Embed Size (px)
Citation preview
D
ue sono i meccanismi immunitariche proteggono l'organismo dal-le infezioni e dal cancro: uno
è la risposta immunitaria mediata dacellule (immunità cellulare) che com-batte funghi e virus e dà inizio al ri-getto di tumori e di tessuti estranei;l'altro è la immunità umorale, efficacecontro le infezioni batteriche e le rein-fezioni virali. Malgrado i due mecca-nismi non siano totalmente indipenden-ti e la loro cooperazione sia a volteimportante, essi sono tuttavia distinti.
La base essenziale di questa divisionedi compiti nel sistema immunitario sitrova in due popolazioni di cellule ori-ginarie del tessuto linfoide, ma che sitrovano anche in altre parti dell'orga-nismo, in particolare nel sangue. Du-rante lo sviluppo, le due linee di cellulelinfoidi sono simili; esse non possonoessere distinte in base a un sempliceesame. Inoltre, tutte e due derivanodagli stessi precursori: le cellule emo-poietiche che danno origine anche anumerosi altri tipi di cellule sanguigne.Malgrado la loro origine comune e lasomiglianza superficiale, queste cellulehanno ruoli diversi nella risposta del-l'organismo verso materiali estranei everso cellule tumorali.
Oltre alla divisione del sistema im-munitario in due classi di cellule lin-foidi, vi è una grande diversità all'in-terno di ciascuna classe. In entrambele classi ogni cellula è in grado di rico-noscere un particolare determinante
antigenico: uno dei gruppi chimici concui sostanze biologiche come le proteinemanifestano la loro identità. Vi sonomilioni di determinanti antigenici e pa-re che vi siano cellule immunologica-mente attive in grado di riconosce cia-scuno di essi.
Le ricerche sul modo in cui, dallaapparente uniformità delle cellule ori-ginarie, si passa alla differenziazionedi vari tipi di cellule possono far me-glio comprendere l'organizzazione e ilfunzionamento del sistema immunità-rio. La conoscenza operativa di questimeccanismi, inoltre, è indispensabileper comprendere il ruolo dei disturbidel sistema immunitario, molti dei qua-li sembrano avere origine da difetti deldifferenziamento cellulare.
Il timo e la borsa di Fabrizio
La duplice natura del sistema immu-nitario fu messa in evidenza da unaserie di esperimenti di Mervil W.Chase e Cari Landsteiner dell'IstitutoRockefeller per la ricerca medica(Landsteiner è più noto per una pre-cedente scoperta: nel 1900 egli definìi gruppi sanguigni A, B e O). Chase eLandsteiner dimostrarono che alcunitipi di reazioni immunitarie potevanoessere trasferiti da un animale all'altrosolo con lo scambio di cellule vive,mentre altri potevano essere trasmessidal siero ematico. Le cellule richiestenel primo tipo di esperimento erano i
linfociti: cellule piccole e non pigmen-tate comprese insieme a numerose altrenel gruppo dei leucociti o globuli bian-chi. Fino a quel momento la funzionedei linfociti era sconosciuta.
Il componente del siero sanguignocapace di trasferire l'immunità consi-ste in molecole proteiche chiamate an-ticorpi che si combinano con sostanzeestranee. Alcuni anni prima era statoscoperto che gli anticorpi appartengo-no a quella parte del siero chiamatafrazione gamma globulinica. Gli anti-corpi sono secreti da plasmacellule cheoriginano dai linfociti, ma non daglistessi linfociti che, come si scoprì piùtardi, sono responsabili dell'immunitàcellulare.
Che la duplice natura funzionale delsistema immunitario potesse avere uncarattere evolutivo fu suggerito dallascoperta di alcune malattie caratteriz-zate da deficienza immunitaria. OdgenC. Bruton, un pediatra del Centro me-dico militare Walter Reed mise per pri-mo in evidenza una disfunzione dell'im-munità umorale in un ragazzo affettoda infezioni batteriche multiple. Lamalattia era caratterizzata da una mam-canza di plasmacellule e da una con-seguente incapacità di fabbricare anti-corpi. I linfociti del sistema immuni-tario cellulare, d'altro canto, erano ab-bondanti e permettevano al paziente diresistere piuttosto bene alle infezionivirali e da funghi. È stato anche sco-perto un quadro opposto di deficienzaimmunitaria. Una disfunzione dell'im-munità cellulare. Gli individui affettida questo disturbo sono vulnerabiliai virus e ai funghi, hanno un numeroinferiore al normale di linfociti, mapossiedono plasmacellule e produconoanticorpi circolanti. Alcuni bambininascono senza linfociti e plasmacellule;senza i due sistemi immunitari essi soc-combono rapidamente alle infezionicausate dai vari microrganismi.
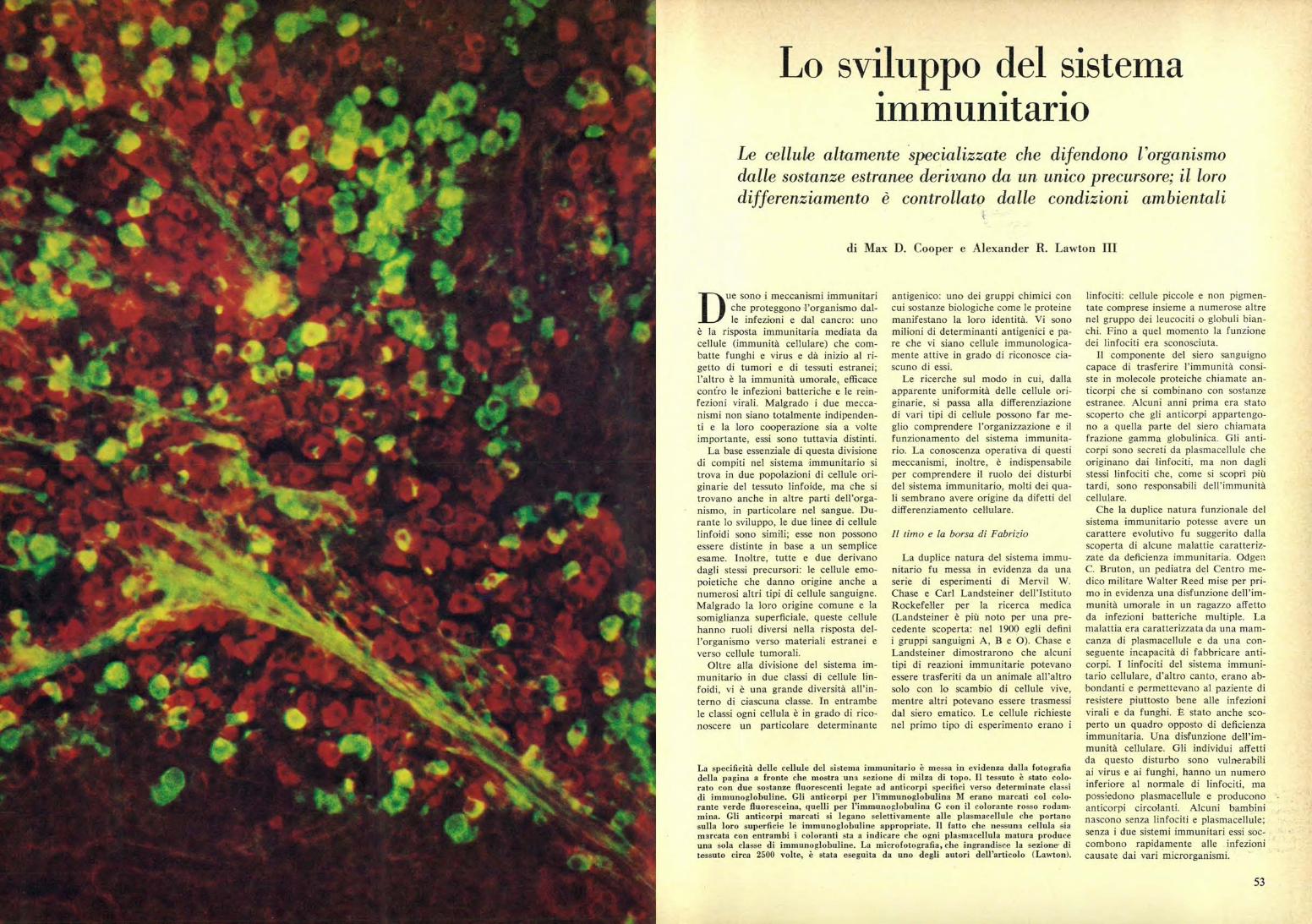
La specificità delle cellule del sistema immunitario è messa in evidenza dalla fotografiadella pagina a fronte che mostra una sezione di milza di topo. Il tessuto è stato colo.rato con due sostanze fluorescenti legate ad anticorpi specifici verso determinate classidi immunoglobuline. Gli anticorpi per l'immunoglobulina M erano marcati col colo-rante verde fluoresceina, quelli per l'immunoglobulina G con il colorante rosso rodam-mina. Gli anticorpi marcati si legano selettivamente alle plasmacellule che portanosulla loro superficie le immunoglobuline appropriate. Il fatto che nessuna cellula siamarcata con entrambi i coloranti sta a indicare che ogni plasmacellula matura produceuna sola classe di immunoglobuline. La microfotografia, che ingrandisce la sezione ditessuto circa 2500 volte, è stata eseguita da uno degli autori dell'articolo (Lawton).
Lo sviluppo del sistemaimmunitario
Le cellule altamente specializzate che difendono l'organismodalle sostanze estranee derivano da un unico precursore; il lorodifferenziamento è controllato dalle condizioni ambientali
di Max D. Cooper e Alexander R. Lawton III
53
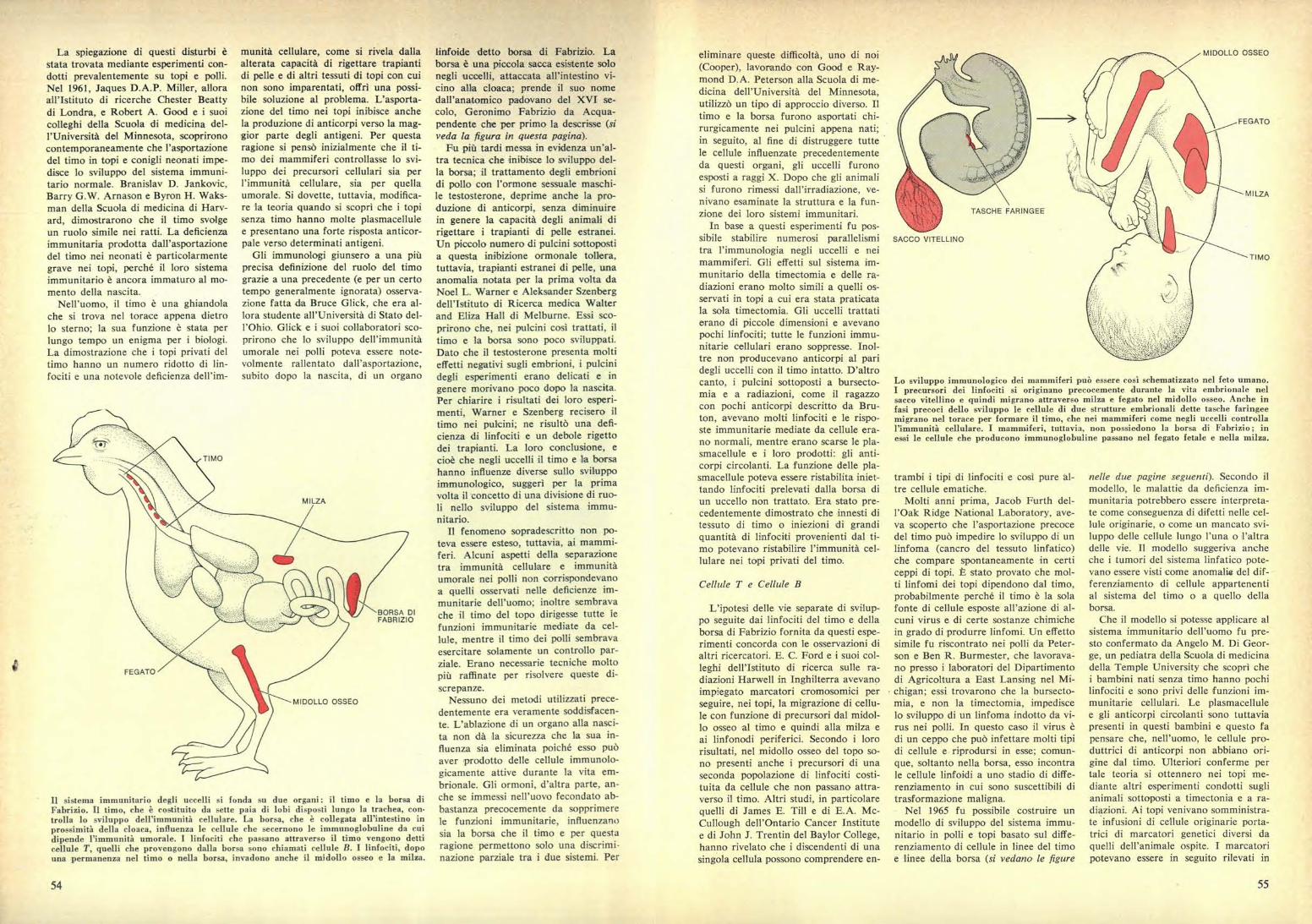
Lo sviluppo immunologico dei mammiferi può essere così schematizzato nel feto umano.I precursori dei linfociti si originano precocemente durante la vita embrionale nelsacco vitellino e quindi migrano attraverso milza e fegato nel midollo osseo. Anche infasi precoci dello sviluppo le cellule di due strutture embrionali dette tasche faringeemigrano nel torace per formare il timo, che nei mammiferi come negli uccelli controllal'immunità cellulare. I mammiferi, tuttavia, non possiedono la borsa di Fabrizio; inessi le cellule che producono immunoglobuline passano nel fegato fetale e nella milza.
