Embed Size (px)
Citation preview

J Mol Cell Cardiol 28, 1271–1277 (1996)
Mild Hypoxia Induces Hypertrophy ofCultured Neonatal Rat Cardiomyocytes:A Possible Endogenous Endothelin-1-mediated MechanismHiroshi Ito, Susumu Adachi, Mimi Tamamori, Hiroyuki Fujisaki,Masato Tanaka, Meihong Lin, Hajime Akimoto, Fumiaki Marumo andMichiaki HiroeSecond Department of Internal Medicine, Tokyo Medical and Dental University, Tokyo, Japan
(Received 7 April 1995, accepted in revised form 12 December 1995)
H. I, S. A, M. T, H. F, M. T, M. L, H. A, F. M M. H. MildHypoxia Induces Hypertrophy of Cultured Neonatal Rat Cardiomyocytes: A Possible Endogenous Endothelin-1-mediated Mechanism. Journal of Molecular and Cellular Cardiology (1996) 28, 1271–1277. Hypoxic or ischemicstresses on cardiomyocytes may cause a variety of compensatory responses including cell hypertrophy. In thisstudy, we examined whether hypoxia induces hypertrophy of cardiomyocytes in vitro and whether hypoxia-induced hypertrophy is inhibited by an endothelin A receptor antagonist (BQ123). Neonatal rat cardiomyocyteswere cultured in 10% O2/85% N2/5% CO2 or 95% N2/5% CO2 to produce a mild or severe hypoxic condition,respectively. Cardiomyocytes exposed to severe hypoxia revealed degenerative morphological changes and adecrease of cell number, suggesting the toxicity of severe hypoxia on cardiomyocytes. In contrast, cardiomyocyteswith mild hypoxia developed hypertrophy; cell surface area of cardiomyocytes as evaluated by an image analysersystem increased by 1.6-fold over control after 48 h. [3H]leucine incorporation into the cells was significantlyincreased by mild hypoxia but decreased by severe hypoxia. mRNA level of skeletal a-actin, a genetic marker ofcardiac hypertrophy, up-regulated after 6–24 h by mild hypoxia. A transient increase of preproET-1 mRNA anda time-dependent increase of ET-1 protein in the culture medium were also observed in cardiomyocytes exposedto mild hypoxia. BQ123 partially inhibited either hypoxia-induced [3H]leucine incorporation or skeletal a-actinmRNA in a dose-dependent manner. These data suggest that mild hypoxia induces hypertrophy of cardiomyocytesand that activation of endogenous ET-1 may, at least in part, mediate this hypertrophic responses as an autocrine/paracrine factor. 1996 Academic Press Limited
K W: Hypoxia; Cardiomyocytes; Hypertrophy; Skeletal a-actin; Endothelin-1.
cardiomyocytes in vitro (Webster et al., 1993). TheIntroductionactivation of these protooncogenes in car-diomyocytes is known to be important for the in-Cardiomyocyte hypertrophy is commonly observed
in ischemic heart diseases such as myocardial in- duction of cardiac hypertrophy (Dunnmon et al.,1990). On the other hand, hypoxic or ischemicfarction (Anversa et al., 1985; Ginzton et al., 1989)
and ischemic cardiomyopathy (Pantely et al., 1984). stresses are known to be a direct trigger of growth-promoting effects in many physiologic and pa-Recently, Webster and coworkers reported that hyp-
oxic stresses mediate an early and sustained in- thologic conditions, such as wound healing (Ni-inikoski et al., 1972) and fibrosis of the skinduction of c-fos and c-jun protooncogenes in
Please address all correspondence to: Hiroshi Ito, Assistant Professor of Medicine, Division of Cardiology, Second Department of InternalMedicine, Tokyo Medical and Dental University, 1-5-45, Yushima, Bunkyo-ku, Tokyo 113, Japan.
