Embed Size (px)
Citation preview

UNIVERSITEIT GENT
FACULTEIT DIERGENEESKUNDE
Academiejaar 2015 - 2016
MOGELIJKE INVLOED VAN MYCOTOXINES OP STAARTNECROSE BIJ BIGGEN
door
Nienke AUPING
Promotoren: Prof. Dr. S. Croubels Onderzoek in het kader van Prof. Dr. D. Maes de Masterproef
© 2016 Nienke Auping


Universiteit Gent, haar werknemers of studenten bieden geen enkele garantie met betrekking tot de juistheid of
volledigheid van de gegevens vervat in deze masterproef, noch dat de inhoud van deze masterproef geen inbreuk
uitmaakt op of aanleiding kan geven tot inbreuken op de rechten van derden.
Universiteit Gent, haar werknemers of studenten aanvaarden geen aansprakelijkheid of verantwoordelijkheid voor
enig gebruik dat door iemand anders wordt gemaakt van de inhoud van de masterproef, noch voor enig
vertrouwen dat wordt gesteld in een advies of informatie vervat in de masterproef.

UNIVERSITEIT GENT
FACULTEIT DIERGENEESKUNDE
Academiejaar 2015 - 2016
MOGELIJKE INVLOED VAN MYCOTOXINES OP STAARTNECROSE BIJ BIGGEN
door
Nienke AUPING
Promotoren: Prof. Dr. S. Croubels Onderzoek in het kader van Prof. Dr. D. Maes de Masterproef
© 2016 Nienke Auping

Inhoudsopgave Samenvatting ....................................................................................................................................... 1
I. Literatuurstudie ............................................................................................................................ 2
1. Inleiding: wat zijn mycotoxines? .............................................................................................. 2
2. Problematiek staartnecrose bij biggen .................................................................................... 6
3. Fusarium mycotoxines en hun toxicokinetiek .......................................................................... 8
3.1. Deoxynivalenol ................................................................................................................ 8
3.2. Zearalenone ..................................................................................................................... 9
3.3. T-2 toxine ....................................................................................................................... 11
4. Wetgeving in voeder .............................................................................................................. 13
5. Analysemethoden .................................................................................................................. 14
II. Onderzoek ................................................................................................................................. 15
1. Inleiding en probleemstelling ................................................................................................. 15
2. Materiaal en methoden .......................................................................................................... 18
2.1. Plasma zeugen en biggen ............................................................................................. 18
2.2. Melkmonsters ................................................................................................................ 19
2.3. Berekeningen en statistiek ............................................................................................ 20
3. Resultaten .............................................................................................................................. 21
3.1. DON-GlcA in zeugenplasma ......................................................................................... 21
3.2. DON-GlcA in biggenplasma .......................................................................................... 24
3.3. Zearalenone en metabolieten in zeugenplasma ........................................................... 24
3.4. Zearalenone en metabolieten in biggenplasma ............................................................ 25
3.5. Correlaties mycotoxines en metabolieten tussen zeugen- en biggenplasma ............... 27
3.5. Mycotoxines in melkmonsters zeugen ........................................................................... 29
4. Discussie ............................................................................................................................... 30
Literatuur ............................................................................................................................................ 33
Bijlagen .............................................................................................................................................. 40
I. Voederanalyse resultaten voor de 12 mycotoxines die werden gedetecteerd in minstens één voeder, zijnde DON, ZEA, T-2, HT-2, NIV, 3-ADON, 15-ADON, AOH, STERIG, FB1, FB2 en ROQ-C. De resultaten zijn weergegeven in functie van het type bedrijf (probleembedrijf - controlebedrijf) ............................................................................................................................... 40
II. Overzicht van de plasmaconcentraties van zeugen (A) en biggen (B) geanalyseerd met HRMS uitgedrukt in chromatografische piek oppervlakte in functie van het type bedrijf (1= probleembedrijf; 0= controlebedrijf) ............................................................................................... 41
III. Overzicht berekende Spearman rank correlaties tussen ZEA en metabolieten in zeugenplasma en ZEA in voeder bij de zeugen ............................................................................ 49
IV. Overzicht berekende Spearman rank correlaties tussen ZEA en metabolieten in biggenplasma en ZEA in voeder bij de zeugen ............................................................................. 50
V. Overzicht berekende Spearman rank correlaties tussen mycotoxines in zeugen- en biggenplasma voor ZEA, α-ZOL, β-ZOL, zearalenol glucuronide (ZEL-GlcA), α-ZAL en β-ZAL .. 51

Samenvatting
Mycotoxines zijn toxines geproduceerd door verschillende schimmels. Het varken is voornamelijk voor
Fusarium mycotoxines gevoeliger dan andere diersoorten. Deze toxines hebben vermoedelijk een
effect op neonatale staartnecrose bij biggen. Naast mycotoxines zijn er nog vele andere factoren die
zouden kunnen bijdragen aan neonatale staartnecrose, zoals infecties en genetica. In een vorig
masterproefonderzoek (Van Neste, 2015) werd aangetoond dat er een positieve associatie is tussen
de aanwezigheid en concentratie van deoxynivalenol in het zeugenvoeder en deoxynivalenol in
zeugenplasma en staartnecrose bij neonatale biggen. Daarbij werden 10 probleembedrijven en 10
controlebedrijven bemonsterd. Per bedrijf werd er bloed genomen van 5 zeugen met ieder 2 biggen,
werden van iedere zeug melkmonsters en enkele urinemonsters genomen, werden de
bedrijfsgegevens verzameld en werden er voederstalen genomen.
In deze studie werden op dezelfde plasmastalen bijkomend de voornaamste fase I en fase II
metabolieten van deoxynivalenol en zearalenone bepaald in zowel zeugen als biggen via UHPLC-
HRMS. Met behulp van deze techniek is aangetoond dat er een significant hogere concentratie van
deoxynivalenol-glucuronide is in het zeugenplasma bij bedrijven met neonatale staartnecrose (11.158
± 7.976 piek oppervlakte) versus controlebedrijven (5.394 ± 4.286 piek oppervlakte); p<0,05). Ook
was er een correlatie tussen deoxynivalenol in het voeder en deoxynivalenol-glucuronide in
zeugenplasma (r=0,547; p<0,05). Ook deoxynivalenol en deoxynivalenol-glucuronide in
zeugenplasma zijn gecorreleerd (r=0,766; p<0,05). Daarnaast werd aangetoond dat deoxynivalenol-
glucuronide een betere biomerker is voor blootstelling van de zeug aan deoxynivalenol dan
deoxynivalenol zelf, vermits deze metaboliet in hogere concentraties voorkomt. Voor deoxynivalenol-
glucuronide in biggenplasma kunnen geen uitspraken worden gedaan wegens het lage aantal
positieve resultaten.
Voor zearalenone en metabolieten werden geen significante verschillen gevonden tussen
probleembedrijven en controlebedrijven, mogelijk wegens het lage aantal positieve resultaten. Wel
lagen de gemiddelde concentraties van zearalenone en metabolieten in biggenplasma hoger bij de
probleembedrijven dan bij de controlebedrijven. De meest frequent gedetecteerde fase II metaboliet
van zearalenone was zearalenol-glucuronide, deze zou bijgevolg een goede biomerker kunnen zijn
voor blootstelling aan zearalenone.
Verder werden er enkele melkstalen van zeugen geanalyseerd met behulp van UPLC-MS/MS. In geen
enkele staal werd zearalenone, α-zearalenol of β-zearalenol teruggevonden. Mogelijk ligt de
kwantificatielimiet te hoog om deze toxinen aan te tonen.
Verder onderzoek is nodig om specifieke oorzaken en risicofactoren voor neonatale staartnecrose bij
biggen aan te tonen.

2
I. Literatuurstudie
1. Inleiding: wat zijn mycotoxines?
Mycotoxines zijn door schimmels geproduceerde substanties die specifieke intoxicatie symptomen
kunnen geven bij mens en dier (Wyllie & Morehouse, 1978). Deze substanties zijn één van de vele
secundaire metabolieten gevormd door schimmels (D’Mello & Macdonald, 1997). Schimmels komen
voor op verschillende levensmiddelen zoals granen, noten en pinda’s (Glenn, 2007; Russell et al.,
2009). Niet alle schimmels produceren toxines en niet alle secundaire metabolieten van schimmels
zijn toxisch (Hussein & Brasel, 2001). De belangrijkste toxine producerende schimmels met hun
mycotoxines zijn opgelijst in tabel 1 (D’Mello & Macdonald, 1997).
Tabel 1: Overzicht van de voornaamste mycotoxine producerende schimmels (naar D’Mello & Macdonald,
1997)
Schimmelsoort Mycotoxine
Aspergillus flavus; A. parasiticus Aflatoxines
A. flavus Cyclopiazonzuur
Fusarium culmorum; F. graminearum; F. sporotrichioides Deoxynivalenol
F. sporotrichioides; F. graminearum; F. poae Diacetoxyscirpenol
Acremonium coenophialum Ergopeptide alkaloïden
F. moniliforme Fumonisinen
A. lolii Lolitrem alkaloïden
A. ochraceus; Penicillium viridicatum; P. cyclopium Ochratoxine A
P. expansum Patuline
Phomopsis leptostromiformis Phomopsinen
Pithomyces chartarum Sporidesmine
F. sporotrichioides; F. poae T-2 toxine
F. culmorum; F. graminearum; F. sporotrichioides Zearalenone
De groei van schimmel en de productie van toxines zijn afhankelijk van vele factoren. Dit zijn o.a. de
soort plant en schimmel, milieufactoren en de wijze van oogsten en opslaan van de gewassen.
Schimmels groeien in vele gevallen op het veld gedurende de ontwikkeling van de plant maar ook
tijdens de oogst, het drogen en de opslag. De belangrijkste factoren voor schimmelgroei zijn de
milieufactoren temperatuur en vocht (CAST, 2003). Iedere species heeft zijn optimale temperatuur
voor groei en toxineproductie (Russell et al., 2009). De relatieve vochtigheid blijkt positief gecorreleerd
met het voorkomen van deoxynivalenol in tarwe in de zomermaanden (Landschoot et al., 2012). Door
de opwarming van de aarde (‘global warming’) zullen gewassen die groeien in een gematigd klimaat
waarschijnlijk meer risico lopen op de contaminatie met mycotoxines omdat bij dit type klimaat
bepaalde schimmels zich gemakkelijk zullen kunnen ontwikkelen. Daarentegen zullen landen met een
koud klimaat de risicotemperatuur allicht niet bereiken. Gebieden met een warm klimaat hebben
mogelijk een verminderd risico op schimmelgroei daar de temperatuur boven de ideale
groeitemperatuur uitstijgt (Russell et al., 2009).

3
Er bestaan verschillende methoden om mycotoxineproductie te verminderen, zowel vóór als na de
oogst. De meest belangrijke preventieve maatregel vóór de oogst is het toepassen van ‘Good
Agriculture Practices (GAP)’, zoals rotatie van verschillende gewassen, juiste grondbewerking en het
correcte gebruik van irrigatie- en bestrijdingsmiddelen. Na de oogst zijn goede opslagcondities en het
gebruik van geschikte voederadditieven van belang. Deze additieven kunnen opgedeeld worden in
mycotoxine binders en mycotoxine modifiers. Mycotoxine binders zijn grote moleculen die
mycotoxines binden of sequesteren en doorheen het gastro-intestinaal systeem passeren. Hieronder
bestaan organische en anorganische binders. Mycotoxine modifiers zorgen voor biodegradatie van
mycotoxines in minder toxische metabolieten. Deze laatste worden onderverdeeld in bacteriën,
schimmels, gisten en enzymen (Devreese et al., 2013).
De belangrijkste vorm van blootstelling aan mycotoxines gebeurt door inname van gecontamineerd
voeder. Daarnaast zijn de huid en inhalatie ook mogelijke blootstellingsroutes (CAST, 2003).
Varkensvoeder bestaat grotendeels uit verschillende granen zoals tarwe, gerst en maïs en daarnaast
nog uit andere gewassen en additieven zoals soja (Solà-Oriol et al., 2013), vitaminen en mineralen
(Hoorens et al., 1973; Noblet et al., 1994; Noblet & Milgen, 2004). In tabel 2 wordt een overzicht
gegeven van de concentraties van verschillende mycotoxines in maïs van 2003 tot 2007 in België,
Duitsland, Frankrijk en Nederland (Van Asselt et al., 2012). In tabel 3 wordt een overzicht gegeven
van de concentratie van verschillende mycotoxines in verschillende landen voor de periode 2004-
2011. In figuur 1, 2 en 3 wordt een overzicht gegeven van het aantal positief geteste stalen (Streit et
al., 2013).
In de Belgische en Franse maïsoogst werd er in 2014 een verhoogd gehalte aan deoxynivalenol en
zearalenone vastgesteld. Deze trend blijkt voor maïs ook aanwezig in Nederland en Duitsland. De
maximale richtwaarden werden evenwel niet overschreden (http://www.bemefa.be/).
De economische verliezen die mycotoxine productie met zich meebrengen, zijn moeilijk te
kwantificeren. De consumptie van gecontamineerd voeder kan leiden tot acute dood, maar zal vaker
resulteren in productievermindering, orgaanbeschadiging, immunosuppressie en afwijkende
vruchtbaarheid (CAST, 2003; Wu, 2007). In deze thesis zal onderzocht worden of er een mogelijke
invloed is van de Fusarium mycotoxines op neonatale staartnecrose bij biggen, door de blootstelling
aan mycotoxines te kwantificeren in plasma, met nadruk op de fase II metabolieten. Dit zijn
metabolieten die gevormd worden in het lichaam van varkens om zodoende de uitscheiding van het
mycotoxine uit het lichaam te bevorderen. Sommige van deze metabolieten kunnen evenwel ook een
toxisch effect hebben.
4
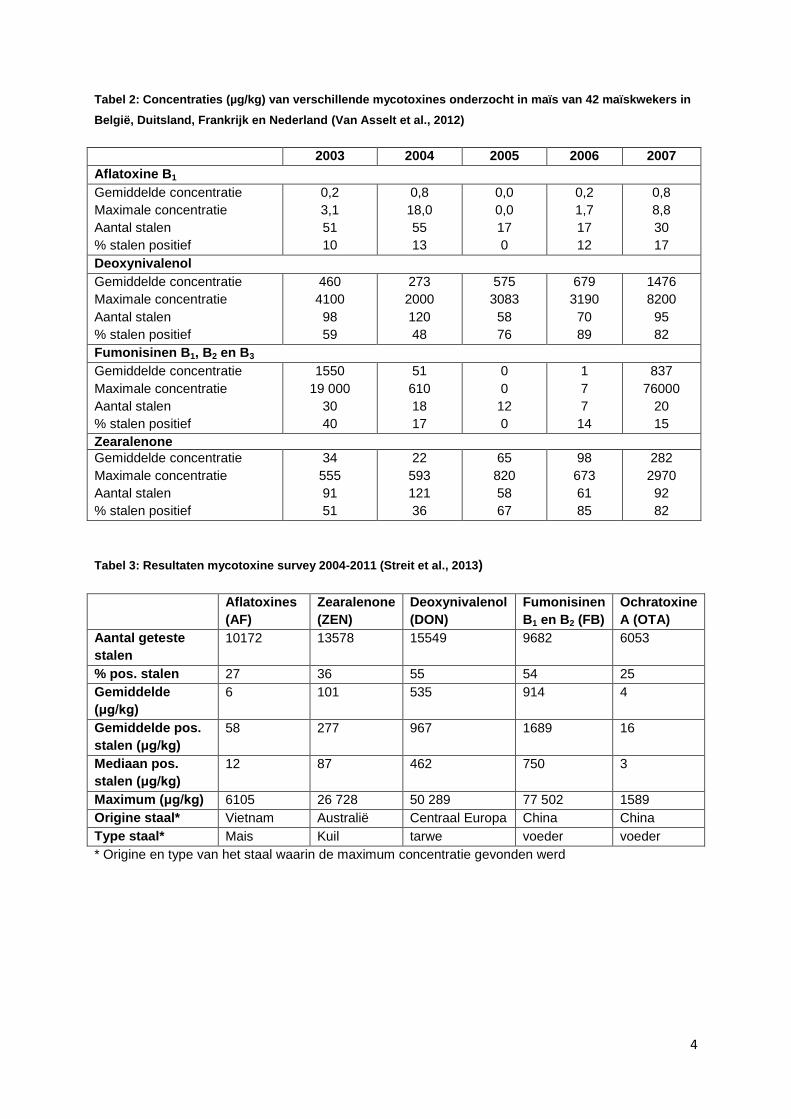
Tabel 2: Concentraties (µg/kg) van verschillende mycotoxines onderzocht in maïs van 42 maïskwekers in
België, Duitsland, Frankrijk en Nederland (Van Asselt et al., 2012)
2003 2004 2005 2006 2007
Aflatoxine B1
Gemiddelde concentratie
Maximale concentratie
Aantal stalen
% stalen positief
0,2
3,1
51
10
0,8
18,0
55
13
0,0
0,0
17
0
0,2
1,7
17
12
0,8
8,8
30
17
Deoxynivalenol
Gemiddelde concentratie
Maximale concentratie
Aantal stalen
% stalen positief
460
4100
98
59
273
2000
120
48
575
3083
58
76
679
3190
70
89
1476
8200
95
82
Fumonisinen B1, B2 en B3
Gemiddelde concentratie
Maximale concentratie
Aantal stalen
% stalen positief
1550
19 000
30
40
51
610
18
17
0
0
12
0
1
7
7
14
837
76000
20
15
Zearalenone
Gemiddelde concentratie
Maximale concentratie
Aantal stalen
% stalen positief
34
555
91
51
22
593
121
36
65
820
58
67
98
673
61
85
282
2970
92
82
Tabel 3: Resultaten mycotoxine survey 2004-2011 (Streit et al., 2013)
Aflatoxines
(AF)
Zearalenone
(ZEN)
Deoxynivalenol
(DON)
Fumonisinen
B1 en B2 (FB)
Ochratoxine
A (OTA)
Aantal geteste
stalen
10172 13578 15549 9682 6053
% pos. stalen 27 36 55 54 25
Gemiddelde
(μg/kg)
6 101 535 914 4
Gemiddelde pos.
stalen (μg/kg)
58 277 967 1689 16
Mediaan pos.
stalen (μg/kg)
12 87 462 750 3
Maximum (μg/kg) 6105 26 728 50 289 77 502 1589
Origine staal* Vietnam Australië Centraal Europa China China
Type staal* Mais Kuil tarwe voeder voeder
* Origine en type van het staal waarin de maximum concentratie gevonden werd
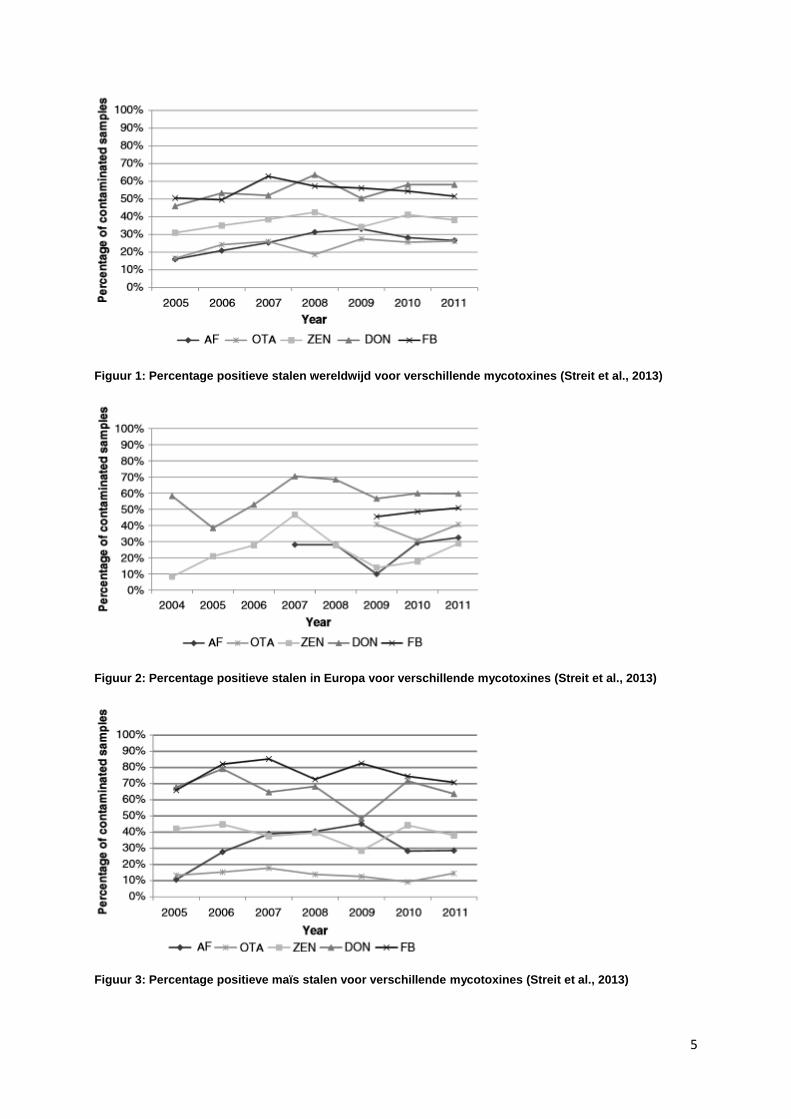
5
Figuur 1: Percentage positieve stalen wereldwijd voor verschillende mycotoxines (Streit et al., 2013)
Figuur 2: Percentage positieve stalen in Europa voor verschillende mycotoxines (Streit et al., 2013)
Figuur 3: Percentage positieve maïs stalen voor verschillende mycotoxines (Streit et al., 2013)

