chịu hạn tốt, trung bình và kém với số mẫu giống tương ứng là 14 mẫu giống (điểm 1 - 3 ), chiếm 14%; 36 giống (điểm 5), chiếm 36%; và 50 mẫu giống (điểm 7 và điểm 9), chiếm 50% tổng số mẫu nghiên cứu (Bảng 2).
Kết quả nảy mầm của các nguồn gen sau khixử lý PEG 6000 (40%)
Không nảy mầm
1010,0
Số mẫu giốngTỷ lệ
Thấp hơn Đ/C(<21.7)
Tương đương hoặccao hơn Đ/C
6464,0
2626,0
100
80
60
40
20
0
Số m
ẫu g
iống
Khả năng chiu hạn
Số mẫu
Tỷ lệ (%)
Mẫu giống điển hình
Tốt (điểm 0-1) 32 32 SĐK 12, SĐK 299
Trung bình ( điểm 3-5) 31 31 SĐK 22, SĐK 537
Kém/rất kém (điểm 7-9) 37 37 SĐK 26, SĐK 523
Tổng cộng 100 100
Kết quả đánh giá khả năng chịu hạn, khả năng phục hồicủa các mẫu giống lúa giai đoạn 3 lá
Số m
ẫu g
iống
100
80
60
40
20
0
Số mẫu giống chịu hạnSố mẫu giống phục hồi
Tốt3236
TB3132
Kém3732

18
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Các mẫu giống được đánh giá chịu hạn khá tốt ở giai đoạn sinh trưởng sinh dưỡng khi hình thái và màu sắc lá của chúng vẫn biểu hiện bình thường hay chỉ bị cuốn nhẹ theo hình chữ V nông là SĐK12, SĐK134, SĐK289, SĐK299, SĐK584, SĐK585, SĐK601, SĐK607, SĐK643, SĐK648, SĐK651, SĐK671, SĐK688, SĐK732.
Bảng 2. Phân nhóm 100 mẫu giống lúa theo mức độ chịu hạn ở giai đoạn sinh trưởng sinh dưỡngtrong nhà lưới, năm 2016 (Hoài Đức, Hà Nội)
Khả năng phục hồi được đánh giá sau 10 ngày cho nước vào theo thang điểm từ 1 - 9 (tương đương với tỷ lệ cây phục hồi từ 100% - 0%) cho thấy, 25/100 mẫu giống phục hồi tốt (điểm 1-3) và 39/100 mẫu giống phục hồi trung bình có thang điểm đánh giá là từ điểm 3,1 - điểm 5 (Hình 3).
Hình 3. Khả năng chịu hạn, phục hồi của 100 mẫu giống lúa ở giai đoạn sinh trưởng,
sinh dưỡng trong nhà lưới năm 2016
Kết quả đánh giá tổng hợp khả năng chịu hạn và khả năng phục hồi cho thấy giữa hai chỉ tiêu này có sự liên quan chặt chẽ. Ở giai đoạn sinh trưởng sinh dưỡng, cả 14 mẫu giống có tính chịu hạn tốt kể trên, cũng nằm trong số mẫu được đánh giá có khả năng phục hồi tốt. Đáng chú ý, một số giống tuy khả năng chịu hạn trung bình nhưng khả năng phục hồi tốt như Tứ thời Thanh Hóa (SĐK 12), Dự Thanh Hóa
(SĐK 134), Tám thơm Thanh Hóa (SĐK 289), Tám đỏ Thanh Hóa (SĐK 299), Mùa Trắng Thanh Hóa (SĐK 585).
3.3.2. Khả năng chịu hạn, khả năng phục hồi của các mẫu giống lúa ở giai đoạn sinh trưởng sinh thực trong nhà lưới
Sau khi có kết quả đánh giá khả năng chịu hạn và khả năng phục hồi ở giai đoạn sinh trưởng sinh dưỡng, tổng số 64 mẫu giống lúa được đánh giá có khả năng phục hồi tốt và trung bình được tiếp tục gây hạn nhân tạo ở giai đoạn sinh trưởng sinh thực.
Kết quả đánh giá đã xác định được 3/64 mẫu giống là SĐK 12 - Tứ thời Thanh Hóa; SĐK 299 - Tám đỏ Thanh Hóa và SĐK 585 - Mùa Trắng Thanh Hóa (đạt 4,6% các giống có khả năng chịu hạn ở giai đoạn sinh trưởng sinh dưỡng) và CH5 - giống đối chứng; có khả năng chịu hạn tốt ở giai đoạn sinh trưởng sinh thực; 25/64 mẫu giống có khả năng chịu hạn trung bình (Hình 4). Như vậy, những giống có khả năng chịu hạn và khả năng phục hồi tốt ở giai đoạn sinh trưởng sinh dưỡng thường có khả năng chịu hạn tốt ở giai đoạn sinh trưởng sinh thực.
Hình 4. Phân nhóm khả năng chịu hạn, khả năng phục hồi của 64 mẫu giống lúa
ở giai đoạn sinh trưởng sinh thực
Sau khi bị gây hạn, các nguồn gen này tiếp tục được đánh giá khả năng phục hồi. Kết quả cho thấy có 4 mẫu giống (Tứ thời Thanh Hóa - SĐK 12; Tám đỏ Thanh Hóa - SĐK 299; Nếp lốc Nghệ An - SĐK 325 và Mùa Trắng Thanh Hóa - SĐK 585) có khả năng phục hồi tốt và 43 nguồn gen phục hồi trung bình sau gây hạn.
Lúa chịu hạn là giống lúa có khả năng duy trì sự phát triển và cho năng suất tương đối ổn định trong điều kiện khô hạn được gọi là lúa chịu hạn. Trong nghiên cứu này, ở giai đoạn sinh trưởng sinh thực, mức suy giảm năng suất của hầu hết các giống đều
Khả năng chiu hạn
Số mẫu
Tỷ lệ (%) Mẫu điển hình
Tốt (điểm 0-1) 14 14 SĐK12, SĐK134
Trung bình(điểm 3-5) 36 36 SĐK13; SĐK106
Kém, rất kém (điểm 7-9) 50 50 SĐK 573; SĐK742
Tổng cộng 100 100
Kết quả đánh giá khả năng chịu hạn, khả năng phục hồigiai đoạn sinh trưởng sinh dưỡng
Số m
ẫu g
iống
100
80
60
40
20
0Tốt1425
TB3639
Kém5036
Số mẫu giống chịu hạnSố mẫu giống phục hồi
Kết quả đánh giá khả năng chịu hạn, khả năng phục hồigiai đoạn sinh trưởng sinh thực
Số m
ẫu g
iống
64
54
44
34
24
14
4
-6Tốt34
TB2543
Kém3617
Số mẫu giống chịu hạnSố mẫu giống phục hồi

19
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
khá nghiêm trọng, chỉ còn 3 mẫu giống SĐK 12 - Tứ thời Thanh Hóa; SĐK 299 - Tám đỏ Thanh Hóa và SĐK 585 - Mùa Trắng Thanh Hóa và đối chứng CH5 là có khả năng chịu hạn sau gây hạn 10 ngày. Đây là thời kỳ cây lúa có nhu cầu về nước rất lớn để tăng sinh khối, tích lũy chất khô vào hạt, do đó rất mẫn cảm với sự khủng hoảng về nước.
Sau khi đánh giá khả năng phục hồi, đã tiếp tục theo dõi, quan sát thấy hiện tượng lá khô héo (điểm 9) xẩy ra ở hầu hết các mẫu giống, ngoại trừ 4 mẫu giống chịu được hạn. Các nguồn gen phục hồi trung bình, bông trỗ chậm hơn và hạt lép nhiều.
Số liệu bảng 3 cho thấy, một số mẫu giống chịu hạn triển vọng đều có thời gian sinh trưởng (TGST)
trong điều kiện gây hạn đều bị kéo dài, dẫn tới kéo dài thời gian trỗ cũng như quá trình chín. Ngoại trừ khối lượng 1000 hạt (P1000) không có sự thay đổi lớn, hầu hết các yếu tố cấu thành năng suất của các mẫu giống lúa trong điều kiện gây hạn đều thấp. Trong đó, số bông/khóm và tỷ lệ hạt chắc/bông có sự chênh lệch lớn nhất, tỷ lệ phần trăm thấp đáng kể. Số hạt/bông của các nguồn gen đều thấp hơn so với đối chứng CH5. Như vậy, hạn đã ảnh hưởng đến hầu hết các yếu tố cấu thành năng suất của các nguồn gen lúa, trong đó làm giảm rõ rệt số bông/khóm, và tỷ lệ hạt chắc/bông. Có thể nói đây là điểm mấu chốt và là nguyên nhân chính làm giảm năng suất lúa trong điều kiện hạn.
Trong điều kiện gây hạn, số bông/khóm của các mẫu giống triển vọng hầu hết đều cao hơn và tương đương so với đối chứng, cao nhất là Nếp Lốc Thanh Hóa, đạt 5,8 bông/khóm. Năng suất lý thuyết các mẫu giống doa động từ 3,55 - 5,04 tấn/ha. Có ba mẫu giống là Tứ Thời Thanh Hóa (SĐK 12), Nếp Lốc Thanh Hóa (SĐK 325), Mùa trắng Thanh Hóa (SĐK 585) cho năng suất lý thuyết cao hơn đối chứng CH5 (4,18 tấn/ha). Cao nhất là Nếp Lốc Thanh Hóa (đạt 5,04 tấn/ha) vượt giống đối chứng CH5 tới 20,5% và thấp nhất Dự Thanh Hóa (đạt 2,93 tấn/ha).
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnTổng số 100 mẫu giống lúa địa phương được thu
thập từ vùng Bắc Trung bộ đã được đánh giá trong điều kiện gây hạn nhân tạo và phân nhóm theo mức độ chịu hạn và khả năng phục hồi ở các giai đoạn nảy mầm, mạ 3 lá, đẻ nhánh, từ phân hóa đòng đến trỗ bông, vào chắc. Bốn mẫu giống chịu hạn qua tất cả các giai đoạn là Tứ thời Thanh Hóa - SĐK 12, Tám đỏ Thanh Hóa - SĐK 299, Nếp Lốc Thanh Hóa - SĐK 325 và Mùa trắng Thanh Hóa - SĐK 585,
trong đó mẫu giống Tứ thời Thanh Hóa (SĐK 12) có khả năng chịu hạn và phục hồi tốt ở cả 4 giai đoạn. Ngoại trừ Nếp Lốc Thanh Hóa - SĐK 325, ba mẫu giống còn lại có năng suất vượt hoặc tương đương đối chứng CH5 (4,18 tấn/ha) trong điều kiện gây hạn nhân tạo.
4.2. Đề nghi Đề nghị sử dụng các mẫu giống có khả năng
chịu hạn, phục hồi tốt phục vụ mục đích khai thác và làm vật liệu khởi đầu cho các nghiên cứu sâu hơn.
TÀI LIỆU THAM KHẢONguyên Hữu Cường, Nguyên Thi Kim Anh, Đinh Thi
Phòng, Lê Thi Muội, Lê Trần Bình, 2003. Mối tương quan giữa làm lượng proline và tính chống chịu hạn ở cây lúa. Tạp chí Công nghệ sinh học, 1(1): 85-95.
Nguyên Thi Thu Hoài, 2005. Nghiên cứu khả năng chịu hạn và mối quan hệ di truyền của một số giống lúa cạn địa phương. Luận văn Thạc sỹ sinh học. Trường Đại học Sư phạm - Đại học Thái Nguyên.
Vũ Tuyên Hoàng, Nguyên Ngọc Ngân, 1992. Một số kết quả nghiên cứu lúa chịu hạn. Kết quả nghiên cứu cây lương thực, thực phẩm (86 - 90). Viện Cây lương
Bảng 3. Năng suất và các yếu tố cấu thành năng suất của một số mẫu giống lúa triển vọng trong điều kiện hạn nhân tạo
SĐK Tên mẫu giống TGST(Ngày)
Số bông/ khóm
Số hạt/ bông
Tỷ lệ hạt chắc (%)
P.1000 hạt (g)
NSLT (tấn/ha)
12 Tứ thời Thanh Hóa 139 5,1 113,3 62,0 32,8 4,66289 Tám thơm Thanh Hóa 156 5,2 110,7 56,3 31,6 3,55299 Tám đỏ Thanh Hóa 152 5,1 114,0 54,6 33,4 3,85325 Nếp Lốc Thanh Hóa 153 5,8 117,0 60,7 33,8 5,04585 Mùa trắng Thanh Hóa 146 5,0 112,2 61,5 33,8 4,20601 Gié Hoa Hà Tĩnh 132 5,3 112,0 59,1 35,0 3,88
14574 CH5 (ĐC) 123 4,9 119,0 56,6 26,1 4,18

20
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
thực và Cây thực phẩm. NXB Nông nghiệp. Hà Nội, trang 47 - 57.
IRRI, 2002. Hệ thống tiêu chuẩn đánh giá nguồn gen lúa (Vũ Văn Liết biên dịch). Trường ĐH Nông nghiệp Hà Nội.
Phạm Anh Tuấn, Nguyên Lan Hoa, Nguyên Thi Minh Nguyệt, Nguyên Bá Ngọc, Nguyên Thi Kim Dung, Nguyên Thi Thanh Thuỷ, 2008. Đánh giá đặc tính chị hạn của một số giống lúa địa phương Việt Nam thông qua phương pháp kiểu hình và ứng dụng chỉ thị phân tử. Tạp chí Nông nghiệp và PTNT, trang 28- 35.
Dorner JW, Cole RJ, Sanders TH and Blankenship PD., 1989. Interrelationship of kernel water activity, soil temperature, maturity, and phytoalexin production in pre-harvest aflatoxin contamination of drought
stressed peanuts. Mycopathologia, 105: 117-128.FAO, 2015. Crops, National Production (FAOSTAT)
Dataset. Food and Agriculture Organization of the United Nations. http://data.fao.org/dataset-d at a- f i l te r ?ent r yId=29920434-4e a2-b e e d-01b832e60609&tab
Fischer K. S., Atlin, G. N., Blum, A., Fukai, S., Lafitte, R. and Mackill, D., 2003. In Breeding Rice for Drought-Prone Environments (pp. 1-4) Philippines: International Rice Research Institute.
Money N. P., 1989. “Osmotic Pressure of Aqueous Polyethylene Glycols: Relationship betweenMolecular Weight and Vapor Pressure Deficit”.
Yoshida S., D. A. Forno, 1971. Laboratory manual for physiological studies of rice. Los Banos, Philippines, International Rice Research Institute (IRRI).
Drought tolerant ability of local rice varieties maintaining at the National Crop Genebank
Luu Quang Huy, Nguyen Thi Ngoc Hue, Vu Linh Chi, Duong Hong Mai, Vu Dang Toan, Bui Thi Thu Huyen,
Ha Minh Loan, Tran Danh SuuAbstractThe study aimed to determine the drought tolerance and restoration ability of 100 local rice varieties collected in the North Central Vietnam by artificial drought method at different growth stages. The results showed that the different varieties responded differently to drought conditions at each growth stage. 100 rice varieties were grouped by the drought tolerance level and restoration ability at the growth stages. Four good drought tolerant varieties (Score 1) were identified at all stages including Tu thoi Thanh Hoa (Acc. No 12), Tam do Thanh Hoa (Acc. No 299), Nep Loc Thanh Hoa (Acc. No 325) and Mua trang Thanh Hoa (Acc. No 585). Except for Tam do Thanh Hoa variety (Acc. No 299), the other three varieties had higher yield or equivalent to CH5 control variety (4.18 tons / ha) under artificial drought conditions.Key words: Rice, local rice, artificial drought, drought tolerance, evaluation, restoration
Ngày nhận bài: 17/6/2017Ngày phản biện: 21/6/2017
Người phản biện: TS. Trần Thị Thu HoàiNgày duyệt đăng: 27/7/2017
1 Trường Đại học Nông lâm Thái Nguyên
NGHIÊN CỨU KHẢ NĂNG SINH TRƯỞNG, PHÁT TRIỂN VÀ NĂNG SUẤT CỦA MỘT SỐ GIỐNG ĐẬU TƯƠNG TRÊN ĐẤT
NƯƠNG RẪY TẠI HUYỆN HOÀNG SU PHÌ, TỈNH HÀ GIANGDương Trung Dũng1
TÓM TẮTHoàng Su Phì là huyện vùng cao của tỉnh Hà Giang, gieo trồng đậu tương chủ yếu trên đất nương rẫy. Người
dân chủ yếu sử dụng giống địa phương năng suất thấp, nên việc đưa giống mới cho năng suất cao vào sản xuất là cần thiết. Nghiên cứu 8 giống đậu tương ĐT22, ĐT31, ĐT51, DT2012, ĐT30, ĐT26, DT2008 và DT84 cho thấy về thời gian sinh trưởng các giống đều thuộc nhóm chín sớm và nhóm chín trung bình. Năng suất thực thu biến động từ 14,74 -24,85 tạ/ha (vụ Hè Thu năm 2015) và từ 13,91- 24,07 tạ/ha (vụ Xuân năm 2016). Ba giống DT2008, ĐT22, ĐT26 cho năng suất cao ở cả hai vụ gieo trồng. Kết quả xây dựng mô hình trình diễn của 3 giống ưu tú năng suất đạt từ 20,63 - 23,59 tạ/ha, trong đó giống DT2008 đạt năng suất trung bình cao nhất (23,59 tạ/ha), giống ĐT22 được người dân ưa thích và lựa chọn.
Từ khóa: Đậu tương, giống, Hoàng Su Phì, năng suất, ưu tú

21
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀCây đậu tương [Glycine max (L.) Merrill] là cây
công nghiệp ngắn ngày có tác dụng rất nhiều mặt và là cây có giá trị kinh tế cao. Cây đậu tương là cây có khả năng cải tạo và bồi dưỡng đất rất tốt do có sự cộng sinh giữa rễ với vi khuẩn nốt sần có khả năng cố định đạm trong không khí làm giàu đạm cho đất. Sau mỗi vụ trồng, đậu tương có thể cố định và bổ sung vào đất từ 60 - 80 kg N/ha (Ngô Thế Dân và ctv., 1999).
Hoàng Su Phì là huyện vùng cao, núi đất nằm ở phía Tây tỉnh Hà Giang có tổng diện tích đất tự nhiên 63.261,82 ha, trong đó: đất nông lâm nghiệp 46.562,31 ha, chiếm 73,6%; đất phi nông nghiệp 1.727,97 ha, chiếm 2,73%; đất chưa sử dụng 14.971,54 ha, chiếm 23,67% ( Chi cục Thống kê tỉnh Hà Giang). Đến năm 2015, diện tích đậu tương cả năm của huyện hơn 5.400 ha, chủ yếu là gieo trồng trên diện tích đất nương rẫy (chiếm trên 70%), trong đó vụ Xuân khoảng 2.600 ha, vụ Hè Thu khoảng 2.800 ha. Cơ cấu giống chủ yếu là giống DT 84 và giống địa phương chiếm khoảng 15% chủ yếu là giống đậu tương hạt vàng và hạt xanh. Năng suất đậu tương của huyện bình quân đạt 15,2 tạ/ha, sản lượng hàng năm đạt trên 8.000 tấn. Theo số liệu thống kê hàng năm thì thu nhập từ cây đậu tương chiếm từ 16 - 18% trong tổng thu nhập kinh tế hộ gia đình (Chi cục Thống kê Hoàng Su Phì, 2016).
Tuy nhiên, do nhiều nguyên nhân về sản xuất nhất là việc đầu tư thâm canh, áp dụng khoa học kỹ thuật chưa hợp lý, công tác giống chưa được chú trọng, số hộ sử dụng giống địa phương đã bị thoái hoá còn chiếm tỷ lệ lớn. Việc quản lý và cung cấp giống còn nhiều bất cập dẫn đến chất lượng giống bị pha tạp, vì vậy chất lượng thương phẩm kém, năng suất bình quân thấp. Mặc dù là huyện có diện tích, năng suất, sản lượng đậu tương lớn nhất toàn tỉnh và cây đậu tương đã được Cấp ủy - Chính quyền huyện xác định là cây trồng phù hợp, có thế mạnh nhưng trong thời gian qua vẫn chưa thực sự trở thành cây trồng chủ lực giúp người nông dân phát triển kinh tế, xóa đói, giảm nghèo và làm giàu.
Xuất phát từ những lý do trên, cần thiết tiến hành nội dung: “Nghiên cứu khả năng sinh trưởng, phát triển và năng suất của một số giống đậu tương trên đất nương rẫy tại huyện Hoàng Su Phì, tỉnh Hà Giang”
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1 Vật liệu nghiên cứu Tiến hành nghiên cứu với 8 giống đậu tương:
ĐT22, ĐT31, ĐT51, DT2012, ĐT30, ĐT26, DT2008 và DT 84 (giống đối chứng).
2.2. Phương pháp nghiên cứu
2.2.1. Phương pháp bố trí thí nghiệm- Các thí nghiệm khảo nghiệm cơ bản, xây dựng
mô hình đều thực hiện theo Quy chuẩn kỹ thuật Quốc gia về khảo nghiệm giá trị canh tác và sử dụng của giống đậu tương QCVN 01-58: 2011/BNNPTNT (Bộ Nông nghiệp và PTNT, 2011).
Thí nghiệm được bố trí theo kiểu khối ngẫu nhiên hoàn chỉnh (RCBD) gồm 8 công thức và 3 lần nhắc lại.
Quy trình kỹ thuật: Áp dụng theo QCVN 01-58: 2011/BNNPTNT của Bộ nông nghiệp và Phát triển nông thôn (Bộ Nông nghiệp và PTNT, 2011).
- Xây dựng mô hình trình diễn: Qui mô: 2,4 ha (0,6 ha/giống, kể cả diện tích đối chứng). Làm đất: cày bừa kỹ, nhặt sạch cỏ dại, lên luống, rạch hàng. Mật độ: 35 cây/m2. Lượng phân bón (cho 1 ha): Phân chuồng: 10 tấn phân chuồng đã ủ hoai. Phân vô cơ: 40 kgN + 60 kg P2O5 + 40 kg K2O.
2.2.2. Các chỉ tiêu theo dõiChỉ tiêu và phương pháp đánh giá: Phương pháp
nghiên cứu các chỉ tiêu sinh trưởng, phát triển của các giống thí nghiệm được tiến hành theo Quy phạm khảo nghiệm giống đậu tương QCVN 01-58: 2011/BNNPTNT của Bộ Nông nghiệp và Phát triển nông thôn (Bộ Nông nghiệp và PTNT, 2011).
2.2.3. Phương pháp xử lý số liệu Các số liệu thu được của các cây/ô chia cho số
cây theo dõi để lấy số liệu trung bình của từng ô rồi tính trung bình của các lần nhắc lại
Số liệu được xử lý theo IRRISTAT 5.0 và Excel 2010.
2.3. Thời gian và đia điểm nghiên cứu - Thời gian nghiên cứu: Từ tháng 6 năm 2015 đến
hết tháng 9 năm 2016. Vụ Xuân trồng từ 25/2- 10/3, vụ Hè Thu trồng từ 25/6 - 10/7. Thực hiện cả 2 xã
- Địa điểm nghiên cứu: xã Chiến Phố và xã Tụ Nhân của huyện Hoàng Su Phì, tỉnh Hà Giang.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Kết quả nghiên cứu khả năng sinh trưởng, phát triển các giống thí nghiệm vụ Hè Thu 2015 và vụ Xuân 2016 tại huyện Hoàng Su Phì, tỉnh Hà Giang
Kết quả nghiên cứu khả năng sinh trưởng, phát triển của các giống, số liệu được trình bày ở bảng 1.

22
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Ở bảng 1 cho thấy tất cả các giống đậu tương thí nghiệm giai đoạn từ gieo đến mọc ở cả 2 vụ, 2 địa điểm thí nghiệm khoảng 4 - 6 ngày. Sau quá trình gieo hạt ở cả vụ Hè Thu 2015 và vụ Xuân 2016 đều gặp điều kiện thời tiết bất thuận.
- Giai đoạn từ gieo đến ra hoa: Ở vụ Hè Thu 2015 và vụ Xuân 2016 thời gian từ gieo đến ra hoa của các giống đậu tương biến động từ 36 - 48 ngày, sự ra hoa giống DT84 đối chứng sớm hơn các giống khác (37 ngày sau gieo hạt ở vụ Hè Thu 2015, 36 ngày sau gieo hạt ở vụ Xuân 2016). Giống DT 2008 thời gian từ gieo đến ra hoa dài nhất (48 ngày sau gieo), tiếp đến là các giống ĐT 30, DT2012, ĐT31, ĐT 26, DT2012, ĐT30, ĐT51, DT84.
- Giai đoạn từ gieo đến chắc xanh: Thời gian từ gieo đến chắc xanh của các giống đậu tương phụ thuộc vào giống và địa điểm trồng. Thời gian từ gieo đến chắc xanh biến động 74 - 89 ngày (vụ Hè Thu 2015), 73 - 89 ngày (vụ Xuân 2016). Các giống tham gia thí nghiệm đều có thời gian từ gieo đến chắc xanh muộn hơn đối chứng 4 - 16 ngày.
- Giai đoạn từ gieo đến chín (thời gian sinh trưởng): Thời gian từ gieo đến chín (TGST) của các giống đậu tương ở vụ Hè Thu 2015 biến động từ 88 - 102 ngày. Các giống thí nghiệm có TGST dài hơn đối chứng: giống DT2008 có tổng TGST dài nhất (102 ngày), dài hơn giống đối chứng 14 ngày. Ở vụ Xuân 2016, thời gian từ gieo đến chín (TGST) biến động từ 86 - 98 ngày, các giống ở vụ Xuân đều có TGST ngắn hơn vụ Hè Thu từ 2 đến 4 ngày. Với thời gian sinh trưởng này các giống đều thuộc nhóm có thời gian sinh trưởng trung bình (phù hợp với kết quả của Cục Trồng trọt, 2006).
3.2. Một số đặc điểm hình thái của các giống đậu tương thí nghiệm
Ở vụ Hè Thu 2015 chiều cao cây biến động 39,2
- 49,5 cm, cao nhất là giống ĐT 30 đạt 49,5 cm cao hơn giống đối chứng DT 84 (42,5 cm) là 7,0 cm, chắc chắn ở mức độ tin cậy 95%; các giống còn lại có chiều cao cây tương đương giống đối chứng. Vụ Xuân 2016, giống ĐT31 có chiều cao cây đạt 35,6 cm thấp hơn giống đối chứng DT84 là 7,9 cm, chắc chắn ở mức độ tin cậy 95%; các giống còn lại có chiều cao cây tương đương giống đối chứng.
- Cành cấp I: Khả năng phân cành cấp I của các giống đậu tương ở vụ Hè Thu năm 2015 có khả năng phân cành nhiều hơn vụ Xuân 2016, biến động từ 2,1 - 3,8 cành, trong đó các giống ĐT31, ĐT51 và ĐT30 có tỷ lệ phân cành tương đương giống đối chứng DT 84; các giống còn lại có tỷ lệ phân cành cao hơn đối chứng, chắc chắn ở mức độ tin cậy 95 %. Ở vụ Xuân 2016 biến động từ 1,6 - 3,2 cành, có 3 giống ĐT31; ĐT30; ĐT51 có tỷ lệ phân cành thấp hơn đối chứng, các giống còn lại đều có khả năng phân cành cấp I tương đương hoặc tốt hơn giống đối chứng, chắc chắn ở mức độ tin cậy 95%.
- Số đốt/thân chính: Ở vụ Hè Thu 2015 các giống có thời gian sinh trưởng dài hơn vụ Xuân 2016 do chênh lệnh biên độ ngày và đêm nên số đốt/thân chính của hầu hết các giống đậu tương cũng lớn hơn, số đốt/thân chính dao động từ 11,0 - 14,7 đốt. Giống ĐT 30 có số đốt/thân chính tương đương giống đối chứng, các giống còn lại có số đốt/thân chính nhiều hơn giống đối chứng chắc chắn ở mức độ tin cậy 95%; trong đó nhiều nhất là giống DT2008 đạt 14,7 đốt, cao hơn giống đối chứng là 3,7 đốt. Ở vụ Xuân 2016 các giống đậu tương thí nghiệm có TGST ngắn hơn nên đều có số đốt/thân chính ít hơn ở vụ Hè Thu 2015 và biến động từ 10,0 - 12,8 đốt. Trong đó giống DT2008 số đốt/thân chính cao nhất đạt 12,8 đốt, cao hơn giống đối chứng (10,0 đốt) 2,8 đốt, chắc chắn ở mức tin cậy 95%.
Bảng 1. Các giai đoạn sinh trưởng, phát triển của các giống thí nghiệm vụ Hè Thu 2015 và vụ Xuân 2016
TT GiốngVụ Hè Thu 2015 (ngày) Vụ Xuân 2016 (ngày)
Ngày mọc
Ngày ra hoa
Ngày chắc xanh
Ngày chín
Ngày mọc
Ngày ra hoa
Ngày chắc xanh
Ngày chín
1 DT 84 (đ/c) 4 37 74 88 4 36 73 862 ĐT 22 4 40 79 91 4 39 77 883 ĐT 31 4 43 81 93 4 42 80 904 ĐT51 4 43 83 93 4 42 80 905 DT2012 5 45 85 98 6 47 84 936 ĐT 30 5 45 85 98 5 45 84 937 ĐT 26 5 43 79 91 4 42 80 888 DT2008 5 48 89 102 6 48 89 98

23
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 2. Đặc điểm hình thái của các giống thí nghiệm
3.3. Tình hình nhiêm sâu hại của các giống đậu tương thí nghiệm
Qua theo dõi cho thấy mật độ sâu cuốn lá ở cả hai vụ Hè Thu 2015 và vụ Xuân 2016 đều phá hại trên tất cả các giống đậu tương và gây hại nặng nhất ở thời kỳ ra hoa. Trong đó giống DT2008 vụ Hè Thu bị nhiễm nhẹ nhất và ĐT31, ĐT26 ở vụ Xuân có tỉ lệ bị hại nhẹ hơn giống đối chứng, chắc chắn ở mức tin cậy 95%. Các giống còn lại có tỉ lệ sâu cuốn lá tương đương giống đối chứng.
Bảng 3. Một số sâu hại chính của các giống đậu tương thí nghiệm
Sâu đục quả gây hại ở cả 2 vụ Hè Thu 2015 và vụ Xuân 2016. Ở vụ Hè Thu, tỷ lệ sâu đục quả gây hại
biến động từ: 0,87 - 2,34%, các giống đậu tương bị sâu đục quả gây hại tương đương giống đối chứng. Ở vụ Xuân 2016, tỷ lệ quả bị hại dao động từ 2,00 - 5,19%, giống DT2012 bị sâu đục quả gây hại tương đương đối chứng. Giống ĐT26, ĐT31 và DT2008 bị sâu đục quả nhẹ hơn giống đối chứng. Các giống còn lại bị sâu đục quả nặng hơn đối chứng, chắc chắn ở mức tin cậy 95%. Kết quả nghiên cứu phù hợp với kết quả nghiên cứu của Dương Trung Dũng (2010).
3.4. Các yếu tố cấu thành năng suất và năng suất của các giống đậu tương
- Số quả chắc/cây: Vụ Hè Thu 2015 có 3 giống ĐT31, ĐT51, ĐT30 có số quả chắc trên cây thấp hơn đối chứng (DT84: 19,43 quả), các giống còn lại có số quả chắc trên cây cao hơn đối chứng, chắc chắn có ý nghĩa ở mức độ tin cậy 95%. Trong đó cao nhất là giống DT2008 đạt 27,9 quả, cao hơn giống đối chứng là 8,47 quả. Ở vụ Xuân năm 2016, số quả chắc trên cây biến động từ 16,97 - 28,03 quả. Giống DT2008 có số quả chắc/cây cao nhất đạt 28,03 quả, tiếp đến là giống ĐT 22 (27,73 quả), cao hơn giống đối chứng lần lượt là 11,06 quả và 10,76 quả, chắc chắn ở mức độ tin cậy 95%. Các giống còn lại đều có số quả chắc trên cây tương đương giống đối chứng.
- Số hạt chắc/ quả: Số hạt chắc/quả của các giống đậu tương biến động từ 2,04 - 2,15 hạt/quả (vụ Hè Thu 2015), 2,00 - 2,11 hạt/quả (vụ Xuân 2016). Tất cả các giống đậu tương đều có số hạt chắc/quả tương đương giống đối chứng, chắc chắn ở mức độ tin cậy 95%.
- Khối lượng 1000 hạt: Ở vụ Hè Thu 2015, P1000 hạt của các giống đậu tương rất khác nhau, biến
GiốngVụ Hè Thu 2015 Vụ Xuân 2016
Chiều cao cây (cm)
Số cành cấp I (cành)
Số đốt/thân chính (đốt)
Chiều cao cây (cm)
Số cành cấp I (cành)
Số đốt/thân chính (đốt)
DT 84(đ/c) 42,5 2,2 11,0 43,5 2,4 10,0ĐT 22 41,4 3,8 14,0 40,5 3,2 12,7ĐT 31 39,2 2,3 12,9 35,6 1,6 11,8ĐT 51 41,7 2,4 13,0 39,3 1,9 11,8DT 2012 45,7 3,6 14,4 42,4 2,4 12,5ĐT 30 49,5 2,1 11,7 44,5 1,6 11,8ĐT 26 41,7 3,6 12,3 39,5 3,1 12,0DT 2008 43,6 3,6 14,7 40,6 3,2 12,8P < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05CV(%) 7,1 9,0 5,4 7,0 8,1 3,8LSD0,05 5,38 0,46 1,22 4,83 0,34 0,79
Giống
Vụ Hè Thu 2015 Vụ Xuân 2016 Sâu
cuốn lá(con/m2)
Sâu đục quả
(% quả bi hại)
Sâu cuốn lá(con/m2)
Sâu đục quả
(% quả bi hại)
DT 84(đ/c) 15,49 1,57 7,65 3,80ĐT 22 15,54 1,28 7,49 5,19ĐT 31 15,99 0,89 5,25 2,47ĐT 51 15,01 1,04 7,79 4,77DT 2012 16,32 0,87 7,58 3,23ĐT 30 15,17 1,14 8,34 4,17ĐT 26 17,49 1,40 6,07 2,00DT 2008 5,73 2,34 6,58 2,56P < 0,05 < 0,05 < 0,05 < 0,05CV(%) 13,5 19,1 10,9 13,3LSD0,05 3,45 1,43 1,35 0,80
24
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
động từ 140,70 - 174,57 g. Vụ Xuân 2016 biến động từ 127,03 - 170,00 g. Trong đó giống ĐT26 và ĐT30 có P1000 hạt tương đương giống đối chứng. Các giống đậu tương còn lại trong vụ Xuân 2016 đều có P1000 hạt thấp hơn đối chứng, chắc chắn có ý nghĩa ở mức độ tin cậy 95%.
- Năng suất lý thuyết (NSLT): Trong điều kiện vụ Hè Thu 2015 và vụ Xuân 2016, NSLT của các giống đậu tương rất khác nhau, biến động 21,24 - 32,62 tạ/ha (vụ Hè Thu 2015), 19,61 - 28,83 tạ/ha (vụ Xuân 2016). Ở vụ Xuân 2016, giống ĐT30, ĐT51, ĐT31 có NSLT tương đương giống đối chứng. Các giống còn lại có NSLT cao hơn giống đối chứng, chắc chắn ở mức tin cậy 95% .
- Năng suất thực thu (NSTT): Vụ Hè Thu 2015, năng suất thực thu của các giống đậu tương biến
động từ 14,74 - 24,85 tạ/ha. Giống ĐT30 có năng suất thực thu thấp hơn giống đối chứng (14,74 tạ), chắc chắn ở mức độ tin cậy 95%. Giống ĐT31 có NSTT tương đương giống đối chứng. Các giống còn lại đều có năng suất thực thu cao hơn giống đối chứng, chắc chắn ở mức độ tin cậy 95%. Trong đó giống DT2008 có NSTT cao nhất đạt 24,85 tạ/ha, cao hơn giống đối chứng là 8,79 tạ/ha. Vụ Xuân 2016, năng suất thực thu của các giống đậu tương từ 13,91- 24,07 tạ/ha. Giống ĐT30 có năng suất thực thu thấp hơn giống đối chứng (ĐT30: 13,91 tạ/ha), chắc chắn ở mức độ tin cậy 95%; giống ĐT31 có NSTT tương đương giống đối chứng. Các giống còn lại (DT2008, ĐT22, ĐT26, ĐT51, DT2012) đều có năng suất thực thu cao hơn giống đối chứng, chắc chắn ở mức tin cậy 99%. Giống DT2008 là giống có NSTT cao nhất đạt 24,07 tạ/ha.
3.5. Kết quả xây dựng mô hình trình diên vụ Hè Thu năm 2016
- Trên cơ sở thực tế, căn cứ vào các đặc tính ưu việt của giống: thời gian sinh trưởng, khả năng chống chịu, các yếu tố cấu thành năng suất, màu sắc hạt của các giống tham gia thí nghiệm, chọn ra 3 giống có triển vọng: DT2008, ĐT22, ĐT26 và giống DT84 (đối chứng) tham gia xây dựng mô hình.
Kết quả xây dựng mô hình giống DT2008: Có 3 hộ gia đình tham gia trồng trên diện tích 0,6 ha, năng suất trung bình đạt 23,59 tạ/ha, cao hơn DT84 là 7,84 tạ/ha (DT84 đạt 15,75 tạ/ha). Trong đó gia đình ông Lù Văn Dợ đạt năng suất cao nhất (25,85 tạ/ha) với diện tích 0,2 ha. Giống ĐT22, có 3 hộ gia đình tham gia trồng
trên diện tích 0,6 ha. Năng suất trung bình đạt 21,38 tạ/ha, cao hơn đối chứng 7,26 tạ/ha (DT84: 14,12 tạ/ha). Trong đó gia đình ông Lù Seo Chang đạt năng suất cao nhất (22,83 tạ/ha) với diện tích 0,2 ha. Giống ĐT26, có 3 hộ gia đình tham gia trồng trên diện tích 0,6 ha. Năng suất trung bình đạt 20,63 tạ/ha, cao hơn đối chứng 8,13 tạ/ha (DT84: 12,5 tạ/ha). Trong đó gia đình Ông Lý Văn Kim đạt năng suất cao nhất (21,34 tạ/ha) với diện tích 0,2 ha. Như vậy, qua xây dựng mô hình trồng đậu tương vụ Hè Thu 2016 tại thôn Võ Thấu Chải, xã Chiến Phố, huyện Hoàng Su Phì cho thấy giống DT2008 đạt năng suất trung bình cao nhất (23,59 tạ/ha), tiếp đến là giống ĐT22 (21,38 tạ/ha) và thấp nhất là giống ĐT26 (20,63 tạ/ha).
Bảng 4. Các yếu tố cấu thành năng suất và năng suất của các giống đậu tương tham gia thí nghiệm vụ Hè Thu 2015, vụ Xuân 2016
Giống
Vụ Hè Thu 2015 Vụ Xuân 2016
Số quả chắc(quả/cây)
Số hạt chắc/quả (hạt)
P1000 hạt (g)
NSLT (tạ/ha)
NSTT (tạ/ha)
Số quả chắc (quả/cây )
Số hạt chắc/quả (hạt)
P1000 hạt (g)
NSLT (tạ/ha)
NSTT (tạ/ha)
DT 84(đ/c) 19,43 2,05 172,67 24,07 16,06 16,97 2,07 165,50 20,37 15,91ĐT 22 26,50 2,15 140,70 28,10 22,85 27,73 2,11 127,03 26,01 20,75ĐT 31 17,40 2,04 171,03 21,24 16,30 17,67 2,09 152,47 19,61 15,47ĐT 51 18,03 2,14 157,20 21,24 17,52 20,03 2,09 147,37 21,54 19,14DT 2012 22,20 2,05 169,30 26,96 21,04 22,83 2,09 153,37 25,60 19,18ĐT 30 18,10 2,04 167,27 21,65 14,74 17,90 2,00 159,33 19,93 13,91ĐT 26 21,57 2,07 174,57 27,25 21,48 20,77 2,06 170,00 25,41 20,26DT 2008 27,90 2,05 162,60 32,62 24,85 28,03 2,02 145,17 28,83 24,07CV(%) 2,3 6,6 1,0 7,2 2,2 9,9 4,0 11,7LSD0,05 0,84 0,24 0,27 0,81 6,2 0,36 10,7 1,26

25
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 5. Kết quả xây dựng mô hình trồng đậu tương
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luận- Thời gian sinh trưởng của các giống đậu tương
ở vụ Xuân 2016 ngắn hơn ở vụ Hè Thu 2015, biến động từ 86 - 98 ngày (vụ Xuân 2016) và 88 - 102 ngày (vụ Hè Thu 2015); với thời gian sinh trưởng này các giống đều thuộc nhóm có thời gian sinh trưởng trung bình.
Năng suất thực thu biến động từ 14,74 -24,85 tạ/ha (vụ Hè Thu 2015) và từ 13,91- 24,07 tạ/ha (vụ Xuân 2016). Ba giống DT 2008, ĐT22, ĐT26 tỏ ra có
NSTT cao nhất ở cả hai vụ gieo trồng, hai địa điểm nghiên cứu và cao hơn giống đối chứng chắc chắn có ý nghĩa từ 5,42 - 11,97 tạ/ha (vụ Hè Thu 2015) và từ 4,35 - 8,16 tạ/ha (vụ Xuân 2016).
- Kết quả xây dựng mô hình trình diễn của 3 giống ưu tú so sánh với giống đối chứng (DT84) cho thấy: Năng suất trung bình của 3 giống đậu tương trong trình diễn đều cao hơn hẳn năng suất của giống đối chứng, đạt từ 20,63 - 23,59 ta/ha, trong đó giống DT2008 đạt năng suất trung bình cao nhất (23,59 tạ/ha).
4.2. Đề nghiCho phép phát triển giống đậu tương DT 2008,
ĐT 22, ĐT 26 vào sản xuất đại trà trong huyện Hoàng Su Phì.
TÀI LIỆU THAM KHẢO Bộ Nông nghiệp và PTNT, 2011. QCVN 01-58:2011/
BNNPTNT. Quy chuẩn kỹ thuật quốc gia về khảo nghiệm giá trị canh tác và sử dụng của giống đậu tương. Hà Nội.
Chi cục thống kê tỉnh Hà Giang. Niên giám thống kê các năm 2005; 2006; 2007; 2008; 2009; 2010.
Cục Trông trọt, 2006. Kết quả điều tra giống 13 cây trồng chủ lực của cả nước giai đoạn 2003 - 2004. Nhà xuất bản Nông nghiệp. Hà Nội, tr 139 - 141.
Ngô Thế Dân, Trần Đình Long, Trần Văn Lài, Đỗ Thi Dung, Phạm Thi Đào, 1999. Cây đậu tương. NXB Nông nghiệp.
Dương Trung Dũng, 2010. Nghiên cứu thực trạng sản xuất và một số giải pháp kỹ thuật nhằm phát triển Đậu tương tại Thái Nguyên. Luận án tiến sỹ nông nghiệp, Trường đại học Nông lâm Thái Nguyên.
TT Hộ tham gia GiốngDiện tích(ha)
Năng suất
(tạ/ha)1 Nông văn Trúc DT 84 (Đ/c) 0,2 15, 752 Nông Văn Đức DT 2008 0,2 21,613 Lù Văn Dợ DT2008 0,2 25,854 Lùng Văn Hai DT 2008 0,2 23,32
Trung bình 23,595 Vàng Văn Minh DT 84 (Đ/c) 0,2 14,126 Tải Seo Chấn ĐT 22 0,2 19,677 Lù Seo Chang ĐT 22 0,2 22,838 Lục Văn Thanh ĐT 22 0,2 21,64
Trung bình 21,389 La thị Dì DT 84 (Đ/c) 0,2 12,5
10 Lý Văn Kim ĐT 26 0,2 21,3411 Lù Thị Seo ĐT 26 0,2 19,4512 Nông Thị Phương ĐT 26 0,2 21,12
Trung bình 20,63
Study on growth, development ability and yield of soybean varieties on upland farm in Hoang Su Phi district, Ha Giang province
Duong Trung DungAbtractsHoang Su Phi is an upland district of Ha Giang province and soybean cultivation is mainly on the upland farm in the mountain. People mostly use low productivity local variety, so application of new variety for high yield is neccesary. Research on 8 soybean varieties including DT22, DT31, DT51, DT2012, DT30, DT26, DT2008 and DT 84 showed that the growth duration of the varieties was belonged to early and medium maturity group. The real yield varried from 14.74 to 24.85 quintals/ha (summer autumn season in 2015) and 13.91 - 24.07 quintals/ha (spring season in 2016). Three soybean varieties such as DT2008, ĐT22, ĐT26 gave high yield in both crop seasons. The results of the demonstration plots of 3 elite varieties showed that the average yield reached from 20.63 to 23.59 quintals/ha, among which, DT2008 variety had the highest average yield (23.59 quintals/ha), DT22 variety is preferred by farmers and selected.Key words: Soybean, variety, Hoang Su Phi, yield, elite
Ngày nhận bài: 6/7/2017Ngày phản biện: 15/7/2017
Người phản biện: PGS.TS. Ninh Thị PhípNgày duyệt đăng: 27/7/2017

26
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀCà phê vối là cây trồng chủ lực trên đất đỏ bazan
vùng cao nguyên Di Linh, tỉnh Lâm Đồng. Theo Cục Thống kê Lâm Đồng (2014), toàn tỉnh có 153.432 ha cà phê trong đó cà phê vối chiếm 95% tổng diện tích. Tình hình sử dụng phân khoáng NPK cũng như phân hữu cơ của nông dân còn nhiều bất cặp về liều lượng và tỷ lệ , nhìn chung lượng phân nông dân bón dao động (kg/ha/năm): 194 - 897 N; 160 - 620 P2O5; 80 - 900 K2O, bình quân tỷ lệ phân NPK là 1,38 : 1 : 0,94. So với mức khuyến cáo cho năng suất 3 tấn/ha của Viện Khoa học kỹ thuật Nông Lâm nghiệp Tây Nguyên trên nền đất đỏ bazan là 250 N; 90 P2O5; 250 K2O (kg/ha/năm) thì nông dân ở Lâm Đồng sử dụng phân bón cho cà phê vối không cân đối về tỷ lệ cũng như liều lượng. Do vậy năng suất cà phê cũng rất biến động (1,5 tấn đến >5 tấn/ha/năm) giữa các hộ trong vùng (Lâm Văn Hà, 2016). Từ những thực trạng nói trên nghiên cứu ảnh hưởng của phân khoáng N, P và phân hữu cơ đến năng suất cà phê vối (Coffea Canephora Pierre) trên đất đỏ bazan vùng cao nguyên Di Linh, tỉnh Lâm Đồng là cần thiết, nhằm xác định được liều lượng phân khoáng N, P kết hợp với phân hữu cơ để đạt năng suất cao và đảm bảo duy trì được độ phì nhiêu đất đáp ứng nhu cầu thâm canh cà phê bền vững trên đất đỏ bazan.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứuCây cà phê vối (Coffea Canephora Pierre) được
nghép giống cao sản TS1 (Trường Sơn 1), độ tuổi 15 năm, với chế độ thâm canh không có cây che bóng.
Phân bón hóa học sử dụng gồm ure (46% N), bón 4 lần trong năm với tỷ lệ (15% mùa khô: tưới nước lần 2; 35% đầu mùa mưa; 25% giữa mùa mưa và 25% cuối mùa mưa); lân nung chảy (16% P2O5), bón 1 lần với tỷ lệ 100% vào đầu mùa mưa; kali clorua (60% K2O), bón 4 lần trong năm (10% tưới nước lần 2; 20% đầu mùa mưa; 35% giữa mùa mưa và 35% cuối mùa mưa). Phân hữu cơ bón gồm (35% heo + 35% gà + 30% vỏ cà phê + chế phẩm vi sinh vật) được trộn chung ủ hoai mục, bón với lượng 10 tấn/ha, thời điểm bón giữa mùa mưa (tháng 6).
2.2. Phương pháp nghiên cứu - Phương pháp bố trí thí nghiệmThí nghiệm gồm 3 yếu tố: Phân đạm, lân và phân
hữu cơ trong đó 4 mức đạm: 250; 320; 390 và 460 kg N/ha; 3 mức lân: 100; 150 và 200 kg P2O5/ha và 2 mức phân hữu cơ: 0 và 10 tấn/ha. Các nghiệm thức được nhắc lại 3 lần. Thí nghiệm được bố trí theo kiểu Lô phụ của lô phụ (Split-Split-Plot design). Diện tich ô nhỏ là 100 m2, tương đương với 9 cây cà phê.
- Các chỉ tiêu theo dõiThu hoạch toàn bộ quả tươi của từng ô thí nghiệm
cân năng suất quả tươi/ô (kg) (T1).Tính năng suất quả tươi/ha (tấn) của từng
nghiệm thức.+ Năng suất (kg) quả tươi/ô thí nghiệm thực thu
chia cho tổng số cây/ô được trung bình năng suất quả tươi/cây (kg).
+ Năng suất bình quân quả tươi/cây ˟ số cây/ha (1100 cây) được năng suất tươi/ha (tấn).
1 Trung tâm Nghiên cứu Đất, Phân bón và Môi trường phía Nam, Viện Thổ nhưỡng Nông hóa
ẢNH HƯỞNG CỦA PHÂN KHOÁNG N, P VÀ PHÂN HỮU CƠ ĐẾN NĂNG SUẤT CÀ PHÊ VỐI (Coffea canephora Pierre)
TRÊN ĐẤT ĐỎ BAZAN VÙNG CAO NGUYÊN DI LINH, TỈNH LÂM ĐỒNGLâm Văn Hà1
TÓM TẮTNghiên cứu ảnh hưởng của phân khoáng N, P và phân hữu cơ đến năng suất cà phê vối (Coffea Canephora Pierre)
được tiến hành trên đất đỏ bazan vùng cao nguyên Di Linh, tỉnh Lâm Đồng từ năm 2012 đến 2015. Thí nghiệm được tiến hành với 4 mức đạm (250, 320, 390 và 460 kg N/ha), 3 mức lân (100, 150, 200 kg P2O5/ha) và 2 mức phân hữu cơ (0 và 10 tấn phân chuồng/ha) với tổng số là 24 nghiệm thức được bố trí theo kiểu Split - Split - Plot, mỗi nghiệm thức được nhắc lại 3 lần. Vườn thí nghiệm với cà phê vối ghép giống cao sản, năng suất bình quân 4,7 tấn/ha. Năng suất cà phê được theo dõi ở năm thứ 2, 3 và 4 của thí nghiệm. Kết quả cho thấy, việc bón phân khoáng N và phân hữu cơ ảnh hưởng đến năng suất cà phê một cách có ý nghĩa ở mức 95%. Với mức bón 10 tấn phân chuồng + 320 kg N - 100 kg P2O5 - 350 kg K2O (ha/năm) cho năng suất cao nhất.
Từ khóa: Phân khoáng N, P; phân hữu cơ; năng suất cà phê vối

27
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
- Tính tỷ lệ quả tươi/nhân khô: + Phơi khô toàn bộ năng suất quả tươi của ô thí
nghiệm đạt độ ẩm 13%, tiến hành tách vỏ và cân lại nhân được năng suất nhân/ô (kg) thí nghiệm (T2).
+ Tính tỷ lệ quả tươi/nhân = T1 (kg)/T2 (kg).+ Năng suất nhân/ô thí nghiệm thực thu chia
cho tổng số cây/ô được trung bình năng suất nhân/cây (kg).
+ Năng suất bình quân nhân/cây ˟ số cây/ha (1100 cây) được năng suất nhân/ha (tấn).
2.3. Thời gian và đia điểm nghiên cứu Vườn thí nghiệm có tọa độ N 11o41’55,3’’, E
108o10’15,6’’ ở xã Đan Phượng, huyện Lâm Hà, tỉnh Lâm Đồng từ năm 2012 đến 2015. Năng suất cà phê được theo dõi từ năm 2013 đến 2015.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Ảnh hưởng của phân hữu cơ đến năng suất cà phê vối sau 4 năm bón liên tiếp
Ảnh hưởng của phân hữu cơ đến năng suất (NS) cà phê: Hình 1, 2 cho thấy sự khác biệt về tỷ lệ quả tươi/nhân, NS quả tươi/ô và NS nhân/ô giữa nghiệm thức có bón và không bón phân hữu cơ qua mùa vụ (2013, 2014 và 2015) là có ý nghĩa thống kê với mức 95%.
Hình 1. Ảnh hưởng của phân hữu cơ đến tỷ lệ quả tươi/nhân
Khi bón phân chuồng với lượng 10 tấn/ha/năm
tỷ lệ quả tươi/nhân của mùa vụ năm 2013 là giảm 2%, mùa vụ năm 2014 giảm 4% và mùa vụ năm 2015 giảm 6% so với không bón phân chuồng. Chứng tỏ bón phân hữu cơ đã làm cho tỷ lệ tươi/nhân giảm ngược lại khi canh tác không có phân hữu cơ thì làm cho tỷ lệ tươi/nhân tăng dần qua các năm.
Qua hình 2 cho thấy, khi bón phân chuồng với lượng 10 tấn/ha/năm đã tăng NS quả tươi/ô trong năm 2013 lên 12%, 2014 là 12% và 2015 là 14% so vơi không bón phân chuồng. Điều này chứng tỏ bón phân hữu cơ đã làm tăng NS quả cà phê tươi. So sánh NS quả tươi/ô giữa các mùa vụ của năm 2013, 2014 và 2015 cho thấy, việc thường xuyên bón phân hữu cơ đã làm cho NS cà phê tươi được duy trì tương đối ổn định so với không bón phân hữu cơ. Khác so với thực tế hiện nay nông dân canh tác cà phê hoàn toàn dùng phân hóa học thì NS không ổn định qua các năm.
Hình 2 cho thấy, khi bón 10 tấn phân chuồng/ha/năm đã làm tăng NS nhân/ô trong năm 2013 là 12%, năm 2014 là 14% và năm 2015 là 18% so với không bón phân chuồng. Điều này chứng tỏ bón phân chuồng đã làm tăng NS cà phê nhân. So sánh NS nhân/ô giữa các mùa vụ của năm 2013, 2014 và 2015 cho thấy, việc thường xuyên bón phân hữu cơ đã làm cho NS cà phê nhân không những được duy trì mà ngày càng tăng lên.
Khi bón 10 tấn phân chuồng/ha/năm qua mùa vụ (2013, 2014 và 2015) đều cho NS quả tươi/ô cũng như NS nhân/ô tăng so với không bón phân chuồng. Sự chênh lệch về NS quả tươi/ô cũng như nhân/ô giữa các mùa vụ (2013, 2014 và 2015) khi có bón phân chuồng là không đáng kể so với không bón phân chuồng. Ở các nghiệm thức không bón phân chuồng NS quả tươi/ha cũng như NS nhân/ha giảm dần qua các mùa vụ. Điều này chứng tỏ rằng canh tác cà phê không có phân hữu cơ sẽ làm cho NS ngày càng giảm đồng nghĩa với sự kém bền vững của vườn cà phê trong quá trình thâm canh. Vì vậy, để thực hiện thâm canh cà phê bền vững thì việc bón bổ sung phân hữu cơ là không thể thiếu.
LSD 5%: 0.13
Tỷ lệ quả tươi/nhânHCO (n = 36) 0 tấn PC/ha
Tỷ lệ quả tươi/nhân(2013)
Tỷ lệ quả tươi/nhân(2014)
Tỷ lệ quả tươi/nhân(2015)
4.74.6
4.8
4.5
4.9
4.4
HC1 (n = 36) 10 tấn PC/ha
LSD 5%: 0.20LSD 5%: 0.15
LSD 5%: 6.7 LSD 5%: 8.1 LSD 5%: 7.8
Năng suất quả tươi kg/ôHCO (n = 36) 0 tấn PC/ha
154.5
194.5
154.9
194.9
152
195.3
NS quả tươi kg/ô(2013)
NS quả tươi kg/ô(2014)
NS quả tươi kg/ô(2015)
HC1 (n = 36) 10 tấn PC/ha
LSD 5%: 1.6LSD 5%: 1.9LSD 5%: 1.7
Năng suất nhân kg/ô
32.9
42.3
32.3
43.3
31
44.4
NS nhân kg/ô(2013)
NS nhân kg/ô(2014)
NS nhân kg/ô(2015)
HCO (n = 36) 0 tấn PC/ha HC1 (n = 36) 10 tấn PC/ha
Hình 2. Ảnh hưởng của phân hữu cơ đến năng suất cà phê (quả tươi/ô và nhân/ô)

28
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
3.2. Ảnh hưởng của phân đạm đến năng suất cà phê - Ảnh hưởng của các mức phân N đến tỷ lệ quả
tươi/nhân: Hình 3 cho thấy, tỷ lệ quả tươi/nhân qua các mùa vụ (2013, 2014 và 2015) ở các mức N khác nhau là có sự khác biệt có ý nghĩa thống kê với mức 95%.
Hình 3. Ảnh hưởng của các mức phân N đến tỷ lệ quả tươi/nhân.
Biến thiên các mức phân N với tỷ lệ quả tươi/nhân qua các mùa vụ: Năm 2013 ở mức N1 là 4,6 và giảm ở mức N2 là 4,4 sau đó tăng dần ở mức N3 là 4,8 và tăng cao nhất ở mức N4 là 4,9; Năm 2014 ở mức N1 là 4,6 giảm với mức N2 là 4,4 và tăng cao nhất ở mức N4 là 4,9; Năm 2015 ở mức N1 là 4,5 giảm ở mức N2 là 4,3 và tăng cao nhất mức N4 là 5,0. So sánh ảnh hưởng của các mức phân N đến tỷ lệ quả tươi/nhân qua các mùa vụ 2013, 2014 không có sự khác biệt nhưng đến mùa vụ 2015 tỷ lệ này thấp ở các mức N1,2,3 và cao ở mức N4. Vậy có thể thấy việc bón phân N ở mức cao trong thời gian dài sẽ làm cho tỷ lệ quả tươi/nhân tăng, mà tỷ lệ này càng nhỏ thì năng suất nhân/ha càng cao. Điều này cũng có thể khẳng định rằng bón phân N hợp lý cho cà phê vối ghép giống cao sản sẽ làm tăng trọng lượng nhân trên quả tươi.
- Ảnh hưởng các mức phân N đến năng suất quả tươi/ô và năng suất nhân/ô.
Bảng 1 cho thấy, ảnh hưởng các mức phân N đến NS quả tươi/ô và NS nhân/ô qua mùa vụ (2013, 2014 và 2015) là có sự khác biệt với ý nghĩa thống kê ở mức 95%.
Phân tích biến thiên các mức phân N đến NS quả tươi/ô qua các mùa vụ (2013, 2014 và 2015) ở hình 4 cho thấy: Mùa vụ năm 2013, 2014 (năm thứ 2 và 3 của thí nghiệm) tăng dần từ mức N1 và đạt cao nhất ở mức N4 (thứ tự 21%, 25%, 27%, 27%). Đều này chứng tỏ bón phân N đã làm tăng trọng lượng quả cà phê tươi. Lý giải cho vấn đề này, có lẻ khi bón phân N với lượng cao, cây hấp thu nhiều NH4
+ gây dư thừa trong tế bào lúc này cây phải hấp thu nhiều nước và tích nước trong tế bào để chống ngộ độc NH4
+ dẫn đến tăng trọng lượng quả tươi. Nhưng sang mùa vụ năm 2015 (năm thứ tư của thí nghiệm) NS quả tươi/ô tăng dần từ mức N1 đến mức N3 và giảm ở mức N4 (tương ứng là 21%, 26%, 27%, 26%). Qua đây cho thấy, ở mùa vụ năm 2015, NS quả tươi/ô ở mức N1 không có sự khác biệt lớn so với mùa vụ
năm 2013 và 2014; ở mức N2 thì NS tăng cao hơn so với mùa vụ năm 2013 và 2014; ở mức mức N3 và N4 giảm đi rõ rệt so với mùa vụ 2013 và 2014. Chứng tỏ bón phân N với liều lượng cao cho cà phê vối giống cao sản trên đất đỏ bazan giai đoạn đầu sẽ làm cho NS quả tươi tăng nhưng về lâu dài là giảm.
Phân tích biến thiên các mức phân N đến NS nhân/ô qua các mùa vụ (2013, 2014 và 2015) ở hình 3 cho thấy, NS đều tăng từ mức N1 đến mức N2 và giảm dần ở mức N3, thấp nhất ở mức N4. Đặc biệt ở mùa vụ năm 2015 ở mức N2 năng suất nhân/ô tăng và giảm ở mức N3, N4 so với mùa vụ năm 2013, 2014 rõ rệt. Kết quả này cũng phù hợp với nghiên cứu của Y Kanin Hdơk và Trình Công Tư (2005), mức bón N lần lượt (200: 300: 400: 500: 600) kg/ha/năm thì năng suất nhân tấn/ha lần lượt (3,0: 3,5: 3,4: 3,3: 3,2). Melke and Ittana (2015), tại Tây Nam Ethiopia NS cà phê tăng dần với mức đạm bón vào và đạt cao nhất ở mức bón 300 kg N/ha/năm sau đó giảm với mức bón cao hơn 450 kg N/ha/năm.
Tỷ lệ quả tươi/nhân 2014
N1 = 250 N/ha(n = 18)
N2 = 320 N/ha(n = 18)
N2 = 320 N/ha(n = 18)
N4 = 460 N/ha(n = 18)
5.2
5
4.8
4.6
4.4
4.2
4
3.8
4.64.5
4.44.3
4.84.7
54.9
Tỷ lệ quả tươi/nhân 2015
Tỷ lệ quả tươi/nhân 2013
Bảng 1. Ảnh hưởng của các mức phân N đến năng suất quả tươi/ô và năng suất nhân/ô.
Ghi chú ý: NS là năng suất.
NT N1 = 250 N/ha (n=18)
N2 = 320 N/ha (n= 18)
N3 = 390 N/ha (n= 18)
N4 = 460 N/ha (n= 18) LSD 5%
NS quả tươi/ô (kg) 2013 147,7 176,3 190,7 191,3 14,4NS quả tươi/ô (kg) 2014 144,9 175,1 185,8 188,7 14,5NS quả tươi/ô (kg) 2015 146,7 178,7 183,0 182,5 14,9
NS nhân/ô (kg) 2013 32,1 40,4 39,7 38,8 3,39NS nhân/ô (kg) 2014 31,5 39,8 38,7 38,5 3,60NS nhân/ô (kg) 2015 32,6 41,9 38,5 36,5 3,85

29
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Hình 4. Ảnh hưởng các mức phân N đến NS quả tươi/ô và NS nhân/ô qua các mùa vụ (2013, 2014 và 2015)
Hình 4 cho thấy, khi bón N tăng thì NS tươi/ô tăng nhưng ngược lại năng suất nhân/ô giảm. Vậy bón tăng dần lượng phân N từ 28% đến 84% đã làm cho NS quả tươi cũng như năng suất nhân của giống cà phê vối cao sản TS1 tăng ở mùa vụ năm 2013, 2014 và 2015. Nhưng về NS nhân khi bón tăng dần
lượng phân N lên 56% và 84% so với mức N1 ở mùa vụ năm 2013, 2014 và 2015 là giảm. Qua đây chứng tỏ năng suất cà phê ổn định khi bón lượng phân N ở mức cân đối (320 kg/ha/năm) và sẽ không ổn định có su hướng giảm dần khi bón N ở liều lượng cao.
N1 = 250 N/ha(n = 18)
N2 = 320 N/ha(n = 18)
N3 = 390 N/ha (n = 18)
N4 = 460 N/ha(n = 18)
NS quả tươi/ô (kg) 2013 NS quả tươi/ô (kg) 2014
NS quả tươi/ô (kg) 2015
N1 = 250 N/ha(n = 18)
N2 = 320 N/ha(n = 18)
N3 = 390 N/ha (n = 18)
N4 = 460 N/ha(n = 18)
NS nhân/ô (kg) 2013 NS nhân/ô (kg) 2014
NS nhân/ô (kg) 2015
3.3. Ảnh hưởng phân lân đến năng suất cà phê vối trên đất đỏ bazan
Ảnh hưởng của các liều lượng phân lân đến tỷ lệ quả tươi/nhân qua các mùa vụ năm (2013, 2014 và 2015) ở bảng 2 cho thấy, là có khác nhau tỷ lệ này thấp nhất ở mức bón lân P1 sau đó tăng dần và cao nhất ở mức bón lân P3. Khi bón tăng lượng phân lân trong mùa vụ 2013, 2014 (năm thứ 2, 3 của thí nghiệm) làm tăng tỷ lệ quả tươi/nhân có ý nghĩa thống kê ở mức 95%, nhưng mùa vụ năm 2015 sự khác biệt không có ý nghĩa.
Phân tích biến thiên các liệu lượng phân lân đến NS quả tươi/ô thí nghiệm: Bảng 3 cho thấy, NS quả tươi/ô và NS nhân/ô ở các mùa vụ (2013, 2014 và 2015) đạt cao nhất ở mức P2 và giảm dần ở mức lân P1và P3. Nhưng sự khác biệt này không có ý nghĩa thống kê với mức 95%. Kết quả này khác với nghiên cứu của De Lima Dias (2015), ở Brazin năng
suất cà phê Arabica tăng dần với mức bón từ 0 - 600 kg P2O5/ha. Nhưng phù hợp với nghiên cứu của Y Kanin Hdơk, Trình Công Tư (2005), trên đất đỏ bazan Tây Nguyên NS cà phê vối tăng dần với mức bón từ 0 - 150 kg P2O5/ha và sau đó không tăng khi tăng mức bón P2O5.
Lý giải cho vấn đề này, có lẽ trên nền đất đỏ bazan giàu lân tổng số việc bón lân trong canh tác cà phê vối chỉ cần với mức 100 kg P2O5/ha/năm đã đáp ứng đủ nhu cầu lân của cây và duy trì độ phì về lân trong đất. Theo Trương Hồng và cộng tác viên (2016), trên nền đất đỏ bazan Tây Nguyên khi không bón lân 1 đến 2 vụ không ảnh hưởng đến NS cà phê so với bón đầy đủ NPK và vẫn duy trì hàm lượng lân dễ tiêu trong đất (>3 mg/100 g đất) đảm bảo nhu cầu của cây cà phê. Theo quan điểm của J. N. Wintgens (2014), lân ít có ý nghĩa đến NS cà phê trừ khi bón với số lượng lớn.
Bảng 2. Ảnh hưởng các liều lượng phân lân đến năng suất (NS) cà phê vối
Ghi chú: TL là tỷ lệ
NT P1 = 100 P2O5/ha (n=24)
P2 = 150 P2O5/ha (n=24)
P3 = 200P2O5/ha(n=24) LSD 5%
TL tươi/nhân 2013 4,5 4,6 4,7 0,15TL tươi/nhân 2014 4,6 4,7 4,8 0,18TL tươi/nhân 2015 4,6 4,7 4,7 NSNS quả tươi/ô 2013 172,2 175,5 175,8 NSNS quả tươi/ô 2014 173,1 175,8 174,2 NSNS quả tươi/ô 2015 171,7 174,4 174,3 NS
NS nhân/ô 2013 37,8 37,4 37,3 NSNS nhân/ô 2014 37,7 37,6 36,7 NSNS nhân/ô 2015 37,4 37,6 37,1 NS

