Embed Size (px)
Citation preview
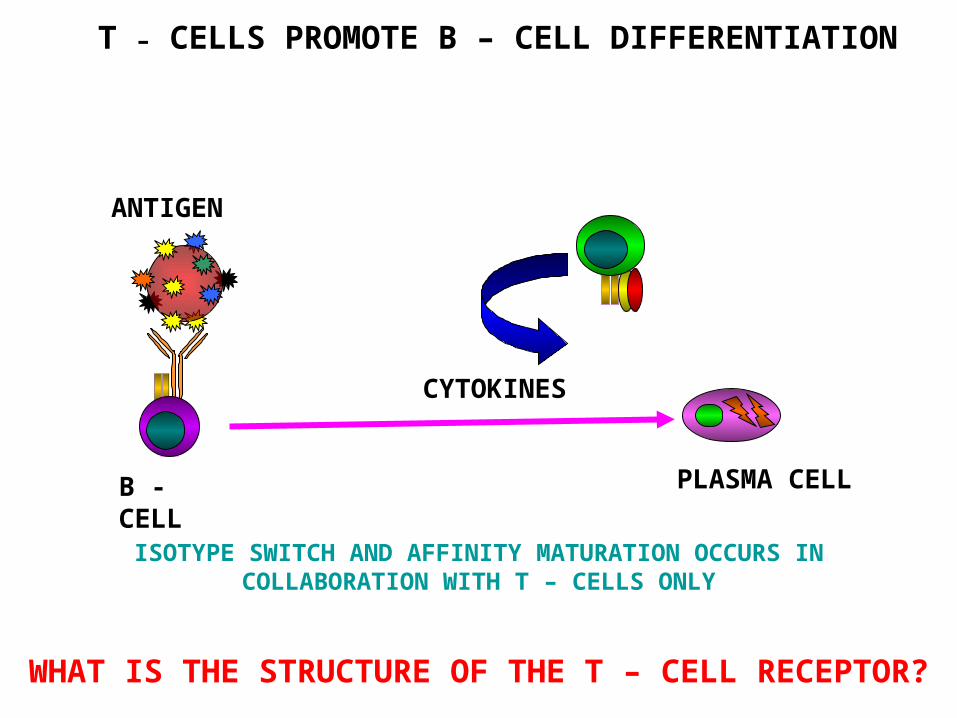
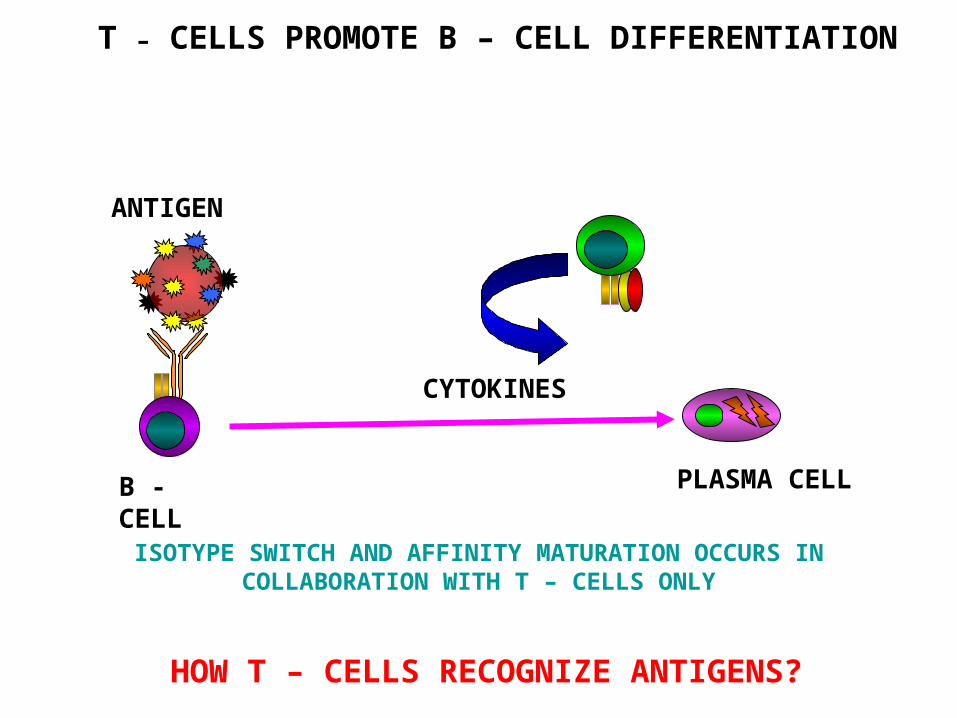
PLASMA CELL
ANTIGEN
CYTOKINES
B -CELL
T – CELLS PROMOTE B – CELL DIFFERENTIATION
ISOTYPE SWITCH AND AFFINITY MATURATION OCCURS IN COLLABORATION WITH T – CELLS ONLY
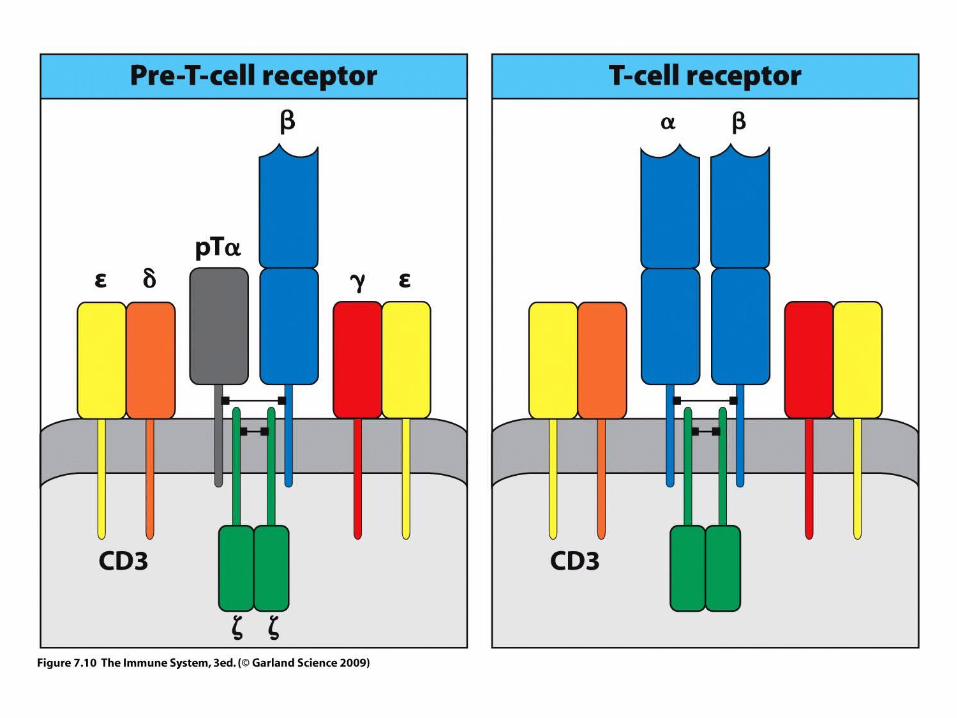
WHAT IS THE STRUCTURE OF THE T – CELL RECEPTOR?
