Embed Size (px)
Citation preview



ANALES
VOL. 24, DICIEMBRE 2011
REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCIA ORIENTAL

© Real Academia de Ciencias Veterinarias de Andalucía Oriental
DIRECCIÓN DE LA REVISTA
RACVAO (Real Academia de Ciencias Veterinarias de Andalucía Oriental)C/ Rector Marín Ocete, 10 • 18014 Granadawww.insacan.org/racvao.ahtml
MAQUETACIÓN:
Gráfi cas la Paz de Torredonjimeno, S.L.www.grafi caslapaz.net DEPÓSITO LEGAL: J-1451-2012
I.S.S.N.: 1130-2534
Imprime: Gráfi cas la Paz de Torredonjimeno, S.L.
VOLUMEN 24, DICIEMBRE DE 2011

ANALESDE LA
REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCIA ORIENTAL
PRESIDENTE DE HONOR:Excmo. Sr. D. Julio Boza López
PRESIDENTE:Excmo. Sr. D. Antonio Marín Garrido
VICEPRESIDENTE:Ilmo. Sr. D. Alberto González Ramón
Sección de Almería
VICEPRESIDENTEIlmo. Sr. D. Eduardo Ruíz Villamor
Sección de Granada
VICEPRESIDENTEIlmo. Sr. D. José Luis Fernández Navarro
Sección de Málaga
SECRETARIO GENERAL:Ilmo. Sr. D. Miguel Molina Galindo
Sección Granada
DIRECTORA DE PUBLICACIONES:Iltma. Sra. Dª Catalina Gómez López
Sección Jaén
CONSEJO DE DIRECCIÓNDE LA REVISTA
La Real Academia de Ciencias Veterinarias de Andalucía Oriental, no se responsabiliza de las opiniones expresadas por los diferentes autores.
Dr. D. Aguilera Tejero, Escolastico
Dra. Dª. Arrazola Saniger, Marcelina
Dr. D. Carrasco Otero, Librado
Dr. D. Contreras Gila, Salvador
Dr.D. Galvez Del Postigo, Antonio
Dr. D. Hernandez Rodriguez, Santiago
Dr. D. Márquez Jiménez, Francisco J.
Dr. D. Moreno Fernández-Caparrós, Luis A.
Dr. D.Palmquist Barrena, Paul
Dr. D. Ros Berruezo, Gaspar
Dr. D. Sánchez de Lollano Prieto, Joaquín
COMITÉ CIENTÍFICO

ÍNDICE
6

ÍNDICE
7
11
13
15
33
45
49
65
75
91
95
ÍNDICE
EDITORIAL ___________________________________________________
ANTONIO GALA. UNA COLABORACIÓN ___________________________
VIDA ACADÉMICA
DISCURSOS DE RECEPCIÓN
CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR (1885-2010): NUEVAS APORTACIONES. LUIS ÁNGEL MORENO FERNÁNDEZ-CAPARRÓS _____________________________
LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES. ANTONIO GÁLVEZ DEL POSTIGO RUIZ _____
CONTESTACIÓN AL DISCURSO DEL ILMO. SR. D. ANTONIO GALVEZ DEL POSTIGO RUIZ. ILTMA. SRA. DRA. Dª. CATALINA GÓMEZ LÓPEZ ______________
LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS. ANTONIO VILLALBA GÓMEZ __________________________________________
CONTESTACION AL DISCURSO EN LA R.A. DE CIENCIAS VETERINARIAS DE ANDALUCIA ORIENTAL DEL ILTMO. SR. D. ANTONIO VILLALBA GÓMEZ. MANUEL MUÑOZ MARTÍN____________________________________________
CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS. LIBRADO CARRASCO OTERO ________________________________________________
CONTESTACIÓN AL DISCURSO DEL ILTMO. SR. DR. D. LIBRADO CARRASCO OTERO. JULIO BOZA LÓPEZ _____________________________
EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL. PEDRO JOSÉ SÁNCHEZ CORDÓN ___________________

ÍNDICE
8
107
111
125
135
145
157
167
193
213
235
CONTESTACIÓN AL DISCURSO DEL ILMO. SR. DR. D. PEDRO JOSÉ SÁNCHEZ CORDÓN. EDUARDO RUIZ VILLAMOR _________________________
TRABAJOS DE INVESTIGACION Y REVISIONES
BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS. Mª JOSÉ GRANDE BURGOS, R. LUCAS, Mª. C. LÓPEZ AGUAYO, R. PÉREZ PULIDO, A. GÁLVEZ _______________
BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN. GASPAR ROS BERRUEZO, CARMEN MARTÍNEZ GRACIÁ Y JOSÉ ANTONIO VALENCIA ARQUES ______________________________
SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E (GENOTIPO 3) EN PERROS DEL SUR DE ESPAÑA. B. HUERTA, C. TARRADAS, M. GONZÁLEZ, B. PERALTA, E. M. MATEU, R. J. ASTORGA, A. CARBONERO, I. LUQUE ___________________________________
EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011. PILAR BARROSO GARCÍA ___________
BASES DE LA RESPUESTA INFLAMATORIA EN LA FORMA RESPIRATORIA DEL PRRS. GÓMEZ-LAGUNA J., RODRÍGUEZ-GÓMEZ I. M., BARRANCO I., QUEREDA J. J., GARCÍA-NICOLÁS O., RAMIS G., PALLARÉS F. J., CARRASCO L. ______________
FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS.F. ROMERO-PALOMO, P.J. SÁNCHEZ CORDÓN, M.A. RISALDE, M. PEDRERA, V. MOLINA, E. RUIZ-VILLAMOR, J.C. GÓMEZ-VILLAMANDOS __________________________
FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA. ESTRELLA AGÜERA BUENDÍA ; F. REQUENA DOMENECH ______
SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES. A. MORENO BOISO, J. HERVÁS RODRÍGUEZ Y F. CHACÓN ___________
GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO (Cervus elaphus hispanicus) A TRAVÉS DE SU DENTICIÓN: REVISIÓN METODOLÓGICA Y TÉCNICAS DE ELECCIÓN. CONCEPCIÓN AZORIT CASAS __

ÍNDICE
9
265
267
269
271
273
275
CIENCIA Y SOCIEDAD:
MEMORIA DEL CICLO “OLORES Y SABORES EN LA DIETA MEDITERRÁNEA”. ANTONIO MARÍN GARRIDO ___________________________________________
MEMORIA DEL CICLO “LA SEGURIDAD ALIMENTARIA EN EL CONTEXTO DE LA SOCIEDAD GLOBALIZADA”. ANTONIO MARÍN GARRIDO _____________
MEMORIA CICLO “ACEITES DE ALMERÍA. PROPIEDADES NUTRICIONALES Y NUEVAS TECNOLOGÍAS”. ANTONIO MARÍN GARRIDO ___________________
OTRAS ACTIVIDADES __________________________________________
OBITUARIO __________________________________________________
NORMAS DE PUBLICACIÓN _____________________________________

ÍNDICE
10

ÍNDICE
11
Coincide esta anualidad con el 250 aniversario de la creación de la Escuela de Veterinaria de Lyon y consecuentemente con el nacimiento de la Veterinaria actual.
Nuestra Corporación ha querido sumarse a esta efeméride mediante la programa-ción de un importante número de actos académicos, así como con su presencia en otros organizados por otras Instituciones , de los que se da cumplida cuenta en este número y que han estado siempre presididos por el logo diseñado para esa conmemoración.
ANALES participa del mismo deseo incorporando a su portada el distintivo elegido para divulgar tan transcendente acontecimiento.
En el número correspondiente a la pasada anualidad ANALES hacia pública su preocupación por la situación económica que se vivía y el incierto futuro que de ella podía derivarse.
De momento la publicación, en su formato actual, está garantizada para la pre-sente anualidad, aún cuando las dudas respecto de su futuro no sean tan halagüeñas. Los costos derivados de su edición, sumados a los devenidos de la distribución de la revista nos obligan a pensar que, al igual que ha ocurrido con otras muchas pu-blicaciones de formato semejante, podrían justifi car que la edición en soporte papel pudiera verse reducida a un mínimo testimonial, utilizando el formato digital para la edición masiva lo que ocasionaría una importante reducción de su coste de pro-ducción y distribución .
Un año más, ANALES hace público agradecimiento a todos aquellos colabora-dores que con su inestimable aportación la hacen posible.
EDITORIAL


13ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
El hombre se engaña: confunde sus efímeras leyes con las más altas, no escritas. Enredado en su propio laberinto, confunde la irresponsabilidad del maltrato a los animales con la falta de sanciones. Hubo un hombre pequeño que los respetó y les hablaba. Fue Francisco de Asís. Cantó a las criaturas con voces fraternales, convencido de que quien se considera a la cabeza de la escala zoológica tiene a su cargo a los seres que ocupan los demás peldaños. Si el hombre es de verdad el administrador de este Planeta, debe cumplir unas graves obligaciones a favor del resto de sus habitantes. Sólo así actuará en su propio favor.
Por eso, me alegraría que proliferasen las sanciones a los amos abandonadores o maltratadores de perros. Y que se enteren de lo que vale un peine los desalmados, que cogen y dejan animales (su nombre viene de ánima, alma) como si fuesen hor-quillas. Sólo falta una consecuencia absolutamente lógica: que lo obtenido con las multas se dedique a favorecer a las Sociedades Protectoras, tan abandonadas por los ayuntamientos como los propios perros. Si así no fuera, habría que multar también a los ayuntamientos, que hacen maldito caso de ellos. Como si no fueran, mucho más que los ediles, tan recientes, criaturas de Dios.
ANTONIO GALA1
1 Escritor, poeta y dramaturgo. Veterinario de Honor de la Universidad de Córdoba.

14

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
15ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
Excmo. Sr. Presidente de la Real Academia.
Conspicuos miembros de la Real Corporación.
Excelentísimas e Ilustrísimas autoridades.
Señoras y Señores
Mis apreciados amigos:
Vaya por delante mi agradecimiento al Presidente y al Cuerpo Académico por permitirme pasar a formar parte como miembro correspondiente de esta prestigiosa Institución. Gracias señor presidente, gracias señoras y señores.
Las Ciencias Veterinarias del tiempo presente y sus Reales Academias están viviendo un momento muy interesante. La nueva concepción del Estado de las Au-tonomías con su desarrollo normativo y el reciente Real Decreto 1160/2010 regulador del Instituto de España van a infl uir decidida y positivamente en nuestras Academias. Un trabajo de ilusión y nuevas motivaciones deben afrontar las Academias de Cien-
CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR (1885-2010): NUEVAS APORTACIONES
LUIS ÁNGEL MORENO FERNÁNDEZ-CAPARRÓS
Discurso de ingreso en la Real Academia de Ciencias Veterinariasde Andalucía Oriental como Académico Correspondiente

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
16
cias Veterinarias distribuidas por todas nuestras autonomías y muy particularmente ésta que está llamada a recorrer un nuevo camino de servicio a la sociedad andaluza. El prestigio de la Real Academia de Ciencias Veterinarias de Andalucía Oriental en el ámbito docente, social y profesional viene avalado por su antigüedad y por las publicaciones de sus bien cuidados Anales.
He decidido que mi preceptivo discurso de ingreso sea en torno a la contribu-ción que las Ciencias Veterinarias en general y la Militar en particular han realizado en entornos científi cos en los que ha tenido que convivir fraternalmente con otros profesionales de las denominadas Ciencias de la Salud.
Precisamente la Veterinaria Militar siempre ha tenido ofi ciales veterinarios en otros sectores de la investigación biosanitaria en paridad con nuestros colegas mé-dicos, farmacéuticos, odontólogos, biólogos, psicólogos y en la actualidad con los enfermeros. Como ejemplo, en el ámbito castrense, citaré a los veterinarios militares que desarrollan su trabajo -y en muchos casos dirigiendo los programas de investi-gación- en la Escuela de Defensa NBQ, el Instituto Tecnológico “La Marañosa”, en el Laboratorio de Investigación Aplicada de Córdoba, Hospitales Militares, Comités de ética y bienestar animal, Escuela Militar de Sanidad e Instituto de Medicina Preventiva Capitán médico Santiago Ramón y Cajal. En este último Centro, que el año pasado cumplió su 125 Aniversario, es donde históricamente se inició el rearme científi co, académico y militar de los nuevos ofi ciales que iban a recorrer los primeros años del siglo XX. Fue precisamente en la antigua Sección de Higiene Veterinaria del antiguo Instituto de Higiene Militar donde se gesta una nueva y moderna veterinaria militar. Creo que más de cien años de convivencia entre facultativos en dicho Centro bien merece la pena para realizar una parada y una refl exión. En esos más de cien años la veterinaria militar experimentó una notable mejora, muchas veces desapercibida por nuestros compañeros de las profesiones hermanas, e incluso por los propios veteri-narios. En las siguientes líneas les relataré dónde, cómo y cuándo se inicia el camino de una nueva veterinaria militar.
LA FORMACIÓN VETERINARIA EN EL INSTITUTO DE HIGIENE MILITAR
Desde 1845, fecha de la constitución del Cuerpo de Veterinaria Militar, faltaba un Centro donde los nuevos ofi ciales veterinarios recibiesen una formación especí-fi ca en veterinaria preventiva y asistencia clínica que les capacitase para su ejercicio profesional en las Unidades hipomóviles. Conseguir que los veterinarios militares tuviesen una formación académica y científi ca adecuada y acorde con el paso del

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
17
tiempo fue una constante preocupación de ilustres jefes de veterinaria militar. En 1851 Fernando Sampedro Guzmán, profesor veterinario de primera clase, catedrático y Mariscal mayor publicó su “Higiene Veterinaria Militar”, libro que sirvió de texto para los ofi ciales veterinarios.
Pero las sucesivas dependencias del Cuerpo de Veterinaria Militar del Arma de Caballería (1845-1857), de Sanidad Militar (1857-1864) y nuevamente del Arma de Ca-ballería (1864-1889), en nada contribuyeron a su mejoría técnica, ni a su organización administrativa y logística-operativa. No son del caso analizar ahora las causas del estado de postración en que se hallaba la veterinaria militar de antaño, pero estaban muy relacio-nadas con la percepción social de la profesión, la estructura castrense, la falta de medios económicos y la constante inestabilidad política de la España decimonónica. A pesar de ello fueron las epizootias que asolaban las cabañas ganaderas y en especial las que afectaban a las yeguadas militares, junto con la aparición de zoonosis tan importantes como la rabia, la tuberculosis, el carbunco y el muermo las que pusieron a los veterinarios en el punto de mira de la sanidad castrense.
Combatir estas enfermedades infecto-contagiosas y parasitarias junto a los es-fuerzos realizados en reproducción equina, con el objetivo de suministrar équidos en buen estado de salud a las Unidades, hizo que los veterinarios militares necesitaran una instrucción y un apoyo técnico de mayor calado para prevenir ciertas enfermedades que, como el muermo (producido por la bacteria Burkholderia mallei), diezmaban la cabaña. Desde el momento en que se descubrió la maleína por los veterinarios rusos Helning y Kalning (éste veterinario militar) en 1890 se llevaron a cabo estudios clíni-cos y experimentales en todos los países más adelantados. Durante sus experiencias Kalning se inocula el muermo y paga con su vida el honor de haber descubierto la maleína. En España los primeros estudios clínicos se realizaron en 1884 por el veteri-nario primero (Capitán) Eusebio Molina Serrano en los caballos de la Guardia Civil de Puerto Rico, y después por Julián Mut Mandilago, veterinario segundo (Teniente) del

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
18
Regimiento de Caballería de Treviño, en Barcelona. Con posterioridad a esta primera época maleínica de las postrimerías del siglo XIX, fueron muchos los veterinarios civiles y militares que realizaron interesantes trabajos con la maleína como prueba di-agnóstica del muermo. Precisamente los veterinarios militares Marcelino Ramírez y Juan Igual Hernández trabajaron intensamente en el desarrollo e intro-ducción de la maleína como prueba diagnóstica. Más de un veterinario militar murió estudiando e investigando la forma de combatir y tratar esta enfer-medad que de forma esporádica sufre el hombre. Se realizaron estudios sobre la melioidosis, enfermedad infectocontagiosa producida en el hombre por un microorganismo muy próximo al agente etiológico del muermo (Burkholderia pseudomallei). Como ya hemos apuntado antes, en 1916 el muermo quitó la vida al veterinario 1º Juan Igual Hernández cuando realizaba nuevos experimentos y observaciones en la En-
fermería de Ganado de la guarnición de Melilla. El Capitán Igual se había consa-grado en cuerpo y alma al estudio de esa enfermedad; son nuestros héroes científi -cos olvidados que dieron a la sociedad lo más preciado que tenían: la vida.
En el IX Congreso de Higiene y Demografía organizado en 1898 por el In-stituto de Higiene Militar los veterinarios militares participaron muy activamente y contribuyeron con numerosas comu-nicaciones, una de ellas fue presentada por Molina Serrano bajo el título de “Maleinización”, en cuya discusión intervino, entre otras personalidades, el mismo Nocard, considerado como uno de los grandes veterinarios franceses especializado en microbiología. En su honor la Nocardiosis lleva su nombre. Con respecto al Tétanos, Roux y Nocard lograron en 1892 una alta inmunización en el caballo, estableciendo así las bases para la sueroterapia, que fue
Coronel Veterinario Eusebio Molina Serrano (1853-1924)

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
19
utilizada por primera vez en el hombre en 1894 por Bazy. Por todo ello desde la creación en 1885 del Laboratorio Histológico e Histoquímico éste comenzó a confi gurar unas nuevas perspectivas de apoyo técnico a los veterinarios. Comprobada la utilidad de la maleína por numerosos ensayos efectuados en todos los países y demostrada en 1898 en la obra premiada sobre “Higiene y Policía sanitaria de las habitaciones del ganado militar” de la que era autor el ya citado Molina, se reglamenta por Real Orden de 10 de mayo de 1899 la organización ofi cial de la maleinización en el Ejército.
Fruto de esta actividad fue la obra “Policía Sanitaria: profi laxis y tratamiento de las enfermedades infecto-contagiosas de los animales domésticos”, del mismo Molina Serrano. Sin embargo la primera reglamentación seria y científi ca de la lucha contra las epidemias del ganado del ejército fue también obra de Molina Serrano al redactar en 1898 el “Proyecto de Ley de Policía Sanitaria de los Animales Domésticos”, base sobre la que se sustentó la primera Ley de Epizootias en España. En 1901 se ini-cian en el Instituto los trabajos de veterinaria, entroncados con la clínica y prevención
en el hombre y colaborando como asesores en todo lo relacionado con las enfermedades infecto contagiosas y parasitarias de los ga-nados del ejército y su posible transmisión al hombre. Sus recomendaciones no fueron puestas en práctica hasta ese mismo año en que por Real Orden de 24 de julio se dictan medidas higiénicas para la conservación y entretenimiento del ganado destinado al servicio de las Unidades. También se apro-baron las instrucciones a las que debían ajustarse las prácticas de las desinfecciones y desinsectaciones de las caballerizas militares así como las medidas higiénicas para evitar infecciones y contagios en el ganado y su transmisión a las tropas. También se dieron instrucciones para luchar contra la durina,
Edmond Nocard (1850-1903)
CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
20
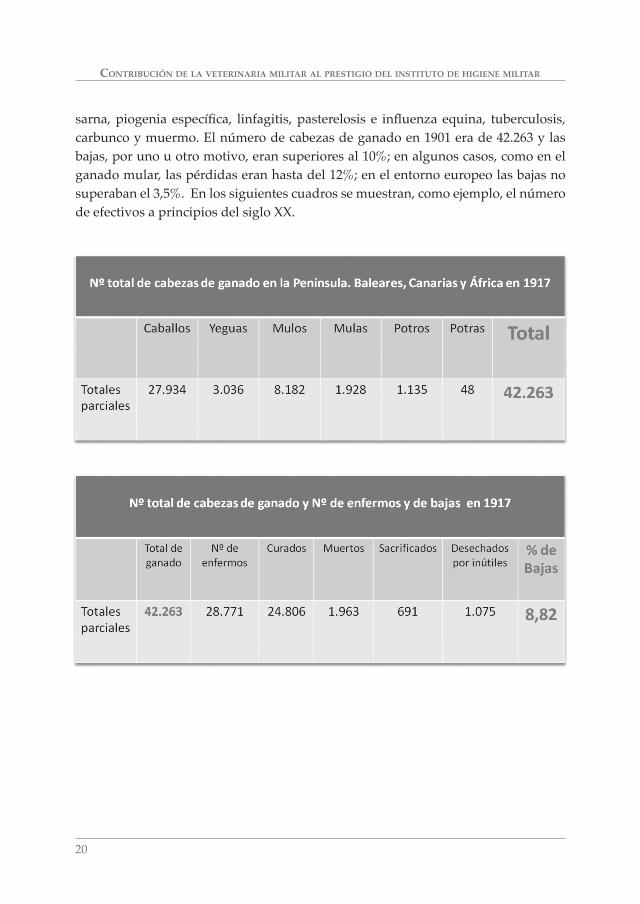
sarna, piogenia específi ca, linfagitis, pasterelosis e infl uenza equina, tuberculosis, carbunco y muermo. El número de cabezas de ganado en 1901 era de 42.263 y las bajas, por uno u otro motivo, eran superiores al 10%; en algunos casos, como en el ganado mular, las pérdidas eran hasta del 12%; en el entorno europeo las bajas no superaban el 3,5%. En los siguientes cuadros se muestran, como ejemplo, el número de efectivos a principios del siglo XX.
CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
21
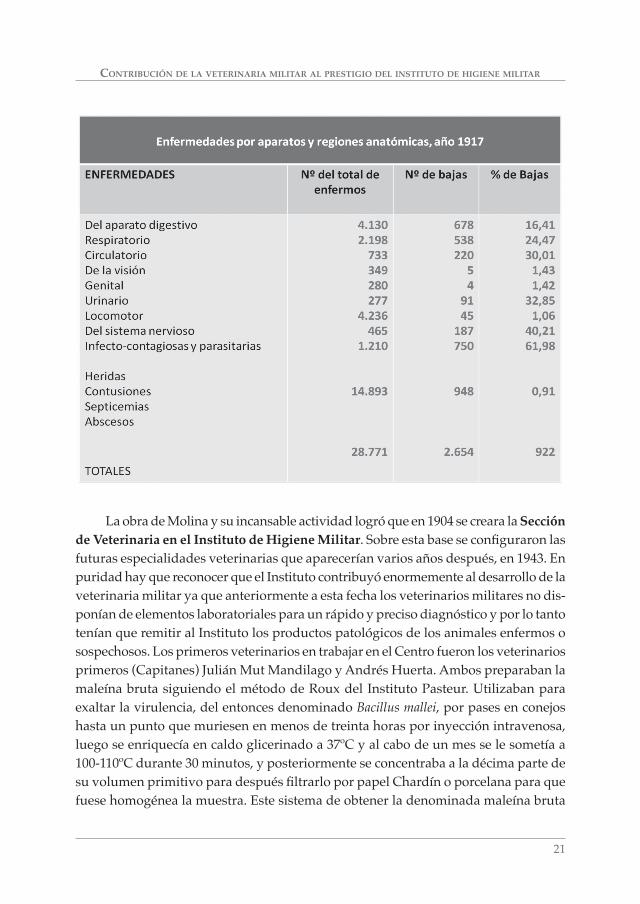
La obra de Molina y su incansable actividad logró que en 1904 se creara la Sección de Veterinaria en el Instituto de Higiene Militar. Sobre esta base se confi guraron las futuras especialidades veterinarias que aparecerían varios años después, en 1943. En puridad hay que reconocer que el Instituto contribuyó enormemente al desarrollo de la veterinaria militar ya que anteriormente a esta fecha los veterinarios militares no dis-ponían de elementos laboratoriales para un rápido y preciso diagnóstico y por lo tanto tenían que remitir al Instituto los productos patológicos de los animales enfermos o sospechosos. Los primeros veterinarios en trabajar en el Centro fueron los veterinarios primeros (Capitanes) Julián Mut Mandilago y Andrés Huerta. Ambos preparaban la maleína bruta siguiendo el método de Roux del Instituto Pasteur. Utilizaban para exaltar la virulencia, del entonces denominado Bacillus mallei, por pases en conejos hasta un punto que muriesen en menos de treinta horas por inyección intravenosa, luego se enriquecía en caldo glicerinado a 37ºC y al cabo de un mes se le sometía a 100-110ºC durante 30 minutos, y posteriormente se concentraba a la décima parte de su volumen primitivo para después fi ltrarlo por papel Chardín o porcelana para que fuese homogénea la muestra. Este sistema de obtener la denominada maleína bruta

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
22
fue el sistema que durante más de sesenta años se siguió practicando en el antiguo Laboratorio y Parque Central de Veterinaria Militar.
En 1907 se dispuso por Real Orden de 25 de abril (C.L. nº68) que los ofi ciales de nuevo ingreso en el Cuerpo de Veterinaria Militar realizasen, en el Instituto de Higiene Militar, un curso de prácticas antes de pasar a prestar servicio a los Regimientos y Yeguadas, ya que el Cuerpo de Veterinaria Militar no disponía de Academia Espe-cial para ello. En 1908 se establecen por Reales Órdenes de 6 de noviembre (Colec-ción Legislativa nº104) y 21 de noviembre de 1910 (Colección Legislativa nº188) los cursos de ampliación de estudios para veterinarios en el citado Instituto y en el 2º Establecimiento de Remonta; estos estudios estaban encaminados a perfeccionar los conocimientos bacteriológicos.
Julián Mut Mandilago, que además de veterinario militar era Licenciado en Filosofía y Letras (en la foto superior vemos a Mut practicando el vendaje ocular que lleva su nombre), fue el encargado en el Instituto de enseñar los últimos avances
de la bacteriología que tanto había prestigiado el químico Pasteur, y los veterinarios franceses Edmon Nocard, Camille Guérin y Gastón Ramón, y en España el veterinario catalán Ramón Turró y Darder, entre otros. La inmu-nización activa en el hombre y en los équidos con toxoide tetánico formolado fue elaborado sistemáticamente desde 1923 por Gastón Ramón y sus colabora-

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
23
dores. En la actualidad se acaba de redactar por el Dr. Galán Torres, veterinario militar jefe del servicio de microbiología e higiene y sanidad ambiental del Centro Militar de Veterinaria, una nueva biografía en español; el libro, prologado por mi persona, recoge aspectos inéditos de esta ilustre fi gura de la medicina veterinaria mundial. De esta sencilla y práctica manera el Instituto comienza a funcionar como Academia Militar y como Centro docente y de apoyo logístico-operativo siendo este el origen de las especialidades veterinarias. En 1909 el Instituto informa sobre la imposibili-dad de fabricar suero antitetánico y que en España no hay ningún laboratorio que lo haga, a pesar de la imperiosa necesidad mostrada por los ofi ciales veterinarios, como consecuencia urgente derivada de las heridas de castración y para atender las lesiones y heridas anfractuosas que se producían en maniobras y operaciones militares. El Dr. Ángel Pulido1, médico y Senador del Reino, es considerado por los veterinarios militares como el político que mejor defendió los intereses de la Veterinaria castrense al comprender de forma temprana (década de 1870) la importancia de la medicina preventiva veterinaria en toda su extensión.
Según un estudio realizado por este político en 1909 el Instituto contaba entonces con cuatro plazas para estabular los caballos siendo insufi ciente este material biológico para atender la producción de suero antitetánico; los establos y cuadras estaban en muy mal estado, no existían espacios para cuarentenas y no había sitio para colocar los animales de experimentación, y para aclarar el estado de postración en que se hallaba la veterinaria militar, añadía:
“La sección de la higiene veterinaria militar, que procura emular con plau-sibles testimonios de hondo saber á la higiene humana, tiene aquí también manifestada su digna representación, y produce, dentro de sus irrisorios y humillantes recursos, sueros y trabajos dignos de estimación. Cerca de mil ampollas de maleína llevaban preparadas en este año para diagnosticar el muermo que sufre el ganado del Ejército, el cual es frecuente.”
1 El Dr. D. Ángel Pulido, Senador del Reino, presentó en 1909 ante la Comisión de Presupuestos el trabajo titulado “La Sanidad Militar, su importancia en la salud del Ejército y en la salud pública. Tras-cendencia de su desenvolvimiento”. Realizó un brillante discurso ante los miembros de la Comisión del que los veterinarios militares se mostraron muy satisfechos. Desde ese instante fue considerado como el “Padre profesional” de la Veterinaria Militar. En 1917 la publicación “Revista de Veterinaria Militar”, le realizó una entrevista como homenaje a su labor política en pro de una mejora técnica y organizativa del Cuerpo de Veterinaria Militar.
Véase nuestro estudio sobre “La revista de veterinaria militar, primera publicación profesional de la veterinaria militar española (1915-1920). Temas de historia de la veterinaria. Coordinador prof. Dr. D. José Manuel Cid Díaz. Servicio de Publicaciones, Universidad de Murcia, pp: 213-240.

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
24
En 1915 por Real Orden Circular de 12 de agosto (Colección Legislativa nº 138) se le asigna al Instituto el cometido de elaborar el suero antitetánico. Esta elaboración se continuó en el antiguo Laboratorio y Parque Central de Veterinaria Militar y en la actualidad en el moderno y modélico Centro Militar de Veterinaria de la Defensa. A partir de esa fecha se han elaborado en las diferentes instalaciones y ubicaciones de los laboratorios veterinarios los siguientes productos biológicos para atender las necesidades de la clínica veterinaria:
• Anatoxina tetánica.• Antivirus.• Maleína diluída.• Maleína bruta.• Suero antiestreptocócico.• Suero anticarbuncoso.• Suero antitetánico.• Suero equino normal.• Vacuna antiabortus equi.• Vacuna anticarbuncosa.• Vacuna antiestreptocócica.• Vacuna antirrábica “Hogyes”.• Vacuna antirrábica “Humeno” para perros.• Vacuna contra el aborto vírico, así como
otras bacterinas y autovacunas.
El día 31 de octubre de 1915 se edita la primera revista profesional de veterinaria militar. Bajo el tí-tulo de “Revista de Veterinaria Militar” se recogieron en sus páginas una interesante e importante activi-dad científi ca, hasta que a fi nales de 1920 dejó de publicarse; algunas de las fotografías que ilustran el texto proceden de ella. En sus diferentes números se recogieron las noticias que generaba el Instituto de Higiene Militar; lo mismo podemos decir de la actividad de los Ateneos de Sanidad Militar (muy especialmente el de Madrid) y de los trabajos origi-nales de los veterinarios militares, todos ellos muy bien redactados y con una metodología científi ca

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
25
que en nada desmerecieron si los comparamos con los actuales. Muchos de ellos contribuyeron a mejorar y modifi car ciertas técnicas quirúrgicas. La microbiología alcanzó notables avances y las enfermedades infectocontagiosas y parasitarias que continuaron siendo el azote de estos motores biológicos, fueron combatidas con me-todología científi ca por parte de los veterinarios militares; las zoonosis de especial emergencia, como la tuberculosis con la práctica de la tuberculinización, la rabia y
la extracción del material nervioso del asta de Ammón y el muermo, con la introducción de la oftalmo-reacción, fueron las enfermedades diana. Como dato anecdótico añadiremos que a los ofi ciales veterinarios no se les autorizó (como a otros sanitarios, en especial los mé-dicos) a asistir como observadores a las ope-raciones militares de la Primera Gran Guerra para comprobar sobre el terreno el grado de organización de los servicios veterinarios de los Ejércitos contendientes, pero de fuentes secundarias sí recogieron numerosos datos de los Hospitales veterinarios, Hospitales de infecciosos, de etapas, enfermerías, Lazaretos, materiales de campaña, centros de carnización y toda la estructura y organización veterinaria. Además, los más inquietos (entre los que des-tacamos ahora a los destinados en el Instituto
de Higiene Militar) ilustraron al resto de los ofi ciales veterinarios con el suministro de datos de la patología más frecuente, brotes epidémicos, distintivos internacionales de la organización veterinaria en campaña y traducciones técnicas y científi cas de las principales revistas, francesas, alemanas e inglesas que se recibían en la Sección Veterinaria del Instituto de Higiene Militar. Uno de los trabajos que se difundió el 31 de octubre de 1915 entre todos los ofi ciales veterinarios fue el de la destrucción de moscas y desinfección de los cadáveres en la zona de combate. En ese mismo año de 1915 la veterinaria internacional incorpora la “Cruz Azul” como distintivo ofi cial de los órganos y estructuras civiles y militares veterinarias. Durante la contienda mun-dial aparecen los primeros “Centros de Carnización” y “Centros de concentración de ganado”, y por primera vez los veterinarios militares tienen que hacerse cargo del saneamiento de los Campamentos y la recogida, retirada y eliminación de los cadáveres de los numerosos équidos y sus híbridos que murieron a miles, unas veces

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
26
municionando a las unidades de Infantería y Artillería, otras transportando víveres y otras evacuando sobre sus lomos a los soldados heridos. La primera Convención de Ginebra de 1864 y la segunda de 1906 no contempló a los veterinarios como personal sanitario sino como contendientes. En 1914, durante la Primera Gran Contienda, se propuso incorporar a los veterinarios como personal sanitario, pero no se logró.
Mientras tanto en el Instituto de Higiene Militar continuaban los trabajos de los dos ofi ciales veterinarios. Andrés Huerta, cuyos artículos de opinión los fi rmaba en la revista de veterinaria militar como “Tahuer”, realizó un interesante trabajo original sobre “La vacunación antimuermosa”, pero los primeros resultados obtenidos acon-sejaron desechar esta vía de investigación. Su vacuna la preparó, según dijo, “…más como entretenimiento que como pretendida solución del asunto, pues no aspiramos a resolver el problema cuya incógnita no parece despejada del todo por los sabios maestros”. La vacuna la preparó siguiendo el método de Mac Kellar, análogo al de Silkman, y desconociendo las dosis que estos aplicaban a los équidos. Huerta utilizó dos lotes de cobayas: uno testigo y otro experimental. En sus conclusiones detalla que para obtener resultados positivos era necesario utilizar una dosis que el consideraba enorme: 150 cc en la primera inyección para un caballo de 300 kilos , y 300 cc en la segunda dosis; fi nalizó Huerta su trabajo diciendo: “La consideración de la enorme cantidad de vacuna necesaria, nos desanimó e hizo que abandonáramos la comenzada serie de experiencias que, quizás, reanu-daremos algún día en otra forma”. Las enseñanzas extraídas durante el desarrollo de las operaciones militares de la primera guerra mundial sobre la sanidad animal hicieron que por Real Orden de 16 de noviembre de 1917 se dispusiese que por el Instituto de Higiene Militar se suministrara a los cuerpos los viales de maleína bruta, que hasta entonces se adquiría en el extranjero. A partir de esa fecha es el propio Instituto quien asume la responsabilidad de su preparación para remitirla a los cuerpos montados, a solicitud de los ofi ciales veterinarios, para practicar el diagnóstico precoz por medio de la oftalmo-reacción. Para comprender el volumen de trabajo y la gravedad de los periódicos brotes epidémicos, que producían unas bajas anuales superiores al 8 por ciento, diremos que el número de semovientes adscritos a las diferentes unidades del ejército ascendía en 1916 a 42.964 cabezas; remontar cada año más de 3.500 cabezas era una importante operación, no exenta de grandes difi cultades. Los países de nuestro entorno tenían unas bajas del 3 al 3,5 por ciento.
La actividad del Instituto quedó claramente demostrada en el “Primer Congreso Nacional de Medicina” de 1917 cuando participó en la “Exposición de Medicina e Higiene” en el Parque del Retiro. El Instituto estuvo presente con una muestra de los productos preparados en sus laboratorios y colecciones fotográfi cas de su actividad; S.M el Rey contempló la fotografía de una yegua a cuyo pie se podía leer este curioso

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
27
rótulo: “Exprimir, 26 años; ha rendido 250 litros de suero antidiftérico valorado en 50.000 pesetas”. Estas abultadas cantidades dinerarias, unidas a la de sueros antitetánicos, vacunas antirrábicas y maleína que anteriormente eran importadas de otros países, generalmente Francia y Alemania, nos dan una muestra de lo que el Instituto, por intermedio de los veterinarios militares, ahorraba al Erario. No tiene nada de extraño que el Veterinario 1º, Manuel Medina, viendo el progreso de la Bacteriología y las nuevas vías de aplicación, se dirigiese al Mando con las siguientes palabras:
“En nuestra cínica veterinaria militar no se obtiene de estas conquistas todo el provecho logrado en otros países; nuestro Instituto de Higiene Militar, que es modelo de Establecimiento de su clase y que realiza una meritísima labor que, quizás, no se sabe apreciar en todo su valor, puede ampliar extraordi-nariamente la utilidad de sus servicios, proporcionando a los veterinarios militares muchos productos de laboratorio de que hoy no se hace uso, o éste es muy limitado, porque ello representa un enorme gasto. Para conseguir este mayor rendimiento, la Sección de Veterinaria del mencionado Instituto debe reorganizarse, aumentando su personal y dotación y encomendándole la obtención de todo género de productos útiles a la profi laxis y curación de las enfermedades del ganado militar y el estudio y comprobación de los im-portantísimos problemas relacionados con esta especialidad del Veterinario. La referida Sección de Veterinaria debe estar formada por un Subinspector de 2ª, un Veterinario Mayor y dos Veterinarios primeros y tener a su cargo la preparación de la maleína, suero equino normal, suero antiestreptocócico para veterinaria, y cuantos productos microbianos sean de reconocida utilidad en la clínica veterinaria, así como la de los diagnósticos bacteriológicos y seroló-gicos de las enfermedades del ganado. A ella debe corresponder el diagnóstico bacteriológico e histológico de la rabia en los animales y la preparación de la vacuna antirrábica”.
En los sucesivos años la Sección preparó dosis de la vacuna antirrábica “Umeno”. Posteriormente, y con el desarrollo y potenciación del Instituto, se constituyeron en 1934 diferentes secciones, una de ellas fue el laboratorio de higiene veterinaria con la misión de elaborar los productos biológicos dedicados a mantener la salud de los animales al servicio del Ejército. En este laboratorio se prepararon los sueros y vacunas como la antitetánica, antiestreptocócica, maleína, vacuna antirrábica, antierisipelótica contra el mal rojo y vacuna estreptobacilar.
También se realizaban los análisis clínicos de las numerosas muestras biológicas remitidas por los ofi ciales veterinarios desde sus enfermerías y botiquines; se con-

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
28
fi rmaban los brotes de sarna y tiña de los équidos y se procesaba toda la anatomía patológica de las piezas obtenidas tras la realización de las necropsias, e incluso las técnicas de aislamiento de asta de Ammón para visualizar los corpúsculos de Negri, patognomónicos de rabia. Los responsables en esa fecha eran: Andrés Huerta López, Subinspector de segunda clase, que actuaba como director de la Sección, auxiliado por los veterinarios mayores José Dornaleteche Zabalza y Victoriano Nieto Magán. En 1967 se crea la Sección de nutrición y alimentación dirigida por el Teniente Coronel Veterinario Arturo López Arruebo (el cual todavía asiste a los anuales ciclos de con-ferencias del Centro Militar de Veterinaria y a las Tertulias Veterinarias del Consejo General de Colegios Veterinarios de España).
Esta Sección tuvo sus orígenes en la donación de equipos de análisis efectuada por los Servicios de Nutrición de Tropas Americanas tras los estudios realizados en 1958 y años siguientes sobre los menús suministrados a un total de 10.727 soldados, quedando demostrado la necesidad de complementar la alimentación del soldado de una forma más racional. Más tarde se hizo cargo de la Sección el Teniente Coro-nel Veterinario Ángel Gonzalo Vitoria, que fue luego General Veterinario, y Manuel Alonso Rodríguez, Doctor en Veterinaria y ambos diplomados en Bromatología e
En el Museo de Veterinaria Militar se exponen los productos biológicos (maleína, sueros, vacunas, autovacunas y bacterinas) elaborados por los veterinarios militares

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
29
Higiene de los Alimentos. Otros Veterinarios que pasaron por esta Sección fueron los Tenien-tes Coroneles Francisco Porta Armendáriz y Alberto Rodríguez Zazo, con dos doctorados, uno en Veterinaria y otro en Medicina; estaban ambos diplomados en microbiología y análisis clínicos veterinarios; Manuel Sánchez Martín y Octavio de Toledo y Ubieto (con dos licenciatu-ras, una en Veterinaria y la otra en Medicina), Diplomados ambos en Bromatología e Higiene de los Alimentos, Comandante Pedro Fernán-dez Domínguez (veterinario y odontólogo), Diplomado en Microbiología e Higiene y Sani-dad Ambiental, y en la actualidad el Teniente Coronel Veterinario Alejandro Palomo Gago, con la misma especialidad; este último, fruto de su experiencia y conocimientos en el campo de la Bromatología fue el autor del manual de “Alimentación y Seguridad Alimentaria” en el ámbito de las Fuerzas Armadas que ha sido publicado en el año 2009 a propuesta e impulso de la Jefatura de Apoyo Veterinario de la Inspección General de Sanidad. La existencia de un animalario, hasta su desaparición y pase a la estructura del Hospital Militar Gómez Ulla, y hoy formando parte de la estructura del Centro Militar de Veterinaria de la Defensa, hizo que la dirección fuese encargada sistemáticamente a los veterinarios militares. Nume-rosas especies animales sirvieron como reactivo biológico (como muy acertadamente lo defi nió Iam Peter), para el diagnóstico y preparación de sueros y vacunas. En el anexo I se recogen todas las vicisitudes por las que ha pasado el antiguo Instituto de Higiene Militar hasta llegar a las modernas instalaciones del actual Centro Militar de Veterinaria de la Defensa.
Y ya para fi nalizar señor presidente, señoras y señores académicos, verdadera-mente podemos concluir, diciendo con satisfacción, que el antiguo Instituto de Higiene Militar contribuyó en grado sobresaliente a la formación de los veterinarios militares, actuando como una Academia Militar de Aplicación de las Ciencias Veterinarias en el ámbito de las Fuerzas Armadas.
He dicho.

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
30
ANEXOI
CUADRO RESUMEN DE LA EVOLUCIÓN DE LA SECCIÓN DE HIGIENE VETERINARIA
Periodo-Fecha Denominación Actividad científi ca y técnica1885-1888 Laboratorio Histológico e
Histoquímico en el Real Monasterio de Nobles
Funciones analíticas y de ayuda al diagnóstico
1888 Instituto Anatómico-Patológico. Comienza a realizar funciones de Academia de Sanidad Militar hasta 1895
1890 Se crea el Instituto Vacunógeno que se instala en el Instituto Anatómico-Patológico, con el que termina fundiéndose.
Se preparan sueros y vacunas
1898 Cambia su denominación a Instituto de Higiene Militar, situado en la calle Pintor Rosales, 10.
Se organiza el IX Congreso de Higiene y Demografía
1904 Se crea la Sección de Veterinaria Militar
Se inician las actividades de diagnóstico laboratorial y de enseñanza de los futuros ofi ciales veterinarios. Se inicia la actividad de preparación de sueros y vacunas
1936-1939 Existen dos Institutos, uno en Madrid y otro en Valladolid. En 1937 comienza a funcionar la “Sección de Veterinaria” dentro del Laboratorio Central de Sanidad. En 1938 el Servicio Veterinario se organizó en tres Agrupaciones con cabeceras en Valladolid, Zaragoza y Sevilla, cada una con un Laboratorio y Parque.
Durante la guerra civil se realizan los análisis microbiológicos, parasitológicos y de alimentos
1939 Por el Ejército Nacional se ocupan las instalaciones de la Jefatura de la Inspección de Veterinaria de la República existente en Barcelona. Una comisión de veterinarios militares se hace cargo del Laboratorio de sueros y vacunas para ganadería que estaba anexo a la Inspección de Veterinaria Militar. Pasa a denominarse Laboratorio Central de Veterinaria Militar, ubicado en Barcelona en la calle Tibidabo.
CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
31
1940 El Laboratorio y todo el material veterinario de los almacenes pasa a Valladolid, constituyéndose el Laboratorio Central y el Parque de Material Veterinario, los cuales se funden en el Laboratorio y Parque Central de Veterinaria Militar
1942 Se traslada a Madrid el Laboratorio y Parque Central de Veterinaria Militar, a la calle Núñez de Balboa 72.
1961 Se traslada, con carácter provisional, el Laboratorio a una de las alas de la Academia de Sanidad Militar en Carabanchel y el Parque se le ubica en unas naves almacén la calle Invencibles nº6, cerca de la Puerta de Toledo.
1981 Se coloca la primera piedra del nuevo Laboratorio y Parque Central de Veterinaria Militar, en la calle Darío Gazapo nº3 en la zona de Campamento. Madrid.
1983 Se traslada el Parque a las nuevas instalaciones
1986 Se traslada el Laboratorio desde la Academia de Sanidad Militar a las nuevas instalaciones de Darío Gazapo (Campamento-Madrid).
1986 Todas las instalaciones veterinarias (excepto la Agrupación de Tropas de Veterinaria Militar de la Reserva General) quedan agrupadas en un nuevo Acuartelamiento recibiendo el nombre de Laboratorio y Parque Central de Veterinaria Militar (LPCVM)
1987 Pasa a denominarse “Centro Militar de Veterinaria” (CEMILVET)
Se le asigna misiones de apoyo logístico general y directo a las Unidades del Ejército de Tierra

CONTRIBUCIÓN DE LA VETERINARIA MILITAR AL PRESTIGIO DEL INSTITUTO DE HIGIENE MILITAR
32
1998-2011 Se integra el Centro Militar de Veterinaria en el Órgano Central del Ministerio de Defensa pasando a denominarse “Centro Militar de Veterinaria de la Defensa” (CEMILVETDEF)Pasa a depender de la Jefatura de Apoyo Veterinario (JAV) de la Inspección General de Sanidad de la Defensa (IGESAN)La veterinaria militar inicia una nueva etapa.
Se le asigna misiones de apoyo logístico general y directo a las Unidades de los Ejércitos de Tierra, Aire, Armada y Órgano Central. Se le dota de laboratorios de bromatología, microbiología y sanidad ambiental, Policlínica Veterinaria, servicio de medicina y cirugía experimental, Unidad de Apoyo Logístico, Escuela Cinológica de la Defensa, Salón de Actos, Aulas, Museo, Biblioteca, Centro de Documentación y un Centro de investigación histórica de veterinaria militar

LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
33ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
Excelentísimo Sr. Presidente de la Real Academia de Ciencias Veterinarias de Anda-lucía Oriental, Excmo. Rector Magnífi co de la Universidad de Jaén, Ilustrísimos señores y señoras Académicos, compañeros y compañeras, querida familia y amigos todos.
En primer lugar, quisiera agradecer a la Real Academia y a sus ilustres miembros su confi anza en mí para este nombramiento, que considero un verdadero privilegio y honor. También quisiera agradecer de un modo especial a D. Antonio Marín su empeño personal en este nombramiento. Tras mi formación como biólogo y espe-cialización como microbiólogo, debo advertir que mi visión del mundo animal es meramente microscópica, valga la paradoja. Por ello me van a permitir que enfoque este discurso desde ese punto de vista, analizando algunos aspectos de las relaciones de los microorganismos con los organismos superiores.
INTRODUCCIÓN
La historia de la humanidad ha estado salpicada por plagas de diversa naturaleza, como recogen numerosos documentos históricos. En muchas de ellas, como la peste o la viruela, las sintomatologías descritas confi rman claramente su etiología microbiana, aunque en muchos otros casos menos evidentes también se sospecha la intervención del
LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
ANTONIO GÁLVEZ DEL POSTIGO RUIZ1
Discurso de Ingreso en la Real Academia de Ciencias Veterinarias de Andalucía Oriental como Académico Correspondiente
1 Catedrático de Microbiología de la Universidad de Jaén

LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
34
látigo fustigador de los microorganismos. Llegado a una situación de equilibrio inesta-ble, como es la actual, entre humanos y microbios, y no siendo mi intención castigar a la audiencia con tintes calamitosos, prefi ero hablaros de los aspectos positivos que nos brindan los microorganismos, esos eternos compañeros de viaje.
Los microorganismos comenzaron a moverse por nuestro planeta hace aproxima-damente 3500 millones de años, de forma que cuando se establecieron los organismos superiores ellos ya conocían a la perfección los secretos de la vida así como los trucos para relacionarse de un modo más provechoso con las nuevas formas evolutivas que iban apareciendo. En la actualidad se asume que las relaciones microbio-hospedador pueden ser de varios tipos, como comensalismo, mutualismo, parasitismo o patoge-nicidad, pudiendo marcar defi nitivamente el estilo de vida del hospedador.
MICROORGANISMOS ASOCIADOS A LOS ANIMALES: EL MICROBIOMA
Las interacciones entre microorganismos y animales presentan una larga historia evolutiva y por consiguiente han tenido un marcado efecto en conformar la vida en nuestro planeta. Para muchos microorganismos somos un sustrato rico en materia orgánica altamente apetecible, un territorio a conquistar, invadir y parasitar hasta su extenuación. Por el contrario, otros microorganismos mucho más modestos, apro-vechan nuestras limitaciones para transformar por completo los componentes del alimento, y eligen como hábitat principal nuestro intestino. Tales microorganismos se han adaptado al ambiente digestivo a lo largo de millones de años, siendo testigos y actores en nuestros cambios evolutivos.
La adaptación a la dieta es uno de los factores principales en la evolución de todas las especies, desde los microorganismos hasta los primates humanos y los no humanos. De hecho, los principales eventos evolutivos en la separación de los huma-nos del resto de primates confl uyen con adaptaciones relacionadas con la dieta. No es sorprendente por tanto encontrar diferencias anatómicas en el tracto digestivo de los vertebrados relacionadas con la calidad y abundancia de alimentos. Si bien el meca-nismo de digestión en vertebrados está conservado en la evolución a lo largo de los diferentes taxones, los sistemas digestivos de las diferentes especies han evolucionado en respuesta a las cualidades estructurales y nutritivas de sus dietas específi cas.
Dado que los microbios del tracto digestivo juegan un papel crítico en la adapta-ción de la especie a una dieta particular proporcionando rutas metabólicas complemen-tarias esenciales, cabe esperar que los microorganismos infl uyan en la evolución del hospedador. Existen, de hecho, numerosas evidencias de que las bacterias comensales

LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
35
o mutualistas obligadas asociadas a insectos son fundamentales para la supervivencia del hospedador no sólo porque le proporcionan nutrientes esenciales, sino también porque le protegen de invasores hostiles. Más aún, el desarrollo normal de diversos animales y plantas requiere la presencia de bacterias simbióticas, como es el caso de los órganos luminosos de la sepia donde se aloja Aliivibrio fi scheri, o las bacterias fi jadoras de nitrógeno en los nódulos de las raíces de las plantas. Lo mismo ocurre en el caso de los mamíferos. Así, por ejemplo, se ha demostrado que el polisacárido producido por Bacteroides fragilis dirige la maduración celular y física durante el desarrollo del sistema inmunológico en ratón, dirigiendo la organogénesis del tejido linfoide e induciendo correcciones de posibles defi ciencias y desequilibrios en las células linfocitarias T. Por otra parte, la leche materna humana contiene oligosacáridos que llegan intactos al intestino grueso donde viven las bifi dobacterias, estimulando su crecimiento y favoreciendo el desarrollo de la incipiente microbiota intestinal del recién nacido. Todo un ejemplo, en defi nitiva, de coevolución y sincronía en el baile de la vida.
Trabajos recientes muestran también cómo el fenotipo metabólico humano está fuertemente infl uenciado por el microbioma del tracto digestivo. Y a su vez, la adaptación del hospedador a una dieta específi ca proporciona a los microorganismos la capacidad de evolucionar y adaptarse a la dieta del hospedador, lo que da como resultado una diversifi cación y co-evolución de ambas especies.
Pero, ¿qué entendemos por microbioma? El término “microbioma” fue propuesto por el premio Nóbel Joshua Lederberg, y se refi ere al conjunto de genomas de nues-tros microbios autóctonos o microbiota. En los estudios realizados en mamíferos, se ha encontrado una relación directa entre la composición de su microbioma intestinal y la dieta, resultando que especies próximas fi logenéticamente también muestran microbiomas más próximos. De los tres grupos estudiados (herbívoros, omnívoros y carnívoros), los primeros mostraron la mayor diversidad microbiana, seguido de omnívoros y carnívoros.
La adaptación a la utilización de vegetales en la dieta fue un hito importante que permitió la expansión de los mamíferos por numerosos hábitats. Se estima que el 80% de los mamíferos extinguidos eran herbívoros. Para poder acceder a los com-plejos carbohidratos de los vegetales, como celulosas y almidones, los mamíferos debieron incrementar la longitud (y a veces el volumen) de sus sistemas digestivos, prolongando el tiempo de digestión para permitir la fermentación microbiana. Esto ocurrió bien al comienzo o al fi nal del tracto digestivo, traduciéndose en diferencias marcadas en las comunidades microbianas de una y otra sección anatómica. Los es-

LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
36
tudios de metagenómica del rumen nos permiten conocer mejor la microbiología y el funcionamiento de este órgano, y a la vez descubrir nuevos genes que podrían tener un enorme interés aplicado, como por ejemplo en la industria para la conversión de materia vegetal en biocombustibles.
EL METAGENOMA HUMANO
Una vez obtenida la secuencia completa del genoma humano, el eminente mi-crobiólogo Julian Davies (Universidad de British Columbia) señaló que descifrar el genoma humano no era sufi ciente para entender nuestra biología, pues hay más de 1000 especies bacterianas viviendo en y sobre nuestro organismo, las cuales afectan a nuestra vida de forma crítica. Julian Davies predijo que este amplio conjunto de bacterias podrían albergar entre dos y cuatro millones de genes nuevos, además de los 30.000 del genoma humano, pudiendo ser que la acción conjunta de ambos (los genes humanos y los microbianos) fuesen determinantes para nuestra salud.
Animados por el éxito de los trabajos de metagenómica a gran escala llevados a cabo por Craig Venter en el mar del Sargaso y por los avances científi cos en secuen-ciación masiva, un grupo investigadores europeos se aventuraron en los estudios de metagenómica humana con el fi n de obtener claves sobre su infl uencia en nuestra salud. En 2005, el Instituto Nacional Francés para la Investigación en Agricultura (INRA) convocó un simposio internacional en París para discutir la investigación del microbioma humano. Esta reunión dio origen al Consorcio Internacional sobre Microbioma Humano, cuya misión era establecer relaciones entre los genes de la microbiota humana y nuestro estado de salud. El proyecto resultante, denominado MetaHIT (Metagenomics of the Human Intestinal Tract) fue fi nanciado por el Séptimo Programa Marco de la Unión Europea y está revolucionando nuestra visión de la mi-crobiota intestinal. Al poco tiempo, el Instituto Nacional de Salud de Estados Unidos puso en marcha en 2007 otro proyecto más ambicioso sobre microbioma humano, enfocado sobre cuatro ambientes diferentes: tracto gastrointestinal, boca, vagina, y piel. Su objetivo es estudiar el microbioma completo de 250 individuos normales y secuenciar 1000 genomas de referencia de las bacterias que se encuentran comúnmente en el tracto intestinal humano, en un periodo de 5 años.
DIVERSIDAD DE LA MICROBIOTA INTESTINAL HUMANA
El metagenoma del tracto digestivo humano es un consorcio complejo formado por miles de millones de microbios, principalmente bacterias, y una proporción mucho

LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
37
menor de hongos, protozoos y virus. Se estima que la microbiota intestinal humana contiene 10 veces más células bacterianas que células tiene nuestro organismo. Lo mismo ocurre con el resto de mamíferos. Así pues, cuando Noé introdujo en el arca una pareja de animales de cada especie, no podía imaginar la cantidad de polizones microbianos que llevaba.
En el primer estudio sobre diversidad microbiana en mucosas y heces de tres individuos sanos se encontraron 395 especies diferentes, en su mayoría Firmicutes (301), seguido de Bacteroidetes (65). El 80% de las especies descritas en este estudio nunca habían sido cultivadas, y el 60% correspondían a nuevas especies. En otro estudio más amplio sobre 124 individuos se encontraron entre 1000 y 1150 especies prevalen-tes, estimándose que cada individuo podría tener al menos 160 especies bacterianas diferentes. Sin embargo, ninguna de las especies mostraba una abundancia relativa superior al 1%, lo que hace difícil defi nir cuál es la microbiota autóctona o enterotipo de cada individuo con la tecnología actual. Más difícil aun es discernir las diferencias existentes a nivel de cepa, lo que sin duda se traduce en diferencias signifi cativas a nivel de funcionalidad. De hecho, aunque el número de enterotipos estables existentes podría ser limitado, la abundancia de especies no se relaciona directamente con las funciones más representadas, como apunta un estudio publicado en el último número de la revista Nature (mes de mayo), en el que se subraya la importancia de realizar análisis funcionales para poder comprender el ecosistema intestinal.
En la actualidad se estima que el microbioma intestinal humano contiene alre-dedor de 150 veces más genes que el propio genoma humano. Dicho de otra forma, sin consideramos la diversidad de especies microbianas presentes en nuestro tracto digestivo y sus genes, podemos decir que somos un 10% humanos y un 90% micro-bios. Este “órgano” esencial (el microbioma intestinal), proporciona al hospedador un aumento considerable en sus capacidades metabólicas, a la vez que le confi ere protección frente a la invasión por patógenos, educa a nuestro sistema inmunitario, y modula el desarrollo del tracto intestinal.
Los estudios de funcionalidad del microbioma humano ponen de manifi esto la complementariedad metabólica que este conjunto de genes aporta a nuestro tracto digestivo. En un estudio reciente se reconstruyeron 11 rutas metabólicas completas, encontrándose una mayor abundancia de las rutas para degradación de carbohidratos, biosíntesis de aminoácidos esenciales y vitaminas. También predominan las rutas de detoxifi cación de compuestos xenobióticos, así como la ruta de metanogénesis, de gran importancia para la retirada de hidrógeno. La retirada de este gas producido durante la fermentación bacteriana en el colon, es fundamental para el procesamien-

LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
38
to efi caz de los polisacáridos ingeridos en la dieta, y para mantener la homeostasis del intestino. Toda una serie de procesos en defi nitiva que nuestro organismo nunca podría llevar a cabo sin la intervención de los microbios.
Resulta también llamativo el estudio sobre microbioma de individuos japoneses, en el que se detectó una mayor abundancia de genes que codifi can para porfi rianasas. Estas son un grupo específi co de enzimas capaces de degradar los porfi ranos presentes en las algas rojas, componente habitual en la dieta de este país. Los genes para estos en-zimas están presentes exclusivamente en microorganismos marinos, pero fueron también adquiridos por una especie bacteriana del intestino denominada Bacteroides plebeius que se ha encontrado solamente en individuos japoneses. Este no es más que un ejemplo de cómo nuestra microbiota se ha adaptado a lo largo de la evolución a nuestros hábitos alimenticios para permitirnos aprovechar mejor los nutrientes que ingerimos. Cabría preguntarse, en un tono jocoso, qué genes ha podido perder o ganar Bacteroides plebeius a lo largo de la evolución de los humanos para adaptarse a nuestro estilo de vida. ¿Te-nemos también una microbiota específi ca para la digestión de alimentos más exquisitos, con especies como Bacteroides aristocraticus, Bacteroides nobile o Bacteroides royale? ¿Podría nuestra microbiota explicar en parte nuestros hábitos alimenticios? ¿Infl uye en nuestra apetencia por determinados alimentos y en el rechazo o intolerancia hacia otros?
MICROBIOTA Y ESTILO DE VIDA
El éxito evolutivo de los humanos se debe en parte a nuestra capacidad para controlar la cantidad y variedad de comida disponible, al menos en los países de-sarrollados, mediante el desarrollo de la ganadería, la agricultura, y los métodos de procesado y conservación de alimentos. Una cuestión clave es conocer cómo infl uyen los diferentes ambientes en que se desarrolla el ser humano y la alimentación de las diferentes civilizaciones en la ecología microbiana del tracto digestivo. Los seres hu-manos actuales estamos adaptados genéticamente al ambiente en que sobrevivieron nuestros ancestros, con un microbioma que codifi ca para funciones metabólicas com-plementaria, como la capacidad de extraer nutrientes y energía a partir de muchos componentes de la dieta. Aunque los hábitos alimenticios son uno de los factores más importantes para el microbioma humano, no está claro cómo ha infl uido la diversi-fi cación de la dieta del hombre moderno en la microbiota del tracto digestivo ni las posibles consecuencias de tales cambios para nuestra salud.
La revolución del periodo Neolítico trajo consigo cambios profundos en la dieta y el estilo de vida de los humanos, derivadas de la introducción de la agricultura y

LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
39
la ganadería. A partir de ese momento, las reservas de alimentos se hicieron más abundantes y constantes, originando una concentración de las poblaciones humanas y posiblemente también las primeras epidemias de enfermedades infecciosas. Se ha postulado que las bacterias del tracto digestivo sufrieron transformaciones intensas durante los cambios sociales y demográfi cos que tuvieron lugar en los primeros asentamientos neolíticos, alcanzando posteriormente un equilibrio más estable que ha perdurado hasta épocas recientes. Sin embargo, durante los siglos 19 y 20 se han producido numerosos cambios en nuestros hábitos de vida. Los países desarrollados han aprendido a controlar las enfermedades infecciosas clásicas mediante una mejora generalizada de las condiciones sanitarias así como mediante el uso de antibióticos y vacunas. Sin embargo, al mismo tiempo se ha producido un incremento considerable de nuevas enfermedades como las alergias y otros desórdenes inmunitarios, o los síndromes de infl amación intestinal. Muchas de estas alteraciones se atribuyen a una interacción incorrecta de la microbiota con el sistema inmunitario.
En el ámbito científi co se reconoce que la microbiota intestinal es esencial para el desarrollo normal y la homeostasis del sistema inmune. La mucosa intestinal es la mayor superfi cie de interacción del organismo con el exterior, y el tejido linfoide asociado es responsable de la tolerancia inmunológica, diferenciando las bacterias comensales que se adhieren de forma normal a la mucosa intestinal de las patógenas, y tolerando también los componentes inmunogénicos de los alimentos. Los estudios científi cos sugieren que la educación de nuestro sistema inmunitario se produce en las primeras etapas de la vida y es esencial para el correcto funcionamiento posterior de nuestro organismo. Sin embargo, los cambios en nuestro estilo de vida parecen estar dando al traste con adaptaciones establecidas a lo largo de miles o incluso millones de años. Nuestro estilo de vida urbanita nos aleja cada vez más de los microorga-nismos con que se educaron nuestros ancestros del Neolítico, pudiendo ser esta la causa de muchos de los confl ictos de salud que padecemos en la actualidad. Estas observaciones han llevado a los investigadores a postular la conocida como hipótesis de la higiene. Paradójicamente, esta hipótesis pretende relacionar la mayor incidencia y gravedad de los procesos alérgicos en el mundo desarrollado con las mejoras de nuestros estándares de higiene y sanidad.
En un afán por defi nir y rescatar la microbiota ancestral de los humanos, y recordando un poco a las cruzadas de Indiana Jones, un estudio reciente comparó el microbioma de niños de una aldea rural de África (en un ambiente que se podría comparar al de los primeros agricultores del Neolítico) con niños del continente eu-ropeo, que consumen dietas representativas de la civilización moderna. Los niños de la muestra Africana, que consumían una dieta rica en fi bra, mostraron unos niveles

LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
40
signifi cativamente superiores de Bacteroidetes y unos niveles de Firmicutes muy infe-riores. Entre los primeros, destacó la abundancia de bacterias que degradan la celulosa y los xilanos presentes en la fi bra vegetal, que por el contrario estaban ausentes en la microbiota de los niños europeos, más habituados a la videoconsola y el bollicao. Estas bacterias tienen la capacidad de producir mayores niveles de ácidos grasos de cadena corta, tales como el butirato, cuyos efectos protectores contra la infl amación intestinal han sido probados. En resumen, la microbiota de los niños africanos está más adaptada a sus hábitos alimenticios específi cos, y les permite obtener un mayor rendimiento energético de la dieta rica en fi bra, a la vez que les proporciona protección frente a las infl amaciones y enfermedades no infecciosas del colon. Por el contrario, la reducción de la riqueza en diversidad microbiana o la pérdida de nuestros antiguos microbios sean posiblemente una de las consecuencia indeseables de la globalización y de nuestros hábitos de consumo hacia una alimentación basada en alimentos gené-ricos, ricos en nutrientes fácilmente asimilables, y libres de microorganismos.
MICROBIOTA Y OBESIDAD
Algunos estudios recientes van todavía un poco más allá y relacionan el des-equilibrio de nuestra microbiota intestinal con la obesidad, otro de los grandes males de nuestro tiempo. Se ha demostrado que la microbiota intestinal es muy dinámica, ya que su composición y su expresión metabólica cambian rápidamente con la dieta. Por ejemplo, la administración de dietas ricas en grasa en ratones provoca cambios en su microbiota que son detectables a las 24 h, incrementando notablemente la po-blación de Firmicutes. De forma sorprendente, cuando esta microbiota se transplanta a ratones gnotobióticos o libres de microbios, aumenta en ellos la acumulación de grasa en hasta un 50% y reduce la sensibilidad a la insulina. Por tanto, es posible hablar de una microbiota obesogénica. Así mismo, en individuos humanos obesos, se observa con frecuencia una microbiota intestinal alterada. En un estudio reciente se han encontrado un conjunto de aproximadamente 500 genes relacionados con la obesidad, que están enriquecidos o ausentes en determinados individuos según su índice de masa corporal. Los genes enriquecidos en individuos obesos provienen ex-clusivamente de dos grupos bacterianos (Actinobacteria y Firmicutes), mientras que los genes más abundantes en individuos delgados pertenecen grupos taxonómicos más variados. A modo de resumen, el estudio concluye que la microbiota de individuos obesos es menos diversa y está enriquecida en genes implicados en el metabolismo de carbohidratos, lípidos y aminoácidos.

LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
41
Existen también datos signifi cativos sobre la infl uencia de la microbiota intesti-nal en la diabetes de tipo 1 y tipo 2. Un estudio reciente indica que los pacientes con diabetes tipo 2 muestran niveles inferiores de una bacteria cuyo nombre requiere un ejercicio intenso de vocalización, denominada Faecalibacterium prausnitzii, bacteria que por el contrario predomina en el intestino de individuos sanos. En consecuencia, se ha propuesto el uso de esta bacteria como probiótico para combatir la resistencia a la insulina.
MEJORA DE NUESTRO ESTADO DE SALUD A TRAVÉS DE LA MICROBIOTA
Los resultados que acabo de comentar nos llevan directamente a otra cuestión que he querido plantear en este discurso: cómo podemos manipular la microbiota intestinal para mejorar nuestro estado de salud. Allá por 1928, el científi co ucraniano Elie Metchnikoff consideró que la mayor longevidad de los habitantes del Cáucaso podría estar relacionada con su elevado consumo de productos lácteos fermentados, los cuales contienen grandes cantidades de bacterias ácido-lácticas. Metchnikoff fue también el primero en proponer la administración de dichas bacterias con fi nes tera-péuticos, para modifi car la fermentación pútrida en el intestino.
La ingestión de bacterias lácticas junto con alimentos fermentados no es algo novedoso, ya que se remonta a varios miles de años en la historia de los humanos, allá por los inicios de la ganadería y la agricultura. En la actualidad consumimos cantidades ingentes de bacterias lácticas a través de los alimentos fermentados. Se estima que el consumo anual de productos fermentados en Europa es de unos 22 kg por habitante, o lo que es lo mismo, un total de 8.5 billones de kg. Si consideramos una carga microbiana de 108 bacterias lácticas por gramo de alimento, tendremos un total de 8.5x1020 bacterias, lo que equivale a 3400 toneladas de biomasa bacteriana. Esos datos no tienen en cuenta el consumo de otros productos fermentados, como vegetales o cárnicos, o el consumo de preparados probióticos. Estos últimos se defi nen como “alimentos a base de microorganismos vivos que benefi cian al animal que los ingiere, mejorando el balance de la microbiota intestinal”. Los probióticos constituyen en la actualidad el 60-70% del mercado total de alimentos funcionales, mercado que se espera vaya en aumento.
Pero, ¿cómo actúan los probióticos? El lanzamiento de esta gama de produc-tos al mercado vino rodeado de grandes expectativas, ya que fueron considerados como panacea para el autocuidado de la salud y la curación de múltiples dolencias y afecciones. Nuestras exigencias para con estas superbacterias no son nada triviales.

LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
42
Las bacterias probióticas deben sobrevivir a la barrea gástrica, implantarse en nues-tra mucosa intestinal compitiendo con el resto de microorganismos del intestino, y ejercer allí una amplia gama de efectos como mejora de la intolerancia a la lactosa, disminución o prevención de diarreas, mejora del tránsito intestinal, modulación de nuestro sistema inmunitario, mejora de determinados procesos infl amatorios intes-tinales y determinados procesos de alergia, reducción de los niveles de colesterol, o incluso disminución del riesgo de padecer determinados tipos de cáncer. Diferentes cepas de lactobacilos y bifi dobacterias, e incluso alguna bacteria entérica y levaduras, compiten en este escenario por demostrar sus bondades para con nuestro organismo.
Si bien algunos de los efectos antes mencionados han sido probados fehacien-temente, en otros casos queda un largo camino por recorrer, con resultados muy variables o poco o nada repetitivos. No obstante, el boom propagandístico sobre los alimentos probióticos ha sido tan exacerbado que la Autoridad Europea en Seguridad Alimentaria (EFSA) y posteriormente la Agencia Española de Seguridad Alimentaria y Nutrición han tomado cartas en el asunto, regulando de un modo estricto la forma en que las industrias pueden hacernos llegar sus recomendaciones e insinuaciones sobre las bondades y milagros de los microorganismos probióticos y sus preparados comerciales.
PROBIÓTICOS PARA ANIMALES
El interés por los probióticos se ha extendido también al campo de la veterinaria, como no podía ser de otro modo. Actualmente existen grandes expectativas sobre la aplicación de probióticos en animales de granja, con fi nes muy diversos como mejorar la asimilación de alimento (y por consiguiente acelerar el proceso de engorde), for-talecer el sistema inmunitario del animal de forma que este se defi enda mejor frente a la infección por patógenos, prevenir la aparición de diarreas y otros trastornos intestinales, o disminuir la colonización por esos patógenos (como Escherichia coli, Salmonella o Campylobacter) que luego originan tantos problemas por su transmisión a lo largo de la cadena alimentaria. Y, por supuesto disminuir el uso de antibióticos, o simplemente como alternativa para hacer frente a las normativas que prohíben de-terminados usos. Los resultados en este campo van desde los claramente alentadores, como el caso de las aves de granja, para las que existen diversos preparados comer-ciales en el mercado, a otros más modestos cuya aplicación en la vida real requiere sin duda de esfuerzos importantes, como por ejemplo los destinados a aumentar la producción de leche en vacas o a combatir la acidosis del rumen.

LA COMPLEJIDAD DE LAS RELACIONES DE LOS MICROORGANISMOS CON ORGANISMOS SUPERIORES
43
No podemos dejar a un lado nuestros animales de compañía, cuya salud pue-de ser tan importante como la nuestra, y con los que mantenemos claros vínculos afectivos. Los probióticos para gatos y perros circulan libremente por el mercado en perfecta armonía, a diferencia de sus destinatarios. Van destinados fundamentalmente a fortalecer el sistema inmunitario del animal, y a reducir los niveles de clostridios y coliformes en heces. Es en este campo donde la publicidad nos hace confundir con más facilidad las diferencias entre efectos probados y posibles, y donde el aspecto económico pasa a menudo a un segundo plano, cosa que las empresas productoras conocen a la perfección.
No quisiera cansar a la audiencia con más contingencias entre el mundo micro-biano y el mundo animal. Muchas gracias por vuestra atención y paciencia, y que vuestra microbiota os sea favorable.
BIBLIOGRAFÍA
Arumugam, M., et al., 2011. Enterotypes of the human gut microbiome. Nature 473(7346):174-180.Cani, P. D., Delzenne, N.M., 2011. The gut microbiome as therapeutic target. Pharmacology and The-
rapeutics 130(2):202-212. De Filippo, C., et al., 2010. Impact of diet in shaping gut microbiota revealed by a comparative study
in children from Europe and rural Africa. Proc Natl Acad Sci U S A 107(33):14691-14696.Hess, M., et al., 2011. Metagenomic discovery of biomass-degrading genes and genomes from cow
rumen. Science 331(6016):463-467.Ley, R.E., et al., 2008. Evolution of mammals and their gut microbes. Science 320(5883): 1647–1651.Mira, A., Pushker, R., Rodríguez-Valera, F., 2006. The Neolithic revolution of bacterial genomes.
TRENDS in Microbiology 14(5): 200-206.Zhu, B., Wang, X., Li, L., 2010. Human gut microbiome: the second genome of human body. Protein
Cell 1(8):718-725.


CONTESTACIÓN AL DISCURSO DE INGRESO DEL ILMO. SR. D. ANTONIO GALVEZ DEL POSTIGO RUIZ
45ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
Excmo Sr. Presidente de la Real Academia de Ciencias Veterinarias de Andalucía Oriental
Excmo. Rector Magnífi co de la Universidad de Jaén
Ilustrísimos Sres y Sras Académicos,
Distinguidas autoridades,
Compañeros y compañeras, Querida familia y
Amigos todos
Con gran satisfacción cumplo el encargo que me ha encomendado este Real Aca-demia de contestar el discurso de ingreso del Prof. Galvez del Postigo, al tiempo que me resulta sumamente grato y un alto honor, darle la bienvenida en nombre de esta Real Corporación. Una persona de tan completa formación en la que confl uyen una esmerada preparación académica, un trabajo infatigable y su pasión investigadora, será un sólido baluarte que prestigiará a esta Academia.
Esta personalidad profesional se refl eja en un extenso y brillante currículo, del que es difícil llevar a cabo una rápida semblanza. Espero disponer de la debida elocuencia.
CONTESTACIÓN AL DISCURSO DE INGRESO DEL ILMO. SR. D. ANTONIO GALVEZ DEL POSTIGO RUIZ
ILTMA. SRA. DRA. Dª. CATALINA GÓMEZ LÓPEZ1
1 Académica de Número de la Real Academia de Ciencias Veterinarias de Andalucía Oriental.

CONTESTACIÓN AL DISCURSO DE INGRESO DEL ILMO. SR. D. ANTONIO GALVEZ DEL POSTIGO RUIZ
46
El Prof. Galvez es licenciado en Ciencias biológicas por la Universidad de Gra-nada y Doctor en Ciencias por la Universidad de Granada. Su actividad docente ha transcurrido por todos los estamentos universitarios, desde becario pre doctoral y profesor asociado en la Universidad de Granada, hasta Profesor titular y Catedrático de la Universidad de Jaén en la Facultad de Ciencias Experimentales, en el Depar-tamento de Ciencias de la Salud en el área de microbiología, cargo que desempeña en la actualidad.
Paralela a esta actividad académica, hay una ingente labor investigadora que se ha centrado muy especialmente en el área de la microbiología y en su aplicación a la Seguridad Alimentaria, participando en más de una quincena de proyectos de investigación fi nanciados en convocatorias públicas tanto nacionales como interna-cionales, en muchos de ellos como investigador principal.
Esta capacidad investigadora ha tenido su refl ejo en la extensa labor divulgativa de sus resultados de investigación que componen un abultado plantel de publicacio-nes. El Prof. Galvez ha publicado más de 170 publicaciones científi cas personales o en colaboración con un amplio equipo de competentes investigadores colaboradores que se han publicado en revistas de alto impacto científi co en su área de conocimiento como Food Microbioly, Meat Science, Journal of Food Protein, Journal of Dairy Scien-ce, Food Chemical Toxicology, etc, y ha publicado en una larga lista de países como Reino Unido, Canada, Francia, EEUU, Checoslovaquia, Alemania, Holanda, India, Australia, Burkina Fasso y por supuesto en España. Los anales de la Real Academia de Ciencias Veterinarias de Andalucía Oriental tiene el honor de contar entre sus colaboradores con el Prof. Gálvez.
La sólida formación científi ca del recipendiario, ha sido aprovechada para la transferencia de conocimientos desde los grupos de investigación al sector productivo participando en 6 proyectos de I+D con empresas como el grupo de leche Pascual o AB-Biotics.
Su formación científi ca se ha consolidado con estancias en diversas Instituciones nacionales y extranjeras, entre las que podemos citar la realizada durante 72 semanas en EEUU en los laboratorios Merck Sharp & Dohme, Res. Labs.
Su inquietud en el campo científi co es admirable y en su currículo nos informa que ha realizado más de 130 contribuciones a Congresos Nacionales e Internaciona-les mediante porters, comunicaciones orales y ponencias en campos como el control de patógenos en los alimentos, antimicrobianos, conservación y degradación de los alimentos, etc. Además, ha dirigido 4 tesinas y 14 tesis doctorales.

CONTESTACIÓN AL DISCURSO DE INGRESO DEL ILMO. SR. D. ANTONIO GALVEZ DEL POSTIGO RUIZ
47
La dimensión académica, investigadora e innovadora del Prof. Gálvez es reco-nocida al ser nombrado Director de Secretariado de Investigación de la Universidad de Jaén.
En fi n, esta sucinta exposición, extraída de su amplio currículo, es más que sufi ciente para comprobar que el Dr. Galvez del Postigo tiene sobrados méritos para su ingreso en esta Real Corporación
Después de haber escuchado el magnífi co trabajo con el recipendiario nos ha recreado y personalmente me ha deleitado, es el momento de cumplir con la norma protocolaria de responder al trabajo expuesto.
Debo reconocer que la lectura previa del discurso del Prof Galvez, además de acercarnos a las más modernas concepciones y teorías acerca de la relaciones fi lo-genéticas que actualmente se discuten en relación con nuestros microorganismos autóctonos, produjo en mi sensaciones agradables que trajeron a mi memoria el tiempo de mi investigación doctoral.
En ella estudie como las bacterias ruminales eran capaces de alimentarse de subproductos inútiles para el hombre (NNP, ácido úrico, materiales fi brosos) pero que ellas, con su maravilloso metabolismo de fermentación, transformaban en AGV y proteínas de alta calidad contenidas en su propia biomasa y que servían de ali-mentos al rumiante que las hospedaba. El moderno término de biosostenibilidad ya fue inventado hace miles de años por unas bacterias que funcionan como fábricas mejoradoras de alimentos.
Aquello fue tan útil y tan mágico que desde entonces quedé totalmente subyu-gada por la capacidad de seducción de las bacterias y desde luego he podido percibir en el discurso leído, que el Prof Galvez ha caído víctima del mismo hechizo.
En similitud con la estrecha simbiosis entre las bacterias ruminales y los herbí-voros, el discurso del prof. Galvez nos ha acercado a la simbiosis de la microbiota y el hombre y las relaciones que comporta para nuestra salud. Nos ha ilustrado sobre el benefi cio sinérgico que el microbioma intestinal proporciona al hombre, aumentando considerablemente sus capacidades metabólicas, o la protección que nos confi ere frente a la invasión por patógenos, a la vez que educa nuestro sistema inmunitario.
Me ha resultado esclarecedor que también en los humanos como en los rumiantes, la fi bra de la dieta es esencial para mantener la microbiota ancestral. Esa fi bra supo-ne una fuente de energía decisiva para mantener en buena salud nuestras bacterias intestinales, lo cual signifi ca efectos positivos para nuestro bienestar.

