Embed Size (px)
Citation preview

ČESKOSLOVENSKÁ
P VĚDECKÁ SP OL EČ N O ST
PRO M Y K O L O G I I
ČESKÁ

Časopis Cs. vědecké společnosti pro mykologii pro šíření znalosti hub po stráncevědecké i praktické
R o č n í k 32 C í s l o l L e d e n 1 9 7 8
V e d o u c í r e d a k t o r : doc. RND r. Zdeněk Urban, DrSc.
R e d a k č n í r a d a : akadem ik Ctibor Blattný, D rSc.; p ro f. Karel Cejp, DrSc.; RND r. Petr Fragner; M UDr. Josef Herink; RND r. Věra Holubová, CSc.; RND r. František Kotlaba, CSc.; ing. Karel Kříž; RND r. Vladim ír Musílek, CSc.; doc. RND r. Jan Nečásek, CSc.; ing. Cyprián Paulech, CSc.; prof. Vladim ír Rypáček, DrSc., člen ko-
resp. Č S A V ; RND r. Miroslav Staněk, CSc.
V ý k o n n ý r e d a k t o r : RND r. Mirko Svrček, CSc.
i 4. sešit 31. ročníku vyše l 25. listopadu 1977
O B S A H
J. V e s e l s k ý , J. K u b i č k a a R. Č u ř í k : Ak tuá ln í poznám ky k otravámmuchomůrkou zelenou — Am anita phalloides (Fr.) L i n k .........................1
M. S v r č e k : N ové nebo méně známé diskomycety. V I I .......................................11Z. P o u z a r : H ypoxylon m acrocarpum Pouz. spec. nov., nová vonná tvrdohouba 19J. S t a n g l a J . V e s e l s k ý : Inocybe descissa (Fr.) Quél. a je j í taxonom ické
umístění. (P říspěvky k poznání vzácnějších vláknic. Část 1 2 .)........................ 22P. F r a g n e r : Kvasinky v lidském m ateriálu u nás a je jich rozlišení. Část I. 32S. Š e b e k : Muchomůrka M aireova — Am an ita m airei F o ley nalezena v Čes
koslovensku 43J. K l á n : Inonotus tam aricis (Pat.) M aire v Řecku, celkové rozšířen í a taxo
nom ické poznám ky k sekci Phym atopilus D o n k .............................................47J. H ř i b a V. R y p á č e k : Růstová reakce dřevokazných hub na přítom
nost smrkového k a l u s u ............................................................................................55i A . A d á m k o v á , M. V á ň o v á a M. L á v i č k a : Rhizopus cohnii jako příčina
m ukormykotického abortu u s k o tu ........................................................................61
R e f e r á t y o l i t e r a t u ř e : D. Ershad, Fungi o f Iran (M. Svrček, str. 63);L. V. L jubarskij a L. N. V as il’jeva, D ěrevorazrušajčije g rib y Dalnego Vosto- ka (F. Kotlaba, str. 63); M. B. Ellis, M ore Dematiaceous Hyphom ycetes (V.
I Holubová-Jechová, str. 10). I. Nuss, Zur Ö kologie der Porlin ge (Z. Pouzar,str. 31); H. D. Thiers, C aliforn ia Mushrooms (V . Čatská, str. 42); E. M ü ller a W. Loe ffle r, M yco logy (V . Čatská, str. 42); D. J. W eber a W. M. Hess, The fungal spore (V. Čatská, str. 46); A . Neuner, B L V Naturführer. P ilze (V. Čatská, str. 54)
P ř í l o h y : č e r n o b í l é t a b u l e : I. K článku: J. Veselský, J. Kubička a B. Čuřík. — II. K článku: J. H řib a V . Rypáček. — III . K článku: A . A dám ková, M. Váňová a M . Lávička. — IV . a V . Inonotus tamaricis (Pat.) Maire.
Č e s k á m y k o l o g i e

ČESKÁ MYKOLOGIEČASOPIS ČESKOSLOVENSKÉ VÉDECKÉ SPOLEČNOSTI PRO MYKOLOGII
ROČNÍK 32 1978 SEŠIT 1
Aktuální poznámky k otravám muchomůrkou zelenou — Amanita phalloides (Fr.) Link
Some Recent Remarks to Death-Cap Poisonings —Amanita phalloides (Fr.) Link
Jaroslav Veselský, J iř í K ub ička a Rom uald Čuřík
Souhrnné pojednání o nynějších znalostech toxinů muchomůrky zelené, o účincích izo lovaných toxinů na organismus laboratorních zv ířa t ve vztahu k alim entárním otravám u lid í celou houbou. Jsou uvedena některá novější pozorování z ČSSR a je diskutováno o léčebných účincích některých látek. Důležitou součástí m edikam entosního léčen í je kyselina thioktová, i když není antidotem toxinů. Zdá se, že je j í léčebné použití v e vysokých dávkách má racionální základy. Podstatnou součástí kom plexní péče je časná očista zažívacího traktu od částic požité houby a zbavení k rve toxinů dříve, než dojde k je jich fixac i v játrech.
A summary dealing w ith the present know ledge of death-cap toxins and o£ the effects o f isolated toxins on laboratory animals in relation to the alim entary poisonings o f man w ith the total fungus. Some new er observations in the ČSSR are stated and m edical e ffects o f some substances are
" discussed. Th ioctic acid seems necessary in the treatment, although it is not
Í a m edicine to counteract a poison. Its m edical use in high doses has a rational basis. Th e use o f cleaning techniques to ensure the elim ination o f muschroom -particles out o f alim entary canal and/or toxins from the blood forth as soon as possible before its fixa tion in the liv e r is substantial in the com plex care.
Roku 1975 napsali Paaso a Harrison v komentáři k první úpěšné léčbě otravy muchomůrkou zelenou v USA, kterou publikoval roku 1972 Finestone se spolupracovníky, že Kubíčkovy návrhy vysokých dávek kyseliny thioktové čili lipoové jsou “ illustrative and encouraging, but as yet unfounded scientifically for clinical treatment“ (1. c. p. 508). Kyselina thioktová resp. lipoová jsou triviáln í názvy dvou sloučenin: kyseliny 6 ,8-dithiooktanové CH2CH,CH— CH,CH,CHoCH2COOH (thiolová forma) a jejího oxidačního pro- I ISH SH
,, duktu, kyseliny 6 ,8-dithiacyklooktanové CH^CHoCH— CH,CH,CH,CHoCOOHI I I
S S(disulfidová forma), jež oDé spolu tvoří redoxní systém. V biochemické literatuře se ujal více název lipoová, popř. lipoát. Návrh léčby vysokými dávkami thioktové kyseliny (Kubička 1963, 1964, 1968) vznikl na základě pozorování, že aktivita sérových enzymů, glutamátoxalacetáttransaminázy (GOT) a gluta- mátpyruváttransaminázy (G PT ) se v průběhu alimentárních otrav u lidí soustavně zvyšuje a empirickým ověřením, že jen velm i vysoké dávky thioktové
1

kyseliny mohou pomoci jaťerním buňkám lidského organismu přežít nejkri- tičtější: fáze otravy. Tento léčebný postup byl od té doby mnohokrát klinicky vyzkoušen a byla publikována řada souhlasných prací z domácích i zahraničních pracovišť (Zulik et al.). Někteří autoři však nemohli potvrdit příznivý efekt z různých důvodů. Podstatným z nich je n e s p r á v n á p ř e d s t a v a0 k y s e l i n ě t h i o k t o v é j a k o a n t i d o t u toxinů muchomůrky zelené (Gulden et Schumacher 1977).
Za spolehlivý důkaz účinnosti nějakého léčiva dnes považujeme jen takové postupy, kdy v sérii pokusů ve srovnání s placebem dosáhneme statisticky významného efektu. Příslušné letální dávky ( L D 5 0 nebo LDjoo) lze stanovit jen na laboratorních zvířatech. Z výsledků lze pak usuzovat na toxicitu u člověka jen spekulativně. Požadavky na zjištění optimální účinnosti nějakého léku u alimentárních otrav muchomůrkou zelenou u lidí jsou mimořádně značné.
Z v í ř e j a k o p o k u s n ý o b j e k t
Tento odstavec je třeba rozdělit na dvě části: na popis pokusů s c e l o u houbou nebo je jím i extrak ty a na pokusy s i z o l o v a n ý m i t o x i n y . Různá zv ířa ta je v í nestejnou rezistenci k muchomůrce zelené. Již v roce 1906 zjis til Ford, že pes je odolnější než člověk. Ještě větší odolnost byla pozorována u králíků, čehož bylo vyu žito s nevelkým úspěchem Lim ousinem k léčebným pokusům v podobě pasty z čerstvých žaludků a mozků králíků. Pozdě ji by lo zjištěno, že schopnost trávení houby zažívacím traktem závisí na enzym ové výbavě zažívacího traktu, která je u většiny zv ířa t odlišná od člověka. Tox in y muchomůrky zelené jsou obvyk le testovány na bílých myších nebo na krysách, ale ukázalo se, že až lOkrát c itlivě jš í (vztaženo na kg váhy) jsou morčata, lidoopi a lid i (T. W ie land 1973). Pokusy s krmením zv ířa t muchomůrkou zelenou nám tedy nemohou poskytnout ani spoleh livé inform ace ani srovnatelné hodnoty pro posouzení účinku toxinů u člověka a lze je brát jen aproxim ativně. Za průměrnou LD u l i d í považu je M oeschlin (1972) m nožství 50 g čerstvé houby s obsahem 4 m g a-amanitinu, 2,5 m g jS-amanitinu a 50 m g faloidinu, což prakticky odpovídá jednom u klobouku dospělé plodnice.
Izolace jednotlivých toxinů, je jich působení ve zv ířec ím experim entu, je jich přem ěny s možností detoxikace m olekuly změnou chem ické stavby, ob jev a n t a m a - n i d u (T. W ieland et al. 1968), pokusy se značeným i tox iny a mnoho dalších cenných poznatků ze školy W ielandů otevře ly netušené možnosti, jichž bude v budoucnu jistě vyu žito i v léčbě. V současné době však nedávají ani ty to cenné poznatky spolehlivou směrnici pro racionální léčen í alim entárních otrav u člověka. Podstatou toxického účinku a - a m a n i t i n u je podle W ielandů (T. et O. W ieland 1972) specifická inhibice nukleoplasm atické R N K -po lym erázy I I eukaryotických buněk s následnou nekrosou buněk ledvinných (“necrosis and nuclear atrophy, especia lly o f the convoluted tubules and the ascending lim b o f the loop o f H en le“ — 1. c. p. 265). A m atox in y jsou desetkrát jedovatějš í než fa lotoxiny. Účinkují pomalu, takže tep rve několikanásobným překročením L D 10o, která je u m yšek 0,5 až 1 mg/kg, se dostavuje sm rtící e fek t za bratší dobu, než 15 hodin. V k lin ice lidských alim entárních otrav nás však nemusí působení am atoxinů na tubuly led vin p ři náležité léčbě příliš znepokojovat. K této problem atice se ještě vrátím e. V roce 1976 na lezli Faulstich a Cochet-M eilhac am atoxiny v nepatrných koncentracích dokonce i v některých jed lých houbách. M nohem více pozornosti si v lidské patologii vyžadu je působení falotoxinů.
F a l o t o x i n y se specifickým účinkem na játra “ “ phallo id in seems to act specifi- ca lly on the liv e r “ — T. et O. W ieland 1972 p. 267) účinkují rych leji, takže L D 10q, která je u m yšek 4—7 mg/kg, usmrtí myšku již p ři dvojnásobném překročení za1 až 2 hodiny. C itu jm e proto alespoň některá důležitá zjištění, týkajíc í se účinku na játra v pokusech na zvířatech. Tak Hess (1956) zjistil, že fa lo id in zasahuje v m ito- chondriích krysích ja ter do aerobní fosforylace na stupni cytochromů dýchacího řetězce. O. W ieland a Szabados pub likovali r. 1968 (Intern. Congr. Clin. Chem. 4 : 59) názornou elektronm ikroskopickou fo togra fii ja tem í buňky u myši. Fotografie byla zhotovena za 30 m inut po otravě čistým fa lo id inem a lze na n í v id ět ve lik é vakuoly v endoplasm atickém retikulu (“the huge vacuoles arising from the tubular m em brane“ ) podobně jako na naší fo togra fii č. 1 , avšak m itochondrie jsou (na rozd íl od našeho nálezu u 271eté ženy) bez m orfo logických změn. F loersheim (1956) podporuje svým i
2
Č e s k á m y k o l o g i e 32 (i) 1978

nálezy hypotézu, že fa lo id in nemusí být sám o sobě tox ický a patrně se m ění v toxický m ětabolit až ja tern ím i enzym y. W ielandové v roce 1972 zdůrazňují, že prim ární účinek fa lo id inu je znám ý daleko méně, než účinek a-amatinu, a že dosud nebyl spoleh livě definován žádný specifický enzym ový systém, k terý by m ěl b ýt fa lo id inem postižen. Pom ocí značeného dem ethylfalo inu zjis tili T. W ieland a spolupracovníci, že 40% jedu růstalo u krysy zachyceno v játrech. N a otázku, proč to jsou právě játra, která jsou intoxikací n e jv íce postižena, přesvědčivě odpovídají Fauser a Faulstich (1973) svým i pokusy na m alých loveckých psech plem ena “B eagle“ : jatern í buňka je in toxi- kováná cestou e n t e r o h e p a t á l n í h o , tedy portáln ího krevn ího oběhu. Větší část parenterálně podaného 14C-methyl-y-am anitinu v dávkách 0,03 až 0,12 mg/kg otrávení psi vym očili, avšak 16% toxinu zůstalo v e žluči a enterohepatální oběh zvyšoval zpětnou resorpcí ce lkový tox ický účinek. Těm ito pokusy autoři zpochybnili terapeutický význam hem odialýz, perfuzí i ve likých krevních transfuzí, pokud nejsou prove- dény zavčas. L ze sice namítnout, že in jekční aplikace jedu neopravňu je k analogiím s peroráln ím podáním, ale je zřejm é, že koncentrace jedů v séru krevn ím nemůže být po je jich požití vyšší než po podání injekčním . N e jn ově ji se uvádí, že detoxikační účinek jm enovaných léčebných postupů lze očekávat jenom v prvn ích 24 až 36 hodinách po požili hub (Bartels; Costantino et Damiia).
P řesvědčivým důkazem k líčové úlohy ja ter a význam ným příspěvkem československých autorů je p rvn í a úspěšné provedení transplantace ja ter u m iniaturních prasat, která byla experim entá lně otrávena extraktem m. z. (Kořístek et al. 1975 cf. Pa lyza 1976.)
K c h e m i c k é s t r u k t u ř e t o x i n ů m u c h o m ů r k y z e l e n é
W ielandové a spolupracovníci (1972) stanovili chemickou strukturu toxinů muchom ůrky zelené. Jsou to b icyk lické polypep tidy se zbytkem tryptofanu a cysteinu; f a l o t o x i n y jsou heptapeptidy, kdežto a m a t o x i n y jsou oktapeptidy. N e jn ově ji se ob jevu jí názory, že ty to tox iny nejsou v houbě volné, ale vázané na m akrom olekuly. T ím je možné vysvě tlit některé rozporné pathofysio logické nálezy. Staron a Courtillot (1975) našli při použití jiných m etod struktury značně kom plikované: cyk lopeptidy jsou podle nich soustředěny kolem polysacharidového jádra. Tyto kom p lexy o ve lké m olekulové hmotnosti, takže neprocházejí dialysačním i membránami, by ly nazvány m y r i a t o x i n y . Izo lované tox iny m. z. jsou ovšem dialysovatelné (Moeschlin 1972) a lze je in v i t r o ještě účinněji e lim in ovat perfuzí přes aktivn í uhlí, avšak v experim entu i n v i v o se nezdařilo ani hem operfuzí udržet na živu krysy, které by ly otráveny faloid inem v dávce 1,5 až 2 mg/kg (Róckel et al. 1976).
Pathofysio logické účinky jednotlivých č is tých , toxinů jsou někdy nekriticky srovnávány s toxickým i účinky celé houby, nebývá přih líženo k rozdílnosti účinku po parenterálním a peroráln ím podání jedů a některým i autory pak b ýva jí nejednou vynášeny unáhlené soudy o účinnosti anebo neúčinnosti některých léčebných postupů u lidí.
N ě k t e r é n a š e p o z n a t k y u a l i m e n t á r n í c h o t r a v l i d í
Ledviny mohou být poškozeny amatoxiny přímo, jak zjistili Wielandové, ale u člověka spíše druhotně ztrátou tekutin a elektrolytů. Projevy selhání ledvin jsou u alimentárních otrav lidí považovány za druhotné (Sicot et al. 1968), takže Dudová se spolupracovníky (1973) po stejných zkušenostech soudí, že lidé neumírají za příznaků amatoxinové smrti, jak se obecně uvádí v literatuře, ale daleko dříve, než se dá histologicky prokázat rozpad buněčných jader s charakteristickou fragmentací jadérka (Fiume et Laschi 1965, cit. ex T. et O. W ieland 1972 p. 269, fig. 7), že tedy umírají f a l o t o x i n o v o u s m r t í . Významným přínosem, který názorně řeší otázku toxického poškození ledviny, je ojedinělé pozorování z CSSR (dosud nepublikované), kdy chlapci, zemřelému v hepatálním komatu na otravu m. z. byla odejmuta ledvina a transplantována jinému. Funkce této ledviny byla po několik let jedna z nejlepších, jichž bylo u nás transplantací dosaženo (J. K.). Nedávno byla u nás v ČSSR provedena poprvé punkční biopsie jater u mladé ženy n a v r c h o l u
VESELSKÝ, KUBICKÁ a CUŘÍK: K OTRAVÁM AM ANITA PHALLOIDES
3

ó s p ě š n ě l é č e n é t ě ž š í a l i m e n t á r n í o t r a v y m. z. (Dudová et al.1973). Elektromikroskopickým vyšetřením, které provedl jeden z nás (R. C.), byly zjištěny v jaterní buňce obrovské mitochondrie s nápadnými parakrysta- lickými inkluzemí (Bednář 1970) a v cytoplasmě zmnožené a rozšířené vaky hladkého endoplasmatického retikula (Cuřík et al. 1976). V iz obr. č. 1. Nález patologických mitochondrií není ovšem specifický jenom pro otravu m. z. Pro náš výklad toxického účinku m. z. u lid í vycházíme z cenných histochemických studií, které uvádí Bednář, že v parakrystalech byl prokázán vysoký obsah cytochromoxidázy a sukcinátdehydrogenázy. Významné je zjištění, že u žádného z našich kontrolovaných pacientů nepřetrvávají chronická poškození jater, dříve častěji popisovaná, což u čtyř z nich ověřili po 120 dnech od otravy kontrolní punkční biopsií jater Dudová et al. 1973.
N á š b i o c h e m i c k ý v ý k l a d p ř í č i n y s m r t i u l i d í
Ze sekčního čili nekroptického materiálu i na materiálu bioptickém (z punkce jater přežívajících osob) a z celého klinického průběhu otravy m. z. u člověka s přihlédnutím k charakteristickému pohybu referenčních hodnot sérových enzymů (SGPT, SGOT a glutamáthydrogenázy GLDH) a nejnověji nálezem parakrystalických inklusí v matrici jaterní buňky (obr. 1) a na základě experimentálních poznatků z citované literatury lze vyvod it tento logický soud:
P ř í č i n o u sm r t i na otravu muchomůrkou zelenou je u člověka porušený metabolizmus kyslíku v jaterních buňkách, a to v samé podstatě dýchání. Enzymová činnost mitochondrií je metabolicky inhibována falotoxiny, čímž je narušen nejen citrátový cyklus, ale i aerobní fosforylace a buňka se „udusí“ . Inhibiční účinek lze vysvětlit analogicky jako akci jiných inhibitorů aerobní fosforylace, např. oligomycinu, který „brzdí u intaktních mitochondrií pevné spřažení dýchání i fosforylace“ (Karlson 1. c. p. 230). Falotoxin tedy účinkuje, podobně jako známé enzymové jedy kysličník uhelnatý nebo kyanidy, na cytochromový systém přenosu elektronů, a to dle Hesse „snad v místě fixace anorganického fosfátu“ a jeho toxický účinek testovaný na krysách lze „v y číslit na základě měření procenta redukce enzymů dýchacího řetězce ve vztahu k spotřebě O2 a koncentraci ADP. Nejde tedy o direktivní účinek na určitý článek dýchacího řetězce“ (Hess 1956). Z pitevního i bioptického materiálu, který jsme studovali — zčásti i elektronmikroskopicky (R. C.) — jsme zjistili, že „udušení“ jaterní buňky nastává u lidí dříve, než se na jadérku objeví charakteristická fragmentace, působená účinkem amatoxinů, tak jak na elek- tronmikroskopické fotografii myšího hepatocytu dokumentují r. 1965 Fiume a Laschi (cit. ex T. et O. Wieland 1972 1. c. p. 269 fig. 7).
P o l o č a s ž i v o t a m i t o c h o n d r i í j e 5— 10 dn í , c o ž n á p a d n ě s o u h l a s í s d o b o u ú m r t í p o p o ž i t í h o u b y u l i d í .
M e c h a n i s m u s p ř e d p o k l á d a n é h o ú č i n k u l i p o á t u
Lipoát (Lipoic acid, Thioktsáure, kyselina a-lipoová, S-lipoová, thioktová) je složkou lipoátového koenzymu, který operuje v několika známých enzymech. Je to předně p y r u v á t d e h y d r o g e n á z a EC 1.2.4.1, člen multienzymového systému účastnícího se oxidace a dekarboxylace pyruvátu na acetylkoenzym A, dále k e t o g l u t a r á t d e h y d r o g e n á z a EC 1.2.4.2, jež pracuje v obdobném multisystému, který oxiduje a dekarboxyluje ketoglutarát na sukcinyl- koenzym A (v citrátovém cyklu). Do této množiny patří ještě 2 - k e t o i s o -
Č e s k á m y k o l o g i e 32 (i) 1978
4

k a p r o n á t d e h y d r o g e n á z a (EC 1.2.4.3) a 2 - k e t o i s o v a l e r á t d e - h y d r o g e n á z a (EC 1.2.4.4.). Krom ě toho operují s lipoátem t éž ' l i p o a m i d d e h y d r o g e n á z a EC 1.6.4.3., jež má flavinový koenzym a l i p o á t a c e t y l t r a n s f e r á z a EC 2.3.1.12, jež váže acetyl na redukovaný lipoát. Vzhledem k předpokládané inhibici enzymové funkce účinkem m. z., měla by otrava houbou charakter akutní „avitaminosy“ , v níž kyselina thioktová má úlohu účinného vitamínu.
Důležitý význam má odevzdávaná volná energie z redoxního potenciálu kyseliny thioktové. Polarografické měření potenciálu ukázalo hodnotu 325 mV (K e B., Biochim, biophys. Acta 25 : 650, 1957), což odpovídá potenciálu N A D +-NADH, který je udáván 320 mV.
Neméně cenné je zjištění, že thioktová kyselina způsobuje vzestup cyto- chromoxidázy, čímž lze vysvětlit podporu regenerace jater. Potvrzením je pozorování na experimentálně ztukovělých játrech: thioktová kyselina brání úbytku glykogenu v játrech, kdežto u kontrolních normálních zvířat působí signifikantní vzestup (Pagliaro L. et Catania A., Patología 45; 117, 1957).
V l i v k y s e l é h o p r o s t ř e d í n a t o x i c i t u A. p h a l l o i d e s
V r. 1803 popsal vratislavský lékař Krocker svůj protilék při otravě muchomůrkou zelenou takto: “Zwiebelsaft mit spiritus vitrio li aethereus (Schwefelsäure) 12— 20 Tropfen stündlich“ — tedy doslova „cibulová šťáva s éterem ( = aether sulphuricus čili jak se dříve říkalo vitrio l) 12 až 20 kapek každou hodinu“ .
Roku 1851 (cit. Bezděk 1905 1. c. p. 101) provedl jakýsi Gérard ve francouzské “Conseil d’hygiène et de la salubrité de la Seine“ pokus na sobě a na členech své rodiny, kdy za komisionálního dohledu a po předchozím určení houby několika členy Francouzské mykologické společnosti pojedl beze škody zelenou muchomůrku, když j i byl před tím máčel 2 hodiny v octové vodě a potom V2 hodiny povařil. „Zpráva ta nedostala se hned do širší veřejnosti, neboť zdravotní rada obávala se, aby lid nechopil se toho prostředku a nedostatečným snad vymočením neb povařením aby přece jen nebyly požity houby, jed ještě obsahující“ (Bezděk 1. c. p. 102).
Theodor Wieland (1968, 1973) uveřejnil svá pozorování, že toxicitu uvedených peptidů lze zrušit účinkem silnějších kyselin. Peptidové vazby se rozštěpí a z falotoxinů tak vznikne nejedovatý s e k o-15-falotoxin, z a-amanitinu neje- dovatý s e k o-15-a-amanitin. Jeden z nás (J. V .) proto naložil čerstvou plodnici m. z. do konzumního octa a odeslal ji k chromatografické detekci amanitinů. T y lze totiž v nekyselém prostředí dokázat již v 0,5 mg čerstvé houby (Palyza1974). Na chromatogramu látek získaných z tohoto relativně značného množství pletiva muchomůrky nebyla však nalezena skvrna odpovídající amanitinů (Palyza in litt.). Naproti tomu totéž množství houby naložené v kyselině 1-askor- bové (Celaskon) mělo při detekci, jak v houbovém pletivu, tak i v tekutině (!), prokazatelný a- i ¡3-amanitin ve zcela obvyklých koncentracích (Palyza 2. IX . 1977 in litt.).
Rovněž nebylo možno detekovat amanitiny v houbě, která již prošla žaludkem a střevem postiženého člověka, ani v obsahu žaludečních a střevních tekutin, resp. výplachů. Této okolnosti bývá používáno k podpoře názoru, že toxiny jsou v té době již dávno resorbovány. Tento názor nemůžeme přijmout, protože v pitevním materiálu dvou zemřelých na masivní otravu m. z. jsme našli v místech přechodu tenkého střeva v tlusté a v ohybech (flexurách)
VESELSKÝ, KUBICKÁ a CURlK: K OTRAVÁM AM ANITA PHALLOIDES
5

tlustého střeva veliké shluky spór m. z. jednak se zachovanými episporiálními valy, jednak v různých stadiích jejich porušení (obr. č. 2). Přihlédneme-li k veliké toxicitě spór m. z., jak experimentálně ověřil r. 1938 Dujarric de la Rívíěre (5 mg jemně rozptýlených spór v sterilním fysiologickém roztoku způ-_ sobilo po injekčním podání uhynutí k r á l í k a do 48 hodin a 30 až 50 mg spór uhynutí m y š í do 24 hodin), nelze považovat účinek toxinů, které se postupně uvolňují z nitra výtrusů, za zanedbatelné, tím spíše, že se tak děje v alkalickém prostředí střevním a až po rozrušení vlastního episporiálního valu, o němž praví Singer (1962 1. c. p. 73) “ it responds chemically to most tests for amidon, it can be dissolved by several chemical substances, such as concentrated HNO3, and less uniformly by NaOH, KO H etc. in a heated con- centrated solution“ .
Reintoxikace jater cestou oběhu vrátnicové žíly (Fauser a Faulstich 197-3) vysvětluje, proč v klinickém průběhu dochází k fázovým nárazům toxických příznaků a proč v cirkulující krvi velkého oběhu krevního je možno nejnovější r a d i o i m u n o l o g i c k o u metodou zjistit toxiny m. z. jenom v prvních 24 hodinách ( “dans la plupart des cas“ ) a jen ve 20 % případů později po požití houby (Costantino et Damia 1977 1. c. p. 2316)! Na základě pozorování svých pacientů soudíme, že u otrávených s žaludeční h y p e r a c i d i t o u probíhá otrava benigněji, než u anacidních, což vyžaduje ověření na větším počtu postižených.
Roku 1976 publikoval francouzský lékař Bastien svá opakovaná autoexperi- mentální pozorování, že na otravy m. z. má příznivý léčebný účinek L - a s k o r - b á t čili vitmain C v injekčních dávkách dvakrát 1 g denně a doložil je úspěšným vyléčením svých 12 pacientů. V této souvislosti není bez zajímavosti, že člověk, primáti a morčata jsou jediní savci, kteří nedovedou z cukrů vytvářet L-askorbovou kyselinu a že to jsou právě morčata a lidi, kteří jsou na rozíl od bílých myšek a krys až lOkrát citlivější (vztaženo na kg váhy) na toxiny m. z. (Th. Wieland 1973). Kyselina askorbová je ovšem jenom relativně silná kyselina, důležité jsou je jí redukční vlastnosti, které se mohou užitečně uplatnit při vzniku aktivních SH-skupin z disulfidových vazeb enzymů a koenzymů (Mass et Butterworth 1. c. p. 72). Touto cestou by mohla zasahovat i do citrá- tového cyklu a dýchacího řetězce tak jako kyselina thioktová. Zajím avé sdělení učinili Rosenberg H. R. et Zulik R. v přednáškovém cyklu 13.—18. IV. 1958 v Division of Biochemical Chemistry of the American Chemical Society, San Francisco (cit. ex Documenta Homburg: Thioctacid p. 7), že kyselina thioktová, podávaná každý druhý den morčatům s experimentální avitaminosou C, má tentýž antiskorbutický účinek jako „suboptimální“ léčení vitaminem C a ž e k o m b i n o v a n é p o d á v á n í l i p o á t u a a s k o r . b á t u účinkuje lépe, než každá substance sama o sobě.
H o d n o c e n í l é č e b n ý c h v ý s l e d k ů u l i d í a j e h o ú s k a l í
Analýza alimentárních otrav muchomůrkou zelenou v materiálu z dřívějších let ukázala, že úmrtnost se pohybovala kolem 50 až 70 % i více (Starkenstein et al. 1929). Antifaloidní sérum v té době ještě oblíbené ztratilo později na významu zjištěním, že toxiny m. z. nemají antigenní vlastnosti. Podstatným přínosem do léčení bylo poznání, že amatoxiny i falotoxiny jsou sice in vitro dialyzovatelné, ale v intoxikovaném organizmu jen velm i krátkou dobu kolují v krvi (Faulstich et Fauser 1973), takže novodobé epurační (krev čistící) technické metody jako dialýza krevní nebo peritoneální, exsanguinační transfuse,
Č e s k á m y k o l o g i e 32 (i) 1978
0

