Embed Size (px)
Citation preview

c.u.biwt..ltf.mpo
UNIVERSIDAD AUT~NOMA MEIROLPOLITANA IZTAPALAPA
DMSI6N DE CIENCIAS BIOLdGICAS Y DE LA SALUD DEPARTAMENTO DE BIOTECNOLOGfA
OBTENCI6N DE MARCADORES GEN&TICOS EN Penicillium chrysogenum UTILIZANDO a T O D O S DE SELECCI6N RACIONAL Y CARACTERIZACI6N PARCIAL DE LAS
MUTANTES.
TESIS Para obtcnrr el grado de
MAESTROENBIOTECNOLOG~A presenta
EDELMIRA(MANZAN0 CUEVAS
M6xic0,D.F. 1998

CONTENIDO.
Lista de abreviaciones. Indice de figuras.
Indice de tablas.
1. Introducción.
I1 Hipótesis.
111. Objetivo.
1V. Generalidades.
1. Clasificacih. 2. Metabolitos ligados y no ligados al crecimiento. 3. Penicilina. 4. Fermentación sólida.
4. l . Definición. 4.2. Ventajas y desventajas.
5.1 . Mutación. 5. Mejoramiento genktico.
V Antecedentes.
111 1v V
1
3
3
4
16
I

VI. Materiales y Métodos.
1. Microorganismos. l . l . Conservación.
2. Medios de cultivo. 3. Condiciones de cultivo.
3.1 Fermentación Líquida. 3.2 Fermentación Sólida.
4. Mutagdnesis y Curva de Sobrevivencia. 5. Obtención de mutantes Auxótrofas a aminoácidos.
5.1 . Procesos de enriquecimiento por filtraciones sucesivas. 5.2. Enzima lítica Novozyme.
6.1. Aislamiento de mutantes resistentes al 5 FOA. 6. Obtención de mutantes Auxótrofos a uridina.
VU. Resultados.
1. Selección de cepas. 2. Aislamiento de auxótrofos.
2. I . Mutaghesis. 2.2. Mutantes Auxótrofos a aminoácidos. 2.3. Mutantes Auxótrofos a uridina.
en mutantes auxótrohs. 3. Evaluaci6n del nivel de producción de penicilina
VI11. Discusión.
I . Selección de cepas. 2. Aislamiento de auxótrofos.
2.1. Mutaghesis. 2.2. Mutantes Auxdtrofos a aminohcidos. 2.3. Mutantes Auxótrofos a uridina.
3. Caracterización de las cepas parentales y auxótrofas. 4. Evaluación del nivel de producción de penicilina
en mutantes aux6trofas.
21
21 21 21 24 24 26 27 28 28 30 31 31
33
33 38 38 42 44
45
54
54 55 55 56 58 59
60
1X. Conclusiones. 63
X. Bibliografia. 64

Lista de abreviaciones.
'AFA " C cm DNA EMS ESP 5FOA FS FL Fig.
h MC MM mg m1 M min MET NTG OMP PdPl PS/PL PDA PEG 'pm S orgfinico SFS SFL SMM TSA UMP uv
-
Acid0 f e d acético.
centímetros. ácido desoxi-ribonucleico. etil metano sulfonato esporas. ácido 5 fluororótico. femcntacih dida. fermentación líquida. figura. gramos. horas. medio completo. medio mínimo. miligramos. mililitros. microgamos. minutos. fenotipo de auxotrofia a metionina. nitroso metil guanidina orotidin monofosfato. producción relativa. productividad relativa. agar papa dextrosa. polietilenglicol. revoluciones por minuto. fenotipo de auxotrofia a fuentes inorgánicas de d e . sistema de fermentación sólida. sistema de fermentación liquida. sólidos de maceración del maíz. agar de soya y tripticaseina. uridin monofosfato. radiación ultravioleta.
grados centígrados.
111

Indice de figuras.
Figura 1 : (pag. 5) Estructura química de las penicilinas.
Figura 2 : (pag. 13) Ruta biosintética de la UMP en hongos filamentosos.
Figura 3 : (pag. 36) Cinéticas de a) producción de penicilina, b)pH y c)humedad de las cepas P2-32 y P2-37 de P. chrysogenum cultivadas en FS a 25 "C y con un flujo de aire de 2.4 V h .
Figura 4 : (pag. 37) Cinacas de a) producción de penicilina, b)pH y c)humedad de l a s cepas P2-32 y P2-37 de P. chrysogenum cultivadas en FL, a 25 "C y 270 rpm.
Figura 5 : (pag. 39) Curva de sobrevivencia. Porciento en función de la irradiación con luz UV y esporas sobrevivientes de la cepa de P. chrysogenum P2-32. Condiciones : 27 ml de suspención acuosa a una concentración de esporas totales de 8 X 1 O6 esp /m], agitación de 36 rpm e intencidad de irradiación UV de 250pW/cm2.
Figura 6 : (pag. 40) Curva de sobrevivencia. Porciento en hción de la irradiación con luz UV y esporas sobrevivientes de la cepa de P. chrysogenum P2-37. Condiciones : 27 ml de suspención acuosa a una concentraci6n de esporas totales de 8 X lo6 esp /ml, agitación de 36 rpm e intencidad de irradiación UV de 250pW/cm2.
Figura 7 : (pag. 41) Comparacibn entre las mutates morfológicas y el porcentaje de sobrevivencia de la cepa de P. chrysogenum P2-37.
Figura 8 : (pag. 48) Cinéticas de producción de penicilina de l a s cepas a) P2-32 y b) P2-37 de P. chrysogenurn tanto de su primera (al inicio del trabajo experimental) como de su segunda caracterización (al caracterizar l a s mutantesauxótrofas aisladas), cultivadas en FS a 25 OC y con un flujo de aire de 2.4 Vh.
IV

Figura 9 : @ag. 49) Cinéticas de produccibn de penicilina de l a s cepas a) P2-32 y b) P2-37 de P. chvsogenum tanto de su primera (al inicio del trabajo experimental) como de su segunda caracterizacibn (al caracterizar l a s mutantesauxbtrofas aisladas), cultivadas en FL a 25 "C y 270 rpm.
Figura 10 : (pag. 50) Cindticas de a) produccidn de penicilina, b)pH y c)humedad de la cepa parental P2-32 y de la mutante auxbtrofa E-32U de Y. chwsogenum cultivadas en FS a 25 "C y con un flujo de aire de 2.4 Vh.
Figura 11 : (pag. 51) Cindticas de a) produccibri de penicilina, b)pH y c)humedad de la cepa parental P2-32 y de la mutante auxdtrofa E-32U de Y. chrysogenurn cultivadas en FL a 25 "C y 270 rpm.
Figura 12 : (pag. 52) Cinéticas de a) produccibn de penicilina, b)pH y c)humedad de la cepa parental P2-37 y de la mutante auxdtrofa E-37M1 de P. chrysogenum cultivadas en FS a 25 "C y con un flujo de aire de 2.4 Vh.
Figura 13 : (pag. 53) Cindticas de a) produccibn de penicilina, b)pH y c)humedad de la cepa parental P2-37 y de la mutante auxbtrofa E-37M1 de Y. chrysogenum cultivadas en FL a 25 "C y 270 rpm.
V

Indice de Tablas.
Tabla 1 : @ag. 30) Suplementación de aminoAcidos al MM para determinar tipo de auxotrofia presente en los mutantes (Gunasekaran 1995).
Tabla 2 : (pag. 33) Comparación de la producción y productividad máxima de penicilina alcanzadas por cinco cepas de P. chrysogenum cultivadas en FS a 25 "C yfl~jodeairede2.4YhyenFLa25~Cy270rpm.
Tabla 3 : (pag. 34) Comparación de los factores de producción y productividad relativa entre l a s cinco cepas de P. chrysogeiurn cultivadas en FS a 25 "C y flujo de airede2.4IihyenFLa25"Cy270rpm.
Tabla 4 : bag. 42) Eficiencia de filtración de tres materiales diferentes.
Tabla S : (pag. 43) Resultados de los procesos de enriquecimiento.
Tabla 6 : (pag. 43) Mutantes aisladas de la cepa P2-37
Tabla 7 : (pag. 44 ) Efecto de tres dosis de 5FOA empleadas para determinar la resistencia de la cepa P2-32 sin mutar de P. chrysogenum, en MM con lactosa y suplementada con 100 pgjd de uridina.
Tabla 8 : (pag. 46) Comparación de la producción y productividad máxima de peniciha alcanzadas entre las cepas de P. chrysogenum parentales P2-32 y P2-37 y entre las mutantes auxbtrofas E-32U y E-37 M 1 cultivadas en FS a 25 "C y flujo de aire de 2.4 vh y en FL a 25 "C y 270 rpm.
Tabla 9 : (pag. 47) Comparación de los factores de producción y productividad relativa entre las cepas de P. chrysogenurn parentales P2-32 y P2-37 y entre las mutantes auxótrohs E-32U y E-37 MI cultivadas en FS a 25 "C y flujo de aire de 2.4I/hyenFLa25"Cy270rpm.

lnhoduccibn
1 Introduccibn.
Después de la segunda guerra mundial el sistema de fermentaci6n dlida (SFS) fue restringido por mucho tiempo a solamente la fermentaci6n de alimentos tradicionales y a la producci6n de algunas enzimas líticas (Nagai, 1979). La producci6n a escala industrial se desarroll6 para fmentaci6n líquida (FL), por medio de la cuál se obtienen productos de importancia tales como : enzimas hidrolíticas, aminoácidos, antibi6ticos y otros productos bajo condiciones perfectamente controladas; esta situaci6n no favoreció el desarrollo de cultivos por fermentación s6lida. Sin embargo, desde hace algunos d o s la fmentaci6n s6lida (FS) ha sido reevaluada, permitiendo su modemizacidn y aplicaci6n en nuevos campos (Nagai, 1979; Abdullah y col., 1985; Oriol y col., 1988a; Barrios y col., 1988; Mudget, 1986; Barrios y Mejía 1996).
El SFS constituye una interesante alternativa para antibi6ticos y otros metabolitos secundarios ya que ha mostrado tener un gran potencial (Barrios y col., 1988), ademh de que los productos generados por este sistema se obtienen m& concentrados y los costos de proceso de purificaci6n son menores (Rivera y col., 199 1).
Se ha desarrollado un SFS en el que se utiliza un soporte inerte (bagazo de caAa de azúcar) impregnado con medio líquido (Banios y col., 1988; Viniegra 1997). Este SFS fue aplicado exitosamente para producir penicilina por Barrios y col. (1988). Los rendimientos obtenidos fueron varias veces mayores a los obtenidos en FL bajo condiciones similares de cultivo, ademils de producirse en tiempos mils cortos.
Debido a que uno de los principales aspectos positivos del SFS es que los metabolitos se producen, generalmente, en mucho mayor cantidad que por el sistema de fermentacidn líquida (SFL) (Barrios y co1.,1988; Barrios y col., 1993) resulta de gran importancia el realizar mayores estudios en cuanto a los niveles de producci6n de la cepa, ya que estos estudios se han realizado con cepas de baja produccidn, que son mh parecidas a las cepas silvestres que a las mutantes hiperproductoras usadas en fermentaciones líquidas industriales.
En un estudio realizado en producci6n de enzimas (Shadcaranand y col.,l992) se llev6 a cabo la adaptaci6n de cepas de Bacillus de alta producci6n desmolladas para FL, en procesos de FS, observhdose discrepancias en el nivel de producci6n de eqzimas en ambos sistemas. Los autores concluyen que no se puede confiar en
1

lntroducci6n
que una cepa de alta producción en FL vaya a expresar este mismo nivel de producción en FS. Puede suponerse que durante los procesos de mejoramiento gendtico para la obtención de cepas especialistas y sobreproductoras para FL, se hayan perdido características o capacidades que permitan un buen desarrollo y rendimiento en FS. Surge entonces la necesidad de programas de mejoramiento y selecci6n de cepas mejor adaptadas al medio sólido (Barrios y Mejía, 1996).
Una alternativa de soluci6n a esta necesidad es la mutagbesis, la cuál ha constituido el metodo empírico tradicional para el mejoramiento de cepas; esta es seguida de una selección por pruebas directas de un gran número de purificaciones. Sin embargo el examen al azar representa un proceso de mucho trabajo debido al gran número de aislamientos que deben realizarse para poder detectar l a s cepas mejoradas.
La cantidad de trabajo puede reducirse significativamente mediante el uso de otros procedimientos que además disminuyen los elementos empincos, tanto en el mejoramiento como en la selección, mediante la aplicación de principios específicos de bioquimica y genbtica. Puede entonces recurrirse a procesos naturales de recombinación genbtica que permite el intercambio de informaci6n de dos organismos con diferentes características. Estos procesos ocurren en diferentes micmrganismos incluyendo la conjugación y transducci6n en bacterias y la heterocariosis y ciclo sexual en hongos.
Aunque todos ellos tienen aplicaciones en el contexto industrial, los metodos miis ampliamente usados en la transferencia de genes para el mejoramiento de cepas incluyen a la reproducci6n parased (en hongos), fusi6n de protoplastos e ingeniería genktica.
Hasta, hace poco la reproduccibn parasexual era pdcticamente el único mdtodo disponible para la cruza de hongos industriales, debido a la ausencia del ciclo sexual en' cepas comerciales. La reproducción parasexual es un proceso que se lleva a cabo lentamente, lo que resulta poco conveniente, sin embargo, con la aplicación de procesos de fusi6n de protoplastos la situaci6n cambió y ahora existen tkcnicas de reproducción basadas en la fusibn de protoplastos para una gran variedad de organismos industriales. El trabajo experimental planteado en esta tesis se desarrolla a partir de la necesidad existente de generar una metodologia que permita la obtenci6n de cepas especializadas para SFS, aspecto importante que ha limitado el desarrollo de esta tdcnica de cultivo.
2

Hidtesis v Objetivo
11 Hip6tesis.
Es posible introducir marcadores gen&icos de auxotrofia en Penicillium chrysogenum sin que estos mutantes modifiquen sensiblemente su producción en fermentacibn sblida y en fermentacibn líquida.
111 Objetivo.
Obtenci6n de marcadores gen&icos en cepas de Penicillium chrysogenum con características complementarias para una eventual obtencibn de recombinantes.
Objetivos Particulares.
1. Evaluacibn de la capacidad de produccibn de penicilina de 5 cepas de Penicillium chrysogenum (P2-32, P2-37, €2-47, P2-2V2, ASP-2A) en fermentación sólida y fermentacibn liquida.
2. Seleccibn de cepas de Penicillium chrysogenum con características complementarias.
3. Mutagdnesis y seleccibn de mutantes : -Auxótrofa a aminottcido. -Auxotrofa a uridina.
4. Evaluacibn d e l nivel de producción de penicilina de las cepas mutantes obtenidas en fmentacibn sblida y en fermentacibn líquida.
3

Generalidades
1V Generalidades.
1. ClasificaciiSn.
El microorganismo utilizado para el desarrollo experimental de esta tesis fue Penicillium chrysogenum, que pertenece a la subdivisión Deuteromycetes. Esta subdivisión comprende una gran cantidad de especies de hongos en las que la reproducción se real& solamente por mecanismos asexuales o parasexuales. Debido a que los Deuteromycetes aparentemente carecen de una fase de reproduccibn sexual, tambidn llamada “perfecta”. por lo común son denominados “hongos imperfectos” o tdcnicamente “Fungi imperfect?’ (Herrera, 1990).
2. Metnbolitos ligados v no ligados al crecimiento.
Los metabolitos generados durante la fase log de crecimiento son esenciales para el crecimiento de las cdlulas. Estos productos incluyen amindcidos, proteínas, vitaminas, polisactuidos y etanol y son llamados metabolitos primarios (Bodie, Ward, 1991).
Los metabolitos secundarios no se encuentran involucrados en el metabolismo celular, no son necesarios para el crecimiento, y sblo son generados cuando algún nutriente .se ha agotado. Es deir la velocidad de crecimiento es limitada por uno o más nutrientes, lo cual resulta en una diferenciacibn bioquírnica en el organismo y consecuentemente son producidos en la fase estacionaria o h e de mantenimiento (Bodie, Ward, 1991). Los metabolitos secundarios incluyen productos que tienen actividad antimicrobiana (antibióticos) y promotores del crecimiento, a s í como otros agentes teraphticos. Especies de Penicillium y otros hongos, producen una gran diversidad de estos metabolitos, identifichndose los siguientes : alcaloides, terpenos, esteroles, p- lacthicos, derivados del Acid0 shikímico y derivados de Acidos grasos (Peberdy, 1985). L a importancia de muchas especies de Penicillium radica en los metabolitos secundarios que producen, de esta f o m la penicilina tiene interés industrial como antibibtico.
3. Penicilina.
Los antibióticos p-lactamicos incluyen las penicilinas, cefalosporinas y cefamicinas, todas ellas útiles m6dicamente como antibibticos. Estos antibibticos se llaman p- lactbicos porque contienen un sistema de anillo p-lacthico (Fig. 1) (Sullivan y
4

Generalidades
Abraham, 1981). Los antibióticos p-lacthmicos actúan por inhibición de la síntesis de peptidoglucanos en las paredes celulares de las eubacterias. El blanco de estos antibióticos es la reacción de transpeptidación que partxipa en la etapa de entrecruzamiento de la biosíntesis de los peptidoglucanos. Debido a que esta reacci6n es única para las bacterias los antibióticos p-lacthicos tienen una alta especificidad y toxicidad relativamente baja. La estructura bhsica de las penicilinas es el bid0 6-aminopenicilhico (6-APA), que consiste en un anillo de tiazolidina con un anillo p-lactsmico condensado (Fig I ) (Sullivan y Abraham, 1981). El 6-APA lleva una mitad acil variable (cadena lateral) en la posición 6.
R - C - NHz
II O / F> CH3 CH3
O
coo- -" Cadena Anillo Anillo laterd B-lacthnico Tiazolidinico
FIG 1 : Estructura química de las penicilinas.
4. Fermentaci6n Wda.
4.1. Definicih.
La fermentacih en estado sólido puede ser definida (Lonsane y col., 1985) como un cultivo microbian0 que se demolla en la superficie y al interior de una matriz s6lida y en ausencia de escurrimiento de líquido. La matriz porosa puede ser un sustrato húmedo o un soporte inerte capaz de absorber los nutrientes disueltos en una
5

Generalidades
solucibn. De esta manera se pueden distinguir dos tipos de FS en funcibn de la naturaleza de la fase sblida utilizada I 11 Cultivo sólido con una fase sustratesoporte. La fase sólida esta constituida de un material que asume simultáneamente la funci6n de soporte y de fuente de nutrientes (Barrios, 1994). 21 Cultivo sblido con un soporte impregnado de un medio líquido. En este tipo de fermentaciones, la fase sblida estd constituida por un soporte inerte que sirve de repertorio de una solucibn nutritiva. Los materiales utilizados son diversos y entre ellos se encuentra el bagacdlo de cafla de azúcar (Barrios y col., 1988). Mudgett (1986) menciona que se puede distinguir a los substratos sblidos como una mezcla de gas líquido &lido en donde la fase acuosa esta íntimamente asociada con las superficies sólidas en varias etapas de la sorción y esta en contacto continuo con una fase gaseosa que proviene de un ambiente externo. Dependiendo del contenido de humedad del sblido, una parte del agua esta firmemente asociada a la superficie sblida, otra parte solo parcialmente asociada y otra existe en estado libre en regiones capilares del &lido. La interfase gas-liquid0 provee de una zona para el intercambio de dibxido de carbono oxígeno y para la transferencia de calor.
4.2. Ventajas y desventajas.
Se presentan algunas ventajas y desventajas de este mbtodo en comparacibn con el SFL. Algunas ventajas del SFS citadas por Mugett (1 986) y Barrios (1 994) incluyen las siguientes : l . Los contenedores de fermentaci6n pueden ser pequeilos debido a que se utiliza poca agua y los substratos obtenidos se encuentran concentrados. 2. No se requieren de tanques para medio semilla, ya que se pueden incluir inoculaciones con esporas. 3. La baja humedad reduce los problemas de contaminación, 4. Las condiciones del crecimiento füngico,son similares a aquellas de sus hdbitats naturales. 5 . La aireacibn es facilitada debido a la porosidad del material. 6. Las producciones obtenidas pueden ser mucho mayores a l a s obtenidas por medio líquido. 7 . Se requiere menor energía de proceso que las correspondientes en FL. 8. Disminucibn de efluentes Uquidos a tratar si el producto se utiliza directamente o el producto se obtiene más concentrado. 9. Muchas veces se hace un uso directo de los sólidos fermentados. 10. Para las fermentaciones tradicionales la microflora del soporte sirve como inbCul0.
6