30
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
3.4. Ảnh hưởng tương hỗ của phân đạm, lân và phân hữu cơ đến năng suất cà phê vối trên đất đỏ bazan sau 4 năm liên tiếp bón phân
Phân tích hỗ tương của phân bón N, P và phân hữu cơ đến NS cà phê trên nền đất đỏ bazan ở cao nguyên Di Linh, Lâm Đồng: Bảng 3 cho thấy, tỷ lệ quả tươi/nhân ở các nghiệm thức mùa vụ (2013, 2014 và 2015) là khác nhau có ý nghĩa thống kê với mức 95%. Mùa vụ năm 2013 tỷ lệ này thấp nhất ở nghiệm thức NT14 (320 N - 100 P2O5 - 350 K2O + 10 tấn phân chuồng/ha) là 4,2 và cao nhất ở nghiệm thức NT4 (460 N - 100 P2O5 - 350 K2O + 0,0 tấn phân chuồng/ha), NT20 (460 N - 150 P2O5 - 350 K2O + 10 tấn phân chuồng/ha), NT24 (460 N - 200 P2O5 - 350 K2O + 10 tấn phân chuồng/ha) là 5,0. Mùa vụ năm 2014 tỷ lệ tươi/nhân thấp ở nghiệm thức NT14, 18, 22 nhìn chung các nghiệm thức này đều có hàm lượng N và phân chuồng là giống nhau chỉ khác ở hàm lượng lân (320 N - 350 K2O + 10 tấn phân chuồng/ha) và cao ở các nghiệm thức NT4, 12, đều có hàm lượng đạm giống nhau chỉ khác ở liều lượng lân (460 N - 350 K2O + 0 tấn phân chuồng/ha/năm). Mùa vụ năm 2015 tỷ lệ tươi/nhân cao ở nghiệm thức NT4, 12 đều có hàm lượng N giống nhau chỉ khác ở hàm lượng lân và thấp ở các nghiệm thức NT14 (320 N - 100 P2O5 - 350 K2O + 10 tấn phân chuồng/ha) là 4,2. Qua đây cho thấy, liều lượng phân N và phân chuồng có ảnh hưởng lớn đến tỷ lệ quả tươi/nhân bất kể liều lượng phân lân. Cùng một giống cà phê và kỹ thuật canh tác như nhau thì tỷ lệ quả tươi/nhân phụ thuộc nhiều vào chế độ dinh dưỡng mà trong đó quan trọng là hàm lượng và tỷ lệ N, P, K. Tỷ lệ quả tươi/nhân ở các mùa vụ (2013, 2014 và 2015) có sự biến thiên không lớn giữa các nghiệm thức, nhìn chung tỷ lệ này thấp ở các nghiệm thức bón kết hợp giữa phân khoáng với phân hữu cơ (NT13 đến NT24) và cao ở các nghiệm thức chỉ bón phân khoáng không có phân hữu cơ (NT1 đến NT12), sự khác biệt này thể hiện rõ nhất ở năm thứ tư của thí nghiệm (2015). Liều lượng phân N càng cao và không có phân chuồng bổ sung trong lâu dài sẽ làm cho tỷ lệ quả tươi/nhân tăng lên.
Kết quả này cũng phù hợp với nghiên cứu của Lê Hồng Lịch (2008) là bón (300 kg N + 100 kg P2O5 + 300 kg K2O/ha/năm - tỷ lệ quả tươi/nhân là 4,36) và Nguyễn Tiến Sĩ (2009) bón (320 kg N + 90 kg P2O5 + 300 kg K2O/ha/năm thì tỷ lệ quả tươi/nhân là 4,4). Nhưng khi tăng hàm lượng N lên thì tỷ lệ này cũng tăng lên và khi bón phối hợp giữa phân khoáng NPK với phân hữu cơ thì tỷ lệ này giảm xuống. Xét tương quan giữa trọng lượng nhân của cà phê với tỷ lệ quả tươi/nhân với hệ số r = 0,67, p <0,05 là mối tương tác chặt.
Xét ảnh hưởng phân khoáng N, P và phân hữu cơ đến NS quả tươi/ô thí nghiệm: Bảng 2 cho thấy, ảnh hưởng của các liều lượng phân khoáng N, P và phân hữu cơ đến NS quả tươi/ô ở các mùa vụ (2013, 2014 và 2015) của các nghiệm thức là có sự khác biệt với ý nghĩa thống kê ở mức 95%. Điều này chứng tỏ liều lượng phân khoáng N, P và phân hữu cơ có ảnh hưởng lớn đến NS tươi của cà phê. Năng suất thay đổi theo liều lượng bón phối hợp giữa 3 yếu tố đạm, lân và phân chuồng ở mùa vụ 2013 cao nhất ở nghiệm thức NT20 (460 N - 200 P2O5 - 350 K2O + 10 tấn phân chuồng/ha) là 217,9 kg quả tươi/ô; 26,63 tấn quả tươi/ha và thấp nhất ở nghiệm thức NT1 (250 N - 100 P2O5 - 350 K2O + 0,0 tấn phân chuồng/ha) là 124,4 kg/ô; 15,20 tấn/ha; Mùa vụ 2014 NS quả tươi/ô cũng như NS quả tươi/ha cao nhất ở nghiệm thức NT20 (460 N - 150 P2O5 - 350 K2O + 10 tấn phân chuồng/ha) là 212,1 kg và thấp nhất ở nghiệm thức NT1 (250 N - 100 P2O5 - 350 K2O + 0,0 tấn phân chuồng/ha) là 132,8 kg; Mùa vụ 2015 năng suất quả tươi/ô cũng như NS quả tươi/ha cao nhất ở nghiệm thức NT24 (460 N - 200 P2O5 - 350 K2O + 10 tấn phân chuồng/ha) là 210,6 kg và thấp nhất ở nghiệm thức NT1 (250 N - 100 P2O5 - 350 K2O + 0,0 tấn phân chuồng/ha) là 124,0 kg. Nhìn chung NS quả tươi/ô cũng như NS quả tươi/ha ở các nghiệm thức bón kết hợp phân khoáng N, P với phân chuồng (NT13 đến NT24) đều cao hơn so với các nghiệm thức chỉ bón phân khoáng N, P (NT1 đến NT 12). So sánh NS quả tươi/ô cũng như quả tươi/ha qua các mùa vụ 2013, 2014 và 2015 ở các nghiệm thức chỉ bón phân khoáng N, P giảm dần, ở các nghiệm thức bón phân khoáng N, P kết hợp với phân chuồng có xu hướng tăng.
Xét ảnh hưởng của phân khoáng N, P và phân hữu cơ đến NS cà phê nhân/ô thí nghiệm và NS cà phê nhân/ha (2013, 2014 và 2015): Đây là sản phẩm cuối cùng đêm lại giá trị thặng dư cho người nông dân, nếu NS cao, chất lượng tốt cộng với đầu tư hợp lý sẽ mang lại hiệu quả kinh tế cho người sản xuất. Bảng 2 cho thấy, ảnh hưởng của các liều lượng phân khoáng N, P và phân hữu cơ đến NS cà phê nhân/ô cũng như nhân/ha qua các mùa vụ (2013, 2014 và 2015) là có sự khác biệt với có ý nghĩa thống kê ở mức 95%.
Mùa vụ 2013 năng suất cà phê nhân/ô cũng như nhân/ha thấp nhất ở nghiệm thức NT1 (250 N - 100 P2O5 - 350 K2O + 0,0 tấn phân chuồng/ha/năm) và cao nhất ở nghiệm thức TN14 (320 N - 100 P2O5 - 350 K2O + 10 tấn phân chuồng/ha/năm); Mùa vụ 2014 năng suất cà phê nhân/ô cũng như nhân/ha thấp nhất ở nghiệm thức NT1 (250 N - 100 P2O5 -

31
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 3. Hỗ tương giữa liều lượng phân khoáng N, P và phân hữu cơ đến các yếu tố cấu thành năng suất và năng suất cà phê vối
NTNăng suất quả tươi/ô (kg) Năng suất nhân/ô (kg) Tỷ lệ quả tươi/nhân2013 2014 2015 2013 2014 2015 2013 2014 2015
NT1 124,4 132,8 124,0 27,6 28,9 27,6 4,5 4,6 4,5NT2 155,1 157,3 152,8 36,1 35,8 34,7 4,3 4,4 4,4NT3 161,6 160,3 159,3 33,7 34,1 32,5 4,8 4,7 4,9NT4 163,3 161,4 158,5 32,7 32,3 31,1 5,0 5,0 5,1NT5 130,7 136,7 129,9 27,8 29,1 27,6 4,7 4,7 4,7NT6 161,5 159,2 157,8 36,7 35,4 35,9 4,4 4,5 4,4NT7 167,1 162,5 162,7 34,8 33,2 33,2 4,8 4,9 4,9NT8 160,5 163,4 161,5 32,8 33,3 32,3 4,9 4,9 5,0NT9 132,4 143,5 124,5 28,2 29,9 27,1 4,7 4,8 4,6
NT10 162,4 159,0 158,4 36,1 34,6 34,4 4,5 4,6 4,6NT11 166,3 160,8 165,9 34,6 32,8 33,2 4,8 4,9 5,0NT12 168,8 162,3 160,2 34,4 32,5 31,4 4,9 5,0 5,1NT13 163,0 159,1 168,3 37,9 37,0 38,3 4,3 4,3 4,4NT14 193,3 197,1 199,7 46,0 45,8 47,5 4,2 4,3 4,2NT15 207,9 205,1 200,3 45,2 43,6 44,5 4,6 4,7 4,5NT16 209,2 211,6 208,5 43,6 43,2 44,4 4,8 4,9 4,7NT17 160,9 166,5 170,5 35,8 36,2 37,9 4,5 4,6 4,5NT18 193,6 197,9 205,1 44,0 46,0 47,7 4,4 4,3 4,3NT19 211,7 208,4 205,6 45,0 44,3 44,7 4,7 4,7 4,6NT20 217,9 212,1 199,3 43,6 44,2 42,4 5,0 4,8 4,7NT21 163,1 167,6 167,9 35,5 36,4 37,3 4,6 4,6 4,5NT22 192,1 195,6 198,5 44,7 45,5 46,2 4,3 4,3 4,3NT23 206,7 199,5 204,4 43,1 41,6 43,5 4,8 4,8 4,7NT24 214,5 205,5 210,6 42,9 41,9 43,9 5,0 4,9 4,8ĐC 168,3 164,6 158,9 35,8 33,6 33,1 4,7 4,9 4,8
LSD0,05 10,2 14,0 6,4 2,6 3,5 1,3 0,24 0,5 0,1CV(%) 12,4 12,5 13,0 13,6 14,5 15,5 5,3 5,8 5,5
350 K2O + 0,0 tấn phân chuồng/ha/năm) và cao nhất ở nghiệm thức TN18 (320 N - 150 P2O5 - 350 K2O + 10 tấn phân chuồng/ha/năm); Mùa vụ 2015 năng suất cà phê nhân/ô cũng như nhân/ha thấp nhất ở nghiệm thức NT9 (250 N - 200 P2O5 - 350 K2O + 0,0 tấn phân chuồng/ha/năm) và cao nhất ở nghiệm thức NT18 (320 N - 150 P2O5 - 350 K2O + 10 tấn phân chuồng/ha/năm).
Nhìn chung NS cà phê ở các nghiệm thức bón kết hợp giữa phân khoáng và phần chuồng (NT13 đến NT24) cao hơn so với các nghiệm thức chỉ bón phân khoáng không có phân chuồng (NT1 đến NT12) và so với đối chứng. Chứng tỏ phân hữu cơ đã ảnh hưởng lớn đến NS cà phê nhân. Ảnh hưởng của việc bón phân khoáng kết hợp với phân hữu cơ thể hiện
rõ nhất ở năm thứ tư của thí nghiệm (2015), các nghiệm thức chỉ bón phân khoáng không có phân hữu cơ NS giảm kể cả các nghiệm thức được cho là bón cân đối NPK (NT2) so với năm 2013 và 2014. Ngược lại các nghiệm thức bón phân khoáng kết hợp với phân chuồng thì năng suất cà phê nhân tăng hơn so với mùa vụ (2013 và 2014). Điều này chứng tỏ bón phân khoáng kết hợp với phân hữu cơ không chỉ duy trì mà còn cải thiện NS trong thâm canh cà phê. Kết quả này cũng phù hợp với nghiên cứu của Hoàng Ngọc Thuận và cộng tác viên ctv (2014), bón vùi 3,5 tấn vỏ cà phê + men vi sinh kết hợp với 300kg N - 100 kg P2O5 - 350 kg K2O cho năng suất cà phê nhân tăng 14% so với công thức chỉ bón NPK.

32
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
IV. KẾT LUẬN VÀ KIẾN NGHỊ
4.1. Kết luậnQua 4 năm liên tiếp thí nghiệm bón phân cho
cà phê vối giống ghép cao sản TS1 trên nền đất đỏ bazan cho thấy, ảnh hưởng của các liều lượng phân N (250 kg; 320 kg; 390 kg và 460 kg) với 10 tấn phân chuồng/ha/năm đến năng suất cà phê là rất lớn bất kể liều lượng phân lân.
Việc bón cân đối liều lượng phân NPK (320 N - 100 P2O5 - 350 K2O kết hợp với 10 tấn phân chuồng ha/năm) cho năng suất cà phê nhân là tối ưu so với các nghiệm thức còn lại.
4.2. Kiến nghiThực hiện bón phân cân đối, bón kết hợp giữa
phân khoáng với phân hữu cơ trong thâm canh cà phê vối là điều cần thiết.
TÀI LIỆU THAM KHẢO Cục Thống kê tỉnh Lâm Đông. Niên giám thống kê Lâm
Đồng 2014.Lâm Văn Hà, 2016. Nghiên cứu thực trạng sử dụng
phân đạm, lâm, kali và lưu huỳnh cho cà phê vối ở tỉnh Lâm Đồng. Tạp chí Nông nghiệp và PTNT, 287 (8): 29-34.
Trương Hông, Trinh Xuân Hông, Nguyên Văn Bộ, Nguyên Văn Phương, Võ Chí Cường, Nguyên Đình Thoảng, 2016. Nghiên cứu hiệu lực trực tiếp, tồn dư và cộng dồn của phân lân trên cây cà phê vối. Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam, 2 (63): 55-57.
Y Kanin H’Dơk và Trình Công Tư, 2005. Nghiên cứu hiệu quả của phân N, P, K đối với cà phê vối kinh doanh trên đất bazan Đắk Lắk. Tạp chí Nông nghiệp và Phát triển Nông thôn, Nhà xuất bản Nông nghiệp.
Lê Hông Lich, 2008. Nghiên cứu sử dụng phân lân hợp lý cho cây cà phê vối giai đoạn kinh doanh trên đất bazan ở Đắk Lắk. Luận án tiến sĩ Nông nghiệp, Trường Đại học Nông nghiệp Hà Nội.
Nguyên Tiến Sĩ, 2009. Nghiên cứu một số tính chất cơ bản của đất phát triển trên đá bazan phục vụ thâm canh cà phê tỉnh Đắk Nông. Luận án tiến sĩ Nông nghiệp, Trường Đại học Nông nghiệp Hà Nội.
Hoàng Ngọc Thuận, Trần Thi Tâm, Đào Trọng Hùng, Nguyên Thi Ngọc Mai, Phạm Thi Nhung, Nguyên Thi Nhiệm, Nguyên Hữu Thành, Vũ Dương Quỳnh và Hô Công Trực, 2014. Nghiên cứu ảnh hưởng việc vùi phụ phẩm nông nghiệp đến tăng năng suất cây trồng, giảm lượng phân khoáng và cải thiện tính chất của đất. Kết quả Nghiên cứu Khoa học và Chuyển giao Công nghệ, quyển 6. Viện Thổ nhưỡng Nông hóa, tr 209 - 222.
Abayneh Melke1 and Fisseha Ittana, 2015. Nutritional Requirement and Management of Arabica Coffee (Coffea arabica L.) in Ethiopia. National and Global Perspectives, American Journal of Experimental Agriculture, 5(5).
De Lima Dias, Neto, Guimarães, Reis, & de Oliveira, 2015. Coffee yield and phosphate nutrition provided to plants by various phosphorus sources and levels. Ciênc. Agrotec, Lavras, Vol.39, No.2: 110-120.
Wintgens J. N., 2014. Coffee: Growing, Processing sustainable production; Wiley-VCH.
Effects of inorganic fertilizer (N, P) and organic fertilizers on coffee Coffea canephora yield in basalt soil, Lam Dong province
Lam Van HaAbstractThe study on the effect of N and P fertilizers and organic fertilizer on Robusta coffee yield was conducted in basaltic red soil in Di Linh plateau of Lam Dong province from 2012 to 2015. The experiment was conducted with four nitrogen doses (250, 320, 390 and 460 kg N/ha), three phosphorus doses (100, 150, 200 kg P2O5/ha) and two levels of organic fertilizer (0 and 10 manure tons/ha) and was designed in a Split-Split-Plot with treatments, three replications. The experiment was conducted in the garden of an intensive high-yielding 15 years-old Robusta coffee (with an average yield of 4.7 tons/ha). Coffee yield was monitored in year 2, 3 and 4 of the experiment. The results showed that N fertilizer and organic fertilizer affected the yield significantly at 95%. The fertilizer application was 10 tons of manure - 320 kg N - 100 P2O5 kg -350 kg K2O (ha/year) gave the highest yield.Key words: Mineral fertilizer N, P; compost; robusta coffee yield
Ngày nhận bài: 9/6/2017Ngày phản biện: 18/6/2017
Người phản biện: TS. Đặng Bá ĐànNgày duyệt đăng: 25/6/2017

33
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀLâm Đồng có 158.944 ha cà phê (Cục Thống kê
tỉnh Lâm Đồng, 2016). Đây là loại cây trồng có nhu cầu dinh dưỡng cao, bao gồm cả các yếu tố đa, trung và vi lượng. Trước đây năng suất còn thấp (trung bình khoảng 2 tấn/ha), các chất khoáng trung và vi lượng được cung cấp chủ yếu bởi đất hoặc một lượng nhỏ thông qua các loại phân bón (hữu cơ, hỗn hợp khoáng) cũng đủ đáp ứng cho nhu cầu của cây. Hiện nay, năng suất cà phê đã tăng cao, nhiều vườn đạt 4 - 5 tấn/ha, tạo nên áp lực cung cấp các chất dinh dưỡng từ đất đặc biệt là các yếu tố trung và vi lượng do không được bổ sung thường xuyên. Tại một số vườn, sau vài năm cho năng suất cao liên tục đã biểu hiện thiếu dinh dưỡng trung và vi lượng.
Polysulphate (Poly) chứa 4 yếu tố dinh dưỡng chính gồm: Kali, canxi, magiê và lưu huỳnh, với hàm lượng các chất dễ tiêu tương ứng 14% K2O, 17% CaO, 6% MgO và 48% SO3, nếu được sử dụng hợp lý, không những có khả năng thay thế một phần kali mà còn bổ sung các yếu tố trung lượng thiết yếu cho cây trồng. Bài viết này đánh giá hiệu quả của phân Poly đối với cây cà phê trên đất nâu đỏ bazan tỉnh Lâm Đồng.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu- Cây cà phê vối 10 năm tuổi trồng thuần, không
che bóng- Phân: Poly (14% K2O, 48% SO3, 6% MgO, 17%
CaO); Urea (46% N); Lân nung chảy (15% P2O5); kali clorua KCl (60% K2O).
- Đất nâu đỏ bazan (Rhodic Ferralsols) với các
tính chất cơ bản như sau: Đất chua, pHKCl: 4,58; OM tổng số: 3,92%; N tổng số: 0,187%; P2O5 dễ tiêu: 6,2 mg/100 g; K2O dễ tiêu: 13,6 mg/100 g; Ca++: 3,5 meq/100 g; Mg++: 2,1 meq/100 g.
2.2. Phương pháp nghiên cứuThí nghiệm gồm 5 công thức tương ứng với các
tổ hợp Poly + KCl khác nhau (kg/ha): Công thức T1: 400 KCl (Đ/C); công thức T2: 200 Poly + 353,3 KCl; công thức T3: 400 Poly + 306,7 KCl; công thức T4: 600 Poly + 260,0 KCl; và công thức T5: 800 Poly + 213,3 KCl. Mỗi tổ hợp chứa 240 K2O. Nền: 259,5 N + 82,5 P2O5 (Bộ Nông nghiệp và PTNT, 2002). Các công thức thí nghiệm được sắp xếp theo kiểu khối ngẫu nhiên đầy đủ (RCB), nhắc lại 4 lần, diện tích ô cơ sở 90 m2 (10 cây cà phê). Phương pháp bón phân và các chăm sóc khác được thực hiện theo 10TCN: 478-2001. Lấy mẫu lá theo Loue’ (1958). Phân tích mẫu theo Viện Thổ nhưỡng Nông hóa (1998).
2.3. Thời gian và đia điểm nghiên cứuNghiên cứu được tiến hành trong 2 năm: 2015
và 2016, tại xã Lộc Thắng, huyện Bảo Lâm, tỉnh Lâm Đồng.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Ảnh hưởng của bón phân Poly đến tích lũy một số nguyên tố trong lá cà phê
Trong lá cà phê trưởng thành bình thường có: 2,8 - 3,5% N; 0,11 - 0,13% P và 2,0 - 2,2% K (Nguyễn Tri Chiêm, 1993). Ngưỡng thích hợp của các yếu tố trung lượng trong lá cà phê là: 0,12 - 0,17% S; 0,6 - 0,9% Ca và 0,20 - 0,26% Mg (Mavolta, 1990). Đối
1 Trung tâm Nghiên cứu Đất, Phân bón và Môi trường Tây Nguyên - Viện Thổ nhưỡng Nông hóa
ĐÁNH GIÁ HIỆU QUẢ CỦA PHÂN POLYSULPHATE ĐỐI VỚI CÀ PHÊ VỐI TRÊN ĐẤT NÂU ĐỎ BAZAN TỈNH LÂM ĐỒNG
Hồ Hữu Huân1, Trình Công Tư1
TÓM TẮTTỉnh Lâm Đồng có 158.944 ha cà phê. Đây là loại cây trồng có nhu cầu dinh dưỡng cao, bao gồm cả các yếu tố đa,
trung và vi lượng. Polysulphate (Poly) là loại phân bón mới chứa 4 yếu tố dinh dưỡng chính gồm kali, canxi, magiê và lưu huỳnh, với hàm lượng các chất dễ tiêu tương ứng 14% K2O, 17% CaO, 6% MgO và 48% SO3. Nhằm đánh giá hiệu lực của loại phân bón này đối với cây cà phê vối trên đất nâu đỏ bazan tỉnh Lâm Đồng, một thí nghiệm gổm 5 mức bón Poly khác nhau (0, 200, 400, 600 và 800 kg/ha) đã được thực hiện trong thời gian 2 năm: 2015 - 2016. Thí nghiệm được bố trí nhắc lại 3 lần theo kiểu khối ngẫu nhiên đầy đủ (RCBD), với kích thước ô cơ sở gồm 20 cây cà phê (180 m2). Kết quả nghiên cứu cho thấy bón Poly giúp ổn định hàm lượng các yếu tố S, Ca, Mg trong lá cà phê ở ngưỡng thích hợp, tăng cường phát triển cành và đốt dự trữ, giảm tỉ lệ rụng, tăng khối lượng và kích thước quả, giảm tỉ lệ tươi/nhân, cải thiện năng suất cà phê. Bón 200 - 800 kg Poly/ha làm tăng năng suất 0,17 - 0,37 tấn cà phê nhân /ha, tương ứng 4,7 - 10,2% so với đối chứng. Công thức cho năng suất và lợi nhuận cao nhất là 400 kg Poly/ha, với 3,95 tấn cà phê nhân/ha, tương ứng 75,0 triệu đồng tiền lãi/ha.
Từ khóa: Cà phê, đất nâu đỏ bazan, năng suất, Poly

34
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
3.2. Ảnh hưởng của bón phân Poly đến sinh trưởng, phát triển của cà phê
Cây cà phê vối có đặc điểm ra không lại hoa trên phần gỗ đã mang quả trong vụ trước. Do vậy các biện pháp tác động nhằm phát triển cành và đốt dự trữ cho vụ sau là khâu kỹ thuật đầu tư hết sức quan trọng, đảm bảo có được năng suất ổn định, tránh hiện tượng mất mùa cách năm. Kết quả thí nghiệm tại bảng 2 cho thấy, qua 6 tháng mùa mưa, cành dự trữ tăng thêm 28,5 - 34,3 cm chiều dài và 4,8 - 6,5 đốt. Trong đó, công thức không bón Poly (T1) chỉ tăng được 28,5 cm chiều dài cành và 4,8 đốt/cành. Bón phân Poly làm tăng khả năng phát triển cành, đốt của cây cà phê, thêm 30,7 - 34,3 cm chiều dài và 5,5 - 6,5 đốt/cành, cao hơn đối chứng 7,7 - 20,4% về chiều dài cành và 14,6 - 35,4% về số đốt. Nhìn chung, mức bón Poly càng cao thì khả năng tăng trưởng cành, đốt càng mạnh. Tuy nhiên, mức bón có ý nghĩa tăng chiều dài và số đốt/cành về mặt thống kê là 400 kg Poly/ha, bón nhiều hơn tuy có làm tăng các chỉ tiêu nói trên nhưng không đáng kể (Bảng 2).
Trong chu trình phát triển của cây cà phê, thường xảy ra hiện tượng rụng quả non vào đầu và giữa mùa mưa. Điều đó có thể do đặc điểm sinh lý của cây, mưa bão, va chạm với dụng cụ sản xuất, hoặc ảnh hưởng của chế độ nước, dinh dưỡng. Mức độ rụng quả cà phê trong thí nghiệm khá cao, biến động trong khoảng 28,1 - 37,1%.
Sử dụng phân Poly, cung cấp đồng thời các yếu tố K, Ca, Mg và S cho cây cà phê, có tác dụng hạn chế
được tình trạng rụng quả. Các công thức thí nghiệm bón 200 - 800 kg Poly/ha làm giảm 3,9 - 9,0% lượng quả rụng so với đối chứng (Bảng 3). Nghiên cứu của Lương Đức Loan và cộng tác viên (1997) cho thấy bón khuyết các yếu tố Ca, Mg, S đã làm tăng 14,9 - 18,9% lượng quả rụng ở cây cà phê (Bảng 3).
Bảng 2. Ảnh hưởng phân Poly đến tăng trưởng cành, đốt trong mùa mưa
Công thức không bón phân Poly có khối lượng trung bình của 100 quả là 92,4 g và thể tích 100 quả là 93,3 cm3. Khối lượng và thể tích 100 quả ở các công thức bón 200 - 800 kg Poly/ha biến động trong khoảng 92,8 - 94,1 g và 93,8 - 94,9 cm3. Như vậy, bón phân Poly có khả năng làm tăng khối lượng và kích thước quả cà phê, tuy nhiên mức tăng đó không đáng kể, chỉ với 0,4 - 1,8% về khối lượng quả và 0,5 - 1,7% về thể tích quả so với đối chứng, không có ý nghĩa về mặt thống kê (Bảng 4).
chiếu kết quả phân tích với ngưỡng trên cho thấy hàm lượng các nguyên tố N, P, K, S, Ca và Mg trong lá trước bón phân nằm ở mức cận thiếu. Đây là thời điểm cần thiết phải bổ sung dinh dưỡng cho cây. Sau bón phân, hàm lượng N, P, K trong lá đáp ứng được yêu cầu để cây cà phê sinh trưởng, phát triển bình thường. Chứng tỏ lượng phân đạm, lân và kali đề nghị của qui trình 10TCN478-2001 (259,5 N + 82,5 P2O5 + 240 K2O) khá thích hợp với cây cà phê vối giai đoạn kinh doanh trên đất nâu đỏ bazan.
Hàm lượng S, Ca và Mg trong lá tiếp tục giảm ở T1 sau bón phân, do cây cà phê không được cung cấp những yếu tố này. Công thức T2 có hàm lượng S, Ca và Mg trong lá cao hơn so với thời điểm trước bón phân, song vẫn dưới ngưỡng yêu cầu, chứng tỏ bón 200 kg Poly/ha chưa giải quyết đủ lượng S, Ca và Mg mà cây cà phê cần. Các công thức T3, T4 và T5 có hàm lượng S, Ca và Mg trong lá sau bón phân nằm trong ngưỡng tối thích, nhờ được cung cấp 400 - 800 kg Poly /ha (Bảng 1).
Bảng 1. Hàm lượng một số nguyên tố trong lá cà phê (%)
Yếu tốTrước bón phân Sau bón phân 20 ngày
T1 T2 T3 T4 T5 T1 T2 T3 T4 T5N 2,66 2,69 2,63 2,64 2,65 3,33 3,35 3,36 3,34 3,31P 0,10 0,10 0,09 0,10 0,09 0,12 0,12 0,13 0,12 0,13K 1,85 1,88 1,86 1,84 1,87 2,11 2,14 2,11 2,09 2,12Ca 0,53 0,55 0,54 0,56 0,51 0,51 0,59 0,64 0,69 0,72Mg 0,18 0,18 0,19 0,18 0,19 0,17 0,19 0,21 0,22 0,24S 0,11 0,10 0,10 0,11 0,10 0,10 0,11 0,12 0,13 0,15
Công thức
Poly (kg/ha)
Tăng trưởng dài cành Đốt tăng/cành
cm % Đốt %T1 (Đ/C) 0 28,5 100,0 4,8 100,0
T2 200 30,7 107,7 5,5 114,6T3 400 32,8 115,1 6,1 127,1T4 600 34,1 119,6 6,5 135,4T5 800 34,3 120,4 6,5 135,4
LSD0,05 1,94 0,53CV(%) 7,7 12,1
35
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 3. Ảnh hưởng phân Poly đến mức độ rụng quả ở cây cà phê
Bảng 4. Ảnh hưởng phân Poly đến trọng lượng và kích thước quả
3.3. Ảnh hưởng của bón phân Poly đến năng suất và hiệu quả sản xuất cà phê
Số liệu ở bảng 5 cho thấy: Công thức có tỉ lệ tươi/nhân cao nhất là đối chứng (không bón Poly), với 4,52 và thấp nhất là bón 800 kg Poly/ha, với 4,22. Giữa lượng Poly được bón và tỉ lệ tươi/nhân có mối quan hệ nghịch. Tuy vậy, về mặt thống kê, khả năng làm giảm tỉ lệ tươi/nhân dừng ở mức 400 kg Poly/ha, bón quá mức này tuy có làm giảm tỉ lệ tươi/nhân nhưng không đáng kể.
Năng suất cà phê giữa các công thức khảo nghiệm rất khác nhau, biến động 3,64 - 4,01 tấn/ha. Công thức có năng suất thấp nhất là T1 (không bón Poly), chỉ với 3,64 tấn nhân/ha. Các công thức T2, T3, T4, T5 có năng suất đạt 3,81- 4,01 tấn cà phê nhân/ha, cao hơn đối chứng không bón Poly 4,7 - 10,2%. Nhìn chung, mức bón Poly càng nhiều thì năng suất càng cao, do cây trồng được cung cấp thêm các yếu tố Ca, Mg, S từ phân Poly. Nghiên cứu trước đây của Bùi Huy Hiền và cộng tác viên (2005) cho thấy bón khuyết các nguyên tố trung lượng (Ca, Mg, S) làm giảm tốc độ sinh trưởng của cây cà phê và giảm 13,88 - 16,27% năng suất so với bón đầy đủ. Tuy vậy,
trong thí nghiệm này mức bón Poly có ý nghĩa làm tăng năng suất cà phê về mặt thống kê là 400 kg Poly/ha, đạt 3,95 tấn cà phê nhân/ha, bón nhiều hơn năng suất tăng không đáng kể.
Bảng 5. Ảnh hưởng phân Poly đến tỷ lệ tươi/nhân và năng suất cà phê
Lợi nhuận thu được từ các công thức thí nghiệm biến động 67,2 - 75,0 triệu đồng/ha. Các công thức có bón Poly cho lợi nhuận 71,5 - 75,0 triệu đồng/ha, cao hơn đối chứng 4,3 - 7,8 triệu đồng/ha.
Như vậy, việc sử dụng phân Poly cho cây cà phê vối giai đoạn kinh doanh trên đất nâu đỏ bazan tỉnh Lâm Đồng không những làm tăng năng suất mà còn có ý nghĩa cải thiện đáng kể hiệu quả kinh tế trong sản xuất. Công thức bón cho lợi nhuận cao nhất là 400 kg Poly/ha, lãi 75,0 triệu đồng/ha (Bảng 6).
Bảng 6. Hiệu quả kinh tế bón phân Poly cho cây cà phê (triệu đồng/ha)
IV. KẾT LUẬN VÀ ĐỀ NGHỊSử dụng phân bón Poly không chỉ có tác dụng
cung cấp K mà còn bổ sung các yếu tố S, Ca, Mg, giúp ổn định hàm lượng các chất dinh dưỡng trong lá cà phê ở ngưỡng cần thiết, tăng cường phát triển cành và đốt dự trữ, giảm tỉ lệ quả rụng, tăng khối lượng và kích thước quả, giảm tỉ lệ tươi/nhân, cải thiện năng suất cà phê. Bón 200 - 800 kg Poly/ha làm tăng năng suất cà phê 0,17 - 0,37 tấn/ha, tương ứng 4,7 - 10,2% so với đối chứng. Công thức cho năng suất và lợi nhuận cao nhất là bón 400 kg Poly/ha, đạt
Công thức Poly (kg/ha)
Tỷ lệ quả rụng (%)
T1 (Đ/C) 0 37,1T2 200 33,2T3 400 29,8T4 600 28,7T5 800 28,1
LSD0,05 1,96CV(%) 11,7
Công thức
Poly kg/ha)
Khối lương 100 quả
Thể tích 100 quả
g % cm3 %T1 (Đ/C) 0 92,4 100,0 93,3 100,0
T2 200 92,8 100,4 93,8 100,5T3 400 93,3 101,0 94,2 101,0T4 600 93,9 101,6 94,7 101,5T5 800 94,1 101,8 94,9 101,7
LSD0,05 1,54 0,90CV(%) 1,0 0,7
Công thức Poly (kg/ha)
Tỉ lệ tươi/nhân
Năng suất nhân
Tấn/ha %
T1 (Đ/C) 0 4,52 3,64 100,0T2 200 4,40 3,81 104,7T3 400 4,28 3,95 108,5T4 600 4,24 3,98 109,3T5 800 4,22 4,01 110,2
LSD0,05 0,115 0,114CV(%) 2,9 11,8
Công thức Poly (kg/ha)
Tổng thu
Tổng chi
Lơi nhuận
T1 (Đ/C) 0 145,6 78,4 67,2T2 200 152,4 80,9 71,5T3 400 158,0 83,0 75,0T4 600 159,2 84,4 74,8T5 800 160,4 85,7 74,7

36
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
3,95 tấn cà phê nhân/ha, tương ứng 75,0 triệu đồng tiền lãi/ha.
Cần tiến hành thêm các nghiên cứu đánh giá ảnh hưởng của Poly đến môi trường đất, nước, làm cơ sở phổ biến sử dụng loại phân bón mới này trong ngành cà phê tỉnh Lâm Đồng nói riêng, vùng Tây Nguyên nói chung.
TÀI LIỆU THAM KHẢO Nguyên Tri Chiêm, 1985. Chuẩn đoán nhu cầu dinh
dưỡng khoáng của cây cà phê để có cơ sở bón phân hợp lý. Kỷ yếu 10 năm nghiên cứu khoa học (1983 -1993). Viện Nghiên cứu Cà phê, trang: 298-311.
Cục Thống kê tỉnh Lâm Đông, 2015. Niên giám thống kê tỉnh Lâm Đồng năm 2014.
Bùi Huy Hiền, Nguyên Đình Thi, Trình Công Tư, 2005. Ảnh hưởng của các chất dinh dưỡng và hiệu lực của các dạng phân lân đối với cà phê vối trên đất đỏ bazan. Kết quả nghiên cứu khoa học Quyển 4. Viện Thổ nhưỡng Nông hóa, NXB Nông nghiệp, trang: 284-296.
Lương Đức Loan, Nguyên Thi Thúy, Bùi Tuấn, Trình Công Tư, 1997. Bón cân đối các yếu tố đa-trung-vi lượng cho cà phê kinh doanh trên đất bazan Tây Nguyên. Tạp chí Nông nghiệp và Công nghiệp Thực phẩm, (9): 379-381.
Viện Thổ nhưỡng Nông hóa, 1998. Sổ tay phân tích đất, nước, phân bón, cây trồng. NXB Nông nghiệp. Hà Nội.
Loue’ A., 1958. The nutrition mineral of robusta coffee and fertilizer on the Ivory Coast.
Effects of polysulphate fertilizer on coffee production on basaltic soil in Lam Dong province
Ho Huu Huan, Trinh Cong TuAbstractLam Dong province has 158,944 hectare of coffee. This crop has a high demand for nutrient, including macro, meso and micro elements. Polysulphate (Poly) is introduced as a new 4-in-1 fertilizer which contains 48.0% SO3, 14.0% K2O, 6.0% MgO and 17.0% CaO. To assess the effect of Poly fertilizer on coffee growth and productivity, a field experiment with 5 treatments of Poly doses (0, 200, 400, 600 and 800 kg ha-1) and 3 replications on basaltic soil in Loc Thang commune, Bao Lam district, Lam Dong province during 2015 - 2016. The results showed that application of Poly increased the development of branch, reduced the rate of dropped fruit and improved the productivity by 0.17 - 0.37 t ha-1, corresponding 4.7 - 10.2% compared to the control treatment. The treatment which had the highest coffee yield (3.95 t ha-1) and economical effectiveness (benefit of 75.0 millions VND ha-1) was with 400 kg Poly ha-1.Key words: Coffee, basaltic soil, productivity, Poly
Ngày nhận bài: 9/6/2017Ngày phản biện: 21/6/2017
Người phản biện: TS. Đặng Bá ĐànNgày duyệt đăng: 25/6/2017
1 Trường Đại học Nông lâm Thái Nguyên2 Viện Khoa học kỹ thuật Nông lâm nghiệp miền núi phía Bắc
NGHIÊN CỨU ẢNH HƯỞNG CỦA MỘT SỐ LOẠI PHÂN HỮU CƠ VI SINH ĐẾN SINH TRƯỞNG, PHÁT TRIỂN GIỐNG CHÈ SHAN TẠI THUẬN CHÂU, SƠN LA
Dương Trung Dũng1, Trần Xuân Hoàng2
TÓM TẮTDiện tích trồng chè Shan ở huyện Thuận Châu, Sơn La ngày càng giảm, nguyên nhân do chưa có quy trình canh
tác hợp lý nên năng suất và chất lượng kém trong nhiều năm gần đây. Tuy nhiên, đến nay vẫn chưa có một nghiên cứu nào về quy trình sản xuất chè Shan theo hướng vừa bảo vệ đất vừa tăng năng suất, chất lượng chè tại huyện Thuận Châu, Sơn La. Để khắc phục tình trạng trên các tác giả đã nghiên cứu ảnh hưởng của một số loại phân hữu cơ vi sinh đến sinh trưởng, phát triển giống chè Shan tại đây. Kết quả cho thấy: Khi bón bổ sung phân hữu cơ vi sinh đã tăng số lứa hái đạt 6,7 - 8 lứa/năm, thời gian trung bình giữa 2 lứa hái dao động từ 28,7 - 36 ngày, mật độ búp từ 506,22 - 536,44 búp/m2, khối lượng búp đạt 0,42 - 0,51 g/búp, năng suất đạt 2,73 - 3,13 tấn/ha. Mặt khác, còn giúp cải thiện đất, tăng độ xốp và hàm lượng mùn trong đất, công thức NTT có độ xốp đạt cao nhất 67,26 - 67,63% và là công thức bón phân tốt nhất.
Từ khóa: Phân hữu cơ vi sinh, chè Shan, sinh trưởng, phát triển

37
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀSơn La là một tỉnh miền núi vùng Tây Bắc, điều
kiện thiên nhiên ưu đãi để phát triển chè đặc sản, trong phải kể đến cao nguyên Mộc Châu, Nà Sản, Thuận Châu và chè nơi đây đã trở thành thương hiệu không chỉ ở trong nước mà cả nước ngoài. Sản lượng chè búp tươi toàn tỉnh đạt 25.722 tấn/năm, năng suất bình quân toàn tỉnh 6,8 tấn/ha, trong đó huyện Thuận Châu năng suất đạt 9,0 tấn/ha (Cục thống kê Sơn La, 2014). Trong những năm gần đây diện tích chè giảm dần, chuyển đổi sang trồng các cây hàng năm khác với lý do cây chè ít được đầu tư chăm sóc và chưa có qui trình nghiên cứu kỹ thuật phát triển nên năng suất thấp (Nguyễn Xuân An, 2006). Hơn nữa, từ trước tới nay chưa có nghiên cứu về quy trình sản xuất chè Shan theo hướng vừa bảo vệ đất vừa tăng năng suất, chất lượng chè tại huyện Thuận Châu, Sơn La.
Xu thế hiện nay, các nhà khoa học đều cho rằng bón phân cho cây trồng nói chung, cây chè nói riêng đều dựa trên nguyên tắc: “Duy trì độ phì sẵn có trong đất là giải pháp dễ dàng và đỡ tốn kém hơn là khôi phục độ phì của đất do hậu quả của việc bón không hợp lý trong thời gian dài”. Đối với đất trồng chè giai đoạn giảm mùn nhiều nhất là 4 - 5 năm trồng mới, do vậy bón phân hữu cơ vi sinh là một biện pháp tốt để bảo vệ đất trồng chè vì phân hữu cơ vi sinh làm tăng hàm lượng mùn trong đất, cải thiện tính chất vật lý đất, mùn lại làm tăng cường hoạt động sinh vật đất, kích thích sự tăng trưởng của cây chè (Đỗ Văn Ngọc, 2006; Đặng Xuân Toàn, 2008; Lê Như Kiểu và ctv., 2011). Do vậy, “Nghiên cứu ảnh hưởng của một số loại phân hữu cơ vi sinh đến sinh trưởng, phát triển giống chè Shan tại Thuận Châu - Sơn La” là rất cần thiết trong giai đoạn hiện nay.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứuGiống chè Shan thời kỳ kinh doanh 5 tuổi; Các
loại phân hữu cơ vi sinh: Sông Gianh, NTT, Quế Lâm và Bình Điền; trong đó, phân bón Sông Gianh (Độ ẩm: 30%; Hữu cơ: 15%; P2O5: 1,5%; axit humic: 2,5%; Trung lượng: Ca, Mg, S; Các chủng vi sinh vật hữu ích: 3 ˟ 106 CFU/g). Phân bón NTT (mùn: 3,5%; N:P:K = 2,5:1:1 ; axit humic 6%; Vi sinh vật hữu hiệu 2 ˟ 106 Cfu/g. N-P2O5(hh)-K2O kết hợp). Phân bón Quế Lâm (thành phần: Hữu cơ = 23%, axit humic=3%, độ ẩm =20%, N:P:K=3:1:1). Phân bón Bình Điền (hữu cơ tinh khiết 18%, hàm lượng axit humic cao, NPK: 2-2-1+TE, nấm đối kháng Trichoderma, vi khuẩn đối kháng Pseudomonas putida).
2.2. Phương pháp nghiên cứu
2.2.1. Phương pháp bố trí thí nghiệm- Nội dung nghiên cứu gồm 5 công thức: CT1:
Bón phân theo quy trình (Nền); CT2: Nền + Phân vi sinh Sông Gianh: 3 tấn/ha; CT3: Nền + Phân hữu cơ vi sinh NTT: 3 tấn/ha; CT4: Nền + Phân hữu cơ vi sinh Quế Lâm: 3 tấn/ha; CT5: Nền + phân hữu cơ vi sinh Bình Điền: 3 tấn/ha.
- Thí nghiệm được bố trí trên nương chè Shan thời kỳ kinh doanh và theo kiểu khối ngẫu nhiên đầy đủ (RCBD), 3 lần nhắc lại. Diện tích mỗi ô thí nghiệm là 54 m2, tổng diện tích thí nghiệm của 2 địa điểm là 2.000m2 (cả bảo vệ). Quy trình nền thí nghiệm: 120 N + 90 P2O5 + 60 K2O bón sâu 6 - 8 cm, giữa hàng lấp kín. Thời gian bón N vào tháng 2, 4, 6, 8, đối với P2O5 bón tháng 2, K2O bón tháng 4. Bón 40 - 20 - 30 - 10 hoặc 40 - 30 - 30% N; 100% P2O5; 60 - 40% K2O.
2.2.2. Các chỉ tiêu theo dõi- Các chỉ tiêu sinh trưởng: Số lứa hái/năm, thời
gian trung bình 1 lứa hái.- Các yếu tố cấu thành năng suất: Mật độ búp
(búp/m2), khối lượng búp (g/búp), năng suất lý thuyết và năng suất thực thu (tấn/ha).
- Theo dõi một số sâu hại chính ở chè: (1) Điều tra mật độ rầy xanh: dùng khay kim loại có kích thước 25 ˟ 20 ˟ 5cm, dưới đáy tráng 1 lớp mỏng dầu, đặt khay dưới gầm, rìa tán chè nghiêng 45oC, so với thân cây, dùng tay đập mạnh trên tán chè 3 đập thẳng góc với khay, sau đó đếm số rầy trên khay. (2) Điều tra bọ trĩ (bọ cánh tơ): Mật độ bọ cánh tơ (hoặc bọ trĩ): điều tra vào buổi sáng. Hái 5 điểm chéo góc, mỗi điểm 20 búp cho vào túi PE đem về phòng đếm số bọ trĩ trên từng búp và phân cấp bị hại. (3) Điều tra bọ xít muỗi: điều tra theo 5 điểm đường chéo, mỗi điểm hái ngẫu nhiên bất kì 20 búp, cho vào túi PE về phòng đếm số búp có vết do bọ xít muỗi hại, tính tỉ lệ % búp bị hại.
- Các chỉ tiêu về lý, sinh học đất: (1) Độ xốp đất được tính bằng công thức sau P(%) = (1 – D/d) ˟ 100, trong đó: P là độ xốp của đất, D là dung trọng đất, d là tỷ trọng đất. (2) Hàm lượng mùn trong đất: Phân tích hàm lượng mùn bằng phương pháp Walkley - Black. (3) Giun đất: Mỗi công thức theo dõi 3 điểm theo đường chéo. Mỗi điểm chọn 1 m2
đất, sau đó dùng nước đổ ngập vào đất cho đến khi nước ngấm toàn bộ xuống đất, sau thời gian khoảng 4 giờ thu nhặt tất cả giun chui từ đất lên và đem cân khối lượng.

38
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
2.2.3. Phương pháp xử lý số liệuSố liệu được xử lý theo IRRISTAT 5.0 và Excel 2010.
2.3. Thời gian và đia điểm nghiên cứuNghiên cứu được thực hiện từ tháng 4/2015 đến
tháng 12/2016 tại xã Phỏng Lái, xã Chiềng Pha, huyện Thuận Châu, tỉnh Sơn La.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Ảnh hưởng của các công thức phân bón hữu cơ vi sinh đến số lứa hái và thời gian trung bình giữa 2 lứa hái chè Shan tại Thuận Châu
Kết quả nghiên cứu ảnh hưởng của các công thức phân bón hữu cơ vi sinh đến số lứa hái và thời gian trung bình giữa 2 lứa hái của giống chè Shan số liệu được trình bày ở bảng 1.
Số liệu ở bảng 1 cho thấy số lứa hái của các công thức CT2, CT3, CT4 và CT5 đạt 6,7 - 8 lứa/năm; ở xã Phỏng Lái và Chiềng Pha có số lứa hái tương đồng nhau. Sự sai khác giữa các công thức thí nghiệm có ý nghĩa ở mức tin cậy 95%. Trong đó, công thức CT3 (bón phân NTT) và CT4 (bón phân Quế Lâm) có số lứa hái cao nhất (8 lứa), công thức CT1 (đối chứng) là thấp nhất (6,7 lứa). Thời gian trung bình giữa 2 lứa hái dao động 28,7 - 36 ngày, công thức CT1 có thời gian dài nhất, công thức CT3 và CT4 có thời gian ngắn nhất và tương đồng xếp hạng với nhau.
Như vậy, việc bón bổ sung phân hữu cơ vi sinh đã ảnh hưởng đến việc rút ngắn thời gian giữa 2 lứa hái và tăng số lứa hái lên so với đối chứng (chỉ bón phân vô cơ). Trong đó CT3 và CT4 là 2 công thức tốt nhất và tương đồng nhau trong tác dụng đến tăng số lứa hái và rút ngắn thời gian giữa 2 lứa hái.
Bảng 1. Ảnh hưởng của phân hữu cơ vi sinh đến số lứa hái, thời gian trung bình giữa 2 lứa hái chè
(4/2015 - 12/2016)
Ghi chú: Bảng 1, 2, 3, 4: (1) PL: xã Phỏng Lái, CP: xã Chiềng Pha; (2) CT1: Bón phân theo quy trình (Nền); CT2: Nền + Phân vi sinh Sông Gianh: 3 tấn/ha; CT3: Nền + Phân hữu cơ vi sinh NTT: 3 tấn/ha; CT4: Nền + Phân hữu cơ vi sinh Quế Lâm: 3 tấn/ha; CT5: Nền + phân hữu cơ vi sinh Bình Điền: 3 tấn/ha; (3) Trong cùng một cột, các giá trị mang chữ cái khác nhau thì khác nhau có ý nghĩa ở mức α = 0,05.
3.2. Ảnh hưởng của phân bón hữu cơ vi sinh đến năng suất chè Shan tại Thuận Châu
Kết quả nghiên cứu ảnh hưởng của các công thức phân bón hữu cơ vi sinh đến năng suất và các yếu tố cấu thành năng suất chè Shan, số liệu được trình bày ở bảng 2.
Công thức
Số lứa hái/năm (lứa/năm)
Thời gian TB của 1 lứa hái (ngày)
Xã Phỏng
Lái
Xã Chiềng
Pha
Xã Phỏng
Lái
Xã Chiềng
Pha
CT1 - Đ/C 6,7b 6,7b 36,0a 35,7a
CT2 7,7a 7,7a 31,7ab 31,7ab
CT3 8,0a 8,0a 28,7b 28,7b
CT4 8,0a 8,0a 28,7b 29,3b
CT5 7,7a 7,7a 31,7ab 31,0ab
CV(%) 5,88 5,88 8,48 8,8
LSD0,05 0,84 0,84 5 5,16
Bảng 2. Ảnh hưởng của phân bón hữu cơ vi sinh đến năng suất và yếu tố cấu thành năng suất chè (4/2015 - 12/2016)
Ghi chú: NSLT: năng suất lý thuyết, NSTT: năng suất thực thu.
Công thứcMật độ búp
(búp/m2)Khối lương búp
(g/búp)NSLT
(tấn/ha)NSTT
(tấn/ha)
PL CP PL CP PL CP PL CP
CT1 - Đ/C 520,09d 506,22c 0,46 0,48 4,00c 3,77d 2,73c 2,60c
CT2 531,11b 514,40bc 0,50 0,46 4,24b 4,12bc 2,90b 2,87b
CT3 536,44a 530,76a 0,51 0,49 4,46a 4,28a 3,13a 3,03a
CT4 524,71c 517,96b 0,46 0,42 4,06c 3,98c 2,90b 2,80b
CT5 529,33b 522,58ab 0,49 0,45 4,33ab 4,16ab 2,83bc 2,73bc
P <0,05 <0,05 >0,05 >0,05 <0,05 <0,05 <0,05 <0,05
CV(%) 0,16 0,55 1,94 2,08 2,84 2,84
LSD0,05 3,19 10,63 0,15 0,16 0,15 0,15

39
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Số liệu ở bảng 2 cho thấy: Mật độ búp của các công thức CT2, CT3, CT4 và CT5 dao động từ 506,22 - 530,76 búp/m2 (Chiềng Pha), 520,09 - 536,44 búp/m2 (Phỏng Lái) đều cao hơn CT1 (đối chứng), trong đó công thức CT3 có mật độ búp cao nhất (530,76 - 536,44 búp/m2) ở cả hai địa điểm nghiên cứu. Tại Phỏng Lái mật độ búp của công thức CT2 và CT5 tương đương nhau, thấp hơn công thức CT3 và cao hơn công thức CT4, CT1 (đối chứng) ở mức tin cậy 99%. Tại Chiềng Pha mật độ búp ở các công thức CT2, CT3, CT4 và CT5 sai khác nhau có ý nghĩa ở mức tin cậy 95%. Như vậy, chứng tỏ ưu thế của phân NTT đối với cây chè cao hơn các loại phân khác. NSLT ở các công thức CT2, CT3, CT4 và CT5 đều cao hơn đối chứng, trong đó công thức CT3 (bón phân NTT) có năng suất lý thuyết đạt (4,28 - 4,46 tấn/ha) ở mức tin cậy 95%.
NSTT ở 2 điểm thí nghiệm có sự xếp hạng các giá trị trung bình của các công thức thí nghiệm giống nhau, theo đó bón phân NTT có năng suất cao nhất (3,13 và 3,03 tấn/ha), thứ 2 là bón phân Sông Gianh và Quế Lâm, tiếp là phân Bình Điền; công thức đối chứng có năng suất thấp nhất ở mức tin cậy 95%. Khối lượng búp của các công thức khi bổ sung phân hữu cơ vi sinh có khối lượng búp cao hơn là không bổ sung (đối chứng). Như vậy, việc bón phân hữu cơ vi sinh rất có ý nghĩa trong tăng năng suất, tăng mật độ búp chè và phân hữu cơ vi sinh NTT là phân có ưu thế nhất.
3.3. Ảnh hưởng của phân hữu cơ vi sinh đến sâu hại chè
Sau khi bón phân hữu cơ vi sinh cho chè Shan kinh doanh tại các địa điểm nghiên cứu xã Phỏng Lái và Chiềng Pha, kết quả điều tra theo dõi một số loại sâu hại chè chính, số liệu được trình bày ở bảng 3.
Bảng 3. Ảnh hưởng của phân bón đến tình hình sâu hại chính (4/2015 - 12/2016)
Số liệu ở bảng 3 cho thấy: Các công thức phân bón hữu cơ vi sinh tại xã Phỏng Lái đều có mật độ bọ trĩ thấp hơn so với đối chứng, trong đó công thức bón phân NTT có mật độ thấp nhất (3 con/búp) ở mức tin cậy 95%. Còn tại xã Chiềng Pha mật độ bọ trĩ ở các công thức thí nghiệm không sai khác nhau. Về mật độ rầy xanh tại xã Phỏng Lái và xã Chiềng Pha ở các công thức thí nghiệm có kết quả tương tự như bọ trĩ. Tỷ lệ bọ xít muỗi gây hại tại xã Phỏng Lái, xã Chiềng Pha ở các công thức phân bón hữu cơ vi sinh đều có tỷ lệ thấp hơn so với đối chứng ở mức tin cậy 95% và công thức phân NTT có tỷ lệ sâu gây hại thấp nhất. Mức độ các loài sâu hại tại xã Chiềng Pha thấp hơn xã Phỏng Lái, có thể thấy điều kiện khí hậu tại Phỏng Lái thuận lợi hơn cho sâu hại phát triển.
3.4. Ảnh hưởng của phân bón hữu cơ vi sinh đến tính chất lý và sinh học đất
Bón phân hữu cơ vi sinh đã làm ảnh hưởng đến các chỉ tiêu lý, sinh học đất là yếu tố giúp tạo nên môi trường đất phù hợp cho cây chè phát triển. Kết quả nghiên cứu ảnh hưởng của phân bón hữu cơ vi sinh đến tính chất lý tính, sinh học (giun đất), số liệu được thể hiện ở bảng 4 và bảng 5.
Bảng 4. Ảnh hưởng của phân hữu cơ vi sinh đến lượng giun đất (g/m2) (4/2015 - 12/2016)
Ghi chú: TTN: Trước thí nghiệm; STN: Sau thí nghiệm.
Số liệu ở bảng 4 cho thấy: Sau bón phân số lượng giun đất ở các công thức CT2, CT3, CT4 và CT5 tăng hơn trước thí nghiệm tương tự ở cả 2 địa điểm và không có sự sai khác, giá trị đạt 24,67 - 32,33 g/m2. Sau thí nghiệm số lượng giun đất ở công thức CT1 (đối chứng) giảm đi từ 24,67 - 29,67 g/m2 tương ứng ở 2 địa điểm. Công thức CT3 có số lượng giun cao nhất đạt 35,67 - 36,67 g/m2. Như vậy, việc bón phân hữu cơ vi sinh tạo nên môi trường đất thích hợp cho giun đất phát triển, trong đó phân NTT tạo môi trường đất thích hợp nhất cho giun đất sinh sống.
Công thức
Bọ trĩ (con/búp)
Rầy xanh (con/khay) Bọ xít muỗi
PL CP PL CP PL CPCT1 - Đ/C 3,78a 2,53 5,38a 4,71 3.51a 1.91a
CT2 3,36b 2,56 4,76b 4,67 2.84b 1.76ab
CT3 3,00c 2,44 4,44b 4,67 2.58b 1.42c
CT4 3,33b 2,42 4,8b 4,53 2.80b 1.69abc
CT5 3,33b 2,44 4,89ab 4,71 2.87b 1.49bc
P <0,05 >0,05 <0,05 >0,05 <0,01 <0,05CV(%) 5,08 5,6 6 10,62LSD0,05 0,32 0,51 0,33 0,33
Công thức
Xã Phỏng Lái Xã Chiềng PhaTTN STN TTN STN
CT1 - Đ/C 26,00 24,67b 30,67 29,67b
CT2 25,67 31,00a 32,33 35,67a
CT3 25,00 35,67a 31,67 36,67a
CT4 24,67 31,00a 32,33 34,33a
CT5 25,00 33,00a 31,33 32,67ab
P >0,05 <0,05 >0,05 <0,05CV(%) 8,14 7,17LSD0,05 4,76 4,56

40
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Số liệu bảng 5 cho thấy: Ở cả 2 địa điểm nghiên cứu thì xã Phỏng Lái có độ xốp và hàm lượng mùn tổng số sau thí nghiệm đều cao hơn xã Chiềng Pha ở tất cả các công thức thí nghiệm.
- Về chỉ tiêu độ xốp đất: Trước thí nghiệm độ xốp đất tại xã Chiềng Pha và Phỏng Lái của các công thức đạt 66,21 - 67,15%, và công thức 4 (phân Quế Lâm) đạt cao nhất (66,84 - 67,15%). Sau thí nghiệm tại xã Chiềng Pha và Phỏng Lái của các công thức thí nghiệm đạt 66,3 - 67,63%, và công thức 3 (phân NTT) đạt cao nhất (67,26 - 67,63%), mức tăng chênh lệch trước và sau thí nghiệm cũng cao nhất (0,46 - 0,52), tiếp đến lần lượt là CT4, CT2 và CT5. Như vậy có thể thấy việc bón phân hữu cơ vi sinh có tác dụng cải thiện độ xốp của đất.
- Về chỉ tiêu hàm lượng mùn tổng số: Trước thí nghiệm tại xã Chiềng Pha và Phỏng Lái đạt 3,63 - 3,95%, trong đó công thức 4 (phân Quế Lâm) tại Phỏng Lái có hàm lượng mùn cao nhất. Sau thí nghiệm tại xã Chiềng Pha và Phỏng Lái có hàm lượng mùn trong đất tăng lên từ 3,74 - 4,25%, trong đó công thức 3 (phân NTT) đạt cao nhất (4,19 - 4,25%), mức tăng chênh lệch so trước thí nghiệm cũng cao nhất (0,33 - 0,35); Tiếp đến lần lượt là CT2, CT5 và CT4; Công thức đối chứng có hàm lượng mùn và mức tăng chênh lệch trước và sau thí nghiệm thấp nhất ở cả 2 địa điểm nghiên cứu. Như vậy, việc bón phân hữu cơ vi sinh có tác dụng lớn trong việc tăng hàm lượng mùn trong đất.
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luận Bón bổ sung phân hữu cơ vi sinh đã tăng số lứa
hái đạt 6,7 - 8 lứa/năm, thời gian trung bình giữa
2 lứa hái từ 28,7 - 36 ngày, mật độ búp đạt 506,22 - 536,44 búp/m2, khối lượng búp đạt 0,42 - 0,51 g/búp, năng suất thực thu đạt 2,73 - 3,13 tấn/ha, trong đó phân NTT là loại phân tốt nhất giúp tăng năng suất chè tối ưu nhất và sâu hại đều gây hại ở mức nhẹ.
Các công thức phân hữu cơ vi sinh có số lượng giun đất đều tăng lên sau khi bón phân, công thức NTT có giá trị cao nhất 35,67 - 36,67 con/m2. Ngoài ra, còn cải thiện đất, tăng độ xốp và hàm lượng mùn trong đất, công thức NTT có độ xốp đạt cao nhất 67,26 - 67,63%.
4.2. Đề nghiĐề nghị áp dụng bón 3 tấn/ha phân hữu cơ NTT
cho vùng sản xuất chè Shan kinh doanh tại xã Phỏng Lái và Chiềng Pha, huyện Thuận Châu, tỉnh Sơn La.
TÀI LIỆU THAM KHẢONguyên Xuân An, 2006. Nghiên cứu khả năng nhân
giống một số giống chè mới bằng phương thức giâm cành và ghép tại Gia Lai - Đắk Lắk. Luận án TS Khoa học Nông nghiệp. ĐHNN1. Hà Nội.
Cục thống kê Sơn La, 2014. Niên giám thông kê tỉnh Sơn La. Nhà xuất bản thống kê.
Lê Như Kiểu, Nguyên Văn Huân, Lê Thi Thanh Thủy, 2011. Nghiên cứu khả năng tổ hợp của các chủng vi sinh vật để sản xuất phân hữu cơ vi sinh đa chức năng cho chè. Tạp chí Khoa học và Công nghệ Nông nghiệp Việt Nam, 47 (2): 133 – 138.
Đỗ Văn Ngọc, 2006. Cây chè Shan vùng cao một cây trồng có lợi thế phát triển ở vùng núi cao miền Bắc Việt Nam. Hội thảo nghiên cứu phát triển chè Shan. Hiệp hội chè Việt Nam. Hà Nội.
Đặng Xuân Toàn, 2008. Báo cáo kết quả khảo nghiệm phân bón qua lá Agro Dream trên cây chè, rau và lúa. Công ty Cổ phần kỹ thuật môi trường.
Ghi chú: TTN: Trước thí nghiệm; STN: Sau thí nghiệm.
Công thức
Độ xốp (%) Mùn tổng số (%)Xã Phỏng Lái Xã Chiềng Pha Xã Phỏng Lái Xã Chiềng Pha
TTN STN Chênh lệch TTN STN Chênh
lệch TTN STN Chênh lệch TTN STN Chênh
lệchCT1 - Đ/C 66,52 66,63 0,11 66,21 66,30 0,09 3,88 4,00 0,12 3,63 3,74 0,11CT2 66,70 67,11 0,41 66,39 66,76 0,37 3,91 4,19 0,28 3,79 4,06 0,27CT3 67,11 67,63 0,52 66,80 67,26 0,46 3,92 4,25 0,33 3,84 4,19 0,35CT4 67,15 67,56 0,41 66,84 67,19 0,35 3,95 4,19 0,24 3,75 3,97 0,22CT5 66,58 66,99 0,41 66,27 66,67 0,40 3,85 4,10 0,25 3,75 4,01 0,26

41
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Effect of microbial organic fertilizer types on growth and development of Shan tea variety in Thuan Chau, Son La
Duong Trung Dung, Tran Xuan HoangAbstractPlanting area of Shan tea in Thuan Chau district, Son La province has been decreasing due to inapropriate cultivation practices and therefore the yield and quality have beeen decreasing in recent years. However, there has not been any study on the procedures of Shan tea production toward protecting soil while increasing tea productivity and quality in Thuan Chau district, Son La province. To solve this situation, the authors studied the effect of microbial organic fertilizer types on growth and development of Shan tea variety in this area. The results showed that supplementing with microbial organic fertilizer, the number of litter picking reached from 6.7 - 8 litters per year, the average time interval between 2 litter picking ranged from 28.7 - 36 days, the density of buds reached 506.22 - 536.44 buds/m2, the weight of buds was recorded at 0.42 - 0.51 g/bud and the yield was of 2.73 - 3.13 tons/ha. On the other hand, applying microbial organic fertilizer could improve the soil as increasing porosity and humus content in the soil; NTT treatment had porosity from 67.26 - 67.63% and it was the best fertilizer treatment.Key words: Microbial organic fertilizer, Shan tea, growth, development
Ngày nhận bài: 6/7/2017Ngày phản biện: 14/7/2017
Người phản biện: PGS.TS. Lê Như KiểuNgày duyệt đăng: 27/7/2017
1 Khoa Công nghệ sinh học, Học viện Nông nghiệp Việt Nam 2 Viện Khoa học Nông nghiệp Việt Nam
NGHIÊN CỨU CHỦNG XẠ KHUẨN CÓ KHẢ NĂNG ĐỐI KHÁNG VỚI VI KHUẨN Erwinia carotovora GÂY BỆNH THỐI NHŨN
TRÊN MỘT SỐ LOẠI CÂY TRỒNGNguyễn Xuân Cảnh1, Nguyễn Thị Khánh1, Phạm Hồng Hiển2
TÓM TẮTNghiên cứu này được thực hiện với mục đích tuyển chọn, nghiên cứu đặc điểm sinh học của chủng xạ khuẩn có
khả năng đối kháng với vi khuẩn Erwinia carotovora gây bệnh thối nhũn trên một số loại cây trồng. Bằng phương pháp khuếch tán trên đĩa thạch đã sàng lọc và xác định được 05 chủng trong số 192 chủng xạ khuẩn có khả năng đối kháng với vi khuẩn Erwinia carotovora. Chủng L2.5 thể hiện hoạt tính kháng khuẩn rất mạnh với đường kính vòng kháng khuẩn đạt 23 mm. Nghiên cứu một số đặc điểm sinh học của chủng này cho thấy chủng L2.5 có khả năng tạo chuỗi bào tử dạng thẳng sau 03 ngày nuôi cấy, không sinh sắc tố tan trên môi trường ISP-6, sinh trưởng tốt ở ngưỡng nhiệt độ 300C- 35°C, pH trung tính và chịu được nồng độ muối thấp dưới 1%. Chủng L2.5 có khả năng sử dụng một số nguồn các bon và ni tơ khác nhau bao gồm sucrose, fructose, cellulose, raffinose, cao thịt bò, pepton và KNO3. Phân tích trình tự 16S rRNA cho thấy chủng L2.5 và chủng Streptomyces psammoticus KP1404 có độ tương đồng cao tới 99%. Kết hợp các đặc điểm hình thái, nuôi cấy, sinh lý, sinh hóa và phân tích sinh học phân tử đã xác định chủng xạ khuẩn L2.5 thuộc vào loài Streptomyces psammoticus.
Từ khóa: Erwinia carotovora, Streptomyces sp., bệnh thối nhũn, xạ khuẩn
I. ĐẶT VẤN ĐỀ Thối nhũn (Soft rot) là một trong số các bệnh
gây thiệt hại lớn cho ngành trồng trọt, bệnh này phổ biến trên toàn thế giới có thể xảy ra trên nhiều đối tượng cây trồng quan trọng như các loại cây họ cải (Brassicaceae), họ cà (Solanaceae) … và một số loại hoa cây cảnh. Triệu chứng của bệnh thối nhũn có thể bắt gặp trong hầu hết các giai đoạn của quá trình sản xuất bao gồm cả trong giai đoạn trên đồng
ruộng, trong vận chuyển, bảo quản và bày bán nông sản (Bhat et al., 2010). Ước tính thiệt hại do bệnh này gây ra vào khoảng 15 - 30% giá trị cây trồng mỗi năm (FAOSTAT data, 2012). Nguyên nhân của bệnh được xác định là do nhiều loài vi khuẩn khác nhau, trong đó Erwinia carotovora (E. carotovora) được xem là tác nhân chính và gây thiệt hại lớn hơn cả (Bhat et al, 2010; Prombelom et al., 2002). Hiện nay chưa có biện pháp nào thực sự hiệu quả để phòng trừ

42
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
bệnh này, mặc dù một số hóa chất đã được sử dụng tuy nhiên chúng cũng bị hạn chế do giá thành cao, ảnh hưởng đến sức khỏe con người và gây ô nhiễm môi trường. Việc sử dụng các vi sinh vật đối kháng để kiểm soát các tác nhân gây bệnh trên cây trồng trong đó có vi khuẩn Erwinia carotovora đang được nghiên cứu và ứng dụng rộng rãi (Abd-El-Khair et al., 2007; Doolotkeldieva et al., 2016; Tao et al., 2011; Xu et al., 1986). Do các vi sinh vật và sản phẩm tự nhiên của chúng là tiềm năng quan trọng cho kiểm soát sinh học bệnh cây mà không gây ảnh hưởng tới môi trường.
Xạ khuẩn (Actinomyces) là tác nhân được nghiên cứu và ứng dụng nhiều nhất trong kiểm soát sinh học (Mitra et al., 2008; Watve et al., 2001). Trên thế giới, các nghiên cứu về xạ khuẩn có khả năng đối kháng với vi khuẩn E.carotovora đã được công bố từ lâu (Kondo et al., 1975). Năm 2005 nhóm nghiên cứu của Zamanian đã xác định loài Streptomyces plicatus có khả năng đối kháng với vi khuẩn E.carotovora subsp.carotovora gây hại cây trồng trong điều kiện in vitro (Zamanian et al., 2005). Khi đã khảo sát hoạt tính của chất kháng sinh neomycin từ chủng Streptomyces fradiae HTP, nhóm nghiên cứu của Tao cũng đã ghi nhận chủng xạ khuẩn này có khả năng đối kháng với E.carotovora subsp. carotovora (Tao et al., 2011). Nghiên cứu này được thực hiện với mong muốn phát hiện và xác định được những chủng xạ khuẩn có hoạt tính kháng vi khuẩn E.carotovora trong điều kiện Việt Nam. Từ đó tìm kiếm các hoạt chất mới, an toàn, hiệu quả và thân thiện với môi trường để ứng dụng trong nông nghiệp nói chung và nghề trồng trọt nói riêng.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứuChủng vi khuẩn Erwinia carotovora subsp.
carotovora ATCC15713 được cung cấp bởi Viện Di truyền Nông nghiệp. Các chủng xạ khuẩn phân lập từ nhiều nguồn khác nhau được lưu trữ tại phòng thí nghiệm Bộ môn Công nghệ Vi sinh, Khoa Công nghệ Sinh học, Học viện Nông nghiệp Việt Nam.
2.2. Phương pháp nghiên cứu
2.2.1. Tuyển chọn các chủng xạ khuẩn có khả năng đối kháng vi khuẩn Erwinia carotovora
Việc sàng lọc và tuyển chọn các chủng xạ khuẩn được thực hiện bằng phương pháp khuếch tán trên đĩa thạch theo các phương pháp đã mô tả chi tiết trong các công bố trước đây (Nguyen et al., 2016;
Nguyễn Xuân Cảnh và ctv., 2016). Xạ khuẩn được cấy đều trên đĩa petri chứa môi trường Gause-1 ở 30oC. Sau 7 ngày nuôi cấy, thỏi thạch xạ khuẩn được cấy vào đĩa petri chứa môi trường MPA (Meat Peptone Agar) đã được cấy trải vi khuẩn, ủ ở 4oC
trong 2 giờ để các hoạt chất từ thỏi thạch khuếch tán vào môi trường, sau đó cho vào tủ nuôi. Đường kính vòng ức chế sinh trưởng được xác định sau một ngày nuôi cấy ở 30oC.
2.2.2. Nghiên cứu một số đặc điểm sinh học của chủng xạ khuẩn đã tuyển chọn (L2.5)
Các nghiên cứu nhằm xác định một số đặc điểm sinh học của chủng xạ khuẩn đã tuyển chọn tiếp tục được thực hiện bao gồm: Xác định hình thái, kích thước khuẩn lạc; Xác định hình thái chuỗi sinh bào tử và bề mặt bào tử; Kiểm tra khả năng sinh sắc tố melanin; Kiểm tra khả năng đồng hóa các nguồn các bon và ni tơ; Khảo sát ảnh hưởng của nhiệt độ, pH, nồng độ NaCl tới sinh trưởng và phát triển. Các nghiên cứu này được thực hiện theo các phương pháp đã mô tả chi tiết trong các công bố trước đây (Nguyen et al., 2016; Nguyễn Xuân Cảnh và ctv., 2016).
2.2.3. Định danh chủng xạ khuẩn L2.5 Các đặc điểm như hình thái khuẩn lạc, màu sắc
khuẩn ty cơ chất, khuẩn ty khí sinh, cuống sinh bào tử và bề mặt bào tử trên môi trường nuôi cấy. So sánh các đặc điểm này với các chủng xạ khuẩn đã biết trong hệ thống phân loại quốc tế (Internatinal Streptomyces Project, ISP) (Shirling and Gottlied, 1966). ADN từ chủng xạ khuẩn đã tuyển chọn được tách chiết theo phương pháp mô tả bởi Marmur (1961). Phản ứng PCR khuếch đại vùng bảo thủ của 16S rRNA với cặp mồi 27F và 1492R lần lượt có trình tự: 5’-AGAGTTTGATCCTGGCTCAG-3’ và 5’- ACGGCTACCTTGTTACGACTT-3’. Sản phẩm PCR được kiểm tra trên gel agarose 1% sau đó gửi đi đọc trình tự tại công ty 1tsBASE (Malaysia). Mức độ tương đồng về trình tự gen mã hóa 16S rRNA của chủng nghiên cứu được so sánh với các chủng đã công bố trên ngân hàng gen thế giới sử dụng công cụ tra cứu Blast (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Phần mềm MEGA6 được dùng để xây dựng cây xác định mối quan hệ di truyền.
2.3. Thời gian và đia điểm nghiên cứuNghiên cứu được thực hiện tại phòng thí nghiệm
Khoa Công nghệ sinh học, Học viện Nông nghiệp Việt Nam từ 2014 - 2016.

