-
1
1
NUDT21 links mitochondrial IPS-1 to RLR-containing stress
granules and 2
activates host antiviral defense 3
4
5
Saeko Aoyama-Ishiwatari1, Tomohiko Okazaki1*, Shun-ichiro
Iemura2†, Tohru 6
Natsume2, Yasushi Okada3,4,5, Yukiko Gotoh1,5 7
8
9 1Graduate School of Pharmaceutical Sciences, The University of
Tokyo, Tokyo 113-10
0033, Japan. 11 2Molecular Profiling Research Center for Drug
Discovery (molprof), National Institute 12
of Advanced Industrial Science and Technology (AIST), Tokyo
135-0064, Japan. 13 3Laboratory for Cell Dynamics Observation,
Center for Biosystems Dynamics Research 14
(BDR), RIKEN, Osaka 565-0874, Japan. 15 4Department of Physics,
Universal Biology Institute (UBI) 16
5International Research Center for Neurointelligence (WPI-IRCN),
The University of 17
Tokyo, Tokyo 113-0033, Japan. 18
19
*Corresponding author: [email protected] 20
21
†Present address: Translational Research Center, Fukushima
Medical University, 22
Fukushima 960-1295, Japan. 23
24
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
2
Summary 25
Viral RNA in the cytoplasm of mammalian host cells is recognized
by retinoic acid–26
inducible protein–I (RIG-I)–like receptors (RLRs), which
localize to cytoplasmic stress 27
granules (SGs). Activated RLRs associate with the mitochondrial
adaptor protein IPS-1, 28
which activates antiviral host defense mechanisms including type
I interferon (IFN) 29
induction. It has remained unclear, however, how RLRs in SGs and
IPS-1 in the 30
mitochondrial outer membrane associate physically and engage in
information transfer. 31
Here we show that NUDT21, an RNA binding protein that regulates
alternative 32
transcript polyadenylation, physically associates with IPS-1 and
mediates its 33
localization to SGs in response to transfection with poly(I:C),
a mimic of viral double-34
stranded RNA. We found that, despite its well-established
function in the nucleus, a 35
fraction of NUDT21 localizes to mitochondria in resting cells
and becomes localized to 36
SGs in response to poly(I:C) transfection. NUDT21 was also found
to be required for 37
efficient type I IFN induction in response to viral infection.
Our results together indicate 38
that NUDT21 links RLRs in SGs to mitochondrial IPS-1 and thereby
activates host 39
defense responses to viral infection. 40
41
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
3
Introduction 42
The innate immune system provides the first line of defense
against viral infection. The 43
initial step of this defense is detection of “non-self” cues
known as pathogen-associated 44
molecular patterns (PAMPs) by specialized sensors, known as
pattern recognition 45
receptors (PRRs), in host cells. Recognition of viral PAMPs by
PRRs results in the 46
activation of a series of mechanisms to combat viral
propagation. In vertebrates, 47
activation of PRRs induces the production of type I interferons
(IFNs) such as IFN-α 48
and IFN-β and the subsequent expression of hundreds of
IFN-stimulated genes (ISGs) 49
that play a major role in restriction of viral replication
within infected cells (1–3). 50
Retinoic acid–inducible protein–I (RIG-I)–like receptors (RLRs)
are a family 51
of PRRs consisting of DEAD box–containing RNA helicases that
recognize viral RNA 52
in the cytoplasm. Among RLRs, RIG-I recognizes RNA molecules
containing a 5'-53
triphosphate group as well as relatively short (2 kb) dsRNAs,
with both types of dsRNA being derived from a wide 56
range of RNA viruses (4–6). The binding of RLRs to such viral
RNAs triggers their 57
interaction with a key antiviral hub protein, IFN-β promoter
stimulator–1 (IPS-1, also 58
known as MAVS, CARDIF, and VISA) (7–10). IPS-1 is anchored to
the mitochondrial 59
outer membrane, and activated IPS-1 forms large prionlike
aggregates on these 60
organelles (11) that in turn activate transcription factors such
as interferon regulatory 61
factor 3 (IRF3), nuclear factor–κB, and activator protein–1,
resulting in the expression 62
of type I IFNs (8,12–16). The pivotal role of IPS-1 in antiviral
responses is exemplified 63
by the finding that IPS-1–deficient mice are more vulnerable to
viral infection than are 64
wild-type (WT) mice (17, 18). The NH2-terminal caspase
activation and recruitment 65
domain (CARD) of RLRs is required for the interaction with IPS-1
and was indeed 66
found to be essential for IFN induction (19). 67
Viral RNAs, RLRs, and RLR-associated proteins have been found to
be 68
localized to stress granules (SGs) in virus-infected cells. SGs
are membraneless 69
structures that are composed of translation-stalled mRNAs and
proteins and in which 70
translation is generally inhibited (20). They are formed in
response to various cellular 71
stresses including viral infection. Viral RNA–mediated
activation of dsRNA-activated 72
protein kinase (PKR) results in the phosphorylation of
eukaryotic initiation factor 2a, 73
translational arrest, and SG nucleation (21). The RNA binding
proteins Ras-GAP SH3 74
domain–binding protein 1 (G3BP1), T cell–restricted
intracellular antigen 1 (TIA1), and 75
TIA1-related protein (TIAR) participate in the formation of SGs
(22, 23), which is 76
thought to involve liquid-liquid phase separation (24). SGs have
been proposed to serve 77
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
4
as a platform for RLR recognition of viral RNA and consequent
activation of antiviral 78
responses, with such SGs also having been termed antiviral SGs
(avSGs) (25). 79
Inhibition of SG formation by depletion of G3BP1 indeed
suppressed type I IFN 80
expression in response to infection with influenza A virus or
Newcastle disease virus 81
(NDV) (25, 26). The fact that many viral factors interfere with
SG formation (27–31) 82
also supports an antiviral function of SGs. 83
The mitochondrial localization of IPS-1 has been thought to be
essential for its 84
function. For instance, deletion of the COOH-terminal
transmembrane (TM) domain of 85
IPS-1 abrogated both its mitochondrial localization and the
induction of type I IFN (8, 86
32). Caspase-mediated cleavage of IPS-1 that results in
detachment of the TM domain 87
also inactivates IPS-1 function (33, 34). Given the
mitochondrial localization of IPS-1, 88
it has remained unclear how RLRs within SGs encounter and
activate IPS-1. Of interest 89
in this regard, a fraction of IPS-1 appears to colocalize with
TIAR, a marker of SGs, in 90
cells infected with viruses or transfected with
polyinosinic-polycytidylic acid 91
[poly(I:C)], a synthetic analog of viral dsRNA (26, 35),
suggesting that viral RNA may 92
induce the localization of IPS-1 to SGs and its association with
RLRs. The mechanism 93
that might underlie such an effect has remained unknown,
however. 94
We here identify nucleoside diphosphate–linked moiety X
(Nudix)–type motif 95
21 (NUDT21) as an IPS-1 interactor. NUDT21 is an RNA binding
protein that 96
constitutes the CFIm complex in nucleus and regulates the choice
of polyadenylation 97
site (PAS) by binding to the UGUA element in the 3' untranslated
region (UTR) of 98
transcripts (36–39). NUDT21-mediated alternative polyadenylation
has been shown to 99
influence various cell fate decision processes as well as
tumorigenesis (40, 41). Virus 100
infection was previously shown to induce genome-wide changes in
PAS selection in 101
host transcripts and was also reported to limit a host gene
expression through 102
NUDT21(42, 43). Unexpectedly, we found that, whereas NUDT21 is
localized mostly 103
to the nucleus, where alternative polyadenylation takes place, a
fraction of NUDT21 is 104
associated with mitochondria in resting cells but also localizes
to SGs in response to 105
poly(I:C) transfection. Moreover, NUDT21 was found to associate
with IPS-1 and to 106
play an important role in its localization to SGs as well as in
the efficient induction of 107
IFN expression in response to poly(I:C) transfection or to
infection with 108
encephalomyocarditis virus (EMCV). Our results thus suggest an
unexpected role for 109
NUDT21 in the recruitment of mitochondrial IPS-1 to SGs and in
the consequent 110
promotion of RLR-mediated activation of IPS-1 and IFN induction
in virus-infected 111
cells. 112
113
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
5
Materials and Methods 114
Cell culture and transfection 115
HeLa S3 and HEK293T cells were maintained in Dulbecco’s modified
Eagle’s medium 116
supplemented with 10% fetal bovine serum and 1%
penicillin-streptomycin. HEK293T 117
cells were transfected with the use of the GeneJuice
Transfection Reagent (Merck 118
Millipore), whereas transfection of HeLa S3 cells with
expression vectors was 119
performed with Lipofectamine 2000 (Thermo Fisher Scientific).
