Embed Size (px)
Citation preview

La relation entre traits d’histoire de vie et répartition des arbres à la lumière du
conservatisme phylogénétique
Xavier Morin & Isabelle Chuine
L’écologie des communautés en devenir : approche intégrative – GDR ComEvol
Montpellier 15 Décembre 2006

Répartition géographique
Biologie
?Webb & Gaston 2003, 2005 Am Nat

Répartition géographique
Biologie
Niche écologique traits
Répartition géographique
Biologie
Niche écologique
Macroécologie
traits
Brown 1995
Blackburn & Gaston 2003

Arbres de la zone tempérée
Amérique du nord Europe
lat N
lat SSurface
Amplitude

0.005-6.60Log Haut.
0.452.04Log Age
0.002-1.81Log Gm
<0.001-3.38St. Succ.
P-valueCoef.Variable
Amplitude = α St. Succ. + β log(Graine m) + γ log(Age) + δ log(Hauteur)
r²=0.30***
118 esp.
Artefact causé par les relations phylogénétiques entre espèces ?
Lien entre traits des espèces et répartition ?

Conservatisme de niche
Peat & Fitter 1994 Biol Rev. Camb. Phil. Soc.
Lord et al. 1995 Am. Nat.
Ackerly & Reich 1999 Am. J. Bot.
Conservatisme d’habitats
Conservatisme de traits
Prinzing et al. 2001 Proc. R. Soc. Lond.
Peterson et al. 1999 Science
Martinez-Mayer et al. 2004 Ecography
Wiens & Donoghue 2004 TREE
Le conservatisme phylogénétique

Peterson et al. 1999 Science
37 paires d’espèces-sœurs isolées depuis 2-10 millions d’années de part et d’autre de l’isthme de Tehuantepec, Mexique
Atthis heloisa
A. ellioti
Peromyscusmelanocarpus
P. zarhynchus
Pereute charopscharops
P. c. nigricans
Le conservatisme de niche

Conservatisme de niche
« Conservatisme »
des répartitions géographiques
« Héritabilité »Jablonski 1987 Science
History of lineagesBrown et al. 1996 Ann. Rev. Ecol. Syst.
Sax 2001 J. of Biogeography
Le conservatisme de niche
Répartition géographique
Biologie
Niche écologique ???
traits des espèces

Jablonski 1987 Science
Héritabilité des répartitions ?
Conservatisme de répartition ?
Pour les arbres…Ricklefs & Latham 1992
Qian & Ricklefs 2004
Peat & Fitter 1994
Résultats contradictoires
Problèmes de méthodes

Différentes méthodes
Espèces d’arbres nord-américains et européens
52 genres
35 familles
19 ordres
7 classes
Angiospermes
Gymnospermes
530 espèces
Conservatisme de traits
Conservatisme de répartition

Masse spécifique de graine
Conservatisme des traits ?
-5
-4
-3
-2
-1
0
1
2
-5 -4 -3 -2 -1 0 1 2-5
-4
-3
-2
-1
0
1
2
-5 -4 -3 -2 -1 0 1 2
Log (masse de graine) esp 1
Log
(mas
se d
e gr
aine
) esp
2
Rééchantillonnage 30 répétitions r² = 0.0001
Régression sib-sib
169 espèces
49 genres
« Modèle nul »
(Smith et al. 2004 Am. Nat.)
r² = 0.77***

