Embed Size (px)
Citation preview

MITOCHONDRIE

Mitoch


Specifický lipid vnitřní mitoch. membrány


Komplexy vnitřní membrány rostlinných mitochondrií

Mitochondrie se dělí „zaškrcením“ podobně jako
plastidy

Vnitřní prstýnek - FtsZ1 a 2 (vzdál. příb. tubulinuGTPdep. polymerace)
a spol. Vnější prstýnek - dynamin (ARC5).

B - na dělení mitoch. se podílejívnitřní i vnější komponenty

Land Plant Mitochondrial Genome Organization
208-2400 kb depending on species
Relatively constant coding but highly variable organization among and even within a species
Physical mapping with overlapping cosmid clones
Entire complexity maps as a single “master circle”
All angiosperms except Brassica hirta have one or more recombination repeats.
Repeats not conserved among speciesDirect and/or inverted orientationsRecombination generated inversions
(inverted repeats)Recombination generated subgenomic
molecules (deletions) (direct repeats), some present at very low copy number (sublimons)
Leads to complex multipartite structures


Recombination across direct repeats leads to deletions
1 23’
4’
1 2 3 4
4’ 3’ 2’ 1’
432’1’

Plant mitochondrial genome organization(modified from Backert et al. Trends Plant Sci 2:478)
> >>>
>>
>
>
>
>>
> >>
>
>

Physical structures of plant mitochondrial DNA(Backert et al. Trends Plant Sci 2:478)
ConclusionsNo discrete size classesCircles including smaller than predictedLinears including longer than predicted
Potential MechanismsRosettesNucleoid complexesRecombination complexes
Long linear molecules and sigma moleculesRolling circle replicationRecombination
Branched linear moleculesRecombination or replication of linear molecules
Small circlesRecombination across short direct repeats

Physical structures of plant mitochondrial DNA(modified from Backert et al. Trends Plant Sci 2:478)

Physical structures of DNA obtained via rolling circle DNA replication(from Freifelder, 1983, Molecular Biology)

Physical structures of DNA obtained viarecombination of linear molecules
(from Freifelder, 1983, Molecular Biology)

Plant mitochondrial genome coding content
In organello protein synthesis indicates 30-50 proteins encoded by most plant mitochondrial genomes
Complete sequence of A. thaliana mitochondrial genome identified 57 genes respiratory complex componentsrRNAs, tRNAs, ribosomal proteinscytochrome c biogenesis
Plant mitochondrial genomes do not encode a complete set of tRNAs mit encoded tRNAs of native (mitochondrial origin)mit encoded tRNAs from imported plastid genomemissing tRNAs are nuclear encoded and imported into mitochondria to
complete the set
42 orfs in A. thaliana mit genome that might be genes
A. thaliana mitochondrial gene density (1 gene per 8 kb) is lower than the nuclear gene density (1 gene per 4-5 kb)!


Mitochondrial regulation of plant nuclear genes Plant mitochondrial respiratory electron transfer chain includes an alternative pathway for electron flow Single subunit alternative oxidase (AOX) – je rezistentní ke kyanidu! A je inhibovánaSlicylhydroxamovou kyselinou (SHAM).
Encoded by a nuclear gene (aox)
Bypasses two of three sites for H+ transfer coupled to ATP synthesis
Transcription of nuclear aox is upregulated when electron flow through the cytochrome pathway is disrupted by the inhibitor antimycin A (AA)

Význam této respirační dráhy je vedle výjimečných případů
produkce tepla (Arum) pravděpodobně v možnosti
„upouštět“ přebytečnou energii a udržet v chodu aerobní
glykolýzu, pentózový a Krebsův cyklus (tj. anabolické fce
mitochondrií).

