Embed Size (px)
Citation preview

1 Microbiologia - Artini
Patogeni e Saprofiti........................................................................................................................................................................................................................................................................................................................................................................................................................................................... 5
Fattori di Virulenza ...............................................................................................................................................................................................................................................................................................................................................................................................................................................................8
Adesione .....................................................................................................................................................................................................8
Produzione di agressine ..............................................................................................................................................................................9
Capsula ......................................................................................................................................................................................................9
Invasività ..................................................................................................................................................................................................9
Tossine ed enzimi degradativi .....................................................................................................................................................................9
Resistenza agli antibiotici ...................................................................................................................................................................... 10
Altri meccanismi ....................................................................................................................................................................................... 10
Regolazione genetica .................................................................................................................................................................................................................................................................................................................................................................................................................................................. 10
La cellula batterica ............................................................................................................................................................................................................................................................................................................................................................................................................................................................... 10
Colorazione di Gram ................................................................................................................................................................................. 11
Capsula .................................................................................................................................................................................................... 11
Pili (fimbriae) ........................................................................................................................................................................................... 11
Genoma batterico ...................................................................................................................................................................................... 11
Regolazione genica nei batteri .................................................................................................................................................................................................... 12
Plasmidi ........................................................................................................................................................................................................................................... 12
Genetica batterica: trasferimento genico orizzontale............................................................................................................................. 12
Trasformazione .............................................................................................................................................................................................................................. 12
Coniugazione................................................................................................................................................................................................................................... 12
Trasduzione..................................................................................................................................................................................................................................... 13
La spora o endospora ................................................................................................................................................................................ 13
Metabolismo batterico .............................................................................................................................................................................................................................................................................................................................................................................................................................................13
Reazioni cataboliche ................................................................................................................................................................................ 13
Reazioni anaerobiche ................................................................................................................................................................................ 14
Biofilm o sociologia dei batteri ........................................................................................................................................................................................................................................................................................................................................................................................................14
Quorum sensing ......................................................................................................................................................................................... 15
Caratteristiche dei batteri nei biofilm nel corpo umano ........................................................................................................................... 16
Un antibiotico è ugualmente efficace su batteri planctonici o sessili (biofilm)? .................................................................................... 17
Biofilm biologici (sedi).............................................................................................................................................................................. 17
Farmaci antibatterici .......................................................................................................................................................................................................................................................................................................................................................................................................................................................18
Antibiotici ................................................................................................................................................................................................ 18
Spettro d’azione ........................................................................................................................................................................................................................... 18
Microbiologia Artini

2 Microbiologia - Artini
Meccanismi d’azione ..................................................................................................................................................................................................................... 18
Chemioterapici...........................................................................................................................................................................................20
Chinoloni......................................................................................................................................................................................................................................... 20
In generale.. ............................................................................................................................................................................................ 21
Resistenza ai farmaci antibatterici ......................................................................................................................................................... 21
Resistenza genotipica .................................................................................................................................................................................................................... 21
Significato biologico della resistenza agli antibiotici ............................................................................................................................................................ 21
Antibiogramma ............................................................................................................................................................................................................................... 21
Classificazione e identificazione dei batteri ...........................................................................................................................................................................................................................................................................................................................................22
La specie................................................................................................................................................................................................... 22
Classificazione......................................................................................................................................................................................... 22
Ceppo ............................................................................................................................................................................................................................................... 22
Identificazione dei batteri ....................................................................................................................................................................... 22
Morfologia delle cellule batteriche.......................................................................................................................................................................................... 23
Disposizione delle cellule ............................................................................................................................................................................................................ 23
Morfologia delle colonie ............................................................................................................................................................................................................. 23
Capacità di metabolizzare substrati particolari .................................................................................................................................................................... 23
Sequenziamento di specie batteriche – informazioni precise ricavate dal genoma.................................................................................... 24
Batteriologia speciale.............................................................................................................................................................................................................................................................................................................................................................................................................................................. 25
Cocchi Gram+ ............................................................................................................................................................................................ 25
Stafilococchi................................................................................................................................................................................................................................... 25
Streptococchi.................................................................................................................................................................................................................................. 27
Pseudomonas .............................................................................................................................................................................................. 30
Pseudomonas aeruginosa............................................................................................................................................................................................................... 31
Enterobatteri ........................................................................................................................................................................................... 32
Fattori di virulenza ...................................................................................................................................................................................................................... 32
Generi più importanti dal punto di vista medico ................................................................................................................................................................... 33
.................................................................................................................................................................................................. 35
Mycobacterium tuberculosis ....................................................................................................................................................................................................... 35
Clostridium ............................................................................................................................................................................................... 39
Clostridium difficile ..................................................................................................................................................................................................................... 39
C. tetani e C.botulinum ............................................................................................................................................................................................................... 41
Legionella ................................................................................................................................................................................................ 41
VIRUS......................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................42

3 Microbiologia - Artini
Ciclo di replicazione virale...................................................................................................................................................................... 43
1. Fase di adsorbimento .............................................................................................................................................................................................................. 43
2. Penetrazione ............................................................................................................................................................................................................................ 43
3. Scapsidamento.......................................................................................................................................................................................................................... 44
4. Replicazione.............................................................................................................................................................................................................................. 44
5-6. Maturazione e liberazione .............................................................................................................................................................................................. 44
Genetica dei virus .................................................................................................................................................................................... 44
Patogenesi virale ..................................................................................................................................................................................... 45
Virus dell’epatite ..................................................................................................................................................................................... 46
Hepatitis A Virus (HAV) ............................................................................................................................................................................................................... 46
Hepatitis B virus (HBV) ................................................................................................................................................................................................................. 46
Hepatitis C virus (HCV) ................................................................................................................................................................................................................ 47
Cirrosi epatica................................................................................................................................................................................................................................ 47
Herpesvirus ............................................................................................................................................................................................... 48
Herpes Smilpex , HSV-1 ............................................................................................................................................................................................................... 48
Herpes Simplex, HSV-2................................................................................................................................................................................................................. 49
Virus Varicella Zoster (VZV)...................................................................................................................................................................................................... 49
Virus di Epstein-Barr (EBV) ......................................................................................................................................................................................................... 49
Citomegalovirus (CMV) ................................................................................................................................................................................................................ 50
Coronavirus .............................................................................................................................................................................................. 50
Infezione da SARS-CoV-2 – dati di maggio 2020 .......................................................................................................................................................... 51
Miceti o funghi........................................................................................................................................................................................................................................................................................................................................................................................................................................................................... 52
Caratteristiche dei miceti......................................................................................................................................................................... 52
Muffe ........................................................................................................................................................................................................ 52
Lieviti ...................................................................................................................................................................................................... 52
Funghi dimorfi........................................................................................................................................................................................... 52
Ciclo vitale e riproduzione ....................................................................................................................................................................... 52
Patologie da miceti .................................................................................................................................................................................. 53
Farmaci antimicotici – diretti contro gli steroli fungini ....................................................................................................................... 53
Farmaci antimicotici- danneggiano la parete cellulare fungina ............................................................................................................. 53
Farmaci antimicotici - inibiscono la sintesi degli acidi nucleici ............................................................................................................. 53
Sterilizzazione e disinfezione.........................................................................................................................................................................................................................................................................................................................................................................................................54
Antisettici ............................................................................................................................................................................................... 54
Disinfezione .............................................................................................................................................................................................. 54
Sterilizzazione ........................................................................................................................................................................................ 55

4 Microbiologia - Artini
Sterilizzazione a gas plasma ....................................................................................................................................................................................................... 55
Sterilizzazione con agenti chimici in forma gassosa .............................................................................................................................................................. 55
Conservanti .................................................................................................................................................................................................................................... 55
Agenti fisici.................................................................................................................................................................................................................................... 55

5 Microbiologia - Artini
Patogeni e Saprofiti Il primo ad osservare organismi microscopici fu Anton van Leeuwenhoek (fine 1600). Utilizzando lo strumento rudimentale (in foto) in cui, il materiale biologico montato sulla punta di un ago, viene osservato attraverso una lente d’ingrandimento fissata nel foro a sinistra. Egli
osservò per la prima volta, quelli che poi chiamò animalicula (animaletti). Un italiano contemporaneo a Leeuwenhoek, Francesco Redi, mise in crisi la teoria della generazione spontanea, che spiegava la comparsa di vermi
nella carne in putrefazione con la prosecuzione della creazione divina nella storia, dimostrando che i vermi compaiono solo se la carne viene lasciata all’aria aperta, contrariamente a quanto accade se la stessa carne
viene chiusa in un barattolo. Dopo un secolo il problema della generazione spontanea continuava ad infervorare i naturalisti e fu dopo la metà del 700 che Lazzaro Spallanzani pubblicò i risultati di un esperimento analogo a quello di Francesco Redi, dimostrando che per far crescere
microrganismi in un brodo di coltura sterilizzato, bisognava lasciar aperta la bottiglia per far entrare i batteri. Infatti, nella bottiglia sigillata il brodo rimaneva sterile. Ma solo dopo un secolo si capì quanto fosse importante il contagio con materiale infetto da batte ri per
la trasmissione delle malattie. Fu infatti Ignaz Semmelweis, un medico che operò a Vienna a metà ‘800, a dimostrare che l’alta mortalità delle donne che avevano partorito in ospedale era causata dalla scarsa igiene dei colleghi medici che andavano in sala parto senza lavare le mani dopo aver
sezionato cadaveri in sala settoria. Con Semmelweis nasce la moderna disciplina dell’igiene e vengono codificate le prime regole per ridurre il contagio. L’epoca di Semmelweis è considerata l’epoca d’oro della m icrobiologia, nel 1800 infatti
furono scoperte e classificate centinaia di specie batteriche e si capì il ruolo che svolgevano nei processi di fermentazione, necessario per produrre cibi e bevande che l’uomo produce da millenni. Negli stessi anni si comprese il ruolo di molti agenti infettivi responsabili di malattie. Louis Pasteur è uno dei più importanti microbiologi operanti in quel periodo e si occupò dei seguenti argomenti (ma
anche di molti altri): • anomalie della fermentazione della birra (1854); • fermentazione del vino e dell'aceto (1861-62); • pasteurizzazione (1862);
• alterazioni del vino di origine fungina o batterica (1863-64); • malattie del baco da seta (1865-70); • colera dei polli (1880);
• carbonchio di bovini, ovini, equini (1881); • rabbia silvestre e sieroterapia.
E come Pasteur, molti altri microbiologi iniziarono a identificare i microrganismi responsabili di molte malattie. Se si osserva la
colonna destra di questa tabella (1.2), si può notare che gli anni di identificazione dei patogeni sono tutti compresi negli ultimi decenni del 1800 e i primi del 1900.
Quelli furono gli anni in cui vennero standardizzate le tecniche di laboratorio necessarie per isolare e far crescere i batteri in co ltura.

6 Microbiologia - Artini
Come si fa:
• Si raccoglie un campione biologico (es. campione urina, espettorato bronchiale o pus) e si semina in un terreno di coltura liquido, che viene lasciato a 37 °C
fino a quando il terreno diviene completamente torbido.
• Quindi si prende un campione dalla coltura e si semina su una piastra di Petri che contiene terreno di coltura gelatinoso grazie all’aggiunta di agar, ponendo cura a
diffondere su tutta la superficie il prelievo per cercare di separare le singole cellule batteriche invisibili sul terreno.
• Dopo 24-48 h, da ogni singola cellula batterica si sviluppa una colonia visibile, formata da cellule tutte
figlie della cellula madre. L’osservazione delle caratteristiche di questa colonia (colore, forma, dimensioni, margini, odori) permette di identificare la specie.
In questo modo si poteva finalmente capire quale fosse il patogeno responsabile della malattia, semplicemente isolandolo da un campione prelevato da un paziente. La maggior parte da un campione biologico ottenuto da un paziente, vengono selezionati tanti microrganismi appartenenti a spe cie
diverse. Come si identifica il batterio responsabile della patologia, in questo caso? Chi è il colpevole (patogeno)? Si deve a Robert Koch, negli ultimi anni del 1800, la standardizzazione dei passaggi necessari per identificare il patogeno responsabile di una malattia, cioè dei criteri che devono essere soddisfatti per stabilire la relazione di causa-effetto che lega un
microrganismo a una malattia. Prima di vedere questi criteri, vediamo come si definisce un patogeno e un saprofita. Si definisce patogeno un microrganismo capace di produrre malattia. Si definisce saprofita un microrganismo che colonizza un altro
organismo derivandone benefici senza causare danno, o addirittura arrecando egli stesso benefici all’ospite. Il protocollo di Koch prevede che vengano soddisfatti tutti questi
passaggi per individuare il patogeno responsabile di una malattia. Da un prelievo di materiale infetto, preso da un soggetto malato, si isola un microrganismo in coltura pura, con le tecniche descritte prima. Un campione della coltura batterica pura, sospesa in
terreno liquido, viene inoculato in un soggetto sano. In questo soggetto sano, deve svilupparsi la malattia con le stesse caratteristiche del soggetto a cui era stato fatto il prelievo iniziale.
E, da un prelievo di materiale infetto da questo soggetto infettato artificialmente, deve essere isolato nuovamente, in coltura pura, lo stesso microrganismo prelevato dal primo animale. Se tutti
questi passaggi sono soddisfatti, questo microrganismo può essere considerato il patogeno responsabile della malattia. Postulati di Koch (1890)
1. Il microrganismo deve essere sempre isolabile nella malattia in esame;
2. Il microrganismo isolabile dall’ospite ammalato deve crescere in coltura PURA; 3. Inoculando microrganismi cresciuti in una cultura pura ad un animale sensibile e sano si deve riprodurre la malattia; 4. Nell’animale infettato sperimentalmente deve essere reperibile nuovamente il microrganismo e quest’ultimo deve essere
coltivabile in coltura pura. Dobbiamo molto a Koch che, con rigore tedesco, definì una volta per tutte le regole per identificare le specie patogene, ma alla luce delle scienze moderne, ci sono molte eccezioni ai postulati che lui ha elaborato. Tra esse:
• lo stesso microrganismo può essere isolato da individui sani e cagionare malattia in altri (1) o Staphiloccocus aureus, responsabile di infezioni importanti che può però essere isolato come saprofita nel naso
di molti soggetti).
• Molti patogeni causano infezioni sub-cliniche (1) o Virus influenzale preso dai componenti della stessa famiglia
• Esistono condizioni di danno a distanza dal sito di replicazione per liberazione di tossine (1) o C. difteriae o C. botulinum, in cui l’intensissima sindrome sintomatologica è dovuta alla ingestione delle tossine senza alcuna
ingestione del batterio
• Produzione di danno per l’innesco di meccanismi immunitari operanti dopo la risoluzione dell’infezione (1, 2)

7 Microbiologia - Artini
• In alcuni microrganismi i geni codificanti per determinanti di patogenicità possono essere alternativamente espressi/repressi (3)
• Alcuni fattori di virulenza sono trasferibili ed espressi da fagi e plasmidi (tutti)
• Microrganismi considerati tradizionalmente saprofiti possono diventare patogeni in particolari condizioni (tutti). Pertanto, è in atto da tempo una revisione profonda dei criteri grazie ai quali ogni singolo microrganismo può essere definito patogeno o saprofita. Restano quindi immutate le categorie generali quali patogenicità e saprofitismo (definiscono i reciproci rapporti tra agente infettante ed ospite) ma si modificano costantemente, sulla base delle nuove scoperte, i criteri che stabiliscono l’attribuzione
di un microrganismo ad una o all’altra delle due categorie. Non è cioè più facile stabilire se un microrganismo sia patogeno o saprofita. Come si classificano allora in saprofiti o patogeni tutte le specie che colonizzano il corpo umano? Si possono classificare in 3 modi diversi:
1. Si distinguono nettamente i saprofiti dai patogeni. È la modalità più antica, quella ai tempi di Koch e di Pasteur. 2. Si distinguono saprofiti da patogeni, ma c’è un terzo gruppo che si può comportare sai da patogeno che da saprofita, a
seconda delle condizioni.
3. Tutte le specie batteriche possono classificarsi da patogeni a saprofiti a seconda delle condizion i in cui si trovano. Personalemnete, il prof. Artini preferisce la classificazione di mezzo (2), perché non trova corretto distinguere nettamente saprofiti da patogeni essendoci delle specie intermedie e, sempre secondo lui, la visione della classificazione 3 è troppo buonista in quanto ci sono alcune specie che si comportano senza dubbio da patogeno e basta per la potenza dei fattori di virulenza che hanno.
Quindi la patogenicità non è più possibile considerarla come una proprietà assoluta dei batteri, ma come una proprietà relativa che dipende dalle condizioni generali dell’ospite umano, dalle difese immunitarie dell’ospite, dalla carica dell’inoculo (dalla quantità di
batteri che tutti insieme colonizzano il corpo umano) e da situazioni iterferenti dopo l’infezione. Negli ultimi decenni, quindi, abbiamo assistito all’enorme evoluazione delle conoscenze scientifiche sulla patogenicità dei batteri, sia
grazie alla disponibilità di nuove tecniche per lo studio della risposta dell’ospite, soprattutto della r isposta immunitaria, sia grazie all’avanzamento delle conoscenze sui fattori di virulenza dei batteri, cioè sulle armi che i batteri utilizzano per conquistare terreno all’interno dell’ospite colonizzato. Da slide: fattori di virulenza dei batteri: evoluzione delle tecniche di microscopia, affinamento delle tecniche colturali e diffusione
dell’uso delle tecniche di biologia molecolare. Risposta dell’ospite: meccanismi della risposta immunitaria innata, meccanismi della risposta specifica. Volendo costruire un grafico che definisca il rapporto tra
il numero o l’aggressività dei fattori di virulenza di un ceppo batterico e la sua capacità di comportarsi da patogeno, si ottiene la retta nel grafico, che all’estremità in basso colloca i saprofiti e in quella in alto vede i
patogeni stretti. In altri temini, più sono i fattori di virulenza più il batterio può comportarsi da patogeno. È importante ricordare che alcuni fattori di virulenza sono
trasferibili, in altri termini un ceppo batterico può acquisisre fattori di virulenza che non possiede, acquisendo dall’ambiente il materiale genetico (le
porzioni di DNA) che codifica per quel fattore di virulenza. Pertanto, in ogni specie, fra i vari ceppi che compongono quella specie, può variare la virulenza a seconda dei fattori di virulenza posseduti da ciascun
ceppo. Ma a prescindere da queste possibili oscillazioni, ogni specie ha un suo grado di patogenicità. Pertanto, a
seconda del numero di fattori di virulenza si possono distinguere specie di saprofiti che in condizioni di immunodepressione si possono comportare da patogeni opportunisti rispetto a specie di patogeni facoltativi, di
patogeni comuni e di patogeni stretti, che sono quelli che hanno fattori di virulenza talmente potenti o in tale numero da potersi comportare solo da patogeni.
Pertanto, a seconda del grado di patogenicità di una specie o un ospite, la relazione ospite-parassita può
essere classificata in:
• Commensalismo: relaizone simbiotica dove un orgnaimso, commensale, tra da essa beneficio, mentre l’altro, l’ospite, non viene né danneggiato né favorito;
saprofiti
patogeniopportunisti
patogenifacoltativi
patogeniopportunisti
0
20
40
60
80
100
120
0
0,2
0,4
0,6
0,8
1
Pat
oge
nic
ità
N. fattori di virulenza
Rapporto tra virulenza e patogenicità
saprofita
patogeno
patogeni stretti
Virulenza

8 Microbiologia - Artini
• Mutualismo: tipo di relazione dalla quale derivano benefici reciproci per entrmbi i partner e dove i membri sono metabolcamente dipendenti l’uno dall’altro;
• Parassitismo: il parassita tra un vantaggio (nutrimento, protezione) a spese dell’ospite, creandogli un danno biologico. Di quali tipi di patologie sono responsabili i microrganismi opportunisti?
In genere, causano malattie in pazienti immunocompromessi, in cui le capacità di difesa sono gravemente ridotte, e sono batteri facenti parte della flora saprofita oppure batteri che sono stati acquisiti dall’ambiente, capaci di sopravviverci, che generalmente colonizzano temporaneamente l’organismo umano senza riuscire a stabilire dei rapporti di danno.
I seguenti sono esempi di alcune specie batteriche che riescono a stabilire infezioni opportunistiche:
• Cute o Staphylococcus aureus, o S. epidermidis, o Propionibacterium acnes
• Intestino o Bacteroides
o Enterobacteriaceae Le caratteristiche delle infezioni sostenute da patogeni stretti sono completamente diverse. Infatti, i patogeni stretti sono responsabili di epidemie perché colpiscono le persone sane, quindi non necessitano un organismo indebolito (co me nel caso delle persone
immunodepresse). Tra le malattie causate da patogeni stretti si hanno: la tubercolosi, la gonorrea, la tularemia, la malaria. L’esito dell’interazione tra batteri e ospite umano dipende da fattori dell’ospite e fattori del micorganismo. Tra i fattori dell’ospite si citano:
1. le barriere anatomiche: l’integrità della cute e delle mucose, il funzionamento delle ciglia delle cellule cigliate negli epiteli,
il flusso di muco che elimina i batteri dalle vie bronchiali, ph basso, pressione ossigeno, presenza di enzimi. 2. Un secondo importantissimo meccanismo di barriere è costituito dalla normale flora saprofita dell’ospite che inibisce la
ploriferazione di microrganismi estranei: per competizione con i nutrienti e recettori cellula ri, per stimolazione continua del
sistema immunitario e con produzione di sostanze antibatteriche (battericide). È stato calcolato che per ogni cellula umana nel nostro organismo sono presenti da 100 a 1000 cellule batteriche che svolgono proprio questo ru olo di difesa dai patogeni.
3. Il terzo meccanismo di difesa è costituito dall’immunità. Nel suo braccio di immunità naturale e di immunità specifica.
Fattori di Virulenza Per quanto rigurada invece i fattori di cui in batterio dispone, per vincere o per difendersi dall’interazione con l’ospite, i più importanti
sono:
• La capacità d aderire a cellule e sueprfici;
• La capacità di invadere le cellule dell’ospite;
• La capcità di diffondere nei tessuti dell’ospite;
• La capacità di acquisire ed utilizzare risorse dell’ospite;
• La capacità di interferire con il sistema immunitario fino ad evadere dalle difese dell’ospite. A tutte queste capacità corrispondono dei fattori di virulenza dei batteri, che sono vere e proprie armi di cui i batteri dispongono per
aggredire l’ospite umano. Esse sono:
• Adesione
• Produzione di agressine
• Capsula
• Invasività
• Tossine ed enzimi degradativi
• Resistenza agli antibiotici
• Altri meccanismi
Per poter colonizzare un ospite, un batterio ha bisogno prima di tutto di aderire ai suoi tessuti, alle sue cellule. L’adesione avviene
ad opera delle adesine, prodotte dalle cellule batteriche, che riconoscono recettori delle cellule eucariotiche dell’ospite. Le adesine possono essere di 2 tipi:
• Fimbriate: le fimbrie sono utilizzate da molte specie GRAM-
• Non-fimbriate: sono proteine di varia struttura che spesso hanno una serie di funzioni. I recettori possono essere espressi:
• Sulla membrana cellulare: glicolipidi, glicoproteine e proteoglicani;
• Nella matrice extracellulare: fibronectina, vitronectina, lamina, plasminogeno, ecc.

9 Microbiologia - Artini
Con il nome di agressine definiamo un intero gruppo di fattori di virulenza che agiscono in modo da ostacolare la risposta immunitaria dell’ospite. Con questo nome definiamo:
• strutture della parete e della capsula batterica che mascherano le adesine (riconosciute dai fagociti professionisti per legare i batteri),
• componenti strutturali che consentono ai batteri patogeni di non essere individuati come non-self grazie a mimetismo molecolare (molecular mimicry). Es. proteina M di S. pyrogenes.
• Sostanze prodotte dai batteri dopo l’adesione per resistere alle difese dell’ospite (matrice extracellulare del biofilm, proteine di superficie, carboidrati, enzimi come la ialuronidasi o la DNAsi, tossine).
Un altro fattore di virulenza importante è la capsula, costituita da una rete di polimeri non strutturali che ricopre un microrganismo (in genre polisaccaridi proteine o miscele di proteine e carboidrati). Essa protegge dalla risposta immunitaria/infiammatoria dell’ospite con i seguenti meccanismi:
• Inibizione di complemento e della fagocitosi;
• Down-regulation della secrezione di citochine;
• Inibizione dell’accumulo di leucociti;
• Induzione di T-suppressor;
• Inibizione della presentazione dell’antigene;
• Inibizione della linfoproliferazione. Alcune capsule mimano polisaccaridi dell’ospite per eludere la risposta immunitaria umorale (anticorpi). Non tutte le cellule batteriche sono capaci di produrre una capsula. Quando acquisiscono il materiale genetico dall’esterno, tramite plasmidi che li rende capaci di produrre una capsula, si dice che le cellule divengono competenti. In una colonia di batteri in crescita,
l’acquisizione della competenza avviene quando la quantità di batteri (densità della popolazione batterica) supera una certa soglia, con un meccanismo che viene definito meccanismo di Quorum sensing.
à L’invasività è un altro fattore di virulenza molto importante, di cui sono capaci alcune specie batteriche, con il quale riescono a sopravvivere all’interno di alcune cellule eucariotiche. I batteri invasivi, infatti, superano le barriere dell’ospite e passano attraverso
le cellule o vi penetrano per nascondersi dalle difese (fagocitosi e aggressione dai fattori del complemento) e diffondere nell’ospite. Alcuni batteri sono capaci di produrre tossine che uccidono i fagociti, altri possono sopravvivere nei fagociti con una serie di strategie quali:
• uscire dal fagosoma prima della fusione con il lisosoma
• impedire la fusione fagosoma-lisosoma
• sciogliere la membrana del fago-lisosoma. Nel seguente elenco si trovano alcune delle specie capaci di invadere le cellule eucariotiche, raggruppate a seconda dei distretti del corpo umano che riescono a invadere.
• Apparato gastroenterico: S. enterica, Y. enterocolitica, S. dysenteriae, E.coli EIEC, L. monocytogenes;
• Cavo orale (es. parodontite): P.gingivalis, P. intermedia, P. nigrescens, A. actinomycetemcomitans;
• Apparato respiratorio: B. pertussis, H. influenzae, S. pneumoniae, P. aeruginosa ;
• Tratto urogenitale (es. cistiti recidividanti resistenti agli antibiotici, perché l’antibiotico spesso esso non riesce ad entrare nella cellula in cui si nasconde il batterio): E. coli, P. mirabilis, N. gonorrhoeae, S. agalactiae, L. monocytogenes;
• Occhio : P. aeruginosa, S. aureus;
• Endotelio: E. coli, N. meningitidis, P. endodontalis, S. aureus, L. monocytogenes.
Un gruppo importante dei fattori di virulenza sono le tossine, che comprendono:
• Esotossine: sono sostanze di origine proteica, secrete da Gram+ e Gram- durante la crescita esponenziale, termolabili, altamente tossiche anche a basse dosi, ed esercitano effetti diversi a seconda della specie batterica che le ha prodotte (neurotrope, pantotrope, enterotossiche, …) tanto che a partire da l qudro di sintomi, si può risalire alla specie batterica che ha infettato il soggetto;
• Endotossine: sono costituenti delle strutture esterne dei Gram- (lipopolisaccaride o LPS) pertanto sono termostabili e hanno sempre lo stesso effetto tossico, qualsiasi sia la specie che le ha prodotte (CID, febbre, ipotensione).
Sia gli enzimi degradativi che le tossine sono prodotti dalla cellula batterica ed estrosi all’esterno. Gli ezimi degradativi sono esoenzimi che distruggono il tessuto connettivo e permettono la penetrazione tissutale. Alcuni esempi sono:
• Ialuronidasi: idrolizza gli acidi ialuronici, i mucopolisaccard acidi presenti nella matrice acellulare del t.c. (S. aureus);
• Pneumolisina: di S. pneumoniae che distrugge le cellule epiteliali ciliate dei polmoni;
• Mucinasi: di H. pylori che degrada la mucosa gastrica. Grazie agli enzimi degradativi alcuni batteri riescono a superare le barriere epitelili, a prenetrare nei tessuti profondi fino a raggiungere il flusso ematico e il circolo linfatico. In questo caso si distinguono due tipi di invasione del circolo ematico:

10 Microbiologia - Artini
• Batteremia: quando i batteri responsabili di processi morbosi localizzati (polmoniti, enteriti) invadono anche se transitoriamente, il circolo ematico.
• Setticemia: patologia estremamente grave, quando si instaura una costante e massiccia presenza di batteri in circolo (e loro prodotti tossici).
Un altro di gruppo importante di fattori di virulenza è quello delle resistenze ai farmaci antibatterici. Nella resistenza genotipica si parla di resistenza dovuta alla presenza
di un gene sul cromosoma batterico o su un plasmide che permette al batterio di essere resistente a un antibiotico o a una famiglia di antibiotici. In questo caso, quindi la resistenza è ceppo specifica, può essere acquisita dall’esterno tramite un plasmide e può essere
espressa o meno, in base alle condizioni in cui si trova il batterio. La resistenza fenotipica, invece, è una genrica resistenza a molte classi di antibiotici, se non a tutte, dovuta d un comportamento genrico
di una specie batterica che la rende resitente agli antibiotici, quindi è specie-specifica e non ceppo-specifica.
La capacità di interferire con i sistemi di difesa operati dal sistema immunitario dell’ospite, costitu isce un altro capitolo dei fattori di virulenza. Fra questi qui vengono citati:
• La capacità di interferire con la lisi batterica mediata dal complemento che può essere prodotta da una serie di meccanismi quali:
o Incapacità di attivare il complemento o Distacco di molecole attivatrici dalla superficie del batterio; o Interruzione della cascata del complemento prima della formazione del complesso C5b-C9
• La digestione enzimatica degli anticorpi o Produzione di proteasi anti-IgA (S. sanguis).
La capcità di formare biofilm, infine, cioè la capacità di formare società batteriche complesse costituisce un fattore di virulenza importante per alcune specie batteriche.
Regolazione genetica Ricordiamo alcuni aspetti di regolazione genetica dei fattori di virulenza. La comparsa di mutazioni è spontanea nei batteri e continua visto che replicano ad altissima velocità, in questo modo possono accumularsi mutazioni che conferiscono per esempio resistenza
agli antibiotici o nuovi comportamenti. Quindi la comparsa di mutazioni determina variabilità genetica. Le isole di patogenicità, invece, sono aree presenti sul cromosoma e spesso veicolate da un batterio all’altro tramite plasmidi, in cui sono raccolti tutti insieme geni che codificano per fattori di virulenza. Queste aree possono essere eliminate dalla cellula o acquisite
dall’ambiente tutte insieme e per questo si chiamano isole (perché isolate) di patogenicità. Dalla slide – Molti fattori di virulenza presenti su plasmidi sono adiacenti e fiancheggiati da sequenze ripetitive o trasposoni. Spesso i fattori di virulenza sono trasferiti insieme su blocchi di materiale genetico mobilizzabile (trasformazione, trasduzione, coniugazione). I blocchi di materiale genetico possono inserirsi in siti caldi del cromosoma batterico (isole di patogenicità) –
La cellula batterica Rispetto alla cellula eucariotica, che è molto complessa per il materiale genetico rinchiuso all’interno del nucleo e al citoplasma popolato da vari organelli, la cellula procariotica è estremamente più semplice e più piccola. Nei batteri il DNA è libero nel citoplasma e viene direttamente
trascritto in RNA messaggero che viene tradotto in proteine. Il citoplasma è rinchiuso all’interno di una membrana citoplasmatica costituita da un doppio strato di fosfolipidi che a sua volta è ricoperta in tutti i batteri da
una parete. Alcune specie batteriche hanno anche intorno una capsula polisaccaridica e alcune specie batteriche possono avere uno o più flagelli che gli permettono di spostarsi.
La parete cellulare è una struttura essenziale in grado di proteggere la cellula dalla lisi osmotica. È costituita da peptidoglicano: un polimero di
Resistenza genotipica (geni sul cromosoma o plasmidi):
• Ceppo specifica
• Acquisibile
• Facoltativa l’espressione Resistenza fenotipica:
• Specie-specifica
• Instabilità naturale o costitutiva di una specie ad un farmaco
• Es. biofilm

11 Microbiologia - Artini
disaccaridi legati da corte catene di amminoacidi (peptidi). La maggior parte dei batteri può essere classificata in due gruppi diversi a seconda di caratteristiche tintoriali della parete che dipendono dalla struttura.
• Nei batteri Gram+, la parete cellulare è spessa (15-80 nm), è costituita da numerosi strati di peptidoglicano associato ad altre
molecole denominate acidi teicoici.
• Nei batteri Gram-, la parete cellulare è relativamente sottile (10 nm) ed è composta da un singolo strato di peptidoglicano circondato da una struttura membranosa chiamata membrana
esterna (outer membrane) che contiene una componente unica lipopolisaccaride (LPS or endotossina) che è tossica.
Per colorare i batteri con il metodo di Gram, cellule batteriche disciolte in terreno di coltura vengono deposte su un vetrino quindi
seccate e fissate. A questo punto si aggiunge il Cristalvioletto per effettuare la colorazione e un fissativo, che in questo caso è il liquido di Lugol, in questo modo tutte
le cellule batteriche assumeranno un colore blu-violetto intenso, profondo. Segue una fase di decolorazione con una soluzione di alcool e acetone, essi essendo liposolubili,
sciolgono la membrana esterna dei Gram-, quindi questi perderanno il colore assunto con la prima colorazione. Questo non avverrà per i Gram+, in cui il Cristalvioletto si è tenacemente fissato al peptidoglicano della parete esterna.
L’aggiunta di un secondo colorante fucsia, colorerà di rosa chiaro i batteri Gram- che hanno perso il primo colorante e non colorerà i Gram+ perché il primo colorante molto più
scuro dominerà sul rosa. Il risultato è quello che si vede nella foto. Questa tecnica mantiene ancora la sua validità perché permette di dividere la maggior parte delle cellule batteriche in due grossi gruppi che hanno delle caratteristiche di comportamento biologico di patogenicità differenti. I Gram+, che hanno una parete più robusta sono più resistenti e sopravvivono meglio in ambiente secco e difficilmente invadono la
cellula eucariotica, d’altra parte sono più patogeni perché hanno la capacità di acquisire più plasmidi e quindi più fattori di virulenza. I Gram- invece hanno una parete più sottile con una membrana esterna e sono più deboli nell’ambiente vivendo preferibilmente in ambienti umidi, sono però più invasivi riescono a penetrare meglio nella cellula eucariotica pur essendo in generale meno patogeni.
Alcune specie batteriche in condizioni particolari secernono polimeri polisaccaridici che si depositano esternamente alla parete formando un rivestimento esterno detto capsula. Questa
rappresenta un fattore di virulenza aggiuntivo che ha le funzioni di adesione ai tessuti dell’ospite, resistenza alla fagocitosi e proprietà antigeniche, cioè permette di distinguere differenti ceppi di batteri sulla base di differenti proprietà antigeniche. Questo in alcuni casi è importante perché quando alcuni ceppi hanno certi tipi di capsula si sa che sono molto più patogeni.
Identificazione di batteri capsulati tramite inchiostro di china: le particelle di carbone non riescono a penetrare la capsula e quindi i batteri appariranno circondati da un alone chiaro.
Molte specie batteriche possono produrre all’occorrenza dei pili che sono appendici proteiche rigide costituite da subunità di pilina. È importante distinguerli in:
• Pili comuni: funzione adesiva, per colonizzare le mucose;
• Pili sessuali: mediano l’adesione fra le ce llule batteriche durante il processo di coniugazione, per il trasferimento del fattore F.
L’organizzazione del genoma batterico è estremamente semplice, esiste un cromosoma batterico costituito da una doppia catena di DNA circolare contenente tutti i geni fondamentali per la vita del batterio, ed esistono degli elementi accessori costituiti da plasmidi. I plasmidi sono degli elementi circolari mobili di DNA che contengono dei geni codificanti per fattori di virulenza e per altre funzioni
utili ma non strettamente necessarie per la sopravvivenza della cellula batterica. Il cromosoma batterico è unico, aploide, circolare, non contiene introni a differenza delle cellule eucariotiche, ha dimensioni da 150 000 a 9 000 000 paia di basi e un gene ogni 1000 paia di basi, non esistono geni codificanti per poliproteine e i geni non sono sovrapposti.

12 Microbiologia - Artini
Regolazione genica nei batteri In ogni batterio l’espressione dei geni viene continuamente modificata per adattare l’attività metabolica al mutare delle condizioni
esterne. Per facilitare questo adattamento, il cromosoma batterico è organizzato in operoni, in cui più geni sono controllati da un solo promotore e sintetizzano proteine correlate.
Plasmidi Il DNA accessorio è invece organizzato nei plasmidi: molecole di DNA circolare bicatenario, indipendenti dal cromosoma batterico, si trovano nel citosol. Contengono geni non essenziali ma utili, perché conferiscono vantaggi alla cellula batterica. Possono codificare
per fattori di virulenza (tossine) o per fattori di resistenza ad antibiotici. Replicano autonomamente. In una cellula possono essere presenti una o più copie e il batterio può eliminarle, al bisogno, quando non sono più necessarie o acquisirli dall’ambiente con vari metodi.
Il trasferimento di DNA accessorio plasmidico da una cellula batterica ad un’altra può avvenire mediante:
• Trasformazione: uccisione di una cellula non distrugge il DNA che mantiene le sue proprietà, nel caso penetri in una nuova cellula batterica.
• Coniugazione: due cellule batteriche entrano in contatto tramite una struttura detta sex pilus che permette il trasferimento di materiale genetico (es. plasmidi).
• Trasduzione (conversione fagica): il trasferimento genetico è mediato da batteriofagi, ovvero virus capaci di infettare batteri. Può essere specializzata o generalizzata.
Trasformazione Mentre le cellule batteriche crescono in un determinato ambiente, alcune cellule possono morire liberando all’esterno il loro DNA (sia
cromosomiale che plasmidico). Le altre cellule invece continuano a crescere e quando raggiungono una certa numerosità acquisiscono la cosiddetta competenza, cioè diventano permeabili al DNA che sta nell’ambiente esterno (eterologo) che può entrare all’interno. In questo modo, le cellule competenti vengono trasformate e acquisiscono i plasmidi delle altre cellule. Tuttavia, la competenza è una condizione transitoria di una popolazione batterica. Batteri competenti: sia Gram+ (Bacillus/Streptococcus) sia
Gram- (Neisseria, Haemophilus)
Coniugazione La coniugazione è un processo attraverso il quale il materiale genetico (plasmidi) di
un batterio viene trasferito da un batterio donatore ad un batterio ricevente previo contatto tra le due cellule. In questo caso la trasmissione unidirezionale di DNA da una cellula donatrice (F+) a una ricevente (F-) avviene attraverso il pilo sessuale. Di solito avviene tra membri della stessa specie o specie affini ma anche tra procarioti e
cellule eucariotiche vegetali, animali e fungine. L’accoppiamento di due cellule batteriche dipende dalla presenza, nel maschio, o assenza, nella femmina, di un plasmide coniugativo (es. plasmide F di E.coli). il plasmide F (fertility) è definito
coniugativo perché contiene tutti i geni necessari per il proprio trasferimento compresa la capacità di produrre pili sessuali e di iniziare la sintesi di DNA. Il plasmide spesso va incorporarsi nel cromosoma della cellula batterica ricevente e nel momento
in cui si scorpora dal cromosoma può portare con sé un frammento del cromosoma. In questo caso la coniugazione con una nuova cellula batterica porterà al trasferimento di una parte della sequenza plasmidica e di una porzione del DNA cromosomiale batterico, causando scambio genico fra batteri.

13 Microbiologia - Artini
Trasduzione Il trasferimento genetico mediante trasduzione è mediato da virus batteriofagi che raccolgono frammenti di DNA e li impacchettano
in particelle virali. Il DNA viene trasferito alle cellule infettate e incorporato nel genoma batterico. Può capitare che il DNA del cromosoma del batterio venga trascinato nel fago che esce da una cellula infettata, in questo caso, questi fagi passando da una cellula A ad una cellula B trasferiranno DNA cromosomico, modificando il corredo genetico de lla cellula B.
La spora rappresenta un fenotipo caratteristico di alcune specie di bacilli Gram+ appartenenti al genere Bacillus e Clostridium. È una forma di differenziamento cellulare che si origina in ambiente di vita sfavorevoli
(scarsamente dotati di acqua e nutrienti). In ambienti sfavorevoli, nella spora si riduce al minimo il citoplasma e viene raddoppiato lo spessore della parete fino a formare una corteccia che rende la cellula batterica completamente impermeabile rispetto all’ambiente esterno.
In condizioni di carenza di acqua e di nutrienti, dalla cellula batterica deriva una spora che può rimanere nell’ambiente anche per centinaia di anni. Quando le condizioni ambientali ritornano ad essere favorevoli alla
replicazione batterica, la spora torna alla forma vegetativa ritornando a un pieno metabolismo e alla replicazione.
Metabolismo batterico Il metabolismo comprende tutte le reazioni chimiche che avvengono all’interno di un organismo vivente e può essere suddiviso in:
• Reazioni cataboliche o degradative: portano alla degradazione delle molecole complesse in molecole più semplici al fine di ricavare sia composti organici che energia;
• Reazioni anaboliche o biosintetiche: processi che portano alla formazione di moleco le complesse a partire da molecole più semplici, chiamate precursori.
Una reazione catabolica fornisce energia ad una reazione anabolica, quindi sono sempre accoppiate. Questo è possibile grazie alla molecola di ATP perché capace di immagazzinare energia dalle razioni cataboliche e rilasciarla per far avvenire quelle anaboliche. Il trasferimento
di energia da un composto ad un altro avviene tramite una serie di reazioni di ossido-riduzione (trasferimento di elettroni).
I microrganismi usano il processo della respirazione cellulare e/o della fermentazione per produrre energia a partire dal glucosio. Entrambi i processi cominciano con la glicolisi o via di Embden-Meyerhof-Parnas (EMP), per poi differenziarsi.
• Nella respirazione cellulare la glicolisi1 è seguita dal ciclo di Krebs o ciclo degli acid i tricarbossilici (TCA) o ciclo dell’acido citrico e il trasporto degli elettroni avviene attraverso il sistema dei citocromi. Resa energetica = 36 molecole ATP
• Nella fermentazione, la glicolisi è seguita dalla conversione dell’acido piruvico in uno o più prodotti, a seconda del substrato e del corredo enzimatico di cui è dotato il microrganismo. Resa energetica = 2ATP, generati dalla glicolisi
Gli organismi aerobi utilizzano O2 come accettore finale degli elettroni, mentre quelli anaerobi possono utilizzare un’altra molecola
inorganica diversa o, di rado, una molecola organica.
• Contemporaneamente alla glicolisi, molti batteri possono utilizzare la via dei pentoso fosfati o shunt dell’esoso monofosfato. Essa utilizza come substrato zuccheri a 5 atomi di carbonio, producendo importanti composti intermedi
1 Processo anaerobio

14 Microbiologia - Artini
utilizzati nella sintesi degli acidi nucleici e di alcuni amminoacidi. Resa energetica = 1 ATP per ogni molecola di glucosio ossidata.
• Alternativamente alla glicolisi, molti batteri possono usare la via di Entner-Doudoroff, che permette ai batteri di metabolizzare glucosio senza ricorrere alle altre vie metaboliche. È un processo che si realizza in presenza id O2 ed è
utilizzata in pochi batteri Gram-. I microrganismi dotati di proteasi, peptidasi, deaminasi e lipasi possono utilizzare per la produzione di energia composti organici alternativi al glucosio.
In base alla capacità o meno di utilizzare l’O2 come accettore finale di elettroni, i batteri sono stati distinti in:
• Aerobi obbligati: l’O2 è essenziale per svolgere la respirazione cellulare;
• Anaerobi obbligati: l’O2 è letale.
• Aerobi-anaerobi facoltativi: la presenza di O2 non è strettamente necessaria, ma la capacita di crescita è notevolmente più rigogliosa in sua presenza;
• Microaerofili: crescono meglio a basse concentrazioni di O2 (2-8%).
Affinché i batteri riescano a portare avanti le vie biosintetiche necessarie alla loro replicazione, essi devono trovarsi in
presenza di fattori ambientali o fisici adeguati. Per ogni specie microbica esiste un intervallo di temperatura che ne consente la crescita e una temperatura in cui la crescita avviene in maniera
ottimale (T.O.). La crescita microbica può verificarsi solo in un ambiente in cui sia presente l’acqua, perché i batteri riescono ad assumere le sostanze nutrizionali solo se presenti in soluzione. Le specie di
batteri ben adattate ad alte concentrazioni saline sono chiamate alofile.
Biofilm o sociologia dei batteri Dobbiamo a Bill Costerton (1934-2012), microbiologo e ricercatore canadese, l’approfondimento
sistematico del ruolo dei biofilm nella patologia e nella fisiologia umana. Tutto iniziò nel 1982 con la descrizione di uno strano caso clinico a Calgary (nel Canada): un uomo di 56 anni fu ricoverato in ospedale per una setticemia comparsa dopo un’infezione purulenta del gomito sostenuta da uno stafilococco aureo.
Dall’emocoltura venne isolato lo stesso stafilococco e sulla base dell’antibiogramma fu instaurata la terapia antibiotica e il paziente guarito fu dimesso. Dopo due settimane lo stesso paziente tornò con la stessa gravissima sintomatologia e dalla coltura di un prelievo di sangue fu prelevato lo stesso ceppo batterico, con lo stesso profilo di sensibilità agli antibiotici, quindi il paziente ripeté la terapia e fu dimesso dopo la
guarigione. Al terzo ricovero effettuato per lo stesso identico quadro sostenuto dallo stesso ceppo batterico, i medici in accordo con il microbiologo Costerton sostituirono il pacemaker nel dubbio che questo fosse infetto. Costerton andò ad osservare la superficie dell’elettrocatetere del pacemaker in microscopia
elettronica e trovò quello che si vede nell’immagine a fianco. La punta dell’elettrocatetere era ricoperta da una massa informe, che ad un ingrandimento maggiore si rivelò essere solcata da profondi solchi e canyon. Ingrandendo ulteriormente, Costerton capì che questa massa era
composta da una quantità enorme di batteri, che ricoprivano la superficie dell’elettrocatetere, né più né meno di ciò che fanno i batteri ambientali nei laghi e nei fiumi. Era noto da tempo infatti che, ovunque vi sia acqua, i batteri aderiscono nell’interfaccia fra il solido ed il liquido e si ricoprono di una matrice extracellulare contenente polisaccaridi, proteine
e DNA. Perché ciò avvenga nel corpo umano, su un dispositivo artificiale, la superficie del dispositivo deve essere prima ricoperta da glicoproteine presenti nel plasma o nel fluido intercellulare (cascata di glicoproteine plasma-derivate: fibronectina, fibrinogeno, laminina,
collagene, albumina, immunoglobuline, mucine, parti di globuli rossi, piastrine, ecc.). I batteri che vengono portati dalla superficie esterna o dal flusso sanguigno, entrano in contatto con questa superficie attivata e aderiscono, passando da una fase reversibile ad una irreversibile mediata da
adesine e fimbrie. Più cellule batteriche possono aggregarsi e avviare la colonizzazione del dispositivo impiantabile. Formando dapprima delle piccole colonie che rapidamente evolvono a formare un biofilm maturo che può essere considerato già formato entro le prime 24 h dall’imp ianto di una protesi artificiale.
Temperatura
Microrganismi intervallo di T (°C) T.O. (°C)
psicrofili 0-25 10
mesofili 20-45 30-37
termofili 45-70 50-55
pH
Microrganismi intervallo di pH
alcalofili >9
neutrofili 6 - <8
acidofili <4

15 Microbiologia - Artini
Nel caso dei biofilm che si formano sulle superfici esterne del corpo umano più specie batteriche possono aderire ad una superficie e rapidamente colonizzarla fino a formare delle colonie mature,
che si ricoprono rapidamente di una matrice extracellulare densa, a formare il biofilm completo. Nelle immagini ottenute con il microscopio a fluorescenza utilizzando la tecnica FISH, che permette di distinguere varie specie batteriche con specifiche sonde fluorescenti, si osserva che specie batteriche diverse si dispongono in un relativo ordine nella placca batterica del cavo orale.
Si vedono in rosa corynebatteri che occupano g li strati più profondi dove scarseggia l’ossigeno ricoperti dagli streptococchi colorati in verde, che si sporgono all’esterno in un ambiente più ricco di ossigeno.
In un biofilm maturo la popolazione batterica cresce fino a formare torri e ammassi di batteri attraversati da pori, canali e stratificazioni che favoriscono il passaggio di ossigeno e nutrienti in entrata e l’eliminazione di anidride carbonica e prodotti metabolici di scarto in uscita. I batteri immersi
in un biofilm comunicano tra di loro per cambiare comportamento quando questo serve ad elaborare risposte rispetto ai cambiamenti dell’ambiente esterno. Questa forma di comunicazione si chiama quorum sensing (tradotto: percepire la numerosità della popolazione). Fasi formazione del biofilm:
1. Formazione di film condizionante il substrato: rapido, influenza l’adesione dei microrganismi pionieri (Es. mucine salivari)
2. Movimento delle cellule batteriche sul substrato: forze convettive, moto browniano,
contatto casuale, flagelli, chemiotassi. 3. Adesione dei batteri al substrato: reversibile tramite forze di Van der Waals,
Coulomb, adesione irreversibile tramite adesine specifiche (EPS) e
coaggregazione e coadesione 4. Sviluppo del biofilm: formazione di microcolonie 5. Maturazione del biofilm: comunicazione intercellulare o quorum sensing.
Le ricerche che hanno condotto alla scoperta del quorum sensing sono state condotte da Bonnie Bassler, studiando un piccolo calamaro che vive lungo le coste degli arcipelaghi dell’Oceano pacifico. Questo calamaro si nasconde sotto la sabbia di giorno ed esce a nutrirsi di notte e per sfuggire ai predatori emette una debole luminescenza che lo rende indistinguibile dalle luci del tappeto
di stelle che ricopre l’oceano. La luce non è emessa dal calamaro, ma da milioni di cellule batteriche di Vibrio fischeri che colonizzano le sue tasche laterali. Al mattino, tali tasche vengono svuotate dal calamaro con una potente contrazione, i pochi batteri rimasti riprendono a crescere e moltiplicarsi e si accendono solo quando hanno raggiunto una certa densità di popolazione il che avviene
solo verso sera. Bonnie Bassler ha riprodotto questo fenomeno osservando al buio una colonia di Vibrio fischeri che cresce in una bottiglia di laboratorio: solo quando i batteri in coltura raggiungono una certa numerosità, si accendono tutti insieme contemporaneamente e la bottiglia si illumina all’istante come una lampadina. Il quorum sensing è il processo con cui i batteri controllano la densità della loro popolazione misurando la concentrazione di piccole
molecole segnale chiamate autoinduttori (AI). Mentre la popolazione di batteri cresce, ogni cellula produce e secerne gli AI nell’ambiente extracellulare, in questo modo ogni momento la concentrazione degli AI esterna è correlata con la densità della popolazione. Monitorando la concentrazione degli AI i batteri si contano e modificano l’espressione di geni bersaglio.
Bonnie Bassler ha identificato gli AI di molte specie batteriche, distinguendo gli AI dei batteri Gram – che hanno una struttura chimica di acil-omoserin-ottoni. Questi si
accumulano nell’ambiente fino a rientrare nelle cellule batteriche dove legano e attivano recettori intracellulari che legano il
promotore di specifici geni, attivandone l’espressione. Nell’immagine si osservano alcuni AI dei Gram-, la struttura a sinistra è
simile, ma la parte a destra è differente. In questo modo gli individui di una specie batterica comunicano tra di loro senza interferire con la comunicazione delle altre
specie. I batteri Gram + comunicano in modo analogo ma gli AI hanno la struttura chimica di piccoli peptidi (oligopeptidi).

16 Microbiologia - Artini
I comportamenti batterici controllati dal quorum sensing sono quelli finalizzati alla sopravvivenza delle cellule batteriche.
Per esempio, nella tabella i batteri in rosso esprimono fimbrie e adesine per aderire ad una superficie, per colonizzarla e formare biofilm quando ci sono condizioni ambientali favorevoli. O al contrario, il distacco di cellule dal singolo
biofilm che tornano al fenotipo planctonico per migrare lontano quando scarseggiano ossigeno e nutrienti. Nelle specie evidenziate in verde, il quorum sensing controlla
l’espressione di fattori di virulenza come tossine, enzimi extracellulari o capsula che vengono prodotti in contemporanea da tutte le cellule batteriche insieme, in
modo da paralizzare le capcità di difesa o di contrattacco da parte dell’ospite umano.
Una forma di comunicazione tra batteri completamente diversa, che si può defin ire da contatto diretto, è stata proposta dopo l’osservazione di queste nanofibre che collegano tra di loro le
cellule di un biofilm. Poiché queste nanofibre possono condurre corrente elettrica, è stato suggerito che le connessioni tra batteri in questo modo possano essere assimilati al passaggio di impulsi nei cercuiti neuronali del SN.
Qui si possono vedere le connessioni mediate da nanofibre fra batteri di un ceppo isolati da un’infezione ossea umana. Quando il corpo umano deve contrastare un patogeno, non si trova solamente davanti a una “truppa” più o meno numerosa da contrastare ma si trova davanti a una comunità organizzata che può diventare imbattibile, perché grazie alla comunicazione mette in
atto strategie complesse adattabili al cambiare delle condizioni esterne e quindi vincenti.
La storia di un biofilm quindi nasce da cellule plantoniche che aderiscono a una sperficie e rapidamente si ricoprono con una matrice
extracellulare di acqua, polisaccaridi, proteine e DNA fino a crescere come ne lla fase 2 dello schema, a formare un biofilm maturo costituito da cellule che hanno acquisito un metabolismo e un profilo differente. Queste cellule si definiscono come cellule in fenotipo sessile. Quando la popolazione batterica nel biofilm è troppo numerosa rispetto ai nutrienti e allo spazio disponibile, con un
meccanismo di comunicazione intercellulare basato su quorum sensing, le cellule più esterne ritornano al fenotipo planctonico per andare in sedi più adatte alla sopravvivenza (fase 3). Pertanto, da un biofilm maturo si staccano sempre cellule superficiali che tornano in fenotipo planctonico per andare a colonizzare siti distanti, formando un nuovo biofilm. Le cellule p lanctoniche quindi
rappresentano solo un fenotipo transitorio, sono cioè cellule in viaggio da un biofilm ad un altro. Per i batteri è molto meglio vivere cicondati da cellule di altre specie commensali sotto la protezione della matrice extracellu lare. Infatti è stato calcolato che in natura il rapporto tra i batteri in forma planctonica/sessile è di 1:1.000 – 1:10.000, quindi le forme sessili sono quelle assolutamente prevalenti in natura.
Pertanto, il biofilm rappresenta la principale forma di organizzazione di vita microbica. La forma planctonica dei batteri può essere considerata una forma fenotipica transitoria dovuta al distacco di cellule dai biofilm per colonizzare nuove superfici.
L’ecosistema biofilm fa acquisire ai batteri proprietà che le cellule planctoniche isolate non possiedono: 1. Resistenza alle difese dell’ospite: fagocitosi, Ig, complemento, ecc. 2. Resistenza alle sostanze antibatteriche naturali (lattoferrina, peptidi ricchi in istidina) ed artificiali (antibiotici).