Generalidades
En lo concerniente a las desventajas, se citan las siguientes : l . La adicibn de agua en Eases tempranas de la fmentacibn, puede incrementar los riesgos de contaminacibn bacteriana. 2. La inoculacih de esporas puede necesitar que se produzcan y siembren asdpticamente. 3. Dificil regulacibn de m e t r o s de cultivo como pH. 4. Falta de conocimientos bhicos y tecnolbgicos; 5 . La dificil adicibn de nutrientes y agentes controladores. 6. Se puede dificultar el recobrar el producto, si éste es absorbido o contaminado con residuos sblidos.
5. Meioramiento pen6tico.
El mejoramiento de una cepa generalmente sigmfica aumentar la productividad de un organismo, aunque también puede ser deseable el manipular características especificas tales como : la habilidad para utilizar diferentes tipos de sustratos que pueden resultar m& baratos, capacidad de esporulacibn etc. Un buen ejemplo en la aplicacibn del mejoramiento de una cepa lo constituye el proceso de fermentacibn de penicilina donde los renhentos de penicilina producidos por P. chrysogenum han sido incrementados aproximadamente hasta unas 500 veces desde su aislamiento original (Rowlands, 1984a).
En general el mejoramiento de cepas de hongos que son utilizados en procesos biotecnolbgicos involucra mutagenesis al azar y una subsecuente seleccibn de las cepas. mutantes con productividades incrementadas. Aunque este tipo de estrategia ha sido provechosa en muchos casos, existen desventajas y limitaciones. La repetici6n de tratamientos mutagbcos puede provocar incrementos en el número de mutaciones que en un inicio no son detectables (daflos en la información gendtica) lo cual puede resultar en la inestabilidad de la cepa y subsecuentemente en la p&dida de una cepa de buena producción ( Swart y col., 1990).
Existen procesos alternativos, bbicamente dos, para aumentar el rendimiento de producto de microorganisnos industriales : recombinacibn e ingeniería gendtica. Cada una presenta diferentes ventajas, pudidndose emplear estas dos técnicas ademh de la mutagénesis en conjunto, para maximizar la produccibn. La recombinacibn gendtica permite la construccibn de cepas con un gran número de diferentes combinaciones de mutaciones que determinan la proporcibn de los productos de interds. Con el desarrollo de las tecnicas de h ibn de protoplastos es
7

Generalidades
relativamente simple el recombinar propiedades de una amplia variedad de microorganismos industriales que no estb bien caracterizados tanto gendtica como bioquímicamente (Baltz ,1986). La ingeniería gen&ica, recibe desde hace algunos d o s atencibn seria para la produccibn de metabolitos füngicos de inter&, lo cud permitirá comprender mejor los mecanismos que controlan la expresibn de los genes involucrados en la biosintesis del antibi6tico (Timberlake y Marshall 1989, Bmedo y col., 1989b).
5.1. Mutacih.
Muchos productos generados por hongos fílamentosos han sido aprovechados a nivel industrial, sin embargo se ha requerido de la implementacih de metodologías que pennitan obtener mejores rendimientos de los productos generados por estos. El metodo esthdar por medio del cud la industria los ha explotado ha sido la mutacibn, seguida de la búsqueda de una cepa mejorada mediante un gran número de aislamientos(Upshall,1992). Sin embargo los aislamientos de mutantes al azar son procedimientos laboriosos debido al gran número de aislamientos que deben llevarse a cabo para a s í detectar la cepa mejorada (Davies, 1964; Rowlands, 1984b). La mutaghesis se define como la induccidn de cambios heredables, mutaciones, en el material genetic0 de cualquier organismo. Estas mutaciones son causadas por cambios ,en el genotipo y pueden ser detectadas como una modificacih fenotipica del organismo (mutante). Las mutaciones pueden ocurrir in vivo espontheamente o despues de la inducci6n por mutsgenos tales como agentes químicos o radiaciones. Las mutaciones pueden a s í mismo ser inducidas in vitro mediante tecnicas de ingeniería genetica (Crueger, 1993). Las mutaciones sufiidas por un organismo varían según las alteraciones presentadas en el genoma, así se distinguen tres clases : 1 .Mutaciones en el genoma, que pueden causar cambios en el número de cromosomas 2. Mutaciones en el cromosoma que pueden causar cambios en el orden de los genes dentro del cromosoma. 3. Mutaciones en el gen o mutaciones puntuales que pueden resultar de cambios en la secuencia de bases en el gen. Las mutaciones en el cromosoma pueden ser inducidas por muthgenos químicos, o
por radiaciones como la luz U.V. o rayos X. Las mutaciones en el gen implican la generacib de nuevos alelos en el mismo. En el caso de macrolesiones se involucran un gran número de nucle6tidos. Por errores de inversidn en la replicaci6r1, pueden ocurrir deficiencias e inserciones, en las que se ven involucrados grandes secuencias de un gen. En microlesiones o mutaciones puntuales una base es substituida por otra , o bien, uno o pocos nucle6tidos son
m

Generalidades
borrados o insertados. Una caracterhtica de las mutaciones puntuales es que éstas pueden revertir (Crueger, 1993). Las mutsiciones puntuales se dividen en dos clases : substituciones de pares de
bases y mutaciones en el marco de lectura. Cuando la substitución de pares de bases ocmen en un gen, produce efectos sobre las proteinas, produciendo ya sea substituciones en aminoficidos, o bien codones de cadenas terminales. Las mutaciones en el marco de lectura consisten en la deleción o adición de un peqwflo número de pares de bases, alterando el marco de l e c t u r a en las etapas de transcripción y traducción. Tanto la radiación como un gran número de agentes ’
químicos pueden producir mutaciones puntuales a bajas dosis de muthgeno. Los cambios mutacionales se llevan a cabo ya sea por errores en la replicación o reparaci6n mediante la incorporaci6n de bases incorrectas en la cadena polinucledtida, o mediante la interacción química directa con una base (Crueger, 1993).
Por otro lado, para el dxito en el mejoramiento de una cepa, se requiere de la selección apropiada tanto del tipo de mutigeno como de la dosis correcta. Para cada tipo de muthgeno a s í como para cada organismo, existen una serie de combinaciones que permiten obtener la m b alta proporción de una clase de mutante en particular entre los sobrevivientes del tratamiento mutaghico, tales como: concentración del muthgeno, tiempo de exposición y condiciones del tratamiento (Queener y col.,
En cuanto a la dosis, debe tomarse en cuenta que altas dosis de mutfigeno usualmente incrementan la posibilidad de producir simulthneamente mutaciones secundarías indeseables (Rowlands, 1984a; Bos, 1987). Un b t o r de importancia que debe considerarse es la facilidad de uso y seguridad del muthgeno. Todos los mutagenos son potencialmente cancerígenos ya que actúan Wando al DNA. La radiacidn UV lejana es indudablemente el muthgeno de mejor conveniencia, ya que puede producir muchos de los tipos de daflos conocidos en el DNA y ficilmente se pueden tomar las precauciones de seguridad necesarias para su uso. De los mutfigaos líquidos como el EMS son mucho m h fficiles de manipular que los de polvo como el NTG. Otra desventaja del NTG es su tendencia a producir mutaciones en clusters cercanamente relacionados, aunque ello puede ser una ventaja para algunos sistemas (Rowlands, 1984a). Por otro lado, es conveniente obtener una curva de sobrevivencia antes de poder determinar l a s condiciones óptimas para la mutag6nesis. Además, para el caso de tratamientos mutagdnicos realizados con luz UV, es conveniente llevarlos a cabo en ausencia de iluminación directa, ademb de cuidar que el crecimiento celular se lleve a cabo en la obscuridad o con l u z roja despuds de una exposicidn a la luz UV, para
1986):
Y

Generolidodes
de esta forma evitar que se lleven a cabo los mecanismos de reparacibn estimulados por la luz visible (Miller, 1972).
A las cblulas mutadas se les debe permitir una etapa de replicacibn del DNA y crecimiento celular para convertir de esta f o m el dafío inducido en el material genbtico en un daÍí0 estable (Qucener y col., 1986; Campuzano y col., 1993). La distincibn mis importante entre los diferentes tipos de mutageno es el tipo de mutacibn que se induce (Rowlands, 1984a), la cual depende de dos factores : El tipo de daÍí0 causado por el mutageno al DNA y la acci6n de los mecanismos de reparacih celular sobre el daÍí0 causado al DNA. Muchos de los muagenos producen msls de un tipo de desarreglo sobre el DNA, aunque el grado con que lo inducen varía con cada uno de ellos. Por ejemplo la radiación lejana de UV (-260 nm) produce una alta proporcih de dímeros de pirimidha mientras que la radiacibn ionizante provoca un alto grado de rompimientos en el cromosoma, en tanto que la EMS y la NTG son agentes alqudantes. Sin embargo la luz UV lejana, suele producir bases hidroxiladas y entrecruzamiento (crosslinking) de las cadenas del DNA. Las rutas de reparacibn del DNA pueden ser por sí mismas no mutagtkicas (libre de errores) o bien mutagbicas, d-endo del mecanismo eflzimsfico involucrado. Las rutas de reparacibn no mutagenicas incluyen a la fotoreactivaci6n, reparacibn por excisibn y la reparaci6n por recombinacibn. La ruta mutagtkica principal y mejor conocida es el mecanismo de reparacibn SOS en Escherichia coli, la cuál se presenta como mecanismo de reparacibn en daflos causados tanto por muagenos químicos como fi sicos incluyéndo la radiacih UV lejana. Otras rutas de reparacibn menos caracterizadas se ven involucradas al inducirse mutaciones mediante EMS o NTG (Rowlands, 1984a).
Estrategias de seleccibn.
Debido a que las especies de hongos filamentosos de importancia industrial son generalmente imperfkctos, se ha recurrido a mbtodos de genbtica parasexual que permiten llevar a cabo procesos de mejoramiento genbtico para lo cuál se hacen necesarias mutaciones gembticas selectivas del tipo de la auxotroíia que permiten recobfar productos de fusibn derivados de cruzas entre dos cepas logrhndose el reconocimiento de la etapa de heterociuibn o de diploidia (Peberdy, 2984).
Los sistemas de complementacibn auxotrbfica son msls diversos debido a la clonaci6n de genes. Muchos de los sistemas de seleccibn positiva, es* basados en la resitencia a una toxina; el primer sistema desarrollado para hongos filamentosos
10

Generalidades
se bas6 en la clonaci6n del gen AmdS de A. nidulans. Este gen permite al hongo utilizar acetamida como imica fuente de carbono ( Upshall, 1992 ).
Otro sistema de seleccidn positiva que resulta particulannente apropiado para hongos flamentosos, estsi basado gen6ticamente en la resistencia de dominancia a los fungicidas. Se han utilizado dos sistemas, uno basado en la resistencia al hgicida benomyl y el otro basado en la resistencia a h oligomicina. El gene de resistencia al benomyl de A. niger ha sido usado para transformar P. chvsogenum (Upshall, 1992). Tambih pueden ser introducidos por trulsformaci6n marcadores selectivos del tipo de la resistencia a antibi6ticos tales como higromicina y fleomicina (Talbot y col., 1988; Pebcrdy, 1991).
Auxotroíia a pirimidha mediante la resistencia al 5 FOA.
Se ha desarrollado un m&odo de seleccibn positiva para la selecci6n directa de mutantes ura 3 am6trofas a uracil, usando 5 AFO, reportado por primera vez en Saccharomyces cerevisiae por Boeke y col. (1984). Se pueden inducir mutaciones en el locus URA 3 de Sacharomyces cerevisiue mediante un sistema de selecci6n positiva. Cepas de levaduras silvestres son incapaces de crecer en medios que contienen 5 AFO, el cuhl es un d o g o de la pirimidina, mientras que los mutantes ura 3 -crecen normalmente. Este tipo de selecci6n puede ser aplicada en una gran variedad de cblulas eucariotas y procariotas, la cuhl esta basada en la p6rdida de actividad de la descarboxilasa orotidin 5 fosf’ato. En experimentos donde se seleccionaron mutantes resistentes al 5 AFO inducidas por radiaci6n UV s6lo un 5- 10 % de los mutantes fueron ura 3 . No se sabe cuantos tipos diferentes de mutantes puedan causar el fenotipo de resistencia; sin embargo los mutantes URA 1, URA 2 y URA 4 son sensibles al 5 AFO mientras que los mutantes URA 3 y URA 5. son resistentes (los mutantes URA 5 son s6lo parcialmente resistentes) (Boeke, 1984). El 5 AFO puede ser muy útil para el aislamiento de mutantes ura 3 - en l e v a d u r a s de inter& indwirial, para las cuales no existen sistemas de transformacibn útiles que permitan obtener marcadores aux6trofos (Boeke, 1984). La biosíntesis del UMP en hongos filmentosos procede del aspamto y del carbamil fosfato a.trav6s del intermediario &ido or6tico al orotidin 5 monofosfato (Fig. 2). Los genes involucrados en la conversi6n del Bcido or6tico a Uh4P se conocen como Pyr F:(OMP pirofosforilasa) y Pyr G ( O W descarboxilasa) en-Aspergiffus. Al parecer el 5 AFO tiene un amplio espectro de accibn, ya que se ha encontrado que inhibe el crecimiento de Saccharomyces, Schizosaccharomyces y Candida, permitiendo el crecimiento de cblulas deficientes en enzimas O W pirofosforilasa u
-
11

Generalidades
O W descarboxilasa. La toxicidad del 5 AFO radica en su conversión a 5 Fluor0 UMP un metabolito que interfiere con la sintesis del DNA . El sistema de selección implementado por Boeke y col. (1984) se ha aplicado exitosamente en otros hongos (Oakley y col., 1987; Smit y Tudzynsky, 1992; Akileswaran y col., 1993). En la mayoría de los casos se encontraron mutantes deficientes en el gen Pyr G y en el gen Pyr F. En el trabajo realizado por Campuzano y col. (1993), se describe el aislamiento, caracterización bioquímica y mapeos de una serie de mutantes de Phycomyces. Los resultados mostraron que los auxótrofos a pirimidina aislados por resistencia al 5FOA caen dentro de dos grupos de complementación, los mutantes deficientes en el gen Pyr F y en el gen Pyr G. La proporción de auxótrofos de pirimidina entre los mutantes resistentes he del 7 %. La mayoría de ellos aproximadamente el 85 % heron Pyr F mientras que el restante 15 % heron mutantes Pyr G. En un trabajo realizado por Diez y col. (1 987) se aislaron auxótrofos a pirimidina de P. chrysogenum de entre mutantes resistentes al 5 AFO, denominhdolos como mutantes Pyr G. No se sabía si todos los mutantes aislados eran deficientes de la enzima OMP descarboxilasa o si había otros tipos diferentes de mutantes que conferian el mismo fenotipo de resistencia. Se mostró por otro lado, que la fiecuencia de mutantes resistentes al 5 AFO fue m& alto en P. chrysogenum que en S. cerevisiae, lo cual sugiere un mayor nivel intrínseco de resistencia al 5 AFO que en las levaduras.
Auxotrofia a aminobidos mediante procesos de enriquecimiento.
Una Mctica frecuente en cepas de inter& industrial (&S, 1987) es la generación de cepas de alto nivel de producción con niveles de sobrevivencia de O. 1-1 % mediante altas dosis de rnweno, incrementando con ello mutaciones indeseables, especialmente cuando se aplican tratamientos mutaghicos recurrentes. Se ha observado que altas dosis de muthgeno producen nuevos arreglos en el cromosoma y mutaciones que no son percibidas que alteran los antecedentes de la información genaca. Se ha observado en Aspergillus nidulans la relación entre la frecuencia del mutante y la sobrevivencia; mosthdose que para diferentes tipos de mutantes, el rendimiento m& alto, se obtiene a bajas dosis de muthgeno (20-50% de sobrevivencia). La fiecuencia de mutantes se incrementa cuando a su vez se incrementa la dosis del mutslgeno, pero se estabiliza y despues decrece a dosis m& altas. NO hay una relación linear simple entre la iiecuencia de mutantes y el logaritmo de la dosis del muageno. Estos estudios dan lugar a elaborar pdcticas
12

HN I
+ O
COOH P O CARBAMIL FOSFATO CARBAMIL ASPARTATO
A I
ASPARTATO ZADP + GLU +P
GLUN +ZATP
coz t O
ENZIMAS
A ........ Sintetasa carbamil fosfato B ........ Transferasa carbamil aspartato C........Dihidm orotasa D ........ Hidrogenasa dihidm orotato E ........ Pirofosforilasa OMP F. ....... Descarboxilasa O W I DIHIDRO OROTATO
N? I
OROTIDIN S-MONO FOSFATO
N
O )-I URIDIN 5-MONO FOSFATO OH OH
OROTATO
FIG 2 : Ruta biosintbtica de la UMP en hongos filamentosos.
13

Generalidades
que resulten en la induccibn de bajos niveles de mutdgeno para obtener un mayor rendimiento de mutanes. Se ha llevado a cabo la búsqueda de metodologias que permitan una seleccibn eficiente de las mutantes deseadas incluyhdose la resistencia a antibibticos a s í como metodos de enriquecimiento utilizando enzimas que degradan la pared celular (Piedra y Hemera, 1976; Sipiczky y Ferenczy, 1978; Delgado y col., 1979). Con procedimientos apropiados de enriquecimiento, pueden llevarse a cabo mejores aislamientos de mutantes auxotrbficas a bajas dosis de muthgeno. El mktodo de enriquecimiento por filtracibn ha probado ser muy eficiente para el aislamiento de mutantes de A. nihfans que requieren una fuente específica de carbono (Bos y col., 1981; Uitzetter y col., 1986), a s í como el enriquecimiento mediante una preparacih de enzima lítica Novozyme 234 (Debets y col., 1989). El procedimiento de enriquecimiento por filtracibn es un metodo cliisico basado en la remocibn selectiva de protbtrofos, demollada originalmente por Fries (1947) para Ophiostoma multiannulatum y subsecuentemente adaptada a Neurospora crassa por Woodward y col. (1954). Bos y col., (1992) indujeron mutantes de A. niger con bajas dosis de muthgeno (luz UV) para eliminar &os en la informacibn general o a h c i o n e s cromosómicas (Bos, 1987), obtenibdose un alto nivel de sobrevivencia y por tanto una baja fiecuencia de mutantes entre los protbtrofos sobrevivientes, Consecuentemente se hizo necesaria una etapa eficiente de enriquecimiento como prerequisito. Miis de 100 mutantes auxbbrofss de A. niger fueron aisladas mediante procesos de enriquecimiento por íiltracibn. Los mutantes obtenidos de esta manera fueron predominantemente auxbtrofos a aminoticidos y dificilmente se encontraron mutantes deficientes a vitaminas. La hcuencia fue baja con promedio de una mutante por experimento. Un aspecto importante relacionado con la baja eficiencia de aislamiento de mutantes es que muchas de las conidias que no germinaron fueron descargadas junto con aquellas que si lo hicieron, debido a que probablemente quedaron atrapadas en la red micelial formada durante la filtracibn o bien debido a la agregacibn de las conidiosporas durante la incubacibn en medio líquido. De tal forma que la mayoría de las mutantes que no germinaron se perdieron durante el enriquecimiento. Para eliminar estas limitaciones, se cuenta con otro metodo de enriquecimiento la utilizacibn de la preparacibn de enzima lítica Novozyme 234. Esta tknica fue origdmente desanollada para el aislamiento de auxbtrofos diploides recombinantes y ha sido descrita con anterioridad por Debets y col. (1989). El principio del procedimiento por enriquecimiento con Novozyme, es que la conidia genninada inmovilizada en medio agar es eliminada selectivamente despues de cierto periodo de incubacibn. Las puntas de Was jbvenes de A. niger son lisadas
14