Mitochondrial regulation of plant nuclear genes NtAI genes (Maxwell et al. Plant J 29:267)
Nuclear genes up-regulated in response to AA, including aox
Seven additional genes identified by differential mRNA display, most associated with stress responsesacc oxidase glutathione S transferaseSar8.2 cysteine proteasepathogen-induced lipase SA-induced glucosyl transferase
Also induced by reactive oxygen species (ROS) (eg H2O2)
Induction is blocked by antioxidants such as flavones
Of all inducers, AA is the most rapid. This implicates mitochondria as the site coordinating ROS signaling in the plant cell

Mitochondrial regulation of plant nuclear genes NtAI genes (Maxwell et al. Plant J 29:267)
Figure 3. Antioxidants lower intracellular ROS levels and inhibit gene induction. (a) Effects of antioxidant addition on AA, H2O2, and SA-dependent accumulation of intracellular ROS in tobacco suspension cells. ROS levels were measured 4 h after AA (5 µm), H2O2 (5 mm) and SA (1 mm) addition with and without preincubationfor 45 min with N-acetylcysteine (25 mm) or flavone (1 mm). Data represent means ± SD for three experiments. (b) Effect of the antioxidant treatment described above on the AA-, H2O2-and SA-dependent expression of Aox1 and the NtAIgenes.
NAC - N-acetylcystein a flavon = antioxidanty


Evidence for the importance of cis-guiding sequences in plant mitochondrial RNA editing
Editing of recombinant or rearranged mitochondrial genesRecombination breakpoint immediately 3’ to an editing site in rice atp6 did not abolish editingRecombination breakpoint seven nucleotides 5’ to an editing site in maize rps12 did abolish editingRecombination breakpoint 21 nucleotides 5’ to an editing site in maize rps12 did not abolish editing
Electroporation of genes into isolated mitochondria, followed by isolation of mitochondrial cDNA
Editing of mutated coxII gene demonstrated sequences from –16 to +6 required for editing
RNA editing



Import bílkovin do mitochondrií

Recognition of proteins destined for mitochondria
• This involves both terminal signal sequences and internal signal patches
• In some cases proteins for import are packaged with chaperones, especially Hsp70, but in other cases it would appear that the fully-folded protein is imported intact.
• There are two major recognition sites on the outer membrane, one recognisingfully folded proteins, the other proteins associated with chaperones

N´term. Lokalizační značka pro import do mitochondrií se nazývá
také presekvence.Často jsou to pozitivně nabité
α-helixy.

Entry of proteins into
mitochondria, general
principles
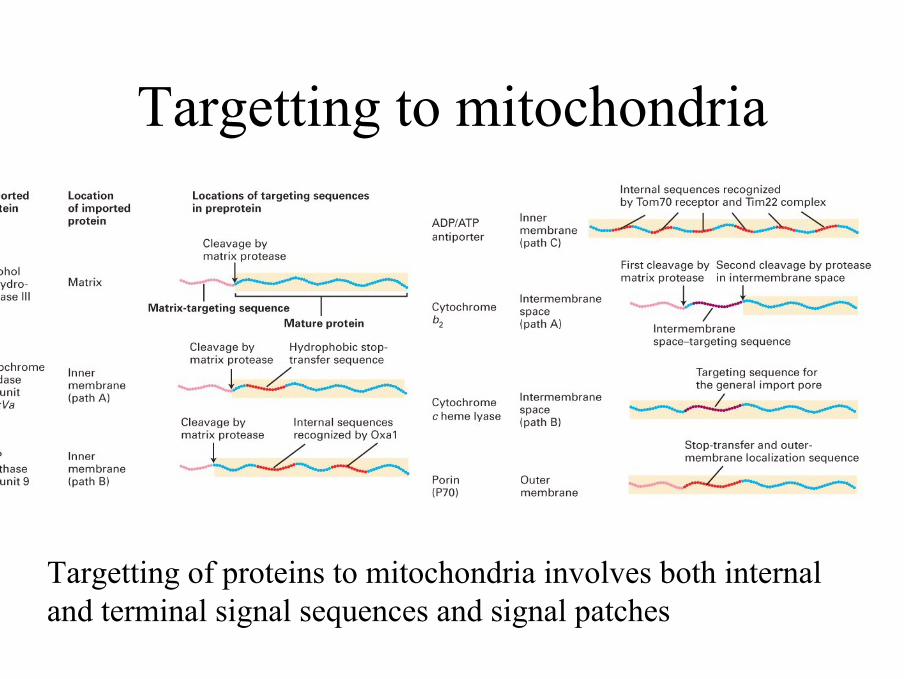
Targetting to mitochondria
Targetting of proteins to mitochondria involves both internal and terminal signal sequences and signal patches