6
2. Problematiek staartnecrose bij biggen
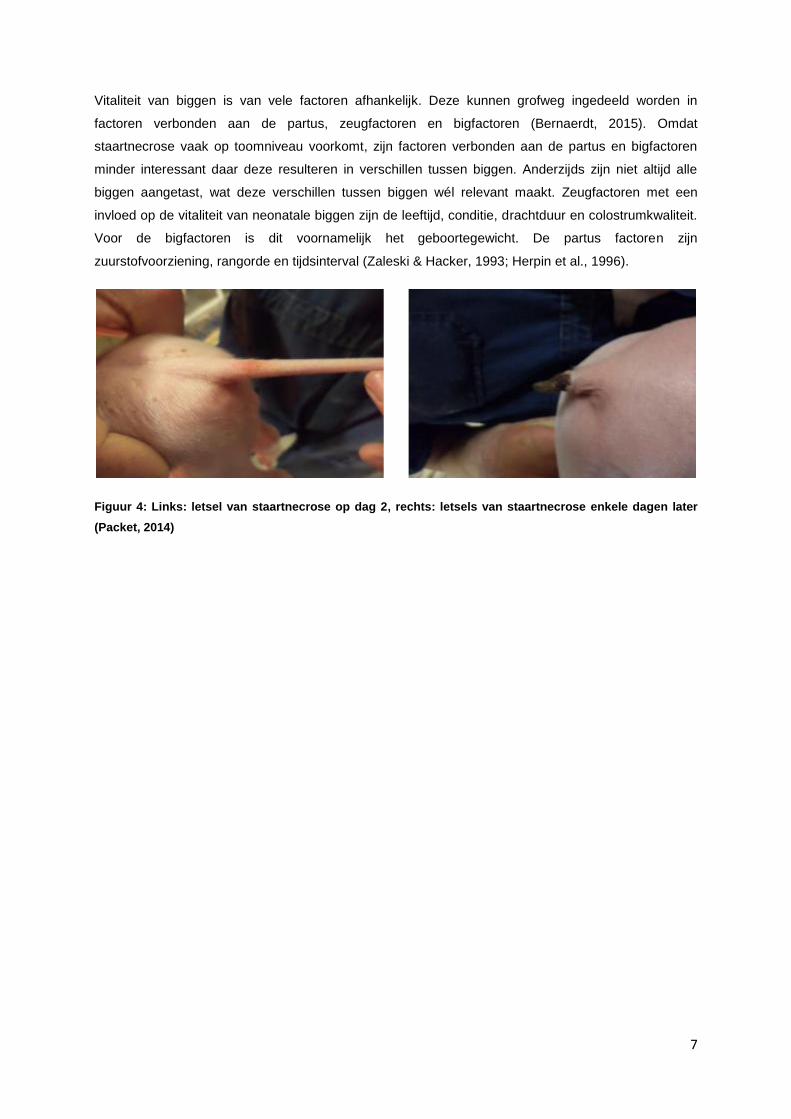
Neonatale staartnecrose, zoals hier onderzocht, begint één cm distaal van het proximale begin van de
staart (figuur 4). Wat het effect hiervan is, is niet gekend. Soms wordt staartnecrose gezien in
combinatie met splayleg en zwakke biggen (Mul et al., 2006).
Over de oorzaken van staartnecrose bij zuigende biggen is weinig gekend in de literatuur. Mogelijk
heeft dit een multifactoriële oorzaak en zijn hierbij o.a. verschillende mycotoxines betrokken.
Aanwezigheid van Zearalenone (ZEA) in het voeder van de zeug kan leiden tot biggen met een rode
uitslag rond de staart en op de rug. Ook kan het een zwelling van de vulva en tepels veroorzaken met
staartnecrose (Dacasto et al., 1995; Mul et al., 2006; Jajić et al., 2010). Daarnaast zou ook
ochratoxine A (OTA), mede door zijn placenta passage, kunnen bijdragen aan het probleem van
neonatale staartnecrose (Fink-Gremmels, 1999).
Ergot alkaloïden kunnen gangreen veroorzaken van de extremiteiten en kunnen betrokken zijn bij
distale staartnecrose (López et al., 1997; Bennet & Klich, 2013). Dit wordt o.a. gezien bij het konijn
(Korn et al., 2014). Bij muizen is gekend dat deoxynivalenol (DON) necrose kan veroorzaken van het
gastro-intestinaal systeem, het beenmerg, het lymfoïde stelsel en de huid (Pestka, 2007; Kalantari &
Moosavi, 2010).
Over de invloed van andere mycotoxines is weinig bekend. Mycotoxicose (DON, ZEA, T-2 toxine) in
het algemeen zou aanleiding kunnen geven tot zwakke biggen, biggen met spreidzit, neonatale
staartnecrose en vulva necrose (Mul et al., 2006).
Packet (2014) beschrijft een opvolging van een bedrijf met neonatale staartnecrose. In dit bedrijf was
er een vermoeden dat mycotoxines een rol zouden spelen. Daarnaast werd er ook een mogelijk
verband gezien met de voederadditieven en de conditie van de zeug aan het eind van de dracht. Ook
werd er PRRSV (porcien reproductief en respiratoir syndroom virus) gevonden in aangetaste biggen.
Nadat er op dit bedrijf tegen PRRS werd gevaccineerd, werd het probleem minder groot, maar was
het daarmee niet opgelost. Als laatste was er een sterke reductie van het aantal aangetaste tomen
nadat er een andere eindbeer werd ingezet. Hieruit kon afgeleid worden dat neonatale staatnecrose
een multifactorieel probleem is, waarbij genetica ook een rol kan spelen. In dit bedrijf bestond de
indruk dat de neonatale staartnecrose vooral voorkwam bij minder vitale biggen. Omdat over dit
onderwerp weinig literatuur beschikbaar is, is niet gekend of deze link er daadwerkelijk is. Wel is
gekend dat ook genetica en het voeder van de zeug belangrijk zijn voor de vitaliteit van pasgeboren
biggen (Edwards, 2002). Ook is er PPRS aangetoond, wat aanleiding kan geven tot zwakke biggen
(Alexopoulos et al., 2005). Mul et al. (2006) onderzochten of de symptomen die in praktijk worden
geassocieerd met mycotoxines mogelijk door PRRS werden veroorzaakt dan wel versterkt. Daarnaast
zouden mycotoxines kunnen zorgen voor immunosuppressie en zo weer kunnen predisponeren voor
PRRS infecties. Andersom zouden infecties, zoals PRRS, voor immunosuppressie kunnen zorgen en
zo kunnen bijdragen aan sterkere klinische verschijnselen van mycotoxicosen (Mul et al., 2006).
7
Vitaliteit van biggen is van vele factoren afhankelijk. Deze kunnen grofweg ingedeeld worden in
factoren verbonden aan de partus, zeugfactoren en bigfactoren (Bernaerdt, 2015). Omdat
staartnecrose vaak op toomniveau voorkomt, zijn factoren verbonden aan de partus en bigfactoren
minder interessant daar deze resulteren in verschillen tussen biggen. Anderzijds zijn niet altijd alle
biggen aangetast, wat deze verschillen tussen biggen wél relevant maakt. Zeugfactoren met een
invloed op de vitaliteit van neonatale biggen zijn de leeftijd, conditie, drachtduur en colostrumkwaliteit.
Voor de bigfactoren is dit voornamelijk het geboortegewicht. De partus factoren zijn
zuurstofvoorziening, rangorde en tijdsinterval (Zaleski & Hacker, 1993; Herpin et al., 1996).
Figuur 4: Links: letsel van staartnecrose op dag 2, rechts: letsels van staartnecrose enkele dagen later
(Packet, 2014)

8
3. Fusarium mycotoxines en hun toxicokinetiek
De drie hieronder besproken mycotoxines, zijnde DON of vomitoxine, T-2 toxine en ZEA, worden alle
geproduceerd door verschillende species van het genus Fusarium (CAST, 2003). Deoxynivalenol en
T-2 toxine behoren tot de groep van de trichothecenen. T-2 toxine behoort tot groep A van de
trichothecenen en DON behoort tot groep B (Nathanail et al., 2015).
Toxicokinetiek bestaat uit absorptie, distributie, metabolisme en excretie (ADME) van een bepaalde
gifstof. De lever is het belangrijkste orgaan voor de metabolisatie van verschillende stoffen. Deze
metabolisatie gebeurt via verschillende pathways die verdeeld kunnen worden in fase I en fase II
reacties. Fase I reacties omvatten hydrolyse, oxidatie en reductie. Fase II reacties omvatten o.a.
glucuronidatie, methylatie en sulfatatie. Fase I metabolieten kunnen substraat zijn voor fase II
reacties. Het doel van deze metabolisatie is om stoffen meer polair en oplosbaar te maken om de
uitscheiding te faciliteren (Riviere, 2011). Hieronder wordt voornamelijk de metabolisatie en excretie
van de Fusarium mycotoxines besproken, daar de metabolieten voor dit onderzoek het meest van
belang zijn.
Bij varkens gebeurt metabolisatie door middel van sulfaat conjugatie in zeer geringe mate ten opzichte
van andere diersoorten (Rathbone & Gurny, 2000; Riviere, 2011). Dit komt omdat varkens over weinig
sulfaat beschikken en daarnaast een specifiek type van het sulfotransferase enzym missen
(Calabrese, 1991). Dit in tegenstelling tot de kip, waar DON-3α-sulfaat de belangrijkste metaboliet is
(Devreese et al., 2015).
3.1. Deoxynivalenol
Metabolieten van DON kunnen worden gevormd door schimmels, dieren, bacteriën en planten
(Maresca, 2013). Voedsel kan gecontamineerd zijn door zowel DON als door zijn metabolieten.
Schimmels vormen voornamelijk geacetyleerde metabolieten zijnde 3-actetyl-DON (3aDON) en 15-
acetyl-DON (15aDON). Deze komen samen met DON voor in verschillende granen. Sommige granen
bezitten de capaciteit om DON te conjugeren tot een minder toxische metaboliet zijnde
deoxynivalenol-3-D-glucopyranoside (D3G) (Berthiller et al., 2013). 15aDON is het meest toxisch,
daarna komen respectievelijk DON, 3aDON en de-epoxy-deoxynivalenol (Broekaert, 2015).
In vivo kan de conjugatie van DON gebeuren met glucuronzuur in het endoplasmatisch reticulum tot
DON-glucuronide (DON-GlcA). Dit is de belangrijkste fase II reactie voor de eliminatie van DON,
zowel bij de mens als bij het varken. Dit gebeurt voornamelijk in de lever en mogelijk ook nog in
andere weefsels zoals longen, gastro-intestinaal systeem, nieren en de huid. DON wordt zowel bij de
mens als het varken voornamelijk uitgescheiden via glucuronide metabolieten (Kollarczik et al., 1994;
Xianai et al., 2007; Turner et al., 2011). Bij de mens domineert de aanwezigheid van DON-15-
glucuronide in de urine over de aanwezigheid van DON-3-glucuronide (Warth et al., 2012). DON-GlcA
is een detoxificatie product van DON, hypothetisch is het minder toxisch omdat het minder goed zou
passen op de 60S subunit van eukaryote ribosomen (Goyarts & Dänicke 2006; Xianai et al., 2007).

9
De microflora van de darm de-epoxideert DON tot de-epoxydeoxynivalenol (DOM-1), voornamelijk in
de caudale darm segmenten, waardoor het zijn cytotoxiciteit verliest (Kollarczik et al., 1994; Xianai et
al., 2007). Niet bij alle varkens is de microflora in staat DON te de-epoxideren. Mogelijk is deze
capaciteit er wel na contact met feces van varkens waarvan de microflora DON wel kan de-epoxideren
(Eriksen et al., 2002). Ook wanneer de microflora de-epoxiderende capaciteit bezit, wordt er geen
DOM-1 gevonden in plasma en urine (Eriksen et al., 2003). Nochtans vonden Razzazi-Fazeli et al.
(2003) wel DOM-1 terug in varkensurine met behulp van High Performance Liquid
Chromatography/Mass Spectrometry (HPLC/MS). Deze metaboliet wordt ook aangetroffen in feces
van varkens (Dänicke et al., 2004). Vorming van DOM-1 lijkt niet een van de belangrijkste detoxificatie
routes (Turner et al., 2010). DOM-1 wordt niet teruggevonden in humane feces (Sundstøl Eriksen &
Pettersson, 2003). Enkele chemische structuren worden weergegeven in figuur 5.
Figuur 5: a) Chemische structuur van DON, 3aDON en 15aDON b) structuur van DOM-1 (Naar Razzazi-
Fazeli et al., 2003)
DON en metabolieten worden voornamelijk uitgescheiden via de urine, een klein deel via de gal en de
rest via de feces. In de urine wordt voornamelijk DON teruggevonden, in feces voornamelijk DOM-1
(Prelusky et al., 1988; Goyarts & Dänicke, 2006; Turner, 2010). Er is zeer weinig DON accumulatie in
weefsels en de eliminatie halfwaardetijd ligt tussen de 3.9 - 5.8 uur (Pestka, 2007).
De ongeboren biggen worden in de baarmoeder al blootgesteld aan DON en in kleine mate aan DOM-
1, voor DON-GlcA zijn geen gegevens bekend. De blootstelling start ten minste vanaf 35 dagen dracht
(Dänicke et al., 2007; Goyarts et al., 2007).
DON heeft geen effect op de melkkwaliteit en wordt in zeer lage hoeveelheden uitgescheiden in de
melk en is daarom mogelijk verwaarloosbaar (Friend et al., 1986).
3.2. Zearalenone
Naast ZEA worden er vele metabolieten teruggevonden in granen. In maïs en tarwe kunnen de
metabolieten α-zearalenol (α-ZOL), β-zearalenol (β-ZOL), zearalenone-4-glucoside, α-zearalenol-4-
glucoside, β-zearalenol-4-glucoside en zearalenone-4-sulfaat worden teruggevonden (De Boevre et
al., 2012). Nathanail et al. (2015) vonden ook nog zearalenone-14-glucoside, zearalenone-14-sulfaat,

10
zearalenone-16-glucoside, α-zearalenol-14-glucoside en β-zearalenol-14-glucoside terug in gerst,
haver en tarwe.
Metabolieten aanwezig op granen kunnen worden geproduceerd door de plant zelf of door schimmels.
Conjugaten gevormd door schimmels zijn o.a. zearalenone-4-sulfate, zearalenone-4-glucoside en
zearalenone-2,4-diglucoside. Conjugaten gevormd door planten zijn o.a. zearalenone-4-glucoside, β-
zearalenol-4-glucoside en α-zearalenol-4-glucoside (Berthiller et al., 2009).
De belangrijkste in vivo gevormde metabolieten bij het varken zijn de glucuronide metabolieten en α-
ZOL. ZEA en zijn metabolieten bekomen hun oestrogene effecten door zich te binden aan de
oestrogeen receptor en daarbij gedragen ze zich als competitieve agonisten/antagonisten van deze
receptor (Shier et al., 2001; Fink-Gremmels & Malekinejad, 2007). Er is een opmerkelijk verschil in
potentie voor deze receptor: ten opzichte van de standaard (diethylstilbestrol) heeft α-ZOL een
potentie van 138%, ZEA 7,4% en β-ZOL 0,5% (Fitzpatrick et al., 1989). In plasma komen zowel ZEA,
α-ZOL, ZEA-glucuronide en α-ZOL-glucuronide voor (Biehl et al., 1993). α-ZOL en β-ZOL kunnen
verder gemetaboliseerd worden naar α-zearalanone, β-zearalanone en zearalanone (Zöllner et al.,
2002). Enkele van deze metabolieten worden weergegeven in figuur 6.
Figuur 6: Zearalenone en metabolieten (Bravin et al., 2009)
De mucosa van de darm van het varken bezit zowel de capaciteit om ZEA te reduceren alsook om te
conjugeren, en deze capaciteit is het grootst in het duodenum en jejunum. De reducerende capaciteit
van de mucosa is echter vele malen kleiner dan die van de lever. Maar omdat de darm een groot
oppervlak heeft, draagt deze voor een belangrijk deel bij aan de reductie. Hier domineert de
aanwezigheid van β-ZOL over α-ZOL. Een groter deel van ZEA wordt geconjugeerd tot ZEA-
glucuronide (Olsen et al., 1987).
In microsomen van de lever van het varken wordt ZEA gehydroxyleerd tot β-ZOL en hoofdzakelijk α-
ZOL. Het varken vormt sneller α-ZOL dan andere diersoorten (Malekinejad et al., 2005). Wanneer
ZEA in heel lage concentraties aanwezig is (≤50 μM) domineert vorming van β-ZOL in de lever. De
glucuronidatie van ZEA, α-ZOL en β-ZOL vindt ook plaats in de lever, deze capaciteit is hoger bij het

11
varken dan bij andere diersoorten. De glucuronidatie van deze 3 bovengenoemde mycotoxines
varieert tussen de 48 en 100% (Malekinejad et al., 2006).
De excretie van ZEA en metabolieten gebeurt voornamelijk via de urine en de gal. Na de galexcretie
worden ZEA en zijn metabolieten deels via de enterohepatische kringloop terug geresorbeerd, wat de
halfwaardetijd verlengt. Het resterende deel wordt uitgescheiden via de feces. In het galvocht komen
voornamelijk de metabolieten α-ZOL-glucuronide en ZEA-glucuronide voor (Biehl et al., 1993). Ook
bijna al het aanwezige ZEA en α-ZOL in plasma en urine is geconjugeerd (Olsen et al., 1985).
Daarnaast worden kleine hoeveelheden van ZEA en metabolieten geïncorporeerd in lever- en
spierweefsel (Zöllner et al., 2002). De eliminatie halfwaardetijd in plasma voor ZEA is 2.63 uur
(Dänicke et al., 2005).
De foetussen worden in de baarmoeder blootgesteld aan ZEA en α-ZOL al vanaf 75 dagen dracht
(Dänicke et al., 2007). Daarnaast worden ze ook via de melk blootgesteld, De samenstelling
gevonden in de melk is ongeveer 85% β-ZOL, 15% α-ZOL en 1% ZEA (Palyusik et al., 1980).
3.3. T-2 toxine
Naast T-2 toxine (T-2) worden er eveneens verschillende metabolieten teruggevonden in granen. In
tarwe, gerst en haver wordt zowel T-2, HT-2 en metabolieten T2-3-glucoside, HT-2-3-glucoside, HT2-
4-glucoside, T2-triol, T2-tetraol, de-epoxy-T2 en de-epoxy-HT-2 teruggevonden (De Boevre et al.,
2012; Lattanzio et al., 2012; Nathanail et al., 2015).
T-2 wordt in vivo snel gemetaboliseerd tot verschillende metabolieten. Dit gebeurt met behulp van
hydrolyse en hydroxylatie en daarnaast nog via de-epoxidatie en conjugatie. De conjugatie van T-2 en
metabolieten gebeurt zeer snel (Li et al., 2011; Sun et al., 2012). De belangrijkste metabolieten
gevonden in weefsels en plasma zijn T-2, HT-2, de-epoxy-HT-2, T-2-triol, de-epoxy-T-2-triol, 3’-
hydroxy-T-2, 3’-hydroxy-HT-2, T-2-tetraol, en de-epoxy-T-2-tetraol (Wu et al., 2010). Enkele structuren
zijn weergegeven in figuur 7.