forsírovaná diuréza (“hyperosmale forcierte Diurese“ , Moeschlin 1. c. p. 453) a nejnověji plazmoforéza (Costantino et Damia 1977) a Changova perfuze krevní přes aktivní uhlí ( “ charcoal haemoperfusion“ ) jsou prakticky účinné jenom v prvních 36 hodinách po požití houby (Bartels 1976, Costantino et Damia 1977). V pozdějších stadiích otravy m. z. se uvedených metod používá obvykle již jenom k odstraňování druhotných metabolických poruch anebo krvácívých stavů, někdy velm i úporných, v souvislosti s portální hypertenzí. Tak poklesla v nejnovějších statistikách úmrtnost na 15 až 10 %■
P r o p ř e ž i t í o t r a v y u č l o v ě k a j e r o z h o d u j í c í :1. M n o ž s t v í p o ž i t é h o u b y . Praktická pozorování ukázala, že požití
4 a více klobouků muchomůrky zelené vede obvykle k nezadržitelné smrti. Jeden náš pacient snědl k obědu a znovu pak ještě k večeři asi po 20 plodnicích m. z. a zemřel ještě za plného vědomí vykrvácením z jícnových varixů již třetího dne otravy. Pacientka, kterou uvádí Moeschlin (1. c. p. 458), snědla v sebevražedném úmyslu 5— 10 kusů syrové m. z. a zemřela šestého dne za příznaků progresivního hypovolemického šoku, prudké jaterní dystrofie a ter- minální hemorragické diatézy. Zvláště tragický byl případ llletého děvčátka v naší sestavě postižených, která si nasbírala a sama na plotýnce opekla a snědla 5 klobouků m. z. Přes intenzivní péči na dokonale vybaveném specializovaném oddělení dívenka umírá pátého dne za příznaků odumření jater, provázeného krátce před smrtí klinickým obrazem nápadného přetlaku v střev- ně-jaterním (portálním) oběhu, tzv. “ caput medusae". Usuzovat z podobných extrémních příhod na účinnost anebo neúčinnost léčebných postupů, jichž bylo použito, bylo by neuvážlivé.
2 . Č a s o v ý ú s e k m e z i p o ž i t í m m. z. a z a h á j e n í m i n t e n z í v n í l é č e b n é p é č e . V prvních 36 hodinách lze při použití citované epurační techniky dosáhnout i nulového procenta úmrtnosti (Costantino et Damia 1977).
3. O d s t r a n ě n í z b y t k ů h u b a s p ó r z e z a ž í v a c í r o u r y je jedním ze základních léčebných zásahů (Dudová et al. 1973 a, 1. c. p. 13). I když vysokými klysmaty se stává bilance ztrát tekutin ještě obtížnější, měli naši nemocní příznaky deplece vodní a elektrolytové pouze první den po přijetí (tj. 2. a 3. den otravy!). Vysoké nálevy je třeba provádět denně tak dlouho, dokud lze mykologickým (sporologickým) vyšetřením čistící tekutiny prokázat v ní spory muchomůrky zelené (Veselský et al. 1973). Stanovisko Moeschlinovo, že výplachy zažívací roury mohou ještě více poškodit pacienta, který před tím delší čas zvracel a měl průjmy (Moeschlin 1. c. p. 459), nemůžeme potvrdit, pokud ovšem jde o pacienta, jemuž je poskytována intenzívní léčebná péče na specializovaných odděleních.
4. V č a s n é z a h á j e n í l é č b y s p o m o c í ú č i n n ý c h l é k ů . Významu kyseliny thioktové ve v e l k ý c h dávkách a pokusu o biochemické vysvětlení mechanismu jejího účinku jako nezbytného vitamínu jsme věnovali podstatnou část svého příspěvku, protože teoretické zdůvodnění jejího využití k léčení otrav m. z. u lidí nebylo dosud nikým provedeno. Podobný účinek má pravděpodobně i podávání askorbátu ve v e l k ý c h dávkách. Roku 1972 upozornil Floersheim na velm i příznivý účinek kombinovaného podávání p e n i c i l i n u G a c y t o c h r o m u c. Roku 1973 doporučila Dudová se spolupracovníky doplnit léčebné postupy o podávání pekařských kvasnic ( d r o ž d í , baker-yeast) v období, kdy pacient nezvrací, v denních dávkách 100 g jako zdroje esenciálních bílkovin, enzymů — jmenovitě klíčově důležité h e x o k i - n á z y — a vitamínů skupiny B. Pacienti, dospělí i děti, snášeli perorální
VESELSKÝ, KUBICKÁ a CUŘIK: K OTRAVÁM AM ANITA PHALLOIDES
7

podávání kvasnic velm i dobře. Injekční formu glukóza-6-fosfátdehydrogenázy (G -6-PDH) čili ATP-D-hexosa-6-fosfotransferázy EC 2.7.1.1. jsme bohužel nedostali.
Jsme přesvědčeni o tom, že Kubíčkovo empirické vyzkoušení a léčebné použití kyseliny thioktové ve vysokých dávkách (300— 900 mg na osobu denně v infuzích) a Bastienovo autoexperimentální ověření ochranného působení velikých dávek vitamínu C (2krát 1 g denně na osobu nitrožilně), ale i Floers- heimem navržené megadávky penicilinu, jehož použili u lidí v dávce 1 000 000 U/kg denně spolu s podáváním 500 mg thioktové kyseliny nejnověji Costantino a Damia, mají ve světle současných biochemických znalostí a k linických pozorování racionální základy. Základem léčebných úspěchů je však dle našich praktických zkušeností dokonalá eliminace houbového pletiva ze zažívací trubice, spolu s obrovským množstvím uvolněných spór, a to z v l á š t ě p e č l i v ě p r o v á d ě n á v o b d o b í , k d y p o s t i ž e n ý j i ž n e z v r a c í a n e m á p r ů j m y (viz obr. 2).
Podrobnou kazuistiku svých souborů spolu s návrhem jednotných léčebných postupů uvedeme v lékařském tisku.
P o d ě k o v á n í
Autoři děkují prof. RNDr. J. K o š t í ř o v i za cenné recenzní připomínky a účinnou pomoc při výkladu některých biochemických pochodů a MUDr. V. P a l y z o v i za laskavé provedení laboratorních analýz.
S u m m a r y
W ith respect to the recent clin ical observations and contemporaneous biochem ical know ledge w e can conclude that Kubicka ’s personal experience concerning the treatm ent o f A m anita phalloides poisoning in humans w ith necessary high doses o f th ioctic acid (300—900 m illigram m es da ily into infusion during 24 hours) and Bastien’s autoexperim ental find ing o f protective action o f ascorbic acid in high doses (2—4 grammes daily ) are rationaly based. Both agents are ab le to in tervene in the various stages o f the liv e r cell m etabolism inhibiting the liv e r dam aging action of toxic agents and are excellen t tolerated, even i f g iven in high doses. The results of the treatm ent depends o f early and repeated elim ination o f toadstool-particles and spores out o f the alim entary tract as soon as possible thus preventing the fatal hepatic coma.
L i t e r a t u r a
B a r t e l s O. (1976): P ilzverg iftungen . Neue Therapienm oglichkeiten. Fortschr. Med. 94 (10): 539-544.
B a s t i e n P . (1976): A propos de m on intoxication volontaire. Documents scientifiques Guigoz 99 : 22 — 28.
B e z d ě k J. (1905): H ouby jed lé a jim podobné jedovaté 2. Pp. 1 — 116, Hranice. C o s t a n t i n o D. et D a m i a G. (1977): L ’in toxication phalloidienne. Résultats
de diverses thérapeutiques chez 47 malades. Nouv. Presse méd. 6 (26): 2315 — 2317. Č u ř í k R., V e s e l s k ý J. et K u b i č k a J. (1976): Jaterní biopsie u ženy na vrcholu
m asivn í o travy muchomůrkou zelenou — Am an ita phalloides. Čas. Lék. čes. 115 (45): 1399-1400.
D u d o v á V., B e n e š J., B l a ž e j V., L i c h n o v s k á M., N i e s 1 a n i k J. et V e s e l s k ý J. (1973a): O trava muchomůrkou zelenou. Pp. 1—47. Ostrava.
D u d o v á V., B l a ž e j V., V e s e l s k ý J., N i e s l a n i k J. et Č u ř í k R. (1973b): Léčba o travy muchomůrkou zelenou na podkladě vlastních k lin ických zkušeností. Čs. Gastroenter. V ý ž iva 27 (8): 543—545.
D u j a r r i c de l a R i v i e ř e R. et G a r n a l P . (1938): Sur la tox ic itě de certains Amanites. Rev. M ycol. 3: 54 — 59.
Č e s k á m y k o l o g i e 32 (i) 1978

F a u l s t i c h H. et C o c h e t - M e i l h a c M. (1976): Am atoxins in ed ib le mushrooms. FEBS Letters Am sterdam 64: 73 — 75.
F a u l s t i c h H. et F a u s e r U. (1973): Untersuchungen zur Frage der Häm odialyse bei der K no llenb lätterp ilzvergiftung. Dtsch. med. Wschr. 98 (47): 2258 — 2259.
F a u s e r U. et F a u l s t i c h H. (1973): Beobachtungen zur Therap ie der K no llen b lätterp ilzvergiftung. Dtsch. med. Wschr. 98 (47): 2259.
F i n e s t o n e A . J., B e r m a n R., W i d m e r B., M a r k o w i t z J. et L a q u e r U. J. (1972): Thioctic acid treatm ent o f acute mushroom poisoning. Pennsylvania M edicine 75: 49 — 51.
F l o e r s h e i m G. L. (1966): Protektion gegen Am anitatoxine. H elv. physiol, phar- macol. A c ta 24: 219 — 228.
F l o e r s h e i m G. L. (1972): Neue Gesichtspunkte zur Therap ie von Vergiftungen durch den K no llen b lä tterp ilz (Am an ita phalloides). Schweiz, med. Wschr. 102 (26): 901-909.
F o r d W . W. (1906): The toxicologica l constitution o f Am anita phalloides. J. pharm, exper. Therap. 8 : 437 — 450.
G u l d e n G. et S c h u m a c h e r T. (1977): G iftsopper og soppforgiftn inger. Pp. 1 — 116. U n iversitestforlaget Oslo.
H e s s B. (1956): Ü ber d ie Hemm ung der oxydativen Phosphorylierung durch Phal- lo id in auf der Cytochrom -Stufe. Biochem. Z. 328: 325 — 327.
K a r l s o n P. (1971): Zák lady biochem ie ed. 8. Pp. 1 — 474. Praha.K o s t í ř J. (1965): Chem ie a fyz ika živých soustav. Pp. 1 — 353. Praha.K o š t í ř J. (1974): B iochem ie. Pp. 1 — 565. Praha.K r o c k e r A . J. (1803): Nom enclator flo rae Silesiae. Manuscriptum cit. ap. Schroeter
J. in Cohn F. (1889) K ryptogam en-F lora von Schlesien 3: P ilze 1: 7 — 8. Breslau.K u b i č k a J. (1963): Neue M öglichkeiten der Behandlung von P ilzverg iftungen durch
den grünen K no llen b lä tterp ilz — Am an ita phalloides. M ykol. M ittb l. H a lle (Saale) 7: 92-94.
K u b i č k a J. (1964): P revence a léčba otrav muchomůrkou zelenou v Jihočeském kra ji. P raktický lékař 44 (18): 702 — 704.
K u b i č k a J. (1968): Zw anzig Jahre K am p f gegen P ilzverg iftungen . Schweiz. Z. P ilz kunde 46 (6): 81-99.
K u b i č k a J. e t A 1 d e r A . E. (1968): Ü ber eine neue Behandlungsmethode der V erg iftung durch den K nollenb lätterp ilz . P raxis 57 (38): 1304 — 1306.
L i m o u s i n H. (1932): Essai de traitem ent des intoxications causées par les champignons vénéneux. Presse méd. 40: 1685 — 1687.
M o e s c h l i n S. (1972): K lin ik und Therap ie der Vergiftungen, ed. 5. Pp. 1 — 534 (Kno llenb lätterp ilzvergiftung p. 452—461). Stuttgart.
M o o s D. W. et B u t t e r w o r t h P. J. (1974): Enzym ology and medicine. Pp. 1—175. P itm an M edical London.
P a a s o E. et H a r r i s o n D. C. (1975): A new look at an old problem : Mushroom poisoning. Am er. J. M edicine 58 (4): 505 — 509.
P a l y z a V. (1974): Schnelle Iden tifiz ierung von Am anitine in P ilzgew eben. Arch. Toxikol. 32: 109-114.
P a l y z a V. (1976): T ox iko log ie muchomůrky, zelené (Am anita phalloides) IV . Léčen í experim entá ln í o travy transplantací jater. M ykol. zpravodaj Brno 20 (2): 57 — 58. [Ref. ex K ořístek V., Černý J. H ök l J., F iku lka J., G regor Z., Peřestý S., Busch K., Böhm F., Štěpánek V., V lastyak J., C ik l M., Ku lhánek V., Pa lyza V. et D vořák E. (1975): Transplantace ja ter jako léčebná m etoda při experim entá ln í otravě A m a- nitou phallo ides u m iniaturních prasat. Rozh ledy v Chirurgii 54 (6): 381 — 386.]
R o c k e l A. , S c h m i d G., R u p p P., K u l t J., H e n n e m a n n H., H e i d l a n d A. et B r a n d A . (1976): Häm operfusion m it A k tivkoh le und Ionenaustauscher bei phallo id in -verg ifteten Ratten. Dtsch. med. Wschr. 101 (11): 418—420.
S i c o t C., B i s m u t h C h., F r e j a v i l l e J. P., R u e f f B., B o i v i n P. e t B e j i - h a m o n J. P. (1968): In téret des dosages répétées de la transaminase glutamo- pyruvique pour prognostic des intoxications phalloidiennes. Rév. fr. Etud. Clin. B iol. 13 (10): 1022-1024.
S i n g e r R. (1962): Th e Agarica les in modern taxonomy, ed. 2. Pp. 1 — 915, W einheim .S t a r k e n s t e i n E., R o s t E. et P o h l J. (1929): Lehrbuch der Toxikologie.
B erlin—Wien.S t a r o n T. et C o u r t i l l o t M. (1975): Quelques données nouvelles sur les toxins
d ’Am an ita v irosa et Am anita phalloides. Bull. Soc. m ycol. Fr. 91: 556.
VESELSKÝ. KUBICKÁ a ČURlK: K OTRAVÁM AM ANITA PHALLOIDES
9

V e s e l s k ý J., D u d o v á V., B l a ž e j V., L i c h n o v s k á M., N i e s l a n i k J. et Č u ř í k R. (1974): The ro le o f a m ycologist in the treatm ent o f Am an ita phalloides intoxication. Čes. M ykol. 28 (2): 106.
W i e l a n d O. et W i e l a n d T. (1972): The toxic peptides o f Am an ita species. In Kadis S., C ieg ler A . et A j l S. J.: M icrob ia l toxins 8 (10): 249 — 280. Acacjemic press N ew Y o rk and London.
W i e l a n d T. (1968): Poisonous principles o f mushrooms o f the genus Am anita.Science 159 (1 M arch): 946 — 952.
W i e l a n d T. (1973): Uber die G ifts to ífe der Gattung Am anita. Z. P ilzkunde 39: 103-112.
W i e l a n d T., L í i b e n G., O t t e n h e y m H., F a e s e l J., d e V r i e s J. X., K o n z W., P r o x A . et S c h m id J. (1968): Antam anid. Seine Entdeckung, Isoliérung, Strukturaufklárung und Synthese. Angew . Chem 80 (6): 209 — 213.
Z u l i k R . , B a k o F., K i s b a n A . (1973): Zur Behandlung der Knolléňb látterpilz- vergiftung. Med. K lin . 68 : 1371 — 1372.
Adresa autorů: M UDr. Jaroslav Veselský, Výškovická 100, 704 00 Ostrava-Zábřeh. M UDr. Jiří Kubička, 39811 P ro tiv ín 202.
M. B. E 11 i s : More Dematiaceous Hyphomycetes. Com m onwealth M ycologica l Institute, K ew , Surrey, England, 1976. 507 stránek, 383 vyobrazen í (perokresby). Cena £ 18.00.
V e ve lm i krátké době p řip ravil M. B. E llis druhou knihu o tm avě zabarvených hyfom ycetech, která úzce navazuje na jeho „Dematiaceous Hyphom ycetes“ z roku 1971 (v iz recenze v České M ykologii, Praha, 28: 64, 1974). A u tor v ní zachovává sýstema- tické uspořádání rodů jím navržené v p rvn ím díle. N ově uvedené rody (celkem 76) jsou systematicky začleněny do d řívějš ího souboru 295 rodů, takže obě knihy tvoří jeden celek, který zahrnuje celkem 371 rodů. Popisy rodů, které jsou uvedeny v p rvé knize, nejsou již opakovány, ale je uveden pouze odkaz. Recenzovaná publikace je ve lm i cenným přínosem, neboť uvádí přesné, m oderní popisy a detailn í vyobrazen í dalších 732 druhů. Autor v této kn ize zahrnul ty rody a druhy, které by ly m ykology od roku 1970 nově popsány a také ty, které un ikly jeho pozornosti při p říp ravě p rvn ího díla. U vád í však jen ty druhy, které sám studoval. Publikace přináší taxony z celého světa. Zcela nově je v ní popsáno 10 rodů, 60 nových druhů a je navrženo 115 nových taxonom ických a nomenklatorických přeřazení, z toho 43 je provedeno F. C. Deightonem. Autor se v tom to svazku podrobněji věnu je některým ekonom icky v ý znamným rodům, jako jsou A ltern a ria , Cercospora, C ladosporium , Drechslera, Ph a e- oram ularia, Ph ia lophora , Spilocaea, Stachybotrys, S tigm ina a V locla d iu m . V úvodní části knihy je k líč na určování rodů, ale týká se jen těch rodů, k teré jsou probírány v tom to svazku. P ro praktické použití v y však bylo vhodnější, kdyby autor tento rodový k líč sestavil pro všechny rody jím pojednávané v obou dílech. V h lavn í části knihy autor popisuje jednotlivé rody a druhy stejným způsobem a ve stejném rozsahu jako v p rvn ím svazku. U vád í nejdů ležitě jš í synonymiku, přesný popis, hostitelské substráty a místa výskytu, a to na základě vlastního výzkumu. V závěru knihy je uveden substrátový a hostitelský index, slovníček m orfo logických term ínů užívaných v rodových a druhových popisech a celkový rejstřík.
Recenzovaná E llisova knižn í publikace je stejně tak vyn ika jíc í jako předcházející z roku 1971 a je neocenitelnou pomůckou pro m ykology na celém světě, k teří se zab ýva jí touto skupinou hub. M. B. E llisovi se podařilo v y tvo ř it m oderní příručku, v které shrnul většinu popsaných a přija tých rodů tm avě zabarvených hyfomycetů. Jeho d ílo je výsledkem více než 301etého intensivního studia v této skupině hub. P roto také zahrnuje mnoho nových poznatků a Zcela nová taxonom ická hodnocení jednotlivých taxonů. Další výzkum y v této skupině im perfektn ích hub budou m oci nejlépe prověřit a ocenit E llisovo d ílo a jeho nové taxonom ické závěry.
V. H olubová -Je chová
Č e s k á m y k o l o g i e 32 (i) 1978
10

N ew or less known Discomycetes. V II.
Nové nebo méně známé diskomycety. VII.
M irk o Svrček
F ive new species o f Discom ycetes from Czechoslovakia and Germ an Dem ocratic Republic are described: Coprobia p ile iform is , C ororie llaria benkertii, Haglundia m agnipilosa, Hymenoscyphus rhy tid iadelph i and Hysterostegiella zelendarkensis. The new records o f Hagludia perelegans and H. sarm entorum from Bohem ia are discussed.
Je popsáno pět nových druhů diskomycetů z územ í ČSSR a N D R : C oprob ia p ile ifo rm is , C oronella ria benkertii, Haglundia magnipilosa, Hym enoscyphus rhytid iadelphi a H ysterostegiella zelendarkensis. Je pojed-
; náno o Hp.glundia, perelegans a H. sarm entorum na podkladě nových nálezů ; Z Čech. * T /1 : I i / ■ jí ' :
Coprobia pileiformis spec. nov.
Apothecia 1— 3 mm diam., plerumque dense gregaria usque confluentia, super- ficialia, absque hypothallo, molliter succoso-carnosa, primům subglobosa vel pyriform ia, apice truncata, disco orbiculari, parvo, immarginato, sed mox late convexo, denique conspecte semigloboso usque obtuse conico, alto, pileiformi, margine saepe irregulariter undulato, superficie granuloso, fulgide aurantiaco vel aurantiaco-luteo; pars in ferior crasse stipitiform iter angustata, obconica vel subcylindracea, 1— 3 mm alta, sanguineo-rubra, obscure rubra vel auran- tiaCo-rubra, extus nuda, minutissime granulata.
Exčipulum textura globulosa, crassum, aurantiaco-coloratum, cellulis globosis, magnis, 20— 40 ^m diam., flavidis, membranis subincrassatis.
Asci 110— 135 X 12— 15 um, cylindracei, apice obtusi, cperculati, non amy- loidei, tenuiter tunicati, deorsum breviter crasseque attenuati, emarginati, octošpori, sporis monostichis. Paraphyses copiosae, crasse cylindraceae, 5— 8 fim crassae, parte basali ramosae, sursum parum usque admodum dilatatae (8— 12 u.m), forma variabili, clavatae, subcapitatae sed etiam obtuse lanceolatae, remote septatae, subhyalinae. Ascosporae 12— 14 X 6,5— 8 l(am, forma irregulari, inaequales, ellipsoideae, ovato-ellipsoideae, ovoideae vel oblongo-ovoideae, pal- lide luteolae, intus granulosae vel nebulosae, tunica subincrassata, perisporio verrucis cyanophilis inaequalibus sparse ornato.
H a b. Ad basim stipitis carposomatis putrescenti Lactarii vellerei (Fr.) Fr. quoque in detrito adiecto (e. g. fragmenta putrida acuum P in i silvestris, caules musei Pleurozii schreberi), in pineto vetusto cum Vaccinio m yrtilli.
L o c a l i t a s t y p i : Bohemoslovakia, Bohemia maridionalis, Rakovické Chalupy prope Mirotice (districtus Písek), in silva “Bořky“ dieta, 17. V III. 1963 leg. M. Svrček (typus, PRM 805292).
The shape of the conspicuously bright colored apothecia, the form of paraphyses and the relatively small, irregularly ornamented ascospores distinguish this new species from the other ones of Coprobia. The ascospores are covered w ith a delicate outer coating consisting o f sparse unequally large, strongly cyanophilic warst or folds. The species is distinct ecologically too, perhaps restricted to the rotting fruit-bodies of some Agaricales. In spite of rather numerous carposomata of Lactarius vellereus occurring on the locality, no other material o f this Coprobia was found and also none in follow ing years (1964— 1977) when I visited again the same place. Thus it seems to be a rare species.
11

Coronellaria benkertii spec. nov.
Apothecia 0,2— 0,5 mm diam., primům suberumpentia, sed mox late sessilia, superficialia, orbicularia, disciformia, disco plano, albido vel pallide sulphureo, extus margineque dense brevissime pilosa, basi subfusca.
Excipulum textura globulosa, cellulis parte basali globosis vel subglobosis, subcrasse tunicatis, usque ad 10 um diam., brunneis, marginem versus cellulis elongatis, oblongis, sat tenuiter tunicatis, pallide fuscis, usque ad 14 X 10 um magnis, margine hyphis 25— 50 X 4— 5 ,um, cylindraceis, apice obtusis, tenuiter tunicatis, plerumque flexuosis, remote septatis, subhyalinis.
i i í t f f. D ü © - f e e n ÍH
/ I Li // / i - JI(Ml "" '>
1. C oprobia p ile ifo rm is Svrček. Ascospores (in Cotton-blue -f- oil-im m ersion), apothecia, ascus, tip o f one ascus, paraphyses (P R M 805292, holotype).
M. Svrček del.
Asci 70— 110 X 11— 14 um, octospori, oblongo-clavati, deorsum sensim stipiti- form iter angustati saepeque arcuati, apice obtusi conspecteque inerassati, poro non amyloideo. Paraphyses forma variabili, sursum sensim dilatatae, cylindra- ceae, 3— 3,5 um crassae, septatae, apice obtusae, non raro irregulares, usque ad 6 dilatatae, sed etiam digitato-angustatae, ascos non superantes, granulis hyalinis sparse incrustatae vel nudae, nonnumquam lateraliter ramosae, cum ramis tenuibus (1,5— 2 (um), flavidae. Ascosporae 17— 20,5 X 4— 4,5 um, partim distichae, anguste cylindraceae vel fusoideo-cylindraceae, basi plerumque angustatae, fere cuneiformes, rectae vel parum curvatae, guttulis binis magnis impletae, rarior eguttulatae, hyalinae, unicellulares, sed postea (an omnes?) 3-septatae.
Č e s k á m y k o l o g i e 32 (i) 1978
12

H a b. Ad. folia emortua Caricis rostratae.L o c a l i t a s t y p i Germania (D. D. R.), “Fresdorfer Moor“ (districtus
Potsdam), in alneto paludoso, 27. V II. 1970 leg. D. Benkert (typus, PRM).Young apothecia o f this noteworthy species develop beneath the epidermis
of fallen leaves of Carex, erumpent, but are sessile very soon on the surface. The marginal hairs are short and blunt, form ing a downy coating, the asci have a conspicuously thicked apex (5— 6 um), thick-walled, provided w ith an inamyloid pore. The large, septate and often encrusted paraphyses are sometimes divided into slender lateral branches. Coronellaria pulicaris Karst., the type species o f the genus, is distinct in the amyloid pore of asci, which are smaller sim ilarly as ascospores. Coronellaria caricinella Karst., the type specimen o f which was revised recently (Müller, Hütter et Schüepp 1970) d iffers in the structure of the excipulum (having long, pointed hairs) and in the shape of ascospores, paraphyses and the amyloid pore o f the asci. According to Nannfeldt (1932), this species is not typical representative o f Coronellaria and must be excluded from it. Coronellaria benkertii, described above, resembles also (e. g. in the shape of its ascospores) species of genera Hysteropezizella Höhn, and Hysterostegiella Höhn., apothecia o f which are permanently and often deeply immersed in plant tissue, and lanceolate paraphyses are mostly much longer than asci.
The new species is dedicated to Dr. D. B e n k e r t (Museum of Natural History o f Humboldt’s University, Berlin), who collected it and sent me for identification, together w ith other interesting Discomycetes.
Haglundia magnipilosa spec. nov.
Apothecia 1 mm diam., late sessilia, disciformia, regulariter orbicularia, anguste marginata, extus brevissime fusco-pilosa, disco obscure cinereo (sicca!), basi hyphis obscure brunneis insidentia.
Excipulum textura globulosa, cellulis 16— 18 um, vel usque ad 25 X 8— 16 fim elongatis, subcrasse tunicatis, pallide brunneolis, margine pilis 50— 70 X X 5— 14 um magnis, clavatis, cylindraceo-clavatis vel subcylindraceis, apice obtusis, unicellularibus vel 2— 3-septatis, subcrasse tunicatis, pallide flavi- dobrunneolis vel cinereobrunneolis. Hyphae medullae 2,5— 4 um crassae, irreguläres, hyalinae, crebre septatae, tenuiter tunicatae. Hyphae and basim excipuli numerosae, 2,5— 5 ¿¡m crassae, obscure brunneae, septatae.
Asci 70— 85 X 6— 7 (um, oblongo-cylindracei, basi sensim angustati, apice subobtusi, poro distincte amyloideo, octospori, sporis distichis. Paraphyses 2,5— 3 um, simpliciter filiformes, cum ascis aequilongae, hyalinae, non oleosae nec guttulatae. Ascosporae 9—10 X 2—2,5 um, anguste cylindraceae, basi angusta- tae, rectae, eguttulatae, unicellulares, hyalinae.
H a b. Ad lignum putridissimum frondosum.
L o c a l i t a s t y p i : Bohemoslovakia, Bohemia centralis, Tobolka prope Beroun, 23. IV. 1944 leg. J. Kubicka (typus, PRM, collectio J. Herink 97/44. ut Pirottaea sp.).
This species differs from Haglundia perelegans Nannf. in its much larger, mostly unicellular (rarely 2-cellular) hairs, as well as by longer and larger asci and larger ascospores. Apothecia are seated on a spare web o f brown- -colored hyphae growing through rotten wood. Haglundia elegantior Graddon (1967) has the ascospores 11— 13,5 X 3— 4 um large.
SVRCEK: NEW OR LESS KNOWN DISCOMYCETES. VII.
13

C E S K A M Y K O L O G IE 32 (1) 1978
Haglundia perelegans Nannf.
Stud.'M brphol. Syst. nicht-lichen. Inoper. Discom. p. 108, fig. 11, tab. 4, fig. 2—4, 1932 S y n . : CfUstula quercina var. carpatica Velen., Mon. Discom. Boh. p. 143. 1934
P iro ttaea g lauco -v irid is Vacek, Ces. M ykol. 4: 44, 1950
This species, originally described by Nannfeldt from Sweden, is not uncommon iri Bohemia. It occurs always on rotten wood o f old stumps o f oak, over- -grown with mosses, preferably in their cavities, but also on the under part of the stumps lying on the ground, sometimes on the walls of the cavities: at
« nflil ih L Milo ft
I I y ' CTI2. 1, Haglundia m agnipilosa Svrcek. One apothecium, ascus w ith paraphysis,. asco
spores, hairs, cells o f the excipulum (PR M , holotype). — 2. Haglundia perelegans Nannf. Ascospores, tw o hairs (P R M 616108). — 3. Haglundia perelegans Nannf. T w o paraphyses, f iv e hairs (PR M 149538, lectotype o f Crustula quercina var. ca rpatica Velen.).
M. Svrcek del.
the basis of oak trunks too. Our collections agree perfectly w ith the Nannf eldt’s original description (1932) as w ell as w ith the exsiccata from Sweden, examined by me. Macroscopically it is striking by its large apothecia (when fresh up to 6 mm diam., PRM 611919), broadly sessile, then flat, on the outer surface covered by short brown hairs. Apothecia are seated on a common subiculum form ing a sparse brown-colored web in their next vicinity. In comparison with the species described by me as Haglundia magnipilosa, the hairs of H. perelegans are much slender, narrow, 60— 90(— 110) X (2,5— )3,5— 5 ¡¿m, 2— 6-septate, often strangulate, sporadically enlarged up to 7 ptm, pale-brown, relatively thin-walled, blunt at the tip, the apical cell lanceolate-tapering or on the contrary narrowly clavate. Asci 55— 75 X 4— 5 /¿m, narrowly clavate, 8-spored, with the stipe mostly emarginate at the base, pore inamyloid. Paraphyses
14

