-
Metabolism
PlasminogenActivator Inhibitor 1 (PAI1) PromotesActin
Cytoskeleton Reorganization andGlycolyticMetabolism in
Triple-Negative Breast CancerBrock A. Humphries1, Johanna M.
Buschhaus1,2, Yu-Chih Chen3,4,5, Henry R. Haley1,Tonela Qyli1,
Benjamin Chiang1, Nathan Shen1, Shrila Rajendran1, Alyssa
Cutter1,Yu-Heng Cheng3, Yu-Ting Chen6, Jason Cong6, Phillip C.
Spinosa7, Euisik Yoon2,3,Kathryn E. Luker1, and Gary D.
Luker1,2,8
Abstract
Migration and invasion of cancer cells constitute funda-mental
processes in tumor progression and metastasis. Migra-tory cancer
cells commonly upregulate expression of plasmin-ogen activator
inhibitor 1 (PAI1), and PAI1 correlates withpoor prognosis in
breast cancer. However, mechanisms bywhich PAI1 promotes migration
of cancer cells remain incom-pletely defined. Here we show that
increased PAI1 drivesrearrangement of the actin cytoskeleton,
mitochondrial frag-mentation, and glycolyticmetabolism in
triple-negative breastcancer (TNBC) cells. In two-dimensional
environments, bothstable expression of PAI1 and treatment with
recombinantPAI1 increased migration, which could be blocked with
thespecific inhibitor tiplaxtinin. PAI1 also promoted invasioninto
the extracellular matrix from coculture spheroids withhuman mammary
fibroblasts in fibrin gels. Elevated cellularPAI1 enhanced
cytoskeletal features associated with migra-tion, actin-rich
migratory structures, and reduced actin stress
fibers. In orthotopic tumor xenografts, we discovered thatTNBC
cells with elevated PAI1 show collagen fibers alignedperpendicular
to the tumor margin, an established marker ofinvasive breast
tumors. Further studies revealed that PAI1activates ERK signaling,
a central regulator of motility, andpromotes mitochondrial
fragmentation. Consistent withknown effects of mitochondrial
fragmentation on metabo-lism,fluorescence lifetime
imagingmicroscopyof endogenousNADH showed that PAI1 promotes
glycolysis in cell-basedassays, orthotopic tumor xenografts, and
lung metastases.Together, these data demonstrate for the first time
that PAI1regulates cancer cell metabolism and suggest targeting
metab-olism to block motility and tumor progression.
Implications: We identified a novel mechanism throughwhich
cancer cells alter their metabolism to promote
tumorprogression.
IntroductionPlasminogen activator inhibitor 1 (PAI1, or serpin
family E
member 1; SERPINE1) is a serine protease inhibitor and the
mostprominent negative regulator of the proteolytic urokinase
plas-minogen activator system. Although originally hypothesized
tohave tumor suppressor effects, cancers typically upregulate
PAI1with higher levels correlating with worse prognosis in breast
and
other malignancies (1–3). Given the strong links between
PAI1levels and prognosis in breast cancer, the American Society
forClinical Oncology (ASCO) recommends analysis of PAI1 levels asa
biomarker for risk assessment and treatment decisions in
lymphnode–negative breast cancers (4).
We identified PAI1 as a key driver of cancer cell migration
andchemotaxis in triple-negative breast cancer (TNBC) using a
high-throughput, single-cell microfluidic device that separates
migra-tory and nonmigratory subpopulations of cancer cells (5).
Thisdevice enables recovery of migratory and nonmigratory
cancercells for subsequent analysis, whichprovides a powerful
approachto identify cells with highest capacity to migrate within a
hetero-geneous bulk population. RNA sequencing revealed PAI1 as
themost upregulated gene inmigratory cells from twodifferent
TNBCcell lines. These data reinforce a recent screening study
thatidentified PAI1 as a key gene driving
epithelial-to-mesenchymaltransition and migration in TNBC (6).
Cells require a large amount of energy to coordinate
cytoskel-etal reorganization and subsequent migration. Therefore,
migrat-ing cancer cells must adopt a metabolic phenotype that
allowsthem to generate energy near sites of rearrangement in the
actincytoskeleton (7). Prior studies indicate cancer cells
typically relyon glycolysis rather than oxidative phosphorylation
to drivemigration and invasion, consistent with studies associating
gly-colysis with more aggressive malignancies (7–9).
Identifying
1Center for Molecular Imaging, Department of Radiology,
University of Michigan,Ann Arbor, Michigan. 2Department of
Biomedical Engineering, University ofMichigan, Ann Arbor, Michigan.
3Department of Electrical Engineering andComputer Science,
University of Michigan, Ann Arbor, Michigan. 4Comprehen-sive Cancer
Center, University of Michigan, Ann Arbor, Michigan.
5ForbesInstitute for Cancer Discovery, University of Michigan, Ann
Arbor, Michigan.6Computer Science Department UCLA, Boelter Hall,
Los Angeles, California.7Department of Chemical Engineering,
University of Michigan, Ann Arbor,Michigan. 8Department of
Microbiology and Immunology, University ofMichigan, Ann Arbor,
Michigan.
Note: Supplementary data for this article are available at
Molecular CancerResearch Online (http://mcr.aacrjournals.org/).
Corresponding Author: Gary D. Luker, University of Michigan, Ann
Arbor, MI.Phone: 734-763-5476; Fax: 734-763-5447; E-mail:
[email protected]
doi: 10.1158/1541-7786.MCR-18-0836
�2019 American Association for Cancer Research.
MolecularCancerResearch
Mol Cancer Res; 17(5) May 20191142
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://crossmark.crossref.org/dialog/?doi=10.1158/1541-7786.MCR-18-0836&domain=pdf&date_stamp=2019-4-13http://mcr.aacrjournals.org/
-
mechanisms promoting glycolytic metabolism in migrating can-cer
cells offers the potential to block this key process in
tumorprogression. Recent studies establishing correlations
betweenPAI1 with obesity and metabolic syndrome suggest PAI1 as
amolecular regulator of metabolism that may translate to
cancerbiology (10, 11). In support of PAI1 as a regulator of
cancermetabolism, secretion of this molecule by fibroblasts
increasedmitochondrial mass in cocultured breast cancer cells (12).
How-ever, a direct relationship between PAI1 and cancer
metabolismremains to be established.
This study investigates mechanisms through which PAI1
drivesmigration and an aggressive phenotype in TNBC, focusing
onactin dynamics andmetabolism. Increasing expression of PAI1
inbreast cancer cells causes dramatic actin reorganization and
drivesfragmentation of mitochondria and glycolytic metabolism in
2Dand 3D culture environments. Overall, this research for the
firsttime establishes PAI1 as a novel regulator of cancer cell
metab-olism and highlights the critical interplay between
metabolismand motility in breast cancer.
Materials and MethodsCell culture
We purchased MDA-MB-231 cells from the ATCC and culturedcells in
DMEM supplemented with 10% FBS and 1% penicillin/streptomycin
(Thermo Fisher Scientific). We obtained SUM159cells from Dr.
Stephen Ethier (now at The Medical University ofSouth Carolina,
Charleston, SC) and cultured cells in F-12 mediasupplemented with
10% FBS, 1% penicillin/streptomycin, 1%glutamine, 5 mg/mL
hydrocortisone, and 1 mg/mL insulin. Weauthenticated all cells by
short tandem repeats analysis andcharacterized cells as free
ofMycoplasma at the initial passage. Weused all cells within 3
months after resuscitation, and we main-tained all cells at 37�C in
a humidified incubator with 5% CO2.
Lentiviral vectorsWe cloned human full-length PAI1 fused to
NanoBiT (Pro-
mega) with a Tev-cleavable linker into the pLVX-Puro
vector(ClonTech; PAI1) and verified products by sequencing. We
pro-duced recombinant lentiviral vectors and transduced target
cells asdescribed previously (13). We first generated cells stably
expres-sing click beetle green luciferase (SUM159-CBG and
MDA-MB-231-CBG) as described previously through selection with
blas-ticidin (14). We subsequently transduced cells with the PAI1
viralvector and used puromycin selection for cells stably
expressingPAI1.We confirmed expressionof PAI1 in these cells
throughqRT-PCR and luminescence of NanoBiT.