120
121
Plasmids and reagents 122
The plasmid pEF-BOS-FLAG-IPS-1 encoding WT human IPS-1 was
kindly provided 123
by M. Yoneyama (Division of Molecular Immunology, Medical
Mycology Research 124
Center, Chiba University, Japan). A full-length cDNA for NUDT21
was amplified by 125
PCR from a mouse cDNA library with the primers 5′-126
GGCAGATCTATGTCTGTGGTGCCGCCCAA-3′ and 5′-127
GGCGAATTCTCAGTTGTATATAAAATTGA-3′ (sense and antisense,
respectively), 128
and was subcloned into either the BglII and EcoRI sites of the
pCS4 vector or the 129
BamHI and EcoRI sites of pcDNA3.1. Human G3BP1 cDNAs with or
without stop 130
codon were amplified by PCR from pN1/G3BP1-iRFP (Okada lab
plasmids, 131
Addgene #129339) with the sense primer 5′-132
GCCAGATCTATGGTGATGGAGAAGCCTAG-3′ and the antisense primers
5′-133
GCCGAATTCGGATCCTTACTGCCGTGGCGC-3′ or 5′-134
GCCGAATTCGGATCCCTGCCGTGGCGCAAG-3′, respectively, and were
135
subcloned into the BamHI and EcoRI sites of pcDNA3. Myc
epitope–tagged full-length 136
human IPS-1 and Myc–IPS-1(ΔTM) cDNAs were amplified by PCR from
a Myc–IPS-1 137
expression vector described previously (44) with the sense
primer 5′-138
ACTGCGGCCGCATGAGCAAAAGCTCATTT-3′ and the antisense primers
5′-139
GCCTCTAGACTAGTGCAGACGCCGCCG-3′ or 5′-140
GCCTCTAGACTAGTCTACCTGGGATGCCA-3, respectively, and were
subcloned 141
into the NotI and XbaI sites of either pcDNA3 or the G3BP1
expression vector. Myc–142
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
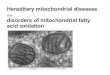
6
IPS-1 (amino acids 1–98), Myc–IPS-1 (amino acids 99–200) and
Myc– IPS-1 (amino 143
acids 201–540) cDNAs were amplified by PCR from a Myc–IPS-1
expression vector 144
with the sense primer 5′-GGCAGATCTATGCCGTTTGCTGAAGACAAGACC-3′
145
and the antisense primer 5′-GGCAGATCTCTACCGAGGCTGGTAGCTCTGGT-3′,
146
the sense primer 5′-GGCAGATCTACCTCGGACCGTCCCCCAGAC-3′ and the
147
antisense primer 5′-GGCAGATCTCTATGTGTCCTGCTCCTGATGCCCGCT-3′,
148
or the sense primer
5′-GGCAGATCTATGGAACTGGGCAGTACCCACACAGCA-149
3′ and the antisense primer
5′-GGCAGATCTCTAGTGCAGACGCCGCCGGT-150
3′, respectively, and were subcloned into the BglII and EcoRI
sites of the pCS4 vector. 151
Poly(I:C) was obtained from GE Healthcare and was introduced
into cells by 152
transfection with Lipofectamine 2000 (Thermo Fisher Scientific).
153
154
Antibodies 155
Antibodies to NUDT21 were obtained from Proteintech; those to
Myc (9E10), to p38, to 156
TIAR, and to TOMM20 were from Santa Cruz Biotechnology; those to
IPS-1 for 157
immunostaining as well as those to phospho-p38, to phospho-JNK,
to cleaved caspase-158
3, to cleaved PARP, and to phospho-IRF3 were from Cell Signaling
Technology; those 159
to FLAG (M2) were from Sigma; those to HA were from Roche; those
to IPS-1 for 160
immunoblot analysis were from Abcam; those to cytochrome c were
from BD 161
Pharmingen; and those to CFIm68 were from Bethyl Laboratories.
Antibodies to RIG-I 162
were kindly provided by M. Yoneyama (Chiba University). 163
164
RNA interference 165
Knockdown of NUDT21 was achieved with the use of Stealth RNA
interference 166
(Thermo Fisher Scientific). HeLa S3 cells were transfected with
siRNA 167
oligonucleotides with the use of the Lipofectamine RNAiMAX
reagent (Thermo Fisher 168
Scientific) and were used for experiments after incubation for
72 h. The siRNA 169
sequences were 5′-UGAACCUCCU-CAGUAUCCAU-AUAUU-3′ and 5′-170
AAUAUAUGGA-UACUGAGGAG-GUUCA-3′ for NUDT21, and a negative
control 171
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
7
siRNA (Thermo Fisher Scientific, catalog no. 12935300) was also
used. 172
173
Mass spectrometry 174
Liquid chromatography and tandem mass spectrometry were
performed as previously 175
described (45). 176
177
Immunoblot analysis 178
Immunoblot analysis was performed as described previously (44).
In brief, cells were 179
lysed with a solution containing 20 mM Tris-HCl (pH 7.5), 150 mM
NaCl, 10 mM -180
glycerophosphate, 5 mM EGTA, 1 mM Na4P2O7, 5 mM NaF, 0.5% Triton
X-100, 1 mM 181
Na3VO4, 1 mM dithiothreitol, and aprotinin (1 mg/ml), cell
lysates were fractionated by 182
SDS-polyacrylamide gel electrophoresis on a 10% gel, and the
separated proteins were 183
transferred to a polyvinylidene difluoride membrane. The
membrane was incubated first 184
with primary antibodies for 24 h at 4°C and then with
horseradish peroxidase–185
conjugated secondary antibodies (GE Healthcare) for 1 h at room
temperature. After a 186
wash with a solution containing 50 mM Tris-HCl (pH 8.0), 150 mM
NaCl, and 0.05% 187
Tween 20, the membrane was processed for detection of peroxidase
activity with 188
chemiluminescence reagents and an Image Quant LAS4000 instrument
(GE 189
Healthcare). 190
191
Co-immunoprecipitation analysis 192
HEK293T cells harvested 20 h after transfection with plasmids
encoding FLAG–IPS-1 193
and Myc-NUDT21 were lysed with a solution containing 20 mM
Tris-HCl (pH 7.5), 194
150 mM NaCl, 10 mM -glycerophosphate, 5 mM EGTA, 1 mM Na4P2O7, 5
mM NaF, 195
0.2% Triton X-100, 1 mM Na3VO4, 1 mM dithiothreitol, and
aprotinin (1 mg/ml). The 196
cell lysates were incubated with rotation at 4°C first for 45
min with antibodies to 197
FLAG and then for 45 min with protein A–Sepharose (GE
Healthcare). The resulting 198
immunoprecipitates were subjected to immunoblot analysis.
199
200
Immunofluorescence microscopy 201
Cells were fixed with 4% formaldehyde for 10 min at 37°C,
permeabilized with 0.2% 202
Triton X-100 in phosphate-buffered saline (PBS) for 5 min, and
incubated for 24 h in 203
PBS containing 2% fetal bovine serum and 2 % bovine serum
albumin (blocking 204
buffer). They were then exposed at room temperature first for 1
h to primary antibodies 205
in blocking buffer and then for 1 h to Alexa Fluor–conjugated
secondary antibodies 206
(Thermo Fisher Scientific) and Hoechst 33342 in blocking buffer.
Moviol were used as 207
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
8
mounting medium. Images were acquired with a TCS SP5 confocal
microscope (Leica) 208
and were processed with Image J (NIH). 209
210
Colocalization analysis and quantification of SG volume 211
Manders M1 and M2 coefficients for colocalization were
calculated with Coloc 2 of 212
Fiji. A statistical significance test was derived by Costes
(46). For the experiment shown 213
in Figure 2C, samples were prepared in the same way as for
immunofluorescence 214
analysis described above, with the exceptions that ProLong
Diamond (Thermo Fisher 215
Scientific) was used as mounting medium and that images were
acquired with a TCS 216
SP8 confocal microscope (Leica). Three-dimensional images were
acquired in order to 217
meet the Nyquist condition (pixel size of 40.6 nm for x and y,
and of 100 nm for z) and 218
were deconvoluted with Huygens software (Scientific Volume
Imaging). Manders M1 219
and M2 for the colocalization of IPS-1 and TOMM20 were then
calculated. For the 220
experiment shown in Figure 2E, samples were again prepared in
the same way as for 221
immunofluorescence analysis with the exception that ProLong
Diamond (Thermo 222
Fisher Scientific) was used as mounting medium.