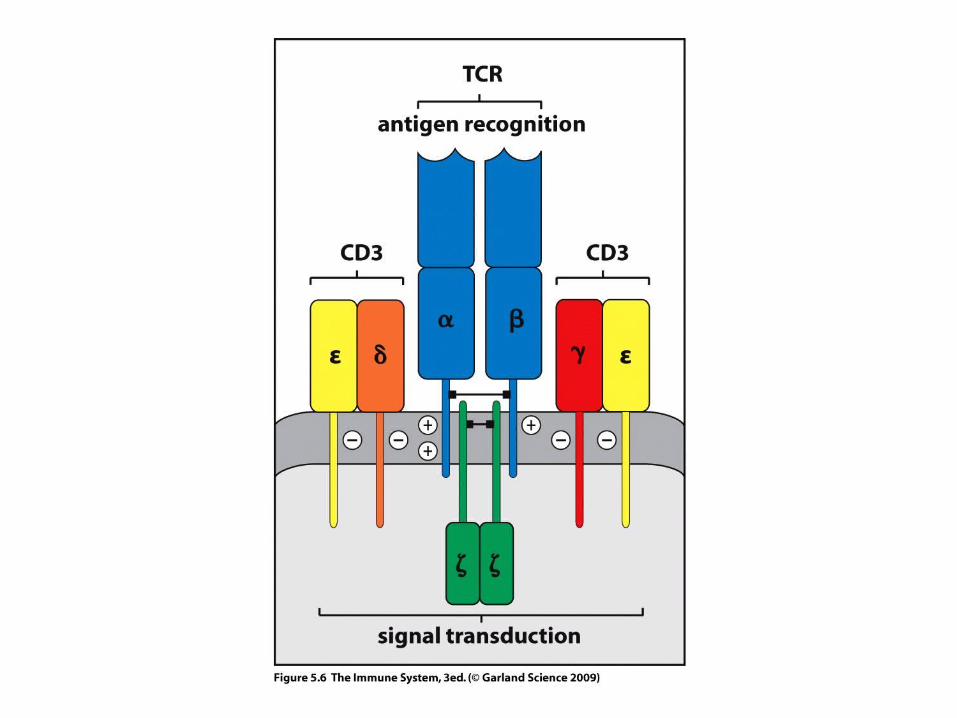
C
mIg H
mIg L
TCR
TCR
TT-SEJT-SEJT
C
VAntigAntigen receptoren receptorTCRTCR
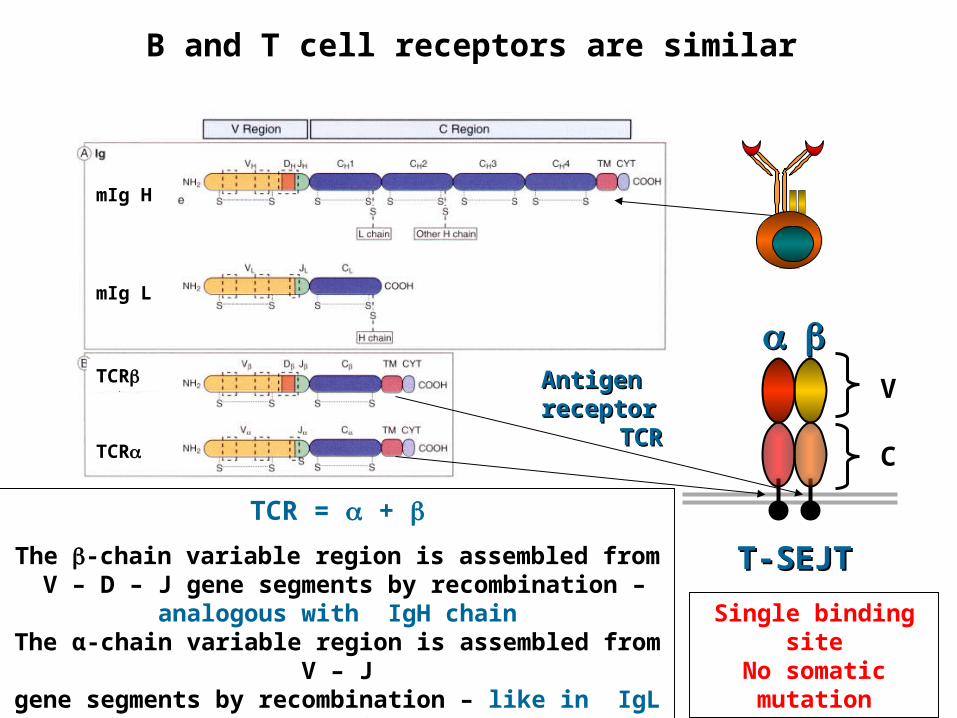
B and T cell receptors are similar
TCR = +
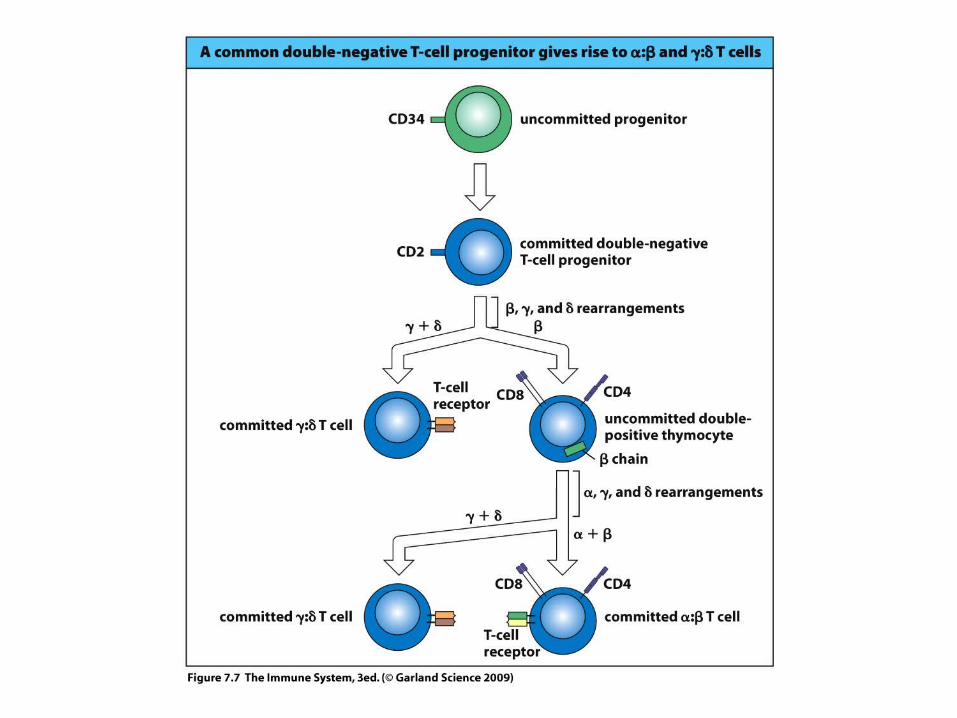
The -chain variable region is assembled from V – D – J gene segments by recombination – analogous with IgH
chainThe α-chain variable region is assembled from V – J gene segments by recombination – like in IgL - chain
Single binding siteNo somatic
mutation
Chr 7
Chr 14
5' 3'
5' 3'
5' 3'
5' 3'
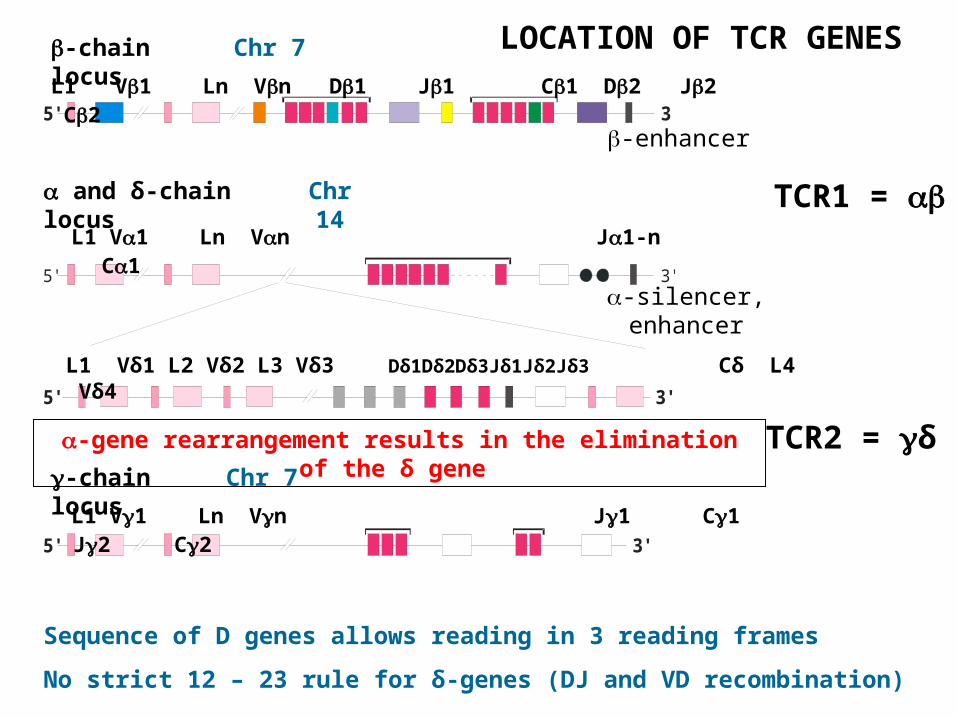
-chain locus
and δ-chain locus
-chain locus
-gene rearrangement results in the elimination of the δ gene
Sequence of D genes allows reading in 3 reading frames
No strict 12 – 23 rule for δ-genes (DJ and VD recombination)
Chr 7
TCR1 =
TCR2 = δ
LOCATION OF TCR GENES
L1 V1 Ln Vn D1 J1 C1 D2 J2 C2
-enhancer
L1 V1 Ln Vn J1 C1 J2 C2
-silencer, enhancer
L1 Vδ1 L2 Vδ2 L3 Vδ3 Dδ1Dδ2Dδ3Jδ1Jδ2Jδ3 Cδ L4 Vδ4
L1 V1 Ln Vn J1-n C1
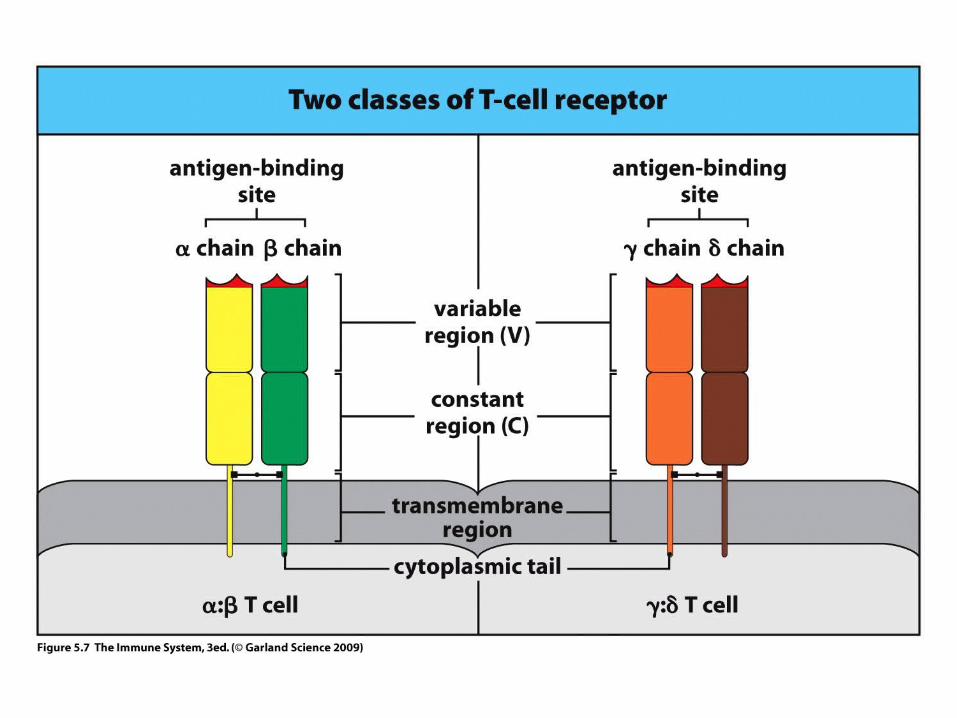
TT-CELL-CELLAntigAntigen receptoren receptor
TCRTCR
VV
CC
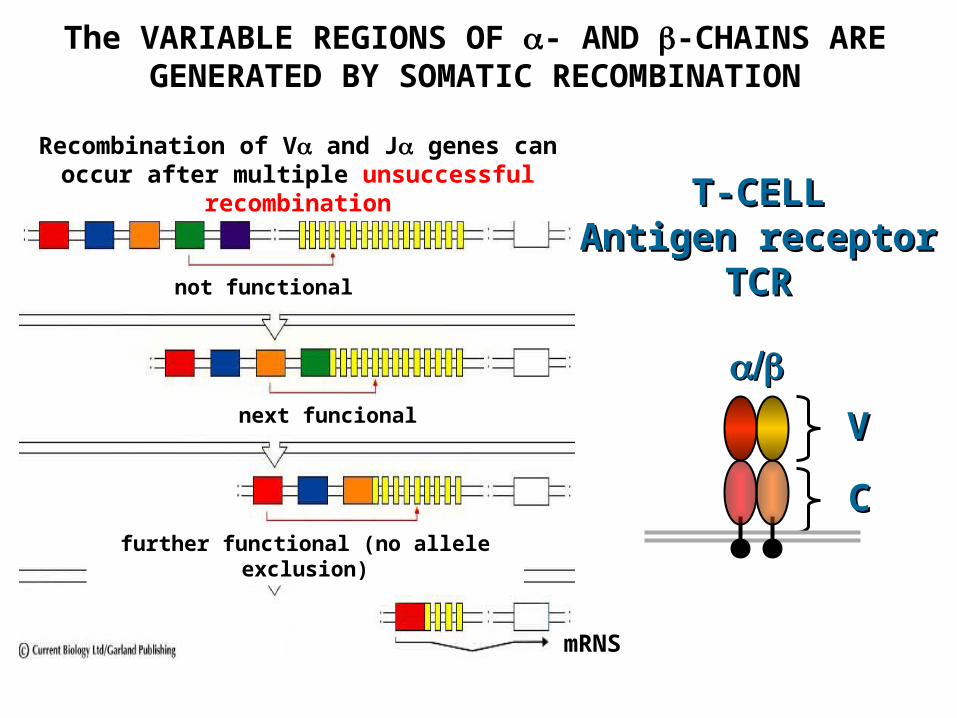
