Embed Size (px)
Citation preview

OOGENESIS

Hembras
Proliferación
Crecimiento
Maduración
DIVISIÓN MITÓTICA DE
OOGONIAS: -Toda la vida (teleósteos, anfibios,
reptiles)
- Fijado antes de la maduración
(elasmobranquios, aves, mamíferos)
-Duplicación ADN, crecimiento
citoplasmático y nuclear
- Oogonia a oocito I (diplotene)
- Reinicio de la Meiosis hasta Metafase II
(excepción erizos que la completan)

Mitosis hasta la semana 20 a 24:
oogonias aproximadamente 7 millones
A partir de 8 a 9 semanas y hasta
6 meses después del nacimiento:
oogonias entran en profase de la
meiosis: oocitos primarios
Reducción oocitaria: al nacer 2 millones
de oocitos primarios y a la pubertad
400000
En humanos

Etapa de crecimiento
Nuclear: - S! de ADN premeíotico
- R! hasta diplonema
- Transcripción de todos los ARN que se utilizarán
y almacenarán en el oocito para su crecimiento.
Citoplasmático: -Acumulación de sustancias de reserva
- Síntesis de gránulos corticales y mitocondrias.
- Formación de microvellosidades
- Síntesis de envolturas vitelinas
Comienza muy temprano y depende del grupo animal. Anfibios: despúes de metamorfosis, aves con la eclosión, ratón en vida
fetal, conejo enseguida del nacimiento, humanos en vida fetal

Estadios de la Profase meiótica en oocitos de anfibios
Estadios de la profase meíotica en oocitos de anfibios
Dumont Duryee Tamaño (μM)
Vitelogénesis y
aspecto Pigmento Meiosis
S! de ac.
Nucleicos G. corticales
Memb.
Vitelina Microvellocidades Ubicación núcleo
1 I 50 (-) transparente (-) cigotene (CS) (-) (-) (-) (-) central
1 II 50-200 (-) transparente (-) paquitene (CO) rADN (-) (-) (-) central
2 III 200-500 (+) opaco-beige premelanosomas diplotene temp.
tARN (+)
(45%),5sARN (+)
(45%),hnARN
(final), 28,18,5s
ARN (final) (+ -) (+ -) (+) central
3 IV 500-750 (++) gris (+)
diplotene max.
(plumulados)
hnARN (++),
28,18,5s ARN
(+++) (++) completa (+) central
4 V 750-850 (+++) polarización diplotene tardío
hnARN (++),
28,18,5s ARN (++) (+) (+) (+) central
5y6 VI 850-1800 (+++) estratificación polarización diplotene final (+ -) (+) (+) (+++) Hemisferio animal
s

18S 28, 5 y 5,8 S 60S
40S

Transcripción génica durante la oogénesis en anfibios
- S! de ARNr 5s (estadio III, antes de la incorporación del vitelo). No se
transcribe a partir del ON. Se acumula en partículas de ribonucleoproteínas
7s y 42s. Se incorpora en partículas de ribosomas de 60s, junto con los
ARNr de 28s y 5.8s.
- ARNr 28s, 18s y 5,8s los productos mas importantes y abundantes s! por
el oocito. Núcleos con gran cantidad de nucleolos, formados durante la
oogénesis (Xenopus 1200 nucleolos). S! estadio IV
- - ARNm, mayor síntesis en estadio IV (cromosomas plumulados). No
activo en S! de proteínas, 80% almacenado en citoplasma como partículas
de ribonucleoproteínas (RNP) para ser usado después de la F!
(histonas, tubulinas, actina, polimerasas, importantes para la S!)
- ARNm, mayor síntesis en estadio IV (cromosomas plumulados). No
activo en S! de proteínas, 80% almacenado en citoplasma como partículas
de ribonucleoproteínas (RNP) para ser usado después de la F!
(histonas, tubulinas, actina, polimerasas, importantes para la S!)
20% es utilizado para el mantenimiento del oocito (FPM, gránulos
corticales, s! MV).

