Embed Size (px)
DESCRIPTION
Origin of diversity Bibliography. Antibody diversity: one enzyme to rule them all Michel C Nussenzweig & Frederick W Alt Nature Medicine 10;1304-5;2004 Immunobiology Janeway et al 6 th ed 2005 (or 7 th ed. 2008). Clonal selection theory. יצירת מאגר של לימפוציטים בשלים. תא אב יחיד מתמיין - PowerPoint PPT Presentation
Citation preview

Origin of diversityBibliography
Antibody diversity: one enzyme to rule them allMichel C Nussenzweig & Frederick W AltNature Medicine 10;1304-5;2004
Immunobiology Janeway et al 6th ed 2005 (or 7th ed. 2008)

Clonal selection theory
Memorycells
B-cells: plasma cellsAb secretion
T cells:T-helper T-killer
תא אב יחיד מתמיין למספר גדול של
לימפוציטים שלכל אחד מהם ספציפיות
שונה
סילוק של לימפוציטיםלא בשלים המגיבים
עם אנטיגנים עצמיים
יצירת מאגר שללימפוציטים בשלים
חלוקה והתמיינות של לימפוציטים
ספציפיים ליצירתקלון של תאים
אפקטוריים

Figure 3-1 part 3 of 3

Figure 3-6
CDR1 CDR1 CDR2CDR2 CDR3 CDR3

Figure 4-2
The chance for successful rearrangement is 33%


Ontogeny of the B Lymphocyte
• Early differentiation– Many stages defined by Ig gene
rearrangements– Progenitor cells
• Hematopoietic stem cells• Pro-B • Pre-B• Immature B

Pathway of B cell Differentiation
Pre-Pro B Pre-B1 Pre-B2 BPro B
Cytoplasmic chain
Heavy chain +surrogate light chain
“Heavy” plus “light” chain
Antigen IndependentAntigen Independent Antigen DependentAntigen Dependent
IgG “Memory” cell
“Plasmablast”
D--->JH VH-->DJH VL-->JL
(חליפית))
Immature
IgM
MatureIgM + IgD

Ontogeny of the B Lymphocyte
• Process of differentiation–Signals that promote survival and
proliferation of early cells• Adhesive interactions with stroma
(non-lymphoid cells that make up marrow matrix)
• Secretion of IL7

Ontogeny of the B Lymphocyte
• Process of differentiation (continued)– Early cells
• Pro-B – heavy chain D gene segment rearranges to J segment
• Pre-B – Heavy chain V gene segment rearranges to join DJ region
– Rearranged VDJ put close to heavy chain constant gene
– Synthesizes a heavy chain

Figure 7-4 part 1 of 3
D-J rearrangement
V-DJ rearrangement

Figure 4-2

Figure 7-6 part 1 of 2

DJ rearrangement on both chromosomes

Ontogeny of the B Lymphocyte
• Pre-B cell – expresses chain as a trans-membrane molecule at cell surface– In conjunction with products of two non-rearranged genes
• Lambda 5 and VpreB– Function as surrogate light chains
– Also in conjunction with additional trans-membrane molecules linked by disulfides
• Ig alpha (CD79a)• Ig beta (CD79b)
– Combination of tran-smembrane molecules (pre-B cell receptor;
pre-BCR)



Ig and Igproteins
Signal Transduction;
1. clonal expansion
2. allelic exclusion
3. light chain rearrangement
heavy chain with surrogate light chains
IL-7 and adhesive interactions between B cells and stromal
cells important for proliferation
IL-7R (IL-7Ra and IL-2 gc chain)
5 KO blocks L chain rearrangementand B cell differentiation It does not block second H-chainrearrangement

Figure 7-18

Sensitive to self antigens

Figure 7-6 part 2 of 2
Sensitive to self antigens


Allelic exclusion

Ontogeny of the B Lymphocyte
• Immature B cells– L chains pair with H chains to form monomeric IgM
then inserted into membrane
– Recognize Ag and respond to it• Long-lasting inactivation instead of expansion and
differentiation
• Interact with self-Ag in bone marrow – inactivate cell– Called negative selection
– Important for development of self-tolerance in B-lineage
» Cells with potential reactivity to self prevented from responding (central tolerance)