TASCHE FARINGEE
SACCO VITELLINO
MIDOLLO OSSEO
MILZA
BORSA DIFABRIZIO
MIDOLLO OSSEO
Il sistema immunitario degli uccelli si fonda su due organi: il timo e la borsa diFabrizio. Il timo, che è costituito da sette paia di lobi disposti lungo la trachea, con•trolla lo sviluppo dell'immunità cellulare. La borsa, che è collegata all'intestino inprossimità della cloaca, influenza le cellule che secernono le immunoglobuline da cuidipende l'immunità umorale. I linfociti che passano attraverso il timo vengono detticellule T, quelli che provengono dalla borsa sono chiamati cellule B. I linfociti, dopouna permanenza nel timo o nella borsa, invadono anche il midollo osseo e la milza.
La spiegazione di questi disturbi èstata trovata mediante esperimenti con-dotti prevalentemente su topi e polli.Nel 1961, Jaques D.A.P. Miller, alloraall'Istituto di ricerche Chester Beattydi Londra, e Robert A. Gooel e i suoicolleghi della Scuola di medicina del-l'Università del Minnesota, scoprironocontemporaneamente che l'asportazionedel timo in topi e conigli neonati impe-disce lo sviluppo del sistema immuni-tario normale. Branislav D. Jankovic,Barry G.W. Arnason e Byron H. Waks-man della Scuola di medicina di Harv-ard, dimostrarono che il timo svolgeun ruolo simile nei ratti. La deficienzaimmunitaria prodotta dall'asportazionedel timo nei neonati è particolarmentegrave nei topi, perché il loro sistemaimmunitario è ancora immaturo al mo-mento della nascita.
Nell'uomo, il timo è una ghiandolache si trova nel torace appena dietrolo sterno; la sua funzione è stata perlungo tempo un enigma per i biologi.La dimostrazione che i topi privati deltimo hanno un numero ridotto di lin-fociti e una notevole deficienza dell'im-
munità cellulare, come si rivela dallaalterata capacità di rigettare trapiantidi pelle e di altri tessuti di topi con cuinon sono imparentati, offrì una possi-bile soluzione al problema. L'asporta-zione del timo nei topi inibisce anchela produzione di anticorpi verso la mag-gior parte degli antigeni. Per questaragione si pensò inizialmente che il ti-mo dei mammiferi controllasse lo svi-luppo dei precursori cellulari sia perl'immunità cellulare, sia per quellaumorale. Si dovette, tuttavia, modifica-re la teoria quando si scoprì che i topisenza timo hanno molte plasmacellulee presentano una forte risposta anticor-pale verso determinati antigeni.
Gli immunologi giunsero a una piùprecisa definizione del ruolo del timograzie a una precedente (e per un certotempo generalmente ignorata) osserva-zione fatta da Bruce Glick, che era al-lora studente all'Università di Stato del-l'Ohio. Glick e i suoi collaboratori sco-prirono che lo sviluppo dell'immunitàumorale nei polli poteva essere note-volmente rallentato dall'asportazione,subito dopo la nascita, di un organo
linfoide detto borsa di Fabrizio. Laborsa è una piccola sacca esistente solonegli uccelli, attaccata all'intestino vi-cino alla cloaca; prende il suo nomedall'anatomico padovano del XVI se-colo, Geronimo Fabrizio da Acqua-pendente che per primo la descrisse (siveda la figura in questa pagina).
Fu più tardi messa in evidenza un'al-tra tecnica che inibisce lo sviluppo del-la borsa; il trattamento degli embrionidi pollo con l'ormone sessuale maschi-le testosterone, deprime anche la pro-duzione di anticorpi, senza diminuirein genere la capacità degli animali dirigettare i trapianti di pelle estranei.Un piccolo numero di pulcini sottopostia questa inibizione ormonale tollera,tuttavia, trapianti estranei di pelle, unaanomalia notata per la prima volta daNoel L. Warner e Aleksander Szenbergdell'Istituto di Ricerca medica Walterand Eliza Hall di Melburne. Essi sco-prirono che, nei pulcini così trattati, iltimo e la borsa sono poco sviluppati.Dato che il testosterone presenta moltieffetti negativi sugli embrioni, i pulcinidegli esperimenti erano delicati e ingenere morivano poco dopo la nascita.Per chiarire i risultati dei loro esperi-menti, Warner e Szenberg recisero iltimo nei pulcini; ne risultò una defi-cienza di linfociti e un debole rigettodei trapianti. La loro conclusione, ecioè che negli uccelli il timo e la borsahanno influenze diverse sullo sviluppoimmunologico, suggerì per la primavolta il concetto di una divisione di ruo-li nello sviluppo del sistema immu-nitario.
Il fenomeno sopradescritto non po-teva essere esteso, tuttavia, ai mammi-feri. Alcuni aspetti della separazionetra immunità cellulare e immunitàumorale nei polli non corrispondevanoa quelli osservati nelle deficienze im-munitarie dell'uomo; inoltre sembravache il timo del topo dirigesse tutte lefunzioni immunitarie mediate da cel-lule, mentre il timo dei polli sembravaesercitare solamente un controllo par-ziale. Erano necessarie tecniche moltopiù raffinate per risolvere queste di-screpanze.
Nessuno dei metodi utilizzati prece-dentemente era veramente soddisfacen-te. L'ablazione di un organo alla nasci-ta non dà la sicurezza che la sua in-fluenza sia eliminata poiché esso puòaver prodotto delle cellule immunolo-gicamente attive durante la vita em-brionale. Gli ormoni, d'altra parte, an-che se immessi nell'uovo fecondato ab-bastanza precocemente da sopprimerele funzioni immunitarie, influenzanosia la borsa che il timo e per questaragione permettono solo una discrimi-nazione parziale tra i due sistemi. Per
eliminare queste difficoltà, uno di noi(Cooper), lavorando con Good e Ray-mond D.A. Peterson alla Scuola di me-dicina dell'Università del Minnesota,utilizzò un tipo di approccio diverso. Iltimo e la borsa furono asportati chi-rurgicamente nei pulcini appena nati;in seguito, al fine di distruggere tuttele cellule influenzate precedentementeda questi organi, gli uccelli furonoesposti a raggi X. Dopo che gli animalisi furono rimessi dall'irradiazione, ve-nivano esaminate la struttura e la fun-zione dei loro sistemi immunitari.
In base a questi esperimenti fu pos-sibile stabilire numerosi parallelismitra l'immunologia negli uccelli e neimammiferi. Gli effetti sul sistema im-munitario della timectomia e delle ra-diazioni erano molto simili a quelli os-servati in topi a cui era stata praticatala sola timectomia. Gli uccelli trattatierano di piccole dimensioni e avevanopochi linfociti; tutte le funzioni immu-nitarie cellulari erano soppresse. Inol-tre non producevano anticorpi al paridegli uccelli con il timo intatto. D'altrocanto, i pulcini sottoposti a bursecto-mia e a radiazioni, come il ragazzocon pochi anticorpi descritto da Bru-ton, avevano molti linfociti e le rispo-ste immunitarie mediate da cellule era-no normali, mentre erano scarse le pla-smacellule e i loro prodotti: gli anti-corpi circolanti. La funzione delle pla-smacellule poteva essere ristabilita iniet-tando linfociti prelevati dalla borsa diun uccello non trattato. Era stato pre-cedentemente dimostrato che innesti ditessuto di timo o iniezioni di grandiquantità di linfociti provenienti dal ti-mo potevano ristabilire l'immunità cel-lulare nei topi privati del timo.
Cellule T e Cellule B
L'ipotesi delle vie separate di svilup-po seguite dai linfociti del timo e dellaborsa di Fabrizio fornita da questi espe-rimenti concorda con le osservazioni dialtri ricercatori. E. C. Ford e i suoi col-leghi dell'Istituto di ricerca sulle ra-diazioni Harwell in Inghilterra avevanoimpiegato marcatori cromosomici perseguire, nei topi, la migrazione di cellu-le con funzione di precursori dal midol-lo osseo al timo e quindi alla milza eai linfonodi periferici. Secondo i lororisultati, nel midollo osseo del topo so-no presenti anche i precursori di unaseconda popolazione di linfociti costi-tuita da cellule che non passano attra-verso il timo. Altri studi, in particolarequelli di James E. Till e di E.A. Mc-Cullough dell'Ontario Cancer Institutee di John J. Trentin del Baylor College,hanno rivelato che i discendenti di unasingola cellula possono comprendere en-
trambi i tipi di linfociti e così puretre cellule ematiche.
Molti anni prima, Jacob Furth del-l'Oak Ridge National Laboratory, ave-va scoperto che l'asportazione precocedel timo può impedire lo sviluppo di unlinfoma (cancro del tessuto linfatico)che compare spontaneamente in certiceppi di topi. È stato provato che mol-ti linfomi dei topi dipendono dal timo,probabilmente perché il timo è la solafonte di cellule esposte all'azione di al-cuni virus e di certe sostanze chimichein grado di produrre linfomi. Un effettosimile fu riscontrato nei polli da Peter-son e Ben R. Burmester, che lavorava-no presso i laboratori del Dipartimentodi Agricoltura a East Lansing nel Mi-chigan; essi trovarono che la bursecto-mia, e non la timectomia, impediscelo sviluppo di un linfoma indotto da vi-rus nei polli. In questo caso il virus èdi un ceppo che può infettare molti tipidi cellule e riprodursi in esse; comun-que, soltanto nella borsa, esso incontrale cellule linfoidi a uno stadio di diffe-renziamento in cui sono suscettibili ditrasformazione maligna.
