Embed Size (px)
Citation preview

Mémoire présenté pour l’obtention du diplôme de
MAGISTER
En Gestion des Ressources Aquatiques
Laboratoire : Réseau de Surveillance Environnementale (L.R.S.E.) /Gestion des Ressources Aquatiques (GeReAq)
Thème :
Présenté par :
Mlle : ROUABHI Ikram Fouzia
Devant la commission du jury :
Président : Mr BOUTIBA ZITOUNI Professeur à l’université d’Oran (Es-Sénia)
Promoteur : Mr ABI-AYAD S-M-El-Amine. Maître de conférences à l’université d’Oran (Es-Sénia)
Examinateur : Mme SENHADJI LAMRI Meriem Y. Professeur à l’université d’Oran (Es-Sénia)
Examinateur : Mr BABA HAMED M. Bey Maître de conférences à l’université d’Oran (Es-Sénia)
Effet du mode de conservation sur la qualité sensorielle et biochimique
des poissons : la sardine commune (Sardina pilchardus), le rouget de
roche (Mullus surmuletus) et le merlan bleu (Micromesistius poutassou)
Année universitaire 2008 -2009

Mémoire présenté pour l’obtention du diplôme de
MAGISTER
En Gestion des Ressources Aquatiques
Laboratoire : Réseau de Surveillance Environnementale (L.R.S.E.) /Gestion des Ressources Aquatiques (GeReAq)
Thème :
Présenté par :
Mlle : ROUABHI Ikram Fouzia
Devant la commission du jury :
Président : Mr BOUTIBA ZITOUNI Professeur à l’université d’Oran (Es-Sénia)
Promoteur : Mr ABI-AYAD S-M-El-Amine. Maître de conférences à l’université d’Oran (Es-Sénia)
Examinateur : Mme SENHADJI LAMRI Meriem Y. Professeur à l’université d’Oran (Es-Sénia)
Examinateur : Mr BABA HAMED M. Bey Maître de conférences à l’université d’Oran (Es-Sénia)
Effet du mode de conservation sur la qualité sensorielle et biochimique
des poissons : la sardine commune (Sardina pilchardus), le rouget de
roche (Mullus surmuletus) et le merlan bleu (Micromesistius poutassou)
Année universitaire 2008 -2009

III
REMERCIEMENTS
C’est un plaisir que de remercier au début d’un tel travail tous ceux qui ont contribué à le rendre possible et agréable. Même si dans mon cas, cette liste peut sembler longue, c’est avec mon enthousiasme le plus vif et le plus sincère et au-delà de tout formalisme que je voudrais rendre mérite à tous ceux qui à leur manière m’ont aidé à mener à bien ce travail.
Je désire alors exprimer ma profonde gratitude Au docteur ABI-AYAD Sidi-Mohammed El-Amine pour avoir accepté de m’encadrer depuis le D.E.S. Je tiens à vous exprimer ma gratitude pour la confiance que vous m’avez témoignée en m’accueillant dans votre laboratoire. Votre rigueur scientifique, votre esprit critique, votre disponibilité et le temps consacré à la correction de mon manuscrit ont permis de mieux structurer et faire progresser rapidement ce travail. Merci de m’avoir permis de trouver, à travers votre personne, que j’espère a quelque peu déteint sur ma personnalité, les valeurs pour lesquelles j’ai emprunté la voie de la Recherche. Ce fut un plaisir de travailler avec vous !
Je tiens à exprimer mes remerciements les plus vifs et les plus sincères aux membres du jury :
• Au professeur BOUTIBA Zitouni, responsable du Laboratoire Réseau de Surveillance Environnementale (L.R.S.E.) pour avoir accepter de présider ce jury.
• Au Professeur SENHADJI LAMRI Meriem qui m’a honoré en acceptant d’évaluer et d’examiner le présent travail.
• Au docteur BABA HAMED Mohamed Bey, chef du département de Biotechnologie, qui a accepté de faire part de ce jury en acceptant d’examiner ce travail, malgré ses nombreuses obligations.
J’adresse mes remerciements les plus sincères à l’agence nationale de développement de recherche en santé (A.N.D.R.S.) pour le financement de ce travail qui rentre dans le cadre d’un projet de recherche portant le n° 03/00/00/07/052.
Je ne saurais oublier le docteur ALI M’HIDI Smail qui a conjugué gentillesse et aide apportée à mon travail lors des dosages des produits de l’oxydation des lipides. Merci pour vos encouragements et votre sympathie.

IV
J’adresse un remerciement particulier à monsieur le docteur TALEB Mohammed Zoheir pour son aide, sa gentillesse et multiples encouragements.
Je dois et j’adresse un remerciement tout particulier, avec toute mon affection et ma reconnaissance, à TOUS mes collègues du laboratoire de Gestion des Ressources Aquatiques (GeReAq) (dans le désordre ...) : El-Batoul, Mohamed, Ayed, Malika, Mouna qui ont rendu ces moments difficiles agréables et Hanane, Lila et Fatima pour leur aide tout au long des expériences et Nabil l’autre « contrôleur » de qualité. (Pardon à celles ou ceux que j’oublie de citer ici...). Sans oublier Adel, Sofiane.
J’adresse mes chaleureux remerciements aux personnes du département de Biotechnologie de l’université d’Oran (Es-sénia) spécialement Abd El Kader et Nacéra.
Et enfin, parce que remercier ne suffit pas, je dédie ce mémoire à ma petite famille

V
A mon père,
pour ses encouragements, ses multiples soutiens
et son affection quotidienne. Merci d’avoir été
présent en toutes circonstances.
Je te dédie ce travail.
A ma mère
A mes sœurs
A mon frère

IV
« Tout ce qui est impossible reste à accomplir »
(JULES VERNE)
« Tous les progrès sont précaires, et la solution d’un
problème nous confronte à un autre problème »
(MARTIN LUTHER KING)

V
LISTE DES ABREVIATIONS
[C] : Concentration
A.G. : Acide gras
A.G.L. : Acide gras libre
A.G.M.I. Acide gras mono-insaturé
A.G.P.I. : Acide gras poly-insaturé
A.G.S. : Acide gras saturé
Amb. : Ambiante
D.A. : Dinar algérien
F.A.O. : Food and Agriculture Organization
GeReAq : Gestion des ressources Aquatiques
I.F.R.E.M.E.R. : Institut Français de Recherche et d’Exploration de la Mer
I.N.R.A. : Institut National de Recherche Agronomique
J : jour
M.D.A. : Malondialdéhyde
M.F. : Matière fraiche
M.S. : Matière sèche
P.C. : Phosphatidylcholine
P.L. : Phospholipide
T.B.A. : Thiobarbituric acid (acide thiobarbiturique)
T.C.A. : Trichlotoacétate
T.G. : Triglycéryde
T° : température
TBA rs : thiobarbituric acid reactive substences (substances réactives à l’acide thiobarbiturique)
Vs : Versus

VI
INDEX DES PHOTOS
Photo 1 : La sardine commune : Sardina pilchardus (Walbaum, 1792)..
Photo 2 : Le rouget de roche : Mullus surmuletus (Linnaeus, 1758).
Photo 3 : Le merlan bleu : Micromesistius poutassou (Risso, 1826).
Photo 4 : Aspect extérieur de la sardine (S. pilchardus) à J1.
Photo 5 : Aspect de la cavité abdominale de la sardine (S. pilchardus) à J1.
Photo 6: Aspect extérieur de la sardine (S. pilchardus) (Lot A) à J2.
Photo 7 : Aspect de la cavité abdominale de la sardine (S. pilchardus) (Lot A) à J3.
Photo 8: Aspect extérieur de la sardine (S. pilchardus) (Lot A) à J5.
Photo 9 : Aspect extérieur de la sardine (S. pilchardus) (Lot B) à J2.
Photo 10 : Aspect extérieur de la sardine (S. pilchardus) (Lot B) à J5.
Photo 11 : Aspect extérieur de la sardine (S. pilchardus) (Lot C) à J30.
Photo 12: Aspect extérieur du rouget de roche (M. surmuletus) à J1.
Photo 13: Aspect extérieur du rouget de roche (M. surmuletus) (Lot A) à J2.
Photo 14: Aspect interne du rouget de roche (M. surmuletus) (Lot B) à J3.
Photo 15: Aspect externe du rouget de roche (M. surmuletus) (Lot B) à J5.
Photo 16: Aspect externe du merlan bleu (M. poutassou) frais.
Photo 17: Aspect externe du merlan bleu (M. poutassou) (Lot A) à J3.
Photo 18: Aspect externe du merlan bleu (M. poutassou) (Lot B) à J5.
Planche 1 : appareillage utilisé lors des procédés expérimentaux.
A : Etuve (Memmert) B : Evaporateur rotatif (Hahn shin Scientific Co, Modèle : HANVAPOR) C : Balance (Pionner, OHAUS) D : Centrifugeuse (Hettich Zentrifugen, Modèle : EBA-20) E : Spectrophotomètre (Genesys) F : Bain-marie (Memmert)
Planche 2 : mise en évidence des réactions substrat/réactif.
A : Réaction des phospholipides avec le réactif de thyocyanate de fer. B : Réaction des hydroperoxydes avec le réactif FOX 2. C : formation d’un complexe rosé entre le M.D.A et le T.B.A.

VII
INDEX DES FIGURES
Figure 1 : Action des différentes phospholipases sur les phospholipides (Bacot, 2004)
Figure 2 : Phase d’initiation de la peroxydation lipidique.
Figure 3 : Phase de propagation de la peroxydation lipidique.
Figure 4 : Mécanisme en chaine de la peroxydation des A.G.P.I. et nature des produits
terminaux formes (exemple de l’acide arachidonique) (Favier 2003).
Figure 5 : Principaux composés naturels ou synthétisé possédant des propriétés antioxydantes (Marc et al., 2004).
Figure 6: Evolution de la teneur en eau (g/100g M.F.) chez la sardine (S. pichardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 7 : Effet du mode de conservation sur la teneur en eau (g/100g M.F.) chez la sardine (S. pilchardus). Les histogrammes portant des lettres ou des chiffres différents sont significativement différents (p < 0,05). Figure 8 : Evolution de la teneur en lipides (g/100g M.F.) chez la sardine (S. pichardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 9 : Effet du mode de conservation sur la teneur en lipides (g/100g M.F.) chez la sardine (S. pilchardus).
Figure 10 : Evolution de la teneur en phospholipides (mg Eq P.C /100g M.F.) chez la sardine (S. pichardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 11 : Effet du mode de conservation sur la teneur en phospholipides (mg Eq P.C /100g M.F.) chez la sardine (S. pilchardus). Les histogrammes portant des lettres, des chiffres ou des symboles différents sont significativement différents (p < 0,05).
Figure 12: Evolution de la concentration en hydroperoxydes (µmoles/g lipide) chez la sardine (S. pichardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 13 : Effet du mode de conservation sur la teneur en hydroperoxydes (µmoles/g lipide)
chez la sardine (S. pilchardus).

VIII
Figure 14: Evolution de la concentration en TBA rs (nmoles MDA/g M.F.) chez la sardine (S. pichardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 15 : Effet du mode de conservation sur la teneur en TBA rs (nmoles MDA/g M.F.) chez la sardine (S. pilchardus). Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 16: Evolution de la teneur en eau (g/100g M.F) dans la chair du rouget de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 17 : Effet du mode de conservation sur la teneur en eau (g/100g M.F.) chez le rouget de roche (M. surmuletus). Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 18: Evolution de la teneur en lipides (g/100g M.F.) chez le rouget de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation.
Figure 19 : Effet du mode de conservation sur la teneur en lipides (g/100g M.F.) chez le rouget de roche (M. surmuletus).
Figure 20: Evolution de la teneur en phospholipides (mg Eq P.C/100g M.F) chez le rouget de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 21 : Effet du mode de conservation sur la teneur en phospholipides (mg Eq P.C/100g M.F) chez le rouget de roche (M. surmuletus).
Figure 22: Evolution de la concentration en hydroperoxydes (µmoles Eq CuOOH/g lipide) chez le rouget de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 23: Effet du mode de conservation sur la teneur en hydroperoxydes (µmoles Eq CuOOH /g lipide) chez le rouget de roche (M. surmuletus). Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 24: Evolution de la concentration en TBA rs (nmoles MDA/g M.F.) dans la chair du
rouget de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation. Les
histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 25: Effet du mode de conservation sur la teneur en TBA rs (nmoles MDA/g M.F.)
chez le rouget de roche (M. surmuletus).

IX
Figure 26: Evolution de la Teneur en eau (g/100g M.F.) dans la chair du merlan bleu (M.
poutassou) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes
portant des lettres différentes sont significativement différents (p < 0,05).
Figure 27 : Effet du mode de conservation sur la teneur en eau (g/100g M.F.), chez le merlan bleu (M. poutassou). Les histogrammes portant des lettres ou des chiffres différents sont significativement différents (p < 0,05).
Figure 28: Evolution de la Teneur en lipides (g/100g M.F.) dans la chair du merlan bleu (M.
poutassou) du lot A, B et C, en fonction de la durée de conservation.
Figure 29 : Effet du mode de conservation sur la teneur en lipides (g/100g M.F.), chez le merlan bleu (M. poutassou).
Figure 30: Evolution de la Teneur en phospholipides (mg Eq P.C/100g M.F.) dans la chair du
merlan bleu (M. poutassou) du lot A, B et C, en fonction de la durée de conservation. Les
histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 31 : Effet du mode de conservation sur la teneur en phospholipides (mg Eq P.C./100g
M.F.), chez le merlan bleu (M. poutassou) le long de la conservation.
Figure 32: Evolution de la concentration en hydroperoxydes (µmoles Eq CuOOH/g lipide) dans la chair du merlan bleu (M. poutassou) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 33 : Effet du mode de conservation sur la concentration en hydroperoxydes (µmoles Eq CuOOH/g lipide) chez le merlan bleu (M. poutassou) le long de la conservation.
Figure 34: Evolution de la concentration en TBA rs (nmoles MDA/g M.F.) dans la chair du merlan bleu (M. poutassou) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 35 : Effet du mode de conservation sur la concentration en TBA rs (nmoles eq MDA/g
lipides) chez le merlan bleu (M. poutassou), le long de la conservation. Les histogrammes
portant des lettres différentes sont significativement différents (p < 0,05).
Figure 36 : Evolution de la Teneur en matière sèche (M.S.) (g/100g M.F.) chez la sardine commune (S. pichardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 37 : Effet du mode de conservation sur la teneur en matière sèche (M.S) (g/100g M.F.) chez la sardine commune (S. pichardus). Les histogrammes portant des lettres ou des chiffres différents sont significativement différents (p < 0,05).

X
Figure 38 : Evolution de la Teneur en matière sèche (M.S.) (g/100g M.F.) chez le rouget de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 39 : Effet du mode de conservation sur la teneur en matière sèche (M.S) (g/100g M.F.) chez le rouget de roche (M. surmuletus). Les histogrammes portant des lettres ou des chiffres différents sont significativement différents (p < 0,05).
Figure 40 : Evolution de la Teneur en matière sèche (M.S.) (g/100g M.F.) chez le merlan bleu (M. poutassou), du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
Figure 41 : Effet du mode de conservation sur la teneur en matière sèche (M.S) (g/100g M.F.) chez le merlan bleu (M. poutassou). Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).

XI
INDEX DES TABLEAU
Tableau 1 : Teneur en lipides et en eau de quelques espèces de poisson. (a : Murray et Burt (1969), b : Poulter et Nicolaides (1985), c : Poulter et Nicolaides (1985).
Tableau 2: Grille d'évaluation organoleptique de l'état de fraîcheur des poissons entiers (Gret, 1993.)
Tableau 3 : niveau d’acceptabilité sensorielle des différents lots des 3 espèces étudiées. Les cases grisées représentent le temps du rejet organoleptique.

XII
TABLE DES MATIERES
INTRODUCTION……………………………… …………………………………… .… 1
ETUDE BIBLIOGRAPHIQUE………………… …………………………………… ... 3
A. Présentation des espèces cibles………………… …………………………… ... 3
I. La sardine commune (Sardina pilchardus)…………………………………. 3
I.1. Position systématique……………………………………………………. 3
I.2. Biologie…………………………………………………………………... 4
II. Le rouget de roche (Mullus surmuletus)……………………………………. 5
II.1. Position systématique…………………………………………………… 5
II.2. Biologie………………………………………………………………….. 6
III. Le merlan bleu (Micromesistius poutassou)……………………………… 7
III.1. Position systématique…………………………………………………… 7
III.2. Biologie………………............................................................................. 8
B. Les lipides et leurs altérations chez les poissons…………………………… ... 9
I. Caractéristiques de la chair du poisson……………………………………... 9
II. Les lipides chez les poissons………………………………………………... 10
II.1. Lipides neutres ou de réserves……………………………………………. 10
II.2. Lipides polaires ou phospholipides………………………………………. 12
III. L’altération du poisson……………………………………………………... 13
III.1. Altérations autolytiques…………………………………………………. 13
III.2. Altérations microbiologiques……………………………………………. 14
III.3. Altérations sensorielles………………………………………………….. 14
III.4. Altérations biochimiques………………………………………………... 15
a. Lipolyse………………………………………………………….... 15
b. Peroxydation lipidique……………………………………………. 16
b.1. Phase d’initiation…………………………………………….. 17
b.2. Phase de propagation……………………………………….... 17
b.3. Phase de terminaison…………………………………………. 18
IV. Les antioxydants……………………………………………………………. 20
IV.1. Antioxydants radicalaires ou primaires………………………………….. 21
IV.2. Antioxydants préventifs ou secondaires……………………………….... 21
IV.3. Antioxydants physiques…………………………………………………. 21

XIII
V. Facteurs influençant l’oxydation des lipides……………………..…………. 21
VI. Méthodes de mesure de l’oxydation des lipides……………………………. 22
VI.1. Mesure des substrats de l’oxydation…………………………………. 22
VI.2. Détermination des diènes conjugués…………………………………. 23
VI.3. Dosage des hydroperoxydes………………………………………….. 23
VI.4. Mesure des produits secondaires……………………………...……… 23
VI.5. Evaluation sensorielle………………………………………...………. 23
PARTIE EXPERIMENTALE………………… ……………………………………… . 25
A. Matériels et Méthodes……………………………………………………… ..... 25
I. Matériel biologique : échantillonnage et conservation……………….. 25
II. L’appréciation sensorielle…..………………………………………… 25
III. Caractéristiques biochimiques……………….………….……………. 26
III.1. Détermination de la teneur en eau…………………………………. 26
III.2. Détermination de la teneur en lipides totaux………………………. 26
• Choix de la méthode…………………………………….... 26
• Principe de la méthode……………………………………. 26
III.3. Détermination de la teneur en phospholipides…………………….… 27
• Choix de la méthode……………………………………… 27
• Principe de la méthode……………………………….…… 27
IV. Détermination du degré d’oxydation des lipides……………………… 27
IV.1. Détermination de la teneur en produits primaires : les hydroperoxydes 28
• Choix de la méthode……………………………………. 28
• Principe de la méthode…………………………………. 28
IV.2. Détermination de la teneur en produits secondaires : les TBA rs……… 28
• Choix de la méthode…………………….……………… 28
• Principe de la méthode…………………………………. 28
V. L’analyse statistique…………...……………………………………… 29
B. Résultats………………………………………… …………………… ..……….. 32
I. La sardine commune (Sardina pilchardus)…………………… .………. 32
I.1. L’analyse sensorielle………………………………………………....... 32
I.2. Caractéristiques biochimiques…………………………………….…… 34

XIV
I.2.1. Teneur en eau…………………………………………………….. 34
a. Effet de la durée de conservation……………………………….. 34
b. Effet du mode de conservation…………………………………. 35
I.2.2. Teneur en lipides……………………………………………………… 36
a. Effet de la durée de conservation……………………………….. 36
b. Effet du mode de conservation…………………………………. 37
I.2.3. Teneur en phospholipides…………………………………………….. 38
a. Effet de la durée de conservation……………………………….. 38
b. Effet du mode de conservation…………………………………. 39
I.3. Evolution du niveau d’oxydation des lipides…………………………… 39
I.3.1. Les hydroperoxydes………………………………………………... 39
a. Effet de la durée de conservation……………………………….. 39
b. Effet du mode de conservation…………………………………. 40
I.3.2. Les TBA rs…………………………………………………………. 41
a. Effet de la durée de conservation……………………………….. 41
b. Effet du mode de conservation…………………………………. 42
II. Le rouget de roche (Mullus surmuletus)………………………………… . 43
II.1. L’analyse sensorielle…………………………………………………….. 43
II.2. Caractéristiques biochimiques…………………………………………... 45
II.2.1. Teneur en eau……………………………………………………… 45
a. Effet de la durée de conservation……………………………….. 45
b. Effet du mode de conservation…………………………………. 46
II.2.2. Teneur en lipides…………………………………………………… 47
a. Effet de la durée de conservation………………………………. 47
b. Effet du mode de conservation…………………………………. 47
II.2.3. Teneur en phospholipides………………………………….……….. 48
a. Effet de la durée de conservation……………………….………. 48
b. Effet du mode de conservation………………………….………. 49
II.3. Evolution du niveau d’oxydation des lipides……………….………... 49
II.3.1. Les hydroperoxydes………………………………………………. 49
a. Effet de la durée de conservation………………………………. 49
b. Effet du mode de conservation…………………………………. 50
II.3.2. Les TBA rs……………………………………………………….. 51

XV
a. Effet de la durée de conservation……………………………….. 51
b. Effet du mode de conservation…………………………………. 51
III. Le merlan bleu (Micromesistius poutassou)…………………………… . 53
III.1. L’analyse sensorielle……………………………………………………… 53
III.2. Caractéristiques biochimiques……………………………………………. 54
III.2.1. Teneur en eau………………………………………………………… 54
a. Effet de la durée de conservation……………………………….. 54
b. Effet du mode de conservation…………………………………. 55
III.2.2. Teneur en lipides……………………………………………………... 56
a. Effet de la durée de conservation……………………………….. 56
b. Effet du mode de conservation…………………………………. 57
III.2.3. Teneur en phospholipides……………………………………...…….. 57
a. Effet de la durée de conservation……………………………….. 57
b. Effet du mode de conservation…………………………………. 58
III.3. Evolution du niveau d’oxydation des lipides………………………….. 59
III.3.1. Les hydroperoxydes………………………………………………... 59
a. Effet de la durée de conservation……………………………….. 59
b. Effet du mode de conservation…………………………………. 60
III.3.2. Les TBA rs………………………………………………………… 61
a. Effet de la durée de conservation………………………………. 61
b. Effet du mode de conservation………………………………… 61
C. Discussion……………………………………… ……………………………… 63
CONCLUSION ET PERSPECTIVES……………………………………………… ... 70
REFERENCES BIBLIOGRAPIQUES……………………………………………… . 71
ANNEXES……………………………………… ……………………………………… 86

XVI
Résumé
Le but de ce travail expérimental est d’étudier l’effet du mode de conservation (à
température ambiante, réfrigérée et de congélation) sur l’altération des lipides (hydrolyse et
oxydation) et la perte des propriétés organoleptiques chez la sardine (Sardina pilchardus), le
rouget de roche (Mullus surmuletus) et le merlan bleu (Micromesistius poutassou) par une
approche biochimique et sensorielle.
L’hydrolyse des lipides (lipides totaux et phospholipides, P.L.), la teneur en produits
primaires (teneur en hydroperoxydes) et en produits secondaires (teneur en TBA rs) de
l’oxydation des lipides ont été déterminés et comparés à une évaluation sensorielle.
L’évolution des paramètres biochimiques ne montre pas de différences significatives (p ≥
0,05) entre la conservation à température ambiante (environ 9° C) et au réfrigérateur (+4°C).
Cependant, ces paramètres montrent un effet significatif en fonction de la durée de
conservation. Au réfrigérateur, les teneurs les plus élevées (p < 0,05) en TBA rs (9350,58,
4193,10 et 6946,83 nmoles eq MDA/g M.F.) sont mesurées à J2, J3 et J5, chez la sardine, le
rouget de roche et le merlan bleu, respectivement. Ces pics mesurés correspondent à une
diminution de la teneur en lipides et en hydroperoxydes ainsi qu’à des niveaux d’altérations
sensorielles très avancés. La conservation au congélateur a un effet préservateur sur le niveau
d’altération des lipides et semble, de ce fait, être le meilleur moyen de conservation du
poisson, s’il n’est pas consommé durant les deux jours qui suivent sa capture.
En général, les altérations lipidiques sont plus élevées et plus rapides chez les poissons
gras (sardine) que chez les poissons semi gras (rouget de roche) et les poissons maigres
(merlan bleu).
Mots-clé : conservation, hydroperoxydes, lipolyse, oxydation des lipides, TBA rs, merlan
bleu, rouget de roche, sardine, Micromesistius poutassou, Mullus surmuletus, Sardina
pilchardus