2— 2,5 jum large, as long as asci butsom etim es exceeding them up to 10 p , blunt at the tip, cylindrical, not enlarged, hyaline. Ascospores 7— 9,5 X X 1,5— 2 biseriate, narrowly fusoid, inequilateral, attenuated towards theirends, straight or very slightly curved, eguttulate, unicellular. Cells of the excipulum on the base 17— 24 ^m diam., elongated towards the margin (up to 15 X 9 (um), brown. Hyphae of the hypothallus 4— 5 um large, reddish- brown, thick-walled, septate.
According to the type collection, revised by me, Crustula quercina var. car- patica, Velen. and Pirottaea glauco-viridis Vacek represent the same species. It seems highly probable, that the species was described already by Feltgen under the name Cenangium lign i var. olivascens Feltgen (1899). This variety was transferred by Le Gal and Mangenot (1958) as a species into the genus M ollisia [as M ollisia olivascens (Feltg.) Le Gal et Mangenot].
M ollisia lign i var. olivascens Feltgen (1899) sensu Grelet is still a different fungus, no doubt belonging to Haglundia too, but its correct name has not yet been solved.
Some o f the exam ined specimens o f Haglundia perelegans: B o h e m i a : Praha, s ilva v iva r ia “ H vězda“ , 14. V I. 1941 leg. J. H erink (holotypus Piro ttaea g lauco-v irid is Vacek, P R M 732232). — K lánovice, s ilva “V idrholec“ , 10. V. 1964, 28. V I. 1964 leg. M. Svrček (P R M 611920, 611919, 611921, 611628). — Sm ržov prope Lom nice nad Lužnicí, ad m arginem piscinae “ D vořiště“ , 6. V II. 1961 leg. M. Svrček (P R M 616108).
S S S R : U craina Transcarpatica (localitas accurata incerta), ad lignum putridum frondosum (? Fagi), V I I I . 1929 leg. A . P ilát, det. J. Velenovský (ut Crustula quercina var. carpatica Velen., lectotypus P R M 149538); the other specimens, P R M 149211 and 149575, are obviously duplicates o f the same gathering. This m ateria l is rather old (or badly dried), but agrees in a ll respects w ith the paratype o f Haglundia perelegartS. \ ■
S w e d e n : Smáland, Fem sjo parish, Dullaberget, on v e ry rotten, fa llen trunk of birch, 13.| V III . 1929 leg. J. A . N ann feld t (Fungi exsiccati suecici etc., No. 191, ut “paratype m ateria l“ , P R M 683036).
Haglundia sarmentorum SvrčekI
Ces. M ýkol. 2 1 : 146, 1967
This, species, described by me from Carpathians (Belanské Tatry Mts., Slovakia) on dead stems of Rubus idaeus, has been collected in Bohemia, too : Praha, Malá Chuchle, in moist pits at the margin of the wood “Chuchelský háj“ , on dead stems o f Rubus fruticosus ly ing on the ground, 30. IV. 1961, leg. M. Svrček (PRM 616109). The collection agrees very w ell with the original description and w ith the type specimen from Slovakia. Apothecia were 0 ,8— 1 mrft diam., sessile, without a subiculum, pale olive-green, at the margin distinctly hairy, marginal hairs pale or whitish, in the lower part shining, on the outer surface of the apothecium dark brown, disc dark olive. Excipulum composed of textura globulosa, cells globose, 3— 9 um diam., relatively thick- -walléd, brown, towards the margin elongated (up to 12 X 4— 7 ^m ) and pale- -colored. Hairs 50— 85 X 3,5— 4 p (above), 5— 7 (um (below), 2— 5-septate, thin-walled, blunt, brownish. Asci 45— 50 X 4,5— 5 urn, 8-spored, narrowly clavate, pore amyloid. Paraphyses 1,5— 2 um thick, not longer than the asci, blunt, hýaline. Ascospores 6— 7(— 8) X 1,5— 1,8(— 2) um, narrowly cuneate, straight, eguttulate, partially biseriate. The species seems to be restricted to Rubus spp.
SVRCEK: NEW OR LESS KNOWN DISCOMYCETES. VII.
15

J p j f elp (n
f * | ' l jjj°’/ j1 i ,;í •«1
3. 1. C oronella ria benkertii Svrček. Asci w ith paraphyses, tip o f one ascus, ascospores (one 3-celled and germ inating), paraphysis, three apothecia, m arginal hairs w ith cells o f the excipulum (PR M , holotype). — 2. Hymenoscyphus rhytid iadelph i Svrček. T w o apothecia, ascospores, cells o f the excipu lum w ith m arginal hyphae, paraphyses, asci (PR M , holotype). — 3. Hysterostegiella zelendarkensis Svrček. T w o apothecia, ascospores, asci w ith paraphyses, tip o f one ascus, m arginal hairs w ith cells o f the excipulum (PR M , holotype).
M. Svrček del.
Č e s k á m y k o l o g i e 32 (i) 1978
16

Hymenoscyphus rhytidiadelphi spec. nov.
Apothecia 0,8— 1 mm diam., solitaria, breviter stipitata, cyathiformia, dein explanata, subcrasse ceracea, margine obsolete denticulata, nuda, tota albida tinctu roseolo, extus glabra, stipite hyalino, nudo.
Excipulum parte basali textura porrecta, ecolorata, hyphis cylindraceis, 6 - 1 0 am crassis instructa, marginem versus cellulis subglobosis vel ellipsoi- deis, 15— 25 X 9— 13 ¡um magnis, strato marginale compacto 70— 90 urn alto, e hyphis paraphysoideis angustis, hyalinis, intus dense guttulatis instructum. Asci 90—110 X 8—10 ^m, cylindracei, deorsum sensim longe stipitiformiter angustati, apice obtuso, subtruncato, incrassato, poro haud amyloideo, octospori, sporis partim distichis. Paraphyses copiosissimae, guttulis impletae, hyalinae, apice 3— 4 um crassae, haud dilatatae vel subdilatatae, rectae. Ascosporae (8,5—) 15,5—17 X 5—6 um, forma magnitudineque valde variabili, plerumque inaequaliter fusoideo-ovoideae usque fusoideae, tenuiter tunicatae, intus subti- liter granulosae, unicellulares, hyalinae.
H a b . Ad folia viva vel pro parte emortua musci Rhytidiadelphus triquetrus.L o c a l i t a s t y p i : Bohemoslovakia, Bohemia meridionalis, Horní Ostro-
vec prope Písek, in silva humida mixta, 9. V III. 1968 leg. M. Svrček (holotypus, PRM).
More collections o f this very characteristic muscicolous discomycete are at present known from Bohemia. It occurs on some larger pleurocarpous mosses, mainly on Rhytidiadelphus triquetrus. Velenovsky’s Helotium procerum Karst. (1934) is the same species, but Karsten’s fungus, according to the revision of the type gathering, made by Dennis (1956, p. 112), is quite distinct. I was not able identify the species described above w ith another species of Helotiales occurring on Bryophyta. The relatively large, broadly fusiform-ovoid ascospores o f rather irregular shape and size seem to be one o f the most striking features of this Hymenoscyphus.
Hysterostegiella zelendarkensis spec. nov.
Apothecia 0,3— 0,8 mm diam., oblonga, ellipsoidea, primum immersa, dein erumpentia, operculo e strato epidermidis dehiscentia, sat carnosa, extus pallide luteo-brunneola, nuda, disco piano, pallide luteolo vel cinereo-cremeo, margine albido, minutissime fimbriato laciniatoque.
Excipulum textura prismatica e cellulis angulatis, isodiametricis usque oblon- gis, 5— 10 ,um diam., subhyalinis, subtenuiter tunicatis, pilis marginalibus 20— 50 X 3— 5,5 Lum, cylindraceis, parte superior! dense incrustatis, granulis usque ad 2 .um altis, unicellularibus vel uniseptatis, tenuiter tunicatis, hyalinis, apice obtusis, non dilatatis. Asci 40— 50 X 4— 5 um, anguste cylindracei, deorsum sensim longe stipitati, apice obtusi, poro amyloideo, octospori. Paraphyses copiosae, lanceolatae, 60— 75 X 4— 5,5 um, sursum sensim acutae, ascos longe superantes guttulatae, nudae, hyalinae. Ascosporae 6—7 X 1,3—1,7 ^m, tenuiter aciculares, cuneatae, rectae vel subrectae, eguttulatae, hyalinae, unicellulares.
H a b. Ad folia emortua Caricis acutiformis.L o c a l i t a s t y p i : Bohemoslovakia, Bohemia meridionalis, Zelendárky
prope Protivín, in pratis uliginosis inter piscinas “ N ový“ et “Starý“ , 10. VI. 1977, leg. M. Svrček (holotypus, PRM).
Défago (1967) in his monograph o f the genera Hysterostegiella Hohn., Hys- teropezizella Hohn. and allies, did not describe any species of Hysterostegiella
♦
SVRCEK: NEW OR LESS KNOWN DISCOMYCETES. VII.
17

occu rrin g on C a re x spp. T h e n ew species d if fe r s in its m in u te , n a r ro w ly ac icu lar ascospores fro m a ll o th er ones. T h e e llip so id a l apoth ec ia a re d eve lo p ed w ith in th e tissue o f dead lea ves som etim es on patches o f d isco lou red ep id erm is , on bo th sides o f le a v e s m os tly so lita ry . T h e p a le y e llo w is h o r g re y -y e llo w is h disc is exp osed b y th e ru p tu re o f th e c o v e r in g ep id e rm is in fo rm o f an opercu lum .
R e f e r e n c e s
D é f a g o G. (1967): Les H ysteropezizella von Höhnel e t leurs form es voisines (Asco- mycetes). Sydow ia 21 (1—6): 1—76.
D e n n i s . R. W . G. (1956) : A revision o f the British Helotiaceae in the Herbarium o f the Royal Botanic Gardens, K ew , w ith notes on related European species. M ycol. Papers 62: 1—216.
F e l t g e n J. (1899): Vorstudien zu einer P ilz -F lo ra des Grossherzogstums Luxem burg.I. Ascomycetes. Pp. 1—417. Luxem burg.
G r a d d o n W . D. (1967): Some new D iscom ycete species. Trans. Br. m ycol. Soc. 50 (1): 9-13.
K a r s t e n P. A . (1869): M onographia Pezizarum fennicarum. Not. Sallsk. Faun.Flor. fenn. 10: 98—206.
L e G a l M. e t M a n g e n o t D. (1958): Contribution à l ’étude des M ollisioïdéee Rev.M ycol. 23 (N. S.), (1): 28-86.
M ü l l e r E., H ü t t e r R. et S c h ü e p p (1964): Ü ber e in ige finnische Discomyceten.Arch. Soc. zool. bot. fenn. Vanam o 18 (3): 189—193.
N a n n f e l d t J. A . (1932): Studien über d ie M orphologie und Systematik der nicht- -lichenisierten Inoperculaten Discomyceten. Uppsala.
S v r č e k M. (1967): Species novae Discomycetum (Helotia les) e montibus Belanské Tatry, Slovakiae. Ces. M ykol. 21: 146—150.
V a c e k V. (1950): P iro ttka šedozelená (P irottaea glauco-virid is sp. n.) Ces. M ykol. 4: 42-44.
V e l e n o v s k ý J. (1934): M onographia Discomycetum Bohemiae. 1—2. Pragae.
Address o f the author: Dr. M irko Svrček, CSc., Národn í muzeum, Sectio m ycologica, Václavské nám. 68, 115 79 Praha 1, Czechoslovakia.
Č e s k á m y k o l o g i e 32 (i) 1978
18

Hypoxylon macrocarpum Pouz. spec. nov., a new fragrant pyrenomycete
Hypoxylon macrocarpum Pouz. spec. nov., nová vonná tvrdohouba
Zdeněk Pouzar
H ypoxy lon m acrocarpum Pouz. spec. nov. d iffers from dose ly related H ypoxy lon fuscum (Pers. ex Hook.) Fr. by larger perithecia, better developed w axy (resinous) layer w h ich is lustrous, and a heavy sweetish smell. It prefers w ood o f A cer, U lm us, Fraxinus, occasionally grow ing also on Car- pinus and Fagus. I t is known from 8 localities in Czechoslovakia and one in Poland.
H ypoxy lon m acrocarpum Pouz. spec. nov. se liš í od ne jb líže příbuzného H ypoxy lon fuscum (Pers. e x Hook.) Fr. větším i perithecii, s ilněji vyvinutou voskovou vrstvou (leskne se), a těžkou, sladkou vůní. Dává přednost dřevu rodů A ce r, U lm us, F raxinus a příležitostně roste také na Carpinus a Fagus.Je znám z osmi loka lit z Československa a z jedné z Polska.
During last four years I have been collecting in forests of both Czechoslovakia and Poland a species of Hypoxylon very close to Hypoxylon fuscum (Pers. ex Hook.) Fr., and not easily distinguishable from it. But after a prolonged study of the problem I am sure that one who is involved in study of taxonomy o f this intricated genus can distinguish this species in every instance. As I failed to identify this fungus w ith any known species (see e. g. M iller 1961 and Martin 1969), I am describing it as a new one — Hypoxylon macrocarpum.
Hypoxylon macrocarpum Pouz. spec. nov.
Stromatibus graveolentibus, v io láceo purpuréis, va lde effusis cum strato ceraceo m etallico nitente, peritheciis 0,5—1,2 m m longis et 0,4—0,7 m m latis, sporis 10—14—(15) X 4,5-5,5 ¿«m.
Holotypus: sylva “ Panónsky há j“ apud C ierna Voda pr. B ratislava (Czechoslovakia, S lovak ia ); ad ramum iacentem F ra x in i angustifoliae subsp. danubialis, 25. X . 1972, leg. Z. Pouzar, P R M 807840.
Stromata at first pulverulent, greenish, soon becoming firm, violet to violetrpurple or vinaceous reddish-purple, finally brownish-purple with age, w idely effused w ith indeterminate margin, at maturity effused-pulvinate, elliptic to irregular (1,4)— 3— 13 cm long, 0,4— 5 cm w ide and 0,8— 1,4 mm thick; perithecia subglobose to ovoid 0,5— 1,2 mm long and 0,4— 0,7 mm broad, from almost free to densely crowded, (stromata w ith densely crowded perithecia are provided w ith a sharply determinate margin); perithecial elevations very prominent to almost absent (prominent in margin o f stroma, less or none at the center). Under the coloured loose, pulverulent surface layer there is abundantly developed the w axy layer which gives the metallic lustrous appearance to the places (fissures or spots) where the coloured layer is discontinuous. Smell o f stroma and especially of the wood infected by mycelium is strong, heavy and sweet reminding somewhat Gaultheria or the core of Phellinus tremulae.
Paraphyses copious, unbranched, thin-walled, 1,5— 2 u m broad, not enlarged above. Asci 120— 140 ¡xm long (w ith stipe 60— 70 um long) cylindric, with amyloid, flat apical ring 2,5— 3 broad. Ascospores 10— 14— (15)X4,5— 5,5 «m ,inaequilaterally ellipsoid to navicular, uniseriate to obliquelly uniseriate, w ith
19

thin hyaline exospore and grayish-black to umber-brown endospore provided by a straight germ-slit running the entire convex part of the spore, in each spore one deBary bubble.
S p e c i m e n s s t u d i e d
C z e c h o s l o v a k i a , Bohem ia: B ranov prope K řivok lá t, ad truncum iacentem Fagi sylvaticae, 9. V II. 1950 leg. M . , Svrček, P R M 688251. — Sub “ Čertova skála“ apud Týřov ice prope K řivok lá t, ad truncum iacentem A ceris campestris, 18. IX . 1977 leg. Z. Pouzar, P R M 807821 (sterilis). — M oravia : S y lva apud “ Zám eček“ prope K rom ěříž (loco reservato), ad truncum iacentem A ceris campestris, 13. IX . 1972, leg. Z. Pouzar, P R M 807832, 807833; ibid. ad ramum iacentem A ceris campestris,13. IX . 1972, leg. Z. Pouzar, P R M 807836. — Montes B ílé Karpaty, sylva v irg inea in monte V elká Javořina supra Strání, ad truncum iacentem A ceris pseudoplatani, 22, V I I I . 1974, leg. Z. Pouzar, P R M 807835, 807839. — S y lva v irg inea “Ranšpurk“ apud Lanžhot, ad truncum iacentem C arp in i betu li, 5. X . 1977, leg. Z. Pouzar, P R M 807822, 807823, 807824, 807825, 807826; ibid. ad truncum iacentem U lm i sp.,5. X. 1977 leg. V. H olubová et Z. Pouzar, P R M 807819. — Sylva v irg inea “ Cahnov“ apud Lanžhot, ad truncum iacentem F ra x in i angustifoliae subsp. danubialis, 4. X . 1977 leg. Z. Pouzar, P R M 807820 (sterilis). — Slovakia: S y lva “Panónsky h á j“ prope Čierna Voda apud V a jn ory ( = D vom ik y ) h. p. Bratislava, ad ramum iacentem F ra x in i angustifoliae subsp. danubialis, 25. X . 1972 leg. Z. Pouzar, P R M 807840 (typus!). — Montes N ízké Tatry, sylva “ Pod Latiborskou holou“ apud K yslá (ni pede mantis); ad truncum iacentem A ceris pseudoplatani, 21. X . 1972 leg. Z. Pouzar, P R M 807828.
P o l o n i a : S y lva v irg inea B ia law ieža apud H ajnów ka,loco reservato “ Pak N aro- dow y“ , quadratum no. 283, ad truncum iacentum U lm i sp., 4. IX . 1973 leg. Z. Pouzar, P R M 807827; ibid. quadratum no. 314, ad truncum iacentem U lm i sp., 4. IX . 1973 leg. Z. Pouzar, P R M 807830, 807838; ibid. 4. IX . 1973 leg. V. H olubová e t Z. Pouzar, P R M 807829 (in societate H ypoxylon is pa p illa ti e t H. ru b ig inos i); ibid. quadratum no. 369, ad truncum iacentem C arp in i betuli, 27. V I I I . 1973. leg. Z. Pouzar, PR M §07831; ibid. quadratum no. 398, ad truncum iacentem U lm i sp., 5. IX . 1973 leg. Z. Pouzar, P R M 807837; ibid. quadratum no. 398, ad truncum iacentem Aceris plata- noidis, 24. V I I I . 1973 leg. V. Holubová, P R M 807834.
Note added in proofs:A n additional collection o f H. m acrocarpum has been found in the herbarium of
National Museum in Prague (P R M ): C z e c h o s l o v a k i a , B ohem ia : M ezn í Louka prope Hřensko, in m onte V ětrovec (450 m s. m .); ad truncum iacentem A ceris pseudoplatani, 5. V II. 1971 leg. M. Svrček (750/71, 751/71), P R M 731424, 731425.
N o t e s
Hypoxylon macrocarpum d iffers from the very closely related Hypoxylon fuscum in these characters:
1. Perithecia are broader (0,4— 0,7 mm) in H. macrocarpum than those ofH. fuscum (0,3— 0,4 mm) — hence the epithet “macrocarpum“. This constant character is the most important one in distinguishing both species. On the other hand the length o f perithecia is variable in both species and cannot be employed in distinguishing both species.
2. The w axy stratum under the coloured surface layer is more strongly developed and when it is naked it is markedly metallic lustrous. The w axy layer is developed in H. fuscum too, but not so strongly and can be seen only when the coloured surface was artificially abraded (rubbed off). In H. macrocarpum the pulverulent coloured surface is so loose that also in untouched places the metallic lustrous layer is shining through small fissures or spots in the coloured layer. Hence no special treatment is needed in H. macrocarpum to make it possible to observe the lustrous w axy layer, whereas in H. fuscum we should abrade carefully the coloured surface first.
3. It produces, when fresh, a heavy smell recalling that of Gaultlieria or the
Č e s k á m y k o l o g i e 32 (i) 1978
20

core of a polypore Phellinus tremulae (Bond.) Bond, et Boris. Hypoxylon fuscum smells sometimes sweetish too, but only very faintly. This is rather a difference of degree of intensity, nevertheless I do not hesitate to include it among characters o f highest importance. Hypoxylon macrocarpum can be identified quite safely directly in forest by this smell even i f barely sterile stromata are at disposal.
4. The surface o f stroma is more v iv id ly coloured than in H. fuscum especially when compared in fresh specimens.
5. H. macrocarpum prefers, w ith one exception, other trees than Hypoxylon fuscum. It grows mostly on wood of Acer, Ulmus and Fraximus (twice collected also on Carpinus and once on Fagus). Hypoxylon fuscum prefers, however, wood o f Alnus, Corylus, Carpinus and Betula, being common also on Fraxinus. The wood of the last genus meets w ell the ecological requirements of both Hypoxylon macrocarpum and H. fuscum. H. macrocarpum was twice collected on Carpinus betulus a common host fo r H. fuscum, nevertheless the stromata w ere quite typical in both collections.
Hypoxylon macrocarpum differs abundantly from H. fuscum even i f both species are very closely related and similar.
Another species which can be confused w ith H. macrocarpum is Hypoxylon vogesiacum (Pers. ex Currey) Sacc. d iffering in its typical variety (var. vogesiacum) by much larger spores (15— 22 um long). The only difficu lty is w ith Hypoxylon vogesiacum var. m icrosporum J. H. Miller, the small-spored variety described from North America, the spores of which are very similar to those of our H. macrocarpum. Nevertheless it differs in less developed perithecial elevations so characteristic for our new species. Also the colour o f H. vogesiacum var. microsporum is not so reddish-purple, but more bluish. The taxonomic status fo r H. vogesiacum var. microsporum J. H. M iller has not yet been settled and requires further investigations.
A somewhat similar fungus is Hypoxylon rubiginosum (Pers. ex Fr.) Fr. too, but it is really not closely related to H. macrocarpum as it belongs to quite another group o f Hypoxylon. The whole group of H. rubiginosum strikingly differs from the group of H. fuscum in its pigments. A simple test can easily be employed in order to distinguish these groups; a small fragment of stroma with surface layer is put on a filter paper and a droplet of 10 % KO H solution let fa ll on it. An orange brown spot is spreading on the paper if a member of H. rubiginosum group is involved while a grayish, bluish-gray, olive green or gray-brown one in the case of a member of H. fuscum group.
Hypoxylon macrocarpum is certainly very close to H. fuscum and those mycologists who are applying a broad species concept may possibly merge it w ith this very variable species. As I am applying a moderately narrow species concept in my work on Hypoxylon I believe that it is an own species. It does not seem to represent a minor species ( “microspecies“ ), as this term is employed in higher plants because it is distinguishable in all instances, especially when studied in fresh specimens.
R e f e r e n c e s
M a r t i n P. (1969): Studies in the Xylariaceae: V. Euhypoxylon. J. South A frican Bot., Pretoria , 35: 149—206.
M i l l e r J. H. (1961): A monograph of the w orld species o f H ypoxylon . Atlanta, pp. 158.
Address o f author: Z. Pouzar, National Museum — Natural H istory Museum, 115 79 Praha 1, Václavské nám. 68.
POUZAR: HYPOXYLON MACROCARPUM SPEC. NOV.
21

Inocybe descissa (Fr.) Quél. und ihre taxonomische Stellung (Beiträge zur Kenntnis seltenerer Inocyben. N r. 12)
Inocybe descissa (Fr.) Quél. a její taxonomické umístění
(Příspěvky k poznání vzácnějších vlákníc. Část 12.)
Johann S tangl und Jaroslav Veselský
Inocybe descissa (Fr.) Quélet im Sinne der klassischen Beschreibungen und Abbildungen in Battarra und Fries w ird von den Verfassern dieses Beitrags als e ine m eist verkannte, jedoch standortgetreue A r t der subxerophilen E ichenwälder und vergleichbaren Parkanlagen m it Eichengruppen und Ahom gew ächsen eingehend behandelt. D ie A r t w ird im System Singers der Sektion In oc ib iu m fLacerae (Fr.) Sacc., F ibrillosae H eim ] als Subsektion Descissae subsect. nov. zugeordnet.
A u toři podrobně popisují na základě vlastních pozorování a v e smyslu původních popisů a vyobrazen í u Battarry a Friese v lákn ici ostřihanou — Inocybe descissa (Fr.) Quél. jako druh m álo poznávaný, avšak věrn ý svému stanovišti, jím ž jsou subxerofiln í doubravy a jim podobné lesoparky se skupinami dubů a javorů. Druh je autory zařazen v systému Singerově do sekce In oc ib iu m [Lacerae (Fr.) Sacc., F ibrillosae H eim ] jako subsekce Descissae subsect. nov.
Bei der Bearbeitung unseres 9. Beitrags (Stangl et Veselský 1976), in dem w ir die mancherorts bezweifelte Inocybe queletii R. Maire et Konrad ex Konrad eingehend behandelt haben, blieb uns eine beträchtliche Zahl von ähnlichen Belegen zurück. Diese Belege stellen einen Übergang zwischen den “ S t i e l n u r a n d e r S t i e l s p i t z e bereiften“ und “S t i e l n i c h t b e r e i f t e n “ Inocyben der Aufschlüsselung von Kühner dar. Da es sich hier um die meist verkannte I. descissa handelt, die von Kühner und Romagnesi (1953) und Kühner (1955) mit Rücksicht auf die verschiedenen Deutungen bei Bresadola Ricken und J. E. Lange aufgegeben wurde, legen w ir unsere reichlich belegten Beobachtungen vor. Sie beruhen auf sorgfältigem Studium aller Originaldiagnosen in Fries und grösstenteils auf eigenen Funden und Terrainskizzen. Von den uns überlassenen Trockenbelegen sind diejenigen aus der Nachlassenschaft des tschechoslowakischen Mykologen F. Šmarda von besonderer Bedeutung, denn sie wurden von ihm auf seinen zuverlässig phytozönologisch und geobotanisch bestimmten Dauerflächen aufgesammelt (F. Šmarda 1972).
Der gemeinsame Nenner aller Funde, die w ir als I. descissa einschliesslich “aurivenia“ , “auricoma“ und “ geophylloides“ bestimmt haben, ist ihr Vorkommen in subxerophillen Eichenwäldern und vergleichbaren Parkanlagen der geobotanischen Assoziation Potentillo-Q uercetum pannonicum K lika und der Vegetationsstufe Carpineto-Quercetum Zlatník, besonders bei Eichen und Haseln oder auf deren abgebrannten Grasplätzen. Es sei noch bemerkt, dass Fries 1857 (1. c. p. 343) ihr Vorkommen “ in silvis acerosis“' erwähnt.
Inocybe descissa (Fr.) Quélet
Bull. Mém. Soc. Emul. M ontbéliard p. 181, 1872; F lore mycol. p. 103, 1888; Schroeter ap. Cohn, K ryptog. Fl. v. Schlesien 3 P ilze 1 p. 586, 1889; Bresadola, Fungi T rid . 2 t. 122, 1892; Iconogr. t. 743 f. 1, 1930; Britzelm ayr, Hymenom. Südbayern p. 234 f. 149 p. 276 f. 366 (Index v. Höhnel, 1904); Massee, Ann. o f Botany 18 p. 478, 1904; Dumée, Essai sur le genre Inocybe p. 30, t. 3 f. 13, 1912; Konrad, Bull. Soc. m ycol. Fr. 45 p. 45, 1929; Heim , Inocybe p. 232, 1931; Pearson, Naturalist 2 (6) p. 128, 1954; P eg le r et Young, K ew Bull. 26 (3) p. 522, 1972.
22

N o n : Ricken, B lä tterp ilze p. 104, 1915 (nom. m ixtum !); Velenovský, Čes. Houby 1 p. 366, 1920 (nom. m ixtum !); J. E. Lange, Fl. Agar. Dan. t. 113 E, 1938 (quod est I. posterula).
N o m e m e s s e n t i a l e a n t i q u u m : Monomyces pileolo discisso J. A . Bat- tarra, Fung. agri A rim inensis Hist. ed. 2, p. 43, t. 18 F, 1759.
B a s i o n y m u m : Agaricus descissus Fries, Epicr. p. 174, 1838, Monogr. 1 p. 343, 1857; Hymen. Europ. p. 233, 1874.
•É¡*
0 l \ n y J /n ) I Z05?) Sfielhaut-
I H*Phen
1. Inocybe descissa (Fr. Quél. var. descissa. — Konturskizze und M ikrom erkm ale.Augsburg, BRD, Siebentischpark, 1. V I. 1974 leg. J. Stangl. PRM .
J. Stangl del.
S y n o n y m a : Agaricus aurivenius Batsch, E lenchi fungorum continuatio prim a p. 137, t. 20 f. 107a, b, 1786; Agaricus descissus form a aurivenia (p ileo fuscescente lúteo-venoso) Fries, Hym. Europ. p. 233, 1874; Agaricus aurivenius Batsch in Fries, Monogr. 1 p. 343 (“ e notis essentialibus A. descisso videtu r subjungendus“ ), 1857; Inocybe descissa (Fr.) Quél. var. aurivenia (Batsch e x Fr.) Quél., Fl. myc. p. 103, 1888; Inocybe aurivenia (Fr.) Ricken, B lätterp ilze p. 104 et 464, 1915. — Agaricus auricomus Batsch, Elenchus p. 75, t. 5 f. 21, ( “p ileo lúteo, lineis fuscis radiatim piloso; stip ite lineari, solido, albo, lam ellis fusco-cinereis“ ), 1783; Agaricus descissus Fr. var. auricomus Batsch, Fries in Epicrisis p. 175, 1838; Hym. Europ. p. 233, 1874; Inocybe descissa (Fr.) Quél. var. auricoma (Batsch e x Fr.) G illet, Hyménomyc. 19 p. 513, 1874; Heim , Inocybe p. 233, t. 20 f. 4, 1931; Inocybe auricoma (Batsch ex Fr.) Ricken,
STANGL et VESELSKÝ: INOCYBE DESCISSA
23

Č E S K Á M Y K O L O G IE 32 (1) 1978
0jJ i ~fZ Sř/rihoní- - H yphen
cm1 a00 o s ()M r\ I * y, ch.u.pt.
1 í M m i??1 i í li \ I 11 A 1 vi 1/i i ' '!U I \ I« ■■I# v
H yphen rom Stíd ^ o m Lo m d /e n o rjsc fjL
2. Inocybe descissa (Fr.) Quél. var. descissa. — a) M ikrom erkm ale des Fundes vom9. V I. 1972, Augsburg, BRD, Siebentischpark, leg. J. Stangl. PR M . — b) M ikrom erkm ale des Fundes vom 5. V I. 1960, K uřim b. Brno, ČSSR, W ald Sibem á, leg. F. Šmarda. PR M . J. Stangl del.
B lätterp ilze p. 104 e t 464, 1915; J. E. Lange, Dansk bot. A rk . 2 (7) p. 37, 1917; Fl. Agar. Dan. t. 113 B, 1938; Velenovský, Čes. Houby 1 p. 374, 1920; Pearson. Naturalist 2 (6) p. 126, 1954; P eg le r et Young, K e w Bull. 26 (3) p. 522, 1972. — Inocybe phaeodisca Kühner in Kühner e t Romagnesi, FI. analyt. p. 227, 1953 (nom. nudum); Kühner, Bull. Soc. nat. Oyonnax 9, suppl. 1 p. 5 e t 75, 1955. — Inocybe phaeodisca Kühner var. geophylloides Kühner, Bull. Soc. nat. Oyonnax 9, suppl. 1 p. 5 et 77, 1955.
24