Nel 1965 fu possibile costruire unmodello di sviluppo del sistema immu-nitario in polli e topi basato sul diffe-renziamento di cellule in linee del timoe linee della borsa (si vedano le figure
nelle due pagine seguenti). Secondo ilmodello, le malattie da deficienza im-munitaria potrebbero essere interpreta-te come conseguenza di difetti nelle cel-lule originarie, o come un mancato svi-luppo delle cellule lungo l'una o l'altradelle vie. Il modello suggeriva ancheche i tumori del sistema linfatico pote-vano essere visti come anomalie del dif-ferenziamento di cellule appartenential sistema del timo o a quello dellaborsa.
Che il modello si potesse applicare alsistema immunitario dell'uomo fu pre-sto confermato da Angelo M. Di Geor-ge, un pediatra della Scuola di medicinadella Tempie University che scoprì chei bambini nati senza timo hanno pochilinfociti e sono privi delle funzioni im-munitarie cellulari. Le plasmacellulee gli anticorpi circolanti sono tuttaviapresenti in questi bambini e questo fapensare che, nell'uomo, le cellule pro-duttrici di anticorpi non abbiano ori-gine dal timo. Ulteriori conferme pertale teoria si ottennero nei topi me-diante altri esperimenti condotti suglianimali sottoposti a timectonia e a ra-diazioni. Ai topi venivano somministra-te infusioni di cellule originarie porta-trici di marcatori genetici diversi daquelli dell'animale ospite. I marcatoripotevano essere in seguito rilevati in
54
55
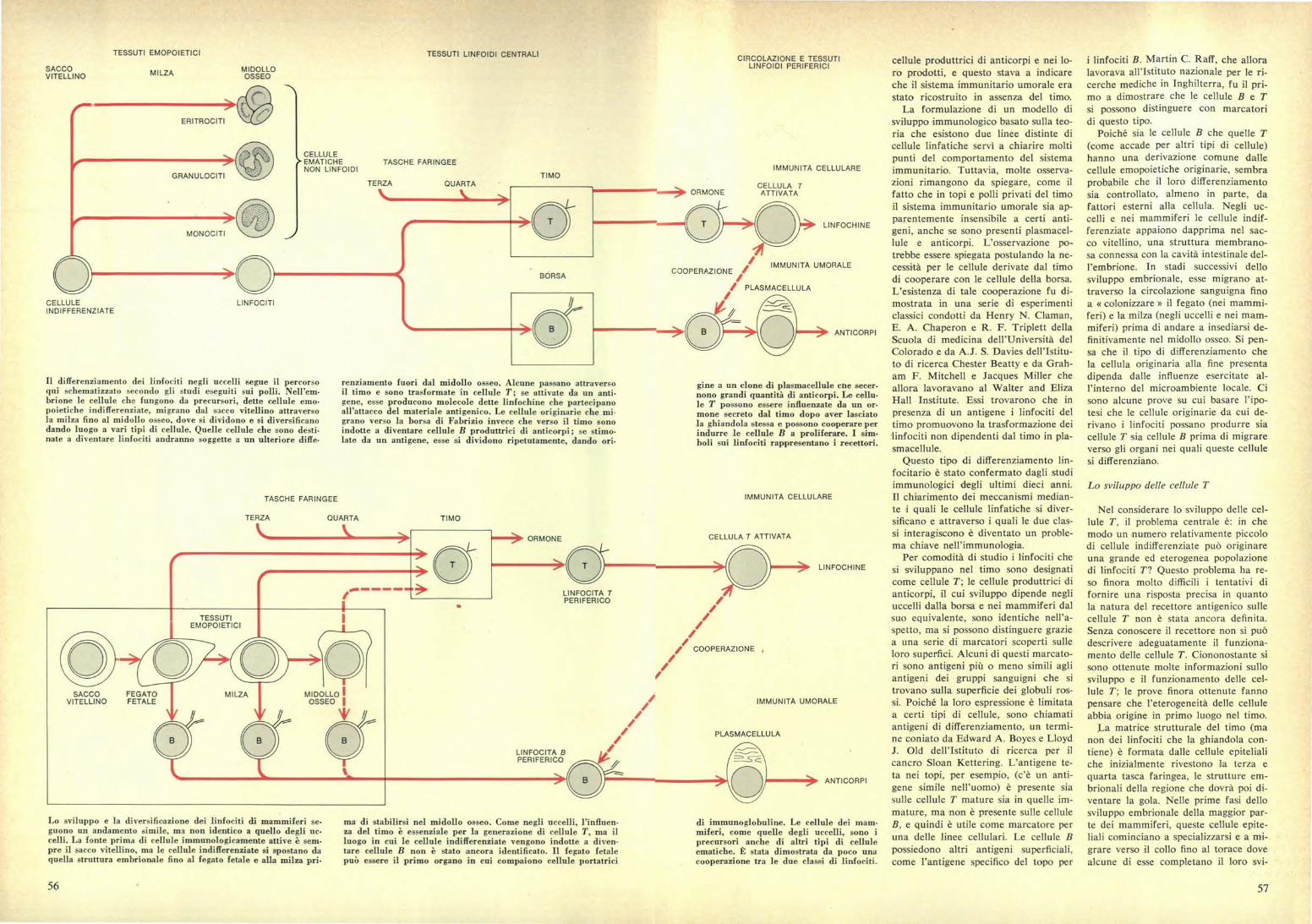
TESSUTI LINFOIDI CENTRALI CIRCOLAZIONE E TESSUTILINFOIDI PERIFERICI
TASCHE FARINGEE~UNITA CELLULARE
TIMOTERZA QUARTA
.•n••> ORMONECELLULA TATTIVATA
IMMUNITÀ UMORALECOOPERAZIONE /
PLASMACELLULA
ANTICORPI
BORSA
nn• LINFOCHINE
renziamento fuori dal midollo osseo. Alcune passano attraversoil timo e sono trasformate in cellule T; se attivate da un anti-gene, esse producono molecole dette linfochine che partecipanoall'attacco del materiale antigenico. Le cellule originarie che mi-grano verso la borsa di Fabrizio invece che verso il timo sonoindotte a diventare cellule B produttrici di anticorpi; se stimo-late da un antigene, esse si dividono ripetutamente, dando ori«
gine a un clone di plasmacellule cne secer-nono grandi quantità di anticorpi. Le cellu«le T possono essere influenzate da un or-mone secreto dal timo dopo aver lasciatola ghiandola stessa e possono cooperare perindurre le cellule B a proliferare. I sim-boli sui linfociti rappresentano i recettori.
ma di stabilirsi nel midollo osseo. Come negliza del timo è essenziale per la generazione diluogo in cui le cellule indifferenziate vengonotare cellule B non è stato ancora identificatopuò essere il primo organo in cui compaiono
uccelli, l'influen-cellule T, ma ilindotte a diven-
. Il fegato fetalecellule portatrici
di immunoglobuline. Le cellule dei mam-miferi, come quelle degli uccelli, sono iprecursori anche di altri tipi di celluleematiche. È stata dimostrata da poco unacooperazione tra le due classi di linfociti.
SACCOVITELLINO MILZA MIDOLLO
OSSEO
ERITROCITI
GRANULOCITI
CELLULE LINFOCITIINDIFFERENZIATE
CELLULE
r
EMATICHENON LINFOIDI
LINFOCITA BPERIFERICO
CELLULA T ATTIVATAORMONE
n~.> LINFOCHINE
LINFOCITA TPERIFERICO
oTESSUTI
EMOPOIETICI
SACCOVITELLINO
FEGATOFETALE
MIDOLLO IOSSEO
MILZA
TASCHE FARINGEE IMMUNITÀ CELLULARE
TERZA QUARTA TIMO
i COOPERAZIONE
e
IMMUNITÀ UMORALE
PLASMACELLULA
ANTICORPI
TESSUTI EMOPOIETICI
Il differenziamento dei linfociti negli uccelli segue il percorsoqui schematizzato secondo gli studi eseguiti sui polli. Nell'ella«brione le cellule che fungono da precursori, dette cellule emo-poietiche indifferenziate, migrano dal sacco vitellino attraversola milza fino al midollo osseo, dove si dividono e si diversificanodando luogo a vari tipi di cellule. Quelle cellule che sono desti-nate a diventare linfociti andranno soggette a un ulteriore diffe«
Lo sviluppo e la diversificazione dei linfociti di mammiferi se-guono un andamento simile, ma non identico a quello degli uc-celli. La fonte prima di cellule immunologicamente attive è sem-pre il sacco vitellino, ma le cellule indifferenziate si spostano daquella struttura embrionale fino al fegato fetale e alla milza pri-
cellule produttrici di anticorpi e nei lo-ro prodotti, e questo stava a indicareche il sistema immunitario umorale erastato ricostruito in assenza del timo.
La formulazione di un modello disviluppo immunologico basato sulla teo-ria che esistono due linee distinte dicellule linfatiche servì a chiarire moltipunti del comportamento del sistemaimmunitario. Tuttavia, molte osserva-zioni rimangono da spiegare, come ilfatto che in topi e polli privati del timoil sistema immunitario umorale sia ap-parentemente insensibile a certi anti-geni, anche se sono presenti plasmacel-lule e anticorpi. L'osservazione po-trebbe essere spiegata postulando la ne-cessità per le cellule derivate dal timodi cooperare con le cellule della borsa.L'esistenza di tale cooperazione fu di-mostrata in una serie di esperimenticlassici condotti da Henry N. Claman,E. A. Chaperon e R. F. Triplett dellaScuola di medicina dell'Università delColorado e da A.J. S. Davies dell'Istitu-to di ricerca Chester Beatty e da Grah-am F. Mitchell e Jacques Miller cheallora lavoravano al Walter and ElizaHall Institute. Essi trovarono che inpresenza di un antigene i linfociti deltimo promuovono la trasformazione deilinfociti non dipendenti dal timo in pla-smacellule.
Questo tipo di differenziamento lin-focitario è stato confermato dagli studiimmunologici degli ultimi dieci anni.Il chiarimento dei meccanismi median-te i quali le cellule linfatiche si diver-sificano e attraverso i quali le due clas-si interagiscono è diventato un proble-ma chiave nell'immunologia.
Per comodità di studio i linfociti chesi sviluppano nel timo sono designaticome cellule T; le cellule produttrici dianticorpi, il cui sviluppo dipende negliuccelli dalla borsa e nei mammiferi dalsuo equivalente, sono identiche nell'a-spetto, ma si possono distinguere graziea una serie di marcatori scoperti sulleloro superfici. Alcuni di questi marcato-ri sono antigeni più o meno simili agliantigeni dei gruppi sanguigni che sitrovano sulla superficie dei globuli ros-si. Poiché la loro espressione è limitataa certi tipi di cellule, sono chiamatiantigeni di differenziamento, un termi-ne coniato da Edward A. Boyes e LloydJ. Old dell'Istituto di ricerca per ilcancro Sloan Kettering. L'antigene te-ta nei topi, per esempio, (c'è un anti-gene simile nell'uomo) è presente siasulle cellule T mature sia in quelle im-mature, ma non è presente sulle celluleB, e quindi è utile come marcatore peruna delle linee cellulari. Le cellule Bpossiedono altri antigeni superficiali,come l'antigene specifico del topo per
i linfociti B. Martin C. Raff, che alloralavorava all'Istituto nazionale per le ri-cerche mediche in Inghilterra, fu il pri-mo a dimostrare che le cellule B e Tsi possono distinguere con marcatoridi questo tipo.
Poiché sia le cellule B che quelle T(come accade per altri tipi di cellule)hanno una derivazione comune dallecellule emopoietiche originarie, sembraprobabile che il loro differenziamentosia controllato, almeno in parte, dafattori esterni alla cellula. Negli uc-celli e nei mammiferi le cellule indif-ferenziate appaiono dapprima nel sac-co vitellino, una struttura membrano-sa connessa con la cavità intestinale del-l'embrione. In stadi successivi dellosviluppo embrionale, esse migrano at-traverso la circolazione sanguigna finoa « colonizzare » il fegato (nei mammi-feri) e la milza (negli uccelli e nei mam-miferi) prima di andare a insediarsi de-finitivamente nel midollo osseo. Si pen-sa che il tipo di differenziamento chela cellula originaria alla fine presentadipenda dalle influenze esercitate al-l'interno del microambiente locale. Cisono alcune prove su cui basare l'ipo-tesi che le cellule originarie da cui de-rivano i linfociti possano produrre siacellule T sia cellule B prima di migrareverso gli organi nei quali queste cellulesi differenziano.