The VARIABLE REGIONS OF - AND -CHAINS ARE GENERATED BY SOMATIC RECOMBINATION
mRNS
not functional
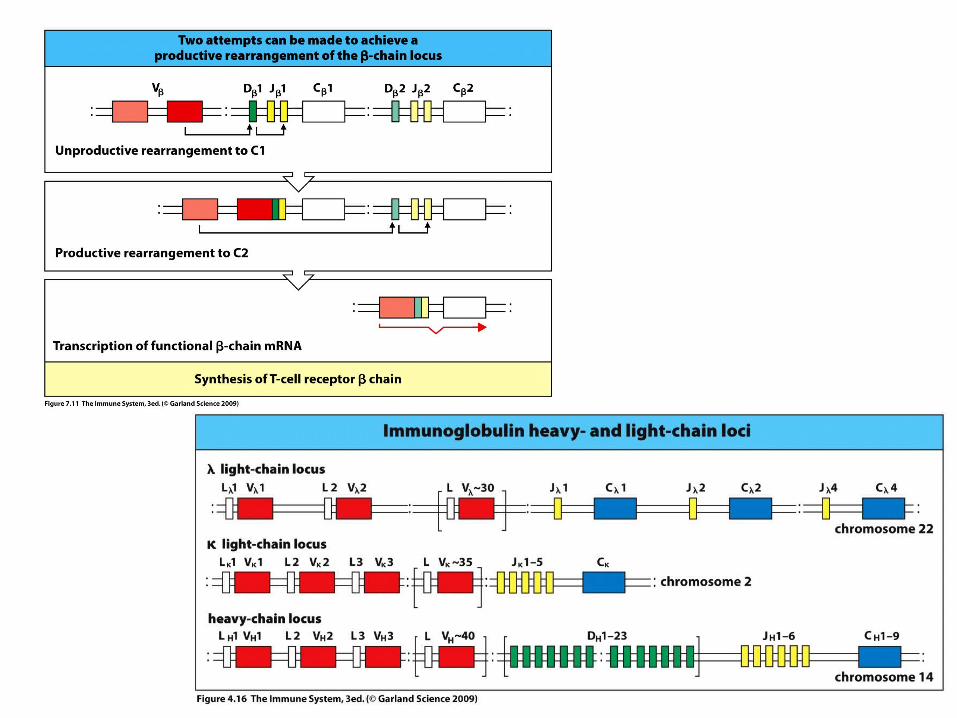
Recombination of V and J genes can occur after multiple unsuccessful recombination
next funcional
further functional (no allele exclusion)
0 10
10
20
20
30
30
40
40
50
50
60
60
70
70
80
80
90
90
100
100
110 120
0 10
10
20
20
30
30
40
40
50
50
60
60
70
70
80
80
90
90
100
100
110 120
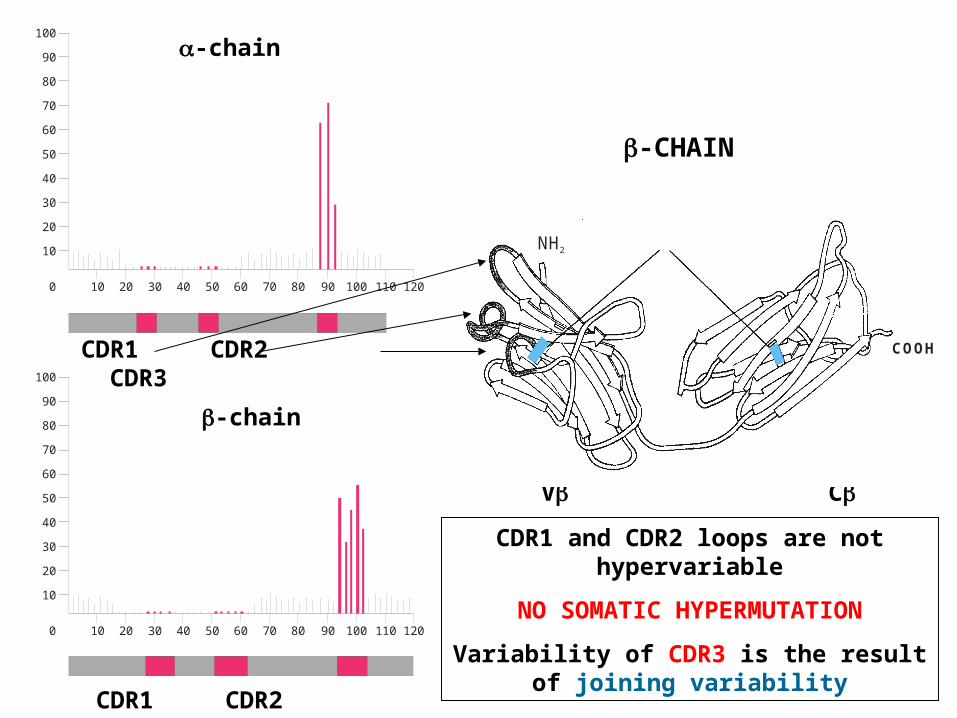
CDR1 CDR2 CDR3
-chain
-chain
CDR1 CDR2 CDR3
V C
-CHAIN
Diszulfid hidak
CDR1 and CDR2 loops are not hypervariable
NO SOMATIC HYPERMUTATION
Variability of CDR3 is the result of joining variability
NH2
COOH
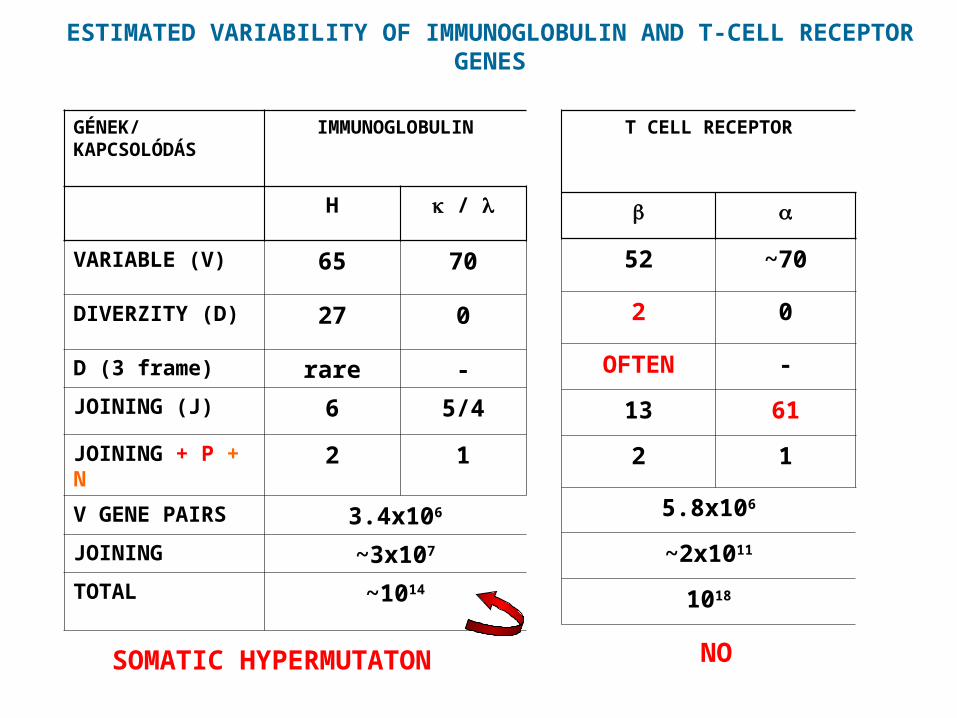
GÉNEK/KAPCSOLÓDÁS
IMMUNOGLOBULIN
H /
VARIABLE (V) 65 70
DIVERZITY (D) 27 0
D (3 frame) rare -
JOINING (J) 6 5/4
JOINING + P + N 2 1
V GENE PAIRS 3.4x106
JOINING ~3x107
TOTAL ~1014
SOMATIC HYPERMUTATON
ESTIMATED VARIABILITY OF IMMUNOGLOBULIN AND T-CELL RECEPTOR GENES
T CELL RECEPTOR
52 ~70
2 0
OFTEN -
13 61
2 1
5.8x106
~2x1011
1018
NO
PLASMA CELL
ANTIGEN
CYTOKINES
B -CELL
T – CELLS PROMOTE B – CELL DIFFERENTIATION
ISOTYPE SWITCH AND AFFINITY MATURATION OCCURS IN COLLABORATION WITH T – CELLS ONLY