Import into mitochondria

Transfer across the Outer membrane
• This involves the TOM (Transport - Outer Membrane) complex which is comprised of at least 8 proteins
• There appear to be three methods for recognisingproteins destined for import.
• Some proteins bind to a TOM20/22 which in turn binds to the universal transporter TOM40
• Others appear require binding to an inner membrane protein OXA1-details under dispute
• Finally yet others bind to TOM70 which presents them to TOM40

Continued• Transfer across the outer membrane does not require
ATP hydrolysis directly but obviously energy is needed as the proteins are moving up a concentration gradient. The energy is probably provided by ATP hydrolysis in binding and releasing Hsp70
• The transfer of proteins destined for the matrix is halted. This may occur soon after the outer membrane is contacted (the so called cis site) but the more important trans site halts translation leaving a length of amino acids projecting into the intermembrane space

The Transfer - Inner membrane complex
• This again is made up of a number of peptides. This complex performs a number of functions
1) It must recognise the signal on peptides projecting from the TOM complex
2) It must guide these through the inner membrane and then pull the rest of the molecule through.
• The matrix targetting signal must now be removed. This is carried out by a special Matrix processing peptidase, sometimes assisted by a second enzyme, the mitochondrial intermediate peptidase
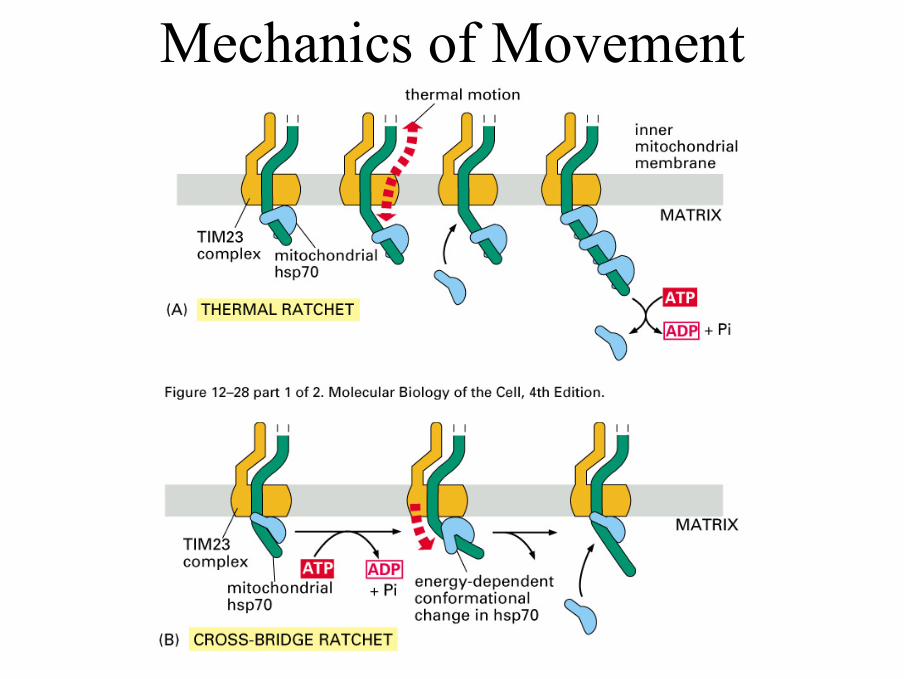
Mechanics of Movement

Contimued• Movement of proteins through the pore depends on the
maintenance of the potential difference across the membrane. This is normally app. 200 mV which is equivalent to 400,000 V/cm
• The key role in dragging the remainder of the proteins through the membrane is played by the mitochondrialform of Hsc70. Transfer requires ATP hydrolysis. Three mechanisms have been suggested. The ratchet mechanism proposes that transfer is by Brownian motion with Hsc70 binding preventing back movement. The molecular motor model proposes that the conformational change of mtHsc70 results in the protein being pulled across the membrane