0022–2828/96/061271+07 $18.00/0 1996 Academic Press Limited1271

H. Ito et al.1272
(Silverstein et al., 1988). Consequently, it can be modifications (Ito et al., 1990). Briefly, the heartsfrom 1- or 2-day-old Wistar rats (Japan Laboratoryhypothesized that hypoxic or ischemic stresses may
exert a hypertrophic effect on cardiomyocytes. How- Animals, Tokyo, Japan) were minced and dis-sociated with 0.1% trypsin. After dispersed cellsever, cardiomyocytes in ischemic heart diseases
may be exposed to many other stresses such as were incubated on 100-mm culture dishes (Falcon,Oxnard, CA, USA) for 60 min at 37°C in 5% CO2mechanical or neurohumoral stimulation. Thus, it
should be clarified whether hypoxic stress itself or incubator, non-attached viable cells were collectedand seeded into 60-mm dishes (2×106 cells/dish)other factors trigger the cardiomyocyte hy-
pertrophic responses. or 12-well plates (8×105 cells/well). Cells werethen incubated in MEM supplemented with 5% calfEndothelin-1 (ET-1) is a 21-amino acid va-
soconstrictor/pressor peptide originally isolated from serum plus cytosine arabinoside (Ara-C) (10−5 )(Wako Pure Chemical Industries, Osaka, Japan) forthe supernatant of cultured porcine aortic endothelial
cells (Yanagisawa et al., 1988). This vasoactive sub- 48 h and then replaced with MEM plus 1% serum24 h before the experiments. The hypoxic conditionstance has been found to induce cardiomyocyte hy-
pertrophy in cultured neonatal rat cardiomyocytes was produced as reported previously (Tanaka et al.,1994). Briefly, cardiomyocytes were placed in an(Ito et al., 1991; Shubetta et al., 1990; Susuki et al.,
1990). Recently, we have shown that cultured rat air-tight desiccator, where air can be completelyreplaced by a gas mixture of 10% O2/85% N2/5%cardiomyocytes express abundant preproET-1 (ppET-
1) transcripts and release mature ET-1 into culture CO2 or 95% N2/5% CO2 to produce mild or severehypoxic conditions, respectively. To measure themedium. Furthermore, we demonstrated that car-
diomyocyte hypertrophy stimulated by angiotensin II oxygen content, the medium cultured with car-diomyocytes was collected by a syringe immediatelyis partially blocked by a receptor antagonist selective
for ETA subtype (Ito et al., 1993a). These in vitro results after 30 min, 6 h and 24 h in mild or severe hypoxia,and P2 was measured using a pH/Blood Gas Ana-have led us to speculate that endogenous ET-1 pro-
duced by cardiomyocytes may be involved in the lyzer (Type 178, Ciba-Corning Diagnostic Co., NY,USA). P2 of the medium reached to 40–55 torr aspathogenesis of cardiac hypertrophy via an au-
tocrine/paracrine mechanism. early as 30 min, which continued up to 24 h inmild hypoxia and of 10–15 torr with severe hypoxiaIn this study, we examined whether hypoxia
induces hypertrophy of cultured rat car- during the time.diomyocytes, and whether hypoxia-induced hy-pertrophy is mediated by endogenous ET-1 as an
Measurement of cell number and surface areaautocrine/paracrine factor.
Cultured cardiomyocytes after normoxic or hypoxicconditions were removed from wells by 100 ll of0.05% trypsin-EDTA (Gibco BRL Products, Gai-Materials and Methodsthersburg, MD, USA), and stained with 0.4% trypanblue (Gibco BRL Products, Gaithersburg, MD, USA)Drugs and cDNAfor 5 min at 37°C. The number of trypan blue-negative cells were counted using a hemocytometer.BQ123 (Ihara et al., 1992) was provided by Banyu
Phase-contrast pictures of cultured car-Pharmaceutical Co. Ltd (Tokyo, Japan). cDNA fordiomyocytes under normoxic or hypoxic conditionsrat ppET-1 was generously provided by Dr M. Masaki(original magnification;×200) were photographed(Kyoto University, Japan). cDNA for rat ribosomalon 35-mm films and digitized into computer im-RNA was from Japanese Cancer Research Resourcesaging. The measurement software (Image, versionBank (Tokyo). cDNA probes for 3′-untranslated1.41, Wetness helix. nih. gov.) loaded on an Appleregion of rat skeletal a-actin (184 base pairs) andcomputer (Apple Computer, Inc., Cupertino, CA,cardiac a-actin (129 base pairs) were synthesizedUSA) was used to measure the surface area ofas previously described (Ito et al., 1993b).cardiomyocytes. The surface area was determinedby the mean value of 50 cells.