CONTESTACIÓN AL DISCURSO DE INGRESO DEL ILMO. SR. D. ANTONIO GALVEZ DEL POSTIGO RUIZ
48
Hace 2500 millones de años, no había rincón de la superfi cie de la Tierra en el que no pulularan las bacterías que ya entonces, habían inventado ingeniosos meca-nismos metabólicos esenciales en la actualidad, como la fermentación, la respiración del azufre, la biosíntesis o la fi jación del nitrógeno, y se las habían ingeniado para buscar soluciones a la contaminación, convirtiendo un terreno de aspecto lunar poblado de cráteres en el fértil planeta en que vivimos. La unidad mas pequeña de vida es un monumento de formas y procesos que no tiene rival en el universo tal y como lo conocemos.
La afi rmación del Dr. Galvez de que somos un 90% microbios y un 10% huma-nos tiene sobrados argumentos. El biólogo británico Richard Dawking en su libro el relojero ciego, escribe una frase que nos debería hacer refl exionar “El mundo esta dividido, fundamentalmente entre bacterías y el resto”. En fi n, siempre creyendo que estabamos en la cima del Universo y resulta que somos un 90% bacterías. Es la maravillosa lección de humildad con la que nos obsequia la naturaleza.
Quede como principal desafío científi co de las refl exiones planteadas por el Prof, Galvez que el conocimiento, cuidado y mejora de nuestra microbiota será esencial para mejorar nuestro estado de salud y el de nuestros animales.
Puesto que la valía personal del nuevo académico no necesita de adornos ba-nales y haciendo de la brevedad una virtud, quisiera terminar ya esta intervención.
Esté seguro el Dr. Galvez de que la Academia lo recibe con gran beneplácito para ella; yo les felicito a ambos; a ella, porque adquiere en este acto un elemento valioso y digno del honor que esto signifi ca y a Ud., porque satisface esa justa y plausible aspiración de ver reconocidos mérito y trabajo.
Para fi nalizar les deseo a todos ustedes que interaccionen correctamente con su microbiota y ello les confi era niveles elevados y permanen tes de salud. Muchas gracias

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
49ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
Excelentísimo Sr. Presidente de la Real Academia de Ciencias Veterinarias de Andalucía Oriental, Excmo. Sr. Presidente del Instituto de Academias de Andalucía, Ilustrísimo Sres. Presidentes de los Ilustres Colegios Ofi ciales de Veterinarios de Gra-nada y Almeria, Sr. Vicepresidente del Ilustre Colegio Ofi cial de Médicos de Málaga, Iltmo. Sr. Alcalde de Benalmádena, Ilustrísimos Señores Académicos, compañeros, familia y amigos todos.
Representa para mi un gran honor y es motivo de un sano orgullo encontrarme hoy, y precisamente en nuestra casa, que es la de todos, rodeado de este apretado nú-cleo de excelentes compañeros y amigos para que, si procede, me deis el visto bueno para ingresar en este egregio colectivo de nuestra Veterinaria Andaluza, al que, en su caso, contribuiré con lo mejor de mí para su engrandecimiento.
A estos efectos, y aunque se merecieran más y mejores palabras, es de rigor que con las pocas y torpes que se me ocurren dé las gracias a nuestro Presidente Antonio Marín Garrido y a Manuel Muñoz, que con su gran liberalidad aceptó de inmediato el encargo de ser mi padrino en este acto que es para mi motivo de especial agrade-cimiento.
Pero antes de entrar en materia y de cambiar el percal por la seda, como tantas veces nos apuntaba nuestro siempre recordado Profesor de Microbiología cuando éramos estudiantes, quiero pediros que me permitáis que con el mayor de los sen-
LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
ANTONIO VILLALBA GÓMEZ
Discurso de ingreso en la Real Academia de Ciencias Veterinariasde Andalucía Oriental como Académico Correspondiente

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
50
timientos fi liales ofrezca este acto a la memoria de mi padre, que de haber vivido, hoy se sentiría orgulloso no sólo de verme titulado y al frente de un colectivo tan importante como el nuestro, sino también, rodeado de lo mejor de nuestra clase, a punto, si obtengo vuestra venia, de ingresar en nuestra Academia.
Y a lo que iba. La Fiesta de los Toros y las Normas que la han regido en las dis-tintas épocas, siempre fueron, como se dice, cogidas de la mano.
Desde la aparición de las Tauromaquias de “Pepe Hillo” y de “Paquiro” hasta nuestros días se han sucedido un sinfín de disposiciones que han ordenado y regu-lado, parcial y a veces deslavazadamente, el desarrollo de los espectáculos taurinos tal y como los conocemos hoy día.
Desde la considerada primera regulación ofi cial del Espectáculo Taurino, el “Reglamento para las funciones de toros en la Plaza de Madrid” aprobado el 30 de junio de 1852 por el Gobernador D. Melchor Ordóñez , donde se ordena por primera vez de forma ofi cial el reconocimiento de los caballos de picar y se habla de las características que tenían que presentar los toros, hasta la publicación del DECRETO 68/2006 de 21 de Marzo, por el que se aprueba el REGLAMENTO TAURINO DE ANDALUCIA, la presencia Veterinaria en los Espectáculos Taurinos ha sido siempre discutida y lo sigue siendo hoy día.
Sin embargo, hay un hecho incuestionable, la permanencia veterinaria, la pre-sencia ininterrumpida del veterinario todas las tardes y en todos los espectáculos a lo largo de más de un siglo y medio.
El veterinario, a lo largo de estos años habrá gozado de mejor o peor imagen, pero a pesar de las controversias esta presencia ininterrumpida ha ido siempre acom-pañada de un incremento de sus cometidos.
No obstante, debemos reconocer que a pesar de nuestra fi rme pero a veces discreta actuación en el desarrollo de nuestras funciones en el espectáculo taurino, algunas generaciones de afi cionados, críticos e incluso autoridades implicadas en la fi esta han considerado e incluso propalado que los veterinarios ocupásemos, en un segundo plano, el lugar que se otorga a técnicos que solventan situaciones y problemas de importancia sin tener en cuenta, unos y otros, que “la tarde memorable que perdura en el recuerdo del buen afi cionado tiene detrás la callada labor de un equipo de Veterinarios”.
Los cambios sufridos por el espectáculo a lo largo de estos años por razones tanto políticas como de la propia evolución de la tauromaquia o debidas a los cambios de sensibilidad del público han condicionado por supuesto la labor del veterinario.

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
51
La suerte de varas quizás haya sido la que más ha cambiado y la que mas polé-micas ha suscitado. La implantación del peto en los caballos de picar en 1928 se debe, entre otras razones, a la decidida voluntad de un veterinario, D. José García Armen-dáriz, Inspector General de Sanidad Veterinaria en el Ministerio de Gobernación.
Aquel peto, sin embargo, era muy distinto al que concluida la Guerra Civil fue preciso introducir, con objeto de proteger mejor la escasa ganadería caballar que había quedado después de aquella triste contienda y que es el mismo que se utiliza en la actualidad ,con las correspondientes reformas.
Los reconocimientos veterinarios, han planteado una problemática distinta en cada época y evolucionan junto a los cambios experimentados por el toro, el criterio de la afi ción, de los ganaderos y, por que no decirlo, de los propios diestros. Ha sido un merito de la profesión veterinaria la rapidez para adaptarse a estos cambios.
La normativa taurina, como expresión de la regulación del espectáculo, ha sido el soporte para las decisiones y actuaciones del veterinario en la plaza de toros, pero también, y hay que reconocerlo, el quebradero de cabeza al tener que lograr su justo cumplimiento.
Nuestros cometidos en los espectáculos taurinos se han visto muchas veces fortalecidos por normativas colaterales, como lo fue en su día la Ley de Epizootias de 1954, o la creación del Libro Genealógico de la Raza de Lidia.
La responsabilidad del veterinario se acrecienta al estar en el centro de una compleja trama en la que se mezclan intereses muy fuertes y diversos de empre-sarios, ganaderos, apoderados, fi guras del toreo, afi cionados, políticos, periodistas etc.; y donde las decisiones hay que tomarlas en poco tiempo y a veces bajo una gran presión mediática.
La competencia se ve ampliada en la continua actitud ante el fraude o el deterioro del espectáculo: Manipulación fraudulenta de las astas, “afeitado”, enmascarar defectos y enfermedades “ dopaje”, o la perdida de calidad del espectáculo por las caídas y falta de fuerzas , todo ello sumado a otras crisis, como las debidas a epizootias, fi ebre aftosa, peste equina o mas recientemente la lengua azul, sin olvidar la aparición de la Encefalopatía Espongiforme Bovina en el año 2000.
Me gustaría, por todo ello, aprovechar esta intervención para hacer una breve reseña histórica de lo que ha sido la actuación veterinaria en los Espectáculos Taurinos en este último siglo y medio.

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
52
Las primeras noticias que se tienen de las actuaciones de nuestros precursores en el tiempo en los espectáculos taurinos, diferentes que eran, naturalmente, a las actuales, se remontan al siglo XVI, y se refi eren sobre todo a las intervenciones clínicas realizadas a los caballos heridos en el transcurso de las celebraciones.
Muchos son los manuales de albeitería pertenecientes a dicho siglo, donde se documentan estas actuaciones. D. Francisco de la Reina, por ejemplo, dedica un capitulo de su” Libro de Albeyteria” a las cornadas sufridas por los caballos en los espectáculos taurinos, haciendo referencia en él a los tipos de herida, la intervención quirúrgica y los posibles pronósticos.
El albéitar Don Pedro García Conde, en su libro “Verdadera Albeytería,” publicado ya en el siglo siguiente, año 1685, nos informa sobre una faceta inédita hasta el mo-mento: “la colaboración entre albéitares y picadores para el diagnóstico de las enfermedades de los caballos, así como la descripción de las evoluciones que debían realizar los jinetes con el caballo y de los signos que mostraban los animales y que facilitaban el diagnóstico que el albéitar debía realizar.”
Pero además de curar las heridas de los caballos y los perros, los albéitares tam-bién se atrevían a aconsejar sobre las cualidades que tenían que presentar los caballos para el toreo y sobre la forma en que era más conveniente que los sujetara el picador.
Las labores de asesoramiento llegaban aún más lejos, hasta el punto de que los albéitares se permitían aconsejar a los contratistas de caballos en las plazas, como lo demuestra de manera indirecta la crítica del matador José Delgado, “Pepe-Hillo”, en su libro “La tauromaquia o arte de torear”, a la actuación de los mismos en los festejos tau-rinos, cuando, al describir las cualidades que a su juicio tenían que reunir los caballos de picar, afi rmaba que: “no se debía tener en cuenta la opinión de los albéitares por parte de los asentistas, sino el dictamen de los picadores; porque no es la sanidad lo que se busca, sino las aptitudes de plaza, que mucho mejor las conocen estos últimos.”
Existen documentos que demuestran que, durante los siglos XVI Y XVII, en el norte de España se curaba a los toros después de los festejos, reservándolos para nuevos espectáculos.
Aunque en la práctica ya existiese una implicación real de los albéitares en los festejos taurinos, hay que esperar sin embargo a la publicación en 1868 del reglamento taurino del Marqués de Villamagna, titulado “ Reglamento para las corridas de toros en Madrid” para que, por primera vez la actuación veterinaria, se viera ofi cialmente regulada en los espectáculos taurinos, recibiendo los profesionales el nombre de

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
53
“revisores veterinarios”, lo cual legitimaba por fi n el trabajo desarrollado con anterio-ridad por el gremio de albéitares y albéitares-herradores en la cura de animales y en el asesoramiento en los festejos y espectáculos taurinos.
El hecho de que el término que apareciese en la normativa taurina del siglo XIX fuera el de veterinario puede deberse al proceso de unifi cación de las diferentes denominaciones existentes en torno a ese único término profesional, el cual se inicia con la Real Resolución de 23 de febrero de 1792 por la que se funda la primera Real Escuela de Veterinaria en Madrid, y se continúa con el Real Decreto de 19 de agosto de 1847 por el que se aprueba el primer Plan de Estudios de la Carrera de Veterinaria, perfectamente reglado ya y con mayor base científi ca.
Desde el primer momento de la presencia veterinaria ofi cialmente regulada en los espectáculos taurinos, su actuación fue cuestionada por los distintos sectores implicados, pero sobre todo, por aquellos cuyos intereses podían amenazar sus de-cisiones sanitarias. Pero lo cierto es que su labor se fue consolidando, seguramente por basarse, como señalan diversos autores, en la propia necesidad- ya expresada por el propio Francisco Montes, Paquiro, en su obra “Tauromaquia completa de 1836” - de contar con la presencia de un hombre que pudiera asesorar al presidente, y al que se debía capacitar para tomar decisiones en determinadas circunstancias.
Un hombre “fi el, competente,” que entre otra misiones tendría que “reconocer el ganado antes de traerlo a la plaza, para ver si tienen los hierros y marcas de las ganaderías a que dice el asentista que pertenecen, para que no engañen al publico, como sucede todos los días anunciando toros de castas acreditadas u oriundas de ellas y corriéndolas luego cuneros. Deberá también este hombre examinar si los toros tienen la edad y fuerza sufi ciente y por ultimo, si la vista y demás requisitos necesarios se hallan como se desean, para desechar los que carezcan de las proporciones oportunas para la lidia. “
Las misiones encomendadas al “fi el”, a lo largo de la historia de la Tauromaquia han pasado casi en su totalidad a ser actuaciones veterinarias.
Antes de la publicación del Reglamento de 1868 ya se venia hablando de que sería conveniente realizar un reconocimiento pericial de las reses que se fuesen a lidiar para evitar los abusos que frecuentemente se cometían.
Prueba de ello es un escrito del empresario de Madrid D. Justo Hernandez, el 17 de Abril de 1850, al Sr. Jefe Superior de Policía , donde se quejaba de estos episodios y solicitaba además la supresión de la suerte de los perros de presa, que los ganaderos consideraban como un castigo, pero que por otra parte el público demandaba cada vez que el toro presentado para la lidia no era totalmente de su agrado.

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
54
No será hasta la considerada primera regulación ofi cial del espectáculo taurino, es decir, el Reglamento para las funciones de toros en la Plaza de Madrid, aprobado el 30 de junio de 1852 por el Gobernador Don Melchor Ordóñez, cuando se ordenará por primera vez de forma ofi cial el reconocimiento de los caballos de picar por la autoridad competente, aunque sin especifi carse el mismo todavía, y se profundizará además en las características que tenían que presentar los toros, señalándose los posibles defectos que no debían mostrar y permitiéndose a la autoridad el rechazo de los animales que no cumpliesen los requisitos.
Todavía, no obstante, no se recogía en este documento legal la realización de un reconocimiento de los toros.
Los reconocimientos, sin embargo, se realizaban, aunque fuese todavía de mane-ra extraofi cial, a pesar de que los responsables de los reconocimientos no estuvieran específi camente nombrados, ni descrita la emisión de los informes derivados de los mismos.
La evidencia de que son los propios veterinarios los encargados de realizarlos, se fundamenta en el recurso interpuesto en Cádiz, en 1866, por el Subdelegado de Sanidad Veterinaria, José María Offarrel O´Connor, donde reclamaba sus honorarios por los reconocimientos practicados a los animales que habían intervenido en los espectáculos taurinos.
Habría que esperar aún hasta el Reglamento para las corridas de toros de Madrid, aprobado el 28 de mayo de 1868 por el Excmo. Sr. Gobernador de la provincia, y fi rmado el 30 de mayo de 1868 por el Marqués de Villamagna, Alcalde-Corregidor de Madrid, para ver por primera vez refl ejada en una regulación ofi cial la presencia y funciones de los veterinarios en los espectáculos taurinos.
Su función, en un primer momento, se dirigió al reconocimiento de perros, caballos y toros, consistiendo en la valoración morfológica de los animales y en su correcta identifi cación.
Así, en cuanto al examen de los caballos, se expresaba que debía considerarse su alzada y fuerza; pero sobre los toros no se señalaba en qué debía consistir su reco-nocimiento, aunque sí se especifi caba que se haría la reseña de ellos, se dibujaría el hierro y se anotaría el orden de salida al ruedo.
En los perros la situación era similar a la de los toros, ya que sólo se indicaba la existencia de su reconocimiento y la obligación de reseñarlos. Igualmente, se desconoce la existencia de un modelo de certifi cado para dichos reconocimientos.

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
55
Con este reglamento se comienza a exigir al veterinario la entrega por escrito del resultado de los reconocimientos, aunque no se indica en el texto si estas certifi -caciones eran o no vinculantes para el presidente del festejo, ni tampoco el número de profesionales que debían realizarlas, ni los honorarios a
devengar por estas actividades. Sí se señala, en cambio, la forma de avisarles para la realización del trabajo y las localidades gratuitas que se les tenían que facilitar.
La presencia veterinaria se consolida en el siguiente “Reglamento para la plaza de toros de Madrid,” fi rmado el 14 de febrero de 1880, siendo Gobernador el Conde de Heredia Spínola, y donde se sigue manteniendo como la actividad principal de los veterinarios el reconocimiento de los animales. En esta ocasión, sin embargo, se amplía a dos el número de los reconocimientos que debían realizarse a los caballos de picar, en los que se obviaba su sanidad, y aparece la fi gura del perito dirimente, la cual tenía que recaer en el Subdelegado Facultativo del Distrito.
Se recoge también por primera vez la obligación de inspeccionar, por parte de los Subdelegados de Veterinaria, las canales y vísceras de los animales lidiados antes de ser derivadas a consumo. Y se contempla además la sanción a los veterinarios en el caso de que hubieran dado por válido algún toro que presentase en la plaza un defecto físico.
En relación con estos nuevos cometidos, desde el primer momento los vete-rinarios van a encontrarse con limitaciones y difi cultades a la hora de certifi car los requisitos de sanidad y utilidad para la lidia de los toros.
Prueba de ello es la forma de redactarse el acta perteneciente a la corrida ex-traordinaria de toros, celebrada el 30 de mayo de 1880,en Madrid, en la que los Sub-delegados de Veterinaria, afi rmaban:
“... hasta donde lo permiten las condiciones especiales de estas reses, al hacerlas pasar de un corral a otro, y a la distancia que han sido vistas, aparecen dotadas de la debida sanidad y utilidad para la lidia que serán objeto...”
A pesar de las limitaciones y las difi cultades que conllevaban los reconocimientos, pronto sus actuaciones se vieron denostadas, cuando el público no quedaba satisfecho con el resultado ofrecido por los toros.
Un artículo publicado el 28 de diciembre de 1885 en la Gaceta Medico-Veterinaria se hacía eco de los ataques a la actuación veterinaria, realizados por críticos taurinos y parte del público, al no quedar contentos con el juego dado por los toros.

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
56
Para contrarrestar estas críticas, y con el fi n de aumentar el crédito de la actua-ción veterinaria ante estos sectores, con el titulo“El reconocimiento de los toros de plaza por los profesores veterinarios en Madrid”, se publico en la misma revista un artículo donde se describían las funciones que realizaban los profesores veterinarios en los espectáculos taurinos en estos términos:
“Para que se vea toda la importancia del trabajo de la inspección de reses bravas y la de los caballos que sirven para la lidia, bastará fi jarse en que el reconocimiento de los toros en vida debe abarcar:
1º. Su sanidad. 2º. Si tienen o no la edad reglamentaria. 3º. Contrastación del hierro de la ganadería, y 4º. Si reúne condiciones para la lidia. En cuanto al reconocimiento de caballos hay que tener en cuenta: 1º. El recuento del número que sea costumbre presentar en cada plaza. 2º. Su clasifi cación. 3º. Apreciar las condiciones que deben tener esos animales para no com-prometer, por su debilidad, enfermedad o resabios, la vida del picador.”
Bajo el título “El reconocimiento facultativo de los toros de lidia” publicado el 15 de mayo de 1891 en la revista “La Veterinaria Contemporánea”, el autor vuelve a refl ejar las situaciones tan complicadas que vivían los veterinarios en la realización de los reconocimientos, al tener que hacerlos con todos los animales juntos en el mismo co-rral, estando durante ese acto rodeados de multitud de gente expresando su opinión y sufriendo vejaciones.
En aplicación de lo dispuesto en los reglamentos taurinos, se venía enjuiciando y sancionando la actuación veterinaria en relación con sus funciones. Las malas con-diciones de los toros durante la lidia, en aquel tiempo, eran causa de sanción para los veterinarios.
El aumento de responsabilidades veterinarias asumidas en las actas de recono-cimiento, no exigidas reglamentariamente, se constata con la nota manuscrita que acompañaba al acta, de la corrida extraordinaria celebrada el 30 de mayo de 1887 en Madrid, con estas palabras:
“La edad con exactitud se hará constar después del reconocimiento de boca, terminada la corrida.”

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
57
El examen “post-mortem” de la boca, sin estar expresamente reglamentado, también se haya refl ejado en otra serie de documentos..
Así, en el acta de reconocimiento perteneciente a la corrida extraordinaria en benefi cio del Hospital Provincial de Madrid celebrada el 31 de mayo de 1888, además de las reseñas de las reses reconocidas con la mención explícita de estar dotadas de la sanidad y utilidad para la lidia hasta donde lo permitían las condiciones de estas reses y las del local donde se efectuaba el reconocimiento, se vuelve a hacer referen-cia a que la edad de los toros reseñados se refl ejará con exactitud en una segunda certifi cación con estas palabras:
“Que los toros reseñados al margen, cuya edad se hará constar con exactitud en segunda certifi cación por el examen de las respectivas mandíbulas...”
Ya en siglo XX Manrique Cantalapiedra en la II Asamblea Nacional Veterinaria celebrada en Madrid en 1907, reclamaba un mayor cometido científi co en los reco-nocimientos de los toros de lidia, pidiendo que en éstos se determinara la sanidad y utilidad para la lidia, obligándose a la entrega de los resultados en actas por triplicado.
El término de sanidad se englobaba en una idea de integridad del organismo, y el de utilidad para la lidia lo unía a la idea de trapío, entendiendo por este último término:
“...desarrollo general conformación ó conjunto que ha de ser lo más armonioso posible, su edad, aparente estado de salud, y la mayor desenvoltura en sus movimientos.”
Además, el examen, según el autor, debía ser tenido en cuenta dentro de las condiciones en las que se realizaban los reconocimientos a distancia de los animales y con todos los toros del festejo en un mismo corral.
Los reconocimientos de los caballos de picar, se consideraban muy defi cientes, ya que solamente se comprobaba la alzada y doma, ignorándose su sanidad, lo que ocasionaba consecuencias, trágicas a banderilleros y espadas en caso de cogidas por la posible transmisión de enfermedades infecciosas a través de los cuernos.
Igualmente, consideraba inadmisible la carencia de unos honorarios dignos a devengar por los servicios veterinarios prestados.
Por último, concluye su disertación señalando el problema de la inspección de la carne de lidia para el consumo humano y el de los nombramientos veterinarios para actuar en los espectáculos taurinos, aspectos que habrían de tener honda repercusión dentro de la profesión a lo largo del siglo XX.

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
58
A principios del siglo XX, debido a la existencia de diferentes reglamentos para las distintas plazas de toros de España, unido a que todavía no se había resuelto de forma general y precisa el desarrollo de las funciones veterinarias, unas reglamen-tadas y otras asumidas simplemente por la costumbre, se hacían necesarios para la profesión veterinaria los recordatorios de los cometidos que se llevaban a cabo en los espectáculos taurinos, así como de las disposiciones que los regulaban, difundidos por lo general en revistas profesionales de la época.
Ejemplo de ello es un artículo publicado en 1915, en la Revista Veterinaria Española, en donde se exponían estas funciones, además de señalarse que sería de aplicación el Reglamento de Madrid en todas aquellas localidades que no tuvieran reglamento propio .
Con la publicación del “Reglamento de las corridas de toros, novillos y becerros” de 1917 volverían a aumentar las funciones de los veterinarios, ya que se incluirá por primera vez un reconocimiento previo de las instalaciones que habían de albergar a los animales vivos y del desolladero; además, también se obligaba al reconocimiento de las canales y vísceras de los toros de lidia.
Se contemplaba la imposición de multas a los Subdelegados de Veterinaria que hubieran dado por útiles toros que no reuniesen las condiciones reglamentarias. Comentario...
En el siguiente “Reglamento ofi cial de las corridas de toros, novillos y becerros que ha de regir en las plazas de primera categoría” de 1923, los reconocimientos veterinarios versarán sobre aspectos similares recogidos en el predecesor, se volverá a exigir un certifi cado de las instalaciones previo al espectáculo, se instaurarán dos reconocimien-tos para los toros y para los caballos de picar y se obligará a la presencia veterinaria en los apartados y enchiqueramiento de los toros. Uno de los subdelegados que hubiese hecho el reconocimiento de las reses, deberá permanecer en el palco de la presidencia, por si esta tuviese que consultarle casos dudosos de inutilidad de las reses.
Con la publicación de la “Real Orden de 2 de mayo de 1925” del Ministerio de la Gobernación, se tratará de organizar, aspectos como el tiempo que debía transcurrir entre los reconocimientos de los animales y el festejo o la forma de abonar los hono-rarios a los Subdelegados encargados del reconocimiento. Se incluía en su articulado, la rotación de todos los veterinarios nombrados en los distintos festejos; contemplán-dose la actuación como dirimente de la Inspección Provincial Pecuaria en el caso de discrepancia en los informes de reconocimiento, tanto de los animales vivos como de sus respectivas canales.

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
59
La inclusión, en la misma orden, de posibles sanciones a los veterinarios causó un gran malestar en la profesión, lo que es recogido en un acta de la Asamblea de la Asociación Nacional Veterinaria Española, de enero de 1926.
En 1930 se publica el nuevo “Reglamento ofi cial para la celebración de espectáculos taurinos y de cuanto se relaciona con los mismos”, aumentándose las responsabilidades asignadas a los veterinarios. Se mantiene la exigencia de la disponibilidad y presencia del veterinario en la plaza, el asesoramiento presidencial y los reconocimientos en vivo de los animales que habían de versar sobre la edad, defensas y utilidad para la lidia, hablándose por primera vez del tipo zootécnico del toro. La comprobación de la edad, peso y defensas se tenía que verifi car en el desolladero.
Aparece la fi gura del perito de parte, veterinario designado por la empresa, ganadero o ambos, que emitiría su informe cuando existiera discrepancia con el dic-tamen de los veterinarios de servicio.
El informe veterinario derivado del reconocimiento de los animales no era vin-culante, pero los veterinarios seguían estando sujetos a posibles sanciones derivadas de su actuación.
La publicación de la “Orden de 10 de febrero de 1953” del Ministerio de la Gober-nación,
volvería a restablecer los artículos del Reglamento taurino de 1930 respecto a los pesos, edad y comprobación de éstos por parte de los veterinarios, a la vez que se les responsabilizaba del examen “post-mortem” de las astas de las reses.
El examen de boca y la comprobación de la edad de los animales por su tabla dentaria efectuada por los veterinarios, fue polémico, ya que podía generar una sanción económica tanto para el ganadero como para el empresario, en caso de demostrarse que las reses no tenían la edad reglamentaria.
Al no existir en esta época el guarismo, ni los certifi cados de nacimiento, se lidiaban como toros animales que eran utreros y de ahí la polémica.
En el Texto refundido del nuevo “Reglamento de Espectáculos Taurinos, de 15 de marzo de 1962”, se reunifi carán las diferentes actuaciones veterinarias en el espec-táculo, tales como el reconocimiento de corrales y chiqueros con certifi cación de la Inspección Provincial de Sanidad, la certifi cación de la sanidad de las reses fi rmada por el veterinario titular de la población donde se ubicase la explotación de origen, los reconocimientos periciales de las reses en la plaza de toros y, por último, el segui-miento laboratorial realizado por los Servicios Veterinarios de la Escuela Nacional de Sanidad por procedimiento iniciado por los veterinarios de plaza ante la sospecha de manipulación de las astas de los toros.

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
60
Los nombramientos se realizaban por la Dirección General de Seguridad en Madrid y por los Gobernadores Civiles en provincias, a propuesta de las Inspecciones de Sanidad Veterinaria.
Se describe la composición del palco presidencial, siendo el veterinario con más antigüedad de los intervinientes en los reconocimientos el que asesore al presidente. Si fuesen varios los festejos a celebrar los veterinarios irán turnándose en el puesto de asesor.
En las corridas de toros y novilladas con picadores se designarán cuatro veterina-rios: dos para el reconocimiento de reses y dos en el de caballos, en las novilladas sin picadores y becerradas se designarán solo dos, y uno en corridas de inferior categoría.
Los reconocimientos versarán sobre la sanidad, edad, peso aparente- en plazas de tercera categoría-, defensas y utilidad para la lidia y, en general, sobre todo lo que el tipo zootécnico del toro requiere.
En los reconocimientos post-morten se comprobará que los toros tienen como mínimo los seis dientes permanentes completamente desarrollados.
Se contempla la posibilidad de sanciones a los veterinarios si dieran por útiles reses que no reunieran las condiciones reglamentarias y por tal motivo fueran de-vueltas a los corrales.
Se hace obligatorio por parte de los veterinarios el examen de las astas, con el fi n de comprobar su posible manipulación.
Se mantenía la precariedad en las condiciones y medios para la realización de los reconocimientos de los toros y caballos de picar. Se presentaron puntualizaciones a los diferentes artículos que afectaban a las funciones veterinarias, principalmente las referentes a la edad de las reses, abogando por la vuelta a la verifi cación de la edad del toro por la dentición y surcos corneales.
Un articulo publicado en la revista “Información Veterinaria” en 1990, bajo el titulo “Reforma del Reglamento Taurino desde el punto de vista veterinario”, D. Jesús Bengoechea, Veterinario del Ayuntamiento de Madrid y de la plaza de toros de las Ventas, volvía a poner de relieve la incomprensión hacia las actuaciones veterinarias, expresándolo de esta forma:
“Algunos estamentos pretenden que los veterinarios desaparezcan de las presidencias, incluso algo mas grave y descabellado, de los reconocimientos. “

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
61
La publicación de la “Ley 10/1991, de 4 de abril, sobre potestades administrativas en materia de espectáculos taurinos”, y los posteriores decretos que la desarrollan, no cambiarían las condiciones en las que se venían realizando los reconocimientos, aña-diendo, sin embargo, nuevas responsabilidades a los veterinarios.
A las ya existentes, se añade la documentación que acompañaba a los animales tras la implantación defi nitiva del Libro de Registro de Toros de Lidia, del Documento de Identifi cación Bovina y demás documentación sanitaria exigida según las condi-ciones epizoóticas del momento.
La primera normativa que deriva de la Ley 10/1991 es el Real Decreto 176/1992 de 28 de febrero, por el que se aprueba el Reglamento de Espectáculos Taurinos.
Las novedades de este Reglamento con respecto a las actuaciones veterinarias son:
- Se permite el arreglo de las defensas en presencia del veterinario cuando estas se hayan deteriorado por accidente.
- El primer reconocimiento previo de las reses se hará con una antelación de 24 horas con respecto al comienzo del festejo.
- Los veterinarios serán designados a por la autoridad competente a pro-puesta del Colegio Ofi cial de Veterinarios de la provincia donde vaya a celebrarse el espectáculo. Para las corridas de toros y novilladas picadas se designaran tres veterinarios y dos para los demás festejos.
- También contempla la toma de muestras biológicas por parte de los veterinarios, en caso de comportamiento sospechoso de los animales durante el desarrollo de la lidia, pero no será hasta la publicación de la Orden de 7 de Julio de 1997, cuando se establezcan los procedimientos y el material necesario para la toma de estas muestras en plazas de primera y segunda categoría.
En 1996 se publica el “Real Decreto 145/1996 que modifi ca y da nueva redacción al Reglamento de Espectáculos Taurinos”.
Su redacción fue motivada, según se expresa en el preámbulo, por no conseguir su predecesor la erradicación de los fraudes en la integridad de las astas.
Aparece el veterinario de parte (de ganaderos y de empresarios), que puede asistir a los reconocimientos previos y post-morten, con el fi n de garantizar el principio de contradicción y que no se produzca indefensión.
En la disposición adicional tercera se indica, por primera vez, la necesidad de garantizar la formación técnica de los veterinarios que actúan en los espectáculos,

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
62
correspondiendo al Consejo General de Colegios Veterinarios o por delegación a los Colegios Provinciales dicha formación, al igual que la habilitación y propuesta de los veterinarios que hayan de ser nombrados por la autoridad competente.
También describe los aspectos sobre los que ha de versar el primer reconoci-miento, expresándose que los veterinarios de servicio emitirán un informe motivado, por escrito y separado, de la falta de características de las reses en razón de la clase espectáculo y de la categoría de la plaza.
Y llegamos fi nalmente al “Decreto 68/2006 de 21 de marzo por el que se aprueba el Reglamento Taurino de Andalucía.”
En su artículo 24 se establecen los requisitos para la designación del equipo veterinario de servicio en los reconocimientos previos y post-morten. Como novedad respecto al Reglamento Nacional, establece que los veterinarios:
- No tienen que tener interés directo de tipo económico, profesional o de parentesco con: ganaderos, empresarios, toreros o representantes de ganaderías que intervengan en el espectáculo.
- Al menos una de las personas nombradas en el equipo veterinario deberá prestar servicio en la Consejeria de Agricultura y Pesca.
En cuanto a las funciones (Articulo 25) las novedades son:
- Asistencia, en su caso, a los señalamientos de las reses en las ganaderías. (Ya lo decía “Paquiro” en su Tauromaquia en 1836, al defi nir al “ hombre fi el”).
- Vigilancia de las normas de bienestar animal, en especial, durante el transporte.
- Asistencia veterinaria a los caballos que se accidenten a consecuencia de su intervención en la lidia.
No quisiera terminar sin recordar a las mujeres veterinarias en este mundo tan machista del toro.
Prueba de ello es el Art.96 del Reglamento de 1917 “Queda prohibido en absoluto tomar parte en la lidia de toros, novillos y becerros, a los menores de dieciséis años y a las mujeres.”
Anteriormente algunos Reglamentos locales, basados en el de Madrid de 1880, ya lo contemplaban, como el de Valencia. “Queda terminantemente prohibido que en la plaza de toros de Valencia puedan tomar parte mujeres en ninguna corrida, sea esta de la clase y categoría que fuese.”

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
63
En el Reglamento Taurino de 1962 se refl ejaba como algo novedoso el permitir a las mujeres torear a caballo, pero no a pie. En su Art. 49, punto c), se decía:
“Queda absolutamente prohibido tomar parte en festejo taurino alguno a mujeres, si bien podrán llevar a efecto la lidia a caballo como rejoneadoras, pero sin echar pie a tierra para rematar la res.”
En ese mismo reglamento no debieron plantearse la posibilidad de la actuación de mujeres veterinarias en las plazas de toros, ya que, al describirse la suerte de rejo-nes en el capítulo XII, Art. 131, en lo relativo al reconocimiento pericial de las reses, se redactaba por primera vez en un reglamento que este examen debía ser realizado por “señores veterinarios”.
La Orden de 10 de agosto de 1974, del Ministerio de la Gobernación, ya permitía la libre participación de las mujeres en los espectáculos taurinos.
La incorporación de la mujer a las funciones veterinarias que se desempeñan en los Espectáculos Taurinos, ha sido lenta, más o menos similar a su incorporación a la propia profesión veterinaria.
La primera solicitud femenina para cursar estudios de veterinaria fue realizada por Juliana Vidal Rodríguez en 1916, pero habría que esperar hasta el año 1925 para que por primera vez en España una mujer obtuviese el titulo de veterinario. Se lla-maba María Cerrato Rodríguez y estaba matriculada de forma libre en la Escuela de Veterinaria de Córdoba.
En 1928 terminó la carrera Justina González Morilla en la Escuela de Veterinaria de León, acabando ese mismo año también la carrera de Medicina, profesión por la que se decantó, y no ejercería como veterinaria.
Pero sería en los años 70 cuando se inicia un ascenso imparable del número de mujeres matriculadas en las Facultades de Veterinaria, llegando a alcanzar en el curso 2005/2006 en la Facultad de Madrid el 75% del total del alumnado matriculado en primer curso.
La primera mujer de que se tiene conocimiento de haber participado como pro-fesional veterinario en Espectáculos Taurinos, reconociendo los animales e incluso asesorando en el Palco Presidencial, fue Vicenta Ferreres Meseguer, actuando en la Plaza de Toros de Málaga en el año 1956. Concluyó los estudios de veterinaria en Zaragoza en 1936. Fue titular en Lorca y desde febrero de 1942 Inspector Municipal Veterinario del Ayuntamiento de Málaga.

LA ACTUACIÓN VETERINARIA EN LOS ESPECTÁCULOS TAURINOS
64
Posteriormente en 1978 fue Angélica Orne Zubiaur la primera mujer veterinaria en actuar en una plaza de primera categoría como la de Bilbao. Y a partir de entonces se van incorporando al resto de plazas de España.
Con lo que llevo dicho, amigos, no es que el tema esté agotado porque, sin duda, tiene la sufi ciente enjundia como para redactar un tratado, pero el cansancio que os haya podido causar este fárrago de fechas, referencias y vicisitudes de nuestra Fiesta relacionadas con su ordenación, me está indicando con voz silenciosa que deje aquí mis palabras.
Sin embargo, no las puedo concluir sin agradecerles su presencia en este acto, que le da el lustre que merece, y la paciente escucha con la que me han obsequiado, y a nuestro Presidente manifestarle mi gratitud por permitirme a partir de ahora formar parte de este selecto Colectivo de compañeros que integran nuestra Academia.
Muchas Gracias.

CONTESTACION AL DISCURSO DE INGRESO DE ANTONIO VILLALBA
65ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
Excmo. Sr. Presidente de la R.A. de Ciencias Veterinarias de A. OrientalIlustres Señoras y Señores Académicos,Amigos todos.
Es la presente una oportunidad que me honra, ocupar esta tribuna para responder a la magnífi ca y apretada exposición que nos ha hecho nuestro recipiendiario con el propósito de dejarnos cumplida constancia de la intervención del veterinario en los espectáculos taurinos incluida como misión primordial en los diferentes Reglamentos que a partir de la mediación del Siglo XIX se fueron estableciendo en nuestro País para la mejor conducción de los espectáculos taurinos. El amplio comentario que nos ha hecho de estos diversos módulos legislativos, al mismo tiempo que ha recreado mis oídos, y me supongo que también los vuestros, por la claridad con que los ha expuesto, ha reverdecido en mi memoria viejos recuerdos que al parecer ya tenía olvidados y que, si me lo permitís, voy a exponerles algunos, no como complemento de sus acertadas palabras que por supuesto no me permiten esta opción, sino sim-plemente como pobre exposición de un anecdotario que podría ser conveniente traer a colación como divertimento circunscrito a nuestro particular ambiente malagueño.
Para ninguno de ustedes es un secreto que las fi estas de toros son, y con mucho, muy anteriores en el tiempo a cualquier patrón legislativo de los que nos ha dictado nuestro nuevo Académico. Es igualmente cierto que la primitiva fi esta era muy di-ferente a la actual, y que evolucionada en sus diversas secuencias a otras formas ha
CONTESTACION AL DISCURSO DE INGRESO EN LA R.A. DE CIENCIAS VETERINARIAS DE ANDALUCIA ORIENTAL DEL ILTMO. SR. D. ANTONIO VILLALBA GÓMEZ
MANUEL MUÑOZ MARTÍN1
1 Académico de Número de la Real Academia de Ciencias Veterinarias de Andalucía Oriental.

CONTESTACION AL DISCURSO DE INGRESO DE ANTONIO VILLALBA
66
venido a concretarse en la actual que conocemos. Pero en un principio, no era más que la lucha, valga la expresión, de un aguerrido y valeroso afi cionado, con una bestia que se batía según sus primitivos y ancestrales instintos defensivos.
Amplío que cualquier tipo de lidia o, para mejor decir, de lucha con una res brava, porque el concepto de lidia taurómaca, al menos para mí, es posterior a estas fechas, la hacían los actuantes para mostrar al público espectador su resistencia, fortaleza y habilidad ante animales de llamativa bravura, pero todavía, como es natural, no controlada, y sin percibir estipendio alguno. Ya Alfonso X el Sabio consideró conve-niente en su completa colección de Las Partidas, legislar que nadie “recibiese precio por lidiar con alguna bestia”, añadiendo que “son infamados los que lidian con vestias bravas por dineros que les den”. Está claro que alude directamente a quienes ya se dedicaban como trabajo o profesión a esta actividad, dentro de estos festejos populares. Pero no es el hecho de luchar con un animal lo que condena nuestro Rey, sino, simple-mente, el hecho de que se hiciera por dinero, “pues de otro modo ganará prez de valiente y esforzado”. Las diferencias con los criterios actuales son, simplemente, abismales. Por estas fechas, para acabar con las reses después de hacer con ellas las peripecias que cada cual podía según su atrevimiento y facultades, fi nalmente eran alanceadas con un venablo, modalidad que parece estaba al alcance de cualquier espectador, incluidas las mujeres. De cualquier forma, este modo de acabar con el animal reque-ría una habilidad especial y ya, un conocimiento especializado de este arte, como el que se dice que mostró Don Pero Niño en Sevilla en un festejo ante el Doliente Rey Enrique III “ansi a pie como a caballo, a donde él lansó muchas hermosas lansas, esperando a los toros, poniéndose en gran peligro con ellos y haciendo golpes de espada tales que todos eran maravillados”.
¿Existían entonces reglas formales a las que sujetar las actuaciones de aquellos atrevidos lidiadores?. Interpreto que no. Lo que sí me parece seguro es que por la mediación del XVI ya no existía la antigua norma de clavar en el suelo, hasta el fi n de la pelea, aquella espada o venablo con que estos luchadores debían acabar con el animal, aguantando mientras las embestidas del toro sin más defensas que sus propias fuerzas y habilidad para evitarlas. Bastante tiempo después, cuando esta actividad empezó a hacerse por dinero, tal forma de mostrar a los espectadores una valentía sin igual quedó derogada. El bravo toreador ya había pensado en su particular integridad, que había de reservar para espectáculos sucesivos. De cualquier modo, en una fi esta no ordenada todavía, era costumbre, debo añadir, que los espectadores lanzaran al toro no sólo ballestas sino cualesquiera elementos punzantes o cortantes que debilitaran sus fuerzas, antes de que el matador acabara defi nitivamente con él. Esta costumbre tan irregular parece que pervivió entre nosotros durante mucho

CONTESTACION AL DISCURSO DE INGRESO DE ANTONIO VILLALBA
67
tiempo, pero cambiando los instrumentos cortantes por otros menos traumatizantes, pero de igual modo perjudiciales a la lidia.
No obstante, y en tanto seamos partidarios, por no tener otras noticias, de que nuestra particular afi ción taurina y sus modalidades partieron de las que nos transmitieron los conquistadores cristianos, habida cuenta de que hasta el momento carecemos de argumentos para demostrar que las pudimos heredar del pueblo árabe, si así fuera, no hemos de perder de vista, y aquí debo recordarlo, que además del carácter lúdico a que antes he hecho referencia, las corridas tuvieron también, con mucha frecuencia, un carácter cuasi religioso, como se deduce de los motivos sobre que se fundamentaban las corridas llamadas votivas, nupciales y funerarias, todas ellas, por su ancestral y supuesta pero vivida y asimilada relación que en pasados tiempos se hizo entre el toro, ser sagrado y principio de vida, y el hombre, que al sacrifi carlo en el ruedo asimilaba de su sangre multitud de sus esencias, entre ellas la virilidad. Votivas eran las corridas que se celebraban en honor de un santo de veneración local, en ciertos momentos de crisis, como pudieran haber sido la aparición de una epidemia, o como pago, valga la expresión, de haberlos librado de ella. Las corridas funerarias, al parecer bastante prodigadas en la encrucijada de los siglos XVI y XVII, se celebraban para cumplir la voluntad de ciertos personajes que las dejaban así dispuestas en su testamento como una manda más, y que necesariamente habían de cumplir sus deudos si no querían verse envueltos después en supuestos disgustos y contrariedades de diversos tipos. Ambas modalidades de festejos, si así podían llamársele, no dejaban de tener un fondo religioso, quizás heredopagano, pero tremendamente supersticioso, en el que la Iglesia no dejada de tener su crematística intervención. Finalmente, las corridas nupciales, que en el medioevo tuvieron gran predicamento, consistían en una especie de encierro, más bien de encerrona, que se hacía de una res vacuna, de bravura contrastada, y de su subsiguiente sacrifi cio que el novio y sus amigos habían de culminar en la puerta de su prometida, después que a través de diversas calles de la localidad la hicieran correr desde la dehesa hasta la puerta de la casa nupcial, donde de las más diversas maneras y con los más insospechados instrumentos acababan con el animal, en cuya sangre, una vez descuartizado, aquélla impregnaba un pañuelo blanco, que en el momento ofrecía al novio como símbolo de su intacta virginidad. Parece ser que la carne del sacrifi cado animal era después consumida en el banquete nupcial, en el que participaba todo el pueblo.
No quisiera que en este breve relato de curiosas efemérides que vengo haciendo gracias a la benevolencia de ustedes, se me pasara algo de una capital importancia íntimamente relacionado con las fi estas de toros que de una manera u otra se desa-rrollaban en las sucesivas plazas de toros que conoció nuestra urbe, que sirvieron

CONTESTACION AL DISCURSO DE INGRESO DE ANTONIO VILLALBA
68
para que nuestros vecinos disfrutaran de este espectáculo, y que alcanzaron, claro está, a conocer los primeros años de la actual de La Malagueta. Solemos decir cuan-do hablamos de esta cuestión, y más referida a aquellos pasados tiempos, que se celebraron tales o cuáles corridas, pero, posiblemente, a la gran mayoría de afi cionados le pase casi del todo desapercibido un importante detalle, y es el de cómo venían a nuestra ciudad los ganados que habían de lidiarse. Cuando este espectáculo no tenía aún la forma que hemos visto nos ha explicado Antonio, deducida de los concretos Reglamentos por que se rigieron, y cuando todavía estas manifestaciones no eran más que meras exhibiciones de fuerza y temeridad ante los vecinos de personas ge-neralmente pertenecientes a la aristocracia, las reses que éstos utilizaban para tales juegos eran de las que los tablajeros, los carniceros, traían para ser sacrifi cadas para el abasto público y que apreciada por éstos la acometividad de algunas de ellas, las ofrecían a aquellos señores para los citados juegos. Es más, me consta que así las cosas, aquellos carniceros ya estimulaban a sus proveedores a que reservaran para sus clientes el ganado que en las dehesas aparentaba la bravura que se exigía para tales funciones. Eran, simplemente, unidades que resaltaban de entre las demás que se criaban para el abasto, por unas facultades ideales y propias para estos juegos. La selección, al estilo de lo que hoy conocemos, entiendo que no existió de principio. Esta fue, al decir de los técnicos, una cuestión que se suscitó después en razón a unas fi nalidades muy concretas: la lidia formal, ya muy evolucionada. Y en tanto esta situación se fue concretando, y antes que se pensara que el recién creado ferrocarril, el primero que vieron los vecinos en nuestra zona, el de Córdoba a Málaga, pudiera ser un medio ideal de transporte de estas reses ¿cómo y de qué forma venían estos ganados hasta nuestra ciudad?. Curiosas por demás debían ser llamémoslas las ce-remonias que confl uían en la conducción de las reses hasta su destino. Por supuesto que el desplazamiento de los animales desde los campos a la ciudad se hacía de siempre a pié, aprovechando las mismas realengas, veredas y coladas destinadas desde tiempo inmemorial al tránsito de los ganados, que perduraron en la memoria popular y que posteriormente fueron califi cadas por la Ley con el nombre de vías pecuarias. Pues bien, a través de estas vías accedía después el ganado que había de lidiarse en nuestra ciudad. Entre éstas, el valle del Rio Guadalmedina era una de las más importantes para los ganados procedentes del Norte e incluso para los que pro-cedían del valle del Guadalquivir, que aún a expensas de dar un gran rodeo, venían a descansar previamente al espectáculo en la parte alta de este río o en dehesas más o menos próximas a la ciudad, como las llamadas del Garabato, situada a la derecha de lo que es hoy la vulgarmente conocida Carretera de Cádiz, o la de Trévenez, en la fi nca de igual nombre, cerca de Campanillas. Los papeles consultados nos dicen que estos traslados duraban entre veinte y veinticinco días para las ganaderías más

CONTESTACION AL DISCURSO DE INGRESO DE ANTONIO VILLALBA
69
lejanas, y que como con mucha frecuencia los festejos se componían de varias corridas, si éstas procedían de ganaderías distintas que se iban incorporando al lote según se hallaban en el camino, era muy natural que los conductores tomaran las oportunas precauciones para evitar las consabidas peleas entre reses que no se conocían. Así, pues, estos diversos encierros procuraban separarlos prudentemente unos de otros, pero no mucho con el objeto de que de haber problemas, los conductores pudieran auxiliarse los unos a los otros. Tales caminatas hacían, naturalmente, que los anima-les perdieran bastante peso, más, muchísimos más, por supuesto, que los que hoy se dejan enclaustrados en los típicos cajones y con los medios de transporte que se utilizan y que, muchísimos más rápidos, no dejan de ser, no obstante, sumamente traumatizantes para los animales. El traslado de una corrida normal precisaba de unos diez o doce bueyes y de un número más o menos igual de vaqueros. Hoy, con el conductor del camión y un ayudante, el asunto queda resuelto plenamente. De aquellos conductores, unos, los más diestros y llamados de estribo, caminaban sobre entrenados caballos a lado y lado de la manada y al fi nal de la misma, y al menos uno, al lado del que abría el contingente, que solía ser el mayoral. Los llamados vaqueros de tropa caminaban igualmente a caballo pero confundidos con las reses, con la mi-sión de evitar las peleas que solían presentarse aún entre reses de un mismo hierro. Vaqueros de cola eran los que cuidaban de arrear las reses que por cualquier motivo se quedaban rezagadas o trataban de salirse de la manada. De los bueyes conductores, uno, que recibía el nombre de portero, más o menos acostumbrado a tomar las puertas una vez llegada la manada a su destino, era el que abría esta particular comitiva que, dicho sea de paso, causaba grandísimo interés en los habitantes de los lugares por donde transitaba. Uno o dos vaqueros, según el volumen del lote en camino, era el encargado de llevar sobre lomos de mulos generalmente, las provisiones, vituallas y enseres propios del personal, que habían de utilizarse en tan largas caminatas, tanto de ida como de regreso al lugar de origen. Como es natural, unos viajes tan largos y en los que se había de pasar por tantos sitios diferentes, con frecuencia debían dar lugar a peripecias de las más diversas clases, unas resueltas de modo favorable y otras no tanto, y que contadas luego por estos hombres a su regreso a la ganadería, habían de ser siempre del agrado de los atentos oyentes sentados junto a la lumbre, en las largas noches de invierno. Una vez las reses en la proximidad de nuestra ciudad, eran conducidas a sitios previamente conocidos, las dehesas que antes he referido, en las que durante algún tiempo se procuraba darles el descanso necesario, no sólo para reponer sus aspeadas pezuñas sino también para que recuperaran parte del peso perdido y rellenaran con éste sus visibles osamentas. Y aquí permanecían hasta la noche antes de la corrida, en que reglamentariamente había de verifi carse su encierro en la plaza. A estos efectos y para evitar cualquier tipo de accidente, la Autoridad

CONTESTACION AL DISCURSO DE INGRESO DE ANTONIO VILLALBA
70
municipal tomaba las precauciones que eran de rigor, pues a lo que parece, la afl uencia de público a este venerable rito taurino para contemplar de cerca las características del ganado, debía ser muy llamativa. Me consta, por ejemplo, que para una corrida que había de celebrarse, todavía en la Plaza Mayor, luego llamada de las Cuatro Calles y, fi nalmente de La Constitución, nuestro entonces Alcalde decía al Subinspector de la Milicia Nacional que “debiendo celebrarse en la noche de este día el encierro de los novillos que han de lidiarse mañana a la tarde, espero de la atención de Vd. se sirva disponer se hallen a las diez de la noche en la Plaza destinada a esta diversión un piquete de veinte hombres de tropa del exercito y otros veinte de la Milicia Nacional”.
Prosiguiendo en mi empeño de referirles diversas curiosidades de añejos tiempos relativas a nuestra Fiesta, y siempre y cuando no les causen cansancio, haciéndome eco de la referencia que nos ha hecho nuestro compañero Antonio del bando que para el mejor orden del espectáculo publicó nuestro entonces gobernador y paisano Melchor Ordóñez Viana Cárdenas en 1º de junio de 1847, no debo dejar de referirles que entre sus peculiaridades disponía que “En cada uno de los cuatro ángulos de la Plaza habrá dos hombres con seis espuertas llenas de tierras y una vacía, con objeto, las primeras, de cubrir en el momento la sangre que arrojen los caballos, y la espuerta vacía será para re-coger en ella las tripas que de aquellos se desprendan. Ambas operaciones se verifi carán sin pérdida de tiempo tan luego como el toro se separe del punto donde la sangre esté, y el mozo que desempeñe con morosidad este servicio sufrirá ocho días de cárcel. Llevará cada uno de estos mozos un palo de media vara de largo, con dos ganchos en la punta para romper y echar en las espuertas las tripas de los caballos. Recalcaba nuestro Gobernador que por cada golpe de tablón que desde el momento del encierro hasta salir a la plaza reciba algún toro, pagará el empresario quinientos reales, y respecto del ganado a lidiar, que no podrá tener ningún toro menos de cinco años cumplidos, ni podrán pasar de ocho: la falta de esta condición será penada por cada uno con quinientos reales de multa. Que en la cuadra habrá cuarenta caballos, sin que ninguno sea de menos alzada de siete cuartas cumplidas; este ganado será reconocido por la autoridad competente el día antes de cada función, a las cuatro de la tarde. Que para evitar la malicia que suele haber en el uso de las banderillas de fuego pretextando equivocaciones por parte de los que han de ponerlas, se previene que éstas sólo se usarán cuando la autoridad que manda la plaza ondee un pañuelo encarnado. La tardanza en la ejecución será penada con cien reales de multa a cada banderillero o tres días de cárcel. Y fi nalmente, que el empresario reservará en los tendidos de sombra sitios sufi cientes donde se coloquen doscientos hombres de tropa que vayan para conservar el orden público, debiendo estar estos con holgura, y además se dará local proporcionado para la banda de música, y serán libres las entradas de guardias civiles y municipales y empleados de seguridad pública”. Las muchas puntualizaciones que se destacan en este conato de reglamento, aunque algunas son tan claras que no

CONTESTACION AL DISCURSO DE INGRESO DE ANTONIO VILLALBA
71
necesitan de comentario, otras merecerían un substancioso estudio, que en este mo-mento no me puedo permitir, es obvio, pero que podíamos dejar para otra ocasión.
El Alcalde de la ciudad entonces, José Freüiller Alcalá Galiano, Abogado de los Tribunales de la Nación, para no ser menos que su paisano el Jefe Político, con la mis-ma fecha que éste publicó un Bando en el que prevenía a los vecinos que “Deseando que el público pueda disfrutar con toda tranquilidad de dichos espectáculos, he determinado se guarden y cumplan las prevenciones siguientes: que para mayor comodidad del público y para evitar los desórdenes y disgustos que son consiguientes a permanecer en el centro de la plaza indistintamente las personas que van a sol y a sombra, teniendo que agolparse en con-fusión en el momento del despejo para adquirir las localidades, que no siempre consiguen, he dispuesto que se verifi que la entrada sólo por entre barreras, pasando cada cual a ocupar su asiento, quedando prohibido desde luego el entrar y pasear en la plaza. Y añadía: no se permite que ningún individuo entre con capa, y el que lo intentare será detenido y expulsado; ni tam-poco con palos, con porras, bastones de hierro o con puños de metal fundido, y todo el que se presente con ellos será detenido y consignado a disposición de la autoridad para que sufra la pena correspondiente; ni tampoco con botellas de vino, de licores o tarros de los mismos, como cualquier efecto de esta especie, y los contraventores serán detenidos y puestos a disposición de la autoridad que manda la plaza, prohibiendo de igual modo que se arroje nada a la misma, incluso los sombreros, aunque sea con el sólo objeto de aplaudir Concluía nuestro Alcalde anunciando que tomadas las medidas oportunas con el fi n de que la diversión no se pueda convertir en disgusto para ninguno de los concurrentes por demasías o excesos, y esperando de este ilustrado público que conservará el buen comportamiento que es indispensable en las grandes reuniones.
De lo que acabo de exponerles se deduce el carácter excesivamente festivo de nuestros vecinos de antaño, del que las autoridades estaban plenamente al corriente y la curiosa y violenta manera de reaccionar ante determinadas secuencias del es-pectáculo que en algún momento no fuera de su agrado, como por ejemplo, del que la empresa de Álvarez les ofreció el 15 de julio de 1849, en el que a causa del pobrí-simo estado de las reses presentadas y el escasísimo juego que dieron, el público, en protesta, arrancó los tablones de los tendidos, tirándolos al ruedo junto con las sillas, agrediendo a los agentes que intentaron de estorbarlo.
No andaríamos muy alejados de la realidad si por nuestra mente cruzara la imagen dantesca del incorrecto y brutal comportamiento de aquella gente, puesta en pie, gritando y lanzando al ruedo, no sabemos si contra las reses de su desagrado o contra los mismos lidiadores, los más insospechados proyectiles, especialmente los envases, siempre de vidrio, de las bebidas que en demasía consumían, según parece

CONTESTACION AL DISCURSO DE INGRESO DE ANTONIO VILLALBA
72
era costumbre entonces, y que en principio a nadie le llamaba la menor atención, así como sombreros, el típico de pleita de juncos, de precio muy asequible, pero que con frecuencia acaba siendo el envoltorio de un proyectil de los prohibidos.
Palos y garrotes. Se dice que con éstos, los asistentes próximos al ruedo solían asustar a las reses dando tremendos golpes sobre las maderas que formaban las barreras, lo que, naturalmente, desquiciaba el comportamiento de los animales y al mismo tiempo los distraía de la atención que debían prestar a los maestros de los di-versos órdenes que intervenían en el festejo. Que no era esta, en general, una manera de protestar, airada, de lo que pudiera pasar en la arena, sino, con mucha frecuen-cia, un argumento para armar alboroto, que es en lo que para muchos de aquellos asistentes consistía su asistencia a la fi esta. Pero si tal actitud, de cara al festejo, tenía una importancia sobresaliente, más la tenía en la opinión de la autoridad, cuando estos argumentos físicos se empleaban de modo exclusivo para dirimir las cuestiones suscitadas en las discusiones propias de los asistentes, partidarios y no partidarios de los diestros que se jugaban la piel sobre el ruedo.
Como la brutal costumbre de arrojar en los tendidos repletos de público y sobre los lidiadores aquellas andanadas de plumas de gallinas, que en el mejor de los casos, a éstos últimos los distraía de su trabajo, con harto peligro para su integridad, y al resto de asistentes a la fi esta los incomodaba con el marcado deseo de armar el ma-yor alboroto posible, como ya antes he apuntado, consecuencia a veces del consumo excesivo de alcohol.
Las capas. La capa era entonces, y hasta no hace mucho, sobre todo en invierno, una prenda de vestir de uso corriente, que a la par que daba un cierto prestigio a los que la usaban, servía, y no hay que hacer muchos aspavientos de ésto, para ocultar ropas de inferior calidad o de gastado uso que necesariamente habían de portar sus dueños. Es curiosa la existencia de los llamados caperos, que durante el día hacían la vida en el cauce del Guadalmedina, entre el puente de La Aurora y el de Santo Domingo, ambos de madera, y que por algunas monedas cubrían con aquella pren-da, para no ser vistos, a quienes tenían necesidad apremiente de exonerar el vientre mientras hacían esta operación. No es que ahora estemos mejor servidos que entonces de mingitorios públicos, pero ante una urgente necesidad, parece que hoy siempre hay a la mano un bar o una cafetería que suple a la referida capa. De rigor es pensar que en las fechas en que estas celebraciones tenían lugar, el pleno estío, tal vestimenta no era la más apropiada para lucir, y mucho menos para soportar la calina a pleno sol en una localidad de este sello. Pero era un argumento más para ocultar a vigilantes, a acomodadores, si es que los había, y demás encargados de guardar el orden público aquellos ya referidos instrumentos llevados expresamente para alborotar.

CONTESTACION AL DISCURSO DE INGRESO DE ANTONIO VILLALBA
73
Pero estas cosas, y muchísimas otras más que podría traer o colación, pero que para evitarles el natural cansancio, o quizás el aburrimiento, es mejor que sigan re-posando en mi memoria, estas cuestiones, repito, con el paso del tiempo se fueron puliendo, lo mismo que los modos de los espectadores, y adaptando cada vez mejor a un festejo que al mismo tiempo evolucionaba a otras formas, y que con mayor o menor precisión acabaron por desembocar en los Reglamentos que tan magnífi camente nos ha recordado nuestro nuevo académico, para terminar en el último, proclamado para nosotros por la Junta de Andalucía y que es el que, con algunos de sus defectos, han de utilizar nuestros compañeros que cumplen su misión en nuestras plazas de toros.
Y termino. Sólo me queda expresar con singular afecto a Antonio Villalba nuestra más cordial enhorabuena por este reconocido mérito a que hoy se ha hecho acreedor, y animarle para que desde este puesto, como hace desde el de Presidente del Colegio que con tanto acierto dirige, contribuya con su demostrada capacidad al deseado engrandecimiento de nuestra Clase. Y a ustedes, amigos todos, darles las más expresivas gracias por la paciencia con que, sin lanzarme tarros, sombreros ni plumas, que sin duda me he merecido, han soportado mis pobres palabras. Gracias.


CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
75ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
“Si conoces al enemigo y a ti mismo,no necesitas temer el resultado
de cientos de batallas”El arte de la guerra. Sun Tzu, 400 AC
En primer lugar quisiera expresar mi agradecimiento más sincero y profundo a los miembros de esta Real Academia de Ciencias Veterinarias de Andalucía Oriental por el honor que me dispensan. Hoy se hace realidad el sueño de pertenecer a esta docta y querida Institución, cuyo principal objetivo es el de difundir y compartir el conocimiento, pero además, me llena de orgullo el hecho de que mi ingreso se realice en mi querida y añorada Granada, la ciudad que me vio crecer como niño y de la que marché siguiendo mi vocación como Veterinario. Una vocación alentada desde peque-ño por mi padre que ejerció nuestra noble profesión en esta ciudad de la Alhambra y del Generalife, que lo acogió, le vio crecer personal y profesionalmente y le brindó la oportunidad de desarrollar no solo nuestra profesión, sino también involucrarse en la investigación y en la actividad colegial, ya que como él decía «somos de donde dicta nuestro corazón».
Mi labor de investigación se ha centrado, principalmente, en el estudio de la patogenia de las enfermedades víricas. Unos estudios que inicié bajo la dirección de mi maestro, Miguel Ángel Sierra Plana, y que me encaminaron a estudiar al macró-fago como una célula fundamental en el establecimiento de la respuesta infl amatoria
CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
LIBRADO CARRASCO OTERO*
* Catedrático de Anatomía y Anatomía Patológica Comparadas. Decano de la Facultad de Veterinaria “Campus de Excelencia Internacional CeiA3”, Universidad de Córdoba.
Discurso de ingreso en la Real Academia de Ciencias Veterinariasde Andalucía Oriental como Académico Correspondiente

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
76
e inmune de las enfermedades, y de cómo algunos virus provocaban la aparición de lesiones vasculares o la muerte de las células vecinas a las infectadas mediante la activación de los macrófagos. Una cuestión que me planteaba la siguiente pregunta ¿por qué una célula defensiva es la responsable de las lesiones más características de algunas enfermedades?, la respuesta a este enigma estaba en que la activación de esa célula desencadenaba la producción de una serie de mediadores, que regulan al organismo dentro de un delicado equilibrio que permite luchar y vencer la infección por diferentes patógenos pero que al mismo tiempo su sobreexpresión puede, inclu-so, conducir a la muerte del individuo. Estos mediadores son conocidos hoy bajo la denominación genérica de citoquinas
INTRODUCCIÓN
Las citoquinas o citocinas (del griego “kyto” célula e “ina” sustancia) son mo-léculas de bajo peso molecular (de 10 a 40 kD) y constituidas por unos 120 a 180 aminoácidos, que poseen la capacidad de modular la función de células y tejidos, y que están producidas, principalmente, por los leucocitos, aunque algunas de ellas también pueden ser secretadas por otros tipos celulares.
En condiciones fi siológicas, las citoquinas no se producen en cantidades sig-nifi cativas, siendo necesaria la activación de las células para que se produzcan en cantidades sufi cientes para ejercer sus efectos biológicos. La mayoría de ellas son se-cretadas al espacio extracelular en forma glicosilada, lo que incrementa su estabilidad y solubilidad. No obstante, algunas citoquinas se pueden acumular en el interior de la célula o adherirse a la membrana celular.
Las citoquinas poseen una vida media muy corta y actúan a muy bajas concen-traciones, del orden de picogramos, mediante la unión a receptores de alta afi nidad que están presentes en la superfi cie de las células. En función del tipo de células sobre las que actúan, se habla de efecto autocrino, cuando actúan sobre las propias células productoras, efecto paracrino, cuando actúan sobre diferentes tipos celulares, o in-cluso pueden tener un efecto endocrino, cuando ejercen su efecto en otros órganos y tejidos, transportadas por circulación sanguínea.
NOMENCLATURA
Los términos con los que se conocen estas moléculas han sido modifi cados desde su descubrimiento por Lindenman en 1957, al detectar la producción del conocido

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
77
actualmente como Interferón gamma en ensayos con cepas de la infl uenza. Inicial-mente, como las citoquinas se identifi caron como la respuesta de los linfocitos frente a determinados antígenos se las denominaron linfoquinas. Cuando posteriormente se comprobó que también eran secretadas por otros tipos celulares se optó por de-nominarlas citoquinas. Dentro del nombre genérico de citoquinas se empleó la no-menclatura de interleuquina (IL) para aquellas moléculas que servían como señales de comunicación entre distintos tipos de leucocitos, numerándose a medida que se descubrían, así nos encontramos desde la IL-1 hasta la IL-43. Sin embargo, y como hemos señalado anteriormente algunas de las citoquinas se identifi caron mediante ensayos «in vitro», por lo que adoptaron el nombre de la función biológica que de-sarrollaban, como es el caso del Interferón (IFN), Factor de Necrosis Tumoral (TNF), Factores Estimuladores de la Formación de Colonias (CSF) o Factor Transformador del Crecimiento (TGF)
Funciones de las citoquinas
Las citoquinas, además de tener un papel fundamental en la respuesta infl ama-toria e inmune, van a estar implicadas en numerosos procesos biológicos como en la hematopoyesis, la embriogénesis y la angiogénesis, así como en diferentes procesos celulares como la mitosis, la diferenciación, la migración o incluso la muerte celular.
Las dos principales características funcionales de las citoquinas son el pleiotro-pismo, ya que una misma citoquina es capaz de ejercer efectos biológicos diferentes al actuar sobre distintos tipos celulares, y la redundancia, ya que varias citoquinas pue-den contribuir al desarrollo del mismo efecto en una célula. Además, las citoquinas pueden actuar en lo que se conoce como cascadas de amplifi cación, estimulando la producción de otras citoquinas de forma consecutiva que amplían sus efectos bio-lógicos, o bien desencadenando un efecto antagonista , regulando de esta forma la respuesta originada o su intensidad. Nos encontramos, por tanto, con un complejo sistema, donde el efecto de una molécula está estrechamente regulado, positiva o negativamente, por otras moléculas del mismo sistema y que, además, cuando falta una citoquina sus funciones pueden ser reemplazadas total o parcialmente por otras.
La expresión y actividad de las citoquinas se puede medir mediante la utiliza-ción de: bioensayos, sobre líneas celulares, inmunoensayos, como el ensayo inmu-noenzimático (ELISA), que permite cuantifi car su concentración en diferentes fl uidos biológicos, y el ELISPOT, que deriva de la técnica anterior, y que permite determinar el número de células productoras, así como mediante citometría de fl ujo, con la que

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
78
es posible no sólo cuantifi car, sino también caracterizar las células productoras, las técnicas de Reacción en Cadena de la Polimerasa (PCR), que nos permite medir los niveles de RNA mensajero que codifi can la producción de una determinada citoquina y las técnicas inmunohistoquímicas, como el Avidin Biotin Peroxidasa (ABC) que nos permiten detectarlas en los tejidos y órganos.
Clasifi cación
Las citoquinas pueden clasifi carse según su estructura, sus receptores o su fun-cionalidad, pero no hay una clasifi cación rígida de las mismas ya que debido a sus características funcionales, no todas las citoquinas que tienen la misma estructura tienen la misma función, y el pueden utilizar diferentes receptores. Actualmente, la clasifi cación más utilizada es la funcional, aunque debemos de tener presente que, debido a que una misma citoquina es capaz de ejercer efectos biológicos diferentes al actuar sobre distintos tipos celulares, se encuadran, según su actividad sea más relevante, en una de la tres principales funciones en las que estas moléculas participan: la respuesta infl amatoria, la respuesta inmune y la hematopoyesis.
Citoquinas que intervienen en la Respuesta Infl amatoria
En la respuesta infl amatoria las citoquinas pueden actuar de dos formas totalmen-te opuestas, ya que mientras que algunas van a favorecer el desarrollo de la misma, las citoquinas pro-infl amatorias, otras van a tener un marcado efecto supresor de la infl amación, las denominadas citoquinas inmunosupresoras (Tabla 1).
Tabla 1: Principales citoquinas que participan en la respuesta infl amatoria
Citoquinas de actividad pro-infl amatoria IL-1
IL-6
TNF-α
TNF-β
IFN-γCitoquinas de actividad inmunosupresora IL-4
IL-10
IL-13
TGF

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
79
1. Citoquinas pro-infl amatorias
Las citoquinas de actividad pro-infl amatoria (Tabla 1) son moléculas producidas, principalmente, por monocitos y macrófagos, activados tras el contacto con un agente patógeno, aunque también pueden ser sintetizadas por otras poblaciones celulares como las células endoteliales o las células dendríticas entre otras.
• La interleuquina 1 (IL-1), de la que existen dos formas, alfa y beta, que, aunque solamente tienen un 25 % de homología, comparten el mismo receptor y ejercen efectos biológicos similares. Es el principal pirógeno endógeno, induciendo la fi ebre, gracias a la producción de prostaglan-dinas, y actúa sobre el SNC, provocando el sueño y la anorexia que se asocian a los procesos infecciosos. Además, esta citoquina induce la liberación de histamina, responsable de la vasodilatación y aumento de la permeabilidad vascular en el lugar de la infl amación, y es un factor quimiotáctico de los leucocitos.
• El Factor de Necrosis Tumoral (TNF) del que también se han descrito dos moléculas, alfa y beta, que tienen una elevada homología y que están estrechamente relacionadas. El TNF-alfa es producido por los monocitos, macrófagos, linfocitos T y B, células natural killer, fi broblastos y mastoci-tos. Está implicado, junto con la IL-1, en el desarrollo de la fi ebre, caquexia y sueño que se instauran en los procesos infecciosos, siendo un potente activador de los monocitos y de los polimorfonucleares neutrófi los. Además, induce la expresión de moléculas de adhesión y estimula la producción de otras citoquinas, como la IL-8, por las células del endotelio vascular, lo que contribuye a la extravasación de linfocitos, neutrófi los y monocitos, y la producción de proteínas de fase aguda mediante la estimulación de la IL-6. Por otra parte, el TNF-beta, es producido, exclu-sivamente, por los linfocitos T activados, aunque se une a los mismos receptores que el TNF-alfa e induce funciones similares. Además, el Factor de Necrosis Tumoral tiene la capacidad de inducir la necrosis de algunos tipos de células tumorales, así como iniciar los fenómenos de apoptosis, o muerte celular programada, en algunas células, al unirse a los receptores de superfi cie que inician la cascada de la vía extrínseca, que es la forma en la que un estímulo extracelular puede inducir a que una célula muera sin originar una respuesta infl amatoria.
• La interleuquina 6 (IL-6) es la responsable de la producción de proteínas de fase aguda por el hígado, proteínas que tienen una función protectora y reguladora del proceso infl amatorio a través de su actividad antipro-

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
80
teasa,. Además, esta citoquina induce en la médula osea la producción de células precursoras de neutrófi los, monocitos y megacariocitos y promueve la diferenciación de linfocitos B hacia células plasmáticas, induciendo la producción de inmunoglobulinas. Pero esta citoquina también es capaz de ejercer una función inmunosupresora ya que va a inducir la síntesis de los antagonistas de los receptores de la IL-1 beta, por lo que esta citoquina no va a encontrar receptores a los que unirse y por lo tanto desarrollar su función pro-infl amatoria
• Para terminar con este grupo de citoquinas, citaremos al interferón (IFN), inicialmente descrito como una proteína producida por las cé-lulas infectadas por algunos virus. Estas moléculas se clasifi can en dos tipos: el Tipo I que incluye el IFN-alfa y el IFN-beta, con propiedades antivirales y antiproliferativas, y el tipo II, que incluye el IFN-gamma, que tiene un efecto inmunomodulador. Mientras que el IFN-alfa es secretado por fi broblastos y algunas células epiteliales, el IFN-gamma es producido fundamentalmente por monocitos y macrófagos, siendo considerado el principal activador de los macrófagos y un desencade-nante de la respuesta inmune celular, al estimular la diferenciación de los linfocitos T a Th1. Sin embargo los interferones tipo I (alfa y beta) también se pueden considerar citoquinas inmunosupresoras debido a su efecto regulador de la producción de citoquinas y a su capacidad antiproliferativa. Efecto antiproliferativo que le confi ere al mismo tiempo una actividad antitumoral
2. Citoquinas inmunosupresoras
El grupo de las citoquinas inmunosupresoras (Tabla 1) incluye a un grupo de moléculas que inhiben el crecimiento celular, suprimen o bloquean la secreción de otras citoquinas. Entre ellas se encuentran algunas citoquinas producidas por los lin-focitos Th 2, como la IL-4, IL-10 e IL-13, que activan a los linfocitos B al mismo tiempo que inhiben la respuesta infl amatoria. Y el Factor Transformador del Crecimiento (TGF-beta) que inhibe el crecimiento y la función de muchos tipos celulares, así como la síntesis de determinadas citoquinas y la actividad citotóxica natural y específi ca.
• La interleuquina 4 (IL-4) es una citoquina que, como se ha mencionado anteriormente, presenta la característica de ejercer efectos biológicos diferentes al actuar sobre distintos tipos celulares. Así, tiene un efecto antiinfl amatorio al inhibir la producción de diferentes mediadores

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
81
infl amatorios de los macrófagos y bloquea la acción de la citoquina IL-1, una de las principales citoquinas pro-infl amatoria. Pero al mismo tiempo promueve el establecimiento de una respuesta inmune de tipo humoral a promover la diferenciación de linfocitos T hacia células de tipo Th2, estimulando de esta forma el crecimiento y diferenciación de los linfocitos B.
• La interleuquina 10 (IL-10) es considerada como la citoquina inmu-nosupresora por excelencia ya que inhibe la síntesis de muchas otras citoquinas como el TNF-alfa, la IL-2 o la IL-12. Además, regula las fun-ciones mediadas por los linfocitos B, induciendo la síntesis de IgG, y por linfocitos T, infl uyendo en el desarrollo de los timocitos y de las células T. Esta citoquina también tiene un papel fundamental en la angiogénesis.
• La interleuquina 13 (IL-13) comparte muchas de sus funciones con la IL-4. Es una citoquina inmunosupresora ya que inhibe, junto con la IL-4 y la IL-10, la producción de diferentes citoquinas infl amatorias por los monocitos/macrófagos. Adicionalmente, esta citoquina puede inducir la proliferación de monocitos y de linfocitos B, y promueve la producción de IgE
• El Factor Transformador del Crecimiento (TGF) del que existen dos tipos, alfa y beta, que no poseen ninguna similitud estructural ni funcional. El TGF-beta, es producido por los linfocitos T, plaquetas y otros muchos tipos celulares, y tiene efectos inmunomoduladores. Además, induce la proliferación de fi broblastos, osteoblastos y células musculares lisas e incrementa la síntesis de proteínas de la matriz extracelular, lo que favorece la curación de las heridas. El TGF-beta también tiene efectos inmunosupresores ya que inhibe tanto el crecimiento de diferentes tipos celulares como la síntesis del IFN-gamma, TNF-alfa, TNF–beta, IL-1, IL-2 e IL-3, inhibiendo también la citotoxicidad tanto natural como específi ca.
Citoquinas que intervienen en la Respuesta Inmune
En la respuesta inmune las citoquinas pueden actuar en el desarrollo de la respuesta inmune inmediata y no especifi ca, conocida como respuesta innata y que está presente en todas las plantas y animales, y en el desarrollo de la respuesta in-mune específi ca, capaz de reconocer al antígeno y por lo tanto de crear una memoria inmunológica que perdura aún cuando el agente patógeno es eliminado, conocida como respuesta inmune adaptativa, y que tan solo está presente en los vertebrados

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
82
1. Citoquinas involucradas en la respuesta inmune innata
En el grupo de las citoquinas que están involucradas en la respuesta inmune innata (Tabla 2) están incluidas las citoquinas producidas, principalmente, por los monocitos y los macrófagos activados de forma inmediata tras el contacto con un agente patógeno, y que por lo tanto están involucradas en el desarrollo de la respuesta infl amatoria. Esta es la razón por la que entre las principales citoquinas que se produ-cen en esta respuesta inmune innata nos encontramos tanto con la IL-1, el TNF-alfa, la IL-6 y el IFN-gamma, que como hemos visto anteriormente también participan en la respuesta infl amatoria, como con la IL-10 que tiene una clara acción inmunosupre-sora. Además, en la respuesta inmune innata van a participar otras citoquinas como:
• La interleuquina 12 (IL-12) que inicialmente se describió como el factor estimulador de las células natural killer. Esta citoquina que es producida por monocitos/macrófagos, células dendríticas y linfocitos B, incrementa la actividad citotóxica de las células natural killer, induce las células LAK (linfocitos asesinos activados por linfoquinas), estimula la producción de IFN-gamma y activa a los linfocitos T citotóxicos.
• La interleuquina 18 (IL-18) posee la misma capacidad de inducción de la producción de IFN-gamma en linfocitos T y células natural killer que la IL-12, aunque es producida por las células de Kupffer y de la glándula adrenal.
Tabla 2: Principales citoquinas que participan en la respuesta inmune
Respuesta inmune innata IL-1
IL-6
IL-10
IL-12
IL-18
TNF-α
IFN-γRespuesta inmune adaptativa IL-2
IL-4
IL-12
IL-15
IL-16

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
83
2. Citoquinas producidas en la respuesta inmune adaptativa
La activación de los linfocitos T y B en reposo para ejercer sus funciones en la respuesta inmune celular y humoral requiere de la participación de determinadas citoquinas (Tabla 2). Así, la presencia de IL-12 promueve la diferenciación hacia lin-focitos Th1, que en colaboración con los macrófagos, están implicados en la respuesta inmune celular. Mientras que la IL-4 condiciona el desarrollo de linfocitos Th2 que promueven la respuesta inmune humoral. Además, los linfocitos T CD8+ se diferen-cian hacia linfocitos T citotóxicos como respuesta a la estimulación antigénica y a la presencia de citoquinas secretadas por otras células, y ejercen su función efectora mediante la secreción de IL-2, IL-16, IFN-gamma y TNF.
• La interleuquina 2 (IL-2) es secretada por linfocitos T CD4+ y CD8+ activados y se describió como factor de crecimiento de células T, ya que es el principal agente que controla su proliferación. Es un factor estimu-lador del crecimiento de linfocitos T , B y natural killer, y promueve la actividad citotóxica mediada por linfocitos T, natural killer, y células LAK (células asesinas activadas por citoquinas). Tras unirse a su receptor en los linfocitos T, activa la secreción de IFN-alfa, IL-3 e IL-4. Ejerce otros muchos efectos sobre el sistema inmune, teniendo un papel esencial en el desarrollo de las respuestas infl amatorias crónicas, tanto humorales como celulares
• La interleuquina 15 (IL-15) comparte la mayoría de sus actividades biológicas con la IL-2, aunque no es producida por los linfocitos Th1, ya que es secretada por una amplia variedad de células, entre las que se encuentran los monocitos, células epiteliales, miocitos y hepatocitos.
• La interleuquina 16 (IL-16) es producida por los linfocitos T CD8+ en respuesta a la estimulación con serotonina o histamina. Originariamente se identifi có como factor quimiotáctico de linfocitos, recibiendo el nombre de linfotactina.
Citoquinas que estimulan la hematopoyesis
Un amplio grupo de citoquinas está conformado por aquellas producidas por las células del estroma de la médula ósea o por linfocitos maduros activados y que van a inducir el crecimiento y/o diferenciación de las células sanguíneas, por lo que algunas reciben el nombre genérico de factores estimuladores de la formación de colonias (CSF)

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
84
• La interleuquina 3 (IL-3) induce la proliferación y diferenciación de pro-genitores hematopoyéticos tempranos de todas las series sanguíneas, es-pecialmente en situaciones de stress que requieren una respuesta rápida.
• La interleuquina 5 (IL-5) es esencial en la proliferación y diferenciación de las células precursoras de los eosinófi los, así como en el mantenimiento de la actividad de los mismos, siendo la responsable de la eosinofi lia que se produce en las infecciones parasitarias. Sobre los linfocitos B actúa incrementando su proliferación y estimulando la producción de IgA.
• La interleuquina 9 (IL-9) tiene un amplio espectro de actividades, entre las que se encuentra la proliferación de los precursores de la serie eritroide, participando también en la respuesta inmune al estimular la proliferación de linfocitos T y la producción de inmunoglobulinas por los linfocitos B.
• La interleuquina 11 (IL-11) estimula la formación de megacariocitos y comparte con la IL-6, la producción de proteínas de fase aguda por el hígado. Además, participa en la respuesta inmune humoral al esti-mular la secreción de inmunoglobulinas por células B en respuestas T-independientes.
• El factor estimulador de la formación de colonias de granulocitos (G-CSF) es producido por fi broblastos, células endoteliales y monocitos en respuesta a estímulos específi cos y provoca la proliferación de los pre-cursores de los granulocitos. La proliferación de los polimorfonucleares neutrófi los que se observa asociada a la infección por diferentes bacterias es debido a que algunos lipopolisacáridos de las paredes bacterianas son un inductor de esta citoquina. Además, esta citoquina actúa como esti-mulador de la fagocitosis y de la citotoxicidad mediada por anticuerpos.
• El factor estimulador de la formación de colonias de macrófagos (M-CSF) está implicado tanto en el desarrollo de las células progenitoras de los macrófagos como en el desarrollo de la placenta, siendo producido también por las células del epitelio uterino en respuesta a los estrógenos.
Los efectos adversos de las citoquinas.
Como hemos visto anteriormente las citoquinas van a tener un importante papel en la regulación de diferentes funciones del organismo. Sin embargo, algunas de ellas debido a su capacidad de ejercer efectos biológicos diferentes al actuar sobre distintos tipos celulares van a tener un efecto adverso sobre el organismo (Tabla 3). Como ejemplos tenemos:

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
85
• La IL-6, que aunque juega un papel muy importante en la respuesta in-fl amatoria y en el establecimiento de la respuesta inmune innata, también puede inducir resorción ósea, atrofi a muscular, anemia y desencadenar una coagulopatía de consumo.
• La IL-4, que juega un papel clave en los fenómenos de inmunosupresión, pero que al mismo tiempo se ha relacionado con el desarrollo de los pro-cesos alérgicos, ya que induce la producción de IgE por los linfocitos B.
Tabla 3: Efectos adversos de las principales citoquinas
IL-1 Fiebre, acidosis metabólica, leucopenia, trombocitopenia, hemorragias, edema pulmonar
IL-4 Desarrollo de procesos alérgicosIL-6 Resorción ósea, atrofi a muscular, anemia, coagulopatía de consumoTNF-α Fiebre, acidosis metabólica, leucopenia, trombocitopenia, hemorragias, edema
pulmonarIFN-γ Efecto sinérgico con endotoxinas bacterianas
Pero además, los efectos adversos de las citoquinas pueden ser el resultado de la respuesta del organismo frente a un patógeno. Así, las citoquinas juegan un papel muy importante en la patogenia del shock que se instaura tras la liberación de exotoxinas o endotoxinas bacterianas en el transcurso de infecciones locales o sistémicas, ya que estas toxinas bacterianas inducen la producción y liberación, entre otros mediadores infl amatorios, del TNF-alfa y de la IL-1beta, que de forma sinérgica estimulan la respuesta infl amatoria aguda. Posteriormente, otras citoquinas, como la IL-4, IL-8 o la IL-10, van a controlar la intensidad de esta respuesta infl amatoria. El desenlace de este equilibrio entre estos dos grupos de citoquinas es lo que determina las manifestaciones clínicas y el desenlace del proceso séptico. Así, si en el proceso séptico predominan los niveles de citoquinas pro-infl amatorias (TNF-alfa, IL-1 beta) se producirá el shock séptico y la muerte del individuo. Ya que el estado de shock, que se instaura en el proceso séptico, es una consecuencia de la alta concentración sérica de las citoquinas pro-infl amatorias, ya que tanto el TNF-alfa como la IL-1 beta, inducen la aparición de la fi ebre, de la acidosis metabólica, de la leucopenia y trom-bocitopenia, así como de las hemorragias y del edema pulmonar. Alteraciones que se han inducido en modelos animales tras la administración de TNF-alfa recombinante. Pero no solo estas citoquinas tiene efectos adversos en los procesos sépticos, ya que otras citoquinas como el IFN-gamma va a incrementar los efectos adversos de las ci-toquinas pro-infl amatorias, al activar a los macrófagos y al estimular la producción de

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
86
anticuerpos contra los polisacáridos de la pared bacteriana, ejerciendo de esta forma un efecto sinérgico con las endotoxinas bacterianas. Por este motivo, se ha ensayado como estrategia terapéutica el bloqueo del IFN-gamma cuando el shock séptico ya está instaurado.
Este efecto adverso producido por el desequilibrio de las citoquinas pro- y anti-infl amatorias, en el que predominan una respuesta exagerada del sistema inmune, es lo que recientemente se ha dado en llamar como “tormenta de citoquinas”, un fenómeno que ha cobrado un gran protagonismo en los recientes brotes infl uenza causados por los virus H1N1 y H5N1, ya que ha sido esta respuesta infl amatoria exagerada la responsable de los cuadros clínicos y de la mortalidad observada en el transcurso de las infecciones.
Otros virus, como el del síndrome respiratorio y reproductivo porcino, van a utilizar la sobreexpresión de citoquinas inmunosupresoras, como la IL-10, como estra-tegia para evadir la respuesta inmune o, como en el caso del virus de la Peste Porcina Africana, van a inducir los fenómenos de apoptosis en los linfocitos, células en las que no se replica, mediante la expresión de TNF alfa, originado de esta forma un estado de inmunosupresión e impidiendo el desarrollo de una correcta respuesta inmune.
Pero no solo se va a producir un efecto adverso como consecuencia del des-equilibrio de los niveles de determinadas citoquinas ya que, además, existen algu-nas citoquinas que se expresan de forma constitutiva en diferentes órganos como el pulmón, hígado, riñón, bazo o la piel, como la denominada factor inhibitorio de la migración del macrófago, y que tiene como función la de contrarrestar las acciones antiinfl amatorias e inmunosupresoras de los glucocorticoides, pero que al mismo tiempo es liberada rápidamente tras la interacción con los lipopolisacáridos (LPS) de las membranas bacterianas, potenciando su efecto como estimulador de la liberación de citoquinas pro-infl amatorias.
Recientemente, se han descritos los denominados «superantígenos», que son proteínas microbianas, como las presentes en algunas exotoxinas estafi locócicas, que son capaces de estimular, sin especifi cidad antigénica, a numerosos linfocitos T CD4+, originando una enfermedad clínica caracterizada por la liberación de gran cantidad de citoquinas, especialmente TNF e IL-2, por los linfocitos T, provocando la instauración de un shock tóxico.
Por último, me gustaría destacar el papel que juegan las citoquinas en las reac-ciones de hipersensibilidad, ya que ellas van a actuar modulando la respuesta infl a-matoria que se desencadena. Una respuesta infl amatoria que va a ser la responsable de la aparición de las lesiones en los tejidos.