TOMs and TIMs
• It is clear that there is no permanent conncetionbetween the two complexes. It is less clear whether there are temporary around the transfer pore connections or whether the sole junction is by the protein in transit. The inner membrane protein TIM54 has a projection which may reach the inner membrane
• It is unclear whether there are permanent contact sites between the inner and outer membrane or whether in real life the outer membranes lie close to the outermost parts of the inner membrane

Supply of the intermembrane space

Further processing
• Proteins are transferred across the TOM/TIM system as extended chains. A variety of chaperone proteins assist in the correct folding following transfer to the matrix
• Chaperones also assist in the assembly of inner membrane components which contain both polypeptides specified by nuclear genes and polypeptides specified by mitochondrial DNA

Import bílkovin do mitochondriíje příbuzný importu do plastidů.



Male sterility, nuclear genes
• MsMs and Msms plants are male fertile• msms plants are male sterile• A pure msms population can not be
produced– Why?

Male sterility, nuclear genes
• MsMs and Msms plants are male fertile• msms plants are male sterile• A pure msms population can not be
produced– Why?– Because msms plants can not be self-pollinated– Pollinate with Msms plants and remove male
fertile plants from the offspring

Cytoplasmic male sterility (CMS)
• Controlled by mitochondrial genes• Maternally inherited• Used for hybrid production in many crops
– Onion, carrot, cabbage– Corn, sorghum, pearl millet, sunflower, sugar
beets

CMS in onion
Allium galanthum CMS male sterile umbel

Restorer genes
• Nuclear genes (Rf) can restore male fertility (One locus restorer system)
– CMS, rfrf is male sterile– CMS, Rfrf is male fertile– CMS, RfRf is male fertile– N, rfrf is male fertile– N, Rfrf is male fertile– N, RfRf is male fertile

Flower phenotypes in carrot
a) Normal (N-cytoplasm, restored CMS plants)b) Brown anther CMS (Sa)c) Petaloid CMS (Sp)

Fertility restoration in maize
From Roger Wise’ research lab. http://www1.iastate.edu/~imagefpc/Subpages/research.html

Simple hybrid with cms and restoration
Maintainer line (B-line)N, rfrf
N1
C1
Large amountsof CMS line
xCMS line (A-line)CMS, rfrf
N1
C1
N1
C1C2x N2
Male line (C-line)N and RfRf
C1
Fertile F1 hybridCMS, Rfrf

Breeding hybrid carrots


Organelle DNA copy number can regulategene expression
Cytoplasmic male sterility (CMS) in Phaseolus vulgaris
CMS = maternally inherited failure to produce or shed functional pollen
Usually results from gain-of-function mutations (novel genes) in the plant mitochondrial genome (probably via recombination).
Mitochondrial CMS gene (orf239) is on a subgenomic molecule
The nuclear gene Fr depresses the copy number of the orf239 subgenome, decreasing the accumulation of orf239 transcripts and preventing the expression of the male sterility trait (Mackenzie and Chase Plant Cell 2:905



PPR=pentatricopeptidrepeat.

Aktivace PCD mitochondriálními signály.A regulated pathway of cell death important in normal development and disease and defense responses in plants and animals
In animals, release of mitochondrial signaling molecules, especially the electron carrier cytochrome c, activates the cell death machinery, which is already in place. This is not regulation of gene expression!A role for mitochondria in plant PCD is becoming evident, although the nature of the signaling molecules is not yet known
Figure 3. Figure 2. Many death signals converge onto mitochondria and are mediated through members of the Bcl-2 protein family called `BH3-only' proteins, such as Bid and Bad. Caspase 8 then cleaves Bid, whose C-terminal fragment (tBid) translocates to mitochondria, where it activates Bax or Bax-like proteins and results in cytochrome-c (cyt c) release. Once in the cytosol, cytochrome c activates caspase 9 by binding to Apaf-1 and dATP. from Desagher and Martinou Trends Cell Biol 10:369

„Dual targeting“ – podvojné adresování.Některé bílkoviny nesou lokalizační N-term.signál
pro lokalizaci jak do mitochondrii, tak do plastidů(příp.peroxisomů). Např. jedna ze 3 NEP
fág.podob. RNApol., AtZn-metalloproteasepro preseq./trans.pept. odštěpování. Jde bud´o
nespecifický trans./pres. peptid či o dva „lokalizační“ peptidy tandemově za sebou.

