Embed Size (px)
DESCRIPTION
fisica
Citation preview



EL LIBR O MUERE CU ANDO LO FO TOCOPI A
AMIGO LECT OR:
La obraqueustedtieneensusmanosposeeungranvalor.En ella, su autorha vertidoconocimientos,experiencia y muchotrabajo.El editorhaprocuradounapresentacióndignadesucontenidoy estáponiendotodosuempe-ñoy recursosparaqueseaampliamente difundida,a través desureddecomerciali-zación.
Al fotocopiarestelibro, el autory el editordejandepercibir lo quecorrespondea lainversión que han realizado y se desalienta la creación denuevas obras.Rechacecualquier ejemplar“pirata”o fotocopiailegal de este libro, puesde lo contrarioestarácontribuyendoal lucro dequienesse aprovechanilegítimamentedel esfuer-zo delautory deleditor.
La reproducciónnoautorizadadeobrasprotegidasporel derechodeautornosóloesundelito,sinoqueatentacontrala creatividady la difusión dela cultura.
Paramayorinformacióncomuníqueseconnosotros:
EL LIBRO MUERE CUANDO LO FOTOCOPIAAMIGO LECTOR:
La obra que usted tiene en sus manos posee un gran valor.En ella, su autor ha vertido conocimientos, experiencia y mucho trabajo. El editor ha procurado una presentación digna de su contenido y está poniendo todo su em-peño y recursos para que sea ampliamente difundida, a través de su red de comer-cialización.
Al fotocopiar este libro, el autor y el editor dejan de percibir lo que corresponde a la inversión que ha realizado y se desalienta la creación de nuevas obras. Rechace cualquier ejemplar “pirata” o fotocopia ilegal de este libro, pues de lo contrario esta-rá contribuyendo al lucro de quienes se aprovechan ilegítimamente del esfuerzo del autor y del editor.
La reproducción no autorizada de obras protegidas por el derecho de autor no sóloes un delito, sino que atenta contra la creatividad y la difusión de la cultura.
Para mayor información comuníquese con nosotros:

Dr. Juan Silva PereyraFacultad de Estudios Superiores Iztacala
Universidad Nacional Autónoma de México
Editor Responsable:Lic. Santiago Viveros FuentesEditorial El Manual Moderno
0205 MP47-30205 MP47-3

Nos interesa su opinión, comuníquese con nosotros:
Editorial El Manual Moderno, S.A. de C.V.,Av. Sonora núm. 206,Col. HipódromoDeleg. Cuauhtémoc06100 México, D.F.
(52-55)52-65-11-00
[email protected]@manualmoderno.com
Métodos en neurociencias cognoscitivas
D.R. © 2011 por Editorial El Manual Moderno,S.A. de C.V.ISBN: 978-607-448-100-6 978-607-448-163-1 Versión Electrónica
Miembro de la Cámara Nacionalde la Industria Editorial Mexicana, Reg. núm. 39
Todos los derechos reservados. Ninguna parte deesta publicación puede ser reproducida, almacenadaen sistema alguno o transmitida por otro medio —electrónico, mecánico, fotocopiador, etcétera— sin permiso previo por escrito de la Editorial.
Director editorial:Dr. Marco Antonio Tovar Sosa
Editora asociada:LCC Tania Uriza Gómez
Diseño de portada:LCS Adriana Durán Arce
Silva Pereyra, Juan Métodos en neurociencias cognoscitivas / Juan Silva Pereyra. -- -- México : Editorial El Manual Moderno, 2011. x i i , 128 p. i l . ; 23 cm. I nc luye b ib l i og ra f í a s e í nd i ce ISBN 978-607-448-100-6 978-607-448-163-1 Versión Electrónica 1. Neurociencias cognoscitivas. 2. Cognición. 3. Neuropsicología. I. t.
612.82-scdd21 Biblioteca Nacional de México

v
Colaboradores . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . viiAgradecimientos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . viiiIntroducción . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ix
Referencias . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xi
Capítulo 1. Métodos de registro electrofisiológico . . . . . . . . . . . . . 1Introducción . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1Métodos electrofisiológicos . . . . . . . . . . . . . . . . . . . . . . . . . . . 2Técnicas de registro . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .7Apéndice: electricidad y circuitos eléctricos básicos . . . . . . . . . . 19Referencias . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
Capítulo 2. El electroencefalograma: medición de la actividad eléctrica cerebral . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
Introducción . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26 Significado de la actividad registrada . . . . . . . . . . . . . . . . . . . 28 Las frecuencias observadas en el EEG . . . . . . . . . . . . . . . . . . . 30 Normalidad y anormalidad. . . . . . . . . . . . . . . . . . . . . . . . . . . 30 Desventajas del EEG convencional para la investigación . . . . . . . .31 Digitalización de la señal electroencefalográfica . . . . . . . . . . . . 32 Análisis espectral . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32 Medidas espectrales y neurometría . . . . . . . . . . . . . . . . . . . . . 34 Alternativas para eliminar el efecto de la referencia . . . . . . . . . . 36 Representación gráfica del EEG cuantitativo . . . . . . . . . . . . . . . 36 Futuro de la electroencefalografía. . . . . . . . . . . . . . . . . . . . . . .37 Conclusión . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38 Referencias . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
Capítulo 3. Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales . . . . . . . . . . . . . . . . . . . . . . . . 41
Introducción . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41 Comparación de los potenciales relacionados con eventos (PRE) con otras técnicas . . . . . . . . . . . . . . . . . . . . . . 42 Los PRE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44Clasificación . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45 Nomenclatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46 Bases fisiológicas de los PRE . . . . . . . . . . . . . . . . . . . . . . . . . 47 ¿Qué son los componentes de los PRE? . . . . . . . . . . . . . . . . . . 47 Breviario de componentes de los PRE . . . . . . . . . . . . . . . . . . . 49 Componentes endógenos . . . . . . . . . . . . . . . . . . . . . . . . . . . 50 Componentes de los PRE relacionados con la respuesta . . . . . . . 55
Contenido

vi
Supuestos básicos para hacer inferencias funcionalesa partir de los PRE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57 Aspectos conceptuales para interpretar datos de los PRE . . . . . . 60 Referencias . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
Capítulo 4. Potenciales relacionados con eventos: técnicas y métodos de registro . . . . . . . . . . . . . . . . . . . . . . . . . 68
Introducción . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68 Elementos para el registro de los PRE . . . . . . . . . . . . . . . . . . . 68 Artefactos y ruido. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 72 Edición del EEG . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .75 Promediación, corrección de línea base, filtrado y suavizado . . . . .77 Medición de la señal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81 Referencias . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84
Capítulo 5. Localización de fuentes de corriente de la actividad eléctrica cerebral: tomografía eléctrica cerebral. . . . . . . . . . . . . 85
Origen de la actividad electromagnética cerebral. . . . . . . . . . . . 85 Solución al problema inverso . . . . . . . . . . . . . . . . . . . . . . . . . 86 Modelos de la cabeza . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88 Posición de los electrodos . . . . . . . . . . . . . . . . . . . . . . . . . . . 90 Corregistro . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91 Señal electroencefalográfica adquirida . . . . . . . . . . . . . . . . . . 91 La Referencia. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 92 Artefactos e interpolación . . . . . . . . . . . . . . . . . . . . . . . . . . . 92 Selección del modelo inverso . . . . . . . . . . . . . . . . . . . . . . . . . 93 Estadística de imágenes . . . . . . . . . . . . . . . . . . . . . . . . . . . . 98 Conclusión . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 102 Referencias . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103
Capítulo 6. Métodos y teorías en neuropsicología:una perspectiva histórica y actual . . . . . . . . . . . . . . . . . . . . . . . 107
Definición y objeto de estudio . . . . . . . . . . . . . . . . . . . . . . . . 107 Aspectos históricos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .108 Métodos actuales en neuropsicología . . . . . . . . . . . . . . . . . . . 117 Nuevos tópicos de interés . . . . . . . . . . . . . . . . . . . . . . . . . . . 121 Conclusión . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 122 Referencias . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123

Colaboradores
Jaime A. Barral Proyecto Neurociencias UIICSE FES Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México, 54090, México. E-mail: [email protected] Capítulo 1Jorge Bernal Hernández Proyecto Neurociencias UIICSE FES Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México, 54090, México. E-mail: [email protected]ítulos 3, 4
Ismael Jiménez Estrada Departamento de Fisiología, Biofísica y Neurociencias, CINVESTAV IPN, México.Capítulo 1Juan Antonio Laville Instituto de Fisiología Celular, Universidad Nacional Autónoma de México, Ciudad Universitaria, México, DF.Capítulo 1Erzsébet Marosi Proyecto Neurociencias UIICSE FES Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México, 54090, México. E-mail: [email protected]ítulo 2Belén Prieto Corona Proyecto Neurociencias UIICSE FES Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México, 54090, México. E-mail: [email protected]ítulos 3, 4Pablo Razgado Laboratorio de Metodología Científica, FES Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México, 54090, México. Capítulo 1Mario Rodríguez Camacho Proyecto Neurociencias UIICSE FES Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México, 54090, México. E-mail: [email protected] Capítulos 3, 4
Helena Romero Romero Proyecto Neurociencias UIICSE FES Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México, 54090, México. E-mail:[email protected]ítulo 4Bertha Segura Alegría Laboratorio de Morfofisiología. FES Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México, 54090, México.Capítulo 1 Juan Silva-Pereyra Proyecto Neurociencias UIICSE FES Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México, 54090, México. E-mail: [email protected] Capítulo 5Guillermina Yáñez-Téllez Proyecto Neurociencias UIICSE FES Iztacala, Universidad Nacional Autónoma de México, Tlalnepantla, Estado de México, 54090, México. E-mail: [email protected] Capítulo 6
vii

viii
Agradecimientos
A VirgiliaJuan Silva Pereyra
A la memoria de mi madre y de mi abuela Mario Rodríguez Camacho
* Los autores agradecen a los programas PAPIIT (DGAPA UNAM) (IN204407: IN213310) y PASPA 2009-2010 (FES Iztacala, UNAM) por los recursos brindados para el desarrollo de las investigaciones en el capítulo 1, asimismo agradecen a los programas PAPIIT (DGAPA UNAM) IN303507 y CONACYT 59066, por el apoyo brindado en los capítulos 3, 4 y 5.

ix
Las neurociencias cognoscitivas son un conjunto de ciencias que se abocan al estudio del cerebro-mente. Ambiciosamente, se han propuesto estudiar procesos que involucran des-de sistemas moleculares y de fisiología del cerebro hasta la cognición y el funcionamien-to de la mente. Aunque pueden conjuntar información proveniente de distintas disciplinas científicas como bioquímica, fisiología, neurología, bioestadística, cibernética, psicología, lingüística, entre otras, no han podido alcanzar una explicación global de dichos fenómenos.
Tratar de entender o explicar el funcionamiento biológico de la mente, implica un complejo proceso interactivo entre el método y la teoría. Además, implica ir y venir por los diversos ni-veles —desde lo micro hasta lo macro—− que son objeto de estudio de estas disciplinas. Des-de hace mucho tiempo, la ciencia positiva se ha encauzado por el camino del superanálisis sin retorno. Para el estudio del acontecer cognoscitivo, se desmenuzan los hechos que van desde lo psíquico hasta lo biológico, sin establecerse puentes entre los diversos niveles. Con este proceder se han mostrado hallazgos sin precedente y se han formulado teorías muy ex-tensas y abarcativas. Sin embargo, aún no son suficientes para poder dar una visión completa del problema. También, con el avance de la tecnología, se hace indispensable una nueva for-ma de pensar acerca del método en las ciencias del comportamiento, la mente y su biología.
La complejidad desconcertante del cerebro-mente no nos deja más alternativa que mejorar nuestros métodos si pretendemos aportar una explicación integral. En las neurocien-cias cognoscitivas existe una diversidad de métodos que dependen de las disciplinas científicas aglomeradas en éstas. Cada método puede proporcionar información de donde se pueden obtener conclusiones en varios niveles. Esto también se debe a que los métodos en las neuro-ciencias tienen una resolución diferente a través de las dimensiones espacial y temporal. Por ejemplo, en el caso de la electrofisiología celular (capítulo 1), se puede tener un registro muy detallado de los sucesos en la dinámica celular ante la manipulación de neurotrasmisores. De esta manera, a partir de los datos obtenidos mediante estos métodos y técnicas para respon-der las preguntas científicas ―aunque tienen una resolución temporal y espacial exquisita―, no se podrá obtener una explicación del funcionamiento de los circuitos neuronales que participan en la memoria, o de los mecanismos neurobiológicos que nos hacen mover una mano.
Las preguntas experimentales y el marco conceptual, al usar estos métodos de registro, se mantienen en el nivel teórico de las vías neuroquímicas que incluyen diversas regiones en el sistema nervioso central y de los canales asociados a los distintos tipos de neurotransmi-sores. Se puede también extrapolar la información y permitirse integrar distintos hallazgos y englobar hasta un nivel de circuitos neurales más complejos. Por desgracia, con estos méto-dos no podemos ir más allá. Dadas sus características, no es posible trabajar en humanos, por lo que se deben diseñar experimentos con procesos que tengan ciertas similitudes con aque-llos que ocurren en los animales. En la figura I se muestra la adaptación de una gráfica que exponía en dos dimensiones (es decir, espacial y temporal) los distintos métodos de las neu-rociencias cognoscitivas (Gazzaniga et al., 1998). En esta nueva representación, agregamos una dimensión que implica diversos niveles de conocimiento científico, la cual toma en con-sideración los alcances de los resultados de esos métodos al construir explicaciones y teorías.
En este caso, los métodos electrofisiológicos descritos en el capítulo 1 tienen alcances muy limitados en esta escala, ya que sólo pueden dar respuesta a preguntas relacionadas con la fisiología celular, la neurofisiología de diversas vías cerebrales y algunos mecanismos
Introducción

PRE y MEGEstimulación
magnética
transcranealRMf
PET
Registro unitario
Patch clamp
Microlesiones
Lesiones
Mente
Procesos
dominio
general
Procesos
dominio
específicoFisiología
celular
Biología
molecular
Cerebro
Mapa
Columna
Capa
Neurona
Dendrita
Sinapsis
Log espacioLog tiempo
7
23 4
56
10-1-2-3
3
2
1
0
-1
-2
-3
-4
Métodos en Neurociencias Cognoscitivas
x
Figura I. Métodos en las neurociencias cognoscitivas en tres dimensiones: a) en el tiempo a escala logarítmica, b) en el espacio a escala logarítmica y c) en los niveles del conocimiento.
fisiológicos de procesos básicos. Podemos saltar de los registros celulares hasta los registros de la actividad electromagnética obtenidos en la superficie de la piel cabelluda (capítulos 2, 3 y 4). Dichos métodos permiten al investigador estudiar en humanos la actividad neuronal ante tareas mentales definidas con precisión. Una de sus características fundamentales es que representan, al menos en apariencia, información directa de los procesos men-tales proveniente de la actividad neuronal. Como se ve en la figura I, hay una excelente re-solución temporal, muy cercana a la resolución que se obtiene con los métodos patch clamp o de registro intracelular, aunque la resolución espacial es extraordinariamente pobre. Asi-mismo, en la dimensión “nivel de explicación teórico”, estos métodos podrían arrojar luz acerca de los mecanismos de procesamiento de dominio específico y de dominio general. El procesamiento de dominio específico se refiere a aquellos modelos de carácter cognos-citivo, sin mucho conocimiento de su correlato neuroanatómico, pero cuyo procesamiento es de información específica. Por ejemplo, cuando hablamos del lenguaje o de la percepción de caras, se da un tipo muy específico de procesamiento cuyos componentes no intervie-nen en otro tipo de procesamiento y podríamos decir que son exclusivos. Entonces se pue-de hablar de cierta clase de encapsulamiento de la información en este tipo de dominio. En contraste, el procesamiento de dominio general involucra sistemas que engloban a los de dominio específico, como la memoria o la atención.

xi
Lo interesante de los métodos electrofisiológicos es que la misma información puede ser analizada de dos formas diferentes: en el dominio del tiempo (capítulos 3 y 4) y en el dominio de la frecuencia (capítulo 2). En los potenciales relacionados con eventos (PRE), cuya información es analizada en el dominio del tiempo, podemos ver con mayor claridad las bondades de la alta resolución temporal a costa de la espacial, lo que impide bajar a un nivel de explicación distinto del de los mecanismos de dominio específico. En el análi-sis de los mecanismos cognoscitivos dados, por ejemplo, durante la comprensión de ora-ciones, los PRE pueden proporcionar mucha información acerca del efecto de nuestras variables respecto del tiempo. Sin embargo, la extracción puntual de la acción de diver-sos circuitos neurales por medio de este método sigue siendo un misterio. Sabemos que los PRE son el resultado de la suma de los potenciales postsinápticos distribuidos a través del volumen conductor, el cerebro. Cuando se analizan las ondas de los PRE, se puede inferir de antemano que probablemente en una de ellas se encontrará la suma e interacción de múltiples regiones del cerebro que actúan en el mismo momento. Muchas veces la sepa-ración temporal de diversos procesadores cognoscitivo-cerebrales es imposible. Como vemos, las explicaciones y teorías que se pueden desprender con el uso de estos métodos se quedan en un nivel tan general que no se puede acceder a los niveles explicativos de la neurofisiología ni de los circuitos neurales.
Por otro lado, la señal electroencefalográfica se puede analizar en términos de la cantidad de energía por frecuencia sacrificando la información de fase. Con el uso de estos métodos se puede ilustrar bastante bien el funcionamiento, y el mal funcionamiento, del sistema en su conjunto. Por ejemplo, cuando se quiere determinar un perfil electroencefalográfico atí-pico relacionado con una enfermedad, o de un grupo de sujetos que realiza una tarea men-tal en particular, se emplea un procedimiento básico estadístico que consiste en determinar diferencias entre la norma y las observaciones. Una forma de hacer estas comparaciones es descomponer los patrones electroencefalográficos en el dominio de la frecuencia. Esto significa transformar los valores de amplitud o voltaje respecto del tiempo que se obtienen del registro electroencefalográfico en sus correspondientes valores de frecuencia, de ma-nera que podamos tener la cantidad de energía para cada banda de frecuencia; con ello, se tienen disponibles patrones “normales” del EEG, cuantitativamente hablando, en esta-do de reposo. La aplicación del criterio se da al compararse los valores de un sujeto con los valores de frecuencia normales en una situación específica. Quizá lo más usual es hacer comparaciones entre los valores de sujetos con alguna enfermedad (p. ej., esquizofrenia, depresión, trastorno bipolar) y los perfiles normales. Así, el criterio estadístico es crucial en la investigación clínica y en la investigación básica.
Con el propósito de mejorar la resolución espacial de los métodos de registro de la acti-vi-dad eléctrica en la superficie de la piel cabelluda, se han desarrollado técnicas matemáti-cas para ubicar la fuente de corriente de la señal magneto o electroencefalográfica (capítulo 5) en la corteza cerebral. Sin embargo, una desventaja básica de estas técnicas es el pro-blema de indeterminación del origen de la fuente de corriente. La localización de las fuen-tes de corriente —o, como también se le conoce, la tomografía electromagnética cerebral (TEC)— puede dar solución única a este problema, aunque en realidad existe una infinidad de ellas que hacen de éste un problema matemáticamente mal planteado. La TEC se basa en la aplicación de una serie de restricciones matemáticas a un modelo espacial de flujo de la corriente en un volumen conductor a partir de la información recogida de la superficie del volumen. Se puede estudiar así una gran variedad de modelos cognoscitivos que antes se analizaban con el registro magnético o eléctrico del cerebro, pero con un correlato directo
Introducción

Métodos en Neurociencias Cognoscitivas
xii
de las regiones cerebrales involucradas o, incluso, de los circuitos neurales en acción. Sin embargo, existe una gran cautela por parte de la mayoría de los investigadores en esta área con respecto a los resultados obtenidos con la TEC en estudios de psicología cognoscitiva, debido a que las fuentes de corriente se localizan, en términos prácticos, como producto de una modelación hecha con métodos de maximización, pero no por evidencias empíri-cas como otros métodos de neuroimagen o como en los resultados de la neuropsicología.
Quizás el método más tradicional dentro del estudio cerebro-mente es la aproximación neuropsicológica (capítulo 6). Aunque desde este método se puede establecer una relación directa entre la conducta y la participación de una estructura cerebral, la variable indepen-diente (p. ej., la ubicación de una lesión cerebral) no puede ser manipulada por el investiga-dor. Las lesiones asociadas con las fallas y deficiencias en el comportamiento son accidentes cuya extensión, ubicación y grado de daño son impredecibles y escapan al control experi-mental. Lo anterior hace verdaderamente complicado el uso de este método, a menos que se trate de un individuo con una lesión muy focalizada y que no pueda ejecutar una tarea muy particular que implique un solo componente cognoscitivo. Aun así, lesiones muy focalizadas, pero que se encuentren entre sustancia gris y blanca, por ejemplo, tendrán repercusiones muy diferentes en términos de conducta. De esta manera, los estudios con una aproxima-ción neuropsicológica pueden decirnos si un área es esencial para alguna función, pero no nos dicen mucho acerca de cómo son los circuitos neuronales que están involucrados en la realización de una tarea en particular. Como podemos darnos cuenta, aunque tenemos una visión bastante global con este método, se pierden los detalles.
Como hemos descrito a lo largo de esta introducción, los diferentes métodos en las neurociencias permiten obtener información a diferentes niveles de lo que se conoce como cerebro-mente, desde la exquisitez del detalle fisiológico con los registros unita-rios hasta lo más global como los procedimientos empleados en la neuropsicología. Es de destacar que, mientras se incrementa la resolución espacial, con facilidad puede perder-se la temporal, pero, independientemente de tal relación, el alcance en el conocimien-to —considerado en la dimensión agregada al gráfico de los métodos— también puede quedar encapsulado a un solo nivel. Es decir, se puede identificar un método que pueda tener una buena resolución hasta en dos dimensiones, pero no más. De esta forma, los estudios que se plantean en las neurociencias no deberían supeditarse a un método en particular, sino más bien a la integración de varios métodos con el fin de tener un pano-rama global, pero sin perder los detalles de la fisiología o la biología molecular. Esto no sólo nos permitiría generar teorías explicativas que vayan más allá de atajar un solo nivel de conocimiento, sino también crear puentes entre los niveles de conocimiento.
En este contexto, esta obra pretende transmitir al lector especialista, al investigador o al estudiante de psicología y al curioso no especialista información pertinente, actual y acce-sible sobre los métodos en las neurociencias cognoscitivas. Un aspecto sobresaliente de esta obra es que se revisa y expone el estado actual del arte de los métodos en las neurociencias cognoscitivas en una selección de las principales direcciones en las que actualmente se trabaja en la investigación de esta vasta área de la ciencia. Lo que se ofrece al lector en esta obra es el resultado del esfuerzo conjunto de un grupo de investigadores mexicanos desta-cados cuyo trabajo se encuentra directamente relacionado con los métodos aquí expuestos.
ReferenciasGazzaniga, M., Ivry, R. y Mangun, G. (1998). Cognitive neuroscience: The biology of mind. Nueva
York: W.W. Norton y Company.

1
Capítulo 1. Métodos de registro electrofisiológico
Jaime Aurelio Barral Caballero, Juan Antonio Laville,Pablo Razgado, Ismael Jiménez Estrada y Bertha Segura Alegría
Resumen En este capítulo se describirán brevemente algunas de las técnicas de registro electrofisiológico más utilizadas en las neurociencias y se hace hincapié en el registro ex-tracelular, ya que la mayor parte de los registros electrofisiológicos utilizados en di-versos estudios clínicos se basan en las técnicas de registro extracelular, tal es el caso del electroencefalograma (EEG), el electromiograma (EMG) y el electrocardiograma (ECG). También se describirán las técnicas de registro intracelular y de parches de mem-brana que permiten realizar experimentos para propósitos específicos, como la transmi-sión sináptica, la medición de potenciales de acción compuestos, el registro en rebanadas, el EMG y el ECG. Al final del capítulo, se incluye un apéndice donde se describen algu-nos conceptos importantes sobre electricidad y electrónica relacionados con el registro electrofisiológico.
Introducción Las células realizan un sinnúmero de intercambios de sustancias e iones entre el
interior y el exterior de la membrana citoplasmática. Establecen también múltiples formas de comunicación inter- e intracelular por medio de receptores colocados en diferentes puntos de la membrana citoplásmica, además de incontables cadenas de señalización intracelular, que en última instancia producen la apertura o el cierre de ca-nales iónicos en la superficie celular. Este flujo de iones a través de la membrana genera corrientes iónicas que son susceptibles de ser medidas con diversas técnicas y disposi-tivos experimentales. Aunque es posible registrar la actividad de las células excitables con diferentes métodos que recurren a trazadores radioactivos o térmicos, lo usual es que la actividad de las células excitables se estudie con métodos electrofisiológicos, in-cluso la medición de los eventos eléctricos rápidos que ocurren a través de la membrana (Keynes y Aidley, 2001).
Las técnicas electrofisiológicas más utilizadas incluyen las de registro extra e in-tracelular, así como los registros en microáreas de membrana en sus diferentes configuraciones. Las técnicas extracelulares por lo general miden los cambios en la densidad de corriente. En el laboratorio, las técnicas extracelulares más utilizadas son el registro unitario, el registro de potenciales de campo y la actividad muscular. En la clínica, así como en la investigación con seres humanos, las técnicas más comunes son el electroencefalograma (EEG), el electrocardiograma (ECG) y el electromiograma (EMG).
Las técnicas de registro intracelular con fijación de voltaje permiten la medición del flujo de corriente a través de la membrana. Los registros con los protocolos de fijación de corriente, también permiten la medición de voltajes que se generan a través de la membrana por la apertura y cierre de canales iónicos y del potencial intracelular de las

Métodos en Neurociencias Cognoscitivas
2
células excitables. Las técnicas de registro electrofisiológico se empezaron a utilizar desde el siglo xviii como se muestra en los trabajos de Luigi Galvani, Alexandro Vol-ta y Alexander Von Humbolt (Kettenman, 1997; Piccolino, 1997, 1998). En el siglo xix, este nuevo campo del conocimiento fue impulsado por los trabajos de Matteucci, Von Helmholtz y Du Bois-Reymond (Del Castillo, 1997), quienes registraron con galvanó-metros algunos eventos electrofisiológicos. Sin embargo, no fue sino hasta el siglo xx, con el desarrollo tecnológico de la década de 1920, cuando aparecieron el oscilosco-pio de rayos catódicos y el amplificador de bulbos, que se pudieron realizar medicio-nes precisas de eventos rápidos (Aidley, 1998). De 1912 a 1926, destacan los trabajos de Adrian, Zotterman, Forbes, Davis, Passer, Lucas, Williams Newcomer y Erlanger. Quizá la innovación que dio origen a las técnicas de registro como las conocemos en la actualidad fue el desarrollo del microelectrodo de registro intracelular, primero utiliza-do en el axón gigante del calamar y después en casi todo tipo de células. No obstante, la técnica de microelectrodos no fue funcional sino hasta finales de la década de 1940 y principios de la de 1950 con los trabajos de Graham, Gerard, Katz, Ling, Hodgkin y Nastuk (Frank, 1986).
Entre 1939 y 1942, con el desarrollo de los microelectrodos de vidrio, los grupos de Cole y Curtis, así como el de Hodking y Huxley, de manera independiente, obtuvieron registros intracelulares utilizando invertebrados como el calamar debido al gran tamaño de sus cé-lulas. Desde la década de 1950 y hasta la fecha, la técnica de registro electrofisiológico se ha logrado gracias al desarrollo tecnológico de amplificadores, estiradores de pipetas, microscopios y, recientemente, al de equipos de cómputo para la obtención, desarrollo y análisis experimentales. Esto ha permitido realizar registros en células cada vez más pe-queñas, incluso en axones y dendritas de algunas de ellas.
Si bien el registro intracelular con microelectrodos permite registrar la actividad eléctrica de células excitables, en particular de neuronas, tiene el defecto de producir una corriente de fuga, o deriva eléctrica, que impide medir corrientes transmembra-nales aisladas como lo habían hecho Hodgkin y Huxley (1952). Fue hasta principios de la década de 1980, cuando Neher y Sakmann (Hamill et al., 1981) desarrollaron una técnica innovadora que incluye el uso de micropipetas no penetrantes, a diferencia de los experimentos con microelectrodos. En ellos, la corriente de fuga disminuye con el tiempo hasta llegar a un valor constante que genera una resistencia óhmica del orden de los gigaohms (GΩ). En este caso, la corriente de fuga generada era varios órdenes de magnitud menor que la del electrodo penetrante. Se obtenía un parche (patch en in-glés), al que se le podía fijar (clamp en inglés) ya sea la corriente o el voltaje. Por esta ra-zón, el nombre de esta técnica es parches de membrana, o simplemente patch clamp. Para ello, Neher y Sakmann desarrollaron una pipeta de vidrio chata que no penetra la célula y que, mediante una suave succión a través de la pipeta de vidrio, puede tomar un parche de la membrana, es decir, una parte de la misma, con el fin de registrar sólo los canales iónicos que en tal parche hayan quedado y poder fijar de manera eficiente en ambos lados del parche el voltaje o la corriente (Hamill et al., 1981). Esto se debe a que los lípidos de la membrana tienen una cierta afinidad con el vidrio de las pipetas; enton-ces, la propia membrana es capaz de sellar en parte el agujero generado por el electrodo.
Métodos electrofisiológicos Las señales eléctricas son fundamentales para la función del sistema nervioso, por lo que es importante determinar las propiedades eléctricas que se propagan a lo largo de

3
Métodos de registro electrofisiológico
las células excitables. Sin embargo, a pesar de la gran diversidad de técnicas de regis-tro y protocolos experimentales, existen algunos elementos comunes en todas ellas.
Electrodos de registro Todas las técnicas electrofisiológicas implican el uso de electrodos que permiten regis-trar la diferencia de voltaje entre dos puntos por medio de un conductor de volumen. Para ello, se utilizan electrodos conectados a amplificadores adecuados a la técnica de re-gistro; uno o varios electrodos funjen como el o los electrodos de registro y su actividad se compara con un electrodo de referencia, de “tierra” o “indiferente”. Los electrodos de referencia no deben ser afectados por el paso de pequeñas corrientes a través de ellos, por lo cual son colocados lo suficientemente lejos de la actividad eléctrica de tal modo que su potencial no cambie significativamente con el tiempo. El potencial en este punto puede ser medido y se define para fines prácticos como “potencial cero”.
Los electrodos más utilizados son los de platino o tungsteno, si sólo se usan para registrar potenciales de acción. Tienen el inconveniente de polarizarse por el paso de corriente eléctrica desde o hacia la solución salina con la cual están en contacto; para evitar esto, se utilizan electrodos de plata con un recubrimiento de cloruro de plata. Se considera que los electrodos que permiten las mediciones más agudas son los electro-dos que contienen calomel (mercurio/cloruro de mercurio) (Keynes y Aidley, 2001). Los electrodos de registro se conectan a las áreas de interés, o bien, al interior de las células por medio de pipetas de vidrio estiradas por calentamiento, y que contienen en su inte-rior soluciones salinas que permiten la conducción de corrientes iónicas.
La forma y el tamaño de los electrodos dependen en gran medida del tipo de re-gistro que se va a realizar. Por ejemplo, los electrodos extracelulares para el registro electroencefalográfico son, por lo general, pequeñas cazoletas de plata de aproximada-mente 10 mm de diámetro que se colocan sobre la piel cabelluda. En cambio, los elec-trodos destinados al registro en rebanadas de cerebro son pequeñas pastillas de cloru-ro de plata comprimida con adaptaciones para la inserción de electrodos de vidrio que entran en contacto con el tejido.
Los electrodos para registro en fijación de voltaje o de corriente son adaptadores de material no conductor con alambres de plata muy delgados que conectan la punta del electrodo de vidrio con los sistemas de amplificación y registro. Los electrodos de vidrio que se utilizan para los registros intra- y extracelular difieren en el diámetro de la boca del electrodo y en la resistencia eléctrica de la punta del mismo. Los electrodos destinados para registro intracelular poseen una punta muy fina (menos de 0.5 μm de diámetro, 20-150 MΩ de resistencia en la punta). Aquí el propósito consiste en penetrar en el inte-rior de la célula con la punta del electrodo sin lesionar la superficie de la membrana ce-lular. Aquellos electrodos que se usan en el registro extracelular en rebanadas tienen la punta “rota” o tienen un gran diámetro (100-200 μm de diámetro, 1-3 MΩ de resistencia en la punta). En este caso el propósito no es penetrar en el interior de una célula, sino obtener un registro de una población de células (por lo común cientos o miles) que se encuentren cerca de la boca del electrodo.
Los electrodos para patch clamp son, por lo general, de puntas redondeadas por calor y de baja resistencia (1-10 μm de diámetro, 2-10 MΩ de resistencia en la punta), aunque no tanta como los usados en el registro extracelular. En este caso, la punta del electrodo se posa sobre la superficie celular, de modo que se puede obtener un registro de la pequeña área de membrana que abarca el electrodo.

Métodos en Neurociencias Cognoscitivas
4
Amplificación electrónica Las células excitables generan diferencias de potencial que pueden ser medidas con auxi-lio de amplificadores diseñados para cada tipo de registro. El uso de sistemas de amplificación es necesario, ya que las señales biológicas son, por lo general, muy pe-queñas. Entonces, la medición electrofisiológica implica la amplificación de señales rá-pidas, pequeñas y que, por lo general, van acompañadas de “ruido” (tanto electrónico como biológico). Es necesario que las mediciones sean precisas, bajas en ruido, estables y confiable (Aidley, 1998).
En general la investigación electrofisiológica realiza mediciones de los cambios de voltaje o del flujo de la corriente respecto del tiempo, ya sea en registro unitario, en po-tenciales de campo, en el cerebro completo o en rebanadas del mismo. El ECG, EMG, EEG y, en general, todas las técnicas comunes en la investigación clínica requieren de la medición de señales de baja amplitud. Los amplificadores para registro extracelular permiten registrar diferencias de voltaje muy pequeñas (20 a 100 μV). Sin embargo, esto conlleva un incremento en el “ruido” electrónico, que son fluctuaciones de vol-taje al azar debidas a diversos factores no relacionadas con la actividad biológica. Para atenuar el ruido electrónico en la medida de lo posible, es necesario contar con amplificadores de bajo nivel de ruido, ya que de otra forma se dificultaría la medición de señales pequeñas. También hay que tomar medidas para que el ruido de la línea de alimentación y del resto del equipo electrónico no se incremente. En algunos disposi-tivos de registro es necesario filtrar las señales recibidas, ya que los electrodos pueden hacer las veces de una antena del ruido electrónico proveniente de equipos cercanos si no está debidamente aterrizada.
Registro electrofisiológico Cuando se usan electrodos de registro en un tejido y se manda algún tipo de estimu-lación con la finalidad de observar la respuesta de las células excitables, se obtiene la medición de una diferencia de voltaje entre el electrodo de registro y el de referencia. Esto se comporta entonces como un potencial registrado en un volumen conductor, cuya propagación puede ser diferente de lo predicho por la teoría de cable (Brinley, 1980; Woodbury, 1965). El registro de un potencial en un punto del volumen conductor respecto de un electrodo de referencia puede ser difícil de interpretar, ya que es compli-cado determinar la localización de las fibras activas debido a que las corrientes se des-pliegan a lo largo de todo el volumen conductor. Además, el tamaño y el curso temporal de la descarga son inciertos, dado que la distancia entre el electrodo de registro y el tejido activo cambia; entonces el potencial también es cambiante respecto de su amplitud y velocidad de propagación.
Las células excitables están rodeadas por fluido intersticial, el cual se convierte en un volumen conductor que rodea a todo el tejido de interés. Esto implica que toda la activi-dad que se genere en ese tejido producirá cambios en el voltaje medido entre los electro-dos de registro y referencia. De no haber flujo de corriente en el tejido de interés, el poten-cial medido entre el electrodo de registro y el de referencia deberá ser isopotencial. Por lo anterior, se establece una relación entre el electrodo de registro y las regiones de las célu-las excitables donde se está generando la actividad, así como con el electrodo de referen-cia. En términos generales, prácticamente todos los tipos de registro electrofisiológico se pueden considerar como registros en un volumen conductor, ya que para fines prácticos el electrodo de referencia está alejado de la zona de actividad bioeléctrica.

5
Métodos de registro electrofisiológico
Cuando las células excitables están en reposo, sus membranas no producen ningún flujo de corriente. Sin embargo, cuando la neurona está activa, se polariza de una mane-ra no uniforme, es decir, que en algún momento el potencial de membrana registrado por el soma puede ser diferente del que se observa en el axón o en las dendritas. Como el po-tencial de acción se propaga a lo largo del axón, ocurren cambios dinámicos en los valores del potencial de membrana en las diferentes regiones de la célula. Entonces, se produ-cen flujos de corriente de una parte de la neurona a otra, que forman un dipolo a tra-vés del fluido extracelular, lo que establece un campo eléctrico (Woodbury, 1965; Brin-ley, 1980; Johnston y Wu, 1995). Supongamos que se colocan los electrodos de estimulación, de registro y de referencia en una cámara de perfusión donde tenemos una rebanada de tejido nervioso y estamos haciendo circular solución salina (figura 1-1). En el momento que el electrodo de regis-tro y el de referencia entran en contacto con la solución salina, se establece un circuito eléctrico, y la diferencia de potencial entre los dos electrodos debe ser cero, ya que en reposo la carga se encuentra uniformemente distribuida.
Supongamos que los potenciales de acción ocupan un periodo finito de tiempo y se propagan a lo largo de las fibras nerviosas con una velocidad también finita, por lo que en un momento dado un potencial de acción podría observarse en cualquier punto a lo largo de la fibra. Esto implica que la variación de un potencial temporal en un punto fijo es equivalente a la variación espacial en un tiempo fijo. En la prác-tica se establece un punto fijo al colocar el electrodo de registro (posición B en la figura 1-1) y medir el curso temporal de los potenciales que se generan. Conceptual-mente, el potencial que se registra en un volumen conductor es a menudo visto como el potencial que pasa o que es “visto por el electrodo”. En general, es más simple considerar al potencial como “congelado en el tiempo” y observar, en diferentes puntos de la fibra, el tránsito de este potencial en diferentes momentos a lo largo del periodo de tiempo.
La membrana establece una diferencia activa en la distribución de cargas entre el interior y el exterior de la célula. El interior de las células es más negativo que el exterior. Sin embargo, cuando los electrodos se encuentran sumergidos en el tejido nervioso y se manda un estímulo de baja intensidad, el cambio de voltaje permite que los canales de sodio sensibles al voltaje se abran y generen una corriente entrante de cargas posi-tivas, por lo que, al menos en ese punto, el interior de las células es positivo respecto del exterior (la polaridad del registro se invierte). El cambio de voltaje también permite que canales de potasio rectificadores lentos se activen con el voltaje y ocurra una salida de cationes, por lo que el interior de las células tiende a repolarizarse. Posteriormente, transcurre un periodo refractario en el que las células no son capaces de responder a la estimulación. El registro extracelular es capaz de detectar estos cambios en el poten-cial; no obstante, la diferencia de potencial medida por los electrodos cambia respecto del tiempo y respecto de la posición relativa en relación con las células que presentan la actividad eléctrica. Este tipo de registro extracelular recibe el nombre de registro po-blacional o potencial de campo.
La membrana establece una diferencia activa en la distribución de cargas entre el interior y el exterior de la célula. El interior de las células es más negativo que el exterior. Sin embargo, cuando los electrodos se encuentran sumergidos en el tejido nervioso y se manda un estímulo de baja intensidad, el cambio de voltaje permite que los canales de sodio sensibles al voltaje se abran y generen una corriente entrante de cargas positivas,

Métodos en Neurociencias Cognoscitivas
6
por lo que, al menos en ese punto, el interior de las células es positivo respecto del ex-terior (la polaridad del registro se invierte). El cambio de voltaje también permite que canales de potasio rectificadores lentos se activen con el voltaje y ocurra una salida de cationes, por lo que el interior de las células tiende a repolarizarse. Posteriormente, transcurre un periodo refractario en el que las células no son capaces de responder a la estimulación. El registro extracelular es capaz de detectar estos cambios en el poten-cial; no obstante, la diferencia de potencial medida por los electrodos cambia respecto del tiempo y respecto de la posición relativa en relación con las células que presentan la actividad eléctrica. Este tipo de registro extracelular recibe el nombre de registro po-blacional o potencial de campo.
Figura 1-1. Curso temporal del registro extracelular en corteza cerebral. En (A), cuando se manda un es-tímulo a la fibra nerviosa, se produce una corriente entrante en esa porción de la célula que se observa como una zona donde las cargas positivas entran a la célula. Entonces, esa parte actúa en ese mo-mento como un pozo o sumidero de corriente, por lo que se observa una negatividad en el registro. Cuando la señal propagada se acerca a (B), el pozo está sobre el electrodo, por lo que el déficit de cargas positivas (que se internalizaron) produce un cambio en el registro que se observa como una negatividad. (C) El estímulo continúa propagándo-se a lo largo de la fibra, por lo que se produce una segunda positividad en el registro.
El potencial de campo registrado extracelularmente es una onda trifásica producida por la diferencia de potencial debida al cambio en la localización de los sumideros y las fuentes de corriente activa. A la zona donde las corrientes fluyen hacia el interior de las células se le denomina sumidero o pozo (corrientes entrantes activas), mientras que a la zona donde salen se le llama fuente (corrientes salientes que cierran el circuito; figura 1-2). En el registro extracelular, las corrientes salientes se observan como deflexiones positivas y las corrientes entrantes, como deflexiones negativas.
Por lo general, la medición con el electrodo se realiza al comparar los voltajes en-tre el electrodo de registro y la tierra de referencia. A la neurona se le puede considerar como un dipolo eléctrico, ya que durante el pico del potencial de acción en la membra-na del soma se da lugar a una corriente activa (sumidero), mientras que en las dendritas se da una corriente pasiva (fuente) que cierra el circuito; ambas son positivas respec-to del soma. Las aferentes sinápticas dan lugar a cambios en estas corrientes. En ese

Sumidero
Sumidero
Fuente
Fuente
Axón Soma Dendritas
B
A
7
Métodos de registro electrofisiológico
sentido las entradas sinápticas excitatorias que dan lugar a una corriente entrante en las dendritas (sumidero) producen en el soma corrientes salientes pasivas (fuente). De-bido a que estas corrientes cambian con el tiempo, es común obtener ondas trifásicas, que representan la propagación del potencial de acción (figura 1-2).
Cuando se registra cerca de los somas neuronales, se observa solamente una onda bifásica que consiste en una espiga negativa seguida de otra positiva. Esto se debe a que el potencial de acción produce una rápida despolarización de la membrana por las co-rrientes entrantes de sodio, así como una rápida repolarización por las corrientes salien-tes de potasio. El componente negativo corresponde a la despolarización de la membra-na del soma por el flujo de corriente entrante. La fase positiva corresponde a la reversión en el flujo de corriente.
Figura 1-2. Fuentes y sumideros. Se pre-senta de manera esquemática la apa-rición de fuentes y sumideros en una célula cualquiera cuando, B) después de generarse un potencial en el soma, la se-ñal llega a las terminales axónicas, o A) después de generarse en el axón y diri-girse al soma.
Técnicas de registro Registro extracelular La técnica de registro extracelular es la precursora de las técnicas de registro electrofisiológico utilizadas hoy día. El registro extracelular mide los cambios de voltaje en el espacio extracelular que rodea a neuronas o axones detectados por microelectrodos de vidrio de baja resistencia. Al introducir el electrodo al tejido, se forma un sello de baja resistencia entre la punta del electrodo y las células que lo rodean, el cual permite detec-tar pequeñas variaciones de voltaje entre el electrodo de registro y las membranas celula-res. En el registro extracelular, el electrodo no rompe ni lesiona las membranas celulares, ni las células son penetradas por los electrodos, de tal modo que el contenido celular no se ve perturbado. La principal ventaja del registro extracelular radica en que es la menos in-vasiva de las técnicas de registro electrofisiológico, lo que permite registros repetidos de la misma célula, población celular o tejido, sin tener que empalar y, en consecuecia, dañar las células registradas. Además, hay muchas situaciones experimentales en las que no es posible introducir microelectrodos en las células, por lo que no es practicable el registro intracelular, o bien, en el curso de la investigación es deseable no lesionar de modo alguno las poblaciones celulares de interés. En esas condiciones, es preferible iniciar las investi-gaciones empleando registros extracelulares. Lo anterior también es válido para realizar
Amplitud (µV)
Intervalo interespiga (ms)
N1
S1 S2
N2
Métodos en Neurociencias Cognoscitivas
8
pruebas farmacológicas con algunos reactivos, pues los datos que arroja el registro extra-celular provienen de cientos o miles de neuronas que disparan simultáneamente, lo que, además, proporciona validez estadística a ese tipo de registros. A nivel celular, la técnica más utilizada es el registro de potenciales de campo, útil en especial para el estudio de la transmisión sináptica. El registro extracelular es también valioso como una técnica de re-gistro no invasiva; es de particular importancia en el diagnóstico clínico, como es el caso del electroencefalograma (capítulo 2), el electrocardiograma y el electromiograma, ya que éstos se rigen bajo los mismos principios electrofisiológicos.
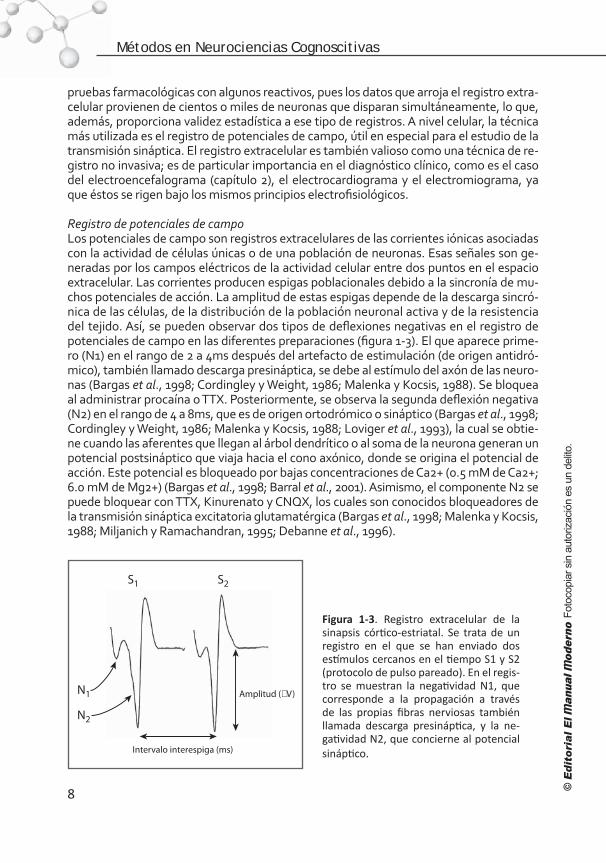
Registro de potenciales de campo Los potenciales de campo son registros extracelulares de las corrientes iónicas asociadas con la actividad de células únicas o de una población de neuronas. Esas señales son ge-neradas por los campos eléctricos de la actividad celular entre dos puntos en el espacio extracelular. Las corrientes producen espigas poblacionales debido a la sincronía de mu-chos potenciales de acción. La amplitud de estas espigas depende de la descarga sincró-nica de las células, de la distribución de la población neuronal activa y de la resistencia del tejido. Así, se pueden observar dos tipos de deflexiones negativas en el registro de potenciales de campo en las diferentes preparaciones (figura 1-3). El que aparece prime-ro (N1) en el rango de 2 a 4ms después del artefacto de estimulación (de origen antidró-mico), también llamado descarga presináptica, se debe al estímulo del axón de las neuro-nas (Bargas et al., 1998; Cordingley y Weight, 1986; Malenka y Kocsis, 1988). Se bloquea al administrar procaína o TTX. Posteriormente, se observa la segunda deflexión negativa (N2) en el rango de 4 a 8ms, que es de origen ortodrómico o sináptico (Bargas et al., 1998; Cordingley y Weight, 1986; Malenka y Kocsis, 1988; Loviger et al., 1993), la cual se obtie-ne cuando las aferentes que llegan al árbol dendrítico o al soma de la neurona generan un potencial postsináptico que viaja hacia el cono axónico, donde se origina el potencial de acción. Este potencial es bloqueado por bajas concentraciones de Ca2+ (0.5 mM de Ca2+; 6.0 mM de Mg2+) (Bargas et al., 1998; Barral et al., 2001). Asimismo, el componente N2 se puede bloquear con TTX, Kinurenato y CNQX, los cuales son conocidos bloqueadores de la transmisión sináptica excitatoria glutamatérgica (Bargas et al., 1998; Malenka y Kocsis, 1988; Miljanich y Ramachandran, 1995; Debanne et al., 1996).
Figura 1-3. Registro extracelular de la sinapsis córtico-estriatal. Se trata de un registro en el que se han enviado dos estímulos cercanos en el tiempo S1 y S2 (protocolo de pulso pareado). En el regis-tro se muestran la negatividad N1, que corresponde a la propagación a través de las propias fibras nerviosas también llamada descarga presináptica, y la ne-gatividad N2, que concierne al potencial sináptico.
-30
-40
-50
-60
-70
-80
-90
-100
-110
-120
-0.8 -0.6 -0.4 -0.2 0.2 0.4 0.60.0
Corriente (nA)
Volta
je (m
V)50 ms
20 m
V
-80 mV
nA0.60.6
-0.6
9
Métodos de registro electrofisiológico
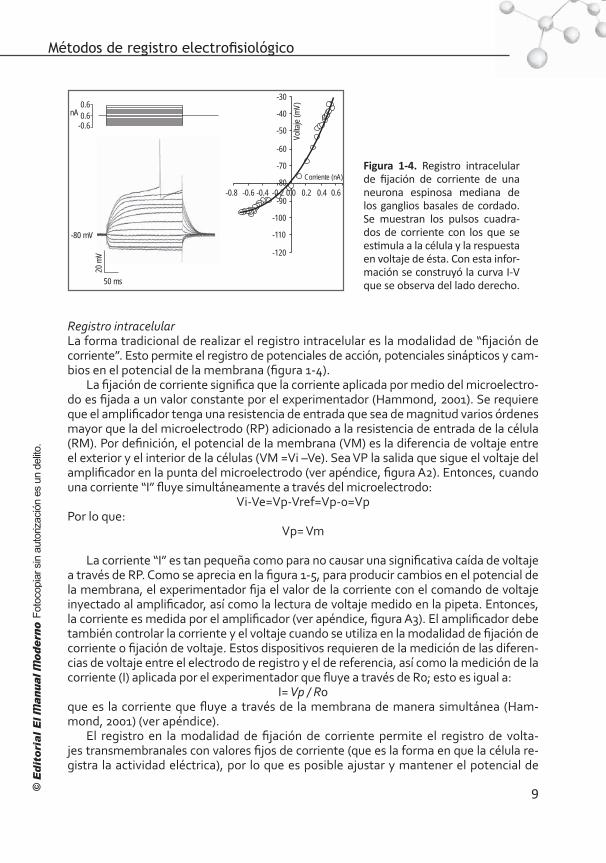
Figura 1-4. Registro intracelular de fijación de corriente de una neurona espinosa mediana de los ganglios basales de cordado. Se muestran los pulsos cuadra-dos de corriente con los que se estimula a la célula y la respuesta en voltaje de ésta. Con esta infor-mación se construyó la curva I-V que se observa del lado derecho.
Registro intracelular La forma tradicional de realizar el registro intracelular es la modalidad de “fijación de corriente”. Esto permite el registro de potenciales de acción, potenciales sinápticos y cam-bios en el potencial de la membrana (figura 1-4).
La fijación de corriente significa que la corriente aplicada por medio del microelectro-do es fijada a un valor constante por el experimentador (Hammond, 2001). Se requiere que el amplificador tenga una resistencia de entrada que sea de magnitud varios órdenes mayor que la del microelectrodo (RP) adicionado a la resistencia de entrada de la célula (RM). Por definición, el potencial de la membrana (VM) es la diferencia de voltaje entre el exterior y el interior de la células (VM =Vi –Ve). Sea VP la salida que sigue el voltaje del amplificador en la punta del microelectrodo (ver apéndice, figura A2). Entonces, cuando una corriente “I” fluye simultáneamente a través del microelectrodo:
Vi-Ve=Vp-Vref=Vp-0=Vp Por lo que:
Vp= Vm
La corriente “I” es tan pequeña como para no causar una significativa caída de voltaje a través de RP. Como se aprecia en la figura 1-5, para producir cambios en el potencial de la membrana, el experimentador fija el valor de la corriente con el comando de voltaje inyectado al amplificador, así como la lectura de voltaje medido en la pipeta. Entonces, la corriente es medida por el amplificador (ver apéndice, figura A3). El amplificador debe también controlar la corriente y el voltaje cuando se utiliza en la modalidad de fijación de corriente o fijación de voltaje. Estos dispositivos requieren de la medición de las diferen-cias de voltaje entre el electrodo de registro y el de referencia, así como la medición de la corriente (I) aplicada por el experimentador que fluye a través de R0; esto es igual a:
I= Vp / R0que es la corriente que fluye a través de la membrana de manera simultánea (Ham-mond, 2001) (ver apéndice).
El registro en la modalidad de fijación de corriente permite el registro de volta-jes transmembranales con valores fijos de corriente (que es la forma en que la célula re-gistra la actividad eléctrica), por lo que es posible ajustar y mantener el potencial de

Métodos en Neurociencias Cognoscitivas
10
membrana en valores determinados por el experimentador durante el registro intra-celular. Sin embargo, el flujo de corriente y el potencial de la membrana cambian con-tinuamente, lo que hace difícil registrar eventos que ocurren en la propia membrana (Aidley, 1998). Entonces, la técnica que se utiliza para medir los cambios en el flujo de corriente transmembranal es la fijación de voltaje. Para ello, se monitorean los cambios de voltaje de la membrana y se inyecta corriente de tal modo que se puede mantener el potencial de la célula en el valor determinado por el experimentador (Aidley, 1998).
El propósito de la técnica de fijación de voltaje consiste en ajustar continuamente el potencial de la membrana (VM) a un voltaje predeterminado por el experimentador (VCMD). Para ello, el potencial de la membrana es continuamente medido y se manda una corriente (I) a través de la membrana de tal modo que el valor de VM quede ajusta-do al comando de voltaje VCMD seleccionado por el experimentador. Básicamente se manejan dos técnicas intracelulares de fijación de voltaje que dependen sobre todo del tamaño de las células que se están registrando: a) con dos electrodos, donde uno de ellos registra el voltaje mientras el otro pasa corriente a través de la membrana y b) con un electrodo, donde a través del mismo se pasa corriente y registra el voltaje. Con el mis-mo electrodo se pueden registrar al mismo tiempo el potencial de la membrana y la corriente que se está enviando a través de ella, lo cual puede ser de manera continua o discontinua, según el amplificador (Hammond, 2001).
Con las técnicas intracelulares, el voltaje puede fijarse en una pequeña área de la membrana. Sin embargo, resulta muy complicado debido a la gran resistencia de los electrodos. Otras técnicas como la voltametría y la amperometría incluyen electrodos sensibles a ciertos iones, ya que miden cambios en los niveles de pequeños iones, neu-rotransmisores en tejidos, en las células o cerca de ellas. Esas técnicas requieren re-gistrar pequeños potenciales después de hacer pasar grandes corrientes a través de ellas. Los electrodos selectivos de iones requieren una entrada diferencial, poca co-rriente de fuga y un seguidor de voltaje de alta impedancia. Estas técnicas electroquí-micas se usan para medir cambios rápidos en la concentración de neurotransmisores y neuromoduladores.
La mayor parte de las técnicas de registro requieren de la aplicación de estímulos eléctricos a las células excitables registradas para generar diferentes tipos de respues-tas. Desde un pequeño choque eléctrico que sincronice una población de neuronas para que disparen al mismo tiempo, hasta elaborados estímulos en forma de rampa, trian-gulares, sinusoidales e, incluso, simulaciones de potenciales de acción simulados por medio de una computadora. Es importante considerar que no todos los estímulos son eléctricos; algunos, como en el electroretinograma, requieren estímulos diferentes, por ejemplo, la luz en diferentes longitudes de onda, mientras que los experimentos de ca-nales dependientes de ligando, como los involucrados en la modulación sináptica, re-quieren diferentes estímulos químicos. Patch clamp La técnica de patch clamp o parches de membrana tiene la gran ventaja de no presentar corrientes de fuga, por lo que es posible realizar fijación de voltaje, lo cual no es posi-ble con el registro intracelular. En el patch clamp, el voltaje o la corriente que fluye a tra-vés de canales iónicos individuales y que contribuye a las corrientes totales de la célula puede ser medido directamente, como se verá más adelante. Esta técnica es equivalen-te al registro intracelular con microelectrodos, pero tiene la ventaja de poderse aplicar a células muy pequeñas o con morfologías que impiden ser registradas de otra forma.