12
Figuur 7: Structuur van T-2 en enkele metabolieten (Lattanzio et al., 2012)
Uitscheiding gebeurt via de gal, feces en voornamelijk via de urine. Via de feces wordt slechts een
minimale hoeveelheid uitgescheiden (Robinson et al., 1979). De belangrijkste ongeconjugeerde
metabolieten in gal en urine zijn 3’-hydroxy-HT-2 en T-2-triol. De belangrijkste geconjugeerde
metabolieten zijn 3’-hydroxy-T-2, neosolaniol, HT-2, 3’-hydroxy-HT-2, T-2-triol, 4-deacetylneosolaniol,
en T-2-tetraol. De meeste metabolieten worden geconjugeerd uitgescheiden, 77% van de
metabolieten in gal en 63% van de metabolieten in urine (Wu et al., 2010). De plasma eliminatie
halfwaardetijd van T-2 is zeer kort en bedraagt slechts 13,8 minuten (Swanson et al., 1987).
Bij de muis (hemochoriale placenta) is beschreven dat T-2 gemakkelijk door de placenta passeert en
schade veroorzaakt aan de foetussen, waaronder thymusatrofie. Bij het varken (epitheliochoriale
placenta) zijn hierover geen gegevens (Holladay et al., 1993; Senger, 2003; Haghighi Poodeh et al.,
2012). T-2 wordt uitgescheiden in de melk van de zeug (Robison et al., 1978).
13
4. Wetgeving in voeder
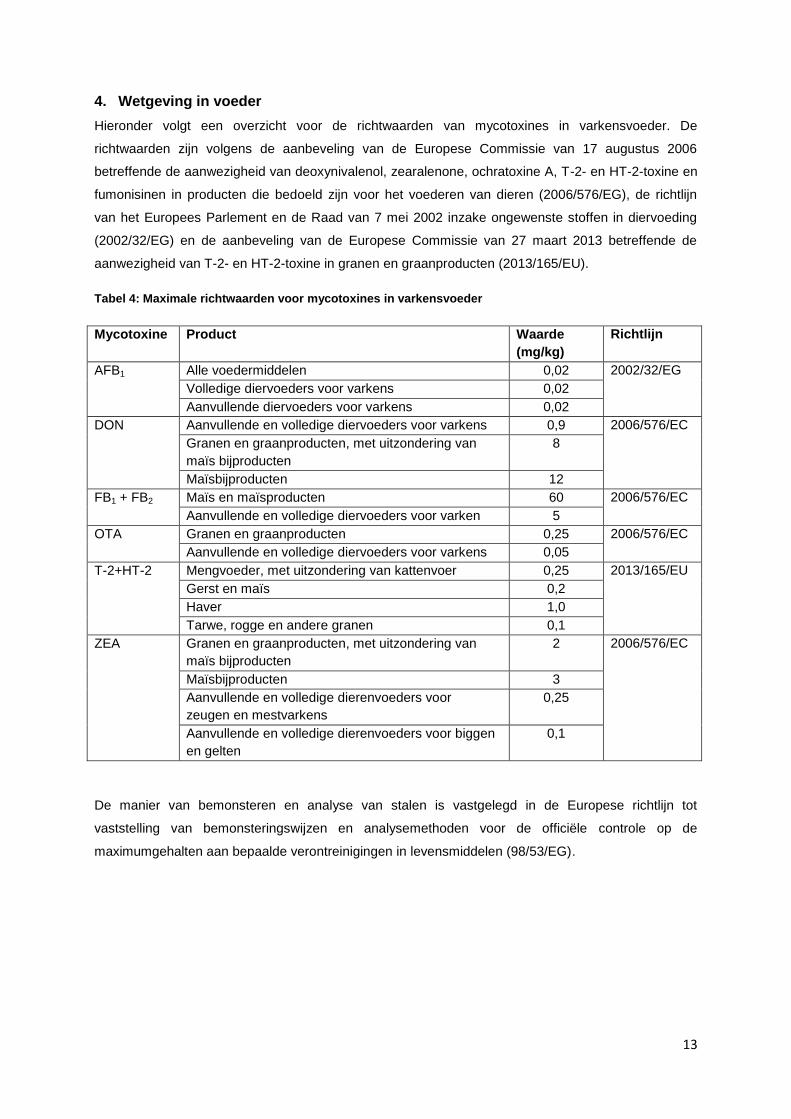
Hieronder volgt een overzicht voor de richtwaarden van mycotoxines in varkensvoeder. De
richtwaarden zijn volgens de aanbeveling van de Europese Commissie van 17 augustus 2006
betreffende de aanwezigheid van deoxynivalenol, zearalenone, ochratoxine A, T-2- en HT-2-toxine en
fumonisinen in producten die bedoeld zijn voor het voederen van dieren (2006/576/EG), de richtlijn
van het Europees Parlement en de Raad van 7 mei 2002 inzake ongewenste stoffen in diervoeding
(2002/32/EG) en de aanbeveling van de Europese Commissie van 27 maart 2013 betreffende de
aanwezigheid van T-2- en HT-2-toxine in granen en graanproducten (2013/165/EU).
Tabel 4: Maximale richtwaarden voor mycotoxines in varkensvoeder
Mycotoxine Product Waarde
(mg/kg)
Richtlijn
AFB1 Alle voedermiddelen 0,02 2002/32/EG
Volledige diervoeders voor varkens 0,02
Aanvullende diervoeders voor varkens 0,02
DON Aanvullende en volledige diervoeders voor varkens 0,9 2006/576/EC
Granen en graanproducten, met uitzondering van
maïs bijproducten
8
Maïsbijproducten 12
FB1 + FB2 Maïs en maïsproducten 60 2006/576/EC
Aanvullende en volledige diervoeders voor varken 5
OTA Granen en graanproducten 0,25 2006/576/EC
Aanvullende en volledige diervoeders voor varkens 0,05
T-2+HT-2 Mengvoeder, met uitzondering van kattenvoer 0,25 2013/165/EU
Gerst en maïs 0,2
Haver 1,0
Tarwe, rogge en andere granen 0,1
ZEA Granen en graanproducten, met uitzondering van
maïs bijproducten
2 2006/576/EC
Maïsbijproducten 3
Aanvullende en volledige dierenvoeders voor
zeugen en mestvarkens
0,25
Aanvullende en volledige dierenvoeders voor biggen
en gelten
0,1
De manier van bemonsteren en analyse van stalen is vastgelegd in de Europese richtlijn tot
vaststelling van bemonsteringswijzen en analysemethoden voor de officiële controle op de
maximumgehalten aan bepaalde verontreinigingen in levensmiddelen (98/53/EG).

14
5. Analysemethoden
Er zijn vele verschillende technieken om mycotoxines in stalen te detecteren en te kwantificeren. De
meeste methoden gebruikt voor het opsporen van mycotoxines vereisen een extractie en zuivering
van het staal. Welke methode hier het best voor gebruikt wordt, is afhankelijk van de structuur van het
toxine en de matrix. Een overzicht wordt gegevens in Turner et al. (2009). Tegenwoordig wordt vooral
massaspectrometrie gebruikt om het toxine te identificeren en kwantificeren (Turner et al., 2009;
Frenrich et al., 2014).
Liquid chromatography tandem mass spectrometry (LC-MS/MS) is een goede bruikbare methode voor
het opsporen van mycotoxines in verschillende matrices zoals voeder (Mol et al., 2008). LC-MS/MS is
een gevoelige methode die kwantificatie toe laat. In dit onderzoek zal gebruik gemaakt worden van
Ultra High Performance Liquid Chromatography - High Resolution Mass Spectrometer (UHPLC-
HRMS) analyse. Dit is een combinatie van vloeistofchromatografie en massa spectrometrie. HRMS
laat detectie toe van analyten in hetzelfde massabereik als LC-MS/MS, maar HRMS heeft geen
beperking in het aantal te detecteren analyten, zelfs indien geen analytische standaard beschikbaar is,
zoals van sommige fase I en II metabolieten (Lacina et al., 2012). Deze methode is een gevoelige,
accurate en precieze methode voor de detectie van mycotoxines (Rubert et al., 2012). Ten opzichte
van LC-MS/MS methode is UHPLC-HRMS meer massa-accuraat door de hoge resolutie.
Chromatografie is een analytische techniek voor het scheiden van 2 of meerdere opgeloste
substanties in een mengsel. Er treedt een verdeling op in 2 fasen, elke substantie heeft een
verschillende affiniteit voor een bepaalde fase. Op deze manier treedt er een scheiding op. Bij HPLC
worden moleculen gescheiden op basis van hun affiniteit voor de kolom en mobiele fase (Bird, 1989).
Bij MS worden de analyten geïoniseerd en gefragmenteerd en vervolgens op basis van massa en
lading gedetecteerd, met deze methode is de detectielimiet zeer laag (Niessen, 1999).
Niet alle analyse methoden kunnen de verschillende metabolieten in voeding en voeders detecteren
(De Boevre et al., 2012). Dit wil zeggen dat naast het parent mycotoxine nog gemaskeerde
mycotoxines in het voeder kunnen voorkomen, zoals bijvoorbeeld DON-glucoside. Doordat in de
meeste gevallen enkel de originele stof wordt gedetecteerd, ligt het totale gehalte aan mycotoxines
mogelijk vele malen hoger (Berthiller et al., 2009). Daarnaast zou bij co-exposure aan verschillende
mycotoxines de effecten van deze toxines synergetisch of additief kunnen werken (McKean et al.,
2006). Daarnaast kan de microbiota in het gastro-intestinaal stelsel DON-glucoside hydrolyseren tot
DON en kan DON op deze manier terug vrijkomen (Broekaert, 2015).

15
II. Onderzoek
1. Inleiding en probleemstelling
Het is bekend dat mycotoxines negatieve effecten kunnen hebben op zowel reproductieve als niet
reproductieve organen bij varkens. Zo beïnvloeden mycotoxines de folliculaire en embryonale groei en
hebben ze effect op o.a. proteïne synthese en het immuunsysteem (Tiemann & Dänicke, 2006). Ook
is het varken voor vele mycotoxines de meeste gevoelige diersoort (Prelusky et al., 1988; Diekman &
Green, 1992). Het doel van dit onderzoek is om een eventuele link tussen mycotoxines, of hun
metabolieten, en staartnecrose aan te tonen met inbegrip van de fase II metabolieten.
Wat het effect is van neonatale staartnecrose op de biggen zelf is ook niet gekend. Soms wordt
staartnecrose gezien in combinatie met splayleg en zwakke biggen (Mul et al., 2006). Of dit altijd het
geval is, is nog niet onderzocht.
Dit onderzoek is een verderzetting van een eerder uitgevoerde masterproef met onderzoek (Van
Neste, 2015). In dit onderzoek werd er reeds gekeken naar de link tussen bepaalde mycotoxines en
neonatale staartnecrose. Hierbij werden er verschillende stalen genomen, maar deze werden niet
allemaal onderzocht. Ook zijn de stalen niet op alle metabolieten onderzocht. Hieronder volgt een
korte samenvatting van de voorafgaande studie (figuur 8).
Figuur 8: Overzicht voorgaand onderzoek, gebaseerd op de masterproef van Van Neste, 2015
Er werden 20 bedrijven geselecteerd, 10 bedrijven met neonatale staartnecrose en 10 controle-
bedrijven. Per bedrijf werden een 5 tal zeugen bemonsterd met telkens ook 2 biggen per zeug. Van de
zeugen werd een melkmonster (2 ml) en een bloedmonster (8 ml) en zo mogelijk een urinemonster

16
(50 ml, mid-stream) genomen. Van de biggen werd steeds 2 ml bloed genomen. Ook werden er voor
ieder bedrijf gegevens verzameld en werd er een voeder- en waterstaal genomen.
De voederstalen werden geanalyseerd voor aflatoxine B1/B2/G1/G2, ochratoxine A, DON, ZEA,
fumonisine B1/B2/B3, T-2, HT-2, nivalenol, 3aDON, 15aDON, diacetoxyscirpenol, fusarenon-X,
neosolaniol, alternariol, alternariol methylether, roquefortine C en sterigmatocystine na vermalen,
homogeniseren en extractie m.b.v. LC-MS/MS. Het plasma van de bloedstalen van de biggen en
zeugen werd na centrifugatie en extractie geanalyseerd voor DON, DOM-1, ZEA, α-/β-ZOL, T-2 en
HT-2 m.b.v. LC-MS/MS. De waterstalen werden geanalyseerd door DGZ. De melkmonsters en
urinemonsters werden niet geanalyseerd. De bekomen gegevens werden geanalyseerd m.b.v. IBM
SPSS statistics 22.
Er werd een significant verschil gezien in productiegetal, dit was lager bij de bedrijven met neonatale
staartnecrose. Maar aangezien vele factoren een invloed kunnen hebben op het productiegetal, kan
dit niet direct worden toegewezen aan de staartnecrose. Bij een toenemende pariteit leek het
percentage aangetaste biggen te stijgen.
Bij de voederanalyse werden 12/23 onderzochte mycotoxines teruggevonden. De resultaten zijn
weergegeven in bijlage I. De concentratie van DON was in de probleembedrijven significant hoger dan
in de controlebedrijven (484,0 ± 212,21 µg.kg-1
vs. 257,4 ± 89,18 µg.kg-1
). De concentratie ZEA was
ook hoger in de probleembedrijven, maar niet significant. T-2 werd slechts in een staal aangetoond, dit
staal was afkomstig van een probleembedrijf. 3aDON en 15aDON werden meer teruggevonden bij
probleembedrijven.
Voor DON werd een significant hogere plasmaconcentratie gevonden bij de zeugen op
probleembedrijven (0,967 ± 0,692 ng.mL-1
vs. 0,510 ± 0,311 ng.mL-1
). Ook werd er een positieve
correlatie gevonden tussen de concentratie van DON in het voeder en de concentratie van DON in
zeugenplasma. In tabel 5 wordt een overzicht gegeven van de gemiddelde concentraties van de
verschillende mycotoxines in biggenplasma. Voor DON werden hogere concentraties gevonden bij
biggen afkomstig van probleembedrijven. Maar dit verschil was niet significant.
Bij de wateranalyse werden geen significante verschillen gevonden tussen probleem- en
controlebedrijven.
Uit bovenstaand onderzoek kan geconcludeerd worden dat DON een rol speelt in de pathogenese van
neonatale staartnecrose bij biggen. Daarnaast is gebleken dat plasma een goede matrix is om DON te
analyseren bij zeugen. Voor ZEA en T-2 kunnen geen conclusies getrokken worden. Mogelijk omdat
enkel de parent mycotoxines DON, ZEA, T-2 en hun fase I metabolieten DOM-1, α-ZOL, β-ZOL en
HT-2 werden geanalyseerd. Daarom zal in de huidige masterproef ook gekeken worden naar de fase
II metabolieten van ZEA en DON. In plaats van de LC-MS/MS detectiemethode zal nu gewerkt worden
met UHPLC-HRMS. Deze techniek heeft geen beperking in het aantal te detecteren analyten. Omdat
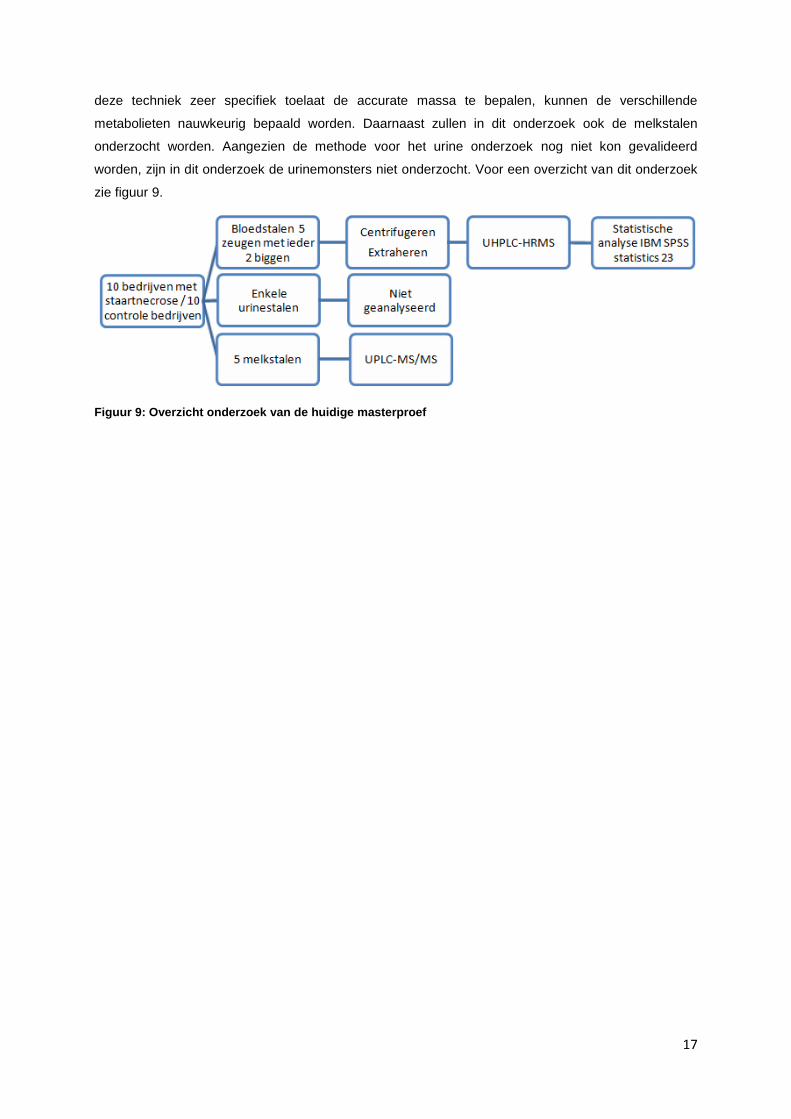
17
deze techniek zeer specifiek toelaat de accurate massa te bepalen, kunnen de verschillende
metabolieten nauwkeurig bepaald worden. Daarnaast zullen in dit onderzoek ook de melkstalen
onderzocht worden. Aangezien de methode voor het urine onderzoek nog niet kon gevalideerd
worden, zijn in dit onderzoek de urinemonsters niet onderzocht. Voor een overzicht van dit onderzoek
zie figuur 9.
Figuur 9: Overzicht onderzoek van de huidige masterproef

18
2. Materiaal en methoden
2.1. Plasma zeugen en biggen
De verschillende staalnamen werden hierboven weergegeven. De bloedstalen werden
gecentrifugeerd (2851g, 10 min, 4 °C) om plasma te verkrijgen. Het plasma werd in hoeveelheden van
250 μL bewaard bij ≤−15 °C.
Er werd 25 μL van de interne standaard (IS) oplossing (100 ng/mL ZEA-13
C18 en DON-13
C15)
toegevoegd aan 250 μL plasma, gevolgd door het toevoegen van acetonitrile (ACN) tot het volume
van 1.0 mL. Daarna werd het staal gehomogeniseerd met behulp van een vortex mixer (10 s) en
gecentrifugeerd (8517g, 10 min, 4 °C). Het supernatant werd drooggedampt met behulp van stikstof
(N2, 40 ± 5 °C). Het droge residu werd opgelost in 200 μL water/methanol (MeOH) (85/15, v/v).
Daarna werd het staal 15 s gehomogeniseerd en gefilterd doorheen een Millex GV-PVDF 0.22 µm
spuitfilter en overgebracht naar een conische autosampler vial. Hiervan werd tweemaal 5 µL
geïnjecteerd in het HRMS instrument, éénmaal ter analyse van DON en de respectievelijke
metabolieten en éénmaal ter analyse van ZEA en metabolieten. Het HPLC-systeem bestond uit een
Accela pomp van het type 1250 en een autosampler. De HPLC werd gekoppeld aan een Exactive
Orbitrap MS toestel.
Voor zowel DON als ZEA gebeurde de chromatografische scheiding aan de hand van een Hypersil
Gold kolom (50 mm × 2.1 mm i.d., dp 1.9 μm) in combinatie met een prekolom van hetzelfde type (10
mm × 2.1 mm i.d., dp 3 μm). Voor DON werden de volgende mobiele fasen gebruikt, UPLC water met
0.3% azijnzuur (A) en UPLC methanol met 0.3% azijnzuur (B). Er werd een gradiënt elutie toegepast:
0-1.0 min: 5% B; 1.0-4.0 min: lineaire gradiënt naar 80% B; 4.0-6.7 min: 80% B; 6.7-7.0: lineaire
gradiënt naar 5% B; 7.0-10.0 min: 5% B. Voor de analyse van ZEA en diens metabolieten werd UPLC
water met 0.01% azijnzuur (A) en UPLC ACN (B) gebruikt met volgende gradiënt: 0-0.5 min: 30% B;
0.5-5.0 min: lineaire gradiënt naar 70% B; 5.0-6.4 min: 70% B; 6.4-6.5 min: lineaire gradiënt naar 30%
B; 6.5-10.0 min: 30% B. De stroomsnelheid voor beide methoden was 300 μL/min en de temperatuur
van de kolom oven en autosampler werden ingesteld op respectievelijk 45 en 5 ° C.
Aangezien bepaalde standaarden van fase II metabolieten niet commercieel beschikbaar zijn, is deze
methode beperkt tot semi-kwantitatieve analyses op basis van de oppervlakte van de
chromatografische piek van de metabolieten.
De volgende mycotoxines werden onderzocht in zowel zeugen- als biggenplasma: DON-GlcA, ZEA,
ZEA-glucuronide, α-ZOL, β-ZOL α-zearalanol (α-ZAL) en β-zearalanol (β-ZAL), zearalenol-
glucuronide, zearalanone (ZAL). Voor bedrijf 1, 2 en 3 zijn de resultaten van DON-GlcA niet in
beschouwing genomen. Doordat deze stalen een extra dooi-vries cyclus hebben doorgemaakt, zijn de
mycotoxines in deze stalen mogelijk gedegradeerd en daardoor niet meer aantoonbaar.
Alle gevonden concentraties zijn uitgedrukt in oppervlakten van de chromatografische pieken.