Generalidades
eficientemente con Novozyme 234, provocando que el citoplasma fluya y se efectúe la muerte celular debido a los bajos valores osmóticos de la solución enzimática. Las paredes conidiales, a s í como las paredes bgicas subapicales, no son susceptibles a la acción de la Novozyme 234. La conidia germinada con una hifa septada y ramificada, por otro lado, no es eliminada apropiadamente ya que a pesar de que las puntas de las hih si son lisadas, no se alcanza a desintegrar todo el citoplasma. La eficiencia de este procedimiento se ve reducido considerablemente si la conidia germinada es sometida a bajas temperaturas por algunas horas, evithdose la lisis por la Novozyme 234. Es esencial por esta &n que el tratamiento enzimihco se realice inmediatamente despues de la incubación. Consecuentemente, solamente las conidiosporas activas metabólicamente con una hifa muy joven pueden ser eliminadas con tratamientos con Novozyme. Otro aspecto de importaucia es la necesidad de incluir una etapa de propagación en MC de las esporas, debido a que la radiacibn con UV afecta su fisiología originando que la germinacih se desincronice, repercutiendo de forma importante en la eficiencia del procsdimiento con Novozyme. Es probable que con altas dosis de mutttgeno se vea alterada por completo la sincronización de la geminación conidial (1 -2 % de sobrevivencia), lo cuil es una pdctica común llevada a cabo por muchos investigadores; en tal caso el enriquecimiento de aux6trofos mediante enzimas líticas no es posible. Es de suponer que el efecto en el enriquecimiento se vea limitado en aquellos procedimientos de enriquecimiento en donde la etapa de propagación es omitida. La sincronizaci6n de la genninación despues del tratamiento mutagénico probablemente pueda mejorar el procedimiento de enriquecimiento. Ademh otra ventaja de la etapa de propagacibn, es la posibilidad de la segregación de alelos mutantes, y la expresibn fenotipica de las mutaciones inducidas. Dependiendo del tipo de mutante deseada, uno o ambos procedimientos pueden determinar un m&odo efectivo para el enriquecimiento de mutantes con una deficiencia metaMlica (Bos y col., 1992).
15

Antecedentes
V Antecedentes.
Fusi6n de protoplastos.
Desde hace algunos dos, existe un gran inter6s en la aplicaci6n de la recombinaci6n gen6tica para la obtenci6n de productos de importancia industrial, tal como los antibi6ticos (Kurth, 1985). Con el descubrimiento del polietilenghcol como mediador en la fusi6n de protoplastos, se cuenta con una herramienta de gran importancia para la recombinaci6n genaca de microorganisnos productores de antibi6ticos.
En un inicio la fusi6n de protoplastos, se UW en ctlulas auimales y en c6lulas de plantas y posteriormente en hongos y bacterias unicelulares. Finalmente el trabajo de Okanishi y col. (1 974) en formacibn de protoplastos, h i 6 n y regeneracibn, aceler6 el uso de esta t 6 c a para la manipulaci6n genaca en Streptomyces (Kurth, 1985).
Recientemente Kazunobu y col. (1996) realizaron la recombinaci6n entre dos levaduras utilizando una cepa de K. marxianus y otra de S. cerevisiae; los autores pretendían conjuntar la mejor capacidad fermentativa de la cepa de S. cerevisiue a 30 oC con el buen crecimiento a altas temperaturas (43 oC) de K. mamiunus usando la tecnica de fusi6n de protoplastos. Obtuvieron un grupo de fusionantes que presentaba características superiores a las cepas parentales en cuanto a crecimiento a alta temperatura y capacidad fermentativa. Sin embargo dste grupo no mantuvo la característica de tennoestabilidad de la cepa de K. marxianus.
Por su parte Ann6 y Peberdy (1 975) utilizaron a P. chrysogenum entre otros hongos, para sus estudios iniciales sobre k i 6 n de protoplastos. Los m6todos y principios importantes de esta metodología han sido discutidos por (Peberdy, 1980). Se ha llevado a cabo exitosamente la fusi6n de protoplastos entre interespecies de P. chrysogenum y Penicillium cyaneo-filvum utilizando marcadores nutricionales (Peberdy y col., 1977).
A s í mismo, se ais16 una progenie hhida de interespecies de P. chrysogenum y P. stoloniferum, P. patulum y P. verrucosum var cyclopium, mediante la induccidn de fusi6n de protoplastos encontrhdose una correlacibn entre la estabilidad de las colonias híbridas y los rendimientos de penicha, ya que los híidos m& estables son los que produjeron los mayores rendimientos (Ann6, 1982).
16

Antecedentes
En ese mismo aflo, Chang y col. (1982), lograron producir híbridos de las especies de P. chrysogenum y Cephalosporium acremonium mediaute fusión de protoplastos. Los autores concluyen que dada la prototrofia y la alteración morfol6gica observada en los productos de fusibn, se sugiere la posibilidad de transferencia de genes entre Cephalosporium y Penicillium.
Posteriormente Ann6 y Peberdy (1985) comentan que la viabilidad de híbridos generados despu6s de un proceso de hión de protoplastos es debida a la commbilidad gen&ica y bioquímica de las especies relacionadas. Y en el sentido inverso, tanto la letalidad como el comportamiento anomal de una progenie híbrida, es consecuencia de las diferencias relacionadas con el cdcter nuclear ylo del citoplasma de las eqpies involucradas en la fusión.
A s í mismo, estos autores (Ann6 y Peberdy 1985), tambib mencionan que se refleja un mayor nivel de compatibilidad entre especiese de Penicillium, que entre otras especies menos relacionadas de Aspergillus o que entre especies de levaduras, debido a las mayores hcuencias de h i 6 n observadas entre cruzas de interespecies de Penicillium.
Se plantea entonces, la viabilidad de la generacih de organismos híiridos estables en donde se realiza la h i 6 n del genoma de las especies involucradas, tanto en especies de Penicillium como en otros microorganismos.
El inicio .en la capacidad de produccih de hííridos entre especies íüngicas se debió al desarrollo de mejores cultivos y m6todos de fusión de protoplastos (Anné y Peberdy, 1975, 1976). Estos estudios permitieron la investigacih de nuevas rutas bioquimicas dadas en un nuevo ambiente celular.
Mos miis tarde, Upez Nieto y col. (1980), valihdose de esta metodología, cruzaron por h ión seis diferentes cepas auxotróficas de P. chrysogenum con la finalidad.de obtener recombinantes con mayores producciones de penicilina que las cepas progenitoras. Los resultados mostraron que los recombinantes obtenidos de una cepa de baja y otra de alta produccibn nunca produjeron tanto como la cepa parental de alta producci6n, sugiriendo que los mecanismos regulatorios de las cepas progenitoras de baja produccih existen en las recombinantes y pueden contramestar la producción eficiente que, existe en la cepa progenitora desregulada. En cambio, los recombinantes obtenidos a partir de cepas de baja producci6n o de cepas no productoras en algunos casos produjeron penicilina en niveles superiores a las de las. cepas parentales; esto puede ser explicado como un rearreglo de la informaci6n
17

Antecedenteg
genetica codificada para la biosíntesis de penicilina, resultando en una cepa mejor productora. Los autores concluyen que para obtener cepas de alta producción por h i 6 n de protoplastos, es recomendable partir de cepas parentales de alta produccih.
La fusidn de protoplastos, se plantea entonces como una metodología para generar cepas de alta producci6n, la cuitl, se puede enfocar al desarrollo de cepas dtamente productoras para FS.
Fermentacih s6lida.
Viniegra Gonziilez (1 989) r e m un ansllisis de las perspectivas y hmitaciones de la FS, concluyendo que este es un proceso tradicional que puede funcionar como una alternativa para la produccih de alimentos (sabores), probi6ticos, aditivos, biopesticidas, enzimas y metabolitos secundarios (antibi6ticos. giberelina etc.). Menciona que hace falta iuvestigaci6n para el desarrollo de productos, ademk que a excepcih del J@n, hay muy poca experiencia en cuanto a la construcci6n y operacibn de plantas industriales que utilizan FS. La investigacih necesaria, debe cubrir aspectos enfocados tanto a las condiciones que afectan 6ste sistema de cultivo, al control de la fermentacih y a la generacibn de cepas especiales para el SFS.
Entre los trabajos enfocados al conocimiento de las condiciones que afectan la FS se encuentran los siguientes :
-Se han llevado a cabo investigaciones sobre cin6ticas de crecimiento de Aspergillus niger en soportes sdidos de bagazo de Caaa impregnados con medio líquido, obmvhdose elevados niveles de crecimiento y demostrbdose la viabilidad del cultivo de hongos filamentosos en FS para la produccib de metabolitos con tecnología de bajo costo (Oriol y col., 1988a).
-Tambi6n se han realizado estudios para determinar la disponibilidad de oxígeno en la fase líquida, lo cuitl permite estudiar la influencia que tienen los flujos de aire y el porciento de materia seca en el medio sobre el coeficiente de transferencia volum6trica de oxígeno (Duran4 1988). A s í mismo se han hecho estudios en donde se observa la influencia de la actividad de agua (aw) sobre el crecimiento de hongos en sustratos dlidos (Oriol, 1988b).
18

Antecedentes
-Se cuenta tambib con trabajos relacionados con la transferencia de calor en FS, en donde mediante modelos matemíiticos, se pretende comprender mejor los fendmenos de transporte que se llevan a cabo en FS, permitiendo al mismo tiempo evaluar varias altemativas de control de temperatura en FS , como lo son por ejemplo flujos de aire e incrementos del contenido de agua (Saucedo y col., 1989).
-Se han efectuado investigaciones en relacidn a factores que afectan el desarrollo de la fmentacidn s6lida. De esta forma se ha deteminado, que ciertas concentraciones de COZ en el ambiente gaseoso pueden ejercer diversos efectos, de acuerdo con el tipo de microorganismo involucrado, en el crecimiento, en la conidiacidn y en la produccih (Desgranges, 1990; Mejía y col. 1996).
-Por su parte Barrios y col. (1 993) consideran que el tamaflo de partícula del soporte dlido, la densidad de empaque y la agitacidn son variables de importancia a considerar en FS. En la producci6n de penicilina, obtienen incrementos del 37 % al utilizar partículas grandes (14mm) de soporte sdlido ( c m de azúcar); a s í mismo, obtuvieron 20% mhs produccidn de penicilina al utilizar altas densidades de empaque (0.35 g/ ml), y observaron que la agitacidn no muestra efectos negativos en la produccidn si se restituye la humedad perdida durante el progreso de la fmentacih.
Por otro lado, tambib existen antecedentes en cuanto a la produccidn de enzimas y metabolitos secundarios, que gracias a un buen control del SFS se han generado resultados exitosos en su produccih; ejemplos de ello son los siguientes :
-Tradicionalmente, se han obtenido enzimas de inter& industrial mediante FL debido a su facilidad de manejo y control. Sin embargo actualmente se han comenzado a producir por FS. Se han efectuado estudios sobre la produccidn microbiana de lipasas tanto por FL como por FS utilizando Penicillium candidurn obsenhdose para &te último sistema de cultivo los m& altos rendimientos y producciones estables (Mufloz y col., 1991).
-Posteriormente Autier y col. (1 993). mencionan que la productividad de pectinasas en procesos de FS es mucho mayor que la productividad obtenida por FL, adem& de que la industria utiliza ambos procesos de cultivo para la produccidn de pectinasas utilizando Aspergillus niger, resultando ser m& apropiado el proceso de FS debido a los mayores rendimientos de pectinestmasa y poligalacturonasa obtenidos.
19

Antecedentes
-Por su parte Barrios G o d e z y col. (1988) obtuvieron rendimientos y productividades muy altos en relacibn con el metodo convencional de FL en la produccidn de penicilina. Estos resultados indican que la FS es una alternativa para la producci6n de metabolites secundarios. Los autores enfatizan que un factor limitante importante para la explotaci6n del potencial de este sistema es la falta de conocimiento de la fisiología fúngica en medio s6lido. A d d de que deben realizarse mayores esfuerzos enfocados al desarrollo de cepas especialistas para este sistema.
En lo referente a la selecci6n de cepas especialistas para FS no se encuentran suficientes trabajos al respecto, de ahí la importancia de este estudio. Una referencia del tema se presenta a continuaci6n :
-Algunos aflos mh tarde este mismo autor (Barrios G o d e z y col., 1993) determin6 la producci6n de penicilina de cinco cepas de P. chrysogenum tanto en FS como en FL. Los resultados mostraron que las cepas de mayor produccibn en FL tambih eran l a s de mayor producci6n en FS. De estas mismas cepas se aislaron mutantes observbdose variaciones importantes en el nivel de produccibn, de tal forma que se observ6 una tendencia inversa a las cepas de las c d e s provenían, es decir, las mutantes m& altamente productoras en FS tienden a ser las menos productoras en FL. Los autores concluyeron que la capacidad de una cepa para producir penicilina en FS no esta directamente relacionada con la capacidad para producir el antibibtico en FL, es decir que algunas cepas expresan mejor su potencial de producci6n en FS, de ahí la necesidad de buscar una metodologia que permita generar cepas especialistas para este sistema.
20

Materiales Y M¿todos
VI Materiales y Mbtodos.
1. Microoraanismos.
Las cepas de P. crhysogenum usadas fueron : P2-32, P2-37, P2-47, P2-2V2, ASP- 2A, todas ellas aisladas por Barrios y col. (1993). Para iniciar los experimentos, se parti6 de stocks de suspensibn de esporas en glicerol al 25 % almacenados a -2O"C, de cada una de las cepas antenomente citadas. Para la realizacibn de los bioensayos de actividad antibiótica se utilizó Bacillus subtifis ATCC 6633 el cual fue mantenido en medio de esporulación a 4 "C.
l.l.Conservaci6n.
Se prepararon stocks de esporas de las cepas seleccionadas, P2-32 y P2-37, a s í como de las mutantes aisladas, en &cero1 al 25 % a -70°C para su conservación a mediano plazo y para su conservación a largo plazo se hizo un preparado de liofilizados de cada una de ellas.
2. Medios de cultivo.
Fermentacibn Sblida.
Se utilizaron como medio de cultivo para el SFS aquellos reportados por Somerson y col. (1961) modificados, donde la composicibn del medio complejo de FS para las cepas P2-32, P2-37, P2-47 Y P2-2V2 en g/l es la siguiente: Lactosa (Bioxon) 110; Glucosa (Bioxon) 14; Caco3 (Baker) 20; K H 2 P 0 4 (Baker) 6; MgSO4 (Baker) 6; sblidos de maceracibn del maíz (S") (donacibn CIBIOSA Saltillo Coahuila [Centro Industrial Bioquimico S.A.]) 70; Bcido fenil-acetic0 (MA) (CIBIOSA) 1.3. El pH se ajustb a 6.5. La composici6n en g/l del medio complejo de FS para la cepa ASPQA es la reportada por Luengo y col. (1980) modificada, siendo la siguiente : Glucosa (Bioxon) 14; Lactosa (Bioxon) 75; Caco3 (Baker) 7.5; P h m a media (CIBIOSA) 30; AFA 1.2. El pH se ajustb a 6.5.
21

Materiales Y Mbtodos
Fermentaci6n Líquida.
Los medios complejos de in&ulo y los medios complejos de producci6n usados, fueron aquellos reportados por Somenon y col. (1961). El medio de inóculo utilizado para las cepas P2-32, P2-37, P2-47 y P2-2V2 en g/l fue la siguiente: Caco3 (Baker) 5; SMM (CIBIOSA) 20; extracto de levadura (Bioxon) 5; melazas de remolacha 26. ajustando el pH entre 6 - 6.5. El medio de produccibn para estas mismas cepas en g/l h e de : Lactosa (Bioxon) 55; Caco3 (Baker) 10; SMM
Ajustando el pH a 6.5. El medio de iu&culo utilizado para la cepa ASP-2A en g/l fue el siguiente (Luengo y col., 1980) : SMM (CIBIOSA) 20; Sacarosa (Bioxon) 20; extracto de levadura (Bioxon) 5; Caco3 (Baker) 5; ajusthndose el pH a 5.7. El medio complejo de producci6n en g/l para dsta misma cepa fue el siguiente : Lactosa (Bioxon) 50; (N& )2 SO4 (Baker) 4; Caco3 (Baker) 5; Pharma media (CIBIOSA) 20; AFA (CIBIOSA) 2.5. Se ajust6 el pH a 6.8.
(CIBIOSA) 35; MgS04 7H20 (Baker) 3; KH2PO4 (Baker) 7; AFA (CIBIOSA) 2.5.
Medio de esporulaci6n para Bacillus subtillis.
El medio de esporulacidn de B. subtillis, tiene la siguiente composici6n en g/ l : peptona (Bioxon) 8; extracto de came (Bioxon) 3; MnC12 (Baker) 10 M. Se ajust6 el pH a 7.2.
Medio para Bioensayo.
Para .la cuantificaci6n de penicilina se realizaron bioensayos, utilizando el medio agar de soya y tripticaseina (TSA) (Biox6n) siendo su composición en g/l : peptona de caseína 17; peptona de soya 3; NaCl 5; K2 Hpo4 2.5; Dextrosa 2.5; Agar bacteriol6gico 10; se ajust6 el pH a 7.3.
Medio de esporulaci6n.
Para las cepas parentales, se empld como medio de esporulaci6n de P. chrysogenum, medio Power cuya composici6n en g/l es la siguiente : Sacarosa (Bioxon) 15; Lactosa (Bioxon) IS; NaNo3 (Baker) 1 ; K2 HPO4 (Baker) 0.25; CuSO4 7H20, (Tecnica quhica) 0.0005; FeCl3 6H2O (Baker) 0.0015; W2m4 (Baker) 0.03; MgS04 7H20 (Baker) 0.275; FeS04 7H20 (Baker) 0.005; Bactopeptona (Difco) 2.5; SMM (CIBIOSA) 0.25; NaCl (Baker) 2.0; Agar bacteriol6gico (Bioxon) 20.0.
22

Materiales Y Mktodos
Medio de esporulación para cepas auxótrofas.
Para llevar a cabo la esporulación de las cepas auxótrofas (Met- y Un -), se empleó medio completo (MC), reportado por Ann6 y col. (1974). La composición en @l es la siguiente : KC1 (T6cnica química) 0.5; MgSO4 7H20 (Baker) 0.5; M2m4 (Baker) 1.0; FeS04 7H20 (Baker) 0.01; SMM 10; Difco Bacto Casitone (Difco) 2.0; Extracto de levadura (Bioxon) 2.0; D-L Metionina (Sigma) 0.05; Sacarosa (Bioxon) 30.0; Agar (Bioxon)l8.0; ajusthdose el pH a 6.0.
Medios de cultivo involucrados en la selecci6n de mutantes aux6trofas a aminoicidos.
Procesos de enriquecimiento.
Se utilizó medio mínimo (MM) para los procesos de íiltración, reportado por Diez y col. (1 987), utilizandose glucosa en lugar de lactosa y sin la adición de agar ya que el medio fue líquido; su composición en g/l : Glucosa (Bioxon) 30; N a N 4 (Baker) 2; K 2 W 4 (Baker) 1; MgS04 7H20 (Baker) 0.5; KC1 (T6cnica química) 0.5; FeSO4 7H20 (Baker) 0.01; se ajustó el pH a 6.5.
Medios para prueba de auxotrofia.,
Para probar la auxotrofia de las cepas se emplearon MM y MC reportados por Diez y col. (1987) y Ann6 y col. (1974) respectivamente. La composición del MM (adicionado de 30 g/l de agar [Bioxon]) y MC ha sido descrita anteriormente.
Medios para defínir tipo de auxotrofia.
Se utiliz6 MM suplementado de aminoAcidos a concentraciones fínales de 200 mg/d (Swart y col., 1990; Bos y col., 1992). Los aminohcidos adicionados al MM como suplemento, se esterilizaron en autoclave a excepción de la metionina la cual se esteriliz6 por filtración a trav6s de una membrana millipore de 0.22 p. Se prepar6 un stock de la metionina a s í esterilizada y se mantuvo almacenado a 4°C.
23