ANOVA emboîtée
Masse de graine
Stade de successionGenre(Famille(Ordre))
Genre 8.7%
Famille 9.2%
Ordre 76.4%
94.3 %
Genre 22.0%
Famille 7.3%
Ordre 44.2%
73.5 %
169 esp
Conservatisme important
Angiospermes Gymnospermes
Genre 43.3%
Famille 4.1% 47.4 %
Genre 42.9%
Famille 6.6% 49.5 %
(Smith et al. 2004 Am. Nat.)
Masse de graine
Stade de succession
Conservatisme des traits ?
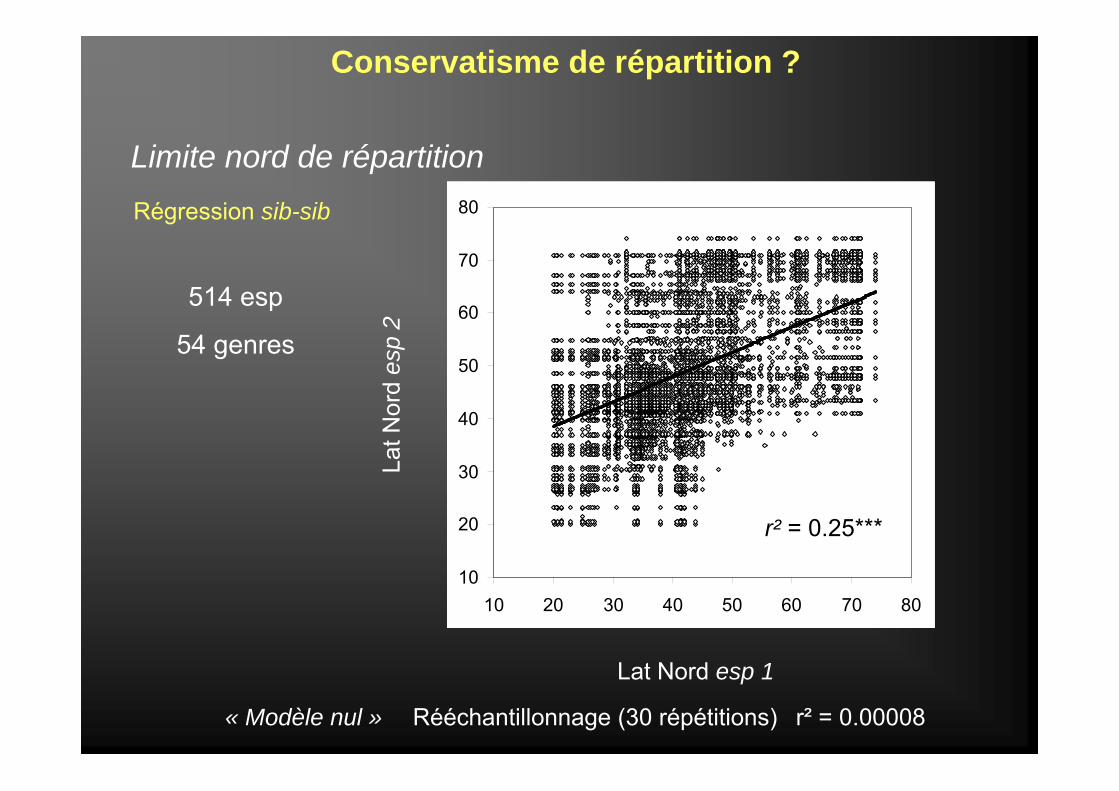
Limite nord de répartition
Conservatisme de répartition ?
Lat Nord esp 1
LatN
ord
esp
2
Rééchantillonnage (30 répétitions) r² = 0.00008
10
20
30
40
50
60
70
80
10 20 30 40 50 60 70 8010
20
30
40
50
60
70
80
10 20 30 40 50 60 70 80
514 esp
54 genres
Régression sib-sib
« Modèle nul »
r² = 0.25***

< 0.001 ns
< 0.001 ns
< 0.0001 ns
< 0.001 nsr² random
0.01***0.15***0.25***0.04***r² obs366514514514n
SurfaceLat SudLat NordAmplitude
Régression sib-sib
30 rép.
lat N
lat SSurface
Amplitude
Conservatisme de répartition ?

Amplitude et surface de répartition
Limites latitudinales
Genre(Famille(Ordre))
23-28 %
43-50 %
530 esp Angiospermes Gymnospermes
15-16 %
25%
Amplitude et surface de répartition
Limites latitudinales
ANOVA emboitée
Conservatisme de répartition ?