A B C D
100 ms
0.1 nA
11
Métodos de registro electrofisiológico
La técnica ilustrada en la figura 1-5 muestra un electrodo de vidrio pulido con una punta de 1 a 2μm de diámetro, el cual se llena con una solución salina isotónica que reproduce, de acuerdo con el interés del experimentador, las concentraciones iónicas externas o internas de la célula en cuestión.
Como se aprecia en la figura, el electrodo se coloca sobre la superficie de la célula, de modo que la resistencia entre el interior del electrodo y el exterior es del orden de decenas a cientos de MΩ. En este momento, se aplica una suave succión a través de la punta del electrodo que produce un contacto estrecho entre el vidrio de la pipeta y la membrana celular, lo que provoca un incremento en la resistencia del electrodo en el orden de los GΩ, llamado gigasello. La formación del gigasello evita la corriente de fuga, que produce una disminución en el ruido electrónico. Esto permite registrar co-rrientes muy pequeñas que fluyen a través de canales iónicos.
Figura 1-5. Técnica de registro de fijación de voltaje en célula entera que muestra la formación del gigasello. (A) Al acercarse el electrodo y adherirse a la membrana se empieza a formar un sello de gran resistencia. (B) En este proceso es común observar pequeñas corrientes sinápticas extracelulares, reflejo de la actividad celular. (C) Una vez que se ha formado el gigasello (cell attach) se obtienen registros de gran resolución. (D) Una suave succión después de formado el gigasello permite la ruptura de la membrana y producir una continuidad entre el electrodo (y el contenido de la pipeta) con el interior celular, por lo que es posible registrar las corrientes totales de la célula (cortesía de José Bargas y Elvira Galarraga).
Actualmente, el registro de patch clamp se ha refinado en diferentes protocolos más o menos generalizados. Una vez que se ha formado el gigasello, se pueden desarrollar di-ferentes modalidades de registro, como se aprecia en la figura 1-6. Cabe destacar que en cada una de ellas se pueden hacer múltiples configuraciones. En general, se consideran las siguientes modalidades:

A B
C
D
E
Célula adherida Célula entera
Interior hacia afuera
Parche perforado
Exterior hacia afuera
Membrana lipídicaCanal iónicoPoros de antibióticoPipeta
Métodos en Neurociencias Cognoscitivas
12
Figura 1-6. Diferentes configuraciones de patch clamp. A) Registro de actividad espontánea de neuronas GABAérgicas en el núcleo supraquiasmático de la rata en la configuración de cell attach. B) Registro de neuronas espinosas en el cuerpo estriado en la configuración who-le cell. C) Registro en la configuración outside out. D) Registro en la configuración inside out. E) registro de patch clamp en la configuración perforated patch de neuronas GABAérgicas en el núcleo supraquiasmático de la rata.
1. Cell attached (célula adherida). La célula queda adherida a la pipeta mediante el parche sin romper la membrana (figura 1-6A). Esta modalidad permite el regis-tro de canales unitarios.
2. Whole cell (célula entera). Cuando está en el modo de célula adherida, se vuelve a succionar a través de la pipeta, de tal forma que se rompe el parche y la pipeta queda en contacto con el interior de la célula, tanto eléctricamente como con la solución de la pipeta, que, por difusión, substituirá a la solución intracelular. En la actualidad, es la más utilizada y substituye a la técnica de registro intracelular (figuras 1-6B, 1-7B y 1-7C).
3. Inside out (el interior hacia fuera). El interior de la membrana plasmática queda expuesta a la solución donde se lleva a cabo el experimento. Ésta fue la técnica original de fijación de voltaje en microáreas de membrana desarrollada para el estudio de las propiedades de los canales iónicos (figura 1-6C).
4. Outside out (el exterior hacia fuera). Después de succionar se separa con rapidez la membrana para que el interior de la membrana plasmática atrape la solución donde se llevaba a cabo el experimento, y se cierra de nuevo para que el exterior de la membrana plasmática quede hacia fuera. Es como hacer una minicélula, pero sin organelos, y se cambia la solución en el interior de la pipeta de acuerdo con el estudio que se desea hacer (figura 1-6D).

13
Métodos de registro electrofisiológico
5. Perforated patch (parche perforado). Se pone en la pipeta algún antibiótico (por ejemplo nistatina) que haga agujeros en la membrana plasmática del parche y así se pueda acceder eléctricamente al interior de la célula. Resulta muy útil en el estudio de cadenas de señalización intracelular, para prevenir que, por la dife-rencia de concentraciones entre el citoplasma y la pipeta de registro, se pueda dializar el interior celular (figura 1-6E).
Las técnicas con el interior o exterior hacia fuera se usan cuando se quiere estudiar, por ejemplo, canales iónicos aislados, la corriente que pasa por un canal unitario o la respuesta intracelular o extracelular de algún receptor que esté en el parche. Por lo ge-neral, se sobreexpresa en células grandes, como los ovocitos de rana, el canal iónico o receptor a estudiar, de tal forma que el parche esté cubierto de estos últimos. La figura 1-6 (B y C) muestra el registro de un canal unitario como ejemplo de esta técnica.
La técnica de patch en la modalidad de célula adherida permite hacer registro extra-celular a nivel unicelular. Es útil cuando se quiere ver la actividad eléctrica de la célula prácticamente intacta o para hacer farmacología y observar el efecto del fármaco sobre la respuesta global de la célula (figura 1-6A). El parche perforado sirve para ver la activi-dad eléctrica de la célula, tal como en el registro de electrodo penetrante, pero sin co-rriente de fuga y mantener intacta la solución intracelular durante un periodo relativa-mente largo (30-60 minutos). Es ideal para estudiar los efectos de fármacos o estímulos sin cambiar nada del metabolismo intracelular (figura 1-6E).
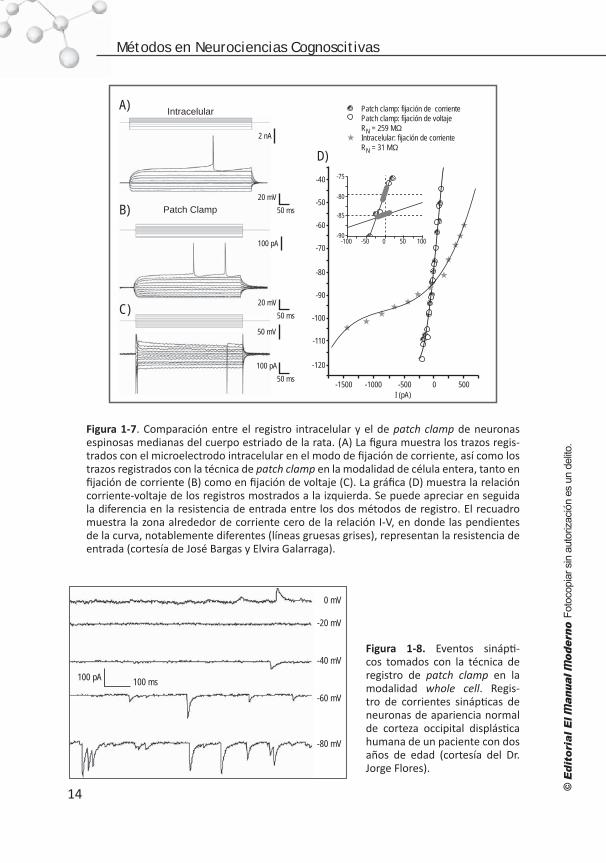
Aunque no fue la primera en usarse, quizá la técnica de registro de patch clamp en la configuración de célula entera es la que más auge ha tenido (figura 1-6B), y es a la que generalmente se refieren los electrofisiólogos cuando dicen que hicieron un regis-tro de patch clamp. En este caso, también se observa la actividad eléctrica de la célula como en el registro de electrodo penetrante sin corriente de fuga. Además, ahora es posible cambiar la solución interna de la célula. En este caso, se puede manipular tanto el exterior como el interior de la célula mientras se observa la respuesta eléctrica de la misma. Aquí se muestran registros de una neurona del neoestriado de rata, en fijación de corriente y en fijación de voltaje; también se presentan diferentes corrientes iónicas estudiadas con fijación de voltaje con la configuración de célula entera (figura 1-7).
Con la técnica de patch clamp ha sido posible medir la resistencia de entrada verda-dera de las diferentes células excitables, lo cual era casi imposible con el electrodo de registro intracelular debido a la corriente de fuga. La relación corriente-voltaje de una neurona medida con registro con electrodo penetrante o con la modalidad de célula en-tera se muestra en la figura 1-7. Por último, pero no menos importante, la modalidad de célula entera ha permitido la observación directa de eventos sinápticos muy pequeños que no eran advertidos con el electrodo penetrante y, ya que es posible fijar el voltaje, se observaron por primera vez las corrientes sinápticas en las neuronas (figura 1-8), así como los efectos de neurotransmisores y fármacos sobre dichas corrientes.
Electromiograma La electromiografía (EMG) es una prueba clínica que se utiliza para evaluar y registrar la actividad eléctrica de los músculos en reposo o activos. La contracción de los múscu-lo se provoca cuando son alcanzados por los potenciales de acción que viajan a través de los nervios motores, provenientes de las neuronas motoras que se localizan en la médula espinal. La interacción funcional entre una neurona motora y el grupo de fibras musculares
A)
B)
D)
C)
Intracelular
Patch Clamp
2 nA
100 pA
50 mV
20 mV50 ms
20 mV50 ms
100 pA50 ms
-40
-50
-60
-70
-80
-90
-100
-110
-120
I (pA)-1500 -1000 -500 0 500
Patch clamp: fijación de corrientePatch clamp: fijación de voltajeRN = 259 MΩIntracelular: fijación de corrienteRN = 31 MΩ
-75
-80
-85
-90-100 -50 0 50 100
0 mV
-20 mV
-40 mV
-60 mV
-80 mV
100 pA 100 ms
Métodos en Neurociencias Cognoscitivas
14
Figura 1-7. Comparación entre el registro intracelular y el de patch clamp de neuronas espinosas medianas del cuerpo estriado de la rata. (A) La figura muestra los trazos regis-trados con el microelectrodo intracelular en el modo de fijación de corriente, así como los trazos registrados con la técnica de patch clamp en la modalidad de célula entera, tanto en fijación de corriente (B) como en fijación de voltaje (C). La gráfica (D) muestra la relación corriente-voltaje de los registros mostrados a la izquierda. Se puede apreciar en seguida la diferencia en la resistencia de entrada entre los dos métodos de registro. El recuadro muestra la zona alrededor de corriente cero de la relación I-V, en donde las pendientes de la curva, notablemente diferentes (líneas gruesas grises), representan la resistencia de entrada (cortesía de José Bargas y Elvira Galarraga).
Figura 1-8. Eventos sinápti-cos tomados con la técnica de registro de patch clamp en la modalidad whole cell. Regis-tro de corrientes sinápticas de neuronas de apariencia normal de corteza occipital displástica humana de un paciente con dos años de edad (cortesía del Dr. Jorge Flores).

15
Métodos de registro electrofisiológico
que inerva se denomina unidad motora. Cuando se activa una unidad motora, el im-pulso nervioso (también denominado potencial de acción) se transmite por el axón de la motoneurona, fibra eferente, hacia el músculo. Cuando llega un potencial de acción a la unión neuromuscular, se libera una sustancia neurotransmisora, la acetilcolina, la cual induce un cambio en la permeabilidad de la membrana de las fibras musculares a ciertos cationes (Na+,K+ yCa2+). Esto produce un cambio de potencial eléctrico en la membrana que se denomina potencial de placa terminal, el cual, a su vez, conduce a la generación de potenciales de acción que se trasmiten a todo lo largo de las fibras mus-culares, lo que provoca que éstas se contraigan. La suma de la actividad eléctrica ge-nerada en todas las fibras musculares de una unidad motora se denomina potencial de unidad motora. La composición de la unidad motora, el número de fibras musculares por unidad motora, el tipo de fibras musculares, entre otros factores, afectan la forma de los potenciales de las unidades motoras en el EMG.
Para llevar a cabo el EMG, se inserta un electrodo de aguja muy fino y estéril (predo-minantemente de acero inoxidable o de platino) a través de la piel en el músculo que se va a analizar. El registro se realiza cuando el músculo se encuentra en reposo o contraí-do. En ocasiones, por el movimiento que realiza el músculo al contraerse, es necesario relocalizar el electrodo para mejorar el registro o para encontrar un mejor sitio de regis-tro. La actividad eléctrica detectada por el electrodo de aguja es transmitida mediante un cable a un receptor-amplificador y éste al electromiógrafo o a un osciloscopio.
La inserción del electrodo de EMG puede ser un poco dolorosa para el paciente y el músculo puede sentirse blando por unos días. Cuando se inserta el electrodo en el músculo se observa cierta actividad eléctrica (figura 1-9), causada por la irritación asociada con la propia inserción del electrodo (“actividad de inserción”). Esta actividad da informa-ción muy importante sobre el estado del músculo y del nervio que lo inerva. Por lo ge-neral, los músculos normales no presentan actividad eléctrica cuando están en reposo; sin embargo, cuando se presenta actividad espontánea anormal, es indicativo de que el músculo o el nervio se encuentran dañados. La actividad eléctrica del músculo se incre-menta en enfermedades del nervio o disminuye en desórdenes musculares prolonga-dos, en donde el tejido muscular es reemplazado por tejido fibroso o por grasa.
Después de colocar el electrodo se solicita al paciente que contraiga el músculo (es decir, que flexione o extienda la articulación en donde se inserte el músculo), o bien, la persona que realiza el EMG orienta el movimiento de la extremidad de la articula-ción del músculo. Cuando el paciente contrae voluntariamente el músculo, aparecen potenciales de unidades motoras en el registro del EMG (figura 1-9). A medida que se incrementa la fuerza de la contracción muscular, se reclutan más grupos de fibras mus-culares y, en consecuencia, se registra mayor actividad electromiográfica. Cuando el músculo está totalmente contraído, aparece un grupo desordenado de potenciales de unidad motora, cuya amplitud y ocurrencia es variable.
La interpretación el EMG no es simple, ya que se requiere del análisis del comien-zo, duración, amplitud y otras características de los patrones de potenciales. La dismi-nución de la amplitud y la duración de los potenciales podrían estar asociadas con en-fermedades musculares que también muestran un reclutamiento más rápido de otras fibras musculares para compensar la debilidad muscular. Por lo general, el EMG se em-plea como ayuda para el diagnóstico de distintas enfermedades que producen debili-dad muscular. A pesar de que el EMG es una prueba del sistema motor, también ayuda en la identificación de anormalidades de los nervios motores o de la médula espinal,
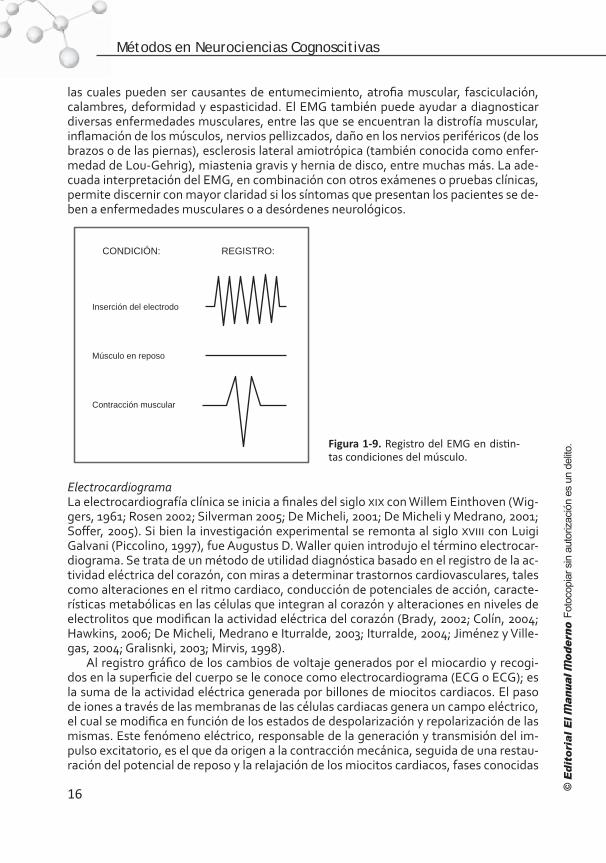
CONDICIÓN: REGISTRO:
Inserción del electrodo
Músculo en reposo
Contracción muscular
Métodos en Neurociencias Cognoscitivas
16
las cuales pueden ser causantes de entumecimiento, atrofia muscular, fasciculación, calambres, deformidad y espasticidad. El EMG también puede ayudar a diagnosticar diversas enfermedades musculares, entre las que se encuentran la distrofía muscular, inflamación de los músculos, nervios pellizcados, daño en los nervios periféricos (de los brazos o de las piernas), esclerosis lateral amiotrópica (también conocida como enfer-medad de Lou-Gehrig), miastenia gravis y hernia de disco, entre muchas más. La ade-cuada interpretación del EMG, en combinación con otros exámenes o pruebas clínicas, permite discernir con mayor claridad si los síntomas que presentan los pacientes se de-ben a enfermedades musculares o a desórdenes neurológicos.
Figura 1-9. Registro del EMG en distin-tas condiciones del músculo.
Electrocardiograma La electrocardiografía clínica se inicia a finales del siglo xix con Willem Einthoven (Wig-gers, 1961; Rosen 2002; Silverman 2005; De Micheli, 2001; De Micheli y Medrano, 2001; Soffer, 2005). Si bien la investigación experimental se remonta al siglo xviii con Luigi Galvani (Piccolino, 1997), fue Augustus D. Waller quien introdujo el término electrocar-diograma. Se trata de un método de utilidad diagnóstica basado en el registro de la ac-tividad eléctrica del corazón, con miras a determinar trastornos cardiovasculares, tales como alteraciones en el ritmo cardiaco, conducción de potenciales de acción, caracte-rísticas metabólicas en las células que integran al corazón y alteraciones en niveles de electrolitos que modifican la actividad eléctrica del corazón (Brady, 2002; Colín, 2004; Hawkins, 2006; De Micheli, Medrano e Iturralde, 2003; Iturralde, 2004; Jiménez y Ville-gas, 2004; Gralisnki, 2003; Mirvis, 1998).
Al registro gráfico de los cambios de voltaje generados por el miocardio y recogi-dos en la superficie del cuerpo se le conoce como electrocardiograma (ECG o ECG); es la suma de la actividad eléctrica generada por billones de miocitos cardiacos. El paso de iones a través de las membranas de las células cardiacas genera un campo eléctrico, el cual se modifica en función de los estados de despolarización y repolarización de las mismas. Este fenómeno eléctrico, responsable de la generación y transmisión del im-pulso excitatorio, es el que da origen a la contracción mecánica, seguida de una restau-ración del potencial de reposo y la relajación de los miocitos cardiacos, fases conocidas
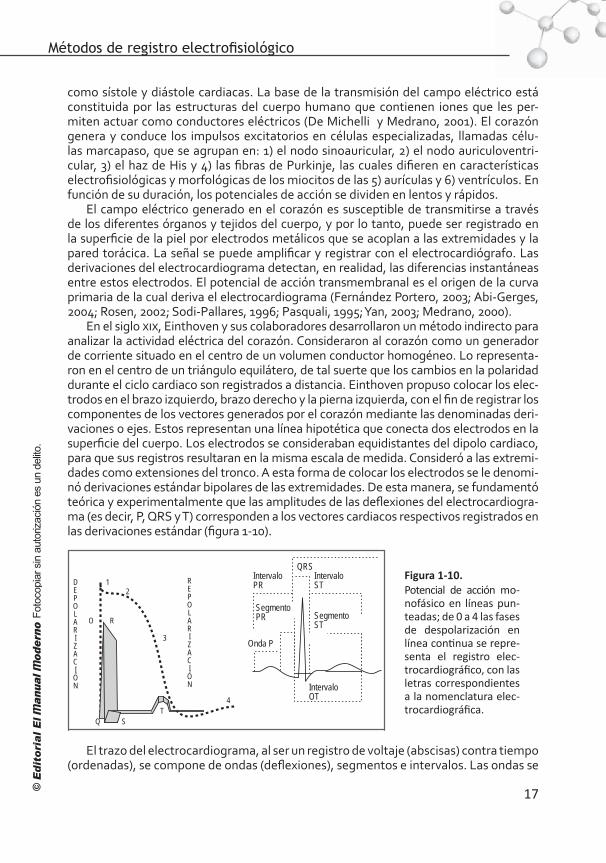
Q ST
4
3
21
RO
REPOLARIZACIÓN
DEPOLARIZACIÓN
IntervaloPR
IntervaloST
SegmentoPR
Onda P
Intervalo OT
SegmentoST
QRS
17
Métodos de registro electrofisiológico
como sístole y diástole cardiacas. La base de la transmisión del campo eléctrico está constituida por las estructuras del cuerpo humano que contienen iones que les per-miten actuar como conductores eléctricos (De Michelli y Medrano, 2001). El corazón genera y conduce los impulsos excitatorios en células especializadas, llamadas célu-las marcapaso, que se agrupan en: 1) el nodo sinoauricular, 2) el nodo auriculoventri-cular, 3) el haz de His y 4) las fibras de Purkinje, las cuales difieren en características electrofisiológicas y morfológicas de los miocitos de las 5) aurículas y 6) ventrículos. En función de su duración, los potenciales de acción se dividen en lentos y rápidos.
El campo eléctrico generado en el corazón es susceptible de transmitirse a través de los diferentes órganos y tejidos del cuerpo, y por lo tanto, puede ser registrado en la superficie de la piel por electrodos metálicos que se acoplan a las extremidades y la pared torácica. La señal se puede amplificar y registrar con el electrocardiógrafo. Las derivaciones del electrocardiograma detectan, en realidad, las diferencias instantáneas entre estos electrodos. El potencial de acción transmembranal es el origen de la curva primaria de la cual deriva el electrocardiograma (Fernández Portero, 2003; Abi-Gerges, 2004; Rosen, 2002; Sodi-Pallares, 1996; Pasquali, 1995; Yan, 2003; Medrano, 2000).
En el siglo xix, Einthoven y sus colaboradores desarrollaron un método indirecto para analizar la actividad eléctrica del corazón. Consideraron al corazón como un generador de corriente situado en el centro de un volumen conductor homogéneo. Lo representa-ron en el centro de un triángulo equilátero, de tal suerte que los cambios en la polaridad durante el ciclo cardiaco son registrados a distancia. Einthoven propuso colocar los elec-trodos en el brazo izquierdo, brazo derecho y la pierna izquierda, con el fin de registrar los componentes de los vectores generados por el corazón mediante las denominadas deri-vaciones o ejes. Estos representan una línea hipotética que conecta dos electrodos en la superficie del cuerpo. Los electrodos se consideraban equidistantes del dipolo cardiaco, para que sus registros resultaran en la misma escala de medida. Consideró a las extremi-dades como extensiones del tronco. A esta forma de colocar los electrodos se le denomi-nó derivaciones estándar bipolares de las extremidades. De esta manera, se fundamentó teórica y experimentalmente que las amplitudes de las deflexiones del electrocardiogra-ma (es decir, P, QRS y T) corresponden a los vectores cardiacos respectivos registrados en las derivaciones estándar (figura 1-10).
Figura 1-10.Potencial de acción mo-nofásico en líneas pun-teadas; de 0 a 4 las fases de despolarización en línea continua se repre-senta el registro elec-trocardiográfico, con las letras correspondientes a la nomenclatura elec-trocardiográfica.
El trazo del electrocardiograma, al ser un registro de voltaje (abscisas) contra tiempo (ordenadas), se compone de ondas (deflexiones), segmentos e intervalos. Las ondas se

Nodo senoauricular
Músculo auricularNodoauriculo-ventricular
Ramas del Haz de HisRamas del Haz de His
Fibras de Purkinje
Músculo ventricular
Células derespuestarápida
Células automáticasde respuestalenta
Métodos en Neurociencias Cognoscitivas
18
denominan P (por presístole) que corresponde a la despolarización auricular; si se siguen ciertas reglas se utilizan las siguientes letras del alfabeto (Q, R, S, T y U) para denominar a las restantes. El punto de referencia es una línea plana por delante de la onda P que se considera isoeléctrica (Kuchar, 1988; Fleming, 1985; Houssay, 1973; Selvester, 1988).
En la figura 1-11 se representa la curva del potencial de acción transmembranal de una fibra muscular ventricular, registrada con un microelectrodo y comparado con el registro de un electrocardiograma. En esta curva se consideran cinco segmentos co-nocidos como fases 0, 1, 2, 3 y 4. En la misma figura se puede apreciar la correlación entre el potencial de acción transmembranal y el electrocardiograma clínico. La mor-fología del QRS-T en el ECG de superficie corresponde, en general, con las diferen-tes fases de los potenciales de acción ventriculares registrados simultáneamente en fibras miocárdicas aisladas. Como se mencionó previamente, la estructura cardiaca con mayor descarga automática de potenciales de acción es el nodo sino-auricular. La progresión ondulatoria de la excitación (es decir, despolarización, inversión de la polaridad y repolarización) genera una diferencia de cargas entre las zonas activadas y las zonas no activadas de las aurículas. Esta diferencia de cargas se puede represen-tar como un vector, con un frente de cargas positivas seguido por cargas negativas, cuya corriente fluye del interior de las zonas inactivas (positivas) hacia el medio circundan-te, para penetrar en las regiones activas (negativas).
Se denomina vector instantáneo a una fuerza direccional en un instante de tiempo, y vector resultante, al efecto de más de un vector que actúa desde un punto común de origen. Dicho vector representa un dipolo, en el que la onda de activación determina la positividad (potencial positivo) en aquellos sitios a los que se aproxima y la negati-vidad (potencial negativo) en aquellos lugares de donde se aleja. Cuando un vector se dirige perpendicularmente a la línea de observación de una derivación, las deflexiones son isodifásicas o isoeléctricas. Por lo tanto, este dipolo queda definido por tres pará-metros: la intensidad o momento del dipolo, la localización y la orientación. La relación entre la dirección de la activación, la orientación del dipolo de corriente y la polaridad de los potenciales es básica en electrocardiografía (Sodi-Pallares, 1996). Los principios generales del análisis vectorial son esenciales para comprender la génesis de los ECG normales y patológicos.
Figura 1-11. Correlación anatómica del ECG con las células de respuesta rápida y las células automáticas.

19
Métodos de registro electrofisiológico
Electroencefalograma Una de las técnicas de registro electrofisiológico más utilizadas en la clínica es, sin duda, el electroencefalograma (EEG), en el cual una diversidad de potenciales mul-tifásicos de la superficie cortical pueden ser registrados al aplicar electrodos en la superficie de la cabeza. La actividad eléctrica que produce el EEG es el producto de flujos de corriente en el espacio extracelular causado por la generación de potenciales de acción y sinápticos en la superficie neural. Entonces se pueden observar oscilacio-nes en el potencial eléctrico entre los electrodos de registro y el de referencia colo-cados en la superficie de la cabeza. Esas oscilaciones difieren en frecuencia y en am-plitud, según el sitio donde están colocados los electrodos de registro y el estado de vigilia del individuo. La utilidad clínica consiste en que las oscilaciones en el registro persisten aun en diferentes estados como la vigilia, el sueño, o bien, en estados alte-rados, como ataques epilépticos o coma. En teoría los cambios en las ondas del EEG podrían servir como un índice directo de cambios en la actividad cerebral. Aunque esto no ha sido alcanzado en su cabalidad, ciertos tipos de actividad en los patrones de las ondas cerebrales en algunos estados de funciones cerebrales alterados se presentan con regularidad. Por ello, el estudio de las ondas cerebrales ha sido, y seguirá siendo, una importante herramienta en el diagnóstico de la medicina clínica como se verá en el capítulo 2.
Perspectivas No cabe duda que a lo largo del siglo xx, sobre todo en la segunda mitad, la técnica de registro electrofisiológico se ha consolidado, tanto en el terreno de la investigación bá-sica como en la clínica. En la actualidad, el registro electrofisiológico se ha convertido en una herramienta indispensable para el estudio del sistema nervioso, ya que permite estudiar el funcionamiento de neuronas completas o disociadas, así como fragmentos de sus membranas celulares. Además, permite estudiar poblaciones neuronales, todo ello sin observar, en la mayoría de los casos, las propias células. Asimismo, el estudio no invasivo de las células cerebrales y de otras células excitables, como las musculares, permite su utilización en la clínica, lo que reditúa en más y mejores diagnósticos de te-jidos que se encuentran en regiones de difícil acceso y que en un momento dado pudie-ran ser muy vulnerables a la manipulación o lesión.
La técnica de registro electrofisiológico constituye un magnífico ejemplo de cómo el avance tecnológico y la experimentación rigurosa proveen al investigador o al especialis-ta de una poderosa técnica para el estudio de las células excitables.
Apéndice: electricidad y circuitos eléctricos básicos Este apéndice busca dar definiciones de algunos conceptos básicos de electrici-dad y circuitos eléctricos para que el lector pueda entender las técnicas de registro electrofisiológico. No pretende ser una guía para enseñar electricidad; para ello, refe-rimos al lector a textos de física e ingeniería (Grob, 1984). Tampoco se pretende que el lector pueda volverse un experto en el equipo usado para el registro electrofisiológico; para eso, se recomienda consultar a los expertos en esta área. Sin embargo, sí presu-pone que el lector tenga nociones elementales de física y se da por hecho cierto cono-cimiento de electricidad y circuitos eléctricos, al menos intuitivo. Se espera que sea de utilidad para entender conceptos que en el texto se hayan dados por supuestos, pero que no todo lector domina.

Métodos en Neurociencias Cognoscitivas
20
Definiciones, conceptos y leyes • Fuerza electromotriz (FEM): fuerza que mueve una carga eléctrica generada por
la diferencia de carga. Se mide en voltios. Un voltio es la diferencia de potencial entre dos puntos, donde se requiere del gasto de un joule de trabajo para mover un coulomb de carga entre esos dos puntos. En sistemas biológicos, la FEM ge-neralmente se debe a la diferencia de concentración de iones entre el interior y el exterior de la célula y se le considera prácticamente invariante en el tiempo.
• Coulomb: cantidad de carga que experimenta la fuerza de un newton en un campo eléctrico de un volt por metro.
• Corriente eléctrica: cantidad de carga que fluye por unidad de tiempo, general-mente a lo largo de un conductor.
• Voltaje: diferencia de carga que existe entre dos puntos. Al igual que la FEM, se mide en voltios, pero con la diferencia de que varía en el tiempo. Esta es la varia-ble para medir en la mayoría de los experimentos electrofisiológicos.
• Capacitancia: Propiedad de los materiales para almacenar carga en presencia de voltaje.
• Leyes de Kirchhoff: se derivan de uno de los principios más utilizados en la física: el principio de conservación. En el caso particular de la electricidad, el principio se utiliza para la conservación de la carga y energía. Las leyes de Kirchhoff no son más que el corolario de la aplicación de dicho principio a los circuitos eléctricos. Estas son dos:
Ley de la corriente: la suma algebraica de todas las corrientes en una inter-sección o nodo es igual a cero (conservación de la carga). Por convención, las corrientes que fluyen hacia el nodo son positivas, mientras que las que se ale-jan de él son negativas. Ley del voltaje: la suma algebraica de todas las fuentes de voltaje o caídas de potencial por elementos pasivos en un circuito cerrado, más conocido como “bucle”, es igual a cero (conservación de la energía). Por convención, la co-rriente positiva produce una caída de potencial al atravesar una resistencia con polaridad (+ hacia -) en la dirección del flujo de corriente.
• Fuente de voltaje: una fuente ideal de voltaje es aquella que mantiene un va-lor de voltaje entre dos puntos sin importar la impedancia que se coloque entre sus terminales de salida. Es útil cuando queremos fijar el voltaje. Es claro que en la vida real tales fuentes no existen, pero se puede llegar a situaciones que se aproximan en rangos de tiempo y voltaje acotados.
• Fuente de corriente: una fuente ideal de corriente es aquella que mantiene la co-rriente sin importar la impedancia puesta entre sus terminales de salida. Ésta es útil cuando queremos fijar la corriente. Es claro que en la vida real tales fuentes no existen, pero se puede llegar a situaciones que se aproximan en rangos de tiem-po y corriente acotados.
• Impedancia: razón de fuerza aplicada a la carga dividida por la cantidad de movi-miento de carga generado. Un caso particular de la impedancia es la ley de Ohm, según la cual la razón de la fuerza aplicada (el voltaje) dividida por la cantidad de movimiento de carga generado (la corriente eléctrica) es constante y se le deno-mina resistencia eléctrica. Una de las propiedades de un sistema eléctrico es la medida de su impedancia para diferentes valores de voltaje o corriente. Relación corriente-voltaje: Esta relación, también llamada curva I-V, se obtiene a partir de

InterfasePreparación biológica Amortiguador Amplificador
Electrodo Pre-amplificadorDispositivo de salida
21
Métodos de registro electrofisiológico
fijar la corriente aplicada (fijación de corriente) a varias intensidades y, enton-ces, determinar la respuesta en voltaje, para así calcular la impedancia a partir de estos valores. La curva I-V también puede obtenerse al fijar el voltaje (fijación de voltaje), observando la corriente generada para diferentes valores del voltaje fijados por el experimentador, y nuevamente calculando la impedancia a partir de los valores obtenidos. Como se puede ver, la relación I-V no es más que la me-dida de la impedancia para diferentes valores de corriente o voltaje. Si la relación corriente voltaje muestra una línea recta, entonces el sistema eléctrico medido cumple con la ley de Ohm:
V=R x I y se dice que tiene un comportamiento lineal u óhmico.
• Impedancia de entrada y resistencia de entrada: la impedancia de entrada es la razón entre la fuerza aplicada y el resultante flujo de energía a través de un sis-tema. No importa cuán complicado sea un sistema; se puede medir su compor-tamiento con un solo parámetro (la impedancia de entrada) nada más aplicando un voltaje y midiendo la corriente resultante (en fijación de voltaje), o aplicando una corriente y observando el voltaje resultante (en fijación de corriente). Esta impe-dancia de entrada puede variar en el tiempo y se distingue de la resistencia de entrada en que esta última es la medida de la impedancia en el estado estacio-nario, es decir, cuando la impedancia de entrada ya no varía en el tiempo. Por lo tanto la impedancia de entrada es función de la frecuencia (variación en el tiem-po), mientras que la resistencia de entrada no lo es.
Medición de señales biológicas Medir señales eléctricas de tejidos biológicos involucra al menos tres diferentes fases. La primera es la interfase o unión entre el flujo de corriente iónica en las soluciones en donde está inmerso el tejido y el flujo de corriente de electrones a través de los alam-bres. Esta fase involucra por lo general algún tipo de electrodo, vidrio y metal para re-gistro extracelular, y vidrio para electrodo intracelular, así como para el patch clamp. La segunda fase de la medición requiere un preamplificador que tenga la función de igua-lar la impedancia del electrodo con la impedancia de entrada del dispositivo medidor o amplificador. La tercera fase requiere del amplificador final donde se lleva a cabo la medición de la señal eléctrica (figura A1).
Figura A1. Medición de señales biológicas. Esquematización de las diferentes fases del registro electrofsiológico.

-
+
Rm
RfRpu
Cp
VcVo
Ag/AgCl
EpuRpuRsEpu
Er
Cm
Cm
Er
Rm
Re
RN
Vo
I
A
B
Métodos en Neurociencias Cognoscitivas
22
La figura A2(A) muestra el circuito generado en el contacto entre un electrodo de registro intracelular, o bien, un electrodo de patch clamp y la célula. Varios problemas sur-gen de esta conexión. El primero es la diferencia de potencial que hay entre dos líquidos con concentraciones iónicas distintas al entrar en contacto (el líquido en el electrodo y el citoplasma de la célula), lo que se denomina potencial de unión. También existe un potencial de unión entre el líquido de la pipeta y el alambre de plata que conduce electrones hasta el dispositivo medidor. Por lo tanto, si queremos medir la diferencia de potencial entre el interior y el exterior de la célula, el dispositivo medidor necesitará de circuitos que compensen estos potenciales de unión.
Figura A2. Circuito generado entre el electrodo de registro y la célula. El diagrama muestra el contacto entre un electrodo de registro in-tracelular, o bien, un electrodo de patch clamp con la célula.
A lo anterior hay que agregar la caída de potencial debida a la resistencia del elec-trodo normalmente denominada resistencia en serie. En la figura A2B, se muestra el circuito equivalente, en donde Re es la resistencia “en serie” del electrodo, y Rn y V0 son la resistencia de entrada y el voltaje asociados con el sistema de medición del voltaje, por ejemplo, un osciloscopio de rayos catódicos.
Valores típicos de Re= 107 ohms y Rn= 106 ohms darían como resultado:
Vo= Rn Vm / Re + Rn = 106 / 1.01 X 108 Vm ≈ 0.01 Vm
Como puede observarse la medida no sería muy confiable para la medición se se-ñales biológicas, por lo que es necesario poner entre el electrodo y el amplificador un preamplificador llamado de seguimiento o almacenamiento, con una resistencia de entrada muy grande y una ganancia unitaria. De este modo, ahora la resistencia del preamplificador se suma a la del electrodo por estar en serie, dominada así por el gran valor de la resistencia del preamplificador. Al aplicar nuevamente los valores a nuestro circuito equivalente (ahora Re=1012 es del orden de la resistencia del preamplificador), da como resultado:
Vo= Rn Vm / Re + Rn = 1012 / 1.0001 X 1012 Vm ≈ Vm

23
Métodos de registro electrofisiológico
Otros problemas generados por el electrodo ―la capacitancia del electrodo, en-tre ellos― se resuelven con circuitos especiales, como circuitos para la corrección de la capacitancia. La figura A3 muestra el circuito básico de registro intracelular con preamplificador, puente y capacitancia negativa. Cuando se pasa corriente a través del electrodo, se genera una caída de potencial a causa de la resistencia en serie de la célula con elelectrodo. Esta caída de potencial puede tergiversar el valor del voltaje que se de-sea medir. La caída del potencial debida a esta resistencia puede ser restada de la señal total al usar un circuito de puente, que resta de la salida total del preamplificador una señal proporcional a la corriente aplicada. La capacitancia del electrodo está en parale-lo con su resistencia y produce un filtro pasa bajas de la señal proveniente de la célula. Esto produce que la fidelidad de la medición no sea especialmente buena si se desean medir cambios de alta frecuencia en el potencial de la célula, lo cual se puede compen-sar al tomar la salida del preamplificador, pasarlo por un capacitor y retroalimentarlo a la entrada del preamplificador. Esto se realiza con un circuito de capacitancia negativa o compensación de la capacitancia.
Figura A3. Circuito básico de registro. Se muestra el diagrama básico de sistema de registro intracelular con preamplifcador, puente y capacitancia negativa.

Métodos en Neurociencias Cognoscitivas
24
ReferenciasAbi-Gerges, N., Philp, K., Pollard, C., Wakefield, I., Hammond. T.G. y Valentin J.P. (2004). Sex
differences in ventricular repolarization: from cardiac electrophysiology to torsades de Piontes. Fundamental and Clinical Pharmacology, 18, 139-151.
Aidley, D. J. (1998). The physiology of excitable cells (4a. ed.). Cambridge: Cambridge University Press.
Bargas, J., Ayala, G. X., Hernández, E. y Galarraga, E. (1998). Ca2+
-Channels involved in neos-triatal glutamatergic transmission. Brain Research Buletin, 45, 521-524.
Barral, J., Poblette, F., Mendoza, E., Pineda, J. C., Galárraga, E. y Bargas, J. (2001). High-affinity inhibition of glutamate release from corticostriatal synapses by ω-agatoxin T. K. European Journal of Pharmacology, 430, 167-173.
Brady, W. (2002). ABC of clinical electrocardiography. London: gbr bmj Publishing Group.Brinley, F. J. (1980). Volume conductor theory. En V. Mountcastle (Ed.), Medical physiology (14a.
ed.). (pp. 290-295). Estados Unidos: C.V. Mosby Co. Colin, L. (2004). El electrocardiograma en el diagnóstico diferencial de las taquicardias con
QRS ancho. Archivos de Cardiología de México, 74 (supl. 1), 44-49. Cordingley, G. E. y Weight, F. F. (1986). Non-cholinergic synaptic excitation in neostriatum:
pharmacological evidence for mediation by glutamate-like transmitter. British Journal of Pharmacology, 88, 847-856.
De Micheli, A. (2001). El centenario del electrocardiógrafo de Einthoven. Archivos de cardiolo-gía de México, 71 (2), 160-166.
De Micheli A., y Medrano, A. (2001). Utilidad del análisis vectorial del electrocardiograma. Archivos de Cardiología de México, 71 (3), 273-240.
De Micheli, A., Medrano, G. e Iturralde, P. (2003). En torno al valor clínico del electrocardiogra-ma. Archivos de Cardiología de México, 73 (1), 38-45.
Debanne, D., Guérineau, M.C., Gähwiler, B.H. y Thompson, S.M. (1996). Paired-pulse facilita-tion and depression at unitary synapses in rat hippocampus: quantal fluctuation affects subsequent release. Journal of Physiology London, 491, 163-176.
Del Castillo, J. (1997). Electrophysiology on the 19th
century. Puerto Rico Health Sciences Journal, 16, 417-427.
Fernández-Portero, P., Castejón, M., Agüera, B., Escribano, B. y Rubio, M. D. (2003). Revisión electrocardiográfica en suidos: Sístole eléctrica auricular, despolarización y
repolarización eléctrica ventricular. Vet. Méx., 34 (4), 343-371. Fleming, J. (1985). Atlas de electrocardiografía práctica (2a. ed.). España: Ediciones Doyma. Frank, R. G. (1986). The Columbian exchange: American physiologist and neuroscience tech-
nics. Fed Proc, 45, 2665-2672. Gralinski, M. (2003). The dog’s role in the preclinical assessment of QT interval prolongation.
Journal of Toxicology. Pathology, 31 (supl.), 11-16. Grob, B. (1984). Electrónica básica (5a. ed). México: McGraw-Hill. Hamill, O. P., Marty, A., Neher, E., Sakmann, B. y Sigworth, F. J. (1981). Improved patch-clamp
techniques for high-resolution current recording from cells and cell-free membrane pat-ches. Pflugers Archiv, 391 (2), 85-100.
Hammond, C. (2001). Cellular and Molecular Neurobiology (2a ed.). Estados Unidos: Academic Press. Hawkins, N. M. (2006). Selecting patients for cardiac resynchronization therapy: electrical or
mechanical dyssynchrony? European Heart Journal, 27, 1270–1281. Hodgkin, A.L., Huxley, A.F. y Katz, B. (1952). Measurements of current voltage relations in the
membrane of the giant axon of Loligo. Journal of Physiology London, 116, 424-448.

25
Métodos de registro electrofisiológico
Houssay, A. B. (1973). Fisiología Humana (4a. ed.). Argentina: El Ateneo. Iturralde, P. (2004). Aplicaciones actuales del electrocardiograma en los síndromes de preexci-
tación. Archivos de cardiología de México, 74 (supl. 1), 84-87. Jiménez, M. y Villegas, J. (2004). Registro del electrocardiograma. Revista Mexicana de
Enfermería Cardiológica, 12 (2), 76-78.Johnston D. y Wu, S. (1995). Foundations of cellular neurophysiology. Estados Unidos: MIT
Press.Kettenman, H. (1997). Alexander von Humbolt and the concept of animal electrici-ty. TINS, 20, 239-242.Keynes, R. D. y Aidley, D. J. (2001). Nerve and muscle (3a. ed.) Reino Unido: Cambridge University Press.
Kuchar L., Thornburn W. y Sammel, N. (1988). Surface electrocardiographic manifestationes of tachyarrhymias: Clues to diagnosis and mechanism. Pace, 11, 61-82.
Lovinger, D. M., y Tyler, E., Fidler, S. y Merritt, A. (1993). Properties of a presynaptic metabotro-pic glutamate receptor in rat neostriatal slices. Journal of Neurophysiology, 69, 1236-1244.
Malenka, R. C. y Kocsis, J. D. (1988). Presynaptic actions of carbachol and adenosine on cor-ticostriatal synaptic transmission studied in vitro. Journal of Neuroscience, 8, 3750-3756.
Medrano, G., De-Michelli, A. y Aranada, A. (2000). ¿Es aún válido el concepto de “salto de onda”? Archivos del Instituto Nacional de Cardiología de México, 70, 19-29.
Miljanich, G. P. y Ramachandran, J. (1995). Antagonist of neural calcium channels: Structure, function and therapeutic implications. Annual Review of Pharmacology and Toxicology, 35, 707-734.
Mirvis, M. (1998). What´s wrong with electrocardiography? Journal of Electrocardiology, 31 (4), 313-316.
Pasquali, G. (1995). Electrocardiografía razonada, teoría y práctica. México: Manual Moderno. Peyronnard, J. M. y Charron, L. (1982). Motor and sensory neurons of the rat sural nerve: a hor-
seradish peroxidase study. Muscle and Nerve, 5, 654-660. Piccolino, M. (1997). Animal electricity and the birth of electrophysiology: The legacy of Luigi
Galvani. Brain Research Bullettin, 46, 381-407. Piccolino, M. (1998). Luigi Galvani and animal electicity: Two centuries after the foundation of
electrophysiology. TINS, 20, 443-448. Rosen, M. (2002). The electrocardiogram 100 years later. Electrical Insights Into Molecular
Messages Circulation, 106, 2173-2179. Selvester, R. (1988). A call to concerted action of change. The recording, display, and reporting
of the routine electrocardiogram. Journal of electrocardiology, 31 (4), 367-370. Silverman M., Grove D. y Upshaw, C. B, Jr. (2006). Why does the heart beat? The discovery of
the electrical system of the heart? Circulation, 113, 2775-2781. Sodi-Pallares, D., Medrano, G., Bisten, A. y Ponce, J. (1996). Electrocardio
grafía clínica, análisis deductivo. México: Méndez Editores.Soffer, A. (2005). Einthoven´s machine, alive and well. Chest, 126 (2), 487-488. Wiggers, C. (1961). Willem Einthoven (1860-1927). Some facets of his life and work. Circulation
Research, 9 (2), 225-234. Woodbury, J.W.O. (1965). Potentials at a volume conductor. En T. Ruch, y H. D. Patton (Eds.),
Physiology and biophysics (19a. ed.). (pp. 85-91). Filadelfia: W.B. Saunders Co. Yan, G., y Martin, J. (2003). Electrocardiographic T wave: a symbol of transmural dispersión
of repolarization in the ventricules. Journal of Cardiac Electrophysiology, 14 (6), 639-640.

26
Capítulo 2. El electroencefalograma:medición de la actividad eléctrica cerebral
Erzsébet Marosi Holczberger
Resumen Este capítulo está destinado a llevar al lector al mundo de la electroencefalografía (EEG), que está lleno de misterios, descubrimientos y retos por resolver. Inicio con una historia breve y trato de presentar en orden cronológico la evolución de esta técnica. Intento aclarar puntos importantes sobre el tema del EEG convencional clínico y llevo al intere-sado hacia temas más recientes, como el EEG computarizado y su papel en la clínica e investigación. Trato de mostrar las líneas futuras, todavía en vías de desarrollo, por las que se conduce esta técnica. Lo anterior ha conducido al cálculo de los generadores de la actividad eléctrica cerebral por distintos modelos físico-matemáticos que buscan so-luciones precisas que permitan identificar las estructuras cerebrales involucradas en la generación del EEG. Además, el desarrollo de esta técnica ha permitido acercamientos más adecuados para entender los porqués psicofisiológicos de la conducta, así como los mecanismos neurofisiológicos implicados en el registro electroencefalográfico.
Introducción En las últimas dos décadas, el electroencefalograma (EEG) se ha convertido en un estu-dio muy importante, no sólo por ser una prueba no invasiva que evalúa con fines clínicos la epilepsia y otras enfermedades del sistema nervioso central, sino también porque es una de las pocas técnicas que usan en forma rutinaria, tanto con humanos como con ani-males, en la investigación. Así, el EEG se ha convertido en el instrumento preferido para evaluar la forma en que el cerebro, animal y humano, maneja la información cognos-citiva, además de proporcionar información sobre el mecanismo neuronal subyacente.
El desarrollo tecnológico ha puesto a la orden de los investigadores en neurocien-cias nuevas técnicas de neuroimagen (—p. ej., tomografía por emisión de positrones (PET), tomografía axial computada (TAC) e imágenes de resonancia magnética fun-cional (RMf)— y, además, amplió las posibilidades del registro electroencefalográfico y magnetoencefalográfico (EEG cuantitativo y MEG). Así, tenemos métodos más precisos y eficientes para el estudio de las funciones superiores mentales. Cada una de estas téc-nicas tiene ventajas y desventajas. Por ejemplo, los estudios de neuroimagen, aunque tienen una alta resolución espacial, tienen una baja resolución temporal y son de alto costo. Por el contrario, el EEG tiene excelente resolución temporal, muestra la actividad ce-rebral en milisegundos de exactitud y es de bajo costo, pero ofrece una baja resolución espacial. Todos estos estudios nos permiten una evaluación más precisa de la actividad cerebral durante la ejecución de diferentes tipos de tareas cognoscitivas, y a la vez un mejor entendimiento de la organización y funcionamiento del cerebro.
Desde el siglo xviii (es decir, los estudios de Volta y Galvani), se sabe que todas las células vivas tienen actividad eléctrica. Su registro, desde aquella época, pasó por varias

27
El Electroencefalograma: medición de la actividad eléctrica cerebral
etapas experimentales. La primera persona que logró registrar un EEG en animales fue Richard Caton, aunque decimos que la época del EEG empezó en 1929, cuando Hans Berger publicó la posibilidad de obtener, sobre la piel cabelluda en humanos, cambios de voltaje representados por ondas sinusoidales constantes, de frecuencia alrededor de 10 Hz (es decir, actividad alfa), que eran sensibles a la apertura y cierre de los ojos. Des-de este tiempo, el EEG está en continuo uso y desarrollo (Swartz y Goldensohn, 1998).
En la primera etapa, alrededor de la década de 1950, los investigadores se dedicaron a perfeccionar la técnica y reunir información sobre la actividad eléctrica cerebral. Se empezó con la estandarización de la colocación de los sensores o electrodos en luga-res específicos, a la que se llamó sistema internacional 10-20 de colocación de los elec-trodos. Dicha estandarización hizo posible que los registros preparados en cualquier parte del mundo sean interpretables para cualquiera y que los electrodos se encuentren sobre la misma estructura cerebral, a pesar de que los sujetos tengan diferentes formas de cabeza. Luego, se procedió con la recolección de la información normativa. Los in-vestigadores se preguntaban ¿cómo es la actividad eléctrica cerebral en gente normal y en pacientes con diversas patologías? Además, ¿qué significado funcional tienen las oscilaciones en el cerebro? Dado que la actividad registrada en el EEG proviene de gran-des poblaciones de neuronas corticales, la relación entre el EEG y el funcionamiento de estos grupos de neuronas específicas o la conducta es todavía desconocida. Los datos normativos recolectados pertenecen al dominio de la estadística, donde se describen los cambios que suele mostrar la mayoría de los sujetos tanto normales como con di-ferentes patologías del sistema nervioso (ciertas frecuencias, amplitudes, morfología de ondas y reacciones a los cambios del medio ambiente). Estos criterios establecidos obviamente no corresponden al 100% de los casos. Siempre existen los falsos positi-vos, es decir, gente con morfología no típica o anormal, pero sin patología alguna; no obstante, en la mayoría de los casos, estos parámetros han permitido establecer los criterios de interpretación del EEG tradicional. De manera paralela, se han realizado investigaciones para entender el origen de estos cambios de voltaje y relacionarlos con la función. Sherrington describió la sinapsis, por lo que en 1932 fue galardonado con el premio Nobel. En su tiempo se supuso que la actividad registrada en el EEG reflejaba potenciales de acción dentro de las neuronas. Sin embargo, gracias a que se puede ob-tener un registro de actividad eléctrica cerebral durante anestesia, aunque no ocurran potenciales de acción dentro de la neurona, pronto se descubrió que los potenciales de acción no pueden dar origen a los ritmos registrados en la superficie. Trabajos posterio-res dejaron claro que, más que los potenciales dentro de las neuronas, son los potenciales extracelulares los que contribuyen al registro electroencefalográfico. Ahora sabemos que la actividad registrada en la superficie se origina en los potenciales postsinápticos excitatorios e inhibitorios sumados. Éstos nacen en las dendritas apicales o en el soma de las neuronas piramidales (Brazier, 1949). Sin embargo, todavía no está clara la fun-cionalidad, el significado de estas oscilaciones y la forma como se relacionan con cier-tas conductas.
Se sabe que el registro de la actividad eléctrica cerebral por medio del electroence-falograma depende del arreglo perpendicular que tienen las neuronas piramidales en la corteza cerebral. Dicho arreglo de la estructura neuronal puede ser representado eléctricamente por un gran número de dipolos colocados verticalmente. La neuronas piramidales de la corteza muestran tal arreglo estructural porque sus axones están co-locados en forma perpendicular a la superficie; son paralelas una con otra.

