Embed Size (px)
Citation preview

12 živa 1/2004www.cas.cz/ziva
Více než 5 000 druhů krásivek (Desmi-diales) patří bezesporu k nejpozoruhod-nějším zástupcům rostlinné říše. V jejichpřípadě skutečně můžeme s plným právemhovořit o pravých rostlinách, přestože jdevětšinou o jednobuněčné mikroorganismy,mezi nimiž i ty největší zástupce zahlédne-me bez mikroskopu jen jako velmi drobnézelené vločky. Přesto jsou krásivky cévna-tým rostlinám — třeba stromům neboorchidejím — v podstatě velmi příbuzné;rozhodně jim jsou daleko bližší než obrov-ské mořské chaluhy, ruduchy a dokoncei než většina dalších zelených řas.
Podle současných pohledů na evoluciautotrofních — fotosyntetizujících — orga-nismů můžeme totiž za rostliny považovatv zásadě zástupce těch skupin, které svéchloroplasty získaly tzv. primární endosym-biózou. Tu si představujeme jako evolučnísymbiotický proces, v němž dávný jedno-buněčný heterotrofní organismus pohltilsinici — tedy vlastně autotrofní bakterii —a natrvalo ji „ubytoval“ ve svých buňkách.Potomky této sinice jsou všechny dnešnírostlinné chloroplasty. Podle současnýchnázorů tento proces proběhl v evolucinezávisle na sobě třikrát. V prvním případěvznikly ruduchy (Rhodophyta), ve druhémmalá skupina Glaucophyta a ve třetím zele-né řasy a všechny další cévnaté rostliny.Pravé rostliny, tzv. podříše Viridiplantae
zahrnuje pouze onen třetí a poslední pří-pad; tedy organismy, které mají v chloro-plastech jako hlavní pigmenty chlorofylya a b a zásobní látky ukládají hlavně veformě škrobu.
Do podříše Viridiplantae taxonomovéřadí všechny zelené řasy — od nejprimitiv-nějších bičíkovců až po parožnatky, dálepak všechny mechorosty a cévnaté rostliny.Ale i v rámci této skupiny lze ještě naléztdvě velké skupiny — oddělení Chlorophy-ta a Streptophyta. Liší se zdánlivě zcelaokrajovým znakem — uspořádáním mikro-tubulů při buněčném dělení, ale zdá se, žeprávě tento znak ukazuje na jejich dávnéevoluční odlišení. Chlorophyta zahrnujívětšinu zelených řas, do oddělení Strepto-phyta patří cévnaté rostliny, mechorostya několik skupin řas k cévnatým rostlinámevolučně nejbližších. A právě jednou z nichje i třída Zygnematophyceae s řádem Des-midiales — krásivky. Jen pro pořádek jetřeba dodat, že všechny ostatní skupiny(chaluhy, rozsivky, zlativky, různobrvky,obrněnky) rostlinami nejsou. Jejich chloro-
plasty pocházejí z tzv. sekundární endo-symbiózy. Při ní došlo k dlouhodobé sym-bióze nějaké dávné rostliny, často zástupceglaukofyt či ruduch, s heterotrofním jedno-buněčným organismem. Tato symbiogene-tická událost se stala v evoluci mnohokráta stále k ní u některých skupin, např. u obr-něnek, často dochází.
Krásivky jsou tedy z evolučního hlediskablízkými příbuznými cévnatých rostlin, byťse zdá, že se z nich, ani z jim podobnýchorganismů cévnaté rostliny v evoluci nevyvi-nuly. Třída Zygnematophyceae se dnes jevíspíše jako vedlejší rostlinná vývojová větev,kterou charakterizuje mnoho unikátníchvlastností a znaků, jež se u žádných jinýchrostlin nevyskytují. V rámci této třídy jednes známo asi 6 000 druhů. Z nich asi 900patří do řádu Zygnematales — spájivky,které mají většinou vláknitou nevětvenoustélku. Zbylých asi 5 100 druhů jsou krásiv-ky. Přestože jsou Zygnematophyceae skupi-nou vývojově izolovanou, můžeme vidět, žerozhodně nejsou zatím evolučně neúspěšné— alespoň co do počtu recentních druhů.
