Embed Size (px)
Citation preview

BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 241, 509–512 (1997)ARTICLE NO. RC977854
Molecular Cloning of a Peptidylglycine a-HydroxylatingMonooxygenase from Sea Anemones
Frank Hauser, Michael Williamson, and Cornelis J. P. Grimmelikhuijzen1
Department of Cell Biology and Anatomy, Zoological Institute, University of Copenhagen,Universitetsparken 15, DK-2100 Copenhagen Ø, Denmark
Received October 30, 1997
tide amidation and, thereby, render them biologicallyCnidarians are the lowest animal group having a active, must play an essential role in the functioning
nervous system. The primitive nervous systems of of the cnidarian nervous system.cnidarians produce large amounts of a variety of neu- In mammals, it has been found that C-terminallyropeptides, of which many or perhaps all are ami-
amidated peptides originate from peptidyl-glycine thatdated at their C terminus. In vertebrates, peptideis converted into peptidyl-amide in two steps. First,amidation is catalyzed by two enzymes acting sequen-peptidyl-glycine is converted into peptidyl-hydroxyg-tially, peptidyl-glycine a-hydroxylating monooxygen-lycine by the enzyme PHM (peptidyl-glycine a-hydrox-ase (PHM) and peptidyl-a-hydroxyglycine a-amidat-ylating monooxygenase), a reaction that requires Cu2/,ing lyase (PAL). In mammals both enzymatic activi-O2 and ascorbic acid [4-7]. Second, peptidyl-hydroxyg-ties are contained within a bifunctional protein thatlycine is cleaved into peptidyl-amide and glyoxylic acidis coded for by a single gene. Using PCR and degener-by the enzyme PAL (peptidyl-a-hydroxyglycine a-ami-ated oligonucleotides derived from conserved regionsdating lyase) [4-9]. By cloning several preprohormonesof PHM, we have now cloned a PHM from the sea
anemone Calliactis parasitica showing 42% amino from cnidarians, we have found that also the cnidarianacid sequence identity with rat PHM. Among the con- amidated neuropeptides originate from a peptidyl-gly-served (identical) amino acid residues are five histi- cine, suggesting that enzymes similar to mammaliandine and one methionine residue, which bind two PHM and PAL must be present in the cnidarian ner-Cu2/ atoms that are essential for PHM activity. No vous system [10-15]. In the present paper, we describecDNA coding for PAL could be identified, suggesting the molecular cloning of such a cnidarian PHM.that sea anemone PAL is coded for by a gene that isdifferent from the sea anemone PHM gene, a situationsimilar to the one found in insects. This is the firstreport on the molecular cloning of a cnidarian PHM.q 1997 Academic Press
Cnidarians are the lowest animal group having anervous system and it was probably within this phylumor a closely related ancestor phylum that nervous sys-tems first evolved [1]. The primitive nervous systemsof cnidarians produce large amounts of a variety ofneuropeptides, which play important roles both in neu-rotransmission and in developmental processes such
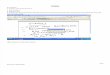
FIG. 1. Schematic representation of the cDNA coding for seaas metamorphosis [2, 3]. All cnidarian neuropeptidesanemone PHM (top) and the positions of the various overlappingisolated so far, carry an amide group at their C termi-PCR clones (bottom). The broad bar in the top figure represents thenus, which is necessary for their biological action. coding region, whereas the two flanking small bars represent the
Therefore, the enzymes that catalyze C-terminal pep- noncoding regions of the cDNA. The black box in the cDNA indicatesthe signal sequence (amino acid positions 1–18 of Fig. 2) and the twogrey boxes represent regions containing conserved histidine clusters(amino acid positions 95–102 and 236–243 of Fig. 2) to which the1 Address correspondence to Prof. C. J. P. Grimmelikhuijzen. Fax:
00 45 - 35 32 12 00. first two degenerate PCR primers were directed.
0006-291X/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
509
AID BBRC 7854 / 6942$$1061 12-02-97 01:24:33 bbrcg AP: BBRC

Vol. 241, No. 2, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 2. cDNA and deduced amino acid sequence of the sea anemone PHM. This figure consists of the compiled data obtained fromsequencing of six PCR clones (Fig. 1). Nucleotides are numbered from 5* to 3* end, and the amino acid residues are numbered startingwith the first ATG in the open reading frame. The translation termination codon is indicated by an asterisk.