Ontogeny of the B Lymphocyte
• Development of self-tolerance– Immature B cell exposed in bone marrow to:
• Self-molecule on surface cells – Apoptosis (deletion)
• Non-cell surface molecule (soluble Ag)– Cell is inactivated but not deleted (anergized)
• Reactivation of VDJ recombinase (receptor editing)– Ig L chain genes undergo secondary rearrangement
» Use unrearranged V or J segments» Generates specificity for non-self Ag – rescued from
inactivation


After gene rearrangements and production of a functional molecule, the cell tests whether its specificity is anti-self
Maintenance of tolerance requires the persistence of antigenbecause self-antigens are always present but foreign antigens are transient

Before clonal deletion of an anti-self B cell, the cell can attempt receptor editing of the light chain
Fig 7.26
Immature T cells continues to rearrange chain until the cell gets positive selection signal. (will eventually die if it does not receive positive selection in a few days). (needs signal to stop “editing”)
ImmatureImmature B cell “edits” light chain if it binds antigen (gets negative selection signal). This could rescue the cell from negative selection (i.e., death). (needs signal to edit)
So, light chain can use repeated rearrangements and can receptor edit. Repeated rearrangements are to make a functional molecule whereas receptor editing is to avoid clonal deletion of anti-self specific B cells.
RAGs expression is still on

Figure 7-18

Allelic exclusion



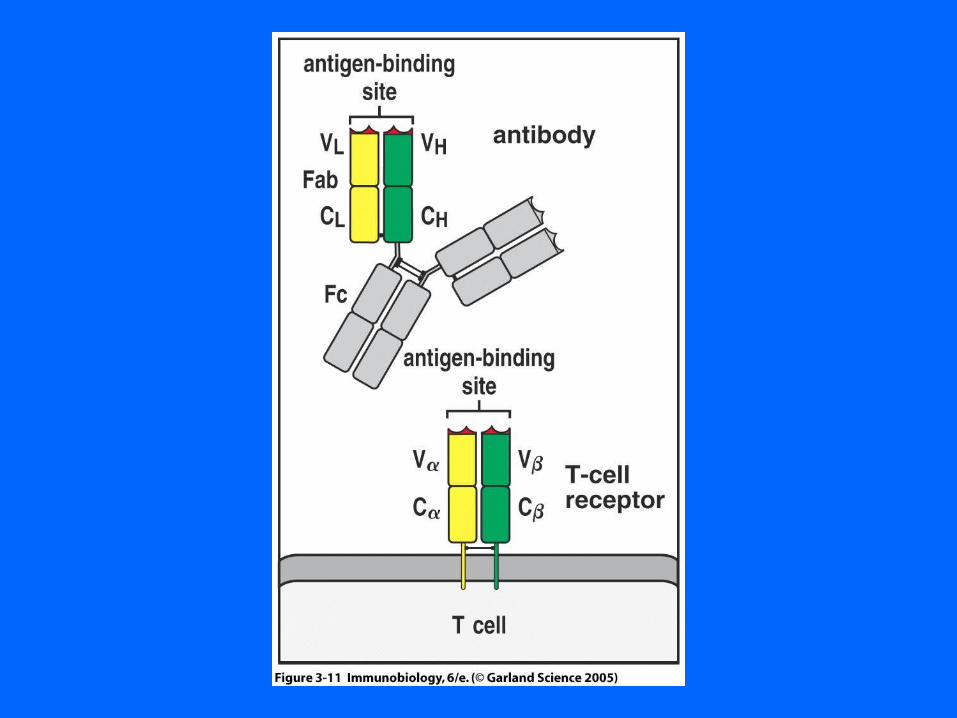
Figure 3-11
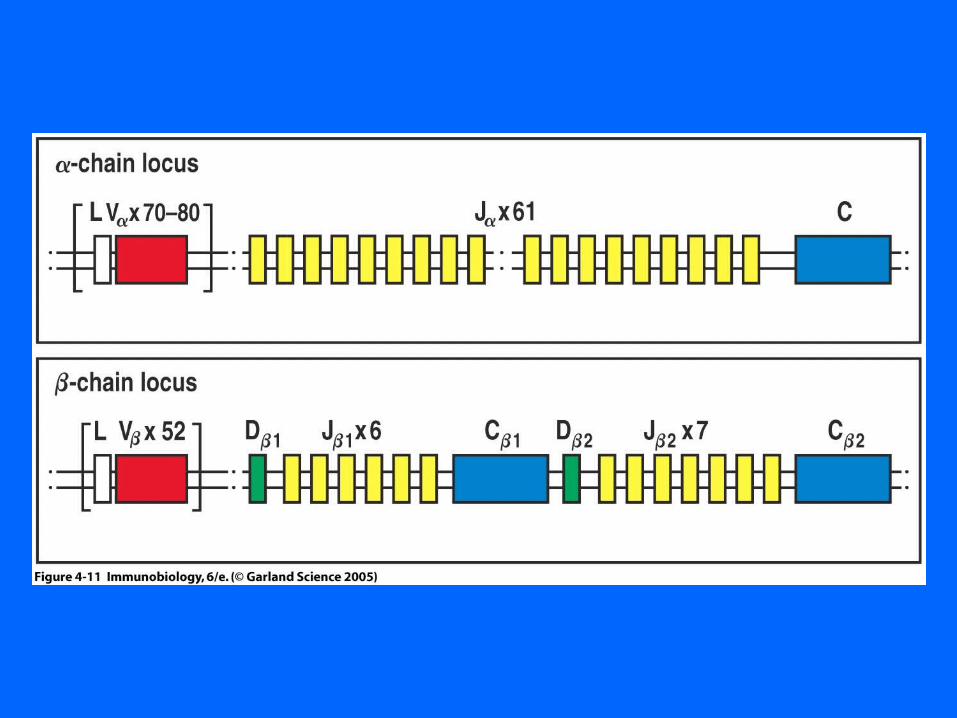