Materiales Y Met&
Medios de cultivo involucrados en La selecci6n de mutantes aux6trofos II
uridina.
Aislamiento de mutantes resistentes al 5 AFO.
Se utilizó MM con lactosa (adicionado con 30 g/l de agar [Bioxon]) como hente de carbono, descrito anteriormente. Este medio se suplementó con 100 pglml de uridina (Sigma) y con 1 mg/ml de 5 AFO (Sigma). El 5 AFO se esterilizó por filtración a traves de ma membrana millipore de 0.22 p.
Medios para prueba de auxotrofia.
Se emplearon MM lactosa y MM lactosa suplementado con 100 pglml de uridina (Sigma) (adicionados con 30 g/l de agar [Bioxon]}.
3. Condiciones de cultivo.
3.1 Fermentaci6n Liquida.
Obtención de suspensidn de esporas.
Se prepararon matraces con 40 ml de medio power, se inocularon con 0.25-0.3 ml de suspención de esporas y se incubaron a 25 OC durante el tiempo necesario para alcanzar la esporulación. Una vez presentada la coloración verde característica de la formación de esporas, estas se suspendieron en agua estbril con tween 80 al 0.2 % con ayuda de m agitador y b a m magn6tica. Se realizó la cuenta de esporas cosechadas en c h a m de Neubauer. La suspensión obtenida se utiliz6 para inocular tanto los medios semilla para FL como para los medios de FS.
La FL requiere de m in6culo en forma de micelio, para lo cuid se preparó medio complejo de semilla para cada una de las cepas. Se inoculó con 2 X 1 O esporadml. Se incubaron matraces con 50 ml de medio a 25 O C y con una agitación de 270 Tm. Este medio se utilizb para la obtencih de micelio necesario para la inoculación del medio complejo de producción.
24

Materioles Y Métdm
Producci6n.
Para la producci6n de penicilina se prepararon matraces con 45 ml de medio complejo de producci6n para cada una de las cepas. Se inocularon con 5 ml de micelio crecido en medio complejo semilla y se incubaron a 25°C y 270 rpm. El tiempo de producci6n varia conforme a la cepa.
Andisis de l a s muestras.
Biomasa (peso seco). Las muestras de FL se filtraron, se lavaron y se secaron en homo a 80 "C hasta peso constante para determinar la cantidad de biomasa. El filtrado se separó antes de los lavados, y se coloc6 en tubos de ensaye; la cantidad de penicilina y pH se determinaron del filtrado.
PH. La medici6n de pH se efectu6 directo del sobrenadante mediante un potencibmetro. Penicilina.
Para la cuantificaci6n de penicilina, se realizaron bioensayos, utilizando B. subtilis ATCC 6633. Esta tknica se basa en la formaci6n de un halo de inhibici6n del crecimiento, cuyo dihetro es proporcional al logaritmo de la concentraci6n de penicilina.
Para poder efectuar el biuensayo se requiri6 obtener una suspensi6n de esporas de B. subtilis, para lo CUAI se inocul6 el medio de esporulacibn indicado anteriormente, con dos, asadas procedentes de una colonia aislada con anterioridad en agar nutritivo. Se incub6 en matraces de 250 ml con 125 ml de medio a 30 O C durante 24 horas con una agitaci6n de 200 rpm. Cuando se obtuvo la suspensibn, se determind su densidad 6ptica a 340 nm y se inocul6 el medio TSA.
Se utilizaron cajas de a d c o de 30 X 30 cm y se colocaron 350 ml de TSA inoculando con 0.2 ml de in6culo por cada 100 ml de medio TSA para una suspensidn de esporas de B. subtillis con densidad 6ptica de l . Una vez gelificado el medio se hicieron pozos con un sacabocados y se llenaron con 60 pl de muestra de fernentach y soluci6n patr6n. Las cajas se incubaron a 35 "C durante 16 horas para la formaci6n de los halos de inhibici6n.
25

Materiales Y Mdtodos
3.2. Fermentacien S6lida.
hetratamiento del soporte.
Se empleó como soporte bagacillo de cafla. El bagacillo se tamiz6 en mallas No. 30 y 50 utilizando únicamente las partículas retenidas entre estas dos mallas.
Para el pretratamiento del bagacillo, se mezcl6 6ste con el 50 % del agua total que se iba a utilizar en el medio y se llevó a 3 Ib/pul? ( 101°C) de presión durante 30 minutos y una vez transcumdo el tiempo de pretratamiento se esterilizó a 15 Ib/pul& (1 2 1 "C) durante 15 minutos.
Condiciones de cultivo.
Al bagacillo de caila tratado se le adicionó el medio apropiado para cada una de las cepas. El medio s6lido conteniendo bagacillo, nutrientes y las esporas (2 X 10 espodml) tiene una humedad inicial de 70 % y 30 % de sólidos. El cultivo se realizó en columnas de vidrio conteniendo 12 g de medio de cultivo con una densidad de empaque de 0.26 dml, sumergidas en bailo de agua a 25°C para el control de tem- y se mantuvo un sistema de aireaci6n a un flujo de aire de 2.4 vh. El flujo de aire se hizo burbujear en agua antes de pasar por cada columna con el fin de humidificar el aire.
M i s i s de muestras.
Para el muestre0 en FS se tomaron dos columnas completas de cada tiempo, y se pesó la cantidad de medio de la c o l u m n a . Del medio se tomó 1 g para la determinacih de pH, 1.5 g para la determinación de penicilina producida y 3 g se pesaron y se secaron para determinar la humedad del medio.
PH. Para la determinación de pH se mezcló 1 g de muestra con 10 ml de agua destilada agitando durante 1 O min. con agitador magn6tico antes de la determinación.
Penicilina. Para la extracción de penicilina, se ped 1 g de muestra y se asladieron 6 m l . de amortiguador de fosfatos (pH 5.5), se agit6 y se ajustó el pH entre 5 - 5.1 5, midiendo la cantidad de &ido o base empleadas para posteriormente tomarlas en cuenta en los cirJculos de dilucih. Se centrifugó a 163 x g durante 20 m i n . y se
26

Materiales Y Metodos
separ15 el sobrenadante, el cual se utiliz6 para determinar la Concentración de penicilina por bioensayo.
Humedad. Para determinar humedad se pesaron 3 g de muestra y se colocaron en papel aluminio previamente puesto a peso constante. Las muestras se colocaron en estufa a 80" C por 24 horas determinhdose su peso final.
4. Mutae6nesis Y Curva de Sobrevivencia.
Siembra y cosecha de esporas.
Se sembraron esporas de P. chrysogenum de las cepas P2-32 y P2-37 con un in6culo de 0.3 ml de suspensi6n de esporas en medio power; una vez esporulado el medio, se realiz6 la cosecha de esporas con tween 80 al 0.2%, agitador y barra magn¿tica. La suspensi6n de esporas a s í obtenida se filtr6 a traves de fibra de vidrio con el fin de eliminar el micelio que pudiera haberse desprendido durante la cosecha de esporas.
Radiacih con luz U.V.
Se realizd una cuenta de esporas de la suspensi6n filtrada y en base a ella se realiz6 una dilucibn para obtener UM concentraci6n de 8 X 1 O espodml. Se colocaron 27 ml de la suspensi6n de esporas a la cmcentraci6n antes mencionada en una caja petri estkril y manteniemdo una agitaci6n de 30 rpm, mediante un agitador magnbtico. Se llev6 a cabo la i1~adiaci6n con luz U.V. aplichdose con una intensidad de 250 pW/cm2 durante 10 min. Se tomaron muestras de l m l a los siguientes tiempos : 0,1,2,3,4,5,6,8 y 10 min. Las muestras irradiadas se colocaron en viales y se conservaron en glicerol al 25% envolvihdose en papel aluminio, para mantenerlas en la obscuridad.
Despues de la mutagfinesis, se llev6 a cabo un periodo de replicaci6n del DNA y crecimiento micelial de las esporas mutadas, para que de esta forma el &o causado en el material genetic0 se estableciera como un dairo estable y se permitiera transferirlos como mutaciones heredables. (Queener y Lively, 1986).
A s í mismo se incluy6 un periodo de sincronizaci6n de la germinaci6n de las conidiosporas, una vez fijada la mutaci6n, ya que de acuerdo a un estudio realizado por Bos y col., (1992), se observ6 que la irradiaci6n con luz U.V. afecta la
21

Materiales Y MPtodos
fisiología de las conidiosporas, reflejándose en una desincronización de su germinación. Para sincronizar la genninación se mantuvo el cultivo de esporas mutadas por un periodo de 7 días en almacenamiento a 4°C (Bos, 1993).
Elaboración de la curva de sobrevivencia.
Se tomó 0.1 ml de las muestras obtenidas para cada una de los tiempos de irradiación, y se realizaron diluciones que abarcaron un intervalo desde 1 X 10" hasta I X 10" . Cada una de l a s diluciones se plaquearon en PDA, por
duplicado (0.1 ml de la dilución por placa) y se incubaron en la obscuridad (envueltas en bolsas negras) a 25 "C durante el tiempo necesario para el crecimiento de las colonias ( 7-9 días). Una vez que surgieron las colonias, se realizó un conteo de las mismas y se generó la curva correspondiente de sobrevivencia.
Cuantiíicación de las mutantes morfológicas.
Las colonias se desarrollaron en PDA para deteminar la viabilidad, y se llevó a cabo el conteo de aquellas colonias que presentaban diferencias morfológicas con respecto a la cepa parental sin mutagenizar.
5.Obtenci6n de mutantes Aux6trofas a amino%cidos.
5.1 Procesos de enriquecimiento por filtraciones sucesivas.
Preparación de inkulo de esporas irradiadas.
Se sembraron 0.3 ml. de la suspensión de esporas irradiadas de la cepa P2-37 en MC, Ilevhdose a cabo su crecimiento en la obscuridad (cubriendo los matraces con papel aluminio). UM vez que el micelio completó la etapa de esporulación, se introdujo un periodo de sincronización de la germinación, almacenando el micelio esporulado a 4°C durante 7 días en la obscuridad (cubriendo los matraces con papel aluminio); Posteriormente se r e m la cosecha de esporas, como ya se mencionó anteriormente. Inmediatamente después se comenzaron los procesos de filtración
28

Materiales Y Mdrodos
Procesos de filtración.
Una vez preparado el inhulo de esporas irradiadas, se inocularon matraces con 50 ml de MM líquido (2.5X105 esp/ml). Se agregaron perlas de ebullición a los matraces para evitar la agregacih del micelio germinado y con ello el atrapamiento de esporas con probabilidad de ser auxótrofas. Los matraces inoculados se incubaron a 25°C y con agitación de 150 rpm durante 60 horas. A intervalos de 30 horas se realizaron filtraciones del MM a travds de fibra de vidrio para remover el micelio que llegó a desarrollarse. El filtrado resultante se centrifugó a 1200 x g por 30 minutos y por decantación se recuperaron las esporas residuales que aún no habían germinado, para posteriormente resuspenderlas en MM nuevo y volver a incubación. Para la última filtración, las esporas no germinadas recuperadas se resuspendieron en agua con tween y se efectuó una cuenta de esporas. Se realizaron las diluciones correspondientes para efectuar el plaque0 de cajas de MC y lograr un crecimiento de alrededor de 300 colonias por caja.
Aislamiento de colonias.
De las esporas residuales no germinadas recuperadas que se desarrollaron en MC, se seleccionaron aqudllas que presentaban un menor tamaílo, ya que se suponía que podían una mayor dificultad para crecer debido a algún desaneglo gendtico provocado por la mutaghesis, que resultara en un tipo de auxotrofia, mientras que aquellas colonias sanas mostrarían un crecimiento vigoroso. Se llevó a cabo el aislamiento de las colonias a s í seleccionadas en M C hasta su esporulación.
Prueba de auxotrofia.
La prueba de la auxotrofia se hizo despuds de impregnar la punta de un palillo con esporas de las colonias aisladas en MC, por picadura directa con el palillo en cajas petri con MM y M C respectivamente. Una vez comprobado que una colonia creció en M C y no lo hizo en M M , es decir que presentó una auxotrofia, entonces se continuó con la detenninación del tipo de auxotrofia, la cual se realizó haciendo crecer la colonia auxótrofa en MM suplementados con los diferentes aminokcidos.
29

Materiales Y Mbtodos
Determinación del tipo de auxotrofia.
Se prepararon 9 cajas petri con "-agar suplementados con una combinación específica de aminoAcidos (ver tabla 1) y a una concentración de 200 mg/ml. Se sembraron por medio de picadura con palillo, esporas de las colonias que presentaron auxotroffa. La siembra de las mismas, se realizó en una posición especifica, mantenihdose el mismo orden en todas las cajas, con el fin de seguir su identificación.
Tabla 1: Suplementacibn de amino4cidos al MM para determinar tipo de auxotrofia presente en los mutantes (Gunasekaran, 1995).
Cuando una colonia crece en alguna de las combinaciones dadas para las cajas 1-5, y lo hace tambib en alguna de las combinaciones dadas para las cajas 6-9, se permite la identificación directa de un Eactor de crecimiento. Por ejemplo, si una colonia crece en la caja 1 y en la 7 requiere como factor de crecimiento histidina; si crece en la caja 3 y tambih lo hace en la 8 entonces requiere triptofano. Si una colonia crece solamente en una de las 9 cajas entonces probablemente requiera más 'de un nutriente.
5.2. Enzima lítica Novozyme.
Se preparó una suspensión de esporas irradiadas de la cepa P2-37 como se mencionó anteriormente. Se sembraron IO4 esporas por caja petri de 5 cm de dibetro conteniendo "-agar (triplicado). Se incubaron a 25 "C durante 29 horas, periodo durante el cual los tubos germinales comienzan a ramificarse, siendo entonces el momento m& propicio para la acción de la enzima. El monitoreo de la genninación se efectuó a travds de un microscopio óptico. Al salir del periodo de incubación, se adicionó 1.5 ml de la enzima Novozyme (enzima lítica de Trichoderma hanianum; Sigma Chemical C.O. L-2265) en cada una de las cajas, en una concentración de 10 mg/ml y se incubaron nuevamente a 25 "C por un periodo de 1 hora. Terminado este se llevó a cabo la remoción de la enzima, seguida de lavados con agua estdril para asegurar su total eliminación. Se incubaron
30

Materiales Y Metodos
nuevamente a 25 "C por 30 horas para dar oportunidad de que las esporas que no germinaron, lo hicieran. Se repitió posteriormente el tratamiento con Novozyme y se incubaron las cajas con la enzima a 25 "C toda la noche. Despubs se realizó la remoción de la enzima y los lavados. Se agregó una pequeila capa de MC a las cajas con 0.8 % de agar y se regresó a incubación a 25 "C por el tiempo necesario para la germinación de las esporas residuales que probablemente eran auxótrofas. Una vez germinadas, se seleccionaron con el criterio autes descrito y se aislaron en MC. El agua de lavado (Id por caja) se recuperó y se centrifugó a 1200 x g durante 30 min para recuperar las esporas que no germinaron y que fueron arrastradas en el agua de lavado. Estas esporas recuperadas se sembraron en MC y una vez germinadas las colonias se seleccionaron las miis pequefías y se aislaron en el mismo medio. La prueba y la determinación de la se realizó a s í mismo como se describe anteriormente.
6, Obtencibn de mutantes Aur6trofos a uridina.
Determinación de la dosis mhs adecuada de 5 AFO para la cepa €2-32.
Para llevar a cabo el aislamiento de mutantes de P. chrysogenum de la cepa P2-32 resistentes al 5 AFO, se implement6 la metodología reportada por Diez y col. (1987). Sin embargo antes de llevarla a cabo, se requirió determinar la concentración de 5 AFO que inhibiera el crecimiento y permitiera el desarrollo de colonias resistentes, en la cepa P2-32. Este proceso se llevó a cabo en la cepa sin mutar con el fin de ,detectar qud tan sensible era al 5 AFO, y posteriormente aplicar la dosis encontrada en la cepa mutada. Se aplicaron tres dosis diferentes de 5 AFO: 0.75 mgd, 1 mdml y 1.25 mg/ml.
6.1. Aislamiento de mutantes resistentes al 5 AFO.
Una vez determinada la concentración de 5 AFO m& adecuada para la cepa P2-32, se preparó el inóculo de esporas irradiadas de la cepa por el metodo descrito anteriormente. Se realizaron las diluciones necesarias y se sembraron por plaque0 1.2 X 1 o6 esporas por caja petri con MM con Iactosa suplementadas con 100 pg / m1 de uridina y con 1 mg / ml de 5 AFO (Diez y col., 1987). Se incubaron a 25 "C durante el tiempo necesario para el crecimiento de l a s colonias (8- 10 días aproximadamente). Se aislaron en MC algunas de las colonias resistentes que lograron crecer a la concentración de 5 AFO empleada. El aislamiento de las
31

Moterioles v Mtodo>
colonias resistentes se llevó a cabo antes de su espodación para evita: contaminación de esporas de colonias adyacentes. De tal forma que se efectuó 1; propagacih de micelio de l a s colonias resitentes en MC.
Prueba de auxotrofia a uridina.
Se probó la auxotrofia especfficamente a uridina en MM con lactosa y MM cor lactosa suplementado con 100 pg / ml de uridma, inoculando esporas de l a s colonia: aisladas por medio de la picadura directa con palillo en estos medios.
32
VI1 Resultados.
1. Seleccih de cenas.
Se caracterizaron 5 cepas de P. chrysogenum: P2-32, P2-37, P2-47, P2-2V2 Y ASP- 2A, las cuales provenían de clones P2 y ASP aislados por Barrios y col. (1993) y que presentaban la característica de ser buenos productores de penicilina en SFS. La caracterización consistió en el desarrollo de fermentaciones tanto por el SFS como por el SFL de las cepas anteriormente mencionadas
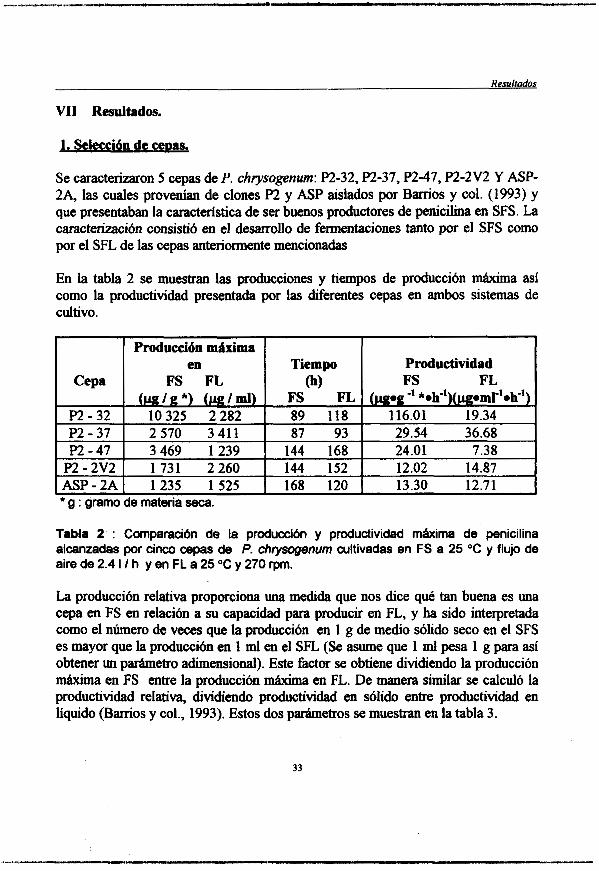
En la tabla 2 se muestran las producciones y tiempos de producción m-a a s í como la productividad presentada por las diferentes cepas en ambos sistemas de cultivo.
Productividad
* g : gramo de materia m.
Tabla 2 : Comparacibn de la produccibn y productividad mkima de penicilina alcanzadas por cinco cepas de P. chrysogenum cultivadas en FS a 25 "C y flujo de aire de 2.4 I I h y en FL a 25 "C y 270 rpm.
La producción relativa proporciona una medida que nos dice qué tan buena es una cepa en FS en relación a su capacidad para producir en FL, y ha sido interpretada como el número de veces que la producción en 1 g de medio sólido seco en el SFS es mayor que la produccith en 1 ml en el SFL (Se asume que 1 ml pesa 1 g para así obtener un p h e t r o adimensional). Este factor se obtiene dividiendo la producción m w a en FS entre la producción -a en FL. De manera similar se calculó la productividad relativa, dividiendo productividad en sólido entre productividad en líquido (Barrios y col., 1993). Estos dos parhetros se muestran en la tabla 3.
33