19
2.2. Melkmonsters
Voor de analyse van mycotoxines in melk werd een methode ontwikkeld aan het ILVO. Deze werd
oorspronkelijk ontwikkeld voor de detectie van mycotoxines in melkvervangers bij kalveren, maar werd
hier ook gebruikt voor zeugenmelk. De volgende mycotoxines werden voor zeugenmelk onderzocht:
ZEA, α-ZOL en β-ZOL.
Voor staalname zie hierboven. De melkstalen van de probleembedrijven 1, 8, 9, 10 en van
controlebedrijven 12 en 18 zijn onderzocht. Wegens tijdgebrek was het niet mogelijk om alle stalen
van alle bedrijven te analyseren. Er is gekozen voor deze stalen omdat op deze ZEA in grootste
hoeveelheid in het voeder werd teruggevonden (8, 9, 10, 12 en 18). Daarnaast zijn ook de stalen van
probleembedrijf 1 geanalyseerd omdat er in bedrijf 1 een aantal zeugen hoge plasmaconcentraties
hadden voor ZEA, α-ZOL of β-ZOL.
Er werd 1.0 mL melk afgevuld in een proefbuis. Dan werd de spike oplossing gemaakt. Zie hieronder,
per bedrijf zijn 5 stalen geanalyseerd m.u.v. bedrijf 1 (3 stalen). Vervolgens werd 2 mL MeOH
toegevoegd en werd er achtereenvolgend 1 min gevortext, 10 min geschud en 10 min gecentrifugeerd
bij 3000 rpm. Het supernatant werd overgebracht in een nieuwe proefbuis. 2 mL ACN/H2O/azijnzuur
(79/20/1) werd toegevoegd, opnieuw werd er 1 min gevortext, 10 min geschud en 10 min
gecentrifugeerd bij 3000 rpm. Het supernatant werd bij het vorige gevoegd. Vervolgens werd er 5 min
gecentrifugeerd bij 3000 rpm. Het supernatant werd overgebracht in een gegradueerde puntbuis. Dit
werd ingedampt tot 1 mL onder N2-stroom bij 40 °C. Vervolgens werd het staal gefilterd (0,22 µm) in
een vial.
Spike oplossingen:
Ijklijn punt 1: blanco biestmelk + 20 µL OPL IS10
Ijklijn punt 2: blanco biestmelk + 20 µL OPL IS10 + 25 µL OPL A0,1
Ijklijn punt 3: blanco biestmelk + 20 µL OPL IS10 + 50 µL OPL A0,1
Ijklijn punt 4: blanco biestmelk + 20 µl OPL IS10 + 75 µL OPL A0,1
Ijklijn punt 5: blanco biestmelk + 20 µL OPL IS10 + 10 µL OPL A1
Ijklijn punt 6: blanco biestmelk + 20 µL OPL IS10 + 15 µL OPL A1
Ijklijn punt 7: blanco biestmelk + 20 µL OPL IS10 + 20 µL OPL A1
Ijklijn punt 8: blanco biestmelk + 20 µL OPL IS10 + 50 µL OPL A1
Staal bedrijf nr. 1: biestmelk staal nr. 1 + 20 µL OPL IS10
Staal bedrijf nr. 8: biestmelk staal nr. 8 + 20 µL OPL IS10
Staal bedrijf nr. 9: biestmelk staal nr. 9 + 20 µL OPL IS10
Staal bedrijf nr. 10: biestmelk staal nr. 10 + 20 µL OPL IS10
Staal bedrijf nr. 12: biestmelk staal nr. 12 + 20 µL OPL IS10
Staal bedrijf nr. 18: biestmelk staal nr. 18 + 20 µL OPL IS10
*OPL A10:[10 ng/ µL] ZEA, α-ZOL, β-ZOL: 3 x 10 µL [1000 ng/ µL] + 970 µl H2O
*OPL A1:[1 ng/ µL] ZEA, α-ZOL, β-ZOL: 100 µL [10ng/ µL] ZEN, α-ZOL, β-ZOL + 900 µl H2O

20
*OPL A0,1:[0,1 ng/ µL] ZEA, α-ZOL, β-ZOL: 100 µL [1 ng/ µL] ZEN, α-ZOL, β-ZOL + 900 µL
H2O
*OPL IS10: [10 ng/ µL] 13
C18-ZEA: 10 µL [1000 ng/ µL] + 990 µL H2O
Chromatografische gradiëntscheiding werd uitgevoerd met een AcquityTM
UHPLC (Waters) systeem
uitgerust met een Acquity CSH Fluoro Phenyl column (1.7 µm, 2.1 x 150 mm, Waters). De mobiele
fase bestond uit een mix van H2O + 0.3% azijnzuur (A) en MeOH + 0.3% azijnzuur (B). Detectie werd
gedaan met een Xevo TQ-S triple quadrupole MS systeem (Waters).
Er werd een ijklijn gemaakt met volgende punten: 0 - 2,5 – 5 – 7,5 – 10 – 15 – 20 – 50 µg/kg van de
componenten (ZEA-13
C18 als IS).
2.3. Berekeningen en statistiek
Verschillen in mycotoxineconcentraties tussen probleem- en controlebedrijven werden statistisch
getoetst (ANOVA). Daarnaast werden verschillende correlaties (Spearman rank, Pearson) onderzocht,
zoals o.a. de correlatie tussen parent toxine en metabolieten. De resultaten van probleembedrijf
nummer 1, 2 en 3 voor DON-GlcA werden uitgesloten daar deze resultaten niet betrouwbaar waren
door een technisch defect. Correlaties kleiner dan 0,3, tussen 0,3 and 0,6 en groter dan 0,6 worden
respectievelijk als zwak, matig en sterk geïnterpreteerd. Er wordt pas aangenomen dat er een
correlatie is wanneer er meer dan 10 paren zijn, tenzij anders vermeld. Een paar bestaat wanneer de
mycotoxines (of metabolieten) waartussen de correlatie wordt berekend beide voorkwamen in het
plasmastaal. De resultaten van Van Neste (2015) zijn eveneens meegenomen in de statistische
berekeningen. Bij deze gegevens waren er veel resultaten onder de detectie- en kwantificatielimiet,
hier is dan steeds de helft van de betreffende waarde genomen voor de statistische berekeningen.
De significantie grens werd vastgelegd op p<0,05. De analyses werden uitgevoerd met IBM SPSS
statistics 23.
21
3. Resultaten
3.1. DON-GlcA in zeugenplasma
In bijlage II bevindt zich een overzicht van de gevonden mycotoxine concentraties in het plasma van
de zeugen. Aangezien er bij het eerste probleembedrijf 6 zeugen met ieder 2 biggen zijn bemonsterd
i.p.v. 5 zeugen is er 1 staal meer. Van de probleembedrijven waren 28/51 stalen positief. Door een
technisch defect zijn de resultaten van de eerste 3 probleembedrijven niet betrouwbaar en zijn om die
reden uitgesloten. Dit maakt dat 28/35 (80%) zeugenplasma stalen positief waren voor DON-GlcA.
Van de controlebedrijven ontbraken 5 stalen, de overige 45/45 (100%) waren allemaal positief voor
DON-GlcA.
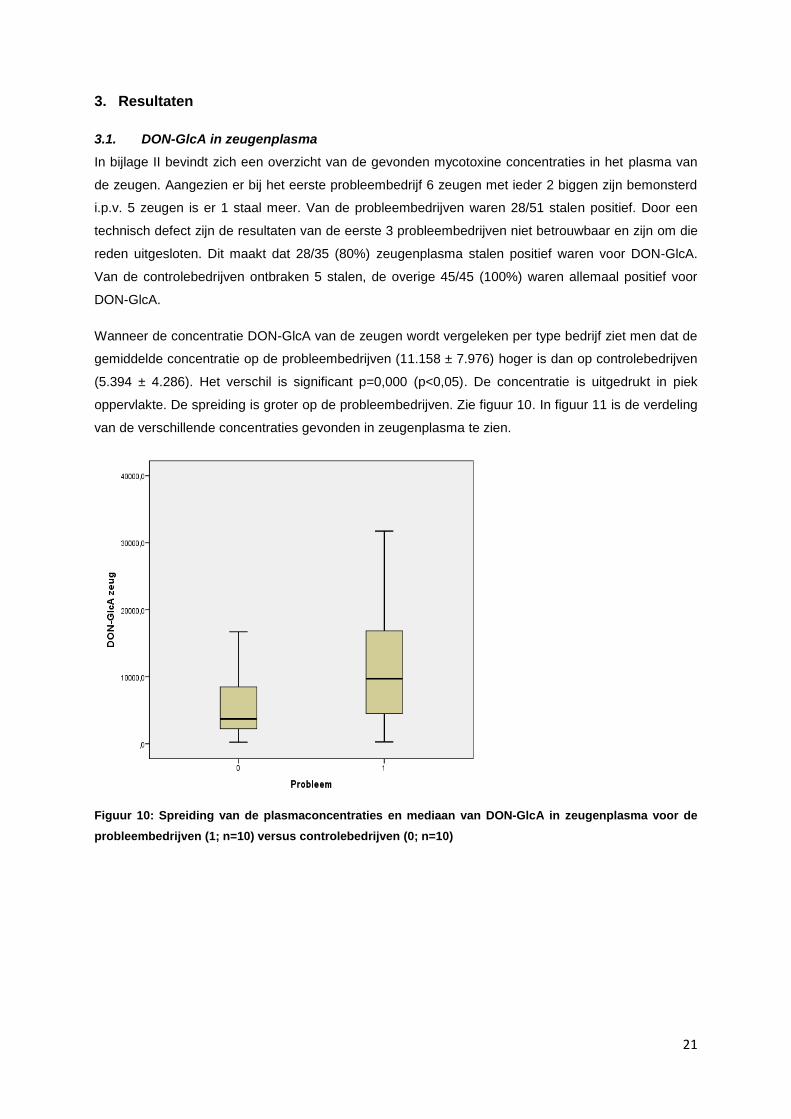
Wanneer de concentratie DON-GlcA van de zeugen wordt vergeleken per type bedrijf ziet men dat de
gemiddelde concentratie op de probleembedrijven (11.158 ± 7.976) hoger is dan op controlebedrijven
(5.394 ± 4.286). Het verschil is significant p=0,000 (p<0,05). De concentratie is uitgedrukt in piek
oppervlakte. De spreiding is groter op de probleembedrijven. Zie figuur 10. In figuur 11 is de verdeling
van de verschillende concentraties gevonden in zeugenplasma te zien.
Figuur 10: Spreiding van de plasmaconcentraties en mediaan van DON-GlcA in zeugenplasma voor de
probleembedrijven (1; n=10) versus controlebedrijven (0; n=10)
22
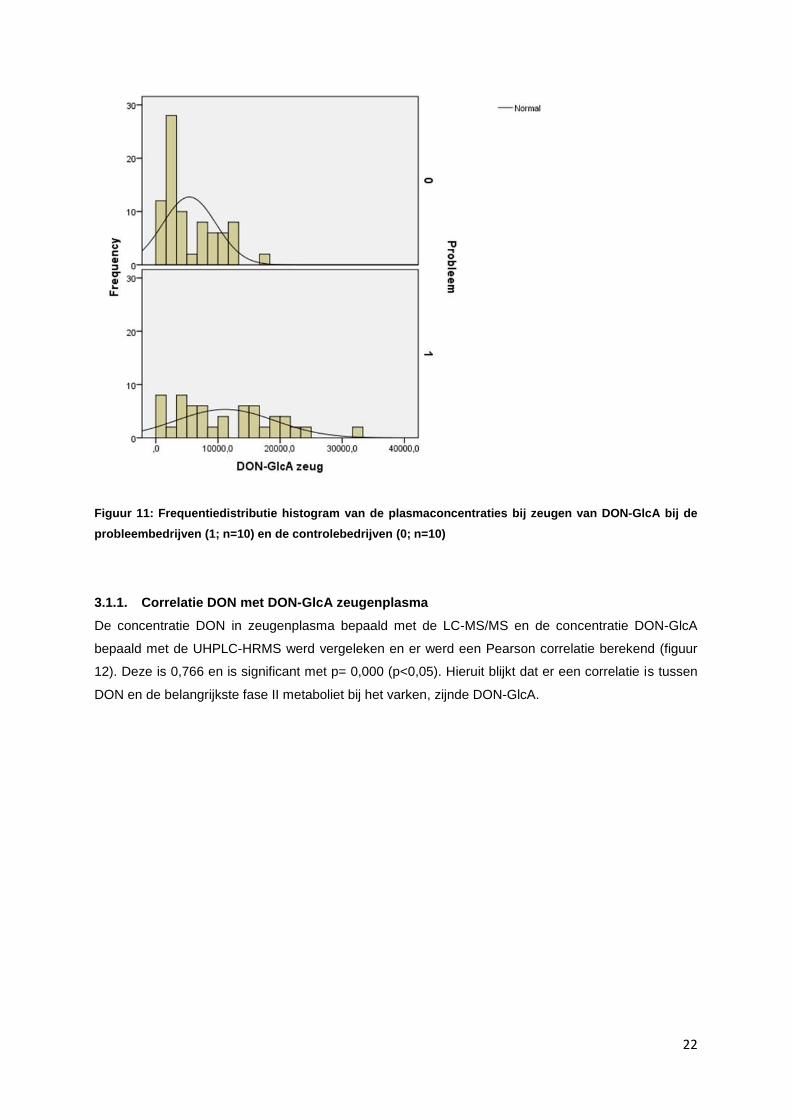
Figuur 11: Frequentiedistributie histogram van de plasmaconcentraties bij zeugen van DON-GlcA bij de
probleembedrijven (1; n=10) en de controlebedrijven (0; n=10)
3.1.1. Correlatie DON met DON-GlcA zeugenplasma
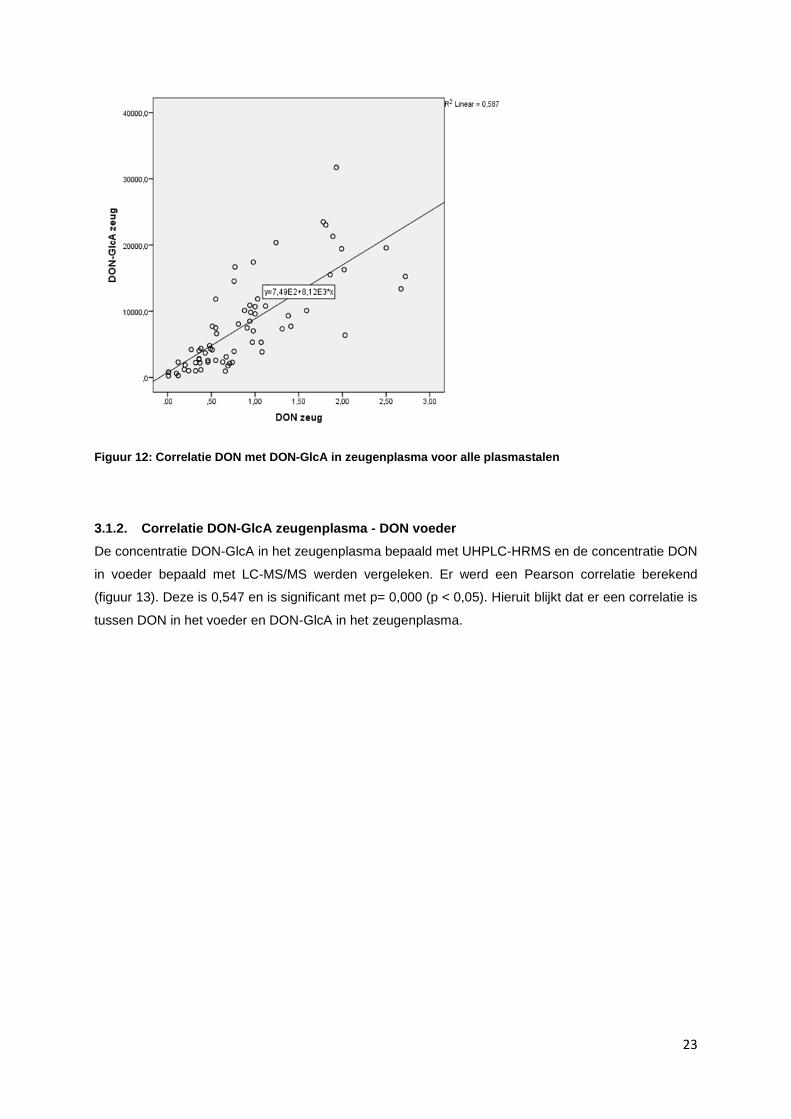
De concentratie DON in zeugenplasma bepaald met de LC-MS/MS en de concentratie DON-GlcA
bepaald met de UHPLC-HRMS werd vergeleken en er werd een Pearson correlatie berekend (figuur
12). Deze is 0,766 en is significant met p= 0,000 (p<0,05). Hieruit blijkt dat er een correlatie is tussen
DON en de belangrijkste fase II metaboliet bij het varken, zijnde DON-GlcA.
23
Figuur 12: Correlatie DON met DON-GlcA in zeugenplasma voor alle plasmastalen
3.1.2. Correlatie DON-GlcA zeugenplasma - DON voeder
De concentratie DON-GlcA in het zeugenplasma bepaald met UHPLC-HRMS en de concentratie DON
in voeder bepaald met LC-MS/MS werden vergeleken. Er werd een Pearson correlatie berekend
(figuur 13). Deze is 0,547 en is significant met p= 0,000 (p < 0,05). Hieruit blijkt dat er een correlatie is
tussen DON in het voeder en DON-GlcA in het zeugenplasma.

24
Figuur 13: Correlatie DON-GlcA in zeugenplasma en DON in voeder van alle plasma- en voederstalen
3.2. DON-GlcA in biggenplasma
In bijlage II bevindt zich een overzicht van de gevonden mycotoxine concentraties in het plasma van
de biggen. Door een technisch defect zijn de resultaten van de stalen van de eerste 3
probleembedrijven niet betrouwbaar. Deze zijn om die reden uitgesloten. Er waren 3/70 (4%) stalen
positief (gemiddelde 115) bij de stalen van de probleembedrijven. Bij de controlebedrijven waren
3/100 (3%) stalen positief (gemiddelde 458). Vanwege het laag aantal positieve stalen voor DON-GlcA
kon er geen vergelijking worden gemaakt in de concentratie van DON-GlcA tussen probleembedrijven
en controlebedrijven.
Verder is er geen correlatie gevonden tussen DON-GlcA zeugenplasma en DON-GlcA biggenplasma.
Ook was er geen correlatie tussen DON in biggenplasma en DON-GlcA in het zeugenplasma.
Eveneens werd er geen correlatie gevonden tussen DON en DON-GlcA in biggenplasma.
3.3. Zearalenone en metabolieten in zeugenplasma
Voor een overzicht van de concentraties van ZEA en metabolieten in zeugenplasma, zie bijlage II. Er
kon geen significant verschil in concentratie van ZEA (of metabolieten) aangetoond worden tussen
probleembedrijven en controlebedrijven door een te laag aantal positieve waarden. Een overzicht
wordt weergeven in tabel 5.
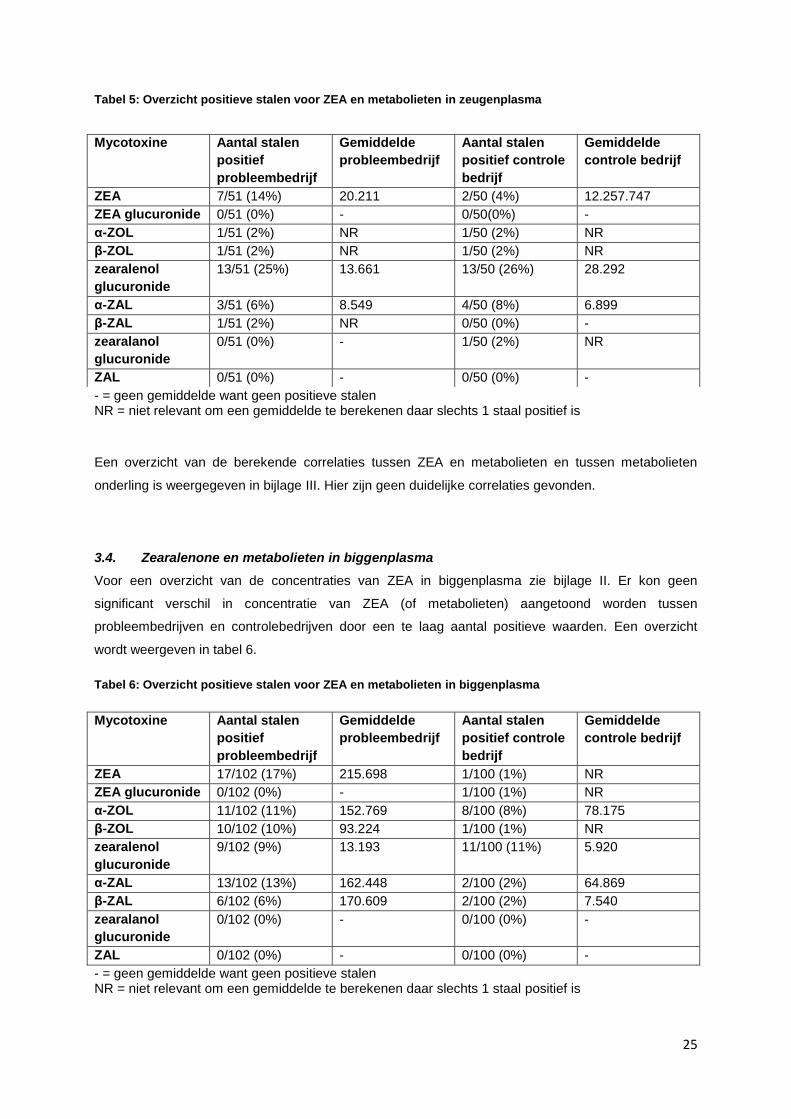
25
Tabel 5: Overzicht positieve stalen voor ZEA en metabolieten in zeugenplasma
- = geen gemiddelde want geen positieve stalen NR = niet relevant om een gemiddelde te berekenen daar slechts 1 staal positief is
Een overzicht van de berekende correlaties tussen ZEA en metabolieten en tussen metabolieten
onderling is weergegeven in bijlage III. Hier zijn geen duidelijke correlaties gevonden.
3.4. Zearalenone en metabolieten in biggenplasma
Voor een overzicht van de concentraties van ZEA in biggenplasma zie bijlage II. Er kon geen
significant verschil in concentratie van ZEA (of metabolieten) aangetoond worden tussen
probleembedrijven en controlebedrijven door een te laag aantal positieve waarden. Een overzicht
wordt weergeven in tabel 6.
Tabel 6: Overzicht positieve stalen voor ZEA en metabolieten in biggenplasma
Mycotoxine Aantal stalen
positief
probleembedrijf
Gemiddelde
probleembedrijf
Aantal stalen
positief controle
bedrijf
Gemiddelde
controle bedrijf
ZEA 17/102 (17%) 215.698 1/100 (1%) NR
ZEA glucuronide 0/102 (0%) - 1/100 (1%) NR
α-ZOL 11/102 (11%) 152.769 8/100 (8%) 78.175
β-ZOL 10/102 (10%) 93.224 1/100 (1%) NR
zearalenol
glucuronide
9/102 (9%) 13.193 11/100 (11%) 5.920
α-ZAL 13/102 (13%) 162.448 2/100 (2%) 64.869
β-ZAL 6/102 (6%) 170.609 2/100 (2%) 7.540
zearalanol
glucuronide
0/102 (0%) - 0/100 (0%) -
ZAL 0/102 (0%) - 0/100 (0%) -
- = geen gemiddelde want geen positieve stalen NR = niet relevant om een gemiddelde te berekenen daar slechts 1 staal positief is
Mycotoxine Aantal stalen
positief
probleembedrijf
Gemiddelde
probleembedrijf
Aantal stalen
positief controle
bedrijf
Gemiddelde
controle bedrijf
ZEA 7/51 (14%) 20.211 2/50 (4%) 12.257.747
ZEA glucuronide 0/51 (0%) - 0/50(0%) -
α-ZOL 1/51 (2%) NR 1/50 (2%) NR
β-ZOL 1/51 (2%) NR 1/50 (2%) NR
zearalenol
glucuronide
13/51 (25%) 13.661
13/50 (26%) 28.292
α-ZAL 3/51 (6%) 8.549 4/50 (8%) 6.899
β-ZAL 1/51 (2%) NR 0/50 (0%) -
zearalanol
glucuronide
0/51 (0%) - 1/50 (2%) NR
ZAL 0/51 (0%) - 0/50 (0%) -
26
Een overzicht van de berekende correlaties tussen ZEA en metabolieten en tussen metabolieten
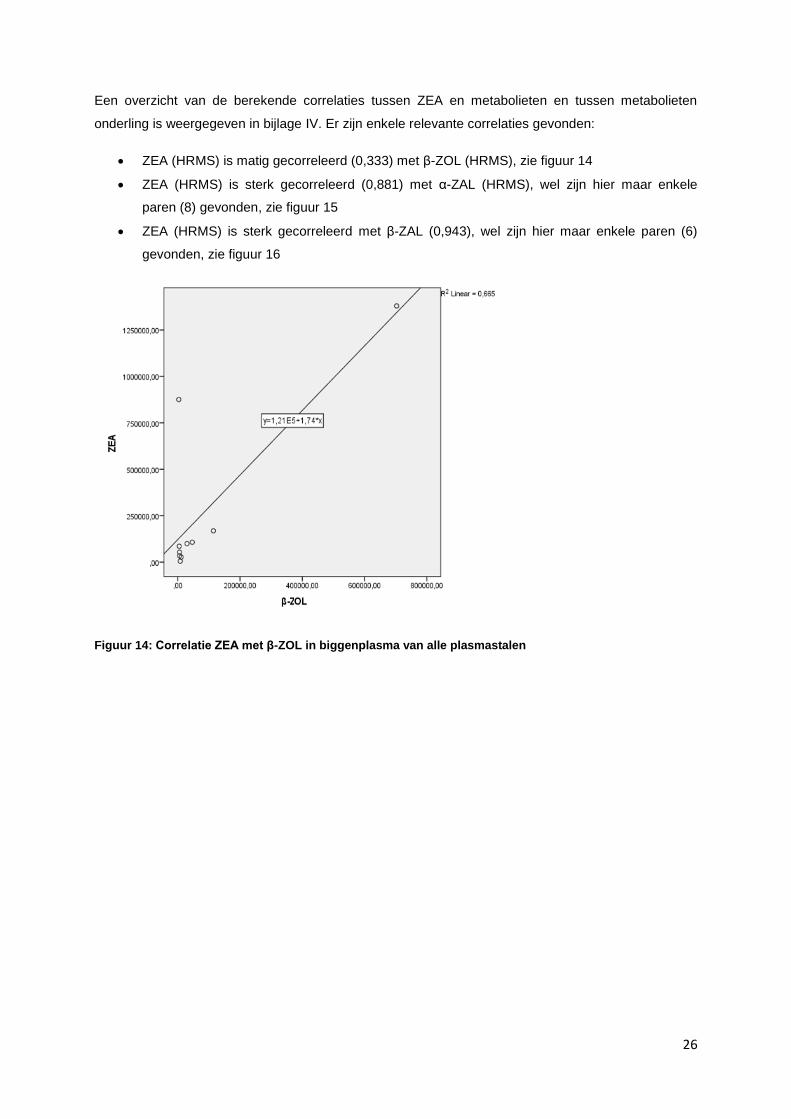
onderling is weergegeven in bijlage IV. Er zijn enkele relevante correlaties gevonden:
ZEA (HRMS) is matig gecorreleerd (0,333) met β-ZOL (HRMS), zie figuur 14
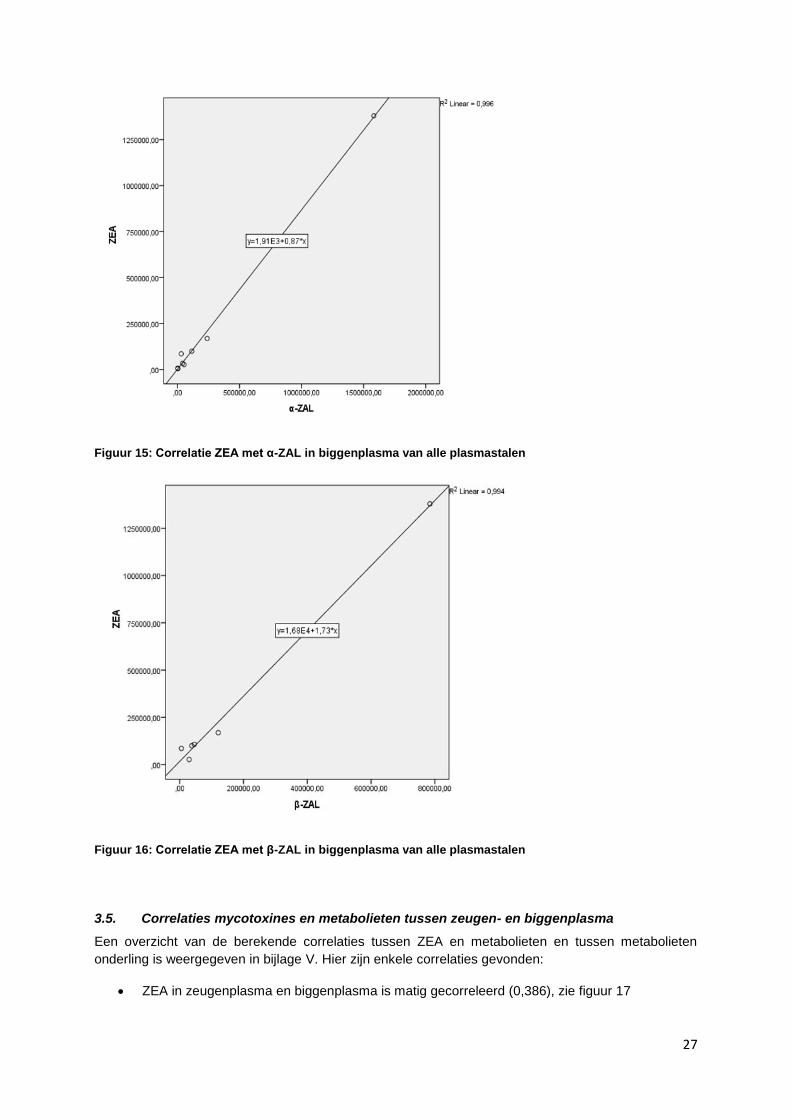
ZEA (HRMS) is sterk gecorreleerd (0,881) met α-ZAL (HRMS), wel zijn hier maar enkele
paren (8) gevonden, zie figuur 15
ZEA (HRMS) is sterk gecorreleerd met β-ZAL (0,943), wel zijn hier maar enkele paren (6)
gevonden, zie figuur 16
Figuur 14: Correlatie ZEA met β-ZOL in biggenplasma van alle plasmastalen
27
Figuur 15: Correlatie ZEA met α-ZAL in biggenplasma van alle plasmastalen
Figuur 16: Correlatie ZEA met β-ZAL in biggenplasma van alle plasmastalen
3.5. Correlaties mycotoxines en metabolieten tussen zeugen- en biggenplasma
Een overzicht van de berekende correlaties tussen ZEA en metabolieten en tussen metabolieten
onderling is weergegeven in bijlage V. Hier zijn enkele correlaties gevonden:
ZEA in zeugenplasma en biggenplasma is matig gecorreleerd (0,386), zie figuur 17