Figure 4-12

Figure 4-15V are mixed with V




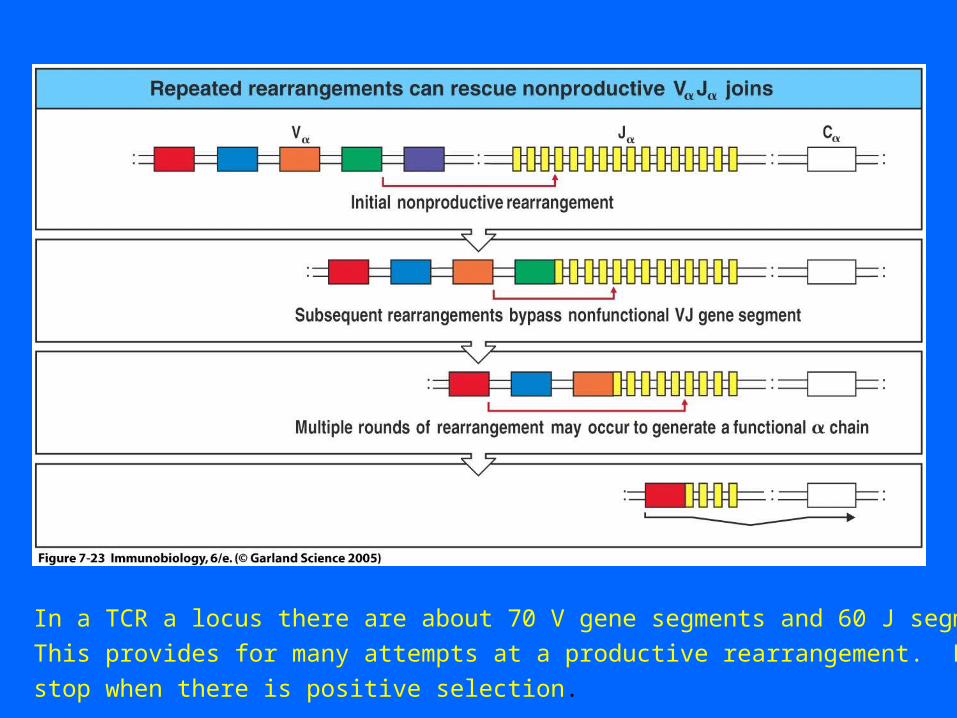
In a TCR a locus there are about 70 V gene segments and 60 J segments.