Corrélogrammes de Moran
Similarité entre deux niveaux taxonomiques
0.4
0.2
0
-0.2
-0.4
S/G G/F F/O O/C
I / I m
ax
0.4
0.2
0
-0.2
-0.4
S/G G/F F/O O/C
I / I m
ax
Angiospermes Gymnospermes
0.4
0.2
0
-0.2
-0.4
S/G G/F F/OI /
I max
0.4
0.2
0
-0.2
-0.4
S/G G/F F/OI /
I max
Surface
Indice de Moran standardisé
Gittleman & Kot 1990 Syst. Zool.
Conservatisme de répartition ?
significatif
non significatif

0.4
0.2
0
-0.2
-0.4
S/G G/F F/O O/C
I / I m
ax
0.4
0.2
0
-0.2
-0.4
S/G G/F F/O O/C
I / I m
ax
0.4
0.2
0
-0.2
-0.4
S/G G/F F/O
I / I m
ax
0.4
0.2
0
-0.2
-0.4
S/G G/F F/O
I / I m
ax
Angiospermes Gymnospermes
Corrélogrammes de Moran
Limite Nord
Conservatisme de répartition ?

Conservatisme de répartition ?
Conservatisme prendre en compte…
Différentes méthodes
Harvey & Pagel 1991
Felsenstein 1985 Am. Nat.
Cheverud et al. 1985 Evolution

A B C D E A B C D E03333
30222
32011
32101
32110
Espèces
A
B
C
D E
A B
C D E1
0
Matrice de distance
phylogénétique
Matrice de corrélation
phylogénétique
00.330.670.671
00.330.6710.67
00.3310.670.67
010.330.330.33
10
00
0
Valeurs de traits pour chaque espèce+
X2 X2
X1 X1
Régressions de traits des espèces (X1 et X2) avec pondération des corrélations
d’après Paradis & Claude 2002 JTB
La méthode GEE (Generalized Estimating Equations)
Prise en compte des distances phylogénétiques
Approche multivariéepossible
0