28
α-ZAL in zeugenplasma en biggenplasma is sterk gecorreleerd (0,973), wel waren er slechts 7
paren, zie figuur 18
Figuur 17: Correlatie ZEA in zeugen- en biggenplasma van alle plasmastalen
Figuur 18: Correlatie α-ZAL in zeugen- en biggenplasma van alle plasmastalen
29
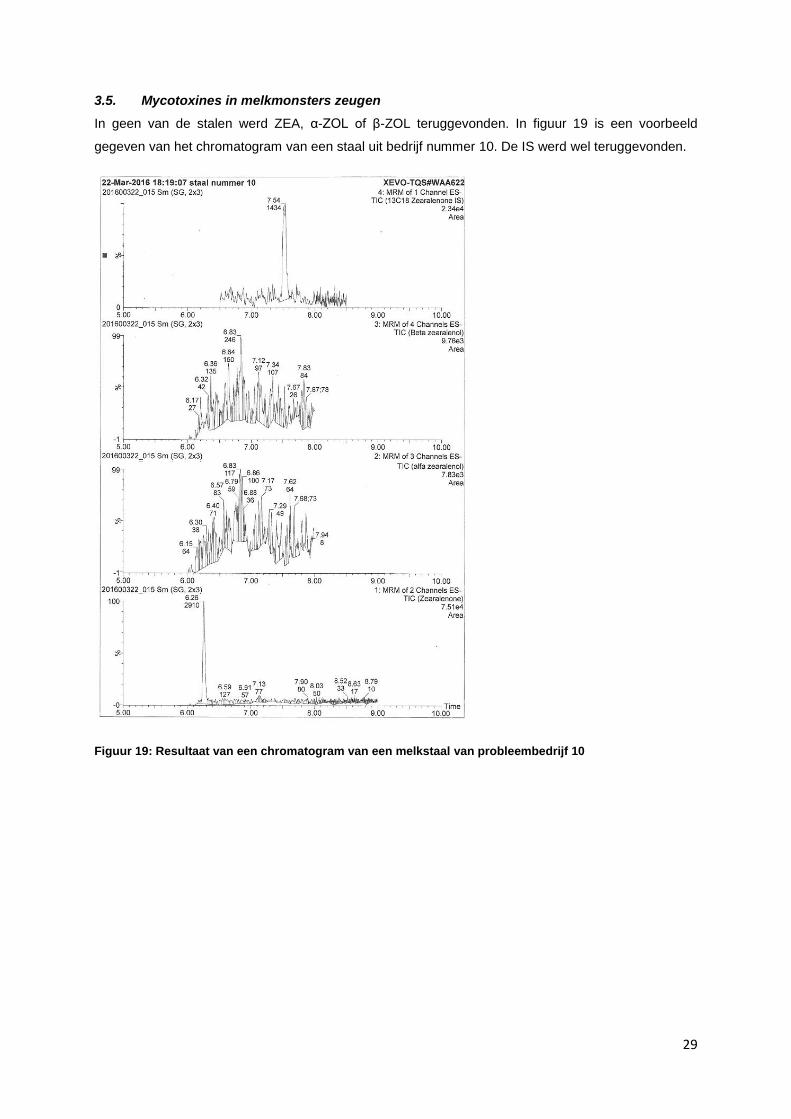
3.5. Mycotoxines in melkmonsters zeugen
In geen van de stalen werd ZEA, α-ZOL of β-ZOL teruggevonden. In figuur 19 is een voorbeeld
gegeven van het chromatogram van een staal uit bedrijf nummer 10. De IS werd wel teruggevonden.
Figuur 19: Resultaat van een chromatogram van een melkstaal van probleembedrijf 10

30
4. Discussie
Het gehalte DON-GlcA is significant hoger op de probleembedrijven (11.158 ± 7.976) versus de
controlebedrijven (5.394 ± 4.286) in zeugenplasma. Eerder is er ook een significant verschil
aangetoond voor DON tussen de probleembedrijven en de controlebedrijven in het plasma van de
zeugen. Daarnaast is er ook een significant verschil gevonden in DON concentratie in het voeder
tussen controle- en probleembedrijven (Van Neste, 2015). Dit was te verwachten, aangezien er een
sterke correlatie is tussen DON voeder en DON zeugenplasma (Van Neste, 2015) en een correlatie
tussen DON voeder en DON-GlcA zeugenplasma. Hieruit kunnen we afleiden dat er een associatie is
tussen DON in het zeugenvoeder en daarmee in het zeugenplasma en neonatale staartnecrose bij
biggen.
Er is bovendien een sterke correlatie (0,766) tussen DON en DON-GlcA in het plasma van de zeug.
DON-GlcA is echter een betere biomerker voor blootstelling aan DON dan DON zelf, daar DON-GlcA
in grotere hoeveelheden wordt teruggevonden. DON-GlcA is ook de belangrijkste metaboliet van DON
bij het varken (Kollarczik et al., 1994; Xianai et al., 2007; Turner et al., 2011). Het is mogelijk om DON-
GlcA te kwantificeren in plaats van de chromatografische piekoppervlakte te hanteren, maar dit is
vanwege de tijdsbeperking van deze masterproef nog niet gedaan.
Bij de biggen werd slechts in een beperkt aantal plasmastalen DON-GlcA gevonden. Mocht dit
onderzoek nog verdergezet of uitgebreid worden, zou het interessant kunnen zijn de stalen van
probleembedrijf 1 tot en met 3 opnieuw te analyseren. Aangezien er bij de biggenstalen in deze
bedrijven met de LC-MS/MS bij enkele biggen wel DON is aangetoond, is hier potentieel ook DON-
GlcA aantoonbaar. Meer positieve stalen zou potentieel kunnen resulteren in aantoonbare verschillen
en bepaalde correlaties. Het feit dat er in deze studie geen correlaties tussen metabolieten of
verschillen tussen controle- en probleembedrijf zijn aangetoond, wil niet zeggen dat deze er niet zijn.
Maar op basis van deze dataset kunnen ze niet worden aangetoond.
Voor ZEA en metabolieten konden geen significante verschillen aangetoond worden tussen probleem-
en controlebedrijven, noch voor zeugen-, noch voor biggenplasma. Bij de gemiddelde concentraties
van ZEA en metabolieten in zeugenplasma zien we dat voor ZEA en zearalenol-glucuronide de
gemiddelde concentratie hoger ligt bij de controlebedrijven. Bij ZEA komt dit doordat er één zeug was
bij de controlebedrijven die enorm hoge waarden had voor ZEA, α-ZOL en β-ZOL. Één enkele
uitschieter is niet relevant. Voor zearalenol-glucuronide ligt de concentratie bij de controlebedrijven
wel veel hoger (28.292 vs. 13.661), waarom is niet gekend. Van de berekende correlaties tussen ZEA
en metabolieten en metabolieten onderling zijn er geen in beschouwing genomen. Een correlatie werd
voor alle combinaties berekend, en leek soms aanwezig, maar op basis van de scatter diagrammen
kon uitgemaakt worden dat deze niet betrouwbaar waren. De meest frequent gedetecteerde
metaboliet van ZEA bij de zeugen in dit onderzoek was zearalenol-glucuronide, dit is voor het varken
een van de meest belangrijkste in vivo gevormde metabolieten (Olsen et al., 1987; Biehl et al., 1993),
en zou bijgevolg ook als biomerker voor DON blootstelling kunnen overwogen worden.

31
Bij de biggen zien we dat voor α-ZOL, α-ZAL, β-ZAL en zearalenol-glucuronide de concentraties
gemiddeld hoger liggen voor probleembedrijven versus controlebedrijven (respectievelijk 152.769 vs.
78.175, 162.448 vs. 64.869, 170.609 vs. 7.540 en 13.193 vs. 5.920). Voor ZEA en β-ZOL was er voor
ieder slechts 1 staal positief bij de controlebedrijven en daarom kunnen we geen gemiddelde
vergelijken. Omdat de waarden niet significant verschillend zijn, kunnen we hieraan geen conclusies
verbinden. Er waren correlaties tussen ZEA en β-ZOL, α-ZAL en β-ZAL. Deze 2 laatstgenoemden
waren wel op basis van minder dan 10 paren, maar omdat het voor beide een sterke correlatie was
nemen we deze toch in beschouwing. Dit betekent dat als ZEA in hogere concentraties aanwezig was,
de metabolieten dit ook waren. De meest frequent gedetecteerde metaboliet van ZEA bij de biggen in
dit onderzoek was eveneens zearalenol-glucuronide, daarnaast werden α-ZOL, α-ZAL en β-ZOL ook
nog regelmatig gevonden. Het is niet gekend of de biggen zelf de metabolieten gevormd hebben of
dat ze deze in utero of via de melk hebben binnengekregen.
Ook zijn er correlaties gevonden tussen ZEA in zeugenplasma en biggenplasma en tussen α-ZAL in
zeugenplasma en biggenplasma. Voor deze laatste waren wel minder dan 10 paren, maar omdat het
een sterke correlatie was nemen we deze eveneens in beschouwing. Uit deze correlaties blijkt dus dat
er een verband is tussen de concentraties van deze mycotoxines in zeugenplasma en biggenplasma.
Mogelijk wordt er bij een hogere plasmaconcentratie van de zeug meer overgedragen naar de big. Of
dit gebeurt in utero, via de melk of via beide wegen is niet gekend. Wel is voor ZEA geweten dat beide
wegen mogelijk zijn (Palyusik et al., 1980; Dänicke et al., 2007).
T-2 toxine en zijn metabolieten zijn niet onderzocht in deze studie. Deze zijn niet geanalyseerd omdat
de eliminatie halfwaardetijd van T-2 vele malen korter is dan van ZEA en DON (respectievelijk 13,8
min, 2.63 u en 3.9 - 5.8 u (Swanson et al., 1987; Dänicke et al., 2005; Pestka, 2007). In Van Neste
(2015) werden geen gehaltes voor T-2 of HT-2 toxine gevonden boven de kwantificatielimiet. Voor een
eventuele verderzetting van het onderzoek zou het mogelijk interessant zijn deze toch te analyseren,
ook al is de kans zeer laag om T-2 terug te vinden. Bijvoorbeeld wanneer er bloed zou worden
genomen van een aantal aangetaste biggen onmiddellijk na het optreden van de symptomen van
staartnecrose.
In de onderzochte melkstalen is geen ZEA, α-ZOL of β-ZOL gevonden, met een kwantificatielimiet
(LOQ) van 2.5 µg/kg. Uit eerder onderzoek is gebleken dat deze toxinen wel via de melk naar de big
overgedragen kunnen worden (Palyusik et al., 1980). Voor de volledigheid van het onderzoek zou het
nuttig zijn om de andere stalen ook te onderzoeken, aangezien enkel de stalen zijn onderzocht die
hoge ZEA concentraties in het voeder dan wel het plasma hadden. Mogelijk ligt de LOQ te hoog om
ZEA en metabolieten terug te vinden. Eventueel zou het interessant zijn om de melk ook te
analyseren voor DON en metabolieten.
Voor een eventueel volgend onderzoek zou het interessant zijn om de mycotoxineconcentratie te
meten in gal (Brezina et al., 2016). Zowel ZEA, DON als T-2 worden uitgescheiden via gal, daarnaast
ondergaat ZEA ook nog een enterohepatische kringloop (Robinson et al., 1979; Biehl et al., 1993;
Goyarts & Dänicke, 2006). Het nadeel hiervan is dat gal enkel post-mortaal te verzamelen is.

32
Wanneer men in de praktijk een zeug wil bemonsteren, kan eventueel een reforme zeug in het
slachthuis bemonsterd worden. Nadeel hiervan is dat dit niet op het moment is dat de staartnecrose
voorkomt. In een experimentele setting kunnen er dieren geëuthanaseerd worden om de mycotoxine
concentratie in gal te bepalen.
Op basis van dit en voorgaand onderzoek kan er een associatie aangetoond worden tussen DON en
neonatale staartnecrose; over ZEA en metabolieten kunnen geen uitspraken gedaan worden. Omdat
dit onderzoek een observationele gevallen-controle studie is, kunnen we dus geen causaal verband
aantonen. Dit komt doordat de vele verschillen in factoren en omstandigheden tussen de bedrijven
onderling niet in beschouwing worden genomen. Om een causaal verband aan te tonen moet idealiter
een experimentele studie worden opgezet. Om het mogelijke effect van DON en ZEA aan te tonen,
zou een onderzoek er als volgt uit kunnen zien: één groep die enkel ZEA toegediend krijgt, één groep
die enkel DON toegediend krijgt, één groep die beide toegediend krijgt en een controlegroep. De
verdere omstandigheden dienen dan voor alle groepen gelijk te zijn. De biggen zouden bij het
vertonen van de symptomen plasma afgenomen kunnen worden en daarna geëuthanaseerd om gal te
verkrijgen. Al zijn dergelijke experimenten ethisch vaak moeilijk te verantwoorden.
We kunnen besluiten dat de problematiek van neonatale staartnecrose multifactorieel is. De resultaten
van deze masterproef tonen aan dat er een associatie is tussen DON in het zeugenvoeder en de
problematiek van staartnecrose bij de biggen. Bovendien is aangetoond dat DON-glucuronide in
plasma een meer geschikte biomerker voor blootstelling aan DON is, t.o.v. DON zelf. Deze fase II
metaboliet kan namelijk in hogere gehaltes teruggevonden worden dan DON, waardoor de kans op
het aantonen van een DON blootstelling bij het levende dier groter is. Dit is zeer nuttig in de
diagnostiek van DON mycotoxicose bij het varken. Tot slot is aangetoond dat de meest frequent
gedetecteerde metaboliet van ZEA bij zowel zeugen als biggen, zearalenol-glucuronide is. Deze fase
II metaboliet in het plasma zou bijgevolg ook een geschikte biomerker voor blootstelling aan ZEA
kunnen zijn.

