Embed Size (px)
Citation preview

[CANCER RESEARCH 58. 1538-1543. April 1. 19<)8|
Overexpressed WAFl/Cipl Renders Glioblastoma Cells Resistant to ChemotherapyAgents l,3-Bis(2-chloroethyl)-l-nitrosourea and Cisplatin1
Sanbao Rúan, M. Fatih Okcu, Jiang Ping Ren, Paul Chiao, Michael Andreeff, Victor Levin, and Wei Zhang2
Departments of Neuro-Oncology ¡S.R.. J. P. R.. V. L., W. Z.J. Hematology ¡M.F. O.. M. A.]. Pediatrics ¡M.F. O.I, Surgical Oncology ¡P.C.¡,and Tumor Biology ¡P.C.. W. Z./.The Universit\ of Texas M. D. Anderson Cancer Center. Houston, Texan 77030
ABSTRACT
Previous studies have shown that the negative cell cycle regulator
WAFl/Cipl is often overexpressed in human gliomas and that WAF1/Cipl overexpression may be a factor in cancer chemoresistance. Weestablished a doxycycline-inducible WAFl/Cipl expression system in two
glioblastoma cell lines and examined the role of WAFl/Cipl in theirresponse to the chemotherapy agents l,3-bis(2-chloroethyl)-l-nitrosourea(BCNU) and ri'.v-diamminedichloroplatinum (cisplatin), in an isogeneic
background. Our results showed that the induction of WAFl/Cipl expression rendered glioma cells resistant to cell death induced by BCNU andcisplatin. Using an in vivo host-cell reactivation DNA repair assay, wedemonstrated that WAFl/Cipl enhances the repair of BCNU-inducedDNA damage. We conclude that WAFl/Cipl allows repair of BCNU- andcisplatin-damaged DNA and protects glioma cells from chemotherapyagent-induced apoptosis. Thus, blocking WAFl/Cipl production or func
tion may serve as a useful chemosensitization regimen for glioma.
INTRODUCTION
Gliomas are the most common primary tumor in the central nervoussystem, and glioblastoma multiforme is the most advanced stage (1).Chemotherapy agents used to treat brain tumors include the alkylatingagent BCNU' (carmustine) and cisplatin (2. 3). Patient response to
chemotherapy, however, is rather poor, and median survival of glio-
blastoma patients is less than 1 year (4). The seriousness of this tumortype is evidenced by its refractory response to current therapy regimens and by the psychosocial and financial costs it inflicts on patients,their families, and society. It is important, therefore, to understand themechanisms for chemoresistance of gliomus and to develop strategiesto improve therapeutic response in glioblastoma patients.
Normal cell proliferation is regulated by essential proteins at checkpoints in the cell cycle. These proteins either stimulate or inhibit cellcycle progression (5, 6). The most studied of the positive regulatorsare CDKs that associate with cyclins and PCNA, forming a kinasecomplex (7).
WAF 1/Cip 1 (also known as p21, Sdi 1, Pic 1. and CAP20) is a targetof p53 transactivation (8) and an inhibitor of a variety of CDKs(9-11). Overexpression of WAFl/Cipl from a transfected expression
vector inhibits the growth of various tumor cells to different degreesin clonogeneic assays (8. 12, 13). The WAFl/Cipl protein alsointeracts with PCNA directly and inhibits DNA replication independent of cyclin-CDK activity (14). Interaction of WAFl/Cipl with
Received 1/6/98: accepted 1/23/98.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Seclion 1734 solely to indicate this fact.
1This work was partially supported by a grant from The University of Texas M. D.
Anderson Cancer Center Physicians Referral Service and by NIH Grants CA67987 andCA55I64.
: To whom requests for reprints should be addressed, at Department of Neuro-
Oncology. Box 316. The University of Texas M. D. Anderson Cancer Center, 1515Holcombe Boulevard. Houston. TX 77030. Phone: (713) 792-3778; Fax: (713) 745-1183;E-mail: l2507vKfulmdacc.mda.uth.tmc.edu.
'The abbreviations used are: BCNU. l.3-bis(2-chloroethyl)-l-nitrosourea; cisplatin,
r/.v-diamniinedichloroplaiinum; CDK. cyclin-dependent kinase; PCNA. proliferating cellnuclear antigen; AMI., acute myelogenous leukemia: AGAT, O^-alkylguanine-DNA
alkyltransferase; ITA. tctracycline (ranscriptional activator.
PCNA may also affect the DNA repair process, because PCNA is animportant factor in DNA repair (15).
To understand the role of WAFl/Cipl in human cancers, wepreviously analyzed the expression of WAFl/Cipl in samples oftumors from patients with AML and from patients with glioblastoma. Samples from 17% of the AML patients studied had elevatedWAFl/Cipl expression, and those patients were four times moreresistant to chemotherapy than patients with low levels of WAF I/Cipl (16). Similarly, WAFl/Cipl was found to be overexpressedin the majority of gliomas, whereas it was expressed at extremelylow levels in normal glial cells ( 17). We hypothesized that elevatedWAFl/Cipl is an important factor in the chemoresistance of gliomas and that WAFl/Cipl repairs DNA damage caused by chemotherapy agents. Several recent publications support this hypothesis. Using a homologous recombination technique, Waldman etal. (18) established a colon cancer cell line that has the WAFl/Ciplgene deleted. The WAF1/Cip I-defective cells were more sensitiveto apoptosis induced by the chemotherapy agent doxorubicin (Ad-
riamycin) than were parental cells (18). McDonald et al. (19)further demonstrated that these W/4/r//C//j/-defective cells were
less efficient in repairing DNA damaged by UV radiation orcisplatin than were parental cells. Along the same lines and incontrast to original speculation (20), WAFl/Cipl did not promotecell death (21) and actually protected certain cancer cells fromapoptosis (22, 23).
In this study, we established a stable doxycycline-inducible WAF I/Cipl expression system in glioblastoma cell lines LN-Z308 and U251.(Doxycycline is a derivative of tetracycline.) This doxycycline turn-on-inducible system (24) allows a target gene to express in cells when
doxycycline is added to the cell culture. Using this inducible isogeneicsystem, we showed that WAFl/Cipl-induced cells were significantlymore resistant than were parental cells to BCNU- and cisplatin-causedcell death. The WAFl/Cipl-induced cells also were better able torepair BCNU-generated DNA damage. Therefore, we identified a
novel mechanism of chemoresistance in gliomas and a new target fortherapeutic chemosensitization regimens.