Métodos en Neurociencias Cognoscitivas
28
Muchas veces atraviesan varias capas corticales y se ramifican de modo abundante en las capas superiores. La posibilidad de registrar su actividad en forma extracelular se basa en el hecho de que los cambios de potenciales dentro de la neurona causan una corriente iónica a través de la membrana en el espacio extracelular. Esto crea un po-tencial de campo positivo en la región de la membrana presináptica y negativo en el líquido extracelular cercano. Las corrientes que llevan iones positivos se llaman fuente o generador y las que llevan iones negativos se llaman sumidero. En el caso de una sinapsis excitatoria, la fuente se encuentra cerca de las dendritas apicales de las células piramida-les en las capas corticales más profundas. Como consecuencia, obtendremos positividad en lo profundidad de la corteza cerebral y negatividad en su superficie, lo que llamamos “un dipolo”. La idea de que hay dipolos en el cerebro fue propuesta por Brazier en 1949; más tarde, se comprobó su existencia experimentalmente. Se registra actividad sobre la piel cabelluda sólo de aquellos campos que son abiertos (corrientes del soma a través del axón y de las dendritas), y de un campo eléctrico cerrado (corrientes dentro del soma) no se obtiene registro de potencial.
La distribución de los potenciales, determinada por fuentes de dipolos múltiples, puede mostrarse en tres dimensiones.Puede calcularse en la superficie por superpo-sición lineal de los potenciales para cada dipolo individual. Los registros superficiales de los campos eléctricos del cerebro indican la actividad de muchos elementos neu-ronales; incluso puede reflejar la actividad de estructuras profundas que se activan simultáneamente.
Significado de la actividad registrada Al hablar del EEG tenemos que dejar claro dos fenómenos básicos que lo componen: las oscilaciones y la sincronización. ¿Qué papel puede tener la oscilación en la función cere-bral? No se sabe con exactitud, pero la vida misma tiene cambios cíclicos en todos sus aspectos: luz-oscuridad, día-noche, sueño-despertar, cambios hormonales y de neuro-trasmisores. Las oscilaciones de cada célula viva pueden ser manifestación de la vida misma. En 1949, experimentos de Moruzzi y Magoun (Steriade et al., 1990) proporcio-naron una idea clara sobre el origen de la ritmicidad en el EEG. Estimularon la forma-ción reticular del tallo cerebral y produjeron así ondas rápidas de bajo voltaje en el re-gistro cortical, acompañadas, por lo general, de alerta conductual. Estudios posteriores con perros revelaron que la reactividad de ritmo alfa dependía de la red tálamo-cortical.
En el cerebro, las oscilaciones se producen y se mantienen por el circuito tálamo-cortical, descrito por Steriade (1993). Los tres componentes de esta gran obra de ingeniería son la corteza cerebral ―con neuronas excitatorias―, la vía tálamo-cortical ―también con neuronas excitatorias― y el núcleo reticular ―con neuronas inhibitorias. La hiperpolari-zación rítmica se debe a los potenciales inhibitorios postsinápticos prolongados, gene-rados en las neuronas reticulares. La hiperpolarización de una célula inhibitoria induce a una activación de las corrientes dependientes del calcio en las neuronas tálamo-corticales de umbral bajo, lo cual excita a la neurona reticular que inhibe el circuito tálamo-cortical y así se mantiene la oscilación. Por ejemplo, durante la percepción visual se excitan las neuronas del núcleo pulvinar, que resulta en diversos ritmos desincronizados de estas células, mientras que los otros núcleos talámicos se mantienen relativamente sin-cronizados. Aferencias del tallo, específicamente del tegmento pedúnculo-pontino y del tegmento laterodorsal, también ejercen efecto sobre el circuito tálamo-cortical, lo que modifica la tonicidad general de esta red.

29
El Electroencefalograma: medición de la actividad eléctrica cerebral
Otro fenómeno crucial en la producción de los ritmos cerebrales es la sincronización. Las oscilaciones en varias frecuencias son la característica prominente del EEG espontá-neo. Cuando grandes poblaciones neuronales descargan al mismo ritmo, probablemente dirigidas por células marcapaso (como directores de orquesta), hablamos de sincroniza-ción. ¿Qué significado pueden tener los ritmos sincronizados a ciertas frecuencias? Exis-te la idea de que la sincronización de las señales optimiza la entrada, es decir, aumenta la eficiencia terminal mientras exista una sincronización de los disparos neurales indivi-duales. Además, se ha planteado la idea de que el patrón de oscilaciones sincronizadas probablemente esté relacionado con ciertos estados psicológicos (Singer, 1993; Nunez, 1981, 1995). Sin embargo, la relación entre los estados psicológicos o resolución de tareas cognoscitivas específicas y las oscilaciones sincronizadas es desconocida, a pesar de ser bastante estudiada, tanto con modelos electrofisiológicos como con simulaciones por computadora.
En particular, las oscilaciones alfa (8-13 Hz) han atraído la atención de los investi-gadores. Debido a la complejidad de la señal del EEG, hasta este momento no se puede distinguir de modo confiable esta actividad del ruido blanco filtrado, pues, según algunos autores (Linkenkaer et al., 2001), la población neuronal subyacente probablemente no obedece a la dinámica de dimensiones bajas. Linkenkaer et al. (2007) demostraron en estudios con gemelos mono y dicigóticos, con la medida de correlación de rangos largos, que las oscilaciones alfa no pueden ser explicadas por cambios causados por variables experimentales, sino más bien reflejan parámetros fisiológicos genéticamen-te determinados que modulan la dinámica intrínseca de las redes neuronales. Nuevas ideas (Fields, 2006) sugieren que la sincronización entre neuronas es la capacidad que hace que el cerebro humano sea superior a los de los animales. La pregunta que todavía no se ha podido responder es ¿cómo coordina el cerebro grandes poblaciones neurona-les en una actividad coherente y para qué? Se ha observado oscilación sincrónica en tejidos in vitro, aun en el caso de que todas las sinapsis estén bloqueadas. Se sospecha, aunque no se sabe por el momento, que la sincronización de las oscilaciones se debe a sustancias neuromoduladoras, o a la glía. Todas éstas son ideas formuladas sobre la función probable de las oscilaciones. Futuras investigaciones deben aclarar la función de la oscilación y la sincronización.
Al hablar del electroencefalograma, debemos mencionar que los cambios de vol-taje en la corteza cerebral son muy pequeños (millonésima parte de un voltio, o sea, un microvoltio), por lo que deben pasar por una serie de amplificadores a fin de poder observarlos. Los amplificadores diferenciales que se usan para la recolección de la señal electroencefalográfica tienen dos entradas de señal y una salida, por lo que muestran la diferencia entre las dos entradas. Este hecho determina que al colocar electrodos en dos áreas activas diferentes (es decir, registro bipolar) no sabemos qué actividad tie-ne una u otra; solamente podemos observar la diferencia entre ellas. La solución para evitar este problema parecía sencilla. Si se coloca un electrodo sobre un área activa y otro en alguna parte diferente del cuerpo que no tiene actividad cerebral (registro mo-nopolar, también llamado registro referencial), el resultante de la diferencia reflejaría la actividad del área activa. Por desgracia, el supuesto de medir sólo el electrodo activo no resulta ser verdadero, ya que no se puede poner electrodos en puntos lejanos a la ca-beza, dado que es más factible que ambos recojan el ruido eléctrico ambiental con ma-yor eficiencia que la actividad cerebral. El electrodo de referencia debe estar arriba del cuello (mandíbula, lóbulos auriculares, mastoides, etc.), aunque en estas partes puede

Métodos en Neurociencias Cognoscitivas
30
colectarse algún tipo de actividad cerebral también. En un intento por eliminar el efecto de la actividad eléctrica de la referencia, nacieron varias soluciones. Primero se colocó un electrodo en un lóbulo auricular, lo que añadió el problema de tener diferentes dis-tancias entre los electrodos activos y la referencia. Esto también eliminó la ventaja de la colocación estandarizada (diferencias proporcionales parecidas entre electrodos). Para resolver este problema, se propuso colocar dos electrodos, uno en cada lóbulo auricular, y conectarlos del hemisferio derecho a la referencia derecha, así como los del hemisferio izquierdo a la referencia izquierda. Esta táctica dificultó las medidas de simetría, ya que las dos referencias afectaron en forma distinta la salida de la información. Ahora el uso de las referencias interconectadas es muy común a pesar de que ya existen programas que calculan el promedio de los valores entrantes y restan este promedio de cada una de los canales de registro. De este modo, la actividad común a todos los electrodos (es decir, lo que captan los electrodos interconectados) se elimina y sólo se consideran los valores que son propios de cada lugar. La eliminación de la referencia promedio es importante, sobre todo cuando se calculan medidas de coherencia, ya que la actividad común a las referencias aumenta en forma dramática la coherencia.
Las frecuencias observadas en el EEG Tradicionalmente, en el EEG se registran oscilaciones con frecuencias de 1 a 30 ciclos por segundo. Esto no significa que el cerebro tenga el límite superior de sus ritmos en 30 Hz; más bien, los aparatos están hechos para usar esta ventana de frecuencias para el registro. El electroencefalógrafo es como el ojo humano: está ajustado para regis-trar ciertas frecuencias y es ciego a otras. Sin embargo, se han reportado registros con equipos modernos en frecuencias altas (200 Hz), cuyo uso rutinario todavía no es muy frecuente (Llinás y Ribary, 1993; Lutzenberger et al., 1995).
Los EEGs tradicionales muestran cambios de voltaje originados en la corteza cerebral durante el tiempo de captación. En los registros que se muestran en la figura 2-1, apare-cen grafoelementos típicos, cuya formación determina las bandas anchas tradiciona-les. Las llamamos bandas anchas porque cada una involucra varias frecuencias. Estas bandas son: a) delta (<4 Hz), b) theta (4.5-7.5 Hz), c) alfa (7.5-13 Hz) y d) beta (14-30 Hz). Aunque algunos autores consideran que la banda beta tiene frecuencias entre 13-20 Hz y de 20 a 30 Hz, otros consideran esta última como gamma.
Dado que estas bandas muestran características parecidas y cambian según los esta-dos de los sujetos (edad, vigilia-sueño, atención-relajación, salud o patología), la división en estas bandas parece lógica, pero no debemos olvidar que son límites arbitrariamente determinados. De hecho, es común observar que las bandas anchas no se comportan con regularidad, por lo que cada vez es más frecuente encontrar estudios con bandas más estrechas (rangos entre 1 o 2 Hz) o divisiones entre bandas (es decir, alfa baja y alfa alta).
Normalidad y anormalidad La normalidad de los registros tradicionales se determina con base en la inspección vi-sual, en la búsqueda de los grafoelementos observados en los registros de la población normal. Las frecuencias observadas dependen del estado psicológico del sujeto (vigilia o sueño, atención o relajación, y de su edad). Al controlar estos factores, el registro es bastante estable, sin mostrar mayores cambios. En la población de adultos sin pato-logía alguna, en vigilia relajada y con los ojos cerrados, se observa actividad alfa en la mayor parte de la cabeza. Los niños más jóvenes tienen actividad lenta de alto voltaje
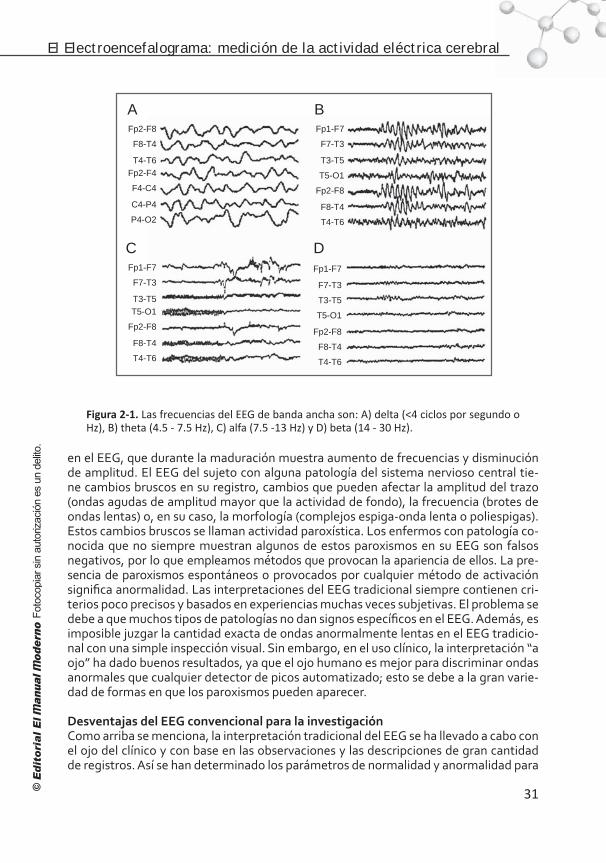
Fp2-F8
F8-T4
T4-T6Fp2-F4
F4-C4
C4-P4
P4-O2
Fp1-F7
F7-T3
T3-T5T5-O1
Fp2-F8
F8-T4
T4-T6
Fp1-F7
F7-T3
T3-T5
T5-O1
Fp2-F8
F8-T4
T4-T6
Fp1-F7
F7-T3
T3-T5
T5-O1
Fp2-F8
F8-T4
T4-T6
A B
C D
31
El Electroencefalograma: medición de la actividad eléctrica cerebral
Figura 2-1. Las frecuencias del EEG de banda ancha son: A) delta (<4 ciclos por segundo o Hz), B) theta (4.5 - 7.5 Hz), C) alfa (7.5 -13 Hz) y D) beta (14 - 30 Hz).
en el EEG, que durante la maduración muestra aumento de frecuencias y disminución de amplitud. El EEG del sujeto con alguna patología del sistema nervioso central tie-ne cambios bruscos en su registro, cambios que pueden afectar la amplitud del trazo (ondas agudas de amplitud mayor que la actividad de fondo), la frecuencia (brotes de ondas lentas) o, en su caso, la morfología (complejos espiga-onda lenta o poliespigas). Estos cambios bruscos se llaman actividad paroxística. Los enfermos con patología co-nocida que no siempre muestran algunos de estos paroxismos en su EEG son falsos negativos, por lo que empleamos métodos que provocan la apariencia de ellos. La pre-sencia de paroxismos espontáneos o provocados por cualquier método de activación significa anormalidad. Las interpretaciones del EEG tradicional siempre contienen cri-terios poco precisos y basados en experiencias muchas veces subjetivas. El problema se debe a que muchos tipos de patologías no dan signos específicos en el EEG. Además, es imposible juzgar la cantidad exacta de ondas anormalmente lentas en el EEG tradicio-nal con una simple inspección visual. Sin embargo, en el uso clínico, la interpretación “a ojo” ha dado buenos resultados, ya que el ojo humano es mejor para discriminar ondas anormales que cualquier detector de picos automatizado; esto se debe a la gran varie-dad de formas en que los paroxismos pueden aparecer.
Desventajas del EEG convencional para la investigación Como arriba se menciona, la interpretación tradicional del EEG se ha llevado a cabo con el ojo del clínico y con base en las observaciones y las descripciones de gran cantidad de registros. Así se han determinado los parámetros de normalidad y anormalidad para
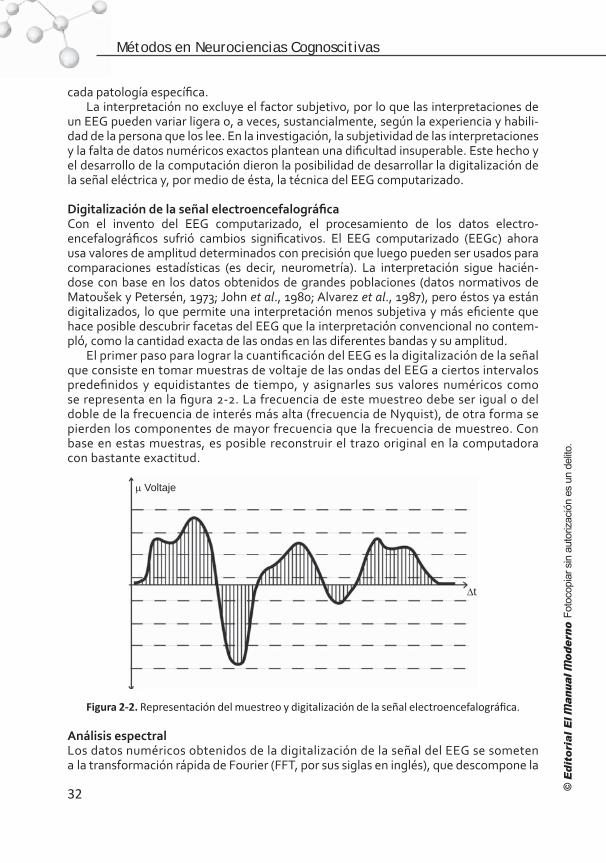
µ Voltaje
∆t
Métodos en Neurociencias Cognoscitivas
32
cada patología específica. La interpretación no excluye el factor subjetivo, por lo que las interpretaciones de
un EEG pueden variar ligera o, a veces, sustancialmente, según la experiencia y habili-dad de la persona que los lee. En la investigación, la subjetividad de las interpretaciones y la falta de datos numéricos exactos plantean una dificultad insuperable. Este hecho y el desarrollo de la computación dieron la posibilidad de desarrollar la digitalización de la señal eléctrica y, por medio de ésta, la técnica del EEG computarizado.
Digitalización de la señal electroencefalográfica Con el invento del EEG computarizado, el procesamiento de los datos electro-encefalográficos sufrió cambios significativos. El EEG computarizado (EEGc) ahora usa valores de amplitud determinados con precisión que luego pueden ser usados para comparaciones estadísticas (es decir, neurometría). La interpretación sigue hacién-dose con base en los datos obtenidos de grandes poblaciones (datos normativos de Matoušek y Petersén, 1973; John et al., 1980; Alvarez et al., 1987), pero éstos ya están digitalizados, lo que permite una interpretación menos subjetiva y más eficiente que hace posible descubrir facetas del EEG que la interpretación convencional no contem-pló, como la cantidad exacta de las ondas en las diferentes bandas y su amplitud.
El primer paso para lograr la cuantificación del EEG es la digitalización de la señal que consiste en tomar muestras de voltaje de las ondas del EEG a ciertos intervalos predefinidos y equidistantes de tiempo, y asignarles sus valores numéricos como se representa en la figura 2-2. La frecuencia de este muestreo debe ser igual o del doble de la frecuencia de interés más alta (frecuencia de Nyquist), de otra forma se pierden los componentes de mayor frecuencia que la frecuencia de muestreo. Con base en estas muestras, es posible reconstruir el trazo original en la computadora con bastante exactitud.
Figura 2-2. Representación del muestreo y digitalización de la señal electroencefalográfica.
Análisis espectral Los datos numéricos obtenidos de la digitalización de la señal del EEG se someten a la transformación rápida de Fourier (FFT, por sus siglas en inglés), que descompone la

33
El Electroencefalograma: medición de la actividad eléctrica cerebral
actividad electroencefalográfica en sus ondas componentes (es decir, series de ondas seno y coseno según las frecuencias requeridas), tal como se muestra en la figura 2-3.
Figura 2-3. Transformación rápida de Fourier del EEG.
La señal original del EEG puede ser reconstruida de modo exacto a partir de los va-lores de las funciones seno y coseno. Al aplicar el operador Fourier a los datos del elec-troencefalograma se pierde la información de fase y sólo se obtienen los componentes de frecuencia por bandas. Para la transformación al dominio de la frecuencia se deben escoger segmentos del EEG que muestren las características especiales del registro en cuestión y que no tengan ningún artefacto (es decir, potencial eléctrico que se sabe de antemano que se no origina en el cerebro). Estos segmentos no llevan la continuidad en el tiempo; son segmentos cortos que se escogen en diferentes tiempos, ya que la aparición de los artefactos o las características especiales que nos interesan ocurren en forma azarosa. Así, las ventanas o segmentos del EEG escogidos para su transforma-ción en el dominio de la frecuencia ya no guardan la relación con el tiempo y el análisis espectral convierte los datos del EEG, que tuvieron relación con el tiempo, en una serie de datos relacionados con la frecuencia. Esto da como resultado la medida de potencia absoluta (el área bajo la curva, medida que refleja la amplitud) en cada banda de fre-cuencia. Esta medida está calculada con base en el promedio de amplitudes contenidas en la totalidad del número de ventanas escogidas para la transformación, por lo que las medidas espectrales muestran las características generales del registro y las peculiari-dades de cada ventana desaparecen.
Esta técnica es robusta cuando los datos no cambian mucho o cuando los cambios fuertes no ocurren en proporción considerable (los paroxismos únicos y repentinos no cambian mucho el promedio). Consecuentemente, el análisis espectral basado en la transformación de Fourier necesita que el EEG sea un proceso estacionario, lo que quiere decir que exista un patrón de ondas que se repita con respecto al tiempo. El EEG es actividad no estacionaria por naturaleza; los estados medidos en el EEG cam-bian con gran frecuencia. Por ejemplo, algunos de los cambios temporales frecuentes son la somnolencia o el sueño o la variación en la atención-relajación. Sin embargo, los modelos estadísticos exigen que los segmentos del EEG sean exactamente del mismo estado, es decir, que el registro sea estacionario para computar los parámetros definidos. En la práctica electroencefalográfica esto es difícil de lograr, pero se obtie-ne una buena aproximación con lapsos cortos de tiempo de análisis, pues se reduce la
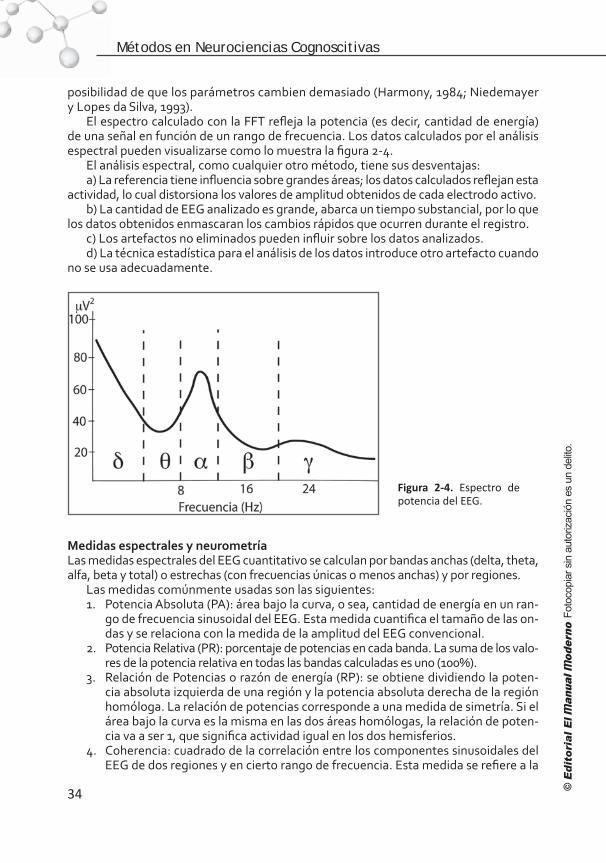
Métodos en Neurociencias Cognoscitivas
34
posibilidad de que los parámetros cambien demasiado (Harmony, 1984; Niedemayer y Lopes da Silva, 1993).
El espectro calculado con la FFT refleja la potencia (es decir, cantidad de energía) de una señal en función de un rango de frecuencia. Los datos calculados por el análisis espectral pueden visualizarse como lo muestra la figura 2-4.
El análisis espectral, como cualquier otro método, tiene sus desventajas: a) La referencia tiene influencia sobre grandes áreas; los datos calculados reflejan esta
actividad, lo cual distorsiona los valores de amplitud obtenidos de cada electrodo activo. b) La cantidad de EEG analizado es grande, abarca un tiempo substancial, por lo que
los datos obtenidos enmascaran los cambios rápidos que ocurren durante el registro. c) Los artefactos no eliminados pueden influir sobre los datos analizados. d) La técnica estadística para el análisis de los datos introduce otro artefacto cuando
no se usa adecuadamente.
Figura 2-4. Espectro de potencia del EEG.
Medidas espectrales y neurometría Las medidas espectrales del EEG cuantitativo se calculan por bandas anchas (delta, theta, alfa, beta y total) o estrechas (con frecuencias únicas o menos anchas) y por regiones.
Las medidas comúnmente usadas son las siguientes: 1. Potencia Absoluta (PA): área bajo la curva, o sea, cantidad de energía en un ran-
go de frecuencia sinusoidal del EEG. Esta medida cuantifica el tamaño de las on-das y se relaciona con la medida de la amplitud del EEG convencional.
2. Potencia Relativa (PR): porcentaje de potencias en cada banda. La suma de los valo-res de la potencia relativa en todas las bandas calculadas es uno (100%).
3. Relación de Potencias o razón de energía (RP): se obtiene dividiendo la poten-cia absoluta izquierda de una región y la potencia absoluta derecha de la región homóloga. La relación de potencias corresponde a una medida de simetría. Si el área bajo la curva es la misma en las dos áreas homólogas, la relación de poten-cia va a ser 1, que significa actividad igual en los dos hemisferios.
4. Coherencia: cuadrado de la correlación entre los componentes sinusoidales del EEG de dos regiones y en cierto rango de frecuencia. Esta medida se refiere a la

35
El Electroencefalograma: medición de la actividad eléctrica cerebral
semejanza de las ondas cerebrales en regiones espacialmente separadas sobre la piel cabelluda. En el caso de que dos áreas desempeñen la misma actividad (consecuentemente sean iguales), la coherencia es 1, y cuando dos áreas están involucradas en actividades completamente diferentes, es 0. Normalmente, la coherencia más alta se obtiene durante las descargas epilépticas. También se ha observado que hay una alta coherencia cuando músicos ensayan composiciones mentalmente. La coherencia de la actividad alfa en las regiones posteriores du-rante el descanso con ojos cerrados suele ser entre 0.5 y 0.8. Esta medida refleja la interconectividad morfológica o funcional, ya que la coherencia de dos áreas le-janas es alta sólo si dichas áreas tienen interconexiones directas o indirectas o si están involucradas en la ejecución de la misma actividad o tarea. La coherencia calculada con la fórmula de Başar (1980) no refleja las diferencias de fase. Dos áreas con actividad igual no necesariamente empiezan esta actividad al mismo momento. La propagación causa un retraso entre la región donde dicha activi-dad inició y aquélla que sigue esta actividad. Este retraso es la información de fase que en el caso de las ondas sinusoidales se expresa en ángulos. El cálculo de fases todavía no es común en los aparatos comerciales. En el caso de la fórmula de Başar, se pierde la información referente a la fase en-tre dos regiones cerebrales, ya que se trabaja con el cuadro de la potencia (es decir, con las matrices espectrales y cross-espectrales). No se sabe en dónde em-pezó y hacia qué rumbo se propaga la actividad, nada más se muestra la simili-tud entre las dos áreas calculadas. Esta es la expresión de coherencia:
Coh(f)2 = Gxy(f)2 / Gxx(f) Gyy(f)
donde Gxx es el espectro de una derivación (p. ej., en el sitio T5), Gyy es el espec-tro de otra derivación (p. ej., en el sitio F8) y Gxy es el espectro cruzado de las dos derivaciones (T5 x F8). Dado que el efecto de la referencia (en caso de registro monopolar o referencial) aumenta sustancialmente las medidas de coherencia por la actividad común y desconocida producida por la referencia, para trabajar con esta medida, es necesario calcular la actividad promedio debajo de todos los electrodos y quitar el valor del promedio de la actividad registrada bajo cada uno de ellos. Así, la actividad común (el efecto de la referencia) se elimina y queda so-lamente el cambio específico de cada lugar.
5. Frecuencia dominante o promedio de frecuencias (FM): medida que muestra el pro-medio de las frecuencias registradas en cada región y por bandas. Refleja los cam-bios sutiles en cada frecuencia que no traspasan los límites de la banda determinada. Estas medidas espectrales obtenidas se organizan en matrices N x p, en donde cada línea es un sujeto (N) y cada columna es una variable o sea medida espectral (p).
Al obtener la matriz espectral, las medidas espectrales se someten a análisis esta-dístico para su evaluación según los intereses específicos de cada estudio. Este proce-so se llama neurometría. La neurometría es un procedimiento basado en las medidas cuantitativas de la actividad eléctrica del cerebro que emplea la estadística para evaluar la integridad anatómica, la maduración y los procesos sensoriales, perceptuales y cog-noscitivos (John, 1977). Dada la flexibilidad del procedimiento, existe una gama amplia de posibilidades para la investigación, ya que es posible comparar los datos de un

Métodos en Neurociencias Cognoscitivas
36
grupo de sujetos con otro, o las medidas de los mismos sujetos durante la aplicación de diferentes tareas o estados.
El análisis estadístico paramétrico exige una distribución normal de los datos. Se supone que las medidas espectrales del EEG de los sujetos normales tienen una dis-tribución normal, suposición que no siempre se cumple. Cuando la distribución no co-rresponde a la curva de distribución normal, tenemos dos opciones: usar estadística no paramétrica o aplicar transformaciones que permiten una distribución normal. Con el fin de lograr o acercar los datos a la distribución normal, John et al. (1980) y Gasser et al. (1982) propusieron las siguientes transformaciones:
Potencia absoluta = ln(x) Potencia relativa = ln(x/1-x) Coherencia (transformación de Fisher) = 0.5 ln(1+x)/(1-x) .
Alternativas para eliminar el efecto de la referencia El uso del EEG computarizado y el desarrollo técnico crean la necesidad de resolver dos problemas básicos de la electroencefalografía:
1 La eliminación del efecto de la referencia, que con su actividad desconoci-da y cambiante reduce significativamente la exactitud que proporciona el EEG computarizado.
2 Encontrar aquellas áreas corticales que son responsables o contribuyen a la acti-vidad registrada sobre la piel cabelluda. El cálculo de la densidad de corriente tiene la ventaja de que elimina el efecto de la referencia. Ésta es la derivada espacial del campo eléctrico medido en μv/cm². La curvatura del campo eléctrico se puede calcular en un lugar particular al sumar el voltaje de 4 a 6 electrodos equidistantes y simétricamente colocados alrededor de un electrodo central. Se calcula el voltaje sumado de los N electrodos y se resta N veces el voltaje en el punto central
L=V1 + V2 + V3 + V4 – 4Vo
donde Vo es el potencial del electrodo central y V1, V2, V3, V4 son los potenciales de los cuatro electrodos vecinos. Este método remueve la importancia del elec-trodo de referencia, mostrando la distribución de voltaje en medidas de flujo de corriente entrando y saliendo en cada lugar de electrodo. Los mapas laplacianos ofrecen una resolución espacial mejor para el cálculo de generadores intracra-neales, aunque necesitan un mayor número de electrodos, lo que es el problema básico de esta técnica. Otra desventaja es que no es posible calcular precisamen-te el voltaje en los electrodos de la periferia.
Representación gráfica del EEG cuantitativo Mapas topográficos Los mapas topográficos son útiles para estudiar la distribución de los potenciales eléctricos sobre la superficie de la cabeza. Es un método de visualización en dos o tres dimensiones.
Las medidas espectrales, cuyos mapas se preparan, se dividen en rangos. A cada ran-go se le asigna algún tipo de color y los colores correspondientes se proyectan en modelos esféricos, en imágenes tomográficas o de resonancia magnética. Hay una gran variedad de representaciones topográficas, algunas de ellas se muestran en la figura 2-5.
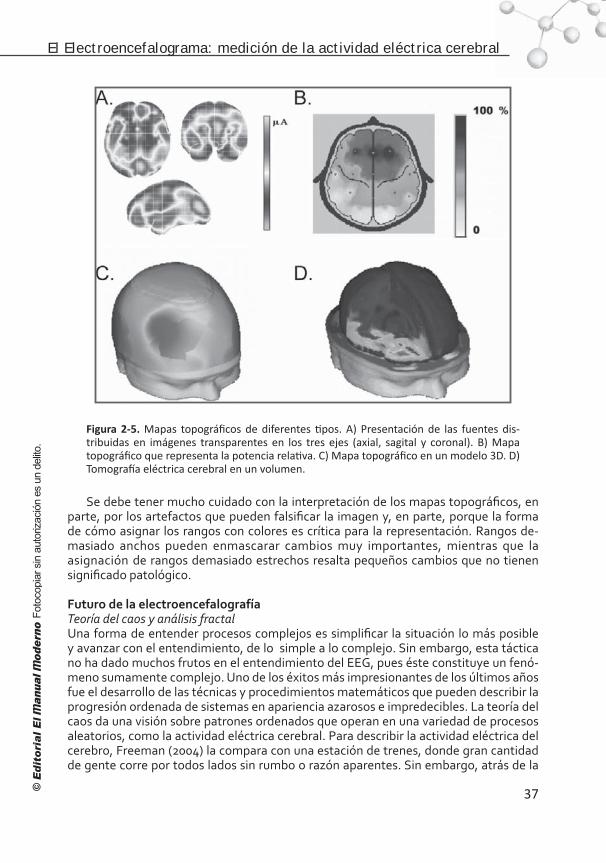
37
El Electroencefalograma: medición de la actividad eléctrica cerebral
Figura 2-5. Mapas topográficos de diferentes tipos. A) Presentación de las fuentes dis-tribuidas en imágenes transparentes en los tres ejes (axial, sagital y coronal). B) Mapa topográfico que representa la potencia relativa. C) Mapa topográfico en un modelo 3D. D) Tomografía eléctrica cerebral en un volumen.
Se debe tener mucho cuidado con la interpretación de los mapas topográficos, en parte, por los artefactos que pueden falsificar la imagen y, en parte, porque la forma de cómo asignar los rangos con colores es crítica para la representación. Rangos de-masiado anchos pueden enmascarar cambios muy importantes, mientras que la asignación de rangos demasiado estrechos resalta pequeños cambios que no tienen significado patológico.
Futuro de la electroencefalografía Teoría del caos y análisis fractal Una forma de entender procesos complejos es simplificar la situación lo más posible y avanzar con el entendimiento, de lo simple a lo complejo. Sin embargo, esta táctica no ha dado muchos frutos en el entendimiento del EEG, pues éste constituye un fenó-meno sumamente complejo. Uno de los éxitos más impresionantes de los últimos años fue el desarrollo de las técnicas y procedimientos matemáticos que pueden describir la progresión ordenada de sistemas en apariencia azarosos e impredecibles. La teoría del caos da una visión sobre patrones ordenados que operan en una variedad de procesos aleatorios, como la actividad eléctrica cerebral. Para describir la actividad eléctrica del cerebro, Freeman (2004) la compara con una estación de trenes, donde gran cantidad de gente corre por todos lados sin rumbo o razón aparentes. Sin embargo, atrás de la

Métodos en Neurociencias Cognoscitivas
38
complejidad de la superficie, hay un orden, cada persona tiene una meta y se apura para alcanzar un tren en particular. La característica principal de estos sistemas es que la condición inicial se puede perturbar con influencias pequeñas sólo en apariencia, pero de consecuencias serias, que hacen que el sistema caótico caiga en un nuevo estado de organización “impredecible”. No obstante, se puede demostrar que este patrón aleato-rio está gobernado por atractores que son propiedades de los sistemas (Kozma y Free-man, 2001). Para el análisis de los datos electroencefalográficos, esta técnica se basa en la dinámica no lineal que examina la complejidad dimensional del EEG, considerándolo como series de tiempo caóticas. La complejidad dimensional se refiere al número de parámetros que contribuyen al comportamiento de las series de tiempo. El caos dis-minuido en el cerebro puede significar patología seria, como el estado de coma o las crisis epilépticas que son de alta coherencia. Para computar los descriptores caóticos de los datos crudos del EEG, es necesario reconstruir un atractor extraño en un espacio de fases multidimensionales. El problema se simplifica mediante el análisis fractal que subdivide el EEG en curvas similares, llamadas dimensión fractal. Usa el algoritmo de Higuchi a fin de transformar los datos del EEG en su dimensión fractal, lo cual permite re-ducir la cantidad de los datos de EEG sin perder información diagnósticamente impor-tante acerca del espectro de las ondas cerebrales (Higuchi, 1988; Ciszewski et al., 1999).
Transformada de ondeletas (wavelet transform) La transformada de ondeletas continua es otro método prometedor y una solución nue-va al problema del análisis de señales de tiempo-frecuencia, como es el caso del EEG. Esta transformación se ha usado para estudiar la información no estacionaria y es una alternativa a la FFT. La idea básica reside en que el EEG tiene formas de ondas repetiti-vas, cuya identificación, descripción y frecuencia de aparición con algoritmos especiales ayuda a describir la totalidad del EEG con gran fidelidad. También se ha utilizado en el estudio de las epilepsias, donde se intenta caracterizar ondas de las etapas ictales, inter y postictales (ictus significa crisis). En este caso, la pregunta básica es, con el propósi-to de predecir crisis epilépticas, si existen ondas preictales varios minutos antes de la aparición de la convulsión (Petrosian et al., 1996) para poder pronosticar su ocurrencia. El problema básico del método es la excesiva exigencia computacional que el proceso requiere. Sin embargo, el desarrollo de potentes computadoras y del uso de algoritmos más rápidos han reducido dramáticamente el tiempo de cómputo requerido, sin com-prometer la exactitud de extracción de información característica. La trasformada de ondeletas usa una estructura de ventanas autoajustadas y de longitud variable que da como resultado una representación en el dominio de tiempo-frecuencia, con una reso-lución variable de los componentes espectrales (Schiff et al., 1994).
Conclusión Con estas nuevas técnicas de análisis de EEG se espera acercarse más a la solución del problema mayor de cómo el cerebro integra los fragmentos de información derivados de grupos de neuronas altamente especializadas a fin de crear una unidad de percepción y pensamiento en forma instantánea. Estos esfuerzos tienen el objetivo de dar al mun-do científico una nueva técnica de evaluación de los registros electroencefalográficos que en un tiempo cercano pueda ayudarnos a entender cómo trabaja el cerebro.

39
El Electroencefalograma: medición de la actividad eléctrica cerebral
ReferenciasÁlvarez, A., Valdés, P. y Pascual, R. (1987). EEG developmental equations confirmed for Cuban
schoolchildren. EEG and Clinical Neurophysiology, 67, 330-332. Başar, E. EEG brain dynamics: Relation between EEG and brain evoked potentials. Nueva York:
Elsevier. Berger, H. (1929). Über das Elektroenkephalogramm des Menschen. Archiv für Psychiatrie und
Nervenkrankheiten, 87, 527-570. Brazier, M.A. (1949). A study of the electric field at the surface of the head. EEG and Clinical
Neurophysiology, 2, 38-52. Ciszewski, J., Klonowski, W., Stepien, R., Jernajczyk, W., Karlinski, A. y Niedzielska, K. (1999).
Application of chaos theory for EEG-signal analysis in patients with seasonal affective di-sorder. Medical and Biological Engineering and Computing, 37 (supl. 1), 359-360.
Fields, R.D. (2006). Beyond the neuron doctrine. Scientific American, 17 (3), 21-27. Freeman, W.J. (2004). Origin, structure, and role of background EEG activity. Clinical
Neurophysiology, 115, 2077-2088. Gasser, T., Bacher, P. y Mocks, J. (1982). Transformation toward the normal distribution of broad
band spectral parameters of the EEG. EEG and Clinical Neurophysiology, 53, 119-124. Harmony, T. (1984). Neurometric assessment of brain dysfunction in neurological patients.
Functional neurosciences (vol. III). Hillsdale: Lawrence Erlbaum. Ass. Higuchi, T. (1988). Approach to an irregular time series on the basis of the fractal theory.
Physica, 31, 277-283. John, E. R., Karmel, B. Z., Corning, W. C., Easton, P., Brown, D., Ahn, H., John, M., Harmony, T.,
Prichep, L., Toro, A., Gerson, I., Bartlett, F., Thatcher, R., Kaye, H., Valdés, P. y Schwartz, E. (1977). Neurometrics. Science, 196, 1393-1410.
John, E. R., Ahn, H., Prichep, L., Trepetin, M., Brown, D. y Kaye, H. (1980). Developmental equations for the EEG. Science, 210, 1255-1258.
Kozma, R. y Freeman, W. J. (2001). Chaotic resonance: methods and applications for robust classification of noisy and variable patterns. International Journal of Bifurcation and Chaos, 10, 2307-2322.
Linkenkaer-Hansen, K., Nikouline, V. V., Palva, J. M. y Ilmoniemi, R. J. (2001). Long-range temporal correlations and scaling behavior in human brain oscillations. Journal of Neuroscience, 21 (4), 370-1377.
Linkenkaer-Hansen, K., Smit, D. J. A., Barkil, A., van Beijsterveldt, T. E. M., Brussaard, A. B., Boomsma, D. I., van Ooyen, A., y de Geus, E. J. C. (2007). Genetic contribution to long rage temporal correlations in ongoing oscillations. Journal of Neuroscience, 27 (59), 13882-13889.
Llinás, R. y Ribary, U. (1993). Coherent 40 Hz oscillation characterizes dream state in humans. Proceedings of the National Academy of Science, 90, 2078-2081.
Lutzenberger, W., Pulvermüller, F., Elbert, T. y Birbaumer, N. (1995). Visual stimulation alters local 40 Hz responses in humans: an EEG study. Neuroscience Letters, 183 (1-2), 39-42.
Matousek, M., y Petersen, I. (1973). Frequency analysis of the EEG in normal children and in normal adolescents. En P. Kellaway y I. Petersen (Eds.). Automation of clinical EEG. (pp. 75-102). Nueva York: Raven Press.
Niedermayer, E. y Lopes da Silva, F. (1993). Electroencephalograpy. Nueva York: Lippincott Williams y Wilkins.
Nuñez, P. L. (1981). Electric Fields of the Brain: The Neurophysics of EEG. Nueva York: Oxford University Press.

Métodos en Neurociencias Cognoscitivas
40
Nuñez, P. L. (1995). Non linear analysis and chaos. En P. L. Núñez (Ed.), Neocortical dynamics and human EEG rhythms. Nueva York: Oxford University Press.
Petrosian, A., Homan, R., Prokhorov, D. y Wunsch, D. C. (1996). Classification of epileptic EEG using neural network and wavelet transform. Spie, 2825, 834-843.
Schiff, S. J., Aldroubi, A., Unser, M. y Sato, S. (1994). Fast Wavelet Transformation of EEG. EEG and Clinical Neurophysiology, 91 (6), 442-455.
Shaker, M. M. (2006). EEG wave’s classifier using wavelet transform and Fourier transform. Intern. Journal of Biomedical Sciences, 1 (2), 1206-1216.
Singer, W. (1993). Synchronization of cortical activity and its putative role in information pro-cessing and learning. Annual Review of Physiology, 55, 349-374.
Steriade, M., Gloor, P., Llinas, R. R., Lopes de Silva, F. H. y Mesulam, M. M. (1990). Report of IFCN Committee on Basic Mechanisms. Basic mechanisms of cerebral rhythmic activities. Electroencephalography and Clinical Neurophysiology, 76, 481–508.
Steriade, M., McCormick, D. A., y Sejnowski, T. J. (1993). Thalamocortical oscillations in the sleeping and aroused brain. Science, 262, 679-685.
Swartz, B. E. y Goldensohn, E. S. (1998). Timeline of the history of EEG and associated fields. EEG and Clinical Neurophysiology, 106 (2), 173-176.

41
Capítulo 3. Potenciales relaciona-dos con eventos (PRE): aspectos básicos y conceptuales
Mario Rodríguez Camacho, Belén Prieto y Jorge Bernal
Resumen El registro de la actividad eléctrica cerebral provocada es una de las técnicas electrofisiológicas más importantes en la comprensión de las bases fisiológicas de los pro-cesos cognoscitivos. El uso extendido de esta técnica reside en que tiene la mayor resolu-ción temporal respecto de todas las demás actualmente utilizadas aunque su resolución espacial es pobre. Los PRE reflejan cambios en la actividad eléctrica relacionados tempo-ralmente con un estímulo que los dispara y que está ligado a un proceso sensorial, percep-tivo, motor o cognoscitivo. Una de sus ventajas es que permite la evaluación en tiempo real de la relación dinámica entre la actividad cerebral y el proceso cognosctivo que se está estudiando; así, proporciona información sobre la temporalidad y secuencia de los procesos cognoscitivos con una gran resolución temporal. El presente capítulo busca ser una introducción al estudio de los PRE: incluye sus aspectos generales, la comparación con otras técnicas de estudio del sistema nervioso, su clasificación y nomenclatura, así como su origen neuronal; presenta una discusión sobre la naturaleza de los componentes de los PRE y un breviario de ellos; hace hincapié en los componentes endógenos y finaliza con una introducción sobre las inferencias que pueden hacerse a partir de los datos obte-nidos con los PRE y las suposiciones que les dan fundamento.
Introducción Los procesos cognoscitivos se refieren a lo que se conoce como percepción, atención, memoria, aprendizaje, comprensión y producción del lenguaje, solución de problemas, creatividad, toma de decisiones y razonamiento. El estudio de estos procesos constitu-ye el objetivo de la psicología cognoscitiva.
Esta disciplina estudia la conducta en situaciones de laboratorio para elaborar con-clusiones sobre los procesos mentales y se relaciona con la neurociencia cognoscitiva, que utiliza diversos métodos para tratar de relacionar la actividad cerebral con las me-didas conductuales (Kellog, 2007).
Es interesante observar cómo a pesar de los grandes avances en la comprensión del sistema nervioso en los niveles celular y molecular, las bases funcionales de los proce-sos cognoscitivos, como la atención, la memoria y el lenguaje, no han alcanzado aún un avance semejante. En parte, esto pudiera deberse a la naturaleza misma de los pro-cesos cognoscitivos, cuya complejidad es objeto de estudio de muchos campos del co-nocimiento, desde la ingeniería a la antropología. Por otra parte, parece ser que los procesos cognoscitivos alcanzan su mayor desarrollo en los seres humanos. Esto limita la aplicación de técnicas invasivas que permitirían un análisis más detallado de la activi-dad cerebral relacionada con dichos procesos.

Métodos en Neurociencias Cognoscitivas
42
Por lo anterior, ha sido conveniente el uso de técnicas no invasivas, como las que registran la actividad eléctrica del sistema nervioso. Una característica fundamental del sistema nervioso es que produce actividad eléctrica y el registro de ésta nos propor-ciona una ventana funcional única que no comparte con otros métodos de estudio, ni siquiera con los sobrevaluados por su alta tecnología y su alto costo. El registro de la actividad eléctrica cerebral es la técnica que permite examinar un secreto celosamente guardado por el sistema nervioso, el de su extraordinaria rapidez para transmitir impul-sos. Así, el registro de los potenciales relacionados con eventos es una de las técnicas electrofisiológicas más importantes para la comprensión de las bases fisiológicas de los procesos cognoscitivos. El uso extendido de esta técnica reside en que tiene la mayor resolución temporal respecto de todas las técnicas actualmente utilizadas, aunque su re-solución espacial es pobre.
Comparación de los PRE con otras técnicasLa mayor parte del conocimiento actual sobre las relaciones entre el cerebro y los pro-cesos cognoscitivos proviene de diferentes métodos: el estudio de pacientes con le-sión cerebral, las técnicas de neuroimagen y las técnicas de registro electrofisiológico. Otros métodos adicionales han incluido la estimulación eléctrica, las intervenciones neuroquirúrgicas y las manipulaciones farmacológicas. Por desgracia, ninguna de estas técnicas refleja de manera clara y detallada los mecanismos neuronales asociados con los procesos cognoscitivos. Cada una de ellas tiene ventajas y desventajas.
Así, los estudios de los déficit cognoscitivos presentes en sujetos lesionados tienen la mayor tradición histórica, como los trabajos de Broca en el lóbulo frontal o los de Harlow en su célebre paciente Phinneas Gage. Sin embargo, esta clase de estudios se complica por toda una serie de factores, entre los que se podrían mencionar la imposibi-lidad de controlar la localización y extensión de una lesión dada, conocer exactamente el tipo y grado de daño neurobiológico causado por diferentes patologías y la influencia de procesos compensatorios que mejoran el funcionamiento del paciente o, incluso, caracterizar con toda exactitud el déficit cognoscitivo-conductual resultante de una lesión específica. En consecuencia, los estudios de lesiones pueden señalar si un área cerebral es esencial para una función, pero no sobre los circuitos o la organización fun-cional que le subyacen (Osterhout, Kim y Kuperberg, 2006; Price, Mummery, Moore, Frakowiak y Friston, 1999).
Las técnicas de neuroimagen —como la tomografía por emisión de positrones (PET) y la resonancia magnética funcional (RMf)— actualmente dominan la escena de los es-tudios funcionales de la cognición. La mayor fortaleza de estas técnicas es su resolución espacial (hasta 1 mm3), ya que proveen un panorama de las estructuras cerebrales in-volucradas en el procesamiento cognoscitivo. No obstante, no hay que perder de vista que estos métodos no miden directamente la actividad cerebral, sino los cambios en el flujo y la oxigenación sanguíneos que se asumen como sinónimos de aquélla. Aunque hay alguna evidencia que apoya este supuesto (Mukamel, Gelbard, Arieli, Hasson, Freid y Malach, 2005), se desconoce bastante sobre el acoplamiento preciso entre actividad neuronal y flujo sanguíneo (Logothetis y Pfeuffer, 2004). Tan importante como lo an-terior es el hecho de que los cambios en el flujo sanguíneo resultantes de un aumento en la actividad cerebral son muy lentos en comparación con la dinámica eléctrica sub-yacente a dicha actividad. En consecuencia, estas técnicas carecen de una adecuada resolución temporal para el estudio de los procesos cognoscitivos, cuya duración se
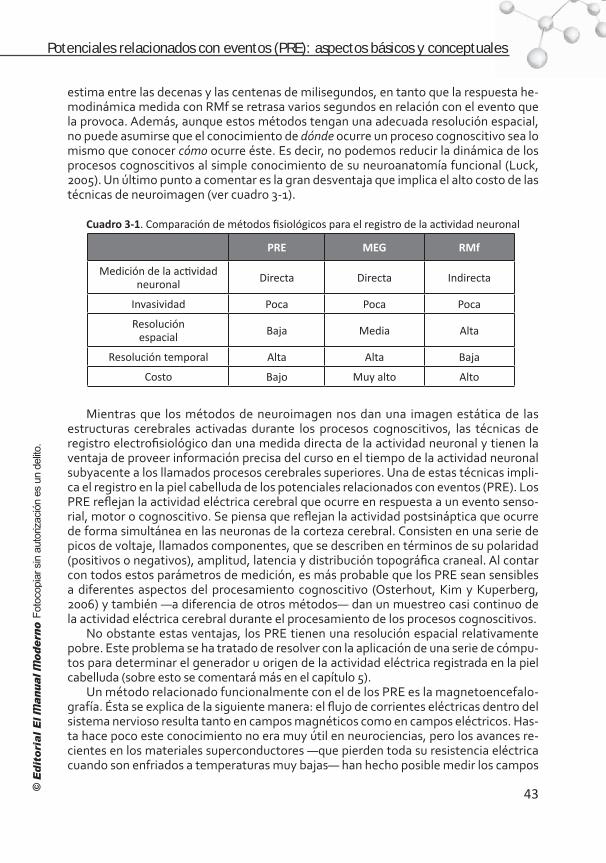
PRE MEG RMf
Medición de la actividad neuronal Directa Directa Indirecta
Invasividad Poca Poca Poca
Resolución espacial Baja Media Alta
Resolución temporal Alta Alta Baja
Costo Bajo Muy alto Alto
43
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
estima entre las decenas y las centenas de milisegundos, en tanto que la respuesta he-modinámica medida con RMf se retrasa varios segundos en relación con el evento que la provoca. Además, aunque estos métodos tengan una adecuada resolución espacial, no puede asumirse que el conocimiento de dónde ocurre un proceso cognoscitivo sea lo mismo que conocer cómo ocurre éste. Es decir, no podemos reducir la dinámica de los procesos cognoscitivos al simple conocimiento de su neuroanatomía funcional (Luck, 2005). Un último punto a comentar es la gran desventaja que implica el alto costo de las técnicas de neuroimagen (ver cuadro 3-1).
Cuadro 3-1. Comparación de métodos fisiológicos para el registro de la actividad neuronal
Mientras que los métodos de neuroimagen nos dan una imagen estática de las estructuras cerebrales activadas durante los procesos cognoscitivos, las técnicas de registro electrofisiológico dan una medida directa de la actividad neuronal y tienen la ventaja de proveer información precisa del curso en el tiempo de la actividad neuronal subyacente a los llamados procesos cerebrales superiores. Una de estas técnicas impli-ca el registro en la piel cabelluda de los potenciales relacionados con eventos (PRE). Los PRE reflejan la actividad eléctrica cerebral que ocurre en respuesta a un evento senso-rial, motor o cognoscitivo. Se piensa que reflejan la actividad postsináptica que ocurre de forma simultánea en las neuronas de la corteza cerebral. Consisten en una serie de picos de voltaje, llamados componentes, que se describen en términos de su polaridad (positivos o negativos), amplitud, latencia y distribución topográfica craneal. Al contar con todos estos parámetros de medición, es más probable que los PRE sean sensibles a diferentes aspectos del procesamiento cognoscitivo (Osterhout, Kim y Kuperberg, 2006) y también —a diferencia de otros métodos— dan un muestreo casi continuo de la actividad eléctrica cerebral durante el procesamiento de los procesos cognoscitivos.
No obstante estas ventajas, los PRE tienen una resolución espacial relativamente pobre. Este problema se ha tratado de resolver con la aplicación de una serie de cómpu-tos para determinar el generador u origen de la actividad eléctrica registrada en la piel cabelluda (sobre esto se comentará más en el capítulo 5).
Un método relacionado funcionalmente con el de los PRE es la magnetoencefalo-grafía. Ésta se explica de la siguiente manera: el flujo de corrientes eléctricas dentro del sistema nervioso resulta tanto en campos magnéticos como en campos eléctricos. Has-ta hace poco este conocimiento no era muy útil en neurociencias, pero los avances re-cientes en los materiales superconductores —que pierden toda su resistencia eléctrica cuando son enfriados a temperaturas muy bajas— han hecho posible medir los campos

Métodos en Neurociencias Cognoscitivas
44
magnéticos neuronales, técnica denominada magnetoencefalograma (MEG), de ma-nera semejante a lo que se hace con el electroencefalograma. La MEG requiere de un dispositivo de interferencia cuántico superconductor (SQUID, por sus siglas en inglés). Tanto el registro de los PRE como de la MEG requieren la presencia de grandes pobla-ciones de neuronas con una configuración geométrica específica (de campos abiertos) y que disparen sincrónicamente. Pero los registros obtenidos con estas técnicas son di-ferentes en los siguientes puntos: 1) mientras que en los PRE se registran las corrientes extracelulares, en la MEG se registran las corrientes intracelulares, 2) los registros de la MEG no son afectados por las propiedades conductoras del cerebro y los tejidos que lo rodean; en contraste, los PRE son distorsionados por la resistencia del cerebro y sus cubiertas, 3) los campos magnéticos se miden en unidades independientes de cualquier sistema de referencia, en tanto que los PRE deben medirse en relación con un voltaje de referencia y 4) los campos magnéticos que se registran desde la superficie del cerebro se generan por corrientes que fluyen tangencialmente a la superficie, por lo que reflejan la actividad neuronal dentro de las fisuras o surcos más que la que se origina en los gi-ros. Por su parte, los PRE se generan por corrientes que fluyen perpendicularmente a la superficie.
Podemos decir que la MEG nos da información sobre la actividad sincrónica de cé-lulas piramidales localizadas en la profundidad de la corteza cerebral, mientras los PRE dan un panorama acerca de la actividad neuronal en respuesta a un estímulo, lo que incluye la actividad de células situadas en la superficie de la corteza. Por último debe mencionarse que la MEG implica tecnología avanzada y de muy alto costo (Hillyard y Picton, 1987; Kutas y Van Petten, 1994) (ver cuadro 3-1).
Los PRESi un par de electrodos se colocan sobre la piel cabelluda y se conectan a un amplificador diferencial, se obtiene un registro que representa las variaciones de voltaje en el tiem-po. Este registro de la actividad eléctrica cerebral espontánea se llama electroencefalo-grama (EEG). Aunque varía de sujeto a sujeto, la amplitud del EEG normal puede estar en el rango de -100 a +100 microvoltios (μV) y su frecuencia de 0.1 a 40 Hz o más.
Podemos definir un segmento (época) del EEG que esté ligado en el tiempo a un es-tímulo presentado si se manifiesta el estímulo a un sujeto mientras se registra su EEG. Es común definir una época que comience 100 ms antes de la presentación del estímulo y que termine 1 000 ms después de su presentación. Dentro de esta época habrá cam-bios de voltaje relacionados específicamente con la respuesta cerebral al estímulo. Son estos cambios de voltaje los que constituyen los potenciales relacionados con eventos (Coles y Rugg, 1995) (ver figura 3-1).
Una definición de los PRE dice que son “cambios en los patrones de voltaje del EEG en curso que están ligados en el tiempo a eventos sensoriales, motores o cognoscitivos” (Hillyard y Picton, 1987). Es decir, los PRE están incluidos en la actividad espontánea o actividad de base del cerebro. Hay que mencionar que la amplitud de un PRE registrado en el cráneo (5-10 microvoltios) es bastante más reducida que la del EEG de base (50-100 microvoltios), por lo que el PRE tiene que ser extraído del EEG mediante promedia-ción. Esto significa que los PRE se obtienen de presentaciones repetidas de estímulos, si se parte de la idea de que la respuesta cerebral a cada estímulo será esencialmente la misma y, al obtener el promedio, se tendrá la actividad eléctrica “más representa-tiva” respecto del estímulo empleado, en tanto que aquella actividad no relacionada

Amplificador
Señal promediadaEstímulo visual
EEG en curso
CZ-
1200 m E E E E E
15
10
5
-100-5
0Mic
rovo
lts
500300100 700 900 1100 ms
45
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
con el procesamiento de éste (“ruido”) tenderá a desaparecer. Lo anterior tiene con-secuencias para considerar el número de estímulos necesarios para obtener un promedio confiable, ya que entre menor amplitud tenga el PRE que se desea estudiar, se necesi-tará un mayor número de ensayos para obtenerlo (Kutas et al., 2006).
Por lo común, los PRE eran conocidos como potenciales evocados (por la traducción directa de evoked potentials) o potenciales provocados. Esto se debió posiblemente a la creencia de que los potenciales reflejaban una actividad cerebral que era provocada de manera exclusiva por la presentación del estímulo y se relacionaba con procesos sen-soriales básicos. Como lo mostró la investigación subsecuente, los potenciales reflejan mucho más que la actividad provocada, lo que llevó al uso del término “potenciales relacionados con eventos” (Coles y Rugg, 1995).
La técnica de los PRE es una de las más útiles para el estudio de las bases fisiológicas de los procesos cognoscitivos. Los PRE reflejan cambios en la actividad eléctrica cere-bral que guardan una relación temporal específica con los estímulos físicos o los proce-sos cognoscitivos que los disparan. Su principal ventaja es que permite la evaluación en tiempo real de la relación dinámica entre la actividad cerebral y el proceso psicológico que se estudia, y proporciona información sobre la cronología y secuenciación de los procesos cognoscitivos con una gran resolución temporal (Rodríguez et al., 2006).
Figura 3-1. Obtención de los PRE en una tarea de atención visual. El trazo superior repre-senta el EEG registrado mientras el sujeto realiza la tarea. Los PRE no pueden visualizarse en el EEG, el cual debe promediarse a partir de muchas presentaciones del estímulo. El trazo inferior representa el PRE visual obtenido al promediar muchas épocas de 1.2 se-gundos de EEG.
Clasificación Exógenos vs endógenos Los PRE consisten de una serie de cambios de voltaje que se visualizan como picos positivos o negativos (es decir, valles) y se presentan en tiempos sucesivos a partir de la presentación del estímulo. A estos picos de voltaje se les suele llamar componentes de los PRE (ver sección “¿Qué son los componentes de los PRE?”). En la figura 3-2(A) se ilustran los PRE obtenidos como respuesta a un estímulo auditivo. Los componentes
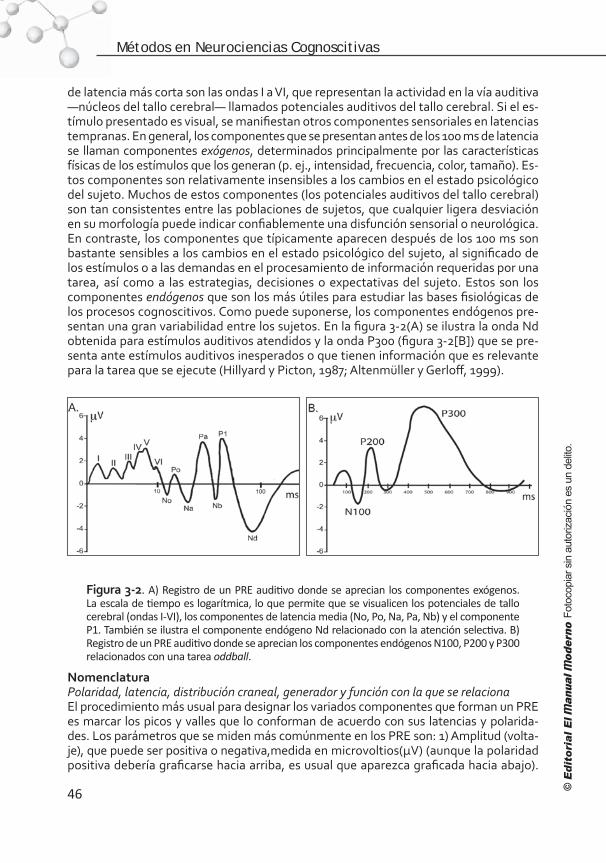
Métodos en Neurociencias Cognoscitivas
46
de latencia más corta son las ondas I a VI, que representan la actividad en la vía auditiva —núcleos del tallo cerebral— llamados potenciales auditivos del tallo cerebral. Si el es-tímulo presentado es visual, se manifiestan otros componentes sensoriales en latencias tempranas. En general, los componentes que se presentan antes de los 100 ms de latencia se llaman componentes exógenos, determinados principalmente por las características físicas de los estímulos que los generan (p. ej., intensidad, frecuencia, color, tamaño). Es-tos componentes son relativamente insensibles a los cambios en el estado psicológico del sujeto. Muchos de estos componentes (los potenciales auditivos del tallo cerebral) son tan consistentes entre las poblaciones de sujetos, que cualquier ligera desviación en su morfología puede indicar confiablemente una disfunción sensorial o neurológica. En contraste, los componentes que típicamente aparecen después de los 100 ms son bastante sensibles a los cambios en el estado psicológico del sujeto, al significado de los estímulos o a las demandas en el procesamiento de información requeridas por una tarea, así como a las estrategias, decisiones o expectativas del sujeto. Estos son los componentes endógenos que son los más útiles para estudiar las bases fisiológicas de los procesos cognoscitivos. Como puede suponerse, los componentes endógenos pre-sentan una gran variabilidad entre los sujetos. En la figura 3-2(A) se ilustra la onda Nd obtenida para estímulos auditivos atendidos y la onda P300 (figura 3-2[B]) que se pre-senta ante estímulos auditivos inesperados o que tienen información que es relevante para la tarea que se ejecute (Hillyard y Picton, 1987; Altenmüller y Gerloff, 1999).
Figura 3-2. A) Registro de un PRE auditivo donde se aprecian los componentes exógenos. La escala de tiempo es logarítmica, lo que permite que se visualicen los potenciales de tallo cerebral (ondas I-VI), los componentes de latencia media (No, Po, Na, Pa, Nb) y el componente P1. También se ilustra el componente endógeno Nd relacionado con la atención selectiva. B) Registro de un PRE auditivo donde se aprecian los componentes endógenos N100, P200 y P300 relacionados con una tarea oddball.
Nomenclatura Polaridad, latencia, distribución craneal, generador y función con la que se relaciona El procedimiento más usual para designar los variados componentes que forman un PRE es marcar los picos y valles que lo conforman de acuerdo con sus latencias y polarida-des. Los parámetros que se miden más comúnmente en los PRE son: 1) Amplitud (volta-je), que puede ser positiva o negativa,medida en microvoltios(μV) (aunque la polaridad positiva debería graficarse hacia arriba, es usual que aparezca graficada hacia abajo).