Am. Nord Europe
Angiospermes
Gymnospermes
PlatanusoccidentalisLiquidambarstyracifluaGleditsiatriacanthosRobiniapseudoacaciaOstryavirginianaCarpinuscarolinianaCarpinusbetulusAlnusrubraBetulalentaBetulapapyriferaBetulaalleghaniensisBetulapendulaBetulananaBetulapubescensAlnusglutinosaAlnusincanaCaryaglabraCaryaillinoensisCaryatomentosaCaryalaciniosaCaryacordiformisFagusgrandifoliaFagussylvaticaCastanopsischrysophyllaLithocarpusdensiflorusQuercusstellataQuercusshumardiiQuercusdouglasiiQuercusnigraQuercusbicolorQuercuslyrataQuercusfalcataQuercusrubraQuercuslaurifoliaQuercuskelloggiiQuercuspetraeaQuercusroburPrunusserotinaUlmusamericanaPrunusaviumSalixnigraSalixbebbianaSalixherbaceaPopulusdeltoidesPopulustremuloidesPopulusbalsamiferaTiliaamericanaTiliacordataAcermacrophyllumAcernegundoAcerpennsylvanicumAcerrubrumAcersaccharinumAcersaccharumAcercircinatumAesculusglabraAcercampestreAcerplatanoidesAcerpseudoplatanusCornusfloridaCornusnuttalliiFraxinuslatifoliaDiospyrosvirginianaFraxinusprofundaFraxinusexcelsiorSassafrasalbidumLiriodendrontulipiferaMagnoliaacuminataMagnoliagrandifloraJuniperusscopulorumSequoiasempervirensSequoiadendrongiganteumThujaplicataChamaecyparislawsonianaJuniperuscommunisAJuniperuscommunisTaxusbaccataAbiesamabilisAbiesbalsameaAbiesconcolorAbiesgrandisAbieslasiocarpaAbiesmagnificaAbiesproceraTsugaheterophyllaTsugacanadensisTsugamertensianaPseudotsugamenziesiiLarixoccidentalisLarixlaricinaPseudotsugamacrocarpaPiceaglaucaPiceaengelmanniiPiceapungensPicearubensPiceasitchensisPiceabrewerianaPinuscembroidesPinusbanksianaPinusclausaPinuscontortaPinusglabraPinusjeffreyiPinusleiophyllaPinuspalustrisPinusponderosaPinusresinosaPinusrigidaPinussabinianaPinusserotinaPinustaedaPinusvirginianaPinuselliottiiPinussylvestrisPinusalbicaulisPinusmonticolaPinusstrobiformisPinusstrobus
Stade de succession
Masse de graine
Hauteur et âge à 1ère
reproduction
118 esp
Amplitude de rép.
Arbres
Analyses corrigées pour l’inertie phylogénétique
GEE
collab. N. Alvarez
PlatanusoccidentalisLiquidambarstyracifluaGleditsiatriacanthosRobiniapseudoacaciaOstryavirginianaCarpinuscarolinianaCarpinusbetulusAlnusrubraBetulalentaBetulapapyriferaBetulaalleghaniensisBetulapendulaBetulananaBetulapubescensAlnusglutinosaAlnusincanaCaryaglabraCaryaillinoensisCaryatomentosaCaryalaciniosaCaryacordiformisFagusgrandifoliaFagussylvaticaCastanopsischrysophyllaLithocarpusdensiflorusQuercusstellataQuercusshumardiiQuercusdouglasiiQuercusnigraQuercusbicolorQuercuslyrataQuercusfalcataQuercusrubraQuercuslaurifoliaQuercuskelloggiiQuercuspetraeaQuercusroburPrunusserotinaUlmusamericanaPrunusaviumSalixnigraSalixbebbianaSalixherbaceaPopulusdeltoidesPopulustremuloidesPopulusbalsamiferaTiliaamericanaTiliacordataAcermacrophyllumAcernegundoAcerpennsylvanicumAcerrubrumAcersaccharinumAcersaccharumAcercircinatumAesculusglabraAcercampestreAcerplatanoidesAcerpseudoplatanusCornusfloridaCornusnuttalliiFraxinuslatifoliaDiospyrosvirginianaFraxinusprofundaFraxinusexcelsiorSassafrasalbidumLiriodendrontulipiferaMagnoliaacuminataMagnoliagrandifloraJuniperusscopulorumSequoiasempervirensSequoiadendrongiganteumThujaplicataChamaecyparislawsonianaJuniperuscommunisAJuniperuscommunisTaxusbaccataAbiesamabilisAbiesbalsameaAbiesconcolorAbiesgrandisAbieslasiocarpaAbiesmagnificaAbiesproceraTsugaheterophyllaTsugacanadensisTsugamertensianaPseudotsugamenziesiiLarixoccidentalisLarixlaricinaPseudotsugamacrocarpaPiceaglaucaPiceaengelmanniiPiceapungensPicearubensPiceasitchensisPiceabrewerianaPinuscembroidesPinusbanksianaPinusclausaPinuscontortaPinusglabraPinusjeffreyiPinusleiophyllaPinuspalustrisPinusponderosaPinusresinosaPinusrigidaPinussabinianaPinusserotinaPinustaedaPinusvirginianaPinuselliottiiPinussylvestrisPinusalbicaulisPinusmonticolaPinusstrobiformisPinusstrobus

<0.0010.849-3.499Log Haut.
0.0171.238-3.173Log Age
0.0020.363-1.238Log Gm
<0.0010.498-4.004St. Succ.
P-valueS. E.Coef.Variable
= α St. Succ. + β log(Graine m) + γ log(Age) + δ log(Hauteur)
Inertie phylogénétique éliminée
r²=0.27***
118 esp.
Amplitude de répartition et traits des espèces
Morin & Chuine 2006 Ecol Lett.
Amplitude de répartition