Nejtypičtějším znakem celé skupiny jeunikátní způsob pohlavního rozmnožová-ní, tzv. spájení. U dvou vegetativních bu-něk se celý protoplast změní na améboidnígametu (což je mimochodem jedinýznámý výskyt améboidní organizace stélkyu zelených řas), obě buňky se k sobě při-blíží, vytvoří se mezi nimi kopulační kaná-lek a jedna z gamet (améb) se přesunekanálkem ke druhé a společně vytvořízygotu (viz obr.). Zygota je většinou kulo-vitá, má velmi často tlustou a strukturova-nou buněčnou stěnu a je schopna přežíti poměrně dlouhou dobu a nepříznivépodmínky (např. zmrznutí), než z ní vyklí-čí nová vegetativní buňka či vlákno.
Krásivky — mikroskopické skvosty našich vod a mokřadů
Jiří Neustupa
Micrasterias thomasiana patří rovněž k druhůmvyskytujícím se v mokřadech střední Evropy,nahoře © Několik buněk různých druhů roduClosterium, dole
Některé krásivky středoevropských mokřadů.Buňky krásivek jsou složeny ze dvou semicel —polobuněk. Nahoře vlevo buňka Micrasterias crux–melitensis © Uprostřed rostoucí buňka Micra-sterias rotata © Dole trichální krásivka Desmi-dium swartzii se spirálně stočeným vláknem
13živa 1/2004 www.cas.cz/ziva
a alpinská rašeliniště v našich sudetskýchpohořích (Krušné hory, Krkonoše, Jizerskéhory), kde dnes kvůli acidifikaci přežívajípouze nejodolnější druhy krásivek. Mezinejcennější lokality s největší biodiverzitoukrásivek v ČR patří např. PR Borkovickáblata na Táborsku či oblast Modravských
V celém životním cyklu se jako u jedinéskupiny v rámci zelených řas nevyskytuježádné bičíkaté stadium.
Krásivky jsou na rozdíl od spájivek větši-nou jednobuněčné, byť několik desítekdruhů je vláknitých. Žijí výhradně ve slad-kých vodách, v sladkovodních mokřadech,vzácně i mimo vodní prostředí (subaeric-ky) ve vlhkých slizových nárostech na ska-lách či kůře stromů a v půdě. Zatím žádnékrásivky nebyly nalezeny v mořích. Jejichbuňka je rozdělena na dvě polobuňky —semicely, které jsou u vývojově pokročilej-ších rodů krásivek odděleny zúženýmmůstkem — isthmem (viz obr.). (Názevřádu Desmidiales pochází z řeckého des-mós, tj. svazek, vazba, spojení; právě podlespojení dvou semicel úzkým buněčnýmmůstkem.) V isthmu se nachází jádro.Každá ze semicel obsahuje jeden neboi větší počet chloroplastů, v nichž se častovyskytují kulovitá tělíska (pyrenoidy) obsa-hující enzym Rubisco, podílející se na tem-nostní fázi fotosyntézy. Krásivky se vedlepohlavního spájení mohou rozmnožovat— a většinou také rozmnožují — nepohlav-ním dělením. Buňky se dělí právě v roviněisthmu; po dělení dorůstá druhá semicelado dospělé velikosti. V přírodě se tak lzečasto setkat s čerstvě rozdělenými jedincis nestejně velkými semicelami (viz obr.).