47
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
Debe tenerse en cuenta que la amplitud se compara a una línea base preestímulo, corto periodo de actividad (es decir, 100 ms) inmediatamente previo a la presentación del estímulo. La amplitud es entonces positiva o negativa respecto de la línea base. 2) La-tencia. Tiempo que transcurre entre la presentación del estímulo y la aparición del pico máximo de un componente de los PRE. La latencia se mide en milisegundos (ms). To-mando en cuenta lo anterior, el nombre P300 de un PRE significa que tiene polaridad positiva y la latencia de su pico máximo es de 300 ms. 3) Distribución topográfica. Lugar del cráneo en el que el componente se registra con mayor amplitud.
Aunque cada PRE debería nombrarse de acuerdo con su generador en el sistema nervioso central y con su papel funcional en el procesamiento de información, la reali-dad es que se desconocen tanto el generador como la función exacta de la mayor parte de los componentes de los PRE. Sin embargo, algunos de ellos pueden nombrarse por su origen, como las ondas más tempranas de los potenciales auditivos (figura 3-2[A]) llamados potenciales del tallo cerebral por originarse en núcleos del tallo cerebral que forman parte de la vía auditiva. Otros componentes reciben su nombre de la función con la que supuestamente se relacionan, como la negatividad de procesamiento (pro-cessing negativity) o la variación contingente negativa (contingent negative variation); o también por su topografía craneal, como la negatividad anterior izquierda (left anterior negativity, LAN) (ver sección “Componentes de los PRE”).
Bases fisiológicas de los PRE En general, se acepta que los PRE reflejan actividad originada dentro del cerebro. Sin embargo, la relación de lo que pasa en el cerebro con lo que puede observarse en el registro craneal aún no se entiende por completo (Coles y Rugg, 1995). No obstante, algunos hechos fisiológicos se aceptan sin controversia: los PRE representan campos eléctricos asociados con la actividad de poblaciones neuronales. Para que un PRE pue-da registrarse en la piel cabelluda, las neuronas que forman parte de la población neuro-nal implicada en un proceso sensorial o cognoscitivo deben activarse sincrónicamente y deben tener una configuración espacial tal que sus campos eléctricos individuales se sumen para formar un dipolo, es decir, un campo con cargas positivas y negativas entre las cuales pueda fluir la corriente. Estas configuraciones espaciales se llaman de campo abierto e implican la alineación de las neuronas en una orientación en paralelo y per-pendicular al cuero cabelludo. Desde el punto de vista de la neurofisiología celular, los PRE reflejan principalmente potenciales postsinápticos que se generan en las células piramidales de la corteza cerebral (Osterhout et al., 2006).
¿Qué son los componentes de los PRE? Hemos mencionado que los picos de los PRE se nombran de acuerdo con su polaridad y latencia (es decir, N100 o P300), que en ocasiones también se designan por su polaridad y su posición ordinal en la onda del PRE (N1, P1, N2, P2) y que otras veces el nombre se designa de acuerdo con su descripción funcional (es decir, la onda negativa de dispari-dad o mismatch negativity, MMN). Ahora bien, debemos distinguir entre los picos de los PRE observados a simple vista y el concepto más abstracto de componente (Donchin et al., 1978; Allison et al., 1986; Kutas et al., 2006).
Se podría pensar que al obtener el PRE uno se enfoca en un pico o valle de la onda resultante que vendría a ser el componente de interés y se determinaría su ampli-tud, su latencia y su distribución topográfica. El mayor impedimento a esta sencilla

Métodos en Neurociencias Cognoscitivas
48
aproximación es el “traslape de componentes”, lo cual se refiere al hecho de que cual-quier onda que observemos en el registro es el resultado de la suma de actividad eléctri-ca, producto de diferentes generadores en el cerebro. Esto se debe a que el cerebro es un medio conductor y la actividad que se genera en un punto en particular se propaga y puede ser detectada en otras localizaciones (Coles y Rugg, 1995).
De acuerdo con lo anterior, un pico de los PRE que encontráramos en una latencia de, digamos, 200 ms podría reflejar no solamente la actividad de un generador cerebral que estuviera activo al máximo en esa latencia, sino la actividad combinada de dos o más generadores que estuviesen activos al máximo antes y después de los 200 ms, y cuyos campos eléctricos se sumaran hasta un máximo en esa latencia. Esta ambigüe-dad en la interpretación de los picos en las formas de onda de los PRE ha llevado a la propuesta de que estos rasgos debieran ser descritos por el término teóricamente neutro “deflexión”, y reservar el término de “componente” a aquellos rasgos de los PRE que puedan atribuirse a la actividad de un generador neuronal específico (Näätänen y Picton, 1987; Coles y Rugg, 1995).
Según Coles y Rugg (1995), para definir el componente, habría un punto de vista “fisiológico” consistente en identificar su generador anatómico dentro del cerebro (Näätänen y Picton, 1987). Ésta es una de las áreas de mayor desarrollo reciente y ha incluido el uso de registros intracraneales en humanos donde la actividad eléctrica re-gistrada directamente en el cerebro se relaciona con la de la piel cabelluda. También se han usado técnicas de neuroimagen que relacionan los PRE con la actividad neuronal localizada por medio de tareas similares con ambas técnicas. Además, se ha aplicado el desarrollo de modelos animales de técnicas de lesión, neuroquímicas y registros úni-cos y múltiples al estudio de sistemas neuronales que pudieran corresponder a los sis-temas generadores de los PRE en humanos. Asimismo, ha habido estudios de casos neurológicos que permiten identificar las relaciones entre las lesiones cerebrales y las distorsiones de los PRE registrados en la piel cabelluda. A pesar de este esfuerzo los procedimientos arriba descritos obtienen información algo indirecta, por lo que una tendencia reciente es la estimación directa de las fuentes generadoras de los PRE por medio de algoritmos matemáticos (ver capítulo 5 de este libro).
Otros investigadores en el campo de los PRE adoptan una aproximación “funcional” para la definición de un componente en términos de la etapa del procesamiento de in-formación con la cual se relaciona (Donchin, 1979; 1981). Así, un componente se define en términos de la función cognoscitiva que se piensa que ejecutan los sistemas cerebrales cuya actividad se registra en el cráneo. El proceso cognoscitivo se puntualiza por la ma-nipulación de variables independientes que afectan el componente (en términos de su amplitud y latencia) y por las relaciones entre el componente y algunas medidas con-ductuales (como la velocidad y exactitud de respuestas). Desde este punto de vista, el principal problema es seleccionar un rasgo específico de la forma de onda del PRE que se relacione con un proceso psicológico específico. Ya que es probable que diferentes operaciones del procesamiento de información ocurran en paralelo, entonces los picos o valles de los PRE registrados en la superficie craneal reflejan más de un proceso. Visto de esta manera, se ha procedido a restar las formas de ondas obtenidas en di-ferentes condiciones experimentales para aislar el componente cuya presencia dis-crimina entre ellas, de modo que lo que es distinto entre dos formas de onda sería el componente de interés y este componente se identificaría con el proceso cognosci-tivo que se cree diferente entre las condiciones. De esta manera, se han identificado

49
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
componentes como la Nd, la negatividad de procesamiento o la MMN (Hillyard y Hansen, 1986; Näätänen, 1992).
Otros intentos para desarrollar métodos de extracción de los componentes han tra-bajado sobre los patrones de covarianza presentes en los conjuntos de datos de los PRE. El más popular de tales métodos es el análisis de componentes principales (ACP) cuyo propósito es identificar las fuentes comunes de varianza en un conjunto de da-tos. Para los PRE, estos datos comprenden los valores que representan las variaciones de voltaje en el tiempo durante la época de registro, las variaciones de voltaje en diferentes localizaciones de electrodos y las variaciones de voltaje en diferentes manipulaciones ex-perimentales (Donchin y Heffley, 1978). Un método de reciente aplicación es el análisis de componentes independientes: ACI (Makeig et al., 1996; Delorme y Makeig, 2004).
Es muy poco probable que los sistemas cerebrales particulares y los procesos cog-noscitivos específicos se activen de manera aislada y, por lo tanto, se necesitan técnicas que permitan identificar con certeza los componentes cuya actividad contribuye a los PRE observados en la superficie craneal. Es importante destacar que aunque hay dife-rentes técnicas para aproximarse al problema (que se mencionan en este apartado), aún no se ha encontrado la solución óptima.
Breviario de componentes de los PRE En esta sección describiremos de manera breve los componentes de los PRE que se reportan comúnmente en la investigación en neurociencia cognoscitiva. Pueden en-contrarse descripciones más extensas en Rugg y Coles, (1995), Regan, (1989) y sobre todo en Hillyard y Picton (1987).
Respuestas sensoriales visuales P1: Potencial positivo que se observa principalmente en los electrodos occipitales
laterales. Se inicia entre los 60-90 ms postestímulo y alcanza un pico máximo entre los 100 y 130 ms. Su latencia varía dependiendo de las características físicas del estímulo, en especial del contraste. Estudios que intentan evaluar el origen de este componente han señalado elementos de la corteza extraestriada dorsal (es decir, giro occipital me-dio y giro fusiforme) como los generadores de este componente (Di Russo et al., 2002). Debido a este posible origen, la P1 es sensible a variaciones en los parámetros de los estímulos visuales. La P1 también es sensible a la dirección de la atención espacial (Hill-yard et al., 1998) y al estado de alertamiento del sujeto (Vogel y Luck, 2000).
N1: Este componente sigue a la P1. Se observa en regiones anteriores y posterio-res. En regiones anteriores tiene su máxima amplitud entre 100-150 ms y parece haber al menos dos componentes N1 posteriores con máximos en 150-200 ms postestímulo, con origen en corteza parietal y corteza occipital lateral, respectivamente. Varios estu-dios han mostrado que la atención espacial influye en estos componentes (ver Hillyard et al., 1998; Mangun, 1995).
N170: Se le llama así al componente que parece ser específico para el procesamiento de caras. Aparece en las regiones occipito-temporales y es de mayor amplitud para los estímulos visuales de caras que se presentan en forma invertida (o sólo se invierten los ojos) que para los estímulos presentados en forma correcta (Bentin, 2002). Eimer (2000) menciona que la N170 se relaciona con los últimos estadios de la codificación estruc-tural de caras, donde las representaciones de las configuraciones globales de la cara se generan para ser utilizadas en procesos subsecuentes del reconocimiento facial. De

Métodos en Neurociencias Cognoscitivas
50
acuerdo con Blaur et al. (2007), la N170 puede ser influenciada por el contenido de la expresión emocional dentro de los estímulos faciales; los estímulos atemorizantes exa-geran la respuesta de la N170. Lo anterior sugiere que este componente se relaciona con un proceso ligado al análisis estructural de las caras que es precedido por el análisis de la expresión emocional.
Respuestas sensoriales auditivas Potenciales auditivos de tallo cerebral (PATC). Componentes que aparecen dentro de los 10 ms postestímulo auditivo. Por lo común se relacionan con diferentes relevos de la vía auditiva. Son útiles para la valoración de la función o patología auditiva en infantes, especialmente en el periodo neonatal. Los PATC son seguidos por los componentes de latencia media que se presentan entre los 10-50 ms (ver figura 3-2) y que probablemen-te se originan del cuerpo geniculado medial y la corteza auditiva primaria.
N1: En forma similar a la N1 visual, la N1 auditiva tiene distintos subcomponentes que incluyen: a) un componente fronto-central con máximo alrededor de los 75 ms; al pa-recer se genera en la corteza auditiva y se relaciona con la atención selectiva temprana (compara el estímulo atendido con el estímulo almacenado en la memoria); b) una onda con máxima amplitud en vértex a los 100 ms de origen desconocido, y c) un componen-te de distribución más lateral con máximo cerca de los 150 ms y que parece generarse en el giro temporal superior (ver resumen por Näätänen y Picton, 1987).
Onda negativa de disparidad o mismatch negativity (MMN). Componente negativo descrito por Näätänen y colaboradores por primera vez en 1978, que aparece entre los 160-220 ms en electrodos centrales de la línea media; este componente se relaciona con la atención involuntaria. La MMN se presenta cuando el sujeto escucha una cadena repetitiva de estímulos idénticos en donde se presentan de forma ocasional estímulos diferentes (es decir, en una secuencia con muchos tonos de 1 000 Hz se presentan tonos ocasionales de 1 200 Hz). Se obtiene al restar los PRE a los estímulos infrecuentes de los frecuentes. La MMN se observa aun cuando el sujeto no utilice los estímulos para una tarea o no ponga atención a ellos (es decir, si está leyendo un libro cuando los estímulos se presenten). Se piensa que la MMN refleja un proceso automático que compara el estí-mulo entrante a una huella en la memoria sensorial del estímulo precedente. A la MMN también se le conoce como N2a.
Componentes endógenos P2. La P200 sigue a la N1 y presenta una topografía fronto-central (Luck, 2005). Aun-que no es claro el proceso cognoscitivo relacionado con P200, Luck y Hillyard (1994) mencionan que en los adultos corresponde a la evaluación del estímulo y a la demanda atencional.
Johnson (1989) encontró que la amplitud de P200 se modifica con la demanda de atención de la tarea que se está ejecutando y Stauder y colaboradores (1993), Taylor y Smith (1995) y Taylor y Kahn (2000) mencionan que la latencia y amplitud de P200 decrementan con la edad, lo que es consistente con otros estudios cognoscitivos de los PRE, en los cuales se muestra que conforme se incrementa la edad, los sujetos se vuel-ven más rápidos o usan estrategias más eficientes para evaluar los estímulos visuales.
N200 o N2: Doscientos milisegundos después de la presentación de un estímulo visual o auditivo, se presenta un componente negativo denominado N200 o N2, aun cuando muchos investigadores han identificado varios componentes diferentes en este rango de

51
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
latencia (ver Luck y Hillyard, 1994; Näätänen y Picton, 1986; Pritchard et al., 1991). El requisito fundamental para la presentación de este componente es que el estímulo se desvíe en alguna forma del contexto precedente. Si el estímulo diferente es un tono no relevante para la tarea, se acompañará de una MMN (N2a), pero si es relevante para la tarea se observará un componente N2 más tardío, llamado N2b (este componente parece covariar con el componente P300 o P3b). Los estímulos diferentes, tanto visuales como auditivos, se acompañan de una N2b más grande sobre sitios centrales para estímu-los auditivos y sobre sitios posteriores para estímulos visuales (Simson et al., 1977). Se piensa que la N2b es un signo del procesamiento de categorización de estímulos.
En la presentación repetitiva de estímulos, la N2a muestra una distribución corti-cal anterior, ya sea por atender conscientemente o por ignorar un estímulo desviado (Näätänen, 1990); la N2b es una negatividad con topografía central, que se observa sólo durante la atención consciente de los estímulos; la N2c presenta una distribución fronto-central en tareas de categorización (Pritchard, Shappell y Brandt, 1991). En los estímulos relevantes para tareas de búsqueda visual con lateralidad específica, se puede observar una N2pc como reflejo de un cambio atencional; este componente tiene una distribu-ción occipito-temporal en la corteza contralateral (Luck, Girelli, Mcdermott, 1997). El componente N2pc no es sensible a la probabilidad y refleja el enfoque de la atención espacial en la localización del blanco (Luck, 2005).
N2b: Refleja un procesamiento voluntario y es provocada cuando los sujetos atien-den selectivamente a estímulos desviados en paradigmas oddball. A diferencia de la MMN, la N2b no se restringe a estímulos auditivos y no refleja específicamente el aban-dono de una colección de estímulos estándar. La N2b es provocada por la desvia-ción de una expectación almacenada mentalmente del estímulo estándar (Sams, Alho y Näätänen, 1983). Se ha sugerido que los generadores de la N2b se encuentran en la corteza temporal superior y en la corteza frontal (Potts, Dien, Hartry-Speiser, McDo-ugal y Tucker, 1998).
P300 o P3: Es el componente más estudiado de los PRE debido a que es de fácil producción y de gran amplitud (de 5 a 20 μV). El paradigma estándar para provocarlo (es decir, paradigma oddball) consiste en una serie de estímulos (visuales o auditivos) de dos clases: frecuentes e infrecuentes. Se requiere que el sujeto responda de algu-na manera a los estímulos infrecuentes. Este componente es una onda positiva que se presenta con mayor predominio en regiones parietales-centrales (ver figura 3-3); su latencia y amplitud pueden variar de acuerdo con:
•Latencia. Va de los 300 hasta los 900 ms (para estímulos auditivos). Esta variabi-lidad de la latencia está controlada por la facilidad con la que un estímulo puede ser categorizado como frecuente o infrecuente. Entre más difícil sea la categori-zación mayor será la latencia. Por esta razón, se ha argumentado que la latencia puede utilizarse como medida de tiempo de evaluación del estímulo (Coles et al., 1995). En consecuencia, cualquier manipulación que posponga la categorización del estímulo debe incrementar la latencia de P300. Dichas manipulaciones inclu-yen aumentar el tiempo requerido para el procesamiento sensorial de bajo nivel o la categorización (Luck, 2005). Se ha probado que la latencia de P300 depende del tiempo requerido para categorizar el estímulo, pero no de los procesos de catego-rización que le siguen; así varios estudios han mostrado que la latencia de P300 no es sensible a la cantidad de tiempo que se requiere para seleccionar y ejecutar una respuesta una vez que el estímulo ha sido categorizado (Kutas et al., 1977).
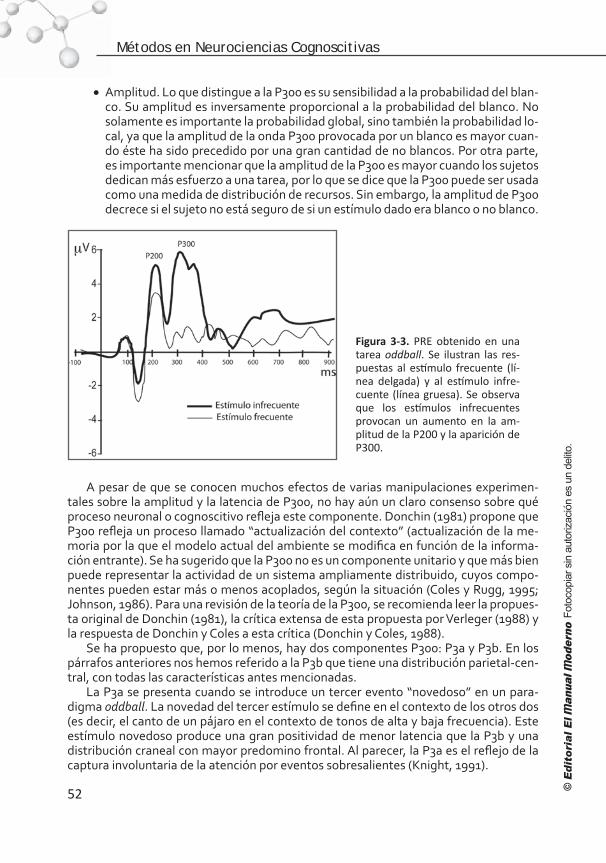
Métodos en Neurociencias Cognoscitivas
52
•Amplitud. Lo que distingue a la P300 es su sensibilidad a la probabilidad del blan-co. Su amplitud es inversamente proporcional a la probabilidad del blanco. No solamente es importante la probabilidad global, sino también la probabilidad lo-cal, ya que la amplitud de la onda P300 provocada por un blanco es mayor cuan-do éste ha sido precedido por una gran cantidad de no blancos. Por otra parte, es importante mencionar que la amplitud de la P300 es mayor cuando los sujetos dedican más esfuerzo a una tarea, por lo que se dice que la P300 puede ser usada como una medida de distribución de recursos. Sin embargo, la amplitud de P300 decrece si el sujeto no está seguro de si un estímulo dado era blanco o no blanco.
Figura 3-3. PRE obtenido en una tarea oddball. Se ilustran las res-puestas al estímulo frecuente (lí-nea delgada) y al estímulo infre-cuente (línea gruesa). Se observa que los estímulos infrecuentes provocan un aumento en la am-plitud de la P200 y la aparición de P300.
A pesar de que se conocen muchos efectos de varias manipulaciones experimen-tales sobre la amplitud y la latencia de P300, no hay aún un claro consenso sobre qué proceso neuronal o cognoscitivo refleja este componente. Donchin (1981) propone que P300 refleja un proceso llamado “actualización del contexto” (actualización de la me-moria por la que el modelo actual del ambiente se modifica en función de la informa-ción entrante). Se ha sugerido que la P300 no es un componente unitario y que más bien puede representar la actividad de un sistema ampliamente distribuido, cuyos compo-nentes pueden estar más o menos acoplados, según la situación (Coles y Rugg, 1995; Johnson, 1986). Para una revisión de la teoría de la P300, se recomienda leer la propues-ta original de Donchin (1981), la crítica extensa de esta propuesta por Verleger (1988) y la respuesta de Donchin y Coles a esta crítica (Donchin y Coles, 1988).
Se ha propuesto que, por lo menos, hay dos componentes P300: P3a y P3b. En los párrafos anteriores nos hemos referido a la P3b que tiene una distribución parietal-cen-tral, con todas las características antes mencionadas.
La P3a se presenta cuando se introduce un tercer evento “novedoso” en un para-digma oddball. La novedad del tercer estímulo se define en el contexto de los otros dos (es decir, el canto de un pájaro en el contexto de tonos de alta y baja frecuencia). Este estímulo novedoso produce una gran positividad de menor latencia que la P3b y una distribución craneal con mayor predomino frontal. Al parecer, la P3a es el reflejo de la captura involuntaria de la atención por eventos sobresalientes (Knight, 1991).
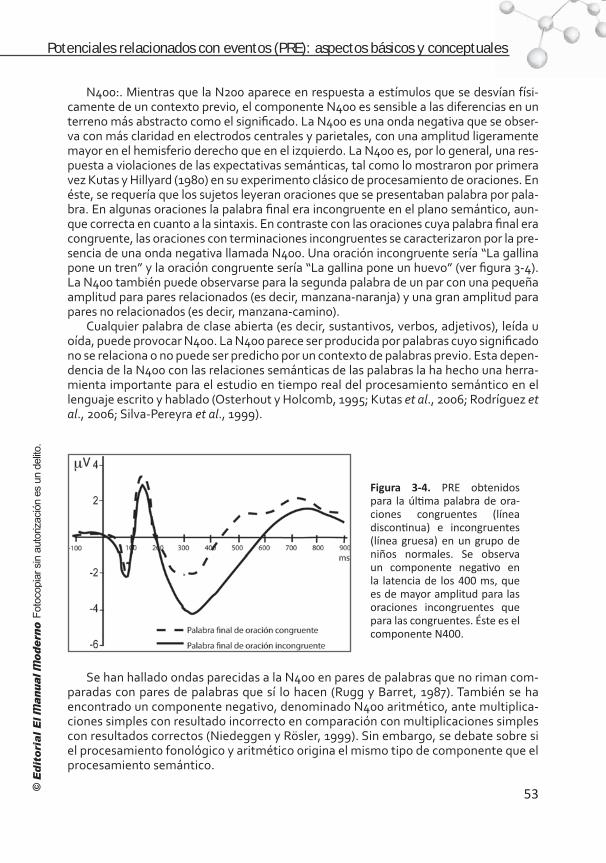
53
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
N400:. Mientras que la N200 aparece en respuesta a estímulos que se desvían físi-camente de un contexto previo, el componente N400 es sensible a las diferencias en un terreno más abstracto como el significado. La N400 es una onda negativa que se obser-va con más claridad en electrodos centrales y parietales, con una amplitud ligeramente mayor en el hemisferio derecho que en el izquierdo. La N400 es, por lo general, una res-puesta a violaciones de las expectativas semánticas, tal como lo mostraron por primera vez Kutas y Hillyard (1980) en su experimento clásico de procesamiento de oraciones. En éste, se requería que los sujetos leyeran oraciones que se presentaban palabra por pala-bra. En algunas oraciones la palabra final era incongruente en el plano semántico, aun-que correcta en cuanto a la sintaxis. En contraste con las oraciones cuya palabra final era congruente, las oraciones con terminaciones incongruentes se caracterizaron por la pre-sencia de una onda negativa llamada N400. Una oración incongruente sería “La gallina pone un tren” y la oración congruente sería “La gallina pone un huevo” (ver figura 3-4). La N400 también puede observarse para la segunda palabra de un par con una pequeña amplitud para pares relacionados (es decir, manzana-naranja) y una gran amplitud para pares no relacionados (es decir, manzana-camino).
Cualquier palabra de clase abierta (es decir, sustantivos, verbos, adjetivos), leída u oída, puede provocar N400. La N400 parece ser producida por palabras cuyo significado no se relaciona o no puede ser predicho por un contexto de palabras previo. Esta depen-dencia de la N400 con las relaciones semánticas de las palabras la ha hecho una herra-mienta importante para el estudio en tiempo real del procesamiento semántico en el lenguaje escrito y hablado (Osterhout y Holcomb, 1995; Kutas et al., 2006; Rodríguez et al., 2006; Silva-Pereyra et al., 1999).
Figura 3-4. PRE obtenidos para la última palabra de ora-ciones congruentes (línea discontinua) e incongruentes (línea gruesa) en un grupo de niños normales. Se observa un componente negativo en la latencia de los 400 ms, que es de mayor amplitud para las oraciones incongruentes que para las congruentes. Éste es el componente N400.
Se han hallado ondas parecidas a la N400 en pares de palabras que no riman com-paradas con pares de palabras que sí lo hacen (Rugg y Barret, 1987). También se ha encontrado un componente negativo, denominado N400 aritmético, ante multiplica-ciones simples con resultado incorrecto en comparación con multiplicaciones simples con resultados correctos (Niedeggen y Rösler, 1999). Sin embargo, se debate sobre si el procesamiento fonológico y aritmético origina el mismo tipo de componente que el procesamiento semántico.
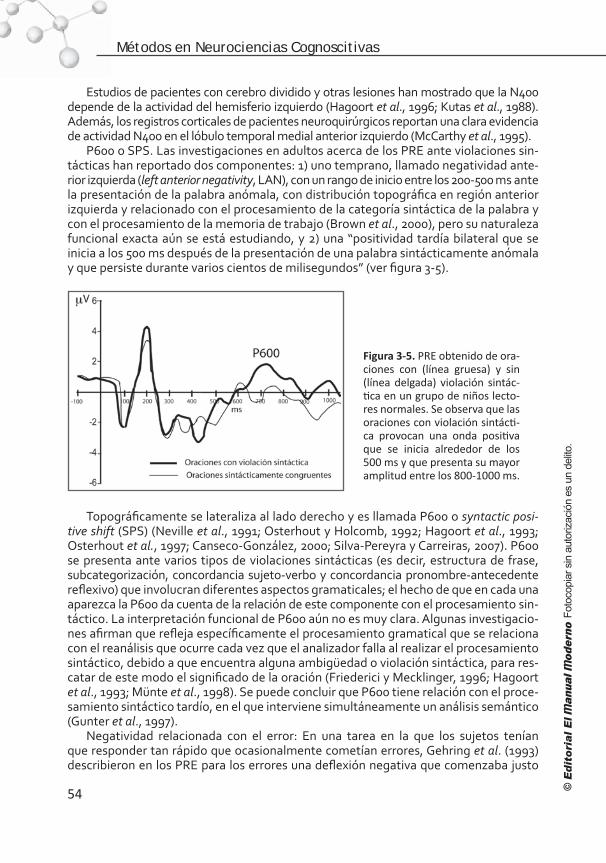
Métodos en Neurociencias Cognoscitivas
54
Estudios de pacientes con cerebro dividido y otras lesiones han mostrado que la N400 depende de la actividad del hemisferio izquierdo (Hagoort et al., 1996; Kutas et al., 1988). Además, los registros corticales de pacientes neuroquirúrgicos reportan una clara evidencia de actividad N400 en el lóbulo temporal medial anterior izquierdo (McCarthy et al., 1995).
P600 o SPS. Las investigaciones en adultos acerca de los PRE ante violaciones sin-tácticas han reportado dos componentes: 1) uno temprano, llamado negatividad ante-rior izquierda (left anterior negativity, LAN), con un rango de inicio entre los 200-500 ms ante la presentación de la palabra anómala, con distribución topográfica en región anterior izquierda y relacionado con el procesamiento de la categoría sintáctica de la palabra y con el procesamiento de la memoria de trabajo (Brown et al., 2000), pero su naturaleza funcional exacta aún se está estudiando, y 2) una “positividad tardía bilateral que se inicia a los 500 ms después de la presentación de una palabra sintácticamente anómala y que persiste durante varios cientos de milisegundos” (ver figura 3-5).
Figura 3-5. PRE obtenido de ora-ciones con (línea gruesa) y sin (línea delgada) violación sintác-tica en un grupo de niños lecto-res normales. Se observa que las oraciones con violación sintácti-ca provocan una onda positiva que se inicia alrededor de los 500 ms y que presenta su mayor amplitud entre los 800-1000 ms.
Topográficamente se lateraliza al lado derecho y es llamada P600 o syntactic posi-tive shift (SPS) (Neville et al., 1991; Osterhout y Holcomb, 1992; Hagoort et al., 1993; Osterhout et al., 1997; Canseco-González, 2000; Silva-Pereyra y Carreiras, 2007). P600 se presenta ante varios tipos de violaciones sintácticas (es decir, estructura de frase, subcategorización, concordancia sujeto-verbo y concordancia pronombre-antecedente reflexivo) que involucran diferentes aspectos gramaticales; el hecho de que en cada una aparezca la P600 da cuenta de la relación de este componente con el procesamiento sin-táctico. La interpretación funcional de P600 aún no es muy clara. Algunas investigacio-nes afirman que refleja específicamente el procesamiento gramatical que se relaciona con el reanálisis que ocurre cada vez que el analizador falla al realizar el procesamiento sintáctico, debido a que encuentra alguna ambigüedad o violación sintáctica, para res-catar de este modo el significado de la oración (Friederici y Mecklinger, 1996; Hagoort et al., 1993; Münte et al., 1998). Se puede concluir que P600 tiene relación con el proce-samiento sintáctico tardío, en el que interviene simultáneamente un análisis semántico (Gunter et al., 1997).
Negatividad relacionada con el error: En una tarea en la que los sujetos tenían que responder tan rápido que ocasionalmente cometían errores, Gehring et al. (1993) describieron en los PRE para los errores una deflexión negativa que comenzaba justo

55
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
después de la respuesta. Esta negatividad presentaba una topografía fronto-central con un pico máximo entre 50 y 150 ms después de la respuesta (Luu et al., 2000; Hajcak et al. 2005). A este componente se le conoce como negatividad relacionada con el error (ERN, por sus siglas en inglés). Estudios recientes han demostrado que la ERN también puede presentarse por retroalimentación negativa después de una respuesta incorrecta o por observar que alguien comete una respuesta incorrecta (Gehring y Willoughby, 2002; van Schie et al., 2004). La interpretación funcional de ERN es que refleja la actividad de un sistema que monitorea las respuestas o es sensible al conflicto entre la respuesta intentada y la respuesta real. La evidencia con otras técnicas de estudio sugiere que estas funciones se originan en la corteza cingulada anterior (Holroyd et al. 2004; Ito et al., 2003; Luck, 2005).
Componentes de los PRE relacionados con la respuesta Potencial de preparación: El potencial de preparación (en alemán Bereitschaftspoten-tial) fue descrito por Kornhuber y Deeke en 1965, en sus estudios de los movimientos voluntarios, cuando se instruía a los sujetos a dar respuestas manuales sin estímulos que las dispararan. Dichas respuestas eran precedidas por un cambio de voltaje nega-tivo, lento, en los electrodos frontales y centrales que empezaba un segundo antes de que el sujeto diera la respuesta física y tenía su máximo en el momento de la respuesta. La topografía del potencial de preparación es contralateral a la mano que va a ser em-pleada para responder.
La porción lateralizada del potencial de preparación se llama potencial de prepara-ción lateralizado (en inglés lateralized readiness potential) y ha sido ampliamente uti-lizado en estudios cognoscitivos por la facilidad con que puede aislarse de otros com-ponentes de los PRE. Así como LPR es contralateral respecto de la mano que responde (mientras otros componentes de los PRE no son lateralizados), se facilita determinar cuándo una manipulación experimental dada afecta la latencia o amplitud de este com-ponente. El LPR se genera, al menos en parte, en la corteza motora (Coles, 1989; Miller et al., 1992) y parece reflejar algún aspecto clave de la preparación de las respuestas motoras. Así, las respuestas son más rápidas cuando LPR es de mayor amplitud en el momento de inicio del estímulo y hay un umbral de la amplitud de la LPR más allá del cual se dispara inexorablemente una respuesta (Gratton et al., 1988; Luck, 2005).
Variación contingente negativa (CNV): Descrita por primera vez por Walter et al. (1964), quienes presentaron pares de estímulos separados por un intervalo de tiempo y establecieron entre ellos una contingencia. En el experimento original, el primer estí-mulo era un clic (estímulo de advertencia) y el segundo una luz titilante (estímulo blanco); se requería que el sujeto oprimiera un botón al estímulo blanco. El intervalo entre el es-tímulo de advertencia y el blanco era de un segundo. Durante el intervalo entre ambos tipos de estímulos se presentaba una onda negativa lenta, con máximo en regiones fronto-centrales. La morfología de la CNV es como de rampa y tiende a alcanzar su máxi-ma negatividad al tiempo en que se presenta el segundo estímulo (Coles y Rugg, 1995). Cuando el periodo de tiempo entre el estímulo de advertencia y el estímulo blanco aumenta a varios segundos, es posible observar que la CNV realmente consiste en una negatividad que sigue al estímulo de advertencia, un retorno a la línea base y luego una negatividad que precede al estímulo blanco. La primera negatividad parece reflejar el procesamiento del estímulo de advertencia y fue llamada onda O por Loveless y Sanford (1974), debido a que relacionaban esta respuesta a un signo de orientación. La segunda negatividad refleja
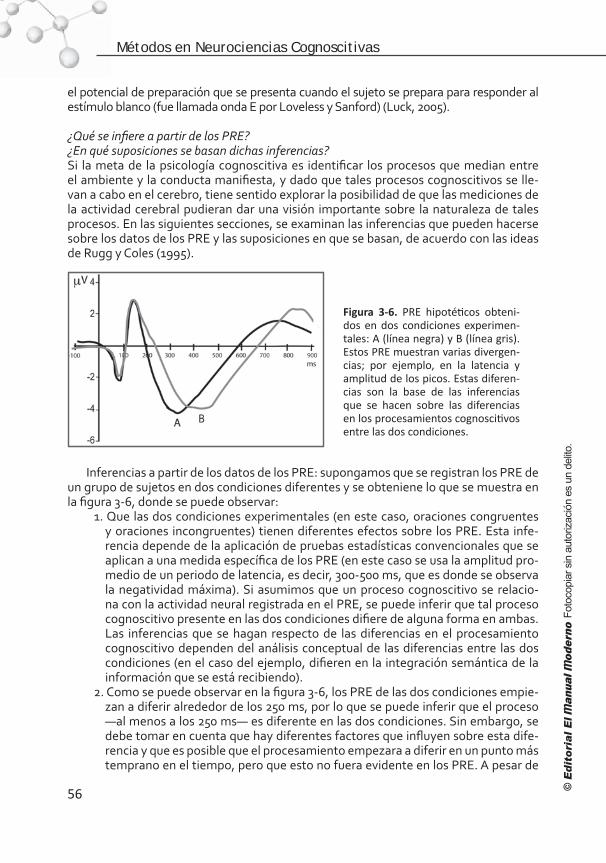
Métodos en Neurociencias Cognoscitivas
56
el potencial de preparación que se presenta cuando el sujeto se prepara para responder al estímulo blanco (fue llamada onda E por Loveless y Sanford) (Luck, 2005).
¿Qué se infiere a partir de los PRE? ¿En qué suposiciones se basan dichas inferencias? Si la meta de la psicología cognoscitiva es identificar los procesos que median entre el ambiente y la conducta manifiesta, y dado que tales procesos cognoscitivos se lle-van a cabo en el cerebro, tiene sentido explorar la posibilidad de que las mediciones de la actividad cerebral pudieran dar una visión importante sobre la naturaleza de tales procesos. En las siguientes secciones, se examinan las inferencias que pueden hacerse sobre los datos de los PRE y las suposiciones en que se basan, de acuerdo con las ideas de Rugg y Coles (1995).
Figura 3-6. PRE hipotéticos obteni-dos en dos condiciones experimen-tales: A (línea negra) y B (línea gris). Estos PRE muestran varias divergen-cias; por ejemplo, en la latencia y amplitud de los picos. Estas diferen-cias son la base de las inferencias que se hacen sobre las diferencias en los procesamientos cognoscitivos entre las dos condiciones.
Inferencias a partir de los datos de los PRE: supongamos que se registran los PRE de un grupo de sujetos en dos condiciones diferentes y se obteniene lo que se muestra en la figura 3-6, donde se puede observar:
1. Que las dos condiciones experimentales (en este caso, oraciones congruentes y oraciones incongruentes) tienen diferentes efectos sobre los PRE. Esta infe-rencia depende de la aplicación de pruebas estadísticas convencionales que se aplican a una medida específica de los PRE (en este caso se usa la amplitud pro-medio de un periodo de latencia, es decir, 300-500 ms, que es donde se observa la negatividad máxima). Si asumimos que un proceso cognoscitivo se relacio-na con la actividad neural registrada en el PRE, se puede inferir que tal proceso cognoscitivo presente en las dos condiciones difiere de alguna forma en ambas. Las inferencias que se hagan respecto de las diferencias en el procesamiento cognoscitivo dependen del análisis conceptual de las diferencias entre las dos condiciones (en el caso del ejemplo, difieren en la integración semántica de la información que se está recibiendo).
2. Como se puede observar en la figura 3-6, los PRE de las dos condiciones empie-zan a diferir alrededor de los 250 ms, por lo que se puede inferir que el proceso —al menos a los 250 ms— es diferente en las dos condiciones. Sin embargo, se debe tomar en cuenta que hay diferentes factores que influyen sobre esta dife-rencia y que es posible que el procesamiento empezara a diferir en un punto más temprano en el tiempo, pero que esto no fuera evidente en los PRE. A pesar de

57
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
ello, el hallazgo de que los PRE comienzan a diferir en un punto particular en el tiempo puede señalar restricciones importantes sobre el curso en el tiempo del proceso cognoscitivo en cuestión. Nuevamente aquí, el análisis conceptual de las diferencias puede aplicarse.
3. Es posible hacer inferencias que busquen evidencia de un procesamiento en pa-ralelo analizando la dimensión espacial de los PRE. Por ejemplo, supongamos que una variable experimental afecta a los PRE en el electrodo Pz, mientras que otra variable provoca, al mismo tiempo, un efecto diferente en los PRE en el electrodo Fz. Tal hallazgo daría buena evidencia de que las variables en cuestión involucrarían sistemas neurales (y por lo tanto, funcionales) diferentes que se su-perponen en el tiempo.
4. Como puede verse en la figura 3-6, los PRE son diferentes en la latencia del pico negativo máximo. La confiabilidad de la diferencia se puede establecer por medio de métodos estadísticos. Si se sabe qué proceso se refleja en este pico ne-gativo, se infiere que las dos condiciones son distintas respecto de este proceso. Mientras que para las inferencias precedentes la especificación de los procesos dependía del análisis conceptual de las diferencias entre las dos condiciones, las inferencias en este punto dependen de la información previa sobre el significado funcional del componente. Estudios anteriores han mostrado que la integración semántica se refleja en la N400, por lo que en el ejemplo usado se infiere que esta integración semántica toma más tiempo en la condición b que en la condición a. Por supuesto, esta inferencia también depende de asumir que todos los otros fac-tores que pudieran influir en la latencia de N400 fueron equivalentes en las dos condiciones.
5. En la figura 3-6, también se observa que la amplitud del pico máximo difiere en ambas condiciones. Esta diferencia se puede evaluar nuevamente por méto-dos estadísticos apropiados. Si sabemos que las variaciones en la amplitud de un componente reflejan las variaciones en el grado en que se activa un proce-so, entonces podemos inferir que las dos condiciones difieren de este respec-to. Al igual que en el punto anterior, esta clase de conocimiento se obtiene por estudios independientes que han explorado tanto las determinantes como las consecuencias de las variaciones de amplitud del PRE en cuestión. Estos estu-dios tienen como objetivo determinar el tipo de variables experimentales cuya manipulación tiene influencia en la amplitud de un componente, así como la descripción de la relación entre la variabilidad de la amplitud y alguna variable conductual del sujeto (tiempos de reacción). Este tipo de datos se usa para ge-nerar una teoría que especifica el proceso cognoscitivo que se relaciona con el componente. En el caso del ejemplo usado, se sabe que la N400 refleja el proce-so de integración semántica. La diferencia de amplitud entre las dos condicio-nes señala que dicho proceso consume más recursos en la condición a que en la condición b.
Supuestos básicos para hacer inferencias funcionales a partir de los PRE Al hablar sobre las inferencias que pueden hacerse a partir de los datos obtenidos con los PRE, debe recordarse que dependen de los supuestos sobre las relaciones entre la actividad cerebral, los procesos cognoscitivos y los PRE. ¿Cuáles son, entonces, los as-pectos más generales que pueden restringir la interpretación funcional de los PRE y

Métodos en Neurociencias Cognoscitivas
58
otros datos fisiológicos (como los obtenidos con las técnicas de neuroimagen)? 1. Establecer una relación entre un sistema neuronal y un proceso cognoscitivo
(mapeo inter-dominio). Cuando se utilizan los PRE para determinar si dos condi-ciones experimentales tienen efectos diferentes en el sistema nervioso, la lógica empleada es bastante clara. Si se encuentran diferencias confiables en los pará-metros de los PRE, esto significa que los procesos que produjeron dichos PRE se asociaron con una actividad neuronal diferente. Sin embargo, ¿cuál es la relación causal de las diferencias en los procesos cognoscitivos a las que dieron origen las manipulaciones experimentales? La respuesta a la pregunta anterior comienza con el siguiente supuesto: cualquier intento de diferenciar estados funcionales distintos con datos fisiológicos requiere de un marco teórico que interrelacio-ne los estados físicos (estados físicos de un sistema neuronal dentro del SNC) con los estados funcionales (procesos cognoscitivos que se asocien al funciona-miento de dicho sistema) del sistema nervioso. Sin este marco teórico, no habría ninguna base para usar alguna de las técnicas fisiológicas para el estudio de los procesos cognoscitivos. El siguiente problema se centra en la cuestión de los crite-rios para decidir cuándo los datos de los PRE nos sugieren procesos funcional-mente distintos, y no el mismo proceso (o procesos) activado en diferentes grados. En general, se asume —a menudo de manera implícita— que las diferen-cias en amplitud de los PRE sin diferencias en la topografía craneal significan va-riaciones en el grado o intensidad con el que uno o varios procesos cognoscitivos se han involucrado en una manipulación experimental. Se supone entonces que diferentes niveles de activación del generador de un PRE indican diferentes nive-les de activación del mismo proceso funcional. De la suposición anterior se des-prende que, cuando dos tratamientos experimentales se asocian con diferencias en la topografía craneal de los PRE, se puede afirmar que dichos tratamien-tos experimentales dieron origen a diferentes patrones de actividad neuronal, por lo que posiblemente se asocien a diferentes procesos funcionales. Sin em-bargo, esto sería una condición necesaria pero no suficiente para concluir que se han identificado procesos funcionalmente distintos. Una razón que apoya esto es que, por el momento, no se sabe qué tan diferentes tienen que ser dos distri-buciones (o qué tan diferente debe ser la localización de los generadores) para justificar la conclusión de que reflejan diferentes procesos cognoscitivos.
2. ¿Qué significan funcionalmente los datos que obtenemos con los PRE? Éste es un tipo más complejo de inferencia a partir de los datos de los PRE. Establecer el significado funcional de los efectos encontrados en un estudio con PRE significa describir la actividad neuronal reflejada en los PRE en términos del pa-pel de dicha actividad en la concreción de un proceso u operación cognosciti-va específica. Aun suponiendo —en forma muy simplificada— que cada proceso cognoscitivo estuviera asociado con un conjunto único de procesos neuronales, establecer la relación entre los dos dominios es un problema formidable consi-derando lo siguiente: 1) Los PRE nos dan información sólo sobre una fracción de toda la actividad neuronal asociada con el procesamiento de un estímulo. De modo que es poco probable que un “PRE característico” de un proceso cognos-citivo fuera equivalente a “un proceso neuronal característico” de dicho proceso. 2) Los intentos para relacionar la actividad neuronal con los procesos cognosci-tivos tienen que confrontar el problema de que las versiones del procesamiento

59
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
de información en una tarea en particular pueden tomar muchas formas diferen-tes. Dichas versiones difieren en sus niveles de análisis y abstracción y, de ma-nera más importante, en sus suposiciones sobre cuestiones básicas, tal como si las versiones funcionales adecuadas sobre el comportamiento fueran posibles sin compromiso con alguna clase de arquitectura “simbólica” del procesamien-to. De esta forma, los intentos para asignar un significado funcional a las varia-ciones en la actividad neuronal relacionadas con una tarea deben, por lo tanto, confrontar los siguientes puntos: 1) Dada la naturaleza de la actividad neuro-nal en cuestión (en el caso de los PRE, cambios sincrónicos en el flujo iónico de grandes poblaciones neuronales), ¿qué nivel de descripción es el más apropia-do para asignar un significado funcional a dicha actividad? 2) ¿Dentro de qué marco debe articularse la descripción? Mientras que las versiones funcionales basadas en distintas arquitecturas de procesamiento son más complementarias que competitivas, parece improbable que todas las versiones funcionalmente plausibles fueran igualmente susceptibles de ser investigadas mediante medi-das fisiológicas, como los PRE.
3. De la correlación a la causalidad. El mayor problema para determinar el significado funcional de los PRE viene de que los estudios de la cognición que se hacen con ellos sólo nos permiten correlacionar, por ejemplo, un proceso cognoscitivo que presun-tamente se activa con un cambio en la amplitud de algún componente de los PRE. Sin embargo, es difícil ver cómo tal relación, por estrecha que sea, podría alguna vez permitirnos concluir que el proceso neuronal responsable de la generación del componente en cuestión también contribuye a la concreción del proceso cognos-citivo en estudio, porque es difícil imaginar cómo un experimento que manipule una variable cognoscitiva pueda diseñarse para poder descartar la posibilidad de que un efecto de los PRE sea una mera consecuencia de la variación en el proceso manipulado y reflejara, más bien, una variación en algún otro proceso contingen-te al que se está manipulando. Este argumento es válido también para otras medi-ciones fisiológicas de la cognición como las técnicas de neuroimagen. Saber que la varianza de un efecto de los PRE está altamente correlacionada con el grado en el que un proceso cognoscitivo específico está involucrado en una tarea puede ser adecuado para muchos propósitos. Por ejemplo, este conocimiento no depende de las bases neuronales, ni del PRE en cuestión, ni del proceso cognoscitivo con el que está asociado. De esta manera, es habitual que los PRE se usen como un índi-ce de procesamiento cognoscitivo en ausencia de medidas conductuales. Sin em-bargo, para otra clase de propósitos, las relaciones de correlación son insuficientes. Considérense, por ejemplo, las aplicaciones cronométricas de los PRE, en las que es necesario conocer la identidad del proceso (o procesos) cognoscitivo reflejado di-rectamente por un efecto de los PRE.
¿Cómo se pueden reemplazar los argumentos de correlación entre los PRE y los procesos cognoscitivos con argumentos de causa-función? En este contexto, es ilus-trativo hacer notar que exactamente la misma dificultad se presenta con otras medidas que podríamos llamar “más directas” sobre la actividad neuronal relacionada con una tarea o un estímulo. Por ejemplo, con las técnicas de registro de neurona única en la corteza visual de monos, se han identificado poblaciones neuronales sensibles de ma-nera específica a una variedad de atributos del estímulo (Zeki, 1993). Pero, como en el