Aire plus grande
Aire plus petite
plus hautes latitudes
taille plus petite
pionnières
graines plus légères
maturité plus précoce
coûts métaboliques bas
plus basses latitudes
taille plus grande
fin de succession
graines plus lourdes
maturité plus tardive
coûts métaboliques forts
Morin & Chuine 2006 Ecol Lett.
Règle de Rapoportst
ress
énergie
Amplitude de répartition et traits des espèces

Jansson & Dynesius 2002 Ann. Rev. Ecol. Syst. Hewitt 2004 Phil. Trans. R. Soc. Lond.Wright et al. 2006 PNAS
Fort taux d’extinction
Taux d’extinction faible
Conservation des taxons
Taxons plus jeunes
Taxons plus vieux
Esp. pionnières
Esp. fin de succ.
Relation entre âge des espèces et répartition

PlatanusoccidentalisLiquidambarstyracifluaGleditsiatriacanthosRobiniapseudoacaciaOstryavirginianaCarpinuscarolinianaCarpinusbetulusAlnusrubraBetulalentaBetulapapyriferaBetulaalleghaniensisBetulapendulaBetulananaBetulapubescensAlnusglutinosaAlnusincanaCaryaglabraCaryaillinoensisCaryatomentosaCaryalaciniosaCaryacordiformisFagusgrandifoliaFagussylvaticaCastanopsischrysophyllaLithocarpusdensiflorusQuercusstellataQuercusshumardiiQuercusdouglasiiQuercusnigraQuercusbicolorQuercuslyrataQuercusfalcataQuercusrubraQuercuslaurifoliaQuercuskelloggiiQuercuspetraeaQuercusroburPrunusserotinaUlmusamericanaPrunusaviumSalixnigraSalixbebbianaSalixherbaceaPopulusdeltoidesPopulustremuloidesPopulusbalsamiferaTiliaamericanaTiliacordataAcermacrophyllumAcernegundoAcerpennsylvanicumAcerrubrumAcersaccharinumAcersaccharumAcercircinatumAesculusglabraAcercampestreAcerplatanoidesAcerpseudoplatanusCornusfloridaCornusnuttalliiFraxinuslatifoliaDiospyrosvirginianaFraxinusprofundaFraxinusexcelsiorSassafrasalbidumLiriodendrontulipiferaMagnoliaacuminataMagnoliagrandifloraJuniperusscopulorumSequoiasempervirensSequoiadendrongiganteumThujaplicataChamaecyparislawsonianaJuniperuscommunisAJuniperuscommunisTaxusbaccataAbiesamabilisAbiesbalsameaAbiesconcolorAbiesgrandisAbieslasiocarpaAbiesmagnificaAbiesproceraTsugaheterophyllaTsugacanadensisTsugamertensianaPseudotsugamenziesiiLarixoccidentalisLarixlaricinaPseudotsugamacrocarpaPiceaglaucaPiceaengelmanniiPiceapungensPicearubensPiceasitchensisPiceabrewerianaPinuscembroidesPinusbanksianaPinusclausaPinuscontortaPinusglabraPinusjeffreyiPinusleiophyllaPinuspalustrisPinusponderosaPinusresinosaPinusrigidaPinussabinianaPinusserotinaPinustaedaPinusvirginianaPinuselliottiiPinussylvestrisPinusalbicaulisPinusmonticolaPinusstrobiformisPinusstrobus
Pionnier
Mil. et fin de succession
P<0.001Pionnier < Mil. et fin
N=234
P=0.0003Relation Age - Amplitude

Répartition géographique
Biologie
Niche écologique
• Macroécologie
Grand nombre d’entités Inertie phylogénétique à considérer
• Lien entre biologie et répartition
espèces
Utilisation pour comprendre le déterminisme de la répartition à
l’échelle spécifique
Nouveaux outils
Données
Conclusion