ARNt: Se acumula en partículas 42s, la s! comienza antes de
la incorporación del vitelo
ARNhn se s! en los cromosomas plumulados. Precursor del
ARNm. Se s! al final del estadio III


¿Qué tipo de ARNm se almacenan?
-Los que codifican proteínas necesarias para la segmentación
(embrión requiere mucha cromatina, hacer membranas celulares,
componentes de citoesqueleto), ciclinas que regulan el ciclo
celular.
-Codifican proteínas que determinan el destino de las células
(por ejemplo ARNm bicoide y nanos, dan información de
posición en Drosophila)

TAA

-Estabilidad del
ARNm
-Le permite salir del
núcleo
-Le permite la
traducción de
proteínas
- Importante
para la unión al
ribosoma.

El transcripto tendrá una región 5’ no traducible (5’UTR), exones, intrones y una 3’UTR.
Modificados antes de salir del núcleo. En el 5’ guanosinas metiladas necesarias para la unión al ribosoma y posterior traducciones. En el 3’ agregado de poliA, adeninas reunidas enzimáticas. AMBAS PROTECCION A EXONUCLEASAS
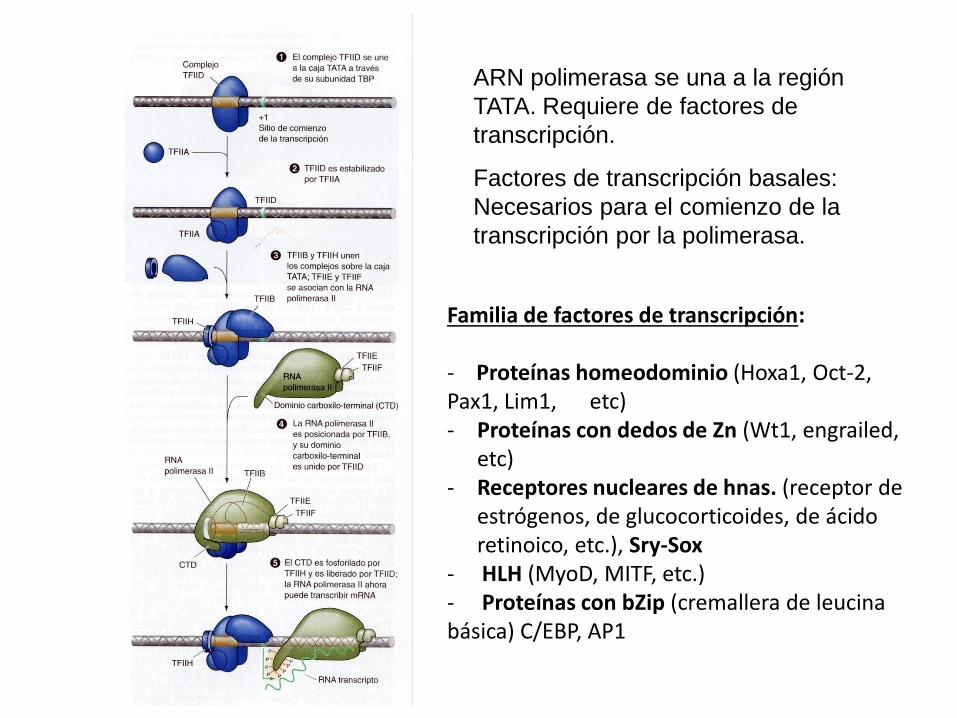
ARN polimerasa se una a la región
TATA. Requiere de factores de
transcripción.
Factores de transcripción basales:
Necesarios para el comienzo de la
transcripción por la polimerasa.
Familia de factores de transcripción: - Proteínas homeodominio (Hoxa1, Oct-2, Pax1, Lim1, etc) - Proteínas con dedos de Zn (Wt1, engrailed,
etc) - Receptores nucleares de hnas. (receptor de
estrógenos, de glucocorticoides, de ácido retinoico, etc.), Sry-Sox
- HLH (MyoD, MITF, etc.) - Proteínas con bZip (cremallera de leucina básica) C/EBP, AP1

¿Cómo se almacenan los ARNm?
- Hipótesis del ARNm disfrazado
- Hipótesis de la cola de poli A