Three-dimensional images were 223
acquired in order to match the Nyquist condition (pixel size of
50 nm for x and y, and of 224
100 nm for z) and were deconvoluted with Huygens software
(Scientific Volume 225
Imaging). Manders M1 coefficient for the colocalization of IPS-1
and TIAR was then 226
calculated. The SG volume per cell was measured with the 3D
object counter 227
(threshold, 25; size filter, 1) of Huygens software in images
for which the background 228
intensity of the cytosol had been subtracted. 229
230
RT and real-time PCR analysis 231
Total RNA was obtained from cells with the use of RNAiso
(TaKaRa). RT was 232
performed with 1 μg of total RNA and ReverTra Ace qPCR RT Master
Mix with gDNA 233
Remover (Toyobo). The resulting cDNA was subjected to real-time
PCR analysis in a 234
Roche LightCycler with the use of a KAPA SYBR Fast qPCR Kit
(Nippon Genetics). 235
The abundance of each target mRNA was normalized by that
glyceraldehyde-3-236
phosphate dehydrogenase (GAPDH) mRNA. The PCR primers (sense and
antisense, 237
respectively) were 5′-ACTCCTCCACCTTTGACGCT-3′ and 5′-238
TCCTCTTGTGCTCTTGCTGG-3′ for human GAPDH, 5′-239
CTGGCTGGAATGAGACTATTGTT- 3′ and 5′-CTTCAGTTTCGGAGGTAACCTG-3′
240
for human IFN-β, 5′-AACCTGAACCTTCCAAAGATGG-3′ and 5′-241
TCTGGCTTGTTCCTCACTACT-3′ for human IL-6, and 5′-242
ATGAGCACTGAAAGCATGATCC-3′ and 5′-GAGGGCTGATTAGAGAGAGGTC-3′
243
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
9
for human TNF-α. 244
245
Reporter gene analysis 246
HEK293T cells seeded in 24-well plates (7.5 × 104 cells/well)
were transiently 247
transfected for 20 h with 8 ng of a reporter plasmid encoding
Renilla luciferase under 248
the control of the human IFN-β gene promoter (p-125-RLuc)
together with 5 ng of an 249
expression plasmid encoding FLAG-tagged full-length IPS-1 and
either 50 or 500 ng of 250
a plasmid for Myc-tagged IPS-1(ΔTM). The activity of Renilla
luciferase in total cell 251
lysates was measured with the use of a Dual-Luciferase Reporter
Assay System 252
(Promega) and was normalized by that of firefly luciferase
derived from 80 ng of a 253
control plasmid. 254
255
Virus infection 256
EMCV were kindly provided by M. Yoneyama (Chiba University). The
cells were 257
incubated in culture medium with 0.1 (+) or 0.2 (++) PFU of EMCV
for 2 h, and then 258
replaced in EMCV-free culture medium for 10 h. 259
260
Statistical analysis 261
Quantitative data are presented as means ± s.e.m. or ± s.d. and
were compared with 262
Student’s t test. A P value of
-
10
Results 265
NUDT21 forms a complex with IPS-1 266
In a search for regulators of IPS-1, we performed
co-immunoprecipitation analysis to 267
identify IPS-1–associated molecules. FLAG epitope–tagged IPS-1
expressed in 268
HEK293T cells was thus immunoprecipitated with antibodies to
FLAG, and the 269
resultant precipitates were analyzed by a highly sensitive
direct nanoflow liquid 270
chromatography–tandem mass spectrometry system. We identified
NUDT21 among the 271
proteins that coprecipitated with FLAG–IPS-1. To confirm this
result, we performed co-272
immunoprecipitation analysis with HEK293T cells expressing
FLAG–IPS-1 and Myc 273
epitope–tagged NUDT21. We found that Myc-NUDT21 coprecipitated
with FLAG–274
IPS-1 (Figure 1A), suggesting that NUDT21 indeed forms a complex
with IPS-1. 275
To examine which domains of IPS-1 interact with NUDT21, we
generated 276
Myc-tagged fragments of human IPS-1 that contain either CARD
(amino acids 1–98), 277
the proline-rich domain (amino acids 99–200), or the
COOH-terminal domain including 278
the TM domain (amino acids 201–540). We found that hemagglutinin
epitope (HA)–279
tagged NUDT21 coprecipitated with Myc–IPS-1 fragments containing
CARD or the 280
proline-rich domain, but not with that containing the
COOH-terminal domain (Figure 281
S1). These results suggested that IPS-1 associates with NUDT21
through its NH2-282
terminal domains including CARD and the proline-rich domain.
283
284
A fraction of NUDT21 associates with mitochondria in resting
cells and localizes to 285
SGs on exposure to cytoplasmic dsRNA 286
These results suggestive of a physical interaction between IPS-1
and NUDT21 were 287
unexpected, given the previously described localization of
NUDT21 in the nucleus and 288
that of IPS-1 to the mitochondrial outer membrane (8).
Immunofluorescence analysis of 289
HeLa S3 cells indeed revealed that most NUDT21 was localized to
the nucleus (Figure 290
1B). However, we detected a fraction of the NUDT21 signals in
the cytoplasm (Figure 291
1B). Knockdown of NUDT21 by transfection of the cells with a
small interfering RNA 292
(siRNA) resulted in a reduction in these cytoplasmic and nuclear
signals, supporting 293
their specificity for NUDT21 (Figure 1B). The NUDT21 signals in
the cytoplasm 294
overlapped with those for the mitochondrial marker cytochrome c
(Figure 1B). By 295
contrast, CFIm68, a cofactor of NUDT21 in its nuclear function,
did not appear to 296
localize to mitochondria (Figure S2). Together, these results
suggested that a fraction of 297
NUDT21 localizes to mitochondria, where it may associate with
IPS-1. 298
We next transfected HeLa S3 cells with the viral dsRNA mimic
poly(I:C) to 299
examine its possible effect on the intracellular distribution of
NUDT21. NUDT21 300
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
11
immunoreactivity was detected as granulelike aggregates in the
cytoplasm at 6 h after 301
poly(I:C) transfection, and these granulelike signals were again
attenuated by NUDT21 302
knockdown (Figure 1C). Moreover, we found that these cytoplasmic
signals of 303
NUDT21 colocalized with the SG marker TIAR (Figure 1C). Indeed,
26.89 ± 3.65% of 304
the area of TIAR-positive foci overlapped with NUDT21
immunoreactivity (Figure 305
1D), suggesting that NUDT21 becomes localized to SGs in the
presence of cytoplasmic 306
dsRNA and consequent activation of the RLR pathway. 307
308
NUDT21 plays an important role in IPS-1 localization to SGs
309
The interaction of NUDT21 with IPS-1 as well as its
mitochondrial localization and its 310
appearance at SGs in response to dsRNA exposure prompted us to
examine whether 311
NUDT21 regulates the localization of IPS-1. Consistent with
previous studies (35, 26), 312
we found that a fraction of IPS-1 colocalized with TIAR in cells
transfected with 313
poly(I:C), whereas most IPS-1 appeared to colocalize with the
mitochondrial import 314
receptor subunit TOM20 homolog (TOMM20) in both control and
poly(I:C)-transfected 315
HeLa S3 cells (Figure 2A). We calculated Manders coefficients
for the colocalization of 316
IPS-1 and TIAR (47). Manders M1 (sum of TIAR signal intensity
overlapping with IPS-317
1 versus total TIAR signal intensity) was significantly
increased by poly(I:C) 318
transfection (Figure 2B), whereas Manders M2 (sum of IPS-1
signal intensity 319
overlapping with TIAR versus total IPS-1 signal intensity) did
not differ significantly 320
between cells with or without poly(I:C) exposure (Figure 2B).
These results suggested 321
that a fraction of IPS-1 becomes localized to SGs in response to
the presence of 322
cytoplasmic dsRNA. We also found the 34.21 ± 5.88% of the area
of TIAR-positive foci 323
overlapped with IPS-1 at 6 h after poly(I:C) transfection,
indicating that IPS-1 is not 324
distributed to all SGs. To examine IPS-1 localization at a
higher resolution, we stained 325
endogenous IPS-1 and TOMM20 at 6 h after poly(I:C) transfection
in HeLa S3 cells 326
expressing a fusion protein of G3BP1 and near-infrared
fluorescent protein (iRFP) and 327
obtained z-stack images deconvoluted with Huygens software. A
fraction of IPS-1 328
signals was found to localize to G3BP1-iRFP foci in
poly(I:C)-transfected cells, 329
whereas TOMM20 was detected exclusively outside of G3BP1-iRFP
foci (Figure 2C). 330
We then examined whether poly(I:C) transfection affects the
colocalization of IPS-1 and 331
TOMM20. Although Manders M1 (sum of IPS-1 signal intensity
overlapping with 332
TOMM20 versus total IPS-1 signal intensity) was not
significantly altered by poly(I:C) 333
transfection, Manders M2 (sum of TOMM20 signal intensity
overlapping with IPS-1 334
versus total TOMM20 signal intensity) was slightly but
significantly reduced in cells 335
exposed to poly(I:C) (Figure 2D), suggesting that a fraction of
IPS-1 separates from 336
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
12
TOMM20 in response to poly(I:C) stimulation. Together, these
results indicated that a 337
fraction of IPS-1, but not of another mitochondrial protein
(TOMM20), becomes 338
localized to SGs in response to the presence of cytoplasmic
dsRNA. 339
We next examined the effect of NUDT21 on the distribution of
IPS-1 in cells 340
transfected with poly(I:C). We thus calculated Manders
coefficients for IPS-1 and TIAR 341
in z-stack images deconvoluted with Huygens software and found
that Manders M1 342
(sum of TIAR signal intensity overlapping with IPS-1 versus
total TIAR signal 343
intensity) was markedly reduced by NUDT21 knockdown (Figure 2E,
F). Importantly, 344
NUDT21 knockdown did not affect the overall (mitochondrial)
distribution of IPS-1 in 345
control (nontransfected) cells (Figure S3), indicating that the
effect of NUDT21 on the 346
localization of IPS-1 was stimulus dependent. By contrast,
NUDT21 knockdown did not 347
significantly alter the volume of TIAR-positive foci (SGs) per
cell (Figure 2G) or the 348
abundance of IPS-1 (Figure S4). We also found that NUDT21
knockdown did not 349
significantly affect the ratio of SG-containing cells (Figure
2H) or RIG-I accumulation 350
at SGs in response to poly(I:C) transfection (Figure 2I).