Hesultados
Produccibn Productividad Cepa Relativa Relativa
P2-47 1 2.8 I 3.25 P2-2v2 I 0.77 0.81
IASP-2AI 1.47 I 1 .O5 I
Tabla 3: Comparacidn de los factores de produccidn y productividad relativa entre las cinco cepas de P. chrysogenum evaluadas.
Los resultados muestran que la cepa de mejor producción de penicilina en el SFS es la cepa P2-32 ya que produjo aproximadamente 4.5 veces mas que las otras cepas evaluadas. Mientras que la mejor producción de penicilina en el SFL lo muestra la cepa P2-37, sin embargo no se observó una diferencia muy marcada (1.4 veces) con respecto a las otras cepas. Los tiempos de producción mAxima son diferentes para cada cepa, observiindose que la que alcanza la mayor producción de penicilina en menor tiempo es la cepa P2-37, esto tanto para FS como para FL (tabla 2).
Las cepas P2-32 y P2-47 presentaron niveles mayores de producción en FS que en FL. Mientras que un comportamiento inverso es mostrado por las cepas P2-37, P2- 2V2 y ASP-2A. La producción y productividad relativas son a s í mismo diferentes para cada cepa, es decir, se confirma que no todas las cepas tienen la misma capacidad para producir en FS. La cepa P2-32 presenta la mayor eficiencia de producción en FS.
Por los datos anteriores se seleccionaron estas dos cepas, para conjuntar en un solo organismo la capacidad de biosíntesis de penicilina mostrada por la P2-37 en FL y las características que penniten una buena producción de penicilina mostrada por la P2-32 en FS. Es decir, una cepa de alto pdpl con una de bajo pdpl.
En la Figura 3 se muestran l a s cín&ícas de producción penicilina, pH y humedad en FS de las cepas seleccionadas. .Se observa que la humedad tiende a aumentar conforme transcurre la fmentacih para ambas cepas. El compommiento de pH muestra una tendencia al incremento durante la fmentacidn. La cepa P2-32 muestra una producción de penicilina superior (1 O 325 pg/g) a la producción mostrada por la P2-37 (2 570 pg/g) para este sistema de fermentación.
34

Resultados
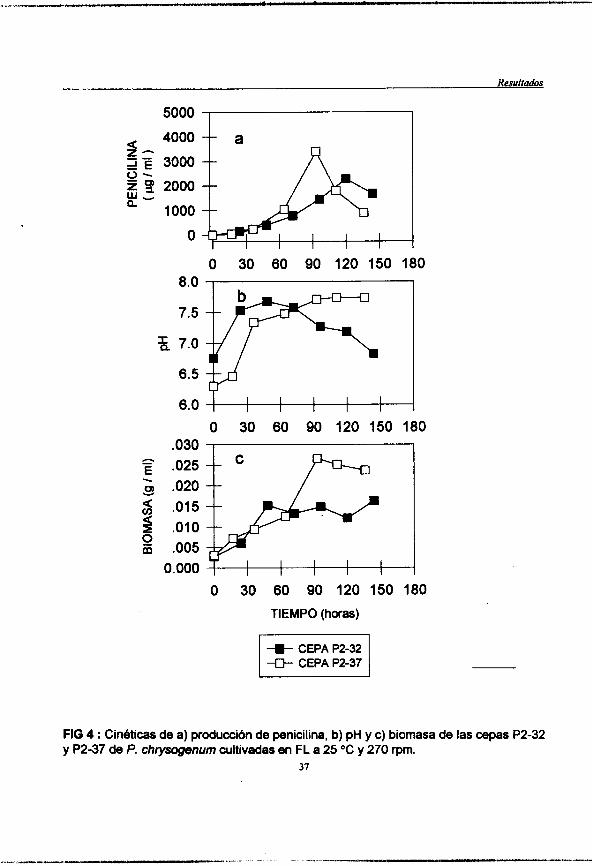
En tanto que en el sistema de FL se observa que la cepa P2-37 creció mejor (0.25 g /ml) que la P2-32 (0.015 g / ml) lo que se ve reflejado en la produccih de penicilina. La P2-37 produjo 2.3 veces m& de penicilina que la cepa P2-32 ( Fig 4). En cuanto al pH, Cste se incrementa en ambos casos hasta l a s 72 horas y observa un decremento posterior para la cepa P2-32 hasta el final de la fermentación.
35

O 30 60 90 120 150 180 8.0
7.5
7.0
6.5
b "
"
6.0 a O 30 60 90 120 150 180
O 30 60 90 120 150 180
TIEMPO (horas)
-C CEPA P2-32 U CEPA P2-37
Fig 3 : Cin6ticas de a) producci6n de penicilina, b) pH y c) humedad de las cepas P2- 32 y P2-37 de P. chrysogenum cultivadas en FS a 25 "C y con un flujo de aire de 2.4 I/h.
36
5000
4000
3000
2000
1 O00
O i i i I I
O 30 60 90 120 150 180
6.5
O 30 60 90 120 150 180 .O30 .O25 C
0.000 ,I O 30 60 90 120 150 180
TIEMPO (horas)
I 1 + CEPA P2-32 U CEPA P2-37
FIG 4 : Cin6ticas de a) produccibn de penicilina, b) pH y c) biomasa de las cepas P2-32 y P2-37 de P. chrysogenum cultivadas en FL a 25 "C y 270 rpm.
31

Resultados
2. Aislamiento de Aux6trofos.
Las cepas seleccionadas como cepas parentales , la P2-32 y la P2-37, no tienen marcadores genbticos, como auxotrofias, por lo que se hizo necesario introducir algún marcador de este tipo, para posteriormente realizar la recombinación genbtica.
Los aminohcidos que intervienen en la ruta de biosíntesis de la penicilina son: valina, cisteína y lisina, por lo que se buscaron marcadores nutricionales diferentes a estos.
2.1. Mutagenesis.
Se generaron las curvas de sobrevivencia para ambas cepas parentales seleccionadas, mediante las cdes , se permite establecer la dosis adecuada para la obtención de sobrevivencia de entre 50-60 %, para cada una de ellas (Fig 5 y Fig 6).
De acuerdo a las curvas de sobrevivencia generadas para ambas cepas, se puede observar, que para que los niveles de sobrevivencia sean de 50 % se requiere de la irradiación de luz U.V. sobre las esporas, por un tiempo de 2.5 minutos para el caso de la cepa P2-37 (Fig 6), mientras que para la cepa P2-32 se requiere un tiempo de 2.1 minutos a l a s condiciones establecidas (Fig 5). Los resultados muestran que la cepa P2-32 es m b sensible a la radiacidn UV al requerir menor tiempo de exposición de la radiación que la cepa P2-37. Ademiis los resultados de cuenta viable observados en la cepa P2-32 fueron menores que los observados para la cepa P2-37 (Fig 5 y Fig 6).
Se re&ó un seguimiento de las mutantes morfológicas aleatoriamente en la cepa P2-37, que surgieron a cada tiempo de la curva de sobrevivencia (Fig 7).
38
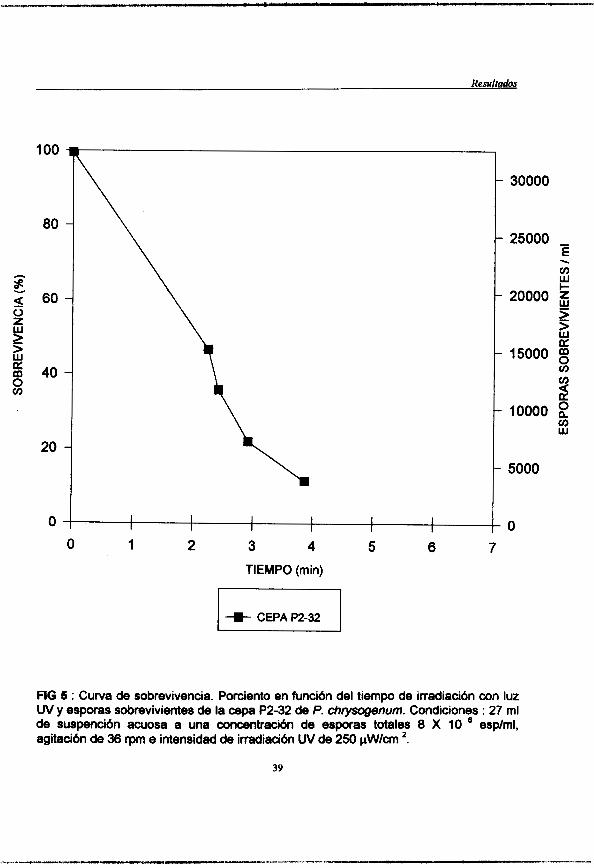
Resultados
1 O 0
80
60
40
20
O I I I I I I
O 1 2 3 4 5 6 7
TIEMPO (min)
-C CEPA P2-32
- 30000
. 25000 - E
3 . 10000 g
u3 W
5000
O
FIG S : Curva de sobrevivencia. Porciento en funci6n del tiempo de irradiaci6n con luz UV y esporas sobrevivientes de la capa P2-32 de P. chrysogenum. Condiciones : 27 ml de suspenci6n acuosa a una comtracitk de esporas totales 8 X 10 esplml, agitacih de 36 rpm e intensidad de irradiaci6n UV de 250 pW/m ’.
39
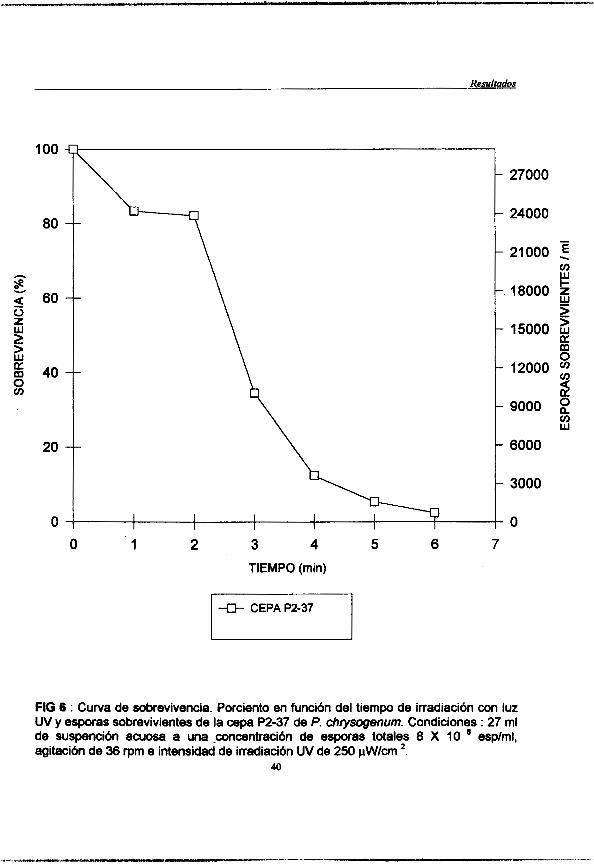
Resultados
1 O 0
80
60
40
20
O O
. 27000
. 24000
- 21000
. 18000
- 15000
- 12000
- 9000
- 6000
- 3000
- 0
1 2 3 4
TIEMPO (min)
5 6 7
L U CEPA P2-37
FIG 6 : Curva de sobrevivencia. Porciento en funcidn del tiempo de irradiacidn con luz UV y esporas sobrevivientes de la cepa P2-37 de P. chrysogenum. Condiciony : 27 m1 de suspenci6n acuosa a una -concentracidn de esporas totales 8 X 10 esplml, agitacibn de 36 rpm e intensidad de irradiaci6n UV de 250 pW/m ’.
40
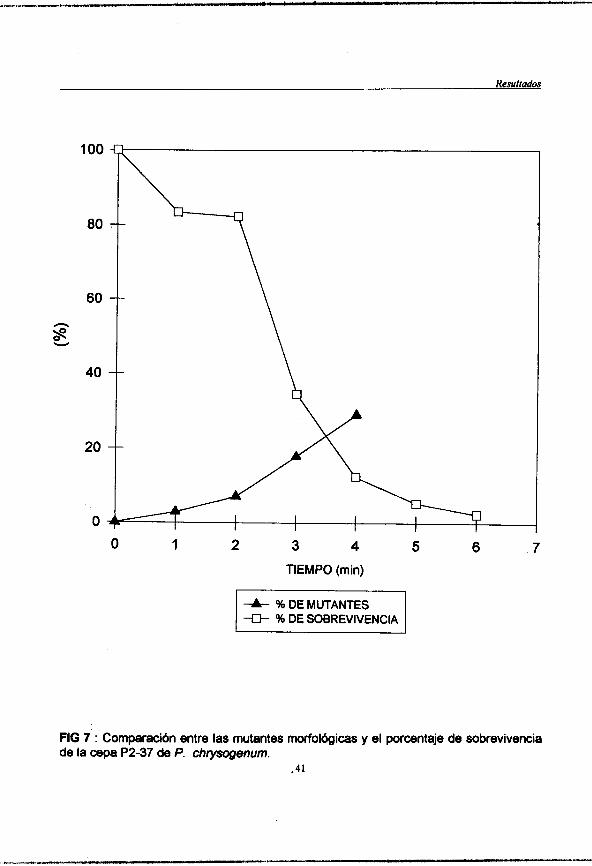
1 O 0
80
60
n 5 40
20
O
O 1 2 3 4
TIEMPO (min)
5 6
-A- % DE MUTANTES U % DE SOBREVIVENCIA
FIG 7 : Comparacibn entre las mutantes morfol6gicas y el porcentaje de sobrevivencia de la cepa P2-37 de P. chrysogmum.
.41

2.2. Mutantes auxbtrofos a rminoicidos.
Se aplicaron 2 procesos de enriquecimiento en la cepa P2-37 para el aislamiento de mutates auxótrofas a aminohcidos. Los fundamentos de estas teCnicas son distintos pero persiguen un mismo íin: recuperar esporas no germinadas, con alta probabilidad de ser auxótrofas, de entre un gran número de ellas que sí pueden hacerlo.
El primer proceso de enriquecimiento empleado consistió en la eliminación paulatina de las esporas germinadas capaces de crecer en MM, mediante filtraciones sucesivas, con objeto de concentrar y recuperar las esporas no germinadas con alta probabilidad de ser auxótrofas (Bos y col., 1981; Uitzetter y col., 1986). Para lo cual se evaluó la eficiencia de algunos medios filtrantes determinando el porcentaje de esporas que lograban atravesarlo, y de 6sta fonna lograr la menor *dida de esporas posible al momento de realizar la remoción del micelio germinado. Se probaron 3 tipos de filtros diferentes y la fibra de vidrio resultó el medio filtrante mh adecuado (Tabla 4). Finalmente se hicieron germinar esas esporas en medios específicos, con la finalidad de probar si eran o no auxótrofis.
Cuenta de Material
Esporas / m1 que atraviesan el filtro filtrante esporas * Porciento de esporas
G W
12.6 X 10 74.8 Fibra de vidrio 7.55 x 10 44.8 Nytal 4.8 X 10 28.0
- "" - * La cuenta de esporas de la suspensibn sin filtrar h e de 16.85 X lo6 Espordml.
Tabla 4: Eficiencia de filtración de 3 materiales diferentes.
El segundo proceso de enriquecimiento empleado fue a base de una preparación de enzima lítica Novozyme 234 (Debets, 1989) basado en la eliminación selectiva de micelio germinado inmovilizado en una base de medio-agar, mediante la degradación de la pared celular. Se realiz6 un seguimiento del progreso de la germinación de las esporas mutadas, por medio de un microscopio óptico, ya que se precisaba llevar a cabo el tratamiento con la enzima lítica en un estadio germinativo específico del hongo. Es decir, la enzima Novozyme 234 sólo actúa sobre
42
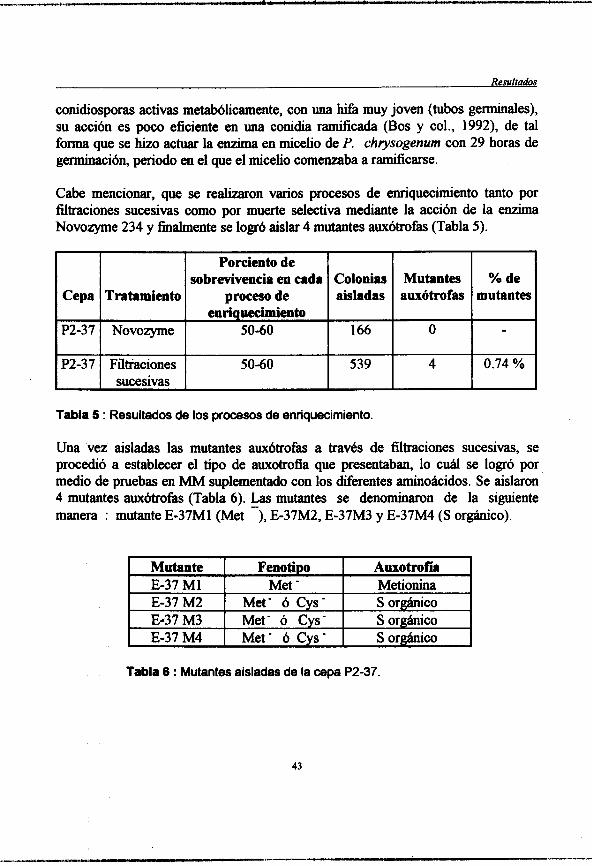
Resultados
conidiosporas activas metabólicamente, con una hifa muy joven (tubos germinales), su acci6n es poco eficiente en una conidia ramificada (Bos y col., 1992), de tal forma que se hizo actuar la enzima en micelio de P. chrysogenum con 29 horas de germinacidn, periodo en el que el micelio comenzaba a ramificarse.
Cabe mencionar, que se realizaron varios procesos de enriquecimiento tanto por filtraciones sucesivas como por muerte selectiva mediante la acci6n de la enzima Novozyme 234 y finalmente se log6 aislar 4 mutantes aux6trofhs (Tabla 5).
Porciento de sobrevivencia en cada Yo de Mutantes Colonias
Cepa
0.74 % 4 539 50-60 Filtiaciones P2-37
- O 166 50-60 Novozyme P2-37
mutantes auxhtrofas aisladas proceso de Tratamiento enriquecimiento
sucesivas
Tabla 5 : Resultados de los procesos de enriquecimiento.
Una vez aisladas las mutantes auxdtrofas a travds de filtraciones sucesivas, se procedió a establecer el tipo de auxotrofia que presentaban, lo cud se logró por medio de pruebas en MM suplementado con los diferentes aminoácidos. Se aislaron 4 mutantes aux6trofas (Tabla 6). Las mutantes se denominaron de la siguiente manera : mutante E-37M1 (Met -), E-37M2, E-37M3 y E-37M4 (S orgánico).
Mutante Auxotrof'ía
E-37 M2
I E-37 M4 I Met 6 Cys- I S orghico I Tabla 6 : Mutantes aisladas de la cepa P2-37.
43

Hesuliados
2.3. Mutantes AuJtrofos a uridina.
Determinación de la dosis mhs adecuada de 5 AFO para la cepa P2-32 sin mutar.
Se encontró que en la cepa P2-32 de P. chrysogenum sin mutagenizar, se inhibió el crecimiento a concentraciones superiores a 0.75 mg/ml de 5 AFO, mientras que se obtuvieron mutantes resistentes a la concentración de 1 y 1.25 m g h l de 5 AFO, sin embargo se decide emplear la dosis de 1 m g / d ya que no se observan diferencias en el número de mutantes resistentes obtenidas (Tabla 7). Se observó una frecuencia de 5 X 1 O para las mutantes espontheas resistentes para ambas concentraciones de 5 AFO (Tabla 7). Se lograron aislar nueve mutantes espontheas, las cuales se probaron en su auxotrofia, sin embargo no se logró aislar ninguna mutante auxótrofa a uridina.
Cepa Auxotrofía Frecuencia de Inkdo Colonias Concentraci6n S AFO
> 300 0.75 caja por caja mglml
a uridina mutaci6n Esporas / resistentes
P2-32 O 5 X 1 0 - 6 6 x 1 0 ’ 3 1
Tabla 7 : Efecto de tres dosis de 5 AFO empleadas para determinar la resistencia de la cepa Pa-32 sin mutar de P. chrysogenum, en MM con lactosa y suplementada con 100 pg/ml de uridina.
Aislamiento de mutantes resistentes al 5 AFO.
Una vez encontrada la dosis de 5 AFO a la cual se aislan mutantes con resistencia a este cOmpuesto, para la cepa €2-32, se sembraron esporas mutadas con UV de esta misma cepa (50-60 % de sobrevivacia) en MM con lactosa con suplementación de 1 m g / d de 5 AFO y 100 pg/d de urih @hez y col., 1987). Se inocularon 1.2 X . 10 esporas por caja, un in6culo mayor del que se había estado aplicando anteriormente (Tabla 7), con el objeto de incrementar el número de colonias resistentes aisladas. La fkecmcia de mutantes resistentes que se observó fue de 5 X 10’’ , lo que llevó al aislamiento de un gran número de colonias resistentes, contando un promedio de 60 colonias resistentes al 5 AFO por caja.
44