17 Microbiologia - Artini
Queste caratteristiche di resistenza dipendono da moltissimi fattori, ma una prima spiegazione alla resistenza agli antibiotici viene sicuramente dalle caratteristiche del
microambiente di un biofilm. Andando dagli strati esterni e superficiali a quelli più profondi troveremo una progressiva riduzione del pH, una riduzione dell’ossigeno e un aumento dell’anidride carbonica. Le cellule in superficie vivono in un ambiente che è favorevole alla replicazione batterica e il metabolismo aerobio; le cellule in
profondità, invece, hanno un metabolismo rallentato e un fenotipo dormiente che conferisce caratteristiche di resistenza agli antibiotici.
èLa risposta è no. Nelle primissime fasi della formazione di un biofilm questo può essere aggredito sia dal sistema immunitario, sia dagli antibiotici, mentre un biofilm maturo sarà insensibile sia agli attori del sistema immunitario sia ai farmaci antibiotici. Il biofilm non viene visto dal sistema immunitario, solo quando si staccano le cellule planctoniche esse inducono la febbre, l’astenia, i brividi,
tutti i sintomi di una sepsi grave e generalizzata. La somministrazione di antibiotici elimina solo le cellule planctoniche circolanti ed eventualmente quelle sulla superficie del biofilm perché entrambe mantengono un fenotipo sensibile agli antibiotici. Le cellule batteriche in profondità non saranno danneggiate dall’antibiotico e sopravviveranno nella matrice. Gli attori delle difese immunitarie
continueranno a rimanere inefficaci nei confronti del biofilm. Quindi, quando si forma un biofilm su un dispositivo artificiale posizionato in profondità, come una protesi vascolare oppure ortopedica, gli antibiotici che dovrebbero essere efficaci, perché testati con l’antibiogramma, in realtà non riescono ad eliminare il
biofilm sia perché non riescono a penetrare negli strati profondi del biofilm sia perché in profondità trovano cellule batteriche con un metabolismo molto rallentato quindi resistenti e, infine, perché in queste cellule si instaura un meccanismo di adattamento tipici dei fenotipi sessili.
Le sedi colonizzate da biofilm fisiologici sono: cute, cavo orale, cavità nasali, colon e mucosa vaginale, ecc. Si noti che le sedi esterne sono abitate dai saprofiti, come già visto nella parte dei saprofiti. Questo significa che in tutte le sedi superficiali in cui i saprofiti formano una barriera all’entrata dei patogeni, lo fanno aggregandosi in biofilm fisiologici costituiti da centinaia di specie saprofite che
si scelgono nel tempo a formare un microbiota complesso di batteri e commensali. La persistenza di questi biofilm fisiologici dipende dalla condizione di non belligeranza e quindi di armonia tra le specie commensali aggregate nel biofilm da un lato e le difese immunitarie dall’altro. La rottura di questo equilibrio, tra il microbiota biofilm e le difese dell’ospite, causa lo squilibrio nella
composizione delle specie del biofilm e l’entrata in scena di specie patogene che altrimenti non sarebbero riuscite ad insediarsi. In questo modo s’instaurano condizioni di infiammazione cronica e di dismicrobismo come la parodontite, colite, sinusite o vagin ite. Quindi i biofilm interessanti per l’uomo si possono classificare in base all’azione che hanno nel corpo umano:
Con l’aumento dell’età media degli italiani, ogni anno, aumentano i soggetti portatori di protesi. Anche i dispositivi temporanei d’infusione di farmaci e soluzioni sono ad altissimo rischio di colonizzazione da biofilm batterico.
Esterno pH O2 CO2
Substrato
Fisiologici
•cute•colon
•bocca•vagina•naso
•ecc.
Patologici
•per alterazione dell'equilibrio con l'ospite di un biofilm fisiologico (parodontite, vaginite, colite, etc.)
•per colonizzazione di una sede profonda normalmente non accessibile a batteri planctonici (impianti dentari, protesi vascolari, protesi ossee, protesi mammarie, ecc.)
•per inalazione di batteri in fase sessuale (polmoniti da Legionella)

18 Microbiologia - Artini
Farmaci antibatterici Un approccio funzionale alla classificazione dei farmaci antibatterici prevede di raggrupparli sulla base del bersaglio della loro azione. Nella figura, si osservano i principali bersagli d’azione degli antibatterici.
Un altro modo di classificare gli agenti antimicrobici è quello di raggruppare da un lato gli antibiotici, che agiscono su bersagli specifici, e dall’altro i disinfettanti
che agiscono su molteplici strutture. I primi agiscono con un meccanismo selettivo, quindi sono efficaci su classi più o meno ristrette di microrganismi, i disinfettanti invece
esercitano un’azione molto generica e non specifica su più strutture, quindi vengono utilizzate per sanificare le superfici del corpo o dell’ambiente umano. Restringendo l’attenzione ai farmaci utilizzati per le
infezioni batteriche (farmaci antibatterici), si distinguono: farmaci di origine naturale (antibiotici) e farmaci ottenuti dall’industria farmaceutica per via sintetica
(chemioterapici).
Le caratteristiche principali degli antibiotici sono:
• Sostanze chimiche a basso peso molecolare (150-5000 Dalton);
• Prodotte da microrganismi;
• A basse concentrazioni sono in grado di inibire la crescita di altri microrganismi (<1 mg/ml). Dal punto di vista del meccanismo d’azione gli antibiotici si dividono in farmaci:
➢ batteriostatici, che inibiscono la crescita dei batteri senza ucciderli;
➢ battericidi, che uccidono le cellule batteriche. Rappresentando in grafico le modalità d’azione di queste due classi di antibiotici, si può osservare che i batteriostatici aggiunti a una coltura di batteri che stavano crescendo in maniera esponenziale bloccano
la replicazione dei batteri ma non li uccidono, quindi andando a contare le cellule batteriche vive rispetto alle cellule totali dopo la somministrazione di un batteriostatico, non si troveranno batteri morti ma solo un arresto nella crescita numerica. Nell’altro grafico, invece, si può osservare che la
somministrazione di un battericida, in una coltura di batteri in crescita esponenziale, riduce progressivamente il numero di cellule vive, come rappresentato dalla curva gialla delle cellule vive. L’efficacia degli antibiotici
batteriostatici dipende dal fatto che il blocco della crescita numerica dei batteri in infezione può aiutare il sistema immunitario ad elaborare una risposta efficace e a riprendere il sopravvento. Si rammenta che alcuni fattori di virulenza vengono espressi da una popolazione batterica, solo dopo che questa ha raggiunto una certa
numerosità e vengono regolati con meccanismi di quorum sensing. È chiaro quindi che contenere la numerosità di una popolazione batterica che altrimenti crescerebbe, serve a limitare l’espressione dei suoi fattori di virulenza e quindi la sua patogenicità, per dare il tempo al sistema immunitario di elaborare una risposta efficace. Le caratteristiche ideali per avere un buon antibiotico sono:
• Non tossicità;
• Scarsa o nulla attività sulla flora commensale;
• Assenza di effetti collaterali;
• Assenza di resistenze.
Spettro d’azione Un antibiotico può essere attivo su tante specie o su un numero limitato di specie. Si definiscono ad ampio spettro gli antibiotici che sono attivi sia sui Gram+ che sui Gram-. Al contrario, gli antibiotici a spettro ristretto sono attivi su un numero limitato di specie, in genere sui Gram+ che hanno caratteristiche di maggior permeabilità alla penetrazione del farmaco.
Meccanismi d’azione Dal punto di vista del meccanismo d’azione, gli antibiotici si possono classificare in 4 gruppi:
• Inibizione della sintesi della parete cellulare
• Lesione delle membrane batteriche
• Inibizione della sintesi proteica
• Inibizione del metabolismo degli acidi nucleici
Figura 1: antibiotici battericidi Figura 2: antibiotici batteriostatici

19 Microbiologia - Artini
Inibenti la sintesi della parete cellulare Hanno un valore storico importante, perché sono i primi ad esser stati utilizzati. Sono in grado di agire sugli enzimi della biosintesi del peptidoglicano determinando il blocco della sintesi della parete cellulare e la conseguente lisi della cellula batterica. Sono attivi
solo in batteri in fase di accrescimento e moltiplicazione, esercitando un’azione di tipo batteriostatico. I principali farmaci appartenenti a questa classe sono:
• β-lattamici (rappresentano la classe più numerosa ed utilizzata);
o si chiamano in questo modo per la presenza dell’anello β-lattamico. Il gruppo include le penicilline e cefalosporine, che prendono il nome dal secondo anello
legato all’anello β-lattamico.
• Bacitracina;
• Vancomicina;
• Cicloserina:
• Novobiocina;
• Ristocetina.
Penicilline In questo gruppo sono inclusi tanti sottogruppi. Ogni nuovo gruppo ha aggiunto altre specie allo spettro d’azione del gruppo
precedente. Quindi le penicilline più recenti hanno uno spettro più ampio.
Penicilline spettro
acido-resistenti:
benzilpenicillina G penicillina v propicillina
azidocillina
streptococchi, bacillus, corinebatteri, actinomiceti, listeria,
neisserie, emofili, clostridi, bacteroides
penicillinasi-RES: meticillina
cloxacillina isoxazolilpenicillina
+stafilococchi
ad ampio spettro: ampicillina e derivati
amoxicillina amidinopenicilline
+stafilococchi, enterobatteri Gram-
ad ampio spettro comprendente P. aeruginosa:
carbenicillina ticarcillina
sulfossipenicilline
cilureidopenicilline
+enterobatteri Gram- e psudomonas aeruginosa
scarsa attività su Gram+
La penicillina è stato il primo antibiotico, isolato da Fleming da colture di Penicillium chrysogenum. Queste molecole sono tutte imparentate chimicamente con la penicillina scoperta da Fleming ma da un decennio oramai vengono ottenute per sintesi chimica.
Cefalosporine Anche le cefalosporine includono farmaci: di I, II e III generazione. Anche in questo caso, i farmaci appartenenti ad ogni generazione hanno ampliato lo spettro d’azione su più specie. Quindi, ogni generazione successiva, ha uno spettro d’azione maggiore.
Cefalosporine spettro
I generazione: cefalotina cefaprina
cefazolina cefacetrile
streptococchi, stafilococchi, bacilli, actinomiceti, corinebatteri, neisserie, E.coli, proteus, klebsiella, shighella, salmonella,
clostridi
II generazione:
cefamandolo cefuroxime cefoxitina
+ branhamella, bordetella, haemophilus, bacteroides
III generazione: cefotaxime
cefoperazone ceftazidime
ceftriaxone
+ enterobacter, serratia, providencia acinobacter, psudomonas

20 Microbiologia - Artini
Attivi sulle membrane batteriche Sono antibiotici battericidi in grado di indebolire la membrana esterna dei batteri Gram-, che perde la sua permeabilità con conseguente fuoriuscita del contenuto cellulare.
Polimixine Sono antibiotici formati da peptidi ciclici. Sono attive solo nei confronti dei Gram-. Le polimixine si legano ai fosfolipidi di membrana dei Gram- distruggendone le proprietà osmotiche. Sono tossiche anche per le cellule eucariotiche dell’ospite quindi possono
essere utilizzati solo per trattamenti topici, sulla cute o sulla mucosa dell’ospite.
Inibenti la sintesi proteica Sono gli antibiotici più largamente usati:
1. Inibiscono le unità ribosomiali bloccando la traduzione
− 30S: tetracicline, aminoglicosidi. Più utilizzati, vengono riservati per i casi in cui la gravità dell’infezione richiede un antibiotico potente anche a costo di rischiare effetti collaterali importanti.
− 50S: macrolidi, cloramfenicolo, lincosamidi. I macrolidi sono privi di effetti collaterali ed utilizzati per questo motivo nella prima infanzia, ma oltre ad essere poco tossici sono anche poco efficaci.
2. Inibiscono le proteine extraribosomiali bloccando l’allungamento e la terminazione proteica (es. acido fusidico). È un gruppo di antibiotici scarsamente usato.
Tetracicline Per questa classe va segnalato il problema degli effetti collaterali. Farmaci utilizzati soprattutto nelle rickettsiosi, ma aggravati nel problema dell’annerimento dei denti nei soggetti che li prendono per periodi prolungati.
Inibenti il metabolismo degli acidi nucleici Interferiscono anche sul metabolismo della cellula dell’organismo ospite , quindi esplicano degli effetti di tossicità importanti, infatti alcuni di questi farmaci (es. rifampicina) vengono utilizzati nelle chemioterapie antitumorali. Scarsa utilizzazione.
− Inibizione della sintesi del DNA: novobiocina, mitomicina, griseofulvina
− Inibizione della sintesi del RNA: rifampicina, acrinomicina, griseofulvina
Rifampicina È un antibiotico ad ampio spettro, usato abbastanza frequentemente, è raramente tossico per alcuni singoli pazienti che dimostrano di avere una selettiva reattività nei confronti della molecola (tossicità selettiva). Si legano alla subunità b della RNA polimerasi batterica.
I chemioterapici sono i farmaci che sono stati disegnati in laboratorio e sintetizzati fin dall’inizio per sintesi chimica. La loro azione antibatterica è selettiva:
• Sulfamidici
• Isonazide (importantissimo per tubercolosi)
• Dapsone
• Chinoloni
• Nitrofurani
Chinoloni Nel gruppo dei chinoloni fluorurati (formula a destra) si trovano alcuni
farmaci che sono costantemente utilizzati nella pratica quotidiana, sia perché esercitano un’attività a spettro molto ampio sia su Gram+ e Gram- sia perché in alcune situazioni sono realmente efficaci e abbastanza
tollerabili.

21 Microbiologia - Artini
Come si può osservare, le cefalosporine prevalgono su tutti, seguite dalle penicilline. Messi insieme questi β-lattamici superano il 50% delle prescrizioni. Tra le grandi classi seguono poi i chinoloni e i macrolidi. Le aminoglicosidi e le tetracicline vengono usati per casi gravi con infezioni importanti in cui il bisogno di attaccare con un farmaco potente il batterio responsabile giustifica il rischio di
effetti collaterali a volte importanti.
Un ceppo batterico è resistente a un farmaco quando si moltiplica in presenza di concentrazioni dello stesso, pari a quelle massime raggiungibili nel corso dell’impiego terapeutico. Questa resistenza può essere:
• naturale (fenotipica): generale insensibilità estesa a tutti gli stipiti di una specie (es. biofilm)
• acquisita (genotipica): assunta ex-novo per modificazione del genoma, quindi tramite mutazioni o acquisizione di geni esterni tramite plasmidi.
Resistenza genotipica La resistenza genotipica ad un antibatterico può dipendere da vari meccanismi. I principali sono:
• produzione di enzimi inattivanti il farmaco
• modificazioni del bersaglio su cui agisce il farmaco
• diminuita permeabilità al farmaco o aumentata eliminazione del farmaco dalla cellula
• sviluppo di una via metabolica alternativa a quella inibita dal farmaco.
Significato biologico della resistenza agli antibiotici Gli antibiotici sono prodotti in natura dai m icrorganismi per competere nell’ecosistema. Anche in ecosistemi lontani dalla civiltà, i
batteri si cimentano quotidianamente con antibiotici (batteriocine) e sviluppano resistenze. L’ambiente urbano comprende ecosistemi ad elevata concentrazione di antibiotici. I meccanismi di resistenza possono selezionarsi autonomamente in un qualsiasi ambiente o giungere da ambienti diversi e qui evolvere a livello di efficienza e regolazione.
Antibiogramma L’antibiogramma è un test in vitro per determinare il grado di resistenza ai più comuni antibiotici di una definita popolazione microbica
isolata dal materiale biologico. Per effettuare l’antibiogramma di un ceppo prelevato da un paziente, si può usare il metodo tradizionale di Kirby Bauer. In accordo con questa tecnica, si preleva un campione batterico da una colonia batterica del ceppo in esame e lo si semina su terreno solido, nella piastra di Petri. Sul terreno, contemporaneamente alla semina dei batter i, vengono posizionati anche dei dischetti di carta assorbente, ciascun dischetto è imbevuto di un differente antibiotico a concentrazioni note.
La piastra viene messa quindi in incubatore e dopo un periodo di tempo prestabilito si può apprezzare una crescita microbica relativamente uniforme su tutta la superficie del terreno, con aloni più o meno ampi intorno ai dischetti. In base all’ampiezza di queste aree di inibizione della crescita batterica, si valuta la suscettibilità del ceppo in esame ad ogni singolo antibiotico testato.
➢ diametro è grande, il microrganismo è sensibile al farmaco; ➢ diametro è piccolo, il batterio presenta una sensibilità intermedia; ➢ diametro è irrisorio o addirittura assente, la specie batterica esaminata è resistente al farmaco.
Per determinare con esattezza la sensibilità batterica, il diametro degli aloni di inibizione viene misurato in mm, paragonando i valori ottenuti con quelli standard per il ceppo batterico. Nella maggior parte dei laboratori ormai tutto è meccanizzato e l’antibiogramma viene effettuato automaticamente, ma anche la procedura delle apparecchiature è basata comunque su questo tipo di principio.
Concentrazione minima inibente in terreno liquido Per valutare meglio la sensibilità agli antibiotici da parte di un ceppo batterico e soprattutto per quantificarla, si deve ricercare la MIC cioè la minima concentrazione di quel farmaco capace di inibire la crescita del ceppo. Per calcolare la MIC di un antibiotico su un
37%
11%14%
17%
3%3%
15%
ANTIBIOTICI E CHEMIOTERAPICI PIÙ USATI
Cefalosporine Macrolidi Chinoloni Penicilline
Aminoglicosidi Tetracicline Altri

22 Microbiologia - Artini
dato ceppo batterico, si fa crescere il ceppo in un terreno liquido fino a portare i batteri ad una densità prestabilita e
standard. Quindi si prepara una serie di provette, in modo che ciascuna contenga uno stesso volume della sospensione batterica per avere la stessa quantità di batteri nello stesso volume di terreno in tutte le provette. A questo
punto si prepara una diluizione scalare dell’antibiotico, secondo un rapporto di 1:2, e si aggiunge l’antibiotico alle varie provette contenenti i batteri in modo che, andando da
sinistra verso destra, ogni provetta contenga la metà del quantitativo di antibiotico della provetta precedente e si lasciano crescere i batteri a crescere nelle provette in
presenza dell’antibiotico per 24h a 37°C. Dopo 24 h, si osserva in quali provette l’antibiotico è riuscito ad inibire la crescita dei batteri: la prima provetta in cui i batteri non sono cresciuti, contiene una concentrazione di antibiotico sufficiente ad inibirne la crescita, quindi quel valore di concentrazione di antibiotico darà la minima concentrazione inibente (MIC). Se si lasciano
le provette in cui i batteri non sono cresciuti per altre 24h a 37°C si troverà con lo stesso metodo la minima concentrazione battericida.
Classificazione e identificazione dei batteri
La specie è l’unità tassonomica fondamentale. Negli organismi superiori, viene definita dalla capacità di produrre progenie fertile (Es. Cavallo + asino = mulo, non fertile) e ciò dipende dalla complessità del corredo genetico degli organismi superiori. I batteri hanno un
genoma molto semplice e non si riproducono per via sessuata, inoltre lo scambio genetico tra individui fenotipicamente diversi non è impossibile. I batteri scambiano informazioni genetiche tra specie e tra unità tassonomiche superio ri (anche tra regni). I batteri differiscono per poche caratteristiche. Ciò nonostante da più di 150 anni, i batteri sono stati divisi in specie, sulla base delle caratteristiche della forma e delle cellule, della
forma delle colonie cresciute in condizioni di laboratorio standardizzate, delle caratteristiche metaboliche e del comportamento. Ne deriva che, poiché i criteri derivano dai dati raccolti con l’osservazione, tali criteri possono essere messi in discussione e modificati man mano che l’evoluzione della scienza permette di osservare i batteri sotto nuovi punti d i vista. Ad esempio, l’esplosione della
genetica molecolare dopo la scoperta del DNA negli anni ’50, ha mostrato affinità impreviste tra specie che invece sembravano distanti. Il risultato è che, ogni tanto, i nomi delle specie vengono cambiati e le classi ficazioni vengono riviste. La definizione di specie batterica è quindi arbitraria e decisa da una commissione internazionale di esperti. Ciò può produrre anomalie.
Il significato delle classificazioni adottate:
• Ceppo: un singolo isolato o la linea (USA 300)
• Tipo: sotto-gruppo di una specie (S. aureus tipo I, II)
• Specie: ceppi correlati (Staphilococcous aureus)
• Genere: specie correlate (Staphilococcus)
• Famiglia: generi correlati (Staphilococcaceae) Quando si isola il batterio da un campione biologico si dice di aver isolato un ceppo batterico, diversi ceppi isolati da campioni
biologici di diversa provenienza costituiscono una specie (es. S.aureus che appartiene al genere staphilococcus cioè all’insieme di specie batteriche che hanno le caratteristiche morfologiche e di comportamento degli stafilococchi.
Ceppo Una specie batterica quindi è costituita da più ceppi differenti. Un ceppo in realtà può essere isolato anche in individu i diversi se è stato trasferito per contagio, in questo caso tutti i membri di un ceppo sono indistinguibili con le tecniche usate per identificarlo. Ceppi
diversi possono differire quando esaminati con metodi adeguati.
Nell’800 sono stati fissati i primi criteri per l’identificazione batterica e nel tempo sono state aggiunti tanti tipi di analisi che hanno
fornito nuovi criteri di classificazione. I passaggi per identificare un batterio sono:
• Esame morfologico 1. Forma 2. Disposizione delle cellule
3. Morfologia delle colonie
• Esame biochimico 4. Struttura della parete batterica 5. Capacità di metabolizzare substrati particolari

23 Microbiologia - Artini
6. Metabolismo
• Altre – caratteristiche peculiari (antibiogramma, tipizzazione fagica, tipizzazione immunologica, etc..)
Morfologia delle cellule batteriche L’esame della morfologia delle cellule batteriche viene effettuato dopo averle colorate con una tecnica che permetta di vederle ed evidenziarne fin dall’inizio alcune caratteristiche. La tecnica di Gram permette di vedere le cellule descrivendone la forma e i rapporti nello spazio e permette di avere una prima e
preziosa informazione sulle caratteristiche della parete batterica. Dopo aver colorato i batteri, si può attribuire loro
un primo nome sulla base della forma delle cellule:
• Sferiche = cocchi
• Bastoncello = bacilli
• A virgola = vibrioni
• A spirale = spirochete
• Bastoncello con estremità rigonfiata = clostridi (dal greco, fuso)
Sulla base di come le cellule si raggruppano, si farà poi un’ulteriore distinzione.
Disposizione delle cellule I cocchi si possono disporre a coppie formando i diplococchi, molti batteri legati a varie polmoniti assumono questa morfologia, altri invece formano catene e si chiamano streptococchi o la forma degli stafilococchi a grappolo.
Morfologia delle colonie Le caratteristiche morfologiche ed organolettiche
(= raccolte con i 5 sensi) di una colonia sono informazioni importanti per l’identificazione dei batteri. L’analisi richiede di valutare l’aspetto generale della colonia, lo spessore, i margini ma
anche l’odore perché alcune specie batteriche emanano un odore caratteristico che può aiutare molto il microbiologo esperto.
A volte è utile valutare la morfologia delle colonie che crescono in fissione, in gelatina nelle provette verticali. Questa modalità di crescita su agar è
detta “a becco di clarino” in brodo e in tubi solidificati per intero con gelatina. In quest’ultimo caso si può avere o non fluidificazione del mezzo per l’attività proteolitica; è importante il modo in cui
evolve la fluidificazione.
Capacità di metabolizzare substrati particolari Col passaggio dal XIX al XX secolo la tecnologia della ricerca si è affinata, permettendo la messa a punto di molte altre tecniche diagnostiche. Quindi nella valutazione delle caratteristiche sono entrate
anche la richiesta di ossigeno, alcune specie hanno bisogno di crescere in atmosfere con poco ossigeno o in totale assenza di ossigeno come gli anaerobi obbligati. Anche la capacità di formare spore, tipica di clostridi e bacilli. La capacità di invadere le cellule eucariotiche e la necessità di
aggiungere alcune forme di nutrienti specifiche nel terreno. Alcune specie infatti hanno bisogno di trovare certi substrati specifici nel terreno di crescita, altre invece crescendo producono prodotti terminali
caratteristici che ne permettono l’identificazione. Per identificare i batteri utilizzando la capacità di metabolizzare substrati particolari si usano delle prove specifiche come:
• Enzimi specifici o Catalasi (Staph+, Strep-): importantissima quando si
ha a che fare con dei cocchi, per identificare se sono streptococchi o stafilococchi, perché a volte la differenza tra le catenelle degli streptococchi e i grappoli degli stafilococchi non è immediatamente evidente.
Caratteristiche colturali Metabolismo
Richiesta di ossigeno (aerobi, anaerobi, microaerofili,
aerotolleranti e anaerobi facoltativi)
Utilizzo di substati specifici come lattosio (SallShilYerl) o citrato
(E.coli -Klebsiella)
Formazione di spore Produzione di determinati
prodotti terminali: prodotti di fermentazione (acidi organici, alcool, ammine,
H2S)
intra-/estracellulari
esigenti/non esigenti

24 Microbiologia - Artini
o Coagulasi (S.aureus+, S.epidermidis-): importante perché permette di distinguere lo stafiloccocco aureo dagli altri stafilococchi. Distinzione importante perché questo è l’unico cocco veramente patogeno che causa infezioni gravi
e pericolose in soggetti perfettamente sani. Gli altri sono saprofiti che possono comportarsi da batteri patogeni opportunisti nei soggetti deboli.
o Ossidasi (neisseria gonorrhoea+) o Ureasi (Proteus+, helicobacter+)
• Antibiogramma
• Tipizzazione fagica
• Profilo acidi grassi
• Immunologiche o Antigeni O-, H-, K-Ag (sierotipizzazione)
o Precipitazione, agglutinazione
Esempio: produzione di catalasi Facilissimo e anche scenografico è il metodo per vedere se un ceppo produce catalasi. La catalasi è un enzima che scinde il perossido di idrogeno (H2O2) in acqua e ossigeno. È un fattore di virulenza importante perché permette agli stafilococchi di
neutralizzare il perossido di idrogeno con cui i macrofagi neutralizzano i batteri fagocitati. Se il ceppo produce la catalasi l’acqua ossigenata aggiunta alla coltura batterica, frigge e fa bollicine per la produzione di ossigeno.
Esempio: produzione di coagulasi La coagulasi agisce sulle proteine del plasma della coagulazione aggregando un coagulo di fibrina intorno al batterio che in questo modo non viene fagocitato.
Test immunologici di sierotipizzazione La sierotipizzazione valuta la presenza di antigeni espressi sulla superficie batterica quindi permette di classificare ceppi batterici
all’interno di una specie. I vantaggi di questa tecnica sono la rapidità, la riproducibilità; in alcuni casi si dimostra una stretta correlazione fra sierotipo, fattori di virulenza e/o evidenza di quadri clinici particolari (es: E.coli O157:H7). La sierotipizzazione viene usata più frequentemente nella tipizzazione di Streptococcus pneumoniae, Legionella pneumophila, Salmonella species, Shigella
species, Haemophilus influenzae, Neisseria meningitidis, Pseudomonas aeruginosa. Gli svantaggi sono: il costo elevato e la non totale disponibilità di antisieri. Il potere discriminatorio per microrganismi appartenenti allo stesso sierogruppo è scarso. Alcuni microrganismi di una determinata specie non sono tipizzabili (Haemophilus influenzae); altri, quali lo stafilococco aureo, non hanno determinanti antigenici sufficientemente vari da essere utilizzati come strumenti di tipizzazione. Quindi è opportuno usare questi test
quando è importante capire se il responsabile di un ’infezione appartiene ad un ceppo particolarmente virulento, per avere una diagnosi completa che permetta di capire quali rischi corre il paziente e come perfezionare la terapia . È importante a volte capire se un ceppo è capace di metabolizzare substrati particolari. Questo può aiutare quando l’identificazione
non è facile. Questo tipo di identificazione è ormai completamente automatizzata e viene affidata ad apparecchiature che leggono la capacità di metabolizzare il substrato specifico misurando il viraggio di colore di un colorante spia, aggiunto nella coltura batterica secondo il principio della cartina tornasole. Purtroppo, in un numero di casi che va da 1 fino a 10%, l’identificazione può non essere
elaborata dal software dell’apparecchio, perché alcuni ceppi possono acquisire capacità metaboliche non tipiche della specie (es. salmonella che fermenta il lattosio, che in genere non succede, viene confusa con un E. coli, che in genere è molto meno aggressivo della salmonella). In questo caso l’identificazione viene affidata ad un microbiologo, che utilizzando tutto il protocollo delle tecniche tradizionali (osservazione macroscopica + microscopica + test enzimatici + caratteristiche sierologiche + caratteristiche genotipiche)
identifica il ceppo, il tutto sempre in stretta collaborazione con i medici del reparto.
Un ultimo accenno alle tecniche più avanzate di identificazione basate sul sequenziamento dei geni, che mutano da una specie
batterica all’altra. Per questo scopo si sequenziano regioni dei geni che codificano per l’rRNA . È il futuro della diagnostica microbiologica, può fornire risposte molto precise in tempi rapidi, almeno per avere un primo orientamento da perfezionare con indagini successive più mirate. Però richiede competenze, laboratori e disponibilità finanziaria che non tutti gli ospedali hanno.
Descritta per la prima volta alla fine degli anni ’70, questa tecnica si è resa disponibile in microbiologia clinica solo recentemente, successivamente alla disponibilità di apparecchiature automatiche. Il sequenziamento a fini identificativi si basa, in genere, sull’amplificazione con primers universali e successiva sequenza del gene che codifica per l’rRNA 16S, presente in tutti i genomi batterici ed estremamente conservato a livello della specie. Altre sequenze geniche utilizzabili possono essere quelle del 23S rDNA,
o quelle di strutture specifiche per determinate specie batteriche. Esempio: geni per la flagellina in Borrelia burgdorferr, i geni sodA, rpoB, hsp60 per stafilococchi coagulasi negativi. La sequenza del 16S rDNA è pari a 1500 bp; quella del 23S rDNA è di 2500 bp . Per l’identificazione di nuove specie batteriche è ovviamente necessario sequenziare l’intero gene, mentre per identificare
microrganismi di specie già descritta è in genere sufficiente sequenziare 500-1000 bp del 16S rDNA.