33
Literatuur
Alexopoulos C., Kritas S.K., Kyriakis C.S., Tzika E., Kyriakis S.C. (2005). Sow performance in
an endemically porcine reproductive and respiratory syndrome (PRRS)-infected farm after
sow vaccination with an attenuated PRRS vaccine. Veterinary Microbiology 111, 151-157.
Bennett J.W., Klich M. (2003). Mycotoxins. Clinical Microbiology Reviews 16, 497-516.
Bernaerdt E. (2015). Vitaliteit van pasgeboren biggen: beïnvloedende factoren, bepaling en
belang. Masterproef Faculteit Diergeneeskunde, Gent, p. 3-6.
Berthiller F., Dall'Asta C., Schuhmacher R., Lemmens M., Adam G., Krska R. (2005). Masked
mycotoxins: Determination of a deoxynivalenol glucoside in artificially and naturally
contaminated wheat by liquid chromatography-tandem mass spectrometry. Journal of
Agricultural and Food Chemistry 4, 3421-3425.
Berthiller F., Schuhmacher R., Adam G., Krska R. (2009). Formation, determination and
significance of masked and other conjugated mycotoxins. Analytical and Bioanalytical
Chemistry 395, 1243-1252.
Biehl M.L., Prelusky D.B., Koritz G.D., Hartin K.E., Buck W.B., Trenholm H.L. (1993). Biliary
excretion and enterohepatic cycling of zearalenone in immature pigs. Toxicology and Applied
Pharmacology 121, 152-159.
Bird I.M. (1989). High performance liquid chromatography - principles and clinical applications.
British Medical Journal 299, 783 -787.
Bravin F., Duca R., C., Balaguer P., Delaforge M. (2009). In vitro cytochrome p450 formation
of a mono-hydroxylated metabolite of zearalenone exhibiting estrogenic activities: possible
occurrence of this metabolite in vivo. International Journal of Molecular sciences 10, 1824-
1837.
Brezina U., Rempe I., Kersten S., Valenta H., Humpf H.U., Dänicke S. (2016). Determination
of zearalenone, deoxynivalenol and metabolites in bile of piglets fed diets with graded levels
of Fusarium toxin contaminated maize. World Mycotoxin Journal 9, 179-193.
Broekaert N. Disposition of 3- and 15-acetyl-deoxynivalenol and deoxynivalenol-3-β-D-
glucoside in broiler chickens and pigs. PhD thesis Faculteit Diergeneeskunde, Gent, p. 50-55.
Calabrese E.J. (1991). Principles of animal extrapolation. Lewis publishers, Chelsea, United
States of America, p. 237.
CAST. (2003). Mycotoxins: Risks in plant, animal, and human systems. Ames IA, Council for
Agricultural Science and Technology, p. 46-66.
Corley R.A., Swanson S.P., Gullo G.J., Johnson L., Beasley V.R., Buck W.B. (1986).
Disposition of T-2 toxin, a trichothecene mycotoxin, in intravascularly dosed swine. Journal of
Agricultural and Food Chemistry 34, 868-875.
Dacasto M., Rolando P., Nachtmann C., Ceppa L., Nebbia C. (1995). Zearalenone
mycotoxicosis in piglets suckling sows fed contaminated grain. Veterinary Human Toxicology
37, 359-361.

34
Dänicke S., Brüssow K.P., Goyarts T., Valenta H., Ueberschär K.H., Tiemann U. (2007). On
the transfer of the fusarium toxins deoxynivalenol (DON) and zearalenone (ZON) from the sow
to the full-term piglet during the last third of gestation. Food and Chemical Toxicology 45,
1565-1574.
Dänicke S., Swiech E., Buraczewska L., Ueberschär K.H. (2005). Kinetics and metabolism of
zearalenone in young female pigs. Journal of Animal Physiology and Animal Nutrition 89, 268-
276.
Dänicke S., Valenta H., Döll S. (2004). On the toxicokinetics and the metabolism of
deoxynivalenol (DON) in the pig. Archives of Animal Nutrition 56, 169-180.
De Boevre M., Di Mavungu J.D., Maene P., Audenaert K., Deforce D., Haesaert G., Eeckhout
M., Callebaut A., Berthiller F., Van Peteghem C., De Saeger S. (2012). Development and
validation of an LC-MS/MS method for the simultaneous determination of deoxynivalenol,
zearalenone, T-2-toxin and some masked metabolites in different cereals and cereal-derived
food. Food Additives and Contaminants: Part A, Chemistry, analysis, control, exposure & risk
assessment 29, 819-835.
Devreese M., Antonissen G., Broekaert N., De Mil T., De Baere S., Vanhaecke L., De Backer
P., Croubels S. (2015) Toxicokinetic study and oral bioavailability of DON in turkey poults, and
comparative biotransformation between broilers and turkeys. World Mycotoxin Journal 8, 533–
539.
Devreese M., De Backer P., Croubels S. (2013). Different methods to counteract mycotoxin
production and its impact on animal health. Vlaams Diergeneeskundig Tijdschrift 82, 181-190.
Diekman M.A., Green M.L. (1992). Mycotoxins and reproduction in domestic livestock. Journal
of Animal Science 70, 1615-1627.
D'Mello J.P.F., Macdonald A.M.C. (1997). Mycotoxins. Animal Feed Science Technology 69,
155–166.
Edwards S.A. (2002). Perinatal mortality in the pig: environmental or physiological solutions?
Livestock Production Science 78, 3-13.
Eriksen G.S., Pettersson H., Johnsen K,. Lindberg J.E. (2002). Transformation of
trichothecenes in ileal digesta and faeces from pigs. Arch Tierernahr Journal 55, 263-274.
Eriksen G.S., Pettersson H., Lindberg J.E. (2003). Absorption, metabolism and excretion of 3-
acetyl DON in pigs. Arch Tierernahr journal 57, 335-345.
Eriksen G.S., Pettersson H. (2003). Lack of de-epoxidation of type B trichothecenes in
incubates with human faeces. Food Additives & Contaminants 6, 579-582.
Fink-Gremmels J. (1991). Mycotoxins: their implications for human and animal health. The
Veterinary Quarterly 21, 115-120.
Fink-Gremmels J., Malekinejad H. (2007). Clinical effects and biochemical mechanisms
associated with exposure to the mycoestrogen zearalenone. Animal Feed Science and
Technology 137, 326–341.
Fitzpatrick D.W., Picken C.A., Murphy L.C., Burm M.M. (1989). Measurement of the relative
binding affinity of zearalenone, alpha-zearalenol and beta-zearalenol for uterine and oviduct

35
estrogen receptors in swine, rats and chickens: an indicator of estrogenic potencies.
Comparative Biochemistry and Physiology 94, 691-694.
Frenich A.G., Romero-González R., Aguilera-Luiz M. (2014). Comprehensive analysis of
toxics (pesticides, veterinary drugs and mycotoxins) in food by UHPLC-MS. Trends in
Analytical Chemistry 63, 158-169.
Friend D.W., Trenholm H.L., Hartin K.E., Prelusky D.B., Thompson B.K. (1986). Effects of
feeding deoxynivalenol (DON)-contaminated wheat diets to pregnant and lactating gilts and on
their progeny. Canadian Journal of Animal Science 66, 229-236.
Glenn A.E. (2007). Mycotoxigenic Fusarium species in animal feed. Animal Feed Science and
Technology 137, 213–240.
Goyarts T., Dänicke S. (2006). Bioavailability of the Fusarium toxin deoxynivalenol (DON)
from naturally contaminated wheat for the pig. Toxicology Letters 163, 171–182.
Goyarts T., Dänicke S., Valenta H., Ueberschär K.H. (2007). Carry-over of Fusarium toxins
(deoxynivalenol and zearalenone) from naturally contaminated wheat to pigs. Food Additives
and Contaminants: Part A 24, 369-380.
Haghighi Poodeh S., Salonurmi T., Nagy I., Koivunen P., Vuoristo J., Räsänen J., Sormunen
R., Vainio S., Savolainen J. (2012). Alcohol-induced premature permeability in mouse
placenta-yolk sac barriers in vivo. Placenta 33, 866-873.
Herpin P., Damon M., Le Dividich J. (2002). Development of thermoregulation and neonatal
survival in pigs. Livestock Production Science 78, 25-45.
Holladay S.D., Blaylock B.L., Comment C.E., Heindel J.J., Luster M.I. (1993). Fetal thymic
atrophy after exposure to T-2 toxin: selectivity for lymphoid progenitor cells. Toxicology and
Applied Pharmacology 121, 8-14.
Hoorens J., Debuyckere M., De Moor A., Maton A., Pensaert M., Vanderplassche M.,
Vanschoubroek F. (1973). Huisvesting, voeding, ziekten van het varken. E. story-scientia
P.V.B.A., België, Gent, p. 495-506.
Hussein H.S., Brasel J. M. (2001). Toxicity, metabolism, and impact of mycotoxins on humans
and animals. Toxicology 167, 101–134.
Jajić I., Jurić V,, Glamočić D., Krstović S. (2010). Occurence of deoxynivalenol and
zearalenone in maize. Contemporary Agriculture 59, 227-233.
Kalantari, H., Moosavi, M. (2010). Review on T-2 Toxin. Jundishapur Journal of Natural
Pharmaceutical Products 5, 26–38.
Kollarczik B., Gareis M., Hanelt M. (1994). In vitro transformation of the Fusarium mycotoxins
deoxynivalenol and zearalenone by the normal gut microflora of pigs. Natural Toxins 2, 105-
110.
Korn A.K., Gross M., Usleber E., Thom N., Köhler K., Erhardt G. (2014). Dietary ergot
alkaloids as a possible cause of tail necrosis in rabbits. Mycotoxin Reasearch 30, 241-2450.
Lacina O., Zachariasova M., Urbanova J., Vaclavikova M., Cajka T. Hajslova J. (2012). Critical
assessment of extraction methods for the simultaneous determination of pesticide residues

36
and mycotoxins in fruits, cereals, spices and oil seeds employing ultra-high performance liquid
chromatography-tandem mass spectrometry. Journal of Chromatography A. 1262, 8-18.
Landschoot S., Waegeman W., Audenaert K., Vandepitte J., Baetens J.M., De Baets B.,
Haesaert G. (2012). An empirical analysis of explanatory variables affecting fusarium head
blight infection and deoxynivalenol content in wheat. Journal of Plant Pathology 94, 135-147.
Lattanzio V.M., Visconti A., Haidukowski M., Pascale M. (2012). Identification and
characterization of new Fusarium masked mycotoxins, T2 and HT2 glycosyl derivatives, in
naturally contaminated wheat and oats by liquid chromatography-high-resolution mass
spectrometry. Journal of Mass Spectrometry 47, 466-475.
Li Y., Wang Z., Beier R.C., Shen J., De Smet D., De Saeger S., Zhang S. (2011). T-2 Toxin, a
trichothecene mycotoxin: Review of toxicity, metabolism, and analytical methods. Journal of
Agricultural and Food Chemistry 59, 3441-3453.
López T.A., Campero C.M., Chayer R., de Hoyos M. (1997). Ergotism and photosensitization
in swine produced by the combined ingestion of Claviceps purpurea sclerotia and Ammi majus
seeds. Journal of Veterinary Diagnostic Investigation 9, 68-71
Malekinejad H., Maas-Bakker R., Fink-Gremmels J. (2006). Species differences in the hepatic
biotransformation of zearalenone. The Veterinary Journal 172, 96–102.
Malekinejad H., Maas-Bakker R.F., Fink-Gremmels J. (2005). Bioactivation of zearalenone by
porcine hepatic biotransformation. Veterinary Research 36, 799-810.
Maresca M. (2013). From the gut to the brain: Journey and pathophysiological effects of the
food-associated Trichothecene mycotoxin deoxynivalenol. Toxins 5, 784-820.
McKean C., Tang L., Tang M., Billam M., Wang Z., Theodorakis C., W., Kendall R., J., Wang
J., S. (2006). Comparative acute and combinative toxicity of aflatoxin B1 and fumonisin B1 in
animals and human cells. Food and Chemical Toxicology 44, 866-876.
Mol H.G.J., Plaza Bolanos P., Zomer P., de Rijk T.C., Stolker A.A.M., Mulder P.P.J. (2008).
Toward a generic extraction method for simultaneous determination of pesticides, mycotoxins,
plant toxins and veterinary drugs in feed and food matrixes. Analytical Chemistry 80 9450-
9459.
Mul M.F., Binnendijk G.P., Kan C.A. (2006). Probleeminventarisatie mycotoxines in de
varkenshouderij: Rapport 02. Wageningen UR Aniomal Science Group, Lelystad, p. 1-16.
Nathanail A.V., Syvähuoko J., Malachová A., Jestoi M., Varga L., Michlmayr H., Adam G.,
Sieviläinen E., Berthiller F., Peltonen K. (2015). Simultaneous determination of major type A
and B trichothecenes, zearalenone and certain modified metabolites in Finnish cereal grains
with a novel liquid chromatography-tandem mass spectrometric method. Analytical and
Bioanalytical Chemistry 407, 4745-4755.
Niessen, W.M.A., Tinke A.P. (1995). Liquid chromatography mass spectrometry - general
principles and instrumentation. Journal of Chromatography A 703, 37-57.
Noblet J., Fortune H., Shi X. S., Dubois S. (1994). Prediction of net energy value of feeds for
growing pigs. Journal of Animal Science 72, 344-354.

37
Noblet J., van Milgen J. (2004). Energy value of pig feeds: Effect of pig body weight and
energy evaluation system. Journal of Animal Science 82, 229-238.
Olsen M., Pettersson H., Sandholm K., Viscontit A., Kiessling K.H. (1987). Metabolism of
zearalenone by sow intestinal mucosa in vitro. Food and Chemical Toxicology 25, 681-683.
Olsen M., Malmlöf K., Pettersson H., Sandholm K., Kiessling K.H. (1985). Plasma and urinary
levels of zearalenone and α-zearalenol in a prepubertal gilt fed zearalenone. Acta
Pharmacologica et Toxicologica 56, 239–243.
Packet S (2014). Neonatale staartnecrose bij biggen. casuïstiek in het kader van de
Masterproef, Gent, p. 3-31.
Palyusik M., Harrach B., Mirocha C.J., Pathre S.V. (1980). Transmission of zearalenone and
zearalenol into porcine milk. Acta Veterinaria Academiae Scientiarium Hungaricae 28, 217-
222.
Pestka J.J. (2007). Deoxynivalenol: Toxicity, mechanisms and animal health risks. Animal
Feed Science and Technology 137, 283–298.
Prelusky D.B., Hartin K.E., Trenholm H.L., Miller J.D. (1988). Pharmacokinetic fate of 14C-
labeled deoxynivalenol in swine. Fundamental and Applied Toxicology 10, 276-86.
Rathbone M.J., Gurny R. (2000). Controlled release veterinary drug delivery: biological and
pharmaceutical considerations, 1st Edition. Elsevier Science, Amsterdam, p 13.
Razzazi-Fazeli E., Böhm J., Jarukamjorn K., Zentek J. (2003). Simultaneous determination of
major B-trichothecenes and the de-epoxy-metabolite of deoxynivalenol in pig urine and maize
using high-performance liquid chromatography-mass spectrometry. Journal of
chromatography B, Analytical technologies 25, 21-33.
Riviere J.E. (2011). Comparative pharmacokinetics: principles, techniques, and applications,
Second Edition. John Wiley & Sons, North Carolina, Raleigh, p. 13-140.
Robison T.S., Mirocha C.J., Kurtz H.J., Behrens J.C., Chi M.S., Weaver G.A., Nystrom S.D.
(1978). Transmission of T- 2 toxin into bovine and porcine milk. Scientific Journal Series of the
Minnesota Agricultural Experiment Station 10, 354.
Rubert J., Dzuman Z., Vaclavikova M., Zachariasova M., Soler C., Hajslova J. (2012).
Analysis of mycotoxins in barley using ultra high liquid chromatography high resolution mass
spectrometry: comparison of efficiency and efficacy of different extraction procedures. Talanta
99, 712-719.
Russell R., Paterson M., Nelson L. (2009). How will climate change affect mycotoxins in food?
Food Research International 43, 1902–1914.
Senger P.L. (2003). Pathways to pregnancy and parturition. Second edition. Current
Conceptions, Inc. Washington, United States of America, p. 304-325.
Shier W.T., Shier A.C., Xie W., Mirocha C.J. (2001). Structure-activity relationships for human
estrogenic activity in zearalenone mycotoxins. Toxicon 39, 1435-8.
Solà-Oriol D., Roura E., Torrallardona D. (2014). Feed preference in pigs: Relationship
between cereal preference and nutrient composition and digestibility. Journal of Animal
Science 92, 220-228.

38
Streit E., Naehrer K., Rodrigues I., Schatzmayr G. (2013). Mycotoxin occurrence in feed and
feed raw materials worldwide: long-term analysis with special focus on Europe and Asia.
Journal of the Science of Food and Agriculture 93, 2692-2899.
Sun L.Y., Li Q., Meng F.G., Fu Y., Zhao Z.J., Wang L.H. (2012) T-2 Toxin Contamination in
grains and selenium concentration in drinking water and grains in Kashin-Beck Disease
endemic areas of Qinghai Province. Biological Trace Element Research 150, 371–375.
Sundstøl Eriksen G., Pettersson H. (2003). Lack of de-epoxidation of type B trichothecenes in
incubates with human faeces. Food Additives and Contaminantes 20, 579-582.
Sundstøl Eriksen G., Pettersson H., Lundh T. (2004). Comparative cytotoxicity of
deoxynivalenol, nivalenol, their acetylated derivatives and de-epoxy metabolites. Food and
Chemical Toxicology 42, 619-624.
Swanson S.P., Rood H.D., Behrens J.C., Sanders P.E. (1987). Preparation and
characterization of the deepoxy trichothecenes: deepoxy HT-2, deepoxy T-2 triol, deepoxy T-2
tetraol, deepoxy 15-monoacetoxyscirpenol, and deepoxy scirpentriol. Applied and
Environmental Microbiology 53, 2821-2826.
Tiemann U., Dänicke S. (2007). In vivo and in vitro effects of the mycotoxins zearalenone and
deoxynivalenol on different non-reproductive and reproductive organs in female pigs: a review.
Food Additives and Contaminantes 24, 306-314.
Turner N.W., Subrahmanyam S., Piletsky S.A. (2009). Analytical methods for determination of
mycotoxins: a review. Analytica Chimica Acta 632,168-180.
Turner P.C., Hopton R.P., White K.L.M., Fisher J., Cade J.E., Wild C.P. (2011). Assessment
of deoxynivalenol metabolite profiles in UK adults. Food and Chemical Toxicology 49, 132-
135.
Turner P.C., Hopton R.P., Lecluse Y., White K.L.M., Fisher J., Lebailly P. (2010).
Determinants of urinary deoxynivalenol and de-epoxy deoxynivalenol in male farmers from
Normandy, France. Journal of Agricultural and Food Chemistry 58, 5206-5212.
Turner, P.C. (2010). Deoxynivalenol and nivalenol occurrence and exposure assessment.
World Mycotoxin Journal 3, 315-321.
Van Asselt E. D., Azambuja W., Moretti A., Kastelein P., De Rijk T. C., Stratakou I., Van Der
Fels-Klerx H. J. (2012). A Dutch field survey on fungal infection and mycotoxin concentrations
in maize. Food additives & contaminants Part A, Chemistry, analysis, control, exposure & risk
assessment 29, 1556-1565.
Van Neste K. (2015). Plasma als matrix voor de diagnose van Fusarium mycotoxicosis bij
varken. Masterproef Faculteit Diergeneeskunde, Gent, p. 18-44.
Warth B., Sulyok M., Fruhmann P., Berthiller F., Schuhmacher R., Hametner C., Adam
G., Fröhlich J., Krska R. (2012). Assessment of human deoxynivalenol exposure using an LC-
MS/MS based biomarker method. Toxicology Letters 20, 85-90.
Wu Q., Dohnal V., Huang L., Kuca K. (2010). Metabolic pathways of trichothecenes. Drug
Metabolism Reviews 42, 250-267.