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
87
Las citoquinas como herramienta terapéutica
La medicina actual se encuentra frente a un incremento de la resistencia de los patógenos frente a los antibióticos, una pérdida de sensibilidad que es debida al uso indiscriminado que se ha hecho en muchos casos de los antibióticos. Adicionalmente, en medicina veterinaria nos encontramos con el handicap que supone los periodos de supresión tras la administración de los antibióticos o la prohibición de determi-nados antibióticos, antiparasitarios y promotores del crecimiento en los animales de producción. Por este motivo, la investigación biomédica ha comenzado a explorar diferentes alternativas, como la utilización de aceites esenciales, con las que estimular y reforzar la respuesta inmune innata y adaptativa. La infl uencia y los efectos que tiene algunas citoquinas sobre el sistema inmune ha sido el motivo de que hoy en día se estén diseñando terapias que incluyen su utilización, son las llamadas “terapias basadas en la respuesta inmunológica”. Sin embargo, estas terapias se están encon-trado con diferentes problemas, algunos de ellos derivados del delicado equilibrio en el que se encuentran las citoquinas en el organismo, pues como hemos mencionado anteriormente, estas se encuentran en muy pequeñas concentraciones y su combina-ción produce diferentes respuestas inmunológicas, así mientras que las dosis bajas de IL-2 estimulan a las células natural killer, las dosis altas e intermitentes de esta misma citoquina estimula la producción de las células CD4+. Por lo que los investigadores se enfrentan al reto de descubrir sus funciones biológicas y de cómo administrarlas para alcanzar la respuesta deseada (Tabla 4). Entre las citoquinas utilizadas y comer-cializadas como agentes terapéuticos hoy en día tenemos:
• El eritropoetin-alfa, que es parte de los tratamientos de las anemias de tipo leve o moderada
• El factor estimulante de las colonias de granulocitos, que es utilizado para prevenir los efectos adversos de los tratamientos frente al VIH o los citomegalovirus, ya que estos virus suelen causar una neutropenia y, por tanto favorecer las infecciones bacterianas secundarias.
• El interferón alfa, es un antiviral de amplio espectro y se ha probado con éxito en el tratamiento de la hepatitis vírica y del Sarcoma de Kaposi, que está provocado por el virus del herpes 8 y cuya aparición está relacionada con el SIDA. Aunque en los ensayos “in vitro” el interferón alfa actúa contra el VIH, los resultados “in vivo” han sido contradictorios. Actual-mente se está estudiando su aplicación en la prevención de la diabetes y en el tratamiento de determinados tumores como los melanomas.
• La interleuquina 12, su utilización se basa en su efecto estimulador de las células CD4+, demostrándose además que si estás células son trata-

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
88
das con esta citoquina responden mejor frente a la infección por el VIH. Además, su efecto estimulador de los linfocitos T ha hecho que se use como potenciador de determinadas vacunas.
Tabla 4: Tratamientos en los que se están utilizando citoquinas
IL-12 Potenciación de vacunasIL-15 Control de la infección por el VIHIFN- α Tratamientos antiviralesG-CSF Tratamiento frente a la neutropeniaEritropoetin- α Tratamiento de anemias leves y moderadas
Actualmente, se está investigando que efectos podría tener en el tratamiento frente a la infección por el Virus de la Inmunodefi ciencia Humana la utilización de diferentes citoquinas como:
• La interleuquina 7. El interés en esta citoquina radica en el hecho de que cuando el número de células CD4+ cae por debajo de los límites nor-males, como en los casos de la infección por el VIH o por el trasplante de médula, se produce un aumento de los niveles de esta citoquina que estimula el timo y la producción de nuevas células CD4+. Sin embargo, esta citoquina también tiene un efecto adverso, ya que se ha demostrado que es capaz de activar al VIH, una activación que haría necesaria su administración junto con otros fármacos contra este virus.
• La interleuquina 15 ya que es un promotor de las células CD8+, que juegan un importante papel en la eliminación de las células infectadas e inhibe la apoptosis de las células activadas inmunológicamente. En este sentido la utilización de esta citoquina se ha asociado con un mejor control de la infección por el VIH.
• El interferón-gamma se ha ensayado en el tratamiento de las infecciones simultaneas de tuberculosis y VIH. Sin embargo los resultados obtenidos indican que esta citoquina tendría un efecto dependiente de la dosis, ya que mientras que en dosis baja si parece que ayuda a controlar la infec-ción por VIH, las dosis altas parecen estimular la replicación del virus.
• La interleuquina 10 ya que, aunque tiene como principal efecto al dis-minución de la respuesta inmune, se ha observado “in vitro” que es capaz de provocar una disminución de la replicación del VIH, por lo que se está ensayando una terapia con esta citoquina con el objetivo de disminuir los niveles de VIH en las personas infectadas por este virus.

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
89
Otro campo de investigación terapeútica de las citoquinas es el de los trasplantes de órganos ya que el rechazo es modulado por la expresión de las citoquinas pro-infl amatorias e inmunomoduladoras, por lo que el bloqueo de de estas citoquinas o de sus receptores podría ser una herramienta extremadamente útil, ya que al no producirse la reacción infl amatoria e inmune, no se produciría el rechazo del tejido.
CONCLUSIÓN
Está claro que si somos capaces de descifrar el lenguaje de las citoquinas, po-dremos comprender como funciona el complejo sistema inmunológico, y por tanto seríamos capaces de controlar sus respuestas, protegiendo y mejorando de esta forma la salud del hombre y de los animales. El conocimiento de la composición y de los mecanismos de activación de los receptores de las citoquinas son de una gran importancia, ya que nos permitiría comprender dos de sus principales características funcionales, la redundancia y el pleiotropismo, y nos permitiría controlar el desenca-denamiento de sus cascadas de amplifi cación y de antagonismo.
BIBLIOGRAFÍA RECOMENDADA
1. Banyer JL, Hamilton NH, Ramshaw IA, Ramsay AJ (2000). Cytokines in innate and adaptive immunity. Rev Immunogenet 2(3):359-373.
2. Barranco I, Gómez-Laguna J, Rodríguez-Gómez IM, Salguero FJ, Pallarés FJ, Carrasco L (2011). Differential Expression of Proinfl ammatory Cytokines in the Lymphoid Organs of Porcine Re-productive and Respiratory Syndrome in Virus-Infected Pigs. Transboundary and Emerging Dis-eases, doi:10.1111/j.1865-1682.2011.01252.x
3. Biron CA, Sen GC (2001). Interferons and other cytokines. In: Fields of Virology, 4th Edit., D Knipe, P Howley, D Griffi n, R Lamb, M Martin, Eds., Lippincott, Williams & Wilkins, Philadel-phia, pp. 321-349.
4. Borden EC, Sen GC, Uze G, Silverman RH, Ransohoff RM, Foster GR, Stark GR (2007). Interfer-ons at age 50: past, current and future impact on biomedicine. Nat Rev Drug Discov. 6, 975–990.
5. Carrasco L, Núñez A, Salguero FJ, Díaz San Segundo F, Sánchez-Cordón PJ, Gómez-Villaman-dos JC, Sierra MA (2002). African swine fever: expression of interleukin-1 alpha and tumour necrosis factor-alpha by pulmonary intravascular macrophages. Journal of Comparative Patholo-gy, 126:194-201.
6. Carrasco L, Núñez A, Sánchez-Cordón PJ, Pedrera M, Fernández de Marco MM, Salguero FJ, Gómez-Villamandos JC (2004). Immunohistochemical detection of the expression of proinfl am-matory cytokines by ovine pulmonary macrophages. Journal of Comparative Pathology, 131:285-293.
7. Cavaillon JM (1994). Cytokines and macrophages. Biomed Pharmacother 48(10):445-453.8. Gómez-Laguna J, Salguero FJ, Barranco I, Pallarés FJ, Rodríguez-Gómez IM, Bernabé A, Carras-
co L (2010). Cytokine expression by macrophages in the lung of pigs infected with the Porcine Reproductive and Respiratory Syndrome virus. Journal of Comparative Pathology, 142:51-60.

CITOQUINAS: DE FIELES ALIADAS A TEMIBLES ENEMIGAS
90
9. Gómez-Laguna J, Salguero FJ, Fernández de Marco M, Pallarés FJ, Bernabé A, Carrasco L (2009). Changes in lymphocyte subsets and cytokines during European Porcine Reproductive and Re-spiratory Syndrome: increased expression of IL-12 and IL-10 and proliferation of CD4-C8high. Viral Inmunology, 22:261-271.
10. Gómez-Laguna J, Salguero FJ, Pallarés FJ, Fernández de Marco M, Barranco I, Rodriguez-Gó-mez I, Bernabé A, Carrasco L (2010). El papel de las citoquinas como mediadores de la respuesta inmune en el Síndrome Reproductivo y Respiratorio Porcino. Tierras, 164:93-98.
11. Gómez-Laguna J, Rodríguez-Gómez IM, Barranco I, Pallarés FJ, Salguero FJ, Carrasco L (2012). Enhanced expression of TGFb protein in lymphoid organs and lung, but not in serum, of pigs infected with a European fi eld isolate of porcine reproductive and respiratory syndrome virus. Veterinary Microbiology, doi:10.1016/j.vetmic.2012.02.003
12. Moore KW, de Waal Malefyt R, Coffman RL, O’Garra A (2001). Interleukin-10 and the interleu-kin-10 receptor. Annual Review of Immunology 19:683-765.
13. Núñez A, Gómez-Villamandos JC, Sánchez-Cordón PJ, Fernández de Marco M, Pedrera M, Sal-guero FJ, Carrasco L (2005). Expression of proinfl ammatory cytokines by hepatic macrophages in acute Classical swine fever. Journal of Comparative Pathology, 133:23-32.
14. Paliard X, Malefi jt RW, De Vries JE, Spits H (1988). Interleukin-4 mediates CD8 induction on human CD4+ T-cell clones. Nature 335:642-644.
15. Pastoret P, Griebel P, Bazin H, Govaerts A (1998). Handbook of Vertebrae Immunology. Acade-mic Press, San Diego.
16. Pestka S, Krause CD, Walter MR (2004). Interferons, interferon-like cytokines, and their recep-tors. Immunological reviews 202:8-32.
17. Sánchez-Cordón PJ, Núñez A, Salguero FJ, Carrasco L, Gómez-Villamandos JC (2004). Proin-fl ammatory cytokines in viral diseases. Current Trends in Immunology, 6:127-137.
18. Tizard, IR (2008). Cell Signaling: Cytokines and Their Receptors. In: Veterinary immunology: an introduction, 8th Edit., Tizard I.R. (eds.), Elsevier Science, Philadelphia, pp. 70-80.
19. Vicek J, Sen G (1996). Interferons and other cytokines. In: Fields’ Virology, Fields B, Knipe D, Howley P (eds.), 3rd ed. Vol. 13, Lippincott–Raven Publishers, Philadelphia. pp. 375–400.
20. Wolf SF, Sieburth D, Sypek J (1994). Interleukin 12: a key modulator of immune function. Stem Cells 12:154-168.

CONTESTACIÓN DISCURSO ILMO. SR. CARRASCO OTERO
91ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
Nos decía Friedrich Nietzsche que la vida está hecha de algunos momentos de gran intensidad, y de innumerables intervalos, y lo mismo sucede con la actividad de estas corporaciones que junto con sus labores cotidianas, vive momentos mágicos de gran intensidad, cuando la Academia se viste de gala y en sesión solemne le da la bienvenida a un nuevo miembro, y máxime en este 2011 declarado “Año Mundial del Veterinario”, conmemorando la fundación de la Escuela Nacional de Veterinaria en Lyon en 1761, hace 250 años por el insigne profesor Claude Bourgelat y por mandato de Luis XV el Rey Sol.
Esta Real Academia de Ciencias Veterinarias de Andalucía Oriental me hizo el honor de designarme para que contestara el discurso de ingreso del Ilmo. Sr. D. Librado Carrasco Otero, y pienso que ello fue debido a la amistad que me unía primero con sus padres D. Librado y Dª Paquita, y posteriormente con nuestro fl amante Académico que también me distingue con su aprecio. Creo sinceramente que otros compañeros de la Corporación, hubiesen glosado mejor que yo su laudatio, y la evidente importancia de su brillante discurso, pero el honor que se me hacía no podía echarlo en puerta ajena por el deber que la amistad obliga, aunque de antemano pensara que ya por mi edad no estaría a la altura de este cometido, en el que seguro no me acompañara la fortuna pero si la buena voluntad.
Como decía Cela en su discurso el “Elogio de la fábula” (1989), no sé dónde puede levantar su aduana la frontera de la vejez, pero por si acaso, me acojo a la sentencia
CONTESTACIÓN AL DISCURSO DE INGRESO DEL ILTMO. SR. DR. D. LIBRADO CARRASCO OTERO
EXCMO. SR. DR. D. JULIO BOZA LÓPEZ1
1 De la Real Academia de Ciencias Veterinarias de Andalucía Oriental.

CONTESTACIÓN DISCURSO ILMO. SR. CARRASCO OTERO
92
de Quevedo todos deseamos llegar a viejos y todos negamos haber llegado ya, pero se bien que no se puede volver la cara a la evidencia, y tampoco ignoro que el calendario es algo inexorable, y es por lo que me dispongo a contestar al Prof. Carrasco Otero, sin dejar el menor hueco a la inspiración ni a la improvisación, por lo que en ésta ocasión me van a permitir que sólo lea unas sentidas palabras como homenaje de acogida.
Como señaló el escéptico tudense Francisco Sánchez en el siglo XVI: “Es inna-to en los hombres querer saber. Pero pocos son los que emprenden el camino de la ciencia, y menos aun los que los que lo alcanzan”, y entre esos pocos esta nuestro conferenciante, como veremos en la sucinta síntesis de su historial profesional y científi co que les he preparado.
Tu paisano Séneca nos pide mesura hasta en el sufrimiento, y el belmontino Baltasar Gracián nos aconseja que seamos breves. Pues bien, mesurada y brevemente, siguiendo estas dos sabias y prudentes sentencias, sólo podré mostrar unas tenues pinceladas de sus muchos y elevados méritos, dejando en el tintero cuestiones impor-tantes que he sido incapaz de extractar, pero al haberlo escuchado todos nos hemos percatado de tu extraordinaria valía.
Nuestro conferenciante nació en Córdoba la próxima capital de la cultura, licen-ciándose en Veterinaria en 1985 y doctorándose en 1988, en la Universidad de esta ciudad. Completo su formación el Prof. Carrasco Otero en la Facultad de Veterinaria de Hannover en Alemania, en el Instituto de Salud Animal de Reino Unido, En la Universidad de Concepción de Chile, y en la Universidad estatal de Kansas en los Estados Unidos.
Es catedrático de la Universidad de Córdoba y decano de su Facultad de Vete-rinaria desde 2006.
Esta diplomado por el Colegio Europeo de Veterinarios Patólogos, y por el Co-legio Europeo de Salud y Producción Porcina, y es también Académico número de la Real Academia Sevillana de Ciencias Veterinarias.
Desde su incorporación como profesor a la Facultad de Veterinaria en 1986, ha desempeñado diferentes puestos docentes en las asignaturas de Citología e Histología,
Anatomía Patológica General y Especial en la Licenciatura de Veterinaria. Ha participado en 12 proyectos de innovación docente, 20 programas de doctorado, y en varias ediciones del Máster de Medicina, Sanidad y Mejora Animal de la Universidad de Córdoba. Junto con ello, ha impartido 75 conferencias dentro de jornadas o cursos de posgrado y especialización, siendo además profesor de varias ediciones del Curso

CONTESTACIÓN DISCURSO ILMO. SR. CARRASCO OTERO
93
Internacional de Enfermedades Exóticas que organiza el Centro de Investigaciones en Sanidad Animal (CISA-INIA).
Es coautor de varios materiales didácticos, entre los que destacan diversos atlas, como el Curso Digital de Enfermedades Infecciosas Porcinas, así como dos capítulos del tratado de Histología Veterinaria. Su activad docente está avalada por la concesión de dos diplomas de excelencia por la Universidad de Córdoba.
Como investigador se ha dedicado al estudio de la patogenia de las enfermedades víricas, participando en 15 proyectos de investigación, destacando sus contribuciones al conocimiento de la Peste porcina africana, Peste clásica, Peste equina africana, y Síndrome reproductivo y respiratorio porcino, Es autor de más de 180 trabajos la mayoría de ellos en publicaciones incluidas en el Journal Citation Repors del Instituto de Ciencias de la Información (ISI). Igualmente ha presentado más de 260 ponencias y comunicaciones a congresos, simposios y jornadas nacionales e internacionales en su especialidad. Su actividad investigadora ha sido acreditada por varios premios concedidos por la European Society of Veterinary Pathology.
También ha dirigido cinco tesis doctorales, dos de ellas con mención europea.
Nuestro nuevo académico es referí de diversas revistas de investigación, y pertenece a diversos comités científi cos, entre el que destacamos ser experto de la Agencia Nacional de Evaluación y Prospectiva del Ministerio de Educación, entre otros nacionales e internacionales.
En el exordio de su discurso titulado “Citoquinas de fi eles aliadas a temibles enemigos” nos ha mostrado la naturaleza de estas moléculas, el tipo de células de quienes proceden tanto en condiciones fi siológicas como patológicas, además de señalarnos las peculiaridades de las distintas citoquinas y de sus capacidades fun-cionales, poniendo de manifi esto el interés actual del tema elegido. En la exposición nos ha señalado las implicaciones de estas moléculas en los procesos biológicos, y las funciones de la diversidad de ellas pre-infl amatorias, inmuno-depresoras, en la respuesta inmune innata y adaptativa, y en aquellas que estimulan la hematopoyesis.
En la argumentación de su discurso nos ha evidenciado a las citoquinas en terapias basadas en la respuesta inmunológica, con sus diferentes problemas conse-cuencia del equilibrio entre las distintas moléculas, y mostrándonos las utilizadas y comercializadas actualmente como agentes terapéuticos. Finalmente en el epílogo nos ha ilustrado sobre las investigaciones que se están efectuando con estas moléculas frente a las enfermedades emergentes, así como un interesante apartado concerniente a las propiedades adversas de las citoquinas, que a la vista de ellos las podríamos

CONTESTACIÓN DISCURSO ILMO. SR. CARRASCO OTERO
94
llamarlas como “el paradigma del bien y del mal”. Termina su intervención presen-tando los objetivos principales de investigación sobre estas moléculas, con la fi nalidad de conocer la efectividad potencial de las mismas en la protección de nuestra salud.
Esta lección magistral sobre estos mediadores moleculares solubles, derivados de los leucocitos y comprometidos con la respuesta inmunitaria celular, ha sido la tarjeta de presentación a esta Real Academia de un destacado docente y prestigioso investigador, que nos ha entusiasmado hasta el extremo que tu viejo compañero te pide que nunca abandones la ilusión con la que nos lo has expuesto, compendio de tu preparación y capacidad de trabajo, que están por encima de todos los elogios.
Termino señalando que es un honor tenerte entre nosotros, deseándote una larga vida académica colmada de aciertos, frente a los embates que siempre aparecen y siempre se estrellan contra el alma de los elegidos, y más en el tiempo inclemente que vivimos, con circunstancias que difi cultan las actividades de estas corporaciones. Pero demos tiempo al tiempo, según decía Cervantes en “La Gitanilla”, que suele ser dulce salida a muchas amargas difi cultades.
Y un deseo más, que desde ahora sean frecuentes tus aportaciones a la Academia para que podamos disfrutar de tu ciencia y tu presencia entre nosotros, y hoy siendo portavoz ocasional de todos, te doy la enhorabuena por esta distinción, así como te felicito y nos felicitamos por tu incorporación a esta Real Corporación.
Muchas gracias por la atención que me han prestado.
He dicho.

EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
95ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
Excelentísimos e Ilustrísimos Señores Académicos, Señoras y Señores. Con su permiso procederé a la lectura del mi discurso de ingreso en la Real Academia de Ciencias Veterinarias de Andalucía Oriental. Pero antes de comenzar quisiera agra-decer a la Real Academia, y a sus ilustres miembros, por la confi anza depositada en mí para este nombramiento. Es para mí un honor y un privilegio formar parte de esta institución, en la que con mi humilde labor espero contribuir para bien de las Ciencias Veterinarias.
Mi discurso versará sobre distintos aspectos históricos y nuevas perspectivas en la investigación de la lengua azul, enfermedad emergente que ha ocupado no pocas portadas en los últimos años, considerada como modelo para el estudio de otras enfermedades víricas animales y humanas transmitidas por vectores. Las experien-cias aprendidas con la lengua azul ponen de manifi esto la posibilidad real de que enfermedades similares, o incluso más nefastas y comunes en latitudes remotas, nos afecten directamente debido a la capacidad de los vectores transmisores de globalizar estos procesos como consecuencia de cambios que aún no alcanzamos a comprender.
Pero antes, quisiera comenzar haciendo una pequeña referencia a una raza de ovejas íntimamente ligada a esta enfermedad, me refi ero a la raza Merina.
EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
PEDRO JOSÉ SÁNCHEZ CORDÓN1
1 Dr. en Veterinaria. Investigador del Programa Ramón y Cajal. Dpto. de Anatomía y Anatomía Patológica Comparadas. Facultad de Veterinaria. Universidad de Córdoba.
Discurso de ingreso en la Real Academia de Ciencias Veterinariasde Andalucía Oriental como Académico Correspondiente

EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
96
Como es bien sabido, la oveja de raza Merina representó un lucrativo negocio durante la Baja Edad Media. Los propietarios de rebaños, en su mayoría nobles terrate-nientes, órdenes militares y monasterios, obtuvieron una serie de ventajas económicas y jurídicas, organizándose entorno al “Honrado Concejo de la Mesta”, asociación de propietarios ganaderos de Castilla creada en 1273 por Alfonso X “El Sabio”.
La Mesta, que controlaría el monopolio de la lana, prohibió la salida de animales de raza Merina, a la vez que estableció fuertes controles sobre las exportaciones de lana a Flandes, Gran Bretaña y otras zonas de Europa entre los siglos XIV y XVI. A excepción de los efectivos que cruzaron nuestras fronteras por cuestiones de estado, o a raíz de concesiones impuestas, la raza Merina permaneció en España, aislada del mundo, hasta el último cuarto del siglo XVIII. En una primera fase, los efectivos ovinos de esta raza se difundieron por Alemania, Francia y Reino Unido. Fue tal su impacto, debido a los ávidos deseos del mundo entero por disponer de estos valiosos ejemplares, productores de la más fi na de las lanas, que recientemente se ha conme-morado el 225 aniversario de dicha salida. En una segunda fase, la raza se expandió por América del Sur, Australia y Sudáfrica, lugares donde encontraron condiciones más propicias para su explotación.
Precisamente en tierras de Sudáfrica, fueron los holandeses quienes en 1652 arrebataron a los portugueses unos pequeños asentamientos situados en el cabo de Buena Esperanza. Estos asentamientos crecieron hasta formar la Colonia del Cabo, constituida en gran parte por granjeros traídos de Holanda y Alemania conocidos como “Boers”, que hicieron prosperar la agricultura y la cría de ganado en la zona. Sin embargo, tras una serie de intentos frustrados, en 1806, serían los ingleses quienes se apoderarían defi nitivamente de la colonia holandesa.
Por caprichos de la Historia, sería en esta colonia británica donde por primera vez de describiría una enfermedad, la lengua azul, en una raza de ovejas, la Merina, originariamente española.
La lengua azul (LA) es una enfermedad históricamente africana y, aunque no se ha descrito en todos los países de África, es posible que sea enzoótica en todo el continente. La LA fue probablemente reconocida tan pronto como las razas de ovejas europeas, y particularmente ovejas de raza Merina, se introdujeron en Sudáfrica. Sin embargo, como enfermedad que afectaba al ganado ovino y bovino, fue descrita por primera vez a fi nales del siglo XVIII por el biólogo francés Francois de Vaillant durante sus viajes al Cabo de Buena Esperanza entre 1781 y 1784. Años más tarde, el Ofi cial Veterinario Jefe de la Colonia del Cabo, Duncan Hutcheon, registró algunas

EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
97
características de la enfermedad en su informe anual de 1880. Pero no sería hasta 1902 cuando publicara en la revista británica “The Veterinary Record” un artículo titulado “fi ebre catarral malárica en ovejas”, donde ya postuló que el agente de la enfermedad podría ser transmitido por insectos hematófagos.
Ese mismo año, el Cirujano Veterinario J. Spreull publicó los primeros resultados obtenidos en sus experimentos con fi ebre catarral malárica en ovejas. Pero no sería hasta 1905 cuando publicara un artículo donde, por primera vez, se describirían con detalle los signos clínicos típicos de esta enfermedad. Entre éstos destacaba la aparición de fi ebre alta en fases iniciales del proceso y la presencia, entre 7 y 10 días después, de lesiones distintivas como una coloración azul oscura de la lengua, consecuencia de la cianosis. A raíz de la observación de estas lesiones, Spreull sugirió el nombre de lengua azul para defi nir a esta enfermedad, término con el que los granjeros Boers se habían referido a la misma durante generaciones.
Poco tiempo después, en 1906, Sir Arnold Theiler, demostró que el agente res-ponsable de la LA era fi ltrable, probablemente un virus estrechamente relacionado con la sangre. Sin embargo, fue el propio Spreull quien demostró que la enfermedad era transmisible a vacas y cabras, así como que la infección en estas especies rumiantes cursaba de forma subclínica.
Pese a no disponer de equipos y técnicas sofi sticadas, los pioneros en la investi-gación de la LA supieron compensar estas defi ciencias con el poder de la observación, el razonamiento lógico, el ingenio y la perseverancia. Determinaron que la LA era una enfermedad más frecuente durante los meses de verano, especialmente después de la temporada de lluvias, y que cesaba de forma súbita tras las primeras heladas. También observaron que la enfermedad era más común en las tierras bajas y pasti-zales de los valles que en las tierras alta, así como que las ovejas estabuladas en los cobertizos durante las noches de verano escapaban a la infección. Establecieron, por tanto, evidencias circunstanciales de que la LA era una enfermedad infecciosa, no contagiosa, causada por un virus que afectaba a rumiantes, en especial a los ovinos, y que era transmitida por insectos, posiblemente mosquitos hematófagos, defi nición que es aceptada hoy en día.
Fue Nieschultz quien, en 1934, llevó a cabo distintos intentos para demostrar la implicación de mosquitos del género Aedes en la transmisión de la LA, auque sin resultados convincentes. Ello llevó a du Toit a considerar a otros insectos hemató-fagos como vectores potenciales del virus, entre los que se incluían mosquitos del género Culicoides, un grupo de insectos abundante en Sudáfrica, especialmente en

EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
98
los meses de mayor incidencia de la enfermedad, que hasta la fecha no habían sido señalados como potenciales vectores de ningún agente patógeno. Para sorpresa de los entomólogos, du Toit demostró en 1944 la implicación de Culicoides imicola en la transmisión del virus de la LA, especie que durante décadas sería considerada como el principal vector del virus, si no el único, hasta que a partir de la década de los 90, distintas investigaciones llevaron a señalar el papel de otras especies de Culicoides en diferentes regiones del mundo. Por tanto, se demostró que la presencia y abundancia del vector, junto a unas condiciones climáticas apropiadas, condicionaría la existencia de la enfermedad y su difusión.
Aunque se sospechaba de la existencia de múltiples cepas del virus, fue Neitz quien en 1948 demostraría, de forma concluyente, su presencia en la naturaleza. Pos-teriormente, sería Howell en 1960 quien, a partir de trabajos previos que permitieron el cultivo del virus, confi rmaría mediante seroneutralización la existencia de distintos serotipos. Hasta la fecha, 26 serotipos han sido reconocidos globalmente, habiéndose aislado el virus en todos los continentes excepto en la Antártida.
Hasta la década de los años cuarenta del siglo XX, la enfermedad estuvo apa-rentemente confi nada en África. Los primeros brotes bien documentados de LA fuera del continente afectaron, en 1943, a ovejas de la isla de Chipre, si bien parece que la enfermedad pudo haber estado presente desde al menos 1924. La intensidad de las formas clínicas observadas y su elevada mortalidad focalizaron la atención sobre esta “nueva enfermedad”. Los brotes aparecidos en años posteriores en Oriente Medio y Turquía sirvieron para reforzar la idea de que una nueva enfermedad estaba emer-giendo, concepto que se vería reforzado por la detección de la misma en los estados norteamericanos de Texas y California en 1952, donde un proceso, que en principio se conoció como “hocico dolorido”, se confi rmaría como causado por el virus de la LA.
En 1956, casi al mismo tiempo que se describió la aparición de la LA en Norte-américa, un importante brote se detectó por primera vez en la Europa continental, concretamente en el sur de Portugal. Dicho brote se extendió rápidamente a España en 1957, continuando en la Península hasta 1960. Un índice de mortalidad elevado, alrededor del 75%, provocó la muerte de unas 180.000 ovejas. Con posterioridad, la enfermedad no volvería a ser descrita en Europa hasta pasados 20 años, cuando de nuevo apareció en las islas griegas de Rodas y Lesbos entre 1979 y 1980.
Durante la década de los años 50 y 60, la LA se expandió por Asia y la India. Sin embargo, en Australia, pese a que el virus se aisló por primera vez en 1977, en mosquitos Culicoides capturados en regiones septentrionales, el país permaneció libre

EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
99
de enfermedad en años posteriores. Dicha ausencia fue debida, en parte, a las restric-ciones impuestas al comercio de animales, así como al hecho de que la mayor parte de las explotaciones ovinas no se ubicaban en regiones del norte y este de Australia. En estas latitudes se identifi caron hasta 8 serotipos distintos, la mayoría poco patógenos, caracterizados por inducir formas asintomáticas de enfermedad.
Por tanto, existen evidencias de una graduación en la virulencia de los serotipos del virus que se localizan en Asia y la India, donde la enfermedad muestra intensas formas clínicas, con respecto a Australia, donde la enfermedad rara vez ha sido ob-servada. Esta situación de aparente expansión de la LA entre la década de los años 40 y 80, fue la razón por la que la enfermedad fue incluida en la antigua Lista A de la Ofi cina Internacional de Epizootías (OIE).
Sin embargo, desde principio de los años 90, ante la ausencia de brotes y formas clínicas de cierta intensidad, muchos científi cos se preguntaron si realmente la LA debería ser considerada una enfermedad emergente. Sus argumentos se basaron en el hecho de que, pese a tratarse de un enfermedad de distribución mundial, su presencia se restringía a ecosistemas relativamente estables, defi nidos por las competencias de vectores Culicoides autóctonos de áreas tropicales y subtropicales, confi nadas normal-mente entre los 35ºS y los 40ºN, auque en ciertas áreas de América y China podían extenderse entorno a los 50ºN. Dichos investigadores postularon que las barreras climáticas y geográfi cas restringirían los movimientos, tanto del los vectores como de los hospedadores de cada ecosistema. De este modo, aquellos virus que fueran introducidos en un ecosistema diferente, se suponía que no sobrevivirían debido a la ausencia de un vector efi ciente. Por tanto, la inclusión de la LA en la citada lista de enfermedades de declaración obligatoria fue tachada de “precipitada y contraria a argumentos razonados”.
Desafortunadamente, “la retrospectiva es el pariente pobre de la previsión”. Así, en plena discusión, en 1998 volvieron a producirse brotes de la enfermedad en cuatro islas griegas. En los siguientes siete años la enfermedad se expandió, afectando a die-ciséis países mediterráneos, incluidos nueve donde nunca antes había sido descrita, y alcanzando latitudes hacia el norte de Europa donde nunca antes había llegado. En 2005, el virus ya había matado entorno a un millón de ovejas, lo que constituyó la mayor epidemia de LA nunca vista hasta entonces.
La expansión europea del virus entre 1998 y 2005 se atribuyó, al menos en parte, a la extensión hacia el norte del hábitat de su principal vector en el Viejo Mundo, Cu-licoides imicola, especie afro-asiática cuya diseminación parece estar ligada al cambio

EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
100
climático. Al igual que en Europa, investigaciones llevadas a cabo en Norteamérica demostraron que durante el periodo de siete años transcurrido desde 1998, diez nuevos serotipos habían penetrado en el sureste de Estados Unidos procedentes de la cuenca caribeña, donde eran considerados enzoóticos. Es posible que junto a Culicoides sono-rensis, vector tradicional del virus en Norteamérica, Culicoides insignis así como otras especies originarias del ecosistema tropical vecino, pudieran actuar como vectores.
Asimismo, en Australia, tras un periodo de 21 años de aparente estabilidad, en-tre 2007 y 2008 se identifi caron dos nuevos serotipos en explotaciones centinelas del norte del país. Además del cambio climático, el predominio de ciertos vientos, como los procedentes del continente africano en el caso de Europa o los vientos monzóni-cos en el caso de Australia, podrían favorecer el transporte de los vectores del virus.
De vuelta al escenario europeo, de entre todos los serotipos identifi cados, el sero-tipo 8 merece una mención aparte. Su aparición y rápida diseminación comenzó en el norte de Europa en 2006, afectando a países del nordeste del continente como Bélgica, Holanda, Luxemburgo, Alemania y norte de Francia. De forma sorprendente, el virus logró sobrevivir al invierno, reapareciendo con fuerza en abril de 2007 en los países afectados. Su expansión prosiguió hacia Reino Unido, República Checa, Dinamarca y la península Escandinava, extendiéndose hasta los 58ºN, el punto más septentrional del mundo en el que se hubiera registrado su presencia. También se extendió hacia el sur, describiéndose brotes en regiones meridionales de Francia hacia fi nales de 2007 y en el norte de España en enero de 2008. En total, se vieron afectadas más de 45.000 explotaciones agrícolas, estimándose pérdidas por valor de unos 150 millones de euros. Por primera vez en la Historia, especies Culicoides Paleárticas autóctonas, y no Culicoides imicola, considerado el principal vector del virus en África y Asia, se vieron implicadas en la transmisión de este serotipo. Dicha implicación parece estar relacionada con el aumento de temperatura que se vive en muchas partes de Europa, donde la efi ciencia de estos vectores aumenta con la misma.
Otro aspecto a destacar del serotipo 8 fue su elevada virulencia, dando lugar a formas clínicas muy intensas que afectaron no sólo al ganado ovino, sino también al ganado vacuno y caprino, así como a rumiantes de vida salvaje, camélidos como llamas y alpacas e incluso carnívoros de zoológico como linces euroasiáticos. Esta cepa de serotipo 8 poseía la capacidad de atravesar la placenta de los rumiantes y provocar infecciones fetales con efectos teratógenos, lo cual era algo inusual para otras cepas de campo en cualquier parte del mundo. De hecho, la capacidad de atravesar la placenta era una propiedad exclusiva de ciertas cepas de laboratorio empleadas en vacunas vivas atenuadas. Curiosamente, en octubre de 2008 un nuevo serotipo,

EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
101
el serotipo 6, fue detectado durante análisis rutinarios en las mismas áreas cercanas a la frontera entre Alemania y Holanda donde, 2 años antes, se detectó el serotipo 8. Poco después, en enero de 2009, otro serotipo inédito, el serotipo 11, fue detectado en Bélgica. Al contrario de lo que ocurría con el resto de serotipos que afectaron a Europa, cuyas rutas de entrada desde Turquía, Chipre y norte de África estaban perfectamente identifi cadas, las rutas de entrada de los serotipos 6, 8 y 11 permanecen sin aclarar. Además, las cepas de los serotipos 6 y 11, mostraron una homología cercana al 100% con prototipos de virus o virus incluidos en vacunas vivas atenuadas procedentes de laboratorios sudafricanos, por lo que la implicación de la mano del hombre en estos brotes, mediante el tráfi co ilegal de vacunas, podría estar en el origen de estos graves hechos.
La expansión del virus en Europa, Norteamérica y Australia, la implicación de especies de vectores nóveles y el incremento de la virulencia de las cepas identifi cadas, sugirieron que importantes cambios se estaban produciendo en la epidemiología de la LA a nivel mundial. Lejos del planteamiento rancio que la consideró como una en-fermedad emergente de interés histórico, la LA se convirtió en una preocupación para científi cos, agricultores, servicios veterinarios y legisladores, surgiendo la pregunta de si esta enfermedad no es sino el anticipo de otros males aún por llegar.
Por otro lado, hay que reconocer, que mientras el problema se mantuvo en paí-ses vecinos a la Unión Europea, o bien restringido en países miembros situados en la cuenca mediterránea como Grecia, Italia, Portugal o España, las autoridades sanitarias europeas y los dirigentes políticos responsables no le prestaron la atención necesaria. Sin embargo, en 2006, con la llegada de la LA al corazón de Europa, dicha actitud cambió radicalmente. La toma de decisiones políticas coordinadas entre los distin-tos gobiernos se acompañó de cuantiosas aportaciones económicas, lo que permitió avanzar considerablemente en distintos aspectos de la enfermedad. En este punto me gustaría aclarar que, cualquier parecido de la evolución de la LA en Europa con la actual crisis económica, que tanto está afectando a estos países mediterráneos, es pura coincidencia, y por supuesto no achacable a la picadura de ningún mosquito.
Sin embargo, la LA es una enfermedad compleja, que aún presenta numerosas cuestiones sin aclarar. Uno de estos aspectos es el concerniente a los mecanismos de diseminación y persistencia del virus, que han permitido su extensión hasta latitudes nunca antes contempladas. Está claro que su presencia está ligada a la del vector Culicoides y a la existencia de unas condiciones climáticas apropiadas para la supervivencia de éste. Pero sólo unas 50 de las 1500 especies de Culicoides que se co-nocen parecen estar implicadas en la transmisión del virus, y es posible que no todas

EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
102
puedan transmitir los 26 serotipos conocidos. Ello puede deberse a la incapacidad de este mosquito de alimentarse sobre rumiantes, o bien a la existencia de lo que se ha dado en llamar “sistemas barrera”, los cuales podrían impedir al virus completar su ciclo en el vector más allá de las células intestinales, y cuyas bases moleculares permanecen sin aclarar.
Otro de los aspectos de la enfermedad sobre el que más se ha especulado, y que más confusión genera entre los investigadores, es el referido a la capacidad de “hibernación” del virus, es decir, como ciertos serotipos en regiones templadas del mundo tales como Sudáfrica, California o Europa, son capaces de desaparecer duran-te el invierno y reaparecer al verano siguiente, sin que se conozcan los mecanismos exactos ni la relativa contribución de hospedadores y vectores. Tanto la actividad de los mosquitos como la replicación del virus en éstos, cesan a bajas temperaturas, lo que interrumpe su transmisión. Pero los brotes se reanudan tras meses de “silencio epidemiológico” cuya duración, normalmente superior a 6 meses, es mayor que la vida media de los mosquitos adultos o que el periodo de viremia de los hospedadores mamíferos. Resulta evidente que el reservorio del virus debe existir durante el perio-do de hibernación, pero su localización, y el mecanismo por el cual se restablece la transmisión entre el vector y el mamífero hospedador, son cuestiones aún por aclarar. Por tanto la gran pregunta es: ¿dónde está el virus durante los meses de invierno, primavera y algunos meses de verano?.
Distintas explicaciones potencialmente plausibles han sido propuestas, entre las que destacan algunas poco probables como la transmisión vertical transovárica del virus en el mosquito Culicoides, lo que daría lugar a una nueva generación de mosquitos portadores, fenómeno que no ha sido documentado hasta la fecha. Tam-bién se ha apuntado la posible existencia de un complicado y, hasta el momento, no demostrado ciclo de hibernación, que involucraría a algunos hospedadores animales no identifi cados como roedores, carnívoros salvajes, pequeños mamíferos o reptiles, los cuales actuarían como reservorios. Con igual suerte, se ha postulado el posible papel de otros vectores de vida más longeva que Culicoides, como ciertos tipos de garrapatas, sin resultados concluyentes.
Otras líneas más probables se han encaminado al estudio de la persistencia del virus en mosquitos Culicoides adultos capaces de prolongar su vida media, hipótesis que podría ser posible, ya que algunos individuos han conseguido sobrevivir durante meses, especialmente cuando las temperaturas invernales no fueron especialmente bajas. Además, se ha demostrado que, al contrario de lo que se pensaba, los hábitos de estos vectores les permiten volar y, en consecuencia, refugiarse en el interior de

EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
103
las instalaciones ganaderas, sobreviviendo así a las bajas temperaturas exteriores. Igualmente, se considera como posible el establecimiento, durante estos meses inver-nales, de “ciclos de transmisión de perfi l bajo” entre rumiantes y Culicoides adultos, lo que favorecería que la enfermedad se mantuviera en algunos animales, que pasarían desapercibidos, hasta la siguiente estación cálida.
Con respecto al papel de los hospedadores, se ha demostrado que la infección del virus puede ser prolongada, pero no persistente. Es conocido que algunos serotipos puede transmitirse de hospedador a hospedador por vía oral a través de la ingestión de calostro. También se ha demostrado que el virus puede transmitirse a través del semen, siendo algunos serotipos, como el serotipo 8, capaces incluso de atravesar la placenta. Esto puede dar lugar al nacimiento de animales virémicos donde la presencia del virus puede ser prolongada pero, igualmente, no persistente. Aunque la importan-cia epidemiológica de estos mecanismos en la transmisión normal de la enfermedad parece ser mínima, su relevancia en la supervivencia del virus podría ser capital.
Algunos trabajos han señalado la infección persistente de los linfocitos T gamma/delta de los hospedadores habituales, como posible lugar de latencia del virus durante la hibernación, de manera que tras las picaduras de los vectores durante el verano y el otoño, se produciría una migración de dichos linfocitos hacia la piel, poniendo así el virus de nuevo a disposición del vector. Dicha teoría no ha sido corroborada ni experimentalmente ni en casos de campo.
Sin embargo, junto a las células endoteliales de los capilares sanguíneos, consi-deradas tradicionalmente como las principales células diana del virus, éste infecta a otras células inmunocompetentes entre las que destacan los monocitos-macrófagos, las células dendríticas así como los linfocitos B y T. Estas células infectadas han sido detectadas principalmente en órganos linfoides, aunque también en multitud de localizaciones orgánicas de los hospedadores. Por tanto, podrían jugar un papel fundamental, no sólo en la diseminación orgánica del virus, sino también en el meca-nismo de hibernación, debiéndose determinar con exactitud los lugares de replicación vírica en animales enfermos, así como los posibles lugares de acantonamiento tras la recuperación clínica de los individuos afectados.
La experiencia acumulada hasta ahora sugiere que, dada la complejidad de la enfermedad, a la hora de afrontar su estudio deberíamos considerar a cada uno de los serotipos como agentes etiológicos individuales, capaces de inducir cuadros clínicos, lesiones, cambios infl amatorios y respuestas inmunes en los hospedadores muy dispares unas de otras. Además, los mecanismos de acción de cada uno de estos

EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
104
serotipos, difi eren considerablemente dependiendo de la especie rumiante y de la raza afectada. Si hacemos una combinación de estas variantes podremos comprobar que nuestros conocimientos sobre la enfermedad son limitados, siendo aún muy largo el camino que nos queda.
Sin duda, cada una de las nuevas aportaciones que se haga en estos campos contribuirá al desarrollo de vacunas más efectivas, las cuales son fundamentales, junto a la restricción de movimientos, para el control de la enfermedad. Sin embargo, la ausencia de inmunidad cruzada entre los 26 serotipos difi culta las estrategias de vacunación, obligando a la aplicación de vacunas específi cas para cada uno de los serotipos.
Fue precisamente Theiler, el primero que apuntó a un virus como posible causa de la LA, el pionero en la aplicación de una vacuna atenuada mediante pasajes seria-dos en ovejas. Más de 50 millones de dosis de esta vacuna fueron usados entre 1907 y 1943, fecha en la que se desestimó su uso, debido a que dicha vacuna monovalente no confería inmunidad frente a otros muchos serotipos circulantes en Sudáfrica.
Precisamente eran vacunas de manufacturación sudafricana las únicas disponi-bles comercialmente cuando, en 1998, aparecieron los nuevos brote de LA en Europa. Entre los inconvenientes de estas “vacunas vivas atenuadas”, con un bajo perfi l de seguridad, destacaban los posibles efectos teratógenos del virus en animales gestantes, la capacidad de replicación del virus vacunal en los animales inmunizados, la apa-rición de viremia y la transmisión del virus por el vector, así como la posibilidad de recombinación entre los virus vacunales y los virus de campo, lo que podría originar virus nuevos, más patógenos o con propiedades antigénicas nuevas. El uso de estas vacunas en Europa, salvo excepciones, fue desechado por las autoridades, ya que en el contexto europeo, las vacunas se requerían no sólo para proteger a los animales, sino también para erradicar la enfermedad, rompiendo con el ciclo de transmisión del virus.
Todo ello llevó a un gran acuerdo entre las autoridades sanitarias y la industria farmacéutica que permitió, en relativamente poco tiempo, el desarrollo de “vacunas inactivadas”, desarrolladas a partir de virus genéticamente inerte, lo que les confería un elevado grado de seguridad y efi cacia. Desde el comienzo de su aplicación en 2005 hasta la actualidad, estas vacunas han constituido la mejor opción disponible, si bien sólo se han desarrollado frente a unos pocos serotipos, siendo escasa la disponibilidad de vacunas polivalentes de este tipo. Además, este tipo de vacunas no permiten la

EVOLUCIÓN HISTÓRICA Y NUEVAS PERSPECTIVAS EN LA INVESTIGACIÓN DE LA LENGUA AZUL
105
diferenciación entre animales infectados y animales vacunados, lo que difi cultaría el movimiento de ganado durante un posible brote.
Esto ha llevado a los investigadores a apostar por un nuevo tipo de vacunas denominadas “VLP (virus-like particle)”. Este tipo de vacunas de última generación, están constituidas por proteínas estructurales víricas, inductoras de la respuesta in-mune, que se ensamblan formando estructuras semejantes al virus, pero que carecen de material genético. Aunque en fase experimental, estas vacunas permitirán protegen frente a multitud de serotipos a la vez, así como diferenciar animales vacunados de infectados, por lo que vislumbran como las vacunas del futuro.
Aunque el paisaje que ha dejado la LA en Europa ha sido desolador, parece ser que el virus también posee una interesantísima utilidad terapéutica. Investigaciones recientes han señalado la capacidad de 5 serotipos del virus para destruir, de manera selectiva, células cancerígenas humanas, lo que abre una puerta de esperanza para el tratamiento de algunos tumores resistentes a la radio y quimioterapia, así como para otros tumores de mal pronóstico.
Y para que la Ciencia avance en estos campos, no existen recetas mágicas. Junto a un mínimo apoyo económico e institucional, los verdaderos equipos de científi cos, es decir, aquellos donde la confi anza y el apoyo entre cada uno de sus miembros va mas allá del laboratorio, sólo necesitan de paciencia, prudencia y perseverancia para alcanzar sus objetivos. Con el trabajo diario todo llega. Por desgracia, a esta receta hay que añadirle también una pizca de desconfi anza. Desconfi anza de aquellos círculos poblados de personajes pseudocientífi cos que buscan el protagonismo mediático, el poder político o simplemente lucrarse con la Ciencia, y que no dudan en utilizar el sacrifi cio y la esperanza de los que les rodean para alcanzar sus objetivos. También hay que recelar de aquellos que en su búsqueda de una gloria pasajera, y desde la más absoluta ignorancia científi ca y/o intelectual, no dudan en desprestigiar a sus compañeros, rompiendo con una máxima que debería prevalecer sobre todas en el seno de cualquier grupo de investigación: “el triunfo de uno es el triunfo de todos”.
Muchas gracias.
Granada, 16 de Diciembre de 2011.


CONTESTACIÓN AL DISCURSO DE INGRESO DEL ILMO. SR. D. PEDRO JOSÉ SÁNCHEZ CORDÓN
107ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
Ilustrísimo Sr. D. Pedro José Sánchez Cordón,
Excelentísimo Sr. Presidente de la Real Academia de Ciencias Veterinarias de Andalucía Oriental,
Excelentísimo Sr. Presidente de Honor de la Real Academia de Ciencias Veteri-narias de Andalucía Oriental,
Excelentísimos e Ilustrísimos Señores Académicos,
Familiares y amigos,
Señoras y Señores.
Agradeciendo, antes de nada, a los presentes por su asistencia, también quiero manifestar mi más sincera gratitud a esta Real Corporación por haberme honrado, permitiéndome efectuar la contestación al Discurso de ingreso del Doctor Sánchez Cordón, al tiempo que, pletórico de felicidad, me resulta sumamente grato darle la bienvenida en nombre de la misma. Tenemos hoy la satisfacción de recibir a un nuevo académico, joven pero de enorme valía, y en lo que a mi respecta, amigo mayúsculo, compañero del alma, de venturas y desventuras.
Pedro, quiero felicitarte por este discurso histórico, elegante y clásico en su de-sarrollo, con el que nos has regalado, y que no voy a valorar desde un punto de vista
CONTESTACIÓN AL DISCURSO DE INGRESO DEL ILMO. SR. DR. D. PEDRO JOSÉ SÁNCHEZ CORDÓN
ILTMO. SR. DR. D. EDUARDO RUIZ VILLAMOR1
1 Académico de número de la Real Academia de Ciencias Veterinarias de Andalucía Oriental.

CONTESTACIÓN AL DISCURSO DE INGRESO DEL ILMO. SR. D. PEDRO JOSÉ SÁNCHEZ CORDÓN
108
científi co, porque, si de eso se tratara, hay personas infi nitamente más cualifi cadas y conocedoras del tema que yo. Pero sí quiero resaltar el acierto en la elección del tema, tanto por su actualidad como por su importancia, sobre todo para los veterinarios y ganaderos de Andalucía. Además, ha sido un discurso de ingreso equilibrado, técnico, y basado en la experiencia científi ca que posees sobre el tema y en el que partiendo de las premisas históricas, nos has dado una visión del presente y futuro de esta enfermedad.
A esta altura del discurso, y aun sin levantar la vista de este papel, que leo para contestar en orden y concierto a tu ingreso en esta Real Corporación, estoy conven-cido que tu tez ya se habrá ruborizado y tu corazón andará como caballo sin freno, respondiendo a lo que en fi siología se denomina “reacción de alarma de Selye”, y que la mayoría de los mortales reconocemos, en estos casos, como vergüenza. Pero ya decía Sigmund Froid que uno puede defenderse de los ataques, pero contra los elogios está indefenso, así es que lo siento por ti, pues tengo que referenciar breve-mente tu currículum y acto seguido reconocer que es un honor y un lujo tenerte en nuestra Academia.
Pedro José Sánchez Cordón, nace en Córdoba a principio de la década de los setenta, y cursará sus primeros estudios en Rute, su pueblo natal, donde el contacto con el campo y con los ganaderos y veterinarios de la zona empezarán a sembrar la semilla que años más tarde habría de germinar.
Realiza los estudios de la Licenciatura en la antigua Facultad de Veterinaria de la Universidad de Córdoba, concluyendo los mismos en el año 1998. Ese mismo año obtendría el Grado de Licenciatura gracias a la realización de una tesina sobre Peste Porcina Clásica Experimental, que dicho sea de paso, me permitió hacer con él las veces de maestro en el laboratorio, y que sería más tarde uno de los capítulos de mi tesis Doctoral. En el año 2003 obtiene el grado de Doctor en Veterinaria, con una califi cación de sobresaliente “cum laude” por unanimidad y recibe el premio extraordinario de doctorado.
Ha sido becario del programa de “Formación de personal investigador y de desarrollo” de la Junta de Andalucía, becario del programa “Acciones de movilidad de investigadores y tecnólogos” del Ministerio de Educación y Ciencia, becario post-doctoral de la Universidad de Córdoba, contratado post-doctoral del Ministerio de Educación y Ciencia, contratado con cargo al programa “Juan de la Cierva” del Minis-terio de Educación y Ciencia, y en la actualidad es Investigador, a tiempo Completo, dentro del programa “Ramón y Cajal” del Ministerio de Ciencia e Innovación.

CONTESTACIÓN AL DISCURSO DE INGRESO DEL ILMO. SR. D. PEDRO JOSÉ SÁNCHEZ CORDÓN
109
Por otra parte, su formación se ha visto notablemente enriquecida con varias estancias en Centros de reconocido prestigio internacional, entre los que cabe destacar el Animal Health and Veterinary Laboratories Agency en Surrey, Reino Unido, y el Institut für pathologie, tierärztlichen hochschule en Hannover, Alemania, entre otros.
Dentro de las líneas de investigación en las que ha trabajado, destacan la patoge-nia de las infecciones víricas y encefalopatías espongiformes transmisibles animales, estudiando las respuestas inmune e infl amatoria. Así, ha participado en 15 proyectos de investigación subvencionados y 4 contratos de investigación de especial relevancia con administraciones nacionales e internacionales; es autor de 42 trabajos publicados en revistas recogidas en el Journal Citation Reports, 23 trabajos publicados en revistas no catalogadas, 4 libros y varios trabajos bibliográfi cos o de divulgación. Asimismo, es autor de más de 150 comunicaciones presentadas en congresos nacionales e inter-nacionales.
Es revisor científi co de varias publicaciones de impacto como Veterinary patholo-gy, Canadian journal of veterinary research, American journal of veterinary research, Research in veterinary science y Current bioactive compounds; además de tener expe-riencia en la organización de actividades de I+D, es evaluador de la Agencia nacional de evaluación y prospectiva (ANEP) y consultor de la FAO, designado como experto en peste porcina africana desde 2009.
Ha dirigido 2 tesinas de licenciatura, 4 trabajos de investigación de fi n de master y 3 tesis doctorales, encontrándose ocupado en la actualidad en otras 6 tesis doctorales en fase de realización.
Respecto a su actividad docente, posee la evaluación positiva para las fi guras de profesor ayudante doctor y profesor contratado doctor, habiendo participado en 8 proyectos de innovación y mejora de la calidad docente. Asimismo, ha impartido docencia universitaria de grado en la universidad de Córdoba y en la universidad complutense de Madrid, así como docencia en cursos de postgrado de la universidad de Córdoba, siendo tutor docente del master en “Medicina, sanidad y mejora animal” y responsable de la asignatura “enfermedades emergentes y exóticas” desde el curso 2008-09. También ha impartido docencia en centros públicos no universitarios como el CISA-INIA (Madrid) y empresas privadas como la Schering-plough animal health.
Por último, su actividad investigadora ha sido distinguida con 2 premios nacio-nales y otros 4 internacionales.
Después de haber presentado esta síntesis curricular, creo que sobran, a la inteli-gencia de los asistentes, las palabras para reconocer que es un honor y un lujo tenerte

CONTESTACIÓN AL DISCURSO DE INGRESO DEL ILMO. SR. D. PEDRO JOSÉ SÁNCHEZ CORDÓN
110
en nuestra Academia. Personalmente me siento orgulloso del camino que has andado, en la medida que representan el tesón y la ilusión que yo también compartí en su día, pero ya sabes que los caminos del Señor son inescrutables y harto difíciles de entender.
Solo me queda felicitarte y también hacer extensiva mi enhorabuena a tu familia mas cercana, a Miriam tu Esposa, y a Maria Teresa y Pedro tus hijos, porque quieras que no, ellos también escriben cada renglón del curriculum vitae, en la medida en que éste es el discurrir de tu vida. Y tampoco quiero olvidarme de tu hermano, ni de tus Padres aquí presentes, que deben sentirse orgullosos de ti, no solo como hijo, sino también como persona, al comprobar que el esfuerzo y la educación invertida rinden tanto fruto.
Y hablando de dar fruto, quiero terminar con estos bellos renglones que escribió José Echegaray: «La ciencia pura es como la soberbia nube de oro y grana que se dilata en Occidente, entre destellos de luz y matices maravillosos: no es ilusión, es resplandor, la her-mosura de la verdad. Pero una nube se eleva, el viento la arrastra sobre los campos y ya toma tintas más oscuras y más severas; es que va a la faena y cambia sus trajes de fi esta, digámoslo así, por la blusa de trabajo. Y entonces se condensa en lluvia, y riega las tierras, y se afana en el terruño, y prepara la futura cosecha, y al fi n dan los hombres el pan nuestro de cada día. Lo que empezó por hermosura para el alma y para la inteligencia, concluye por ser alimento para la pobre vida corporal».
Espero y deseo, querido amigo, que el esfuerzo realizado y la valía ya demostrada te rindan fi nalmente el provecho que mereces, y que tu próxima partida en busca del progreso personal y familiar no sea un adiós sino solo un hasta luego.
He dicho.

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
111ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
RESUMEN
Las bacteriocinas, así como las cepas bacterianas que las producen, han sido ampliamente estudiadas para la conservación de los alimentos por métodos naturales (bioconservación). En productos cárnicos (abarcando toda la gama, desde la materia prima hasta productos elaborados) se han desarrollado numerosos estudios empleando bacteriocinas como nisina, pediocinas y otras, bien solas o aplicadas con otros antimicrobianos (como ácidos orgánicos o agentes que-lantes) o tratamientos (como el calor, las altas presiones o la luz pulsada) para la inactivación de bacterias patógenas o alterantes. Los resultados de muchos de estos tratamientos son bastante prometedores, como por ejemplo el empleo de bacteriocinas como recubrimientos protectores en salchichas. Las cepas de bacterias lácticas seleccionadas por su capacidad productora de bacteriocinas ofrecen buenas perspectivas como cultivos protectores, pero sobre todo como cultivos iniciadores en la elaboración de productos cárnicos fermentados, mejorando la se-guridad microbiológica de los mismos. El empleo de cultivos iniciadores bacteriocinogénicos con propiedades funcionales defi nidas abre nuevas vías para la innovación y el desarrollo de nuevos productos cárnicos fermentados.
Palabras clave: bacteriocinas, productos cárnicos, bioconservación
ABSTRACT
Bacteriocins as well as their producer strains have been widely investigated for appli-cation in natural food preservation processes (biopreservation). Numerous studies have been carried out on application of bacteriocins such as nisin, pediocin and others in different types
BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
Mª JOSÉ GRANDE BURGOS, R. LUCAS, Mª. C. LÓPEZ AGUAYO, R. PÉREZ PULIDO, A. GÁLVEZ1
1 Área de Microbiología. Dpto. de Ciencias de la Salud. Facultad de Ciencias Experimentales. Uni-versidad de Jaén. Campus Las Lagunillas, s/n. 23071-Jaén. E-mail: [email protected]

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
112
of meat products, either singly or in combination with other antimicrobials (such as organic acids or chelating agents) or treatments (such as heat, pulsed light or high hydrostatic pres-sure). Many of the results reported are highly promising, like for example the application of bacteriocins in edible coatings for sausages. Bacteriocin-producing lactic acid bacteria are also interesting, in view of their possible applications as protective cultures or as starter cultures in fermented meat products, increasing the product safety. Bacteriocin-producing starter cultures with additional functional properties open new possibilities for innovation and development of new fermented meat products.
Keywords: bacteriocins, meat products, biopreservation
INTRODUCCIÓN
Las bacteriocinas son proteínas o péptidos antimicrobianos de síntesis ribosómica (Jack et al., 1995). Debido a su relación con los alimentos, las bacterias del ácido láctico (BAL) son el grupo de bacterias productoras de bacteriocinas más estudiado hasta el momento y donde más bacteriocinas se han descrito. Las bacteriocinas producidas por las BAL se pueden clasifi car en cuatro clases: I, lantibióticos; II, péptidos termoestables no modifi cados; III, proteínas termolábiles (bacteriolisinas) y IV, péptidos cíclicos (Nes et al, 2007). La mayoría de estas clases incluyen también varias subclases en función de la estructura de la bacteriocina.
Las bacteriocinas de las BAL y sus cepas productoras pueden ser utilizadas para mejorar la calidad y conservación de los alimentos (Thomas et al., 2000; Gálvez et al., 2007; Gálvez et al., 2008; Settani y Corsetti, 2008). (i) son reconocidas generalmente como sustancias seguras, (ii) no son activas ni tóxicas frente a células eucariotas, (iii) son inactivadas por enzimas proteolíticos, (iv) son en general estables en amplios intervalos de pH y temperatura, (v) presentan un modo de acción bactericida frente a muchas bacterias patógenas y/o alterantes de los alimentos, (vi) poseen un mecanismo de acción bactericida actuando sobre la membrana citoplasmática bacteriana, lo cual signifi ca que por lo general no presentan resistencia cruzada con los antibióticos de uso clínico, ya que actúan sobre dianas celulares diferentes y (vii) sus determinantes genéticos están mayoritariamente localizados en plásmidos. Los plásmidos pueden ser transferidos mediante manipulación genética y de forma natural a cepas de mayor interés industrial. Otras bacteriocinas como las producidas por bacterias coliformes (colicinas) o algunas bacteriocinas producidas por bacilos formadores de endosporas también pueden ser interesantes para la bioconservación. Sin embargo, se conoce muy poco sobre su función en sistemas alimentarios en comparación con las bacteriocinas producidas por las BAL.

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
113
Uno de los principales objetivos en la adición de bacteriocinas es reducir la carga microbiana de bacterias patógenas transmitidas por los alimentos. Muchas bacteriocinas producidas por las BAL tienen actividad bactericida frente a bacterias Gram-positivas patógenas o toxicogénicas, tales como Listeria monocytogenes, Bacillus cereus, Clostridium botulinum o Staphylococcus aureus. Algunas bacteriocinas también puede inhibir a bacterias Gram-negativas, especialmente cuando se combinan con otros agentes antimicrobianos como veremos a lo largo de esta revisión. La actividad inhibidora de las bacteriocinas también puede ser utilizada frente a la proliferación de bacterias patógenas durante el almacenado de los alimentos. El procesado y almacenamiento de alimentos en condiciones de refrigeración da lugar a nuevas oportunidades para la proliferación de bacterias psicrotrofas, tales como L. mono-cytogenes. En ausencia de microbiota competitiva (como ocurre en muchos alimentos procesados mediante tratamientos térmicos ), la proliferación de microorganismos procedentes de contaminación cruzada (como L. monocytogenes) se ve claramente facilitada, a menos que se empleen barreras adicionales que impidan su crecimiento. Algunos microorganismos (como L. monocytogenes) presentan la capacidad de crecer en condiciones de refrigeración. Por otra parte, las interrupciones accidentales de la cadena de frío también aumentan los riesgos de proliferación de bacterias en alimentos refrigerados. Por tanto, la incorporación de bacteriocinas en alimentos refrigerados, puede proporcionar una protección extra frente a la proliferación de diversos tipos de microorganismos, bien sean patógenos o alterantes.
CONTROL DE BACTERIAS PATÓGENAS Y ALTERANTES EN CARNE CRUDA
Las bacteriocinas han sido ensayadas en la conservación de carne cruda, o bien en combinación con otros antimicrobianos para la descontaminación y/o inhibición del crecimiento bacteriano en carne fresca almacenada (Tabla 1). Se han aplicado en forma de lavado, pulverización o inmersión en soluciones de bacteriocina sola o en combinación con otros agentes antimicrobianos para potenciar su actividad. Para incrementar la efi cacia de los tratamiento, y también para evitar la contaminación cruzada, la carne cruda tratada ha sido congelada, envasada en diferentes condiciones atmosféricas tales como el envasado al vacío, atmósferas modifi cadas, o envasado activo con agentes secuestradores de O2 o sistemas generadores de CO2 (Coma, 2008; McMillin, 2008 ). También se han utilizado métodos adicionales como la irradiación en dosis bajas, descontaminación de la superfi cie con luz UV o las altas presiones (Aymerich et al., 2008). Todos estos tratamientos durante el procesado actúan sobre la microbiota inicial y pueden actuar sinérgicamente con las bacteriocinas para aumentar

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
114
la seguridad y vida útil del alimento. Aunque los productos de carne cruda son pro-cesados antes de su consumo mediante tratamientos que normalmente destruyen las bacterias patógenas, éstos pueden ser una fuente considerable de contaminación cru-zada e incluso en algunos casos pueden producir toxinas microbianas termoestables.
La nisina ha sido ampliamente estudiada en la conservación de carne cruda (Thomas et al., 2000). Sin embargo, la aplicación de nisina en carne presenta algunas limitaciones derivadas de su baja solubilidad a pH próximo a neutralidad, interacción con los fosfolípidos y la inactivación por el glutatión (Thomas et al., 2000; Stergiou et al., 2006). Sin embargo, en la descontaminación de la superfi cie de carne cruda los mejores resultados se obtuvieron combinando nisina junto con otros agentes anti-microbianos tales como ácidos orgánicos, quelantes, lisozima, envasado al vacío, o envasado en atmósfera modifi cada, antes de la elaboración y envasado el producto, produciéndose un incremento en la inactivación de L. monocytogenes, Brochothrix thermosphacta y Escherichia coli O157:H7. El tratamiento de carne cruda con pediocinas (especialmente pediocina PA-1/Ach) también puede inhibir el crecimiento de bacte-rias Gram-positivas (por ejemplo, B. thermosphacta) y/o reducir las poblaciones de L. monocytogenes y Clostr. perfringens (Rodríguez et al, 2002; Nieto-Lozano et al., 2006). Otras bacteriocinas tales como sakacinas, carnobacteriocinas, bifi docinas, lactocinas, lactococcinas o pentocinas muestran una actividad variada frente a bacterias pató-genas o alterantes en carne o volatería crudas (Aymerich et al, 2008;. Gálvez et al, 2008.). En carne picada, la combinación de bacteriocinas junto con aceites esenciales procedentes de plantas (empleados a baja concentración para paliar el sabor demasiado intenso de algunos de ellos) está siendo considerada como una forma de aumentar la inactivación de L. monocytogenes, y también para inhibir a Salmonella Enteritidis (Solomakos et al., 2008; Govaris et al., 2010).
Una forma interesante de incrementar la efectividad de las bacteriocinas en carne cruda ha sido mediante inmovilización en sustratos (como perlas, liposomas, recubrimientos o películas). La nisina (sola o en combinaciones con ácido cítrico, EDTA y Tween 80) incorporada en una gran variedad de sustratos (tales como geles de alginato cálcico, revestimientos de agar, películas palmitoladas de alginato, clo-ruro de polivinilo, polietileno de baja densidad o nylon) mostró una gran actividad inhibitoria frente a bacterias como L. monocytogenes, B. thermosphacta, S. aureus o S. Typhimurium en carne cruda refrigerada (Chen y Hoover, 2003; Aymerich et al., 2008; Gálvez et al., 2007, 2008). La combinación del almacenamiento a temperatura próxima al punto de congelación junto con el empleo de envases antimicrobianos ha demostrado ser efi caz en la mejora de la calidad microbiológica de piezas de carne, inhibiendo a BAL, carnobacterias y B. thermosphacta (Ercolini et al., 2010).

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
115
Dado que muchas BAL asociadas con productos cárnicos pueden crecer a tem-peraturas de refrigeración, aquellas cepas productoras de bacteriocinas carentes de efectos adversos sobre la carne pueden ser seleccionadas como cultivos protectores para la conservación de carne cruda (Tabla 1). En trabajos anteriores se ha demostrado la efi cacia de cepas de Lactobacillus sakei y Lactobacillus curvatus productoras de bacte-riocinas en la inhibición de L. monocytogenes o B. thermosphacta en productos cárnicos crudos (Castellano et al., 2008). Las cepas de Lb. sakei productoras de inhibidores de tipo bacteriocina retrasaron la aparición de hinchazón en los envases provocada por Clostridium estertheticum e inhibieron el crecimiento de Campylobacter jejuni en carne de vacuno (Jones et al., 2009). En carne de pollo, el cultivo protector de L. fermentum ACA-DC179 consiguió disminuir el crecimiento de S. enteritidis (Maragkoudakis et al., 2009). La carne de vacuno fresca inoculada con Lb. curvatus CRL705 como cultivo protector mostró un aumento neto de los aminoácidos libres, debido a la actividad complementaria de las proteasas bacterianas sobre las proteínas sarcoplásmicas de la carne, por lo que se propuso que este cultivo protector podría contribuir a la ma-duración de la carne y a la vez mejorar su vida útil (Fadda et al., 2008).
CONTROL DE BACTERIAS PATÓGENAS Y ALTERANTES EN PRODUCTOS CÁRNICOS COCIDOS
Los productos de carne cocida se comercializan como alimentos listos para consumo. Puede tratarse de piezas de carne enteras, pero lo más común es que se trate de productos elaborados mediante mezcla y picado de trozos secundarios de carne, grasa, órganos animales, o sangre con otros ingredientes, seguido por el relleno/moldeo y cocción. El proceso de cocción inactiva la microbiota natural, lo que facilita el camino para el crecimiento de microorganismos contaminantes tras la etapa de procesado. Los valores de pH de la mayoría de los productos cárnicos cocidos son compatibles con el crecimiento de bacterias patógenas y alterantes, que pueden proliferar a temperaturas de refrigeración durante la vida útil del producto. Algunos de estos productos también pueden ser sometidos a procesos adicionales tales como corte, retirada de recubrimientos, y envasado, lo que aumenta los riesgos de contaminación cruzada (Murphy et al., 2005). Las preparaciones de bacteriocinas como pediocina o nisina ofrecen grandes oportunidades de mercado como barreras frente a patógenos y bacterias alterantes en productos cárnicos cocidos. Los principales estudios realizados están enfocados a la adición de preparaciones de bacteriocina a las suspensiones de carne antes del proceso de calentamiento, aplicación de bacteriocinas en superfi cie antes del envasado, o la aplicación de películas o recubrimientos que contienen bacteriocinas.

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
116
Algunas cepas de BAL (sobre todo Lactobacillus y Leuconostoc) son consideradas como el principal grupo de bacterias alterantes en varios tipos de carne envasada a vacío, provocando cambios sensoriales típicos tales como agriado, producción de gas, SH2 o limo (Korkeala et al., 1988; Björkroth y Korkeala., 1997). Estas BAL recontaminan rápidamente los productos cárnicos durante su manipulación y loncheado (Lucke, 2000). Las enterocinas A y B inhiben la producción de limo por Lb. sakei en lonchas de jamón de cerdo cocido envasadas a vacío, y la nisina previene la formación de limo por Leuconostoc carnosum (Hugas et al., 2003; Aymerich et al., 2008). Cuando las bacteriocinas anteriores y la pediocina PA-1/AcH se incorporaron mezcladas en el jamón cocido en combinación con un tratamiento por alta presión hidrostática, sólo la nisina evitó la reaparición de L. carnosum. El tratamiento combinado con alta presión y nisina aumentó la inactivación de E. coli y evitó su reaparición durante el almace-namiento (Garriga y col., 2002). La nisina también fue la bacteriocina que causó una mayor disminución de los estafi lococos, pero sin embargo no impidió el crecimiento de L. monocytogenes (mientras que enterocinas como sakacina y pediocina si lo hi-cieron). Estos resultados muestran la efi cacia variable de las diferentes bacteriocinas dependiendo de las bacterias diana y las condiciones de procesamiento del alimento.
La efi cacia de la nisina y la lacticina 3147 contra bacterias Gram-positivas y Gram-negativas en embutidos (tales como los de carne de cerdo, jamón cocido o salchichas bologna) aumentó en combinación con la adición de ácidos orgánicos, lisozima, y EDTA. Del mismo modo, las combinaciones de pediocina con diacetato de sodio o lactato de sodio incrementaron la inhibición de L. monocytogenes en salchichas de Frankfurt o de L. monocytogenes y Yersinia enterocolitica en trozos de carne de ave cocidos almacenados en atmósfera modifi cada a 3,5 º C (O´Sullivan et al., 2002; Chen y Hoover, 2003; Aymerich et al., 2008; Gálvez et al., 2008). Estos resultados facilita-ron el camino para la aplicación de recubrimientos envases tratados o activados con mezclas de sustancias antimicrobianas en productos cárnicos listos para el consumo (Coma, 2008). Las películas activadas con bacteriocinas pueden ser muy apropiadas para productos cárnicos cocidos, actuando como barreras frente a la contaminación externa del producto elaborado. Entre los distintos tipos de recubrimientos comestibles que han sido ensayados en productos envasados al vacío (tales como salchichas o jamón cocido) los mejores resultados se obtuvieron para recubrimientos que contenían nisina en combinación con otros antimicrobianos durante el almacenado en frío del producto. Un estudio reciente demostró que las películas de celulosa que contienen pediocina PA-1/AcH retrasan el crecimiento de Listeria en el jamón cocido envasado a vacío almacenado a 12 º C simulando temperaturas abusivas como las que pueden darse en los supermercados (Santiago-Silva et al., 2009).