The pLenti.PGK.LifeAct-GFP.W vector was a gift from
RustyLansford (Addgene plasmid # 51010; University of
SouthernCalifornia, Los Angeles, CA). We transduced wild-type and
PAI1cells with LifeAct-GFP viral vector and sorted cells by flow
cyto-metry to obtain populations with homogeneous expression
ofLifeAct-GFP. For in vivo fluorescence lifetime imaging
microscopy(FLIM) studies, we transduced wild-type and PAI1 cells
withmCherry Nuc-FUW viral vector and sorted cells by flow
cytometrytoobtainapopulationexpressingnuclearmCherry.
For3Dspheres,we transduced human mammary fibroblasts (HMF,
providedby Daniel Hayes, University of Michigan, Ann Arbor, MI)
withmCherry viral vector and sorted for stable cells by flow
cytometry.
The pLentiCMV Puro DEST ERK KTRClover was a gift fromMarkus
Covert (Addgene plasmid # 59150; ref. 15). We replaced
mClover fluorescent protein with mCitrine and added a
nuclearH2B-mCherry and a puromycin selection marker through
P2Alinker sequences using the NEB HiFi DNA Assembly Kit (NewEngland
BioLabs). This vector allows us to visualize ERK activa-tion via a
nucleocytoplasmic shuttling event of the mCitrine ERKreporter,
while mCherry demarks the nucleus. We cloned theconstruct into the
Piggyback transposon vector (Systems Bios-ciences) and transfected
cells using FuGENE HD (Promega). Oneweek after transfection, we
treated cells with puromycin to iden-tify stable integrants and
confirmed expression by fluorescencefrom mCitrine and mCherry.
qRT-PCRTo analyze levels of PAI1, we performed qRT-PCR for
PAI1
and GAPDH using SYBR Green detection as described previous-ly
(16). Primers for PAI1 were 50-CGCAACGTGGTTTTCTC-30
and50-CATGCCCTTGTCATCAATC-30 and for GAPDH were
50-GA-AGGTGAAGGTCGGAGT-30 and 50-GAAGATGGTGATGGGATT-TC-30.
Whole transcriptome next-generation sequencingWe performed whole
transcriptome next-generation sequenc-
ing as described previously (17). We deposited these data as
GEOaccession number GSE125802.
Bioluminescence growth, migration, and cell adhesion assaysWe
analyzed effects of PAI1 on cell growth using biolumines-
cence imaging for CBG with medium binning and 30-secondexposure
as described previously (14).
We used our previously published microfluidic device
andwound-healing assays to verify PAI1 as a regulator of cell
migra-tion. We performed microfluidic migration assays and
imagedcells as described previously (17). For wound-healing assays,
weseeded 1� 105 cells into 35-mm dishes and allowed cells to
formconfluent monolayers before creating a linear scratch with a
200-mL pipette tip. We washed dishes once with PBS and then
addedfreshmediumcontaining the proliferation inhibitorMitomycinC(1
mg/mL). When indicated, we also added vehicle, the PAI1inhibitor
tiplaxtinin (5 mmol/L) (Selleckchem), or recombinantPAI1 (rPAI1,
Sigma-Aldrich, 40 nmol/L) at the time of wounding.Wemeasured the
size of thewound in eachmonolayer of cells at 0and 17 hours. We
used the following formula to calculate woundclosure over time:
(1�(woundwidth at 17 hours/woundwidth at0 hours)) � 100.
We also analyzed the ability of PAI1 tomodulate cell adhesion.We
seeded 1� 105 HMFs expressingmCherry into a 24-well plate48 hours
before the assay. After 48 hours, we seeded 2.5 � 105MDA-MB-231 and
SUM159 wild-type or PAI1 LifeAct-GFP–expressing cells onto the HMFs
and incubated at 37�C for 15minutes. We removed nonadherent cells
with PBS and visualizedadherent cells by fluorescence imaging. We
present data as thenumber of adherent cells to total number of
cells seeded relative tomatched wild-type cells.
Immunofluorescence stainingTo visualize the actin cytoskeleton,
we performed immunoflu-
orescence staining. We seeded 2.5 � 104 cells on glass cover
slipsand incubated overnight.Wefixed cellswith 4%formaldehyde for15
minutes at room temperature and then washed three timeswith PBS for
5minutes each.We permeabilized cells with ice-cold100% methanol for
10 minutes at �20�C; washed with PBS for
PAI1 Drives TNBC Cytoskeletal Reorganization and Glycolysis
www.aacrjournals.org Mol Cancer Res; 17(5) May 2019 1143
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
5 minutes; and blocked with 10% goat serum for 1 hour at
roomtemperature. Next, we incubated cells for 1 hour at room
tem-perature with an antibody against phosphorylated
paxillin(p-paxillin, Cell Signaling Technology) in 5% goat serum.
Afterincubation, we washed cells with PBS 3 times for 5 minutes
eachand costained with Alexa Fluor 488–conjugated secondary
anti-body (Jackson ImmunoResearch Laboratory) for p-paxillin
andconjugated Texas Red-X phalloidin (Thermo Fisher Scientific)
foractin for 1 hour at room temperature in the dark. We washed
theslides andmountedwithmedium containing an anti-fade reagentand
DAPI (ProLong Gold, Thermo Fisher Scientific). We acquiredimages on
an Olympus IX73 microscope with a DP80 CCDcamera (Olympus) and we
analyzed epifluorescence images withcellSens software (Olympus) and
stress fibers with in-houseMATLAB code.
Three-dimensional assaysWe formed spheroids as described
previously (14), coculturing
MDA-MB-231 or SUM159 wild-type or PAI1 LifeAct-GFP–expres-sing
cells with human mammary fibroblasts (HMF) expressingmCherry. We
embedded spheroids or single cells (1� 106/mL) infibrin gels (final
concentration 4 mg/mL fibrin, 2.5 U/mL throm-bin, and 0.01 U/mL
aprotinin; ref. 18) and imaged actin andfibroblasts by two-photon
microscopy. We analyzed the perim-eter of spheroids using in-house
MATLAB code. We mechanicallydissociated spheroids after 24 hours by
pipetting up and down 5times with a micropipette in the bottom of a
96-well plate. Weenumerated dissociated cells by fluorescence
microscopy.
Kinase translocation reporterTo quantify activation of ERK by
PAI1, we used cells stably
expressing a validated kinase translocation reporter (KTR) for
thiskinase. This reporter utilizes a known downstream substrate
ofERK to drive reversible translocation of the reporter into and
outof the nucleus (15).We seeded 1.2� 105MDA-MB-231 and 8.5�104
SUM159 wild-type cells containing the KTR for ERK and anuclear
marker (H2B-mCherry) on to 35-mm dishes with a 20-mm glass bottom
(Cellvis) in FluoroBrite DMEMmedia (ThermoFisher Scientific,
A1896701). After 2 days, we changedmedium tolow (1%) serum in
Fluorobrite DMEM and added tiplaxtinin (5mmol/L)where indicated.
The next day,we treated cellswith eitherrPAI1 (40nmol/L) or vehicle
control and acquired images at timeslisted in thefigure.We analyzed
images for activationof ERKusingcustom MATLAB code.
FLIM and metabolic analysisWe used FLIM to quantify metabolic
differences between wild-
type and PAI1 cells based on endogenous fluorescence fromNADH as
described previously (ISS FastFLIM; refs. 19, 20). Forboth NADH and
mCherry, we used 740 nm excitation, 25� NIRcorrected objective,
512�512matrix, 15% laser power, 4ms dwelltime, and 2.5� electronic
zoom (Olympus FVMPE-RS uprightmicroscopewith Spectra-Physics
InsightDSþ laser).We separatedlight emitted from NADH and mCherry
with band pass filters of410–460 nm and 545–645 nm,
respectively.
We exported modulation lifetime and direct counts (DC,intensity)
data as ".tiff" files from ISS VistaVision into MATLABalongside
red-channel intensity data obtained by two-photonmicroscopy from
the Olympus software for analysis. We filteredimages to remove all
data points with NADH lifetimes less than0.5 ns and greater than 5
ns, as well as points with DC's less than
15 as per themanufacturer's instructions.We created amask
usingmCherry tomark locations of cancer cells in images. We
analyzedNADH lifetimes of pixelswithin themask for each cell
andpresentlifetimes on images based on a pseudo color scale.