39
Wu X., Murphy P., Cunnick J., Hendrich S. (2007). Synthesis and characterization of
deoxynivalenol glucuronide: its comparative immunotoxicity with deoxynivalenol. Food and
Chemical Toxicology 45, 1846-1855.
Wyllie T.D., Morehouse L.G. (1978). Mycotox fungi, mycotoxins, mycotoxicoses. Marcel
Dekker inc., USA, New York, p. 100-140.
Zaleski H.M., Hacker R.R. (1993). Variables related to the progress of parturition and
probability of stillbirth in swine. Canadian Veterinary Journal 34, 109-113.
Zöllner P., Jodlbauer J., Kleinova M., Kahlbacher H., Kuhn T. (2002). Concentration levels of
zearalenone and its metabolites in urine, muscle tissue, and liver samples of pigs fed with
mycotoxin-contaminated oats. Journal of Agricultural and Food Chemistry 24, 2494-2501.
Wetten
- Aanbeveling van de Commissie van 17 augustus 2006 betreffende de aanwezigheid van
deoxynivalenol, zearalenone, ochratoxine A, T-2- en HT-2-toxine en fumonisinen in producten
die bedoeld zijn voor het voederen van dieren (2006/576/EG).
- De aanbeveling van de Commissie van 27 maart 2013 betreffende de aanwezigheid van T-2-
en HT-2-toxine in granen en graanproducten.
- De richtlijn 2002/32/EG van het Europees Parlement en de Raad van 7 mei 2002 inzake
ongewenste stoffen in diervoeding.
- Europese richtlijn tot vaststelling van bemonsteringswijzen en analysemethoden voor de
officiële controle op de maximumgehalten aan bepaalde verontreinigingen in levensmiddelen
(98/53/EG).
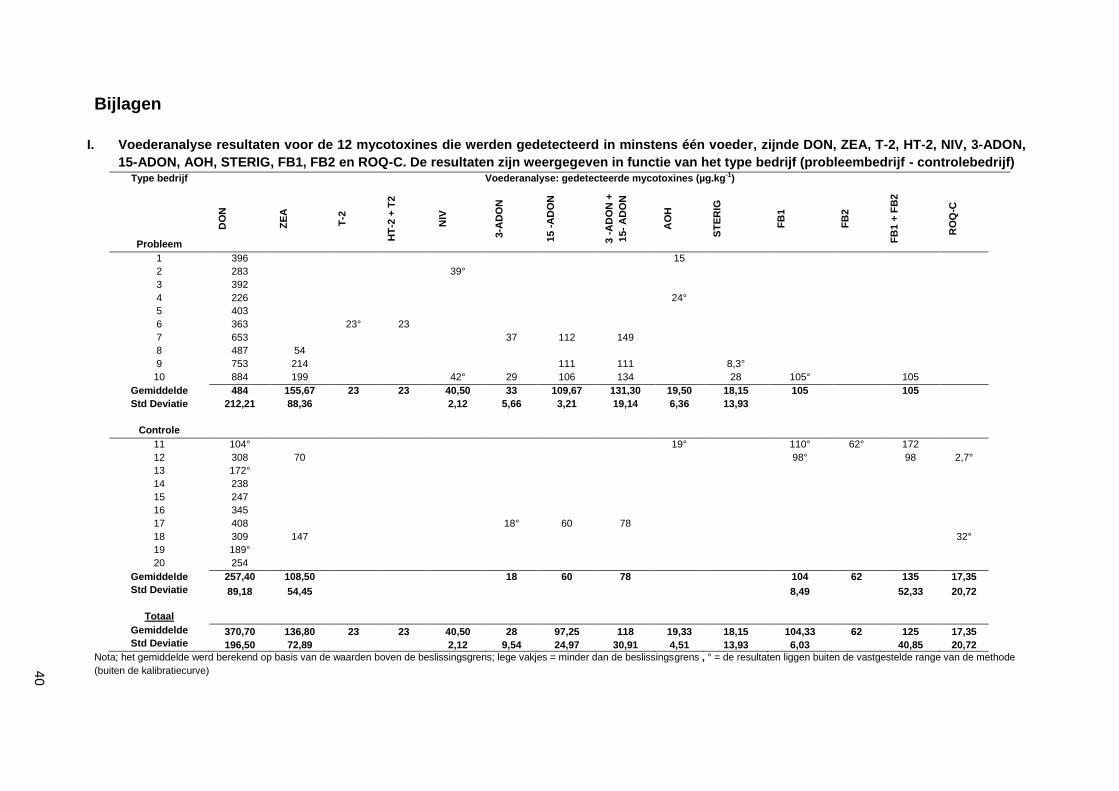
Bijlagen
I. Voederanalyse resultaten voor de 12 mycotoxines die werden gedetecteerd in minstens één voeder, zijnde DON, ZEA, T-2, HT-2, NIV, 3-ADON,
15-ADON, AOH, STERIG, FB1, FB2 en ROQ-C. De resultaten zijn weergegeven in functie van het type bedrijf (probleembedrijf - controlebedrijf) Type bedrijf Voederanalyse: gedetecteerde mycotoxines (µg.kg-1)
Probleem
DO
N
ZE
A
T-2
HT
-2 +
T2
NIV
3-A
DO
N
15 -
AD
ON
3 -
AD
ON
+
15-
AD
ON
AO
H
ST
ER
IG
FB
1
FB
2
FB
1 +
FB
2
RO
Q-C
1 396 15
2 283 39°
3 392
4 226 24°
5 403
6 363 23° 23
7 653 37 112 149
8 487 54
9 753 214 111 111 8,3°
10 884 199 42° 29 106 134 28 105° 105
Gemiddelde 484 155,67 23 23 40,50 33 109,67 131,30 19,50 18,15 105 105
Std Deviatie 212,21 88,36 2,12 5,66 3,21 19,14 6,36 13,93
Controle
11 104° 19° 110° 62° 172
12 308 70 98° 98 2,7°
13 172°
14 238
15 247
16 345
17 408 18° 60 78
18 309 147 32°
19 189°
20 254
Gemiddelde 257,40 108,50 18 60 78 104 62 135 17,35
Std Deviatie 89,18 54,45 8,49 52,33 20,72
Totaal
Gemiddelde 370,70 136,80 23 23 40,50 28 97,25 118 19,33 18,15 104,33 62 125 17,35 Std Deviatie 196,50 72,89 2,12 9,54 24,97 30,91 4,51 13,93 6,03 40,85 20,72
Nota; het gemiddelde werd berekend op basis van de waarden boven de beslissingsgrens; lege vakjes = minder dan de beslissingsgrens , ° = de resultaten liggen buiten de vastgestelde range van de methode
(buiten de kalibratiecurve)
40
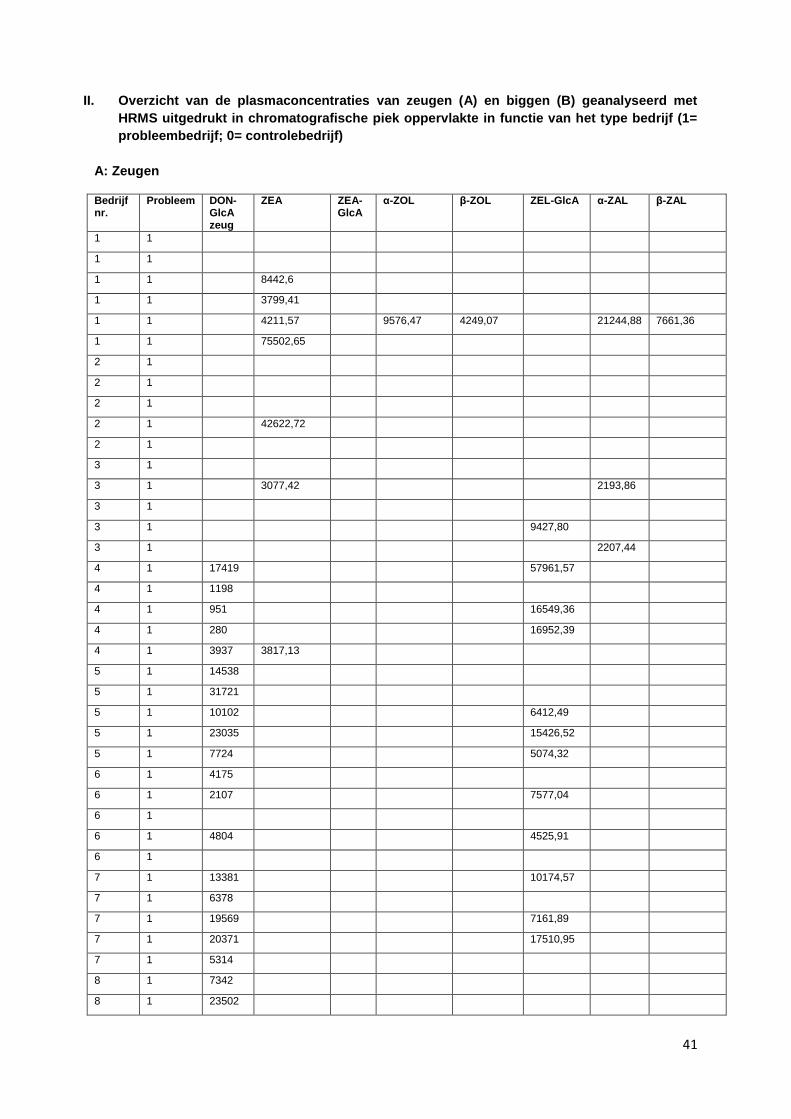
41
II. Overzicht van de plasmaconcentraties van zeugen (A) en biggen (B) geanalyseerd met
HRMS uitgedrukt in chromatografische piek oppervlakte in functie van het type bedrijf (1=
probleembedrijf; 0= controlebedrijf)
A: Zeugen
Bedrijf nr.
Probleem DON-GlcA zeug
ZEA ZEA-GlcA
α-ZOL β-ZOL ZEL-GlcA α-ZAL β-ZAL
1 1
1 1
1 1 8442,6
1 1 3799,41
1 1 4211,57 9576,47 4249,07 21244,88 7661,36
1 1 75502,65
2 1
2 1
2 1
2 1 42622,72
2 1
3 1
3 1 3077,42 2193,86
3 1
3 1 9427,80
3 1 2207,44
4 1 17419 57961,57
4 1 1198
4 1 951 16549,36
4 1 280 16952,39
4 1 3937 3817,13
5 1 14538
5 1 31721
5 1 10102 6412,49
5 1 23035 15426,52
5 1 7724 5074,32
6 1 4175
6 1 2107 7577,04
6 1
6 1 4804 4525,91
6 1
7 1 13381 10174,57
7 1 6378
7 1 19569 7161,89
7 1 20371 17510,95
7 1 5314
8 1 7342
8 1 23502
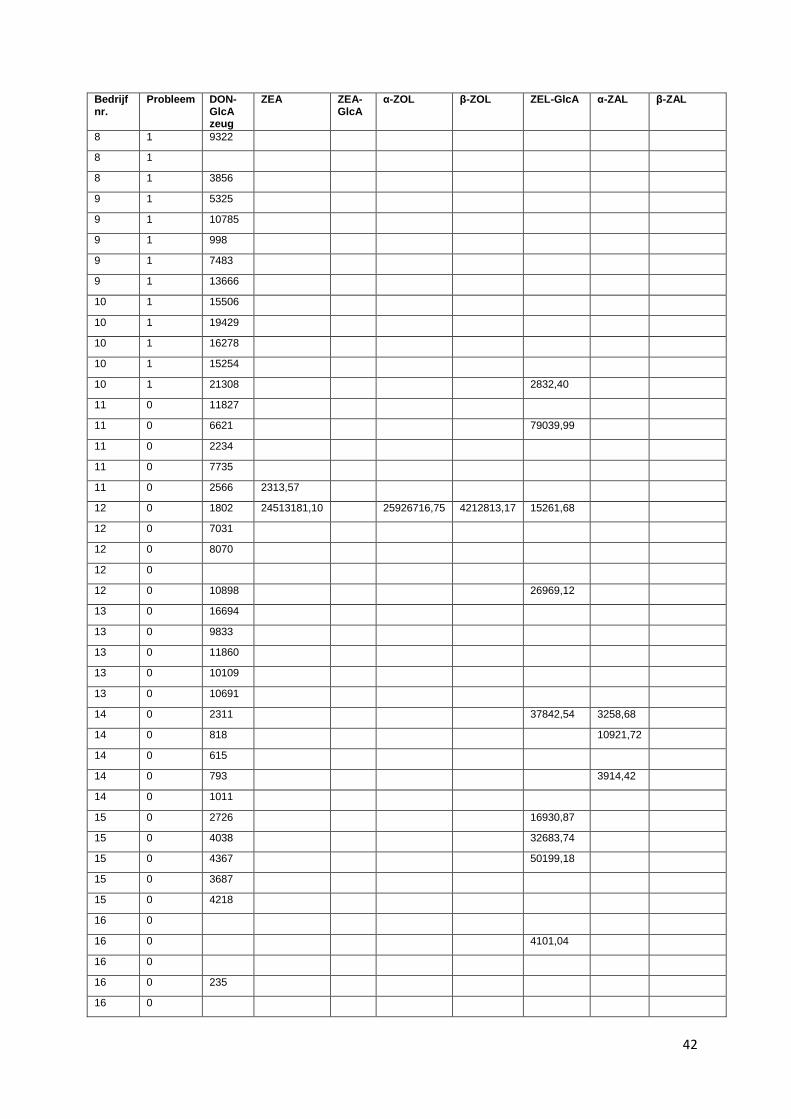
42
Bedrijf nr.
Probleem DON-GlcA zeug
ZEA ZEA-GlcA
α-ZOL β-ZOL ZEL-GlcA α-ZAL β-ZAL
8 1 9322
8 1
8 1 3856
9 1 5325
9 1 10785
9 1 998
9 1 7483
9 1 13666
10 1 15506
10 1 19429
10 1 16278
10 1 15254
10 1 21308 2832,40
11 0 11827
11 0 6621 79039,99
11 0 2234
11 0 7735
11 0 2566 2313,57
12 0 1802 24513181,10 25926716,75 4212813,17 15261,68
12 0 7031
12 0 8070
12 0
12 0 10898 26969,12
13 0 16694
13 0 9833
13 0 11860
13 0 10109
13 0 10691
14 0 2311 37842,54 3258,68
14 0 818 10921,72
14 0 615
14 0 793 3914,42
14 0 1011
15 0 2726 16930,87
15 0 4038 32683,74
15 0 4367 50199,18
15 0 3687
15 0 4218
16 0
16 0 4101,04
16 0
16 0 235
16 0
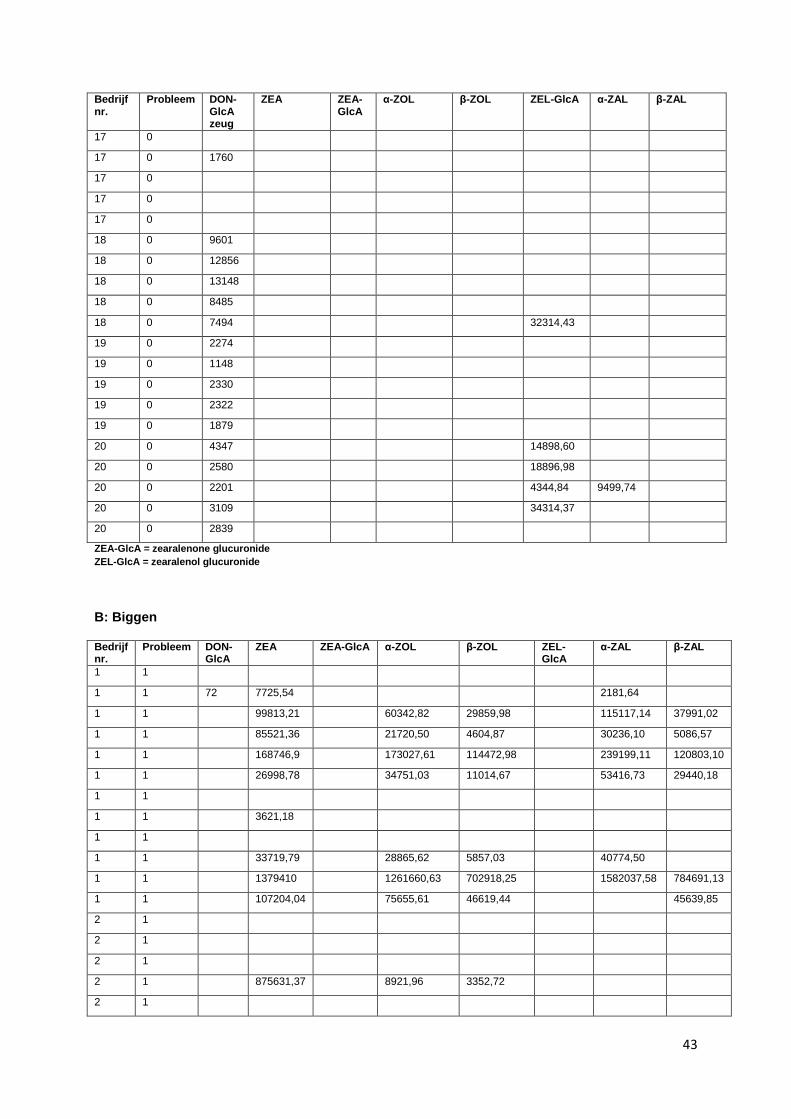
43
Bedrijf nr.
Probleem DON-GlcA zeug
ZEA ZEA-GlcA
α-ZOL β-ZOL ZEL-GlcA α-ZAL β-ZAL
17 0
17 0 1760
17 0
17 0
17 0
18 0 9601
18 0 12856
18 0 13148
18 0 8485
18 0 7494 32314,43
19 0 2274
19 0 1148
19 0 2330
19 0 2322
19 0 1879
20 0 4347 14898,60
20 0 2580 18896,98
20 0 2201 4344,84 9499,74
20 0 3109 34314,37
20 0 2839
ZEA-GlcA = zearalenone glucuronide
ZEL-GlcA = zearalenol glucuronide
B: Biggen
Bedrijf nr.
Probleem DON-GlcA
ZEA ZEA-GlcA α-ZOL β-ZOL ZEL-GlcA
α-ZAL β-ZAL
1 1
1 1 72 7725,54 2181,64
1 1 99813,21 60342,82 29859,98 115117,14 37991,02
1 1 85521,36 21720,50 4604,87 30236,10 5086,57
1 1 168746,9 173027,61 114472,98 239199,11 120803,10
1 1 26998,78 34751,03 11014,67 53416,73 29440,18
1 1
1 1 3621,18
1 1
1 1 33719,79 28865,62 5857,03 40774,50
1 1 1379410 1261660,63 702918,25 1582037,58 784691,13
1 1 107204,04 75655,61 46619,44 45639,85
2 1
2 1
2 1
2 1 875631,37 8921,96 3352,72
2 1
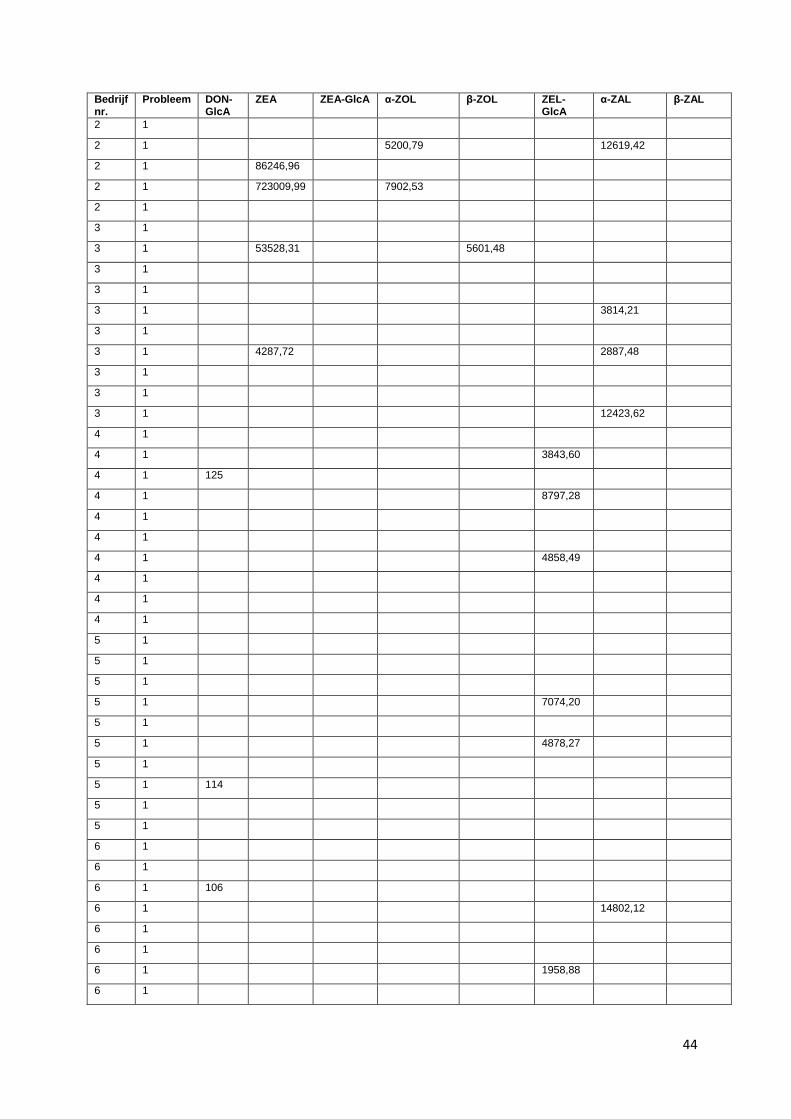
44
Bedrijf nr.
Probleem DON-GlcA
ZEA ZEA-GlcA α-ZOL β-ZOL ZEL-GlcA
α-ZAL β-ZAL
2 1
2 1 5200,79 12619,42
2 1 86246,96
2 1 723009,99 7902,53
2 1
3 1
3 1 53528,31 5601,48
3 1
3 1
3 1 3814,21
3 1
3 1 4287,72 2887,48
3 1
3 1
3 1 12423,62
4 1
4 1 3843,60
4 1 125
4 1 8797,28
4 1
4 1
4 1 4858,49
4 1
4 1
4 1
5 1
5 1
5 1
5 1 7074,20
5 1
5 1 4878,27
5 1
5 1 114
5 1
5 1
6 1
6 1
6 1 106
6 1 14802,12
6 1
6 1
6 1 1958,88
6 1

45
Bedrijf nr.
Probleem DON-GlcA
ZEA ZEA-GlcA α-ZOL β-ZOL ZEL-GlcA
α-ZAL β-ZAL
6 1 4514,74
6 1
7 1 3924,24
7 1 2317,86
7 1
7 1 49053,20
7 1
7 1 33760,40
7 1
7 1
7 1 2473,16
7 1
8 1
8 1
8 1
8 1
8 1
8 1
8 1
8 1
8 1
8 1
9 1
9 1
9 1
9 1
9 1
9 1
9 1
9 1
9 1
9 1
10 1
10 1
10 1
10 1 2408,04
10 1
10 1
10 1
10 1
10 1 4995,20 7934,40
10 1
11 0
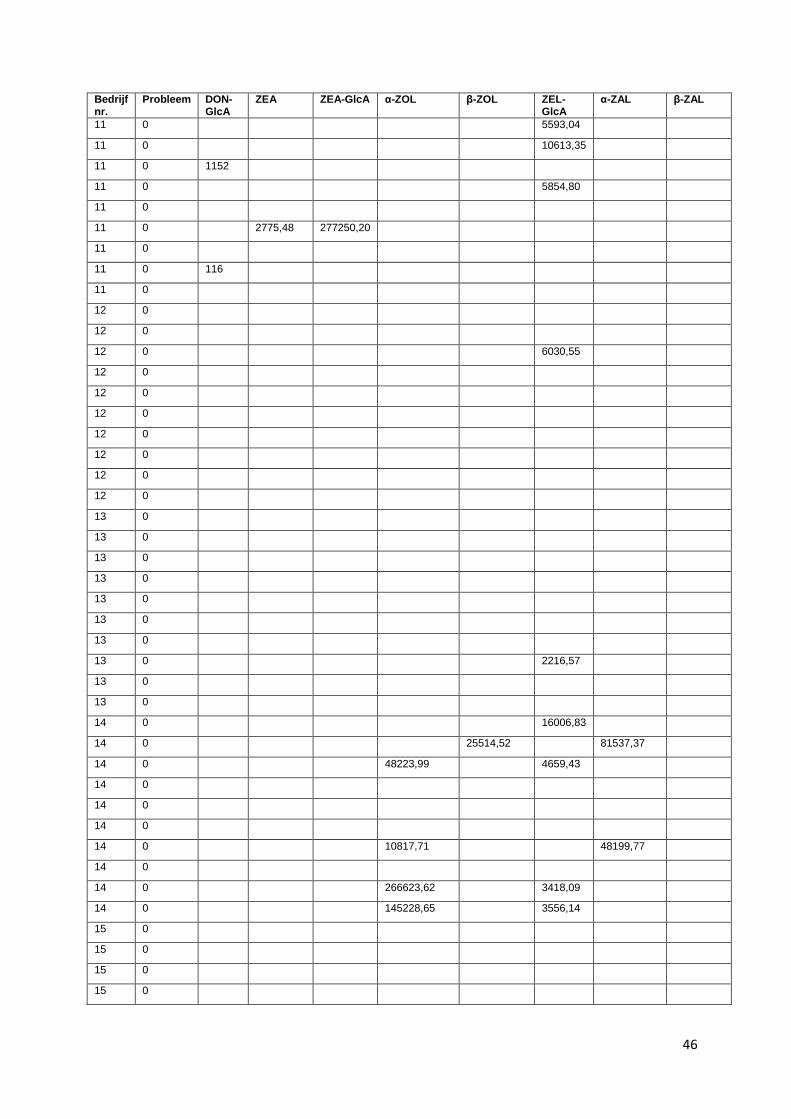
46
Bedrijf nr.
Probleem DON-GlcA
ZEA ZEA-GlcA α-ZOL β-ZOL ZEL-GlcA
α-ZAL β-ZAL
11 0 5593,04
11 0 10613,35
11 0 1152
11 0 5854,80
11 0
11 0 2775,48 277250,20
11 0
11 0 116
11 0
12 0
12 0
12 0 6030,55
12 0
12 0
12 0
12 0
12 0
12 0
12 0
13 0
13 0
13 0
13 0
13 0
13 0
13 0
13 0 2216,57
13 0
13 0
14 0 16006,83
14 0 25514,52 81537,37
14 0 48223,99 4659,43
14 0
14 0
14 0
14 0 10817,71 48199,77
14 0
14 0 266623,62 3418,09
14 0 145228,65 3556,14
15 0
15 0
15 0
15 0