MATERIALS AND METHODS
Cells and Reagents. U25I glioblastoma cells were purchased from theAmerican Type Culture Collection (Rockville, MD). LN-Z308 glioblastoma
cells were generously provided by Dr. Erwin Van Meir (University Hospital.
Lausanne, Switzerland). A mutant p53 is present in U251 cells (25), and nop53 is expressed in LN-Z308 cells (26). Transfeclion experiments were done
using the Superfest method (Qiagen, Inc., Chatsworth, CA). which routinelyachieves a 30-40% transfection efficiency.
BCNU, cisplatin. and doxorubicin (Adriamycin) were obtained from TheUniversity of Texas M. D. Anderson Cancer Center Pharmacy and weremanufactured by Bristol-Myers Squibb Co. (Princeton, NJ). Cell viability was
measured by trypan blue exclusion assay. Harvested cells were mixed with anequal volume of trypan blue dye. Dead cells absorbed the blue dye and
appeared as blue cells under light microscopy. To quantitate cell viability,250-300 cells were counted, and viability was calculated as the percentage of
live (bright) cells in the total cell population.
1538
on June 6, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
WAFI/Cipl CONFERS CHKMORKSISTANCE
A. U251 clone 1 clone2 clone3 clone4
B. LN-Z308Dox:
actin
clone 1 clone2 clone3 clone4 clones cloneo
WAFI/Cipl «»^«
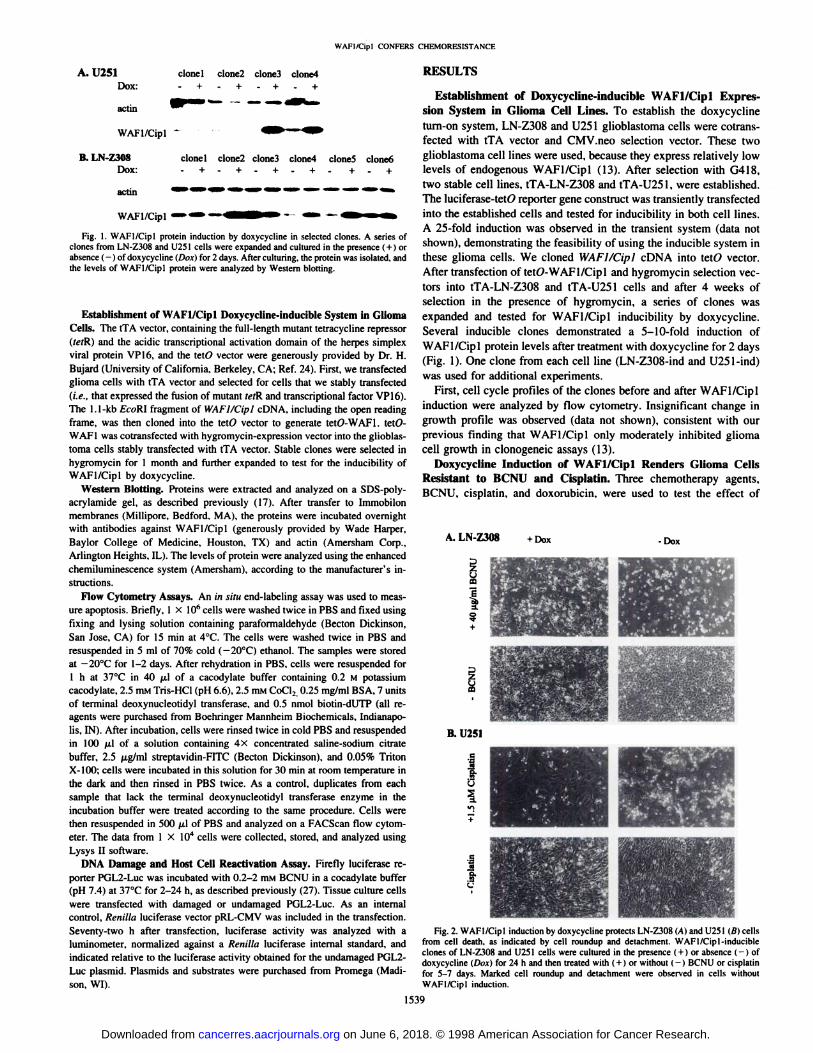
Fig. 1. WAFI/Cipl protein induction by doxycydine in selected clones. A series ofclones from LN-Z308 and U251 cells were expanded and cultured in the presence (+ ) orabsence (—¿�)of doxycydine (Dox) for 2 days. After culturing. the protein was isolated, andthe levels of WAFI/Cipl protein were analyzed by Western blotting.
Establishment of WAFl/Cipl Doxycycline-inducible System in GliomaCells. The tTA vector, containing the full-length mutant tetracycline repressor
(tetR) and the acidic transcriptiona! activation domain of the herpes simplexviral protein VP 16. and the tetO vector were generously provided by Dr. H.Bujard (University of California. Berkeley. CA: Ref. 24). First, we transfectedglioma cells with tTA vector and selected for cells that we stably transfected
(i.e., that expressed the fusion of mutant rerR and transcriptional factor VP 16).The 1.1-kb EcoRl fragment of WAFI/Cipl cDNA. including the open readingframe, was then cloned into the tetO vector to generate tetO-WAFl. tetO-WAF1 was cotransfected with hygromycin-expression vector into the glioblas-
toma cells stably transfected with tTA vector. Stable clones were selected inhygromycin for 1 month and further expanded to test for the inducibility ofWAFl/Cipl by doxycydine.