■ S T A N G L et V E S E L S K Ý : IN O C YB E DESCISSA
B e m e r k u n g e n z u r T a x o n o m i e
Es sei hier bemerkt, dass wir, auf die ältesten Beschreibungen in Battarra■ und Fries fussend und mit voller Berücksichtigung die subspezifische Glie-
derung in Fries betreffs der absolut unlösbar gefassten Taxone A. aurivenius und A. auricomus, w ie in Batsch, so auch in Fries, absichtlich auf eine Glie- derung der I. descissa auf Varietäten oder Formen verzichten. Die ausführlich lesbare und m it einer glänzenden Federzeichnung versehene Beschreibung
H ¿ ‘f f f V Aťw? SheloLertml \ [
I 3. Inocybe descissa (Fr.) Quél. var. geophylloides ex K ü h n er . — Konturskizze undI M ikrom erkm ale. Inzigkofen, BRD, auf Auw aldboden , 24. IX . 1974 leg. J. Stangl.■ M. 1031. J. Stangl del.
I Battarras hat Fries als Agaricus descissus übernommen und das ganz eindeutigeI Merkmal “ stipite apice albo-pulverulento“ zugegeben. Es stellte sich aber■ heraus, dass gerade das Merkmal “ Stiel nur an der Spitze bereift“ zuverlässigI erst nach exakter Untersuchung, insbesondre bei angewachsenen bzw. ange-I hefteten Lamellen, festgestellt werden kann. W ir haben an eigenem MaterialI wiederholt beobachtet, dass das erwähnte Merkmal an einzelnen Fruchtkörpern,I die am selben Ort und Stelle aufgesammelt wurden, sehr variabel sein kann
und auch übersehen wird. Erst bei mikroskopischer Untersuchung können an der Stielspitze vereinzelt Kaulozystiden, öfters aber reichlich Dermato- pseudozystiden ohne Metuloiden nachgewiesen werden.
25

do m
vOTvy¡¡I 11 W c h .u f í v í u
nMkft/ n A. \A O I ) | ) \\ ¡ I \ )
\ A A nÍ\Kau/ozysHden VÍ|) 'rom S t/el o b e n \ j
4. Inocybe descissa (Fr.) Quél. var. geophylloides ex Kühner. — M ikrom erkm ale des Fundes vom 10. X . 1960, K uřim b. Brno, CSSR, W a ld Zlobice, leg. F. Smarda. PRM .
J. Stangl del.
Inocybe phaeodisca Kühner muss als ein Taxonomisches Synonymum zur I. descissa (Fr.) Quél. gestellt werden. Selbst Kühner gibt zum Abschluss seiner Beschreibung folgende Notiz bei: “ Il s’agit certainement du I. descissa Fr., tel que le figure Bresadola (le. 743,1); il n’est pas impossible, qu’il s’agisse du descissa de Fries lui-même (Monogr.), bien que le stipe soit plein“ . (Kühner1. c. p. 76). Da Kühner merkwürdigerweise seine Inocybe phaeodisca der Gruppe
Č e s k á m y k o l o g i e 32 (i) 1978
26

n i c h t S t i e l b e r e i f t e n Inocyben zugeordnet hat, muss man bei genauem Suchen und Auffinden von Kaulozystiden ohne Ergebnis aufgeben. Überdies betont Kühner keine Stielbereifung: “ stipe... non pruineux dans la moitié ou le tiers supérieur“ (1. c. p. 79), als das Unterscheidungsmerkmal zu I. geo- phylla, aber bei der Beschreibung gibt er wörtlich folgendes an:
bei 1. phaeodisca: “ Stipe . . . passant au sommet à une furfuration blanche . . .“ H{1. c. p. 76) also “ S t ie l. . . an der Spitze in weisse Kleien übergehend“ ;
I ' i I■ 5. Inocybe descissa (Fr.) Quél. var. auricom a (Batsch ex Fr.) Helm . — M ikrom erkm aleI m it au ffä llig aufgeblasenen Hymenialzystiden. Ostrava, CSSR, Halde “ H rabůvka“ ,I auf e iner Brandstelle am Rand des Laubwaldes, 30. V I I I . 1970. leg. J. Veselský.■ J. Stangl del.
I bei var. geophylloides: “ S t ip e ... pruiné ou villeux-tomenteux à l ’extrémeI sommet seulement (1. c. p. 78), also “ S t ie l. . . nur an der äussersten SpitzeI bereift oder zottig“ .I Die I. descissa var. brunneoatra Heim und var. microspora (J. E. Lange) Heim ||||I entziehen sich ausser kleineren Abweichungen der Grundmerkmale schon■ weitgehend der hier beschriebenen Ökologie (in unserem Material) und w irI betrachten sie mit Dennis-Orton-Hora übereinstimmend, als gute Arten, dieI gemeinsam mit 1. descissa (Fr.) Quél. eine selbständige Subsektion in derI Sektion Nr. 6 in Singers Agaricales vorstellen.
I D i a g n o s i s l a t i n a s u b s e c t i o n i s n o v a e :I Descissae Stangl et Veselský subsect. nov. — Characteres sectionis N o 6 In ocib iu m■ im Singer, Agarica les ed. 3 p. 573, 1975. P ileus et pigm enta sine umbris violaceis;I stipes apice pruinosus, raris cum dermatocystidiis, sed m ultis dermatopseudocystidiis.
I — Grundmerkmale der Subsektion Descissae Stangl et Veselský subsect. nov.:Hut und Pigmente nicht violettlich; Stiel nur an der Spitze bereift. Dermato- zystiden (typische Metuloiden) selten, Dermatopseudozystiden reichlich vorhanden.
H STANGL et VESELSKÝ: INOCYBE DESCISSA
27
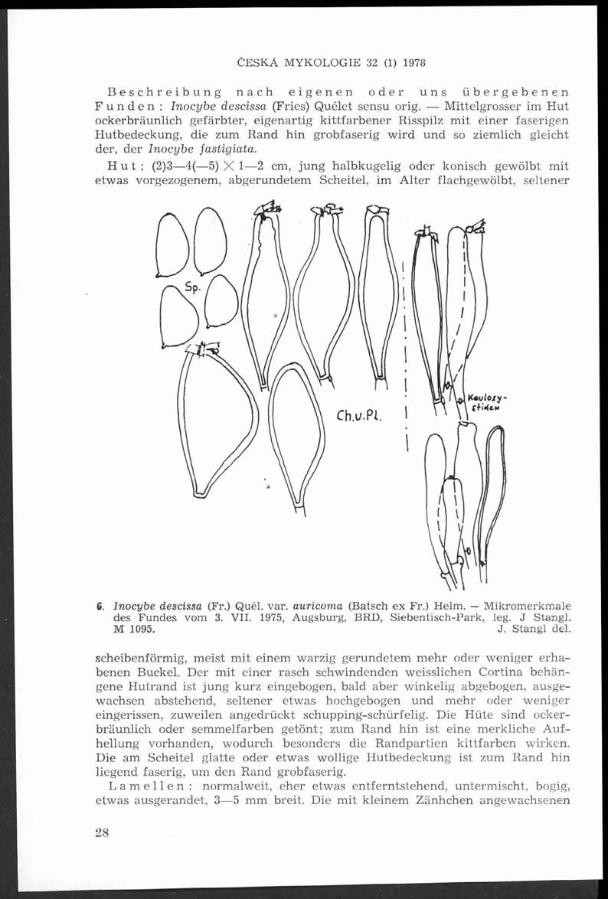
B e s c h r e i b u n g n a c h e i g e n e n o d e r u n s ü b e r g e b e n e n F u n d e n : Inocybe descissa (Fries) Quélet sensu orig. — Mittelgrosser im Hut ockerbräunlich gefärbter, eigenartig kittfarbener Risspilz mit einer faserigen Hutbedeckung, die zum Rand hin grobfaserig w ird und so ziemlich gleicht der, der Inocybe fastigiata.
H u t : (2)3— 4(— 5) X 1— 2 cm, jung halbkugelig oder konisch gewölbt mit etwas vorgezogenem, abgerundetem Scheitel, im A lter flachgewölbt, seltener
l y '#¥6. Inocybe descissa (Fr.) Quél. var. auricom a (Batsch ex Fr.) Helm . — M ikrom erkm ale
des Fundes vom 3. V II. 1975, Augsburg, BRD, Siebentisch-Park, leg. J Stangl. M 1095. J. Stangl del.
scheibenförmig, meist mit einem warzig gerundetem mehr oder weniger erhabenen Buckel. Der mit einer rasch schwindenden weisslichen Cortina behän- gene Hutrand ist jung kurz eingebogen, bald aber w inkelig abgebogen, ausgewachsen abstehend, seltener etwas hochgebogen und mehr oder weniger eingerissen, zuweilen angedrückt schupping-schürfelig. Die Hüte sind ockerbräunlich oder semmelfarben getönt; zum Rand hin ist eine merkliche A u fhellung vorhanden, wodurch besonders die Randpartien kittfarben wirken. Die am Scheitel glatte oder etwas wollige Hutbedeckung ist zum Rand hin liegend faserig, um den Rand grobfaserig.
L a m e l l e n : normalweit, eher etwas entferntstehend, untermischt, bogig, etwas ausgerandet, 3— 5 mm breit. D ie mit kleinem Zänhchen angewachsenen
Č e s k á m y k o l o g i e 32 (i) 1978
28

Lamellen sind jung hellbeige gefärbt und werden alt lichtbraun; seltener ist schwacher Olivstich wahrnehmbar. Die glatten, seltener etwas wolligen Lamellenschneiden sind durchgehend weisslich bewimpert.
S t i e l : 2— 5(— 6 ) X (0,3)04,— 0,6(— 0,7) cm, rundlich, vorwiegend gleichdick, etwas verbogen, jung ausgestopft-voll, alt hohl werdend, mit einer gleichdicken, seltener minimal verdickten Basis. Die zartholzfarbenen Stiele sind fein liegend befasert und tragen zuweilen im oberen Stieldrittel ein kleines faseriges, oft nur angedeutetes Kränzchen. Unter dem Lamellenansatz kann am Stiel durchaus eine schmale bereifte, bis beschürfelte Zone vorhanden sein.
F l e i s c h : im Hut weisslich, 1— 2 mm dick, im Stiel faserig holzfarbig bis lieb testbraun, faserig brechend; im Geruch etwas modrig-mehlig, kaum auffällig.
S p o r e n s t a u b : tabakbraun (Moser B 10, D 10).B a s i d i e n : 30— 35 X 9— 10 ^m, vorwiegend mit 4 Sterigmen.B a s i d i o s p o r e n : (8)9— 10,(— 12— 14) X 5,5— 6— 6,5 ^m, länglichoval,
selten etwas deformiert. Es sei hier erwähnt, dass Heim (1. c. p. 236) bei seinerI. descissa var. m a crosp o ra , die Sporendimensionen 10—13(14—17,5) X 5,2—6,5 (—7,5) um feststellte. Der Sporenquotient schwankt abermals um 1,90—1,95. Die Sporenwänden sehen im Lichtmikroskop etwas hebelomaartig rauhwarzig aus und an die Stirps F lo ccu lo sa Heim auffällig erinnernd. Cf. unseren 10. Beitrag (1977) mit Farbtafel Nr. 91.
C h e i l o - u. P l e u r o z y s t i d e n : (40)50— 70 X (13)15— 20(— 24!) u m mit bis zu 2,5 um dicken, in NH/tOH kaum gilbenden Wänden.
K a u l o z y s t i d e n : dünnwandig, mit Kristallen nur ausnahmsweise und vereinzelt im Bereich des Lamellenansatzes zu finden. Vorwiegend ist der Stieloberteil m it langgestreckten, in der M itte etwas blasig auf getriebenen 50— 70— 80 um langen und 8— 15(— 20) um breiten Flockenhyphen (Dermato- pseudozystiden), die reichlich Schnallen tragen, besetzt.
M ö g l i c h e V e r w e c h s l u n g e n
Die im Terrain sehr ähnliche und auch im System nahe stehende A rt ist die I. le p to cy s tis Atk., bei der auch vereinzelt Kaulozystiden zu finden sind (A. Einhellinger in litt.). Fast unmöglich ist die I. descissa bloss nach Exsikkaten zu beurteilen. Erhebliche Unterschiede in der Sporenlänge und Zystidenbreite, aber auch in Kaulozystiden an Stielspitzen wurden festgestellt. Hellere Formen der I. d eg lu ben s (Fr.) Gill, und I. s u b tig r in a Kühner könnten damit vertauscht werden ohne alle Merkmale nicht kritisch zu erwägen. Über die I. q u e le t i i haben w ir in unserem 9. Beitrag (1976) berichtet und die I. g r ise ov e la ta Kühner w ird im vorangehenden 11. Beitrag behandelt.
D a s u n t e r s u c h t e M a t e r i a l
a) D ie als 1. descissa Fr. ss. Bresadola bestimmten Belege:K u rim bei Brno, Landkr. Südmähren, CSSR, W a ld “ S ibem ä“ . 330 m, subxerophiler
E ichenwald der Assoziation Po ten tillo -Q u e rce tu m pannonicum K lika , mässiger Podsolboden au f Granodiorit, Beobachtungsquadrat Nr. 1: 5. V I. 1960 (P R M 803160);I I . V I. 1960 (P R M 803147); 20. V I I I . 1960 (PR M 803148) leg. F. Smarda.
b) D ie als I. descissa Fries f. auriven ia Fr. ss. Heim f = 1. aurivenia (Fr.) Ricken 1915] aufgesam m elten Belege:
K u fim bei Brno, Landkr. Südmähren, CSSR, W ald “Z lob ice“ , 330 m, subxerophiler E ichenwald der Assoziation P o ten tillo -Q u e rce tu m pannonicum K lika, mässig ver- g leyter W aldboden der brünner Eruptivmasse (A p lit) m it angewehtem Löss, Beo-
STANGL et VESELSKY: INOCYBE DESCISSA
29

bachtungsquadrat Nr. 10: 25. IX . 1960 (P R M 803149); 16. X . 1960 (P R M 803150) leg. F. Šmarda.
c) D ie als I. descissa Fr. var. auricoma (Batsch ex Fr.) H eim [ = I. auricoma (Batsch ex Fr.) R icken 1915] aufgesam m elten Belege:
Ostrava, Landkr. Nordm ähren, ČSSR, Hochofenschlackenhalde “ H rabůvka“ im A llu v ia lgeb ie t des Flusses Ostravica, z. T. w ildbewachsen, z. T. als Laubw ald reku ltiv iert, besonders B irken und Kanada-Pappeln , Schwarzpappeln, vere in zelt d ie F laum -Eiche (Q. pubescens W illd .) angepflanzt, auf e iner a lten m it frischen Gras bewachsenen Brandstelle am Rand des Laubw aldes; 27. V I I I . 1970 (P R M 803151),30. V II I . 1970 (P R M 803152) leg. J. Veselský. - Augsburg, BRD, Siebentischpark, Dauerfläche Nr. 1, bei Ahorn, 3. V II. 1975 leg. J. Stangl (M 1095); Siebentischstrasse, bei Eichen und Buchen an Löschwegrand im Strassengraben, 17. V I. 1976 leg. J. Stangl (Herb. J. Veselský).
d) D ie als I. phaeodisca Kühner au f gesam m elten Belege:Augsburg, BRD, Siebentischpark, Dauerfläche Nr. 2 (cf. J. Stangl 1965), k iesiger
Boden im kurzen Gras bei Eichen: 4. V I. 1970 (Herb. J. Stangl), 10. V I. 1971 (Herb. J. Stangl), 25. V I. 1971 (P R M 803153), 9. V I. 1972 (P R M 803154), 11. V I. 1973 (P R M 803155), I. V I. 1974 (P R M 803156), 14. V I. 1975 (P R M 803157) und 21. V I. 1975 (Herb. J. Veselský) leg. J. Stangl.
A n m e r k u n g : D er a llererste Fund au f dieser Loka litä t vom 30. V. 1966 w urde als Inocybe auricoma (Batsch ex Fr.) Lge. erk lärt und als Beleg ins Herbarium M unter d. Num m er 254 hinterlegt.
e) D ie als I. phaeodisca Kühner var. geophylloides Kühner aufgesam m elten und bestimmten Belege:
K u řim bei Brno, Landkr. Südmähren, ČSSR, W ald “Z lob ice“ , 330 m, subxerophiler E ichenwald der Assoziation Potentillo-Quercetum pannonicum K lika , Beobachtungsquadrat N r. 5: 11. IX . 1960 (P R M 803148) und 10. X . 1960 (P R M 803159) leg. F. Smarda, det. J. Veselský. — Inzigofen, Landkr. Siegmaringen, BRD, auf Auw aldboden bei Laub-u. Nadelbäumen: 24. IX . 1974 leg. J. Stangl (M 1031).
D a n k s a g u n g
Für gern gew ährte H ilfe und w ertvo lle taxonom ische H inw eise sind w ir dem H errn Z. Pouzar zu grossem Dank verpflichtet.
iL i t e r a t u r
B a t s c h A . J. G. (1783): Elenchus fungorum. Halae.B a t s c h A . J. G. (1786): E lenchi fungorum continuatio prim a, Halae.B a t t a r r a J. A . (1759): Fungorum agri Arim inensis historia. E d itio secunda. Fa-
ventiae.B r e s a d o l a G. (1881—1892): Fungi T ridentin i nov i v e l nondum delineati descripti e t
iconibus illustrati. Tridenti.B r e s a d o l a G. (1930): Iconographia m ycologica 15. M ilano.B r i t z e l m a y r M. (1879—1897): Hym enom yceten aus Südbayem . Pp. 1 — 390,
t. 1 — 761 (Index von v. Höhnel, 1906). Augsburg.F r i e s E. M. (1838): Epicrisis systematis m ycologici. Upsaliae e t Lundae.F r i e s E. M. (1857): M anographia Hym enom ycetum Sueciae. Upsaliae.F r i e s E. M. (1874) : Hym enom ycetes Europaei. Upsaliae.H e i m R. (1931): L e genre Inocybe. Paris.K o n r a d P. (1929) : Inocybe eutheles (Berk, e t Br.) Saccardo et espèces voisines.
Bull. Soc. mycol. France 45: 41—46.K ü h n e r R. (1955): Com plém ents à la F lo re analytique V. Inocybe leiosporés
cystidiés. Bull. Soc. nat. Oyonnax 9, suppl. 1: 1—95.L a n g e J. E. (1917): Studies in the Agarics o f Denm ark II I . Pluteus, Collybia, Inocybe.
Dansk bot. Ark . 2 (7): 23-48.L a n g e J. E. (1938): F lora Agaric ina Danica 3. Copenhagen.M a s s e e G. (1904): A monograph o f the genus Inocybe Karsten. Annals o f Botany
18 (71): 459-504.P e a r s o n A . A . (1954): The genus Inocybe. Naturalist 2 (6): 117—140.P e g 1 e r D. N. e t Y o u n g T. W. K . (1972): Basidiospore form in the british species
o f Inocybe. K e w Bull. 26 (3): 499-537.Q u é l e t L. (1872—1875): Les champignons du Jura et des Vosges. M ontbéliard.
Č e s k á m y k o l o g i e 32 u ) 1978
30

■ Q u é l e t L. (1888): F lore m ycologique de la France et des pays lim itrophes. Paris.■ R i c k e n A . (1915): D ie B lätterp ilze (Agaricaceae) Deutschlands und der angren-■ zenden Länder besonders Österreichs und der Schweiz 1—2. Leipzig.■ S c h r o e t e r J . (1889): P ilze. Erste H älfte . Cohn F.: K ryptogam en-F lora von Schle-■ sien 3. Breslau.H S i n g e r R. (1975): The Agarica les in m odern taxonom y. Ed. 3, Vaduz.■ S t a n g l J. (1965): Zur Kenntnis der P ilzvegetation in Parkanlagen. P ilze in den■ Siebentischanlagen. Zeitschr. f. P ilzkude 31: 85—100.■ S t a n g l J. et V e s e l s k ý J. (1971: Beitrag zur Kenntnis der selteneren Inocybe-■ -Arten. Ces. M ykol. 25 (1): 1—9.■ S t a n g l J. et V e s e l s k ý J. (1976): B eiträge zur Kenntnis seltenerer Inocyben■ Nr. 9: Inocybe queletii R. M aire e t Konrad. Ces. M ykol. 30 (3—4): 176—180.■ S m a r d a F . (1972): D ie P ilzgesellschaften ein iger Laubw älder Mährens. A cta Sc.■ nat. Brno, 6 (6): 1—53.■ V e l e n o v s k ý J . (1920): České houby 1. Praha.
■ A n sch rift der Autoren : Johann Stangl, von der Tannstraße 48, 8900 Augsburg.■ BRD. — Jaroslav Veselský, Dr. med., Výškovická 100, 704 00 Ostrava, ČSSR.
■ I n g o N u s s : Zur Ökologie der Porlinge. Bibliotheca M ycologica vol. 45, pp. 1—258. I V yda l J. Cram er v Gantner Verlag, Vaduz. Cena DM 50.
■ K n iha je výsledkem původní badatelské práce autora o ekologii chorošů, k terý ji I vypracoval jako disertaci na universitě v západním Berlíně. Podtitu l práce „Výzkum y■ sporulace některých chorošů a průzkum na nich nalezených brouků“ nám nepodává I ještě úplný obraz o nových a za jím avých zjištěních, která v této knize najdeme. Je to I skutečně přík ladná disertace, kdy autor se snažil na pom ěrně m além m ateriálu pro- I zkoumat nejrůznější otázky, které by m ohly m ít význam ať již pro ekologii chorošů■ nebo pro je jich taxonom ii. H lavn í část práce pojednává o vlastním výzkum u sporulace I některých chorošů (celkem 27 druhů). Tato otázka není nová a zabývali se j í v m inu- I losti např. Orloš, Parm asto a jin í. Je známo, že plodnice mnoha druhů chorošů jsou■ po dlouhé období v roce úplně steriln í a v určitém, někdy dosti krátkém období na-■ stane sporulace, během které p lodnice vyprodukuje m nožství výtrusů. Au tor přichází I se zajím avou aparaturou, která reg istru je vypadáván í výrusů z plodnice přím o v te- I rénu v určitých časových intervalech. V ýsledky jsou pak zpracovány v grafech a ta- I bulkách. Věnova l pozornost i m orfo log ii výtrusů, kterou v některých případech studoval I pom ocí rastrovacího elektronového mikroskopu. Práci uzavírá seznam brouků, které I na choroších zjistil.I Celkově m ožno uzavřít, že práce přináší tyto důležité podněty:I 1. Rozdělen í chorošů podle sezónního sporulačního rytmu.I 2. P lodn ice produkují krom ě norm álních výtrusů i tak zvané “proterospory“ (v h lav-■ ním textu zvané “ protospory“ — toto je však zadaný term ín), výtrusy m alé v e li-I kosti a většinou s tenčí blanou. O b jev těchto proterospor je neobyčejně za jím avýI a vysvětlova lo by se tím ve lm i mnoho zejm éna v některých kra jně podivných pří-I pádech variab ility . N icm éně z hlediska obecné sporologie má toto zjištěn í ještěI další dosah, k terý však autor v práci nevzpom íná. Je totiž nyní zřejm é, že basidio-I m ycety mohou produkovat vlastně dva typy anomálnícch basidiospor: jednak ty,I k teré m a jí b lány tenčí a jsou drobnější, tedy depauperátní basidiospory zvané Nus-I sem proterospory, a na druhé straně basidiospory s extém ně tlustou blanou, kteréI vzácně pozoru jem e zejm éna u čeled i Tricholomataceae a které bychom m ohli na-I zvat “ sklerospory“ . Nussovo zjištěn í proderospor bude třeba ještě h louběji prozkou-I mat, protože by to m ohlo vnést světlo jak do m orfologických, tak i do fy logenetic-I kých otázek.I 3, Au tor potvrd il d řívě jš í výzkum y o skladbě různých broucích společenstev žijíc ích I u různých druhů chorošů, často se liš ících i podle stáří plodnice:I Je to bezesporu jedna z nejzdařilejších prací o b io logii velkých hub posledních let,I cenná zejm éna původním m yšlenkovým i m etodickým přínosem.I Zdeněk Pouzar
I STANGL et VESELSKÝ: INOCYBE DESCISSA

Kvasinky v lidském materiálu u nás a jejich rozlišení. Část I
Yeasts in Human Material in our Country and their Differentiation.
Part I
P e t r F ra g n e r
Uveden diagnostický systém k určování kvasinek, vyskytu jíc ích se v lid ském m ateriá lu na našem území. Jsou v něm shrnuty naše zkušenosti, získané na v íce než 15 000 kulturách během posledních 20 let. Pod le našich postupů je možno s dobrou přesností určit asi 98% kvasinek za 48 hodin nebo dříve, za předpokladu, že vycházím e z čistých kultur.
K líč je založen v p rvé řadě na a) asim ilaci glukózy, galaktózy, sacharózy, m altózy a laktózy, b ) asim ilaci kaliumnitrátu, c) m akroskopickém a m ikroskopickém vzhledu kultur. V některých skupinách je použito ještě auxano- gramů dalších zdrojů uhlíku, zymogram ů atd.
P rvn í část publikace (I) obsahuje odstavce: Uvod, M ateriá l a metody, Laboratorn í práce a ž ivné půdy, Přeh led druhů, Výskyt a rozšíření, K líč. Další části budou obsahovat podrobné charakteristiky druhů, diskusi, závěr a literaturu. ______________
A diagnostic system fo r the determ ination o f yeasts occurring in human m ateria l on our territory is presented. I t summarizes our experiences gained w ith m ore than 15 000 cultures during the last 20 years. Accord ing to our procedures it is possible to fa ir ly exactly determ ine approxim ate ly 98% o f yeasts w ith in 48 hours o r earlier, presum ing that pure cultures are at hand.
The key is based in firs t place on: a ) glucose, galactose, sucrose, m altose and lactose assimilation, b) potassium nitrate assim ilation, c) macroscopic and m icroscopic appearance o f the cultures. For some cultures also auxano- grams o f further carbon sources, zymograms, etc. are used.
The first part o f the publication (Part I) contains the fo llow in g paragraphs: Introduction, M ateria l and Methods, Laboratory Studies and N utrien t Media, Survey o f the Species, Incidence and Distribution, K ey. The fo llow in g parts w il l contain detailed characteristics o i the species, discussion, conclusion and literature.
Ú v o d
Přesné, botanické určování kvasinek v plném rozsahu (Lodderová et al. 1970) je nesnadné. Vyžaduje příliš mnoho laboratorní práce, vzácných a čistých chemikálií a především mnoho času. V oblasti lékařské mykologie včasná diagnóza mnohdy rozhoduje o životě nemocného a urychlení laboratorní diagnostiky je proto úkolem prvořadým. V minulosti celá řada autorů (včetně nás) vypracovala nejrůznější, dílčí rychlé metody, z nichž některých se ještě používá. V současné době jde spíše o zhotovení celých diagnostických systémů, často využívajících mikrometod a poloautomatických přístrojů, s vyhlídkou na úplnou automatizaci a se zapojením počítačů v budoucnosti. O spolehlivosti některých z nich diskutují například Bowman et Ahearn (1976) a Segalová et A je llo (1976).
Spolehlivost těchto systémů je závislá nejen na reprodukovatelnosti biochemických reakcí a na jejich indikátorech, ale především na dokonalosti a kapacitě určovacích klíčů. Klíče, zahrnující jen „klin icky významné“ kvasinky a nikoliv také současně nejčastější náhodné nálezy a kontaminaci z prostředí, mají — pro svoji nepřirozenou jednoduchost — malou naději pro použití v praxi. Náhodné nálezy z prostředí budou však z velké části geograficky odlišné a proto univerzální, celosvětový klíč nebude možný.
Každé zjednodušení problematiky tak složité nevyhnutelně povede k celé řadě omylů. Bude záležet na jejich únosnosti. Budoucí systém musí bezpodmí-
32

H nečně rozlišit druhy kvasinek patogenních či oportunistických od těch, jejichžH škodlivost pro člověka nebyla prokázána anebo se zatím nepředpokládá. Roz-■ lišení neškodných kvasinek navzájem není již pro lékařskou mykologii pod-H statné.■ Jedinou možnou cestou k řešení — podle našeho názoru — je shromáždit■ všechny kvasinky v těch kulturálních formách, které se vyskytují v lidskémH materiálu z určité oblasti. Na vlastních kulturách zjistit jejich hlavní diferen-H ciálně diagnostické znaky a začlenit je do určovacího klíče. Použít metodiky■ co nejjednodušší, a nejrychlejší, i za cenu některých nepřesností.■ Výsledkem této cesty je následující návrh diagnostického systému, podleH něhož lze určit čisté kultury kvasinek z lidského materiálu během 48 hodin.
■ M a t e r i á l a m e t o d y
■ Práce je založena na souhrnu charakteristik kvasinek, které jsme izolovali■ z různých chorobných projevů (také nemykotického původu) pacientů ve Stře-I dočeském kraji a v hlavním městě Praze během posledních dvaceti let. Za tutoI dobu prošlo našima rukama více než 15 000 kultur. Do roku 1976 jsme je■ určovali podle monografie Lodderové a Kreger-van R ijové (1952) a podle někte-■ rých dalších, menších prací, později podle nového vydání publikace Lodderové■ et al. (1970). Poněvadž toto nové vydání přineslo četné změny, bylo nutno naše■ starší údaje přehodnotit a asi 300 kmenů (udržovaných v Castellaniho vodních■ kulturách) přeurčit. Nové izoláty byly hodnoceny již podle nových metodik.
■ L a b o r a t o r n í p r á c e a ž i v n é p ů d y
■ P r i m o k u l t u r y a č i s t á k u l t u r a . Prim okultury jsm e získávali pěstováním■ na Sabouraudově glukózovém agaru s aneurinem s přím ěsí chloram fenikolu (0,1 gI na 1 litr), k terý odstranil (pochopitelně jen citlivou ) bakteriá ln í kontaminaci. P ře-■ čištění bylo provedeno na Sabouraudově glukózovém agaru s aneurinem v PetrihoI m iskách běžnou, bakteriologickou technikou očkováním kličkou do izolovaných kolo-I nií. Za čistou kulturu se považu je ta, která dává izo lované kolonie stejného vzhledu.I D vakrát opakovaný postup je považován za dostačující. Skrytá bakteriá lní konta-H m inace je pom ěrně vzácná. Z jedné, typické, izo lované kolonie, dostatečně vzdálenéI od ostatních, vyočku jem e kličkou na šikm ý Sabouraudův g lukózový agar s aneurinemI ve zkumavce, pom nožím e a s touto kulturou (případně s dalším i je jím i subkultu-I ram i) p rovád ím e všechny zkoušky.I I n k u b a č n í t e p l o t a je v e všech případech 24 °C.I S a b o u r a u d ů v g l u k ó z o v ý a g a r s a n e u r i n e m : Aneurin HC1 0,05,I baktopepton Spofa 10,0, glukóza subst. Spofa 40,0, agar (podle jakosti, druhu a stáří)I 15,0 až 30,0, destilovaná voda 1000,0; pH, které bývá kolem 5,8—6,2, neupravujeme.I Rozehřejem e 30—35 m inut v průběžné páře (m nožství 3 litry rozehřívám e 45 minut),I dokonale promícháme, roz lijem e do zkum avek nebo do baněk a sterilizu jem e v prů-I béžné páře (100 °C ) 2 hodiny. Zkum avky šikm o položíme. Půda v Erlenm eyerovýchI baňkách nám slouží jako zásobní a k rozléván í do Perih o misek. Baňky či zku-■ m avky se ztuhlým a dokonale vych lad lým agarem m ůžeme přechovávat v chladničceI při asi + 5 °C.I Ž i v n é p ů d y v P e t r i h o m i s k á c h p řip ravu jem e tak, že zásobní půduI v E rlenm eyerově baňce rozehřejem e na vodn í lázn i a rozlijem e do sterilních PetrihoI misek. A g a r v miskách, ztuhlý a zcela studený, osušíme v sušárně 15—20 minut přiI 80 °C : do vyh řá té sušárny um ístím e ved le sebe otevřené m isky s agarem, každouI na svo je víčko, se zvláštn í pozorností, abychom je nekontam inovali. P ro 15—20I m inutách setřepem e zbytky kondenzační vod y s víček, m isky uzavřem e a nechámeI schladnout v íčkem dolů na pracovním stole.I C u k r o v é a u x a n o g r a m y nám ukazují, k terý cukr (nebo jin ý zdroj uhlíku)I je kultura schopna asim ilovat. Příslušnou živnou půdu (v iz dále) v Petrih o miskáchI osušíme (v iz výše) a na spodní straně m isek derm ografem vyznačím e výseče pro
H F R A G N E R : K V A S IN K Y V L ID S K É M M A T E R IÁ L U . I.
I 33