XVII
Abstract
The effect of the storage mode on the hydrolytic and oxidative rancidity development and
the quality loss was studied at different temperature (ambient, chilled and freezed storage) and
performed by a biochemical and sensorial indices. These experiments were conducted on the
sardine (Sardina pilchardus), the red mullet (Mullus surmuletus) and the blue whiting
(Micromesistius poutassou).
The lipid hydrolysis (lipid and phospholipids), the primary (peroxyde value, P.V) and the
secondary (TBA rs index) lipid oxidation products were determinate and compared to the
sensory assessment. Most of lipid damage indices did not show significant differences (p ≥
0.05) between the different modes of conservation, whatever the studied specie. However,
theses parameters show a significant effect (p < 0.05) with the storage time, in both storage
means. The results show that the TBA rs index reaches its highest values (9350.58, 4193.10
and 6946.83 nmoles eq MDA/g F.M., respectively) measured at D2, D3 and D5, respectively
(in the sardine, the red mullet and the blue whiting, respectively). These highest values
correspond to the decrease of the lipid hydrolysis and the peroxide values and the highest
levels of the sensorial alteration. The freezing storage has a preserving effect on the lipid
damage and the sensorial alteration. Therefore, it seems to be the best mean of storage if the
fish is not consumed during the two days following its capture.
Mainly, the fatty species (sardine) present a higher and a faster lipid alteration than the
leaner ones.
Key-words: conservation, hydroperoxydes values, lipid hydrolysis, lipid oxidation, TBA rs
index, blue whiting, red mullet, sardine, Micromesistius poutassou, Mullus surmuletus,
Sardina pilchardus

XVIII
ملخص
طریقة الحفاظ على درجة حرارة (مفعول الھذف من ھذا العمل التجریبي ھو دراسةوفقدان الممتلكات )تحلیل و األكسدة ( على تخریب اللیبیدات ) عادیة، باردة أو درجة التثلیج
Micromesistiusو Mullus surmuletus و Sardina pilchardus النوعیة عند حوثpoutassou.
كمیة ( كمیة المواد األولیة ) الفوسفولیبیداتاللیبیدات الخامة و ( تحلیل اللیبیدات األكسدة اللیبیدات قدرت وقورنت ) TBA rs كمیة ( و المواد الثانویة) الھیدروبیروكسیدات
.للسمك) إحساسي( لتقدیر خارجي
بین (p ≥ 0,05) تطویر العوامل البیوكیمائیة ال تظھر أي مفعول أي تغیرات حساسة
لكن °C°4+)( ودرجة المبرد) C°9تقریبا ( الحفاظ على السمك على درجة حرارة عادیة د )p < 0,05( نالحظ أن ھذه العوامل تظھر مفعول حساس بالنسبة لمدة الحفاظ في المبرّ
(+4°C) ،لكن نالحظ أن ھذه العوامل تظھر مفعول حساس (p > 0.05) بالنسبة لمدة حفاظ TBA rs المؤشر (p > 0,05) ألكثرا المرتفعةفي المبرد كمیات
(91350.58, 4193,10, 6946.83 n moles eq MDA/g M.F.) ثم قیاسھا خاللھذه . التوالي على M .poutassouوS. pilchardus, M. surmuletus ، عند5، 2،3الیوم
الھیدروبیروكسیدات و تناسب أیضا اللیبیدات و الكمیات المرتفغة تتناسب مع انخفاض كمیة .انخفاض شدید في النوعیة الخارجیة للسمك
یمارس مفعول إیجابي على تخریب اللیبیدات و یمثل (C°18-) الحفاظ في درجة التثلیج .على السمك إن لم یستھلك خالل الیومین التابعین للصید طریقة للحفاظ أفضل
مقارنة مع السمك(S.pilchardus) الدسم عموما التخریب اللیبیدي مرتفع عند السمك
. (M.poutassou) و السمك الرقیق (M.surmuletus) النصف الدسم
، ھیدرو بیروكسیدات ، تحلیل اللیبید ، أكسدة اللیبید ، مؤشر حفاظ: الكلمات المفتاحیة
TBArs Micromesistius poutassou, , Mullus surmuletus, Sardina pilchardu

Introduction
1
INTRODUCTION
La qualité et la sécurité sont des aspects centraux du commerce du poisson sur les marchés
locaux et internationaux. Les consommateurs se préoccupent de plus en plus des questions de
sécurité et de qualité, et font pression pour que celles-ci soient assurées dans les produits de
la pêche.
La mer méditerranéenne est connue par sa richesse en produits marins. Selon la direction
de la pêche et des ressources halieutiques de la wilaya d'El Tarf, la production de poissons, en
Algérie, a enregistré en 2008 une augmentation de l'ordre de 74 % par rapport à l'année
précédente. Une pêche «miraculeuse» qui a atteint 5028 tonnes incluant toutes les variétés de
poissons et de crustacés, alors que la production de l'année 2007 n'était que de 2884 tonnes.
Néanmoins, le secteur de la pêche souffre d’un problème important qui est le gaspillage des
poissons du à la mauvaise gestion et à une conservation insuffisante.
Le poisson représente non seulement un produit de grande valeur socio-économique mais
aussi un aliment de haute valeur nutritionnelle vue sa richesse exceptionnelle en éléments
nutritifs essentiels (protéines, lipides, vitamines liposolubles, éléments minéraux, …). De
plus, il est particulièrement apprécié pour sa haute teneur en acides gras poly insaturés
(Ackman, 1989) dont les effets bénéfiques sur la santé humaine ne sont plus à démontrer
(Kinsella, 1987 ; Ackman et Ratnayake, 1990). Cependant, cette grande qualité représente
aussi l’un des principaux problèmes liés à sa conservation. En effet, durant cette dernière, la
qualité du poisson décline suite à l’action de différents facteurs comme les acides gras
hautement insaturés qui représentent le substrat préférentiel de l’oxydation des lipides
(Pearson et al., 1977 ; Pigott et Tucker, 1987). De plus, c’est une denrée rapidement
périssable, en particulier dans les zones méditerranéennes et tropicales où les techniques de
réfrigération n’existent pas toujours. La qualité du poisson se dégrade rapidement après la
capture. Sous les températures ambiantes, il s’altère en moins de 12 heures (Mazorra-
Manzano et al., 2000).
Le présent travail a un objectif essentiellement pratique : il consiste en l'examen critique
des méthodes susceptibles de renseigner le plus exactement possible sur l'état de fraîcheur ou
le degré d'altération des poissons. En général, il n'est certes pas expédient de recourir à des
recherches de laboratoire pour reconnaître si un lot de poissons est propre à la consommation ;
l'observation des caractères organoleptiques peut suffire. Mais il existe tout de même des cas

Introduction
2
où il serait utile de disposer de techniques purement objectives qui définissent la condition
hygiénique du poisson de façon suffisamment rapide et précise.
Afin de disposer d’outils permettant de limiter le développement des réactions d’oxydation
des lipides responsables de l’altération de la chair et de ce fait la baisse de la qualité du
poisson. Il est question, dans ce travail, de suivre l’évolution temporelle de la qualité
organoleptique et sensorielle du poisson, ainsi que de déterminer les étapes critiques de
conservation à différents modes.
Pour ce faire, 3 modes de conservation sont testés à savoir, la température ambiante,
réfrigérée et de congélation. De plus, la qualité du produit est abordée de différents points de
vue. En effet, nous avons suivi la dynamique des composés biochimiques comme l’eau, les
lipides et les phospholipides, l’évolution de l’oxydation des lipides par la mesure d’un produit
primaire (hydroperoxydes) et d’un produit secondaire (TBA rs) ainsi qu’une évaluation
sensorielle afin de compléter les analyses biochimiques et de vérifier la potentielle corrélation
entre les méthodes biochimiques et l’analyse sensorielle.
Ce travail expérimental est effectué sur 3 espèces différentes de poisson. La sardine
commune (Sardina pilchardus), un poisson gras, le rouget de roche (Mullus surmuletus),
poisson semi gras et le merlan bleu (Micromesistius poutassou) ou faux merlan, poisson
maigre. Toutes les espèces étudiées sont largement consommées en Algérie.
En améliorant les méthodes de conservation de certaines espèces de poisson, un aliment de
haute qualité en protéines et en lipides peut être mis à disposition du consommateur algérien
sur une échelle de temps plus prolongée.

Etude bibliographique
3
ETUDE BIBLIOGRAPHIQUE
A. Présentation des espèces cibles I. La sardine commune (Sardina pilchardus)
I.1. Position systématique
Embranchement : Vertebrés
Sous embranchement : Gnatostomes
Super classe : Poissons
Classe : Ostéichtyens
Sous classe : Téléostéens
Super ordre : Clupéiformes
Ordre : Clupéoides
Famille : Clupéidés
Genre : Sardina
Espèce : Sardina pilchardus (Walbaum, 1792)
Photo 1 : La sardine commune : Sardina pilchardus (Walbaum, 1792).
RO
UA
BHI, 2009

Etude bibliographique
4
I.2. Biologie
La sardine commune (Sardina pilchardus) (photo 1) est un poisson pélagique vivant dans
les eaux côtières et jusqu'à 120 m de profondeur. Elle évolue en Atlantique Nord-Est, de la
Norvège à l’Ecosse jusqu’au Sénégal et en Méditerranée (Forest, 2001). La sardine vit en
bancs parfois importants, près de la surface la nuit et plus en profondeur le jour. Sa taille
moyenne est de 10-20 cm avec une taille maximale de 25 cm. Elle fraie toute l’année avec
une période de ponte variant en fonction de la répartition géographique (Dumay, 2006). C’est
une espèce planctonophage. Les jeunes se nourrissent de phytoplanctons ainsi que d’œufs et
de larves de petits crustacés. Les adultes consomment surtout des crustacés planctoniques
(copépodes), mais également différentes larves présentes dans le zooplancton (crabes,
ophiures, …).
La sardine est un poisson gras qui possède un grand intérêt nutritionnel. En effet, c’est
l’un des poissons les plus riches en lipides et particulièrement en acides gras de la famille des
(n-3) (20 à 30% des acides gras totaux). Elle présente, également, un excellent profile
protéique, ce qu’il lui a permis d’être classée parmi les 11 espèces de poisson possédant les
meilleures recommandations nutritionnelles par la société américaine du cœur (American
Heart Association). Les autres espèces étant le maquereau (Scomber scombrus), le hareng de
l’Atlantique (Harengus clupea) et du Pacifique (Clupea pallasii), la truite de rivière (Salmo
trutta lacustris), le saumon de l’Atlantique (Salmo salar), le saumon royal (Oncorhynchus
tshawytscha), le saumon rouge, l’anchois (Engrolis encrasicholus), la morue noire (Gadus
ogac) et le tassergal (Pomatomus saltatrix) (Sidhu, 2003).
La pêche à la sardine est une activité fortement influencée par les conditions
hydrologiques. En effet, la température agit directement sur les migrations ainsi que sur
l’importance et la localisation des concentrations de sardines et, donc sur leur accessibilité
aux flottilles de pêche (Forest, 2001). La sardine est pêchée à la bolinche et de plus en plus, au
chalut pélagique. Elle est consommée fraîche, salée, parfois fumée mais principalement en
conserve. Figure emblématique de la pêche en Algérie, la sardine est une espèce largement
consommée. En effet, elle jouit d’excellents apports nutritionnels ainsi que d’un prix
raisonnable, convenable à toutes les bourses. Cependant, nous assistons récemment à une
diminution des débarquements accompagnée par une flambée des prix atteignant jusqu’à 500
D.A. le kilogramme.

Etude bibliographique
5
II. Rouget de roche (Mullus surmuletus)
II.1. Position systématique
Embranchement : Chordés
Sous-embranchement : Vertébrés
Super-classe : Poissons
Classe : Osteichtyens
Sous-classe : Actinoptérigiens
Super ordre : Téléostéens
Ordre : Perciformes
Sous-ordre : Percoidés
Famille : Mullidés
Genre : Mullus
Espece : Mullus surmuletus (Linnaeus, 1758).
Photo 2 : Le rouget de roche : Mullus surmuletus (Linnaeus, 1758).
.
1ère dorsale rayée 3 Bandes jaunes horizontales
Ecailles suborbitaires
RO
UA
BHI, 2009

Etude bibliographique
6
II.2. Biologie
Le rouget de roche (Mullus surmuletus) (photo 2) est un poisson benthique que l’on
retrouve dans l’océan Atlantique depuis les côtes de Norvège jusqu’aux côtes ouest africaines
(Whitehead et al., 1986) et dans tout le bassin méditerranéen. Cette espèce est caractérisée
par une tête haute et courte avec deux longs barbillons sous le menton. La bouche n’atteint
pas le niveau antérieur du bord de l’œil. Sous l’orbite de l’œil, deux grandes écailles (écailles
suborbitaires) sont présentes. La première nageoire dorsale montre des points ou rayures
foncés. La coloration du dos et des flancs va de rougeâtre à rouge écarlate. Trois bandes
jaunes horizontales sont visibles au niveau inférieur des flancs (Fage, 1909 ; Desbrosses, 1935
et 1936 ; Bougis, 1952 ; Quéro 1984 ; Hureau, 1986 ; Bauchot, 1987 ; Quéro et Vayne, 1997).
À partir de 19 cm, tous les rougets barbets de roche sont matures. Le rouget barbet de
roche pond à la fin du printemps. La période de ponte est comprise entre le mois d’avril et le
mois de juillet. C’est un poisson carnivore (Labropoulou et al., 1997) et euryphage (Mamuris
et al., 1998) dont les proies principales sont endogées ou épigées. Il ne faut pas confondre le
rouget barbet de roche (Mullus surmuletus) avec le rouget de vase (Mullus barbatus), espèce
avec laquelle il a longtemps était confondu.
Les statistiques mondiales des pêches (F.A.O.) de 1950 à nos jours montrent que les
captures mondiales sont passées de 800 tonnes à 14 500 tonnes en 50 ans. (Mahé et al., 2005).

Etude bibliographique
7
III. Le merlan bleu : Micromesistius poutassou (Risso, 1826)
III.1. Position systématique
Embranchement : Chordés
Sous-embranchement : Vertébrés
Super-classe : Poissons
Classe : Osteichtyens
Sous-classe : Actinoptérigiens
Super ordre : Téléostéens
Ordre : Gadiformes
Famille : Gadidés
Genre : Micromesistius
Espece : Micromesistius poutassou (Risso, 1826)
Photo 3 : Le merlan bleu : Micromesistius poutassou (Risso, 1826).
RO
UA
BHI, 2009

Etude bibliographique
8
III.2. Biologie
Le merlan bleu Micromesistius poutassou (Risso, 1826), aussi appelé " poutassou " par les
pêcheurs ou faux merlan en Algérie (photo 3), est un Gadidé océanique bathypélagique dont
les populations vivent essentiellement au-dessus des fonds du sommet du talus continental
(Hureau et al., 1973), du nord de la Norvège au Maroc (cap Bojador) ainsi que dans le bassin
occidental de la Méditerranée (Maurin, 1968; Robles, 1970). Très abondant au large des côtes
algériennes au-dessous de 150 m sur fonds meubles (Bauchot, 1987). Il forme des bancs
importants, particulièrement denses en été, se déplaçant vers la surface pendant la nuit. La
reproduction est observée à partir du mois de février, au-dessus du bord du talus continental,
entre 10 et 30 m du fond.
Micromesistius poutassou est un poisson prédateur euryphage dont l'alimentation se
compose essentiellement de crustacés plus ou moins pélagiques (Euphausiacés et
Amphipodes), rarement de poissons. Les jeunes merlans bleus, d'une taille inférieure à 200
mm, se nourrissent de petites proies : Copépodes planctoniques et jeunes Euphausiacés. Les
individus plus âgés consomment préférentiellement des proies plus volumineuses telles que
les Euphausiacés Meganuctiphanes noruegica et Nyctiphanes couchii, espèces
caractéristiques du sommet du talus atlantique (Sorbe, 1980 ; MacPherson, 1978).
Malgré des captures au chalut parfois très importantes, ce poisson est actuellement peu
exploité, au niveau mondial, car il est peu apprécié du public du fait de sa fragilité.
Cependant, l'appauvrissement constant de pêche des fonds du plateau continental entraînera
peut-être, dans un avenir plus ou moins proche, les professionnels de la pêche à s'intéresser
plus activement à cette espèce de poisson. Contrairement aux tendances mondiales, le merlan
bleu est très apprécié en Algérie. En effet, c’est un poisson qui coute très cher, dont le prix
peut excéder 700 D.A.

9
B. Les lipides et leurs altérations chez les poissons
La maîtrise de la qualité des produits est l’un des enjeux actuels de la filière aquatique.
Parmi les critères de qualité, la maîtrise des propriétés organoleptiques et biochimiques du
poisson sont d’une importance capitale. En effet, la texture, la qualité nutritive et gustative de
la chair des poissons dépendent principalement des caractéristiques des composants
chimiques et de leur organisation structurale. Par ailleurs, les lipides du tissu musculaire, dont
la teneur et la dégradation représentent un facteur clé de la qualité de la chair, peuvent
affecter les qualités texturales, organoleptiques et nutritives de cette dernière.
I. Caractéristiques de la chair des poissons
La chair de poisson est, d'un point de vue nutritionnel, un produit carné, et présente donc
des qualités nutritionnelles proches de la viande. Elle contient en moyenne 70 à 80 % d’eau,
16 à 22 % de protéines, peu de glycogène (moins de 1% en général) et des lipides en quantité
très variable allant de 1 à 20 % selon les espèces et leur alimentation (Medale, 2005). La
teneur en protéines de la chair de poisson varie peu d'une espèce à l'autre. Elle augmente
progressivement lors de la croissance pour se stabiliser à une valeur proche de 20% (Médale
et al., 2003 ; Lefevre et Bugeon, 2008). L'apport en micronutriments (caroténoïdes, vitamines,
minéraux et oligo-éléments) de la chair de poissons varie fortement d'une espèce à l'autre en
fonction de leur alimentation et de leur milieu de vie, mais peut être particulièrement
intéressant pour certains éléments comme le phosphore (Médale et al., 2003 ; Médale, 2004).
La chair de poisson se différencie de celle des autres animaux élevés par l'organisation
structurale des muscles qui la constituent et par ses composants. Elle est composée de deux
principaux types de muscles qui se distinguent par la nature des fibres qui les composent
majoritairement, à savoir :
• Le muscle brun, de type oxydatif
Caractéristique des téléostéens, il est généralement présent sous forme d’une fine couche
située sous la peau ; il est plus abondant sur les flancs du poisson. Sa proportion dans la chair
varie d’une espèce à l’autre. Ce muscle participe au déplacement du poisson ce qui explique
sa forte vascularisation. De plus, sa réserve lipidique est élevée pouvant représenter jusqu'à
30% du poids (Bendiksen et Jobling, 2003).
• Le muscle blanc, de type glycolytique C’est le plus important
quantitativement puisqu’il représente jusqu’à 50% de la masse corporelle du poisson. Le

10
muscle blanc contient davantage de protéines que le muscle rouge mais moins de lipides et
de glycogène (Bendiksen et Jobling, 2003).
II. Les lipides chez les poissons
Les lipides sont des constituants cellulaires fondamentaux. Ce sont des molécules
organiques insolubles dans l’eau et solubles dans les solvants organiques tels que l’éther, le
chloroforme, l’hexane ou le benzène (Oumansour, 2001). Les lipides forment une classe très
hétérogène d’un point de vue fonctionnel : triglycéride (réserve énergétique), sphingomyéline
(constituant du tissu nerveux) et qui ont la caractéristique commune d’être constitués d’acides
acycliques à longues chaînes linéaires, dénommés acides gras.
Chez les poissons, la teneur et la composition en lipides varient avec l’âge, le cycle sexuel
et les facteurs environnementaux tels que la température et la salinité de l’eau (Wood et
Lister, 1973 ; Gandemer, 1997). Elle est également sous le contrôle de facteurs nutritionnels
(teneur en lipides alimentaires) (Médale et al., 2003), et de facteurs génétiques (Quillet et al.,
2005).
Les lipides de poissons sont caractérisés par un haut degré d’insaturation. Ils incluent
jusqu’à 40% d’acides gras poly insaturés (A.G.P.I.) (14 à 22 atomes de carbone) (Medale,
2005), alors que la graisse des mammifères contient rarement plus de deux doubles liaisons
par molécule d’acide gras avec une prédominance des A.G.P.I. de la série (n-6) et (n-9) (Abi-
ayad, 1998). En effet, la teneur en A.G.P.I. n-3 de la chair de poisson varie entre 15 et 36%
alors qu’elle n’est que de 1% chez le porc, 2 % chez le bœuf et 4 % chez le poulet (Gandemer,
1992). Cette richesse est liée à la forte concentration en A.G.P.I. (n-3) dans la chaîne
alimentaire aquatique (Sargent et al., 1989).
Les lipides sont présents dans les muscles des poissons sous deux formes :
II.1. Lipides neutres ou lipides de réserve Essentiellement représentés par des triglycérides (T.G.) qui sont hautement insaturés
comparés aux animaux d’élevage terrestres. En effet, les dépôts gras du poisson contiennent
plusieurs acides gras avec cinq ou six doubles liaisons (Girard et Paquotte, 2003).
La composition en acides gras des triglycérides de la chair est le reflet de celle des lipides
alimentaires (Henderson et Tocher, 1987; Corraze et Kaushik, 1999). En effet, c’est la
résultante d’un mélange de lipides alimentaire et de lipides endogènes (synthèse de novo et/ou
élongation et désaturation) (Abi-ayad, 1998). Ces T.G. qui représentent les réserves
lipidiques chez les poissons, permettent de classer ces derniers en (tableau 1):

11
• Poissons maigres : dont la teneur en lipides dans le muscle est inférieure à 1% et
déposent massivement les lipides dans le tissu hépatique (jusqu'à 75% du poids du foie) ex. la
morue (Gadus morhua), l’églefin (Melanigrammus aeglefinus) ou le turbot (Psetta Maxima).
• Des poissons gras : chez qui la teneur en lipides dans le muscle est supérieure à 15%,
tels que le hareng (Clupea harengus), le maquereau (Scomber scombrus) ou l’anguille
européenne (Anguilla anguilla).
• Des poissons « intermédiaires » ou semi- gras : qui déposent les lipides dans le
muscle mais aussi dans d’autres sites tels que le tissu adipeux périviscéral comme c’est le cas
pour les salmonidés par exemple (Henderson et Tocher, 1987; Sheridan, 1988 et 1994;
Ackman, 1995).
Tableau 1 : Teneur en lipides et en eau de quelques espèces de poisson. (a : Murray et Burt (1969), b : Poulter et Nicolaides (1985a), c : Poulter et Nicolaides (1985b).
Espèce Nom scientifique Eau (%)
Lipides (%) source
Merlan bleu Micromesistius poutassou 79-80 1,9-3 a
Cabillaud Gadus morhua 78-83 0,1-0,9 a
Anguille Anguilla anguilla 60-71 8-31 a
Hareng Clupea harengus 60-80 0,4-22 a Carrelet Pleuronectes platessa 81 1,1-3,6 a
Saumon Salmo salar 67-77 0,3-14 a
Truite Salmo trutta 70-79 1,2-10,8 a
Thon Thunnus sp 71 4,1 a
Langoustine Nephrops norvegicus 77 0,6-2 a
Pjerrey Basilichthys bornariensis 80 0,7-3,6 b
Carpe Cyprinus carpio 81,6 2,1 b
Sabalo Prochylodus platensis 67 4,3 c
Pacu Colossoma macropomum 67,1 18 c
Tambaqui Colossoma brachypomum 69,3 15,6 c
Chincuina Pseudoplatystoma tigrinum 70,8 8,9 c
Corvina Plagioscion squamosissimus 67,9 5,9 c
Bagré Ageneiosus spp. 79 3,7 c

12
En général, la teneur en lipides des filets de poissons maigres est basse et stable alors que
la teneur en lipides des poissons gras est extrêmement variable. Cependant la variation du
pourcentage (%) de graisse se reflète dans le pourcentage d’eau. En effet, la graisse et l’eau
constituent environ 80 % du filet (Rodriguez et al., 2003).
II.2. Lipides polaires ou phospholipides (P.L.)
Les phospholipides sont des composants majeurs des membranes cellulaires. Leur teneur et
leur composition sont relativement constantes. Les phospholipides représentent moins de 1%
du poids du muscle. Ils sont principalement composés de phosphatidylcholine (50 à 60 % des
phospholipides) et de phosphatidyl éthanolamine (20 à 30% des phospholipides) (Aursand et
al., 1994). Ils se caractérisent par une grande richesse (jusqu'à 60%) en A.G.P.I. à longue
chaîne, avec une prépondérance de l'acide eicosapentaénoïque (E.P.A. ou 20:5 (n-3)) et de
l'acide docosahéxaénoïque (D.H.A. ou 22:6 (n- 3)) (Henderson et Tocher, 1987; Ingemansson
et al., 1991). Ces acides gras permettent de maintenir la fluidité membranaire, même à basse
température. En effet, Kiessling et al. (2001) a montré que les acides gras polyinsaturés
(A.G.P.I.) entrent dans la composition des lipides polaires, tandis que les acides gras saturés
(A.G.S.) et monoinsaturés (A.G.M.I.) sont représentatifs des lipides neutres.
En effet, parmi toutes les classes de lipides, les phospholipides se révèlent être les plus
actifs. Certains phospholipides caractéristiques de bactéries possèdent une activité anti-
bactérienne (Tamehiro et al. - 2002). Les phospholipides possèdent également des actions
cytotoxiques et anti-prolifératives, leur donnant un rôle potentiel dans la lutte contre le cancer.
En effet, les phospholipides jouent des rôles divers dans le métabolisme cellulaire, notamment
du fait de leur participation dans la composition de la membrane cellulaire (Dumay, 2006).
Les phospholipides jouent aussi un rôle en aquaculture. Des études menées conjointement
entre l’I.F.R.E.M.E.R. et l’I.N.R.A. ont montré que les phospholipides sont des constituants
essentiels des aliments pour larves de poissons (Dumay, 2006). En effet, ils constituent de très
bons vecteurs pour apporter aux larves de poissons les acides gras essentiels à leur
développement. De plus, une alimentation des larves de poissons enrichie en phospholipides
induit une mortalité plus basse et une prise de poids plus importante (Cahu et Zambonino
Infante - 2001; Cahu et al. - 2003).