25 Microbiologia - Artini
Batteriologia speciale Non imparare a memoria la classificazione e tutte le caratteristiche di un batterio, ma focalizzarsi sulle caratteristiche che lo distinguono dagli altri e lo rendono importante per la patogenesi. A volte si possono dedurre informazioni utili considerando anche solo la morfologia: un cocco è Gram+ (solo Neissere sono Gram-), asporigeno (solo bacilli e clostridi producono spore), immobile (solo alcuni enterococchi hanno flagelli), non produce endotossine
(perché sono prodotte solo dai Gram-), e quindi molto probabilmente produrrà esotossine. Tutti i batteri d’interesse medico sono generalmente aerobi/anaerobi facoltativi (tranne Brucelle che sono aerobi obbligati) e pochi gruppi anaerobi obbligati (es. clostridi).
+I cocchi Gram+ che costituiscono l’enorme maggioranza dei cocchi, si dividono in: aerobi ed anaerobi. Le specie di anaerobi sono raramente responsabili di ascessi profondi. Tra gli aerobi si distinguono gli stafilococchi dagli streptococchi. Entrambi includono molte
specie batteriche responsabili di infezioni diffuse ed importanti. Il test della catalasi, visto in precedenza, serve per distinguere gli stafilococchi, che producono catalasi, dagli streptococchi. Se il test è risultato positivo, si eseguirà il test della coagulasi per capire se si è di fronte ad uno stafilococco aureo (patogeno) o uno dei tanti saprofiti coagulasi negativi.
Stafilococchi In generale quindi gli stafilococchi sono Gram+ e risultano positivi al test della catalasi, in coltura liquida si raggruppano nelle
caratteristiche strutture a grappolo, sono immobili e possono vivere in presenza di abbondante o scarso ossigeno. Infine, resistono ad altre concentrazioni di cloruro di sodio (sono alofili), proprietà importante che permette loro di colonizzare la pelle ma anche di contaminare i cibi conservati nel sale. Sono componenti della flora commensale (cute e naso).
Le specie principali patogene per l’uomo sono:
• S. aureus
• S. epidermidis
• S. haemolyticus
• S. lugdunensis
• S. saprophyticus
Staphylococcus aureus Nella foto affianco si possono osservare le caratteristiche
morfologiche fondamentali dello stafilococco aureo. In piastra di Petri, su terreno solido contenente sangue, si vedono le tipiche colonie grandi, bianche e traslucide con alone di emolisi, perché
lo stafilococco aureo lisa i globuli rossi. In un terreno contenente sale e il mannitolo (zucchero particolare), lo stafilococco aureo riesce a crescere e le colonie assumono un caratteristico colore
giallo-oro, perché la digestione del mannitolo abbassa il pH facendo virare al giallo un colorante inserito apposta perché varia colore quando si abbassa il pH.
Comportamenti strutturali e antigenici Lo stafilococco aureo può comportarsi come un’importante patogeno, anche se in anni recenti è stato isolato come commensale nelle cavità nasali di un 20-30% di portatori sani. Alcuni fattori di virulenza dello stafilococco aureo interferiscono molto efficacemente con le difese immunitarie dell’ospite, tra queste:

26 Microbiologia - Artini
• La capsula (11 sierotipi: V e VIII associati a patologie umane);
• Proteine superficiali di adesione: legate al peptidoglicano. o Proteina A, contenuta nella parete cellulare, si lega al frammento Fc delle immunoglobuline di tipo IgG, esplicando
un’azione anticomplementare e antifagocitaria. Non si trova nei coagulasi negativi. o Fibronectin binfing protein A e B
• Discreta capacità di formare biofilm
• Catalasi
• Coagulasi
• Esotossine (pericolose nella prima infanzia)
o Leucocidina e stafilotossina: 5 tossine citolitiche (α, β, γ, δ e leucocidina di Panton-Valetine) che uccidono i globuli bianchi.
o Tossina esfoliativa ET-A, ET-B: sindrome della cute scottata (malattia di Ritter) nella quale la tossina raggiunge lo
strato granuloso dell’epidermide determinandone lo scollamento. o Enterotossina: causa enterocolite (almeno 6 tipi A-F), termostabili e resistenti agli enzimi dell’intestino. Bastano 25
mg di enterotossina perché essa sia efficace a manifestarsi come patogena. o TSST1, tossina della sindrome da shock tossico: causa febbre, per liberazione di IL-1, shock, eruzione cutanea
desquamativa, ipotensione, disfunzioni a livello di organi, morte.
• Enzimi degradativi (usati per diffondere nei tessuti profondi) o Ialuronidasi: fattore di diffusione in quanto rompe l’acido ialuronico presente nel tessuto connettivo. o Lipasi: consente l’idrolisi dei lipidi consentendo al microrganismo di vivere nelle zone sebacee.
o Emolisine (α, β, γ): tossine che lisano gli eritrociti e danneggiano le piastrine. Le infezioni sostenute dall’aureo iniziano sempre con un’infezione localizzata in sede cutanea o mucosa e si propagano in profondità
grazie all’azione combinata del gran numero di fattori di virulenza descritti sopra.
Patogenesi A seconda da quanti fattori di virulenza sono posseduti dal ceppo che causa
l’infezione, si può avere un livello di infezioni e quindi di aggressione e gravità dell’infezione molto differenti. Si distingueranno quindi:
• Infezioni superficiali, limitate alla cute o alle mucose o Mastite, ferite o Follicoliti, foruncoli/favi, impetigine, idrosadenite suppurativa
• Infezioni profonde localizzate o Infezione Osteoarticolari
• Infezioni disseminate o Infezioni locali con effetti a distanza o Sindrome della cute scottata o Sindrome Shok tossico (associata o non alle mestruazioni)
• Infezione sistemica, diffuse a tutto il corpo o ad un distretto importante o Batteriemia/setticemia → endocardite, meningite, osteomielite
o Infezione polmonare
• Intossicazione alimentare, dovute ad ingestione di cibo solo contaminato dall’esotossina enterica (o enterotossina) che causa un’improvvisa e importante enterite diarroica. È un caso nettamente distinto, perché il quadro clinico è dovuto all’ingestione della tossina ma non all’infezione del tessuto da parte del batterio, quindi in questo caso non si somministrano antibiotici.
Patologie • Polmonite
• Osteomielite
• Artrite settica
• Setticemia
• Endocardite
• Ascessi/foruncoli
• Altre infezioni cutanee Questo è uno dei più comuni patogeni nelle infezioni ospedaliere e di comunità.
È importante ricordare che ormai quasi tutti i ceppi acquisiti in infezioni ospedaliere sono resistenti a molti antibiotici, quindi queste infezioni possono diventare quadri clinici molto temibili.
Le resistenze ben note sono:
• MRSA: ceppi con elevata resistenza alla Meticillina, vecchio antibiotico non utilizzato da anni che viene

27 Microbiologia - Artini
utilizzato nell’antibiogramma come spia di resistenza ad un ampio ventaglio di antibiotici differenti che comprende praticamente tutti i tipi di antibiotici con anello β-lattamico. Fino a qualche anno fa la Vancomicina era l’unico antibiotico
attivo su questi ceppi Meticillina resistenti, ma ormai si stanno diffondendo ceppi resistenti anche alla Vancomicina.
• VISA: ceppi con resistenza intermedia alla Vancomicina
• VRSA: ceppi con elevata resistenza alla Vancomicina
Infezioni cutanee
Favo: infezione localizzata con molto pus che colpisce più
foruncoli contigui confluenti e scende in profondità, invadendo spesso il torrente circolatorio, con febbre e brividi.
Impetigine: tipica nei bambini, coinvolge soprattutto il volto e gli arti, parte da una singola lesione purulenta che viene trasferita gradualemnte per contatto in siti adiacenti nella pelle
Sindrome della cute scottata: colpisce prevalentemente bambini o adulti immunocompromessi, è dovuta ad un’esotossina esfoliativa che causa lo scollamento del derma
dall’epidermide.
Stafilococchi coagulasi negativi (CNS) Sono saprofiti commensali ubiquitari poco virulenti: si trovano sulla cute e sulle mucose dell’uomo e possono comportarsi occasionalmente da patogeni opportunisti perché riescono a causare infezioni in soggetti immunodepressi o perché riescono a
colonizzare una protesi profonda o un dispositivo impiantabile temporaneo (es. catetere venoso centrale), formando biofilm. La loro diffusione li rende patogeni in conseguenza di quasi tutte le manovre chirurgiche che mandano in circolo questi batteri. Freq uenti sono infatti: le endocarditi, le infezioni localizzate nel peritoneo o nei reni in soggetti drenati, infezioni di protesi o valvole cardiache.
Presentano diffusa farmaco-resistenza, rivolta in molti casi a più farmaci. Questi batteri non producono coagulasi, non fanno alone emolitico in terreni contenente sangue (non emolizzano) e non fermentano il mannitolo. Le specie più frequenti sono:
• Staphylococcus epidermidis → il principale saprofita della cute che causa infezioni su protesi artificiali, valvole cardiache, cateteri, osteomieliti, etc.
• Staphylococcus saprophyticus → responsabile soprattutto di infezioni del tratto urinario
Terapia S. aureus è resistente a molti antibiotici e la velocità con cui sviluppa resistenza è preoccupante.
• Penicilline e cefalosporine non sono più efficaci,
• β-lattamine protette
• glicopeptidi: vancomicina e teicoplanina
• emergono ceppi resistenti Molti coagulasi negativi invece sono resistenti a tutti gli antibiotici conosciuti e rappresentano un problema per immunocomp romessi e degenti in terapia intensiva.
Streptococchi La caratteristica morfologica degli streptococchi è la disposizione in
catenelle, soprattutto quando sono in terreno liquido, oppure in coppie, come si osserva frequentemente nel responsabile della polmonite batterica (Strep pneumoniae, diplococco lanceolato). Quindi sono Gram positivi appartenenti a una parte della flora microbica
saprofita, sono negativi al test della catalasi, sono aerobi ma possono facoltivamente adattarsi a condizioni di carenza di ossigeno, spesso sono capsulati e includono alcune specie di patogeni franchi.

28 Microbiologia - Artini
Classificazione Per l’identificazione e la classificazione si riparte dalla divisione dei cocchi Gram+ in base al test della catalasi. Con un test negativo si hanno gli streptococchi,
mentre è positivo per gli stafilococchi perché possiedono questo enzima capace di neutralizzare il perossido di idrogeno dei macrofagi. Gli streptococchi vengono fatti crescere in terreno solido contenente sangue, per
verificare se il ceppo in esame ha proprietà emolitiche. Quindi gli streptococchi sono classificati in base a:
• tipizzazione sierologica 2specificità sierologica (gruppi di Lancefield: da A a W)
• tipo di emolisi su agar sangue o α emolisi – Streptococcus pneumoniae, S. viridans e alcuni S.
faecalis– Causano un’emolisi incompleta delle emazie
alterando l’emoglobina che assume un colore verdastro, per questo motivo questi streptococchi sono chiamati viridanti.
o β emolisi – Streptococcus pyogenes, S. haemolyticus – Causano un’emolisi completa delle emazie, con
formazione di un alone chiaro e netto intorno alle colonie. o γ emolisi – S.faecalis, non patogeni – non lisano le emazie e non causano modifiche nel terreno in agar sangue.
• Reazioni biochimiche Ma dal punto di vista pratico-clinico conviene ragionare distinguendo:
• Streptococchi β emolitici → classificati secondo Lancefield in tipi diversi a seconda del polisaccaride C della parete: o 18 tipi diversi (da A a V) o Di maggior rilievo clinico: S.pyogenes di gruppo A e S.agalactiae di gruppo B
• Streptococchi α emolitici e γ emolitici → classificati per caratteristiche biochimiche
• Streptococchi enterici (enterococchi)
Fattori di virulenza Ciò che rende alcuni streptococchi particolarmente temibili sono i fattori di virulenza.
Streptococcus pyogenes È uno streptococco che produce emolisi β in agar sangue ed appartiene al gruppo A di Lancefield. È la specie clinicamente più importante di questo genere, uno dei patogeni più diffusi. Un inoculo di scarsa entità riesce ad invadere la cute o mucose
apparentemente intatte, causando infezioni rapidamente progressive. Vi rimane nell’ambiente, quindi pazienti infetti o portatori sani. La trasmissione avviene tramite aerosol-respiratorio oppure con lesioni della pelle tramite contatto diretto. Può colonizzare l’orofaringe, senza dare malattia, quindi ci sono molti portatori sani.
Fattori di virulenza: • Strutture legate alla parete che favoriscono l’adesione alle cellule dei tessuti umani e impediscono la fagocitosi:
o Capsula costituita da acido ialuronico – attività antifagocitaria
2 Ricerca di antigeni nella parete batterica che permettono di classificare un ceppo in vari gruppi secondo la classificazione di Lancefield

29 Microbiologia - Artini
o Proteina M (antigene superficiale) – 80 tipi sierologicamente diversi, responsabili di malattia reumatica (classe I) e glomerulonefriti (classe I e II).
• Esotossine o Streptolisina S – è responsabile dell’emolisi quindi può lisare eritrociti, leucociti e piastrine.
o Streptolisina O – distrugge le membrane cellulari, compare presto (titolo anti-streptolisinico) o Esotossine eritrogeniche pirogene (SPE) – responsabile dell’esantema, nella scarlattina, e delle forme più acute.
Sono antigeniche e stimolano la produzione di anticorpi neutralizzanti.
• Enzimi che digeriscono componenti della matrice interstiziale favorendo l’invasione dei tessuti (Ialuronidasi, streptochinasi , DNAsi).
Patogenesi e manifestazioni cliniche Le manifestazioni cliniche di questo batterio sono tante e diverse a seconda di quanti e quali fattori di virulenza sono presenti nel ceppo batterico isolato da un paziente.
Un primo gruppo è costituito dalle malattie di tipo suppurativo. Con questo termine s’intende manifestazioni acute, immediatamente successive al contagio e veloci, localizzate nella sede colonizzata dai batteri dove c’è produzione di pus abbondante. Suppurativo infatti significa con produzione di pus, cioè globuli bianchi morti. Quindi si possono avere infezioni:
• localizzate a livello di gola e naso quando il contagio è avvenuto per via aerea tramite l’aerosol di un altro individuo infetto o Faringite (faringe, tossile) → scarlattina o Otite (orecchio medio) o Sinusite (seni nasali)
• Della cute, quando l’infezione si diffonde a livello cutaneo. Le Infezioni suppurative dei tessuti molli sono dovute a una colonizzazione iniziale della cute seguita dalla penetrazione del batterio nei tessuti superficiali e profondi, in seguito a lesioni
cutanee che causano quadri diffusi. o Impetigine (Piodermite) o Erisipela o Fascite necronizzante, causata da ceppi definiti “mangia carne” perché distruggono e decompongono alcuni tessuti
profondi fino ai piani muscolari in un tempo rapidissimo. Naturalmente sia dalle sedi orofaringee che da quelle cutanee i batteri possono penetrare fino invadere i vasi sanguigni disseminandosi in circolo fino a causare la setticemia.
Il secondo gruppo è quello delle manifestazioni cliniche non suppurative dovute ad un danno tardivo, causato su tessuti distanti dalla sede dell’infezione. In questo caso, il meccanismo della malattia è totalmente diverso da quello delle malattie suppurative perché il danno non è dovuto all’aggressione diretta da parte dei batteri ma dal coinvolgimento di tipo infiammatorio dovuto all’accumulo di
immunocomplessi circolanti, come avviene nell’artrite e nella glomerulonefrite, oppure ad una reazione di tipo autoimmune dovuta al fatto che anticorpi rivolti ad antigeni dello streptococco legano antigeni delle valvole cardiache scatenan do una reazione infiammatoria e immunitaria contro i tessuti del cuore. Quindi si ha:
• Febbre reumatica: o Cuore: pancardite o Articolazioni o Noduli sottocutanei
• Glomerulonefrite acuta o Infiammazione dei glomeruli
o Edema o Ipertensione o Ematuria e proteinuria
Diagnosi La diagnosi viene fatta mediante isolamento in terreni arricchiti con sangue, in cui si dovrà vedere l’emolisi β. Si può aggiungere l’esecuzione di test sierologici per identificare i gruppi secondo la classificazione di Lancefield (precipitazione, agglutinazione, immunofluorescenza) ma una diagnosi più veloce può essere posta usando sonde tecniche di biologia molecolare che identificano
sequenze del genoma possedute esclusivamente dallo S.pyogenes.
Un metodo per differenziare lo streptococco β-emolitico di gruppo A dagli altri β-emolitici (B, C, G) può essere attuato testando la sensibilità alla bacitracina.
Streptococcus agalactiae È β-emolitico ma fa parte del gruppo B. Esso fa parte della flora saprofita della vagina, uretra, retto e occasionalmente faringe ma sta diventando sempre più spesso responsabile di infezioni urinarie multiresistenti ma anche meningiti, endocarditi, sepsi, etc,
soprattutto in soggetti anziani o con patologie. La comparsa di ceppi multiresistenti di S.agalactiae sta trasformando questa specie innocua in un patogeno occasionale. Aumenta il rischio di sepsi post partum in portatrici. Infezioni gravi da S. di gruppo B si verificano nei neonati che si infettano in utero,
durante il parto o nella madre/in ospedale.

30 Microbiologia - Artini
Streptococchi viridanti o α-emolitici Essi in agar sangue emolizzano parzialmente i globuli rossi, quindi le colonie e il terreno circostante si tingono di verde. Nei nostri
distretti esterni (vie respiratorie, vie genitourinarie, intestino, etc.) questi batteri costituiscono una quota importante della flora saprofita, ma alcune specie possono comportarsi da patogeni franchi se riescono a guadagnare gli strati più interni. Sono esigenti, richiedono supplementi del sangue e cofattori, spesso microaerofili e la maggior parte non ha un antigene specifico. Sono la causa
più comune di endocardite subacute. Vengono associati a infezioni suppurative addominali e carie dentale. Inizialmente molto sensibili a penicillina e derivati, sono diventati moderatamente/altamente resistenti.
Streptococcus pneumoniae Nella colorazione di Gram di lato si osservano le macchie rosse, che sono polimorfonucleati neutrofili coinvolti nella risposta immunitaria, e i batteri scuri sono S.pneumoniae. Sono aggregati questi batteri tipicamente in brevi
catenelle o in coppie (come in figura) e in questo caso le due cellule hanno la forma di punta di freccia rivolta verso l’esterno. Le cellule sono:
• immobili e capsulate, come si può intuire dall’alone chiaro che si nota nel cerchio azzurro,
• negative al test della catalasi, come tutti gli streptococchi
• in agar sangue producono un alone di emolisi α, cioè di colore verdastro.
Lo S.pneumoniae:
• È presente nel tratto respiratorio di circa la metà della popolazione.
• Se invade il tratto respiratorio inferiore può causare polmonite e avviene spesso in seguito ad infezione virale (infezioni virali favorenti).
• È causa del 95% di tutte le polmoniti batteriche.
• È responsabile di sinusiti, otiti medie, meningiti, betteriemie. La colonia che forma S.pneumoniae è ampio-mucoide, caratteristica tipica dei batteri che producono capsula. Essa rappresenta il principale fattore di virulenza di questa specie, capace di inibire la fagocitosi dei batteri dai macrofagi. Gli antigeni capsulari permettono la classificazione di questi ceppi in moltissimi tipi, ma non sono classificabili secondo Lancefield. Diagnosi
L’identificazione con siero ematico degli antigeni capsulari è usata per la diagnosi, naturalmente dopo l’esame microscopico diretto dei batteri di campioni biologici di pus o espettorato bronchiale. Per la dimostrazione della capsula di fa la reazione di quellung, in cui la capsula si rigonfia dopo che i batteri sono stati cimentati con anticorpi contro i suoi antigeni polisaccaridici.
Sensibilità alla bile e all’optochina. Terapia
Anche per questa specie vengono usati antibiotici β-lattamici per la terapia, ma stanno aumentando i ceppi resistenti. La maggior parte dei ceppi di S.pneumoniae sono sensibili alla penicillina G. Lo sviluppo di resistenza è al 5-10% (MIC>2). Moderatamente resistenti 20% (MIC = 0,1-1).
Vaccino antipneumococco È un vaccino coniugato 3e basato sull’inoculazione di un mix di polisaccaridi capsulari ottenuti da vari (13-23) tipi di pneumococchi. Sono consigliati soprattutto per la III età perché lo pneumococco è ancora di gran lunga il principale responsabile di polmoniti che
nella III età possono risultare fatali. Ce ne sono due tipi:
• Il 13-valente è protettivo nei confronti di 7 sottotipi responsabili della quasi totalità dei casi di meningite e sepsi da pneumococco. È il vaccino più indicato in tutte le età, compresa la prima infanzia (>2 anni).
• Il 23-valente, utilizzabile sono nei bambini di oltre 2 anni d’età e negli adulti.
Pseudomonas aeruginosa è una specie appartenente al genere pseudomonas, che letteralmente significa falsamente uno perché le cellule di questi bacilli Gram- si presentano in coppie talmente tanto ravvicinate da sembrare ingannevolmente un solo batterio. Gli Pseudomonas sono microrganismi ubiquitari, si trovano nel suolo, nella materia organica in
decomposizione, nelle piante e nell’acqua, pertanto non necessitano di terreni arricchiti per la coltivazione in vitro. Questa vasta distribuzione ambientale è possibile grazie alle semplicissime esigenze di crescita. Infatti, crescono bene in un ampio intervallo di temperature (4-42 °C). Non
possono essere definiti saprofiti perché non risiedono tra i commensali del microbioma presente sul corpo umano, ma le caratteristiche di adattabilità e le scarse esigenze nutritive unite alla notevole resistenza ai disinfettanti e antibiotici permettono a questi batteri di comportarsi da patogeni occasionali nei soggetti immunodepressi o indeboliti e nei portatori di endoprotesi artificiali.
Caratteristiche generali:
• Bacilli Gram-, aerobi, mobili (flagelli).
3 Legato cioè ad una proteina per aumentarne l’efficacia

31 Microbiologia - Artini
• Ossidasi-positivi.
• Non fermentanti; utilizzano solo alcuni carboidrati (glucosio, ribosio e gluconato) durante i processi metabolici ossidativi.
• Alcuni ceppi appaiono mucoidi per un’abbondante capsula polisaccaridica formata da alginato
Pseudomonas aeruginosa È la specie più frequentemente coinvolta in patologia umana. Presenta lunghi flagelli raccolti in un polo, che permettono a q uesto
bacillo Grma- di spostarsi, e pili che circondano il corpo cellulare. Questo non è un patogeno stretto ma è responsabile di infezioni che in genere vengono contratte in ambiente ospedaliero e comunque sono contratte da soggetti che abbiano condizioni di debolezza che predispongono all’infezione. Sono responsabili di infezioni nosocomiali e sono resistenti a molti antibiotici e chemioterapici. Alcuni ceoppi producono il colorante fluorescente detto fluoresceina. La fluorescenza al buio sotto luce UV permette l’identi ficazione
precoce di P.aeruginosa, sia in coltura che nei tessuti infettati.
Alcuni ceppi producono piocianina e pioverdina, pigmenti solubile in acqua, che danno agli essudati una leggera sfumatura blu -
verde.
Fattori di virulenza: Componenti strutturali
Sono quelle di definizione dei fattori di virulenza, quindi favoriscono l’adesione. Tra di essi:
• Pili e flagelli, permettono l’attacco alle cellule epiteliale dell’ospite.
• Capsula polisaccaridica di alginato, favorisce l’adesione alla cellula ospite in pazienti affetti da malattie respiratorie e possiede attività antifagocitaria.
• Lipopolisaccaride (LPS). Enzimi Gli enzimi vengono prodotti per degradare i componenti del tessuto interstiziale e per facilitare la progressione dei batteri in profondità:
• Elastasi: degradano l’elastina con danno al parenchima polmonare e lesioni emorragiche
• Fosfolipasi C: degradano lipidi e lectina facilitando la distruzione dei tessuti
• Proteasi alcalina: contribuisce alla distruzione dei tessuti e alla diffusione
• Piocianina: catalizza la produzione di superossido e di perossido di idrogeno, forme tossiche dell’ossigeno . Tossine
• Endotossina (lipide A-LPS): media i diversi effetti biologici della sindrome setticemica
• Esotossina A: inibisce la sintesi proteica nelle cellule eucariote
• Citotossina (Leucocidina): citotossica per le membrane eucariotiche perché impedisce ai leucociti di agire, produce lesioni microvascolari polmonari.
Antibioticoresistenza
È un batterio intrinsecamente resistente a molti antibiotici e può acquisire geni oppure mutare, durante la terapia, originando cappi ancor più resistenti. Si distinguono classicamente, per questo batterio, 3 forme di antibioticoresistenza:
• Intrinseca: dovuta a scarsa capacità di movimento degli antibiotici attraverso i pori della membrana esterna e rapida eliminazione dalle pompe di efflusso. (resistenza fenotipica)
• Acquisita: trasferimento orizzontale di geni di resistenza plasmidici o comparsa di mutazioni. (resistenza genotipica)
• Adattativa: indotta quando è esposto a stimoli ambientali, formazione di biofilm e produzione di β-lattamasi che inattivano
antibiotici β-lattamici. (resistenza fenotipica)
Flagelli unipolari
foto al SEM Pili

32 Microbiologia - Artini
Infezioni È un patogeno opportunista un microrganismo che generalmente provoca infezioni umane in particolari condizioni (traumi, ustioni, impianti protesici, individui immunocompromessi) e dopo antibiotici. Può causare infezioni cutanee, osteoarticolari, delle basse vie
respiratorie, urinarie oculari, auricolari, vascolari. In tutti questi distretti deve essere presente un qualche fattore pred isponente per causare l’infezione.
• Infezioni cutanee: le più frequenti avvengono su cute ustionata. Il microrganismo colonizza la superficie nuda delle ustioni provocando danno vascolare localizzato, necrosi tissutale e batteriemia.
• Infezioni osteoarticolari: si possono verificare come complicanza infettiva di interventi chirurgici o più spesso come conseguenza della diffusione ematogena del batterio in soggetti che fanno uso di sostanze stupefacenti o in pazienti
diabetici.
• Infezioni polmonari: possono causare quadri che variano da una colonizzazione delle basse vie respiratorie o una tracheobronchite benigna a broncopolmoniti necronizzanti. Si manifestano in soggetti con cause predisponenti locali (pazienti intubati nell’unità intensiva) o generali (soggetti immunodepressi). I soggetti di fibrosi cistica presentano un’anomalia genetica che causa il malfunzionamento delle pompe del cloro in tutte le cellule del corpo umano, da cui deriva
la produzione di secrezioni e tenaci in tutte le mucose. Il muco bronchiale denso predispone alla colonizzazione delle vie aeree di molte specie batteriche tra cui prevale P.aeruginosa. Sono pazienti che muoiono presto per infezioni croniche recidivanti e intrattabili per via aerea.
• Infezioni urinarie: si riscontrano principalmente in pazienti con catetere urinario. Pielonefrite come conseguenza di un’infezione ascendente delle basse vie urinarie; pielonefriti ed ascessi perirenali per via ematogena negli ustionati.
• Infezioni oculari: sono complicanze post-operatorie e sono rappresentate da cheratiti ulcerose. Si verificano in seguito ad esposizione allo P.aeruginosa presente nell’acqua contaminata dopo un trauma della cornea (abrasioni dovute a lenti a contatto, graffi alla superficie dell’occhio, ulcere corneali).
• Infezioni auricolari: otiti dell’orecchio esterno e otiti croniche dell’orecchio medio, dovute a prolungata esposizione a liquidi abbondantemente contaminati. L’infezione si può estendere ai tessuti sottostanti, può danneggiare nervi cranici e le ossa e può risultare letale.
• Batteriemia ed endocardite: la batteriemia non è distinguibile da quella causata da altri batteri Gram-, tuttavia il grado di mortalità tra i pazienti affetti è più alto. La batteriemia si verifica in pazienti affetti da neutropenia, diabete, ustioni diffuse.
L’endocardite è più comune in soggetti che fanno uso di stupefacenti.
La famiglia delle Enterobacteriaceae comprende moltissime specie di batteri Gram- che vivono come saprofiti commensali
nell’intestino dell’uomo (coliformi) e di altri animali, ma sono microrganismi ubiquitari. Le caratteristiche comuni di questo genere sono:
• Bacilli Gram-, asporigeni
• la possibilità di movimento per le specie che esprimono flagelli peritrichi,
• la presenza di pili che permettono l’adesione alla mucosa intestinale,
• crescono su terreni selettivi (MacConkey agar) e non selettivi (agar sangue).
• la possibilità di vivere sia in presenza che in carenza di ossigeno e in questo caso la possibilità di fermentare il glucosio e altri polisaccaridi con produzione di acidi e gas.
Tutte le enterobacteriaceae posseggono 3 tipi di antigeni presenti sulla loro superficie che permettono la loro identificazione: ➢ Antigene O: porzione polisaccaridica del lipopolisaccaride (LPS), esistono oltre 150
differenti antigeni somatici O.
➢ Antigene H (flagellari): è rappresentato dalle proteine flagellari; ne esistono oltre 50 tipi differenti.
➢ Antigeni K (capsulari): all’esterno degli antigeni O in alcune enterobacte riaceae, alcuni
sono polisaccaridi mentre altri sono proteine; esistono oltre 100 differenti antigeni K.
Fattori di virulenza • Endotossina (lipide A del LPS)
• Capsula, protegge i microrganismi dalla fagocitosi
• Variazione di fase: gli antigeni capsulare K e flagellare H possono essere alternativamente espressi o non espressi proteggendo così il batterio dalla morte cellulare anticorpo-mediata.
• Sistemi di secrezione di tipo III: siringa molecolare che inietta i fattori di virulenza nella cellula
• Sequestro dei fattori di crescita: composti chelatanti il ferro, siderofori che sequestrano il ferro dai globuli rossi ma anche emolisine che agiscono sui globuli rossi.