To confirm our FLIM data, we performed metabolic flux anal-ysis
to measure the glycolytic capacity of cells by measuring
theextracellular acidification rate (ECAR) of cells using a
SeahorseBioscience XFe96 Extracellular Flux Analyzer and XF Cell
EnergyPhenotype test kit (Agilent) as described previously
(19).
Mouse xenograft implantationThe University of Michigan
Institutional Animal Care and Use
Committee approved all animal procedures (protocol00006795). The
animals used in this study received humane carein compliance with
the principles of laboratory animal careformulated by the National
Society for Medical Research andGuide for theCare andUse of
Laboratory Animals prepared by theNational Academy of Sciences and
published by the NIH(Bethesda, MD; publication no NIH 85-23,
revised 1996). Weestablished orthotopic tumor xenografts in the
fourth inguinalmammary fat pads of 17–21-week-old female NSG mice
(TheJackson Laboratory; ref. 16), implanting 2� 105MDA-MB-231
orSUM159 wild-type or PAI1 cells that express LifeAct-GFP.
Wequantified tumor growth and metastasis by bioluminescenceimaging
as described previously (13). We euthanized mice 7–8 weeks after
injection (n ¼ 5 mice in each group) and thenimmediately visualized
collagen and actin by two-photonmicros-copy. We used an 880 nm
excitation wavelength and collectedsecond harmonic signal from
fibrillary collagen and emissionfrom LifeAct-GFP with 410–460 nm
and 495–540 nm filters,respectively (21). For FLIM experiments, we
implanted 2 � 105MDA-MB-231 or SUM159 wild-type or PAI1 cells
expressingmCherry Nuc-FUW to demark nuclei of cancer cells. Six
weeksafter injection, we euthanized mice, removed the tumor
andlungs, and immediately imaged metabolism of cancer cells
inorthotopic tumors or lung metastases ex vivo by FLIM. We notethat
previous research demonstrates NADH/FAD metabolism ofcancer cells
maintained in intact tumors and organs ex vivoremains the same as
in vivo for up to eight hours (22, 23). Tocalculate numbers of
cells in an image, we used FIJI (24) tobinarize and analyze numbers
of nuclei in a region. We thenpooled and plotted the density from
each image.
MATLAB analysisMATLAB programs used to measure the kinase
translocation
reporter (KTR), collagen directionality, stress fiber density,
andspheroids are available through a Material Transfer
Agreement(MTA).
Statistical analysisWe used a nonparametric Mann–Whitney U test
for compar-
isons of cell migration and motility with a significance level
of0.05 considered statistically significant. For sphere formation
andother experiments, we used two-tailed, unpaired Student t
tests.We prepared bar graphs (mean valuesþ SD or SEM as denoted
infigure legends) and box plots andwhiskers using GraphPad Prism7
orOrigin 9.0. For box plots andwhiskers, the bottom and top ofa box
define the first and third quartiles, and the band inside thebox
marks the second quartile (the median). The ends of thewhiskers
represent the 5th percentile and the 95th percentiles,respectively.
The "þ" inside the box indicates the mean, dots
Humphries et al.
Mol Cancer Res; 17(5) May 2019 Molecular Cancer Research1144
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
outside thebox andwhiskers indicate outliers, and the "x" refers
tothe maximum and minimum of all data.
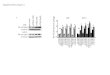
ResultsPAI1 enhances cell migration
We recently used our migration-based microfluidic device
toidentify critical positive and negative regulators of cell
migration(ref. 17; Fig. 1A and B). RNA-sequencing data revealed
plasmin-ogen activator inhibitor 1 (PAI1) as one of the most
highlyupregulated genes (�8-fold relative to nonmigratory cells, P
<0.0001) in both highly migratory SUM159 and
MDA-MB-231triple-negative breast cancer (TNBC) cells. To
investigate effects ofPAI1 on migration and motility of TNBC cells
in vitro and in vivo,we stably expressed PAI1 in SUM159 and
MDA-MB-231 cells.qRT-PCR analysis revealed higher expression of
PAI1 in stablytransduced SUM159 andMDA-MB-231 PAI1 cells
comparedwithwild-type control (Supplementary Fig. S1). RNA
sequencing withgene set enrichment analysis (GSEA) identified
significant asso-ciations with cell movement and organization of
the cytoskele-ton/cell projections in PAI1 cells (Table 1),
confirming the effectof PAI1 on cell migration. Stable expression
of PAI1 significantlyincreased migration of both SUM159 and
MDA-MB-231 cell inour migration device (Fig. 1C and D) and a
wound-healing assay(Fig. 1E and F). Adding exogenous recombinant
active PAI1
(rPAI1) also significantly increased migration of wild-type
cellsfrom both cell lines, and a PAI1-specific inhibitor,
tiplaxtinin,significantly reduced cellmigration (Fig. 1E and F;
SupplementaryFig. S1). Together, these data show that elevated
expression ofPAI1 enhances migration.
PAI1 promotes actin cytoskeleton reorganization and aninvasive
phenotype
Cell migration critically depends on the actin
cytoskeleton.Lamellipodia, membrane ruffles, filopodia, and stress
fibers con-stitute the most important and studied actin-based
structures formotility. A lamellipodium and membrane ruffles demark
actin-rich sheet-like projections of a motile cell, while a
filopodium
Figure 1.
PAI1 enhances migration of TNBCcells. A, Schematic of the
migrationdevice. We initially loaded cells intothe left and right
chambers of thedevice in serum-free media. Aftercells adhered, we
loadedmediacontaining a chemoattractant(serum) into the middle
channel tostimulate migration for 24 hours.B,We used trypsin to
selectivelyrecover cells from the middlechannel for further
analyses. C andD, Graphs show box and whiskerplot summaries (top)
and frequencydistributions (bottom) for migrationof SUM159 (C) and
MDA-MB-231 (D)WT and PAI1 cells toward serum inthe device (n¼ 600
cells per group;��� , P < 0.0001). E and F, Summaryof
percentages of wound closurepresented as meanþ SD (n¼ 9).� , P <
0.05; �� , P < 0.01, as comparedwithWT control cells.
Table 1. Relevant GO biological processes of PAI1-expressing
cells based onGSEA
GO Biological Process P
Biological adhesion 2.60E�31Movement of cell or subcellular
component 4.43E�30Homophilic cell adhesion via plasmamembrane
adhesionmolecules 2.22E�25Regulation of intracellular signal
transduction 2.49E�25Cell projection organization 3.66E�25Cell–cell
adhesion via plasma membrane adhesion molecules
9.20E�25Cytoskeleton organization 1.11E�24Positive regulation of
gene expression 2.28E�23
PAI1 Drives TNBC Cytoskeletal Reorganization and Glycolysis
www.aacrjournals.org Mol Cancer Res; 17(5) May 2019 1145
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
Humphries et al.
Mol Cancer Res; 17(5) May 2019 Molecular Cancer Research1146
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
defines a finger-like projection extending from a
lamellipodium.Stress fibers form from contractile actomyosin
bundles. Effects ofPAI1 on each of these actin-based structures
remain poorlydefined. Phalloidin staining revealed dramatic
reorganization ofthe actin cytoskeleton in PAI1 cells compared with
wild type.Stable expression of PAI1 or addition of rPAI to cells
increased theformation of actin-rich migratory structures
(lamellipodia andmembrane ruffles; Figs. 2A and 3A) and reduced
stress fiberdensity (stress fiber area divided by total cell size;
SupplementaryFig. S2) compared with control cells. In addition, we
found thatPAI1 increases, and tiplaxtinin reduces, the aspect ratio
(length/width) of cells (Figs. 2B and 3B), which suggests PAI1
supports amore mesenchymal morphology associated with cell
migration.