Collectively, these results 351
suggested that NUDT21 plays a role in IPS-1 localization to SGs
in response to the 352
presence of cytoplasmic dsRNA, whereas it does not overtly
affect SG formation or 353
RIG-I accumulation at SGs. 354
355
NUDT21 is required for antiviral responses induced by
cytoplasmic dsRNA 356
The interaction between RLRs and IPS-1 triggers activation of
downstream signaling 357
pathways in response to virus infection (7–10). Given that our
data implicated NUDT21 358
in localization of IPS-1 to SGs, which contain RLRs, we next
asked whether NUDT21 359
contributes to antiviral responses mediated by RLRs and IPS-1
such as the induction of 360
IFN-β and the proinflammatory cytokines interleukin-6 (IL-6) and
tumor necrosis 361
factor–α (TNF-α) in HeLa S3 cells transfected with poly(I:C). We
found that NUDT21 362
knockdown markedly suppressed the increase in the amount of
IFN-β mRNA induced 363
by poly(I:C) stimulation (Figure 3A). The up-regulation of IL-6
and TNF-α mRNAs in 364
response to poly(I:C) transfection was also significantly
attenuated by depletion of 365
NUDT21 (Figure 3B, C). These results thus suggested that NUDT21
is necessary for 366
the efficient induction of IFN-β and proinflammatory cytokines
in response to the 367
presence of cytoplasmic dsRNA. 368
We next examined whether NUDT21 mediates activation of the
transcription 369
factor IRF3 or the mitogen-activated protein kinases (MAPKs) p38
and c-Jun NH2-370
terminal kinase (JNK), all of which are essential factors for
activation of the promoters 371
of IFN-β and inflammatory cytokine genes in response to
cytoplasmic dsRNA (15, 48). 372
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
13
Knockdown of NUDT21 suppressed the increase in the abundance of
phosphorylated 373
(activated) forms of IRF3 as well as of p38 and JNK induced by
poly(I:C) transfection 374
(Figure 3D), suggesting that NUDT21 promotes the activation of
these signaling 375
molecules that is essential for the transcription of IFN-β and
inflammatory cytokine 376
genes. In addition, NUDT21 knockdown suppressed the cleavage of
caspase-3 and 377
poly(ADP-ribose) polymerase (PARP) induced by poly(I:C)
transfection (Figure 3D), 378
suggesting that NUDT21 also plays a key role in the induction of
caspase activation, 379
and perhaps cell death, in response to the presence of
cytoplasmic dsRNA. Together, 380
these results indicated that NUDT21 mediates antiviral cellular
responses induced by 381
the presence of cytoplasmic dsRNA. 382
383
NUDT21 mediates type I IFN induction in response to virus
infection 384
To investigate further the role of NUDT21 in antiviral responses
triggered by RLRs and 385
IPS-1, we asked whether NUDT21 knockdown affects type I IFN
induction in cells 386
infected with viruses. To this end, we studied EMCV, whose RNAs
released into the 387
cytoplasm are recognized predominantly by the RLRs MDA5 (6). We
found that 388
NUDT21 knockdown significantly suppressed the EMCV-induced
phosphorylation of 389
IRF3 and increase in IFN-β mRNA abundance (Figure 4A, B).
Together, these results 390
implicated NUDT21 in the optimal induction of IFN-β in response
to viral infection 391
392
Forced localization of IPS-1 to SGs promotes type I IFN
induction in response to 393
cytoplasmic dsRNA 394
Disruption of SGs has been shown to attenuate IFN induction by
dsRNA (25, 30), 395
although SGs appear to be dispensable for antiviral responses in
some instances (49). 396
We therefore investigated whether forced targeting of IPS-1 to
SGs might enhance IFN 397
induction in poly(I:C)-transfected cells. We first constructed
an expression vector for a 398
fusion protein containing the SG protein G3BP1 and Myc-tagged
IPS-1. However, 399
expression of G3BP1-Myc-IPS-1 resulted in formation of abnormal
aggregates within 400
HeLa S3 cells that appeared to include mitochondria as shown by
TOMM20 401
immunostaining (Figure S5). To avoid such mitochondrial
aggregation, we generated a 402
fusion protein containing G3BP1 and a Myc-tagged form of IPS-1
that lacks the 403
COOH-terminal TM domain (Figure 5A). We found that this
G3BP1-Myc-IPS-1(ΔTM) 404
fusion protein was present mostly in the cytosol of HeLa S3
cells in the absence of 405
poly(I:C) transfection, but preferentially localized to SGs in
the presence of poly(I:C) 406
(Figure 5B). We then measured the level of IFN-β mRNA in cells
expressing G3BP1, 407
Myc-tagged IPS-1(ΔTM), or G3BP1-Myc-IPS-1(ΔTM) at 9 h after
poly(I:C) 408
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
14
transfection (Figure 5C, D). Forced expression of G3BP1 resulted
in slight enhancement 409
of the increase in the amount of IFN-β mRNA induced by poly(I:C)
transfection, 410
consistent with a previous observation (50). We also found that
expression of Myc-411
tagged IPS-1(ΔTM) slightly enhanced the increase in IFN-β mRNA
abundance induced 412
by poly(I:C). Expression of G3BP1-Myc-IPS-1(ΔTM) resulted in a
markedly greater 413
increase in the level of IFN-β mRNA in poly(I:C)-transfected
cells compared with that 414
induced by G3BP1 or Myc-tagged IPS-1(ΔTM) (Figure 5D).
Importantly, the amount of 415
G3BP1-Myc-IPS-1(ΔTM) in the cells was markedly lower than that
of Myc–416
IPS(1ΔTM). These results together suggested that the
localization of IPS-1 to SGs 417
enhances IFN induction in the presence of cytoplasmic dsRNA.
418
419
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
15
Discussion 420
In the innate immune system, the site of receptor-ligand
interaction is often dissociated 421
from that of signal transduction, likely in order to prevent
aberrant activation of 422
antipathogen programs in the absence of infection (51). However,
these distinct 423
compartments must encounter each other soon after ligand
detection. We have now 424
identified NUDT21 as a link between RLR-containing stress
granules and 425
mitochondrial IPS-1 as well as an essential mediator of
antiviral responses. Our study 426
has therefore unveiled the existence of a cellular machinery
that links the site of 427
pathogen recognition and that of antiviral signal transduction.
428
What is the mechanism by which NUDT21 mediates type I IFN
production? 429
Although we cannot exclude the possibility that NUDT21 regulates
type I IFN 430
expression through alternative polyadenylation in the nucleus,
we propose a model 431
whereby NUDT21 promotes antiviral responses by recruiting IPS-1
to antiviral SGs on 432
the basis of the following observations: (1) Both mass
spectrometry and co-433
immunoprecipitation analyses indicated that NUDT21 forms a
complex with 434
mitochondrial IPS-1. (2) A fraction of NUDT21 was found to be
localized to 435
mitochondria in resting cells and became localized to SGs in
response to poly(I:C) 436
transfection. (3) Another component of the CFIm complex, CFIm68,
appeared to be 437
localized only in the nucleus, not being detected in the
cytoplasm, suggesting that 438
NUDT21 has a cytoplasmic function independent of the CFIm
complex. (4) 439
Knockdown of NUDT21 suppressed the change in the localization of
IPS-1 from 440
mitochondria to SGs, but not the formation of SGs, in response
to poly(I:C) stimulation. 441
Further studies are required to understand in more detail how
NUDT21 regulates IPS-1 442
localization in response to poly(I:C) transfection. 443
Caspase-mediated IPS-1 cleavage at a juxtamembrane site has been
proposed 444
to inactivate the function of IPS-1 (33, 34). However, an IPS-1
mutant lacking the TM 445
domain was found to form prionlike fibers (“seeds”) that could
convert native IPS-1 446
into functional aggregates (11). The role of IPS-1 cleavage
therefore remains 447
controversial. We found that expression of an IPS-1(ΔTM) mutant
significantly 448
enhanced the activity of the IFN-β gene promoter only in the
presence of full-length 449
IPS-1 (Figure S6). Moreover, our results showed that forced
localization of IPS-1(ΔTM) 450
to SGs enhanced type I IFN induction by poly(I:C) transfection.