Cell cultureNorthern blot analysis
Primary culture of neonatal rat cardiomyocyteswas prepared by the method originally described Total RNA of neonatal rat cardiomyocytes with or
without hypoxic stimulation was isolated by theby Simpson (Simpson et al., 1982) with minor
Cardiomyocyte Hypertrophy by Mild Hypoxia 1273
EDTA). 32P-labeled cDNA probes were prepared bythe random primer method (Feinberg et al., 1983).The membranes (Magnagraph nylon; Micron Sep-arations Inc., Westborough, MA, USA) were washedtwice with 5×SSPE/10% SDS at room temperature,once each for 15 min with 2×SSPE/10% SDS at37°C and 0.2×SSPE/10% SDS at 37°C. Auto-radiography was performed on a Fuji RX film withan intensifying screen at −80°C. Autoradiogramswere quantified using BAS 2000 (Fuji Film Cor-poration, Tokyo, Japan). Quantitative data of ET-1mRNA from three independent experiments werenormalized to 18S ribosomal RNA expression andwere statistically analysed.
Radioimmunoassay
Cardiomyocytes (4×105 cells per well) were cul-tured in 12-well plates (Falcon, Oxnard, CA, USA),with 0.5 ml of MEM supplemented with 1% calfserum. After 6-h incubation in normoxic or mildhypoxic conditions, ET-1-like immunoreactivity (ET-1-LI) was determined by a specific radio-immunoassay (RIA) as previously described (Ito etal., 1993b); the antibody used cross-reacted fullywith ET-1, 2% with big ET-1, but <0.1% with ET-2 and ET-3.
Incorporation of [3H]leucine
Protein synthesis by cardiomyocytes was evaluatedby incorporating [3H]leucine into the cells. Car-diomyocytes plated on 12-well plates (2×105 cells/well) were incubated for 4,12 or 24 h under nor-moxic or hypoxic conditions. Four hours before theend of the incubation period, 0.5 lCi [3H]leucinewas added to the culture medium. Cultures were
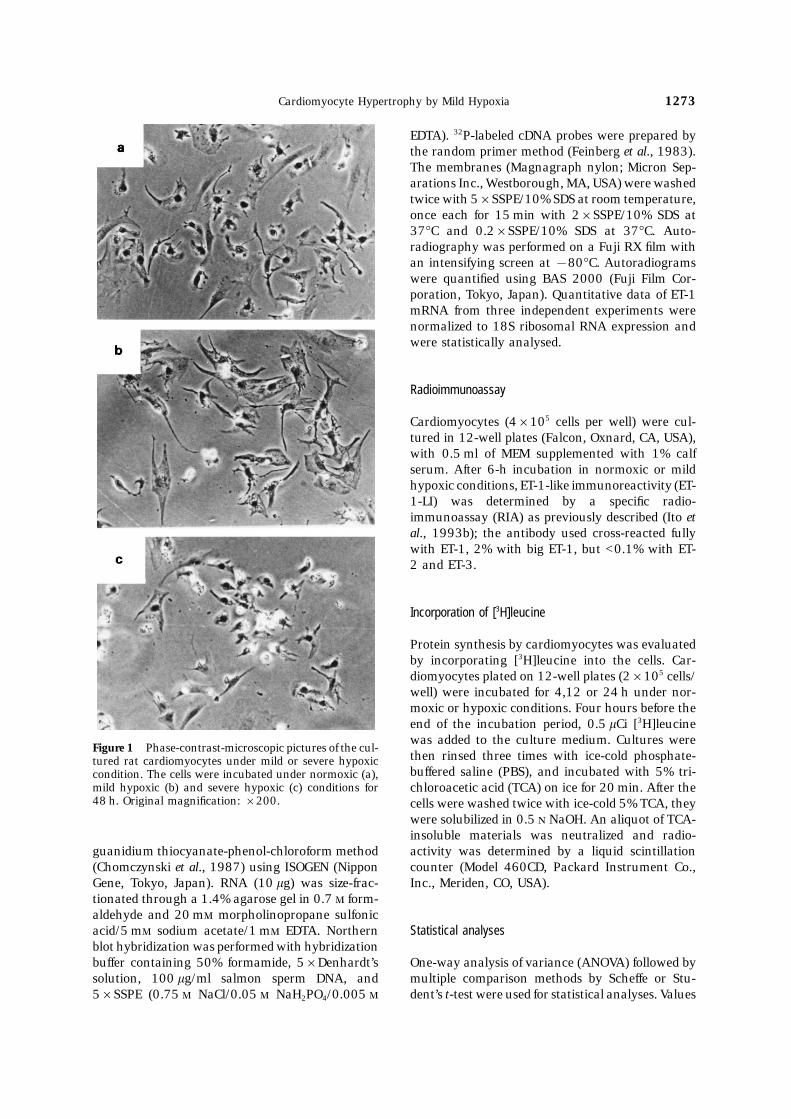
Figure 1 Phase-contrast-microscopic pictures of the cul-then rinsed three times with ice-cold phosphate-tured rat cardiomyocytes under mild or severe hypoxicbuffered saline (PBS), and incubated with 5% tri-condition. The cells were incubated under normoxic (a),