43
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Tuyển chọn chủng xạ khuẩn có khả năng đối kháng với vi khuẩn Erwinia carotovora
Trong quá trình nuôi cấy xạ khuẩn sẽ tiết vào môi trường một số các hợp chất có hoạt tính sinh học khác nhau, vì vậy phương pháp khuếch tán trên đĩa thạch được sử dụng để tuyển chọn và đánh giá hoạt tính kháng vi khuẩn của các chủng xạ khuẩn. Tiến hành nuôi cấy các chủng xạ khuẩn trên môi trường Gause-1, chủng vi khuẩn kiểm định Erwinia carotovora subsp. carotovora ATCC15713 được nuôi trên môi trường MPA, việc sàng lọc các chủng xạ khuẩn có hoạt tính được thực hiện như mô tả trong nội dung phương pháp. Sau quá trình sàng lọc, 05 trong số 192 chủng xạ khuẩn nghiên cứu được xác định có khả năng đối kháng với vi khuẩn Erwinia carotovora (Bảng 1).
Trong số 05 chủng xạ khuẩn có hoạt tính thì chủng L2.5 thể hiện khả năng đối kháng mạnh nhất với đường kính vòng vô khuẩn là 21 mm khi sử dụng thỏi thạch và 23 mm khi sử dụng giếng thạch (Hình 1). Một số công bố trên thế giới đã tìm ra các chủng xạ khuẩn có khả năng đối kháng với vi khuẩn Erwinia carotovora gây bệnh thối nhũn trên
cây trồng (Doolotkeldieva et al., 2016; Zamanian et al., 2005). So sánh với những kết quả trước đây, nhận thấy rằng 02 chủng L2.4 và L2.5 có hoạt tính tương đối mạnh. Hơn thế nữa trong một nghiên cứu trước đây, chủng L2.5 đã được xác định là có hoạt tính mạnh đối với nấm Rhzoctonia solani gây bệnh khô vằn trên lúa (Nguyễn Hoài Nam và ctv., 2015). Chính vì thế chủng xạ khuẩn L2.5 có khả năng sinh ra một số hoạt chất có hoạt tính sinh học khác nhau và có tiềm năng để phát triển ứng dụng.
Bảng 1. Hoạt tính kháng vi khuẩn Erwinia carotovora của một số chủng xạ khuẩn được xác định bằng
phương pháp khuếch tán trên đĩa thạch
Hình 1. Hoạt tính kháng vi khuẩn Erwinia carotovora của chủng xạ khuẩn L2.5 được xác định bằng phương pháp khuếch tán sử dụng thỏi thạch (A) và phương pháp khuếch tán sử dụng giếng thạch (B)
Chủng xạ khuẩn
Đường kính vòng vô khuẩn (D-d, mm)
Khuếch tán qua thỏi thạch
Khuếch tán qua giếng thạch
C4.1 11±2 5±2L2.4 17±2 14±2L2.5 21±2 23±2
L3.10 6±2 4±2X 12±2 10±2
A B
3.2. Đặc điểm sinh học của chủng xạ khuẩn L2.5
3.2.1. Đặc điểm hình tháiCăn cứ đầu tiên thường được sử dụng để nghiên
cứu đặc điểm sinh học và phân loại xạ khuẩn là các đặc điểm hình thái (Miyadoh et al., 2016). Chủng xạ khuẩn L2.5 được nuôi trên các môi trường khác nhau và quan sát các đặc điểm màu sắc, kích thước, hình dạng của khuẩn lạc. Trên môi trường Gause-1 khi nuôi ở 30°C sau 05 ngày khuẩn lạc chủng L2.5 có dạng tròn, bề mặt thô ráp, màu xám với viền
khuẩn lạc màu nhạt hơn. Màu sắc khuẩn lạc thay đổi theo thời gian nuôi cấy, ở 1-2 ngày nuôi cấy đầu khuẩn lạc màu trắng, khi nuôi cấy được 4 - 5 ngày thì khuẩn lạc chuyển sang màu nâu xám với đường kính khoảng 1,5 - 2 mm. Tiếp theo tiến hành các nghiên cứu xác định hình dạng cuống sinh bào tử, chuỗi bào tử và bề mặt bào tử của chủng L2.5. Chủng L2.5 được nuôi trên môi trường Gause-1 ở 30°C, tiến hành làm tiêu bản và quan sát sơ bộ hình thái sợi xạ khuẩn bằng kính hiển vi quang học ở độ phóng đại 1000 lần, kết quả cho thấy sau 72 h nuôi

44
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
cấy chủng xạ khuẩn L2.5 bắt đầu hình thành bào tử. Các bào tử được sắp xếp thành từng chuỗi thẳng và dài, trên mỗi chuỗi chính có sự phân nhánh. Sau 90 h nuôi cấy các bào tử bắt đầu phân cắt tách rời khỏi chuỗi và phát tán vào môi trường. Việc xác định chính xác hơn hình thái chuỗi bào tử và bề mặt bào tử là điều kiện quan trọng nhằm phân nhóm xạ khuẩn, tiếp tục quan sát hình thái chuỗi sinh bào tử và bề mặt bào tử của chủng xạ khuẩn L2.5 dưới kính
hiện vi điện tử quét (SEM). Ở độ phóng đại 2500 lần có thể dễ dàng quan sát thấy chuỗi bào tử dạng thẳng, trên mỗi chuỗi chính có khoảng 30-50 bào tử, ở một số chuỗi chính xuất hiện sự phân nhánh, các nhánh này cũng có dạng thẳng mỗi nhánh có khoảng 08-15 bào tử (Hình 2 A). Bào tử chủng L2.5 có dạng hình bầu dục, trơn, kích thước dao động từ 1,2 - 1,6 ˟ 6,0 - 7,0 µm (Hình 2 B). B
Hình 2. Hình thái chuỗi sinh bào tử và bề mặt bào tử của chủng L2.5 dưới kính hiển vi điện tử quét (SEM) ở độ phóng đại 2.500 lần (A) và 35.000 lần (B)
3.2.2. Khả năng hình thành sắc tố MelaninRất nhiều chủng xạ khuẩn được xác định là có khả
năng sinh sắc tố melanin trong quá trình nuôi cấy, vì thế đây cũng là một trong những tiêu chí trong việc phân nhóm xạ khuẩn (Shirling and Gottlied, 1966). Chủng L2.5 được nuôi cấy trên môi trường ISP-6 ở 30°C và quan sát sự thay đổi màu sắc môi trường trong 21 ngày nuôi cấy. Kết quả cho thấy sau 21 ngày nuôi cấy màu của môi trường không có sự thay đổi đáng kể, điều này chứng tỏ chủng L2.5 không có khả năng sinh sắc tố melanin.
3.2.3. Khả năng sử dụng các nguồn đường và ni tơ của chủng L2.5
Theo Shirling và Gottlieb (1966), khả năng đồng hóa và sử dụng các nguồn các bon khác nhau được đánh giá theo 04 mức. Chủng L2.5 được nuôi cấy trên môi trường ISP-9 có bổ sung các nguồn đường khác nhau sau đó kiểm tra khả năng phát triển của chủng này. Kết quả cho thấy chủng L2.5 có khả năng đồng hóa tốt các nguồn đường như sucrose, fructose, cellulose, raffinose nhưng không đồng hóa các nguồn đường D-xylose, L-arabinose và inositol (Bảng 2). Tiếp đó nghiên cứu xác định khả năng sử dụng các nguồn ni tơ khác nhau của chủng L2.5 cũng đã thực hiện. Chủng này được nuôi trên môi trường Starch Nitrate với nguồn NaNO3 được thay thế bằng các nguồn ni tơ khác nhau như mô tả trong phần phương pháp. Kết quả cho thấy chủng L2.5 có khả năng sử dụng nguồn ni tơ từ các nguồn khác nhau như cao thịt bò, pepton, KNO3 (Bảng 2). Kết quả khảo sát này
đã cung cấp thông tin quan trọng làm căn cứ để tiến hành phân loại xạ khuẩn theo hệ thống ISP, đồng thời cung cấp thông tin về dinh dưỡng của chủng L2.5 phục vụ quá trình lên men sau này.
Bảng 2. Khả năng sử dụng các nguồn cacbon và ni tơ khác nhau của chủng L2.5
Ghi chú: (++) Chủng L2.5 có khả năng sử dụng tốt nguồn các bon hoặc ni tơ; (+) Chủng L2.5 có khả năng sử dụng nguồn các bon hoặc ni tơ; (±) Không xác định được khả năng sử dụng nguồn các bon hoặc ni tơ cho chủng L2.5; (-) Chủng L2.5 không có khả năng sử dụng nguồn các bon hoặc ni tơ
Nguôn cacbon
Khả năng phát triển của chủng L2.5 sau 05 ngày nuôi cấy
Nguôn nitrogen
Khả năng phát triển của chủng L2.5 sau 05 ngày nuôi cấy
Sucrose + NaNO3 ++R-Hannose ± KNO3 ++
Cellulose ++ Cao thịt bò ++
Fructose ++ NH4Cl -L-arabinose - Pepton ++
Raffinose + (NH4)2SO4 -D-xylose - NH4NO3 ±Inositol -
D-manitol ±

45
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
3.2.3. Khả năng thích nghi với một số điều kiện môi trường của chủng L2.5
Điều kiện môi trường có ảnh hưởng rất lớn đến quá trình trao đổi chất và sinh trưởng của các loài vi sinh vật. Các loài vi sinh vật khác nhau sẽ có khả năng thích nghi với các điều kiện môi trường khác nhau. Nhằm mục đích cung cấp thông tin về điều kiện nuôi cấy của chủng L2.5 phục vụ các nghiên cứu sau này các nghiên cứu khảo sát các yếu tố môi trường đến sự sinh trưởng và phát triển của chủng xạ khuẩn này đã được thực hiện. Chủng xạ khuẩn L2.5 được nuôi trên trên môi trường Gause-1 ở các điều kiện nhiệt độ, pH và các nồng độ muối khác nhau. Khả năng sinh trưởng và phát triển của chủng này sau 05 ngày nuôi cấy đã được kiểm tra, kết quả được tổng hợp trong bảng 2. Kết quả cho thấy chủng xạ khuẩn L2.5 có khả năng năng phát triển tốt trong điều kiện nhiệt độ từ 30°C - 35°C, thích hợp với môi trường trung tính hoặc hơi kiềm với khoảng pH từ 6 - 8 và nồng độ NaCl 1%. So với các nghiên cứu trước đây, nhận thấy chủng L2.5 có khả năng thích ứng với môi trường tương đối kém đặc biệt là khả năng chịu muối, do vậy có thể xếp chủng xạ khuẩn này vào nhóm có khả năng chịu muối kém.
3.3. Đinh danh chủng xạ khuẩn L2.5Sử dụng phương pháp sinh học phân tử dựa trên
việc so sánh độ tương đồng của đoạn gen 16S rARN của chủng L2.5 với các chủng xạ khuẩn đã công bố trên ngân hàng gen để định danh chủng xạ khuẩn
này. ADN tổng số của chủng xạ khuẩn L2.5 được tách chiết theo phương pháp của Marmur (1961). Phản ứng PCR được thực hiện với cặp mồi 27F và 1492R nhằm khuếch đại đoạn gen 16S rARN của chủng L2.5. Sản phẩm PCR được kiểm tra trên gel agarose 1%, kết quả điện di cho thấy có 01 băng ADN kích thước khoảng 1500 bp phù hợp với kích thước lý thuyết có thể đạt được khi nhân gen bằng cặp mồi này (Hình 3).
Hình 3. Kết quả điện di sản phẩm PCR trên gel agarose 1%, marker 1kb
Sau khi kiểm tra sản phẩm PCR được gửi tới công ty 1stBASE (Malaysia) để tinh sạch và giải trình tự . Tiến hành so sánh trình tự thu được với các trình tự khác trên ngân hàng gen nhờ công cụ blast, xây dựng cây phân loại cho chủng L2.5 bằng phần mềm MEGA6. Kết quả được thể hiện ở hình 4.
Hình 4. Cây phân loại dựa trên trình tự 16S rARN của chủng xạ khuẩn L2.5
99
51
21
18
31
34
98
28
18
2165
99 l2.5S. psammoticus KP1404
S. aureofaciens
S. sayamaensis NBRC
S. aureofaciens xsd08158S. aureofaciens NBRC13178
S. kaniharaensis NBRC13790
S. aureofaciens NBRC12843
S. psammoticus CSSP729
99
S. aureofaciens NBRC3187
S. aureofaciens NBRC12594
S. rimosus MJM8796
S. aburaviensis NBRC12830
S. avellaneus xsd08159
S. aburaviensis CSSP531
S. aureofaciens NBRC13183
S. aburaviensis S-66
S. aureofaciens NBRC3712
Marker
1500 pb
L2.5

46
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Kết quả xây dựng cây phả hệ dựa trên trình tự gen 16S rRNA cho thấy chủng L2.5 nằm ở cùng nhánh với chủng Streptomyces psammoticus KP1404 với giá trị bootstrap 99. Giá trị bootstrap này nằm khoảng trong tin cậy, cùng với kết quả căn trình tự nucleotide cho thấy mức tương đồng của 2 trình tự 16S rRNA của chủng L2.5 với Streptomyces psammoticus KP1404 là 99%, hoàn toàn đảm bảo mức tin cậy về mối quan hệ loài. Vì vậy có thể khẳng định chủng L2.5 là Streptomyces psammoticus và được đặt tên là Streptomyces psammoticus L2.5.
IV. KẾT LUẬNTừ 192 chủng xạ khuẩn phân lập đã xác định
được 05 chủng có khả năng đối kháng lại vi khuẩn Erwinia carotovora gây bệnh thối nhũn trên cây trồng, trong đó chủng L2.5 có hoạt tính mạnh nhất.
Đã thực hiện nghiên cứu các đặc điểm sinh học của chủng xạ khuẩn L2.5 bao gồm đặc điểm về hình thái, nuôi cấy, sinh lý và sinh hóa.
Sử dụng phương pháp sinh học phân tử để định danh chủng xạ khuẩn L2.5 cho thấy chủng xạ khuẩn này là loài là Streptomyces psammoticus.
TÀI LIỆU THAM KHẢONguyên Xuân Cảnh, Hô Tú Cường, Nguyên Thi
Đinh, Phạm Thi Hiếu, 2016. Nghiên cứu chủng xạ khuẩn có khả năng đối kháng với vi khuẩn Vibrio parahaemolyticus gây bệnh trên tôm. Tạp chí Khoa học Nông nghiệp Việt Nam 14(11): 1809-1816.
Nguyên Hoài Nam, Nguyên Minh Trang, Đặng Phú Hoàng, Nguyên Văn Hùng, Nguyên Xuân Cảnh, Tống Văn Hải, Nguyên Đức Bách, 2015. Sàng lọc xạ khuẩn Actinomycestes sp. Có khả năng đối kháng với nấm gây bệnh khô vằn lúa Rhzoctonia solani. Tạp chí Khoa học và Phát triển 13(8): 1474-1480.
Abd-El-Khair, H. and Karima, H.E.H., 2007. Application of Some Bactericides and Bioagents for Controlling the Soft Rot Disease in Potato. Research Journal of Agricultural and Biological Science, 3: 463-473.
Bhat K.A., Masoodi S.D., Bhat N.A., Ahmad M., Zargar M.Y., Mir S.A., and Ashraf B. M., 2010. Studies on the Effect of Temperature on the Development of Soft Rot of Cabbage (Brassica oleracea var. Capitata) Caused by Erwinia carotovora sub Sp. Carotovora. J Phytol Vol. 2; 64-67.
Doolotkeldieva, T., Bobusheva, S. and Suleymankisi, A., 2016. Biological Control of Erwinia carotovora
ssp. carotovora by Streptomyces Species. Advances in Microbiology 6: 104-114.
FAOSTAT data, 2012. The Top 5 Potato Producing Countries; Potato World, Commodity report.
Kondo H., Honke T., Hasegawa R., Shimoda T., Nakamura S., 1975. Isolation of maltotetraose from Streptomyces as an antibiotic against Erwinia carotovora. Journal of Antibiotic (Tokyo) 28(2): 157-60.
Miyadoh S., Tsuchizaki N., Ishikawa J., Hotta K., 2016. Digital Atlas of Actinomycete. The Society for Actinomycetes Japan, Asakura Co.
Mitra, A., S.C. Santra and J. Mukharjee., 2008. Distribution of actinomycetes, the antagonistic behavior and the Physio-chemical characteristic of the world’s lagest tidal mangrove forest. Applied Microbial Biotechnology 80: 685 - 695.
Nguyen X.C., Phan Thi T.T., Tran T.T.H., 2016. Isolation and identification of an actinomycete strain with biocontrol effect against Xanthomonas oryzae pv. oryzae causing bacterial blight disease in rice. Vietnam Journal of Agriculture Science 14(10): 1564 -1572.
Shirling, E.B. & Gottlieb D., 1966. Methods for characterization of Streptomyces species. International Journal of Systematic Bacteriology 16, 313 - 340.
Perombelon M.C.M., van der Wolf J.M., 2002. Methods for the detection and quantification of Erwinia carotovora subsp. atroseptica (Pectobacterium carotovorum subsp. atrosepticum) on potatoes:a laboratory manual. Scottish Crop Research Institute Occasional Publication No.10, Revised Version 2002.
Tao K., Fan J., Shi G., Zhang X., Zhao H., Hou T., 2011. In vivo and in vitro antibacterial activity of neomycin against plant pathogenic bacteria. Academic Journals, 6(34): 6829-6834.
Xu, G.W. and Gross, D.C., 1986. Selection of Fluorescent Pseudomonads Antagonistic to Erwinia carotovora and Suppressive of Potato Seed Piece Decay. Phytopathology 76: 414-422.
Watve M. G., Tickoo R., Jog M. M., Bhole B. D., 2001. How many antibiotics are produced by the genus Streptomyces? Archives of Microbiology, 176(5): 386 - 390.
Zamanian S., Shahidi Bonjar G.H., Saadoun I., 2005. First report of antibacterial properties of a new (strain 101) against Erwinia carotovora from Iran. Biotechnology, 4: 114-120.

47
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Characterization of actinomyces strain with bioactivity against Erwinia carotovora causing soft rot disease on some crops
Nguyen Xuan Canh, Nguyen Thi Khanh, Pham Hong HienAbstractIn this study experiments were performed to screen and identify the actinomyces trains that were capable of antagonism to Erwinia carotovora causing the soft rot disease on plants. Using the agar diffusion plate method, 05 strains that were capable of antagonism to Erwinia carotovora were obtained. The strain number L2.5 had strongest activity with a diameter of 23 mm clear zone of bacteria. The L2.5 strain was capable to produce the straight spore chains after 03 days of culture, non-induced the soluble pigments on ISP-6 medium, growing well at temperatures between 30 - 35 °C and neutral pH, and adapting to low salt concentration medium. It could be used some carbon and nitrogen sources including sucrose, fructose, cellulose, raffinose, meat extract, peptone and KNO3. Results of 16S rRNA sequence analysis showed that strain L2.5 had a similarity of 99% comparing to Streptomyces psammoticus KP1404. Based on morphology, culture, physiological, biochemical characteristics and molecular biological analyzes we have identified the strain L2.5 belonging to Streptomyces psammoticus species.Key words: Actinomyces, Erwinia carotovora, Streptomyces sp., soft rot
Ngày nhận bài: 7/6/2017Ngày phản biện: 14/6/2017
Người phản biện: TS. Trần Thị Mỹ HạnhNgày duyệt đăng: 25/6/2017
1 Khoa Công nghệ Sinh học, Học viện Nông nghiệp Việt Nam2 Viện Khoa học Nông nghiệp Việt Nam
ẢNH HƯỞNG CỦA ĐIỀU KIỆN NUÔI CẤY, MÔI TRƯỜNG DINH DƯỠNG ĐẾN KHẢ NĂNG SINH TRƯỞNG CỦA CHỦNG XẠ KHUẨN GL30
Nguyễn Văn Giang1, Đinh Văn Lợi1, Phạm Hồng Hiển2
TÓM TẮTMục đích của thí nghiệm này là khảo sát ảnh hưởng của trạng thái nuôi (tĩnh, lắc) và các nguồn carbon, nitơ,
nhiệt độ, pH môi trường nuôi cấy tới khả năng sinh trưởng và khả năng kháng khuẩn của chủng xạ khuẩn GL30. Chủng xạ khuẩn GL30 đối kháng mạnh nhất với 3 chủng vi sinh vật gây bệnh là Bacillus cereus ATCC-11778, Escherichia coli VTCC-B-482, Staphylococcus aureus VTCC-B-658. Khi nuôi chủng xạ khuẩn GL30 trong bình tam giác (V=250 ml) với các nguồn carbon và nitơ khác nhau, giá trị pH môi trường nuôi ban đầu từ 3 - 11, tại các nhiệt độ (25, 30, 40, 500C), kết quả cho thấy hoạt tính kháng vi khuẩn Staphylococcus aureus VTCC-B-658 của chủng GL30 đạt cao nhất sau 4 ngày nuôi cấy ở điều kiện lắc. Chủng GL30 sinh tổng hợp chất kháng khuẩn và sinh trưởng tốt tại pH 6 - 8, nhiệt độ 300C, lắc 150 vòng/phút, thể tích dịch nuôi cấy/thể tích môi trường là 20%. Bột ngô 2%, pepton 2% là nguồn cacrbon và nitơ tốt nhất cho chủng GL30 sinh trưởng và sinh chất kháng khuẩn.
Từ khóa: Trạng thái nuôi cấy, môi trường dinh dưỡng, khả năng đối kháng, Streptomyces
I. ĐẶT VẤN ĐỀÔ nhiễm môi trường sống đang tác động trực tiếp
đến sức khỏe con người, đặc biệt làm gia tăng rất nhiều bệnh truyền nhiễm do vi sinh vật gây ra. Hơn nữa, các vi sinh vật gây hại không ngừng biến đổi để thích nghi với môi trường và có khả năng kháng lại chất diệt khuẩn. Điều này đã đặt ra thách thức trong nghiên cứu phát hiện và phân tích những loại hợp chất kháng khuẩn mới. Trong số các vi sinh vật sinh chất kháng khuẩn đã biết, xạ khuẩn, đặc biệt là các loài thuộc chi Streptomyces, đóng vai trò hàng đầu và được xem là nguồn sản xuất chất kháng khuẩn nhiều nhất (Intra et al., 2011). Tuy nhiên, hàm lượng
kháng khuẩn thu được phụ thuộc rất nhiều vào điều kiện nuôi cấy và môi trường dinh dưỡng (Gesheva et al., 2004).
Quá trình tổng hợp các hợp chất trao đổi chất thứ cấp chịu tác động rất lớn bởi thành phần môi trường (nguồn C, N và P), tốc độ sinh trưởng, nhiệt độ, pH và tốc độ cung cấp ôxi (Sánchez et al., 2010). Trong đó, C và N có vai trò quan trọng trong quá trình nuôi cấy vi sinh vật, chúng cung cấp năng lượng và các tiền chất để vi sinh vật trao đổi chất, tạo sinh khối và sinh tổng hợp các hợp chất trao đổi thứ chất (Wang et al., 2010). Trong nghiên cứu này, một số chủng xạ khuẩn có khả năng sinh chất diệt khuẩn đã được

48
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
phân lập và tuyển chọn tại địa bàn huyện Gia Lâm. Ảnh hưởng của các điều kiện nuôi cấy đã được khảo sát nhằm cải thiện mức độ sinh tổng hợp chất kháng khuẩn từ xạ khuẩn. Đây là cơ sở cho nghiên cứu thu nhận chất kháng khuẩn, hướng tới sản xuất thuốc trong tương lai.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứuCác mẫu đất được thu thập ở độ sâu từ 5 - 10
cm tại một số khu vực thuộc huyện Gia Lâm - Hà Nội. Chủng vi sinh vật kiểm định, bao gồm Bacillus cereus ATCC-11778, Escherichia coli VTCC-B-482, Staphylococcus aureus VTCC-B-658, Salmonella typhimurium VTCC-B-480, Shigella flexneri VTCC-B-479, Listeria monocytogenes được cung cấp từ khoa Công nghệ Sinh học, Học viện Nông nghiệp Việt Nam.
Môi trường nuôi xạ khuẩn: Môi trường Gause I (g/l): tinh bột tan 20 g, KNO3 1 g, MgSO4.7H2O 0,5 g, K2HPO4 0,5 g, FeSO4 0,01 g, NaCl 0,5 g, agar 20 g, pH 7,0 - 7,2. Môi trường Starch Nitrate Broth (g/l): starch, 20 g; NaNO3 2 g; K2HPO4 1 g; MgSO4 0,5 g; KCl, 100 g; 1ml dung dịch muối kim loại vết, gồm FeSO4. 7H2O (0,1 g/100 ml); MnCl2.4H2O (0,1 g/100 ml); ZnSO4.7H2O (0,1 g/100 ml), pH 7,1 ± 0,2 at 25°C.
2.2. Phương pháp nghiên cứu- Phương pháp phân lập và đánh giá khả năng
sinh kháng khuẩn: Chủng xạ khuẩn được phân lập trên môi trường Gause I theo phương pháp của Vinogradkii (1952) (trích theo Nguyễn Thành Đạt, 2000). Khả năng đối kháng của chủng xạ khuẩn với vi sinh vật kiểm định được tiến hành dựa theo phương pháp của Nguyễn Lân Dũng và cộng tác viên (1997).
- Phương pháp khảo sát một số đặc điểm sinh học chủng xạ khuẩn: Đặc điểm hình thái của chủng xạ khuẩn được xác định dựa trên các đặc điểm nuôi cấy, bao gồm màu sắc của khuẩn ty khí sinh, màu sắc của khuẩn ty cơ chất, khả năng sinh sắc tố tan (Tresner and Backus, 1963) và sự hình thành sắc tố melanin trên hệ thống môi trường ISP.
- Phương pháp khảo sát ảnh hưởng của nhiệt độ, pH, tỷ lệ dịch nuôi cấy/thể tích bình: Xạ khuẩn được nuôi trong môi trường nhân giống cấp 1 (w/v): 1% yeast extract; 1% dextrose dạng lỏng (pH 7,0). Hút 10ml dịch nuôi bổ sung vào 100 ml môi trường Starch Nitrate Broth (SNB) (pH 6,8 - 7,0) chứa trong bình tam giác (V= 250 ml). Các bình này được đặt
ở hai trạng thái tĩnh và lắc 150 vòng/phút, với dải nhiệt độ 25 - 50 oC và pH 3 - 11. Sau 3, 4, 5, 6, 7, 9 ngày nuôi tiến hành xác định khả năng sinh chất kháng khuẩn (Nguyễn Lân Dũng và ctv., 1997) và sinh khối khô (Oskay, 2011). Ảnh hưởng của tỷ lệ dịch nuôi cấy chủng xạ khuẩn/thể tích bình được xác định dựa theo nghiên cứu trước của Nguyễn Văn Hiếu và cộng tác viên (2012). Môi trường SNB được bổ sung vào bình tam giác 250ml với thể tích tương ứng là 5, 10, 15, 20, 25, 30, 35, 40% thể tích bình. Tiếp theo, 10% giống cấp 1 được bổ sung và nuôi ở trạng thái nhiệt độ, pH tối ưu. Trọng lượng sinh khối khô và hoạt tính kháng khuẩn được xác định vào ngày nuôi cấy tối ưu.
- Phương pháp khảo sát ảnh hưởng của nguồn C và N: Xạ khuẩn được nuôi khởi động 3 ngày trong môi trường nhân giống cấp 1 dạng lỏng (w/v): 1% yeast extract; 1% dextrose, pH 7. Dịch nuôi cấy sau đó được chuyển sang môi trường SNB với các nguồn C và N khác nhau. Hàm lượng tinh bột được thay thế bằng các nguồn đường, như bột gạo, bột đao, bột ngô với tỷ lệ thay thế 2%. Môi trường chứa 2% tinh bột là đối chứng dương, môi trường không bổ sung nguồn C là đối chứng âm. Tiếp theo, NaNO3 trong môi trường SNB được thay thế bằng các nguồn N khác, như peptone, cao thịt bò, (NH4)2SO4, KNO3, urea, bột đậu tương, bã đậu và bột canola. Môi trường chứa 0,2% NaNO3 là đối chứng dương, môi trường không bổ sung nguồn N là đối chứng âm. Hoạt tính kháng khuẩn được xác định bởi đường kính vòng đối kháng với Staphylococcus aureus VTCC-B-658.
2.3. Thời gian và đia điểm và nghiên cứuNghiên cứu được thực hiện tại Khoa Công nghệ
sinh học, Học viện Nông nghiệp Việt Nam từ tháng 3 đến tháng 12 năm 2014.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Kết quả phân lập và tuyển chọn các chủng xạ khuẩn thu thập
Từ các mẫu đất thu thập tại địa bàn Gia Lâm - Hà Nội đã phân lập được các chủng xạ khuẩn với đặc điểm hình thái và màu sắc khuẩn lạc khác nhau trên môi trường Gause I. Các chủng này sau đó đã được kiểm tra khả năng kháng vi sinh vật kiểm định bằng hai phương pháp thỏi thạch và khuếch tán đĩa thạch. Kết quả thu được 4 chủng có hoạt tính mạnh với đường kính vòng kháng khuẩn từ 2 - 18 mm (Bảng 1). Chủng GL30 có biểu hiện hoạt tính mạnh nhất nên được lựa chọn để tiến hành các thí nghiệm tiếp theo.

49
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Khuẩn lạc của chủng GL30 có kích thước từ 4 - 6 mm, bề mặt khuẩn lạc khô, lồi ở tâm, hình phóng xạ và được bao phủ bởi nhiều lớp khuẩn ty khí sinh màu nâu trắng. Khuẩn ty cơ chất có màu vàng nhạt sau 7 ngày nuôi cấy. Bên cạnh đó, khuẩn ty của GL30 là khí sinh dạng thẳng, phân nhánh, không bị đứt, cuống sinh bào tử thẳng, bào tử có bề mặt trơn, kích thước khoảng 0,7 - 1.37 ˟ 0,6 - 0,9 µm (Hình 1). Màu sắc hệ sợi khí sinh và cơ chất của chủng GL30 khi nuôi cấy trên các môi trường hầu như không có sự thay đổi lớn. Sau 21 ngày, màu sắc khuẩn lạc hầu như không thay đổi. Trên hệ thống môi trường Gause - I, ISP, sau thời gian 7, 14 và 21 ngày nuôi cấy, chủng GL30 có màu xám, xám trắng, trắng. Đặc biệt, khảo sát khả năng sinh sắc tố melanin trên môi trường ISP 6, không có sự thay đổi màu rõ rệt qua các ngày nuôi cấy được ghi nhận. Kết quả cho thấy, chủng xạ khuẩn GL30 không có khả năng sinh sắc tố melanin.
Hình 1. Hình thái chuỗi bào tử của chủng xạ khuẩn GL30 (A ˟ 1800); (B ˟ 25000)
3.2. Kết quả khảo sát ảnh hưởng của điều kiện nuôi cấy đến sinh trưởng và sinh tổng hơp chất kháng khuẩn của chủng xạ khuẩn GL30
Khi nuôi ở 30oC, GL30 sinh chất kháng khuẩn mạnh nhất với đường kính vòng đối kháng S. aureus VTCC-B-658 lên tới 23 mm. Ở 25 và 400C, chủng GL30 cho kích thước vòng kháng khuẩn lần lượt là 18 và 2 mm (Hình 2). Chủng GL30 không sinh trưởng tại 50oC. Như vậy, dải nhiệt độ từ 25 - 300C thích hợp cho chủng xạ khuẩn GL30 sinh trưởng và sinh chất kháng khuẩn. Trước đó, Sunita Bundale và cộng tác
viên (2015) cũng đã kết luận rằng nhiệt độ 300C là thích hợp nhất với chủng xạ khuẩn Streptomyces sp. phân lập từ đất.
Hình 2. Ảnh hưởng của nhiệt độ đến khả năng sinh trưởng và sinh kháng khuẩn của chủng GL 30
Chủng GL30 sau 3 - 9 ngày nuôi ở trạng thái lắc và tĩnh đã có biểu hiện sai khác rõ rệt về khả năng sinh chất kháng khuẩn và khối lượng sinh khối khô. Khi nuôi lắc, hoạt tính của chủng GL30 xuất hiện ngay từ ngày nuôi cấy thứ 3, sau 4 ngày đường kính vòng đối kháng với S. aureus VTCC-B-658 đạt 26 mm. Chủng GL30 sinh trưởng cực đại với lượng sinh khối khô lên tới 0,51 g/100 ml sau 7 ngày nuôi (Hình 3), sau đó giảm dần từ ngày thứ 9. Các chủng xạ khuẩn trong nghiên cứu của Sunita Bundale và cộng tác viên (2015) cũng biểu hiện tính kháng khuẩn mạnh nhất sau 4 ngày nuôi và duy trì trong khoảng 10 ngày.
Hình 3. Ảnh hưởng của thời gian nuôi cấy đến sinh trưởng và khả năng sinh chất
kháng khuẩn của chủng GL30
Khi được nuôi trong môi trường với dải pH từ 5 - 11 (Hình 4.A), chủng GL30 có thể sinh trưởng và biểu hiện hoạt tính kháng S. aureus VTCC-B-658 tương đối mạnh (đường kính vòng kháng khuẩn từ
Bảng 1. Hoạt tính kháng vi sinh vật của 4 chủng xạ khuẩn được tuyển chọn
Ghi chú: I - phương pháp thỏi thạch; II - phương pháp khuếch tán đĩa thạch
Chủng
Hiệu quả ức chế vi sinh vật kiểm đinh(đường kính vòng kháng khuẩn, mm)
E. coli S. aureus S. typhimurium S. flexneri L.
monocytogenes B. cereus
I II I II I II I II I II I IIVS15 11 10 10 10 4 7 8 10 4 4 4 5VS18 5 5 13 17 2 4 9 13 5 5 3 3VS28 5 4 16 20 3 5 14 16 4 5 4 6GL30 9 10 15 18 5 6 12 12 6 6 2 2
25Nhiệt độ (oC)
: Khối lượng: Đường kính
Đườ
ng k
ính
vòng
khán
g kh
uẩn
(mm
)
Khối
lượn
g si
nhkh
ối k
hô (g
/100
ml)
30 40 50
0,4
0,2
0
30
20100

50
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
19 - 23 mm). Trước đó, Ababutain và cộng tác viên (2013) đã khẳng định xạ khuẩn có thể sinh trưởng tốt tại các giá trị pH này. Tại pH 5, chủng xạ khuẩn GL30 sinh trưởng chậm, tuy nhiên hoạt tính đối kháng mạnh, đường kính vòng kháng khuẩn đạt 23 mm. Tại pH 6 - 8, sinh khối của chủng này cao nhất từ 0,30 -
0,33 g/100 ml, đường kính vòng kháng khuẩn đạt 21 - 22 mm. Khi tiếp tục tăng pH, hoạt tính đối kháng của chủng GL30 không tăng và có khuynh hướng giảm. Kết quả này cũng đã được ghi nhận trước đó trên các nhóm xạ khuẩn khác (Oskay, 2011; Nguyễn Văn Hiếu và ctv., 2012; Sunita Bundale et al., 2015).
Bên cạnh đó, nuôi chủng xạ khuẩn GL30 trên môi trường SNB và thay đổi thể tích môi trường lên men đã ghi nhận sự ảnh hưởng đến khuếch tán ôxi không khí vào môi trường nuôi. Kết quả thí nghiệm cho thấy với lượng môi trường lên men chiếm 20% thể tích bình sẽ cho hiệu quả tổng hợp chất kháng khuẩn cao nhất, vòng kháng khuẩn tương ứng với chủng S. aureus VTCC-B-658 là 28 mm (Hình 4.B). Kết quả này có sự sai khác so với ghi nhận trước đó của Nguyễn Văn Hiếu và cộng tác viên (2012) khi đánh giá ảnh hưởng của độ thông khí tới hoạt tính kháng khuẩn của chủng xạ khuẩn HLD 3.16.
3.3. Kết quả khảo sát ảnh hưởng của nguôn C và NTrong số các nguồn C thí nghiệm, bột ngô và
tinh bột là 2 nguồn hydratcacbon thích hợp nhất cho chủng GL30 sinh trưởng và sinh chất kháng khuẩn. Đặc biệt, khi nuôi trong môi trường có bột ngô, kích thước đường kính vòng kháng khuẩn của chủng GL30 lên tới 26 mm (Hình 5.A). Bột ngô chứa các thành phần protein, lipit, tinh bột, đường, chất xơ ở vỏ nội nhũ và cả trong phôi, vì vậy nó là nguồn chất dinh dưỡng thích hợp cho vi sinh vật sinh trưởng. Gần đây, nhóm nghiên cứu của Silva cũng kết luận bột ngô và cao nấm men thích hợp với chủng xạ khuẩn No. 01 (Silva et al., 2012).
Hình 4. Ảnh hưởng pH môi trường (A) và độ thông khí (B) đến khả năng sinh trưởng, sinh tổng hợp chất kháng khuẩn của chủng xạ khuẩn GL30
Hình 5. Ảnh hưởng của các nguồn carbon (A) và các nguồn nitơ (B) đến hoạt tính kháng khuẩn của chủng GL30
Trên các môi trường với các nguồn N vô cơ và hữu cơ khác nhau, chủng GL30 đều có thể sinh trưởng và biểu hiện hoạt tính kháng khuẩn. Trong đó, pepton và NaNO3 thích hợp cho chủng GL30 sinh tổng hợp chất kháng khuẩn với kích thước vòng kháng khuẩn lớn nhất, trong đó nguồn pepton cho kích thước lên đến 27 mm (Hình 5.B). Kết quả này cũng được ghi nhận trong một số nghiên cứu trước đó (Oskay,
2011; Nguyễn Văn Hiếu và ctv., 2012) trong các thí nghiệm trên đối tượng Streptomyces sp. KGG32 và HLD 3.16. Tuy nhiên Gao và cộng tác viên (2009) lại công bố nguồn nitơ tốt nhất cho chủng xạ khuẩn Streptomyces avermitilis 14-12A là cao nấm men. Nghiên cứu này sẽ được tiếp tục nhằm nghiên cứu khả năng thu nhận chất kháng khuẩn, hướng tới sản xuất thuốc trong tương lai.

51
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
IV. KẾT LUẬNĐã phân lập được 4 chủng xạ khuẩn từ mẫu đất
thu tập tại địa bàn Gia Lâm - Hà Nội. Trong đó, chủng xạ khuẩn GL30 có hoạt tính kháng khuẩn cao, biểu hiện ở ngày nuôi cấy thứ 3 và đạt cực đại sau 4 ngày nuôi trong điều kiện nuôi lắc 150 vòng/phút tại nhiệt độ 30oC, pH 6 - 8.
Chủng GL30 sinh trưởng tốt ở ngưỡng nhiệt độ từ 25 - 400C, tốt nhất tại 30 oC, pH 6 - 8, thể tích dịch nuôi cấy/thể tích bình là 20%, sinh khối cực đại đạt từ 0,30 - 0,33 g/100 ml, đường kính vòng kháng khuẩn đạt 21 - 22 mm.
Bột ngô và pepton đã được xác định là nguồn C và N thích hợp nhất cho chủng xạ khuẩn GL30 phát triển. Hai nguồn này có tiềm năng thay thế cho tinh bột và NaNO3 có trong môi trường Starch Nitrate Broth.
TÀI LIỆU THAM KHẢONguyên Lân Dũng, Phạm Văn Ty, Nguyên Đình
Quyến, 1997. Vi sinh vật học - tập II. Nhà xuất bản Đại học và Trung học chuyên nghiệp. Hà Nội.
Nguyên Thành Đạt, 2000. Sinh học vi sinh vật, Nhà xuất bản Giáo dục.
Nguyên Văn Hiếu, Nguyên Phương Nhuệ, Vũ Thi Hạnh Nguyên, Phan Thi Hông Thảo, Phạm Thanh Huyền, Phí Quyết Tiến, Lê Gia Hy, 2012. Nghiên cứu chủng xạ khuẩn HLD 3.16 có hoạt tính kháng khuẩn phân lập từ vùng ven bờ biển Việt Nam. Tạp chí Khoa học và Công nghệ, 50(5): 581-593.
Ababutain, I.M., Aziz, Z.K.A., AL-Meshhen, N.A., 2013. Optimization of environmental and nutritional conditions to improve growth and antibiotic productions by Streptomyces sp. isolated from Saudi Arabia soil. Int Res J Microbiol, 4(8): 179-187.
Da Silva, I.R., Martins, M.K., Carvalho, C.M., de Azevedo, J.L., de Lima Procópio, R.E., 2012. The effect of varying culture conditions on the production
of antibiotics by Streptomyces spp., isolated from the Amazonian soil. Ferment Technol, 1(105): 1-3.
Gao, H., Liu, M., Liu, J., Dai, H., Zhou, X., Liu, X., Zhuo, Y., Zhang, W., Zhang, L., 2009. Medium optimization for the production of avermectin B1a by Streptomyces avermitilis 14-12A using response surface methodology. Bioresour Technol, 100(17): 4012-4016.
Gesheva, V., Ivanova, V., Gesheva, R., 2004. Effects of nutrients on the production of AK-111-81 macrolide antibiotic by Streptomyces hygroscopicus. Microbiological, 160(3): 243-248.
Intra, B., Mungsuntisuk, I., Nihira, T., Igarashi, Y., Panbangred, W., 2011. Indentification of actinomyces from plant rhizospheric soils with inhibitory activity against Colletotrichum spp, the causative agent of anthracnose disease. BMC Research Notes, 4: 98.
Oskay M., 2011. Effects of some environmental conditions on biomass and antimicrobial metabolite production by Streptomyces sp., KGG32. Int J Agric Biol, 13(3): 317-324.
Sánchez, S., Chavez, A. , Forero, A., Garcıa-Huante, Y., Romero, A., Sánchez, M., Rocha, D., Sánchez, B., Avalos, M., Guzman-Trampe, S., Rodrıguez-Sanoja, R., Langley, E., Ruiz, B., 2010. Carbon source regulation of antibiotic production. J. Antibiot., 63(8): 442-459.
Sunita, B., Begde, D., Nashikkar, N., Kadam, T., Upadhyay, A., 2015. Optimization of Culture conditions for production of bioactive metabolites by Streptomyces spp. isolated from soil. Adv Microbiol, 5(6): 441-451.
Tresner H. D., Backus E. J., 1963. System of color wheels for streptomycete taxonomy. Applied microbiology, 11(4): 335-338.
Wang, X., Huang, L., Kang, Z., Buchenauer, H., Gao, X., 2010. Optimization of the fermentation process of actinomycete strain Hhs.015T. J. Biomed Biotechnol, 2010: 1-10.
Effects of cultural and nutritional conditions on growth of new isolated GL30 Streptomyces strain
Nguyen Van Giang, Dinh Van Loi, Pham Hong HienAbstractThe purpose of the present study was to survey the effects of some cultural conditions, including temperature, pH, different resources of C and N, the culture medium / flack’s volume ratio, cultural status (static and shacking) on growth and antibacterial metabolite production of Streptomyces GL30 strain, isolated from different soil samples. Strain GL30 exhibited antagonistic ability to three pathogens: Bacillus cereus ATCC-11778, Escherichia coli VTCC-B-482 and Staphylococcus aureus VTCC-B-658. Antibacterial activity of Streptomyces GL30 strain was evaluated based on the diameter of inhibition zone of pathogenic microorganisms. The results showed that the antibacterial activity of GL30 strain reached highest after 4 days of growth under shacking conditions, the temperature of 30 oC, pH 6-8 and culture media/ flack’s volume was 20%. Cornstarch of 2%, peptone 2% was the best carbon and nitrogen source for growth and antibacterial production of strains GL30. The results also showed that strain GL30 was a potential soil microorganism with antimicrobial activity and may be used for biotechnological purposesKey words: Cultural conditions, nutrient conditions, antimicrobial activity, Streptomyces
Ngày nhận bài: 10/7/2017Ngày phản biện: 13/7/2017
Người phản biện: TS. Nguyễn Xuân Cảnh Ngày duyệt đăng: 27/7/2017

52
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀĐông trùng hạ thảo (Cordyceps militaris), là một
loại dược liệu quý có nhiều tác dụng chữa bệnh, tốt cho sức khoẻ và phù hợp với nhiều lứa tuổi. Cordycepin và adenosine là hai hợp chất quan trọng nhất dùng để đánh giá chất lượng nguyên liệu này. Cordycepin chỉ có trong đông trùng hạ thảo, đóng vai trò quan trọng trong việc hỗ trị điều trị ung thư (Fan et al., 2012, Peter et al., 2008), chống viêm (Shashidhar et al., 2013), bảo vệ gan, phổi (Bueriz et al., 2005), bảo vệ thận và giảm chấn thương lách do tiểu đường (Ma et al., 2015). Adenosine có tác dụng chống viêm, tác dụng tốt trên phổi, hệ tim mạch, tuần hoàn và thần kinh trung ương (Hori et al., 1991).
Đông trùng hạ thảo gồm phần quả thể và phần giá thể sau thu hoạch (đế). Phần quả thể đang được quan tâm rất nhiều trong khi đế dù chứa nhiều hoạt chất sinh học nhưng chưa được sử dụng hiệu quả.
Trong đế vẫn còn cơ chất môi trường giàu dinh dưỡng và hoạt chất. Lượng hoạt chất phụ thuộc nhiều yếu tố trong đó có môi trường nuôi cấy và điều kiện xử lý nhiệt. Đế khô mới chỉ được ứng dụng để trích ly lấy hoạt chất hay bổ sung vào các sản phẩm như cháo, rượu hay ủ uống như trà, tuy nhiên điều kiện ngâm chiết như thế nào để đạt được hàm lượng hoạt chất cao nhất cũng chưa được nghiên cứu nhiều. Trong nghiên cứu này, ảnh hưởng của hai môi trường nuôi cấy tạo đế: môi trường bán tổng hợp (MT1) và môi trường tự nhiên (MT2); chế độ gia nhiệt khô (sấy) và nhiệt ẩm (ngâm chiết) tới sự biến đổi lượng hoạt chất và tính chất cảm quan
của sản phẩm sau sấy, của dịch chiết được quan tâm nhằm đề xuất điều kiện tác động phù hợp sao cho hàm lượng hoạt chất cordycepin và adenosine được giữ lại nhiều nhất và chất lượng cảm quan tốt nhất. Đây cũng là cơ sở để khuyến cáo những ứng dụng về sau.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP
2.1. Vật liệu nghiên cứuĐế đông trùng hạ thảo nuôi trên môi trường
MT1 gồm thành phần chính là: Gạo lứt, peptone, glucose, vi lượng, nhộng tằm, trứng, nước cốt dừa, khoai tây, vitamin…; Môi trường MT2 gồm thành phần chính là gạo lứt, nhộng tằm, trứng, nước cốt dừa, khoai tây được cung cấp bởi Viện Công nghệ sinh học - Viện Hàn lâm Khoa học và Công nghệ Việt Nam và Khoa Công nghệ sinh học - Học viện Nông nghiệp Việt Nam. Đế tươi sau khi thu về được tiến hành thí nghiệm trong vòng 24 giờ.
2.2. Phương pháp nghiên cứu
2.2.1. Phương pháp bố trí thí nghiệm- Thí nghiệm 1: Ảnh hưởng của nhiệt độ sấy tới
hàm lượng hoạt chất và chất lượng cảm quan của đế đông trùng hạ thảo. Yếu tố phi thí nghiệm: Đế đông trùng hạ thảo nuôi ở môi trường MT1 và MT2. Yếu tố thí nghiệm: Nhiệt độ sấy đối lưu lần lượt là 50, 60, 70, 80°C. Sấy cho tới khối lượng không đổi. Chỉ tiêu theo dõi: Độ ẩm, lượng hoạt chất và cảm quan.
- Thí nghiệm 2: Ảnh hưởng của chế độ ngâm chiết tới hàm lượng hoạt chất và chất lượng cảm quan của dịch chiết đông trùng hạ thảo. Đế đông
1 Khoa Công nghệ thực phẩm, Học viện Nông nghiệp Việt Nam2 Viện Công nghệ sinh học, Viện Hàn lâm Khoa học và Công nghệ Việt Nam
ẢNH HƯỞNG CỦA NHIỆT ĐỘ SẤY VÀ CHẾ ĐỘ NGÂM CHIẾT TỚI SỰ BIẾN ĐỔI THÀNH PHẦN HOÁ HỌC VÀ CẢM QUAN CỦA GIÁ THỂ SAU THU HOẠCH ĐÔNG TRÙNG HẠ THẢO
Nguyễn Thị Thanh Thuỷ1, Phí Quyết Tiến2
TÓM TẮTĐông trùng hạ thảo (ĐTHT) có tác dụng tăng cường sức khoẻ, chống ung thư, chống viêm nhờ một số hoạt chất
sinh học như adenosine và cordycepin chứa trong nó. Thành phần này thay đổi tùy thuộc vào nhiều yếu tố trong đó có môi trường nuôi cấy và phương thức xử lý nhiệt. Ngoài quả thể là thành phần chính được thu nhận, giá thể sau thu hoạch (đế) của ĐTHT hiện mới chỉ được sấy khô, sử dụng ở dạng thô. Nghiên cứu này nhằm tìm ra được nhiệt độ sấy khô và nhiệt độ ngâm chiết tới sự biến đổi hoạt chất và tính chất cảm quan. Hai loại giá thể sau thu hoạch sử dụng được nuôi trên môi trường bán tổng hợp (MT1) có hàm lượng adenosine và cordycepin lần lượt là 0,34 mg/g; 2,34 mg/g và môi trường tự nhiên (MT2) với hàm lượng hai hoạt chất này đạt 0,36 mg/g; 2,71 mg/g. Sấy ở 70°C cho hoạt chất thu được là cao nhất với điểm cảm quan xếp loại khá. Chế độ ngâm chiết cho hàm lượng hoạt chất và cảm quan tốt nhất là 90°C - 15 phút và 90°C- 20 phút cho đế MT1; 95°C -10 phút cho đế MT2.
Từ khoá: Adenosine, cordycepin, đế, đông trùng hạ thảo, giá thể sau thu hoạch

53
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
trùng hạ thảo sau khi sấy khô ở nhiệt độ thích hợp, được nghiền thành bột và rây ở kích cỡ sàng 0,5mm để đạt được độ đồng đều. Bột đế được ngâm trong nước nóng với các nhiệt độ và thời gian khác nhau. Yếu tố phi thí nghiệm: Lượng bột đế/nước là 1g/100 ml (tính theo chất khô tuyệt đối). Yếu tố thí nghiệm: Chế độ ngâm với nhiệt độ lần lượt là 80, 85, 90, 95, 100°C với thời gian lần lượt là 5, 10, 15, 20, 25 phút. Chỉ tiêu theo dõi: Hàm lượng các hoạt chất và chất lượng cảm quan của dịch chiết.
2.2.2. Phương pháp phân tích- Xác định độ ẩm bằng phương pháp sấy đến khối
lượng không đổi. - Xác định hàm lượng protein bằng phương pháp
Kjeldahl.- Xác định hàm lượng chất béo bằng phương
pháp Soxhlet. - Xác định hàm lượng polysaccharide bằng
phương pháp so màu. Hàm lượng polysaccharide trong mẫu thí nghiệm được xác định và tính toán theo công thức và dựa trên bảng chuẩn (Dubois, 1956; Mo, 2013).
- Xác định hàm lượng cordycepin và adenosine bằng sắc khí lỏng hiệu năng cao (HPLC). Sử dụng hệ thống sắc ký lỏng HPLC-Shimadzu để xác định hàm lượng adenosine và cordycepin. Phần mềm LC- Solution dùng để xử lý peak. Cột C18 pha đảo (4,6 mm × 150 mm). Detector UV-Vis bước sóng 250 nm, nhiệt độ cột 30oC, tốc độ dòng 1,0 ml/phút, thể tích bơm vào 10 µL. Thành phần các pha động gồm nước (A) và methanol (B). Từ giá trị diện tích peak đo được ta tính được hàm lượng (mg/g).
2.2.3. Phương pháp đánh giá chất lượng cảm quan theo TCVN 3215-79
Đông trùng hạ thảo trong đề tài được đánh giá cảm quan theo TCVN 3215-79 cho hai nhóm thí nghiệm, nhóm 1 là nhóm sản phẩm thu được sau khi sấy khô ở các điều kiện khác nhau và nhóm 2 là nhóm sản phẩm thu được sau khi ngâm 1g trong 100ml nước nóng ở các điều kiện nhiệt độ khác nhau. Do chưa có các tiêu chuẩn trước và hệ số trọng lượng cho các chỉ tiêu về cảm quan đối với sản phẩm này nên dựa trên thực tế, hệ số trọng lượng được xếp thứ tự là màu - mùi - vị cho sản phẩm là 1,2 - 1,4 - 1,4. Mẫu nào mặc dù có tổng điểm cao nhưng nếu có chỉ tiêu có điểm sau khi nhân hệ số trọng lượng <2,8 thì mẫu đó coi như không đạt yêu cầu.
2.2.4. Phương pháp xử lý số liệuCác thí nghiệm được lặp lại 3 lần. Kết quả nghiên
cứu được xử lý và lấy số liệu trung bình theo lý thuyết thống kê sinh học trên phần mềm Excel.
2.3. Thời gian và đia điểm nghiên cứuNghiên cứu được thực hiện từ tháng 6 năm 2016
đến tháng 2 năm 2017 tại Viện công nghệ sinh học - Viện Hàn lâm Khoa học và Công nghệ Việt Nam và Khoa Công nghệ thực phẩm - Học viện Nông nghiệp Việt Nam.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Xác đinh thành phần hoá học trong đế đông trùng hạ thảo nuôi cấy trên hai môi trường khác nhau
Ngoài các hoạt chất sinh học trong đó có adenosin và cordycepin là điểm làm nên chất lượng của đông trùng hạ thảo thì nguyên liệu này cũng chứa các thành phần hoá học cơ bản bao gồm protein, lipid, polysaccharid... hàm lượng các chất cơ bản trong đế đông trùng hạ thảo dùng trong nghiên cứu được thể hiện trong bảng 1.
Kết quả bảng 1 cho thấy, hàm lượng các chất cơ bản và hoạt chất của đế đông trùng hạ thảo nuôi trên môi trường MT1 và môi trường MT2 có sự khác nhau tuy nhiên không nhiều. Hàm lượng polysaccharide trong đế đông trùng hạ thảo ở môi trường MT2 thấp hơn so với cùng hàm lượng này ở môi trường MT1. Hàm lượng lipid, protein thì ngược lại, hàm lượng lipid trong đế nuôi ở MT2 cao hơn so với cùng hàm lượng này trong đế nuôi ở MT1, sự chênh lệch lần lượt là 0,76% và 1,11%. Đối với hai hoạt chất adenosine và cordycepin thì các chất này có hàm lượng cao hơn ở đế nuôi trên MT2, đạt lần lượt là 0,36 và 2,71 mg/g.
Bảng 1. Thành phần hoá học và hoạt chất trong đế đông trùng hạ thảo tươi (% CK)
So sánh với kết quả của nghiên cứu khác thì kết quả trên đây cho hàm lượng của các hoạt chất thấp hơn so với công bố của Đài Loan. Điều đó có thể
Mẫu
Chỉ tiêu
Đế đươc nuôi trong môi trường
MT1
Đế đươc nuôi trong môi trường
MT2Chất khô (%) 20,22 ± 0,06 17,71 ± 0,02Protein (%) 12,18 ± 0,06 13,29± 0,12Lipid (% ) 3,78 ± 0,06 4,54 ± 0,02Polysaccharide (mg/ml) 7,87 ± 0,11 7,56 ± 0,10Adenosine (mg/g) 0,34 ± 0,01 0,36 ± 0,01Cordycepin (mg/g) 2,34 ± 0,07 2,71 ± 0,02

54
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
dễ hiểu vì nguyên liệu trong nghiên cứu này là đế còn nghiên cứu của Yu (2016) trên quả thể chứa 0,91 mg/g adenosine và 9,49 mg/g cordycepin. Tuy nhiên, hàm lượng cordycepin lại đạt gần hàm lượng cordycepin có trong quả thể đông trùng hạ thảo được Huang (2009) công bố với lượng cordycepin là 2,65 mg/g.
3.2. Ảnh hưởng của nhiệt độ sấy đến chất lương đế đông trùng hạ thảo
3.2.1. Ảnh hưởng của nhiệt độ sấy tới adenosine và cordycepin trong đế ĐTHT
Hoạt chất adenosine và cordycepin là hai hoạt chất quý quan trọng nhất trong quả thể cũng như trong đế đông trùng hạ thảo. Ảnh hưởng của nhiệt độ sấy tới hàm lượng hoạt chất được thể hiện trong
bảng 2.Thời gian kết thúc quá trình sấy cả đế MT1 và
MT2 đạt khối lượng không đổi dựa trên độ ẩm và nhiệt độ sấy ban đầu cho thấy tổng thời gian khi sấy đế ở 50, 60, 70, 80°C lần lượt là 136, 104, 72, 48 ±1 giờ (số liệu không trình bày).
Kết quả cho thấy, khi sấy ở nhiệt độ 50°C, 60°C và 70°C thì hàm lượng các chất chỉ giảm rất nhẹ, sau đó hàm lượng giảm hơn một chút khi sấy ở nhiệt độ 80°C. Hàm lượng adenosine và cordycepin ở đế đông trùng hạ thảo được nuôi ở cả 2 môi trường cao nhất khi sấy ở nhiệt độ 70°C lần lượt là 0,32 mg/g và 2,31 mg/g đối với đế nuôi ở MT1 và 0,31 mg/g và 2,61 mg/g đối với đế nuôi ở MT2. Từ đó, nhiệt độ sấy đế được đề xuất 70°C.
Nhìn chung khi sấy ở các nhiệt độ khác nhau thì hàm lượng các hoạt chất trong đế đông trùng hạ thảo không bị thay đổi nhiều, trong nghiên cứu của Li (2015) đã đưa ra kết luận cordycepin bền với nhiệt độ cao, còn theo nghiên cứu của Fu (2008) lại kết luận rằng adenosine nhạy cảm với nhiệt độ cao, tuy nhiên đó là khi xử lý với nhiệt độ trên 100°C.
3.2.3. Ảnh hưởng của nhiệt độ sấy tới chất lượng cảm quan của đế ĐTHT
Chất lượng của đế đông trùng hạ thảo được tạo nên bởi các chất có hoạt tính sinh học và chất lượng
cảm quan. Kết quả đánh giá cảm quan đế đông trùng hạ thảo sấy khô được thể hiện trên bảng 3 với các tiêu chí chấm điểm và hệ số quan trọng được nêu trong phần 2.3.3.
Kết quả cảm quan đế đông trùng hạ thảo ở nhiệt độ sấy khác nhau có sự khác nhau về chất lượng cảm quan. Có thể thấy rõ rằng, đế đông trùng hạ thảo MT1 sấy 70°C có điểm cảm quan cao hơn so với các mẫu còn lại, còn đế nuôi trên MT2 đạt được kết quả khá khi sấy ở cả 60 và 70°C. Kết hợp với kết quả về hoạt chất, lựa chọn mẫu sấy 70°C đem tiếp tục ngâm chiết ở chế độ nhiệt độ, thời gian khác nhau.
Bảng 2. Ảnh hưởng của nhiệt độ sấy tới lượng hoạt chất adenosine và cordycepin
Nhiệt độ sấy (°C)
Adenosin (mg/g) Cordycepin (mg/g)MT1 MT2 MT1 MT2
50 0,33 ± 0,01 0,34 ± 0,01 2,33 ± 0,29 2,66 ± 0,0460 0,32 ± 0,03 0,33 ± 0,01 2,32 ± 0,46 2,63± 0,0270 0,32 ± 0,05 0,32 ± 0,01 2,31± 0,01 2,61 ± 0,0280 0,30± 0,04 0,29 ± 0,01 2,26 ± 0,02 2,52 ± 0,02
Bảng 3. Ảnh hưởng của nhiệt độ sấy tới chất lượng cảm quan của đế ĐTHT
Ghi chú: * Điểm đã được tính theo hệ số trọng lượng
Mẫu sấy Màu* Mùi* Vi* Tổng điểm Xếp loại
MT1
Mẫu 50°C 4,05 4,55 4,90 13,50 ± 0,43 Trung bìnhMẫu 60°C 4,35 5,08 5,08 14,50 ± 0,42 Trùng bìnhMẫu 70°C 5,40 6,13 6,13 17,65 ± 0,42 KháMẫu 80°C 4,35 5,08 5,08 14,50 ± 0,42 Trung bình
MT2
Mẫu 50°C 4,05 4,38 4,90 13,33 ± 0,43 Trung bìnhMẫu 60°C 4,95 5,25 4,90 15,10 ± 0,19 KháMẫu 70°C 5,40 6,30 5,95 17,65 ± 0,45 KháMẫu 80°C 4,20 4,90 4,90 14,00 ± 0,40 Trung bình

55
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Hình 1. Ảnh hưởng của chế độ ngâm tới hàm lượng adenosin và cordycepin (mg/g)
3.3. Ảnh hưởng của chế độ ngâm tới sự biến đối chất lương và tính chất cảm quan của đế đông trùng hạ thảo
Đế sấy thu từ thí nghiệm trên được chọn làm tiếp cho ngâm chiết. 1 gam bột đế được ngâm với 100ml nước nóng ở các chế độ nhiệt độ và thời gian khác nhau. Sở dĩ có sự lựa chọn này vì mục đích tiếp theo dự kiến sẽ áp dụng cho các sản phẩm trà, trà túi lọc,
trà hòa tan.
3.3.1. Ảnh hưởng của chế độ ngâm tới lượng hoạt chất của đế ĐTHT
Đế được nuôi ở cả 2 môi trường sau khi ngâm chiết ở các chế độ khác nhau có cho hàm lượng adenosine và hàm lượng cordycepin tương đối gần nhau. Kết quả được thể hiện qua hình 1.
Dịch chiết từ đế đông trùng hạ thảo được nuôi trên MT1, có hàm lượng adenosine và cordycepin tăng dần khi nhiệt độ trích ly tăng từ 80°C lên 90°C, sau đó giảm dần ở nhiệt độ trích ly là 95°C và 100°C. Trích ly ở các khoảng thời gian khác nhau, nhiệt độ được cố định, nhìn chung hàm lượng adenosine và cordycepin tăng với thời gian trích ly kéo dài từ 5 phút đến 20 phút. Về hàm lượng adenosine, sau khi kéo dài thời gian trích ly đến 25 phút thì hàm lượng adenosine bắt đầu giảm. Trong nghiên cứu của Li (2015) khi tác động ở 100°C cũng đã chỉ ra rằng trong đông trùng hạ thảo thành phần adenosine nhạy cảm với nhiệt độ, việc tác động nhiệt trong thời gian kéo dài hơn 20 phút sẽ làm giảm lượng hoạt chất này. Đối với hàm lượng cordycepin, khi kéo dài thời gian trích ly đến 25 phút hàm lượng này có giảm nhẹ, riêng chỉ có công thức trích ly trong 25 phút tại 80°C, hàm lượng cordycepin vẫn tăng, điều này cũng phù hợp với kết quả nghiên cứu của Wu
(2014), kết quả này đã chỉ ra rằng với nhiệt độ 80°C việc xử lý nhiệt có thể kéo dài 60 phút cũng sẽ không ảnh hưởng tới sự giải phóng cordycepin của nguyên liệu. Có 3 công thức cho hàm lượng adenosine và cordycepin cao nhất đó là 85°C - 20 phút; 90°C - 15 phút và 90°C - 20 phút.
Dịch chiết từ đế đông trùng hạ thảo được nuôi trên MT2, hàm lượng adenosine và cordycepin tăng dần khi nhiệt độ trích ly tăng, so với việc trích ly đế nuôi trên MT1 thì khi nhiệt độ lên đến 95°C hàm lượng hai hoạt chất này vẫn tiếp tục tăng và chỉ giảm khi nhiệt độ trích ly lên đến 100°C. Ba công thức cho hàm lượng adenosine và cordycepin cao nhất trong dịch chiết của đế nuôi trên MT2 lần lượt là 90oC - 15 phút, 95°C - 10 phút và 95°C - 15 phút.
Do vậy, các công thức 85°C - 20 phút; 90°C - 15 phút, 90°C - 20 phút (của MT1) và 90°C - 15 phút, 95°C - 10 phút, 95°C - 15 phút (của MT2) sẽ được sử dụng tiếp theo cho phần đánh giá cảm quan.