Métodos en Neurociencias Cognoscitivas
60
caso de los PRE y las medidas conductuales, las correlaciones entre la actividad de la neurona única y los atributos del estímulo no pueden más que sugerir las funciones pro-bables llevadas a cabo por una población neuronal dada. Para enfrentar directamente las preguntas funcionales en este caso, es necesario buscar evidencia de otras fuentes, como pueden ser los efectos de las lesiones, la microestimulación o las manipulaciones farmacológicas.
El ejemplo anterior ilustra que: 1) el problema de elucidar el significado funcional de un correlato fisiológico del procesamiento de información por el sistema nervioso no es de ninguna manera exclusivo de los PRE. 2) Para ir más allá de las inferencias de corre-lación sobre los PRE y los procesos cognoscitivos, es necesario probar directamente la existencia de relaciones causales entre la actividad fisiológica que se mide y el proceso cognoscitivo con el que ésta se correlaciona manipulando al sistema neuronal que se supone responsable de un efecto particular en los PRE, para observar las consecuencias funcionales de dicha manipulación.
La conclusión de la discusión precedente es que un entendimiento completo del significado funcional de los PRE implica identificar los procesos y estructuras neurona-les que los generan. Entonces, será posible determinar las consecuencias funcionales de los cambios en la actividad neuronal que se refleja directamente en una onda de los PRE. Esto también permitiría integrar por completo la investigación con los PRE con el conocimiento vasto, y siempre en expansión, de las bases neurales de la cognición y conocer íntegramente el papel funcional de diferentes estructuras neuronales para documentar hipótesis sobre el significado funcional de los diferentes efectos obser-vados en los PRE.
El conocimiento de los generadores de los PRE, necesario para completar esta inte-gración, puede provenir de diferentes fuentes. Los avances recientes en este campo se exponen en el capítulo 5 de este libro. En forma complementaria, se mencionan tam-bién los estudios sistemáticos de los efectos de lesiones cerebrales focales, que pueden dar información sobre el papel de diferentes regiones cerebrales en la modulación y la generación de los PRE, y permitir que las hipótesis específicas sobre la relación entre los efectos de los PRE y los procesos cognoscitivos puedan probarse (Rugg, 1992). Por ejemplo, supongamos que el componente N400 de los PRE refleja los procesos cog-noscitivos necesarios para la comprensión normal del lenguaje, entonces, una lesión cerebral, suficiente para causar una disfunción de estos procesos, debería asociarse con un N400 anormal. Si esta relación no se encontrara, se negaría la hipótesis que liga N400 con la comprensión del lenguaje, sin importar que hubiera altas correlaciones en-tre la amplitud de N400 y las medidas conductuales de comprensión bajo condiciones normales.
Se concluye que para establecer el significado funcional de los PRE relacionados con la cognición, se requiere la identificación tanto de sus correlatos cognoscitivos como de sus orígenes neuronales. A pesar de los problemas discutidos en esta sección, la técnica de los PRE ya ha hecho una contribución significativa en varias áreas de la investigación en la cognición humana.
Aspectos conceptuales para interpretar datos de los PRE 1. ¿Qué refleja exactamente un PRE? Los PRE reflejan no uno sino muchos efectos
causados por el procesamiento de un estímulo efectuado por el sistema nervio-so: cuando analizamos un PRE es conveniente recordar que en él se representa

61
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
actividad neuronal que puede tener distintos generadores cerebrales y que se agrega en el tiempo y el espacio. Esto origina la superposición (overlapping) de componentes. Por ejemplo, cuando el sujeto atiende a una tarea visual supone-mos que la principal área cerebral que se activa es la corteza visual. Sin embargo, se sabe que simultáneamente pueden estar activas otras áreas, como la corteza auditiva o la frontal, cuya actividad se agrega a la de la corteza visual que estamos registrando.Por lo tanto, concluir que un componente de los PRE tiene su origen únicamente en un conjunto restringido de neuronas corticales puede ser inexacto. Es necesario co-nocer con precisión los generadores de un componente de los PRE para obtener la información del área cerebral que con mayor probabilidad provoca el PRE que se re-gistra en la piel cabelluda. Para tratar de caracterizar de manera individual las formas de onda que componen un PRE, existen métodos que descomponen las formas de onda, como pueden ser las aproximaciones de la estadística multivariada (análisis de componentes principales) (Donchin y Heffley, 1978) o el uso de filtros analógicos o digitales que atenúan actividad eléctrica con frecuencias mayores o menores a la del PRE de interés (Rugg y Coles, 1995).En cambio, hay actividad originada en el cerebro como respuesta al procesa-miento de un estímulo que nunca es registrada en la piel cabelluda. Si conside-ramos lo expuesto hasta aquí, es fácil imaginar que hay poblaciones neuronales en el cerebro involucradas en un determinado proceso cognoscitivo, pero que no tienen una configuración de “campo abierto”, o cuya actividad no está suficientemente sincronizada para generar un campo eléctrico y que, por ello, no pueden registrarse como PRE.
2. La distribución topográfica de un PRE no equivale a su generador. Es muy importan-te aclarar que la distribución topográfica craneal de un componente de los PRE no implica que dicho componente sea generado en ese sitio del cerebro. Dado que cual-quier PRE registrado en la piel cabelluda puede provenir de más de una configuración de generadores (fuentes), es muy limitada la inferencia que podemos hacer sobre la localización de un generador basados únicamente en la distribución topográfica cra-neal del PRE. La máxima amplitud de un PRE no necesariamente es registrada en los electrodos más cercanos a su generador. Por esta razón, son necesarios los métodos de localización de las fuentes que se detallan en el capítulo 5. Para obtener conclusiones sólidas sobre los generadores de los PRE que regis-tramos en la superficie craneal, se han construido modelos matemáticos ba-sados en las propiedades conductoras del cerebro y los generadores (fuentes) responsables del campo eléctrico registrado en la superficie (PRE). Algunos de estos modelos asumen un conductor de forma esférica con capas de diferente conductividad para el cerebro, el líquido cerebro-espinal, las membranas que lo rodean (meninges), el cráneo y la piel cabelluda. Para obtener información adi-cional sobre los generadores de los PRE, se han utilizado otras técnicas como los registros de profundidad, las mediciones de los campos magnéticos (MEG), los estudios de lesiones y las técnicas de neuroimagen.
3. ¿Por qué cuando no hay diferencias de amplitud en los PRE entre dos condicio-nes no significa que la actividad cerebral que las originó sea exactamente igual? Si para dos condiciones hay una diferencia confiable entre los PRE en función de

Métodos en Neurociencias Cognoscitivas
62
una manipulación experimental, esto quiere decir que tal manipulación dio ori-gen a una actividad cerebral distinta para cada condición. Sin embargo, la au-sencia de esa diferencia no permite hacer la conclusión opuesta, es decir, que la manipulación experimental no resultó en una actividad eléctrica distinta para cada condición. Esto puede deberse a que, a pesar de las diferencias en la ac-tividad cerebral, éstas pudieron ser de una amplitud muy pequeña como para detectarse en la piel cabelluda, o pudieron haberse generado en una zona con una configuración no favorable para la generación de potenciales de campo. Por lo tanto, no se pueden extraer conclusiones definitivas con base en resultados nulos. Así, para extraer conclusiones sólidas basadas en un efecto nulo, en los PRE no es suficiente determinar que dos condiciones experimentales resulten en PREs sin diferencias significativas.
4. ¿Es la amplitud de los PRE equivalente a la intensidad de un proceso cognosciti-vo? La amplitud (voltaje) que un componente de los PRE presenta no es igual a la “fuerza” de un proceso cognoscitivo dado. Cuando se hacen comparaciones en-tre dos o más condiciones experimentales y se obtienen diferencias de amplitud en algún componente, conviene recordar que dichas diferencias pueden presen-tarse sin cambios en la fuerza (eléctrica) de la actividad neural subyacente. Con respecto a esto, es preciso tener en cuenta que la promediación asume homoge-neidad a lo largo de los ensayos, sobre todo en la latencia en que se presenta el PRE en cada ensayo, lo cual es muy difícil que ocurra, sobre todo para los compo-nentes endógenos. Por ello, las diferencias de amplitud se pueden presentar en realidad como consecuencia de las diferencias en la latencia de ocurrencia de la señal (latency jitter), o —como es frecuente en la práctica— porque hay diferente proporción de ensayos para cada condición.
5. ¿Puede interpretarse la polaridad de un PRE en términos de los procesos cognosci-tivos en estudio? La polaridad con la que se presenta una forma de onda de los PRE no tiene un significado particular. La polaridad de un PRE depende de la localiza-ción de los electrodos de referencia, de la línea base contra la cual el efecto del PRE es comparado y de la localización y orientación de los generadores intracerebrales de los PRE. También la polaridad puede variar por razones neurofisiológicas: que el impulso nervioso de entrada a la neurona sea excitatorio o inhibitorio, o sea, que este impulso se reciba por sinapsis distales o proximales al cuerpo celular de la neurona en cuestión. La resultante de éstas y otras variables determina la polari-dad de un PRE registrado en la piel cabelluda, y no las variables cognoscitivas a las que supuestamente podría asociarse esta polaridad.
6. Correlación frente a causalidad. Todas las inferencias que provienen de los estu-dios de neuroimagen (FMRI, PET, SPECT, etc.) y de los PRE son correlativas. La actividad neural registrada en estos estudios no necesariamente es causada por el proceso cognoscitivo en cuestión. Puede ser incidental o colateral a dicho pro-ceso. Para estudiar la causa de un proceso cognoscitivo, hay que interferir con la actividad neuronalal en la que supuestamente está basado el proceso y estudiar las consecuencias de ello.

63
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
Referencias
Allison, T., Wood, C. C. y McCarthy, G.M. (1986). The central nervous system. En M. G. H. Coles, E. Donchin y S. W. Porges (Eds.), Psychophysiology: Systems Processes and applications (pp. 5-25). Nueva York: Guilford.
Altenmüller, E. y Gerloff, C. (1999). Psychophysiology and the EEG. En E. Niedermeyer y F. Lopes da Silva (Eds.), Electroencephalography: Basic principles, clinical applications and re-lated fields (pp. 637-655). Nueva York: Lippincott Williams and Wilkins.
Bentin, C. (2002). Domain specificity versus expertise: factors influencing distinct processing of faces. Cognition, 83 (1), 1-29.
Blaur, V. C., Maurer, U., Tottenham, N., McCandliss, B. D. (2007). The face specific N170 com-ponent is modulated by emotional facial expression. Behavioral and Brain Functions, 3, 7.
Brown, C., Hagoort, P. y Kutas, M. (2000). Postlexical integration processes during language comprehension: Evidence from brain-imaging research. En M. Gazzaniga (Ed.), The New Cognitive Neuroscience (pp. 881-895). Cambridge: mit Press.
Canseco-González, E. (2000). Using the recording of event-related brain potentials. En Y., Shapiro, L. y Swinney, D. (Eds.), The study of sentence processing. Grodzinsky, language and the brain: representation and processing (pp. 229-266). Nueva York: Academic Press.
Coles, M. G. H. (1989). Modern mind-brain reading: psychophysiology, physiology and cogni-tion. Psychophysiology, 26, 251-269.
Coles, M. G. H. y Rugg, M. D. (1995). Event-related potentials: An introduction. En M. Rugg y M. Coles (Eds.), Electrophysiology of mind: event-related brain potentials and cognition (pp.1-26). Oxford: Oxford University Press.
Coles, M. G. H., Smid, H., Scheffers, M. K., y Otten, L. J. (1995). Mental chronometry and the study of human information processing. En M. Rugg y M. Coles (Eds.), Electrophysiology of mind: event-related brain potentials and cognition (pp. 86-131). Oxford: Oxford University Press.
Delorme, A. y Makeig, S. (2004). EEGlab: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. Journal of Neuroscience Methods, 134, 9-21.
Di Russo, F., Martínez, A., Sereno, M.I., Pitzalis, S. y Hillyard, S. A. (2002). Cortical sources of the early components of the visual evoked potential. Human Brain Mapping, 15, 95-111.
Donchin, E. (1979). Event-related brain potentials: A tool in the study of human infor-mation processing. En H. Begleiter (Ed.), Evoked potentials and behavior (pp.13-75). Nueva York: Plenum.
Donchin, E. (1981). Surprise!...Surprise? Psychophysiology, 18, 493-513.Donchin, E. y Coles, M. G. H (1988). Is the P300 component a manifestation of context upda-
ting? Behavioral and Brain Sciences, 11, 355-372. Donchin, E. y Heffley, E. (1978). Multivariate analysis of event-related potential data: A tutorial
review. En D. Otto (Ed.), Multidisciplinary perspectives in event-related potential research (pp. 555-572). Washington: Goverment Printing Office.
Donchin, E., Ritter, W. y McCallum, C. (1978). Cognitive psychophysiology: The endogenous components of the ERP. En E. Callaway, P. Tueting y S.H. Koslow (Eds.), Brain event-re-lated potentials in man (pp. 349-411). Nueva York: Academic.
Eimer, M. (2000). The face-specific N170 component reflects late stages in the structural enco-ding of faces. Neuroreport, jul 14, 11 (10), 2319-2324.

Métodos en Neurociencias Cognoscitivas
64
Friederici, A. y Mecklinger, A. (1996). Syntactic parsing as revealed by brain responses: First-pass and second-pass parsing processes. Journal of Psycholinguistic Research, 25 (1), 157-176.
Gehring, W. J. y Willoughvy, A. R. (2002). The medial frontal cortex and the rapid processing of monetary games and losses. Science, 295, 2279-2282.
Gehring, W. J., Goss, B., Coles, M. G. H., Meyer, D. E. y Donchin, E. (1993). A neural system for error-detection and compensation. Psychological Science, 4, 385-390.
Gratton, G., Coles, M. G. H., Sirevaag, E. J., Eriksen, C. W., y Donchin, E. (1988). Pre and post-stimulus activation of response channels: A psychophysiological analysis. Journal of Experimental Psychology: Human Perception and Performance, 14, 331-344.
Gunter, T., Stowe, L. y Mulder, G. (1997). When syntax meets semantics. Psychophysiology, 34, 660-676.
Hacjak, G., Muser, J., Yeung, N. y Simons, R. (2005). On the ERN and the significance of errors. Psychophysiology, 42, 151-160.
Hagoort, P., Brown, C. y Groothusen, J. (1993). The syntactic positive shift as a ERP measure of syntactic processing. Language and cognitive processes 8, 439-484.
Hagoort, P., Brown, C. M. y Swaab, T. Y. (1996). Lexical-semantic event-related potential effects in patients with left hemisphere lesions and aphasia and patients with right hemisphere lesions wihout aphasia. Brain, 119, 627-649.
Hillyard, S. A. y Hansen, J. C. (1986). Attention: electrophysiological approaches. En M. G. H. Coles, E. Donchin y S. W. Porges (Eds.), Psychophysiology: systems processes and applica-tions (pp. 227-243). Nueva York: Guilford.
Hillyard, S. A. y Picton, T. W. (1987). Electrophysiology of cognition. En F. Plum (Ed.), Handbook of physiology: Section I. the nervous system (vol.5). Higher functions of the brain (part II). (pp.519-584). Bethesda: Waverly Press.
Hillyard, S. A., Vogel, E. K. y Luck, S. J. (1998). Sensory game control (amplification) as a mechanism of selected attention: Electrophysiological and neuroimaging evidence. Philosophical Transactions of the Royal Society: Biological Sciences, 353, 1257-1270.
Holroyd, C. B., Niewenhuis, S., Yeung, N., Nystrom, L., Mars, R. B., Coles, M. G. H. y Cohen, J. D. (2004). Dorsal anterior cingulate cortex shows fMRI response to internal and external errors signals. Nature Neuroscience, 7, 497-498.
Ito, S., Stuthorn, V., Brown, J. W. y Schall, J. D. (2003). Performance monitoring by the anterior cingulate cortex during saccade countermanding. Science, 302, 120-122.
Johnson, R., Jr. (1986). A triarchic model of P300 amplitude. Psychophysiology, 23, 367-384. Johnson, R., Jr. (1989). Developmental evidence for modality dependent P300 generators: a
normative study. Psychophysiology, 26, 651-667.Kellog, R. (2007). Fundamentals of cognitive psychology (p. 3) California: Sage Publications.Knight, R.T. (1991). Evoked potential studies of attention capacity in human frontal lobe le-
sions. En H. S. Levyn, H. M. Eisenberg y A. L. Benton (Eds.), Frontal lobe function and dis-function (pp. 139-153). Nueva York: Oxford University Press.
Kutas, M. y Hillyard, S. A. (1980). Reading senseless sentences: Brain potentials reflect seman-tic incongruity. Science, 207, 203-205.
Kutas, M. y van Petten, C. (1994). Psycholinguistics electrified. Event-related brain potential in-vestigations. En M. A. Gernsbacher (Ed.), Handbook of psycholinguistics (pp. 83-143). San Diego: Academic Press.
Kutas, M., Hillyard, S. A., y Gazzaniga, M. (1988). Processing of semantic anomaly by right and left hemispheres of commissurotomized patients. Brain, 111, 553-576.

65
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
Kutas, M., McCarthy, G. y Donchin, E. (1977). Augmenting mental chronometry: The P300 as a measure of stimulus evaluation time. Science, 197, 792-795.
Kutas, M., van Petten, C. K. y Kluender, R. (2006). Psycholinguistics electrified II (1994-2005). En M. A. Gernsbacher y M. Traxler (Eds.), Handbook of psycholinguistics, 2nd edition (p.659-724) Nueva York: Elsevier Press. .
Logothetis, N. y Pfeuffer, J. (2004). On the nature of the BOLD fMRI contrast mechanism. Magnetic Resonace Imaging, 22 (10), 1517-1531.
Loveless, M. E. y Sanford, A. J. (1974). Effects of age on the contingent negative variation and pre-paratory sets in a reaction-time task. Journal of Gerontology, 29, 52-63.
Luck, S. J. (2005). An introduction to the event-related potential technique. Cambridge: MIT Press.
Luck, S. J. y Hillyard, S. (1994). Electrophysiological correlates of feature detection during vi-sual search. Psychophysiology, 31, 291-308.
Luck, S. J., Girelli, M., Mcdermott, M. T. y Michelle, A. (1997). Bridging the gap between monkey neurophysiology and human perception: An ambiguity resolution theory of visual selective attention. Cognitive Psychology, 33, 64-87.
Luu, P., Collins, P. y Tucker, D. M. (2000). Mood, personality and self-monitoring: Negative effect and emotionality in relation to frontal lobe mechanisms of error monitoring. Journal of Experimental Psychology: General, 129, 43-60.
Makeig, S., Bell, A. J., Jung, T. P. y Sejnowski, T. J. (1996). Independent component analysis of electroencephalographic data. En D. Touretzky, M. Mozer y M. Hasselmo (Eds.), Advances in neural information processing systems (vol. 8), (pp. 145-151). Cambridge: MIT Press.
Mangun, G. (1995). Neural mechanisms of visual selective attention. Psychophysiology, 32, 4-18. McCarthy, G., Nobre, A., Bentin, S. y Spencer, D. (1995). Language-related field potentials
in the anterior-medial temporal lobe: Intracranial distribution and neural generators. Journal of Neuroscience, 15, 1080-1089.
Miller, J., Riehle, A. y Requin, J. (1992). Effects of preliminary perceptual output on neuronal ac-tivity of the primary motor cortex. Journal of Experimental Psychology: Human Perception and Performance, 18, 1121-1138.
Mukamel, R., Gelbard, H., Arieli, A., Hasson, U., Freid, I. y Malach, R. (2005). Coupling between neuronal firing, field potentials and fMRI in human auditory cortex. Science, 309, 951-954.
Münte, T., Heinze, H., Matzke, M., Wieringa, B. y Johannes, S. (1998). Brain potentials and syntactic violations revisited: no evidence for specificity of the syntactic positive shift. Neuropsychologia, 36 (3), 217-226.
Näätänen, R. (1990). The role of attention in auditory information processing as revealed by event-related potentials and other brain measures of cognitive function. Behavioral and Brain Sciences, 13, 201-288.
Näätänen, R. (1992). Attention and brain function. Hillsdale: Erlbaum. Näätänen, R y Picton, T. (1986). N2 and automatic versus controlled processes. En W. McCallum,
R. Zappoli y F. Denoth (Eds.), Cerebral psychophysiology: studies in event-related poten-tials (pp. 169-186). Amsterdam: Elsevier.
Näätänen, R y Picton, T. (1987). The N1 wave of the human electric and magnetic response to sound: A review and an analysis of the component structure. Psychophysiology, 24, 375-425.
Neville, H., Nicol, J., Barss, A., Forster, K. y Garrett, M. (1991). Syntactically based sentence processing classes: Evidence from event-related brain potentials. Journal of Cognitive Neuroscience, 3, 151-165.

Métodos en Neurociencias Cognoscitivas
66
Niedeggen, M. y Rösler, F. (1999). N400 effects reflect activation spread during arithmetic fact retrieval. Psychological Science, 10, 271-276.
Osterhout, L. y Holcomb, P. (1992). Event-related potentials elicited by syntactic anomaly. Journal of Memory and Language, 31, 785-806.
Osterhout, L. y Holcomb, P. (1995) Event-related potentials and language comprehension. En M. Rugg y M. Coles (Eds.), Electrophysiology of mind: event-related brain potentials and cogni-tion (pp. 171-209. Oxford: Oxford University Press.
Osterhout, L., Kim, A. y Kuperberg, G. (2006). The neurobiology of sentence comprehension. En M. Spivey, M. Joannisse y K. Macrae (Eds.), The Cambridge handbook of psycholinguis-tics (pp. 295-317). Cambridge: Cambridge University Press.
Osterhout, L., McLaughlin, J. y Bersick, M. (1997). Event-related brain potentials and human language. Trends in Cognitive Sciences, 1 (6), 203-209.
Potts, G., Dien, J., Hartry-Speiser, A., McDougal, L. y Tucker, D. (1998). Dense sensor array topography of the event-related potential to task-relevant auditory stimuli. Electroencephalography and Clinical Neurophysiology, 106, 444-456.
Price, C., Mummery, C., Moore, C., Frakowiak, R. y Friston, K. (1999). Delineating necessary and sufficient neural system with functional imaging studies of neuropsychological patients. Journal of Cognitive Neuroscience, 11 (4), 371-382.
Pritchard, W., Shappell, S. y Brandt, M. (1991). Psychophysiology N200/ N400: a review and classification scheme. En J. Jennings, P. Ackles y M. Coles (Eds.), Advances in psychophy-siology (vol. 4), (pp. 43-106). London: Jessica Kingsley.
Reagan, D. (1989). Human brain electrophysiology: evoked potentials and evoked magnetic fields in science and medicine. Nueva York: Elsevier. Rodríguez, M., Prieto, B., Bernal, J., Marosi, E., Yáñez, G., Harmony, T., Silva-Pereyra, J., et al.
(2006). Language event-related potentials in poor readers. En S. Randall (Ed.), Learning di-sabilities new research (pp. 187-217). Estados Unidos: Nova Publisher.
Rugg, M. (1992). Event-related potentials in clinical neuropsychology. En J. Crawford, W. Mckinlay y D. Parker (Eds.), Handbook of neuropsychological assessment (pp. 393-412). Hillsdale: Erlbaum.
Rugg, M., y Barret, S. (1987). Event-related potentials and the interaction between orthogra-phic and phonological information in a rhyme-judgment taks. Brain and Language, 32, 336-361.
Rugg, M. y Coles, M. (1995). The ERP and cognitive psychology: Conceptual issues. En M. Rugg y M. Coles (Eds.), Electrophysiology of mind: event-related brain potentials and cognition. Nueva York: Oxford University Press.
Sams, M., Alho, K. y Näätänen, R. (1983). Sequential effects on the ERP in discriminating two stimuli. Biological Psychology, 17, 41-58.
Silva-Pereyra, J., y Carreiras, M. (2007). An ERP study of agreement features in Spanish. Brain Research, 1185, 201-211.
Silva-Pereyra, J., Harmony, T., Villanueva, G., Fernández, T., Rodríguez, M., Galán, L., et al. (1999). N400 and lexical decisions: Automatic or controlled processing? Clinical Neurophysiology, 110, 813-824.
Simson, R., Vaughan, H. y Ritter, W. (1977). The scalp topography of potentials in auditory and visual discrimination task. Electroencephalography and Clinical Neurophysiology, 42, 528-535.
Stauder, J., Molenaar, P. y van der Molen, M. (1993). Scalp topography of event-related bra-in potentials and cognitive transition during childhood. Child Development, 64, 769-788.

67
Potenciales relacionados con eventos (PRE): aspectos básicos y conceptuales
Taylor, M. y Khan, S. (2000). Top-down modulation of early selective attention processes in chil-dren. International Journal of Psychophysiology, 37 (2), 135-147.
Taylor, M. y Smith, M. (1995). Age related ERP changes to verbal and nonverbal memory tasks. Journal of Psychophysiology, 9, 283–297.
Van Schie, H., Mars, R., Coles, M. y Bekkering, H. (2004). Modulation of activity in medial fron-tal and motor cortices during error observation. Nature Neuroscience, 7, 549–554.
Verleger, R. (1988). On the utility of P3 latency as an index of mental chronometry. Psychophysilogy, 34, 131–156.
Vogel, E. y Luck, S. (2000). The visual N1 component as an index of a discrimination process. Psychophysiology, 37, 123–190.
Walter, W., Cooper, R., Aldridge, V., McCallum, W. y Winter, A. (1964). Contingent negative va-riation: An electric sign of sensorimotor association and expectancy in the human brain. Nature, 428, 748-751.
Zeki, S. (1993). A vision of the brain. Oxford: Blackwell Scientific.

68
Capítulo 4. Potenciales relaciona-dos con eventos: técnicas y méto-dos de registro
Jorge Bernal, Mario Rodríguez Camacho, Belén Prieto yHelena Romero
Resumen En este capítulo se tratan las bases del registro de los potenciales relacionados con even-tos (PRE) y los aspectos técnicos sobre la promediación, uso de filtros, rechazo y correc-ción de artefactos que se utilizan para obtenerlos. Asimismo, se revisan los temas rela-cionados con la medición de las latencias y amplitudes de los componentes de los PRE.
Introducción El registro de la actividad eléctrica cerebral provocada es una de las técnicas electrofisiológicas más importantes para estudiar las bases fisiológicas de los procesos cognoscitivos. El uso extendido de esta técnica reside en que tiene mayor resolución temporal respecto de otras técnicas actualmente utilizadas, aunque su resolución es-pacial es pobre.
Los PRE reflejan cambios en la actividad eléctrica relacionados temporalmente con un estímulo que los dispara, ligado a un proceso sensorial, perceptivo, motor o cognos-citivo. Una de sus ventajas es que permite evaluar en tiempo real la relación dinámica entre la actividad cerebral y el proceso cognoscitivo que se está estudiando, por lo que proporciona información precisa sobre la temporalidad y secuencia de los procesos cognoscitivos. El presente capítulo pretende ser una introducción a las bases técnicas de los PRE; incluye los principios básicos de su registro y los aspectos relacionados con la promediación, uso de filtros, rechazo y corrección de artefactos. Asimismo, se tratan los temas de medición de latencia y amplitud de los componentes de los PRE.
Elementos para el registro de los PRE Electrodos
Los electrodos conducen los potenciales eléctricos del paciente al electroencefaló-grafo, por lo que es importante que sean buenos conductores eléctricos. Además, es ne-cesario usar una pasta conductora para conectar el electrodo a la piel. A lo largo de la histo-ria de la electroencefalografía, se han usado electrodos de diferentes clases de metales, los cuales difieren en su conductancia, que es la capacidad para conducir la electricidad. En la actualidad, probablemente los electrodos que más se usan son los de estaño, plata-cloruro de plata y los de plata-cloruro de plata que están sinterizados.Estos últimos tienen la ventaja de que, además de su alta conductancia, requieren de bajo mantenimiento.
Electrodos activos y de referencia El registro de la actividad electroencefalográfica (actividad eléctrica cerebral espontá-nea) se hace con el amplificador diferencial, el cual utiliza un electrodo activo (A), un

69
Potenciales relacionados con eventos técnicas y métodos de registro
electrodo de referencia (R) (que puede estar en alguno de los sitios que se suelen usar para tal efecto: lóbulos de las orejas, protuberancia mastoidea, etc.) y un electrodo co-nectado a tierra (T) (conectado habitualmente en alguna parte de la cabeza). De esta manera, el registro electroencefalográfico es resultado de la diferencia de voltajes en-tre el electrodo activo y el electrodo de tierra (A-T) menos el voltaje registrado entre electrodo de referencia y el electrodo de tierra (R-T). Dado que técnicamente el ruido eléctrico registrado en el circuito de tierra es el mismo que el registrado en A-T y R-T, al hacer la sustracción este ruido será eliminado.
La selección del sitio del electrodo de referencia más adecuado se hace consideran-do los lugares eléctricamente más neutrales, aunque en la práctica, habrá una actividad eléctrica, por mínima que sea, en cualquier lugar que se escoja para ello, por lo que no hay razón para pensar que un sitio puede ser mejor que otro. Lo recomendable es que la selección del sitio se base en que éste sea confortable para el paciente, que no registre actividad eléctrica generada en un solo hemisferio cerebral (es decir, poner como refe-rencia sólo un mastoides o sólo un lóbulo de la oreja) y, por último, que siempre se es-coja el mismo sitio en los experimentos que un mismo investigador efectúa, sobre todo si éstos están relacionados. Lo anterior es importante porque el potencial registrado en un sitio varía en función de la actividad registrada en el electrodo de referencia. También es una buena idea seleccionar el mismo sitio de referencia que otros investigadores usan y trabajan en la misma temática, toda vez que esto permite hacer comparaciones entre experimentos (Luck, 2005).
Para evitar que la actividad de un solo hemisferio influya en el registro, por lo ge-neral se emplean los electrodos de referencia físicamente cortocircuitados. Éste es el método que se ha empleado y se sigue empleando en muchos laboratorios, por lo que su uso está por completo justificado. No obstante, de acuerdo con Luck (2005), las referencias cortocircuitadas distorsionan la distribución de voltajes sobre la piel cabelluda y reducen las asimetrías en los PRE. Aunque existen otros métodos que se han usado para resolver el problema de la referencia, como la referencia promedio de todos los electrodos registrados o la referencia de otro electrodo activo, Luck (2005) recomienda el método descrito por Nunez (1981), que consiste en combinar matemáti-camente los sitios de referencia izquierdo y derecho mediante un promedio de ambas señales y, después, tomar este resultado como referencia para el electrodo activo. Por fortuna, los equipos actuales permiten el registro del EEG de los electrodos de referencia como un canal independiente, lo que posibilita aplicar los métodos que se recomiendan.
Convertidor analógico-digital El siguiente elemento necesario para tener un registro adecuado del EEG es el con-vertidor analógico-digital. Éste es un dispositivo electrónico que, como su nombre lo indica, digitaliza la señal eléctrica que recibe en forma analógica después de que ha sido amplificada, lo que permite que los datos se puedan procesar con la computadora. Así, la señal que llega en forma continua a los amplificadores se convierte en una secuencia de valores discretos que se aproximan a la señal original. Cada punto se expresa como un par de números, en donde uno de ellos representa el voltaje y el otro el tiempo en que ocurre. Esta información es almacenada para su posterior análisis.
Aunque en la práctica se podría decir que la señal digitalizada es igual a la señal con-tinua que recibe el convertidor analógico-digital, en realidad no es así, pues cuando se captura un punto de la señal continua, pasa un intervalo de tiempo hasta que se captura

Métodos en Neurociencias Cognoscitivas
70
un nuevo punto. Si el intervalo entre punto y punto es suficientemente corto se podrán obtener señales digitalizadas muy parecidas a la original, pero si se alarga aparecerán distorsiones que serán mayores a medida que se alarga dicho intervalo. Estas distorsio-nes dan origen a lo que se ha llamado “fenómeno ALIAS”, que consiste en componentes de la señal digitalizada que no aparecen en la señal original. Los puntos que se capturan secuencialmente se denominan muestras; al tiempo que transcurre entre la captura de una muestra y otra se denomina periodo de muestreo, y al número de muestras que se capturan en un segundo se le llama ritmo de muestreo, el cual se mide en hertzios (Hz). Por ejemplo, cuando se habla de un ritmo de muestreo de 500 Hz significa que se están capturando 500 muestras en un segundo, lo que nos permite calcular que cada muestra se está tomando en un periodo de muestreo de 2 milisegundos. El ritmo de muestreo puede fijarse en los equipos de EEG, dentro sus limitaciones.
Con base en lo anterior, cabe preguntarse ¿cuál es el ritmo de muestreo al que debe registrarse el electroencefalograma? De acuerdo con el teorema de Nyquist, la infor-mación contenida en una señal continua se puede capturar en forma digital a un ritmo de muestreo de, al menos, el doble de la frecuencia más alta de la señal que se está cap-turando. En el caso particular del EEG, las ondas de interés se encuentran en el rango de 0-100 Hz, pero, según el interés del investigador, el rango puede ser menor, por lo que es necesario fijar el rango de frecuencias que se quiere amplificar para determinar el ritmo de muestreo. Para seleccionarlo, se recurre a los filtros pasa baja y pasa alta de frecuencia, los cuales determinan las frecuencias que habrán de amplificarse. Un filtro pasa baja determina la frecuencia máxima que se amplificará. Por ejemplo, un filtro pasa baja de 100 Hz amplificará todas las frecuencias menores a 100 Hz; por otra parte, un filtro pasa alta de 0.5 Hz amplificará todas las frecuencias que se encuentren por arriba de esa frecuencia. Es habitual que para un registro simple de EEG se escoja un filtro pasa baja de 30 Hz y uno pasa alta de 0.3 o 0.5 Hz, ya que las frecuencias de interés clínico se encuentran en este rango. En el caso de la investigación de los componen-tes cognoscitivos (endógenos) de los PRE, los filtros pasa alta se pueden fijar en un ran-go entre 30 y 100 Hz, según se trate de componentes tempranos o tardíos, y dejar las frecuencias más altas para los primeros componentes y las más bajas para los últimos.
La precisión con que se midan los voltajes dependerá de la resolución del convertidor, expresada en el número de bits. En la actualidad los convertidores más usados tienen 12 bits de resolución, o sea que pueden asignar 212 (4 096) valores de voltaje. Existen otros con una resolución mayor, como el de 16 bits, que puede asignar hasta 65 536 va-lores. De esta forma, según el valor mínimo y máximo de voltaje que acepte un conver-tidor, el valor mayor se asigna al valor máximo de voltaje que acepta y el valor cero, al mínimo. A todos los voltajes que excedan el valor mínimo se les asignará el mismo valor y a los que excedan el máximo se les asignará también el valor máximo. Evidentemente un convertidor con mayor cantidad de bits podrá medir con precisión valores mayores de voltaje que uno con menor resolución.
Por otra parte, la selección de la ganancia es un factor importante a considerar, ya que si se selecciona una ganancia que produzca valores que excedan a los que acepta el convertidor, la señal se distorsionará. Así, es conveniente escoger una ganancia en que los valores más extremos del EEG se ajusten a los valores máximos y mínimos permitidos por el convertidor, para aprovechar óptimamente su resolución. En general, la mayoría de los sistemas de registro trabaja bien con ganancias en el rango de 1 000 a 50 000.

71
Potenciales relacionados con eventos técnicas y métodos de registro
Filtros Como ya se mencionó, los filtros son un elemento de suma importancia en el registro y en la edición de la actividad eléctrica cerebral. El filtrado se hace, por lo regular, me-diante circuitos electrónicos incorporados a los amplificadores o interpuestos entre el amplificador y otro dispositivo de registro, como el convertidor analógico-digital. Este tipo de filtro electrónico se llama filtro analógico por trabajar con señales continuas y difiere del filtro digital, entre otras cosas, en que este último se aplica a señales discre-tas o representadas numéricamente (Edgar, Stewart y Millar, 2005). El filtrado se aplica al EEG durante la adquisición del mismo, o fuera de línea (offline) antes y después del proceso de promediación.
Aunque es absolutamente necesario usar filtros durante la adquisición del EEG, así como aplicarlos a registros ya adquiridos offline, hay que tener en cuenta que el filtrado puede distorsionar con severidad los PRE sin que sea necesariamente obvio. Por ejemplo, el uso de filtros puede cambiar el inicio y duración de un componente de los PRE, puede hacer que formas de onda monofásicas aparezcan como multifásicas, puede inducir osci-laciones artificiales o interferir con la localización de generadores de un PRE.
Los filtros más utilizados en la investigación con PRE se usan para atenuar rangos específicos de frecuencias. Los filtros que se usan fuera de línea son los digitales, ya que dependen de la aplicación de funciones matemáticas a la señal eléctrica en cuestión. Los filtros digitales obedecen los mismos principios que los filtros analógicos que, como ya se dijo, se encuentran instalados en los amplificadores y dependen de circuitos elec-trónicos que filtran la señal eléctrica. Este tipo de filtros tiene varias ventajas sobre los filtros analógicos, como el hecho de que la señal se puede guardar tal como se registró, para ser analizada después con filtros digitales de diferentes características. Además, los filtros digitales se pueden construir de tal modo que no produzcan ninguna distor-sión de fase o de frecuencia, a diferencia de los filtros analógicos, que sí las producen.
El uso de filtros es necesario principalmente por dos razones (Luck, 2005): 1.Por la conversión analógico-digital de la señal que se está adquiriendo. De
acuerdo con el teorema de Nyquist, es posible convertir una señal analógica continua, como el EEG, en un conjunto de muestras discretas sin perder infor-mación, siempre y cuando la tasa de digitalización sea de al menos el doble que la frecuencia más alta que contenga la señal que se está digitalizando. Por otra parte, este teorema también establece que, si la señal original contiene fre-cuencias que sean de más del doble de la tasa de digitalización, estas muy altas frecuencias aparecerán como artefactos de baja frecuencia en los datos digita-lizados. En consecuencia, los amplificadores para EEG contienen filtros para suprimir frecuencias altas; por lo general están configurados para atenuar fre-cuencias más altas que la mitad de la tasa de muestreo. Por ejemplo, en un expe-rimento en el que se analicen los PRE endógenos, la tasa de digitalización podría ser 250 Hz (se toma un punto cada 4 ms) y, por lo tanto, se filtrarían todas las fre-cuencias por arriba de 125 Hz.
2. La segunda meta principal del filtrado es la reducción del ruido, lo cual es bastan-te complicado. La idea básica es que el EEG se compone de señal más algún ruido. Algo de este ruido es suficientemente diferente en frecuencia de la señal y pue-de suprimirse atenuando ciertas frecuencias. Sin embargo, a medida que las fre-cuencias de la señal y las del ruido sean más similares, será más difícil suprimir el ruido sin distorsionar de manera significativa la señal. Por ejemplo, las ondas alfa

Métodos en Neurociencias Cognoscitivas
72
pueden ser una fuente representativa de ruido en el análisis de los PRE, pero de-bido a que su frecuencia está alrededor de los 10 Hz, es difícil filtrarlas sin distor-sionar la forma de onda de los PRE. Además de usar los filtros para suprimir altas frecuencias, los filtros también se usan para suprimir frecuencias muy bajas. El uso más común de tales filtros es remover cambios de voltaje muy lentos de origen no neural durante el proceso de adquisición de datos. Artefactos como los producidos por el sudor y cambios en la impedancia de los electrodos pueden llevar a cambios lentos sostenidos en la línea base de la señal de EEG y, usualmente, estos cambios lentos de voltaje se eliminan al filtrar aquellas frecuencias más bajas de 0.01 Hz. Esto es importante cuando se quiere obtener registros de pacientes clínicos o de niños, porque los movimientos de la cabeza y el cuerpo son una causa común de estos cambios de voltaje y, si no son eliminados, pueden saturar el amplificador, lo que implicaría perder datos. Aun si no saturan el amplificador, pueden causar distorsiones en las formas de ondas de los PRE debido a que se trata de voltajes muy grandes.
Los filtros se describen usualmente en términos de su habilidad para suprimir o dejar pasar varias frecuencias diferentes. Los tipos más comunes de filtros son: filtros pasa baja, que atenúan las frecuencias altas y dejan pasar las frecuencias bajas; filtros pasa alta, que atenúan las frecuencias bajas y dejan pasar las frecuencias al-tas; filtros pasa banda, que suprimen tanto las frecuencias bajas como las altas y dejan pasar solamente un rango intermedio de frecuencias, y filtros notch, que atenúan un rango estrecho de frecuencias, tales como los filtros para 60Hz; estos filtros dejan pasar todas las demás frecuencias.
Artefactos y ruido Es muy frecuente que la señal electroencefalográfica se vea distorsionada por seña-les eléctricas que se registran al mismo tiempo que el PRE, pero que no provienen del cerebro, sino de fuentes externas a éste, ya sean de otras partes del organismo, del equipo de registro o de otras partes del laboratorio. En este último caso, la causa más común es la línea de corriente eléctrica que alimenta al laboratorio, la cual puede ser de 50 o 60 Hz según el país en que se encuentre. No obstante, los equipos de registro cuentan con filtros llamados notch, utilizados para filtrar estas frecuencias. Los artefac-tos provenientes de los componentes del equipo de registro pueden originarse en los amplificadores, en el convertidor analógico digital o incluso en los monitores. Dentro de los artefactos provenientes de otras partes del organismo tenemos los de los ojos, sea por movimientos o por parpadeo, los artefactos musculares, los que surgen por aumento en la resistencia debida a varios factores relacionados con el electrodo y la superficie de contacto con la piel cabelluda, por el electrocardiograma (ECG), el pulso o por movimientos del cuerpo. Así, se puede decir que las ondas eléctricas registradas en cada presentación de estímulo están compuestas por tres elementos: el PRE, el ruido proveniente de la actividad de fondo o EEG y los artefactos. Basados en esto, Talsma y Woldorff (2005) proponen la siguiente definición de un artefacto: los artefactos son actividad eléctrica de cualquier fuente que pueda ser registrada por el equipo de EEG, no originada en fuentes cerebrales; los artefactos se pueden distinguir con claridad de la actividad de fondo o son suficientemente grandes como para modificar la forma ver-dadera del PRE.

73
Potenciales relacionados con eventos técnicas y métodos de registro
A menudo, los artefactos provenientes de los amplificadores y del convertidor ana-lógico-digital se observan en forma de ondas muy lentas, con una tendencia lineal en los registros, producto de la saturación de los amplificadores o del rango de digita-lización. Dicha saturación se produce cuando los amplificadores o convertidores alcan-zan su límite en el rango de amplificación o de digitalización, punto en el cual la señal se recorta y en consecuencia aparece como una línea plana. Estos artefactos son comunes cuando el registro se realiza sin un filtro pasa alta, como ocurre en los registros de co-rriente directa (DC por sus siglas en Inglés, que no presenta oscilaciones de voltaje). Los diversos equipos contienen elementos que permiten que los amplificadores regresen a su línea base para continuar con un registro libre de estos artefactos.
Otra fuente común de ruido proviene de la radiación que emite el monitor. Este tipo de interferencia se puede notar por la presencia en el registro de un pulso con un in-tervalo correspondiente al ritmo de barrido de la pantalla, el cual es de 16.667 ms de duración para un ritmo de refresh de 60 Hz. Luck (2005) recomienda que si se detecta que el monitor provoca ruido en el registro, lo mejor es usar una cámara de Faraday para el monitor. No obstante, es necesario mencionar que en la actualidad los moni-tores modernos emiten mucho menos radiación, por lo que son más apropiados para un sistema de registro. La empresa Neuroscan (2003) recomienda que, dado que la radiación de los monitores es mayor por los lados que por enfrente, se trate de cambiar la orientación del monitor hasta disminuir la interferencia.
Como se mencionó antes, entre los artefactos producidos por otras partes del orga-nismo, se encuentran los que tienen su origen en el ojo, ya sea por movimientos ocu-lares o por el parpadeo. El conocimiento de estos artefactos es importante porque se registran con una frecuencia y características similares a los PRE. En términos genera-les, estos artefactos se originan porque el globo ocular funciona como un dipolo, con carga positiva y negativa en cualquiera de los dos lados, que produce campos eléctricos fluctuantes que se propagan en la piel cabelluda en sentido anteroposterior (Coles y Rugg, 1995). Ambos movimientos pueden detectarse con un registro bipolar, colocan-do un electrodo por encima de un ojo, referido a otro situado en la comisura externa del mismo ojo. Más aún, se puede distinguir entre los dos tipos de movimientos si se coloca un electrodo en la parte superior referido a otro colocado en la parte inferior para la detección del parpadeo, y se ponen electrodos en las comisuras externas de cada ojo, para la detección de los movimientos horizontales. Con estos arreglos de electrodos es muy fácil detectar a simple vista este tipo de artefactos, localizados de forma muy mar-cada en la región frontal y fronto-temporal, por lo que pueden eliminarse del registro los segmentos de EEG contaminados con este tipo de actividad o, de preferencia, se pueden usar los algoritmos de reducción de artefacto ocular que contienen los sistemas comerciales de registro, con lo que se evita la pérdida de estos segmentos.
También son frecuentes los artefactos musculares en los registros de EEG, debido a la contracción de los músculos de la cara. Por fortuna, su frecuencia de onda está por arri-ba de la que presentan la mayoría de los PRE, por lo que se pueden filtrar con facilidad, o se pueden evitar con ejercicios de relajación. Por ejemplo, en los experimentos en neurociencias que utilizan los PRE como técnica, las ondas de interés se encuentran en un rango de frecuencia que va de 0.01 a 30 Hz, mientras que la contracción muscular produce una actividad electromiográfica que está por arriba de los 100 Hz, por lo que un filtro que suprima estas frecuencias será suficiente para eliminar este artefacto sin causar cambios importantes en el PRE.

Métodos en Neurociencias Cognoscitivas
74
Los artefactos causados por un aumento de la resistencia en la conexión entre el electrodo de registro y la piel de la piel cabelluda (o impedancia de la piel) pueden de-berse a varios factores. Como lo describe Luck (2005), la resistencia en un circuito eléc-trico se refiere a la oposición que presentan los elementos de dicho circuito al paso de la corriente directa, que se mueve de un polo a otro. La impedancia se refiere a la resistencia que presenta un circuito eléctrico al paso de la corriente alterna (es decir, que cambia de polaridad, negativa o positiva, en la unidad de tiempo) entre un polo y otro. Esta impedancia (resistencia) está formada por la inductancia, la capacitancia y la resistencia de los componentes del circuito. En los registros electroencefalográficos, el circuito que registra dicha actividad está formado por el electrodo (con una conduc-tancia determinada), la pasta o gel conductor y la piel. Estos tres elementos pueden funcionar como un capacitor que atenúa la trasmisión de las ondas de baja frecuencia. Así, dado que la actividad electroencefalográfica registrada por los electrodos consiste en oscilaciones de voltaje, es más apropiado hablar de impedancia y no de resistencia.
La impedancia se mide en unidades llamadas Ohms (Ω) o, con mayor frecuencia, en miles de ohms (KΩ). Los valores que se manejan por lo general en la electroencefalo-grafía son menores a los 10 KΩ, pero se ha comprobado que los valores menores a 5 KΩ son los que permiten mejores registros, y son los más reportados en la literatura. Una de las causas de alta impedancia puede ser que entre el electrodo y la piel haya materia que dificulte el paso de la corriente hacia el electrodo. Dicha materia está formada por células muertas de la piel, suciedad y grasa, las cuales tienen que ser removidas para lograr un buen contacto. Si la impedancia no se mantiene dentro de estos límites, en-tonces se afecta el modo común de rechazo de los amplificadores, esto es, su capacidad para reducir el registro del ruido eléctrico, lo que da por resultado registros contaminados. Por fortuna, un factor que ayuda a suprimir el ruido, además de la correcta limpieza del área de contacto entre el electrodo y la piel, es el uso de amplificadores con una impe-dancia de entrada muy alta.
Otro efecto de la alta impedancia en los electrodos de registro es el aumento de los potenciales de la piel. Éstos se refieren al potencial constante que existe entre la superficie de la piel y sus capas más profundas, pero que varía con los cambios en la resistencia de la piel. Se producen cuando el sujeto suda o se mueve y genera un cambio en la posición del electrodo y, con ello, en la resistencia, lo que resulta en un cambio en el potencial. Se ha demostrado que con impedancias bajas de la piel se reducen en gran medida estos potenciales (Luck, 2005).
La actividad electrocardiográfica es otro artefacto bioleléctrico que se registra con cierta frecuencia en el EEG. A menudo esta actividad puede registrarse con varios de los componentes del ECG, sobre todo en la región posterior izquierda. Por fortuna, los sistemas de registro también contienen algoritmos que permiten reducir este tipo de artefactos. De igual forma, el pulso sanguíneo puede registrarse en el EEG con cierta frecuencia. Se identifica como pequeñas ondas rítmicas, rápidas y triangulares, regis-tradas, por lo general, en electrodos cercanos a alguna arteria. Estos artefactos se pre-sentan durante todo el registro y no se pueden remover, por lo que en nuestro labora-torio hemos optado por eliminar el registro de los electrodos contaminados con estos artefactos.
Finalmente, los artefactos producidos por el movimiento de diferentes partes del cuer-po también son fáciles de distinguir del resto de la actividad electroencefalográfica. A me-nudo puede detectarse la parte del cuerpo que mueve el sujeto, por lo que, en la mayoría

75
Potenciales relacionados con eventos técnicas y métodos de registro
de los casos, basta con instruirlo para que se deje de mover. Sin embargo, una vez que se ha registrado algún artefacto de este tipo, es necesario rechazar los segmentos de EEG contaminados.
Edición del EEG Rechazo y corrección de artefactos Rechazo y corrección de artefactos se refieren al proceso que se lleva a cabo para eli-minar la actividad eléctrica registrada en el EEG, cuyo origen no es cerebral, o para disminuir la influencia de este tipo de artefactos en el registro. Como ya se dijo, estos artefactos pueden provenir de muchas fuentes, como el parpadeo, los movimientos oculares, los potenciales de piel, el pulso cardiaco, los movimientos respiratorios, la actividad muscular, el voltaje de la línea de corriente eléctrica, etc. Por lo regular, el voltaje que produce dichos artefactos es mayor que el de los PRE, por lo que pueden provocar que la relación señal-ruido sea pequeña. Además, puede suceder que algunos artefactos ocurran con más frecuencia ante una situación que en otra, como podría su-ceder con un mayor parpadeo en una condición experimental que en otra, lo cual puede alterar la amplitud de los PRE.
Por lo tanto, es de primordial importancia proceder a la “limpieza” de los registros de EEG, ya sea eliminando los segmentos “contaminados” con los artefactos o redu-ciendo su influencia en el registro a partir de los algoritmos que proporcionan los siste-mas computarizados disponibles en el mercado, los cuales pueden contener algoritmos para reducción del artefacto ocular y de las ondas del ECG o sólo del primero.
Reducción de artefactos oculares En general, uno de los artefactos más frecuentes en los registros electroencefalográfi-cos es el de los movimientos oculares, que afectan principalmente a los electrodos fron-tales y temporales. Una manera de eliminarlos es rechazar las épocas o segmentos de EEG contaminados con ellos. Sin embargo, en la mayoría de los casos, esto equivaldría al rechazo de prácticamente todo el registro. Dado que la mayoría de los sujetos pre-senta movimientos oculares, sería necesario registrar un gran número de sujetos para obtener unos cuantos registros libres de artefacto ocular; aun así, probablemen-te permanecerían otros artefactos. Por ello, se han diseñado algoritmos que reducen la influencia de estos artefactos a tal punto, que la pérdida de datos por esta causa se ha reducido de manera notable. Aunque hay varios métodos para realizar esta reduc-ción que, en general, se basan en la sustracción de la parte del EEG contaminada con electrooculograma del EEG. De estos métodos, los más confiables son aquellos que emplean análisis de regresión en combinación con el promedio de los artefactos. Quizá los métodos más confiables son los que parten de la selección, por parte del experimen-tador, de los segmentos contaminados y que los algoritmos toman como un patrón a partir de los cuales harán los cálculos necesarios para disminuir el artefacto. Estos mé-todos han resultado muy confiables, como se observa en la figura 4-1.
Eliminación de otro tipo de artefactos Con frecuencia, los artefactos en los registros electroencefalográficos son evidentes, porque su tamaño sobrepasa notablemente al del EEG. Por ello, un sistema compu-tarizado de detección de artefactos y de rechazo de los segmentos contaminados con aquéllos suele realizar este trabajo una vez que se detecta una amplitud mínima,
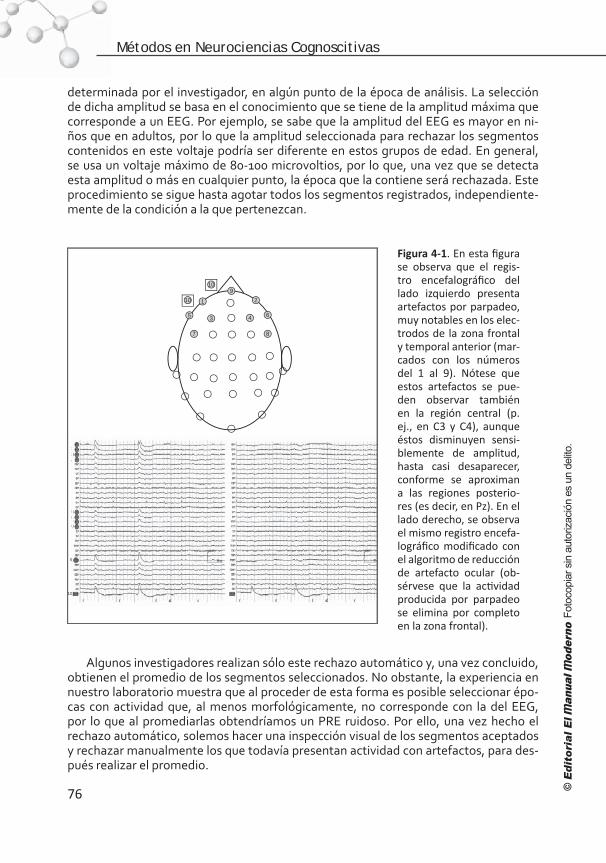
8
10
5
9
7
3 4
2
6
110
1234
5678
9
10
111122333344
5555667778888
99
00
Métodos en Neurociencias Cognoscitivas
76
determinada por el investigador, en algún punto de la época de análisis. La selección de dicha amplitud se basa en el conocimiento que se tiene de la amplitud máxima que corresponde a un EEG. Por ejemplo, se sabe que la amplitud del EEG es mayor en ni-ños que en adultos, por lo que la amplitud seleccionada para rechazar los segmentos contenidos en este voltaje podría ser diferente en estos grupos de edad. En general, se usa un voltaje máximo de 80-100 microvoltios, por lo que, una vez que se detecta esta amplitud o más en cualquier punto, la época que la contiene será rechazada. Este procedimiento se sigue hasta agotar todos los segmentos registrados, independiente-mente de la condición a la que pertenezcan.
Figura 4-1. En esta figura se observa que el regis-tro encefalográfico del lado izquierdo presenta artefactos por parpadeo, muy notables en los elec-trodos de la zona frontal y temporal anterior (mar-cados con los números del 1 al 9). Nótese que estos artefactos se pue-den observar también en la región central (p. ej., en C3 y C4), aunque éstos disminuyen sensi-blemente de amplitud, hasta casi desaparecer, conforme se aproximan a las regiones posterio-res (es decir, en Pz). En el lado derecho, se observa el mismo registro encefa-lográfico modificado con el algoritmo de reducción de artefacto ocular (ob-sérvese que la actividad producida por parpadeo se elimina por completo en la zona frontal).
Algunos investigadores realizan sólo este rechazo automático y, una vez concluido, obtienen el promedio de los segmentos seleccionados. No obstante, la experiencia en nuestro laboratorio muestra que al proceder de esta forma es posible seleccionar épo-cas con actividad que, al menos morfológicamente, no corresponde con la del EEG, por lo que al promediarlas obtendríamos un PRE ruidoso. Por ello, una vez hecho el rechazo automático, solemos hacer una inspección visual de los segmentos aceptados y rechazar manualmente los que todavía presentan actividad con artefactos, para des-pués realizar el promedio.