mild hypoxic (b) and severe hypoxic (c) conditions for chloroacetic acid (TCA) on ice for 20 min. After the48 h. Original magnification: ×200. cells were washed twice with ice-cold 5% TCA, they
were solubilized in 0.5 NaOH. An aliquot of TCA-insoluble materials was neutralized and radio-activity was determined by a liquid scintillationguanidium thiocyanate-phenol-chloroform method
(Chomczynski et al., 1987) using ISOGEN (Nippon counter (Model 460CD, Packard Instrument Co.,Inc., Meriden, CO, USA).Gene, Tokyo, Japan). RNA (10 lg) was size-frac-
tionated through a 1.4% agarose gel in 0.7 form-aldehyde and 20 m morpholinopropane sulfonicacid/5 m sodium acetate/1 m EDTA. Northern Statistical analysesblot hybridization was performed with hybridizationbuffer containing 50% formamide, 5×Denhardt’s One-way analysis of variance (ANOVA) followed by
multiple comparison methods by Scheffe or Stu-solution, 100 lg/ml salmon sperm DNA, and5×SSPE (0.75 NaCl/0.05 NaH2PO4/0.005 dent’s t-test were used for statistical analyses. Values

H. Ito et al.1274
Table 1 Effects of severe or mild hypoxia on [3H]leucine incorporation and number of cardiomyocytes
Leucine incorporation Cell number/well Leucine incorporation/cellsHours (cpm/well) (×100) (cpm/1000 cells)
4 control 6387±159 127±2.4 495±12severe hypoxia 5859±195 127±6.6 461±15mild hypoxia 6566±275 124±6.5 529±22
24 control 5939±100 119±5.0 499±9severe hypoxia 4165±136∗† 89±4.0∗† 468±21∗†mild hypoxia 7233±302∗† 121±9.1 538±27∗†
∗ P<0.05 v 24-h control, † P<0.05 v 4-h control.
hypertrophy was developed under hypoxic con-ditions (Fig. 1); the cell surface area of car-diomyocytes (as evaluated by an image analysersystem) was increased by 1.6-fold over the controlafter 48 h. [3H]leucine incorporation into cells wassignificantly increased by mild hypoxia (Table 1).The mRNA level of skeletal a-actin, a genetic markerof cardiac hypertrophy, was up-regulated undermild hypoxic conditions for 6–24 h (Fig. 2).