jednotlivé cukry. Zkoušenou kulturu m asivně naočkujem e (3krát křížem ) kličkou po celém povrchu agaru. P o naočkování nasypem e na povrch agaru v určených výsečích m alé hrom ádky cukrů, případně dalších zdrojů uhlíku. Inkubujem e víčkem vzhůru. P op rvé odečítám e po 24 hodinách, podruhé po 48. Jestliže je kultura schopna látku asim ilovat, ob jev í se růst v m ístech nasypání hromádky. V jedné m isce o průměru 10 cm lze provést pět zkoušek různých cukrů současně.
Jako zdroje uhlíku užívám e: glukózu (Glucosum Spofa subst. Čsl. 3), galaktózu (D-Galaktosa Spofa-Léčiva, diagnostikům), sacharózu (Sacharosa p. a. Lachem a), maltózu (M altose fů r biochem ische Zw ecke M erck), laktózu (Lactosa p. a. Lachem a), rafinózu (Raffinose, Pentahydrat, fů r die Bakterio log ie M erck), inositol (M eso-inosit p ro bakto puriss. Lachem a), D -xylózu (D- X ylosa purum Spofa-Léčiva, diagnostikům), r ib ito l (Adon it čistý Lachem a), celobiózu (Cellotoiosa čistá Lachem a), L-rham nózu (L-Rham nosa čistá Lachem a), rozpustný škrob (Škrob rozpustný p. a. pod le Leuliera, Lachema, některé šarže jsou m álo vhodné), m elezitózu (D -M elezitose d ihydrate puriss. bact. K och-L ight), trehalózu (aa — Trehalosa čistá Lachem a), D -m anitol (D -M annit Lachem a — Chemapol), ery trito l (m eso-Erythrit fů r biochem ische Zw ecke M erck) a případně další.
P ů d a p r o c u k r o v é a u x a n o g r a m y : ( NH4)2SO/, — 10,0, K H 2P 0 4 — 2,0, M gS 0 4 — 1,0, kvasničný extrak t D ifco 3,0 (nebo kvasničný autolyzát Spofa 4,0), agar (podle jakosti, druhu a stáří) 30,0 až 60,0, dest. voda ad 2000,0. Rozehřejem e, rozp l- n ím e do baněk a sterilizu jem e v průběžné páře (100 °C ) 2 hodiny.
D u s í k o v é a u x a n o g r a m y nám ukazují, k terý zdroj dusíku je kultura schopna asim ilovat. P ro naše účely zcela postačuje zkouška asim ilace dusičnanu draselného K N O 3 a jako kontro ly síranu amonného (N H 4)2S 0 4. Používám e k tomu dva druhy šikmých půd v e zkumavkách, které očkujem e zkoušenou kulturou lehkým tahem k ličky (n iko liv masivně). Odečítáme po 24 a 48 hodinách. Všechny kultury musí růst na půdě kontrolní (amoniumsulfát), a le na půdě zkušební (kalium nitrát) jen ty, které jsou schopny ho asim ilovat.
P ů d y p r o d u s í k o v é a u x a n o g r a m y .Půda kontrolní: (N H <)2SO/, — 8,0, K H ;P O ( — 2,0, M gS 0 4 — 1,0, glukóza subst. Spofa
40,0, agar (podle jakosti) 30,0 až 60,0, dest. voda ad 2000,0.Půda zkušební: K N O 3 — 6,0, K H 2P 0 4 — 2,0, MgSO/, — 1,0, glukóza subst. Spofa 40,0,
agar (podle jakosti) 30,0 až 60,0, dest. voda ad 2000,0.Obě půdy samostatně rozehřejem e, rozp ln ím e po 200 m l do baněk a sterilizu jem e
v průběžné páře (při 100 °C ) 2 hodiny.Pod le potřeby ty to zásobní půdy rozehřejem e na vodn í lázni, na 200 m l přidám e
1 m l roztoku vitam ínů (v iz dále), rozm íchám e a asepticky rozlijem e do sterilních zkumavek, které šikmo položíme. Obě půdy v e zkumavkách (s v itam ín y ) lze dále skladovat v chladničce.
Abychom dosáhli výsledků opravdu zřetelných, musíme používat k přípravě obou půd čistého agaru (např. drcený, argentinský „chubutagar“ vyh ovu je ). M éně kva litn í řasový agar vyčistím e opakovaným propíráním . N a odvážené m nožství (50—60 g) řasového agaru v baňce na lijem e 2000 m l destilované vody, protřepem e a ponecháme při laboratorn í tep lotě do druhého dne. Druhý den vodu s lijem e a odm ěřím e je jí m nožství. N a lijem e dalších 2000 m l destilované vod y a další den opět s lijem e a odm ěříme. Properem e tak celkem třik rá t 2000 m l destilované vod y a agarem pohlcenouyodu odečtem e z celkového množství, udaného v receptech.
R o z t o k v i t a m í n ů : D -biotin 0,02 mg, calcium pantothenicum 4 mg, inositol 20 mg, n iacin (P P ) 4 mg, acidum p-am inóbenzoicum 2 mg, p yridox in hydrochlor. (B 6) 4 mg, aneurin HC1 (B l) 4 mg, r ibo fla v in (B2) 2 mg, dest. voda 100 ml. Rozpustíme, filtru jem e Seitzovým E K filtrem a uchovávám e v chladničce p ři asi + 5 °C.
P ro snadnější navážení D-biotinu postupujem e takto: navážím e 2 m g a rozpustím e v 10 m l dest. vod y ; odtud přenesem e 1 m l (tj. 0,2 m g) do dalších 9 m l dest. vody, prom íchám e a odtud přenesem e 1 m l (tj. 0,02 m g) do roztoku ostatních vitam ínů. Pak tep rve sterilizu jem e filtrací.
Z y m o g r a m y ( z j i š ť o v á n í k v a s n ý c h s c h o p n o s t í ) . Používám e řady te kutých půd, obsahujících různé cukry (v iz dále). Půdy v e steriln ích zkumavkách, v m nožství asi 2—4 m l očkujem e kličkou ze zkoušené kultury na šikmém Sabouraudově agaru. Odečítám e po 24 a 48 hodinách. S ledujem e změnu m odrozelené barvy půdy do žlutá.
P ů d a p r o z y m o g r a m y ( z j i š ť o v á n í k v a s n ý c h s c h o p n o s t í ) : K vasničný extrak t D ifco 4,0 g, Na2H PO ,5 — 2,5 g, K H 2P 0 4 0,5 g, dest. voda 2000 ml.
Č e s k á m y k o l o g i e 32 (i) 1978
34

Rozpustím e za m írného zahřátí, obarvím e roztokem brom thym olové m odři (asi 6—7 m l nasyceného, alkoholického roztoku) a na každých 300 m l tohoto základního roztoku v samostatných baňkách přidám e 6 g jednoho z cukrů (glukóza, galaktóza, sacharóza, maltóza, laktóza a ra finóza; náš určovací k líč je založen na zymogram u pouze tř í cukrů: glukózy, sacharózy a laktózy). P o rozpuštění rozlijem e do zásobních zkum avek (po asi 10—15 m l), označím e pod le druhu cukru a sterilizu jem e v průběžné páře (při 100 °C ) 2 hodiny. P o vych ladnutí uchovávám e v chladničce p ři asi + 5 °C.
T yto půdy m ůžeme p řip ravovat též s p lynovkam i (vrstva půdy asi 1 cm nad p ly- novkou) a sledovat tvorbu plynu. P lyn se však vy tvá ř í obvyk le až později.
M i k r o s k o p i c k ý p r e p a r á t z k u l t u r y . Z okraje kolonie opatrné vy jm em e kličkou nebo skalpem m alou část i s kouskem agaru, položím e na podložní sklo a stiskneme krycím sklíčkem tak, aby se m ateriá l co nejdokonaleji rozprostřel.
M i k r o k u l t u r a n a p o d l o ž n í m s k l e . Pasteurovou pipetou rozlijem e trochu rozehřáté, steriln í agarové půdy na obyčejné, podložní sklo, které je umístěné v Petriho m isce na trojhránku z ohnuté skleněné tyčky nebo trubičky. N a dno nale- jem e trochu steriln í vody, aby kultura nevysychala. Povrch ztuhlého agaru na podložn ím skle naočkujem e dvěm a tahy kličkou. M isku uzavřem e a inkubujem e víčkem vzhůru. P řed m ikroskopováním vy jm em e sk líčko s m ikrokulturou z Petrih o misky, otřem e spodní stranu a um ístím e na stolek mikroskopu. Proh líž ím e jen suchými, slabším i ob jektivy.
Tro jhránek a podložní sklo sterilizu jem e v uzavřené m isce horkovzdušně. M ám e připraveno vžd y několik desítek steriln ích souprav.
C h l a m y d o s p o r y C. albicans prokazujem e v m ikrokulturách (v iz výše ) na žlučovém agaru (v iz dále), po 18—20 hodinách. V e sporných případech odečítám e ještě jednou po 48 hodinách. Inkubační teplota nesmí přestoupit 24 ° C !
Ž l u č o v ý a g a r p r o t v o r b u c h l a m y d o s p o r C . albicans: F el tauri siccum (Spofa ) 5,0, agar (podle jakosti) 5,0 až 12,5, dest. voda 500,0. Rozehřejem e, rozlijem e do zásobních zkum avek a sterilizu jem e v průběžné páře (při 100 °C ) 2 hodiny. Ztuhlou půdu uchovávám e v chladničce při asi -f- 5 °C.
S t í n o v ý o b r a z n a v í č k u P e t r i h o m i s e k m ůžeme pozorovat u druhů rodu Sporobolom yces, čerstvě izolovaných. Ku ltu ry na Sabouraudově agaru inkubujem e vžd y v íčkem dolů. P o 7 dnech v id ím e na vn itřn í stěně v íčka m isky zrcadlový, „ fo togra fick ý “ obraz kolonií, tvořený prachem balistospor, které by ly vystřeleny z kolonií. Některé, zv láště sb írkové kultury schopnost vystře lovat balistospory ztrácejí a je jich určení bývá ve lice obtížné.
M o ž n o s t i u r y c h l e n í n ě k t e r ý c h l a b o r a t o r n í c h p r a c í . Časové údaje o urychlen í laboratorn í diagnostiky kvasinek mohou být předm ětem mnoha nedorozumění. Často se zam ěňuje doba nutná k získání prim okultury z infekčního m ateriálu s dobou potřebnou k je jím u určení.
Pod le našich zkušeností postačují k záchytu nečisté kultury z in fekčního m ateriálu3—5 dny, m axim álně 7 dní. Rych lost určení záleží nejen na charakteru zachycené kultury, a le také na pohotovosti a zkušenostech laboratorn ího pracovníka. Barva kolon ií je často nápadná již v prim okulturáeh: můžeme rozeznat např. Candida pu l- cherrim a od druhů rodů Rhodotoru la a (nebo) Sporobolom yces.
G eo trich u m candidum pro nápadný tvar kolonie rozeznáme od trichosporonú a k ověřen í stačí nativn í preparát. Z kolon ií porcelánově bílých a poloku lovitého tvaru m ůžem e již z nečisté prim okultury naočkovat žlučový agar v m ikrokulturách, abychom prokázali chlam ydospory C. albicans. V těchto a podobných případech — při vyu žití všech možností — známe (předběžný) výsledek často již během 3—5 dnů od doručení vzorku do laboratoře.
Auxanogram ů, zym ogram ů a případně dalších znaků (podle dále uvedených charakteristik ) užívám e tam, kde m ožnosti rych lejš ího určení b y ly vyčerpány a pochopitelně vždy, kdy se nechceme spokojit s určením jen předběžným nebo rámcovým .
Pro zhotovování auxanogramů a zym ogram ů potřebujem e čistou kulturu a proto přečišťování kvasinek do izolovaných kolon ií považu jem e za samozřejmé. P ř i určován í je výhodné m ít je u ruky, poněvadž vzh led izo lovaných kolon ií často mnoho napoví: Slouží nám také k příp ravě nativních preparátů. Je možné z nich vyočkovat po 4—7 dnech. Pom nožení na šikmém agaru 1—2 dny je nezbytné.
Časová bilance vypadá pak asi takto: prim okultury 3—7 dní, přečištění do izo lovaných kolon ií 4—7 dní, pom nožení na šikmém agaru 1—2 dny, v lastn í určení 2 dny; celkem 10—18 dní.
Zhodnocení význam u a spoleh livosti některých ze zm íněných metod je uvedeno dále v diskusi.
FRAGNER: K V A S IN K Y V LIDSKÉM M ATER IÁLU . I.
35

Č e s k á m y k o l o g i e 32 ( i ) i9?8
P ř e h l e d d r u h ů , k t e r é z a h r n u j e m e d o n a š í s e s t a v y
V elm i často se vyskytu jíc í:
1. Candida albicans (Robin) Berkhout 1923,2. Candida parapsilosis (Ashf.) Langeron et Ta lice 1932,3. Candida trop ica lis (Cast.) Berkhout 1923,4. Torulopsis candida (Saito) Lodder 1934,5. Torulopsis glabrata (Anderson) Lodder e t de V ries 1939.Často se vyskytu jíc í:
6. Candida claussenii Lodder et K reger-van R ij 1952,7. Candida krusei (Cast.) Berkhout 1923,8. Candida lam bica (L indner et Genoud) van Uden e t Buckley 1970,9. Candida pellicu losa R edaelli 1925,
10. Candida pseudotropicalis (Cast.) Basgal 1931,11. Candida stellatoidea (Jones e t M artin ) Langeron et Guerra 1939,12. Candida u tilis (Henneberg) Lodder e t K reger-van R ij 1952,13. Candida valida (Leberle ) van Uden et Buckley 1970,14. G eotrichu m candidum L in k 1809 ex Persoon 1822,15. K loeckera apiculata (Reess emend. K lócker) Janke 1928,16. Rhodotoru la rubra (Dem m e) Lodder 1934,17. Saccharomyces cerevisiae Hansen 1883,18. Torulopsis inconspicua Lodder e t K reger-van R ij 1952,19. Torulopsis norvegica Reiersól 1958,20. Torulopsis sphaerica (H am m er e t Cordes) Lodder 1934,21. Trichosporon capitatum Diddens e t Lodder 1942,22. T rich osporon cutaneum (de Beurm., Gougerot et Vaucher) Ota 1926.
M éně často se vyskytu jíc í:23. Candida bo id in ii R am irez 1953,24. Candida freyschusii Buckley e t van Uden 1968,25. Candida kefyr (B eijerinck ) van Uden et Buckley 1970,2#. Candida lusitaniae van Uden et do Carmo-Sousa 1959,27. Candida m esenterica (G eiger) Diddens et Lodder 1942,28. Candida pu lch errim a (L indner) W indisch 1940,Í9. Candida zeylanoides (Cast.) Langeron et Guerra 1938,30. C ryptococcus neoform ans (Sanfelice) V u illem in 1901,31. C ryptococcus albidus (Saito) Skinner 1947 var. albidus,32. Cryptococcus albidus (Saito) Skinner var. d iffluens (Zach) P h a ff e t F e ll 1970,33. Cryptococcus lau ren tii (K u ffera th ) Skinner var. flavescens (Saito) Lodder e t K re
ger-van R ij 1952,34. K luyverom yces bulgaricus (Santa M aria ) van der W a lt 1970,35. Rhodotoru la g lu tin is (Fres.) Harrison 1928,36. R hodotoru la m inuta (Saito) Harrison 1928 var. m inuta,37. R hodotoru la pallida Lodder 1934,38. Saccharomyces bayanus Saccardo 1895,39. Saccharomyces globosus O sterw alder 1924,40. Saccharomyces m icroellipsodes Osterw alder 1924,41. Saccharomyces rosei (Gu illierm ond) Lodder e t K reger-van R ij 1952,42. Saccharomyces unisporus Jorgensen 1909,43. Sporobolom yces sa lm on icolor (F ischer e t Brebeck) K lu yver et van N ie l 1925,44. Sporobolom yces roseus K lu yve r et van N ie l 1925,45. Torulopsis h o lm ii (Jorgensen) Lodder 1934,46. Torulopsis magnoliae Lodder e t K reger-van R ij 1952,47. Torulopsis stellata (K roem er et Krum bholtz) Lodder 1932.
Jména jsou uvedena pod le Lodderové e t al. (1970) a jsou platná podle m ezinárodního botanického kódu. Z několika m ožných názvů (zvláště to p latí o některých perfektních stadiích) vyb írám e zde ty, s n im iž se v lékařské m ykologii setkávám e nejčastěji. Jména perfektních stadií a některých význam nějších synonym jsou u vedena dále v podrobnějších charakteristikách.
G eotrichu m candidum není kvasinka; poněvadž bývá často zam ěňováno s druhy rodu Trichosporon a běžně se nás vyskytuje, zařazu jem e je do naší sestavy.
3«

V ý s k y t a r o z š í ř e n í
Candida albicans se vyskytu je v nejrůznějších vzorcích lidského původu nepo- H chybně ze všech kvasinek nejčastěji. N ěkteré jin é kvasinky nalézám e převážně jen■ v určitých vzorcích. N a přík lad Candida parapsilosis bývá přítom na v onychom ykó-■ zách rukou, paronychiích prstů rukou a v otom ykózách stejně často a možná i častěji
než C. albicans, někdy spolu s ní v e smíšených infekcích. Naproti tomu v jiném m ateriálu bývá rela tivně vzácná a představuje spíše náhodné nálezy. G eotrichum
■ candidum a druhy rodu Saccharomyces b ýva jí časté v e stolici a v e sputu, přičem ž p rvn í může vyvo lá va t střevn í geotrichózy.
N ěkteré kvasinky u člověka pocházejí zcela evidentně z jeho pracovního a životn ího prostředí a z potravy. P ro to je nalézám e na povrchu kůže (po kontaktu s in fikova
l i ným i předm ěty), v e sputu a v dutině ústní (jestliže b y ly vdechnuty nebo pojídány),v e stolici (když spolu s potravou prošly žaludkem do střeva). Tak na přík lad T r ich o - sporon cutaneum se hojně vyskytu je v prostředí m lékárenských výroben a na kůži zaměstnanců, G. candidum v tvarohu, Saccharomyces cerevisiae v droždí a pivu,Cryptococcus neojorm ans a jiné druhy kryptokoků v holubím trusu, Candida krusei p ři kysání zelí, C. pseudotropica lis a C. kefyr v kefíru a v jiných m léčných výrob
c i cích atd.H Frekvence nálezů jednotlivých druhů závisí tedy především na tom, jaké vzorky
a jakého původu laboratoř vyšetřu je. P ro to výše uvedené rozdělení podle častého H výskytu je jen přibližné. Některé údaje o původu kultur jsou uvedeny dále v podrobu j nějších charakteristikách druhů.
I K l í č
■ Náš diagnostický klíč je založen v prvé řadě na asimilaci pěti základních■ cukrů (glukózy, galaktózy, sacharózy, maltózy a laktózy) a podle jejich kom-■ binací je rozdělen do deseti skupin. Druhým, spolehlivým znakem je asimilace■ kaliumnitrátu. Poněvadž zmíněných deset skupin je co do počtu druhů různě■ obsažných, je v některých skupinách zapotřebí různého počtu dalších diagnos-■ tických znaků. To znamená, že rozlišení druhů je v některé skupině snadnější,■ v jiné méně snadné a pracovně náročnější. Za předpokladu, že všechny potřebné■ zkoušky byly provedeny současně, lze kulturu určit do 48 hodin. Z ekonomic-■ kého hlediska (šetření čistými chemikáliemi z dovozu) je někdy výhodnějšíI přistoupit k dalším zkouškám teprve po zařazení kultury do některé z deseti■ skupin. Může to však představovat zdržení výsledku o 24 až 48 hodin.I Jako dalších znaků je zde v některých případech použito: 1. asimilace rafi-I nózy, inositolu, trehalózy, D-xylózy, celobiózy. melezitózy, L-rhamnózy. erytri- 9■ tolu, rozpustného škrobu a ribitolu, 2. kvašení glukózy, sacharózy a laktózy,I 3. tvorby chlamydospor na žlučovém agaru, 4. vzhledu a zbarvení kolonií,I 5. mikroskopického obrazu v nativním preparátu z agarové kultury, 6. tvorbyI stínového obrazu z vystřelených balistospor na víčku Petriho misek, 7. schop-I nosti růst při 37 °C.
■ I. Asim ilace glukózy, galaktózy, sacharózy, maltózy a laktózy . skupina A■ II. Asimilace glukózy, galaktózy, sacharózy a maltózy. Laktóza není asimi-I l o v á n a .............................................................................................skupina BI III. Asimilace glukózy, sacharózy, maltózy a laktózy. Galaktóza není asimi-I lo v á n a .............................................................................................skupina CI IV. Asim ilace glukózy, galaktózy, sacharózy a laktózy. Maltóza není asimi-I lo v á n a ............................................................................................ skupina DI V. Asimilace glukózy, sacharózy a maltózy. Galaktóza a laktóza nejsou asi-I m i l o v á n y ...................................................................................... skupina E
VI. Asimilace glukózy, galaktózy a sacharózy. Maltóza a laktóza nejsou asim ilovány ..................................................................................... skupina F
H FRAGNER: K V A S IN K Y V LIDSKÉM M ATERIÁLU . I.
37

VII. Asimilace glukózy, galaktózy a maltózy. Sacharóza a laktóza nejsou asi-m i l o v á n y ......................................................................................... skupina G
V III. Asimilace glukózy a sacharózy. Galaktóza, maltóza a laktóza nejsou asi-m i l o v á n y ......................................................................................... skupina H
IX . Asimilace glukózy a galaktózy (někdy slabá). Sacharóza, maltóza a laktózanejsou a s im i lo v á n y ......................................................................skupina I H
X. Asimilace glukózy. Galaktóza, sacharóza, maltóza a laktóza nejsou asi- Hm i l o v á n y ......................................................................................... skupina J H
S k u p i n a A H
1 a. Kaliumnitrát je asimilován. Inositol je asimilován: HCryptococcus albidus var. albidus H
b. Kaliumnitrát není a s im ilo v á n ..................................................................2 H
2 a. Vlákna se rozpadají v artrospory, často uspořádané cik-cak: HTrichosporon cutaneum H
b. Vlákna, pokud se vůbec vytvářejí, se nerozpadají v artrospory . . 3 H
3 a. Laktóza je kvašena: Torulopsis sphaerica Hb. Laktóza není k v a š e n a .....................................................................................4 H
4 a. Inositol je asimilován: Crytococcus laurentii var. flavescens Hb. Inositol není asimilován: Torulopsis candida H
S k u p i n a B H
1 a. Kaliumnitrát je as im ilován .............................................................................12 Hb. Kaliumnitrát není a s im ilo v á n ........................................................................2 B
2 a. Kolonie jsou silně růžově až světle červeně zbarveny karotenoidním Ipigmentem, který nedifunduje do půdy: ■
Rhodotorula rubra Ib. Kolonie jsou silně purpurově červeně až hnědočerveně zbarveny pul- I
cheriminem. který difunduje do půdy: ICandida pulcherrim a I
c. Kolonie nejsou nápadněji zb a rven y ................................................................. 3 I
3 a. Vlákna se rozpadají v artrospory, často uspořádané cik-cak: ITrichosporon cutaneum ■
b. Vlákna, pokud se vytvářejí, se nerozpadají v artrospory . . . . 4 I
4. a. Inositol je asim ilován: Cryptococcus neoformans Ib. Inositol není asim ilován .....................................................................................5 I
5 a. Rafinóza je asim ilována.....................................................................................6 Ib. Rafinóza není a s im ilo v á n a .............................................................................. 7 I
6 a. D-xylóza, lib ito l a celobióza jsou asimilovány: ITorulopsis candida I
b. D-xylóza, ribitol a celobióza nejsou asimilovány: ISaccharomyces cerevisiae I
7 a. L-rhamnóza je asimilována: Candida lusitaniae Ib. L-rhamnóza není a s im ilo v á n a ........................................................................8 I
Č e s k á m y k o l o g i e 32 (i) 1978 H
38 I

8 a. Škrob je asim ilován ........................................................................................... 9b. Škrob není a s im ilo v á n ................................................................................... 11
9 a. Na žlučovém agaru se vytvářejí typické chlamydospory:Candida albicans
b. Na žlučovém agaru se chlamydospory nevytvářejí anebo nejsou typické 10
10 a. Sacharóza je kvašena: Candida tropicalisb. Sacharóza není kvašena: Candida claussenii
11 a. Celobióza je asimilována: Candida pulcherrim ab. Celobióza není asimilována: Candida parapsilosis
12 a. Kolonie jsou zbarveny výrazně růžově nebo o ra n žo v ě .............................. 14b. Kolonie nejsou zbarveny růžově nebo oranžově............................................13
13 a. Inositol je asimilován: Cryptococcus albidus var. diffluensb. Inositol není asimilován: Candida pelliculosa
14 a. Na víčku Petriho misky stínový obraz z vystřelených balistospor. Ku ltury růžové s nádechem oranžovým:
Sporobolomyces roseus b. Na víčku Petriho misky není stínový obraz z vystřelených balistospor.
Kultury růžové až červené, bez oranžového nádechu:Rhodotorula glutinis
S k u p i n a C
Kaliumnitrát je asimilován. Inositol je asimilován:Cryptococcus albidus var. albidus
S k u p i n a D
Kaliumnitrát není asimilován. Inositol není asimilován:1 a. Melezitóza je asimilována: Torulopsis sphaerica
b. Melezitóza není asim ilována.............................................................................. 2
2 a. D-xylóza je asim ilována.....................................................................................3b. D-xylóza není asimilována: Candida kefyr
3 a. Celobióza je asimilována: Candida pseudotropicalisb. Celobióza není asimilována: Kluyveromyces bulgaricus
S k u p i n a E
1 a. Kaliumnitrát je a s im ilován .............................................................................. 5b. Kaliumnitrát není a s im ilo v á n ........................................................................2
2 a. Erytritol je asimilován, D-xylóza a L-rhamnóza nikoliv:Candida mesenterica
b. Erytritol není a s im i lo v á n .............................................................................. 3
3 a. D-xylóza je a s im i lo v á n a ..............................................................................4b. D-xylóza není asimilována: Saccharomyces bayanus
4 a. L-rhamnóza, celobióza a melezitóza jsou asimilovány, škrob nikoliv:Candida freyschussii
FRAGNER: K V A S IN K Y V LIDSKÉM M ATERIÁLU . I.
39

b. Škrob je asimilován, L-rhamnóza, celobióza a melezitóza nikoliv:Candida albicans (vzácná forma)
5 a. Kolonie jsou zbarveny výrazně růžově nebo oranžově. Na víčku Petriho misky stínový obraz z vystřelenýcsh balistospor:
Sporobolomyces roseus b. Kolonie nejsou nápadně zbarveny. Na víčku misky se nevytváří obraz
z ba lis tospor.........................................................................................................6
6 a. Inositol je asimilován: Cryptococcus albidus var. diffluensb. Inositol není asim ilován ..................................................................................... 7
7 a. Erytritol a škrob jsou asimilovány:Candida pelliculosa
b. Erytritol a škrob nejsou asimilovány:Candida utilis
S k u p i n a F
1 a. Kaliumnitrát je asimilován: Torulopsis magnoliaeb. Kaliumnitrát není a s im ilo v á n ........................................................................2
2 a. Kolonie zbarveny světle růžově nebo cihlově červeně karotenoidnímpigmentem. Rafinóza není asim ilována...........................................................3
b. Kolonie nejsou zbarveny růžově nebo červeně karetonoidním pigmentem. Rafinóza je a s im i lo v á n a ........................................................................4
3 a. Melezitóza je asimilována: Rhodotorula minutab. Melezitóza není asimilována: Rhodotorula pallida
4 a. Buňky pravidelně kulovité nebo subglobózní:Saccharomyces m icroellipsodes
b. Buňky kulovité, oválné a dlouze oválné v téže kultuře současně:Torulopsis holm ii
S k u p i n a G
Kaliumnitrát není asimilován: Candida stellatoidea
S k u p i n a H
1 a. Kolonie jsou nápadně růžové. Na víčku Petriho misky stinový obrazz vystřelených balistospor. Kaliumnitrát je asimilován:
Sporobolomyces salmonicolor b. Kolonie nejsou nápadně růžové. Na víčku misky se nevytváří stínový
obraz z balistospor. Kaliumnitrát není asim ilován .......................................2
2 a. D-manitol a trehalóza jsou asimilovány:Saccharomyces rosei
b. D-manitol a trehalóza nejsou asimilovány:Torulopsis stellata
S k u p i n a IKaliumnitrát není asimilován:
1 a. Kolonie zřetelně vláknitého charakteru, alespoň na okrajích. Vláknase rozpadají na a r t r o s p o ry .............................................................................. 2
Č e s k á m y k o l o g i e 32 (i) 1978
40

H b. Kolonie nemají vláknitý charakter. Vlákna, pokud se vytvářejí, se■ nerozpadají na a r t r o s p o r y ................................................................................3
■ 2 a. Kolonie rychleji rostoucí, ploché a nízké, připomínající jemnou krajku,jemně zrnité, jakoby moukou poprášené. Artrospory 3— 8 um široké. Blas
a i tospory nepřítomny. Asimilace galaktózy silná:Geotrichum candidum
■ b. Kolonie pomaleji rostoucí, mírně vyvýšené až polokulovité, uprostřed■ někdy vmáčklé, jemně zrnité nebo chmýřité. Artrospory 2,5— 3,5 ^m ši-
roké. Blastospory často v typických hlavičkách. Asimilace galaktózy vět- šinou slabá: Trichosporon capitatum
3 a. Buňky kulovité, oválné a válcovité, 2— 6,5— 8,5 X 4— 11 ^m, nejčastějikolem 4,5— 6,5 um. Válcovité někdy uspořádány za sebou jako náznak primitivního pseudomycelia : Saccharomyces globosus
b. Buňky kulovité, vejčité a krátce oválné, 2— 4,5 X 2— 6,5 um, nejčastějikolem 4 um. Pseudomycelium se nevytváří:
Saccharomyces unisporus
H S k u p i n a J
H 1 a. Kaliumnitrát je asim ilován .............................................................................. 9H b. Kaliumnitrát není a s im ilo v á n .............................................................2
H 2 a. Buňky nápadně citronovitého tvaru s oběma konci zašpičatělými nebo■ protáhlými. Převážně bipolární pučení. Celobióza je asimilována:■ Kloeckera apiculata■ b. Buňky nejsou nápadně citronovitého tvaru, nepřevažuje bipolární■ pučení. Celobióza není asim ilována.................................................................. 3
■ 3 a. D-manitol je asimilován. Dále asimilovány (pomaleji) trehalóza a■ ribitol: Candida zeylanoides■ b. D-manitol není a s im ilován ................................................................... 4
I 4 a. Trehalóza je asimilována. Kolonie vždy hladké a lesklé. Pseudomyce-I lium se nikdy nevytváří. Glukóza je kvašena:I Torulopsis glabrataI b. Trehalóza není asim ilována................................................................... 5
I 5 a. D-xylóza je asimilována: Candida lambicaI b. D-xylóza není as im ilován a .........................................................................6
I 6 a. Glukóza je kvašena ........................................................................7 HI b. Glukóza není k v a š e n a .......................................................................... 8
I 7 a. Růst při 37 °C vždy velm i dobrý: Candida kruseiI b. Růst při 37 °C žádný nebo slabý: Candida valida
I 8 a. Růst při 37 °C vždy velm i dobrý. Pseudomycelium se nevytváří nebo jeprimitivní, složené jen z řetízků buněk:
Torulopsis inconspicua b. Růst při 37 ° žádný nebo slabý. Pseudomycelium z dlouhých, válcovitých
buněk často bývá hojně vyvinuté:Candida valida
FRAGNER: K V A S IN K Y V LIDSKÉM M ATER IÁLU . I.
41