To study actin dynamics in living cells, we stably
expressedLifeAct-GFP, a 17-amino-acid peptide that binds to
filamentousactin (F-actin) and does not interfere with actin
dynamics (25), inbothwild-type andPAI1 cells. To confirmour
phalloidin staining,we imaged SUM159PAI1 and control cells
expressing LifeAct-GFPas theymigrated through ourmicrofluidic
device. Consistent withour previous data, we found that PAI1 cells
had greater actin-richmigratory structures compared with control
cells (SupplementaryVideo S1 and S2). Next, we uniformly embedded
single cellsexpressing LifeAct-GFP into fibrin gels and allowed
them toproliferate. Fibrin promotes adhesion and migration of cells
andprovides structural integrity for breast tumors (26). Aggregates
ofSUM159 PAI1 cells displayed a more invasive phenotype withmore
actin-rich structures extending into the surrounding gel(Fig. 2C).
These projections contained concentrated actin-basedleading edges,
suggesting an actin-richmigratory structure such aslamellipodia or
membrane ruffles. We also noted that SUM159PAI cells formed smaller
aggregates than wild-type cells. MDA-MB-231 cells did not form
aggregates in gels, independent of PAI1expression. However,
wild-type MDA-MB-231 cells did formgrape-like nonadherent
structures, whereas PAI1 cells formedadherent structures that
displayed actin-based projections(Fig. 3C).
We next formed spheroids using LifeAct-GFP cells andhuman
mammary fibroblasts (HMF) and embedded spheroidsinto fibrin gels to
investigate effects of PAI1 on cancer cells in aphysiologically
relevant, coculture 3D model. While HMFsshowed similar phenotypes
between groups, wild-type Life-Act-GFP cells displayed a compact,
spherical shape with adefined edge (Fig. 2D and 3D). By comparison,
both SUM159and MDA-MB-231 PAI1 cells demonstrated greater
invasioninto the adjacent fibrin gel (Figs. 2D and 3D). We also
discov-ered that cancer cells near the edge of PAI1 spheroids
showedmore actin-rich migratory structures, suggesting actin
reorga-
nization and enhanced motility. Using custom MATLAB codewe
identified the margins of cancer cells in spheroids. Relativeto
spheroids with control cells, PAI1 spheroids contained
moreprojections from a spheroid, and these projections
extendedfarther from the spheroid core into the adjacent gel (Figs.
2Dand 3D). Overall, these data establish that PAI1 promotes
actincytoskeleton reorganization to display a more migratory
phe-notype in 2D and 3D settings.
PAI1 enhances cellular adhesionIn addition to cell migration,
GSEA also revealed multiple sets
with associations to cell adhesion or attachment (Table 1).
Wenoted that cancer cells invading from a spheroid into the
geltracked closely with HMF cells, suggesting that PAI1
promotesintercellular adhesion. To test this hypothesis, we
mechanicallydissociated coculture spheroids of cancer cells andHMFs
and thenenumerated single cancer cells. We found more single
wild-typecancer cells following mechanical disruption of spheroids,
con-sistent with reduced adhesion relative to cancer cells with
elevatedPAI1 (Supplementary Fig. S3).We also identified greater
adhesionof PAI1 cancer cells to HMFs in monolayer culture
(Supplemen-tary Fig. S3). Together, these data demonstrate that
PAI1promotesintercellular adhesion of breast cancer cells.
To investigate mechanisms through which PAI1 promotesadhesion,
we focused on focal adhesions, structures of the actincytoskeleton
that form anchor points to the extracellular matrix(ECM).
Immunofluorescence staining for paxillin, a protein com-ponent of
focal adhesions, showed reduced numbers of focaladhesions in breast
cancer cells with elevated PAI1 relative tocontrol (Supplementary
Fig. S4). This result suggests that PAI1increases adhesion of
breast cancer cells independent of focaladhesions.
PAI1 activates ERK signaling and alters
mitochondrialmorphology
MAPK signaling through ERK defines one prominent pathwaythat
promotes reorganization of actin, turnover of focal adhe-sions, and
cell motility (27). We investigated to what extent PAI1activates
ERK in single cells using a kinase translocation reporter(KTR)
system (15). Using cells stably expressing the ERK KTR, wefound
that exogenous addition of rPAI1 to both wild-typeSUM159 and
MDA-MB-231 cells increased kinase activity of ERKat 20 minutes
(Fig. 4A; Supplementary Fig. S5). In addition,tiplaxtinin inhibited
activation of ERK by rPAI1 (SupplementaryFig. S5). Activation of
ERK provides one potential mechanismthrough which PAI drives cell
migration.
Figure 2.Stable expression of PAI1 promotes actin cytoskeletal
reorganization and amore invasive phenotype in SUM159 TNBC cells.
A, Representative overlaid images ofimmunofluorescence staining of
phalloidin (red) and nuclear DAPI (blue) ofWT cells; WT cells
treated with rPAI1 (40 nmol/L); PAI1 cells; and PAI1 cells
treatedwith tiplaxtinin (5 mmol/L; left). Arrows point to
actin-rich migratory structures. Right, Frequency distribution of
number of actin-rich migratory structures percell for each group
(n¼ 50 cells). B, Box plot and whiskers for the aspect ratio
(length/width) of cells in each group demonstrates a more elongated
morphologyof PAI1 cells (n¼ 50 cells). ��� , P < 0.0001. C,
Representative images of spheres formed from single cells of
SUM159WT and PAI1 cells that express LifeAct-GFP(left). Arrows
point to projections. Right, Analysis of the mean number of
projections per spheroidþ SD (top) and the mean circumference of
spheroidsþ SD ineach group (bottom; n¼ 7 spheroids per group). � ,
P < 0.05; �� , P < 0.01. D, Representative images of
spheroids composed of SUM159WT or PAI1 cells thatexpress
LifeAct-GFP (green) and humanmammary fibroblasts (HMFs, red, i).
Arrows in zoomed picture point to actin-rich migratory structures.
Example imagesthat define the perimeter of cancer cells in a
spheroid (green line), the center of the spheroid (red dot), and
the spheroid core (red circle, circle from the center ofthe
spheroid with a radius of the distance from the center to the
closest point on the perimeter) for each of the groups (ii).
Frequency distribution of the distanceof points on the perimeter of
the spheroid from the spheroid core (n¼ 5 spheroids, iii). Graph
showing the mean number of points on the perimeterþSD that
falloutside of the spheroid core (n¼ 5 spheroids; iv). � , P <
0.05. These data show that spheroids made from cells that stably
express PAI1 havemore projections,farther projections, and amore
invasive phenotype than spheroids fromWT cells.
PAI1 Drives TNBC Cytoskeletal Reorganization and Glycolysis
www.aacrjournals.org Mol Cancer Res; 17(5) May 2019 1147
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
Humphries et al.
Mol Cancer Res; 17(5) May 2019 Molecular Cancer Research1148
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
Recent studies show that ERK promotes mitochondrial fis-sion to
drive tumor growth (28) and cellular reprogram-ming (29), and more
migratory cancer cells frequently exhibitgreater fission of
mitochondria. Staining with MitoTrackerGreen showed greater
fragmentation of mitochondria, indica-tive of mitochondrial
fission, in both SUM159- and MDA-MB-231-PAI cells. Treatment of
wild-type parental cells with rPAI1also shifted mitochondria from
more fused, elongated struc-tures to a more fragmented morphology,
characteristic offission (Supplementary Fig. S6).
PAI1 promotes glycolytic metabolism in vitroHaving established
that PAI1 promotes mitochondrial fission,
we next investigated effects of PAI1 on cellmetabolism in
vitro.Weused FLIM of fluorescence from endogenous NADH to
measurerelative glycolytic versus oxidative metabolism in single
TNBCcells (30). Cells predominantly relying on glycolysis have
greateramounts of free NADH, which exhibits a shorter lifetime
thanprotein-bound NADH in cells utilizing oxidative
phosphoryla-tion (OXPHOS; ref. 31). We determined that PAI1
promotesglycolytic metabolism in both SUM159 and MDA-MB-231 cellsin
culture as shown by a shorter lifetime for fluorescence fromNADH
(Fig. 4B). We also found that PAI1 promotes glycolyticmetabolism in
cells seeded infibrin gels (Fig. 4C). Consistentwithour FLIM data,
we found that PAI1 increased the extracellularacidification rate
(ECAR) in both SUM159 and MDA-MB-231cells (Supplementary Fig. S7),
suggesting an increase in glycolyticcapacity in cells that express
PAI1. Together, these data demon-strate that PAI1 shifts cellular
metabolism to glycolysis.