77
Potenciales relacionados con eventos técnicas y métodos de registro
Promediación, corrección de línea base, filtrado y suavizado Promediación Los PRE se extraen del electroencefalograma, el cual por lo regular tiene mayor voltaje que los PRE. Salvo en casos raros, estos potenciales no se pueden observar a simple vista. Por tal motivo, se tiene que implementar un método para extraer dichos poten-ciales; de ellos, el más usado es el de la promediación. Este método descansa en dos supuestos fundamentales: 1) que la actividad neuronal asociada a la presentación de un evento en particular (un estímulo, una respuesta, etc.) siempre es la misma y 2) que la actividad neuronal no asociada al evento registrado en el EEG representa ruido que varía al azar de ensayo a ensayo. No obstante que estos dos supuestos no siempre se cumplen, al tomar en cuenta que el PRE es sólo el promedio de los segmentos, no re-presenta ningún problema para la interpretación de la amplitud de los componentes de los PRE (Luck, 2005).
En la actualidad, la mayoría de laboratorios obtienen los PRE a partir de un registro continuo de EEG, al mismo tiempo que se estimula a los sujetos. Para promediar la ac-tividad electroencefalográfica relacionada con los eventos que se presentan, trátese de estímulos o respuestas, es necesario saber con precisión el momento en que se presen-tan dichos estímulos o respuestas en el registro continuo. En la mayoría de los equipos, esto se logra poniendo marcas que indican el comienzo de presentación del even-to y su categorización (figura 4-2(A). De esta forma, es posible separar la actividad electroencefalográfica asociada antes y después al evento de interés del registro con-tinuo. Estos trazos de EEG constituyen las épocas de registro, que de manera habitual se componen de un intervalo de tiempo antes del evento y otro que se mide a partir de que comienza el evento en cuestión. Cuando el evento es un estímulo, es común que el primer intervalo tenga una duración de 100-200 ms (intervalo preestímulo), mientras que el intervalo, que comienza con el inicio del estímulo, tiene una duración variable pero suele ser de 1 a 1.5 seg (figura 4-2(B).
Una vez seleccionadas las épocas de registro que se consideran útiles, se procede a hacer la promediación punto por punto y electrodo por electrodo. La actividad eléctrica relacionada con el estímulo permanece, mientras que la actividad no relacionada tien-de a cero. El promedio se obtiene de la suma algebraica de cada punto dividida por el número de épocas, lo que da como resultado final el PRE (figuras 4-2(B) y 4-2(C). Con este procedimiento, el PRE se va haciendo más nítido a medida que se promedian más épocas, es decir, a medida que aumenta el número de épocas promediadas el ruido se reduce y, por tanto, se acerca más a cero, mientras que el PRE se hace más patente (figura 4-2(C).
Así, puede decirse que la cantidad de ruido presente en un PRE está en función del número de ensayos que se promedian. Se sabe que el ruido se reduce en función de la raíz cuadrada del número de épocas promediadas y, dado que —en teoría— la señal permanece constante en el proceso de promediación, la relación señal-ruido (S-R) se incrementa en función del número de épocas promediadas (Litt y Fisher, 1990).
Por lo anterior se enfatiza la importancia de obtener los PRE con un número suficiente de épocas (segmentos). Aun en poblaciones clínicas o en niños, en donde esto puede resultar difícil, debe hacerse lo posible por acercarse a las veinte épocas o más. También es importante que, en un experimento, las diferentes categorías del es-tímulo sean promediadas con un número igual o muy semejante de segmentos, ya que si difieren en esto, será más probable que los PRE obtenidos tengan diferente relación
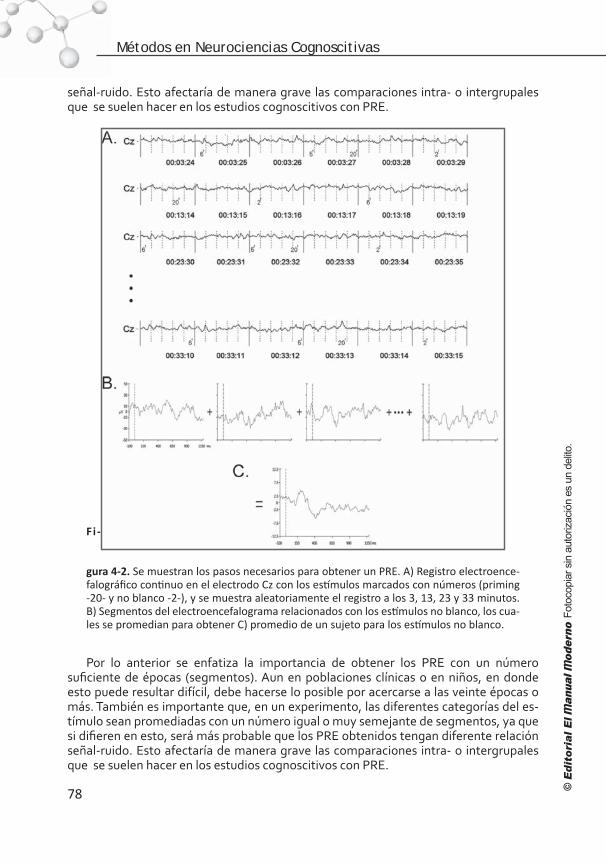
Métodos en Neurociencias Cognoscitivas
78
señal-ruido. Esto afectaría de manera grave las comparaciones intra- o intergrupales que se suelen hacer en los estudios cognoscitivos con PRE.
Fi-
gura 4-2. Se muestran los pasos necesarios para obtener un PRE. A) Registro electroence-falográfico continuo en el electrodo Cz con los estímulos marcados con números (priming -20- y no blanco -2-), y se muestra aleatoriamente el registro a los 3, 13, 23 y 33 minutos. B) Segmentos del electroencefalograma relacionados con los estímulos no blanco, los cua-les se promedian para obtener C) promedio de un sujeto para los estímulos no blanco.
Por lo anterior se enfatiza la importancia de obtener los PRE con un número suficiente de épocas (segmentos). Aun en poblaciones clínicas o en niños, en donde esto puede resultar difícil, debe hacerse lo posible por acercarse a las veinte épocas o más. También es importante que, en un experimento, las diferentes categorías del es-tímulo sean promediadas con un número igual o muy semejante de segmentos, ya que si difieren en esto, será más probable que los PRE obtenidos tengan diferente relación señal-ruido. Esto afectaría de manera grave las comparaciones intra- o intergrupales que se suelen hacer en los estudios cognoscitivos con PRE.

79
Potenciales relacionados con eventos técnicas y métodos de registro
Promediación sincronizada con las respuestas Otra forma de obtener el PRE es mediante la sincronización de la respuesta del indi-viduo con el EEG en cada ensayo. Para lograr esto, sólo se toma como referencia el punto donde respondió el sujeto considerando un tiempo hacia atrás y hacia adelante de dicho punto para conformar las épocas de registro y, después, promediar punto por punto, como cuando se sincroniza con los estímulos.
Corrección de línea base Una vez que se ha obtenido un PRE, es necesario establecer el punto a partir del cual se realizarán las mediciones de las amplitudes, lo cual es un paso crítico para determinar de modo correcto el tamaño de los picos y valles de los PRE. Dicho punto de referencia es llamado línea base y es el promedio de las amplitudes de los puntos considerados en el tiempo preestímulo de la época de registro, que suele ser de 100-200 ms. De esta manera, la medición de las amplitudes tiene como base el escalamiento de todo el PRE con respec-to al tiempo preestímulo cuyo promedio es igual a 0μv. Los programas de edición de los PRE de los sistemas comerciales calculan la amplitud de cada punto del PRE y lo grafican restando el valor de dicho promedio a cada punto del PRE (ver figura 4-3A y 4-3A’).
Este procedimiento hace sensibles al ruido las mediciones de las amplitudes, de modo que, si el ruido del tiempo preestímulo es diferente entre condiciones, pueden resultar diferencias entre ellas. Además, las amplitudes varían en función del tamaño del tiempo preestímulo, ya que, entre más cortos, aportan mayor cantidad de ruido (Handy, 2005). No obstante, el supuesto que existe para considerar el periodo preestí-mulo como línea base es que éste no resulta afectado por la presentación del estímulo, es decir, que es un tiempo en el que el individuo está en reposo. Aunque esta presunción no siempre es del todo cierta, esta forma parece ser la más recomendable para obtener mediciones de amplitud más o menos confiables.
Filtrado y suavizado Una vez que se han obtenido los PRE, uno puede darse cuenta de que contienen fre-cuencias que más bien pueden deberse a artefactos, producidas ya sea por los filtros analógicos o por el ruido con el que se hizo el registro. Aunque siempre es mejor tomar todas las precauciones para obtener registros “limpios”, en la práctica esto no siempre es posible, por lo que es necesario someter las señales a procedimientos de filtrado digital para atenuar la señal no deseada.
Una de las grandes ventajas del filtrado digital es que puede hacerse fuera de línea, es decir, una vez que se hizo el registro de EEG. Éste se puede revisar y filtrar según el interés en eliminar algún tipo de frecuencia. Además, existen filtros digitales como los llamados filtros de respuesta al impulso finita (FIR, por sus siglas en inglés), que son filtros en el dominio del tiempo que se pueden diseñar para no producir distorsión de fase, como lo hacen los filtros analógicos de modo inevitable (Gotman, 1990). Hay tres tipos de filtros digitales: a) con respuesta al impulso finita, b) con respuesta al im-pulso infinita y c) en el dominio de la frecuencia usando la transformación rápida de Fourier (FFT, por sus siglas en inglés). De éstos, el más simple es el filtro FIR, que ade-más tiene la ventaja de que se aplica con mayor facilidad a señales lentas, comparado con los filtros basados en la FFT (Fisch, 1999). En nuestra experiencia, los FIR producen menos distorsión en los PRE, por lo que a continuación se menciona de forma general su cálculo (ver Fisch, 1999).
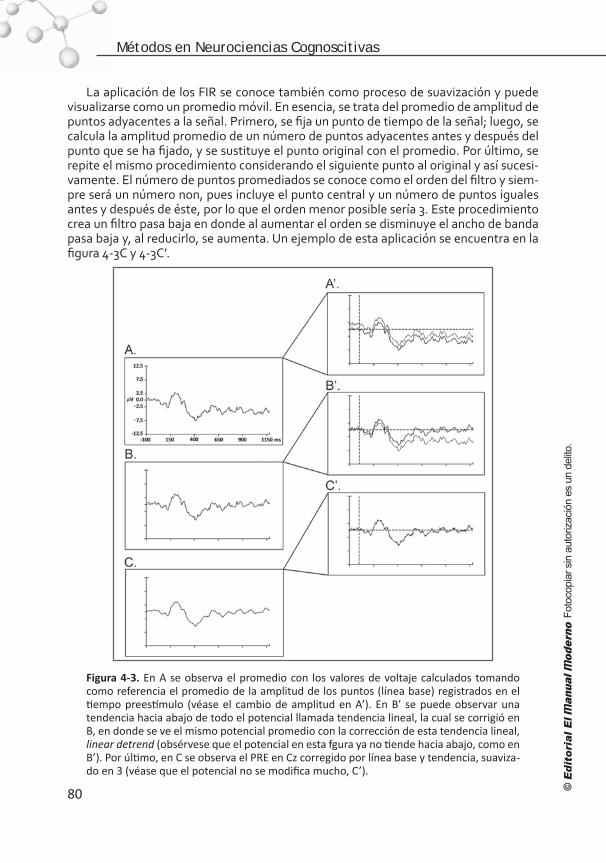
Métodos en Neurociencias Cognoscitivas
80
La aplicación de los FIR se conoce también como proceso de suavización y puede visualizarse como un promedio móvil. En esencia, se trata del promedio de amplitud de puntos adyacentes a la señal. Primero, se fija un punto de tiempo de la señal; luego, se calcula la amplitud promedio de un número de puntos adyacentes antes y después del punto que se ha fijado, y se sustituye el punto original con el promedio. Por último, se repite el mismo procedimiento considerando el siguiente punto al original y así sucesi-vamente. El número de puntos promediados se conoce como el orden del filtro y siem-pre será un número non, pues incluye el punto central y un número de puntos iguales antes y después de éste, por lo que el orden menor posible sería 3. Este procedimiento crea un filtro pasa baja en donde al aumentar el orden se disminuye el ancho de banda pasa baja y, al reducirlo, se aumenta. Un ejemplo de esta aplicación se encuentra en la figura 4-3C y 4-3C’.
Figura 4-3. En A se observa el promedio con los valores de voltaje calculados tomando como referencia el promedio de la amplitud de los puntos (línea base) registrados en el tiempo preestímulo (véase el cambio de amplitud en A’). En B’ se puede observar una tendencia hacia abajo de todo el potencial llamada tendencia lineal, la cual se corrigió en B, en donde se ve el mismo potencial promedio con la corrección de esta tendencia lineal, linear detrend (obsérvese que el potencial en esta fgura ya no tiende hacia abajo, como en B’). Por último, en C se observa el PRE en Cz corregido por línea base y tendencia, suaviza-do en 3 (véase que el potencial no se modifica mucho, C’).

81
Potenciales relacionados con eventos técnicas y métodos de registro
Técnica de promediación para PREs superpuestos Cuando una respuesta neuronal a un estímulo se presenta antes de que haya finalizado la respuesta producida ante un estímulo previo, ambas respuestas pueden verse distor-sionadas debido a que se superponen, como es el caso en los estímulos presentados en una sucesión rápida (con menos de un segundo entre estímulos). Hay varias maneras de resolver este problema (ver Woldorff 1988), pero sólo nos referiremos a una de ellas: considérese un experimento donde se presentan dos estímulos, que pueden ser de la misma modalidad sensorial o no, a intervalos de tiempo cortos, variables o fijos, como sucede por ejemplo en los experimentos de inhibición conductual, donde la apari-ción sorpresiva de un estímulo es señal para inhibir una respuesta. Para ello, se presen-ta a los sujetos un primer estímulo al que tienen que responder (estímulo blanco). A éste le puede seguir o no, al azar, un segundo estímulo que puede presentarse a diferentes intervalos de tiempo y que condiciona que los sujetos inhiban su respuesta. Dado que los estímulos que condicionan la inhibición se pueden presentar a intervalos cortos, el PRE que producen —y que es el de interés— se traslapa con el que produjo el estímulo blanco, por lo que es necesario quitar el efecto del traslape.
Tomando en cuenta que en la situación experimental se presenta a los sujetos un estímulo visual seguido a los 400 ms por un estímulo auditivo, la forma en que hemos procedido se describe a continuación (figura 4-4):
1 Se obtienen las épocas del EEG sincronizadas con el estímulo visual, más las épo-cas en las que aparece el estímulo auditivo. En este caso, tomamos un segmento del EEG donde aparece el estímulo auditivo 400 ms después del estímulo visual, más el tiempo considerado para analizar el segundo estímulo (en este caso 1100 ms contando el tiempo preestímulo), por lo que la época queda de 1 600 ms, to-mando en cuenta 100 ms de tiempo preestímulo antes del visual (figura 4-4(A).
2 Se obtienen las épocas del EEG sincronizadas con el estímulo visual en la condi-ción en la que no le sigue ningún estímulo auditivo, considerando el tiempo pre-estímulo (100 ms) más el tiempo programado para analizar el segundo estímulo, por lo que la época queda de 1 600 ms y se procede a promediar los segmentos para obtener el PRE (ver figura 4-4(B).
3 Se hace una diferencia entre el PRE auditivo-PRE visual: esto tiene el objetivo de restar la actividad provocada por el estímulo visual un poco antes (100 ms) de que se presente el estímulo auditivo y durante toda la época de este último (figura 4-4(C). Como puede observarse en la figura 4-4(D), este método resulta útil para quitar el efecto de sobrelapamiento entre los PRE. Nótese, por ejemplo, que los picos positivos son de mayor amplitud y los negativos de menor magni-tud antes de quitar dicho efecto.
Medición de la señal Amplitud
La mayoría de las investigaciones que utilizan los PRE realizan mediciones tanto de amplitud como de latencia. En el primer caso, pueden utilizarse las mediciones de los picos (positivos o negativos) o las amplitudes promedio en una ventana de tiempo. Para medir un pico de los PRE, por lo general se designa un intervalo de tiempo (basado en conocimientos teóricos o empíricos), en el cual se buscará la amplitud máxima. Sin em-bargo, esta forma de medición puede llevar a errores, pues la amplitud máxima en el intervalo designado no necesariamente podría corresponder a un pico. Luck (2005)
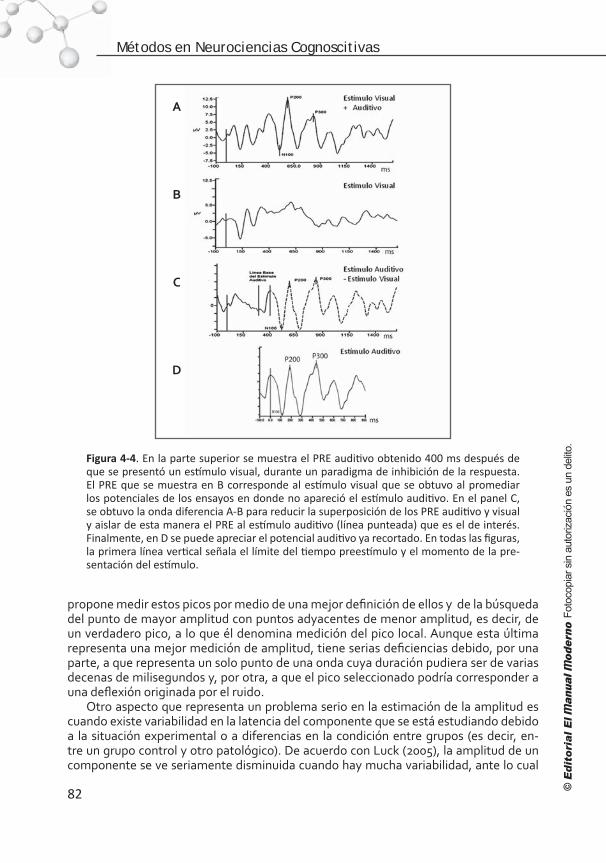
A
B
C
D
Métodos en Neurociencias Cognoscitivas
82
Figura 4-4. En la parte superior se muestra el PRE auditivo obtenido 400 ms después de que se presentó un estímulo visual, durante un paradigma de inhibición de la respuesta. El PRE que se muestra en B corresponde al estímulo visual que se obtuvo al promediar los potenciales de los ensayos en donde no apareció el estímulo auditivo. En el panel C, se obtuvo la onda diferencia A-B para reducir la superposición de los PRE auditivo y visual y aislar de esta manera el PRE al estímulo auditivo (línea punteada) que es el de interés. Finalmente, en D se puede apreciar el potencial auditivo ya recortado. En todas las figuras, la primera línea vertical señala el límite del tiempo preestímulo y el momento de la pre-sentación del estímulo.
propone medir estos picos por medio de una mejor definición de ellos y de la búsqueda del punto de mayor amplitud con puntos adyacentes de menor amplitud, es decir, de un verdadero pico, a lo que él denomina medición del pico local. Aunque esta última representa una mejor medición de amplitud, tiene serias deficiencias debido, por una parte, a que representa un solo punto de una onda cuya duración pudiera ser de varias decenas de milisegundos y, por otra, a que el pico seleccionado podría corresponder a una deflexión originada por el ruido.
Otro aspecto que representa un problema serio en la estimación de la amplitud es cuando existe variabilidad en la latencia del componente que se está estudiando debido a la situación experimental o a diferencias en la condición entre grupos (es decir, en-tre un grupo control y otro patológico). De acuerdo con Luck (2005), la amplitud de un componente se ve seriamente disminuida cuando hay mucha variabilidad, ante lo cual

83
Potenciales relacionados con eventos técnicas y métodos de registro
recomienda ampliamente la medición del área bajo la curva o la estimación de la latencia basada en esta misma medida, la que se obtiene al encontrar el punto de la curva que la divide en dos porciones iguales.
Uno de los problemas que se presentan con los PRE es que debido a que los picos de sus componentes pueden variar en latencia de un ensayo a otro, una vez que se obtiene el promedio, la amplitud de estos componentes puede verse modificada en relación con que si se hubiera medido en los ensayos individuales; este mismo efecto se puede observar cuando la latencia cambia entre sujetos y los PRE se promedian para obtener el gran promedio. Así la amplitud del pico disminuirá a medida que aumenta la variabili-dad en la latencia entre ensayos o entre sujetos en los grandes promedios. De esta for-ma se podría llegar a conclusiones erróneas al considerar las diferencias en la amplitud de un determinado componente en función de la condición o el grupo.
Una mejor aproximación para la medición de la amplitud de los componentes de los PRE es la amplitud promedio del intervalo de tiempo seleccionado, que, como su nombre lo indica, es el promedio de las amplitudes de los puntos del intervalo. Ésta es la técnica más empleada en la literatura y se usa con algunas variantes, una de las cuales puede consistir en encontrar un pico y definir el intervalo de tiempo con igual cantidad de puntos a la izquierda y a la derecha del punto de mayor amplitud.
No obstante, Luck (2005) considera que una de las maneras más apropiadas para eliminar el efecto de la variabilidad de la latencia en la amplitud de los componentes es medir el área bajo la curva, ya que se considera que el promedio de las áreas bajo la curva de un número de ensayos es equivalente al área bajo la curva del PRE obtenido de esos mismos ensayos, independientemente de la latencia del componente en cada uno.
Otra forma de medir la amplitud de los PRE es la llamada amplitud pico a pico. Ésta consiste en la medición de la amplitud que hay entre un pico y un valle adyacente en el PRE. La ventaja que tiene esta medición es que no la afecta el ruido de la línea base. Sin embargo, tiene el serio defecto de que, si el punto de referencia es influido por la manipulación experimental o si no refleja el mismo proceso subyacente como el com-ponente de interés, entonces no sirve como un punto de referencia válido para medir las amplitudes. En estas circunstancias, sería prácticamente imposible determinar si, en el caso de que existieran diferencias entre condiciones en las mediciones pico a pico, éstas se deben a un efecto en el pico de interés o si el cambio fue en el punto de referencia (Handy, 2005). Es quizá por esta razón que en la actualidad casi ningún estu-dio utiliza esta medida.
Latencia La medición de la latencia es un aspecto muy importante en los experimentos que miden tiempos de procesamiento de la información y toman la latencia del componen-te de interés como una medida de dichos tiempos. Por lo general, la forma de calcular la latencia es anotar el tiempo en que aparece el punto de mayor amplitud en un intervalo de tiempo seleccionado o, para mayor confiabilidad, como ya se mencionó, anotar la latencia del pico local. Este último procedimiento es el más empleado y está sujeto a las mismas limitaciones que las medidas de amplitud. No obstante, como sucede con la amplitud, la estimación de la latencia basada en la medida del área bajo la curva pa-rece tener ventajas. Este método consiste en calcular el punto en el tiempo que divide el área en dos mitades iguales, que constituiría la latencia del componente. Esta medición tiene una sola desventaja y es que funciona cuando el intervalo de tiempo utilizado para

Métodos en Neurociencias Cognoscitivas
84
medir el área abarca el rango de latencias de un solo componente y no hay la contribu-ción de múltiples componentes superponiéndose entre sí.
Otra medida utilizadapara estimar los tiempos de procesamiento de la información mediante los PRE es el momento en el que comienza a aparecer un componente que, se sabe o se presume, está relacionado con un tipo de procesamiento. Esta medición llamada latencia de inicio está justificada en tanto puede variar entre condiciones sin que esto necesariamente se refleje en una diferencia en la latencia del pico (Handy, 2005). Aunque es un procedimiento difícil de llevar a cabo, sobre todo en PRE muy rui-dosos, Luck (2005) enumera una serie de métodos que pueden usarse para determinar la latencia de inicio de los componentes. Recomienda que es necesario remover la posi-ble superposición de distintos componentes para aislar el que sea de interés, lo cual se puede llevar a cabo al obtener la onda diferencia entre dos condiciones determinadas.
Una vez que se han realizado las mediciones de amplitud y de latencia y se han consi-derado las limitaciones señaladas, el siguiente paso es la comparación entre condiciones o entre grupos, lo cual nos remite a consideraciones estadísticas.
Referencias
Coles, M. G. H. y Rugg, M. D. (1995). Event-related potentials: an introduction. En M. Rugg y M. Coles (Eds.), Electrophysiology of mind: event-related brain potentials and cognition (pp. 1-26). Oxford: Oxford University Press.
Edgar, J. C., Stewart, J. L. y Miller, G. A. (2005). Digital filters in ERP research. En T. C. Handy (Ed.), Event-related potentials: a methods handbook (pp. 85-114). Cambridge: mit Press.
Fisch, B. J. (1999). Fisch and Spehlmann’s EEG primer: Basic principles of digital and analog EEG (3a. ed.). Nueva York: Elsevier.
Gotman, J. (1990). The use of computers in analysis and display of EEG and evoked potentials. En D. D. Daly y T. A. Pedley (Eds.), Current practice of clinical electroencephalography (pp. 51-84). Nueva York: Raven Press.
Handy, T.C. (2005). Basic principles of ERP quantification. En T. C. Handy (Ed.), Event-related potentials: a methods handbook (pp. 33-56). Cambridge: mit Press.
Litt, B. y Fisher, R. S. (1990). EEG engineering principles. En D. D. Daly y T. A. Pedley (Eds.), Current practice of clinical electroencephalography (pp. 1-28). Nueva York: Raven Press.
Luck, S. J. (2005). An introduction to the event-related potential technique. Cambridge: mit Press. Scan 4.3 (vol II). (2003). Nunez, P. L. (1981). Electric fields of the brain. Nueva York: Oxford University Press. Talsma, D. y Woldorff, M. G. (2005). Methods for the estimation and removal of artifacts and
overlap in ERP waveforms. En T. C. Handy (Ed.), Event-related potentials: a methods han-dbook (pp. 115-148). Cambridge: mit Press.
Woldorff, M. G. (1988). Adjacent response overlap during the erp averaging process and a tech-nique (Adjar) for its estimation and removal. Psychophysiology, 25, 490.

85
Capítulo 5. Localización defuentes de corriente de la actividad eléctrica cerebral: tomografía eléctrica cerebral
Juan Silva Pereyra
Resumen La localización de las fuentes de corriente de la actividad electromagnética cerebral es un método de análisis de la señal electrofisiológica que sólo tiene tres décadas de historia en la investigación científica. Para llevar a cabo dicho método es necesario con-siderar varios pasos que implican la combinación de varios tipos de información como la espacial, geométrica, del campo eléctrico y de la electrofisiológica adquirida durante una tarea cognoscitiva. Cada paso debe ser considerado con cuidado y seleccionado sobre la base de la información que uno quiere obtener. En este capítulo, analizaremos el procedimiento que precede a la estimación de una fuente y cómo el resultado de ésta puede integrarse con una imagen de resonancia magnética de la cabeza.Además, presentaremos análisis estadísticos de imágenes con las fuentes localizadas, en diseños experimentales tan complejos como los empleados en la resonancia magnética funcio-nal (RMf). Este capítulo se centra en los registros del electroencefalograma, aunque también esto es aplicable a los magnetoencefalográficos. Gran parte de los esfuerzos se dirigen al uso de la información en el dominio del tiempo (como en el caso de los po-tenciales relacionados con eventos, PRE); sin embargo, también es posible la aplicación de este método en el domino de la frecuencia.
Origen de la actividad electromagnética cerebral Es difícil pensar en un sistema tan complejo como el cerebro y no considerar redes neu-ronales distribuidas implicadas en su funcionamiento (Mesulam, 1998). Los meca-nismos básicos de interacción entre diferentes módulos de estas redes neuronales son procesos de antero y retroalimentación por inhibición y excitación. El funcionamiento del sistema nervioso central se ha estudiado no sólo al analizar los mecanismos básicos de la estructura o neuroanatomía cerebral, sino también en relación con su funcionamiento. De ahí que el principal objetivo de los métodos y técnicas de neuroimagen, como la reso-nancia magnética funcional (RMf) y la tomografía por emisión de positrones (PET, por sus siglas en inglés), es ubicar en el cerebro tales módulos funcionales implicados en una tarea cognoscitiva específica (Cabeza y Nyberg, 2000). Sin embargo, estos métodos no son los mejores para proporcionar información directa acerca de los procesos cognosci-tivos involucrados en la realización de una tarea; pues aunque tienen una alta resolución espacial, cuentan con una pobre resolución temporal, además de proveer información indirecta acerca de la activación neuronal al evaluar más el flujo sanguíneo y la desoxige-

Métodos en Neurociencias Cognoscitivas
86
nación de la sangre por ciertas regiones cerebrales durante la realización de una tarea. Hay métodos que directamente evalúan la actividad neuronal, al medir la actividad
eléctrica postsináptica, y se usan para investigar las redes neuronales con una alta re-solución temporal, aunque con una pobre resolución espacial. El electroencefalograma (EEG) y el magnetoencefalograma (MEG) son métodos que miden la actividad electro-magnética de asambleas neuronales en una escala temporal inferior a los milisegundos. Se dice que estos métodos son los que tienen mejores propiedades para el estudio de procesos cognoscitivos, así como para el análisis de las interacciones entre módulos de una red neuronal, procesos en serie o en paralelo y mecanismos de retroalimentación. Como arriba se mencionó, estos métodos enfrentan el problema de la pobre resolución espacial, pues existe una ambigüedad biofísica del origen de la fuente de corriente den-tro del cerebro.
Como se menciona en los capítulos 1 y 2 de este libro, las fuentes de corriente en el cerebro que pueden ser detectables por el EEG dependen de la organización de las neu-ronas en campos abiertos. Les llamaremos campos abiertos a aquellos campos eléctri-cos cuyo producto final no es cero. Estos dependerán de la organización o arreglo de las neuronas. En núcleos subcorticales puede haber un arreglo desordenado de las neuronas, por lo que no se puede representar el flujo de la corriente como un dipolo. Existen dife-rencias importantes en el registro de la actividad cerebral entre el EEG y el MEG. Mien-tras el electroencefalograma puede capturar la actividad eléctrica postsináptica de los grupos neuronales que están organizados de manera perpendicular con respecto a la superficie de la piel cabelluda (es decir, asambleas neuronales que se encuentran en las áreas como los giros cerebrales), el magnetoencefalograma sólo puede registrar la ac-tividad que proviene de regiones profundas o cisuras del cerebro, puesto que las neuronas están organizadas ahí en forma tangencial a la superficie (figura 5-1). Los algoritmos de maximización espacial, como los cálculos de densidad de corriente, pueden ayudar a identificar la topografia de la actividad cerebral (Nunez, 1981).
Otra condición fundamental para colectar la actividad eléctrica cerebral es la sincro-nía de grandes grupos de neuronas. Si las asambleas neuronales disparan en diferentes instantes de tiempo, la suma del campo eléctrico con respecto del tiempo se aproxi-mará a cero y es muy poco probable que se detecte actividad eléctrica ante un proceso cognoscitivo. De esta manera, la detección de la actividad eléctrica con sensores en la superficie de la piel cabelluda estará condicionada por la geometría de la asamblea neuronal y por la sincronía en los disparos.
Solución al problema inverso Desde mediados del siglo xix, Helmholtz descubrió que no es posible determinar el ori-gen de una fuente de corriente dentro de una esfera a partir de las mediciones del vol-taje en la superficie de la misma. Una multiplicidad de configuraciones de fuentes puede generar el mismo patrón en la superficie del volumen conductor (en este caso el cerebro). También lo inverso, una multiplicidad de configuraciones registradas en la superficie de la cabeza pueden producirse por el mismo patrón de configuración de fuentes dentro del cerebro. El análisis de la distribución del campo eléctrico es un método precursor para la localización de fuentes, aunque esto no provee información concluyente acerca de la lo-calización y distribución de las mismas. La única manera de localizar las fuentes eléctricas en el cerebro es por medio de la solución del llamado problema inverso. Éste puede resol-verse si se introducen supuestos de la generación de la señal EEG y, con esto, se restringe
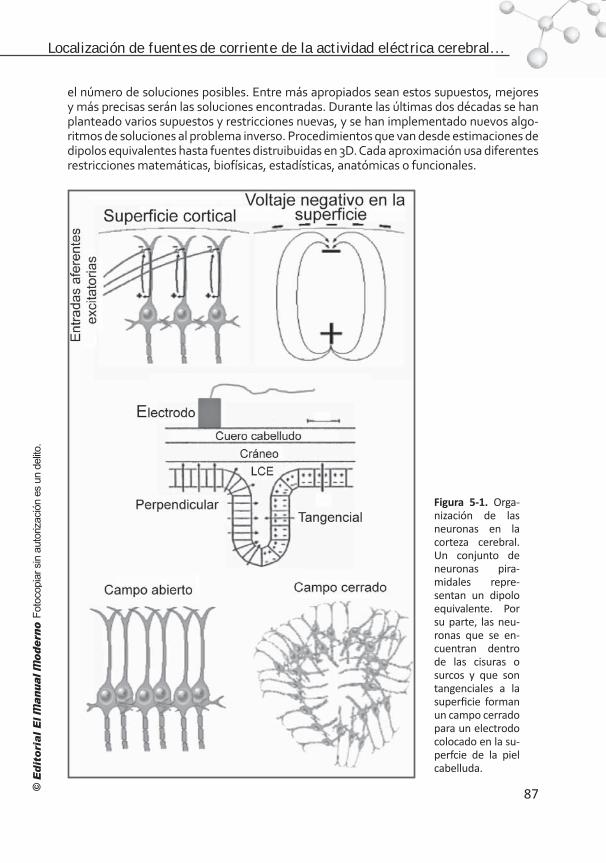
87
Localización de fuentes de corriente de la actividad eléctrica cerebral...
el número de soluciones posibles. Entre más apropiados sean estos supuestos, mejores y más precisas serán las soluciones encontradas. Durante las últimas dos décadas se han planteado varios supuestos y restricciones nuevas, y se han implementado nuevos algo-ritmos de soluciones al problema inverso. Procedimientos que van desde estimaciones de dipolos equivalentes hasta fuentes distruibuidas en 3D. Cada aproximación usa diferentes restricciones matemáticas, biofísicas, estadísticas, anatómicas o funcionales.
Figura 5-1. Orga-nización de las neuronas en la corteza cerebral. Un conjunto de neuronas pira-midales repre-sentan un dipolo equivalente. Por su parte, las neu-ronas que se en-cuentran dentro de las cisuras o surcos y que son tangenciales a la superficie forman un campo cerrado para un electrodo colocado en la su-perfcie de la piel cabelluda.

Métodos en Neurociencias Cognoscitivas
88
El procedimiento para el cálculo y proyección de las soluciones al problema inverso es largo y laborioso. Como en el caso de los estudios de resonancia magnética funcional (RMF), en la tomografía eléctrica cerebral también se debe hacer un preprocesamiento de una imagen estructural de RM, lo que nos permitirá construir nuestro modelo de la cabeza. En este modelo podremos calcular las fuentes de corriente y nos permitirá también proyectar sus soluciones. El modelo de la cabeza tendrá toda la información estructural (es decir, posición en el volumen de las posibles ubicaciones de las fuentes, así como la información sobre sus conductividades) que se complementará con la in-formación de la posición de los electrodos y la señal electrofisiológica.
La posición espacial (en un plano cartesiano) de cada uno de los electrodos de regis-tro durante la recolección de los datos electrofisiológicos se sobreimpondrá y ajustará al modelo de la cabeza seleccionado. Un paso previo al cálculo de las fuentes y que consume una gran cantidad de tiempo de cómputo es el cálculo del lead field (LF). El LF viene a ser un campo vectorial que concretamente representa una propiedad del es-pacio, debido a la cual una carga eléctrica puntual sufrirá los efectos de una fuerza. Un campo eléctrico estático puede ser representado con un campo vectorial o con líneas vectoriales (líneas de campo).
Las líneas vectoriales se utilizan para crear una visualización del campo. Aunque se puedan trazar en un papel en dos dimensiones, también es posible representarlas en un espacio tridimensional. En realidad, existen infinitas líneas de campo; sin embargo, se representan sólo unas pocas por claridad. En el caso del LF para la localización de fuen-tes en el cerebro, es necesario hacer una reconstrucción tridimensional del volumen del cerebro con pequeños cubos a los cuales llamaremos vóxeles (es decir, en realidad este término parte de píxel, que está en dos dimensiones, a tres dimensiones). Así para el cálculo del LF se considerará el paso de la corriente a través de cada vóxel.
La señal electrofisiológica colectada durante una tarea cognoscitiva es otra parte impor-tante de los datos que, con el LF calculado, servirá para el cálculo de las fuentes.
Modelos de la cabeza La selección del modelo de la cabeza es el inicio del largo procedimiento para la locali-zación de fuentes de corriente con datos electrofisiológicos. El modelo de la cabeza determina la manera en que las fuentes, localizadas en ciertas regiones del cerebro, producen las mediciones sobre la piel cabelluda. Esto incluye las propiedades electro-magnéticas (es decir, permeabilidad y conductividad) y las propiedades geométricas (es decir, forma) de un volumen dentro del cual se calculan las soluciones inversas. Como se mencionó en la sección anterior, estas propiedades se expresan matemáticamente en la matriz LF.
El modelo de la cabeza más simple y más usado en la actualidad es el modelo esférico (ver figura 5-2). Los modelos esféricos de la cabeza con conductividades homogéneas permiten una solución analítica del problema anterógrado, fácil y rápida de calcular. Sin embargo, nuestra cabeza no es esférica y su conductividad eléctrica no es espacialmen-te uniforme. Si se incorporan diferentes parámetros de conductividad en un modelo de esferas concéntricas (Berg y Scherg, 1994) puede mejorarse la precisión en la localiza-ción de la fuente. Varios estudios han mostrado que se puede conseguir una adecuada matriz LF si se usa un modelo de volumen conductor con geometría real (Cuffin, 1996). Sin embargo, la mayoría de estos estudios se han hecho para la localización de un solo dipolo y no hay evidencias con modelos de soluciones inversas distribuidas.
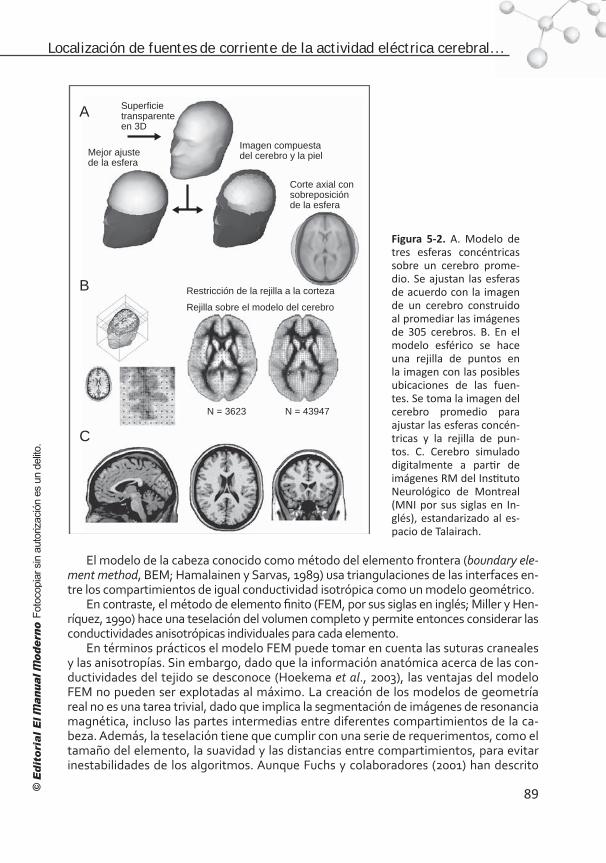
Superficietransparenteen 3D
Mejor ajustede la esfera
Imagen compuestadel cerebro y la piel
Corte axial consobreposiciónde la esfera
Restricción de la rejilla a la corteza
Rejilla sobre el modelo del cerebro
N = 3623 N = 43947
A
B
C
89
Localización de fuentes de corriente de la actividad eléctrica cerebral...
Figura 5-2. A. Modelo de tres esferas concéntricas sobre un cerebro prome-dio. Se ajustan las esferas de acuerdo con la imagen de un cerebro construido al promediar las imágenes de 305 cerebros. B. En el modelo esférico se hace una rejilla de puntos en la imagen con las posibles ubicaciones de las fuen-tes. Se toma la imagen del cerebro promedio para ajustar las esferas concén-tricas y la rejilla de pun-tos. C. Cerebro simulado digitalmente a partir de imágenes RM del Instituto Neurológico de Montreal (MNI por sus siglas en In-glés), estandarizado al es-pacio de Talairach.
El modelo de la cabeza conocido como método del elemento frontera (boundary ele-ment method, BEM; Hamalainen y Sarvas, 1989) usa triangulaciones de las interfaces en-tre los compartimientos de igual conductividad isotrópica como un modelo geométrico.
En contraste, el método de elemento finito (FEM, por sus siglas en inglés; Miller y Hen-ríquez, 1990) hace una teselación del volumen completo y permite entonces considerar las conductividades anisotrópicas individuales para cada elemento.
En términos prácticos el modelo FEM puede tomar en cuenta las suturas craneales y las anisotropías. Sin embargo, dado que la información anatómica acerca de las con-ductividades del tejido se desconoce (Hoekema et al., 2003), las ventajas del modelo FEM no pueden ser explotadas al máximo. La creación de los modelos de geometría real no es una tarea trivial, dado que implica la segmentación de imágenes de resonancia magnética, incluso las partes intermedias entre diferentes compartimientos de la ca-beza. Además, la teselación tiene que cumplir con una serie de requerimentos, como el tamaño del elemento, la suavidad y las distancias entre compartimientos, para evitar inestabilidades de los algoritmos. Aunque Fuchs y colaboradores (2001) han descrito

Métodos en Neurociencias Cognoscitivas
90
un procedimiento automático que calcula el LF usando un modelo completo realista en una computadora personal y toma algunos minutos, la facilidad en el cómputo de los modelos esféricos hace que éstos todavía sean los más empleados para el cálculo del LF. Esto se ve con claridad en la mayoría de los paquetes de cómputo que usan ajuste de dipolos.
Posición de los electrodos Número de electrodos y su posición El número de electrodos es un factor muy importante dentro del procedimiento de la localización de las fuentes de corriente. Los electrodos son pequeños sensores que co-lectan la actividad eléctrica y se colocan sobre la piel cabelluda. Los primeros registros electroencefalográficos usaban pocos electrodos dado que sólo buscaban actividad pa-tológica. Desde entonces, ya se colocaban en una posición estándar llamada sistema internacional 10-20 (ver capítulo 2). Al incrementarse la capacidad de los computadores y permitir su graficación digital, se posibilitó el aumento del número de canales y por tanto el número de electrodos en los registros. Se mantuvo la misma posición estándar y sólo se cambió primero a un sistema 10-10 para que pudieran colocarse más de estos sensores.
En teoría, cuanto mayor número de medidas espaciales se tengan, mejor resolución topográfica. Primero, es necesario considerar que las distancias entre electrodos deben ser, al menos, de 2 a 3 cm para evitar distorsiones en el potencial espacial (Gevins et al., 1990). Para saber qué distribución de los sensores y cuántos de ellos habría que usar para no distorsionar las imágenes tomográficas, varios autores han realizado simulaciones y han aplicado a éstas varios métodos de localización de fuentes. Se ha demostrado que los registros de 31 electrodos son insuficientes para una adecuada localización, mientras que la diferencia entre 63 y 123 electrodos es mínima (Michel et al. 2004a).
Los procedimientos de localización de fuentes también requieren que la distribu-ción de los electrodos sea homogénea sobre la piel cabelluda, dado que la posición no equidistante de los sensores o la mayor densidad de electrodos en alguna región puede provocar una distorsión muy grande a la hora de calcular la fuente. Los estudios de lo-calización de fuentes de corriente del EEG en registros convencionales de 21 electrodos deben de interpretarse con cautela aunque se han tenido buenos resultados (Fernán-dez et al., 2000; Silva-Pereyra et al., 2003).
Posición de los electrodos estándar y por sujeto En la actualidad, se emplean gorros elásticos con la posición estándar de los electro-
dos, de manera que al estirarse para amoldarse a un tamaño de cabeza en particular, se conserva la distancia entre los electrodos y su distribución.
En tal caso, las medidas de algunos puntos clave sobre la piel cabelluda podrían ser suficientes para la reconstrucción de las posiciones del resto de los electrodos (Le et al., 1998). Hasta el momento no es posible tomar la posición de los electrodos durante el registro del EEG al mismo tiempo que se adquieren las imágenes de RM. Por ello, resulta práctico tomar las medidas en un sistema cartesiano con un digitalizador en 3D. El sistema digitalizador espacial crea, por lo regular, un campo magnético alrededor de una referencia que a su vez es colocada en el borde del gorro con los electrodos. Una pluma metálica va cortando el campo magnético y tomando una medida de sus coor-denadas cartesianas.

91
Localización de fuentes de corriente de la actividad eléctrica cerebral...
Este procedimiento provee una medición de la posición de los electrodos en el mo-mento en que se toma el registro de EEG, de manera que muestra las variaciones interin-dividuales en la colocación de los electrodos (Towle et al., 1993). Otro procedimiento útil, aunque aún no usado, sería la reconstrucción de una imagen 3D por múltiples fotografías instantáneas en diferentes ángulos. Este procedimiento de reconstrucción de una imagen 3D por múltiples fotografías instantáneas se ha empleado en algunas películas de acción.
Además de los procedimientos arriba mencionados, se pueden usar sistemas de coor-denadas estándares con las posiciones de los electrodos dependiendo del arreglo que se haya usado en la colecta de los datos electrofisiológicos. Estos sistemas están ajustados a una imagen digital tridimensional de la cabeza (ésta puede ser el cerebro simulado del Instituto Neurológico de Montreal, MNI, estandarizado y puesto en el espacio de Talaira-ch). Si se usa un gorro de 32 canales, por ejemplo, se podrá emplear un sistema de coor-denadas estándar para los 32 canales.
Corregistro Este paso requiere que las posiciones de los electrodos concuerden con la superficie de la piel cabelluda definida por la imagen de RM, tal vez con el uso de alguna transformación (es decir, rotación o traslación). Estos parámetros se obtienen por lo regular al medir marcas comunes durante la adquisición del EEG y de las imágenes de RM. Se ubican mar-cas visibles en las imágenes de RM sobre la piel que corresponden a la posición de los electrodos y de algunas otras marcas, como las fiduciarias (es decir, nasion, inion, puntos periauriculares y vertex). Para que esto aparezca en la imagen RM, se puede hacer con una cápsula de vitamina k para cada punto.
De manera similar, estas marcas en la cabeza se miden durante el registro del EEG con el digitalizador de la ubicación espacial de los electrodos. De esta información se pueden determinar los parámetros de transformación para proyectar las fuentes del EEG en las coordenadas anátomicas y mostrarlas en las imágenes de RM (Scherg et al., 1999). Como se mencionó antes, cuando no es factible tener las imágenes de RM estruc-tural por cada sujeto y hacer el registro digital espacial de cada electrodo con las marcas fiduciarias en cada sujeto, es posible usar un modelo general estándar de RM estructural (es decir, el cerebro MNI del instituto Neurológico de Montreal) y asumir un sistema de coordenadas estándar. En este caso, los parámetros predefinidos de traslación se usan para cotejar el EEG con el espacio de la RM. Se ignoran, entonces, las diferencias indivi-duales en el tamaño de la cabeza y de las posiciones de los electrodos, lo que permite una limitada precisión con respecto del detalle anatómico de la localización de las fuentes.
Señal electroencefalográfica adquirida El registro de la actividad eléctrica cerebral que se empleará para la localización de fuentes se debe llevar a cabo con el mayor número de electrodos posible. Se debe cui-dar que la tarea cognoscitiva en cuestión evoque los componentes que se piensa anali-zar de acuerdo con el diseño experimental previamente planeado. Así, debemos consi-derar un registro de la actividad eléctrica extraordinariamente limpio (es decir, libre de artefactos), dado que los algoritmos empleados para la localización de fuentes son muy sensibles a los cambios bruscos de amplitud o de frecuencia que se ven en la actividad eléctrica que no es propia del cerebro. Muchos de los parámetros para la adquisición de la señal electroencefalográfica se han detallado ampliamente en los capítulos 1, 2 y 4 de este libro, por lo que no nos detendremos en ello.

Métodos en Neurociencias Cognoscitivas
92
La señal del EEG se puede analizar en el dominio del tiempo o en el dominio de la frecuencia. Para el cálculo de las fuentes en estos dos dominos se deben seguir proce-dimientos diferentes, por lo que es importante destacar algunos puntos del análisis en el dominio de la frecuencia.
Muchos investigadores han estado interesados en la localización de fuentes de las diferentes bandas de frecuencia electroencefalográficas, dado que el análisis espec-tral del EEG provee información importante de patologías relacionadas con el sueño, epilepsia, enfermedades psiquiátricas y el abuso de sustancias psicotrópicas. Como se menciona en el capítulo 2, una forma de uso del registro multicanal del EEG que se ha he-cho muy popular es el análisis topográfico de las bandas de frecuencia para cuantificar estados cerebrales normales y patológicos (es decir, el llamado EEG cuantitativo o computarizado, EEGc; Duffy et al., 1981; John et al., 1977). El paso siguiente sería inten-tar localizar las fuentes de dichos ritmos del EEG. Por desgracia, los procedimientos de localización de fuentes no se pueden aplicar directamente a los mapas de “potencia” debido a que los mapas representan el cuadrado de los valores y, así, la información de polaridad dada por los ángulos de fase se pierde. Además, los mapas de potencia son dependientes de la referencia (Lehmann, 1987). Sin embargo, hay métodos que permi-ten el análisis de fuentes en el dominio de la frecuencia.
Se ha demostrado que, sobre la base de las matrices de covarianza, hay varios mé-todos directos de localización de fuentes en el dominio de la frecuencia (Bosch-Bayard et al,2001).
La Referencia La referencia siempre ha sido fuente de debate en la electrofisiología, sobre todo cuan-do se vierte esa información en datos topográficos. Este problema se ha tomado como una desventaja mayor en el EEG que en el MEG (Williamson et al., 1991). La referencia tiene una fuerte influencia en el análisis de las ondas de los PRE, pero en la actualidad la referencia es por completo irrelevante para el análisis topográfico y para la locali-zación de fuentes, en tanto se considere dentro del modelo. Esto se debe a que la configuración de la distribución topográfica es independiente del electrodo de referencia. Es interesante que ésta corresponde matemáticamente al cálculo de la referencia promedio del potencial superficial (Lehmann, 1987). Así, la referencia promedio se usa de modo automático en el cálculo de las fuentes.
Artefactos e interpolación Un importante punto a considerar es cuando hay canales contaminados con artefac-tos a lo largo del registro (es decir, canales que muestran un mal funcionamiento del amplificador, registro de actividad muscular o quizá una gran impedancia), se usan go-rros con un mínimo número de electrodos y se quiere aplicar los procedimientos de lo-calización de fuentes. En principio, estos “malos” electrodos pueden nada más quitarse del análisis; sin embargo, un procedimiento mejor es interpolarlos sobretodo si se está haciendo un análisis de grupo donde los datos deben tener la misma configuración de electrodos. Hay dos procedmientos de interpolación ampliamente usados: el vecino más cercano y el spline (Fletcher et al., 1998). En el método del vecino más cercano, los valores “incógnita” se computan como un promedio pesado del potencial de los elec-trodos vecinos, cuyos pesos son dependientes de la distancia euclidiana entre el sitio a estimar y su vecino. En el caso del método spline del plano delgado (Perrin et al., 1987),

Z
F
X
Y
θ
93
Localización de fuentes de corriente de la actividad eléctrica cerebral...
se minimiza la energía circundante de una hoja infinitamente plana que pasa a través de los datos dados. El spline esférico (Hassaina et al., 1994) usa la mínima energía circun-dante en la superficie deformada de una esfera para pasar a través de un número finito de puntos conocidos. En general, las interpolaciones spline se comportan mejor que las interpolaciones por el método del vecino más cercano.
Selección del modelo inverso Soluciones dipolares El supuesto básico subyacente al modelo dipolar es que un pequeño número de fuentes de corriente en el cerebro pueden modelar de manera adecuada las medidas de los po-tenciales en la superficie. Para garantizar una solución única, el número de parámetros desconocidos tiene que ser menor o igual al número de mediciones independientes (es decir, electrodos). En la figura 5-3, se representa el número de parámetros o incógnitas en el modelo dipolar para el cálculo de las fuentes. La posición del dipolo en el volu-men involucra 3 parámetros (x, y, z), la dirección del mismo 2 (ángulos theta y alfa). Muchos de estos parámetros se pueden “fijar” y así se reduce la complejidad en las ecuaciones del modelo. Por ejemplo, si se ubican a priori fuentes en una región dado el conocimiento de que las áreas cerebrales involucradas durante la realización de una tarea estarán activas, entonces al menos se quitan tres parámetros de los seis que hay que estimar (es decir, x, y, z).
Se puede encontrar la mejor ubicación de un número limitado de fuentes al com-putar el mapa del potencial eléctrico de la supeficie con cierto modelo anterógrado (es decir, cómo un dipolo en una posición y orientación determinada propaga la señal sobre la piel cabelluda) y después se compara con el mapa del potencial medido. Esta compa-ración suele basarse en el cálculo del promedio del error cuadrado entre los dos mapas. La solución con un error cuadrado mínimo se acepta como la mejor explicación de la medición (así se usa el término estimación de fuentes por mínimos cuadrados).
Figura 5-3. Solución dipolar. Deben considerarse seis pa-rámetros para calcularse. La posición del dipolo en el vo-lumen conductor (x, y, z), la orientación del dipolo (theta y alfa) y la fuerza del mismo.