Figure 2 Effects of mild hypoxia on steady-state level ofmRNA for skeletal a-actin and preproendothelin-1 in ratcardiomyocytes. Cells were incubated under mild hypoxic Mild hypoxia increases production of endothelin-1 andconditions for indicated times. Northern blot hy- preproendothelin-1 mRNA in cardiomyocytesbridization (10 lg of total RNA/each lane) was performedas described in the text. Hybridization was performed
A transient increase of ppET-1 mRNA, peaking atusing [32P]-labeled rat skeletal a-actin and pre-proendothelin-1 (ET-1) cDNA as probes. Ethidium brom- 1 h (412±52% over control, n=3), was observedide-stained 18S-ribosomal RNA bands are shown to in cardiomyocytes in mild hypoxia (Fig. 2), in-indicate equal loading of RNA. Similar results were ob- dicating the expression of ET-1 in cardiomyocytestained from three independent cultures.
preceding the induction of hypertrophy. ET-1-likeimmunoreactivity in the medium cultured withcardiomyocytes in mild hypoxia was significantlywere expressed as mean±... A P value of lesshigher than that in normoxic condition at 6 hthan 0.05 was considered significant.(129±26 fmol/106 cells v 42±14 fmol/106 cells,n=4, P<0.01) and at 24 h (239±64 fmol/106 cellsv 73±18 fmol/106 cells, n=4, P<0.01).
Results
Severe hypoxia is cytotoxic to cardiomyocytes
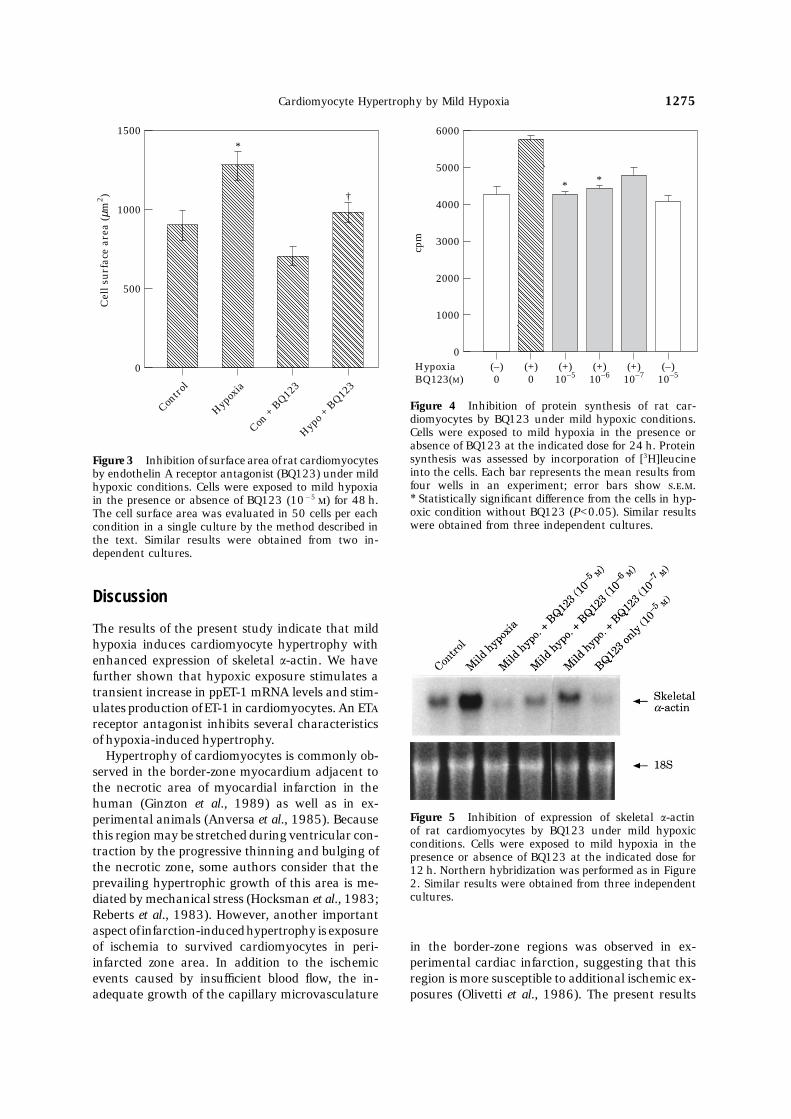
BQ123 partially inhibits cardiomyocyte hypertrophyCardiomyocytes exposed to severe hypoxia exhibited induced by mild hypoxiadegenerative morphological changes (Fig. 1). Fur-thermore, protein synthesis (as evaluated by [3H] We next examined effects of an ETA receptor ant-leucine incorporation) and a number of the cells agonist, BQ123, on hypertrophic cardiomyocyteswere decreased in cardiomyocytes cultured under induced by mild hypoxia. As shown in Figure 3,severe hypoxic conditions (Table 1). BQ123 (10−5 ) partially inhibited increases in the
surface area of cardiomyocytes stimulated by mildhypoxia at 48 h. [3H]leucine incorporation was alsoinhibited under the mild hypoxic conditions byMild hypoxia induces hypertrophy of cardiomyocytesBQ123 at 24 h in a dose-dependent manner (Fig.4). Further, BQ123 inhibited skeletal a-actin mRNAIn contrast to the overt toxic effects of severe hyp-
oxia, mild hypoxia did not induce any cytotoxicity in in cardiomyocytes induced by mild hypoxia for 12 h(Fig. 5).cultured cardiomyocytes. Moreover, cardiomyocyte
Cardiomyocyte Hypertrophy by Mild Hypoxia 1275
6000
0
cpm
5000
1000
*
2000
3000
4000
(–)0
(+)0
(+)10–5
(+)10–6
(+)10–7
(–)10–5
*
HypoxiaBQ123(M)
Figure 4 Inhibition of protein synthesis of rat car-diomyocytes by BQ123 under mild hypoxic conditions.Cells were exposed to mild hypoxia in the presence or
1500
0
Contro
l
Cel
l su
rfac
e ar
ea (
µm2 )
500
1000
Hypox
ia
Con +
BQ12
3
Hypo +
BQ12
3
*
†