PAI1 orthotopic tumors are smaller, but show a more
invasivephenotype
To extend our studies to an in vivo setting, we
orthotopicallyimplanted SUM159 or MDA-MB-231 wild-type or PAI1
cells thatexpress LifeAct-GFP into the fourth mammary fat pad of
NSGmice. Consistent with our data showing that PAI1 cells form
fewer(Supplementary Fig. S8) and smaller tumor spheres than
wild-type cells (Figs. 2C and 3C), mice injected with PAI1 TNBC
cellsproduced significantly smaller tumors (Fig. 5A). Even though
wesaw significantly smaller orthotopic tumors, we did not
observeany differences in growth of PAI1 versus wild-type cells in
stan-dard culture (Supplementary Fig. S8). Interestingly, we
foundreduced density of cancer cells in PAI1 tumors (number of
cancercells divided by total tumor area; Fig. 5B and C). PAI1 TNBC
cellsalso displayed more actin-rich migratory structures than
cancercells in wild-type tumors (Fig. 5B). In addition to imaging
cancercells, we analyzed the architecture of peritumoral collagen
bysecond harmonic two-photonmicroscopy. Alignment of collagenfibers
perpendicular to the margin of a tumor defines a moreinvasive
phenotype with poor prognosis in breast cancer (32).
Tumors formed from SUM159- and MDA-MB-231-PAI cellsshowed
greater alignment of collagen perpendicular to the tumormargin than
wild-type, consistent with a more locally invasivephenotype (Fig.
5B and D; Supplementary Fig. S9). We note thatboth bioluminescent
and two-photon imaging revealed no dif-ferences in metastasis
between PAI and control tumors (data notshown), potentially because
we euthanized mice at a relativelylate time point (8 weeks) after
implantation to facilitate imagingby two-photon microscopy.
PAI1 promotes glycolytic metabolism in a mouse model ofbreast
cancer
To determine effects of PAI on metabolism of cancer cellsin
vivo, we used FLIM to quantify glycolysis versus OXPHOS
inorthotopic tumors and lung metastases produced by SUM159
orMDA-MB-231 wild-type or PAI1 cells. For these studies, we
usedcancer cells expressing nuclearmCherry to avoid interference
fromGFP in detecting NADH and distinguish disseminated cancercells.
Unlike studies that recover cancer cells from tumors andanalyze
metabolism of bulk populations of dissociated cancercells in
standard culture, FLIM allows quantitative analysis ofmetabolism in
single cancer cells in intact tumors andmetastases.We analyzed mice
six weeks after injection. Both SUM159 andMDA-MB-231 PAI1 cells in
orthotopic tumors displayed moreglycolytic metabolism (Fig. 6A and
B). In addition to orthotopictumors, we discovered enhanced
glycolysis inMDA-MB-231 PAI1cells in lung metastases (Fig. 6C). We
were unable to detectSUM159 metastases independent of PAI1
expression at thetime of imaging. These data establish that PAI1
promotes glyco-lytic metabolism at both the orthotopic tumor and
the lungmetastatic site.
DiscussionDeveloping more effective treatments to block local
invasion
and metastasis in cancer requires mechanistic understanding
ofpathways driving cancer cell migration. While well-established
asa prognostic and predictive biomarker in breast cancer (1),
target-ingPAI1 for cancer therapy remains challenging due to
limitationsof existing drugs and incomplete understanding of
functions ofPAI1 in tumor progression (33). While other studies
have iden-tified PAI1 as an important regulator of actin dynamics
and cellmigration (34–36), mechanisms for how PAI1 promotes
cancercell migration and tumor progression remain
incompletelydefined. Here we make the unique discovery that PAI1
activatesERK, connecting PAI1 signaling to glycolytic metabolism
andcytoskeletal reorganization.
Building on our discovery that highly migratory TNBC
cellsexpress high levels of PAI1, this study established that PAI1
drivescell migration by promoting cytoskeletal reorganization
and
Figure 3.Stable expression of PAI1 promotes actin cytoskeletal
reorganization and amore invasive phenotype in MDA-MB-231 TNBC
cells. A, Representative overlaidimages of immunofluorescence
staining of phalloidin (red) and nuclear DAPI (blue) ofWT cells; WT
cells treated with rPAI1 (40 nmol/L); PAI1 cells; and PAI1
cellstreated with tiplaxtinin (5 mmol/L; left). Arrows point to
actin-rich migratory structures. Right, Frequency distribution of
number of actin-rich migratorystructures per cell for each group
(n¼ 50 cells). B, Box plot and whiskers for the aspect ratio
(length/width) of cells in each group demonstrates more
elongatedmorphology of PAI1 cells (n¼ 50 cells). � , P < 0.05;
��, P < 0.01; ��� , P < 0.0001. C, Representative images of
spheres formed from single cells of MDA-MB-231 WTand PAI1 cells
that express LifeAct-GFP. D, Representative images of spheroids
composed of MDA-MB-231 WT or PAI1 cells that express LifeAct-GFP
(green) andhumanmammary fibroblasts (HMFs, red; i). Arrows in
zoomed picture point to actin-rich migratory structures. Example
images that define the perimeter ofcancer cells in a spheroid
(green line), the center of the spheroid (red dot), and the
spheroid core (red circle) as defined in Fig. 2 (ii). Frequency
distribution of thedistance of points on the perimeter of the
spheroid from the spheroid core (n� 9 spheroids; iii). Graph
showing the mean number of points on the perimeterþSD that fall
outside of the spheroid core (n� 9 spheroids; iv; � , P <
0.05).
PAI1 Drives TNBC Cytoskeletal Reorganization and Glycolysis
www.aacrjournals.org Mol Cancer Res; 17(5) May 2019 1149
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
Figure 4.
PAI1 activates ERK signaling and promotes glycolytic metabolism
in TNBC cells. A, Representative fluorescence images of ERK KTR
(yellow) and nuclear H2B-mCherry (red) in SUM159 and MDA-MB-231 WT
cells at the initial timepoint (0 minutes) and 20minutes after
addition of rPAI1 (40 nmol/L) showing translocationof yellow
fluorescence out of the nucleus (darker nucleus, ERK activation;
left). Right, frequency distribution of cytoplasmic-to-nuclear
ratio (CNR) of SUM159 (n> 350 cells) and MDA-MB-231 (n > 670
cells) WT cells at the initial timepoint (0minutes) and 20minutes
after addition of rPAI1 (n¼ 2). A shift to the rightdemonstrates
activation of ERK signaling. B, Frequency distributions for NADH
lifetime and representative FLIM images of SUM159 and MDA-MB-231 WT
andPAI1 cells in 2D (B, SUM159: n� 12, and MDA-MB-231: n� 9) and in
3D fibrin gels (C, SUM159: n� 39, and MDA-MB-231: n� 36). Both
SUM159 and MDA-MB-231PAI1 cells shift toward glycolytic metabolism
as defined by a shorter NADH lifetime in 2D and 3D
environments.
Humphries et al.
Mol Cancer Res; 17(5) May 2019 Molecular Cancer Research1150
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
glycolytic metabolism. Recent studies show migrating cancercells
utilize energy generated through glycolysis rather thanOXPHOS (7,
8), suggesting that elevated glycolysis mediated byPAI1drives
cellmigration.Mechanistically, weobserved that PAI1activates
several processes that potentially link cell migration
andglycolytic metabolism. Cells with high expression of PAI1
(i)activate ERK; (ii) exhibit fragmented mitochondria; and
(iii)utilize glycolysis to a greater extent than control breast
cancercells in environments from 2D cultures to metastases in
mice.Overall, our data suggest anovelmetabolic regulatory function
forPAI1 in migration and tumor progression.
Dysregulation of cell migration during cancer progressionshapes
the metastatic capacity of cancer cells. Cell migration isthe
result of amulti-step process driven by dynamic changes in the
actin cytoskeleton. Consistent with other studies (37, 38),
wefound that PAI1 expression reduces stress fiber density
whileincreasing the formation of actin-rich migratory structures
instandard cell culture. We also found that cells expressing
PAI1displayed reduced focal adhesions as determined by
paxillinstaining. Paxillin is a key component of integrin
signaling, andphosphorylation of paxillin is required for
integrin-mediatedreorganization of the actin cytoskeleton. Previous
work demon-strated that phosphorylation of paxillin by ERK promotes
trans-location of paxillin from focal adhesions to the cytosol,
resultingin disassembly of focal adhesions (39, 40). Therefore, one
expla-nation for reduced focal adhesions in cells expressing
PAI1maybethrough activation of ERK. Although cells displayed a
moremigratory phenotype in culture conditions, breast cancer
cells
Figure 5.