Resultados
Finalmente, se lograron aislar 128 colonias resistentes al 5 AFO, y mediante pruebas en MM y MM suplementado con uridma, se encontró una mutante auxótrofa a uridina. La mutante aislada se denominó E-32U.
3. Evrluacihn del nivel de Droduccith de Denrcdma en mutantes auxdtrofas. . ..
Es necesario hacer menci6n de que muchas de las investigaciones que involucran el trabajo con microorganismos marcados genéticamente han recibido tratamientos mutaghicos que generan niveles de sobrevivencia muy bajos, menores al 10 %, ylo bien tratamientos con mutitgenos químicos.
Sin embargo, a pesar de que los niveles de sobrevivencia generados en las cepas seleccionadas fueron de entre 50-60 % para prevenir daílos indeseables en el material genético, aún a s í es necesario evaluar el nivel de producción de penicilina en las mutantes auxótrofas aisladas y asegurar de esta forma que la producción de penicilina no ha sido a f e c t a d a . Esto se observd al realizar una comparación entre las cineticas de produccidn de penicilina de las mutantes y las cepas parentales.
En la tabla 8 se muestran las producciones máximas, los tiempos de producción mkimos y la productividad de las cepas parentales P2-32 y P2-37 en una segunda caracterización llevada a cabo, a s í como de las mutantes auxótrofas E-32U y E- 37M1 e n , cultivos de FS y FL. Las producciones y productividades relativas se muestran en la Tabla 9.
En la segunda caracterizacidn de las cepas parentales, se observa una reducción en la produccih de penicilina en el SFL con respecto a la primera tanto en la cepa P2- 32 como en la P2-37 (tabla 2 y tabla 8) y (Fig 9); un comportamiento similar es presentado en la cepa P2-32 en el SFS, mientras que la cepa P2-37 por lo contrario, incrementa su producción en este mismo sistema (tabla 2 y tabla 8) y (Fig 8 ).
La produccih relativa de la cepa P2-32 (tabla 9) disminuye considerablemente en su segunda evaluacih con respecto a la primera (tabla 3 y tabla 9), mientras que la P2- 37, la aumenta tambih de forma importante (tabla 3 y tabla 9).
Al llevar a cabo una comparación entre las mutantes y las cepas parentales en su última evaluacih (tabla 8), se observa una disminución de la producción en el SFS tanto para la cepa P2-32 como en la P2-37, presentando la mutante E-32U una disminución del 40 %, en tanto la E-37M1, muestra una disminución del 8%. Un comportamiento inverso se pone de manifiesto en el SFL en donde un aumento de
45

entre 25-30 YO en la producción es observado para ambas mutantes. Se presenta entonces una dismiuución de la eficiencia de producción en sólido en ambas mutantes aisladas, la E-32U y la E-37M1.
En la Fig. 10 se muestran las cindticas de producción de penicilina, pH y humedad de la mutante E-32U comparada con la cepa parental P2-32 en el SFS. La Fig. 11 muestra las cindticas de producción de penicilina, pH y biomasa de las mismas cepas pero en el SFL. En la Fig. 12, se muestran a s í mismo las cindticas de producción de penicilina, pH y humedad de la mutante E-37M1 comparada con la cepa parental P2-37 en el SFS. En tanto que la Fig 13 muestra las cinkticas de producción de penicilina, pH y biomasa de las mismas cepas para el SFL.
Se observa en la Fig. 10 que la humedad se incrementa gradualmente conforme transcurre la fermentación tanto para la cepa P2-32 como para la awótrofa E-32 U. El comportamiento general del pH para ambas cepas es de un incremento gradual hasta el final de la fmentación. En la Fig. 1 1 la biomasa de ambas cepas se incrementa, en tanto que el pH pra la cepa P2-32 aumenta para luego permanecer estable; el pH de la awótrofb E-32U disminuye despuds de las 60 horas de fermentación para luego incrementarse de forma gradual hasta el h a l del cultivo.
En la Fig 12 el comportamiento de la humedad en las cepas P2-37 y E-37 M1 es de un incremento gradual hasta el final de la fennentacih al igual que el pH. A s í mismo, en la Fig. 13 la biomasa tambikn se incrementa de forma gradual hasta el final de la fermentacibn. Se observa el mismo comportamiento en el pH para la cepa P2-37 mientras que el pH de la'awótrofb E-37 M1 decrece despuds de las 80 horas de fermentación y hasta el final de la fermentación.
46
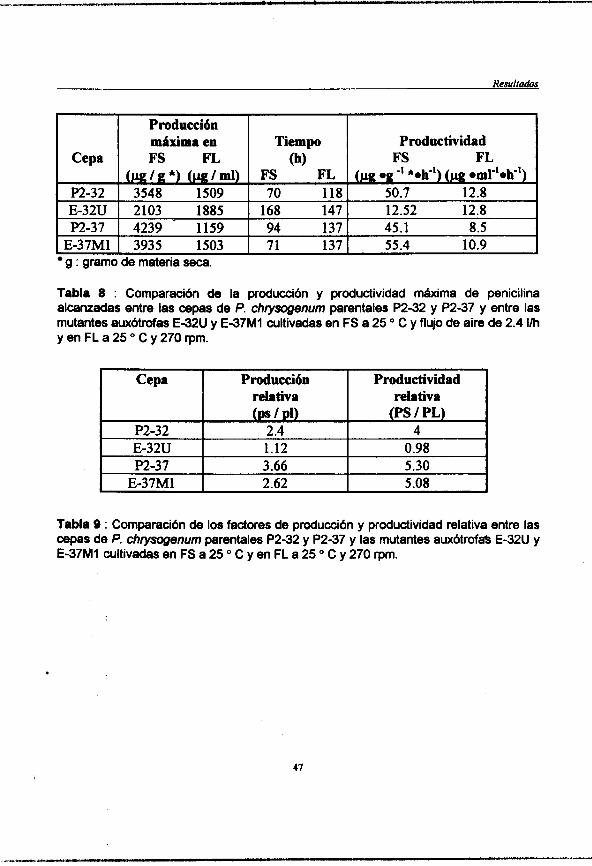
Cepa Cepa
¡ P2-32 P2-32 E-32U
E-3 7M 1
E-32U P2-37
E-3 7M 1
Producci6n mixima en FS FL
( M / E * ) (M!/ml) 3548 1509
Producci6n mixima en FS FL
( M / E * ) (M!/ml) 3548 1509 2103 1885 2103 1885 4239 1159 3935 1503 4239 1159 3935 1503
Tiempo Productividad (h) FS FL
FS FL (pg og *oh") (pg om1"oh") ~
70 118 50.7 12.8
Tiempo Productividad (h) FS FL
FS FL (pg og *oh") (pg om1"oh") ~
70 118 50.7 12.8 168 147
55.4 10.9 - 71 137 45.1 8.5 94 137 12.52 12.8
~~~
168 147
55.4 10.9 - 71 137 45.1 8.5 94 137 12.52 12.8
~~~
g : gramo de materia s e c a .
Tabla 8 : Comparacidn de la producci6n y productividad h i m a de penicilina alcanzadas entre las cepas de f. chrvsogenum parentales P2-32 y P2-37 y entre las mutantes awdtrofas E-32U y E37M1 cultivadas en FS a 25 o C y flup de aire de 2.4 Ilh yenFLa25OCy270rpm.
Produccidn Productividad I relativa I relativa I (ps I PI) (Ps / PL)
P2-32 4 2.4 I E-32U I 1.12 I 0.98 I
P2-37 5.08 2.62 E-37M1 5.30 3.66
Tabla 9 : Comparacidn de los factores de producci6n y productividad relativa entre las cepas de f. chrvsogenum parentales P2-32 y P2-37 y las mutantes aw6trofas E-32U y E-37M1 cultivadas en FS a 25 O C y en FL a 25 O C y 270 rpm.
41
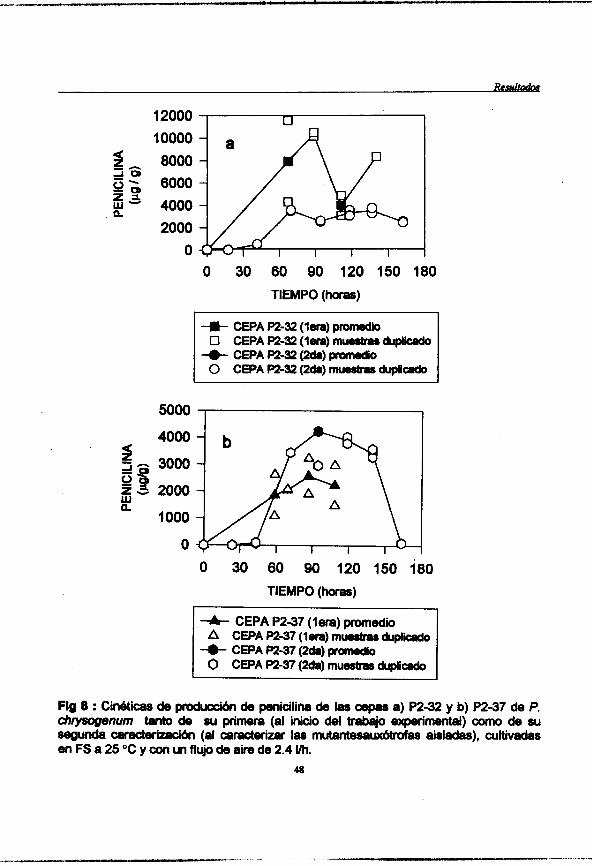
O 30 60 90 120 150 180
TIEMPO (horas)
O CEPA P2-32 (lm) numstms duplicado -0- CEPA P2-32 (a) -0
O CEPA P2-32 (w) duplicpdo
9
3- ag p.
4000
3000
2000
1 O00
O O 30 60 90 120 150 180
TIEMPO (horas)
A CEPA P2-37 (le@ muostmr duplicado
O CEPA m-37 (2da) mueabas dupticlldo CEPA P2-37 (2dr) pronndio
Fig 8 : CiMtiCas de producci6n de penicilina de las cepas a) P2-32 y b) P2-37 de P. dvysogenum tantu de su primera (al inicio del trabajo experimental) como de su segunda - (al cprpcterjzar las mutant- aisladas), a r l t i i s enFSa25°Cyconunflupdeairede2.4Uh.
. .
48
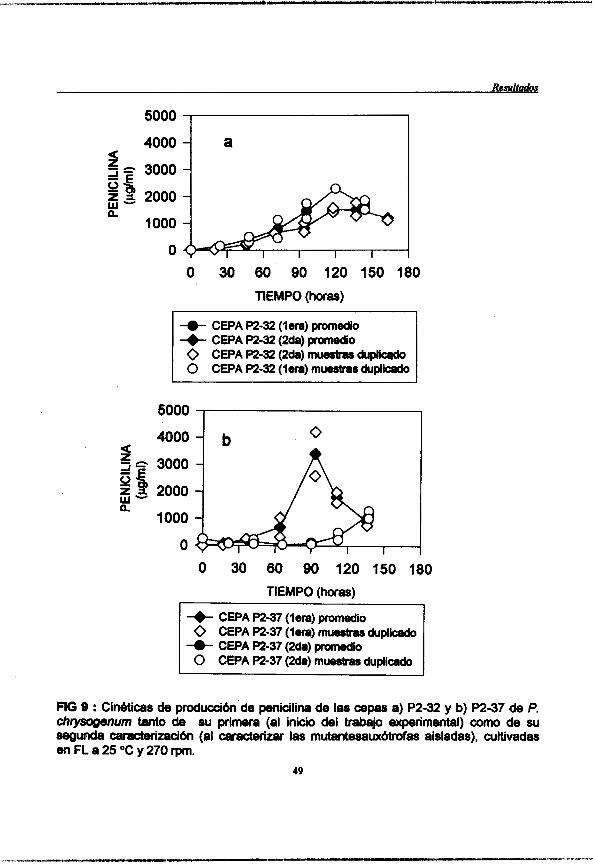
5000
4000
3 3000 0 3 n E 2000
1 O00
O
O 30 60 90 120 150 180 TIEMPO (horas)
a
u- P
+ CEPA P2-32 (lera) promedio -e- CEPA P2-32 (2da) promedio 0 CEPA P2-32 (2da) rnueslm duplicKk O CEPA P2-32 (lefa) muestras duplicado
4000
3000
2000
1 O00
O
O
O 30 60 90 120 150 180
L. TIEMPO (horas)
CEPA P2-37 (lera) promedio
+ CEPA P2-37 (2da) promedio 0 CEPA P2-37 (l-) duplicado
O CEPA P2-37 (2da) muestras duplicado 1 FIG 9 : Cin6ticas de p r o d u o c i 6 n ' d e penicilina de las cepas a) P2-32 y b) P2-37 de P. ahfysogenum tanto de su primera (al inicio del trabajo experimental) como de su segunda camcterizaci6n (al c8ract6rizar las mutantesaw6trofas aisladas), cultivadas enFLa25OCy2701pm.
49
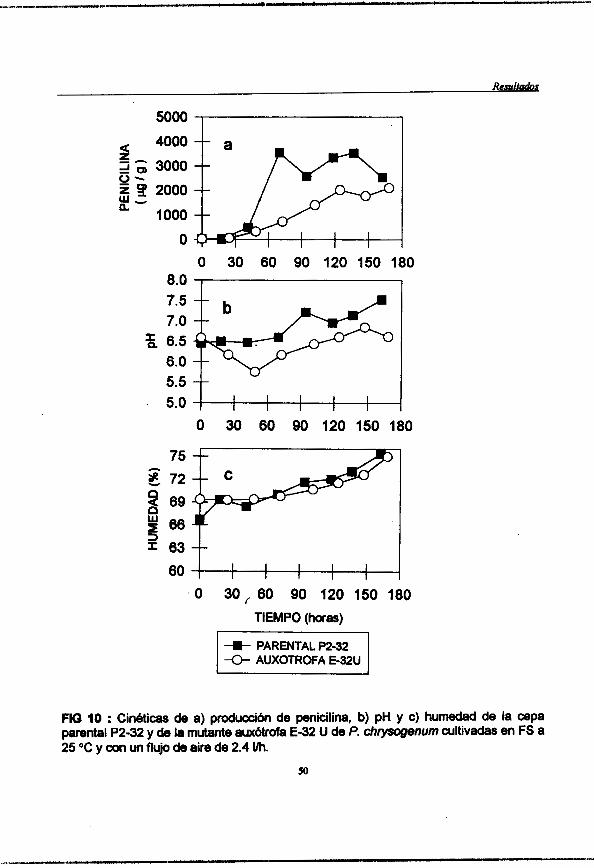
O 30 60 90 120 150 180 8.0 I I
5.5 " U
. 5.0 I I I I I
O 30 60 90 120 150 180
60 6 3 L H " l ' O 30 ~ 60 90 120 150 180
TIEMPO (horas)
-0- AUXOTROFA E-32U
FIG 10 : Cin6ticas de a) produccibn de penicilina, b) pH y c) humedad de la cepa parental P2-32 y de la mutante aux&ofa E-32 U de P. chfysogenum cultivadas en FS a 25°Cyconunflupdeairede2.4Vh.
M
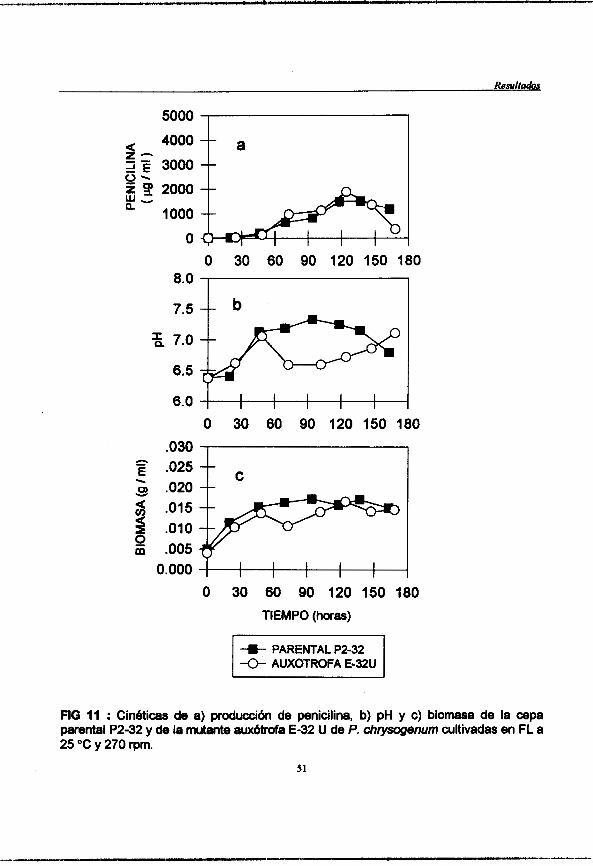
"
Resultados
5000 I 4000
3000
2000
1 O00
O
-- a "
"
"
O 30 60 90 120 150 180 8.0 '11
6.0 O 30 60 90 120 150 180
.O30
.O25 a
0.000 .~
O 30 60 90 120 150 180 TIEMPO (horas)
PARENTAL P2-32 -0- AUXOTROFA E-32U
FIG 11 : Cineticas de a) prduccih de penicilina, b) pH y c) biomasa de la cepa parental P2-32 y de la mutante awdtrofa E32 U de P. chrysogenum cultivadas en FL a 25 "C y 270 rpm.
51

O 30 60 90 120 150 180 8.0
O 30 60 90 120 150 180
6 3 L " H - 60 O 30 60 90 120 150 180
TIEMPO (horas)
U PARENTAL P2-37 -@- AUXOTROFA E-37-M1
FIG 12 : Cineticas de a) produccidn de penicilina, b) pH y c) humedad de la cepa parental P2-37 y de la mutante Bw6fTofB E37Mlde P. chrysogenum cultivadas en FS a 25 "C y con un flujo de aire de 2.4 I/h.
52
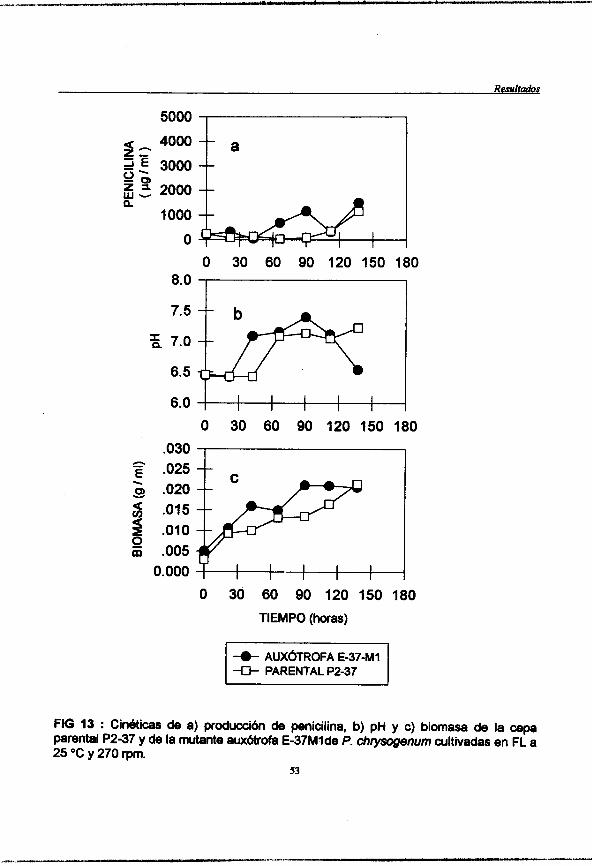
5000
v W - = 3 2000 n
looo I O
O 30 60 90 120 150 180
8.0
O 30 60 90 120 150 180
I
O 30 60 90 120 150 180
TIEMPO (horas)
-e AUX~TROFA ~ - 3 7 ~ -0- PARENTAL P2-37