Western Blotting. Proteins were extracted and analyzed on a SDS-poly-
acrylamide gel, as described previously (17). After transfer to Immobilonmembranes (Millipore, Bedford. MA), the proteins were incubated overnightwith antibodies against WAFl/Cipl (generously provided by Wade Harper.Baylor College of Medicine. Houston, TX) and actin (Amersham Corp..Arlington Heights. IL). The levels of protein were analyzed using the enhancedchemiluminescence system (Amersham), according to the manufacturer's in
structions.Flow Cytometry Assays. An in situ end-labeling assay was used to meas
ure apopto.sis. Briefly, 1 X IO6cells were washed twice in PBS and fixed using
fixing and lysing solution containing paraformaldehyde (Becton Dickinson.San Jose, CA) for 15 min at 4°C.The cells were washed twice in PBS andresuspended in 5 ml of 70% cold (—20°C)ethanol. The samples were storedat —¿�20°Cfor 1-2 days. After rehydration in PBS. cells were resuspended forl h at 37°C in 40 /xl of a cacodylate buffer containing 0.2 M potassium
cacodylate, 2.5 mM Tris-HCl (pH 6.6), 2.5 mM CoCU 0.25 mg/ml BSA. 7 unitsof terminal deoxynucleotidyl transferase, and 0.5 nmol biotin-dUTP (all re
agents were purchased from Boehringer Mannheim Biochemicals, Indianapolis, IN). After incubation, cells were rinsed twice in cold PBS and resuspendedin 100 ¡uof a solution containing 4X concentrated saline-sodium citratebuffer. 2.5 fig/ml streptavidin-FITC (Becton Dickinson), and 0.05% TritonX-IOO: cells were incubated in this solution for 30 min at room temperature in
the dark and then rinsed in PBS twice. As a control, duplicates from eachsample that lack the terminal deoxynucleotidyl transferase enzyme in theincubation buffer were treated according to the same procedure. Cells werethen resuspended in 500 ¡úof PBS and analyzed on a FACScan flow cytom-eter. The data from 1 X IO4 cells were collected, stored, and analyzed using
Lysys II software.DNA Damage and Host Cell Reactivation Assay. Firefly luciferase re
porter PGL2-Luc was incubated with 0.2-2 mM BCNU in a cocadylate buffer(pH 7.4) at 37°Cfor 2-24 h, as described previously (27). Tissue culture cells
were transfected with damaged or undamaged PGL2-Luc. As an internalcontrol, Renilla luciferase vector pRL-CMV was included in the transfection.
Seventy-two h after transfection, luciferase activity was analyzed with a
luminometer. normalized against a Renilla luciferase internal standard, andindicated relative to the luciferase activity obtained for the undamaged PGL2-
Luc plasmid. Plasmids and substrates were purchased from Promega (Madison, WI).
RESULTS
Establishment of Doxycycline-inducible WAFl/Cipl Expression System in Glioma Cell Lines. To establish the doxycydineturn-on system, LN-Z308 and U251 glioblastoma cells were cotrans
fected with tTA vector and CMV.neo selection vector. These twoglioblastoma cell lines were used, because they express relatively lowlevels of endogenous WAFl/Cipl (13). After selection with G418,two stable cell lines. tTA-LN-Z308 and ITA-U251. were established.The luciferase-tetO reporter gene construct was transiently transfected
into the established cells and tested for inducibility in both cell lines.A 25-fold induction was observed in the transient system (data not
shown), demonstrating the feasibility of using the inducible system inthese glioma cells. We cloned WAFI/Cipl cDNA into tetO vector.After transfection of tetO-WAFl/Cipl and hygromycin selection vectors into ITA-LN-Z308 and ITA-U251 cells and after 4 weeks of
selection in the presence of hygromycin. a series of clones wasexpanded and tested for WAFl/Cipl inducibility by doxycydine.Several inducible clones demonstrated a 5-10-fold induction of
WAFl/Cipl protein levels after treatment with doxycydine for 2 days(Fig. 1). One clone from each cell line (LN-Z308-ind and U251-ind)
was used for additional experiments.First, cell cycle profiles of the clones before and after WAFl/Cipl
induction were analyzed by flow cytometry. Insignificant change ingrowth profile was observed (data not shown), consistent with ourprevious finding that WAFl/Cipl only moderately inhibited gliomacell growth in clonogeneic assays (13).
Doxycydine Induction of WAFl/Cipl Renders Glioma CellsResistant to BCNU and Cisplatin. Three chemotherapy agents.BCNU. cisplatin. and doxorubicin. were used to test the effect of
A. LN-Z308 +DOX -Dox
B. U251
U5
Fig. 2. WAFl/Cipl induction by doxycydine protects LN-/30H (Ai and 11251(«Icellsfrom cell death, as indicated by cell roundup and detachment. WAFl/Cipl-inducibleclones of LN-Z308 and U251 cells were cultured in the presence (+ ) or absence (-) ofdoxycydine (Di«)for 24 h and ihen treated with (+ 1or without (-) BCNU or cisplatinfor 5-7 days. Marked cell roundup and detachment were observed in cells withoutWAFl/Cipl induction.