This provides for many attempts at a productive rearrangement. Rearrangements
stop when there is positive selection.
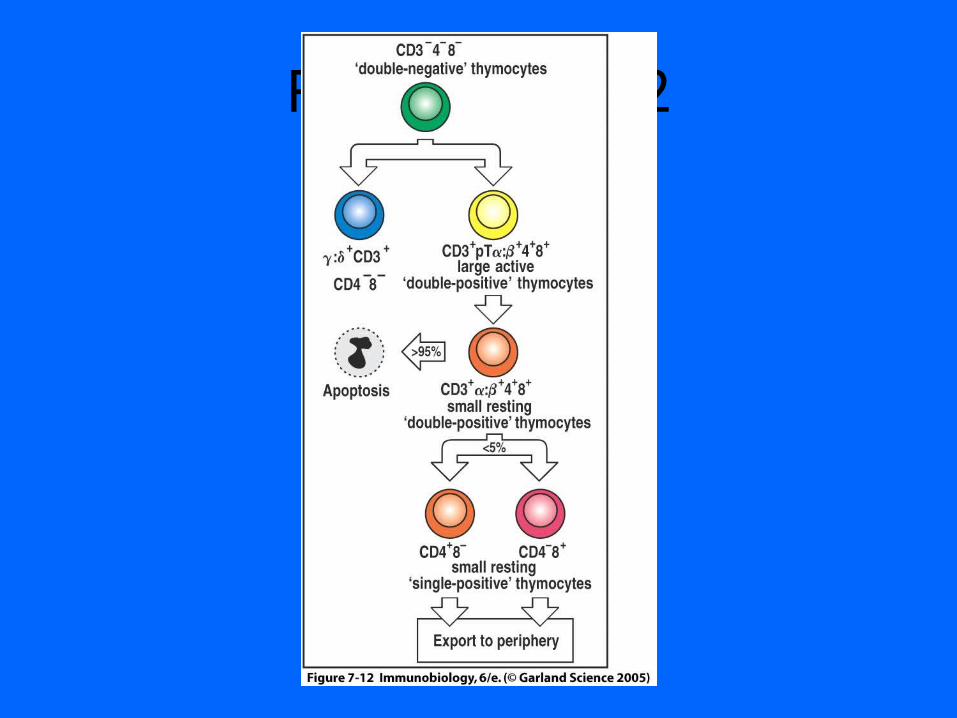
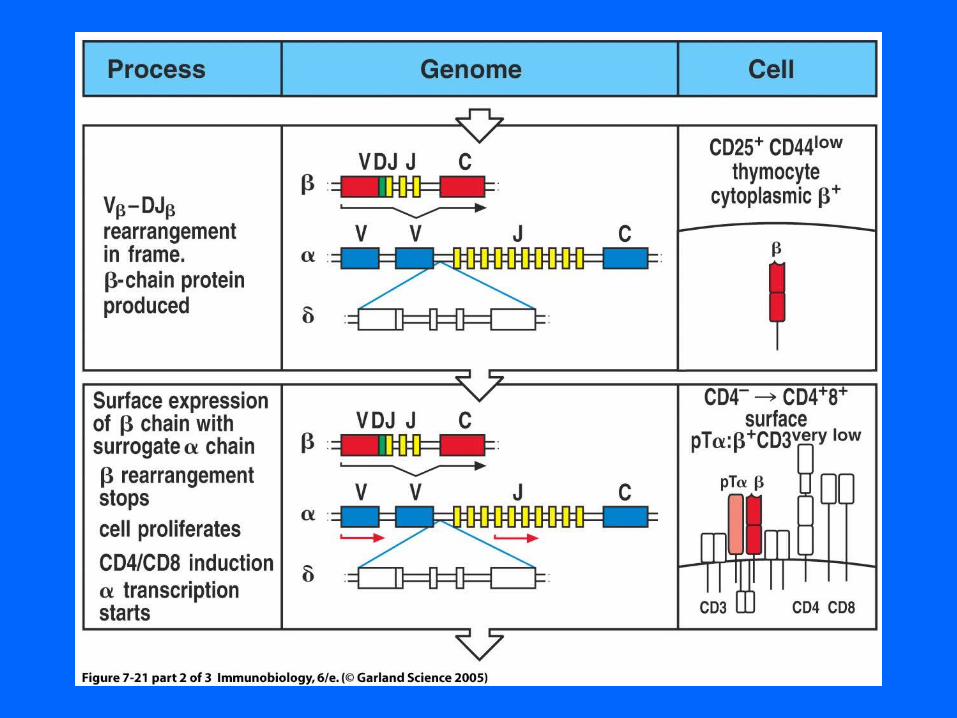
Figure 7-12