47
Bedrijf nr.
Probleem DON-GlcA
ZEA ZEA-GlcA α-ZOL β-ZOL ZEL-GlcA
α-ZAL β-ZAL
15 0
15 0 10408,16
15 0
15 0
15 0
15 0
16 0
16 0
16 0
16 0
16 0
16 0
16 0
16 0
16 0
16 0
17 0
17 0
17 0
17 0
17 0
17 0 21063,66
17 0
17 0
17 0
17 0
18 0
18 0
18 0
18 0
18 0
18 0
18 0
18 0 2452,08
18 0
18 0
19 0 4506,91 4671,95
19 0
19 0
19 0
19 0
19 0 107
19 0
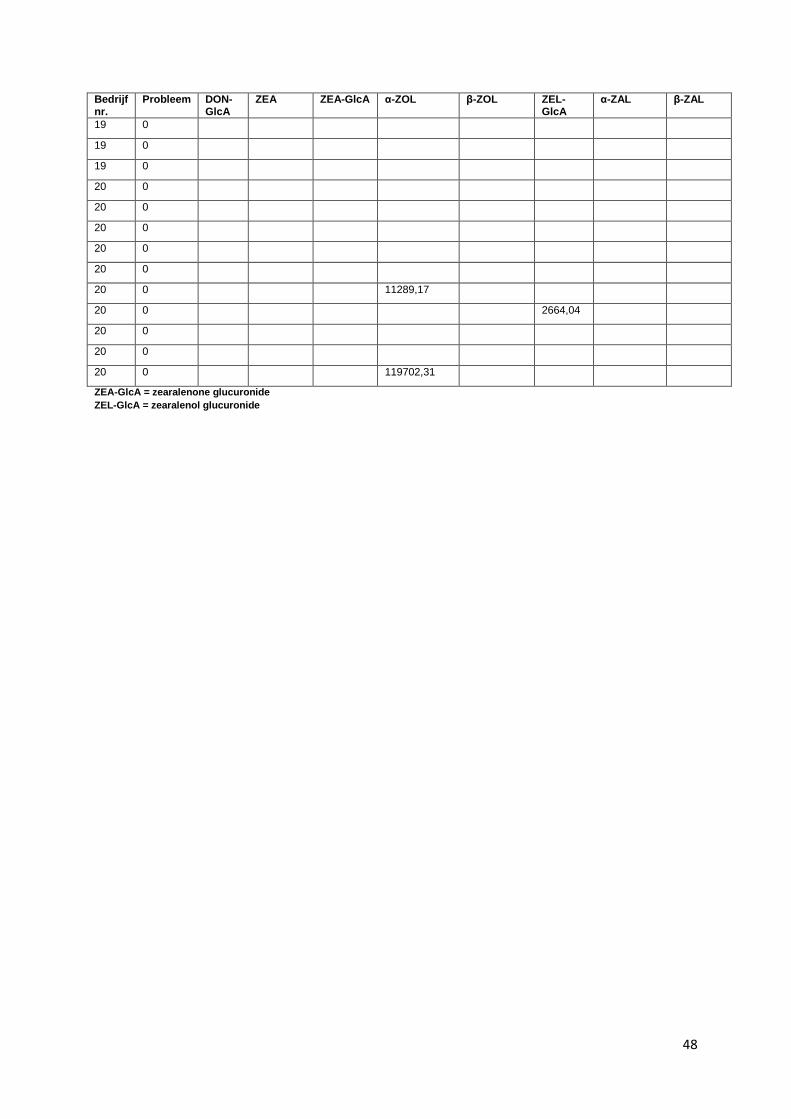
48
Bedrijf nr.
Probleem DON-GlcA
ZEA ZEA-GlcA α-ZOL β-ZOL ZEL-GlcA
α-ZAL β-ZAL
19 0
19 0
19 0
20 0
20 0
20 0
20 0
20 0
20 0 11289,17
20 0 2664,04
20 0
20 0
20 0 119702,31
ZEA-GlcA = zearalenone glucuronide
ZEL-GlcA = zearalenol glucuronide
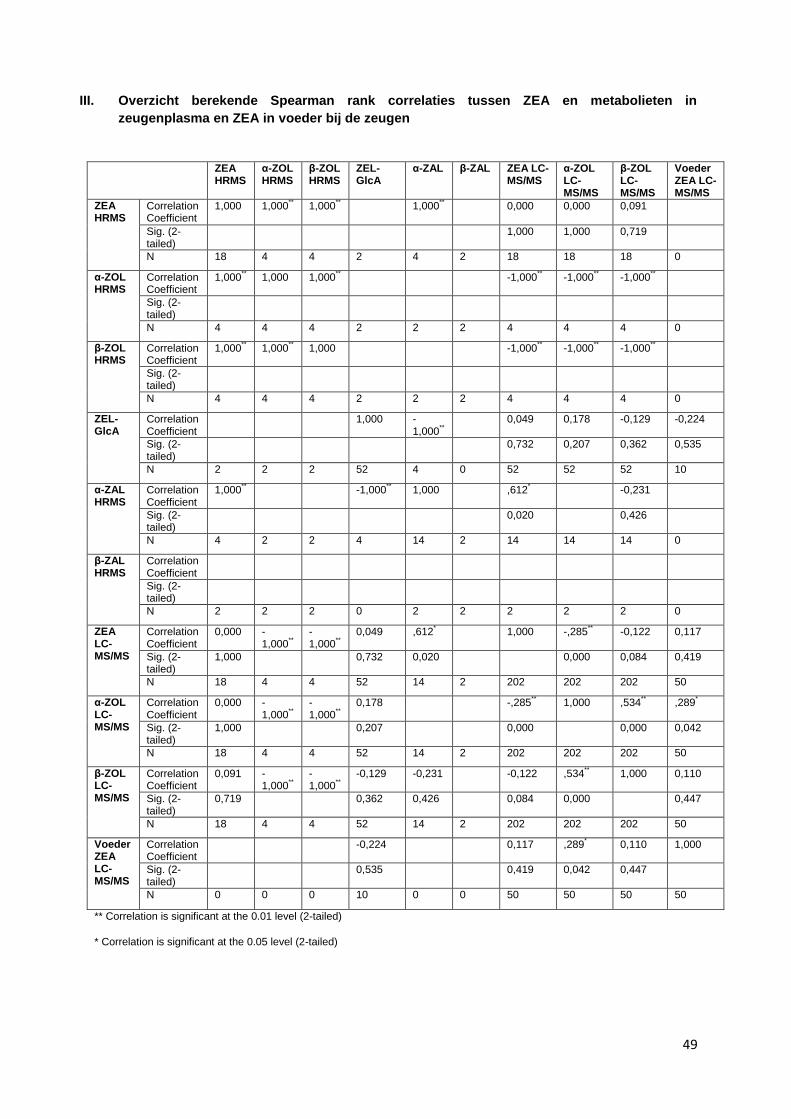
49
III. Overzicht berekende Spearman rank correlaties tussen ZEA en metabolieten in
zeugenplasma en ZEA in voeder bij de zeugen
ZEA HRMS
α-ZOL HRMS
β-ZOL HRMS
ZEL-GlcA
α-ZAL β-ZAL ZEA LC-MS/MS
α-ZOL LC-MS/MS
β-ZOL LC-MS/MS
Voeder ZEA LC-MS/MS
ZEA HRMS
Correlation Coefficient
1,000 1,000** 1,000
** 1,000
** 0,000 0,000 0,091
Sig. (2-tailed)
1,000 1,000 0,719
N 18 4 4 2 4 2 18 18 18 0
α-ZOL HRMS
Correlation Coefficient
1,000** 1,000 1,000
** -1,000
** -1,000
** -1,000
**
Sig. (2-tailed)
N 4 4 4 2 2 2 4 4 4 0
β-ZOL HRMS
Correlation Coefficient
1,000** 1,000
** 1,000 -1,000
** -1,000
** -1,000
**
Sig. (2-tailed)
N 4 4 4 2 2 2 4 4 4 0
ZEL-GlcA
Correlation Coefficient
1,000 -1,000
**
0,049 0,178 -0,129 -0,224
Sig. (2-tailed)
0,732 0,207 0,362 0,535
N 2 2 2 52 4 0 52 52 52 10
α-ZAL HRMS
Correlation Coefficient
1,000** -1,000
** 1,000 ,612
* -0,231
Sig. (2-tailed)
0,020 0,426
N 4 2 2 4 14 2 14 14 14 0
β-ZAL HRMS
Correlation Coefficient
Sig. (2-tailed)
N 2 2 2 0 2 2 2 2 2 0
ZEA LC-MS/MS
Correlation Coefficient
0,000 -1,000
**
-1,000
**
0,049 ,612* 1,000 -,285
** -0,122 0,117
Sig. (2-tailed)
1,000 0,732 0,020 0,000 0,084 0,419
N 18 4 4 52 14 2 202 202 202 50
α-ZOL LC-MS/MS
Correlation Coefficient
0,000 -1,000
**
-1,000
**
0,178 -,285** 1,000 ,534
** ,289
*
Sig. (2-tailed)
1,000 0,207 0,000 0,000 0,042
N 18 4 4 52 14 2 202 202 202 50
β-ZOL LC-MS/MS
Correlation Coefficient
0,091 -1,000
**
-1,000
**
-0,129 -0,231 -0,122 ,534** 1,000 0,110
Sig. (2-tailed)
0,719 0,362 0,426 0,084 0,000 0,447
N 18 4 4 52 14 2 202 202 202 50
Voeder ZEA LC-MS/MS
Correlation Coefficient
-0,224 0,117 ,289* 0,110 1,000
Sig. (2-tailed)
0,535 0,419 0,042 0,447
N 0 0 0 10 0 0 50 50 50 50
** Correlation is significant at the 0.01 level (2-tailed)
* Correlation is significant at the 0.05 level (2-tailed)
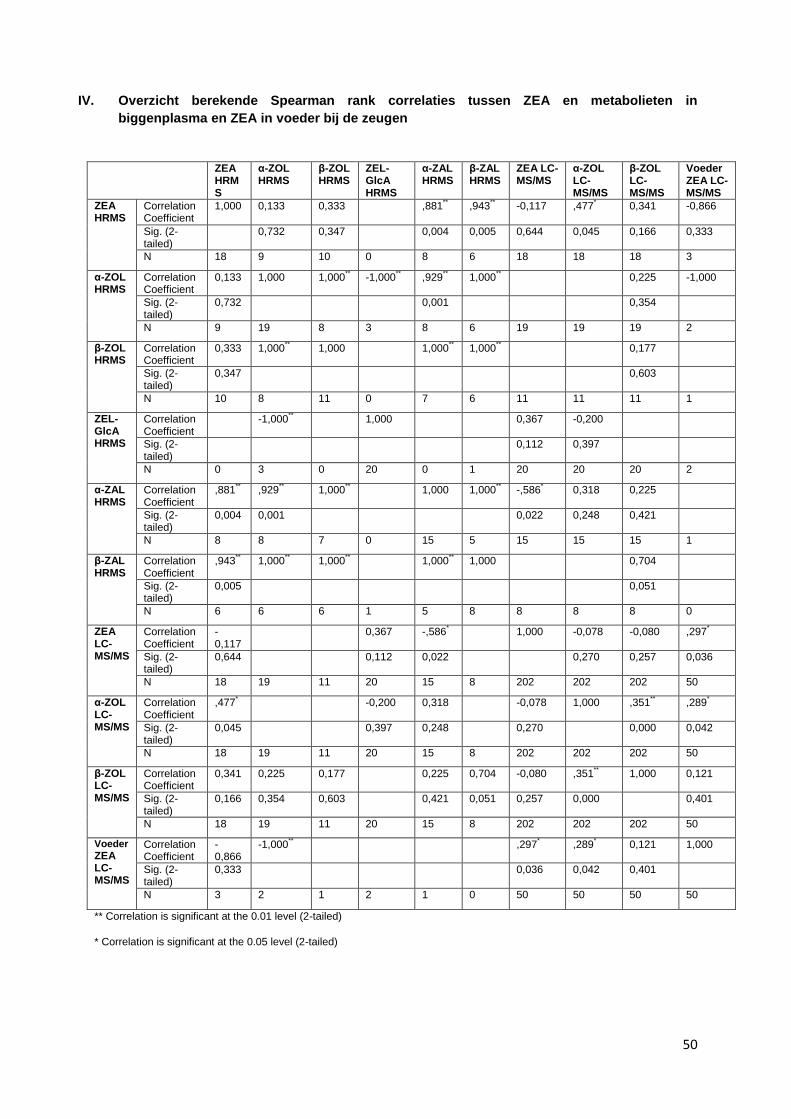
50
IV. Overzicht berekende Spearman rank correlaties tussen ZEA en metabolieten in
biggenplasma en ZEA in voeder bij de zeugen
ZEA HRMS
α-ZOL HRMS
β-ZOL HRMS
ZEL-GlcA HRMS
α-ZAL HRMS
β-ZAL HRMS
ZEA LC-MS/MS
α-ZOL LC-MS/MS
β-ZOL LC-MS/MS
Voeder ZEA LC-MS/MS
ZEA HRMS
Correlation Coefficient
1,000 0,133 0,333 ,881** ,943
** -0,117 ,477
* 0,341 -0,866
Sig. (2-tailed)
0,732 0,347 0,004 0,005 0,644 0,045 0,166 0,333
N 18 9 10 0 8 6 18 18 18 3
α-ZOL HRMS
Correlation Coefficient
0,133 1,000 1,000** -1,000
** ,929
** 1,000
** 0,225 -1,000
Sig. (2-tailed)
0,732 0,001 0,354
N 9 19 8 3 8 6 19 19 19 2
β-ZOL HRMS
Correlation Coefficient
0,333 1,000** 1,000 1,000
** 1,000
** 0,177
Sig. (2-tailed)
0,347 0,603
N 10 8 11 0 7 6 11 11 11 1
ZEL-GlcA HRMS
Correlation Coefficient
-1,000** 1,000 0,367 -0,200
Sig. (2-tailed)
0,112 0,397
N 0 3 0 20 0 1 20 20 20 2
α-ZAL HRMS
Correlation Coefficient
,881** ,929
** 1,000
** 1,000 1,000
** -,586
* 0,318 0,225
Sig. (2-tailed)
0,004 0,001 0,022 0,248 0,421
N 8 8 7 0 15 5 15 15 15 1
β-ZAL HRMS
Correlation Coefficient
,943** 1,000
** 1,000
** 1,000
** 1,000 0,704
Sig. (2-tailed)
0,005 0,051
N 6 6 6 1 5 8 8 8 8 0
ZEA LC-MS/MS
Correlation Coefficient
-0,117
0,367 -,586* 1,000 -0,078 -0,080 ,297
*
Sig. (2-tailed)
0,644 0,112 0,022 0,270 0,257 0,036
N 18 19 11 20 15 8 202 202 202 50
α-ZOL LC-MS/MS
Correlation Coefficient
,477* -0,200 0,318 -0,078 1,000 ,351
** ,289
*
Sig. (2-tailed)
0,045 0,397 0,248 0,270 0,000 0,042
N 18 19 11 20 15 8 202 202 202 50
β-ZOL LC-MS/MS
Correlation Coefficient
0,341 0,225 0,177 0,225 0,704 -0,080 ,351** 1,000 0,121
Sig. (2-tailed)
0,166 0,354 0,603 0,421 0,051 0,257 0,000 0,401
N 18 19 11 20 15 8 202 202 202 50
Voeder ZEA LC-MS/MS
Correlation Coefficient
-0,866
-1,000** ,297
* ,289
* 0,121 1,000
Sig. (2-tailed)
0,333 0,036 0,042 0,401
N 3 2 1 2 1 0 50 50 50 50
** Correlation is significant at the 0.01 level (2-tailed)
* Correlation is significant at the 0.05 level (2-tailed)
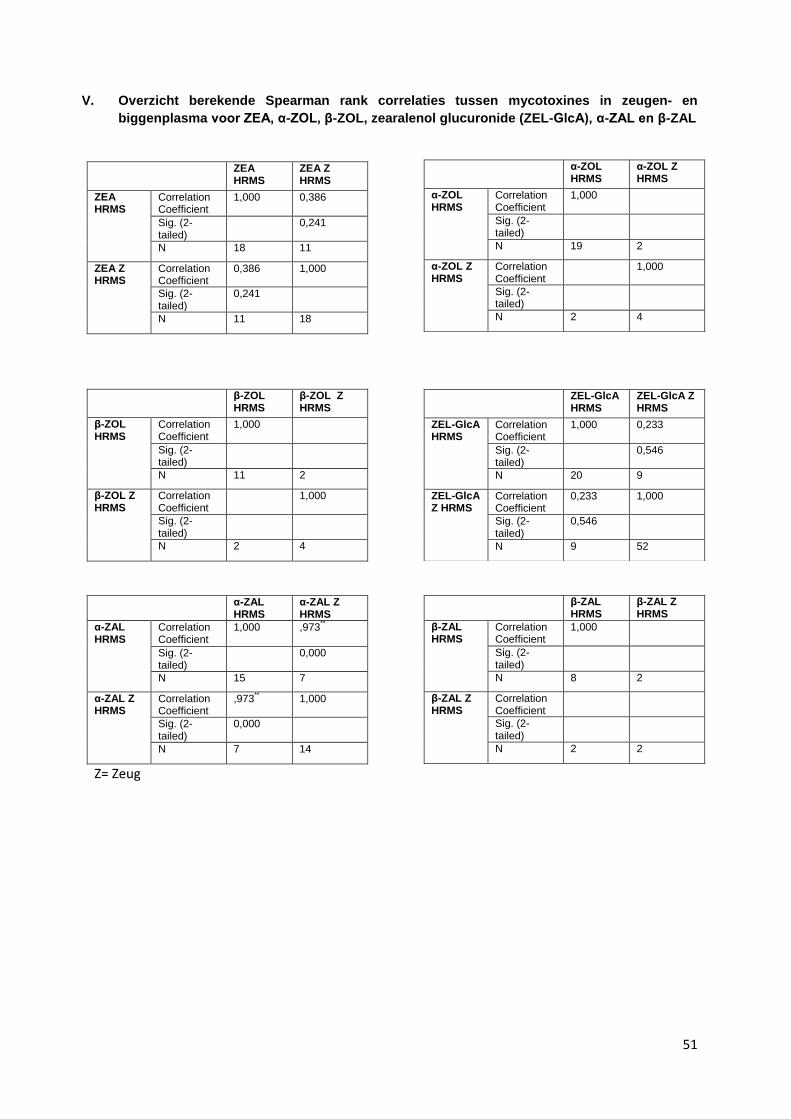
51
V. Overzicht berekende Spearman rank correlaties tussen mycotoxines in zeugen- en
biggenplasma voor ZEA, α-ZOL, β-ZOL, zearalenol glucuronide (ZEL-GlcA), α-ZAL en β-ZAL
ZEA HRMS
ZEA Z HRMS
ZEA HRMS
Correlation Coefficient
1,000 0,386
Sig. (2-tailed)
0,241
N 18 11
ZEA Z HRMS
Correlation Coefficient
0,386 1,000
Sig. (2-tailed)
0,241
N 11 18
β-ZOL HRMS
β-ZOL Z HRMS
β-ZOL HRMS
Correlation Coefficient
1,000
Sig. (2-tailed)
N 11 2
β-ZOL Z HRMS
Correlation Coefficient
1,000
Sig. (2-tailed)
N 2 4
α-ZAL HRMS
α-ZAL Z HRMS
α-ZAL HRMS
Correlation Coefficient
1,000 ,973**
Sig. (2-tailed)
0,000
N 15 7
α-ZAL Z HRMS
Correlation Coefficient
,973** 1,000
Sig. (2-tailed)
0,000
N 7 14
Z= Zeug
α-ZOL HRMS
α-ZOL Z HRMS
α-ZOL HRMS
Correlation Coefficient
1,000
Sig. (2-tailed)
N 19 2
α-ZOL Z HRMS
Correlation Coefficient
1,000
Sig. (2-tailed)
N 2 4
ZEL-GlcA HRMS
ZEL-GlcA Z HRMS
ZEL-GlcA HRMS
Correlation Coefficient
1,000 0,233
Sig. (2-tailed)
0,546
N 20 9
ZEL-GlcA Z HRMS
Correlation Coefficient
0,233 1,000
Sig. (2-tailed)
0,546
N 9 52
β-ZAL HRMS
β-ZAL Z HRMS
β-ZAL HRMS
Correlation Coefficient
1,000
Sig. (2-tailed)
N 8 2
β-ZAL Z HRMS
Correlation Coefficient
Sig. (2-tailed)
N 2 2





