1539
on June 6, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
WAFI/Cipl CONFERS CHEMORESISTANCE
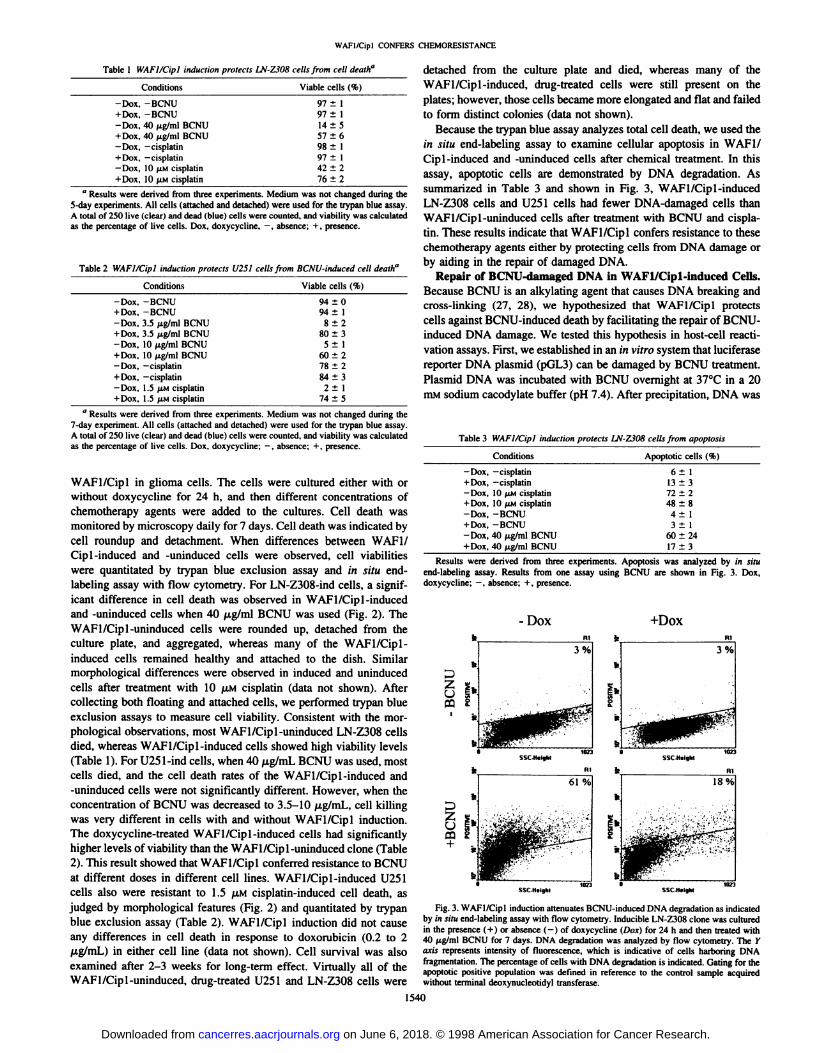
Table 1 WAFl/dpl induction protect!. LN-ZJOS cells from cell death"
-Dox,+DOX,-Dox,+DOX,-Dox,+DOX,-Dox,+DOX,Conditions-BCNU-BCNU40
/Ag/mlBCNU40/Ag/mlBCNU-cisplatin—
cisplatin10
/AMcisplatin10fiM cisplatinViable
cells(%)97971457989742115611276
2" Results were derived from three experiments. Medium was not changed during the
5-day experiments. All cells (attached and detached) were used for the trypan blue assay.A total of 250 live (clear) and dead (blue) cells were counted, and viability was calculatedas the percentage of live cells. Dox, doxycycline, -, absence; +, presence.
Table 2 WAFI/Cipl induction protects U25I cells from BCNU-induced ceil death"
-Dox,+DOX,-Dox,+DOX.-Dox,+DOX.-Dox,+DOX,-Dox,+DOX,Conditions-BCNU-BCNU3.5
/Ag/mlBCNU3.5/Ag/mlBCNU10
/Ag/mlBCNU10/Ag/mlBCNU—
cisplatin-cisplatin1.5
/AMcisplatin1.5 /AMcisplatinViable
cells(%)94
±094±18±280
±35±1M±278±284±32±
174±5
" Results were derived from three experiments. Medium was not changed during the7-day experiment. All cells (attached and detached) were used for the trypan blue assay.A total of 250 live (clear) and dead (blue) cells were counted, and viability was calculatedas the percentage of live cells. Dox. doxycycline; —¿�,absence; + , presence.
WAFI/Cipl in glioma cells. The cells were cultured either with orwithout doxycycline for 24 h, and then different concentrations ofchemotherapy agents were added to the cultures. Cell death wasmonitored by microscopy daily for 7 days. Cell death was indicated bycell roundup and detachment. When differences between WAF1/Cip 1-induced and -uninduced cells were observed, cell viabilitieswere quantitated by trypan blue exclusion assay and in situ end-labeling assay with flow cytometry. For LN-Z308-ind cells, a significant difference in cell death was observed in WAFI/Cipl-inducedand -uninduced cells when 40 pig/ml BCNU was used (Fig. 2). TheWAFl/Cipl-uninduced cells were rounded up, detached from theculture plate, and aggregated, whereas many of the WAFl/Cipl-
induced cells remained healthy and attached to the dish. Similarmorphological differences were observed in induced and uninducedcells after treatment with 10 ¡JLMcisplatin (data not shown). Aftercollecting both floating and attached cells, we performed trypan blueexclusion assays to measure cell viability. Consistent with the morphological observations, most WAFl/Cipl-uninduced LN-Z308 cellsdied, whereas WAFI/Cipl-induced cells showed high viability levels(Table 1). For U251-ind cells, when 40 /xg/mL BCNU was used, mostcells died, and the cell death rates of the WAFl/Cipl-induced and-uninduced cells were not significantly different. However, when theconcentration of BCNU was decreased to 3.5-10 /j,g/mL, cell killing
was very different in cells with and without WAFI/Cipl induction.The doxycycline-treated WAFl/Cipl-induced cells had significantlyhigher levels of viability than the WAFl/Cipl-uninduced clone (Table
2). This result showed that WAFI/Cipl conferred resistance to BCNUat different doses in different cell lines. WAFl/Cipl-induced U251cells also were resistant to 1.5 fiM cisplatin-induced cell death, as
judged by morphological features (Fig. 2) and quantitated by trypanblue exclusion assay (Table 2). WAFI/Cipl induction did not causeany differences in cell death in response to doxorubicin (0.2 to 2fxg/mL) in either cell line (data not shown). Cell survival was alsoexamined after 2-3 weeks for long-term effect. Virtually all of theWAFl/Cipl-uninduced, drug-treated U251 and LN-Z308 cells were
detached from the culture plate and died, whereas many of theWAFl/Cipl-induced, drug-treated cells were still present on the
plates; however, those cells became more elongated and flat and failedto form distinct colonies (data not shown).
Because the trypan blue assay analyzes total cell death, we used thein situ end-labeling assay to examine cellular apoptosis in WAF1/Cip 1-induced and -uninduced cells after chemical treatment. In this
assay, apoptotic cells are demonstrated by DNA degradation. Assummarized in Table 3 and shown in Fig. 3, WAFl/Cipl-inducedLN-Z308 cells and U251 cells had fewer DNA-damaged cells thanWAFl/Cipl-uninduced cells after treatment with BCNU and cispla
tin. These results indicate that WAFI/Cipl confers resistance to thesechemotherapy agents either by protecting cells from DNA damage orby aiding in the repair of damaged DNA.
Repair of BCNU-damaged DNA in WAFl/Cipl-induced Cells.