Lo sviluppo delle cellule T
Nel considerare lo sviluppo delle cel-lule T, il problema centrale è: in chemodo un numero relativamente piccolodi cellule indifferenziate può originareuna grande ed eterogenea popolazionedi linfociti T? Questo problema ha re-so finora molto difficili i tentativi difornire una risposta precisa in quantola natura del recettore antigenico sullecellule T non è stata ancora definita.Senza conoscere il recettore non si puòdescrivere adeguatamente il funziona-mento delle cellule T. Ciononostante sisono ottenute molte informazioni sullosviluppo e il funzionamento delle cel-lule T; le prove finora ottenute fannopensare che l'eterogeneità delle celluleabbia origine in primo luogo nel timo.
La matrice strutturale del timo (manon dei linfociti che la ghiandola con-tiene) è formata dalle cellule epitelialiche inizialmente rivestono la terza equarta tasca faringea, le strutture em-brionali della regione che dovrà poi di-ventare la gola. Nelle prime fasi dellosviluppo embrionale della maggior par-te dei mammiferi, queste cellule epite-liali cominciano a specializzarsi e a mi-grare verso il collo fino al torace dovealcune di esse completano il loro svi-
56
57
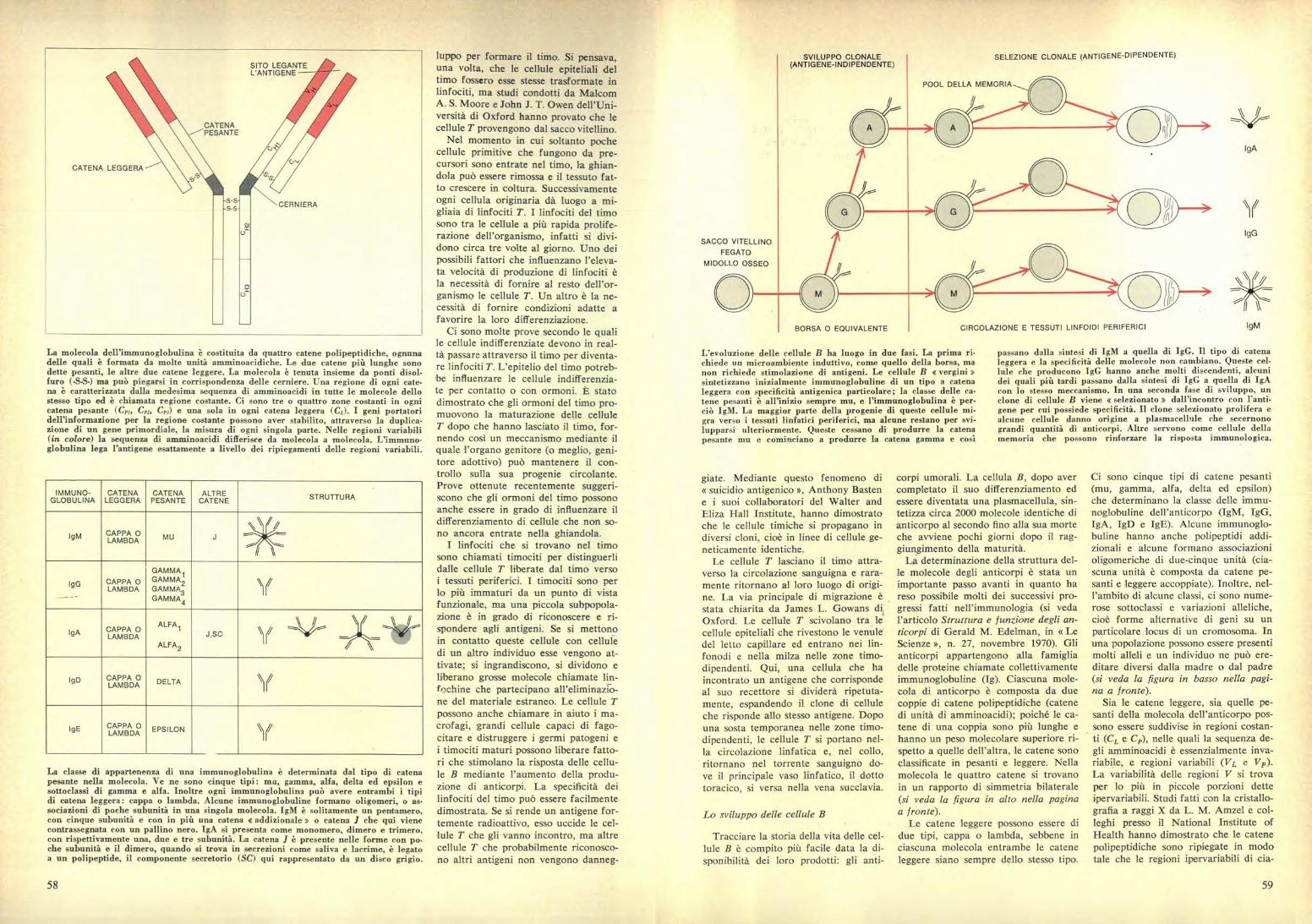
L'evoluzione delle cellule B ha luogo in due fasi. La prima ri-chiede un microambiente induttivo, come quello della borsa, manon richiede stimolazione di antigeni. Le cellule B e vergini »sintetizzano inizialmente immunoglobuline di un tipo a catenaleggera con specificità antigenica particolare; la classe delle ca-tene pesanti è all'inizio sempre mu, e Pimmunoglobulina è per-ciò IgM. La maggior parte della progenie di queste cellule mi-gra verso i tessuti linfatici periferici, ma alcune restano per svi.lupparsi ulteriormente. Queste cessano di produrre la catenapesante mu e cominciano a produrre la catena gamma e così
passano dalla sintesi di IgM a quella di IgG. Il tipo di catenaleggera e la specificità delle molecole non cambiano. Queste cel-lule che producono IgG hanno anche molti discendenti, alcunidei quali più tardi passano dalla sintesi di IgG a quella di IgAcon lo stesso meccanismo. In una seconda fase di sviluppo, unclone di cellule B viene « selezionato » dall'incontro con Vanti-gene per cui possiede specificità. Il clone selezionato prolifera ealcune cellule danno origine a plasmacellule che secernonograndi quantità di anticorpi. Altre servono come cellule dellamemoria che possono rinforzare la risposta immunologica.
SELEZIONE CLONALE (ANTIGENE-DIPENDENTE)SVILUPPO CLONALE(ANTIGENE-INDIPENDENTE)
POOL DELLA MEMORIA
SACCO VITELLINOFEGATO
MIDOLLO OSSEO
BORSA O EQUIVALENTE CIRCOLAZIONE E TESSUTI LINFOIDI PERIFERICI
IgA
IgG
IgM
SITO LEGANTEL'ANTIGENE
CATENA LEGGERA
CERNIERA
CATENAPESANTE
La molecola dell'immunoglobulina è costituita da quattro catene polipeptidiche, ognunadelle quali è formata da molte unità amminoacidiche. Le due catene più lunghe sonodette pesanti, le altre due catene leggere. La molecola è tenuta insieme da ponti disol-furo (-S-S-) ma può piegarsi in corrispondenza delle cerniere. Una regione di ogni cate-na è caratterizzata dalla medesima sequenza di amminoacidi in tutte le molecole dellostesso tipo ed è chiamata regione costante. Ci sono tre o quattro zone costanti in ognicatena pesante ( Cpl, CP2, Cp3) e una sola in ogni catena leggera (C L). I geni portatoridell'informazione per la regione costante possono aver stabilito, attraverso la duplica-zione di un gene primordiale, la misura di ogni singola parte. Nelle regioni variabili(in colore) la sequenza di amminoacidi differisce da molecola a molecola. L'immuno-globulina lega l'antigene esattamente a livello dei ripiegamenti delle regioni variabili.
IMMUNO-GLOBULINA
CATENALEGGERA
CATENAPESANTE
ALTRECATENE STRUTTURA
lgM CAPPA OLAMBDA MU J
\ /
\
IgG
---
CAPPA OLAMBDA
GAMMA 1GAMMA2GAMMA3GAMMA 4
) (
IgA CAPPA OLAMBDA
ALFA 1
ALFA 2
J,SCA___ -.-„\F
— —)( — / \
IgD CAPPA OLAMBDA DELTA
)(
IgE CAPPA OLAMBDA EPSILON
)(
La classe di appartenenza di una immunoglobulina è determinata dal tipo di catenapesante nella molecola. Ve ne sono cinque tipi: me, gamma, alfa, delta ed epsilon esottoclassi di gamma e alfa. Inoltre ogni immunoglobulina può avere entrambi i tipidi catena leggera: cappa o lambda. Alcune immunoglobuline formano oligomeri, o as-sociazioni di poche subunità in una singola molecola. IgM è solitamente un pentamero,con cinque subunità e con in più una catena « addizionale» o catena J che qui vienecontrassegnata con un pallino nero. IgA si presenta come monomero, dimero e trimero,con rispettivamente una, due e tre subunità. La catena J è presente nelle forme con po-che subunità e il dimero, quando si trova in secrezioni come saliva e lacrime, è legatoa un polipeptide, il componente secretorio (SC) qui rappresentato da un disco grigio.
luppo per formare il timo. Si pensava,una volta, che le cellule epiteliali deltimo fossero esse stesse trasformate inlinfociti, ma studi condotti da MalcomA. S. Moore e John J. T. Owen dell'Uni-versità di Oxford hanno provato che lecellule T provengono dal sacco vitellino.
Nel momento in cui soltanto pochecellule primitive che fungono da pre-cursori sono entrate nel timo, la ghian-dola può essere rimossa e il tessuto fat-to crescere in coltura. Successivamenteogni cellula originaria dà luogo a mi-gliaia di linfociti T. I linfociti del timosono tra le cellule a più rapida prolife-razione dell'organismo, infatti si divi-dono circa tre volte al giorno. Uno deipossibili fattori che influenzano l'eleva-ta velocità di produzione di linfociti èla necessità di fornire al resto dell'or-ganismo le cellule T. Un altro è la ne-cessità di fornire condizioni adatte afavorire la loro differenziazione.
Ci sono molte prove secondo le qualile cellule indifferenziate devono in real-tà passare attraverso il timo per diventa-re linfociti T. L'epitelio del timo potreb-be influenzare le cellule indifferenzia-te per contatto o con ormoni. È statodimostrato che gli ormoni del timo pro-muovono la maturazione delle celluleT dopo che hanno lasciato il timo, for-nendo così un meccanismo mediante ilquale l'organo genitore (o meglio, geni-tore adottivo) può mantenere il con-trollo sulla sua progenie circolante.Prove ottenute recentemente suggeri-scono che gli ormoni del timo possonoanche essere in grado di influenzare ildifferenziamento di cellule che non so-no ancora entrate nella ghiandola.
I linfociti che si trovano nel timosono chiamati timociti per distinguerlidalle cellule T liberate dal timo versoi tessuti periferici. I timociti sono perlo più immaturi da un punto di vistafunzionale, ma una piccola subpopola-zione è in grado di riconoscere e ri-spondere agli antigeni. Se si mettonoin contatto queste cellule con celluledi un altro individuo esse vengono at-tivate; si ingrandiscono, si dividono eliberano grosse molecole chiamate lin-fechine che partecipano all'eliminazio-ne del materiale estraneo. Le cellule Tpossono anche chiamare in aiuto i ma-crofagi, grandi cellule capaci di fago-citare e distruggere i germi patogeni ei timociti maturi possono liberare fatto-ri che stimolano la risposta delle cellu-le B mediante l'aumento della produ-zione di anticorpi. La specificità deilinfociti del timo può essere facilmentedimostrata. Se si rende un antigene for-temente radioattivo, esso uccide le cel-lule T che gli vanno incontro, ma altrecellule T che probabilmente riconosco-no altri antigeni non vengono danneg-
giate. Mediante questo fenomeno di« suicidio antigenico », Anthony Bastene i suoi collaboratori del Walter andEliza Hall Institute, hanno dimostratoche le cellule timiche si propagano indiversi cloni, cioè in linee di cellule ge-neticamente identiche.