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
117
La aplicación de bacteriocinas sobre la superfi cie del producto en combinación con tratamientos posteriores al envasado es otro aspecto de interés. La aplicación de bacteriocina seguida de la aplicación de un tratamiento térmico al producto enva-sado puede proporcionar una combinación efi caz para controlar a L. monocytogenes en productos tales como salchichas o mortadela de pavo, como en el caso de los estudios realizados con pediocina, nisina, nisina-lisozima o combinaciones de estas bacteriocinas con lactato sódico/diacetato sódico (Chen et al., 2004b; Mangalassary et al., 2008). En salchichas, la aplicación de un baño de Nisaplina seguido por la ex-posición a la luz pulsada (9,4 J/cm2) redujo la población de Listeria innocua e inhibió el crecimiento microbiano durante el almacenamiento del producto bajo refrigeración (Uesugi y Moraru, 2009). Debido a que la aplicación de la luz pulsada está aceptada para la descontaminación de las superfi cies de los alimentos, el tratamiento com-binado podría ser aplicado como una etapa de post-procesamiento para reducir la contaminación superfi cial y aumentar la seguridad de los productos cárnicos listos para el consumo. La aplicación de tratamientos con bajas dosis de irradiación sobre salchichas previamente tratadas con pediocina o sobre carne de pavo lista para con-sumo envasada al vacío recubierta con una película de pectina-nisina aumentó la inactivación de L. monocytogenes en gran medida y redujo también la proliferación de supervivientes durante el almacenamiento (Chen et al., 2004a; Jin et al., 2009). Los resultados mostraron que el tratamiento combinado podría servir para prevenir la listeriosis asociada a la contaminación post-proceso de los alimentos, a la vez que se reducían las dosis de radiación y el impacto en la calidad del producto.
La aplicación de películas activadas con bacteriocina en combinación con altas presiones es otra alternativa para incrementar la inactivación de los microorganismos en productos cárnicos listos para el consumo (Aymerich et al., 2008). En jamón cocido envasado en películas activadas con enterocinas, el tratamiento combinado con altas presiones redujo los recuentos de L. monocytogenes y también inhibió la recuperación de las listerias viables durante condiciones de abuso de temperatura (Marcos et al., 2008). La aplicación de tratamientos combinados de nisina con altas presiones fue efi caz para lograr la ausencia de Salmonella en muestras de 25 g de jamón cocido loncheado almacenado en condiciones de refrigeración (Jofré et al., 2008).
Las BAL productoras de bacteriocinas podrían ser utilizadas como cultivos pro-tectores en productos cárnicos ligeramente procesados o cocidos sin llegar a alterar sus propiedades organolépticas, consiguiendo a la vez una inhibición de las BAL alterantes y de Listeria (Hugas et al. 1998; Lucke, 2000; Chen y Hoover, 2003; Aymerich et al., 2008; Gálvez et al., 2008). La inoculación de cepas productoras de sakacinas, pediocinas,

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
118
leucocinas, plantaricinas, enterocinas, bavaricinas y curvaticinas en productos cárni-cos procesados ha demostrado ser efi caz en la inhibición del crecimiento de Listeria y en algunos casos la formación de limo (Aymerich et al., 1998; Hugas et al., 1998). Existen diferentes cultivos de BAL en el mercado que son utilizados como cultivos iniciadores o bioprotectores con el objeto de mejorar la seguridad microbiológica de productos cárnicos ligeramente procesados o precocinados (Aymerich et al., 2008).
APLICACIONES EN PRODUCTOS CÁRNICOS FERMENTADOS
Las preparaciones de bacteriocina se pueden añadir a la masa cárnica para la inactivación de patógenos en productos cárnicos fermentados. La disminución de pH que se alcanza en embutidos en comparación con las carnes frescas puede aumentar la solubilidad de algunas bacteriocinas como la nisina, y probablemente su actividad antimicrobiana también. Las bacteriocinas como la nisina, enterocinas (CCM 4231, A, B y AS-48) o leucocinas mejoró la reducción de L. monocytogenes y de S. aureus en productos fermentados (Rodríguez et al., 2002; Chen y Hoover., 2003; Aymerich et al., 2008; Gálvez et al., 2008). El empleo de bacteriocinas puede ser una barrera interesante para la inactivación de microorganismos en embutidos ligeramente fermentados, en los que su pH más elevado y el mayor contenido en agua pueden facilitar la super-vivencia y proliferación de ciertas bacterias patógenas.
Las BAL juegan un papel importante en la fermentación de productos cárnicos. Por lo tanto, las cepas productoras de bacteriocinas podrían ser empleadas como cul-tivos iniciadores activos frente a patógenos como Listeria monocytogenes (Työppönen et al., 2003; Leroy et al., 2006; Aymerich et al., 2008). Diversas cepas de Lactobacillus productoras de bacteriocinas (principalmente Lb. sakei y Lb. curvatus, Lb. rhamnosus y Lb. plantarum) han demostrado tener efectos inhibidores frente a Listeria en fermen-taciones de salchichas o salami, dependiendo en gran medida de la cepa y el tipo de carne (Erkkila et al., 2001; Leroy et al., 2005b; Dicks et al., 2004; Benkerroum et al., 2005; Todorov et al., 2007) (Tabla 1). La cepa Lb. sakei CTC 494 (productora de sakaci-na K) es un cultivo iniciador interesante con actividad frente a Listeria, siendo capaz de inhibir con éxito a L. monocytogenes en embutidos fermentados al estilo español y alemán (Aymerich et al., 2008) o reducir las poblaciones de Listeria en salchichas belgas, salami italiano y salami Cacciatore (Ravyst et al., 2008). La efi cacia de Lb. sakei está infl uenciada por factores ambientales tales como ingredientes, sal, grasa, concen-traciones de nitrito, nivel de acidifi cación y temperatura (Leroy et al., 2006). Debido a que Lb. sakei y Lb. curvatus pueden hidrolizar las proteínas musculares sarcoplásmicas y, en menor medida, las proteínas miofi brilares, pueden contribuir a la generación de

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
119
péptidos pequeños y aminoácidos que contribuyen como potenciadores del sabor o como precursores directos de otros compuestos de sabor durante la maduración de los embutidos fermentados (Leroy et al., 2006). El estudio de estos procesos podría dar lugar a nuevas aplicaciones de cultivos iniciadores de nueva generación con propiedades funcionales de interés industrial o nutricional (Leroy et al., 2006).
Los pediococos productores de bacteriocina pueden reducir las poblaciones de L. monocytogenes en productos cárnicos fermentados (Amézquita y Brashears, 2002; Rodríguez et al., 2002; Aymerich et al., 2008). Los pediococcos son preferidos como cultivos iniciadores (en lugar de los lactobacilos) en ciertos productos, por ejemplo en embutidos al estilo americano fermentados a temperaturas más altas. Las cepas productoras de Pediocina PA-1 no inhiben a bacterias que son importantes para la fermentación de los embutidos como estafi lococos y micrococos (González y Kunka, 1987). Algunas cepas de lactococos productoras de nisina aisladas de embutidos fer-mentados también se han propuesto como cultivos adjuntos para mejorar la seguridad microbiológica de los productos cárnicos fermentados elaborados en condiciones higiénicas defi cientes, tales como en productos tradicionales caseros (Noonpakdee et al., 2003).
Los enterococos son a menudo parte de la microbiota normal en las fermenta-ciones de productos cárnicos y han demostrado tener actividad frente a Listeria y S. aureus en productos fermentados (Foulquié Moreno et al., 2003; Aymerich et al., 2008; Gálvez et al., 2008). Sin embargo, su aplicación en alimentos da lugar a controversia debido a su potencial como patógenos oportunistas. En este aspecto, la cepa de Ente-rococcus faecalis CECT7121 (productora de la enterocina de amplio espectro MR99) es interesante porque carece de factores de virulencia como los genes para producción de hemolisina y de gelatinasa y además no produce aminas biógenas (Sparo et al., 2008). Los embutidos fermentados inoculados con la cepa CECT7121 presentaron una disminución del recuento de células viables de Staphylococcus, enterobacterias y otros cocos gram-positivos al fi nal de la fermentación y ausencia de viables de S. aureus y enterobacterias al fi nal del proceso.
Los estafi lococos y micrococos también pueden ser explotados como fuente de sustancias antibacterianas aplicables en la fermentación de embutidos. El gen que codifi ca para la lisostafi na (una endopeptidasa que rompe específi camente los en-laces glicina-glicina en los puentes transversales de la pared celular de S. aureus) se puede introducir en lactobacilos y posteriormente utilizarlos como cultivo iniciador para evitar el crecimiento de S. aureus en productos cárnicos (Cavadini et al., 1998). Staphylococcus xylosus aislado de salchicha produce una sustancia antimicrobiana que
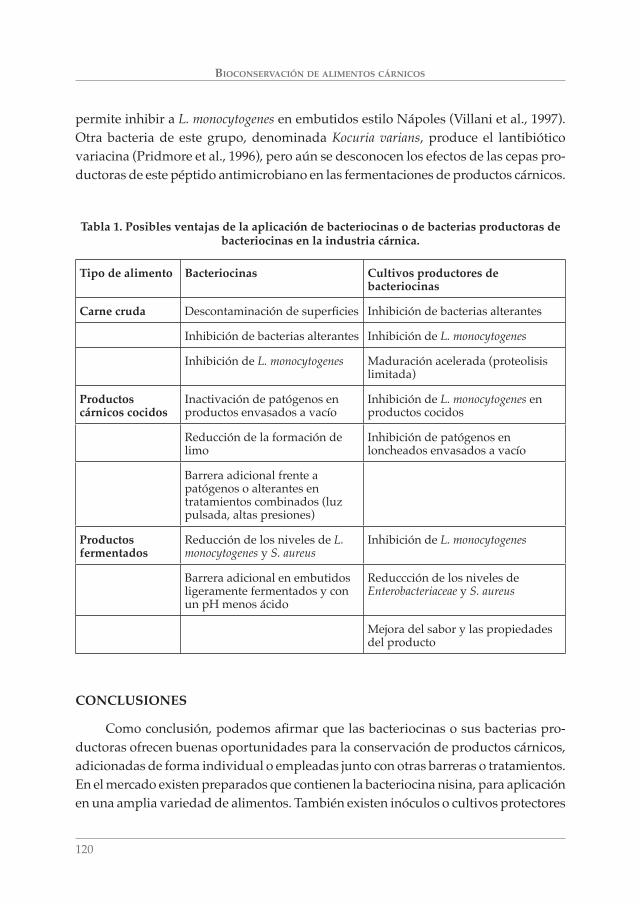
BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
120
permite inhibir a L. monocytogenes en embutidos estilo Nápoles (Villani et al., 1997). Otra bacteria de este grupo, denominada Kocuria varians, produce el lantibiótico variacina (Pridmore et al., 1996), pero aún se desconocen los efectos de las cepas pro-ductoras de este péptido antimicrobiano en las fermentaciones de productos cárnicos.
Tabla 1. Posibles ventajas de la aplicación de bacteriocinas o de bacterias productoras de bacteriocinas en la industria cárnica.
Tipo de alimento Bacteriocinas Cultivos productores de bacteriocinas
Carne cruda Descontaminación de superfi cies Inhibición de bacterias alterantes
Inhibición de bacterias alterantes Inhibición de L. monocytogenes
Inhibición de L. monocytogenes Maduración acelerada (proteolisis limitada)
Productos cárnicos cocidos
Inactivación de patógenos en productos envasados a vacío
Inhibición de L. monocytogenes en productos cocidos
Reducción de la formación de limo
Inhibición de patógenos en loncheados envasados a vacío
Barrera adicional frente a patógenos o alterantes en tratamientos combinados (luz pulsada, altas presiones)
Productos fermentados
Reducción de los niveles de L. monocytogenes y S. aureus
Inhibición de L. monocytogenes
Barrera adicional en embutidos ligeramente fermentados y con un pH menos ácido
Reduccción de los niveles de Enterobacteriaceae y S. aureus
Mejora del sabor y las propiedades del producto
CONCLUSIONES
Como conclusión, podemos afi rmar que las bacteriocinas o sus bacterias pro-ductoras ofrecen buenas oportunidades para la conservación de productos cárnicos, adicionadas de forma individual o empleadas junto con otras barreras o tratamientos. En el mercado existen preparados que contienen la bacteriocina nisina, para aplicación en una amplia variedad de alimentos. También existen inóculos o cultivos protectores

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
121
a base de bacterias lácticas que son empleados por la industria cárnica en la elabora-ción de alimentos fermentados. El empleo de cultivos iniciadores con propiedades funcionales (incluyendo su carácter probiótico) es un aspecto poco estudiado aun, y constituye un campo interesante de investigación para el desarrollo de productos más seguros, enriquecidos en sabor de forma más natural, o con propiedades funcionales.
BIBLIOGRAFÍA
Amezquita A, Brashears MM (2002) Competitive inhibition of Listeria monocytogenes in ready-to-eat meat products by lactic acid bacteria. J Food Prot 65:316-325.
Aymerich MT, Hugas M, Monfort JM (1998) Review: bacteriocinogenic lactic acid bacteria associa-ted with meat products. Food Sci Technol Int 4:141-158.
Aymerich T, Picouet PA, Monfort JM (2008) Decontamination technologies for meat products. Meat Sci 78:114-129.
Benkerroum N, Daoudi A, Hamraoui T et al (2005) Lyophilized preparations of bacteriocinogenic Lactobacillus curvatus and Lactococcus lactis subsp. lactis as potential protective adjuncts to con-trol Listeria monocytogenes in dry-fermented sausages. J Appl Microbiol 98:56-63.
Björkroth J, Korkeala H (1997) Ropy slime-producing Lactobacillus sake strains possess a strong com-petitive ability against a commercial biopreservative. Int J Food Microbiol 38:117–123.
Castellano P, Belfi ore C, Fadda S et al (2008) A review of bacteriocinogenic lactic acid bacteria used as bioprotective cultures in fresh meat produced in Argentina. Meat Sci 79:483–499.
Cavadini C, Hertel C, Hammes WP (1998) Application of lysostaphin-producing lactobacilli to con-trol staphylococcal food poisoning in meat products. J Food Prot 61:419–424.
Chen H, Hoover DG (2003) Bacteriocins and their food applications. Comp Rev Food Sci Food Safety 2:82-100.
Chen CM, Sebranek JG, Dickson JS et al (2004a) Combining pediocin with postpackaging irradia-tion for control of Listeria monocytogenes on frankfurters. J Food Prot 67:1866-1875.
Chen CM, Sebranek JG, Dickson JS et al (2004b) Combining pediocin (ALTA 2341) with postpac-kaging thermal pasteurization for control of Listeria monocytogenes on frankfurters. J Food Prot 67:1855-1865.
Coma V (2008) Bioactive packaging technologies for extended shelf life of meat-based products. Meat Sci 78:90–103.
Dicks LMT, Mellett FD, Hoffman LC (2004) Use of bacteriocin-producing starter cultures of Lac-tobacillus plantarum and Lactobacillus curvatus in production of ostrich meat salami. Meat Sci 66:703-708.
Ercolini D, Ferrocino I, La Storia A et al (2010) Development of spoilage microbiota in beef stored in nisin activated packaging. Food Microbiol 27:137–143.
Erkkilä S, Suihko ML, Eerola S et al (2001) Dry sausage fermented by Lactobacillus rhamnosus strains. Int J Food Microbiol 64:205-210.
Fadda S, Chambon C, Champomier-Vergès MC et al (2008) Lactobacillus role during conditioning of refrigerated and vacuum-packaged Argentinean meat. Meat Sci 79:603–610.
Foulquié Moreno MR, Rea MC, Cogan TM et al (2003) Applicability of a bacteriocin-producing En-terococcus faecium as a co-culture in Cheddar cheese manufacture. Int J Food Microbiol 81: 73-84.
Gálvez A, Abriouel H, López RL et al (2007) Bacteriocin-based strategies for food biopreservation. Int J Food Microbiol 120:51-70.

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
122
Galvez A, Lopez RL, Abriouel H et al (2008) Application of bacteriocins in the control of foodborne pathogenic and spoilage bacteria. Crit Rev Biotechnol 28:125-152.
Garriga M, Aymerich MT, Costa S et al (2002) Bactericidal synergism through bacteriocins and high pressure in a meat model system during storage. Food Microbiol 19:509-518.
Gonzalez CF, Kunka BS (1987) Plasmid-associated bacteriocin production and sucrose fermentation in Pediococcus acidilactici. Appl Environ Microbiol 53:2534-2538.
Govaris A, Solomakos N, Pexara A et al (2010) The antimicrobial effect of oregano essential oil, nisin and their combination against Salmonella Enteritidis in minced sheep meat during refrigerated storage. Int J Food Microbiol 137: 175-180.
Hugas M, Pagés F, Garriga M et al (1998). Application of the bacteriocinogenic Lactobacillus sakei CTC494 to prevent growth of Listeria in fresh and cooked meat products packaged with differ-ent atmospheres. Food Microbiol 15: 639-650.
Hugas M, Garriga M, Aymerich MT (2003) Functionalty of enterococci in meat products. Int J Food Microbiol 88:223-233.
Jack RW, Tagg JR. Ray B (1995) Bacteriocins of Gram positive bacteria. Microbiol Reviews 59: 171–200.
Jin T, Liu L, Sommers CH et al (2009) Radiation sensitization and postirradiation proliferation of Listeria monocytogenes on ready-to-eat deli meat in the presence of pectin-nisin fi lms. J Food Prot 72:644-649.
Jofré A, Aymerich T, Garriga M et al (2008) Assessment of the effectiveness of antimicrobial pac-kaging combined with high pressure to control Salmonella sp. in cooked ham. Food Control 19:634-638.
Jones RJ, Zagorec M, Brightwell G et al (2009) Inhibition by Lactobacillus sakei of other species in the fl ora of vacuum packaged raw meats during prolonged storage. Food Microbiol 26:876–881.
Korkeala H, Suortti T, Măkelă P (1988) Ropy slime formation in vacuum-packed cooked meat pro-ducts caused by homofermentative lactobacilli and a Leuconostoc species. Int J Food Microbiol 7:339–347.
Leroy F, Lievens K, De Vuyst L (2005) Interactions of meat-associated bacteriocin-producing Lac-tobacilli with Listeria innocua under stringent sausage fermentation conditions. J Food Prot 68:2078-2084.
Leroy F, Verluyten J, De Vuyst L (2006) Functional meat starter cultures for improved sausage fer-mentation. Int J Food Microbiol 106: 270-285.
Lücke FK (2000) Utilization of microbes to process and preserve meat. Meat Sci 56:105-115.Mangalassary S, Han I, Rieck J et al (2008) Effect of combining nisin and/or lysozyme with in-pac-
kage pasteurization for control of Listeria monocytogenes in ready-to-eat turkey bologna during refrigerated storage. Food Microbiol 25:866–870.
Maragkoudakis PA, Mountzouris KC, Psyrras D et al (2009) Functional properties of novel protec-tive lactic acid bacteria and application in raw chicken meat against Listeria monocytogenes and Salmonella enteritidis. Int J Food Microbiol 130:219–226.
McMillin KW (2008) Where is MAP Going? A review and future potential of modifi ed atmosphere packaging for meat. Meat Sci 80:43–65.
Murphy RY, Hanson RE, Feze N et al (2005) Eradicating Listeria monocytogenes from fully cooked franks by using an integrated pasteurization-packaging system. J Food Prot 68: 507–511.
Nes IF, Yoon S, Diep DB (2007) Ribosomally synthesized antimicrobial peptides (bacteriocins) in lactic acid bacteria: A review. Food Sci Biotechnol 5: 675–690.
Nieto-Lozano JC, Reguera-Useros JI, Peláez-Martínez MC et al (2006) Effect of a bacteriocin pro-duced by Pediococcus acidilactici against Listeria monocytogenes and Clostridium perfringens on Spanish raw meat. Meat Sci 72:57-61.

BIOCONSERVACIÓN DE ALIMENTOS CÁRNICOS
123
Noonpakdee W, Santivarangkna C, Jumriangrit P et al (2003) Isolation of nisin-producing Lactococ-cus lactis WNC 20 strain from nham, a traditional Thai fermented sausage. Int J Food Microbiol 81:137-45.
O’Sullivan L, Ross RP, Hill C (2002) Potential of bacteriocin-producing lactic acid bacteria for im-provements in food safety and quality. Biochimie 84:593-604.
Pridmore D, Rekhif N, Pittet AC et al (1996) Variacin, a new lanthionine-containing bacteriocin produced by Micrococcus varians: comparison to lacticin 481 of Lactococcus lactis. Appl Environ Microbiol 62:1799-802.
Ravyts F, Barbuti S, Frustoli MA et al (2008) Competitiveness and antibacterial potential of bacte-riocin-producing starter cultures in different types of fermented sausages. J Food Prot 71:1817-1827.
Rodríguez JM, Martinez MI, Kok J (2002) Pediocin PA-1, a wide-spectrum bacteriocin from lactic acid bacteria. Crit Rev Food Sci Nutr 42:91-121.
Settanni L, Corsetti A (2008) Application of bacteriocins in vegetable food biopreservation. Int J Food Microbiol 121:123-138.
Solomakos N, Govaris A, Koidis P et al (2008) The antimicrobial effect of thyme essential oil, nisin, and their combination against Listeria monocytogenes in minced beef during refrigerated storage. Food Microbiol 25:120-127.
Sparo M, Nuñez GG, Castro M et al (2008) Characteristics of an environmental strain, Enterococcus faecalis CECT7121, and its effects as additive on craft dry-fermented sausages. Food Microbiol 25:607-615.
Stergiou VA, Thomas LV, Adams MR (2006) Interactions of nisin with glutathione in a model pro-tein system and meat. J Food Prot 69:951-956.
Thomas LV, Clarkson MR, Delves-Broughton J (2000) Nisin. In: A,S, Naidu, ed., pp. 463-524. Natural Food Antimicrobial Systems. CRC-Press, FL.
Todorov SD, Koep KSC, Van Reenen CA et al (2007) Production of salami from beef, horse, mutton, Blesbok (Damaliscus dorcas phillipsi) and Springbok (Antidorcas marsupialis) with bacterioci-nogenic strains of Lactobacillus plantarum and Lactobacillus curvatus. Meat Sci 77:405-412.
Työppönen S, Petäjä E, Mattila-Sandholm T (2003) Bioprotectives and probiotics for dry sausages. Int J Food Microbiol 83:233-244.
Uesugi AR, Moraru CI (2009) Reduction of Listeria on Ready-to-Eat sausages after exposure to a combination of pulsed light and Nisin. J Food Prot 72:347–353.
Villani F, Sannino L, Moschetti G et al (1997) Partial characterization of an antagonistic substance produced by Staphylococcus xylosus 1E and determination of the effectiveness of the producer strain to inhibit Listeria monocytogenes in Italian sausages. Food Microbiol 14:555–566.


BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN
125ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
RESUMEN
Los ácidos grasos de cadena corta (AGCC) son ácidos monocarboxílicos con una cadena de uno a seis átomos de carbono saturados. Son los principales aniones del colon. Se presen-tan como ácidos libres en el tracto gastrointestinal de los mamíferos, siendo el producto fi nal de la digestión microbiana de los carbohidratos. Los índices de producción microbiana y de absorción son tan altos que los AGCC son la mayor fuente de energía (70-80%) en mamíferos como la vaca. La contribución de los AGCC a los requerimientos del metabolismo basal de los animales monogástricos depende de factores como la concentración y las cantidades totales de los mismos, índices de producción y absorción y la capacidad relativa del segmento intes-tinal considerando el peso del cuerpo del individuo. Además de su papel energético, van a intervenir en diversos procesos como el transporte de agua y electrolitos en el colon. Debido a su carácter hidrofílico, los ratios y mecanismos de absorción de estos ácidos son claramente diferenciables de los ácidos grasos de cadena larga liposolubles. En este artículo se hace un repaso a la funcionalidad de los ácidos grasos de cadena corta y sus mecanismos de absorción por difusión pasiva como por transporte activo.
Palabras clave: ácido grasos de cadena corta (AGCC), biodisponibilidad, difusión pasiva, transporte activo.
ABSTRACT
Short chain fatty acids (SCFA) are monocarboxylic acids with a chain from one to six carbon atoms saturated. Are the major anions of the colon. Present as free acids in the gas-
BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN
GASPAR ROS BERRUEZO1, 2, CARMEN MARTÍNEZ GRACIÁ Y JOSÉ ANTONIO VALENCIA ARQUES
1 Catedrático de Nutrición y Bromatología. Universidad de Murcia. 2 Académico de Número de la Real Academia de Ciencias Veterinaria de Andalucía Oriental. [email protected]

BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN
126
trointestinal tract of mammals, the fi nal product of microbial digestion of carbohydrates. The rates of microbial production and absorption are so high that the SCFA are the major source of energy (70-80%) in mammals such as cows. The contribution of SCFA to the requirements of basal metabolism of monogastric animals depends on factors such as concentration and the total amounts thereof, production and absorption rates and the relative ability of the intestinal segment considering the individual’s body weight.
Besides its role energy will participate in various processes such as transport of water and electrolytes in the colon. Due to their hydrophilic nature, the rates and mechanisms of absorp-tion of these acids are clearly distinguishable from long chain fatty acids liposoblubles. This article gives an overview of the functionality of short chain fatty acids and their mechanisms of uptake by passive diffusion and active transport.
Key words: Short chain fatty acid (SCFA), bioavailability, passive diffusion, active transport.
MICROBIOTA Y PRODUCCIÓN DE AGCC
El intestino del ser humano contiene una sociedad compleja, dinámica y diversa de bacterias no patógenas. El intestino grueso de los mamíferos, donde se incluye al hombre, contiene un gran número de microorganismos (Simon y Gorbach, 1984). Estos microorganismos son capaces de digerir azúcares y fi bras además de un amplio rango de otros polisacáridos como las celulosas, hemicelulosas y pectinas (Halliwell, 1961; Keys et al., 1969). Bajo condiciones normales los productos fi nales de toda la digestión de los carbohidratos son principalmente tres: acetato, propionato y butirato (Leng, 1970) además de gases como el hidrogeno, el dióxido de carbono y metano (Wolin, 1975). Las concentraciones de AGCC son normalmente encontradas en pe-queñas proporciones, excepto si la dieta es rica en azucares (Leontowicz et al., 1979). La proteolisis que llevan a cabo los microorganismos también va a proporcionar AGCC. Los AGCC también llamados ácidos grasos volátiles (Tabla 1), contribuyen al mantenimiento de las funciones del intestino grueso, a prevenir patologías a través de sus actuaciones en la luz intestinal y sobre la musculatura y vasos del colon y a través de su metabolismo por los colonocitos. Parece ser que el butirato en particular juega un papel importante en el mantenimiento de la población de colonocitos sanos.
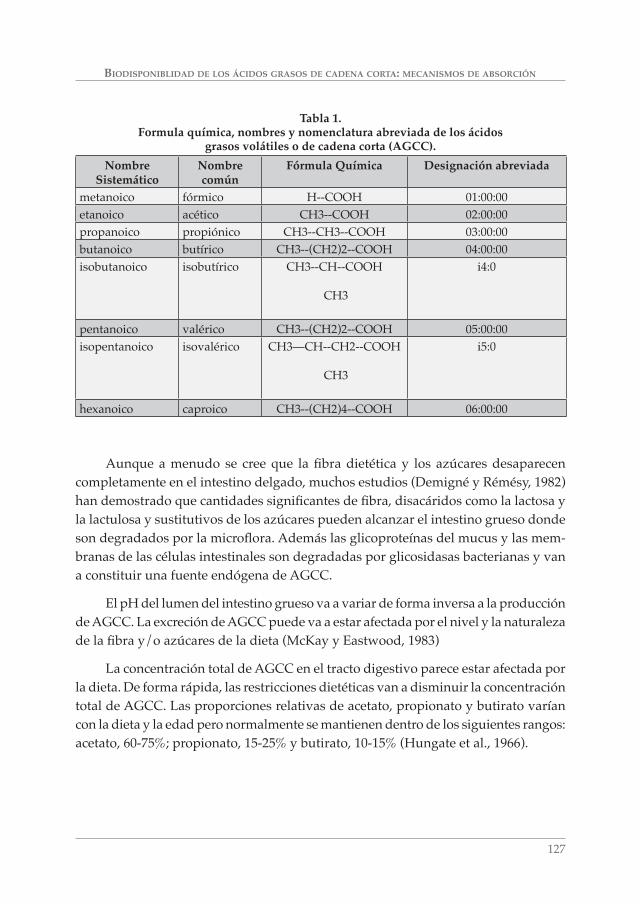
BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN
127
Tabla 1.Formula química, nombres y nomenclatura abreviada de los ácidos
grasos volátiles o de cadena corta (AGCC).Nombre
SistemáticoNombre común
Fórmula Química Designación abreviada
metanoico fórmico H--COOH 01:00:00etanoico acético CH3--COOH 02:00:00propanoico propiónico CH3--CH3--COOH 03:00:00butanoico butírico CH3--(CH2)2--COOH 04:00:00isobutanoico isobutírico CH3--CH--COOH
CH3
i4:0
pentanoico valérico CH3--(CH2)2--COOH 05:00:00isopentanoico isovalérico CH3—CH--CH2--COOH
CH3
i5:0
hexanoico caproico CH3--(CH2)4--COOH 06:00:00
Aunque a menudo se cree que la fi bra dietética y los azúcares desaparecen completamente en el intestino delgado, muchos estudios (Demigné y Rémésy, 1982) han demostrado que cantidades signifi cantes de fi bra, disacáridos como la lactosa y la lactulosa y sustitutivos de los azúcares pueden alcanzar el intestino grueso donde son degradados por la microfl ora. Además las glicoproteínas del mucus y las mem-branas de las células intestinales son degradadas por glicosidasas bacterianas y van a constituir una fuente endógena de AGCC.
El pH del lumen del intestino grueso va a variar de forma inversa a la producción de AGCC. La excreción de AGCC puede va a estar afectada por el nivel y la naturaleza de la fi bra y/o azúcares de la dieta (McKay y Eastwood, 1983)
La concentración total de AGCC en el tracto digestivo parece estar afectada por la dieta. De forma rápida, las restricciones dietéticas van a disminuir la concentración total de AGCC. Las proporciones relativas de acetato, propionato y butirato varían con la dieta y la edad pero normalmente se mantienen dentro de los siguientes rangos: acetato, 60-75%; propionato, 15-25% y butirato, 10-15% (Hungate et al., 1966).

BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN
128
ABSORCIÓN DE LOS AGCC.
La característica físico-química esencial de los AGCC que los distinguen de los ácidos grasos de cadena larga es su gran solubilidad en un medio acuoso. Por ejemplo, a un pH de 7.4 se ha observado que el ácido butírico era 25,000 veces más soluble en agua que el ácido palmítico (Westergaard y Dietschy, 1976). los ácidos grasos de cadena larga necesitan ser solubilizados en los contenidos del lumen a través de una mezcla de sales biliares para su posterior absorción. Por el contrario, los AGCC al ser monómeros en la fase acuosa luminal pueden ser absorbidos por cualquier segmento del tracto digestivo de los mamíferos, por lo que la digestibilidad de los AGCC es total. La absorción de los AGCC no esta infl uenciada por la presencia de micelas de sales biliares en el medio. Como resultado de las diferencias en la solubilidad de los ácidos grasos en relación a la longitud de su cadena, se observó que las vías de absorción de los AGCC y los ácidos de cadena larga difi eren de forma muy signifi cante. Frente a los ácidos grasos de cadena larga que son transportados a través de una proteína específi ca del borde epitelial al retículo endoplasmático, activando derivados de las acil-CoA, siendo reesterifi cados a triacilgliceroles y fi nalmente pasando a la linfa intes-tinal en forma de quilomicrones (Brindley, 1984), los AGCC se difunden libremente a través del citosol de la mucosa y entrando en la sangre venosa mientras manteniendo su forma de ácido graso libre. Sólo se produce un mínimo transporte cuando pasan al sistema linfático. Bugaut (1987) sugirieron que la absorción de los AGCC esta en parte controlada por el citoesqueleto. Pero sin embargo, no se ha visto si la interac-ción AGCC-citoesqueleto puede estar conectada con la bien conocida infl uencia del butirato en la elaboración del citoesqueleto celular (Prasad, 1980). Además, los AGCC no son transportados al retículo endoplasmático donde se produce la reesterifi cación de los ácidos grasos (Brindley, 1974). Es más, la acil-CoA sintetasa, la cual es necesa-ria para la activación de los ácidos grasos antes de su reesterifi cación y la cual esta localizada en la fracción microsomal del enterocito, no presenta actividad hacia los AGCC como se ha demostrado in vitro en el cerdo (Ailhaud et al., 1962), el gato y la cobaya (Brindley y Hübscher, 1966).
Aunque los términos de absorción y de transporte son usados para describir el fl ujo o transferencia de AGCC a través de la mucosa del tracto digestivo desde el lu-men a la sangre, se puede hacer una distinción entre el ratio de absorción el cual esta relacionado con la cantidad de AGCC que deja el lumen para entrar en la mucosa y el ratio de transporte el cual esta relacionado con la cantidad de AGCC que aparece en sangre. Como consecuencia del amplio metabolismo de los AGCC el ratio de transporte puede ser considerablemente más bajo que el ratio de absorción especialmente cuando la capacidad del epitelio para metabolizar AGCC no esta saturada (Stevens, 1970).

BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN
129
INFLUENCIA DE LA LONGITUD DE LA CADENA Y EL PH.
Los AGCC son absorbidos a un ratio el cual aumenta con la longitud de la ca-dena (Oshio y Thata, 1984). Se ha demostrado que en un lumen con un pH bajo, la absorción de los AGCC es más rápida (Rübsamen y Engelhardt, 1978).
MECANISMOS DE ABSORCIÓN.
Difusión Pasiva.
Difusión pasiva. A través de la capa de agua y la membrana celular.
La absorción de AGCC se pensaba que ocurría únicamente por difusión pasiva (Naupert y Rommel, 1975; Mottaz y Worbe, 1977). Al igual que un ácido graso es absorbido de forma pasiva, su transporte a través del epitelio intestinal está deter-minado por su difusión a través de una barrera que consiste en una capa de agua adyacente a las microvellosidades de la membrana y su permeabilidad con respecto a esta membrana (Thomson y Dietschy, 1981). El grosor de la capa de agua es de aproximadamente 600μm en yeyuno humano y depende de la integridad de la mucosa. En el tracto gastrointestinal la mucosa pude participar en la formación de la capa de agua, un hecho que está basado en que la capa de agua y la capa de moco segregado por las células calciformes son similares en grosor (Lukie, 1977). Aunque se piensa que la mucosidad intestinal tiene un papel en el proceso de transporte, su papel como barrera de difusión de los AGCC no ha sido demostrado (Rübsamen y Hörnicke, 1982). Frente a la capa de agua de la mucosa de la membrana celular, cuyo grosor es relativamente pequeño, nos encontramos con una membrana liposoluble formada por lípidos polares, llamados fosfolípidos y colesterol, como principales componentes de esta (Schwarz et al., 1985). Westergaard y Dierschy (1974) demostraron que la mem-brana celular lipídica, y no la capa acuosa, es la que realmente infl uye en el transporte pasivo de los AGCC hidrosolubles. Las diferencias entre la concentraciones de AGCC absorbidos observadas entre distintas especies de roedores puede estar relacionada a diferencias inherentes en las propiedades de las microvellosidades de la membrana (Thomson et al., 1983). Como señalaron Diamon y Wright (1969) las variaciones de los ratios de permeabilidad entre los diferentes ácidos grasos absorbidos por mecanismos pasivos pueden ser explicadas debido a fuerzas intermoleculares que determinan la relación entre las interacciones de soluto:agua y soluto:lípido. Cuando se ha incubado pequeños discos de con muestras de mucosa intestinal se ha observado que el ratio de AGCC absorbido es normalmente más alto de los esperado en la extrapolación de la relación lineal entre N (número de átomos de carbono para el ac. Caprílico, ac.

BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN
130
Cáprico y ac. Láurico) y el ln Jd/D (Jd: ratio de absorción y D: coefi ciente de difusión libre de cada ácido graso). Este fenómeno fue visto usando diferentes técnicas in vi-tro y los diferentes ratios de removido en la fase de digestión donde las muestras de intestino fueron incubadas. Esto también ocurre en animales sometidos a diferentes condiciones fi siológicas, farmacológicas y nutricionales.
Los ratios de absorción de AGCC in vivo aumentan más o menos proporcional-mente con el gradiente de concentración entre el lumen y la sangre portal o simple-mente con la concentración luminal de AGCC como se ha observado en el retículo de los rumiantes, intestino delgado y colon. El fl ujo sanguíneo infl uye en el gradiente de concentración de nutrientes y en su tasa de absorción. Se ha visto que la absorción de AGCC estimula el fl ujo sanguíneo en los vasos de la panza, del colon del perro y el ciego de las ratas a diferentes niveles según el animal. En cambio, la vasodilatación incrementa ligeramente la absorción de butirato en el intestino delgado de los gatos. Además, en el índice de absorción de AGCC no varía por la acción de inhibidores metabólicos y no han sido demostradas interacciones de competitividad (Myers et al., 1967).
Forma Cargada Positivamente.- Como regla general los compuestos hidrosolubles no ionizados pasan a través de la membrana celular lipídica pero sus partes ionizadas no lo hacen al igual ocurre con los AGCC como vemos en los experimentos con cepos eléctricos in vitro (Argenzio et al., 1977). El índice de absorción relativa de un electrolito débil depende no solo de la liposolubilidad relativa del compuesto no ionizado sino que también lo hace de su pK (Schanker et al., 1958). La pK de los AGCC es cercana a 4.8 y el grado de ionización del ácido aumenta con el pH y es aproximadamente 100/1 con un pH 7.0. En condiciones fi siológicas normales, el pH de los contenidos del canal alimentario es mayor que 5.0 a excepción del estomago e incluso algunas veces mas alto que 7.0, por ejemplo en animales en hibernación (Rémésy y Demig-né, 1976). Es posible que los AGCC puedan ser absorbidos incluso en un pH neutro (Rémésy y Demigné, 1976).
Un mecanismo propuesto para la protonación luminal de aniones de AGGC está basado en un proceso de transporte mediado por proteína transportadora que encontramos en todas partes y que cambia Na+ por H+ y la cual podría estar loca-lizada en una de las capas de lípidos de la membrana celular (Vigne et al., 1985). El intercambio de protones de la célula a través de la membrana luminal por el sistema electroneutral de intercambio Na+-H+ puede mantener altos índices de transporte de AGCC en la forma no ionizada. Ahora se ha establecido que la absorción de AGCC está muy conectada con el transporte de agua y electrolitos (Schultz, 1981). El esti-

BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN
131
mulo producido por la absorción de AGCC con respecto a la absorción de Na+ ha sido demostrado en la cabra, la rata, el cerdo, el ternero, la oveja y en la cobaya. Este fenómeno también se ha visto que tiene lugar en el colón humano no parece ocurrir en el intestino delgado de los mismos (Roediger y Moore, 1981). Por otra parte, se ha observado un efecto inhibitorio de los AGCC en el transporte de Na+ en el intestino grueso equino (Argenzio et al.,1977). Estas discrepancias pueden estar causadas en parte por diferentes pHs de la fase luminal durante los experimentos, ya que es la forma disociada de los AGCC la que infl uyen en la absorción de sodio. Lo que es más, el aumento del metabolismo de los AGCC, que pude ser una fuente de energía para activar el transporte de Na+, probablemente varía a lo largo del tracto digestivo y según las especies, y además puede infl uir en el transporte de Na+. Al contrario, se ha demostrado que el Na+ produce una estimulación de la absorción de AGCC en el intestino de rata (Umesaki et al., 1979) o una disminución por la sustitución de Na+ en el ciego y el colon de conejos y en el intestino delgado de ratas (Vernay, 1986). Hay que decir que también se ha encontrado una ausencia de correlación entre la cantidad neta de transporte de Na+ y el transporte de AGCC. La unión indirecta entre la absorción de Na+ y la de AGCC a través de mecanismos simples de intercambio de Na+-H+ no se ha sido ratifi cado en los resultados obtenidos en pruebas realizadas en distintos segmentos del tracto digestivo de varias especies y por distintos investigadores. Este mecanismo por sí solo no puede ser el responsable de los altos índices de absorción de AGCC, ya que cuando se suprime el transporte neto de Na+, los AGCC se siguen absorbiendo de forma moderada (Vernay, 1986). El intercambio de Na+-H+ fue medido por Ruppin et al. (1980) y el resultado fue que no prevalecía en el colón humano. Sin embargo, la unión de AGCC y sodio es un poderoso mecanismo para llevar agua fuera del lumen del intestino grueso. De esta manera, podemos considerar a la producción y absorción de AGCC como un mecanismo antidiarreico (Vernay, 1986).
Modelo de los Tres Compartimentos.- Hasta ahora, habíamos considerado el tejido epitelial como una membrana simple que separaba dos compartimentos, por ejemplo, el lumen y la sangre. Sin embargo, el tejido está constituido por un epitelio columnar o estratifi cado y su pH intracelular que es aproximadamente 7.0 (Kurtin y Charney, 1984) pude ser considerado como un tercer compartimento cuando estudiamos la difusión de los ácidos grasos. El ratio de difusión transepitelial variaría con el pH y con los gradientes electroquímicos entre el epitelio del compartimento y las dos soluciones que están en contacto con él (Jackson et al., 1981). Un modelo aplicable al transporte epitelial de electrolitos débiles. Consiste en dos barreras colocadas en serie separadas por un microcompartimento en cuyo interior el pH es menor que en la fase de agitación. Para mejorar el transporte total de un ácido débil de una fase de agitación a la otra en

BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN
132
contra de gradiente del potencial electroquímico, una barrera debe ser relativamente permeable a las formas ionizadas y no ionizadas. Este sistema de tres compartimen-tos ha sido estudiado en detalle por Stevens (1970), quien ha observado in vitro en el epitelio de la panza un transporte de acetato en dirección sangre-lumen en contra de su gradiente de concentración y sin una diferencia de pH entre las soluciones en que esta bañado el epitelio. Para concretar más sus datos experimentales, los autores propusieron un modelo hipotético de transporte de AGCC, donde la membrana que da al lumen podría ser permeable tanto a AGCC disociados como no disociados, por lo que, en consecuencia, la membrana relativamente impermeable a los aniones podría ser la que estuviera en el lado de la sangre. Esta propuesta permitiría la captación rápida por parte del tejido de AGCC sin la disminución del pH intracelular, el cual debe acompañar a la absorción de AGCC protonado solo. Una absorción de CO2 por parte del epitelio y un suministro de CO2 que proviene de la oxidación de AGCC con una difusión asociada de HCO3
- del tejido hacia el lumen podría ser una fuente de H+ para protonar AGCC- y se tendría en cuenta para la relación entre la absorción de AGCC y la acumulación intraluminal de HCO3
-. Es más, Stevens y sus compañeros resumieron que el índice de difusión de los AGCC podrían ser modifi cado por su metabolismo intraepitelial. El catabolismo de un ácido incrementa su gradiente de concentración entre el lumen y el compartimento de la mucosa y, más allá, incremen-ta su índice de absorción por el lumen. Paralelamente, el catabolismo disminuye el gradiente de concentración del ácido entre el compartimento del tejido y la sangre, además de disminuir su transporte hacia esta última.
TRANSPORTE ACTIVO.
El transporte de muchas sustancias hidrofílicas se supone que se lleva a cabo a través de transportadores específi cos de cada sustrato. Un transporte mediado por estos transportadores que dependen de energía para su funcionamiento es conocido con el nombre de transporte activo. En este caso, el sustrato puede ser absorbido en contra gradiente químico o en contra de gradiente electroquímico. La saturación de movimientos, la dependencia de de sodio y la inhibición del sustrato de competición también caracterizan a este tipo de transporte (Kimmich, 1990). Algunas de las caracte-rísticas de la absorción por transportadores se han observado en relación a los AGCC. De esta manera, el movimiento de AGCC en contra de gradiente de concentración y la reducción de su transferencia por la ausencia de glucosa e incluso su supresión en condiciones anaeróbicas se han observado in vitro en paredes de intestino de ratas (Naupert y Rommel, 1975). Una inhibición del transporte de AGCC en presencia de

BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN
133
inhibidores metabólicos como el 2,4-dinitrofenol o el phlorrhizin (Naupert y Rommel, 1975) y, a la inversa, las pruebas por adicción de una azúcar metabolizable como la glucosa también se han descrito (Barry et al., 1966). La Ouabaína, inhibidor bien co-nocido de la bomba de Na+, reduce la desaparición de AGCC en los colonocitos del conejo (Vernay, 1986) y de la cobaya (Wirthensohn y Engelhardt, 1981) pero no tiene efectos en pruebas in vitro de absorción de propionato en el yeyuno de la rata (Naupert y Rommel, 1975) y de acetato en el colon de rata (Umesaki et al., 1997).
En resumen, el transporte de AGCC parece ser principalmente debido a una difusión pasiva no iónica. Quizá pueda ser considerado como una especie de “trans-porte activo” ya que se requiere energía para los mecanismos de regulación del pH.
REFERENCIA BIBLIOGRÁFICAS
Ailhaud G., Sarda L., Desnuelle P. Formation of hydroxamates of long chain fatty acids by a subce-llular fraction of intestinal mucosa. Biochim Biophys Acta. 1962, 21 (59): 261–272.
Argenzio R.A., Southworth M., Lowe J.E., Stevens C.E. Interrelationiship of Na, HCO3 and volatile fatty acid transport by equine large intestine. Am J Physiol 1977; 233: E469-78.
Brindley D.N. Fatty acid and triacylglycerol metabolism. Br J Clin Pract Suppl. 1984; 31: 58-62.Brindley D.N. The intracellular phase of fat absorption. Biomembranes. 1974; 4B (0): 621-71.Brindley, D.N., Hubscher, G. The effect of chain length on the activation and subsequent incorpo-
ration of fatty acids into glycerides by the small intestinal mucosa. Biochim. Biophy Acta, 1966; 125, 92.
Bugaut M. Occurrence, absorption and metabolism of shortchainfattyacids in the digestive tract of mammals. Comparative Biochemistry and Physiology Part B: Comparative Biochemistry. 1987; 86 (3): 439–472
Demigne C, Remesy C. Infl uence of unrefi ned potato starch on cecal fermentations and volatile fatty acid absorption in rats. J Nutr. 1982 Dec; 112 (12): 2227-34.
Diamond, J.M., E.M. Wright. Biological membranes. The physical basis of ion and non-electrolyte selectivity. Annu. Rev. Biochem. 1969; 31:581
Halliwell G. The action of cellulolytic enzymes from Myrothecium verrucaria. Biochem J. 1961 Apr; 79: 185-92.
Hungate R.E., Smith W., Clarke R.T. Suitability of butyl rubber stoppers for closing anaerobic roll culture tubes. J Bacteriol. 1966 Feb; 91 (2): 908-9.
Jackson M.J., Tai C.Y., Steane J.E. Weak electrolyte permeation in alimentary epithelia. Am J Physiol. 1981, Mar; 240 (3): G191-8.
Keys JE Jr, Van Soest PJ, Young EP. Comparative study of the digestibility of forage cellulose and hemicellulose in ruminants and nonruminants. J Anim Sci. 1969 Jul; 29 (1): 11-5.
Kimmich G.A. Membrane potentials and the mechanism of intestinal Na(+)-dependent sugar transport. J Membr Biol. 1990; Mar; 114 (1): 1–27.
Kurtin P., Charney A.N. Intestinal ion transport and intracellular pH during acute respiratory alka-losis and acidosis. Am J Physiol. 1984, Jul; 247 (1 Pt 1): G24-31.
Leng RA. Glucose synthesis in ruminants. Adv Vet Sci Comp Med. 1970; 14: 209-60.Leontowicz H, Barej W, Kulasek G, Chomyszyn M. Diminishing rumen butyrogenesis in bulls and
sheep fed sugar beets. Ann Rech Vet. 1979; 10 (2-3): 454-6.

BIODISPONIBLIDAD DE LOS ÁCIDOS GRASOS DE CADENA CORTA: MECANISMOS DE ABSORCIÓN
134
Lukie, B.E. Mod. Prob. Paediatr. 1977; 19, 46-54McKay L.F., Eastwood M.A. The infl uence of dietary fi bre on caecal metabolism in the rat. Br J Nutr.
1983; Nov; 50 (3): 679-84Mottaz P, Worbe JF. Transfer of volatile fatty acids in rat isolated cecum wall. C R Seances Soc Biol
Fil. 1977; 171 (2): 375-80.Myers, L.L., Jackson, H.D., Packett, L.V. Absorption of volatile fatty acids from the cecum of sheep.
Journal of Animal Science, 1967; 26, 1450-1458.Naupert C., Rommel K. Absorption of Short and Medium Chain Fatty Acids in the Jejunum of the
Rat. Z. Klin. Chem. Klin. Biochem. 1975; 13., 8.553-562Oshio S., Tahata I. Absorption of dissociated volatile fatty acids through the rumen wall of sheep.
Can. J. Anim. Sci. 1984 64 (Suppl.):1 67–168Pradas, K.N. Butyric acid: A small fatty acid with diverse biological functions. Life Sciences, 1980
27, 1351-1358.Remesy C., Demigne C. Partition and absorption of volatile fatty acids in the alimentary canal of the
rat. Ann Rech Vet. 1976; 7 (1): 39-55.Rubsamen K., Engelhardt W.v. Bicarbonate secretion and solute absorption in forestomach of the
llama. Am. J. Physiol. 1978 235: 1–6.Rübsamen K., Hörnicke H. Infl uence of osmolality, short chain fatty acids and deoxycholic acid on
mucus secretion in the rat colon. Pfl ugers Arch. 1982; Dec; 395 (4): 306-11Schanker, L.S., Tocco, D.J., Brodie, B.B., and Hogben, C.A.M. Absorption of drugs from the rat small
intestines. J. Pharmacol. Exp. Ther. 1958; 123:81–88Schultz, S.G. Federation Proceedings, 1981; 40, 2408-241 1Schwarz S.M., Hostetler B., Ling S., Mone M., Watkins J.B. Intestinal membrane lipid composition
and fl uidity during development in the rat. Am J Physiol. 1985; Feb; 248 (2 Pt 1): G200-7Simon G.L., Gorbach S.L. Intestinal fl ora in health and disease. Gastroenterology. 1984 Jan; 86 (1):
174-93.Stevens, C.E. Fatty acid transport through the rumen epithelium. En: Physiology of Digestion and
Metabolism in the Ruminant, ed. PHILLIPSON, A.T., 1970 pp. 10-1 12. Oriel Press, Newcastle on Tyne.
Thomson A.B.R., Dietschy J.M. Intestinal lipid absorption: major extracellular and intracellular events. In Physiology of the Gastrointestinal Tract, 1981; vol 2, pp 1147-1220 (Johnson LR, ed), Raven Press, New York.
Umesaki, Y., Yajima, T., Yokokura, T. & Mutai, M. PJugers Archiv. 1979; 379, 4 3 4 8 . Umesaki, Y., Okada, Y., Imaoka, H., Setoyama, H., and Matsumoto, S. Interaction between epithelial
cells and bacteria, normal and pathogenic. Science 1997; 276: 964-965Vernay, M. Colonic absorption of inorganic ions and volatile fatty acids in the rabbit. Comp. Bio-
chem. Physiol. 1986; 83A, 775–784.Vigne P., Jean T., Barbry P., Frelin C., Fine L.G., Lazdunski M. [ 3H]Ethylpropylamiloride, a ligand
to analyze the properties of the Na+/H+ exchange system in the membranes of normal and hypertrophied kidneys. J Biol Chem. 1985, Nov 15; 260 (26): 14120-5.
Westergaard H., Dietschy J.M. Normal mechanisms of fat absorption and derangements induced by various gastrointestinal diseases. Med Clin North Am. 1974 Nov; 58 (6): 1413-27.
Wirthensohn K., von Engelhardt W. Uptake of sodium and short-chain fatty acids into colonic epithelial cells. Gastroenterol. Clin. Biol. 1981; 5, 125A
Wolin MJ. Metabolic interactions among intestinal microorganisms. Am J Clin Nutr. 1974 Nov; 27 (11): 1320-8.

SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E...
135ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
RESUMEN
En este trabajo se ha determinado la seroprevalencia de IgG frente al genotipo 3 del Virus de la Hepatitis E (VHE) en una muestra representativa de perros de una provincia del sur de España, y se han analizado los posibles factores de riesgo asociados a la seropositividad. Un total de 296 muestras de suero fueron analizadas mediante ELISA. Todas las muestras con una densidad óptica positiva, cerca del punto de corte, fueron confi rmados por Western Blot. La seroprevalencia global fue del 15,5% (IC del 95%, 11,42-19,66%). El análisis de los diferentes factores relacionados con el animal y con el ambiente demuestran que existe una mayor sero-prevalencia en perros machos (OR 2.07, P = 0,026), y una correlación entre la seropositividad y el aumento de la edad de los animales, así como chequeos sanitarios infrecuentes (P < 0,05). Estos resultados muestran claras similitudes con los obtenidos en la población humana. Todas las muestras seropositivas (n = 46) fueron sometidas a técnicas de RT-PCR, para la detección del genoma vírico en el suero, pero en todos los casos fue negativo. Por este motivo, conside-ramos que se requieren más estudios que aclaren el papel desempeñado por los perros como reservorios de VHE humano.
Palabras clave: virus de la hepatitis E, factores de riesgo en perros.
SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E (GENOTIPO 3) EN PERROS DEL SUR DE ESPAÑA
B HUERTA1, C TARRADAS1, M GONZÁLEZ1, B PERALTA2, E. M. MATEU2, RJ ASTORGA1, A CARBONERO1, I LUQUE1,*
1Dpto. Sanidad Animal. Universidad de Córdoba. Campus Universitario de Rabanales, Campus de Excelencia Internacional CeiA3, 14071-Córdoba
2Centre de Recerca en Sanitat Animal (CReSA). Edifi ci CReSA. Campus UAB. 08193-Bellaterra (Barcelona)*Inmaculada Luque Moreno, Correo electrónico [email protected].

SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E...
136
SUMMARY
The seroprevalence of IgG against genotype 3 of hepatitis E virus (HEV) in dogs in a province of Spain and the possible associated risk factors were determined. A total of 296 serum samples were analyzed by ELISA. All samples with a positive Optical Density, near to cut-off point, were confi rmed by Western Blot. Overall seroprevalence was 15.5% (95% CI, 11.42-19.66%). Analysis of different animal- and management-related factors shown a higher seroprevalence in male dogs (OR 2.07, P = 0.026) and a slight correlation between seropositi-vity and both age and infrequent health checks (P < 0.05). These results show clear similarities to those described for humans. Seropositive samples (n = 46) were tested for HEV-RNA using a semi-nested PCR, but the viral genome was not detected in any case, for this reason further studies are required to clarify the role played by animals as reservoirs for human HEV.
Key Words: Hepatitis E virus, Risk factors in dogs
INTRODUCCIÓN
La hepatitis E es un tipo de hepatitis viral de transmisión entérica, producida por un Hepevirus (familia Hepeviridae) que afecta al hombre, siendo considerada una enfermedad emergente en países desarrollados (Balayan 1997, Farovov y col 1996, Ruiz-Moreno y col 2000; Buti y col 2006, López-Izquierdo y col 2007). Hasta hace relativamente pocos años, la enfermedad se había descrito solo de forma esporádica en personas que habían viajado a países infectados y en drogodependientes. No obstante, en los últimos años ha ido aumentando considerablemente la existencia de casos autóctonos, con tasas de seroprevalencia que han alcanzado en algunas zonas el 11% (Mateos y col 1998, Buti y col 2006, Galiana y col 2008, Krüttgen y col 2011), parece ser que el aumento progre-sivo de esta zoonosis está asociado directamente a fenómenos de globalización, tales como la proliferación del turismo a zonas afectadas y la elevada inmigración en todos los países desarrollados (López-Izquierdo y col, 2007).Por otro lado y desde el punto de vista etiológico cuando se estudia la secuencia genética del virus responsable de la infección se comprueba que existen cuatro genotipos (Mushahwar 2008, Okamoto 2007, Zhang y col 2008). Los genotipos 1 y 2 responsables de la infección en el hombre en países de climas tropicales, con pobres condiciones sanitarias, y los genotipos 3 y 4 aislados de cerdos y aves, responsables también de casos esporádicos de hepatitis E en el hombre pero siempre en países desarrollados (Meng 2010).
Una de las cuestiones que aún quedan por responder, a pesar del número de trabajos publicados, es cuál es el principal reservorio y fuente de infección para el hombre. En países tropicales y subtropicales, un importante reservorio lo constituyen las aguas residuales, de donde se han realizado aislamientos genéticamente similares al virus de la hepatitis en el hombre (Pina y col 2000). Los estudios realizados para

SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E...
137
valorar el posible papel de los animales como reservorio biológico, han permitido poner en evidencia la presencia de anticuerpos en un número elevado de especies animales, como cerdos, roedores, terneros, cabras, gatos, ciervos, caballos, palomas, perros y aves (Kuno y col 2003, de Deus y col 2008, Peralta y col 2008, Zhang y col 2008). No obstante, hasta la fecha, el virus sólo se ha aislado a partir de aves y cerdos, tanto domésticos como silvestres, en distintos estadios de producción (Meng, 2010; Mushahwar, 2008; Zhang et al., 2008). A pesar de todos los estudios realizados, no se ha podido demostrar el verdadero papel de los animales domésticos en la transmi-sión y contagio del virus de la Hepatitis E al hombre, si bien algunos investigadores han puesto de manifi esto que las personas en contacto con animales tienen un riesgo mayor de padecer la infección (Galiana y col 2008, Kuniholm y col 2009, Meng 2010) y por lo tanto se puede sospechar que los animales domésticos pueden jugar un papel fundamental en el ciclo epidemiológico de esta enfermedad.
El principal objetivo de este trabajo es intentar defi nir el papel de los perros en el ciclo epidemiológico de esta zoonosis emergente. Para ello, se ha determinado la prevalencia de anticuerpos específi cos frente al virus de la hepatitis E en la población canina de una provincia del sur de España y los posibles factores de riesgo asociados a la seropositividad de esta especie. También se realizaron pruebas genéticas para detectar la presencia del virus en el suero.
MATERIAL Y MÉTODOS
Población en estudio
Para defi nir la población en estudio se utilizaron los datos del Registro Andaluz de Identifi cación de Animales (RAIA), con una población estimada de 100308 perros. La recogida de muestras se ha realizado siguiendo la distribución en cuatro distritos sanitarios, estratifi cando la población de referencia según su procedencia (Figura 1). El tamaño de la muestra (n = 296 animales, fracción de muestreo <0,05) fue calculada para una prevalencia esperada del 24 por ciento (Peralta y col 2006), un error del 5 por ciento y un nivel de confi anza del 95 por ciento, utilizando el programa informático de epidemiología veterinaria WinEpiscope 2.0. (Ignacio de Blas, Facultad de Veterinaria, Universidad de Zaragoza, 2006).
Durante un periodo de seis meses (enero a junio), los veterinarios responsables de las clínicas participantes obtuvieron las muestras de suero de animales que asistían a consulta veterinaria, siendo congelados a -20ºC hasta su uso. Al mismo tiempo, rellenaron un cuestionario epidemiológico completo, en el que se recogieron datos

SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E...
138
específi cos y antecedentes epidemiológicos de los individuos analizados, con el objetivo de comparar grupos seropositivos y seronegativos e identifi car los posibles factores de riesgo asociados a la infección con el virus HEP. Se recogieron las siguien-tes variables: sexo, edad, hábitat urbano/rural, convivencia con otros animales (perros, gatos, cerdos, rumiantes, aves, équidos y especies exóticas), alimentación (piensos comerciales exclusivamente o alimentación combinada con comida casera) y chequeos sanitarios a los que eran sometidos los animales al año.
Estudio serológico
Los sueros para la detección de anticuerpos anti-HEV fueron analizados mediante una técnica ELISA (Arankalle y col 2001, Vitral y col 2005, Peralta y col 2009), sobre placas de 96 pocillos (Nunc, Ref. 269620) antigenadas con la ORF2 de la cepa paquis-taní SAR55, diluida en 50 mM tampón carbonato-bicarbonato a pH 9.6; utilizando como conjugado anticuerpos policlonales de cabra IgG (H+L) anti-perro (Cultek, S.L) y realizando la lectura a 450 nm (Seminati y col 2007). Aquellas muestras con una densidad óptica cercana al punto de corte (0,300) fueron sometidas a una técnica de Western Blot, siguiendo metodologías descritas previamente (Peralta y col 2009).
Técnicas de PCR
Todas las muestras de suero positivas fueron analizadas mediante técnicas de RT-PCR, tras la extracción del ARN a partir de una muestra de 100 μl suero con Tri-Zol (Invitrogen, SA). Los primers específi cos utilizados para la prueba fueron F4340: 5´-CTDTTYGGCCCNTGGTTCCG-3´ y R4690: 5´-CCATRTTCCARACDGTRTTCC-3´para la primera amplifi cación y para la amplifi cación semi-nested se utilizaron los primers F4340: 5´-CTDTTYGGCCCNTGGTTCCG-3´ y R4670: 5´-CANARNAGGGT-GCCVGGCTC-3´ (Peralta y col 2006). Los productos esperados de la amplifi cación tenían un tamaño de 350 y 330 para la primera y segunda amplifi cación, respectiva-mente. En cada prueba se utilizó una muestra de un cerdo positivo al genotipo 3 del virus como control positivo y como control negativo se utilizaron muestras de agua.
Análisis estadístico
Se calculó la seroprevalencia de la enfermedad y sus intervalos de confi anza (95% CIs). Para realizar el análisis estadístico, la variable edad fue categorizada en cuartiles

SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E...
139
(Tabla 1). La asociación entre las variables categóricas y la respuesta inmune frente a la infección fue determinada mediante el test de Chi-cuadrado (χ²) o el test de Fisher. Un valor de P < 0,05 fue considerado estadísticamente signifi cativo. El grado de la asociación fue establecido mediante el cálculo de la Odds Ratio (OR) y sus corres-pondientes intervalos de confi anza (95% CI) para variables nominales, y mediante el coefi ciente Kendall´s tau-c para variables ordinales. Los cálculos estadísticos se han realizado utilizando el programa SPSS v.15 para Windows.
RESULTADOS Y DISCUSIÓN
Esta ampliamente demostrado que la Hepatitis E es una zoonosis emergente y que la especie porcina actúa como reservorio y fuente de infección para el hombre (Baechlein y col 2010), sin embargo algunos investigadores afi rman que los anima-les de compañía también pueden intervenir en el contagio al hombre (Peralta y col 2006, 2009). No obstante, hasta la fecha no se ha demostrado el verdadero papel de estos animales en el ciclo epidemiológico de la enfermedad ni los factores de riesgo asociados. En este trabajo nos propusimos realizar una encuesta epidemiológica sobre una muestra representativa de la población canina de la provincia de Cádiz, situada al sur de España, con un clima mediterráneo oceánico de la costa atlántica. Esta provincia está dividida en cuatro distritos sanitarios (Figura 1), divisiones que se establecen ofi cialmente para la organización de la atención primaria en el ámbito de la salud pública en España.
Se han analizado un total de 296 sueros caninos, mediante técnicas de ELISA, obteniendo unos valores de seroprevalencia de Hepatitis E (HEV) del 15,5% (95% CI: 11,38-19.62%) (tabla 1), resultados ligeramente inferiores a los obtenidos en Barcelona, situada en la zona noreste de España, con un clima típico mediterráneo (Peralta y col 2006). En estudios epidemiológicos realizados en poblaciones humanas de nuestro país se han obtenido valores de seroprevalencia del 11,6%, tasas muy similares a las encontradas en la población canina encuestada en nuestro estudio (Galiana y col 2008). En otros países con climas tropicales y subtropicales como India, Vietnam y Brasil, se han obtenido valores de prevalencia que oscilan entre el 7 y el 27% (Tien y col 1997, Arankalle y col 2001, Vitral y col 2005).
Se han intentado establecer los posibles factores de riesgo asociados a la seropo-sitividad. En primer lugar se determinó la relación con el distrito sanitario; no obstante, aunque existían zonas con niveles de seroprevalencia más elevada, como la obtenida en el Distrito Bahía de Cádiz (21,6%), las diferencias no han sido signifi cativas (P =

SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E...
140
0,1255) (Tabla 1). En relación con el sexo, de los 296 perros incluidos en el estudio, 142 eran machos y 154 hembras, observando una asociación signifi cativa (P = 0,026) entre la seropositividad y el sexo macho (20,4% machos positivos vs. 11% hembras positivas), con valor de OR de 2,07 (95% CI: 1,08-3,95), es decir es dos veces más probable que un macho sea seropositivo a que lo sea una hembra. Estos resultados coinciden con los obtenidos en la población humana, donde las diferencias son mucho más evidentes, de forma que el riesgo de infección es doce veces mayor en el hombre que en la mujer (Galiana y col 2008, Kuniholm y col 2009).
Otra de las variables estudiadas fue la edad, se analizaron animales que oscila-ban entre los 4 meses y los 15 años, que se agruparon en cuartiles para el estudio estadístico (Tabla 1). Se ha observado un incremento de prevalencia de anticuerpos frente al virus de la hepatitis E a medida aumenta la edad del animal (Kendall´s-tau c = 0,062 P = 0,04). Algunos investigadores aseguran que la seropositividad frente al virus de la hepatitis E en diferentes especies, como aves, cerdos y humanos, depende en gran medida de la edad del individuo, ya que generalmente se obtienen siempre prevalencias más altas en grupos de animales adultos que en individuos jóvenes (Meng y col 1999, Huang y col 2002, Buti y col 2006), resultados que coinciden con los obtenidos en nuestro estudio.
En relación con las condiciones medioambientales, la mayoría de los animales procedían de áreas urbanas (Tabla 1), sin embargo la seroprevalencia encontrada fue muy similar a la obtenida en perros que vivían en áreas rurales (16,8% vs. 12,8%, P = 0.421). Estos resultados difi eren de los obtenidos en estudios realizados sobre la población humana, donde la seroprevalencia ha sido mayor en zonas urbanas (Pina y col 2000, Buti y col 2006). No hemos encontrado asociación entre la seropositividad y otras variables como la convivencia con animales, incluyendo perros, gatos, suidos, rumiantes, aves, équidos y otras especies exóticas (P > 0.05) y la alimentación.
Uno de los indicadores más importantes para defi nir el nivel habitual de cui-dados y medidas preventivas de los dueños sobre sus mascotas es la frecuencia de chequeos sanitarios, los resultados de nuestra encuesta epidemiológica demuestran que más del 80% de los perros visitaban al veterinario al menos una vez al año y como se refl eja en la tabla 1 la prevalencia de anticuerpos anti-HEV era mayor entre los animales sometidos a escaso control sanitario (P = 0,027). Por último, en la encuesta epidemiológica realizada se determinó la presencia de sintomatología derivada de una alteración hepática, que pudiera relacionarse con la infección por HEV. De los 46 animales seropositivos, dieciocho (6.08%) mostraron síntomas compatibles con una alteración hepática (vómitos, diarrea) y tres dolor abdominal, pero sólo un animal presentó aumento de las enzimas hepáticas.

SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E...
141
Según los resultados obtenidos, podemos afi rmar que los perros machos pre-sentan el doble de probabilidad (P = 0.026) de poseer anticuerpos IgG frente al virus de la Hepatitis E, también se ha encontrado una correlación entre la seroprevalencia y otros factores como la edad y los chequeos sanitarios infrecuentes (P < 0.05), estos factores también han estado asociados a la infección en algunas poblaciones humanas (Mushahwar 2008, Kunihlom y col 2009), por lo tanto se puede sugerir que existe una gran similitud entre los factores de riesgo asociados a la presencia de anticuerpos anti-Hepatitis E en el hombre y en el perro.
Por último, para demostrar que el perro puede actuar como reservorio de la in-fección para el hombre, resulta imprescindible poner en evidencia el virus. Todas las muestras serológicamente positivas fueron analizadas genéticamente mediante RT-PCR para detectar la presencia del ARN-HEV, siendo los 46 animales a negativos a las técnicas moleculares, resultados similares a los obtenidos anteriormente por Liu y col (2009) y Galiana y col (2008) en perros y humanos respectivamente. Ahora nos tendríamos que plantear la siguiente cuestión ¿Por qué presentan algunos animales anticuerpos IgG frente al virus de la hepatitis E y no se detecta ninguna partícula vírica en su suero? En primer lugar estos resultados pueden ser debidos a que el virus permanezca durante un tiempo limitado en sangre y sea eliminado del organismo (Billam y col 2005, de Deus y col 2008), o bien que tenga un fuerte tropismo por algunos órganos parenquimatosos, como el hígado, donde queda acantonado dando lugar o no a manifestaciones clínicas. Independientemente de la causa, los resultados obtenidos justifi can sin duda la nece-sidad de continuar los estudios sobre la patogenia y la epidemiología de esta infección en la especie canina, para esclarecer su posible papel como reservorio del virus.
AGRADECIMIENTOS
Este estudio ha sido fi nanciado por el Ilustre Colegio Ofi cial de Veterinarios de Cádiz (España). Los autores quieren agradecer a todos los responsables de las clínicas veterinarias de Cádiz y a Federico Vilaplana presidente del Colegio de Veterinarios de dicha provincia, su estrecha colaboración y disposición para la recogida y envío de muestras, material imprescindible para la realización de este trabajo.
REFERENCIAS
Arankalle VA, MV Joshi, AM Kulkarni, SS Gandhe, LP Chobe, SS Rautmare, AC Mishra, VS Pad-bidri. 2001. Prevalence of anti-hepatitis E virus antibodies in different Indian animal species. J Viral Hepat 8, 223-227.

SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E...
142
Baechlein C, A Schielke, R Johne, RG Ulrich, W Baumgaertner, B Grummer. 2010. Prevalence of Hepatitis E virus-specifi c antibodies in sera of German domestic pigs estimated by using different assays. Vet Microbiol 144, 187-191
Billam P, FF Huang, ZF Sun, FW Pierson, RB Duncan, F Elvinger, DK Guenette , TE Toth, XJ Meng. 2005. Systematic pathogenesis and replication of avian hepatitis E virus in specifi c-pathogen-free adult chickens. J Virol 79, 3429-3437.
Buti M, A Domínguez, P Plans, T Jardí, M Sxhaper, J Espuñes, N Cardeñosa, F Rodríguez-Frías, R Esteban, A Plasència, L Salleras. 2006. Community-Based Seroepidemiological Survey of Hepa-titis E Virus Infection in Catalonia, Spain. Clin Vaccine Immunol 13, 1328-1332.
de Deus N, M Casas, B Peralta, M Nofrarías, S Pina, M Martín, J Segalés. 2008. Hepatitis E virus infec-tion dynamics and organic distribution in naturally infected pigs in a farrow-to-fi nish farm. Vet Microbiol 132, 19-28.
Galiana C, S Fernández-Barredo, A García, MT Gómez, M Pérez-Gracia. 2008. Occupational expo-sure to hepatitis E virus (HEV) in swine workers. Am J Trop Med Hyg 78, 1012-1015.
Huang FF, G Haqshenas, HL Shivapreasad, DK Guenette, PR Woolcok, CT Larsen, FW Pierson, F Elvinger, TE Toth, XJ Meng. 2002. Heterogeneity and seroprevalence of a newly identifi ed avian hepatitis E virus from chickens in the United States. J ClinMicrobiol 40, 4197-4202.
Krüttgen A, S Scheithauer, M Häusler, M Kleines. 2011. First report of an autochthonous hepatitis E virus genotype 3 infection in a 5 month old female child in Germany. J Clin Virol 50. 175-176
Kunihlom MH, RH Purcell, GM Mcquillan, RE Engle, A Wasley, KE Nelson. 2009. Epidemiology of hepatitis E virus in the United States: results from the third national health and nutrition examination survey, 1988-1994. J Infect Dis 200, 48-56.
Liu J, W Zhang, Q Shen, S Yang, F Huang, P Li, X Guo, Z Yang, L Cui, J Zhu, X Hua. 2009. Prevalence of antibody to hepatitis E virus among pet dogs in the Jiang-Zhe area of China. Scandinavian J Infect Dis 41, 291-295.
Meng XJ, S Dea, RE Engle, R Friendship, YS Lyoo, T Sirinarumitr, K Urairong, D Wang, D Wong, D Yoo, Y Zhang, RH Purcell, SU Emerson SU. 1999. Prevalence of antibodies to the hepatitis E virus in pigs from countries where hepatitis E is common or is rare in the human population. J Med Virol 59, 297-302.
Meng XJ. 2010. Hepatitis E virus: Animal reservoirs and zoonotic risk. Vet Microbiol 140, 256-65.Mushahwar IK. 2008. Hepatitis E virus: molecular virology, clinical features, diagnosis, transmis-
sion, epidemiology, and prevention. J Med Virol 80, 646-658.Peralta B, M Martín, E Mateu. 2006. Detección de IgG e IgM contra el virus de la Hepatitis E en la
población canina de Barcelona. Avedila 36, 2-6.Peralta B, M Casas, N de Deus, M Martín, A Ortuño, E Pérez-Martín, S Pina, E Mateu. 2009. Anti-
HEV antibodies in domestic animal species and rodents from Spain using a genotype 3-based ELISA. Vet Microbiol 137, 66-73.
Pina S, M Buti, M Cotrina, J Piella, R Girones. 2000. HEV identifi ed in serum from humans with acute hepatitis and in sewage of animal origin in Spain. J Hepatol 33, 826-833.
Tien NT, HT Clayson, HB Khiem, PK Sac, AL Corwin, KS Myint, DW Vaught. 1997. Detection of immunoglobulin G to hepatitis E virus among several animal species in Vietnam. Am J Trop Med Hyg 57, 211.
Vitral CL, MA Pinto, LL Lewis-Ximenez, YE Khudyakov, DR do Santos, AM Gaspar AM. 2005. Se-rological evidence of hepatitis E virus infection in different animal species from the Southeast of Brazil. Memórias do Instituto Oswaldo Cruz 100, 117-122.
Zhang W, Q Shen, J Mou, G Gong, Z Yang, L Cui, J Zhu, G Ju, X Hua. 2008. Hepatitis E virus infec-tion among domestic animals in eastern China. Zoonoses Public Health 55, 291-298.
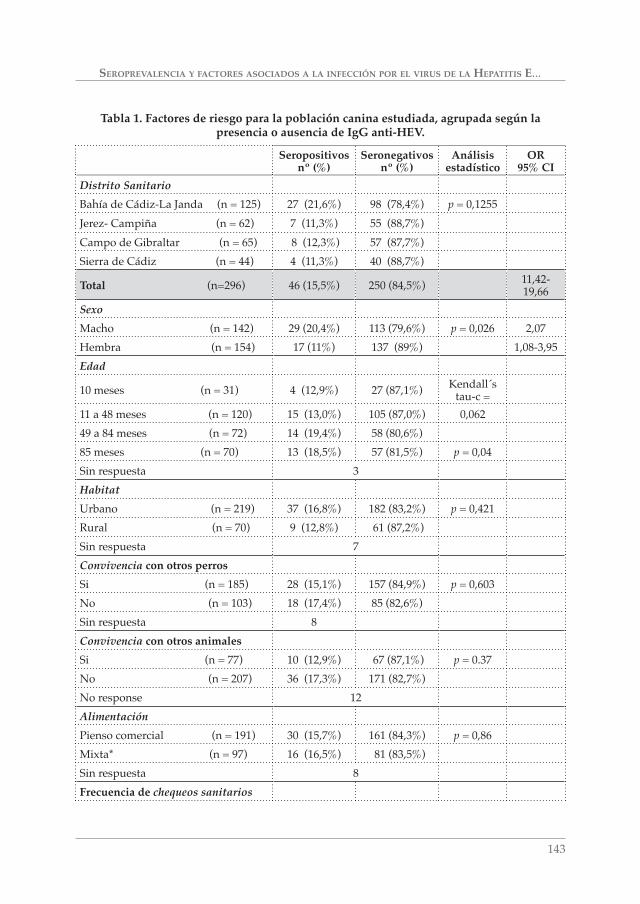
SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E...
143
Tabla 1. Factores de riesgo para la población canina estudiada, agrupada según la presencia o ausencia de IgG anti-HEV.
Seropositivosnº (%)
Seronegativosnº (%)
Análisis estadístico
OR95% CI
Distrito Sanitario Bahía de Cádiz-La Janda (n = 125) 27 (21,6%) 98 (78,4%) p = 0,1255 Jerez- Campiña (n = 62) 7 (11,3%) 55 (88,7%) Campo de Gibraltar (n = 65) 8 (12,3%) 57 (87,7%) Sierra de Cádiz (n = 44) 4 (11,3%) 40 (88,7%)
Total (n=296) 46 (15,5%) 250 (84,5%) 11,42-19,66
Sexo Macho (n = 142) 29 (20,4%) 113 (79,6%) p = 0,026 2,07Hembra (n = 154) 17 (11%) 137 (89%) 1,08-3,95Edad
10 meses (n = 31) 4 (12,9%) 27 (87,1%) Kendall´s tau-c =
11 a 48 meses (n = 120) 15 (13,0%) 105 (87,0%) 0,062 49 a 84 meses (n = 72) 14 (19,4%) 58 (80,6%) 85 meses (n = 70) 13 (18,5%) 57 (81,5%) p = 0,04 Sin respuesta 3 Habitat Urbano (n = 219) 37 (16,8%) 182 (83,2%) p = 0,421 Rural (n = 70) 9 (12,8%) 61 (87,2%) Sin respuesta 7 Convivencia con otros perros Si (n = 185) 28 (15,1%) 157 (84,9%) p = 0,603 No (n = 103) 18 (17,4%) 85 (82,6%) Sin respuesta 8 Convivencia con otros animales Si (n = 77) 10 (12,9%) 67 (87,1%) p = 0.37 No (n = 207) 36 (17,3%) 171 (82,7%) No response 12 Alimentación Pienso comercial (n = 191) 30 (15,7%) 161 (84,3%) p = 0,86 Mixta* (n = 97) 16 (16,5%) 81 (83,5%) Sin respuesta 8 Frecuencia de chequeos sanitarios
SEROPREVALENCIA Y FACTORES ASOCIADOS A LA INFECCIÓN POR EL VIRUS DE LA HEPATITIS E...
144
Una vez al año (n = 236) 41 (17,3%) 195 (82,7%) Kendall´s tau-c =
Dos veces al año (n = 17) 2 (11,7%) 15 (88,3%) - 0,027 ≥Tres veces al año (n = 18) 2 (11,1%) 16 (88,9%) p = 0,027 Sin respuesta 25
*Régimen de alimentación mixta de pienso comercial con comida hecha en casa.
Figura 1. Número de perros censados en cada distrito sanitario de Cádiz (RAIA)

EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
145ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
RESUMEN
En la provincia de Almería la brucelosis ha presentado, a lo largo de los años, tasas de incidencia elevadas en humanos y alta prevalencia en ganado ovino y caprino. El objetivo de este estudio es describir la evolución de las tasas de brucelosis humana en la provincia de Al-mería, las características de los casos declarados en el periodo 2004-2011 y valorar la infl uencia de las actuaciones llevadas a cabo tras la priorización como problema de salud en la provincia, en el III Plan Andaluz de Salud.
Metodología: Estudio descriptivo de variables de persona, lugar y tiempo de los casos de brucelosis y tasas en la provincia de Almería en el período 2004-2011.
Resultados: Se notifi caron 150 casos de brucelosis en humanos. El año con mayor tasa correspondió al 2004 con 7,9 casos por 100000 habitantes y el de menor tasa el 2011 con 0,85. Un 74% fueron hombres. La media de edad fue de 44,8 años. Un 13,3% correspondió a población inmigrante. En el 54% de los casos el mecanismo de transmisión fue el contacto. Un 60,2% ejercía alguna profesión considerada de riesgo de contraer la enfermedad.
Conclusiones: En la provincia de Almería se ha producido un descenso de las tasas de brucelosis en los últimos años, coincidiendo con la puesta en marcha de programas de preven-ción y control. Se considera que la priorización de la brucelosis como un problema de Salud Pública en la provincia en el III Plan Andaluz de Salud ha contribuido a este descenso de tasas.
Palabras clave: Brucelosis, mecanismos de transmisión, programas de prevención y control, Plan Andaluz de Salud.
EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
EVOLUTION OF RATES OF BRUCELLOSIS IN THE PROVINCE OF ALMERIA, IN THE PERIOD 2004-2011
PILAR BARROSO GARCÍA1, 2
1 Servicio de Salud Pública de la Delegación Provincial de Salud de Almería. Email: [email protected]
2 Académico correspondiente de la Real Academia de Ciencias Veterinarias de Andalucía Oriental.

EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
146
SUMMARY
In the province of Almeria brucellosis has presented over the years high incidence rates in humans and high prevalence in sheep and goats. The aim of this study is to describe the evolution of rates of human brucellosis in the province of Almeria, characteristics of reported cases in the period 2004-2011 and assess the infl uence of the actions carried out after prioritizing health problem in the province, in the III Andalusian Health Plan.
Methodology: Descriptive study of variables of person, place and time of brucellosis cases and rates in the province of Almeria in the period 2004-2011.
Results: There were 150 cases of brucellosis in humans. The year 2004 corresponded to the highest rate with 7.9 cases per 100000 population and the lowest rate in 2011 with 0.85. A 74% were men. The mean age was 44.8 years. A 13.3% were immigrants. In 54% of cases the mechanism of transmission was through contact. 60.2% practiced a profession considered at risk of contracting the disease.
Conclusions: In the province of Almeria brucellosis rates has decreased in recent years, coinciding with the implementation of prevention and control programs. It is considered that the prioritization of brucellosis as a public health problem in the province in the III Andalusian Health Plan has contributed to this decline in rates.
Keywords: Brucellosis, mechanisms of transmission, prevention and control programs, Andalusian Health Plan.
INTRODUCCIÓN
La brucelosis continúa siendo un problema de Salud Pública en distintos países del Norte de África, Este de Europa, India y Latinoamérica (1,2). Para países del sudeste europeo y el mediterráneo su control se considera un reto prioritario (3-5). Algunos que ya estaban declarados libres de enfermedad, como Bulgaria que así lo estableció en la década de los 50, han vuelto a detectar casos, tanto en animales como en humanos (6). Este resurgir de la enfermedad ha sido debido a distintos motivos, como las guerras en la mitad Este de Europa que han reducido los programas de vigilancia y control, así como al transporte de animales no controlados a través de fronteras abiertas (1,3).
La OMS considera que la brucelosis tiene serias implicaciones para la salud, particularmente en segmentos menos favorecidos de la población y en regiones con insufi ciente atención sanitaria. También tiene muchas repercusiones en el ganado. Los programas de control y erradicación representan costes importantes, tanto en pruebas diagnósticas, como en la vacunación de ganado y en la compensación a los ganaderos por el sacrifi cio de animales (3).

EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
147
A nivel mundial se puede decir que existe un predominio de la infección humana por B. melitensis, y por ello la brucelosis tiene un fuerte vínculo con el ganado ovino y caprino, principal reservorio y fuente de infección (3). Aunque hay excepciones como en Argentina en el que se ha identifi cado mayor prevalencia de B. suis (2).
Diversos estudios han puesto de manifi esto la necesidad de organizar estrate-gias de intervención para el control de la enfermedad, desde medidas de educación sanitaria hasta vacunación de animales (4). Destacando la necesidad de fomentar actuaciones intersectoriales en las que intervengan a la vez médicos, veterinarios, servicios de Salud Pública, otros servicios gubernamentales y asociaciones no gu-bernamentales (3,7-10)
En España la incidencia de brucelosis en humanos ha presentado un descenso continuado desde 1984 (11), aunque en algunas comunidades autónomas como Cata-luña o Andalucía hay provincias en las que el número de casos es mayor que la media de su entorno (12,13). Algunos casos se declaran en forma de brotes epidémicos (14-16). Desde su incorporación a la Unión Europea en 1986 se ha realizado un notable esfuerzo para ordenar las políticas sanitarias tanto en materia de higienización de la leche y sus productos (17-19) como en la más específi ca de control y erradicación de la brucelosis animal (20-23).
En la provincia de Almería la brucelosis ha presentado tasas de incidencia ele-vadas en humanos y alta prevalencia en ganado ovino y caprino (24,25). Diversos estudios han descrito la importancia de la enfermedad en dicha provincia así como las características de la misma en un largo período de tiempo (24-28). Se ha ido produciendo un descenso de tasas en los últimos años, aunque se han mantenido superiores a la media de Andalucía y España. Algunos de los casos declarados se han relacionado en forma de brotes epidémicos (29).
Con el fi n de controlar la enfermedad se pusieron en marcha en la década de los 90 distintos programas específi cos, basados en la vigilancia epidemiológica y la educación sanitaria, así mismo, se intensifi caron las actuaciones en el ganado (25).
Aún así estas medidas no parecían lo sufi cientemente efi caces para controlar las tasas en humanos, de ahí que la morbilidad por brucelosis fuera uno de los problemas de salud priorizados en el III Plan Andaluz de Salud provincializado en Almería (30). En él se plantearon tres estrategias. Por un lado, la mejora de la Vigilancia Epide-miológica; por otro, establecer una coordinación intersectorial: salud, agricultura y entidades locales; y por último, campañas de educación sanitaria dirigidas a población general y profesionales de riesgo. Se haría un seguimiento exhaustivo de cada caso de

EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
148
enfermedad, para ello los distritos sanitarios comunicarían los casos de brucelosis en humanos con fuente de infección conocida a la Ofi cina Comarcal Agraria. Por parte de Agricultura se llevaría a cabo investigación de la fuente de infección. Así mismo, las Entidades Locales controlarían la venta ambulante. En el año 2005 se establecie-ron reuniones provinciales con profesionales representantes de todas las Ofi cinas Comarcales Agrarias, Distritos Sanitarios, Delegación de Salud y de Agricultura, con un seguimiento de casos notifi cados y actuaciones llevadas a cabo para controlar la fuente de infección y con presentación de resultados de las campañas de saneamiento llevadas a cabo por Agricultura.
El objetivo de este estudio es describir la evolución de las tasas de brucelosis en la provincia de Almería, las características de los casos en el periodo 2004-2011 y valorar la infl uencia de las actuaciones llevadas a cabo tras la priorización como problema de salud en la provincia en el III Plan Andaluz de Salud.
MATERIAL Y MÉTODOS
Se trata de un estudio descriptivo de los casos de enfermedad en la provincia de Almería, en el período 2004-2011, notifi cados al Sistema de Vigilancia Epidemiológica por los profesionales de centros de Atención Primaria y Hospitales. Además de los casos declarados, como fuentes de información complementaria se han utilizado, la notifi cación microbiológica y el Conjunto Mínimo Básico de Datos Hospitalario (CMBD) de los hospitales, estos datos se aportan de forma periódica o continua y se van incorporando al Sistema de Vigilancia una vez que se ha depurado la información, quedando todo ello registrado en el Sistema Integrado de Alertas.
Se han estudiado variables de persona, lugar y tiempo. Calculando frecuencias y porcentajes para las variables cualitativas y medias y rango para variables cuan-titativas. Se han calculado las tasas de la enfermedad para la provincia y distritos sanitarios, utilizando datos de los censos de Almería obtenidos a partir del INE. La provincia de Almería está dividida en tres distritos sanitarios: Levante Alto Alman-zora, Almería y Poniente de Almería.
RESULTADOS
En el periodo 2004-2011 se notifi caron 150 casos de enfermedad en humanos. El año con mayor tasa correspondió al 2004 con 7,9 casos por 100000 habitantes, superio-res a la de Andalucía (3,2) y España (1,5). Y el de menor tasa el 2011 con 0,85. Figura 1.
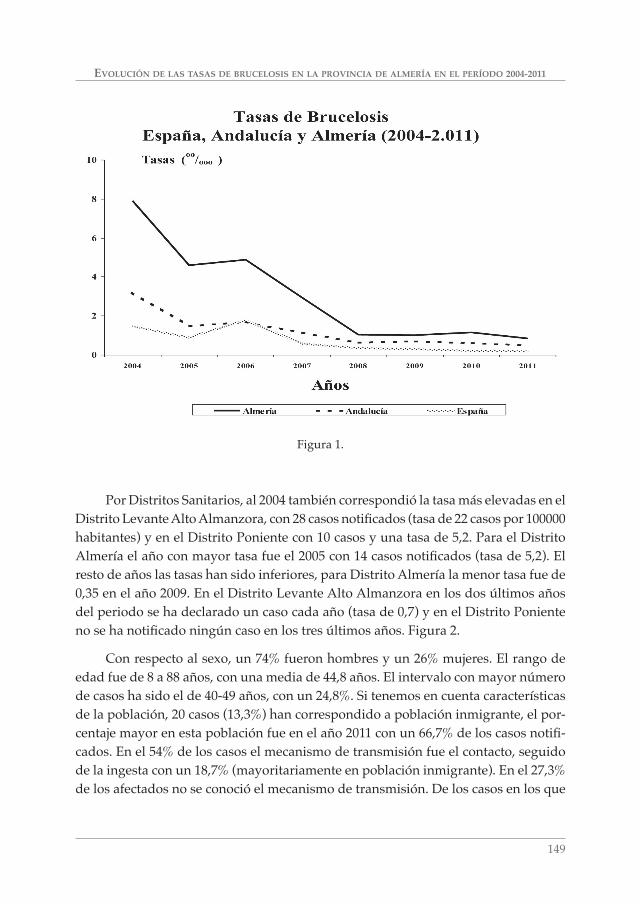
EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
149
Figura 1.
Por Distritos Sanitarios, al 2004 también correspondió la tasa más elevadas en el Distrito Levante Alto Almanzora, con 28 casos notifi cados (tasa de 22 casos por 100000 habitantes) y en el Distrito Poniente con 10 casos y una tasa de 5,2. Para el Distrito Almería el año con mayor tasa fue el 2005 con 14 casos notifi cados (tasa de 5,2). El resto de años las tasas han sido inferiores, para Distrito Almería la menor tasa fue de 0,35 en el año 2009. En el Distrito Levante Alto Almanzora en los dos últimos años del periodo se ha declarado un caso cada año (tasa de 0,7) y en el Distrito Poniente no se ha notifi cado ningún caso en los tres últimos años. Figura 2.
Con respecto al sexo, un 74% fueron hombres y un 26% mujeres. El rango de edad fue de 8 a 88 años, con una media de 44,8 años. El intervalo con mayor número de casos ha sido el de 40-49 años, con un 24,8%. Si tenemos en cuenta características de la población, 20 casos (13,3%) han correspondido a población inmigrante, el por-centaje mayor en esta población fue en el año 2011 con un 66,7% de los casos notifi -cados. En el 54% de los casos el mecanismo de transmisión fue el contacto, seguido de la ingesta con un 18,7% (mayoritariamente en población inmigrante). En el 27,3% de los afectados no se conoció el mecanismo de transmisión. De los casos en los que
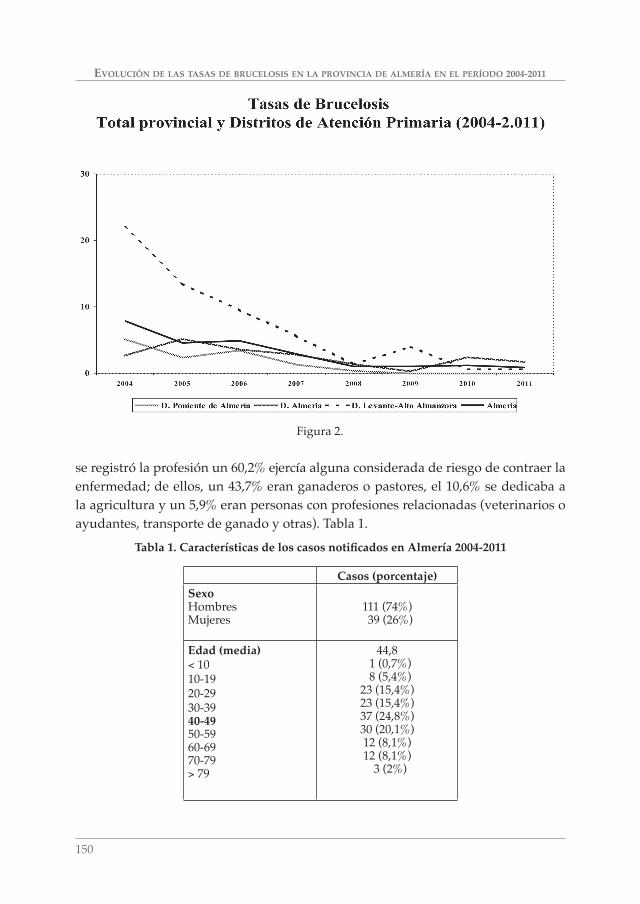
EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
150
se registró la profesión un 60,2% ejercía alguna considerada de riesgo de contraer la enfermedad; de ellos, un 43,7% eran ganaderos o pastores, el 10,6% se dedicaba a la agricultura y un 5,9% eran personas con profesiones relacionadas (veterinarios o ayudantes, transporte de ganado y otras). Tabla 1.
Tabla 1. Características de los casos notifi cados en Almería 2004-2011
Casos (porcentaje)SexoHombresMujeres
111 (74%) 39 (26%)
Edad (media)< 1010-1920-2930-3940-4950-5960-6970-79> 79
44,8 1 (0,7%) 8 (5,4%)23 (15,4%)23 (15,4%)37 (24,8%)30 (20,1%)12 (8,1%)12 (8,1%) 3 (2%)
Figura 2.
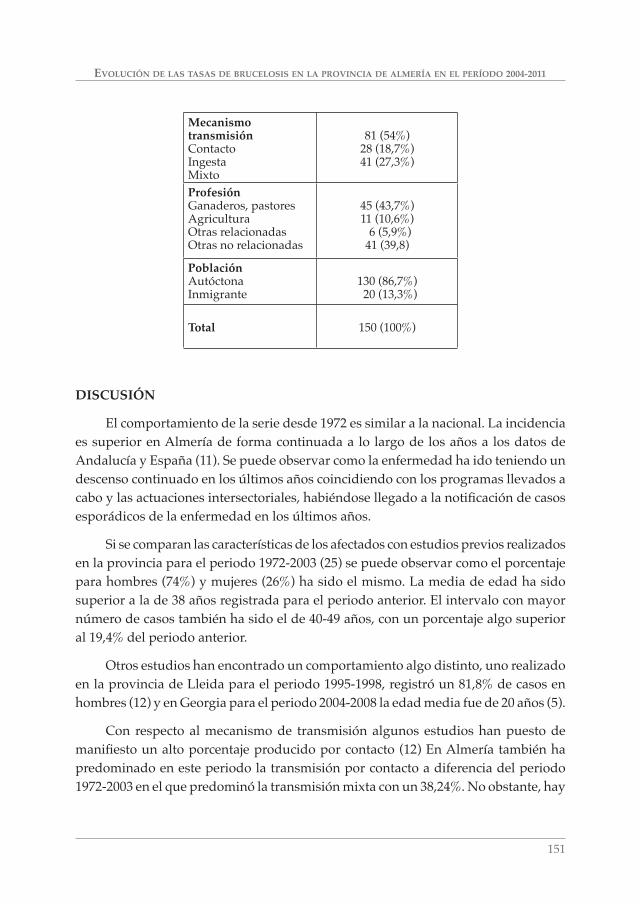
EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
151
Mecanismo transmisiónContactoIngestaMixto
81 (54%)28 (18,7%)41 (27,3%)
ProfesiónGanaderos, pastoresAgriculturaOtras relacionadasOtras no relacionadas
45 (43,7%)11 (10,6%) 6 (5,9%)41 (39,8)
PoblaciónAutóctonaInmigrante
130 (86,7%) 20 (13,3%)
Total 150 (100%)
DISCUSIÓN
El comportamiento de la serie desde 1972 es similar a la nacional. La incidencia es superior en Almería de forma continuada a lo largo de los años a los datos de Andalucía y España (11). Se puede observar como la enfermedad ha ido teniendo un descenso continuado en los últimos años coincidiendo con los programas llevados a cabo y las actuaciones intersectoriales, habiéndose llegado a la notifi cación de casos esporádicos de la enfermedad en los últimos años.
Si se comparan las características de los afectados con estudios previos realizados en la provincia para el periodo 1972-2003 (25) se puede observar como el porcentaje para hombres (74%) y mujeres (26%) ha sido el mismo. La media de edad ha sido superior a la de 38 años registrada para el periodo anterior. El intervalo con mayor número de casos también ha sido el de 40-49 años, con un porcentaje algo superior al 19,4% del periodo anterior.
Otros estudios han encontrado un comportamiento algo distinto, uno realizado en la provincia de Lleida para el periodo 1995-1998, registró un 81,8% de casos en hombres (12) y en Georgia para el periodo 2004-2008 la edad media fue de 20 años (5).
Con respecto al mecanismo de transmisión algunos estudios han puesto de manifi esto un alto porcentaje producido por contacto (12) En Almería también ha predominado en este periodo la transmisión por contacto a diferencia del periodo 1972-2003 en el que predominó la transmisión mixta con un 38,24%. No obstante, hay

EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
152
que tener en cuenta que ya se había detectado un cambio de tendencia a partir del año 1990 en la provincia de Almería con predominio de la transmisión por contacto (25,27).
El porcentaje de casos que ejercía profesión considerada de riesgo es similar al periodo anterior (61% en 1972-2003), aunque inferior al 71% comunicado en otros estudios (12). Sin embargo, en este periodo el mayor porcentaje corresponde a la pro-fesión de ganadero o pastor con un 43,7%, mientras que en el periodo anterior había sido la agricultura. Otros autores también han identifi cado más casos en personas que cuidan ganado (5).
En este estudio se ha identifi cado un porcentaje importante de casos en población inmigrante, siendo éste mayor en el año 2011. En este sentido hay que tener en cuenta que Almería en su condición de frontera Norte-Sur y por su reclamo laboral, acogió un incremento inmigratorio que comenzó a principios de 1980 y alcanzó sus niveles más altos en la primera década del siglo XXI. La mayoría de estos casos se ha debido a consumo de leche no higienizada. En otras zonas de España también se han decla-rado casos en población inmigrante. En la ciudad de Elche que presenta en general tasas inferiores a la media nacional, se notifi có un brote con nueve afectados, en dos familias de inmigrantes marroquíes, todos ellos habían tomado leche cruda de cabra no pasteurizada (31). Estos datos hacen ver la necesidad de llevar a cabo medidas de educación sanitaria en esta población para evitar casos de estas características.
Algunos autores ponen de manifi esto que no haya estudios que relacionen y vinculen datos de brucelosis humana y animal con el fi n de poder realizar una pre-dicción del riesgo humano (32). En Almería, sí se había realizado un estudio para el periodo 1990-1998, comparando datos de brucelosis humana y animal, encontrándose que tanto en la dispersión como en la prevalencia para ganado, los porcentajes más elevados correspondían al Distrito Poniente, que a su vez presentaba las menores tasas en humanos. Sin embargo, estos porcentajes eran inferiores para el Distrito Levante Alto Almanzora que presentaba las mayores tasas en humanos (24,25). Aunque con cifras inferiores este comportamiento fue similar en estudios posteriores. Según datos aportados por la Delegación de Agricultura en las reuniones de coordinación para el año 2006, se registró un 26,2% de dispersión y un 2,8% de prevalencia en Poniente; y un 6,3% de dispersión y 0,4% de prevalencia en Alto Almanzora. Sin embargo, si se tenía en cuenta la distribución del ganado, el distrito con mayor censo ganadero (52%) y explotaciones ganaderas (60%) correspondía al Levante Alto Almanzora que ha mantenido las tasas más elevadas en humanos (24,25).
Con respecto a la brucelosis en otros animales, un estudio realizado en España en el periodo 1999 y 2009, encuentra que la prevalencia mayor corresponde a jabalís

EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
153
salvajes oscilando entre 25 y 46%, sobre todo en el Sur y Centro de España, lo que puede suponer una amenaza para los cerdos domésticos, no habiéndose identifi cado como reservorio a los salvajes rumiantes (33).
En cuanto al control y erradicación de la enfermedad, en la última conferencia internacional sobre brucelosis llevada a cabo en Macedonia se insistió en la necesidad de que se desarrollaran programas con actuaciones bien planifi cadas y coordinadas, y que la FAO y la OMS desarrollara un proceso de evaluación (3).
En este sentido hay que decir que se están desarrollando actuaciones en distintos países, hay países como Mongolia donde se ha puesto de manifi esto los benefi cios de las intervenciones con vacunación en animales (34). En otros se ha puesto de manifi esto un mayor descenso de tasas cuando se combinan medidas de educación sanitaria y vacunación. En un estudio llevado a cabo en Grecia se registró una disminución de tasas tras la intervención con medidas de educación sanitaria (a 10,3 casos por 1000 habitantes), aunque el descenso fue aún mayor cuando se combinaron además progra-mas de vacunación en animales, llegando a descender a 0,3 casos por 1000 habitantes (4). En un estudio realizado anteriormente para el periodo 1972 a 1998 en la provincia de Almería (26), en el que se comparaban las tasas medias de brucelosis humana en tres períodos marcados por la puesta en marcha de programas de prevención, se obtuvo en el total provincial un descenso signifi cativo en la tasa media en los tres períodos estudiados. Los datos aportados en el trabajo actual indican un descenso importante del número de casos, habiéndose llegado a la notifi cación de casos esporádicos de la enfermedad en los últimos años.
Se considera que el hecho de haber priorizado la enfermedad como un problema de Salud Pública en el III Plan Andaluz de Salud (30) también ha infl uido de manera muy positiva. El III Plan Andaluz de Salud se publicó para el periodo 2005-2008. En su provincialización se priorizaron algunos problemas de Salud Pública. En la provincia de Almeria uno de ellos fue la brucelosis. Como ya se ha indicado, desde la década de los 90 se habian puesto en marcha distintos programas de prevención y control, no llegándose a conseguir una disminución de tasas a cifras cercanas a las que se notifi caban en el ámbito nacional o a la media andaluza, manteniendo la pro-vincia cifras superiores (35). Con las actuaciones planteadas en el plan se intensifi có el seguimiento de los casos de enfermedad, y se daba conocimiento además de todas las actuaciones contempladas en los programas de control y erradicación llevados a cabo en animales. Las reuniones periódicas mantenidas en la Delegación de Salud, con representación de profesionales de Salud y de Agricultura permitieron compartir información y llevar a cabo actuaciones de manera rápida y efi caz.

EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
154
Como conclusión se puede decir que en la provincia de Almería se ha producido un descenso de las tasas de brucelosis en los últimos años, coincidiendo con la puesta en marcha de programas de prevención y control. Se considera que la priorización de la brucelosis como un problema de Salud Pública en la provincia, en el III Plan Andaluz de Salud ha contribuido a este descenso de tasas.
BIBLIOGRAFÍA
1. Gwida M, Al Dahouk S, Melzer F , Rösler U, Neubauer H, Tomaso H. Brucellosis – Regionally Emerging Zoonotic Disease?. Croat Med J 2010; 51: 289–95.
2. Lucero NE, Ayala SM, Escobar GI, Jacob NR. Brucella isolated in humans and animals in Latin America from 1968 to 2006. Epidemiol Infect 2008; 136: 496–503.
3. Donev DM. Brucellosis as Priority Public Health Challenge in South Eastern European Coun-tries. Croat Med J 2010; 51: 283–4.
4. Jelastopulu E, Bikas C, Petropoulos C, Leotsinidis M. Incidence of human brucellosis in a rural area in Western Greece after the implementation of a vaccination programme against animal brucellosis. BMC Public Health 2008; 8: 241.
5. Akhvlediani T, Clark DV, Chubabria G, Zenaishvili O, Hepburn MJ. The changing pattern of human brucellosis: clinical manifestations, epidemiology, and treatment outcomes over three decades in Georgia . BMC Infect Dis 2010; 10: 346.
6. Russo G, Pasquali P, Nenova R, Alexandrov T, Ralchev S, Vullo V et al. Reemergence of Human and Animal Brucellosis, Bulgaria. Emerg Infect Dis 2009; 15: 314–16.
7. Mainar Jaime RC, Vazquez Boland JA. Associations of veterinary services and farmer character-istics with the prevalences of brucellosis and border disease in small ruminants in Spain. Prev Vet Med 1999; 40 (3-4): 193-205.
8. Economides P. Control of zoonosis in Cyprus. Rev Sci Tech 2000; 19 (3): 725-734.9. Benkirane A. Surveillance epidemiologique et prophylaxie de la brucellose des ruminants:
l´exemple de la region Afrique du Nord et Proche-Orient. Rev Sci Tech 2001; 20 (3): 757-767. 10. Lithg Pereira PL, Mainar Jaime RC, Alvarez Sanchez MA, Rojo Vazquez FA. Evaluation of of-
fi cial eradication campaigns data for investigating small ruminant brucellosis in the province of Leon, Spain. Prev Vet Med 2001; 51 (3-4): 215-225.
11. Sánchez Serrano LP, Ordóñez Banegas P, Díaz García MO, Torres Frías A. Human and animal incidence of brucellosis declining in Spain. Eurosurveillance Weekly 2005; 10.
12. Serra Alvarez J, Godoy García P. Incidencia, etiología y epidemiología de la brucelosis en un área rural de la provincia de Lleida. Rev Esp Salud Pública 2000; 74: 45-53.
13. García León J, Bernal M ,Niebla P, Delgado E, Gallardo V, Carmona JC. Cambio del patrón epi-demiológico de la brucelosis en Andalucía. Gac Sanit. 1999; 13: 9165.
14. Rodríguez Valín ME et al. La brucelosis como enfermedad profesional: estudio de un brote de transmisión aérea en un matadero. Rev. Esp. Salud Publica 2001;75:159-170.
15. Méndez Martínez C, Páez Jiménez A, Cortés Blanco M, Salmoral Chamizo E, Mohedano Mohe-dano E, Plata C et al. Brucellosis outbreak due to un pasteurized raw goat cheese in Andalucia (Spain), January-March 2002: Euro Surveill 2003; 8: 164-168.
16. Castell Monsalve J, Gutiérrez Ávila G, Ruiz Valdepeñas MA. Tres brotes de brucelosis inves-tigados en un año de vigilancia de salud laboral en Ciudad Real. Gac Sanit 2009; 23: 562-563.
17. Diario Ofi cial de las Comunidades Europeas. Directiva 92/46/CEE relativa a la comercializa-ción de leche y productos lácteos. DOCE núm. L 268 de 14/09/1992.

EVOLUCIÓN DE LAS TASAS DE BRUCELOSIS EN LA PROVINCIA DE ALMERÍA EN EL PERÍODO 2004-2011
155
18. Boletín Ofi cial del Estado. Real Decreto 1679 /1994 por el que se establecen las condiciones sani-tarias aplicables a la producción y comercialización de leche cruda, leche tratada térmicamente y productos lácteos. BOE núm. 229, 24/09/1994.
19. Boletín Ofi cial del Estado. Real Decreto 402/1996 por el que se modifi ca el Real Decreto 1679/1994. BOE núm. 85, 08/04/1996.
20. Boletín Ofi cial del Estado. Real Decreto 2611/1996 por el que se regulan los programas naciona-les de erradicación de enfermedades de los animales. BOE núm.307, 21/12/1996.
21. Boletín Ofi cial del Estado. Real Decreto 1047/2003, por el que se modifi ca el Real Decreto 2611/1996, de 20 de diciembre, por el que se regulan los programas nacionales de erradicación de enfermedades de los animales. BOE núm.216, 09/09/2003.
22. Boletín Ofi cial del Estado. Real Decreto 51/2004, por el que se modifi ca el Real Decreto 2611/1996, de 20 de diciembre, por el que se regulan los programas nacionales de erradicación de enfermedades de los animales. BOE núm.17, 20/01/2004.
23. Boletín Ofi cial de la Junta de Andalucía. Orden de 29 de noviembre de 2004, por la que se desa-rrollan las normas de ejecución de los programas nacionales de vigilancia, prevención, control y erradicación de las enfermedades de los animales en Andalucía. BOJA núm. 241, 13/12/2004.
24. Barroso García P, Rodríguez-Contreras Pelayo R, Parrón Carreño T, Luna del Castillo JD. Es-tudio comparativo de brucelosis humana y en ganado en la provincia de Almería 1990-1998. Centro de Salud 2002; 10: 396-400.
25. Barroso García P. La brucelosis en la provincia de Almería, aspectos históricos y epidemiológi-cos. Anales de la Real Academia de Ciencias Veterinarias de Andalucía Oriental 2004; 17: 13-26.
26. Barroso García P, Rodríguez-Contreras Pelayo R, Parrón Carreño T, Ocaña Riola R. Evolución de las tasas de brucelosis en la provincia de Almería durante el período 1972-1998. Medicina de Familia (And) 2001; 2: 50-53.
27. Barroso García P, Parrón Carreño T, Rodríguez-Contreras Pelayo R. Estudio descriptivo de la brucelosis en la provincia de Almería. Evolución de los mecanismos de transmisión. Medicina de Familia (And) 2001; 2: 46-52.
28. Barroso García P, Rodríguez-Contreras Pelayo R, Gil Extremera B, Maldonado Martín A, Gui-jarro Huertas G, Martín Salguero A, Parrón Carreño T. Estudio de 1595 casos de brucelosis en la provincia de Almería (1972-1998) según datos epidemiológicos de declaración de la enferme-dad. Rev Clín Esp 2002; 202(11): 577-82.
29. Barroso García P. Brotes de brucelosis en el Distrito Sanitario Levante Alto Almanzora en el periodo 2003-2006. Dirección General de Salud Pública y Participación. Consejería de Salud de la Junta de Andalucía. Sistema de Vigilancia Epidemiológica de Andalucía (SVEA) 2007; 12.
30. Almería. Priorización e implantación del III Plan Andaluz de Salud. 2005-2008. Consejería de Salud de la Junta de Andalucia, 2005.
31. Ramos JM, Bernal E, Esguevillas T, Lopez-García P, Gaztambide MS, Gutiérrez F. Non-imported brucellosis outbreak from unpasteurized raw milk in Moroccan immigrants in Spain. Epide-miol Infect 2008; 136: 1552–55.
32. Scotch M, Odofi n L, Rabinowitz P. Linkages between animal and human health sentinel data. BMC Vet Res 2009; 5: 15.
33. Muñoz PM, Boadella M, Arnal MC, de Miguel MJ, Revilla M, Martínez D et al. Spatial distribu-tion and risk factors of Brucellosis in Iberian wild ungulates. BMC Infect Dis 2010; 10: 46.
34. Zinsstag J, Schelling E, Roth F, Bonfoh B, de Savigny D, Tanner M. Human Benefi ts of Animal Interventions for Zoonosis Control. Emerg Infect Dis 2007; 13: 527–31.
35. Barroso García P, de Dios Luna del Castillo J, Rodríguez-Contreras Pelayo R, Parrón Carreño T, Durán Ferrer M. Brucelosis en Almería: su evolución en el período 1972-2002, en relación con intervenciones intersectoriales y nivel socioeconómico. Investig Clin 2007; 10: 96-106.


BASES DE LA RESPUESTA INFLAMATORIA EN LA FORMA RESPIRATORIA DEL PRRS
157ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
RESUMEN
El Síndrome Reproductivo y Respiratorio Porcino (PRRS) es una enfermedad de distri-bución mundial que causa graves pérdidas económicas al sector porcino. Este virus no sólo es importante como agente causal del PRRS sino también por su participación en el desarrollo del Complejo Respiratorio Porcino. Su interacción con las defensas pulmonares, la alteración de la respuesta inmune y su persistencia en los órganos linfoides conlleva a que los cerdos tengan difi cultades para luchar contra la enfermedad.
Palabras clave: PRRS, macrófago, citoquinas, proteinas de fase aguda
ABSTRACT
Porcine Reproductive and Respiratory Syndrome (PRRS) is considered as the most eco-nomically important disease of the modern swine industry. The importance of this virus lies in not only being the causative agent of PRRSV, but also due to its implication in the onset of the Porcine Respiratory Disease Complex. The interaction of the virus with pulmonary defenses, the impairment of the immune response as well as the viral persistence in lymphoid organs make overcoming the disease diffi cult to infected pigs.
Key words: PRRS, macrophage, cytokines, acute phase proteins
BASES DE LA RESPUESTA INFLAMATORIA EN LA FORMA RESPIRATORIA DEL PRRS
GÓMEZ-LAGUNA J.1, RODRÍGUEZ-GÓMEZ I. M.2, BARRANCO I.2, QUEREDA J. J.3, GARCÍA-NICOLÁS O.4, RAMIS G.5, PALLARÉS F. J.4, CARRASCO L.2
1 Dpto. de I+D+i, CICAP, Pozoblanco, Córdoba, España.2 Dpto. de Anatomía y Anatomía Patológica Comparadas, Facultad de Veterinaria, Universidad de
Córdoba, Córdoba, España.3 Dpto. de Biotecnología Microbiana, Centro Nacional de Biotecnología, Consejo Superior de Inves-
tigaciones Científi cas (CSIC), Campus de Cantoblanco, Madrid, España4 Dpto. de Anatomía y Anatomía Patológica Comparadas.5 Dpto. de Producción Animal, Facultad de Veterinaria, Universidad de Murcia, Murcia, España;
e-mail: [email protected]

BASES DE LA RESPUESTA INFLAMATORIA EN LA FORMA RESPIRATORIA DEL PRRS
158
INTRODUCCIÓN
El Síndrome Reproductivo y Respiratorio Porcino (PRRS; del inglés, Porcine Reproductive and Respiratory Syndrome) es una enfermedad causada por un arterivirus conocido por el mismo nombre del síndrome (virus del PRRS, PRRSV), distinguién-dose actualmente dos genotipos del PRRSV, el europeo o tipo I y el americano o tipo II, que presentan importantes diferencias antigénicas y patogénicas entre sí. La forma respiratoria de la enfermedad afecta principalmente a cerdos en transición y cebo, causando una neumonía de tipo intersticial que induce tanto difi cultad respiratoria (distrés respiratorio) como mayor susceptibilidad a otros patógenos.
La mayor susceptibilidad que tienen los cerdos que desarrollan la forma respi-ratoria del PRRS a sufrir infecciones secundarias por otros patógenos, es lo que ha motivado que esta forma de la enfermedad haya cobrado una gran importancia, al ser uno de los principales patógenos implicados en el desarrollo del complejo respirato-rio porcino (CRP). El CRP se considera como una enfermedad multifactorial en cuya patogenia tienen un papel destacado distintos factores: i) la ausencia en las explota-ciones de medidas de manejo adecuadas; ii) la ausencia o descuido en las medidas de bioseguridad; y, iii) la presencia de patógenos del tracto respiratorio (Tabla 1). Los principales agentes infecciosos implicados en el desarrollo del CRP son tanto víricos, como el circovirus porcino tipo 2 (PCV-2) o el PRRS, como bacterianos, como Myco-plasma hyopneumoniae, los cuales van a estar implicados en una mayor susceptibilidad de los animales a infecciones respiratorias por patógenos secundarios, dando lugar a un cuadro respiratorio grave en las naves de transición y cebo, con el consecuente aumento del número de bajas y la merma en los parámetros productivos. Además, la concomitancia de distintos patógenos (primarios y secundarios) agrava el cuadro respiratorio presente en las explotaciones, así, la neumonía intersticial que aparece durante el PRRS se hace más intensa y duradera cuando existe una coinfección con Mycoplasma hyopneumoniae. Por otro lado, diversos estudios han demostrado que los cerdos infectados por el PRRSV presentan una mayor susceptibilidad a infecciones secundarias por otras bacterias como Streptococcus suis, Haemophilus parasuis, Myco-plasma hyopneumoniae, Actinobacillus pleuropneumoniae, y Salmonella spp., y otros virus, como el de la infl uenza porcina (Tabla 2).
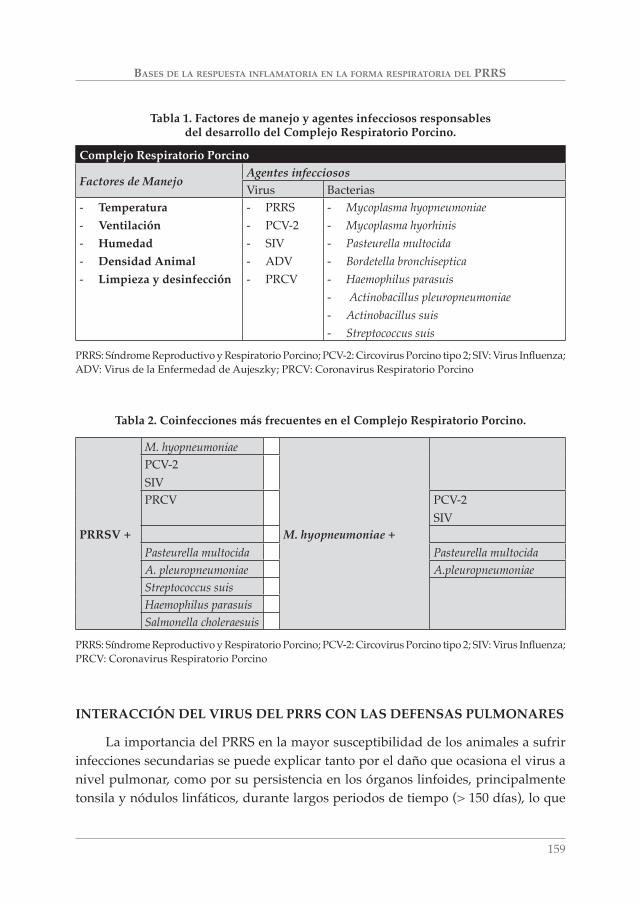
BASES DE LA RESPUESTA INFLAMATORIA EN LA FORMA RESPIRATORIA DEL PRRS
159
Tabla 1. Factores de manejo y agentes infecciosos responsables del desarrollo del Complejo Respiratorio Porcino.
Complejo Respiratorio Porcino
Factores de ManejoAgentes infecciososVirus Bacterias
- Temperatura - Ventilación - Humedad - Densidad Animal - Limpieza y desinfección
- PRRS - PCV-2 - SIV - ADV - PRCV
- Mycoplasma hyopneumoniae - Mycoplasma hyorhinis - Pasteurella multocida - Bordetella bronchiseptica - Haemophilus parasuis - Actinobacillus pleuropneumoniae - Actinobacillus suis - Streptococcus suis
PRRS: Síndrome Reproductivo y Respiratorio Porcino; PCV-2: Circovirus Porcino tipo 2; SIV: Virus Infl uenza; ADV: Virus de la Enfermedad de Aujeszky; PRCV: Coronavirus Respiratorio Porcino
Tabla 2. Coinfecciones más frecuentes en el Complejo Respiratorio Porcino.
PRRSV +
M. hyopneumoniae
M. hyopneumoniae +
PCV-2SIVPRCV PCV-2
SIV
Pasteurella multocida Pasteurella multocidaA. pleuropneumoniae A.pleuropneumoniaeStreptococcus suisHaemophilus parasuisSalmonella choleraesuis
PRRS: Síndrome Reproductivo y Respiratorio Porcino; PCV-2: Circovirus Porcino tipo 2; SIV: Virus Infl uenza; PRCV: Coronavirus Respiratorio Porcino
INTERACCIÓN DEL VIRUS DEL PRRS CON LAS DEFENSAS PULMONARES
La importancia del PRRS en la mayor susceptibilidad de los animales a sufrir infecciones secundarias se puede explicar tanto por el daño que ocasiona el virus a nivel pulmonar, como por su persistencia en los órganos linfoides, principalmente tonsila y nódulos linfáticos, durante largos periodos de tiempo (> 150 días), lo que

BASES DE LA RESPUESTA INFLAMATORIA EN LA FORMA RESPIRATORIA DEL PRRS
160
está íntimamente relacionado tanto con la interacción de este virus con las barreras de defensa del aparato respiratorio, como con el fallo en la instauración de una respuesta inmune adecuada por parte del organismo frente a este virus.
A continuación vamos a describir brevemente la interacción del PRRSV con las principales barreras de defensa del aparato respiratorio:
- La primera línea de defensa está constituida por el epitelio de las vías aéreas, un epitelio pseudoestratifi cado cilíndrico ciliado. Este epitelio se caracteriza por la presencia de células caliciformes y de células ciliadas, lo que le permite capturar las partículas vehiculadas por el aire (como son los patógenos respiratorios) así como, la eliminación de las mismas mediante el movimiento de los cilios. Los animales infectados por el PRRSV sufren la destrucción del sistema mucociliar, lo que impide que se pueda producir tanto la captura como la eliminación de aquellas partículas y/o microorganismos vehiculados por el aire, lo que facilita la progresión de otros patógenos tanto primarios como secundarios.
- La siguiente línea de defensa está representada por el macrófago alveolar porcino (MAP), el cual se encarga, principalmente, de fagocitar y eliminar todas aquellas sustancias extrañas al organismo y que, vehiculadas por el aire inspirado, son capaces de llegar hasta el alveolo. Este macrófago es considerado como la principal célula diana de replicación del PRRSV, el cual al replicarse en los mismos provoca un efecto sobre varias de sus funciones, entre las que se encuentra la fagocitosis, que es la herramienta que tiene este macrófago para capturar y eliminar a los patógenos res-piratorios. Además, este virus puede inducir la muerte de estas células, lo que se traduce en una disminución, no sólo de sus funciones sino también de su número y, por tanto, de este mecanismo de defensa, lo que facilita la colonización del alveolo por aquellos patógenos que llegan a alcanzarlo.
- En tercer lugar, y no sólo a nivel pulmonar, sino en la mayoría de los órganos que componen el sistema inmune, el PRRSV es capaz de inducir la apoptosis (o muerte celular programada), tanto de linfocitos como de macrófagos, impidiendo de esta forma el desarrollo de una respuesta inmune efi caz.

BASES DE LA RESPUESTA INFLAMATORIA EN LA FORMA RESPIRATORIA DEL PRRS
161
INTERACCIÓN DEL PRRSV CON LA RESPUESTA DE FASE AGUDA
La respuesta de fase aguda (RFA) surge como consecuencia de la alteración de la homeostasis normal por diversos estímulos como pueden ser la infección, el desarrollo de un proceso infl amatorio, una situación de estrés, un traumatismo o cu-alquier tipo de daño tisular. Esta RFA se desencadena por la síntesis y liberación de citoquinas proinfl amatorias, como la interleuquina 1 (IL-1), la interleuquina 6 (IL-6) y el factor de necrosis tumoral alfa (TNF-α), en el lugar de la lesión. Estas citoquinas, que se liberan al torrente sanguíneo, llegan al hígado e inducen la producción de las proteínas de fase aguda (PFA) por los hepatocitos. Las PFA se clasifi can como “positivas” o “negativas” en función del aumento o disminución de su concentración sérica, respectivamente. La haptoglobina (Hp), proteína C-reactiva (CRP), amiloide A sérica (AAS), y la “Pig-Major acute phase protein” (Pig-MAP) se consideran como las principales PFA en el cerdo. En general se acepta que las PFA son inductores de una reacción proinfl amatoria y de la fi ebre, pero su sobreexpresión puede llevar también a una respuesta anti-infl amatoria. Por lo tanto, las PFA son utilizadas en la actualidad como posibles marcadores biológicos en el control del bienestar animal y del estatus sanitario, tanto a nivel de explotaciones como a nivel individual. Por otra parte, las PFA también se están utilizando para determinar la virulencia de diferentes aislados de bacterias o virus, o la efi cacia de las vacunas.
El PRRSV se caracteriza, a diferencia de la mayoría de los patógenos, por inducir una pobre producción de citoquinas proinfl amatorias a lo largo de la infección, lo que sería una de las razones por las que no se produce ni una sintomatología respiratoria evidente ni alteraciones sistémicas como la aparición de un cuadro febril, por lo que en los animales inoculados experimentalmente con el PRRSV contrasta la ausencia de un incremento de la temperatura rectal con el desarrollo de una intensa neumonía intersticial.
Estudios previos sobre la patogenia de otras enfermedades respiratorias han sugerido que, durante las infecciones pulmonares, la respuesta infl amatoria puede activarse de manera efi ciente a nivel local en el pulmón, mientras que la concentración sérica de las citoquinas proinfl amatorias puede no presentar cambios signifi cativos. En este sentido, nuestro grupo de investigación ha demostrado un aumento signifi cati-vo en la expresión in situ de IL-1α, IL-6 y TNF-α en el pulmón de cerdos infectados con un aislado europeo del PRRSV, existiendo una correlación signifi cativa entre la expresión tisular de estas citoquinas y el desarrollo de la neumonía intersticial típica de esta enfermedad. Por otra parte, se sabe que la infección por el PRRSV induce una modulación de la respuesta inmune evitando el establecimiento de una respuesta

BASES DE LA RESPUESTA INFLAMATORIA EN LA FORMA RESPIRATORIA DEL PRRS
162
inmune efi ciente en el animal infectado. Por lo tanto, la falta de cambios signifi cativos en la concentración sérica de las citoquinas proinfl amatorias puede apuntar a una estrategia del virus para escapar de la respuesta inmune del huésped, previniendo así la activación de los hepatocitos para inducir una RFA efi ciente.
La Hp es una PFA que modula la respuesta inmune por medio de interacciones complejas con diferentes mediadores. Estudios previos han demostrado un aumento en la concentración sérica de Hp coincidiendo con el aumento en la expresión de IL-6 y TNF-α, de forma que estas citoquinas podrían inducir una leve RFA a partir de la cual se indujera la síntesis de Hp. No obstante, también se ha observado un aumento de Hp relacionado con un aumento de la citoquina anti-infl amatoria IL-10, lo que se podría originar a través de la interacción con el receptor CD163. El receptor CD163 actúa eliminando los complejos hemoglobina-Hp en circulación, de manera que disminuye la hemoglobina disponible para otros patógenos. De este modo, se ha descrito igualmente como la interacción de la Hp con el receptor CD163 da lugar a la liberación de mediadores anti-infl amatorios como la IL-10. Estos resultados apuntan a que la Hp jugaría un papel importante en la patogenia de la enfermedad, estando involucrada esta PFA en la modulación de la respuesta inmune, probablemente a través de la inducción de ciertas citoquinas.
Recientemente se ha llevado a cabo una clasifi cación de los aislados del PRRSV en función de si son inductores de TNF-α, de IL-10 o de ninguna de estas citoquinas. El TNF-α juega un importante papel en la respuesta infl amatoria, ya que esta citoquina puede actuar como una citoquina antiviral, protegiendo a las células de la infección por determinados virus, o eliminando de forma selectiva a las células infectadas por virus de una forma independiente al interferón. Así, se ha descrito que la adición de TNF-α recombinante porcino es capaz de inhibir la replicación in vitro del PRRSV, observándose igualmente una reducción en la expresión de esta citoquina tras la infección de macrófagos alveolares porcinos con el PRRSV. La pobre expresión de TNF-α observada en las infecciones experimentales con algunas cepas del PRRSV, puede apuntar a un mecanismo de evasión del virus para evitar la respuesta inmune del huésped, evitando así la inducción de una eliminación efi ciente del PRRSV.
Por su parte, la IL-10 es una citoquina conocida como inmunomoduladora, ya que es capaz de desarrollar, entre otras, acciones anti-infl amatorias al impedir una síntesis adecuada de citoquinas proinfl amatorias. En este sentido, se ha propuesto que esta citoquina puede desempeñar un papel importante en la patogenia de la enfer-medad al inhibir la síntesis de interferón gamma (IFN-γ), una de cuyas funciones es la de impedir la replicación vírica. De este modo, al igual que la IL-10 puede impedir

BASES DE LA RESPUESTA INFLAMATORIA EN LA FORMA RESPIRATORIA DEL PRRS
163
la producción de IFN-γ, esta citoquina, la IL-10, podría estar también impidiendo el desarrollo de una respuesta proinfl amatoria más marcada en los animales infecta-dos por el PRRSV, lo que se traduciría en una respuesta sistémica a la enfermedad, al producirse un aumento de la concentración de las citoquinas proinfl amatorias en circulación.
La menor expresión de citoquinas proinfl amatorias en la infección por el PRRSV, con respecto a otras infecciones víricas, también podría estar relacionada con la ex-presión de niveles más bajos de PFA, ya que las citoquinas proinfl amatorias actúan como inductoras de la producción de PFA por los hepatocitos. En un estudio llevado a cabo por nuestro grupo de investigación, se observó una expresión temprana de Hp y Pig-MAP, mientras que la concentración sérica de CRP y AAS mostró un aumento más tardío. La CRP y la AAS son PFA que desarrollan varias funciones biológicas relacionadas con la respuesta inmune. La CRP participa en la activación del com-plemento y opsonización, e induce la producción de citoquinas por los macrófagos, mientras que la AAS es quimiotáctica para monocitos, linfocitos T y leucocitos po-limorfonucleares. Por lo tanto, el aumento tardío observado en ambas PFA puede contribuir al establecimiento de una respuesta inmune inefi ciente por parte de los animales infectados.
Recientemente nuestro grupo de investigación ha demostrado que tanto la saliva como el jugo de carne representan unas muestras biológicas muy útiles para la medición y monitorización de las PFA a lo largo de la infección con el PRRSV, pre-sentando importantes ventajas respecto al suero, al permitir una toma de muestras menos invasiva y de fácil acceso en el caso de la saliva, disminuyendo tanto el estrés de los animales, como del tiempo empleado para el muestreo de los mismos, ya que en los casos de jugo de carne, la toma de muestra en la cadena de sacrifi cio es mucho más rápida y cómoda que la toma de sangre.
CONCLUSIÓN
A modo de conclusión nos gustaría destacar que el PRRSV es capaz de desa-rrollar diversos mecanismos que conllevan una alteración de la respuesta inmune a nivel local, como es la destrucción del sistema mucociliar y la disminución de las funciones y del número de los macrófagos alveolares, y la inducción de apoptosis de células inmunes competentes (como linfocitos y macrófagos). Todos estos mecanis-mos impiden que el animal que está infectado por el PRRSV pueda establecer una respuesta inmune efi caz, lo que junto a la disminución en la actividad bactericida de

BASES DE LA RESPUESTA INFLAMATORIA EN LA FORMA RESPIRATORIA DEL PRRS
164
los macrófagos podría explicar la persistencia de este virus en el animal, así como, el aumento en la prevalencia de infecciones secundarias concomitantes, las cuales se observan con frecuencia en el CRP.
Por otro lado, los estudios realizados sobre el papel de las PFA en el PRRS seña-lan que la Hp y la Pig-MAP serían las más útiles para la monitorización de las fases tempranas de la infección por este virus. Además, el PRRSV sería capaz de modular la respuesta inmune al inducir una mayor expresión de Hp en las fases tempranas, y a una expresión tardía y errática de TNF-α, CRP y AAS, lo que haría viable el esta-blecimiento de una viremia prolongada, al evitar una eliminación efi caz de este virus.
AGRADECIMIENTOS
Este trabajo ha sido fi nanciado por el Ministerio de Educación y Ciencia, número de proyectos AGL2006-04146/GAN y AGL2009-12438/GAN.
BIBLIOGRAFIA RECOMENDADA
Asai T, Mori M, Okada M, Uruno K, Yazawa S, Shibata I. Elevated serum haptoglobin in pigs in-fected with porcine reproductive and respiratory syndrome virus. Vet Immunol Immunopathol 1999; 70:143–148.
Barranco I, Gómez-Laguna J, Rodríguez-Gómez IM, Salguero FJ, Pallarés FJ, Carrasco L. Differen-tial Expression of Proinfl ammatory Cytokines in the Lymphoid Organs of Porcine Reproduc-tive and Respiratory Syndrome Virus-Infected Pigs. Transbound Emerg Dis 2011 Aug 16. doi: 10.1111/j.1865-1682.2011.01252.x.
Barranco I, Gómez-Laguna J, Rodríguez-Gómez IM, Salguero FJ, Pallarés FJ, Bernabé A, Carrasco L. Immunohistochemical detection of extrinsic and intrinsic mediators of apoptosis in porcine paraffi n-embedded tissues. Vet Immunol Immunopathol 2011; 139 (2-4):210-216.
Darwich L, Díaz I, Mateu E. Certainties, doubts and hypotheses in porcine reproductive and respi-ratory syndrome virus immunobiology. Virus Res 2010; 154(1-2):123-132.
Díaz I, Darwich L, Pappaterra G, Pujols J, Mateu E. Immune responses of pigs after experimental infection with a European strain of porcine reproductive and respiratory syndrome virus. J Gen Virol 2005; 86(Pt 7): 1943-1951.
Gimeno M, Darwich L, Diaz I, de la Torre E, Pujols J, Martín M, Inumaru S, Cano E, Domingo M, Montoya M, Mateu E. Cytokine profi les and phenotype regulation of antigen presenting cells by genotype-I porcine reproductive and respiratory syndrome virus isolates. Vet Res 2011; 42(1): 9.
Gómez-Laguna J, Rodríguez-Gómez IM, Barranco I, Pallarés FJ, Salguero FJ, Carrasco L. Enhanced expression of TGF protein in lymphoid organs and lung, but not in serum, of pigs infected with a European fi eld isolate of porcine reproductive and respiratory syndrome virus. Vet Microbiol 2012; 158 (1-2):187-193.
Gómez-Laguna J, Salguero FJ, Pallarés FJ, Rodríguez-Gómez IM, Barranco I, Carrasco L. Acute phase proteins as biomarkers in animal health and welfare. Book chapter of Acute phase pro-

BASES DE LA RESPUESTA INFLAMATORIA EN LA FORMA RESPIRATORIA DEL PRRS
165
teins as early non-specifi c biomarkers of human and veterinary diseases. Intech. April, 2011. Chapter 12, 259-298. ISBN 978-953-307-873-1.
Gómez-Laguna J, Gutiérrez A, Pallarés FJ, Salguero FJ, Cerón JJ, Carrasco L. Haptoglobin and C-re-active protein as biomarkers in the serum, saliva and meat juice of pigs experimentally infected with porcine reproductive and respiratory syndrome virus. Vet J 2010; 185(1):83-87.
Gómez-Laguna J, Salguero FJ, Barranco I, Pallarés FJ, Rodríguez-Gómez IM, Bernabé A, Carrasco L. Cytokine expression by macrophages in the lung of pigs infected with the porcine reproductive and respiratory syndrome virus. J Comp Pathol 2010; 142(1):51-60.
Gómez-Laguna J, Salguero FJ, Pallarés FJ, Fernández de Marco M, Barranco I, Cerón JJ, Martínez-Subiela S, Van Reeth K, Carrasco L. Acute phase response in porcine reproductive and respira-tory syndrome virus infection. Comp Immunol Microbiol Infect Dis 2010; 33(6):e51-e58.
Gómez-Laguna J, Salguero FJ, De Marco MF, Pallarés FJ, Bernabé A, Carrasco L. Changes in lym-phocyte subsets and cytokines during European porcine reproductive and respiratory syn-drome: increased expression of IL-12 and IL-10 and proliferation of CD4(-)CD8(high). Viral Immunol 2009, 22(4):261-271.
Gutiérrez AM, Martínez-Subiela S, Soler L, Pallarés FJ, Cerón JJ. Use of saliva for haptoglobin and C-reactive protein quantifi cations in porcine respiratory and reproductive syndrome affected pigs in fi eld conditions. Vet Immunol Immunopathol 2009; 132(2-4):218-23.
Martínez-Lobo FJ, Díez-Fuertes F, Segalés J, García-Artiga C, Simarro I, Castro JM, Prieto C. Com-parative pathogenicity of type 1 and type 2 isolates of porcine reproductive and respirato-ry syndrome virus (PRRSV) in a young pig infection model. Vet Microbiol 2011; 154(1-2):58-68.
Parra MD, Fuentes P, Tecles F, Martínez-Subiela S, Martínez JS, Muñoz A, Cerón JJ. Porcine acute phase protein concentrations in different diseases in fi eld conditions. J Vet Med B Infect Dis Vet Public Health 2006; 53(10):488-93.
Qiao S, Feng L, Bao D, Guo J, Wan B, Xiao Z, Yang S, Zhang G. Porcine reproductive and respira-tory syndrome virus and bacterial endotoxin act in synergy to amplify the infl ammatory re-sponse of infected macrophages. Vet Microbiol 2011; 149(1-2):213-20.
Renukaradhya GJ, Alekseev K, Jung K, Fang Y, Saif LJ. Porcine reproductive and respiratory syn-drome virus-induced immunosuppression exacerbates the infl ammatory response to por-cine respiratory coronavirus in pigs. Viral Immunol 2010; 23(5):457-66.
Rodríguez-Gómez IM, Gómez-Laguna J, Barranco I, García-Nicolás O, Pallarés FJ, Ramis G, Carras-co L. El virus del Síndrome Reproductivo y Respiratorio Porcino como llave de entrada al Com-plejo Respiratorio Porcino. Tierras, Otoño 2011. Vol 184, 42-45.
Van Gucht S, van Reeth K, Pensaert M. Interaction between porcine reproductive-respiratory syn-drome virus and bacterial endotoxin in the lungs of pigs: potentiation of cytokine production and respiratory disease. J Clin Microbiol 2003; 41(3):960-6.


FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
167ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
RESUMEN
Las células dendríticas (CDs) son leucocitos que juegan un importante papel tanto en la inmunidad innata como en la adaptativa, siendo las células presentadoras de antígeno más potentes que existen y con la capacidad única de activar linfocitos T colaboradores que no han tenido contacto antigénico previo. No sólo son importantes en la regulación de respuestas inmunógenas efectivas, sino también en la inducción de fenómenos de tolerancia inmunológi-ca, necesarios para evitar la aparición de procesos autoinmunes. Tras la captación y procesa-miento de los antígenos, las CDs se dirigen principalmente a los órganos linfoides, donde se produce la presentación de antígenos a los linfocitos T, proceso en el cual no sólo es necesaria la interacción del CMH-II con el receptor de linfocitos T sino también la interacción de otras moléculas accesorias como las moléculas coestimuladoras y las moléculas de adhesión. Las CDs son células de una gran plasticidad funcional y que van a caracterizarse por su diferente localización en el organismo, su estado de madurez y su origen. En este último aspecto, cabe destacar las importantes diferencias funcionales entre las células dendríticas foliculares (CDF) de origen estromal y las clásicas CDs de origen hematopoyético, que van a derivar a su vez de progenitores mieloides o linfoides y que van a clasifi carse de manera distinta según la especie estudiada.
FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
F. ROMERO-PALOMO1*, P.J. SÁNCHEZ CORDÓN1, M.A. RISALDE1, M. PEDRERA1, V. MOLINA1, E. RUIZ-VILLAMOR2, J.C. GÓMEZ-VILLAMANDOS1
1Departamento de Anatomía y Anatomía Patológica Comparadas, Facultad de Veterinaria, Univer-sidad de Córdoba-Campus de Excelencia Internacional Agroalimentario (ceiA3), Edifi cio Sanidad Animal, Campus de Rabanales, 14014, Córdoba, España.
2Laboratorio Central de Veterinaria de Santa Fe, Camino del Jau s/n, 18320, Santa Fe, Granada, España*E-mail: [email protected]

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
168
1. ANTECEDENTES
La primera vez que se observaron células con morfología dendrítica, presentes en la piel data de 1868, siendo Paul Langerhans, un estudiante alemán de medicina, el responsable del hallazgo [1]. En aquel momento se pensó que estas células formaban parte del sistema nervioso, cuando en realidad se trataba de las actualmente cono-cidas como células dendríticas epidérmicas o células de Langerhans. A pesar de su descubrimiento, no se consiguió averiguar la verdadera naturaleza y función de estas células, que permanecieron como un misterio durante más de 100 años.
En la teoría de la selección clonal propuesta por Frank Macfarlane Burnet en 1957 se postulaba que los linfocitos proliferan en respuesta a antígenos sólo si el antígeno se une a su receptor [2], pero esta teoría no explicaba cómo se presentaba el antígeno desencadenante de la respuesta inmune. Ralph M. Steinman, que en esta época continuaba con sus estudios de medicina, se interesó por estas cuestiones y se dedicó a investigar qué agente posibilitaba la presentación de antígeno para iniciar la respuesta inmunitaria linfocítica, ya que observó que añadiendo antígenos específi cos a los linfocitos no se conseguía una respuesta inmune primaria.
Robert Mishell y Richard Dutton publicaron en 1966 un estudio en el que consiguieron desencadenar por primera vez una respuesta primaria de anticuerpos in vitro añadiendo glóbulos rojos de oveja a una suspensión de células de bazo de ratón [3]. En este caso, dicha suspensión de células no sólo contenía poblaciones de linfocitos, como se había hecho anteriormente, lo que llevo a la conclusión en trabajos posteriores [4] de que dicha respuesta primaria de anticuerpos sólo se produce en presencia de una variedad de células accesorias del bazo, cuyo principal componente son los macrófagos.
En 1970 Ralph M. Steinman comenzó a trabajar junto con Zanvil A. Cohn en estas células accesorias de bazo de ratón y descubrieron mediante microscopía de contraste de fases una escasa población de células con múltiples ramifi caciones, móviles y ricas en mitocondrias. Dada su peculiar morfología, ambos investigadores utilizaron por primera vez en 1973 el término “células dendríticas” (CDs) para referirse a estas células [5-6]. Durante los años 70, la mayoría de los inmunólogos consideraron a los macrófagos como la principal célula presentadora de antígenos (CPA) en el sistema inmune, siendo poco aceptada la hipótesis de Steinman sobre el papel clave de las CDs en la generación de la respuesta inmune. Steinman y otros investigadores car-acterizaron las proteínas expresadas en la superfi cie de las CD [7], que fueron clave para determinar la función de las CDs. Uno de los hallazgos más signifi cativos fue la elevada expresión de proteínas del complejo mayor de histocompatibilidad (CMH),

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
169
que más tarde demostraron ser necesarias para la presentación de antígenos a los linfocitos T. Tras una serie de experimentos basados en la reacción linfocitaria mixta (MLR), Steinman pudo demostrar que las CDs eran entre 100 y 1000 veces más efi caces para iniciar la respuesta inmunitaria que las células genéricas del bazo [8].
Dado que las CDs son tan escasas, los estudios iniciales fueron difíciles y no fue hasta los años 80 cuando se aceptó que las CDs son “CPA profesionales”. Estudios posteriores describieron los procesos de maduración de las CDs, mediante los cuales, las CDs inmaduras capturan antígenos en tejidos periféricos para transformarse en efi cientes iniciadores de la respuesta inmune [9].
La escasez de CDs supuso un impedimento para su estudio, por lo que los in-vestigadores descubrieron en los años 90 cómo generar in vitro grandes cantidades de CDs a partir de precursores de médula ósea o de monocitos, eliminando la ardua tarea de purifi carlas a partir de órganos linfoides [10-11]. Distintas terapias basadas en la aplicación de CDs obtenidas in vitro están siendo objeto de numerosas investigaciones.
2. FUNCION DE LAS CÉLULAS DENDRÍTICAS:
2.1.) Células presentadoras de antígenos (CPAs)
Las CDs son células especializadas del sistema inmune conocidas principalmente por su papel como CPAs a los linfocitos T, lo que permite el establecimiento de una respuesta inmune apropiada. Pero no son las CDs las únicas que desempeñan esta función presentadora de antígenos, ni tampoco esta presentación es realizada a un solo tipo de linfocito T [12]. Para que tenga lugar este proceso de presentación, es necesaria la presencia de una molécula denominada complejo mayor de histocom-patibilidad (CMH). En humanos, el CMH es denominado “sistema HLA” (Human leukocyte antigen), ya que estas proteínas se descubrieron como antígenos presentes en los leucocitos. Los receptores de linfocitos T (TCR - T cell receptor) son incapaces de reconocer antígenos intactos (como hacen los linfocitos B), por lo que necesitan que el antígeno sea procesado por una CPA y que ésta lo presente en su superfi cie como fragmentos unidos al CMH. Existen 2 tipos de moléculas CMH, denominadas CMH tipo I y tipo II. La primera se expresa en la mayoría de células nucleadas, que van a presentar antígenos intracelulares a los linfocitos T citotóxicos (CTL o CD8+), como es el caso de las nuevas proteínas sintetizadas en las células infectadas por virus. Una vez activados, los linfocitos T citotóxicos pueden destruir directamente la célula diana. Las moléculas CMH tipo II, sin embargo, se localizan principalmente en linfocitos B, macrófagos y células dendríticas, que son responsables de la presentación a linfocitos

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
170
T colaboradores (Th o CD4+) de antígenos extracelulares que han sido captados por la CPA mediante fagocitosis o por endocitosis mediada por receptores. Estos linfocitos T CD4, al activarse se transforman en potentes reguladores de la respuesta inmune [13]. Las CDs presentan una particularidad ausente en los macrófagos denominada “presentación cruzada” mediante la cual, antígenos no replicativos, normalmente procesados por la vía exógena y presentados en el contexto de moléculas CMH-II, son presentados por moléculas CMH-I con la consiguiente inducción de respuestas de linfocitos T CD8+ [14].
La mayoría de células nucleadas del organismo pueden actuar como CPA por su capacidad de presentar antígenos a linfocitos T CD8+ por medio de las moléculas CMH-I; sin embargo, este término es a menudo utilizado para referirse únicamente a las células capaces de presentar antígenos a los linfocitos T CD4, es decir, las que expresan CMH-II. Existen células que en determinadas ocasiones pueden activarse y expresar CMH-II, como es el caso de algunas células epiteliales (endotelio vascular, epiteliales del timo), siendo por tanto consideradas como “CPA no profesionales”. Por otro lado, las CPA que expresan CMH-II mejor defi nidas son las CDs, los fagoci-tos mononucleares y los linfocitos B, también denominadas “CPA profesionales”, siendo su capacidad fagocítica y procesadora de antígenos muy superior a la de cualquier otro tipo celular. Dentro de estas últimas, los macrófagos y linfocitos B presentan antígenos a linfocitos T colaboradores activos, es decir, linfocitos que han tenido un contacto previo con el antígeno. Las CDs en cambio, además de presentar antígenos a linfocitos T colaboradores activos, tienen la habilidad única de inducir respuestas inmunes primarias mediante la activación de linfocitos T vírgenes [15]. Las CDs expresan además una cantidad de complejos CMH-II-péptido muy superior a la expresada por linfocitos B o monocitos, siendo las CPAs más potentes de todo el sistema inmune [16-17].
2.2.) Circulación de las CDs e interacción con los linfocitos T
Las CDs tienen su origen en la médula ósea, donde las células madre se diferen-cian y migran como precursores de CDs hacia la sangre. Desde allí, las CDs inmadu-ras buscan los tejidos en los que actúan como células centinela, vigilando la posible entrada de patógenos invasores, a los cuales capturan, procesándolos en fragmentos antigénicos [13]. Una vez que se ha capturado el patógeno, la DC inmadura recibe señales de activación, que inician su maduración y migración a los órganos linfoides secundarios donde presentan los antígenos procesados a los linfocitos T vírgenes para la inducción de una respuesta inmune específi ca frente a esos antígenos. La

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
171
maduración y la migración de las CDs están minuciosamente dirigidas por diversas quimiocinas y moléculas de adhesión [18-19]. Una vez en las áreas T de los nódulos linfáticos, las quimiocinas atraen a los linfocitos T vírgenes hacia las CDs, permitiendo que se establezca la interacción entre la CD y el linfocito T (ver Figura 1). En esta interacción o sinapsis inmunológica intervienen diversas moléculas: En primer lugar, se establece la “señal 1” resultado de la interacción entre la molécula CMH-II de la CD, cargada con el antígeno, y el receptor del linfocito T. Las CDs expresan además distintas moléculas coestimuladoras que incluyen miembros de la familia B7 como CD80 (B7-1) y CD86 (B7-2), que pueden interactuar con CD28 y CD152 (CTLA4) presentes en el linfocito T [20], así como miembros de la familia TNF, como es el caso de CD40, cuyo ligando en el linfocito T es CD154 [21]. Esta segunda señal (“señal 2”) generada por las moléculas coestimuladoras, presentes también en otras CPA [22], es necesaria para la correcta activación de los linfocitos T. Dicho de otro modo, la señal 2 debe acompañar a la señal 1 para que se produzca inmunidad, ya que la señal 1 en ausencia de coestimulación se asocia con la inducción de tolerancia [23]. Hay que matizar que esta segunda señal inmunógena es resultado de la interacción con CD28 en el linfocito T, ya que si se produce con CTLA4, tendrá lugar una inactivación del linfocito, es decir, una respuesta tolerogénica. Existen además interacciones en las que intervienen moléculas de adhesión intercelular, que proporcionan estabilidad a la unión, como es el caso de CD58 (LFA-3) y CD54 (ICAM-I) presentes en la CD, cuyos correceptores en el linfocito T son LFA-2 y LFA-1 respectivamente. Se han identifi cado nuevas moléculas de la superfi cie celular de las CDs que pueden contribuir en su función, como es el caso de las lectinas de tipo C [24], que regulan muchas funciones implicadas en el establecimiento de la inmunidad innata y adaptativa. Algunas de estas moléculas pueden no solo reconocer patógenos sino también regular la interac-ción celular con los linfocitos T, como es el caso de CD209 (DC-SIGN) [25].
Figura 1. Principales moléculas participantes en la interacción entre Células dendríticas y linfocitos T. CD (grupo de diferenciación), CTLA-4 (Antígeno de linfocito T citotóxico- 4), LFA (Antígeno asociado a la función linfocítica), ICAM (Molécula de adhesión intercelular). DC-SIGN (Molécula de adhesión intercelular no asociada a integrina, específi ca de células dendríticas)

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
172
2.3.) Células dendríticas e Inmunidad innata:
Las CDs son conocidas por el importante papel que juegan enlazando la inmu-nidad innata y la adaptativa [26-27]. La respuesta inmune innata limita la infección y activa a la CPA para desencadenar la inmunidad adaptativa, que incrementa la especifi cidad y crea memoria inmunológica. Existen varios mecanismos que llevan a la activación innata de las CDs, compartiendo todos ellos un nexo de unión con las infecciones [28]. Las células del sistema inmune, incluyendo las CDs poseen los denominados receptores de reconocimiento de patrones moleculares (PRR - Pattern-recognition receptor) que como su propio nombre indica, reconocen distintos patrones moleculares asociados a patógenos (PAMP - Pathogen-associated molecular pattern) presentes en virus, bacterias, hongos y protozoos, como pueden ser su material ge-nético, lipopolisacáridos (LPS), etc. Los PRR mejor estudiados son los Receptores de tipo Toll (TLR - Toll-like Receptors), los cuales tienen un importante papel en la biología de las CDs [29], aunque se ha visto que el repertorio de TLRs entre los tipos de CDs no es el mismo entre distintas especies como es el caso de ratones y humanos [27, 29]. Es posible hacer una distinción entre las distintas vías de activación de CDs, que pueden ser dependientes o independientes de los PAMPs. La activación dependiente de PAMP puede producirse de manera directa por contacto con PAMPs, o bien de manera indirecta mediada por citoquinas producidas por otros tipos celulares que han contactado con el patógeno. La activación independiente de PAMP se produce en respuesta a moléculas propias del organismo o alteraciones del medio interno, como las proteínas de choque térmico [27-28]. Además de los TLRs, responsables de señali-zaciones intracelulares, existen otros PRR incluidos en la familia de lectinas de tipo C [24, 30], que están siendo objeto de numerosos estudios debido a su presencia en las CDs (DEC205, langerina, DC-SIGN, BDCA-2, etc.). La activación de las CDs implica que éstas secreten distintas citoquinas proinfl amatorias implicadas en la defensa del hospedador [31]. El mejor ejemplo de ello son las CDs plasmacitoides, las cuales son conocidas por ser grandes productoras de INF de tipo 1, de gran importancia en la inmunidad innata frente a los virus [32-34].
2.4.) Células dendríticas e Inmunidad adaptativa:
Las CDs han demostrado ser de vital importancia no sólo para la inducción de respuestas inmunes primarias (inmunidad innata), sino también para la regulación del tipo de respuesta inmune de linfocitos T, que va a ser específi ca del antígeno que procesa y presenta. En este sentido, las CDs van a desencadenar respuestas de linfocitos T colaboradores de tipo 1 (Th1) y de tipo 2 (Th2) [31, 35]. Existe la denominada “señal

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
173
3” para referirse precisamente a estas señales que dan las CDs a los linfocitos T para que estos se diferencien en células efectoras como Linfocitos Th1, Th2 o citotóxicos [36]. La IL-12 es un ejemplo de mediador que proporciona una señal 3 inductora de linfocitos Th1 o CTL [37].
Existe mucha controversia acerca de si los distintos tipos de inmunidad son producidos por distintos tipos de CDs en respuesta a diferentes PAMPs o si una sola subpoblación de CDs tiene el potencial sufi ciente de producir distintas citoquinas dependiendo del estímulo activador [28]. Parece ser que las subpoblaciones están en cierto modo especializadas para inducir distintos tipos de inmunidad, pero conservan-do una plasticidad sufi ciente para ajustar su respuesta a las señales de los patógenos.
2.5.) Células dendríticas y tolerancia inmunológica
Las células dendríticas no solo están relacionadas con la activación de linfocitos T para desencadenar respuestas inmunes adaptativas sino que también están impli-cadas en la inducción de tolerancia inmunológica [38-39], de gran importancia para evitar que el cuerpo produzca un ataque inmune contra antígenos inocuos, incluidos los de los tejidos, células o proteínas del propio organismo. Dependiendo de dónde se produzcan estos fenómenos de tolerancia, hablamos de tolerancia central o de tolerancia periférica. La tolerancia central tiene lugar en el timo, donde se produce no sólo un proceso de selección positiva de aquellos linfocitos T que no reconocen antígenos propios, sino también un proceso de selección negativa mediado por las CDs de la médula, en el que se destruyen aquellos linfocitos T que reconocen complejos CMH-péptidos con alta afi nidad [40]. Dado que algunos linfocitos T pueden esquivar el primer proceso de tolerancia, que además existen otros antígenos propios que no están presentes en el timo, o que otros surgen más tarde en la vida, las células den-dríticas también participan en los fenómenos de tolerancia periférica que restringen su actividad [41]. Estos mecanismos incluyen la muerte, anergia o supresión activa de linfocitos T, para lo cual las CDs pueden inducir linfocitos T reguladores (Treg) a nivel periférico [42]. Fallos en el mantenimiento de la tolerancia inmune pueden conducir a la aparición de enfermedades autoinmunes al igual que un exceso de tolerancia puede crear un ambiente permisivo para agentes infecciosos crónicos, como el VIH. Tradicionalmente se ha considerado que las CDs fenotípicamente inmaduras son las tolerogénicas y que las fenotípicamente maduras son las CDs inmunógenas. Sin embargo, observaciones recientes muestran que las CDs fenotípicamente maduras no siempre promocionan inmunidad de linfocitos T sino que pueden de hecho inducir tolerancia [23, 43]. Ver apartado de tipos de CDs según madurez.

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
174
3. TIPOS DE CÉLULAS DENDRÍTICAS:
Las CDs van a poder clasifi carse en base a distintos criterios como su localización en el organismo, su estado de madurez o si origen.
3.1.) Según su localización en el organismo:
Las CDs presentan un nivel de heterogeneidad que no solo se refl eja fenotípica-mente o en sus distintos orígenes sino también en las diferentes denominaciones que reciben según su localización anatómica. Las CDs circulan en sangre como células precursoras mieloides o linfoides, representando aproximadamente el 1% de las células mononucleares de sangre periférica (PBMCs). En los tejidos no linfoides, a nivel de la piel se encuentran las CDs epidermales, también conocidas como “Células de Langerhans” (CL) [44-45], que contienen unos estructuras intracitoplasmáticas de gran tamaño llamadas gránulos de Birbeck, mientras que en la dermis existen las “CDs dermales” [46], las cuales pertenecen a una subpoblación más amplia de “CDs intersticiales” [47], las cuales se presentan a nivel de la mayoría de órganos, incluyendo hígado, riñón, corazón y otros tejidos conectivos. Las “CDs asociadas a superfi cies mucosas” se encuentran en la mucosa de la cavidad oral, de tracto intestinal y tracto respiratorio. Estas poblaciones de células dendríticas presentes en tejidos no linfoides actúan como células centinela captando antígenos en las barreras de entrada al orga-nismo. Una vez que se han captado los antígenos, las CDs migran hacia los órganos linfoides donde llevan a cabo la presentación de antígeno a los linfocitos T para que éstos sean activados. Aunque el término CL se usa principalmente para referirse a las CDs de la epidermis, este término se ha extendido a las CDs presentes en todos los epitelios estratifi cados [44]. En los tejidos linfoides, el centro germinal, que es el microambiente que permite la generación de linfocitos B de memoria, también contiene “Células Dendríticas Foliculares” (CDF) y “CDs de los centros germinales” (CDCG). Las CDCG son potentes CPA para los linfocitos T [48-49]. Por el contrario, las CDF tienen la peculiar capacidad de captar antígenos en forma de complejos inmunes durante largos periodos de tiempo y promover la activación y selección de linfocitos B de los CG [50-51]. Dadas las importantes diferencias fenotípicas y funcionales con el resto de CDs, las CDF serán estudiadas con mayor detalle en el apartado de CDs de origen mesenquimal. Las CDs que han capturado antígenos y que han migrado desde la piel, otros tejidos intersticiales no linfoides, y de superfi cies mucosas hacia linfa aferente son reconocidas como “CDs de linfa aferente”, también llamadas “Cé-lulas veladas o veliformes” [52-53], que deben su nombre a los procesos en forma de velo que presentan en la superfi cie. Estas células migrantes representan una fase

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
175
intermedia entre las células de Langerhans (CDs inmaduras por excelencia) y las “CDs Interdigitantes” (CDI) [54] en las que se transforman. Estas últimas están presentes principalmente en las áreas T de los órganos linfoides secundarios, presentan un alto grado de maduración y pueden iniciar respuestas inmunes por activación de linfocitos T vírgenes. Al contrario de lo que sucede con las CL, las “CDs del timo” (CD tímicas) parecen ser células no migrantes, que son generadas en el timo, donde completan su ciclo de vida [40]. Por esto, es más probable que sólo se encuentren y presenten antígenos propios, estando así implicadas en la selección negativa de linfocitos T. Ver apartado de células dendríticas y tolerancia inmune.
3.2.) Según su madurez:
Ya se ha visto anteriormente que las CDs tienen la capacidad de captar antígenos extracelulares, procesarlos en distintos compartimentos de su citoplasma y asociarlos con moléculas CMH-II de su superfi cie para que sean presentados a los linfocitos T. Sin embargo, no todas las CDs tienen la misma capacidad para realizar por igual estas tres funciones, lo cual va a depender de su estado de maduración.
Distintos trabajos en los que se comparaban células de Langerhans aisladas en fresco (“inmaduras”) y CL procedentes de suspensiones epidermales posteriormente cultivadas in vitro (“maduradas”) revelaron sus distintas capacidades de captación y procesamiento antigénico y de estimulación de linfocitos T [9, 55]. De manera muy general y a modo de defi nición, las CDs inmaduras se caracterizan por presentar una elevada capacidad fagocítica y de procesamiento antigénico, localizarse princi-palmente en regiones periféricas del organismo como piel y mucosas y presentar una menor cantidad de moléculas CMH-II y de moléculas coestimuladoras. De manera opuesta, las CDs maduras se dirigen a las zonas T de los órganos linfoides secun-darios, donde queda refl ejada su capacidad para la presentación de antígenos a los linfocitos T, siendo su actividad fagocítica más limitada. Además sobreexpresan la molécula CMH-II superfi cialmente (y no en el citoplasma como en las inmaduras) y moléculas coestimuladoras [13]. Estas defi niciones se ajustan al modelo clásico en el que las CDs inmaduras presentes en piel y mucosas, maduran durante su migración a los nódulos linfáticos, disminuyendo su capacidad de captación de antígenos y aumentando su capacidad de estimular a los linfocitos T [56]. Sin embargo, existen numerosas excepciones y matices, como el hecho de que las CDs en estado inmaduro no solo están en piel y mucosas sino también en otros lugares como la sangre o los propios tejidos linfoides secundarios.

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
176
Además de cambios en la expresión de distintas moléculas de superfi cie, las CDs también experimentan cambios de forma muy signifi cativos que van a depen-der del estado en el que se encuentren. Cuando se encuentran en sangre o linfa en forma de células migrantes o de precursores de CDs, su morfología es básicamente redondeada. Sin embargo, cuando se trata de CDs inmaduras existentes en regiones superfi ciales del organismo (principalmente CLs), su superfi cie presenta numerosas prolongaciones para facilitarles un mayor contacto con los antígenos. De manera si-milar, las CDs maduras de las áreas T también presentan numerosas prolongaciones citoplasmáticas, pero en este caso para proporcionarles una mayor superfi cie con la que contactar con los linfocitos T [57-58].
Las CDs inmaduras disponen de distintos mecanismos para la captación de antí-genos. En primer lugar, pueden captar partículas o microorganismos por fagocitosis. En segundo lugar, pueden formar grandes vesículas en las que se toman muestras de fl uido extracelular y solutos, mediante un proceso denominado macropinocitosis. Por último, expresan elevados niveles de receptores que median endocitosis por ad-sorción, incluidos en la familia de Lectinas tipo C [24], como es el caso de DEC-205 [59]. Gracias a esta variedad de mecanismos de captación antigénica, las CDs realizan una presentación tan efi ciente que concentraciones tremendamente inferiores a las necesarias empleadas por otras CPA son sufi cientes [60].
La mayoría de proteínas que entran por vía endocítica en los macrófagos son degradadas en aminoácidos en los lisosomas, lugar en el que existen muy pocas molé-culas CMH-II. Las CDs en cambio se caracterizan por la presencia de “compartimentos ricos en CMH tipo II” (MIICs), que son muy abundantes en las CDs inmaduras. Du-rante la maduración de las CDs, los MIICs se convierten en vesículas no lisosómicas que descargan sus complejos péptido-CMH en la superfi cie [13]. La sobreexpresión en superfi cie de CMH-II y de moléculas coestimuladoras son indicadores fenotípicos de CDs maduras, pero existen otras moléculas como CD83 y CD208 (DC-LAMP) que se conocen por ser marcadores exclusivos de CDs maduras [61-63], aunque los datos obtenidos provienen únicamente de estudios realizados en humanos y ratones.
Reis e Sousa [23] analiza una serie de conceptos erróneos a cerca del paradigma de la maduración de las CDs, incluyendo los siguientes: 1) No todas las CDs de los órganos linfoides secundarios proceden de la periferia ya que gran parte de las CDs presentes en el bazo y nódulos linfáticos proceden de progenitores sanguíneos, 2) No todas las CDs de los órganos linfoides secundarios son maduras, sino que hay muchas (sobre todo las derivadas de progenitores sanguíneos) que están en estado inmaduro y 3) Dado que la expresión superfi cial de elevados niveles de moléculas CMH-II,

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
177
CD40, CD80, CD83 y CD86 se correlaciona normalmente con la habilidad de activar a linfocitos T, se asume de manera generalizada que las CDs que son maduras por criterios fenotípicos, son también maduras funcionalmente, es decir, inmunógenas. Sin embargo, observaciones recientes muestran que las CDs fenotípicamente madu-ras no siempre promocionan inmunidad de linfocitos T sino que pueden de hecho inducir tolerancia [23, 43].
3.3.) Según su origen:
3.3.1.) De origen hematopoyético:
El origen de las CDs ha sido siempre un tema de interés entre los investigadores. Hoy día se sabe que las CDs se originan a partir de precursores hematopoyéticos, a excepción de las CDF, que se verán en el apartado siguiente. Las CDs de origen he-matopoyético, se dividen a su vez en células dendríticas procedentes de progenitores linfoides [64] y procedentes de progenitores mieloides [65]. Las CDs siguen varias vías hematopoyéticas de diferenciación y maduración, y las múltiples y heterogéneas subpoblaciones de CDs varían en la expresión de marcadores de superfi cie [66-67]. La diversidad de funciones de las CDs en la regulación del sistema inmune (respuestas inmunes innatas, adaptativas (Th1-Th2), tolerancia inmune, producción de linfocitos T reguladores, etc.) refl ejan las heterogéneas subpoblaciones con diferente origen y plasticidad funcional.
La clasifi cación de las distintas subpoblaciones de CDs ha sido objeto de es-tudio en distintas revisiones bibliográfi cas de gran complejidad [67-69], por lo que se intentará dar a continuación unas nociones básicas y conceptos importantes que hay que tener en cuenta. Hasta el momento no se ha identifi cado un marcador es-pecífi co de líneas de CDs, y las subpoblaciones de CDs están por tanto actualmente defi nidas como líneas de células CMH-II + en combinación con varios marcadores celulares de superfi cie [69]. Cuando se intenta clasifi car las CDs en base a su origen y funcionalidad, se hace uso de una amplia terminología que a veces puede ser muy complicada y ambigua. Este es el caso de acepciones como “mieloides”, “linfoides”, “convencionales”, “plasmacitoides”, “CDs tipo1”, “CDs tipo2” “IPCs (CDs produc-toras de IFN)”, complicándose todo aún más debido a la diferente subdivisión que se hace de las CDs cuando se habla de modelos murinos o de humanos, que son las dos especies más estudiadas (Figuras 2 y 3). En algunas fuentes se habla en humanos de CDs tipo 1 y CDs tipo 2 para referirse a las CDs convencionales y plasmacitoides respectivamente [57]. En otros textos diferencian CDs tipo 1 y CDs tipo 2 para refe-

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
178
rirse a CDs inductoras de una respuesta Th1 y Th2 respectivamente [28, 70]. Estas denominaciones se basan principalmente en la habilidad de las CDs de origen mo-nocítico y plasmacitoides para inducir respuestas Th1 y Th2 respectivamente [71], sin embargo, se ha visto posteriormente que ambos tipos de CDs (Convencionales y plasmacitoides) son muy fl exibles, pudiendo dar respuestas Th1 y Th2 [33, 72]. Aun así, es frecuente seguir encontrando la denominación CDs tipo 1 y tipo 2 para referirse a las CDs convencionales y plasmacitoides respectivamente [67]. Otra distinción es la que separa las CDs convencionales en CD mieloides tipo I y CDs mieloides tipo 2 según la expresión de marcadores como BDCA-1 (CD1c) y BDCA-3 (CD141) respec-tivamente. Por todo ello, la forma más común de clasifi car a las CDs es aquella que las divide en convencionales (de origen principalmente mieloide) y en plasmacitoides (de origen principalmente linfoide).
Ratones:
Aunque esta revisión no está enfocada al estudio de las CDs en modelos muri-nos, es importante explicar brevemente las grandes diferencias con otras especies ya que muchas de las grandes conclusiones que se obtienen sobre la biología de las CDs proceden de estudios hechos en ratones.
La existencia de subpoblaciones de CDs se describió por primera vez en ratones, mediante marcadores que se utilizarían para los linfocitos T, estableciéndose dos gru-pos principales de CDs, CD8+CD4- y CD8-CD4+ [73]. Este hallazgo de que algunas CDs murinas expresan niveles de CD4 casi tan altos como los de los linfocitos T ahora concuerda con las CDs humanas, que también expresan CD4 [74-75]. Sin embargo, la expresión de altos niveles de CD8 no es una característica de las CDs humanas [75]. Es importante matizar que en el contexto de las CDs de ratón, hablamos de la molécula CD8 como un homodímero αα, y no como el heterodímero αβ presente en los linfocitos T. Es por ello frecuente encontrar dicha molécula denominada como “CD8α murina” para hacer hincapié en esa diferencia. Se puede generalizar entonces que las moléculas CD4 y CD8α no son de utilidad en la diferenciación de las subpoblaciones de CDs humanas, ya que todas son CD4+CD8α-, mientras que las subpoblaciones de CDs de ratón van a presentar distintas combinaciones (CD4+/-CD8α+/-). Hasta hace poco, las “CDs linfoides” eran conocidas como CDs CD8α+ y las “CDs mieloides” como CDs CD8α- pero este concepto se considera actualmente incorrecto [76-77], ya que las subpoblaciones de CDs han resultado ser mucho más complejas de lo que se pensaba [67]. A pesar de todo, la discriminación entre subpoblaciones de CDs basada en la expresión diferencial de CD8 sigue siendo bastante útil en términos prácticos.
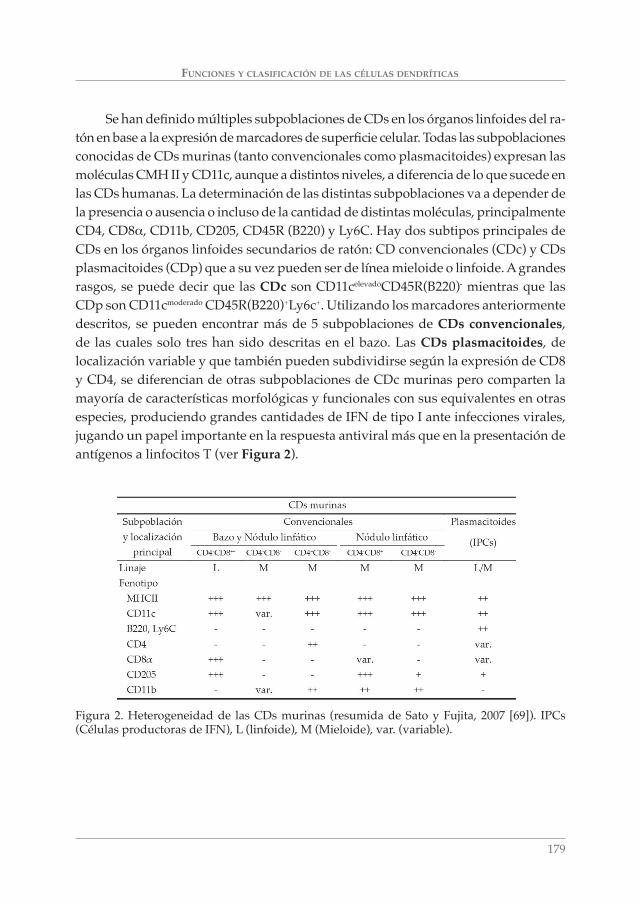
FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
179
Se han defi nido múltiples subpoblaciones de CDs en los órganos linfoides del ra-tón en base a la expresión de marcadores de superfi cie celular. Todas las subpoblaciones conocidas de CDs murinas (tanto convencionales como plasmacitoides) expresan las moléculas CMH II y CD11c, aunque a distintos niveles, a diferencia de lo que sucede en las CDs humanas. La determinación de las distintas subpoblaciones va a depender de la presencia o ausencia o incluso de la cantidad de distintas moléculas, principalmente CD4, CD8α, CD11b, CD205, CD45R (B220) y Ly6C. Hay dos subtipos principales de CDs en los órganos linfoides secundarios de ratón: CD convencionales (CDc) y CDs plasmacitoides (CDp) que a su vez pueden ser de línea mieloide o linfoide. A grandes rasgos, se puede decir que las CDc son CD11celevadoCD45R(B220)- mientras que las CDp son CD11cmoderado CD45R(B220)+Ly6c+. Utilizando los marcadores anteriormente descritos, se pueden encontrar más de 5 subpoblaciones de CDs convencionales, de las cuales solo tres han sido descritas en el bazo. Las CDs plasmacitoides, de localización variable y que también pueden subdividirse según la expresión de CD8 y CD4, se diferencian de otras subpoblaciones de CDc murinas pero comparten la mayoría de características morfológicas y funcionales con sus equivalentes en otras especies, produciendo grandes cantidades de IFN de tipo I ante infecciones virales, jugando un papel importante en la respuesta antiviral más que en la presentación de antígenos a linfocitos T (ver Figura 2).
Figura 2. Heterogeneidad de las CDs murinas (resumida de Sato y Fujita, 2007 [69]). IPCs (Células productoras de IFN), L (linfoide), M (Mieloide), var. (variable).