33 Microbiologia - Artini
Generi più importanti dal punto di vista medico Dal punto di vista della capacità di produrre malattia, le varie specie di enterobacteriaceae possono essere
divise in 3 gruppi:
• Patogeni stretti: specie che sono sempre associate a patologie dell’uomo (Salmonella spp., Shighella spp., Yersinia pestis).
• Saprofiti: sono i membri della normale flora commensale, che possono comportarsi occasionalmente da patogeni in soggetti denutriti o immunodepressi (Escherichia coli, Klebsiella pneumoniae, Proteus mirabillis). In questi casi si parla di infezioni opportunistiche.
• Saprofiti trasformati in patogeni stretti: specie che sono innocue ma diventano aggressive in seguito all’acquisizione di DNA accessorio plasmidico codificante per fattori di virulenza importanti. Esempio: E.coli EPEC, che pur appartenendo a una specie commensale saprofita, riescono a fare gravi danni nella mucosa dei soggetti infettati.
Escherichia coli Sono bacilli Gram-, asporigeni che possono avere la capsula, possono essere mobili quando esprimono il flagello e sono anaerobi facoltativi (fermentano glucosio e lattosio). Crescono bene nelle condizioni di temperatura e di pH dei d istretti del corpo umano che colonizzano (37°C e pH = 7-7,4). L’identificazione è relativamente semplice e può essere fatta su dive rsi tipi
di terreni non selettivi e selettivi. È uno dei principali commensali della flora saprofita intestinale, ma alcuni ceppi sono provvisti di fattori di virulenza importanti presenti sul genoma accessorio plasmidico. Questi ceppi sono considerati dei temibili patogeni.
Classificazione La classificazione in sierotipi è effettuata sugli antigeni O e sulla base degli antigeni H e K, ha permesso di identificare alcuni ceppi patogeni ben caratterizzati e responsabili di quadri molto gravi diffusi soprattu tto nei paesi in via di sviluppo. Essa è basata quindi sui fattori di virulenza:
➢ Fattori implicati nell’adesione alle cellule, come le fimbrie
➢ Produzione di esotossine ➢ Fattori presenti su elementi genetici mobili.
ENTEROTOSSIGENI
ETEC Intestino tenue
Diarrea del viaggiatore, diarrea infantile,
vomito, crampi, nausea febbre
Enterotossine (plasmidi) ipersecrezione
di fluidi ed elettroliti LT-I, LT-II, ST1, ST-II
ENTEROINVASIVI
EIEC intestino crasso
Malattia nei paesi in via di sviluppo,
febbre, crampi, diarrea, feci con sangue
Invasione e distruzione delle cellule
epiteliali del colon plasmide pINV
ENTEROPATOGENI
EPEC Intestino tenue
Diarrea infantile con febbre, nausea,
vomito e feci non sanguinolente
Adesione e distruzione delle cellule
epiteliali. L’adesione mediata da una proteina: intimina
ENTEROEMORRAGICI EHEC
Intestino crasso
Coliti emorragiche, diarrea acquosa e sanguinolenta, poca febbre
Inibizione della sintesi proteica mediata da tossine citotossiche (SLTI, SLTII,
codificate da batteriofagi lisogeni
ENTEROAGGREGANTI EAggEC
Intestino tenue
Diarrea infantile in paesi in via di sviluppo, diarrea acquosa persistente
con vomito, disidratazione e febbre
Adesione aggregante plasmide mediata che previene l’assorbimento dei fluidi
Le caratteristiche comuni sono sottolineate nella tabella: tutti causano sindromi gastroenteriche acute con diarrea e febbre. Molti ceppi colpiscono i bambini soprattutto se malnutriti.
Salmonelle Caratteristiche:
• Bacilli Gram-,
• aerobi-anaerobi facoltativi,
• sporigeni,
• capsulati,
• dotati di molti flagelli peritrichi,
• non fermentanti,
• crescono facilmente in comuni terreni di coltura,
• sono inattivati da cottura ebollizione, pastorizzazione e irradiazione;
• la struttura antigenica è complessa (>2000 sierotipi): o Ag H, flagellari, proteici, inattivati dal calore
Figura 3: ceppo EPEC che hanno distrutto i villi intestinali della mucosa del colon e contraggono rapporti intimi di legame con i monconi di alcuni microvilli

34 Microbiologia - Artini
o Ag O, somatici, lipopolisaccaridici, nella parete, termostabili o Ag K (Vi), nella capsula, glicoproteici, principale Ag Vi.
Le principali specie patogene per l’uomo sono: S.typhi, S.paratyphi, S.typhimurium, S.choleraesuis. Le prime due fortemente adattate all’uomo e non patogene per gli animali, sono responsabili dei quadri più gravi, soprattutto S.typhi. Le altre due invece sono fortemente adattate agli animali ma quando infettano l’uomo possono causare malattia, sono le salmonellosi. Fatta eccezione per S.typhi, responsabile della febbre tifoide (quadro clinico più grave), la carica infettante deve essere 10 5-108
quindi alta ed è contrastata dall’acidità gastrica, la normale flora microbica intestinale, l’immunità locale intestinale sono le difese dell’ospite. La trasmissione avviene tramite ingestione di cibi contaminati (pollame, uova, prodotti caseari) oppure per via diretta oro -fecale nei
bambini. È più frequente nei mesi estivi e autunnali e la diffusione è ubiqu itaria in tutto il globo, ma soprattutto nelle zone a basso standard igienico. I serbatoi son rappresentati da animali (S.typhiumurium, S.cholereaesuis, etc) e l’uomo (S.tiphy e paratyphi). Il trattamento prevede la somministrazione di antibiotici e la reintegrazione idrosalina che vengono abbondantemente perduti con la
diarrea.
Klebsiella È sempre stato considerato un saprofita commensale delle prime vie aeree respiratorie e del tratto intestinale, ma ad oggi è considerato un patogeno opportunista responsabile di molte infezioni ospedaliere in soggetti immunocompromessi, infezioni difficili
da curare per l’enorme diffusione di ceppi resistenti agli antibiotici. Ha caratteristiche simili alle altre enterobacteriaceae da cui si distingue per l’immobilità e per un’abbondante capsula polisaccaridica che è causa del tipico aspetto mucoso delle colonie e della virulenza del microrganismo.
Klebsiella pneumonie è la specie responsabile di: polmoniti contratte in ambiente ospedaliero, infezioni del tratto urinario, otiti, sinusiti, meningiti.
Vibrio cholerae È il patogeno responsabile del Colera, quadro clinico di gastroenterite simile a quelli visti fino ad ora. In realtà esso app artiene alla
famiglia delle vibrionaceae, quindi batteri differenti dagli enterobatteri visti finora. La morfologia è comunque bastoncellare ma tipicamente ricurva a virgola, da cui il termine vibrio. È un Gram-, mobile per un flagello polare, anaerobio facoltativo con scarse richieste nutrizionali (→ facile coltivazione), cresce a pH = 8,5-9 in presenza di NaCL 6% (alofilo). Questa ultima caratteristica lo
rende particolarmente resistente in molti ambienti, inclusa l’acqua di mare. Naturalmente è endemico in paesi a basso standard igienico-sanitario ed è responsabile di epidemie. La trasmissione è dovuta all’eliminazione di batteri con le feci dei malati che contaminano acque dolci o salate che a loro volta
contaminano cibi assunti dai soggetti sani. I ceppi responsabili del colera classico sono i ceppi di gruppo O1 e O139, che esprimono la loro patogenicità grazie a potent i fattori di virulenza di cui sono dotati. Tra questi:
• Tossina colerica → ipersecrezione di elettroliti e acqua
• Pilo, adesina → adesione alle cellule mucose
• Flagello → mobilità
• Siderofori → cattura ferro
Patogenesi e anatomia patologica Vibrio cholerae è patogeno solo per l’uomo e, se ingerito con il cibo, basta un numero di batteri molto basso per il contagio (10 2-104)
perché il cibo tampona l’acidità gastrica che danneggerebbe i batteri. Se invece il veicolo contaminato è l’acqua, la carica infettante è pari a 108-1010 batteri ingeriti. I batteri non arrivano mai al torrente circolatorio, quindi non è un’infezione invasiva, ma si ferma solo al tratto gastrointestinale dove i batteri aderiscono ai microvilli delle cellule epiteliali mediante i pili, si moltiplicano e liberano la tossina colerica.
Tossina colerica In seguito alla colonizzazione della mucosa, i vibrioni iniziano a secernere una tossina che viene internalizzata dalle cellule intestinali e causa la
perdita di ioni dalle cellule verso il lume intestinale. Ne segue una perdita massiva di acqua per osmosi nella stessa direzione che produce una diarrea imponente. Il primo presidio medico infatti in questi pazienti è l’infusione immediata e massiccia di liquidi e di sali per ricostruire il
patrimonio idrosalino, contemporaneamente viene avviata la terapia antibiotica. Il meccanismo d’azione specifico della tossina è il seguente:
• La tossina colerica si lega ad un recettore delle cellule dell’epitelio
• La tossina internalizzata stimla la produzione di cAMP
• cAMP attiva i canali ionici di membrana
• si ha massiccia perdita di ioni NA+, Cl-, K+ nel lume intestinale
• perdita massiva di acqua per osmosi.

35 Microbiologia - Artini
Vaccino orale È disponibile un vaccino inattivato, costituito da una sospensione di vibrioni del colera uccisi. Il vaccino è efficace contro le infezioni
sostenute da Vibrio cholerae di gruppo O1, ma non per il gruppo O139 o altre specie di vibrione.
I batteri appartenenti a questo genere si classificano in base a:
• acido resistenza;
• elevata concentrazione di G:C (61-71%) nel loro DNA;
• presenza di acidi micolici nella parete La parete cellulare di questi batteri infatti è provvista, dalla parte più esterna, di uno spesso strato di lipidi detti anche cere costituiti in gran parte da acidi micolici e arab inogalattani che
conferiscono caratteristiche di resistenza agli acidi, ai detergenti, ai comuni disinfettanti nonché un’estrema lentezza nella crescita e un’elevata capacità di evocare una risposta immunitaria da parte dell’ospite (antigenicità elevata). Tutte caratteristiche che causano una robusta
risposta infiammatoria e immunitaria; per altro spesso incapace di eliminare il patogeno e la combinazione della robustezza della risposta e della persistenza dello stimolo antigenico è la causa del danno di organo.
Mycobacterium tuberculosis Mycobacterium tuberculosis è il principale responsabile della tubercolosi nell’uomo. Le sue caratteristiche sono le seguenti:
• appare al microscopio come un bastoncello immobile
• debolmente Gram+ ma in realtà non può essere considerato né positivo né negativo alla colorazione di Gram, viene quindi ben identificato con una tecnica di colorazione che prevede un passaggio di decolorazione con una soluzione acida che danneggia tutti gli altri batteri ed evidenzia solo i Mycobatteri che sono acido resistenti grazie alle cere della parete. Questa
è la colorazione di Ziehl-Neelsen.
• È aerobio obbligato
• Asporigeno
• Non produce tossine
• Mesofilo
• Tempo di duplicazione cellulare molto lento: 15-20 h.
• È un batterio piuttosto loffio rispetto ai comuni patogeni ed è praticamente privo della maggior parte dei fattori di virulenza ma ha una combinazione di caratteristiche particolari che lo rendono sfuggente rispetto alle difese immunitarie e lo
trasformano in uno dei principali patogeni tra i più diffusi del globo. Mycobacterium tuberculosis riesce a sopravvivere nei macrofagi, bloccando i sistemi con cui queste cellule digeriscono ed eliminano i patogeni, sopravvivendo quindi e replicando in tutta tranquillità nel fagosoma. Interferisce quindi con la fagocitosi:
o Reagisce con vari ligandi per promuovere la fagocitosi
o Previene la fusione tra fagosoma e lisosoma o Usa il vacuolo fagocitario per sopravvivere e replicare
Tubercolosi La tubercolosi è una malattia infettiva acuta e cronica che non è causata da una sola specie batterica ma dalle tre specie raggruppate nel Mycobacterium tuberculosis complex:
• Mycobacterium hominis (bac. Di Koch),
• Mycobacterium africanum (si trova prevalentemente in Africa),
• Mycobacterium bovis (colpisce prevalentemente i bovini). Sono specie molto simili tra loro e probabilmente provengono da un ceppo ancestrale comune. In prima battuta è quasi sempre colpito l’apparato respiratorio e i linfonodi ad esso collegati, ma in caso di cronicizzazione e diffusione della malattia possono essere colpiti praticamente tutti gli organi.
Contagio Il contagio avviene quasi sempre da parte di un individuo con malattia polmonare in fase attiva (tubercolosi polmonare aperta) ed è dovuto al passaggio di goccioline di espettorato, quindi per via aerea. Il batterio può essere trasferito anche: con la polvere ambientale in cui vi resiste anche per settimane, per via alimentare (m.bovis dal latte di bovini infetti), per via cutanea con materiale infetto (urine
o pus eliminato da focolaio di tubercolosi ossea). In ogni caso il contagio è più probabile se è alta la carica di batteri ingeriti e se il ceppo è virulento, ma anche se il soggetto presenta le difese immunitarie ridotte.
Meccanismo dell’infezione Questo batterio non dispone di forti fattori di virulenza, tipici degli altri patogeni, ma è circondato da uno spesso strato di cere che facilitano l’entrata nei macrofagi, bloccano i sistemi che dovrebbero distruggerlo e lo rendono resistente ad alcune classi di antibiotici. Infine, la replicazione lentissima non attiva in prima battuta una forte risposta immunitaria e riduce l’efficacia degli antibiotici, dando
ai batteri tutto il tempo di nascondersi nei macrofagi. Tutto ciò blocca la fagocitosi da parte dei macrofagi, rallenta la risposta immunitaria adattativa o specifica e attiva la risposta d’ipersensibilità di IV tipo, che tipicamente viene evocata dai microrganismi che si nascondono nelle cellule umane. Quindi il sistema immunitario viene attivato ma tardivamente, aumentando il rischio che prevalga

36 Microbiologia - Artini
il danno tissutale sulla capacità di eliminazione del patogeno. Perciò la tubercolosi è una patologia dovuta principalmente a lla risposta dell’ospite all’infezione.
Patogenesi I micobatteri sono inalati nelle vie aeree e negli alveoli vengono fagocitati dai macrofagi che attivano la risposta innata chiamando nuovi macrofagi, i polimorfonucleati neutrofili. Ma i batteri riescono a sopravvivere e a replicare nei macro fagi che iniziano a morire ed accumularsi. Nel giro di pochi giorni viene montata anche la risposta adattativa o specifica, quindi partecipano alla risposta anche
i linfociti T e B che concorrono a formare un granuloma che conterrà macrofagi necrotici al centro, circondati da linfociti B e T e intorno al granuloma si forma un vallo fibroso che tende a circoscrivere e a limitare il “teatro della battaglia” e l’esito dipenderà da una serie di fattori tra cui la virulenza del ceppo, la quantità di batteri e l’efficienza della risposta immunitaria. Se il ceppo è poco
virulento, se i batteri da eliminare sono pochi e il sistema immunitario dell’ospite è efficiente, tutti i batteri saranno uccisi e il granuloma tenderà lentamente verso una progressiva fibrosi e la calcificazione. Se invece i batteri sono troppi per essere tutti uccisi e il sistema immunitario è poco efficiente, come avviene per esempio nei soggetti HIV-positivi, il granuloma resterà in attività, continuerà a
richiamare cellule del sistema immunitario e tenderà a crescere mantenendo al centro un’area necrotica di cellule morte in cui i micobatteri proliferano e non vengono attaccati. Il granuloma può rimanere in queste condizioni di latenza per mesi o molti a nni e in caso di indebolimento delle difese immunitarie, il granuloma può riprendere a crescere facendo aumentare i macro fagi morti che si accumulano al centro del granuloma sciogliendosi a formare una sostanza detta caseosa. Granulomi adiacenti possono liquefarsi e
fondersi man mano che crescono e può capitare che quelli adiacenti ad un bronco si aprano nel suo lume, liberando bacilli tubercolari nell’essudato bronchiale. Questi bacilli vanno a disseminare l’infezione in altre aree del polmone, ma vengono anche eliminat i all’esterno con il respiro e possono andare a contagiare nuovi individui.
Questa fase di attivazione della malattia (cerchiata in rosso) può avvenire dopo pochi mesi dal contagio oppure può comparire anche dopo anni o decenni in soggetti che attraversano un momento di indebolimento delle difese immunitarie.
Quindi dalla risposta adattativa ci sono 3 possibili evoluzioni della malattia:
• in verde, è rappresentata la guarigione, quindi quando tutti i batteri vengono uccisi e il granuloma viene siggillato con la calcificazione. Questa avviene per >90% dei soggetti contagiati.
• In blu, la latenza. La latenza di un’infezione clinicamente inapparente, quando nel granuloma persistono micobatteri in vita, ed è una fase che può durare anche decenni.
• In rosso, la malattia attiva (<10% si soggetti contagiati). Quando l’equilibrio di forze tra il patogeno e l’ospite si sposta a favore dei micobatteri e i granulomi riprendono a crescere nel parenchima polmonare fino a svuotare nei bronchi il proprio contenuto necrotico e infetto.
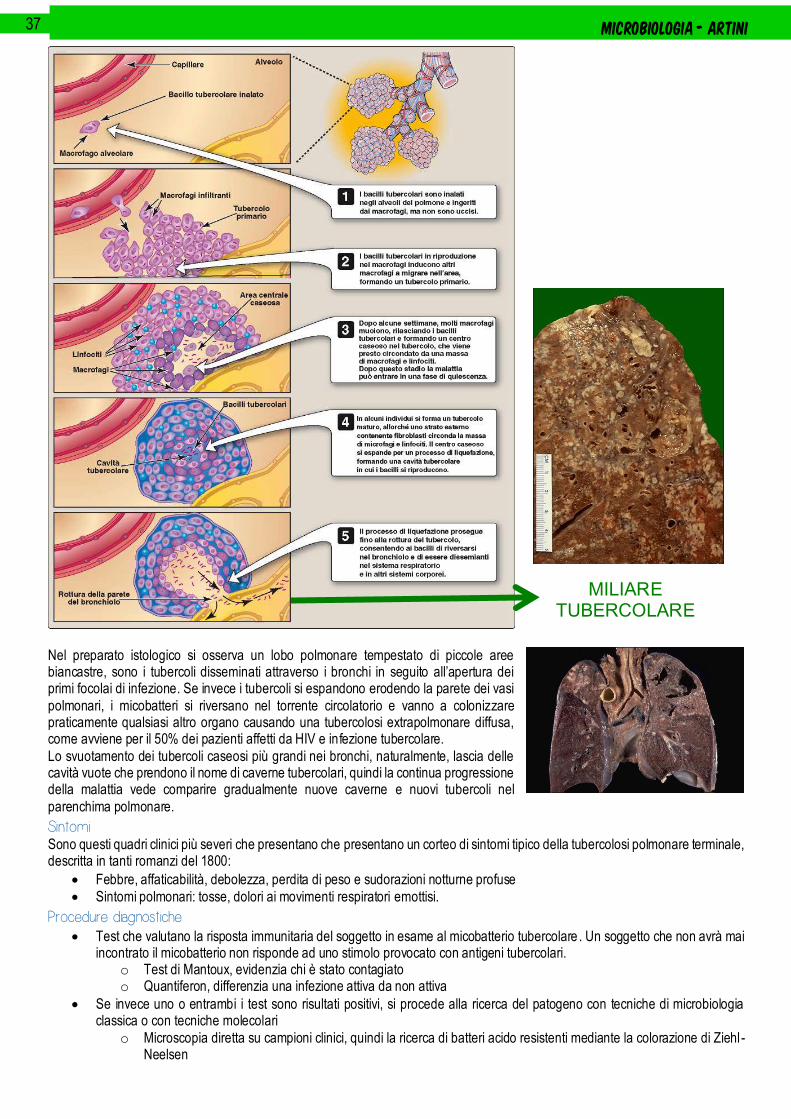
37 Microbiologia - Artini
Nel preparato istologico si osserva un lobo polmonare tempestato di piccole aree biancastre, sono i tubercoli disseminati attraverso i bronchi in seguito all’apertura dei primi focolai di infezione. Se invece i tubercoli si espandono erodendo la parete dei vasi
polmonari, i micobatteri si riversano nel torrente circolatorio e vanno a colonizzare praticamente qualsiasi altro organo causando una tubercolosi extrapolmonare diffusa, come avviene per il 50% dei pazienti affetti da HIV e infezione tubercolare.
Lo svuotamento dei tubercoli caseosi più grandi nei bronchi, naturalmente, lascia delle cavità vuote che prendono il nome di caverne tubercolari, quindi la continua progressione della malattia vede comparire gradualmente nuove caverne e nuovi tubercoli nel
parenchima polmonare.
Sintomi Sono questi quadri clinici più severi che presentano che presentano un corteo di sintomi tipico della tubercolosi polmonare terminale, descritta in tanti romanzi del 1800:
• Febbre, affaticabilità, debolezza, perdita di peso e sudorazioni notturne profuse
• Sintomi polmonari: tosse, dolori ai movimenti respiratori emottisi.
Procedure diagnostiche • Test che valutano la risposta immunitaria del soggetto in esame al micobatterio tubercolare . Un soggetto che non avrà mai
incontrato il micobatterio non risponde ad uno stimolo provocato con antigeni tubercolari. o Test di Mantoux, evidenzia chi è stato contagiato o Quantiferon, differenzia una infezione attiva da non attiva
• Se invece uno o entrambi i test sono risultati positivi, si procede alla ricerca del patogeno con tecniche di microbiologia classica o con tecniche molecolari
o Microscopia diretta su campioni clinici, quindi la ricerca di batteri acido resistenti mediante la colorazione di Ziehl -Neelsen
MILIARE TUBERCOLARE

38 Microbiologia - Artini
o Test molecolari basati su acidi nucleici per l’identificazione delle specie micobatteriche (PCR) o Isolamento in coltura
o Antibiogramma per definire la sensibilità ai farmaci
Intradermoreazione di Mantoux Il test di Mantoux o test cutaneo alla tuberco lina valuta la risposta immunitaria ritardata da ipersensibilità in seguito all’inoculazione
intradermica di un derivato di proteine purificate dal Mycobacterium tuberculosis. Si inoculano le proteine del batterio sotto la cute del braccio ed entro 2-3 giorni si misura in mm la dimensione del pomfo che si è formato in seguito allo stimolo dell’antigene. Più è largo il pomfo più è probabile che l’individuo sia stato contagiato recentemente dal micobatterio tubercolare:
• 5mm: soggetti a stretto contatto con pazienti affetti da TBC attiva, pazienti immunodepressi (HIV+), Rx torace compatibile con gli esiti di TBC;
• 10mm: altri casi. Il problema è che questo test può risultare positivo anche in soggetti vaccinati.
Test Quantiferon È un test più sensibile e specifico per la diagnosi di tubercolosi polmonare attiva e per individuare un’infezione latente di tipo tubercolare nei soggetti maggiormente a rischio. Questo
test misura la quantità di interferone γ (γ-IFN) prodotto dai linfociti T e dai monociti di un soggetto in risposta ad una stimolazione con antigeni del micobatterio tubercolare. Un’elevata
produzione di γ-IFN suggerisce uno stato di attivazione specifica del sistema immunitario e quindi la presenza del micobatterio nel corpo del soggetto in esame. In pratica si effettua un prelievo di sangue venoso che viene lasciato per 24 h in contatto con gli antigeni del micobatterio tubercolare. Se il paziente ha in atto un’infezione anche latente, i linfociti T e i
monociti verranno attivati nella provetta e produrranno citochine infiammatorie, tra cui γ-IFN.
Il giorno successivo si fa il dosaggio del γ-IFN nella provetta utilizzando un metodo basato su anticorpi che riconoscono e legano specificatamente la citochina ed emettono luminescenza
quando avviene il legame. Più è alta la quantità di γ-IFN trovato, maggiore è l’attivazione dell’organismo, quindi maggiore è la probabilità di un’infezione tubercolare attiva operante nel paziente. Questo test può affiancare o può sostituire l’intradermoreazione di Mantoux nella diagnosi di tubercolosi.
Microscopia diretta Una volta accertata la presenza di una risposta immunitaria si può procedere ad effettuare i test microbiologici. Prima si esegue la microscopia diretta sui campioni biologici, in genere nell’espettorato bronchiale, utilizzando strisci allestiti da l sedimento di campioni che vengono colorati con la colorazione di Ziehl-Neelsen. È fondamentale ricordare che il numero di bacilli tubercolari rilevati correla
fortemente con il rischio di trasmissione interumana. Ogni tipo di campione biologico deve essere raccolto secondo un protocollo ben preciso, codificato da tempo secondo accordi internazionali. La modalità di raccolta deve avvenire in contenitori monouso di plastica sterili, con tappo a vi te, non devono essere
utilizzati fissativi o conservanti, deve essere eseguita prima dell’inizio della terapia antimicobatterica ed i campioni raccolti vanno inviati in laboratorio il prima possibile (mezz’ora) entro le 24 h dal prelievo. Tra i vari campioni si trovano:
• Escreato, 3 campioni in giorni diversi;
• Espettorato indotto con aerosol di soluzione salina ipertonica;
• Aspirato gastrico nei pazienti che non espettorano neanche con aerosol;
• Broncoaspirati e lavaggi in corso di broncoscopia a fibre ottiche;
• Urine, 3 campioni della prima urina del mattino (mitto intermedio);
• Sangue, il campione va raccolto in provette eparinate (in immunodepressi);
• Liquor, raccogliere in provette sterili almeno 2 ml;
• Tessuti ed altri liquidi corporei vanno raccolti in contenitori sterili senza fissativo;
• Feci, il campione (≥1g) va raccolto in contenitori di plastica trasparente e inviato in laboratorio allestendo una sospensione omogenea in soluzione fisiologica (Come per i parassiti fecali) – da EVITARE perché presenta il rischio di risultati inattendibili.
Test molecolari su DNA (PCR) Esse permettono di amplificare enormemente quantitativi infinitesimali di DNA batterico. Questi test, che sono più rapidi, de vono essere eseguiti contestualmente alle tecniche di identificazione della microbiologia tradizionale. La sua utilità clinica è data
dall’identificazione diretta del campione. In campioni respiratori con esame microscopico negativo deve essere considerato co n molta cautela l’esito di test molecolari, e sempre nel contesto del sospetto clinico.
Isolamento in coltura La diagnosi definitiva in realtà richiede anche l’isolamento in coltura di Mycobacterium tuberculosis complex, che deve essere fatta in terreni specifici. La crescita in piastra richiede tempi molto più lunghi (3-6 settimane), ma l’adozione di tecniche di coltura in terreno liquido ha dimezzato questi tempi (7-14 giorni). Nonostante ciò, i terreni all’uovo come Loewenstein-Jensen e il terreno di Petragnani,
vanno comunque impiegati poiché consentono la crescita di alcuni ceppi di Mycobacterium tuberculosis complex e di alcune specie non tubercolari che non riescono a svilupparsi negli altri terreni.