On the basis of these 451
observations, we propose a two-step model: In the early stage of
virus infection, the 452
caspase-cleaved form of IPS-1 cooperates with full-length IPS-1
to form large 453
aggregates on the mitochondrial surface that associate with RLRs
in SGs and thereby 454
elicit antiviral responses. In the late stage, caspases cleave
the remaining intact IPS-1 455
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
16
molecules and thereby terminate IPS-1–mediated immune responses.
This model may 456
reconcile the apparent discrepancy in the effects of
caspase-mediated cleavage on IPS-1 457
function mentioned above. 458
The precise nature of the physical interaction between SGs and
IPS-1 459
aggregates remains unclear. Antiviral SGs are membraneless
organelles that are formed 460
by liquid-liquid phase separation within the cytoplasm. We
observed that a fraction of 461
IPS-1 appeared to form fiberlike structures rather than being
evenly distributed within 462
SGs (Figure 2A). This observation and the previous findings on
the prionlike 463
aggregation of IPS-1 (11) suggest that IPS-1 aggregates may form
a subcompartment 464
within SGs, perhaps by undergoing a liquid-to-solid phase
transition (which typically 465
underlies the formation of solid prionlike aggregates or
crystals), or that they may lie 466
adjacent to SGs and associate with RIG-I in SGs at the
interface. In these two cases, the 467
presence of NUDT21 may facilitate functional solidification of
IPS-1 in SGs or 468
functional association between RIG-I in SGs and IPS-1
aggregates, respectively. 469
Lysine-63–linked polyubiquitination by the ubiquitin ligase
TRIM25 has been 470
implicated in the activation of RIG-I and subsequent formation
of large IPS-1 471
aggregates (52, 53, 11). It would thus be of interest to examine
the relation between 472
such TRIM25-mediated polyubiquitination and NUDT21 in the
formation of IPS-1 473
aggregates and their association with SGs. Of note, RIG-I has
been shown to associate 474
exclusively with either TRIM25 or IPS-1, with the two complexes
being localized to 475
distinct compartments (54). Together with our observation that
IPS-1 is not distributed 476
among all SGs in a cell, this finding suggests that RIG-I might
shift from the TRIM25 477
compartment to the IPS-1 compartment in association with IPS-1
activation (54) and 478
that NUDT21 may facilitate this transition. 479
In closing, we have here demonstrated the cytoplasmic
localization of 480
NUDT21 and its unexpected role in regulation of antiviral
proteins in the cytoplasm in 481
addition to its well-established localization to nucleus and
function in alternative 482
polyadenylation. We therefore propose that NUDT21 may function
in broader biological 483
contexts, at least at mitochondria and SGs, than anticipated,
and our results provide a 484
basis for the development of a new target for clinical
intervention in viral propagation. 485
486
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
17
Acknowledgments 487
We thank M. Okajima and K. Takechi (Graduate School of
Pharmaceutical Sciences, 488
The University of Tokyo) for technical assistance; M. Yoneyama
(Chiba University) for 489
providing the pEF-BOS-FLAG-IPS-1 plasmid, antibodies to RIG-I,
and ECMV; and 490
laboratory members for discussion. 491
492
Author Contributions 493
S.A.I. performed experiments and analyzed data. T.N. and S.I.
conducted the mass 494
spectrometric analysis. Y.O. provided the pN1/G3BP1-iRFP plasmid
and technical 495
assistance for imaging analysis. S.A.I., Y.G., and T.O.
conceived the study and wrote the 496
manuscript. 497
498
Funding 499
This work was supported by a Grant-in-Aid from the Ministry of
Education, Culture, 500
Sports, Science, and Technology (MEXT) of Japan; by Core
Research for Evolutionary 501
Science and Technology of the Japan Science and Technology
Agency; by research 502
fellowships from the Japan Society for the Promotion of Science
(JSPS) and the Global 503
Centers of Excellence Program (Integrative Life Science Based on
the Study of 504
Biosignaling Mechanisms) of MEXT; by the Graduate Program for
Leaders in Life 505
Innovation, The University of Tokyo Life Innovation Leading
Graduate School, of 506
MEXT; and by JSPS KAKENHI grants, JP18gm0610013, JP15H05773,
JP16H06481, 507
JP16H06279, and JP16H06479 to Y.G., JP16K19149 and JP18K07168 to
T.O., and 508
JP16H06280 to Y.O., and JP15J10794 to S.A.I.. 509
510
Declaration of Interests 511
The authors declare no competing interests. 512
513
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
18
References 514
1. Stetson, D.B., and R. Medzhitov. 2006. Type I interferons in
host defense. 515
Immunity. 25: 373–381. 516
2. Schneider, W.M., M.D. Chevillotte, and C.M. Rice. 2014.
Interferon-stimulated 517
genes: a complex web of host defenses. Annu. Rev. Immunol. 32:
513–545. 518
3. Akira, S., S. Uematsu, and O. Takeuchi. 2006. Pathogen
recognition and innate 519
immunity. Cell. 124: 783–801. 520
4. Yoneyama, M., M. Kikuchi, T. Natsukawa, N. Shinobu, T.
Imaizumi, M. Miyagishi, 521
K. Taira, S. Akira, and T. Fujita. 2004. The RNA helicase RIG-I
has an essential 522
function in double-stranded RNA-induced innate antiviral
responses. Nat Immunol. 523
5: 730–737. 524
5. Gitlin, L., W. Barchet, S. Gilfillan, M. Cella, B. Beutler,
R.A. Flavell, M.S. 525
Diamond, and M. Colonna. 2006. Essential role of mda-5 in type I
IFN responses to 526
polyriboinosinic: polyribocytidylic acid and
encephalomyocarditis picornavirus. 527
Proc Natl Acad Sci USA. 103: 8459–8464. 528
6. Kato, H., O. Takeuchi, S. Sato, M. Yoneyama, M. Yamamoto, K.
Matsui, S. 529
Uematsu, A. Jung, T. Kawai, K.J. Ishii, O. Yamaguchi, K. Otsu,
T. Tsujimura, C.S. 530
Koh, C. Reis e Sousa, Y. Matsuura, T. Fujita, and S. Akira.
2006. Differential roles 531
of MDA5 and RIG-I helicases in the recognition of RNA viruses.
Nature. 441: 101–532
105. 533
7. Kawai, T., K. Takahashi, S. Sato, C. Coban, H. Kumar, H.
Kato, K.J. Ishii, O. 534
Takeuchi, and S. Akira. 2005. IPS-1, an adaptor triggering
RIG-I- and Mda5-535
mediated type I interferon induction. Nat Immunol. 6: 981–988.
536
8. Seth, R.B., L. Sun, C.K. Ea, and Z.J. Chen. 2005.
Identification and 537
characterization of MAVS, a mitochondrial antiviral signaling
protein that activates 538
NF-κB and IRF3. Cell. 122: 669–682. 539
9. Meylan, E., J. Curran, K. Hofmann, D. Moradpour, M. Binder,
R. Bartenschlager, 540
and J. Tschopp. 2005. Cardif is an adaptor protein in the RIG-I
antiviral pathway 541
and is targeted by hepatitis C virus. Nature. 437: 1167–1172.
542
10. Xu, L.G., Y.Y. Wang, K.J. Han, L.Y. Li, Z. Zhai, and H.B.
Shu. 2005. VISA is an 543
adapter protein required for virus-triggered IFN-β signaling.
Mol Cell. 19: 727–544
740. 545
11. Hou, F., L. Sun, H. Zheng, B. Skaug, Q.X. Jiang, and Z.J.
Chen. 2011. MAVS 546
forms functional prion-like aggregates to activate and propagate
antiviral innate 547
immune response. Cell. 146: 448–461. 548
12. Fitzgerald, K.A., S.M. Mcwhirter, K.L. Faia, D.C. Rowe, E.
Latz, D.T. Golenbock, 549
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
19
A.J. Coyle, S.M. Liao, and T. Maniatis. 2003. IKKε and TBK1 are
essential 550
components of the IRF3 signaling pathway. Nat Immunol. 4:
491–496. 551
13. Chu, W.M., D. Ostertag, Z.W. Li, L. Chang, Y. Chen, Y. Hu,
B. Williams, J. 552
Perrault, and M. Karin. 1999. JNK2 and IKKβ are required for
activating the innate 553
response to viral infection. Immunity. 11: 721–731. 554
14. Du, W., and T. Maniatis. 1992. An ATF / CREB binding site is
required for virus 555
induction of the human interferon beta gene. Proc Natl Acad Sci
USA. 89: 2150–556
2154. 557
15. Honda, K., A. Takaoka, and T. Taniguchi. 2006. Type I
inteferon gene induction by 558
the interferon regulatory factor family of transcription
factors. Immunity. 25: 349–559
360. 560
16. Okazaki, T., M. Higuchi, K. Takeda, K. Iwatsuki-Horimoto, M.
Kiso, M. Miyagishi, 561
H. Yanai, A. Kato, M. Yoneyama, T. Fujita, T. Taniguchi, Y.
Kawaoka, H. Ichijo, 562
and Y. Gotoh. 2015. The ASK family kinases differentially
mediate induction of 563
type I interferon and apoptosis during the antiviral response.