13
III. L’altération du poisson
Par définition, l’altération d’un produit alimentaire est la dégradation ou la diminution
constante de sa qualité c'est-à-dire de sa fraîcheur. La décomposition étant l’étape ultime de
l’altération.
Les altérations les plus importantes atteignent surtout les muscles. A l’état post mortem,
l'arrêt de la circulation sanguine va priver le muscle d'un apport en oxygène et en molécules
énergétiques. Dans un premier temps, le muscle va mobiliser ses substances de réserve. Avec
l'épuisement de ces réserves, les fibres musculaires sont dans l'incapacité de se relaxer: le
muscle va durcir, sa qualité diminue. Toutefois, en raison de la composition et de la structure
particulière du muscle de poisson, la rigidité puis l'attendrissement interviennent plus
rapidement que dans le cas des viandes. Le poisson est ainsi rapidement exposé aux réactions
d'altération et au développement microbien. C'est l'un des produits animaux les plus difficiles
à conserver (Aubourg, 2007).
Après la mort du poisson, les processus biochimiques se développent plus intensément que
chez les animaux terrestres. Cela est expliqué par le fait que l’activité optimale des enzymes
est proche de la température normale de l’animal vivant, condition qui est gardée aussi après
la mort du poisson. De plus, la chair du poisson offre de meilleures conditions pour le
développement des processus autolytiques et le développement de la flore microbienne
d’altération.
La chair de poisson s’altère plus rapidement que la viande des autres animaux d’élevage en
raison de:
ü la teneur très élevée en eau,
ü la quantité réduite du tissu conjonctif,
ü la concentration importante d’azote extractible,
ü la présence de lipides fortement insaturés.
Cette altération peut être de type microbiologique, sensoriel, autolytique ou biochimique.
III.1. Altérations autolytiques
L’altération autolytique est largement représentée par la dégradation de l’A.T.P. Il a été
démontré que les produits issus de la dégradation de l’A.T.P. ont pour conséquence l’arrière
gout amer du poisson altéré (Hughes & Jones, 1966). De même, elle est reliée au

14
ramollissement post mortem du poisson et à l’éclatement de la cavité abdominale sous l’effet
des enzymes protéolytiques tissulaires (cathepsines, calpaines et collagénases) (F.A.O., 2003).
III.2. Altérations microbiologiques
En condition normale, la chair du poisson est stérile (Kyrana & lougrovois, 2002).
Cependant, la peau, les branchies et les viscères renferment une flore commensale plus ou
moins abondante (Bourgeois et al., 1996).
A l’état post mortem et suite à l’autolyse, les enzymes digestives détruisent la barrière
intestinale et de ce fait permettent la dissémination des germes. Ces microorganismes sont de
nature psychrotrophes, ce qui explique leur action même à basse température.
Ce type d’altération peut aboutir à la formation d’un produit toxique : l’histamine, par
décarboxylation de l’histidine (Bourgeois et Leveau, 1991).
III.3. Altérations sensorielles
L’altération sensorielle varie considérablement en fonction de l’espèce et du mode de
conservation (F.A.O., 1999).
Après capture, les caractéristiques organoleptiques et sensorielles du poisson se modifient
comme suit :
ü l’œil saillant et clair s’altère en devenant opaque, brumeux et par la suite blanchâtre,
ü les branchies rouges deviennent rose fade et passent ensuite au gris et au brun,
ü l’anus fermé s’ouvre avant de devenir béant,
ü la chair ferme, élastique et blanche se gélifie et finit par se ramollir,
ü les écailles et la peau passe de l’état brillant à la décoloration pour devenir terne et le
mucus devient opalescent.
L’évaluation sensorielle de la fraicheur du poisson est un outil de mesure immédiat, rapide
et précis (Eymard, 2003). De plus, elle permet de détecter à travers un examen visuel les
imperfections sur le corps du poisson (Eymard, 2003) et de détecter et d’identifier les odeurs
issues des dégradations lipidiques difficilement mesurables par des méthodes biochimiques
(Frankel , 1998 ; Eymard, 2003).
L’analyse sensorielle, permet d’évaluer la qualité du poisson telle qu’elle est appréhendée
par le consommateur.

15
III.4. Altérations biochimiques
a. Lipolyse
La lipolyse est l’un des principaux mécanismes de dégradation des lipides post mortem qui
permet la libération des A.G.P.I. (Shewfelt, 1981 ; Eymard, 2003). C’est un phénomène
enzymatique qui se déroule dans la chair crue au cours de sa maturation ou conservation. Elle
n'est pas observée dans la viande cuite car la cuisson dénature les enzymes lipolytiques. La
lipolyse est catalysée par des enzymes spécifiques : les lipases et les phospholipases (Van Der
Bosch, 1980) qui présentent un pH d'activité optimale basique (Alasnier, 1996). Chez les
poissons, il s’agit de la phospholipase C (PLC) et de la phospholipase A2 (PLA2). Les
phospholipases A1 et D étant des voies mineures (Bacot, 2004) (figure 1). Ces enzymes
hydrolysent les liaisons esters des glycérides et libèrent à partir des triglycérides (T.G.) des
acides gras (A.G.L.), des monoglycérides (M.G.) et des diglycérides (D.G.) (Eymard, 2003)
Figure 1 : Action des différentes phospholipases sur les phospholipides (Bacot, 2004)
PLA2 R1 COO- + lysoPL.
PLA2 R2 COO- + lysoPL.
CH2 O CO R1
CH O CO R2
O
CH2 O P O X
O
PLD Phosphatidate + XOH
PLC D .A.G. + X-P
DAG lipase
M.A.G. + R1- COO-
MAG lipase
Glycerol – R-COO-

16
L’activité lipolytique, bien que faible, persiste lors de la conservation à l’état congelé.
Ainsi, des acides gras sont libérés après au moins 300 jours de conservation à -18°C des filets
et de la chair du merlu blanc du cap (Merluccius capensis) (De Koning et Mol, 1990).
Ces réactions de lipolyse induisent une dégradation du produit. De plus, les acides gras
libérés interagissent avec les protéines favorisant leur dénaturation et conduisant à l’altération
du poisson (Dyer et Fraser, 1959).
b. La peroxydation lipidique
Dans les conditions normales (poisson vivant), la peroxydation lipidique existe. C’est un
processus physiologique naturel et continu, indispensable à la synthèse des prostaglandines,
des leucotriènes, à la leucocytose, à la phagocytose et aux remaniements des membranes
cellulaires. C’est la peroxydation lipidique enzymatique (Marcel, 2002).
En condition post mortem, on parle de peroxydation lipidique non enzymatique ou
spontanée. C’est un processus oxydatif d’altération des lipides portant essentiellement sur les
A.G.P.I. En effet, il est établi que les A.G.P.I. sont des cibles privilégiées en raison de leurs
oxygènes bisallyliques facilement oxydables (Servais, 2004). De plus, les acides gras saturés
(A.G.S.) ne s’oxydent qu’à des températures supérieures à 60°C, tandis que les acides gras
polyinsaturés s’oxydent même lors de l’entreposage des aliments à l’état congelé (Grandjean
2001, Favier 2003). Par ailleurs, les triglycérides sont peu réactifs parce qu'ils contiennent peu
d'acides gras polyinsaturés et que ces acides gras ne contiennent que 2 ou 3 doubles liaisons.
De plus, regroupés dans des gouttelettes, ils n'ont que peu de contact avec les catalyseurs de
l'oxydation qui sont localisés dans la phase aqueuse des cellules adipeuses et musculaires. De
ce fait, ils sont peu atteints par l'oxydation.
En fonction de l’agent initiateur, on classe l’oxydation des lipides en 3 types :
• l’auto-oxydation catalysée par la température, les ions métalliques et les radicaux
libres ;
• la photo-oxydation, initiée par la lumière en présence de photosensibilisateurs ;
• l’oxydation enzymatique initiée par la présence des enzymes d'oxydation.
Il s’agit d’une cascade de réactions radicalaires organisée en 3 phases successives (Frankel,
1984 ; Halliwell, 1990a):

17
b.1. Phase d’initiation
Elle consiste en la soustraction, par un radical libre, d’un atome d’hydrogène à un
groupement méthylène (-CH2-) de la chaine acylée de l’A.G.P.I. Elle conduit à la formation
d’un radical libre de type lipoyle (figure 2), qui se stabilise par un remaniement électronique
aboutissant à la formation de 2 diènes conjugués (Murray et al. 2002, Bacot, 2004 ; Coulon,
2004).
Figure 2 : Phase d’initiation de la peroxydation lipidique.
b.2. Phase de propagation
C’est une phase explosive d’amplification, durant laquelle il y’a formation d’un radical
peroxyle qui réagit à son tour avec une autre molécule d’A.G.P.I. pour former des
hydroperoxydes qui sont très instables et se décomposent en présence de métaux ionisés ou de
structure héminique (figure 3). Ce processus aboutit à la formation de radicaux alkoxyles,
alkyloperoxyles ainsi qu’à des alcanes et aldéhydes (Sevanian et Hoschstein, 1985 ; Penfield
et Campbell, 1990 ; Chaveron, 1999).
Figure 3 : Phase de propagation de la peroxydation lipidique.
Comme alternative à cette voie, le radical peroxyle, après évolution en un peroxyde
cyclique et coupure de la molécule, peut libérer différents aldéhydes hautement cytotoxiques
et mutagènes, tels que le malondialdéhyde (M.D.A.) et le 4-hydroxynonenal (4-H.N.A.). Ces
derniers peuvent réagir avec l’ADN ou des protéines et provoquent des lésions structurales et
Agent initiateur
A.G.P.I. Radical lipoyle
Lumière, chaleur, catalyseur
Radical libre Radical peroxyle Hydroperoxydes
+ O2 R-H

18
fonctionnelles (Frei, 1994 ; Clausse, 2001 ; Favier, 2003). Un tel processus amplifie
notablement le phénomène de peroxydation lipidique (Clausse, 2001 ; Hill’s 2001 ; Delattre
et al. 2005).
Il est admis que chaque radical libre soit à l’origine d’une centaine de molécules
d’hydroperoxydes avant que ne survienne la phase d’arrêt de la peroxydation lipidique
(Coulon, 2004). Cette étape de propagation peut se reproduire jusqu’à épuisement des
A.G.P.I. et/ou de l’oxygène (chabaud, 2007).
b.3. Phase de terminaison
Elle correspond à diverses réactions de couplage entre deux espèces radicalaires formant
des produits non radicalaires et stables qui terminent la chaine de réactions. Globalement, ce
processus conduit à des hydrocarbures, des aldéhydes, des cétones, des acides, des esters, des
peracides, des peroxydes, mais aussi à des produits de polymérisation (figure 4). Sous son
effet, l’aliment perd de sa qualité nutritionnelle ou organoleptique (rancissement, changement
de couleur…) (Chabaud, 2007).

19
Figure 4 : Mécanisme en chaine de la peroxydation des A.G.P.I. et nature des produits terminaux formes (exemple de l’acide arachidonique) (Favier 2003).
PRODUITS TERMINAUX
endoperoxydes
Hydrooperoxydes ROOH
REACTION EN CHAINE RADICALAIRE
Arachidonate
Radical peroxyle
Radical arachidonique
Radical diène conjugué
Acide arachidonique
Malondialdéhyde
Esoprostane
Hydroxynonénal Pentane
Ethane

20
IV. Les antioxydants
Le maintien d’un niveau non toxique des radicaux libres est assuré par des systèmes
antioxydants. Un antioxydant, dans sa définition la plus large, est défini comme étant une
substance capable, à concentration relativement faible, d’entrer en compétition avec d’autres
substrats oxydables et ainsi retarder ou empêcher leur oxydation (Halliwell, 1990b ; Servais,
2004). Ces antioxydants peuvent être de nature enzymatique (superoxyde dismutase, catalase,
glutathion peroxydase) ou non enzymatique (vit E, vit C) (Mates et al., 1999 ; Comhair et
Erzurums 2002 ; Droge, 2002), naturels ou de synthèse (figure 5).
Figure 5 : Principaux composés naturels ou synthétisés possédant des propriétés
antioxydantes (Chabaud, 2007).
Vitamine ξ : α-tocophérol
Flavone
(lutéoline)
Flavone
(quercétine)
Flavonoide
(courmarine)
Flavonoide
(Ac. cinnamique)
Β- carotène
lycopène
Vitamine C : Ac. ascorbique

21
Les antioxydants susceptibles de protéger les lipides de l’oxydation peuvent être répartis
selon leur mode d’action en:
IV.1. Antioxydants radicalaires ou primaires
Ils permettent l’interruption de la chaîne autocatalytique. Ils fonctionnent comme des
pièges aux radicaux libres. (polyphénols). On peut citer parmi cette famille d’antioxydants les
composés naturels phénoliques (tocophérols), des composés synthétiques tels que le B.H.T.
(butylhdroxytoluene), les caroténoïdes, vitamine C, etc. … (Kortenska et al., 2002).
IV.2. Antioxydants préventifs ou secondaires
Ils inhibent la production des radicaux libres. Ce sont des substances qui interceptent les
radicaux propagateurs de la peroxydation lipidique et retardent la peroxydation ou
interceptent les espèces responsables de l’initiation de la lipoperoxydation. Ils interrompent
directement la chaîne de lipopéroxydation (α-tocophérol) ou participent indirectement à cette
interruption (acide ascorbique, polyphénols). (Buettner, 1993 ; Leger, 2006).
IV.3. Antioxydants physiques
Dans cette catégorie, on trouve les procédés de protection physique : teneur en oxygène,
humidité relative, température, lumière. Les aliments sont donc protégés par des « barrières
adéquates » souvent apportées par les emballages et les conditions de stockage.
Les agents synergiques améliorent le pouvoir antioxydant d’autres composés. Il peut s’agir
d’acides (ascorbique, lactique, tartrique ou phosphorique) ou de sels (de potassium, de sodium
ou de calcium) (Cheriot, 2007).
V. Facteurs influençant l’oxydation des lipides
L’oxydation des lipides est un facteur qui limite considérablement la durée de vie des
denrées alimentaires. On dénombre un certain nombre de facteurs susceptibles de favoriser
cette oxydation (Mahmoud et Benkakaa, 2007). Ces facteurs peuvent être de nature
intrinsèque ou bien des facteurs externes (Génot et al., 2003) parmi lesquels, on peut citer :
• la nature et le degré de dispersion des lipides dans l’aliment,
• le type métabolique des fibres musculaires, • la présence dans l’aliment d’agents antioxydants et notamment de vitamine E,
• l’activité de l’eau (A w) qui influence aussi l’activité catalytique des métaux,

22
• la présence de substances prooxydantes (hème, ions ou enzymes),
• la pression partielle en oxygène,
• la structure des lipides et le taux d’A.G.P.I. qu’ils renferment,
• le pH, la lumière et la température.
L’agent initiateur est l’élément clé qui provoque l’enchainement des réactions radicalaires
de la peroxydation lipidique. Les métaux de transition jouent un rôle capital dans la genèse
des radicaux libres (Love, 1980). Au sein des systèmes biologiques, ils sont largement
représentés par le fer et le cuivre (Eymard, 2003). De même, la température et le pH l’initient
en raison de leur influence sur la solubilité de l’oxygène (Fernandez et Mackie,
1987 ;Gandemer et al., 1990 ; Ngah et al., 1993).
VI. Méthodes de mesure de l’oxydation des lipides
VI.1. Mesure des substrats de l’oxydation
Lors des études de cinétique d’oxydation des lipides, l’état d’avancement de la réaction
peut être évalué par la mise en évidence de la disparition des substrats de l’oxydation.
L’étude de la consommation d’oxygène permet de suivre les phases d’initiation et de
propagation de la réaction. Ces méthodes sont manométriques (mesure de la pression partielle
en oxygène), polarographiques (mesure de la consommation d’oxygène) ou
chromatographiques ou gravimétriques par mesure de l’augmentation du poids consécutive à
la fixation d’oxygène (Eymard, 2003).
L’analyse des acides gras est réalisée après extraction des lipides, méthylation des acides
gras et chromatographie en phase gazeuse. La difficulté consiste à extraire quantitativement la
matière grasse et à minimiser les pertes au niveau des réactions de méthanolyse (Berset et
Cuvelier, 1996).
Les phospholipides (P.L.) sont de nature, très riches en A.G.P.I., caractère qui fait d’eux
les premiers substrats de la peroxydation lipidique. Le dosage des P.L. permet de renseigner
sur l’état d’avancement de la peroxydation des lipides.
VI.2. Mesure des produits primaires
Les produits primaires de l’oxydation des lipides peuvent être analysés à l’aide de
nombreuses techniques présentant de grandes différences au niveau de leur sensibilité, leur
facilité d’utilisation, et la nature de la matrice. Même si elles présentent l’intérêt d’être simple,

23
beaucoup d’entre elles ne sont pas forcément adaptées à des systèmes plus complexes tels que
le poisson et ses produits transformés (Eymard, 2003).
a. Détermination des diènes conjugués Les produits primaires de l’oxydation des lipides contenant des doubles liaisons
conjuguées peuvent être mesurés en UV (Klein, 1970 ; Corongiu et Banni, 1994). Ces diènes
conjugués absorbent à 232-233 nm et les triènes conjugués à 268 nm. Ils peuvent être
déterminés par mesure de l’absorbance à ces longueurs d’onde. Cette méthode est rapide,
mais peu spécifique (Gray, 1978).
b. Dosage des hydroperoxydes Les hydroperoxydes sont des produits intermédiaires instables qui sont rapidement
dégradés pour donner des composés hydroxylés et carbonylés. Les concentrations en
hydroperoxydes mesurées correspondent en réalité à la différence entre formation et
décomposition des peroxydes. Les concentrations en hydroperoxydes peuvent être
déterminées à l’aide de nombreuses méthodes à adapter en fonction du substrat étudié. Deux
groupes de méthodes peuvent être distingués : les méthodes analytiques permettant de
déterminer la concentration en hydroperoxydes et les techniques chromatographiques
permettant d’identifier et de quantifier la nature et les teneurs en hydroperoxydes spécifiques
(Dobarganes et Valesco, 2002).
VI.3. Mesure des produits secondaires
Deux méthodes colorimétriques sont employées couramment pour les doser : l’indice de p-
anisidine et le test à l’acide thiobarbiturique.
La détermination de l’indice de para-anisidine repose sur le principe qu’en milieu acide, la
p-anisidine donne un complexe coloré en jaune avec des diénals conjugués (Eymard, 2003).
Le test à l’acide 2-thiobarbiturique repose sur la formation d’un complexe coloré rose
résultant de la réaction entre une molécule de malonaldéhyde et deux molécules d’acide 2-
thiobarbiturique. Le complexe coloré formé absorbe à 532-535 nm. De nombreuses
adaptations de cette méthode ont été proposées (Vincke, 1970 ; Guillén-Sans et Guzmàn-
Chozas, 1998 ; Wang et al., 2002).

24
VI.4. Evaluation sensorielle
L’évaluation sensorielle est souvent considérée comme une bonne méthode d’évaluation de
l’oxydation des produits. C’est une discipline scientifique qui consiste à mesurer, analyser et
interpréter les réactions et les caractéristiques d’aliments ou de matières perçues par les sens
de la vue, de l’odorat, du goût et du toucher. Les méthodes sensorielles, souvent considérées
comme subjectives, ne remplacent pas les mesures instrumentales, mais les complètent.

Matériels et Méthodes
25
ETUDE EXPERIMENTALE
A. Matériels et Méthodes I. Matériel biologique : échantillonnage et conservation
Afin d’obtenir du poisson frais, les poissons sont ramenés immédiatement après capture,
du port d’Oran, pour la sardine commune (Sardina pilchardus, 18.24 ± 3,69 g) et le rouget de
roche (Mullus surmuletus, 75,89 ± 20,34 g) et du port de Béni-Saf pour le merlan bleu
(Micromesistius poutassou, 33,51 ± 18,41 g). Le travail expérimental a lieu au sein du
laboratoire de Gestion des ressources aquatiques (Ge.Re.Aq.), au département de
Biotechnologie.
Au total, 60 spécimens pour chaque espèce sont utilisés, à raison de 5 individus par espèce
et par mode de conservation. Les trois espèces de poissons sont répartis selon le mode de
conservation en trois lots comme suit :
Lot A : poissons entiers (non éviscérés), conservés à T° ambiante la journée, au
réfrigérateur la nuit.
Lot B : poissons entiers (non éviscérés), conservés au réfrigérateur (+ 4°C) 24h/24h.
Lot C : poisson entiers (non éviscérés), conservés au congélateur (-18°C).
Les analyses sont effectuées à J1-J2-J3-J5 et éventuellement à J7- J9 (pour les poissons
conservés à T° ambiante et réfrigérés) puis à J15 et J30 pour les poissons congelés.
II. L’appréciation sensorielle
Le suivi de l’évolution de la qualité d’un point de vue sensoriel et organoleptique s’est fait
selon la grille d’évaluation organoleptique de l’état de fraicheur des poissons entiers
préconisée par Gret (1993) (annexe 1). Cette grille prend en charge l’évaluation des poissons
conservés à T° ambiante ou basse. Ainsi, selon cette grille le poisson est classé en 4 catégories
(A, B, C et D) qui correspondent soit à un poisson d’une qualité excellente, bonne, acceptable
ou mauvaise, respectivement. Pour cela, nous procédons à l’évaluation du poisson à travers
l’aspect couleur (peau, écailles et branchies), texture (chair, colonne vertébrale et péritoine) et
odeur (branchies, peau et cavité abdominale).
Cette appréciation est élaborée afin de déterminer le temps de rejet qui correspond au
moment où le poisson est impropre à la consommation.