9 a. Erytritol a ribitol jsou asimilovány: HCandida boidinii H
b. Erytritol a ribitol nejsou asimilovány: ITorulopsis norvegica. H
Upozorňujeme, že náš klíč (jako ostatně každý) je pouze vodítkem: k určení Hje nezbytně nutná konfrontace znaků neznámé kultury s podrobnějšími popisy, Huvedenými dále v „Charakteristice druhů“ . H
Pokračování. H
Adresa autora: RND r. P e tr Fragner M ykologické odd. Hygien ické stanice Středo- Hčeského K N V , Apolinářská 4, 128 00 Praha 2. U
H. D. T h i e r s : California Mushrooms. A F ield Guide to the Boletes. — 261 stran, HC ollier-M acm illan Publ. H afner Press, N ew Y ork — London 1975. Cena £ 9.50. H
Tato kniha je výtečnou m onografií nejen pro odborníky, a le i pro m ykology ama- Itéry. V ysvě tlivky technických termínů, určovací klíče, popisy a ilustrace um ožní i za- Ičátečníkům určit h řibovité houby v terénu bez použití mikroskopu. Zvláštností této Iknihy jsou m ikrofiše 54 barevných fo togra fií nejdůležitějších druhů. I
P rvn í část této knihy popisuje některé dů ležitější strukturální a anatom ické rysy Ihřibovitých hub z taxonom ického hlediska, jako je stavba plodnic, barva masy Ivýtrusů i jednotlivých spor, typ buněk vyskytu jíc ích se běžně v hymeniu, typ lupenů Im ikroskopická struktura kutikulárních vnějších vrstev klobouku, schopnost vytvá ře t Im ykorrhizn í společenstva s některým i strom y a reakce různých částí plodnic Is rozličným chem ickým reagens. V obsáhlé části knihy „Taxonom ické k líče a popisy Idruhů“ jsou tyto vlastnosti popsány u rodů hřibovitých hub vyskytu jících se v K a lifo r- In ii: Gastroboletus, Suillus, Pu lveroboletus, Gyroporus, Leccinum , Ty lop ilus a Boletus. I
A čk o liv název této knihy je „K a liforn ské houby“ , taxonom ické k líče a druhový Ipopis umožní identifikovat i hřibovité houby v jiných zemích, tedy i u nás. I
Vlasta Čatská ■
E. M ü l l e r a W. L o e f f l e r : Mycology. A n Outline fo r Science and M edical IStudents. — 306 stran, G eorg Th iem e Publ., Stuttgart 1976. I
Recensovaná kniha je anglickým překladem původního něm eckého vydán í publi- Ikovaného v letech 1968 a 1971. I
V kapitole „Ekologické aspekty m ykologie“ jsou uvedeny podm ínky pro růst a akti- Iv itu hub. Jsou zde zahrnuty m in im áln í požadavky pro růst, houbové asociace jako Isymbiosa, Parasitismus a saprofytismus, specifické požadavky jako živ iny, teplota, Isvětlo a jiné faktory. V kapitole „K van tita tivn í aspekty m ykologie“ autoři upozorňují Ina to, že houby v půdě tvoř í převážnou část m ikrobiáln í biomasy, p ři čemž stanovení Ihoubové biom asy je ve lm i obtížné. K rom ě toho některé organické látky jako celulosa, I
■ chitin, lignin, arom atické a jin é lá tky b ýva jí rozkládány převážně houbami. IV kapitole „M orfo log ie “ jsou uvedeny popisy a obrázky vegetativn ího i fertiln ího Istadia hub. U ltrastruktury jader, m itochondrií, ribosomů a buněčných stěn jsou po- Ipsány v kapitole „U ltrastruktura“ . V dalších kapitolách „M etabolism us“ a „G enetika“ Ijsou uvedeny m etabolické cesty, vyu žíván í živin, mutace, heterokaryosis a parasexua- Ilita. Houby na jedné straně mohou být vyu žívány v potravinářství, nižší houby mohou Ipřisp ívat k m ineralizaci některých organických látek, řada vyšších hub je jedlá, ně- Ikteré houby jsou producenty antibiotik, na druhé straně však houby způsobují one- Imocnění jak rostlin, tak i zv ířa t a člověka. O těchto činnostech hub je psáno v kapito le I„A p likovan á m ykologie“ . Nejobsáh lejš í kapitolou je „K las ifikace hub“ obsahující Inomenklaturu, klíče, popisy a obrázky nejdů ležitějších a nejrozšířenějších hub. I
Tato kniha je určena především studentům přírodních věd i m edicíny a zejm éna Ipracovníkům v aplikované m ykologii. I
Vlasta Čatská I
č e s k á m y k o l o g i e 32 (i) 1978 I
42 I

Muchomůrka Maireova — Amanita mairei Foley nalezena v Československu
Maire’scher Streifling — Amanita mairei Foley in ČSSR gefunden
Svatop luk Šebek
V článku je popsán nález vzácné muchomůrky M a ireovy (A m anita m aire i F o ley ) z příbuzenstva muchomůrky pošvaté [A m an ita vaginata (Bull. ex Fr.) V itt.] u Poděbrad a u Loučené (okr. Nym burk.) Současně je probráno je jich vzá jem né odlišení. ______________
In dem Beitrag ist ein fů r Tschechoslovakei neuer Fund des M aire'schen S tre iflin gs (A m a n ita m a ire i Foley) aus der Verw andschaft des Grauen Stre iflings [A m a n ita vaginata (Bull. ex Fr.) V itt.] bei Poděbrady und Loučeň (K re is Nym burk, Bez. M ittelbohm en) gleichzeitig m it der D ifferenzierung beider A rten beschrieben.
Muchomůrka pošvatá — Amanita vaginata (Bull. ex Fr.) Vitt., hojná houba našich lesů od nížin až do horského pásma, je současně nejproměnlivějším druhem muchomůrek, charakterizovaným především barevnou variabilitou. Veselý (1934) popisuje 27 forem, o nichž uvádí, že jsou „z největší části nestálé a proto systematicky těžko ohraničitelné. Některé však jsou dosti konstantní a mají snad i cenu var i e t . . a je přesvědčen, že „z největší části jsou systematicky bezvýznamné“ (1. c., str. 49).
Kritická revize „forem “ z okruhu muchomůrky pošvaté, která je nanejvýš žádoucí, by jistě přehodnotila taxonomickou hodnotu řady z nich. O tom, že i od tak zdánlivě dobře známého druhu, jako je A. vaginata, lze odlišit řadu hodnotných a konstantních taxonomů, svědčí už několik nových druhů z jejího okruhu, zjištěných a popsaných v Evropě v posledních letech. Patří k nim i význačný druh, zjištěný spolehlivě i na našem území, jemuž jsou věnovány následující řádky.
Už v r. 1965 jsem u Poděbrad zjistil plodnice muchomůrky ze sekce Am anito- psis Roze, které se nápadně podobaly muchomůrce pošvaté Amanita vaginata (Bull. ex Fr.) Vitt., ale v některých makroznacích (např. rýhování klobouku a jeho barva, tvar a síla třeně) se od jejího popisu lišily. Protože jsem tehdy nemohl tyto plodnice spolehlivě určit, pořídil jsem jejich kresbu a popis, a celou záležitost jsem odložil na pozdější příležitost. Upozorněn zanedlouho potom článkem C. Base (1967) o nálezu druhu Amanita argentea Huijsman v Teutoburském lese, usuzoval jsem na možnost totožnosti mých poděbradských nálezů s touto houbou, popsanou z franc. Jury, a začal jsem po ní cílevědomě pátrat. Teprve nový nález podobných plodnic v roce 1970 na jiné lokalitě mi potvrdil správnost mého předchozího určení. (Později byla tato houba ztotožněna se severoafrickým druhem Amanita mairei Foley, popsaným v r. 1949; jeho jméno je tedy prioritní.) V následujícím podávám popis muchomůrky M aireovy (Amanita m airei Foley), pořízený podle nálezů z níže uvedených loka lit:
Klobouk v mládí slabě vyklenutý, v dospělosti plochý, 5,5— 7 cm v pr., na okraji krátce jemně rýhovaný, stříbřitě šedý, na vrcholu tmavší, na okraji světle šedobéžový, suchý, hladký, s dužninou poměrně masitou, na okraji ztenčenou, u některých plodnic s 1— 3 většími nepravidelnými špinavě bílými útržky pochvy. Lupeny velm i husté, volné nebo jen krátce připojené, široké, bílé až světle šedokrémové, na ostří bílé, s četnými nepravidelně rozmístěnými
43

A m anita m a ire i F o ley — dvě dospělé plodnice a výtrusy. — Z w e i re ife Fruchtkorpem und Spořen. S. Šebek del.
lamellulami. Třeň 8— 10 cm dlouhý, 1,5 cm široký, v horní části a na bázi poněkud rozšířený, bez prstenu, bílý nebo našedlý, jemně vláknitý, v horní části drobně bíle vločkovitý, vatovitě vycpaný. Dužnina bílá, neměnná, křehká, bez chuti a vůně. Pochva kožovitá, silnostěnná, špinavě bílá, téměř přitisklá, jen v horní okrajové části mírně odstávající, 2,5— 3,5 cm vys. Výtrusný prach bílý. Výtrusy elipsoidní, bezbarvé, silnostěnné, s velkou olejovou kapkou, ne- amyloidní, 11— 13,5 X 8— 9 «m vel.
Hab.: Poděbrady, na ok ra ji go lfovéh o hřiště pod topoly, 2 exem pláře 16. V I. 1965 leg. S. Šebek; tam též 6 ex. 9. IX . 1967 leg. M ir. K rom bholz; tamtéž, 3 ex. 15. V I. 1968 a 3. IX . 1969 leg. S. Šebek; tamtéž, 22. V I. 1970 leg. M ir. K rom bholz. — Loučen, sub- xero filn í doubrava při levé straně silnice na M cely, na ok ra ji lesní cesty pod duby, 1 ex. 10. IX . 1970 leg. S. Šebek. Doklad předán do herbáře M uzea v Poděbradech.
Č e s k á m y k o l o g i e 32 (i) 1978
44

H Še b e k : a m a n i t a m a i r e i v Cs s r
H V evropské literatuře je tato muchomůrka známa jako Amanita argentea;■ pod tímto jménem ji popsal v r. 1959 H. S. C. Huijsman ze stř. Francie a tak ji■ také později interpretuje Parrot (1965) a Bas (1967). Priorita však patří jménu■ Amanita m airei, pod nímž ji popsal H. Foley (1949) ze sev. A friky. Protože■ podle originálního popisu (Foley, 1949) i podle podrobného popisu Malengonova■ a Bertaultova (1970) se tato houba v makroznacích prakticky neliší od Huijs-■ manova popisu druhu Amanita argentea a udávaná velikost výtrusů [u Foleye
a Malengon-Bertaulta 11,5— 13,5 X 7,6— 8,3(— 9,3) jj.m, u Huijsmana 10,5— 12 (— 13,8) X 8,6— 10(— 11,5) ^m, u Parrota 10— 14 X 8,5— 11,5 fim, u Base 11,5— 14 X 8,5— 11 (¡m] se patrně pohybuje v hranicích přípustného rozmezí, lze oba druhy ztotožnit, jak správně připouští už Malengon-Bertault (1970 : 71).
Am anita mairei Foley je dobrý druh, který by mohl být na první pohledzaměněn s běžnou Amanita vaginata; od ní se však liší některými význačnýmiznaky. Především je to robustnější postava plodnic s velm i plochým kloboukem bez terminálního hrbolku a nápadnou stříbřitě šedou barvou pokožky. Amanita vaginata má klobouk tence masitý, obyčejně s hrbolkem, a je jí barva, ponej- více šedavá až šedohnědá, varíruje od bílé přes nažloutlou až po temně hnědou. Velm i charakteristické je také krátké rýhování klobouku, které je omezeno jen asi na 5— 8 mm širokou okrajovou zónu (podle Base 0,1— 0,2 R), zatímco rýhy na okraji klobouku muchomůrky pošvaté zasahují mnohem dále k jeho středu (podle Base 0,3— 0,4 R). V e srovnání s dlouhým úzkým nahoře ztenčeným
H třeném muchomůrky pošvaté je třeň muchomůrky M aireovy relativně kratší■ a širší a nahoře se rozšiřuje. Nápadný rozdíl je také v umístění báze třené ve■ vztahu k lemu na vnitřní straně volvy, rozlišitelný už v primordiálním stadiu■ plodnic, zřetelně patrný zejména na průřezu dospělých plodnic včetně volvy.■ Lem na vnitřní straně vo lvy je u muchomůrky M aireovy značně zakrnělý a bázeH jejího třené je tedy blízko jeho horního okraje; u muchomůrky pošvaté je■ naopak lem na vnitřní straně vo lvy dobře vyvinut a poměrně vysoký, takže■ se zdá, že báze jejího třeně je umístěna mnohem hlouběji (srv. Huijsman, 1959,■ fig. 2 na str. 19). Charakterem vela generále se tedy Amanita mairei vývojově■ blíží muchomůrce císařce — Am anita caesarea (Scop. ex Fr.) Pers. ex Schw,I Pokud se týká rozdílů v mikroznacích jsou nejvíce patrny mezi široce elipsoid-■ nimi sporami Amanita m airei a mezi kulovitými sporami u Amanita vaginata.■ Z chemických reakcí upozorňuje Bas (1967) na fenolovou, při níž dochází■ u Amanita mairei po ± 1 minutě na dužnině a lupenech k vínově červenému■ až cihlově červenému zbarvení.
I Am anita mairei Foley byla popsána z Alžíru, kde na své původní lokalitěH v okolí Birmandreis u Icosie roste v říjnu—listopadu v písčité půdě pod PinusI halepensis a v pistáciových macchiích. Malengon-Bertault (1970) j i udávajíI z Maroka ze světlých lesů, macchií a houštin v okolí Mesety, Tangeru (podI Pinus pinea), Rabatu a El-Khatouatu (pod Quercus suber).■ Evropské lokality této houby se od severoafrických ekologicky značně liší.■ V Evropě byla tato vzácná muchomůrka sbírána (jako Amanita argentea) po-I prvé Huijsmanem (1959) ve stř. Francii (Doubs, okolí Montbéliardu, jednotlivě
pod listnáči); později udává ji Parrot (1960) z listnatého lesa u Dancharia v Nízkých Pyrenejích v již. Francii. V r. 1966 ji nalezl Huijsman ve Švýcarsku u Neuenburgu (v písčité půdě pod buky a duby nedaleko Neuenburského jezera) (Bas, 1967). V r. 1967 byla tato houba zjištěna i na území NSR, kde ji sbíralC. Bas v Beller Holz u Bad Meinbergu vých. od Teutoburského lesa ve vých. Westfálsku (ve smíšeném lese s duby a buky a roztroušenými smrky, olšemi
45

a kleny, 25. 9. 1965). Evropské lokality (včetně obou československých) před-■ stavují vesměs listnaté (nebo smíšené) lesy.|| Z Československa nebyl tento význačný druh dosud uváděn, i když není po-H chyby o tom, že při podrobnějším mykofloristickém průzkumu budou jistě
zjištěna jeho další naleziště.
■ L i t e r a t u r a ^ B
■ B a s C. (1967): Am anita argentea Huijsman am Teutoburger Walcl geíunden. W est- ^ BH falische P ilzb r ie fe 6: 125—129, 19G7.B F o l e y H. (1949): Une Am an ite nord aíricaine nouvelle: Am anita M airei, n. s. Foley. ^ BB Trav. bot. déd. Mém. R. M aire (Mém. Soc. Hist. nat. A fr . du N ord 11: 117—120).
H u i j s m a n H. S. C. (1959): Deux Am anites méconnues. Bull. Soc. m ycol. France ^ B75: 14-32. ■
M a l e n g o n G. et B e r t a u l t R. (1970): F loře des Champignons supérieurs de M aroc I. Rabat.
B P a r r o t A. G. (1960): Am anites du sud-ouest de la France. Centr. Rech. Etud. Sci.B iarritz.
H V e s e l ý R. (1934): Am anita-Muchom ůrka. In : K av in a K . et P ilá t A., A tla s hub evrop-ských. 1. Praha.
Adresa autora: Svatopluk Šebek, Boleslavská 481/30, 288 00 Nym burk.
B B D. J. W e b e r a W. M. H e s s (edit.): The Fungal Spore. Form and Function. — H895 stran, A W iley—Interscience Publ., N ew Y o rk — London — Sydney — Toronto 1976. ■Cena $ 33.30. ■
Tato kniha byla vydána na základě druhého m ezinárodního symposia ,.H ou bové BM g s p o ry konaném v roce 1974 v B irm ingham u. V knize je shrnuto vše, co je známo B■ o ultrastruktuře a b iochem ii houbových spor. K n iha je rozdělena na pět sekcí. P rvn í Bffifjg sekce „Dorm ance a aktivace houbových sp o r1 obsahuje tři kapitoly, v nichž jsou po- BH B psány spory v k lidovém stadiu, inh ib itory a aktivátory k líčen í houbových spor. V druhé H9SS| sekci „K líčen í houbových spor“ se šesti kapitolam i je popsán metabolismus a struktura B■51 buněčné stěny v průběhu k líčen í houbových spor. Nejobsáhlejší sekce ,.Tvar a funkce B
houbovývých spor s osmi kapitolam i se zabývá resistentním i strukturami u M yxom y ce - IH tes, tvarem a funkcí spor u Chytrid iom ycetes, Zygom ycetes, Coelom ycetes, Basidioray- H
cetes Ascom ycetes a tvarem a funkcí kon idií im perfektn ích hub. V sekci pracovního Izasedání v kapitole „Fosiln í houbové spory“ je diskutováno o paleoekologii, paleo- ■geografii a fylogenesi typů houbových spor. I
H H Tato kniha je nejen zdrojem in form ací vědeckých studií špičkových pracovníků Iv tomto oboru, zdrojem literárn ího přehledu, a le zejm éna zdrojem názorů o budoucím ■
n zam ěření a směru studia houbových spor. Účastníky symposia bylo navrženo, aby příští Isymposium se zabývalo především fys io log ií a biochem ii houbových spor, biochem ií Ia povrchovou strukturou spor, procesy sporogenese včetně biochem ických studií a stu- I
B jlW d ií elektronové m ikroskopie a sporulací. Značný počet elektronových m ikro fo togra fií Iobsahuje již tato kniha. I
B B Doporučit lze tuto knihu vědeckým pracovníkům nejen m ykologům, ale též botani-B frům i m ikrobiologům.
Vlasta Čatská
Č e s k á m y k o l o g i e 32 (i) 1978
46

Inonotus tamaricis (Pat.) M aire in Greece, its general distribution and taxonomic notes to the section Phymatopilus Donk
Inonotus tamaricis (Pat.) Maire v Itecku, celkové rozšíření a taxonomické poznámkyk sekci Phymatopilus Donk
Jaroslav K lá n
Inonotus tam aricis (Pat.) M aire, which has been known up to the present tim e from three localities in Greece, is described and its area o f distribution is g iven including several taxonom ical notes. The taxonom ic position of fu rther species o f the section Phym atop ilus Donk (I.rh ea d es , I. d ryo- philus and I. pseudohispidus) is discussed.
Je uveden popis choroše Inonotus tam aricis (Pat.) M aire, k terý je v ftecku dosud znám pouze na 3 lokalitách, jeho celkový areál a taxonom ické poznámky. Je diskutováno taxonom ické postavení dalších zástupců sekce Phym atop ilus Donk (I . rheades, I. dryophilus a I. pseudohispidus).
The Greek mycoflora has been very little studied and only incomplete information are sometimes found scattered in various journals. Contributions dealing with macromycetes were published by Landerer (1859), Heufler (1868), Diapou- lis (1939), Serejanni (1939), Maire et Politis (1940), Petrak (1943), Serejanni et Demetriades (1951), Pantidou et Watling (1970), Příhoda (1970), Pantidou (1973) and Moravec (1974).
Inonotus tamaricis (Pat.) Maire, a characteristic polypore o f the Mediterranean area, was recorded for the first time in Greece by M aire et Politis (1940) as Polyporus tamaricis (Pat.) Sacc. et D. Sacc. (locality Phaliron, Attiki, on Ta- m arix sp.). Pantidou (1973) inserted this fungus in her list o f Greek fungi. This polypore was collected for the second time in the island of Corfu (Sidari, on Tam arix sp., leg. D. A. Reid 28 May 1966, K ). The latter finding has not been published till now. The third locality was disclosed in the spa of Agía Triás, 25 km southwards from Solune (Thessaloniki); 14 fruit bodies at various stage of development were collected on a living tamarisk tree (Tam arix sp., leg. J. Klán, 7 July 1975, PRM 756258, PRC).
Inonotus tamaricis (Pat.) M aire ap. M aire et Wern. 1937
Syn .: Xanthochrous tam aricis Pat. 1904Polyporus tam aricis (Pat.) Sacc. et D. Sacc: 1905X anthochrous rheades (Pers.) Pat. ssp. tam aricis (Pat.) Bourd. et Galz. 1928
Icon.: Polyporus corruscans Fr. sensu Bres., Iconographia m ycologica 1931, tab. 981 Polyporus rheades Pers. sensu Bres., Fungi Tridentin i I I 1892, tab. 136
Description
Fruit bodies solitary, annual, attached laterally to the substrate, 3—8 X 4—9 X 2—4 cm large, soft and fleshy when young, later fragile.
Pileus convex to hemispherical, densely velvety on surface, later glabrous, yellow-rusty to rusty-brown, becoming blackish when old. Pileus-margin blunt, more and less glabrous, lighter in colour.
Context spongy-fibrous, fragile when dry, mostly yellowish when young, but rusty w ith d i s t i n c t y e l l o w z o n e s o n m a t u r i t y . The zonation is less apparent in old fruit bodies, and the rusty colour prevails. The zones of the context are formed by alternating fascicles of yellow and rusty hyphae.
47

Table 1. Taxonomic characters within
■xr Dimensiones o f spores , „ .Name x ± 3 x s , a; ± s +E +Empim
I . rheades 5.2— 5.7— 6.2 X 5.71 ± 3x0.053 5.71 ± 0.41 1.25— 1.71 1.43X 3.1— 4.0— 4.3 3.95 ± 3x0.033 3.95 ± 0.25
I . dryophilus 6.9— 7.4— 8.2 X 7.44 ± 3x0.052 7.44 ± 0.41 1.17— 1.60 1.32X 5.2— 5.6— 6.2 5.60 ± 3x0.047 5.60 ± 0.37
I . tamaricis 6.2— 6.8— 7.3 x 6.79 ± 3 x 0.050 6.79 ± 0.41 1.09— 1.45 1.30X 4.6— 5.2— 5.7 5.21 ± 3x0.030 5.21 ± 0.25
In young fruit bodies and at the margin o f adult ones, yellow to rust-coloured, septate, slightly branched hyphae are present, 5—8 ym broad, w ith walls 0.5—0.8 thick. In adult fruit bodies are slightly branched hyphae mainly around the core, which are rusty, septate, 6—10 ^m broad, w ith thick walls 1.5—4.5 dum. The context o f old fruit bodies consists mostly of thick-walled hyphae. Hyphal system monomitic, w ith no clamps. Setae absent.
In the place in which the fruit body is attached to the substrate, a d i s t i n c t s u b g l o b o s e g r a n u l a r c o r e i s p r e s e n t . The core is relatively small (1—3 cm in diam.), occupying 1/5 to 1/3 o f the fruit body; it consists of thin-walled septate hyphae which are brightly yellow, yellow to rusty in colour. In addition, there are some thick-walled hyphae, which are rusty, densely intertwined, form ing hard, b l a c k a n d l u s t r o u s g r a i n s .
Tubes are up to 0.5—2 cm long, yellow-rusty, rust-brown, dark-brown tobrownich-black. Pores are rounded, later angular, fringed, 0.3—0.7 ¿¡m in diam., usually 2—3(4) per mm.
Spores, 6.2—6.8—7.3 X 4.6—5.2—5.7 kum*, (measured on dry material in lacto- phenol and in M elzer’s solution), ellipsoidal, thick-walled, yellow to yellow-rusty, finely granular (Fig. la).
Distribution and ecology
The species grows m ain ly on liv in g tam arisk (T a m a rix sp. div.). It has been found up to the present tim e in these countries:P o r t u g a l : P into-Lopes 1953: 214S p a i n : H eim 1933: 61; P ila t 1936—42: 563; R yvarden 1972: 141; [.Catalogo m ico-
log ico del Pais Vasco Munibe, 1973: 57; — Baquio, V izcaya; — San Sebastian, Guipuzcoa; — Casa de Campo, M adrid ; Parque de Donana, H u elva ; in Herb. Botanic Garden, M adrid ; F. D. Calonge private inform ation]
F r a n c e : Bourdot et G alzin 1928: 637; — Toulon, leg. Joachim , M ay 1937, P R M487431
I t a l y : Saccardo 1905: 111; Bresadola 1892: 30; P ila t 1936—42: 563; — L ivorno , leg.G. A rcangeli Aug. 1902 in Saccardo, M yc. ital. No. 1203; Sardinia: leg. Carvara29 June 1900, rev. F. Kotlaba 10 M ay 1973, W 5930
Y u g o s l a v i a : C roatia: — Starigrad-Paklen ica, leg. F. K o tlaba 20 July 1966, 8 July 1968, P R M 770390, 770396; - Seline pr. Zadar, leg. F. K otlaba 6 July 1968, P R M 770407; — inter Stara K u la et Seline pr. Zadar, leg. F. K o tlaba 5 July
* The fo llow in g dimensions are g iven by the authors: Patou illard (1904) 8—9 X X 6 cm, Bourdot et G alzin (1928) 7—8—(9) X 4.5—5—(6) «m , Bresadola (1927—33) 7 -8 X 4.5—5.5 um, P ila t (1936-42) 7-8.5 X 4.5-5.5 u m, Bondarcev (1953) 6.5-7.5 X X (4)—4.5—5.5 urn. P eg le r (1964b) 7.2-9.5 X 5 -7 (8.5 X 6) um.
CESKA M YKO LO G IE 32 (1) 1978
48

■ K L Á N : IN O N O TU S T A M A R IC IS IN GREECE
H Inonotus sect. Phymatopilus Donk
H ... r . Auxiliary featuresDimensions of granular J ,Colour or context ° Dimensions HostCOre o f fruitbody
rusty-yellow to rusty 0.5— 1.5 cm 2— 5 X 3— 10 X Populus sp. div.■ 1/5 to 1/2 o f fruitbody 1— 3.5 cm living or dead■ (Fagus, Salix )■ yellow, yellow-rusty 3— 7 cm 5— 10 X 6— 16 X Quercus sp. div.■ to rusty 1 /2 to 2/3 of fruitbody 3— 8 cm living
distinctly yellow 1— 3 cm 3— 8 X 4— 9 X Tamarix sp. div.and rustily zoned 1/5 to 1/3 o f fruitbody 2— 4 cm living
+E = length width o f spores+Em = mean length H- mean width o f spores
■ 1968, 6 July 1968, P R M 770402, 770401; - Split, leg. M. Bertosi 30 Jan. 1967,■ P R M 647027; - Sp lit-M eje, leg. F. K otlaba 12 July 1968, P R M 770399; - Slano■ prope Dubrovnik, leg. E. G eorg ijev ič Sept. 1971, P R M 717046; — insula Hvar,■ leg. J. Veselský 24 M ay 1969, PR M 674135, 674136; M onte N egro : — Kotor, in■ ripa “ Marsala T ita “ , leg. F. K otlaba 26 M ay 1976, PR M 795924; — Titograd, in■ horto publico ante deversorium “ Crna G ora“ , leg. F. Kotlaba 26 M ay 1976, PR M■ 795926, (the localities w ere summarized by Tortič et Kotlaba, 1976)
I 1. Inonotus tam aricis (Pat.) M aireI a — spores, b — hypha w ith thin w a ll o f context, c — hypha w ith thick w a ll o fI context, d — hypha w ith th ick w a ll o f granular core
I B u l g a r i a : — Sozopol pr. Burgas, leg. F. Kotlaba 22 July 1974, P R M 741831; —I Burgas, leg. F. K otlaba 22 July 1974, P R M 741767; — S liančev briag ap. Nesebar,I leg. F. Kotlaba 20 July 1974, P R M 741761; — Nesebar ap. Burgas, leg. F. KotlabaI 17 July 1974, P R M 741809I S o v i e t U n i o n : Ukrainian S. S. R : Bondarcev 1953: 337; Zerova 1972: 157;I Georgian S. S. R .: Bondarcev 1953: 337; Kazakh S. S. R .: Švarcman 1964: 396I S y r i a : — ad opp. Sawra, leg. F. N ábělek 12 Dec. 1909, det. V. Litschauer, rev. F.I Kotlaba 10 M ay 1973, W 926, W 6633
A l g e r i a : Patou illard 1897: 20; 1904: 51 M a r o c c o : M aire et W erner 1937: 84
49

The list giving the localities of Inonotus tamaricis (Pat.) Maire shows that the species has a mediterranean-oriental distributional pattern (cf. the map on p. 51). The localities in Kazakh S. S. R. in the Soviet Union (Svarcman 1964) may possibly be an eastern exclave of the territory, but it can not be excluded that some other species was involved. However the author had not possibility to study the pertinent specimen.
The three localities in Greece known till now indicate an insufficient investigation of the Greek flora. Inonotus tamaricis (Pat.) Maire w ill probably be found in other localities of the seashore wherever the host tree occurs.
2. Inonotus tam aricis (Pat.) M aireCross section o f fru it bodye — granular core, f — zonation o f the context, g — tubes
Taxonomical notes
Bondarceva (Sinadskij et Bondarceva 1956) described a form : Inonotus tamaricis f. corneus M. Bond, growing on Tam arix ramosissima Pal. and T. palasii Desv. in Uzbek S.S.R., U.S.S.R. (Karakalpacka region). This form differs from the type one by the absence of the granular core, horny consistency of the trama, velvety surface of the pileus, short tubes, minute pores, and dimensions of hyphae and spores. On account o f the absence of the granular core, it probably was not I. tamaricis (Pat.) Maire at all. Pegler (1964a) described a new species Inonotus subhispidus Pegler et Reid, fo r which the absence of the granular core is also characteristic. As synonyms of this species, he mentiones Polyporus subhispidus Lloyd 1924 (found on trunk of a species of the genus Tam arix L.) and Inonotus tamaricis f. corneus M. Bond. Inonotus subhispidus has been known up to the present time from India, Pakistan and Uzbek S.S.R. (U.S.S.R.).
Inonotus tamaricis is closely allied to Inonotus dryophilus (Berk.) Murrill, I. rheades (Pers.) Bond, et Sing, and I. pseudohispidus Kravtz. et Schwarzman. These species belong to the section Phymatopilus Donk 1974, fo r which is typical the presence of the granular core in the place in which the fruit body adheres to the substrate. Furthermore, the setae in the hymenium and the thick-walled yellow to yellow-rusty spores are absent. Bondarcev (1953) distinguished fo r the first time the above mentioned taxa as species. Bourdot et Galzin (1925,
CESKA M YKO LO G IE 32 (1) 1978
50