Figure 4-2


Mechanisms contributing to generation of primary antibody diversity in humans
6000120200VxJx(D)H-L chain associations Hxk 1,200,000
Hxl 720,000

Levels of Regulation of Ig Gene Expression & GOD
1. V-(D)-J rearrangement
2. Class switch recombination (CSR)
3. Somatic hypermutation (SH)
4. Receptor editing\revision

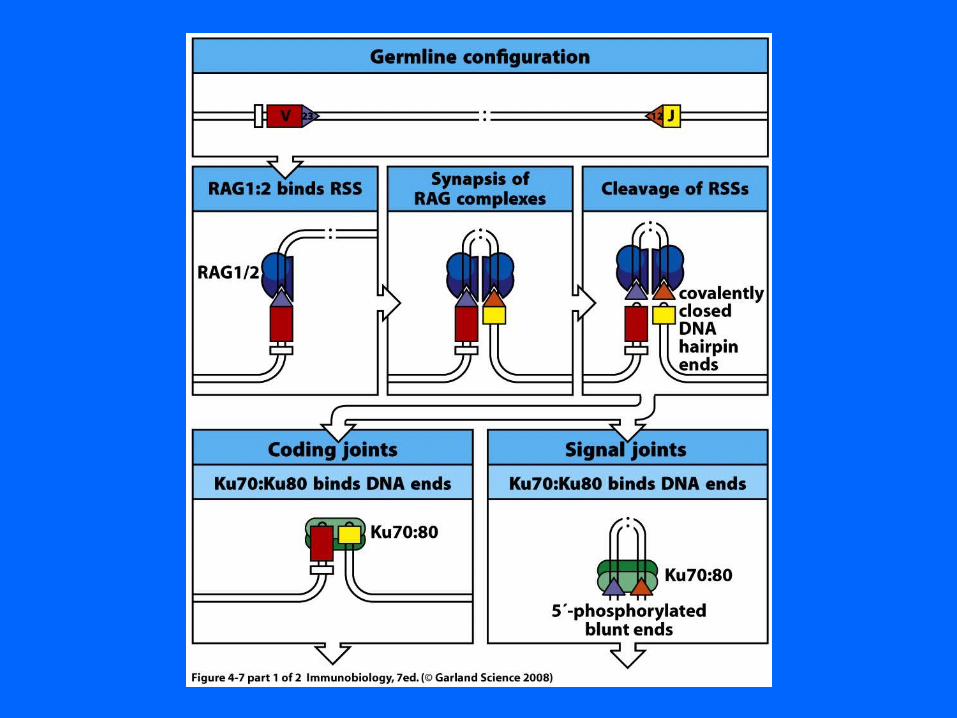
V(D)J Recombination : RAG
RAG 1 & 2 required
RAG 1 or RAG 2 K.O. miceAlso SCID in Man
No VDJ recombination (B or T-cells)No dsDNA breaks

RAG function
• RAG 1 & 2 transfected into fibroblasts + synthetic V-D-J substrate
Recombines exons
Transcription control In synthetic substrate transfected into cell lines and mice, deletion of any of promoters or enhancers blocks rearrangement

Transcription Controls V(D)J Recombination
P Pi Pi
vD J Ei 3’E
μCμ
Gene construct to test control elements: Deletion of any of Promoters (P or Pi) or Enhancers (Ei or 3’Eμ blocks recombination.
Lp



P N P
V JVJ1 TGG GAT T AGCVJ2 TGG GA AGCVJ3 TGG GA CCG AT AGCVJ4 TGG GAT TGG GCVJ5 TGG GA AT AT AGCVJ6 TGG GAT A T T AGCVJ7 TGG GAT TT AT AT AGC
Vg6: TGG GAT A cactcta…………
Jg1: …………cactgtg AT AGC
Nucleotide sequences of TCR V-gamma6 / J-gamma1 junction

Generation of diversity
Mechanism of action of RAG1 & RAG2
N addition by TdT=Terminal deoxynucleotidyl transferase


RSS
RSS
5’ N-O-P-O-N 3’ 3’ N-O-P-O-N 5’



Antibody protein sequence is altered in several independent ways
•Regulation of membrane vs secreted IgM RNA splicing
•(Pre-immune) membrane IgM/IgD co-expression RNA splicing AID dependent (activation induced cytidine deaminase):•Affinity maturation Somatic hyper-mutation
•Isotype switch: Switch to IgG, IgA, IgE antibody H-chain C exons
•Gene conversion

Note: there are exons encoding the membrane and secreted forms of each of the antibody heavy chains.