Matériels et Méthodes
26
Par ailleurs, cette évaluation nous permettra de mettre en évidence la potentielle relation
entre l’appréciation sensorielle et les méthodes analytiques employées lors de ce travail
expérimental. Pour cela, les mêmes poissons sont rapidement inspectés sensoriellement puis
utilisés pour les analyses biochimiques
III. Caractéristiques biochimiques
III.1. Détermination de la teneur en eau
De 4 à 10 g d’échantillon (M1) sont pesés (balance : Pionner, OHAUS, capacité de 200 g,
précision de 0,01 g) puis placé à l’étuve (Memmert) (figure : A, planche 1) pendant une nuit à
110 °C. Après refroidissement, la matière sèche obtenue (M2) est pesée. Les analyses sont
effectuées sur les cinq individus.
ü La teneur en eau des échantillons est calculée selon la formule suivante :
Teneur en eau (g/100g) =
ü La proportion de matière sèche est calculée selon la formule :
Matière sèche (MS) (g/100g) =
III.2. Détermination de la teneur en lipides totaux
• Choix de la méthode
Il existe plusieurs méthodes d’extraction des lipides dont les plus couramment utilisées
celles de Blight et Dyer (1959) et celle Folch et al. (1957). La méthode utilisée lors de ce
travail expérimental est celle Folch et al. (1957). Cette dernière présente l’avantage d’être
rapide (nécessitant une seule séquence d’homogénéisation) et utilisable quelle que soit la
concentration en lipides de l’échantillon, sans une adaptation préalable du volume de solvant
d’extraction (Eymard, 2003).
• Principe
Cette technique repose sur le principe d’une extraction à froid des lipides par un solvant
composé de chloroforme et de méthanol (2 / 1 ; v/v). Le chloroforme extrait les lipides et le
méthanol dissocie les lipides des autres constituants membranaires. L’addition d’une solution
aqueuse de NaCl à 0,58 % (p/v) favorise l’obtention d’un système biphasique. En effet, elle
permet la séparation entre 2 phases ; la supérieure, phase méthanolique, contient le méthanol
�M1 − M2�x 100� 1
M2x 100� 1

Matériels et Méthodes
27
et l’eau et l’inférieure, chloroformique, contient les lipides. A l’interface, se trouvent
majoritairement des protéines (Dumay, 2006).
La phase chloroformique inferieure est récupérée dans un ballon et évaporée à 39°C sous
vide à l’aide d’un évaporateur rotatif (Hahn shin Scientific Co, Modèle : HANVAPOR)
(figure : B, planche 1).
La quantification de la teneur en lipides est réalisée par la pesée du ballon vide et après
l’évaporation (balance Pionner, OHAUS d’une capacité de 210 g et d’une précision de
0,0001) (figure : C, planche 1). La teneur en lipides est exprimée en g de lipides pour 100 g
d’échantillon matière fraiche (M.F.). Les résultats sont également exprimés par rapport à la
matière sèche (g/100 g M.S.).
III.3. Détermination de la teneur en phospholipides
• Choix de la méthode
La méthode utilisée pour la détermination de la teneur en phospholipides est celle
préconisée par Stewart (1980).
• Principe de la méthode Cette technique, colorimétrique, est basée sur le principe de formation d’un complexe entre
les phospholipides de l’échantillon et le ferrocyanate d’ammonium (réactif associé). Le
réactif, composé de chlorure de fer III et de thiocyanate d’ammonium, va se complexer avec
le phosphore contenu dans les phospholipides et donner une coloration plus ou moins intense
suivant la quantité de phosphore présente dans l’échantillon (figure A, planche 2).Les lectures
sont réalisées à 488 nm au spéctrophotometre (Genesys) (figure : E, planche 1).
Une gamme étalon est effectuée avec la phosphatidylcholine, pour des quantités allant de 0
à 200 µg par tube. Les résultats sont obtenus par comparaison avec la droite d’étalonnage et
exprimés en mg équivalent de phosphatidylcholine (mg Eq PC) par g de lipide (mg Eq. PC/g
lipides) ou par kg d’échantillon (mg Eq. PC/kg M.F. ou mg Eq. PC/kg MS).
IV. Détermination du degré d’oxydation des lipides
Pour déterminer le degré d’oxydation des lipides, nous nous sommes intéressés à un
produit primaire (hydroperoxydes) et un produit secodaire (les TBA rs : thiobarbiturique
reactifs substances).

Matériels et Méthodes
28
IV.1. Détermination de la teneur en produits primaires: les hydroperoxydes • Choix de la méthode
Il existe beaucoup de méthodes permettant le dosage des hydroperoxydes (Berset et
Cuvelier, 1996 ; Frankel, 1990, 1998). La méthode, colorimétrique, utilisée lors de la présente
étude, est celle proposée par Hermes-Lima et al. (1995) et modifiée par Eymard et Génot
(2003). Elle présente l’avantage de travailler directement sur l’échantillon, sans passer
préalablement par une extraction des lipides pouvant modifier la teneur initiale en
hydroperoxydes (Eymard, 2003).
• Principe de la méthode
En condition d’acidité favorable, les hydroperoxydes extraits de l’échantillon par le
méthanol, oxydent le Fe2+ (présent dans le réactif) en Fe3+. Ce dernier forme un complexe
coloré avec le xylénol orange (figure B, planche 2). Ce complexe coloré possède un
maximum d’absorption à 560 nm.
Une gamme étalon d’hydropéroxydes de cumène (CuOOH) dans le méthanol est réalisée
pour des concentrations variant de 0 à 20 μM. Les concentrations en hydroperoxydes dans les
échantillons sont obtenus comparaison avec la droite d’étalonnage et sont exprimées en
équivalents d’hydroperoxydes de cumène (Eq CuOOH) puis rapportés en μmoles Eq
CuOOH/g lipides ou en mmoles Eq CuOOH/kg M.F. ou mmoles Eq CuOOH/kg M.S.
IV.2. Détermination de la teneur en produits secondaires: les TBA rs
• Choix de la méthode
La méthode utilisée fait appel à l’acide thiobarbiturique (TBA), elle a été mise au point par
de Salih et al. (1987) et Bostoglou et al. (1994) et modifiée par (Genot, 1996). C’est une
technique simple, rapide et qui peut être utilisée en analyses de routines et à échelle
industrielle.
• Principe de la méthode
Après extraction des TBA rs par l’acide trichloroacétique (T.C.A.) il y’a formation d’un
complexe rose entre le TBA et le malondialdéhyde (M.D.A.) (figure C, planche 2) dont
l’absorption maximale est à λ = 532 nm. le M.D.A. n’est pas le seul produit secondaire issu
de l’oxydation des acides gras polyinsaturés (A.G.P.I.) à réagir avec l’acide thiobarbiturique
(T.B.A.). De ce fait, il est plus approprié de parler de substances réactives au T.B.A. (TBA
rs).

Matériels et Méthodes
29
La concentration en M.D.A., exprimée en nmoles de M.D.A. /g de chair, est calculée à
partir de la formule suivante :
C (nmoles de MDA/g) = D.O. x 104 x volume TCA x 0,641 x dilutions (1/2) x
(poids de l’échantillon)-1
V. L’analyse statistique
Les différents dosages sont effectués sur 5 spécimens (n=5).Dans tous les cas, les
statistiques descriptives (moyennes ± écarts types) sont utilisées pour décrire l’ensemble des
résultats.
Avant toutes analyses statistiques, l’homogénéité des variances des variances a été
vérifiée. Lorsque les variances sont homogènes, l’analyse consiste en un test paramétrique
d’Anova 1 (p< 0,05). Lorsqu’elles ne sont pas homogènes, l’analyse consiste en un test non
paramétrique de Kruskal- Wallis.
Pour déterminer si les différences, entre les moyennes obtenues, sont significatives, nous
utilisons le test de Ficher (P.L.S.D) dans le cas de l’Anova et celui de Mann-Withney dans le
cas de Kruskal- Wallis.

Matériels et Méthodes
30
Planche 1
Planche 1 : appareillage utilisé lors des procédés expérimentaux.
A : Etuve (Memmert) B : Evaporateur rotatif (Hahn shin Scientific Co, Modèle : HANVAPOR)
C : Balance (Pionner, OHAUS) D : Centrifugeuse (Hettich Zentrifugen, Modèle : EBA-20)
E : Spectrophotomètre (Genesys)
F : Bain-marie (Memmert)
A B
C D
E F
Rouabhi, 2009
Rouabhi, 2009
Rouabhi, 2009
Rouabhi, 2009
Rouabhi, 2009
Rouabhi, 2009

Matériels et Méthodes
31
Planche 2
Planche 2 : mise en évidence des réactions substrat/réactif.
A : Réaction des phospholipides avec le réactif de thyocyanate de fer. B : Réaction des hydroperoxydes avec le réactif FOX 2. C : formation d’un complexe rosé entre le M.D.A et le T.B.A.
Rouabhi, 2009
Rouabhi, 2009
Rouabhi, 2009
A
B
C

Résultats
32
B. Résultats
I. La sardine commune (Sardina pilchardus)
I.1. L’analyse sensorielle
A J1, la sardine (S. pilchardus) est caractérisée par une fine odeur d’algues. Le corps
présente un aspect brillant et des écailles bien ancrées, avec présence d’un mucus transparent
de consistance visqueuse. L’œil est convexe et brillant avec une cornée translucide (photo 4).
La paroi abdominale est intacte. Après ouverture du poisson, nous remarquons que les viscères
sont intactes et bien distinctes (photo 5). La chair est ferme et élastique avec une nette
différenciation entre le muscle brun et le muscle blanc.
Photo 4 : Aspect extérieur de la sardine (S. pilchardus) à J1.
Après deux jours (J2) de conservation à température ambiante, nous remarquons que la
sardine maintien un bon niveau de qualité. Les premiers signes d’altérations concernent
l’aspect externe du poisson. En effet, la peau perd de son éclat, des plaques de sang
apparaissent rapidement sous l’opercule et l’œil (photo 6). A J3, l’odeur du poisson s’intensifie
et devient aigre et rance. Nous remarquons une opacification continue de l’œil qui devient
laiteux et épais s’enfonçant complètement (concave). Les viscères éclatent et leur contenu
liquide se dissémine dans la chair lui procurant un aspect mou et pâteux (photo 7). Le poisson
perd de sa fermeté et se détache difficilement de la colonne vertébrale. A J5, la peau devient
totalement terne, dépourvue d’écailles et totalement recouverte d’un liquide visqueux jaunâtre
(photo 8).
Rouabhi, 2009
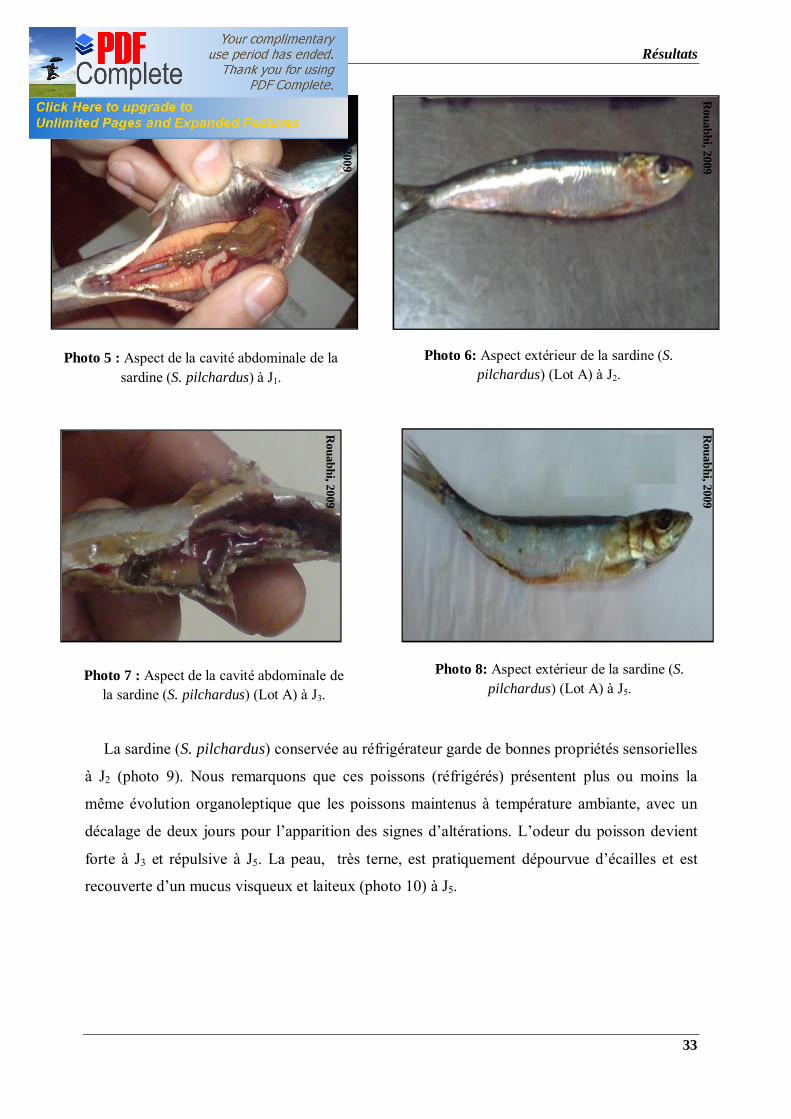
Résultats
33
La sardine (S. pilchardus) conservée au réfrigérateur garde de bonnes propriétés sensorielles
à J2 (photo 9). Nous remarquons que ces poissons (réfrigérés) présentent plus ou moins la
même évolution organoleptique que les poissons maintenus à température ambiante, avec un
décalage de deux jours pour l’apparition des signes d’altérations. L’odeur du poisson devient
forte à J3 et répulsive à J5. La peau, très terne, est pratiquement dépourvue d’écailles et est
recouverte d’un mucus visqueux et laiteux (photo 10) à J5.
Photo 5 : Aspect de la cavité abdominale de la sardine (S. pilchardus) à J1.
Photo 7 : Aspect de la cavité abdominale de la sardine (S. pilchardus) (Lot A) à J3.
Photo 8: Aspect extérieur de la sardine (S. pilchardus) (Lot A) à J5.
Photo 6: Aspect extérieur de la sardine (S. pilchardus) (Lot A) à J2.
Rouabhi, 2009
Rouabhi, 2009
Rouabhi, 2009
Rouabhi, 2009

Résultats
34
.
La sardine (S. pilchardus) conservée au congélateur présente une odeur neutre que ce soit à
J15 ou J30. Par ailleurs, le poisson garde pratiquement les mêmes caractéristiques de J1 (photo
11).
I.2. Caractéristiques biochimiques
I.2.1 Teneur en eau
a. Effet de la durée de conservation
Durant la conservation à température ambiante et au réfrigérateur (+ 4°C), la teneur en eau
du muscle de la sardine (S. pilchardus) diminue jusqu’à la fin des expériences (figure 6). Dans
les deux modes, cette baisse n’est significative (p < 0,05) qu’à partir de J3. En terme de valeur
absolues, la teneur en eau du muscle passe de 76,07± 1,12 g/100 g M.F. à J1 (poisson frais) à
68,09 ± 3,18 g/100 g M.F. à J5 pour les poissons conservés à température ambiante (lot A) et à
52,71 ± 7,68 g/100 g M.F. à J9 pour les poissons réfrigérés (lot B).
Photo 10 : Aspect extérieur de la sardine (S. pilchardus) (Lot B) à J5.
Photo 9 : Aspect extérieur de la sardine (S. pilchardus) (Lot B) à J2.
Photo 11 : Aspect extérieur de la sardine (S. pilchardus) (Lot C) à J30.
Rouabhi, 2009
Rouabhi, 2009
Rouabhi, 2009

Résultats
35
0
10
20
30
40
50
60
70
80
90
j1 j2 j3 j5 j7 j9 j15 j30
Teneur en eau (g/100g M.F.)
Durée (jours)
lot Alot Blot C
La teneur en eau de la sardine conservée au congélateur reste stable dans le temps (p ≥ 0,05)
à J1, J15 et J30.
Figure 6: Evolution de la teneur en eau (g/100g M.F.) chez la sardine (S. pilchardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
b. Effet du mode de conservation
L’analyse statistique Anova 1 montre un effet significatif du mode de conservation sur la
teneur en eau chez la sardine à J2 et J3 en faveur de la réfrigération. En effet, nous remarquons
que cette teneur est plus élevée chez la sardine conservée au réfrigérateur (lot B) que chez
celles conservées à température ambiante (lot A) (figure 7).
a a
b
ab
b
b c
d
a a
: T° amb. : T° +4°C : T° -18°C
b b

Résultats
36
Figure 7 : Effet du mode de conservation sur la teneur en eau (g/100g M.F) chez la sardine (S. pilchardus). Les histogrammes portant des lettres ou des chiffres différents sont significativement différents (p < 0,05).
I.2.2. Lipides
a. Effet de la durée de conservation
Durant la conservation à température ambiante, Anova 1 ne révèle aucune différence
significative (p ≥ 0,05) dans la teneur lipidique chez la sardine. Par contre, en termes de
valeurs absolues, nous remarquons que cette teneur diminue progressivement jusqu’à J5 où elle
atteint une teneur minimale estimée à 1,95 ± 0,52 g/100g M.F. (figure 8).
En ce qui concerne les poissons réfrigérés (lot B), nous remarquons que la teneur en lipide
diminue de manière significative et progressive (p < 0,05) au cours de la conservation. En effet,
cette dernière passe de 3,72 ± 2,92 g/100g M.F. à J1 à 1,66 ± 0,41 g/100g M.F. à J7.
La teneur en lipides dans les muscles des sardines (S. pilchardus) congelées (lot C) est
caractérisée par des fluctuations non significatives (p ≥ 0,05) (figure 8). Par ailleurs, en terme
de valeurs absolues, les lipides atteignent une valeur minimale (2,46 ± 0,48 g/100g M.F.) à J15
puis augment (4,24 ± 1,15 g/100g M.F.) après 1 mois de congélation (J30).
62
64
66
68
70
72
74
76
78
j2 j3 j5
Teneur en eau (g/100g M.F.)
Durée (jours)
Lot A
Lot Ba
b
1
2
: T° amb.
: T° +4°C

Résultats
37
Figure 8 : Evolution de la teneur en lipides (g/100g M.F.) chez la sardine (S. pilchardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
b. Effet du mode de conservation
L’analyse statistique (Anova 1) ne décèle aucun effet significatif (p ≥ 0,05) du mode de
conservation sur la teneur en lipides chez la sardine le long de ses conservations respectives
(figure 9).
Figure 9 : Effet du mode de conservation sur la teneur en lipides (g/100g M.F.) chez la sardine (S.
pilchardus).
0
1
2
3
4
5
6
7
j1 j2 j3 j5 j7 j15 j30
Teneur en lipides (g/100g M.F.).
Durée (Jours).
Lot A
lot B
Lot C
00,5
11,5
22,5
33,5
44,5
j2 j3 j5
Teneur en lipides (g/100g M.F.)
Durée (jours)
Lot A
lot B
a
ab ab
b b
: T° amb. : T° +4°C : T° -18°C
: T° amb. : T° +4°C

Résultats
38
I.2.3. Teneur en phospholipides (P.L.)
a. Effet de la durée de conservation
L’analyse de la variance à 1 critère de classification montre un effet significatif (p < 0,05)
du temps sur la teneur en P.L. chez la sardine conservée à température ambiante (lot A). En
effet, la teneur initiale en P.L. (678,52 ± 323,57 mg Eq P.C/100g M.F.) diminue
significativement au bout de 24 h de conservation puis se stabilise entre J2 et J3 (382 mg Eq
P.C/100g M.F.) et atteint une valeur minimale de 260,52 ± 52,74 mg Eq P.C/100g M.F. à J5
(figure 10).
En ce qui concerne la sardine du lot B (réfrigérée), la teneur en P.L. fluctue de manière
significative (p < 0,05). En effet, cette dernière passe de 678,52 ± 323,57 mg Eq P.C/100g
M.F. à J1 à 173,72 ± 28,72 mg Eq P.C/100g M.F. à J7 (figure 10). Par ailleurs, nous observons
une brusque augmentation à J5.
Chez la sardine congelée (lot C), nous remarquons que la teneur en P.L. diminue
significativement (p < 0,05) au cours du temps. En effet, la teneur en P.L. est de 462,22±
82,78 mg Eq P.C/100g M.F. à J15 puis diminue et atteint une valeur minimale de 173,77 ±
49,57 mg Eq P.C/100g M.F. après un mois de conservation (J30) (figure 10).
Figure 10 : Evolution de la teneur en phospholipides (mg Eq P.C /100g M.F.) chez la sardine (S. pilchardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05)
0
200
400
600
800
1000
1200
j1 j2 j3 j5 j7 j15 j30
P.L. (mg Eq P.C /100g M.F)
Durée (jours)
Lot A
Lot B
Lot Ca a a
b
bcd
b
b
b
ab
bc
b
c
: T° amb.
: T° +4°C
: T° -18°C

Résultats
39
b. Effet du mode de conservation
En comparant les modes de conservations (température ambiante Vs réfrigérée), l’Anova 1
montre des différences significatives (p < 0,05) dans la teneur en P.L. (figure 11).
Figure 11 : Effet du mode de conservation sur la teneur en phospholipides (mg Eq P.C./ 100g M.F.) chez la sardine (S. pilchardus). Les histogrammes portant des lettres, des chiffres ou des symboles différents sont significativement différents (p < 0,05).
I.3. Evolution du niveau d’oxydation des lipides
I.3.1. Les hydroperoxydes
a. Effet de la durée de conservation
A température ambiante et réfrigérée, Anova 1 montre un effet significatif de la
conservation sur la teneur en hydroperoxydes du muscle des poissons (p < 0,05). En effet, à
température ambiante, la teneur en hydroperoxydes augmente progressivement et
significativement (p < 0,05) au cours des 3 jours de conservation, passant ainsi de 7,37 ± 4,14
µmoles/g lipide à J1 à 31,51± 13,07 µmoles Eq CuOOH/g lipide à J3 (figure 12). Au
réfrigérateur, la teneur en hydroperoxydes augmente de manière significative (p < 0,05) puis se
stabilise jusqu’à la fin des expériences (J7) (25,91 ± 12,21 µmoles Eq CuOOH/g lipide) (figure
12).
Lors de la congélation, la concentration en hydroperoxydes fluctue de manière significative
(p < 0,05). En effet, les hydroperoxydes augmentent significativement à J15 puis diminuent
après 1 mois de conservation où ils atteignent une concentration comparable à celle mesurée à
J1 (figure 12).
0
100
200
300
400
500
600
700
800
900
j2 j3 j5
P.L. (mg Eq P.C./100 g M.F.
Durée (jours)
Lot A
Lot B
α
β
a
b
1
2
: T° amb : T° +4°C

Résultats
40
0
5
10
15
20
25
30
35
40
45
50
j1 j2 j3 j5 j7 j15 j30
[C]hydroperoxydes
(µmoles Eq CuOOH/g lipide)
Durée (Jours)
Lot A
Lot B
Lot C
Figure 12: Evolution de la concentration en hydroperoxydes (µmoles Eq CuOOH/g lipide) chez la sardine (S. pilchardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
b. Effet du mode de conservation
L’analyse statistique Anova 1 ne révèle aucun effet significatif (p ≥ 0,05) du mode de
conservation sur la teneur en hydroperoxydes, chez la sardine (figure 13).
Figure 13 : Effet du mode de conservation sur la teneur en hydroperoxydes (µmoles Eq CuOOH/g
lipide) chez la sardine (S. pilchardus).
0
5
10
15
20
25
30
35
40
45
j2 j3
[C]hydroperoxydes
(µmoles eq CUOOH/g lipide)
Durée (jours).
Lot A
Lot B
a a a
b
b
c
c c
c
b
bc
: T° amb.
: T° +4°C
: T° -18°C
: T° amb
: T° +4°C

Résultats
41
0
10000
20000
30000
40000
50000
60000
j1 j2 j3 j5 j7 j9 j15 j30
[C]TBA rs (nmoles MDA/g M.F.)
Durée (Jours).
Lot A
Lot B
Lot C
I.3.2. Les TBA rs
a. Effet de la durée de conservation
Fraîche, la sardine présente une teneur en TBA rs extrêmement faible (396,62 ± 329,29
nmoles MDA/ g de M.F.).
Au cours de la conservation à température ambiante, la concentration en TBA rs augmente
significativement (p < 0,05) au bout de 24 h (J2) puis se stabilise jusqu’à J5. A J7, les TBA rs
augmentent et atteignent un maximum puis diminuent significativement à J9 (15 807± 6 886,26
nmoles MDA/g M.F.) (figure 14).
La concentration en TBA rs de la sardine conservée au réfrigérateur (lot B) augmente
significativement et progressivement (p < 0,05) jusqu’à J7 où elle présente une concentration
maximale de 32 883,3 ± 15 736,17 nmoles MDA/g M.F. puis diminue de manière significative
(p < 0,05) à J9 (figure 10).
Concernant la sardine congelée (lot C), l’analyse de la variance à 1 critère de classification
montre des différences significatives (p < 0,05) dans la concentration en TBA rs. En effet, cette
dernière augmente de manière continue, durant la conservation pour atteindre un maximum à
J30 (figure 14).
Figure 14: Evolution de la concentration en TBA rs (nmoles MDA/g M.F.) chez la sardine (S. pilchardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
b
a a a b
c b
c
d
c
bc bc
bd
bc b
: T° amb.
: T° +4°C
: T° -18°C