HOW T – CELLS RECOGNIZE ANTIGENS?
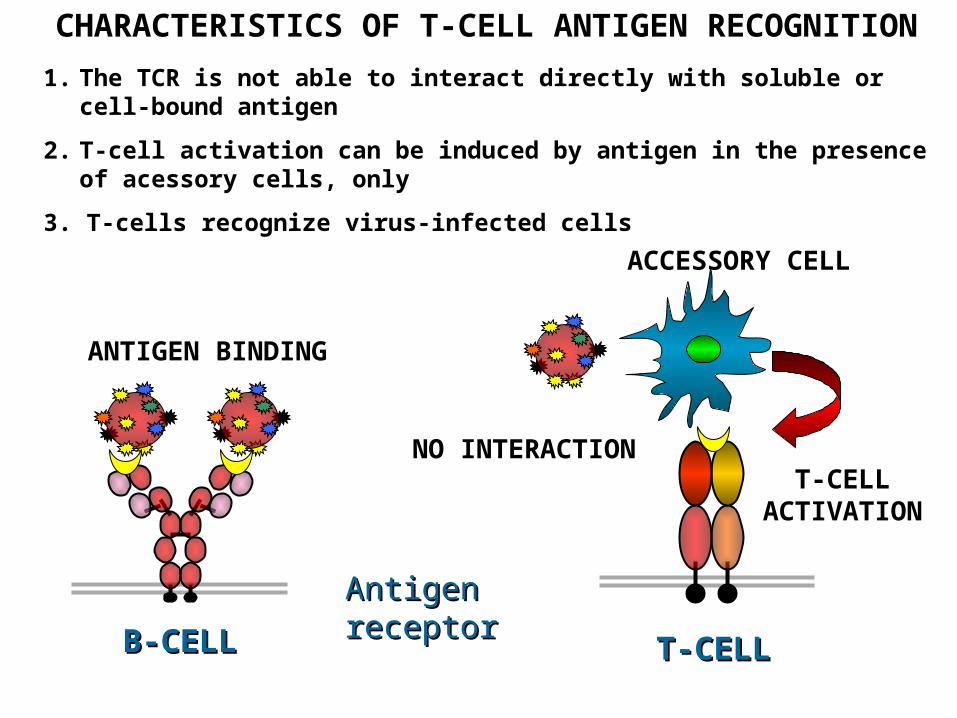
ANTIGEN BINDING
NO INTERACTION
ACCESSORY CELL
T-CELL ACTIVATION
AntigeAntigen receptorn receptor
TT-CELL-CELL B-CELLB-CELL
CHARACTERISTICS OF T-CELL ANTIGEN RECOGNITION
1. The TCR is not able to interact directly with soluble or cell-bound antigen
2. T-cell activation can be induced by antigen in the presence of acessory cells, only
3. T-cells recognize virus-infected cells
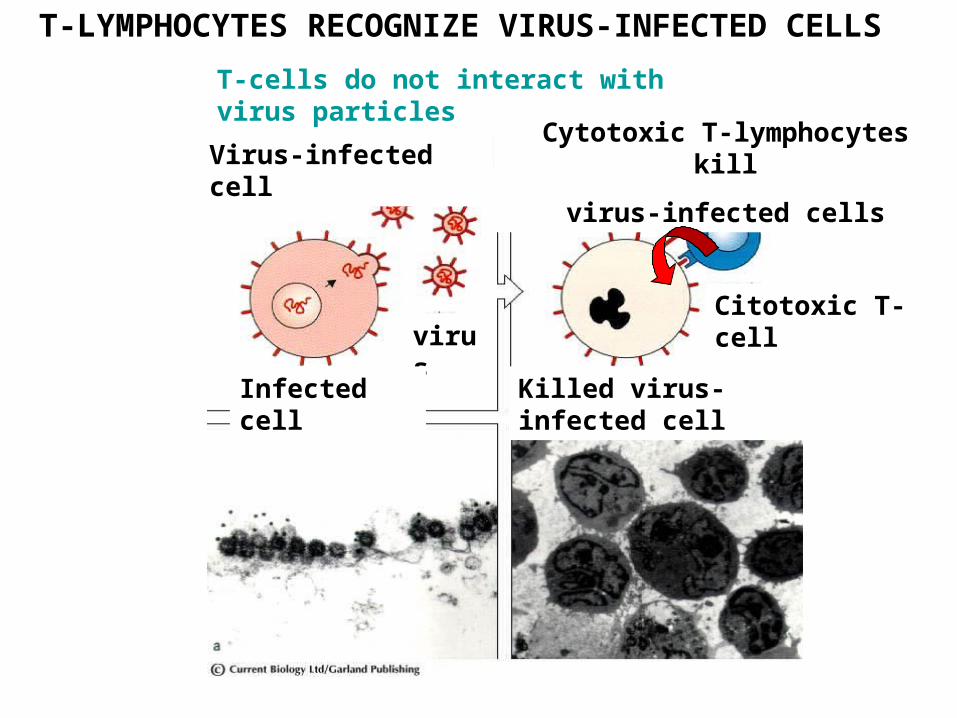
T-LYMPHOCYTES RECOGNIZE VIRUS-INFECTED CELLS
Virus-infected cellCytotoxic T-lymphocytes kill
virus-infected cells
virus
Killed virus-infected cellInfected cell
Citotoxic T-cell
T-cells do not interact with virus particles
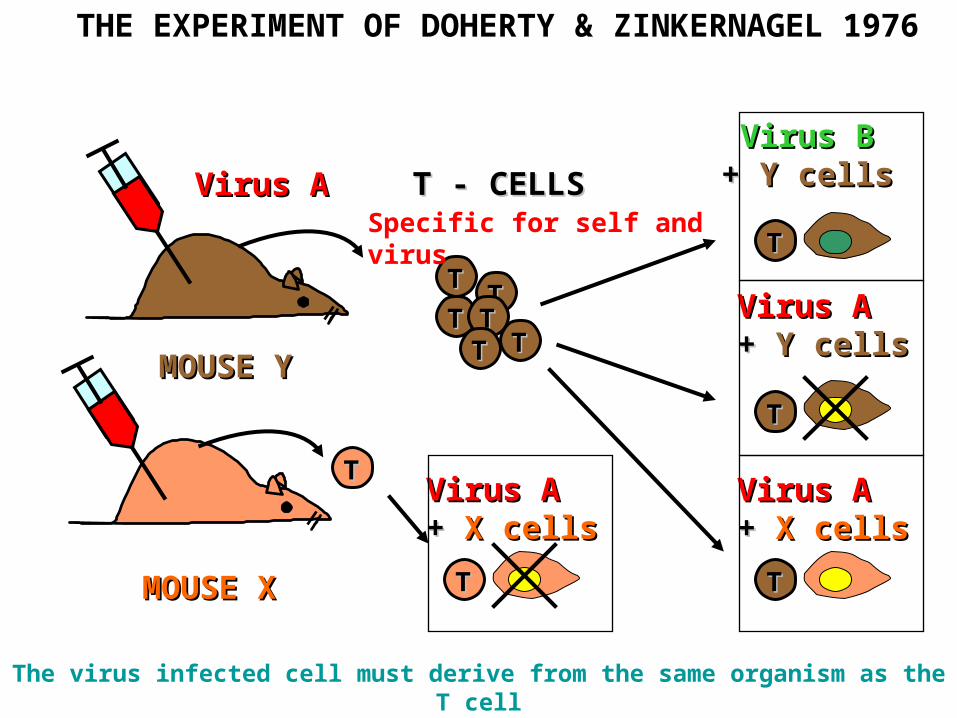
MMOUSEOUSE YY
Virus AVirus A T -T - CELLS CELLS
TT
TTTT
TTTT TT
Virus BVirus B+ + YY cellscells
TT
Virus AVirus A+ + YY cellscells
TT
MMOUSEOUSE X X
Virus AVirus A+ + X X cellscells
TT
Virus AVirus A+ + X X cellscells
TT
TT
THE EXPERIMENT OF DOHERTY & ZINKERNAGEL 1976
The virus infected cell must derive from the same organism as the T cell
Specific for self and virus

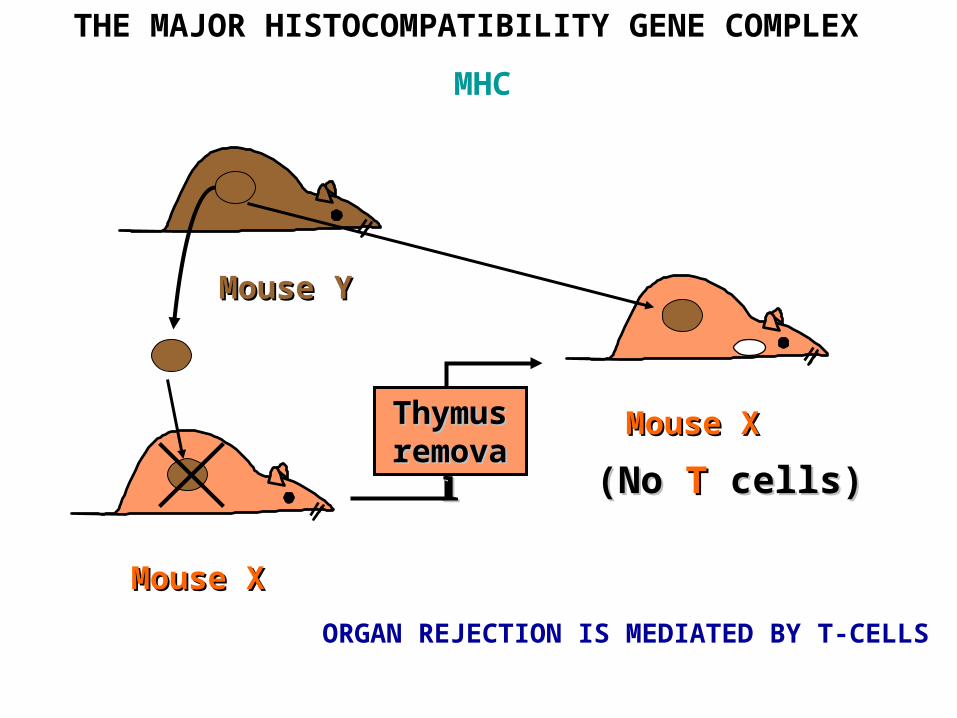