- Hipótesis del ARNm disfrazado
Elemento de unión para la
poliadenilación citoplasmática
Factor específico de
poliadenilación y segmentación
Proteína de unión al
PoliA
Por proteínas de modo que los
ribosomas no pueden unirse al
ARNm F! Cambios
iónicos que
desestabilizan la
unión
anfibios
progesterona
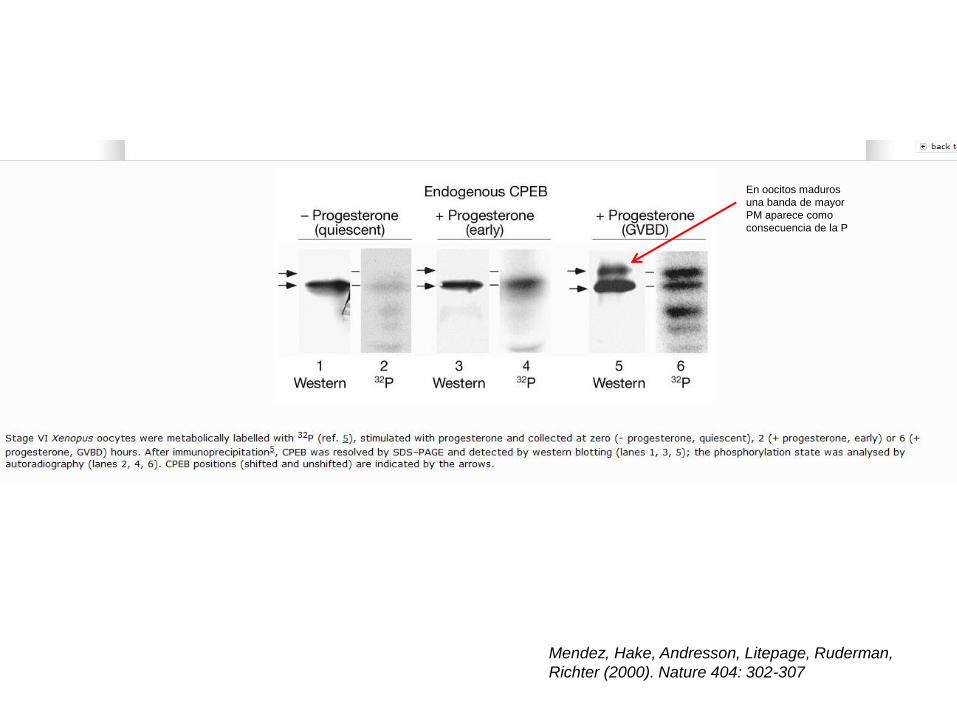
Mendez, Hake, Andresson, Litepage, Ruderman,
Richter (2000). Nature 404: 302-307
En oocitos maduros
una banda de mayor
PM aparece como
consecuencia de la P

Hipótesis de la cola de poli A
Secuencia AATAAA necesaria para la poliadenilación.
Esta región ( UTR 3’) sería la responsable del tamaño de
la cola de poliA. Depende de la presencia de CPEB. Esta
región es también responsable de la localización del
mensajero.
Cola larga, vida corta. Cola corta, vida larga.

Oocitos de Xenopus
- Cuando se rompe la VG y se
reinicia la R! el núcleo libera factores
de desadenilación.
- Los ARNm sin CPEB son
desadenilados, los que la tienen se
poliadenilan.

Otras estrategias:
-En algunos los RNA mensajeros son estabilizados en momentos específicos
en células específicas (Ej. Caseína en glándula mamaria de 1 hora a 28,5 por
prolactina).
- Oocito de un gusano X mRNA sin casquete 5’ metilado , en la F! una
metiltransferasa completa la formación del casquete y entonces se puede
traducir.
- En Drosophila la proteína Smaug (proteína inhibitoria) se une al 3’ UTR de
ARNm de nanos e impide su traducción hasta la fecundación. También en
Drosophila bicoide se une a una región del 3’ UTR y como masquina bloquea el
inicio de la traducción.
- En C. elegans transcripción de ARNm antisentido para uno de sus propios
mensajeros. Hoy se ve que sería un mecanismo conservado.