Résultats
42
b. Effet du mode de conservation
De manière générale, les deux lots (A et B) présentent les mêmes fluctuations au cours de
leurs conservations respectives. Cependant l’analyse statistique Anova 1 montre une différence
significative (p < 0,05) dans la concentration en TBA rs à J7 entre la sardine conservée à T°
ambiante (Lot A) et réfrigérée (Lot B) (figure 15).
Figure 15 : Effet du mode de conservation sur la teneur en TBA rs (nmoles MDA/g M.F.) chez la sardine (S. pilchardus). Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
0
10000
20000
30000
40000
50000
60000
j2 j3 j5 j7 j9
[C]TBA rs (nmoles MDA/g M.F.)
Durée (jours)
Lot A
Lot B
b
a
: T° amb.
: T° +4°C

Résultats
43
II. Le rouget de roche (Mullus surmuletus)
II.1. L’analyse sensorielle
Au laboratoire (J1), le rouget (Mullus sumuletus) est en phase de rigor mortis (rigidité
cadavérique). En effet, le poisson est caractérisé par un corps d’aspect rigide. La peau est
brillante avec une coloration très vive est présentant des écailles qui lui adhèrent complètement.
Le mucus qui recouvre la peau est de faible consistance, transparent et inodore. L’œil est
convexe (bombé) avec une grande cornée transparente et brillante (photo 12). A l’ouverture du
poisson, nous remarquons que les organes internes sont intacts. La chair est très ferme. La
colonne vertébrale est solidement rattachée à la chair.
Lors du 2ème jour d conservation à température ambiante, les spécimens présentent un aspect
terne avec une décoloration partielle. Les écailles n’adhèrent plus à la peau, pour la plupart des
individus (photo 13). De plus, le mucus qui recouvre le poisson est plus visqueux et légèrement
laiteux. Les trois bandes jaunes latérales commencent à s’estomper. Par ailleurs, on perçoit une
odeur intense caractérisée par une certaine acidité. L’œil s’aplatit et devient opaque. Le flanc se
détériore rapidement et la cavité abdominale est éclatée chez tous les individus. La chair est
complètement recouverte d’un liquide visqueux brun provenant de l’éclatement des viscères.
De plus, la chair est molle, de faible élasticité et difficilement séparable de la colonne
vertébrale et de la peau.
Photo 12: Aspect extérieur du rouget de roche (M. surmuletus) à J1.
Rouabhi, 2009

Résultats
44
A J3, le poisson est complètement décoloré. L’œil, concave et opaque, présente une cornée
laiteuse. De plus, le poisson est très desséché. La chair, recouverte d’une couche cireuse,
ramolli et pâteuse. La forte odeur dégagée par les individus de ce lot (lot A) à J3, nous met dans
l’incapacité de prolonger la conservation.
Pour les poissons conservés au réfrigérateur, l’odeur de rance commence à se dégager à J2.
De même, la peau commence à se ternir et à perdre ses écailles. On observe, aussi, l’éclatement
de la cavité abdominale chez la majorité des individus à J2 et la chair perd de son élasticité. A
J3, l’odeur perçue est de plus en plus forte. La couleur du mucus qui recouvre la peau est très
foncée. La décoloration est très avancée. A l’ouverture du poisson (lot B), les organes internes
sont indifférenciables. La chair est molle (flasque), pâteuse, d’élasticité réduite et recouverte
d’une couche cireuse brune (photo 14).
Photo 13: Aspect extérieur du rouget de roche (M. surmuletus) (Lot A) à J2.
Photo 14: Aspect interne du rouget de roche (M. surmuletus) (Lot B) à J3.
Rouabhi, 2009
Rouabhi, 2009

Résultats
45
A J5, le poisson est en stade d’altération très avancée aussi appelé stade de putréfaction. L’odeur est très repoussante. L’œil est complètement enfoncé et opaque. La peau, complètement dépourvue d’écailles, est recouverte d’un mucus de forte odeur (photo 15).
Le rouget de roche du lot C (congelé) présente de bonnes qualités sensorielles que se soit à
J15 ou à J30. La peau est marquée par un léger ternissement, mais les écailles y adhèrent
complètement. Le mucus est très fluide et translucide. L’œil est toujours bombé et brillant.
L’odeur est typique du poisson frais (ça sent le poisson !). La chair est ferme, fibreuse mais
légèrement mouillée.
II.2. Caractéristiques biochimiques
II.1. Teneur en eau
a. Effet de la durée de conservation
La teneur moyenne en eau chez le rouget de roche frais est de 74,92 ± 1,88 g/100g M.F.
Au cours de la conservation à température ambiante et réfrigérée, la teneur en eau fluctue
significativement (p < 0,05) après deux jours de conservation puis atteint une teneur
comparable à celle mesurée à J1 à J3 (figure 16).
Chez les individus du lot C (congelé), nous remarquons que la teneur en eau reste stable (p ≥
0,05) le long de la congélation.
Photo 15: Aspect externe du rouget de roche (M. surmuletus) (Lot B) à J5.
Rouabhi, 2009

Résultats
46
0
20
40
60
80
100
120
j1 j2 j3 j15 j30
Teneur en eau (g/100g M.F.)
Durée (Jours)
Lot A
Lot B
Lot C
Figure 16: Evolution de la teneur en eau (g/100g M.F) dans la chair du rouget de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
b. Effet du mode de conservation
Lors de la comparaison entre la conservation à température ambiante et réfrigérée (lot A et
lot B, respectivement), aucune différence significative (p ≥ 0,05) n’est décelée à J2. Cependant,
à J3, Anova 1 montre que la teneur en eau de la chair des individus du lot A est
significativement inferieure (p < 0,05) à celle des individus du lot B (figure 17).
Figure 17 : Effet du mode de conservation sur la teneur en eau (g/100g M.F.) chez le rouget de roche (M. surmuletus). Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
0
10
20
30
40
50
60
70
80
90
100
j2 j3
Teneur en eau (g/100g M.F.)
Durée (jours)
Lot A
Lot B
b ab a a
b
ab
a
b
: T° amb.
: T° +4°C
: T° -18°C
: T° amb.
: T° +4°C

Résultats
47
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
j1 j2 j3 j15 j30
Teneur en lipides (g/100g M.F.).
Durée (Jours).
Lot A
Lot B
Lot C
II.2.2. Teneur en lipides
a. Effet de la durée de conservation
L’analyse de la variance à 1 critère de classification (Anova 1) ne révèle aucune différence
significative (p ≥ 0,05) dans la teneur en lipides au cours de la conservation. En terme de
valeurs absolues, la teneur en lipides du muscle du rouget de roche diminue le long de la
conservation (figure 18).
Figure 18: Evolution de la teneur en lipides (g/100g M.F.) chez le rouget de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation.
b. Effet du mode de conservation
L’Anova 1 ne révèle aucune différence significative (p ≥ 0,05) du mode de conservation sur
la dynamique des lipides (figure 19).
Figure 19 : Effet du mode de conservation sur la teneur en lipides (g/100g M.F.) chez le rouget de roche (M. surmuletus).
00,5
11,5
22,5
33,5
4
j2 j3
Teneur en lipides (g/100g M.F.)
Durée (jours)
Lot ALot B
: T° amb.
: T° +4°C
: T° -18°C
: T° amb. : T° +4°C

Résultats
48
0
200
400
600
800
1000
1200
1400
j1 j2 j3 j15 j30
P.L. (mg Eq PC/100g M.F.).
Durée (Jours).
Lot A
Lot B
Lot C
II.2.3. Teneur en phospholipides (P.L.)
a. Effet de la durée de conservation
Au 1er jour d’expérience (J1), la chair du rouget de roche (M. surmuletus) présente une
teneur en P.L. de l’ordre de 643, 82 ± 506,68 mg Eq P.C/100g M.F.
Au cours de la conservation à température ambiante, la teneur en P.L. diminue
significativement (p < 0,05) et atteint son minimum à J3. Cette dernière passe de 452,02 ±
164,82 à 74,62 ± 33,73 mg Eq P.C/100g M.F. de J2 à J3 respectivement (figure 20).
Lors de la conservation au réfrigérateur (lot B), nous remarquons que la teneur en P.L.
diminue brusquement et atteint son minimum (p < 0,05) à J2 puis se stabilise jusqu’à J3 (figure
20).
Concernant le lot de rouget de roche (M. surmuletus) congelé, la teneur en phospholipides
diminue significativement (p < 0,05) à J15 puis se stabilise jusqu’à la fin des expériences.
Figure 20: Evolution de la teneur en phospholipides (mg Eq P.C/100g M.F) chez le rouget de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
a a
ab
b
b b
b b
a
: T° amb
: T° +4°C
: T° -18°C

Résultats
49
b. Effet du mode de conservation
Anova 1 ne montre aucun effet significatif (p ≥ 0,05) du mode de conservation sur la teneur
en phospholipides chez le rouget de roche (figure 21).
Figure 21 : Effet du mode de conservation sur la teneur en phospholipides (mg Eq P.C/100g M.F) chez le rouget de roche (M. surmuletus).
II.3. Evolution du niveau d’oxydation des lipides
II.3.1. Les hydroperoxydes
a. Effet de la durée de conservation
Quel que soit le mode de conservation (température ambiante ou réfrigérée), la teneur en
hydroperoxydes du rouget de roche, fluctue de manière significative (p < 0,05). En effet, cette
teneur augmente significativement et atteint son maximum à J3 où elle présente des teneurs
maximales puis diminue fortement à J5, atteignant ainsi les teneurs minimales en fin
d’expériences (figure 14).
Concernant le lot de rouget de roche congelé (lot C), nous remarquons que la concentration
en hydroperoxydes diminue significativement (p < 0,05) à J 15 puis se stabilise (p ≥ 0,05)
jusqu’à la fin de l’expérience (J30).
0
100
200
300
400
500
600
700
j2 j3
P.L. (mg Eq PC/100g M.F.).
Durée (jours)
Lot A
Lot B
: T° amb.
: T° +4°C

Résultats
50
0
0,005
0,01
0,015
0,02
0,025
0,03
0,035
0,04
0,045
j1 j2 j3 j5 j15 j30
[C]Hydroperoxydes
(µmoles Eq CuOOH/g lipide).
Durée (Jours).
Lot A
Lot B
Lot C
Figure 22: Evolution de la concentration en hydroperoxydes (µmoles Eq CuOOH/g lipide) chez le rouget de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
b. Effet du mode de conservation
A J2, l’Anova 1 montre un effet significatif (p < 0,05) du mode de conservation sur la teneur
en hydroperoxydes chez le rouget de roche, en faveur de la conservation au réfrigérateur. A J3
et J5, aucune différence significative (p ≥ 0,05) n’est décelée. Par ailleurs, en terme de valeurs
absolues, la teneur en hydroperoxydes est plus élevée lors de la conservation à température
ambiante qu’au réfrigérateur (figure 23).
Figure 23: Effet du mode de conservation sur la teneur en hydroperoxydes (µmoles Eq CuOOH/g lipide) chez le rouget de roche (M. surmuletus). Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
0
0,005
0,01
0,015
0,02
0,025
0,03
0,035
0,04
0,045
j2 j3 j5
[C]Hydroperoxydes
(µmoles Eq CuOOH/g lipide).
Durée (jour)
Lot A
Lot B
b
b
b
b
a a a a
ab c
c
a
b
: T° amb.
: T° +4°C
: T° -18°C
: T° amb. : T° +4°C

Résultats
51
0
2000
4000
6000
8000
10000
12000
14000
j1 j2 j3 j5 j15 j30
[C]TBA rs (nmole eq
MDA/g M.F.)
Durée (jours)
Lot A
Lot B
Lot C
II.3.2. Les TBA rs
a. Effet de la durée de conservation
A J1, la concentration en TBA rs du muscle rouget (lot A, B et C) est de 2 807,58 ± 858,56
nmoles MDA/g M.F.
Quelle que soit le mode de conservation (à température ambiante ou réfrigérée), nous
remarquons que la concentration en TBA rs fluctue de manière significative (p < 0,05). En
effet, nous observons qu’elle atteint sa valeur minimale à J2 puis augmente significativement à
J3 et rediminue à J5 (figure 24).
Concernant le rouget de roche congelé (lot C), on observe une augmentation significative (p
< 0,05) à J15 puis les TBA rs restent stable (p ≥ 0,05) jusqu’à J30 (figure 15).
Figure 24: Evolution de la concentration en TBA rs (nmoles MDA/g M.F.) dans la chair du rouget
de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes
portant des lettres différentes sont significativement différents (p < 0,05).
b. Effet du mode de conservation
L’analyse statistique Anova 1 ne montre aucun effet significatif (p ≥ 0,05) du mode de
conservation sur la concentration en TBA rs chez le rouget de roche. Par contre, en terme de
valeurs absolues, nous remarquons l’effet bénéfique de la réfrigération sur cette teneur. En
effet, les teneurs en TBA rs mesurées à J2 et J3 sont plus faibles chez les poissons réfrigérés que
ceux maintenus à température ambiante. De plus, en vue des valeurs mesurées à J5, nous
ad
c
d
ad a
b b
c
d
b
b
: T° amb.
: T° +4°C
: T° -18°C

Résultats
52
remarquons que la synthèse des TBA rs est plus avancée chez le rouget conservé à température
ambiante que celui qui est réfrigéré (figure 25).
Figure 25: Effet du mode de conservation sur la teneur en TBA rs (nmoles MDA/g M.F.) chez le
rouget de roche (M. surmuletus).
0
1000
2000
3000
4000
5000
6000
7000
j2 j3 j5
[C]TBA rs (nmole eq
MDA/g M.F.)
Durée (jours)
Lot A
Lot B
: T° amb.
: T° +4°C

Résultats
53
III. Le merlan bleu (Micromesistius poutassou)
III.1. L’analyse sensorielle
En début d’expériences (J1), le merlan bleu (M. poutassou) est caractérisé par une peau
d’aspect rugueux et plus ou moins brillante. L’œil est convexe, brillant et translucide. La cavité
abdominale est intacte chez la majorité des individus (photo 16). Par ailleurs, le poisson n’est
caractérisé par aucune odeur. A l’ouverture du poisson, nous constatons que les organes
internes sont intacts. La chair est ferme, élastique et de couleur très claire.
Durant la conservation à T° ambiante (Lot A), le poisson se ternit, la peau se décolle, l’œil
s’enfonce et devient opaque. Tous les individus de ce lot ont la paroi abdominale éclatée à J2
(photo 17). Par ailleurs, nous remarquons que l’odeur du poisson évolue rapidement et devient
très intense à J3 où l’on note, aussi, une chair est très collante, pâteuse, desséchée et foncée. Il
est difficile de séparer la chair de la peau ou de la colonne vertébrale.
Photo 17: Aspect externe du merlan bleu (M. poutassou) (Lot A) à J3.
Rouabhi, 2009
Photo 16: Aspect externe du merlan bleu (M. poutassou) frais.
Rouabhi, 2009

Résultats
54
Pour le merlan bleu (M. poutassou) conservé au réfrigérateur (Lot B), nous remarquons un
ternissement de la peau à J2. Le mucus qui couvre cette dernière est très consistant. Par
ailleurs, la chair commence à se ramollir
En général, les signes d’altérations s’intensifient en fonction du temps. Ainsi, à J5 le poisson
présente une odeur repoussante, un aspect terne, une décoloration marquée et un desséchement
avancé (photo 18).
Le lot C (congelé) garde des propriétés sensorielles comparables à celle du J 1, que se soit à
J15 ou J30. Le poisson ne dégage aucune odeur particulière (neutre). Par ailleurs, la chair,
toujours ferme et élastique, est imbibée d’eau.
III.2. Caractéristiques biochimiques
III.2.1. Teneur en eau
a. Effet de la durée de conservation
Durant la conservation à T° ambiante, la teneur en eau diminue significativement (p < 0,05)
jusqu’à J5 où elle atteint sa teneur minimale en passant par des valeurs intermédiaires à J2 et J3
(figure 26).
Durant la conservation au réfrigérateur, la teneur en eau reste stable (p ≥ 0,05) les 3 premiers
jours puis diminue brusquement à J5 où elle atteint sa teneur minimale puis regagne une valeur
intermédiaire à J7.
Photo 18: Aspect externe du merlan bleu (M. poutassou) (Lot B) à J5.
Rouabhi, 2009

Résultats
55
65
70
75
80
85
90
j1 j2 j3 j5 j7 j15 j30
Teneur en eau (g/100g M.F.).
Durée (jours).
Lot A
Lot B
Lot C
Les poissons congelés (lot C) présentent une augmentation significative (p < 0,05) de la
teneur en eau. En effet, cette dernière atteint une teneur maximale de 83,77 ± 3,12 g/100g M.F.
(figure 26) après un mois de congélation.
Figure 26: Evolution de la Teneur en eau (g/100g M.F.) dans la chair du merlan bleu (M. poutassou)
du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres
différentes sont significativement différents (p < 0,05).
b. Effet du mode de conservation
L’analyse statistique (Anova 1) ne montre aucun effet significatif (p ≥ 0,05) du mode de
conservation sur la teneur en eau chez le merlan bleu à J2. Cependant, en termes de valeurs
absolues, nous remarquons que cette teneur est plus élevée chez les poissons conservés au
réfrigérateur que ceux conservés à température ambiante. A J3 et J5, la teneur en eau chez les
poissons conservés au réfrigérateur est significativement plus élevée (p < 0,05) que celle
mesurée chez les poissons conservés à température ambiante. (figure 27).
a ab
b
a ab
b b
a a
a
b
ab
: T° amb
: T° +4°C
: T° -18°C

Résultats
56
00,5
11,5
22,5
33,5
44,5
5
j1 j2 j3 j5 j7 j15 j30
Teneur en lipides (g/100g M.F.).
Durée (Jours).
Lot A
Lot B
Lot C
Figure 27 : Effet du mode de conservation sur la teneur en eau (g/100g M.F.), chez le merlan bleu (M. poutassou). Les histogrammes portant des lettres ou des chiffres différents sont significativement différents (p < 0,05).
III.2.2. Teneur en lipides
a. Effet de la durée de conservation
A l’état frais, la chair du merlan bleu (M. poutassou) présente une teneur en lipides de 2,26
± 1,31 g/100g M.F.
Quel que soit le mode de conservation (à température ambiante, réfrigérée ou de
congélation), l’analyse statistique Anova 1 ne montre pas de différences significatives (p ≥
0,05) dans la teneur lipidique au cours de la conservation (figure 28).
Figure 28: Evolution de la Teneur en lipides (g/100g M.F.) dans la chair du merlan bleu (M.
poutassou) du lot A, B et C, en fonction de la durée de conservation.
68
70
72
74
76
78
80
82
j2 j3 j5
Teneur en eau (g/100g M.F)
Durée (jours)
Lot ALot Bb
2 a 1
: T° amb. : T° +4°C
: T° amb.
: T° +4°C
: T° -18°C

Résultats
57
b. Effet du mode de conservation
L’Anova 1 ne montre aucun effet significatif (p ≥ 0,05) du mode de conservation sur la
teneur en lipides du merlan bleu. Par contre, en terme de valeurs absolues, nous remarquons
que les poissons réfrigérés présentent des teneurs en lipides supérieures à celles mesurées chez
les individus maintenus à température ambiante (figure 29).
Figure 29 : Effet du mode de conservation sur la teneur en lipides (g/100g M.F.), chez le merlan bleu (M. poutassou).
III.2.3. Teneur en phospholipides (P.L.)
a. Effet de la durée de conservation
A J1, la teneur en P.L. du muscle du merlan bleu (M. poutassou) est de 699,62 ± 199,87 mg
Eq P.C./100g M.F. Durant la conservation à T° ambiante, cette teneur reste stable (p ≥ 0,05)
pendant 24 heures puis diminue, par la suite, significativement (p < 0,05) à J3.pour atteindre sa
teneur minimale à J7 (p < 0,05) (figure 30).
Concernant le rouget de roche réfrigéré (lot B), nous remarquons que la teneur en P.L.
diminue progressivement (p < 0,05) le long de la conservation. En termes de valeurs absolues,
la teneur en P.L. atteint une valeur minimale de 202,52 ± 29,39 mg Eq P.C./100g M.F. à J7
(figure 30).
00,5
11,5
22,5
33,5
44,5
5
j2 j3 j5
Teneur en lipides (g/100g M.F.)
Durée (jours)
Lot A
Lot B
: T° amb.
: T° +4°C

Résultats
58
0
200
400
600
800
1000
1200
j1 j2 j3 j5 j7 j15 j30
Concentration en P.L. (mg Eq
P.C./100 g M.F.)
Durée (jours)
Lot A
Lot B
Lot C
Figure 30: Evolution de la Teneur en phospholipides (mg Eq P.C/100g M.F.) dans la chair du
merlan bleu (M. poutassou) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes
portant des lettres différentes sont significativement différents (p < 0,05).
Chez le merlan bleu congelé (lot C), nous remarquons que la teneur en P.L. diminue (p <
0,05) et atteint sa valeur minimale à J15 puis augmente (p < 0,05) et regagne une teneur
comparable à celle mesurée à J1, après un mois de congélation (J30) (figure 30).
b. Effet du mode de conservation
L’analyse statistique Anova 1 ne montre aucun effet significatif (p ≥ 0,05) du mode de
conservation sur la teneur en P.L. du merlan bleu. Par contre, en terme de valeurs absolues,
nous remarquons que l’hydrolyse des phospholipides est plus élevée chez les poissons
conservés à température ambiante que chez ceux maintenus au réfrigérateur (figure 31).
a a
d
b c
a
b
b c
a
a
b
: T° amb
: T° +4°C
: T° -18°C

Résultats
59
Figure 31 : Effet du mode de conservation sur la teneur en phospholipides (mg Eq P.C./100g M.F.), chez le merlan bleu (M. poutassou) le long de la conservation.
III.3. Evolution du niveau d’oxydation des lipides
III.3.1. Teneur en hydroperoxydes
a. Effet de la durée de conservation
A l’état initial (J1), le merlan bleu (M. poutassou) présente une concentration en
hydroperoxydes de 3,38 ± 1,85µmoles Eq CuOOH/g lipide. Lors de la conservation à T°
ambiante, nous observons une augmentation significative (p < 0,05) de la concentration en
hydroperoxydes. En effet, cette dernière atteint une teneur maximale de 8,76 38 ± 3,11 µmoles
Eq CuOOH/g lipide à J5 (figure 32).
Durant la conservation au réfrigérateur (lot B), la concentration en hydroperoxydes reste
stable (p ≥ 0,05) entre J1 et J3. A J5, nous observons une augmentation brusque (p < 0,05) de
cette teneur qui se stabilise jusqu’à J7 (figure 32).
Concernant le merlan bleu du lot C (congelé), la concentration en hydroperoxydes reste
stable (p ≥ 0,05) le long de la conservation (figure 32).
0
200
400
600
800
1000
1200
j2 j3 j5
Teneur en P.L. (mg Eq P.C./100 gM.F.)
Durée (jours)
Lot A
Lot B
: T° amb.
: T° +4°C

Résultats
60
0
2
4
6
8
10
12
14
j1 j2 j3 j5 j7 j15 j30
[C]hydroperoxydes (µmoles Eq Cu OOH/g lipide)
Durée (Jours).
Lot A
Lot B
Lot C
Figure 32: Evolution de la concentration en hydroperoxydes (µmoles Eq CuOOH/g lipide) dans la chair du merlan bleu (M. poutassou) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
b. Effet du mode de conservation
L’analyse statistique ne montre aucun effet significatif (p ≥ 0,05) du mode de conservation
sur la concentration en hydroperoxydes chez le merlan bleu (figure 33).
Figure 33 : Effet du mode de conservation sur la concentration en hydroperoxydes (µmoles Eq Cu OOH/g lipide) chez le merlan bleu (M. poutassou) le long de la conservation.
0
2
4
6
8
10
12
14
j2 j3 j5
[C]hydroperoxydes (µmoles Eq Cu OOH/g lipide)
Durée (jours)
Lot A
Lot B
a a a
a
b b
ab a
b
: T° amb.
: T° +4°C
: T° -18°C
: T° amb.
: T° +4°C

