Embed Size (px)
Citation preview

OSMOREGULATION ET EXCRETION AZOTEE
RELATION AVEC LE MILIEU DE VIE

OSMOREGULATION

Osmo-régulation ?
Organisme vivant principalement constituée d’eau
Nécessité de maintenir constant le volume d’eau et la concentration des différents solutés => constance du milieu intérieur nécessaire aux fonctions vitales
Différence de concentrations entre milieu extérieur et milieu intérieur ont tendances a s’estompé par : Diffusion
osmose
Pour limiter ces perturbations : diminuer la perméabilité
diminue le gradient de [C] entre l’intérieur et l’extérieur

Osmorégulation fonction physiologique qui maintient constant le gradient de [C] entre milieu intérieur et environnement
L’osmorégulation est associé à d’autres fonctions physiologiques, principalement à l’excrétion
Excrétion assuré par le rein qui est aussi un organe important dans l’osmorégulation
Plusieurs autres organes impliqués dans l’osmorégulation : Intestin (absorption eau et sels)
Branchies (échanges actifs d’ions)
Épiderme des amphibiens (mouvements d’eau et transport de Na+)
Glandes a sels
(glandes sudoripares, appareil respiratoire pulmonaire)

Diffusion (diffusion simple)
O2 et CO2, petites molécules non chargés => diffuse librement
H20 et Urée diffusent + ou – facilement
Autres molécules : diffusion très lente mais pas impossible (notamment au niveau des surfaces d’échanges)

Osmose = diffusion de l’eau
Osmose = mouvement d’eau entre 2 solutions de concentrations ioniques différentes et séparés par une membrane biologique semi-perméable.
Pression exercé pour empêcher ce mouvement d’eau = pression osmotique
Pression osmotique ne dépend que du nombre de particule et pas de leur taille ou de leur nature

Osmolarité
Correspond à la concentration osmotique d’une solution
Dépend du nombre de particules dissoutes
Osmolarité d’une solution contenant un non-éléctrolyte (saccharose, urée) est équivalente à la concentration molaire
Pour un electrolyte comme NaCl, osmolarité supérieure à la molarité mais pas la double car dissociation du NaCl n’est pas totale
Eau de mer : 1000 mOsm/L
Eau douce : 0,1 à 10 mOsm/L

Osmoconforme et osmorégulateur

mortalité
mortalité
osmolarité de l’eau de mer (mOsmol/l)
osmolarité du MI
(mOsmol/l)
osmolarité de l’eau de mer (mOsmol/l)
osmolarité du MI
(mOsmol/l)
mortalité
Deux types d’osmoconformes : les sténohalins et les euryhalins
Euryhalin : grandes tolérances aux variations de salinités Sténohalin : pas ou peu de tolérance au variations de salinités Termes indépendants du milieu de vie (sténohalin marin ou sténohalin d’eau douce)

osmorégulateur partiel osmorégulateur
osmoconforme

ISO, HYPO ET HYPEROSMOTIQUE
1000 mOsm/L
10 mOsm/L
1000 mOsm/L
300 mOsm/L
300 mOsm/L
300 mOsm/L

ISOTONIQUE OU ISOOSMOTIQUE ? Isotonique = cellule dans un milieu ne gonfle
pas et ne se rétracte pas (GR dans solution de NaCl à 150 mmol/L
Si GR placé dans une solution isoosmotique d’urée gonflent et éclatent => urée traverse rapidement la membrane pour équilibré de part et d’autre, puis entrée d’eau


OSMOREGULATION EN MILIEU MARIN

SELACIENS MARINS
Sélaciens sont isoosmotiques (légèrement hyperosmotique en fait) par rapport à l’eau de mer (1000 mosm/L)
Osmolarité élevé grâce à grande quantité d’urée dans le milieu intérieur
[Urée] 100 x + élevé que chez mammifère => urée est réabsorbée par le rein
Autre composé organique = TMAO
Urée effet néfaste => déstabilise les protéines. TMAO à effet inverse

SELACIENS
Forte osmolarité : pas de perte d’eau (même léger gain pour formation de l’urine)
Entrée de sels par les branchies (Na, Cl, SO4, Mg) et alimentation : élimination Na et Cl par glandes spéciales = glande rectale (=glande a sel)
Autres lieux d’éliminations des ions Cl- et Na+ => branchies via transport actif (cf téléostéens marins)

GLANDE RECTALE
Cotransport Na-K/2 Cl- au niveau basolatérale
Canal Cl- du coté apicale (ressemblance CFTR)
Sécrétion sous contrôle de l’AMPc : contrôle hormonal ?