Krásivky se v přírodě vyskytují hlavněv prostředí s nižším pH, tedy v kyselejšíchvodách. Největší druhové rozmanitostidosahují v prostředí pH 4,5–6,5. Nejtypič-tějším biotopem krásivek jsou bentickáa nárostová společenstva, tedy dna tůněk,mikrobiální porosty na ponořených kame-nech nebo rostlinách. Řada z nich také pro-dukuje velké množství slizu a vytváří sli-zovité kolonie. Některé menší druhy sevyskytují i v planktonu řek, přehradníchnádrží nebo čistších rybníků. Centrum bio-diverzity krásivek ovšem jednoznačně ležív biotopech s malým množstvím živin, tedyhlavně organických látek, dusíku a fosforu.Eutrofizace vodního prostředí (zvyšováníkoncentrace živin), která v našich podmín-kách probíhá převážně činností člověka,populace krásivek decimuje a vede častok jejich lokálnímu vyhynutí. Dánský hydro-biolog Nygaard dokonce na základě počtudruhů sinic, centrických rozsivek a kokál-ních zelených řas (tedy skupin žijících vět-šinou v eutrofním prostředí) ku počtudruhů krásivek vytvořil obecný index bio-indikace eutrofie vodních ekosystémů.
Příkladem škodlivého vlivu eutrofizacezpůsobené lidskou činností je jihočeskáNPR Řežabinec. V posledních 30 letechtam z tůní a mokřadů vymizela více nežpolovina druhů krásivek, včetně některýchvelmi vzácných taxonů, které nebyly zná-my z žádné jiné středoevropské lokality.Jiným vážným rizikem likvidujícím spole-čenstva krásivek v naší přírodě je acidifi-kace. Ta se projevuje hlavně v horskýchrašelinných ekosystémech, kde vlivemkyselých dešťů může dojít k dlouhodobé-mu snížení pH, což vede k vyhynutí valnéčásti mikrobiálních druhů, včetně krásivek.Příkladem mohou být horská subalpinská
Zygota krásivky Micrasterias rotata se dvěmaprázdnými buňkami (vlevo). Foto S. Nováková© Dvě dceřiné buňky spojené po proběhlémnepohlavním dělení převážně severoamerickékrásivky Xanthidium antilopaeum var. polyma-zum. Snímky J. Neustupy, pokud není uvedenojinak
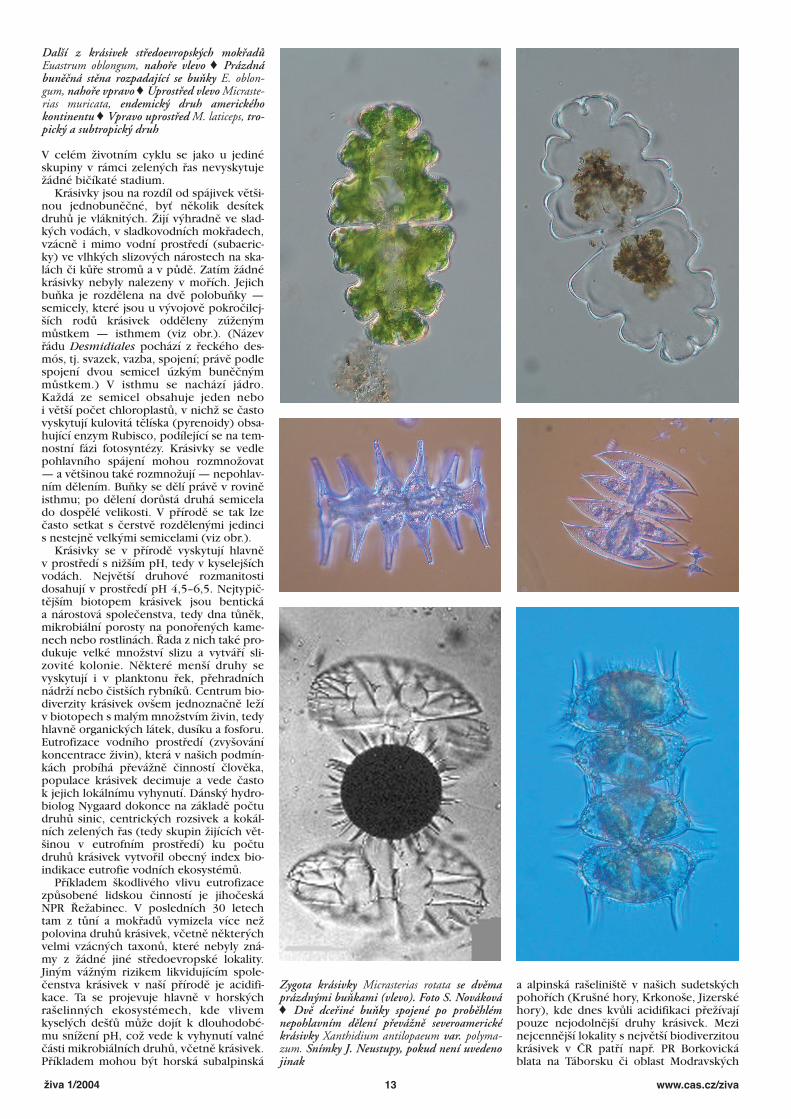
Další z krásivek středoevropských mokřadůEuastrum oblongum, nahoře vlevo © Prázdnábuněčná stěna rozpadající se buňky E. oblon-gum, nahoře vpravo © Uprostřed vlevo Micraste-rias muricata, endemický druh americkéhokontinentu © Vpravo uprostřed M. laticeps, tro-pický a subtropický druh

14 živa 1/2004www.cas.cz/ziva