56
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Từ các kết quả trên cho thấy, đối với đế nuôi trên cả 2 môi trường bán tổng hợp và tự nhiên thì nhiệt độ 100°C không phải là nhiệt độ thích hợp để trích ly được hai hoạt chất adenosine và cordycepin, thêm vào đó đây cũng là khoảng nhiệt độ khá cao, có thể dẫn tới biến tính các hoạt chất và 5 phút là khoảng thời gian quá ngắn để trích ly được các hoạt chất từ đế đông trùng hạ thảo. Điều kiện trích ly cho hàm lượng hai hoạt chất cao nhất là 90°C - 20 phút cho đế nuôi trên MT1 và 95°C - 10 phút cho đế nuôi trên MT2.
3.3.2. Ảnh hưởng của chế độ ngâm tới chất lượng cảm quan của dịch chiết đế ĐTHT
Nhiệt độ và thời gian chiết mẫu cũng làm cho chất lượng cảm quan thay đổi, màu sắc, mùi, vị cũng biến đổi theo. Từ kết quả phân tích hoạt chất ở trên, những mẫu có hàm lượng aenosine và cordycepin
cao nhất được sử dụng để đánh giá cảm quan.Dựa vào bảng 4 có thể thấy được chất lượng cảm
quan của dịch chiết đế đông trùng hạ thảo nuôi trên MT1 đạt mức khá với các công thức 90°C, 15 phút và 90°C, 20 phút. Như vậy, dịch chiết tại 90°C trong 20 phút đều đạt chất lượng về cảm quan cũng như hàm lượng các hoạt chất. Với dịch chiết từ đế nuôi trong MT2, khi ngâm ở 95°C trong 10 phút thì chiết ra được hàm lượng hoạt chất và cho kết quả cảm quan tốt nhất.
Các sản phẩm trà đông trùng hạ thảo hiện nay trên thị trường có trà đông trùng hạ thảo atiso, trà đông trùng hạ thảo ô long, trà đông trùng hạ thảo Biofun,… tuy trong thành phần đều ghi có chứa đông trùng hạ thảo nhưng khối lượng và việc sử dụng quả thể hay đế cũng không được nói rõ, vì vậy rất khó có thể so sánh được sản phẩm của nghiên cứu với các sản phẩm hiện đang có trên thị trường.
IV. KẾT LUẬNĐế đông trùng hạ thảo được nuôi trên hai môi
trường bán tổng hợp MT1 hay tự nhiên và MT2 sau thu hoạch sử dụng trong nghiên cứu có thành phần hoạt chất chênh lệch nhau không nhiều. Nhiệt độ sấy có ảnh hưởng đến hoạt chất của đế đông trùng hạ thảo. Hàm lượng hoạt chất cao nhất và chất lượng cảm quan đạt tốt nhất khi sấy đế ở nhiệt độ 70°C. Chế độ ngâm đế thích hợp để chiết được hàm lượng hoạt chất nhiều và đạt chất lượng cảm quan đạt loại khá là ở nhiệt độ 90°C trong 20 phút với đế nuôi trong môi trường MT1 và 95°C trong 10 phút với đế nuôi trong môi trường MT2. Kết quả này bước đầu cũng có những khuyến cáo cho việc sử dụng nhiệt khô, nhiệt ẩm để thu nhận hoạt chất trong đế đông trùng một cách có hiệu quả. Kết quả cảm quan ở mức khá cũng cho phép nguyên liệu này ứng dụng dễ dàng khi sử dụng đơn lẻ hay kết hợp với nguyên liệu khác.
LỜI CẢM ƠNNghiên cứu này có sử dụng trang thiết bị của
Phòng Thí nghiệm trọng điểm Công nghệ Gen
Quốc gia và Phòng thí nghiệm trung tâm Khoa học thực phẩm - Học viện Nông nghiệp Việt Nam. Công trình được thực hiện nhờ kinh phí của đề tài mã số: VAST02.05.16-17 Cấp Viện Hàn lâm Khoa học và Công nghệ Việt Nam.
TÀI LIỆU THAM KHẢOBueriz E.J, Bauer B.A, Osmundson T.W., Motley T.J.,
2005. The traditional Chinese medicine Cordyceps sinensis and its effects on the apoptotic homeostatic. Journal of Ethnopharmacology, 96:19-29.
Dubois M., Gilles K. A., Hamilton J. K., 1956. Colorimetric method for determination of sugars and related substances. Anal Chem. 1956;28:350e356.
Fan B. and Zhu H., 2012. Cordycepin: pharmacological properties and their relevant mechanisms. TANG Humanitas medicine, 141-147.
Fu J. and Li Y., 2008. The sample pretreatment methods for the determination of adenosine in Cordyceps mycelium powder. Journal of Jiangnan University. 7(1):96-98.
Hori M. and Kitakaze M., 1991. Adenosine, the heart and coronary Circulation. Hypertension, 18:565-574.
Bảng 4. Ảnh hưởng của chế độ ngâm tới chất lượng dịch chiết
Ghi chú: * Điểm đã được tính theo hệ số trọng lượng
Mẫu sấy Màu* Mùi* Vi* Tổng điểm Xếp loại
MT1
85°C, 20 phút 4,50 5,08 5,08 14,65 ± 0,33 Trung bình90°C, 15 phút 5,40 6,30 5,78 17,48 ± 0,45 Khá90°C, 20 phút 4,95 5,43 5,78 16,15 ± 0,41 Khá
MT2
90°C, 15 phút 4,35 5,08 5,08 14,50 ± 0,42 Trung bình95°C, 10 phút 5,35 6,48 6,13 17,85 ± 0,63 Khá95°C, 15 phút 4,80 5,25 5,08 15,13 ± 0,23 Trung bình

57
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Huang L., Li Q., Chen Y., Wang X. and Zhou X., 2009. Determination and analysis of cordycepin and adenosine in the products of Cordyceps spp., African Journal of Microbiology Research 3(12):957-961.
Li J., Guan M., Li Y., 2015. Effects of cooking on the contents of adenosine and cordycepin in Cordyceps militaris. Procedia Engineering, 102:485-491.
Ma L., Zhang S. and Du M., 2015. Cordycepin from Cordyceps militaris prevents hyperglycemia in alloxan. Induced diabetic mice Nutrition research, 35:431-39.
Mo M., Hu S., Xu X., Ma Z., Ni Y., Wei Y, Nie J., 2013. Optimization of extraction technology of polysaccharide of Tricholom giganteum. Pharmacology & Pharmacy, 4:1-5.
Peter C.K. Cheung, 2008. Mushrooms as functional foods. A John Wiley & Sons Inc, USA.
Shashidhar M.G., Giridhar P., Udaya Sankar K., Mahohar B., 2013. Bioactive principles from Cordyceps sinensis: A potent food supplement - A review, Journal of Functional foods, 5(3):1013-1030.
Wu P., Tao Z., Liu H, Jiang G., Ma C., Wang C., Geng D., 2014. Effects of heat on the biological activity of wild Cordyceps sinensis, Journal of Traditional Chinese Medical Sciences, 2:32-38.
Yu S. H., Dubey N. K., Li W. S., Liu M. C., Chiang H. S., Leu S. J. and Deng W. P., 2016. Cordyceps militaris treatment preserves renal function in type 2 diabetic nephropathy mice, PLoS One, 11(11), [e0166342]. DOI: 10.1371/journal.pone.0166342.
Effect of drying and extraction temperature on variation of bioactive compound and sensory properties of spent Cordyceps militaris substrate
Nguyen Thi Thanh Thuy, Phi Quyet TienAbstractCordyceps militaris has an effect for enhancing health, anti-cancer, anti-inflammatory due to some bioactive compounds including adenosine and cordycepin. These compounds vary depending on many factors such as culture media, heat treatment method etc. Apart from the fruiting body being the main parts to be harvested, the spent of Cordyceps militaris substrate is now only dried, used in raw form. This research aims to find the effect of drying and extraction temperatures on the change of bioactive substances and sensory properties. The two types of spent Cordyceps militaris were grown on semi-synthetic media (MT1) with the adenosine of 0.34 mg/g; cordycepin of 2.34 mg/g and on the natural media (MT2), with these two active compounds at 0.36 mg/g; 2.71 mg/g, respectively. The results showed that the obtained bioactive compound was highest with a good sensory point at the drying temperature of 70°C. The extraction conditions indicated the best bioactive substances content and the best sensory points were at 90°C in 15 min and 90°C in 20 min for MT1; 95°C in 10 min for MT2.Key words: Spent Cordyceps militaris substrate, adenosine, cordycepin
Ngày nhận bài: 9/7/2017Ngày phản biện: 13/7/2017
Người phản biện: TS. Nguyễn Xuân CảnhNgày duyệt đăng: 27/7/2017
1Trường Đại học Kinh tế Kỹ thuật Công nghiệp
NGHIÊN CỨU HOẠT TÍNH KHÁNG KHUẨN CỦA MỘT SỐ LOẠI TINH DẦUNguyễn Thị Mai Hương1, Hồ Tuấn Anh1
TÓM TẮTBài báo trình bày nghiên cứu khả năng kháng khuẩn của 5 loại tinh dầu: Tinh dầu hương nhu (Ocimum
gratissimum), tinh dầu quế (Cinnamomum loureiri), tinh dầu húng quế (Ocimum basilicum), tinh dầu bạc hà (Mentha arvensis), tinh dầu nghệ vàng (Curcuma longa) đối với các loài vi khuẩn B. subtilis, B. cereus, S. aureus, E. coli, S. typhimurium, P. putida, L. damsella so sánh đối chứng dương với 2 loại kháng sinh là gentamycin và streptomycin. Nghiên cứu bằng phương pháp khuyếch tán đĩa thạch cho thấy các loại tinh dầu đều có khả năng kháng khuẩn, trong đó tinh dầu quế thể hiện khả năng cao nhất. Đã xác định được nồng độ ức chế tối thiểu (MIC) và thời gian diệt khuẩn của tinh dầu quế đối với B. cereus tương ứng là 0,25% và 10 phút, đối với E. Coli là 0,5% và 20 phút. Theo tỉ lệ nồng độ diệt khuẩn tối thiểu/nồng độ ức chế tối thiểu (MBC/MIC) đã xác định được tinh dầu quế là chất diệt khuẩn, tinh dầu bạc hà là chất kìm khuẩn đối với B. cereus và E. coli. Nghiên cứu đã chỉ ra hiệu quả của một số loại tinh dầu có tác dụng tương đương các chất kháng sinh.
Từ khóa: Tinh dầu, kháng vi sinh vật, vòng kháng khuẩn, nồng độ ức chế tối thiểu, nồng độ diệt khuẩn tối thiểu

58
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀViệt Nam với điều kiện khí hậu nhiệt đới rất
thuận lợi cho sự sinh trưởng của đa dạng các loài thực vật có chứa tinh dầu. Tinh dầu được biết đến từ lâu là hương liệu sử dụng trong các lĩnh vực thực phẩm, mỹ phẩm.
Ngày nay, hiện tượng kháng kháng sinh ngày càng trở nên phổ biến, kháng sinh không còn là liều thuốc vạn năng như khi mới tìm thấy. Tổ chức Y tế Thế giới (WHO) đã báo động về nguy cơ kháng kháng sinh đối với nhiều loại vi sinh vật gây bệnh (Nguyễn Văn Kính và ctv., 2010). Nhiều hướng nghiên cứu khác nhau đã được thực hiện để giảm thiểu sử dụng kháng sinh, trong đó sử dụng tinh dầu là một phương pháp tuy không mới nhưng đã đem lại nhiều kết quả khả quan.
Khả năng kháng khuẩn của một số tinh dầu phụ thuộc vào nhiều yếu tố như thành phần, nồng độ, thời gian tiếp xúc với tinh dầu và chủng vi sinh vật. Sự tăng trưởng của vi sinh vật kháng hoặc đa kháng kháng sinh có thể bị ức chế bởi một số loại tinh dầu. Tinh dầu họ cam quýt, cây oải hương, cây bạc hà, cây bách xù, cây chè, cây húng tây và cây khuynh diệp có hiệu quả đặc biệt chống lại vi khuẩn S. aureus kháng methiciline (MRSA) (Tohidpour et al., 2010) và các khuẩn cầu ruột kháng vancomycine (ERV) (Fisher, 2009).
Nghiên cứu hoạt tính kháng khuẩn của một số loại tinh dầu như hương nhu, quế, bạc hà, nghệ vàng, húng quế có mục tiêu làm phong phú thêm tính ứng dụng của tinh dầu trong bảo quản thực phẩm hoặc trong y tế.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu- 5 loại tinh dầu: Tinh dầu hương nhu (Ocimum
gratissimum), tinh dầu quế (Cinnamomum loureiri), tinh dầu húng quế (Ocimum basilicum), tinh dầu bạc hà (Mentha arvensis), tinh dầu nghệ vàng (Curcuma longa) được chưng cất bằng phương pháp lôi cuốn hơi nước (Đỗ Tất Lợi, 2006).
- Sử dụng các vi khuẩn Bacillus subtilis, Bacillus cereus, Staphylococcus aureus, Escherichia coli, Salmonella typhimurium, Pseudomonas putida, Listonella damsella từ bộ sưu tập giống của Viện Công nghệ sinh học - Viện Hàn lâm Khoa học và Công nghệ Việt Nam.
- Kháng sinh gentamycin 50 μg/ml và streptomycin 50 μg/ml vô trùng làm đối chứng dương.
2.2. Phương pháp nghiên cứu
2.2.1. Phương pháp khuếch tán trên thạch sử dụng đĩa giấy
Các chủng vi khuẩn được hoạt hóa trong môi trường lỏng MPA và nuôi ở 30oC trong thời gian 24 h. Sau đó vi khuẩn được pha loãng trong nước muối sinh lý 0,9%. Mật độ vi sinh vật khoảng 108 CFU/ml bằng phương pháp so sánh độ đục với ống chuẩn 0,5 McFarland (108 CFU/ml) hoặc tiến hành đo quang ở λ=500 nm, DO= 0,125. Lấy 100 µl mỗi chủng cấy trải lên đĩa petri và lấy 20 µl từ 5 loại tinh dầu đưa lên tấm giấy lọc vô trùng có đường kính 0,5 cm.
2.2.2. Xác định nồng độ ức chế tối thiểu (Minimum inhibitory concentration - MIC) và nồng độ diệt khuẩn tối thiểu (Minimum bactericidal concentration - MBC)
MIC là nồng độ nhỏ nhất của tinh dầu mà tại đó nó ức chế sự phát triển của vi khuẩn. MBC là nồng độ thấp nhất của tinh dầu mà tại đó 99,9% lượng vi khuẩn bị tiêu diệt.
- Tinh dầu được pha loãng trong nước có bổ sung thêm 0,1% v/v Tween 20.
- Các chủng vi khuẩn được hoạt hóa trong môi trường lỏng MPA và nuôi ở 30oC trong 24 h.
- Cấy vi sinh vật vào ống có bổ sung tinh dầu ở nồng độ khác nhau. Ống đối chứng không bổ sung tinh dầu. Lấy 100 µl mỗi chủng cấy trải lên đĩa petri môi trường MPA (không có tinh dầu). Sau 24 h nuôi cấy ở nhiệt độ 300C, đánh giá sự phát triển của vi sinh vật bằng cách xác định số lượng khuẩn lạc phát triển trên môi trường đặc.
Khả năng ức chế được tính theo công thức:% ức chế = (1 – T/C) ˟ 100
Trong đó: T là CFU/ml mẫu thử nghiệm; C là CFU/ml đối chứng.
2.2.3. Xác định thời gian ức chế sinh trưởng của tinh dầu quế và bạc hà đối với B. cereus và E. coli
- Lấy 500 µl giống vi khuẩn đã được hoạt hóa trong môi trường lỏng cho vào 2 ống eppendorf trộn đều với 100 µl tinh dầu có nồng độ ức chế tối thiểu. Sau các khoảng thời gian tương ứng: 0’, 5’, 10’, 20’, 30’, 40’, 50’, 60’, 90’ dùng pipette hút 20 µl dịch cấy trên đĩa thạch môi trường đặc. Sau 24 h nuôi cấy ở 300C xác định thời gian diệt khuẩn bằng phương pháp đếm khuẩn lạc.
2.2.4. Phương pháp xử lý số liệuSự sai khác giữa các công thức thí nghiệm được
xác định bằng phân tích phương sai ANOVA với

59
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
sự hỗ trợ của phần mềm R. Thí nghiệm được bố trí theo kiểu khối đầy đủ hoàn toàn ngẫu nhiên với 3 lần lặp lại mỗi công thức thí nghiệm.
2.4. Thời gian và đia điểm nghiên cứuCác thí nghiệm được tiến hành từ 10/2016 -
3/2017 tại phòng thí nghiệm Khoa Công nghệ thực phẩm, Trường Đại học Kinh tế Kỹ thuật Công nghiệp.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Xác đinh hoạt tính kháng khuẩn của một số tinh dầu
Kích thước vòng kháng khuẩn thu được khi thực hiện các thí nghiệm theo mô tả trong 2.2.1 nhằm đánh giá hoạt tính kháng khuẩn của 5 loại tinh dầu và 2 loại kháng sinh đối với 7 chủng vi khuẩn được thể hiện trong bảng 1 và hình 1.
Hình 1. Kích thước vòng kháng khuẩn của 5 loại tinh dầu đối với 7 chủng vi khuẩn
Hoạt tính kháng vi sinh vật của tinh dầu được đánh giá trong mối liên quan với đường kính vòng kháng khuẩn. Có thể phân loại độ mẫn cảm của tinh dầu đối với vi sinh vật dựa vào kích thước vòng kháng khuẩn như sau (Zanil, 2000): Không mẫn cảm (-) : D < 8 mm; Mẫn cảm (+) : D = 9 - 12 mm; Rất mẫn cảm (++): D = 13 -18 mm; Cực kỳ mẫn cảm (+++): D > 18 mm.
Kết quả đạt được cho thấy, tất cả 5 loại tinh dầu đều có hoạt tính kháng khuẩn với 7 chủng vi khuẩn nghiên cứu. Tinh dầu hương nhu Ocimum gratissimum ức chế tất cả các loại vi khuẩn với đường
kính vòng kháng khuẩn nằm trong khoảng 17 - 25 mm. Chủng S. Typhimurium kém nhạy hơn (D =17 mm) so với 5 chủng còn lại.
Tinh dầu húng quế Ocimum basilicum đặc biệt nhạy cảm với vi khuẩn Gram (+) B. subtilis với đường kính vòng kháng khuẩn D = 27 mm. Với các vi khuẩn còn lại, đường kính vòng kháng khuẩn dao động trong khoảng D = 3 - 18 mm.
Tinh dầu bạc hà Mentha arvensis có giá trị kích thước vòng kháng khuẩn D nằm trong khoảng 14 - 30 mm. Giá trị kích thước vòng kháng khuẩn lớn nhất là đối với vi khuẩn S. aureus và nhỏ nhất là vi khuẩn S. typhimurium.
Tinh dầu nghệ vàng Curcuma longa có đường kính vòng kháng khuẩn 15 - 22 mm.
Tinh dầu quế có vòng kháng khuẩn 25 - 46 mm đối với tất cả các chủng. Vòng kháng khuẩn của Gram (+) B. cereus đạt 46 mm trong khi đối với Gram (-) L. damsella là 25 mm (Hình 2). Kết quả đạt được trong thí nghiệm này có tính chất tương đồng với một số công bố trên thế giới (Sikkema, J., 1994 và Burt, S., 2004).
Kết quả trên hình 2 cho thấy, kích thước vòng kháng khuẩn của tinh dầu Cinnamomum loureiri với các loại vi khuẩn Gram (+) lớn hơn so với vi khuẩn Gram (-). Tuy nhiên cũng có công bố rằng, vi khuẩn Gram (-) A. hydrophila đặc biệt nhạy cảm với hoạt tính kháng khuẩn của tinh dầu (Wan et al., 1998).
Bảng 1. Kích thước vòng kháng khuẩn của 5 loại tinh dầu đối với 7 chủng vi khuẩn
Ghi chú: Giá trị trung bình ± độ lệch chuẩn và giá trị phương sai p< 0,05
Chủng vi khuẩn
Kích thước vòng kháng khuẩn, D (mm)Ocimum
gratissimumCinnamomum
loureiriOcimum basilicum
Curcuma longa
Mentha arvensis
Gentamycin 50µg/ml
Streptomycin 50µg/ml
B. subtilis 25 ± 0,01 36 ±0,05 27 ±0,05 24 ±0,11 21 ±0,15 27 ±0,14 30 ±0,01B. cereus 22 ±0,03 46 ±0,02 18 ±0,19 22 ±0,17 27 ±0,02 25 ±0,02 16 ±0,23S. aureus 20 ±0,11 42 ±0,08 15 ±0,01 20 ±0,13 30 ±0,03 26 ±0,01 29 ±0,08E. coli 20 ±0,03 32 ±0,14 10 ±0,03 17 ±0,02 21 ±0,18 24 ±0,01 30 ±0,01S. typhimurium 17 ±0,01 30 ±0,09 3 ±0,01 15 ±0,02 14 ±0,21 23 ±0,17 20 ±0,03P. putida 18 ±0,05 27 ±0,21 12 ±0,11 19 ±0,16 22 ±0,19 18 ±0,03 20 ±0,13L. damsella 21 ±0,01 25 ±0,05 15 ±0,14 16 ±00,1 25 ±0,12 16 ±0,01 19 ±0,07
B. subtilus
L. damsella
P. putida
S. Typhimurium E. coli
S. aureus
B. cereus
Ocimum gratissimum
Cinnamomum loureiri
Ocimum basilicum
Curcuma longa
Mentha arvensis
504540353025201510
50

60
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Hình 2. So sánh kích thước vòng kháng khuẩn của tinh dầu quế Cinnamomum loureiri
với 4 vi khuẩn Gram (-) và 3 vi khuẩn Gram (+)
Vi khuẩn Bacillus cereus phân bố nhiều trong tự nhiên, nhiễm vào các loại thức ăn qua đêm hay trữ lạnh lâu, thường gây ngộ độc thực phẩm. E. coli cũng dễ dàng phân lập được từ các mẫu thực phẩm. Sự hiện diện của E. Coli trong mẫu chỉ thị khả năng có sự hiện diện của các vi sinh vật gây bệnh khác trong thực phẩm.Các nghiên cứu tiếp theo có mục đích đánh giá khả năng kháng khuẩn của 5 loại tinh dầu đối với 2 loài vi khuẩn này. So sánh vòng kháng khuẩn của 5 loại tinh dầu đối với các khuẩn B. cereus và E. coli kết quả thể hiện trên hình 3 và 4.
Hình 3. So sánh kích thước vòng kháng khuẩn đối với vi khuẩn B.cereus và E.coli.
Hình 4. Hình ảnh vòng kháng khuẩn của 5 loại tinh dầu đối với vi khuẩn
1: Tinh dầu hương nhu (Ocimum gratissimum); 2: Tinh dầu quế (Cinnamomum loureiri); 3; Tinh dầu nghệ vàng (Curcuma longa); 4: Tinh dầu bạc hà (Mentha arvensis); 5: Tinh dầu húng quế (Ocimum basilicum)
Kết quả thu được trên hình 3 và 4 cho thấy kích thước vòng kháng khuẩn của 5 loại tinh dầu đối với vi khuẩn B. cereus lớn hơn so với vi khuẩn E. coli. Kết quả thí nghiệm đã tiến hành có tính tương đồng với kết quả của một số nghiên cứu đã công bố (Cimanga et al., 2002; Delaquis et al., 2002; Pintore et al., 2002; Harpaz et al., 2003).
Kháng sinh steptomycin 50μg/ml nhạy với tất cả các loại vi khuẩn, kích thước vòng kháng khuẩn 16 - 30 mm, cao nhất là đối với B. subtilis và E. coli. Đối với trường hợp gentamycin 50 μg/ml kích thước vòng kháng khuẩn nằm trong khoảng 16 - 27 mm. Như vậy 5 loại tinh dầu và 2 loại kháng sinh đều có khả năng kháng khuẩn cao trong đó tinh dầu quế Cinnamomum loureiri cao hơn (Hình 5). Kết quả đạt được cho thấy, việc nghiên cứu, sử dụng tinh dầu để tiêu diệt vi khuẩn gây bệnh trong thực phẩm hoặc thay thế chất kháng sinh là khả quan.
Hình 5. Khả năng kháng khuẩn của 5 loại tinh dầu và 2 kháng sinh đối với vi khuẩn
Theo hình 5, phần diện tích kháng khuẩn của tinh dầu quế Cinnamomum loureiri và tinh dầu bạc hà Mentha arvensis lớn hơn các tinh dầu còn lại, tương ứng với khả năng kháng khuẩn cao nhất của 2 loại tinh dầu này. Trong các thí nghiệm tiếp theo 2 loại tinh dầu quế và bạc hà được nghiên cứu xác định nồng độ ức chế tối thiểu (MIC) và nồng độ diệt khuẩn tối thiểu (MBC) đối với 2 loài vi khuẩn là B. cereus và E. coli.
3.2. Xác đinh nông độ ức chế tối thiểu và nông độ diệt khuẩn tối thiểu của tinh dầu quế (Cinnamomum loureiri) và tinh dầu bạc hà (Mentha arvensis)
Đánh giá ảnh hưởng của nồng độ tinh dầu đến khả năng kháng khuẩn, các thí nghiệm được thực hiện theo 2.2.2. Kết quả xác định số lượng khuẩn lạc tương đương số lượng tế bào sống khi đưa vào canh trường vi sinh vật các nồng độ tinh dầu khác nhau thể hiện trên bảng 2.
Vi khuẩn
Kích t
hước
vòng
khán
g khu
ẩn (m
m)
50
45
40
35
30
25
20
15
10
5
0
B. subtilus
B. cereus
S. aureusE. coli
S. Typhimurium
P. putida
L. damsella
Kích
thướ
c vòn
g khá
ng kh
uẩn (
mm)
50
45
40
35
30
25
20
15
10
5
0
Ocimumgratissimum
Cinnamomumloureiri
Ocimumbasilicum
Tinh dầu
B. cereus E. coli
Curcuma longa Menthaarvensis
B.cereus E. coli
Ocimumgratissimum
Streptomycin50µg
Cinnamomumloureiri
Ocimumbasilicum
Gentamycin50µg
Menthaarvensis
B. subtilus
B. cereus
S. aureus
S. Typhimurium
P. putida
L. damsella
E. coli
Curcuma longa
5040302010
0

61
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 2. Khả năng kháng khuẩn của tinh dầu quế (Cinnamomum loureiri)
Kết quả đạt được trong bảng 2 cho thấy độ mẫn cảm với tinh dầu quế (Cinnamomum loureiri) của B. cereus cao hơn E. coli.
Tỉ lệ MBC/MIC được dùng để xác định hoạt tính kìm khuẩn hoặc là diệt khuẩn của một tinh dầu. Nếu giá trị này nhỏ hơn 4 (MBC/MIC ≤ 4), tinh dầu được coi là một chất diệt khuẩn. Nếu MBC/MIC > 4, tinh dầu được xác định là một chất kìm khuẩn. MIC và MBC của tinh dầu quế với 2 loài vi khuẩn thể hiện trên bảng 3.
Bảng 3. MIC và MBC của tinh dầu quế Cinnamomum loureiri của vi khuẩn
Kết quả bảng 3 cho thấy tinh dầu quế Cinnamomum loureiri là chất diệt khuẩn đối với cả 2 loài vi khuẩn.
Nghiên cứu khả năng kháng khuẩn của tinh dầu bạc hà (Mentha arvensis), các thí nghiệm được thực hiện theo 2.2.2. Kết quả được thể hiện trên bảng 4.
Bảng 4. Khả năng kháng khuẩn của tinh dầu bạc hà (Mentha arvensis)
Kết quả thu được từ các bảng 3 và 4 cho thấy độ mẫn cảm của B. cereus và E. coli với tinh dầu bạc hà cao hơn so với tinh dầu quế. Nồng độ tinh dầu bạc hà ức chế hoàn toàn vi khuẩn B. cereus là 1,5% và 4,5% với E. coli. MIC và MBC của tinh dầu bạc hà với 2 vi khuẩn nghiên cứu thể hiện trên bảng 5.
Bảng 5. Kết quả xác định MIC và MBC tinh dầu bạc hà (Mentha arvensis)
Với tỉ lệ MBC/MIC thu được cho thấy tinh dầu bạc hà là chất kìm khuẩn.
3.3. Xác đinh thời gian ức chế sinh trưởng của tinh dầu quế và bạc hà đối với B. cereus và E. coli
Để xác định thời gian tối thiểu tinh dầu có thể tiêu diệt hoàn toàn 2 vi khuẩn nghiên cứu, thí nghiệm được tiến hành như mục 2.2.3. Các chủng vi khuẩn được nuôi trong môi trường lỏng đến mật độ 108 CFU/ml, bổ sung tinh dầu quế và bạc hà nồng độ 0,25% cho B. cereusvà 0,5% cho E. coli. Kết quả xác định số khuẩn lạc phát triển thể hiện trên bảng 6.
Bảng 6. Ảnh hưởng của thời gian phơi nhiễm lên khả năngkháng khuẩn của tinh dầu
Kết quả đạt được trên bảng 6 cho thấy tinh dầu quế ức chế hoàn toàn với B. cereus sau thời gian 10 phút và với E. coli là 20 phút. Độ mẫn cảm của tinh dầu bạc hà với vi khuẩn thấp hơn so với tinh dầu quế. Tinh dầu bạc hà ức chế hoàn toàn B. cereus sau 40 phút và với E. coli thời gian ức chế hoàn toàn là 90 phút.
Vi khuẩnNông độ tinh dầu
(%)
Mật độ tế bào
(CFU/ml)
% ức chế
Bacillus cereus0 2,8 ˟ 103
0,25 1,4 ˟ 102 95 0,5 0 100
E. coli
0 1,42 ˟ 103
0,5 9,3 ˟ 102 34,5 1,0 5,8 ˟ 102 59,15 1,5 1,3 ˟ 102 90,84 2 0 100
Vi khuẩn MIC MBC MBC/MICB. cereus 0,25 0,5 2E. coli 0,5 2,0 4
Vi khuẩnNông độ tinh dầu,
(%)
Mật độ tế bào
(CFU/ml)% ức chế
B. cereus
0 3,6 ˟ 103
0,25 9,8 ˟ 102 72,77 0,5 3,1 ˟ 102 91,391,0 1,1 ˟ 102 96,941,5 0 100
E. coli
0 4,4 ˟ 103
0,5 3,2 ˟ 103 27,27 2,5 9,7 ˟ 102 77,953 7,3 ˟ 102 83,41 4 1,7 ˟ 102 96,14
4,5 0 100
Vi khuẩn MIC MBC MBC/MICB. cereus 0,25 1,5 6E. coli 0,5 4,5 9
Tinh dầu
VSV (CFU/ml)
TG (phút)
B. cereus103
E. coli103
Quế0 2,08 3,36
10 0 0,220 0 0
Bạc hà
0 2,52 5,3610 1,94 3,5620 1,1 1,4830 0,2 1,640 0,12 1,1250 0 0,8460 0 0,490 0 0

62
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
IV. KẾT LUẬN Quá trình thực nghiệm cho thấy khả năng kháng
khuẩn của một số tinh dầu phụ thuộc vào nhiều yếu tố như chủng vi sinh vật, nồng độ, thời gian tiếp xúc. Kết quả cho thấy các tinh dầu quế, tinh dầu bạc hà có khả năng kháng khuẩn tốt nhất, cao hơn kháng sinh đối chứng. Nồng độ ức chế tối thiểu của tinh dầu quế với B. cereus là 0,25% và với E. coli là 0,5%. Tỉ lệ MBC/MIC thu được đã chỉ ra rằng, tinh dầu quế là chất diệt khuẩn và tinh dầu bạc hà là chất kìm khuẩn đối với B. cereus và E. coli.
Kết quả của các nghiên cứu cho thấy khả năng ứng dụng các hợp chất thiên nhiên có hoạt tính sinh học như tinh dầu thay thế cho kháng sinh.
TÀI LIỆU THAM KHẢONguyên Văn Kính và nhóm Nghiên cứu quốc gia của
GARP- VN, 2010. Phân tích thực trạng: sử dụng kháng sinh và kháng kháng sinh tại Việt Nam p. 2-6.
Đỗ Tất Lơi, 2006. Những cây thuốc và vị thuốc Việt Nam. NXB Y học.
Burt, S., 2004. Essential oils: their antibacterial properties and potential applications in foods-a review. Int. J. Food Microbiol, (94): 223-253.
Fisher, K., Phillips, C., 2009. The ecology, epidemiology and virulence of Enterococcus. Microbiology, 155: 1749-1757.
Harpaz, S., Glatman, L., Drabkin, V., Gelman, A., 2003. Effects of herbal essential oils used to extend the shelf life of fresh water reared Asian sea bass fish (Lates calcarifer). Journal of Food Protection, 66 (3): 410-417.
Pintore, G., Usai, M., Bradesi, P., Juliano, C., Boatto, G., Tomi, F., Chessa, M., Cerri, R., Casanova, J., 2002. Chemical composition and antimicrobial activity of Rosmarinus officinalis L. oils from Sardinia and Corsica. Flavour and Fragrance Journal, 17: 15-19.
Sikkema, J., Bont J.A.M., Poolman, B., 1994. Interactions of cyclic hydrocarbons with biological membranes. J. Biol. Chem, 269: 8022-8028.
Tohidpour, A., Sattari, M., Omidbaigi, R., Yadegar, A., Nazemi, J., 2010. Antibacterial effect of essential oils from two medicinal plants against Methicillin-resistant Staphylococcus aureus (MRSA). Phytomedicine, 17: 142-145.
Wan, J., Wilcock, A., Coventry, M.J., 1998. The effect of essential oils of basil on the growth of Aeromonas hydrophila and Pseudomonas fluorescens. J. Appl. Microbio l, 84: 152-158 .
Zanil, A., Junior, C., 2000. Biological screening of Brazilian meditional plants. Braz J. Sci, 95: 367-373.
Study on the antimicrobial activity of essential oilsNguyen Thi Mai Huong, Ho Tuan Anh
AbstractThis article presents research on the antimicrobial properties of five essential oils from Ocimum gratissimum, Cinnamomum loureiri, Ocimum basilicum, Mentha arvensis, Curcuma longa for B. subtilis, B. cereus, S. aureus, E. coli S. typhimurium, P. putida, L. damsella compared with two antibiotics, gentamycin and streptomycin. Disc diffusion method was established so that all essential oils had antimicrobial ability, in which Cinnamomum loureiri showed the highest ability. It was determined that the minimum inhibitory concentration (MIC) and bactericidal time of Cinnamomum loureiri oil for B. cereus were 0.25% and 10 minutes, respectively, and for E. coli were 0.5%, 20 minutes, respectively. Based on the minimum bactericidal concentration/minimum inhibitory concentration ratio (MBC / MIC), Cinnamomum loureiri oil was determined as a disinfectant, and Mentha arvensisas as a bacteriocin for B. cereus and E. coli. This study showed that some essential oils indicated a comparable effectiveness as an antibiotics.Key words: Essential oil, antimicrobial, antibacterial ring, minimum inhibitory concentration, minimum bactericidal concentration
Ngày nhận bài: 2/7/2017Ngày phản biện: 15/7/2017
Người phản biện: PGS. TS. Nguyễn Thị Minh TúNgày duyệt đăng: 27/7/2017

63
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀValidamycin A (Val-A) là một chất kháng sinh
nhóm aminoglycoside có hoạt tính kháng nấm được sinh tổng hợp chủ yếu bởi Streptomyces hygroscopicus var. limoneus (Liao et al., 2009; Iwasa et al., 1971) và S. hygroscopicus subsp. jinggangensis 5008 (Shen, 1988; Zhou et al., 2014). Nhờ hiệu quả phòng trừ nấm cao cũng như an toàn cho con người và động vật, nên Val-A trở thành một trong những kháng sinh quan trọng nhất và được sử dụng rộng rãi để trừ bệnh đốm vằn hại lúa, ngô, bệnh đốm lá và thân lúa, ngô do Rhizoctonia solani, R. oryzae và Sclerotium oryzae-sativa gây nên. Ngoài ra, val-A còn trừ bệnh thối củ, thối rễ khoai tây, bông, cà chua, nấm hồng trên cây cao su và nhiều loại rau do nấm R. solani gây nên (Wang et al., 2001). Val-A là chất kìm hãm yếu (IC50= 10-3 M) các enzyme thủy phân đường khác như maltase, isomaltase và saccharase trong ruột lợn. Trong R. solani, kháng sinh được vận chuyển nhanh đến sợi nấm và được glycosidase thủy phân thành validoxylamine A, đây là chất kìm hãm trehalase của nấm mạnh cả ở in vitro và in vivo (Kameda et al., 1975). Trong tế bào nấm bệnh, trehalase xúc tác chuyển hóa trehalose thành đường glucose - nguồn năng lượng cho nấm phát triển. Do đó, dưới tác động của val-A, tế bào nấm sẽ tạo nhánh bất thường, bị khô dần và chết do không có khả năng tổng hợp glucose để sinh trưởng (Demain, 1974). Các chủng S.hygroscopicus có hoạt tính tổng hợp kháng sinh rất yếu, không mang tính ổn định và bền vững cao để hạn chế hình thành các sản phẩm phụ, rút ngắn thời gian lên men và giảm chi phí sản xuất. Do vậy, nghiên cứu này trình bày kết quả gây
đột biến ở chủng S. hygroscopicus 11405 dưới tác động của các tác nhân như tia UV, chất gây đột biến MNNG và lựa chọn những biến chủng có hoạt tính cao cho lên men sinh tổng hợp Val-A.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứuChủng xạ khuẩn S. hygroscopicus 11405 và chủng
nấm kiểm định Rhizoctonia solani VP nhận được từ Bộ sưu tập giống vi sinh vật của Phòng Công nghệ lên men, Viện Công nghệ sinh học, Viện Hàn lâm Khoa học và Công nghệ Việt Nam.
Môi trường ISP2 (g/l): Cao nấm men 4; cao malt 10; dextrose 4; thạch 20 pH 7,3, nước cất 1000 ml. Môi trường lên men cơ bản FM3 (g/l): Bột ngô 90,0; bột đậu tương 40,0; cao nấm men 5,0; KH2PO4 1,0; pH 7,0; nước cất 1000 ml. Môi trường xác định hoạt tính kháng sinh A1 (g/l): Saccharose 1,0; cao thịt 5,0; pepton 10,0; thạch 8,0; pH 7,0, nước cất 1000 ml.
2.2. Phương pháp nghiên cứu
2.2.1. Xác định hoạt tính kháng sinh (HTKS)Theo phương pháp màng ngược của Iwasa và cộng
tác viên (1971). Hoạt tính Val-A được xác định bằng phương pháp phân tích sắc ký lỏng cao áp (HPLC) tại Trung tâm Kỹ thuật I, Tổng cục Tiêu chuẩn Đo lường Chất lượng, Bộ Khoa học và Công nghệ.
2.2.2. Đánh giá sự biến động tự nhiên về HTKS của các khuẩn lạc (KL)
Đầu tiên xác định HTKS trung bình (X) của các khuẩn lạc, rồi từ đó xác định độ lệch chuẩn:
δ = √∑(Xi –X)2/(n-1)
1 Viện Công nghệ sinh học, Viện Hàn lâm Khoa học và Công nghệ Việt Nam2 Khoa Công nghệ thực phẩm, Học viện Nông nghiệp Việt Nam
NÂNG CAO KHẢ NĂNG SINH TỔNG HỢP VALIDAMYCIN-A TỪ CHỦNG Streptomyces hygroscopicus 11405 BẰNG ĐỘT BIẾN TẾ BÀO TRẦN
Vũ Thị Hạnh Nguyên1, Nguyễn Thị Thanh Thủy2, Phí Quyết Tiến1
TÓM TẮTValidamycin A (Val-A) là chất kháng sinh thuộc nhóm aminoglycoside được sinh tổng hợp bởi chủng Streptomyces
hygroscopicus. Mục đích nghiên cứu này là tạo tế bào trần và gây đột biến tế bào trần (TBT) chủng S. hygroscopicus 11405 bằng tia UV và N-metyl-N’-nitro-N-nitrosoguanidin (MNNG) để nâng cao hiệu suất chủng sản xuất. TBT S. hygroscopicus 11405 được tạo tốt nhất khi phát triển ở đầu pha logarit có bổ sung 0,4% glycin và xử lý với lysozyme 1 mg/ml ở 37°C, 60 phút. Đột biến TBT chủng 11405 bằng tia UV không cho kết quả tốt, tuy nhiên gây đột biến bằng MNNG với nồng độ 0,1 mg/ml trong 30 - 40 phút tạo được tỷ lệ đột biến cao nhất. Sau khi xử lý đột biến TBT đã thu được 109 biến chủng được phân thành 5 typ màu, trong đó typ 1, 5 chiếm đa số, đạt tới 38,5 - 49,6%. Áp dụng quy trình gây đột biến bằng MNNG đã tạo được biến chủng S. hygroscopicus 11405-115 có hoạt tính Val-A cao hơn chủng gốc 70%. Thời điểm sinh tổng hợp kháng sinh cao nhất của biến chủng này là sau 48 giờ lên men đạt 6,87 mg/ml.
Từ khóa: Đột biến, Rhizoctonia solani, Streptomyces hygroscopicus, tế bào trần, validamycin-A

64
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Xi là HTKS của 1 khuẩn lạc có HTKS lớn hơn X+2δ là những biến chủng dương, còn nhỏ hơn X-2δ là những biến chủng âm.
2.2.3. Thu nhận và phục hồi TBT từ xạ khuẩn Theo phương pháp của Hopwood và cộng tác
viên, 1985.
2.2.4. Gây đột biến bằng cách chiếu tia UV Lấy bào tử từ môi trường nghèo dinh dưỡng, sau
đó tạo TBT và hoạt hóa trên môi trường giàu để các bào tử chuyển dần sang giai đoạn tiền nảy mầm (8 - 10 giờ). Bào tử tiền nảy mầm được chiếu tia UV với công suất của đèn là 30 W, bước sóng 2537 A° (260 nm), cường độ chiếu sáng 8x103 ergs/cm2/giây, thời gian chiếu thay đổi từ 5 - 80 giây. Sau đó pha loãng và cấy gạt trên môi trường ISP2.
2.2.5. Gây đột biến bằng MNNG Dịch TBT được xử lý bằng hóa chất gây đột biến
MNNG (ở các nồng độ 0; 0,001; 0,01; 0,05; 0,1; 0,3; 0,5 và 1,0 mg/ml) trong đệm TM pH = 8,0 (0,05 M Tris pH 8,0; 0,05 M maleic acid) để tạo tế bào đột biến. Đem dịch có chứa tế bào đột biến ly tâm, thu tế bào và pha loãng bằng đệm TM đến nồng độ 10-5 và cấy gạt trên môi trường ISP2, nuôi ở 30°C trong 3-4 ngày. Đếm số khuẩn lạc, quan sát sự biến đổi mầu sắc khuẩn lạc, khuẩn ty cơ chất và xác định hoạt tính kháng sinh (Kim et al., 1983).
2.2.6. Xây dựng đường cong sống sótĐồ thị sống sót biểu diễn mối quan hệ giữa tỷ lệ
% tế bào sống sót với thời gian xử lý. Sau khi nuôi cấy đếm số khuẩn lạc ở các độ pha loãng, thời gian chiếu tia UV, xử lý với MNNG và đĩa kiểm chứng. Từ đó, xác định được tỷ lệ % tế bào sống sót (% SS) ở các thời gian chiếu tia UV hoặc xử lý với MNNG khác nhau theo công thức (Kim et al., 1983):
% SS = NmNo
˟ 100 %
Trong đó Nm: số khuẩn lạc sống sót sau đột biến; No: số khuẩn lạc ở mẫu kiểm chứng.
2.2.7. Xác định sự biến đổi màu sắc khuẩn ty khí sinh (KTKS) và khuẩn ty cơ chất (KTCC)
Các khuẩn lạc của các biến chủng được nuôi cấy trên môi trường ISP2 và màu sắc KTKS và KTCC được so sánh theo bảng màu của Tresner và Backus, 1963.
2.2.8. Động thái quá trình lên menTiến hành lên men trong bình tam giác 500 ml
với pH ban đầu là 6,0, ở 37°C trong 120 giờ. Theo dõi sự biến động của sinh khối và HTKS theo thời gian, cứ 12 giờ lấy mẫu 1 lần để kiểm tra.
2.2.9. Phương pháp xử lý số liệuCác thí nghiệm được lặp lại 3 lần. Kết quả nghiên
cứu được xử lý và lấy số liệu trung bình theo lý thuyết thống kê sinh học trên phần mềm Excel.
2.3. Thời gian và đia điểm nghiên cứu- Thời gian nghiên cứu: Từ năm 2015 đến
năm 2016.- Địa điểm nghiên cứu: Phương pháp phân tích
sắc ký lỏng cao áp (HPLC) thực hiện tại Trung tâm Kỹ thuật I, Tổng cục Tiêu chuẩn Đo lường Chất lượng, Bộ Khoa học và Công nghệ. Các thí nghiệm khác đều được thực hiện tại phòng Công nghệ lên men, Viện Công nghệ sinh học.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Biến động tự nhiên về hoạt tính kháng sinh của chủng S. hygroscopicus 11405
Một trong những tiêu chí chọn chủng sản xuất là chọn các khuẩn lạc có hoạt tính cao do biến động tự nhiên của chúng trong các ống giống theo chiều hướng giảm khả năng sinh kháng sinh (Lê Gia Hy, 1994). Kết quả lựa chọn ngẫu nhiên 76 khuẩn lạc của chủng S. hygroscopicus 11405 sau thời gian bảo quản 3 tháng và xác định hoạt tính kháng sinh của chúng được được trình bày trên các hình 1 và 2. Qua tính toán cho thấy, đường kính vòng ức chế trung bình là 18,8 mm có độ lệch chuẩn: δ = 3,6. Như vậy, các khuẩn lạc có đường kính vòng ức chế trên
+ 2 δ (26,0 mm) là chủng có HTKS cao hơn chủng gốc và chủng có đường kính vòng ức chế nhở hơn
- 2 δ (11,6 mm) là chủng có HTKS thấp hơn chủng gốc S. hygroscopicus 11405 (Hình 1).
Hình 1. Kết quả sàng lọc tự nhiên về hoạt tính kháng nấm R. solani của chủng S. Hygroscopicus 11405

65
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Hình 2. Biến động tự nhiên về hoạt tính kháng nấm R. solani của chủng S. hygroscopicus 11405
sau 3 tháng bảo quản
Chủng S. hygroscopicus 11405 có hoạt tính kháng sinh chưa cao và không ổn định. Chủng S. hygroscopicus 11405 trước khi được bảo quản có khả năng ức chế nấm R. solani với đường kính vòng ức chế dao động trong khoảng 18 - 21 mm. Sau 3 tháng, có 7,9% số khuẩn lạc cấy tách ra giảm hoạt tính kháng nấm và có 6,5% số khuẩn lạc có hoạt tính
kháng nấm cao hơn chủng gốc (Hình 2). Trong 6,5% khuẩn lạc này chọn ra một khuẩn lạc có hoạt tính mạnh nhất để gây đột biến nhân tạo.
3.2. Nâng cao khả năng sinh tổng hơp Val-A bằng kỹ thuật đột biến TBT
3.2.1. Tạo TBT từ chủng S. hygroscopicus 11405Để thu được tế bào trần, chủng 11405 được nuôi
trong môi trường FM3 dịch thể, sinh khối sau đó được xử lý với lysozyme. Xử lý bằng lysozyme với nồng độ 1 mg/ml từ 30 đến 120 phút cho thấy, sau 30 phút các đoạn khuẩn ty bị cắt, TBT cũng được tạo thành, sau 60 phút lượng TBT tạo thành rất nhiều, sau 90 phút số lượng TBT ít đi, có thể một số TBT đã bắt đầu bị phá hủy. Số lượng TBT từ khuẩn ty ở 48 giờ nuôi cấy tạo thành cao nhất, quá trình diễn ra khá nhanh và có hiệu quả nhất (Hình 3). Kết quả cho thấy quá trình tạo TBT hiệu quả nhất khi giống đang phát triển ở đầu pha logarit tức là khoảng 48 giờ nuôi cấy, trong môi trường có bổ sung 0,4% glycin, nồng độ lysozyme và thời gian xử lý tế bào thích hợp nhất là 1 mg/ml ở 37ºC và 60 - 70 phút.
Hình 3. Ảnh hưởng của thời gian nuôi cấy tới khả năng tạo thành TBT của chủng S. hygroscopicus 11405
Hình 4. Đường cong sống sót của chủng S. hygroscopicus 11405 sau khi xử lý UV
Thời gian (giờ)
Số lư
ợng
TBT/
ml (
x105 )
1816141210
86420
24 36 48 60 72
0
20
40
60
80
100
120
0 20 40 60 80
Tỷ lệ
sống
sót (
%)
Thời gian xử lý (giây)
50
45
40
35
30
25
20
15
10
5
0(10-13) (14-17) (18-21) (22-24) (25-29)
Tỷ lệ
chủ
ng (%
)
Đường kính vòng ức chế (mm)
3.2.2. Ảnh hưởng của thời gian chiếu UV và hàm lượng chất MNNG lên khả năng sống sót của chủng S. hygroscopicus 11405
Kết quả tỷ lệ sống sót của TBT chủng S. hygroscopicus 11405 sau khi chiếu UV và số liệu thu được sau xử lý với MNNG cho thấy: Khi chiếu UV (Hình 4), thời gian chiếu càng lâu thì tỷ lệ sống sót càng giảm. Chiếu UV trên 70 giây thì không còn khuẩn lạc nào phát triển. Theo tài liệu được công bố (Adrio and Demain, 2008), ở độ sống sót từ 1-10% sẽ cho xác suất chủng bị đột biến cao nhất. Dựa vào đồ thị sống sót có thể xác định được thời gian chiếu UV lên TBT thích hợp nhất là 40 đến 50 giây. Tác động gây đột biến của tia UV chủ yếu là do acid nucleic và các hợp chất trong tế bào hấp thụ. Sự hấp thụ càng mạnh thì khả năng đột biến càng cao. Acid nucleic
hấp thụ bước sóng 260 nm cao nhất nên bước sóng này có hiệu quả gây đột biến nhiều nhất. Tia UV có thể gây nên các đột biến gen và các đột biến cấu trúc nhiễm sắc thể, gây biến đổi quang oxy hoá các gốc purin và pirimidin. Tác động của tia UV làm cho adenin bị biến thành hypoxanthin, dẫn đến những thay đổi hoá học của DNA.
Xử lý bào tử trần với MNNG ở nồng độ và thời gian khác nhau cho thấy, nồng độ tác động càng cao và thời gian ủ càng dài thì tỉ lệ sống sót của tế bào trần càng giảm. Bảng 1 thể hiện kết quả xử lý với MNNG trong 1 phút. Khi nồng độ MNNG càng cao thì tỷ lệ sống của TBT càng giảm, khi tăng lượng MNNG lên 0,01 và 0,05 mg/ml tỷ lệ sống sót lần lượt 35,2 và 26,1%, còn khi xử lý ở nồng độ 0,1 mg/ml MNNG tỷ lệ sống sót đạt 4,9% và TBT không thể tồn tại ở nồng độ 1 mg/ml trở lên.

66
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 1. Ảnh hưởng của nồng độ xử lý MNNG lên khả năng sống sót TBT
của chủng S. hygroscopicus 11405
Bảng 2. Ảnh hưởng của thời gian xử lý 0,1 mg/ml MNNG lên khả năng sống sót TBT
của chủng S. hygroscopicus 11405
Do vậy, hàm lượng chất gây đột biến MNNG cần sử dụng với mức 0,1 mg/ml và sau 40 phút xử lý TBT (Bảng 2) sẽ cho số khuẩn lạc sống sót trong khoảng 1 - 10% và khả năng gây đột biến là cao nhất. Đây cũng là nồng độ thích hợp cho đột biến TBT của chủng Micromonospora rosaria (Kim et al., 1983).
3.2.3. Sự biến động hình thái và hoạt tính kháng sinh của chủng S. hygroscopicus 11405 sau đột biến
Chủng S. hygroscopicus 11405 gốc có đặc điểm trên môi trường ISP2: bề mặt khuẩn lạc khô, xù xì, xẻ thùy, mầu trắng vàng và có hoạt tính Val-A (đường kính vòng kháng R. solani) là 15 mm. Sau đột biến bằng tia UV và chất gây đột biến thu được 109 chủng S. hygroscopicus 11405 phân thành 5 typ theo màu sắc của khuẩn lạc và khuẩn ty cơ chất và phân thành 4 nhóm theo khả năng sinh tổng hợp kháng sinh. Trong đó typ 5 nhiều nhất chiếm 49,6% KTCC màu vàng và KTKS màu trắng giống với chủng gốc, sau đó là typ 1 chiếm 38,5% KTCC màu đỏ và KTKS màu trắng, nhỏ nhất là typ 4 chiếm 0,9% KTKS và KTCC đều màu vàng, cuối cùng là typ 2 và 3 có KTCC đỏ,
vàng và KTKS đều là xám, lần lượt chiếm 7,3 và 3,7% (số liệu không trình bày). Điều này đã khẳng định tác động của UV và MNNG đến quá trình hình thành màu sắc của KTCC và KTKS. Về hoạt tính kháng sinh thì nhóm 1 gồm biến chủng có hoạt tính cao hơn chủng gốc chiếm số lượng khá lớn là 23, chiếm 21,1%. Nhóm 2 gồm biến chủng có hoạt tính tương đương chủng gốc là 15, chiếm 4,58%. Còn lại là biến chủng (nhóm 3) có hoạt tính nhỏ hơn chủng gốc. Đặc biệt có hai biến chủng (nhóm 4) mất hẳn khả năng sinh kháng sinh (số liệu không trình bày).
Bảng 3. Kết quả lên men các biến chủng của S. hygroscopicus 11405 sau khi đột biến
Đường kính vòng kháng nấm trung bình của các lần đo = 15,1 mm. Độ lệch chuẩn δ = 2,34. Các khuẩn lạc có đường kính vòng vô khuẩn lớn hơn + 2δ (D >19,8 mm) là những biến chủng dương tính, ngược lại các khuẩn lạc có đường kính vòng vô khuẩn nhỏ hơn - 2δ (D < 10,4 mm) là những biến chủng âm tính. Theo đồ thị phân bố hoạt tính kháng sinh với các tác nhân gây đột biến khác nhau có 6 khuẩn lạc là biến chủng dương tính, nghĩa là tần suất xuất hiện đột biến không cao. Khi chiếu tia UV lên TBT đã không thu được chủng nào cho hoạt tính cao (số liệu không trình bày). Sử dụng chất gây đột biến MNNG có tác dụng nhiều hơn. So với chủng gốc 11405 (hàm lượng Val-A là 3,65 mg/ml), các biến chủng có hoạt tính kháng sinh cao hơn (Bảng 3), trong đó biến chủng S. hygroscopicus 11405-115 có hoạt tính cao nhất đạt 6,2 mg/ml, tăng 170%. Như vậy, gây đột biến TBT bằng chất MNNG cho kết quả tốt nhất cũng có thể là do MNNG là gây nên các đột biến khử amin đối với base của DNA. Ngoài ra, MNNG có thể tác dụng làm đứt mối liên kết hydro trong các sợi DNA đang phân chia, tạo ra một loại biến đổi khác như kiểu cấu trúc lại nhiễm sắc thể (Kim et al., 1983).
Nông độ MNNG (mg/ml)
Tỷ lệ sống sót của TBT (%)
0 1000,001 69,50,01 35,20,05 26,10,1 4,90,3 0,540,5 0,151,0 0
Thời gian xử lý (phút) Tỷ lệ sống sót của TBT (%)
0 10010 23,620 17,530 8,240 4,350 0,5960 0,0870 0
Chủng xạ khuẩn
HTKS theo Dimitrieva
(mg/ml)
Tỷ lệ HTKS (%)
ĐC-chủng 11405 3,65 100
11405-1A 6,1 16711405-25A 5,11 14011405-70B 5,58 15311405-115 6,2 17011405-233 5,84 16011405-232 5,18 142

67
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
3.3. Ảnh hưởng của thời gian lên men đến khả năng sinh Val-A của biến chủng S. hygroscopicus 11405-115
Kéo dài thời gian lên men không đồng nghĩa với tăng hoạt tính kháng sinh mà ngược lại, có thể dẫn đến hình thành các chất độc, gây ức chế quá trình sinh tổng hợp kháng sinh (Nguyễn Văn Cách, 2004). Lựa chọn thời gian lên men thích hợp không những có thể rút ngắn thời gian lên men, giảm chi phí sản xuất mà còn giúp thu hồi kháng sinh có hoạt tính cao nhất.
Hình 5. Động thái quá trình lên men sinh tổng hợp Val-A của biến chủng 11405-115
Biến chủng S. hygroscopicus 11405-115 sinh trưởng cực đại sau 60 giờ lên men (đạt 7,6 mg sinh khối ướt/ml) và sinh tổng hợp Val-A cao nhất sau 48 giờ, đạt 6,9 mg/ml. Sau thời gian này, nếu kéo dài thời gian lên men, hoạt tính Val-A không tăng mà có xu hướng giảm nhẹ (Hình 5). Ở thời điểm 96 giờ, hoạt tính kháng sinh còn lại 6,2 mg/ml. Như vậy, thời điểm thích hợp nhất để thu hồi kháng sinh là sau 48 giờ lên men. Đây là thời điểm sớm hơn so với nhiều nghiên cứu khác lựa chọn khi lên men sinh tổng hợp Val-A (Iwasa et al., 1970; Liao et al., 2009).
3.4. Xác đinh hàm lương Val-A của dich lên men thô bằng sắc ký lỏng cao áp (HPLC)
Dịch lên men thu được từ biến chủng S. hygroscopicus 11405-115 nuôi trên môi trường FM3 ở điều kiện lên men thích hợp (37oC; pH 7,0; tỷ lệ tiếp giống 10%) sau 48 giờ, được xác định bằng phương pháp sắc ký lỏng cao áp.
Kết quả hình 6 cho thấy, dịch lên men có chứa Val-A do thời gian lưu (retention time) của hoạt chất của dịch lên men với Val-A chuẩn có độ tương đồng là 15,4 và 15,7 phút. Căn cứ vào độ rộng của đỉnh peak cho thấy, dịch lên men thô từ chủng S. hygroscopicus 11405-115 chứa hàm lượng val-A là 0,687%, tương ứng với 6,87 mg/ml.
Sinh khối9876543210
0 12 24 36 48 60 72 84 96 108Thời gian (giờ)
Hàm
lượn
g V
al -
A v
à si
nhkh
ối ư
ớt (m
g/m
L)
Hàm lượng Val - A
IV. KẾT LUẬNNâng cao hoạt tính kháng sinh bằng kỹ thuật tạo,
phục hồi TBT từ chủng xạ khuẩn S. hygroscopicus 11405 và gây đột biến bằng MNNG, đã chọn lọc được biến chủng S. hygroscopicus 11405-115 có hoạt tính kháng sinh cao hơn 70% so với chủng gốc. Thời điểm sinh tổng hợp kháng sinh cao nhất của biến chủng này trong môi trường lên men trên bình tam giác sau 48 giờ đạt 6,87 mg/ml. Biến chủng nhận
được đáp ứng mục tiêu nghiên cứu là có hoạt tính kháng sinh ổn định trong quá trình bảo quản và lên men validamycin, thời điểm thu nhận Val-A rút ngắn hơn so với một số công bố khác (sau 48 giờ lên men). Kết quả thu được khả quan và thuận lợi cho nghiên cứu tiếp theo để sản xuất Val-A, định hướng tạo sản phẩm thuốc bảo vệ thực vật an toàn và hiệu quả, dùng điều trị bệnh cây trồng do nấm R. solani gây ra, đáp ứng nhu cầu rất lớn trong nông nghiệp.
Hình 6. Sắc đồ HPLC của val-chuẩn (a) và val-A có trong dịch lên men của biến chủng S. hygroscopicus 11405-115 (b)
buV
250000V
alid
amyc
in
150000
200000
100000
50000
0
0 5 10 15 20 25 30min
uV200000
Val
idam
ycin
150000
100000
50000
0
0 5 10 15 20 25 30min
a

68
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
LỜI CẢM ƠNNhóm nghiên cứu cảm ơn sự hỗ trợ kinh phí
của Công ty cổ phần Thuốc sát trùng Việt Nam - Bộ Công thương và sự hỗ trợ trang thiết bị của Phòng thí nghiệm trọng điểm công nghệ gen - Viện Công nghệ sinh học, Viện Hàn lâm Khoa học Công nghệ Việt Nam.
TÀI LIỆU THAM KHẢONguyên Văn Cách, 2004. Công nghệ lên men các chất
kháng sinh. Nhà xuất bản Khoa học Kỹ thuật. Hà Nội.Lê Gia Hy, 1994. Nghiên cứu xạ khuẩn thuộc chi
Streptomyces sinh chất kháng sinh chống nấm gây bệnh đạo ôn và thối cổ rễ phân lập ở Việt Nam. Luận án phó tiến sĩ sinh học. Hà Nội.
Adrio J. L., Demain A. L., 2008. Strain improvement for production of pharmaceuticals and other microbial metabolites by fermentation, Progress in Drug Research. Birkhauser Verlag AG, Basel, Switzerland, 65: 251-290.
Demain, A. L., 1974. How do antibiotic-producing microorganisms avoid suicide. Annuals of the NewYork Academy of Sciences, 235: 601-612.
Hopwood D. A., Bibb M. J., Chater K. F, Kieser T., Bruton C. J., Kieser H. M., Lydiate D. J., Smith C. P., Ward J. M., Norwich H. S., 1985. Genetic manipulation of Streptomyces: A laboratory manual. The John Innes Foundation, Paperback,. ISBN 07084 0036 0: 35-41.
Iwasa T., Higashide E., Yamamoto H., Shibata M., 1971. Studies on validamycins, new antibiotics.III. Bioassay methods for the determination of validamycins. Jantibiot, 24 (2): 114-118.
Kameda Y., Horii S., Yamano T., 1975. Microbial transformation of validamycins. The journal of antibiotic, 28: 298-306.
Kim K. S., Cho N. Y., Pai H. S., Ryu D. D. Y., 1983. Mutagenesis of Micromonospora rosaria by using protoplasts and mycelia fragments. Applied and Environmental Microbiology, 46(3): 689-693.
Liao Y., Wei Z. H., Bai L., Deng Z., Zhong J. J., 2009. Effect of fermentation temperature on validamycin A production by Streptomyces hygroscopicus 5006. Journal of Biotechnology, 142: 271-274.
Shen Y., 1988. Research and development of agro-antibiotic, validamycin. Chinese journal of antibiotics, 6: 58-61.
Tresner H. D., Backus E. J., 1963. System of color wheels for Streptomycete taxonomy. Applied Microbiology, 11: 335-338.
Wang S., Chen S., Yu Z., 2001. Selection of nitrogen sources and proportion of carbon and nitrogen of jinggangmycin of Streptomyces hygroscopicus. Journal of Huazhong Agricultural: 06.
Zhou T. C., Kim B. G., Zhong J. J., 2014. Enhanced production of validamycin A in Streptomyces hygroscopicus 5008 by engineering validamycin biosynthetic gene cluster. Appl Microbiol Biotechnol, 98 (18): 7911-7922.
Enhancement of Validamycin-A productivity of Streptomyces hygroscopicus 11405 by protoplast mutagenesis
Vu Thi Hanh Nguyen, Nguyen Thi Thanh Thuy, Phi Quyet TienAbstractValidamycin A (Val-A) is an anti-fungal aminoglycoside antibiotic produced by Streptomyces hygroscopicus. The present work aims to enhance the antibiotic productivity of the actinomycete strain S. Hygroscopicus 11405 by protoplasts mutagenesis UV and N-methyl-N’-nitro-N-nitrosoguanidine (MNNG). The obtained results showed that a maximal yield of protoplast was obtained within 60 - 70 min at 37°C after adding lysozyme at a concentration of 1 mg/ml to mycelium in the logarithmic phase of growth (48 hours cultivation) in medium containing 0.4% (w/v) glycine. The suitable conditions for MNNG treatment on the protoplasts was established as MNNG of 0.1 mg/ml and treatment time of 30-40 min. Resulting variants by this method expressed antibiotic activity higher than that by UV treament. After mutation, 109 resulting variants with 5 types of color were collected, of which type 1 and type 5 were 38.5 - 49.6%. Among the positive variants, the Val-A productivity of the mutant strain S. hygroscopicus 11405-115 was highest and enhanced 70% compared to the original one. The maximal Val-A activity of the strain S. hygroscopicus 11405-115 reached 6.87 mg/ml after 48 hours of cultivation in shaking flasks. Key words: Anti-fungal, mutant, protoplast, Rhizoctonia solani, Streptomyces hygroscopicus, validamycin-A
Ngày nhận bài: 06/7/2017Ngày phản biện: 10/7/2017
Người phản biện: TS. Trần Danh SửuNgày duyệt đăng: 27/7/2017

69
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀNhững năm gần đây, ngành nuôi cá rô phi
(Oreochromis niloticus) trên thế giới đang gặp phải nhiều dịch bệnh do liên cầu khuẩn Streptococcus gây ra, trong số đó phải kể đến như bệnh Streptococcis do Streptococcus agalactiae (Sun et al., 2016). Để khống chế Streptococcus spp. trong các ao nuôi cá rất nhiều liệu pháp sử dụng thuốc kháng khuẩn (erythromycin, oxytetracyclin, doxycyclin, sulfadiazin và amoxicilin) đã được áp dụng. Việc lạm dụng thuốc kháng khuẩn không những gây ra sự tồn dư thuốc trong cơ thể cá, ảnh hưởng tới chất lượng cá, mà còn dẫn đến hiện tượng kháng chất kháng khuẩn ở S. agalactiae. Nhiều nghiên cứu cho thấy lạm dụng kháng sinh đã dẫn tới hiện tượng “nhờn thuốc” và chính vì thế nhiều quốc gia trên thế giới đã ban hành quy định cấm và hạn chế nhiều loại kháng sinh trong nuôi trồng thủy sản (Cabello, 2006).
Xạ khuẩn được biết đến là nhóm vi sinh vật phân bố rộng rãi trong tự nhiên và có khả năng sinh nhiều hợp chất chất kháng khuẩn có khả năng kháng lại các vi khuẩn, vi nâm gây bệnh. Gần 80% kháng sinh trên thế giới được biết đến có nguồn gốc từ xạ khuẩn, chủ yếu là chi Streptomyces và Micromonospora (Lima et al., 2012). Nghiên cứu này được thực hiện nhằm tuyển chọn và đánh giá khả năng kháng vi khuẩn S. agalactiae của chủng xạ khuẩn HT1. Từ đó, chủng xạ khuẩn được tuyển chọn sẽ được sử dụng như nhân tố sinh học hữu hiệu nhằm kiểm soát và góp phần giảm thiểu dịch bệnh trên cá rô phi.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu Chủng liên cầu khuẩn S. agalactiae và các chủng
xạ khuẩn được Khoa Thủy sản và Bộ môn Công nghệ vi sinh - Khoa Công nghệ sinh học - Học viện Nông nghiệp Việt Nam cung cấp. Các môi trường được sử dụng trong nghiên cứu: Môi trường LB, hệ thống môi trường Intermational Streptomyces Project (ISP) từ ISP I đến ISP 7.
2.2. Phương pháp nghiên cứu- Hoạt tính kháng khuẩn của chủng xạ khuẩn:
Các chủng xạ khuẩn được kiểm tra khả năng kháng S. agalactiae bằng phương pháp khuếch tán trên đĩa thạch (agar disk-diffusion) (Balouiri et al., 2016). Các chủng xạ khuẩn được nuôi trong môi trường ISP4 lỏng, 7 ngày, vi khuẩn Streptococcus agalactiae được nuôi lắc trên môi trường LB lỏng trong vòng 24 h. Hút 0.1ml dịch nuôi vi khuẩn và cấy chang trên môi trường LB đặc. Dùng dụng cụ đục thạch có đường kính 7mm đã được vô trùng đục các thỏi thạch và lấy chúng ra. Hút 0.1ml dịch nuôi cấy xạ khuẩn đã được nuôi lỏng lắc trên môi trường ISP4 từ 5 - 7 ngày trước đó nhỏ vào các lỗ thạch đã được chuẩn bị. Để đĩa thử hoạt tính trong tủ lạnh ở 40C từ 4h để chất kháng sinh khuếch tán vào môi trường, sau đó chuyển ra tủ 300C. Sau 24h quan sát và đo vòng kháng khuẩn. Hoạt tính kháng khuẩn được tính theo công thức: D-d (mm), trong đó D - đường kính vòng vô khuẩn, d - đường kính lỗ thạch.
- Phương pháp xác định đặc điểm sinh học của xạ khuẩn: Chủng xạ khuẩn được nuôi trên hệ thống môi trường ISP (International Streptomyces Project) (Shirling and Gottlieb, 1966) để xác định đặc điểm hình thái, màu sắc khuẩn lạc, sắc tố tan và khả năng hình thành sắc tố melanin.
1 Khoa Công nghệ Sinh học, Học viện Nông nghiệp Việt Nam2 Bộ môn Sinh học Phân tử, Viện Di truyền Nông nghiệp
KHẢO SÁT MỘT SỐ ĐẶC ĐIỂM CỦA CHỦNG XẠ KHUẨN HT1 CÓ KHẢ NĂNG KHÁNG VI KHUẨN Streptococcus agalactiae GÂY BỆNH TRÊN CÁ RÔ PHI
Nguyễn Văn Giang1, Chu Đức Hà2, Nguyễn Thị Thu1
TÓM TẮTBài báo này trình bày một số kết quả về hoạt tính kháng khuẩn Streptococcus agalactiae và đặc điểm sinh học của
chủng xạ khuẩn HT1. Chủng xạ khuẩn HT1 có hoạt tính kháng S.agalctiae gây bệnh trên cá rô phi mạnh nhất với vòng kháng khuẩn đạt 21 mm sau 24 giờ nuôi. Khuẩn lạc của xạ khuẩn HT1 có kích thước từ 4 - 6 mm, hình tròn, bề mặt khô, màu xám trắng sau 21 ngày nuôi trên môi trường ISP4, cuống sinh bào tử dạng hơi lượn sóng, phân nhánh, chuỗi sinh bào tử có dạng xoắn lò xo. Chủng xạ khuẩn HT1 sinh trưởng tốt nhất khi được nuôi lỏng, lắc 200 vòng/phút, nhiệt độ là 30oC với pH 7, lượng tiếp giống ở mức 3%, thể tích môi trường lên men/thể tích bình nuôi là 10%. Nguồn cacbon và nitơ thích hợp cho sinh trưởng và sinh chất kháng khuẩn của chủng HT1 là xylose, pepton và KNO3 với đường kính vòng kháng khuẩn tương ứng đạt 23,24 và 24,67 mm.
Từ khóa: Hoạt tính kháng khuẩn, Streptococcus agalactiae, xạ khuẩn, đặc điểm sinh học, cá rô phi