TELEOSTEENS MARINS
Osmotiquement + dilué que l’eau => poisson perd constamment de l’eau
- beaucoup par les branchies
- un peu par l’urine
- pertes par tégument sont négligeables
- Diffusion de sels a travers les branchies

Pertes d’eau compensé par absorption d’eau de mer => absorption d’ions (Na et Cl) supplémentaires
élimination active de ces ions grâce à transport actif au niveau des branchies
Élimination de ces ions par les reins impossible (urine isoosmotique) mais élimination des ions sulfates et magnésium

FONCTIONNEMENT DES BRANCHIES
Complexe de cellule a chlorure formé d’une cellule à chlorure mère (ou cellule a chlorure différenciés) et d’une cellule à chlorure accessoire (cellule fille)

Eau douce
Eau de mer Grande densité de mitochondrie : fournissent ATP nécessaire au transport actif
Pompage par pompe NaK
Cl suit par canal chlorure

Fonctionnement cellule à chlorure
Jonction lâche permet le passage des ions Na+ selon gradient de [C]
Si pas de cellule accessoire => uniquement jonction serrée : pas d’élimination des ions Na+ car pompe Na/K localisé uniquement dans membrane baso-latérale
Si pompe Na/K sur pole basal => dépendant du K+ pour excrétion Na+ !
Jonction lâche

Reptiles et oiseaux marins
Pas d’entrée de sels par le tégument ou les surfaces respiratoires (poumons). Le risque de surcharge en sels est uniquement dû à l’ingestion d’eau de mer et d’aliments salés.
L’excès de sels ne peut pas être éliminé par les reins : urine isotonique (reptiles) ou faiblement hypertonique (oiseaux).
élimination du sel assurée par des glandes à sel

Les glandes à sel possèdent des cellules sécrétrices dont le fonctionnement est très semblable à celui des cellules à chlorure des branchies de poissons

animaux tendance à la perte d’eau par osmose
concentration du LEC/milieu
concentration de
l’urine/sang
Téléostéens marins + hypotonique isotonique
Sélaciens (requins,
raies) 0 isotonique isotonique
Reptiles marins
0 hypotonique isotonique
Oiseaux marins
0 hypotonique faiblement
hypertonique
Mammifères marins
0 hypotonique fortement
hypertonique
boit de l’eau de mer
sécrétion branchiale
ne boit pas
sécrétion G.rectale
boit de l’eau de mer larmes salées boit de l’eau de mer
sécrétions nasales salées
ne boit pas urine très hyperton.
Bilan

OSMOREGULATION EN EAU DOUCE

TELEOSTEENS D’EAU DOUCE
Osmotiquement + concentré que l’eau => poisson gagne constamment de l’eau par les branchies

gains d’eau compensé par urine abondante => pertes d’ions (Na et Cl)
Également pertes ions pas branchies
Transport actif de ces ions grâce à transport actif au niveau des branchies
Élimination de ces ions par les reins impossibles mais élimination des ions sulfates et magnésium

FONCTIONNEMENT CELLULE A CHLORURE
La pompe à protons augmente la ddp transmembranaire apicale, ce qui favorise l’influx de Na+ par diffusion. La pompe Na/K basale expulse ensuite Na+ vers le sang.

Bilan

Changement de milieu au cours de la vie : poissons migrateurs
alevin
tacon
smolt
saumon adulte
oeufs

Répartition des saumons en France Trajet des saumons vers leur zone d’engraissement

MIGRATION EAU DOUCE/EAU DE MER
Ex du Saumon de l’Atlantique (Salmo salar)
Le tacon (jeune saumon eau douce) subit une smoltification pour former le smolt (jeune saumon vivant dans l’estuaire : phase transitoire d’adaptation)
Smoltification s’accompagne de grands changements morphologiques, biochimiques et physiologiques
Modification au niveau branchiale => activité Na/K ATPasique augmente fortement
Espèce anadromique
Également espèce catadromique (anguille)