THE MAJOR HISTOCOMPATIBILITY GENE COMPLEX
MHC
MMouseouse XXThymus Thymus removalremoval
((No No TT cellscells))
MMouseouse YY
MMouseouse X X
ORGAN REJECTION IS MEDIATED BY T-CELLS

MMouseouse X X ((YY))MMouseouse YY
MMouseouse YY and the congenic and the congenic MMouse ouse XX((YY)) carry carry an identical an identical MHCMHC gene locus gene locus
TT-cells recognize -cells recognize pproducts of MHC genes as sroducts of MHC genes as seelf or non-selflf or non-self
CONGENIC MICE SHARE COMMON MHC GENES
If any cell of an individual starts to produce foreign (viral or bacterial) or abnormal If any cell of an individual starts to produce foreign (viral or bacterial) or abnormal (tumor associated) proteins, the T-cells recognize these antigen presenting cells (tumor associated) proteins, the T-cells recognize these antigen presenting cells
as altered self cells and respond against themas altered self cells and respond against them
HISTOCOMPATIBILITY IS DETERMINED BY GENES OF THE MHC

MMouseouse X XMMouseouse YY
TThe immune response to protein antighe immune response to protein antigeens is also dependent on MHC ns is also dependent on MHC genesgenes
Protein antigens are tProtein antigens are taaken up from the environment by phagocytic ken up from the environment by phagocytic cells and via MHC proteins present for T-lymphocytes cells and via MHC proteins present for T-lymphocytes
THE MAJOR HISTOCOMPATIBILITY GENE COMPLEX AND THE RESPONE TO PROTEIN ANTIGENS
Antigen
IMMUNE RESPONSE NO IMMUNE RESPONSE
Antigen

T-LYMPHOCYTES RECOGNIZE ANTIGEN-DERIVED PROTEIN FRAGMENTS (PEPTIDES) EXPRESSED ON THE SURFACE OF SELF
ANTIGEN PRESENTING CELLS
VIRUS-INFECTED CELLS ARE RECOGNIZED BY T-LYMPHOCYTES IN MHC-DEPENDENT MANNER
TISSUE TRANSPLANTATION IS RESTRICTED BY MHC MOLECULES
THE IMMUNE RESPONSE TO PROTEIN ANTIGENS IS REGULATED BY INDIVIDUALLY POLYMORPHIC MHC GENES

ANTIGEN PRESENTING CELLSANTIGEN PRESENTING CELLS
Synthesize antigens – endogenous antigensendogenous antigens (virus, tumor)
Internalize antigens – exogenous antigensexogenous antigens (any protein)
Degrade protein antigens to peptides – processingprocessing
Protein – derived peptides are presented by MHC (HLA) membrane proteins – antigen presentationantigen presentation