70
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
- Ảnh hưởng của điều kiện nuôi: Chủng HT1 được nhân giống trên môi trường nhân giống cấp 1 (Yeast extract 10 g/l, dextrose 10 g/l, pH 6,8) sau đó được tiếp giống vào môi trường ISP4 chứa trong bình tam giác (V = 250 ml) lần lượt khảo sát ảnh hưởng của thời gian và trạng thái nuôi cấy (chủng xạ khuẩn được nuôi ở hai trạng thái lắc (200 vòng/phút) và tĩnh, nồng độ tiếp giống (2 - 10% v/v), nhiệt độ (25, 30, 35, 40 và 500C), pH môi trường (từ 3 đến 11), thể tích môi trường nuôi/thể tích bình: 10, 15, 20, 25, 30, 35, 40 và 45%; nguồn cacbon (glucose, sucrose, maltose, xylose, lactose, fructose, tinh bột) và nguồn nitơ (pepton, cao nấm men, urea, NaNO3, KNO3, NH4NO3, (NH4)2SO4). Hoạt tính kháng khuẩn được xác định bằng phương pháp khuếch tán đĩa thạch.
- Phương pháp xác định sinh khối khô: Chủng xạ khuẩn HT1 được nuôi trên môi trường: Yeast extract 10 g, dextrose 10 g, 1000 ml nước cất, pH 6.8 ở 300C. Sau 3 ngày nuôi, hút 10 ml dịch nuôi bổ sung vào 100ml môi trường ISP4 lỏng đựng trong bình có thể tích 250 ml. Sau đó nuôi các bình ở 300C trên hai trạng thái tĩnh và lắc 200 vòng/phút. Toàn bộ lượng mẫu nuôi được lọc qua giấy lọc Whatmann No 1, lượng sinh khối ướt được sấy khô trong không khí nóng. Lượng sinh khối khô được tính bằng sự chênh lệch giữa trọng lượng giấy lọc có sinh khối và trọng lượng giấy lọc đã cân trước đó (Bhavana et al., 2014).
2.3. Thời gian và đia điểm nghiên cứuNghiên cứu được thực hiện từ tháng 2/2016 -
tháng 2/2017 tại phòng thí nghiệm Bộ môn Công nghệ vi sinh, Khoa Công nghệ sinh học, Học viện Nông nghiệp Việt Nam.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Xác đinh đặc điểm sinh học của chủng xạ khuẩn có khả năng kháng vi khuẩn S. agalactiae
Từ 90 chủng xạ khuẩn được lưu giữ trong phòng thí nghiệm sau khi sàng lọc dựa trên khả năng kháng vi khuẩn S.agalactiae, đã chọn được chủng xạ khuẩn HT1 có khả năng kháng vi khuẩn S. agalactiae mạnh nhất với đường kính vòng kháng khuẩn đạt 21mm sau 24 giờ (Hình 1A). Chủng HT1 sau đó được cấy sang hệ thống môi trường ISP để quan sát đặc điểm hình thái và một số đặc tính sinh học cơ bản.
Màu sắc khuẩn lạc của chủng xạ khuẩn HT1 khi nuôi trên các môi trường ISP (từ ISP1 đến ISP7) có sự khác nhau. Khuẩn ty khí sinh của chủng HT1 có
màu xám ở các môi trường ISP1, 2, 3 và 7, trong khi các khuẩn ti này có màu trắng khi nuôi trên môi trường ISP4 và 5. Khuẩn ty cơ chất của chủng HT1 có màu xám vàng khi nuôi trên môi trường ISP1, màu vàng trên môi trường ISP2, 4 và 6, màu trắng trên môi trường ISP5 và màu xám khi nuôi trên môi trường ISP7. Sự phát triển của chủng xạ khuẩn HT1 trên môi trường ISP không làm thay đổi màu sắc vùng thạch quanh khuẩn lạc, chứng tỏ chủng HT1 không có khả năng sinh sắc tố melanin, khuẩn lạc điển hình có kích thước từ 4 - 6 mm, hình tròn, bề mặt khô, màu xám trắng sau 21 ngày nuôi (Hình 1B). Quan sát dưới kính hiển vi quang học ở độ phóng đại 1000, chủng HT1 có cuống sinh bào tử dạng hơi lượn sóng, phân nhánh, chuỗi sinh bào tử có dạng xoắn lò xo (Hình 1C).
Hình 1. Sàng lọc (A), quan sát khuẩn lạc (B) sau 7 ngày nuôi trên môi trường ISP4 và xác định hình thái chuỗi bào tử (C, 1000x) của chủng xạ khuẩn HT1
3.2. Kết quả theo dõi ảnh hưởng của các điều kiện lên men đến khả năng sinh trưởng và sinh chất kháng khuẩn của chủng xạ khuẩn HT1
Hoạt tính kháng khuẩn của xạ khuẩn nói chung chịu tác động của môi trường nuôi (lên men) khác nhau. Như trình bày ở hình 2, khả năng sinh chất kháng khuẩn và khối lượng sinh khối khô có sự sai khác rõ rệt ở các điều kiện nuôi khác nhau. Trong đó, chủng HT1 có khả năng sinh trưởng và sinh chất kháng khuẩn mạnh khi được nuôi trong môi trường ISP4 lỏng, lắc 200 vòng/phút. Khả năng sinh hoạt tính kháng khuẩn được ghi nhận từ ngày thứ 3 và kéo dài đến ngày thứ 9, đạt cực đại sau 4 ngày với kích thước vòng kháng khuẩn đạt 31,33 mm.
A B
Môi trườngISP
C

71
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Với mục đích tìm hiểu ảnh hưởng của mật độ giống đến khả năng hình thành chất kháng khuẩn, nồng độ tiếp giống cũng được khảo sát. Trong dải nồng độ tiếp giống từ 2; 3; 4; 5; 6; 7; 8; 9 và 10%, chủng HT1 có khả năng sinh chất kháng khuẩn cao nhất ở ngưỡng nồng độ tiếp giống 3% với đường kính vòng kháng khuẩn đạt 22,3 mm. Trước đó, Song và cộng tác viên (2012) đã cho rằng bổ sung lượng giống là 5% và thể tích môi trường là 75 ml/trên thể tích bình tam giác (250 ml) thì hoạt tính kháng khuẩn của S. felleus đạt tối ưu.
Hình 2. Ảnh hưởng của thời gian và trạng thái nuôi cấy đến khả năng sinh chất kháng khuẩn
của chủng HT1
Bên cạnh đó, ảnh hưởng của nhiệt độ và pH đến sinh trưởng của chủng xạ khuẩn HT1 cũng được quan tâm trong nghiên cứu này. Tiến hành thử nghiệm dải nhiệt độ từ 25, 30, 35, 40 và 50oC cho thấy, HT1 sinh trưởng tốt nhất ở ngưỡng 30oC, khi nhiệt độ đạt ngưỡng 50oC, HT1 mất hoàn toàn khả năng đối kháng vi khuẩn S. agalactiae. Naik và cộng tác viên (2015) cũng cho rằng ở nhiệt độ 500C, chủng xạ khuẩn S. zaomyceticus có khả năng sinh trưởng nhưng lại không có khả năng sinh chất kháng khuẩn. Khi nuôi cấy HT1 trong điều kiện lỏng, lắc 200 vòng/phút, ở dải pH 4; 5; 6; 7; 8; 9; 10 và 11, nhiệt độ 300C, sinh khối khô của chủng HT1 đạt cao nhất (2,5 - 3,3 mg/100ml) và có khả năng sinh kháng khuẩn mạnh nhất (đường kính vòng kháng khuẩn từ 12,6 đến 22,3 mm) ở ngưỡng pH trung tính (từ 6 - 8). Lê Thị Thanh Xuân và Tăng Thị Chính (2007) cũng kết luận xạ khuẩn thuộc nhóm vi sinh vật phát triển tốt ở môi trường trung tính hoặc hơi kiềm.
Chủng HT1 được tiếp giống với lượng 3% thể tích giống được nuôi vào bình có dung tích 250 ml chứa môi trường ISP4, pH = 7 với tỷ lệ dịch nuôi/thể tích bình là 10; 15; 20; 25; 30; 35; 40 và 45%. Nuôi các bình này ở 300C với tốc độ lắc 200 vòng/phút, sau 4 ngày nuôi đem đi kiểm tra khả năng kháng vi khuẩn S. agalactiae. Thay đổi thể tích môi trường
lên men dẫn đến sự thay đổi lượng ôxi không khí được khuếch tán vào môi trường, vì thế ảnh hưởng tới sự phát triển của vi sinh vật. Trên cơ sở đánh giá hoạt tính kháng S. agalactiae, kết quả thí nghiệm cho thấy, với lượng môi trường lên men chiếm 10% thể tích bình, hiệu quả tổng hợp chất kháng khuẩn tốt nhất, tương ứng với vòng kháng S. agalactiae là 23,67 mm.
Như vậy, chủng xạ khuẩn HT1 có thể sinh trưởng và có hoạt tính sinh kháng khuẩn trong dải nhiệt độ 25 - 40oC, pH từ 4 - 11, tối ưu ở 30oC, pH 6 - 8. Chủng xạ khuẩn HT1 thích hợp trong nuôi cấy lỏng, lắc. Với lượng tiếp giống ở mức 3%, lượng môi trường lên men chiếm 10% thể tích bình, xạ khuẩn HT1 sinh trưởng tốt và có hoạt tính kháng khuẩn mạnh nhất.
3.3. Ảnh hưởng của nguôn cacbon và nitơ đến khả năng sinh chất kháng khuẩn của chủng HT1
Trong môi trường nuôi cấy vi sinh vật luôn cần thiết phải cung cấp nguồn carbon và nitơ cho quá trình trao đổi chất và giải phóng năng lượng. Trong nghiên cứu này, nguồn carbon trong môi trường ISP4 được thay thể với khối lượng tương đương bởi tinh bột, maltose, fructose, xylose, glucose và sucrose, trong khi 2 nguồn nitơ đã được tiến hành đánh giá, bao gồm (i) nguồn N vô cơ là (NH4)2SO4, NaNO3, KNO3, NH4NO3 và (NH4)2CO và (ii) nguồn nitơ hữu cơ là cao nấm men và pepton. Kết quả đánh giá khả năng sinh trưởng và khả năng sinh kháng khuẩn được thể hiện ở hình 3.
Chủng HT1 có khả năng sinh trưởng và sinh chất kháng khuẩn trên nhiều nguồn C khác nhau. Trên môi trường có bổ sung xylose, chủng HT1 có khả năng sinh chất kháng khuẩn mạnh nhất với đường kính vòng kháng khuẩn đạt 23 mm. Kết quả này cũng khá tương đồng với kết quả nghiên cứu của Bundale và cộng tác viên công bố trước đó. Các tác giả này cho rằng xylose là một nguồn carbon tốt cho sinh trưởng và sinh chất kháng khuẩn của chủng xạ khuẩn R5 (Bundale et al., 2015).
Cũng như carbon, nitơ là là thành phần nguyên liệu cho sự tổng hợp các sản phẩm của tế bào, ngoài ra các hợp chất chứa nitơ còn giúp tế bào thực hiện quá trình trao đổi chất và điều hòa quá trình chuyển hóa. Tuy nhiên, các dạng khác nhau của nguồn nitơ đã có ảnh hưởng tới khả năng sử dụng của vi sinh vật. Chủng HT1 có khả năng sinh trưởng và sinh chất kháng khuẩn khi nuôi trong môi trường chứa các nguồn nitơ khác nhau (Hình 3). Pepton và KNO3
: Vòng kháng khuẩn nuôi tĩnh: Vòng kháng khuẩn nuôi lắc
Thời gian nuôi cấy (ngày)
35
30
25
20
15
10
5
03 4 5 6 7 8 9
6
5
4
3
2
1
0
Đư
ờng
kính
vòn
g kh
áng
khuẩ
n (m
m)
Khố
i lư
ợng
sinh
khố
i khô
(mg/
100m
l)
: Sinh khối khô nuôi lắc: Sinh khối khô nuôi tĩnh

72
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
là 2 nguồn nitơ giúp xạ khuẩn HT1 có khả năng sinh chất kháng khuẩn mạnh nhất, đường kính vòng kháng khuẩn lần lượt là 24 và 24,67 mm. Kết quả này tuy không cao bằng hoạt tính kháng khuẩn của chính chủng HT1 khi nuôi trên môi trường ISP4 nhưng rất hữu ích trong điều kiện lên men công nghiệp, khi
cần phải sử dụng các nguồn carbon, nitơ sẵn dễ tìm và không đắt. Như vậy, xylose, pepton và KNO3 là nguồn carbon và nitơ thay thế thích hợp giúp chủng xạ khuẩn HT1 sinh trưởng và có khả năng sinh chất kháng khuẩn S. agalactiae mạnh nhất.
IV. KẾT LUẬN- Đã tuyển chọn được chủng xạ khuẩn HT1 có
hoạt tính kháng S. agalactiae mạnh nhất. Chủng HT1 không có khả năng sinh sắc tố melanin trên môi trường ISP, khuẩn lạc chủng HT1 có kích thước từ 4 - 6 mm, hình tròn, bề mặt khô, màu xám trắng trên môi trường ISP sau 21 ngày nuôi cấy. Cuống sinh bào tử của chủng HT1 có dạng hơi lượn sóng, phân nhánh, chuỗi sinh bào tử có dạng xoắn lò xo.
- Các điều kiện thích hợp cho chủng xạ khuẩn HT1 sinh trưởng tốt: nhiệt độ 30oC, pH trung tính (6 - 8), nuôi cấy lỏng, lắc 200 vòng/phút, lượng tiếp giống 3%, thể tích môi trường nuôi/thể tích bình nuôi là 10%.
- Chủng xạ khuẩn HT1 sinh trưởng tốt và có hoạt tính kháng S. agalactiae trong môi trường có nguồn carbon là xylose, nguồn nitơ là Pepton và KNO3 với đường kính vòng kháng khuẩn đạt tương ứng là 23, 24 và 24,67 mm.
TÀI LIỆU THAM KHẢONguyên Xuân Cảnh, Hô Tú Cường, Nguyên Thi
Đinh, Phạm Thi Hiếu, 2016. Nghiên cứu chủng xạ khuẩn có khả năng đối kháng với vi khuẩn Vibrio parahaemolyticus gây bệnh trên tôm. Tạp chí Khoa học Nông nghiệp Việt Nam, 14(11): 1809-1816.
Lê Thi Thanh Xuân, Tăng Thi Chính, 2007. Ảnh hưởng của các điều kiện lên men lên khả năng sinh chất kháng khuẩn kháng nấm Fusarium oxysporum của hai chủng xạ khuẩn Streptomyces cyaneogriceus HD54 và Streptomyces hygroscopicus HD58. Tạp chí Sinh học, 29(1): 89-94.
Balouiri, M., Sadiki, M., Ibnsouda, S.K., 2016. Methods for in vitro evaluating antimicrobial activity: A review. J Pharm Anal, 6: 71-79.
Bhavana, M., Talluri, V.P., Kumar, K.S., Rajagopal, S.V., 2014. Optimization of culture conditions of Streptomyces carpaticus (MTCC-11062) for the production of antimicrobial compound. Int J Pharm Pharm Sci, 6(8): 281-285.
Bundale, S., Begde, D., Nashikkar, N., Kadam, T., Upadhyay, A., 2015. Optimization of culture conditions for production of bioactive metabolites by Streptomyces spp. isolated from soil. Adv Microbiol, 5(6): 11.
Cabello F.C., 2006. Heavy use of prophylactic antibiotics in aquaculture: a growing problem for human and animal health and for the environment. Environ. Microbiol., 8: 1137-1144.
Lima, R.E., Silva, I.R., Martins, M.K., Azevedo, J.L., Araújo, J.M., 2012. Antibiotics produced by Streptomyces. Braz J Infect Dis, 16: 466-471.
Shirling, E.B. and Gottlieb, D., 1966. Methods for characterization of Streptomyces sp. Int. J. Syst. Bacteriol., 16: 313-340.
Song, Q., Huang, Y., Yang, H., 2012. Optimization of fermentation conditions for antibiotic production by actinomycetes YJ1 strain against Sclerotinia sclerotiorum. J Agri Sci, 4(7): 95-102.
Sun, J., Fang, W., Ke, B., He, D., Liang, Y., Ning, D., Tan, H., Peng, H., Wang, Y., Ma, Y., Ke, C., Deng, X., 2016. Inapparent Streptococcus agalactiae infection in adult/commercial tilapia. Sci Rep, 6: 26319.
Đườ
ng k
ính
vòng
khá
ngkh
uẩn
(mm
)
Khôn
g bổ
sun
g C
1% ti
nh b
ột
Mal
tose
Fruc
tose
Xylo
se
Glu
cose
Lact
ose
Sucr
ose
Khối
lượn
g si
nh k
hối k
hô(m
g/10
0ml)
3025201510
50
: Vòng kháng khuẩn: Sinh khối khô
876543210
Đườ
ng k
ính
vòng
khá
ngkh
uẩn
(mm
)
Khôn
g bổ
sun
g N
(NH
4) 2SO4
NaN
O3
KNO
3
NH
4NO
3
(NH
4) 2CO
Khôn
g bổ
sun
g N
Cao
nấm
men
Pept
on
Khối
lượn
g si
nh k
hối k
hô(m
g/10
0ml)
3025201510
50
Vô cơ Hữu cơ 5
4
3
2
1
0
Hình 3. Ảnh hưởng của nguồn cacbon và nitơ đến khả năng sinh trưởng và sinh chất kháng khuẩn của chủng HT1 khi nuôi lắc 200 vòng/phút ở nhiệt độ 300C

73
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Biological characterization of HT1 strain of Streptomyces with potential antimicrobial activity against Streptococcus agalactiae causing disease on tilapia
Nguyen Van Giang, Chu Duc Ha, Nguyen Thi ThuAbstractThe article presents the biological characteristics and antimicrobial activity of actinomyces strain HT1 against Streptococcus agalactiae causing disease on tilapia. The ‘HT1’ colonies were observed to be circular, diameter 4 - 6 mm, dried surface with grayish white color, aerial hyphae are long, straight branching with a chain of spherical spore on ISP 4 medium after 21 day of cultivation. The favourable conditions for growth and antimicrobial activity of the strain HT1 were established: temperature of 30oC, pH (6 - 8), shaking speed of 200 rpm, the inoculums size of 3% (v/v), volume of medium in shaking flask of 10% (v/v). Our results also revealed that ‘HT1’ strain could grow well on the media containing xylose (C resource), peptone and KNO3 (N resource) with clear zone of inhibition against S. agalactiae 23, 24 and 24.67 mm, respectively.Key words: Antimicrobial activity, biological characteristics, Streptococcus agalactiae, actinomyces, tilapia
Ngày nhận bài: 10/6/2017Ngày phản biện: 20/6/2017
Người phản biện: TS. Đinh Trường SơnNgày duyệt đăng: 25/6/2017
1 Khoa Công nghệ thực phẩm, Học viện Nông nghiệp Việt Nam2 Khoa Công nghệ thực phẩm, Trường Đại học Kinh tế Kỹ thuật Công nghiệp
TỐI ƯU HÓA ĐIỀU KIỆN TỰ PHÂN TẾ BÀO NẤM MEN BIA THẢI Nguyễn Thị Thanh Thủy1, Hồ Tuấn Anh2
TÓM TẮTNấm men bia thải sau khi xử lý được tối ưu hóa các điều kiện tự phân theo thiết kế Box-Behnken. Hàm số kì
vọng đơn Deringer được sử dụng để tối ưu hoá các yếu tố đầu ra. Kết quả cho thấy, khả năng phân giải bã men bia phụ thuộc nhiều vào yếu tố đơn lẻ, quan trọng nhất là nhiệt độ, thời gian và pH. Sự kết hợp giữa các yếu tố với nhau ít hoặc không làm thay đổi kết quả ở mức ý nghĩa ngoại trừ tương tác giữa pH với thời gian gây giảm các chất hòa tan. Với điều kiện tối ưu bã nấm men : nước là 1 : 3, tốc độ khuấy 30 vòng/phút, nhiệt độ tự phân 52oC, pH 5,8, thời gian tự phân 22h, tỷ lệ protein chuyển hóa đến dạng amin tự do là 41,3%, tỷ lệ protein chuyển hóa đến dạng hòa tan là 73,6%, và tỷ lệ chuyển hóa chất khô từ sinh khối vào dịch chiết nấm men là 52,1%. Giá trị kỳ vọng đạt cho cả ba hàm mục tiêu là 94,3%.
Từ khóa: Nấm men bia, tối ưu hóa, tự phân, mô hình toán học, thiết kế thí nghiệm
I. ĐẶT VẤN ĐỀTự phân (autolysis) là quá trình xảy ra tự nhiên,
khi tế bào kết thúc chu kỳ phát triển và chuyển sang pha suy vong, lúc đó các enzyme nội bào sẽ được tiết ra phân hủy tế bào (Hasan and Huseyin, 2007). Mặc dù tạo ra sản phẩm với hiệu suất thu hồi cao, nhưng phương pháp phân giải tế bào bằng acid hiện nay ít được sử dụng rộng rãi do chi phí đầu tư ban đầu cao, ngoài ra có khả năng tạo ra các sản phẩm phụ gây độc như mono và dichloropropanol (Chae et al., 2001). Cho đến nay việc sử dụng các enzyme như glucanase, protease, nuclease và deaminase để phân giải nấm men đang được phổ biến (Chung et al., 1999; Chae et al., 2001; Kim et al., 2001). Tuy nhiên, kinh phí sử dụng cho phương pháp này còn cao. Phân giải tế bào nấm men bia thải ứng dụng
trong sản xuất thức ăn chăn nuôi thì tự phân là kinh tế nhất, có tính khả thi cao khi áp dụng ở quy mô công nghiệp.
Theo các tài liệu đã công bố và kết quả nghiên cứu của nhóm đề tài, quá trình tự phân nấm men bia thải chịu ảnh hưởng của nhiệt độ, pH, thời gian, tốc độ khuấy, tỷ lệ phối trộn sinh khối nấm men với nước (Kenji, 2002; Иванова et al., 1989).
Quá trình tự phân của nấm men không chỉ chịu ảnh hưởng của các đơn yếu tố mà còn là sự kết hợp của các yếu tố với nhau. Việc thực hiện tối ưu hóa điều kiện tự phân nấm men nhằm xác định được mức độ của từng yếu tố ảnh hưởng cũng như sự tương tác giữa các yếu tố với nhau là cần thiết để nâng cao hiệu quả của quá trình tự phân.

74
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứuNấm men bia thải thu từ Công ty TNHH Một
thành viên Bia Rượu Eresson, khu công nghiệp Quang Minh, Mê Linh, Hà Nội. Trong quá trình tiền xử lý, nấm men thải được rửa và tách đắng bằng nước, dung dịch H3PO4 0,01N và NaCl 0,05% với tỷ lệ 3: 1 so với sinh khối, nhiệt độ 0 - 50C, thời gian lắng 50 phút. Dịch nấm men được đưa qua sàng loại bỏ các cặn thô, lọc vắt nhằm loại bỏ nước tự do để thu được nấm men ở dạng đặc, màu kem đồng nhất, tỷ lệ tế bào sống 92 - 95%, (Hồ Tuấn Anh, 2016). Nấm men vắt có hàm lượng chất khô tuyệt đối 20,50%, hàm lượng protein và các thành phần chứa nitơ chiếm 54,72% so với chất khô tuyệt đối. Tỷ lệ protein chuyển hóa đến FAN là 4,1%, tỷ lệ protein chuyển hóa đến dạng hòa tan là 14,2%.
2.2. Phương pháp nghiên cứu
2.2.1. Phương pháp bố trí thí nghiệm tối ưu hóa điều kiện tự phân nấm men bia thải
Để xác định điều kiện tối ưu cho quá trình tự phân của nấm men, trong nghiên cứu này 5 yếu tố chính được xác định là tỷ lệ bã nấm men : nước (X1); tốc độ khuấy (vòng/ phút) (X2); nhiệt độ tự phân (oC) (X3); pH (X4); thời gian tự phân (giờ) (X5). Giá trị biến thiên của 5 yếu tố được xác định dựa trên nghiên cứu ảnh hưởng các yếu tố đến quá trình tự phân nấm men, cụ thể như sau: Tỷ lệ sinh khối nấm men : nước: 1 : 2; 1 : 3; 1 : 4, tốc độ khuấy: 25; 30; 35 vòng /phút, nhiệt độ tự phân: 42; 47; 52oC, pH: 5,0; 5,4; 5,8, thời gian tự phân: 14; 18; 22 h. Các điều kiện tự phân trước và sau mã hóa được bố trí theo thiết kế Box-Behnken (Bảng 1). Mô hình toán học sử dụng để xác định giá trị tối ưu cho các yếu tố có dạng như sau:
Yi = fi(X1, X2,…., Xk) hay Y = Bo + B1X1 + B2X2 + B3X3 + B4X4 + B5X5+ B6X1X2 + B7X1X3 + B8X1X4 + B9X1X5 + B10X2X3 + B11X2X4 + B12X2X5 + B13X3X4 + B14X3X5 + B15X4X5 + B16X1
2 + B17X22 + B18X3
2 + B19X42
+ B20X52
Trong đó: Bo, B1, B2, B3, B4, B5, B6, B7, B8, B9, B10, B11, B12, B13, B14, B15, B16, B17, B18, B19, B20 : là hệ số hồi quy. X1, X2, X3, X4, X5: là các yếu tố thí nghiệm cần tối ưu.
Yi: Là các hàm mục tiêu bao gồm: tỷ lệ protein chuyển hóa đến dạng amin tự do (FAN) (y1), tỷ lệ protein chuyển hóa đến dạng hòa tan không kết tủa sau ly tâm (y2), và tỷ lệ chuyển hóa chất khô từ sinh khối vào dịch chiết nấm men (y3).
Hàm số kì vọng đơn Deringer được sử dụng để tối ưu hoá các yếu tố đầu ra (tỷ lệ protein chuyển hóa đến dạng amin tự do (FAN), tỷ lệ protein chuyển hóa đến dạng hòa tan không kết tủa sau ly tâm, và tỷ lệ chuyển hóa chất khô từ sinh khối vào dịch chiết nấm men). Hàm số kì vọng này thay đổi phụ thuộc vào điểm kì vọng, dao động từ 0 (không mong muốn) đến 1 (mong muốn), hàm này có thể đạt giá trị lớn nhất hoặc nhỏ nhất, tuỳ thuộc vào mục đích thí nghiệm. Phương trình hàm số kì vọng có dạng như sau:
Trong đó y i,min và y i,max là mức tối thiểu, mức tối đa tương ứng của các biến độc lập, đây là giá trị nhỏ nhất và lớn nhất của điểm ưa thích cho các chỉ tiêu chất lượng. Các điểm kỳ vọng đơn sau đó được kết hợp với nhau tạo thành kỳ vọng tổng thể (D) và được tính theo công thức dưới, trong đó ri thể hiện hệ số trọng lượng cho mỗi chỉ tiêu. Trong nghiên cứu này ri được cố định là 1.
2.2.2. Phương pháp phân tíchXác định hàm lượng chất khô bằng phương pháp
sấy đối lưu ở 60ºC đến khối lượng không đổi. Xác định hàm lượng FAN của dịch chiết nấm men
theo phương pháp ninhydrin của Wylie & Johnson, đo màu ở bước sóng 570 nm, mô tả trong mục 8.10 của EBC (Analitica - EBC, 2005).
Xác định hàm lượng protein tổng số và protein hòa tan theo phương pháp của Kjeldahl, mô tả trong 8.9.1 của EBC (Analitica - EBC, 2005).
2.2.3. Phương pháp xử lý số liệuSố liệu thu nhận được xử lý bằng phần mềm
Excel 2010 và JMP version 13.
2.3. Thời gian và đia điểm nghiên cứuNghiên cứu được thực hiện từ tháng 7 năm 2016
đến tháng 2 năm 2017 tại Khoa Công nghệ thực phẩm - Học viện Nông nghiệp Việt Nam.

75
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 1. Thiết kế thí nghiệm tối ưu hóa quá trình tự phân
Công thức
Trước mã hóa Sau mã hóaTỷ lệ
bã nấm men:nước
Tốc độ khuấy
(vòng/ phút)
Nhiệt độ tự phân (oC) pH
Thời gian tự phân
(h)X1 X2 X3 X4 X5
1 1:3 30 42 5,4 14 0 0 -1 0 -12 1:2 25 47 5,4 18 -1 -1 0 0 03 1:3 30 47 5,4 18 0 0 0 0 04 1:4 30 47 5,0 18 1 0 0 -1 05 1:2 35 47 5,4 18 -1 1 0 0 06 1:3 35 52 5,4 18 0 1 1 0 07 1:3 30 52 5,4 14 0 0 1 0 -18 1:3 25 42 5,4 18 0 -1 -1 0 09 1:3 30 52 5,8 18 0 0 1 1 0
10 1:3 35 47 5,0 18 0 1 0 -1 011 1:3 35 47 5,4 22 0 1 0 0 112 1:3 30 42 5,8 18 0 0 -1 1 013 1:4 35 47 5,4 18 1 1 0 0 014 1:2 30 47 5,4 22 -1 0 0 0 115 1:4 30 47 5,4 14 1 0 0 0 -116 1:3 25 47 5,8 18 0 -1 0 1 017 1:3 30 47 5,0 22 0 0 0 -1 118 1:2 30 47 5,8 18 -1 0 0 1 019 1:3 25 47 5,4 22 0 -1 0 0 120 1:3 25 47 5,4 14 0 -1 0 0 -121 1:2 30 42 5,4 18 -1 0 -1 0 022 1:3 30 47 5,8 14 0 0 0 1 -123 1:3 30 42 5,4 22 0 0 -1 0 124 1:3 30 47 5,0 14 0 0 0 -1 -125 1:3 35 42 5,4 18 0 1 -1 0 026 1:2 30 52 5,4 18 -1 0 1 0 027 1:4 30 52 5,4 18 1 0 1 0 028 1:3 25 52 5,4 18 0 -1 1 0 029 1:4 30 42 5,4 18 1 0 -1 0 030 1:3 35 47 5,8 18 0 1 0 1 031 1:3 30 47 5,4 18 0 0 0 0 032 1:4 30 47 5,8 18 1 0 0 1 033 1:2 30 47 5,0 18 -1 0 0 -1 034 1:3 30 47 5,4 18 0 0 0 0 035 1:3 30 52 5,0 18 0 0 1 -1 036 1:2 30 47 5,4 14 -1 0 0 0 -137 1:4 30 47 5,4 22 1 0 0 0 138 1:3 30 42 5,0 18 0 0 -1 -1 039 1:3 35 47 5,4 14 0 1 0 0 -140 1:3 30 52 5,4 22 0 0 1 0 141 1:4 25 47 5,4 18 1 -1 0 0 042 1:3 30 47 5,8 22 0 0 0 1 143 1:3 25 47 5,0 18 0 -1 0 -1 0

76
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
III. KẾT QUẢ VÀ THẢO LUẬNSau khi có bảng mã hóa các công thức, tiến hành
bố trí thí nghiệm như thiết kế của mô hình, kết quả thu được trong bảng 2.
Kết quả cho thấy: đối với hàm y1, tỷ lệ protein chuyển hóa đến FAN dao động trong khoảng 39,6 - 41,2%; hàm y2, tỷ lệ protein chuyển hóa tới dạng hòa tan thu được trong khoảng 70,7 - 73,5%; hàm y3, tỉ lệ chất khô chuyển hóa vào dạng hòa tan thu được trong khoảng 49,0 - 51,8%. Tuy nhiên, để tìm ra được phương trình toán học cho các hàm mong đợi, lựa chọn được điểm tối ưu theo mong muốn, các số liệu đã có được xử lý tối ưu trong phần mềm JMP.
Bảng 2. Hiệu quả tự phân của tế bào nấm men ở các điều kiện khác nhau
Công thức
y1 (%)
y2 (%)
y3 (%)
Công thức
y1 (%)
y2 (%)
y3 (%)
1 39,7 70,9 49,0 23 40,4 72,3 50,42 40,3 71,8 50,1 24 40,9 71,7 50,23 40,4 72,1 50,4 25 39,2 71,1 49,24 39,9 71,9 50,2 26 40,5 71,4 49,85 40,5 71,8 50,2 27 40,4 72,5 50,86 40,7 73,0 51,4 28 40,6 73,3 51,67 39,9 72,5 50,6 29 40,3 72,8 51,08 40,1 71,2 49,4 30 40,2 71,6 50,29 41,1 73,5 51,8 31 41,2 72,8 51,2
10 40,0 71,6 50,0 32 40,4 72,0 50,311 41,2 72,6 51,0 33 41,1 73,1 51,412 40,9 71,9 50,2 34 39,7 71,1 49,413 40,6 72,2 50,6 35 40,4 72,1 50,414 40,9 72,1 50,6 36 39,9 72,3 50,615 39,9 72,1 50,2 37 39,7 71,3 49,416 40,8 72,6 50,8 38 41,1 72,9 51,417 40,7 72,3 50,8 39 39,7 70,7 49,018 40,9 72,3 50,6 40 40,0 71,8 49,819 40,8 72,4 50,8 41 41,1 73,3 51,820 39,6 71,6 49,6 42 40,3 72,4 50,621 40,2 70,9 49,2 43 40,9 72,6 51,122 39,7 70,9 49,0
Với 5 yếu tố đầu vào là tỷ lệ bã nấm men : nước (X1), tốc độ khuấy (vòng/ phút) (X2), nhiệt độ tự phân (oC) (X3), pH (X4), thời gian tự phân (X5), 3 yếu tố đầu ra là tỷ lệ protein chuyển hóa đến dạng amin tự do (FAN) (y1), tỷ lệ protein chuyển hóa đến dạng hòa tan không kết tủa sau ly tâm (y2), và tỷ lệ chuyển hóa chất khô từ sinh khối vào dịch chiết nấm men
(y3) phần mềm cho thấy được mức độ ảnh hưởng của các yếu tố đầu vào tới chất lượng dịch tự phân của nấm men. Kết quả được thể hiện trong bảng 3.
Từ bảng 3 có thể thấy quá trình tự phân của nấm men chịu ảnh hưởng của cả 5 yếu tố đầu vào. Tuy nhiên ảnh hưởng lớn nhất là nhiệt độ tự phân với mức quan trọng 17,8; tiếp đến là thời gian tự phân và pH môi trường tự phân có tầm ảnh hưởng gần tương tự nhau với mức quan trọng 14,8 và 14,2. Tốc độ khuấy ảnh hưởng yếu hơn khi mức quan trọng chỉ còn 10,5. Tỷ lệ nấm men : nước là yếu tố có mức độ ảnh hưởng nhỏ nhất đến chất lượng của dịch sau tự phân khi mức quan trọng chỉ có 5,6.
Bên cạnh sự ảnh hưởng của các đơn yếu tố, hiệu quả tự phân của nấm men còn chịu tác động của tương tác giữa pH và thời gian tự phân, tuy nhiên chỉ ở mức độ tương đối nhẹ thể hiện ở mức quan trọng chỉ dừng lại ở 3,5.
Để thấy rõ được tương tác của các yếu tố đến từng hàm mục tiêu, kết quả phân tích hệ số hồi quy của các yếu tố thí nghiệm được thể hiện trong bảng 4.
Từ kết quả ước lượng hệ số hồi quy, phương trình mô tả mối tương quan giữa tỷ lệ protein chuyển hóa đến dạng amin tự do (FAN) và các yếu tố thí nghiệm là:
y1 = 40,4 + 0,06875X1 + 0,18125X2 + 0,1125X3 + 0,5375X4 + 0,575X5 - 0,25X4X5
Phương trình mô tả mối tương quan giữa tỷ lệ protein chuyển hóa đến dạng hòa tan không kết tủa sau ly tâm và các yếu tố thí nghiệm là:
y2 = 70,566667 + 0,35625X1 + 0,0625X2 + 0,80625X3 + 0,54375X4 + 0,39375X5 - 0,225X4X5
Phương trình mô tả mối tương quan giữa tỷ lệ chuyển hóa chất khô từ sinh khối vào dịch chiết nấm men và các yếu tố thí nghiệm như sau:
y3 = 48,866667 + 0,36875X1 + 0,13125X2 + 0,7875X3 + 0,54375X4 + 0,59375X5
_ 0,225X4X5
Với kết quả bảng 4, ảnh hưởng của các đơn yếu tố, từ X1 đến X5 đến 3 hàm mục tiêu đều có ý nghĩa với P<0,05. Sự tương tác giữa các yếu tố với nhau hầu như không có ý nghĩa trừ sự tương tác giữa X4*X5. Hai yếu tố pH và thời gian có ảnh hưởng đến ti lệ các chất hòa tan thu được ở mức ý nghĩa, tuy nhiên sự ảnh hưởng này là tiêu cực. Nó không làm tăng tỉ lệ chất hòa tan mà làm biến đổi ngược lại.
Để xác định giá trị tối ưu của các yếu tố thí nghiệm, điểm kỳ vọng sẽ được tối đa hóa khi sử dụng đồ thị dự đoán hiệu suất tự phân, kết quả được thể hiện trên hình 1.
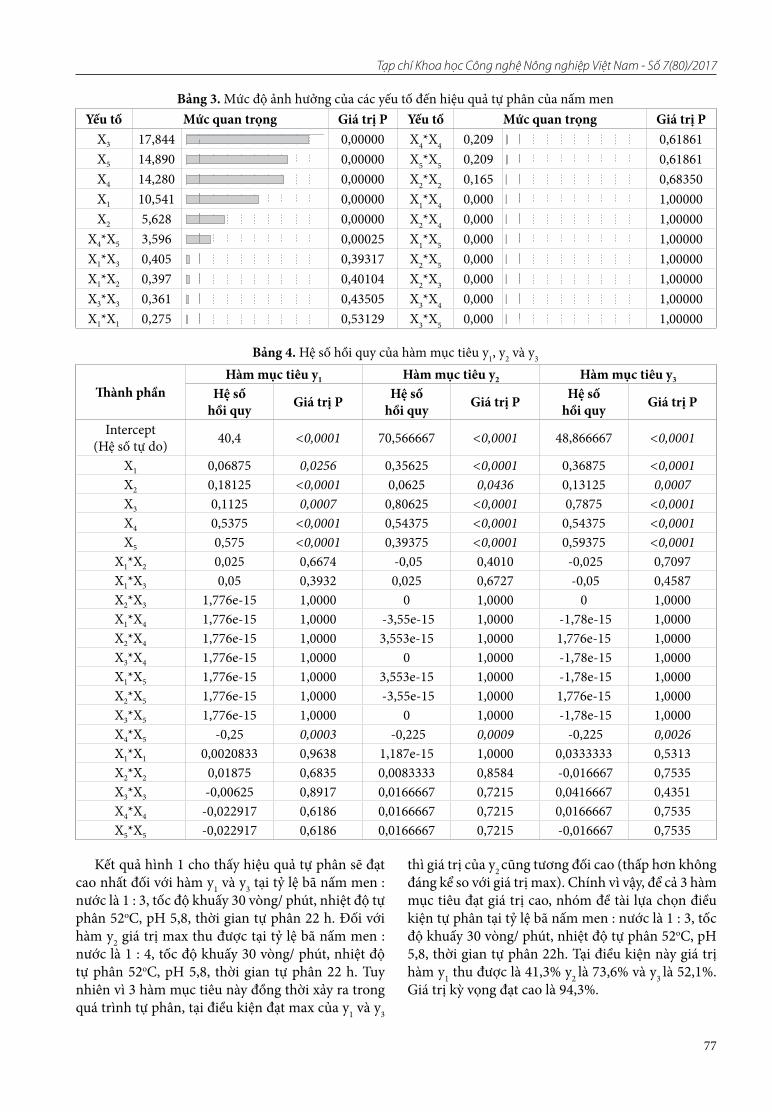
77
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 4. Hệ số hồi quy của hàm mục tiêu y1, y2 và y3
Bảng 3. Mức độ ảnh hưởng của các yếu tố đến hiệu quả tự phân của nấm men
Thành phầnHàm mục tiêu y1 Hàm mục tiêu y2 Hàm mục tiêu y3
Hệ số hôi quy Giá tri P Hệ số
hôi quy Giá tri P Hệ số hôi quy Giá tri P
Intercept (Hệ số tự do) 40,4 <0,0001 70,566667 <0,0001 48,866667 <0,0001
X1 0,06875 0,0256 0,35625 <0,0001 0,36875 <0,0001X2 0,18125 <0,0001 0,0625 0,0436 0,13125 0,0007X3 0,1125 0,0007 0,80625 <0,0001 0,7875 <0,0001X4 0,5375 <0,0001 0,54375 <0,0001 0,54375 <0,0001X5 0,575 <0,0001 0,39375 <0,0001 0,59375 <0,0001
X1*X2 0,025 0,6674 -0,05 0,4010 -0,025 0,7097X1*X3 0,05 0,3932 0,025 0,6727 -0,05 0,4587X2*X3 1,776e-15 1,0000 0 1,0000 0 1,0000X1*X4 1,776e-15 1,0000 -3,55e-15 1,0000 -1,78e-15 1,0000X2*X4 1,776e-15 1,0000 3,553e-15 1,0000 1,776e-15 1,0000X3*X4 1,776e-15 1,0000 0 1,0000 -1,78e-15 1,0000X1*X5 1,776e-15 1,0000 3,553e-15 1,0000 -1,78e-15 1,0000X2*X5 1,776e-15 1,0000 -3,55e-15 1,0000 1,776e-15 1,0000X3*X5 1,776e-15 1,0000 0 1,0000 -1,78e-15 1,0000X4*X5 -0,25 0,0003 -0,225 0,0009 -0,225 0,0026X1*X1 0,0020833 0,9638 1,187e-15 1,0000 0,0333333 0,5313X2*X2 0,01875 0,6835 0,0083333 0,8584 -0,016667 0,7535X3*X3 -0,00625 0,8917 0,0166667 0,7215 0,0416667 0,4351X4*X4 -0,022917 0,6186 0,0166667 0,7215 0,0166667 0,7535X5*X5 -0,022917 0,6186 0,0166667 0,7215 -0,016667 0,7535
Yếu tố Mức quan trọng Giá tri P Yếu tố Mức quan trọng Giá tri PX3 17,844 0,00000 X4*X4 0,209 0,61861X5 14,890 0,00000 X5*X5 0,209 0,61861X4 14,280 0,00000 X2*X2 0,165 0,68350X1 10,541 0,00000 X1*X4 0,000 1,00000X2 5,628 0,00000 X2*X4 0,000 1,00000
X4*X5 3,596 0,00025 X1*X5 0,000 1,00000X1*X3 0,405 0,39317 X2*X5 0,000 1,00000X1*X2 0,397 0,40104 X2*X3 0,000 1,00000X3*X3 0,361 0,43505 X3*X4 0,000 1,00000X1*X1 0,275 0,53129 X3*X5 0,000 1,00000
Kết quả hình 1 cho thấy hiệu quả tự phân sẽ đạt cao nhất đối với hàm y1 và y3 tại tỷ lệ bã nấm men : nước là 1 : 3, tốc độ khuấy 30 vòng/ phút, nhiệt độ tự phân 52oC, pH 5,8, thời gian tự phân 22 h. Đối với hàm y2 giá trị max thu được tại tỷ lệ bã nấm men : nước là 1 : 4, tốc độ khuấy 30 vòng/ phút, nhiệt độ tự phân 52oC, pH 5,8, thời gian tự phân 22 h. Tuy nhiên vì 3 hàm mục tiêu này đồng thời xảy ra trong quá trình tự phân, tại điều kiện đạt max của y1 và y3
thì giá trị của y2 cũng tương đối cao (thấp hơn không đáng kể so với giá trị max). Chính vì vậy, để cả 3 hàm mục tiêu đạt giá trị cao, nhóm đề tài lựa chọn điều kiện tự phân tại tỷ lệ bã nấm men : nước là 1 : 3, tốc độ khuấy 30 vòng/ phút, nhiệt độ tự phân 52oC, pH 5,8, thời gian tự phân 22h. Tại điều kiện này giá trị hàm y1 thu được là 41,3% y2 là 73,6% và y3 là 52,1%. Giá trị kỳ vọng đạt cao là 94,3%.

78
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
IV. KẾT LUẬNTối ưu hóa các điều kiện tự phân theo thiết kế
Box-Behnken cho thấy quá trình tự phân của nấm men chịu ảnh hưởng chủ yếu bởi các yếu tố riêng rẽ mà ít chịu ảnh hưởng bởi sự kết hợp giữa nhiều yếu tố. Yếu tố ảnh hưởng lớn nhất là nhiệt độ, tiếp theo là thời gian, pH. Chịu ảnh hưởng ít là tỉ lệ bã nấm men: nước và tốc độ khuấy.
Tỷ lệ protein chuyển hóa đến dạng amin tự do là 41,3%, tỷ lệ protein chuyển hóa đến dạng hòa tan là 73,6% và tỷ lệ chuyển hóa chất khô từ sinh khối vào dịch chiết nấm men là 52,1% trong điều kiện tối ưu tỷ lệ bã nấm men : nước là 1 : 3, tốc độ khuấy 30 vòng/phút, nhiệt độ tự phân 52oC, pH 5,8, thời gian tự phân 22 h. Tuy nhiên, tùy thuộc vào mong muốn sản phẩm đầu ra cần chỉ tiêu gì vượt trội mà lựa chọn các điều kiện tối ưu dựa trên kết quả mô hình đã tìm được.
LỜI CẢM ƠNNghiên cứu được hỗ trợ kinh phí của Sở Khoa
học và Công nghệ Hà Nội trong khuôn khổ đề tài “Nghiên cứu ứng dụng công nghệ sinh học trong tái chế men bia thải làm thức ăn chăn nuôi cho địa bàn Hà Nội”.
TÀI LIỆU THAM KHẢO Hô Tuấn Anh, 2016. Nghiên cứu phương pháp tiền xử
lý nấm men bia thải. Hội thảo khoa học toàn quốc 2016. “Tiến bộ kỹ thuật thực phẩm và kỹ thuật sinh học: Từ nghiên cứu đến sản xuất, 10 - 11/10/2016,
Trường Đại học Bách khoa Hà Nội.”. Nhà xuất bản Bách khoa Hà Nội, 253-261.
Chae, H. J., Joo, H., In, M. J., 2001. Utilization of brewer yeast cells for the production of food grade extract. Part 1: effect of different enzymatic treatments on solid and protein recovery and flavor characteristic. Bioresource Tech.,76: 253-258.
Chung, Y., Chae, H. J., Kim, D. C., Oh, N. S., Park, M. J., Lee, Y. S., In, M. J., 1999. Selection of comercial proteolytic enzymes for the production of brewer yeast extract. Food Eng. progr., 3: 159-163.
EBC (European Brewery Convention), 2005. Analitica - EBC. Copyright Fachverlag Hans Carl, Nurnberg. Printed in Germany by Fahner Druck GmbH, Lauf a.d. Pegnitz. ISBN 3-418-00759-7.
Hasan T., Huseyin E., 2007. Utisation of spent brewer yeast for yeast extract production by autolysis: the effect of temperature. Food and Bioproducts processing, 86:317-321.
Kenji Satake, 2002. Tận dụng men thừa trong các nhà máy bia. Hội thảo Công nghệ xử lý chất thải và tận dụng nấm men trong ngành sản xuất bia 13/3/2002, TP. Hồ Chí Minh. Viện Nghiên cứu Bia, Nước giải khát (RIB), 11-16.
Kim D. C., Chae H. J., Kim D. C., Oh N. S., In M. J., 2001. Effect of cell lytic enzymes on the production of yeast extract. J. Kor. Soc. Agri. Che. Biotechnol. 44: 273-275.
Иванова, Т. Ат. Колева, Н. Люцканов, 1989, Получаване, характеристика и приложение на автолизати от пивни дрожди, Научни трудове на ВИХВП, том 36, свитък 2, 129-140.
Hình 1. Điều kiện tự phân nấm men tối ưu
Tỷ lệ nấmmen:nước
0
-1
-0.5
-0.5
-0.5
-0.5
-0.50.5
0.5
0.5
0.5
0.5
0.5
0.25
0.750 0 0 0 0 01 11 1 1 1-1 -1 -1 -1
Tốc độkhuấy
0Nhiệt độtự phân
11
phThời giantự phân
1
Desirability
Des
irabi
lity
Y3Y2
Y141.32292
73.63542
52.10833
[41.0646,41.5812]
4140.5
39.5
73.5
72.5
71.5
70.5
51.5
50.5
49.5
48.549
50
1
00.25
0.750.5
51
72
71
52
73
3974
40
[73.3727,73.8982]
[51.81,52.4066]
0.943304

79
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Optimization of autolysis conditions for waste brewer’s yeast Nguyen Thi Thanh Thuy, Ho Tuan Anh AbstractTreated brewers’ yeast is optimized for the autolysis conditions followed the Box-Behnken design. Derringer’s desirability function is used for optimization of output factors. The results showed that the autolysis ability of waste beer yeast depended on various factors, but the most important ones were temperature, composting time and pH. The combined factors had very little or insignificant impact on the result, except the interaction between pH and composting time caused reducing the dissolved substances. Under the optimum condition the ratio of yeast: water at 1: 3, stirring speed at 30 rpm, temperature at 52ºC, pH at 5.8 and composting time in 22h, the percentage of protein conveted to free amino nitrogen, of the protein transformed to dissolved form, and of the dry matter transformed into extract were 41.3; 73.6 and 52.1%, respectively. The desirability value for all three target function was 94.3%. Keywords: Waste beer yeast, optimization, autolysis, mathematical model, experimental design
Ngày nhận bài: 5/7/2017Ngày phản biện: 10/7/2017
Người phản biện: TS. Nguyễn Xuân CảnhNgày duyệt đăng: 27/7/2017
1 Khoa Công nghệ thực phẩm, Học viện Nông nghiệp Việt Nam2 Trường Đại học Kinh tế Kỹ thuật Công nghiệp
TUYỂN CHỌN CHỦNG VI KHUẨN LACTIC CÓ KHẢ NĂNGSINH ENZYME β - GALACTOSIDASE CHỊU AXIT (pH 2 - 3)
Nguyễn Hoàng Anh1, Hồ Tuấn Anh2
TÓM TẮT Trong nghiên cứu này, 82/265 chủng vi khuẩn lactic được xác định là sinh enzyme β -galactosidase bằng phương
pháp nuôi cấy trên môi trường thạch có bổ sung X-gal. Trong đó, chủng RGH7.1, RGH6.1, RGH8.8 sinh enzyme β-galactosidase ngoại bào có hoạt độ cao nhất tương ứng là 685,95 U/L, 498,92 U/L và 492,23 U/L. β-galactosidase của ba chủng này đều có hoạt độ cao ở pH 2 và 3 với hoạt độ tương đối tương ứng dao động từ 74,32 - 83,16%, 86,49 - 93,24%. Hoạt độ của β-galactosidase của ba chủng này còn trên 50% sau 4 giờ ủ ở pH 2 và 3, trong đó chủng RGH 7.1 có độ bền với pH 2 và pH 3 tốt nhất, hoạt độ còn lại sau 4 giờ ủ tương ứng là 50,01% và 65,14%. Kết quả của nghiên cứu này chỉ ra rằng enzyme β-galactosidase ngoại bào từ chủng RGH 7.1 có tiềm năng lớn ứng dụng trong công nghiệp chế biến sữa không lactose, cũng như viên nang uống chứa enzyme β-galactosidase bền ở pH 2 và 3 cho người không dung nạp lactose
Từ khóa: Vi khuẩn lactic, β-galactosidase, bền pH
I. ĐẶT VẤN ĐỀEnzyme β-galactosidase còn được gọi là lactase,
là enzyme xúc tác cho quá trình thủy phân lactose thành glucose và galactose (Davail et al.,1994). Nhờ khả năng phân giải lactose mà β-galactosidase được sử dụng nhiều trong công nghiệp sản xuất các sản phẩm từ sữa để giảm hàm lượng lactose, một thành phần chính trong sữa mà phần lớn người trưởng thành dung nạp rất kém và đặc biệt một số ít người lớn kể cả trẻ sơ sinh có dị ứng mạnh với đường lactose (Järvelä et al., 2009). β-galactosidase được bổ sung vào quá trình sản xuất bơ sữa để tránh sự kết tinh lactose và tăng độ ngọt của sản phẩm, cải thiện các chức năng của các sản phẩm từ sữa. Ngoài ra, trong y dược chúng được sử dụng hỗ trợ tiêu hóa cho những người có khả năng hấp thụ lactose kém
(Nakayama and Amachi, 1999). Khả năng chịu pH acid đóng vai trò quan trọng trong quá trình thủy phân lactose ở các sản phẩm chế biến và sản phẩm sữa lên men. β-galactosidase có khả năng hoạt động ở pH acid trong sản xuất sữa chua và phomat sẽ làm tăng quá trình acid hóa, làm giảm khả năng đông đặc của sữa chua và tăng tốc độ phát triển cấu trúc và hương vị cho phomat (Parmjit S Panesar et al., 2010). β-galactosidase có thể tìm thấy ở động vật, thực vật, nấm men, nấm mốc và vi khuẩn (Lê Xuân Phương, 2001). Trong đó, enzyme thu nhận từ vi khuẩn ưu việt hơn cả vì vi khuẩn có sinh khối nhỏ, sinh sản nhanh, nhưng tỉ lệ enzyme trong tế bào lớn. Mặt khác, môi trường dinh dưỡng để nuôi cấy vi khuẩn lại rẻ tiền, dễ kiếm nên quy trình sản xuất chế phẩm enzyme khá dễ dàng, hiệu suất thu hồi cao và ít tốn

80
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
kém (Ngô Xuân Mạnh và ctv., 2005). Trong các loài vi khuẩn được nghiên cứu và sản xuất β-galactosidase, vi khuẩn lactic đang rất được quan tâm vì ngoài các ưu điểm trên vi khuẩn lactic còn có những đặc tính rất cần thiết như: Là vi khuẩn rất an toàn trong thực phẩm; được sử dụng thường xuyên trong thực phẩm của con người (sữa chua, dưa muối,...); dễ tìm kiếm hơn so với nhiều loài vi khuẩn khác; dễ dàng tìm được β-galactosidase. Cho đến nay, đã có một số nghiên cứu về enzyme β-galactosidase từ một số vi khuẩn lactic (Splechtna et al., 2006; Hsu et al., 2005). Tuy nhiên, các nghiên cứu về enzyme β-galactosidase từ vi khuẩn lactic có hoạt tính cao và bền ở pH axit thấp vẫn còn rất hạn chế.
Từ những cơ sở trên, nghiên cứu này được thực hiện với mục tiêu tuyển chọn được chủng lactic sinh enzyme β-galactosidase bền acid (pH 2 và 3), làm cơ sở cho ứng dụng vào việc chế biến các sản phẩm từ sữa, các viên nang enzyme uống cho người không dung nạp được lactose.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứuBộ sưu tập 265 chủng vi khuẩn lactic đã được
phân lập có trong ngân hàng chủng giống của Khoa Công nghệ thực phẩm, Học viện Nông nghiệp Việt Nam được sử dụng để tuyển chọn chủng sinh enzyme β-galactosidase có hoạt độ cao và bền ở pH 2 và pH 3. Các chủng có nguồn gốc từ sữa bò tại các nhà máy sữa, sữa nguyên chất lên men, bắp cải muối chua, cải bẹ muối chua, ruột gà hồ, ruột gà ri, nước thải chuồng bò, ống nước thải nhà máy sữa, đất xung quanh chuồng bò ở các vùng khác nhau.
2.2. Phương pháp nghiên cứu
2.2.1 Phương pháp tuyển chọn chủng vi khuẩn lactic sinh enzyme β-galactosidase
- Chủng vi khuẩn lactic sinh enzyme β-galactosidase được tuyển chọn bằng phương pháp nuôi cấy trên môi trường thạch MRS có bổ sung X-gal theo Toru Nakayama và Teruo Amachi (1996) được tóm tắt như sau:
Nguyên tắc: β-galactosidase thuỷ phân X-gal hình thành sản phẩm kết tủa màu xanh da trời kiểu indigo. Vì vậy, vi khuẩn dương tính với enzyme này sẽ tạo khuẩn lạc màu xanh khi nuôi cấy trên môi trường đĩa thạch bổ sung chỉ thị X-gal và lactose.
- Xác định khả năng sinh enzyme β-galactosidase bằng phương pháp cấy chấm điểm trên đĩa thạch. Môi trường MRS được hấp vô trùng và làm nguội, sau đó bổ sung 60 µl X-gal nồng độ 20 mg/ml và 10 µl IPTG 0,1M trên mỗi đĩa, chang đều. Đĩa cấy vi sinh
vật được đưa vào tủ nuôi cấy ở nhiệt độ 37oC, trong điều kiện tối. Kiểm tra kết quả sau 24 h, 48 h và 72 h.
2.2.2. Phương pháp nuôi cấy chủng vi khuẩn lactic để thu enzyme β-galactosidase
Chủng vi khuẩn lactic được nuôi cấy trong môi trường MRS lỏng có bổ sung lactose được tiến hành theo phương pháp của Nguyễn Lân Dũng và cộng tác viên (1978) và được tóm tắt như sau: Tiếp giống 1% từ ống nghiệm đã hoạt hóa vào bình tam giác chứa môi trường MRS có bổ sung 1% đường lactose đã hấp khử trùng ở 121oC trong 15 phút và nuôi cấy ở 37 oC, lắc 200 vòng/phút trong 24h. Tiến hành li tâm 6000 vòng/phút ở 4oC trong 20 phút, gạn lấy phần dịch nổi, cô đặc 10 lần bằng màng cô đặc có kích thước 10 kDa được sử dụng như là dịch enzyme ngoại bào, phần sinh khối được sử dụng để phá vỡ tế bào thu dịch enzyme nội bào như mô tả bởi Nguyen và cộng tác viên (2012). Dịch enzyme nội bào và ngoại bào được bảo quản ở 4oC để xác định hoạt độ, ảnh hưởng của pH đến hoạt độ và độ bền của enzyme.
2.2.3. Xác định hoạt độ của β-galactosidase ngoại bào và nội bào
Hoạt độ của β-galactosidase xác định bằng cách sử dụng o-nitrophenyl-β-D-galactopyranoside (oNPG) làm cơ chất như mô tả bởi Nguyen và cộng tác viên (2006), mô tả như sau: phản ứng enzyme - cơ chất được bắt đầu bằng cách thêm 20 μL mẫu enzyme được chuẩn bị như ở mục 2.2.2 vào 480 μL 22 mM oNPG pha trong đệm 50 mM phosphate pH 6,5, sau đó ủ trong 10 phút ở 30oC, tốc độ lắc 600 vòng/phút. Phản ứng được dừng bằng cách cho thêm 750 μl 0,4 M Na2CO3, o-nitrophenol (oNP) tạo ra được xác định bằng cách đo độ hấp thụ ở 420 nm. Một đơn vị hoạt độ của β-galactosidase được định nghĩa là lượng enzyme giải phóng 1 μmol oNP / 1phút trong các điều kiện phản ứng như trên.
2.2.4. Ảnh hưởng của pH 2, pH 3 đến hoạt độ và độ bền của enzyme β-galactosidase
Ảnh hưởng của pH 2, pH 3 đến độ bền của enzyme β-galactosidase được xác định theo phương pháp của Nguyen và cộng tác viên (2012), như sau: dịch enzyme β-galactosidase thô được ủ trong đệm Britton robinson ở pH 2 và pH 3, nhiệt độ 30ºC. Tại các thời gian ủ khác nhau, enzyme lấy ra để xác định hoạt độ còn lại (theo phương pháp mô tả ở mục 2.2.3). Độ bền pH của enzyme được xác định bằng cách so sánh % hoạt độ còn lại của enzyme tại thời điểm đo với thời điểm bắt đầu ủ.
2.3. Thời gian và đia điểm nghiên cứu Các thí nghiệm được tiến hành từ tháng 1/2017 -

81
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
tháng 5/2017 tại phòng thí nghiệm Trung tâm Khoa học và công nghệ thực phẩm, Khoa Công nghệ thực phẩm, Học viện Nông nghiệp Việt Nam.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Kết quả sàng lọc các chủng lactic có khả năng sinh enzyme β-galactosidase
Thực hiện theo 2.2.1, các chủng vi khuẩn lactic được sử dụng để tuyển chọn chủng sinh enzyme β-galactosidase bằng phương pháp nuôi cấy trên đĩa thạch có bổ sung X-gal. Kết quả thể hiện màu xanh trên đĩa thạch (minh họa ở hình 1) chỉ ra rằng 82/265 chủng vi khuẩn lactic có khả năng sinh enzyme β-galactosidase (Bảng 1).
Bảng 1. Kết quả xác định khả năng sinh β-galactosidase của các chủng vi khuẩn lactic trên môi
trường thạch bổ sung X-gal
Hình 1. Kết quả kiểm tra khả năng sinh enzyme β-galactosidase của chủng vi khuẩn lactic RGH 7.1
trên môi trường có chứa X-gal
Các chủng RGH 6.11, RGH7.1, RGH8.8, BE1.9, BE3.2, BA3.2, ON3 xuất hiện màu xanh đậm và sớm nhất sau 24 giờ nuôi cấy được sử dụng để xác định hoạt tính enzyme β–galactosidase nội bào và ngoại bào.
3.2. Xác đinh hoạt độ của enzyme β-galactosidase của vi khuẩn lactic
Kết quả xác định hoạt độ β-galactosidase của 7 chủng vi khuẩn lactic xuất hiện màu xanh đậm sau 24 giờ nuôi cấy được thể hiện ở bảng 2.
Bảng 2. Hoạt độ β-galactosidase ngoại bào của 7 chủng vi khuẩn lactic
Kết quả bảng 2 cho thấy: Cả 7 chủng đều có hoạt tính enzyme nội bào và ngoại bào. Tuy nhiên, hoạt tính của enzyme ngoại bào cao hơn nhiều lần so với nội bào, trong đó hoạt độ enzyme ngoại bào của chủng RGH7.1, RGH6.11, RGH8.8 cao nhất tương ứng là 685,95 U/L, 498,92 U/L và 492,23 U/L. Kết quả này tương đồng với kết quả của Maurya và Padalia., 2016, khi nghiên cứu sản xuất β-galactosidase ngoại bào từ vi khuẩn Lactobacillus acidophilus, hoạt độ của enzyme là 671 U/L. Các chủng còn lại có hoạt độ enzyme ngoại bào dao động trong khoảng từ 50,43 U/L - 170,7 U/L. Hoạt độ của enzyme nội bào thấp, chỉ từ 17,06 U/L - 46,32 U/L.
Các chủng RGH7.1, RGH6.11, RGH8.8 được tuyển chọn để sử dụng cho những nghiên cứu tiếp theo.
3.3. Xác đinh ảnh hưởng của pH 2 và 3 tới hoạt độ, độ bền của β-galactosidase từ chủng vi khuẩn lactic
3.3.1. Xác định ảnh hưởng của pH 2, pH 3 đến hoạt độ của enzyme
Các chủng RGH7.1, RGH6.11, RGH8.83 có hoạt độ enzyme cao được dùng để xác định ảnh hưởng của pH 2 và pH 3 đến hoạt độ của enzyme. Kết quả thể hiện ở bảng 3.
STT Nguôn phân lập
Kí hiệu mẫu
Tổng số chủng vi
khuẩn lactic
Tổng số chủng có khả năng sinh β-galactosidase
1 Sữa bò tại nhà máy sữa ST 26 9
2 Sữa nguyên chất lên men SB 33 9
3 Bắp cải muối chua BA 33 16
4 Cải bẹ muối chua BE 23 15
5 Ruột gà hồ RGH 29 96 Ruột gà ri RGD 20 3
7 Nước thải chuồng bò NT 39 9
8Ống nước thải nhà máy sữa
ON 29 9
9Đất xung quanh chuồng bò
Đ 33 3
STT Tên chủng
Hoạt độ của
enzyme ngoại bào
(U/L)
Hoạt độ của
enzyme nội bào(U/L)
Tổng hoạt độ(U/L)
1 BE1,9 121,3 46,32 167,62
2 BE3.2 75,9 18,69 94,593 BA3.2 170,7 36,42 207,124 RGH6.11 498,92 23,56 522,485 RGH7.1 685,95 34,85 720,806 RGH8.8 492,23 45,89 538,127 ON3 50,43 17,06 67,49

82
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 3. Kết quả xác định ảnh hưởng của pH 2, pH 3 đến hoạt độ của enzyme
Kết quả thể hiện cho thấy cả ba chủng RGH6.1, RGH8.8, RGH7.1 đều có hoạt tính cao ở pH 2, pH 3 với hoạt độ enzyme tương đối tương ứng dao động từ 74,32 - 83,16%, 86,49 - 93,24%. Trong đó, chủng RGH8.8 có hoạt độ tương đối cao nhất ở pH 2 và pH 3 tương ứng là 83,16% và 93,24%.
3.3.2. Xác định độ bền của β-galactosidase tại pH 2, pH 3
Các kết quả nghiên cứu độ bền ở pH 2 và pH 3 của enzyme β-galactosidase được thể hiện ở hình 2. Phân tích các giá trị đạt được cho thấy, enzyme β-galactosidase của chủng RGH6.11, RGH7.1, RGH.8.8 đều bền sau 4 giờ ở pH 2 và pH 3. Sau 1 giờ đầu thì hoạt độ của enzyme β-galactosidase ở pH 2 và pH 3 vẫn còn khá cao tương ứng là 78,84 - 84,21% và 81,20 - 90,02%. Sau 2 giờ thì hoạt độ tương đối của enzyme ở pH 2 và pH 3 còn lại tương ứng là 52,32 - 62,26% và 69,57 - 82,68%. Sau 3 giờ hoạt độ tương đối của enzyme ở pH 2 và pH 3 giảm không nhiều so với ở thời điểm 2 giờ, tương ứng là 52,45 - 57,74%. Sau 4 giờ thì hoạt độ tương đối của enzyme ở pH 2 và pH 3 còn lại tương ứng là 50,12 - 54,16% và 51,05 - 65,14%.
Theo công bố khoa học của Bộ Y tế, khả năng tiêu hóa thức ăn trong dạ dày từ 3 đến 4 giờ. Sau khoảng thời gian 4 giờ enzyme β-galactosidase của chủng RGH7.1 vẫn duy trì hoạt độ 65,14%, ở pH 3 và 50,01% ở pH 2 so với ban đầu. Kết quả của nghiên cứu này chỉ ra rằng có thể sử dụng chủng vi khuẩn lactic RGH7.1 để nghiên cứu, sản xuất enzyme β-galactosidase, ứng dụng enzyme trong sản xuất thực phẩm không lactose.
Hình 2. Độ bền của enzyme β-galactosidase ở pH 2 và pH 3
IV. KẾT LUẬNĐã tuyển chọn được chủng vi khuẩn lactic RGH7.1
phân lập từ ruột gà hồ sinh enzyme β-galactosidase ngoại bào cao (685,95 U/L). Enzyme này có hoạt độ cao và bền ở pH 2 và pH 3, sau 4 giờ ủ ở pH 2 và pH 3 hoạt độ tương đối của enzyme còn lại tương ứng là 50,01% và 65,14%. Kết quả này chỉ ra rằng enzyme beta-galactosidase từ chủng vi khuẩn Lactic RGH7.1 có tiềm năng ứng dụng trong chế biến sữa và các sản phẩm từ sữa, các viên nang enzyme uống cho người không dung nạp được lactose.
LỜI CẢM ƠNNhóm tác giả xin trân trọng cảm ơn Dự án Việt
Bỉ, Học viện Nông nghiệp Việt Nam đã tài trợ kinh phí để thực hiện nghiên cứu này.
TÀI LIỆU THAM KHẢONguyên Lân Dũng, Đoàn Xuân Mươu, Nguyên Phùng
Tiến, Đặng Đức Trạch, Phạm Văn Ty, 1978. Một số phương pháp nghiên cứu vi sinh vật học, Tập 3. NXB Khoa học và Kĩ thuật Hà Nội.
Ngô Xuân Mạnh, Võ Nhân Hậu, Nguyên Thi Tú, 2005. Nghiên cứu các điều kiện tối ưu cho việc thu nhận α-amylase chịu nhiệt từ vi khuẩn Bacillus lichenifomis. Tạp chí Khoa học Kỹ thuật nông nghiệp, 5(1): 412-416.
Lê Xuân Phương, 2001. Vi sinh công nghiệp. Nhà xuất bản Xây dựng.
Nakayama, T. and T. Amachi, 1999. Beta-galactosidase, Enzymology. In Encyclopedia of Bioprocess Technology: Fermentation, Biocatalysis, and Bioseparation; Flickinger, M. C., Drew, S. W., Eds.; John Willey: New York: 1291-1305.
Davail. S., Feller. G., Narinx. E., Gerday. C., 1994, Cold adaptation of proteins. Purification, characterization, and sequence of the heat-labile subtilisin from the antarctic psychrophile Bacillus TA41. Biol Chem, 269 (26): 17448-17453.
0
120
100
80
60
40
20
0
Hoạ
t độ
tươn
g đố
i (%
)
Thời gian (h)1 2 3
RGH6.11 pH2
RGH6.11 pH3
RGH7.1 pH2
RGH7.1 pH3
RGH8.8 pH2
RGH8.8 pH3
4
Tên chủng
Hoạt độ/Hoạt độ
tương đối
pH
pH=6,5 pH=2 pH=3
RGH6.11
Hoạt độ (U/L) 498,92 376,7 431,5
Hoạt độ tương đối (%) 100 75,5 86,49
RGH7.1
Hoạt độ (U/L) 685,95 509,80 626,34
Hoạt độ tương đối (%) 100 74,32 91,31
RGH8.8
Hoạt độ (U/L) 492,23 409,3 459
Hoạt độ tương đối (%) 100 83,16 93,24