Because BCNU is an alkylating agent that causes DNA breaking andcross-linking (27, 28), we hypothesized that WAFI/Cipl protectscells against BCNU-induced death by facilitating the repair of BCNU-induced DNA damage. We tested this hypothesis in host-cell reacti
vation assays. First, we established in an in vitro system that luciferasereponer DNA plasmid (pGL3) can be damaged by BCNU treatment.Plasmid DNA was incubated with BCNU overnight at 37°Cin a 20
mM sodium cacodylate buffer (pH 7.4). After precipitation, DNA was
Table 3 WAFI/Cipl induction protects LN-2308 cells from apoptosis
Conditions-Dox,-cisplatin+
DOX,—¿�cisplatin—Dox. 10 /AMcisplatin+DOX, 10 /AMcisplatin-Dox,
-BCNU+DOX.-BCNU-Dox,
40/Ag/mlBCNU+
DOX,40 /Ag/ml BCNUApoptotic
cells(%)6
±113±372
±248±84±13
±160±2417
±3
Results were derived from three experiments. Apoptosis was analyzed by in situend-labeling assay. Results from one assay using BCNU are shown in Fig. 3. Dox,doxycycline; —¿�,absence; + , presence.
-Dox +DOX
§1*.CQ £
3%
18%
SSCHilghl
Fig. 3. WAFI/Cipl induction attenuates BCNU-induced DNA degradation as indicatedby in situ end-labeling assay with flow cytometry. Inducible LN-Z308 clone was culturedin the presence (+ ) or absence (-) of doxycycline (Dox) for 24 h and then treated with40 /Ag/mlBCNU for 7 days. DNA degradation was analyzed by flow cytometry. The Yaxis represents intensity of fluorescence, which is indicative of cells harboring DNAfragmentation. The percentage of cells with DNA degradation is indicated. Gating for theapoptolic positive population was defined in reference to the control sample acquiredwithout terminal deoxynucleotidyl transferase.
1540
on June 6, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

WAFl/Cipl CONFERS CHEMORESISTANCE
BCNU (mM) 0 0.2 0.5 0.75 1.0 2.0
Fig. 4. DNA damage by BCNU. Two ¿igof luciferase reporter plasmid DNA wereincubated with BCNU at the concentrations indicated for 24 h at 37°Cin 20 mM sodium
cacodylate buffer (pH 7.4). The DNA was then precipitated with ethanol and run on a l%agarose gel.
analyzed on an agarose gel. As seen in Fig. 4, BCNU treatment causeda dose-dependent conversion of normal supercoiled DNA (form I) to
nicked forms of DNA (forms II and III). A concentration of 2 mMBCNU caused such severe DNA damage that supercoiled DNA (formI) disappeared. DNA damage induced by cisplatin was also examinedusing this in vitro assay, but we failed to detect DNA damage as seenwith BCNU treatment.
The luciferase reporter plasmids, which were treated with 0.2, 0.75,and 2 mM BCNU, were transfected into the WAFl/Cipl-inducible
U251 clone after the cells were incubated either with or withoutdoxycycline for 24 h. The undamaged Renilla luciferase plasmid wascotransfected as an internal control for transfection efficiency. Thetransfected cells were incubated for an additional 48 h. The firefly andRenilla luciferase activities were then determined on two substratesusing a luminometer, and the ratios of firefly luciferase (from theBCNU-treated plasmid):/?«!///«luciferase (from the control, untreated
plasmid) were calculated (Table 4). It is evident that luciferase activityderived from the BCNU-treated plasmid decreased as the BCNU
concentration used to treat the plasmids increased, which was consistent with the extent of DNA damage (Fig. 4). The firefly:/?«!///«luciferase ratios of transfected WAFl/Cipl-induced and -uninduced
cells were compared to determine whether WAFl/Cipl contributes tothe repair of damaged firefly luciferase reporter plasmid. DNA damage from 2 mM BCNU was so severe that no DNA repair in WAF1/Cip 1-induced cells could be observed (ratios from WAFl/Cipl-in
duced cells are equivalent to ratios from control cells). In contrast,2-fold DNA repair was detected in WAFl/Cipl-induced U251 cellswhen 0.2 or 0.75 mM BCNU-treated firefly luciferase plasmids were
transfected (i.e., twice as much firefly luciferase activity was detectedwhen damaged DNA was transfected into WAFl/Cipl-induced cells
as in control cells).
DISCUSSION
Primary anaplastic glial neoplasms are notoriously refractory totherapy. Although surgery, radiotherapy, and chemotherapy can prolong survival, these treatments usually have a poor therapeutic indexand high toxicity. Multiple mechanisms may account for the refractory nature of gliomas. In the case of resistance to alkylating agents,the nitrosourea can first form an alkyl substitution at the O1' positionof guanine and then cross-link DNA. AGAT can repair O6-alkylgua-
nine adducts and thereby confer resistance to tumor cells treated with
alkylating agents (29, 30).Here, we demonstrated a novel mechanism of drug resistance in
gliomas. The central player of this mechanism is WAFl/Cipl, thepreviously known functions of which include inhibition of CDKs andDNA replication. We demonstrated that WAFl/Cipl can also repairDNA damage and confer to glioma cells chemoresistance to thealkylating agents BCNU and cisplatin.
This study stemmed from two previous studies. Our clinical correlation study of WAFl/Cipl in human AML first revealed that WAF1/Cipl positively contributes to the chemoresistance of these cancercells (16). We then showed that WAFl/Cipl overexpression constitutes a phenotype of human gliomas (17), which are notorious for theirresistance to chemotherapy. Accumulating evidence supports thisthesis. In a recent study. WAFl/Cipl was deleted from a colon cancercell line and WAFl/Cipl -negative cells were found to be moresensitive to doxorubicin-induced apoptosis (18). Along the same lines,
WAFl/Cipl was found to protect cells from apoptosis during celldifferentiation (22, 23, 31, 32).