Sci Signal. 8: ra78. 564
17. Kumar, H., T. Kawai, H. Kato, S. Sato, K. Takahashi, C.
Coban, M. Yamamoto, S. 565
Uematsu, K.J. Ishii, O. Takeuchi, and S. Akira. 2006. Essential
role of IPS-1 in 566
innate immune responses against RNA viruses. J Exp Med. 203:
1795–1803. 567
18. Sun, Q., L. Sun, H.H. Liu, X. Chen, R.B. Seth, J. Forman,
and Z.J. Chen. 2006. The 568
specific and essential role of MAVS in antiviral innate immune
responses. 569
Immunity. 24: 633–642. 570
19. Yoneyama, M., M. Kikuchi, K. Matsumoto, T. Imaizumi, M.
Miyagishi, K. Taira, E. 571
Foy, Y.M. Loo, M. Gale Jr., S. Akira, S. Yonehara, A. Kato, and
T. Fujita. 2005. 572
Shared and unique functions of the DExD/H-Box helicases RIG-I,
MDA5, and 573
LGP2 in antiviral innate immunity. J Immunol. 175: 2851–2858.
574
20. Buchan, J.R., and R. Parker. 2009. Eukaryotic stress
granules : the ins and outs of 575
translation. Mol Cell. 36: 932–941. 576
21. McCormick, C., and D.A. Khaperskyy. 2017. Translation
inhibition and stress 577
granules in the antiviral immune response. Nat Rev Immunol. 17:
647-660. 578
22. Kedersha, N.L., M. Gupta, W. Li, I. Miller, and P. Anderson.
1999. RNA-binding 579
proteins TIA-1 and TIAR link the phosphorylation of eIF-2α to
the assembly of 580
mammalian stress granules. J Cell Biol. 147: 1431–1442. 581
23. Tourrière, H., K. Chebli, L. Zekri, B. Courselaud, J.M.
Blanchard, E. and Bertrand, 582
J. Tazi. 2003. The RasGAP-associated endoribonuclease G3BP
assembles stress 583
granules. J Cell Biol. 160: 823–831. 584
24. Boeynaems, S., S. Alberti, N.L. Fawzi, T. Mittag, M.
Polymenidou, F. Rousseau, J. 585
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
20
Schymkowitz, J. Shorter, B. Wolozin, L. Van Den Bosch, P. Tompa,
and M. 586
Fuxreiter. 2018. Protein Phase Sparation: A New Phase in Cell
Biology. Trends Cell 587
Biol. 28: 420–435. 588
25. Onomoto, K., M. Jogi, J.S. Yoo, R. Narita, S. Morimoto, A.
Takemura, S. 589
Sambhara, A. Kawaguchi, S. Osari, K. Nagata, T. Matsumiya, H.
Namiki, M. 590
Yoneyama, and T. Fujita. 2012. Critical role of an antiviral
stress granule containing 591
RIG-I and PKR in viral detection and innate immunity. PLoS One.
7: e43031. 592
26. Oh, S.W., K. Onomoto, M. Wakimoto, K. Onoguchi, F. Ishidate,
T. Fujiwara, M. 593
Yoneyama, H. Kato, and T. Fujita. 2016. Leader-Containing
Uncapped Viral 594
Transcript Activates RIG-I in Antiviral Stress Granules. PLoS
Pathog. 12: 595
e1005444. 596
27. White, J.P., A.M. Cardenas, W.E. Marissen, and R.E. Lloyd.
2007. Inhibition of 597
cytoplasmic mRNA stress granule formation by a viral proteinase.
Cell Host 598
Microbe. 2: 295–305. 599
28. Khaperskyy, D.A., T.F. Hatchette, and C. McCormick. 2012.
Influenza A virus 600
inhibits cytoplasmic stress granule formation. FASEB J. 26:
1629–1639. 601
29. Borghese, F., and T. Michiels. 2011. The leader protein of
cardioviruses inhibits 602
stress granule assembly. J Virol. 85: 9614–9622. 603
30. Ng, C.S., M. Jogi, J.S. Yoo, K. Onomoto, S. Koike, T.
Iwasaki, M. Yoneyama, H. 604
Kato, and T. Fujita. 2013. Encephalomyocarditis virus disrupts
stress granules, the 605
critical platform for triggering antiviral innate immune
responses. J Virol. 87: 606
9511–9522. 607
31. Finnen, R.L., M. Zhu, J. Li, D. Romo, and B.W. Banfield.
2016. Herpes Simplex 608
Virus 2 Virion Host Shutoff Endoribonuclease Activity Is
Required To Disrupt 609
Stress Granule Formation. J Virol. 90: 7943–7955. 610
32. Li, X.D., L. Sun, R.B. Seth, G. Pineda, and Z.J. Chen. 2005.
Hepatitis C virus 611
protease NS3/4A cleaves mitochondrial antiviral signaling
protein off the 612
mitochondria to evade innate immunity. Proc Natl Acad Sci USA.
102: 17717–613
17722. 614
33. Rebsamen, M., E. Meylan, J. Curran, and J. Tschopp. 2008.
The antiviral adaptor 615
proteins Cardif and Trif are processed and inactivated by
caspases. Cell Death 616
Differ. 15: 1804–1811. 617
34. Ning, X., Y. Wang, M. Jing, M. Sha, M. Lv, P. Gao, R. Zhang,
X. Huang, J.M. 618
Feng, and Z. Jiang. 2019. Apoptotic Caspases Suppress Type I
Interferon 619
Production via the Cleavage of cGAS, MAVS, and IRF3. Mol Cell.
74: 19-31.e7. 620
35. Zhang, Peifen, Y. Li, J. Xia, J. He, J. Pu, J. Xie, S. Wu,
L. Feng, X. Huang, and 621
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
21
Ping Zhang. 2014. IPS-1 plays an essential role in dsRNA-induced
stress granule 622
formation by interacting with PKR and promoting its activation.
J Cell Sci. 127: 623
2471–2482. 624
36. Brown, K.M., and G.M. Gilmartin. 2003. A mechanism for the
regulation of pre-625
mRNA 3’ processing by human cleavage factor I m. Mol Cell. 12:
1467–1476. 626
37. Di Giammartino, D.C., K. Nishida, and J.L. Manley. 2011.
Mechanisms and 627
consequences of alternative polyadenylation. Mol Cell. 43:
853–866. 628
38. Yang, Q., M. Coseno, G.M. Gilmartin, and S. Doublié. 2011.
Crystal structure of a 629
human cleavage factor CFI(m)25 / CFI(m)68 / RNA complex provides
an insight 630
into poly (A) site recognition and RNA looping. Structure. 19:
368–377. 631
39. Elkon, R., A.P. Ugalde, and R. Agami. 2013. Alternative
cleavage and 632
polyadenylation: extent, regulation and function. Nat Rev Genet.
14: 496–506. 633
40. Masamha, C.P., Z. Xia, J. Yang, T.R. Albrecht, M. Li, A.B.
Shyu, W. Li, and E.J. 634
Wagner. 2014. CFIm25 links alternative polyadenylation to
glioblastoma tumour 635
suppression. Nature. 510: 412–416. 636
41. Brumbaugh, J., B. Di Stefano, X. Wang, M. Borkent, E.
Forouzmand, K.J. Clowers, 637
F. Ji, B.A. Schwarz, M. Kalocsay, S.J. Elledge, Y. Chen, R.I.
Sadreyev, S.P. Gygi, 638
G. Hu, Y. Shi, and K. Hochedlinger. 2018. Nudt21 Controls Cell
Fate by 639
Connecting Alternative Polyadenylation to Chromatin Signaling.
Cell. 172: 106-640
120.e21. 641
42. Jia, X., S. Yuan, Y. Wang, Y. Fu, Yong Ge, Yutong Ge, X.
Lan, Y. Feng, F. Qiu, P. 642
Li, S. Chen, and A. Xu. 2017. The role of alternative
polyadenylation in the 643
antiviral innate immune response. Nat Commun. 8: 14605. 644
43. Gaucherand, L., B.K. Porter, R.E. Levene, E.L. Price, S.K.
Schmaling, C.H. 645
Rycroft, Y. Kevorkian, C. McCormick, D.A. Khaperskyy, and M.M.