Adresování do organel je ovšem silně závislé na
diferenciačním/vývojovém stavu buňky – tj. (ne)-přítomnosti
trans-regul. bílkovin.

PEROXISOMY

PeroxisomyA-asociované s ER
B-barvené diaminobenzidinem
(kataláza)

Peroxisomal Variability
Extreme ranges Humanhepatocytes
Shape Spherical, ovoid, tubular, Ovoidsquare, triangular
Internal Amorphous, AmorphousStructure paracrystalline
Size 0.1-2.0 microns 0.5 microns
No. per cell 1-1000s 100s
% vol of cell 0.1-80% 1%

1. Lipid -oxidation (VLCFA only in mammals)(acyl-CoA-oxidase, bi/tri-functional enzyme, acyl-CoA-thiolase)
2. Steroid side chain oxidation / bile acid formation
3. Dolichol and cholesterol synthesis
4. Plasmalogen (ether lipid) synthesis(acyl-CoA:dihydroxyacetone phosphate acyltransferase,acyl-CoA:dihydroxyacetone phosphate synthase)
5. Miscellaneous H2O2-generating oxidations(urate oxidase, D-amino acid oxidase, L--hydroxy acid oxidase)
6. H2O2 catabolism(catalase)
7. Glyoxylate detoxification(alanine:glyoxylate aminotransferase)
8. Glyoxylate cycle (glyoxysomes, yeast peroxisomes)(isocitrate lyase, malate synthase)
9. Photorespiration (green plants)(glycolate oxidase, serine:glyoxylate aminotransferase)
10. Glycolysis (Kinetoplastids)
LIPIDMETABOLISM
H2O2METABOLISM
GLYOXYLATEMETABOLISM


Only somemembrane proteins(Pex3p & pex19p)travel via ER
Other membrane proteinsimported like matrix proteins

Import of Proteins into Peroxisomes
• Peroxisomes are a type of microbody. Microbodies are cell organelles bounded by a single membrane and are used for a variety if different processes. For example peroxisomescontain enzymes which produce hydrogen peroxide (and have the means for destroying it). In addition plants have glyoxysomes which contain the enzymes of the glyoxylate cycle and yeasts have a variety of microbodies including ones involved in methanol oxidation.
• PEROXINY – proteiny biogeneze peroxisomů.

Import – „peroxisome targetingsignal“ = PTS
PTS1 – Ser-Lys-Leu na C´ není odštěpován
PTS2 – N´odštěpovaný. Do matrix.

PEROXISOMAL TARGETING SEQUENCES (PTSs)
Type 1 (PTS1)
1 Carboxy-terminal2 Small (3 amino acids)3 Consensus sequence = S-K-L4 No known structure5 Not cleaved following import
Type 2 (PTS2)
1 Amino-terminal or near the amino terminus2 Medium sized (~9 amino acids)3 Consensus = (X)n-R-L-(X)5-H/Q-L4 No known structure5 Cleaved following import (in mammals)
PTS1s are much more common than PTS2s.

Targeting to peroxisomes
• Fortunately this is nice and simple. Two major targetting signals PTS1R and PTS2R have been identified. The PTS1R signal is a carboxy-terminal tripeptide SKL.
• The mechanism of PTS1R appears to involve binding of the SKL sequence to Pex5. This then interacts with a peroxisomal membrane protein called Pex14 forming a channel. It is not clear whether Pex5 and the protein move together across the channel or whether the imported protein is “pushed” through.
• In the peroxisome the protein is released and Pex5 is recycled with the help of Pex2, 10, and 12.

Podmínky importu• ATP hydrolysis is required for import • Import into peroxisomes does not require unfolding of
the protein chain - even gold particles conjugated to a peroxisomal protein are imported.
• Hsp70 is however needed and becomes bound to the exterior of the peroxisome.
• .