Adaptations de l’osmorégulation chez le saumon lors du passage
eau douce eau de mer
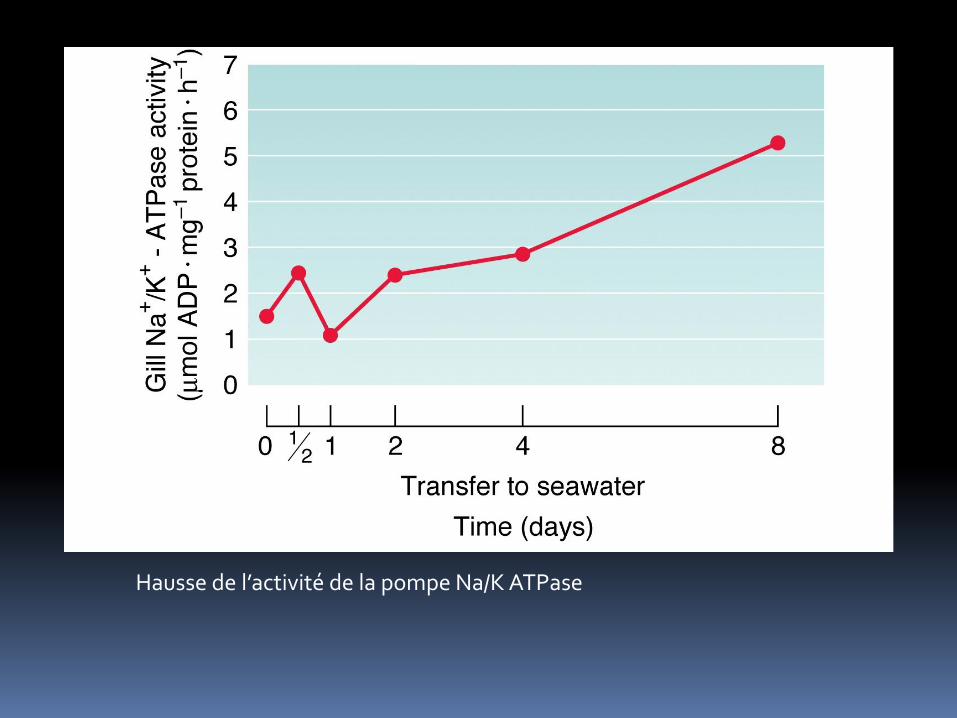
Hausse de l’activité de la pompe Na/K ATPase

1. l’augmentation de la natrémie stimule la
sécrétion de cortisol par les glandes
surrénales et d’hormone de croissance
par l’hypophyse.
2. Ces hormones induisent une du nombre
de cellules à chlorure dans les branchies
et la synthèse de pompes Na/K.
3. En conséquence, la sécrétion de NaCL du
sang vers l’eau de mer
progressivement et la natrémie
redevient normale (au bout d’une
semaine environ.
4. Parallèlement, la synthèse de pompes à
protons dans les cellules épithéliales,
d’où chute de l’absorption de Na+ par les
branchies (mécanisme mal connu).

1) Les jonctions intercellulaires
entre les cellules à chlorure et
les cellules épithéliales
deviennent plus imperméables,
d’où de l’efflux de Na+ par la
voie paracellulaire.
2) La prolactine sécrétée par
l’hypophyse provoque une
diminution du nombre de
cellules à chlorure, d’où du
rejet actif de NaCl dans l’eau.
3) Le nombre de pompes à
protons dans les cellules
épithéliales, d’où de
l’absorption active de Na+ par
les branchies.

OSMOREGULATION EN MILIEU AERIEN Principale problème : déshydratation
Différente adaptation physiologique, morphologique et comportementale permettent de lutter contre la perte d’eau en milieu aérien

EVAPORATION
Evaporation augmente avec la température
Evaporation définit par la pression de vapeur d’H2O (PH20)
Si l’air contient déjà de la vapeur d’eau, différence de PH2O entre surface du liquide et air est plus faible : évaporation plus réduite
Évaporation modifie température de la surface d’évaporation
Mouvement de l’air => renouvellement de l’air saturée en H2O => augmente évaporation