83
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Hsu C., Davoodi-Semiromi Y., Colee J.C., Culpepper T., Dahl W.J., Mai V., Christman M.C., Langkamp-Henken B., 2005. Galactooligosaccharide supplementation reduces stress-induced gastrointestinal dysfunction and days of cold or flu: a randomized, double-blind, controlled trial in healthy university students. Am J Clin Nutr, 93 (6): 1305-1311.
Järvelä, I., Torniainen, S., Kolho, K.L., 2009. Molecular genetics of human lactase deficiencies. Annals of Medicine, 41 (8): 568-575.
Maurya, K and Padalia, U., 2016, Production of Beta-Galactosidase from Lactic Acid Bacteria. International Journal of Engineering Science and Computing, 6 (10): 2674-2676.
Nguyen, H A., Nguyen, T H., Kren, V., Eijsink, V G H., Haltrich, D., Peterbauer, C K., 2012. Heterologous Expression and Characterization of an N-Acetyl-β-D-hexosaminidase from Lactococcus lactis ssp. lactis
IL1403. J. Agricultural and food chemistry, 60 (12): 3275-3281.
Nguyen, TH., Splechtna, B., Steinböck, M., Kneifel, W., Lettner, H.P., Kulbe, K.D., Haltrich, D., 2006. Purification and Characterization of Two Novel β-Galactosidases from Lactobacillus reuteri. J. Agricultural and food chemistry, 54: 4989-4998.
Parmjit S, P., Shweta, K., Reeba, P., 2010. Protential Applications of Ininiobilized β-galactosidase in Food Prossesing Industries. Enzyme Research: 1-16.
Splechtna B, Nguyen T, Zehetner R, Lettner HP, Lorenz W, Haltrich D., 2007. Process development for the production of prebiotic galacto-oligosaccharides from lactose using β-galactosidase from Lactobacillus sp.. Biotechnol J, 2: 480-485.
Toru Nakayama, Teruo Amachi, 1996. Beta-galactosidase enzymology Encyclopaedia of Food science, food technology and nutrition, 3: 1291-1305.
Selection of lactic acid bacteria producing acidic tolerant enzyme β-galactosidase (pH 2 - 3)
Nguyen Hoang Anh, Ho Tuan AnhAbstract In this study, 82/265 strains of lactic acid bacteria were determined to produce β-galactosidase by using agar plate supplemented with X-gal. Of which, strains RGH7.1, RGH6.1, RGH8.8 produced extracellular β-galactosidase with the highest enzyme activity, 685.95 U/L, 498.92 U/L and 492.23 U/L, respectively. β-galactosidase of these three strains had high activity at pH 2 and 3 with their relative activity was from 74.32 - 83.16%, 86.49 - 93.24%. In addition, residual enzyme activity was still remained over 50% after 4 hours of incubation at pH 2 and 3. Among them, β-galactosidase from strain RGH7.1 was the best in terms of stability at pH 2 and 3 after 4 hours of incubation; residual enzyme activity was remained 50.01% and 65.14%, respectively. Results of this research indicated that extracellular β-galactosidase of strain RGH7.1 was promising one which could be applied in free lactose milk production and capsules containing of β-galactosidase with stability at with pH 2 and 3 for intolerant lactose people.Key words: Lactic acid bacteria, β-galactosidase, pH stability
Ngày nhận bài: 29/6/2017Ngày phản biện: 5/7/2017
Người phản biện: TS. Nguyễn Xuân CảnhNgày duyệt đăng: 27/7/2017
1 Viện Bảo vệ thực vật; 2 Trung tâm Ứng dụng Khoa học và Công nghệ Nam Định 3 Cục Ứng dụng và Phát triển Công nghệ
CẢI TIẾN CÔNG NGHỆ SẢN XUẤT CHẾ PHẨM NẤM XANH Metarhizium anisopliae ĐỂ PHÒNG TRỪ RẦY NÂU HẠI LÚA
Trần Văn Huy1, Phạm Văn Nhạ1, Nguyễn Thị Nga1, Nguyễn Mạnh Cường2, Vũ Xuân Trung2, Phạm Việt Hồng3,
Lê Thị Thu Hiền3, Nguyễn Trường Phi3
TÓM TẮTChế phẩm sinh học có nguồn gốc từ nấm xanh Metarhizium anisopliae có nhiều tiềm năng trong phòng trừ rầy
nâu hại lúa. Tuy nhiên, để phát triển thương mại hóa trên thị trường, sản phẩm cần phải cải tiến về tạo dạng sử dụng. Trên cơ sở nghiên cứu kỹ thuật tách bào tử tinh bằng hộp rây với mắt rây 200 µm, bổ sung bi sắt kích thước 10 mm, đã thu được lượng bào tử tinh lớn và ít tạp chất và đã phối trộn bào tử tinh với chất phụ gia PG1 tạo dạng bột thấm nước. Chế phẩm dạng mới bao gồm bào tử tinh khiết cộng chất phụ gia PG1, có hàm lượng bào tử cao trên 1010 bt/g,

84
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀRầy nâu (Nilaparvata lugens) là một trong những
đối tượng gây hại nguy hiểm trên cây lúa ở nước ta (Trần Văn Huy và ctv., 2012). Phòng trừ bằng thuốc hóa học trong một thời gian dài đã hủy diệt thiên địch, thúc đẩy hình thành tính kháng thuốc dẫn đến nguy cơ rầy nâu bùng phát thành dịch. Rầy nâu sống trong hệ sinh thái ruộng lúa, có độ ẩm cao nên chúng bị nhiều loài nấm ký sinh gây chết tự nhiên. Trong các loài nấm ký sinh thì loài nấm xanh Metarhizium anisoplie có nhiều tiềm năng nhân nuôi sản xuất chế phẩm sinh học để phòng trừ rầy nâu (Jin S. F et al., 2008; Lê Văn Trịnh và ctv., 2008; Nguyễn Thị Lộc, 2009). Trong giai đoạn năm 2014 - 2016, Trung tâm Ứng dụng Khoa học và Công nghệ Nam Định đã phối hợp với Viện Bảo vệ thực vật nghiên cứu thực hiện dự án “Hoàn thiện công nghệ sản xuất, ứng dụng chế phẩm sinh học trừ rầy nâu hại lúa tại Nam Định”. Chế phẩm đã mang lại những hiệu quả nhất định: Làm giảm tần suất sử dụng thuốc trừ sâu hóa học, an toàn với người sử dụng và thiên địch và ít hình thành tính kháng thuốc dẫn đến hiệu quả trừ rầy nâu bền vững (Nguyễn Mạnh Cường, 2016). Tuy nhiên để phát triển sản phẩm và thương mại hóa trên thị trường thì chế phẩm có một số hạn chế: Dạng chế phẩm bao gồm bào tử và cơ chất gạo gây ra một số khó khăn trong sử dụng như phải lọc bào tử trước khi đưa vào bình phun. Chế phẩm có khối lượng lớn tốn kém vận chuyển và kho bãi, thời gian bảo quản ngắn 3 tháng. Để khắc phục vấn đề này, trong khuôn khổ thực hiện nhiệm vụ “Trình diễn, kết nối cung - cầu và xúc tiến thương mại hóa công nghệ năm 2016”, Trung tâm Ứng dụng tiến bộ KH và CN Nam Định đã được Cục Ứng dụng và Phát triển công nghệ phối hợp cùng chuyên gia triển khai cải tiến công nghệ sản xuất chế phẩm nấm xanh bằng kỹ thuật tách bào tử tinh và phối trộn với chất phụ gia để tăng thời gian bảo quản.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu- Chế phẩm nấm xanh Metarhizium anisopliae.- Chất phụ gia PG1.
2.2. Phương pháp nghiên cứu
2.2.1. Phương pháp tách lọc bào tử tạo dạng chế phẩm từ sinh khối nấm xanh
- Thí nghiệm xác định kích thước mắt rây bào tử
thích hợp: Thí nghiệm được bố trí với 4 công thức, tương ứng với 4 loại mắt rây có kích thước khác nhau: 150, 200, 300 và 450 µm. Số lượng nấm lắc trên một mẻ là 5 kg và thời gian lắc là 5 phút/mẻ. Mỗi công thức bố trí 3 lần nhắc lại.
- Thí nghiệm bổ sung bi sắt vào hộp rây bào tử: Thí nghiệm được bố trí 4 công thức tương ứng với 4 loại bi sắt có đường kính khác nhau: 5, 10, 15 và 20 mm. Số lượng nấm lắc trên một mẻ là 5 kg và thời gian lắc là 5 phút/mẻ. Mỗi công thức bố trí 3 lần nhắc lại.
- Phương pháp đánh giá khả năng sống của bào tử sau khi phối trộn với chất phụ gia PG1 với tỷ lệ 10% bào tử tinh và 90% chất phụ gia PG1: Đánh giá khả năng sống của bào tử nấm xanh với chất PG1 ở các mốc thời gian: 1, 2, 3, 4, 6, 9, 12 tháng bằng phương pháp kiểm tra sự hình thành khuẩn lạc của bào tử trên môi trường N1: Lấy mẫu 1gr chế phẩm pha loãng ở 10-5 và cấy 0,1ml dịch nên đĩa môi trường nuôi cấy N1 hoặc đặt ở 28oC trong 5 ngày để đếm số lượng khuẩn lạc nấm.
2.2.2. Phương pháp thử nghiệm chế phẩm trên đồng ruộng
Thí nghiệm đánh giá hiệu lực của chế phẩm trừ rầy nâu hại lúa trên đồng ruộng tiến hành tại Nam Định được bố trí với 4 công thức (CT): CT 1 chế phẩm nấm xanh dạng hạt (chưa cải tiến), CT 2 chế phẩm nấm xanh dạng bột thấm nước (cải tiến), CT 3 thuốc trừ sâu Bassa 50EC và CT 4 đối chứng không phun. Mỗi công thức 3 lần nhắc lại, mỗi lần nhắc lại là 40m2. Chỉ tiêu theo dõi: Mật độ rầy nâu tại các công thức trước và sau 7, 10 ngày xử lý.
2.2.3. Phương pháp xử lý số liệu - Các tham số thống kê cơ bản xử lý bằng phần
mềm Microsoft Excel 2007.- Đánh giá sự sai khác các chỉ tiêu bằng phần
mềm Statistix 9.0
2.3. Thời gian và đia điểm nghiên cứu - Thời gian: Từ tháng 1 đến tháng 6 năm 2017.- Trung tâm Ứng dụng tiến bộ KHCN Nam Định.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Kết quả nghiên cứu tách lọc bào tử, tạo dạng chế phẩm từ sinh khối nấm xanh
Theo quy trình cũ, dạng chế phẩm nấm xanh là
liều lượng sử dụng thấp 500 g/ha. Chế phẩm dễ sử dụng, có thể pha trực tiếp vào trong bình phun, không phải tách lọc, không gây tắc bình. Hiệu lực phòng trừ rầy nâu trên đồng ruộng của chế phẩm đạt 72,8%. Thời gian bảo quản chế phẩm kéo dài trên 12 tháng.
Từ khóa: Rầy nâu, nấm xanh Metarhizium anisopliae, bào tử tinh, chất phụ gia PG1

85
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
dạng hạt thô bao gồm hạt gạo và bào tử nấm. Dạng chế phẩm này gây khó khăn cho việc bảo quản, vận chuyển và sử dụng do khối lượng chế phẩm lớn, tốn diện tích bảo quản, vận chuyển. Đặc biệt khi sử dụng phải tiến hành bước tách lọc bào tử ra khỏi hạt gạo nên gây tâm lý ngại dùng của người dân. Để khắc phục nhược điểm này đã tiến hành tách bào tử tinh ra khỏi cơ chất gạo bằng máy rây bào tử. Để thu được lượng bào tử lớn và ít tạp chất đã thử nghiệm các kích cỡ mắt rây khác nhau. Kết quả thử nghiệm các loại rây bào tử được trình bày trong bảng 1. Kết quả thử nghiệm cho thấy khối lượng bào tử thu được tỷ lệ thuận với kích thước mắt rây tuy nhiên ở công thức mắt rây là 300 và 450 cho lượng tạp chất lớn. Đối với kích thước mắt rây là 150 µm cho lượng bào tử 42,4 gam /kg ít hơn so với mắt rây có kích thước 200 µm (51,2 g). Vì vậy kích thước thích hợp để rây bào tử là 200 µm (Bảng 1).
Bảng 1. Ảnh hưởng của kích cỡ mắt rây đến hiệu suất thu bào tử nấm xanh Metarhizium anisopliae
(Viện Bảo vệ thực vật, 2016)
Ghi chú: Bảng 1, 2, 4: Các chữ cái khác nhau trong cùng một cột biểu thị sự sai khác có ý nghĩa 95%.
Để thu được lượng bào tử cao hơn, đã bổ sung bi sắt vào hộp rây, kết quả thử nghiệm kích thước bi sắt bổ sung vào hộp rây được trình bày trong bảng 2. Kết quả thử nghiệm cho thấy khi bổ sung bi sắt vào hộp rây thì lượng bào tử thu được là cao hơn. Trong các loại bi thì loại có được kính là 10mm cho khối lượng bào tử (62,6 g) tuy thấp hơn 2 loại bi có đường kính 15 mm và 20 mm nhưng với lượng tạp chất ít. Vì vậy đã lựa chọn loại bi kích thước 10 mm để bổ sung vào hộp lồng rây bào tử.
Để tăng thời gian bảo quản bào tử nấm xanh, đã phối trộn bào tử tinh với chất phụ gia PG1. Đánh giá thời gian bảo quản bào tử nấm sau khi phối trộn được trình bày trong bảng 3. Kết quả đánh giá khả năng sống của bào tử nấm xanh sau các mốc thời gian kiểm tra cho thấy, sau khi phối trộn với chất
phụ gia PG1 thì thời gian sống của bào tử nấm nâng lên rõ rệt. Tại công thức phối trộn, sau 9 tháng lượng bào tử sống đạt 2,8 ˟ 109 Cfu/g, sau 12 tháng đạt 1,0 ˟ 109 Cfu/g, trong khi ở dạng cũ lượng bào tử giảm rất nhanh sau 3 tháng còn 1,2 ˟ 108 Cfu/g, sau 6 tháng số lượng bào tử sống chỉ còn 1,2 ˟ 106 Cfu/g và sau 9 tháng lượng bào tử sống còn lại rất thấp.
Bảng 2. Ảnh hưởng của kích thước bi bổ sung vào trong hộp rây bào tử đến hiệu suất thu bào tử
nấm xanh (Viện Bảo vệ thực vật, 2016)
Bảng 3. Kết quả thử nghiệm khả năng sống của bào tử của nấm xanh trong chất phụ gia ở điều kiện nhiệt độ
phòng thí nghiệm (Viện Bảo vệ thực vật, 2016)
Ghi chú: (-) Lượng bào tử sống còn lại rất thấp < 103Cfu/g, không ổn định
Từ kết quả cải tiến công nghệ, đề tài đã sản xuất chế phẩm dạng bột thấm nước bao gồm tử tinh và chất phụ gia. Kết quả thử nghiệm hiệu lực của chế phẩm dạng bộn thấm nước trên đồng ruộng (Bảng 4) cho thấy, chế phẩm sản xuất theo công nghệ mới cho hiệu quả phòng trừ đạt 72,8% sau 10 ngày phun ở liều lượng 0,5 kg/ha, cao hơn chế phẩm dạng cũ (hiệu lực đạt 69,1%) với liều lượng là 30 kg/ha. Như vậy chế phẩm dạng mới cho hiệu quả phòng trừ cao hơn với liều lượng sử dụng ít hơn.
Công thức
Kích cỡ mắt rây (µm)
Lương bào tử thu đươc/kg nấm /5 phút (g)
I 150 42,4cII 200 51,2bIII 300 58,1aIV 450 60,4a
CV(%) 2,43LSD0,05 2,57
Công thức Mắt rây
Đường kính bi sắt
(mm)
Lương bào tử thu đươc / kg nấm/5
phút (g) I 200 µm 5 mm 55,3 dII 200 µm 10 mm 62,6c III 200 µm 15 mm 65,4ab IV 200 µm 20 mm 67,5 a
CV(%) 3,58LSD0,05 4,48
Thời gian bảo quản
Bào tử bám trên cơ chất gạo (Cfu/g)
Bào tử tinh phối trộn với chất phụ gia PG1(Cfu/g)
Ban đầu 1,0 ˟ 109 2,0 ˟ 1010 Sau 1 tháng 8,8 ˟ 108 1,8 ˟ 1010 Sau 2 tháng 2,5 ˟ 108 1,5 ˟ 1010 Sau 3 tháng 1,2 ˟ 108 1,1 ˟ 1010 Sau 4 tháng 3,1 ˟ 107 9,2 ˟ 109 Sau 6 tháng 1,2 ˟ 106 5,2 ˟ 109 Sau 9 tháng - 2,8 ˟ 109
Sau 12 tháng - 1,0 ˟ 109

86
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Như vậy sau quá trình nghiên cứu thử nghiệm đã cải tiến và hoàn thiện được quy trình sản xuất để sản phẩm có thể thương mại hóa trên thị trường với các ưu điểm cụ thể được trình bày trong bảng 5.
Bảng 5. Đánh giá ưu điểm của chế phẩm nấm xanh Metarhizium anisopliae sau cải tiến công nghệ
(Viện Bảo vệ thực vật, 2016)
Ưu điểm của công nghệ sau cải tiến là: Dạng chế phẩm là dạng bột thấm nước, bào tử tinh khiết cộng chất phụ gia, hàm lượng bào tử cao 1010 bt/g, liều lượng sử dụng thấp 0,5kg /ha, thời gian bảo quản dài, dễ sử dụng, pha trực tiếp vào trong bình phun.
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luận- Đã tạo dạng chế phẩm dạng bột thấm nước, bào
tử tinh khiết cộng chất phụ gia PG1, hàm lượng bào tử cao trên 1010 bt/g, liều lượng sử dụng chế phẩm thấp 500g/ha, chế phẩm dễ sử dụng trên đồng ruộng.
- Đã nâng cao được thời gian bảo quản chế phẩm nấm xanh trên 12 tháng trong điều kiện nhiệt độ phòng. Hiệu lực phòng trừ rầy nâu trên đồng ruộng của chế phẩm đạt 72,8%.
4.2. Đề nghiSản xuất thử nghiệm chế phẩm nấm xanh dạng
bột thấm nước để trừ rầy nâu trên đồng ruộng theo hướng bền vững.
TÀI LIỆU THAM KHẢONguyên Mạnh Cường, 2016. Báo cáo kết quả dự án sản
xuất thử nghiệm: Hoàn thiện công nghệ sản xuất, ứng dụng chế phẩm sinh học trừ rầy nâu hại lúa tại Nam Định.
Trần Văn Huy, Nguyên Thi Chúc Quỳnh, Vũ Thi Hiên, Phạm Thi Minh Thắng, Phùng Quang Tùng, 2012. Một số đặc điểm hình thái sinh học và khả năng ký sinh của nấm Paecilomyces javanicus đối với rầy nâu và rầy lưng trắng hại lúa. Tạp chí Bảo vệ thực vật, Số 3/2012.
Nguyên Thi Lộc, 2009. Kết quả ứng dụng chế phẩm sinh học Metarhizum anisopliae và Beauveria bassiana trừ sâu hại cây trồng tại đồng bằng sông Cửu Long. Kỷ yếu hội thảo định hướng phát triển ứng dụng BPSH trong phòng chống dịch hại cây trồng. Sóc Trăng, tháng 6/2009, Tr. 90- 98.
Lê Văn Trinh, 2008. Kết quả nghiên cứu và sản xuất chế phẩm nấm có ích phòng trừ rầy nâu hại lúa bền vững. Hội nghị KH hàng năm Viện Bảo vệ thực vật. 16 Tr.
Jin S. F., Feng M. G. , Chen J. Q., 2008. Selection of global Metarhizium isolates for the control of the rice pest Nilaparvata lugens (Homoptera: Delphacidae). Institute of Microbiology, College of Life Science, Zhejiang University, Hangzhou, Zhejiang 310058, People’s Republic of China.
Bảng 4. Kết quả thử hiệu lực phòng trừ rầy nâu trên đồng ruộng của chế phẩm nấm xanh được sản xuất theo công nghệ cải tiến (Nam Định)
Công thức thí nghiệm
Liều lương phun/1ha
Mật độ rầy nâu trước phun
(con/m2)
Hiệu lực phòng trừ rầy nâu sau phun7 ngày 10 ngày
Mật độ rầy nâu (con/m2)
Hiệu lực (%)
Mật độ rầy nâu (con/m2)
Hiệu lực (%)
Nấm xanh(CN cũ ) 20 kg 549,3 357,7b 55,9 484,3b 69,1
Nấm xanh (CN mới) 0,5 kg 562,3 341,7b 58,9 437,3c 72,8
Bassa50EC 1,5 lít 526,3 198,7c 74,5 495,3b 67,1Đối chứng Không phun 524,6 775,3a - 1498,7a -
CV(%) 2,80 2,25LSD0,05 23,4 32,7
Các chỉ tiêu Công nghệ cũ Công nghệ cải tiến
Hàm lượng bào tử 1 ˟ 109 bt/g 2 ˟ 1010 bt/g
Khả năng bảo quản 6 tháng 12 tháng
Dạng sử dụng Dạng hạt: Bào tử + Cơ chất gạo
Bột thấm nước: Bào tử tinh + chất phụ gia PG1
Liều lượng sử dụng 20kg/ha 0,5 kg/ha
Hiệu lực phòng trừ rầy 70,0% 72,8%

87
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Improving production technology of ometar Metarhizium anisopliae preparation for brown backed rice plant hopper prevention
Tran Van Huy, Pham Van Nha, Nguyen Thi Nga, Nguyen Manh Cuong, Vu Xuan Trung, Pham Viet Hong,
Le Thi Thu Hien, Nguyen Truong PhiAbstract Bioproduct of Metarhizium anisopliae shows its great potential in controlling the brown plant hopper. However, the characteristic of the final product is still need to be improved for the better commercialization. A number of pure spores was obtained based on the spore splitting technique by using screen with mesh size of 200 µm. The preparation pervious powder included pure spores with more than 1010 bt/g and PG1 addition agent, and the powder dose used was low by 500 g/ha. The preparation pervious powder can be directly dissolved inside the sprayer without any addition refinement. The 72.8% preventing effect of the preparation was achieved, furthermore, the new form of the product offered a great extension in preservation time which made the preparation keeping use within a year. Key words: Brown planthopper, Metarhizium anisopliae, spore, PG1 addition agent
Ngày nhận bài: 1/7/2017Ngày phản biện: 6/7/2017
Người phản biện: TS. Trịnh Xuân HoạtNgày duyệt đăng: 27/7/2017
1 Viện Bảo vệ thực vật, 2 Trường Đại học Nông Lâm Thái Nguyên
KẾT QUẢ GIÁM ĐỊNH VÀ MỘT SỐ ĐẶC ĐIỂM CỦA NẤM Botrytis cinerea Pers. GÂY BỆNH THỐI XÁM TRÊN HOA THƯỢC DƯỢC (Dahia pinnata Cav.) TẠI VIỆT NAM
Mai Văn Quân1, Dương Thị Nguyên2
TÓM TẮTBotrytis cinerea là tác nhân gây bệnh thối xám trên nhiều loại cây trồng khác nhau tại Việt Nam. Trong nghiên
cứu này, 15 mẫu nấm phân lập từ cây hoa thược dược (Dahia pinnata Cav.) bị bệnh thối xám trồng tại Đồng bằng sông Hồng và Trung du miền núi phía Bắc. Kết quả phân loại dựa vào đặc điểm và kích thước bào tử đã ghi nhận tất cả 15 nguồn nấm đều thuộc nấm Botrytis cinerea. Phản ứng PCR với cặp primer đặc hiệu C729+/C729- đã khuếch đại sản phẩm PCR với kích thước khoảng 730 bp từ tất cả các nguồn nấm. Kết quả giải trình tự gen và phân tích cây phả hệ đã khẳng định nấm B. cinerea Pers. là tác nhân gây bệnh thối xám trên hoa thược dược. Kết quả nghiên cứu đặc điểm sinh học của nấm B. cinerea Pers. trên các loại môi trường, điều kiện nhiệt độ và thời gian chiếu sáng khác nhau cho thấy có sự khác nhau về khả năng phát triển của sợi nấm, sản sinh bào tử và hình thành hạch nấm. Trong 4 loại môi trường, nấm phát triển mạnh nhất trên môi trường BĐ; tuy nhiên, nấm hình thành nhiều loại hạch nấm lớn sau 3 ngày trên môi trường Czapek. Trên môi trường PDA nấm chỉ hình thành bào tử ở nhiệt độ 15oC sau 8 ngày nuôi cấy. Nấm phát triển tốt ở điều kiện tối hoàn toàn và 12 giờ sáng xen kẽ 12 giờ tối. Nấm chỉ sản sinh bào tử trong điều kiện sáng hoàn toàn.
Từ khóa: Botrytis cinerea, bệnh thối xám, Dahia pinnata Cav.
I. ĐẶT VẤN ĐỀNấm Botrytis cinerea Pers. là một trong những
loài nấm gây hại trên nhiều loại hoa, rau, cây ăn quả ôn đới, cây dược liệu ở các vùng khác nhau trên thế giới và Việt Nam. Tại Việt Nam, nấm B. cinerea đã phát sinh và gây hại nghiêm trọng tại các vùng trồng hoa, cây ăn quả ôn đới và cây thực phẩm tại vùng Trung du miền núi phía Bắc, Đồng bằng sông Hồng và Đà Lạt - Lâm Đồng. Bệnh chủ yếu gây hại trên quả, cuống quả, đài hoa, cánh hoa và lá làm cho các bộ phận này bị thối và xuất hiện lớp mốc màu xám bao phủ trên bề mặt. Nấm B. cinerea là một đối
tượng gây hại quan trọng trên các loài hoa, nấm gây thối hoa, lá làm giảm năng suất, chất lượng của các loài hoa hồng, cúc, thược dược (Đặng Vũ Thị Thanh và ctv., 2007, 2010; Mai Văn Quân và ctv., 2016).
Trong thời gian gần đây, các nghiên cứu về phổ ký chủ, đặc điểm sinh học, quy luật phát sinh gây hại và biện pháp phòng trừ bệnh thối xám do nấm B. cinerea gây ra đã được Viện Bảo vệ thực vật tiến hành. Bài báo này phản ánh một số kết quả nghiên cứu về nấm B. cinerea gây bệnh thối xám hại hoa thược dược (Dahia pinnata Cav.) tại vùng Đồng bằng sông Hồng và Trung du miền núi phía Bắc.

88
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu- Mẫu hoa thược dược có biểu hiện triệu chứng
bệnh thối xám được thu thập trong năm 2017 tại một số vùng Đồng bằng sông Hồng và Trung du miền núi phía Bắc.
- Môi trường phân lập thông thường bao gồm: môi trường Water Agar (WA), Bột Đậu (BĐ), PDA, Cà rốt (CR) và Czapek. Các loại hóa chất phục vụ chiết suất ADN, chạy PCR và giải trình tự gen.
2.2. Phương pháp nghiên cứu- Điều tra, thu thập và phân lập tác nhân gây bệnh
thối xám trên thược dược theo phương pháp điều tra phát hiện bệnh cây của Viện Bảo vệ thực vật (1997).
- Chiết suất ADN tổng số bằng phương pháp CTAB (Cetyl Trimethyl Ammonium Bromide) theo mô tả của Doyle & Doyle (1990). Phản ứng PCR để xác định nấm B. cinerea theo Rigotti và cộng tác viên (2002) với cặp primer C729+(5’-AGCTCGAGAGAGATCTCTGA-3’)/C729-(5’-CTGCAATGTTCTGCGTGGAA-3’).
Sản phẩm PCR được tinh sạch từ agarose gel sử dụng QIAquick PCR Purifcation Kit (Qiagen, Đức) và được giải trình tự gen trực tiếp cả hai chiều bằng máy ABI3100 tại Hàn Quốc sử dụng BigDye Terminator 3.1 Kit (Applied Biotech). Trình tự các mẫu được so sánh với Ngân hàng Gen bằng phần mềm trực tuyến http://blast.ncbi.nlm.nih.gov/Blast.cgi; Cây phả hệ xây dựng theo phương pháp Neighbor-joining với khoảng cách di truyền giữa các chuỗi được xác định dựa trên mô hình thay thế Kimura hai tham số, giá trị thống kê bootstrap (%) với 1000 lần lặp lại trong phần mềm MEGA 6.0 (Tamura et al., 2013).
- Nghiên cứu khả năng phát triển của nấm B. cinerea (1) trên môi trường PDA, Cà rốt, Bột đậu, Czapek; (2) các mức nhiệt độ 10, 15, 20, 25 và 30oC trên môi trường PDA; (3) các điều kiện chiếu sáng hoàn toàn, tối hoàn toàn, 12 giờ sáng 12 giờ tối ở nhiệt độ 20oC trên môi trường PDA. Các nguồn nấm B. cinerea trong nghiên cứu được làm thuần bằng đơn bào tử. Theo dõi hình thái và màu sắc tản nấm, thời gian hình thành bào tử và hạch nấm trên môi trường.
- Phương pháp xử lý số liệu: Số liệu được xử lý bằng phần mềm Excell 2007, Statistics 9.0.
2.3. Thời gian và đia điểm nghiên cứuThí nghiệm được tiến hành tại phòng thí nghiệm
của Viện Bảo vệ thực vật trong năm 2017.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Triệu chứng bệnh trên cây hoa thươc dươcNấm gây hại trên các bộ phận của cây, đặc biệt là
phần non như cánh hoa, nụ hoa, lá non. Trên hoa, bệnh hại trên cánh hoa làm cho cánh hoa khô lại, trên bề mặt có lớp nấm màu xám (Hình 1). Nấm xâm nhập vào nụ hoa làm cho nụ hoa bị thối không hình thành được hoa. Gặp điều kiện mưa phùn và ẩm độ cao bộ phận bị bệnh phủ lớp mốc màu xám tro. Cành bào tử mọc đơn lẻ, thẳng, đa bào, cành phân nhánh ngắn, trên đỉnh cành hơi phình to thành hình cầu, có đính các núm nhỏ. Cành không màu hoặc có màu nâu nhạt. Bào tử đính trên các núm nhỏ, hình trứng hay hình bầu dục, đơn bào, không màu kích thước bào tử nấm trên môi trường PDA 9,86 ˟ 5,92 µm (Hình 1).
Hình 1. Triệu chứng điển hình của bệnh thối xám trên cây hoa thược dược tại Hà Nội
(A). Bông hoa bị nhiễm bệnh thối xám (hình bên trái); bông hoa không bị nhiễm bệnh (hình bên phải).
(B). Cành bào tử phân sinh và bào tử nấm gây bệnh thối xám (độ phóng đại 40 lần).
3.2. Xác đinh nấm B. cinerea gây bệnh thối xám trên hoa thươc dươc bằng phương pháp PCR
DNA tổng số của các mẫu nấm gây bệnh thối xám trên cây hoa thược dược được chiết suất phục vụ phản ứng PCR sử dụng cặp primer C729+/C729-. Kích thước sản phẩm PCR của tất cả các mẫu nấm có kích thước khoảng 730 bp. Tất cả các sản phẩm PCR đều được giải trình tự gen trực tiếp cả hai chiều bằng primer C729+/C729-. Trình tự gen của tất cả các mẫu thu được đều đồng nhất. So sánh với Ngân hàng Gen, trình tự DNA của sản phẩm PCR trùng với các gen mã hóa của nấm Botrytis cinerea gây bệnh.
Cây phả hệ được xây dựng dựa trên 16 trình tự đoạn gen của 11 loài nấm Botrytis sp. (bao gồm B. fabae, B. eucalypti, B. tulipae, B. squamosa, B. elliptica, B. prunorum, B. byssoidea, B. fabiopsis, B. convoluta, B. aclada và B. cinerea) khác nhau từ Ngân hàng Gen.
Chủng nấm Bo.HN06 thu trên cây hoa thược dược tại vùng Hà Nội đã cùng với 2 đại diện của loài nấm B. cinerea có mã số Ngân hàng Gen AJ422103 (gây bệnh thối xám trên cây dâu tây tại Thụy Sĩ) và KU936083 (từ Ấn Độ) tạo thành một nhánh riêng biệt so với các loài Botrytis khác trên cây phả hệ (Hình 2).
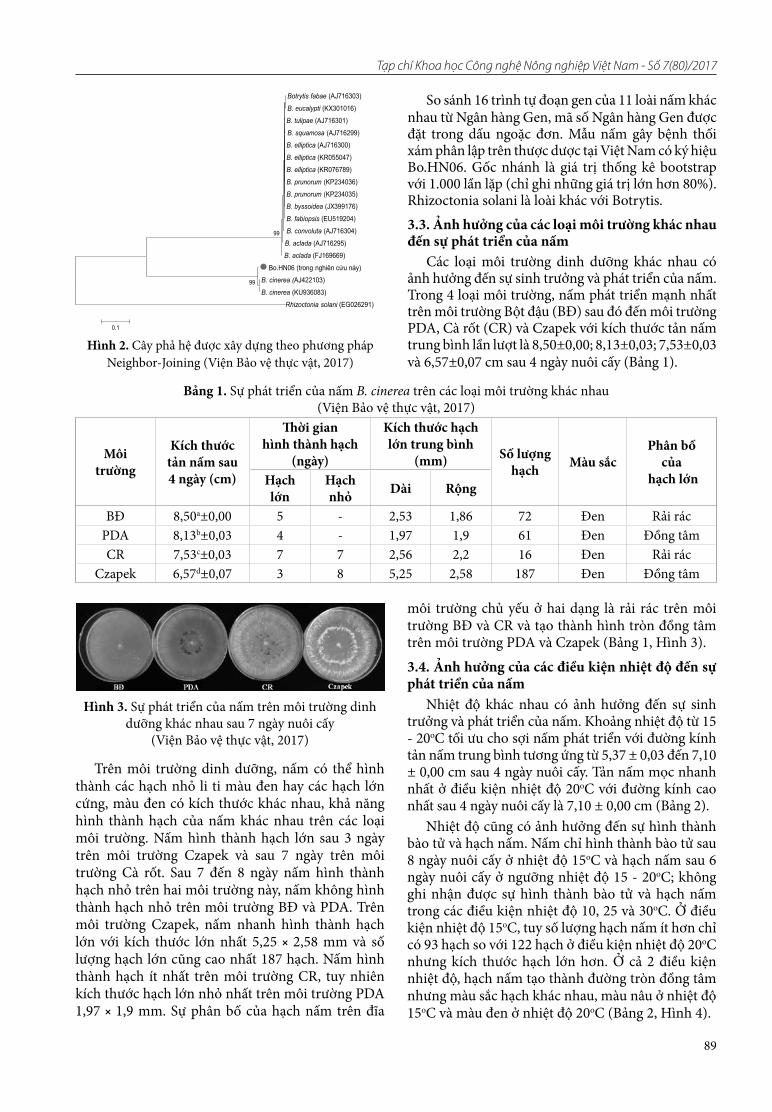
89
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Hình 2. Cây phả hệ được xây dựng theo phương pháp Neighbor-Joining (Viện Bảo vệ thực vật, 2017)
So sánh 16 trình tự đoạn gen của 11 loài nấm khác nhau từ Ngân hàng Gen, mã số Ngân hàng Gen được đặt trong dấu ngoặc đơn. Mẫu nấm gây bệnh thối xám phân lập trên thược dược tại Việt Nam có ký hiệu Bo.HN06. Gốc nhánh là giá trị thống kê bootstrap với 1.000 lần lặp (chỉ ghi những giá trị lớn hơn 80%). Rhizoctonia solani là loài khác với Botrytis.3.3. Ảnh hưởng của các loại môi trường khác nhau đến sự phát triển của nấm
Các loại môi trường dinh dưỡng khác nhau có ảnh hưởng đến sự sinh trưởng và phát triển của nấm. Trong 4 loại môi trường, nấm phát triển mạnh nhất trên môi trường Bột đậu (BĐ) sau đó đến môi trường PDA, Cà rốt (CR) và Czapek với kích thước tản nấm trung bình lần lượt là 8,50±0,00; 8,13±0,03; 7,53±0,03 và 6,57±0,07 cm sau 4 ngày nuôi cấy (Bảng 1).
Botrytis fabae (AJ716303)
B. eucalypti (KX301016)
B. tulipae (AJ716301)
B. squamosa (AJ716299)
B. elliptica (AJ716300)
B. elliptica (KR055047)
B. elliptica (KR076789)
B. prunorum (KP234036)
B. prunorum (KP234035)
B. byssoidea (JX399176)
B. fabiopsis (EU519204)
B. convoluta (AJ716304)
B. aclada (AJ716295)
B. aclada (FJ169669)
Bo.HN06 (trong nghiên cứu này)
B. cinerea (AJ422103)
B. cinerea (KU936083)
Rhizoctonia solani (EG026291)
99
99
0.1
Bảng 1. Sự phát triển của nấm B. cinerea trên các loại môi trường khác nhau (Viện Bảo vệ thực vật, 2017)
Môi trường
Kích thước tản nấm sau 4 ngày (cm)
Thời gian hình thành hạch
(ngày)
Kích thước hạch lớn trung bình
(mm) Số lương hạch Màu sắc
Phân bố của
hạch lớnHạch lớn
Hạch nhỏ Dài Rộng
BĐ 8,50a±0,00 5 - 2,53 1,86 72 Đen Rải rácPDA 8,13b±0,03 4 - 1,97 1,9 61 Đen Đồng tâmCR 7,53c±0,03 7 7 2,56 2,2 16 Đen Rải rác
Czapek 6,57d±0,07 3 8 5,25 2,58 187 Đen Đồng tâm
Hình 3. Sự phát triển của nấm trên môi trường dinh dưỡng khác nhau sau 7 ngày nuôi cấy
(Viện Bảo vệ thực vật, 2017)
Trên môi trường dinh dưỡng, nấm có thể hình thành các hạch nhỏ li ti màu đen hay các hạch lớn cứng, màu đen có kích thước khác nhau, khả năng hình thành hạch của nấm khác nhau trên các loại môi trường. Nấm hình thành hạch lớn sau 3 ngày trên môi trường Czapek và sau 7 ngày trên môi trường Cà rốt. Sau 7 đến 8 ngày nấm hình thành hạch nhỏ trên hai môi trường này, nấm không hình thành hạch nhỏ trên môi trường BĐ và PDA. Trên môi trường Czapek, nấm nhanh hình thành hạch lớn với kích thước lớn nhất 5,25 ˟ 2,58 mm và số lượng hạch lớn cũng cao nhất 187 hạch. Nấm hình thành hạch ít nhất trên môi trường CR, tuy nhiên kích thước hạch lớn nhỏ nhất trên môi trường PDA 1,97 ˟ 1,9 mm. Sự phân bố của hạch nấm trên đĩa
môi trường chủ yếu ở hai dạng là rải rác trên môi trường BĐ và CR và tạo thành hình tròn đồng tâm trên môi trường PDA và Czapek (Bảng 1, Hình 3).
3.4. Ảnh hưởng của các điều kiện nhiệt độ đến sự phát triển của nấm
Nhiệt độ khác nhau có ảnh hưởng đến sự sinh trưởng và phát triển của nấm. Khoảng nhiệt độ từ 15 - 20oC tối ưu cho sợi nấm phát triển với đường kính tản nấm trung bình tương ứng từ 5,37 ± 0,03 đến 7,10 ± 0,00 cm sau 4 ngày nuôi cấy. Tản nấm mọc nhanh nhất ở điều kiện nhiệt độ 20oC với đường kính cao nhất sau 4 ngày nuôi cấy là 7,10 ± 0,00 cm (Bảng 2).
Nhiệt độ cũng có ảnh hưởng đến sự hình thành bào tử và hạch nấm. Nấm chỉ hình thành bào tử sau 8 ngày nuôi cấy ở nhiệt độ 15oC và hạch nấm sau 6 ngày nuôi cấy ở ngưỡng nhiệt độ 15 - 20oC; không ghi nhận được sự hình thành bào tử và hạch nấm trong các điều kiện nhiệt độ 10, 25 và 30oC. Ở điều kiện nhiệt độ 15oC, tuy số lượng hạch nấm ít hơn chỉ có 93 hạch so với 122 hạch ở điều kiện nhiệt độ 20oC nhưng kích thước hạch lớn hơn. Ở cả 2 điều kiện nhiệt độ, hạch nấm tạo thành đường tròn đồng tâm nhưng màu sắc hạch khác nhau, màu nâu ở nhiệt độ 15oC và màu đen ở nhiệt độ 20oC (Bảng 2, Hình 4).

90
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Hình 4. Sự phát triển của nấm ở các điều kiện nhiệt độ khác nhau sau 14 ngày nuôi cấy
(Viện Bảo vệ thực vật, 2017)
3.5. Ảnh hưởng của các điều kiện chiếu sáng đến sự phát triển của nấm
Thời gian chiếu sáng khác nhau có ảnh hưởng đến sự sinh trưởng và phát triển của nấm. Kích thước tản nấm đạt 6,87 ± 0,03; 7,27 ± 0,03 và 7,63 ± 0,03 cm tương ứng với các điều kiện chiếu sáng hoàn toàn, tối hoàn toàn và 12 giờ sáng xen kẽ 12 giờ tối sau 4 ngày nuôi cấy. Đặc biệt ở điều kiện chiếu sáng 12 giờ sáng xen kẽ 12 giờ tối, nấm phát triển theo nhịp điệu sinh trưởng. Trong ba điều kiện chiếu sáng, nấm chỉ hình thành bào tử nấm trong điều kiện chiếu sáng hoàn toàn. Nấm sản sinh ra hạch lớn sau 6 - 8 ngày nuôi cấy với kích thước hạch khác nhau lần lượt 3,4 ˟ 3,02; 2,45 ˟ 1,50 và 2,80 ˟ 1,48 mm, nấm không hình thành hạch nhỏ. Số lượng hạch lớn cao nhất ở điều kiện tối hoàn toàn, tiếp đến là sáng hoàn toàn và cuối cùng là 12 giờ sáng xen kẽ 12 giờ tối. Hạch nấm đều có màu đen nhưng sự phân bố của hạch nấm trên bề mặt đĩa môi trường khác nhau gồm 2
dạng là hình thành rải rác ở điều kiện 12 giờ chiếu sáng, và tạo thành hình tròn đồng tâm ở 2 điều kiện còn lại (Bảng 3, Hình 5).
Hình 5. Sự phát triển của nấm trong các điều kiện chiếu sáng khác nhau (Viện Bảo vệ thực vật, 2017)
IV. KẾT LUẬN - Bệnh thối xám trên cây hoa thược dược do nấm
Botrytis cinerea gây ra, nấm gây hại trên các bộ phận non như cánh hoa, nụ hoa trong vụ Đông Xuân và xuân tại các vùng Trung du miền núi phía Bắc, Đồng bằng sông Hồng.
- Trong 4 loại môi trường, nấm phát triển mạnh nhất trên môi trường bột đậu. Tuy nhiên nấm hình thành hạch lớn sau 3 ngày trên môi trường Czapek và có kích thước và số lượng hạch lớn nhất, nấm không hình thành hạch nhỏ trên môi trường BĐ và PDA.
- Trên môi trường PDA nấm chỉ hình thành bào tử ở nhiệt độ 15oC sau 8 ngày nuôi cấy. Nấm hình thành hạch lớn ở mức nhiệt độ 15 - 20oC sau 6 ngày nuôi cấy và hạch phân bố dạng đồng tâm.
- Nấm phát triển tốt ở điều kiện tối hoàn toàn và 12 giờ sáng xen kẽ 12 giờ tối. Nấm phát triển theo nhịp điệu sinh trưởng ở điều kiện 12 giờ sáng xen kẽ 12 giờ tối. Nấm chỉ hình thành bào tử trong điều kiện sáng hoàn toàn. Hạch nấm hình thành sau 6 ngày nuôi cấy ở tất cả các điều kiện chiếu sáng.
Bảng 2. Sự phát triển của nấm B. cinerea ở các điều kiện nhiệt độ khác nhau (Viện Bảo vệ thực vật, 2017)
Ghi chú: BT: bào tử
Nhiệt độ (oC)
Kích thước tản nấm
sau 4 ngày (cm)
Thời gian hình
thành BT (ngày)
Thời gian hình thành hạch
(ngày)
Kích thước hạch lớn trung bình
(mm)Số
lương hạch
Màu sắc
Phân bố hạch lớn
Hạch lớn
Hạch nhỏ Dài Rộng
10 1,67e±0,03 - - - - - - - -15 5,37b±0,03 8 6 - 3,70 2,53 93 Nâu Đồng tâm20 7,10a±0,00 - 6 - 3,17 1,77 122 Đen Đồng tâm25 3,53c±0,03 - - - - - - - -30 1,97d±0,03 - - - - - - - -

91
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 3. Sự phát triển của nấm B. cinerea trong các điều kiện chiếu sáng khác nhau (Viện Bảo vệ thực vật, 2017)
Ghi chú: BT: bào tử
TÀI LIỆU THAM KHẢOMai Văn Quân, Trinh Xuân Hoạt, Đặng Vũ Thi Thanh,
Trần Thi chi, Hà Văn Dũng, Lê Thi Thanh Thuỷ, Nguyên Công Thành, 2016. Một số kết quả nghiên cứu về nấm Botrytis cinerea Pers. gây bệnh thối xám trên cây trồng. Tạp chí Bảo vệ thực vật, 6: 37-41.
Đặng Vũ Thi Thanh, Vũ Duy Hiện, Mai Văn Quân, 2007. Nghiên cứu đặc điểm sinh học của nấm Botrytis cinerea gây bệnh thối xám trên đào, hoa hồng, hoa lily ở vùng Sa Pa, Lào Cai. Những nghiên cứu cơ bản trong khoa học sự sống. NXB Nông nghiệp. Hà Nội, tr 370-380.
Đặng Vũ Thi Thanh, Vũ Duy Hiện, Mai Văn Quân, 2010. Nghiên cứu phổ ký chủ của nấm Botrytis
cinerea Pers. gây bệnh thối xám trên cây trồng. Tạp chí Bảo vệ thực vật, 1: 8-9.
Viện Bảo vệ thực vât, 1997. Phương pháp nghiên cứu bảo vệ thực vật tập 1. NXB Nông nghiệp. Hà Nội, tr 46-57,
Rigotti S., Gindro K., Richter H., Viret, O., 2002. Characterization of molecular markers for specific and sensitive detection of Botrytis cinerea Pers.: Fr. in strawberry (Fragaria x ananass Duch.) using PCR. FEMS Microbiology Letters, 209 (2): 169-174.
Tamura, K., Stecher, G., Peterson, D., Filipski, A., Kumar, S., 2013. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Molecular Biology and Evolution, 30: 2725-2729.
Điều kiện chiếu sáng
Kích thước tản nấm sau 4 ngày (cm)
Nhip điệu sinh
trưởng
Thời gian hình
thành BT (ngày)
Thời gian hình thành hạch (ngày)
Kích thước hạch lớn
(cm)Số
lương hạch
Màu sắc
Phân bố hạch lớn
Hạch lớn
Hạch nhỏ Dài Rộng
12 giờ sáng xen kẽ 12 giờ tối 7,63a±0,03 Có - 8 - 2,80 1,48 32 Đen Đồng tâm
Sáng hoàn toàn 6,87c±0,03 Không 8 7 - 3,40 3,02 55 Đen Rải rác
Tối hoàn toàn 7,27b±0,03 Không - 6 - 2,45 1,50 212 Đen Đồng tâm
Identification and biological characteristics of Botrytis cinerea Pers. causing gray mold on dahlia (Dahia pinnata Cav.) in Vietnam
Mai Van Quan, Duong Thi NguyenAbstractBotrytis cinerea is an important pathogen that causes gray mold different crops in Vietnam. In present study, a total of 15 isolates were isolated from dahlia (Dahia pinnata Cav.) in Northern midland mountainous and the Red River delta. The morphological characterization was based on conidiophore and conidial length; and the results indicated that all isolates belonged to morph species Botrytis cinerea Pers. PCR with specific primer pair C729+/C729- amplified DNA fragments of about 730 bp from all isolates. The DNA sequencing and phylogenetic analysis confirmed that B. cinerea was the causal agent of gray mold disease on dahlia. Using medium plate culture method, the effect of various culture conditions on mycelium growth, sporulation, sclerotia formation of dahlia B. cinerea was detected. Among 4 media, the mycelium cultured on BĐ medium growed fastest with the production of gray mycelium and dense colonies; however, the highest number of big sclerotia was formed on Czapek 3 days after incubation. The optimum temperature for mycelium growth and sporulation of dahlia B. cinerea was 15oC on PDA medium. The optimum lighting conditions for mycelium growth was fluorescent light with alternating cycles of 12 hours light and 12 hours darkness; and the continous light was optimum condition for sporulation of the fungus.Key words: Botrytis cinerea, gray mold, biological characteristics, Dahia pinnata Cav.
Ngày nhận bài: 9/7/2017Ngày phản biện: 13/7/2017
Người phản biện: TS. Hà Minh ThanhNgày duyệt đăng: 27/7/2017

92
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀCây nghệ có tên khoa học là Curcuma longa L.
thuộc họ gừng (Zingiberaceae), là cây trồng lấy củ có điều kiện sinh trưởng và phát triển rất phù hợp ở những vùng khí hậu nóng ẩm như các nước thuộc khu vực châu Á và Thái Bình Dương. Trong đó, Việt Nam có nguồn sản xuất nghệ phong phú, phân bố ở nhiều tỉnh thành như Vĩnh Phúc, Hải Dương, Đắk Lắk, Nghệ An, Quảng Trị, Quảng Nam… Về giá trị sử dụng, củ nghệ từ lâu đã được sử dụng rộng rãi làm gia vị, chất bảo quản và chất tạo màu trong thực phẩm (Đào Hùng Cường và ctv., 2008). Ngoài ra, trong y học, do hoạt tính sinh học của curcumin, nghệ có tác dụng kìm hãm sự phát triển của tế bào ung thư và điều trị nhiều căn bệnh viên khớp, giải độc gan, loét dạ dày, tá tràng... (Anand et al., 2007). Sản phẩm tinh bột nghệ được tiêu thụ phần lớn trên thị trường hiện nay được sản xuất theo phương pháp truyền thống thô sơ, lạc hậu. Đặc biệt, công đoạn sấy thường phơi tinh bột trực tiếp bằng ánh nắng mặt trời hay sử dụng khói lò, than, củi đốt để làm khô dẫn đến khó khăn trong kiểm soát độ ẩm, tổn thất curcumin, không đảm bảo chất lượng vệ sinh an toàn thực phẩm (Lê Văn Hoàng và ctv., 2007). Hiện nay, đã có một số công trình nghiên cứu về ảnh hưởng của yếu tố kỹ thuật sấy bơm nhiệt trên một số sản phẩm có giá trị dinh dưỡng. Nghiên cứu khảo sát ảnh hưởng yếu tố kỹ thuật sấy bơm nhiệt trên ớt đã đưa ra kết luận: Sấy ớt ở nhiệt độ 55˚C, độ ẩm giảm từ 87,6 % xuống còn 8% trong 22 giờ (Vũ Minh Tâm và Nguyễn Đình Kiên, 2009). Kết quả công bố sấy bơm nhiệt kiểu thùng quay đối với cà rốt ở nhiệt độ 45˚C, vận tốc tác nhân sấy 2,5 m/s, thì độ ẩm còn 10% trong thời gian sấy 6 giờ (Võ Duy Mạnh và Lê Chí Hùng, 2011). Tuy nhiên, các công bố hiện nay về tác động của sấy bơm nhiệt đối với chất lượng sản phẩm tinh bột nghệ còn hạn chế.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu Củ nghệ tươi được thu mua tại thôn Bản Sơn
(làng Cùa), xã Cam Nghĩa, huyện Cam Lộ, tỉnh Quảng Trị. Nghệ được thu hoạch sau 8 - 9 tháng kể từ khi ngày gieo trồng.
2.2. Phương pháp nghiên cứu
2.2.1. Phương pháp bố trí thí nghiệmTừ nguyên liệu nghệ tươi để thu được bột nghệ
ướt phục vụ thí nghiệm, thực hiện theo quy trình sau: Nghệ tươi à rửa sạch à nghiền à ly tâm tách bã à ly tâm tách nước à bột nghệ ướt.
Mẫu thí nghiệm được bố trí sấy ở các nhiệt độ 45ºC; 50ºC; 55ºC và 60ºC cố định trong thời gian 11 giờ. Sau khi chọn được nhiệt độ sấy thích hợp, tiến hành khảo sát ảnh hưởng của thời gian sấy. Mẫu được sấy ở các thời gian lần lượt là 2; 4; 6; 8; 10 và 12 giờ. Dịch tinh bột nghệ ướt trước khi sấy có hàm lượng nước 40 ÷ 45%, diện tích khay sấy 540 ˟ 740 ˟ 20 mm, bề dày vậy liệu 0,5 cm. Sản phẩm tinh bột nghệ, sau khi sấy được tiến hành xác định các chỉ tiêu chất lượng như: độ ẩm, hàm lượng curcumin, đánh giá chất lượng cảm quan (màu sắc, mùi, vị).
2.2.2. Phương pháp phân tíchĐộ ẩm được xác định bằng phương pháp sấy đến
khối lượng không đổi theo TCVN 4415:1987 (Bộ Khoa học và Công nghệ, 1987). Hàm lượng tinh bột được xác định theo TCVN 9936:2013 (Bộ Khoa học và Công nghệ, 2013). Hàm lượng cellulose được xác định theo TCVN 4590:1988 (Bộ Khoa học và Công nghệ, 1988). Hàm lượng protein được xác định bằng phương pháp Kjeldahl theo TCVN 9936:2013 (Bộ Khoa học và Công nghệ, 2013). Hàm lượng lipid tổng số được xác định theo TCVN 8137:2009
1 Trường Đại học Nông Lâm, Đại học Huế
NGHIÊN CỨU ẢNH HƯỞNG CỦA CHẾ ĐỘ SẤY (NHIỆT ĐỘ, THỜI GIAN) BẰNG PHƯƠNG PHÁP SẤY BƠM NHIỆT ĐẾN SẢN PHẨM TINH BỘT NGHỆ
Nguyễn Văn Toản1, Nguyễn Văn Huế1
TÓM TẮTNghiên cứu tiến hành xác định ảnh hưởng của chế độ sấy (nhiệt độ, thời gian) bằng phương pháp sấy bơm nhiệt
đến chất lượng sản phẩm tinh bột nghệ nhằm tạo ra sản phẩm có chất lượng cao từ củ nghệ tươi. Khảo sát chế độ sấy được tiến hành ở các nhiệt độ: 45oC; 50oC; 55oC; 60oC và thời gian: 2; 4; 6; 8; 10 và 12 giờ cho các mẫu dịch tinh bột nghệ có hàm lượng nước 40 ÷ 45%, diện tích khay sấy 540 ˟ 740 ˟ 20 mm, bề dày vật liệu 0,5 cm. Kết quả nghiên cứu cho thấy với chế độ sấy ở nhiệt độ 55oC trong thời gian 10 giờ cho sản phẩm tinh bột nghệ có chất lượng cao nhất, tương ứng với hàm lượng curcumin 1,02%, độ ẩm 8,46% cùng với các chỉ tiêu cảm quan là tốt nhất.
Từ khóa: Củ nghệ, tinh bột nghệ, thời gian sấy, nhiệt độ sấy, hàm lượng curcumin, độ ẩm, chất lượng cảm quan

93
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
(Bộ Khoa học và Công nghệ, 2009). Định lượng hàm lượng tinh dầu bằng phương pháp chưng cất lôi cuốn hơi nước theo TCVN 7039-2002 (Bộ Khoa học và Công nghệ, 2002). Định lượng hàm lượng curcumin bằng phương pháp sắc ký lỏng hiệu cao năng (HPLC) (Phạm Thị Phương và ctv., 2014). Đánh giá cảm quan bằng phương pháp cho điểm thị hiếu theo thang điểm Hedonic (Hà Duyên Tư, 2006).
2.2.3. Phương pháp xử lý số liệu Các thí nghiệm được lặp lại 3 lần. Kết quả thí
nghiệm được phân tích phương sai ANOVA và kiểm định LSD (5%) để so sánh sự khác biệt trung bình giữa các nghiệm thức. Các phân tích thống kê được xử lý trên phần mềm IBM SPSS 20.
2.3. Thời gian và đia điểm nghiên cứuNghiên cứu được tiến hành từ 1/2015 đến 6/2017.
Củ nghệ ngay sau thu hoạch được vận chuyển về phòng thí nghiệm thuộc Bộ môn Công nghệ thực phẩm, Khoa Cơ khí - Công nghệ, Trường Đại học Nông Lâm, Đại học Huế để xử lý và chế biến.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Kết quả xác đinh hàm lương một số thành phần hóa học của củ nghệ vàng
Bảng 1. Thành phần hoá học chính của củ nghệ vàng
Kết quả phân tích ở bảng 1 cho thấy: Hàm lượng nước trong củ nghệ tươi tương đối cao (chiếm 79,82%) sẽ thúc đẩy các quá trình sinh lý, sinh hóa diễn ra mạnh mẽ và làm tăng tổn thất chất dinh dưỡng. Hàm lượng tinh bột của củ nghệ chiếm 14,67%, điều này có ý nghĩa quan trọng trong sản xuất tinh bột nghệ. Hàm lượng tinh bột càng cao thì hiệu suất thu hồi càng lớn. Thành phần cần được quan tâm nhất trong củ nghệ là curcumin với hàm lượng tương đối cao (0,63%). Bên cạnh đó, curcumin có vai trò quyết định màu sắc cho sản phẩm tinh bột nghệ sau khi sấy. Vì vậy, trong quá trình sản xuất và chế biến người ta quan tâm đến
điều chỉnh các thông số kỹ thuật làm hạn chế mức thấp nhất tổn thất curcumin trong sản phẩm. Ngoài ra, trong củ nghệ có chứa một số thành phần có hàm lượng tương đối thấp nhưng nó mang lại giá trị dinh dưỡng cao như: protein 0,71%; lipid 0,73%, cellulose 2,371% và tinh dầu 1,96%.
3.2. Khảo sát ảnh hưởng của nhiệt độ sấy đến chất lương sản phẩm tinh bột nghệ
3.2.1. Ảnh hưởng nhiệt độ sấy đến độ ẩm sản phẩmNhiệt độ sấy là một yếu tố rất quan trọng trong
quá trình sấy các loại nông sản khác nhau. Ảnh hưởng của nhiệt độ đến vấn đề “tổn thất” hoạt chất sinh học - curcumin, ảnh hưởng đến mùi vị của sản phẩm cũng như hiệu suất sản xuất. Kết quả thực nghiệm thu được từ đồ thị hình 1 cho thấy: Độ ẩm của sản phẩm thu được ở các nhiệt độ sấy tỷ lệ nghịch với nhiệt độ sấy. Độ ẩm sản phẩm thấp nhất (w = 9,89%) tại nhiệt độ sấy 60˚C và cao nhất (w = 15,51%) ở nhiệt độ 45˚C. Tại nhiệt độ 50˚C và 55˚C sau thời gian sấy 11 giờ có độ ẩm thu được của sản phẩm (tương ứng 13,46% và 10,39% ). Kết quả này cũng phù hợp với nguyên lý quá trình sấy: Nhiệt độ càng cao thì khả năng truyền nhiệt của tác nhân không khí nóng vào nguyên liệu sẽ càng nhanh. Do đó, hàm ẩm trên bề mặt vật liệu sấy sẽ bốc hơi nhanh hơn so với nhiệt độ thấp (Lê Bạch Tuyết, 1996). Đối chiếu với tiêu chuẩn công bố của Việt Nam về các sản phẩm chế biến từ tinh bột yêu cầu duy trì độ ẩm của sản phẩm ≤13% (TCVN 10546:2014). Do vậy, nhiệt độ sấy 55˚C ÷60˚C là phù hợp trong nghiên cứu này.
Hình 1. Ảnh hưởng của nhiệt độ sấy đến độ ẩm tinh bột nghệ.
Ghi chú: Hình 1, 2, 3, 4; Bảng 2, 3: Các chữ cái khác nhau thể hiện sự sai khác có ý nghĩa về mặt thống kê với α= 0,05.
STT Thành phần Đơn vi Hàm lương1 Hàm lượng nước % 79,82±0,522 Tinh bột % 14,67±0,163 Protein % 0,71± 0,034 Lipid % 0,73 ±0,125 Tinh dầu % 1,96± 0,076 Cellulose % 2,37±0,017 Curcumin % 0,63±0,01
18
16
14
12
10
8
6
4
2
045
15,51a
13,46b
10,39c9,89c
50 55 60
Độ
ẩm (%
)
Nhiệt độ sấy (oC)

94
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
3.2.2. Ảnh hưởng nhiệt độ sấy đến hàm lượng curcumin
Kết quả nghiên cứu ảnh hưởng nhiệt độ sấy đến hàm lượng curcumin trong sản phẩm tinh bột nghệ được trình bày ở đồ thị hình 2.
Hình 2. Ảnh hưởng nhiệt độ sấy đến hàm lượng curcumin trong tinh bột nghệ
Từ số liệu thu được cho thấy: Khi nhiệt độ tăng thì hàm lượng curcumin có trong tinh bột nghệ sau sấy tăng theo: Mẫu tinh bột nghệ sấy ở nhiệt độ 45˚C có hàm lượng curcumin 0,88%; mẫu sấy ở nhiệt độ 50˚C có hàm lượng curcumin 0,97%; mẫu sấy ở nhiệt độ 55˚C có hàm lượng curcumin 1,01% và mẫu tinh bột nghệ sấy ở nhiệt độ 60˚C có hàm lượng curcumin 1,03%. Theo kết quả công bố của Ravindram (2007) và Lê Văn Hoàng (2007) thì nhiệt độ sấy khô nghệ phù hợp nhất là không vượt quá 60˚C và hàm lượng curcumin tổn thất trong quá trình sấy là thấp nhất. Kết quả thực nghiệm hoàn toàn phù hợp với công bố này. Như vậy, khi sấy ở nhiệt độ 55˚C cho sản phẩm tinh bột nghệ có hàm lượng curcumin đạt giá trị tương đối cao (1,01%).
3.2.3. Ảnh hưởng nhiệt độ sấy đến chất lượng cảm quan của sản phẩm
Kết quả đánh giá chất lượng cảm quan của sản phẩm tinh bột nghệ được thể hiện trong bảng 2.
Bảng 2. Ảnh hưởng nhiệt độ sấy đến chất lượng cảm quan sản phẩm tinh bột nghệ
Trong các mẫu sấy, chi tiêu cảm quan của sản phẩm tinh bột nghệ có sự khác nhau rõ rệt giữa các nhiệt độ sấy. Khi ở nhiệt độ 50˚C cho màu sắc có mức độ yêu thích cao nhất so với các nhiệt độ
sấy còn lại. Nhiệt độ sấy 60˚C có điểm màu thấp nhất (5,38). Nguyên nhân là do quá trình oxy hóa phenolic dưới sự xúc tác oxy tạo thành các các sắc tố màu không mong muốn (Prathapan et al., 2009). Mùi, vị cũng là chỉ tiêu quan trọng trong đánh giá chất lượng của tinh bột nghệ. Thông qua quá trình đánh giá cảm quan cho thấy các mẫu sấy bơm nhiệt có mùi, vị được mọi người yêu thích; đặc biệt, mẫu sấy 55˚C có điểm mùi, vị cao nhất, tương ứng 8,17 và 7,77. Khi nhiệt độ sấy thấp, độ ẩm trong sản phẩm cao, các tinh dầu và acid nhựa chưa phân hủy hết sẽ ảnh hưởng trực tiếp đến mùi và vị của sản phẩm; do đó, kết quả cảm quan mùi và vị của mẫu sấy 45˚C và 50˚C có điểm thấp. Mẫu được sấy ở nhiệt độ 55˚C có chỉ tiêu màu sắc tương đối thích; tuy nhiên, chỉ tiêu mùi và vị được đánh giá mức độ yêu thích cao nhất. Do đó, khi sấy tinh bột nghệ ở nhiệt độ 55˚C cho sản phẩm được người tiêu dùng đánh giá tốt nhất. Tóm lại, có thể kết luận rằng mẫu sấy ở 55˚C là mẫu đạt yêu cầu thủy phần của sản phẩm cũng như chất lượng cảm quan cao nhất, bên cạnh đó còn hạn chế tổn thất curcumin. Như vậy, nhiệt độ sấy thích hợp nhất là 55˚C. Kết quả này hoàn toàn phù hợp với công bố nghiên cứu của Vũ Minh Tâm và Nguyễn Đình Kiên (2009) khi sấy ớt bằng máy sấy bơm nhiệt ở nhiệt độ 55˚C cho sản phẩm chất lượng cao, hạn chế phân hủy các chất dinh dưỡng có trong sản phẩm.
3.3. Ảnh hưởng thời gian sấy đến chất lương sản phẩm tinh bột nghệ
3.3.1. Ảnh hưởng thời gian sấy đến độ ẩm sản phẩm tinh bột nghệ
Khảo sát ảnh hưởng của thời gian sấy (ở nhiệt độ sấy 55˚C) đến độ ẩm của sản phẩm tinh bột nghệ và thu được kết quả ở đồ thị hình 3.
Hình 3. Ảnh hưởng thời gian sấy đến độ ẩm sản phẩm tinh bột nghệ
Kết quả thu được từ hình 3 cho thấy: Trong quá trình sấy, độ ẩm của các mẫu giảm dần theo thời gian sấy. Độ ẩm giảm nhanh chóng từ khi bắt đầu
Nhiệt độ (˚C)
Chỉ tiêuMàu sắc Mùi Vi
45 6,40b 5,43c 4 ,80c
50 7,23a 5,93c 5,97b
55 6,57b 8,17a 7,77a
60 5,83c 6,50b 5,60b
Độ
ẩm (%
)
Thời gian sấy (h)
4540353025201510
50
0
40,74a38,73b
25,23c
16,35d
10,39e
8,46f 7,88f
2 4 6 8 10 12
Nhiệt độ sấy (oC)
Hàm
lượn
gC
urcu
min
(%)
45
0,88c 0,97b 1,01a 1,03a1.401.201.000.800.600.400.200.00
50 55 60