Cévnaté rostliny (Tracheophyta) před-stavují skupinu vyšších rostlin, u nichžv ontogenetickém vývoji převažuje sporofyt(nepohlavní, většinou diploidní generaceprodukující spory) nad gametofytem (po-hlavní, většinou haploidní generace produ-kující gamety). Pro fylogenetický vývoj cév-natých rostlin je charakteristický postupnýústup gametofytu až k jeho úplné závislostina sporofytu, jak je tomu u rostlin, kterédosáhly vývojového stupně semennosti(Spermatophyta). Sporofyt cévnatých rost-lin (kormus) se rozlišuje na vegetativní a ge-nerativní orgány, které jsou tvořeny systémypravých pletiv. Jednotlivými orgány probí-hají vodivá pletiva vytvářející vodivý (vasku-lární) systém rostliny.
Evoluce cévnatých rostlin je úzce spjatas přechodem rostlin z vodního prostředí nasouš. Za předky suchozemských tracheofytse považují semiakvatické zelené řasy, nej-pravděpodobněji z příbuzenstva r. Coleo-chaete nebo z příbuzenstva obdobnýchhypotetických vyhynulých typů. Nejstaršídosud známé suchozemské rostliny, jež lzesnad již považovat za rostliny cévnaté, dolo-žené nálezy spor a makrofosilí (úlomky telo-mů, tj. koncových prýtů nesoucích sporan-
gium), jsou zástupci r. Cooksonia (odděleníProtracheophyta) ze středního siluru (420mil. let). Kolonizace souše tracheofyty bylaspojena s mnoha anatomicko–morfologic-kými a fyziologickými adaptacemi na zcelanové ekologické podmínky prostředí: atmo-sférické sucho, přímé sluneční záření, odliš-ný způsob příjmu uhlíku, minerálních láteka vody, zvýšené působení gravitace (rostlinyjiž nebyly nadnášeny vztlakovou silou vody)a rozmnožování v odlišných podmínkách(Živa 2003, 4: 153–156). K upevnění rostlink substrátu a čerpání živin sloužily nejstar-ším suchozemským cévnatým rostlinámspecializované orgány telomového původu,tzv. rhizomoidy. Povrch rostlin kryla pokož-ka (epidermis) s kutikulou (tenká vrstvanepropustná pro vodu). Tyto povrchovéstruktury bránily vysychání rostlin a sloužilyk ochraně před slunečním zářením. V epi-dermis byly již diferencovány průduchy(stomata), které zajišťovaly regulovanoutranspiraci a tím i řízené hospodařenís vodou (cévnaté rostliny jsou až na výjimkyhomoiohydrické = hydrostabilní). Transpi-race také vytvářela hnací sílu vzestupnéhotranspiračního proudu a zajišťovala účin-nou termoregulaci (ochlazování) pletiv.