PAI1 reduces orthotopic tumorgrowth and promotes
collagenalignment. A, Area-under-the-curvephoton flux for
orthotopic tumorgrowth of SUM159 and MDA-MB-231WT and PAI1 cells.
Graphs showmeanþ SEM (n� 6 tumors;� , P < 0.05; �� , P <
0.01).B, Representative overlaid imagesof tumors (green) and
collagen(gray) from SUM159 or MDA-MB-231WT or PAI1 cells that
expressLifeAct-GFP. Tumors from PAI1expressing cells demonstrate
morealigned collagen fibers. C,Graphsdemonstrate reduced cell
density intumors from PAI1 cells relative toWT (SUM159: n > 30
z-axis sections� 20 mm in depth, and MDA-MB-231: n¼ 36 z-axis
sections� 20 mmin depth). Graphs showmeanþSEM. � , P < 0.05; ���
, P < 0.0001.D, Frequency distribution ofcollagen alignment
angle revealsgreater alignment of collagen fibersperpendicular to
margins of tumorsfrom PAI1 cells. Graphs show theorientation of the
collagen fibersrelative to the tumor (n� 5tumors).
PAI1 Drives TNBC Cytoskeletal Reorganization and Glycolysis
www.aacrjournals.org Mol Cancer Res; 17(5) May 2019 1151
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
with elevated expression of PAI1 exhibited greater adhesion
toHMFs. Apparent discordance between greater migration andadhesion
may arise from measuring these processes in 2D and
3D environments, respectively. Furthermore in orthotopictumors,
we found that PAI1 promotes alignment of tumor-asso-ciated collagen
perpendicular to the margin of a tumor, a
Figure 6.
PAI1 promotes glycolytic metabolism in both orthotopic tumor
andmetastatic sites. Left, Frequency distributions of NADH lifetime
for SUM159 (A) or MDA-MB-231 (B) WT control and PAI1 cells in an
orthotopic tumor (n� 5 tumors) and MDA-MB-231 WT and PAI1 cells in
lung metastatic sites (C; n� 3 lungs, n� 15metastatic sites per
group). Decreased lifetime of NADH fluorescence in PAI1 cells
indicates increased glycolysis in both the orthotopic tumor and
lungmetastases. Right, Representative fluorescence (red) and FLIM
images of cells in orthotopic tumor and lung metastatic sites.
Humphries et al.
Mol Cancer Res; 17(5) May 2019 Molecular Cancer Research1152
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
signature associated with tumor invasion and poor progno-sis
(32). These data suggest that PAI1 enhances migration notonly
through effects on cancer cells, but also on the surroundingtumor
microenvironment.
Reprogramming of cellular metabolism represents a
hallmarkfeature of cancer with malignant cells frequently shifting
toglycolytic metabolism (41). Breast cancers upregulate
glycolyticenzymes relative to normal breast tissue and benign
breastlesions, and glycolytic metabolism correlates with worse
out-comes (42, 43). Mechanisms underlying a switch from oxidativeto
glycolytic metabolism in cancer cells remain incompletelydefined.
Here, we discovered a previously unknown function ofPAI1 to promote
glycolysis in TNBC cells, a metabolic switchlinked to cell
migration (7). Consistent with a previous studyshowing
mitochondrial effects of PAI1 (12), we found that PAI1promotes
mitochondrial fission in TNBC cells. Mitochondrialfission promotes
a shift to glycolytic metabolism in both normaland malignant
settings (44–46), linking changes in mitochon-drialmorphology to
effects of PAI1 onmetabolic reprogramming.ERK signaling drives
mitochondrial fission (28, 29), suggestingactivation of this kinase
as a mechanism through which PAI1promotes mitochondrial fission and
glycolysis. In addition, wefound that PAI1 cells retain their
enhanced glycolytic phenotypefollowing colonization to lung. This
contrasts with other studiesdemonstrating that breast cancer cells
engage OXPHOS to metas-tasize to the lung (47, 48). Differences
among studies may be dueto the use of human breast cancer cells in
our current study versusmurine breast cancer cells in prior
publications. In addition, wequantified metabolism in single cells
in situ using FLIM of endog-enous NADH (49, 50), while studies
showing OXPHOS in lungmetastases relied on profiles of gene
expression and cell cultureassays with populations of cancer cells
selected for enhanced lungmetastasis. Our data establish that PAI1
drives glycolysis in cell-based assays, orthotopic xenografts, and
lungmetastases, suggest-ing that PAI1 glycolytic reprogramming as
an integral function ofPAI1 in breast cancer.
In summary, this study not only provides the first evidence
thatPAI1 promotes glycolytic metabolism but also suggests an
under-lying mechanism mediated by ERK and known effects of this
kinase on mitochondrial morphology. Our data support
recentresearch proposing that highly migratory cells rely on
energyobtained primarily through enhanced glycolysis for
migrationand chemotaxis (7, 8). Data from our study provide a
strongrationale for targeting cancer cell metabolism as a
potentialtherapeutic approach not only for aggressive
andhighlymigratorycancers, such as TNBC, but also for patients with
elevated levels ofPAI1 in primary tumors and/or circulating tumor
cells.
Disclosure of Potential Conflicts of InterestG.D. Luker is a
consultant/advisory board member for Polyphor. No poten-
tial conflicts of interest were disclosed by the other
authors.
Authors' ContributionsConception and design: B.A. Humphries, E.
Yoon, K.E. Luker, G.D. LukerDevelopment of methodology: B.A.
Humphries, J.M. Buschhaus, Y.-C. Chen,B. Chiang, Y.-H. Cheng, E.
Yoon, K.E. Luker, G.D. LukerAcquisition of data (provided animals,
acquired and managed patients,provided facilities, etc.): B.A.
Humphries, J.M. Buschhaus, Y.-C. Chen,H.R. Haley, T. Qyli, B.
Chiang, N. Shen, A. Cutter, K.E. Luker, G.D. LukerAnalysis and
interpretation of data (e.g., statistical analysis,
biostatistics,computational analysis): B.A. Humphries, J.M.
Buschhaus, Y.-C. Chen,H.R. Haley, T. Qyli, B. Chiang, S. Rajendran,
A. Cutter, J. Cong, P.C. Spinosa,K.E. Luker, G.D. LukerWriting,
review, and/or revision of the manuscript: B.A. Humphries,J.M.
Buschhaus, Y.-T. Chen, K.E. Luker, G.D. LukerAdministrative,
technical, or material support (i.e., reporting or organizingdata,
constructing databases): Y.-T. Chen, E. Yoon, K.E. Luker, G.D.
LukerStudy supervision: E. Yoon, K.E. Luker, G.D. Luker
AcknowledgmentsThe authors acknowledge funding from NIH grants
R01CA196018,
R01CA195655, U01CA210152, and R37CA222563. This study was
supportedby NIH grants R01CA196018, R01CA195655, U01CA210152,
andR37CA222563.
The costs of publication of this articlewere defrayed inpart by
the payment ofpage charges. This article must therefore be hereby
marked advertisement inaccordance with 18 U.S.C. Section 1734
solely to indicate this fact.
Received August 7, 2018; revised October 22, 2018; accepted
January 29,2019; published first February 4, 2019.
References1. Duffy MJ, McGowan PM, Harbeck N, Thomssen C,
Schmitt M. uPA and
PAI-1 as biomarkers in breast cancer: validated for clinical use
in level-of-evidence-1 studies. Breast Cancer Res 2014;16:428.
2. Chambers SK, Ivins CM, Carcangiu ML. Plasminogen activator
inhib-itor-1 is an independent poor prognostic factor for survival
inadvanced stage epithelial ovarian cancer patients. Int J Cancer
1998;79:449–54.