¿En que momento se traducen los ARNm almacenados por el
oocito?
-Durante la maduración
-Después de la Fecundación
-En el desarrollo temprano
Ej. Huevos de erizo de mar con
actinomicina D se desarrollan hasta
blástula

Cambios citoplasmáticos
-Almacenamiento de ARNs
-Aumento de mitocondrias, pueden reunirse alrededor del
núcleo (anillos de Balbiani)
-Formación de gránulos corticales
-Acumulación de sustancias de reserva (lípidos, proteínas,
glucógeno)
-Formación de microvellosidades en la membrana
citoplasmática

Formación de gránulos corticales: S! a partir del Golgi.
- En erizos de mar, peces, anuros, moluscos bivalvos, algunos
anélidos, mamíferos (hamster, conejo, hombre).
-Ausente en urodelos, aves, moluscos, gasterópodos, rata, cobayo.
-Membrana limitante y un gránulo central, denso, homogeneo,
constituido por mucopolisacáridos sulfatados y enzimas
proteolíticas o proteasas.
Almacenamiento de sustancias de reserva: Gránulos de vitelo o
plaquetas vitelinas.
-Componente principal: lipofosfoproteínas o vitelogeninas y en
menor grado grasas neutras.
- vitelo graso o proteico.

Fosvitina + lipovitelina Lipofosfoproteína o
vitelogeninas
(principal componente del
vitelo) Viaja parcialmente
fosforilada
Soluble en agua
En mitocondrias de oocito de rana:
Fosvitina parc. Fosforilada + ATP
Fosvitina totalmente
fosforilada + ADP proteinquinasa
Mg++
Fosvitina
lipovitelina
plaqueta
En sangre

Vitelo (plaqueta vitelinas, gránulos de vitelo, esferas o glóbulos de vitelo)
Fosvitina (PM 40000, 8,4% de P)
Lipovitelina (PM 400000,
17,5% de lípidos)
Red hexagonal
central
Material amorfo
Glucógeno: en gránulos pequeños
Lípidos: en lipocondrios

Al final de la vitelogénesis…..
Ribosomas,
glucógeno,
vesículas
lipídicas, Ret.
endoplasmático
corteza
Gránulos
corticales,
pigmento,
mitocondrias
Plaquetas vitelinas

Anfibios Rana pipiens y Rhinella arenarum, 3
años
Acumulación
de vitelo
previtelogénesis vitelogénesis






Maduración
-Desde el reinicio de la Meiosis (diplonema de la primera %)
hasta el oocito en Metafase II.
-Dentro del ovario (mamíferos, erizos de mar)
-Fuera del ovario (anfibios). Liberación del corpúsculo polar en
oviducto o antes de entrar.
-Comienza con la ruptura de la vesícula germinal (RVG),
seguida por la condensación de los cromosomas, formación del
huso, y continuación de la R! hasta Metafase II.

Maduración del oocito en Xenopus
Gonadotrofinas
6 hs. Post Progesterona: Ruptura de la VG, retracción de microvellosidades,
desintegración de nucleolos, cromosomas se contraen y migran al PA.
Completa I % y ovulación.
(cél. Foliculares)
(activa cascada de P)

P34 fosforila: -H1 (condensación de cromosomas)
-lamininas (proteínas sobre la cara interna de la MN)
-Proteína nucleolar (desensamble de nucleolo)
-ARN pol (evitaría la transcripción durante la % celular)
Estadio VI: -p34 sin P y no unida a ciclina.
-p34P + ciclina: Pre FPM -Treonina 161 P por una protein quinasa
serina treonina: CAK. Fosforilación activante.
-Treonina 14, tirosina 15 por medio de la
proteinquinasa dependiente de AMPc (PKA). Fosforilaciones inhibitorias.
Treonina 161
Treonina 14 Tirosina 15
P34 ciclina

Progesterona
ARNm c-mos
CPEB
Proteina c-mos
Ca2+
Disminución de AMPc
Disminución de PKA
Pre FPM:
ciclina +p34
cdc25
FPM
Ca2+
CSF
gonadotrofinas
(de depósitos
internos)
(Desfosforila
-ción de tyr
15 y thr 14)
(vuelve a los
depósitos por
aumento de FPM)

(Anti CPEB)

C-mos y cinasa 2
dependiente de
ciclina (cdk2)
- cdk2
-
C-mos

Células foliculares del oocito:
-Involucradas en la síntesis de estructuras no celulares que rodean
al oocito (Membranas secundarias)
-Sintesis de hormonas esteroideas (17ßestradiol y progesterona)
-Permite el pasaje de ciertos componentes hacia el oocito
-Uniones GAP entre ellas y entre ellas y el oocito.
Membranas que envuelven al oocito:
Primarias: s! por el propio oocito
Constituída por proteínas fibrosas
Moluscos, insectos, equinodermos, algunos peces, aves, erizos de mar,
mamíferos (ZP1, ZP2, ZP3)
Secundarias: S! por las foliculares, proteicas
Corión de insectos y algunos peces
Terciarias: Secretadas por el oviducto.
Gelatina de anfibios, aves (albúmina, ms. Cáscara, cáscara), elasmobranquios