39 Microbiologia - Artini
Sensibilità ai farmaci È attualmente raccomandato che tutti i nuovi isolati di Mycobacterium tuberculosis complex siano saggiati nei confronti dei farmaci
antitubercolari di prima scelta (isoniazide, rifampicina, pirazinamide, etambutolo e streptomicina). Una forma di Tb resistente ai farmaci particolarmente pericolosa è la Mdr-Tb (multidrug resistant) provocata da batteri resistenti almeno ai due medicinali di prima linea anti Tb più potenti, l’isoniazide e la rifampicina. La Mdr-Tb va quindi curata necessariamente
con farmaci di seconda linea. Secondo l’OMS, la Mdr-Tb è ormai presente praticamente in ogni area del mondo e costituisce uno dei problemi più importanti nel controllo e trattamento della Tb. L’antibiogramma dovrebbe essere ripetuto anche quando le colture continuano ad essere positive dopo tre mesi di terapia.
Contagio - Epidemiologia L’uomo è l’unico serbatoio naturale dell’infezione ed è stimato dall’OMS che circa un terzo della popolazione mondiale sia
infettata da M.tuberculosis. Naturalmente la diffusione non è omogenea su tutta la terra ma prevale nei paesi più poveri e a basso standard igienico-sanitario dell’Africa sub-
sahariana, dell’Asia e del Sud America. È noto che anche in Italia da alcuni anni è in atto un importante ritorno dell’infezione e ogni
giorno aumenta il numero di nuovi casi diagnosticati. Questo è dovuto a tante cause tra cui l’arrivo di profughi ed immigranti
provenienti da Paesi ad alta endemia, ma anche l’aumento di soggetti italiani con riduzione delle difese immunitarie.
Vaccino (BCG) La vaccinazione antitubercolare viene praticata inoculando batteri vivi di un ceppo attenuato, quindi privato della virulenza tipica dei ceppi selvatici. Il vaccino è consigliato per chi convive con soggetti affetti da tubercolosi, a chi vive in aree ad alta endemia e a chi svolge attività lavorative che espongono al rischio di contagio. Poiché le cellule batteriche del preparato vaccinale sono vi ve, la
vaccinazione è sconsigliata in tutti i soggetti che hanno le difese immunitarie ridotte, quindi gli immunodepressi per qualsiasi causa.
Il genere Clostridium include un vasto insieme eterogeneo di bacilli Gram+, anaerobi e sporigeni. Patogeni importanti come C.tetani
e C.botulinum sono ben conosciuti ma hanno ormai un valore più che altro storico, invece la patologia provocata da C.difficile si è diffusa negli ultimi anni come complicanza infettiva dell’uso di antibiotici ad ampio spettro, sia negli ospedali che nella popolazione. Si può dire che l’uso massiccio di antibiotici è stata quindi la causa di diffusione delle affezioni intestinali da C.difficile.
Clostridium difficile Fino alla metà degli anni ’70, l’importanza clinica di questo batterio non era stata ben valutata, il microrganismo veniva isolato
raramente in colture di materiale fecale e il suo ruolo nelle enteriti dell’uomo era sconosciuto. Tuttavia, numerosi studi hanno dimostrato chiaramente che questo clostridio è responsabile di malattie gastrointestinali associate agli antibiotici, che vanno dalla diarrea autolimitante alla colite pseudomembranosa grave e a volte potenzialmente fatale. Ad oggi è considerato un patogeno
importante, responsabile di episodi di diarree recidivanti e coliti che si manifestano più spesso in ambito ospedaliero. I fattori di rischio
Stima di incidenza nel 2015 Fonte: OMS
Categorie a rischio nei paesi occidentali:
• Senza fissa dimora • alcolisti • Tossicodipendenti • Detenuti • HIV positivi

40 Microbiologia - Artini
noti sono 2: l’assunzione recente di antibiotici e il ricovero in ambiente ospedaliero, anche se quest’ultimo è stato ridimen sionato recentemente.
C.difficile è un bacillo Gram+, anaerobio e sporigeno che esprime la propria virulenza producendo 2 esotossine:
• Tossina A (enterotossina): stimola l’infiltrazione di polimorfonucleati neutrofili e il rilascio di citochine
• Tossina B (citotossina): distruzione del citoscheletro per depolimerizzazione dell’actina. Tra gli altri fattori di virulenza di questo batterio ci sono: proteasi, fimbrie, flagelli e capsula. I ceppi presentano vari abilità nella virulenza. Le spore sono abbondantemente presenti nell’ambiente e il rischio di contagio è alto sia nell’uomo che negli animali. È noto che esistono molti portatori asintomatici di C.difficile, quindi la semplice colonizzazione del corpo umano non è sufficiente ad
esprimere la malattia, quindi non è un patogeno stretto. In ospedale aumenta la concentrazione di soggetti portatori ma soprattutto di ricoverati con infezione attiva, inoltre le corsie ospedaliere sono spesso contaminate dalle spore che sono resistenti e molto difficili da eliminare. È noto che il rischio di contagio aumenta con la lunghezza della degenza ed è in rapporto con condizioni di stress fisico e psichico del paziente che possono
verificarsi durante il ricovero. Quindi un ricovero lungo, stressante e con lunghi cicli di terapia antibiotica sono condizioni predisponenti per il passaggio da una condizione di colonizzazione intestinale a quello di infezione, prima, e di malattia conclamata poi. Per scatenare la malattia devono
concorrere fattori favorenti sia dal microrganismo che dal paziente. Da parte del microrganismo sono fattori favorenti: un’alta carica infettante, alta patogenicità del singolo ceppo che deve essere capace di produrre le tossine A e B. Da parte dell’ospite son o fattori favorenti: l’età avanzata, lunga terapia antibiotica e condizioni di debilitazione. I sintomi compaiono sempre in soggetti ricoverati o recentemente dimessi dall’ospedale che abbiano assunto o stiano ancora
assumendo antibiotici. Il quadro clinico può essere di 3 tipi:
• Diarrea acuta, si sviluppa 5-10 giorni dopo l’inizio della terapia antibiotica con cefalosporine o fluorochilonuri, comunque in genere è breve, autolimitante ma può prolungarsi nel tempo.
• Colite
• Colite pseudomembranosa, rara. È la forma più grave d’infezione sostenuta da C.difficile con diarrea abbondante, crampi addominali, febbre e la presenza di pseudomembrane alla colonscopia, ovvero
delle placche biancastre che spiccano sul tessuto infiammato e arrossato della mucosa sottostante. Le complicanze possono essere perforazione, megacolon tossico e sepsi.
Le pseudomembrane si formano quando la citotossina B danneggia le cellule
della mucosa, generando uno stato d’infiammazione locale che richiama dai vasi sanguigni sottostanti i monociti e neutrofili che migrano nel lume intestinale formando con la fibrina una matrice mucoide densa che ricopre le
aree di mucosa intestinale danneggiata.
Patogenesi Se durante il ricovero il paziente viene contaminato dalle spore e contemporaneamente assume grandi quantitativi di antibioti ci ad ampio spettro, la flora intestinale saprofita viene fortemente ridotta dagli antibiotici e vengono alterate le condizioni di equilibrio tra le
specie commensali del colon. In questa condizione di fragilità del microbioma intestinale, le spore del C.difficile tornano allo stato vegetativo e la nuova specie cresce rapidamente fino a prevalere sulla flora saprofita. Quindi l’intestino è rapidamente invaso dai clostridi. Nel giro di poco tempo, le esotossine prodotte dal batterio danneggeranno i tessuti: l’esotossina A causa la diarrea e
l’esotossina B danneggia l’epitelio fino a formare le pseudomembrane con il meccanismo descritto sopra.
Diagnosi diretta La diagnosi microbiologica di infezione da C.difficile va cercata sempre in un contesto clinico di diarrea, febbre e colite insorte dopo
una terapia con antibiotici ad ampio spettro. Si parte dall’isolamento e identificazione del ceppo ma è fondamentale dimostrare la presenza delle tossine A e B. Per questo scopo, sono disponibili tecniche diverse che presentano difficoltà e costi different i ma la sola dimostrazione della presenza del clostridio è insufficiente. Tra queste tecniche:
• Citotossicità fecale specifica su colture cellulari (gold standard)
• Ricerca antigeni: tossina A, tossina B, altri antigeni
• Metofi biomolecolari (tossine, 16SrRNA)
Terapia Premesso che la sospensione dell’antibiotico ad ampio spettro, che ha alterato l’equilibrio del microbioma dell’intestino, spesso
migliora il quadro. In casi in cui il quadro dovesse essere molto grave, si possono prescrivere altri antibiotici che hanno un’azione più mirata come vancomicina e metronidazolo. Le raccomandazioni per la gestione del paziente vengono organizzate in modo da formare l’acronimo SIGHT:
• Suspect that a case may be infective where there is no clear alternative cause of diarrhoea;
• Isolate the patient in a side room, consulting with the Infection Control Team (ICT);
• Gloves and aprons must be used for all contacts with the patient and their environment;
• Hand washing with soap before and after each contact with the patient and the patient’s environment;
• Test the stool for toxin immediately.

41 Microbiologia - Artini
C. tetani e C.botulinum Sono entrambi bacilli Gram+, anaerobi stretti quindi le loro forme vegetative muoiono rapidamente quando sono esposte all’ossigeno
e sono sporigeni, quindi il contagio avviene sempre tramite le spore presenti nell’ambiente. I ceppi patogeni di C.tetani producono tetanospasmina, neurotossina trasportata dalla sede d’infezione a specifici recettori situati sulla superficie di neuroni motori. La tossina viene internalizzata e trasportata lungo l’assone fino al copro dei neuroni situati nel
midollo spinale, dove inattiva i sistemi che controllano il rilascio di neurotrasmettitori portando a un’attività sregolata delle sinapsi eccitatorie con una conseguente paralisi spastica. La paralisi spastica è il sintomo caratteristico del tetano, può essere localizzata ma più frequentemente generalizzata con alcuni segni tipici come:
• Riso sardonico dovuto allo spasmo dei muscoli facciali
• Spasmo persistente di tutti i muscoli dorsali (opistotono) Questi quadri clinici sono rarissimi nei Paesi in cui è diffuso l’uso estensivo della vaccinazione e per la possibilità di somministrare
siero antitetanico ai soggetti potenzialmente esposti al contagio perché si sono feriti con oggetti esposti da tempo all’ambiente esterno. Anche il C.botulinum esprime la sua virulenza grazie ad una esotossina che però deve essere assunta con il cibo, quindi questa è una tossinfezione alimentare dovuta all’ingestione di cibo contaminato dalla tossina. Per contenere la tossina, il cibo deve essere
contaminato dalle spore di clostridio, diffusissime nell’ambiente, e dev’essere confezionato sottovuoto o comunque in modo da permettere condizioni di assenza di ossigeno per permettere alle spore di tornare alla forma vege tativa. All’apertura del barattolo, i batteri muoiono immediatamente ma la tossina che ha contaminato il cibo persiste e ne basta una quantità infinitesimale che leghi i
recettori presenti sulla superficie dei neuroni motori per bloccare il rilascio di ace tilcolina nelle sinapsi periferiche. Ne deriva una progressiva paralisi flaccida che ha caratteristiche opposte a quella spastica del tetano e può portare a morte per paralisi dei muscoli respiratori.
Legionella è un batterio che produce patologia nell’uomo grazie alle opportunità e nuovi habitat offerti dall’evoluzione tecnologica. Si chiama così perché il microrganismo fu identificato in seguito ad un’epidemia di polmonite che colpì i veterani ritornati dal Vietnam (legionari) riuniti per una convention a Philadelphia del 1976. Si scoprì poi che tutti i veterani colpiti dalla polmonite erano stati
contagiati da un batterio, mai isolato precedentemente, che fu trovato anche nell’impianto di condizionamento dell’edificio in cui si era tenuta la convention. Le legionelle sono bacilli Gram- molto sottili, di dimensioni molto variabili (pleomorfismo), aerobi obbligati, mobili (uno o più flagelli),
asporigeni. Per la coltura sono esigenti perché richiedono terreni addizionati (L -cisteina e ferro). Presenta la parete cellulare contenente acidi grassi a catena ramificata, in genere assenti nei batteri Gram-. La specie Legionella pneumophila di sierogruppo 1 è responsabile del 90% dei casi di polmonite.
Le legionelle sono diffusissime in natura, ma sempre in associazione con raccolte di acqua come laghi, fiumi, sorgenti termali tiepide, falde idriche e ambienti umidi in generale. Da questi serbatoi naturali le Legionelle colonizzano ambienti idrici artificiali costruiti dall’uomo: impianti per innaffiare, torri di raffreddamento industriali, sanitari domestici incluso le docce, impianti termali, fontane ornamentali, impianti di condizionamento dell’aria, etc. La nebulizzazione dell’acqua contaminata dalle Legionelle, viene trasportata
nell’aria e gli aerosol contaminati possono entrare nell’apparato respiratorio degli individui presenti in quell’ambiente portando le Legionelle alle vie aeree superiori fino a giungere agli alveoli e ai polmoni. Tutti questi impianti quindi possono essere colonizzati da biofilm microbici ed è noto che le Legionelle sono ottimi formatori di biofilm.
Un moderato riscaldamento dell’acqua, come avviene in estate, è la causa di un aumento della crescita di microrganismi e la nebulizzazione causa la diffusione nell’ambiente umano. Una volta che Legionella pneumophila penetra nell’ospite, per inalazione di aerosol contaminati nelle vie respiratorie, i batteri
raggiungono i polmoni e vengono fagocitati dai macrofagi alveolari. In questa sede riescono ad evadere le difese del sistema immunitario perché hanno caratteristiche che altri patogeni delle vie aeree non possiedono. Quando un microrganismo (non Legionella) riesce ad attraversare tutte le barriere delle prime vie aeree fino ad arrivare agli alveoli, viene fagocitato dai macrofagi alveolari, digerito e rapidamente eliminato. In questo modo i macrofagi che stabilmente stazionano
negli alveoli polmonari impediscono l’insorgenza di infezioni eliminando prontamente tutti i microrganismi che riescono ad ar rivarvi. La Legionella invece ha due modi per sfuggire a questo tipo di difesa:
• Il primo meccanismo dipende dalla capacità di questo batterio di sopravvivere e replicare nelle cellule umane, soprattutto nei macrofagi. Una volta fagocitato, infatti, questo batterio impedisce la fusione del fagosoma con il lisosoma e replica attivamente nel fagolisosoma. Il macrofago reagisce producendo un’enorme quantità di citochine chemochine
infiammatorie, ma alla fine lisa e le Legionelle che si erano replicate al suo interno vengono liberate e possono entrare in altri macrofagi o nelle cellule dell’epitelio alveolare. Il risultato è un’enorme attivazione dell’infiammazione locale ma senza l’eliminazione dei batteri, quindi un duplice danno. La legionella, infatti, sopravvive e replica sia nell’ambiente naturale
all’interno di microrganismi unicellulari eucarioti, come l’ameba, sia nel corpo umano e in particolare nei macrofagi ma anche in altri tipi cellulari. Questa capacità impedisce al corpo umano di eliminare il batterio aumenta moltissimo l’infiammazione a livello polmonare.
• Il secondo meccanismo di difesa delle Legionelle dipende dalla loro enorme capacità di formare biofilm negli impianti idrici contaminati, quindi è molto probabile che piccoli frammenti di biofilm vengano nebulizzati nell’aria come aerosol
contaminato. Quando un frammento di biofilm di Legionella riesce ad arrivare fino all’alveolo, non viene visto dai macrofagi, quindi i batteri restano indisturbati nell’alveolo e replicano lentamente all’interno del biofilm, continuando a crescere

42 Microbiologia - Artini
indisturbati perché le difese immunitarie non sono attivate, occupando in questo modo gli spazi negli alveoli e liberando cellule planctoniche che possono nuovamente invadere le cellule eucariotiche dell’alveolo.
Esistono alcune condizioni dell’ospite che favoriscono l’infezione da Legionella e la sua piena espressione: età avanzata, sesso maschile, alcolismo, tabagismo, viaggi frequenti, bronchiti croniche, insufficienza renale cronica, diabete, trapianti, neoplasie. Focolai sporadici compaiono tutto l’anno ma piccole epidemie si verificano soprattutto d’estate e d’autunno, quando aumenta la temperatura. Esistono due forme della malattia:
• Forma più leggera, ha le caratteristiche di una malattia febbrile autolimitante con febbre, brividi, mialgia e cefalea;
• Forma più grave, polmonite detta “malattia del legionario) dove compaiono bruscamente astenia, cefalea, dolori muscolari nonché febbre, tosse secca, dolore toracico, fame d’aria a cui possono associarsi anche sintomi d’interessamento intestinale come diarrea, nausea, vomito, dolori addominali e sintomi del sistema nervoso centrale come stato confusionale e agitazione.
Diagnosi Per la diagnosi:
• dimostrazione delle Legionelle in colture su terreni selettivi, metodo migliore ma richiede molto tempo perché la crescita è lenta;
• sierologia (IgG e IgM sieriche);
• Ricerca di antigeni nelle urine: facile esecuzione, positivizzazione precoce, positivizzazione anche dopo 60 gg, solo sierotipo 1
• Ricerca dei genomi per amplificazione genica (PCR)
Terapia Essendo Legionella un microrganismo intracellulare è necessario utilizzare antibiotici con una buona penetrazione intracellulare:
macrolidi, chinoloni, rifampicina, tetracicline, trimetoprim-sulfometoxazolo.
VIRUS I virus parassitano cellule eucariotiche o procariotiche e utilizzano la loro energia e i loro sistemi enzimatici per riprodursi, quindi
sono dei parassiti obbligati. Replicano, ma non sono affatto autonomi. Sono particelle organizzate in una struttura regolare che partecipa delle proprietà della materia vivente, ma sono obbligati ad usare le cellule per replicare. Forse è possibile considerarli delle porzioni di cellule contenenti informazioni genetica che migrano tra una cellula e l’altra, trasferendo materiale genetico.
La caratteristica comune è che sono parassiti intracellulari obbligati. Passano da:
• una fase intracellulare, in cui replicano sfruttando il macchinario delle cellule infettate,
• ad una fase extracellulare, dove la particella virale completa è libera e infettante nei fluidi corporei dell’ospite. Le particelle virali complete o virioni sono tipiche della sola fase extracellulare. Il virione contiene sempre un core, formato da un acido nucleico e da alcune proteine ad esso associate. Questo genoma è circondato da un capside proteico che lo avvolge e lo protegge. Il nucleocapisde è il termine per indicare il core e il capside assieme. In alcune specie virali il capside è circondato da un mantello o envelope di
natura lipoproteica, costituito da un doppio strato di fosfolipidi da cui sporgono proteine virali. Lo spazio tra capside ed envelope si chiama tegumento e può contenere altre proteine virali con funzione enzimatica.
Tutti i virus possiedono materiale genetico, che può essere costituito da DNA o RNA a doppia o singola catena. La quantità di materiale genetico è estremamente variabile da una specie virale all’altra e può contenere da 4 -5 geni fino a 100 geni nei virus più grandi. Per economia di spazio, i geni sono spesso sovrapposti e possono essere tradotti con splicing multipli.
Le proteine codificate dal genoma virale sono classificate in:
• Proteine funzionali: enzimi necessari per la replicazione virale o Enzimi che permettono la replica dell’acido nucleico (RNA o DNA polimerasi) o Proteine associate agli acidi nucleici o Proteine che interferiscono con funzioni e strutture della cellula infettata
• Proteine strutturali: vengono incorporate nelle particelle virali nascenti per formare il capside o l’envelope oppure entrano nello spazio del tegumento.
Virus nudo (senza envelope) Virus con envelope
Sono più resistenti alle alte temperature, agli acidi, ai detergenti
e all’essicamento. Resistono meglio nell’intestino dell’ospite. Conseguenze: il virus si diffonde più facilmente nell’ambiente, perché resistono meglio sugli oggetti, nella polvere e sulle mani.
Replicano lasciando la cellula infettata in seguito alla sua lisi.
Meno resistenti ad acidi, detergenti, essicamento e calore. Non
sopravvivono nel tratto gastro-intestinale. Conseguenze: deve rimanere in ambiente umido, si diffonde con gocce d’acqua, secrezioni, trasfusioni di sangue e trapianti
d’organo. Sono quindi più fragili e meno resistenti nell’ambiente. Escono dalla superficie della cellula infettata per gemmazione,
quindi senza danneggiarla.

43 Microbiologia - Artini
Essendo un parassita intracellulare obbligato, un virus può replicare solo in una cellula ospite che permetta la completa espressione
del suo genoma e la produzione di nuove particelle virali. I passaggi fondamentali per questo scopo sono:
• L’adesione del virus alla cellula ospite e l’introduzione del suo genoma .
• Il genoma deve essere in grado di esprimersi e di utilizzare l’apparato sintetico della cellula per la sintesi dei costituenti virali.
• Questi costituenti si combineranno in modo da formare nuovi virioni.
• I virioni prodotti, liberati dalla cellula estenderanno l’infezione ad altre cellule dell’ospite.
• La replicazione virale può avvenire solo in cellule sensibili e permissibili, capaci di far esprimere completamente il genoma virale.
Per permettere la replicazione virale, la cellula ospite deve possedere due caratteristiche:
• Sensibilità: strettamente condizionata dalla presenza di adeguati recettori cellulari che permettono l’attacco del virus alla cellula ospite (adsorbimento).
• Permissività: dipende da tutti i meccanismi molecolari e cellulari che permettono al completa trascrizione del genoma virale e la sintesi delle proteine virali.
La cellula eucariotica può essere:
• Non sensibile → il virus non entra
• Sensibile ma non permissiva → o Infezione abortiva: in alcuni casi l’infezione può non completarsi. Ciò si verifica quando il virus infettante è difettivo
(mancano parti essenziali del genoma) o quando la cellula consente solo l’espressione di alcuni geni virali, portando all’arresto dell’infezione.
o Infezione restrittiva: quando le cellule sono permissive solo in una certa fase del loro ciclo vitale.
o Entrambe le infezioni possono consentire la persistenza del virus all’interno della cellula ospite generando una condizione di latenza virale. In questo caso, il genoma virale può integrarsi in quello della cellula ospite, nascondendosi in forma episomale anche per anni. La replicazione virale potrà ripartire in un qualsiasi momento in cui nella cellula tornino condizioni ambientali favorevoli.
• Sensibile e permissiva → infezione produttiva: infezione di una cellula permissiva da parte di un virione completo che porta alla produzione di nuovi virioni in numero variabile a seconda del tipo di virus o di cellula.
Per convenzione, il ciclo di replicazione virale è diviso in 6 fasi:
1. Adsorbimento 2. Penetrazione 3. Scapsidazione 4. Replicazione
5. Maturazione 6. Liberazione
In caso di latenza dell’infezione, il periodo compreso tra la
penetrazione del virus nella cellula ospite e l’uscita dei nuovi virioni, viene definito periodo di eclissi, non essendo il virus più replicabile come entità morfologica nella cellula. Il periodo di eclissi può durare
da giorni fino ad anni, per i virus che integrano il proprio genoma nei cromosomi della cellula ospite.
1. Fase di adsorbimento È la fase in cui il virus deve aderire alla cellula eucariotica con un meccanismo di riconoscimento specifico e recettoriale. Questa fase richiede un’interazione stretta tra gli antirecettori presenti sul virus e i recettori della membrana citoplasmatica del la cellula da infettare. Il legame avviene tra cariche complementari tra gli antirecettori del virus e i recettori della cellula ospite. La natura chimica
di molti antirecettori è stata descritta, in genere sono glicoproteine in virus con envelope e proteine semplici nei virus nudi. Questa è la fase fondamentale che stabilisce il tipo di malattia che segue l’infezione da una determinata specie virale, perché possono essere infettate solo le cellule che esprimono i recettori riconosciuti dagli antirecettori di quel virus. In genere i recet tori della cellula ospite
che permettono l’adesione virale sono espressi sulla membrana cellulare che svolge una funzione fisiologica ben precisa che non ha nulla a che fare con l’infezione virale. Sono i virus che si sono adattati esprimendo antirecettori specifici che permettono loro di utilizzare i recettori cellulari come porta d’ingresso alle cellule eucariotiche. Questa fase non richiede dispendio energetico da parte della cellula eucariotica che partecipa passivamente.
2. Penetrazione In questa fase la cellula partecipa attivamente spesso anche con consumo di energia. Il virus può entrare per:
• Traslocazione: quando viene trasportato tutto intero da molecole di membrana ed entra come tale nel citoplasma (virus nudi);
ADSORBIMENTO
PENETRAZIONEPENETRAZIONESINTESI VIRALESINTESI VIRALE
ASSEMBLAGGIOASSEMBLAGGIO(MATURAZIONE)
TRASCRIZIONE
REPLICAZIONE
USCITA
SCAPSIDAZIONESCAPSIDAZIONE TRADUZIONE

44 Microbiologia - Artini
• Endocitosi: quando viene portato ne l citoplasma attraverso l’invaginazione della membrana che lo accoglie nel vacuolo fagocitario (virus con envelope, virus nudi);
• Fusione: quando la membrana che costituisce l’envelope virale si fonde con la membrana citoplasmatica liberando il nucleocapside nel citoplasma (virus con envelope).
3. Scapsidamento Il genoma viene liberato dal capisde. Le proteine del capside possono disaggregarsi spontaneamente oppure possono essere digerite da proteasi o enzimi lisosomiali che si riversano nel fagolisosoma.
4. Replicazione solo se la cellula è anche permissiva viene subito avviata la replicazione del virus, che ovviamente richiede la sintesi di nuove
proteine virali e nuovi genomi. Le proteine virali neosintetizzate si distinguono in:
• Proteine precoci: di natura enzimatica, sono sintetizzate per prime ed intervengono nella replicazione del genoma virale;
• Proteine tardive: sintetizzate dopo la replicazione del genoma virale, soprattutto strutturali. Contemporaneamente si avvia la produzione di nuovi genomi, questa fase è attivata a scapito della salute della cellula perché in generale causa una riduzione della sintesi di acidi nucleici della cellula a favore della sintesi di acidi nucleici utilizzat i dal virus. Nella cellula infettata, infatti, l’espressione dei geni virali è in competizione con l’espressione dei geni cellulari, per prendere il sopravvento i virus producono mRNA modificati in modo che vengano favoriti nel processo di traduzione o messaggeri che hanno evoluto
meccanismi capaci di bloccare o limitare le sintesi macromolecolari della cellula ospite. Le sedi e le modalità di duplicazione saranno diverse a seconda se il virus sia costituito da un genoma a RNA o DNA. I meccanismi di replicazione del genoma virale sono complessi e variano tra le specie, basta ricordare che poiché la cellula
eucariotica non possiede enzimi capaci di sintetizzare mRNA a partire da una molecola di RNA. Né possiede enzimi che trascrivono il DNA a partire dall’RNA, solo i virus che possiedono genoma a DNA e siano in grado di raggiungere il nucleo della cellula infettata possono usare il solo sistema di trascrizione della cellula a proprio vantaggio. Tutti gli altri devono necessariamente possedere polimerasi virus-specifiche con caratteristiche necessarie a seconda del tipo di genoma e della sede di replicazione.
Dal punto di vista della regolazione dell’espressione genica è importante ricordare che la cellula eucariotica è predisposta per la traduzione di messaggi monocistronici, in cui a un gene corrisponde una sola proteina. Quindi alcuni RNA messaggeri virali che trascrivono più di un gene, cioè codificano per più di una proteina, vengono tradotti in una poliproteina che viene poi tagliata nelle
varie proteine funzionali del virus. Nella cellula infettata l’espressione dei gen i virali è in competizione con i geni cellulari. Per poter avere il sopravvento, i virus o producono mRNA che sono favoriti nel processo di traduzione, o hanno evoluto meccanismi in grado di bloccare o limitare le sintesi macromolecolari della cellula ospite.
5-6. Maturazione e liberazione Una volta sintetizzati i genomi e le proteine, questi componenti
devono associarsi in modo da costituire i nuovi virioni. Le nuove particelle virali devono fuoriuscire dalle cellule che le hanno prodotte per poter infettare altre cellule. Prima si formano i nucleocapsidi per aggregazione delle proteine
del capside intorno al genoma virale. Per i virus nudi la formazione di nucleocapsidi maturi porta alla liberazione di questi per lisi dalla cellula, che muore liberando la progenie virale. Per i virus dotati
di envelope, le proteine di superficie vengono portate dal citoplasma della cellula infettata verso la membrana; nella zona della membrana cellulare in cui is sono affollate le proteine della superficie del virus, i virioni si formano per gemmazione verso
l’esterno e in genere senza portare a morte la cellula. Quando un virus patogeno per l’uomo si diffonde per contagio nella popolaz ione umana, i soggetti contagiati che guariscono sviluppano una immunità persistente di quel virus e non potranno essere contagiati nuovamente. Poniamo il caso che una mutazione
apparsa casualmente in un virus, conferisca ad esso un vantaggio e venga trasferita alla progenie virale. Quando questi virus mutati infettano soggetti già immuni al virus progenitore, lo stato di immunitizzazione non li protegge più. Questo avviene soprattutto se la nuova mutazione ha determinato un cambiamento di sequenza delle proteine di superficie del virus. In questo caso, i nuovi virioni
non verranno più riconosciuti dal sistema immunitario dei soggetti immuni che erano stati infettati prima che comparisse la mutazione e di conseguenza si diffonderà una nuova ondata di infezioni trasmesse per contagio.
I virus sono in continua evoluzione, grazie alla comparsa di nuove caratteristiche geniche (mutazioni ma non solo) e alla loro
stabilizzazione nella popolazione. Le nuove caratteristiche compaiono casualmente durante la replicazione del genoma. Se il cambiamento non peggiora le capacità di replicazione, si potrà trasmettere alla progenie virale e stabilizzare nella popolazione. Esistono vari tipi di mutazioni che compaiono con meccanismi differenti e producono conseguenze diverse.
• Mutazioni minori: Il primo tipo è rappresentato dalle mutazioni puntiformi che compaiono più frequentemente nei virus a RNA. Esempio del virus influenzale: in esso compaiono ogni anno varianti del ceppo circolante, con mutazioni nelle due proteine di superficie che mediano l’adesione alle cellule dell’ospite: l’emoagglutinina (HA) e la neuraminidasi (NA). La
5.- 6. MATURAZIONE E LIBERAZIONE
Una volta sintetizzati i genomi e le proteine, questi componenti devono
associarsi in modo da costituire i nuovi virioni
Le nuove particelle virali devono fuoriuscire dalle cellule che le hanno prodotte
per poter infettare altre cellule
Biosintesi degli alfavirus Liberazione del virus influenzale
LIBERAZIONE DI VIRUS CON ENVELOPE (I)