Probabilité de maturationdes fruits
Le modèle PHENOFIT
Succès reproducteur
Probabilité de présence
Survie
Tj
Paramètres ajustés
SénescenceSénescenceFloraisonFloraisonDébourrementDébourrement MaturationMaturation
Energie disponible depuis la floraison
Dommages du gel sur les fleurs
Dommages du gel sur les feuilles
Dommages par le gel
ETP
W
Survie à la sécheresse
Survie aux températures
extrêmes
Observations phénologiques
Modèles phénologiques
Tj Pm
Tj
Chuine & Beaubien 2001 Ecol. Lett.(modif. Morin & Chuine 2005 GCB)

Succès reproducteur
Probabilité de présence
Survie
Tj
Paramètres ajustés
Probabilité de maturationdes fruits
Energie disponible depuis la floraison
Dommages du gel sur les fleurs
Dommages du gel sur les feuilles
Dommages par le gelDommages par le gel
ETP
W
Survie à la sécheresse
Survie aux températures
extrêmes
Observations phénologiques
Modèles phénologiques
Tj Pm
Tj
SénescenceSénescenceFloraisonFloraisonDébourrementDébourrement MaturationMaturation
Le modèle PHENOFIT

110°0'0"O
110°0'0"O
100°0'0"O
100°0'0"O
90°0'0"O
90°0'0"O
80°0'0"O
80°0'0"O
70°0'0"O
70°0'0"O
60°0'0"O
60°0'0"O
23°0
'0"N
23°0
'0"N
33°0
'0"N
33°0
'0"N
43°0
'0"N
43°0
'0"N
53°0
'0"N
53°0
'0"N
Acer saccharumAcer circinatumAcer grandidentatum
Acer macrophylumAcer glabrum
Le modèle PHENOFIT
…conservatismeMorin et al. soumis

Merci
Nadir Alvarez
Julien Claude
Georges Kunstler
Denis Vile
Cyrille Violle
Bernhard Schmid
Stephan Hattenshwiler
Jean-Luc Dupouey
Emmanuel Paradis
Jacques Roy


La règle de Rapoport
Oiseaux
Mammifères
Arbres
…
(Gaston 2003)
Rapoport (1982)
Stevens (1989)
Gradient de richesse spécifique
Gradient latitudinal de la taille des aires de répartition

Variabilité climatique
Compétition
Gradient d’énergie
Taux d’extinction différentiel
Contraintes géographiques
Plusieurs hypothèses…
La règle de Rapoport
Gradient latitudinal de la taille des aires de répartition

Hypothèse de variabilité climatique
Stevens (1989)
Gradient latitudinal de variabilité climatique
Tps
Tps
Tolérance Taille d’aire

Hypothèse de compétition
Pianka (1989)
Gradient latitudinal de compétition
Gradient latitudinal de richesse spécifique
Gradient latitudinal de compétition
Gradient latitudinal de taille d’aire de répartition

Hypothèse du gradient d’énergie
Wright (1983)
Gradient latitudinal d’énergie
Gradient de taille d’aire de répartition des espèces
Tolérance aux contraintes énergétiques
Bonn et al.(2004)

éner
gie
var. clim.
Aptitudes compétitives -> coût en tolérance aux conditions abiotiques stressantes Tolérance au stress
Pouvoir compétitif
tolérance au stress
Espèces à forte
tolérance
Faibles compétitrices Espèces à
faible tolérance
Fortes compétitrices
pouv. compétitif
Pouv. Comp.
Echelle locale
Echelle continentale
Nord
Sud
Largeur de niche ≈ taille d’aire
Largeur de niche / Pouvoir compétitif
Gradients énergie / climatPatrons latitudinaux de taille d’aire de rép. et de pouvoir compétitif des espèces
taille d’aire

Vérification de la loi de Rapoport
P<0.001
P<0.001
P<0.001
169 esp. 65 esp.Taille d’aire
Gradient latitudinal de taille d’aire de répartition