• ANFIBIOS (4 glicoproteínas) 38 kDa < PM < 120 kDa • MAMIFEROS (Zp3 - Zp2 – Zp1)
* Unión a espermatozoide
* Inducción de la reacción acrosómica
* Prevención de la polispermia
LA ENVOLTURA VITELINA
FUNCIONES:


INSECTOS: TIPOS DE OVARIOLA
Sin cél nutricias,
solo foliculares.
Cromosomas
plumulados
Blátidos
Con células nutricias con
cromosomas politénicos
Dípteros
Drosophila
Hemípteros,
coleópteros

Locusta migratoria: ortóptero
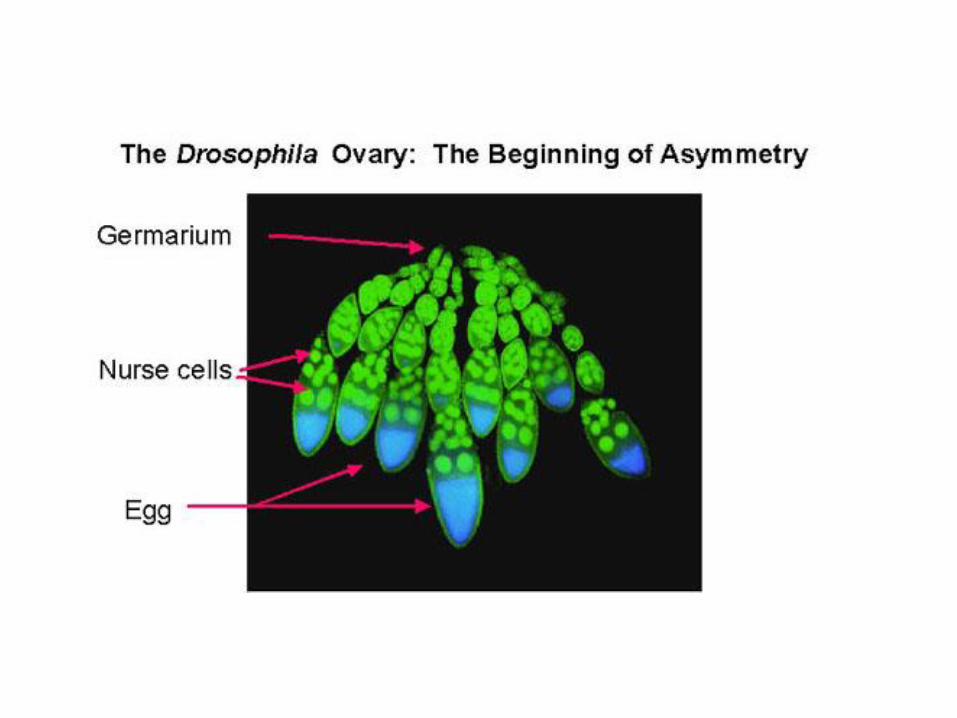

cistocitos