DiscvSibn
VI11 Discusibn.
1. Seleccibn de cmas,
De acuerdo con la hipdtesis, en donde se establece que para lograr generar cepas especialistas en el SFS se requiere partir de cepas que cuenten con características deseables complementarias para este sistema de fermentacih, y que a traves del proceso de recombinaci6n gen&ica estas lleguen a remine en un s610 organismo recombinante, la mejor opcih en cuanto a complementaci6n de caracteristicas deseables la presenta la pareja de cepas P2-32 y P2-37, ya que por una parte, la cepa P2-32 presenta un alto potencial de producci6n de penicilina en el SFL (pdpl bajo). En tanto que la cepa F?2-37, desarrolla una alta eficiencia de producci6n en el SFS (ps/pl alto). Esta eleccitm esta sustentada en evidencias experimentales gen- por.Banios y col. (1993). Los autores evaluaron mutantcs espont8neas, aisladas de cepas altamente produaoras en FL y encontraron que algunas de &stas mutantes expresaron mejor su potencial de producci6n en el SFS que las otras. Ellos determinaron esta habilidad, mediante el dculo de su pdpl, de tal modo que las cepas con una elevada prodwi&n del antibibtico en FS mostrabau un pdpl alto, y tendían a expresar u118 menor producci6n en FL, mientras que por lo contrario, las cepas con una baja producci6n del antibi6tico en FS mostrabau un pdpl bajo y tench a expresar una myor produCci6n en FL. La posible interpretacih que dau a estos resultados es que las cepas con bajas producciones en FL poseen ciertas características que, aunque no esth directamente relacionadas con la biosíntesis d e l antibi6tic0, son Útiles para su crecimiento y producci6n en el SFS.
Por tanta, se sugiere que la cepa P2-37 posee alguna o algunas características, que le pennite expresar mejor su potencial de produccih en FS que la P2-32; mientras que la cepa P2-32 esta expresando, probablemente, otras cmcterkticas que le permiten mostrar una buena producci6n en FL.
Las características de in- mostradas independientemente por las cepas P2-32 y P2-37 penniten sugerir la existencia de dos grupos de genes independientes entre sí, que codifican para cada una de estas dos características deseables, en donde el grupo de genes que participa en la gencracih de informach para una alta eficiencia de producci6n (pdpl) codifica amcterísticas útiles para el crecimiento y producci6n del hongo en el SFS y que no aportan ningún beneficio para el SFL. Mientras que el otro grupo de genes contiene la informacih relacionada de biosintesis necesaria para codificar un alto potencial de prodwci6n expresado en el SFL. ‘ D e esta forma, al fusionarse las cepas y generarse recombinaci6n genetica de
54

Discusidn
cada una de ellas, podrian conjuntarse ambos grupos de genes, es decir, ambas características en un solo organismo, de tal forma que le permitiera desarrollarse eficientemente en FS y expresar un potencial de producci6n aún .mayor que el presentado por las cepas parentales de las d e s proceden.
2. Aislamiento de AusuMrofos.
2.1 Mutag6nesis.
Los marcadores que se buscaron en la cepa P2-37 fueron auxotrofias a aminohidos como leucina, metionina y prolina (Ann6 y col., 1975; Peberdy y col., 1977, Ann& 1982) y en la cepa P2-32, aux6trofos a uridina (Dw y col., 1987). La elecci6n de este tipo de marcadofes para cada una de las cepas, fue alwtoria, de este modo, pudo haberse realizado de forma invertida, es decir, buscar auxotrofias a aminobidos en la cepa P2-32 y motrofia a uridina en la p2-37. En el caso de los aw6trofos a aminohcidos, se buscaron estos ya que, add de no intervenir en la ruta de biosintesis de penicilina (valina, cisteína, lisina), otros autores que han realizado trabajos similares con P.chrysogenum han obtenido este tipo de marcadores; la importancia de aislar marcadores similares radicq en que si se genera un seguimiento en los trabajos por parte de otros investigadores, se genera mhs informacih que puede ser útil 6 aplicable en ciertas íhas, de tal forma que con los resultados generados & este estudio, tambib se tenga acceso a dar seguimiento a trabajos posteriores relacionados, e inclusive encontrar infonnacih adicional igualmente importaute. Ahora bien, para el caso del aux6trofo a uridiua se busc6 este tipo de mutaute por la ventaja que representaria si fuera un mutante pyr G (que c&ca para la enzima descdmilasa orotidin 5- monofosfato, OMP - descarboxilasa) debido a que estos mutantes tienen aplicabilidd en el Brea de ingeniería gen&ica, donde este gen es marcador de los principales vehículos moleculares para hongos. De tal modo que se llevaron a cabo mutaciunes de las cepas parentelas mediante radiacih con luz U.V., debido a que presenta ciertas ventajas sobre algunos otros tipos de mutageno (Rowlands, 1984a). Se ha 0b-o que mutantes aux6troík de Asprgiffus niger obtenidas por tratamiento con hitrosoguauidina mostraron un crecimiento lento y una esporulaci6n pobre en contraste con mutantes similares obtenidas con tratamiento de luz U.V., en base a lo cual se decidi6 no ub.lliaar mutagenos químicos, sino inducir la mutacib en las cepas paratales mediante la aplicacih de luz U.V. Se obmyaron diferencias entre las curvas de sobrevivencia de las cepas P2-32 y P2- 37. La cepa P2-32 requiri6 de menos tiempo para generar niveles de sobrevivencia
55

Discwidn
del 50 %, en tanto que la cepa P2-37 necesit6 m& tiemp para generar los mismos resultados. De acuerdo a referencias bibliogrhficas, se tiene que la coloraci6n de las esporas determiaa la resistencia a la radiaci6n U.V. @os y col., 1992; Loera 1994) de esta forma esporas lncgras de A. niger requieren de una intencidad de radiaci6n U.V..de 700 pw/m2 mientras que esporas mtts claras como las de P. chrysogenurn quieren de 250 pW/cm2 . Las esporas de ambas cepas la P2-32 y la P2-37 son verdes y es posible que se presente alguna diferencia en pigrnentaci6n de poca intensidad entre ellas sin embargo no es apreciable a simple vista. Esto podria explicar la mayor sensibilidad de la cepa P2-32. El porcentaje de mutantes morfol6gicas se consider6 como un estimador que pennitiera comprobar que realmente se habfan inducido cambios a nivel de material gdt ico y particuhmmte se consideraba impartante que estas mutaciones se presen&ran a los niveles de sobrevivencia de inter&, es decir, a niveles entre 50- 60%.
2.2. Muturfes Aux6trafos a amindcidos.
El utilizar bajas dosis de mutageno para cancelar aberraciones crmos6micas, ' implica el obtener una alta proporci6n de sobrevivientes y una baja fiemencia de
mutantes entre los prat6trofbs sobrevivientes por lo que se requiere una etapa eficiente de enriqueciemiento (Bos y col., 1992).
El proceso de enriquecimiento por filtraciones sucesivas result6 ser eficiente para el aislamiento de mutantes que se encuentran bloqueados en alguna vía metab6lica, en este 08so se aislaron mWantes Met y S org8niC0, a dosis bajas de mutAgeno (50-60%), logr&dose el aislamiento de las cepas E-37M1, E-37M2 E-37M3 y E-37 M4 a partir de la cepa P2-37. La mutante E-37 M1 (Met -) fue incapaz de metabolizar metionina, por lo que se hizo necesario la suplementaci6n de este aminoacido en el medio de cultivo, es decir se afect6 la ruta de biosíntesis de la metimina. Mientras que las mutantes, E-37M2, E-37M3 y E-37 M4 fueron aux6trofhs a S - orghiao, es decir presentaron la incapacidad de asimilar d e inorghico, por lo que el suministro de una fuente orghca del mismo se hizo necesario, siendo la metionha o bien la cisteína aquellos aminohidos que podían proporcionar esta h t e orghmca de d e . Trabajos anteriores realizados en este sentido (Bos, 1992), mostraron que al aplicar el mhdo de filtraciones sucesivas, permiti6 el aislamiento de una gran variedad de mutant-, deficientes en la shtesis de aminoscidos, así como de mutantes que requieren adenina ademas de m pequeflo número de mutaates deficientes en vi-. Encontraron que la aplicaci6n de este m&& deriva en una baja
m
- -

En el proceso de exmiquecimiento mediante la aplicacih de la enzima lítica Novozyme, se hizo un seguimiento de la m i 6 n con la ayuda del microscopio 6ptico, pretendiendo cacontrar un detemido de los tubos germinales. En trabajos similares @ebets y col., 1989), se reporta que la accih de la enzima litica es mhs eficiente cuando el tamaflo de los tubos gennmales es de 20 .veces el tamafko del dihetro de la mpora. En el caso de P.chrysogenum, se monitor& la gembci6n COnfoRne tramamía el tiempo de incubacib, y cuando el tubo germinal c o d a con un tamafko aproximado de de 3-5 veces el dihelro de la espora, se dio inicio una compleja dcacih genpndo por tanto varios tubos genninales en varias direcciones con longdudes diferentes, de tal forma que se decidi6realizarlaadicihdelaenzjmaalptesentarseunaestructuracomplejaen cuanto a forma ( d m c i h ) actuando asi en el micelio joven y poco ramificado; esto se llev6 a acabo a llas 29 horas de incubacih. Debido a las observaciones encontradas en el monitoreo de la germinacibn para el caso de P. chrysogenum y bajo las condiciones experimentales reportadas en este estudio, se tiene que el valor promedio del tamaflo de los tubos gemmales que pmnite las condiciones msS propicias para la adici6n de la enzima Novozime es de 1Oveceseldiametrod&laespora.Estevalorseencuentrapordebajodelreportado en literatura, el cud era a su vez el mhs propicio para un microorganismo diferente (A. niger) y bajo condiuiones tambih diférentes.
El proceso de enriquecimiento con la enzima lítica Novozyme se implement6 en una etapa posterior al de fil@ciones SUCeSiV8S, de tal forma que no se log6 la aplicaci6n de forma importante de esta metodologh, y se suspendi6 al encontrarse las mutautes requeridas en-, por procesos de m i o n e s sucesivas. De este modo no resulta posible el poder evaluar la eficacia del m&&. Sin embargo la aplicabilidad de esta metodología fue así mismo discutida por Bos y col. (1992) encontrando nuevos tipos de mutrmtes con requerimientos a amimicidos, a vitaminas o nucle6tidos. Ademhs de que el número de mutantes independientes por experimento y el aislamiento de nuevos tipos de mutantes, mostraron que el emiqwximiento por medio de enzimas líticas es mucho mhs eficiente que el metodo de enriquecimiento por filtraciones sucesivas.

Disncsibn
2.3. Mutantes Aux6trafos a uridina.
Esta metodología se fundamenta en el aislamiento de mutautes resistentes al compuesto 5 AFO, las cuales son Mcientes en enzimas OMP descarboxilaza, por lo que la ruta de biosíntesis de la UMP precursor de las bases pirimidicas se bloquea, y con ello la &tesis del ADN, en consecuencia se requiere el summstm de uridina en el medio para permitir el crecimiento del organismo.
. .
La mutante aux&rofh a uridina se ais16 en MM lactosa en lugar de glucosa. La glucosa es m represor de la biosíntesis de penicilina y la lactosa no lo es; en el transcursodela~~6~alacabarselaghcosaseactivalaacci6ndela~- galactosidasa y desdobla a la lactwa en sus unidades : glucosa y galactosa. La glucosa entonces se nrttnrnrstra poco a poco evitando la represih en la prduccih de penicilina. Si se adiuionara glucosa en el MM habría la posibilidad de aislar una mutante que no sintetioe la p - w s i d a s a , es decir, una p-galactosidasa - y esa mutante no es de inter& ya que se requiere el desdoblamiento de la lactosa.
. .
Para el aislamiento de mutantes resistentes al 5 AFO se requiere de por lo menos una etapa de crecimiento y esporulacih del stock on@ de esporas mutagemadas, ya que despds de la mutaghsis, las esporas se encuenimn daWas debido al tratamiento de irradiacih por lo que tienden a manifestar bajas expectativas de vida. Es por ello que una etapa de geminaci6n de las esporas sobrevivientes tan prcmto como sea posible, resultani ventajoso para la obtencih de esporas saludables con el mismo porcentaje de mutantes que el stock o r i g d de esporas mutagentzadas (Campuzano, 1993).
Se requiri6 encontrar h dosis de 5 AFO mhs adecuada para la cepa P2-32 que permitiera el aislamiento de colonias resistentes a este compuesto. Este proceso se implement6 en la cepa sin mutar, debido a que se necesitaba detemhr que tan sensible era la cepa al compuesto t6xico. Adem& se permitia contar con el aislamiento de mutautes espontaneas que mautenían la probabilidad de contar con la auxotmíla a widiua. La proporci6n de auxwfos a piximidina de entre las colonias resistentes al 5 AFO aisladas de la cepa mutagenizada P2-32, en este caso fue de 1 de entre 128 , es decir, el : 0.78%. La pr'porci6n encontrada en trabajos similares (Diez y col., 1987) ha sido superior, repdmdo porcentajes de hasta 10 %; cabe aclarar que el tratamiento mutaghico8 ha sido realizado con muthgenos quimicos adem& de haber generado bajos niveles de sobrevivencia de entre 10-20 % p e z y col., 1987). A s í mismo, Campuzmo (1993), usando la misma metodologia en Phycomyces
S8

Discusih
blakesleeanus, report6 porcentajes de hasta 7 % utilizando de igual fonna mutagenos quhcos y niveles de sobrevivencia aún menores (517%). Sin embargo a pesar de lo anterior, el proceso de aislamiento de mutantes a piximidina es eficiente y se lo@ aislar la mutante E-32U a partir de la cepa P2-32. En la mayoría de los casos en donde se han obtenido aux6trofos a pirimidina por medio de la resistencia al 5 AFO, se hau realizado paralelamente caLacferizBciones bioquimicas de estas mutantes y se ha encontrado que en la mayoría de los casos los mutautes auxWfos a pirimidina esth altedos en el gen pyr G (que &ca para la enzima desmboxilam ddin 5- monofosfato, OMP - descarboxilasa) y en el gen pyr F (que codiíica para la enzima orotato fosforibosil trausf- OPR- T d m ) (Oakley y col., 1987; Smit y Tudzynsky, 1992; Akileswaran y col., 1993). Por su parte Diez y col. (1987) para determinar si la mutaci6n de una cepa de P. chrysogenum aux6troíh a pirimidina obtenida por resistencia al 5 AFO, se encontraba en el gen pyr G, realipiron mediciones de la actividad enzhitica de la OMP d e s c a r b o ~ los resubdos mostraron la carencia de actividad enzimstica de la OMP desmboxilasa por lo que concluyen que la mutante era un aux6trofo pyr G. Ademh conkmu sus resultados al complementar la mutaci6n con el uso de los plfismidos pDJBl y pDJB2 los cuales portaban el gen pyr 4 de Neurospora crassu (gen que codiíica para la enzima OMP descarboxdasa). Recupemon la actividad de la OMP descarboxilasa en la mutante aux6troh a pirimidha, lo cud indica que el gen pyr 4 de Neurospora crassa se puede expresar en P. chrysogenum y que la enzimaOMP~~lasacodificadaporestegcnesfimcionalenlarutade biosíntesis de la pirimidina en P. chrysogenwn. Se considera importante dar seguimiento a estudios relacionados con la identificaci6n de la mutaci6n del aux6trofoauridinaE32U,yaquesisetratademmutantepyrG,resultariade utilidad en estudios de Ingenieria gcn&ica. Actualmente es m hecho que el gen 41 F de P. chrysogenum ya ha sido clonado y es posible construir vehículos m o l e c h s con ese d o r (J.F. Martin comunicaci6n personal).
3. Cancteha6n de las cenas nanntrlcs Y aur6trofas.
En la caracterizaci6n efectuada de las cepas parentales y aux6troh, se consideran las cineticas de pH, humedad y biomasa como que proporcionan infOrmac6n acerca de la femmtaci6n. Así el pH cambia y se basiíica COnfoRne transcurre la fennentaai6n debido a que la concentraci6n del kid0 fd adtico (AFA) presente en el medio de cultivo disminuye para in- en la ruta m w l i c a de biosíutesis de penicilina, adem& puede ser indicador de posibles
59

contaminacones del cultivo. Por otro lado, se considera que el contenido de agua, la humedad, aumenta en la FS durante el cultivo y de esta forma se puede inferir el comportamiento en cuanto al crecimiento del hongo. Ademis la humedad se establece como un control que permite adjudicar la prochrccibn de penicilina al microorganismo en sí y no a cambios en la humedad ya que se ha comprobado que la humedad inicial en intervalos de 60-78 % no afecta el tiempo de inicio de la produccibn pero si time una importante influencia en la producci6n de penicih, encontrandose que se obtiene una producci6n msvrima utilizando 70 y 73 % de humedad inicial (Barrio$ y col., 1988). En las amchimciona efectuadas tanto en las cepas parentales P2-32 y E-37 como en las aux6trob E-32 U y E 37 M1, las fermentaciones comenzaron con humedades iniciales de entre 68-71% para de esta fmma proporcionar las condiciones de humedad m& hvorables y obtener los niveles mhximos de producci6n de penicilina. Por tanto .los cambios obseavados en las producciones de penicilina generados por las diferentes cepas, son atriluibles al microorganismo ya que no se observan variaciones importantes en lo que a humedad inicial se refiere.
4. Evaluaciijn del niVd D I W ~ U C C ~ ~ I ~ de m muta ntes auxijtrofas. . .. De acuerdo a los resultados obtenidos de la primera y segunda caracterizacib de las cepas parentales €2-32 y P2-37 en cuanto al nivel de produccih de penicilina, se observa que la produccibn disminuye en ambos sistemas de cultivo a excepcibn de la P2-37, la cuhl presenta un incremento importaote en la producci6n de penicilina en el SFS durante su segada caracterizacih (Tabla 2 y Tabla 8) y (Fig 8 y Fig 9). El comportamiento anterior puede sue un cierto grado de inestabilidad genbtica p r e s e n & , e n las cepas ylo bien una variacibn clonal de las esporas empleadas, es decir, heterogeneidad a los niveles de produccib generados.cabe mencionar que tanto las condiciones del proceso de caracterizaci6n de las cepas y el equipo empleado, fueron los mismos en la primera y segunda wa&rhci6n. Los cambios observados en prodw;ci6n pueden ser consecuencia del manejo de inadecuados sistemas de conservacibn que causaran un ambiente propicio para permitir alteraciones en el genoma. J% este estudio, la conservaci6n efectuada a lo largo! de casi 12 meses h e en un sistema de glicerol al 25 % a -70°C. Se sugiere entonces. considerar sistemas alternativos de conservacibn que aseguren estabilidad de las características presentes en una cepa. Estos resultados sefíalan la necesidad de ham estudios de estabilidad gen&ica como parte de la caracterizaci6n de las cepas parentales.
60

Discusibn
Ahora bien, si se observan las cineticas de produccih de penicilina, de la cepa parental F2-32 y de la mutante aux6trofi E-32U en FL (Fig. ll), se tiene que esta, pr$cticamente es igual en ambas cepas. Es decir, el proceso de mutagenesis no afect6 a la mutante en su dwmpeiío en FL. En cambio al observar las cineticas de producci6n de peniciliua de las mismas cepas pero en FS (Fig. lo), se detecta claramente, que el prowso de mutagbsis &st6 fúnciones en la mutante E-32U que s610 son necesarias para producir en FS, ya que se observa una pdrdida importante en la producci6n maxima de penicilina (del orden del 40 %) (Tabla 8).
Es poco probable que la auxotrofh a uridina sea la responsable de este fenotipo en la mutante ya que se han obtenido aux6trofos a midina en otro tipo de microorganisnos por la via de la resistencia al 5 AFO sin alterar otras características, ademhs se ha &&ado que en la mayoría de los casos las mutantes aux6troh a uridina estan alterados en el gen pyr G y en el gen pyr F. Lo mhs probable es que haya otra(@ mutacion(es); en este caso el otro gen mutado sería un gen que pennite una mayor pmducci6n en FS. La existencia de esta mutante, da margen a realizar estudios posteriores entbcados a lajdentiiicaci6n del gen que pennite una mejor produccih en FS. El llevar a cabo este tipo de estudios sería de gran importancia ya que se reconoceda el gen (o los genes) activador de la producci6n de penicilina en FS. Estos estudios contemplarían la búsqueda del gen en una genoteca que complemente esta mutaci6n. Cabe mencionar que ademas de esta impdante disminucih en la producci6n del antibidtico, se presume un deesajuste general de la fisiología de la mutante E-32U, debido a que present6 deficiencia en la formaci6n de esporas. Esto hizo necesario realizar modificacioaes en la metodologia respectiva a la evaluaci6n de la produccibn en el SFS ya que se inocul6 con micelio y no con esporas como se hace habiwente; esto se llev6 a cabo tanto en la cepa parental como en la mutante con la halidad de estandarizar los resultados, y poder efectuar comparaciones entre las producciones de penicilina registradas para ambas cepas. Esta modificaci6n en la metodología, introduce ' u n a variable no considerada en la primera evaluaci6n de la cepa parental, ya que esta se e f d utilizando un inoculo de esporas y no de micelio. De esta fbrma se podrim generar cambios en caracteristicas como el pdpl. La deficiencia en esporulaci6n de la mutante E-32U indica que el proceso de mutacih aplicado, m&c6 i n f o m i h a nivel genaco que afect6 la formacih de esporas; este evento, nu era deseable, sin embargo la caracteristica de deficiencia en esporulacih permite amtar adem& con un marcador de tipo morfol6gico en esta mutante, . lo cusll facilitadia el proceso de aislamiento de recombinantes.
61