95
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
sấy (w = 40,74%) đến 6 giờ sấy (w = 16,35%) giảm 40,13%. Tuy nhiên, từ kết quả thu được cũng cho thấy, mẫu được sấy trong thời gian 10 giờ và sấy 12 giờ thì độ ẩm giảm ở mức thấp và không có sự khác biệt khi xử lý ANOVA với mức ý nghĩa 5%.
3.3.2. Ảnh hưởng thời gian sấy đến hàm lượng curcumin
Phân tích ảnh hưởng thời gian sấy đến hàm lượng curcumin trong sản phẩm tinh bột nghệ có kết quả được thể hiện ở đồ thị hình 4.
Hình 4. Ảnh hưởng thời gian sấy đến hàm lượng curcumin
Số liệu cho thấy: Thời gian sấy càng dài thì hàm lượng curcumin càng tăng. Nguyên liệu trước khi đem vào sấy có độ ẩm 40,74% với hàm lượng curumin 0,43%. Tuy nhiên, sau 12 giờ sấy, hàm lượng curumin tăng lên với giá trị xác định là 1,04%. Kết quả này có thể giải thích như sau: Thời gian sấy càng dài, độ ẩm thu được trong sản phẩm càng giảm nên nồng độ chất khô thu được của sản phẩm sẽ tăng. Xử lý thống kê về hàm lượng curcumin trên biến động ANOVA và so sánh giá trị trung bình của các mẫu sấy với mức ý nghĩa α = 0,05, cho thấy hàm lượng curcumin của hai mẫu 10 giờ và 12 giờ không sai khác về mặt thống kê (5%). Để tăng hiệu quả kinh tế, tiết kiệm chi phí trong sản xuất, thời gian sấy thích hợp nhất được chọn là 10 giờ.
3.3.3. Ảnh hưởng thời gian đến chất lượng cảm quan sản phẩm tinh bột nghệ
Cảm quan là chỉ tiêu quan trọng góp phần đánh giá chất lượng của sản phẩm. Chỉ tiêu mùi, vị và màu sắc bị ảnh hưởng bởi thời gian sấy. Kết quả đánh giá cảm quan sản phẩm được trình bày trong bảng 3.
Trong các mẫu, chỉ tiêu cảm quan của sản phẩm đã có sự thay đổi rõ rệt ở các thời gian sấy khác nhau. Mức độ yêu thích sản phẩm tinh bột nghệ tăng dần trong thời gian sấy. Mẫu có điểm số cao nhất ở thời gian sấy 10 giờ với điểm số các chỉ tiêu đều trên 7 (thích). Các mẫu có điểm số thấp nhất là mẫu sấy thời gian 2 giờ; 4 giờ và 6 giờ với số điểm cảm quan
trung bình dưới 4 (tương đối không thích). Tuy nhiên, nếu kéo dài thời gian sấy, độ ẩm thấp, sản phẩm bị cháy có màu sắc không mong muốn.
Bảng 3. Ảnh hưởng thời gian sấy đến chất lượng cảm quan của sản phẩm
Từ kết quả thu được, đã lựa chọn được chế độ sấy bằng phương pháp bơm nhiệt ở nhiệt độ 550C, thời gian sấy 10 giờ là thích hợp nhất trong quá trình sản xuất tinh bột nghệ từ củ nghệ tươi. Ở chế độ lựa chọn này, sản phẩm tinh bột nghệ có độ ẩm ≤13%, các chỉ tiêu chất lượng cảm quan tốt nhất và hàm lượng curcumin đạt được ở mức cao (1,02%).
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnĐã xác định được chế độ sấy (nhiệt độ và thời
gian) phù hợp nhất với mục đích độ ẩm của tinh bột nghệ (8,46%) ≤13%, chất lượng cảm quan sản phẩm tốt nhất và hàm lượng curcumin đạt được ở mức cao (1,02%) là: Nhiệt độ sấy 550C và thời gian sấy 10 giờ.
4.2. Đề nghi Áp dụng kết quả thu được để tiếp tục nghiên cứu
và hoàn thiện quy trình sản xuất tinh bột nghệ trên quy mô công nghiệp.
TÀI LIỆU THAM KHẢOBộ Khoa học và Công nghệ, 2002. TCVN 7039:2002.
Gia vị và thảo mộc - xác định hàm lượng dầu dễ bay hơi (phương pháp chưng cất bằng hơi nước).
Bộ Khoa học và Công nghệ, 2009. TCVN 8137:2009. Thịt và sản phẩm thịt - xác định hàm lượng chất béo tự do.
Bộ Khoa học và Công nghệ, 2013. TCVN 9936:2013. Tinh bột và sản phẩm tinh bột - xác định hàm lượng nitơ bằng phương pháp Kjeldahl - Phương pháp chuẩn độ.
Bộ Khoa học và Công nghệ, 2014. TCVN 10546:2014. Quy định Tinh bột sắn- sản phẩm từ tinh bột.
Bộ Nông nghiệp và PTNT, 2005. Tuyển tập tiêu chuẩn nông nghiệp Việt Nam, Tập VI: Tiêu chuẩn rau quả (xác định độ ẩm theo TCVN 4415: 1987; xác định
Thời gian sấy (giờ)
Hàm
lượn
g C
urcu
min
(%) 1,2
1
0,8
0,6
0,4
0,2
00
0,43f
0,55e0,62d
0,76c
0,89b
1,02a 1,04a
2 4 6 8 10 12
Thời gian (giờ)
Chỉ tiêuMàu sắc Mùi Vi
2 3,13e 2,67e 2,53e
4 3,83d 3,53d 3,10e
6 4,90c 4,80c 4,53d
8 6,30b 5,93b 6,03c
10 7,13a 7,40a 7,53a
12 6,13b 6,93a 6,80b

96
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
hàm lượng Cellulose theo TCVN 4590:1988; xác định hàm lượng protein theo TCVN 9936:2013). Hà Nội.
Đào Hùng Cường, Đỗ Thi Thúy Vân, Nguyên Đình Anh, 2008. Nghiên cứu ứng dụng curcumin trong phối màu thực phẩm. Tạp chí Hóa học và Ứng dụng, số 7 (79):150-156.
Lê Văn Hoàng, Đào Hùng Cường, Nguyên Đình Anh, 2007. Nghiên cứu ảnh hưởng điều kiện sấy đến hàm lượng curcumin của củ nghệ vàng, Tạp chí Hóa học và Ứng dụng, số 7(67):48-49.
Võ Duy Mạnh, Lê Chí Hùng, 2011. Nghiên cứu sấy cà rốt bằng sấy bơm nhiệt kiểu thùng quay. Tạp chí Khoa học, Trường Đại học Cần Thơ, 20b: 209-216.
Phạm Thi Phương, Nguyên Quỳnh Ngọc, Phạm Thi Thanh Hương, Đào Văn Đôn, Nguyên Thi Thanh Phương, 2014. Nghiên cứu định lượng đồng thời curcumin và piperrin bằng phương pháp sắc ký lỏng hiệu năng cao. Tạp chí Y - Dược, số 6: 7-12.
Vũ Minh Tâm, Nguyên Đình Kiên, 2009. Nghiên cứu quy trình công nghệ sấy ớt tính toán, thiết kế máy sấy
bơm nhiệt năng suất 200kg/mẻ. Luận văn tốt nghiệp, Trường Đại học Nông Lâm Thành Phố Hồ Chí Minh.
Lê Bạch Tuyết, 1996. Các quá trình công nghệ cơ bản trong sản xuất thực phẩm. Nhà xuất bản Giáo dục. Hà Nội.
Hà Duyên Tư, 2006. Kỹ thuật phân tích cảm quan thực phẩm. Nhà xuất bản Khoa học và Kỹ thuật. Hà Nội.
Anand P, Kunnumakkara AB, Newman RA, Aggarwal BB, 2007. Bioavailability of curcumin: problems and promises. Molecular Pharmaceutics, 4(6):807-818.
Prathapan A., M. Lukhman, C. Arumughan, A. Sundaresan and K. G. Raghu, 2009. Effect of heat treatment on curcuminoid, colour value and total polyphenols of fresh turmeric rhizome. International Journal of Food Science and Technology, 44: 1438-1444.
Ravindram, P. N., Babu Nirmal, K. Sivaraman, 2007. Turmeric - The Genus Curcuma. Medicinal and Aromatic Plants-Industrial Profiles, Volume 45. CRC Press. Taylor & Francis Group.
Effect of drying regime (temperature, duration) by using heat pump drying technology on quality of refined turmeric powder
Nguyen Van Toan, Nguyen Van Hue AbstractThis research studied effect of drying regime (temperature, duration) by heat pump drying technology on quality of refined turmeric powder. The experiment was carried out with 4 treatments of temperatures as 45oC; 50oC; 55oC; 60oC, in 2; 4; 6; 8; 10 and 12 hours for wet refined tumeric powder with humidity content of 40 ÷ 45%; the dimension of the trays was 540 x 740 x 20 mm, the thickness of the tumeric powder was 0.5 cm. The results showed that the most suitable temperature of drying was 55oC for 10 hours making the best quality of refined tumeric powder with humidity of 8.46%; the curcumin content of 1.02% and the best quality of sensory indicators. Key words: Turmeric, refined tumeric powder, drying duration, drying temperature, curcumin content, humidity, sensory quality
Ngày nhận bài: 1/7/2017Ngày phản biện: 10/7/2017
Người phản biện: PGS. TS. Hoàng Thị Lệ HằngNgày duyệt đăng: 27/7/2017
1 Trường Đại học Nông Lâm, Đại học Huế
2 Chi cục An toàn vệ sinh Thực phẩm thành phố Đà Nẵng
NGHIÊN CỨU ẢNH HƯỞNG CỦA AMINOETHOXYVINYLGLYCINE (AVG) KẾT HỢP XỬ LÝ NƯỚC NÓNG ĐẾN QUÁ TRÌNH
CHÍN QUẢ BƠ (BOOTH7) SAU THU HOẠCH Nguyễn Văn Toản1, Nguyễn Hồng Phúc2
TÓM TẮTNghiên cứu sử dụng chất kìm hãm quá trình sinh tổng hợp ethylene (AVG) kết hợp xử lý nước nóng (nhiệt độ
500C, thời gian 10 phút) đến quá trình chín quả bơ được thực hiện với mục đích kiểm soát làm chậm quá trình chín và kéo dài thời gian bảo quản sau thu hoạch quả bơ (BOOTH7). Kết quả cho thấy quả bơ được xử lý với AVG làm cho hoạt lực của ACC oxydase, cường độ hô hấp và cường độ sản sinh ethylene thấp hơn so với đối chứng. Trong 4 nồng độ (320 ppm; 370 ppm; 420 ppm; 470 ppm) được khảo sát, nồng độ xử lý AVG 420 ppm làm cho đỉnh hô hấp và

97
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀBơ (Persea americana) là cây ăn quả nhiệt đới và
cận nhiệt đới nổi tiếng với trái cây chứa hàm lượng chất béo có giá trị dinh dưỡng rất cao. Ở Việt Nam, bơ Booth7 được xem như giống bơ nhiệt đới thích hợp với đặc điểm khí hậu do khả năng sinh trưởng và phát triển mạnh, thu hoạch trái vụ và cho năng suất cao. Bơ là loại quả hô hấp đột biến nên khi có dấu hiệu chín thì chín rất nhanh và chóng hỏng, làm giảm chất lượng quả. Quá trình chín này do sự sản sinh hormone thực vật - ethylene, giữ vai trò quan trọng trong việc tạo ra hô hấp đột biến và kích thích quá trình chín của quả (Nguyễn Văn Toản, 2011). Hiện nay, trên thế giới cũng như ở Việt Nam đã tìm ra được nhiều phương pháp bảo quản khác nhau để kéo dài thời gian bảo quản bơ; trong đó có phương pháp sử dụng aminoethoxyvinylglycine (AVG). AVG là hợp chất có hoạt tính sinh học và tác động kìm hãm hoạt lực của enzyme ACC synthase làm hàm lượng ACC (chất tiền ethylene) tạo thành thấp (Nguyễn Văn Toản và ctv., 2010). Tác giả Burhan Ozturk (2012) đã xử lý AVG trên quả mận với nồng độ 100 và 200 mg/l trước 2 tuần thu hoạch, kết hợp với bảo quản lạnh. Tác giả Salvatore (2010) đã phun AVG với nồng độ 125 mg/l hoặc 250 mg/l 2 tuần trước thu hoạch quả lê. Kết quả cho thấy AVG làm chậm quá trình chín, kéo dài thời gian bảo quản sau thu hoạch và nâng cao chất lượng quả tươi. Tuy nhiên, ở Việt Nam, đến nay chưa có công trình công bố về tác động của AVG đến quá trình chín của quả bơ Booth7. Do đó, việc nghiên cứu và xác định được nồng độ AVG thích hợp cho mục tiêu trên là nội dung chủ yếu cần đạt được.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu - Quả bơ (Booth7) thuộc giống bơ sáp, được thu
hái từ vườn ươm của Công ty trách nhiệm hữu hạn Trịnh Mười, tỉnh Đắk Lắk. Bơ được thu hoạch sau 8 - 9 tháng kể từ khi nở hoa. Vào thời điềm này, vỏ quả có màu xanh lục đậm, có độ bóng sáng, trạng thái quả cứng và khi lắc không phát ra tiếng. Phương pháp lấy mẫu thực hiện theo TCVN 9017:2011.
- Chế phẩm Aminoethoxyvinylglycine (AVG), tên thương mại là Retain, ở dạng bột, hòa tan trong nước, được mua từ Valent BioSciences corp (Úc). Thùng carton loại 3 lớp được sản xuất tại Việt Nam. Bao bì LDPE có chiều dày 25 µm được sản xuất tại Việt Nam. 1-Aminocyclopropane-1-carboxylic acid (ACC), cơ chất tiền ethylene, được mua từ công ty Sigma-Aldrich (Mỹ). Tricine là chất đệm, được mua từ công ty Merk KgaA (Đức). Maltose được mua từ công ty Prolabo (Đan Mạch).
2.2. Phương pháp nghiên cứu
2.2.1. Phương pháp phân tích Cường độ hô hấp được xác định theo phương
pháp đo kín, sử dụng máy ICA 250 (Anh) để đo lượng CO2 (Nguyễn Văn Toản, 2011). Cường độ sản sinh ethylene được xác định trên máy đo ethylene ICA 56 do hãng Dual Analyser, Nhật Bản sản xuất (Nguyễn Văn Toản, 2011). Hoạt lực của enzyme 1- Aminocyclopropane - 1- carboxylate oxydase (ACCO) được xác định theo phương pháp cải tiến của Moya - Léon và John (1995). Hàm lượng lipid tổng số được xác định theo TCVN 8137:2009. Hàm lượng vitamin C được xác định theo TCVN 4715-1989. Xác định hao hụt khối lượng tự nhiên bằng phương pháp cân (sử dụng cân kỹ thuật Sartorius - Đức).
2.2.2. Phương pháp bố trí thí nghiệmThí nghiệm được tiến hành theo sơ đồ sau: Quả
bơ → Thu hoạch → Lựa chọn → Xử lý nước nóng (nhiệt độ 500C, thời gian 10 phút)→ Xử lý AVG [ở các nồng độ: 320 ppm; 370 ppm; 420 ppm; 470 ppm và ĐC (không xử lý AVG)] → Bao gói → Bảo quản. Thí nghiệm được bố trí hoàn toàn ngẫu nhiên, mỗi thí nghiệm được thực hiện với 3 lần lặp, các mẫu có khối lượng 200 kg và tiến hành xử lý nước nóng bằng phương pháp nhúng. Sau đó, các mẫu được bao gói bằng bao bì LDPE 25µm và bảo quản ở cùng điều kiện (t0 = 80C, φkk= 80 - 85%) (Mongy - El Abd et al., 2009). Tiến hành phân tích các chỉ tiêu chất lượng cũng như tỷ lệ hư hỏng của các mẫu với tần suất 3 ngày/lần. Quá trình theo dõi kết thúc khi mẫu hư hỏng hoàn toàn.
sản sinh ethylene xuất hiện muộn hơn, quả bơ chín chậm hơn 9 ngày so với đối chứng. Đồng thời, nghiên cứu cũng xác định được một số chỉ tiêu về chất lượng của quả bơ sau 30 ngày bảo quản ở điều kiện xử lý (AVG 420 ppm, nhiệt độ 500C, thời gian 10 phút): Hàm lượng vitamin C 7,45 mg%; hàm lượng lipid tổng số là 14,28%; hao hụt khối lượng 3,14%; cường độ sản sinh ethylene 23,29 (μl C2H4.kg-1.h-1); cường độ hô hấp 38,96 (ml CO2.kg-1.h-1); hàm lượng ACC oxydase 9,01 (nmol C2H4.g
-1.h-1). Từ khóa: Cường độ hô hấp, cường độ sản sinh ethylene, hoạt lực ACC oxydase, nhiệt độ thấp, xử lý nước nóng,
xử lý AVG
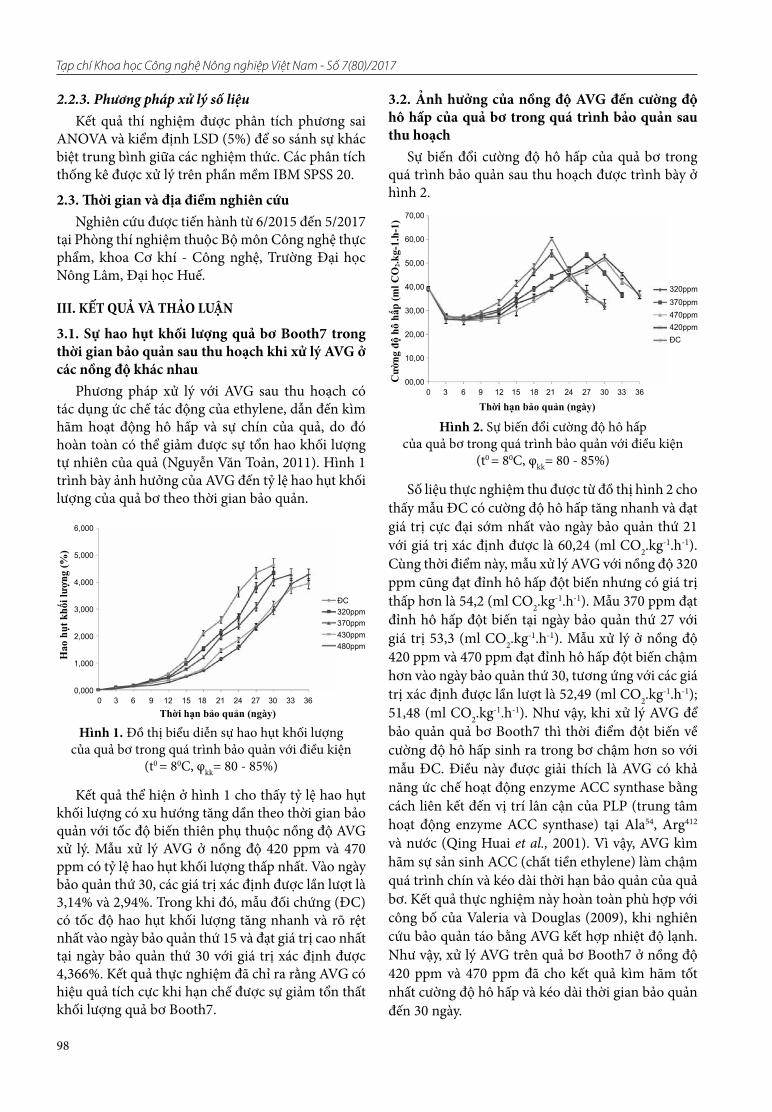
98
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
2.2.3. Phương pháp xử lý số liệuKết quả thí nghiệm được phân tích phương sai
ANOVA và kiểm định LSD (5%) để so sánh sự khác biệt trung bình giữa các nghiệm thức. Các phân tích thống kê được xử lý trên phần mềm IBM SPSS 20.
2.3. Thời gian và đia điểm nghiên cứuNghiên cứu được tiến hành từ 6/2015 đến 5/2017
tại Phòng thí nghiệm thuộc Bộ môn Công nghệ thực phẩm, khoa Cơ khí - Công nghệ, Trường Đại học Nông Lâm, Đại học Huế.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Sự hao hụt khối lương quả bơ Booth7 trong thời gian bảo quản sau thu hoạch khi xử lý AVG ở các nông độ khác nhau
Phương pháp xử lý với AVG sau thu hoạch có tác dụng ức chế tác động của ethylene, dẫn đến kìm hãm hoạt động hô hấp và sự chín của quả, do đó hoàn toàn có thể giảm được sự tổn hao khối lượng tự nhiên của quả (Nguyễn Văn Toản, 2011). Hình 1 trình bày ảnh hưởng của AVG đến tỷ lệ hao hụt khối lượng của quả bơ theo thời gian bảo quản.
Hình 1. Đồ thị biểu diễn sự hao hụt khối lượng của quả bơ trong quá trình bảo quản với điều kiện
(t0 = 80C, φkk= 80 - 85%)
Kết quả thể hiện ở hình 1 cho thấy tỷ lệ hao hụt khối lượng có xu hướng tăng dần theo thời gian bảo quản với tốc độ biến thiên phụ thuộc nồng độ AVG xử lý. Mẫu xử lý AVG ở nồng độ 420 ppm và 470 ppm có tỷ lệ hao hụt khối lượng thấp nhất. Vào ngày bảo quản thứ 30, các giá trị xác định được lần lượt là 3,14% và 2,94%. Trong khi đó, mẫu đối chứng (ĐC) có tốc độ hao hụt khối lượng tăng nhanh và rõ rệt nhất vào ngày bảo quản thứ 15 và đạt giá trị cao nhất tại ngày bảo quản thứ 30 với giá trị xác định được 4,366%. Kết quả thực nghiệm đã chỉ ra rằng AVG có hiệu quả tích cực khi hạn chế được sự giảm tổn thất khối lượng quả bơ Booth7.
3.2. Ảnh hưởng của nông độ AVG đến cường độ hô hấp của quả bơ trong quá trình bảo quản sau thu hoạch
Sự biến đổi cường độ hô hấp của quả bơ trong quá trình bảo quản sau thu hoạch được trình bày ở hình 2.
Hình 2. Sự biến đổi cường độ hô hấp của quả bơ trong quá trình bảo quản với điều kiện
(t0 = 80C, φkk= 80 - 85%)
Số liệu thực nghiệm thu được từ đồ thị hình 2 cho thấy mẫu ĐC có cường độ hô hấp tăng nhanh và đạt giá trị cực đại sớm nhất vào ngày bảo quản thứ 21 với giá trị xác định được là 60,24 (ml CO2.kg-1.h-1). Cùng thời điểm này, mẫu xử lý AVG với nồng độ 320 ppm cũng đạt đỉnh hô hấp đột biến nhưng có giá trị thấp hơn là 54,2 (ml CO2.kg-1.h-1). Mẫu 370 ppm đạt đỉnh hô hấp đột biến tại ngày bảo quản thứ 27 với giá trị 53,3 (ml CO2.kg-1.h-1). Mẫu xử lý ở nồng độ 420 ppm và 470 ppm đạt đỉnh hô hấp đột biến chậm hơn vào ngày bảo quản thứ 30, tương ứng với các giá trị xác định được lần lượt là 52,49 (ml CO2.kg-1.h-1); 51,48 (ml CO2.kg-1.h-1). Như vậy, khi xử lý AVG để bảo quản quả bơ Booth7 thì thời điểm đột biến về cường độ hô hấp sinh ra trong bơ chậm hơn so với mẫu ĐC. Điều này được giải thích là AVG có khả năng ức chế hoạt động enzyme ACC synthase bằng cách liên kết đến vị trí lân cận của PLP (trung tâm hoạt động enzyme ACC synthase) tại Ala54, Arg412 và nước (Qing Huai et al., 2001). Vì vậy, AVG kìm hãm sự sản sinh ACC (chất tiền ethylene) làm chậm quá trình chín và kéo dài thời hạn bảo quản của quả bơ. Kết quả thực nghiệm này hoàn toàn phù hợp với công bố của Valeria và Douglas (2009), khi nghiên cứu bảo quản táo bằng AVG kết hợp nhiệt độ lạnh. Như vậy, xử lý AVG trên quả bơ Booth7 ở nồng độ 420 ppm và 470 ppm đã cho kết quả kìm hãm tốt nhất cường độ hô hấp và kéo dài thời gian bảo quản đến 30 ngày.
Hao
hụt
khố
i lượ
ng (%
)
6,000
5,000
4,000
3,000
2,000
1,000
0,0000 3 6 9 12 15 18 21 24 27 30 33 36
ĐC320ppm370ppm430ppm480ppm
Thời hạn bảo quản (ngày)
Thời hạn bảo quản (ngày)C
ường
độ
hô h
ấp (m
l CO
2.kg-
1.h-
1)0 3 6 9 12 15 18 21 24 27 30 33 36
70,00
60,00
50,00
40,00
30,00
20,00
10,00
00,00
ĐC
320ppm370ppm470ppm420ppm

99
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
3.3. Sự biến thiên cường độ sản sinh ethylene của quả bơ Booth7 khi xử lý AVG ở các nông độ khác nhau
Hình 3. Sự biến thiên cường độ sản sinh ethylene trong quá trình bảo quản với điều kiện
(t0 = 80C, φkk= 80 - 85%)
Kết quả thu được từ đồ thị hình 3 cho thấy cường độ sản sinh ethylene của tất cả các mẫu có xử lý và không xử lý AVG đều tăng chậm, tương đối đồng đều trong 12 ngày bảo quản đầu tiên, sau đó có xu hướng tăng nhanh dần và đạt đến đỉnh đột biến về ethylene ở các thời điểm khác nhau. Quy luật này hoàn toàn không mẫu thuẫn với kết quả nghiên cứu trước đó của các tác giả Penelope và Roger (1980), khi tiến hành nghiên cứu tác động của AVG đến quá trình sinh tổng hợp ethylene. Trong đó, mẫu 420 ppm và 470 ppm có cường độ sản sinh ethylene duy trì ở hàm lượng thấp nhất, đạt đỉnh đột biến ethylene đến chậm nhất vào ngày bảo quản thứ 30 với giá trị xác định được là 41,01 (μl C2H4.kg-1.h-1); 39,46 (μl C2H4.kg-1.h-1). Kết quả nghiên cứu cũng hoàn toàn tương đồng với kết quả của tác giả Qing Huai (2001), khi chỉ ra rằng AVG là chất ức chế quá trình sinh tổng hợp ethylene bằng con đường tác động trực tiếp, kìm hãm enzyme ACC synthase, ngăn cản sự hình thành ACC nên cường độ sản sinh ethylene được hạn chế. Chính vì vậy, hàm lượng ethylene nội sinh tạo thành thấp và kéo dài được thời hạn bảo quản bơ sau thu hoạch. Như vậy, xử lý AVG trên quả bơ Booth7 ở nồng độ 420 ppm và 470 ppm đã hạn chế được sự sản sinh ethylene và kéo dài được thời gian bảo quản quả bơ sau thu hoạch đến 30 ngày.
3.4. Ảnh hưởng của nông độ AVG đến hàm lương lipid tổng số trong quá trình bảo quản
Hàm lượng lipid tổng số là một trong những chỉ tiêu chất lượng phản ánh giá trị dinh dưỡng của quả bơ. Sự biến thiên hàm lượng lipid tổng số trong thời gian bảo quản bơ ứng với thời gian xử lý AVG khác nhau được thể hiện ở đồ thị hình 4.
Hình 4. Sự thay đổi hàm lượng lipid quả bơ Booth7 trong thời gian bảo quản với điều kiện
(t0 = 80C, φkk= 80 - 85%)
Kết quả thực nghiệm trên đồ thị hình 4 cho thấy mẫu ĐC có hàm lượng lipid tăng nhanh nhất và đạt giá trị cao vào ngày bảo quản thứ 21 với giá trị 14,01%; cùng thời điểm này thì mẫu xử lý 320 ppm cũng đạt giá trị lớn nhất là 14,11%. Trong khi đó, mẫu 370 ppm; 420 ppm và 470 ppm có sự biến động hàm lượng lipid diễn ra chậm hơn so với hai mẫu (ĐC và 320 ppm). Vào ngày bảo quản thứ 30, hai mẫu 420 ppm và 470 ppm cho hàm lượng lipid cao nhất với các giá trị lần lượt là 14,28% và 14,35%. Hàm lượng lipid đạt giá trị cực đại cùng thời điểm bơ đạt đến trạng thái chín, sau đó bắt đầu giảm dần làm giảm chất lượng quả bơ. Kết quả thực nghiệm này không mâu thuẫn với kết quả nghiên cứu tác giả Yoshio và Louis (1968) về những biến đổi hàm lượng lipid trong quá trình bảo quản bơ sau thu hoạch. Trong nghiên cứu này đã xác định được nồng độ 420 ppm và 470 ppm cho hiệu quả rõ ràng nhất.
3.5. Ảnh hưởng của nông độ dung dich AVG xử lý đến sự biến thiên hoạt lực ACC oxydase trong quá trình bảo quản quả bơ Booth7
Theo báo cáo của Kende (1993), ACC oxydase là một enzyme có hoạt tính sinh học và tác dụng xúc tác cho sự chuyển hóa ACC thành ethylene. Sự thay đổi hoạt lực ACC oxydase trong quá trình bảo quản khi xử lý AVG trên quả bơ Booth7 sau thu hoạch được thể hiện ở hình 5.
Hình 5. Ảnh hưởng xử lý nồng độ AVG đến hoạt lực ACC oxydase trên quả bơ Booth7 với điều kiện bảo quản (t0 = 80C, φkk= 80 - 85%)
Cườn
g độ s
ản si
nh E
thyle
ne (µ
l C2H
4.kg-1
.h-1)
70,000
60,000
50,000
40,000
30,000
20,000
10,000
0,0000 3 6 9 12 15 18 21 24 27 30 33 36
Thời hạn bảo quản (ngày)
ĐC
320ppm370ppm470ppm420ppm
Hàm
lượn
g lip
id (%
)
ĐC
320ppm370ppm420ppm470ppm
Thời hạn bảo quản (ngày)0 3 6 9 12 15 18 21 24 27 30 33 36
18,00
16,00
14,00
12,00
10,00
8,00
6,00
4,00
2,00
0,00
Thời hạn bảo quản (ngày)0 3 6 9 12 15 18 21 24 27 30 33 36
ĐC
320ppm370ppm420ppm470ppm
Hoạt l
ực enz
yme A
CCO (
nmol C
2H 4.g-1
.h-1) 25,000
20,000
15,000
10,000
5,000
0,000

100
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Kết quả thu được biểu thị từ hình 5 cho thấy hoạt lực ACC oxydase ở các mẫu xử lý với nồng độ AVG khác nhau, thì có sự biến động hoàn toàn không giống nhau. Mẫu xử lý 420 ppm và 470 ppm có hoạt lực ACC oxydase tăng chậm hơn so với các mẫu còn lại; thời điểm đạt giá trị cực đại cũng muộn hơn vào ngày bảo quản thứ 27 với các giá trị xác định được lần lượt là: 15,95 (nmol C2H4.g
-1.h-1); 15,53 (nmol C2H4.g
-1.h-1). Điều này cho thấy AVG kìm hãm đáng kể hoạt lực ACC oxydase trong thời gian bảo quản bơ Booth7. Nguyên nhân là do AVG khóa hoạt động của enzyme ACC synthase, ức chế sự tạo thành ACC, làm giảm lượng cơ chất tham gia trong quá trình oxy hóa tạo thành ethylene. Chính vì vậy, dễ dàng nhận thấy hoạt lực ACC oxydase của các mẫu có xử lý AVG sẽ bị kìm hãm và tăng chậm hơn so với mẫu ĐC (Qing Huai et al., 2001). Kết quả thực nghiệm thu được hoàn toàn phù hợp với công bố của các tác giả Baker và cộng tác viên (1982) khi cho rằng AVG có tác dụng kìm hãm hoạt lực enzyme ACC oxydase trong quá trình sinh tổng hợp ethylene trên quả bơ sau thu hoạch.
3.6. Sự thay đổi hàm lương vitamin C trên quả bơ Booth7 trong quá trình bảo quản sau thu hoạch
Sự thay đổi hàm lượng vitamin C được trong quá trình bảo quản được trình bày ở hình 6. Kết quả thực nghiệm thu được từ đồ thì hình 6 cho thấy thời gian bảo quản càng dài thì hàm lượng vitamin C càng giảm. Điều này có thể giải thích là hàm lượng vitamin C giảm theo thời gian bảo quản khi quả bơ chín, các biến đổi hóa học bắt đầu diễn ra mạnh mẽ, protopectin và hemicelluloses bị thủy phân làm cho thịt bơ mềm, cấu trúc tế bào lỏng lẻo, khi đó oxy bên ngoài dễ xâm nhập bên trong cấu trúc quả, thúc đẩy quá trình oxy hóa vitamin C. Mặt khác, vitamin C là một hợp chất hữu cơ nên nó tham gia vào quá trình hô hấp trong quá trình chín quả (Tôn Nữ Minh Nguyệt và ctv., 2009). Kết quả nghiên cứu này hoàn toàn phù hợp với kết quả do Ahmed Dorria và cộng tác viên (2010) đưa ra khi nghiên cứu mối quan hệ giữa các chỉ tiêu chất lượng trong quá trình chín quả bơ sau thu hoạch.
Qua những biến đổi hàm lượng vitamin C thu được trong quá trình nghiên cứu, nhận thấy AVG có tác dụng hiệu quả khi giảm tổn thất hàm lượng vitamin C. Trong đó, mẫu xử lý 420 ppm và 470 ppm có hàm lượng vitamin C tổn thất ở mức thấp, duy trì đến ngày bảo quản thứ 30 và giữ được giá trị dinh dưỡng quả bở Booth7 tốt nhất sau thu hoạch với các giá trị xác định lần lươt là 9,15 mg%; 9,45 mg%. Xử lý thống kê về hao hụt hàm lượng vitamin C trên
biến động ANOVA và so sánh giá trị trung bình của các mẫu bảo quản với mức ý nghĩa α = 0,05, cho thấy hàm lượng vitamin C của hai mẫu 420 ppm và 470 ppm vào ngày bảo quản thứ 30, sai khác không có ý nghĩa ở mức 5%. Vì vậy, kết hợp với xem xét về hiệu quả kinh tế, nồng độ AVG 420 ppm được lựa chọn nhằm kéo dài thời gian bảo quản và ổn định chất lượng cho quả bơ sau thu hoạch.
Hình 6. Sự thay đổi hàm lượng vitamin C quả bơ Booth7 theo thời gian bảo quản với điều kiện
(t0 = 80C, φkk= 80 - 85%)
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luận- Đã xác định được nồng độ AVG phù hợp nhất
với mục đích kìm hãm hoạt lực ACC oxydase, cường độ hô hấp, cường độ sản sinh ethylene và kéo dài thời gian bảo quản của quả bơ là 420 ppm.
- Đã xác định được các thông số công nghệ về cường độ sản sinh ethylene, cường độ hô hấp, hoạt lực ACC oxydase của mẫu đối chứng (mẫu ĐC) và mẫu có xử lý AVG (420 ppm) tại ngày bảo quản thứ 21 sau thu hoạch: Cường độ sản sinh ethylene của mẫu ĐC: 55,29 (μl C2H4.kg-1.h-1); trong khi đó, mẫu xử lý 420 ppm có giá trị: 23,29 (μl C2H4.kg-1.h-1); cường độ hô hấp mẫu ĐC: 60,24 (ml CO2.kg-1.h-1); mẫu 420 ppm có cường độ hô hấp thấp chỉ 38,96 (ml CO2.kg-1.h-1); hàm lượng ACC oxydase mẫu ĐC: 17,99 (nmol C2H4.g
-1.h-1); tuy nhiên, mẫu có xử 420 ppm AVG có giá trị xác định được: 9,01 (nmol C2H4.g
-1.h-1). - Đã xác định được các thông số kỹ thuật chính
khi sử dụng AVG nhằm kéo dài thời gian bảo quản quả bơ sau thu hoạch lên đến 30 ngày, cụ thể: Nồng độ AVG xử lý là 420 ppm, nhiệt độ xử lý nước nóng là 500C, thời gian xử lý nước nóng 10 phút, nhiệt độ môi trường bảo quản 80C, độ ẩm môi trường bảo quản 80 - 85%.
4.2. Đề nghi Áp dụng kết quả thu được để tiếp tục nghiên cứu
và hoàn thiện quy trình bảo bơ tươi sau thu hoạch cho mục đích xuất khẩu.
Thời hạn bảo quản (ngày)0 3 6 9 12 15 18 21 24 27 30 33 36
ĐC320ppm370ppm
420ppm470ppm
Hàm
lượng
vitam
in C (
mg%
)
25,00
20,00
15,00
10,00
5,00
0,00

101
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
TÀI LIỆU THAM KHẢOBộ Khoa học và Công nghệ, 1989. TCVN 4715-1989.
Đồ hộp rau quả - Phương pháp xác định hàm lượng vitamin C (axit ascobic).
Bộ Khoa học và Công nghệ, 2011. TCVN 9017:2011. Tiêu chuẩn Quốc gia - Quả tươi - Phương pháp lấy mẫu trên vườn sản xuất.
Tôn Nữ Minh Nguyệt, Lê Văn Việt Mẫn, Trần Thi Thu Hà, 2009. Công nghệ chế biến rau trái (Tập 1: Nguyên liệu và công nghệ bảo quản sau thu hoạch). Nhà xuất bản Đại học Quốc gia TP. Hồ Chí Minh.
Nguyên Văn Toản, 2011. Điều tiết quá trình sinh tổng hợp etylen nhằm kéo dài thời gian chín sau thu hoạch của quả chuối tiêu. Luận án tiến sĩ kỹ thuật. Đà Nẵng.
Nguyên Văn Toản, Lê Văn Luận, Trương Thi Minh Hạnh, Lê Thi Liên Thanh và Đỗ Chí Thinh, 2010. Ảnh hưởng của các loại bao bì kết hợp phun chất kháng ethylene (AVG) ở giai đoạn cận thu hoạch đến quá trình sinh tổng hợp ethylene của chuối tiêu. Tạp chí khoa học, Đại học Huế, (63):129-139.
Ahmed Dorria M, Yoursef Aml R.M, Hassan H S. A, 2010. Relationship between electrical conductivity, softening and color of Fuerte avocado fruits during ripening. Agriculture and Biology Journal of North America., 1: 878-885.
Baker J.E., Anderson J.D., Adams D.O., Apelbaum A., Lieberman M, 1982. Biosynthesis of ethylene from methionine in aminoethoxyvinylglycine- resistant avocado tissue. Plant Physiology., 69: 93-97.
Burhan Ozturk, Emine Kucuke, Sedat Karaman, Yacup Ozkan, 2012. The effects of cold storage and aminoethoxylvinylglycine (AVG) on bioactive compounds of plum fruit (Prunus Salicina Lindell cv. ‘Black Amber’). Postharvest Biology and Technology, 72: 35-41.
Kende H., 1993. Ethylene biosynthesis. Annu. Rev. Plant Physio., Plant Mol. Biol., 44: 283-307.
Mongy - El Abd, B. Abu-Aziz, Fayek M. Ahmed; Dorria M. Ahmed and Aml R. Yousef, 2009. Utilization of hot water treatments for reducing external damage and maintain quality of Hass avocado fruits. Journal of Agriculture and Biological Sciences., 5(6): 1046-1053.
Moya-León, M.A., John, P, 1995. Activity of 1-aminocyclopropane-1- carboxylate (ACC) oxidase (ethylene-forming enzyme) in the pulp and peel of ripening bananas. J. Hort. Sci., 69: 243-250.
Penelope S. Ness, Roger J. Romani, 1980. Effects of aminoethoxyvinylglycine and countereffects of ethylene on ripening of bartlett pear fruits. Plant Physiology, 65(2): 372-376.
Qing Huai, Yuanhong Xia, Yongquan Chen, Brian Callahan, Ning Li, Hengming Ke, 2001. Crystal structures of 1 - Aminocyclopropane - 1 - carboxylate (ACC) Synthase in complex with Aminoethoxyvinylglycine and Pyridoxal - 5’ - Phosphate provide new insight into catalytic mechanisms. The Journal of Biological Chemistry, 276(41): 38210-38216.
Salvatore D’Aquino , Mario Schirra, Maria Giovanna Molinu, Marco Tedde, Amedeo Palma, 2010. Preharvest aminoethoxyvinylglycine treatments reduce internal browning and prolong the shelf-life of early ripening pears. Scientia Horticulturae, 125: 353-360.
Valeria Sigal Escalada, Douglas D. Archbold, 2009. Preharvest Aminoethoxyvinylglycine Plus Postharvest Heat Treatments Influence Apple Fruit Ripening after Cold Storage. HortScience, 44(6): 1637-1640.
Yoshio Kikuta, Louis C. Erickson, 1968. Seasonal changes of avocado lipids during fruit development and storage.Califonia Avocado Society Yearbook., 52:102-108.
Effect of aminoethoxyvinylglycine combining with hot water treatment on ripen time of postharvest avocado Booth7
Nguyen Van Toan, Nguyen Hong PhucAbstractThe study used an inhibitor of ethylene biosynthesis (AVG) in combination with hot water treatment (temperature of 50 0C; in 10 mins) to prolong the shelf life of postharvesting avocado fruits (BOOTH7). The results showed that avocado fruits treated with AVG had lower activity of ACC oxydase, respiratory intensity and ethylene production than that of the control. Among 4 studied concentrations (320 ppm; 370 ppm; 420 ppm; 470 ppm), the AVG treatment concentration of 420 ppm showed that the peak of respiration and ethylene production appeared more slowly in comparison to the control and the ripen time was 9 days longer than the control. The results also showed that the avocado fruits treated with AVG 420 ppm after 30 days of preservation had the best quality with indicators (AVG 420 ppm, temperature of 50 0C; in 10 mins): Vitamin C content of 7.45 mg%; total lipid content of 14.28%; weight loss of 3.14%; ethylene production rate of 23.29 (μl C2H4.kg -1.h -1); respiration rate of 38.96 (ml CO2.kg-1.h-1); the content of ACC oxydase of 9.01 (nmol C2H4.g-1.h-1).Key words: Respiration rate, ethylene production rate, ACC oxydase activity, low temperature, hot water treatment, AVG treatmentNgày nhận bài: 1/7/2017Ngày phản biện: 14/7/2017
Người phản biện: TS. Nguyễn Văn PhongNgày duyệt đăng: 27/7/2017

102
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀ Tam thất hoang (Panax stipuleanatus H. T. Tsai
et K. M. Feng) là một trong 12 loài thuộc chi Panax - họ nhân sâm (Araliaceae) đã được các nhà khoa học trên thế giới nghiên cứu. Ở Việt Nam loài này chỉ xuất hiện ở huyện Sa Pa (núi Hàm Rồng - thị trấn Sa Pa và xã Tả Phìn) và huyện Bát Xát (xã Trung Lèng Hồ), tỉnh Lào Cai. Tam thất hoang là loại cây ưa bóng, ưa ẩm thích hợp với nền khí hậu quanh năm mát mẻ. Đây là nguồn gen quý, có giá trị, thuộc nhóm rất nguy cấp (CR) trong sách đỏ Việt Nam 2007 và nằm trong nhóm IIa của Nghị Định 32-CP.
Thành phần hóa học chính của Tam thất hoang là các saponin triterpen khung olean (Trần Công Luận và ctv., 2009). Tam thất hoang đã được chứng minh có các tác dụng: Giúp lưu thông tuần hoàn máu, giảm lượng Cholesterol trong máu, hạ đường huyết, kích thích hệ miễn dịch, ức chế vi khuẩn và siêu vi khuẩn, chống viêm tấy giảm đau… (Nguyễn Thị Thu Hương và ctv., 2009).
Nghiên cứu dược lý cho thấy cắn chiết ethanol thân rễ và rễ củ của loài Tam thất hoang có khả năng giảm stress, saponin toàn phần có tác dụng chống oxy hóa. Ngoài ra các polyacetylen có tác dụng độc với một số dòng tế bào ung thư (Liang C, 2010, 2013).
Do là cây thuốc quý, hiếm gặp trong tự nhiên, giá bán cao nên Tam thất hoang thường xuyên bị tìm kiếm khai thác dẫn đến suy giảm nghiêm trọng số lượng cá thể và quần thể, nhiều nơi trong rừng tự nhiên hầu như không còn nguồn gen quý này. Mặc dù là một dược liệu quí nhưng cho đến nay những công trình nghiên cứu trong nước về loài này còn ít. Để góp phần hoàn thiện các nghiên cứu về loài cây
này, nhóm nghiên cứu đã tiến hành khảo sát các chỉ tiêu để xây dựng tiêu chuẩn cở sở cho dược liệu Tam thất hoang.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứuThân rễ và rễ củ Tam thất hoang thu hái tại Vườn
Quốc gia Hoàng Liên.
2.2. Hóa chất - thuốc thửCác dung môi, hóa chất sử dụng đều đạt tiêu
chuẩn hóa chất tinh khiết phân tích (PA). Các dung môi dùng cho phân tích sắc ký (TLC, HPLC) đều được mua của hãng Merck (hoặc tương đương). Chất chuẩn acid oleanolic (AO) của hãng Sigma-Aldrich, độ tinh khiết đạt 98,0%.
2.3. Phương pháp nghiên cứuXây dựng tiêu chuẩn dược liệu Tam thất hoang
dựa trên các chỉ tiêu chung được qui định tại dược điển Việt Nam gồm: Mô tả, soi bột, vi phẫu, độ ẩm dược liệu, tro toàn phần, định tính, định lượng,… (Bộ Y tế, 2009).
- Mô tả: Mô tả bằng cảm quan, kiểm tra kích thước bằng cách đo.
- Vi phẫu, bột: Phương pháp hiển vi, thử theo Dược điển Việt Nam IV (DĐVN IV), phụ lục 12.18.
- Hàm ẩm: Phương pháp cân, thử theo DĐVN IV, Phụ lục 9.6
- Độ tro: Thử theo DĐVN IV, Phụ lục 9.8.- Định tính: Acid oleanolic bằng phương pháp
sắc ký lớp mỏng. Bản mỏng: Silica gel 60F254 (Merck); Dung môi khai triển: n-hexan/ethyl acetat (4/1).
1 Vườn Quốc Gia Hoàng Liên; 2 Viện Dược liệu
NGHIÊN CỨU XÂY DỰNG TIÊU CHUẨN CƠ SỞ DƯỢC LIỆU TAM THẤT HOANGNguyễn Quang Vĩnh1, Nguyễn Thị Phương2, Bùi Tuấn Anh1,
Trần Văn Tú1, Vũ Thị Hải1, Lê Huy Công1
TÓM TẮTTiêu chuẩn cơ sở dược liệu Tam thất hoang được xây dựng bao gồm: Mô tả, vi phẫu, soi bột, độ ẩm, tro toàn
phần, tạp chất khác, định tính và định lượng. Kết quả nghiên cứu cho thấy dược liệu Tam thất hoang thường nhiều đốt, cong ngoằn ngoèo, dài 3 - 10 cm, đường kính 0,3 - 1,0 cm; Mặt ngoài màu nâu hay màu vàng xám, có những vết nhăn dọc, mảnh, mùi thơm nhẹ đặc trưng, vị đắng, hơi ngọt. Vi phẫu thân thấy có lớp bần gồm từ 7 - 10 lớp tế bào hình chữ nhật, xếp chồng lên nhau, đều đặn; Mô mềm vỏ chứa nhiều hạt tinh bột, rải rác có những ống tiết chứa chất nhựa và tinh thể calci oxalat hình cầu gai; Libe - gỗ xếp thành từng bó hướng vào tâm; Mạch gỗ rất ít. Bột có màu nâu nhạt, vị thơm, vị hơi ngọt; Hạt tinh bột hình tròn, hình chuông hay hình nhiều cạnh. Hàm ẩm không quá 13 %, độ tro không quá 8 %, định tính dược liệu phải có acid oleanolic và hàm lượng AO không được thấp hơn 1%. Nghiên cứu này góp phần kiểm soát tốt chất lượng và nâng cao giá trị của dược liệu Tam thất hoang.
Từ khóa: Tam thất hoang, tiêu chuẩn cơ sở dược liệu, mô tả, vi phẫu

103
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
+ Dung dịch thử: Cân khoảng 1,0 g rễ tam thất hoang đã xay nhỏ cho vào bình cầu dung tích 50 ml, có nút nhám, thêm 20 ml dung môi methanol chứa acid chlohydric 2% (TT), đun hồi lưu cách thủy ở nhiệt độ 70oC trong 1 giờ. Để nguội bình cầu về nhiệt độ phòng, lọc lấy dịch chiết. Cô dịch chiết đến cắn, hòa tan cắn bằng 5 ml methanol (TT), lọc, thu được dung dịch thử.
+ Dung dịch đối chiếu 1: Dung dịch acid oleanolic 0,5 mg/ml trong methanol (TT).
+ Dung dịch đối chiếu 2: Lấy 1,0 g bột rễ tam thất hoang đối chiếu, tiến hành chiết như dung dịch thử.
+ Cách tiến hành: Chấm riêng biệt lên bản mỏng 10 μl mỗi dung dịch thử và các dung dịch đối chiếu. Sau khi khai triển, bản mỏng được để bay hết dung môi ở nhiệt độ phòng. Phun thuốc thử dung dịch acid sulfuric 10% trong ethanol, sấy bản mỏng ở 105oC đến khi xuất hiện rõ các vết.
- Định lượng: Định lượng acid oleanolic (AO) bằng phương pháp sắc ký lỏng cao áp hiệu năng cao (HPLC).
+ Hệ thống, thiết bị và điều kiện phân tích HPLCHệ thống HPLC (Shimadzu, Nhật Bản), gồm:
bơm LC-20AD, bộ tiêm mẫu tự động SIL-20AHT, detector UV-VIS, phần mềm Labsolution để truy xuất hình ảnh và số liệu trên máy HPLC. Phần mềm Excel được sử dụng để tính toán kết quả.
Điều kiện phân tích HPLC: cột pha đảo Ascentis C18 (250 mm x 4,6 mm; 5 µm), detector UV-VIS (bước sóng 205 nm); pha động là nước và acetonitril (tỷ lệ 20 – 80), chế độ rửa giải đẳng dòng; tốc độ rửa giải là 0,5 ml/phút; thể tích mẫu tiêm vào cột là 10 μl.
+ Chuẩn bị mẫuMẫu chuẩn AO: cân chính xác 5,0 mg chất chuẩn
AO, hòa tan trong 5,00 ml methanol. Các dung dịch chuẩn AO có nồng độ nhỏ hơn được chuẩn bị bằng cách pha loãng dung dịch trên bằng methanol.
Mẫu thử: Cân chính xác khoảng 2,0 g mẫu thử (đã xay nhỏ và xác định độ ẩm) cho vào bình cầu 250 ml, thêm khoảng 100 ml methanol chứa acid chlohydric 2% , chiết hồi lưu cách thủy trong 2 giờ. Để nguội về nhiệt độ phòng. Lọc lấy dịch chiết, cô dịch chiết đến cắn. Hòa cắn bằng 50 ml methanol, lọc vào bình định mức 50 ml, định mức vừa đủ bằng methanol. Lọc qua màng lọc cellulose acetat 0,45 µm trước khi tiêm vào cột.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Mô tả dươc liệu Tam thất hoangThân rễ thường nhiều đốt, cong ngoằn ngoèo, ít
khi có hình trụ thẳng, dài 3 - 10 cm, đường kính 0,3 - 1,0 cm. Mặt ngoài màu nâu hay màu vàng xám, có những vết nhăn dọc, mảnh; những vết vân ngang nổi rõ chia thân rễ thành nhiều đốt, đặc biệt có nhiều sẹo do thân khí sinh hàng năm tàn lụi để lại. Thể chất cứng chắc, giòn, dễ bẻ, mặt bẻ lởm chởm, màu xám nhạt. Mùi thơm nhẹ đặc trưng, vị đắng, hơi ngọt (Hình 1).
Hình 1. Hình ảnh dược liệu Tam thất hoang
3.2. Vi phẫu dươc liệu Tam thất hoangQuan sát từ ngoài vào trong thấy có: Lớp bần
gồm từ 7 - 10 lớp tế bào hình chữ nhật, xếp chồng lên nhau, đều đặn. Mô mềm vỏ gồm tế bào có khích thước và hình dạng khác nhau, chứa nhiều hạt tinh bột, rải rác có chứa những ống tiết chứa chất nhựa và tinh thể calci oxalat hình cầu gai. Libe - gỗ xếp thành từng bó hướng vào tâm, libe nằm ngoài, gỗ nằm trong, các bó libe-gỗ cách nhau bởi những tia ruột, gồm nhiều tế bào (bên trong có chứa tinh bột). Mạch gỗ rất ít (Hình 2).
Hình 2. Vi phẫu cắt ngang rễ Tam thất hoang1. Bần, 2. Mô mềm, 3. Tinh thể canxi oxalat,
4. Bó libe-gỗ, 5. Tia ruột, 6. Ống tiết, 7. Libe , 8. Gỗ
3.3. Đặc điểm bột dươc liệu Tam thất hoangBột có màu nâu nhạt, vị thơm, vị hơi ngọt. Quan
sát dưới kính hiển vi thấy có: mảnh bần là những tế bào hình chữ nhật hay nhiều cạnh, thành dày.

104
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Mảnh mô mềm gồm tế bào hình nhiều cạnh, hoặc tròn, thành mỏng, có chứa hạt tinh bột và nhiều tinh thể calci oxalat hình cầu gai. Đôi khi thấy ống tiết trong có chất tiết màu vàng nâu. Hạt tinh bột hình tròn, hình chuông hay hình nhiều cạnh. Mảnh mạch mạng, mạch vạch, mô dày (Hình 3).
Hình 3. Bột dược liệu Tam thất hoang1- Mảnh bần; 2- Mảnh mô mềm; 3- Mô dày;
4,5- Tinh thể canxi oxalat; 6,7,8,9- Hạt tinh bột; 10- Mảnh mạch mạng; 11- Mạch vạch; 12- Ống tiết
3.4. Kết quả kiểm tra hàm ẩm và độ tro của các mẫu dươc liệu Tam thất hoang
Từ kết quả bảng 1 cho thấy hàm ẩm và độ tro của các mẫu Tam thất hoang lần lượt là 10,1 ± 0,9; 11, 0 ± 0,8; 12,4 ± 1,0; 6.06 ± 0,15; 5.56 ± 0,40; 4.98 ± 0,17.
Tiêu chuẩn Tam thất hoang đề xuất hàm ẩm không quá 13,0 % và độ tro không quá 8%.
Tiêu chuẩn cơ sở Tam thất hoang qui định tạp chất không quá 1 %.
Bảng 1. Hàm ẩm và độ tro của các mẫu dược liệu Tam thất hoang
3.5. Đinh tính acid oleanolic bằng phương pháp sắc ký lớp mỏng
Tiến hành định tính acid oleanolic bằng phương pháp sắc ký lớp mỏng. Kết quả được trình bày tại hình 4.
Hình 4. Sắc ký đồ TLC định tính Tam thất hoang1, 2: Mẫu tam thất hoang;
3: Chất đối chiếu acid oleanolic
Kết quả cho thấy sắc ký đồ TLC mẫu thủ cho các vết tách rõ ràng, trong đo có 01 vết Rf khoảng 0,5 có cùng màu hồng và cùng Rf với vết của mẫu đối chiếu AO, chứng tỏ lựa chọn định tính AO là thích hợp.
3.6. Hàm lương AO trong các mẫu dươc liệu Tam thất hoang
Kết quả định lượng AO cho thấy, trên sắc đồ HPLC của các mẫu thử đều có pic cùng thời gian lưu với pic của AO và hàm lượng AO tổng số của các mẫu tam thất đều trên 1 %, trung bình khoảng 1,3 %, vì vậy tiêu chuẩn Tam thất hoang đề xuất hàm lượng acid olealoic không được thấp hơn 1 %.
Bảng 2. Kết quả hàm lượng AO trong các mẫu Tam thất hoang
Hình 5. Sắc ký đồ HPLC định lượng acid oleanolic trong Tam thất hoang
1, 2, 3: Dược liệu Tam thất hoang; 4: Mẫu chuẩn acid oleanolic
TT Mẫu đánh giá Hàm ẩm (% ) Độ tro
1 Mẫu 1 10,1 ± 0,9 6,06 ± 0,152 Mẫu 2 11, 0 ± 0,8 5,56 ± 0,403 Mẫu 3 12,4 ± 1,0 4,98 ± 0,17
Trung bình 11,16 % 5,53 %
TT Mẫu đánh giá Hàm lương % AO 1 Mẫu 1 1,54 ± 0,092 Mẫu 2 1,12 ± 0,083 Mẫu 3 1,32 ± 0,08
1
2
3
uV(x1,000,000)
2.00
1.75
1.50
1.25
1.00
0.75
0.50
0.25
0.00
-0.25
-0.50
-0.75
5.0 10.0 15.0 20.0 25.0 30.0 35.0 40.0 45.0 50.0 min

105
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
IV. KẾT LUẬN Nghiên cứu đã xây dựng được các tiêu chí trong
tiêu chuẩn của dược liệu Tam thất hoang gồm các chỉ tiêu: mô tả, hàm ẩm, độ tro, định tính, định lượng. Kết quả nghiên cứu cũng đã bước đầu đề xuất được giới hạn cần đạt trong tiêu chuẩn dược liệu Tam thất hoang: hàm ẩm không quá 13%, độ tro không quá 8%, định tính dược liệu phải có acid oleanolic và hàm lượng AO không được thấp hơn 1%. Chỉ tiêu mô tả, vi phẫu, bột phải giống như mô tả. Việc tiêu chuẩn hóa dược liệu Tam thất hoang sẽ giúp kiểm soát tốt hơn chất lượng của dược liệu này trên thị trường cũng như trong trồng trọt, thu hái, góp phần nâng cao giá trị của dược liệu này.
TÀI LIỆU THAM KHẢOBộ Y Tế, 2009. Dược điển Việt Nam IV. Nhà xuất bản Y
học, tr 348-349.
Nguyên Thi Thu Hương, Trần Công Luận, Nguyên Tập, 2009. Nghiên cứu một số tác dụng dược lý của Tam thất hoang Panax stipuleanatus Tsai et Feng, họ Araliaceae. Tạp chí Dược liệu, 14(2): 99-103.
Trần Công Luận, Lưu Thảo Nguyên, Nguyên Tập, 2009. Nghiên cứu thành phần hóa học của hai loài Sâm vũ diệp và Tam thất hoang. Tạp chí Dược liệu, 14(1): 17-23.
Liang C, 2010. Oleanane-type triterpenoids from Panax stipuleanatus and their anticancer activities. Bioorganic & Medicinal Chemistry Letters, 20: 7110-7115.
Liang C, 2013. Oleanane-triterpenoids from Panax stipuleanatus inhibit NF-kB. Journal of Ginseng Research, 37(1): 74-79.
Establishment of baseline standards for medicinal herb (Panax stipuleanatus) Nguyen Quang Vinh, Nguyen Thi Phuong,
Bui Tuan Anh, Tran Van Tu,Vu Thi Hai, Le Huy CongAbstractThe baseline standards for medicinal herb (Panax stipuleanatus) were established based on: Morpological characterization, microsurgery, powder, moisture, total ash, impurities. The research results showed that Panax stipuleanatus was curvy, scattered into several installments, each 3-10 cm long, 0.3-1.0 cm diameter. The outer surface was brown or yellowish-gray, with slim lines, fragments, slight odor, bitter taste, and slightly sweet taste. Vascular surgery consisted of 7 to 10 layers in rectangular shapes, stacked over equally. Soft shell contained many starch granules, occasionally appeared tubes containing the resin and crystalline calcium oxalate which is spherical hemisphere. Libe - the wood packs in a bunch directed to the core; Very few wood circuit. Powder was light brown, tasting slight sweet. Grain starch was round or multi-edged. Moisture content is not more than 13%, ash content is not more than 8%, qualitative pharmaceutical materials must contain oleanolic acid and AO content is not less than 1%. This research contributes to good quality control and enhances the value of Panax stipuleanatus.Key words: Panax stipuleanatus, medicinal baseline standards, characterization, microsurgery
Ngày nhận bài: 18/6/2017Ngày phản biện: 29/6/2017
Người phản biện: PGS. TS Nguyễn Thị Bích ThuNgày duyệt đăng: 27/7/2017
1 Vườn Quốc gia Hoàng Liên, 2 Viện Dược liệu
NGHIÊN CỨU XÂY DỰNG TIÊU CHUẨN CƠ SỞ DƯỢC LIỆU HOÀNG LIÊN Ô RÔNguyễn Quang Vĩnh1, Nguyễn Thị Phương2, Bùi Tuấn Anh1,
Trần Văn Tú1, Vũ Thị Hải1, Lê Huy Công1
TÓM TẮTTiêu chuẩn cơ sở cho dược liệu Hoàng liên ô rô được xây dựng gồm: Mô tả, vi phẫu, soi bột, độ ẩm, tro toàn phần.
Kết quả nghiên cứu cho thấy dược liệu Hoàng liên ô rô, thân hình trụ, màu vàng nhạt đường kính khoảng 0,5-2 cm, vỏ thân bề mặt sần sùi. Vi phẫu thân thấy có lớp bần gồm 1 - 2 lớp tế bào thành dày hình thoi, hình đa giác, xếp đều nhau, chứa nhiều đám sợi và đám tế bào mô cứng; Vùng libe dày; Tia ruột xuyên từ libe vào đến hết mạch gỗ, trên tia ruột vùng libe chứa các tinh thể canxi oxalat hình thoi, hình lăng trụ. Mô mềm ruột chiếm phần lớn thiết diện của thân, gồm những tế bào hình đa giác thành dày đã hóa cellulose một phần. Hàm ẩm không quá 12 %, độ tro không quá 8 %, định tính dược liệu phải có berberin chlorid và hàm lượng berberin chlorid không được thấp hơn 0,5%. Nghiên cứu này góp phần kiểm soát tốt chất lượng và nâng cao giá trị của dược liệu Hoàng liên ô rô.
Từ khóa: Hoàng liên ô rô, tiêu chuẩn cơ sở dược liệu, mô tả, vi phẫu

106
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀHoàng liên ô rô có tên khoa học là Mahonia
nepalensis DC, còn có tên gọi khác là Thích hoàng liên, Hoàng bá gai, phân bố ở vùng ôn đới ấm hoặc cận nhiệt đới châu Á, bao gồm Trung Quốc, Ấn Độ và một số nước ở vùng Trung Á. Ở Việt Nam, Hoàng liên ô rô là cây thuốc thuộc diện quý hiếm, thường có ở các tỉnh như Hà Giang, Cao Bằng, Lào Cai (Đỗ Tất Lợi, 2001). Bộ phận dùng của cây Hoàng liên ô rô là cả lá, thân, rễ và quả. Theo y học cổ truyền Trung Quốc, Hoàng liên ô rô có vị đắng, tính mát, có tác dụng thanh nhiệt ở phế, vị, can, thận. Quả lợi tiểu và làm dịu kích thích. Ở Việt Nam, dược liệu Hoàng liên ô rô vốn được biết đến là một vị thuốc dùng để chữa các bệnh như kiết lỵ, tiêu chảy, viêm ruột, ăn uống không tiêu, vàng da, đau mắt, mẩn ngứa, ho lao (Bộ Giáo dục-Đào tạo và Bộ Y tế, 2006; Đỗ Huy Bích và ctv., 2006). Về thành phần hóa học, thành phần hoá học của dược liệu Hoàng liên ô rô chủ yếu là các alkaloid như berberin, palmatin, berbamin, jatrozzhizin, magnoflorin, oxyacanthin,… (Cao Thùy Hương và ctv., 2009; Shion-Jane Lin et al., 1996).). Trong đó, berberin là thành phần chính, chiếm từ 0,35 đến 2,5 % trong bộ phận thân cây Hoàng liên ô rô. Dược liệu này là nguồn nguyên liệu chính được dùng để sản xuất hợp chất berberin chlorid.
Trong tự nhiên, Hoàng liên ô rô (Mahonia nepalensis DC) mọc ở vùng núi Hoàng Liên Sơn (Sa Pa), là loài cây thuốc có nhiều giá trị, công dụng tốt và đang có nguy cơ tuyệt chủng cao. Mặc dù là một dược liệu quí, nhưng cho tới nay Hoàng liên ô rô vẫn chưa được đưa vào Dược điển Việt Nam. Vì vậy, để góp phần hoàn thiện các nghiên cứu về loài cây này nghiên cứu xây dựng tiêu chuẩn cở sở cho dược liệu Hoàng liên ô rô được tiến hành.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứuCác mẫu dược liệu thân Hoàng liên ô rô thu hái
tại vườn Quốc gia Hoàng Liên.
2.2. Hóa chất-thuốc thửCác dung môi, hóa chất sử dụng đều đạt tiêu
chuẩn hóa chất tinh khiết phân tích (PA). Các dung môi dùng cho phân tích sắc ký (TLC, HPLC) đều được mua của hãng Merck (hoặc tương đương). Chất chuẩn berberin clorid và palmatin clorid của hãng Sigma-Aldrich, độ tinh khiết đạt 98,0%.
2.3. Phương pháp nghiên cứuXây dựng tiêu chuẩn dược liệu Hoàng liên ô rô
dựa trên các chỉ tiêu chung được qui định tại dược điển Việt Nam gồm: Mô tả, soi bột, vi phẫu, độ ẩm dược liệu, tro toàn phần, định tính, định lượng,… (Bộ Y tế, 2009).
- Mô tả: Mô tả bằng cảm quan, kiểm tra kích thước bằng cách đo.
- Vi phẫu, bột: Phương pháp hiển vi, thử theo DĐVN IV, phụ lục 12.18.
- Hàm ẩm: Phương pháp cân, thử theo DĐVN IV, phụ lục 9.6
- Độ tro: Thử theo DĐVN IV, Phụ lục 9.8.- Định tính: Berberin, palmatin bằng phương
pháp sắc ký lớp mỏng. + Bản mỏng: Silica gel 60F254 (Merck); Dung môi
khai triển:n-Butanol:acid acetic:nước (7:1:2).+ Dung dịch thử: Cân khoảng 0,2 g thân, rễ
Hoàng liên ô rô đã xay nhỏ cho vào bình cầu dung tích 50 ml, có nút nhám, thêm 20 ml dung môi methanol chứa acid chlohydric 2% (TT), đun hồi lưu cách thủy ở nhiệt độ 70oC trong 1 giờ. Để nguội bình cầu về nhiệt độ phòng, lọc lấy dịch chiết. Cô dịch chiết đến cắn, hòa tan cắn bằng 5 ml methanol (TT), lọc, thu được dung dịch thử.
+ Dung dịch đối chiếu 1: Dung dịch berberin chlorid 0,05 mg/ml trong methanol (TT). Dung dịch palmatin chlorid 0,05 mg/ml trong methanol (TT).
+ Dung dịch đối chiếu 2: Cân khoảng 0,2 (g) Hoàng liên ô rô (mẫu chuẩn), tiến hành chiết như dung dịch thử.
+ Cách tiến hành: Chấm riêng biệt lên bản mỏng 10 μl mỗi dung dịch thử và các dung dịch đối chiếu. Sau khi khai triển, bản mỏng được để bay hết dung môi ở nhiệt độ phòng. Quan sát bản mỏng dưới đèn tử ngoại UV 366 nm.
- Định lượng: Định lượng berberin chlorid bằng phương pháp sắc ký lỏng
+ Tiêm riêng biệt các dung dịch chuẩn, tiến hành sắc ký, ghi nhận sắc ký đồ thời gian lưu và diện tích pic của pic berberin chlorid tương ứng. Thiết lập phương trình hồi quy tuyến tính mô tả sự phụ thuộc giữa nồng độ dung dịch chuẩn berberin chlorid (mg/ml) và diện tích pic tương ứng theo phương trình y = ax + b.
+ Tiêm dung dịch thử, tiến hành sắc ký, ghi nhận sắc ký đồ thời gian lưu và diện tích pic của pic có cùng thời gian lưu với pic của berberin chlorid chuẩn. Tính nồng độ C (mg/ml) berberin chlorid

107
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
trong dung dịch thử (mg/ml) dựa trên phương trình hồi quy µl.
- Hàm lượng (%) berberin chlorid có trong mẫu thử (tính theo khối lượng khô kiệt) được tính theo công thức:
X (%) = C ˟ 50 ˟ 100 ˟ P1000 ˟ m ˟ (100 – b)
Trong đó: C là nồng độ berberin clorid trong dung dịch mẫu thử (mg/ml); m: khối lượng của mẫu thử đem phân tích (g) ; b: độ ẩm của mẫu thử (%); P: độ tinh khiết của berberin clorid chuẩn (%).
2.4. Thời gian và đia điểm nghiên cứu Nghiên cứu thực hiện được thực hiện từ tháng
8/2017 đến tháng 11/2017 tại Vườn Quốc gia Hoàng Liên và Viện Dược liệu.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Mô tả dươc liệu Hoàng liên ô rôThân hình trụ, màu vàng nhạt đường kính
khoảng 0,5 - 2 cm, vỏ thân bề mặt sần sùi, mang vế tích lá còn sót lại.
Hình 1. Hình ảnh dược liệu Hoàng liên ô rô
3.2. Vi phẫu dươc liệu Hoàng liên ô rôVi phẫu thân: Thiết diện cắt ngang thân Hoàng
liên ô rô hình tròn, quan sát từ ngoài vào trong gồm: Bần gồm 1 - 2 lớp tế bào thành dày, xếp đều nhau. Lớp bần thứ cấp gồm nhiều tế bào hình thoi, hình đa giác kích thước không đều xếp sát nhau, chứa nhiều đám sợi và đám tế bào mô cứng. Mô mềm vỏ là những tế bào thành mỏng chứa nhiều đám sợi, và đám tế bào cứng. Vùng libe dày, gồm những tế bào thành mỏng hơi bị ép bẹp. Tia ruột xuyên từ libe vào đến hết mạch gỗ, trên tia ruột vùng libe chứa các tinh thể canxi oxalat hình thoi, hình lăng trụ. Mô mềm ruột chiếm phần lớn thiết diện của thân, gồm những tế bào hình đa giác thành dày đã hóa cellulose một phần.
Hình 2. Vi phẫu cắt ngang thân Hoàng liên ô rô1. Bần, 2. Đám sợi, 3. Libe, 4. Tia ruột, 5. Tinh thể
canxi oxalat hình lăng trụ, 6. Gỗ, 7. Mô mềm ruột
3.3. Đặc điểm bột dươc liệu Hoàng liên ô rôBột màu vàng hay vàng nâu, không mùi, vị hơi
đắng. Quan sát dưới kính hiển vi thấy có: Mô mềm là những tế bào hình chữ nhật hay hình đa giác, thành mỏng. Rất nhiều tế bào cứng có dạng hình thoi màu vàng, thành dày, có ống trao đổi rõ. Sợi tập trung thành từng đám, màu vàng hay màu nâu nhạt. Tinh thể calci oxalat hình khối hay hình lăng trụ nằm rải rác.
Hình 3. Bột dược liệu Hoàng liên ô rô1, 2: Tinh thể calci oxalat; 3, 4, 5: Tế bào cứng; 6, 9: Bó
sợi; 7,8: Mô mềm
3.4. Kết quả kiểm tra hàm ẩm của các mẫu dươc liệu Hoàng liên ô rô
Tiến hành xác định độ ẩm của các mẫu dược liệu Hoàng liên ô rô khác nhau kết quả được trình bày ở bảng 1.
123
56
7