Résultats
61
0
5000
10000
15000
20000
25000
30000
j1 j2 j3 j5 j7 j15 j30
[C]TBA rs (nmoles MDA/g M.F.)
Durée (Jours)
Lot A
Lot B
Lot C
III.3.2. Les TBA rs
a. Effet du mode de conservation
A l’état initial (J1), la concentration en TBA rs est de 3 870,04 ± 7 133,24 nmoles MDA / g
M.F. Lors de la conservation à T° ambiante et au réfrigérateur, nous observons que la
concentration en TBA rs diminue significativement à J2 où elle atteint sa teneur minimale (lot
A et B) (p <0,05). A partir de J3, la concentration en TBA rs augmente significativement (p <
0,05) et atteint une concentration maximale de 6 946,83 ± 1 164,05 nmoles MDA/g M.F. à J5
(figure 34).
La concentration en TBA rs du lot B (réfrigéré) fluctue de manière non significative durant
les 5 premiers jours de conservation (J1-J5). A J7, nous notons une augmentation significative (p
< 0,05) de la concentration en TBA rs (figure 34).
Pendant la conservation au congélateur, aucune différence significative dans la
concentration en TBA rs n’est décelée (p < 0,05). En termes de valeurs absolues, la
concentration en TBA rs diminue à J15 puis augmente à nouveau après 1 mois de congélation
(figure 34).
Figure 34: Evolution de la concentration en TBA rs (nmoles MDA/g M.F.) dans la chair du merlan bleu (M. poutassou) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
a abc
a a
ab
b
b
abc c
: T° amb.
: T° +4°C
: T° -18°C
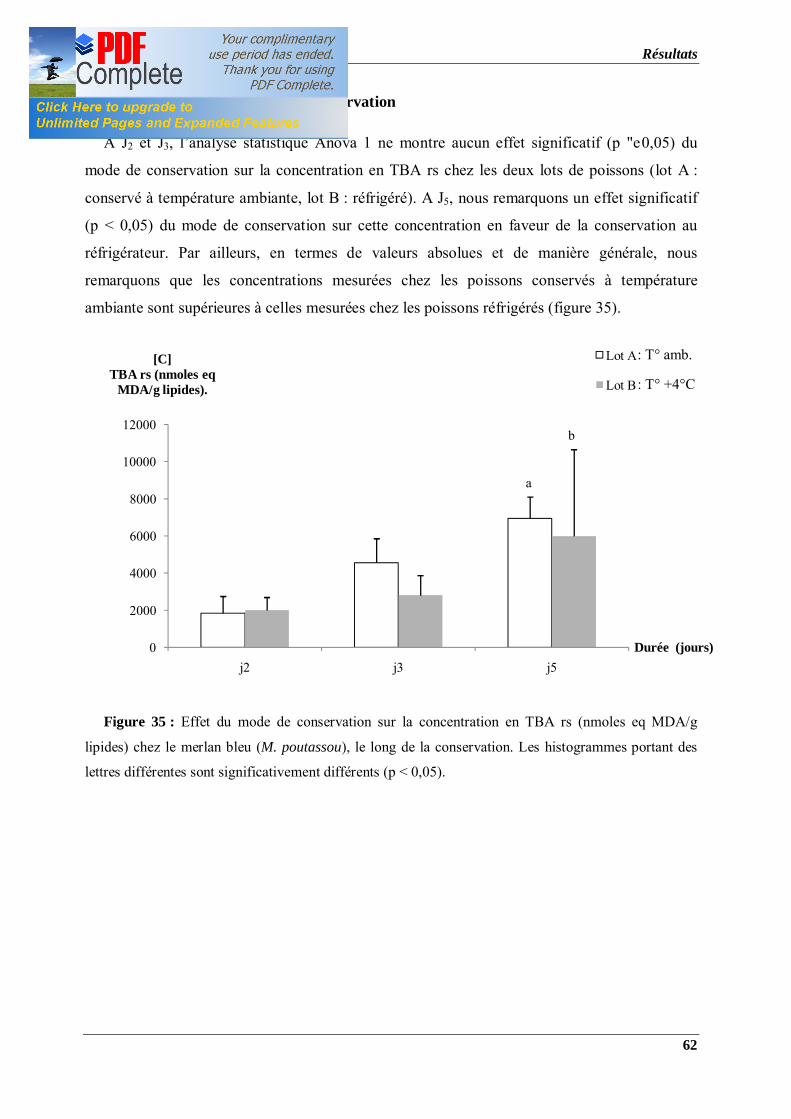
Résultats
62
b. Effet du mode de conservation
A J2 et J3, l’analyse statistique Anova 1 ne montre aucun effet significatif (p ≥ 0,05) du
mode de conservation sur la concentration en TBA rs chez les deux lots de poissons (lot A :
conservé à température ambiante, lot B : réfrigéré). A J5, nous remarquons un effet significatif
(p < 0,05) du mode de conservation sur cette concentration en faveur de la conservation au
réfrigérateur. Par ailleurs, en termes de valeurs absolues et de manière générale, nous
remarquons que les concentrations mesurées chez les poissons conservés à température
ambiante sont supérieures à celles mesurées chez les poissons réfrigérés (figure 35).
Figure 35 : Effet du mode de conservation sur la concentration en TBA rs (nmoles eq MDA/g
lipides) chez le merlan bleu (M. poutassou), le long de la conservation. Les histogrammes portant des
lettres différentes sont significativement différents (p < 0,05).
0
2000
4000
6000
8000
10000
12000
j2 j3 j5
[C]TBA rs (nmoles eq
MDA/g lipides).
Durée (jours)
Lot A
Lot B
b
a
: T° amb.
: T° +4°C

Discussion
63
C. Discussion
Durant la conservation, la qualité du poisson décroit sous l’effet de plusieurs facteurs
(Cheftel et Cheftel, 1976 ; Aubourg et al., 2005). Par ailleurs l’hydrolyse et l’oxydation des
lipides sont les principaux facteurs incriminés (Pearson et al, 1977 ; Miyashita et Takagi,
1986 ; Pigott et Tucker, 1987 ; Han et Liston, 1988 ; Hsieh et Kinsella, 1989a ; German,
1992).
Chez les poissons, les modifications sensorielles interviennent très rapidement dès
l’installation de la rigor mortis. En effet, la qualité du poisson décline rapidement à partir de
ce stade. Les températures basses ont un effet préservateur sur le maintien de la qualité
organoleptique quelle que soit l’espèce considérée (sardine, rouget de roche ou merlan bleu).
Contrairement au mode de conservation à température ambiante, la conservation au
réfrigérateur semble retarder l’apparition et l’évolution des signes d’altérations. En effet, à
température ambiante, le temps de rejet organoleptique est de 24 heures, 24 heures et 3 jours
de conservation, pour la sardine, le rouget de roche et le merlan bleu, respectivement. Alors
qu’au réfrigérateur, ils deviennent impropres à la consommation après 3, 3 et 5 jours de
conservation, respectivement. A partir de ces résultats, il en ressort que le poisson maigre se
conserve mieux que le poisson gras. En effet, Afilal et al. (2006) a démontré qu’à 30°C, le
Dicentrarchus punctatus se conserve jusqu’à 36 heures alors que la sardine (Sardina
pilchardus) ne peut être conservée au-delà de 7 heures. Par ailleurs, la congélation maintient
les poissons à un bon niveau de qualité organoleptique.
Durant la conservation, le poisson présente des odeurs de rance de plus en plus fortes. Ces
dernières traduisent des altérations biochimiques comme la dégradation de l’ATP (Hughes et
Jones 1966). Dans les cas les plus avancés d’altérations ou putréfaction (à partir de J3, pour la
sardine et le rouget de roche et à partir de J5, pour le merlan bleu), les odeurs de rance se
transforment en odeurs soufrées suite à la prolifération de microorganismes producteurs
d’acide sulfurique (Gram, 1989 ; Jorgensen et al., 2000). De plus, cette putréfaction est reliée
à l’abondance des lipides qui sont reliés à la formation de produits aromatiques tels que le
propanol, le pentanol et l’éthanol (Bennour et al., 1991). Donc, plus le poisson est gras plus il
s’altère rapidement. Chez les petits pélagiques comme la sardine (Sardina pilchardus), les
principaux responsables des mauvaises odeurs sont les produits carbonylés, la triméthylamine
n’état pas très bien développée chez ces poissons (Eymard, 2003).

Discussion
64
Le poisson ne bénéficie pas d’un temps de maturation post mortem. La chair est
suffisamment tendre dès le départ. Au cours de la conservation, cette dernière se ramollie et
perd de son élasticité. La texture de la chair dépend de plusieurs facteurs biologiques
intrinsèques comme la densité des fibres musculaires, la teneur en lipides et en collagène
(Sigurgrsladottir et al., 1999 ; Olafsdottir et al., 2004). En effet, la chair de poisson connait
une activité protéolytique très importante (Aubourg et al., 2004). De plus, le collagène du
poisson est moins résistant, moins réticulé et présente une température de fusion plus basse
que celle de la viande (Eymard, 2003). Par ailleurs, le changement de texture serait,
principalement, du à l’interaction des chaines peptidiques avec les produits d’oxydation des
lipides comme les aldéhydes (Pokorny, 1981 ; Sikorski et Kolakowska, 1994).
La sardine se dessèche et perd de sa forme longitudinale au cours de la conservation. De
plus c’est un poisson gras qui présente une forte proportion de muscle brun. Or ce dernier peut
rétrécir jusqu’à 52 % alors que le muscle blanc ne perd que 15 % de sa longueur (Penfield et
campbell, 1990). Nos résultats concordent avec ceux d’autres travaux ayant comparé
l’évolution sensorielle chez un poisson maigre et un autre gras (Perrez villareal et Pozo, 1990,
Ruis capilas et Moral, 2001). Aubourg (1998a) a conclut que l’altération sensorielle du merlan
bleu (Micromesistius poutassou) est moins rapide que celle de la sardine (Sardina
pilchardus). Le rouget de roche (M. surmuletus), bien que d’excellente qualité à J1, se
détériore très rapidement que se soit à température ambiante ou réfrigérée.
Par ailleurs, il a été prouvé que les poissons de tailles moyennes (merlan, merlan bleu et
turbot) ont une durée d’acceptabilité sensorielle longue (Perez villareal et Pozo, 1990 ; Ruiz
capillas et Moral, 2001 ; Aubourg, 2005), contrairement aux petits pélagiques comme la
sardine ou le saurel (Trachurus trachurus) (Nunes et al., 1992 ; Aubourg, 2001). En général,
la durée de conservation est affectée par le niveau d’altération initial et la température de
conservation (Church, 1998).
La teneur en eau est inversement proportionnelle à celle des lipides (Picklet, 1987 ;
Aubourg, 2007). En effet, les poissons maigres présentent une teneur en eau supérieure à celle
des poissons gras (Aubourg et al., 1998a ; Aubourg, Sotelo et Martin, 1998b ; Aubourg et
Medina, 1999a). En accord avec les résultats d’Eymard (2003), nous remarquons que la
teneur en eau de la sardine est inferieure à celle du pomfret atlantique (Brama brama) et du
merlan bleu (Micromesistius poutassou). La faible teneur en eau, mesurée chez le rouget de

Discussion
65
roche, est probablement du aux variations interindividuelles (Aubourg, 2001) ou saisonnières
(Ozyurt, 2005).
Durant la conservation à température ambiante ou réfrigérée, les tissus des poissons
perdent leur intégrité (Eymard, 2003) et la membrane cytoplasmique perd sa fonction de
barrière. De ce fait, l’accroissement de la concentration des solutés dans l’espace
extracellulaire provoque, par osmose, l’extraction de l’eau en dehors de la cellule puis son
évaporation. Cette perte peut être, aussi, du à une dégradation des protéines. En effet, la
dégradation des protéines aboutit à la diminution de la capacité de rétention d’eau par la
fraction myofibrillaire (Seet et Brown, 1983 ; Castrillon et al., 1996 ; Rodriguez et al., 2008).
La teneur en eau chez la sardine et le rouget de roche congelés reste stable jusqu’à un mois
de conservation. Par contre et comme le souligne Aubourg (1993, 1998b) nous remarquons
que le merlan bleu est caractérisé par une forte augmentation de la teneur en eau, qui peut être
attribuée à la diffusion de l’eau au sein des structures musculaires.
Les variations dans la teneur en lipides au sein d’un même lot issu de la même capture
reflètent l’hétérogénéité de la matière première. Elles résulteraient de variations inter
individus liées aux variations de poids, de taille, de sexe, de statut physiologique et d’âge
(Aubourg, 2001 ; Eymard, 2003). Frais, la teneur en lipides est de 2,26 % chez le merlan
bleu, 2,62 % chez le rouget de roche et de 3,72 % chez sardine. La teneur en lipides des
poissons gras (sardine commune) est caractérisée par une grande élasticité. En effet, la sardine
péchée près des côtes portugaises varie considérablement au cours de l’année. Elle atteint 18
% au mois de septembre et octobre (saison abondante en nourriture), et environ 1,2 % au mois
de mars et avril (saison de frai) (Bandarra et al., 1997). Par ailleurs, nous remarquons que la
teneur en P.L est relativement comparable chez les trois espèces de poissons et de l’ordre de
600 mg Eq P.C./ 100 g M.F.). Ceci est la preuve que les P.L., en tant que constituants
fonctionnels, restent stables chez les trois espèces étudiées. Quelle que soit l’espèce
considérée, nous remarquons que la teneur en lipides et en P.L. diminuent au cours de la
conservation. Nous remarquons, aussi, que l’hydrolyse des lipides et P.L. est plus importante
chez les spécimens conservés à température ambiante qu’au réfrigérateur. En effet, à
température ambiante, nous assistons à des cassures des liaisons faibles (Chan, 1987). Les
légères fluctuations observées sont dû aux variations inter individus au sein de chaque espèce
(Aubourg, 2001). La teneur en lipide et en P.L. du merlan bleu et le rouget de roche restent
stables, durant la congélation. Par contre, chez la sardine, nous assistons à une forte

Discussion
66
diminution des P.L. tandis que la teneur en lipides reste comparable à celle mesurée en début
d’expérience. D’après Hwang et Regenstein (1993) et Aubourg (1998a), l’activité des lipases
diminue après 7 jours de congélation, alors que celle des phospholipases persiste au-delà de
15 jours. En effet, à l’état congelé, l’hydrolyse des P.L. est le principal facteur de dégradation
des lipides (Ohshima et al.,1985 ; Eymard, 2003). Rossel et al. (1989) montrent que 70 % des
A.G. libres proviennent de l’hydrolyse des P.L. Cette hydrolyse est catalysée par des enzymes
endogènes qui restent fonctionnels jusqu’à 300 jours de congélation (Eymard, 2003). Bien
que l’abaissement des températures réduise la vitesse d’oxydation, l’augmentation de la
concentration des solutés due à la congélation entraine une accélération de ce processus. Il en
résulte que la lipolyse peut devenir le facteur limitant de la durée de congélation. Ceci est
particulièrement vrai chez les poissons gras (Ozyurt et al., 2007).
Les acides gras libérés au cours de l’hydrolyse des lipides représentent le substrat
préférentiel de la peroxydation lipidique (Howell, 1995 ; Careche et Tejada, 2002). Quelle
que soit l’espèce considérée (sardine, rouget de roche, merlan bleu), nous remarquons que la
formation des hydroperoxydes est déjà initiée à l’état frais. En effet, dès la mort de l’animal
nous assistons à une baisse du pH qui représente un agent initiateur puissant de l’oxydation
des lipides. Par ailleurs, nous remarquons que cette teneur initiale est plus importante chez la
sardine (poisson gars) que chez le merlan bleu (poisson maigre). Chez le rouget de roche, la
teneur en hydroperoxydes est extrêmement réduite. Elle est expliquée par le fait que le
poisson est en rigor mortis à l’arrivée au laboratoire. D’après les résultats obtenus et en
accord avec Rodriguez (2008), il semble que la conservation à température réfrigérée ne soit
pas suffisante pour inhiber ou ralentir la formation des hydroperoxydes.
Durant la conservation à température ambiante ou au réfrigérateur, la sardine est
caractérisée par une augmentation considérable de la concentration en hydroperoxydes durant
les premières 24 heures. En effet, chez les petits pélagiques, la formation des hydroperoxydes
est favorisée par les premières heures de conservation (Eymard et Génot., 2003). Par ailleurs,
il semble que cette période corresponde à la phase de propagation qui se prolonge jusqu’à J5.
Chez la sardine, cette phase est longue en raison du pH post mortem qui est inferieur à 6, qui
favorise la présence des formes actives de myoglobine (puissant catalyseur de la
lipoperoxydation). En effet, les réactions d’oxydations catalysées par la myoglobine sont pH
dépendantes (Baron et Anderson, 2002), de même que celles catalysées par les ions
métalliques (Eymard, 2003). Après cinq jours de conservation, cette teneur diminue en raison
de la décomposition des hydroperoxydes en molécules plus petites (Aubourg, 1999b).

Discussion
67
Chez le rouget de roche, nous remarquons que la teneur en hydroperoxydes est au
maximum de sa propagation à J3. Contrairement aux autres espèces, la phase de propagation
est brève. Ainsi, après cinq jours de conservation, la teneur en hydroperoxydes est très
réduite. Chez le merlan bleu, nous remarquons que les teneurs en hydroperoxydes sont
inferieures à celles mesurées chez la sardine. En effet, le merlan contient une quantité en
lipides moindre et de ce fait, une quantité de substrat pour la peroxydation lipidique moindre.
La phase de propagation est plus longue (5 jours) que chez les autres espèces. Au-delà, nous
assistons à une diminution de leur concentration. D’après Aubourg (2001), cette baisse est du
à leur interaction à des constituants biologiques ou à leur décomposition en produits
secondaires. La conservation au réfrigérateur ne semble pas ralentir la formation des TBA rs,
comparée à la conservation à température ambiante.
Chez la sardine congelée, la cinétique de formation des hydroperoxydes suit son schéma
classique. A savoir une augmentation à J15 (phase de propagation) et une diminution à j30
(transformation en produits secondaires). Chez le rouget de roche et le merlan bleu, l’effet
préservateur des basses températures est bien établi. En effet, nous remarquons que la teneur
en hydroperoxydes se stabilise après 15 jours chez le rouget de roche et reste stable dès le
début de l’expérience chez le merlan bleu. A partir de ces résultats nous pouvons conclure que
la cinétique de formation des hydroperoxydes varie considérablement en fonction de la teneur
en lipides du poisson.
La nature des TBA rs et leurs proportions relatives dépendent de l’A.G. oxydé (Eymard,
2003). Les produits secondaires contribuent à une perte de saveur et à la genèse de
groupements toxiques fréquemment associés à la rancidité (Chow, 1980 ; Halliwell et Chirico,
1993). De plus, les produits carbonylés, issus de la fission des aldéhydes et des
hydroperoxydes, peuvent réagir avec les groupements amines conduisant à une baisse de la
valeur nutritive de la chair (Nielson et al., 1985, Hidalgo et al., 1992), à la production
d’odeurs désagréables (F.A.O., 1992, 1994) et à la perte de saveur (Pearson et al., 1977 ;
Pokorny, 1981 ; Hseih et Kinsella, 1989b ; Harris, 1994). La production des TBA rs suit la
même tendance que ce soit chez les poissons conservés à température ambiante ou réfrigérée.
Par ailleurs, cette concentration reste plus importante chez la sardine (poisson gras) que chez
les autres poissons étudiés (rouget de roche et merlan bleu). En accord avec les résultats
d’Aubourg et al (1998a, 1998b, 2004), la production de TBA rs est plus importante chez la
sardine que chez le merlan bleu, lors de la congélation.

Discussion
68
La sardine doit être consommée fraîche et ne doit être conservée au-delà d’une journée à
température ambiante et de 2 jours au réfrigérateur. La congélation n’est pas conseiller pour
cette espèce. En effet, chez les petits pélagiques comme la sardine, l’hydrolyse et l’oxydations
des lipides persistent (Aubourg et al., 1998a ; Undeland et al., 1998 ; Saeed et Howell, 2002)
et contribuent à la dégradation du poisson (Jacobsen, 1999). Par ailleurs, la sardine contient
une grande quantité de lipides et substance pro- oxydantes (Decker et Hultin, 1990 ; Saeed et
Howell, 2001) et doit de ce fait être conservée entière et non sous forme de filets pour limiter
la surface de contact avec les agents initiateurs extérieurs (Santos et Regenstein, 1990 ;
Aubourg et al., 2004).
Lors de la conservation à température ambiante, la grande altération lipidique et les fortes
variations organoleptiques affectent considérablement la qualité du poisson.par conséquent,
ce mode de conservation est à éviter. Les conservations au réfrigérateur et au congélateur sont
très utilisées pour retenir le poisson à un bon niveau de qualité (Madrid et al., 1994 ;
Erickson, 1997). Cependant beaucoup d’études montrent que la qualité du poisson décroit lors
de durées prolongées de réfrigération (Shewfelt, 1981 ; Bennour, 1991 ; El Merrakchi et al.,
1990 ; Verma et al., 1995). Par conséquent, si le poisson n’est pas consommé durant les deux
jours qui suivent sa capture, il est préférable de le congeler (Erickson, 1987 ; Pigott et Tucker,
1987). L’effet bénéfique des basses températures de congélation contre l’altération des
lipides est largement connu (Aubourg, 1999a et b).
Le suivi de l’oxydation des lipides est un outil très important dans l’évaluation de la qualité
du poisson (Kim et Labella, 1983 ; Melton, 1983). Cependant le problème relié à leur
utilisation est du au fait que l’oxydation des lipides prend la forme d’une cloche (Eymard,
2003). Donc on ne sait pas toujours dans quel intervalle on est situé par rapport au maximum
de propagation. De plus, les produits de l’oxydation des lipides interagissent rapidement avec
des constituants biologiques qui ne sont pas sensibles aux dosages utilisés (Aubourg et al.,
2007 ; Rapeepan et al, 2009). D’après Ozyurt et al. (2005), le suivi d’un substrat de
l’oxydation (P.L ou acides gras libres) et le dosage des TBA rs est une combinaison pertinente
pour l’évaluation de la qualité du poisson. En accord avec Eymard (2003) et Aubourg (1998a,
2001, 2004), nous pensons que l’analyse sensorielle, très pertinente, complétée par le dosage
des TBA rs constituent un moyen très précis. En effet, parmi tous les paramètres
biochimiques, les TBA rs sont les plus liés à la formation d’odeurs de rance (Olafsdottir et al.,
1997).

Discussion
69
Durant le présent travail, le problème lié à l’hétérogénéité de la matière première est
soulevé. En effet, l’utilisation de 5 individus et le dosage de 10 échantillons (par espèce) n’est
pas suffisant pour s’affranchir des variations inter individuelles. Aubourg (1999a, 2004) a
également travaillé sur 5 individus et est parvenu à la même remarque. On pourrait, par
conséquent, nous interroger sur le nombre de spécimens à analyser pour aboutir à un indice de
qualité représentatif de la fraicheur réelle du poisson. Cependant, ayant travaillé sur 10
individus de saurel (Trachurus trachurus), Eymard (2003) trouve que ce nombre ne suffit
toujours pas.

Conclusion et perspectives
70
Conclusion et perspectives
L’évolution de l’hydrolyse et l’oxydation des lipides est influencée par deux facteurs, à
savoir, l’espèce et le mode de conservation. Concernant l’espèce, il a été prouvé que le
poisson gras présente une altération des lipides plus élevées par rapport au poisson maigre
(Aubourg, 2004). En effet, au cours de notre travail, nous sommes arrivés à la même
conclusion à savoir : la sardine s’altère le plus rapidement tandis que merlan bleu se garde le
mieux. Quant au mode de conservation, il s’avère que la congélation représente le meilleur
moyen pour conserver un bon niveau de qualité, si le poisson n’est pas consommé dans les
deux jours qui suivent sa capture.
A l’issu de ce travail, nous pensons qu’il serait très intéressant de compléter nos résultats
par le dosage des composés volatiles afin de déterminer et de suivre la cinétique de formation
des composés responsables des mauvaises odeurs. Lors d’une étude portant sur la perception
des odeurs des produits de la mer, Prost et al. (2002) a montré la relation entre l’évolution de
l’odeur du produit et la nature et l’évolution des concentrations des produits volatiles. En
effet, le dosage de triméthylamine corrèle bien avec l’analyse sensorielle (Eymard, 2003). De
plus, l’oxydation des lipides pourrait être contrôlée par la régulation du pH en incorporant du
carbonate de calcium à la chair. Procédé déjà utilisé à l’échelle industrielle pour la
transformation du poisson.
Pour conclure, nous pouvons dire qu’à une époque où les notions de qualité et de
traçabilité prennent de l’importance, des normes de qualités, à l’échelle locale, devrait être
mises en place. De plus, ces dernières doivent être continuellement actualisées par rapport aux
nouvelles techniques adaptées à ces matrices (produits de la mer, notamment les poissons).
L’assurance de produits aquatiques seins et de bonne qualité reste un challenge à relever en
Algérie.