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
180
Humanos y otras especies:
Al igual que en las CDs de ratón, las CDs humanas también comprenden múltiples subpoblaciones en cuanto a la expresión de numerosos marcadores super-fi ciales, aunque parece que están más relacionados con la madurez o funcionalidad que con su origen. Las CDs humanas también se defi nen por ser CMH II +, aunque todas ellas expresan CD4 y carecen de CD8. Además, se dividen en subpoblaciones de CDs convencionales, de linaje mieloide (CD11c+) y CDs plasmacitoides, de linaje linfoide CD11c-, a diferencia de las CDs de ratón, que expresan en su totalidad CD11c independientemente de su origen.
En sangre periférica de humanos se han descrito dos subpoblaciones de CDs mieloides convencionales denominadas como CDm1 (CD4+CD1a+CD11c+↑BDCA-1+) y CDm2 (CD4+CD1a-CD11c+↓BDCA-3+) [78-80]. Ambas subpoblaciones expresan ade-más CD13, CD33, CD45RO y CD116 pero otras moléculas como CD2, CD11b, CD32 o CD64 se restringen solo a CDm1. Las subpoblaciones CDm1 y CDm2 parecen ser los precursores directos de las CL y de las CD intersticiales respectivamente [78]. Estas CDs convencionales pueden dar lugar a respuestas de tipo Th1 o Th2 dependiendo del entorno infl amatorio existente [35].
Las CDs plasmacitoides son reconocidas en humanos como subpoblaciones de CDs únicas de origen linfoide. Estas células se identifi caron por primera vez en áreas paracorticales de nódulos linfáticos reactivos con una morfología similar a la de las células plasmáticas y con marcadores comunes a linfocitos T y monocitos. Se observó también que estas células producían grandes cantidades de IFN de tipo 1 en respuesta a ciertos virus y que en determinadas condiciones de cultivo (IL3, CD40L) se transformaban en células de morfología estrellada y con capacidad para activar linfocitos T vírgenes. Por todo ello, los linfocitos T plasmacitoides, monocitos plasma-citoides o Células productoras de IFN naturales resultaron ser el mismo tipo celular denominado CD plasmacitoide [33, 81-82]. Las CDp humanas se identifi can como subpoblaciones CMHII+ CD4+ CD45RA+ IL-3Rα(CD123)+ ILT3+ ILT1– CD11c- [33] con ausencia de otros marcadores mieloides observados en CDc y con dos marcadores adicionales, BDCA-2 (CD303) y BDCA-4 (CD304), restringidos a CDp humanas en sangre periférica y médula ósea [79]. Ver Figura 3.
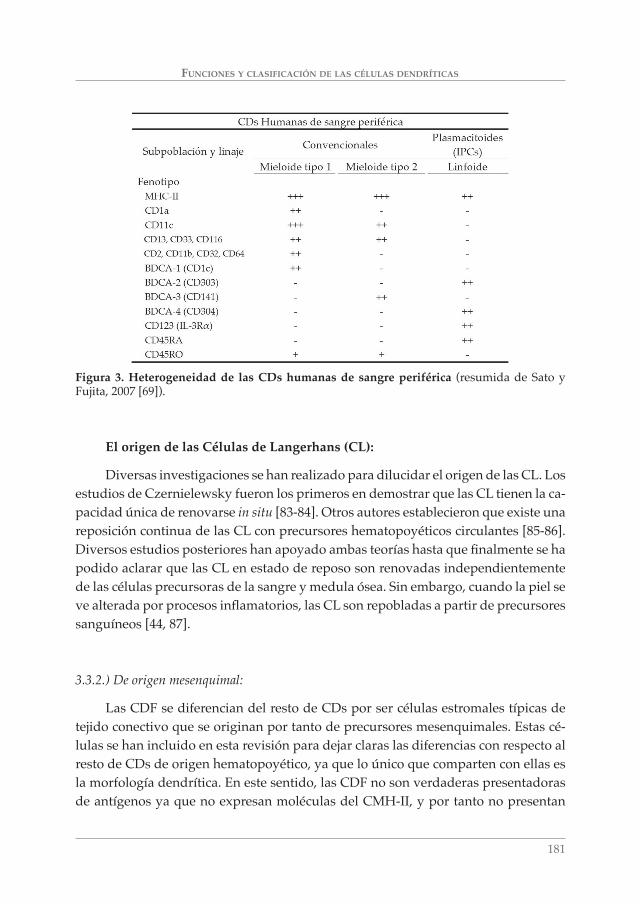
FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
181
Figura 3. Heterogeneidad de las CDs humanas de sangre periférica (resumida de Sato y Fujita, 2007 [69]).
El origen de las Células de Langerhans (CL):
Diversas investigaciones se han realizado para dilucidar el origen de las CL. Los estudios de Czernielewsky fueron los primeros en demostrar que las CL tienen la ca-pacidad única de renovarse in situ [83-84]. Otros autores establecieron que existe una reposición continua de las CL con precursores hematopoyéticos circulantes [85-86]. Diversos estudios posteriores han apoyado ambas teorías hasta que fi nalmente se ha podido aclarar que las CL en estado de reposo son renovadas independientemente de las células precursoras de la sangre y medula ósea. Sin embargo, cuando la piel se ve alterada por procesos infl amatorios, las CL son repobladas a partir de precursores sanguíneos [44, 87].
3.3.2.) De origen mesenquimal:
Las CDF se diferencian del resto de CDs por ser células estromales típicas de tejido conectivo que se originan por tanto de precursores mesenquimales. Estas cé-lulas se han incluido en esta revisión para dejar claras las diferencias con respecto al resto de CDs de origen hematopoyético, ya que lo único que comparten con ellas es la morfología dendrítica. En este sentido, las CDF no son verdaderas presentadoras de antígenos ya que no expresan moléculas del CMH-II, y por tanto no presentan

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
182
antígenos procesados a los linfocitos T CD4+ [88]. En su lugar, presentan antígenos completos en forma de complejos inmunes a los linfocitos B, ayudando a su activación o maduración selectiva y contribuyendo en la formación de los centros germinales y de células plasmáticas y de linfocitos B de memoria [89]. Siempre que se hable del término CDs en esta revisión, nos referiremos exclusivamente a las de origen hematopoyético, utilizando la denominación completa de CDF cuando queramos referirnos a estas últimas.
Relación con la reacción de los centros germinales.
A pesar de sus diferencias funcionales, las CDF guardan cierta relación con los antígenos presentados por las CDs, que queda refl ejada durante la reacción de los centros germinales (CG) [90]. La formación de los CG (ver Figura 4) comienza cuando las CDs presentes en las áreas T, que muestran antígenos (Ags) en su superfi cie, activan linfocitos T CD4 específi cos, que proliferan y maduran a células efectoras capaces de activar linfocitos B específi cos para esos Ags. Una vez activados por los linfocitos T CD4, los linfocitos B proliferan para formar un foco primario de linfocitos B específi cos frente al Ag en cuestión, pudiendo a continuación seguir dos vías de diferenciación. Algunos linfocitos B del foco primario persisten en el área T, diferenciándose a células plasmáticas y secretando anticuerpos (Acs) (que darán lugar a los complejos inmunes que penetran en el folículo linfoide (FL) y son captados por las CDF). Sin embargo, otros linfocitos B activos migran desde el foco primario y penetran en los FL primarios cercanos, siendo estos los responsables de la verdadera reacción de los CG. Una vez en el folículos, estos linfocitos B activos comienzan a proliferar rápidamente (expansión monoclonal). Esta expansión clonal a la que se ven sometidos los linfocitos B (denomi-nados centroblastos) se ve acompañada a su vez por diversas mutaciones en el ADN que codifi ca los fragmentos Fab de sus BCR (receptores de linfocito B). Este proceso, denominado hipermutación somática, da lugar a la formación de anticuerpos de di-ferentes afi nidades a la del antígeno original, generándose así una gran diversidad de clones en el CG. Debido a este proceso de mutación, algunos linfocitos B obtienen una mayor afi nidad en sus BCR mientras que otros perderán afi nidad. Es incluso posible que aparezcan nuevas especifi cidades de BCRs durante estas mutaciones aleatorias [91]. Ésta progenie de linfocitos B o centroblastos migra de la zona oscura en la que han proliferado a la zona clara de los CG, donde pierden la capacidad mitótica y comienzan a exponer sus anticuerpos en la superfi cie, siendo denominados en esta fase como centrocitos. En la zona clara, los centrocitos sufren un proceso de selección mediado por las CDF. Los linfocitos B cuya mutación ha dado lugar a una unión más

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
183
fuerte con los antígenos presentados por las CDF (mediante receptores CR y FcR que se verán posteriormente) serán seleccionados para sobrevivir mientras que los lin-focitos B cuya mutación reduce su afi nidad por los antígenos morirán por apoptosis [92]. Los linfocitos B funcionales tienen ahora que interactuar con linfocitos Th para recibir señales fi nales de diferenciación que determinan su transformación en células plasmáticas secretoras de anticuerpos o en linfocitos B de memoria [93]. Además de la previa mutación de los genes de Fab, la diferenciación fi nal implica cambios de isotipo en los BCR de los linfocitos B [51, 94-95]. Los linfocitos B inactivos recirculan por los FL primarios sin apenas establecer interacciones con las CDF, mientras que los linfocitos B que han sido activados son los que sufren la hipermutación somática y la posterior selección mediada por CDF. Estudios recientes han demostrado que las CDF ayudan a mantener los FL primarios como nichos exclusivos de linfocitos B y que desempeñan un papel crítico en la retención de células dentro de los CG [96].
Figura 4. Reacción de los centros germinales.
Expresión de moléculas en las CDF
En relación con toda esta maquinaria, las CDF expresan varias moléculas para la presentación de antígenos. Los icosomas (del inglés Iccosome “Immunecomplex coated bodies”) son estructuras formadas por complejos antígeno-anticuerpo presentes en la

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
184
superfi cie de las CDF [97], que son liberadas de sus dendritas y rápidamente captadas por los linfocitos B de los CGs. Este mecanismo permite a los linfocitos B de los CGs obtener antígenos intactos que son procesados y presentados a linfocitos T. Estructu-ralmente, las prolongaciones dendríticas pueden tanto tener una apariencia fi liforme como formar cadenas de icosomas [50, 97]. La superfi cie de la membrana plasmática de las CDF está por tanto recubierta de moléculas que les permiten atrapar y retener antígenos no procesados en forma de complejos inmunes (Ag-Ac, Ag-Complemento o Ag-Ac-Compl). Entre estas moléculas destacan los receptores de los fragmentos Fc de anticuerpos, como FcγRIIb (CD32) y FcεRII (CD23), y los receptores de com-plemento, como CR2 (CD21) y CR1 (CD35). Algunas de estas moléculas pueden ser utilizadas para identifi car CDF, pero la mayoría no son completamente específi cas. Por ejemplo, CD21 se expresa en gran cantidad en las CDF, pero también, aunque en menor cantidad, en linfocitos B, y CD35 se expresa en muchos otros tipos celulares, incluyendo linfocitos B, neutrófi los y eritrocitos [98].
Además de para presentar los antígenos en forma de complejos inmunes, las CDF también expresan otras moléculas relacionadas con la atracción de linfocitos B como la quimiocina CXCL13, también denominada como BCA-1 (B cells attracting chemokine-1), que se une a CXCR5 (BLR1) en los linfocitos B [99]. La secreción de BCA-1 no está restringida a las CDF, existiendo evidencias de que otros tipos celula-res pueden secretar esta quimiocina incluso en mayor cantidad. En la interacción de los linfocitos B con las CDF intervienen además las moléculas de adhesión LFA-1/ICAM-1 y VLA-4/VCAM-1. La expresión de ICAM-1 y VCAM-1 es típica en las CDF, pero de ninguna manera específi ca [91].
Origen de las CDF
Durante mucho tiempo ha existido una gran controversia sobre el origen de las CDF, pero parece que su origen estromal ha quedado claro con numerosas evi-dencias, como es el hecho de compartir distintos marcadores con los fi broblastos [100-101], carecer del marcador leucocitario común (CD45) o presentar un conjunto único de moléculas en su superfi cie [51]. Como se ha visto, las CDF se localizan ex-clusivamente en los FL primarios, así como en la zona clara de los FL secundarios, siendo muy escasas en la zona oscura. En este contexto, es importante recordar que en los FLs también existen las denominadas CDCG, cuyo origen es hematopoyético (CD4+CD11c+CD3-) [48-49, 51, 95].

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
185
4. CÉLULAS DENDRITICAS EN LA DIARREA VIRICA BOVINA (DVB)
Pocos son los trabajos en los que se ha estudiado el papel de las células dendríti-cas en la patogenia de la DVB. En algunos estudios in vivo se ha visto la detección del virus en células de morfología dendrítica, localizadas principalmente en el interior de los folículos linfoides, en áreas interfoliculares y en timo, mediante técnicas inmu-nohistoquímicas [102-108]. También ha sido posible detectar mediante hibridación in situ la presencia de células infectadas de morfología dendrítica en el interior de los folículos linfoides [109]
En otros estudios se ha estudiado el potencial como CPAs de los monocitos aislados a partir de sangre de animales infectados persistentemente con el DVB, sugiriendo dicho estudio que las CPA no ven alterada su capacidad de presentación antigénica [110]. Glew y colaboradores [111] estudiaron in vitro el efecto del virus de la DVB sobre monocitos y CDs derivadas de monocitos, concluyendo que ambos tipos celulares eran susceptibles a la infección por el virus. En este estudio también se observó que los monocitos veían afectada su capacidad de presentación antigénica y que eran susceptibles al efecto citopático del virus, no observándose dichas altera-ciones en el caso de las CDs.
Estas conclusiones proceden de estudios in vitro, siendo necesarios estudios in vivo en los que se caractericen e identifi quen las localizaciones y variaciones cuan-titativas de las CDs en tejidos de animales infectados con el virus de la DVB, para poder llegar así a un mejor entendimiento del papel de las células dendríticas en la patogenia de la enfermedad.
AGRADECIMIENTOS.
Este trabajo ha sido fi nanciado por la Consejería de Innovación y Ciencia de la Junta de Andalucía a través del proyecto de excelencia P09-AGR-4671 y por el Minis-terio de Ciencia e Innovación (MICINN) a través del proyecto AGL2009-13174-C02-01. Fernando Romero es benefi ciario de una beca-contrato fi nanciada por el proyecto de excelencia P09-AGR-4671 de la Junta de Andalucía. Pedro José Sánchez Cordón es benefi ciario de un contrato dentro del “Programa Ramón y Cajal” del Ministerio de Ciencia e Innovación, (España).

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
186
BIBLIOGRAFÍA:
[1]Langerhans P. Uber die nerven der menschlichen haut. Archives of Pathological Anatomy. 44: 325-37 (1868).
[2]Burnet FM. A modifi cation of Jerne’s theory of antibody production using the concept of clonal selection. Austr. J. Sci. 20: 67-69 (1957).
[3]Mishell RI, Dutton RW. Immunization of normal mouse spleen cell suspensions in vitro. Science. 153: 1004-6 (1966).
[4]Mosier DE. A requirement for two cell types for antibody formation in vitro. Science. 158: 1573-5 (1967).
[5]Steinman RM, Cohn ZA. Identifi cation of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J. Exp. Med. 137: 1142-62 (1973).
[6]Steinman RM, Cohn ZA. Identifi cation of a novel cell type in peripheral lymphoid organs of mice. II. Functional properties in vitro. J. Exp. Med. 139: 380-97 (1974).
[7]Steinman RM, Kaplan G, Witmer MD, Cohn ZA. Identifi cation of a novel cell type in peripheral lymphoid organs of mice. V. Purifi cation of spleen dendritic cells, new surface markers, and maintenance in vitro. J. Exp. Med. 149: 1-16 (1979).
[8]Steinman RM, Witmer MD. Lymphoid dendritic cells are potent stimulators of the primary mi-xed leukocyte reaction in mice. Proc. Natl. Acad. Sci. U. S. A. 75: 5132-6 (1978).
[9]Schuler G, Steinman RM. Murine epidermal Langerhans cells mature into potent immunostimu-latory dendritic cells in vitro. J. Exp. Med. 161: 526-46 (1985).
[10]Inaba K, Inaba M, Romani N, Aya H, Deguchi M, Ikehara S, Muramatsu S, Steinman RM. Ge-neration of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor. J. Exp. Med. 176: 1693-702 (1992).
[11]Romani N, Reider D, Heuer M, Ebner S, Kampgen E, Eibl B, Niederwieser D, Schuler G. Gene-ration of mature dendritic cells from human blood. An improved method with special regard to clinical applicability. J. Immunol. Methods. 196: 137-51 (1996).
[12]Knight SC, Stagg AJ. Antigen-presenting cell types. Curr. Opin. Immunol. 5: 374-82 (1993). [13]Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 392: 245-52
(1998). [14]Belz GT, Carbone FR, Heath WR. Cross-presentation of antigens by dendritic cells. Crit. Rev.
Immunol. 22: 439-48 (2002). [15]Hart DN. Dendritic cells: unique leukocyte populations which control the primary immune
response. Blood. 90: 3245-87 (1997). [16]Inaba K, Pack M, Inaba M, Sakuta H, Isdell F, Steinman RM. High levels of a major histocompa-
tibility complex II-self peptide complex on dendritic cells from the T cell areas of lymph nodes. J. Exp. Med. 186: 665-72 (1997).
[17]Guery JC, Adorini L. Dendritic cells are the most effi cient in presenting endogenous naturally processed self-epitopes to class II-restricted T cells. J. Immunol. 154: 536-44 (1995).
[18]Sallusto F, Lanzavecchia A. Mobilizing dendritic cells for tolerance, priming, and chronic in-fl ammation. J. Exp. Med. 189: 611-4 (1999).
[19]Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, Pulendran B, Palucka K. Immu-nobiology of dendritic cells. Annu. Rev. Immunol. 18: 767-811 (2000).
[20]Orabona C, Grohmann U, Belladonna ML, Fallarino F, Vacca C, Bianchi R, Bozza S, Volpi C, Salomon BL, Fioretti MC, Romani L, Puccetti P. CD28 induces immunostimulatory signals in dendritic cells via CD80 and CD86. Nat Immunol. 5: 1134-42 (2004).
[21]Ma DY, Clark EA. The role of CD40 and CD154/CD40L in dendritic cells. Semin. Immunol. 21: 265-72 (2009).

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
187
[22]Johnson JG, Jenkins MK. Co-stimulatory functions of antigen-presenting cells. J. Invest. Derma-tol. 99: 62S-65S (1992).
[23]Reis e Sousa C. Dendritic cells in a mature age. Nat Rev Immunol. 6: 476-83 (2006). [24]Figdor CG, van Kooyk Y, Adema GJ. C-type lectin receptors on dendritic cells and Langerhans
cells. Nat Rev Immunol. 2: 77-84 (2002). [25]Geijtenbeek TB, Engering A, Van Kooyk Y. DC-SIGN, a C-type lectin on dendritic cells that un-
veils many aspects of dendritic cell biology. J. Leukoc. Biol. 71: 921-31 (2002). [26]Clark GJ, Angel N, Kato M, Lopez JA, MacDonald K, Vuckovic S, Hart DN. The role of dendritic
cells in the innate immune system. Microbes Infect. 2: 257-72 (2000). [27]Reis e Sousa C. Activation of dendritic cells: translating innate into adaptive immunity. Curr.
Opin. Immunol. 16: 21-5 (2004). [28]Reis e Sousa C. Dendritic cells as sensors of infection. Immunity. 14: 495-8 (2001). [29]Reis e Sousa C. Toll-like receptors and dendritic cells: for whom the bug tolls. Semin. Immunol.
16: 27-34 (2004). [30]Kanazawa N. Dendritic cell immunoreceptors: C-type lectin receptors for pattern-recognition
and signaling on antigen-presenting cells. J. Dermatol. Sci. 45: 77-86 (2007). [31]Moser M, Murphy KM. Dendritic cell regulation of TH1-TH2 development. Nature immunology.
1: 199-205 (2000). [32]McKenna K, Beignon AS, Bhardwaj N. Plasmacytoid dendritic cells: linking innate and adapti-
ve immunity. J. Virol. 79: 17-27 (2005). [33]Colonna M, Trinchieri G, Liu YJ. Plasmacytoid dendritic cells in immunity. Nat Immunol. 5:
1219-26 (2004). [34]Diebold SS, Montoya M, Unger H, Alexopoulou L, Roy P, Haswell LE, Al-Shamkhani A, Flavell
R, Borrow P, Reis e Sousa C. Viral infection switches non-plasmacytoid dendritic cells into high interferon producers. Nature. 424: 324-8 (2003).
[35]Maldonado-Lopez R, Moser M. Dendritic cell subsets and the regulation of Th1/Th2 responses. Semin. Immunol. 13: 275-82 (2001).
[36]Kalinski P, Hilkens CM, Wierenga EA, Kapsenberg ML. T-cell priming by type-1 and type-2 polarized dendritic cells: the concept of a third signal. Immunol. Today. 20: 561-7 (1999).
[37]Trinchieri G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat Rev Immunol. 3: 133-46 (2003).
[38]Huang FP, MacPherson GG. Continuing education of the immune system--dendritic cells, im-mune regulation and tolerance. Curr Mol Med. 1: 457-68 (2001).
[39]Steinman RM, Hawiger D, Nussenzweig MC. Tolerogenic dendritic cells. Annu. Rev. Immunol. 21: 685-711 (2003).
[40]Ardavin C. Thymic dendritic cells. Immunol. Today. 18: 350-61 (1997). [41]Steinman RM, Nussenzweig MC. Avoiding horror autotoxicus: the importance of dendritic
cells in peripheral T cell tolerance. Proc. Natl. Acad. Sci. U. S. A. 99: 351-8 (2002). [42]Roncarolo MG, Levings MK, Traversari C. Differentiation of T regulatory cells by immature
dendritic cells. J. Exp. Med. 193: F5-9 (2001). [43]Lutz MB, Schuler G. Immature, semi-mature and fully mature dendritic cells: which signals
induce tolerance or immunity? Trends Immunol. 23: 445-9 (2002). [44]Merad M, Ginhoux F, Collin M. Origin, homeostasis and function of Langerhans cells and other
langerin-expressing dendritic cells. Nat Rev Immunol. 8: 935-47 (2008). [45]Valladeau J, Ravel O, Dezutter-Dambuyant C, Moore K, Kleijmeer M, Liu Y, Duvert-Frances
V, Vincent C, Schmitt D, Davoust J, Caux C, Lebecque S, Saeland S. Langerin, a novel C-type lectin specifi c to Langerhans cells, is an endocytic receptor that induces the formation of Birbeck granules. Immunity. 12: 71-81 (2000).

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
188
[46]Poulin LF, Henri S, de Bovis B, Devilard E, Kissenpfennig A, Malissen B. The dermis contains langerin+ dendritic cells that develop and function independently of epidermal Langerhans cells. J. Exp. Med. 204: 3119-31 (2007).
[47]Hart DN, McKenzie JL. Interstitial dendritic cells. Int. Rev. Immunol. 6: 127-38 (1990). [48]Grouard G, Durand I, Filgueira L, Banchereau J, Liu YJ. Dendritic cells capable of stimulating T
cells in germinal centres. Nature. 384: 364-7 (1996). [49]Goval JJ, Greimers R, Boniver J, de Leval L. Germinal center dendritic cells express more ICAM-
1 than extrafollicular dendritic cells and ICAM-1/LFA-1 interactions are involved in the capa-city of dendritic cells to induce PBMCs proliferation. J. Histochem. Cytochem. 54: 75-84 (2006).
[50]Tew JG, Kosco MH, Burton GF, Szakal AK. Follicular dendritic cells as accessory cells. Immunol. Rev. 117: 185-211 (1990).
[51]Liu YJ, Grouard G, de Bouteiller O, Banchereau J. Follicular dendritic cells and germinal cen-ters. Int. Rev. Cytol. 166: 139-79 (1996).
[52]Howard CJ, Hope JC. Dendritic cells, implications on function from studies of the afferent lym-ph veiled cell. Vet. Immunol. Immunopathol. 77: 1-13 (2000).
[53]Knight SC. Veiled cells--”dendritic cells” of the peripheral lymph. Immunobiology. 168: 349-61 (1984).
[54]Steinman RM, Pack M, Inaba K. Dendritic cells in the T-cell areas of lymphoid organs. Immunol. Rev. 156: 25-37 (1997).
[55]Romani N, Koide S, Crowley M, Witmer-Pack M, Livingstone AM, Fathman CG, Inaba K, Stein-man RM. Presentation of exogenous protein antigens by dendritic cells to T cell clones. Intact protein is presented best by immature, epidermal Langerhans cells. J. Exp. Med. 169: 1169-78 (1989).
[56]Steinman RM. The dendritic cell system and its role in immunogenicity. Annu. Rev. Immunol. 9: 271-96 (1991).
[57]Grayson MH, Holtzman MJ. Emerging role of dendritic cells in respiratory viral infection. J Mol Med (Berl). 85: 1057-68 (2007).
[58]Shutt DC, Daniels KJ, Carolan EJ, Hill AC, Soll DR. Changes in the motility, morphology, and F-actin architecture of human dendritic cells in an in vitro model of dendritic cell development. Cell Motil. Cytoskeleton. 46: 200-21 (2000).
[59]Jiang W, Swiggard WJ, Heufl er C, Peng M, Mirza A, Steinman RM, Nussenzweig MC. The re-ceptor DEC-205 expressed by dendritic cells and thymic epithelial cells is involved in antigen processing. Nature. 375: 151-5 (1995).
[60]Trombetta ES, Mellman I. Cell biology of antigen processing in vitro and in vivo. Annu. Rev. Immunol. 23: 975-1028 (2005).
[61]Lechmann M, Berchtold S, Hauber J, Steinkasserer A. CD83 on dendritic cells: more than just a marker for maturation. Trends Immunol. 23: 273-5 (2002).
[62]Cao W, Lee SH, Lu J. CD83 is preformed inside monocytes, macrophages and dendritic cells, but it is only stably expressed on activated dendritic cells. Biochem. J. 385: 85-93 (2005).
[63]de Saint-Vis B, Vincent J, Vandenabeele S, Vanbervliet B, Pin JJ, Ait-Yahia S, Patel S, Mattei MG, Banchereau J, Zurawski S, Davoust J, Caux C, Lebecque S. A novel lysosome-associated mem-brane glycoprotein, DC-LAMP, induced upon DC maturation, is transiently expressed in MHC class II compartment. Immunity. 9: 325-36 (1998).
[64]Ardavin C, Wu L, Li CL, Shortman K. Thymic dendritic cells and T cells develop simultaneously in the thymus from a common precursor population. Nature. 362: 761-3 (1993).
[65]Inaba K, Inaba M, Deguchi M, Hagi K, Yasumizu R, Ikehara S, Muramatsu S, Steinman RM. Granulocytes, macrophages, and dendritic cells arise from a common major histocompatibility complex class II-negative progenitor in mouse bone marrow. Proc. Natl. Acad. Sci. U. S. A. 90: 3038-42 (1993).

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
189
[66]Liu YJ. Dendritic cell subsets and lineages, and their functions in innate and adaptive immunity. Cell. 106: 259-62 (2001).
[67]Shortman K, Liu YJ. Mouse and human dendritic cell subtypes. Nat Rev Immunol. 2: 151-61 (2002).
[68]Pulendran B, Banchereau J, Maraskovsky E, Maliszewski C. Modulating the immune response with dendritic cells and their growth factors. Trends Immunol. 22: 41-7 (2001).
[69]Sato K, Fujita S. Dendritic cells: nature and classifi cation. Allergol Int. 56: 183-91 (2007). [70]Tizard IR. 2009. Inmunología Veterinaria. 8th ed. Elsevier. Barcelona. pp94.[71]Rissoan MC, Soumelis V, Kadowaki N, Grouard G, Briere F, de Waal Malefyt R, Liu YJ. Recipro-
cal control of T helper cell and dendritic cell differentiation. Science. 283: 1183-6 (1999). [72]Cella M, Facchetti F, Lanzavecchia A, Colonna M. Plasmacytoid dendritic cells activated by
infl uenza virus and CD40L drive a potent TH1 polarization. Nat Immunol. 1: 305-10 (2000). [73]Vremec D, Zorbas M, Scollay R, Saunders DJ, Ardavin CF, Wu L, Shortman K. The surface
phenotype of dendritic cells purifi ed from mouse thymus and spleen: investigation of the CD8 expression by a subpopulation of dendritic cells. J. Exp. Med. 176: 47-58 (1992).
[74]Sotzik F, Rosenberg Y, Boyd AW, Honeyman M, Metcalf D, Scollay R, Wu L, Shortman K. As-sessment of CD4 expression by early T precursor cells and by dendritic cells in the human thymus. J. Immunol. 152: 3370-7 (1994).
[75]Winkel K, Sotzik F, Vremec D, Cameron PU, Shortman K. CD4 and CD8 expression by human and mouse thymic dendritic cells. Immunol. Lett. 40: 93-9 (1994).
[76]Wu L, D’Amico A, Hochrein H, O’Keeffe M, Shortman K, Lucas K. Development of thymic and splenic dendritic cell populations from different hemopoietic precursors. Blood. 98: 3376-82 (2001).
[77]Manz MG, Traver D, Miyamoto T, Weissman IL, Akashi K. Dendritic cell potentials of early lymphoid and myeloid progenitors. Blood. 97: 3333-41 (2001).
[78]Ito T, Inaba M, Inaba K, Toki J, Sogo S, Iguchi T, Adachi Y, Yamaguchi K, Amakawa R, Valladeau J, Saeland S, Fukuhara S, Ikehara S. A CD1a+/CD11c+ subset of human blood dendritic cells is a direct precursor of Langerhans cells. J. Immunol. 163: 1409-19 (1999).
[79]Dzionek A, Fuchs A, Schmidt P, Cremer S, Zysk M, Miltenyi S, Buck DW, Schmitz J. BDCA-2, BDCA-3, and BDCA-4: three markers for distinct subsets of dendritic cells in human peripheral blood. J. Immunol. 165: 6037-46 (2000).
[80]Narbutt J, Lesiak A, Zak-Prelich M, Wozniacka A, Sysa-Jedrzejowska A, Tybura M, Robak T, Smolewski P. The distribution of peripheral blood dendritic cells assayed by a new panel of anti-BDCA monoclonal antibodies in healthy representatives of the polish population. Cell Mol Biol Lett. 9: 497-509 (2004).
[81]Dzionek A, Inagaki Y, Okawa K, Nagafune J, Rock J, Sohma Y, Winkels G, Zysk M, Yamagu-chi Y, Schmitz J. Plasmacytoid dendritic cells: from specifi c surface markers to specifi c cellular functions. Hum. Immunol. 63: 1133-48 (2002).
[82]Grouard G, Rissoan MC, Filgueira L, Durand I, Banchereau J, Liu YJ. The enigmatic plasma-cytoid T cells develop into dendritic cells with interleukin (IL)-3 and CD40-ligand. J. Exp. Med. 185: 1101-11 (1997).
[83]Czernielewski J, Vaigot P, Prunieras M. Epidermal Langerhans cells--a cycling cell population. J. Invest. Dermatol. 84: 424-6 (1985).
[84]Czernielewski JM, Demarchez M. Further evidence for the self-reproducing capacity of Langer-hans cells in human skin. J. Invest. Dermatol. 88: 17-20 (1987).
[85]Katz SI, Tamaki K, Sachs DH. Epidermal Langerhans cells are derived from cells originating in bone marrow. Nature. 282: 324-6 (1979).

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
190
[86]Frelinger JG, Hood L, Hill S, Frelinger JA. Mouse epidermal Ia molecules have a bone marrow origin. Nature. 282: 321-3 (1979).
[87]Merad M, Manz MG, Karsunky H, Wagers A, Peters W, Charo I, Weissman IL, Cyster JG, Engle-man EG. Langerhans cells renew in the skin throughout life under steady-state conditions. Nat Immunol. 3: 1135-41 (2002).
[88]Abbas AK, Lichtman AH. 2003. Inmunología Celular y Molecular. 5th ed. Elsevier. p206.[89]Tew JG, Wu J, Fakher M, Szakal AK, Qin D. Follicular dendritic cells: beyond the necessity of
T-cell help. Trends Immunol. 22: 361-7 (2001). [90]MacLennan IC. Germinal centers. Annu. Rev. Immunol. 12: 117-39 (1994). [91]van Nierop K, de Groot C. Human follicular dendritic cells: function, origin and development.
Semin. Immunol. 14: 251-7 (2002). [92]Lindhout E, de Groot C. Follicular dendritic cells and apoptosis: life and death in the germinal
centre. Histochem. J. 27: 167-83 (1995). [93]Thorbecke GJ, Amin AR, Tsiagbe VK. Biology of germinal centers in lymphoid tissue. FASEB J.
8: 832-40 (1994). [94]Liu YJ, Malisan F, de Bouteiller O, Guret C, Lebecque S, Banchereau J, Mills FC, Max EE, Mar-
tinez-Valdez H. Within germinal centers, isotype switching of immunoglobulin genes occurs after the onset of somatic mutation. Immunity. 4: 241-50 (1996).
[95]Dubois B, Barthelemy C, Durand I, Liu YJ, Caux C, Briere F. Toward a role of dendritic cells in the germinal center reaction: triggering of B cell proliferation and isotype switching. J. Immunol. 162: 3428-36 (1999).
[96]Wang X, Cho B, Suzuki K, Xu Y, Green JA, An J, Cyster JG. Follicular dendritic cells help esta-blish follicle identity and promote B cell retention in germinal centers. J. Exp. Med. (2011).
[97]Terashima K, Dobashi M, Maeda K, Imai Y. Follicular dendritic cell and iccosomes in germinal center reactions. Semin. Immunol. 4: 267-74 (1992).
[98]Kosco-Vilbois MH. Are follicular dendritic cells really good for nothing? Nat Rev Immunol. 3: 764-9 (2003).
[99]Cyster JG, Ansel KM, Reif K, Ekland EH, Hyman PL, Tang HL, Luther SA, Ngo VN. Follicular stromal cells and lymphocyte homing to follicles. Immunol. Rev. 176: 181-93 (2000).
[100]Lee IY, Choe J. Human follicular dendritic cells and fi broblasts share the 3C8 antigen. Biochem. Biophys. Res. Commun. 304: 701-7 (2003).
[101]Bofi ll M, Akbar AN, Amlot PL. Follicular dendritic cells share a membrane-bound protein with fi broblasts. J. Pathol. 191: 217-26 (2000).
[102]Odeon AC, Kelling CL, Marshall DJ, Estela ES, Dubovi EJ, Donis RO. Experimental infection of calves with bovine viral diarrhea virus genotype II (NY-93). J. Vet. Diagn. Invest. 11: 221-8 (1999).
[103]Stoffregen B, Bolin SR, Ridpath JF, Pohlenz J. Morphologic lesions in type 2 BVDV infections experimentally induced by strain BVDV2-1373 recovered from a fi eld case. Vet. Microbiol. 77: 157-62 (2000).
[104]Raya AI, Gomez-Villamandos JC, Sanchez-Cordon PJ, Bautista MJ. Virus Distribution and Role of Thymic Macrophages During Experimental Infection With Noncytopathogenic Bovine Viral Diarrhea Virus Type 1. Vet. Pathol. (2011).
[105]Liebler EM, Kusters C, Pohlenz JF. Experimental mucosal disease in cattle: changes of lym-phocyte subpopulations in Peyer’s patches and in lymphoid nodules of large intestine. Vet. Immunol. Immunopathol. 48: 233-48 (1995).
[106]Liebler-Tenorio EM, Ridpath JF, Neill JD. Distribution of viral antigen and development of le-sions after experimental infection of calves with a BVDV 2 strain of low virulence. J. Vet. Diagn. Invest. 15: 221-32 (2003).

FUNCIONES Y CLASIFICACIÓN DE LAS CÉLULAS DENDRÍTICAS
191
[107]Liebler-Tenorio EM, Ridpath JE, Neill JD. Distribution of viral antigen and tissue lesions in persistent and acute infection with the homologous strain of noncytopathic bovine viral diarr-hea virus. J. Vet. Diagn. Invest. 16: 388-96 (2004).
[108]Pedrera M, Sanchez-Cordon PJ, Romero-Trevejo JL, Risalde MA, Greiser-Wilke I, Nunez A, Gomez-Villamandos JC. Morphological changes and virus distribution in the ileum of colos-trum-deprived calves inoculated with non-cytopathic bovine viral diarrhoea virus genotype-1. J. Comp. Pathol. 141: 52-62 (2009).
[109]Collins ME, Desport M, Brownlie J. Bovine viral diarrhea virus quasispecies during persistent infection. Virology. 259: 85-98 (1999).
[110]Glew EJ, Howard CJ. Antigen-presenting cells from calves persistently infected with bovine viral diarrhoea virus, a member of the Flaviviridae, are not compromised in their ability to pre-sent viral antigen. J. Gen. Virol. 82: 1677-85 (2001).
[111]Glew EJ, Carr BV, Brackenbury LS, Hope JC, Charleston B, Howard CJ. Differential effects of bovine viral diarrhoea virus on monocytes and dendritic cells. J. Gen. Virol. 84: 1771-80 (2003).


FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
193ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
RESUMEN
Con este trabajo se persigue aportar nuevas ideas sobre la fi siopatología del ejercicio en el toro bravo, siendo el objetivo principal, valorar los cambios metabólicos que se presentan de forma secundaria a las lesiones producidas durante la lidia como factores limitantes del rendi-miento físico. Se obtuvieron muestras de sangre antes y después de la lidia y biopsias musculares después de la lidia de 30 toros de 4-5 años de edad pertenecientes a ganaderías de las provincias de Córdoba, Sevilla y Badajoz. La sangre fue separada en dos fracciones: una para el estudio de sangre entera y otra para el estudio bioquímico del plasma. Se utilizó Sysmex- F-820. Se estudió el hemograma [recuento de glóbulos rojos, concentración de hemoglobina, valor hematocrito e índices volumétricos [volumen corpuscular medio (VCM), hemoglobina corpuscular media (HCM), concentración de hemoglobina corpuscular media (CHCM)] y el leucograma [recuento de glóbulos blancos, linfocitos, neutrofi los, cociente neutrófi los /linfocitos (N/L)]. En plasma se investigó ácido láctico con el analizador Analox, Champion P-LM5, cortisol con un analizador automático Inmulite, potasio mediante fotómetro Corning 480 y por espectrofotometría glucosa proteínas plasmáticas totales y las actividades enzimáticas de creatin quinasa (CK), aspartato aminotransferasa (AST ó GOT), lactato deshidrogenasa (LDH),. Biopsias musculares: las biopsias musculares fueron extraídas del músculo glúteo medio derecho a una profundidad absoluta de 5 cm. Las biopsias se extrajeron dentro de los cinco minutos posteriores a la fi nalización de la lidia. En estas biopsias se estudiaron glucógeno, concentración de lactato y pH. Se realizó una Valora-ción del Rendimiento Físico para lo cual se confeccionó una fi cha de valoración del mismo para cada animal durante la lidia. Estas notas las tomó siempre el mismo observador. Los resultados presentados hacen referencia a los valores medidos antes y después de la lidia El eritrograma mostró diferencias signifi cativas en los glóbulos rojos y CHCM de p ≤ 0.05 (*). Asimismo, en los parámetros bioquímicos plasmáticos medidos se encontraron diferencias signifi cativas de:
FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
ESTRELLA AGÜERA BUENDÍA1 ; F. REQUENA DOMENECH2
1 Profesora Titular. Departamento Biología Celular, Fisiología e Inmunología. Facultad de Veterinaria de Córdoba. Grupo Investigación AGR-019..
2 Profesor Asociado. Facultad de Veterinaria. Director Gerente de Gen-Ova Veterinaria, S.L. Grupo de Investigación AGR-019.

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
194
-p ≤ 0.05 (*) en cortisol, potasio y AST; p ≤ 0.01 (**) en LDH y proteínas plasmáticas totales, p ≤ 0.001 (***) en CK y lactato. No se obtuvieron diferencias signifi cativas en la glucosa aunque si un aumento en los valores medios de la misma después de la lidia. En cuanto al rendimiento físico demostrado durante la lidia, según la fi cha de valoración del observador, se observó que los que tenían un rendimiento alto presentaban altos niveles de cortisol. Se ha concluido que se pueden considerar factores limitantes la disminución del número de glóbulos rojos, lesión muscular, aumento del gasto cardiaco por alteración del potasio e incapacidad motora debida a la depleción de glucógeno y acidosis muscular.
Palabras Clave: toro de lidia, fi siopatología, ejercicio, cambios metabólicos
SUMMARY
This work seeks to contribute new ideas on the physiopathology of exercise in the fi ght-ing bull, the main objective being to assess the metabolic changes presented secondarily to the lesions produced during the fi ght as limiting factors of physical performance. Blood samples were obtained before and after the fi ght, and muscle biopsies after the fi ght, from 30 bulls 4-5 years old belonging to stock farms in the provinces of Córdoba, Seville and Badajoz. The blood was divided into two fractions: one for the study of whole blood and the other for a biochemi-cal study of the plasma. Sysmex-F-820 was used. The haemogram was studied [red blood cell count, haemoglobin concentration, haematocrit value] and volumetric indices[mean corpus-cle volume (MCV), mean corpuscular haemoglobin (MCH), mean corpuscular haemoglobin concentration (MCHC)]and the leucogram [white blood cell count, lymphocytes, neutrophyls, neutrophyl/lymphocyte quotient (N/L)]. In plasma, lactic acid was investigated with the Analox analyzer, Champion P-LM5, cortisol with an automatic Inmulite analyzer, potassium by means of a Corning 480 photometer, and, by spectrophotometry, glucose, total plasmatic proteins and the enzymatic activities of creatine kinase (CK), aminotransferase aspartate (AST or GOT), and lactate dehydrogenase (LDH). Muscle biopsies: the muscle biopsies were extracted from the middle right gluteus muscle at an absolute depth of 5 cm. The extraction was carried out within the 5 minutes after the end of the bullfi ght. In these biopsies, glucogen, lactate concentration and pH were studied. A Valuation of Physical Performance was carried out for which a valuation card was made for each animal during the fi ght. These notes were always taken by the same observer. The results presented refer to the values measured before and after the fi ght. The erythrogram showed signifi cant differences in the red blood cells and MCHC of p ≤ 0.05 (*). Similarly, in the plasmatic biochemical parameters measured, signifi cant differences were found of : -p ≤ 0.05 (*) in cortisol, potassium and AST, p ≤ 0.01 (**) in LDH and total plasmatic proteins, and p ≤ 0.001 (***) in CK and lactate. No signifi cant differences were obtained in glucose although there was an increase in its mean values after the fi ght. With regard to the physical performance demonstrated during the fi ght, it was observed that those bulls which performed well presented high levels of cortisol. It was concluded that the factors which can be considered as being limiting ones are: a diminution in the number of red blood cells, muscle lesions, an increase in cardiac output due to an alteration in potassium, and a motor incapacity due to glucogen depletion and muscle acidosis.
Key words: fi ghting bull, physiopathology, exercise, metabolic changes ,

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
195
INTRODUCCIÓN
La importancia económica y social de la que gozan los festejos taurinos en nuestro país, Portugal, Sur de Francia y otros países Iberoamericanos se refl eja en el número creciente de corridas de toros y otros eventos relacionados, así como el incremento en el número de espectadores. Sin embargo, los animales no siempre entregan lo esperado ni favorecen el lucimiento de los espadas, con la consiguiente decepción del público.
Actualmente, el problema generalizado que nos encontramos en las corridas de toros es que en la mayoría de ellas falla el elemento fundamental, el toro. Es frecuente observar al animal que sale a la plaza con síntomas evidentes de fatiga, con una falta de acometividad total, debido a un agotamiento físico extremo, que aparece mucho antes de lo que debiera.
El diccionario de la Real Academia Española defi ne al estrés como “situación de un individuo, o de alguno de sus órganos o aparatos, que, por exigir de ellos un rendimiento superior al normal, los pone en riesgo próximo de enfermar”. No obstante, también se ha dicho que desde el punto de vista de la biología y la psicología el estrés se defi ne como “cualquier tensión o interferencia que altera el funcionamiento de un organismo”. Tanto el hombre como los animales responden al estrés físico y psicológico con una serie de respuestas que, generalmente, recuerdan una activación del sistema nervioso autónomo. Si el estrés es muy fuerte, o las defensas orgánicas inadecuadas, se puede producir una alteración psicosomática.
En la respuesta al estrés parecen existir tres etapas. En la primera, de alerta, el organismo reconoce la situación anormal y se prepara para la acción, ya sea de agresión o de fuga. En la segunda, de resistencia, el organismo repara cualquier daño causado por la fase anterior, pero si el estrés persiste el cuerpo permanece alerta y no puede reparar los daños. En la tercera, que se inicia al continuar la resistencia y recibe el nombre de etapa de agotamiento, las consecuencias pueden incluso llegar a ser fatales.
Es indudable que el toro bravo es sometido a numerosas situaciones desconocidas tales como el embarque, el transporte, el desencajonamiento y el apartado de los toros en las plazas correspondientes. Estas circunstancias ocasionan estrés psíquico en los ani-males produciendo cambios valorables en los niveles de algunas variables sanguíneas, que refl ejan alteraciones del bienestar animal y que, indirectamente, afectan al compor-tamiento del toro en el ruedo. La lidia es un ejercicio intenso y agotador para él, con el agravante de las lesiones producidas durante la suerte de varas lo que implica estrés físico para el animal que está realizando ejercicio a la vez que hay una gran pérdida de sangre y una serie de trastornos que suponen una situación traumática para el animal.

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
196
Igualmente, es posible evaluar el nivel de estrés metabólico en el trauma durante la lidia mediante la valoración de diferentes parámetros fi siológicos, tanto hemato-lógicos como musculares: hemograma, leucograma, glucosa, proteínas plasmáticas, potasio, cortisol, enzimas plasmáticas, lactato plasmático y muscular, glucógeno muscular y pH muscular. En todo tipo de lesión se van a presentar de forma secun-daria una serie de cambios fi siopatológicos que son considerados en conjunto como la respuesta metabólica al trauma (1) y pueden ser considerados factores limitantes del rendimiento físico para el toro durante la lidia.
La fi nalidad de la respuesta metabólica es producir la energía necesaria que permita sostener las funciones vitales durante la lidia y la aparición de un proceso infl amatorio consecuente con las lesiones ya que después de la faena, por la caracterís-ticas de la misma, no hay que contemplar la reparación tisular. Para esto es necesaria la movilización de sustratos hacia áreas con elevadas demandas energéticas como son el hígado y el músculo lesionado. La respuesta del organismo, que inicialmente constituye un mecanismo de defensa ante el trauma, dependiendo de su severidad y duración, puede llegar a convertirse en un factor muy limitante para que el toro realice el ejercicio que se le exige durante la lidia. Todos estos cambios metabólicos son inducidos y mantenidos por mediadores de la infl amación.
Con este trabajo se persigue aportar nuevas ideas sobre la fi siopatología del ejercicio en el toro bravo, siendo el objetivo principal, valorar los cambios metabólicos que se presentan de forma secundaria a las lesiones producidas durante la lidia como factores limitantes del rendimiento físico.
MATERIAL Y MÉTODOS
En la presente investigación se han estudiado un total de 30 toros de 4-5 años pertenecientes a ganaderías de las provincias de Córdoba, Sevilla y Badajoz. El trabajo experimental se realizó en el momento anterior al embarque y en las distintas plazas de toros después de la corrida. Se obtuvieron muestras de sangre antes y después de la lidia y biopsias musculares después de la lidia.
Muestras de sangre: para las muestras de sangre se utilizaron agujas, porta-tubos y tubos de vacío Venojet 10 ml, unos con EDTA-3K y otros con heparina-litio. Antes de la lidia se obtuvieron por punción en la vena coxígea, previa desinfección de la zona, aprovechando la inmovilización del animal en el cajón de curas o mueco, el día anterior a la lidia e incluso el mismo día, antes del embarque de las reses en el camión que las transportaría a la plaza. Inmediatamente después de la lidia, las muestras

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
197
se obtuvieron en el desolladero de las distintas plazas de toros en el momento del degollado de los toros.
La sangre fue separada en dos fracciones: una en tubos con EDTA-3K para el estudio de sangre entera y otra en tubos con heparina-litio para el estudio bioquímico del plasma. Dentro de los cinco minutos posteriores a la toma de muestras la sangre se centrifugó durante 5 minutos a 3.000 r.p.m. para conseguir el plasma. La sangre se analizó antes de 48 horas de su recogida y el plasma fue almacenado a -20C para su posterior análisis.
El análisis de sangre se hizo mediante un contador celular semiautomático, mo-delo Sysmex- F-820. Se estudió el hemograma [recuento de glóbulos rojos, concentra-ción de hemoglobina, valor hematocrito e índices volumétricos [volumen corpuscular medio (VCM), hemoglobina corpuscular media (HCM), concentración de hemoglobina corpuscular media (CHCM)] y el leucograma [recuento de glóbulos blancos, linfocitos, neutrofi los, cociente neutrófi los /linfocitos (N/L)]. En plasma se investigó ácido láctico con el analizador Analox, Champion P-LM5, cortisol con un analizador automático Inmulite, potasio mediante fotómetro Corning 480 y por espectrofotometría glucosa proteínas plasmáticas totales y las actividades enzimáticas de creatin quinasa (CK), aspartato aminotransferasa (AST ó GOT), lactato deshidrogenasa (LDH),.
Biopsias musculares: las biopsias musculares fueron extraídas del músculo glúteo medio derecho a una profundidad absoluta de 5 cm., aproximadamente a 5 cm. en sentido dorso caudal a la tuberosidad coxal, en la línea que une esta tuberosidad con la tuberosidad isquiática. Se utilizó una aguja de biopsia muscular Bergström modifi cada (2). Todas las biopsias musculares fueron obtenidas por la misma per-sona para estandarizar la toma de muestras. Tras su recogida fueron introducidas en viales criogénicos procediéndose a su congelación en nitrógeno líquido para su posterior análisis. Las biopsias se extrajeron dentro de los cinco minutos posteriores a la fi nalización de la lidia. En estas biopsias se estudiaron glucógeno, concentración de lactato y pH.
Se realizó una Valoración del Rendimiento Físico para lo cual se confeccionó una fi cha de valoración del mismo para cada animal durante la lidia. Estas notas las tomó siempre el mismo observador. En ella se diferenciaron tres secciones relacionadas con la aptitud del animal en cada una de las etapas de la lidia, caídas y claudicaciones, y al concluir la lidia se tuvo en cuenta la fuerza demostrada, ejercicio realizado y fatiga observada. Se anotaron las particularidades ocurridas en la lidia de cada uno. Con todas estas características se hicieron tres grupos de rendimiento físico A, B y C según fue alto, bajo o muy bajo respectivamente. Se correlacionaron los incrementos

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
198
medios de los parámetros medidos con la capacidad física demostrada en la lidia por los distintos grupos de animales surgidos de la clasifi cación anterior.
En cuanto al estudio estadístico, en primer lugar se procedió al cálculo de los estadísticos básicos para cada uno de los parámetros antes y después de la lidia. Se realizó un ANOVA para observar diferencias antes y después de la lidia en los parámetros hemáticos, plasmáticos y musculares, así como los cambios metabólicos que se presentan de forma secundaria a las lesiones producidas durante la corrida.
RESULTADOS
El eritrograma de los toros antes y después de la lidia se presenta en la tabla I, encontrándose diferencias signifi cativas en los glóbulos rojos y CHCM de p ≤ 0.05 (*).
La tabla II refl eja los parámetros bioquímicos plasmáticos medidos antes y des-pués de la lidia encontrándose diferencias signifi cativas de:
- p ≤ 0.05 (*) en cortisol, potasio y AST; - p ≤ 0.01 (**) en LDH y proteínas plasmáticas totales - p ≤ 0.001 (***) en CK y lactato
No se obtuvieron diferencias signifi cativas en la glucosa aunque si un aumento en los valores medios de la misma después de la lidia.
En la tabla III se exponen los valores medios de y desviación estándar de las con-centraciones de glucógeno, lactato y pH en músculo glúteo medio después de la lidia.
En la tabla IV se muestra la correlación existentes entre el incremento medio de cortisol con respecto a los grupos de rendimiento físico, aumentando directamente proporcional.
El gráfi co 1 se representa los valores medios de linfocitos y neutrófi los aprecián-dose una inversión de la fórmula leucocitaria.
En el gráfi co 2 se aprecian la relación del cortisol y glucosa donde se puede valorar la situación estresante después de la lidia y la hiperglucemia asociada al aumento de cortisol.
El gráfi co 3 clasifi ca a los toros en grupos de rendimiento físico demostrado durante la lidia, según la fi cha de valoración del observador.
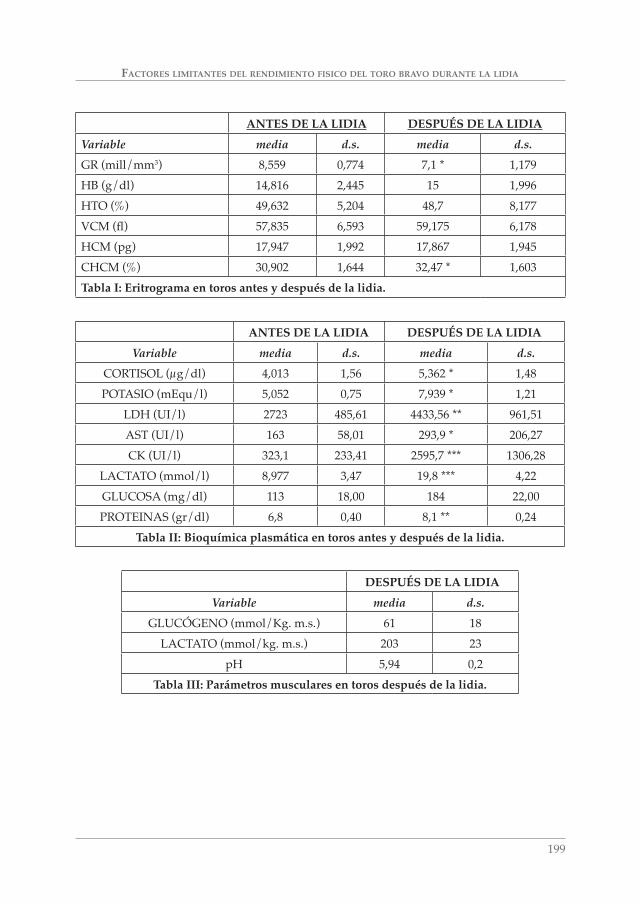
FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
199
ANTES DE LA LIDIA DESPUÉS DE LA LIDIAVariable media d.s. media d.s.GR (mill/mm3) 8,559 0,774 7,1 * 1,179HB (g/dl) 14,816 2,445 15 1,996HTO (%) 49,632 5,204 48,7 8,177VCM (fl ) 57,835 6,593 59,175 6,178HCM (pg) 17,947 1,992 17,867 1,945CHCM (%) 30,902 1,644 32,47 * 1,603Tabla I: Eritrograma en toros antes y después de la lidia.
ANTES DE LA LIDIA DESPUÉS DE LA LIDIAVariable media d.s. media d.s.
CORTISOL (μg/dl) 4,013 1,56 5,362 * 1,48POTASIO (mEqu/l) 5,052 0,75 7,939 * 1,21
LDH (UI/l) 2723 485,61 4433,56 ** 961,51AST (UI/l) 163 58,01 293,9 * 206,27CK (UI/l) 323,1 233,41 2595,7 *** 1306,28
LACTATO (mmol/l) 8,977 3,47 19,8 *** 4,22GLUCOSA (mg/dl) 113 18,00 184 22,00PROTEINAS (gr/dl) 6,8 0,40 8,1 ** 0,24
Tabla II: Bioquímica plasmática en toros antes y después de la lidia.
DESPUÉS DE LA LIDIAVariable media d.s.
GLUCÓGENO (mmol/Kg. m.s.) 61 18LACTATO (mmol/kg. m.s.) 203 23
pH 5,94 0,2Tabla III: Parámetros musculares en toros después de la lidia.

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
200
RENDIMIENTO FÍSICO A LA LIDIAVariable MUY BAJO BAJO ALTOCORTISOL (μg/dl) 0,640 1,347 1,995
Tabla IV: Relación entre los incrementos medios de cortisol en cada grupo de rendimiento físico.
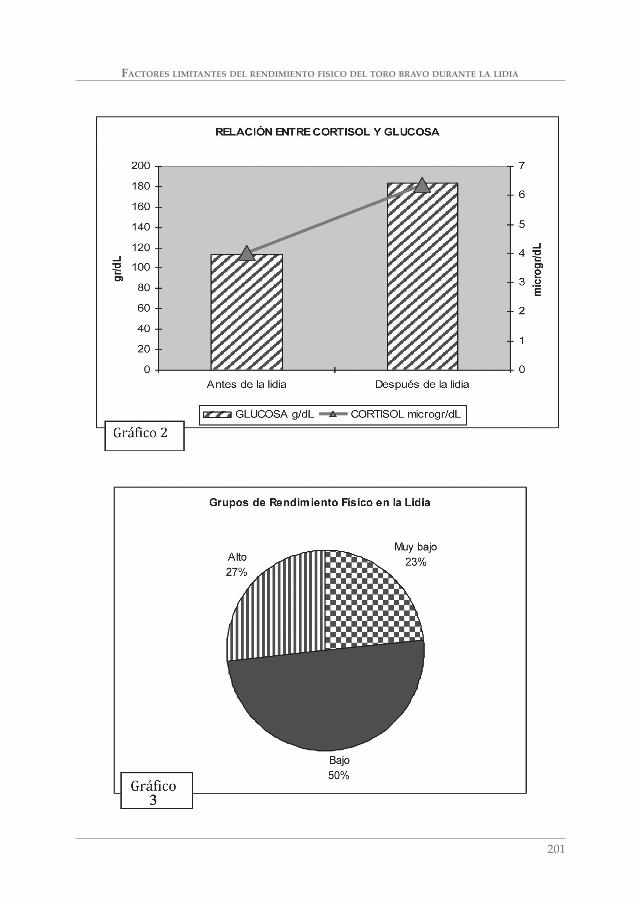
FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
201

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
202
DISCUSIÓNLos mecanismos que inician, regulan y sostienen la respuesta metabólica al
trauma siguen defi niéndose en términos de tres componentes: 1) respuesta neuroen-docrina en la que interviene hipotálamo, hipófi sis y glándula adrenal; 2) reacción metabólica que se deriva de las hormonas producidas en el eje anterior y sus efectos en órganos como el páncreas, el hígado y el músculo esquelético principalmente y 3) cambios hematológicos.
1. RESPUESTA NEUROENDOCRINA:
Los toros bravos acusan el estrés de manejo mucho más que otras razas de bo-vinos. Este animal pasa su vida en el campo, sin realizar apenas ejercicio físico, de manera que son escasos los estímulos externos que vienen a perturbar su tranquilidad.
El caballo ha conseguido el califi cativo de “atleta” por su elevado potencial físico que le proporciona atributos de velocidad, fuerza y resistencia como pocas especies dentro del reino animal. Durante siglos, los caballos de deporte han sido selecciona-dos para realizar un elevado rendimiento durante el ejercicio, variando éste según las modalidades ecuestres (3). Sin embargo, La selección del vacuno bravo comprende aspectos genealógicos, morfológicos y funcionales, que quedan supeditados a la comprobación de la descendencia. No se han tenido en cuenta factores fi siológicos que contribuyan a un rendimiento atlético superior como son la capacidad aerobia, propiedades musculoesqueléticas, cardiovasculares, biomecánicas, concentración de hemoglobina, intercambio de gases y heredabilidad de todas estas variables. En el transcurso de la lidia, este animal realiza una actividad física intensa e intermitente, con una duración aproximada de 20 minutos, en los que se espera que este rumiante actúe como un verdadero atleta.
El síndrome de adaptación al estrés puede ser el responsable de las alteraciones hormonales y bioquímicas que padece el animal (4). El transporte, el desencajona-miento, el apartado y la lidia suponen para el toro una situación estresante que se va a incrementar en la plaza con las percepciones visuales, lesiones producidas durante la faena, estímulos dolorosos y otras sensaciones, que unidas al ejercicio físico al que se le somete, desencadena una rápida respuesta hormonal para intentar adaptarse a la nueva situación. El cortisol tiene muchas funciones en el metabolismo corporal que incluyen estimulación de la gluconeogénesis, incremento de la proteólisis y de la síntesis de alanina, sensibilización del tejido adiposo a la acción de las hormonas lipolíticas, efectos antinfl amatorios y de resistencia a la insulina (5). Específi camente

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
203
en el hígado, el cortisol inhibe la vía colateral de fosfato pentosa, la acción de la in-sulina y de varias enzimas reguladoras de la glucólisis; además facilita la captación de aminoácidos; promueve la síntesis y actividad de varias enzimas reguladoras de la gluconeogénesis y potencia las acciones de glucagón y adrenalina (1) En músculo estriado no ejerce efecto directo en el metabolismo de la glucosa, aunque inhibe la captación de glucosa mediada por insulina. Además, disminuye la captación e incre-menta la liberación de aminoácidos (1). En resumen, esta hormona es importante para la conservación de los niveles normales de glucosa durante las situaciones de estrés, como es la lidia, y para incrementar la disponibilidad de sustratos hacia el músculo y para la gluconeogénesis hepática. Debido a la inhibición de la acción de la insulina y potenciación de la acción del glucagón que ejerce el cortisol, en estos toros los valores de glucosa han aumentado desde 113 mg/dl (antes de la lidia) hasta 184 mg/dl (des-pués de la lidia), lo que demuestra la acción hiperglucemiante del cortisol (gráfi co 2).
Los valores de cortisol plasmático obtenidos antes de la lidia de estos toros es-taban dentro de los límites expuestos como normales por la mayoría de los autores (6,7,8). Es obvio, que los valores hallados en el presente trabajo como basales estén algo elevados debido a la reacción de estrés que desencadena el inmovilizar al toro bravo para la extracción de sangre antes de la lidia. Después de la lidia se evidenció un fuerte incremento de los niveles de cortisol como se aprecia en la tabla III. También se obtuvieron incrementos de cortisol directamente proporcionales al rendimiento o capacidad física mostrada por los animales durante la lidia (tabla IV). Estos datos coinciden con los presentados por otros autores (7, 9) que encontraron valores de cortisol elevados en animales que más ejercicio habían realizado. La hiperventilación producida por el ejercicio conlleva a un aumento de la presión parcial de CO2 y una hipoxia en el tejido nervioso, lo que desencadena una rápida respuesta hormonal con la consiguiente liberación de cortisol (10,11). Es posible que en el ejercicio que realiza el toro durante la lidia se produzca una hipoxia del tejido nervioso debido a la forma de embestir tras la muleta con la cabeza agachada y describiendo semicircunferencias cerradas, generándose una fatiga considerable, a lo que hay que unir la cantidad de sangre que pierde durante la lidia. Esto explicaría que los incrementos en los niveles plasmáticos de cortisol sean más acentuados en los toros después de la lidia que antes de la misma. Se ha descrito (12) que los ejercicios de intensidad superior al 60% de la capacidad aerobia máxima provocan una elevación de los niveles plasmáticos de cortisol de forma considerable. Parece ser que, debido a la intensidad y duración del ejercicio que realiza el toro, sobrepasa este umbral la mayoría de las veces.
Muchos tipos de traumatismos se caracterizan por incremento de la secreción cortisol, lo cual guarda correlación con la intensidad de la lesión (1). Se piensa que

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
204
el cortisol es un mediador importante de la respuesta metabólica al traumatismo, esto se ha comprobado porque los animales con ausencia de glándulas adrenales tienen una respuesta pobre cuando se encuentran sometidos a estrés. Se piensa que la importancia del cortisol durante la respuesta al estrés radica en que modifi ca el metabolismo de la glucosa poniendo a disposición del cerebro y músculo mayor cantidad de este sustrato energético, facilitando la acción de las catecolaminas y previniendo una reacción exagerada del sistema inmune a las lesiones. En el toro bravo la liberación de cortisol puede estar relacionada con la colocación de la puya y banderillas que lesionan los músculos del cuello y el dorso provocando una serie de fenómenos como hiperglucemia, alteraciones hidroelectrolíticas, cambios neuroen-docrinos, hipertermia y cambios hemodinámicos que se conoce como un síndrome de respuesta infl amatoria sistémica (SRIS). Por tanto, a partir de la suerte de varas, el toro presenta lesión muscular y hemorragia profusa que inducen a disminución de la perfusión tisular (hipoxia), hipovolemia y dolor, siendo estos dos últimos los factores principales que desencadenan las respuestas neuroendocrinas a las lesiones. Todo ello tiene que hacerlo compatible con el ejercicio y demostrar un alto rendimiento físico que es lo que se espera de él en la plaza.
2. RESPUESTA METABÓLICA
La respuesta metabólica muscular a la lidia en el presente trabajo muestra un marcado carácter glucolítico, observándose concentraciones bajas de glucógeno, junto con niveles altos de lactato y pH bajo, lo que sugiere que la glucogenolisis es la vía más importante de liberación de energía durante las situaciones estresantes que ocurren durante la lidia coincidiendo con autores de la bibliografía consultada (13).
El acúmulo de glucógeno en el músculo está supeditado a muchos factores, sien-do quizá los más relevantes la dieta y el nivel de entrenamiento (14). Una alimentación rica en glúcidos y lípidos favorece el almacenamiento de glucógeno, propiciando su uso como fuente energética durante un esfuerzo (15).
Varios motivos podrían proponerse para explicar este fuerte consumo de glucó-geno en respuesta a la lidia obtenido en este trabajo, tal y como se refl eja en la tabla III. En primer lugar, no se puede obviar el efecto metabólico del estrés psíquico. Un segundo motivo de la importante depleción glucogénica podría ser la duración de la lidia. De los 20 minutos que dura la misma, el toro realiza ejercicio sólo 387 segundos, lo que supone 1/3 del total. Las actividades físicas de duración prolongada dependen energéticamente tanto de los glúcidos como de los lípidos. En opinión de los autores,

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
205
los lípidos desempeñarían un papel bastante secundario durante la lidia. Los mo-vimientos respiratorios y jadeos del toro bravo refl ejarían un alto grado de hipoxia, circunstancia que se ha comprobado tras la determinación de los niveles de lactato en el músculo (tabla III). Ello lleva a suponer que el consumo de oxígeno (VO2) no alcanzaría una magnitud sufi ciente para metabolizar los lípidos.
En tercer lugar la disminución de los fosfatos de alta energía podría ser con-secuencia del metabolismo anaerobio derivado de hipoperfusión y la hipoxia o del efecto quimiotáctico de las células infl amatorias. La disminución de los fosfatos de alta energía durante la lidia, igual que en cualquier lesión suele ocurrir en el hígado y los riñones antes que en el miocardio y músculo esquelético (1), pudiendo ser la causa de la incapacidad motora que presentan algunos toros en los espectáculos taurinos.
Otro resultado sorprendente ha sido el reducido valor del pH muscular, bastante por debajo de los límites fi siológicos (pH medio de 5.94±0.2). Ciertas investigaciones realizadas en atletas humanos han propuesto un pH de 6.5 como el límite inferior para una actividad muscular razonable (16, 17). Además, estudios realizado en ca-ballos en caballos, se hallaron concentraciones de lactato de 149 y 204 mmol/kg p.s., correspondientes a pHs de 6.5 y 6.3 respectivamente. En este trabajo en el toro bravo, a pesar de encontrar niveles de lactato similares (203 mmol/kg p.s), el pH muscular fue sustancialmente inferior. No existe una explicación para esta circunstancia, ya que se desconoce la actividad tamponadora del músculo bovino. Esta viene determinada por las tasas de fosfocreatina y sobre todo por el sistema bicarbonato, el cual se sabe que se incrementa con el entrenamiento. Además, los sprinters de élite presentan ma-yores concentraciones de lactato e inferiores de bicarbonato tras un ejercicio máximo, refl ejando el potencial buffer para tamponar los hidrogeniones, de manera que el músculo muestra un pH superior al de los atletas menos exitosos (18, 19, 20), lo que indica que el toro no se encuentra entre las especies atléticas.
Quizás el pH inferior del músculo del toro bravo en respuesta a niveles de lactato similares al caballo resalte la selección ejercida sobre esta última especie, encauzada para el deporte. Además, estos datos provienen del caballo Pura Sangre Inglés, raza conocida por su importancia en carreras de velocidad. En los bovinos, por el contrario, se ha realizado una selección hacia la producción cárnica o láctea, o más exactamen-te, en el caso particular del toro bravo, con relación a unos criterios morfológicos y comportamentales, sin tener en cuenta criterios de su potencial físico.
Sin duda, la acidosis también contribuye a la aparición de fatiga muscular durante la lidia, ya que el descenso de pH va asociado a una disminución de la capacidad

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
206
respiratoria de la fi bra muscular, con efectos directos sobre la actividad del aparato contráctil (21).
El ácido láctico es producido por la glucólisis anaerobia. Los cambios en el ritmo de producción de lactato refl ejan las variaciones en el grado al cual el metabolismo anaerobio contribuye a la producción de energía. Ante una situación de ejercicio intenso, son las fi bras musculares de contracción rápida y baja capacidad oxidativa (tipo II) las que mayoritariamente entran en funcionamiento. Hay que tener en cuenta que la concentración plasmática de lactato aumenta en muchas lesiones de manera correlacionada con la gravedad de éstas. La acumulación del metabolito en el toro explica, en parte, su acidosis progresiva y se deriva del metabolismo anaerobio de los tejidos isquémicos. Durante la lidia es posible que el toro sufra el efecto negativo del ácido láctico, dando lugar a la fatiga muscular. El mecanismo de acción de este metabolito es indirecto a partir de la liberación de protones, produciendo un descenso importante del pH, tal como se observado en este estudio, que inhibe la actividad de enzimas como fosfofructoquinasa (enzima glucolítica) y la miosín-ATP-asa (22).
El exceso de lactato no se debe confundir con sus valores séricos totales, dado que aquél es la cantidad de lactato sanguíneo que hace que aumente la razón de lactato/piruvato por arriba de lo normal (1).
Se ha demostrado que la actividad de las enzimas plasmáticas se incrementa después de alteraciones musculares o ejercicio intenso. Las elevaciones de las acti-vidades enzimáticas de CK, LDH y AST son superiores signifi cativamente (tabla II) a los valores descritos en la bibliografía para el ganado vacuno pero coinciden con los valores expuestos por Purroy y col., (1) que indicarían un grado de lesión mus-cular. La CK es indicadora de la severidad del ejercicio, mientras que la AST puede ser indicadora del sobreentrenamiento. La LDH además de indicar lesión muscular es clave en el metabolismo anaerobio. Se han encontrado diferencias signifi cativas superiores al 99.9% al comparar la actividad enzimática de CK antes y después de la lidia que es el principal marcador de daño muscular (Tabla II). La lidia supone un ejercicio intenso para el animal si consideramos el aumento de enzimas provocado por la hipoxia muscular que se produce por la escasa preparación física de los toros para el ejercicio y como se ha mencionado anteriormente, por la disminución del riego tisular como consecuencia de la hemorragia. Esto conlleva a un incremento de la generación de radicales libres y especies reactivas del oxígeno y del nitrógeno (12, 23), lo que unido a una reducida capacidad para eliminarlos, por la baja concentración de antioxidantes como GSH-Px, expondría a los animales a sufrir más fácilmente lesiones en la membrana celular y en las mitocondrias (24). En un estudio llevado a

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
207
cabo por Agüera y cols., (14), expusieron que los biomarcadores antioxidantes pueden mejorarse en el toro bravo si se somete a éste a un periodo de entrenamiento de 24 semanas, lo que evitaría estas posibles lesiones de la membrana celular provocada por el ejercicio físico y mejorando su rendimiento en la plaza.
El ejercicio realizado durante la lidia provoca liberación del potasio desde la fi bra muscular. El potasio, una vez fuera, actúa como un potente vasodilatador para asegurar la hipertermia en el músculo activo (25, 26, 27). El incremento de la con-centración de potasio en plasma durante el ejercicio máximo puede ser asignado, en parte, al incremento extracelular, el cual se acompaña de acidemia y liberación de K+ a través de la fi bra muscular ejercitada, concomitante con una pérdida de sudor (28, 2). El descenso del potasio intracelular desencadena una pérdida del control de la membrana sarcolémica por el organismo.
Todo traumatismo origina cambios rápidos del volumen extracelular funcional, volumen circulante efectivo, osmolalidad extracelular y composición electrolítica, que resulta en la estimulación del sistema neuroendocrino. A su vez, la respuesta de éste induce alteraciones de las funciones renal y circulatoria encaminadas a mejorar el esta-do hidrosalino. Estas alteraciones serán más acentuadas cuanto más grave sea lesión.
El exceso de potasio encontrado en plasma esta investigación induce a pensar que, la fatiga de toro bravo durante la lidia puede deberse, en parte, a un exceso de potasio en el líquido extracelular, que hace que el corazón se dilate al extremo y quede fl ácido, disminuyendo la frecuencia cardiaca, ya que baja el potencial de reposo de las fi bras del miocardio, lo que debilita progresivamente la contracción del corazón (29).
3. CAMBIOS HEMATOLOGICOS
Los fundamentos fi siológicos de los valores hematológicos están, sobre todo, en el papel fundamental que juega la hemoglobina en la cadena del transporte de oxígeno. La capacidad del transporte de oxígeno de la sangre depende del volumen de células rojas del cuerpo (30). Este volumen se expresa con diferentes parámetros: número de glóbulos rojos/ml de sangre; la concentración de hemoglobina (gr/dl); el hematocrito (%).
El análisis hematológico se halla infl uenciado por numerosos factores extrínse-cos e intrínsecos del animal. En consecuencia los cambios hematológicos ocurridos durante la lidia divergen en función del estrés, de las lesiones y del ejercicio llevado a cabo por el animal durante la corrida.

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
208
En la lidia, como en todo tipo de ejercicio hay un incremento en la fuerza de bombeo del corazón y una redistribución del fl ujo sanguíneo que incrementa dicho fl ujo a través del músculo esquelético. Estos ajusten proveen aumento del aporte de oxígeno para la producción aeróbica de energía y mantener la respuesta del organis-mo al ejercicio (31).
Aunque después del trabajo realizado por el toro cabría esperar un aumento del número de glóbulos rojos como consecuencia de la esplenocontracción por el ejercicio, los valores que se presentan el la tabla I demuestran la disminución del número de glóbulos rojos, como consecuencia de la pérdida de sangre sufrida por estos animales. La respuesta neuroendocrina clásica a la hemorragia trata de mantener la perfusión para el corazón y el cerebro, a veces a expensas de sacrifi car otros órganos y sistemas. Esta vasoconstricción selectiva se presenta por acción de mecanismos mediados por vía central (32), lo que puede afectar al aporte de oxígeno al sistema muscular del toro tan necesario a la hora de realizar el ejercicio. La activación de la respuesta infl amatoria a la hemorragia y a la lesión tisular es un componente importante de la fi siopatología del shock hemorrágico.
Se descarta que la lidia provoque una hemólisis fi siológica por el esfuerzo intenso que supone ya que los valores que muestran en la tabla I no varían signifi cativamente antes y después de la lidia. En cuanto al hematocrito se obtiene un ligero descenso debido a la hemorragia, al contrario de lo que se esperaba, ya que el toro manifi esta deshidratación. Esto se detecta por el aumento signifi cativo de las proteínas plas-máticas totales tal y como se muestra en la tabla II, pues es uno de los parámetros fi siológicos más constantes de la bioquímica sanguínea.
Los índices volumétricos se mantienen constantes apreciándose diferencias signifi cativas sólo del 95% en el CHCM porque la hemoglobina se mantiene y dis-minuye el número de glóbulos rojos debido a la hemorragia profusa que mantiene el toro durante toda la faena.
El leucograma del toro bravo exige una interpretación cuidadosa debido a que existen ciertos factores que pueden modifi carlo (33). La fórmula leucocitaria del bo-vino es tipo linfoide. En esta investigación se describe la fórmula leucocitaria de tipo neutrofílico coincidiendo con otros autores consultados (9). Se han descrito depresión en la función de los linfocitos y cambios en la subpoblación de linfocitos en animales y humanos después de traumatismo o shock hemorrágico (34). Esta neutrofi lia (gráfi co 1) puede ser debida al estrés, al ejercicio y a la respuesta infl amatoria sistémica que incluyen las lesiones por isquemia-reperfusión y la activación de neutrófi los. La dis-

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
209
minución del fl ujo sanguíneo por los capilares con isquemia prolongada con ausencia de restitución del fl ujo y reperfusión puede iniciar la activación de los neutrófi los en los toros con shock hemorrágico, es decir los lidiados. Durante el ejercicio disminuye el cociente neutrófi los/linfocitos por la liberación de estos a la circulación (35, 21).
Rendimiento físico
El rendimiento físico se evaluó tal y como se describe en el material y métodos. El gráfi co 3 representa la clasifi cación de los toros en grupos de rendimiento según la capacidad física mostrada durante la lidia.
No se apreció una correlación signifi cativa, en los parámetros estudiados, excepto para el cortisol en el que se apreció un aumento de los incrementos medios de éste tras la lidia, directamente proporcional al rendimiento físico observado, es decir, los toros que hicieron más ejercicio tuvieron un incremento de cortisol más alto como se puede apreciar en la tabla IV.
CONCLUSIONES
Como conclusiones a esta investigación, se exponen como factores limitantes del toro durante la lidia los siguientes:
- Disminución de la volemia y del número de glóbulos rojos, y con ello disminución de la capacidad de transporte de oxígeno, debido a la he-morragia, lo que provoca una vasoconstricción selectiva para mantener el aporte a órganos vitales como cerebro y corazón, afectando al sistema muscular y, por tanto el rendimiento físico durante la lidia.
- La lesión muscular que provoca dolor, hipoperfusión y el desencadena-miento de los mediadores de la infl amación lo que da lugar al síndrome de respuesta infl amatoria sistémica.
- La deshidratación del animal por la realización de ejercicio y acentuada por la hemorragia.
- La fatiga del toro bravo durante la lidia puede deberse el aumento de los niveles de potasio, el cual provoca alteraciones funcionales del corazón, a la vez que se requiere de éste un aumento del gasto cardíaco durante el ejercicio.