absence of BQ123 at the indicated dose for 24 h. Proteinsynthesis was assessed by incorporation of [3H]leucineFigure 3 Inhibition of surface area of rat cardiomyocytesinto the cells. Each bar represents the mean results fromby endothelin A receptor antagonist (BQ123) under mildfour wells in an experiment; error bars show ...hypoxic conditions. Cells were exposed to mild hypoxia∗ Statistically significant difference from the cells in hyp-in the presence or absence of BQ123 (10−5 ) for 48 h.oxic condition without BQ123 (P<0.05). Similar resultsThe cell surface area was evaluated in 50 cells per eachwere obtained from three independent cultures.condition in a single culture by the method described in
the text. Similar results were obtained from two in-dependent cultures.
Discussion
The results of the present study indicate that mildhypoxia induces cardiomyocyte hypertrophy withenhanced expression of skeletal a-actin. We havefurther shown that hypoxic exposure stimulates atransient increase in ppET-1 mRNA levels and stim-ulates production of ET-1 in cardiomyocytes. An ET
receptor antagonist inhibits several characteristicsof hypoxia-induced hypertrophy.
Hypertrophy of cardiomyocytes is commonly ob-served in the border-zone myocardium adjacent tothe necrotic area of myocardial infarction in thehuman (Ginzton et al., 1989) as well as in ex-
Figure 5 Inhibition of expression of skeletal a-actinperimental animals (Anversa et al., 1985). Becauseof rat cardiomyocytes by BQ123 under mild hypoxicthis region may be stretched during ventricular con-conditions. Cells were exposed to mild hypoxia in the
traction by the progressive thinning and bulging of presence or absence of BQ123 at the indicated dose forthe necrotic zone, some authors consider that the 12 h. Northern hybridization was performed as in Figureprevailing hypertrophic growth of this area is me- 2. Similar results were obtained from three independent
cultures.diated by mechanical stress (Hocksman et al., 1983;Reberts et al., 1983). However, another importantaspect of infarction-induced hypertrophy is exposureof ischemia to survived cardiomyocytes in peri- in the border-zone regions was observed in ex-
perimental cardiac infarction, suggesting that thisinfarcted zone area. In addition to the ischemicevents caused by insufficient blood flow, the in- region is more susceptible to additional ischemic ex-
posures (Olivetti et al., 1986). The present resultsadequate growth of the capillary microvasculature

H. Ito et al.1276
that hypertrophy of cultured cardiomyocytes under The question of whether ppET-1 gene expressionmay be stimulated by hypoxic stress alone or sub-mild hypoxic conditions, which are free from the
effect of mechanical stresses, indicate that mild hyp- sequent reoxygenation might be raised. In our study,ET-1 mRNA induction was detectable immediatelyoxia itself can induce cardiomyocyte hypertrophy.