H K L A N : IN O N O TU S T A M A R IC IS IN GREECE
1928) hold Inonotus rheades for a species and I. dryophilus and I. tamaricis for its subspecies. Pilat (1936—1942) considers these taxa even to be mere forms of the species I. rheades.
■ Another species of an unclear taxonomic status which requires further study is Inonotus pseudohispidus Kravtz. et Schwarzman described in 1950 by B.I. Kravcew and S. R. Svarcman from Kazakh S.S.R. (U.S.S.R.). Except for the data available from the literature for I. pseudohispidus no further information
I 3. The general distribution o f Inonotus tam aricis.
■ could be given here. The definition of the species by Pegler (1964b) and Svarc-I man (1964) is very unclear. Pegler (1964b) gives the chief distinguishing features■ in which it differs from I. tamaricis as “Pileus surface glabrescent but otherwiseI unchanged on drying or w ith age“ , growth on Populus (Acer, Fraxinus), where-I as in I. tamaricis, Pegler mentiones “Pileus surface becoming black and brittleI on drying or with age“ , growth on Tamarix. Svarcman (1964: 402) describes■ the colour of the pileus as cinnamon-brown, later (i. e. w ith age) rusty black!I There virtually remains only one distinguishing feature mentioned by bothI authors, namely the different host trees, in I. tamaricis the liv ing Tam arix sp.I div. and in I. pseudohispidus Populus, Acer and Fraxinus. The occurrence onI different host trees cannot be considered as the chief distinguishing feature.I From the study on the rather extensive material of the three species of theI section Phymatopilus Donk in the Herbarium of the National Museum in PragueI (PRM ), it is possible to make several conclusions about their taxonomical value.I Inonotus rheades (Pers.) Bond, et Sing. [ = Polyporus rheades Pers., Xantho-I chrous rheades (Pers.) Pat.], a species w ith a clear taxonomical definition mayI be quite reliably distinguished from the other species b y t h e s i z e o f t h eI s p o r e s . These are generally smaller and neither in their width nor in theirI length reach the extreme values o f both allied species. The recorded values of
5.2—5.7—6.2 X 3.1—4.0—4.3 (um agree w ith the values mentioned by other authors
51

as Bourdot et Galzin (1925, 1928) 5—6 X 3.5—4.5 um, Pilát (1936—1942), forma B rheades (Pers.) Pil. 5.5—6 X 4—4.5 um, Bondarcev (1953) 5—6 X 3.5—4 um,R Kotlaba et Pouzar (1969) 5—6.7 X 3.4—4.5 ^m. Kotlaba et Pouzar later studied
the type material of Persoon and gave the follow ing measurements 5.6—6.7 X X (4.4—)3.9—4.6 um (Kotlaba et Pouzar 1970). Pegler (1964b) mentiones the spores to be somewhat longer i. e. 5—7 ^m. The distinction of I. rheades from the other species on the basis of the spore size was confirmed by statistical evaluation (Table 1); the significance of the differences in the average values among the sets of 60 spores measured from five different localities was Hproved. Thus the chief character distinguishing I. rheades from I. tamaricis and HI. dryophilus is the size of the spores. The other features which have rather an Hauxiliary importance are indicated in Table 1. Kotlaba et Pouzar (1969) and HTortic et Kotlaba (1976) named several of them. H
Inonotus dryophilus (Berk.) Murrill ] = Polyporus corruscans Fr., X antho- Hchrous rheades ssp. corruscans (Fr.) Bourd. et Galz., Inonotus rheades f. cor- Hruscans (Fr.) Pil.] is closely allied to I. tamaricis. The distinction o f the two Hspecies on the basis of the host trees, i. e. I. dryophilus on living Quercus sp. Hdiv. and I. tamaricis on living Tam arix sp. div., is possible, but does not fu lly Hmeet the requirement of an exact taxonomic approach. Pegler (1964b) and Do- Hmaňski (1975) indicate erroneously that both species can be distinguished on Hthe basis of the thickness of the spore-wall: spores of I. dryophilus are thin- H-walled contrary to thick-walled ones of I. tamaricis. However, on the basis of Hour observations, the spores of I. dryophilus are distinctly thick-walled (aboud H1 jum ); Bondarcev’s drawing is quite right (Bondarcev 1953: 334, Fig. 76), the Isame being reported by Kotlaba (1973). According to our observations, there Hare two important distinguishing characters in which these two species differ, Hi. e. t h e c o l o u r o f t h e c o n t e x t and t h e s i z e o f t h e s p o r e s . The Hcontext of I. dryophilus is yellow, yellow-rusty to rusty, in dependence on the Hage of fruit bodies. Old fruit bodies, often sterile, having entirely rusty-colour- Hed context. The context of I. tamaricis is r u s t y w i t h y e l l o w z o n e s I(the zonation in old fruit bodies being evanescent, while the rusty colour pre- Ivails). Similar zones have never been observed in I. dryophilus. The context o f HI. tamaricis is formed of two types of hyphae: yellow to yellow-rusty thin ■walled (wall approx. 0.5—0.8 ^m thick) and rusty, thick walled (wall approx. I0.7—2 (um thick). The first type of hyphae prevails around the core, whereas Ithe other is predominant towards the distal part of the fruit body. I
A further character, which distinguishes I. dryophilus from I. tamaricis is ■the spore size. The respective values in I. dryophilus are 6.9—7.4—8.2 X 5.2— I—5.6—6.2 um, in I. tamaricis 6.2—6.8—7.3 X 4.6—5.2—5.7 um. The mentioned Ispecies cannot be distinguished reliably according to such kind o f record so Icommonly employed. From the statistical elaboration (according to the methods Iof Hrubý 1950; Nečásek 1950) of the dimensions of the sets of 60 spores I(measured in lactophenol, oil immersion 1250 X) from five different localities, Ithe follow ing values have been obtained: x ± 3 X sx, x + s, E, Em; for I. dryo- Iphilus, length 7.44 + 3 X 0.052, width 5.60 ± 3 X 0,047; for I. tamaricis, length I6.79 + 3 X 0.050, width 5.21 ± 3 X 0.030 (Table 1). The difference between Ithe average values of length and width ascertained by the t-test was significant I(tuš = 9.03, tug = 6,96). For the statistical evaluation the adequate value was Ifound to be N = 20. Thus the size of the spores is statistically a significant Idiacritical feature for the distinction of the two species. The other distinguish- Iing features, which rather belong to auxiliary ones, are summarized in Table 1. I
Č e s k á m y k o l o g i e 32 (i) 1978 H
52

K L Á N : IN O N O TU S T A M A R IC IS IN GREECE
A c k n o w l e d g e m e n t s
I w ish to thank Professor M. E. Pantidou (Athens) fo r the generous provision of uneasily ava ilab le literature and fo r her kind inform ation, Dr D. N. P eg le r (K ew )fo r his advice about one o f the tw o localities in G reece and D r F. D. Calonge (M adrid) fo r in form ing me about the localities in Spain. M any thanks are expressed to Dr F. Kotlaba fo r his notes from the H erbarium at V ienna and fo r his interest in m y work. Special thanks are g iven to D r Z. Pouzar fo r the reading o f the manu- srip l and fo r his valuable comments.
L i t e r a t u r a
B o n d a r c e v A . C. (1953): T ru tovy je griby jevropejskoj časti SSSR i Kavkaza.M oskva—Leningrad.
B r e s a d o l a J. (1892): Fungi Tridentin i. 2. Tridenti.B r e s a d o l a J. (1927—33): Iconographia m ycologica. M ilano.B o u r d o t H. et G a l z i n A . (1925): Hym énom ycětes de France (X I. Porés). Bull.
Soc. mycol. France, Paris, 41: 98—255.B o u r d o t H. et G a l z i n A . (1928): Hym énom ycětes de France. Sceaux 1927. D i a p o u l i s Ch. A . (1939): Contribution to the study o f the fungi o f P ilion . Rep.
physical. Sei. 5: 33—39. [In Greek]D o m a n s k i S. (1975): Basidiomycetes (Podstaw czaki)-Aphyllophora les (Bez-
blaszkowe). In : M ala flo ra grzybów . W arszaw a—Kraków .D o n k M. A . (1974): Check list o f European polypores. Am sterdam —London. H e i m R. (1933): Treballs Mus. Ciences Nat. Bare. 15: 61. [sec. F. D. Calonge
(M adrid ) — private in form ation]H e u f i e r L. J. (1868): Specimen florae cryptogam ae šeptem insularum V I. Fungi.
Verh zool.-bot. Ges. W ien, 18: 427—428.H r u b ý K . (1950): V ariab ilita a korelace v b iologii. Rozpr. 2. tř. Ces. Akad., Praha,
60 (17): 1-100.J a a p O. (1916): B eiträge zur Kenntnis der P ilz e Dalmatiens. Ann. mycol., Berlin,
14: 1-44.K o t l a b a F. (1973): O dvou vzácných rezavcích na dubech v Československu: Ino
notus dryadeus (Pers. ex Fr.) M u rrill a I. dryophilus (Berk.) M urrill. Ces. M y kol., Praha, 27: 69-83.
K o t l a b a F. et P o u z a r Z. (1969): Inonotus rheades (Pers.) Bond, et Sing. — Re- zavec skořicový. Ces. Mykol.,, Praha, 23: 163—170.
K o t l a b a F. et P o u z a r Z. (1970): Revision o f the original m ateria l o f Phel- linus sulphurascens Pil., Xanthochrous glomeratus ssp. heinrichii P il. and P o ly porus rheades Pers. (Hym enochaetaceae). Ces. M ykol., Praha, 24: 146—152.
L ä n d e r e r X . (1858): Botanische Notizen aus Griechenland I. Ü ber d ie in G rie chenland vorkom m enden Schwämme. Flora, Regensburg, 41: 675—683.
M a i r e R. et W e r n e r R. G. (1937): Fungi Maroccani. Catalogue raisonné des Champignons connus jusqu’ici au Maroc. M émor. Soc. Sei. nat. Maroc, Rabat- -Paris, 45: 1—147.
M a i r e R. et P o l i t i s J. (1940): Fungi H ellenici. Catalogue raisonné des champignons connus jusq ’ici en Grěce. Actes Inst. bot. Univ. Athénes 1: 27—179.
M o r a v e c J. (1974): Severa l operculate Discomycetes from G reece and remarks on the genus Scutellin ia (Cooke) Lamb, emend. L e Gal. Ces. M ykol., Praha, 28: 19-25.
N e č á s e k J. (1950): On the variab ility oi the spores of higher Fungi. Stud, bot..čechosl., Praha, 11 (1—2): 49—64.
P a n t i d o u M. E. (1973): Fungus — host index fo r Greece. K iphissia—Athens.P a n t i d o u M. E. et W a t l i n g R. (1970): A contribution to the study o f the
Boletaceae-Suilloideae. Notes Roy. bot. Garden, Edinburgh, 30: 207—237. P a t o u i l l a r d N. (1897): Catalogue des champignons de la Tunisie. Bull. Soc.
mycol. France, Paris, 13: 197—216.P a t o u i l l a r d N. (1904): Champignons algéro-tunisiens nouveaux ou peu connus.
Bull. Soc. m ycol. France, Paris, 20: 51—54.P e g l e r D. N. (1964a): N ew species o f Inonotus (Polyporaceae). Trans, brit. m y
col. Soc., Cambridge, 47: 167—173.P e g l e r D. N. (1964b): A survey o f the genus Inonotus (Polyporaceae). Trans, brit.
mycol. Soc., Cambridge, 47: 175—195.
53

P e t r á k F. (1943): Fungi. In : Hechinger K . H., f il. : F lora Aegaea, Denkschr. Akad.Wiss. W ien, Cl. math.-nátur., 105/1: 10—15.
P i l á t A . (1936—42): Polyporaceae — Houby chorošovité. In : K av in a K . et A . P i- lát: A tlas hub evropských. 3. Praha.
P i n t o - L o p e s (1953): Po lyporaceae de Portugal. Revisao das coleccoes Portu- guesas. Revista Fac. Sci. Univ. Lisboa IIA . Ser. C, 3: 157—237.
P ř í h o d a A . (1970): Battarea stevenni (L ib .) Fr. v Řecku. Ces. M ykol., Praha, 24:40-43.
R y v a r d e n L. (1972) : Studies on the Aphyllophorales o f the Canary Islands w ith a note on the genus Perenniporia Murr. Norw . J. Bot., Oslo, 19: 139—144.
S a c c a r d o P. A . (1905) : Sylloge fungorum omnium hucusque cognitorum. Pavii. S e r e j a n n i J. A . (1939) : Catalogue com menté des champignons rencontrés sur
les plantes cultivées en Grèce. Ann. Inst. Phytopath. Benaki, 3: 9—14.S e r e j a n n i J. A. et D e m e t r i a d e s S. D. (1951): Catalogue commenté No. 2
des champignons et baoteries rencontrés sur ies plantes cultivées en Grèce.Ann. Inst. Phytopath. Benaki, 5: 5—11.
S i n a d s k i j J. V. et B o n d a r c e v a M. A . (1956): M alo izvestn yje tru tovik i na Populus i Tam arix i ich značen ije v K ara-Kalpakskoj A.S.S.R. Bot. 2um .,M oskva—Leningrad, 41: 1177—1183.
S v a r c m a n S. R. (1964) : Auriculariales, Trem ellales, Dacrym ycetales, Exobasi- diales, Aphyllophorales. In : F lora sporovych rasténij Kazachstana. 4. A lm a-A ta .
T o r t i é M. et K o t l a b a F. (1976) : A handful o f polypores, rare or not previously published from Yugoslavia. A c ta bot. Croat., Zagreb, 35: 217—231.
Z é r o v a M. J., R a d z i e v s k i j G. G. et S e v č e n k o S. V. (1972): Bazydio- m icety. — In : Z erov D. K. (éd.): Vyznačnyk g ryb iv U krajiny. 5 (1), K y jiv .
Address o f the author: RND r. Jaroslav K lán, Departm ent o f Botany, Charles Un i-versity, 128 01 Praha 2, Benátská 2, Czechoslovakia H
A. N e u n e r : B L V Naturführer. Pilze. A ile w ich tige P ilze nach Farbfotos bestim - Hmen. — 143 stran, B L V Verlagsgesellschaft, München — Bern — W ien 1975. H
Tato kniha patří m ezi populární knihy, je jich ž význam nelze přehlížet. M ůže slou- Hžit nejen m ykologům amatérům, ale i vědeckým pracovníkům jiných oborů k získání Hobecnějšího přehledu a znalostí o houbách rostoucích v lesích. Kapesn í vydán í této ■knihy s barevně perfektn ím i obrázky a krátkým popisem 91 druhů hub vyskytu jících Hse přím o na stanovišti se zdůrazněním některých zvláštních znaků jako je např. Hspodní část klobouku, umožňuje sběratelům hub přím é použití v přírodě. K lepšímu Hsrovnání a vyloučení možnosti zám ěny jsou obvyk le společně na jedné straně v y obra- Izeny houby jed lé s jedovatým i dvojn íky. P ro tě jš í strana vždy obsahuje krátký popis Inejdůležitějších znaků jako je tvar, velikost, barva, chuť a vůně hub a u hub s m ož- ■ností záměny popis rozlišovacích znaků. I
Autor této knihy — vedoucí nejstarší a největší m ykologické poradny N SR v M n i- Ichově — poukazuje na to, že většina hub rostoucích v lesích je jedlých, zatímco počet nepoživatelných nebo jedovatých je m alý. A kdo zná zejm éna houby smrtelně jedovaté, zbavu je se rizika při požíván í lesních hub.
P ro řadu lid í je sběr lesních hub koníčkem a zdravým sportem ; s pom ocí tohoto atlasu hub lze získat nejen základní znalosti o houbách, ale i větší jistotu při určování zejm éna hub jedlých a je jich jedovatých dvojníků.
Vlasta Čatská
Č e s k á m y k o l o g i e 32 (i) 1978
54

■ The Growth Response of Wood-Destroying Fungi to the■ Presence of Spruce Callus I
H Růstová reakce dřevokazných hub na přítomnost smrkového kalusu
J iř í H řib et V la d im ír Rypáček*)
Interaction between spruce tissue culture and selected species o f wood- -destroying fungi, attacking either liv ing spruces or their dead wood, was studied. The experim ents w ere carried out on Brow n and Law rence m odified medium. The fungi responded to the presence o f tissue culture either by grow th inhibition o f d iffe ren t intensity or by grow th stimulation. The grow th response o f fungi to the presence o f tissue culture indicates the degree o f their aggressivity.
H B yla studována interakce m ezi tkáňovou kulturou smrku a vybraným idruhy dřevokazných hub, které napadají buď ž ivé sm rky nebo je jich m rtvé
H dřevo. Pokusy by ly prováděny na m odifikované půdě podle B row na a HH Lawrence. Použité houby reagova ly na přítom nost tkáňové kultury buď
různě mohutnou inh ibicí růstu nebo jeho stimulací. Pod le toho, jak houby reagují na přítom nost tkáňové kultury, lze usuzovat na stupeň je jich agre-
■ siv ity. I
I n t r o d u c t i o n
Detailed studies have been carried out concerning interactions between viruses, pathogenic bacteria and fungous parasites o f culture plants on the one hand and tissue cultures on the other till this time. A survey of these problems was presented, for example, by Ingram (1973). Interaction between tissue cultures of trees and wood-destroying fungi have not been studied up to now. The first experimental statements on this problem were reported by Hřib
■ and Rypáček (1976). IH The present paper describes the growth response of selected species of wood-■ -destroying fungi, which are specialized on living spruces or their dead wood,■ to the presence o f spruce callus.
■ M a t e r i a l a n d m e t h o d s
■ The culture o f spruce callus was derived from the hypocotyl o f a P icea excelsa■ seedling in 1974 and is maintained by passaging on Brown and Law rence B-24■ m odified m edium (W inton 1972) w ith the addition o f 5 m g . I -1 a-napthylacetic acidH and 0.1 m g . l - ’ benzylam inopurine.■ T h e cultures o f wood-destroying fungi w ere obtained from the Department o f P lant■ B iology, J. E. Purkyně U n iversity Brno. W ith regard to the fact that the most■ intensive w ood decay is caused by isolates from the same wood species (Santra and■ N andi 1975) those species w ere used w hich w ere isolated from samplings on liv in gI spruces (each species is presented w ith the strain number): 172 Heterobasidion■ annosus (Fr.) Bref., 17 Fom itopsis p in ico la (Sw. ex Fr.), P. Karst., 144 LaetioporusI tu lphureus (Bull, ex Fr.) Bond, e t Singer, 68 Phellinus abietis (P. Karst.) P ilá t and■ 92 Phaeolus schw ein itz ii (Fr.) Pat. For the comparison tw o exclusively saprophyticI species w ere used: 3 Conio-phora puteana (Fr.) P. Karst, and 4 P o ria v a illa n tii (DC.
ex Fr.) Sacc.The grow th response o f m ycelium to the presence o f spruce callus was investigated
in P e tri dishes on the same medium, which the callus has been m aintained on.For the inoculation the fungus m ycelium was taken w ith m alt-agar substrate o f about 1 X 1 cm in size from the m argin o f the culture. Th e inoculum o f the fungus was put in the m idd le o f the dish. The callus used fo r inoculation was grow n fo r three
*) Botanical Institute o f the Czechoslovak Academ y o f Sciences, Department of Forest B iology, Poříč í 3b, 603 00 Brno, CSSR.
55

Table 1 — The growth response o f selected wood-destroying
Variant aSpecies Acceleration ( + ) Temporary Permanent Attacko f the fungus Retardation (— ) inhibition zono inhibition zone o f callus
o f mycelium growth towards
callusdays yes/no yes/no yes/no
P. abietis — 1 to— 4 yes yes noP. schweinilzii — 8 to— 12 no yes noH. annosus + 7 to +10 yes no yesL. sulphureus — 18 yes no yes/■'. pinicola 0 to + 1 no no yesC. puteana 0 to — 1 no no yesP. vaillantii — 1 to — 2 no no yes
weeks from the last passage. Its inoculum was put at the margin o f the dish in the distance o f about 2.5 cm from the inoculum o f the fungus.
The experim ent was arranged in th ree fo llow in g variants: The fungus was ino- culated simultaneously w ith the callus (a), 14 days (b ), and 23 days (c ) a fter the callus inoculation. The m ixed cultures w ere incubated at 22 ± 1 °C in the dark. Each variant w as established in three repeatings. The cultures w ere photographed in tw o days’ intervals, the grow th o f m ycelium being measured towards the callus and opposite to it. H
R e s u l t s a n d d i s c u s s i o n H
When compared the growth rate of mycelium towards callus and opposite Hto it (Table 1), it was found that, in the majority of cases, the callus caused Hthe growth retardation, regardless of the fact if the callus was inoculated Hsimultaneously with the fungus or if the fungus was inoculated later. An Hexception is represented by H . annosus, the growth of which was accelerated Hsignificantly in all three variants and all repeatings towards the callus (Plate I, HPhoto 1 to 3, Fig. Id ); it reached the callus by 6 to 12 days earlier than the Isame distance to the opposite side. Acceleration of the mycelium growth H
|H towards the callus was found also in the fungus F. p in ico la , but only by one Iday and in one case when the fungus and the callus were inoculated simul- Itaneously. I
The majority of fungi used attacked the callus. An exception is represented Iby P . a b ie tis and P. s ch w e in itz ii which did not grow into the callus. In these Icases an inhibition zone was formed between the callus and the mycelium, Iwhich persisted for the whole time of the experiment (36 days). The width I
SSi of this permanent inhibition zone varied from 1 to 13 mm; the widest inhibition IHR zone was found from 1 to 3 mm and from 9 to 13 mm in P. a b ie tis and IH j P. s ch w e in itz ii, respectively. The permanent inhibition zone was formed when I
the mycelium reached the callus (Plate I., Photo 4 to 6, Fig. la ); such a response I■ was exhibited by P. s ch w e in itz ii in the variants a, b and by P. a b ie tis in the■ variants b, c. In two cases the growth of the fungus was stopped at a certain
distance from the callus for a certain time, then it continued to be stopped permanently near the callus (Fig. lb ). In the course of the mycelium growth
HHj towards the callus a temporary inhibition zone was formed first, which wasin P. a b ie tis in the variant a from 5 to 8 mm wide and lasted from 4 to 6 days,
Č e s k á m y k o l o g i e 32 (i) 1978
56

■ H Ř IB et R Y P Á Č E K : TH E G R O W TH RESPONSE OF W O O D -D E S TR O Y IN G F U N G I
■ F ungi to the presence o f spruce callus
■ Variant b■ Acceleration ( + ) Temporary Permanent Attack of callus
Retardation (— ) inhibition zone inhibition zoneo f mycelium growth
■ towards callus■ days yes/no yes/no yes/no
— 4 to — 8 no yes no— 2 to — 13 no yes no
+ 6 yes no yes— 15 to — 19 yes no yes
0 no no yes— 4 no no yes— 5 no no yes
Continued on the following page
I in P. schweinitzii in the variant c it was from 5 to 10 mm and lasted from 6■ to 8 days. Nevertheless, this temporary inhibition was overcome by both the■ fungi and their growth was stopped definitively close by the callus.■ The temporary stopping of growth can also be found in the fungi which■ attack callus (Fig. lc). This response was exhibited by L. sulphureus in all■ variants. In the variant a its growth was stopped at the distance of 4 to 7 mm■ from the callus for 6 to 10 days, in the variant b at the distance of 4 to 7 mm■ for 6 to 12 days and in the variant c at the distance of 2 to 9 mm for 8 to 12I days. The attack of the callus by the fungi followed after overcoming thisI temporary inhibition (Plate I, Photo 7 to 9). In the fungus H. annosus, whichI was stimulated in growth by the presence of the callus in all variants, this■ response was found only in one case in the variant a; the mycelium growthI was stopped at the distance of 11 mm from the callus for 4 days and thenI continued more intensively towards the callus which was attacked by theI fugus at once.I Nevertheless, the exclusively saprophytic fungi (C. puteana and P. vaillantii)I attacked the callus, too. They grew towards the callus either at the same rateI as to the opposite side (Fig. 2a), or their growth was retarded towards theI callus by 5 days at the most (Fig. 2b). The presence of the callus never caused,I however, a temporary stopping of their growth. I f the callus was inoculatedI earlier by 18 or 23 days, the growth retardation of the fungus towards theI callus was more remakrable than in the case when the fungus and callus wereI inoculated simultaneously. Similar growth response to the presence of spruceI callus was also exhibited by the fungus F. pinicola. In the majority of casesI its growth rate towards the callus was the same as that to the opposite side,I or its growth towards the callus was either accelerated or retarded only byI one day.I Nevertheless, saprophytic wood-destroying fungi do not attack in nature theI wood of living trees. I f they are sometimes found on living trees, then theyI occur only on dead fragments of the branches or on wounds of trunks whereI the wood has died away. For the preventions against infection of living treesI with saprophytic fungi should be searched elsewhere. They are probably

Continuation from the previous page
Variant cSpecies Acceleration (-j-) Temporary Permanent Attacko f the fungus Retardation (— ) inhibition zone inhibition zone o f callus
of mycelium growth towards callus
days yes/no yes/no yes/no
P . abietis — 10 to — 17 no yes noP . schweinitzii — 18 to — 19 yes yes noH. annosus + 7 to -4-12 no no yesL . sulphureus — 17 to — 23 yas no yesF. pinicola 0 to — 1 no no yesC. puteana — 2 to — 5 no no yesP . vaillantii — 2 to — 3 no no yes
localized in the cambial region, i. e. in the youngest sapwood, bast and in the Hyoungest periderms (Vikstrom 1975). H
The infection of the tree w ith wood-destroying fungi depends upon the level Io f natural antibiotics (Findlay 1966). In spite o f the fact that from our first Hexperiments no generally accepted conclusions can be drawn, they indicate Hthat the callus keeps antibiotical properties only against some species of wood- I-destroying fungi which attack living tress. H. annosus and L. sulphureus Hattack the callus. They are especially aggressive species which attack not only Hthe heartwood, bult also the liv ing sapwood and reach the cambium finally. IThe resistance against wood-destroying fungi increases not only from the base Ito the top of the tree, but also from its middle part to the periphery (Rao, IRao 1975, Domahski 1953). This resistance can be overcome by the both species. HIn addition to it, the infection does not penetrate into the tree only through Ithe wounded spots, but also through the intact bark o f the roots, as it was Ifound, for example by Woeste (1956), Peek et al. (1972) in H. annosus. I
The species P. abietis and P. schweinitzii do not attack the callus and they Istop their growth at a certain distance from it. These species never attack the Iliving sapwood. They are specialized only on dead inner wood, penetrating Iinto it through wounds or open knots but they never penetrate the sapwood. IEven though the seedlings can also be infected with these species, the disease Iremains in the latent stage for a long time to be manifested later. These species Ican be found especially in the old-aged stands. I
The resistance o f living trees against wood-destroying fungi is a rather Icomplicated problem. It depends not only upon climatic and trophic conditions Iunder which the tree or the whole stand grows and upon its age and vita lity I(Manka 1960, Gibbs 1967, 1968, and others), but also upon the degree of the Iaggressivity of the fungus and the amount of infection. Essentially the spread Io f the fungous infection depends also on the interaction w ith living tissue Iof the host and its metabolites. To solve the interaction between the fungus Iand the tree, the study of growth responses exhibited by the fungi to the Ipresence of its callus appears to be one of the prospective methods. I
Č e s k á m y k o l o g i e 32 (i) 1978 H
58 I

■ K L A N : IN O N O TU S T A M A R IC IS IN GREECE
I callus■ Z O 1-------- 1--------1-------- 1--------1--------1--------1--------1--------
■ mm ^ ______________ I
I mmH 2 5 ____ 1___ 1______ 1___ L__—1___1___ 1-----I callus■ Z d 1--------1------- 1-------- 1--------1--------1--------1--------1--------■ mm ________________________ --
I mm2 5 ___ 1___ 1__^ ___ 1___ 1___ 1___ 1___ 1___
I ~ r callus■ A D 1--------1--------1--------1--------1------ 1-------------— i--------■ m m _________________________^
I mm25 i \ i i i i i i___
I 2 5 ----- t----- 1----- \----- 1--------- —i---- 1----- 1-----I mm
I mm ‘—I 2 5 1______ 1______1______ 1______ 1______ 1--------1 r~i>—
I 4 8 12 16 20 24 28 32 II daysI 1. The types o f grow th responses o f the fungi attacking the wood o f liv in g spruce to
the presence o f spruce callus.a) P. schw einitzii, inoculated 18 days a fter callus inoculation,b) the same species inoculated 23 days a fter callus inoculation,c) L. sulphureus and d) H. annosus, both species inoculated 23 days a fter callu* inoculation. The grow th o f the colony tow ards the callus is indicated up and the grow th to the opposite side dow n from the zero point.