Le cellule T lasciano il timo attra-verso la circolazione sanguigna e rara-mente ritornano al loro luogo di origi-ne. La via principale di migrazione èstata chiarita da James L. Gowans diOxford. Le cellule T scivolano tra lecellule epiteliali che rivestono le venuledel letto capillare ed entrano nei lin-fonodi e nella milza nelle zone timo-dipendenti. Qui, una cellula che haincontrato un antigene che corrispondeal suo recettore si dividerà ripetuta-mente, espandendo il clone di celluleche risponde allo stesso antigene. Dopouna sosta temporanea nelle zone timo-dipendenti, le cellule T si portano nel-la circolazione linfatica e, nel collo,ritornano nel torrente sanguigno do-ve il principale vaso linfatico, il dottotoracico, si versa nella vena succlavia.
Lo sviluppo delle cellule B
Tracciare la storia della vita delle cel-lule B è compito più facile data la di-sponibilità dei loro prodotti: gli anti-
corpi umorali. La cellula B, dopo avercompletato il suo differenziamento edessere diventata una plasmacellula, sin-tetizza circa 2000 molecole identiche dianticorpo al secondo fino alla sua morteche avviene pochi giorni dopo il rag-giungimento della maturità.
La determinazione della struttura del-le molecole degli anticorpi è stata unimportante passo avanti in quanto hareso possibile molti dei successivi pro-gressi fatti nell'immunologia (si vedal'articolo Struttura e funzione degli an-ticorpi di Gerald M. Edelman, in « LeScienze », n. 27, novembre 1970). Glianticorpi appartengono alla famigliadelle proteine chiamate collettivamenteimmunoglobuline (Ig). Ciascuna mole-cola di anticorpo è composta da duecoppie di catene polipeptidiche (catenedi unità di amminoacidi); poiché le ca-tene di una coppia sono più lunghe ehanno un peso molecolare superiore ri-spetto a quelle dell'altra, le catene sonoclassificate in pesanti e leggere. Nellamolecola le quattro catene si trovanoin un rapporto di simmetria bilaterale(si veda la figura in alto nella paginaa fronte).
Le catene leggere possono essere didue tipi, cappa o lambda, sebbene inciascuna molecola entrambe le cateneleggere siano sempre dello stesso tipo.
Ci sono cinque tipi di catene pesanti(mu, gamma, alfa, delta ed epsilon)che determinano la classe delle immu-noglobuline dell'anticorpo (IgM, IgG,IgA, IgD e IgE). Alcune immunoglo-buline hanno anche polipeptidi addi-zionali e alcune formano associazionioligomeriche di due-cinque unità (cia-scuna unità è composta da catene pe-santi e leggere accoppiate). Inoltre, nel-l'ambito di alcune classi, ci sono nume-rose sottoclassi e variazioni alleliche,cioè forme alternative di geni su unparticolare locus di un cromosoma. Inuna popolazione possono essere presentimolti alleli e un individuo ne può ere-ditare diversi dalla madre o dal padre(si veda la figura in basso nella pagi-na a fronte).
Sia le catene leggere, sia quelle pe-santi della molecola dell'anticorpo pos-sono essere suddivise in regioni costan-ti (CL e Ce), nelle quali la sequenza de-gli amminoacidi è essenzialmente inva-riabile, e regioni variabili (VL e Ve).La variabilità delle regioni V si trovaper lo più in piccole porzioni detteipervariabili. Studi fatti con la cristallo-grafia a raggi X da L. M. Amzel e col-leghi presso il National Institute ofHealth hanno dimostrato che le catenepolipeptidiche sono ripiegate in modotale che le regioni ipervariabili di cia-
58
59
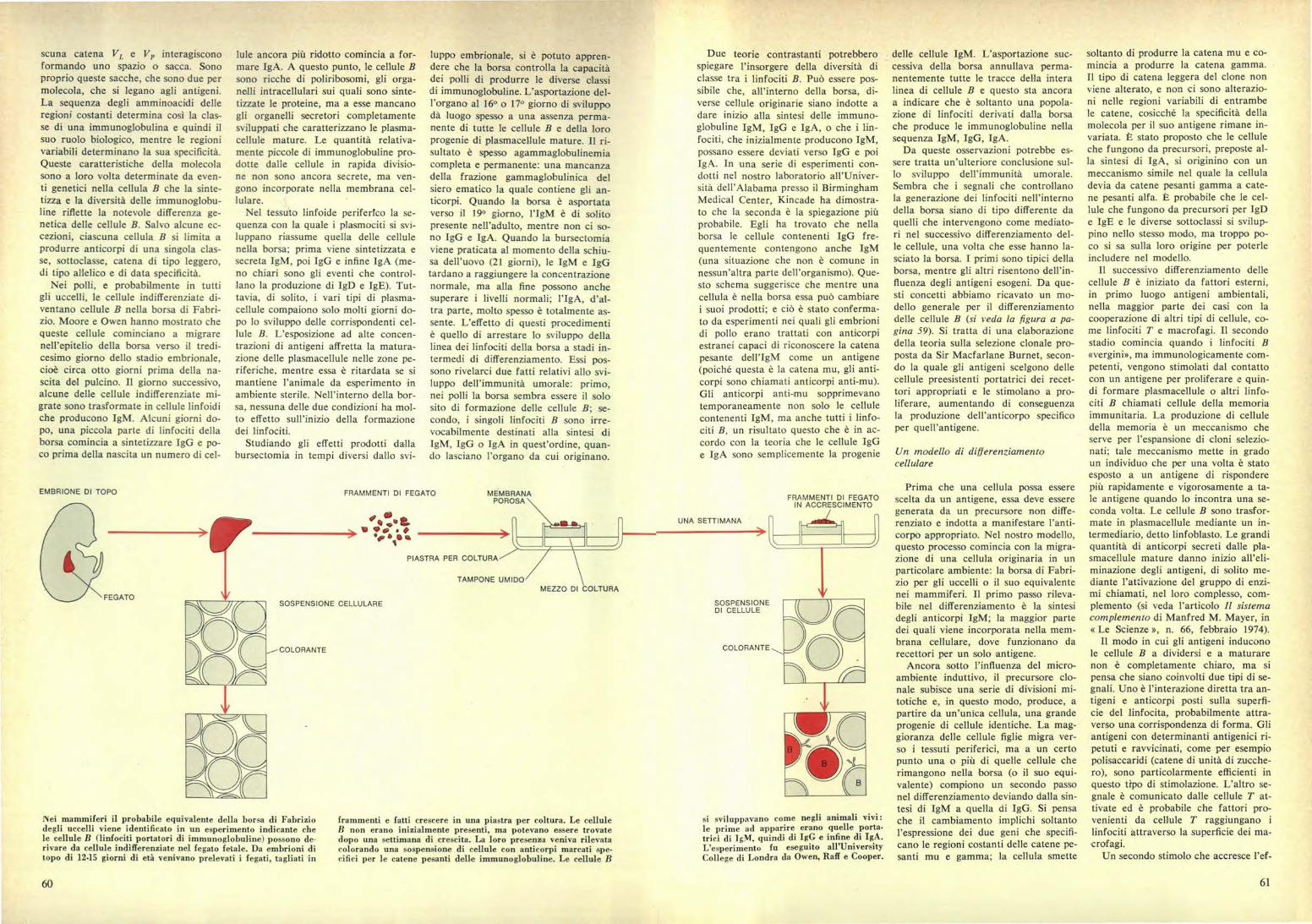
INei mammiferi il probabile equivalente della borsa di Fabriziodegli uccelli viene identificato in un esperimento indicante chele cellule B (linfociti portatori di immunoglobuline) possono de-rivare da cellule indifferenziate nel fegato fetale. Da embrioni ditopo di 12-15 giorni di età venivano prelevati i fegati, tagliati in
frammenti e fatti crescere in una piastra per coltura. Le celluleB non erano inizialmente presenti, ma potevano essere trovatedopo una settimana di crescita. La loro presenzi veniva rilevatacolorando una sospensione di cellule con anticorpi marcati spe.cifici per le catene pesanti delle immunoglobuline. Le cellule B
EMBRIONE DI TOPO
,-COLORANTE
FEGATO
FRAMMENTI DI FEGATO MEMBRANAPOROSA
o ‘-')e su a0 • p
PIASTRA PER COLTURA
TAMPONE UMIDOMEZZO DI COLTURA
SOSPENSIONE CELLULARE
FRAMMENTI DI FEGATOIN ACCRESCIMENTO
UNA SETTIMANA
SOSPENSIONEDI CELLULE
COLORANTE
scuna catena VL e Vp interagisconoformando uno spazio o sacca. Sonoproprio queste sacche, che sono due permolecola, che si legano agli antigeni.La sequenza degli amminoacidi delleregioni costanti determina così la clas-se di una immunoglobulina e quindi ilsuo ruolo biologico, mentre le regionivariabili determinano la sua specificità.Queste caratteristiche della molecolasono a loro volta determinate da even-ti genetici nella cellula B che la sinte-tizza e la diversità delle immunoglobu-line riflette la notevole differenza ge-netica delle cellule B. Salvo alcune ec-cezioni, ciascuna cellula B si limita aprodurre anticorpi di una singola clas-se, sottoclasse, catena di tipo leggero,di tipo allelico e di data specificità.
Nei polli, e probabilmente in tuttigli uccelli, le cellule indifferenziate di-ventano cellule B nella borsa di Fabri-zio. Moore e Owen hanno mostrato chequeste cellule cominciano a migrarenell'epitelio della borsa verso il tredi-cesimo giorno dello stadio embrionale,cioè circa otto giorni prima della na-scita del pulcino. Il giorno successivo,alcune delle cellule indifferenziate mi-grate sono trasformate in cellule linfoidiche producono IgM. Alcuni giorni do-po, una piccola parte di linfociti dellaborsa comincia a sintetizzare IgG e po-co prima della nascita un numero di cel-
lule ancora più ridotto comincia a for-mare IgA. A questo punto, le cellule Bsono ricche di poliribosomi, gli orga-nelli intracellulari sui quali sono sinte-tizzate le proteine, ma a esse mancanogli organelli secretori completamentesviluppati che caratterizzano le plasma-cellule mature. Le quantità relativa-mente piccole di immunoglobuline pro-dotte dalle cellule in rapida divisio-ne non sono ancora secrete, ma ven-gono incorporate nella membrana cel-lulare.
Nel tessuto linfoide periferIco la se-quenza con la quale i plasmociti si svi-luppano riassume quella delle cellulenella borsa; prima viene sintetizzata esecreta IgM, poi IgG e infine IgA (me-no chiari sono gli eventi che control-•ano la produzione di IgD e IgE). Tut-tavia, di solito, i vari tipi di plasma-cellule compaiono solo molti giorni do-po lo sviluppo delle corrispondenti cel-lule B. L'esposizione ad alte concen-trazioni di antigeni affretta la matura-zione delle plasmacellule nelle zone pe-riferiche, mentre essa è ritardata se simantiene l'animale da esperimento inambiente sterile. Nell'interno della bor-sa, nessuna delle due condizioni ha mol-to effetto sull'inizio della formazionedei linfociti.