45 Microbiologia - Artini
comparsa di una o più mutazioni dei geni di queste due proteine, determina la diffusione di una nuova variante del ceppo che avrà caratteristiche di virulenza e patogenicità non dissimili dall’anno precedente ma avrà caratteristiche antigeniche
sufficientemente diverse da riuscire a scavalcare le difese immunitarie dei soggetti che già avevano contratto l’influenza nella stagione precedente. Quindi molte persone saranno contagiate nuovamente pur sviluppando una malattia con caratteristiche attenuate
• Mutazioni maggiori: mutazioni che possono comparire in un ospite che venga infettato contemporaneamente da due specie virali diverse ma geneticamente simili. Quando una cellula di questo ospite viene coinfettata da entrambi i virus, può
avvenire che i due virus replichino contemporaneamente e che avvengano degli errori o nella fase di trascrizione dei genomi o nella fase di impacchettamento delle particelle virali nascenti. Il risultato è il mescolamento di parte del loro patrimonio genetico e la comparsa di un nuovo ceppo virale completamente inedito che ha ereditato caratteristiche da entrambi i ceppi
virali da cui è derivato. Questo tipo di rimescolamento del patrimonio genetico da due specie simili può avvenire fondamentalmente in due modi diversi:
o Ricombinazione del materiale genetico: scambio di segmenti di materiale genetico tra virus diversi ma correlati e avviene con modalità differenti ma fondamentalmente sempre dovute ad errori degli enzimi che sorvegliano la
sintesi di nuovi genomi come le polimerasi. Nei virus a DNA avviene la rottura e risaldatura di segmenti neosintetizzati; mentre nei virus a RNA la RNA-polimerasi durante la sintesi del nuovo filamento salta da un filamento stampo ad uno omologo appartenente all’a ltro virus. Il risultato sarà un nuovo RNA chimera, diverso da
quello dei due virus che avevano infettato la cellula. o Riassortimento: quando nella stessa cellula si incontrano e replicano due virus a RNA con genoma segmentato,
cioè diviso in tanti segmenti staccati e distinti. Nel momento dell’assemblaggio delle nuove particelle virali, è possibile che segmenti omologhi dei due virus vadano a segregarsi nei capsidi in maniera disordinata. Durante
l’assemblaggio e maturazione di questi segmenti vengono riassortiti diversamente nella progenie virale. È il meccanismo alla base della variabilità antigenica che genera le epidemie influenzali.
Il meccanismo del riassortimento genico è tipico delle zoonosi, cioè delle infezioni sostenute da un virus capace di contagiare sia
l’uomo che un animale che possa entrare in contatto con l’uomo. In questo caso, l’animale funge da serbatoio virale nei periodi in cui si fermano i contagi tra essere umani e lo stesso animale può diventare la sede del riassortimento di geni in caso di coinfezione da più virus. Il maiale, ad esempio, può essere infettato con la stessa efficienza sia dal virus dell’influenza degli uccelli che da ceppi
virali dell’influenza umana. Quando questo avviene, si può avere un riassortimento tra i geni dei due virus e i geni del virus aviario possono andare a finire nel virus dell’influenza umana. In questo caso l’uomo è contagiato tramite il maiale, da un ceppo con caratteristiche antigeniche nuove e profondamente diverse e quindi seguirà un’ondata di influenza altamente contagiosa, perché non esistono individui immunizzati contro quel ceppo nuovo e potenzialmente molto pericolosa perché il virus potrebbe contenere fattori
di virulenza nuovi, presi dal virus aviario. Questo avviene in condizioni di scarsissima igiene e di convivenza molto stretta tra uomo, uccelli e maiali., come avviene nelle campagne della Cina meridionale. Le mutazioni geniche minori, che compaiono ogni anno, causano la comparsa delle varianti epidemiche, perlopiù annuali, e dovuta
a una lenta e progressiva deriva (drift) del patrimonio genetico del ceppo prevalente. Le mutazioni geniche maggiori compaiono più raramente, all’incirca ogni 10 anni, causano la comparsa delle varianti pandemiche dovute ad un brusco cambiamento (shift) nel patrimonio genetico del ceppo.
Nel rapporto tra virus e uomo, entrambi cercano di ottenere un vantaggio cercando di sopravvivere. Il virus è un parassita che opera sabotando i meccanismi cellulari dell’ospite ed utilizzando le sue riserve d’energia. L’ospite reagisce sviluppando le difese immunitarie e il virus sfugge ad esse mutando in continuazione. Tra le tante mutazioni che compaiono nel virus, la sele zione naturale
favorisce quelle che non uccidono l’ospite. Infatti, un virus troppo patogeno danneggerebbe sé stesso se uccidesse i suoi ospiti. D’altra parte i meccanismi dell’infiammazione locale, messi in atto dall’ospite, per arginare la diffusione del l’infezione fra le cellule, danneggiano i tessuti umani. Il quadro della malattia generato da ogni virus risulta dal bilanciamento di questi due fattori:
• Azione diretta dannosa del virus alle cellule infettate
• La risposta infiammatoria dell’ospite infettato per cercare di eliminare il virus, che spesso causa danni maggiori Comunque il danno all’ospite dipende anche dal grado di patogenicità del ceppo, cioè da quanti e quali fattori di virulenza possiede
il ceppo infettante:
• Capacità replicativa;
• Capacità di diffondere nell’ospite o di passare da un ospite ad un altro;
• Capacità di eludere le difese dell’ospite;
• Capacità di produrre effetti tossici. Per essere quantificata la virulenza si possono usare parametri come:
• Dose Letale 50 (DL 50): dose infettante che uccide il 50% degli animali inoculati;
• Dose infettante 50 (DI 50): dose che infetta 50% delle colture cellulari Alcune definizioni importanti:
• Ceppo virulento: causa una malattia evidente o più grave di uno meno virulento
• Ceppo avirulentato: non causa malattia (usato per vaccini)
• Ceppo attenuato: non causa malattia o la causa molto ridotta (usato per vaccini).

46 Microbiologia - Artini
Alcuni virus possono causare infezioni persistente portando ad un’epatite cronica che può evolvere in cirrosi ed eventualmente causare il cancro del
fegato. Nella tabella sono riassunte informazioni fondamentali sui virus dell’epatite:
• nella riga in alto ci sono l’elenco dei principali virus responsabili dell’epatite
• La tabella è divisa in azzurro e arancione per raggruppare i virus a seconda della modalità di trasmissione
o Epatite A ed E sono trasmesse per via oro-fecale, perché il virus è eliminato con le feci e tramite esse
viene tramesso ai soggetti che lo assumono per via orale con acqua o cibi contaminati.
o Epatite V, C e Delta sono tramesse per via parenterale cioè attraverso il sangue, quindi è necessario che il sangue di un soggetto portatore dell’infezione entri in contatto con il sangue di un oggetto sano. In passato questo avveniva per trasfusione di una sacca non controllata, adesso può avvenire in seguito ad una puntura con uno strumento contaminato. Più
raramente può essere trasmesso per via sessuale, attraverso le mucose genitali. Il virus dell’epatite A ed E non cronicizzano mai e causano solo infezioni acute che si risolvono spontaneamente in pochi mesi con nessun potenziale oncogeno, al contrario il virus dell’epatite B, C e Delta possono dare infezioni croniche che potenzialmente durano
per tutta la vita se non curate e possono condurre alla comparsa di epatocarcinoma. La diagnosi di infezione viene fatta ricercando antigeni virali o anticorpi specif ici nel sangue e viene confermata l’identificazione dell’acido nucleico del virus sempre nel sangue. Sono disponibili i vaccini solo per i virus dell’epatite A e B. Il primo è suggerito per coloro che viaggiano in zone ad alta endemia,
mentre il secondo è obbligatorio in Italia da molti anni e copri tutti i nati a partire dal 1979.
Hepatitis A Virus (HAV) Il nome della famiglia è Picornavirus, quindi è un virus molto piccolo, resistente ai detergenti, agli acidi e a una temperatura di 60 °C. Potendo sopravvivere per diversi mesi sia in acqua dolce che salata. Non ha un envelope, ma solo un piccolo capside icosaedrico formato da proteine virali. Il genoma è costituito da una singola catena lineare di RNA polarità positiva che funziona immediatamente
da mRNA nelle cellule infettate, senza il bisogno di generare trascritti. Il genoma codifica per più di 10 proteine, di cui alcun e sono proteine strutturali e altre proteine non strutturali. Quando il virus, che è stato ingerito con cibo o acqua contaminata, arriva al fegato si manifesta l’infezione acuta in seguito ad un periodo di latenza che dura 2-8 settimane. Questa fase acuta esita nella guarigione nel 99,5% dei soggetti, mentre ad un 0,5% può
comparire un quadro di epatite fulminante con un danno epatico talmente esteso da far rischiare la morte. Quando il quadro di epatite acuta è sintomatico, non obbligatorio, compaiono i sintomi classici dell’epatite: febbre, astenia, nausea, anoressia, dolore addominale, urine scure e feci chiare.
Hepatitis B virus (HBV) Questo virus è endemico nel mediterraneo come l’HAV, ma è molto più
pericoloso. HBV appartiene alla famiglia dell’Hepadnavirus (virus epatici a DNA), è piccolo con un genoma circolare a doppia catena incompleta, in cui la catena a polarità negativa è completa mentre la
catena complementare a polarità positiva non è completa nei virioni circolanti ma viene completata solo quando il genoma virale entra nel nucleo delle cellule infettate. Questo DNA codifica per 4 mRNA che vengono tradotti in proteine strutturali e non. Nel virione circolante il
DNA è circondato da un capside icosaedrico formato da proteine virali, circondato a sua volta da un envelope fosfolipidico contenente le proteine di superficie del virus.
La replicazione di HBV è unica: pur essendo un virus a DNA, HBV replica mediante un intermedio ad RNA utilizzando una trascrittasi inversa virale, quindi solo dal punto di vista della replicazione questo virus si comporta come un retrovirus. Dopo la penetrazione nella
cellula, il capside rilascia il genoma nel nucleo, dove il DNA virale a doppio filamento incompleto, viene completato per formare la doppia
a = nelle feci, b = nel siero

47 Microbiologia - Artini
catena circolare. A questo punto nel nucleo, vengono trascritti un mRNA lungo che codifica per l’intero genoma e gli mRNA che verranno tradotti nelle proteine dei virioni nascenti. Tutti questi RNA vengono portati nel citoplasma della cellula infettata, dove
l’mRNA maggiore viene impacchettato nei nucleocapsidi nascenti insieme alla trascrittasi inversa. Essa sintetizza sia la prima catena a polarità negativa sia una parte della catena di DNA complementare. A questo punto, il capside si riveste con una porzione di membrana di RE contenente le proteine virali di superfice e il virus esce dalla cellula senza danneggiarla, per andare ad infettare altre cellule.
La storia clinica delle infezioni da HBV è articolata e può prendere diverse direzioni:
• il virus raggiunge il fegato tramite il sangue ed inizia a replicare nelle cellule epatiche entro 3 giorni dal momento del contagio, ma i sintomi possono passare inosservati fino a 6 settimane. Si è in piena epatite acuta, quando la risposta immunitaria cellulo-mediata fa morire le cellule epatiche producendo gli stessi sintomi dell’epatite A.
o Se la risposta immunitaria è tempestiva e robusta, la malattia evolve verso la guarigione, come avviene in ~80% dei casi.
o Se la risposta immunitaria ritarda ed è scarsa, la malattia evolve verso la cronicizzazione (~20% dei casi). L’epatite cronica non trattata persiste per tutta la vita, danneggiando progressivamente il fegato. In ~20% dei portatori
d’infezione cronica, compare un quadro di epatite cronica attiva che evolve in cirrosi. ~3% dei soggetti cirrotici ogni anno sviluppa epatocarcinoma. Questa storia clinica può essere rallentata o più raramente interrotta con le terapie antivirali.
Hepatitis C virus (HCV) Il HCV causa un’infezione con alcune caratteristiche simili a quelle dell’HBV pur
essendo un virus diverso. È un flavivirus (famiglia Flaviviridae) e si trasmette anch’esso per via ematica e più difficilmente per via sessuale. Il genoma è un unico filamento di RNA polarità positiva che codifica per 10 proteine strutturali ed enzimatiche. Nel virione circolante l’RNA genomico è contenuto in un capside di
proteine virali, avvolta in un pericapside che contiene glicoproteine virali. La replicazione di HCV si svolge tutta nel citoplasma. Il virus entra nell’epatocita utilizzando vari tipi di recettore. Quindi il genoma si libera dall’envelope e dal capside.
Nel citoplasma inizia subito la traduzione delle proteine virali sull’RNA genomico che funge da mRNA e contemporaneamente inizia la replicazione dell’RNA ad opera di una polimerasi virale. Le cellule eucariotiche non possiedono una RNA polimerasi
RNA dipendente. Quindi si assemblano i nuovi capsidi intorno agli RNA genomici appena sintetizzati a formare le nuove particelle virali. Le particelle virali nascenti escono dagli epatociti per gemmazione senza danneggiare gli epatociti infettati. Infatti, il danno epatico è immuno-mediato, cioè dovuto all’eliminazione di epatociti infettati da parte dei lin fociti T citotossici.
Sulla base di differenze nella sequenza genica dei ceppi virali isolati, il HCV è stato diviso in 6 genotipi che sono distribuiti geograficamente in maniera diseguale (guardare tabella). I genotipi hanno una diversa aggressività come il genotipo 1b che in genere causa quadri clinici più aggressivi e meno sensibili alle
terapie virali. Il quadro clinico dell’epatite da HCV ricalca quello da HBV ma con un ribaltamento delle percentuali di cronicizzazione. Si passa quindi da una fase di epatite acuta, quasi sempre asintomatica, ma al contrario di HBV, solo il 20-30% dei casi va spontaneamente incontro a guarigione, mentre più dei 2/3 cronicizzano con un 20% di casi cronicizzati che evolvono in cirrosi ed in alcuni casi fino
all’epatocarcinoma. L’epatite da HCV è più subdola, perché la fase acuta dell’infezione è quasi sempre asintomatica e la cronicizzazione e le sue brutte conseguenze compaiono in 2 pazienti su 3.
Cirrosi epatica È un quadro di completo sovvertimento dell’architettura epatica, causata dal portarsi di un insulto di qualsiasi natura sul parenchima epatico capace di
produrre infiammazione cronica, morte delle cellule, rigenerazione del tessuto epatico danneggiato e riparazione dell’insulto con la fibrosi. La classica triade della cirrosi è. necrosi degli epatociti, fibrosi a partenza dai tranci infiammatori e rigenerazione del tessuto epatico. Il risultato è una
formazione di noduli di tessuto di rigenerazione epatica strozzati dai tralci di fibrosi. Nel tempo l’architettura epatica viene completamente sovvertita, con una riduzione del volume totale del fegato, una riduzione del tessuto epatico
funzionante che viene lentamente sostituito da tessuto cicatriziale fibroso (tralci bianchi). È proprio l’infiammazione protratta la principale causa della progressione verso l’epatocarcinoma
primitivo.
Regione Genotipo prevalente
Europa, Nord America,
Giappone
Genotipo 1° e 1b (genotipo 2 e 3
meno comuni)
SudEst Asia Genotipo 3
Egitto e
Medioriente, Africa Centrale
Genotipo 4
Sud Africa Genotipo 5
Asia Genotipo 6

48 Microbiologia - Artini
Sono tipicamente suddivisi in 3 sottofamiglie:
α - herpesvirus
Herpes simplex virus (HSV-z e HSV-2)
Virus Varicella Zoster (VZV)
• Spettro d’ospite ampio
• Ciclo riproduttivo breve
• Portano alla completa distruzione delle cellule infettate (citolisi)
• Nell’ospite infezione o Litica in cellule epiteliali
(epidermide e mucosa) o Latente in cellule neuronali
(neurotropismo α)
β - herpesvirus
Citomegalovirus umano (HCMV)
Roseolovirus (HHV-6 e HHV7)
• Spettro d’ospite più ristretto
• Ciclo riproduttivo lento
• Nell’ospite infezione o Litica in cellule linfoidi o Latente nelle ghiandole, cellule
linforeticolari, fegato e altri tessuti
• No neurotropismo
γ - herpesvirus
Virus Epstein Barr (EBV)
Herpesvirus umano 8 (HHV-8)
• Spettro d’ospite molto ristretto
• Ciclo riproduttivo lento
• Nell’ospite infezione latente di linfociti B e T
• No neurotropismo
Andando dall’alto verso il basso, si restringe lo spettro di ospiti infettabili, il ciclo riprodu ttivo si fa più lento ed aumenta la tendenza
ad instaurare rapporti di latenza intracellulare del virus. In generale, gli herpesvirus sono virus piuttosto grandi, hanno per genoma una doppia catena lineare di DNA contenuto in un capside icosaedrico a sua volta avvolto da un envelope di derivazione cellulare che contiene molte glicoproteine codificate dal genoma virale.
Nel tegumento sono contenute proteine ed enzimi virali che favoriscono l’inizio della replicazione. La caratteristica fondamentale di questi virus è la capacità di rimanere latenti in specifici tipi cellulari dell’ospite, dopo l’infezione primaria. In queste condizioni di latenza, il virus esprime pochissimi geni virali, nascondendosi dalle difese immunitarie. In risposta a stimoli di varia natura, il virus si può riattivare tornando al ciclo litico. È caratteristico proprio di queste specie virali la possibilità di passare dal ciclo litico alle condizioni
di latenza e viceversa, a seconda delle condizioni dell’ospite. Dopo che un herpesvirus ha riconosciuto la sua cellula ospite, tramite i recettori di membrana, l’envelope si fonde con la membrana plasmatica della cellula e rilascia il nucleocapside nel citoplasma ove migra lungo i microtubuli, verso il nucleo nel quale viene
rilasciato il genoma virale. Nel nucleo vengono immediatamente trascritti i geni delle proteine precocissime che regolano la trascrizione genica e il controllo del metabolismo cellulare. È il momento in cui il virus prende il controllo della cellula ospite. A seconda se le condizioni metaboliche della cellula sono favorevoli o meno alla replicazione virale, vengono prese due direzioni
alternative:
• Se la cellula è permissiva, vengono espressi i geni precoci e tardivi del virus che permettono la replicazione virale e l’assemblaggio dei nuovi virioni che vengono portati attraverso le strutture del citoplasma verso la superficie della
cellula ospite per essere rilasciati da questa.
• Se la cellula non è permissiva, il virus non replica ma rimane nel nucleo in condizioni di latenza, fino a quando un cambiamento nelle condizioni di salute ed equilibrio dell’ospite non porteranno la cellula in condizioni di
permissività. In questo caso il virus passa dalla latenza alla replicazione attiva.
Herpes Smilpex , HSV-1 Si tramette per contatto con la saliva o il fluido delle vescicole cutanee. In genere causa solo infezioni orali ed è estremamente diffuso visto che il 90% della popolazione possiede anticorpi contro questo virus. ¼ di queste hanno riaccensioni periodiche della
replicazione virale. Le cellule epiteliali dell’epidermide o delle mucose sono la sede dell’infezione litica sia nell’infezione primaria che nelle riaccensioni di malattia, quindi queste cellule muoiono per rilasciare le nuove particelle virali. Nelle cellule dei gangli nervosi sensitivi il virus rimane latente per tutta la vita dell’ospite. La sede più frequente della prima infezione è il cavo orale ove può

49 Microbiologia - Artini
manifestarsi con una gengivostomatite che può anche essere molto grave. Quando invece si ha una riaccensione di malattia, questa si manifesta con il classico herpes labiale.
Dopo aver replicato nelle cellule epiteliali sede dell’infezione, le cellule muoiono liberando i virioni che entrano nell’assone delle fibre nervose sensitive locali ove migrano fino al corpo cellulare del neurone situato nel ganglio sensitivo. Nel nucleo di questo neurone il virus rimane latente anche
per molti anni, fino a quando, in risposta a stimoli locali o a stimoli sistemici, si ha una riattivazione del virus latente che riprende a replicare e migra nuovamente lungo lo stesso assone andando a raggiungere la regione di cute
o di mucosa ove si era verificata l’infezione inizia le. Qui i virioni infettano nuovamente le cellule epiteliali, dove la replicazione è nuovamente litica quindi compare di nuovo il danno del tessuto. Occasionalmente, in condizioni
di grave deficit dell’immunità, il virus può risalire lungo le vie nervose f ino al SNC dando una gravissima encefalite. In genere il virus rimane latente nel ganglio del nervo trigemino e in caso di affaticamento o calo delle difese immunitarie si riattiva migrando a ritroso
lungo gli assoni che aveva percorso per arrivare al ganglio. Per questo motivo la recidiva di herpes labiale compare sempre nello stesso punto (sede del primo contagio).
Herpes Simplex, HSV-2 Ha caratteristiche molto simili a HSV-1 ma si verifica prevalentemente a livello genitale. Il contagio avviene per via sessuale o all’atto
della nascita per i nati da madri con infezione attiva al momento del parto. Anche questo ciclo compie un ciclo replicativo litico nelle cellule epiteliali, mentre nelle cellule nervose dei gangli sensitivi rimane latente. Se il contagio avviene in età adulta, la malattia si manifesta con la comparsa di papule che si trasformano in vescicole che po i ulcerano. Le lesioni mucose o cutanee saranno simili ma attenuate nell’infezione ricorrente che può comparire dopo fasi di latenza
più o meno lunghe. Se il contagio avviene al momento della nascita, la malattia sarà devastante con un’infezione dissemina ta agli organi viscerali e un’encefalite. Sono sempre quadi molto gravi che lasciano gravi danni neurologici anche in caso di guarigione.
Virus Varicella Zoster (VZV) Via di trasmissione: aerosol respiratorio o fluido delle vescicole.
È un α-herpesvirus, come gli herpes simplex, quindi ha in comune con essi: la capacità di instaurare infezioni latenti nei neuroni, manifestazioni ricorrenti, ruolo dell’immunità cellulo-mediata, le lesioni vescicolari caratteristiche.
L’infezione primaria è più frequente nell’infanzia ed è più spesso asintomatica, quando è sintomatica si manifesta con il quadro clinico tipico della varicella. Nella varicella i virioni che hanno infettato le prime vie respiratorie diffondono nei linfonodi regionali della gola, in cui replicano per poi riversarsi nelle vie linfatiche
andando a contaminare il fegato e la milza. In questi organi si ha una nuova ondata di replicazione cui segue una seconda ondata di viremia che porta i virioni a infettare la cute
provocando il tipico esantema della varicella. Entro un paio di settimane la malattia si risolve spontaneamente ma il virus può penetrare nei neuroni cutanei, migrando verso i gangli nervosi dove rimangono latenti per anni potenzialmente per tutta la
vita. Dopo l’infezione primaria sintomatica come nella varicella o asintomatica, l’infezione può rimanere latente nei neuroni dei
gangli sensoriali per molti anni e manifestarsi clinicamente in occasione di un indebolimento, di uno stress psicofisico o di una riduzione delle difese immunitarie. Questo tipo di infezione ricorrente è più frequente nell’adulto.
In genere l’infezione ricorrente colpisce una sola radice nervosa e si manifesta sul dermatomero (singolo ganglio spinale sensitivo) corrispondente. La comparsa dell’esantema
è sempre preceduta da un dolore intenso e da un arrossamento della cute.
Virus di Epstein-Barr (EBV) È il virus responsabile della mononucleosi infettiva. La via di trasmissione è rappresentata dalla saliva (chiamata anche malattia del bacio). La prevalenza dell’infezione della popolazione è molto alta, ma molto individui non sanno di aver contratto l’infezione perché
è più frequente il contagio in età infantile e nei bambini l’infezione è asintomatica. Nelle cellule epiteliali di bocca e fa ringe, l’infezione
3 Fasi dell’esantema:
Vescicole contenenti fluido con virioni infettanti
Opacizzazione successiva delle vescicole, quando esse
diventano purulente per un’infezione batterica
Essiccazione delle vescicole sostituite da croste