Références bibliographiques
71
REFERENCES BIBLIOGRAPHIQUES
Abi-ayad S. M., E.-A., 1998- Etude expérimentale de la biologie de la reproduction de la
perche fluviatile (Perca fluviatilis). Effet de la composition en acides gras de la série (n-3)
de l’alimentation des géniteurs sur la qualité des œufs et des larves. Thèse de doctorat,
faculté des sciences, Université de Liège, Belgique, 147p.
Ackman R., 1989- Fatty acids. In R.G. Ackman, marine biogenic lipids,.fats and oils. 1: 103-
137). Boca Raton, FL: CRC press.
Ackman R. & Ratnayake W., 1990- Chemical and analytical aspects of assuring an effective
supply of omega – 3 fatty acid to the cpnsummer. In omega-3 fatty acids in Health and
disease, Lees R. & Karel M. (Ed.), pp 215-233. Marcel Decker, Inc., New York and Basel.
Ackman R.G., 1995. In “ Fish and Fishery Products ”, ed. Ruiter A., pp 117-156, CAB
International, Oxon UK.
Afilal M.A., Daoudi H., Jdaini S., Asehraou A. & Bouali A., 2006- Study of the histamine
production in the red flesh fish (Sardina pilchardus) and a with flesh fish (Dicentrarchus
ounctatus). Turkish journal of fisheries and aquatic sciences. 6 :43-48.
Alanasier C., 1996- Évolution post mortem des lipides musculaires et des activités
lipolytiques chez le lapin. Thèse de Doctorat, Université de Bordeaux I. 165 p.
Aubourg, S. P., 1993- Review: interaction of malondialdehyde with biological molecules-
new trends about reactivity and significance. International Journal of Food Science and
Technology. 28: 323–335.
Aubourg S., Medina I. & Gallardo J., 1998a- Quality assessment of blue whiting
(Micromesistius poutassou) during chilled storage by monitoring lipid damages. Journal of
the Agricultural and Food Chemistry. 46: 3662–3666.
Aubourg S., Sotelo C. & Perez-Martin R., 1998b- Assessment of quality changes in frozen
sardine (Sardina pilchardus) by fluorescence detection. Journal of the American Oil
Chemists’ Society. 75: 575–580.
Aubourg S., & Medina I., 1999a- Influence of storage time and temperature on lipid
deterioration during cod (Gadus morhua) and haddock (Melanogrammus aeglefinus)
frozen storage. Journal of the Science of Food and Agriculture.79: 1943–1948.

Références bibliographiques
72
Aubourg S., 1999b- Review: recent advances in assessment of marine lipid oxidation by
using fluorescence. Journal of the American Oil Chemists’ Society. 76: 409-419.
Aubourg S., 2001- Damage detection in horse mackerel (Trachurus trachurus) during chilled
storage. Journal of the American Oil Chemists’ Society. 78: 857–862.
Aubourg S.P., Rodriguez A. & Gallardo J.M., 2004- Rancidity development during frozen
storage of mackerel (Scomber scombrus): effect of catching season and commercial
presentation. Eur. J. Lipid Sci. Technol. 107: 316-323.
Aubourg S, Carmen Pineiro , Jose M. Gallardo & Jorge Barros-Velazquez, 2005-
Biochemical changes and quality loss during chilled storage of farmed turbot (Psetta
maxima), Food Chemistry. 90: 445–452
Aubourg S., Quitral V., Larraın A., Rodrıguez A., Gomez J., Maier L., & Vinagre J.,
2007- Autolytic degradation and microbiological activity in farmed Coho salmon
(Oncorhynchus kisutch) during chilled storage. Food Chemistry. 104: 369-375.
Aursand M., Bleivik B., Rainuzzo J.R., Jorgensen L. & Mohr V., 1994.- J. Sci. Food
Agric. 64: 239-248.
Bacot S., 2004- Caractérisation d’un nouvel aldéhyde plaquettaire (4- Hydroxydodecadienal)
et formation d’adduits de Mickael avec les phospholipides à l’éthanolamine. Thèse de
doctorat. Université De Lyon. p187.
Bandarra, N.M., Batista, I., Nunes, M.L., Empis, J.M. and Christie, W., 1997- Seasonal
changes in lipid composition of sardine (Sardina pilchardus). Journal of Food Science. 62:
40-42.
Baron, C.P. & Andersen, H.J., 2002- Myoglobin-induced lipid oxidation. A review.
Journal of Agricultural and Food Chemistry. 50: 3887-3897.
Bauchot, M.L., 1987- Poissons osseux : 891-1421. In W. Fischer, M.L. Bauchot et M.
Schneider (eds). Fiches FAO d’identification pour les besoins de la pêche (rev.1).
Mediterranee et mer Noire. Zone de peche 37. Vol. II. Commission des Communautes
Europeennes, F.A.O., Rome.
Bendiksen E.A. & Jobling M., 2003- Fish Physiol. Biochem. 29:133-140.
Bennour M., El Marrakchi A., Bouchriti N., Hamama A. & El Ouadaa M., 1991-
Chemical and microbiogical assessments of mackerel (Scomber scombrus) stored in ice. J.
Food Pro. 54: 789-792.

Références bibliographiques
73
Berset C. & Cuvelier M. E., 1996- Methods of estimating the degree of lipid oxidation
and of measuring antioxidizing power. Sciences des Aliments. Vol. 16, pp 219-245.
Bligh E.G. and Dyer W.J.,1959- A rapid method of total lipid extraction and purification.
Canadian Journal of Biochemistry and Physiology. 37: 911-917.
Bostoglou N.A., Fletouris D.J., Papageorgiou G.E., Vassilopoulos V.N., Mantis A.J. &
Trakatellis A.G., 1994- Rapid sensitive and specific thiobarbituric acid method for
measuring lipid peroxidation in animal tissue, food and feedstuff samples. Journal of
Agricultural and Food Chemistry.42 : 1931-1937.
Bougis, P., 1952- Recherches biométriques sur les rougets (Mullus barbatus L. et Mullus
surmuletus L.). Archives de Zoologie Expérimentale et Générale. 89 (2): 57-174
Bourgeois, C.M. & Leveau, J.Y., 1991- Techniques d’analyse et de contrôle dans les
industries agro-alimentaires. Le contrôle microbiologique, Lavoisier, Apria.
Bourgeois, C.M. Mescle, G.F. & Zucca, J., 1996- Microbiologie alimentaire. Aspect
microbiologique de la sécurité et de la qualité des aliments. Londres, Paris.
Buettner G.R., 1993- The pecking order of free radicals and antioxidants: lipid peroxidation,
alpha-tocopherol and ascorbate. Arch Biochem Biophys. 300: 535-43.
Cahu C. & Zambonino Infante J. L., 2001 - Substitution of live food by formulated diets in
marine fish larvae. Aquaculture. 2000: 161-80.
Cahu C., Zambonino Infante J. L., & Takeuchi T., 2003 - Nutritional components
affecting skeletal development in fish larvae. Aquaculture. 227: 245-58.
Careche M. & Tejada M., 2002- The effect of neutral and oxidized lipids on functionality in
hake (Merlucciuc merluccius): A dimethylamine and formaldehyde forming species during
frozen storage. Food Chemistry. 36: 113-128.
Castrillon, A., Alvarez-Pontes, E., Garcıa, M., & Navarro, P., 1996- Influence of frozen
storage and defrosting on the chemical and nutritional quality of sardine (Clupea
pilchardus). Journal of the Science of Food and Agriculture. 70 : 29-34.
Chabaud M., 2007 – Utilisation des antioxydants en hépatologie chez les carnivores
domestiques. Thèse de doctorat. Université Claude Bernard, Lyon I. 119pp.
Chan H., 1987- Autooxidation of unsaturated lipids; Chan H., Eds. Academic press: New
York.

Références bibliographiques
74
Chaveron H., 1999- Molécules toxiques. Molécules formées au cours des réactions
d’oxydation ed. Introduction à la toxicologie nutritionnelle. Londres : Tec Et Doc, pp 175-
83.
Cheftel J. & Cheftel H., 1976- Introduction to the biochemistry and technology of foods.
Editorial acribia, Zaragoza.
Cheriot S. 2007- Role des produits de la réaction de Maillard dans l’inhibition de l’oxydation
enzymatique des phenols et des lipides. Thèse de doctorat. Université de Paris, Agro Paris
Tech. Pp239.
Chow, K.W., 1980- Storage problems of feedstuffs. FAO/UNDP Training Course in Fish
Feed Technology, College of Fisheries, University ofWashington, Seattle, 9 October– 15
December 1978, pp. 216–224. ADCP/REP/SO/ll.
Church, N., 1998- MAP fish and crustaceans-sensory enhancement. Food Science and
Technology Today. 12(2) : 73–83.
Clausse F., 2001- Radicaux libres et molécules à activité antioxydante. Thèse de doctorat
veterinaire, Faculte de medecine, Creteil, 92p.
Comhair S.A. & Erzumus C., 2002 – Antioxidant responses to oxidant mediated long
diseases. Am. J. Physiol. 283: 246-255.
Corongiu F.P. & Banni S., 1994- Detection of conjugated dienes by second derivative
ultraviolet spectrophotometry. Methods in Enzymology 233 303-313.
Corraze G., Kaushik S.J., 1999- O.C.L. 6: 111-115.
Coulon L., 2004- Effet d’un hydroperoxyde lipidique et des LDL oxydés sur les enzymes
impliqués dans la libération de l’acide arachidonique des phospholipides plaquettaires.
Thèse de doctorat. Université de Lyon. 194 pp.
De Koning A.J. & Mol T.H., 1990- Rates of free fatty acid formation from phospholipids
and neutral lipids in frozen cape hake ( Merlussius spp ) mince at various temperatures.
Journal of the Science of Food and Agriculture. 50: 391-398.
Decker E.A. & Hultin H.O., 1990- Nonenzymic Catalysts of Lipid Oxidation in Mackerel
Ordinary Muscle. Journal of Food Science. 55 : 951-953.
Delattre J., Beaudeux J.-L., Bonnefont & Rousselot D., 2005- Radicaux libres et stress
oxydant : aspects biologiques et pathologiques. Lavoisier, Paris, 549p.

Références bibliographiques
75
Desbrosses, P., 1935- Contribution a la connaissance de la biologie du rouget barbet en
Atlantique Nord (III). Mullus barbatus (rond) surmuletus (Fage). Mode septentrional Fage.
Rev. Trav. Off. Pêch. marit. 8(4) : 351-376.
Desbrosses, P., 1936- Contribution a la connaissance de la biologie du rouget barbet en
Atlantique nord (IV). Rev. Trav. Inst. (Scient. Tech.) Pêch. Marit. 92 : 329-399.
Direction des pêches et des ressources halieutiques de la Wilaya d’El Tarf, 2008.
Dobarganes, M.C. and Velasco, J. , 2002- Analysis of lipid hydroperoxides. European
Journal of Lipid Science and Technology. 104: 420-428.
Droge W., 2002 - Free radicals in the physiological control of cell function. Physiol. Rev. 82:
47-95.
Dumay J., 2006 – Extraction des lipides en voie aqueuse par bioréacteur enzymatique
combiné à l’ultrafiltration: Application à la valorisation de co-produits de poisson (Sardina
pilchardus). Thèse de doctorat de l’université de Nantes. 318 pp.
Dyer, W.J. & Fraser, D.I., 1959- Proteins in Fish Muscle. 13. Lipid Hydrolysis. Journal of
Fisheries Research Board of Canada. 16: 43-52.
El Marrakchi A., Bouchriti N., Bennour N., Hmama A. & Tagafit H., 1990- Sensory,
chemical and microbiological assessment of moroccan sardines (Sardina pilchardus)
stored in ice. J. Food Prot. 53: 600-605.
Erickson M., 1997- Lipid oxidation : flavor and nutritional quality deterioration in frozen
foods. In: Quality in frozen foods. Eds. Erickson M. & Hung Y. C. Chapman & Hall, New
York (U.S.A.). pp 141-173.
Eymard S., 2003 - Mise en évidence et suivi de l'oxydation des lipides au cours de la
conservation et de la transformation du chinchard (Trachurus trachurus): choix des
procédés. Thèse de doctorat de l'Université de Nantes, 143 pp.
Eymard S. & Genot C., 2003- A modified xylenol orange method to evaluate formation of
lipid hydroperoxides during storage and processing of small pelagic fish. European
Journal of Lipid and science technology.105: 497-501.
F.A.O., 1994- Food and Agriculture Organization of the United Nations inform Fshery
statistics, yearbook 1992. (82: 187-188) Rome (Italy)
F.A.O., 1999- The State of Food and Agriculture 1998. F.A.O. Agric. Ser. 26 F.A.O., fisheries department, 2003 - Review of the state of world fishery resources.
Technical paper, 457.

Références bibliographiques
76
Fage, L., 1909- Etude de la variation chez le Rouget (Mullus barbatus L., M. surmuletus L.)."
Archives de Zoologie Expérimentale et Générale. 1(5) : 55p.
Favier A., 2003- Le stress oxydant : Intérêt conceptuel et expérimental dans la
compréhension des mécanismes des maladies et potentiel thérapeutique. Act. Chim. 56:
108-115.
Fernandez S.J. & Mackie I., 1987- Comparative rates of spoilage of fillets and whole fish
during storage of haddock (Melanogrammus aeglefinus) and herring (Clupea harengus) as
determined by the formation of non-volatile and volatile amines. Int. J. Food Sci. Technol.,
22: 385-390.
Folch J., Lees M. & Sloane Stanley G.H.S., 1957- A simple method for the isolation and
purification of total lipides from animal tissues. Journal of Biological Chemistry. 226 :
497-509.
Forest A., 2001 - Ressources halieutiques hors quotas du Nord Est Atlantique: bilan des
connaissances et analyse de scénarios d'évolution de la gestion. Ifremer Eds, tome 2: 215
pp.
Frankel E.N., 1984- Lipid oxidation: Mechanism, products and biological significance. J
Am Oil Chem Soc. 61: 1908-1917.
Frankel, E.N., 1998 - Lipid oxidation. The Oily Press.10. Dundee, Scotland. 10.
Frankel E.N., Neff W.E. & Weisleder D., 1990- Determination of methyl linoleate
hydroperoxides by 13C nuclear magnetic resonance spectroscopy. Methods in Enzymology
186 :380-387.
Frei B., 1994- Reactive oxygen species and antioxidant vitamins: mechanisms of action.
Am. J. Med. 97 (3A) :5S-13S.
Gandemer G., Pichou D., Bouguennec B., Caritez J.C., Bergé P., Briand E. & Legault
C., 1990- Influence du système d'élevage et du génotype sur la composition chimique et
les qualités organoleptiques du muscle long dorsal chez le porc. Journées Rech Porc.
France. 22 :101-10.
Gandemer G., 1992- Les cahiers de l’ENSBANA. 8 : 25-48.
Gandmer G., 1997- Phospholipides, lipolyse, oxydation et flaveur des produits carnés.
OCL. Vol. 4, pp 1-7

Références bibliographiques
77
Genot C., 1996- Some factors influencing TBA test. Report of diet-ox project (AIRIII-CT
92-1577).
Genot C., Meynier A., Riaublanc A. & Chobert J.M., 2003- ProteinAlterations Due to to
Lipid Oxidation in Multiphase Systems. In Lipid oxidation pathways, Kamal-Eldin A.
(Ed.), AOACS Press Champaign, 265-292.
German, J.B., 1992- Lipid oxidation in fish tissue. enzymatic initiation via lipoxygenase.
Journal of Agricultural and Food Chemistry. 33 : 680-683.
Girard S., Pacquotte P., 2003- Cah. Nutr. Diet. 38, 1: 17-28.
Gram L., 1989- Identification, characterization and inhibition of bacteria isolated from
tropical fish. PhD thesis. Royal Veterinary and Agricultural University. Denmark
Granjean D., 2001- Le stress oxydatif cellulaire chez le chien: conséquences et prévention
nutritionnelle. Bull. Acad. Vet. De France. 154, (3suppl.):49-61.
Gray J.I., 1978- Measurement of lipid oxidation: a review. Journal of the American Oil
Chemists' Society. 55: 539-545.
Gret F., 1993- conserver et protéger l’aliment. www.es-sahel.com
Guillén-Sans R. & Guzmàn-Chozas G., 1998- The thiobarbituric acid (TBA) reaction in
foods: A review. Critical Reviews in Food science and Nutrition. 38 : 315-330.
Halliwell B. & Gutteridge J.M., 1990a – Role of free radicals and catalytic metal anions in
human diseases: an overview. Methods. Enzymol. 186: 1-85.
Halliwell B., 1990b- How to characterize a biological antioxidant. Free Radic Res Commun.
13: 1-32.
Halliwell B.& Chirico S., 1993- Lipid-peroxidation-its mechanism, measurement, and
significance. Am. J. Clin. Nutr. 57: 715–725.
Han T. J. & Liston J., 1988- Correlation between lipid peroxidation and phospholipid
hydrolysis in frozen fish muscle. J. Food. Sci. 53: 1917-1918.
Harris P., 1994- Rancidity in fish. In: Rancidity in foods. Eds. Allen J., Hamilton R.
Chapman & Hall, London (UK), pp 256-272.
Henderson R.J., Tocher D.R., 1987- Prog. Lipid Res. 26: 281-347.
Hermes-Lima M., Willmore W.G. &Storey K.B., 1995- Quantification of lipid
peroxidation in tissue extracts based on Fe(III) xylenol orange complex formation. Free
Radical Biology & Medicine. 19: 271-280.

Références bibliographiques
78
Hidalgo J., Zamora R. & Giron J., 1992- Grasas Acietes. 28: 43-97/100. In. Aubourg S.P.,
1998a.
Hill’s, 2001- Nutrition et Protection Contre les Lésions Oxydatives. Hill’s Pet Nutrition. 40p.
Howell N., 1995- Interaction of proteins with small molecules. In: Ingedient interactions
Effects on food quality. Ed. Gaonkar A. & Dekker M., New York (U.S.A.), pp-269-289.
Hsieh R.J. & Kinsella J.E., 1989b- Lipoxygenase Generation of Specific Volatile Flavor
Carbonyl Compounds in Fish Tissues. Journal of Agricultural and Food Chemisttry. 37:
279-286.
Hsieh, R.J. and Kinsella, J.E., 1989a- Oxidation of polyunsaturated fatty acids:
mechanisms, products, and inhibition with emphasis on fish. Advances in Food and
Nutrition Research.33: 233-341.
Hughes, R.B. & Jones, N.R., 1966 - Measurement of hypoxantine concentration in canned
herring as an index of the freshness of the raw material, with a comment on the flavor
relations. J. Sea. Food Agree.17: 434-436.
Hureau J.C., 1986- Mullidae. In Whitehead, P. J. P., Bauchot M.-L., Hureau, J.-C., Nielsen
J. & E. Tortonese (1986). Fishes of the North-eastern Atlantic and the Mediterranean. Les
Presses de l’Unesco. II : 877-882.
Hureau J-C., 1973 - Biologie comparée de quelques poissons antarctiques (Nototheniidae).
Bull. Inst. océanogc. Monaco. 68: 1391- 244.
Hwang, K.T. & Regeinstein, J.M., 1993- Characteristics of mackerel mince lipid hydrolysis.
Journal of Food Science. 58: 79-83.
Ingemansson T., Olsson N.U., Herslöf B.G., Ekstrand B., 1991- J. Sci. Food Agric. 57:
443-447.
Jacobsen C., 1999- Sensory impact of lipid oxidation in a complex food systems. Fett/Lipid.
101: 484-492.
Jorgensen L., Huss H. & Dalgard P., 2000- The effect of biogenic amine production by
single bacteria cultures and metabolisis on cold-smoked salmon. Journal of Applied
Microbiology. 56: 920-934.
Kiessling, A., Pickova, J., Johansson, L., Asgard, T., Storebakken, T. & Kiessling, K-H.,
2001- Changes in fatty acid composition in muscle and adipose tissue of farmed rainbow
trout (Oncorhynchus mykiss) in relation to ration and age. Food. Chem. 73: 271-284.

Références bibliographiques
79
Kim R. & Labella F., 1987- Comparison of analytical methods for monitoring autooxidation
profiles of authentic lipids. J. lipid. Res. 28: 1110-1117.
Kinsella J. 1987 - Dietery fats and cardiovascular disease. In Seafoods and fish oils in
human health and disease. Lees R. & Karel M. (Ed.), p 1-23. Marcel Decker, Inc. New
York and Basel.
Klein R.A., 1970- The detection of oxidation in liposome preparations. Biochemistry and
Biophysic Acta. 210 :486-489.
Kortenska V. D., Yanishlieva N. V., Kasaikina O. T., Totzeva I. R., Boneva M. I. &
Russina I. F., 2002- Phenol antioxidant efficiency in various lipid substrates containing
hydroxy compounds. Eur. J. Lipid Sci. Technol. 104: 513-519
Labropoulou M., Machias, A. Tsimenides, N. & Eleftheriou A., 1997- Feeding habits and
ontogenetic shift of the striped red mullet, Mullus surmuletus Linnaeus, 1758. Fish. Res.
31 : 257-267.
Lefevre F., Cos I. & Bugeon J., 2008- Déterminisme biologique de la qualité des poissons.
Poster 12èmes JSMTV.
Leger C. L., 2006 - Anti-oxydants d’origine alimentaire : diversité, modes d’action anti-
oxydante, interactions. OCL VOL. 13 N° 1 JANVIER-FEVRIER 2006.
Love R.M., 1980 -The Chemical Biology of Fishes. Vol.2. Academic Press London; 1968-
1977.
Macpherson, C., 1978 - Regimen alimentario de Micromesistius poutassou (Risso, 1810) y
Gadiculus argenteus argenteus GUICHENOT, 1850 (Pisces, Gadidae) en el Mediterraneo
occidental. — Inu. Pesq. 42 (2): 305-316.
Madrid A., Madrid J. & Madrid R., 1994- Technologia del pescado y productos derivados.
Madrid A. Vicente, Ediciones y Mundi- Prensa Libros, S.A., Madrid (Spain), pp 45-103.
Mahé, K., Destombes, A., Coppin, F., Koubbi, P., Vaz, S., Le Roy, D. & Carpentier, A.,
2005- Le rouget barbet de roche Mullus surmuletus (L. 1758) en Manche orientale et mer
du Nord. Rapport de Contrat IFREMER/CRPMEM Nord-Pas-de-Calais,187p.
Mahmoud, S & Benkakaa, K., 2007- Effet du mode de conservation sur l’oxydation des
lipides ; concentration en malodialdéhyde (TBARS) chez Sardina pilchardus (Walbaum,
1792) et Merluccius merluccius (Linnaeus, 1758). Mémoire d’ingéniorat. Univ. D’Oran,
Es Sénia. 29p.

Références bibliographiques
80
Mamuris, Z., Apostolidis, A.P. & Triantaphyllidis, C., 1998- Genetic protein variation in
red mullet (Mullus barbatus) and striped mullet (M. surmuletus) populations from the
Mediterranean sea. Mar. Biol. 130 : 353-360.
Marcel J., 2002 – les troubles de la reperfusion : Perspectives thérapeutiques, les lazéroides.
Thèse de doctorat. Université Claude Bernard, Lyon I. 128pp.
Mates J.M., Perez-Comez C. & Nunez de Castro, 1999 – Antioxidant enzymes and human
diseases. Clin. Biochem. 32 : 595-603.
Maurin, C., 1968 - Ecologie ichthyologique des fonds chalutables atlantiques (de la baie
ibéro-marocaine à la Mauritanie) et de la Méditerranée occidentale. — Reu. Trau. Inst.
Pèches Macit. 32 (1), 147 p.
Mazorra-Manzano M.A., PachecoAguilar R., Diaz-Rojas E.I., & Lugo-Sanchez H.E.,
2000- Post mortem changes in black skipjack muscle during storage in ice. J. Food Sci. 65:
774779.
Medale F., Lefevre F., Corraze G., 2003- Cahier de Nutrition et Diététique. 38: 37 44.
Medale F., 2004- V.P.C., Sp. Issue 10èmes JSMTV, 87-93.
Medale F., 2005- Caractéristiques nutritionnelles des poissons et facteurs de variations
Aquaculture. 79 : 87-93.
Melton S.L., 1983 – Methodology for following lipid oxidation in muscle foods. Food
Technol. 37: 105-111,116.
Miyashita K. & Takagi T, 1986- Study on the oxidative rate and prooxidant activity of free
fatty acids. J. Am. Oil Chem. Soc. 63: 1380-1384.
Murray R. K., Granner D. K., & al., 2002- Biochimie de Harper. 25eme edition
américaine revue et mise a jour. Traducteur : Cohen, P., Presses de l’Université de Laval,
933p.
Murray C.K., Burt J.M. & Shewan K.H., 2000- The microbial spoilage of fish with
special reference to the role of psychrotrophs. In: Russell, A.D. and R. Fuller (eds.) Cold
tolerant microbes in spoilage and the environment, Academic Press, 117-136.
Murray J. & Burt G.R., 1969- The composition of fish. J. Fish Biol. 9: 329-334.