Studiando gli effetti prodotti dallabursectomia in tempi diversi dallo svi-
luppo embrionale, si è potuto appren-dere che la borsa controlla la capacitàdei polli di produrre le diverse classidi immunoglobuline. L'asportazione del-l'organo al 16° o 17° giorno di sviluppodà luogo spesso a una assenza perma-nente di tutte le cellule B e della loroprogenie di plasmacellule mature. Il ri-sultato è spesso agammaglobulinemiacompleta e permanente: una mancanzadella frazione gammaglobulinica delsiero ematico la quale contiene gli an-ticorpi. Quando la borsa è asportataverso il 19° giorno, l'IgM è di solitopresente nell'adulto, mentre non ci so-no IgG e IgA. Quando la bursectomiaviene praticata al momento della schiu-sa dell'uovo (21 giorni), le IgM e IgGtardano a raggiungere la concentrazionenormale, ma alla fine possono anchesuperare i livelli normali; l'IgA, d'al-tra parte, molto spesso è totalmente as-sente. L'effetto di questi procedimentiè quello di arrestare lo sviluppo dellalinea dei linfociti della borsa a stadi in-termedi di differenziamento. Essi pos-sono rivelarci due fatti relativi allo svi-luppo dell'immunità umorale: primo,nei polli la borsa sembra essere il solosito di formazione delle cellule B; se-condo, i singoli linfociti B sono irre-vocabilmente destinati alla sintesi diIgM, IgG o IgA in quest'ordine, quan-do lasciano l'organo da cui originano.
Due teorie contrastanti potrebberospiegare l'insorgere della diversità diclasse tra i linfociti B. Può essere pos-sibile che, all'interno della borsa, di-verse cellule originarie siano indotte adare inizio alla sintesi delle immuno-globuline IgM, IgG e IgA, o che i lin-fociti, che inizialmente producono IgM,possano essere deviati verso IgG e poiIgA. In una serie di esperimenti con-dotti nel nostro laboratorio all'Univer-sità dell'Alabama presso il BirminghamMedical Center, Kincade ha dimostra-to che la seconda è la spiegazione piùprobabile. Egli ha trovato che nellaborsa le cellule contenenti IgG fre-quentemente contengono anche IgM(una situazione che non è comune innessun'altra parte dell'organismo). Que-sto schema suggerisce che mentre unacellula è nella borsa essa può cambiarei suoi prodotti; e ciò è stato conferma-to da esperimenti nei quali gli embrionidi pollo erano trattati con anticorpiestranei capaci di riconoscere la catenapesante dell'IgM come un antigene(poiché questa è la catena mu, gli anti-corpi sono chiamati anticorpi anti-mu).Gli anticorpi anti-mu sopprimevanotemporaneamente non solo le cellulecontenenti IgM, ma anche tutti i linfo-citi B, un risultato questo che è in ac-cordo con la teoria che le cellule IgGe IgA sono semplicemente la progenie
si sviluppavanosviluppavano come negli animali vivi:le prime ad apparire erano quelle porta-trici di IgM. quindi di IgG e infine di IgA.L'esperimento fu eseguito all'UniversityCollege di Londra da Owen, Raff e Cooper.
delle cellule IgM. L'asportazione suc-cessiva della borsa annullava perma-nentemente tutte le tracce della interalinea di cellule B e questo sta ancoraa indicare che è soltanto una popola-zione di linfociti derivati dalla borsache produce le immunoglobuline nellasequenza IgM, IgG, IgA.
Da queste osservazioni potrebbe es-sere tratta un'ulteriore conclusione sul-lo sviluppo dell'immunità umorale.Sembra che i segnali che controllanola generazione dei linfociti nell'internodella borsa siano di tipo differente daquelli che intervengono come mediato-ri nel successivo differenziamento del-le cellule, una volta che esse hanno la-sciato la borsa. I primi sono tipici dellaborsa, mentre gli altri risentono dell'in-fluenza degli antigeni esogeni. Da que-sti concetti abbiamo ricavato un mo-dello generale per il differenziamentodelle cellule B (si veda la figura a pa-gina 59). Si tratta di una elaborazionedella teoria sulla selezione clonale pro-posta da Sir Macfarlane Burnet, secon-do la quale gli antigeni scelgono dellecellule preesistenti portatrici dei recet-tori appropriati e le stimolano a pro-liferare, aumentando di conseguenzala produzione dell'anticorpo specificoper quell'antigene.
Un modello di differenziamentocellulare
Prima che una cellula possa esserescelta da un antigene, essa deve esseregenerata da un precursore non diffe-renziato e indotta a manifestare l'anti-corpo appropriato. Nel nostro modello,questo processo comincia con la migra-zione di una cellula originaria in unparticolare ambiente: la borsa di Fabri-zio per gli uccelli o il suo equivalentenei mammiferi. Il primo passo rileva-bile nel differenziamento è la sintesidegli anticorpi IgM; la maggior partedei quali viene incorporata nella mem-brana cellulare, dove funzionano darecettori per un solo antigene.
Ancora sotto l'influenza del micro-ambiente induttivo, il precursore clo-nale subisce una serie di divisioni mi-totiche e, in questo modo, produce, apartire da un'unica cellula, una grandeprogenie di cellule identiche. La mag-gioranza delle cellule figlie migra ver-so i tessuti periferici, ma a un certopunto una o più di quelle cellule cherimangono nella borsa (o il suo equi-valente) compiono un secondo passonel differenziamento deviando dalla sin-tesi di IgM a quella di IgG. Si pensache il cambiamento implichi soltantol'espressione dei due geni che specifi-cano le regioni costanti delle catene pe-santi mu e gamma; la cellula smette
soltanto di produrre la catena mu e co-mincia a produrre la catena gamma.Il tipo di catena leggera del clone nonviene alterato, e non ci sono alterazio-ni nelle regioni variabili di entrambele catene, cosicché la specificità dellamolecola per il suo antigene rimane in-variata. È stato proposto che le celluleche fungono da precursori, preposte al-la sintesi di IgA, si originino con unmeccanismo simile nel quale la celluladevia da catene pesanti gamma a cate-ne pesanti alfa. È probabile che le cel-lule che fungono da precursori per IgDe IgE e le diverse sottoclassi si svilup-pino nello stesso modo, ma troppo po-co si sa sulla loro origine per poterleincludere nel modello.
Il successivo differenziamento dellecellule B è iniziato da fattori esterni,in primo luogo antigeni ambientali,nella maggior parte dei casi con lacooperazione di altri tipi di cellule, co-me linfociti T e macrofagi. Il secondostadio comincia quando i linfociti B«vergini», ma immunologicamente com-petenti, vengono stimolati dal contattocon un antigene per proliferare e quin-di formare plasmacellule o altri linfo-citi B chiamati cellule della memoriaimmunitaria. La produzione di celluledella memoria è un meccanismo cheserve per l'espansione di cloni selezio-nati; tale meccanismo mette in gradoun individuo che per una volta è statoesposto a un antigene di risponderepiù rapidamente e vigorosamente a ta-le antigene quando lo incontra una se-conda volta. Le cellule B sono trasfor-mate in plasmacellule mediante un in-termediario, detto linfoblasto. Le grandiquantità di anticorpi secreti dalle pla-smacellule mature danno inizio all'eli-minazione degli antigeni, di solito me-diante l'atZivazione del gruppo di enzi-mi chiamati, nel loro complesso, com-plemento (si veda l'articolo Il sistemacomplemento di Manfred M. Mayer, in«Le Scienze », n. 66, febbraio 1974).
Il modo in cui gli antigeni induconole cellule B a dividersi e a maturarenon è completamente chiaro, ma sipensa che siano coinvolti due tipi di se-gnali. Uno è l'interazione diretta tra an-tigeni e anticorpi posti sulla superfi-cie del linfocita, probabilmente attra-verso una corrispondenza di forma. Gliantigeni con determinanti antigenici ri-petuti e ravvicinati, come per esempiopolisaccaridi (catene di unità di zucche-ro), sono particolarmente efficienti inquesto tipo di stimolazione. L'altro se-gnale è comunicato dalle cellule T at-tivate ed è probabile che fattori pro-venienti da cellule T raggiungano ilinfociti attraverso la superficie dei ma-crofagi.
Un secondo stimolo che accresce l'ef-
60
61
CAPPA
LAMBDA
PESANTE
ci
C=MMIE1CIZMia
111===MIIIIMI
CAPPA
LAMBDA
PESANTE
(::7:1=1•~1=,--
r CAPPA
LAMBDA
PESANTE
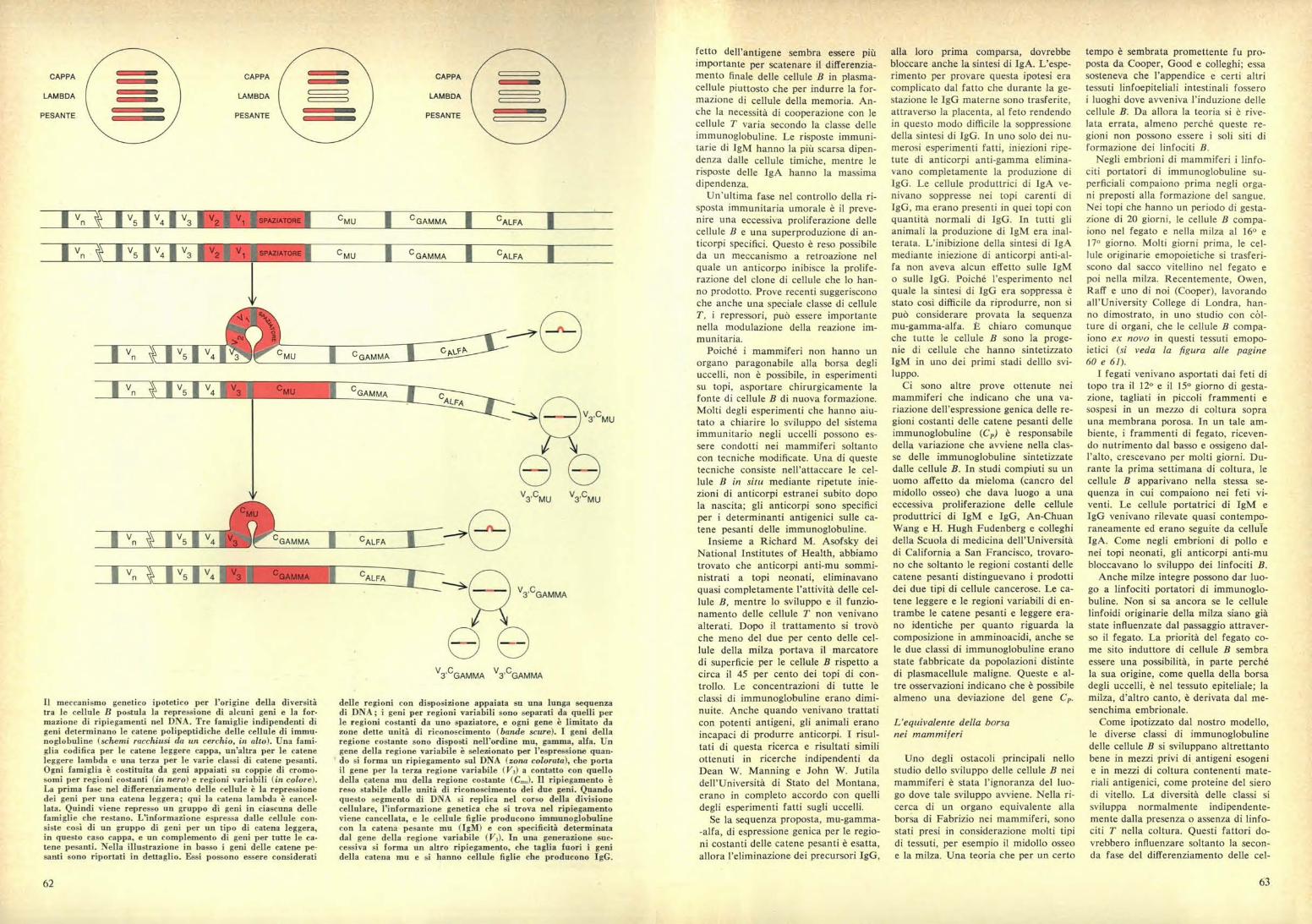
delle regioni con disposizione appaiata su una lunga sequenzadi DNA; i geni per regioni variabili sono separati da quelli perle regioni costanti da uno spaziatore, e ogni gene è limitato dazone dette unità di riconoscimento (bande scure). I geni dellaregione costante sono disposti nell'ordine mu, gamma, alfa. Ungene della regione variabile è selezionato per l'espressione quan-do si forma un ripiegamento sul DNA (zona colorata), che portail gene per la terza regione variabile (1/3) a contatto con quellodella catena mu della regione costante (Cm.). Il ripiegamento èreso stabile dalle unità di riconoscimento dei due geni. Quandoquesto segmento di DNA si replica nel corso della divisionecellulare, l'informazione genetica che si trova nel ripiegamentoviene cancellata, e le cellule figlie producono immunoglobulinecon la catena pesante mu (IgM) e con specificità determinatadal gene della regione variabile (V 3). In una generazione suc-cessiva si forma un altro ripiegamento, che taglia fuori i genidella catena mu e si hanno cellule figlie che producono IgG.