of each primer, 0.2 mM of all four dNTPs and 2.5 units of Taq polymer-MATERIALS AND METHODSase (Boehringer Mannheim). Thermal cycling parameters were 35 cy-cles of the following step program: 947C for 1 min, 487C for 1 min,Poly(A/) RNA from Calliactis parasitica was prepared as described727C for 1 min. The PCR products were separated on a 1% agarosein [10]. Single-stranded cDNA was synthesized using random primersgel, isolated (Qiaquick extraction kit, Qiagen), subcloned into pCR2.1and a RT-PCR kit from Stratagene. For PCR, two degenerate primers,(Invitrogen) and sequenced (Sequenase version 2.0 DNA sequencingsense CAC/TCAC/TATGC/TTIC/TTITA/TC/TGGITG (coding for thekit; U.S. Biochemical Corp.). Each DNA sequence was sequenced sev-conserved amino acid sequence HHMLLF/YGC; Drosophila aminoeral times in different PCR clones (Fig. 1).acid positions 95-102 of Fig. 3), and antisense TGIGTA/GTGIA/GC/
TICG/T A/GA/TAIGCA/GAA (coding for the conserved amino acid se- 5*/3 *- RACE. All reactions of the 5*/3*- RACE were carried outfollowing the protocol and using the chemicals of the 5*/3*- RACE kitquence FAYRVHTH; Drosophila amino acid positions 236-243 of Fig.
3) were used. The reaction mixture consisted of 50ml of 1 1 Taq buffer from Boehringer Mannheim. cDNA was prepared as above. For 5*-RACE the sense primer GATTAAAAGCTTCTTTCTGTGG and the(Boehringer Mannheim) containing 2.5 mM MgCl2, 0.1 mg cDNA, 2mM
510
AID BBRC 7854 / 6942$$1061 12-02-97 01:24:33 bbrcg AP: BBRC

Vol. 241, No. 2, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 3. Amino acid comparison of the PHMs from Calliactis parasitica (CP1), Drosophila melanogaster (DRO), Xenopus laevis (XL1)and rat (RAT). Amino acid residues that are identical between the sea anemone PHM and at least one of the other PHMs are boxed. Thefive histidine and one methionine residue that form complexes with two Cu2/ atoms are indicated by asterisks and printed in boldface type.The eight conserved cysteine residues that form four putative cysteine-cysteine bridges are marked with closed circles and also printed inboldface type. The data from Drosophila, Xenopus and rat are from [19–22, 24].
nested sense primer GGAATTGCAGCCCGCTCTCTGT, and for 3*- we were able to clone PCR products from the sea anem-RACE the antisense primer TGGCTGCAATTTTAGTTCTAAC and the one Calliactis parasitica coding for a partial sequence ofnested antisense primer ACAGAGAGCGGGCTGCAATTCC were used. a cnidarian PHM (clones pCP3 and pCP5; Fig.1). After
5*/3*- RACE, we combined the cDNA sequences of allRESULTS PCR clones and obtained the complete cDNA for the cnid-
arian PHM (Figs. 1, 2). The coding region of this cDNAUsing oligonucleotide probes coding for regions that is 1092 nucleotides long. It is preceded by an untrans-
were conserved between various vertebrate PHMs and lated 5* region of 140 bp and followed by a 330 bp trailercontaining the first 13 nucleotides of the poly(A/) tail.Drosophila PHM (see MATERIALS AND METHODS),
511
AID BBRC 7854 / 6942$$1061 12-02-97 01:24:33 bbrcg AP: BBRC

Vol. 241, No. 2, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
The cDNA codes for a protein that is 364 amino acid ACKNOWLEDGMENTSresidues long (Fig. 2). The start codon is followed by a
We thank Dr. Paul Taghert (Washington University, Medicalhydrophobic signal sequence that is probably cleavedSchool, St. Louis, MO) for making the Drosophila PHM sequenceoff at Gly-18 [16].available to us prior to publication, Christina Færch-Jensen for tech-
Fig. 3 shows an alignment of the PHMs from Calli- nical assistance, Astrid Juel Jensen for typing the manuscript, andactis, Drosophila, Xenopus and rat. When we compared the Danish Natural Science Research Council for financial support.the central portion of the cnidarian PHM (between Leu-52 and Pro-347 of the CP1 sequence of Fig. 3) with the REFERENCEScorresponding regions of the rat, Xenopus and Drosoph-
1. Mackie, G. O. (1990) Amer. Zool. 30, 907–920.ila PHMs, we found 42% amino acid sequence identity2. Grimmelikhuijzen, C. J. P., Leviev, I., and Carstensen, K. (1996)with rat PHM, 46% with Xenopus PHM and 46% with
Int. Rev. Cytol. 167, 37–89.Drosophila PHM (Fig. 3). Considerably less identity3. Leitz, T., Morand, K., and Mann, M. (1994) Develop. Biol. 163,(27%) exists when the central portion of cnidarian PHM
440–446.was compared with the corresponding region of rat do-4. Bradbury, A. F., Finnie, M. D. A., and Smyth, D. G. (1982) Na-pamine b-hydroxylase, an enzyme that is structurally
ture 298, 686–688.related to PHM [17].