50 Microbiologia - Artini
è produttiva, mentre i linfociti B sono la sede di infezione latente, che potenzialmente persiste per tutta la vita. Si distinguono 3 sindromi cliniche nelle infezioni da EBV:
• Sindrome acuta – mononucleosi infettiva positiva ad anticorpi eterofili
• Sindrome cronica – malattia cronica ricorrente ciclica (fatigue sindrome, febbricola, cefalea)
• Sindrome cronica – disordini linfoproliferativi Il percorso della malattia acuta inizia con l’infezione dell’orofaringe. L’infezione delle cellule epiteliali causa una faringite e la diffusione di nuovi virioni nella saliva e quindi il contagio di nuovi soggetti. Contemporaneamente vengono infettati i linfociti B loca li che vengono
spinti verso la proliferazione incontrollata e l’immortalizzazione, di conseguenza vengono attivati i linfociti T che si attivano per limitare la crescita dei linfociti B e promuovono la latenza del virus. L’abnorme attivazione dei linfociti T causa un’abbondante presenza di forme atipiche nel sangue causa anche la colonizzazione di milza, fegato e linfonodi da parte di un numero enorme di linfociti T.
Questa è la causa di alcuni sintomi caratteristici di questa malattia: l’importante splenomegalia4 che può portare alla rottura della milza, l’epatomegalia e l’ingrossamento dei linfonodi prevalentemente a livello della nuca e del collo. La risoluzione dell’infezione e molti dei sintomi derivano quindi dai linfociti T.
Il quadro tipico della mononucleosi infettiva si manifesta dopo 4-7 settimane (periodo di incubazione lungo) e include sempre la febbre, la faringite, l’ingrossamento dei linfonodi della nuca e del collo. Al quadro clinico acuto di mononucleosi, si possono aggiungere raramente episodi recidivanti di stanchezza cronica, febbricol a, cefalea e mal di gola o saltuariamente la comparsa di episodi di proliferazione incontrollata dei linfociti B che negli anni possono
esitare in linfomi e in altri tumori delle linee cellulari linfatiche.
Citomegalovirus (CMV) Via di trasmissione: sangue, saliva, latte, secrezioni, trapianti. È un virus altamente diffuso nella popolazione, essendo stati trovati sedi di infezione nel 70% degli adulti sani. Riesce a resistere per un tempo indefinito nell’organismo contagiato, dove rimane latente anche molto a lungo, ma nei soggetti sani difficilmente causa malattia. Deve essere considerato quindi un agente opportunistico,
capace di produrre malattia solo in soggetti immunocompromessi. Solo nei linfociti T e nei macrofagi resta latente mentre l’infezione produttiva avviene nelle cellule epiteliali e altri tipi ce llulari. L’immunità cellulo-mediata dei linfociti T è fondamentale per la risoluzione della malattia ed il mantenimento della latenza dei virus. Quindi la comparsa di malattia grave e recidive, deriva sempre da una forte riduzione dell’immunità cellulo-mediata. Nei soggetti immunocompetenti non compare malattia, sono tutti portatori asintomatici. La
malattia può invece comparire nei neonati da madre sieropositiva, sia in infezione congenita per passaggio transplacentare de l virus sia in infezione perinatale, per assunzione con il colostro o il latte dopo la nascita. La malattia può comparire nei soggetti immunocompromessi adulti o affetti da AIDS, dove compare una malattia sintomatica multicentrica in cui possono essere colpiti tutti
gli organi perché il recettore cellulare cui si lega questo virus è ubiquitario (sta su tutte le cellule). I soggetti a rischio sono quindi: neonati da madre con infezione, individui sessualmente attivi, riceventi di sangue o organi, ustionati e immunocompromessi.
Sono una vasta famiglia di virus noti da anni che causano malattie che vanno dal comune raffreddore a malattie più gravi come la sindrome respiratoria medio-orientale (MERS) e la sindrome respiratoria acuta grave (SARS). Il virione di dimensioni medie include per genoma una singola catena lineare di RNA a polarità positiva che quindi nelle cellule dell’ospite può funzionare direttamente da mRNA per la produzione di proteine. Il genoma è complessato con la proteina N a formare un core o capside che si avvolge a spirale
su sé stesso. Un’envelope o pericapside circonda tutto. Le principali proteine sono:
• la nucleoproteina N, che forma il core elicoidale con RNA;
• la proteina E1: proteina transmembrana dell’envelope
• la proteina E2: spike (spuntoni), lega il recettore cellulare e fonde virus con la cellula permissiva
• la proteina L: l’RNA polimerasi RNA dipendente necessaria per la replicazione del virus;
• la proteina H1: emoagglutinina Le caratteristiche del comportamento biologico sono:
• resistenza ai detergenti grazie a corona di glicoproteine nell’envelope
• la maggior parte replica a 35°C (solo nelle vie aeree)
• la maggior parte causano raffreddore comune (~10% dei raffreddori)
• SARS-COV1, MERS-COV e SARS-COV2 replicano a 37°C, quindi nelle vie aeree più profonde → polmoniti Il recettore delle cellule permissive è il ACE2 (Angiotensinconverting enzyme), espresso nelle vie aeree ma soprattutto nelle cellule epiteliali degli alveoli. Viene espresso anche in altri organi come il tratto intestinale, rene e cuore, quin di anche queste sedi il virus
potrebbe in teoria entrare ed infettare. La proteina E2 lega il recettore ACE2 e poi fonde il virus con la membrana della cel lula epiteliale. Tutta la replicazione si svolge nel citoplasma. Una volta entrato nella cellula tram ite un endosoma, l’RNA virale viene liberato nel citoplasma dove è tradotto dai ribosomi in un’unica poliproteina, che viene scissa in varie proteine, che vanno a comporre
la RNA polimerasi RNA dipendente che trascrive il genoma virale a polarità positiva in un’RNA complementare a polarità negativa che a sua volta è usato come stampo per produrre due tipi di RNA. Prima vengono sintetizzati una serie di RNA positivi subgen omici di varie taglie, che verranno poi tradotti in proteine strutturali del virus nel RER della cellula. Quindi vengono trascritte tante copie del genoma completo, sempre a polarità positiva, che andranno a costituire il corredo genetico dei virioni nascenti. Quando sono pronte
4 Aumento di volume della milza, oltre le normali dimensioni
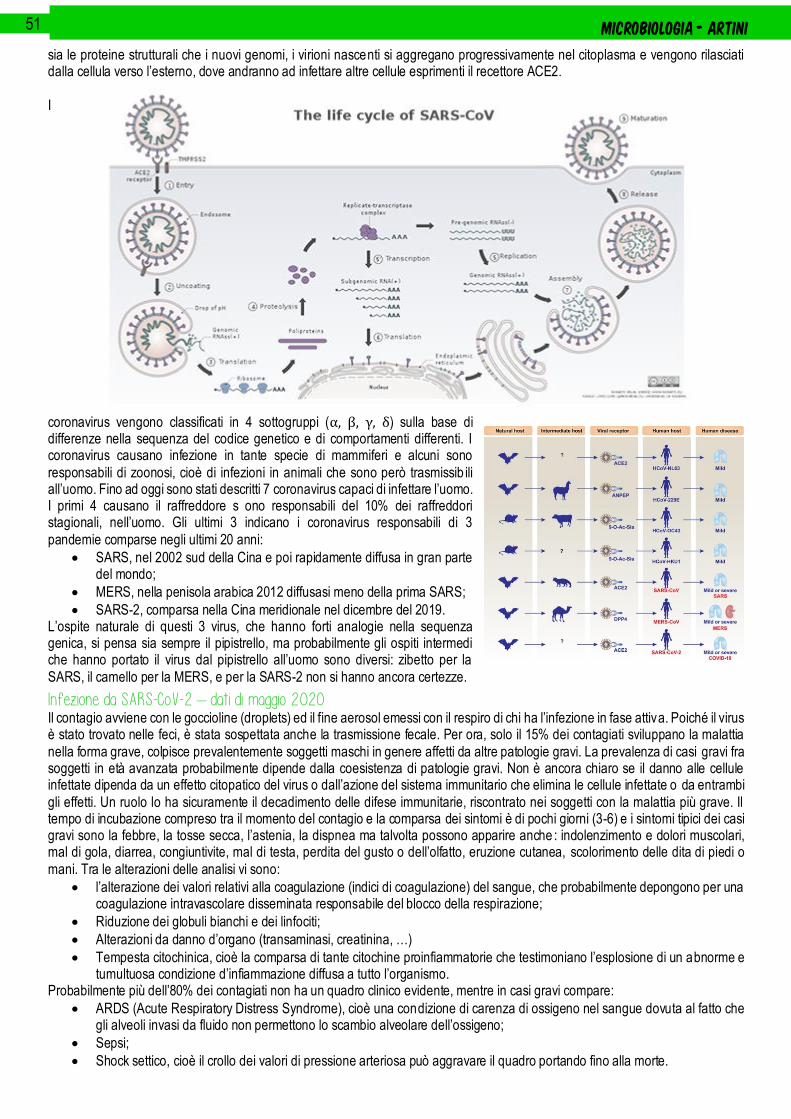
51 Microbiologia - Artini
sia le proteine strutturali che i nuovi genomi, i virioni nascenti si aggregano progressivamente nel citoplasma e vengono rilasciati dalla cellula verso l’esterno, dove andranno ad infettare altre cellule esprimenti il recettore ACE2.
I
coronavirus vengono classificati in 4 sottogruppi (α, β, γ, δ) sulla base di differenze nella sequenza del codice genetico e di comportamenti differenti. I coronavirus causano infezione in tante specie di mammiferi e alcuni sono
responsabili di zoonosi, cioè di infezioni in animali che sono però trasmissib ili all’uomo. Fino ad oggi sono stati descritti 7 coronavirus capaci di infettare l’uomo. I primi 4 causano il raffreddore s ono responsabili del 10% dei raffreddori stagionali, nell’uomo. Gli ultimi 3 indicano i coronavirus responsabili di 3
pandemie comparse negli ultimi 20 anni:
• SARS, nel 2002 sud della Cina e poi rapidamente diffusa in gran parte del mondo;
• MERS, nella penisola arabica 2012 diffusasi meno della prima SARS;
• SARS-2, comparsa nella Cina meridionale nel dicembre del 2019. L’ospite naturale di questi 3 virus, che hanno forti analogie nella sequenza genica, si pensa sia sempre il pipistrello, ma probabilmente gli ospiti intermedi che hanno portato il virus dal pipistrello all’uomo sono diversi: zibetto per la
SARS, il camello per la MERS, e per la SARS-2 non si hanno ancora certezze.
Infezione da SARS-CoV-2 – dati di maggio 2020 Il contagio avviene con le goccioline (droplets) ed il fine aerosol emessi con il respiro di chi ha l’infezione in fase attiva. Poiché il virus è stato trovato nelle feci, è stata sospettata anche la trasmissione fecale. Per ora, solo il 15% dei contagiati sviluppano la malattia
nella forma grave, colpisce prevalentemente soggetti maschi in genere affetti da altre patologie gravi. La prevalenza di casi gravi fra soggetti in età avanzata probabilmente dipende dalla coesistenza di patologie gravi. Non è ancora chiaro se il danno alle cellule infettate dipenda da un effetto citopatico del virus o dall’azione del sistema immunitario che elimina le cellule infettate o da entrambi
gli effetti. Un ruolo lo ha sicuramente il decadimento delle difese immunitarie, riscontrato nei soggetti con la malattia più grave. Il tempo di incubazione compreso tra il momento del contagio e la comparsa dei sintomi è di pochi giorni (3-6) e i sintomi tipici dei casi gravi sono la febbre, la tosse secca, l’astenia, la dispnea ma talvolta possono apparire anche : indolenzimento e dolori muscolari, mal di gola, diarrea, congiuntivite, mal di testa, perdita del gusto o dell’olfatto, eruzione cutanea, scolorimento delle dita di piedi o
mani. Tra le alterazioni delle analisi vi sono:
• l’alterazione dei valori relativi alla coagulazione (indici di coagulazione) del sangue, che probabilmente depongono per una coagulazione intravascolare disseminata responsabile del blocco della respirazione;
• Riduzione dei globuli bianchi e dei linfociti;
• Alterazioni da danno d’organo (transaminasi, creatinina, …)
• Tempesta citochinica, cioè la comparsa di tante citochine proinfiammatorie che testimoniano l’esplosione di un abnorme e tumultuosa condizione d’infiammazione diffusa a tutto l’organismo.
Probabilmente più dell’80% dei contagiati non ha un quadro clinico evidente, mentre in casi gravi compare:
• ARDS (Acute Respiratory Distress Syndrome), cioè una condizione di carenza di ossigeno nel sangue dovuta al fatto che gli alveoli invasi da fluido non permettono lo scambio alveolare dell’ossigeno;
• Sepsi;
• Shock settico, cioè il crollo dei valori di pressione arteriosa può aggravare il quadro portando fino alla morte.

52 Microbiologia - Artini
La diagnosi dei soggetti sintomatici può essere posta sulla base del quadro clinico e confermata dalla dimostrazione del virus direttamente nel tampone faringeo e dalla ricerca degli anticorpi mediante un test ELISA. Le terapie per ora sono solo di supporto,
quindi non dirette all’eliminazione del virus ma ad aiutare il paziente a superare la fase critica. Tra esse ci sono:
• la ventilazione assistita che in molti casi non serve più, forse perché i pazienti vengono ricoverati subito e perché si incomincia immediatamente la correzione dello sbilanciamento della coagulazione dei vasi sanguigni e il reintegro dei liquidi e i farmaci antinfiammatori.
• Fluidi
• Antibiotici per superinfezioni batteriche Tra i trial sperimentali si ha un cocktail di antivirali, anticorpi monoclonali e farmaci sperimentali. Sono in studio anche varie formulazioni di vaccini.
Miceti o funghi I miceti sono microrganismi eucarioti. Appartengono alla categoria dei miceti: lieviti, muffe e funghi eduli. I funghi e le muffe svolgono
un ruolo fondamentale nella catena alimentare perché sono i principali decompositori delle parti dure dei tessuti vegetali. I funghi eduli vengono utilizzati come alimento nell’uomo.
Possono presentarsi come microrganismi uni- o pluricellulari. Il corpo o tallo ha un’organizzazione pluricellulare nelle muffe e nei
funghi eduli, mentre nei lieviti è unicellulare. La struttura cellulare è tipicamente eucariotica. Il citoplasma è racchiuso dal plasmalemma (membrana di glicoproteine, fosfolipidi ed ergosterolo). All’esterno di essa si trova la tunica, una parete cellulare rigida. Alcuni lieviti possiedono una capsula di natura polisaccaridica.
Il metabolismo è di tipo chemioeterotrofo, in quanto i funghi utilizzano composti organici come fonte di energia e di carbonio. Generalmente, sono aerobi o aerobi facoltativi, solo alcuni funghi che vivono nell’intestino degli animali sono anaerobi. I funghi crescono meglio ad un pH ~5 e resistono maggiormente alla pressione osmotica, infatti sopportano alte concentrazioni di zucchero
o sale. Riescono a crescere su substrati con un basso contenuto di azoto e di umidità, condizioni non favorevoli per la crescita dei batteri. Per queste caratteristiche è più semplice l’isolamento colturale.
Sono miceti con un tallo filamentoso (micelio) costituito da ife fittamente intrecciate. Le ife sono lunghe filamento di cellule unite fra
loro e si distinguono in:
• Cenocitiche: muffe le cui ife appaiono come singole cellule estremamente allungate dotate di molteplici nuclei;
• Settate: sono presenti delle pareti trasversali (Setti) che le dividono in singole unità similcellulari mononucleate o polinucleate. Nei setti sono spesso presenti dei pori che permettono la continuità citoplasmatica fra cellule adiacenti.
Nel micelio si distinguono due parti funzionali: ➢ Micelio aereo: svolge una funzione replicativa. Si proietta al di sopra della superficie del substrato sul quale cresce la muffa
e produce le spore che ne rappresentano la forma riproduttiva ➢ Micelio vegetativo: svolge una funzione nutritiva. Le ife vegetative crescono aderenti al substrato su cui la muffa si moltiplica
favorendo l’assorbimento dei nutrienti.
I lieviti sono miceti non filamentosi che hanno un tallo unicellulare. La loro forma è tipicamente ovale o arrotondata. Vengono dette anche blastospore perché si riproducono per gemmazione. Se le blastospore vanno incontro a gemmazioni successive senza staccarsi, si forma una corta catena di gemme chiamata pseudoifa.
Dal punto di vista metabolico si comportano come anaerobi facoltativi.
Sono i miceti ceh pososno presentarsi alternativamente sotto forma di l ieviti o muffe al variare delle condizioni ambientali. Si
presentano sotto forma di ife quando crescono nell’ambiente, mentre nella fase parassitaria tissutale si presentano come blastospore. Nei funghi il dismorfismo è temperatura dipendente: a 37 °C si comportano da lieviti e a 25 °C da muffe.
I miceti si riproduzione esclusivamente mediante spore. Le spore possono avere origine sessuale o asessuale. I due tipi di riproduzione si alternano nel ciclo vitale di uno stesso micete. Le spore fungine sono una vera e propria forma riproduttiva che non manifesta l’estrema tolleranza al calore e la longevità delle spore batteriche.
La riproduzione asessuata (funghi imperfetti) può avvenire mediante: ➢ ife non specializzate con produzione di tallospore ➢ ife aree specializzate per tale scopo e si produrranno dei conidi.
Le spore asessuate sono prodotte mediante mitosi e conseguente divisione cellulare senza fusione di nuclei cellulari. Quindi, quando
germinano, le spore diventano organismi geneticamente identici al micete che le ha generate. I tipi di spore asessuate sono: ➢ artospore, tallospore che si formano in seguito a frammentazione di un’ifa settata in singole cellule; ➢ blastospore, tallospore che si formano da gemme che protrudono alla superficie della cellula madre;

53 Microbiologia - Artini
➢ clamidospore, tallospore a parete spessa che derivano dall’arrotondamento ed espansione di un segmento ifale o di una blastospora terminale o intercalare in una pseudoifa;
➢ conidiospore (o conidi), spore unicellulari o pluricellulari prodotte dall’estremità di un’ifa specializzata detta conidioforo. ➢ Sprorangiospore, conidiospore che si formano all’interno di un sacco (sporangio) portato all’estremità di un’ifa aerea
(sporangioforo). La riproduzione sessuata (funghi perfetti) prevede la formazione di spore con un processo divisibile in 3 fasi:
1. La plasmogamia, in cui un nucleo aploide di una cellula donatrice penetra nel citoplasma di una cellula ricevente; 2. La cariogamia, che consiste nella fusione dei nuclei delle due cellule coinvolte con formazione di uno zigote diploide; 3. La meiosi.
In base alle modalità con cui si espletano queste tre fasi i miceti vengono distinti in: zigomiceti, ascomiceti e basidiomiceti.
Un’infezione fungina è chiamata micosi. Per molti miceti saprofiti il parassitismo è facoltativo, transitorio o accidentale. Alcuni miceti
vivono come commensali sulla superficie delle mucose dell’uomo. Il commensalismo fungino può essere assimilato a un parassitismo ben tollerato. Il micete si sviluppa a spese del suo ospite senza causargli un danno apprezzabile ma senza dargli alcun vantaggio. Quando l’equilibrio tra replicazione del micete e i meccanismi di difesa dell’ospite si spezza, il micete può prendere il sopravvento comportandosi da patogeno. In questo caso si utilizza il termine micosi opportunistiche. La malattia può essere favorita da una
serie di fattori, come l’alterazione della normale flora microbica dopo terapia antibiotica ad ambio spettro, la deabilitazione dell’ospite in seguito a particolari terapie (Steroidi, radioterapia o chemioterapia) o alterazioni delle difese del sistema immunitario (AIDS). In un ospite gravemente immunodepresso, debilitato o traumatizzato anche un innocuo micete ambientale può diventare patogeno e
causare una micosi opportunistica. Esistono anche miceti parassiti obbligati, che vivono solo a spese di uno o più ospiti. Nell’uomo, i miceti cheratinofili antropofili sono parassiti obbligati che non hanno una vita saprofitica. In genere, essi sono responsabili di infezioni croniche (a lungo decorso) poiché crescono lentamente e in sedi non sottoposte al controllo del sistema immunitario.
Le micosi vengono classificate in 4 gruppi in funzione del grado di interessamento dei tessuti e delle modalità di ingresso nell’ospite: ➢ Micosi sistemiche: sono infezioni profonde che colpiscono diversi tessuti e organi. Queste infezioni hanno inizio
tipicamente nei polmoni e la trasmissione avviene per inalazione di spore presenti nell’ambiente. Dalla prima sede polmonare per diffusione ematica l’infezione può poi interessare altri tessuti. Esempio: criptococcosi.
➢ Micosi sottocutanee: sono infezioni che interessano il derma e sono causate generalmente da funghi saprofiti che vivono nel terreno o sulla vegetazione. La sporotritrae è un’infezione sottocutanea, a cui sono esposti giardinieri e agricoltori, che si contrae per impianto diretto di spore o frammenti di micelio a seguito di una ferita cutanea anche di lieve entità.
➢ Micosi cutanee: interessano esclusivamente l’epidermide e i suoi annessi (capelli, unghie e peli). I dermatofiti sono il più tipico esempio di miceti che causano questo tipo di infezioni. Essi si trasmettono da uomo a uomo o da animale a uomo tramite contatto diretto o indiretto con peli o squame epidermiche infette. I dermatofiti sono in grado di degradare la cheratina
presente negli strati crnei della cute e annessi utilizzando l’enzima cheratinasi. ➢ Micosi superficiali: interessano solo la superficie dell’epidermide e dei peli. Queste infezioni sono prevalenti nei climi
tropicali.
La membrana citoplasmatica dei miceti è caratterizzata dalla presenza di ergosterolo. L’inibizione della biosintesi dell’ergosterolo è alla base della tossicità selettiva di molti antimicotici, fra i quali: polieni, azoli e allilamine. Quando questa biosintesi è interrotta, la membrana diventa eccessivamente permeabile, portando alla morte della cellula.
L’amfotericina B è il più utilizzato fra i componenti della famiglia degli antimicotici polienici per la terapia delle infezioni fungine sistemiche. LA tossicità di questo farmaco, soprattutto a carico dei reni, è un fattore che ne limita fortemente l’uso. La somministrazione in dorma di liposomi è in grado di moderare la tossicità.
I primi azoli utilizzati furono gli imidazoli, come il clotrimazolo e il miconazolo, che sono utilizzati per applicazioni topiche per il trattamento di micosi cutanee e per le infezioni vaginali da lieviti. Il ketoconazolo possiede uno spettro d ’azione estremamente ampio e può essere somministrato per via orale come alternativa meno tossica in caso di infezioni sistemiche. Le preparazioni topiche di questo farmaco sono utilizzate nella cura delle dermatomicosi. L’utilizzo di tale farmaco è stato ridotto drasticamente in seguito
all’introduzione dei triazolici. I farmaci di questo gruppo (fluconazolo e itraconazolo) sono meno tossici e sono idrosolubili, quindi la somministrazione è più semplice ed efficace in caso di infezioni sistemiche. Un nuovo antimicotico ad ampio spettro è il voriconazolo e si pensa possa rimpiazzare l’amfotericina B nella terapia delle micosi del
SNC grazie alla sua capacità di penetrare la barriera emato-encefalica. Le allilamine bloccano la biosintesi dell’ergosterolo a uno stadio precoce, inibendo l’enzima squalene epossidasi. La terbinafina e la naftina sono allilamine frequentemente utilizzate in caso di resistenze ad antimicotici azolici.
Le echinocandine inibiscono la sintesi dei glucani compoennti caratteristici della parete cellulare dei funghi, inducendo la produzione di una parete cellulare incompleta e quindi la lisi della cellula. L’unica molecola attualmente in commergio è il caspofungin.
La flucitosina interferisce con la biosintesi dell’RNA e quindi con la sintesi proteica. La sua tossicità selettiva deriva dalla capacità della cellula fungina di confertire la flucitosina in 5-fluorouracile, che viene incorporato nell’RNA e determina un blocco della sintesi

54 Microbiologia - Artini
proteica. Le cellule dei mammiferi non possiedono un enzima capace di convertire il farmaco nella forma attiva. Ha uno spettro di attività ristretto e la sua tossicità per i reni e il midollo osseo ne limita ulteriormente l’uso.
Sterilizzazione e disinfezione Quando si vuole ridurre o eliminare la presenza di batteri, virus, funghi o alghe ma anche spore (le più difficili da eliminare) si effettua una disinfezione o una sterilizzazione.
Definizioni importanti:
• Sterilizzazione: elimina ogni forma di vita, è un processo che rende sterile e con il quale si uccidono anche le spore. Mai s i può pensare di sterilizzare una superficie o un tessuto del corpo umano.
• Disinfezione: riduce la carica di microrganismi, uccidendo batteri e miceti o inattivando i virus; non sempre elimina le spore.
• Antisettico: sostanza che si oppone alla crescita o all’azione dei microrganismi, sia uccidendo o inibendo la crescita. Il termine è di solito usato per indicare la detersione ferita o per la superficie corporea.
• Sanificazione: comprende la pulizia e la disinfezione. Si usano composti che riducono i contaminanti microbici a livelli di sicurezza come determinato da requisiti di sanità pubblica.
• “-cida”: uccide i batteri o inattiva i virus
• “-statico”: arresta la crescita dei batteri o inibisce la replicazione del virus Per bloccare l’efficacia batteriostatica o battericida di un composto chimico possono essere utilizzate la MIC (minima concentrazione inibente) oppure il metodo di Kirby-Bauer. A seconda della sede in cui possono essere utilizzati gli agenti chimici per ridurre la
crescita microbica possono essere definiti come:
• Antisettici → antisepsi
• Disinfettanti → disinfezione
• Agenti per sterilizzare → sterilizzazione
• Conservanti → conservazione dei cibi
• Antibiotici sintetici → agenti chemioterapici
• Antibiotici
Si parla di antisepsi quando si vuole ridurre drasticamente la carica microbica sulla pelle o sulle mucose, senza eliminazione completa
dei microrganismi che in queste sedi non può essere raggiunta. Per l’antisepsi si usano agenti chimici innocui e non tossici per l’uomo, tra essi: mercurio, nitrato di argento, soluzioni iodate, alcoli, detergenti. Solo per uso esterno. La prima forma di antisepsi passa per il lavaggio accurato delle mani con detergenti. La clorexidina è meno efficace di quanto dichiarato perché alcuni batteri Gram- sono resistenti. Comunque clorexidina e soluzioni iodate sono i presidi più utilizzati per lo
scrub chirurgico, il lavaggio accurato di chi entra in sala operatoria. L’alcol rientra nei principali antisettici anche se alla lunga secca la pelle. L’uso dei guanti ovviamente è sempre consiglia to ed è necessario quando si lavora su agocannule, flebo, dispositivi intravenosi o intrarteriosi.
• Lozioni e saponi: lavaggio delle mani per contatto con pazienti non infetti di 15-30 sec. È una forma di antisepsi debole, infatti batteri e virus possono rimanere sulla pelle.
• Clorexidina 2% 15-30 sec. – contatto con pazienti infetti o con i relativi fluidi corporei. Svantaggio: allergia di alcuni individui, in questi casi usare iodio, saponi antibatterici o alcol. Vantaggio: la sua attività antisettica persiste dopo l’asciugatura.
• Clorexidina 4% 6 min – scrub chirurgico
• Iodio – scrub chirurgico; spesso dopo lavaggio con alcol
• Alcol – molto efficace ma non uccide le spore; secca la pelle e l’attività non persiste dopo l’asciugatura
• Uso dei guanti, da cambiare tra un paziente e l’altro, riduce l’eventualità di disseminare l’infezione
Agenti chimici che uccidono i microrganismi ma non necessariamente le loro spore, ma non sono sicuri per l’applicazione su tessuto vivente infatti si usano su
oggetti inanimati come tavoli, pavimenti, strumenti, etc. Tra di essi ci sono: il coloro, ipoclorito, composti clorinati, liscivia, solfato di rame, composti dell’ammonio quaternario.
L’etanolo, l’isopropanolo, la tintura di iodio e il nitrato d0’argento sono i p iù comuni antisettici. L’alcol esercita la sua funzione antisettica in modo migliore quando è diluito in acqua al 70%.
I composti fenolici e i composti di ammonio quaternario possono essere usati come antisettici a bassa concentrazione e come disinfettanti ad alta concentrazione.

55 Microbiologia - Artini
Può essere ottenuta con agenti chimici attivi contro diverse forme di vita con uno o più dei meccanismi seguenti:
• Ossidazione della cellula
• Modificazione della permeabilità cellulare
• Lisi della cellula
• Denaturazione delle proteine
• Inattivazione di enzimi Sono agenti chimici utilizzati nella sterilizzazione:
• Gas di diossido di cloro: usato per decontaminare edifici che hanno subito un attacco con antrace, è ossidante e uccide le spore;
• Acido peracetico: soluzione liquida ossidante. Rappresenta un metodo di elezione per la sterilizzazione di strumenti sensibili al calore, è limitato alla strumentazione che deve essere usata immediatamente (oggetti umidi);
• Plasma di perossido di idrogeno: ossidante, si attiva con il microonde per aumentare la concentrazione dei radicali di ossigeno che hanno azione “-cida”.
Sterilizzazione a gas plasma È una nube di elettroni, ioni, radicali liberi, molecole dissociate che agisce mediante generazione di radiazioni ionizzanti, prevalentemente UV. Per la sterilizzazione si usa soprattutto il gas plasma di perossido di idrogeno. È una sterilizzazione a freddo, particolarmente utile per lo strumentario adatto al calore o sensibile all’umidità (es. carta o tessuti). È molto usato negli ospedali e
non lascia residui, quindi è privo di tossicità per gli addetti. Il ciclo dura 75’ a 45°C, in ambiente secco.
Sterilizzazione con agenti chimici in forma gassosa Per ottenere chimicamente un efficiente sterilizzazione si possono usare gas altamente tossici, in contenitori dedicati in cui vengono eliminate tutte le forme di vita.
• La glutaraldeide è uno sterilizzante chimico a freddo, è in soluzione acquosa attivata resa alcalina ed elimina anche le spore batteriche. È tossica soprattutto per contatto.
• L’ossido di etilene viene erogato in una piccola camera a gas dedicata. Denatura le proteine di membrana ed è battericida anche su spore se somministrato per 8-12 h a 50°C. Bisogna porre particolarmente attenzione perché viene assorbito dagli
oggetti che non devono essere manipolati per 48 h.
Conservanti Sono agenti chimici usati per inibire la crescita dei microrganismi soprattutto nei cibi. Devono essere continuamente present i per
essere efficaci e quindi dovrebbero essere privi di tossicità. Alcuni esempi sono: propinato di calcio, benzoato di sodio, sali di ammonio quaternario, nitrati, diossido di zolfo.
Agenti fisici Temperatura È il sistema più utilizzato. L’efficienza di un trattamento al calore dipende dal tipo di calore (umido o secco), dal tempo di esposizione
e dalla temperatura raggiunta. In generale, i risultati migliori si ottengono con temperature più alte e tempi di esposizioni più lunghi. I tentativi di sterilizzazione con acqua bollente non sono efficaci perché alla pressione di 1atm la temperatura non può superare i 100 °C e a questa temperatura sopravvivono alcuni batteri termofili.

56 Microbiologia - Artini
Invece, il vapore sotto pressione in autoclave garantisce una sterilizzazione efficacie perché in queste condizioni si denatu rano le proteine microbiche. L’unico limiti è che non si possono sterilizzare miscele contenenti anticorpi, vitamine, antibiotici o zuccheri che
sono labili al calore. È il metodo più pratico, economico ed efficace usato in tutti gli ospedali e laboratori. In autoclave la temperatura viene portata a 121 °C utilizzando una pressione di 1atm per 15 min. Non è un sistema utilizzabile per materiale biologico degradabile o per gli strumenti taglienti perché nel tempo ne smussa i margini. Uccide ogni cosa tranne alcuni termofili estremi. Con il calore a secco si possono sterilizzare contenitori chiusi, gelatine oleose e petrolio che non sono penetrati dal vapore, ma il
tempo necessario per ottenere la sterilizzazione è molto più lungo di quello richiesto dall’autoclave. In genere sono necessa rie 2 h a una temperatura di 160 °C o solo 1 h per una temperatura di 170 °C. nella stufa o forno Pasteur possono essere sterilizzati bisturi o altri taglienti senza che perdano l’affilatura della lama.
Tindalizzazione È l’unico metodo fisico che garantisce l’eliminazione delle spore batteriche usato anche per gli alimenti. Questa tecnica prevede di alternare 30 min di riscaldamento a temperatura moderatamente alta (60-100 °C), seguito da un giorno di incubazione a 37 °C. Il
processo è ripetuto per almeno 3 volte. Ad ogni fase di riscaldamento, a 100 °C vengono uccise le forme vegetative che potrebbero essersi rigenerate dalle spore durante la precedente incubazione di 30 min a 37 °C. In questo modo vengono eliminate tutte le spore.
Radiazioni È un sistema di sterilizzazione per strumenti e superfici. Le onde ad alta energia causano la perdita di elettroni che producono
composti reattivi capaci di uccidere i microrganismi grazie al danneggiamento del DNA e delle proteine. Le radiazioni usate sono:
UV, raggi-X e raggi-γ. Le radiazioni ultraviolette sono utilizzate per ridurre la carica batterica negli ambienti, come le sale operatorie dove vengono accesi di notte o quando non c’è nessuno. Esse distruggono il DNA e ne inibisce la replicazione e inducono la formazione di composti sfavorenti nell’habitat batterico come ozono o perossido di idrogeno. L’efficienza non è completa e questo sistema è spesso
sopravalutato, serve solo ad abbattere la carica e non ad eliminare completamente i microrganismi. Le radiazioni ionizzanti sono utili la sterilizzazione di fili per sutura chirurgici, strumenti per la chirurgia, tubi e dispositivi medici e
chirurgici in materiale plastico come le siringhe. Tra queste radiazioni ci sono i raggi-β o i raggi-γ. Anche in questo caso, le radiazioni causano emissioni di elettroni che danneggiano profondamente le componenti delle cellule batteriche e dei virus.



























![2. Microbiologia - Aula 01 [Introdução à Microbiologia]](https://img.pdfslide.tips/doc/110x75/55cf9336550346f57b9cd187/2-microbiologia-aula-01-introduaao-a-microbiologia.jpg)