Métodos en Neurociencias Cognoscitivas
94
Dado que la búsqueda exhaustiva de una posible ubicación y orientación de las fuentes en el espacio de soluciones es prácticamente imposible, se utilizan métodos de optimización no lineales basados en algoritmos de búsqueda dirigida (Uutela et al., 1998). Obviamente, la complejidad de los algoritmos de búsqueda dirigida se in-crementa con el número de dipolos en el modelo. Así, el máximo número de fuentes independientes, que pueden localizarse en un instante de tiempo, es menor que lo que teóricamente es posible. Entre todas las soluciones compatibles con los datos, la solu-ción mínima global es ligeramente mejor que las otras.
Con el objeto de incrementar el número de dipolos que pudieran ajustarse a un mo-delo, Scherg y sus colaboradores propusieron incorporar el tiempo en el procedimento de ajuste de dipolos (Scherg y von Cramon, 1986). Esta técnica de análisis espacio-tempo-ral de múltiples fuentes (implementada en el software BESA) fija la localización de los dipolos sobre un intervalo de tiempo determinado y entonces usa el bloque completo de datos en el ajuste de mínimos cuadrados. El ajuste resulta entonces de la modu-lación variante en el tiempo de la amplitud de cada uno de esos dipolos. Como todos los modelos dipolares, el aspecto crucial en el modelo espacio-temporal es asumir el número correcto de dipolos. Se proponen dos aproximaciones en el BESA (Scherg et al., 1999): 1) El periodo completo es analizado una primera vez con un incremento en el número de fuentes. Se añaden tantas nuevas fuentes conforme la varianza explicada incremente. 2) El periodo es analizado secuencialmente y los nuevos dipolos se añaden para cada ventana de tiempo adicional si más actividad aún no es explicada. La cuestión clave que se debe resolver es obtener el número correcto de fuentes. Lo más frecuente es la elección basada en el conocimiento fisiológico. Por ejemplo, estudios de poten-ciales relacionados con eventos intracraneales (Blanke et al., 1999) han indicado una rápida activación de diferentes áreas cerebrales fuera de la corteza visual en latencias muy tempranas (Michel et al., 2004b). Así, no siempre está justificado el rechazar dipo-los que no están dentro de las áreas esperadas o que explican una pequeña cantidad de varianza adicional.
Muchos estudios se han propuesto definir el número de dipolos basándose en otros estudios funcionales de neuroimagen, como la PET o la RMf (Foxe et al., 2003). Sin em-bargo, pueden esperarse diferencias entre las dos medidas debido a múltiples razones. Por ejemplo, la activación en la RMf puede verse en las áreas llamadas de “campo ce-rrado” (es decir, áreas donde el patrón de actividad eléctrica neuronal es cero y, por lo tanto, en el EEG no se puede registrar nada de la actividad dada en esos núcleos). Por otro lado, las fuentes del EEG pueden encontrarse sin su correspondiente correlato de aumento de la activación en la RMf debido a su suceptibilidad a los artefactos o, más importante, al umbral de significancia de los datos de RMf, donde las actividades son muy débiles y duran muy poco para alcanzar significación (para una discusión sobre el asunto vea Vitacco et al., 2002). Son más prometedoras las aproximaciones donde se modelan independientes las fuentes del EEG y del MEG y donde la comparación con la RMf se usa para seleccionar las más probables distribuciones de dipolos entre las posibles (Ahlfors et al., 1999). Una tercera alternativa para definir el número de fuen-tes activas sería emplear una técnica usada en la tecnología de radares con la que se puede aislar la señal del ruido. El método, llamado clasificación de la señal múltiple (MUSIC, por sus siglas en inglés), se basa en una descomposición del eigenvalor de los datos para identificar los componentes subyacentes en series de tiempo (Mosher et al., 1992). El volumen completo del cerebro es escaneado para aquellas localizaciones de

95
Localización de fuentes de corriente de la actividad eléctrica cerebral...
fuentes que contribuyen al espacio señal. Una vez encontradas estas fuentes, los cursos de tiempo de sus momentos se determinan de la misma manera que se hace en el aná-lisis de fuentes múltiples espacio-temporal descrito arriba.
Soluciones distribuidas En vista de que hay varios problemas para la determinación del número de fuentes en los modelos dipolares, existen métodos que no necesitan supuestos a priori para la de-terminación del número de fuentes y su ubicación. Sin embargo, estos métodos depen-den de varias restricciones al modelo para su funcionamiento. Estos métodos, llamados modelos de fuentes distribuidas, se basan en la reconstrucción de la actividad eléctrica cerebral en cada punto de una rejilla en tres dimensiones, donde el número de puntos en la rejilla es mucho mayor que el número de puntos de medición en la superficie. Cada punto de la rejilla se considera un punto posible de solución de una fuente de corriente, de manera que no hay supuestos a priori del número de dipolos en el cerebro (Pascual-Marqui, 1999). En estos modelos distribuidos, la densidad de corriente se estima a lo largo de la superficie cortical con cerca de 10000 dipolos equivalentes o fuentes regio-nales (figura 5-4).
Figura 5-4. Soluciones inversas distribuidas en diferentes mode-los de corteza cerebral. Las fuen-tes distribuidas no son puntuales como en el caso de los modelos dipolares, sino que representan “manchas” extensas cuyo centro alcanza el máximo de la densidad de la corriente.
Si la orientación se mantiene fija de acuerdo con el supuesto de fuentes radiales en la corteza, se llama modelo con restricción de orientación cortical. Si sólo la localización es una restricción (es decir, por lo general se usa la frontera sustancia gris-blanca), entonces el modelo será de restricción a la superficie cortical. En este caso, dipolos regionales se ubican en la superficie de la corteza triangulada. La tarea para resolver es en-contrar una única configuración de la actividad en estos puntos de solución de la rejilla que expliquen las medidas en la superficie. Como se dijo al principio de este capítulo, por desgracia, un número infinito de distribuciones de fuentes de corriente dentro de la rejilla 3D puede llevar a exactamente el mismo mapa de distribución del potencial en la supercie de la piel cabelluda. Esto significa que el problema inverso es indeterminado. La naturaleza indeterminada del problema inverso necesita de la aplicación de diferen-tes supuestos con el objetivo de identificar la solución más probable. Las soluciones inversas distribuidas que han sido propuestas en la literatura difieren sólo en la elección

Métodos en Neurociencias Cognoscitivas
96
e implementación de restricciones al número infinito de soluciones. Algunas de estas restricciones son puramente matemáticas, otras incorporan conocimiento fisiológico o biofísico o incorporan hallazgos de otros métodos de imagen estructural y funcional. Es importante enfatizar que la validez de las restricciones a priori define la validez de la solución inversa. A nivel conceptual, cada punto en el espacio de soluciones puede ser concebido como un dipolo equivalente. Sin embargo, en contraste con las soluciones de dipolos equivalentes, los dipolos de las soluciones distribuidas tienen una posición fija. Sólo las orientaciones y la fuerza de los vectores dipolares varían. En consecuencia, las ecuaciones que describen las soluciones inversas son lineales, lo que significa que, para la estimación de la solución, una matriz puede construirse de manera lineal en relación con los datos medidos. Esto implica que las soluciones inversas lineales dis-tribuidas tienen la propiedad de que la estimación de la corriente en un volumen re-produce de modo exacto los datos medidos u observados. En la práctica, los parámetros de regularización se introducen usualmente para explicar el ruido en los datos, y así la distribución de la fuente resultante no explica los datos por completo. Si el parámetro de regularización se basa propiamente en la estimación del ruido, la parte no explicada de los datos debería, de hecho, corresponder a este ruido. El operador de regularización provee estabilidad a la solución, de manera que pequeñas variaciones en los datos no conllevan grandes variaciones en la configuración de las fuentes.
Mínima norma La estimación general para una distribución de fuentes en un modelo cerebral tridi-mensional en la ausencia de cualquier información a priori es la mínima norma (MN) (Hamalainen y Illmonemi, 1984). Este método sólo asume que la distribución de la co-rriente en 3D debe tener la mínima intensidad a lo largo del volumen conductor. Así, el método da una única solución en el sentido de que sólo una combinación de fuentes intracraneales puede tener la intensidad total más baja y, al mismo tiempo, ajustarse de modo exacto a los datos. Sin embargo, la restricción de que la intensidad global deba ser tan baja como sea posible no es necesariamente válida en términos fisiológicos. Por ejemplo, no hay prueba de que la solución con la segunda más baja amplitud global sea, en efecto, la correcta. El algoritmo empuja a soluciones que dan una fuerte activa-ción a un gran número de puntos de solución, esto es, favorece patrones de activación débiles y localizados. En consecuencia, el algoritmo mínima norma favorece las solu-ciones con fuentes superficiales, dado que se requiere menos actividad para la solución en los puntos superficiales para dar cierta distribución superficial del voltaje. Por lo tan-to, las fuentes profundas se proyectan incorrectamente en la superficie, lo que permite interpretaciones incorrectas.
Mínima norma ponderada Se han propuesto diferentes estrategias de ponderación para compensar la tendencia de favorecer las soluciones superficiales con el método de mínima norma. En la solución PROMS propuesta por Greenblatt (1993), la matriz de covarianza se usa para construir una función de ponderación dentro del espacio de las fuentes. En el algoritmo FOCUSS (por sus siglas en inglés), Gorodnitsky y colaboradores (1995) propusieron cambiar ite-rativamente el peso de acuerdo con las soluciones estimadas en el paso previo, lo que permite una solución no lineal. Grave de Peralta y Gonzalez Andino (1998) propusieron imponer restricciones físicas, por ejemplo, que las corrientes estuvieran limitadas por el

97
Localización de fuentes de corriente de la actividad eléctrica cerebral...
volumen cerebral y, así, los componentes radiales deberían ir a cero cuando se aproxi-maran a la superficie del cerebro (solución norma mínima ponderada radialmente, RWMN, por sus siglas en inglés). Mientras estas diferentes estrategias de ponderación solucionan el problema del algoritmo MN que restringe las soluciones a la superficie, se tiene que tener en mente que estas ponderaciones se basan en operaciones puramente matemáticas, sin ninguna base teórica fisiológica que pudiera justificar la selección de la ponderación.
LORETAOtras restricciones que pueden agregarse a la ponderación es la que usa la tomografía de baja resolución (LORETA por sus siglas en inglés; Pascual-Marqui et al., 1994). La restricción particular en este método es que se selecciona la solución con la distribución espacial más “suave” dada por la minimización del laplaciano de las fuentes (es decir, una medida de la robustez espacial). El razonamiento fisiológico debajo de esta restric-ción es que la actividad de neuronas y sus vecinas están altamente correlacionadas, por lo que la actividad cerebral es el reflejo de asambleas neurales.
Mientras que el supuesto de solución suave puede ser básicamente correcto, se ha criticado que la distancia entre los puntos de la solución y la limitada resolución espacial de los registros de EEG permitan una escala espacial donde tales correlaciones no se pueden esperar. De hecho, áreas muy diferentes a nivel funcional pueden estar muy cerca anatómicamente. Sin considerarse tales distinciones anatómicas de modo ex-plícito, el argumento de correlación entre zonas muy próximas, como justificación del algoritmo de LORETA, debe tomarse con cautela. En consecuencia, y dado el supuesto de correlación sobre largas distancias, LORETA provee por lo general soluciones muy suaves, que incluso incluyen a ambos hemisferios o diferentes lóbulos (Trujillo-Barreto et al., 2004).
VARETA La tomografía de resolución variable (VARETA, por sus siglas en inglés; Valdés-Sosa et al., 1996; Bosch et al., 2001) es una técnica también desarrollada recientemen-te para estimar soluciones distribuidas discretas tipo spline (Riera et al., 1997), pero que puede estimar tanto fuentes discretas como distribuidas con igual precisión. Esta técnica impone diferentes cantidades de suavización espacial para diferentes tipos de generadores seleccionados por un procedimiento de búsqueda guiados por los mismos datos. VARETA se ha empleado para localizar fuentes de los potenciales auditivos y su precisión ha sido validada con otros métodos de localización de fuentes como el BESA (Picton et al., 1999).
LAURA El promedio autorregresivo local (LAURA, por sus siglas en inglés) es un método de soluciones inversas distribuidas que incorpora leyes biofísicas a sus restricciones en el algoritmo de mínima norma. De acuerdo con la teoría electromagnética, la fuerza de la fuente decae con la inversa de la distancia cúbica para los campos vectoriales y con la inversa del cuadrado de la distancia para los potenciales de campo. El método asume que la actividad caerá de acuerdo con estas leyes físicas cuando uno se mueva lejos de la fuente. LAURA integra esta ley en términos de un promedio autorregresivo local con coeficientes que dependen de las distancias entre los puntos de solución (Grave de

Métodos en Neurociencias Cognoscitivas
98
Peralta et al., 2001). El promedio autorregresivo determina así la actividad para cada punto que no puede ser estimado únicamente desde los datos. En consecuencia, la ac-tividad en un punto depende de dos contribuciones: una fijada por las leyes biofísicas y la otra libre para ser determinada por los datos. Se pueden incorporar los detalles ana-tómicos en este nivel del modelamiento al variar los coeficientes de regresión.
Aproximaciones bayesianas En la aproximación bayesiana se incorpora un método estadístico que introduce in-formación a priori en la estimación de las fuentes. Incluso si las implementaciones no lineales resultan en problemas matemáticos complejos, éste es probablemente el mé-todo más prometedor, ya que permite una solución por una descripción más detallada anatómica y funcional a priori. Los tipos de información a priori que se pueden incor-porar en esta aproximación incluyen información de la corriente neural (Schmidt et al., 1999), la naturaleza focal escasa de las fuentes (Phillips et al., 1997), restricciones com-binadas espacio-temporales (Baillet y Garnero, 1997), así como también las estrategias para penalizar fuentes fantasmas (Trujillo-Barreto et al., 2004).
Posprocesamiento de las imágenes Visualización de las soluciones inversas en imágenes RM De manera similar a cuando se hace el corregistro entre el modelo de la cabeza y la po-sición de los electrodos, un prerrequisito para poder definir los resultados de las solucio-nes inversas dentro de la imagen de resonancia magnética estructural es el corregistro entre esas dos modalidades de imagen (es decir, el espacio EEG y el de la RM). La última meta de las imágenes de fuentes del EEG es la localización de dichas fuentes en estruc-turas cerebrales definidas, de manera que la comparación directa con otros métodos de imagen, con estudios de lesiones o con registros intracraneales pueda hacerse. Otro objetivo importante es tener las múltiples imágenes de las fuentes para llevar a cabo análisis estadísticos, como se hace en otros métodos de neuroimagen. La mayoría del software libre y comercial disponible provee esta posibilidad, ya que se aplican sobre todo con propósitos de visualización (Aubert, 2001). Con estas herramientas gráficas, pueden generarse mapas tridimensionales con los tres planos ortogonales y el tras-lapamiento de imágenes a color con los valores máximos de las fuentes. Recientemente, esta información ha sido usada para definir las coordenadas de las fuentes en términos de las áreas de Brodmann (o en las coordenadas de Talairach) y plantear conclusiones acerca de estructuras anatomofuncionales que están activas. La precisión de estos si-tios depende de manera crucial de dos aspectos: el modelo de la cabeza seleccionado al inicio para computar la solución inversa y la solución inversa en sí misma.
Estadística de imágenes En los estudios clínicos con imágenes de fuentes del EEG, es común aplicar los algo-ritmos de localización a los datos de un solo paciente. La mayoría de los estudios ex-perimentales limitan la localización de las fuentes sólo a datos promedio de un grupo. Lo anterior es con el objetivo de ilustrar los resultados en cierta ventana de tiempo o su secuencia de activación. En este caso no se provee de la replicabilidad estadística de estas fuentes o su correspondencia a la real localización de las fuentes en el individuo. Si se aplica el algoritmo de localización de fuentes a cada individuo en un estudio, es factible hacer un análisis estadístico con esos datos. Cuando se usan los algoritmos de

99
Localización de fuentes de corriente de la actividad eléctrica cerebral...
localización de dipolos únicos, por ejemplo, es posible aplicar pruebas estadísticas con respecto a un cambio en la ubicación del mismo dentro del plano cartesiano. Esto per-mite obtener conclusiones acerca de los posibles cambios de ubicación del dipolo en una o varias direcciones (Michel et al., 1995). Incluso si un solo dipolo en sí mismo puede o no representar las fuentes en el caso de patrones de activación distribuida, un cambio estadísticamente significativo del “centro de gravedad” de todas las fuentes, debido a una manipulación experimental o debido a cierta droga o enfermedad, puede proveer indicaciones de un cambio en la distribución de la actividad global dentro del cerebro. Con el uso de las soluciones inversas distribuidas, se pueden aplicar análisis estadísti-cos similares a los propuestos para otro tipo de imágenes topográficas funcionales. En los estudios de RMf, el método llamado mapeo estadístico paramétrico (SPM, por sus siglas en inglés) es el procedimiento estadístico más empleado. Después de la norma-lización espacial de las imágenes individuales de RMf en un cerebro estándar, se cons-truye un modelo estadístico dependiente del diseño experimental, donde se establecen los factores, como en el caso de un ANOVA, pero con una serie de volúmenes. Para las comparaciones planeadas, se hace una serie de pruebas t entre dos condiciones expe-rimentales, por ejemplo, por cada vóxel o región de interés para determinar las áreas donde hay diferencias significativas. Varios estudios han aplicado un procedimiento estadístico similar al anterior a los mapas tomográficos derivados de las soluciones in-versas distribuidas.
Al resolver el problema inverso por un modelo general de cabeza y usar el mismo espacio de solución para todos los sujetos, es posible hacer múltiples pruebas t para cada vóxel del espacio de solución (Silva-Pereyra et al., 2003). Así, como en el análisis de la RMf, queda una cuestión importante concerniente al ajuste de los resultados estadís-ticos a un apropiado criterio de significancia. La corrección por múltiples pruebas debe estar basada en el número de medidas independientes y no en el número de vóxeles (Murray et al., 2004).
El análisis estadístico de los mapas de soluciones inversas es sencillo. Tiene muchas ventajas comparado con la presentación de un mapa de solución inversa de un gran promedio de grupo. Primero, los datos de un grupo promediado pueden estar domina-dos por la fuente máxima que es idéntica en dos condiciones experimentales (es decir, las fuentes occipitales en los componentes visuales tempranos). En consecuencia, las débiles pero consistentes diferencias en otras áreas tienen el riesgo de ser ignoradas por el umbral. Segundo, las soluciones inversas distribuidas pueden producir fuentes es-purias, en especial cuando se aplican a datos ruidosos. Sin embargo, tales fuentes es-purias, por lo general, no sobreviven al análisis estadístico debido a su inconsistencia a través de los sujetos. Así, la cuestión no resuelta y delicada de la regularización puede ser minimizada por el análisis estadístico cuando se calculan las soluciones inversas dis-tribuidas. Finalmente, con datos apareados, la información relevante se apoya en las diferencias intrasujetos más que en los promedios separados sobre las condiciones. El análsis estadístico de la solución inversa revelaría diferencias en áreas cerebrales que no son visibles en las imágenes promedio de cada una de las condiciones experimentales.
Cuando se usa un procedimiento estadístico no paramétrico, el supuesto de nor-malidad no es necesario y los niveles de significación pueden calcularse mediante la distribución empírica de los datos. Se han propuesto métodos de cómputo intensivo basados en principios de permutación como una alternativa estadística en la prueba de diferencias entre potenciales relacionados con eventos (Galán et al., 1997) y en mapas

Métodos en Neurociencias Cognoscitivas
100
volumétricos (Blair y Karniski, 1994). Esta alternativa de análisis estadístico se ha usado previamente para la evaluación de procesos cognoscitivos por medio de imágenes del VARETA en el dominio de la frecuencia (Fernández et al., 2000) y con potenciales rela-cionados con eventos (Silva-Pereyra et al., 2003). Este método tiene las siguientes ven-tajas: las pruebas son libres de distribución, no se requieren supuestos de una estructu-ra de correlación subyacente y proveen valores de probabilidad exactos para cualquier número de sujetos, puntos en el tiempo y sitios de registro.
Podríamos describir el proceso de análisis estadístico mediante esta alternativa de la siguiente manera. Supongamos que tenemos dos grupos de sujetos, uno con trastornos del aprendizaje y el otro control. Ambos grupos llevaron a cabo una tarea de atención sostenida y a todos los sujetos se les registraron sus potenciales relacionados con la res-puesta a un estímulo “blanco”. Se identifica un componente de los PRE asociado con la actualización de la memoria de trabajo, conocido como P300 como se menciona en el capítulo 3 de este libro. Se calculan las fuentes de dicho componente para cada uno de los sujetos de ambos grupos. Lo que interesaría en este supuesto estudio es saber si las regiones cerebrales involucradas en la generación del componente P300, ante una tarea de atención sostenida, son diferentes entre sujetos con trastornos del aprendizaje y nor-males. En términos estrictamente estadísticos, nuestra hipótesis Nula sería: la media del grupo 1 es igual a la del grupo 2. Para contrastarla primero es necesario hacer una matriz de ts o comparaciones vóxel a vóxel entre los dos grupos. A este primer acercamiento le llamaremos matriz de las ts originales (figura 5-5).
Figura 5-5. Cálculo de la matriz de ts original sin la primera permutación.
En un siguiente paso del análisis, se permutan las imágenes-rebanada entre grupos y se consideran a todos los sujetos. Cada permutación lleva consigo una comparación vóxel a vóxel, lo que genera un nuevo conjunto de valores ts. Así, en la segunda permu-tación, por ejemplo, se tiene una nueva matriz de ts, como se muestra en la figura 5-6.
Con base en las múltiples matrices de ts por cada permutación, es posible construir la distribución t empírica como se muestra en la figura 5-7.

# de
vóxeles
Frec
uenc
ia
Valores t
# de
vóxeles
Frec
uenc
ia
Valores t
Nivel de
alfa
101
Localización de fuentes de corriente de la actividad eléctrica cerebral...
La distribución estimada por las técnicas de permutación puede entonces usarse para determinar los niveles de significación que controlan el error de juicio experimen-tal (es decir, probabilidad de falsear el rechazo de la hipótesis nula cuando es verdadera en todo el experimento) para las comparaciones simultáneas univariadas. Así, uno pue-de probar de manera simultánea diferencias en las neuroimágenes para todos los pun-tos en el espacio y evitar que se infle el alfa o el error tipo I (ver Blair y Karniski, 1994). Un estadígrafo multivariado puede emplearse para resumir y probar diferencias entre neuroimágenes asociadas con dos condiciones y puede obtenerse del máximo valor de todos los estadísticos univariados. Éste es el máximo de las pruebas t calculadas entre dos conjuntos de datos para todos los vóxeles tmax.
Figura 5-6. Subsecuentes permutaciones y estimaciones de las ts para cada permutación.
Una vez comparados los valores t originales en la distribución empírica previamen-te calculada, se podrán obtener los valores de t que pasan un umbral de significación.
Figura 5-7. Distribución empírica de las ts de comparación entre grupos. Se fija el nivel de alfa como región de rechazo de la hipótesis nula. Es factible tener el valor de probabilidad asociado a los valores de t originales.
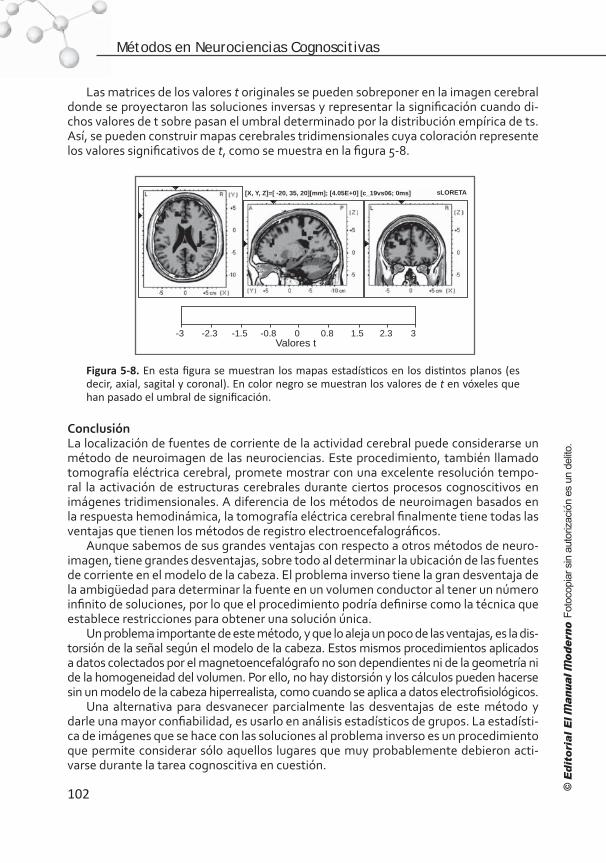
Valores t-2.3 -1.5 -0.8 0 0.8 1.5 2.3 3-3
sLORETA[X, Y, Z]=[ -20, 35, 20][mm]; [4.05E+0] [c_19vs06; 0ms]
Métodos en Neurociencias Cognoscitivas
102
Las matrices de los valores t originales se pueden sobreponer en la imagen cerebral donde se proyectaron las soluciones inversas y representar la significación cuando di-chos valores de t sobre pasan el umbral determinado por la distribución empírica de ts. Así, se pueden construir mapas cerebrales tridimensionales cuya coloración represente los valores significativos de t, como se muestra en la figura 5-8.
Figura 5-8. En esta figura se muestran los mapas estadísticos en los distintos planos (es decir, axial, sagital y coronal). En color negro se muestran los valores de t en vóxeles que han pasado el umbral de significación.
Conclusión La localización de fuentes de corriente de la actividad cerebral puede considerarse un método de neuroimagen de las neurociencias. Este procedimiento, también llamado tomografía eléctrica cerebral, promete mostrar con una excelente resolución tempo-ral la activación de estructuras cerebrales durante ciertos procesos cognoscitivos en imágenes tridimensionales. A diferencia de los métodos de neuroimagen basados en la respuesta hemodinámica, la tomografía eléctrica cerebral finalmente tiene todas las ventajas que tienen los métodos de registro electroencefalográficos.
Aunque sabemos de sus grandes ventajas con respecto a otros métodos de neuro-imagen, tiene grandes desventajas, sobre todo al determinar la ubicación de las fuentes de corriente en el modelo de la cabeza. El problema inverso tiene la gran desventaja de la ambigüedad para determinar la fuente en un volumen conductor al tener un número infinito de soluciones, por lo que el procedimiento podría definirse como la técnica que establece restricciones para obtener una solución única.
Un problema importante de este método, y que lo aleja un poco de las ventajas, es la dis-torsión de la señal según el modelo de la cabeza. Estos mismos procedimientos aplicados a datos colectados por el magnetoencefalógrafo no son dependientes ni de la geometría ni de la homogeneidad del volumen. Por ello, no hay distorsión y los cálculos pueden hacerse sin un modelo de la cabeza hiperrealista, como cuando se aplica a datos electrofisiológicos.
Una alternativa para desvanecer parcialmente las desventajas de este método y darle una mayor confiabilidad, es usarlo en análisis estadísticos de grupos. La estadísti-ca de imágenes que se hace con las soluciones al problema inverso es un procedimiento que permite considerar sólo aquellos lugares que muy probablemente debieron acti-varse durante la tarea cognoscitiva en cuestión.

103
Localización de fuentes de corriente de la actividad eléctrica cerebral...
Referencias
Ahlfors, S. P., Simpson, G. V., Dale, A. M., Belliveau, J. W., Liu, A. K., Korvenoja, A., Virtanen, J., Huotilainen, M., Tootell, R. B., Aronen, H. J. y Ilmoniemi, R. J. (1999). Spatiotemporal acti-vity of a cortical network for processing visual motion revealed by MEG and fMRI. Journal of Neurophysiology, 82, 2545-2555.
Aubert, E. (2001). Síntesis y visualización de imágenes de la actividad eléctrica cerebral. Tesis para obtener el grado de Doctor en Ciencias. La Habana, Cuba.
Baillet, S. y Garnero, L. A. (1997). Bayesian approach to introducing anatomofunctional priors in the EEG/MEG inverse problem. IEEE Transaction of Biomedical Engineering, 44, 374-385.
Barnes, G. R. y Hillebrand, A. (2003). Statistical flattening of MEG beamformer images. Human Brain Mapping, 18, 1-12.
Berg, P. y Scherg, M. (1994). A fast method for forward computation of multiple-shell spherical head models. Electroencephalography Clinical Neurophysiology, 90, 58–64.
Blanke, O., Morand, S., Thut, G., Michel, C.M., Spinelli, L., Landis, T. y Seeck, M. (1999). Visual activity in the human frontal eye field. Neuroreport, 10, 925-930.
Bosch-Bayard, J., Valdés-Sosa, P., Virues-Alba, T., Aubert-Vázquez, E., John, E. R., Harmony, T., Riera-Díaz, J. y Trujillo-Barreto, N. (2001). 3D statistical parametric mapping of EEG source spectra by means of variable resolution electromagnetic tomography (VARETA). Clinical Electroencephalography, 32, 47-61.
Cabeza, R. y Nyberg, L. (2000). Imaging cognition. II. An empirical review of 275 pet and fMRI studies. Journal of Cognitive Neuroscience (review), 12, 1-47.
Cuffin, N.B. (1996). EEG localization accuracy improvements using realistically shaped head mo-dels. IEEE Transaction of Biomedical Engineering, 43, 299-303.
Duffy, F. H., Bartels, P. H. y Burchfiel, J. L. (1981). Significance probability mapping: an aid in the topographic analysis of brain electrical activity. Electroencephalography Clinical Neurophysiology, 51, 455-462.
Fernández, T., Harmony T., Silva-Pereyra J., Fernández-Bouzas, A., Gersenowies, J., Galán, L., Carbonell, F., Marosi, E., Otero, G. y Valdés, S. (2000). Specific EEG frequencies at specific brain areas and performance. Neuroreport, 11, 2663-2668.
Fletcher, E. M., Kussmaul, C. L. y Mangun, G. (1998). Estimation of interpolation errors in scalp topographic mapping. Electroencephalography Clinical Neurophysiology, 1996, 422-434.
Foxe, J. J., McCourt, M. E. y Javitt, D. C. (2003). Right hemisphere control of visuospatial atten-tion: line-bisection judgments evaluated with high-density electrical mapping and source analysis. Neuroimage, 19, 710-726.
Fuchs, M., Wagner, M. y Kastner, J. (2001). Boundary element method volume conductor mo-dels for EEG source reconstruction. Clinical Neurophysiology, 112, 1400-1407.
Galán, L., Biscay, R., Rodríguez, J. L., Pérez-Ávalo, M. C. y Rodríguez, R. (1997). Testing topo-graphic differences between event related brain potentials by using non-parametric com-binations of permutation tests. Electroencephalography Clinical Neurophysiology, 102, 240-247.
Gevins, A. S., Brickett, P., Costales, B., Le, J. y Reutter, B. (1990). Beyond topographic mapping: towards functional–anatomical imaging with 124channel EEGs and 3-D MRIs (Review). Brain Topography, 3, 53-64.
Gorodnitsky, I. F., George, J. S. y Rao, B. D. (1995). Neuromagnetic source imaging with FOCUSS: a recursive weighted minimum norm algorithm. Electroencephalography Clinical Neurophysiology, 231-251.

Métodos en Neurociencias Cognoscitivas
104
Grave de Peralta, R., González, S., Lantz, G., Michel, C. M. y Landis, T. (2001). Noninvasive lo-calization of electromagnetic epileptic activity. I. Method descriptions and simulations. Brain Topography, 14, 131-137.
Grave de Peralta, R. y González, S. (1998). A critical analysis of linear inverse solutions. IEEE Transaction of Biomedical Engineering, 45, 440-8.
Greenblatt, R. (1993). Probabilistic reconstruction of multiple sources in the neuroelectromag-netic inverse problem. Inverse Problems, 9, 271-284.
Hamalainen, M. S. e Ilmoniemi, R. J. (1984). Interpreting measured magnetic fields of the brain: estimates of current distributions. Technical report TKKFA559. Helsinki: Helsinki University of Technology, Espoo.
Hamalainen, M. y Sarvas, J. (1989). Realistic conductor geometry model of the human head for interpretation of neuromagnetic data. IEEE Transaction of Biomedical Engineering, 36, 165-171.
Hassaina, F., Medina, V., Donadey, A. y Langevin, F. (1994). Scalp potential and current densi-ty mapping with an enhanced spherical spline interpolation. Medical Program Technology, 20, 23-230.
Hoekema, R., Wieneke, G. H., Leijten, F. S., van Veelen, C. W., van Rijen, P. C., Huiskamp, G. J., Ansems, J. y van Huffelen, A. C. (2003). Measurement of the conductivity of skull, tempo-rarily removed during epilepsy surgery. Brain Topography, 16, 29-38.
John, E. R., Karmel, B. Z., Corning, W. C., Easton, P., Brown, D., Ahn, H., John, M., Harmony, T., Prichep, L., Toro, A., Gerson, I., Bartlett, F., Thatcher, F., Kaye, H., Valdés, P. y Schwartz, E. (1977). Neurometrics. Science, 196, 1393–1410.
Le, J., Lu, M., Pellouchoud, E. y Gevins, A. (1998). A rapid method for determining standard 10/10 electrode positions for high resolution EEG studies. Electroencephalography Clinical Neurophysiology, 106, 554-558.
Lehmann, D. (1987). Principles of spatial analysis. En A. Gevins y A. Remond (Eds.), Handbook of electroencephalography and clinical neurophysiology (vol. 1). Methods of analysis of bra-in electrical and magnetic signals (pp. 309-354). Amsterdam: Elsevier.
Mesulam, M. M. (1998). From sensation to cognition (Review). Brain, 121, 1013-1052. Michel, C. M., Lehmann, D., Henggeler, B. y Brandeis, D. (1992). Localization of the sources of
EEG delta, theta, alpha and beta frequency bands using the FFT dipole approximation. Electroencephalography Clinical Neurophysiology, 82, 38-44.
Michel, C. M., Murray, M., Lantz, G., González, S., Spinelli, L. y Grave de Peralta, R. (2004a). EEG source imaging. Clinical Neurophysiology, 115, 2195-2222
Michel, C. M., Pascual-Marqui, R. D., Strik, W. K., Koenig, T. y Lehmann, D. (1995). Frequency domain source localization shows state-dependent diazepam effects in 47-channel EEG. Journal of Neural Transmission General Section, 99, 157-171.
Michel, C. M., Seeck, M., y Murray, M. M. (2004b). The speed of visual cognition. En M. Hallett, L. Phillips, D. Schomer, J. Mark (Eds.), Advances in Clinical Neurophysiology (Supplements to Clinical Neurophysiology) (vol.54) (pp. 437-445). The Netherlands: Elsevier. .
Miller, C. E. y Henríquez, C. S. (1990). Finite element analysis of bioelectric phenomena (Review). Critical Reviews in Biomedical Engineering, 18, 207-233.
Mosher, J. C., Lewis, P. S. y Leahy, R. M. (1992). Multiple dipole modeling and localization from spatio-temporal MEG data. IEEE Transaction of Biomedical Engineering, 39, 541-557.
Murray, M. M., Michel, C. M., Grave de Peralta, R., Ortigue, S., Brunet, D., González S., y Schnider, A. (2004). Rapid discrimination of visual and multisensory memories revealed by electrical neuroimaging. Neuroimage, 21, 125-135.

105
Localización de fuentes de corriente de la actividad eléctrica cerebral...
Nunez, P. L. (1981). Electric fields in the brain: the neurophysics of EEG. Nueva York: Oxford University Press.
Pascual-Marqui, R. D. (1999). Review of methods for solving the EEG inverse problem. International Journal of Bioelectromagnetism, 1, 75-86.
Pascual-Marqui, R. D. (2002). Standardized low resolution brain electromagnetic tomography (sLORETA): technical details. Methods and Findings in Experimental Clinical Pharmacology, 24D, 5-12.
Pascual-Marqui, R. D. y Lehmann, D. (1993). Comparison of topographic maps and the reference electrode: comments on two papers by Desmedt and collaborators. Electroencephalography Clinical Neurophysiology, 88 (530-531), 534-536.
Pascual-Marqui, R. D., Michel, C. M. y Lehmann, D. (1994). Low resolution electromagnetic to-mography: a new method to localize electrical activity in the brain. International Journal of Psychophysiology, 18, 49-65.
Perrin, F., Pernier, J., Bertrand, O., Girad, M. H. y Echalier, J. F. (1987). Mapping of scalp poten-tials by surface spline interpolation. Electroencephalography Clinical Neurophysiology, 66, 75-81.
Perrin, F., Pernier, J., Bertrand, O. y Echallier, J. F. (1989). Spherical splines for scalp poten-tial and current density mapping. Electroencephalography Clinical Neurophysiology, 72, 184-189.
Phillips, J. W., Leahy, R. M. y Mosher, J. C. (1997). MEG-based imaging of focal neuronal current sources. IEEE Transaction of Biomedical Engineering, 16, 338-348.
Picton, T., Alain, C., Woods, D., John, M., Sherg, M., Valdés-Sosa, P., Bosch-Bayard, J. y Trujillo, N. (1999) Intracerebral sources of human auditory-evoked potentials. Audiology and Neuro-otology, 4, 64-79.
Robinson, S. E., y Vrba, J. (1999). Functional neuroimaging by synthetic aperture magnetome-try (SAM). En T. Yoshimoto, M. Kotani, S. Kuriki, H. Karibe y N. Nakasoto (Eds.), Recent advances in biomagnetism (pp. 302-305). Senai: Tohoku University Press.
Rodin, E., Rodin, M., Boyer, R. y Thompson, J. (1997). Displaying electroencephalographic di-pole sources on magnetic resonance images. Journal of Neuroimaging, 7, 106-10.
Scherg, M. y von Cramon, D. (1986). Evoked dipole source potentials of the human auditory cor-tex. Electroencephalography Clinical Neurophysiology, 65, 344-60.
Scherg, M., Bast, T. y Berg, P. (1999). Multiple source analysis of interictal spikes: goals, requi-rements, and clinical value (Review). Journal Clinical Neurophysiology 16, 214-24.
Schmidt, D. M., George, J. S. y Wood, C. C. (1999). Bayesian inference applied to the electro-magnetic inverse problem. Human Brain Mapping, 7, 195-212.
Silva-Pereyra, J., Rivera-Gaxiola, M., Aubert, E., Bosch, J., Galán, L. y Salazar, A. (2003). N400 during lexical decision task: A current source localization study. Clinical Neurophysiology, 114, 2469-248.
Towle, V. L., Bolanos, J., Suárez, D., Tan, K., Grzeszczuk, R., Levin, D.N., Cakmur, R., Frank, S. A. y Spire, J. P. (1993). The spatial location of EEG electrodes: locating the best-fitting sphe-re relative to cortical anatomy. Electroencephalography Clinical Neurophysiology 86, 1-6.
Trujillo-Barreto, N. J., Aubert-Vazquez, E. y Valdés-Sosa P. (2004). Bayesian model averaging in EEG/MEG imaging. NeuroImage, 21, 1300-1319.
Uutela, K., Hamalainen, M. y Salmelin, R. (1998). Global optimization in the localization of neu-romagnetic sources. IEEE Transaction of Biomedical Engineering, 45, 716-723.
Valdés, P., Bosch, J., Grave de Peralta, R., Hernández, J., Riera, J., Pascual-Marqui, R. y Biscay, R. (1992). Frequency domain models of the EEG. Brain Topography 4, 309-319.

Métodos en Neurociencias Cognoscitivas
106
Valdés P, Marti F, Garcia F, Casanova R (1996): Variable resolution electro-magnetic tomogra-phy. Proceedings of The Tenth International Conference on Biomagnetism, BIOMAG’96, Santa Fe, New Mexico.
Vitacco, D., Brandeis, D., Pascual-Marqui, R. y Martin, E. (2002). Correspondence of event-re-lated potential tomography and functional magnetic resonance imaging during language processing. Human Brain Mapping, 17, 4-12.
Vrba, J. y Robinson, S. E. (2001). Signal processing in magnetoencephalography (Review). Methods, 25, 249-271.
Wang, M. Y., Maurer, C. R., Fitzpatrick, J. M. y Maciunas, R. J. (1996). An automatic technique for finding and localizing externally attached markers in CT and MR volume images of the head. IEEE Transaction of Biomedical Engineerin, 43, 627-637.
Williamson, S. J., Lu, Z. L., Karron, D. y Kaufman, L. (1991). Advantages and limitations of mag-netic source imaging (Review). Brain Topography, 4, 169-180.

107
Capítulo 6. Métodos y teorías en neuropsicología: una perspectiva histórica y actual
María Guillermina Yáñez-Téllez
Resumen El objetivo de este trabajo es plantear un panorama histórico y actual de las teorías y métodos que han buscado respuesta a uno de los problemas fundamentales en neu-ropsicología: la relación entre el cerebro y el comportamiento. Para ello, se parte de una revisión histórica con el fin de valorar cómo ha sido la evolución de las diferentes aproximaciones que han dado paso a las concepciones más recientes, con las cuales se ha podido avanzar en el conocimiento de esta compleja relación. En la primera parte, se sigue un orden cronológico para estudiar los enfoques pioneros en la época antigua acerca de si las funciones estaban localizadas en el cerebro o en el corazón, el dualis-mo, los primeros estudios de correlaciones clínicopatológicas y el desarrollo de las ideas localizacionistas y antilocalizacionistas. En la segunda parte, se tratan las concepciones teóricas actuales acerca de la organización de las funciones mentales en el cerebro, los conceptos de sistema funcional complejo y redes neurales, y los más recientes aportes teóricos y metodológicos de la neuropsicología cognitiva. En el tercer apartado, se re-sumen los principales métodos en neuropsicología, como el estudio de la lesión cere-bral, el método psicométrico y la neuropsicología experimental. Por último, se señala la importancia de las teorías y métodos que aporta la neuropsicología a las neurociencias cognoscitivas, en particular, lo insustituible que resulta el estudio de pacientes con le-sión cerebral y el enfoque psicométrico para entender la relación cerebro-conducta.
Definición y objeto de estudio Benton (1988) define la neuropsicología humana como una disciplina que investiga, en un nivel empírico y científico, las interrelaciones del cerebro con la mente y la conducta. Considera que es una disciplina en la que confluyen diversos campos de estudio, como neurología y psicología, neuroanatomía y neurofisiología, y neuroquímica y neurofarma-cología. Para Lezak, Howieson y Loring (2004), la neuropsicología clínica es una ciencia aplicada interesada en la expresión de la conducta de la disfunción cerebral. En un sen-tido similar, Kolb y Whishaw (2003) consideran que la neuropsicología es el estudio de la relación entre la función cerebral humana y la conducta. Aunque obtiene información de muchas disciplinas, se enfoca básicamente en el desarrollo de una ciencia de la conducta humana basada en la función del cerebro humano. La diferencia fundamental entre la primera definición y las otras dos radica, quizá, en la connotación dada al término con-ducta, ya que los segundos autores se refieren a la conducta en un sentido muy amplio, para representar las funciones cognoscitivas, las emociones y el comportamiento. Le-zak, Howieson y Loring (2004) estiman que la neuropsicología es una disciplina valiosa y

Métodos en Neurociencias Cognoscitivas
108
reconocida que establece un puente entre las neurociencias y las ciencias de la conducta y se caracteriza por una metodología distintiva y un amplio rango de aplicaciones.
Aspectos históricos Los aspectos históricos en neuropsicología tienen que ver con el problema de la loca-lización de las funciones en la corteza cerebral. Sobre este tema, Luria —en su libro Las funciones corticales superiores del hombre (1977, trabajo publicado originalmente en 1969)— menciona cómo la solución a dicho problema no sólo ha dependido de los medios técnicos disponibles en una determinada época histórica, sino también de los conceptos teóricos acerca de los procesos psíquicos que predominaban en cada periodo histórico. La vigencia de tal afirmación es incuestionable, dado que en neuropsicología son igualmente importantes tanto la teoría disponible acerca de las funciones menta-les, su organización y la forma en que se relacionan con el funcionamiento cerebral, como la disposición de los medios técnicos para su estudio. De aquí que una revisión del desarrollo histórico de estos dos aspectos clave resulta relevante para ubicar el momento actual de desarrollo en esta disciplina. Aunque esta revisión no pretende ser exhaustiva, sí enfatiza aquellos hechos históricos que fueron moldeando los conceptos acerca de la relación mente-cerebro, hasta llegar a los que son de más amplia aceptación en los ámbitos clínico y de investigación de la neuropsicología actual.
Época antigua: hipótesis del cerebro e hipótesis del corazón Para Kolb y Whishaw (2003), desde la Grecia antigua pueden delimitarse dos hipótesis en relación con la localización de las funciones mentales: la de su localización en el cere-bro, a la cual le llaman hipótesis del cerebro, y la idea de que el corazón era el sustento de dichas funciones, a la cual le denominan hipótesis del corazón.
En relación con la hipótesis del cerebro, a Alcamaeon de Crotón, estudiante de Pi-tágoras, se le atribuye haber hecho la afirmación, en el siglo v a. C., de que el cerebro era el asiento físico de la mente, con base en sus observaciones de pacientes con daño cerebral. En esta misma categoría puede incluirse Hipócrates, a quien se le atribuyen diversas descripciones de alteraciones del lenguaje en casos de algunas enfermedades fatales. García-Albea y García-Albea (2006) citan un párrafo del tratado de Hipócrates, Sobre la enfermedad sagrada, poco difundido y que, por su actualidad, parece sorpren-dente para el momento histórico en que vivió:
Conviene que la gente sepa que nuestros placeres, gozos, risas y juegos no proceden de otro lugar sino del cerebro y lo mismo las penas y las amarguras, sinsabores y llantos. Y por él precisamente razonamos e intuimos, y vemos y oímos, y distinguimos lo feo, lo bello, lo malo, lo agradable y lo desagradable […] también por su causa enloquecemos y deliramos, y se nos presentan espasmos y terrores, unos de noche y otros de día, e insom-nios e inoportunos desvaríos, preocupaciones inmotivadas y estados de ignorancia de las circunstancias reales, y extrañezas. Y todas estas cosas las padecemos a partir del cere-bro, cuando éste no está sano, sino que se pone más caliente de lo natural, o bien más frío, o más seco, o sufre alguna otra afección contraria a su naturaleza.
Otro de los pensadores que pueden identificarse con la hipótesis cerebral es Platón, quien desarrolló el concepto de un alma tripartita, con su parte racional en el cerebro.
Entre todos estos pensadores, en quienes prevalecía la hipótesis del cerebro, impe-raba la idea de que éste era propiamente un empaque o envoltura para la sustancia real

109
Métodos y teorías en neuropsicología: una perspectiva histórica y actual
del pensamiento, el fluido cerebroespinal, por lo que los ventrículos cerebrales eran la parte más importante del cerebro. No obstante, para algunos de ellos como Galeno, el tejido cerebral también era importante (citado en Feinberg y Farah, 2000). Galeno observaba los cambios conductuales en los gladiadores romanos heridos y señalaba que no sólo el daño cerebral alteraba la conducta, sino también que los nervios de los órganos de los sentidos iban al cerebro y no al corazón.
Kolb y Whishaw (2003) mencionan a Empédocles de Acragas (427-347 a. C.) como el partidario más antiguo del que se tiene registro de localización de los procesos men-tales en el corazón. Dentro de esta misma hipótesis, Aristóteles (384-322 a. C.) supuso que dado que el corazón era caliente y activo, era la fuente de los procesos mentales, mientras que el cerebro, debido a que era frío e inerte, servía como un radiador para enfriar la sangre.
Edad media En la Edad Media en Europa, los ventrículos continuaban siendo el foco de teorías que relacionaban la mente con el cerebro. Los ventrículos anteriores se asociaron con la percepción; los medios, con la razón, y los posteriores, con la memoria (Pagel, citado en Feinberg y Farah, 2000).
Dualismo de Descartes Durante el Renacimiento, Descartes reemplazó el concepto platónico de un alma tri-partita por el de un alma única e indivisible, a la cual llamó mente. Descartes descri-be la mente como inmaterial y sin extensión espacial, diferente del cuerpo. El cuerpo operaba de acuerdo con los principios de una máquina, pero era la mente la que deci-día qué movimientos debía hacer la máquina. La glándula pineal era el vehículo por el cual la mente influía en el cuerpo. Descartes negaba alguna forma de identidad entre la mente y el cerebro, por lo cual se le conoce como impulsor del dualismo. De acuerdo con esta postura, la mente y el cuerpo estaban separados, aunque podían interactuar. Kolb y Whishaw (2003) consideran que con el dualismo de Descartes se inicia lo que actualmente se conoce como el problema mente-cuerpo. Desde esta perspectiva, es de suponer que no representaba un problema la localización de funciones, ya que la mente no era divisible y era inmaterial.
Localizacionismo y antilocalizacionismo A finales del siglo xviii y principios del xix, Franz Josef Gall y Johann Spurzheim formu-laron una teoría general de la localización cerebral, conocida actualmente como freno-logía. Gall identificó 27 facultades humanas básicas y las asoció con centros cerebrales particulares, que a su vez afectaban la forma del cráneo en el sentido de mayor promi-nencia o depresión de la zona en cuestión. Debido a esta postura Gall es conocido como el pionero del localizacionismo.
Por su parte, se considera a Jean-Pierre Flourens uno de los detractores de la teoría de Gall, como pionero del antilocalizacionismo. Al parecer, la principal preocupación de Flourens era de índole religiosa, ya que la teoría de Gall dividía el alma y el cerebro en partes funcionalmente distintas, mientras que Flourens consideraba que el alma era unitaria. Flourens llevó a cabo una investigación en la cual provocaba extensas lesiones en diversas especies de aves para demostrar la equipotencialidad de la cor-teza. Observó que, pasado cierto tiempo, a partir de la lesión había una recuperación

Métodos en Neurociencias Cognoscitivas
110
del comportamiento, independientemente de la parte destruida. Llegó a la conclu-sión de que la corteza actúa como un todo homogéneo.
Según Feinberg y Farah (2000), fue el populismo de Gall y los ataques con prue-bas empíricas de Flourens los que provocaron que el localizacionismo quedara fuera de los intereses de los científicos de principios del siglo xix. Así, cuando Jean-Baptiste Bouillaud, en 1825, presentó una serie de casos clínicos de pérdida del habla posterio-res a lesiones frontales, su trabajo fue ignorado. De acuerdo con Bouillaud (citado en Hécaen y Dubois, 1983), “investigaciones clínicas tienden a demostrar que la pérdida de la palabra corresponde a la lesión de los lóbulos anteriores del cerebro, y a confirmar la opinión de Gall relativa a la localización del lenguaje articulado”.
A pesar de sus detractores, el enorme interés en la localización de las funciones mentales ya se estaba gestando y fue en este tiempo que Marc-Dax notó la asociación entre daño al hemisferio izquierdo, hemiplejia derecha y afasia, con base en su examen de 40 pacientes en un periodo de veinte años. Su artículo fue escrito en 1836, pero no se publicó en ese momento; sólo distribuyó copias entre sus amigos y colegas.
De acuerdo con Feinberg y Farah (2000), fue hasta 1861 que en este campo se re-conoció al localizacionismo con una mente más abierta. En este año, en la Sociedad Antropológica de París se llevó a cabo una serie de debates entre Pierre Gratiolet, que argumentaba a favor del holismo o el equipotencialismo, y Ernest Aubertin (yerno de Bouillaud), que estaba a favor del localizacionismo. Aubertin describió el caso de su pa-ciente, quien al ser tocado con una espátula en los lóbulos “anteriores” tuvo un cese del habla sin pérdida de consciencia. Dicho paciente había sido intervenido quirúrgicamen-te con una remoción del hueso frontal después de un intento de suicidio.
Aunque todos estos investigadores hicieron su contribución para la mejor compren-sión de la localización de las funciones, muchos autores coinciden en señalar a Broca como el iniciador de la historia moderna de la neuropsicología. En 1861, Broca llevó a cabo los primeros intentos de correlaciones clínico-patológicas al presentar ante la Socie-dad Antropológica de París el caso de un paciente con pérdida del lenguaje expresivo, a lo que llamó originalmente “afemia” (aunque posteriormente se denominó afasia, término que ha prevalecido hasta nuestros días). En una autopsia, Broca detectó una lesión en el tercio posterior de la circunvolución frontal inferior del hemisferio izquierdo (aunque también se ha documentado que la lesión era mucho más extensa). Este hecho, junto con otros hallazgos, le permitió manifestar que el lenguaje articulado se localizaba en un área del cerebro demarcada con claridad, a la que se denominó en su momento “centro de las imágenes motoras de las palabras”, y que actualmente se conoce con el epónimo de área de Broca. Según Luria (1977), esta propuesta de Broca se considera prototípica de una postura localizacionista, en el sentido de que considera “la posibilidad de la localización directa de los procesos mentales complejos en áreas circunscritas de la corteza cerebral”. Las aportaciones de Broca son importantes por dos razones: 1) por primera vez había sido “localizada” una función mental compleja en una zona específica del córtex y esta localización descansaba sobre una base de hechos clínicos, 2) se mostró por primera vez la radical diferencia entre las funciones de los hemisferios cerebrales derecho e izquierdo y se identificó en el hemisferio izquierdo las funciones más importantes del lenguaje. En resumen, las contribuciones de Broca consistieron en haber innovado con el método de análisis clínico patológico al hacer una descripción minuciosa de los síntomas de su paciente y relacionarlos con los hallazgos patológicos, y ser uno de los pioneros del locali-zacionismo al haber planteado como un hecho la lateralización del lenguaje.