MHC molecules present both self and non-self protein MHC molecules present both self and non-self protein – derived peptides– derived peptides
MHC class I molecules are expressed in all nucleated MHC class I molecules are expressed in all nucleated cellscells
MHC class II molecules are expressed by professional MHC class II molecules are expressed by professional antigen presenting cellsantigen presenting cells
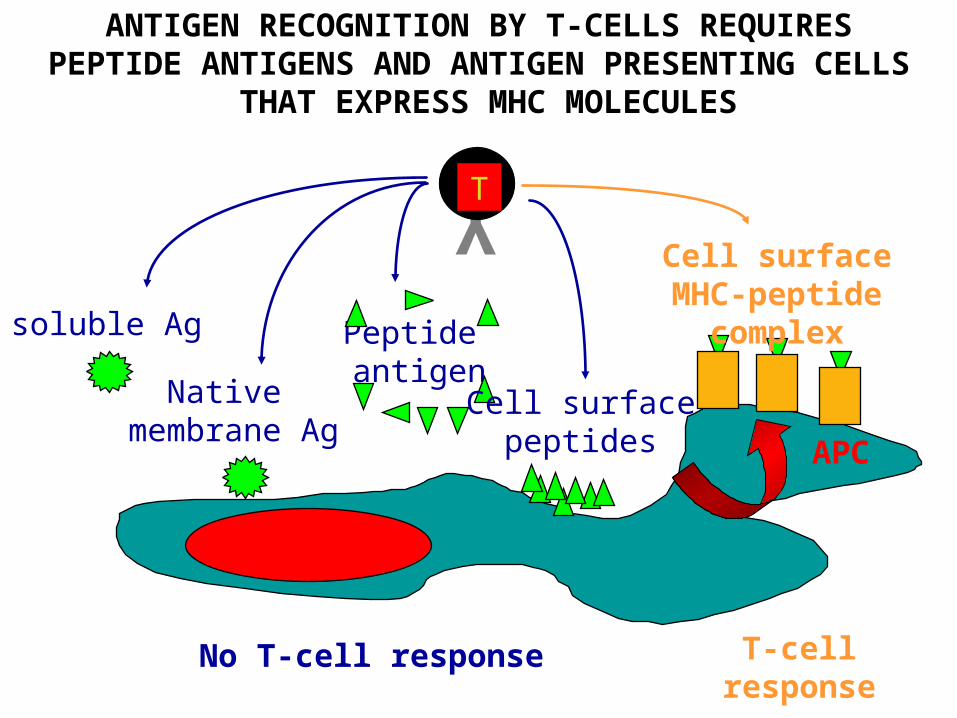
ANTIGEN RECOGNITION BY T-CELLS REQUIRESPEPTIDE ANTIGENS AND ANTIGEN PRESENTING CELLS
THAT EXPRESS MHC MOLECULES
YT
No T-cell response
soluble Ag
Native membrane Ag
Peptide antigen
Cell surface MHC-peptide complex
T-cell response
Cell surfacepeptides APC

PROFESSIONAL ANTIGEN PRESENTING CELLS
Express MHC class I and class II proteins in the cell membrane
Express co-stimulatory molecules (CD40, B7)
B cells – specialized for soluble proteins, toxins
ADAPTIVE
Macrophages – extracellular pathogens (bacteria, yeast)
Dendritic cells – viruses, apoptotic cells
INNATE
T-lymphocytes with αβ TCR recognize MHC – peptide complexes expressed on the surface of professional antigen presenting cells (APC) APC)
T-cell recognition requires the physical contact of APC and T cellT-cell recognition requires the physical contact of APC and T cell