Peroxisome matrix and membrane
proteins enter by different
routes
Tj. nejsou peroxisomy

Mitochondrial Peroxisomalprotein import protein import
Type Post-translational Post-translational(matrix proteins)
Co-translational(some membrane proteins)
Route Direct from cytosol Direct from cytosol (matrix & (matrix proteins)memb proteins) Via ER
(some memb proteins)
Structures Unfolded monomers Folded oligomersimported

Mitochondrial Peroxisomalprotein import protein import
Targeting N-terminal C-terminal (PTS1s)sequences N-terminal (PTS2s)
Cleaved Not cleaved (PTS1s)Cleaved (some PTS2s)
+ charged amphiphilic No obvious structurealpha helices
Receptors TOM20/22 Pex5p (for PTS1s) TOM37/70 Pex7p (for PTS2s)
Location of Mitochondrial Cytosol, perox memb,Receptors memb perox matrix
Molecular Hsp70 (cyto & matrix) None knownChaperones Hsp60 (matrix)


Komunikace kompartmentů

Fotorespirace

Olejová tělíska – vznik a mobilizace.

ACP=Acyl carrier. Prot.


Oleosiny stabilizujicípovrch olejových tělísek pvolněných z tapetamají důležitou rolipři interakci/rozpoznánípylu a blizny.
Glyoxysomy a olejová tělíska.

Mobilizace zásobních tuků a glukoneogeneze.
Rostliny dokážoupřevádět tuky na cukry.
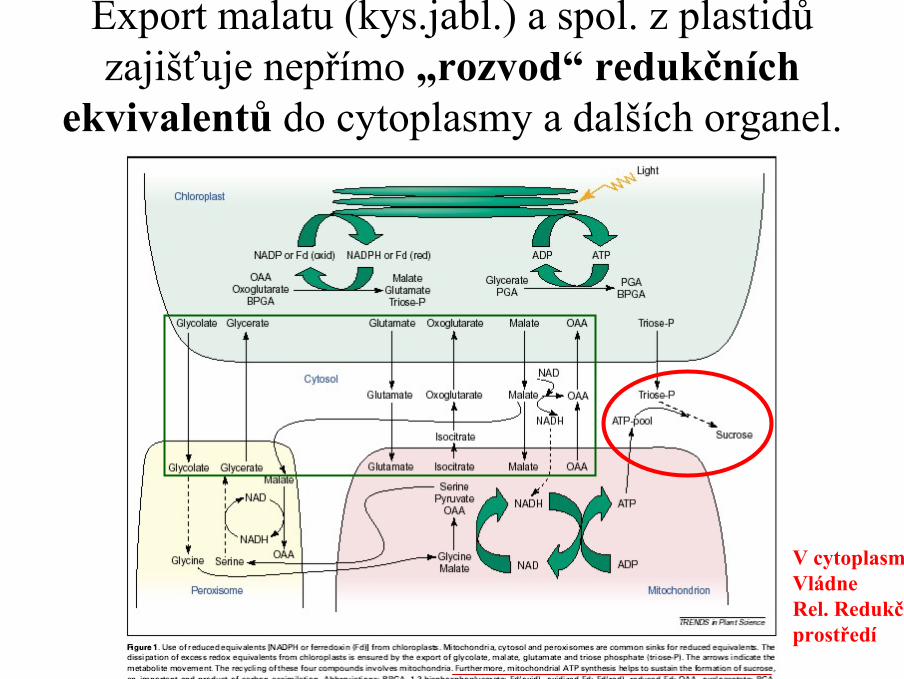
Export malatu (kys.jabl.) a spol. z plastidů zajišťuje nepřímo „rozvod“ redukčních
ekvivalentů do cytoplasmy a dalších organel.
V cytoplasměVládneRel. Redukčníprostředí

Shrnutí metabolické komunikace organel

Rostliny mutanta ječmene albostrians, kompenzují nefunkční mutantní plastidy
aktivací mitochondrií.





















![2 rostl bunka.ppt [Režim kompatibility]kfrserver.natur.cuni.cz/studium/prednasky/anatomie/anatomie/2_rostl_bunka.pdf · vakuola Rostlinná buňka je eukaryotní buňkou se základními](https://img.pdfslide.tips/doc/110x75/5cd69dce88c99300748dfdc4/2-rostl-bunkappt-rezim-kompatibility-vakuola-rostlinna-bunka-je-eukaryotni.jpg)