In this study, we formally tested our hypothesis that WAFl/Ciplcontributes to chemoresistance in glioma cells. To obtain a definitive answer, we established a doxycycline-inducible WAFl/Cipl
expression system to test our hypothesis in an isogeneic background. Using inducible clones of two glioma cell lines, we foundthat WAFl/Cipl-induced cells are more resistant to BCNU- andcisplatin-induced cell death than are WAFl/Cipl-uninduced cells.Our host-cell reactivation assays showed that WAFl/Cipl confers
this resistance at least in part by enhancing DNA repair ability.Therefore, these results provided direct evidence that WAFl/Ciplcontributes to glioma chemoresistance, which explains our initialobservation of elevated WAFl/Cipl expression in tumor cells frompatients with primary gliomas. The AGAT is known to provide amechanism for gliomas to resist BCNU (29, 30). It is possible thatWAFl/Cipl affects AGAT expression and by this means conferschemoresistance. It would be interesting to investigate AGATactivity after WAFl/Cipl induction in glioma cells. However,alteration of AGAT expression may not be the mechanism ofWAFl/Cipl-conferred chemoresistance. because WAFl/Cipl also
renders cells resistant to cisplatin. whereas AGAT does not (33).Nevertheless, considering the finding that WAFl/Cipl is involvedin nuclear factor «B-dependent gene expression through p300 (34),
it is conceivable that WAFl/Cipl is involved in the expression ofchemoresistance genes. Because the p53 gene is defective in bothU251 and LN-Z308 cells, the p53 is not involved in this process.
Our study showed that the effect of WAFl/Cipl on the response ofcells to chemotherapy agents is not universal. WAFl/Cipl inductiondid not affect the response of the cells to doxorubicin. Failure ofWAFl/Cipl to affect doxorubicin-induced cell death was also re
ported in a recent study with a colorectal carcinoma cell line (35). Incontrast. WAFl/Cipl was observed to protect a different colon cancercell line from doxorubicin (18). This difference may reflect thatdoxorubicin works on different pathways in different types of cancercells and that WAFl/Cipl does not cause a general resistance tocell-killing agents. Although doxorubicin is not used for glioma
treatment, investigation of the effect of this agent on gliomas mayprovide additional clues to the mechanism of WAF1/Cipl-associated
chemoresistance.
Table 4 Repair of BCNU-damaged DNA in WAFI/dp I-induced U251 cells"
Ratio of Firefly luciferase from BCNU-treated plasmid:Renilla luciferase from undamaged control plasmid
0.2mM BCNU 0.75mM BCNU 2.0HIMBCNU
Doxycycline treated, 50 ±10 25 ±IWAFl/Cipl induced
Control cells 30 ±1 13 ±I
4 ±1
4± I13Results were derived from three experiments. The numbers shown are the ratios of
firefly luciferase activity (from the BCNU-damaged plasmid): Renilla luciferase activity(from Ihe control undamaged plasmid). Higher number indicates the presence of morefunctional firefly luciferase plasmid in cells. Two-fold higher values were seen inWAFl/Cipl-induced cells than in cells without WAFl/Cipl induction when 0.2 and 0.75mM BCNU-treated plasmids were transfected.
1541
on June 6, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

WAFI/Cipl CONFERS CHEMORESISTANCE
One mechanism that our results can exclude is a cell cycle-dependent event, for the following reasons: (a) BCNU is a cycle-indepen
dent cytotoxic agent, yet WAFI/Cipl induction had a profound protective effect; (b) WAFI/Cipl-induction did not provide protectionagainst doxorubicin-induced cell killing, which occurs predominantly
in replicating cells; and (r) WAFI/Cipl induction had little effect oncell cycle profiles of the glioma clones during the course of ourexperiments.
Our DNA repair studies demonstrated that WAFI/Cipl provideschemoresistance at least partially through playing an active role inDNA repair. This conclusion is supported by a finding by McDonald el ai. (19), who found that WAFI/Cipl-deleted colon
cancer cells had less DNA repair ability. These investigatorsfurther showed that the role of WAFI/Cipl in DNA repair isdependent on its interaction with PCNA: deletion of the PCNA-
interaction domain from WAFI/Cipl abolished its contribution toDNA repair. Thus, finding agents that disrupt the interaction ofWAFI/Cipl and PCNA as a means of sensitizing glioma cells tochemotherapy agents may be an attractive strategy. Another, perhaps more direct, strategy is to down-regulate WAFI/Cipl expres
sion by delivery of antisense WAFI/Cipl, which is currently understudy in our laboratory.
We should point out that our findings and those of McDonald et al.,which were derived from in vivo experiments, appear to be contrary toconclusions concerning the role of WAFI/Cipl in DNA replicationreached by other groups. Using in vitro purified WAFI/Cipl proteinin a reconstitution assay. Li et ai. (36) and Shivji et ai. (37) reportedthat WAFI/Cipl did not inhibit DNA excision repair. Conversely, ina similar in vitro study by Pan et al. (38). WAFI/Cipl was found toinhibit DNA excision repair. One reason that different results wereobtained from those in vitro experiments is that added recombinantWAFI/Cipl may be unstable to form specific complexes. In ourrecent preliminary experiments, we isolated nuclear proteins fromWAFI/Cipl-induced and -uninduced cells and performed in vitro
DNA repair assay. We failed to detect a marked difference in DNArepair (data not shown), consistent with the notion that the putativeWAFI/Cipl DNA repair complexes are not stable. Nevertheless,results from in vivo assays appear to support a positive role forWAFI/Cipl in DNA repair.
It would be interesting to identify two groups of glioma patientswho express low and high levels of WAFI/Cipl and investigatewhether the low-WAFl/Cipl expressors are relatively more respon
sive to therapy. Initial results suggest that this pattern may hold forpatients with anaplastic astrocytomas. In this subgroup, the patientswhose cancer cells had p53 mutations had low levels of WAFI/Ciplexpression (17). These patients were more responsive to chemotherapy regimens, which often included BCNU and cisplatin (39). However, the presence of p53 mutations may also play a part in theresponse of astrocytomas to chemotherapy-induced apoptosis. This
hypothesis was begotten in breast cancer studies (40) and supportedby our early findings that />5.?-defective gliomas were more sensitiveto pentoxifyMine-enhanced radiation-induced apoptosis (41). How
ever, the low expression of WAFI/Cipl may be the mediator for thiseffect of mutant p53. From a therapeutic point of view, WAFI/Cipl,which is located downstream from p53, may be a better target forintervention.