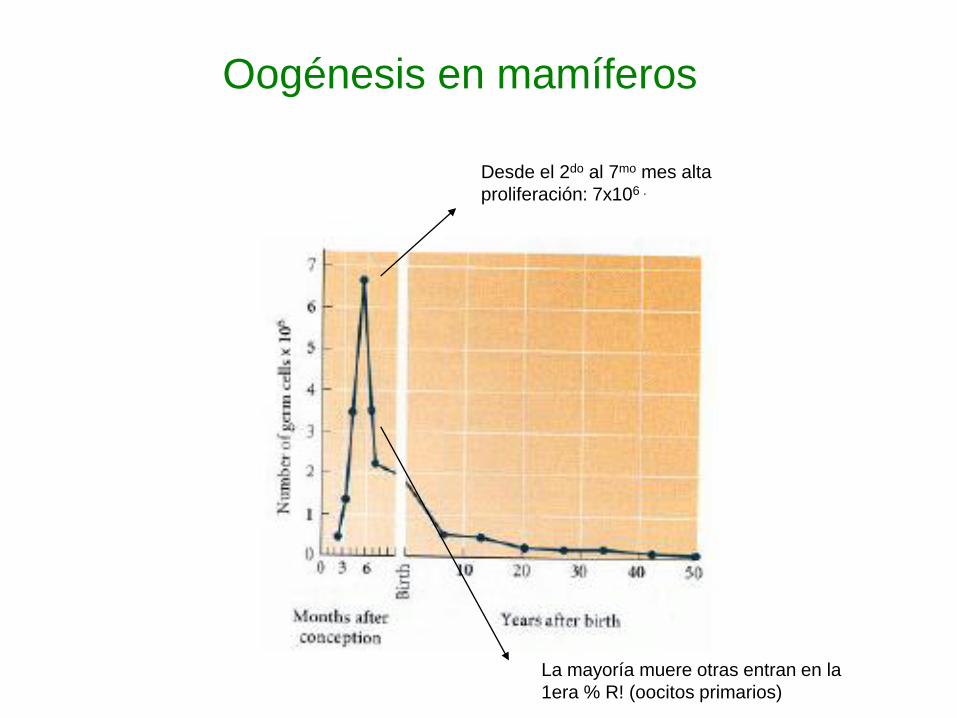
Oogénesis en mamíferos
Desde el 2do al 7mo mes alta
proliferación: 7x106 .
La mayoría muere otras entran en la
1era % R! (oocitos primarios)

CGP
oogonia
++Oogonias
M!
Oocitos primarios
-Folículos primordiales
-Folículo primario (5/6 meses de
gestación)
-Folículos secundarios (GDF9)
Folículos antrales
TGF-ß2,
VEGF,
leptina, FGF2
(Detenidos en 1era Profase R!)
Oogénesis en mamíferos

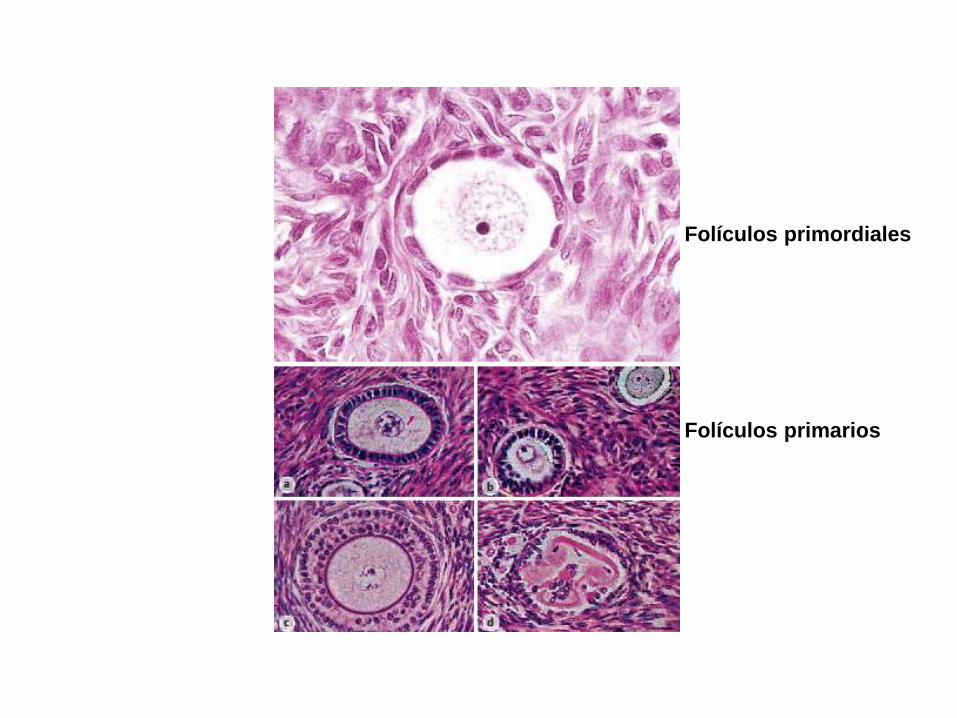
Folículos primordiales
Folículos primarios