108
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 1. Kết quả xác định độ ẩm của các mẫu Hoàng liên ô rô
Độ ẩm các mẫu dược liệu Hoàng liên ô rô trung bình 8,66% (Bảng 1).
Tiêu chuẩn đề xuất: Hàm ẩm cho Hoàng liên ô rô không quá 12,0%.
3.5. Kết quả kiểm tra tro toàn phần trong các mẫu dươc liệu Hoàng liên ô rô
Tiến hành xác định tro toàn phần của các mẫu dược liệu ở 2 mẫu khác nhau kết quả được trình bày tại bảng 2.
Bảng 2. Kết quả xác định tro toàn phần của các mẫu Hoàng liên ô rô
Theo kết quả ở bảng 2, độ tro toàn phần của dược liệu Hoàng liên ô rô nằm trong khoảng từ 2,06 % đến 3,98 %.
Tiêu chuẩn đề xuất: Hoàng liên ô rô qui định độ tro toàn phần không quá 8,0%.
3.6. Tạp chấtTạp chất lẫn trong dược liệu Hoàng liên ô rô gồm
tất cả các chất ngoài qui định của dược liệu đó như: các bộ phận khác của cây như lá, rễ,..
Xác định tạp chất của mẫu dược liệu, kết quả được trình bày tại bảng 3.
Bảng 3. Kết quả xác định tạp chất của các mẫu Hoàng liên ô rô
Tạp chất trong các mẫu Hoàng liên ô rô chủ yếu là lá và chiếm không nhiều trung bình khoảng 0,13% (Bảng 3).
Tiêu chuẩn đề xuất: Chuẩn cơ sở Hoàng liên ô rô qui định tạp chất không quá 1%.
3.7. Đinh tính Berberin, palmatin bằng phương pháp sắc ký lớp mỏng
Tiến hành định tính Berberin, palmatin bằng phương pháp sắc ký lớp mỏng. Kết quả được trình bày tại hình 4.
Hình 4. Sắc ký đồ TLC định tính dược liệu Hoàng liên ô rô
Ký hiệu: Ber: berberin chlorid; Pal: palmatin chlorid; 1,2,3: Mẫu Hoàng liên ô rô thử, 4: Mẫu Hoàng liên ô rô chuẩn. Sắc ký đồ quan sát dưới đèn tử ngoại bước sóng UV 366 nm
Từ kết quả thu được trên hình 4, ta thấy các vết trên sắc ký đồ TLC của các mẫu dược liệu Hoàng liên ô rô tách nhau rõ ràng. Đặc biệt, khi quan sát dưới UV 366 nm, các vết hiện màu vàng đặc trưng tách rời nhau, các vết này có cùng màu sắc và Rf trùng với màu sắc và Rf của các chất đối chiếu Berberin chlorid (Rf = 0,35) và Palmatin chlorid (Rf = 0,30). Cả hai hợp chất này đều là những alkaloid đặc trưng cho dược liệu Hoàng liên ô rô. Chứng tỏ lựa chọn định tính Berberin, palmatin là thích hợp.
3.8. Đinh lương berberin chlorid bằng phương pháp sắc ký lỏng
Kết quả định lượng thành phần berberin clorid trong các mẫu Hoàng liên ô rô được trình bày tại bảng 4.
Bảng 4. Kết quả hàm lượng berberin chlorid trong các mẫu Hoàng liên ô rô
Bảng 4 cho thấy hàm lượng berberin trong các mẫu thử trung bình 0,54%.
Tiêu chuẩn đề xuất: Hàm lượng berberin chlorid trong dược liệu Hoàng liên ô rô không thấp hơn 0,5% tính theo dược liệu khô kiệt.
TT Tên mẫu Hàm ẩm (% )1 Mẫu 1 8,2 ± 0,82 Mẫu 2 9, 0 ± 0,7
Trung bình 8,66
TT Tên mẫu Tro toàn phần (%)1 Mẫu 1 2.06 ± 0,52 Mẫu 2 3,15 ± 0,40
Trung bình 3,06
TT Tên mẫu Hàm lương % tạp chất1 Mẫu 1 0,102 Mẫu 2 0,10
Trung bình 0,10
TT Tên mẫu Hàm lương % berberin chlorid
1 Mẫu 1 0,52 ± 0,122 Mẫu 2 0,57 ± 0,08
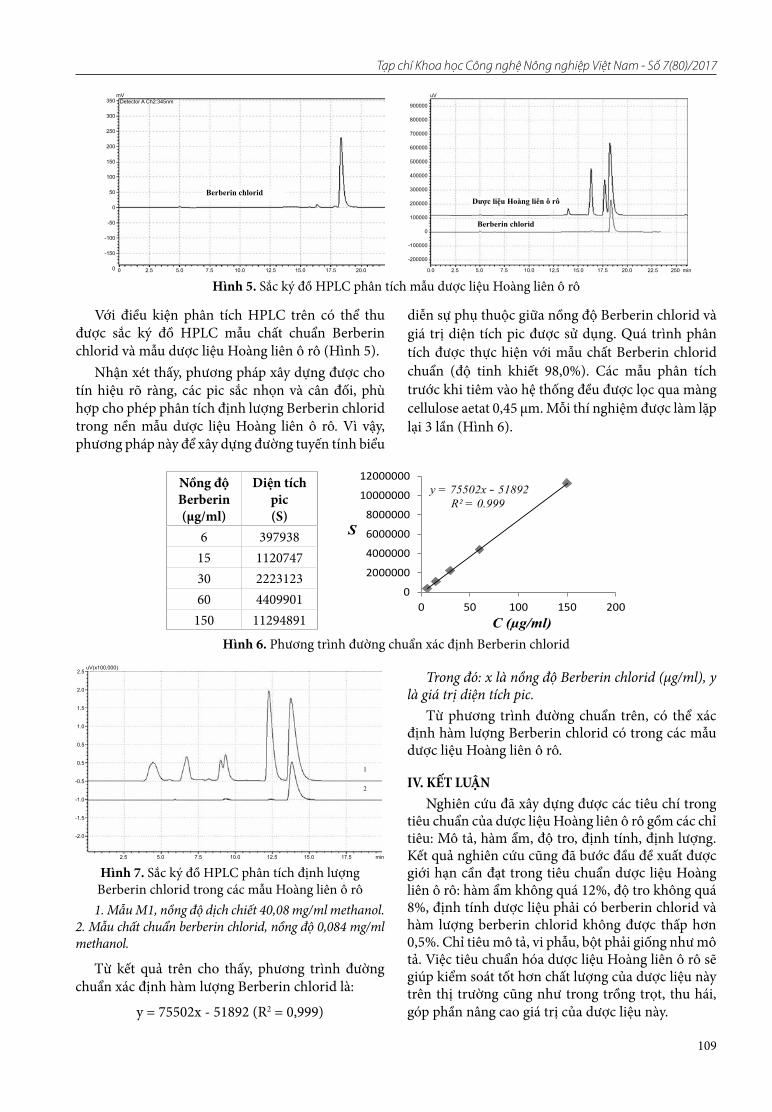
109
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Hình 6. Phương trình đường chuẩn xác định Berberin chlorid
Hình 5. Sắc ký đồ HPLC phân tích mẫu dược liệu Hoàng liên ô rô
Với điều kiện phân tích HPLC trên có thể thu được sắc ký đồ HPLC mẫu chất chuẩn Berberin chlorid và mẫu dược liệu Hoàng liên ô rô (Hình 5).
Nhận xét thấy, phương pháp xây dựng được cho tín hiệu rõ ràng, các pic sắc nhọn và cân đối, phù hợp cho phép phân tích định lượng Berberin chlorid trong nền mẫu dược liệu Hoàng liên ô rô. Vì vậy, phương pháp này để xây dựng đường tuyến tính biểu
diễn sự phụ thuộc giữa nồng độ Berberin chlorid và giá trị diện tích pic được sử dụng. Quá trình phân tích được thực hiện với mẫu chất Berberin chlorid chuẩn (độ tinh khiết 98,0%). Các mẫu phân tích trước khi tiêm vào hệ thống đều được lọc qua màng cellulose aetat 0,45 μm. Mỗi thí nghiệm được làm lặp lại 3 lần (Hình 6).
y = 75502x - 51892R² = 0.999
0
2000000
4000000
6000000
8000000
10000000
12000000
0 50 100 150 200
S
C (µg/ml)
Nông độ Berberin (µg/ml)
Diện tích pic (S)
6 39793815 112074730 222312360 4409901
150 11294891
Hình 7. Sắc ký đồ HPLC phân tích định lượng Berberin chlorid trong các mẫu Hoàng liên ô rô 1. Mẫu M1, nồng độ dịch chiết 40,08 mg/ml methanol.
2. Mẫu chất chuẩn berberin chlorid, nồng độ 0,084 mg/ml methanol.
Từ kết quả trên cho thấy, phương trình đường chuẩn xác định hàm lượng Berberin chlorid là:
y = 75502x - 51892 (R2 = 0,999)
Trong đó: x là nồng độ Berberin chlorid (µg/ml), y là giá trị diện tích pic.
Từ phương trình đường chuẩn trên, có thể xác định hàm lượng Berberin chlorid có trong các mẫu dược liệu Hoàng liên ô rô.
IV. KẾT LUẬN Nghiên cứu đã xây dựng được các tiêu chí trong
tiêu chuẩn của dược liệu Hoàng liên ô rô gồm các chỉ tiêu: Mô tả, hàm ẩm, độ tro, định tính, định lượng. Kết quả nghiên cứu cũng đã bước đầu đề xuất được giới hạn cần đạt trong tiêu chuẩn dược liệu Hoàng liên ô rô: hàm ẩm không quá 12%, độ tro không quá 8%, định tính dược liệu phải có berberin chlorid và hàm lượng berberin chlorid không được thấp hơn 0,5%. Chỉ tiêu mô tả, vi phẫu, bột phải giống như mô tả. Việc tiêu chuẩn hóa dược liệu Hoàng liên ô rô sẽ giúp kiểm soát tốt hơn chất lượng của dược liệu này trên thị trường cũng như trong trồng trọt, thu hái, góp phần nâng cao giá trị của dược liệu này.
350
300
250
200
150
100
50
0
-50
-100
-150
0
mVDetector A Ch2:345nm
Berberin chlorid
0 2.5 5.0 7.5 10.0 12.5 15.0 17.5 20.0
uV
Berberin chlorid
Dược liệu Hoàng liên ô rô
0.0 2.5 5.0 7.5 10.0 12.5 15.0 17.5 20.0 22.5 250 min
900000
800000
700000
600000
500000
400000
300000
200000
100000
0
-100000
-200000
2.5
2.0
1.5
1.0
0.5
0.5
-0.5
-1.0
-1.5
-2.0
uV(x100,000)
1
2
2.5 5.0 7.5 10.0 12.5 15.0 17.5 min

110
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
TÀI LIỆU THAM KHẢOĐỗ Huy Bích, Đặng Quang Chung, Bùi Xuân Chương,
Nguyên Thương Dong, Đỗ Trung Đàm, Phạm Văn Hiển, Vũ Ngọc Lộ, Phạm Duy Mai, Phạm Kim Mãn, Đoàn Thi Nhu, Nguyên Tập, Trần Toàn, 2006. Cây thuốc và động vật làm thuốc ở Việt Nam, tập I. NXB Khoa học và kỹ thuật, tr. 956.
Bộ Giáo dục - Đào tạo và Bộ Y tế, 2006. Nghiên cứu thuốc từ thảo dược. Nhà xuất bản Khoa học và Kỹ thuật, tr. 33-111.
Bộ Y tế, 2009. Dược điển Việt Nam IV. Nhà xuất bản Y học, trang 783.
Cao Thùy Hương, Phan Văn Đệ, Trần Công Luận, 2009. Nghiên cứu đặc điểm vi học và thành phần hóa học trong cây Hoàng liên ô rô Mahonia nepalensis DC. Tạp chí Dược liệu, 14(2): 99-103.
Đỗ Tất Lơi, 2001. Những cây thuốc và vị thuốc Việt Nam. NXB Y học, trang 192.
Shion-Jane Lin, Hshinn-Hshiung Tseng, Kuo-Ching Wen, 1996. Tsi-Tee Suen, Determination of gentiopicroside, mangiferin, palmatine, berberine, baicalin, wogonin and glycyrrhizin in the traditional Chinese medicinal preparation Sann-Joong-Kuey-Jian-Tang by high-performance liquid chromatography. Journal of Chromatography A, 730: 17-23.
Establishment of baseline standards for medicinal herb (Mahonia nepalesis DC)Nguyen Quang Vinh, Nguyen Thi Phuong, Bui Tuan Anh,
Tran Van Tu, Vu Thi Hai, Le Huy CongAbstractThe baseline standards of this medicinal herb were established based on: Morpological characterization, microsurgery, powder, moisture, total ash, impurities. The research results showed that medicinal herb (Mahonia nepalesis DC) had cylinder shape, light orange color with the diameter of about 0.5-2 cm, rough bark. Vascular microsurgery indicated that the vascular containing 1-2 layers of dense polygons, multi-edged packed together, with fibrous tissue and hard tissue. The libe area was thick and the intestine rooted from libe heading to the end of the pulleys, the libre intestinal flux containing calcium oxalate pronged which are prismatic and crystals shapes. The soft tissue of the intestine occupied most of the body’s surface, consisting of thick polygonal cells that partially encode cellulose. Moisture content is not greater than 12%, ash content is not higher than 8%, qualitative medicinal products must contain berberine chlorid and the content of berberine chlorid should not be lower than 0.5%. This research contributes to good quality control and enhances the value of Mahonia nepalesis DC.Key words: Mahonia nepalesis DC, medicinal baseline standards, characterization, microsurgery
Ngày nhận bài: 18/6/2017Ngày phản biện: 29/6/2017
Người phản biện: PGS. TS. Nguyễn Thị Bích ThuNgày duyệt đăng: 27/7/2017
I. ĐẶT VẤN ĐỀNhãn có tên khoa học là Dimocarpus longan,
là loài cây ăn quả có giá trị dinh dưỡng cao. Trong 100 g cùi nhãn tươi cung cấp 48 kcal, có chứa 10,9 g
carbohydrate, 58 mg vitamin C, ngoài ra còn có vitamin B1, B2, B3, canxi, sắt, kẽm, magie... đều là những chất cần thiết cho nhu cầu dinh dưỡng của con người (Nguyễn Công Khẩn và ctv., 2007).
1 Học viện Nông nghiệp Việt Nam
BƯỚC ĐẦU SỬ DỤNG NANO BẠC TRONG PHÒNG TRỪ BỆNH THỐI NÂU DO VI KHUẨN Gluconobacter frateurii GÂY RA TRÊN QUẢ NHÃN
Đồng Huy Giới1
TÓM TẮTNghiên cứu này đã tiến hành đánh giá khả năng ức chế vi khuẩn Gluconobacter frateurii và khả năng phòng trừ
bệnh thối nâu trên quả nhãn của dung dịch nano bạc. Kết quả đã xác định được: (i) Nồng độ nano bạc 10 ppm (với thời gian tiếp xúc 45 phút) và 7,5 ppm (với thời gian tiếp xúc 60 phút và 75 phút) cho hiệu quả ức chế vi khuẩn Gluconobacter frateurii đạt 100%; (ii) Nồng độ nano bạc 20 ppm cho hiệu quả phòng trừ bệnh thối nâu trên quả nhãn là tốt nhất, không có dấu hiệu của bệnh ở cả 2 phương pháp phòng và trừ bệnh sau 48 giờ lây nhiễm.
Từ khóa: Vi khuẩn Gluconobacter frateurii, bệnh thối nâu trên quả nhãn, nano bạc

111
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Cũng giống như những cây trồng khác, nhãn thường bị nhiều loại bệnh hại tấn công, trong đó bệnh thối nâu do vi khuẩn thuộc chi Gluconobacter gây ra là một loại bệnh phổ biến và làm ảnh hưởng nghiêm trọng đến chất lượng, mẫu mã và năng suất của quả. Hà Viết Cường và Trần Thị Định (2016) cũng đã ghi nhận vi khuẩn Gluconobacter oxydans là nguyên nhân chính gây ra bệnh thối nâu trên quả vải.
Biện pháp phổ biến đang được sử dụng để phòng trừ bệnh thối nâu trên nhãn là dùng thuốc bảo vệ thực vật, tuy nhiên hiệu quả thu được không cao. Bên cạnh đó, việc lạm dụng thuốc bảo vệ thực vật sẽ làm ảnh hưởng đến chất lượng quả nhãn, ảnh hưởng đến sức khỏe người tiêu dùng và làm ô nhiễm môi trường, đặc biệt dễ để lại dư lượng trong sản phẩm, ảnh hưởng đến xuất khẩu mặt hàng quả nhãn tươi ra các thị trường trên thế giới, đặc biệt là các thị trường lớn như EU, Nhật, Mỹ… (Trần Thế Tục, 2002).
Trong những năm gần đây, chế phẩm nano đang được sử dụng ngày càng nhiều trong trồng trọt giúp làm tăng năng suất, chất lượng nông sản, đảm bảo sự phát triển một nền nông nghiệp sạch, an toàn, hiệu quả và thân thiện với môi trường (Rostami and Shahsavar, 2012). Mặt khác, nhiều nghiên cứu đã chứng minh rằng vật liệu nano mà đặc biệt là nano bạc có khả năng diệt khuẩn một cách hiệu quả (Shokri et al., 2015). Chính vì vậy, việc nghiên cứu sử dụng nano bạc trong phòng trừ bệnh ở thực vật sẽ mang lại một giải pháp mới an toàn với con người, hiệu quả và thân thiện với môi trường.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu- Vi khuẩn Gluconobacter frateurii được cung cấp
bởi Trung tâm Bệnh cây nhiệt đới, Học viện Nông nghiệp Việt Nam.
- Chế phẩm nano bạc kích thước 15 - 20 nm được điều chế tại Bộ môn Sinh học, khoa Công nghệ sinh học, Học viện Nông nghiệp Việt Nam.
3.3. Phương pháp nghiên cứu
2.2.1. Phương pháp đánh giá khả năng ức chế vi khuẩn Gluconobacter frateurii của dung dịch nano bạc trong điều kiện in vitro
- Trộn vi khuẩn G. frateurii với dung dịch nano bạc ở 5 nồng độ khác nhau (2,5; 5,0; 7,5; 10,0 và 12,5 ppm) với 3 mốc thời gian là 45 phút, 60 phút và 75 phút. Cấy 50µl hỗn hợp dung dịch vi khuẩn-nano bạc trên môi trường LB đặc, nuôi ở 280C, sau
24 giờ đánh giá khả năng ức vi khuẩn G. frateurii của dung dịch nano bạc theo công thức Antibacterial efficacy (ABE):
ABE (%) = (Vc – Vt)/Vc ˟ 100%Trong đó: ABE (%) là hiệu quả ức chế tính theo %;
Vc: là số lượng khuẩn lạc trung bình ở công thức không bổ sung nano bạc; Vt: là số khuẩn lạc trung bình ở công thức thí nghiệm.
- Thí nghiệm lặp lại 3 lần, mỗi lần 3 đĩa/công thức.- Môi trường Luria Bertani (LB) đặc: Thành phần
gồm 10g/l Pepton, 5g/l cao nấm men, 10g/l NaCl, 20g/l agar, pH = 7. Hòa tan thành phần, chỉnh pH theo yêu cầu sau đó hấp khử trùng ở nhiệt độ 1210C, áp suất 1atm trong 15 phút, để nguội đến 50 - 600C, đổ vào đĩa petri.
2.2.2. Phương pháp đánh giá khả năng phòng trừ bệnh thối nâu trên quả nhãn do vi khuẩn Gluconobacter frateurii của chế phẩm nano bạc trong điều kiện in vitroa) Thí nghiệm đánh giá khả năng phòng bệnh
Chọn quả nhãn khỏe, không bị bệnh, rửa sạch dưới vòi nước trong vòng 1 - 2 phút để loại bỏ bớt bụi bẩn và nguồn bệnh nếu có. Rửa lại với nước cất vô trùng. Ngâm trong cồn 70o trong 30 giây, sau đó để khô tự nhiên. Đục 9 lỗ nhỏ trên quả nhãn bằng kim đã hấp khử trùng, sau đó ngâm quả nhãn vào dung dịch nano bạc, lắc đều trong thời gian 5 phút, vớt ra cho vào hộp vô trùng. Nhỏ 20 μl dịch khuẩn (nồng độ khuẩn 106 CFU) lên quả nhãn (chỗ đã đục lỗ), để ở nhiệt độ phòng (28 - 320C) và theo dõi sự phát triển của bệnh (màu sắc và kích thước vết bệnh) trên vỏ quả sau 48 giờ.b) Thí nghiệm đánh giá khả năng trừ bệnh
Chọn quả nhãn khỏe, không bị bệnh, rửa sạch dưới vòi nước trong vòng 1 - 2 phút để loại bỏ bớt bụi bẩn và nguồn bệnh nếu có. Rửa lại với nước cất vô trùng. Ngâm trong cồn 70o trong 30 giây, sau đó để khô tự nhiên. Đục 9 lỗ nhỏ trên quả nhãn bằng kim đã hấp khử trùng, sau đó nhỏ 20 μl dịch khuẩn (nồng độ khuẩn 106 CFU) lên quả nhãn (chỗ đã đục lỗ), để ở nhiệt độ phòng trong 1 giờ. Ngâm quả nhãn vào dung dịch nano bạc, lắc đều trong thời gian 5 phút, sau đó cho vào hộp vô trùng và để trong điều kiện nhiệt độ phòng. Theo dõi sự phát triển của bệnh trên quả sau 48 giờ.
2.3. Đia điểm và thời gian nghiên cứuNghiên cứu được thực hiện từ tháng 1 đến tháng
12 năm 2016 tại Khoa Công nghệ sinh học - Học viện Nông nghiệp Việt Nam.

112
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Kết quả đánh giá khả năng ức chế vi khuẩn Gluconobacter frateurii của dung dich nano bạc bằng phương pháp cấy trộn trực tiếp
Trong thí nghiệm này, dịch khuẩn được trộn trực
tiếp với dung dịch nano bạc ở các nồng độ 2,5 ppm; 5 ppm; 7,5 ppm; 10 ppm và 12,5 ppm trong 45 phút, 60 phút và 75 phút. Sau đó, dung dịch vi khuẩn được cấy lên môi trường LB đặc, nuôi ở 280C sau 24 giờ tiến hành quan sát và đếm số khuẩn lạc. Kết quả được trình bày ở bảng 1, hình 1 và hình 2.
Từ kết quả bảng 1 cho thấy, dung dịch nano bạc ở tất cả các nồng độ thí nghiệm đều có khả năng ức chế vi khuẩn G. frateurii gây thối nâu trên nhãn. Với thời gian xử lý 45 phút, hiệu quả ức chế đạt thấp nhất là 61,92% (nồng độ nano bạc là 2,5 ppm) và hiệu quả ức chế đạt 100% khi nồng độ nano bạc từ 10 ppm trở lên. Với thời gian xử lý là 60 phút và 75 phút, hiệu quả ức chế đạt thấp nhất tương ứng là 77,59% và 79,19% (nồng độ nano bạc là 2,5 ppm) và hiệu quả ức chế đạt 100% khi nồng độ nano bạc từ 7,5 ppm trở lên, không có sự sai khác về hiệu quả ức chế vi khuẩn khi xử lí ở thời gian 60 phút và 75 phút. Kết quả thu được khá tương đồng với kết quả nghiên cứu của Siddhartha và cộng tác viên (2007) khi tiến hành thí nghiệm đánh giá khả năng ức chế 4 loại vi khuẩn là E. coli, E. coli kháng ampicillin, Staphylococcus aureus và Salmonella typhus của dung dịch nano bạc với các nồng độ 5 ppm, 10 ppm, 25 ppm và thời gian tiếp xúc 60 phút. Kết quả nghiên cứu chỉ ra rằng khả năng ức chế của nano bạc với E. coli là 60% ở nồng độ 5 ppm, 90% ở nồng độ 10 ppm và 100% ở nồng độ 25 ppm. Với E. coli kháng ampicillin và Salmonella typhus với nồng độ nano bạc từ 10 ppm cho hiệu quả ức chế là 70 - 75% và trên 25 ppm cho hiệu quả ức chế đạt 100%. Fatemeh và cộng tác viên (2013) cũng đã thử nghiệm ảnh hưởng của thời gian tiếp xúc đến khả năng ức chế vi khuẩn của nano bạc. Các tác giả tiến hành với các nồng độ nano bạc là 100 ppm, 200 ppm và 300 ppm
trên 2 loại vi khuẩn là Pseudomonas aeruginosa và Staphylococcus aureus trong các khoảng thời gian xử lý là 5 phút, 15 phút, 30 phút và 60 phút. Kết quả đối với Pseudomonas aeruginosa, với thời gian xử lý là 5 phút số lượng khuẩn lạc mọc trên đĩa ở cả 3 nồng độ đều trên 35 khuẩn lạc, nhưng khi tăng thời gian xử lý lên 15 phút, 30 phút, 60 phút thì cho kết quả 0 khuẩn lạc mọc. Còn với Staphylococcus aureus với thời gian xử lý là 5 phút số lượng khuẩn lạc ở cả ba nồng độ dao động từ 12 - 14 khuẩn lạc, tuy nhiên phải tăng thời gian xử lý lên 60 phút mới cho hiệu suất diệt khuẩn là 100%. Nói cách khác, thời gian xử lý cũng ảnh hưởng đến khả năng kháng khuẩn của nano bạc, trong đó xử lý với thời gian 60 phút sẽ cho hiệu quả vượt trội.
Hình 1. Kết quả sau 24 giờ nuôi cấy khi cho vi khuẩn Gluconobacter frateurii tiếp xúc với nano bạc trong 45 phút
Ghi chú: Hình 1 và 2: ĐC- : nước cất; CT1: 2,5ppm; CT2: 5ppm; CT3: 7,5ppm; CT4: 10ppm; CT5: 12,5ppm; ĐC+: kháng sinh Streptomycin 10 ppm.
Bảng 1. Khả năng ức chế vi khuẩn Gluconobacter frateurii của dung dịch nano bạc bằng phương pháp cấy trộn trực tiếp
Ghi chú: ĐC (-): nước cất; ĐC (+): kháng sinh Streptomycin 10 ppm; các chữ cái khác nhau trong cùng một cột biểu thị sự sai khác có ý nghĩa 95%.
Nông độ nano bạc
(ppm)
Thời gian tiếp xúc45 phút 60 phút 75 phút
Số lượng khuẩn lạc
Hiệu quả ức chế (%)
Số lượng khuẩn lạc
Hiệu quả ức chế (%)
Số lượng khuẩn lạc
Hiệu quả ức chế (%)
ĐC(-) 93,67± 4,22a 0 96,67± 3,56a 0 97,67±4,22a 02,5 35,67 ± 1,78b 61,92 21,67± 1,56b 77,59 20,33± 1,56b 79,195 15,33 ± 1,78c 83,63 8,00 ± 0,67c 91,72 7,00 ± 0,67c 92,83
7,5 6,67 ± 1,11d 92,88 0 100 0 10010 0 100 0 100 0 100
12,5 0 100 0 100 0 100ĐC (+) 0 100 0 100 0 100LSD0,05 6,23 7,31 8,16

113
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Hình 2. Kết quả sau 24 giờ nuôi cấy khi cho vi khuẩn Gluconobacter frateurii tiếp xúc với nano bạc trong 60 phút
3.2. Kết quả đánh giá khả năng phòng trừ bệnh thối nâu trên quả nhãn do vi khuẩn Gluconobacter frateurii gây ra của chế phẩm nano bạc trong điều kiện in vitro
Căn cứ vào kết quả thí nghiệm đánh giá khả năng ức chế vi khuẩn G. frateurii của nano bạc trong điều kiện in vitro, tiến hành đánh giá khả năng phòng trừ bệnh thối nâu trên quả nhãn của dung dịch nano bạc ở các nồng độ 5 ppm, 10 ppm, 15 ppm và 20 ppm. Kết quả thu được thể hiện ở hình 3, 4 và bảng 2.
Hình 3. Kết quả đánh giá khả năng phòng bệnh thối nâu trên nhãn của dung dịch nano bạc
theo dõi sau 48 h lây nhiễmGhi chú: ĐC: Quả nhãn không lây nhiễm vi khuẩn
G. Frateurii
Hình 4. Kết quả thí nghiệm đánh giá khả năng trừ bệnh thối nâu trên nhãn của dung dịch nano bạc
theo dõi sau 48 h lây nhiễmGhi chú: ĐC: Quả nhãn không lây nhiễm vi khuẩn
G. frateurii
Bảng 2. Kích thước vết bệnh trên vỏ quả nhãn sau 48 h lây nhiễm
Từ kết quả quan sát sự biểu hiện bệnh và kích thước vết bệnh cho thấy: Tất cả các nồng độ nano trong thí nghiệm đều làm giảm mức độ biểu hiện bệnh so với công thức không sử dụng nano. Biểu hiện cụ thể là cùi nhãn ít bị tổn thương hơn, vết bệnh trên vỏ quả nhãn nhạt màu hơn và kích thước vết bệnh nhỏ hơn. Đặc biệt, ở cả hai phương pháp phòng và trừ bệnh, khi xử lí nano bạc với nồng độ 20 ppm thì không thấy bất kỳ dấu hiệu nào của bệnh thối nâu, cùi nhãn và vỏ quả nhãn hầu như không thay đổi sau 48 giờ lây nhiễm.
IV. KẾT LUẬN Ở thí nghiệm đánh giá khả năng ức chế vi khuẩn
Gluconobacter frateurii của dung dịch nano bạc trong điều kiện in vitro, tất cả các nồng độ nano bạc trong thí nghiệm đều có khả năng ức chế vi khuẩn Gluconobacter frateurii gây bệnh thối nâu trên nhãn. Hiệu quả ức chế vi khuẩn đạt 100% ở nồng độ 10 ppm (thời gian tiếp xúc 45 phút) và 7,5 ppm (thời gian tiếp xúc 60 phút và 75 phút).
Nồng độ 20 ppm nano bạc cho hiệu quả phòng trừ bệnh thối nâu trên quả nhãn là tốt nhất, không quan sát thấy sự xuất hiện của bệnh sau 48 giờ lây nhiễm cả trên vỏ quả và thịt quả bên trong.
LỜI CẢM ƠNTác giả xin chân thành cảm ơn PGS. TS. Hà Viết
Cường, Trung tâm Bệnh cây nhiệt đới, Học viện Nông nghiệp Việt Nam đã cung cấp nguồn vi khuẩn Gluconobacter frateurii cho nghiên cứu này.
TÀI LIỆU THAM KHẢOHà Viết Cường, Trần Thi Đinh, 2016. Xác định loài
nấm mốc và vi khuẩn gây bệnh sau thu hoạch trên vải và phương pháp phòng trừ. Tạp chí Khoa học Nông nghiệp Việt Nam, 14 (4): 635-644.
0 ppm 5 ppm 10 ppm
15 ppm 20 ppm ĐC
0 ppm 5 ppm 10 ppm
15 ppm 20 ppm ĐC
Nông độ nano bạc
(ppm)
Kích thước vết bệnh (cm)
Thí nghiệm phòng bệnh
Thí nghiệm trừ bệnh
0 1,13 1,25
5 0,63 0,92
10 0,36 0,77
15 0,12 0,46
20 0,0 0,0

114
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Nguyên Công Khẩn, Nguyên Thi Lâm, Hà Thi Anh Đào, Lê Hông Dũng, Lê Bạch Mai, Nguyên Văn Sĩ, 2007. Bảng thành phần thực phẩm Việt Nam. Nhà xuất bản Y học.
Trần Thế Tục, 2002. Hỏi đáp về nhãn - vải. Nhà xuất bản Nông nghiệp.
Fatemeh Ahmadi, Sara Abolghasemi, Najme Parhizgari, Fatemeh Moradpour, 2013. Effect of Silver Nanoparticles on Common Bacteria in Hospital Surfaces. Jundishapur Journal Mirobiol, Vol. 6, No. 3: 209-214.
Rostami A.A. and Shahsavar A., 2012. Nano-Silver Particles the in vitro Contaminations of Olive
‘Mission’ Explant. Asian Journal of Plant Science, Vol. 8, No. 7: 505-509.
Shokri S., A. Babaei, M. Ahmadian, M.M. Arab, S. Hessami, 2015. The effects of different concentrations of nano - silver on elimination of bacterial contaminations and phenolic exudation of rose (Rosa hybrida L.) in vitro culture. International Society for Horticultural Science,Vol. 3, No.1: 50-54.
Siddhartha S., Tanmay B., Arnab R., Gajendra S., P. Ramachandrarao and Debabrata D., 2007. Characterization of enhanced antibacterial effects of novel silver nanoparticles. Nanotechnology, Vol. 18, No. 22: 1-9.
Preliminary results of using nano silver in controlling brown rot (Gluconobacter frateurii) on longan fruit
Dong Huy Gioi
AbstractIn this study, we have evaluated the inhibition effect of silver nanoparticles on Gluconobacter frateurii bacteria and the ability to prevent brown rot on the longan fruit of silver nanoparticles. The results showed that: (i) silver nanoparticles at a concentration of 10 ppm (with a treatment time of 45 minutes) and 7.5 ppm (with a treatment time of 60 minutes and 75 minutes) achieved 100% inhibitory effect on Gluconobacter frateurii bacteria; (ii) The concentration of 20 ppm nano silver provided the best disease control for brown rot on the longan fruit. No disease appeared after 48 hours of infection.Key words: Gluconobacter frateurii bacteria, brown rot on the longan fruit, silver nanoparticles
Ngày nhận bài: 6/7/2017Người phản biện: TS. Lê Ngọc Anh
Ngày phản biện: 12/7/2017Ngày duyệt đăng: 27/7/2017
1 Bộ môn Kỹ thuật nuôi thủy sản nước ngọt, Khoa Thủy sản, Trường Đại học Cần Thơ
ẢNH HƯỞNG CỦA THỨC ĂN ĐẾN SỰ THÀNH THỤC CỦA CÁ HEO (Botia modesta Bleeker, 1865)
Nguyễn Thanh Hiệu1, Dương Nhựt Long1, Lam Mỹ Lan1
TÓM TẮT Thí nghiệm nuôi vỗ thành thục cá heo được thực hiện với 4 nghiệm thức (NT) thức ăn NT1: tép trấu; NT2: cá
tạp biển, NT3: 50% cá tạp biển + 50% thức ăn công nghiệp 40% đạm và NT4 là thức ăn công nghiệp 40% đạm. Sau 4 tháng nuôi vỗ kết quả đạt được như sau: Nhiệt độ, pH và ôxy trong ao nuôi vỗ luôn nằm trong khoảng thích hợp để cá thành thục sinh dục. Hệ số thành thục của cá cái (2,7 ± 0,99%) cao nhất ở NT1 và khác biệt có ý nghĩa (P<0,05) so với hệ số thành thục của cá cái (1,44 ± 0,84%) ở NT4. Tỷ lệ cá thành thục của NT1 cao nhất (66,67 ± 19,52) và thấp nhất là NT4 (44,44 ± 8,61). Sức sinh sản của cá heo ở NT1 và NT2 khác biệt có ý nghĩa (P<0,05) so với NT3 và NT4. Hàm lượng Vitellogenine (Vg) trong buồng trứng giai đoạn IV ở nghiệm thức NT1, NT2 và NT3 khác biệt có ý nghĩa (P<0,05) so với NT4. Hàm lượng protein trong cơ thấp nhất ở tháng 4 (2,37 - 4,87 mg protein/g mẫu tươi). Protein trong gan lớn nhất (6,88 - 10,85 mg protein/g mẫu tươi) ở tháng 4. Protein trong TSD ở tháng 4 khác biệt không có ý nghĩa (P>0,05) giữa các nghiệm thức.
Từ khóa: Cá heo, thành thục, loại thức ăn khác nhau

115
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
I. ĐẶT VẤN ĐỀTheo Rainboth (1996) cá heo (Botia modesta
Bleeker, 1865) là 1 trong 8 loài cá nước ngọt thuộc giống Botia phân bố khá phổ biến trong các lưu vực của vùng hạ nguồn sông Cửu Long như Thái Lan, Lào, Campuchia và Đồng Bằng Sông Cửu Long - Việt Nam. Hiện nay cá heo là loài rất có giá trị kinh tế và được nhiều nuôi quan tâm. Mặc dù cá heo là loài có giá trị kinh tế nhưng nguồn lợi cá heo thương phẩm đang tiêu thụ trên thị trường chủ yếu là được đánh bắt từ tự nhiên (Trương Thủ Khoa và Trần Thị Thu Hương, 1993). Bước đầu đã nghiên cứu thành công về sản xuất giống cá heo (Nguyễn Thanh Hiệu và ctv., 2015). Để hoàn thiện qui trình kỹ thuật sản xuất giống cá heo thì việc nghiên cứu nuôi vỗ thành thục sinh dục cá heo nhằm xác định loại thức ăn thích hợp trong nuôi vỗ, làm cơ sở khoa học cho việc nghiên cứu kỹ thuật sản xuất giống cá heo là cần thiết.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cứu Cá heo bố mẹ có khối lượng trung 24,96 ±
2,37 g/con.Giai lưới, cân, nhiệt kế, thau, xô, kéo, hóa chất.Thức ăn công nghiệp, tép trấu, cá nục tròn.
2.2. Phương pháp nghiên cứu
2.2.1. Thí nghiệm nuôi vỗ cá heo với các loại thức ăn khác nhaua) Hệ thống thí nghiệm
Thí nghiệm nuôi vỗ thành thục sinh dục cá heo được bố trí trong giai luới có diện tích là 1 m2/giai (theo qui cách 1 ˟ 1 ˟ 2 m) đặt trong ao đất tại Trại Cá Thực Nghiệm Khoa Thủy Sản - Trường Đại Học Cần Thơ. Ao nuôi vỗ cá heo có diện tích khoảng 600 m2, mực nước trong ao dao động từ 1 - 1,2 m. Ao trước khi bố trí nuôi vỗ được cải tạo như sên vét lớp bùn đáy, bón vôi, diệt tạp, phơi ao và lọc nước vào ao.
Hình 1. Hệ thống thí nghiệm (A) và cá heo bố mẹ (B)
b) Cá thí nghiệmCá heo bố mẹ để bố trí thí nghiệm có khối lượng
trung bình khoảng 24,96 ± 2,37 g/con được mua từ các bè nuôi ở huyện An Phú, tỉnh An Giang. Cá được thuần dưỡng 60 ngày, trong giai (2 ˟ 2 ˟ 1,5 m) đặt trong ao đất, bằng thức ăn công nghiệp trước khi bố trí thí nghiệm là 60 ngày. Mật độ nuôi vỗ là 1,5 kg/m3 và tỷ lệ đực cái là 1 : 1.c) Bố trí thí nghiệm
Thí nghiệm được bố trí hoàn toàn ngẫu nhiên với 4 nghiệm thức (NT) thức ăn khác nhau, mỗi nghiệm thức được lập lại 3 lần (Bảng 1).d) Thức ăn nuôi vỗ
Tép trấu Macrobrachium lanchesteri, cá nục tròn (Decapterus punctatus) được mua từ các chợ ở thành phố Cần Thơ tiến hành loại bỏ cá tạp, chất vẫn. Cá
nục tròn, tép trấu được rửa bằng nước máy từ 2 - 3 lần và được băm nhỏ trước khi cho ăn. Thức ăn viên hiệu Aquafish có hàm lượng đạm 40% được trộn với cá tạp xay theo Tỷ lệ 1 : 1 (khối lượng tươi) trước khi cho ăn (Bảng 2).
Bảng 1. Thí nghiệm nuôi vỗ cá heo với các loại thức ăn khác nhau
Nghiệm thức (NT)
Mật độ (kg/m3) Thức ăn
NT1 1,5 Tép trấu
NT2 1,5 Cá biển (Cá nục tròn)
NT3 1,5 Thức ăn công nghiệp (TACN)
NT4 1,5 50% cá tạp biển + 50% TACN

116
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
Bảng 2. Thành phần hóa học của thức ăn nuôi vỗ cá heo (tính theo khối lượng khô)
e) Chăm sóc cá nuôiCá được cho ăn thỏa mãn theo nhu cầu (1 - 3%
khối lượng/ngày) và được cho ăn 2 lần/ngày, sáng lúc 7 giờ và chiều lúc 17 giờ. Để tăng cường ôxy cho cá trong các giai thí nghiệm được lắp đặt hệ thống sục khí. Nước trong ao nuôi vỗ được thay đổi 2 lần/tháng, mỗi lần thay 30% nước trong ao.
2.2.2. Các chỉ tiêu theo dõi - Các chỉ tiêu về môi trường nước: Một số chỉ tiêu
theo dõi về môi trường nước bao gồm nhiệt độ, ôxy, pH được kiểm tra 2 lần/ngày (7h00 và 14h00) bằng máy hiệu YSI 95 do Mỹ sản xuất.
- Các chỉ tiêu về sinh sản cá: Định kỳ 1 lần/tháng thu ngẫu nhiên mỗi nghiệm thức 21 con cá heo bố mẹ, để mổ và xác định các chỉ tiêu sinh sản.
+ Xác định sự thành thục sinh dục (gonadosomatic index - GSI). GSI của cá heo được theo dõi từng tháng và dựa vào công thức của Biswas (1993).
GSI (%) = (GW/BW) ˟ 100Trong đó: GW: là khối lượng tuyến sinh dục cá
(Gonad Weight, g); BW: là khối lượng toàn thân cá (Body Weight, g)
+ Xác định sự thành thục của cá theo thời gianĐo đường kính trứng bằng kính lúp điện tử có
trắc vi thị kính và xác định các giai đoạn của tế bào trứng theo (Pravdin, 1973) và kết hợp với phương pháp của Nikolsky (1963) áp dụng cho nghiên cứu bậc thang thành thục sinh dục trong điều kiện ngoài trời.
Tỷ lệ thành thục (%): 100 ˟ (số cá thành thục)/(số cá được khảo sát)
Độ béo Fulton (F) được xác định theo công thức:
F = PLo3
Độ béo Clark (C) được xác định theo công thức:
C = PoLo3
Trong đó: Po: Khối lượng cá bỏ nội quan; P: Khối lượng của cá; Lo: Chiều dài chuẩn của cá.
+ Xác định sức sinh sản tuyệt đối và sức sinh sản tương đối
Sức sinh sản tuyệt đối (Absolute Fecundity, AF) là số lượng trứng trong buồng trứng của cá cái và được tính theo công thức của Banegal (1967).
AF (trứng/cá thể cái) = n ˟ G/gTrong đó: AF: Sức sinh sản tuyệt đối; G: Khối
lượng buồng trứng (g); g: khối lượng 01 mẫu trứng được lấy đại diện để đếm (g); n: Số lượng trứng có trong 1 mẫu (trứng).
Sức sinh sản tương đối (Relative Fecundity, RF) được xác định bằng số lượng trứng tính trên một đơn vị khối lượng của cá (gram)
- Một số chỉ tiêu sinh hóa liên quan đến sự thành thục sinh dục của cá
Thu mẫu sinh hóa: Định kỳ 30 ngày/lần thu mẫu máu, cơ, gan và tuyến sinh dục (TSD) dùng để phân tích các chỉ tiêu sinh hóa như Vitellogennin (Vg) và hàm lượng protein trong cơ, gan và TSD được phân tích theo phương pháp Lowry và công tác viên, (1951) sử dụng Albumine bovine (BSA, Sigma) làm đường chuẩn.
- Phương pháp phân tích các chỉ tiêu sinh hóa:Đối với cơ, gan và TSD: tiến hành lấy 0,3 g cơ hoặc
gan và sau đó cho vào 3 ml dung dịch phosphate 50 mM KH2PO4/K2HHPO4 50 mM (pH 7,5) để nghiền mẫu. Tiếp theo đem ly tâm 10.000 vòng/phút (10 phút, 4 0C). Sau đó lấy phần nước ở trên đem trữ tủ âm (-800C) đến khi phân tích.
Đối với mẫu máu: Máu được thu từ động mạch lưng bằng kim tiêm, với thể tích từ 0,5 - 1ml rồi cho vào ependoff và sau đó ly tâm lạnh ở 4 oC trong vòng 6 phút (6.000 vòng/phút) sử dụng huyết tương để đo hàm lượng Vg, từ đó xác định mối quan hệ giữa hàm lượng Vg trong huyết tương với các giai đoạn phát triển của buồng trứng. Hàm lượng Vg được xác định bằng phương pháp so màu quang phổ (theo phương pháp ALKALI - LABILE PHOSPHATE dựa trên đường chuẩn phosphate Standard và so màu ở bước sóng 660 nm).
Công thức tính hàm lượng Vg:
Thức ănThành phần (%)
Đạm Béo Tro Ẩm độ
Tép trấu 65,72 9,13 16,65 76,34Cá nục tròn 62,34 17,39 15,82 64,4550% cá nục + 50% TACN 40% Cp 52,3 13,35 12,96 40,30
Thức ăn công nghiệp 40% Cp 42,32 9,55 10,46 11,27
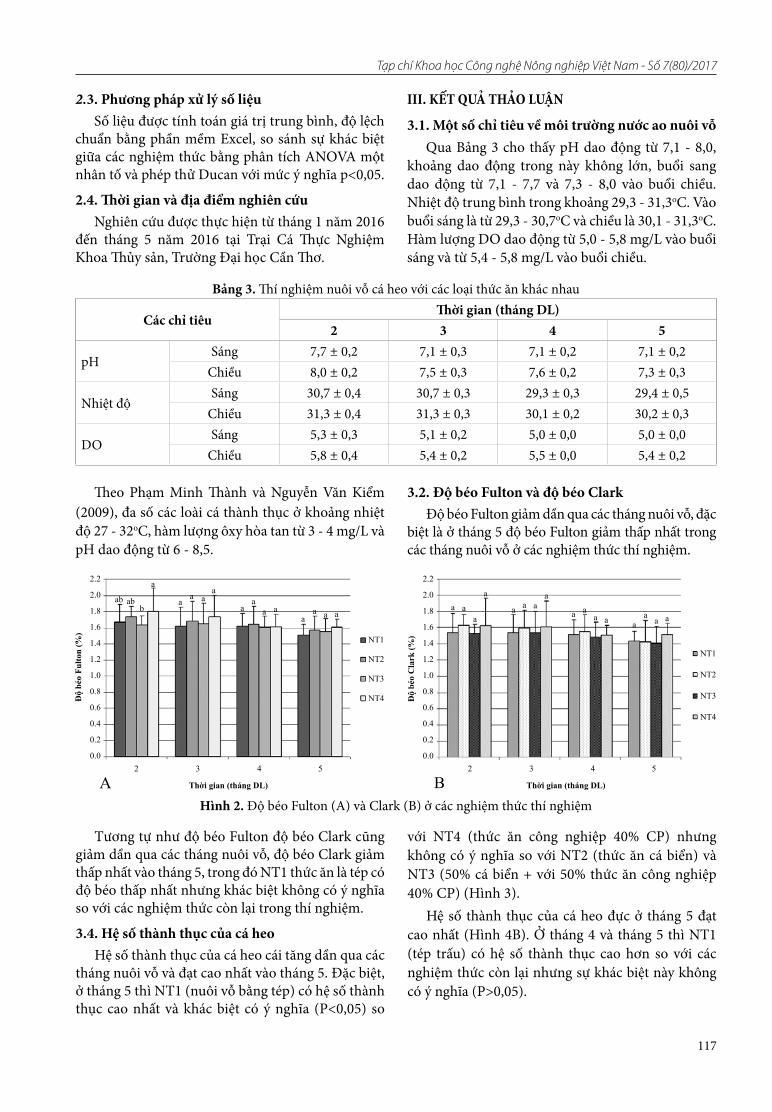
117
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
2.3. Phương pháp xử lý số liệuSố liệu được tính toán giá trị trung bình, độ lệch
chuẩn bằng phần mềm Excel, so sánh sự khác biệt giữa các nghiệm thức bằng phân tích ANOVA một nhân tố và phép thử Ducan với mức ý nghĩa p<0,05.
2.4. Thời gian và đia điểm nghiên cứuNghiên cứu được thực hiện từ tháng 1 năm 2016
đến tháng 5 năm 2016 tại Trại Cá Thực Nghiệm Khoa Thủy sản, Trường Đại học Cần Thơ.
III. KẾT QUẢ THẢO LUẬN
3.1. Một số chỉ tiêu về môi trường nước ao nuôi vỗQua Bảng 3 cho thấy pH dao động từ 7,1 - 8,0,
khoảng dao động trong này không lớn, buổi sang dao động từ 7,1 - 7,7 và 7,3 - 8,0 vào buổi chiều. Nhiệt độ trung bình trong khoảng 29,3 - 31,3oC. Vào buổi sáng là từ 29,3 - 30,7oC và chiều là 30,1 - 31,3oC. Hàm lượng DO dao động từ 5,0 - 5,8 mg/L vào buổi sáng và từ 5,4 - 5,8 mg/L vào buổi chiều.
Bảng 3. Thí nghiệm nuôi vỗ cá heo với các loại thức ăn khác nhau
Hình 2. Độ béo Fulton (A) và Clark (B) ở các nghiệm thức thí nghiệm
Theo Phạm Minh Thành và Nguyễn Văn Kiểm (2009), đa số các loài cá thành thục ở khoảng nhiệt độ 27 - 32oC, hàm lượng ôxy hòa tan từ 3 - 4 mg/L và pH dao động từ 6 - 8,5.
3.2. Độ béo Fulton và độ béo ClarkĐộ béo Fulton giảm dần qua các tháng nuôi vỗ, đặc
biệt là ở tháng 5 độ béo Fulton giảm thấp nhất trong các tháng nuôi vỗ ở các nghiệm thức thí nghiệm.
Các chỉ tiêuThời gian (tháng DL)
2 3 4 5
pHSáng 7,7 ± 0,2 7,1 ± 0,3 7,1 ± 0,2 7,1 ± 0,2
Chiều 8,0 ± 0,2 7,5 ± 0,3 7,6 ± 0,2 7,3 ± 0,3
Nhiệt độSáng 30,7 ± 0,4 30,7 ± 0,3 29,3 ± 0,3 29,4 ± 0,5
Chiều 31,3 ± 0,4 31,3 ± 0,3 30,1 ± 0,2 30,2 ± 0,3
DOSáng 5,3 ± 0,3 5,1 ± 0,2 5,0 ± 0,0 5,0 ± 0,0
Chiều 5,8 ± 0,4 5,4 ± 0,2 5,5 ± 0,0 5,4 ± 0,2
Tương tự như độ béo Fulton độ béo Clark cũng giảm dần qua các tháng nuôi vỗ, độ béo Clark giảm thấp nhất vào tháng 5, trong đó NT1 thức ăn là tép có độ béo thấp nhất nhưng khác biệt không có ý nghĩa so với các nghiệm thức còn lại trong thí nghiệm.
3.4. Hệ số thành thục của cá heoHệ số thành thục của cá heo cái tăng dần qua các
tháng nuôi vỗ và đạt cao nhất vào tháng 5. Đặc biệt, ở tháng 5 thì NT1 (nuôi vỗ bằng tép) có hệ số thành thục cao nhất và khác biệt có ý nghĩa (P<0,05) so
với NT4 (thức ăn công nghiệp 40% CP) nhưng không có ý nghĩa so với NT2 (thức ăn cá biển) và NT3 (50% cá biển + với 50% thức ăn công nghiệp 40% CP) (Hình 3).
Hệ số thành thục của cá heo đực ở tháng 5 đạt cao nhất (Hình 4B). Ở tháng 4 và tháng 5 thì NT1 (tép trấu) có hệ số thành thục cao hơn so với các nghiệm thức còn lại nhưng sự khác biệt này không có ý nghĩa (P>0,05).
Độ
béo
Fulto
n (%
)
2.2
2.0
1.8
1.6
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.0
a
bab ab a
a aa
aa
a aa
a a a
2 3 4 5
NT1
NT2
NT3
NT4
Thời gian (tháng DL)A
Độ
béo
Cla
rk (%
)
Thời gian (tháng DL)B
NT1
NT2
NT3
NT4
2 3 4 5
2.2
2.0
1.8
1.6
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.0
aaa
aaaaa
aaaa
a
aaa

118
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
3.5. Tỷ lệ thành thục của cá heo Tỷ lệ cá heo thành thục tăng dần qua các tháng
nuôi vỗ, ở tháng 4 Tỷ lệ cá heo thành thục dao động từ 27,8 – 52.4%. Ở tháng 5 Tỷ lệ cá thành thục tăng
cao và dao động từ 44,44 – 66.67%, trong đó NT1 (tép trấu) có Tỷ lệ thành thục cao nhất (66.67±19.52%) và khác biệt có ý nghĩa (P,0,05) so với NT4 (thức ăn công nghiệp 40% CP) là 44.44±8.61% (Bảng 4).
Hình 3. Độ béo Fulton (A) và Clark (B) ở các nghiệm thức thí nghiệm
Bảng 4. Tỷ lệ thành thục (%) của cá qua các tháng nuôi vỗ
Bảng 5. Sức sinh sản của cá heo ở các nghiệm thức thí nghiệm
Ghi chú: Bảng 4, 5: Số liệu trong bảng là số liệu trung bình ± độ lệch chuẩn, các số liệu trong cùng một cột có chữ cái khác nhau thì khác biệt có ý nghĩa (P<0,05).
Từ kết quả trên cho thấy thức ăn là tép cho kết quả tốt nhất trong nuôi vỗ thành thục sinh dục cá heo. Ngoài ra, thì cá tạp biển và 50% cá tạp biển + 50% thức ăn công nghiệp 40% CP hoàn toàn có thể thay thế tép trong nuôi vỗ thành thục cá heo.
3.6. Sức sinh sản của cá heo Qua bảng 5 cho thấy sức sinh sản tuyệt đối (AF)
và sức sinh sản tương đối (RF) của cá heo cái ở các
nghiệm thức thí nghiệm có sự khác biệt. Trong đó, nghiệm thức nuôi vỗ bằng tép (NT1) có sức sinh sản cao nhất và khác biệt có ý nghĩa (P<0,05) so với nghiệm thức nuôi vỗ bằng thức ăn công nghiệp 40% CP (NT4), nhưng khác biệt không có ý nghĩa (P>0,05) so với nghiệm thức nuôi vỗ bằng cá tạp biển (NT2) và nghiệm thức 50% cá tạp biển kết hợp với 50% thức ăn công nghiệp (NT3).
Nghiệm thức (NT)Thời gian (tháng DL)
2 3 4 5NT1 23,8±14,7b 38,1±7.4a 52,4±32,2a 66,7±19,5a
NT2 33,3±14,7a 33,3±14,7a 47,6±7,38a 61,9±14,7a
NT3 23,8±19,5b 33,3±14,9a 44,4±8,61a 61,1±8,61a
NT4 19,1±14,7bc 23,8±19,5b 27,8±22,2b 44,4±8,61b
Từ kết quả ở bảng 5 cho thấy, thức ăn nuôi vỗ có ảnh hưởng rất lớn đến sự phát triển của buồng trứng, mặc dù khối lượng cá cái ở các nghiệm thức khác biệt không có ý nghĩa (P>0,05) nhưng khối
lượng buồng trứng ở nghiệm thức nuôi vỗ bằng thức ăn có nguồn gốc động vật (NT1, NT2 và NT3) khác biệt có ý nghĩa (P<0,05) so với nghiệm thức nuôi vỗ bằng thức ăn công nghiệp (NT4). Cá heo là loài có
Nghiệm thức(NT)
Khối lương thân (g)
Khối lương buông trứng (g)
AF(trứng/cá thể)
RF(trứng/kg)
NT1 31,2±3,4a 1,447±0,67a 3.920±1.809a 124.547±51.056a
NT2 30,5±3,6a 1,269±0,54a 3.437±1.454a 111.535±41.685a
NT3 31,2±1,7a 1,085±0,62ab 2.939±1.674ab 95.526±55.517a
NT4 29,9±5,5a 0,531±0,48b 1.438±1.319b 44.314±35.199b
Hệ
số th
ành
thục
cá
cái (
%)
Thời gian (tháng DL)A
NT1
NT2
NT3
NT4
2 3 4 5
3.3
3.0
2.7
2.4
2.1
1.8
1.5
1.2
0.9
0.6
0.3
0.0
aa
a a
a aa
a
aa
a
a
aab
ab
b
Hệ
số th
ành
thục
cá
đực
(%)
Thời gian (tháng DL)B
NT1
NT2
NT3
NT4
2 3 4 5
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0.0
a
a a a a
a
aa
aa a
a
a
aa
a

119
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
tính ăn động vật, thức ăn bao gồm: nhuyễn thể, giáp xác, giun, mùn bã hữu cơ… (Nguyễn Thanh Hiệu và ctv., 2014) là những thức ăn được cá ưa thích. Vì vậy, trong thí nghiệm nuôi vỗ với các loại thức ăn khác nhau, thức ăn có nguồn gốc tươi sống sẽ giúp cá có hệ số thành thục, Tỷ lệ thành thục và sức sinh sản cao hơn so với thức ăn công nghiệp.
3.7. Một số chỉ tiêu sinh hóa liên quan đến sự thành thục sinh dục của cá heo
3.7.1. Hàm lượng Vitellogenin (Vg) trong máu ứng với các giai đoạn phát triển của tuyến sinh dục
Qua thời gian nuôi vỗ cá heo với các loại thức
ăn khác nhau, đã cho thấy sự khác biệt ở các giai đoạn phát triển của buồng trứng, hàm lượng Vg ở các nghiệm thức dao động từ 0,86 - 5,82 µg ALP/mg protein, thấp nhất ở giai đoạn I (dao động từ 0,86 - 3,13 µg ALP/mg protein) và cao nhất ở giai đoạn III (dao động từ 3,71 - 4,83 µg ALP/mg protein). Khi buồng trứng ở giai đoạn III sự khác biệt về hàm lượng Vg ở các nghiệm thức là không có ý nghĩa (P>0,05). Trong khi đó, buồng trứng ở giai đoạn IV sự khác biệt về thức ăn dẫn đến hàm lượng Vg ở các nghiệm thức nuôi vỗ bằng thức ăn có nguồn gốc tươi sống (NT1, NT2 và NT3) khác biệt có ý nghĩa (P< 0,05) so với nghiệm thức ăn công nghiệp (NT4).
Vitellogenin bắt đầu tăng dần và đạt cao nhất khi buồng trứng ở giai đoạn III và sau đó giảm dần ở giai đoạn IV và thấp nhất khi buồng trứng ở giai đoạn I (Hình 5A). Hàm lượng Vg của tinh sào ở các nghiệm thức khác nhau dao động từ 0,16 - 4,77 µg ALP/mg protein, hàm lượng Vg đạt cao nhất khi tinh sào ở giai đoạn III dao động từ 3,08 - 4,77 µg ALP/ml protein và thấp nhất khi tinh sào ở giai đoạn II dao động từ 0,16 - 3,97 µg ALP/mg protein. Hàm lượng Vg trong tinh sào cá heo ở giai đoạn IV giữa các nghiệm thức nuôi vỗ bằng thức ăn có nguồn gốc tươi sống (NT1, NT2 và NT3) khác biệt có ý nghĩa (P<0,05) so với nghiệm thức thức ăn công nghiệp (NT4) (Hình 5B).
3.7.2. Hàm lượng protein trong cơ, gan và tuyến sinh dục của cá heo
Hàm lượng protein trong cơ của cá heo tăng
cao nhất vào tháng 2 (dao động từ 5,64 - 8,20 mg protein/g mẫu tươi) ở các nghiệm thức thí nghiệm và sự khác biệt giữa các nghiệm thức là không có ý nghĩa (P>0,05). Từ tháng 3 đến tháng 5 thì hàm lượng protein trong cơ giảm dần, ở tháng 4 hàm lượng protein trong cơ có sự khác biệt giữa các nghiệm thức với nhau, NT1 (tép trấu) và NT2 (cá tạp biển) khác biệt có ý nghĩa (P<0,05) so với NT3 (50% cá biển + 50% thức ăn công nghiệp 40% Cp) và NT4 (thức ăn công nghiệp 40% Cp).
Ở tháng 2 hàm lượng protein trong gan của cá đạt thấp nhất (3,85 - 5,58 mg protein/g mẫu tươi) ở các nghiệm thức thí nghiệm, do mới nuôi vỗ nên hàm lượng protein trong gan lúc này còn thấp. Hàm lượng protein trong gan của cá heo bắt đầu tăng từ tháng 3 và đạt cao nhất vào tháng 4. Hàm lượng protein trong gan ở tháng 5 (dao động từ 5,97 - 8,15
Hình 4. Hàm lượng Vg trong máu cá cái (A) và cá đực (B) ứng với các giai đoạn của TSD
Hình 5. Hàm lượng protein trong cơ (A), gan (B) và TSD (C) của cá heo
Vg
(µgA
LP/
mg
prot
ein)
Giai đoạn TSD cáiA
NT1
NT2
NT3
NT4
I II III IV
9
8
7
6
5
4
3
2
1
0
a
a
a
a aa
a
a a
a
a
c
ab
b
abV
g (µ
gAL
P/m
g pr
otei
n)
Giai đoạn TSD đựcB
NT1
NT2
NT3
NT4
I II III IV
9
8
7
6
5
4
3
2
1
0
aa
a
aa
a
aa
a
aa a
a
bb
A
NT1
NT2
NT3
NT4
18
16
14
12
10
8
6
4
2
0
Thời gian (tháng DL)
Prot
ein
tron
g cơ
(mg/
g)
2 3 4 5
a
aa
a
aa a a
bb
a
a aa
a
a
B
NT1
NT2
NT3
NT4
18
16
14
12
10
8
6
4
2
0
Prot
ein
tron
g ga
n (m
g/g)
Thời gian (tháng DL)
2 3 4 5
a
aa
a
a
a
a
b
b b
bab
abab
ab ab
C
NT1
NT2
NT3
NT4
18
16
14
12
10
8
6
4
2
0
Prot
ein
tron
g T
SD (m
g/g)
Thời gian (tháng DL)
2 3 4 5
aa
a
aa
aa
ab
ab
ab
a
aa
b
b
c

120
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 7(80)/2017
mg protein/g mẫu tươi) thấp hơn so với tháng 4 (động từ 6,88 - 10,85 mg protein/g mẫu tươi).
Hàm lượng protein trong tuyến sinh dục (TSD) của cá heo tăng cao vào tháng 2 và khác biệt không có ý nghĩa (P>0,05) ở các nghiệm thức. Ở tháng 3 hàm lượng protein ở nghiệm thức nuôi vỗ bằng tép (NT1) thấp nhất và khác biệt có ý nghĩa (P<0,05) so với các nghiệm thức còn lại.
IV. KẾT LUẬN VÀ ĐỀ XUẤT
4.1. Kết luận- Thức ăn tép trấu (NT1) cá có hệ số thành thục
cao nhất (cá cái: 2,7% và cá đực: 0,47%) vào tháng 5. Hệ số thành thục cá cái NT1 cao nhất và khác biệt có ý nghĩa (P<0,05) so với các nghiệm thức còn lại.
- Tỷ lệ thành thục của cá heo NT1 (tép trấu) cao nhất (66.67±19.52%) và khác biệt có ý nghĩa (P<0,05) so với NT4 (44.44±8.61%) thức ăn công nghiệp 40% đạm.
- Thức ăn tép trấu (NT1) và cá tạp biển (NT2) khác biệt có ý nghĩa (P<0,05) so với nghiệm thức chế biến (NT3) và thức ăn công nghiệp 40% đạm (NT4).
- Hàm lượng Vg trong buồng trứng giai đoạn IV ở NT1 (tép trấu), NT2 (cá biển) và NT3 (50% cá biển + 50% thức ăn công nghiệp 40% CP) khác biệt có ý nghĩa (P<0,05) so với nghiệm thức thức ăn công nghiệp 40% CP.
4.2. Đề xuất Tiếp tục nghiên cứu nuôi vỗ thành thục sinh dục
cá heo ở các tháng tiếp theo để làm cơ sở khoa học hoàn thiện qui trình kỹ thuật sản xuất giống cá heo.
TÀI LIỆU THAM KHẢONguyên Thanh Hiệu, Dương Nhựt Long và Lam Mỹ
Lan, 2014. Nghiên cứu một số đặc điểm sinh học của cá heo. Tạp chí Khoa học. Trường Đại học Cần Thơ, 264 - 272.
Nguyên Thanh Hiệu, Dương Nhựt Long và Lương Công Tâm, 2015. Nghiên cứu nuôi vỗ thành thục và kỹ thuật sản xuất giống nhân tạo cá heo (Botia modesta Bleeker, 1865). Báo cáo tổng kết đề tài cấp trường năm 2015. Trường Đại học Cần Thơ.
Trương Thủ Khoa và Trần Thi Thu Hương, 1993. Định loại cá nước ngọt vùng Đồng bằng sông Cửu Long. Trường Đại học Cần Thơ, 360 trang.
Ofxakun và N.A. Buskaia, 1968. Xác định các giai đoạn phát dục và nghiên cứu chu kỳ sinh dục của cá. Bản dịch từ tiếng Nga của Lê Thanh Lựu và Trần Mai Hiên. Nhà xuất bản Nông nghiệp Hà Nội 1982, 46 trang.
Phạm Minh Thành và Nguyên Văn Kiểm, 2009. Sản xuất cá giống. Khoa Thủy sản. Trường Đại học Cần Thơ, 160 trang.
Bagenal T. B., 1967. A short review of the fish fecundity. The Biological Basic of Freshwater Fish Production pp. 89 - 111.
Biswas, S.P., 1993. In Fish Biology. South Asia Publishers. New Delhi. 157 trang.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall R.J., 1951. Protein measurement with the Folin phenol reagent. J. Biol Chem, 193: 265 - 275.
Nikolsky. G. V, 1963. Biology of fish. Academic press. London. 352 p.
Pravdin, I. F, 1973. Hướng dẫn nghiên cứu cá. NXB Khoa học và kỹ thuật Hà Nội. Người dịch Phạm Minh Giang. 276 trang.
Rainboth, W. J, 1996. Fishes of the Cambodian Mekong. FAO Species Identification Field Guide for fishery Purposes. FAO, Rome, 265 p.
Effect of different feed types on sexual maturity of redtail botia Botia modesta Bleeker, 1865)
Nguyen Thanh Hieu, Duong Nhut Long, Lam My LanAbstractThe experiment was carried out with 4 treatments (NT) of NT1 feed: small shrimp; NT2: trash fish, NT3: 50% marine trash fish + 50% commercial pellet feed 40% protein and NT4 are 100% commercial pellet feed 40% protein. After 4 months, the results showed that temperature, pH and oxygen in the culture ponds were always within the appropriate range for maturity stage. Maturation coefficient ratios of female (2.7±0.99%) was highest in NT1 and the difference was significant at P<0.05 compared with coefficient ratios of female 1.44 ± 0.84% in NT4. In treatment NT1, the highest maturity rate was obtained (66.67 ± 19.52%), and the lowest one was in NT4 (44.44 ± 8.61%). The fecundity in NT1 and in NT2 were significantly different (P<0.05) compared to NT3 and NT4. The level of Vitellogenin (Vg) in stage IV ovary in treatments NT1, NT2 and NT3 were significantly different (P <0.05) compared with NT4. The protein content in muscle was lowest in April (2.37 - 4.87). The highest protein in liver (6.88 to 10.85 mg protein/g fresh sample) was in April. Key words: Redtail Botia, sexual maturity, different feed types
Ngày nhận bài: 22/6/2017Ngày phản biện: 27/6/2017
Người phản biện: TS. Lê Quốc ViệtNgày duyệt đăng: 27/7/2017





