ACKNOWLEDGMENTS
We thank Kathryn Carnes and Gerne Zhang for their editorial assistancewith the manuscript.
REFERENCES1. Ringertz, N. "Grading" of gliomas. Acia Palhol. Microbio!. Scand., 27: 51-64, 1950.
2. Friedman, H. S., Colvin, O. M., Skapek, S. X.. Ludeman, S. M., Elion, G. B., Schold.S. C., Jacobsen, P. F.. Muhlbaier, L. H., and Bigner. D. D. Expérimentalchemotherapy of human medulloblastoma cell lines and transplantable xenografts with bifunc-tional alkylating agents. Cancer Res., 48: 4189-4195, 1988.
3. Steward, D. J. The role of chemotherapy in the treatment of gliomas in adults. CancerTreat. Rev., 16: 129-160, 1989.
4. Conrad, C. A., Milosavlijevic. V. P., and Yung, W. K. A. Advances in chemotherapyfor adult primary brain tumors. Neurosurg. Clin. North Am., 13: 795-812, 1995.
5. Sherr. C. J. Mammalian Gl cyclins. Cell, 73: 1059-1065, 1993.
6. Hunter. T., and Pines. J. Cyclins and cancer II: cyclin D and CDK inhibitors come ofage. Cell, 79: 573-582, 1994.
7. Zhang. H.. Xiong. Y., and Beach, D. Proliferating cell nuclear antigen and p21 arecomponents of multiple cell cycle kinase complexes. Mol. Cell. Biol., 4: 897-906,
1993.8. El-Deiry, W. S., Tokino, T.. Velculescu, V. E., Levy, D. B., Parsons. R., Trent. J. M..
Lin, D., Mercer, W. E., Kinzler, K. W., and Vogelstein, B. WAF1. a potentialmediator of p53 tumor suppression. Cell, 75: 817-825, 1993.
9. Harper. J. W., Adami, G. R., Wei, N., Keuomarsi, K., and Elledge. S. J. TheWAFI/Cipl Cdk-interacting protein Cipl is a potent inhibitor of G l cyclin-dependentkinases. Cell, 75: 805-816. 1993.
10. Noda. A., Ning. Y., Venable, S. F., Pereira-Smith, O. M., and Smith, J. R. Cloning ofsenescent cell-derived inhibitors of DNA synthesis using an expression screen. Exp.Cell Res.. 211: 90-98, 1994.
11. Xiong, Y., Hannon, G. J.. Zhang. H., Casso, D., Kobayashi, R., and Beach, D. p21 isa universal inhibitor of cyclin kinases. Nature (Lond.), 366: 701-704, 1993.
12. Zhang, W. Grasso, L., McClain, C. D., Gambel, A. M., Cha, Y., Travali, S.,Deisseroth, A. B., and Mercer, W. E. p53-independent induction of WAFI/Cipl inhuman leukemia cells is correlated with growth arrest accompanying monocyte/macrophage differentiation. Cancer Res., 55; 668-674, 1995.
13. Jung, J. M., Li, H., Kobayashi. T., KyriÃsis,A., Bruner, J., Langford, L., Levin, V.,and Zhang, W. The inhibition of human glioblastoma cell growth by WAFl/Cipl canbe attenuated by mutant p53. Cell Growth Diff., 6: 909-913, 1995.
14. Waga, S., Hannon, G. J., Beach, D., and Stillman, B. The WAFl/Cipl inhibitor ofcyclin-dependent kinases controls DNA replication by interaction with PCNA. Nature(Lond.), 369: 574-577, 1994.
15. Umar, A., Buermeyer. A. B., Simon, J. A.. Thomas, D. C., Clark, A. B., Liskay,R. M., and Kunkel, T. A. Requirement for PCNA in DNA mismatch repair at a steppreceding DNA resynthesis. Cell, 87: 65-73, 1996.
16. Zhang, W.. Kornblau, S. M.. Kobayashi, T., Gambel, A., McClain, C. D., Claxton, D.,and Deisseroth, A. B. High levels of constitutive WAFl/Cipl protein are associatedwith chemoresistance in acute myelogenous leukemia. Clin. Cancer Res.. /: 1051-
1057. 1995.17. Jung. J. M., Bruner. J. M., Rúan.S.-b., Langford. L., Kyritsis, A. P.. Kobayashi, T.,
Levin, V. A., and Zhang. W. Increased levels of p21WAFI/c'pl in human brain tumors.
Oncogene, //: 2021-2028, 1995.
18. Waldman. T.. Lengauer. C.. Kinzler, K. W., and Vogel.stein, B. Uncoupling of Sphase and mitosis induced by anticancer agents in cells lacking WAFl/Cipl. Nature(Lond.). 381: 713-716, 1996.
19. McDonald. E. R.. Wu, G. S.. Waldman. T.. and El-Deiry, W. S. Repair defect in p21WAF1/CIP1 -/- human cancer cells. Cancer Res., 56: 2250-2255, 1996.
20. El-Deiry, W. S.. Harper, J. W., O'Connor, P. M., Velculescu, U. E., Cannman, S. E.,
Jackman, J., Pietenpal, J. A., Burrett, M., Hill, D. E., Worry, Y., Winman, K. G.,Mercer, W. E., Kastan, M. B., Kohn, K. W., Elledge. S. J., Krizler, K. W., andVogel.stein, B. WAF-1/CIP1 is induced in p53-mediated G, arrest and apoptosis.Cancer Res.. 54: 1169-1174. 1994.
21. Kobayashi, T., Consoli. U., Andreeff, M., Shiku, H., Deisseroth, A. B., and Zhang, W.Activation of WAFl/Cipl by a temperature sensitive mutant of human p53 does notlead to apoptosis. Oncogene, II: 2311-2316, 1995.
22. Gorospe, M., Cirielli, C., Wang. X., Seth. P., Capogrossi, M. C., and Holbrook, N. J.p21 (Waf 1/Cipl ) protects against p53-mediated apoptosis of human melanoma cells.Oncogene, 14: 929-935, 1997.