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
210
- La fuerte depleción de glucógeno muscular se asocia al grado de hipoxia tisular, la hipoperfusión, el efecto quimiotáctico de las células infl ama-torias provocando incapacidad motora de la fi bra.
- La acidosis muscular es consecuencia de la vía anaerobia que sigue el glucógeno para la obtención de energía, con el gran acúmulo de lactato que ello supone, y la consiguiente disminución del pH muscular, lo que afecta a la actividad del aparato contráctil del músculo.
BIBLIOGRAFÍA
1. Schwartz S. I. (1990). Principios de Cirugía. Respuestas endocrinas y metabólicas a la lesión. Edi-torial Interamericana. Quinta Edición; 1-59
2. Agüera E. I. , Rubio M. D. , Agüera S. , Escribano B. M. , Castejón F. (1995). Blood parameters and heart rate response to training in Andalusian horses. J. Physiol. Bioch. 51 (2): 55-64.
3. López Rivero J. L. (1990). Anatomía, fi siología y bioquímica del músculo equino. En: Equino: Aspectos de cría y clínica. Consejo General de Colegios de Veterinaria de España. Publex Studio Edición.
4. Agüera E. I. , Rubio M. D. , Vivo R. , Escribano B. M. , Muñoz A. , Villafuerte J. L. , Castejón, F. (1998). Adaptaciones fi siológicas a la lidia en el toro bravo. Parámetros plasmáticos y muscula-res. Vet. Méx. 29 (4): 399-403.
5. Weissman, Ch. (1990). The metabolic response to stress: An overview and update; Anesthesiol-ogy. 73: 308-327
6. Jones S. , Calkins C. , Johnson R. , Dikeman M. (1998). The effects of trombolones acetate on cor-tisol release and growth in steers and bulls. Mp. Univ. Neb. Agric.
7. Gascón M. , Marca M. C. , Purroy A. , García- Belenguer S. , Gonzálz J. M. (1990). Actividad de la LDH y sus isoenzimas en toros bravos después de la lidia. Med. Vet. Vol 7 (10).
8. Castro J. M. (1992). Estudio de la capacidad de adaptación de la raza de lidia a diferentes prácti-cas de manejo. Tesis Doctoral. Facultad de Veterinaria. León.
9. Purroy A. , García- Belenguer S. , González J. M. , Barberán M. (1992). Lesions musculaires et activités enzymatiques chez les bovines de combat Ann. Rech. Vet. 23: 59-62.
10. Galbo H. (1983). Hormonal and metabolic adaptation to exercise. Georg Thieme Velag, Stuttgar-tpp: 30-34; 46-50; 62-85.
11. Fernández J. J. , Diego A. M. , Fernández V. J. (1992). Hormonas y ejercicio. En: Fisiología de la actividad física y del deporte. Ed. Interamerica. McGraw-Hill. 1ª edición. Pp: 95-128.
12. Davies KJA. 1982. Free radicals and tissue damage produced by exercise. Biochem Biophys Res Commun 107:1198-12-05.
13. Agüera E. I. , Muñoz A. , Castejón F. , Essén-Gustavsson B. (2001-a). Skeletal muscle fi bre cha-racteristics in young and old bulls and metabolic response after a bullfi ght. J. Vet. Med. A. 48: 313-319.
14. Agüera E. I. , Escribano B. M. , Rubio M. D. , De Miguel R. , Requena, F. , Tovar, P. (2005). Valo-ración de biomarcadores oxidantes y antioxidantes en toro bravo sometido a un programa de entrenamiento. VII Symposium del toro de lidia. Zafra, Badajoz.
15. Pagan J. P. , Essén-Gustavsson B. , Lindholm A. , Thornton, J. (1987). The effect of dietary energy source on exercise performance in Standardbred horses. En: Equine Exercise Physiology 2. (Gil-lespie, J. R. ; Robinson, N. E. , Eds. ). ICEEP Publications. Davis. CA. pp: 686-700.

FACTORES LIMITANTES DEL RENDIMIENTO FISICO DEL TORO BRAVO DURANTE LA LIDIA
211
16. Sahlin K. , Harris R. C. , Hultman, E. (1975). Creatine kinase equilibrium and lactate content compared with muscle pH in tissue samples obtained after isometric contration. Biochem. J. 152: 173-180.
17. Sahlin K. , Harris R. C. , Hultman E. (1976). Lactate content and pH in muscle samples obtained after dynamic exercise. Pfl ügers Arch. 367: 143-149.
18. Svedenhag J. , Sjödin B. (1984). Maximal and submaximal oxygen uptakes and blood lactate levels in elite middle- and long-distance runners. Int. J. Sports Med. 5: 255-261.
19. Heck H. , Mader A. , Hess G. , Mucke S. , Muller R. , Hollmann W. (1985). Justifi cation of the 4-mmol/l lactate threshold. Int. J. Sports Med. 6: 117-130.
20. Harkins J. D. , Beadle R. E. , Kamerling S. G. (1993). The correlation of running ability and physi-ological variables in thoroughbred racehorses. Equine Vet. J. 25(1): 53-60.
21. Snow D. H. , Valberg S. J. (1994). Muscle anatomy, physiology and adaptations to exercise and training. En: The Athletic Horse: principles and practice of equine sports medicine (Hodgson, D. R. ; Rose, R. J. , Eds. ). W. B. Saunders Company. Philadelphia. pp: 145-179
23. Jackson MJ, Edwards RH, Symons MC. (1985). Electron spin resonance studies of intact mam-malian skeletal muscle. Biochem Biophys. Acta 847:185-190.
24. Rice D. , Kennedy S. (1988) Vitamin E: function and effects of defi ciency. Br. Vet. J. 144: 482-496. 25. Coffman, J. R. (1983). Proceedings of 28th Congress of the American Association of Equine
Practtioners. Las Vegas, pp:15. 26. Carlson G. P. (1987). Hematology and body fl uids in equine athlete. A review. En: Equine Exerci-
se Physiology 2. J. R. Gillespie and N. R. Robinson Eds. ICEEP Publications. Davis, Ca. 393-452. 27. Monreal, L. , Lavin S. , Viñas (1990). Fisiopatología del ejercicio en el caballo: La carrera o el
esfuerzo de velocidad. Med. Vet. 7 (12): 647-665. 28. Cohen N. D. , Rousset A. J. , Lumsden J. H. , Cohen A. C. , Grift E. , Lewis C. (1993). Alterations of
fl uids and electrolyte balance in Thoroughbred race horses following strenuous exercise during training. Can. J. Vet. Res. 57: 9-13.
29. Guyton A. C. , Hall J. E. (2006). Tratado de Fisiología Médica. 10ª Edición. MacGraw-Hill Inte-ramericana.
30. Persson S. G. B. (1983). Evaluation of exercise tolerante and fi tness in the performance horses. En: Equine Exercise Physiology. (Snow, D. H. , Persson, S. G. B, Rose, R. J. Eds. ). Granta Edi-tions. Cambridge. Pp: 441-457.
31. Shen W. (1995) Nitric oxide production and NO synthase gene expression contribute to vascular regulation during exercise. Med. Sci. Sporst. Exerc. 27(8): 1125-1134.
32. Peitzman A. (1995) Hemorrhagic shock. Curr. Probl. Surg. 32(11): 925-1002 33. Agüera E. I, Santisteban R. , Villafuerte J. L. , Escribano M. D. , Rubio, M. D. (2001-b) Estudio del
eritrograma y leucograma en el toro bravo. Med. Vet. Vol 18 (5): 430-435. 34. Tyburski J. (1994). Regional differences in lymphocite function following resuscitated hemor-
ragic shock. J. Trauma. 37(3): 469-47235. Rossdale P. D. , Buguez P. N. , Cash R. S. G. (1982). Changes in blood neutrophil/linphocite ratio
related to adrenocortical function in the horsse. Equine Vet. J. 14: 293-298


SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
213ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
RESUMEN
La ecografía es una técnica de diagnóstico por imagen segura, no invasiva y que no requiere una preparación excesiva del animal. Se utiliza para estudiar tejidos blandos, permi-tiendo la valoración del tamaño, forma, situación y estructura de los mismos. La ecografía o ultrasonografía se basa en los ultrasonidos generados en un dispositivo llamado transductor, compuesto por cristales piezoeléctricos. La Histopatología constituye un medio diagnóstico especializado, complementario de otras técnicas diagnósticas como la ecografía y que, en la mayoría de los casos, resulta esencial en cuanto a establecer un diagnóstico defi nitivo en muchos procesos patológicos. La toma de biopsias para realizar un estudio histopatológico requiere una acción agresiva “in vivo” sobre el animal pudiendo servirse de la técnica ecográfi ca para realizar la toma de muestras de los diferentes órganos internos.El objetivo de este trabajo es el establecimiento de una correlación directa entre la imagen ultrasónica de órganos normales y patológicos procedentes de animales eutanasiados o de extirpaciones quirúrgicas de rutina, con la imagen macroscópica e histopatológica de esos mismos órganos.
Palabras clave: Ecografía, histopatología, biopsia ecoguiada.
ABSTRACT
Ultrasound is a diagnostic imaging technique safe, non invasive and does not require excessive grooming of the animal. Is used to study soft tissue, allowing the assessment of the size, shape, location and structure of the data. Ultrasound sonography, based on the ultrasound
SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
DR. A. MORENO BOISO1, DR. J. HERVÁS RODRÍGUEZ2 Y DR. F. CHACÓN2
1 Hospital Veterinario Alhaurín el Grande. Málaga.2 Histolab Veterinaria. Fuengirola. Málaga

SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
214
generated in a device called a transducer, comprising crystals piezoeléctricos.La constitutes a diagnostic Histopathology specialized complementary to other diagnostic techniques such as ultrasound and that, in most cases, it is essential as for a defi nitive diagnosis in many disease processes. The biopsy for histopathological study requires aggressive action “in vivo” on the animal can use ultrasound technique for sampling of the different organs internos.El aim of this study is to establish a direct correlation between the ultrasound image of normal and pathological organs from animals euthanized or routine surgical excision with macroscopic images and histopathologyf these same organs.
Key words: Ultrasound, histopathology, ultrasound-guided biopsy.
INTRODUCCIÓN
La ecografía es una técnica de diagnóstico por imagen segura, no invasiva y que no requiere una preparación excesiva del animal. Se utiliza para estudiar tejidos blandos, permitiendo la valoración del tamaño, forma, situación y estructura de los mismos. La ecografía o ultrasonografía se basa en los ultrasonidos generados en un dispositivo llamado transductor, compuesto por cristales piezoeléctricos. Las ondas de sonido viajan a través de los tejidos y son devueltas en forma de ecos, que se refl ejan en la pantalla en forma de puntos de luz de intensidad variable. Los transductores en veterinaria utilizan frecuencias de 2.5 a 7.5 MHz. Cuanto mayor es la frecuencia, menor es la profundidad de penetración pero mayor es la resolución o defi nición de la imagen. Generalmente se utilizan 3 MHz en perros de razas gigantes; 5 MHz en razas medianas y pequeñas; 7.5 MHz en perros toy y gatos y 9 MHz en ecografía de ojo.
La imagen ecográfi ca que está formada por puntos de luz de diferente brillo, corresponden a una intensidad variable. Esto se llama ecogenicidad. Se defi ne:
- Hiperecogénico o hiperecoico: corresponde a una mayor refl exión de sonidos. Los puntos en la pantalla aparecen blancos (imágenes de gas y hueso).
- Hipoecogénico o hipoecoico: se ven diferentes tonalidades de gris según la cantidad de ecos producidos. Se corresponden con tejidos blandos.
- Anecogénicos: ausencia total de ecos por una transmisión completa de ultrasonidos. Se ve en negro la pantalla. Se corresponde con líquidos.
Asimismo, se han de tener en cuenta imágenes que no corresponden a la realidad, pero que aparecen con cierta frecuencia en la ecografía:
- Sombra acústica: se forma detrás de una estructura que bloquea el paso de ultrasonidos, por ejemplo un cálculo.
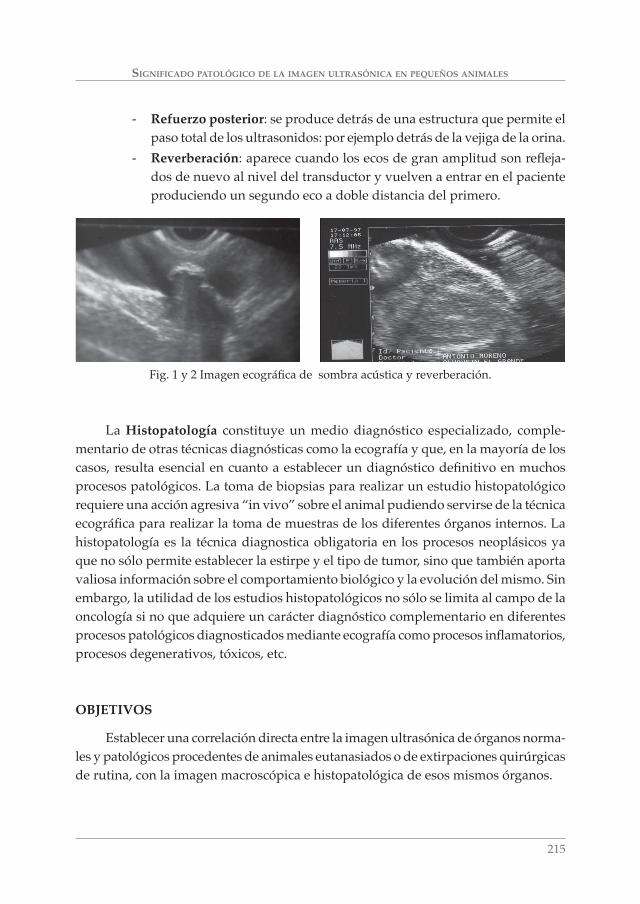
SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
215
- Refuerzo posterior: se produce detrás de una estructura que permite el paso total de los ultrasonidos: por ejemplo detrás de la vejiga de la orina.
- Reverberación: aparece cuando los ecos de gran amplitud son refl eja-dos de nuevo al nivel del transductor y vuelven a entrar en el paciente produciendo un segundo eco a doble distancia del primero.
Fig. 1 y 2 Imagen ecográfi ca de sombra acústica y reverberación.
La Histopatología constituye un medio diagnóstico especializado, comple-mentario de otras técnicas diagnósticas como la ecografía y que, en la mayoría de los casos, resulta esencial en cuanto a establecer un diagnóstico defi nitivo en muchos procesos patológicos. La toma de biopsias para realizar un estudio histopatológico requiere una acción agresiva “in vivo” sobre el animal pudiendo servirse de la técnica ecográfi ca para realizar la toma de muestras de los diferentes órganos internos. La histopatología es la técnica diagnostica obligatoria en los procesos neoplásicos ya que no sólo permite establecer la estirpe y el tipo de tumor, sino que también aporta valiosa información sobre el comportamiento biológico y la evolución del mismo. Sin embargo, la utilidad de los estudios histopatológicos no sólo se limita al campo de la oncología si no que adquiere un carácter diagnóstico complementario en diferentes procesos patológicos diagnosticados mediante ecografía como procesos infl amatorios, procesos degenerativos, tóxicos, etc.
OBJETIVOS
Establecer una correlación directa entre la imagen ultrasónica de órganos norma-les y patológicos procedentes de animales eutanasiados o de extirpaciones quirúrgicas de rutina, con la imagen macroscópica e histopatológica de esos mismos órganos.

SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
216
MATERIAL Y MÉTODOS
En la elaboración de las imágenes ultrasónicas se trabajó con el sistema “FAZONE CB “ de la marca FUJIFILM , provisto con una sonda miniconvex, de 3 a 9 megaherzios .con tecnología “ zonareR que permite imágenes de alta resolución sobre la base de casos clínicos rutinarios del Hospital Veterinario Alhaurín el Grande en Alhaurin el Grande (Málaga)
Para el estudio histopatológico, los tejidos se proce-saron según las técnicas rutinarias en microscopía óptica utilizando un procesador de tejidos Histokinette 2000 y un dispensador de parafi na Histotap Plus. Se realizaron cortes de 4 μm de grosor en los bloques de parafi na ob-tenidos con un microtomo 2025 RM que, tras su fi jación mediante calor, fueron teñidos con la técnica rutinaria en histopatología de Hematoxilina-Eosina, así como con técnicas especiales de tinción como P.A.S., Ziehl-Nielsen, Giemsa, Plata o azul de toluidina.
RESULTADOS
A continuación se relacionan los órganos estudia-dos, tanto en su forma ecográfi ca como sus alteraciones macro y microscópicas.
HÍGADO
Para el estudio ecográfi co del hígado se sitúa el animal en decúbito supino y se coloca el transductor orientado cranealmente inmediatamente caudal a la apófi sis xifoides. El hígado es uniformemente hipoecogénico con respecto al bazo e igual o ligeramente hiperecogénico con respecto a la corteza renal. Presenta una granulación más grosera que la del bazo, pudiéndose observar la vesícula biliar, la vena cava caudal, la vena porta y los vasos intrahepáticos mayores. Los ecos producidos por la cápsula se ven como una fi na línea en las áreas de contacto entre dos lóbulos. La pared de la cava tiene una mayor ecogenicidad que la porta, lo cual es un signo diferencial.
Entre las alteraciones del parénquima hepático encontramos:
• Neoplasias: de distinto tamaño y ecogenicidad. Suele ser hipoecogénicas o de ecogenicidad mixta, deformando el contorno cuando son de gran tamaño. La
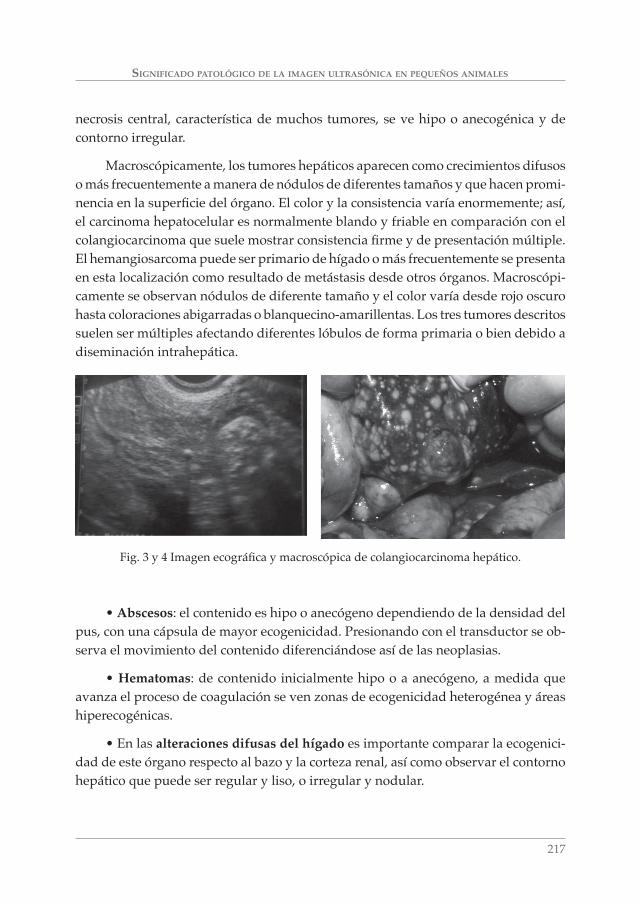
SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
217
necrosis central, característica de muchos tumores, se ve hipo o anecogénica y de contorno irregular.
Macroscópicamente, los tumores hepáticos aparecen como crecimientos difusos o más frecuentemente a manera de nódulos de diferentes tamaños y que hacen promi-nencia en la superfi cie del órgano. El color y la consistencia varía enormemente; así, el carcinoma hepatocelular es normalmente blando y friable en comparación con el colangiocarcinoma que suele mostrar consistencia fi rme y de presentación múltiple. El hemangiosarcoma puede ser primario de hígado o más frecuentemente se presenta en esta localización como resultado de metástasis desde otros órganos. Macroscópi-camente se observan nódulos de diferente tamaño y el color varía desde rojo oscuro hasta coloraciones abigarradas o blanquecino-amarillentas. Los tres tumores descritos suelen ser múltiples afectando diferentes lóbulos de forma primaria o bien debido a diseminación intrahepática.
Fig. 3 y 4 Imagen ecográfi ca y macroscópica de colangiocarcinoma hepático.
• Abscesos: el contenido es hipo o anecógeno dependiendo de la densidad del pus, con una cápsula de mayor ecogenicidad. Presionando con el transductor se ob-serva el movimiento del contenido diferenciándose así de las neoplasias.
• Hematomas: de contenido inicialmente hipo o a anecógeno, a medida que avanza el proceso de coagulación se ven zonas de ecogenicidad heterogénea y áreas hiperecogénicas.
• En las alteraciones difusas del hígado es importante comparar la ecogenici-dad de este órgano respecto al bazo y la corteza renal, así como observar el contorno hepático que puede ser regular y liso, o irregular y nodular.
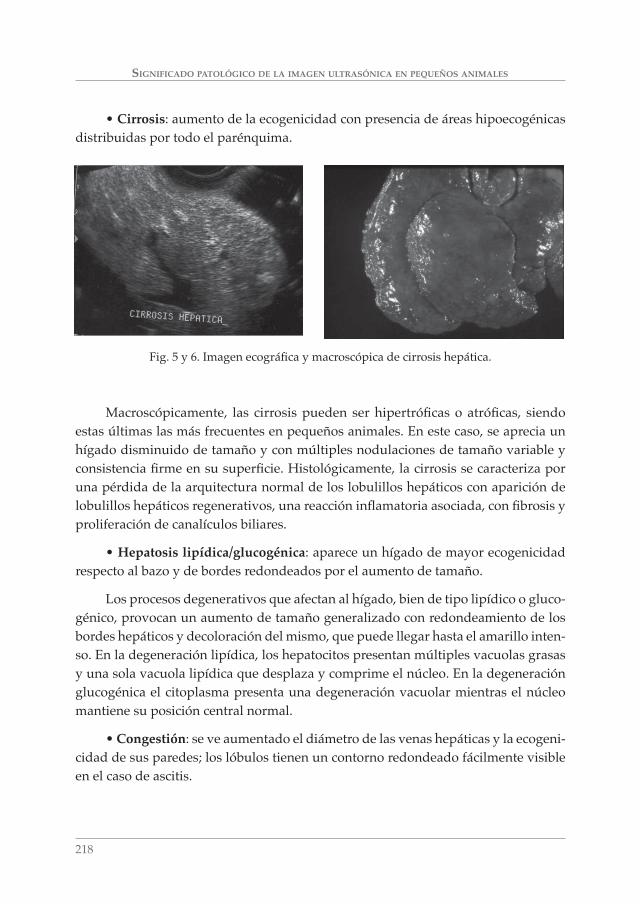
SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
218
• Cirrosis: aumento de la ecogenicidad con presencia de áreas hipoecogénicas distribuidas por todo el parénquima.
Fig. 5 y 6. Imagen ecográfi ca y macroscópica de cirrosis hepática.
Macroscópicamente, las cirrosis pueden ser hipertrófi cas o atrófi cas, siendo estas últimas las más frecuentes en pequeños animales. En este caso, se aprecia un hígado disminuido de tamaño y con múltiples nodulaciones de tamaño variable y consistencia fi rme en su superfi cie. Histológicamente, la cirrosis se caracteriza por una pérdida de la arquitectura normal de los lobulillos hepáticos con aparición de lobulillos hepáticos regenerativos, una reacción infl amatoria asociada, con fi brosis y proliferación de canalículos biliares.
• Hepatosis lipídica/glucogénica: aparece un hígado de mayor ecogenicidad respecto al bazo y de bordes redondeados por el aumento de tamaño.
Los procesos degenerativos que afectan al hígado, bien de tipo lipídico o gluco-génico, provocan un aumento de tamaño generalizado con redondeamiento de los bordes hepáticos y decoloración del mismo, que puede llegar hasta el amarillo inten-so. En la degeneración lipídica, los hepatocitos presentan múltiples vacuolas grasas y una sola vacuola lipídica que desplaza y comprime el núcleo. En la degeneración glucogénica el citoplasma presenta una degeneración vacuolar mientras el núcleo mantiene su posición central normal.
• Congestión: se ve aumentado el diámetro de las venas hepáticas y la ecogeni-cidad de sus paredes; los lóbulos tienen un contorno redondeado fácilmente visible en el caso de ascitis.

SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
219
Macroscópicamente, el hígado aparece aumentado de tamaño, de color rojo oscuro y, al corte, fl uye abundante sangre. Microscópicamente se aprecia un acúmulo de eritrocitos tanto en los sinusoides hepáticos como en los vasos de mayor calibre. Congestión hepática se asocia a multitud de procesos patológicos que van desde el shock anafi láctico hasta procesos endotóxicos.
• En los casos de comunicación porto-sistémica lo más llamativo es el escaso tamaño del hígado; la ecogenicidad es similar a la del bazo y no se observa la porta.
• Hepatitis crónica necrotizante: se observan múltiples focos hipo o anecógenos de contorno irregular que confl uyen unos con otros. El resto del parénquima se ve hiperecogénico debido a la fi brosis.
Otros procesos donde se puede apreciar un aumento difuso del hígado son las hepatitis multifocales del tipo de la hepatitis granulomatosa multifocal que se pro-duce en la leishmaniosis canina, la hepatozoonosis canina o en la hepatitis vírica del perro. Todos estos procesos son fácilmente diagnosticados histológicamente mediante la visualización del agente etiológico en cuestión (formas amastigotes de Leishmania sp. en el citoplasma de macrófagos, macro y microesquizontes de Hepatozoon canis o cuerpos de inclusión intranucleares y basófi los en el caso de la hepatitis vírica canina, respectivamente).
Vesícula biliar
Se localiza en la región paramedial derecha, entre el lóbulo medial derecho y el lóbulo cuadrado. La vesícula biliar aparece con una estructura ovalada y el contenido anecógeno o hiperecogénico. La bilis concentrada es hipoecogénica con preferencia de elementos corpusculares, los cuales sedimentan en reposo (barro biliar); esto pue-de considerarse como un hallazgo ecográfi co normal. El tamaño de la vesícula va a depender del momento de la ingesta.
• El aumento de tamaño se observa en casos de anorexia prolongada y/u obs-trucciones biliares extrahepáticas.
• Los cálculos biliares también son visibles ecográfi camente, apareciendo la som-bra acústica dorsal. Incluso es posible el movimiento de ésta, cambiando de posición al animal. Hay que diferenciarlo de los artefactos producidos por superposiciones del sistema gastrointestinal.
Los colelitos suelen mostrar gran variación de tamaño y consistencia, producien-do en muchos casos un aumento de grosor de la pared de la vesícula biliar debido al desarrollo de una colecistitis crónica de tipo irritativo.

SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
220
• Las infl amaciones y el edema de la pared de la vesícula son fácilmente detecta-bles y en medicina humana, se relacionan con proceso de colecistitis, con insufi ciencia cardiaca congestiva y con insufi ciencia renal, entre otros procesos patológicos en los cuales aparece la estructura trilaminar de la pared.
• Neoplasias de la vesícula biliar. Se encuentran asociados a la pared, presen-tando menos ecogenicidad que los cálculos, y no producen sombras acústicas.
Las neoplasias en este órgano son poco frecuentes, habiéndose descrito papilo-mas, adenomas y, menos frecuentemente, carcinomas asociándose el origen de estas neoplasias al padecimiento de una colelitiasis. El carcinoma invade la pared de la vesícula biliar extendiéndose en el parénquima hepático adyacente habiéndose citado matástasis a ganglios linfáticos regionales pero no a órganos internos.
Tracto gastrointestinal
La ultrasonografía no es el método más indicado, debido a la abundante presen-cia de gas que difi culta la visión, usándose otros métodos diagnósticos alternativos, como la radiología o la endoscopia.
Se recomienda la introducción de líquido mediante una sonda gástrica para vi-sualizar la luz así como las paredes del estómago y primeras porciones del intestino. Con el estudio ecográfi co se pueden valorar la estructura y motilidad, distinguiéndose en la pared cinco capas que se suceden como capas de ecogenicidad opuesta.
Como patologías distinguimos:
• Neoplasias: se ven como estructuras hipoecogénicas de tamaño variable y generalmente bien delimitadas.
Entre las neoplasias más frecuentes que afectan al tubo digestivo encontramos el adenoma y el adenocarcinoma de la mucosa del tubo gastrointestinal y, con especial incidencia, el pólipo colorectal que aparece como una masa pedunculada que puede evidenciarse por la abertura anal. Otra neoplasia muy frecuente y sobre todo en gatos y perros jóvenes es el linfoma alimentario que se presenta como un engrosamiento segmental del intestino con afectación del mesenterio y ganglios linfáticos regionales. Este tumor tiene pronóstico reservado y una evolución muy desfavorable con escaso periodo de supervivencia desde su diagnóstico histopatológico.
• Cuerpos extraños: muy difíciles de ver, podemos encontrarlos caudalmente a zonas de acúmulo de líquido.
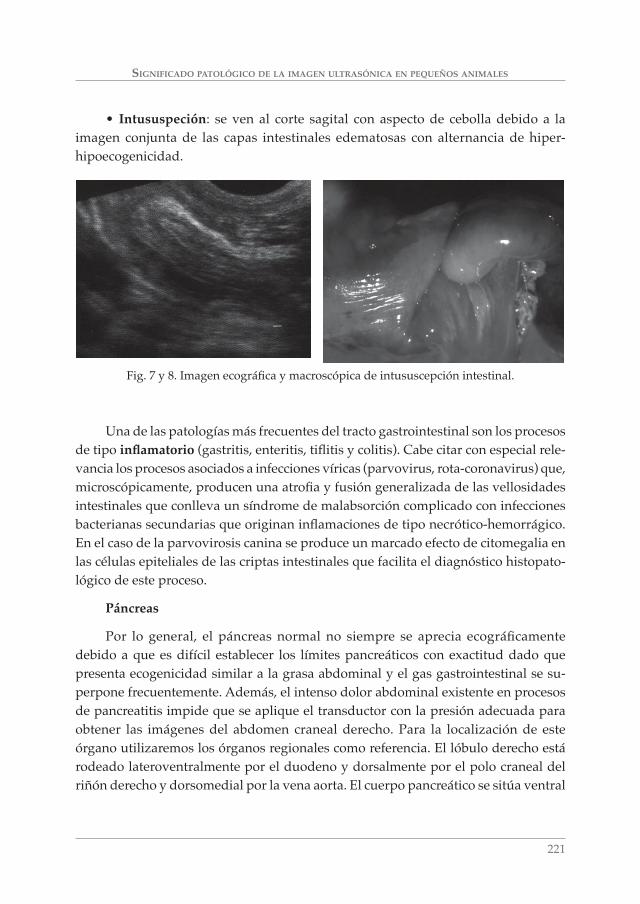
SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
221
• Intususpeción: se ven al corte sagital con aspecto de cebolla debido a la imagen conjunta de las capas intestinales edematosas con alternancia de hiper-hipoecogenicidad.
Fig. 7 y 8. Imagen ecográfi ca y macroscópica de intususcepción intestinal.
Una de las patologías más frecuentes del tracto gastrointestinal son los procesos de tipo infl amatorio (gastritis, enteritis, tifl itis y colitis). Cabe citar con especial rele-vancia los procesos asociados a infecciones víricas (parvovirus, rota-coronavirus) que, microscópicamente, producen una atrofi a y fusión generalizada de las vellosidades intestinales que conlleva un síndrome de malabsorción complicado con infecciones bacterianas secundarias que originan infl amaciones de tipo necrótico-hemorrágico. En el caso de la parvovirosis canina se produce un marcado efecto de citomegalia en las células epiteliales de las criptas intestinales que facilita el diagnóstico histopato-lógico de este proceso.
Páncreas
Por lo general, el páncreas normal no siempre se aprecia ecográfi camente debido a que es difícil establecer los límites pancreáticos con exactitud dado que presenta ecogenicidad similar a la grasa abdominal y el gas gastrointestinal se su-perpone frecuentemente. Además, el intenso dolor abdominal existente en procesos de pancreatitis impide que se aplique el transductor con la presión adecuada para obtener las imágenes del abdomen craneal derecho. Para la localización de este órgano utilizaremos los órganos regionales como referencia. El lóbulo derecho está rodeado lateroventralmente por el duodeno y dorsalmente por el polo craneal del riñón derecho y dorsomedial por la vena aorta. El cuerpo pancreático se sitúa ventral

SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
222
a la vena porta y el lóbulo pancreático izquierdo entre el bazo, el riñón izquierdo y la curvatura mayor del estómago.
Las patologías más comúnmente detectadas son: pancreatitis, neoplasias, abscesos y pseudoquistes. Se han reportado casos bibliográfi cos de dilataciones de conducto pancreático (Lamb C.R. 1989). En la mayoría de las patologías pancreáticas, excepto las neoplasias, se observan formaciones hipoecogénicas. En la detección de neoplasias se observan formaciones nodulares hiperecogénicas de 1 a 2 centímetros.
La patología más frecuente en este órgano es la necrosis pancreática aguda o pancreatitis necrotizante originada tras una liberación de enzimas pancreáticas que conlleva una destrucción progresiva del parénquima pancreático. Microscópicamente se caracteriza por una necrosis de los acinos pancreáticos, edema intersticial, necrosis de estructuras vasculares con fuerte reacción infl amatoria asociada junto a imágenes de necrosis enzimática de la grasa.
En los procesos pseudotumorales y tumorales que afectan al páncreas se descri-be la hiperplasia nodular, el adenoma y el adenocarcinoma pancreático. Este último aparece en forma de nódulos solitarios o múltiples de coloración abigarrada, con área central necrótica frecuentemente calcifi cada. Histológicamente, se describe un patrón tubular, acinar o de crecimiento sólido indiferenciado siendo siempre el grado de atípica celular y el índice mitótico elevados. El adenocarcinoma pancreático se considera una neoplasia altamente agresiva que invade localmente y metastatiza a órganos internos.
Bazo
El estudio ecográfi co del bazo lo comenzamos a nivel del abdomen craneal izquierdo, a partir del cual se extiende hacia el lado derecho y caudal. Estudiamos situación, tamaño y forma del parénquima esplénico. La situación y extensión se realiza mediante cortes longitudinales y sagitales. La superfi cie del bazo normal es continua y lisa, excepto la región del hilio. La cápsula es una capa hiperecogénica muy visible en casos de ascitis. El parénquima es hipoecogénico, homogéneo y de grosor variable, apareciendo los vasos esplénicos anecógenos.
Alteraciones patológicas: esplenomegalia difusa con aumento del grosor y del grano, desituación del bazo (torsiones de estómago) y ruptura esplénica apreciándose una interrupción anecógena del parénquima esplénico asociado a hemoperitoneo. Sin embargo, la patología más frecuente en bazo son las neoplasias. Los abscesos esplénicos son anecógenos, con partículas fl otando en el interior del mismo.
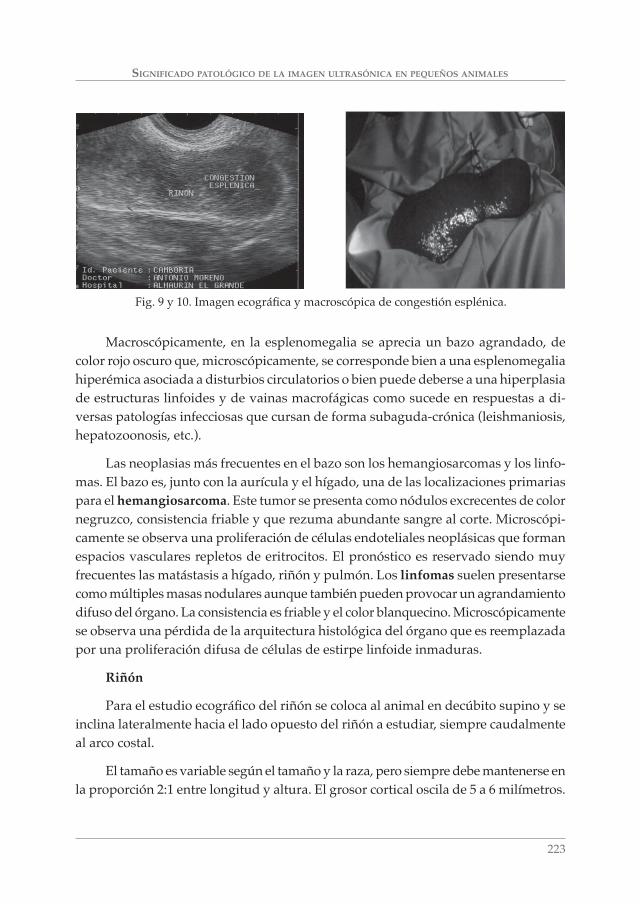
SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
223
Macroscópicamente, en la esplenomegalia se aprecia un bazo agrandado, de color rojo oscuro que, microscópicamente, se corresponde bien a una esplenomegalia hiperémica asociada a disturbios circulatorios o bien puede deberse a una hiperplasia de estructuras linfoides y de vainas macrofágicas como sucede en respuestas a di-versas patologías infecciosas que cursan de forma subaguda-crónica (leishmaniosis, hepatozoonosis, etc.).
Las neoplasias más frecuentes en el bazo son los hemangiosarcomas y los linfo-mas. El bazo es, junto con la aurícula y el hígado, una de las localizaciones primarias para el hemangiosarcoma. Este tumor se presenta como nódulos excrecentes de color negruzco, consistencia friable y que rezuma abundante sangre al corte. Microscópi-camente se observa una proliferación de células endoteliales neoplásicas que forman espacios vasculares repletos de eritrocitos. El pronóstico es reservado siendo muy frecuentes las matástasis a hígado, riñón y pulmón. Los linfomas suelen presentarse como múltiples masas nodulares aunque también pueden provocar un agrandamiento difuso del órgano. La consistencia es friable y el color blanquecino. Microscópicamente se observa una pérdida de la arquitectura histológica del órgano que es reemplazada por una proliferación difusa de células de estirpe linfoide inmaduras.
Riñón
Para el estudio ecográfi co del riñón se coloca al animal en decúbito supino y se inclina lateralmente hacia el lado opuesto del riñón a estudiar, siempre caudalmente al arco costal.
El tamaño es variable según el tamaño y la raza, pero siempre debe mantenerse en la proporción 2:1 entre longitud y altura. El grosor cortical oscila de 5 a 6 milímetros.
Fig. 9 y 10. Imagen ecográfi ca y macroscópica de congestión esplénica.

SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
224
La anchura a nivel del hilio es siempre mayor que la altura. La cápsula se observa marcadamente hiperecogénica debajo de la cual vemos la corteza hipoecogénica que envuelve a una médula hipo o anecogénica. La pelvis presenta una marcada hipoeco-genicidad debido a su contenido en grasa. Es importante comparar la corteza renal con el parénquima hepático y esplénico, para poder detectar si existe un aumento o disminución de la ecogenicidad.
Como patologías renales podemos citar quistes renales, riñones poliquísticos, riñón fruncido, cálculos, neoplasias, hidronefrosis, abscesos y calcifi caciones.
• Riñón fruncido: observamos una disminución del tamaño renal y mucha difi cultad para diferenciar corteza, médula y pelvis. Está no diferenciación de la eco-genicidad del parénquima renal es común a los casos de nefritis, en cuya patología se observa además un incremento en la ecogenicidad total.
El término clínico de riñón fruncido se corresponde macroscópicamente con un riñón disminuido de tamaño y color pálido que presenta numerosas cicatrices en la superfi cie externa y adherencias con la cápsula renal lo que difi culta su decapsula-ción. Microscópicamente se corresponde a un proceso crónico caracterizado por una glomerulonefritis de tipo membranoso-esclerosante y/o nefritis intersticial crónica de tipo linfoplasmocitario con fenómenos más o menos intensos de fi brosis.
• Dilatación de la pelvis renal: se ve el uréter y una pelvis hipoecogénica debido al acúmulo de líquido. Puede evolucionar a una hidronefrosis con un aumento del área anecógena central.
• Quistes renales: se ven zonas circulares anecógenas sin una pared diferen-ciada, situadas en la periferia. En el riñón poliquístico se observan las estructuras anecógenas en forma de celdillas.
La enfermedad quística en el riñón incluye diversas patologías caracterizadas por la presencia de una o más cavidades quísticas en el parénquima renal. Estas se pueden originar durante la organogénesis y se asocian a displasia renal o bien como consecuencia de lesiones obstructivas dando lugar a los quistes por retención ad-quiridos. Los quistes suelen localizarse en la corteza renal, su pared es clara u opaca y el contenido acuoso. Microscópicamente los quistes aparecen delimitados por un epitelio simple aplanado siendo la mayoría uniloculares.
• Neoplasias: se suelen observar estructuras anecógenas que en la mayoría de los casos deforman el contorno renal.

SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
225
Las neoplasias renales son poco frecuentes describiéndose el adenoma y el ade-nocarcinoma. El nefroblastoma es un tumor de animales jóvenes que suele observarse en un polo del riñón de forma única y con superfi cie externa lobulada.
• Cálculos: se observan estructuras muy hiperecogénicas de contornos irregu-lares con marcada sombra acústica. Las calcifi caciones se observan como estructuras hiperecogénicas irregulares de tamaño y situación variables, pero carecen de sombra acústica.
Glándulas adrenales
La ecografía de las glándulas adrenales no siempre es fácil, debido a su peque-ño tamaño y a su situación poco accesible. Se encuentran craneomedial o craneal a los riñones, localizándose la glándula derecha entre el riñón y la cava caudal, y la izquierda entre el riñón y la aorta. Frecuencia empleada: 7.5 MHz.
Patologías: cuando se sospecha alteración de las glándulas adrenales (S. de Cushing), aparecen agrandadas e hipoecogénicas. En algunos casos pueden aparecer áreas de calcifi cación con sombras acústicas y, en caso de neoplasias, aparece aumento de tamaño y ecogenicidad mixta.
Entre los tumores de la corteza adrenal se citan los adenomas y los adenocar-cinomas. Neoplasias de la corteza adrenal o hiperplasia adrenocortical bilateral son causas del síndrome de Cushing junto al adenoma corticotropo de la adenohipófi sis y medicación iatrogénica.
Vejiga de la orina
El estudio de la vejiga debe hacerse con ésta bien distendida para su correcta evaluación, posicionando el transductor cranealmente al pubis, con el animal en de-cúbito supino. Con cortes longitudinales y transversales, valoramos tamaño, forma, situación y pared de ésta. Presenta una estructura redondeada u ovoidal, totalmente anecógena y sin elementos corpusculares. La pared ventral (más cerca del transductor), es muy difícil de valorar, debido a su escasa resolución a ese nivel.
Patologías más frecuentes:
• Los cálculos son visibles ecográfi camente, independientemente de su tamaño y composición, observándose como estructuras hiperecogénicas con sombra acústica.
• En cistitis aparece aumento del grosor de la pared trilaminar con irregulari-dades en las capas, y aparecen puntos en suspensión hipoecogénicos bien defi nidos.

SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
226
En cistitis hemorrágicas se observan los coágulos como estructuras hipoecogénicas de contorno irregular, liso, y adheridas a la pared.
La mayoría de las cistitis son de etiología bacteriana siendo la vía de entrada la uretra. La cistitis enfi sematosa se describe en perros y gatos con diabetes mellitus estando originada por la fermentación de azúcares a cargo de las bacterias implicadas. En las cistitis hemorrágicas existe ulceración de la mucosa, hemorragias y edema de la pared pudiendo en ocasiones observar cuerpos de inclusión intracitoplasmáticos de tinción acidófi la en las células epiteliales de la mucosa típicos del moquillo canino. Las cistitis crónicas se caracterizan por un engrosamiento irregular de la mucosa del órgano y suele asociarse a cálculos vesicales persistentes que, en ocasiones, pueden llegar a formar proyecciones polipoides (cistitis crónica polipoide), proceso en el que el estudio histopatológico es esencial para diferenciarlo de los procesos neoplásicos.
• Las neoplasias producen imágenes ecográfi cas muy variadas. Deben estudiar-se con la vejiga muy distendida, son más ecogénicas que los coágulos y su aspecto es muy variable, siendo normalmente nódulos con base de implantación o, en otros casos, estructuras irregulares.
Entre las neoplasias más frecuentes se citan el papiloma de células transicionales de presentación solitaria o múltiple y el adenoma y el adenocarcinoma del epitelio de transición. Entre los tumores mesenquimales más frecuentes están los derivados de la capa muscular.
• En la rotura vesical observamos una vejiga escasamente distendida, incluso después de forzar la diuresis; además va en aumento la cantidad de líquido libre en abdomen.
Próstata
Valoramos situación, tamaño, forma, así como la uretra intraprostática. Depen-diendo de la raza y edad del paciente, la encontramos distinta (a mayor edad, mayor tamaño y peso de ésta). Dicha glándula se va desplazando cranealmente, posicionando en abdomen. Su morfología también varía con la edad. En cachorros es alargada; en perros de año y medio, es redondeada y, a partir de los dos años y medio de edad, es claramente bilobulada, acentuándose el surco ventral a lo largo de los años. Para su localización se coloca al paciente en decúbito supino y en situación paraprepucial, caudalmente a la vejiga de la orina.
• En las prostatitis aparece ecogenicidad aumentada, con un ligero aumento del tamaño en procesos crónicos, mientras que en procesos agudos hay una disminución de la ecogenicidad y un ligero aumento de tamaño.
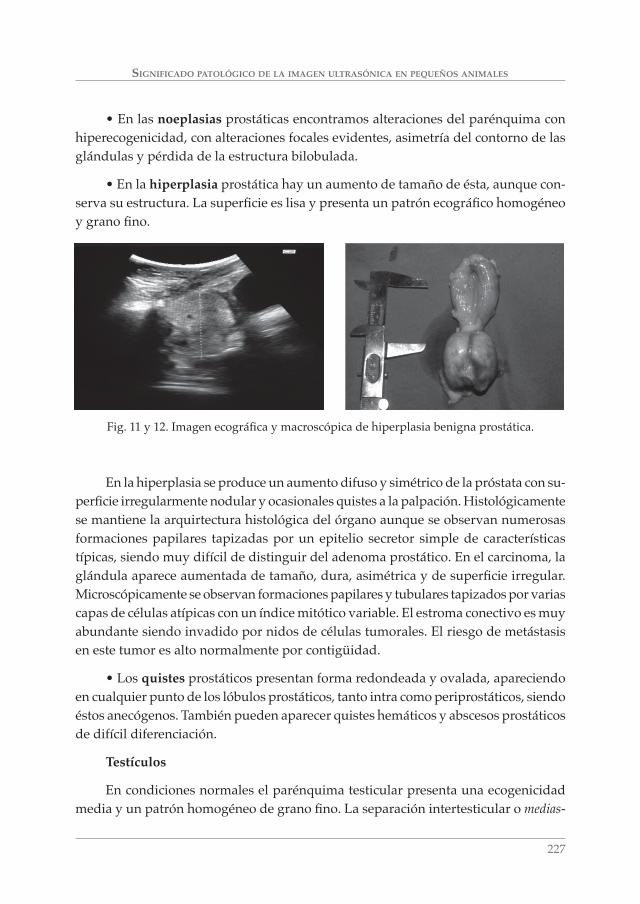
SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
227
• En las noeplasias prostáticas encontramos alteraciones del parénquima con hiperecogenicidad, con alteraciones focales evidentes, asimetría del contorno de las glándulas y pérdida de la estructura bilobulada.
• En la hiperplasia prostática hay un aumento de tamaño de ésta, aunque con-serva su estructura. La superfi cie es lisa y presenta un patrón ecográfi co homogéneo y grano fi no.
Fig. 11 y 12. Imagen ecográfi ca y macroscópica de hiperplasia benigna prostática.
En la hiperplasia se produce un aumento difuso y simétrico de la próstata con su-perfi cie irregularmente nodular y ocasionales quistes a la palpación. Histológicamente se mantiene la arquirtectura histológica del órgano aunque se observan numerosas formaciones papilares tapizadas por un epitelio secretor simple de características típicas, siendo muy difícil de distinguir del adenoma prostático. En el carcinoma, la glándula aparece aumentada de tamaño, dura, asimétrica y de superfi cie irregular. Microscópicamente se observan formaciones papilares y tubulares tapizados por varias capas de células atípicas con un índice mitótico variable. El estroma conectivo es muy abundante siendo invadido por nidos de células tumorales. El riesgo de metástasis en este tumor es alto normalmente por contigüidad.
• Los quistes prostáticos presentan forma redondeada y ovalada, apareciendo en cualquier punto de los lóbulos prostáticos, tanto intra como periprostáticos, siendo éstos anecógenos. También pueden aparecer quistes hemáticos y abscesos prostáticos de difícil diferenciación.
Testículos
En condiciones normales el parénquima testicular presenta una ecogenicidad media y un patrón homogéneo de grano fi no. La separación intertesticular o medias-
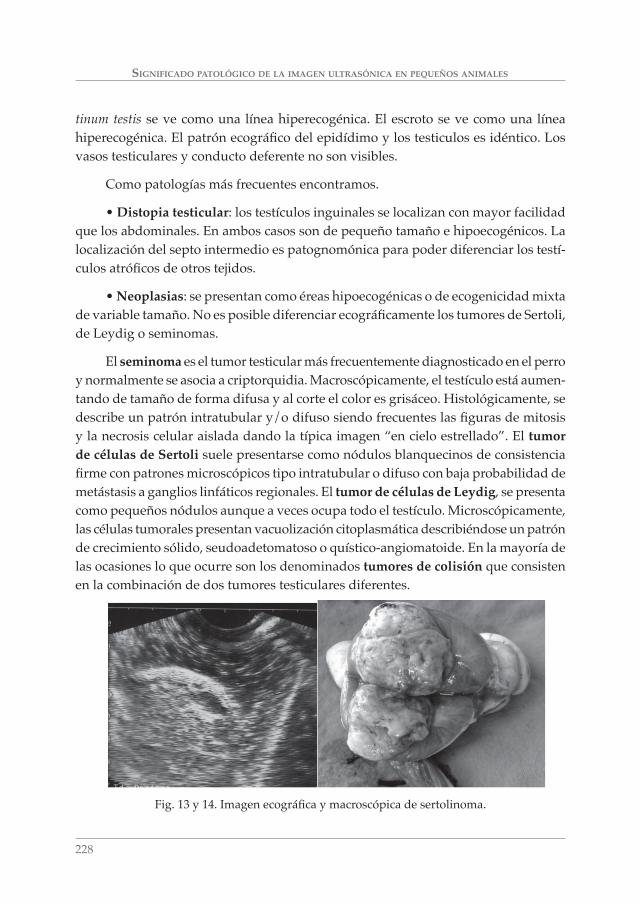
SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
228
tinum testis se ve como una línea hiperecogénica. El escroto se ve como una línea hiperecogénica. El patrón ecográfi co del epidídimo y los testiculos es idéntico. Los vasos testiculares y conducto deferente no son visibles.
Como patologías más frecuentes encontramos.
• Distopia testicular: los testículos inguinales se localizan con mayor facilidad que los abdominales. En ambos casos son de pequeño tamaño e hipoecogénicos. La localización del septo intermedio es patognomónica para poder diferenciar los testí-culos atrófi cos de otros tejidos.
• Neoplasias: se presentan como éreas hipoecogénicas o de ecogenicidad mixta de variable tamaño. No es posible diferenciar ecográfi camente los tumores de Sertoli, de Leydig o seminomas.
El seminoma es el tumor testicular más frecuentemente diagnosticado en el perro y normalmente se asocia a criptorquidia. Macroscópicamente, el testículo está aumen-tando de tamaño de forma difusa y al corte el color es grisáceo. Histológicamente, se describe un patrón intratubular y/o difuso siendo frecuentes las fi guras de mitosis y la necrosis celular aislada dando la típica imagen “en cielo estrellado”. El tumor de células de Sertoli suele presentarse como nódulos blanquecinos de consistencia fi rme con patrones microscópicos tipo intratubular o difuso con baja probabilidad de metástasis a ganglios linfáticos regionales. El tumor de células de Leydig, se presenta como pequeños nódulos aunque a veces ocupa todo el testículo. Microscópicamente, las células tumorales presentan vacuolización citoplasmática describiéndose un patrón de crecimiento sólido, seudoadetomatoso o quístico-angiomatoide. En la mayoría de las ocasiones lo que ocurre son los denominados tumores de colisión que consisten en la combinación de dos tumores testiculares diferentes.
Fig. 13 y 14. Imagen ecográfi ca y macroscópica de sertolinoma.
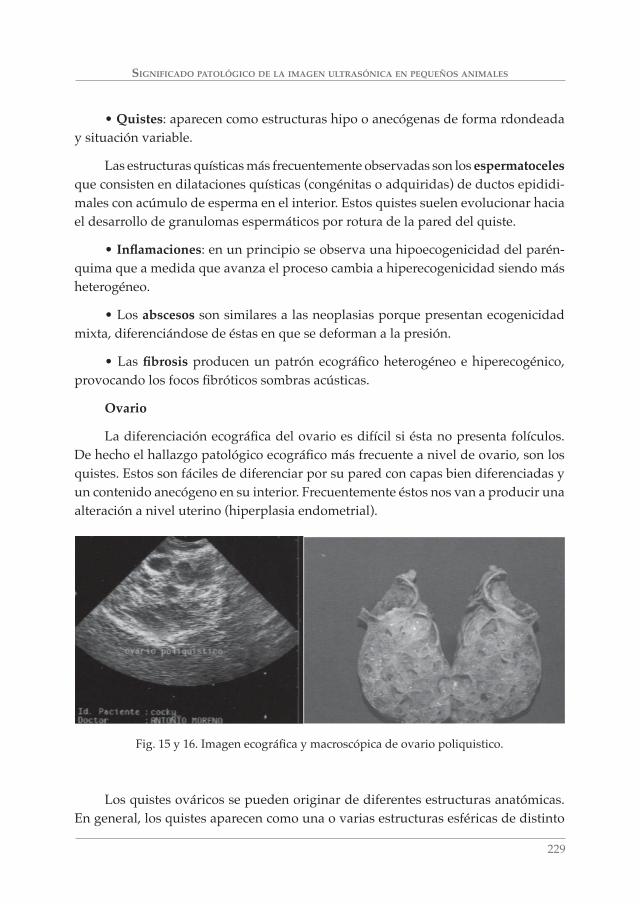
SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
229
• Quistes: aparecen como estructuras hipo o anecógenas de forma rdondeada y situación variable.
Las estructuras quísticas más frecuentemente observadas son los espermatoceles que consisten en dilataciones quísticas (congénitas o adquiridas) de ductos epididi-males con acúmulo de esperma en el interior. Estos quistes suelen evolucionar hacia el desarrollo de granulomas espermáticos por rotura de la pared del quiste.
• Infl amaciones: en un principio se observa una hipoecogenicidad del parén-quima que a medida que avanza el proceso cambia a hiperecogenicidad siendo más heterogéneo.
• Los abscesos son similares a las neoplasias porque presentan ecogenicidad mixta, diferenciándose de éstas en que se deforman a la presión.
• Las fi brosis producen un patrón ecográfi co heterogéneo e hiperecogénico, provocando los focos fi bróticos sombras acústicas.
Ovario
La diferenciación ecográfi ca del ovario es difícil si ésta no presenta folículos. De hecho el hallazgo patológico ecográfi co más frecuente a nivel de ovario, son los quistes. Estos son fáciles de diferenciar por su pared con capas bien diferenciadas y un contenido anecógeno en su interior. Frecuentemente éstos nos van a producir una alteración a nivel uterino (hiperplasia endometrial).
Fig. 15 y 16. Imagen ecográfi ca y macroscópica de ovario poliquistico.
Los quistes ováricos se pueden originar de diferentes estructuras anatómicas. En general, los quistes aparecen como una o varias estructuras esféricas de distinto

SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
230
tamaño haciendo prominencia en la superfi cie del ovario/s afectado/s, la pared es delgada y normalmente transparente. Al corte, los quistes foliculares presentan un líquido fl uido y generalmente ambarino. Miroscópicamente aparecen revestidos internamente por células de la granulosa que se disponen en un número variable de capas con núcleos picnóticos, algunas desprendidas en la luz. El cuerpo lúteo quístico aparece como una estructura prominente de color rojo oscuro y con estigma de ovulación que al corte sale un líquido acuoso transparente. Los quistes de inclu-sión son formaciones quísticas del epitelio de superfi cie ovárico que se originan por pinzamiento e invaginación del mismo después de la ovulación o de un trumatismo manual del ovario.
Los cuerpos lúteos persistentes aparecen promientes, la superfi cie de corte presenta color variable, normalmente con un tono naranja oscuro e histológicamente, se trata de un cuerpo lúteo en fase de degeneración con ligero engrosamiento de los tabiques conjuntivos y paredes vasculares.
Las neoplasias son formaciones sólidas, irregulares y con aumento de ecoge-nicidad. Con más frecuencia son carcinomas ováricos y, con menos frecuencia, los mesoteliomas.
Entre las neoplasias ováricas se encuentran los tumores derivados del epitelio de superfi cie (adenoma papilar, adenocarcinoma papilar, cistadenomas y cistadeno-carcinomas). Al corte se observan cavidades quísticas y áreas más o menos sólidas. Entre los tumores gonadoestromales se describen los tumores de células de la granu-losa, el tecoma y el luteoma que son hormonalmente activos en cuanto a secreción de esteroides. El tumor de células de la granulosa es normalmente unilateral, al corte la consistencia es fi rme observándose áreas sólidas y quísticas y la superfi cie tiene color banco-grisáceo o amarillenta.
Útero
El útero de los animales de compañía es el órgano abdominal más sencillo y rápido de identifi car cuando éste se encuentra alterado. El órgano de referencia para su localización es la vejiga. La imagen ecográfi ca permite confi rmar o descartar una gestación; el número de embriones no siempre puede ser determinado. También es posible valorar el desarrollo del embrión, los latidos cardiacos y movimientos fetales, así como, la situación y tamaño de los fetos. La disminución de la frecuencia cardiaca a la mitad de la de la madre, es considerada como señal de aborto, debiendo tomar medidas. Antes de 20 días es posible ver las vesículas embrionarias, que son formaciones redondeadas y anecógenas y una zona hiperecogénica en la periferia
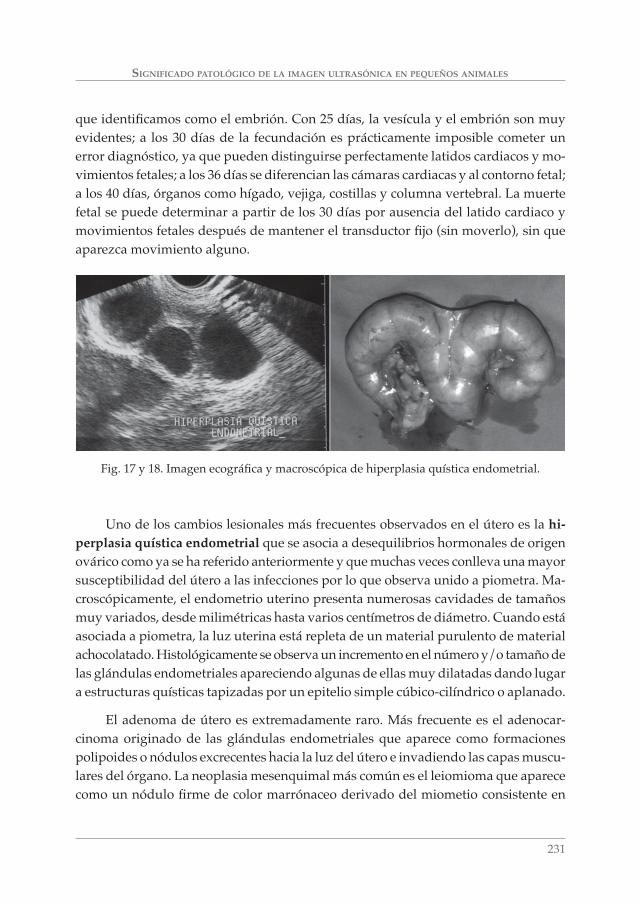
SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
231
que identifi camos como el embrión. Con 25 días, la vesícula y el embrión son muy evidentes; a los 30 días de la fecundación es prácticamente imposible cometer un error diagnóstico, ya que pueden distinguirse perfectamente latidos cardiacos y mo-vimientos fetales; a los 36 días se diferencian las cámaras cardiacas y al contorno fetal; a los 40 días, órganos como hígado, vejiga, costillas y columna vertebral. La muerte fetal se puede determinar a partir de los 30 días por ausencia del latido cardiaco y movimientos fetales después de mantener el transductor fi jo (sin moverlo), sin que aparezca movimiento alguno.
Fig. 17 y 18. Imagen ecográfi ca y macroscópica de hiperplasia quística endometrial.
Uno de los cambios lesionales más frecuentes observados en el útero es la hi-perplasia quística endometrial que se asocia a desequilibrios hormonales de origen ovárico como ya se ha referido anteriormente y que muchas veces conlleva una mayor susceptibilidad del útero a las infecciones por lo que observa unido a piometra. Ma-croscópicamente, el endometrio uterino presenta numerosas cavidades de tamaños muy variados, desde milimétricas hasta varios centímetros de diámetro. Cuando está asociada a piometra, la luz uterina está repleta de un material purulento de material achocolatado. Histológicamente se observa un incremento en el número y/o tamaño de las glándulas endometriales apareciendo algunas de ellas muy dilatadas dando lugar a estructuras quísticas tapizadas por un epitelio simple cúbico-cilíndrico o aplanado.
El adenoma de útero es extremadamente raro. Más frecuente es el adenocar-cinoma originado de las glándulas endometriales que aparece como formaciones polipoides o nódulos excrecentes hacia la luz del útero e invadiendo las capas muscu-lares del órgano. La neoplasia mesenquimal más común es el leiomioma que aparece como un nódulo fi rme de color marrónaceo derivado del miometio consistente en

SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
232
una proliferación de fi bras musculares lisas que se disponen en bandas o paquetes orientadas en las distintas direcciones del espacio con atipia celular e índice mitótico bajos. Leiomiosarcomas y fi brosarcomas también se presentan aunque con menor frecuencia. En ambos casos el índice mitótico puede llegar a ser elevado, el grado de atipia celular es alto y la posibilidad de recidivas y/o metástasis a distancia también es elevada.
CONCLUSIONES
1.- La ecografía es una técnica diagnóstica no invasiva de valor diagnóstico, cuanto no orientativo, en la patología de los animales de compañía, aunque requiere una amplia experiencia para su interpretación.
2.- La biopsia ecoguiada resulta de gran utilidad para la realización de estudios histopatológicos de órganos internos.
3.- Los estudios histopatológicos son de gran valor a la hora de establecer una correlación positiva entre las imágenes ecográfi cas y sus patologías correspondientes.
4.- Por tanto, ecografía e histopatología son dos técnicas complementarias de gran utilidad e interrelación para el estudio de la patología en pequeños animales.
BIBLIOGRAFÍA
Chacón-M. De Lara F., Hervás J. et al. (1996). Intestinal smooth muscle hiperplasia in a goat. J. Vet. Diagn. Invest. 8:390-392.
Dellmann H.D. (1995). Histología Veterinaria. Ed. Acribia. Hervás J. et al. (1995). Acute fatal hepatozoonosis in a puppy. Vet. Rec. 137, 518-519. Hervás J. et al. (1996). Pathological study of visceral leishmaniasis in a jackal. Vet. Rec. 139, 293-295. Henry, C.J. ( 2007.) Transitional Cell Carcinoma. Proceedings of the World Small Animal Veterinary
Association. Sydney, Australia Holzworth J. (1987). Diseases of the Cat. Ed. Saunders Company. México. Jubb KVF., Kennedy PC. And Palmer N. (1992) pathology of Domestic Animals. Academic Press.
London. Moreno Boiso A. , Chacon M. de Lara G-Ripoll, Hervas Rodriguez (1997). Signifi cado patológico
de la imagen ultrasónica en pequeños animales. X Convención Nacional. (Libro de Memorias) pp: 81-89. Octubre 1997, Jalisco (México).
Moreno Boiso, A 1999 et J. Lopez Fernandez,Sanchez Isarria M.A, Chacon M. de Lara G-Ripoll, Hervas Rodriguez l. Manual práctico de ecografía comparada en pequeños animales.ed.colegio Veterinarios de Malaga.
Moreno Boiso A.A., Hervas Rodriguez J. Chacon M de Lara G-Ripoll F. Lopez Fernandez J. M Guer-rero Rueda M. J. Simon Salazar (2003). Manual de endocrinologia .1ª Ed. Intervet Mexico S.A.

SIGNIFICADO PATOLÓGICO DE LA IMAGEN ULTRASÓNICA EN PEQUEÑOS ANIMALES
233
Moreno Boiso A.A., Illera del Portal J.C, Silvan Granado G., Illera del Portal J.C. ( 2011). Endo-crinología de Pequeños Animales de la Fisiología a la clínica Ediciones LID Madrid( España
Mutsaers, AJ; Widmer, WR; Knapp, DW (2003). Canine transitional cell carcinoma. 2003. J Vet. Intern Med., vol. 17, pp.136-144.
Nyland & Mattoon (2006). Diagnostico ecografi co en pequeños animales Ed. multimedica veteri-naria.
O’Brien, RT y Autran de Morais, H (2007 ). Enfermedades no neoplásicas del bazo. En: Ettinger, SJ, Feldman, EC: Tratado de medicina interna veterinaria: enfermedades del perro y el gato (6ª ed). Elsevier. pp.1945-1951
Paclikova K, P Kohout, M Vlasin. (2006). Diagnostic possibilities in the management of canine prostatic disorders. Veterinarni Medicina 51, 1-13.
Penninck, D, D’Anjou M.A (2010). Atlas de ecografi a en pequeños animales multimedica ediciones veterinarias Barcelona España
Poirier, VJ; Forrest, LJ; Adams, WM y col (2004). Piroxicam, mitoxantrone, and coarse fraction radiotherapy for the treatment of transitonal cell carcinoma of the bladder in 10 dogs: a pilot study. 2004. JAAHA vol. 40, p. 131-136
Rodríguez-Franco, F. (2007). Neoplasias digestivas en el perro. 7º Congreso Bayer2007.Tamada H., Kawate N., Inaba T., Kuwamura M., Maeda M., Kajikawa T., Sawada T.,( 2005). Adeno-
myosis with sever infl ammation in the uterine cervix in a dog. Can. Vet. J. 46, 333-334.Thamm, DH (2007). Hemangiosarcoma. En: Withrow, SJ; Vail, DM (ed). Small animal clinical
oncology (4ª ed). Saunders, Missouri. pp. 785-795.Wilkerson, M.J., Dolce, K., Koopman, T. y col.,( 2005). Lineage differentiation of canine lymphoma/
leukemias and aberrant expression of CD molecules. Vet. Immunol. Immunopathol. 106, 179-196.


GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
235ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
RESUMEN
La determinación de la edad de los ciervos cazados es fundamental e imprescindible para valorar tanto la calidad individual como la calidad de la población en su conjunto y pone en perspectiva cuestiones sobre gestión y manejo. La determinación de la edad es la mejor fuente de información para evaluar los resultados de la gestión desarrollada, monitorizar el cumplimiento de los objetivos marcados a medio y largo plazo, y valorar las consecuencias de prácticas de manejo anteriores. También constituye una información básica imprescindible para realizar predicciones sobre el futuro demográfi co de las poblaciones a gestionar, bien sea para incrementar su densidad o para limitarla. Y además, es necesario conocer la edad con el mayor detalle y precisión posibles cuando se investigan temas como crecimiento corporal, re-producción, supervivencia o longevidad. Este trabajo recopila los resultados más destacados de la puesta a punto de diversos métodos y técnicas de determinación de edad en el ciervo ibérico (Cervus elaphus hispanicus) del sur de España, a partir del estudio de ciervos cazados en Sierra Morena Oriental y de edad conocida. Se revisan antecedentes, y fundamentos metodológicos haciendo especial referencia a las técnicas esqueletocronológicas es decir, al uso de marcas de crecimiento en tejidos dentales evidenciadas mediante histología convencional o petrografía. Se discuten y explican las limitaciones y errores intrínsecos a cada método a modo de guía práctica para veterinarios, biólogos y gestores interesados.
Palabras clave: Determinación de edad, Ciervo ibérico, Cervus elaphus hispanicus, sur de España, erupción dental, primer molar, líneas de reposo, cemento dental, esqueletocronología.
GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIER-VO IBÉRICO (Cervus elaphus hispanicus) A TRAVÉS DE SU DENTICIÓN: REVISIÓN METODOLÓGICA Y TÉCNICAS DE ELECCIÓN
CONCEPCIÓN AZORIT CASAS1
1 Área de Zoología del Departamento de Biología Animal, Vegetal y Ecología. Facultad de Ciencias Experimentales. Universidad de Jaén, 23071, España. E-mail: [email protected]

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
236
ABSTRACT:
Aging of hunted deer is essential to evaluate both the individual quality and the quality of the population as a whole, and puts into perspective questions about management and handling. The age determination is the best source of information to evaluate the results of the management undertaken, to monitor the achievement of objectives developed both medium and long term and to evaluate the consequences of the practices of previous management. It also constitutes as basic essential information to make predictions about the demographic future of the populations to manage, whether to increase their density or to limit it. It is also necessary to know the age with the greatest detail and precision as possible when subjects such as body growth, condition, reproduction, survival or longevity are investigated.
This paper collects the most outstanding results of various methods and techniques for determining the age of the Iberian Deer (Cervus elaphus hispanicus) in the south of Spain, from the study of deer hunted in
Oriental Sierra Morena and of a known age. Background is reviewed and methodological fundamentals with special reference to skeletochronology, the use of marks of growth in dental tissues evident through conventional histology or petrography. The limitation and intrinsic error to each method are discussed and explained as a kind of practical guide for interested veterinary, biologists and managers.
Keyword: aging, Iberian red deer, Cervus elaphus hispanicus, south Spain, teeth, fi rst molar, rest lines, skeletochronology.
1. INTRODUCCIÓN:
El ciervo ibérico (Cervus elaphus hispanicus) es la especie de caza mayor más emblemática de la Península Ibérica y su gestión supone una preocupación mayor por motivos tanto medioambientales como económicos. Una de las herramientas más valiosas para el aprovechamiento sostenible de la especie es el conocimiento de su pirámide y dinámica poblacional. Así, necesitamos resolver interrogantes en relación a la densidad, la tasa de renovación de la población, o la determinación de la edad de los individuos que forman parte de ella. De hecho, la proporción entre los distintos grupos de edad es uno de los factores más infl uyentes en la calidad ecológica de una comunidad animal de manera que sin la presencia de un número adecuado de ciervos maduros, o una proporción de edades adecuada, no se asegura un compor-tamiento social equilibrado. Por todo ello, para algunos autores el conocimiento de la tasa de edades es incluso más importante que el conocimiento del número total de individuos (1, 2).
La correcta determinación de la edad es también clave cuando queremos com-parar poblaciones sometidas a diferentes condiciones ambientales, o para monitorizar

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
237
los resultados de actuaciones de gestión dirigidas a mejorar las cualidades físicas o cinegéticas de una especie en un hábitat concreto. Ningún carácter estructural o fi siológico cuantifi cable es independiente de la edad. Ni el estado reproductivo, ni el peso corporal, ni tampoco el tamaño de la cornamenta pueden ser valorados correctamente sin conocer la edad. Aunque cornamentas bien desarrolladas pueden ser consideradas indicadoras de una buena condición corporal, la calidad cinegética de dos poblaciones de ciervos no puede ser comparada enfrentando únicamente el tamaño de las cornamentas de los ciervos cazados sin referenciar la edad de los mismos (3,4). En la práctica, la determinación de la edad de los animales cazados es fundamental e imprescindible para valorar la calidad individual de los ciervos así como la calidad de la población en su conjunto y pone en perspectiva cuestiones de manejo y gestión. La determinación de la edad es la mejor fuente de información para determinar el éxito del tipo de gestión desarrollada en cada fi nca, para valorar el cumplimiento de los objetivos marcados a medio y largo plazo, o las consecuencias de prácticas de manejo anteriores. También constituye una información básica necesaria para realizar predicciones sobre el futuro demográfi co de las poblaciones a gestionar, bien sea para incrementar su densidad o para limitarla (5). Y además, es necesario conocer la edad con el mayor detalle y precisión posibles cuando se investigan temas como crecimiento corporal, reproducción, supervivencia o longevidad.
La edad se conocería con precisión si los animales se marcaran al nacer. Sin em-bargo, esto no suele ocurrir por lo que, desde siempre se ha descrito y utilizado una gran cantidad de métodos de datación tanto en animales domésticos como silvestres (1, 6, 7, 8, 9, 10, 11, 12, 13). Concretamente en cérvidos, los métodos tradicionales de determinación de edad se han basado en cambios en la conformación corporal, colo-ración del pelaje, cambios en la cornamenta y cambios en las características dentales como el recambio y desgaste dentarios, ampliamente utilizados desde la antigüedad. A todos estos métodos se les ha llamado métodos cinegéticos (14) y en la mayoría de los casos aportan información sobre el grupo de edad al que pertenece el animal, aunque a veces sólo es posible la distinción entre crías, jóvenes y viejos. Los basados en la dentición han sido siempre más fi ables, pero una vez completada la erupción de toda la dentición permanente hay que recurrir al desgaste dental o a métodos que usan marcas de crecimiento periódicas presentes en tejidos duros. A éstos se les ha llamado métodos esqueletocronológicos (15), y son métodos de elección cuando se requiere conocer la edad con el mayor detalle y precisión posibles ya que se consideran capaces de informar sobre la edad absoluta del animal (8, 10, 16).
Muchos autores llaman la atención sobre la importancia de la exactitud y precisión en la determinación de la edad ya que errores mínimos pueden ocasionar fallos im-

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
238
portantes en la interpretación de las tablas de vida y dar lugar a la toma de decisiones incorrectas en la gestión, con consecuencias graves en la población y su hábitat (17, 18). La existencia de diferencias en la fi abilidad de la datación según procedimientos y dientes utilizados (19), subespecies, ambientes o poblacionales (20, 21) hace necesaria la validación de cada método siempre que se emplea por primera vez en una nueva especie o subespecie. Así, en el caso del ciervo ibérico la utilización de criterios propios obtenidos a partir de estudios en nuestras latitudes resulta especialmente indicada. Pero, aunque son muchos los estudios referidos a la datación de ciervos centroeuro-peos (22, 23, 24, 25, 26, 27, 28, 29, entre otros), en la Península Ibérica estos estudios han sido más escasos y recientes. La mayoría basados en criterios morfológicos (30, 31) o en criterios dentales como erupción o desgaste dental (32, 33, 34, 35). El uso de las marcas de crecimiento en el cemento o la dentina de incisivos no se inicia hasta principios de los años 90 en nuestro país (36, 37), y no es hasta fi nales de esa década cuando se realizan estudios detallados sobre la validez de las técnicas esqueletocro-nológicas y su fi abilidad según piezas dentales, tejidos y tipo de muestras (38). A partir de entonces se establecen procedimientos estandarizados para el recuento e interpretación de las citadas marcas periódicas en ciervos mediterráneos (39).
Este trabajo recopila los resultados más destacados de la puesta a punto de diferentes técnicas de determinación de edad del ciervo ibérico en el sur de España, partiendo del estudio de ciervos cazados así como de ciervos de edad conocida. Se revisan antecedentes, fundamentos metodológicos y procedimientos, haciendo especial referencia a las técnicas esqueletocronológicas, es decir, al uso de marcas de crecimiento periódico evidenciadas en tejidos duros mediante histología convencional o petrografía. Se aportan resultados inéditos sobre la exactitud de la datación usando preparaciones de molares sin descalcifi car en comparación con las histológicas de dientes descalcifi cados. Se discuten limitaciones y errores intrínsecos a cada método, así como las ventajas de los métodos preferentes según los casos a modo de guía práctica para veterinarios, biólogos y gestores interesados.
2. MATERIAL Y MÉTODOS:
Los trabajos revisados estudiaron un total de 558 ciervos de ambos sexos cazados entre 1993-97 en monterías y cazas selectivas de Sierra Morena Oriental (Jaén). Más recientemente, para investigar la validez de procedimientos petrográfi cos de forma comparada con procedimientos histológicos, se estudiaron 207 ciervos cazados entre 2007-2008. A partir de necropsias de campo se obtuvieron sus mandíbulas para el posterior estudio dental. Se dispuso de 3 ciervos adultos de edad conocida nacidos en

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
239
distintas fi ncas de Sierra Morena de Jaén que fueron marcados en su primer año de vida (un macho llamado Bartolo de 9 años y dos hembras Jara y Rosa de 5 y 13 años, respectivamente), además de 20 crías de la granja experimental de la Universidad de Castilla la Mancha en Albacete.
2.1. Erupción y reemplazamiento
Para determinar el patrón y tiempo de erupción y reemplazamiento dentarios se estudió la dentición de 20 crías nacidas en cautividad, en la granja experimental antes citada, mediante procedimientos manuales durante jornadas rutinarias de manejo, además de 158 ciervos (56 machos, 87 hembras y 15 de sexo no registrado) con dentición incompleta (erupción dentaria en curso) de los que se conocía la fecha exacta de muerte y el mes de nacimiento. La edad se determinó con metodología y simbología descritas previamente (32, 33, 40, 41).
2.2. Marcas de crecimiento periódicas en tejidos dentales
Las marcas de crecimiento parecen ser el refl ejo de un acúmulo de tejido que tiene lugar de forma más rápida e intensa en un periodo y que cesa o se reduce en otro momento del año. El efecto de este acúmulo diferencial es la aparición de capas alternas de distinto grosor (unas anchas y otras más estrechas), y de distinto aspecto y densidad óptica, que en realidad indican distintas fases de la odontogénesis, ce-mentogénesis u osteogénesis. Actualmente está aceptado que las capas anchas que aparecen en el cemento dental (u otros tejidos) son las que resultan de periodos de rápido crecimiento (42), y las marcas estrechas, a las que muchos autores llaman lí-neas de detención de crecimiento o de reposo, resultan de periodos de depósito lento (43, 44). Pero el desconocimiento de las causas que provocan estas marcas, así como las variaciones detectadas en cuanto a la época de crecimiento o reposo según el área geográfi ca o la subespecie (16), así como la existencia de diferencias en la fi abilidad de la datación según procedimientos, dientes (19), subespecies, ambientes o pobla-cionales (20, 21) hace necesaria su validación para el ciervo ibérico (38) y para ello un protocolo de trabajo con una serie de pasos concretos:
1º. Confi rmar la formación de marcas periódicas en dientes de ciervo ibérico y averiguar el tipo de preparación, diente y zona con marcas más visibles y fácilmente interpretables para la determinación de la edad (45), 2º.- Averiguar la periodicidad anual en la formación de cada marca así como la época de mayor crecimiento y reposo

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
240
(47), 3º.- Determinar la edad de aparición de las primeras marcas formadas en cada diente (46), 4º.- Establecer un método estandarizado para su recuento e interpretación que evite en lo posible problemas de subjetividad (39) y 5º.- Estimar la precisión y fi abilidad de la datación, así como el error que se puede cometer al determinar la edad de un ciervo usando un procedimiento u otro y fi nalmente valorar técnicas, métodos y dientes preferentes (39).
Para evidenciar en detalle las marcas de crecimiento en diferentes tipos de tejidos y zonas de cada diente se realizaron preparaciones histológicas de un total de 815 dientes (521 incisivos, 222 molares y 72 caninos) pertenecientes a 521 ciervos machos y hembras de entre 6 y 15 años de edad. Se utilizó un microtomo convencional Lei-ca para muestras descalcifi cadas. El procedimiento se describe en trabajos previos (45). Brevemente: 1) extracción de dientes de la mandíbula, fi jación y preservación de los dientes en una solución de formol al 10 %, 2) descalcifi cación de los dientes con una solución comercial de ácido hypoclorico y polivinil-prolidona al 14% (TBD-® Shandon) durante tiempo variable según el tipo de pieza dental, 3) inclusión en parafi na y cortes de 4-5 μm con microtomo convencional en diferentes orientaciones y zonas del diente, 4) tinción de rutina con Hematoxylina-Eosina, y 5) observación con microscopio de luz transmitida (Leica Leitz LaborluxS) y/o polarizada (Nikon E800 FL-DIC) conectados en ambos casos a una cámara color Sony 3CCD DXC-950P y ordenador Pentium 200 MMX.
Con el fi n de comparar la efi cacia y precisión de las técnicas histológicas con-vencionales con las petrográfi cas, más rápidas y versátiles que no requieren descal-cifi cación dental previa, se realizaron cortes mediante procedimientos petrográfi cos de los M1 de 207 ciervos de los que también se realizaron preparaciones histológicas con microtomo convencional. Se utilizaron dos procedimientos petrográfi cos para la preparación de molares: A) Preparación de láminas delgado-pulidas y B) Obtención de secciones gruesas pulidas, mediante cortes de molares también sin descalcifi car usando un procedimiento simplifi cado del anterior. Las láminas delgado-pulidas del M1 se realizaron según lo descrito (46). Brevemente: 1) corte longitudinal con una cortadora tipo Diamant boart, y selección de corona y almohadilla interradicular, 2) esmerilado en placas de vídrio con carborundum, 3) adherencia de la preparación a placa de vidrio con ayuda de una placa calentadora y pegamento de dos componentes tipo epoxy mounting media tipo microtec, 4) varios cortes con cortadora Petrothin (thin sectioning system de Buehler, USA) hasta un grosor de 50-20 μm, 5) pulido con paño de diamante en máquina Planopol-V, 6) observación con lupa (Leica Zoom tm 2000 con luz refl ejada.