From those results, it may be presumed that car- after exposure to the mild hypoxia without re-oxygenation procedure, suggesting that reperfusion-diomyocytes in ischemic heart diseases may develop
hypertrophy in response to hypoxia by a mechanism mediated effects are not required for ET-1 inductionby mild hypoxia in vitro. This notion is compatibleindependent of mechanical stresses.
Locally-produced humoral factors may play an im- with the observation made in the Webster’s paper(Webster et al., 1993) which demonstrated that in-portant role in the initiation and maintenance of
cardiac hypertrophy. We have recently reported that duction of fos and jun was initiated within the 1sthour of hypoxia, well before metabolic, structural orcultured rat cardiomyocytes express abundant ppET-
1 mRNA and release mature ET-1 into the culture functional damages were detectable in the cells.medium. Further, we showed that ANG-II-inducedcardiomyocyte hypertrophy is partially inhibited byETA receptor antagonist as well as antisense oli- Acknowledgementsgonucleotides against ppET-1 mRNA. We concludedfrom the previous data that ET-1 is an extracellular This study was supported in part by Grant-in-Aid
for General Scientific Research for the Ministry ofmediator of ANG II-induced cardiomyocyte hy-pertrophy (Ito et al., 1993a). In the present study, Education, Japan, and a grant from the Research
Group of Molecular Cardiology, Japan.ppET-1 mRNA, as well as ET-1 like immuno-reactivity, was also up-regulated by exposure to mildhypoxia. Furthermore, hypertrophy of cardio-myocytes and its enhanced expression of skeletal a- Referencesactin was inhibited dose-dependently by ETA receptor
A P, L AV, L V, G G, 1985. Leftantagonist, BQ123. These data support our hy-ventricular failure induced by myocardial infarction.pothesis that mild hypoxia-induced hypertrophyI. Myocyte hypertrophy. Am J Physiol 17: H876–H882.
may be, at least in part, mediated by endogenous C P, S N, 1987. Single-step methodET-1 produced by cardiomyocytes via an autocrine/ of RNA isolation by acid quanidinium thiocyanate-
phenol-chloroform extraction. Anal Biochem 162: 156–paracrine mechanism. In our previous immuno-160.cytochemical studies, our cultures of “cardio-
F AP, V BA, 1983. Technique for radio-myocytes” contain approximately 5% of fibroblasts,labeling DNA restriction endonuclease fragments to
and we found that cardiac fibroblasts also produce high specific activity. Anal Biochem 132: 6–13.ET-1 (Fujisaki et al., 1995). Thus our results of pro- F H, I H, H Y, T M, H M, L
M, A S, A H, M F, H M, 1985.duction of ET-1 by cultured “cardiomyocytes” mayNatriuretic peptides inhibit angiotensin II-induced pro-be partly contributed by a small fraction of cardiacliferation of rat cardiac fibroblasts by blocking en-fibroblasts. In our study, the concentration of ET-1-dothelin-1 gene expression. J Clin Invest 96:
like immunoreactivity was lower than the effective 1059–1065.dose of ET-1 required to induce hypertrophy. How- G LE, C R, R DC, L MM, 1989.
Functional significance of hypertrophy of the non-ever, it is presumed that ET-1 may work through aninfarcted myocardium after myocardial infarction inautocrine/paracrine mechanism and that there mayhumans. Circulation 80: 816–822.be a higher local concentration, though this has not
H JS, B BH, 1983. Pathogenesis of leftyet been proven. It still remained to be clarified how ventrilar aneurysms: an experimental study in the ratthe mild hypoxia can cause an “anaerobic” situation, model. Am J Cardiol 50: 83–88.