LES AMPHIBIENS
Perte d’eau importante par la peau => obligation de vivre près de l’eau, dans un habitat humide Dans l’eau, se comporte comme les téléostéens d’eau douce (urine abondante et diluée, absorption des ions au niveau de la peau) Adaptation au milieu désertique :
• Grenouille du désert Australien : estivation dans des terriers profonds durant la secheresse, ne sorte qu’en cas de pluie • Fabrique une urine très diluée et abondante (30% de la masse) • Certaines grenouilles secrètent une substance cireuse via des glandes cutanés qui imperméabilisent la peau • D’autres sécrètent acide urique plutôt qu’urée pour économie d’eau (cf cours excrétion azotée) • En cas de stress hydrique il peut y avoir accumulation d’urée (hausse osmolarité) •Résistance à la déshydratation : supporte perte H2O très importante

AMPHIBIENS Perte d’eau importante par la peau
=> obligation de vivre près de l’eau, dans un habitat humide
Dans l’eau, se comporte comme les téléostéens d’eau douce (urine abondante et diluée, absorption des ions au niveau de la peau)
Adaptation au milieu désertique : • Grenouille du désert Australien :
estivation dans des terriers profonds durant la sécheresse, ne sorte qu’en cas de pluie
• Fabrique une urine très diluée et abondante (30% de la masse)
• Certaines grenouilles secrètent une substance cireuse via des glandes cutanés qui imperméabilisent la peau
• D’autres sécrètent acide urique plutôt qu’urée pour économie d’eau (cf cours excrétion azotée)
• En cas de stress hydrique il peut y avoir accumulation d’urée (hausse osmolarité)
• Résistance à la déshydratation : supporte perte H2O très importante

INSECTES
Réduction des pertes d’eau cuticulaires. La cuticule des insectes est très imperméable à l’eau, non pas à cause de la chitine, mais de la cire qui la recouvre. Les pertes d’eau brusquement pour une t°C qui coïncide avec le point de fusion de la cire.

Réduction pertes d’eau
réduction des pertes d’eau dans les excréments et l’urine. Les insectes rejettent des excréments déshydratés grâce à la réabsorption d’eau par la paroi du rectum.
Fermeture des stigmates et ouverture quand CO2 trop élevé

REPTILES
Evaporation cutanée (limité par écailles) plus importante que évaporation respiratoire
Relation entre évaporation totale (cutanée et respiratoire) et le milieu de vie => imperméabilité de la peau varie
Économie d’eau : sécrétion d’acide urique (idem pour les oiseaux)

MAMMIFERES
Les mammifères terrestres peuvent
lutter contre la déshydratation en
produisant une urine plus concentrée
que le plasma ( avec vertébrés
inférieurs).
La réabsorption de l’eau par le canal collecteur fait intervenir 2 facteurs :
- l’ADH qui rend ce canal perméable à l’eau
- le gradient osmotique cortico-médullaire.

Les mammifères : mécanismes de la concentration de l’urine
La branche descendante de l’anse de
Henlé est perméable à l’eau et
imperméable aux solutés. C’est l’inverse
pour la branche ascendante.
Le transport actif de NaCl le long de la
branche ascendante l’osmolarité du
liquide interstitiel, ce qui permet la
réabsorption d’eau par osmose, le long
de la branche descendante.
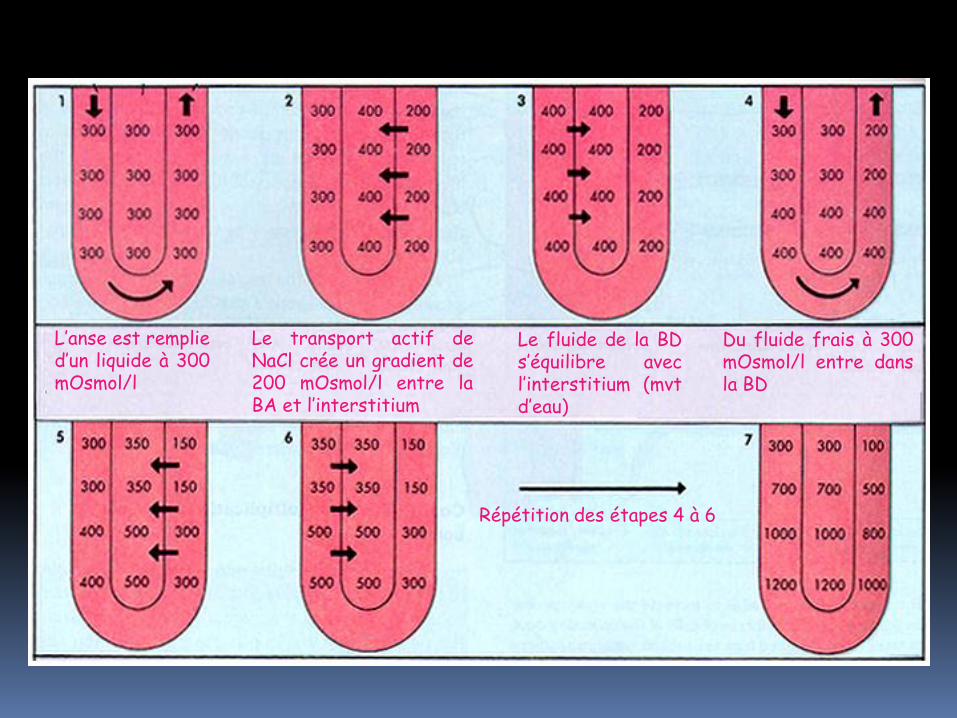
L’anse est remplie d’un liquide à 300 mOsmol/l
Le transport actif de NaCl crée un gradient de 200 mOsmol/l entre la BA et l’interstitium
Le fluide de la BD s’équilibre avec l’interstitium (mvt d’eau)
Du fluide frais à 300 mOsmol/l entre dans la BD
Répétition des étapes 4 à 6