Gaglia. 2019. 646
The Influenza A Virus Endoribonuclease PA-X Usurps Host mRNA
Processing 647
Machinery to Limit Host Gene Expression. Cell Rep. 27:
776-792.e7. 648
44. Okazaki, T., M. Higuchi, and Y. Gotoh. 2013. Mitochondrial
localization of the 649
antiviral signaling adaptor IPS-1 is important for its induction
of caspase activation. 650
Genes Cells. 18: 493–501. 651
45. Natsume, T., Y. Yamauchi, H. Nakayama, T. Shinkawa, M.
Yanagida, N. Takahashi, 652
and T. Isobe. 2002. A direct nanoflow liquid chromatography -
tandem mass 653
spectrometry system for interaction proteomics. Anal Chem. 74:
4725–4733. 654
46. Costes, S.V., D. Daelemans, E.H. Cho, Z. Dobbin, G.
Pavlakis, and S. Lockett. 655
2004. Automatic and quantitative measurement of protein-protein
colocalization in 656
live cells. Biophys J. 86: 3993–4003. 657
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
22
47. Manders, E.M.M., F.J. Verbeek, and J.A. Aten. 1993.
Measurement of co-658
localization of objects in dual-colour confocal images. J.
Microsc. 169: 375–382. 659
48. Mikkelsen, S.S., S.B. Jensen, S. Chiliveru, J. Melchjorsen,
I. Julkunen, M. Gaestel, 660
J.S. Arthur, R.A. Flavell, S. Ghosh, and S.R. Paludan. 2009.
RIG-I-mediated 661
activation of p38 MAPK is essential for viral induction of
interferon and activation 662
of dendritic cells: dependence on TRAF2 and TAK1. J Biol Chem.
284: 10774–663
10782. 664
49. Langereis, M.A., Q. Feng, and F.J. van Kuppeveld. 2013. MDA5
localizes to stress 665
granules, but this localization is not required for the
induction of type I interferon. J 666
Virol. 87: 6314–6325. 667
50. Kim, S.S., L. Sze, C. Liu, and K.P. Lam. 2019. The stress
granule protein G3BP1 668
binds viral dsRNA and RIG-I to enhance interferon-β response. J
Biol Chem. 294: 669
6430–6438. 670
51. Kagan, J.C. 2012. Signaling organelles of the innate immune
system. Cell. 151: 671
1168–1178.. 672
52. Gack, M.U., Y.C. Shin, C.H. Joo, T. Urano, C. Liang, L. Sun,
O. Takeuchi, S. Akira, 673
Z. Chen, S. Inoue, and J.U. Jung. 2007. TRIM25 RING-finger E3
ubiquitin ligase is 674
essential for RIG-I-mediated antiviral activity. Nature. 446:
916–920. 675
53. Jiang, X., L.N. Kinch, C.A. Brautigam, X. Chen, F. Du, N.V.
Grishin, and Z.J. 676
Chen. 2012. Ubiquitin-induced oligomerization of the RNA sensors
RIG-I and 677
MDA5 activates antiviral innate immune response. Immunity. 36:
959–973. 678
54. Sánchez-Aparicio, M.T., J. Ayllón, A. Leo-Macias, T. Wolff,
and A. García-Sastre. 679
2017. Subcellular Localizations of RIG-I, TRIM25, and MAVS
Complexes. J Virol. 680
91: e01155-16. 681
682
683
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
23
Figure Legends 684
Figure 1. A fraction of NUDT21 localizes to mitochondria in
resting cells and 685
contributes to SGs in the cytoplasm in response to poly(I:C)
transfection. 686
(A) Extracts of HEK293T cells transiently transfected with
expression vectors for 687
FLAG-tagged IPS1 and Myc-tagged NUDT21 were subjected to
immunoprecipitation 688
(IP) with antibodies to FLAG, and the resulting precipitates as
well as the original cell 689
extracts (Total) were subjected to immunoblot analysis with
antibodies to FLAG and to 690
Myc. The asterisk indicates a nonspecific band. Data are
representative of three 691
independent experiments. 692
(B) Immunofluorescence analysis of HeLa S3 cells transfected
with control or NUDT21 693
siRNAs. The cells were stained with antibodies to NUDT21 and to
cytochrome c as 694
well as with Hoechst 33342. The boxed regions in the upper
panels are shown at higher 695
magnification in the corresponding middle panels. Scale bars, 5
μm or 2 μm (higher 696
magnification images). Data are representative of three
independent experiments. 697
(C) Immunofluorescence analysis of control and NUDT21-depleted
HeLa S3 cells at 6 698
h after transfection with poly(I:C) (0.25 μg/ml) or mock
transfection. The cells were 699
stained with antibodies to NUDT21 and to TIAR as well as with
Hoechst 33342. Scale 700
bars, 5 μm. Data are representative of three independent
experiments. 701
(D) Quantification of the NUDT21-positive area of TIAR-positive
foci per cell in 702
images similar to those in (C). Data are means ± s.e.m. from
three independent 703
experiments (control, n = 16; NUDT21 knockdown, n = 13). ***P
< 0.005 (Student’s t 704
test). 705
706
Figure 2. NUDT21 knockdown impairs the localization of IPS-1 to
SGs. 707
(A) Immunofluorescence analysis of HeLa S3 cells at 6 h after
transfection with 708
poly(I:C) (0.25 μg/ml) or mock transfection. The cells were
stained with antibodies to 709
IPS-1, to TOMM20, and to TIAR as well as with Hoechst 33342. The
boxed regions of 710
the middle panels are shown at higher magnification in the
corresponding lower panels. 711
Arrowheads indicate SGs. Scale bars, 5 μm or 2 μm (higher
magnification images). 712
Data are representative of three independent experiments.
713
(B) Colocalization (Manders M1 and M2) between IPS-1 and TIAR
was determined for 714
images as in (A). Manders M1 is the sum of TIAR signal intensity
overlapping with 715
IPS-1 versus total TIAR signal intensity, whereas Manders M2 is
the sum of IPS-1 716
signal intensity overlapping with TIAR versus total IPS-1 signal
intensity. Data are 717
means ± s.e.m. from three independent experiments (mock, n = 5;
poly(I:C), n = 11). 718
**P < 0.01 (Student’s t test). 719
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
24
(C) HeLa S3 cells transiently expressing iRFP-fused G3BP1 were
subjected to 720
immunofluorescence staining with antibodies to IPS-1 and to
TOMM20 at 6 h after 721
transfection with poly(I:C) (0.25 μg/ml) or mock transfection.
The fluorescence of iRFP 722
was monitored directly. The z-stack images were deconvoluted
with Huygens software. 723
The boxed regions of the left images are shown at higher
magnification in those on the 724
right. Scale bars, 5 μm or 2 μm (higher magnification images).
Line-scan analysis of the 725
relative fluorescence intensity of IPS-1, TOMM20, and G3BP1-iRFP
along the white 726
broken lines (6 μm) in the enlarged images is also shown. Data
are representative of 727
three independent experiments. 728
(D) Colocalization (Manders M1 and M2) between IPS-1 and TOMM20
was 729
determined for images as in (C). Manders M1 is the sum of IPS-1
signal intensity 730
overlapping with TOMM20 versus total IPS-1 signal intensity, and
Manders M2 is the 731
sum of TOMM20 signal intensity overlapping with IPS-1 versus
total TOMM20 signal 732
intensity. Data are means ± s.e.m. from three independent
experiments (mock, n = 12; 733
poly(I:C), n = 18). *P < 0.05 (Student’s t test). 734
(E) Immunofluorescence analysis of HeLa S3 cells expressing
control or NUDT21 735
siRNAs at 6 h after transfection with poly(I:C) (0.25 μg/ml).
The cells were stained with 736
antibodies to IPS-1, to TOMM20, and to TIAR. The z-stack images
were deconvoluted 737
with Huygens software. The boxed region of the upper image of
each pair is shown at 738
higher magnification in the lower image. Arrowheads indicate
SGs. Scale bars, 5 μm or 739
2 μm (higher magnification images). Data are representative of
three independent 740
experiments. 741
(F) Colocalization (Manders M1, sum of TIAR signal intensity
overlapping with IPS-1 742
versus total TIAR signal intensity) between IPS-1 and TIAR was
determined for images 743
as in (E). Data are means ± s.e.m. of three independent
experiments (control, n = 10; 744
NUDT21 knockdown, n = 10). ****P < 0.001 (Student’s t test).
745
(G) Quantification of SG volume per cell for images as in (E).
Data are means ± s.e.m. 746
of three independent experiments (control, n = 10; NUDT21
knockdown, n = 10). The P 747
value was determined with Student’s t test. 748
(H) Quantification of SG-containing cells. HeLa S3 cells
expressing control or 749
NUDT21 siRNAs were subjected to immunofluorescence staining for
TIAR in order to 750
determine the proportion of SG-containing cells at 6 h after
transfection with poly(I:C) 751
(0.25 μg/ml) or mock transfection. Data are means ± s.e.m. of
three independent 752
experiments. The P value was determined with Student’s t test.