Cél granulosa
Zona pelúcida
Memb. basal
Cél tecales
antro
Vacuola de
Call-Exner
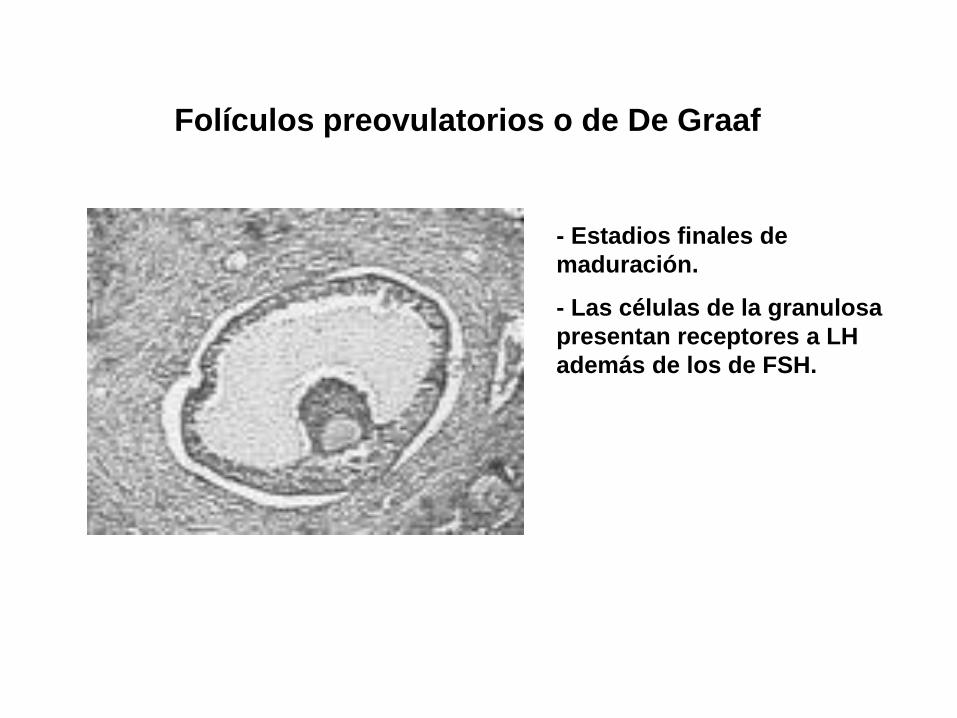
Folículos preovulatorios o de De Graaf
- Estadios finales de
maduración.
- Las células de la granulosa
presentan receptores a LH
además de los de FSH.
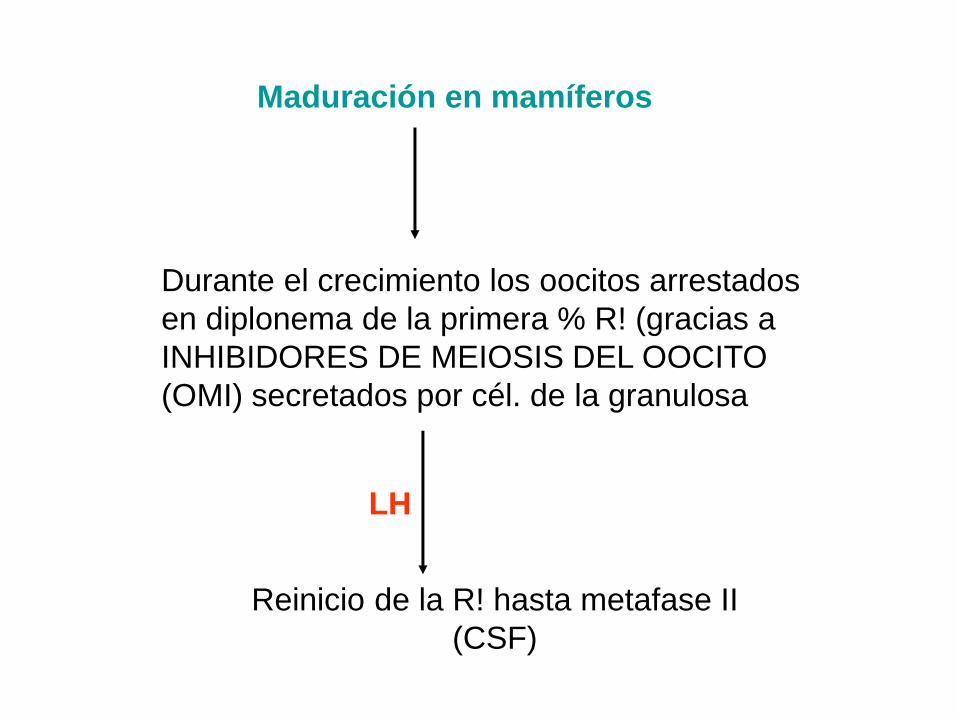
Maduración en mamíferos
Durante el crecimiento los oocitos arrestados
en diplonema de la primera % R! (gracias a
INHIBIDORES DE MEIOSIS DEL OOCITO
(OMI) secretados por cél. de la granulosa
LH
Reinicio de la R! hasta metafase II
(CSF)