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
241
Para una correcta interpretación de las marcas de crecimiento es necesario conocer su periodicidad de formación (anual, semestral, bianual, etc.) y a qué edad del animal, aparece la primera marca en cada diente y tejido. La periodicidad fue determinada a partir del cemento dental de 82 de ciervos de más de 4 años de edad cazados a lo largo de todas las estaciones del año. Este estudio se basó en el registro de la situación y grosor de la última capa de cemento observado en la almohadilla interradicular del primer molar (M1). Para la edad de aparición de las primeras marcas se analizaron dientes (tanto incisivos, como molares y caninos) en distinto grado de erupción para después establecer un procedimiento estandarizado de recuento e interpretación de las marcas para la datación (39, 47).
Para la valoración de la exactitud y precisión del método se realizaron prepara-ciones histológicas convencionales de 64 incisivos y 55 molares de 61 ciervos de 4-44 meses de edad (39). De 20 de estos ciervos se estudiaron sus molares tanto a partir de preparaciones petrográfi cas como histológicas.
2.3. Desgaste dental:
El estudio cuantitativo del desgaste dental en incisivos y molares se desarrolló a partir de 136 ciervos entre machos y hembras según lo descrito en trabajos previos (35) y utilizando medidas similares a las de otros trabajos de referencia (48). La edad de estos animales oscilo entre los 5 y los 163 meses de edad y fue estimada a partir de marcas en el cemento celular del primer molar. Los análisis estadísticos se realizaron con el paquete estadísitico SAS (1992) y se llevaron a cabo análisis de varianza así como procedimientos GLM para determinar infl uencia de factores como sexo, densi-dad, gestión, climatología o hábitat. Se estableció un patrón de desgaste que permite establecer amplias clases de edad en ciervos del área de estudio.
3. RESULTADOS Y DISCUSIÓN:
3.1. Patrón de erupción y reemplazamiento dentarios
La dentición de un ciervo adulto consta de 4 incisivos inferiores, 1 canino supe-rior, 3 premolares superiores y 3 inferiores, y 3 molares superiores y 3 inferiores, ya que en el proceso evolutivo los rumiantes han perdido los incisivos superiores y el canino inferior se ha transformado en un incisivo más, aproximándose al resto de los incisivos inferiores. Como en la mayoría de los artiodáctilos (excepto en los suidos),

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
242
en el ciervo también se ha desaparecido el primer premolar por lo que los premola-res existentes son en realidad el 2°, el 3° y el 4° aunque aquí los llamemos primero, segundo y tercero, respectivamente.
Todos los mamíferos placentarios presentan dientes de leche o deciduos que son sustituidos por los permanentes o defi nitivos en momentos concretos de la vida del animal, siendo los molares los únicos dientes que no tienen un predecesor de leche. La aparición y pérdida de dientes de leche y la erupción de dientes permanentes ocurren en cada especie siguiendo un orden determinado y en periodos cronológicos relativamente independientes del estado fi siológico del animal (1). Mientras que en una misma especie la secuencia de erupción suele permanecer constante, el momento de erupción puede verse alterado por factores genéticos o ambientales (40). El patrón de erupción y reemplazamiento dentario del ciervo centroeuropeo, se conoce detalla-damente gracias a los trabajos de investigadores alemanes (27, 49, 50), ingleses (22, 23, 51) o franceses (52, 53), pero en España existen pocos estudios al respecto, a excepción de los realizados en el norte de la Península (32), incompletos por no disponer de animales menores de 7-8 meses de edad ni informar sobre el reemplazamiento de incisivos, o los de ciervos del sur (33). Las variaciones en el momento y secuencia de erupción de los dientes permanentes del ciervo ibérico se muestran en la Tabla 1 comparado con lo publicado en otras regiones para otras subespecies de ciervo.
En la Figura 1 se muestran 5 mandíbulas que pueden ser utilizadas como patrón dental de otras tantas clases de edad, con diferentes fases de erupción dental donde la dentición de leche va siendo sustituida por dentición permanente. El ciervo ibérico a los 2-3 meses de edad presenta una dentición constituida por dientes de leche (ver Figura 1. 0a) y en ocasiones muestran premolares de leche aún en erupción. El primer diente permanente en emerger a través de la encía es el primer Molar (M1). Empieza a aparecer a los 3-4 meses de edad, no termina de erupcionar hasta los 5 meses y se hace plenamente funcional (cuando su cara oclusal contacta con la del molar superior en la masticación) a los 6 meses de edad (Figura 1. 1a). En ciervos de las montañas Cantábricas, especimenes de 7-8 meses muestran un patrón de desgaste en la superfi cie de masticación del M1 coincidente con lo observado en nuestros ciervos a la misma edad (32). El tiempo de erupción del primer molar en nuestros ciervos coincide con el rango establecido para ciervos de Escocia (51), pero es posterior a lo publicado para ciervos alemanes (27, 49) en los que la erupción se establece a los 4 meses de edad. Estas diferencias pueden deberse a las diferencias en nomenclatura y consideración o no de distintos grados de erupción o funcionalidad dental.

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
243
Tabla 1.- Erupción de la dentición permanente del Ciervo (Cervus elaphus)
Orden y mes de erupción de los dientes permanentes Presente estudio para Cervus elaphus hispanicus en la mitad sur de la Península Ibérica
M1 I1 M2 I2 C I3 I4 Premolares M3 3ª Cúspide Autores6 14-15 15-16 17-18 25-26 27-30 31-32 37-44 (33)
Otras subespecies y latitudesM1 M2 I1 I2 C I3 I4 M3 P2 P3 P1 Autores
5-12 12-14 15 15-17 15-18 18-20 21-23 24-27 27-30 27-30 27-30 (53)4 12 14 16 17 19 19 21 25 25 25 (49)4 12 14 15 16 19 19 21 25 25 25 (27)5 13 - - - - - 26 - - - (29)
4-6 12-15 12-15 19 21 - 21-28 33 21-28 21-28 21-28 (51)- - 15 - - - - - - 24 - (16)
Figura 1.- Fases de la erupción de dientes permanentes en el ciervo ibérico. Letras minúsculas para los dientes de leche y mayúsculas para dientes defi nitivos y premolares numerados según su orden de aparición en la mandíbula, en vez de considerar su lugar
desde el punto de vista evolutivo.

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
244
En el ciervo ibérico el primer incisivo permanente (I1) se observa totalmente erupcionado y sin desgaste dental a los 14-15 meses, coincidiendo con un segundo molar (M2) que no ha terminado aún de emerger, y es a los 15-16 meses cuando esta muela aparece erupcionada totalmente. El segundo incisivo (I2) completa su erupción a los 17-18 meses igual que el canino permanente. Un carácter muy distintivo del tercer premolar de leche son sus tres cúspides. La Figura 1. 2a muestra la dentición típica de los animales llamados varetos o primalas (en el caso de las hembras) que al año y medio tienen casi siempre la mitad de los incisivos de leche y la mitad permanentes, y las dos primeras muelas totalmente funcionales. A los 27-30 meses aparecen los premolares totalmente erupcionados siendo visibles sus cúspides sobre la mandíbula a los 25-26 meses de edad. El estado de erupción del tercer premolar descrito a la edad de 20 meses para ciervos del norte de la Península Ibérica (32) coincide con lo observado en Sierra Morena. De los estudios con subespecies norteñas, los resultados que más se asemejan a los nuestros son en los que el I1 erupciona cuando el animal tiene aproximadamente 15 meses y el último premolar cuando tiene unos 24 (16, 51). La erupción del tercer molar en ciervos alemanes tiene lugar antes que en el ciervo ibérico según algunos autores como se observa en la Tabla 1.
Aunque Eidmann ya en 1932 (27) señala dicha erupción también a los 29-31 meses de edad coincidiendo con nuestras observaciones. Las diferencias pueden deberse a la consideración o no de la erupción de la tercera cúspide de este molar. En el ciervo ibérico, el tercer molar es el último diente permanente que aparece erupcionando totalmente sus primeras dos cúspides a la edad de 31-32 meses mientras que la ter-cera cúspide sólo es visible si eliminamos los tejidos de la encía (ver Figura 1. 3a). Esta tercera cúspide completa su erupción desde los 37 a los 44 meses, momento en el que presenta coloración oscura pero sin tener todavía desgaste en su cara oclusal (Figura 1. 4a).
Como en el ciervo ibérico la erupción de todos los dientes permanentes, inclu-yendo la tercera cúspide del tercer molar, se produce de forma completa a los 37-44 meses de vida del animal, es posible determinar la edad con bastante fi abilidad y detalle hasta los tres años y medio. A partir de entonces tenemos que utilizar otros criterios como el desgaste dental o la esqueletocronología.
3.1.1. Retraso en la erupción de dientes permanentes
Un hallazgo importante en nuestros ciervos ha sido la detección de un porcentaje signifi cativamente alto de casos en los que el reemplazamiento de algunos dientes

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
245
permanentes aparece retrasado (33). Cerca de un 20% de los animales presentaron retraso en la erupción de los incisivos permanentes. Concretamente un 18.75% de los ciervos mostraron retraso en la erupción del I1, apareciendo además animales menores de 18 meses con la primera y segunda muelas totalmente funcionales pero aún con todos los incisivos de leche. Esto puede evidenciar diferencias en el desarrollo general provocado por carencias nutricionales como consecuencia de factores ambientales o poblacionales adversos ya que estos animales son prácticamente el grupo de los ca-zados en 1994 y 1995, que nacieron muy tarde (fi nales de julio-agosto) en años malos de extrema sequía. Por el contrario, los animales cazados en 1997 presentaron a la misma edad los dos primeros incisivos permanentes totalmente erupcionados (Figura 1. 2a), además de mandíbulas más grandes y cornamentas de mayor calidad (3, 33).
Autores ingleses (51) encuentra que algunos ciervos raquíticos mantienen todos sus incisivos de leche aún teniendo ya dos molares y comenta que algunos indivi-duos de dos años y medio, con tres molares, mantienen premolares de leche. Este dato también ha sido constatado en un 8,33% de las hembras de Sierra Morena. La determinación del momento de erupción dental así como conocer estas variaciones específi cas y locales, es interesante no sólo para determinar la edad correctamente, sino para interpretar las marcas de crecimiento en los distintos tejidos dentales en los estudios esqueletocronológicos (9, 10, 16).
Estos retrasos en la erupción dental condicionan la aparición de las primeras marcas de crecimiento y pueden provocar errores de interpretación.
3.2. Utilidad del desgaste dental en la determinación de la edad
Los resultados en los estudios del desgaste dental en el ciervo ibérico indican diferencias de desgaste también entre sexos, destacando un mayor desgaste en los molares de los machos, probablemente debido a diferencias en el uso del hábitat y al tipo de alimento consumido (34, 35). Además los ciervos de fi ncas pequeñas y con mayor densidad presentaron mayor desgaste, infl uyendo también el tipo de gestión. Debido a estos factores de variación el desgaste es un método que necesariamente ha de ser tomado con precaución en la determinación de la edad y siempre habiendo descrito previamente un patrón de desgaste propio de una población en un hábitat determinado. Si en la Figura 1 se muestra el tipo de dentición que tienen los ciervos desde que nacen hasta que termina la erupción de todos los dientes permanentes, la Figura 2 describe el desgaste dental desde los 4 hasta los 13 años de edad. La Figura 2. 4b (vista oclusal de la mandíbula 4a en la Figura 1) muestra el desgaste dental a los

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
246
3-4 años de edad. En las mandíbulas de 4,5-6,5 años de edad (Figura 2. 5b), entre otros detalles podemos observar cómo la dentina expuesta en la cara oclusal de los molares dobla la anchura del grosor del esmalte, todavía son evidentes los infundíbulos de todos los molares. En el M2 se muestra evidente la línea de la corona (LC= línea de separación entre corona y raíz) y la dentina del metacono contacta con la del protoco-no. En las mandíbulas de los ciervos de 5,5-8,5 años (Figura 2. 6b) han desaparecido los infundíbulos de la cara oclusal del M1, y en la M3 es evidente la línea de la corona (LC) pero la dentina del hipocono y paracono aun no contactada. En las mandíbulas de ciervos de 7,5-13,5 años prácticamente han desaparecido los infundíbulos de los molares M1 y M2 y el del hipoconulo del M3 (Figura 2. 7b).
En Sierra Morena Oriental utilizando el reemplazamiento dentario y el patrón de desgaste dental conjuntamente es posible establecer 7 clases de edad, y aunque existe un mayor solapamiento y una menor precisión y fi abilidad a medida que aumenta la edad del animal, el método es óptimo para el conocimiento de la tasa de edades en estudios generales de población.
Figura 2.- Patrón de desgaste dental de ciervos de 4 a 13 años de edad. (LC= línea de la corona que aparece en M2 a partir de los 4 y en M3 a
partir de los 6 años de edad).

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
247
3.3. Interpretación y uso de marcas de crecimiento periódicas en tejidos dentales
3.3.1. Consideraciones generales sobre tejidos dentales y esqueletocronología
El término “esqueletocronología”, utilizado con más frecuencia por autores franceses, fue propuesto por primera vez por Castanet et al. en 1977 (15) para designar una técnica que venía siendo utilizada desde las primeras décadas del siglo XX para la determinación de la edad en vertebrados. Esta técnica está basada en la observación de marcas naturales repetitivas, de manera estacional, inscritas en distintos tejidos esqueléticos en crecimiento. El fenómeno de aparición de marcas o zonas de creci-miento estacionales, o anuales ha sido documentado en una gran variedad de tejidos duros como: tejidos dentales, hueso dermal del cráneo, vértebras, tejido perióstico de alrededor de los huesos, escamas, espinas y otolitos de peces, entre otros. Estas marcas o estructuras incrementales pueden ser estudiadas en cortes histológicos, en secciones gruesas pulidas o láminas delgado-pulidas, observadas con luz transmitida, polarizada o refl ejada. Los primeros trabajos de determinación de la edad con méto-dos esqueletocronológicas fueron los de Clerc (1927) y Harris (1927) (54) basados en estructuras incrementales observadas en hueso. Los primeros estudios en los que se utilizaron dientes fueron realizados por investigadores alemanes que usaron marcas de crecimiento anuales en la dentina de los incisivos de ciervo (Cervus elaphus) (Eid-mann 1932) (27). A partir de estos trabajos y de los realizados para la determinación de la edad en pinnípedos (55, 56), se extiende el uso de esta técnica, tanto en verte-brados acuáticos como en terrestres, con gran auge en los años 60 y 70 para estudios de dinámica de población y manejo de ungulados silvestres.
Los primeros tejidos utilizados fueron el hueso y la dentina, pero es el cemento dental el preferido para la determinación de la edad a partir de los trabajos en incisivos de alce (Alces alces) (57), en oso (Ursus americanus) (58) y en el primer molar de ciervo (22, 23, 51). Esto se debe a que al contrario que el hueso, rara vez el cemento esta sujeto a algún tipo de remodelado. En cuanto a la dentina, las marcas de crecimiento utilizadas son las formadas en la dentina secundaria que crece hacia la cavidad pulpar rellenándola, por lo que su uso se limita al momento en el que la citada cavidad de la pulpa dental se completa dejándose de formar nuevas capas de dentina.
El cemento es un tejido parecido al hueso y a la dentina, formado por una matriz orgánica en la que posteriormente se depositan sales inorgánicas, principalmente fos-fato cálcico (hydroxiapatita). La base orgánica del cemento está compuesta casi en su totalidad por colágeno, pero también contiene un número de proteínas no colagénicas entre las que destacan los glucosaminoglicanos y proteoglicanos, aunque la mayoría

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
248
de las proteínas no colagénicas del cemento no se conocen bien. El cemento dental se localiza alrededor de las raíces de los dientes de todos los mamíferos, pero también puede estar presente en la corona dental de molares de algunos herbívoros con alto grado de hipsodontia como es el caso del caballo y en la corona de algunos roedores o algunos mamíferos marinos. El colágeno del cemento tiene dos orígenes: intrínseco y extrínseco. El intrínseco está sintetizado por los cementoblastos, que se diferencian a partir de las células mesenquimales del ligamento o membrana periodontal y de-positan cemento sobre la superfi cie de la dentina a nivel de la raíz, y el extrínseco, mayoritario en determinados tipos de cemento, que es sintetizado por fi broblastos, en el mismo ligamento periodontal en grandes haces que son los conocidos como fi bras de Sharpey. Estas fi bras extrínsecas en haces llegan a mineralizarse en la ma-triz del cemento sirviendo de anclaje de la raíz del diente al ligamento o membrana periodontal. La función del cemento parece ser la de fi jación y sujeción del diente al alveolo dental, aunque el crecimiento continuo del cemento a lo largo de toda la vida puede también estar compensando la pérdida de altura del diente por el desgaste de la corona.
Todo el cemento que envuelve a las raíces dentales no es igual, sino que existen varios tipos con distintas características y localización. El primer cemento que se forma alrededor de la raíz de los incisivos, formando una primera capa distinta a las capas de cemento que se depositan después, se llama cemento intermediario o hialino. Está constituido por una banda estrecha (10-20 μm en la mayoría de las especies) de tejido calcifi cado situado entre la línea de separación de la dentina y los otros tipos de cemento que se depositan después. Algunas veces se confunde con la capa granular de Tomes de la dentina. Los otros dos tipos de cemento dental más abundantes son el cemento acelular que se encuentra en los laterales de la raíz de los dientes y el cemento celular que se encuentra en la zona apical de la raíz y en la almohadilla de cemento que se forma entre las raíces de los molares. El cemento celular se deposita más rápidamente que el acelular y se caracteriza por la presencia de cementoblastos que han quedado atrapados dentro de la matriz de cemento. Estas células pueden permanecer con vida durante algún tiempo como cementocitos en un estado de reposo, pero como el cemento no está nutrido por vasos o capilares sanguíneos, estas células mueren y dejan huecos o lagunas que dan a este cemento un aspecto característico y fácil de identifi car. El cemento celular suele ser mas ancho y tiene más cantidad de colágeno intrínseco pero pocas o prácticamente ninguna fi bra de Sharpey. Al contra-rio, el cemento acelular se caracteriza por que los cementoblastos, debido a que es depositado relativamente despacio, permanecen en la superfi cie del cemento y no llegan a ser incluidos o embebidos por el tejido. De esta manera, el cemento acelular

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
249
no incorpora cementocitos y no tiene lagunas como en el caso del cemento celular. Además, el cemento acelular normalmente contiene una gran variedad de colágeno extrínseco (fi bras de Sharpey) y tiene relativamente pocas fi bras de colágeno intrínseco.
Se ha comprobado la existencia de diferencias importantes en cuanto al proceso y la tasa de cementogénesis entre los distintos tipos de cemento, pero el mecanismo molecular mediante el que se produce su formación todavía es desconocido. Tampo-co se conocen las causas de que tanto el cemento celular como el acelular, así como el resto de tejidos en los que aparecen marcas estacionales, tengan un crecimiento periódico que se refl eja en la aparición de capas alternas que pueden ser utilizadas para la determinación de la edad.
3.3.2. Dientes, tejidos y métodos para identifi car marcas de crecimiento en el ciervo ibérico
En el ciervo ibérico las marcas de crecimiento son observadas tanto en dentina como en cemento de incisivos, molares y caninos como se muestra en las Figuras 3 y 4. Los caninos fueron los menos apropiados para la identifi cación y distinción de marcas de crecimiento ya que aunque éstas fueron evidentes en dentina, no fue siempre así en el cemento radicular. El tejido de elección fue el cemento dental, al igual que en estudios precedentes ya que la interpretación de las marcas en la dentina fue muy difícil en todas las piezas dentales, principalmente por que son mas fi nas aparecen muy próximas unas a otras y en grupos (Figuras 3d-e) lo que complica distinguir las primarias de las secundarias o las marcas incrementales iniciales formadas en la dentina (46).
Las zonas de elección son: el cemento celular de la zona interradicular del primer molar (Figuras 3b, 3g, 4b-h) y el cemento acelular de la raíz de los incisivos (Figuras 3a, 3c, 3f, 3i, 4a) ambos observados a través de cortes longitudinales, debido a que sobre todo de incisivos, se evidencia una mayor región dental permitiendo confi rmar la continuidad de líneas de reposo y marcas de crecimiento a lo largo del diente. Se diferencian mejor las líneas primarias y se hace posible la identifi cación de la línea de separación entre dentina y cemento (Figuras 3i, 4a). La mayoría de las muestras de incisivos observadas con luz transmitida (Figura 3c) o polarizada (Figura 4i) mostraron fi bras de Sharpey con orientación constante siempre perpendicular a la línea de sepa-ración entre dentina y cemento, al contrario de lo encontrado en otros estudios (79).
En relación al tipo de preparación, son los cortes histológicos obtenidos con microtomo convencional los óptimos. Permiten la observación de mayor cantidad
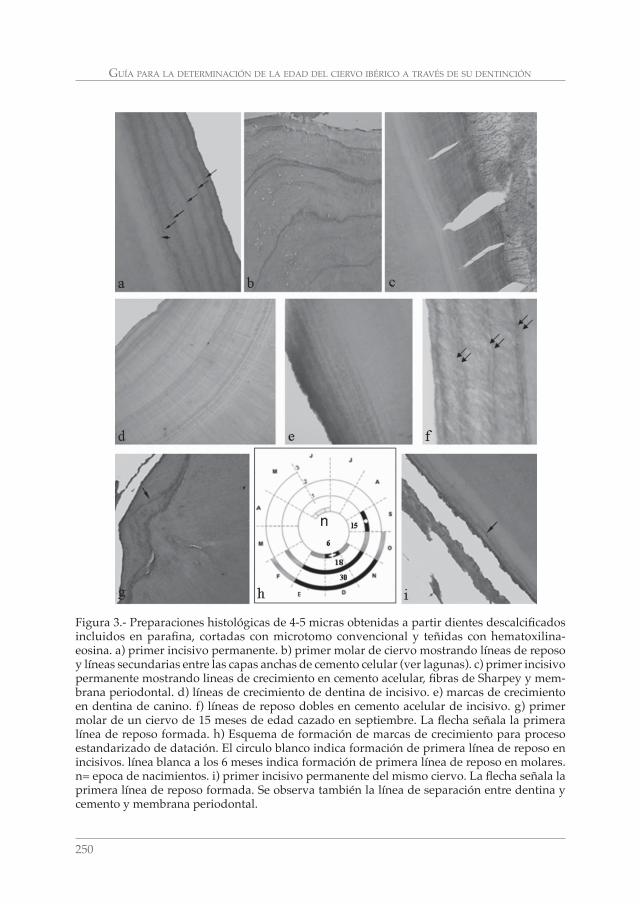
GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
250
Figura 3.- Preparaciones histológicas de 4-5 micras obtenidas a partir dientes descalcifi cados incluidos en parafi na, cortadas con microtomo convencional y teñidas con hematoxilina-eosina. a) primer incisivo permanente. b) primer molar de ciervo mostrando líneas de reposo y líneas secundarias entre las capas anchas de cemento celular (ver lagunas). c) primer incisivo permanente mostrando lineas de crecimiento en cemento acelular, fi bras de Sharpey y mem-brana periodontal. d) líneas de crecimiento de dentina de incisivo. e) marcas de crecimiento en dentina de canino. f) líneas de reposo dobles en cemento acelular de incisivo. g) primer molar de un ciervo de 15 meses de edad cazado en septiembre. La fl echa señala la primera línea de reposo formada. h) Esquema de formación de marcas de crecimiento para proceso estandarizado de datación. El circulo blanco indica formación de primera línea de reposo en incisivos. línea blanca a los 6 meses indica formación de primera línea de reposo en molares. n= epoca de nacimientos. i) primer incisivo permanente del mismo ciervo. La fl echa señala la primera línea de reposo formada. Se observa también la línea de separación entre dentina y cemento y membrana periodontal.
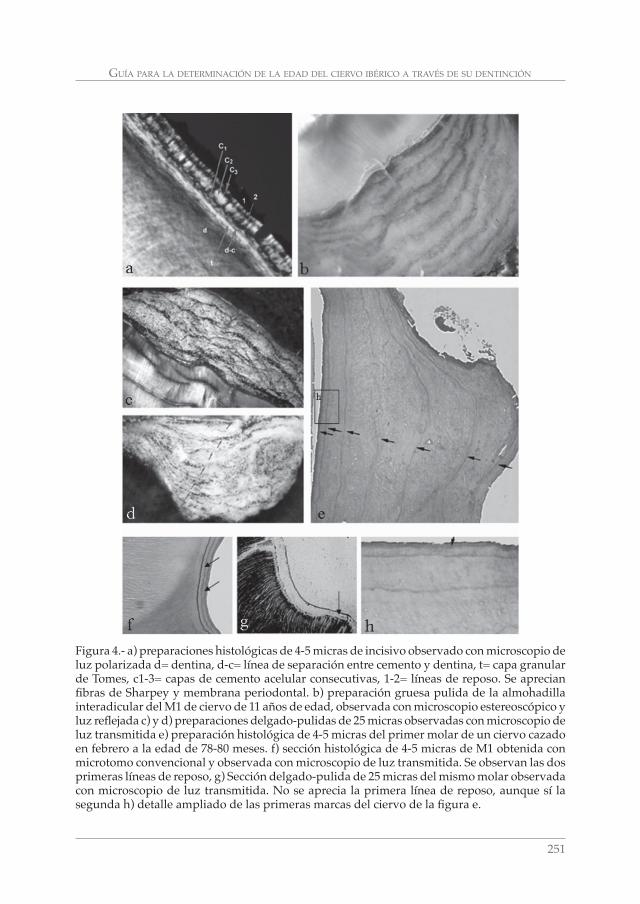
GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
251
Figura 4.- a) preparaciones histológicas de 4-5 micras de incisivo observado con microscopio de luz polarizada d= dentina, d-c= línea de separación entre cemento y dentina, t= capa granular de Tomes, c1-3= capas de cemento acelular consecutivas, 1-2= líneas de reposo. Se aprecian fi bras de Sharpey y membrana periodontal. b) preparación gruesa pulida de la almohadilla interadicular del M1 de ciervo de 11 años de edad, observada con microscopio estereoscópico y luz refl ejada c) y d) preparaciones delgado-pulidas de 25 micras observadas con microscopio de luz transmitida e) preparación histológica de 4-5 micras del primer molar de un ciervo cazado en febrero a la edad de 78-80 meses. f) sección histológica de 4-5 micras de M1 obtenida con microtomo convencional y observada con microscopio de luz transmitida. Se observan las dos primeras líneas de reposo, g) Sección delgado-pulida de 25 micras del mismo molar observada con microscopio de luz transmitida. No se aprecia la primera línea de reposo, aunque sí la segunda h) detalle ampliado de las primeras marcas del ciervo de la fi gura e.

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
252
de detalles de la histología dental, facilitan la identifi cación fi able de las primeras marcas de crecimiento y reposo así como de la línea de separación entre dentina y cemento tanto en molares como en incisivos, y en los incisivos incluso la identifi cación de la capa granular de Tomes en la dentina. Sin embargo, en las secciones pulidas es mucho más difícil distinguir las primeras marcas, así como otros detalles impor-tantes. Las Figuras 4f y 4g muestran el mismo molar preparado mediante histología convencional y petrografía.
En cualquier caso, en casi todas las preparaciones se pueden observar marcas secundarias o accesorias. Algunas muestras de molares mostraron líneas accesorias en el interior de capas anchas de cemento celular (Figura 3b) que han sido relacio-nadas con el debilitamiento físico que sufren algunos machos en el periodo de celo (51), pero que han sido también descritas en hembras (59). En la región apical de los incisivos aparece gran cantidad de líneas accesorias como característica del cemento celular de esta zona (14, 60) difi cultando el recuento e interpretación a este nivel. En algunos ciervos ocasionalmente también se han observado líneas dobles en el cemento acelular de incisivos (Figura 3f) similares a las marcas irregulares descritas en otras subespecies (61). Y en animales muy viejos las líneas aparecen muy juntas y más difícilmente cuantifi cables a la vez que las primeras capas de cemento pueden desaparecer debido a la erosión dental por el proceso de masticación tanto en incisivos como en molares. La identifi cación de líneas accesorias, secundarias, falsas, dobles o suplementarias es esencial para una determinación correcta de la edad.
3.3.3. Periodicidad en la formación de marcas de crecimiento
Las marcas de crecimiento parecen ser el resultado de fl uctuaciones en la tasa y naturaleza de la deposición de tejidos calcifi cados que refl ejan diferentes eventos de la vida del animal. Aunque las causas que las originan han sido objeto de un amplio debate en la literatura, éstas aún no se han determinado satisfactoriamente. Algunos autores consideran que tales marcas tanto en tejidos dentales como en hueso pueden estar relacionadas con cambios estacionales de clima, alimentación, cambios endocrinos y/o cambios regionales (1). Muchos otros opinan que las líneas estrechas corresponden a una restricción en la dieta (8, 24, 62, 63, 64, 65), pudiendo estar también infl uidas por cambios en el fotoperiodo (66, 67). Pero lo cierto es que los procesos fi siológicos para su desarrollo, así como su regulación son casi desconocidos. Debido al desconocimiento de las causas y mecanismos que las ocasionan es necesario determinar su periodicidad, el número de incrementos y líneas de reposo que se producen por año, así como la época de formación de casa tipo de marca para su correcta interpretación.

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
253
En algunos ungulados tropicales, como es el caso del ciervo de cola blanca ve-nezolano (Odocoileus virginianus venz.) que es capaz de reproducirse todo el año, las marcas de crecimiento pueden formarse a intervalos variables de seis meses a más de un año (68). En peces (69, 70, 71), algunos anfi bios urodelos (72, 73) y algunos mamíferos marinos (74) aparecen dos ciclos de crecimiento al año. Pero en la mayoría de los mamíferos terrestres tanto carnívoros (75) como bóvidos y cérvidos (8, 57, 76, 77,) aparece un ciclo de crecimiento anual, formándose cada año una capa ancha de crecimiento y otra estrecha de reposo. No obstante, existen amplias variaciones en cuanto a las épocas correspondientes a los periodos de crecimiento o reposo dependiendo de la localización geográfi ca y de la subespecie (16). Así, en los ciervos de Escocia el crecimiento se observa en verano-otoño y las paradas de crecimiento en invierno-primavera mostrándose las líneas de reposo o detención de crecimiento principalmente desde enero hasta abril (51). En Noruega y Dinamarca las marcas fi nas de detención de crecimiento se observan también en primavera desde marzo a mayo (78).
En el ciervo ibérico de Sierra Morena el cemento interradicular del primer molar presenta dos marcas por año, una amplia formada en primavera-verano cuyo desarrollo termina en agosto-septiembre y una estrecha que se forma durante otoño-invierno parece que como resultado de un “reposo” en el crecimiento del cemento dental. Las línea de reposo o detención de crecimiento aparece con mas frecuencia desde noviembre a enero, con un pico de formación en diciembre (47). Estas líneas aparecen también antes de enero en ciervos franceses (14). Nuestras observaciones también coinciden con lo observado en otros ungulados del área Mediterránea (79) que muestran líneas estrechas en invierno (diciembre) y anchas en verano. En ciervos de Nueva Zelanda las líneas de reposo se desarrollan desde julio a octubre coincidiendo también con el invierno del hemisferio sur (24).
En el área Mediterránea no es el invierno la época más desfavorable desde el punto de vista trófi co por lo que, la aparición de líneas de reposo en invierno y no a fi nales de verano (época limitante) puede estar apoyando las teorías en relación a cambios endocrinos relacionados con el fotoperiodo como infl uyentes en la formación de las citadas marcas (66, 67).
3.3.4. Las primeras marcas de crecimiento y reposo y procedimiento estandarizado de datación.
En los caninos no fue posible determinar el momento de aparición de las primeras marcas de crecimiento periódicas a pesar de disponer de material sufi ciente de edad conocida, debido a la gran variabilidad encontrada entre individuos. Sólo fue posible

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
254
determinar con exactitud la edad de aparición de las primeras marcas en el cemento de incisivos y molares, por lo que los caninos, a pesar de ser piezas accesibles y fáciles de obtener incluso en animales vivos, no resultaron ser esqueletocronológicamente útiles para datar ciervos ibéricos.
Como se describe en trabajos previos (39) en todas las preparaciones histológicas de incisivos permanentes (I1) de ciervos menores de 15 meses se observa una capa de cemento hialino de anchura variable, mientras que a los 15 meses, coincidiendo con el fi nal de la erupción del diente, se observa una línea fi na a la que se ha llamado línea de oclusión justo después de la capa de cemento hialino. También en otros artio-dáctilos ha sido descrita esta línea oclusal incluso antes de que el incisivo hubiera completado el cierre de la raíz dental (79). En los incisivos de ciervos de 18 meses de edad, después de la línea de separación entre dentina y cemento, aparece la capa de cemento hialino, después la línea de oclusión y a continuación una segunda capa de cemento de grosor variable. A partir de los 18 meses la segunda línea de reposo aparece coincidiendo con la época de detención del crecimiento en noviembre-enero. En la siguiente primavera-verano se forma otra capa de cemento acelular adicional y así sucesivamente. En los molares (M1) la primera capa de crecimiento en el cemento inte-rradicular se desarrolla durante la estación de crecimiento primavera-verano cuando el molar está en fase de erupción a través de la encía, mientras que la primera línea de detención de crecimiento se forma durante el primer invierno, justo cuando el ciervo tiene 6 meses y el molar se hace funcional, es decir cuando choca con su homólogo del maxilar. A esta línea que corresponde con el periodo de menor crecimiento en el cemento dental (noviembre-enero) también la podríamos llamar línea oclusal ya que también coincide con la oclusión molar. La segunda capa de crecimiento se deposita durante la siguiente primavera-verano y la siguiente línea fi na de reposo durante el siguiente otoño-invierno y así sucesivamente.
En síntesis, las primeras marcas de crecimiento anchas se forman tanto en incisi-vos como en molares durante la erupción dental y la primera línea estrecha de reposo o detención de crecimiento aparece a los 6 meses en molares (M1) y a los 15 meses en incisivos (I1) coincidiendo con la oclusión dental. La segunda línea de reposo aparece tanto en molares como en incisivos a los 18 meses de edad coincidiendo con la de-tención del crecimiento en otoño-invierno, y cada línea fi na de reposo de crecimiento supone un año más de vida del animal (39, 47). Las Figuras 3g y 3i, correspondientes a preparaciones histológicas del primer molar e incisivo de un ciervo de 15 meses de edad, donde se observan estas primeras marcas en detalle. La determinación de la edad puede realizarse contando las capas anchas o las líneas fi nas tanto en inci-sivos como en molares. Pero es más fácil el recuento de las líneas de reposo. Como

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
255
se esquematiza en la Figura 3h un ciervo que haya nacido en mayo-junio y cuyos dientes molares e incisivos hayan erupcionado según el patrón habitual a los 6 y 15 meses respectivamente, presentarán dos líneas fi nas a los 18 meses, 3 a los 30 meses y así sucesivamente. Cada línea sucesiva indica un año más en la vida del ciervo. En consecuencia, la edad en años de un ciervo se estima a partir del número de líneas de reposo menos 0,5 años.
También es posible estimar la edad en meses conociendo el mes de muerte y asumiendo una hipotética época de nacimiento, que en el área de estudio habitual-mente va de mayo a junio (ver Figura 4f por ejemplo). Pero cuando el mes de muerte es desconocido deducir la edad en meses es más difícil, especialmente en ciervos cazados entre febrero y octubre, ya que puede haber una diferencia de 6 meses entre dos animales que muestren el mismo número de líneas de reposo por que éstas tienen lugar después de septiembre, entre noviembre y enero como sabemos. En estos casos el análisis de la anchura de la última capa de cemento formada puede resultar útil. Por ejemplo, un ciervo muerto en octubre mostrará una capa ancha de cemento en el borde mientras que otro muerto en enero o febrero mostrará una última línea de reposo muy próxima a una última capa de cemento muy delgada que crecerá durante los siguientes 9 meses hasta la formación de la siguiente línea de reposo. En general, esto es fácil de observar en molares con capas de cemento celular más anchas que las del cemento acelular de incisivos.
3.3.5. Fiabilidad, precisión, errores asociados y limitaciones
Según algunos autores (8, 10, 80) la ventaja de la esqueletocronología en com-paración con otras técnicas es que, una vez validada sobre una especie concreta, constituye un método absoluto de determinación de la edad individual. Esta técnica es considerada una de las herramientas disponibles más útiles para la investigación y manejo de la vida silvestre (16). Uno de sus principales valores en investigación es poder ser utilizada como referencia en la evaluación de otros métodos de estimación de edad (81), y también tiene una aplicación interesante en arqueología (79, 82, 83, 84). Sin embargo, se ha reconocido la existencia de un error asociado al método y la necesidad de estimar la frecuencia y la magnitud de este error (85, 86). Además, el grado de precisión de esta técnica no es siempre el mismo, varía según los animales y según procedimientos. Este método es más preciso para mamíferos que viven varios años o incluso varias décadas, que se reproducen una vez durante cierto periodo del año (por ejemplo los ungulados, carnívoros y pinnípedos), y pierde precisión para determinar la edad en pequeños mamíferos que viven 1-2 años y se reproducen varias
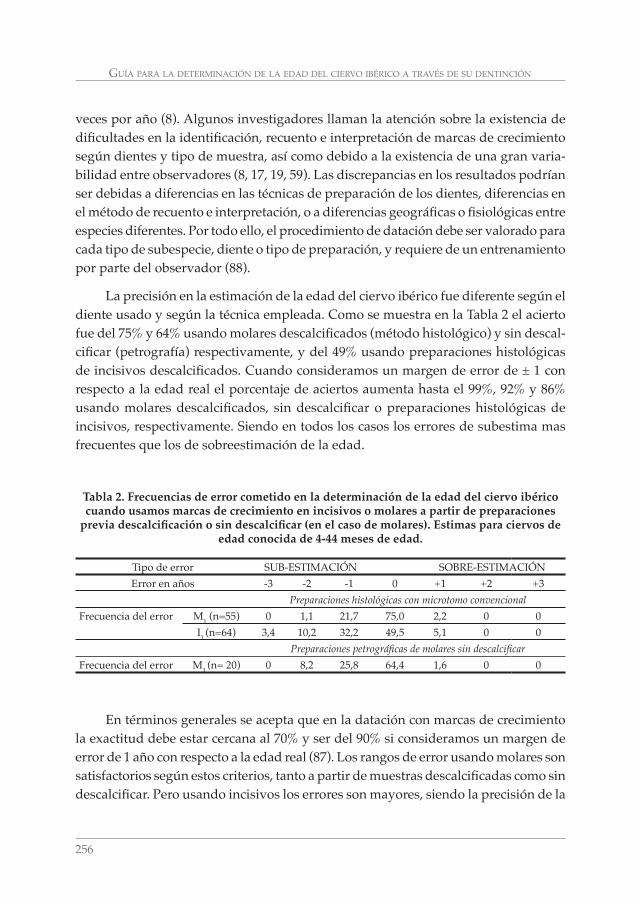
GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
256
veces por año (8). Algunos investigadores llaman la atención sobre la existencia de difi cultades en la identifi cación, recuento e interpretación de marcas de crecimiento según dientes y tipo de muestra, así como debido a la existencia de una gran varia-bilidad entre observadores (8, 17, 19, 59). Las discrepancias en los resultados podrían ser debidas a diferencias en las técnicas de preparación de los dientes, diferencias en el método de recuento e interpretación, o a diferencias geográfi cas o fi siológicas entre especies diferentes. Por todo ello, el procedimiento de datación debe ser valorado para cada tipo de subespecie, diente o tipo de preparación, y requiere de un entrenamiento por parte del observador (88).
La precisión en la estimación de la edad del ciervo ibérico fue diferente según el diente usado y según la técnica empleada. Como se muestra en la Tabla 2 el acierto fue del 75% y 64% usando molares descalcifi cados (método histológico) y sin descal-cifi car (petrografía) respectivamente, y del 49% usando preparaciones histológicas de incisivos descalcifi cados. Cuando consideramos un margen de error de ± 1 con respecto a la edad real el porcentaje de aciertos aumenta hasta el 99%, 92% y 86% usando molares descalcifi cados, sin descalcifi car o preparaciones histológicas de incisivos, respectivamente. Siendo en todos los casos los errores de subestima mas frecuentes que los de sobreestimación de la edad.
Tabla 2. Frecuencias de error cometido en la determinación de la edad del ciervo ibérico cuando usamos marcas de crecimiento en incisivos o molares a partir de preparaciones
previa descalcifi cación o sin descalcifi car (en el caso de molares). Estimas para ciervos de edad conocida de 4-44 meses de edad.
Tipo de error SUB-ESTIMACIÓN SOBRE-ESTIMACIÓNError en años -3 -2 -1 0 +1 +2 +3
Preparaciones histológicas con microtomo convencionalFrecuencia del error M1 (n=55) 0 1,1 21,7 75,0 2,2 0 0
I1 (n=64) 3,4 10,2 32,2 49,5 5,1 0 0Preparaciones petrográfi cas de molares sin descalcifi car
Frecuencia del error M1 (n= 20) 0 8,2 25,8 64,4 1,6 0 0
En términos generales se acepta que en la datación con marcas de crecimiento la exactitud debe estar cercana al 70% y ser del 90% si consideramos un margen de error de 1 año con respecto a la edad real (87). Los rangos de error usando molares son satisfactorios según estos criterios, tanto a partir de muestras descalcifi cadas como sin descalcifi car. Pero usando incisivos los errores son mayores, siendo la precisión de la

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
257
estima inferior a lo observado en estudios similares (88, 89, 90). El uso de incisivos está en el límite de lo aceptable ya que además, se han producido errores de subestima de hasta 3 años, no detectados usando molares. Este error podría ser incluso mayor si el estudio hubiera incluido animales mayores de 44 meses de edad, ya que en animales muy viejos las primeras líneas aparecen muy juntas y son más difícilmente cuantifi ca-bles a la vez que las primeras capas de cemento pueden quedar compactadas debido a la erosión dental o a la presión por el proceso de masticación (27).
Las variaciones de exactitud y precisión entre dientes son debidas a diferencias en el número de líneas o marcas contabilizadas. De todos los ciervos en los que se comparó el número de marcas tanto en incisivos como en molares, el 41% presentó menos marcas en I1. A pesar de que los incisivos fueron cortados con microtomo con-vencional previa descalcifi cación, que permite una mejor visualización de su histología en detalle, en muchos de ellos no fue posible identifi car la primera línea de reposo durante el segundo año de vida del animal. Esto puede ser debido al retraso en la erupción dental de manera que si incisivos permanentes erupcionan después de los 18 meses de edad se formará una única línea de reposo en lugar de dos. El retraso en la erupción del I1 constituye una fuente de error importante en el área de estudio ya que se ha detectado en un 18.75% de los ciervos estudiados retraso en la erupción del (3.1.1.), y difícil de corregir en un procedimiento estandarizado. Una vez completado el reemplazamiento dentario no es posible saber si un animal adulto ha tardado más o menos en mudar sus dientes de leche.
Las discrepancias entre molares parecen ser debidas a diferencias entre méto-dos de preparación de muestras. El cemento celular de la almohadilla interradicular observado a partir de preparaciones histológicas (4-5 micras de grosor), muestra un mayor número de líneas de reposo siendo visibles las primaras marcas formadas así como más fáciles de distinguir líneas accesorias o irregulares. Cuando observamos secciones de esta misma zona de los molares en preparaciones petrográfi cas de grosor superior a 15 micras, las líneas de reposo son más difíciles de identifi car, especial-mente las primeras marcas formadas (ver Figura 3e-h). Incluso se ha observado que en preparaciones petrográfi cas las marcas depositadas durante los tres primeros años de vida son más difíciles de distinguir que las que se forman con posterioridad (45, 46, 85). La primera línea de reposo que tiene lugar durante el primer año de vida en el ciervo ibérico puede no ser detectada a partir de preparaciones delgado-pulidas sin descalcifi car (ver Figura 4f y 4g). Pero este tipo de diferencias no son difíciles de corregir a través de un proceso estandarizado de recuento. La precisión de la estima de a edad en mamíferos mediante recuento de líneas o marcas en el cemento dental

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
258
puede ser mejorada si tenemos un método estandarizado y un trabajo previo de va-lidación a modo de referencia para la especie y/o población objeto de estudio (91).
Cuando usamos molares descalcifi cados (3.3.4) al número de líneas contadas se les resta 0,5 para determinar la edad en años, pero cuando usamos preparaciones petrográfi cas es necesario sumar de 1 a 2 al número total de líneas contadas. Sólo se resta 0,5 cuando la preparación obtenida es de mucha calidad y con certeza se puede observar la primera línea formada. De hecho, todos aquellos autores que usan secciones de molares sin descalcifi car (24, 51) o usan microtomo de congelación (60) encuentran que la primera marca de reposo es observada durante el segundo invierno de la vida del animal. La primera línea de reposo detectada por la mayoría de los autores que usan este tipo de preparaciones corresponde a la segunda línea de reposo observada usando preparaciones histológicas de piezas descalcifi cadas, por ello estos autores añaden 1 al número de marcas contabilizadas para obtener la edad correcta (51).
Las preparaciones delgado-pulidas de molares son rápidas de realizar, menos costosas que las histológicas y más conservativas. Por todo ello, se recomienda su uso cuando la cantidad de animales a datar es grande, pero requieren de un proceso de entrenamiento por parte del observador para una correcta interpretación.
Los investigadores y/o gestores deben ser conscientes del error que se comente con cada método y la elección del método de datación ha de ir en función del grado de precisión que requieran los objetivos de gestión o investigación marcados. Cuando se requiere información rápida y determinar los grupos de edad durante trabajos de campo la combinación de desgaste y reemplazamiento ha sido utilizada satisfacto-riamente (92, 93, 94).
4. CONCLUSIONES
El establecimiento de protocolos estandarizados para cada especie y población y un entrenamiento previo del observador en todos los métodos de datación reduce errores por subjetividad. Los errores cometidos en la datación deben ser sólo los propios o intrínsecos a cada método.
La erupción dental permite determinar la edad hasta los tres años y medio y una combinación de desgaste y erupción dental establecer al menos 7 clases de edad útiles en estudios de población.

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
259
En cuanto a métodos esqueletocronológicos se aconseja usar marcas de creci-miento y reposo en el cemento celular interradicular del primer molar inferior en detrimento del uso de incisivos.
Con muestras de molares (M1) previa descalcifi cación la edad en años se estima restando 0,5 años al número total de líneas de reposo contadas en el cemento celular interradicular. La fi abilidad es la máxima registrada, del 75% (99% considerando un error de ± 1 año).
Para conseguir una precisión similar usando preparaciones delgado-pulidas de molares sin descalcifi car se requiere un ajuste consecuente a la calidad de la prepara-ción. Según los casos es necesario sumar de 1 a 2 años al número de líneas de reposo contadas. Sólo se resta 0,5 cuando la primera marca es ciertamente visible.
AGRADECIMIENTOS:
A Julian Garde López-Brea y Tomás Landete de la Universidad de Castilla la Mancha, a propietarios, gestores y guardas de fi ncas cinegéticas en Sierra Morena Oriental como El Friscalejo, El Morron de las Mujeres, El Mencho, La Alameda, Valquemado o Lugar Nuevo,…entre otras. A taxidermistas como Alfonso Blanco y Juan Pleités, a empresas como Monterías Cencerra y Monterías Chamocho. A los Departamentos de Mineralogía de la Universidad de Granada, Geología de la Uni-versidad de Jaén, Anatomía y Anatomía Patológica Comparadas de la Universidad de Córdoba y a la Estación Biológica de Doñana, especialmente a: Alfonso, Aniceto Méndez, Antonio Piedra y Ernesto García. También a Mario Martínez Pulido, José Hervás, Rafael Carrasco y en especial Mohamed Analla, Joaquín Muñoz-Cobo quien orientó los trabajos, y a G.A. Klevezal, G. Matson y B. Mitchell quienes los revisaron y mejoraron, o facilitaron importante información inicial.
5. BIBLIOGRAFÍA
1. Bourlière, F., Spitz, F. 1975. Les critères d’áge chez les Mamifères. In: Problèmes D’écologie: La démographie des populations de Vertébrés. Pp: 53-76. Eds. M. Lamotte & F. Bourliere. Masson et Cie. París.
2. Riney, T. 1982. Study and Management of large Mammals. Eds. John Wiley and sons. New York.3. Azorit, C., Analla, M., Carrasco, R., Muñoz-Cobo, J. 2002a. Infl uence of age and environment on
antler traits in Spanish red deer. Zeitschrift für Jagdwissenschaft 48: 137-144.4. Azorit, C., Analla, M., Carrasco, R.J., Carrasco, A., Muñoz-Cobo, J. 2002b. Astas, esqueleto y edad
del ciervo de Sierra Morena Oriental: estudio de correlación. Anales de Biología 24: 169-174.

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
260
5. Johnston, D.H., Joachim, D.G., Bachmann, P., Kardong, K.V., Stewart, R.A., Dix, L.M., Strickland, M.A., Watt, I.D. 1987. Aging furbearer using tooth structure and biomarkers. In: Wild furbearer Management and Conservation in North America Ontario, Canada. Pp: 228-243. Eds. Novak, M., Baker, J.A., Obbard, M.E., Malloch, B.
6. Aparicio, G. 1946. Fenotipología Animal. Ministerio de Agricultura, Madrid.7. Aparicio, G. 1956. Exterior de los animales domésticos. Morfología externa. Imp. Moderna. Cór-
doba. 8. Klevezal, G.A., Kleinenberg, S.E. 1967. Age determination of mammals from annual layers in
teeth and bones. Translation by Israel Program for Scientifi c Translations, Jerusalem, 1969, No. 5433.
9. Morris, P. 1972. A review of mammalian age determination methods. Mammal Review 2: 69-104. 10. Morris, P. 1978. The use of teeth for estimating the age of wild mammals. Pp: 483-494 in Butler,
P.M., Joysey, K.A. editors. Development, function and evolution of teeth. Academic Press, Lon-don, United Kingdom.
11. Larson, J.S., Taber, R.D. 1980. Criteria of sex an age. In: Wildlife Management Techniques Man-ual. 4th edition. Ed. Schemitz S.D. Wildlife Society. Washington D.C.
12. Sáenz de Buruaga, M., Lucio, A.J., Purroy, F.J. 1991. Reconocimiento de sexo y edad en especies cinegéticas. Diputación Foral de Álava, Gobierno Vasco.
13. Dimmick, R.W., Pelton, M.R. 1994. Criteria of sex and age. In: Research and management tech-niques for widlife and habitats. Pp: 169-214. Ed. Bookhout, T.A. Fifth Edition. The Wildlife Society, Bethesda, MD. 740 pp.
14. Quéré, J.P., Pascal, M. 1983. Comparaison de divers critères de détermination de l’âge individuel chez le cerf élaphe (Cervus elaphus) de France. Annales des Sciences Naturelles-Zoologie 13: 235-252.
15. Castanet, J., Meunier, F., Ricqles, A. 1977. Lénregistrement de la croisance cyclique par le tissu osseux chez les vertébrés poikilothermes: données comparatives et essai de synthèse. Bulletin Biologique de la France et de la Belgique 111: 183-202.
16. Grue, H., Jensen, B. 1979. Review of the formation of growth lines in tooth cementum of terres-trial mammals. Danish Review of Game Biology 11: 1-48.
17. Roseberry, J.L. 1980. Age determination of white-tailed deer in the Midwest. Methods and prob-lems. White-tailed deer population management in the north central states. North Central Sec-tion of the Wildlife Society, Midwest Fish and Wildlife Conference 41: 73-82.
18. Vincent, J.P., Angibault, J.M., Bideau, E., Gaillard, J.M. 1994. Le problème de la détermination de l’âge: Une source d’erreur négligée dans le calcul des tables de vie transversales. Mammalia 58: 293-299.
19. McCullough, D.R. 1996. Failure of the tooth cementum aging technique with reduced popula-tion density of deer. Wildlife Society Bulletin 24:722-724.
20. Harris, S. 1978. Age determination in the red fox (Vulpes vulpes): an evaluation of technique ef-fi ciency as applied to a sample of suburban foxes. Journal of Zoology 184: 91-118.
21. Schemnitz, S.D. (ed.) 1980. Wildlife Management Techniques Manual. 4ª ed. The Widlife Society. Washington. 575 p.
22. Mitchell, B. 1963. Determination of age in Scottish red deer from growth layers in dental cemen-tum. Nature, London 198: 350-351.
23. Lowe, V.P.W. 1967. Teeth as indicators of age with special reference to red deer (Cervus elaphus) of known age from Rhum. Journal of Zoology London 152: 137-152.
24. Douglas, M.J.W. 1970. Dental cement layers as criteria of age for deer in New Zealand with em-phasis on red deer, Cervus elaphus. New Zealand Journal of Science 13: 352-358.

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
261
25. Almasan, H.A., Rieck, W. 1970. Untersuchungen der Zahnstrucktur zur Altersbestimmung beim Rotwild (Cervus elaphus L.). Zeitschrift für Jagdwissenschaft 18: 222-224.
26. Ueckermann, E., Scholz, H. 1976. Vergleich der ersatzdentinbildung im 1. Scheidezahn und der zementzonenbildung im 1. Molar mit dem abnutzungsgrad der backenzähne im Unterkiefer beim Rothirsch (Cervus elaphus L., 1958). Zeitschrift für Jagdwissenschaft 22: 65-74.
27. Müller-Using, D. 1981. Rotwildalter-Merkblatt. Seventh edition. D. J. V., Bonn, Germany. 28. Godawa, J. 1989. Age determination in the Red deer. Acta Theriologica 34(28): 381-384.29. Brown, W.A., Chapman, N.G. 1991. The dentition of red deer (Cervus elaphus) a scoring scheme
to assess age from wear of the permanent molariform teeth. Journal of Zoology 224: 519-536.30. Costa, L. 1991. Ordenación y gestión de la caza mayor. En: Manual de ordenación y gestión
cinegética. Pp: 259-296. Ed. Alvarado Corrales. IFEBA, Badajoz.31. Arenas, A., Perea, A. 1993. El Ciervo en Sierra Morena. Eds. A. Arenas y A. Perea. Facultad de
Veterinaria, Universidad de Córdoba. Córdoba.32. Mariezkurrena, K. 1983. Contribución al conocimiento del desarrollo de la dentición y el es-
queleto postcraneal de Cervus elaphus. Munibe 35: 149-202.33. Azorit, C., Analla, M., Carrasco, R., Calvo, J.A., Muñoz-Cobo, J. 2002c. Teeth eruption pattern in
red deer (Cervus elaphus hispanicus) from southern Spain. Anales de Biología 24: 193-200. 34. Azorit, C., Analla, M., Carrasco, R., Muñoz-Cobo, J. 2003a. Determinación de la edad por des-
gaste dental en el ciervo ibérico (Cervus elaphus hispanicus, Erxleben, 1777) (Mammalia, Cervi-dae). Boletín de la Real Sociedad Española de Historia Natural (Sec. Biol.) 98(1-4): 123-134.
35. Azorit, C., M. Analla, R. Carrasco, and J. Muñoz-Cobo. 2003b. Efecto del sexo, densidad y año de muerte, sobre el desgaste dental del ciervo ibérico (Cervus elaphus hispanicus, Erxleben, 1777) (Mammalia, Cervidae) en Sierra Morena, España. Boletín de la Real Sociedad Española de His-toria Natural (Sec. Biol.) 98(1-4): 115-122.
36. Braza, F., Soriguer, R.C., San José, C., Delibes, J.R., Aragón S., Fandos, P., León, L. 1994. Métodos para el estudio y manejo de cérvidos. Junta de Andalucía, Sevilla.
37. Soriguer, R.C., Fandos, P., Bernáldez, E., Delibes, J.R. 1994. El ciervo en Andalucía. Eds. Soriguer, R.C., Fandos, P., Bernaldez, E., Delibes, J.R. Consejería de Agricultura y Pesca, Sevilla.
38. Azorit, C. 1999. Estudio de la edad y su aplicación a la gestión cinegética del ciervo en Sierra Morena Oriental. Tesis doctoral. Universidad de Jaén. 280 p.
39. Azorit, C., Muñoz-Cobo, J., Hervás, J., Analla, M. 2004. Aging through growth marks in teeth of Spanish red deer. Wildlife Society Bulletin 32(3): 702-710.
40. Attwell, C.A.M. 1980. Age determination of the blue wildebeest Connochaetes taurinus in Zulu-land. South African Journal of Zoology/Suid Afrikaanse Tydskrif vir Dierkunde 15(3): 121-130.
41. Moore, N.P., Cahill, J.P., Kelly, P.F., Hayden, T.J. 1995. An assessment of fi ve methods of age de-termination in an enclosed population of fallow deer (Dama dama). Biology and Environment: Proceedings of the Royal Irish Academy 95B: 27-34.
42. Phillipson, S., Lindskog, S, Flock, A. 1990. Mineralization of and cementoblast attachement to original and reparative cementum. Scandanavian Journal of Dental Research 98:295-300.
43. Davis, W.L. 1986. Oral Histology. Ed. W.B. Saunders. Philadelphia, PA.44. Jones, S.J. 1981. Dental Tissues. In: Dental Anatomy and Embriology. Pp 66-209. Ed. Osborne,
J.W. Blackwell Scientifi c Publications, Oxford.45. Azorit, C., Hervas, J. Analla, M. Carrasco, R., Muñoz-Cobo, J. 2002d. Histological thin-sections:
a method for the microscopic study of teeth in Spanish red deer (Cervus elaphus hispanicus). Anatomia Histologia Embryologia (Journal of Veterinary Medicine, Series C) 31:224-227.
46. Azorit, C., Analla, M., Hervas, J., Carrasco, R., Muñoz-Cobo, J. 2002e. Growth marks observa-tion: preferential techniques and teeth for ageing of Spanish red deer (Cervus elaphus hispanicus). Anatomia Histologia Embryologia (Journal of Veterinary Medicine, Series C) 31:303-307.

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
262
47. Azorit, C., Muñoz-Cobo, J., Analla, M. 2002f. Seasonal deposition of cementum in fi rst lower molars from Cervus elaphus hispanicus. Mammalian Biology 67:1-3.
48. Kierdorf, U., Becher, J. 1997. Mineralization and wear of mandibular fi rst molars in red deer (Cervus elaphus) of known age. Journal of Zoology London 241: 135-143.
49. Wagenknecht, E. 1980. Der Rothirsch. A. Ziemsen Verlag, Wittenberg Lutherstadt.50. Drescher, H. 1989. Das Rotwild. Landbuch-Verlag GmbH. Hannover.51. Mitchell, B. 1967. Growth layers in dental cement for determining the age of red deer (Cervus
elaphus L). Journal of Animal Ecology 36: 279-293. 52. Lotze, K. 1968. Comment juger un cerf?. Ed. Gerfaut, Paris.53. ONC (Offi ce National de la Chasse) 1984. Le Cerf d´Europe. Paris.54. Castanet, J., Smirina, E. 1990. Introductión to the skeletochronological method in amphibians
and reptiles. Annales des Sciences Naturelles-Zoologie 11: 191-196.55. Scheffer, V.B. 1950. Growth layers on the teeth of Pinnipedia as an indication of age. Science
112(2907): 309-311.56. Laws, R. 1952. A new method of age determination for mammals. Nature, Lon. 169(4310): 972-
974.57. Sergeant, D.E., Pimlott, D.H. 1959. Age determination in moose from sectioned incisor teeth.
The Journal of Wildlife Management 23(3): 315-321.58. Rausch, R.L. 1961. Notes on the Black Bear, (Ursus americanus, Pallas), in Alaska, with Particular
Reference to Dention and Growth. Zeitschrift für Säugetierkunde 2(26): 77-107.59. Lockard, G.R. 1972. Further studies of dental annuli for aging white-tailed deer. The Journal of
Wildlife Management 36: 46-55. 60. Ohtaishi, N., Kaji, K., Miura, S, Wu, J. 1990. Age determination of the white-lipped deer (Cervus
albirostris) by dental cementum and molar wear. Journal of Mammal Society, Japan 15: 15-24. 61. Rice, L. 1980. Infl uences of irregular dental cementum layers on aging deer incisors. The Journal
of Wildlife Management 44: 266-268. 62. Ransom, A.B. 1966. Determining the age of white-tailed deer from layers in cementum of mo-
lars. The Journal of Wildlife Management 30: 197-199.63. Boozer, R.B. 1969. Cementum annuli versus tooth wear aging of the white-tailed deer of Ala-
bama. Dissertation, Auburn University, Auburn, Alabama, USA. 64. Miller, F.L. 1974. Age determination of caribou by annulations in dental cementum. The Journal
of Wildlife Management 38: 47-53. 65. Hass, A. 1977. Contribution a l’étude des appositions de cément dentaire au niveau des molaires
chez Cervus elaphus L. These nº32. Université Louis Pasteur Strasbourg I Faculté Chirugie den-taire.
66. Saxon, A., Higham, C.F.W. 1968. Identifi cation and interpretation of growth rings in the second-ary dental cementum of Ovis aries L. Nature 219(5154): 634-635.
67. Stallibrass, S. 1982. The use of cement layers for absolute ageing of mammalian teeth: a selective review of the literature, with suggestions for further studies and alternative applications. In Ageing and sexing animal bones from archeological sites (eds Wilson, B., Grigson, C., Payne, S.) Pp. 109-126. BAR British Series 109.
68. Brokx, P.A. 1972. Age determination of Venezuelan white-tailed deer. The Journal of Wildlife Management 36: 1060-1067.
69. Meunier, F.J., Lecomte, F., Rojas-Beltrán, R. 1985. Mise en evidence de doubles cycles annuels de croissance sur le squelette de quelques teleostes de Guyane. Bulletin de le Société Zoologique de France 110(3): 285-289.
70. Lecomte, F., Meunier, F.J., Rojas-Beltrán, R. 1985. Mise en évidence d’un double cicle de crois-sance annuel chez un Silure de Guyane, Arius couma (Val., 1839) (Teleostei, Siluriforme, Ariidae)

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
263
à partir de l’étude squeletochronologique des épines des nageoires. Comptes Rendus de l’Aca-démie des Sciences Paris 300: 181-184.
71. Lecomte, F., Meunier, F.J., Rojas-Beltrán, R. 1986. Données préliminaires sur la croisance de deux téléostées de Guyane, Ariun proops (Ariidae, Siluriformes) et Leporinus friderici (Anostomi-dae, Characoidei). Cybium 10: 121-134.
72. Caetano, M.H., Castanet, J., Francillon, H. 1985. Determinatión de l’âge de Triturus marmoratus (Latreille 1800) du Parc National de Peneda Gerês (Portugal) par squeletochronologie. Amphib-ia-Reptilia 6: 117-132.
73. Caetano, M.H., Castanet, J. 1987. Experimental data on bone growth and age in Triturus marmo-ratus. 4th Meeting of the European Society of Herpetology. Nijmegen 1987: 87-90.
74. Klevezal, G.A., Myrick, A.C. 1984. Marks in tooth dentine of female dolphins (Genus stenella) as indicators of parturition. Journal of Mammalogy 65(1): 103-110.
75. Zapata, S.C., García Perea, R., Beltrán, J.F., Ferreras, P., Delibes, M. 1997. Age determination of iberian lynx (Lynx pardina) using canine radiograph and cementum annuli enumeration. Zeitschrift für Säugetierkunde 62: 1-5.
76. Novarowski, 1965. Cemental deposition as an age criterion in bison, and the relation of incisor wear, eye-lens weight, and dressed bison carcass weight to age. Canadian Journal of Zoology 43: 173-178.
77. Klevezal, G.A., Pucek, M. 1987. Growth layers in tooth cement and dentine of european bison and its hybrids with domestic cattle. Acta Theriologica 32(9): 115-128.
78. Ahlen, I. 1965. Studies on the red deer (Cervus elaphus L.) in Scandinavia. III Ecological investiga-tions. Viltrevy 3: 177-376.
79. Lieberman, D.E. 1994. The biological basis for seasonal increments in dental cementum and their application to archaeological research. Journal of Archaeological Science 21: 525-539.
80. Pascal, M., Castanet, J. 1978. Méthodes de détermination de l’âge chez le chat haret des îles Kerguelen. Terre et Vie 32: 529-555.
81. Mitchell, B., Youngson, R.W. 1969. Teeth and age in Scottish Red Deer a practical guide to age assessment. Ed.: Red Deer Commission. Inverness.
82. Bourque, B.J.K., Morris, K., Spiess, A. 1978. Determining the season of death of mammal death of mammal teeth from archeological sites: A new sectioning technique. Science 199(4328): 530-531.
83. Lieberman D.E., Meadow, R.H. 1992. The biology of cementum increments (with an archaeo-logical application). Mammal Review 22: 58-77.
84. Sanchez-Villagra, M.R. 2010. Suture closure as a paradigm to study late growth in recent and fossil mammals: a case study with giant deer and dwarf deer skulls. Journal of Vertebrate Pale-ontology 30(6): 1895–1898.
85. Gasaway, W.C., Harkness, D.B., Rausch, R.A. 1978. Accuracy of moose age determinations from incisor cementum layers. The Journal of Wildlife Management 42: 558-563.
86. Dapson, R.W. 1980. Guidelines for statistical usage in age-estimation technics. The Journal of Wildlife Management 44: 541-548.
87. Matson, G. 1981. Workbook for cementum analysis. Matson’s, Milltown, Montana, USA. 88. Hamlin, K.L., Pac, D.F., Sime, C.A., Desiml, R.M., Desek, G.L. 2000. Evaluating the accuracy of
ages obtained by 2 methods for Montana ungulates. The Journal of Wildlife Management 64: 441-449.
89. Rolandsen, C.M., Solberg, E.J., Heim, M., Holmstrøm, F., Solem, M.I., Sæther, B.E. 2008. Accura-cy and repeatability of moose (Alces alces) age as estimated from dental cement layers. European Journal of Wildlife Research 54: 6-14.

GUÍA PARA LA DETERMINACIÓN DE LA EDAD DEL CIERVO IBÉRICO A TRAVÉS DE SU DENTINCIÓN
264
90. Asmus, J., Weckerly, F.W. 2011 Evaluating precision of cementum annuli analysis for aging mule deer from southern California. The Journal of Wildlife Management 75(5): 1194-1199
91. Fancy, S.G. 1980. Preparation of mammalian teeth for age determination by cementum layers: a review. Wildlife Society Bulletin 8: 242-248.
92. Carranza, J., Mateos, C., Alarcos, S., Sanchez-Prieto, C.B., Valencia, J. 2008. Sex-specifi c strategies of dentine depletion in red deer. Biological Journal of the Linnean Society 93: 487-497.
93. Jacobson, H.A., Reiner, R.J. 1989. Estimating age of white-tailed deer: tooth wear versus ce-mentum annuli. Proceeding of Annual Conference. Southeast Association of Fish and Wildlife Agencies 43: 286-291.
94. Hall, G.P., Murray, P.J., Byrne, M.J. 2012. Is Tooth Wear a Reliable Means of Aging Wild Euro-pean Fallow Deer in Tasmania, Australia? Wildlife Society Bulletin 36(1): 124-129.

265ANALES - VOL. 24 (1) - DIC. 2011 - REAL ACADEMIA DE CIENCIAS VETERINARIAS DE ANDALUCÍA ORIENTAL
MEMORIA DEL CICLO
OLORES Y SABORES EN LA DIETA MEDITERRANEA
ANTONIO MARÍN GARRIDO
Con la colaboración de Académicos de esta Corporación, así como de personal docente perteneciente a las universidades de Almería, Córdoba y Jaén, todos ellos en-cuadrados en Departamentos de Nutrición y tecnología de alimentos, se han celebrado seis conferencias utilizando como sede las correspondientes a los Colegios Ofi ciales de Veterinarios de las cuatro provincias que conforman nuestro ámbito territorial, cedidos comos siempre de forma gratuita por sus respectivas Juntas de Gobierno.
El acto inaugural tenía lugar el 23 de noviembre en la sección de Málaga con una conferencia titulada “Cibaria mediterránea: trepidancia de Olores y sabores” a cargo de D. Pablo Amate López, Periodista especializado en gastronomía, Director del Curso de Gastronomía. Enología y Nutrición de la Universidad de Granada y Académico Correspondiente de esta Real Academjia de Ciencias Veterinarias de An-dalucía Oriental, en la que el ponente hizo un amplio y detallado recorrido por los principales alimentos que integran la dieta de los paises ribereños del Mediterráneo con especial énfasis en los producidos en nuestros país.
El dia 30 de noviembrte, en Almería se celebraban en la misma sesión dos conferencias dictadas la primera de ellas por el Dr. D. Julio Boza López, Profesor de investigación del C.S.I.C., Académico fundador y Presidente de Honor de la Acade-mia, bajo el Titulo: “Historia de la dieta Mediterránea y sus aportaciones a la salud“, constituyó una lección magistral sobre los orígenes y efectos benefi ciosos que su consumo ha producido en la especie humana.

266
Seguidamente, con una conferencia titulada “ Frutas y hortalizas: un placer muy saludable “, la Dra. Dª. Maria del Mar Rebolloso, profesora Titular de la Universidad de Almería dedicada su ponencia a un detallado recorrido por ese amplio grupo de frutas y verduras que constituyen ingredientes básicos de la dieta mediterranea y que contribuyen decididamente a la mejora de la salud de sus consumidores.
El día 1 de diciembre, en Jaén, tenía lugar una nueva jornada del ciclo con la participación de los Dres. D. Rafael Moreno López. Catedrático de la Universidad de Córdoba y D. Rafael Gómez Díaz, profesor Titular de la misma Universidad, con las ponencias “ Embutidos: el sabor de la necesidad, el aroma de la casualidad “ y “ Quesos: saberes y sabores “ respectivamente.
Tras un documentadísimo recorrido por sus respectivas ponencias, en las que pusieron de manifi esto las virtudes de este grupo de alimentos que constituyen un importante aporte nutricional, con independencia del deleite gastronómico que para muchos supone su consumo.
Al fi nalizar las ponencias, un numeroso grupo de entre los asistentes al acto, bajo la dirección del Dr. Gómez Díaz, participaban en una lección teorico-practica de iniciación a la cata de quesos, de acuerdo con criterios normalizados para este fi n.
Días mas tarde, el 13 de diciembre, en Granada, tenía lugar la conferencia de clausura del curso con la conferencia dictada por el Profesor Dr.D. José Juan Gaforio, Profesor Titular de la Universidad de Jaen y Director de Citoliva y Comisionado por la Universidad de Jaén para el Centro de Estudios avanzados en olivar y aceitre de oliva, con su ponencia “ El aceite de oliva virgen: sabor y salud “ en la que destaca-ría las diferencias fundamentales entre los aceites de oliva virgen y otros aceites de consumo habitual, destacando las virtudes del primero y haciendo especial énfasis en las aportadas por el consumo del oliva virgen elaborado con la variedad “picual” que constituyen la base fundamental de los trabajos de investigación que viene rea-lizando desde hace años y fruto de los cuales son los reconocimientos de sus efectos benefi ciosos en la salud de los consumidores.
El acto fue clausurado por el Presidente de la Academia, coordinador del ciclo, con la asistencia del Vicepresidente de la Sección de Granada.

267
MEMORIA DEL CICLO
LA SEGURIDAD ALIMENTARIA EN EL CONTEXTO DE LA SOCIEDAD GLOBALIZADA
ANTONIO MARÍN GARRIDO
Coordinado por el Presidente de la Corporación y siguiendo el mismo criterio que viene empleándose desde hace unos años, el día 25 de noviembre daba comien-zo el ciclo de referencia programado de forma que sus sesiones serían distribuidas entre las cuatros provincias de la Andalucía Oriental, con la valiosa participación de la Cátedra de Microbiología de la Universidad de Jaén que aportaba la totalidad de los ponentes del Ciclo los que, a través de sus respectivas intervenciones pondrían de manifi esto la importancia que sobre la calidad y seguridad alimentarias tienen los trabajos de investigación que vienen realizando, encaminados a la mejora de la producción, procesado y conservación de los alimentos; puntos críticos de mayor interés en la lucha hacia una mejora de la seguridad alimentaria, así como el im-portante papel que en esta tarea desempeñan tanto la microbiótica intestinal, como una alimentación variada y rica en elementos que contribuyan a la disminución de episodios de disbiosis..
La ponencia de apertura, en Málaga, sería dictada por el Dr. D. Antonio Gálvez del Postigo, Catedrático de Microbiología y Coordinador del Master Ofi cial Avances en Seguridad alimentaria de la Universidad de Jaén, con el título: “Nuevas tecnolo-gías para la conservación de alimentos”, seguida con gran interés por los asistentes a los que el sugestivo título de la ponencia había congregado en el salón de actos del Colegio Ofi cial de Veterinarios de aquella provincia.
La segunda sesión se celebraba en Jaén, el 19 de diciembre, con una ponencia titulada “Resistencia a antibióticos y biocidas en la cadena alimentaria” a cago de

268
la Dra. Dª Elena Ortega Morante, profesora titular de la Cátedra de Microbiología y del Master Avances en seguridad alimentaría de la UJA; una disertación en la que destacaría de forma especial su documentadísima exposición sobre los distintos me-canismos de resistencia a los antimicrobianos desarrollados por las bacterias.
En Granada se cerraba el Ciclo, en sesión doble, en la que las Dras, Dª. Hikmate Abriouel Hayani y Dª. Magdalena Martinez Cañamero, profesoras titulares de la Cáte-dra de Microbiología y del Master Avances en seguridad alimentaria de la Universidad de Jaén, desarrollarían respectivamente las ponencias “Las bacterias lácticas y sus bacteriocinas en la mejora de la calidad y seguridad alimentaria” y “Probioticos y macrobiótica intestinal: otra forma de protección frente a los riesgos alimentarios”.
El Ciclo era clausurado por el Presidente de la Academia, asistido por el Vice-presidente de la Sección Provincial de Granada.

269
MEMORIA DEL CICLO
MEMORIA CICLO ACEITES DE ALMERIA. PROPIEDADES NUTRICIONALES Y NUEVAS TECNOLOGÍAS
ANTONIO MARÍN GARRIDO
Organizado por la Real Academia de Ciencias Veterinarias del Andalucía Orien-tal y el Instituto de Estudios Almerienses de la Diputación Provincial de Almería en colaboración con la Almazara Olivar del Desierto S.L Castillo de Tabernas, Bodegas Perfer de Uleila del Campo, Industrias Cárnicas Sierra de María SL, “Los Amadeos” y el Patronato para el Centro Asociado a la UNED en Almería, se han celebrado unas Jor-nadas sobre “Aceites de Almería, propiedades Nutricionales y Nuevas Tecnologías ”.
La sesión teórica tuvo lugar el 20 de octubre de 2011 en el Salón de Actos el Centro de la “UNED” en Almería; el Dr. D. Jose Luis Gaforio Martínez, Profesor Titular del Área de Inmunología del Departamento de Ciencias de la Salud de la Universidad de Jaén, realizó una conferencia sobre “Potencial elaboración de aceites de oliva singu-lares diferenciados por sus propiedades saludables”; seguidamente el Dr. D. Jose Luis Guil Guerrero, Catedrático del Área de Tecnología de Alimentos de la Universidad de Almería, impartió una conferencia sobre “La Tecnología Agroalimentaria en el contexto socioeconómico de la provincia de Almería”, fi nalizando la sesión teórica con una conferencia a cargo de D. Antonio Jesús Casimiro Andújar, Profesor Titular de Actividad Física e Higiene de la Universidad de Almería. que disertó sobre el “Consumo de productos autóctonos en los hábitos alimenticios y deportivos”. Ac-tuando como moderador el Dr. D. Tomás Martínez Moya, Académico numerario de la Real Academia de Ciencias Veterinarias de Andalucía Oriental.
La sesión práctica, con visitas guiadas, se desarrolló en la Almazara Olivar del Desierto ( Tabernas ) y en las Bodegas Perfer ( Uleila del Campo ), el 22 de octubre.

270
En dicha sesión se degustaron productos de las citadas empresas y de las Industrias Cárnicas Sierra de María SL, “Los Amadeos” de la localidad de María.
La exposición se realizó el día 20 de diciembre en el Salón de Actos de la “UNED” en Almería, en presencia de diversas autoridades y del Director del Instituto de Es-tudios Almerienses.
Estas Jornadas, reconocidas de interés Científi co-Sanitario por la Consejería de Salud de la Junta de Andalucía, y coordinadas por D. Alberto González Ramón, Vicepresidente y Académico de número de la RACVAO y por D. Juan J. Espinosa Moya, miembro del Departamento de Ciencia y Tecnología del Instituto de Estudios Almerienses, han contribuido a plantear un foro de conocimiento y discusión sobre las propiedades y aplicaciones en el campo de la Salud de los productos almerienses y la contribución de las nuevas tecnologías en la mejora de su calidad.

271
OTRAS ACTIVIDADES
La Academia, a través de su Presidente, ha participado en actos organizados por diferentes Instituciones de entre los que deben destacarse:
- Conferencia en el Instituto de Estudios Giennenses: “La Veterinaria espa-ñola: síntesis de un largo camino“, organizado por el Seminario Médico del Instituto y celebrada en Jaén el día 14 de abril.
- Ponencia “La Real Academia de Ciencias Veterinarias de Andalucía Oriental: una aproximación a su Historia” dictada en el I Encuentro Científi co de la Asociación Iberoamericana de Academias de Ciencias Veterinarias cele-brada en Madrid los dias del 9-12 de mayo.
- Ponencia: “El laboratorio de control y la seguridad alimentaria”, dictada en la Universidad de Verano “Intendente Olavide” de La Carolina (Jaén).
- Graduación de la Promoción 2006-2011 de la Facultad de Veterinaria de Córdoba, acto en el que era distinguido con la entrega de una placa en reconocimiento a su colaboración con dicha Facultad.
- Imposición de la Medalla de Colegiado de Honor del Colegio Ofi cial de Veterinarios de Almería, al Presidente de la Real Academia de Ciencias Veterinarias de Andalucía Oriental, con ocasión de su presencia en el Acto de entrega del Premio de Investigación “Francisco Fernández”.

272
- Acto de presentación del libro “Rafael Pérez del Alamo. 150 aniversario de la Revolución de Loja“ editado por la Fundación Ibn al-Jatib y en el que se recopilan 6 trabajos relacionados con Pérez del Alamo entre los que fi gura el publicado en ANALES de esta Cor-poración (año 2006 ) bajo el titulo “Una aproximación hacia Rafael Pérez del Álamo: el Albéitar Caudillo“ fi rmado por J. Mollinedo Gómez-Zorrilla y Marín Garrido, A. celebrado en Loja (Granada) el dia 30 de septiembre.

273
OBITUARIO
El pasado día 24 de junio de 2011, a la edad de 101 años, fallecía en Almería el Iltmo.Sr. D. Francisco Colomer Luque, Veterinario almeriense, quien durante su dilatada vida mantuvo hasta el fi nal un envidiable vínculo con la profesión y con esta Real Academia de Ciencias Veterinarias de Andalucía Oriental de la que era Académico Fundador.
ANALES, al tiempo que hace cumplido recuerdo a su memoria, se une al dolor de todos sus deudos y muy especialmente a sus hijos Africa y Eugenio a los que desde esta página desea trasladar y compartir su pesar por tan sentida pérdida.

274

275
NORMAS DE PUBLICACIÓN
Los ANALES se constituyen en el medio de difusión de la vida académica y de las actividades científi cas de esta Real Corporación, en cumplimiento de uno de sus objetivos fundamentales: el fomento y difusión de los estudios e investigaciones en las Ciencias Veterinarias.
ARTÍCULOS CIENTÍFICOS
Sólo se aceptarán los trabajos que no hayan sido editados previamente en otras publicaciones, de cualquier naturaleza o contenido editorial por lo que todo envío deberá estar acompañado de la oportuna declaración acreditativa de esta circuns-tancia.
El texto deberá ser redactado en español, con estilo conciso e impersonal, e in-cluirá un resumen bilingüe (Español/Inglés), cuya extensión recomendada será de 10-15 líneas, así como las palabras clave/keywords que correspondan .
Siempre que sea posible, los trabajos deberán adaptarse a la siguiente estructura: Introducción, métodos, resultados, discusión/conclusiones, notas y referencias.
Las referencias bibliográfi cas se incluirán al fi nal del texto, según el orden de aparición o cita, y con numeración correlativa. La citas o llamadas se identifi caran presentando el número de orden entre paréntesis.
En cada referencia bibliográfi ca se detallará el nombre del autor/es, el título del artículo o capítulo, nombre de la revista, libro o publicación, número del volumen, números de la primera y última paginas y año de publicación.
Aunque no hay un límite de extensión, se recomienda un máximo de 15 páginas para los artículos y trabajos de investigación y 20 para las revisiones.
El documento deberá guardarse en formato doc de Microsoft Word. Con inde-pendencia de su presencia en el documento y a efectos de una adecuada ubicación, los gráfi cos, esquemas, diseños o imágenes, serán guardadas en archivo aparte en formato JPEG, EPS, TIFF o PICT, con una resolución de 300 ppp (dpi). Ambos archivos, debi-damente identifi cados, serán remitidos a la dirección de correo: [email protected] con solicitud de acuse de recibo para confi rmar su correcta recepción.

276
Así mismo, adjunto al trabajo enviado, se remitirá un archivo en el que fi gurarán el nombre y apellidos del autor/es, dirección completa, cargos y lugar de trabajo con los que deseen fi gurar en la publicación, así como el teléfono y dirección de correo electrónico de contacto.
El autor/es cede sus derechos a la Real Academia de Ciencias Veterinarias de Andalucia Oriental para que esta pueda facilitar el acceso libre y gratuito de los textos completos de sus publicaciones tanto desde su propia web, como a través de los repositorios con los que ANALES tenga establecidos convenios y que, sin ánimo de lucro, realicen la difusión de sus contenidos por medio de Internet, autorizando la adaptación de los mismos, si así fuese preciso, a las exigencias del formato que estos repositorios pudieran precisar. facilitando su visualización, reproducción y/o almacenamiento en soporte digital o papel, exclusivamente para uso privado con fi nes de estudio y/o investigación, sin que en ningún caso puedan ser usados con fi nes comerciales o lucrativos.
La titularidad de los derechos morales y de explotación de la propiedad inte-lectual de los trabajos objeto de esta cesión pertenecen a sus respectivos autores y a la Real Academia de Ciencias Veterinarias de Andalucía Oriental.
Esta Corporación no garantiza ni asume responsabilidades por la forma en que los usuarios hagan uso posterior de sus contenidos.
Los trabajos no aceptados por los órganos competentes de la revista serán de-vueltos a sus autores previa solicitud escrita de los interesados.
Toda la correspondencia se realizará vía electrónico a la dirección de correo
administració[email protected],
o por correo postal a
Real Academia de Ciencias Veterinarias de Andalucía Oriental.c/ Rector Marín Ocete, 10. C.P. 18014. GRANADA.
La incorporación de los trabajos a esta publicación supone la aceptación de los autores a las condiciones establecidas en las presentes instrucciones.

277
