l’osmolarité maximale de l’interstitium est de 1200 mOsmol/l (4 fois plus que celle du plasma. NaCl ne contribue à cette valeur que pour 50% environ ; les 50% restants étant dus à de l’urée.

La plus grande partie du néphron est
imperméable à l’urée (trait épais bleu
sur le schéma). Au fur et à mesure que
l’eau est réabsorbée le long du canal
collecteur, la concentration en urée .
La partie terminale du canal collecteur
(pointillés) est perméable à l’urée qui
diffuse alors vers l’interstitium. Cette
diffusion contribue à la très forte
osmolarité de la partie la plus profonde
de la médulla.

castor
lapin
rat des sables

Bilan osmorégulation

EXCRETION AZOTEE

Excrétion : élimination des déchets de la nutrition cellulaire et leur retour dans le milieu extérieur Étendue a élimination des substances en excès (eau, ions divers)
Organes impliqués très divers
Plusieurs rôles pour ces organes notamment régulation de l’équilibre hydrominéral et de l’osmolarité
Élimination du CO2 : appareil respiratoire

Dégradation des protéines produit Aa puis NH4+ hautement toxique : Perturbation du pH
intracellulaire Inhibition des mouvements
ioniques transmembranaires Au niveau mitochondrial :
abolition du gradient de H+
Dégradation des acides nucléiques produit acide urique ou allantoine qui peuvent être transformé en urée

Excrétion de l’azote sous différente forme :
Ammoniotélie => excrétion d’ammoniaque
Uréotélie => excrétion d’urée
Uricotélie => excrétion d’acide urique
DEPEND DE LA DISPONIBILITE EN EAU

Ammoniaque nécessite 500mL H20/g NH4+ = Élimination directe essentiellement par espèces aquatiques
Élimination indirecte : transformation en un composé moins toxique comme l’urée ou l’acide urique
Permet une économie d’eau (50mL H20/g d’urée et 10 mL/ d’acide urique)


Milieu de + en + sec : => passage ammoniotélie à uréotélie puis uricotélie

CHANGEMENT DE MILIEU
Passage d’une excrétion d’ammoniaque à une excrétion d’urée durant la métamorphose chez les amphibiens

CHANGEMENT D’ACTIVITE
Gastéropodes pulmonés terrestres :
- escargots actifs = urée
- Escargots inactifs = acide urique
=>Gastéropodes aquatiques en zone de marée :
- marée basse : acide urique et urée
- Marée haute : uricolyse => libération d’allantoïne et de NH3