En la Tabla 9 se observan los pdpl de las mutantes aux6lrohs aisladas. Estos mantienen la caractedstica de ser uno menor que el otro (E 32U pslpl = l . 12; E 37M1 pdpl = 2.62), lo que permite una c o m p l d e d a d entre ambas es decir, en trabajos posteriores en donde el objetivo sea efectuar h i 6 n de protoplastos, se contaria con una cepa que aporta una alta eficiencia de prodwi6n en medio &lido (alto pdpl) y otra que aporta una alta p r b i 6 n de penicilina (alta produccih en medio líquido y bajo pdpl). Es deseable que las diferencias entre pdpl de ambas cepas sean grandes para poder mtcrprctar los resultados de una forma m& sencilla. Es decir, una ves realizada la c m entre ambas cepas se generanin recombinantes, y entre ellos pueden encontram algunos en los que se haya conjuntado tanto la característica de eficiencia en Alido (pdpl alto) y la de alta produccibn, de tal forma que si la diferencia entre pdpl de ambas cepas es grande, la producclbn de penicilina del recombinante se incremcntarA de tal forma que sea mucho m& alta que aquella mostmh por las cepas parentales @S = [pel alto] X [alta produccih de penicilina]) . Por lo que se genera la sugerencia, para trabajos fbturos, de mutagenizar nuevamente a la cepa E-32U con la fínalidad de aumentar su producci6n de penicilina en FL y de esta forma disminuir su pdpl y generar una mayor diferencia con el pslpl de la cepa E-37 M1 . Ello contemplaria comprobar mediante pruebas en discos de agar, primero, que la mutante conserva la auxotrofia y segundo, que la producci6n de penicilina en FL efectivamente se incremente, para de esta forma generar una mayor diferencia entre los pdpl de las mutantes que se sometan a estudios de mwmbinaci6n gen6tica.
62

IX Conclusiones.
Se realiz6 la evaluaci6n de producci6n de penicilina de cinco cepas de P. chrysogenum tanto por FS como por FL y se enwntr6 que la cepa de mejor producci6n de penicilina en FS fue la cepa P2-32 (10 325 &g), mientras que la de mejor producci6n del antibi6tico en FL fue la P2-37 (3 41 1 pdd). De las observaciones anterims, se llega a la selecci6n de las cepas la P2-32 y la P2-37 como cepas parentales.
Se obtuvieron las curvas de sobrevivhcia de las cepas de P. chrysogenum P2-32 y P2-37, y se detemh6 que para garan- una sobrevivencia de SO%, se deben irradiarlasesporasdelacepaP2-37porunti~pode2.5min.mien~quelacepa p2-32 requiere un tiempo de 2.1 min a una intensidad de irradiacibn de luz UV de 250 pW/cm2, bajo las condiciones experimentales establecidas.
Los procesos de enriquecimiento por filtraciones sucesivas heron eficientes para encontrar mutantes Met - y Met - 6 Cis - con bajas dosis de mutageno (50-60 % de sobrevivencia), logrando el aislamiento de las mutantes E-37 M1 (Met -), E-37 M 2 , E-37 M3, E-37 M4 (W últimas Met - 6 Cis - ) orig.madas de la cepa parental P2- 37 con un porciento de mutantes de 0.74%.
Se logr6.obtener y aislar mutantes de P. chrysogenum de la cepa €2-32 resistentes a 1, mg/ml del compuesto 5 AFO con una frecuencia de 5 X lo5. Y a partir de las mutantes resistentes se efectu6 el aislamiento de una mutante aux6troh a uridina la E-32 U a partir de la cepa parental P2-32 con un porciento de mutantes de 0.78 %, empleando la metodologia propuesta por Diez y col., (1987).
Se llevaron a cabo dos evaluaciones en c k t o a la produccih de penicilina de las cepas parentales P2-32 y P2-37, una al inicio de la experimentaci6n y otra al l i d , así como una evaluacib de la producci6n de penicilina de las mutantes aux6trofas E-32 U y E-37 M1, en cultivos llevados a cabo por FS y por FL. Los resultados sugieren.un cierto @o de inestabilidad en las cepas parentales dada la diferencia en producci6n de penicilina de ambas cepas en la primera y segunda caracterizaci6n re-.
En base a la hi@sis plsmteada, se concluye que no h e posible introducir marcadores gentiticos en P.chrysogenum y obtener mutantes aux6trofas sin modificar sensiblemente la producci6n en FS, no siendo así para FL en donde los niveles de producci6n no se encontraron modi6cados.
63

X Bibliografh.
Abdullah, A.L., Tengerdy, R.P., Murphy, V.G. (1985). Optimization of solid substrate fermentation of wheat straw. Biotechnology and Bioengineering, a , 2 0 - 27.
Akileswarau, L., Alic, M., Clark, E.K., Homick, J.L., Gold, M.H. (1 993). Isolation and transformation of uracil auxotrophs of the lignindegradmg Basidiomycete Phanerochaete chrysosprium. Cum Genet, a, 35 1-356.
M, J., Eyssen, H., and De S o m e r , P. (1974). Fonnation and regeneration of Penicillium cbrysogenuum protoplast. Arch. Microbiol. B, 159-166.
Anne, J., and Peberdy, J.F. (1975). Conditions for induced fusion of hgal protoplasts in polyethylene glycol solutions. Arch. Microbiol. m, 201-205.
Ann&, J., and Pebdy, J.F. (1976). Induced fusion of fuugd protoplasts following treatment with polyethylene glycol. Journal of General Microbiology,B, 4 13-4 17.
And, J. (1982). Comparison of penicillins produced by inter-species hibrids fkom Penicillium chrysogenum. Eur. J. Appl. Microbiol Biotechnol, U, 41 4 6 .
Annb, J., and Peberdy, J.F. (1985). Protoplast fusion and interspecies bibridization in penicillium. En : Fungal protoplasts. Aplications in biochemistry and genetics. Edited by Peberdy, J.F., and Ferenczy, L. M a r c e l Mer, hc. New Yo&, pp.259- 227.
Antier, P., Minjares, A., Roussos, S., Raimbadt, M., Viniegra, g.g., (1993). Pecthnases hipexproducing mutants of Aspeqgillus niger C28B25 for solid state fermentaton of coffe pulp. Enzyme microb technol, B, 254-260.
B a l e R.H. (1986). Strain improvement : introduction. En : Mauual of Industrial Microbiology and Biotccbnology, E d s . Demain, A.L., Solomon N.A., Washington, American Society for Mimbiology, pp. 154.
64

Baredo, J.L., Solingen, V.P., Diez, B., Álvarez, E., Cantoral, J.M., Kattervilder, A., Smaal, E.B., Groenen, M.A.M., Veenstra, A.E., Martín, J.F. (1989b). Cloning and characterization of the acyl-coenyme A : 6-aminipenicilliauic-acid acyltransferase gene of Penicillium chrysogenum. Gene, 83.291-300.
Barrios, G.J., Tomasini, A., Viniegra, G.G., y Upez, L. (1988). Penicillin production by solid state fermentation. Biotechnology Letters, (1 l), 793-798.
Barrios, G.J., Castillo, T., Mejía, A. (1993). Development of high penicillin producing strains for solid state fermentation. Biotech. A&. 2,525-537.
Banios, G.J., G o d e z , H., Mejía, A. (1993). Effect of particle size, packing density and agitation om penicillin production in solid state fermentation. Biotech. A h . 11,539-547.
Barrios, G.J. (1994). Roducci6n de metabolites secundarios por fennentaci6n s6lida. Tesis de doctorado U N A M .
Banios, G.J., y Mejía, A. (1996). Production of secondary metabolites by solid-state fermentation. Biotechnology annual review, (2), 85- 12 l .
Boeke, J.D., La Croute, F., Fink, G.R. (1984). A positive selection for mutants lacking orotidine-5-phosphate descarboxilasc activity in yeast : 5-fluoro-orotic acid resistance. Mol. Gen. Genet, 197,345-346.
BOS, C. J., SMorst, M., Visstr, J., and Roberts, C.C. (198 1). A third unlinked gene controlling the pyruvate dehydrogenase complex in Aspergillus nichrlrmdr. Journal of Bacteriology, =(2), 594-599.
Bos, C.J. (1987). Inductm and isolation of mutants in fimgi at low mutagen doses. Cum. Genet., 12,471-474.
Bos, C.J., Debets, A.J.M., Nachtegaal, H., Slakhorst, S.M., and Swart, K. (1992). Isolation of auxotrophx mutants of Aspergillus niger by filtration enrichment and lyt~c enzymes. Cum. Genet., 21,117-120.
Bos, C.J., Slakhorst, S.M., Debets, A.J.M., Swart, K. (1993). Linkage group analysis in Aspergillus niger. Appl. Microbiol. Biotechnol, 38,742-745.
65

Campuzano, V., Valle, P., Vicente, J.I., Eslava, A.P., Álwez, M.I. (1993). Isolation, characteridon and mapping of pirimidhe auxotrophs of Phycomyces blaResleeanus. Curr. Genet.., 5 15-519.
Chang, L.T., Terasaka, D.T., and Elander, R.P. (1982). Protoplast hion in industrial hgi. Dev. I . . Microbio1.,23.21-29.
Crueger, A. (1993). Mutagenesis. En : Classical Genetics. Genetic Fmdamentals and Genetic Engineeriug. Biotechnology. Edited by Rehm, H.J. and Reed G. in coopefatcm with pnhle?, A., and Stadler P. VCH publishers Inc., N.Y. (USA), pp. 7-45.
Davies, O.L. (1964). Screening for improved mutants in antibiotic research. Biometrics pp. 576-591.
Debets, A.J.M., Swart, K., and Bos, C. J. (1 989). Mtotic mapping in linkage group V of Aspergillus niger based on selection of awcotrophw recombinants by novozym emichment. Can. J. Micmbiol, 35.982-988.
Delgado, J.M., Herrera, L.S., P ~ z , C., and Lbpez, R. (1979). Optmmtion on the selection of awcotmphc mutauts in Candida utilis by snail enzyme treatment Can. J. Micmbiol, 2i, 486-490.
hgraslges, C., and Dunand, A. (1990). Effect of pC@ on growth, conidiation, and etlzyme production in solid state culture on Aspergillus niger and Trichodenna viride TS. Enzyme Microb. Technol., l2,546-551.
Diez, B., Álvarez, E., Cantoral, J.M., Banedo, J.L., and Mart4 J.F. (1987). Selection and caracterization of pyr G mutants of Penicillium crysogenum lacking orotidine-5-phosphate &scarboxylast and complementation by the pyr 4 gene of Neumspqra crussa. Cum Genet., 12,277-282.
Diez, B., Gutierrez, S., B m d o , J.L., Solingen, P.V., Voort, V.L.H.M., Martin, J.F. (1990). The cluster of penicillin biosinthtic genes. Identification and characterization of the pbc AB gene encoding the a-aminoadqyl-cysteinyl-We synthetase and linkage to the pbcc and penDE genes. J. Biol. Chem,, 265, 16358- 16365.
66

Durand, A., Pichon, P., Desgranges, C. (1988). Aproaches to KL a measurements in solid state fermentation. Biotechnology Techniques, 2 (1 ), 1 1 - 16.
Fries, N. (1947). Experiments with difhmt methods of isolating physiological mutations of filamentous hgi. Nmre, 1 5 9 , 1 9 9 . Gunaseh, P. (1995). Bacterial survival against UV irradiation and mutagenesis. En : Laboratory manual in microbiology. New age intemational limeted publishen PP. 78-81.
Herrera, T., Viloa, M. (1990). El reino de los hongos. Micología bstsica y aplicada. UNAM Mexico D.F. Fondo de cultura econ6mica.hpreso en rndxico lera. edicih
Kanmobu, S., Yan, W., Kishida, M., and Sakai, T. (1996). Breeding a fermentative yeast at high temperature using protoplast fusion. Journal of fennentation and Bioengineering, 81 (2), 104-108.
Kurth, R., and Demttin, A.L. (1985).The impact of the new genetics on antibiotic . production. Biotechnology of industrial antibiotics. pp. 781-789.
Loera, C.O. (1994). Caracterizacih de mutautes de Aspergillus niger L sobreproductoras de pectinasas. Tesis de maestría UAMI.
Lasane, B.K., Ghildyal, N.P., Budiatman, S., Ramalaishua, S.V. (1985). Engineering aspects of solid-state fennentation. Enzyme Microbiol. Technol., 2, 258-265.
L6pez, N.M.J., García, I.A., and Martín, J.F. (1980). Penicillin production by recombinant strains of Penicillium chrysogenum obteined by fusion of protoplasts. En : C. Vt5zina y Ka Sin@ Editores. Advances in biotechnology, vol. m, Pergamon Press., Canadh, pp. 57-61.
Luengo, J.M., Revilla, G., Upez, M. J., Villanueva, J.R., Martin, J.F. (1 980). Inhibition and repression of homocitrate synthase by lysine m Penicillium chrysogenum. J. Bacteriol., m, 869-876.
67

Biblioaoflcr
Mejia, A., Tomasini, A, y Barrios, G.J. (1997). Application of on-line measurement of oxygen uptake and carbon dioxide evolution to Penicillin production in solid state fermentation. En : Advances in solid state fermentation. Edited by S. Roussos, B.K. Lonsane, M. Raimbault y G.G. Viniegra. IUuwer Academic Publishers. pp. 417-425.
Miller, J. (1972). ultraviolet light mutagenesis. En : Experiments in molecular genetics. Ed. Cold SpringHarbor U.S.A. pp. 121-122.
Mudgett, R.E. (1986). Solid state fmentation. En : Manual of industrial microbiology and biotechnology. Demain, a.l., Solomon, n.a., editores, American Society for microbiology, Washington, pp. 66-84.
Mufíoz, R.G., Valencia, T. J.R., Sgnchez, S., and Fm6s A. (1991). Production of microbial lipases in a salid state fermentation system. Biotechnology Letters, 13(4), 277-280.
Nagai, S. (1979). Control of solid state cultivation. Proc. GL4M-K pp.413-424.
Oakley, B.R., Rinchart, J.E., Mitchell, B.L., Wey, C.E., Cmona, C., Gray, G.L., May, G.S. (1987). Gene, 61,385-399.
0- M., Swuki, K., and Umezawa, H. (1974). Formation and reversion of streptomycete p r o t o p l a s t s : cultural condition and morphological studies. J. Gen. Microbiol, 80.389-400.
Oriol, e., Schettino, B., Viniegra, G.G., Raimbault, M. (1988a). Solid-state culture of Aspergillus niger on support. J. Ferment. Technol., 66 (l), 57-62.
Oriol,’ E., Raimbault, M., Roussos, S., Viniegra, G.G. (1988b). Water and water activity in the solid state fmentation of cassava starch by Aspergillus niger. Appl. Microbiol. Biotechnol., 22,498-503.
Peberdy, J.F., Eyssen, H., and h 6 , J. (1977). Interespecific hybridization between Penicillium chrysogenum and Penicillium cyanofilvum following protoplast fusion. Molec. Gen. Genet., 157,281-284.
Peberdy, J.F. (1980). Ratoplast fusion, a tool for genetic manipulation and breeding in industrial microorganisms. Enzyme Microb. Technol., 2,23-29.

Peberdy, J.F. (1984). h e t i c recombination in fungi following protoplast h i o n and transformation. pp. 559-581.
Peberdy, J.F. (1985). The biology of Penicillium. En : Biology of Industrial Microorganisms. Demain, A., and Solomon, N. editores, The Benjamdcummings Publishing Company, pp. 407-43 l .
Peberdy, J.F. (1991). Fungal Protoplasts. En : More gene manipulations in hgi. Benuett, J. W., and Lasaae, L., editores, by Academic Press, Inc, pp. 307-3 18.
Piedra, D., and Herr- L.S. (1976). Selection of Auxotrophic mutants in Saccharomyces cereviciae by snail enzyme digestion method. Folia Microbiol, 2, 337-341.
Queener, S.W., Lively, D.H. (1986). Screening and selection for strain improvement. En : Manual of industrid microbiology and biotechnology. Demain, a.l., Solomon, n.a., editores, American Society for microbiology, Washington, pp. 155-169.
Rivera, M.G., Tinm, V.J.R., Sbchez, S. y Fads, A. (1991). Production of microbial lipases in a solid state fermentation system. Biotechnology Letters, u (4), 277-280.
Rowlands, R.T. (1984a). Industrial strain improvement : mutagenesis and random screening pmcedwes. E w e Microb. Technol., & 3-10.
Rowlands, R.T. (1984b). Indudal strain improvement : rational ~ e e n s and genetic recombination. Enzyme Microb. Technol., 6,290-300.
Saucedo, G.C., Gutierrez, M.R., Bacquet, G., Raimbault, M. (1990). Heat transfer simulation in solid substrate fezmentation. Biotechnology and Bioengineering, S, 802-808.
Shankaranand, M. V., Ramesb, B.K., Lonsane. (1 992). Idiosyncrasies of solid-state- fermentation systems in the biosynthesis of metabolites by some bacterial and hgal cultures. Process Biochemistry, z 3 3 - 3 6 .
Sipiczki, .M., and F e m ~ z y , L. (1 979). Enzymic methods for enrichment of íimgal mutants. Enrichment of Schizosaccharomyces p m b e mutants. Mutation Research, a 163-173.
69

Smit, R., Tudzynsk, P. (1992). Efficient transformation of Claviceps purpurea using pyrimidine amatrophic mutants : cloning of the O W decarboxilase gene. Mol. Gen. Genet., 234,297-305.
Somerson, N.L., Demain, AL., Nunheimer, T.D. (1961). Reversal of lysine inhibition of Penicillin production by a-amino adipic acid. Arch. Biochem., 93,238- 241.
Sullivan, J.O., and Abnaham, E.P. (1981). Biosynthesis of p-lactam antibiotics. En : Antibiotics. Edited by Corcaran, J.W. Springer-verlag, New Yo&. vol. 4.
Swart, K., Vondervoort, J.I.V., Witteveen, C.F.B., Visser, J. (1990). Genetic localization of a series of genes a f € t t g glucose oxidase levels in Aspergillus niger. Curr. Genet., u, 435-439.
Talbot, N.J., Coddington, A., Roberts, I.N., and Oliver, R.P. (1 988). Diploid construction by protoplast fusion in Fulvia fitlva synonym Cladosoporium fulvum : genetic analysis of an &perfkt fungal plant pathogen. Curr. Genet., 14,567-572.
Timberlake, W.E., hkshall, M.A. (1989). Genetic engineering of filamentous fungi. Science, 244, 1313-1317.
Uitzetter, J.H.A.A., Bos, C.J., and Viseer, J. (1986). Characterization ofAspergillus niditlans mutants in carbon metabolism isolated after D-galacturonate emichment. Journal of general micmbiology, 132,1167-1 172.
Upshall, A. (1992). The application of molecular genetic methods to filamentous fungi. En : Handbook af applied mycology. Fungal Biotechnology. Richard, P.A., Dilip, K., Elander,Mukerji, K.G., eds., Marcel Mer, Inc. New York, 4,81-93.
Viniegra, G.G. (1989). Perspectives and limitatibns of solid fermentation in Mexico. Ed. by Raimbault ORSTOM.
Viniegra, ' G.G. (1997). Solid state fermentation : Deíinition, characteristics, limitations and monitoring. En : Advances in solid state fermentation. Edited by S. Roussos, B.K. Lonsane, M. Raimbault y G.G. Viniegra. Kluwer Academic Publishers. pp 5-22.

Woodward, W., Zeeuw, J.R., and Adrian, M. (1954). The separation and isolation of particular biochemid mutants of Neurospom by differential germination of conidia, followed by Gltration and selective plating. Proc. N.A.S., 192-200.
71





