Relation traits – répartition
Répartition
Biologietraits
Conservatisme de niche
Inertie phylogénétique ?
Illustration…

Reliée à l’écologie
Reliée à la phylogénie
Non expliquée
a b c d

P<0.001
20 30 40 50 60 70
0.0
0.2
0.4
0.6
0.8
1.0
Latitude (°N)20 30 40 50 60 70
0.0
0.2
0.4
0.6
0.8
1.0
Latitude (°N)20 30 40 50 60 70
0.0
0.2
0.4
0.6
0.8
1.0
Latitude (°N)20 30 40 50 60 70
0.0
0.2
0.4
0.6
0.8
1.0
Latitude (°N)20 30 40 50 60 70
0.0
0.2
0.4
0.6
0.8
1.0
Latitude (°N)20 30 40 50 60 70
0.0
0.2
0.4
0.6
0.8
1.0
Latitude (°N)20 30 40 50 60 70
0.0
0.2
0.4
0.6
0.8
1.0
Latitude (°N)
- Effet Rapoport vérifié
- Relation entre latitude et probabilité de présence des pionniers
Partie 3 Approche intégrative des déterminants des patrons de répartition
Morin & Chuine 2006
Prop
orti
on d
’esp
èces
pi
onni
ères

Gradient de taille d’aire de répartition
Gradient énergétique
Pouvoir compétitif
Gradient de variabilité climatique
Partie 3 Approche intégrative des déterminants des patrons de répartition
Bilan

Différentes méthodes pour estimer l’inertie phylogénétique
Similarité de répartition entre espèces congénériques
Similarité de répartition dans la hiérarchie taxonomique
Régressions sib-sib Comparaison avec assemblages aléatoires
Corrélogrammes de Moran
ANOVA emboîtée

0.4
0.2
0
-0.2
-0.4
S/G G/F F/OI /
I max
0.4
0.2
0
-0.2
-0.4
S/G G/F F/OI /
I max
0.4
0.2
0
-0.2
-0.4
S/G G/F F/O O/C
I / I m
ax
0.4
0.2
0
-0.2
-0.4
S/G G/F F/O O/C
I / I m
ax
Corrélogrammes de Moran
Angiospermes Gymnospermes
Taille de répartition

0.4
0.2
0
-0.2
-0.4
S/G G/F F/O
I / I m
ax
0.4
0.2
0
-0.2
-0.4
S/G G/F F/O
I / I m
ax
0.4
0.2
0
-0.2
-0.4
S/G G/F F/O O/C
I / I m
ax
0.4
0.2
0
-0.2
-0.4
S/G G/F F/O O/C
I / I m
ax
Corrélogrammes de Moran
Angiospermes Gymnospermes
Latitude Sud

0.5
0
-0.5
S/G G/F F/O O/C
I / I m
ax
1
0.5
0
-0.5
S/G G/F F/O O/C
I / I m
ax
1
Angiospermes Gymnospermes
Masse graine
Stade succ.
S/G G/F F/O
0.5
0
-0.5
I / I m
ax
S/G G/F F/O
0.5
0
-0.5
I / I m
ax
S/G G/F F/O O/C
0.5
0
-0.5
I / I m
ax
1
S/G G/F F/O O/C
0.5
0
-0.5
I / I m
ax
1
S/G G/F F/O
0.5
0
-0.5
I / I m
ax
S/G G/F F/O
0.5
0
-0.5
I / I m
ax

Masse de graine
Stade de succession
Hauteur à maturité
Age à 1ère reproduction
Tolérance à l’ombre Baker 1972Coomes & Grubb 2003
Longévité Westoby et al. 1992
Pouvoir compétitif juvénile
Survie juvénile
Baker 1972
Walters 2000
Phénologie
Intégrateur de nombreux traits
Bazzaz 1979
Grime 1979, 2001
Acquisition de ressources
Succès reproducteur
Westoby 1998
Nakashizuka 2001
Loehle 2000
Nakashizuka 2001





