LES APPAREILS EXCRETEURS ET LEURS FONCTIONNEMENTS

FILTER OU SECRETER ?
Excrétion doit éliminer certains composés et en conservés d’autres
Le plus souvent cette élimination se fait par Filtration, qui va former une urine primaire
Le plus souvent des mécanismes de Réabsorption sélective et de Sécrétion vont modifier cette urine primaire pour former l’urine définitive
Filtration à un cout important puisqu’il faut réabsorber activement toute les substance nécessaires (eau, nutriments, etc…)
Sécrétion active a l’extérieur des produits azotés moins coûteuse que filtration et Réabsorption
Avantage : grande adaptabilité à des régimes alimentaires et habitats divers
Toutes nouvelles substances va être filtré et éliminé si elle n’est pas réabsorbée (ex : médicaments)
Sécrétion nécessite système de transport pour chaque molécule à éliminé
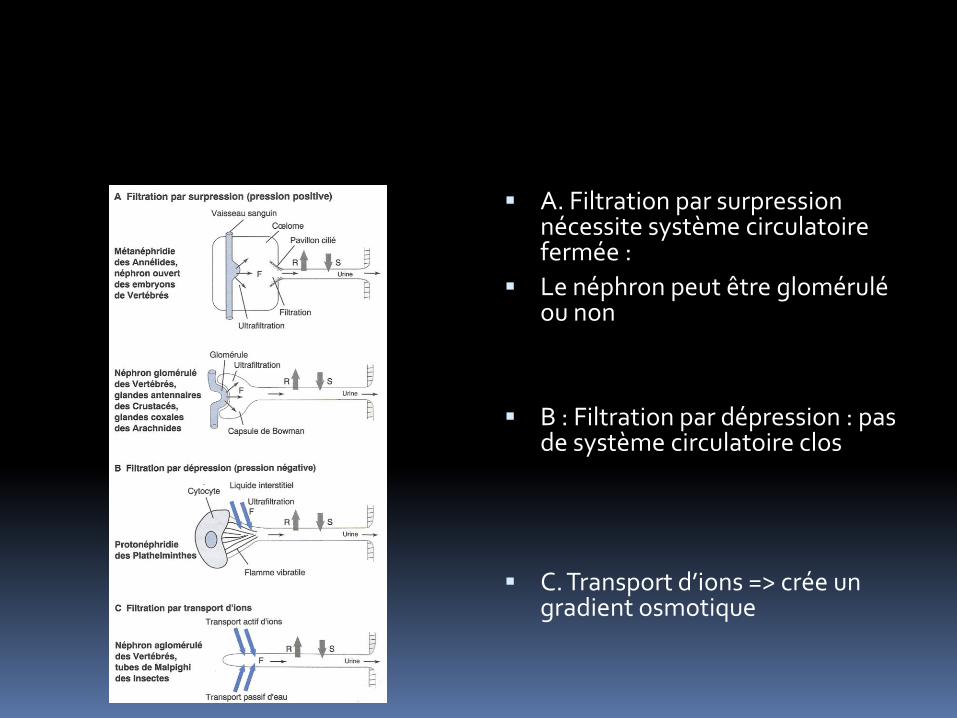
A. Filtration par surpression nécessite système circulatoire fermée :
Le néphron peut être glomérulé ou non
B : Filtration par dépression : pas de système circulatoire clos
C. Transport d’ions => crée un gradient osmotique

LE REIN

LE NEPHRON

Vaisseaux sanguins au niveau des tubules contournés permettent la réabsorption et la sécrétion => modification de l’urine primaire
VASCULARISATION DU NEPHRON

CAPSULE DE BOWMAN ET GLOMERULE DE MALPIGHI

T C P
podocyte
filtrat glomérulaire
feuillet externe de la capsule
artériole afférente
artériole efférente