111
Métodos y teorías en neuropsicología: una perspectiva histórica y actual
Los trabajos de Broca dieron lugar a otra serie de estudios que descubrieron rela-ciones entre signos clínicos y áreas lesionadas. Así, se propuso la existencia de diversos “centros” que sustentaban diversas funciones. En 1869, Bastian observó que había pacientes que tenían déficit no sólo en la articulación de las palabras, sino también en la memoria para las palabras. Bastian propuso, además, la existencia de un centro visual y uno auditivo para la palabra y uno cinestésico para la mano y la lengua. Afirmó que estos centros estaban conectados y que el lenguaje era procesado por el cerebro en diferen-tes formas en cada uno de estos centros. Las lesiones en estos centros podían producir distintos síndromes, según el aspecto del procesamiento que estaba alterado.
Wernicke (1874) notó que las lesiones de la porción posterior de la región temporal superior producían un tipo de afasia en el cual la comprensión era muy deficiente. Pensó que este centro auditivo contenía las imágenes de los sonidos de las palabras y que este centro estaba conectado con el área de Broca por medio de una comisura, por lo que el daño en esta comisura podría desconectar el área de las imágenes de los sonidos del área de las imágenes para los movimientos del habla. De acuerdo con este esquema, se podían explicar la afasia motora, de conducción y la afasia sensorial. Lichtheim (1885), reelaborando las ideas de Wernicke, propuso un esquema complejo para explicar los mecanismos que subyacen a siete tipos de trastornos del habla y del lenguaje.
De acuerdo con Heilman y Valenstein (2003), Bastian, Wernicke y Lichtheim de-mostraron que las conductas complejas pueden ser fraccionadas en componentes modu-lares y desarrollaron modelos de procesamiento de la información por medio de los cuales estos componentes interactúan para producir conductas complejas, como el habla.
Por otra parte, a la postura antilocalizacionista, que ya había perfilado Flourens desde principios del siglo xix, vinieron a sumarse otra serie de estudios tanto anatómicos como fisiológicos, de acuerdo con los cuales se corroboraba la idea de que las alteraciones en la conducta no se podían atribuir a lesiones en determinadas áreas del cerebro, ya que “el grado de alteración estaba en proporción directa con la masa del cerebro eliminada y que, con respecto a las funciones complejas, las áreas diferentes de la corteza de los hemisferios cerebrales eran, de hecho, equivalentes”. Destacan, como representantes más influyentes en esta corriente, John Hughlings Jackson, Henry Head, Kurt Goldstein y Karl Lashley.
John Hughlings Jackson (1878), contemporáneo de Broca, formuló varias hipótesis que impugnaban el localizacionismo estrecho de su época. A su entender, la función tiene una compleja organización “vertical”, representada primeramente en un nivel “inferior” (espinal o del tronco cerebral); aparece representada otra vez (re-represen-tada) a un nivel “medio” de los sistemas motores o sensoriales de la corteza cerebral, y vuelve a aparecer, por tercera vez (re-re-representada), en un nivel “superior” en los lóbulos frontales del cerebro. Así, cada función realizada por el sistema nervioso cen-tral no era el resultado del trabajo de un grupo limitado de células, sino de un todo organizado de modo jerárquico e interactivo en alto grado. De acuerdo con Jackson, la localización del síntoma, la pérdida de una u otra función, que acompaña a la lesión sufrida en un área limitada del sistema nervioso central, de ninguna manera puede ser identificada con la localización de la función. Así, sólo las lesiones, pero no las funcio-nes, son localizables en el cerebro.
Henry Head (1926) estudió las alteraciones complejas del habla mediante la descrip-ción de los trastornos en algunos aspectos del acto verbal y atribuyó estos fenómenos a lesiones en áreas extensas de la corteza cerebral. Para explicar estos hechos, se remitió

Métodos en Neurociencias Cognoscitivas
112
a un factor general de la “vigilancia” como el principio de explicación definitiva (Luria, 1977). Kurt Goldstein formuló otros principios generales como la “actitud abstracta” o la “conducta categórica”, las cuales resultan alteradas en todos los casos de lesiones cere-brales. Goldstein distinguió entre la “periferia” de la corteza, la cual conserva el principio de localización en su estructura, y “la parte central” de la corteza, la cual, a diferencia de la primera, es “equipotencial”. Las lesiones de la “corteza periférica” conducen a la alte-ración de los “medios” de la actividad psíquica, pero dejan intacta “la actitud abstracta”. La lesión en la “parte central” de la corteza provoca cambios profundos en la “actitud abstracta” y la “conducta categorial”, subordinándose a la “acción de la masa”: cuanto mayor sea la parte de la sustancia cerebral afectada por la lesión, tanto más sufre la for-mación de las “estructuras dinámicas” complejas y tanto menos diferenciadas resultarán las relaciones de la “estructura” y “el fondo” que, a juicio de Goldstein, constituyen la base neurológica de esta compleja “conducta categorial”.
Lashley (1929, citado en Luria, 1977) extirpó diferentes áreas del cerebro en ratas y observó los cambios de su conducta en un laberinto. Llegó a la conclusión de que las alteraciones en su conducta no podían atribuirse a lesiones en distintas áreas del cere-bro, que el grado de alteración de sus hábitos estaba en relación directa con la masa de cerebro eliminada y que, con respecto a las funciones complejas, las áreas corticales de los hemisferios cerebrales eran equivalentes.
De acuerdo con Luria (1977), las posturas opuestas en relación con la localización de las funciones psíquicas en la corteza cerebral —la localizacionista y la antilocalizacionis-ta— hicieron su aportación al desarrollo de la teoría científica de la organización funcio-nal de la corteza cerebral. Los localizacionistas descubrieron en la masa aparentemente homogénea del cerebro aquellas áreas que tienen un valor específico para desarrollar las diferentes formas de la actividad psíquica. Elaboraron los conceptos pioneros acerca del trabajo diferenciado del cerebro. Los antilocalizacionistas colocaron en primer pla-no la tesis de la elevada plasticidad de los tejidos cerebrales, la capacidad del substrato cerebral para el reestablecimiento de las funciones y promovieron la idea de la organi-zación jerárquica de las funciones.
Posturas teóricas recientes acerca del problema de la localización de las funciones mentales en el cerebro.
Concepto de sistema funcional complejo Las posturas extremas descritas en el apartado anterior, localizacionista y equipoten-cialista, fueron sustituidas con los nuevos avances del conocimiento por concepciones que proponían que la complejidad de los fenómenos psicológicos no podía explicarse de manera convincente con ninguna de éstas. Una alternativa teórica a estas dos visio-nes opuestas es la que postula Luria (1974, 1977), quien derivó sus planteamientos de observaciones del estudio de pacientes con daño en las funciones mentales durante la Segunda Guerra Mundial y en la posguerra.
Su principal contribución al conocimiento de la organización de las funciones men-tales es el concepto de sistema funcional complejo. Para Luria, las funciones superio-res no pueden ser vistas como el producto directo de un órgano o grupo de células, sino las funciones mentales, como el lenguaje, la memoria, la actividad práxica, etc., deben verse como sistemas funcionales complejos, que, por lo tanto, no pueden loca-lizarse en áreas limitadas del cerebro. El concepto de sistema funcional complejo apli-cado a las funciones mentales, significa que éstas no son producto directo de un grupo

113
Métodos y teorías en neuropsicología: una perspectiva histórica y actual
de células, “no pueden localizarse en zonas restringidas del córtex, o en grupos celu-lares aislados, sino que deben de estar organizadas en sistemas de zonas que trabajan concertadamente, cada una de las cuales ejerce su papel dentro del sistema funcional complejo, y que pueden estar situadas en áreas del cerebro completamente diferen-tes, y a menudo muy distantes” (Luria, 1974). Estos sistemas funcionales complejos no son estáticos, en la medida en que cambian durante el desarrollo del niño y en pe-riodos subsecuentes. Para ejemplificar el carácter dinámico de los sistemas funcionales complejos durante el desarrollo, Luria (1974) pone el ejemplo de la escritura de un niño. Cuando el infante está aprendiendo lo hace por medio de la memorización de la forma gráfica de cada letra y de una cadena de impulsos motores aislados, cada uno de los cua-les es responsable de la ejecución de cada uno de los elementos de la estructura gráfica. Sin embargo, con la práctica, la escritura se convierte en una “melodía cinética”, la cual ya no necesita la memorización de la forma visual ni los impulsos motores individuales.
Contemplada la actividad mental como un sistema funcional complejo, la lesión de cada una de las zonas o áreas participantes puede conducir a la desintegración de todo el sistema funcional. Por lo tanto, la pérdida de una función particular o sínto-ma no es indicativo de su localización. Dado el concepto de sistema funcional, Luria (1974) propone que el objetivo no es “localizar” los procesos psicológicos superiores del hombre en áreas limitadas del córtex, sino averiguar, mediante un cuidadoso aná-lisis, qué grupos de zonas de trabajo concertado del cerebro son responsables de la ejecución de la actividad mental compleja; qué contribución aporta cada una de estas zonas al sistema funcional complejo, y cómo cambia la relación entre estas partes de trabajo concertado del cerebro durante la actividad mental compleja en las distintas etapas de su desarrollo.
El método que propone Luria para el estudio de las funciones superiores y su re-lación con el cerebro consiste en estudiar la estructura de los defectos observados y cualificar los síntomas, ya que sólo identificando el factor básico que yace tras el sín-toma observado, será posible extraer conclusiones referentes a la localización del foco que permanece en la base del defecto. Desde esta perspectiva, la “localización de un foco” no coincide con el de “localización de una función”, por lo que, para extraer con-clusiones acerca de la localización de un sistema funcional, el síndrome debe ser some-tido a un análisis estructural complejo. Luria propone que la metodología para evaluar los trastornos neuropsicológicos se base en el análisis de errores; afirma que no es tan importante saber si un paciente puede o no realizar una prueba, sino analizar el tipo de error producido por el paciente y de qué forma repercute en otras habilidades cognos-citivas; a esto se denomina cualificación del síntoma.
La cualificación del síntoma es sólo el primer paso en el análisis de la organización cerebral de los procesos mentales. Para que los análisis sean fiables, el siguiente paso es pasar a la descripción del complejo sintomático completo o lo que Luria llama análisis sindromático de cambios de la conducta que aparecen en lesiones locales del cerebro. Este análisis se basa en el hecho de que cada área del cerebro implicada en un sistema funcional introduce su propio factor particular esencial para su realización, y la exclu-sión de este factor hace imposible la normal actuación de este sistema funcional.
Dado que la actividad de todo sistema funcional complejo se efectúa con la parti-cipación de diferentes estructuras cerebrales que trabajan en conjunto y que cada una de ellas aporta su contribución específica al sistema, el sistema funcional —como un todo— puede ser alterado por la lesión de un gran número de zonas y también por

Métodos en Neurociencias Cognoscitivas
114
lesiones en diferente localización. Así, para Luria, cada área del cerebro implicada en un sistema funcional aporta su propio factor particular esencial para su realización y la exclusión de este factor hace imposible la normal actuación de este sistema funcional. Así, el neuropsicólogo debe asegurarse de qué factores están involucrados en una ac-tividad mental particular y qué estructuras cerebrales constituyen su base neuronal.
De acuerdo con Luria, en la investigación neuropsicológica se encuentra gran abun-dancia de hechos que demuestran que todo foco patológico local que aparece en el córtex altera la correcta realización de algunos procesos psicológicos mientras deja a otros intactos o, en otras palabras, que todo foco local da lugar a lo que se ha llamado “principio de la doble disociación de la función”. Un cuidadoso análisis del síndrome y las observaciones de la “doble disociación” que aparecen en lesiones locales del cerebro pueden aportar una gran contribución al análisis estructural de los procesos psicológi-cos en sí mismos. Para Luria, el uso correcto del método del análisis sindrómico sólo es posible si se resiste a la tentación de buscar la localización directa de los procesos mentales en el córtex y se reemplaza por el análisis de cómo es alterada la actividad mental en diferentes lesiones del cerebro y qué factores son introducidos dentro de la estructura y formas complejas de actividad mental por cada sistema cerebral.
Concepto de redes neurales Una visión actual que, desde nuestro punto de vista, mantiene algunas de las ideas bá-sicas de Luria es la conceptualización de Mesulam acerca de la organización de las fun-ciones cognoscitivas (1981). Mesulam postula una organización cerebral en redes y la aplica al estudio de la atención. Resume su postura en los siguientes principios:
• Los componentes de una sola función compleja están representados en lugares distintos, pero interconectados, que constituyen una red integrada para la función.
• Las áreas corticales individuales contienen el substrato nervioso para compo-nentes de diversas funciones complejas y pueden, sin embargo, pertenecer a va-rias redes parcialmente superpuestas.
• Las lesiones confinadas en una sola zona cortical tienen la probabilidad de dar lugar a déficit múltiples.
• Las alteraciones graves y duraderas de una función compleja individual requieren, generalmente, la afectación de varios componentes de la red funcional relevante.
• La misma función compleja puede alterarse como consecuencia de una lesión en una o varias áreas corticales, cada una de las cuales es un componente de la red integrada para tal función.
Esta postura de Mesulam se mantiene vigente, como puede verse en muchos tra-bajos de neurociencia cognoscitiva enfocados en estudiar los componentes de una red que originan un determinado proceso y a encontrar los correlatos neuroanatómicos de dichos componentes.
Principales aportaciones de la neuropsicología cognoscitiva En las últimas dos décadas, ha surgido una nueva aproximación teórica y metodológica para el estudio de las funciones cognoscitivas en pacientes con daño cerebral: la neurop-sicología cognoscitiva. Esta nueva aproximación se denomina así debido a que surgió de la fusión de dos aproximaciones que, hasta la década de 1970, estuvieron aisladas: la neu-ropsicología y la psicología cognoscitiva (Feinberg y Farah, 2000). Este enfoque novedo-so ha tenido un impacto en la forma de conceptualizar las funciones cognoscitivas, en la

115
Métodos y teorías en neuropsicología: una perspectiva histórica y actual
forma de hacer neuropsicología clínica (diagnóstico y rehabilitación), en los métodos de investigación y en las neurociencias cognoscitivas en general. A raíz de este enfoque, han emergido nuevos tópicos de interés y ha habido un debate en los objetivos de la disciplina.
Desde el punto de vista teórico, uno de los conceptos fundamentales en neuropsico-logía cognoscitiva es el de la disociación de funciones. Este concepto permite afirmar que dos funciones en apariencia emparentadas pueden estar disociadas funcional y anatómi-camente, tanto en un individuo sano como en uno enfermo. Sin embargo, sólo en este último caso se pueden probar algunas hipótesis al respecto, aunque recientemente tam-bién se han puesto a prueba paradigmas experimentales para probar algunas hipótesis de disociación de funciones en individuos sanos. Ellis y Young (1988) explican la disocia-ción de funciones en términos de que si un paciente presenta una actuación deficiente en la realización de una tarea 1, pero ejecuta con normalidad una tarea 2, se podría afirmar que existe una disociación entre las dos tareas, por ejemplo, no poder leer pseudopala-bras (tarea 1), pero ser capaz de leer palabras reales (tarea 2). Una pseudopalabra es una construcción que se parece a una palabra, que sigue las reglas fonotácticas del idioma, pero que no es una palabra real y por tanto no tiene un significado.
Sin embargo, de acuerdo con Ellis y Young (1988), podría argumentarse que tal dis-crepancia en la ejecución pudiera deberse a que el nivel de dificultad es distinto entre la tarea 1 y la 2. Por lo tanto, para confirmar el supuesto de que la tarea 1 y 2 son in-dependientes sería necesario encontrar en otro paciente el patrón opuesto, es decir, un paciente que ejecute con normalidad la tarea 1 (leer pseudopalabras), pero tenga dificultad en la tarea 2 (leer palabras reales). En este último caso, se habla de una do-ble disociación y el supuesto de la independencia de ambas funciones se ve fortalecido. En otras palabras, la disociación de funciones significa que hay módulos (elementos encargados de cierta función) que están alterados mientras otros están intactos.
Otro concepto clave en este enfoque es la modularidad de las funciones cognosciti-vas, es decir, la suposición de que las funciones cognoscitivas están organizadas en módu-los independientes. Según la hipótesis de la modularidad, “nuestra vida mental es posible gracias a la actividad orquestada de múltiples procesadores cognoscitivos o módulos” (Ellis y Young, 1988). Cada módulo se ocupa de su propia forma de procesamiento, inde-pendientemente de la actividad de aquellos módulos con los que no se halla en comuni-cación directa. Estos módulos son también distintos dentro del cerebro, de tal forma, que las lesiones cerebrales pueden afectar el funcionamiento de algunos módulos, mientras dejan intactos otros.
De acuerdo con Fodor (1986), las propiedades de los módulos son: 1) encapsula-miento informativo, que se refiere a que un módulo puede realizar su propia forma de procesamiento con total ignorancia o aislamiento de los procesos que se producen en otros lugares del sistema cognoscitivo, y 2) especificidad de dominio, que significa que cada módulo acepta sólo un tipo particular de aferencia (visual, auditiva, etc.).
Para Coltheart (2001), es necesario asumir que los módulos funcionales son también anatómicamente modulares, lo que les permite ser susceptibles a disociaciones des-pués de daño cerebral. El carácter obligatorio e innato de los módulos son propiedades postuladas por Fodor que no han sido aceptadas por completo. El carácter obligatorio se refiere a que la actividad de los módulos no se puede detener, está fuera del control voluntario y, si se produce el input apropiado, el módulo llevará a cabo su propio tipo de procesamiento, lo desee o no el propietario de este módulo. El carácter innato se refiere a que los módulos están genéticamente determinados.

Métodos en Neurociencias Cognoscitivas
116
También resulta controvertida la postura de Fodor de que mientras los procesos de entrada relacionados con la percepción del mundo externo —y probablemente los proce-sos de salida vinculados con el control de la acción sobre el mundo— son modulares, pue-den existir también componentes centrales de la mente, como la abstracción y la solución de problemas, cuya organización no es modular.
Otros supuestos de la neuropsicología cognoscitiva señalados por Ellis y Young (1988) son el del isomorfismo o especificidad neurológica, la transparencia y la sus-tractividad, los cuales son indispensables para entender las alteraciones en las funcio-nes cognoscitivas. Según el primero de estos supuestos, hay cierta correspondencia entre la organización de la mente y la organización del cerebro (Shallice, 1981). La transparencia indica que es posible deducir la naturaleza de la alteración subyacen-te de una función a partir del patrón de capacidades preservadas y alteradas. Este proceso se apoya en el supuesto de sustractividad, según el cual el desempeño de un paciente con una lesión cerebral se explica en términos de los componentes del sis-tema cognoscitivo normal intacto menos los componentes perdidos como resultado de la lesión.
La neuropsicología cognoscitiva, desde el punto de vista metodológico, también tiene diferencias con la neuropsicología tradicional. Mientras la neuropsicología tradi-cional estudia la localización y organización funcional de habilidades que dependen de una serie de procesos cognoscitivos componentes, como la lectura, el lenguaje, la memo-ria, etc., la neuropsicología cognoscitiva empezó a estudiar precisamente los procesos componentes de estas habilidades o procesos. En el caso de pacientes, la neuropsi-cología tradicional estudia la asociación de síntomas en síndromes, como la afasia de Broca, donde confluyen una gran diversidad de signos y síntomas como agramatis-mo, dificultades articulatorias, anomia, falta de fluidez verbal, etc. En cambio, en la neuropsicología cognoscitiva se privilegia el análisis del síntoma, en términos de alteración de algún módulo o componente del procesamiento cognoscitivo normal. De acuerdo con esta aproximación, es más probable que sean estos componentes, más que las habilidades o procesos, los que puedan ser localizables en el tejido cerebral.
Otra diferencia metodológica de la neuropsicología cognoscitiva con la neuropsico-logía tradicional es que, mientras esta última privilegia el estudio de grupos de pacien-tes con lesiones similares, la neuropsicología cognoscitiva se interesa en el estudio de casos particulares de importancia teórica (Caramazza, 1986), en el estudio de grupos pequeños seleccionados después de una evaluación con base en perfiles de ejecución muy similares (Denes, Semenza y Bisiacchi, 1988) y en el estudio de individuos normales en experimentos de laboratorio.
Con todo este contenido teórico y metodológico, la neuropsicología cognoscitiva tie-ne como objetivos, de acuerdo con Ellis y Young (1988): 1) explicar los patrones de las funciones cognoscitivas afectadas o intactas que se pueden observar en los pacientes con lesiones cerebrales, en términos de alteración de uno o más componentes de una teoría o modelo del funcionamiento cognoscitivo normal, y 2) extraer conclusiones sobre los pro-cesos cognoscitivos intactos y normales a partir de los patrones de habilidades afectadas e intactas observadas en pacientes con lesiones cerebrales. Para McCloskey (2001), los objetivos de los neuropsicólogos cognoscitivos son tres: 1) entender el funcionamiento y la estructura del sistema cognoscitivo normal, 2) explorar la localización de funciones cognoscitivas en el cerebro y 3) tener una mejor comprensión del déficit per se, como una base para el diagnóstico y tratamiento.

117
Métodos y teorías en neuropsicología: una perspectiva histórica y actual
Los objetivos de la neuropsicología cognoscitiva resultan muy controvertidos. Por ejemplo, para Coltheart (2001) la neuropsicología cognoscitiva está más interesada en la mente que en el cerebro y no es un tipo de neuropsicología, sino una rama de la psi-cología cognoscitiva. Como puede entenderse, esta aproximación no pretende única-mente explicar los casos en que el sistema ha resultado alterado como consecuencia de una lesión cerebral, sino también conocer cómo está funcionando un sistema dado en un individuo normal. Estos objetivos también difieren radicalmente de los objetivos de la neuropsicología tradicional, en la cual se pretende el estudio de síndromes y su correla-ción con el sitio de una lesión.
Una de las diferencias fundamentales de la neuropsicología cognoscitiva de la dé-cada de 1980 respecto de las aproximaciones más recientes, de la década 1990 y de los primeros años de este siglo, es que, mientras en la primera era más importante poner a prueba los modelos cognoscitivos mediante el estudio exhaustivo de casos particula-res, en la aproximación más reciente se enfatizan dos métodos fundamentales: tanto el estudio de pacientes como los estudios con neuroimagen funcional. Para Feinberg y Farah (2000), los primeros estudios de neuroimagen funcional pueden ser considerados como estudios de “calibración”, ya que los investigadores buscaban confirmar princi-pios bien establecidos de neuroanatomía funcional con el uso de nuevas técnicas. En la medida que la neuroimagen funcional ha madurado, los investigadores empiezan a formularse nuevas preguntas de investigación, para las cuales no hay respuestas pre-vias disponibles. En esta última fase de desarrollo, las teorías y métodos de la psicolo-gía cognoscitiva son importantes debido a que especifican los componentes de procesos cognoscitivos involucrados en llevar a cabo una tarea compleja y proporcionan los me-dios para aislarlos experimentalmente.
Métodos actuales en neuropsicología Método de lesión Como puede verse en la revisión histórica de la neuropsicología, el método de la lesión ce-rebral en humanos como recurso para conocer la relación cerebro-conducta ha pre-valecido en todas las etapas históricas y es quizá el que más identifica y diferencia la disciplina de otras aproximaciones. Lo que ha cambiado en particular son los conceptos y teorías a partir de los cuales se interpretan los hallazgos clínicos y los medios técnicos para el estudio de las funciones y de las lesiones. Es decir, para el localizacionismo de finales del siglo xix, el objetivo era encontrar los “centros” para una función determina-da, mientras que en el equipotencialismo se pretende encontrar una función más ge-nérica que pueda explicar los cambios encontrados en un paciente con lesión cerebral, como la “actitud abstracta” de Golstein. Desde la postura de Luria, con su concepto de sistema funcional complejo, el objetivo consistiría en encontrar el factor deficitario. Para los neuropsicólogos cognoscitivos, la interpretación se hace con la suposición de módulos de un sistema cognoscitivo deficitario o dañado. Con los modelos de redes, la región de daño cerebral se estudia como parte de una red a gran escala de sitios corti-cales y subcorticales que operan en conjunto para producir una función particular (Da-masio y Damasio, 2000). Dada la estructura teórica de cómo están constituidas dichas redes y llevan a cabo una función particular, una lesión es una prueba para examinar una hipótesis específica. Así, una lesión permite determinar si el daño a un componente de la red, responsable de una función determinada, altera la conducta de acuerdo con las predicciones hechas por la teoría que dice cómo está constituida una red. Para Damasio

Métodos en Neurociencias Cognoscitivas
118
y Damasio (2000), las lesiones son un medio para apoyar o rechazar la teoría del funcio-namiento cerebral normal.
Aunque con sus diferencias teóricas, se puede afirmar que el método de lesión tie-ne como objetivo predecir, según el sitio de la lesión, el patrón de alteración en algún aspecto de una ejecución cognoscitiva o conductual y, a la inversa, predecir el sitio de lesión a partir de la sintomatología clínica. En relación con este último aspecto, también se hace uso de métodos estadísticos como el análisis de clusters para analizar si los pa-cientes agrupados con base en lesiones cerebrales bien delimitadas y caracterizadas pueden ser discriminados a partir de la sintomatología clínica. En el caso de los síndro-mes afásicos, Kertesz (1979) encuentra una gran coincidencia de los sitios de lesión con la taxonomía y localización de los síndromes afásicos ya conocidos.
Para Heilman y Valenstein (2003), no es lo mismo hacer las correlaciones de cambio conductual y lesión cerebral que intentar deducir, a partir de los efectos conductuales de una lesión, los mecanismos normales de la función cerebral, dado que el tejido cerebral remanente puede reaccionar de modo adverso o compensar la pérdida de la función cau-sada por la lesión y así añadir o minimizar el déficit conductual. Las lesiones pueden tam-bién producir cambios en la conducta por liberar otras áreas cerebrales de la facilitación o la inhibición. Así, puede ser difícil distinguir entre los efectos conductuales causados por una interrupción del procesamiento normal que ocurre en el área dañada y los efectos debidos a alteraciones menos específicas de la función en otras áreas del cerebro.
Deben tenerse en cuenta diversos factores en los estudios de localización de fun-ciones. Uno de ellos es el efecto de la recuperación, ya que un déficit temprano puede relacionarse con edema, reacción celular, isquemia transitoria, etc., por lo que puede haber una gran recuperación espontánea. Por su parte, el déficit crónico se relaciona no sólo con la pérdida de la función, sino con cambios compensatorios en todo el cerebro, o áreas homólogas o vecinas, durante estadios subsecuentes de recuperación (Kertesz, 1979). El efecto de la etiología también debe tomarse en cuenta, ya que algunas lesiones producen más efectos a distancia que otras, a lo cual se le denomina diasquisis. Tales cambios metabólicos o fisiológicos pueden no ser detectables por métodos neuropato-lógicos y contribuir así a una sobreestimación de la función del área lesionada (Heilman y Valenstein, 2003). Finalmente, variables del individuo como edad, sexo, escolaridad, lateralidad y variaciones anatómicas también pueden influir en los resultados.
Los efectos no específicos de una lesión, como la diasquisis o efectos de acción de masa, pueden excluirse como determinantes principales de conducta anormal por el uso de lesiones “control”. Si lesiones de tamaño comparable en otras áreas cerebrales no producen efectos conductuales similares, entonces los efectos no se pueden atribuir a causas no específicas.
Así, el método de lesión es insustituible y ha generado una gran cantidad de infor-mación del cerebro y las funciones mentales. Kertesz (1983) propone algunos principios de aceptabilidad de la evidencia de lesión en la localización de una función. Así, el ejer-cicio de un área se relaciona con la lesión si:
• El mismo déficit funcional siempre sigue a la lesión. • El mismo déficit no es producido por otras lesiones independientes. • El déficit es medido por métodos estandarizados y significativos. La localización
de la lesión es determinada con objetividad y exactitud. • Variables biológicas, como tiempo de inicio, edad, etiología y otras, son controladas. • El autor considera que estas situaciones, aunque nunca satisfechas con plenitud,
deben ser siempre las metas.

119
Métodos y teorías en neuropsicología: una perspectiva histórica y actual
Estudios experimentales Una limitación del método de lesión cerebral es de carácter ético, pues una lesión no puede ser provocada en humanos y es muy complicado encontrar patrones de daño semejante en diferentes individuos. Tal limitación se supera con el estudio de lesión en animales, es decir, se crean modelos experimentales en los cuales se diseñan pa-radigmas que pretenden ser análogos a los aplicados a seres humanos y de mane-ra propositiva se lesionan áreas cerebrales circunscritas en algunos animales, con el objetivo de probar que una determinada función o elemento de una red funcional se correlaciona con una determinada estructura. Un ejemplo de ello son los experi-mentos resumidos por Squire (1992) para el estudio de la memoria. La limitación en este caso deriva de las distancias filogenéticas entre las especies y, por lo tanto, de la menor complejidad estructural y funcional del sistema nervioso de los mamíferos en los cuales se investiga.
Por otra parte, los estudios experimentales en humanos también han contri-buido al conocimiento en neuropsicología. A diferencia de los estudios con anima-les, los estudios con humanos tienen la ventaja de permitir valorar funciones que son específicamente humanas, como el lenguaje. Así, se han diseñado diferentes paradig-mas y manipulaciones experimentales para probar modelos de diversos aspectos del lenguaje; o subtipos de memoria, como la memoria de trabajo (Baddeley, 1986) y la memoria implícita (Tulving, Schacter y Stara, 1982).
Método psicométrico Para Lezak, Howieson y Loring (2004), una importante aportación de la neuropsicología a las neurociencias cognoscitivas es la aplicación del enfoque psicométrico en el diag-nóstico neuropsicológico y en la investigación. De acuerdo con estos mismos autores, aunque las pruebas psicológicas en un principio se usaron para evaluar la “inteligencia” en el área educativa y después en el reclutamiento de personal, en la actualidad ya son parte de la disciplina y de la práctica clínica. Tales pruebas psicológicas se aplican en el ámbi-to de la neuropsicología clínica, tanto en su forma estandarizada para población normal como con algunas modificaciones o adaptaciones para tratar de discernir, de manera más específica, por qué un paciente puede tener dificultad en resolver alguna tarea, más que centrarse sólo en si la puede llevar a cabo o no. Aunque originalmente se utilizaron pruebas psicométricas diseñadas para otros fines, el crecimiento de pruebas neuropsico-lógicas diseñadas ex profeso para su aplicación en la neuropsicología clínica es abundante.
El uso de pruebas estandarizadas es de suma importancia para los estudios de grupo y los análisis estadísticos tienen un enorme impacto en la neuropsicología por permitir calcular con precisión la probabilidad de que el rendimiento en una tarea determinada, de un individuo con alguna lesión o condición particular, se dé a un nivel inferior que el de un grupo control, lo cual contribuye a la comprensión de los procesos psicológicos. Esto, a su vez, proporciona una base para la evaluación neuropsicológica clínica y, además, proporciona un conjunto de procedimientos que pueden ser bastante útiles en el diag-nóstico y la rehabilitación.
Las diferentes ediciones del libro Neuropsychological assessment de Lezak (1976, 1983, 1995) y Lezak, Howieson y Loring (2004), y de A compendium of neuropsychologi-cal tests de Spreen y Strauss (1991, 1998) son un ejemplo del impactante desarrollo que ha tenido la evaluación neuropsicológica del adulto; asimismo, el libro de Baron (2004) es un ejemplo del creciente interés en la evaluación neuropsicológica del niño.

Métodos en Neurociencias Cognoscitivas
120
Tramontana y Hooper (1988) consideran que en la historia de la evaluación neuropsi-cológica en niños pueden distinguirse cuatro estadios de desarrollo, distintos, aunque superpuestos, que se pueden aplicar a la evaluación de los adultos. El primer estadio es denominado aproximación de la prueba singular, la cual dominó el campo de la neuropsi-cología infantil de mediados de la década de 1940 a mediados de la de 1960, y se caracte-rizó por el uso de mediciones generales para diagnosticar daño cerebral u “organicidad”. Un ejemplo es el Test Gestáltico Visomotor de Bender (Koppitz, 1964). Esta aproximación se orientó por la idea de que el daño cerebral se manifiesta en una forma unitaria, ya sea en forma de una pérdida de las habilidades de abstracción, habilidades perceptivo-moto-ras o algunas otras. La meta era diferenciar niños con daño cerebral de los normales y se pensó que una medida cerebral bien elegida podía desempeñar tal objetivo.
El segundo estadio, llamado Batería de Tests-especificación de la Lesión, se ca-racteriza por el uso de baterías de pruebas que cubren un amplio rango de funciones para clasificar niños con daño cerebral y niños normales. La batería neuropsicológica de Halstead-Reitan (Boll, 1986) es la más significativa de este periodo. En esta fase, se hace hincapié en la detección y localización de las lesiones cerebrales. La aproximación, ya sea con pruebas singulares o baterías de pruebas, es empírica y ateórica, encamina-da a obtener puntos de corte y reglas de inferencia para el propósito de maximizar la tasa de diagnóstico categorial (con daño cerebral o normales).
La siguiente fase es la del estadio del perfil funcional de la evaluación neuropsico-lógica de Tramontana y Hooper (1988). El énfasis se desplazó al papel de la evaluación neuropsicológica para especificar el defecto conductual de las lesiones cerebrales. La meta fue diferenciar entre habilidades intactas y alteradas, y entre fortalezas y debili-dades funcionales. La evaluación neuropsicológica tiene aquí el papel de determinar el efecto mental y conductual del daño cerebral, identificar deficiencias funcionales para planear el tratamiento y evaluación del cambio subsecuente.
El cuarto estadio es el de la validez ecológica; la meta es evaluar el funcionamiento actual del individuo en relación con su capacidad para llevar a cabo tareas importantes de la vida diaria. El énfasis se pone no sólo en la descripción, sino en hacer los señalamientos necesarios en cuanto a tratamiento y medio ambiente que puedan ser más favorables.
Quizá se pueda agregar a esta descripción de Tramontana una fase más, que se ca-racterizaría ya no sólo por el uso de pruebas estandarizadas, sino por el diseño de prue-bas más individualizadas y tareas de laboratorio cada vez más específicas para poner a prueba los modelos cognoscitivos mediante la disociación de funciones.
Una innovación reciente en la evaluación neuropsicológica es el uso de versiones automatizadas de pruebas neuropsicológicas clásicas, como la prueba de tarjetas de Wisconsin, la prueba de Stroop, etc. También se han desarrollado métodos de evalua-ción nuevos, que desde su origen se diseñan en versión automatizada, aunque incorpo-ran algunos de los principios teóricos que subyacen a algunas pruebas neuropsicológi-cas. Las versiones automatizadas tienen la ventaja de ahorrar tiempo por ser la máquina la que solicita la información al paciente. Otra ventaja es que permiten la calificación exacta de los aciertos, errores y tiempos de reacción y de ejecución total de la prueba y, por otra parte, la comparación casi de manera automática de los resultados de un determinado individuo con una población de referencia. Esta forma también permite al evaluador centrarse más en la observación cualitativa del desempeño del paciente mientras la máquina hace el registro de la ejecución. Tales métodos pueden ser, so-bre todo, de utilidad con pacientes jóvenes, quienes previamente han tenido una cierta

121
Métodos y teorías en neuropsicología: una perspectiva histórica y actual
familiarización con las computadoras, ya que los pacientes ancianos pueden tener ma-yor dificultad para familiarizarse con el método de respuesta, más que con la tarea en sí misma. Una alternativa para esta dificultad ha sido la utilización de touch screens, las cuales no requieren del empleo de teclas, cajas de respuesta o el mouse. Una desventa-ja de estos métodos es que pueden ser útiles para explorar algunas funciones (es decir, atención visual), pero no otras (es decir, praxias del vestir).
Un ejemplo de un sistema de evaluación automatizado es el sistema automated neuropsychological assessment metrics (ANAM), el cual originalmente fue desarrollado para evaluar a individuos sanos (Reeves et al., 1992). Sin embargo, también se ha uti-lizado en el área clínica y su principal utilidad es que se trata de un instrumento de es-crutinio para pacientes en riesgo de alteración neurocognoscitiva, ya que no es práctico ni económico evaluar a cada paciente en riesgo cognoscitivo con una batería neuropsi-cológica completa. Los hallazgos obtenidos al evaluar varias poblaciones de pacientes (lupus eritematoso, esclerosis múltiple, enfermedad de Parkinson, migraña, etc.) indi-can que las medidas de la ANAM son sensibles a cambios cognoscitivos asociados con trastornos neurológicos.
Otro ejemplo de baterías automatizadas es la Cambridge neuropsychological automa-ted battery (CANTAB) (De Luca et al., 2003). Esta batería de pruebas neurocognoscitivas evalúa con eficiencia las funciones ejecutivas, memoria de trabajo, planeación, inhibi-ción de respuestas, supervisión o monitorización de las mismas y flexibilidad cognosci-tiva, todas asociadas con el funcionamiento del lóbulo frontal. Dentro del campo de la neuropsicología, hay una necesidad imperante de contar con baterías neuropsicológicas que se puedan aplicar en tiempos reducidos y sean capaces de identificar con precisión pacientes con problemas neurocognoscitivos (Kane et al., 2006) para, en su caso, llevar a cabo una evaluación más completa. Ésta es la ventaja que se busca con la automatiza-ción de las pruebas de evaluación neuropsicológica.
Estudios de grupo y estudios de caso Aunque los estudios de grupo son un enorme avance en neuropsicología, los estudios de caso único han venido a ser revalorados por varias razones. En primer lugar, ciertos trastornos son tan raros, que formar un grupo considerable tomaría varios años. Ade-más, en trastornos severos como las afasias, los cuales pueden ser detectados sin mé-todos estadísticos, el sujeto aislado proporciona una vívida y detallada descripción del deterioro que con frecuencia se pierde con el análisis de los datos del grupo. Más aún, una serie de estudios de caso proporciona información que se pierde fácilmente en el curso de un análisis de grupo en torno a la variabilidad de los síntomas presentes entre los individuos que comparten cierta sintomatología o áreas comunes de anormalidad cerebral. La variabilidad se obscurece, o incluso se pierde, en los datos que expresan la función en términos de promedios grupales.
Nuevos tópicos de interés Después de muchos años en que los neuropsicólogos estuvieron centrados en el diag-nóstico clínico, en años recientes ha crecido el interés por la intervención. Lezak (1995) considera que los campos de aplicación de la neuropsicología han variado a través del tiem-po y en la actualidad se centran más en el campo clínico, esto es, en el manejo y tra-tamiento del paciente, y la evaluación como una forma de diagnóstico inicial y de seguimiento de los cambios.

Métodos en Neurociencias Cognoscitivas
122
El interés actual en la rehabilitación se enfoca en conocer qué tanto un paciente, con secuelas cognoscitivas a consecuencia de un daño neurológico, puede beneficiarse o no de un programa de rehabilitación neuropsicológica, cuáles son los mejores enfoques teóricos y las más adecuadas estrategias terapéuticas, en qué momento de las fases de una enfermedad es más conveniente intervenir y cuánto debe durar un determinado programa. Otro aspecto que resulta interesante es la inclusión de algunos avances tec-nológicos para la compensación de funciones en pacientes. Un ejemplo de ello pueden ser las agendas electrónicas para pacientes con demencia o con amnesia.
Mientras hasta hace pocos años el neuropsicólogo estaba únicamente interesado en el estudio de las funciones mentales de individuos con lesiones cerebrales bien conocidas, desde hace ya dos décadas ha crecido el interés por el estudio de los trastor-nos del desarrollo, para los que se presume y se investiga una posible causa biológica, pero para quienes no ha sido totalmente determinada la posible anomalía estructural. Por otra parte, también son objeto de muchos estudios neuropsicológicos algunas en-fermedades neuropsiquiátricas, como esquizofrenia, depresión, trastornos bipolares, trastornos obsesivo-compulsivos y trastornos de la alimentación.
Conclusión Apreciar la historia de la neuropsicología, sus principales corrientes y enfoques, permi-te ubicarnos en la actualidad con la consciencia de quien sabe de dónde parte y hacia dónde se dirige. Ubicarnos en perspectiva en relación con los conceptos y enfoques más trascendentes en la disciplina puede ser importante para no caer involuntariamente en posturas teóricas que ya han sido rebasadas y, en su lugar, retomar aquellas de mayor actualidad y más prometedoras para el avance de la neuropsicología y de la neurocien-cia cognoscitiva.
Las aportaciones de la neuropsicología a la neurociencia cognoscitiva son incuestio-nables, ya que mientras el avance de las técnicas de neuroimagen y neurofisiológicas son elementos determinantes para estudiar la organización de las funciones mentales, el estudio de las mismas no tendría sentido sin los elementos teóricos y conceptuales que articulan y dan sentido a toda la investigación en este rubro. Los modelos teóricos y los conceptos de la neuropsicología permiten articular de manera coherente los ha-llazgos, plantear hipótesis y desarrollar paradigmas para el avance de las neurociencias cognoscitivas.
El método del estudio de pacientes es insustituible. Los estudios de series de casos y la descripción de síndromes neuropsicológicos, y la descripción minuciosa de casos úni-cos han arrojado mucha luz acerca de los procesos cognoscitivos. De ahí que el estudio del paciente con lesión cerebral sigue siendo la esencia de la neuropsicología.
Es posible que con investigaciones futuras en neurociencias se logre avanzar en el estudio de la compleja relación cerebro-conducta. No obstante, a la fecha, tal y como mencionan Feinberg y Farah (2000), la mayoría de los estudios de neuroimagen se ha dedicado a comprobar las relaciones que ya habían predicho los estudios clínicos.
Otra de las aportaciones de la neuropsicología es el uso de las pruebas psicológi-cas, las cuales, aunque no son la única herramienta para el estudio de las funciones cognoscitivas, son cada vez más específicas y mejor estandarizadas para proporcionar resultados más confiables. Al principio, se utilizaron varios instrumentos derivados directamente de la psicología; no obstante, dentro de la propia disciplina se han de-sarrollado una gran cantidad de pruebas que permiten estudiar las diversas funciones

123
Métodos y teorías en neuropsicología: una perspectiva histórica y actual
que pudieran estar afectadas en un paciente. El empleo de la computadora ha permi-tido que algunas funciones se puedan evaluar de una manera más precisa mediante el registro exacto de tiempos de reacción o tipos de error, aunque en neuropsicología la observación del desempeño del paciente es insustituible; asimismo la neuropsicología se ha interesado en la rehabilitación de los pacientes y la reintegración de los mismos a su medio escolar, laboral, familiar y social.
Finalmente, se puede concluir que en neurociencias cognoscitivas los métodos son complementarios y no contrapuestos. Los hallazgos obtenidos por un método, como pudiera ser el de lesión, son complementados por los derivados de estudios experi-mentales con humanos, con modelos animales, electrofisiológicos o de neuroimagen funcional. La comprobación acerca de una determinada relación cerebro-conducta con más de uno de estos métodos contribuye a hacer más sólido el conocimiento en neuro-ciencias cognoscitivas. De esta forma, aunque todos los métodos, incluido el neuropsi-cológico, tienen ventajas y desventajas, es deseable que los trabajos en esta área sean multidisciplinarios para poder disminuir las desventajas de los mismos.
Referencias
Baddeley, A. D. (1986). Working memory. Londres-Nueva York: Oxford University Press. Baron, I. (2004) Behavioral Assessment. En Neuropsychological evaluation of the child (cap.2).
Nueva York: Oxford University Press. Caramazza, A. (1986). On drawing inferences about the structure of normal cognitive systems
from the análisis of patterns of impaired performance: the case for single patient studies. Brain and Cognition, 5, 41-66.
Coltheart, M. (2001). Assumptions and methods in cognitive neuropsychology. En B. Rapp (Ed.), The handbook of cognitive neuropsychology: what deficits reveal about the human mind (pp. 3-22). Estados Unidos: Psychology Press Taylor y Francis Group.
Damasio, H. y Damasio, A. R. (2000). The lesion method in cognitive neuroscience. En M. J. Farah y T. E. Feinberg (Eds.), Patient-based approaches to cognitive neuroscience (pp. 21-34). Cambridge: mit Press.
De Luca, C., Wood, S., Anderson, V., Buchanan, J., Proffitt, T., Mahony, K. y Pantelis, C. (2003). Normative data from the CANTAB I: Development of executive function over the lifes-pan. Journal of Experimental Neuropsychology, 25, 242-254.
Denes, G., Semenza, C. y Bisiacchi, P. (1988). Perspectivas on cognitive neuropsychology. Reino Unido: Lawrence Erlbaum Associates Ltd.

Métodos en Neurociencias Cognoscitivas
124
Ellis, A. W. y Young, A. W. (1988). Human cognitive neuropsychology. Londres: Lea. Feinberg, T. E. y Farah, J. (2000). A historical perspective on cognitive neuroscience. En M.
J. Farah y T. E. Feinberg (Eds.), Patient-based approaches to cognitive neuroscience (pp. 3-20). Cambridge: mit Press.
Fodor, J. A. (1983). The modularity of mind. Cambridge: mit Press. García-Albea, R. E. y García-Albea M. J. (2006). Cerebro, mente y síntoma. Revista de
Neurología, 42 (7), 439-443. Hécaen, H. y Dubois, J. (1983). El nacimiento de la neuropsicología del lenguaje. México: FCE. Heilman, K. M. y Valenstein, E. (2003). Clinical neuropsychology. Nueva York: Oxford University Press. Kertesz, A. (1979). Aphasia and associated disorders. Nueva York: Grune y Stratton. Kertesz, A. (1983). Localization in neuropsychology. Estados Unidos: Academic Press. Kolb, B. y Whishaw, I. Q. (2003). Fundamentals of human neuropsychology. Nueva York: Worth
Publishers. Lezak, M. D. (1983). Neuropsychological assessment (2a ed.). Nueva York: Oxford University
Press. Lezak, M. D. (1995). Neuropsychological assessment (3a ed.). Nueva York: Oxford University
Press. Lezak, M. D., Howieson, D.B. y Loring, D.W. (2004). Neuropsychological assessment (4a ed.).
Nueva York: Oxford University Press. Luria, A. R. (1974). El cerebro en acción. Barcelona: Fontanella. Luria, A. R. (1977). Las funciones corticales superiores del hombre (C. Pérez, Trad.). La Habana:
Orbe. (Trabajo original publicado en 1969). McCarthy, R. A. y Warrington, E. K. (1990). Cognitive neuropsychology. Reino Unido: Academic
Press. McCloskey, M. (2001). The future of cognitive neuropsychology. En B. Rapp (Ed.), The handbook of
cognitive neuropsychology: what deficits reveal about the human mind (pp. 593-610). Estados Unidos: Psychology Press Taylor y Francis Group.
Mesulam, M. (1981). A cortical network for directed attention and unilateral neglect. Annals of Neurology, 10 (4), 309-325.
Spreen, O. y Strauss, E. (1991). A compendium of neuropsychological tests. Nueva York: Oxford University Press.
Spreen, O. y Strauss, E. (1998). A compendium of neuropsychological test: Administration, norms and commentary. Nueva York: Oxford University Press.
Squire, L.R. (1992). Memory and the hippocampus: A synthesis from findings with rats, monkeys and humans. Psychological Review, 99, 195-231.
Tramontana, M. G. y Hooper, S. (1988). Child neuropsychological assessment. En M. G. Tramontana y S. R. Hooper (Eds.), Assessment issues in child neuropsychology (pp. 3-40). Nueva York: Plenum Press.
Tulving, E., Schacter, D. L. y Starck, H. (1982). Priming effects in wordfragment completion are independent of recognition memory. Journal of Experimental Psychology: Human Learning and Memory, 8, 336-342.

AActividad(es)
cerebral, 29eléctrica cerebral, 41, 85
espontánea, 68fuentes de corriente de, 85
electroencefalográfica, 68, 74electromagnética, 86
cerebral, 85global dentro del cerebro, 99mental, 113neuronal, 61práxica, 112
Afasia, 110de Broca, 116de conducción, 111motora, 111sensorial, 111
Afemia, 110Agramatismo, 116Algoritmo
de LORETA, 97FOCUSS, 96
Amnesia, 122Amperometría, 10Amplitud, 46, 52
medición de la señal, 81Análisis
de clusters, 118de componentes principales (ACP), 49espacio-temporal, 94estadístico, 100
paramétrico, 36fractal, 37sindromático, 113
Anomia, 116Antilocalizacionismo, 109Aprendizaje, 41Aproximaciones bayesianas, 98Áreas
corticales individuales, 114de Broca, 110de Brodmann, 98
Artefactos, 72, 92eliminación de otro tipo de, 75oculares, 75
reducción de, 75rechazo y corrección de, 75
Atención, 41Autopsia, 110Axón, 5BBandas anchas, 30
alfa, 30beta, 30delta, 30gamma, 30theta, 30
Bateríade Tests-especificación de la
lesión, 120neuropsicológica de Halstead-
Reitan, 120Boundary element method (BEM), 89CCálculo de la matriz, 100Cámara de Faraday, 73
Cambridge neuropsychological automated battery (CANTAB), 121
Campo eléctrico, 5, 17medido, 36
Capacitancia, 20Célula(s)
adherida, 12automáticas, 18cardiacas, 16de respuesta rápida, 18entera, 12excitables, 1, 2marcapaso, 17, 29piramidales, 28, 44
Cerebro, 27número de dipolos en, 95
Choque eléctrico, 10Ciencias de la conducta, 108Circuito(s)
básico de registro, 23eléctricos básicos, 19tálamo-cortical, 28
Comprensión, 41Conducta
categórica, 112de disfunción cerebral, 107
Convertidor analógico-digital, 69Coordenadas de Talairach, 98Corazón, 17Corregistro, 91Corriente eléctrica, 20Córtex, 113Corteza
auditiva, 50cerebral, 6, 27, 43, 111, 112periférica, 112
Coulomb, 20Creatividad, 41Cualificación del síntoma, 113DDaño
cerebral, 109neurobiológico, 42neurológico, 122
Déficit cognoscitivo-conductual, 42Deflexiones
isodifásicas, 18isoeléctricas, 18
Demencia, 122Dendritas, 7
apicales, 28
Depresión, 122Diasquisis, 118 Dipolo, 28Distrofía muscular, 16Dualismo, 109
de Descartes, 109EElectrocardiografía clínica, 16Electrocardiograma (ECG), 1, 16, 72Electrodo(s), 68, 90
activos, 68conectado a tierra, 69de calomel, 3de estimulación, 5de platino, 3de referencia, 5, 68de registro, 3, 5de tungsteno, 3estándar, 90interconectados, 30número de, 90occipitales, 49para patch clamp, 3por sujeto, 90posición de, 90selectivos de iones, 10
Electroencefalografía (EEG), 26, 70, 92, 94
computarizado (EEGc), 32convencional para la
investigación, 31cuantitativo, 26, 34, 36
representación gráfica del, 36desventajas del, 31espectro de potencia del, 34espontáneo, 29frecuencias observadas en, 30medidas espectrales del, 34para eliminar el efecto de
referencia, 36significado de actividad
registrada, 28sincronizadas, 81tradicional, 27, 30transformación rápida de
Fourier del, 33Electroencefalógrafo, 30Electroencefalograma (EEG), 1, 19,
44, 70, 86medición de actividad
eléctrica, 26Electromiografía, 13Electromiograma (EMG), 1, 13Encapsulamiento informativo, 115Enfermedad(es)
de Lou-Gehrig, 16de Parkinson, 121musculares, 16psiquiátricas, 92
Epilepsia, 92
125
Índice

Equipotencialismo, 110, 117Error de juicio experimental, 101Esclerosis
lateral amiotrópica, 16múltiple, 121
Esquizofrenia, 122Estadística de imágenes, 98Estados psicológicos, 29Estímulo(s)
auditivos, 51blanco, 100
Estudios con PRE, 58experimentales, 119
Evaluación neuropsicológica clínica, 119
Eventos sinápticos, 14Evoked potentials, 45FFenómeno ALIAS, 70Fibras de Purkinje, 17Filtro(s), 71
analógicos, 71de respuesta al impulso finita
(FIR), 79digitales, 71notch, 72
Fórmula de Basar, 35Frecuencia de transformación
rápida de Fourier (FFT), 79Fuente
de corriente, 20de voltaje, 20
Fuerza electromotriz (FEM), 20Función cerebral, 28
humana, 107GGalvanómetros, 2Gigasello, 11H Habilidades cognoscitivas, 113Haz de His, 17Hemisferio cerebral, 69Hertzios (Hz), 70Hiperpolarización, 28Hipótesis
del cerebro, 108del corazón, 108
Holismo, 110IImágenes
del VARETA, 100de resonancia magnética,
posprocesamiento de, 98visualización de soluciones inversas en, 98
Impedancia, 20, 74de entrada y resistencia de
entrada, 21Investigación
científica, 85electrofisiológica, 4neuropsicológica, 114
JJuicio experimental, 101LLatencia, 51
medición de la señal, 83Lateralized readiness potential (LPR), 55Lead field (LF), 88Left anterior negativity (LAN), 54Lenguaje, 41, 110, 112
articulado, 110Lesión cerebral, 117Ley(es)
de Kirchhoff, 20de la corriente, 20del voltaje, 20
Línea(s)base corrección de, 79de campo, 88vectoriales, 88
Localizacionismo, 109Lupus eritematoso, 121MMagnetoencefalograma (MEG),
44, 86Mapa(s)
cerebrales tridimensionales, 102de potencia, 92del potencial eléctrico, 93topográficos, 36, 37
Mapeo estadístico paramétrico (SPM), 99interdominio, 58
Medición de actividad eléctrica, 26de campos magnéticos (MEG),
61, 94de señales biológicas, 21
Memoria, 41, 112de trabajo, 119implícita, 119
Memorización, 113Mente, 109Método(s)
actual en neuropsicología, 117de lesión, 117de neuroimagen, 98, 102de registro,
electroencefalográficos, 102electrofisiológico, 1
del elemento frontera, 89electrofisiológicos, 2en neuropsicología, 107neuropatológico, 118psicométrico, 119spline del plano delgado, 92
Microelectrodo, 9
de registro intracelular, 2de vidrio, 2
Microvoltios, 46Migraña, 121Milisegundos (ms), 47Mínima norma, 96
ponderada, 96Mismatch negativity (MMN), 50Modelo(s)
anterógrado, 93cerebral tridimensional, 96con restricción de orientación
cortical, 95de esferas concéntricas, 88de fuentes distribuidas, 95de la cabeza, 88de tres esferas concéntricas, 89dipolar, 93inverso, 93
Motoneurona, 15NNegatividad relacionada con el error (ERN), 55Neuroanatomía, 107
funcional, 117Neurociencia, 108
cognoscitiva, 41, 122Neurofarmacología, 107Neurofisiología, 107Neurología, 107Neurometría, 32, 34, 35Neuromoduladores, 10Neurona(s), 27
en campos abiertos, 86organización en corteza
cerebral, 87piramidales, 27reticulares, 28tálamo-corticales, 28
Neuropsicología, 122clínica, 107, 115cognoscitiva, 114, 117infantil, 120método actual en, 117tradicional, 116
Neuropsicólogo, 114cognoscitivo, 117
Neuroquímica, 107Neuroquirúrgicas, 42Neuroscan, 73Neurotransmisores, 10Nodo
auriculoventricular, 17sinoauricular, 17
Notch, 72OOhms, 74Onda(s)
cerebrales, 35negativa de disparidad, 50
Métodos en Neurociencias Cognoscitivas
126

P300, 52Ondeletas, 38Organización de neuronas en
corteza cerebral, 87Osciloscopio de rayos catódicos, 2PParche
de membrana, 2perforado, 13
Patch clamp, 2, 10Perforated patch, 13Piel cabelluda, 28, 86Potencia
absoluta (PA), 34relativa (PR), 34
Potencial(es)auditivos de tallo cerebral
(PATC), 50de acción, 15negativo, 18positivo, 18provocados, 45relacionados con eventos
(PRE), 41, 45, 68aspectos conceptuales para interpretar datos, 60auditivo, 46breviario de componentes de, 49característico, 58comparación con otras técnicas, 42componentes de, 47, 55elementos para el registro de, 68en humanos, 48endógenos, 45exógenos, 45filtrado y suavizado, 79hipotéticos, 56inferencias funcionales a partir de, 57muy ruidosos, 84nomenclatura, 46registrados en piel cabelluda, 48relacionados con la respuesta, 55ruidoso, 76superpuestos, 81traslape de componentes, 48
Principio de doble disociación de la función, 114
Procesadores cognoscitivos, 115Procesamiento
aritmético, 53cognoscitivo, 42fonológico, 53semántico, 53
Proceso
cognoscitivo, 58, 62psicológicos superiores, 113
Promediación, 77sincronizada con respuestas, 79
Promedio autorregresivo local (LAURA), 97
Prueba(s) de Stroop, 120de tarjetas de Wisconsin, 120neuropsicológicas, 119, 120psicométricas, 119
Pseudopalabra, 115Psicología, 107
cognoscitiva, 56, 117RRazón de energía (RP), 34Razonamiento, 41Red(es)
neurales, 114neuronales, 29tálamo-cortical, 28
Registro(s) de actividad neuronal, 43de patch clamp, 12de potenciales de campo, 8electroencefalográfico
continuo en el electrodo, 78electrofisiológico, 1, 4, 19extracelular, 1, 5, 7intracelular, 9monopolar, 29referencial, 29
Rehabilitación, 122neuropsicológica, 122
Resonancia magnética, 88funcional (RMf), 26, 42, 85
Respuestas sensoriales auditivas, 50visuales, 49
SSeñal electroencefalográfica, 32
adquirida, 91digitalización de, 32
Sistema(s) automated neuropsychological
assessment metrics (ANAM), 121
cerebral, 114 particular, 49
cognoscitivo, 115normal, 116
de evaluación automatizado, 121funcional, 113
complejo, 112, 113nervioso, 58
central, 47neuronal, 58
Software BESA, 94Solución
norma mínima ponderada
radialmente, 97PROMS, 96
Somas, 28neuronales, 7
Spline, 97esférico, 93
Suicidio, 110Superposición (overlapping), 61Sustancias psicotrópicas, 92Syntactic positive shift (SPS), 54TTallo cerebral, 28Técnica(s)
de análisis espacio-temporal, 94de microelectrodos, 2de neuroimagen, 42, 58, 59, 122de patch clamp, 10de promediación para PRE
superpuestos, 81de registro, 7
electrofisiológico, 1intracelular, 1
electrofisiológicas, 1extracelulares, 1intracelulares, 10
Teoría de cable, 4de Gall, 109del caos, 37electromagnética, 97en neuropsicología, 107
Tomografía axial computada (TAC), 26de baja resolución (LORETA), 97de resolución variable
(VARETA), 97eléctrica cerebral, 85, 88, 102por emisión de positrones
(PET), 26, 42, 85Topografía de la actividad
cerebral, 86Trastorno(s)
bipolares, 122cardiovasculares, 16de la alimentación, 122del desarrollo, 122del habla, 111del lenguaje, 111neuropsicológicos, 113obsesivo-compulsivos, 122
VVCMD (voltaje predeterminado
por experimentador), 10Vida mental, 115Voltaje, 20Voltametría, 10Vóxeles, 88
tmax, 101WWavelet transform, 38
127
Índice

Esta obra ha sido publicada porEditorial El Manual Moderno, S.A. de C.V.,
y se han terminado los trabajos de laprimera edición el 31 de agosto de 2011
en los talleres de Grafiscanner, S.A. de C.V.,Bolívar 455- L, Col. Obrera,
Del. Cuauhtémoc, C.P. 06800,México, D.F.
1a edición, 2011
•