Références bibliographiques
81
Naganawa, H., & Ochi, K., 2002 - Bacilysocin, a novel phospholipid antibiotic produced by
Bacillus subtilis 168. Antimicrob. Agents Chemother. 46: 315-20.
Ngah E., Génot C., Meynier A., Viau M. & Gandemer G. 1993- Phospholipid oxidation in
Turkey precooked meat. 39th Int Congress Meat Sci Technol, Proceeding on disks, 5,
August 1-8, Calgary, Canada.
Nielson H, Hurrell R & Finot H., 1985- Br J. Nut. 53: 57-86.
Nunes, M., Batista, I., & Morao de Campos, R., 1992- Physical, chemical and sensory
analysis of sardine (Sardina pilchardus) stored in ice. Journal of the Science of Food and
Agriculture. 59: 37–43.
Ohshima T., Wada S & Koimusi C., 1985 – Accumulation of lyso-form phospholipids in
several species of fish flesh during storage at -5°C. Bull. Jpn. Sci. Fish. 51: 971-986.
Olafsdottir G., Martinsdottir E., Oehlenschlager J.Dalgaard P., Jensen B., Undeland I.,
Mackie I., Hanahan J., Nielsen G. & Nielsen H., 1997- Methods to evaluate fish
freshness in research and industry. Trends Food Sci. Technol. 8 : 258265.
Olafsdottir G., Nesvadba, P., di Natale, C., Careche, M., Oehlenschlager, J., &
Tryggvadottir, S. & al., 2004- Multisensor for fish quality determination. Trends in Food
Science and Technology. 15 :258-265.
Oumansour Y.F., 2001- Etude de l’altération des huiles de poissons après décongélation.
Effet micro-ondes. Diplôme de Magister. Université M’Hammed Bouguerra, Boumerdes,
95 pp.
Ozyurt G., Ozogul Y., Ozogul F., Kuley E. & Polat A., 2005- Freshness assessment of
European eel (Anguilla Anguilla) by sensory, chemical and microbiological methods. Food
Chemistry. 92: 745-751.
Ozyurt, G.n Polat, A. & Tokur, B., 2007- Chemical and sensory changes in frozen ()18 _C)
wild sea bass (Dicentrarchus labrax) captured at different fishing seasons. International
Journal of Food Science and Technology. 42: 887–893.
Pearson A.M., Love J.D. & Shortland F.D., 1977- Warmed-over-flavor in meat, poultry
and fish. Adv Food Res. 23: 1-74.

Références bibliographiques
82
Penfield M.P. & Campbell A.M., 1990- Fats and their lipids constituents. Chemical
reactions of lipids eds. Experimental food science, 3e ed. San Diego: Academic Press, pp
340 7.
Perez-Villarreal, B., & Pozo, R., 1990- Chemical composition and ice spoilage of albacore
(Thunnus alalunga). Journal of Food Science. 55 : 678–682.
Piclet, G., 1987- Le poisson aliment. Composition et intérêt nutritionnel. Cahiers de
Nutrition et Diététique, XXII, 317–335.
Pigott G. & Tucker B., 1987- Science opens new horizons for marine lipids in human
nutrition. Food Rev. Int. 3: 105-138.
Pokorny J., 1981- Browning from lipid-protein interactions. Progress in Food Nutrition and
Science. 5: 421-428.
Poulter N.H. & Nicolaides L., 1985a- Studies of the iced storage characteristics and
composition of a variety of Bolivian freshwater fish. 1. Altiplano fish. J. Food Technol. 20:
437-449.
Poulter, N.H. & Nicolaides L., 1985b- Studies of the iced storage characteristics and
composition of a variety of Bolivian freshwater fish. 2. Parana and Amazon Basins fish. J.
Food Technol. 20: 451-465.
Prost, C., Hallier, A., Cardinal, M., Courcoux, P., 2002- Contribution of sensory and
olfactometry analysis to discriminate the aroma of different species of seafood products
analysed at three states of freshness. Proceedings of the 10th Weurman Flavour Research
Symposium, Le Queté J.L. & Etievaut P.X. (Eds), Lavoisier Publisher, 24-28 june 2002,
Beaune, France.
Quero, J.C. & Vayne, J.J., 1997- Les poissons de mer des pêches françaises. IFREMER, Ed.
Delachaux & Niestlé, 304p.
Quillet E., Le Guillou S., Aubin J. & Fauconneau B., 2005- Aquaculture. 245: 49-61.
Rapeepan Laohabanjong , Chutima Tantikitti , Soottawat Benjakul , Kidchakan
Supamattaya & Mali Boonyaratpalin , 2009- Lipid oxidation in fish meal stored under
different conditions on growth, feed efficiency and hepatopancreatic cells of black tiger
shrimp (Penaeus monodon). Aquaculture. 286 : 283–289
Robles, R., 1970 - La bacaladilla Micromesistius poutassou (Risso, 1810). — Bol. Inst. Esp.
Ocean. 142: 16 p.

Références bibliographiques
83
Rodrıguez A., Carriles N., Cruz J.M. & Aubourg S.P., 2008- Changes in the flesh of
cooked farmed salmon (Oncorhynchus kisutch) with previous storage in slurry ice (_1.5
_C). LWT - Food Science and Technology. 41: 1726-1732.
Rodrıguez, O., Barros-Velazquez, J., Ojea, A., Pineiro, C., Gallardo, J. M., & Aubourg,
S., 2003- Effect of chilled storage in flow ice on the microbial quality and shelf life of
farmed turbot (P. maxima), isolation and identification of major proteolytic bacteria. In:
Proceedings first joint trans-atlantic fisheries technology conference, TAFT 2003. 33rd
WEFTA and 48th AFTC meetings, 11–14 June 2003, Reykjavik, Iceland (pp. 73–74).
Rossell, J. B., 1989- Measurement of rancidity. In J. C. Allen & R. J. Hamilton (Eds.),
Rancidity in foods (pp. 23–52). New York: Elsevier.
Ruiz-Capillas, C., & Moral, A., 2001- Correlation between biochemical and sensory quality
indices in hake stored in ice. Food Research International. 34: 441–447.
Saeed S. & Howell N. K., 2001- 12-Lipoxygenase activity in the muscle tissue of atlantic
mackerel (scomber scombrus) and its prevention by antioxidants. Journal of the science of
Food and Agriculture. 81 :745-750.
Saeed S. & Howell N., 2002- Effect of lipid oxidation and frozen storage on the muscle
proteins of Atlantic mackerel (Scomber scombrus). J. Scie. Food.Agric. 82: 579-586.
Salih A.M., Smith D.M., Price J.F. & Dawson L.E. , 1987- Modified extraction 2
thiobarbituric acid method for measuring lipid oxidation in poutry. Poultry science. 66
:1483-1488.
Santos S. & Regenstein J., 1990- Effects of vacuum packaging, glazing and erythorbic acid
on the shelf-life of frozen white hake and mackerel. J. Food Sci. 55: 64-70.
Sargent J., Henderson R.J. & Tocher D.R., 1989- In "Fish Nutrition", pp 153-218. ed.
Halver J.E., Academic Press, INC.
Seet S., & Brown D., 1983- Nutritional quality of raw, precooked and canned albacore tuna
(Thunnus alalunga). Journal of Food Science. 48 : 288-289.
Servais S., 2004- altérations mitochondriales et stress oxydant pulmonaire en réponse à
l’ozone : effets de l’âge et d’une supplémentassions en Oméga 3. Thèse de doctorat. 127
pp. Université Claude Bernard-Lyon I. Lyon, France.
Sevanian A. & Hochstein L., 1985 - Mechanisms and consequences of lipid peroxidation in
biological systems. Ann. Rev. nutr. 5: 365-390.

Références bibliographiques
84
Sheridan, M.A., 1988- Lipid dynamics in Fish : aspects of absorption, transportation,
deposition and mobilization. Comp.Biochem.Physiol. 90B: 676-690.
Sheridan M.A., 1994- Comp. Biochem. Physiol. 107 B: 495-508.
Shewfelt, R.L. 1981- Fish muscle lipolysis-A review. Journal of food Biochemistry. 5: 79-
100.
Sidhu K.S., 2003 - Health benefits and potential risks related to consumption of fish or fish
oil. Regul. Toxicol. Pharm. 38: 336-44.
Sigurgisladottir S., Hafsteinsson, H., Jonsson, A., Nortvedt, R., Thomasses, M., &
Torrisen, O., 1999- Textural properties of raw salmon fillets as related to sampling
method. Journal of Food Science. 64: 99-104.
Sikorski Z., & Kolakowska A., 1994- Changes in protein in frozen stored fish. In Z.
Sikorski, B. Sun Pan, & F. Shahidi (Eds.), Seafood proteins (pp. 99e112). New York,
USA: Chapman and Hall.
Sorbe J-C., 1980 - Régime alimentaire de Micromesistius poutassou (RISSO, 1826) dans le
sud du golfe de Gasgones, Rev. Trav. Inst. Pèches marit. 3: 245-255.
Stewart J.C., 1980- Colorimetric determination of phospholipids with ammonium
ferrothiocyanate. Analytical biochemistry. 104: 10-14.
Tamehiro, N., Okamoto-Hosoya, Y., Okamoto, S., Ubukata, M., Hamada, M. &
Trakatellis A.G., 1994- Rapid sensitive and specific thiobarbituric acid method for
measuring lipid peroxidation in animal tissue, food and feedstuff samples. Journal of
Agricultural and Food Chemistry. 42 :1931-1937.
Undeland B., Ekstrand H. & Lingnert I., 1998- Lipid oxidation in minced herring (Clupea
harengus) during frozen storage. Effect of washing and precooking. J. Agric. Food Chem.
46: 2319-2338.
Undeland I., Hultin H.O. & Richards M.P., 2002- An aqueous fraction of cod muscle
inhibits hemoglobin-mediated oxidation of cod muscle membrane lipids. 32nd Annual
WEFTA Meeting, May 13th-15th, Galway (Ireland) 90
Van Der Boch H., 1980- Intracellular phospholipases A. Biochim Biophys Acta. 604: 191-
246.
Verma J., Srikar L, Sudhakara N. & Sarma J., 1995- Effects of frozen storage on lipid
freshness parameters and some functional properties of oil sardine (Sardina pilchardus)
mince. Food Research International. 28:87-90.

Références bibliographiques
85
Vincke W., 1970- Direct determination of the thiobarbituric acid value in thrichloroacetic
acid extracts of fish as a measure of oxidative rancidity. Fette Seifen Anstrichm. 72: 1084-
1096.
Wang B., Pace R.D., Dessai A.P. Bovell-Benjamin A. & Phillips,B., 2002- Modified
Extraction Method for Determining 2-Thiobarbituric Acid Values in Meat with Increased
Specificity and Simplicity. Journal of Food Science. 67: 2833-2836.
Whitehead, P.J.P., Bauchot, M.L., Hureau, J.C., Nielson, J. & Tortonese, T., 1986-
Fishes of the North-eastern Atlantic and the Mediterranean. UNESCO, Paris., Vol.I-II :
1473p.
Wood J.D. & Lister D., 1973- The fatty acid and phospholipid composition of Longissimus
dorsi from Pietrain and Large White pigs. J Sci Food Agric. 24: 449-56.
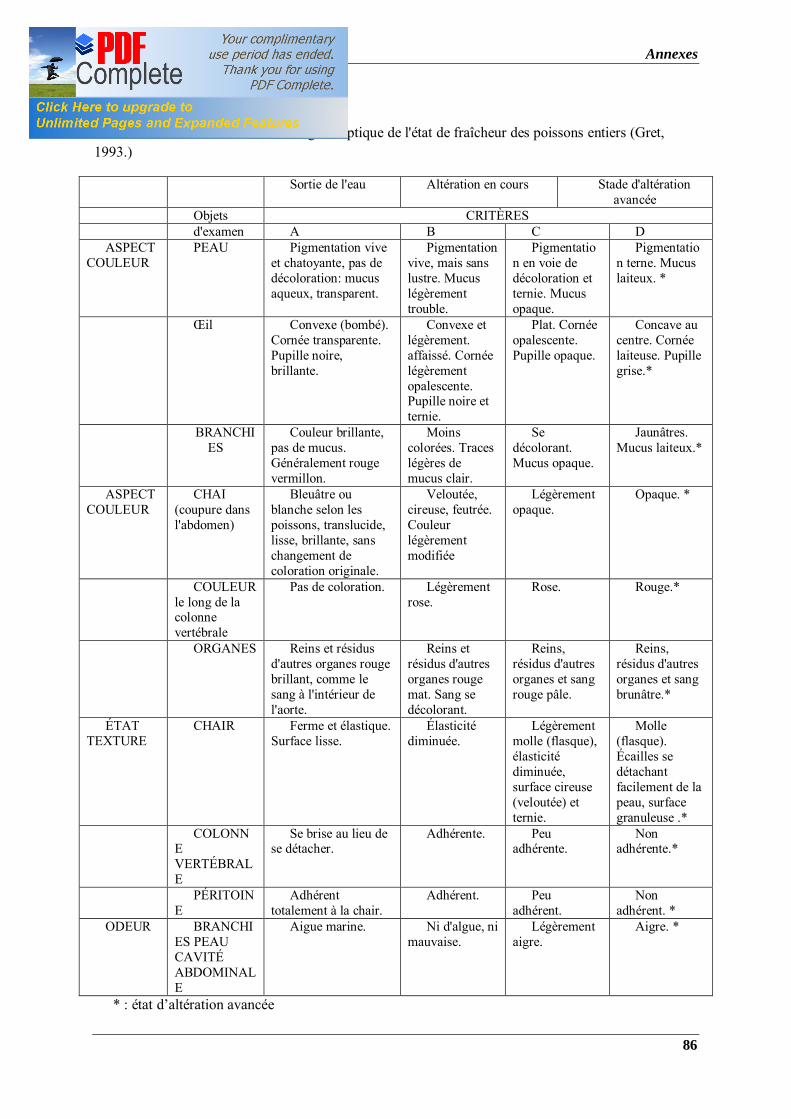
Annexes
86
ANNEXE 1
Tableau 2: Grille d'évaluation organoleptique de l'état de fraîcheur des poissons entiers (Gret, 1993.)
Sortie de l'eau Altération en cours Stade d'altération avancée
Objets CRITÈRES d'examen A B C D ASPECT
COULEUR PEAU Pigmentation vive
et chatoyante, pas de décoloration: mucus aqueux, transparent.
Pigmentation vive, mais sans lustre. Mucus légèrement trouble.
Pigmentation en voie de décoloration et ternie. Mucus opaque.
Pigmentation terne. Mucus laiteux. *
Œil Convexe (bombé). Cornée transparente. Pupille noire, brillante.
Convexe et légèrement. affaissé. Cornée légèrement opalescente. Pupille noire et ternie.
Plat. Cornée opalescente. Pupille opaque.
Concave au centre. Cornée laiteuse. Pupille grise.*
BRANCHIES
Couleur brillante, pas de mucus. Généralement rouge vermillon.
Moins colorées. Traces légères de mucus clair.
Se décolorant. Mucus opaque.
Jaunâtres. Mucus laiteux.*
ASPECT COULEUR
CHAI (coupure dans l'abdomen)
Bleuâtre ou blanche selon les poissons, translucide, lisse, brillante, sans changement de coloration originale.
Veloutée, cireuse, feutrée. Couleur légèrement modifiée
Légèrement opaque.
Opaque. *
COULEUR le long de la colonne vertébrale
Pas de coloration. Légèrement rose.
Rose. Rouge.*
ORGANES Reins et résidus d'autres organes rouge brillant, comme le sang à l'intérieur de l'aorte.
Reins et résidus d'autres organes rouge mat. Sang se décolorant.
Reins, résidus d'autres organes et sang rouge pâle.
Reins, résidus d'autres organes et sang brunâtre.*
ÉTAT TEXTURE
CHAIR Ferme et élastique. Surface lisse.
Élasticité diminuée.
Légèrement molle (flasque), élasticité diminuée, surface cireuse (veloutée) et ternie.
Molle (flasque). Écailles se détachant facilement de la peau, surface granuleuse .*
COLONNE VERTÉBRALE
Se brise au lieu de se détacher.
Adhérente. Peu adhérente.
Non adhérente.*
PÉRITOINE
Adhérent totalement à la chair.
Adhérent. Peu adhérent.
Non adhérent. *
ODEUR BRANCHIES PEAU CAVITÉ ABDOMINALE
Aigue marine. Ni d'algue, ni mauvaise.
Légèrement aigre.
Aigre. *
* : état d’altération avancée

Annexes
87
ANNEXE 2
Tableau 3 : niveau d’acceptabilité sensorielle des différents lots des 3 espèces étudiées.
Les cases grisées représentent le temps du rejet organoleptique.
Mode de
conservation
Temps de conservation (jours)
1 2 3 5 7 9 15 30
La sardine
(S. pichardus)
Lot A. A C D D
Lot B A B C D D D
Lot C A B B
Le rouget
de roche (M.
surmuletus)
Lot A. A B D D
Lot B A B D D
Lot C A B B
Le merlan
bleu (M.
poutassou)
Lot A B C D D
Lot B B B C D D
Lot C B B B

Annexes
88
0
10
20
30
40
50
60
j1 j2 j3 j5 j7 j9 j15 j30
Teneur en M.S. (g/100g M.F.)
Durée (jours)
Lot A
Lot B
Lot C
ANNEXE 3 : L’évolution de la teneur en matière sèche (M.S.) en fonction de la durée et du mode de conservation chez les trois espèces de poissons étudiées.
I. La sardine commune (Sardina pilchardus) a. Effet de la durée de conservation
Figure 36 : Evolution de la Teneur en matière sèche (M.S.) (g/100g M.F.) chez la sardine commune (S. pichardus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
b. Effet du mode de conservation
Figure 37 : Effet du mode de conservation sur la teneur en matière sèche (M.S) chez la sardine commune (S. pichardus) (g/100g M.F.). Les histogrammes portant des lettres ou des chiffres différents sont significativement différents (p < 0,05).
0
5
10
15
20
25
30
35
40
j2 j3 j5
Teneur en M.S. (g/100g M.F.)
Durée (jours)
Lot ALot B
a a a a
b ab
b b c
d
a b 1 2
: T° amb.
: T° +4°C
: T° -18°C
: T° amb.
: T° +4°C

Annexes
89
0
10
20
30
40
50
60
j1 j2 j3 j15 j30
Teneur en M.S. (g/100g M.F.)
Durée (jours)
Lot A
Lot B
Lot C
II. Le rouget de roche (Mullus surmuletus) a. Effet de la durée de conservation
Figure 38 : Evolution de la Teneur en matière sèche (M.S.) (g/100g M.F.) chez le rouget de roche (M. surmuletus) du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
b. Effet du mode de conservation
Figure 39 : Effet du mode de conservation sur la teneur en matière sèche (M.S) (g/100g M.F.) chez le rouget de roche (M. surmuletus). Les histogrammes portant des lettres ou des chiffres différents sont significativement différents (p < 0,05).
0
5
10
15
20
25
30
35
40
j2 j3
Teneur en M.S. (g/100g M.F.)
Durée (jours)
Lot A
Lot B
a a a b
b
a a
a
b 1
2
: T° amb.
: T° +4°C
: T° -18°C
: T° amb : T° +4°C
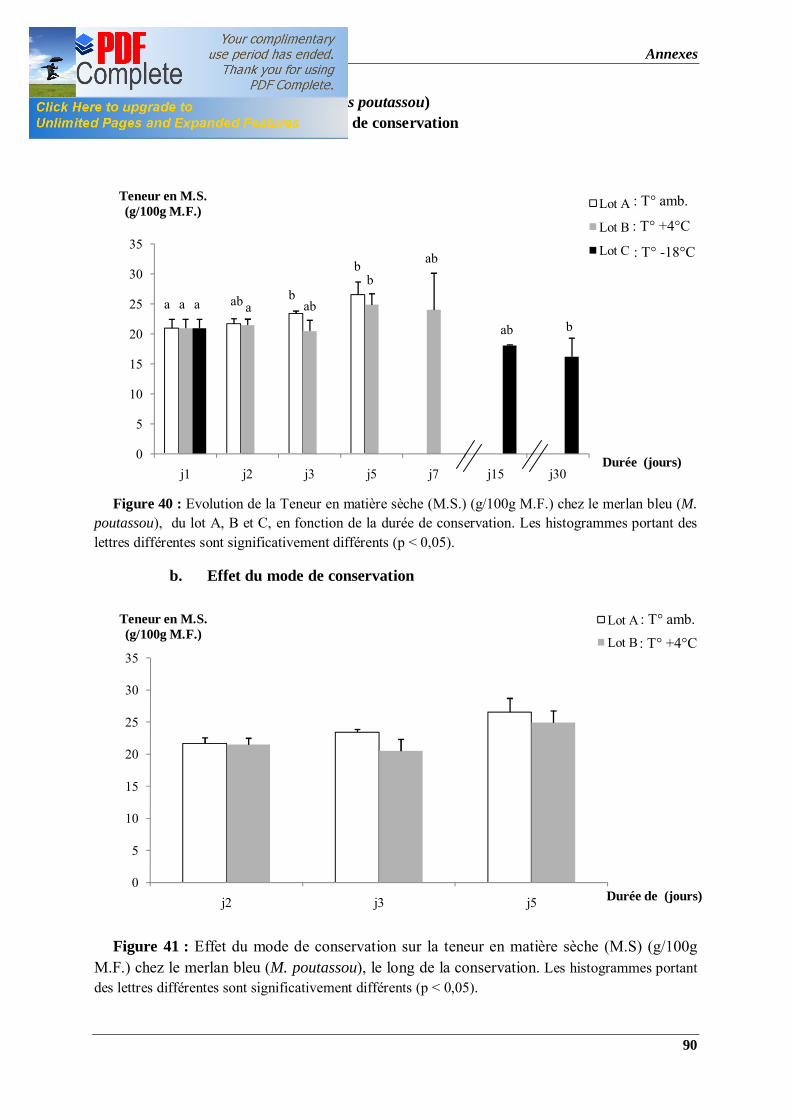
Annexes
90
0
5
10
15
20
25
30
35
j1 j2 j3 j5 j7 j15 j30
Teneur en M.S. (g/100g M.F.)
Durée (jours)
Lot A
Lot B
Lot C
III. Le merlan bleu (Micromesistius poutassou) a. Effet de la durée de conservation
Figure 40 : Evolution de la Teneur en matière sèche (M.S.) (g/100g M.F.) chez le merlan bleu (M. poutassou), du lot A, B et C, en fonction de la durée de conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
b. Effet du mode de conservation
Figure 41 : Effet du mode de conservation sur la teneur en matière sèche (M.S) (g/100g M.F.) chez le merlan bleu (M. poutassou), le long de la conservation. Les histogrammes portant des lettres différentes sont significativement différents (p < 0,05).
0
5
10
15
20
25
30
35
j2 j3 j5
Teneur en M.S. (g/100g M.F.)
Durée de (jours)
Lot A
Lot B
a a a
ab b a ab b
ab
b b
ab
: T° amb.
: T° +4°C
: T° -18°C
: T° amb. : T° +4°C

Annexes
91
ANNEXE 4 : Structure des principaux phospholipides.

Annexes
93
ANNEXE 5 : Schéma des voies catalytiques de l’oxydation de l’acide linolénique en 13-
hydroperoxydes et/ou hydroperoxydes par la lipoxygenase (LOX) (Cheriot, 2007).
Elimination d’un hydrogène
Réarrangem
ent radicalaire
Insertion de l’oxygène
13-hydroperoxyde
9 -hydroperoxyde






































![[EXPERTISE ENVIRONNEMENTALE]](https://img.pdfslide.tips/doc/110x75/62c928a6e75d163779556d2c/expertise-environnementale.jpg)