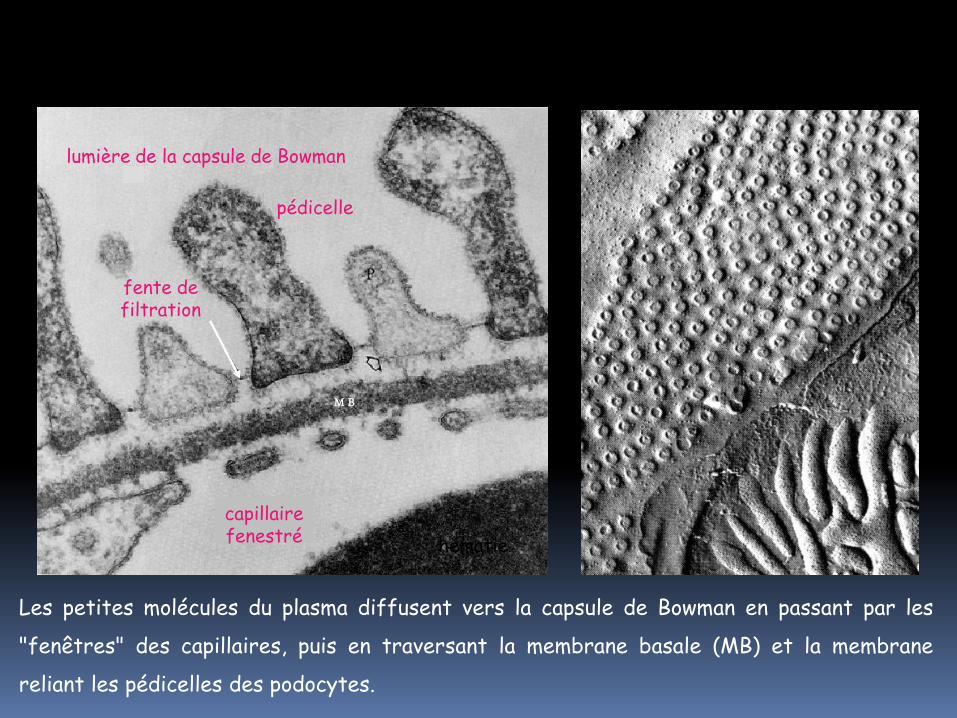
Les petites molécules du plasma diffusent vers la capsule de Bowman en passant par les
"fenêtres" des capillaires, puis en traversant la membrane basale (MB) et la membrane
reliant les pédicelles des podocytes.
lumière de la capsule de Bowman
capillaire fenestré
M B
hématie
fente de filtration
pédicelle

Capillaire fenestré : ne retient
que les cellules sanguines
Lame basale : retient les
grosses protéines
Membrane des fentes de filtration :
retient les petites protéines
fente de filtration
pédicelle

LA MEMBRANE DE FILTRATION
Membrane basale = glycoprotéine anionique => repoussent charge négative donc Protéine plasmatique Fenestration empêche passage des GR

Résistance dans les artérioles glomérulaires : -artériole afférente : protège glomérules des grandes fluctuation de la PA -Artériole efférente : augmente la P hydrostatique dans les glomérules
Baisse de 15% de la pression glomérulaire fait cesser la filtration !! Nécessite une régulation ! En modifiant résistance des artérioles afférentes, variation de la PA supporté de 80 à 180 mmHg

Débit de filtration glomérulaire est de 125 mL/min On utilise inuline : faible PM, ni sécrété ni absorbé Déterminer par la clearance rénal : vol de sang qui contient la quantité d’une substance rejetée en 1 min Clearance Na+ : 2mL/min
Trois forces sont impliquées
dans la filtration :
1 = la pression sanguine
glomérulaire, liée au pompage
cardiaque (55 mm Hg)
2 = la pression capsulaire (15
mm Hg)
3 = la pression oncotique du
plasma, liée aux protéines
plasmatiques (30 mm Hg)
Bilan : pression de filtration
= 55 – (30 + 15) = 10 mm Hg.


TRANSPORT DU NA+

Transport passif des ions négatifs (gradient éléctrique causé par Na+) Transport passif d’eau (cf infra) Transport passif par gradient de [C] Transport actif secondaire (SGLT par ex)
TRANSPORT D’AUTRES SUBSTANCES

TCP : réabsorption la plus importante Anse de Henlé : réabsorption d’eau dans la partie descendante et d’ions dans la partie ascendante TCP et TRC : réabsorption d’urée (et d’eau)

Na+ Glu Na+ H+
Na+ K+
ATP ADP
lumière du TCP
capillaire péritubulaire
H2O
Le transport actif du sodium entre la
lumière tubulaire et le milieu interstitiel
tend à créer un gradient osmotique entre
les deux faces de l’épithélium tubulaire.
L’eau est entraînée par osmose du liquide
tubulaire vers le liquide interstitiel et le
sang.
Les mouvements passifs de l’eau sont
donc provoqués par le transport actif de
sodium (" l’eau suit le sodium ").
Le néphron des mammifères: mécanismes de la réabsorption obligatoire

Rôle de l’ADH
Réabsorption facultative de
l’eau est conditionnée par 2
facteurs :
la sécrétion d’une hormone
(l’ADH) qui rend les cellules du
canal collecteur perméables à
l’eau.
l’existence d’une différence
de concentration entre la
lumière tubulaire et le milieu
interstitiel. canal
collecteur
réabs. H2O
volume
plasma
osmolarité
plasma ADH
neurones à ADH hypothalamiques
terminaisons dans
l’hypophyse
trains de PAs