TCR
APC
MHC
TCR TCR
APC
MHC
APC
MHC
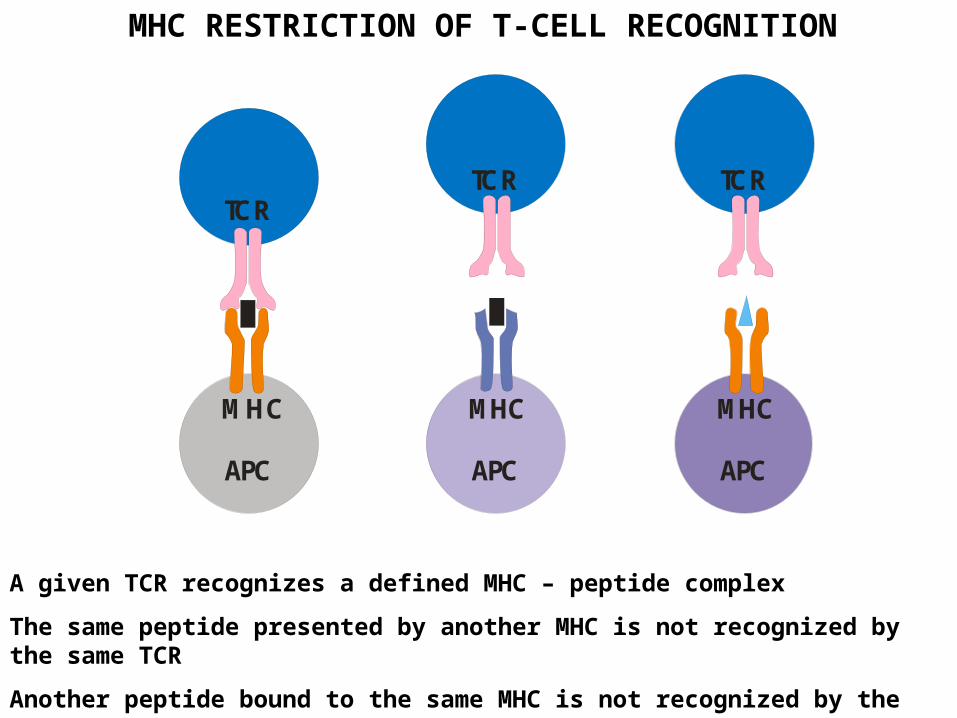
MHC RESTRICTION OF T-CELL RECOGNITION
A given TCR recognizes a defined MHC – peptide complex
The same peptide presented by another MHC is not recognized by the same TCR
Another peptide bound to the same MHC is not recognized by the same TCR
MHC
TCR
CD3
APC
s s
ss
ss
s
ss
V V
C C
s
α βss
ss
ss
ss
CD3
s s
ε δ ε γζ ζ
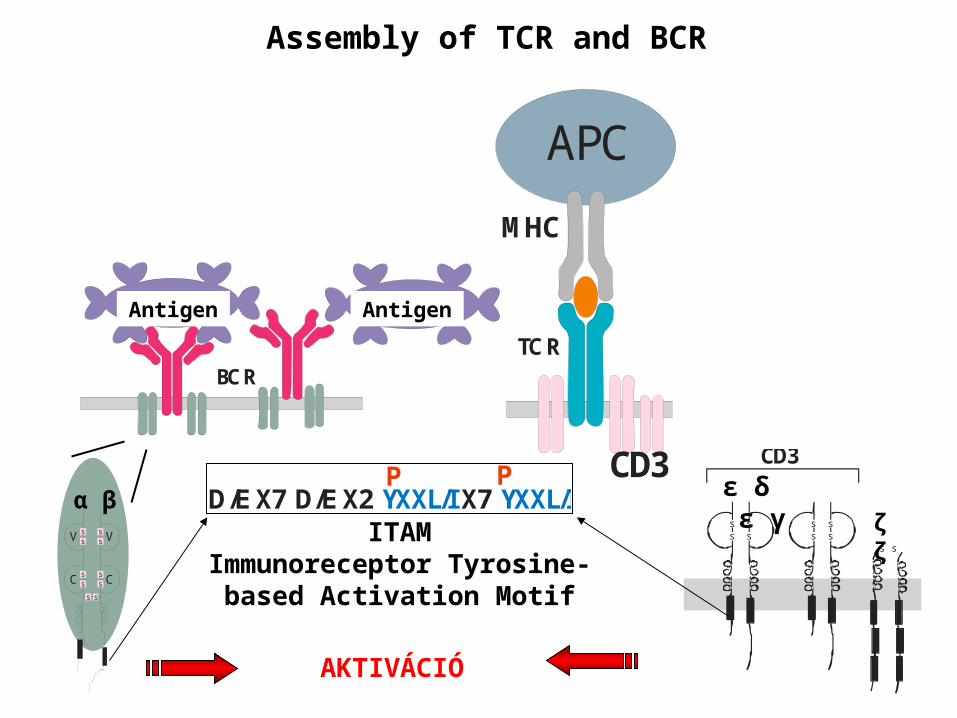
D/E X7 D/E X2 X7 YXXL/I YXXL/IP P
ITAMImmunoreceptor Tyrosine-based
Activation Motif
AKTIVÁCIÓ
Assembly of TCR and BCR
Antigen
BCR
AntigenAntigen Antigen
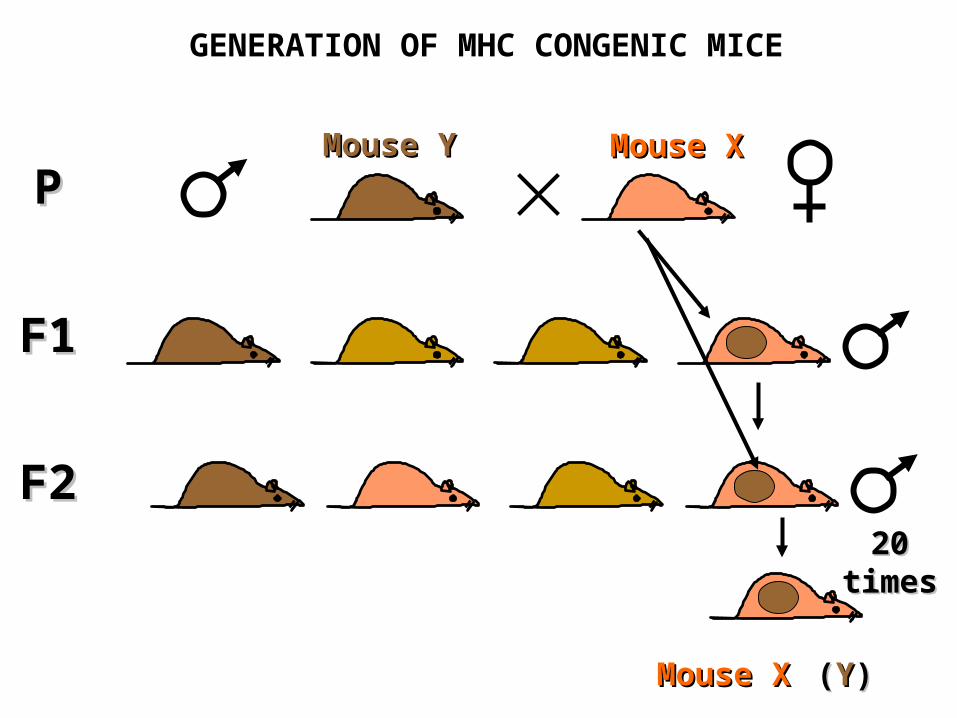
F1F1
PPMMouseouse X XMMouseouse Y Y
F2F2
GENERATION OF MHC CONGENIC MICE
20 times20 times
MMouseouse X X ((YY))



![Fish Cytokines and Immune Response - IntechOpen · Cytokines can modulate immune responses through an autocrine or paracrine manner upon binding to their corresponding receptors [44]](https://img.pdfslide.tips/doc/110x75/5ec39d0259d16726d441c877/fish-cytokines-and-immune-response-intechopen-cytokines-can-modulate-immune-responses.jpg)

