753
(I) Immunofluorescence analysis of control and NUDT21-knockdown
HeLa S3 cells at 754
6 h after transfection with poly(I:C) (0.25 μg/ml) or mock
transfection. The cells were 755
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
25
stained with antibodies to RIG-I and to TIAR as well as with
Hoechst 33342. Scale 756
bars, 5 μm. Data are representative of three independent
experiments. 757
758
Figure 3. NUDT21 plays a key role in the induction of type I IFN
and caspase 759
activation in response to poly(I:C) transfection. 760
(A–C) Quantitative reverse transcription and polymerase chain
reaction (RT-PCR) 761
analysis of IFN-β, IL-6, and TNF-α mRNAs, respectively, in HeLa
S3 cells transiently 762
expressing control or NUDT21 siRNAs at 9 h after transfection
with poly(I:C) (0.25 763
μg/ml) or mock transfection. Data are expressed relative to the
corresponding amount of 764
GAPDH mRNA and are means ± s.e.m. from four independent
experiments. **P < 765
0.01, ****P < 0.001 (Student’s t test). 766
(D) Immunoblot analysis of phosphorylated (p-) forms of IRF3,
p38 MAPK, and JNK 767
as well as of cleaved forms of caspase-3 and PARP in control or
NUDT21-knockdwon 768
HeLa S3 cells at 9 h after transfection with poly(I:C) [0.25 (+)
or 2.5 (++) μg/ml] or 769
mock transfection. The amounts of NUDT21 and total p38 were
examined as controls 770
for knockdown efficiency and protein loading, respectively.
Asterisks indicate 771
nonspecific bands. Data are representative of three independent
experiments. 772
773
Figure 4. NUDT21 is required for the optimal induction of type I
IFN in response to 774
EMCV or NDV infection. 775
(A) Quantitative RT-PCR analysis of IFN-β mRNA in HeLa S3 cells
transiently 776
expressing control or NUDT21 siRNAs at 12 h after infection with
0.2 plaque-forming 777
units (PFU) of EMCV. Data are means ± s.e.m. from four
independent experiments. *P 778
< 0.05 (Student’s t test). 779
(B) Immunoblot analysis of phosphorylated IRF3, NUDT21, and p38
in control or 780
NUDT21-knockdown HeLa S3 cells infected with 0.1 (+) or 0.2 (++)
PFU of EMCV 781
for 12 h. The asterisk indicates nonspecific bands. Data are
representative of three 782
independent experiments. 783
784
Figure 5. Forced localization of IPS-1 to SGs enhances type I
IFN induction in 785
response to poly(I:C) transfection. 786
(A) Schematic representation of human IPS-1, Myc-tagged
IPS-1(ΔTM), and G3BP1-787
Myc-IPS-1(ΔTM). 788
(B) Immunofluorescence analysis of HeLa S3 cells transiently
expressing G3BP1-Myc-789
IPS-1(ΔTM) at 6 h after transfection with poly(I:C) (0.25 μg/ml)
or mock transfection. 790
The cells were stained with antibodies to Myc, to TIAR, and to
TOMM20. Scale bars, 5 791
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
26
μm. Data are representative of three independent experiments.
792
(C) Immunoblot analysis with antibodies to Myc of HeLa S3 cells
transiently 793
expressing Myc–IPS-1(ΔTM) or G3BP1-Myc-IPS-1(ΔTM) at 9 h after
transfection with 794
poly(I:C) (0.25 μg/ml) or mock transfection. Black and blue
arrowheads indicate Myc–795
IPS-1(ΔTM) and G3BP1-Myc-IPS-1(ΔTM), respectively. Data are
representative of 796
three independent experiments. 797
(D) Quantitative RT-PCR analysis of IFN-β mRNA in HeLa S3 cells
transiently 798
expressing G3BP1, Myc–IPS-1(ΔTM), or G3BP1-Myc-IPS-1(ΔTM) at 9 h
after 799
transfection with poly(I:C) (0.25 μg/ml) or mock transfection.
Data are means ± s.e.m. 800
from four independent experiments. *P < 0.05, **P < 0.01,
****P < 0.001 (Student’s t 801
test). 802
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
Con
trol
siR
NA
NU
DT2
1si
RN
A
Poly(I:C)
-
-
+
+
Hoechst NUDT21 TIAR
HoechstNUDT21
TIARC
HoechstNUDT21
(low gain)NUDT21
(high gain) Cytochrome c
ControlsiRNA
NUDT21siRNA
HoechstNUDT21
Cytochrome cB
D
FLAG
Myc
FLAG
Myc
IP: FLAG
Total
FLAG-IPS-1Myc-NUDT21
+ +++
--
A
*
26.590
90 kDa
26.5
Figure 1
0
20
40
60
80
100
ControlsiRNA
NUDT21siRNA
NU
DT2
1+ a
rea/
TIA
R+
area
(%)
***
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
C
D
B
*Colocalization
TOMM20 and IPS-1
050
100150200
0 2 4 6
050
100150200
0 2 4 6Distanc
Rel
ativ
e In
tens
ityR
elat
ive
Inte
nsity
Distanc
TOMM20IPS-1
G3BP1-iRFP
-
+
Figure 2
F
Man
ders
M2
- +0
0.20.40.60.81.0
Con
trol
siR
NA
NU
DT2
1si
RN
A
-
-
+
+
Hoechst RIG-I TIAR
HoechstRIG-ITIAR
I
H
ControlsiRNA
NUDT21siRNA
P = 0.287
0
304050
2010
Volu
e of
TIA
R fo
ci (µ
³
0
10
20
30
ControlsiRNA
NUDT21siRNA
SG+
+ - +-
P = 0.522
P = 0.543
GColocalizationTIAR and IPS-1
A
IPS-1TOMM20
TIARIPS-1 TIAR
ControlsiRNA
NUDT21siRNA
TOMM20E
Man
ders
M1
- +0
0.20.40.60.81.0
Man
ders M
1
0
0.2
0.4
0.6
0.8
1.0 ****
ControlsiRNA
NUDT21siRNA
00.20.40.60.81.0
Man
ders
M1
+-0
0.20.40.60.81.0
+-
Man
ders
M2**
P = 0.993
ColocalizationTIAR and IPS-1
Hoechst IPS-1 TOMM20 TIAR
HoechstIPS-1
TOMM20
HoechstIPS-1TIAR
-
+Deconvoluted
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
D
p-IRF3
p-p38
p-JNK
Cleavedcaspase-3
CleavedPARP
NUDT21
p38
ControlsiRNA
NUDT21siRNA
- + - +Poly(I:C)
*++ ++
*
*
Rel
ativ
em
RN
A a
mou
nt
- + - +Poly(I:C)
ControlsiRNA
NUDT21siRNA
****
1.2
1.0
0.8
0.6
0.4
0.2
0
1.4
Rel
ativ
e IL
-6m
RN
A a
mou
nt- + - +Poly(I:C)
ControlsiRNA
NUDT21siRNA
**
1.2
1.0
0.8
0.6
0.4
0.2
0
1.4
Rel
ativ
em
RN
A a
mou
nt
- + - +Poly(I:C)
ControlsiRNA
NUDT21siRNA
**1.2
1.0
0.8
0.6
0.4
0.2
0
1.4
A B C
48.5 kDa
90
36.5
36.5
48.5
*26.5
26.5
Figure 3
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
p-IRF3
NUDT21
p38
EMCV
ControlsiRNA
NUDT21siRNA
- + - +++ ++
A B
*
Rel
ativ
em
RN
A a
mou
nt
EMCV - - ++
*
ControlsiRNA
NUDT21siRNA
3.0
2.5
2.0
1.5
1.0
0.5
0
36.5
26.5
48.5 kDa
Figure 4
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597
-
Rela
tive
mRN
A am
ount
Control G3BP1 Myc-IPS-1(ΔTM)
G3BP1-Myc-IPS-1 (ΔTM)
- - ++
**3.0
2.5
2.0
1.5
1.0
0.5
0- - ++
P = 0.113
Poly(I:C)
****
*
A
B
C D
Myc
(long exposure)
p38
Myc-IPS-1(ΔTM)
G3BP1-Myc-
IPS-1 (ΔTM)- - ++Poly(I:C)
MycTIAR
TOMM20TOMM20Myc TIARPoly(I:C)
-
+
IPS-1 (full length)
Myc-IPS-1 (ΔTM)
G3BP1-Myc-IPS-1 (ΔTM) 4291
CARDG3BP1 Myc
TM5401
D429
CARD
4291CARDMyc
G3B
P1-M
yc-
IPS-
1 ( Δ
TM)
48.558
90110 kDa
48.558
90110
36.5
Myc
Figure 5
(which was not certified by peer review) is the author/funder.
All rights reserved. No reuse allowed without permission. The
copyright holder for this preprintthis version posted April 10,
2020. ; https://doi.org/10.1101/2020.04.09.033597doi: bioRxiv
preprint
https://doi.org/10.1101/2020.04.09.033597