ADH et aquaporines
Rôle de ADH sur adressage des aquaporines 2 à la surface des cellules des tubes collecteurs (capes 2006)

ADH

Osmolarité passe de 300 à 1200 mOsm/L grâce à Na+ et à urée
CONCENTRATION DE L’URINE


Rein produire Rénine : pas vraiment une hormone : plutôt enzyme Rénine produite par les cellules justaglomérulaires (cf infra) Rénine produite en cas de baisse de la PA (ce qui rend difficile la filtration rénal) Egalement innervation par le système nerveux autonomes => libéré en cas de stress
LA RENINE


REGULATION ENDOCRINE DE LA FONCTION RENAL
Fait intervenir plusieurs hormones qui agissent sur la réabsorption rénal d’eau ou de solutés : -ADH, Hormone antidiurétique (AVP) : réabsorption d’eau
-Système rénine/angiotensine/aldostérone : réabsorption de Na+
-ANF, auricular natriurectic Factor (ou Peptide) : Facteur natriurétique auriculaire, élimination de Na+

FACTEUR NATRIURETIQUE AURICULAIRE
Synthétisé au niveau des oreillettes, => mécanorécepteur : Distension auriculaire provoque synthèse ANF Provoque la sécrétion de Na+ pour favoriser la baisse de la Pression artérielle
Voie de signalisation = GMPc
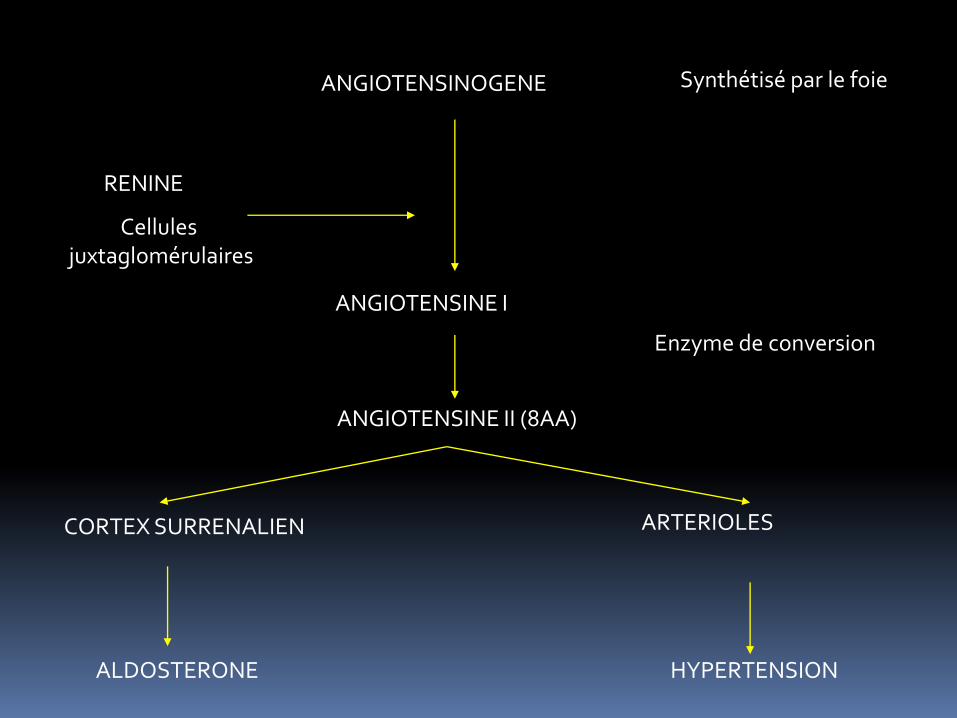
ANGIOTENSINOGENE
RENINE
ANGIOTENSINE I
ANGIOTENSINE II (8AA)
Enzyme de conversion
CORTEX SURRENALIEN ARTERIOLES
HYPERTENSION ALDOSTERONE
Synthétisé par le foie
Cellules juxtaglomérulaires

Aldostérone augmente la réabsorption du Na+ et augmente la sécrétion de K+ Réabsorption de Na+ par transport actif primaire via SGLT1 ou SGLT2

ADH augmente la réabsorption d’eau => Également sensation de soif






