I M, N K, S T, F T, T S,or how the mild hypoxic conditions cause differentK S, F T, I K, N M, Yreactions from those by severe hypoxia in car-M, 1992. Biological profiles of highly potent noveldiomyocyte hypertrophy. In our preliminary study,endothelin antagonists selective for the ETA receptor.
ET-1 mRNA was also upregulated by severe hypoxia Life Sciences 50: 247–255.in a similar manner to that by mild hypoxia (Ito, H. I H, H Y, H M, T M, A S, T
T, N M, T K, M F, 1991. Endothelin-unpublished data), suggesting that cardiomyocytes1 induces hypertrophy with enhanced expression ofmay be exposed to similar “anaerobic” stimulationmuscle-specific genes in cultured neonatal rat car-by mild and severe hypoxia. Thus, it can be spec-diomyocytes. Circ Res 69: 209–215.
ulated that “toxicity” by severe hypoxia may prevent I H, H Y, A S, T M, T M, Kthe cardiomyocyte hypertrophy even in the existence A, N A, M F, H M, 1993a. Endothelin-
1 is an autocrine/paracrine factor in the mechanismof the stimulation.

Cardiomyocyte Hypertrophy by Mild Hypoxia 1277
of angiotensis II-induced hypertrophy in cultures rat ventricular myocytes. A paracrine mechanism for myo-cardial cell hypertrophy. J Biol Chem 265: 20555–cardiomyocytes. J Clin Invest 92: 398–403.
I H, H M, H Y, T M, A S, S 20562.S JL, S VD, M J TA, F V,M, K A, N A, M F, 1993b. Insulin-like
growth factor-I induces hypertrophy with enhanced 1988. Cutaneus hypoxia in patients with systemicsclerosis (scleroderma). Arch Dermatol 124: 1379–expression of muscle specific genes in cultured rat
cardiomyocytes. Circulation 87: 1715–1721. 1382.S P, S S, 1982. Differentiation of rat myocytesI H, M SC, B ME, A H, T S,
W R, G R, L G, K L, T FM, in single cell cultures with and without proliferationnonmyocardial cells. Circ Res 50: 101–116.1990. Doxorubicin selectively inhibits muscle gene
expression in cardiac muscle cells in vitro and in vivo. S T, H H, M Y, 1990. Endothelin stimulateshypertrophy and contractility of neonatal rat cardiacProc Natl Acad Sci USA 87: 4275–4279.
N J, H TK, D JE, 1972. Oxygen supply myocytes in a serum-free medium. FEBS Lett 268:149–151.in healing tissue. Am J Surg 123: 247–252.
O G, R R, B C, G G, A P, 1986. T M, I H, A S, A H, N T,K T, M F, H M, 1994. HypoxiaResponse of the border zone to myocardial infarction
in rats. Am J Pathol 125: 476–483. induces apoptosis with enhanced expression of Fasantigen mRNA in cultured neonatal rat car-P GA, B JD, 1984. Ischemic car-
diomyopathy. Prog Cardiovasc Diseases 27: 95–114. diomyocytes. Circ Res 75: 426–433.W KA, D DJ, B NH, 1993. InductionR CS, ML D, B E, M PR,
K RA, 1983. Topographic changes in the left and nuclear accumulation of Fos and Jun proto-onco-genes in hypoxic cardiac myocytes. J Biol Chem 268:ventricle after experimentally induced myocardial in-
farction in the rat. Am J Cardiol 51: 872–876. 16852–16858.Y M, K H, K S, T Y, K-S HE, MD PM, H AN, K
KU, G CC, B JH, C KR, 1990. En- M, M Y, Y Y, G K, M T,1988. A novel potent vasoconstrictor peptide produceddothelin induction of inositol phospholipid hydrolysis,
sarcomere assembly, and cardiac gene expression in by vascular endothelial cells. Nature 332: 411–415.





