Prior to activation, B cells express two forms of antibody as membrane receptors, IgM and IgD. On any given cell, the antigen specificity is identical.
This is accomplished by differential RNA splicing.
IgMIgD


Low affinityIgM
High affinityIgG
IgM
Antibody protein sequence and bioactivity changes during the immune response

Figure 4-9Somatic hypermutation introduces diversity BCR (not in TCR)
Mutationssilent Phe ttc ttt
neutral Lys aaaArg aga
PositiveGlu gaaGly gga
deleterious Ser tcaStop taa
Affinity maturation
Selection of clones withthe highest affinity

Characteristics of Somatic Mutation only in B cells and not in T cells
1. Occurs at high rates: 10 -4 -10 -3 /bp/generation.2. Occurs by untemplated single base substitutions.3. Restricted to a brief period of B cell differentiation.4. Restricted to the rearranged V region and its immediate flanking sequences.5. Occurs in germinal centers with T cell help.6. Occurs throughout the V region but more frequently in RGYW (A/G G C/T A/T) motifs.
BioEssays 20:227–234, 1998

The mutation DomainThe mutation Domain
• Mutations largely confined to variable regionsMutations largely confined to variable regions• Occur over a 1-2 kB region around the rearranged VJ Occur over a 1-2 kB region around the rearranged VJ
gene segmentgene segment• Found predominantly in CDRs but due to selectionFound predominantly in CDRs but due to selection

Patricia J. Gearhart and Richard D. Wood
Hyper-mutations spread in the V(D)J region


Figure 3-6
CDR1 CDR1 CDR2CDR2 CDR3 CDR3

Requirements for recruiting SHMRequirements for recruiting SHM(Somatic Hyper Mutation)(Somatic Hyper Mutation)
• In kappa locus both intronic and 3’ enhancer In kappa locus both intronic and 3’ enhancer elements are requiredelements are required
• Transcription is required, mutations decrease with Transcription is required, mutations decrease with distance from the promoterdistance from the promoter
• Expression of AID (activation-induced cytidine Expression of AID (activation-induced cytidine deaminase) is essentialdeaminase) is essential

Activation Induced cytidine Deaminase (AID) induces somatic hypermutation
AID
G
C
A
T
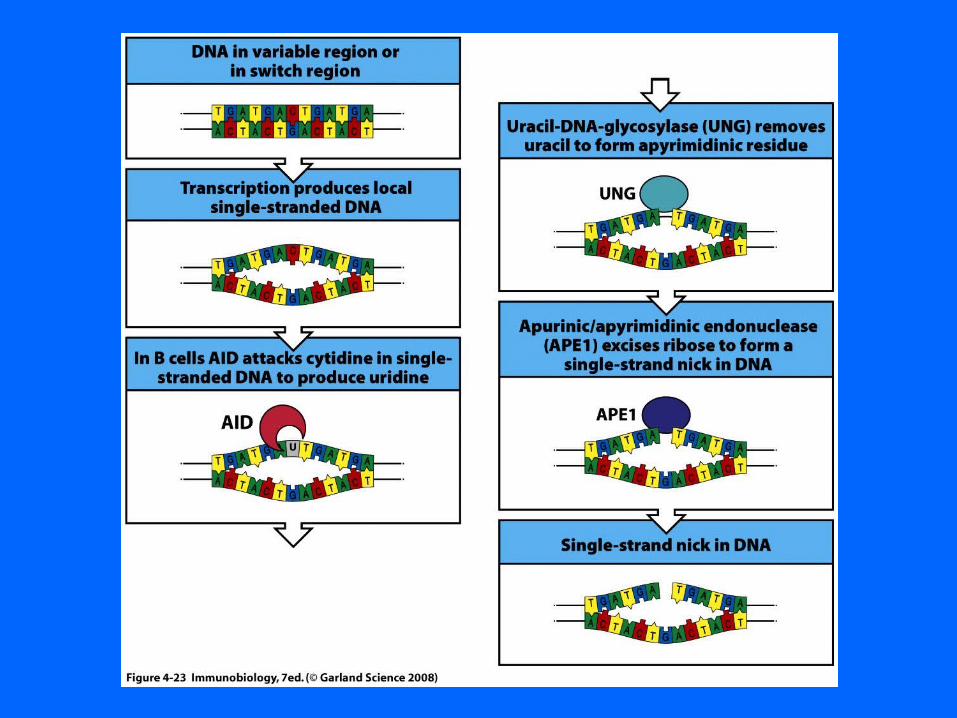