Jednou ze zásadních anatomických adap-tací, které umožnily cévnatým rostlinámosídlit rozsáhlé oblasti pevnin (včetně ob-lastí aridních) ve velmi rozmanitých život-ních formách, byl vznik specializovanýchvodivých pletiv. Tato pletiva zajišťovala v rost-lině transport vody, anorganických a orga-nických látek na delší vzdálenosti. Vytvořilataké zpevňující mechanický skelet umožňu-jící rostlinám odolávat povětrnostním vli-vům. V evoluci mechanických pletiv bylavysoce významná syntéza ligninu, kterýimpregnoval celulózní buněčné stěny pů-vodních tenkostěnných parenchymatickýchpletiv (především dřevních elementů).
Následující text podává stručnou charak-teristiku dřevních a lýkových vodivých ele-mentů. Upřesnění použitých anatomickýchtermínů lze nalézt v Základním slovníkurostlinné anatomie I–VI (Živa 2001, 1–6).
Cévní svazky
Vodivá pletiva vytvářejí v rostlinnýchorgánech cévní svazky (fasciculi vasorum),které sestávají z části dřevní (xylém,hadrom, část vazální), vedoucí vzestupnýtranspirační proud, a části lýkové (floém,leptom, část kribrální), vedoucí sestupnýasimilační proud. Cévní svazky jsou tvořeny
Systémy vodivých pletivcévnatých rostlin
Vladimír Vinter, Michaela Sedlářová
Vodivé elementy xylému. 1— tracheidy proto-xylému brukve zesílené kruhovitě a šroubovitě,2 — schodovitě zesílená tracheida metaxylémuhasivky orličí, 3 — dvůrkaté tracheidy deutero-xylému jehličnanů na radiálním řezu (abie-toidní typ), 4 — schodovitě perforovaná tra-chea metaxylému u hasivky orličí, 5 — síťovitězesílená trachea deuteroxylému tykve, 6 —síťovitě zesílená trachea metaxylému kukuřice,7 — schodovitě perforovaná deska tracheje deu-teroxylému dubu
slatí v NP Šumava. Na těchto lokalitách jemožné v jediné rašelinné tůňce při jedinémodběru nalézt až 50 druhů krásivek. Cel-kem je ze střední Evropy známo téměř 600druhů.
Globální centrum evoluce krásivek ležíovšem v tropech. Z tropických mokřadůjsou popsány téměř dvě třetiny dnes zná-mých druhů. Díky jejich atraktivnímu vzhle-du jsou v současnosti jednou z nejprozkou-manějších skupin. Můžeme se domnívat, ženezbývá snad více než 20 % neobjevenýchdruhů. Díky dobré znalosti rozšíření jednot-livých druhů se podařilo vymezit i několikbiogeografických areálů s typickou flórou
krásivek. Více než 10 % druhů se vyskytujejen na jednom kontinentu. Velmi bohatouflóru krásivek má indomalajská a australskáoblast i tropická část Jižní Ameriky. Směremk pólům rozmanitost druhů ubývá, ale Ark-tida je daleko bohatší než Antarktida, kdekvůli geografické izolaci a extrémním pod-mínkám žije jen 8 nejotužilejších druhů.
Krásivky jsou opravdu velmi krásnýmirostlinami. Symetrické a komplikovaněstrukturované tvary mnoha rodů a druhůje řadí k nejpozoruhodnějším příkladůmbuněčných morfologií vůbec. Zástupci r.Micrasterias jsou v poslední době využívá-ni jako modelové organismy pro studium
buněčné morfogeneze. Tvar buněk je dánzpůsobem růstu buněčné stěny. Její onto-genetický vznik se v průběhu růstu buněkřídí především provazci cytoskeletárníchmikrotubulů, které přinášejí materiál prosyntézu a ovlivňují její odlišnou rychlostrůstu buněčné stěny v různých částechbuňky. V současnosti neexistuje žádná uce-lená teorie, která by vysvětlovala případnýadaptivní význam takto složitých buněč-ných tvarů v evoluci. Krásivky se v každémpřípadě vedle např. květů orchidejí, kresebmotýlích křídel či tvarů a ornamentací hlu-bokomořských plžů řadí mezi nejkrásnějšíjevy v živé přírodě vůbec.





