V5 i ,14 i v, i v, V1 SPAZIATORI C M U C GAMMA I CALFA
IV5 I V4 I V3
V2V1 SPAZIATORE i C M U C GAMMA I cALFA
Il meccanismo genetico ipotetico per l'origine della diversitàtra le cellule B postula la repressione di alcuni geni e la for-mazione di ripiegamenti nel DNA. Tre famiglie indipendenti digeni determinano le catene polipeptidiche delle cellule di immu-noglobuline (schemi racchiusi da un cerchio, in alto). Una fami•glia codifica per le catene leggere cappa, un'altra per le cateneleggere lambda e una terza per le varie classi di catene pesanti.Ogni famiglia è costituita da geni appaiati su coppie di cromo-somi per regioni costanti (in nero) e regioni variabili (in colore).La prima fase nel differenziamento delle cellule è la repressionedei geni per una catena leggera; qui la catena lambda è cancel-lata. Quindi viene represso un gruppo di geni in ciascuna dellefamiglie che restano. L'informazione espressa dalle cellule con-siste così di un gruppo di geni per un tipo di catena leggera,in questo caso cappa, e un complemento di geni per tutte le ca-tene pesanti. Nella illustrazione in basso i geni delle catene pe-santi sono riportati in dettaglio. Essi possono essere considerati
M U
e eV C V
3' MU 3C' MU
/ V5
vn I
c ALFA
3c
GAMMA
eV3' C GAMMA V3'CGAMMA
C GAMMA
GAMMA
I Vn
n 7 IV
fetto dell'antigene sembra essere piùimportante per scatenare il differenzia-mento finale delle cellule B in plasma-cellule piuttosto che per indurre la for-mazione di cellule della memoria. An-che la necessità di cooperazione con lecellule T varia secondo la classe delleimmunoglobuline. Le risposte immuni-tarie di IgM hanno la più scarsa dipen-denza dalle cellule timiche, mentre lerisposte delle IgA hanno la massimadipendenza.
Un'ultima fase nel controllo della ri-sposta immunitaria umorale è il preve-nire una eccessiva proliferazione dellecellule B e una superproduzione di an-ticorpi specifici. Questo è reso possibileda un meccanismo a retroazione nelquale un anticorpo inibisce la prolife-razione del clone di cellule che lo han-no prodotto. Prove recenti suggerisconoche anche una speciale classe di celluleT, i repressori, può essere importantenella modulazione della reazione im-munitaria.
Poiché i mammiferi non hanno unorgano paragonabile alla borsa degliuccelli, non è possibile, in esperimentisu topi, asportare chirurgicamente lafonte di cellule B di nuova formazione.Molti degli esperimenti che hanno aiu-tato a chiarire lo sviluppo del sistemaimmunitario negli uccelli possono es-sere condotti nei mammiferi soltantocon tecniche modificate. Una di questetecniche consiste nell'attaccare le cel-lule B in situ mediante ripetute inie-zioni di anticorpi estranei subito dopola nascita; gli anticorpi sono specificiper i determinanti antigenici sulle ca-tene pesanti delle immunoglobuline.
Insieme a Richard M. Asofsky deiNational Institutes of Health, abbiamotrovato che anticorpi anti-mu sommi-nistrati a topi neonati, eliminavanoquasi completamente l'attività delle cel-lule B, mentre lo sviluppo e il funzio-namento delle cellule T non venivanoalterati. Dopo il trattamento si trovòche meno del due per cento delle cel-lule della milza portava il marcatoredi superficie per le cellule B rispetto acirca il 45 per cento dei topi di con-trollo. Le concentrazioni di tutte leclassi di immunoglobuline erano dimi-nuite. Anche quando venivano trattaticon potenti antigeni, gli animali eranoincapaci di produrre anticorpi. I risul-tati di questa ricerca e risultati similiottenuti in ricerche indipendenti daDean W. Manning e John W. Jutiladell'Università di Stato del Montana,erano in completo accordo con quellidegli esperimenti fatti sugli uccelli.
Se la sequenza proposta, mu-gamma--alfa, di espressione genica per le regio-ni costanti delle catene pesanti è esatta,allora l'eliminazione dei precursori IgG,
alla loro prima comparsa, dovrebbebloccare anche la sintesi di IgA. L'espe-rimento per provare questa ipotesi eracomplicato dal fatto che durante la ge-stazione le IgG materne sono trasferite,attraverso la placenta, al feto rendendoin questo modo difficile la soppressionedella sintesi di IgG. In uno solo dei nu-merosi esperimenti fatti, iniezioni ripe-tute di anticorpi anti-gamma elimina-vano completamente la produzione diIgG. Le cellule produttrici di IgA ve-nivano soppresse nei topi carenti diIgG, ma erano presenti in quei topi conquantità normali di IgG. In tutti glianimali la produzione di IgM era inal-terata. L'inibizione della sintesi di IgAmediante iniezione di anticorpi anti-al-fa non aveva alcun effetto sulle IgMo sulle IgG. Poiché l'esperimento nelquale la sintesi di IgG era soppressa èstato così difficile da riprodurre, non sipuò considerare provata la sequenzamu-gamma-alfa. È chiaro comunqueche tutte le cellule B sono la proge-nie di cellule che hanno sintetizzatoIgM in uno dei primi stadi delllo svi-luppo.
Ci sono altre prove ottenute neimammiferi che indicano che una va-riazione dell'espressione genica delle re-gioni costanti delle catene pesanti delleimmunoglobuline (Ce) è responsabiledella variazione che avviene nella clas-se delle immunoglobuline sintetizzatedalle cellule B. In studi compiuti su unuomo affetto da mieloma (cancro delmidollo osseo) che dava luogo a unaeccessiva proliferazione delle celluleproduttrici di IgM e IgG, An-ChuanWang e H. Hugh Fudenberg e colleghidella Scuola di medicina dell'Universitàdì California a San Francisco, trovaro-no che soltanto le regioni costanti dellecatene pesanti distinguevano i prodottidei due tipi di cellule cancerose. Le ca-tene leggere e le regioni variabili di en-trambe le catene pesanti e leggere era-no identiche per quanto riguarda lacomposizione in amminoacidi, anche sele due classi di immunoglobuline eranostate fabbricate da popolazioni distintedi plasmacellule maligne. Queste e al-tre osservazioni indicano che è possibilealmeno una deviazione del gene Ci,.
L'equivalente della borsanei mammiferi
Uno degli ostacoli principali nellostudio dello sviluppo delle cellule B neimammiferi è stata l'ignoranza del luo-go dove tale sviluppo avviene. Nella ri-cerca di un organo equivalente allaborsa di Fabrizio nei mammiferi, sonostati presi in considerazione molti tipidi tessuti, per esempio il midollo osseoe la milza. Una teoria che per un certo
tempo è sembrata promettente fu pro-posta da Cooper, Good e colleghi; essasosteneva che l'appendice e certi altritessuti linfoepiteliali intestinali fosseroi luoghi dove avveniva l'induzione dellecellule B. Da allora la teoria si è rive-lata errata, almeno perché queste re-gioni non possono essere i soli siti diformazione dei linfociti B.
Negli embrioni di mammiferi i linfo-citi portatori di immunoglobuline su-perficiali compaiono prima negli orga-ni preposti alla formazione del sangue.Nei topi che hanno un periodo di gesta-zione di 20 giorni, le cellule B compa-iono nel fegato e nella milza al 16° e17° giorno. Molti giorni prima, le cel-lule originarie emopoietiche si trasferi-scono dal sacco vitellino nel fegato epoi nella milza. Recentemente, Owen,Raff e uno di noi (Cooper), lavorandoall'University College di Londra, han-no dimostrato, in uno studio con còl-ture di organi, che le cellule B compa-iono ex novo in questi tessuti emopo-ietici (si veda la figura alle pagine60 e 61).
I fegati venivano asportati dai feti ditopo tra il 12° e il 15° giorno di gesta-zione, tagliati in piccoli frammenti esospesi in un mezzo di coltura soprauna membrana porosa. In un tale am-biente, i frammenti di fegato, riceven-do nutrimento dal basso e ossigeno dal-l'alto, crescevano per molti giorni. Du-rante la prima settimana di coltura, lecellule B apparivano nella stessa se-quenza in cui compaiono nei feti vi-venti. Le cellule portatrici di IgM eIgG venivano rilevate quasi contempo-raneamente ed erano seguite da celluleIgA. Come negli embrioni di pollo enei topi neonati, gli anticorpi anti-mubloccavano lo sviluppo dei linfociti B.
Anche milze integre possono dar luo-go a linfociti portatori di immunoglo-buline. Non si sa ancora se le cellulelinfoidi originarie della milza siano giàstate influenzate dal passaggio attraver-so il fegato. La priorità del fegato co-me sito induttore di cellule B sembraessere una possibilità, in parte perchéla sua origine, come quella della borsadegli uccelli, è nel tessuto epiteliale; lamilza, d'altro canto, è derivata dal me-senchima embrionale.
Come ipotizzato dal nostro modello,le diverse classi di immunoglobulinedelle cellule B si sviluppano altrettantobene in mezzi privi di antigeni esogenie in mezzi di coltura contenenti mate-riali antigenici, come proteine del sierodi vitello. La diversità delle classi sisviluppa normalmente indipendente-mente dalla presenza o assenza di linfo-citi T nella coltura. Questi fattori do-vrebbero influenzare soltanto la secon-da fase del differenziamento delle cel-
62
63

Le malattie caratterizzate da deficienza immunitaria possono es-sere considerate come difetti del differenziamento dei linfocitie dei tessuti linfatici. La mancanza delle cellule B e T suggeri-sce una mancanza di precursori dei linfociti e si può ovviaretrapiantando cellule indifferenziate (1) dal fegato fetale o dalmidollo osseo. Individui nati senza timo (2) sono privi dell'im-munità cellulare; le funzioni di questo sistema possono essereripristinate trapiantando un timo fetale. Il mancato sviluppo del-le cellule indifferenziate in linfociti B (3) è una malattia conge-
nita legata al sesso, descritta per la prima volta da Ogden C.Bruton; deriva forse da un difetto del luogo di induzione dellecellule B, probabilmente il fegato fetale. In altre alterazioni del-la produzione di anticorpi, le cellule B sono presenti ma nonsono stimolate da antigeni a dividersi e a svilupparsi in plasma-cellule mature. L'arresto del differenziamento può essere assolu-to, e portare perciò a una assenza di tutte le classi di immuno-globuline (4); può interessare le cellule che producono IgG e IgA(5) o può essere limitato alle sole cellule che producono IgA (6).