23. Gorospe, M., Wang. X., Guyton, K. Z., and Holbrook, N. J. Protective role ofp2l(Wafl/Cipl ) against prostaglandin A2-mediated apoptosis of human colorectalcarcinoma cells. Mol. Cell. Biol., 16: 6654-6660, 1996.
24. Gossen, M., Freundlieb, S., Bender, G., Muller, G., Jillen, W., and Bujard, H.Transcriptional activation by tetracyclines in mammalian cells. Science (WashingtonDC), 268: 1766-1769, 1995.
25. von Deimling, A., Eibi, R. H.. Ohgaki, H., Louis, D. N., von Ammon, K., Peterson.I.. Kleihues, P., Chung. R. Y.. Wiestler, O. D., and Seizinger, B. R. p53 mutations areassociated with 17p allelic loss in grade II and grade III astrocytoma. Cancer Res., 52:2987-2990, 1992.
26. Van Meir, E. C., Kikuchi, U.. Tada, M., Li, H., Diserens, A-C., Wojcik, B. E.,Huang, H-J. S.. Friedmann, T., de Tribolet. N., and Cavenee, W. K. Analysis of
the p53 gene and its expression in human glioblastoma cells. Cancer Res., 54:649-652, 1994.
27. Srivenugopal, K. S., and Ali-Osman F. Stimulation and inhibition of l,3-bis(2-chloroethyl)-l-nitrosourea-induced strand breaks and interstrand cross-linking in Col
El plasmid deoxyribonucleic acid by polyamines and inorganic cations. Biochem.Pharmacol.. 40: 473-479, 1990.
28. Gralla, J. D., Dwight. S. S., and Poljak, L G. Formation of blocking lesions inidentical DNA sequences by the nitrosourea and platinum classes of anticancer drugs.Cancer Res.. 47: 5092-5096, 1987.
1542
on June 6, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

WAFI/Cipl CONFERS CHEMORESISTANCE
29. Beith, J., Hartley. J.. Darling. J.. and Souhami, R. DNA interstrand cross-linking and
cytotoxicity induced by chloroethylnitrosourea and cisplatin in human glioma celllines which vary in cellular concentration of O6-alkylguanine-DNA alkyltransferase.
Br. J. Cancer. 75: 500-505. 1997.30. Hunter, K. J.. Deen. D. F., Pellarin, M., and Marlon, L. J. Effect of a-difluoro-
methylornithine on l,3-bis(2-chloroethyl)-l-nitrosourea and rij-diamminedichloro-platinum(ll) cytotoxicity, DNA interstrand cross-linking, and growth in human braintumor cell lines in vitro. Cancer Res., JO: 2769-2772, 1990.
31. Poluha, W., Poluha, D. K., Chang, B., Crosbie, N. E., Schonhoff. C. M., Kilpatrick, D. L.,and Ross. A. H. The cyclin-dependent kinase inhibitor p21WAF1is required for survival
of differentiating neuroblastoma cells. Mol. Cell. Biol., 16: 1335-1341, 1996.32. Wang J., and Walsh, K. Resistance to apoptosis conferred by Cdk inhibitors during
myocyte differentiation. Science (Washington DC). 273: 359-361. 1996.33. Dolan, M. E.. Mitchell, R. B.. Mummen. C.. Moschel. R. C., and Pegg. A. E. Effect
of O6 benzylguanine analogues on sensitivity of human tumor cells to the cytoxic
effects of alkylating agents. Cancer Res., 51: 3367-3372, 1991.34. Perkins, N. D. Regulation of NF-KB by cyclin-dependent kinases associated with the
p300 coactivator. Science (Washington DC), 275: 523-527, 1997.
35. Sheikh, M. S.. Chen. Y. Q.. Smith. M. L.. and Fornace. A. J.. Jr. Role ofp2iw.fi/cipi/sdii ¡ncel| deam an(J DNA repair as stu(jie(i usmg a tetracycline-
inducible system in p53-deficient cells. Oncogene. 14: 1875-1882, 1997.
36. Li. R., Waga. S.. Hannon, G. J.. Beech. D.. and Stillman, B. Differential effects by theWAFl/Cipl CDK inhibilor on PCNA-dependenl DNA replication and repair. Nature(Lond.). 371: 534-537, 1994.
37. Shivji. M. K. K., Grey. S. J.. Strausfeld. U. P.. Wood. R. D., and Blow. J. J. Ciplinhibits DNA replication but not PCNA-dependent nucleotide excision-repair. Curr.Biol., 12: 1062-1068 1994.
38. Pan, Z-Q., Reardon, J. T., Li. L., Flores-Roas. H., Legerski. R., Sanear, A., andHurwitz, J. Inhibition of nucleotide excision repair by the cyclin-dependenl kinaseinhibitor p21. J. Biol. Chem., 270: 22008-22016. 1995.
39. Kyritsis. T.. Bondy. M. L., Hess, K. R., Cunningham. J. E., Zhu, D.. Amos. C. J.,Yung, W. K. A., Levin, V. A., and Bruner. J. M. Prognostic significance ofp53 immunoreactivity in patients with glioma. Clin. Cancer Res., /: 1617-1622,
1995.40. Fan, S-J., Smith. M. L., Rivet. D. J.. II. Duba, D.. Zhan. Q.. Kohn. K. W., Hornace.
A. J., Jr.. and O'Connor, P. M. Disruption of p53 function sensitizes breast
cancer MCF-7 cells to cisplatin and pentoxifylline. Cancer Res., 55: 1649-1654,
1995.41. Consoli, U., Li, H.. Huang, P., Kobayashi, T., Andreeff. M., Levin. V., and Zhang. W.
Enhancement of apoptosis in irradiated glioblastoma cells by pentoxifylline. Oncol.Rep.. 4: 673-678. 1997.
1543
on June 6, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1998;58:1538-1543. Cancer Res Sanbao Ruan, M. Fatih Okcu, Jiang Ping Ren, et al. 1,3-Bis(2-chloroethyl)-1-nitrosourea and CisplatinResistant to Chemotherapy Agents Overexpressed WAF1/Cip1 Renders Glioblastoma Cells
Updated version
http://cancerres.aacrjournals.org/content/58/7/1538
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/58/7/1538To request permission to re-use all or part of this article, use this link
on June 6, 2018. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from