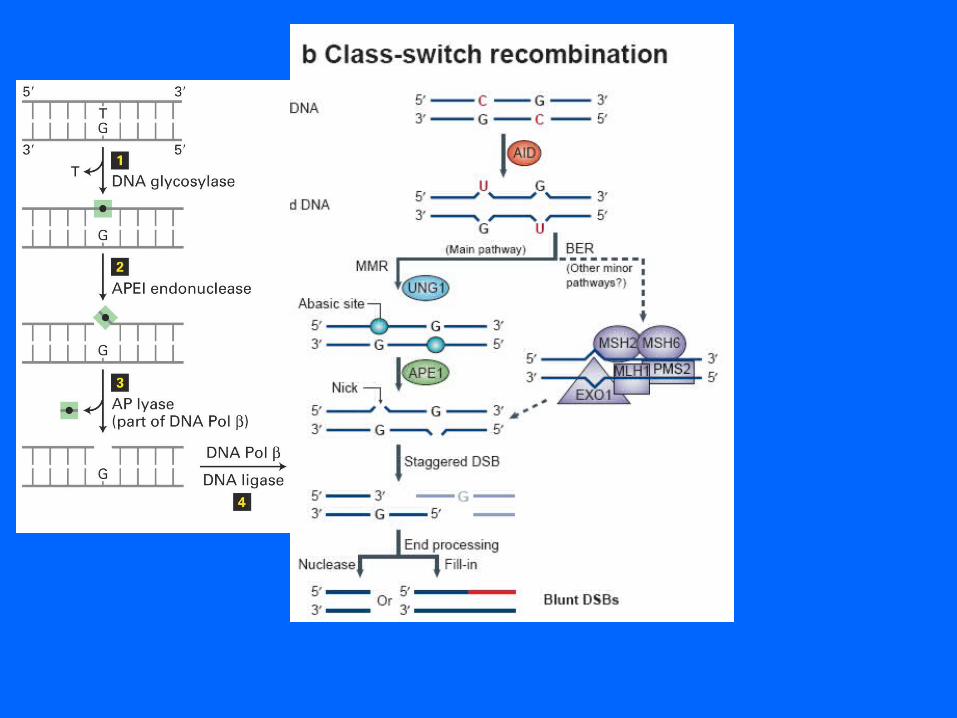
Base excision repair of G-T mismatch
removes base leaves sugar backbone

Mismatch excision repair of newly replicated DNA


Ig Class Switch Recombination (CSR)

Figure 4-21

.
CSR occurs by an intrachromosomal deletional recombination between switch (S) region sequences located upstream of the constant region genes. S region sequences consist of tandem repeats of short (20–80 bp) consensus elements, extending from 2 to 10 kb in length, and recombination can occur at any site within the S regions. The process is thought to be initiated by the creation of double-strand breaks within the S regions, consistent with the ability to detect the deleted DNA as a circle. Although the different S regions have short sequence elements in common (e.g., GGGGT, GGGCT, or GAGCT), they differ too much to undergo homologous recombination and CSR is thought to occur by a type of nonhomologous end joining (NHEJ).

Class Switch Recombination
• Transcription in switch region required
• Cytokine directs transcription to I exon promoter
• RNA/DNA hybrid of product flags target site for recombination

Ig Class Switch Recombination (CSR)Initiated by CD40L:CD40 + appropriate cytokine
Cytokine IgG1 IgG2a IgG2b IgG3 IgA IgE
IL-4 + +IL-5 +IFN + +TGF + +

VDJ joining creates heavy chain variable region domain
VDa

Figure 9-8

Ig Class Switch Recombination
Evidence:
• CSR blocked by deletion of :-– Splice site– I exon promoter – I exon
• RNA/DNA hybrids identified

Ig Class Switch Recombination
• RAG 1 & 2 NOT required
• CSR requires:-– Activation-induced cytidine deaminase (AID,
mice & hu.)– MSH2 (K.O. mice)



Ig Class Switch Recombination
• AID initiates staggered dsDNA breaks
• Cut by endonuclease
• Breaks are repaired by error-prone DNA pol.