Aumento de pulsos por estradiol y
progesterona
+
-
- Por folículo dominante
2 días 200 pg/ml
Secretado por cuerpo luteo
+
Testo y Androstenodiona precursores de
Estradiol
+
Provocan retroalimentación negativa sobre
FSH
Aumento por caída de
estradiol y progesterona
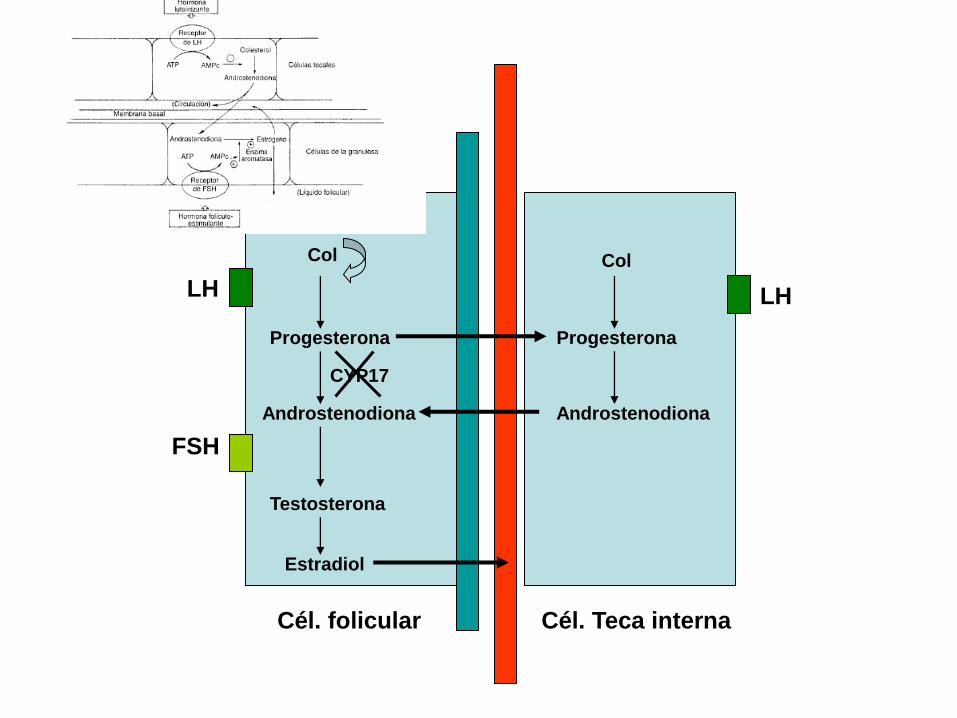
Cél. folicular Cél. Teca interna
Col
Progesterona
Androstenodiona
Testosterona
Estradiol
Col
Progesterona
Androstenodiona
LH
FSH
LH
CYP17


LH
Neutraliza la acción
del péptido Inhibidor
de la Maduración: Se
desarresta la R!
Estimula la síntesis de
Progesterona: aumento
de actividad
proteolítica, debilita la
pared
Respuesta
Pseudoinflamatoria:
síntesis local de
prostaglandinas,
leucotrienos, tromboxanos FSH
Estimula la producción
de
glucosaminoglucanos
que mucifican el
entorno, dispersando
al cumulus oosforo.
Induce a enzimas proteolíticas que catalizan
la degradación de la pared folicular.
Estradiol Pérdida de integridad
del folículo
ovulación