3. Nekarda H, Schmitt M, Ulm K, Wenninger A, Vogelsang H, Becker
K, et al.Prognostic impact of urokinase-type plasminogen activator
and its inhib-itor PAI-1 in completely resected gastric cancer.
Cancer Res 1994;54:2900–7.
4. Harbeck N, Schmitt M, Meisner C, Friedel C, Untch M, Schmidt
M, et al.Ten-year analysis of the prospective multicentre Chemo-N0
trial validatesAmerican Society of ClinicalOncology
(ASCO)-recommended biomarkersuPA and PAI-1 for therapy decision
making in node-negative breast cancerpatients. Eur J Cancer
2013;49:1825–35.
5. Chen YC, Allen SG, Ingram PN, Buckanovich R, Merajver SD,
Yoon E.Single-cell migration chip for chemotaxis-based microfluidic
selection ofheterogeneous cell populations. Sci Rep
2015;5:9980.
6. Rangel R, Lee SC,Hon-KimBanK,Guzman-Rojas L,MannMB,Newberg
JY,et al. Transposon mutagenesis identifies genes that cooperate
with mutant
Pten in breast cancer progression. Proc Natl Acad Sci U S A
2016;113:E7749–58.
7. Shiraishi T, Verdone JE, Huang J, Kahlert UD, Hernandez JR,
Torga G, et al.Glycolysis is the primary bioenergetic pathway for
cell motilityand cytoskeletal remodeling in human prostate and
breast cancer cells.Oncotarget 2015;6:130–43.
8. Beckner ME, Stracke ML, Liotta LA, Schiffmann E. Glycolysis
as a primaryenergy source in tumor cell chemotaxis. J Natl Cancer
Inst 1990;82:1836–40.
9. Gatenby RA, Gillies RJ. Why do cancers have high aerobic
glycolysis?Nat Rev Cancer 2004;4:891–9.
10. Alessi MC, Poggi M, Juhan-Vague I. Plaminogen activator
inhibitor-1,adipose tissue and insulin resistance. Curr Opin
Lipidol 2007;18:240–5.
11. Alessi MC, Juhan-Vague I. PAI-1 and the metabolic syndrome:
links,causes, and consequences. Arterioscler Thromb Vasc Biol
2006;26:2200–7.
12. Castello-Cros R, Bonuccelli G, Molchansky A, Capozza F,
Witkiewicz AK,Birbe RC, et al. Matrix remodeling stimulates stromal
autophagy, "fueling"cancer cell mitochondrial metabolism
andmetastasis. Cell Cycle 2011;10:2021–34.
www.aacrjournals.org Mol Cancer Res; 17(5) May 2019 1153
PAI1 Drives TNBC Cytoskeletal Reorganization and Glycolysis
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
13. Smith MC, Luker KE, Garbow JR, Prior JL, Jackson E,
Piwnica-Worms D,et al. CXCR4 regulates growth of both primary
andmetastatic breast cancer.Cancer Res 2004;64:8604–12.
14. Cavnar SP, Rickelmann AD, Meguiar KF, Xiao A, Dosch J, Leung
BM, et al.Modeling selective elimination of quiescent cancer cells
from bone mar-row. Neoplasia 2015;17:625–33.
15. Regot S, Hughey JJ, Bajar BT, Carrasco S, Covert MW.
High-sensitivitymeasurements of multiple kinase activities in live
single cells. Cell 2014;157:1724–34.
16. Luker KE, Mihalko LA, Schmidt BT, Lewin SA, Ray P, Shcherbo
D, et al. Invivo imaging of ligand receptor binding with Gaussia
luciferase comple-mentation. Nat Med 2011;18:172–7.
17. Chen YC, Humphries B, Brien R, Gibbons AE, Chen YT, Qyli T,
et al.Functional isolation of tumor-initiating cells using
microfluidic-basedmigration identifies phosphatidylserine
decarboxylase as a key regulator.Sci Rep 2018;8:244.
18. Zhao H, Ma L, Zhou J, Mao Z, Gao C, Shen J. Fabrication and
physical andbiological properties of fibrin gel derived from human
plasma.Biomed Mater 2008;3:015001.
19. Cavnar SP, Xiao A, Gibbons AE, Rickelmann AD, Neely T, Luker
KE, et al.Imaging sensitivity of quiescent cancer cells to
metabolic perturbations inbone marrow spheroids. Tomography
2016;2:146–57.
20. LuoM, Shang L, BrooksMD, Jiagge E, Zhu Y, Buschhaus JM, et
al. Targetingbreast cancer stem cell state equilibrium through
modulation of redoxsignaling. Cell Metab 2018;28:69–86.
21. Ranjit S, Dvornikov A, StakicM,Hong SH, LeviM, Evans RM, et
al. Imagingfibrosis and separating collagens using second harmonic
generation andphasor approach to fluorescence lifetime imaging. Sci
Rep 2015;5:13378.
22. Walsh AJ, Poole KM, Duvall CL, Skala MC. Ex vivo optical
metabolicmeasurements from cultured tissue reflect in vivo tissue
status. J BiomedOpt 2012;17:116015.
23. Walsh AJ, Cook RS, Arteaga CL, Skala MC. Optical metabolic
imaging oflive tissue cultures. SPIE Proceedings
2013;8588:858820.
24. Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair
M, Pietzsch T,et al. Fiji: an open-source platform for
biological-image analysis.Nat Methods 2012;9:676–82.
25. Riedl J, Crevenna AH, Kessenbrock K, Yu JH, Neukirchen D,
Bista M, et al.Lifeact: a versatile marker to visualize F-actin.
Nat Methods 2008;5:605–7.
26. Costantini V, Zacharski LR, Memoli VA, Kisiel W, Kudryk BJ,
Rousseau SM.Fibrinogen deposition without thrombin generation in
primary humanbreast cancer tissue. Cancer Res 1991;51:349–53.
27. Huang C, Jacobson K, Schaller MD. MAP kinases and cell
migration. J CellSci 2004;117:4619–28.
28. Kashatus JA, Nascimento A, Myers LJ, Sher A, Byrne FL, Hoehn
KL, et al.Erk2phosphorylationofDrp1promotesmitochondrial fission
andMAPK-driven tumor growth. Mol Cell 2015;57:537–51.
29. Prieto J, Leon M, Ponsoda X, Sendra R, Bort R,
Ferrer-Lorente R, et al. EarlyERK1/2 activation promotes
DRP1-dependent mitochondrial fission nec-essary for cell
reprogramming. Nat Commun 2016;7:11124.
30. Sun Y, Day RN, Periasamy A. Investigating protein-protein
interactions inliving cells using fluorescence lifetime imaging
microscopy. Nat Protoc2011;6:1324–40.
31. Stringari C,Nourse JL, Flanagan LA,Gratton E. Phasor
fluorescence lifetimemicroscopy of free and protein-bound NADH
reveals neural stem celldifferentiation potential. PLoS One
2012;7:e48014.
32. ConklinMW, Eickhoff JC, RichingKM, Pehlke CA, Eliceiri KW,
ProvenzanoPP, et al. Aligned collagen is a prognostic signature for
survival in humanbreast carcinoma. Am J Pathol
2011;178:1221–32.
33. Placencio VR, DeClerck YA. Plasminogen activator inhibitor-1
in cancer:rationale and insight for future therapeutic testing.
Cancer Res 2015;75:2969–74.
34. Durand MK, Bodker JS, Christensen A, Dupont DM, Hansen M,
Jensen JK,et al. Plasminogen activator inhibitor-I and tumour
growth, invasion, andmetastasis. Thromb Haemost 2004;91:438–49.
35. Gramling MW, Church FC. Plasminogen activator inhibitor-1 is
an aggre-gate response factor with pleiotropic effects on cell
signaling in vasculardisease and the tumor microenvironment. Thromb
Res 2010;125:377–81.
36. Czekay RP,Wilkins-Port CE,Higgins SP, Freytag J, Overstreet
JM, Klein RM,et al. PAI-1: an integrator of cell signaling and
migration. Int J Cell Biol2011;2011:562481.
37. Degryse B, Neels JG, Czekay RP, Aertgeerts K, Kamikubo Y,
Loskutoff DJ.The low density lipoprotein receptor-related protein
is a motogenicreceptor for plasminogen activator inhibitor-1. J
Biol Chem 2004;279:22595–604.