I n m II 25 4 8 12 4 8 12 16 Idays I
2. The types o f the grow th responses o f saprophytic fungi to the presence o f spruce Hcallus. ■a) C. puteana, inoculated simultaneously w ith the callus, Hb) the same species, inoculated 23 days a fter callus inoculation. The grow th o f the Hfungal colony towards the callus and opposite to it is indicated in the same w ay Has in Fig. 1. H
R e f e r e n c e s H
D o m a ñ s k i S. (1953): Badania nad przyczyham i powstawania posuszu v starszych Bdrzewostanach sosnowych w W ielkopolsk im Parku N arodow ym w Ludw ikow ie. BPrace Inst. Bad. Lesných, No. 93. B
F i n d l a y W. P. K . (1966): Ecology o f w ood-destroying and w ood-inhabiting fungi. BHolz u. Organismen. Berlin. B
G i b b s J. N. (1967): The role of host v igou r in the susceptibility o f Pinus to Fomes Bannosus. Ann. Bot. 31: 803—815. B
G i b b s J. N. (1968): Resin and the resistance o f Conifers to Fom es annosus. Ann. BBot. 32: 649-665. B
■ H ř i b J. et R y p á č e k V. (1976): Using tissue cultures of wood species fo r the study Bo f the substrate specific ity o f wood-destroying fungi. Internát. Symp. “ App lication Ho f plant exp lantate cultures in breed ing“ Olomouc, CSSR. Sept. 6—11, 1976. I
I n g r a m D. S. (1973): G row th o f p lant parasites in tissue culture. In H. E. Street I(Ed.): P lant tissue and cell culture. O xford—Edinburgh—Melbourne. I
M a ñ k a K . (1960): F itopatología lesna. W arszawa. IP e e k R. D., L i e s e W., P a r a m e s w a r a n N. (1972): In fektion und Abbau der m
W urzelrinde von Fichte durch Fom es annosus. Eur. J. For. Pathol. 2: 104—115. IR a o B. S , R a o R. V. (1975): Variation in decay resistance of Betula pubescens Ehrh. I
against Polyporus vers ico lor. Current Sci. 44: 864—865. IS a n t r a S., N a n d i B. (1975): Strain variations among Fomes darissim um L loyd I
attacking d ifferen t host wood. A cta Soc. Bot. Polon iae 44: 317—321. IV i k s t r o m C. (1975): Fungal decomposition of sapwood and heartwood o f European I
aspen, Populus trém ula L. Eur. J. For. Pathol. 5: 349—356. IW i n t o n L. L. (1972): Callus and cell culture o f Douglas fir. Forest Sci. 18: 151—154. IW o e s t e U. (1956): Anatom ische Untersuchungen über die In fektionsw ege ein iger m
■ W urzelp ilze. Phytopath. Z. 26: 225—272. I
Č e s k á m y k o l o g i e 32 (i) 1978 H
60 I

H Rhizopus cohnii jako příčina mukormykotického abortu u skotu
Rhizopus cohnii as the case of mucormycoíic abortion in cattle
H A lena Adám ková, M a rie Váňová a M iros lav Lávička
■ V článku je popsán m ykotický abortus u skotu, způsobený druhem R h i-■ zopus coh n ii Berlese e t De Ton i (M ucora les). Práce je doplněna popisem
a fo togra fiem i kultury. Dále je uvedena fotodokum entace hyf v nativním preparátu z obsahu slezu (žaludku) zmetka a histologického nálezu hyf v placentě. ______________
■ A case of m ucormycotic abortion caused by Rhizopus cohn ii Berlese etDe Toni (M ucora les ) is described in cattle. The paper is documented bydescription and photos o f the culture. There are also photos o f hyphae onn a tive preparation from the abomasum content (stomach) of the aborted foetus and from the histological find ing o f hyphae in the placenta.
Mykotické aborty skotu, způsobené zástupci řádu Mucorales jsou známé již H delší dobu, i když nejsou tak hojné, jako aborty vyvolané jiným i houbami.■ Např. Moreira — Jacob (1956) udává jako příčinu abortu také druhy Absidia■ ramosa (Lindt) Lendn., A. corym bifera (Cohn in Lichth.) Sacc. et Trotter, M ucorH pusillus Lindt, Rhizopus arrhizus Fischer a R. cohnii Berlese et De Toni.H Austwick a Venn (1957) druhy Absidia ramosa (Lindt) Lendn., M ucor sp. a■ Rhizopus sp. Ainsworth a Austwick (1959) Absidia ramosa (Lindt) Lendn.,■ A. corym bifera (Cohn in Lichth.) Sacc. et Trotter, M ucor pusillus Lindt, M or-■ tierella polycephala Coemans, Rhizopus arrhizus Fischer, R. cohnii Berlese et■ De Toni a R. microsporus van Tiegh. Dvořák o Otčenášek (1975) uvádějí ve■ svém přehledu mykóz jako příčiny abortů z řádu Mucorales druhy Absidia■ corymbifera, A. ramosa, M ortierella polycephala, M ucor pusillus, Rhizopus■ arrhizus a R. oryzae Went et Prinsen Geerlings.■ Na pracovišti ÚSVÚ byly z orgánů zmetků během času vyizolovány a na■ Katedře botaniky určeny druhy Absidia corymbifera, A. ramosa a M ucor■ pusillus.I V roce 1975 se nám podařilo vyizolovat houbu, která byla zjištěna jakoH příčina abortu krávy, zmetající v době březosti sedmi měsíců po předchozíI léčbě akutní intoxikace z krmivá. Mykotický abortus byl diagnostikován naI základě pozitivního mikroskopického nálezu v nativním preparátu z obsahu■ slezu zmetka, kultivačního průkazu houby v obsahu slezu a v plicích zmetkaI a v placentě, jakož i histologického nálezu v kotyledonech placenty.I Makroskopicky bylo pozorováno zduření kotyledonů placenty, špinavě šedéI barvy s povrchovým rozpadem. V orgánech zmetka nebyly zjištěny makrosko-I picky podstatné změny. Histologicky jsme zjišťovali v kotyledonech placenty■ výraznou trombotizaci cév a nekrotický rozpad povrchové třetiny kotyledonů,■ ohraničený demarkační zónou zánětlivé inflitrace. Barvením podle GrocottaI byla pak nalézána v trombotizovaných cévách a nekrotické tkáni četná větvenáI vlákna, nejčastěji široká 8— 11 «m . V plicích zmetka jsme nenalezli žádnouI orgánovou afekci, avšak barvením podle Grocotta jsme v bronších zjistili kulo-I vité a oválné útvary specificky se barvící.I K mykologickému vyšetření byly použity vzorky z plic, obsahu slezu a pla
centy. Každý vrozek byl kultivován při teplotě + 24 °C a + 3 7 °C na Petriho miskách s živným i půdami Czapek-Doxovým agarem a Sabouraudovým agarem. Na obou živných půdách inokulovaných materiálem z plic, obsahu slezu a placenty byl růst při + 24 °C řídký a při + 37 °C hojný. S pomocí literatury, pře-
61

devším Zycha a kol. (1969) a Pidopličko a M ilko (1971) byl patogen určen jako HRhizopus cohnii Berlese et De Toni.
P o p i s p l í s n ě : HKolonie na Sabouraudově agaru při + 37 °C za tři dny vyplňuje celou H
Petriho misku, tvoří vatovité mycelium, zpočátku bílé, později šedočerné. HStolony slabě vyvinuté. Rhizoidy zřetelné. Sporangia černá ve zralosti, H50— 110 tu široká. Kolumely většinou válcovité nebo obráceně hruškovité, málo- Hkdy kulovité, 29— 65 «m , nejčastěji asi 40 ¿¿m široké, s miskovitou apofýzou Ha zřetelným límečkem. Sporangispory kulovité, někdy nepravidelné, slabě ovál- Hné, 3,8— 6 jum, nejčastěji asi 4,5 um, hladké. Chlamydospory v myceliu četné, Hrůzného tvaru i velikosti. Zygospory nezjištěny. H
Kultura je uložena ve Sbírce kultur plísní a hub na Katedře botaniky UK, Hpod číslem 1509. H
Z našeho území je Rhizopus cohnii Berlese et De Toni uváděn (1972 a 1973) HFragnerem a kol. jako původce mykotického onemocnění vepře. H
L i t e r a t u r a H
A i n s w o r t h G. C. et A u s t w i c k P. K . C. (1959): Fungal diseases o f animals. HCom m onwealth Agricu ltu ra l Bureaux, Farnham Royal Books, England. H
A u s t w i c k P. K. C. et V e n n J. A . J. (1957): Routine investigation into m ycotic Habortion. Vet. Rec. 69: 488—491. H
D v o ř á k J. et O t č e n á š e k M. (1975): K etio logii m ykóz skotu. Veterinářství 3: I127-128. ■
F r a g n e r P., V í t o v e c J. et V l a d í k P. (1972): Rhizopus cohnii v m ukorm ykóze Ivepře a diskuse o podobných rhizopech. Ces. M ykol. 26: 167—178. B
F r a g n e r P., V í t o v e c J., V l a d í k P. et P r o k š C. (1973): L iv e r disease in hog Icaused by Rhizopus cohnii. Mycopath. M ycol. appl. 49: 249—254. B
M o r e i r a - J a c o b M. V. e t N. v a n U d e n M. D. (1956): M ycotic abortion in cattle. IA čase record and a rev iew o f the relevan t literatuře. Brit. Vet. J. 112: 453—461. H
P i d o p l i č k o N. M. et M i l k o A . A . (1971): A tlas mukoralnych gribov. Pp. 1—115. HA N USSR, K iev . ■
Z y c h a H., S i e p m a n n R. et L i n n e m a n n G. 1969): Mucorales. Pp. 1—355, IJ. Cramer, Lehre. I
A dresy autorů: Dr. A . Adám ková, M VD r. M. Lávička, Ústřední státní veterinárn í Iústav, Praha 6-Lysolaje; Dr. M. Váňová, K atedra botaniky Univ. K arlovy , IBenátská 2, 128 01 Praha 2. I
Č e s k á m y k o l o g i e 32 (i) 1978 H
I 62 I

L i t e r a t u r a
D. E r s h a d : Fungi o f Iran. M in istry o f Agricu ltu re and Natural Resources.Department o f Botany, publication No. 10. Teheran 1977. Pp. 1—277.
K n ih a je katalogem všech skupin hub dosud zjištěných na území Iránu. Autor, vědecký pracovník Výzkum ného ústavu rostlinných chorob v Teheranu, v ní podchytil většinu publikovaných údajů a navíc p řipo jil některé v lastn í nálezy. Výzkum íránské m ykoflo ry prováděli v m inulosti (v letech 1960 až 1941) výhradně zahraniční badatelé a teprve v posledních několika desetiletích se začala v Íránu rozv íje t v lastn í v ý zkumná činnost. O poznání íránských m ikrom ycetů z taxonom ického hlediska se n e jv íce zasloužil F. P e t r á k , k terý uveře jn il o nich řadu význam ných příspěvků h lavně v Sydow ii, z íránských m ykologů později také E. E s f a n d i a r i , s ním ž Petrák úzce spolupracoval. Petrák zpracoval také m ateriál, získaný známým ra kouským botanikem K. H. Rech ingerem během jeho četných výzkum ných cest po Iránu.
V katalogu D. E r s h a d a jsou jednotlivé druhy uvedeny podle hostitelů, seřazených abecedně podle rodů a druhů. Celkem je takto zaregistrováno 1356 druhů, z toho je 466 basidiomycetů, 313 askomycetů a 512 deuteromycetů, zbytek připadá na jiné skupiny. U každého taxonu jsou uvedeny lokality s odkazy na příslušné literárn í pramsny. Práce je doplněna literaturou, která byla podkladem pro rešerši, a abecedním seznamem druhů hub. V e lm i užitečný je seznam lokalit a přiložená mapka Iránu s vyznačeným i lokalitam i. S výjim kou stručného souhrnu a rejstříku je celý text v angličtině.
V současné době pracuje v Íránu skupina fytopathologů a m ykologů (D. Ershad, M. Saber, G. Scharif, A . Bambadian, A . Gh. Ebrahim i, J. Fatemi, F. Fallahyan, N. Zalpoor atd.). Je nutno ocenit záslužnou práci autora této publikace, D. Ershada, která podává prvn í souborný přeh led o dosavadních znalostech m ykoflory této i po m ykologické stránce tak pozoruhodné a v mnoha skupinách hub dosud málo prozkoumané země.
M irk o Svrček
L. V. L j u b a r s k i j , L. N. V a s i I j e v a : Děrevorazrušajčije griby Dalnego Vostoka.Novosib irsk 1975. Pp. 1—164, 53 tab. Cena 2 ruble 28 kopějek.
K n ih a zahrnuje v zestručněné form ě většinu materiálů, které za dobu 40 let soustavné vědeckovýzkum né práce na Dálném východě SSSR nashromáždil fytopatolog a m ykolog L. V. L j u b a r s k i j (1903—1968). Ten však zem řel,*) aniž rukopis dokončil; to učinila L. N. V a s i l j e v a z dálněvýchodního vědeckého střediska A k a demie věd SSSR, specialistka na lupenaté houby (L jubarskij se zabýval h lavně houbami nelupenatým i, především choroši).
Obsah knihy je krom ě úvodu, předm luvy, závěru, seznamu literatury, indexů druhů a vysvětlivek k tabulím rozdělen do šesti kapitol: 1. Obecné údaje o dřevokazných houbách (p. 6—16). 2. Ekologicko-geografická charakteristika (p. 17—36). 3. Hospodářský význam dřevokazných hub (p. 37—64). 4. Druhy dřevokazných hub, je jich rozšíření a specializace na Dálném východě (abecední seznam více než 300 druhů ve form ě přehledných tabulek s údaji o hostitelských dřevinách a rozšířením — p. 65—85). 5. K líče k určování hlavn ích a některých za jím avých druhů (p. 86—99). 6. Popisy hub vyvo láva jíc ích hniloby kmenů a kořenů živých dřevin (p. 100—148). Tato poslední — bohužel dost krátká — kapitola je pro nás nejza jím avějš í, neboť krom ě popisů obyčejných hub zahrnuje podrobné popisy i takových druhů, které nerostou v Evropě. Patří m ezi ně Cryptoporus volvatus, Phellinus van in ii, P. baum ii, Pelloporus scaurus a Leucophellinus irpicoides. N ěkteré z těchto a řada dalších, uvedených bez popisu jen v tabulkách, roste též v Severn í Am erice, neboť flóra východoasijská a západoamerická má mnoho společných prvků (v m inulých geologických dobách tyto pevn iny spolu souvisely). M ezi uváděným i ohňovci je i Phellinus van in ii L jub. 1962 (p. 125), který je totožný s oním záhadným ohňovcem sbíraným K ravcevem r. 1928 na Dálném východě („distr. A m u r“ ) (v iz Ces. Mykol., Praha, 30: 17—19. 1976) určený a publikovaný A . P ilá tem jako P. torulosus a později jako P. everhartii. Sdělila m i to krátce po v y jit í m ého článku v dopise právě L. N. V asiljeva .
Taxonom ie a nomenklatura hub v knize zahrnutých je dosti konzervativn í a vychází z kn ihy A . S. Bondarceva (1953). Tak např, dobrý druh Phellinus chrysolom a (Fr.) Donk = P. abietis (P. Karst.) P il. je považován jen za odrůdu ohňovce borového —
*) Ž ivotop is L. V. Ljubarského a jeho b ib liogra fii u veře jn il před 7 lety D. V. Sokolov (M ikol. i Fitopat., Leningrad, 4: 93—96, 1970).
63

Phellinus p in i var. abietis (P. Karst.) P il. (p. 100); Hapalopilus fib rillosus (p. 112)B patří podle moderních názorů do rodu Pycnopore llus a jeho správné jm éno je P.
fulgens (Fr.) Donk; Fom itopsis odoratissima Bond. (p. 129) je identický s H aploporus I odorus (Sommert.) Sing.; taxonom icky naprosto neudržitelné je rozeznávat tři odrůdy
Daedaleopsis confragosa — var. confragosa, var. bu llia rd ii a var. rubescens (p. 140),B přičem ž jsou navíc všude navrhovány nové kombinace bez uvedení basionymů, takžeB nom enklatorickv nejsou platně publikované. V knize je zachyceno ve lm i zajím avé
spektrum substrátů jednotlivých druhů hub, které význam ně doplňuje údaje známé ^ BB z ostatních částí Sovětského svazu. Evidentní je m alá znalost zejm éna západní novější ^ B
m ykologické literatury, což nepřízn ivě ov livňu je kvalitu práce: v seznamu literatury ^ Bje citováno pouze 12 západních autorů (včetně m onografie chorošů A . Piláta, bohužel ^ Bvšak n ikoliv jeho příspěvky o houbách východní Asie ), z čehož nejm ladší z nich je ^ Bpráce v íce než 20 let stará. ^ B
Na 53 křídových tabulích jsou silně retušované fotogra fie celkem 67 druhů dřevních ^ Bhub a je třeba s povděkem kvitovat, že reprodukce není většinou špatná. To je ve lm i ^ Bcenný přínos, neboť m ezi vyobrazeným i druhy jsou krom ě již výše uvedených další ^ Bvelice za jím avé nebo vzácné druhy (které bohužel nem ají podrobný popis a jsou za- Hchyceny jen v přehledných tabulkách), např. Fom itopsis maackiae, F. insularis, F. bu -
I cho ltz ii, Lenzites acuta, Daedaleopsis d ickinsii, Cystid iophorus merulioides, M yce lep to -don m uraschinskyi, M . Ijubarski aj. (některé zase s nově navrhovaným i kombinacemi, avšak bez basionymů — včetně nově navrhované kom binace Daedaleopsis tr ico lo r, Hkterou provedli Bondarcev a Singer již r. 1941!). H
Kniha o dřevokazných houbách Dálného východu SSSR je určena především pra- Hcovnikům v lesním hospodářství a dřevařském průmyslu a najde tedy jistě široké Huplatnění, a to nejen na lesnatém Dálném východě. P ro m ykology by však bylo v í- Htanější, kdyby zahrnovala mnohem více popisů druhů vyskytu jících se na Dálném Hvýchodě a chybějících jinde v SSSR. H
František K otlaba H
Č E S K Á M Y K O L O G IE — V ydává Čs. vědecká společnost pro m ykologii v Academ ii, Inakladatelství ČSAV , Vodičkova 40, 112 29 Praha 1. — Redakce: Václavské nám. 68, I115 79 Praha 1, tel: 261441—5. T iskne: Státní tiskárna, n. p., závod 4, Sám ova 12, 10146 IPraha 10. — O bjednávky a předplatné přijím á PNS, admin. odbor tisku, Jindřišská 14, B125 05 Praha 1. L ze také objednat u každého poštovního úřadu nebo doručovatele. BCena jednoho čísla Kčs 8,—, roční předplatné (4 sešity) Kčs 32,—. (T yto ceny jsou Bplatné pouze pro Československo.) — Sole agents fo r a ll western countries w ith the Bexception o f the Germ an Federal Republic and W est B erlin JO H N B E N JA M IN S B. V., BA m steld ijk 44, Am sterdam (Z.), Holland. Orders from the G. F. R. and W est B erlin Bshould be sent to Kubon & Sagner, P. O. Box 68, 8000 München 34, or to any other Bsubscription agency in the G. F. R. Annual subscription: Vol. 31, 1977 (4 issues) Dutch ■Glds. 62.- (DM 53.-). B
Toto číslo vyšlo v lednu 1978. I
© Academ ia, Praha 1978. I
64 I

1. Abnorm áln í m itochondrie ja tern í buňky s parakrystalickou inkluzí; ja tern í biopsie u 271eté ženy 14. den po otravě A. phalloides. — A n abnorm al mitochondrion, of the hepatocyte w ith paracrystalline inclusion; liv e r biopsy o f fem ale (age 27) the 14th day a fter A. phalloides poisoning. (40 000X)
Electron micrograph R. Čuřík
< * '' IK '■ • ’ c M C * I
m é ' ' 1 * ^ V I
i ř . v. ■ ■ f W • I
p . ¡ - > ' Is f e J t é ě ě l.^ á 4 ^ . > v j *
2. Spory muchomůrky zelené v střevn ím obsahu 291eté ženy 5. den těžké otravy; barveno d le M elzera. — Death-cap spores from the intestinal lavage o f fem ale (age 29) 5 days a fter heavy poisoning; smear M elzer stain. (1500X)
L igh t m icrograph R. Čuřík
VESELSKÝ, KU B IC K Á A Č U ftíK : K O TR AVÁM A M A N IT A PHALLOIDES I.

H R lB ET R Y P Á Č E K : THE G R O W TH RESPONSE OB' WOOD - D E S TR O YIN G ]F U N G I I
II. I
I
II
'SN^^HHPBM HM M ^HH^Ktig 11 ' K H I > f /š ýí n.—-íž«8řiajffT8SiaEMB™ nB«B B B » w M M 3 ! JB M F v H I ''¡¡p il
i m ^ K Ê / il h ss lsB H h . i . w
i*wB| B ^ I ^ R gt J I
J iù to H W L j^flflH
Pla te I. -"*■ f lG row th responses o f the colonies of wood-destroying fungi to the presence o f spruce Hcallus. HH. annosus inoculated 23 days a fter callus inoculation : Photo 1 : situation a fter 18 days; HPhoto 2: situation a fte r 22 days, Photo 3: situation a fter 30 days fo llow in g fungus Hinoculation. P. schw ein itz ii inoculated simultaneously w ith the callus: Photo 4: Hsituation a fte r 8 days; Photo 5: situation a fter 11 days; Photo 6: situation a fte r H18 days fo llow in g fungus inoculation. L. sulphurea inoculated 23 days a fter callus inoculation: Photo 7: situation a fter 10 days. Photo 8: situation a fter 14 days; P h o to 9: Ssituation a fte r 28 days fo llow in g fungus inoculation.

K L Á N : IN O N O TU S T A M A R IC IS IN GREECE III. I
1. Inonotus tam aricis (Pat.) M aireFru it body grow ing on liv in g Ta m a rix sp. 1,3X- H(Greece, Solune, 7 July 1975)
Photo J. Klán f l
ém Ij S g k . . . « -4 F :■ ■ ; * , ■ > I
2. Fru it body grow ing on liv in g T a m a rix gallica. 0,5X- (Yugoslavia, “ Crna G ora“ , 26 M ay 1976)

I K L Á N : IN O N O TU S T A M A R IC IS IN GREECE IV .
H 3. Young fru it body grow ing on Ta m a rix gallica. 3 Y .■ (Yugoslavia, Starigrad, 8. July 1968)H Photo F. Kotlaba
Ps-.v.. -m ĚĚĚm - ~
li«' ^- ‘ 4'’
'^ÉÉÉi •'?
j| | l ^ * ^tSeťi ' "’ í
M m ... '|> k '^ i1 r *V^W-rf. IpfljM h W*1 \"_iJg',l
m H -/v j. r '. ^ j n r ► . nv- ^s M fe ^
4. Cross section of fruit body. 1,5X- (Bulgaria. Nesebar, 10 July 1975)
^ ^ B Photo F. Kotlaba

-•■' • ».'*» '-*• J* ♦ »• * 9 ■ W3íh.-~:' /" ^ í % sKSm r MWmTíifni 1
j - >! ■ 1 js
'0 S \ * • ' ’S• i ., m « & :C 8 k /í b B (''"' p"" d '
Rhizopus cohnii Berlese et De Toni — a — v nativním preparátu obsahu zmetka, 250 X — in foetal stomach contents, 250 X — b — v kotyledonech placenty. Barveno podle Grocotta, 400 X — in cotyledon of placenta. Grocott stain, 400 X — c — sporan- g io fo ry se sporangii, kolumelami a rhizoidy v kultuře, 150 X — sporangiophores with sporangia, collumellae and rhizoids in the vultures, 150 X — d — detail kolumel, 850 X — detaile of columellae, 850 X.
Foto A. Adámková a M. Lávička
AD ÁM KO VÁ, V ÁŇ O V Á A LÁ V IČ K A ; RHIZOPUS COHNII V.

V zh ledem k tomu, že většina autorů zasílá redakci rukopisy fo rm álně n e v y h o v u j í c í , u veře jňu jem e nékteré nejdů ležltějS Í zásady pro úpravu rukopisů (jinak odkazu jem e na podrob- inějš i sm ěrn ice u veře jněné v 1. čísle České m ykolog ie , roň. 16, 1962).
■ 1. Č lánek začíná českým nadpisem , pod n im i je p ř e k l a d n á z v u n a d p i s u v někteř í rém ze světových jazyků , a to v tém že, jím ž je psán abstrakt a případně souhrn na konci článku.H Pod n im následuje plné k ř e s t n í j m é n o a p ř í j m e n í a u t o r a (autorů), bez akade-■ m ických titulů. N a konci článku, za citovanou literaturu, nutno uvést adresu autora (včetně PSC).
2. VSechny původní práce musí b ý t doplněny k rá tkým úvodním souhrnem — a b s t r a k t e m■ v č e s k é a n ě k t e r é s v ě t o v é ř e č i . Rozsah abstraktu, ve kterém m ají bý t výstižně a
stručně charakterizovány výs ledk y a přinos po jednání, nesm í přesahovat 1S řádek strojopisu.3. U dů ležitých a význam ných studií doporuču jem e p ř ip o jit (krom ě abstraktu, k terý Je pouze
■ in form ativn í) podrobnější c izo jazyčný souhrn; jeh o rozsah není omezen.K rom ě toho se p ř ijím a jí články psané ce le c izo jazyčně, s českým podtitu lem , doplněné českým
■ abstraktem a popřípadě 1 souhrnem.4. V lastn i rukopis, tj. strojop is (30 řádek po 60 úhozech na stránku o n e jvýše s S přek lepy
nebo Škrty a vp isy na stránku) musl být psán o b y č e j n ý m z p ů s o b e m . Zásadně není■ přípustné psaní autorských Jmen vel. p ísm eny, prok ládán í nebo podtrhován í s lov čí ce lých v í t
atd. To , co chce autor zdůraznit, sm í provést v rukopise p o u z e t u ž k o u (podtrhne přeruš í Sovanou čarou). VeSkerou typografickou úpravu provád í výhradně redakce. Tužkou m ůže autor
po straně rukopisu označit, co má být vysázeno petitem .3. C itace litera tu ry : každý autor s úplnou literárn í citaci je na s a m o s t a t n é m ř á d k u .
Je-li od jednoho autora uváděno v íce citovaných prací, jeh o jm éno se vždy znovu celé vyp isu je i s citací zkratky časopisu, která se opaku je (nepoužívám e „lb id em “ ). Za p říjm en ím následuje (bez čárky ) zkratka křestn ího Jména, pak v závorce letopočet práce, za závorkou dvojtečka a za ní úplná (nezkrácená) citace názvu po jednán í nebo kn ihy. P o tečce za názvem místo, kdekniha vyšla, nebo zkrácená citace časopisu. Jména dvou autorů spoju jem e l a t i n s k o u
H s p o j k o u „e t* a tří č i v íce autorů čárkam i; Jen m ezi posledním i dvěm a Je spojka „ e t “ .H 6. N ázvy časopisů používám e v m e z i n á r o d n ě s m l u v e n ý c h z k r a t k á c h . Jejich
seznam u nás dosud souborně nevyšel, jako vzo r lze však používa t zkratek period ik z 1. svazku H F lory CSR - G asterom ycetes, z posledních ročn íků České m yko log ie , z Lom ského Soupisu cizo-H zem ských period ik (l»5S—lis t ) nebo z botanické b ib liog ra fie Futák-D om ln: B ib liograU a k fló re■ CSR (1960), kde Je 1 stručný výk lad o zkratkách časopisů a b ib liog ra fii vůbec.H 7. Po zkratce časopisu nebo po citaci kn ihy následuje ročn ík nebo d íl kn ihy vždy jen■ a r a b s k ý m i č i i l i c e m i a bez vyp isován í zkratek (roč., tom., Band., vo l., etc.) a přesnáH citace stránek. C islo ročn íku nebo svazku je od citace stránek odděleno dvo ečkou. U Jednodil-■ ných kn ih p líem e m ísto čís lice 1: pouze p. (” pagina, stránka).H 6. P ř i uváděn i dat sběru apod. píSeme m ě s í c e zásadně ř í m s k ý m i č í s l i c e m i (2. V I.).I «. VSechny d r u h o v é názvy zač ína jí zásadně m a l ý m p í s m e n e m (napf. Sclerotln leH vese lý l), 1 kd yž je druh po jm enován po některém badateli.H 10. U pozorňu jem e autory, aby se ve svých příspěvcích p řid ržova li posledního vydán i N o -■ m e n k l a t o r i c k ý c h p r a v i d e l (v iz J. H olub; M ezinárodní kód botanické nom enklatury■ 1966; Zp rávy Cs. bot. Spol. 3 , Příl. 1, 1968). Jde především o uvádění typů u nové popi-I sovaných taxonů, o přesnou citaci basionym ů u nově publikovaných kom binací apod.I 11. Ilustračn í m ateriá l (kresby, fo tog ra fie ) k článkům číslu jte průběžně u každého článkuI zvláSt arabským i čís licem i (bez zkratek obr., Abb lld . apod.) v tom pořadí, v Jakém má být■ uveřejněn.■ 12. Separáty se tisknou na účet autora. Na sloupcové korektuře autor sdělí, žádá-li a ja k ý■ počet separátů (n e jvýše však 70 kusů).■ 13. N evyžádan é rukopisy včetně příloh a tabulí se nevracejí.H 14. Přednostně se o tisku jí p říspěvky členů Československé vědecké společnosti pro m ykolog ii.I P ř i c itaci herbářových dokladů uvádějte zásadné m ezinárodní zkratky všech herbářů (IndexI herbariorum 1956):
I B R A — S lovenské národně múzeum, BratislavaI B R N M — Bot. odd. M oravského muzea, Brno■ BRNS - Ú střední fy tokaran ténn i laboratoř pří O stf. kontr. a zkué. úst. zemSd., Brn®I B R N U — Katedra bo tan iky přírod , fak . J. E. Purkyně, BrnoI O P — Bot. odd. S lezského muzea, OpavaI PR M — Národn í m uzeum , m yko log ícké oddělení, PrahaI PRC - Katedra bo tan iky přírod , fak . K a r lo vy univ., Praha
■ Soukrom é herbáře necitu jem e n ikdy zkratkou, nýbrž příjm en ím m ajite le , n »p ř. herb.■ J. Herink, herb. F. Šm arda apod. Podobně u herbářů ústavů, k teré nem ají m ezinárodní zkratku.
Rukopisy neodpovída jíc í výše u vedeným zásadám budou vráceny výkonným redaktorem zpět autorům k přepracován í, aniž budou projednány redakční radou.
Retfakc* ia io p itu Ctnkd m y k o lo g ii
H Upozorněni p H ip ěn te ltm České mykologie

4ti 23 » I
The journal of the Czechoslovak Scientific Society for Mycology, formed for the Iadvancement of scientific and practical knowledge of the Fungi I
V o l . 32 P a r t i J a n u a r y 1 978 I
C h i e f E d i t o r : Doc. RNDr. Zdeněk Urban, DrSc. I
E d i t o r i a l C o m m i t t e e : Academician Ctibor Blattný, DrSc.; Professor Karel ICejp, DrSc.; RNDr. Petr Fragner; M UDr. Josef Herink; RNDr. Věra Holubová, CSc.; IRNDr. František Kotlaba, CSc.; Ing. Karel Kříž; RNDr. Vladim ir Musilek, CSc.; Doc. IRNDr. Jan Nečásek, CSc.; Ing. Cyprián Paulech, CSc.; Professor Vladim ir Rypáček, I
DrSc.; RNDr. Miroslav Staněk, CSc. I
E d i t o r i a l S e c r e t a r y : RNDr. Mirko Svrček, CSc. I
Part 4 of the 31th volume was published on the 25th November 1977 I
C O N T E N T S I
J. V e s e l s k ý , J. K u b i č k a et R. C u ř í k : Some recent remarks to death-cap Ipoisonings — Amanita phalloides (Fr.) L i n k ................................................... 1 I
M. S v r č e k : New or less known Discomycetes. V II .................................................. 11 IZ. P o u z a r : Hypoxylon macrocarpum Pouz. spec, nov., a new fragrant pyre- I
n o m y c e t e ....................................................................................................................... 19 IJ. S t a n g l et J. V e s e l s k ý : Inocybe descissa (Fr.) Quél. und ihre taxonomi- I
sehe Stellung. (Beiträge zur Kenntnis seltenerer Inocyben. Nr. 12) . . . 22P. F r a g n e r : Yeasts in human material in our country and their differentia
tion. Part I. ...........................................................................................................32S. S e b e k : Maire’scher Streif ling — Amanita mairei Foley in CSSR gefunden 43 J. K l á n : Inonotus tamaricis Pat.) Maire in Greece, its general distribution
and taxonomic notes to the section Phymatopilus D o n k ..................................... 47J. H ř i b et V. R y p á č e k : The growth response of wood-destroying fungi to
the presence of spruce c a l l u s ..................................................................................55A. A d á m k o v á , M. V á ň o v á et M. L á v i č k a : Rhizopus cohnii as the case
of mucormycotic abortion in c a t t le ........................................................................... 61
R e f e r e n c e s ......................................................................................... 10,31,42,46,54
W i t h b l a c k a n d w h i t e p h o t o g r a p h s : I. To the article: J. Veselský,J. Kubička and B. Cuřík. — II. To the article: J. Hřib and V. Rypáček. —III. To the article: A. Adámková, M. Váňová and M. Lávička. — IV. and V.Inonotus tamaricis (Pat.) Maire.
Cena čísla 8,— Kčs
Č e s k á m y k o l o g i e I





