Class Switch Recombination
• DNA breaks at switch sites are staggered
• Breaks are repaired by error-prone DNA pol. & ligation

Class Switch Recombination
Evidence:-
• AID- - hu. & AID or MSH2 K.O. mice No CSR
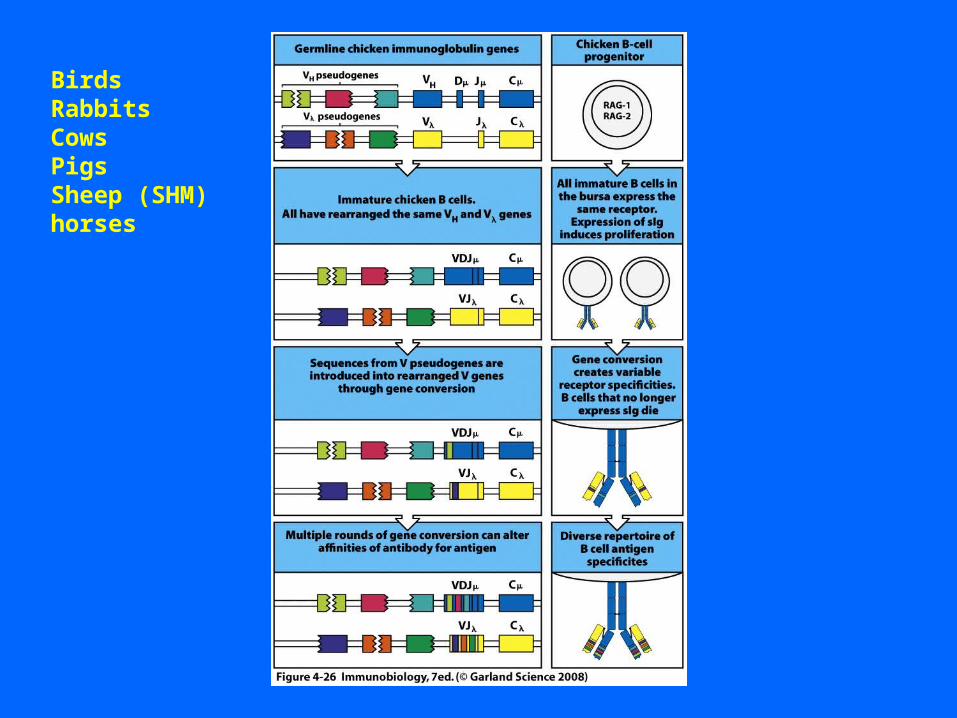
BirdsRabbitsCowsPigsSheep (SHM)horses


Figure 4-13

AID converts C to U

Somatic Hypermutation (SHM) & Class Switch Recombination (CSR)
Activation of B lymphocytes by antigen and costimulatory signals, usually from T lymphocytes, initiates two processes of antibody diversification. Somatic hypermutation (SHM) introduces mutations in the variable region genes, which, in conjunction with antigen selection, generates antibodies with increased affinity.
The second process, class switch recombination (CSR), enables B Cells to diversify the constant region and thereby the effector function of the antibody molecule, while maintaining the same antigen-binding domain.
These two mechanisms have several shared features but both are poorly understood

Activation-induced cytidine deaminase (AID) is essential for both SHM and CSRThe data indicate its role is to convert dC to dU residues within variable genes and S regions. Resolution of the dU residues could introduce mutations that are characteristic both of SHM and of segments surrounding S–S junctions. Furthermore, removal of the dU residues by the base excision repair (BER) pathway could introduce the DNA breaks necessary to initiate CSR. This has not been demonstrated; however, CSR is 90% reduced in B cells from mice deficient in the BER enzyme uracil DNA glycosylase (UNG) that excises dU residues, and even more severely reduced in some patients with hyper IgM syndrome caused by deleterious mutations in UNG.

Class Switch Recombination
Cytokines direct transcription of I-S-CH which initiates CSR