38. Vial D, McKeown-Longo PJ. PAI1 stimulates assembly of the
fibronectinmatrix in osteosarcoma cells through crosstalk between
the alphavbeta5and alpha5beta1 integrins. J Cell Sci 2008;121(Pt
10):1661–70.
39. Ishibe S, Joly D, Zhu X, Cantley LG.
Phosphorylation-dependent paxillin-ERK association mediates
hepatocytes growth factor-stimulated epithelialmorphogenesis. Mol
Cell 2003;12:1275–85.
40. WebbDJ, Donais K,Whitmore LA, Thomas SM, Turner CE, Parsons
JT, et al.FAK-Src signalling through paxillin, ERK and MLCK
regulates adhesiondisassembly. Nat Cell Biol 2004;6:154–61.
41. Pavlova NN, Thompson CB. The emerging hallmarks of cancer
metabo-lism. Cell Metab 2016;23:27–47.
42. HennipmanA, Smits J, vanHouwelingen JC, RijksenG,Neyt JP,
VanUnnikJA, et al. Glycolytic enzymes in breast cancer, benign
breast disease andnormal breast tissue. Tumour Biol
1987;8:251–63.
43. Peng F, Wang JH, Fan WJ, Meng YT, Li MM, Li TT, et al.
Glycolysisgatekeeper PDK1 reprograms breast cancer stem cells under
hypoxia.Oncogene 2018;37:1062–74.
44. Guido C, Whitaker-Menezes D, Lin Z, Pestell RG, Howell A,
Zimmers TA,et al. Mitochondrial fission induces glycolytic
reprogramming in cancer-associated myofibroblasts, driving stromal
lactate production, and earlytumor growth. Oncotarget
2012;3:798–810.
45. Hagenbuchner J, Kuznetsov AV, Obexer P, Ausserlechner MJ.
BIRC5/Survivin enhances aerobic glycolysis and drug resistance by
altered regu-lation of the mitochondrial fusion/fission machinery.
Oncogene 2013;32:4748–57.
46. Salabei JK, Hill BG. Mitochondrial fission induced by
platelet-derivedgrowth factor regulates vascular smooth muscle cell
bioenergetics and cellproliferation. Redox Biol 2013;1:542–51.
47. Dupuy F, Tabaries S, Andrzejewski S, Dong Z, Blagih J, Annis
MG, et al.PDK1-dependent metabolic reprogramming dictates
metastatic potentialin breast cancer. Cell Metab
2015;22:577–89.
48. Andrzejewski S, Klimcakova E, Johnson RM, Tabaries S, Annis
MG,McGuirk S, et al. PGC-1a promotes breast cancer metastasis and
confersbioenergetic flexibility against metabolic drugs. Cell Metab
2017;26:1–10.
49. Skala MC, Riching KM, Bird DK, Gendron-Fitzpatrick A,
Eickhoff J, EliceiriKW, et al. In vivo multiphoton fluorescence
lifetime imaging of protein-bound and free nicotinamide adenine
dinucleotide in normal and pre-cancerous epithelia. J Biomed Opt
2007;12:024014.
50. Skala MC, Riching KM, Gendron-Fitzpatrick A, Eickhoff J,
Eliceiri KW,White JG, et al. In vivo multiphoton microscopy of NADH
and FAD redoxstates, fluorescence lifetimes, and cellular
morphology in precancerousepithelia. Proc Natl Acad Sci U S A
2007;104:19494–9.
Mol Cancer Res; 17(5) May 2019 Molecular Cancer Research1154
Humphries et al.
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/
-
2019;17:1142-1154. Published OnlineFirst February 4, 2019.Mol
Cancer Res Brock A. Humphries, Johanna M. Buschhaus, Yu-Chih Chen,
et al. Triple-Negative Breast CancerCytoskeleton Reorganization and
Glycolytic Metabolism in Plasminogen Activator Inhibitor 1 (PAI1)
Promotes Actin
Updated version
10.1158/1541-7786.MCR-18-0836doi:
Access the most recent version of this article at:
Material
Supplementary
http://mcr.aacrjournals.org/content/suppl/2019/02/13/1541-7786.MCR-18-0836.DC1
Access the most recent supplemental material at:
Cited articles
http://mcr.aacrjournals.org/content/17/5/1142.full#ref-list-1
This article cites 50 articles, 10 of which you can access for
free at:
Citing articles
http://mcr.aacrjournals.org/content/17/5/1142.full#related-urls
This article has been cited by 3 HighWire-hosted articles.
Access the articles at:
E-mail alerts related to this article or journal.Sign up to
receive free email-alerts
Subscriptions
Reprints and
[email protected]
To order reprints of this article or to subscribe to the
journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take
you to the Copyright Clearance Center's (CCC)
.http://mcr.aacrjournals.org/content/17/5/1142To request
permission to re-use all or part of this article, use this link
on July 5, 2021. © 2019 American Association for Cancer
Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst February 4, 2019; DOI:
10.1158/1541-7786.MCR-18-0836
http://mcr.aacrjournals.org/lookup/doi/10.1158/1541-7786.MCR-18-0836http://mcr.aacrjournals.org/content/suppl/2019/02/13/1541-7786.MCR-18-0836.DC1http://mcr.aacrjournals.org/content/17/5/1142.full#ref-list-1http://mcr.aacrjournals.org/content/17/5/1142.full#related-urlshttp://mcr.aacrjournals.org/cgi/alertsmailto:[email protected]://mcr.aacrjournals.org/content/17/5/1142http://mcr.aacrjournals.org/
/ColorImageDict > /JPEG2000ColorACSImageDict >
/JPEG2000ColorImageDict > /AntiAliasGrayImages false
/CropGrayImages false /GrayImageMinResolution 200
/GrayImageMinResolutionPolicy /Warning /DownsampleGrayImages true
/GrayImageDownsampleType /Bicubic /GrayImageResolution 300
/GrayImageDepth -1 /GrayImageMinDownsampleDepth 2
/GrayImageDownsampleThreshold 1.50000 /EncodeGrayImages true
/GrayImageFilter /DCTEncode /AutoFilterGrayImages true
/GrayImageAutoFilterStrategy /JPEG /GrayACSImageDict >
/GrayImageDict > /JPEG2000GrayACSImageDict >
/JPEG2000GrayImageDict > /AntiAliasMonoImages false
/CropMonoImages false /MonoImageMinResolution 600
/MonoImageMinResolutionPolicy /Warning /DownsampleMonoImages true
/MonoImageDownsampleType /Bicubic /MonoImageResolution 900
/MonoImageDepth -1 /MonoImageDownsampleThreshold 1.50000
/EncodeMonoImages true /MonoImageFilter /CCITTFaxEncode
/MonoImageDict > /AllowPSXObjects false /CheckCompliance [ /None
] /PDFX1aCheck false /PDFX3Check false /PDFXCompliantPDFOnly false
/PDFXNoTrimBoxError true /PDFXTrimBoxToMediaBoxOffset [ 0.00000
0.00000 0.00000 0.00000 ] /PDFXSetBleedBoxToMediaBox true
/PDFXBleedBoxToTrimBoxOffset [ 0.00000 0.00000 0.00000 0.00000 ]
/PDFXOutputIntentProfile (None) /PDFXOutputConditionIdentifier ()
/PDFXOutputCondition () /PDFXRegistryName () /PDFXTrapped
/False
/CreateJDFFile false /Description > /Namespace [ (Adobe)
(Common) (1.0) ] /OtherNamespaces [ > /FormElements false
/GenerateStructure false /IncludeBookmarks false /IncludeHyperlinks
false /IncludeInteractive false /IncludeLayers false
/IncludeProfiles false /MarksOffset 18 /MarksWeight 0.250000
/MultimediaHandling /UseObjectSettings /Namespace [ (Adobe)
(CreativeSuite) (2.0) ] /PDFXOutputIntentProfileSelector /NA
/PageMarksFile /RomanDefault /PreserveEditing true
/UntaggedCMYKHandling /LeaveUntagged /UntaggedRGBHandling
/LeaveUntagged /UseDocumentBleed false >> > ]>>
setdistillerparams> setpagedevice