FEGATO FETALE IMMUNITÀ UMORALE
TIMO
> ORMONE CELLULA T ATTIVATA
TERZA QUARTA
LINFOCHINE
CELLULAINDIFFERENZIATA
PLASMACELLULA
> IgM
ANTICORPI
IgA
IgG
TASCHE FARINGEE IMMUNITA CELLULARE
lule B: quello che accade in risposta aun antigene.
Joan L. Press e Norman R. Klin-man della Scuola di medicina dell'Uni-versità di Pennsylvania hanno dimo-strato che i precursori delle cellule por-tatrici di anticorpi contro uno specificoantigene compaiono con frequenza con-frontabile nella milza di topi fetalie topi maturi. Poiché l'antigene nonverrebbe incontrato nella vita fetale, iloro dati sono in favore della ipotesiche la diversità clonale nelle cellule Binsorga indipendentemente dal contat-to con gli antigeni. Patricia G. Speare i suoi colleghi della Università Ro-ckefeller sono arrivati a una simileconclusione con esperimenti in cui im-piegavano una tecnica diversa per rile-vare linfociti B reattivi verso l'antigenenei feti e negli animali adulti. Per oranon si può affermare con certezza chegli antigeni « intrinseci » (come gli an-tigeni dell'istocompatibilità che sono ipiù importanti determinanti dell'auto-riconoscimento) non abbiano alcun ruo-lo nella ontogenesi delle cellule B, maci sono prove in abbondanza per sug-gerire che lo sviluppo della diversitàdei linfociti B non sia una risposta acontatti casuali con antigeni provenien-ti dall'ambiente esterno.
La genetica dell'immunità
Come risulta implicito dagli esperi-menti sopra descritti, ci sono tutte leragioni per pensare che il differenzia-mento primario delle cellule in una po-polazione diversa di cellule B e di altrecellule segua un programma ordinatocodificato dai geni, e che il programmaoperi senza intervento dell'ambienteesterno. Per questo motivo è essenzialeche una spiegazione dello sviluppo im-munologico sia basata su principi ge-netici.
Nella maggior parte delle proteine,un solo gene codifica tutte le informa-zioni per la sintesi di una singola cate-na polipeptidica; questo non è il casodelle immunoglobuline, come WilliamJ. Dreyer e J. Claude Bennett dell'Isti-tuto di tecnologia della California han-no suggerito. Sia gli studi sull'eredita-rietà sia quelli sulle sequenze am-minoacidiche di immunoglobuline sug-geriscono che le regioni costanti e va-riabili di ciascuna catena polipeptidicasiano definite da geni diversi e che cisiano infatti tre famiglie distinte di ge-ni strutturali per le molecole degli an-ticorpi. Ogni famiglia è composta da uncerto numero di geni disposti a coppieche codificano per le regioni viariabili
di catene pesanti o leggere e, in regio-ni vicine sullo stesso cromosoma, daun numero minore di geni per le regio-ni costanti. La prima famiglia determi-na le catene leggere cappa, la secondale catene leggere lambda e la terza lediverse classi e sottoclassi di catene pe-santi. I geni all'interno di ogni fami-glia sono legati, ma le tre famiglie nonlo sono; esse sono probabilmente loca-lizzate in cromosomi distinti.
I cromosomi sono, naturalmente, ac-coppiati, ma viene espressa soltantol'informazione che parte da un cromo-soma di ogni coppia. La repressione diun gruppo di geni in ogni coppia è ne-cessaria se il modello deve servire aspiegare l'osservato comportamento del-le plasmacellule; persino in un indivi-duo eterozigote per un marcatore ge-netico su una immunoglobulina ogniplasmacellula esprime soltanto uno de-gli alleli: un fenomeno chiamato esclu-sione allelica.
La prima fase della costruzione diuna immunoglobulina in base a questoprogramma genetico è quindi la repres-sione funzionale dei geni dell'immuno-globulina, su uno dei cromosomi accop-piati, portatori di ciascuna famiglia ge-nica. Il meccanismo deve poi sceglierequale famiglia di catene leggere, cap-
pa o lambda, deve essere espressa. Ri-mangono poi due famiglie di geni: unaper le catene leggere e una per le ca-tene pesanti; quest'ultima determina laclasse della immunoglobulina.
Il passo successivo è la selezione diun particolare gruppo di geni della re-gione variabile, sia per le catene pesan-ti, sia per quelle leggere; queste deter-mineranno insieme la specificità del-l'anticorpo. Il numero dei geni variabiliereditari rimane materia di controver-sia, ma la maggior parte dei ricercatoriè d'accordo nel ritenere che esso debbaessere molto elevato, o addirittura enor-me. I sostenitori della prima teoria, det-ta la teoria della linea germinale, affer-mano che tutti i geni della regione va-riabile necessari sono ereditari. Altrisostengono che molta della variabilitàdella molecola è generata da mutazionisomatiche o ricombinazioni di geni, nelqual caso essi non sarebbero trasmessialla progenie. È stato dimostrato chei geni che specificano le regioni varia-bili e costanti sono uniti sul DNA delnucleo cellulare e che un solo messag-gio portato dall'RNA viene trascrittodal DNA e impiegato per dirigere lasintesi di ciascuna catena di polipepti-di nella molecola dell'anticorpo.
Sono stati proposti molti modelli perl'unione dei geni delle regioni costantie variabili. Uno di tali mcdelli che po-trebbe spiegare sia l'unione dei geni Ve C sia la deviazione successiva dauna classe di immunoglobuline all'al-tra, ipotizza che i geni Cp siano legatinell'ordine mu, gamma, alfa. Ripiega-menti formati sul DNA escluderebberosuccessivamente ciascuno di questi ge-ni; quando il DNA ripiegato viene tra-scritto o replicato, l'informazione nelripiegamento sarebbe cancellata e diconseguenza sarebbero creati cloni dicellule che producono le singole classidi anticorpi. Lo stesso meccanismo po-trebbe spiegare la selezione di un geneper le regioni variabili da un pool dimolti geni, determinando in questo mo-do la specificità della molecola (si vedala figura a pagina 62).
Un metodo potenziale per sceglieretra le varie teorie di unione e selezionedei geni può essere quello di studiarele apparenti eccezioni alla regola cheuna - singola cellula B passa produrresoltanto una classe di anticorpi allavolta. Benvenuto Pernis dell'Istituto diimmunologia di Basilea ha dimostratoche le cellule B di coniglio produttricidi IgG possono avere IgM alla loro su-perficie; insieme a David S. Rowe dellaboratorio di Losanna dell'Organiz-zazione Mondiale della Sanità ha an-che dimostrato che i linfociti B umanipossono produrre IgM e IgD contem-poraneamente.
Le scoperte di Pernis nei conigli po-trebbero essere spiegate dalla presenzadi una molecola di messaggero a vitalunga per le catene mu, che potrebberimanere attiva dopo che la trascrizio-ne del DNA si è spostata dal gene perla catena mu a quello per la catenagamma. Il secondo caso è molto diffi-cile da spiegare e forse rappresenta unesempio di trascrizione simultanea digeni per gli anticorpi che appartengonoa classi diverse, ma hanno la stessa spe-cificità. Pernis e Maxime Seligmanndell'ospedale Saint Louis di Parigi han-no scoperto in un paziente con altera-zione maligna del linfocita B moleco-le di IgM e IgD sulla superficie cellu-lare che hanno attività comune controIgD. L'espressione della specificità ver-so lo stesso antigene può voler signifi-care che regioni variabili delle catenepesanti di entrambe le immunoglobu-line sono identiche. Se si potesse dimo-strare inequivocabilmente che lo stessogene Vp può essere trascritto simulta-neamente a due geni Ci,, il modello quipresentato sarebbe escluso in favore diuno che prende in considerazione piùdi una coppia di ciascun gene Vp.
Sviluppo anormale
La conoscenza del meccanismo di dif-ferenziamento delle cellule linfoidi per-mette di chiarire anche aspetti dellemalattie del sistema immunitario e inalcuni casi può rivelare fino a che pun-to il sistema sia alterato. I marcatoriche possono essere impiegati per rive-lare i linfociti B, per esempio, sonostati di aiuto nel mostrare che la mag-gior parte dei ragazzi affetti dal morbodi Bruton (deficienza congenita legataal sesso di plasmacellule e anticorpi cir-colanti) sono praticamente privi di cel-lule B a tutti gli stadi di differenzia-mento. L'osservazione suggerisce chela malattia può essere causata da un di-fetto nell'organo che genera cellule B,forse il fegato fetale. D'altra parte, ipazienti nei quali si sviluppa una caren-za di plasmacellule e anticorpi in unostadio successivo della vita hanno spes-so un numero normale di linfociti Bportatori di immunoglobuline di tuttele classi. In questo caso, sembra che ildifetto compaia in stadi successivi deldifferenziamento e impedisca la matu-razione delle cellule B in plasmacellule.
In altri individui con sistema immu-nitario carente è arrestato lo sviluppodi una o più classi, ma non di tutte leclassi di cellule produttrici di anticorpi.Nel caso di carenza di IgA isolata,una condizione nella quale manca sol-tanto quella classe di immunoglobuline,le cellule B portatrici di IgA si trovanoquasi sempre nel sangue. Il difetto è
presente in una persona su cinquecen-to di origine europea. L.Y. Frank Wudel nostro laboratorio ha trovato chele cellule B di alcuni pazienti con ca-renza di IgA possono essere stimolatea produrre e a secernere anticorpi fa-cendole crescere in una coltura con unestratto di fitolacca, una pianta che con-tiene molte sostanze che influenzano ilcomportamento delle cellule linfoidi.L'estratto di fitolacca può stimolare an-che il differenziamento di linfociti inalcuni individui che presentano caren-ze in tutte le classi di immunoglobuline.Tali scoperte fanno sperare che in fu-turo sia possibile intervenire su alcunealterazioni immunitarie letali. Bambininati senza timo sono già stati trattaticon successo mediante trapianti di ti-mo e neonati privi di cellule T e Bsono stati curati immunologicamentemediante trapianti di midollo osseo efegato fetale.
Mediante marcatori specifici per cel-lule TeBe possibile identificare le cel-lule coinvolte in tumori del sistema lin-fatico. Alcuni casi di leucemia linfoci-taria acuta, per esempio, sembrano es-sere dovuti ad alterazioni delle celluleT. La proliferazione maligna di celluleB si manifesta a diversi stadi del diffe-renziamento. I mielomi sono caratte-rizzati da una crescita incontrollata dicloni di plasmacellule mature, mentrei linfociti B sono implicati nella leuce-mia linfocitaria e nel linfoma di Bur-kitt. Le macroglobulinemie di Wald-enstriim, così chiamate da Jan Wald-enstróm del Generai Hospital di Malmòin Svezia, rappresentano un accumulodi cellule in transizione da linfociti Ba cellule che secernono anticorpi.
Il linfoma indotto da virus delle cel-lule della borsa nei polli assomiglia allinfoma di Burkitt nell'uomo, una ma-lattia maligna che sembra anch'essacausata da virus. I linfociti tumoraliproducono invariabilmente IgM, indi-pendentemente dalla loro localizzazionenell'organismo. Sembra quindi plausibi-le che il virus che provoca il tumoretrasformi le cellule durante il differen-ziamento clonale, interrompendo l'abi-tuale corso dell'espressione genica nelmomento in cui le cellule dovrebbero,come al solito, passare dalla sintesi diIgM a quella di IgG. Poiché sembrache le cellule siano suscettibili agli ef-fetti oncogeni del virus soltanto in unostadio particolare del differenziamento,forse la degenerazione del tumore di-pende dalla regione del genoma dellacellula ospite.
Qualunque sia il valore di questeipotesi, esse offrono idee potenzialmenteimportanti che possono essere perfezio-nate con ulteriori ricerche sui processidi sviluppo del sistema immunitario.
64
65





