5. Perkins, S. N., Husten, E. J., and Eipper, B. A. (1990) Biochem.Five histidine and one methonine residue in the cen- Biophys. Res. Commun. 171, 926–932.tral portion of PHM are supposed to form a complex 6. Bradbury, A. F., and Smyth, D. G. (1991) Trends Biochem. Sci.with two Cu2/ atoms, which are essential for the PHM 16, 112–115.reaction [18, 19]. These six residues (indicated by aster- 7. Eipper, B. A., Stoffers, D. A., and Mains, R. E. (1992) Ann. Rev.
Neurosci. 15, 57–85.isks in Fig. 3) are conserved between the cnidarian8. Katopodis, A. G., Ping, D., and May, S. W. (1990) Biochemistryand the other three PHMs. Furthermore, the three-
29, 6115–6120.dimensional structure of rat PHM is fixed with the help9. Katopodis, A. G., Ping, D., Smith, C. E., and May, S. W. (1991)of five cysteine-cysteine bridges [19]. Eight of these ten
Biochemistry 30, 6189–6194.cysteine residues are conserved between rat and Dro-10. Darmer, D., Schmutzler, C., Diekhoff, D., and Grimmelikhuijzen,sophila PHM [19, 20]. These eight conserved cysteine
C. J. P. (1991) Proc.Natl. Acad. Sci. USA 88, 2555–2559.residues (marked with filled circles in Fig. 3) are also11. Schmutzler, C., Darmer, D., Diekhoff, D., and Grimmelikhuijzen,present in the cnidarian PHM. C. J. P. (1992) J. Biol.Chem. 267, 22534–22541.12. Schmutzler, C., Diekhoff, D., and Grimmelikhuijzen, C. J. P.
(1994) Biochem. J. 299, 431–436.DISCUSSION13. Reinscheid, R. K., and Grimmelikhuijzen, C. J. P. (1994) J. Neu-
rochem. 62, 1214–1222.This is the first report describing the molecular clon- 14. Leviev, I., and Grimmelikhuijzen, C. J. P. (1995) Proc. Natl.
ing of a PHM in lower invertebrates. In mammals, both Acad. Sci. USA. 92, 11647–11651.PHM and PAL are contained within a bifunctional pro- 15. Leviev, I., Williamson, M., and Grimmelikhuijzen, C. J. P. (1997)
J. Neurochem. 68, 1319–1325.tein PAM (peptidyl a-amidating monooxygenase) that16. Heijne von, G. (1986) Nucleic Acids Res. 14, 4683–4690.is coded for by a single gene, where the PHM coding
exons are located at the 5* end of those coding for PAL 17. Southan, C, and Kruse, L. I. (1989) FEBS Lett. 255, 116–120.[7, 21, 22]. In the tetraploid claw frog Xenopus laevis, 18. Eipper, B. A., Quon, A. S. W., Mains, R. E., Boswell, J. S., and
Blackburn, N. J. (1995) Biochemistry 34, 2857–2865.there are two closely-related genes coding for PAM [23,19. Kolhekar, A. S., Keutmann, H. T., Mains, R. E., Quon, A. S. W.,24]. In the fruitfly Drosophila melanogaster, however,
and Eipper, B. A. (1997) Biochemistry 36, 10901–10909.a single PHM occurs, and no bifunctional PAM. On the20. Kolhekar, A. S., Roberts, M. S., Jiang, N., Johnson, R. C., Mains,genomic level, one gene for PHM could be cloned, but
R. E., Eipper, B. A., and Taghert, P. H. (1997) J. Neurosci. 17,PAL appears to be encoded by a separate and yet un- 1363–1376.identified gene [20]. 21. Kato, I., Yonekura, H., Tajima, M., Yanagi, M., Yamamoto, H.,
The situation in cnidarians appears to be similar to and Okamoto, H. (1990) Biochem. Biopys. Res. Commun. 172,197–203.the one found in insects. We were unable to find PCR
clones after 5*/3 *- RACE that were longer than the ones 22. Quafik, L., Stoffers, D. A., Campbell, T. A., Johnson, R. C.,Bloomquist, B. T., Mains, R. E., and Eipper, B. A. (1992) Mol.given in Fig. 1, thereby leaving no space for a PALEndocrinol. 6, 1571–1584.message. This strongly suggests that PHM and PAL
23. Mizuno, K., Ohsuye, K., Wada, Y, Fuchimura, K., Tanaka, S.,are coded for by different genes in cnidarians. Thus,and Matsuo, H. (1987) Biochem. Biophys. Res. Commun. 148,PHM and PAL appear to have originated as separate 546–552.
genes and only later in evolution these genes appear 24. Ohsuye, K., Kitano, K., Wada, Y., Fuchimura, K., Tanaka, S.,to have fused forming the single gene coding for bifunc- Mizuno, K., and Matsuo, H. (1988) Biochem. Biophys. Res. Com-
mun. 150, 1275–1281.tional PAM that we know from vertebrates.
512
AID BBRC 7854 / 6942$$1061 12-02-97 01:24:33 bbrcg AP: BBRC